Изобретение относится к биологии, медицине и биотехнологии и касается разработки способа оценки пролиферативного состояния лимфоидных клеток человека иммуноцитохимическим методом по характеру экспрессии С-концевого фрагмента ядрышкового белка В23/нуклеофозмина (В23).
Ядрышко является важнейшим структурным элементом клеток человека, принимающим участие в процессах пролиферации, клеточного старения и апоптоза. Поэтому идентификации белков, входящих в его состав, придается очень большое значение. Одним из подходов к изучению состояния ядрышка и его белковых компонентов является иммуноцитохимическое выявление входящих в его состав белков с помощью высокоспецифичных моноклональных антител (МКА) и антисывороток.
Белок В23 (м.м. 37-40 кДа) является одним из основных белков ядрышка с полифункциональной направленностью, включающей контроль биогенеза рибосом, трансформацию и пролиферацию клеток, стабильность генома и канцерогенез (Chan РК et al., J Biol Chem., 1986, 5; 261(4):1868-72). В то же время ранее нами было показано, белок В23 является более ранним маркером активации клеток к пролиферации, чем общепринятый маркер пролиферации Ki-67 (Булычева Т.И. и др. Цитология, 2000, №3, т.42 (10), с.944-954; Владимирова Н.М. и др., Биол. мембраны, 2004, 21, 94-101; Булычева и др., Terra Med, Лаб. диагностика, 2006, №2, стр.16-21; Булычева и др., Патент РФ на изобретение №2310201 от 10.11.2007). Поэтому определение экспрессии В23 имеет важное значение для ранней диагностики злокачественных гематологических и онкологических заболеваний, а также при мониторинге больных.
Известно что белок В23 рассматривается в последнее время многими исследователями в качестве раннего маркера рака (Ulanet DB et al, 2003,. PNAS, 100, 12361-12366; Lim M.Y. et al., 2006, Cancer Detect. Prev., 30, 481-490; Grisendy S etal, 2006, Nature, Rev. 6, 493-505; Falini В. et al., 2007, Haematologica, 92, 519-532;
Gmmmit CG et al., 2008, J. Biol. Chem., 283, 23326-23332). При этом обнаружено, что в опухолевых клетках его экспрессия возрастает более чем в 20 раз.
К настоящему времени установлено, что белок В23 имеет две изоформы, причем изоформа В23.1 локализована преимущественно в ядрышке, а изоформа В23.2 - преимущественно в нуклеоплазме. Обе изоформы функционируют как в мономерной (N-концевая последовательность), так и в олигомерной (С-концевая последовательность) форме (Сауткина Е.Н. и др., 2008, Приклад, биохим. и микробиол., т.44, №3, 287-295).
Все МКА к В23, имеющиеся к настоящему времени, как полученные нами впервые МКА ВСА-РС штамма ЗС9 (Малашенко О.С.и др. Патент РФ на изобретение №2145634/13 от 20.02.2000), так и коммерческие образцы моноклональных и поликлональных антител к В23, появившиеся после 2004 г. (Sigma, Sunta Cruz, США и др.), маркируют при цитохимических исследованиях только цельную молекулу В23 с молекулярной массой 37-40 кДа.
До недавнего времени специфических антител, распознающих отдельно С- и N- концевые фрагменты белка В23 в мире не существовало. В 2008 году в Институте биоорганической химии им. академиков М.М.Шемякина и Ю.А.Овчинникова РАН (ИБХ РАН) получены новые, не имеющие аналогов в мире, противопептидные антитела, селективно выявляющие в иммуноблоттинге мономерные и олигомерные формы белка В23 (Шалгунов B.C. и др. Биоорган. химия, 2009, т.35. №6, 799-807). При этом было выявлено, что антитела к N-концевому фрагменту полипептидной цепи (19-36) общему для всех форм белка В23 при иммунохимическом исследовании (в иммуноблоттинге) с лизатами перевиваемых опухолевых клеток HeLa и RAMOS взаимодействуют только с мономерами белка В23 с мол. массой - 38-40 кДа (Короев Д.О. и др., Заявка на патент РФ №2009112515 от 06.04.2009). В то же время антитела к С-концевому участку полипептидной цепи (283-294), характерному только для изоформы В23.1, выявили на иммуноблотах в лизатах вышеназванных клеток только олигомеры белка с мол. массой 210-230 кДа (Короев Д.О и др., Заявка на патент РФ №2009112519 от 06.04.2009).
Однако в иммуноблотинге можно определить лишь наличие и количество каждой из форм белка В23, выявляемых с помощью полученных антител, а установить локализацию, характер распределения и экспрессии каждого белка можно лишь при визуальной оценке реакции на цельных клетках под микроскопом. С этой целью нами были проведены предварительные иммуноцитохимические исследования полученных в ИБХ антител (любезно предоставленных нам к.х.н., с.н.с.ИБХ РАН Владимировой Н.М.) с лимфоцитами здоровых лиц и опухолевыми клетками RAMOS и HeLa для выявления локализации каждой из форм и характера их экспрессии в клетках в реакции непрямой иммунофлюоресценции.
Было обнаружено, что антитела к N-концевому фрагменту выявляют белок, локализованный в виде многочисленных мелких фокусов средней интенсивности в цитоплазме и нуклеоплазме свежевыделенных лимфоцитов и культуральных клеток, а к С-концевому фрагменту - в основном в ядрышке клетки в виде одного фокуса средней интенсивности в интактных лимфоцитах донора и многочисленных фокусов свечения в культивируемых опухолевых клетках (Шалгунов B.C. и др., Биоорган, химия, 2009, т.35, №6, С.799-807).
Учитывая полученные результаты необходимо было выяснить, какие изменения претерпевает каждая из выявляемых форм белка В23 в процессе пролиферации и какая из них наиболее точно отражает состояние пролиферативной активности клеток, играя доминирующую роль в отнесении цельного белка В23 к маркерам пролиферации.
Для этого был использован классический метод искусственной стимуляции интактных лимфоцитов периферической крови здоровых лиц к пролиферации с помощью фитогемагглютинина (ФГА), применяемый для изучения изменений в содержании и локализации основных белков ядра и ядрышка в процессе пролиферации (Булычева и др. Цитология, 2000, 3, 42 (10) 944-954, Малышева М.В. и др. «Иммунология», 2010, 1,13-17, Galand et al, Leukemia, 1995, 9(6), 1075-1084; Lopez et al., Cytometry, 1991, 12 (I): 42-49 и др.). Этот метод позволяет определить пролиферативное состояние клеток по включению в них 3Н тимидина. В результате четко установлено, что лимфоциты вступают в раннюю S-фазу клеточного цикла через 24 часа после их стимуляции ФГА, а максимум включения изотопа наблюдается через 48-72 ч. после их активации, при этом процент меченых (т.е. пролиферирующих) клеток возрастет в несколько раз по сравнению с нестимулированными (интактными) лимфоцитами (Артеменко Е.Г. автореф. канд. дис., М., 2004, Булычева и др. Цитология, 2000, 3, 42 (10) 944-954, Дергунова и др, Иммунология 2003, 24 №4. 205-208).
Проведенные ранее в этом методе исследования экспрессии цельного белка В23 в лимфоцитах человека позволили выявить, что в процессе нарастания пролиферативной активности клеток, обусловленной стимуляцией их ФГА, происходит как его количественное увеличение (в иммуноблотах), так и усиление экспрессии в ядрышках клеток (при иммуноцитохимических исследованиях). Это позволило отнести белок В23 к числу маркеров пролиферации (Малашенко О.С. и др., Патент РФ №2145634 от 20.02.2000, Артеменко Е.Г. автореф. канд. дис., М., 2004, Булычева и др. Цитология, 2000, 3, 42 (10) 944-954, Dergunova et al., Immunology letters, 2002, 83 pp.67-72).
Однако выяснить какая из форм белка В23 наиболее точно отражает состояние пролиферативной активности клетки и может являться истинным маркером пролиферации, стало возможным лишь при создании специфических антител к отдельным его фрагментам. Было необходимо оценить информативность изменения экспрессии отдельно С- и N-концевых фрагментов белка В23, отражающих его олигомерную и мономерную формы, при визуальной оценке клеток в реакции непрямой иммунофлюоресценции при стимуляции лимфоцитов к пролиферации с помощью ФГА.
В результате проведенных исследований обнаружено, что в процессе нарастания пролиферативной активности клеток основные изменения касаются белка, выявляемого антителами к С-концевому фрагменту белка В23, что позволяет считать его доминирующим, т.е. определяющим маркером пролиферации внутри цельного белка В23, чему и посвящена данная заявка.
В качестве аналогов предлагаемого способа определения пролиферативной активности лимфоидных клеток человека по экспрессии одного из фрагментов ядрышкового белка В23 можно рассматривать лишь выявление цельного белка В23 в клетках в процессе пролиферации в иммуноцитохимических и иммунохимических методах:
1. Выявление экспрессии белка В23 с помощью полученных нами ранее МКА ВСА-РС, секретируемых гибридомным клоном 3С9 (Малашенко О.С. и др., Патент РФ №2145634 от 20.02.2000) в процессе ФГА-стимуляции лимфоцитов человека в реакции непрямой иммунофлюоресценции (Артеменко Е.Г. автореф. канд. дисс., М., 2004, Булычева и др. Цитология, 2000, 3, 42 (10) 944-954, Дергунова и др, Иммунология 2003, 24 №4. 205-208). В этих работах четко показано, что белок В23 как структурный элемент ядрышка уже присутствует в интактных лимфоцитах до начала стимуляции их с ФГА. Однако после инкубации с ФГА экспрессия его постепенно нарастает, достигая максимума через 48-72 часа от начала стимуляции, т.е когда большинство лимфоидных клеток находится в S-периоде клеточного цикла, что подтверждено как параллельным исследованием в этих клетках ядерного антигена Кi-67, так и уровнем включения изотопа 3Н-тимидина (Булычева и др. Цитология, 2000, 3, 42 (10) 944-954, Дергунова и др, Иммунология 2003, 24 №4. 205-208., Dergunova et al., Immunology letters, 2002, 83 pp.67-72).
2. Увеличение содержания белка В23 в процессе усиления пролиферативной активности лимфоцитов под влиянием ФГА-стимуляции было подтверждено также в иммунохимическом методе по ширине белковой полосы, выявляемой на иммуноблотах (Булычева и др., Цитология, 2000, 3, 42 (10) 944-954, Дергунова и др., Иммунология 2003, 24 №4. 205-208, Dergunova et al., Immunology letters, 2002, 83 pp.67-72). При этом было обнаружено, что увеличение содержания белка В23 в лимфоидных клетках начинается уже через 16 часов от начала инкубации их с ФГА, т.е. еще в G1 периоде, в отличие от Ki-67, появляющегося только после 24 часов стимуляции, что соответствует S-периоду клеточного цикла.
3. Феномен повышения экспрессии белка В23 при усилении пролиферативной активности клеток, присущий большинству опухолевых клеток, был использован в клинической практике для выявления различий при диагностике лимфопролиферативных заболеваний. С помощью иммунопероксдидазной реакции, позволяющей избежать маркирования интактных лимфоцитов по белку В23, удалось выявить, что при заболеваниях с более злокачественным течением и наличием наибольшего количества пролиферирующих опухолевых клеток экспрессия белка В23 в клетках достигает максимальных значений (Артеменко Е.Г. Автореф. канд. дисс. М., 2004), что свидетельствует о диагностическом значении В23 в оценке пролиферативного состояния клеток.
4. Обнаружение в иммуноблотах более раннего количественного нарастания белка В23 при искусственной стимуляции лимфоцитов к пролиферации с помощью ФГА оказалось информативным для раннего выявления или прогнозирования наступающего прогрессирования опухолевого заболевания (рецидива). Выявленный при мониторинге больных феномен количественного нарастания белка В23 в лизатах лимфоидных клеток до начала экспрессии белка Ki-67, свидетельствующий о начале активации клеток к пролиферации, явился основой для разработки способа прогнозирования прогрессирования хронического лимфолейкоза по количеству белка В23 (Булычева Т.И. и др. «Terra Medica», Лабораторная диагностика, №2 (10) 2006, стр.16-21. Булычева Т.И. и др., Патент №2310201 от 10.11.2007).
Однако все перечисленные аналоги, доказывающие участие белка В23 в процессе пролиферации клеток, касаются лишь цельного белка В23 и не позволяют провести дифференцированное изучение его отдельных изоформ. Как уже указывалось, все существующие в мире до настоящего времени антитела к белку В23 способны выявлять лишь цельную молекулу белка В23 (с мол. массой 37-40 кДа), хотя некоторые из них и обозначаются как направленные к С- и N-фрагментам белка.
Выявить дифференцированное участие С- и N-концевых фрагментов белка В23 в процессе пролиферации стало возможным лишь с появлением антител к пептидным цепям белка В23, характерным для мономерной и олигомерной форм белка (Короев Д.О. и др., Заявка на патент РФ №2009112515 от 06.04.2009, Короев Д.О и др., Заявка на патент РФ №2009112519 от 06.04.2009).
Прототипом данного способа можно считать выявление мономерных и олигомерных форм белка В23 в культивируемых опухолевых клетках методом иммуноблоттинга с помощью полученных поликлональных антител к синтезированным пентодам, входящим в состав N- и С-концевых последовательностей белка В23 (Шалгунов B.C. и др. Биоорган, химия, 2009, т.35. №6, 799-807). В этом методе выявлены различия между мономерной и олигомерной формами белка В23 по их молекулярной массе (38-40 и 210-230 кДa, соответственно). Предварительное определение локализации каждой из форм белка в клетке, проведенное нами при иммуноцитохимическом исследовании с опухолевыми клетками RAMOS и HeLa, позволило выявить, что антитела к N-концевому фрагменту белка В23 окрашивают нуклеоплазму и цитоплазму, а к С-концевому фрагменту - ядрышки (Шалгунов B.C. и др. Биоорган, химия, 2009, т.35. №6, 799-807). Эти данные свидетельствовали о том, что полученные антитела способны визуализировать локализацию мономерной и олигомерной форм белка В23, что позволило проводить в дальнейшем дифференцированный анализ их участия в биологических процессах, в том числе в процессе пролиферации.
Полученные нами результаты иммуноцитохимических исследований динамики экспрессии отдельных форм белка В23 в лимфоцитах, искусственно стимулированных к пролиферации, позволили выявить наиболее динамичную форму этого белка, объективно отражающую пролиферативное состояние клеток. Эти данные и легли в основу предлагаемого нового способа оценки пролиферативного состояния лимфоидных клеток человека по экспрессии С-концевого фрагмента белка В23.
Способ осуществляли следующим образом.
I этап. Выделение лимфоцитов из периферической крови здоровых лиц
Лимфоциты выделяли из венозной крови 9 здоровых лиц (в 5 опытах) с помощью градиентного центрифугирования на фиколле (d=l,077 г/см3, Pharmacia, Швеция), дважды промывали средой RPMI 1640 и помещали в среду RPMI 1640, содержащую 10% пуловой сыворотки человека АВ (IY) группы, 0.01 М буфера HEPES, глутамин и гентамицин. В полученной взвеси подсчитывали общее количество клеток в камере Горяева и делили на две равные порции: 1-я порция исследовалась в качестве контроля (без добавления стимулятора пролиферации - ФГА), 2-я порция была опытной (с добавлением стимулятора пролиферации - ФГА). Вся работа проводилась в стерильных условиях.
II этап. Стимуляция лимфоцитов к пролиферации
Для стимуляции лимфоцитов к пролиферации в опытные порции добавляли 50 мкг/мл ФГА (Difco, США) и разливали для культивирования при 37°С в СO2 инкубаторе в 3 пластиковых матраса объемом 5.0 мл из расчета 1×10 лимфоцитов в 1 мл среды для исследования на разных сроках инкубации: через 24, 48 и 72 часа. В качестве контроля исследовали лимфоциты, которые культивировали в тех же условиях без ФГА, в том числе и выделенные до начала инкубации (в G0 периоде).
III этап. Иммуноцитохимическое исследование лимфоцитов в реакции непрямой иммунофлюоресценции
Отмытую от культуральной среды суспензию лимфоцитов каждого срока культивирования раскапывали на стекла с лунками, предварительно обработанными 0,1% поли-Е-лизином (Serva, Германия), инкубируя в течение 30 мин во влажной камере. За счет этого достигалось прочное прикрепление клеток к поверхности стекла. Для получения наилучшей визуализации нами были опробированы следующие методы фиксации:
- фиксация абсолютным ацетоном 10 мин при -20°С;
- фиксация абсолютным метанолом 10 мин при -20°С;
- фиксация 2%-ным параформальдегидом на PBS при комнатной температуре 20 мин, затем обработка 0,5%-ным раствором Тритона Х-100 на PBS 5 мин;
- обработка 0,5%-ным Тритоном Х-100 на PBS 5 мин, затем фиксация 2%-ным параформальдегидом на PBS 20 мин при комнатной температуре.
Наилучшей оказалась фиксация 2%-ным параформальдегидом на PBS при комнатной температуре в течение 20 мин, с последующей обработкой 0,5%-ным раствором Тритона Х-100 на PBS в течение 5 мин при комнатной температуре. После фиксации клетки отмывали в PBS 3 раза по 5 мин и инкубировали в течение 30 минут при комнатной температуре во влажной камере со следующими антителами:
- анти-3С9 к белку В23 (ГНЦ РАМН, Россия) - в разведении 1:50,
- анти-В23 (Sigma, США) - в разведении 1:1000,
- анти-К1-67 (Sigma, США) - в разведении 1:20,
- антитела к N-фрагменту В23 (ИБХ РАН, Россия) - в разведении 1:50,
- антитела к С-фрагменту В23 (ИБХ РАН, Россия) - в разведении 1:50,
МКА анти-В23 и анти-3С9 были взяты в качестве контроля - аналогов, выявляющих цельный белок В23, для сравнения их с его N- и С-концевыми фрагментами.
Антиген Ki-67 исследовали в качестве общепринятого маркера пролиферации, отсутствующего в интактных лимфоцитах, находящихся в фазе G0 клеточного цикла, но появляющегося с нарастающей экспрессией в процессе пролиферации, достигающей максимума в S-фазе клеточного цикла (Самойлова и др, Клин. лаб. диагн., 2003, 11, 35-39; Gerdes et al., Int. J. Cancer, 1983 15, 31(1):13-20; Endl et al., Exp.Cell Res. 2000; 257(2): 231-237; Leile R. J., Acta Histochemica, Supplementband, 1990, 39: 109-124; MacCallum et al., Exp.Cell Res. 1999; 252(1): 186-198; Hadar et al., Pathol. Oncol. Res., 2005; 11(1): 45-49).
После отмывания в трех сменах PBS по 10 мин клетки для люминисцентного окрашивания результатов реакции инкубировали с вторичными антителами: козьей сывороткой против иммуноглобулинов мыши, меченной ФИТЦ (Сорбент, Россия) в разведении 1:80 для МКА анти-3С9, анти-В23 и Ki-67, а для выявления N- и С-концевых фрагментов (кроличьих антисывороток) метили антителами к иммуноглобулинам кролика, конъюгированными с ФИТЦ («Медгамал», Россия) при комнатной температуре в течение 30 мин. После инкубации с вторичными антителами клетки отмывали в трех сменах PBS по 10 мин при тех же условиях. В качестве контроля для исключения прямого иммуномечения (окрашивания) клеток вторичными антителами вместо исследуемых первичных МКА использовали PBS. После отмывания клетки заключали в мовиол (Calbiochem, Швейцария).
Полученные препараты изучали с помощью люминесцентного микроскопа "Axiophot" (Carl Zeiss, Германия), используя объективы х100/1,3 и окуляры х10. Захват изображения производился с помощью цифровой камеры Delta Pix.
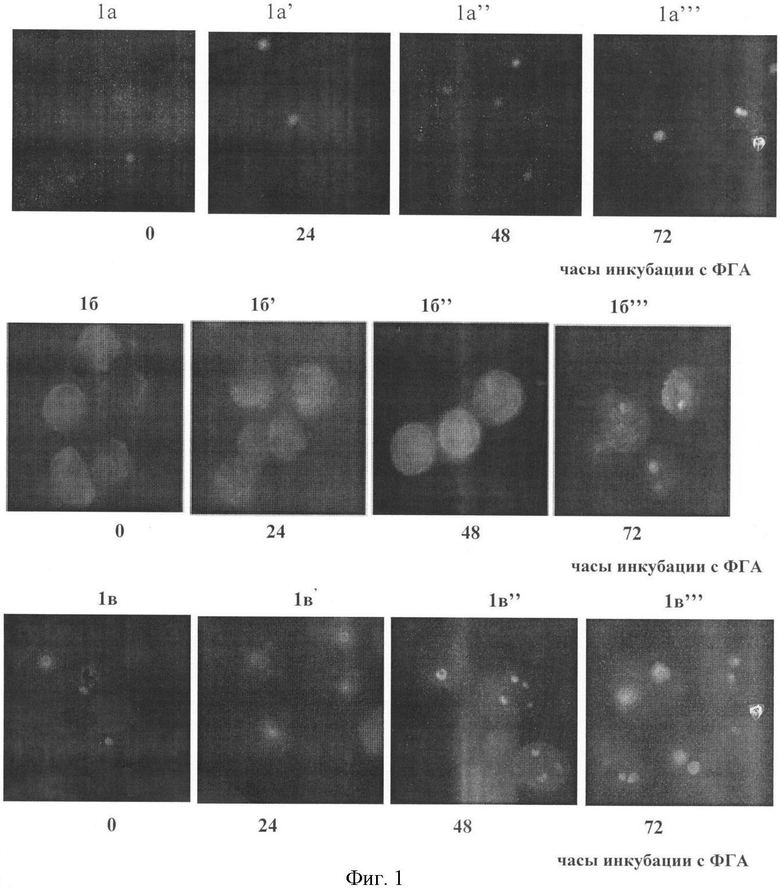
В результате проведенных исследований получены следующие данные, которые в сокращенном виде приведены в таблице и представлены на фото (фиг.1. «Локализация и характер экспрессии цельного белка В23 и его N- и С-концевых фрагментов в процессе ФГА-стимуляции лимфоцитов»).
До начала ФГА-стимуляции
- Ki-67 практически отсутствовал в интактных лимфоцитах, т.е. менее 1% клеток окрашивалось антителами к Ki-67.
- Характер окрашивания интактных лимфоцитов клеток МКА анти-3С9 и анти-В23 приблизительно совпадал: антитела окрашивали ядрышки, в среднем, 65% клеток (от 50 до 95%), при этом ядрышки имели незначительные размеры и не более одного в клетке (фиг.1, 1а).
- Антитела к N-концевому фрагменту белка В23 окрашивали диффузно цитоплазму и нуклеоплазму у подавляющего большинства (более 90%) клеток, не фокусируясь при этом в ядрышках клеток (фиг.1, 1б).
- Антитела к С-концевому фрагменту белка В23 в отличие от N-концевого фрагмента четко окрашивали ядрышки, в среднем, в 60% клеток, но не более одного в клетке, при этом слабое диффузное окрашивание наблюдалось и в цитоплазме (фиг.1, 1в).
Через 24 часов стимуляции ФГА
- Ki-67 выявлялся, в среднем, в ядрах 5% лимфоидных клеток.
- Локализация фокусов окрашивания клеток МКА анти-3С9 и анти-В23 совпадала с интактными лимфоцитами. Степень интенсивности окрашивания несколько увеличилась, количество окрашенных клеток увеличивалось не более чем на 5% (фиг.1, la').
- Антитела к N-концевому фрагменту окрашивали диффузно цитоплазму и нуклеоплазму в таком же количестве (более 90%) клеток, не фокусируясь при этом в ядрышках клеток, но по сравнению с интактными лимфоцитами интенсивность свечения в лимфоцитах сильно увеличилась (фиг.1, 1б').
- Антитела к С-концевому фрагменту окрашивали, в среднем, около 80% клеток, при этом в 15% клеток появлялось 2-3 ядрышка (фиг. 1, 1в'). Ядрышки (даже единичные) увеличивались в размерах. Интенсивность свечения нарастала по сравнению с интактными лимфоцитами, при этом слабое диффузное окрашивание наблюдалось и в цитоплазме.
Через 48 часов стимуляции ФГА
- Ki-67 выявлялся, в среднем, в ядрах 45% лимфоцитов.
- Локализация фокусов окрашивания клеток МКА анти-3С9 и анти-В23 совпадала с интактными лимфоцитами, но интенсивность свечения и количество окрашенных клеток увеличились: антитела окрашивали ядрышки, в среднем, 75% клеток (от 65 до 95%) клеток. Если в клетках, окрашенных антителами анти-3С9, ядрышки были не очень четкие и наблюдалось окрашивание нуклеоплазмы (фиг.1, 1а"), то при окрашивании клеток антителами анти-В23 ядрышки выявлялись очень четко, количество их увеличивалось по сравнению с интактными лимфоцитами.
- Антитела к N-концевому фрагменту окрашивали диффузно цитоплазму и нуклеоплазму в 95% клеток, не фокусируясь при этом в ядрышках клеток, но по сравнению с 24 часами инкубации интенсивность свечения в лимфоцитах значительно возросла (фиг. 1, 1б").
- Антитела к С-концевому фрагменту окрашивали, в среднем, около 80% клеток, из них в 65% клеток окрашивалось более одного (2-3) ядрышка, причем количество окрашенных ядрышек доходило до 4 (фиг.1, 1в"). Ядрышки увеличивались в размерах. Интенсивность свечения нарастала, по сравнению с лимфоцитами после 24 часов инкубации, при этом слабое диффузное окрашивание наблюдалось и в цитоплазме. Через 72 часа стимуляции ФГА
- Ki-67 выявлялся, в среднем, в ядрах 74% лимфоцитов.
- Локализация фокусов окрашивания клеток МКА анти-3С9 и анти-В23 не изменялась. Яркое и четкое свечение ядрышек наблюдалось, в среднем, в 85% клеток (фиг.1, la'"), причем интенсивность и четкость свечения и МКА анти-3С9 и анти-В23 была ниже, чем у С-концевого фрагмента. Подсвет цитоплазмы незначителен.
- N-концевой фрагмент - интенсивность свечения очень высокая, при этом в 7% клеток можно различить появившиеся ядрышки, хотя и не очень ярко окрашенные (фиг.1, 1б'"). Свечение большей частью локализовалось в нуклеоплазме, хотя в 35% клеток продолжала светить и цитоплазма. Общее количество окрашенных клеток составило 95%.
- С-концевой фрагмент - общее количество окрашенных клеток, в среднем, составило 90%. Лишь в 15% клеток светит 1 точечное ядрышко, в остальных клетках количество ядрышек варьирует от 2 до 5. По сравнению с 48 часами от начала ФГА-стимуляции четкость очертания ядрышек уменьшилась, а интенсивность экспрессии усилилась, при этом ядрышки сильно увеличены в размерах (фиг.1, 1в'").
Как уже указывалось, в контрольных пробах исследовали лимфоциты, находящиеся в тех же условиях и сроках культивирования, но без добавления ФГА. На протяжении всего срока культивирования локализация и распределение в клетках выявляемых антигенов практически не изменялось по сравнению с исходными показателями (то есть была сходной с фото на фиг.1(1а, 1б, 1в), но интенсивность свечения падала в связи с постепенной гибелью клеток.
Как видно из описания приведенных результатов, данные, полученные при исследовании контрольных антител, совпадали с общепризнанными данными по локализации и нарастанию экспрессии белков Ki-67 и В23 в процессе ФГА-стимуляции лимфоцитов. Это позволило достоверно оценить результаты реакции с антителами к N- и С-концевым фрагментам В23.
Полученные данные свидетельствуют о том, что антитела к N-концевому фрагменту В23 выявляют белок, локализующийся в цитоплазме и нуклеоплазме подавляющего большинства клеток вне зависимости от фазы пролиферативной активности клеток. Лишь на максимуме пролиферации (через 72 часа от начала стимуляции) в единичных клетках появляется ядрышковая локализация этого фрагмента. В то же время антитела к С-концевому фрагменту выявляют белок, находящийся в ядрышках интактных клеток в виде одного светящегося фокуса и изменяющийся в процессе нарастания их пролиферативной активности, коррелируя с данными контрольных сывороток (к К.1-67, анти-В23 и анти-3С9). Эти изменения выражались как в увеличении количества маркируемых ядрышек (с 1 до 4-5) в пролиферирующих лимфоцитах, так и в повышении уровня экспрессии (с + до +++) и размеров самих ядрышек по мере увеличения сроков инкубации с ФГА.
Таким образом, с помощью иммуноцитохимического метода исследования были выявлены не только различия в локализации и характере распределения концевых фрагментов белка В23, характерных для моно- и олигомерных форм, но и различное участие их в процессе пролиферации. Выявлено, что изменения касаются, в основном, С-концевых фрагментов белка В23 в ядрышковом аппарате клеток, т.е. его олигомерной формы. Это подтверждается увеличением количества маркированных анти-С-антителами ядрышек с одного (в состоянии G0 в интактных лимфоцитах) до 4-5 на пике пролиферации клеток (через 72 часа от начала ФГА-стимуляции), т.е. в S-периоде клеточного цикла, с увеличением их размеров и интенсивности экспрессии.
Полученные данные позволяют пересмотреть молекулярно-биологические свойства и роль цельной молекулы белка В23 в пролиферативном процессе, выявив его более индифферентную (мономерную) и активную (олигомерную) форму, принимающую непосредственное участие и претерпевающую четко видимые изменения в процессе пролиферации. В связи с этим можно сделать вывод о том, что характер экспрессии именно С-фрагмента белка В23 определяет пролиферативное состояние клеток, что позволяет назвать С-фрагмент белка В23 - маркером пролиферации.
Пример 1
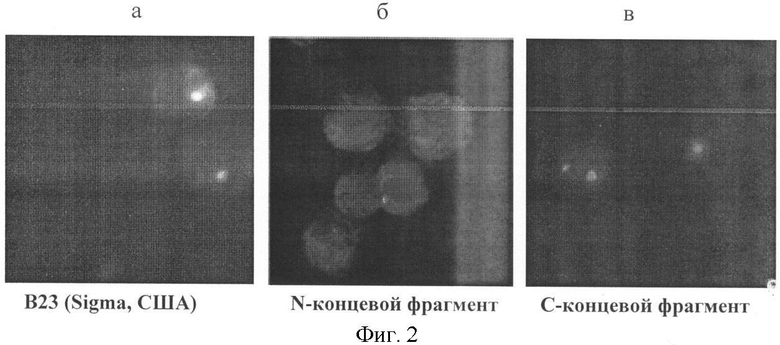
В лабораторию на исследование доставлена периферическая кровь больного Т., 65 лет, с диагнозом: хронический лимфолейкоз, плановое обследование (срок заболевания 6 лет). При исследовании пролиферативной активности лимфоцитов в реакции непрямой иммунофлуоресценции с МКА Ki-67 выявлено 6% антиген-позитивных клеток (в норме 0-1%). На фиг. 2 показана локализация цельного белка В23 и его N- и С-концевых фрагментов в клетках этого больного. При исследовании его лимфоцитов с МКА анти-В23 - в 80% клеток выявлен 1 фокус свечения в ядрышке (фиг.2, 2а), антитела к N-концевым фрагментам в 90% клеток диффузно окрашивали цитоплазму и нуклеоплазму (фиг.2, 2б), антитела к С-концевым фрагментам в 85% клеток окрашивали 1 ядрышко, а в 6% клеток - 2 ядрышка (фиг.2, 2в). Заподозрено начало прогрессирования заболевания. Рекомендовано повторное исследование через 3 мес для мониторинга опухолевого процесса и возможной коррекционной терапии.
Пример 2
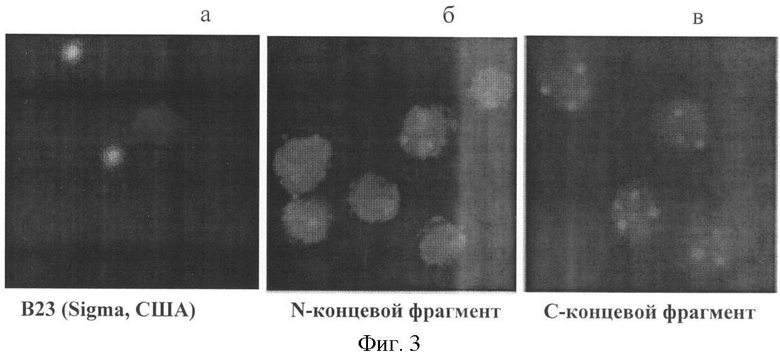
В лабораторию на исследование доставлена периферическая кровь больного Б., 83 лет, для подтверждения диагноза: В-крупноклеточная лимфосаркома высокой степени злокачественности. При исследовании клеток выявлено 65% Ki-67-позитивных клеток, с МКА анти-В23 - в 75% клеток светились ядрышки (фиг.3, 3а). При исследовании клеток с антителами к N-концевым фрагментам на фоне яркого диффузного окрашивания цитоплазмы и нуклеоплазмы, наблюдаемом в 90% клеток, в 5% из них были различимы окрашиваемые антителами 2-3 ядрышка, что подтверждало крайне высокую степень пролиферативной активности (фиг.3, 3б). Антитела к С-концевым фрагментам окрашивали 3-4 ядрышка в 90% клеток, причем ядрышки были увеличены в размерах, что также свидетельствовало о максимальной пролиферативной активности клеток (фиг.3, 3в). Больному срочно была назначена противоопухолевая терапия. Локализация и характер экспрессии цельной молекулы белка В23 и его N- и С-концевых фрагментов в клетках больного Б. представлены на фиг.3.
Приведенные примеры демонстрируют информативность визуализации результатов иммуноцитохимического исследования клеток больных с антителами к N- и С-концевым фрагментам белка В23 и возможность их использования в клинико-лабораторной практике для более точной оценки пролиферативного состояния клеток. Это имеет важное значение для ранней диагностики и прогнозирования злокачественных гематологических и опухолевых заболеваний. Учитывая, что оригинальные антитела к N- и С-фрагментам В23 получены отечественными авторами (в ИБХ РАН), способ иммуноцитохимической оценки пролиферативного состояния клеток по характеру экспрессии С-концевого фрагмента белка В23 в клетках может быть в ближайшее время рекомендован к внедрению в широкую медицинскую практику.
Литература
1. Артеменко Е.Г. «Иммуноцитохимический анализ ядерных антигенов лимфоцитов человека, выявляемых с помощью моноклональных антител», автореферат диссертации на соискание ученой степени кандидата биологический наук, Москва, 2004.
2. Булычева Т.И., Дейнеко Н.Л., Самойлова Р.С., Артеменко Е.Г., Зацепина О.В. "Способ прогнозирования прогрессирования хронического лимфолейкоза." Патент РФ на изобретение №2310201 от 10.11.2007.
3. Булычева Т.И., Артеменко Е.Г., Дергунова Н.Н., Дудник О.А., Шпакова А.П., Малашенко О.С., Зацепина О.В. "Анализ пролиферативной активности клеток с помощью новых моноклональных антител к ядрышковому белку В23/нуклеофозмину». Цитология, 2000, №3, т.42 (10), 944-954.
4. Булычева Т.И., Дейнеко Н.Л., Артеменко Е.Г., Самойлова Р.С., Зацепина О.В. «Диагностическое значение ядрышкового белка В23-нуклеофозмина при хронических лимфопролиферативных заболеваниях», «Terra Medica», Лабораторная диагностика,2006, №2 (10), 16-21.
5. Владимирова Н.М., Сауткина Е.Н., Табданов Е.Б., «Идентификация и характеристика мономерных и олигомерных изоформ ядрыкового белка В23/нуклеофозмина в клетках Hela», M., Биологические мембраны, 2004, №21,94-101.
6. Дергунова Н.Н. Шпакова А.П. Гемждан Э.Г. Булычева Т.И. Дейнеко Н.Л. Митогенное и токсическое действие фитогемагглютинина на лимфоциты периферической крови в культуре. Иммунология, 2003, том 24, №4. С.205-208.
7. Короев Д.О., Шалгунов B.C., Владимирова Н.М., Лобанова Н.В., Волкова Т.Д., Вольпина О.М. Пептид, стимулирующий образование специфических антител против мономерной формы нуклеофозмина в опухолевых клетках. Заявка на патент РФ на изобретение №2009112515 от 06.04.2009.
8. Короев Д.О., Шалгунов B.C., Владимирова Н.М., Лобанова Н.В., Волкова Т.Д., Вольпина О.М. Пептид, стимулирующий образование специфических антител против олигомерной формы нуклеофозмина в опухолевых клетках. Заявка на патент РФ на изобретение №2009112519 от 06.04.2009.
9. Малашенко О.С., Булычева Т.И., Дудник О.А. «Штамм культивируемых клеток мышиной гибридомы, используемый для получения моноклональных антител к ядрышковому антигену, ассоциированному с клеточной пролиферацией». Патент РФ №2145634 от 20.02.2000.
10. Малышева М.В., Дейнеко Н.Л., Булычева Т.И., Зацепина О.В. и др. «Сравнительный анализ экспрессии ключевых белков ядрышка в лимфоцитах периферической крови здоровых доноров, активированных к пролиферации in vitro», Иммунология (2010), №1, Т.31, с.13-17.
11. Сауткина Е.Н., Потапенко Н.А., Булычева Т.И., Владимирова Н.М., «Выделение белка В23/нуклеофозмина из ядер клеток HeLa», Прикладная биохимия и микробиология, 2008, т.44, №3, 287-295.
12. Самойлова Р.С., Булычева Т.И. Иммунофенотипирование в диагностике хронических лимфоидных заболеваний. Клиническая лабораторная диагностика, 2003, 11, 35-39.
13. Шалгунов B.C., Лобанова Н.В., Булычева Т.И., Дейнеко Н.Л., Волкова Т.Д., Филатова М.П., Камынина А.В., Ким Я.С., Владимирова Н.М., Короев Д.О., Ахидова Е.В., Вольпина О.М. «Антитела к синтетическим фрагментам нуклеофозмина для специфического выявления его мономерных и олигомерных форм». Биоорган, химия, 2009, т.35, №6, С.799-807.
14. Chan PK, Aldrich M, Cook RG, Busch H. Ammo acid sequence of protein B23 phosphorylation site. J Biol Chem. 1986 Feb 5;261(4):1868-72.
15. Dergunova N.N., Bulycheva T.I., Artemenko E.G., Shpakova A.P., Pegova A.N., Gemijan E.G., Dudnik О.А., Zatsepina O.V., Malashenko O.S.. A major nucleolar protein B23 as a marker of proliferation activity of human peripheral lymphocytes. Immunology Letters, 2002, 83, 67-72.
16. Endl E., Gerdes J. The Ki-67 protein: fascinating forms and an unknown function. //Exp.Cell Res. 2000; 257(2): 231-237.
17. Falini В, Nicoletti I, Bolli N, Martelli MP, Liso A, Gorello P, Mandelli F, Mecucci C, and Martelli MF «Translocations and mutations involving the nucleophosmin (NPM1) gene in lymphomas and leukemias», Haematologica, 2007, 92, 519-532.
18. Galand P, Del Bino G, Morret M, Capel P, Degraef C, Fokan D, Feremans W. PCNA immunopositivity index as a substitute to ЗН-thymidine pulse-labeling index (TLI) in methanol-fixed human lymphocytes. Leukemia. 1995 Jun;9(6): 1075-84.
19. Gerdes J, Schwab U, Lemke H, Stein H. Production of a mouse monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. IntJCancer., 1983 Jan, 15;31(1):13-20.
20. Grisendy S., Meccuci C., Falini B. et al. «Nucleophosmin and cancer», 2006, Nature, Rev., 6493-505.
21. Grummit CG, Townsley FM, Johnson CM, Warren AJ, and Bycroft M., «Sructural Consequences of Nucleophosmin Mutations in Acute Myeloid Leukemia», J. Biol. Chem, 2008, 283, 23326-23332.
22. Hadar Т, Shvero J, Yaniv E, Ram E, Shvili I, Koren R. Expression ofp53, Ki-67 and Bcl-2 in parathyroid adenoma and residual normal tissue. Pathol Oncol Res., 2005:11(1):45-49.
23. Lelle R. J. In situ determination of the Ki -67 growth fraction (Ki -67 GF) in human tumors (studies in breast cancer). Acta Histochemica/ Supplementband, 199Q, 39:109-124.
24. Lopez F, Belloc F, Lacombe F, Dumain P, Reiffers J, Bernard P, Boisseau MR. Modalities of synthesis of Ki67 antigen during the stimulation of lymphocytes. Cytometry, 1991;12(1):42-9.
25. MacCallum D.E., Hall P.A. Biochemical characterization of pKi67 with the identification of a mitotic-specific form associated with hyperphosphorylation and altered DNA binding. // Exp.Cell Res., 1999; 252(1): 186-198.
26. Lim M.J., X. Wang, Nucleophosmin and human cancer, Cancer Detect. Prev., 30, 481-490.
27. Ulanet DB, Torbenson M, Dang CV, Carciola-Rosen C, and Rosen A. «Unique conformation of cancer autoantigen B23 in hepatoma: A mechanism for specificity in the autoimmune response», PNAS, 2003, 100, 12361-12366.

Изобретение относится к области биотехнологии, конкретно к способам оценки пролиферативного состояния лимфоцитов, и может быть использовано в диагностике. Способ иммуноцитохимической оценки пролиферативного состояния лимфоцитов по характеру экспрессии С-концевого фрагмента белка В23/нуклеофозмина в реакции непрямой иммунофлюоресценции предусматривает визуальную оценку в клетках количества и размеров светящихся ядрышек, маркируемых специфическим противопептидным антителом к С-концевому фрагменту белка В23/нуклеофозмина, выявляющим только олигомерную форму белка В23.1 с молекулярной массой 210-230 кДа. При этом показателем интактных лимфоцитов, находящихся в фазе G0 клеточного цикла, является наличие в ядре клетки не более одного светящегося ядрышка, выявляемого указанным антителом к С-фрагменту В23, а в процессе пролиферативной активности лимфоцитов, вызванной стимуляцией их ФГА, через 24 часа от начала стимуляции в лимфоцитах выявляется 2-3 светящихся ядрышка, а через 48-72 часа от начала стимуляции, когда большинство лимфоцитов находится на пике пролиферативной активности в S-периоде клеточного цикла, в клетках выявляется до 4-5 светящихся ядрышек, более крупного размера и большей интенсивности свечения от + до +++, по сравнению с исходными показателями. Изобретение позволяет провести раннюю диагностику злокачественных гематологических и онкологических заболеваний. 3 ил., 1 табл.
Способ иммуноцитохимической оценки пролиферативного состояния лимфоцитов по характеру экспрессии С-концевого фрагмента белка В23/нуклеофозмина в реакции непрямой иммунофлюоресценции, предусматривающий визуальную оценку в клетках количества и размеров светящихся ядрышек, маркируемых специфическим противопептидным антителом к С-концевому фрагменту белка В23/нуклеофозмина, выявляющим только олигомерную форму белка В23.1 с молекулярной массой 210-230 кDа, где показателем интактных лимфоцитов, находящихся в фазе G0 клеточного цикла, является наличие в ядре клетки не более одного светящегося ядрышка, выявляемого указанным антителом к С-фрагменту В23, а в процессе пролиферативной активности лимфоцитов, вызванной стимуляцией их фитогемагглютинином (ФГА), через 24 ч от начала стимуляции в лимфоцитах выявляется 2-3 светящихся ядрышка, а через 48-72 ч от начала стимуляции, когда большинство лимфоцитов находится на пике пролиферативной активности в S-периоде клеточного цикла, в клетках выявляется до 4-5 светящихся ядрышек более крупного размера и большей интенсивности свечения от + до +++ по сравнению с исходными показателями.
| ШАЛГУНОВ B.C | |||
| и др | |||
| Антитела к синтетическим фрагментам нуклеофозмина для специфического выявления его мономерных и олигомерных форм, Биоорганическая химия, 2009, т.35, №6, с.799-807 | |||
| БУЛЫЧЕВА Т.И | |||
| и др | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |

