РОДСТВЕННАЯ ЗАЯВКА
Данная заявка испрашивает приоритет и преимущество предварительной заявки США серийного № 60/830831, поданной 14 июля 2006 года, описание которой приведено здесь в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способам получения гетерологичных рекомбинантных белков, производимых в клеточной культуре. Изобретение включает способы выделения и очистки повторно свернутых рекомбинантных белков, которые были произведены в прокариотических клетках-хозяевах и присутствуют в этих клетках, как правило, в периплазматическом или внутриклеточном пространстве. Рекомбинантные белки, произведенные в прокариотических клетках-хозяевах, также можно обнаружить в виде растворимых белков или в виде смеси растворимых и нерастворимых белков.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Много относящихся к терапии рекомбинантных белков производится множеством организмов-хозяев. Большинство белков можно экспрессировать в их нативной форме в эукариотических клетках, таких как клетки CHO. Культура клеток животных, как правило, требует продолжительных времен роста для достижения максимальной плотности клеток и, в конечном счете, достигает меньшей плотности, чем культуры прокариотических клеток (Cleland, J. (1993) ACS Symposium Series 526, Protein Folding: In Vivo and In Vitro, American Chemical Society). Кроме того, культуры клеток животных зачастую требуют дорогостоящих сред, содержащих компоненты роста, которые могут препятствовать очистке целевого белка. Системы экспрессии на основе бактерий-хозяев предоставляют рентабельную альтернативу получения в промышленном масштабе рекомбинантных белков. Существуют многочисленные патенты США по общей бактериальной экспрессии рекомбинантных белков, включая патенты США № 4565785, 4673641, 4795706 и 4710473. Основным преимуществом способа получения является возможность легко отделять продукт от клеточных компонентов путем центрифугирования или микрофильтрации. Смотри, например, Kipriyanov and Little, (1999) Molecular Biotechnology, 12:173-201; и Skerra and Pluckthun, (1988) Science, 240:1038-1040.
Однако бактериальные системы экспрессии, такие как E. coli, не имеют клеточных механизмов, способствующих правильному рефолдингу белков, и, как правило, не приводят к секреции крупных белков в культуральную среду. Рекомбинантные белки, экспрессированные в бактериальных клетках-хозяевах, зачастую обнаруживаются в виде телец включения, состоящих из плотных масс частично свернутого и неправильно свернутого восстановленного белка. Смотри, например, Baneyx, (1999) Current Opin. Biotechnology 10:411-421; и Villaverde and Carrio, (2003) Biotech. Letts. 25:1385-1395. Белки также можно экспрессировать без образования телец включения. Смотри, например, Id. Как правило, рекомбинантный белок в тельцах включения в основном неактивен.
Кроме того, рефолдинг часто приводит к образованию неправильно свернутых и соединенных дисульфидными связями димеров, тримеров и мультимеров (Morris et al., (1990) Biochem. J., 268:803-806; Toren et al., (1988) Anal. Biochem., 169:287-299). Этот феномен ассоциации является довольно обычным при рефолдинге белков, особенно при более высоких концентрациях белка, и, как представляется, часто включает ассоциацию посредством гидрофобного взаимодействия частично свернутых интермедиатов (Cleland and Wang, (1990) Biochemistry. 29:11072-11078).
Неправильное сворачивание происходит либо в клетке при ферментации, либо в ходе процедуры выделения. Белки, выделенные из периплазматического или внутриклеточного пространства, необходимо растворить и растворимый белок повторно свернуть в нативное состояние. Смотри, например, Rudolph, Renaturation of Recombinant, Disulfide-Bonded Proteins From «Inclusion Bodies» в Modern Methods in Protein - and Nucleic Acid Research (Walter de Gruyter New York, 1990), p. 149-172. Способы рефолдинга белков in vitro в правильную, биологически активную конформацию являются существенными для получения функциональных белков. Обычная последующая обработка белков, выделяемых из телец включения, включает растворение телец включения при высокой концентрации денатурирующего средства, такого как мочевина, с последующим разведением денатурирующего средства, чтобы дать возможность произойти рефолдингу (смотри патенты США № 4512922, 4511502 и 4511503). Смотри также, например, Rudolph and Lilie, (1996) FASEB J. 10:49-56; Fischer et al., (1993), Biotechnology and Bioengineering, 41:3-13; Misawa & Kumagai, (1999) Biopolymers 51:297-307; и Clark, (1998) Current Opinion in Biotechnology, 9:157-163; и Tsumoto et al., (2003) Protein Expression and Purification 28:1-8. Такие способы выделения рассматриваются как повсеместно применимые, с незначительными модификациями, для выделения биологически активных рекомбинантных белков из телец включения. Эти способы были применены для гепаринсвязывающего белка (HBP), такого как VEGF (Siemeister et al. (1996), выше). Этими способами добиваются элиминации случайных дисульфидных связей перед переводом рекомбинантного белка в его биологически активную конформацию посредством других его стабилизирующих сил, и можно не удалять неправильно свернутые интермедиаты, получать гомогенные популяции правильно свернутого продукта или получать достаточные количества правильно свернутого продукта.
Обращенные мицеллы или ионообменную хроматографию использовали для облегчения рефолдинга денатурированных белков путем заключения отдельного белка внутри мицелл или выделения их на смоле, и затем удаляя денатурирующее средство (Hagen et al., (1990) Biotechnol. Bioeng. 35:966-975; Creighton (1985) в Protein Structure Folding and Design (Oxender, DX. Ed.), p.249-251, New York: Alan R. Liss, Inc.). Эти способы были пригодны для предотвращения агрегации белка и облегчения правильного рефолдинга. Для изменения скорости или степени рефолдинга осуществляли специфичный по отношению к конформации рефолдинг с лигандами и антителами к нативной структуре белка (Cleland and Wang, (1993), в Biotechnology, (Rehm H. J., and Reed G. Eds.), p.528-555, New York, VCH). Например, креатинкиназу повторно сворачивали в присутствии антител к нативной структуре (Morris et al., (1987) Biochem. J. 248:53-57). В дополнение к антителам для усиления рефолдинга были использованы лиганды и кофакторы. Эти молекулы могут преимущественно взаимодействовать со сворачивающимся белком после образования нативного белка. Таким образом, равновесие фолдинга можно «сдвинуть» к нативному состоянию. Например, скорость рефолдинга феррицитохрома была увеличена при помощи внешнего лиганда к осевому положению железа гема (Brems and Stellwagon, (1983) J. Biol. Chem. 258:3655-3661). Для облегчения белкового фолдинга также были использованы белки-шапероны и катализаторы фолдинга. Смотри, например, Baneyx, (1999) Current Opinion in Biotechnology, 10:411-421; & Carrio & Villaverde, (2003) FEBS Letters 537:215-221. Впрочем, эти способы не всегда эффективны или достаточны для количественного выхода белкового продукта.
Существует необходимость в новых и более эффективных способах фолдинга и/или выделения рекомбинантных белков из культуры клеток-хозяев, например, для эффективного и экономичного получения рекомбинантных белков в культуре бактериальных клеток. Эти новые и более эффективные способы обеспечивают улучшенное выделение высокоочищенного, биологически активного, правильно повторно свернутого белка и, в общем, применимы для получения белков в промышленном масштабе. Изобретение относится к этим и другим потребностям, как будет ясно при ознакомлении со следующим описанием.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к способу выделения и очистки повторно свернутых рекомбинантных белков из культуры клеток. В частности, изобретение относится к способу выделения рекомбинантного белка из прокариотических клеток-хозяев, например бактериальных клеток. Способы по изобретению широко применимы для рекомбинантных белков. В некоторых вариантах осуществления рекомбинантный белок представляет собой фактор роста, например фактор роста эндотелия сосудов (VEGF). В одном из вариантов осуществления фактор роста представляет собой VEGF165.
В одном из вариантов осуществления способ включает: (a) выделение указанного рекомбинантного белка из культуры прокариотических клеток; (b) растворение указанного белка в первом буферном растворе, pH более 9, содержащем первое хаотропное средство; (c) рефолдинг указанного растворенного белка во втором буферном растворе, pH>9, но ≤11, содержащем второе хаотропное средство, два или более восстанавливающих средств, и добавление воздуха или кислорода в течение такого времени и при таких условиях, при которых происходит рефолдинг рекомбинантного белка; и (d) выделение указанного повторно свернутого рекомбинантного белка. В одном из вариантов осуществления первый буферный раствор и/или второй буферный раствор дополнительно содержит аргинин. В одном из вариантов осуществления первый буферный раствор в конечной концентрации содержит 1 M мочевину, 300 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, pH 11. В другом варианте осуществления первый буферный раствор содержит в конечной концентрации 1 M мочевину, 300 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, pH 11. В одном из вариантов осуществления второй буферный раствор содержит два или более восстанавливающих средств, например DTT и цистеин. В одном из вариантов осуществления второй буферный раствор в конечной концентрации содержит 1 M мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, pH 10. В другом варианте осуществления второй буферный раствор содержит в конечной концентрации 1 M мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, pH 10.
В одном из вариантов осуществления способ включает: (a) выделение указанного рекомбинантного белка из культуры прокариотических клеток; (b) растворение и рефолдинг указанного белка в комбинированном буферном растворе, pH>9, но ≤11, с добавлением воздуха или кислорода; и (c) выделение указанного рекомбинантного белка. В одном из вариантов осуществления комбинированный буферный раствор в конечной концентрации содержит 1 M мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, pH 10. В другом варианте осуществления комбинированный буферный раствор в конечной концентрации содержит 1 M мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, pH 10.
Кислород или воздух для реакции рефолдинга можно предоставить посредством источника воздуха или источника обогащенного кислородом сжатого газа. В одном из вариантов осуществления применяют kLa 0,004 мин-1, например ту, которая соответствует скорости перемешивания 200-400 об/мин и скорости барботирования 0,3 куб. см/мин/л во флаконе на 2,5 л, содержащем импеллер морского типа. В других вариантах осуществления для получения повторно свернутого белка применяют kLa = 0,01 мин-1 или 0,1 мин-1.
Растворение и/или рефолдинг можно проводить при различных температурах. В одном из вариантов осуществления температура инкубации для растворения и/или рефолдинга представляет собой комнатную температуру. Время инкубации может варьировать согласно тому, как происходит восстановление и рефолдинг рекомбинантного белка. В одном из вариантов осуществления рекомбинантный белок инкубируют в первом буферном растворе в течение, по меньшей мере, 1 часа или от 1 до 2 часов. В одном из вариантов осуществления растворенный белок инкубируют во втором буферном растворе в течение от приблизительно 3 до 24 часов. В одном из вариантов осуществления выделенный рекомбинантный белок инкубируют в комбинированном буферном растворе в течение от 3 до 24 часов.
Кроме того, изобретение относится к процессам и способам рефолдинга рекомбинантных белков либо самим по себе, либо в связи с выделением рекомбинантного белка, как описано здесь. В конкретном варианте осуществления способы очистки включают осветление раствора, содержащего рекомбинантный белок, и приведение указанного повторно свернутого рекомбинантного белка в осветленном растворе в контакт с носителем комбинированного типа, катионным хроматографическим носителем, первым хроматографическим носителем для гидрофобного взаимодействия и, необязательно, вторым гидрофобным хроматографическим носителем или ионообменным носителем; и селективное выделение или элюирование повторно свернутого рекомбинантного белка с каждого носителя. В одном из вариантов осуществления осветление раствора включает добавление детергента до конечной концентрации 1%, доведение pH до приблизительно 8,5-9,5, инкубирование раствора в течение от 1 до 10 часов при 25-30°C, центрифугирование раствора; и фильтрование жидкости, полученной на стадии центрифугирования. В одном из вариантов осуществления pH составляет приблизительно 8,7. В другом варианте осуществления pH составляет приблизительно 9,0. Предполагается, что стадии для стадий выделения можно проводить в любом порядке, например последовательно или изменяя порядок хроматографических носителей. В некоторых вариантах осуществления изобретения предоставлены способы выделения и очистки повторно свернутых рекомбинантных белков из клеточной культуры производственного или промышленного масштаба.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 иллюстрирует пример исследования с течением времени процесса рефолдинга, как описано здесь, который оценивали путем хроматографии rpHPLC.
Фиг. 2 иллюстрирует хроматограмму для VEGF, полученную при нанесении на колонку CaptoMMCTM бактериального штамма W3110. Колонку уравновешивают 25 мМ HEPES, pH 9,0. VEGF элюируют с колонки MMC изократически в 1 M аргинине/25 мМ HEPES, pH 6-9.
Фиг. 3 иллюстрирует хроматограмму для VEGF, полученную при нанесении на колонку SPHP бактериального штамма W3110. Например, колонку SPHP уравновешивают 50 мМ HEPES, pH 7,5. Колонку элюируют с использованием линейного градиента с 0,0-1,2 M ацетата натрия в 50 мМ HEPES, pH 7,5, в 1 объеме колонки. Элюант контролируют при 280 нм. Белок выделяют из фракций с наибольшим поглощением при 280 нм (макс OD при ~42 мСм/см), которые обычно содержат >90% VEGF, их собирают для дальнейшей обработки.
Фиг. 4 иллюстрирует хроматограмму для VEGF, полученную при нанесении на колонку HiPropyl бактериального штамма W3110.
Фиг. 5 иллюстрирует хроматограмму для VEGF, полученную при нанесении на колонку с фенилсефарозой бактериального штамма W3110.
Фиг. 6 иллюстрирует эффект мочевины и аргинина на условия рефолдинга.
Фиг. 7 иллюстрирует эффект N2 на стабилизацию повторно свернутого пула вплоть до 48 часов в виде оценки с помощью rpHPLC с течением времени, когда N2 добавляли через 6 часов после начала рефолдинга.
Фиг. 8 иллюстрирует эффект варьирования скорости воздушного барботирования на условия рефолдинга.
Фиг. 9 иллюстрирует анализ на колонке, где для анализа повторно свернутого VEGF применяют катионообменную HPLC.
Фиг. 10 иллюстрирует аминокислотную последовательность VEGF165 с отмеченными дисульфидными связями.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Как применяют здесь, «полипептид» относится, как правило, к пептидам и белкам из любого источника клеток, имеющим более чем приблизительно десять аминокислот. «Гетерологичные» полипептиды представляют собой полипептиды, чужеродные используемой клетке-хозяину, такие как белок человека, вырабатываемый в E. coli. Несмотря на то что гетерологичный полипептид может быть прокариотическим или эукариотическим, он является предпочтительно эукариотическим, более предпочтительно полипептидом млекопитающего и более предпочтительно человека. В некоторых вариантах осуществления изобретения его получают рекомбинантно (например, рекомбинантный полипептид или рекомбинантный белок).
Примеры полипептидов млекопитающего включают молекулы, такие как, например, фактор роста; гепаринсвязывающий фактор роста; фактор роста эндотелия сосудов (VEGF), например VEGF-A (изоформы), VEGF-B, VEGF-C и VEGF-D; рецептор и антитело к VEGF, такие как rhuFab V2 и бевацизумаб, ранибизумаб; антитело к рецепторам VEGF; реннин; гормон роста, включая гормон роста человека (hGH); бычий гормон роста; рилизинг-фактор гормона роста; гормон паращитовидной железы; тиреотропный гормон; липопротеины; 1-антитрипсин; А-цепь инсулина; B-цепь инсулина; проинсулин; тромбопоэтин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; рецепторы гормона роста; высвобождающий белок для гормона роста (GHRP); LIV-1 (EP 1263780); TRAIL; факторы свертывания крови, такие как фактор VIIIC, фактор IX, тканевой фактор и фактор фон Виллебранда; фактор VIII; B домен фактора VIII; противотканевой фактор; факторы, препятствующие свертыванию крови, такие как белок C; предсердный натрийуретический фактор; сурфактант легких; активатор плазминогена, такой как урокиназа или мочевой или тканевой активатор плазминогена человека (t-PA) и их варианты; бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухоли альфа и бета; антитела к домену(ам) ErbB2, энкефалиназа; сывороточный альбумин, такой как сывороточный альбумин человека; ингибирующее вещество Мюллера; A-цепь релаксина; B-цепь релаксина; прорелаксин; гонадотропин-ассоциированный пептид мыши; микробный белок, такой как бета-лактамаза; ДНКаза; ингибин; активин; рецепторы для гормонов или факторов роста; интегрин; белок A или D; ревматоидные факторы; нейротрофический фактор, такой как нейротрофический фактор мозгового происхождения (BDNF), нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6), или фактор роста нервов, такой как NGF; кардиотрофины (фактор кардиальной гипертрофии), такие как кардиотрофин-1 (CT-1); фактор роста тромбоцитов (PDGF) (A, B, C или D); фактор роста фибробластов, такой как aFGF и bFGF; фактор роста эпидермиса (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулиноподобный фактор роста I и II (IGF-I и IGF-II) и их рецепторы, такие как IGFBP-1-IGFBP-6; des(1-3)-IGF-I (IGF-I мозга), белки, связывающие инсулиноподобный фактор роста; белки CD, такие как CD-3, CD-4, CD-8 и CD-19; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; белок морфогенеза костей (BMP); интерферон, такой как интерферон-альфа, -бета и -гамма; сывороточный альбумин, такой как сывороточный альбумин человека (HSA) или бычий сывороточный альбумин (BSA); колониестимулирующие факторы (CSF), например M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например IL-1-IL-10; антитело анти-HER-2; лиганд Apo2; супероксиддисмутаза; анти-CD20; херегулин, анти-IgE, анти-CD11a, анти-CD18; фактор некроза опухолей (TNF) и антитела к нему, рецептор TNF и соответствующие антитела, IgG к TNF-рецептору, факторы, связанные с рецептором TNF (TRAF), и их ингибиторы, T-клеточные рецепторы; поверхностные мембранные белки; стимулятор гемолиза; анти-TGF, такие как анти-TGF-бета; анти-активин; анти-ингибин; антитела анти-Fas; ингибитор лиганда Apo-2; рецептор Apo-2; Apo-3; апоптические факторы; Ced-4; DcR3; рецептор смерти и агонистические антитела (DR4, DR5); лимфотоксин (LT); пролактин; рецептор пролактина; белки SOB; WISP (wnt-индуцированные секретированные белки); анти-NGF; ДНКаза; антиген гепатита; антиген простого герпеса; лептин; белок Toll, лиганды TIE, CD40 и анти-CD40, иммуноадгезины, субтилизин, фактор роста гепатоцитов (HGF), тромбопоэтин (TPO); простатоспецифичный раковый антиген (PSCA); вирусный антиген, такой как, например, часть оболочки вируса СПИДа; транспортные белки; домашние рецепторы; адрессины; регуляторные белки; антитела; и фрагменты любых вышеперечисленных полипептидов. Термин «антитело» применяется здесь в самом широком смысле и, в частности, охватывает интактные моноклональные антитела, поликлональные антитела, поливалентные антитела (например, бивалентные антитела), образованные, по меньшей мере, из двух интактных антител, и фрагменты антител, до тех пор пока они проявляют требуемую биологическую активность. В некоторых вариантах осуществления изобретения рекомбинантный полипептид представляет собой фактор роста. В одном из вариантов осуществления рекомбинантный полипептид является полипептидом VEGF млекопитающего. В другом варианте осуществления рекомбинантный полипептид представляет собой VEGF человека (например, VEGF165). В одном из вариантов осуществления рекомбинантный полипептид не является ангиостатином. В одном из вариантов осуществления рекомбинантный полипептид не является IGF-1.
Как применяют здесь, «фактор роста эндотелия сосудов» или «VEGF» относится к фактору роста млекопитающего, полученному первоначально из гипофизарных фолликулярных клеток быка, обладающему аминокислотной последовательностью, описанной в Castor, C. W., et al., (1991) Methods in Enzymol. 198:391-405, вместе с его функциональными производными, обладающими качественной биологической активностью соответствующего нативного VEGF, включая в качестве неограничивающих примеров аминокислотную последовательность VEGF человека, как сообщено в Houck et al., (1991) Mol. Endocrin. 5:1806-1814. Смотри также Leung et al. (1989) Science. 246:1306, Robinson & Stringer, (2001) Journal of Cell Science. 144(5):853-865, патент США № 5332671. Его также обозначают как VEGF-A. Другие члены семейства отмечены буквенным символом в конце VEGF, например VEGF-B, VEGF-C или VEGF-D. Преобладающая форма VEGF, или VEGF-A, представляет собой 165-аминокислотный гомодимер, обладающий шестнадцатью остатками цистеина, которые образуют 7 внутримолекулярных дисульфидных связей и две межмолекулярные дисульфидные связи. Альтернативный сплайсинг принимает участие в образовании множественных полипептидов VEGF человека, состоящих из 121, 145, 165, 189 и 206 аминокислот, при этом вариант VEGF121 не содержит гепаринсвязывающий домен других вариантов. Все изоформы VEGF имеют часть общего аминоконцевого домена, но отличаются длиной карбоксиконцевой части молекулы. Предпочтительная активная форма VEGF, VEGF165, содержит дисульфидные связи между аминокислотными остатками Cys26-Cys68; Cys57-Cys104; Cys61-Cys102; Cys117-Cys135; Cys120-Cys137; Cys139-Cys158; Cys146-Cys160 в каждом мономере. Смотри Фиг. 10. Смотри также, например, Keck et al., (1997) Archives of Biochemistry and Biophysics 344(1):103-113. Молекула VEGF165 состоит из двух доменов: аминоконцевого рецепторсвязывающего домена (аминокислоты 1-110 связанного дисульфидными связями гомодимера) и карбоксиконцевого гепаринсвязывающего домена (остатки 111-165). Смотри, например, Keyt et al., (1996) J. Biol. Chem., 271(13):7788-7795. В некоторых вариантах осуществления изобретения выделенный и очищенный VEGF165 не гликозилирован по остатку 75 (Asn). Смотри, например, Yang et al., (1998) Journal of Pharm. & Experimental Therapeutics, 284:103-110. В некоторых вариантах осуществления изобретения выделенный и очищенный VEGF165 по существу не диамидирован по остатку Asn10. В некоторых вариантах осуществления изобретения выделенный и очищенный VEGF165 представляет собой смесь диамидированного (по остатку Asn10) и недиамидированного белка, как правило, с преобладанием недиамидированного белка. Поскольку VEGF165 представляет собой гомодимер, диаминирование может иметь место на одной или обеих полипептидных цепях.
Термин «гепаринсвязывающий белок», как применяют здесь, относится к полипептиду, способному связывать гепарин (как определено здесь). Определение включает зрелую, пре-, препро- и проформы нативных и рекомбинантно получаемых гепаринсвязывающих белков. Типичными примерами гепаринсвязывающих белков являются «гепаринсвязывающие факторы роста», включающие в качестве неограничивающих примеров фактор роста эпидермиса (EGF), фактор роста тромбоцитов (PDGF), основный фактор роста фибробластов (bFGF), кислотный фактор роста фибробластов (aFGF), фактор роста эндотелия сосудов (VEGF), фактор роста гепатоцитов (HGF) (также известный как рассеивающий фактор, SF) и фактор роста нервов (NGF), IL-8, и т.д.
«Гепарин» (также обозначаемый как гепариновая кислота) представляет собой гетерогенную группу высокосульфатируемых, анионных мукополисахаридов с прямой цепью, называемых гликозаминогликанами. Хотя другие сахара могут присутствовать, основными сахарами в гепарине являются: α-L-идуроновая кислота-2-сульфат, 2-дезокси-2-сульфамино-α-глюкоза-6-сульфат, β-D-глюкуроновая кислота, 2-ацетамидо-2-дезокси-α-D-глюкоза и L-идуроновая кислота. Эти и, необязательно, другие сахара присоединены при помощи гликозидных связей, образуя полимеры различных размеров. Благодаря наличию у него ковалентно связанных сульфатных и карбоксильных кислотных групп гепарин является сильно кислым. Молекулярная масса гепарина варьирует от приблизительно 3000 до приблизительно 20000 дальтон, в зависимости от источника и способа определения. Нативный гепарин является компонентом различных тканей, особенно печени и легкого, и клеток молочной железы у некоторых видов млекопитающих. Гепарин и соли гепарина (гепарин-натрий) коммерчески доступны и используются, главным образом, в виде антикоагулянтов в различных клинических ситуациях.
Как применяют здесь, «правильно свернутый» или «биологически активный» VEGF или другой рекомбинантный белок и т.п. относится к молекуле с биологически активной конформацией. Квалифицированный специалист признает, что неправильно свернутые интермедиаты и интермедиаты с неправильными дисульфидными связями могут обладать биологической активностью. В таком случае правильно свернутый или биологически активный VEGF или рекомбинантный белок соответствует нативному характеру фолдинга VEGF (описанного выше) или другого рекомбинантного белка. Например, правильно свернутый VEGF обладает отмеченными выше дисульфидными парами, в дополнение к двум межмолекулярным дисульфидным связям в димерной молекуле, однако другие интермедиаты могут быть получены при помощи бактериальной клеточной культуры. Для правильно свернутого VEGF имеют место две межмолекулярные дисульфидные связи между теми же остатками, Cys51 и Cys60, каждого мономера. Смотри, например, WO98/16551. Биологические активности VEGF включают в качестве неограничивающих примеров активацию сосудистой проницаемости, стимулирование роста эндотелиальных клеток сосудов, связывание с рецептором VEGF, связывание и передачу сигналов через рецептор VEGF (смотри, например, Keyt et al., (1996) Journal of Biological Chemistry, 271(10):5638-5646), индуцирование ангиогенеза и т.д.
Термины «очищенный» или «чистый рекомбинантный белок» и т.п. относятся к материалу, свободному от веществ, которые в норме сопровождают его, как обнаружено при его рекомбинантном получении и особенно в прокариотической или бактериальной культуре клеток. Поэтому термины относятся к рекомбинантному белку, который свободен от загрязнений ДНК, белками клетки-хозяина или другими молекулами, связанными с его окружением in situ. Термины относятся к степени чистоты, которая составляет, по меньшей мере, приблизительно 75%, по меньшей мере, приблизительно 80%, по меньшей мере, приблизительно 85%, по меньшей мере, приблизительно 90%, по меньшей мере, приблизительно 95% или, по меньшей мере, приблизительно 98% или более.
Термины «тельца включения» или «преломляющие тельца» относятся к плотным внутриклеточным массам представляющих интерес агрегированных полипептидов, которые составляют значительную часть общего клеточного белка, включая все клеточные компоненты. В некоторых случаях, но не во всех случаях, эти агрегаты полипептидов можно обнаружить в виде ярких пятен, видимых внутри очертания клеток под фазово-контрастным микроскопом при увеличении ниже чем 1000-кратное.
Как применяют здесь, термин «неправильно свернутый» белок относится к преципитированным или агрегированным полипептидам, которые содержатся в преломляющих тельцах. Как применяют здесь, «нерастворимый» или «неправильно свернутый» VEGF или другой рекомбинантный белок относится к преципитированному или агрегированному VEGF или рекомбинантному белку, который содержится внутри периплазмы или внутриклеточного пространства прокариотических клеток-хозяев, или иным способом ассоциированному с прокариотической клеткой-хозяином и принимает биологически неактивную конформацию с неправильно образованными или несформированными дисульфидными связями. Нерастворимый рекомбинантный белок, как правило, но необязательно, содержится в преломляющих тельцах, т.е. он может или не может быть визуализирован в фазово-контрастный микроскоп.
Как применяют здесь, «хаотропное средство» относится к соединению, которое в соответствующей концентрации в водном растворе способно изменить пространственную конфигурацию или конформацию полипептидов посредством изменений на их поверхности таким образом, чтобы сделать полипептид растворимым в водной среде. Изменения могут происходить путем изменения, например, характера гидратации окружающего растворителя или взаимодействия растворителя с поверхностью. Концентрация хаотропного средства будет непосредственно влиять на его силу и эффективность. Сильно денатурирующий хаотропный раствор содержит хаотропное средство в высоких концентрациях, которое в растворе будет эффективно разворачивать присутствующий в растворе полипептид, эффективно элиминируя вторичную структуру белков. Разворачивание должно быть относительно обширным, но обратимым. Умеренно денатурирующий хаотропный раствор содержит хаотропное средство, которое в достаточных концентрациях в растворе обеспечивает частичное сворачивание полипептида из любой нарушенной конформации, принятой полипептидом, через интермедиаты, растворимые в растворе, в пространственную конформацию, в которой он существует при функционировании в активной форме при эндогенных или гомологичных физиологических условиях. Примеры хаотропных средств включают гидрохлорид гуанидина, мочевину и гидроксиды, такие как гидроксид натрия или калия. Хаотропные средства включают сочетания этих средств, такие как смеси гидроксида с мочевиной или гидрохлоридом гуанидина.
Как применяют здесь, «восстанавливающее средство» обозначает соединение, которое в подходящей концентрации в водном растворе сохраняет свободные сульфгидрильные группы таким образом, что интра- или межмолекулярные дисульфидные связи химически разрываются. Репрезентативные примеры подходящих восстанавливающих средств включают дитиотреитол (DTT), дитиоэритрит (DTE), бета-меркаптоэтанол (BME), цистеин, цистамин, тиогликолят, глутатион и борогидрид натрия.
Как применяют здесь, «буферный раствор» обозначает раствор, который устойчив к изменениям pH путем действия своих спаренных компонентов кислота-основание.
«Бактерии» для целей настоящего описания включают эубактерии и архебактерии. В некоторых вариантах осуществления изобретения эубактерии, включая грамположительные и грамотрицательные бактерии, применяют в описанных здесь способах и процессах. В одном из вариантов осуществления изобретения применяют грамотрицательные бактерии, например Enterobacteriaceae. Примеры бактерий, принадлежащих к Enterobacteriaceae, включают Escherichia, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, Serratia и Shigella. Другие типы подходящих бактерий включают Azotobacter, Pseudomonas, Rhizobia, Vitreoscilla и Paracoccus. В одном из вариантов осуществления изобретения применяют E. coli. Подходящие хозяева E. coli включают E. coli W3110 (ATCC 27325), E. coli 294 (ATCC 31446), E. coli B и E. coli X1776 (ATCC 31537). Эти примеры являются скорее иллюстративными, чем ограничивающими, и W3110 представляет собой один из примеров. Можно также использовать мутантные клетки любой из вышеупомянутых бактерий. Конечно, необходимо отбирать подходящие бактерии, принимая во внимание воспроизводимость репликона в клетках бактерии. Например, виды E. coli, Serratia или Salmonella можно соответствующим образом использовать в качестве хозяина, в котором используют в качестве источника репликона хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Смотри далее относительно примеров подходящих бактериальных клеток-хозяев.
Как применяют здесь, выражения «клетка», «клеточная линия», «штамм» и «клеточная культура» используют взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичную из рассматриваемых клеток и культуры, полученные из нее безотносительно к количеству пассажей. Также понятно, что все потомство не может являться в точности идентичными по содержанию ДНК вследствие преднамеренных или случайных мутаций. Включается мутантное потомство, которое обладает той же самой функцией или биологической активностью, как и те, поиск которых осуществляли в исходной трансформированной клетке. Там, где предполагаются различные обозначения, это станет ясно из контекста.
Комбинированный тип колонки обозначает колонку со смолой, которая обладает как катионообменными свойствами, так и гидрофобными взаимодействиями.
Рекомбинантные белки
Рекомбинантные белки, например факторы роста, такие как кислый фактор роста фибробластов, основный фактор роста фибробластов и фактор роста эндотелия сосудов, были выделены и очищены из ряда источников, включая бактерии (Salter D.H. et al., (1996) Labor. Invest. 74(2):546-556 (VEGF); Siemeister et al., (1996) Biochem. Biophys. Res. Commun. 222(2):249-55 (VEGF); Cao et al., (1996) J. Biol. Chem. 261(6):3154-62 (VEGF); Yang et al., (1994) Gaoiishu Tongxun. 4:28-31 (VEGF); Anspach et al., (1995) J. Chromatogr. A 711(1):129-139 (aFGF и bFGF); Gaulandris (1994) J Cell. Physiol. 161(1):149-59 (bFGF); Estape and Rinas (1996) Biotech. Tech. 10(7):481-484 (bFGF); McDonald et al., (1995) FASEB J. 9(3):A410 (bFGF)). Например, преобладающая активная форма VEGF представляет собой гомодимер двух 165-аминокислотных полипептидов (VEGF-165). В этой структуре каждая субъединица содержит 7 пар внутрицепочечных дисульфидных связей и две дополнительные пары, которые образуют ковалентную связь двух субъединиц (Ferrara et al., (1991) J. Cell. Biochem. 47:211-218). Нативная конформация включает сильно основный домен, который, как показано, легко связывается с гепарином (Ferrara et al. (1991), выше). В то время как ковалентная димеризация VEGF необходима для эффективного связывания рецептора и биологической активности (Potgens et al., (1994) J. Biol. Chem. 269:32879-32885; Claffey et al., (1995) Biochim. et Biophys. Acta 1246:1-9), бактериальный продукт потенциально содержит несколько неправильно свернутых интермедиатов и интермедиатов с неправильно сформированными дисульфидными связями. Предложены способы, которые пригодны при выделении, очистке и реактивации белков, которые образуются в клетках-хозяевах в форме «преломляющих телец», а также растворимых белков.
Выделение рекомбинантного белка
Нерастворимый, неправильно свернутый рекомбинантный белок выделяют из прокариотических клеток-хозяев, экспрессирующих белок с помощью любого из ряда общепринятых в области способов. Например, нерастворимый рекомбинантный белок выделяют в подходящем буфере для выделения, помещая клетки в буфер с подходящей ионной силой для растворения большинства белков хозяина, но в котором рассматриваемый белок по существу нерастворим, или разрушая клетки таким образом, чтобы высвободить тельца включения или белок из периплазматического или внутриклеточного пространства и сделать их доступными для выделения, например, центрифугированием. Этот способ хорошо известен и описан, например, в патенте США № 4511503. В Kleid et al. описана очистка преломляющих телец путем гомогенизации с последующим центрифугированием (Kleid et al., (1984) в Developments in Industrial Microbiology, (Society for Industrial Microbiology, Arlington, VA) 25:217-235). Смотри также, например, Fischer et al., (1993) Biotechnology and Bioengineering 41:3-13.
В патенте США № 5410026 описан типичный способ выделения белка из телец включения, и он суммирован, как описано далее. Прокариотические клетки суспендируют в подходящем буфере. В основном буфер состоит из буферного вещества, подходящего для забуферивания pH между 5 и 9 или приблизительно 6-8, и соли. Любая подходящая соль, включая NaCl, пригодна для поддержания достаточной ионной силы в буферном растворе. В основном используют ионную силу приблизительно 0,01-2 M или от 0,1 до 0,2 M. Клетки, при суспендировании в этом буфере, разрушают или лизируют с использованием общепринятых способов, таких как, например, механические способы, например гомогенизатор (пресс Manton-Gaulin, Microfluidizer или Niro-Soavi), французский пресс, шаровая мельница или звуковой осциллятор, или путем химических или ферментативных способов.
Примеры химических или ферментативных способов разрушения клеток включают сферопластирование, которое влечет использование лизоцима для лизирования бактериальной стенки (H. Neu et al., (1964) Biochem. Biophys. Res. Comm., 17:215), и осмотический шок, который предусматривает обработку живых клеток гипертоническим раствором и промывку гипотонической холодной водой для высвобождения полипептидов (H. Neu et al., 1965 J. Biol. Chem. 240(9):3685-3692). Обработку ультразвуком, как правило, используют для разрушения бактерий, содержащихся в объемах среды для ферментации аналитических масштабов. При более крупных масштабах в основном используют гомогенизацию при высоком давлении.
После того как клетки разрушены, суспензию обычно центрифугируют при низкой скорости, как правило, например, в районе от 500 до 15000×g, например в одном из вариантов осуществления изобретения используют приблизительно 12000×g, в общепринятой центрифуге в течение времени, достаточном для осаждения по существу всего нерастворимого белка. Такие периоды времени можно легко определить, и они зависят от центрифугируемого объема, а также от конструкции центрифуги. В основном от приблизительно 10 минут до 0,5 часов достаточно для осаждения нерастворимого белка. В одном из вариантов осуществления суспензию центрифугируют при 12000×g в течение 10 минут.
Полученный осадок содержит по существу всю фракцию нерастворимого белка. Если процесс разрушения клеток не закончен, то осадок может также содержать интактные клетки или фрагменты разрушенных клеток. Завершение разрушения клеток можно анализировать путем ресуспендирования осадка в небольшом количестве того же буферного раствора и исследования суспензии в фазово-контрастный микроскоп. Присутствие фрагментов разрушенных клеток или целых клеток указывает на то, что необходима дополнительная обработка ультразвуком или другие способы разрушения для удаления фрагментов или клеток и ассоциированных непреломляющих полипептидов. После такого дополнительного разрушения, при необходимости, суспензию снова центрифугируют и осадок выделяют, ресуспендируют и повторно исследуют. Процесс повторяют до тех пор, пока визуальное исследование не выявит отсутствие фрагментов разрушенных клеток в осажденном материале, или до тех пор, пока дополнительная обработка не сможет уменьшить размер полученного осадка.
Вышеупомянутый процесс можно применять там, где нерастворимый белок или является внутриклеточным, или находится в периплазматическом пространстве. В одном из вариантов осуществления изобретения условия, приведенные здесь для выделения рекомбинантного белка, относятся к тельцам включения, агрегированным в периплазматическом пространстве или внутриклеточном пространстве, и в особенности относятся к VEGF. Однако способы и процедуры предполагается применять для рекомбинантных белков в основном с незначительными модификациями, как отмечено по ходу следующего текста. В некоторых вариантах осуществления изобретения способы и процедуры применяют для получения, рефолдинга и очистки рекомбинантного белка в производственном или промышленном масштабе.
В одном из вариантов осуществления выделенный рекомбинантный белок в осадке инкубируют в первом буферном растворе, достаточном по существу для растворения рекомбинантного белка. Эта инкубация имеет место в условиях концентрации, времени инкубации и температуры инкубации, которые будут обеспечивать растворение требуемого количества или по существу всего рекомбинантного белка.
Первый буферный раствор содержит буферное вещество, подходящее для поддержания диапазона pH буфера, по меньшей мере, приблизительно 9 или более, где обычный диапазон составляет 9-11. В одном из вариантов осуществления pH для VEGF составляет pH 11. Примеры подходящих буферов, которые будут обеспечивать pH внутри этого последнего диапазона, включают ТРИС (трис[гидроксиметил]аминометан), HEPPS (N-[2-гидроксиэтил]пиперазин-N'-[3-пропансульфоновая кислота]), CAPSO (3-[циклогексиламино]-2-гидрокси-1-пропансульфоновая кислота), AMP (2-амино-2-метил-1-пропанол), CAPS (3-[циклогексиламино]-1-пропансульфоновая кислота), CHES (2-[N-циклогексиламино]этансульфоновая кислота), аргинин, лизин и борат натрия. В одном из вариантов осуществления буфер в данной работе включает CHES и аргинин при pH приблизительно 11. В другом варианте осуществления буфер в данной работе включает Трис и аргинин при pH приблизительно 11. В одном из вариантов осуществления буфер в данной работе включает CHES при pH приблизительно 11. В другом варианте осуществления буфер в данной работе включает Трис при pH приблизительно 11. В некоторых вариантах осуществления первый буферный раствор содержит хаотропное средство.
Хаотропные средства, подходящие для осуществления на практике этого изобретения, включают, например, мочевину и соли гуанидина или тиоцианата, например мочевину, гидрохлорид гуанидина, тиоцианат натрия и т.д. Количество хаотропного средства, которое должно присутствовать в буфере, представляет собой количество, достаточное для разворачивания рекомбинантного белка в растворе. В некоторых вариантах осуществления изобретения хаотроп присутствует из расчета приблизительно между 0,5-5 моль/л. В одном из вариантов осуществления изобретения хаотропное средство представляет собой мочевину из расчета приблизительно 1 M.
Концентрация белка в буферном растворе должна быть такой, при которой белок будет по существу растворенным при определении по оптической плотности. Точное количество для применения будет зависеть, например, от концентраций и типов других ингредиентов в буферном растворе в частности, концентрации белка, хаотропного средства и pH буфера. В одном из вариантов осуществления изобретения концентрация рекомбинантного белка находится в диапазоне 0,5-5,5 мг на мл или 1,5-5,0 мг/мл. Растворение в основном осуществляют при приблизительно 0-45°C, или приблизительно 2-40°C, или приблизительно 20-40°C, или приблизительно 23-37°C, или приблизительно 25-37°C, или приблизительно 25°C в течение, по меньшей мере, от приблизительно одного до 24 часов. В основном, на температуру явным образом не влияют уровни соли, восстанавливающего средства и хаотропного средства. В некоторых вариантах осуществления растворение осуществляют при атмосферном давлении.
Измерение степени растворения в буферном растворе можно определить и соответствующим образом осуществить, например, определением мутности, анализом фракционирования между супернатантом и осадком после центрифугирования, в восстановленных гелях SDS-PAGE, анализом белка (например, набором для анализа белка Bio-Rad) или путем HPLC.
Необязательно, разрушенные клетки не центрифугируют, но разводят, например, 1:4, 1:6, 1:8 во втором описанном здесь буферном растворе (буфер для рефолдинга). Эта инкубация имеет место в условиях концентрации, времени инкубации и температуры инкубации, которые будут обеспечивать растворение и рефолдинг рекомбинантного белка. В одном из вариантов осуществления приблизительно 30% или более рекомбинантного белка растворено и повторно свернуто.
Рефолдинг рекомбинантных белков
После растворения полипептида или, альтернативно, разрушения клеток, его помещают во второй буферный раствор или растворяют в нем, где раствор содержит, по меньшей мере, одно восстанавливающее средство и хаотропное средство, в концентрации, которая обеспечивает рефолдинг рекомбинантного белка, вместе с добавлением воздуха или кислорода, например, с использованием постоянного коэффициента волюметрического массообмена kLa = 0,004-0,1 мин-1 (например, для флакона на 2,5 л с импеллером морского типа, скорость барботирования воздухом составляет 0,3-10 куб. см/мин/л, 0,3-3 куб. см/мин/л, или 1 куб. см/мин/л, или 25 куб. см/мин/л со скоростью перемешивания 200-400 об/мин). Кислород или воздух для реакций рефолдинга можно обеспечить с помощью источника воздуха или обогащенного кислородом источника сжиженного газа. Эффективность массообмена из газовой фазы в жидкую фазу контролируют путем перемешивания, барботирования и повышения давления и ее выражают в виде коэффициента волюметрического массообмена, kLa. Смотри, например, Blanch, & Clark, Biochemical engineering, Marcel Dekker, New York, 1997; и Aunins, & Henzler, Aeration in cell culture bioreactors, в Biotechnology: A multi-volume comprehensive treatise, G. Stephanopoulos, Ed., Weinheim, New York, 1993, p. 219-281. В одном из вариантов осуществления используют kLa 0,004 мин-1, что соответствует скорости перемешивания 200-400 об/мин и скорости барботирования 0,3 куб. см/мин/л во флаконе на 2,5 л, содержащем импеллер морского типа. В других вариантах осуществления используют kLa = 0,01 мин-1 или 0,1 мин-1 для получения правильно свернутого белка.
В некоторых вариантах осуществления изобретения второй буферный раствор содержит два или более восстанавливающих средства. Полипептид можно разбавить буфером для рефолдинга, например, по меньшей мере, в пять раз, или, по меньшей мере, приблизительно в десять раз, или приблизительно в 20 раз, или приблизительно в 40 раз. Условия этой второй инкубации растворимого, развернутого белка, как правило, будут такими, чтобы происходил рефолдинг требуемого количества или существенный или полный рефолдинг белка. Точные условия будут зависеть, например, от pH буфера и типов и концентраций присутствующих хаотропных и восстанавливающих средств. Температура инкубации, как правило, составляет приблизительно 0-40°C, и инкубацию, как правило, будут проводить, по меньшей мере, от приблизительно 1 часа до приблизительно 48 часов для осуществления рефолдинга. В некоторых вариантах осуществления реакцию осуществляют, например, при приблизительно 0-45°C, или приблизительно 2-40°C, или приблизительно 20-40°C, или приблизительно 23-37°C, или приблизительно 25-37°C, или приблизительно 25°C, в течение, по меньшей мере, приблизительно 3 часов, в течение, по меньшей мере, приблизительно 10 часов, или между приблизительно 3 и 30 часами, или между приблизительно 3 и 24 часами. В некоторых вариантах осуществления реакцию осуществляют при атмосферном давлении.
Второй буферный раствор содержит буферное вещество, подходящее для поддержания диапазона pH буфера, по меньшей мере, приблизительно 9 или более чем 9, где типичный диапазон составляет 9-11, хаотропное средство и, по меньшей мере, одно восстанавливающее средство. В некоторых вариантах осуществления второй буферный раствор содержит два или более восстанавливающих средства. В одном из вариантов осуществления pH для VEGF составляет pH 10. Примеры подходящих буферов, которые будут обеспечивать pH внутри этого последнего диапазона, включают ТРИС (трис[гидроксиметил]аминометан), HEPPS (N-[2-гидроксиэтил]пиперазин-N'-[3-пропан-сульфоновая кислота]), CAPSO (3-[циклогексиламино]-2-гидрокси-1-пропансульфоновая кислота), AMP (2-амино-2-метил-1-пропанол), CAPS (3-[циклогексиламино]-1-пропансульфоновая кислота), CHES (2-[N-циклогексиламино]этансульфоновая кислота), аргинин, лизин и борат натрия. В одном из вариантов осуществления второй буферный раствор в данной работе содержит CHES и аргинин при pH приблизительно 10 (приблизительно в конечной концентрации 10 мМ и 100 мМ соответственно), с двумя или более восстанавливающими средствами и, по меньшей мере, одним хаотропным средством. В другом варианте осуществления второй буферный раствор в данной работе содержит Трис и аргинин при pH приблизительно 10 (приблизительно в конечной концентрации 10 мМ и 100 мМ соответственно), с двумя или более восстанавливающими средствами и, по меньшей мере, одним хаотропным средством.
Аргинин (или другая положительно заряженная аминокислота), например L-аргинин/HCl, может присутствовать в первом буферном растворе и втором буферном растворе. В некоторых вариантах осуществления изобретения концентрация аргинина составляет, например, приблизительно 50-500 мМ, приблизительно 75-300 мМ, или приблизительно 100-300 мМ, или приблизительно 100 мМ, или 300 мМ в конечной концентрации и т.д. В некоторых вариантах осуществления изобретения белок находится в первом буферном растворе при pH более чем 9 и с 0,5-3 M мочевиной, 50-500 мМ аргинином и 5 мМ EDTA в конечной концентрации. В одном из вариантов осуществления используют 10 мМ CHES в конечной концентрации. В другом варианте осуществления используют 10 мМ Трис в конечной концентрации. В одном из вариантов осуществления первый буферный раствор содержит в конечной концентрации 1 M мочевину, 300 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, pH 11. В другом варианте осуществления первый буферный раствор содержит в конечной концентрации 1 M мочевину, 300 мМ аргинин, 10 мМ Трис, 5 мМ EDTA, pH 11. В некоторых вариантах осуществления изобретения белок находится во втором буферном растворе (буферный раствор для рефолдинга) при pH>9, но <11, содержащем в конечной концентрации 0,5-3 M мочевину, 50-500 мМ аргинин, 0,25-1 мМ DTT, 5-20 мМ цистеин и 2-10 мМ EDTA. В одном из вариантов осуществления используют 10 мМ CHES в конечной концентрации. В другом варианте осуществления используют 10 мМ Трис в конечной концентрации. В одном из вариантов осуществления белок находится в буферном растворе для рефолдинга в конечной концентрации с 1 M мочевиной, 15 мМ цистеином, 2 мМ DTT, 100 мМ аргинином, 10 мМ CHES, 5 мМ EDTA, pH 9-10. В другом варианте осуществления белок находится в буферном растворе для рефолдинга в конечной концентрации с 1 M мочевиной, 15 мМ цистеином, 0,5-2 мМ DTT, 100 мМ аргинином, 10 мМ Трис, 5 мМ EDTA, pH 9-10.
Как отмечено, раствор также содержит, по меньшей мере, одно восстанавливающее средство. Примеры подходящих восстанавливающих средств включают в качестве неограничивающих примеров дитиотреитол (DTT), β-меркаптоэтанол (BME), цистеин, DTE и т.д. Количество восстанавливающего средства, присутствующего в буфере, будет зависеть в основном от типа восстанавливающего средства и хаотропного средства, типа и pH используемого буфера, количества кислорода, поглощенного раствором или введенного в него, и концентрации белка в буфере. Например, восстанавливающее средство соответствующим образом отбирают из средств, описанных выше, в диапазоне концентрации от приблизительно 0,5 до приблизительно 20 мМ для цистеина, 0,25-3,0 мМ для DTT (например, 0,5-2 мМ DTT) и менее чем приблизительно 0,2 мМ для BME. В одном из вариантов осуществления изобретения используют два или более восстанавливающих средства, например DTT из расчета приблизительно 0,5-2 мМ и от 0,5 до приблизительно 20 мМ цистеина. Поскольку DTT и BME можно использовать в связи с представленными здесь процедурами для рекомбинантных белков в общем случае, сочетание цистеина из расчета приблизительно 15 мМ и DTT, как описано выше, представляет собой пример для выделения VEGF. В одном из вариантов осуществления восстанавливающее средство представляет собой DTT из расчета приблизительно 2 мМ, с 15 мМ цистеином, в конечной концентрации. В другом варианте осуществления восстанавливающее средство представляет собой DTT из расчета приблизительно 0,5 мМ, с 15 мМ цистеином, в конечной концентрации.
Второй буферный раствор содержит, по меньшей мере, одно хаотропное средство в такой концентрации, чтобы происходил рефолдинг рекомбинантного белка. Как правило, хаотроп присутствует из расчета приблизительно между 0,5 и 2 моль/л в конечной концентрации. В одном из вариантов осуществления изобретения хаотропное средство в данной работе представляет собой мочевину из расчета приблизительно 0,5-2 M, 0,5-2 M или из расчета приблизительно 1 M в конечной концентрации. В другом варианте осуществления изобретения хаотропное средство представляет собой гидрохлорид гуанидина из расчета приблизительно 0,1-1 M в конечной концентрации.
Буфер для рефолдинга может необязательно содержать дополнительные вещества, такие как любое из множества неионных детергентов, таких как ТРИТОН™ X-100, НОНИДЕТ™ P-40, ряд ТВИН™ и ряд БРИДЖ™. Неионный детергент присутствует из расчета приблизительно между 0,01% и 1,0% в конечной концентрации. В одном из примеров концентрации неионного детергента составляют между приблизительно 0,025% и 0,05% или приблизительно 0,05% в конечной концентрации.
Степень рефолдинга соответствующим образом определяют путем анализа жидкостной хроматографией при высоком давлении (HPLC) с использованием, например, хроматографической колонки rpHPLC, катионообменной HPLC (гелевой колонки SP-5PW TSK, Tosoh Bioscience LLC) или другой подходящей гепарин-аффинной колонки. Увеличение размера правильно свернутого рекомбинантного пика в катионообменном анализе HPLC или гепаринсвязывающем анализе HPLC прямо коррелирует с возрастанием количеств свернутого, биологически активного рекомбинантного белка, присутствующего в буфере. Инкубацию осуществляют с тем, чтобы максимально увеличить отношение правильно свернутого рекомбинантного белка к выделенному неправильно свернутому рекомбинантному белку, как определено анализом rpHPLC.
В одном из вариантов осуществления качество и количество правильно свернутого VEGF оценивают с использованием гепаринсвязывающего анализа. Образцы, содержащие разведенный рекомбинантный белок, наносят, например, на колонку Heparin-5PW (7,5×75 мМ, Tosoh Biosciences LLC, Tokyo, Japan) или другую подходящую гепарин-аффинную колонку. Например, колонку Heparin-5PW уравновешивают 10 мМ фосфата натрия, pH 7,4, содержащим 0,15 M хлорида натрия. При скорости потока 1 мл/мин или 2 мл/мин колонку элюируют с использованием линейного градиента из расчета 0,15-2 M хлорида натрия в 10 мМ фосфата натрия, pH 7,4, в течение 10 минут. Элюант контролируют при 280 нм. В одном из вариантов осуществления белок выделяют в одиночном пике, соответствующем биологически активному правильно повторно свернутому VEGF. В одном из вариантов осуществления изобретения анализ для определения правильно повторно свернутого VEGF представляет собой RPHPLC. Дисульфидные связи можно, необязательно, подтверждать путем пептидного картирования. Круговой дихроизм можно также использовать для определения 2- и 3-мерной структуры/фолдинга.
В одном из вариантов осуществления растворение и рефолдинг выполняют в одну стадию. После получения осадка из разрушенных клеток, его помещают в описанный выше второй буферный раствор или разводят им (в этом случае озаглавленном как комбинированный буферный раствор). Полипептид можно разводить комбинированным буферным раствором, например, по меньшей мере, в пять раз, или, по меньшей мере, приблизительно в 10 раз, или приблизительно в 20 раз, или приблизительно в 40 раз. Условия этой инкубации осадка, как правило, будут такими, чтобы происходили растворение и рефолдинг требуемого количества или существенные или полные растворение и рефолдинг белка с добавлением воздуха или кислорода. В одном из вариантов осуществления используют kLa 0,004 мин-1, соответствующую скорости перемешивания 200-400 об/мин и скорости барботирования 0,3 куб. см/мин/л во флаконе на 2,5 л, содержащем импеллер морского типа. В других вариантах осуществления используют kLa = 0,01 мин-1 или 0,1 мин-1 для получения правильно свернутого белка. Точные условия будут зависеть, например, от pH буфера и типов и концентраций присутствующих хаотропного и восстанавливающего средств. Температура инкубации составляет, как правило, приблизительно 0-40°C, и инкубацию будут, как правило, проводить в течение, по меньшей мере, от приблизительно 1 до 48 часов, чтобы осуществить растворение и рефолдинг. Реакцию осуществляют, например, при приблизительно 0-45°C, или приблизительно 2-40°C, или приблизительно 20-40°C, или приблизительно 23-37°C, или приблизительно 25-37°C, или приблизительно 25°C, в течение, по меньшей мере, приблизительно 3 часов, в течение, по меньшей мере, приблизительно 10 часов, или между приблизительно 3 и 48 часами, или между приблизительно 3 и 30 часами. В некоторых вариантах осуществления реакцию осуществляют при атмосферной температуре.
Выделение и очистка рекомбинантных белков
Хотя при выделении и очистке рекомбинантного белка можно применять разнообразные способы и известные процедуры для разделения таких белков, как, например, фракционирование с помощью соли и растворителя, поглощение коллоидными веществами, гель-фильтрация, ионообменная хроматография, аффинная хроматография, иммуноаффинная хроматография, электрофорез и жидкостная хроматография при высоком давлении (HPLC), описан пример стадии осветления и многостадийной хроматографической процедуры. Стадия осветления включает добавление детергента до конечной концентрации 1% (например, тритон-X-100), доведение pH до приблизительно 8,5-9,5 (или приблизительно 8,7, или приблизительно 9), инкубирование раствора в течение от 1 до 10 часов при 25-30°C, центрифугирование раствора и фильтрование жидкости, полученной со стадии центрифугирования. Многостадийная хроматографическая процедура включает приведение указанного повторно свернутого рекомбинантного белка в контакт со смолой комбинированного типа, катионным хроматографическим носителем, первым носителем для гидрофобной хроматографии и, необязательно, вторым носителем для гидрофобной хроматографии или ионообменным носителем; и селективное выделение или элюирование рекомбинантного белка с каждого носителя. Предусмотрено, что стадии любой из процедур можно осуществлять в любом порядке. В одном из вариантов осуществления изобретения стадии выполняют последовательно.
Подходящая первая стадия дальнейшего выделения и очистки рекомбинантного белка обычно обеспечивает концентрирование рекомбинантного белка и уменьшение объема образца. Например, вторая описанная выше стадия инкубации может приводить к значительному возрастанию объема выделенного рекомбинантного белка и сопутствующему разбавлению белка буфером для рефолдинга. Подходящие первые хроматографические носители обеспечивают уменьшение объема выделенного рекомбинантного белка и могут успешно обеспечивать некоторую очистку белка от нежелательных загрязняющих белков. Подходящие первые хроматографические стадии включают хроматографические носители, с которых можно элюировать и наносить непосредственно на второй хроматографический носитель.
Иллюстративные первые хроматографические носители включают в качестве неограничивающих примеров смолу комбинированного типа (например, CaptoMMC™, GE Healthcare или MEP Hypercel, Pall Corporation), гидроксиапатитные хроматографические носители, например CHT керамического типа I и типа II (формально известного как MacroPrep ceramic), Bio-Gel HT, Bio-Gel HTP, Biorad, Hercules, CA и т.д.; металло-хелатирующие хроматографические носители, состоящие из инертной смолы с иммобилизованными ионами металла, такого как медь, никель и т.д.; а также не содержащие производных силикагели. В одном из вариантов осуществления изобретения первые хроматографические носители для очистки и выделения VEGF представляют собой смешанные ионообменные хроматографические носители. Элюцию с первого хроматографического носителя осуществляют согласно общепринятым в данной области способам. Подходящие условия элюции и буферы будут облегчать нанесение элюируемого рекомбинантного белка непосредственно на катионный хроматографический носитель, как описано ниже.
Разнообразные анионные компоненты можно присоединить к матрицам для получения катионных носителей для хроматографии. Анионные компоненты включают карбоксиметильные, сульфоэтильные группы, сульфопропильные группы, фосфат и сульфонат (S). Целлюлозные ионообменные смолы, такие как SE52, SE53, SE92, CM32, CM52, CM92, P11, DE23, DE32, DE52, EXPRESS ION™ S и EXPRESS ION™ C, доступны от Whatman LTD, Maidstone Kent U.K. Ионообменники на основе SEPHADEX™ и SEPHAROSE™ и поперечносшитые также известны под торговыми названиями CM SEPHADEX™ C-25, CM SEPHADEX™ C-50 и SP SEPHADEX™ C-25, SP SEPHADEX™ C-50 и SP-SEPHAROSE™ High Performance, SP-SEPHAROSE™-XL, SP-SEPHAROSE™ Fast Flow, CM-SEPHAROSE™ Fast Flow и CM-SEPHAROSE™ CL-6B, все доступны от Pharmacia AB. Примеры ионообменников для осуществления на практике изобретения включают в качестве неограничивающих примеров, например, ионообменники под торговыми названиями MACROPREP™, такие как, например, носитель MACROPREP™ S, носитель MACROPREP™ High S и носитель MACROPREP™ CM от BioRad, Hercules, CA.
Элюцию с катионных хроматографических носителей, как правило, осуществляют путем увеличения концентраций соли. Вследствие того что элюция с ионных колонок включает добавление соли и вследствие того что, как упомянуто здесь, HIC повышается в концентрации соли, необязательно используют введение стадии HIC вслед за ионной стадией или другими стадиями с применением соли. В одном из вариантов осуществления изобретения катионообменная хроматографическая стадия предшествует, по меньшей мере, стадии HIC, например первому хроматографическому носителю для гидрофобного взаимодействия и/или второму гидрофобному взаимодействию.
Гидрофобные колонки можно использовать при очистке рекомбинантного белка, например, на 2-й, 3-й и/или 4-й стадии очистки. Гидрофобная хроматография хорошо известна в данной области и основана на взаимодействии гидрофобных частей молекулы, взаимодействующих с гидрофобными лигандами, присоединенными к «хроматографическим носителям». Гидрофобный лиганд, присоединенный к матрице, по-разному обозначают как хроматографический носитель для HIC, гель для HIC или колонка для HIC и т.п. Более того, принимают во внимание, что сила взаимодействия между белком и колонкой для HIC является не только функцией отношения неполярных поверхностей на белке к полярным, но также распределения неполярных поверхностей.
При изготовлении колонок для HIC можно использовать ряд матриц. Наиболее широко используют агарозу, хотя можно использовать смолы на основе кремнеземных и органических полимеров. Пригодные гидрофобные лиганды включают в качестве неограничивающих примеров алкильные группы, содержащие от приблизительно 2 до приблизительно 10 углеродных атомов, такие как бутильные, пропильные, или октильные, или арильные группы, такие как фенил. Общепринятые носители для HIC для гелей и колонок можно коммерчески получить от поставщиков, таких как Pharmacia, Uppsala, Sweden, под торговыми названиями butyl-SEPHAROSE™, butyl-SEPHAROSE™-Fast Flow, phenyl-SEPHAROSE™ CL-4B, octyl SEPHAROSE™ FF и phenyl SEPHAROSE™ FF, Tosoh Corporation, Tokyo, Japan под торговыми названиями TOYOPEARL™ butyl 650M (Fractogel TSK Butyl-650) или TSK-GEL phenyl 5PW.
Плотность лиганда является важным параметром потому, что она влияет не только на силу взаимодействия белка, но также и на емкость колонки. Плотность лиганда коммерчески доступных фенильных или октилфенильных гелей составляет порядка 5-40 мкмоль/мл слоя геля. Емкость геля является функцией конкретного рассматриваемого белка, а также pH, температуры и концентрации соли, но, как правило, можно ожидать, что она окажется в диапазоне 3-20 мг/мл геля.
Выбор конкретного геля может определить квалифицированный специалист. В общем случае сила взаимодействия белка и лиганда для HIC возрастает с длиной цепи алкильных лигандов, но лиганды, обладающие от приблизительно 4 до приблизительно 8 углеродных атомов, подходят для большинства разделений. Фенильная группа обладает приблизительно такой же гидрофобностью, что и пентильная группа, хотя избирательность может отличаться благодаря возможности пи-пи-взаимодействия с ароматическими группами белка.
Адсорбция белка на колонке для HIC облегчается посредством высокой концентрации соли, но фактическая концентрация может различаться в широком диапазоне в зависимости от природы белка и конкретного выбранного лиганда для HIC. В общем случае пригодна концентрация соли между приблизительно 1 и 4 M.
Элюцию с носителя для HIC, или постадийно или в форме градиента, можно осуществлять множеством способов, таких как a) путем изменения концентрации соли, b) путем изменения полярности растворителя или c) путем добавления детергентов. При уменьшении концентраций соли адсорбированные белки элюируют в порядке возрастания гидрофобности. На изменения полярности можно влиять путем добавлений растворителей, таких как этиленгликоль или изопропанол, таким образом, уменьшая силу гидрофобных взаимодействий. Детергенты функционируют в качестве вытеснителей белков, и их преимущественно используют в связи с очисткой мембранных белков.
Примеры способов для очистки VEGF описаны ниже в данной работе, например смотри примеры IV и V.
Экспрессия рекомбинантного белка
В кратком изложении, экспрессирующие векторы, способные к автономной репликации и экспрессии белка по отношению к геному прокариотической клетки-хозяина, вводят в клетку-хозяина. Конструирование подходящих экспрессирующих векторов хорошо известно в данной области, включая нуклеотидные последовательности рекомбинантных белков, описанных здесь. Смотри, например, Sambrook et al., Molecular Cloning, A Laboratory Manual., Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001); Ausubel et al., Short Protocols in Molecular Biology, Current Protocols John Wiley and Sons (New Jersey) (2002); и Baneyx, (1999) Current Opinion in Biotechnology, 10:411-421. Подходящие прокариотические клетки, включая бактерии, экспрессирующие векторы коммерчески доступны, например, от American Type Culture Collection (ATCC), Rockville, Maryland. Способы крупномасштабного выращивания прокариотических клеток и, особенно, культуры бактериальных клеток хорошо известны в данной области, и эти способы можно использовать в контексте изобретения.
Например, прокариотические клетки-хозяева трансфицируют экспрессирующими или клонирующими векторами, кодирующими интересующий рекомбинантный белок, и культивируют в общепринятой питательной среде, модифицированной соответствующим образом для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности. Нуклеиновая кислота, кодирующая интересующий полипептид, представляет собой соответствующую РНК, кДНК или геномную ДНК из любого источника, при условии, что она кодирует интересующий(е) полипептид(ы). Хорошо известны способы для отбора подходящей нуклеиновой кислоты для экспрессии гетерологичных полипептидов (включая их варианты) в микробных хозяевах. Молекулы нуклеиновой кислоты, кодирующей полипептид, получают множеством способов, известных в данной области. Например, ДНК, кодирующую VEGF, выделяют и секвенируют, например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с геном, кодирующим VEGF.
Гетерологичную нуклеиновую кислоту (например, кДНК или геномную ДНК) соответствующим образом вставляют в реплицирующийся вектор для экспрессии в микроорганизме под управлением подходящего промотора. Многие векторы доступны для этой цели, и выбор подходящего вектора в основном будет зависеть от размера нуклеиновой кислоты, которую вставляют в вектор, и конкретной клетки-хозяина, которую трансформируют вектором. Каждый вектор содержит различные компоненты в зависимости от конкретной клетки-хозяина, с которой он совместим. В зависимости от конкретного типа хозяина компоненты вектора, как правило, включают в качестве неограничивающих примеров один или несколько из следующего: сигнальную последовательность, точку начала репликации, один или несколько маркерных генов, промотор и последовательность терминации транскрипции.
В общем случае плазмидные векторы, содержащие репликон и управляющие последовательности, которые получают из видов, совместимых с клеткой-хозяином, используют в связи с микробными хозяевами. Вектор исходно несет участок репликации, а также маркерные последовательности, которые способны обеспечить отбор по фенотипу трансформированных клеток. Например, E. coli в основном трансформируют с использованием pBR322, плазмиды, полученной из вида E. coli (смотри, например, Bolivar et al., (1977) Gene, 2:95). pBR322 содержит гены устойчивости к ампициллину и тетрациклину и, таким образом, предоставляет простые средства для идентификации трансформированных клеток. Плазмида pBR322, или другая бактериальная плазмида, или фаг также, как правило, содержит, или модифицирована так, чтобы содержать, промоторы, которые может использовать хозяин для экспрессии селективных маркерных генов.
(i) Сигнальная последовательность
Полипептиды по изобретению можно получать рекомбинантным способом не только непосредственно, но также в виде полипептида, слитого с гетерологичным полипептидом, который представляет собой в основном сигнальную последовательность, или другим полипептидом, обладающим специфическим участком расщепления на N-конце зрелого белка или полипептида. Выбранная гетерологичная сигнальная последовательность в основном представляет собой последовательность, которая узнается и процессируется (т.е. расщепляется сигнальной пептидазой) клеткой-хозяином. Для прокариотических клеток-хозяев, которые не распознают и процессируют сигнальную последовательность нативного полипептида, сигнальную последовательность замещают на выбранную прокариотическую сигнальную последовательность, например, из группы лидерных последовательностей щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II.
(ii) Компонент точки начала репликации
Экспрессирущие векторы содержат последовательность нуклеиновой кислоты, которая позволяет вектору реплицироваться в одной или нескольких выбранных клетках-хозяевах. Такие последовательности хорошо известны для множества микробов. Точка начала репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий, таких как E. coli.
(iii) Компонент селективного гена
Экспрессирующие векторы, как правило, содержат селективный ген, также называемый селективным маркером. Этот ген кодирует белок, необходимый для выживания или роста трансформированных клеток-хозяев, растущих в селективной культуральной среде. Клетки-хозяева, не трансформированные вектором, содержащим селективный ген, не будут выживать в культуральной среде. Этот селективный маркер выбирают из генетических маркеров, как применяют и определяют по этому изобретению. Типичные селективные гены кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют ауксотрофные недостаточности, отличные от тех, которые вызываются присутствием генетического(их) маркера(ов), или (c) обеспечивают необходимыми питательными веществами, которые не доступны в комплексных средах, например ген, кодирующий D-аланинрацемазу в Bacilli.
В одном из примеров схемы селекции используют средство, которое вызывает остановку роста клетки-хозяина. В этом случае те клетки, которые успешно трансформированы интересующей нуклеиновой кислотой, вырабатывают полипептид, придающий устойчивость к средству, и, таким образом, выживают в режиме селекции. В примерах такой доминантной селекции используют средства неомицин (Southern et al., (1982) J. Molec. Appl. Genet. 1:327), микофеноловую кислоту (Mulligan et al., (1980) Science 209:1422) или гигромицин (Sugden et al., (1985) Mol. Cell. Biol., 5:410-413). В этих трех приведенных выше примерах используют бактериальные гены под эукариотическим контролем для придания устойчивости к подходящему средству G418 или неомицину (генетицин), xgpt (микофеноловая кислота) или гигромицину соответственно.
(iv) Компонент промотора
Экспрессирующий вектор для получения интересующего рекомбинантного белка содержит подходящий промотор, который распознается организмом хозяина и функционально связан с нуклеиновой кислотой, кодирующей интересующий полипептид. Промоторы, подходящие для использования в прокариотических хозяевах, включают промоторную систему бета-лактамазы и лактозы (Chang et al., (1978) Nature, 275:615; Goeddel et al., (1979) Nature, 281:544), промоторную систему арабинозы (Guzman et al., (1992) J. Bacteriol. 174:7716-7728), промоторную систему щелочной фосфатазы, триптофана (trp) (Goeddel, (1980) Nucleic Acids Res. 8:4057 и EP 36776) и гибридные промоторы, такие как tac-промотор (deBoer et al., (1983) Proc. Natl. Acad. Sci. USA, 80:21-25). Однако подходят и другие известные бактериальные промоторы. Их нуклеотидные последовательности опубликованы, таким образом, позволяя квалифицированному специалисту функционально лигировать их с ДНК, кодирующей интересующий полипептид (Siebenlist et al., (1980) Cell, 20:269) с использованием линкеров или адаптеров для обеспечения любыми необходимыми участками рестрикции. Смотри также, например, Sambrook et al., выше; и Ausubel et al., выше.
Промоторы для использования в бактериальных системах также, как правило, содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей интересующий полипептид. Промотор можно выделить из ДНК бактериального источника путем расщепления ферментами рестрикции и вставить в вектор, содержащий требуемую ДНК.
(v) Конструирование и анализ векторов
При конструировании подходящих векторов, содержащих один или несколько из вышеперечисленных компонентов, применяют общепринятые способы лигирования. Выделенные плазмиды или фрагменты ДНК расщепляют, подготавливают и повторно лигируют в форме, требуемой для получения необходимых плазмид.
Для анализа подтверждения правильных последовательностей в сконструированных плазмидах, смеси после лигирования используют для трансформации E. coli K12, штамм 294 (ATCC 31446), или других штаммов и успешные трансформанты отбирают по устойчивости к ампициллину или тетрациклину, там где это подходит. Плазмиды получают из трансформантов, анализируют путем расщепления эндонуклеазами рестрикции и/или секвенируют способом по Sanger et al., (1977) Proc. Natl. Acad. Sci. USA. 74:5463-5467 или Messing et al., (1981) Nucleic Acids Res. 9:309, или способом по Maxam et al., (1980) Methods in Enzymology, 65:499. Смотри также, например, Sambrook et al., выше; и Ausubel et al., выше.
Нуклеиновую кислоту, кодирующую интересующий рекомбинантный белок, вводят в клетки-хозяева. В основном это осуществляют путем трансформации клеток-хозяев вышеописанными экспрессирующими векторами и культивированием в общепринятой питательной среде, модифицированной так, чтобы подходить для индукции различных промоторов.
Культивирование клеток-хозяев
Подходящие прокариотические клетки для осуществления на практике изобретения хорошо известны в данной области. В основном используют клетки-хозяева, которые экспрессируют рекомбинантный белок в большом количестве в форме телец включения или в периплазматическом или внутриклеточном пространстве. Подходящие прокариоты включают бактерии, например эубактерии, такие как грамотрицательные или грамположительные организмы, например E. coli, Bacilli, такие как B. subtilis, виды Pseudomonas, такие как P. aeruginosa, Salmonella typhimurium или Serratia marcescens. Один из примеров хозяина E. coli представляет собой E. coli 294 (ATCC 31446). Также подходят другие штаммы, такие как E. coli B, E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Эти примеры являются скорее иллюстративными, чем ограничивающими. Штамм W3110 представляет собой типичного хозяина, вследствие того что он является общепринятым штаммом-хозяином для ферментаций продуктов рекомбинантных ДНК. В одном из аспектов изобретения клетка-хозяин должна секретировать минимальные количества протеолитических ферментов. Например, штамм W3110 можно модифицировать для получения генетической мутации в генах, кодирующих белки, примеры таких хозяев, включая E. coli W3110, штаммы 1A2, 27A7, 27B4 и 27C7, описаны в патенте США № 5410026, опубликованном 25 апреля 1995. Например, штамм для получения VEGF представляет собой штамм E.coli W3110, обладающий генотипом tonAΔ ptr3 phoAΔE15 Δ(argF-lac)169 degP41 ilvg, обозначенный 49B3. Смотри также, например, таблицу, находящуюся на страницах 23-24 WO2004/092393.
Прокариотические клетки, используемые для получения интересующего рекомбинантного белка, выращивают в средах, известных в данной области и подходящих для культивирования отобранных клеток-хозяев, включая среды, в общем описанные в Sambrook et al., Molecular Cloning, A Laboratory Manual., Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001). Среды, которые подходят для бактерий, включают в качестве неограничивающих примеров среду AP5, питательный бульон, среду Лурия-Бертани (LB), минимальную среду Нейдхардта и минимальную или полную среду C.R.A.P., плюс необходимые питательные добавки. В некоторых вариантах осуществления среды также содержат селективное вещество, выбранное на основе конструирования экспрессирующего вектора, для обеспечения селективного роста прокариотических клеток, содержащих экспрессирующий вектор. Например, ампициллин добавляют в среды для роста клеток, экспрессирующих ген устойчивости к ампициллину. Любые необходимые добавки помимо источников углерода, азота и неорганического фосфата также можно включать в подходящих концентрациях при введении по отдельности или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно культуральная среда может содержать одно или несколько восстанавливающих средств, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритрита и дитиотреитола.
Примеры подходящих сред приведены в патентах США No. 5304472 и 5342763. Среда с ограниченным содержанием фосфата C.R.A.P. состоит из 3,57 г (NH4)2(SO4), 0,71 г цитрата Na-2H2O, 1,07 г KCl, 5,36 г дрожжевого экстракта (сертифицированного), 5,36 г HycaseSF™-Sheffield, где pH доведен KOH до 7,3, объем доведен до 872 мл деионизованной H2O и проавтоклавировано; охлаждено до 55°C и добавлено 110 мл 1 M MOPS pH 7,3, 11 мл 50% глюкозы, 7 мл 1 M MgSO4. Затем можно добавить карбенициллин для индукции культуры в концентрации 50 мкг/мл.
Прокариотические клетки-хозяева культивируют при подходящих температурах. Для роста E. coli, например, температура варьирует, например, от приблизительно 20°C до приблизительно 39°C, или от приблизительно 25°C до приблизительно 37°C, или при приблизительно 30°C.
Там, где используют промотор щелочной фосфатазы, клетки E. coli, используемые для получения интересующего полипептида по этому изобретению, культивируют в подходящей среде, в которой промотор щелочной фосфатазы может быть частично или полностью индуцирован, как в общем виде описано, например, в Sambrook et al., Molecular Cloning, A Laboratory Manual., Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001). Культивирование никогда не может иметь места в отсутствие неорганического фосфата или при истощенных уровнях фосфата. Сначала среда содержит неорганический фосфат в количестве, превышающем уровень индукции синтеза белка и достаточном для роста бактерий. По мере роста клеток и утилизации фосфата они снижают уровень фосфата в среде, таким образом, вызывая индукцию синтеза полипептида.
Если промотор представляет собой индуцируемый промотор, то для получения индукции в основном клетки культивируют до достижения некоторой оптической плотности, например, A550 приблизительно 200 с использованием процесса с высокой плотностью клеток, при котором начало индукции инициируется (например, добавлением индуктора, истощением компонента среды и т.д.) для индукции экспрессии гена, кодирующего интересующий полипептид.
Любые необходимые добавки можно также включать в подходящих концентрациях, которые должны быть известны специалисту в данной области, при внесении по отдельности или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. pH среды может представлять собой любой pH из приблизительно 5-9, что в основном зависит от организма хозяина. Для E. coli pH составляет, например, от приблизительно 6,8 до приблизительно 7,4 или приблизительно 7,0.
Применение рекомбинантных белков
Для таким образом полученного полипептида можно получить композицию с фармацевтически приемлемым носителем и использовать для различных диагностических, терапевтических или других применений, известных для таких молекул. Например, описанные здесь белки можно использовать в иммуноанализах, таких как ферментные иммуноанализы.
Также предусмотрены терапевтические применения рекомбинантных белков, полученных с использованием описанных здесь способов. Например, фактор роста или гормон, например VEGF, можно применять при желании для усиления роста. Например, VEGF можно использовать для стимуляции заживления ран, например острой раны (например, ожога, хирургической раны, обычной раны и т.д.) или хронической раны (например, диабетической язвы, язвы вследствие давления, пролежня, венозной язвы и т.д.), для стимуляции роста волос, для стимуляции роста ткани и заживления и т.д.
Терапевтические композиции рекомбинантных белков получают для хранения путем смешивания молекулы, например полипептида, обладающего требуемой степенью чистоты с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 18th edition, Osol, A. Ed. (1995)) в форме лиофилизированных композиций или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие, как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие вещества, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как ТВИН™, ПЛЮРОНИКИ™ или полиэтиленгликоль (PEG).
В некоторых вариантах осуществления используемые композиции для введения in vivo являются стерильными. Это легко осуществить фильтрованием через стерильные мембраны для фильтрования. Рекомбинантные белки можно хранить в лиофилизированной форме, или в виде водного раствора, или в форме геля. pH препаратов рекомбинантного белка может составлять, например, от приблизительно 4 до 8 (в одном из вариантов осуществления pH 5,0), хотя более высокие или низкие значения pH могут также подходить в некоторых случаях. Будет понятно, что использование некоторых эксципиентов, носителей или стабилизаторов может приводить к образованию солей рекомбинантного белка.
Способ введения полипептида находится в согласии с известными способами, например, местным введением, инъекцией или инфузией внутривенным, внутрибрюшинным, интрацеребральным, внутримышечным, интраокулярным, внутриартериальным или внутриочаговым способами, или с помощью системы замедленного высвобождения, как отмечено ниже. Полипептид можно вводить непрерывно инфузией или болюсной инъекцией.
В основном при заживлении ран для рекомбинантного белка получают композицию для сайт-специфичной доставки. При местном применении рекомбинантный белок соответствующим образом комбинируют с другими ингредиентами, такими как носители и/или адъюванты. Отсутствуют ограничения на природу таких других ингредиентов, за исключением того, что они должны быть фармацевтически приемлемыми и эффективными для своего предполагаемого введения и они не могут в значительной степени снижать активность активных ингредиентов композиции. Примеры подходящих носителей включают мази, кремы, гели, спреи или суспензии, с очищенным коллагеном или без него. Композициями также можно пропитывать стерильные повязки, трансдермальные пластыри, пластыри и бандажи, необязательно в жидкой или полужидкой форме.
При получении гелевой композиции рекомбинантный белок, получаемый в жидкой композиции, можно смешивать с эффективным количеством водорастворимого полисахарида или синтетического полимера, такого как полиэтиленгликоль, с образованием геля подходящей вязкости для местного применения. Полисахарид, который можно использовать, включает, например, производные целлюлозы, такие как этерифицированные производные целлюлозы, включая алкилцеллюлозы, гидроксиалкилцеллюлозы и алкилгидроксиалкилцеллюлозы, например метилцеллюлозу, гидроксиэтилцеллюлозу, карбоксиметилцеллюлозу, гидроксипропилметилцеллюлозу и гидроксипропилцеллюлозу; крахмал и фракционированный крахмал; агар; альгиновую кислоту и альгинаты; гуммиарабик; пуллулан; агарозу; каррагинан; декстраны; декстрины; фруктаны; инулин; маннаны; ксиланы; арабинаны; хитозаны; гликогены; глюканы и синтетические биополимеры; а также камеди, такие как ксантановая камедь; гуаровая камедь; камедь плодоворожкового дерева; гуммиарабик; камедь трагаканта и камедь карайи; и их производные и смеси. В некоторых вариантах осуществления изобретения гелеобразующее вещество в данной работе представляет собой вещество, которое, например, инертно по отношению к биологическим системам, нетоксично, просто в получении и/или не является слишком текучим или вязким и не будет дестабилизировать рекомбинантный белок, который находится внутри него.
В некоторых вариантах осуществления изобретения полисахарид представляет собой этерифицированное производное целлюлозы, в другом варианте осуществления он хорошо определен, очищен и находится в перечне USP, например производные метилцеллюлозы и гидроксиалкилцеллюлозы, такие как гидроксипропилцеллюлоза, гидроксиэтилцеллюлоза и гидроксипропилметилцеллюлоза. В одном из вариантов осуществления метилцеллюлоза является полисахаридом.
Полиэтиленгликоль, пригодный для гелеобразования, представляет собой, как правило, смесь низко- и высокомолекулярных полиэтиленгликолей для получения подходящей вязкости. Например, смесь полиэтиленгликоля молекулярной массы 400-600 с полиэтиленгликолем 1500 может являться эффективной для этой цели при смешивании в правильном соотношении до получения пасты.
Предполагается, что термин «водорастворимый» применительно к полисахаридам и полиэтиленгликолям включает коллоидные растворы и дисперсии. В общем случае растворимость производных целлюлозы определяют по степени замещения эфирных групп, и стабилизирующие производные, пригодные в данной работе, должны обладать достаточным количеством таких эфирных групп на звено ангидроглюкозы в целлюлозной цепи, чтобы сделать производные водорастворимыми. Степень замещения эфиром, по меньшей мере, 0,35 эфирных групп на звено ангидроглюкозы, как правило, является достаточной. Кроме того, производные целлюлозы могут находиться в форме солей щелочных металлов, например солей Li, Na, K или Cs.
Например, если метилцеллюлозу используют в геле, то она содержит приблизительно 2-5%, или приблизительно 3%, или приблизительно 4%, или приблизительно 5% геля, и рекомбинантный белок находится в количестве приблизительно 300-1000 мг на мл геля.
Активные ингредиенты можно также заключать в полученные микрокапсулы, например, способами коацервации или межповерхностной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидную систему доставки лекарственного средства (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences 18th edition, Osol, A. Ed. (1995). Смотри также Johnson et al., Nat. Med. 2:795-799 (1996); Yasuda, Biomed. Ther. 27:1221-1223 (1993); Hora et al., Bio/Technology, 8:755-758 (1990); Cleland, «Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems» в Vaccine Design: Subunit and Adjuvant Approach, Powell and Newman, eds, (Plenum Press: New York, 1995), p. 439-462; WO 97/03692, WO 96/40072, WO 96/07399 и патент США № 5654010.
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих полипептид по изобретению, где матрицы находятся в виде оформленных продуктов, например пленок или микрокапсул. Примеры матриц для замедленного высвобождения включают полимерные сложные эфиры, гидрогели (например, поли-2-гидроксиэтилметакрилат или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагающийся этиленвинилацетат, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, образованные сополимером молочной кислоты-гликолевой кислоты и леупролид-ацетатом), полимер полимолочной-гликолевой кислоты (PLGA) и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этиленвинилацетат и молочная кислота-гликолевая кислота, обеспечивают высвобождение молекул в течение 100 суток, некоторые гидрогели высвобождают белки в течение более коротких периодов времени. Если инкапсулированные белки остаются в организме в течение длительного времени, то они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к утрате биологической активности и возможным изменениям иммуногенности. Можно разработать рациональные стратегии для стабилизации в зависимости от вовлеченного механизма. Например, если выявлено, что механизм агрегации представляет собой образование межмолекулярной S-S-связи посредством тио-дисульфидного обмена, стабилизации можно достичь модификацией сульфгидрильных остатков, лиофилизацией из кислотных растворов, контролированием содержания влаги, использованием подходящих добавок и разработкой специфических композиций полимерных матриц.
Композиции с замедленным высвобождением полипептида также включают полипептиды, заключенные в липосомы. Липосомы, содержащие белок, получают по сути известными способами: DE 3218121; Epstein et al., (1985) Proc. Natl. Acad. Sci. USA, 82:3688-3692; Hwang et al., (1980) Proc. Natl. Acad. Sci. USA, 77:40304034; EP 52322; EP 36676; EP 88046; EP 143949; EP 142641; японская патентная заявка 83-118008; патенты США № 4485045 и 4544545 и EP 102324. Обычно липосомы представляют собой небольшую (около 200-800 ангстрем) однослойную структуру, в которой содержание липидов составляет более чем приблизительно 30 мол.% холестерина, где выбранное соотношение приспосабливают для наиболее эффективной терапии полипептидом.
Эффективное количество рекомбинантного белка для применения в терапии будет зависеть, например, от целей терапии, способа введения и состояния пациента. Соответственно, необходимо для терапевта, если потребуется, подбирать дозировку и изменять способ введения для получения наиболее благоприятного терапевтического эффекта. Типичная суточная дозировка может варьировать от приблизительно 1 мкг/кг вплоть до 10 мг/кг или более, в зависимости от вышеупомянутых факторов. В основном лечащий врач будет вводить полипептид до тех пор, пока дозировка не достигнет величины, которая приводит к требуемому эффекту. Пациенту можно также вводить полипептид под наблюдением лечащего врача. За прогрессом этого лечения легко наблюдать с помощью общепринятых анализов.
Следующие примеры представлены для иллюстрации, а не для ограничения.
ПРИМЕРЫ
ПРИМЕР 1: Растворение и рефолдинг рекомбинантного VEGF человека, экспрессированного в Escherichia coli
Способы
Плазмида для экспрессии VEGF 165. Плазмиду pVEGF171 сконструировали для экспрессии VEGF165 человека (смотри, например, Leung et al., (1989) Science, 246:1306-1309) в периплазме E. coli. Транскрипцию кодирующей последовательности VEGF помещали под непосредственное управление промотора щелочной фосфатазы (AP) (смотри, например, Kikuchi et al., (1981) Nucleic Acids Research, 9:5671-8), а последовательности, необходимые для инициации трансляции, обеспечила область Шайна-Дальгарно trp (смотри, например, Yanofsky et al., (1981) Nucleic Acids Research, 9:6647-68). Кодирующую последовательность VEGF сливали ниже сигнальной последовательности бактериального термостабильного энтеротоксина II (STII) (смотри, например, Lee et al., (1983) Infect. Immun. 42:264-8; Picken et al., (1983) Infect. Immun. 42:269-75) для последующей секреции в периплазму E. coli. Модификации кодонов в сигнальной последовательности STII обеспечивают отрегулированный уровень трансляции, который дает оптимальный уровень накопления VEGF в периплазме (смотри, например, Simmons and Yansura, (1996) Nature Biotechnology, 14:629-34). Терминатор транскрипции лямбда (смотри, например, Scholtissek and Grosse, (1987) Nucleic Acids Research 15:3185) размещен ниже кодона терминации трансляции VEGF. Точка начала репликации и гены устойчивости как к ампициллину, так и к тетрациклину получены из плазмиды pBR322. Смотри, например, Bolivar et al., (1977) Gene 2:95-113.
Гомогенизация клеток и получение преломляющих телец . Массу целых клеток из клеток Escherichia coli, продуцирующих рекомбинантный белок, гомогенизировали микрофлуидизатором или Niro Soavi при давлениях, превышающих 8000 фунт/кв. дюйм. Гомогенат разводили 1:1 160 мМ MgSO4, 0,0375 декстран-сульфатом и 1% Тритоном X-100 перед собиранием осадка центрифугированием (центрифуга BTUX, Alfa Laval., Швеция).
Растворение и рефолдинг . Осадок (например, 1 грамм) суспендировали в 4 объемах (например, 4 мл) буфера для растворения: 1 M мочевина/300 мМ аргинин, 10 мМ Трис или CHES, 5 мМ EDTA, pH 11, в конечной концентрации (4 л/кг осадка). Суспензию тщательно перемешивали в течение 1-2 часов при комнатной температуре (15-30°C). Рефолдинг инициировали добавлением 3 объемов (1:4 об/об) буфера на объем буфера для растворения, что приводит к конечной концентрации буфера для рефолдинга в виде 1 M мочевины, 15 мМ цистеина, 0,5-2 мМ DTT, 100 мМ аргинина, 10 мМ Трис или CHES, 5 мМ EDTA, pH 9-10. Смесь перемешивали с добавлением воздуха или кислорода при постоянном коэффициенте волюметрического массообмена kLa (например, для барботирования воздухом во флаконе на 2,5 л kLa = от 0,004 до 0,01 мин-1, скорость барботирования составляет 0,3-3 куб. см/мин/л, и скорость перемешивания составляет 200-400 об/мин) в течение 6-24 часов при комнатной температуре. Смотри фиг. 1 для рефолдинга с течением времени. Необязательно VEGF можно стабилизировать в буфере для рефолдинга добавлением азота (например, 0,3-3 куб. см/мин/л для емкости на 2,5 л) вместо воздуха после 6 часов. Смотри фиг. 7. Фолдинг контролировали по SDS-PAGE, катионообменной хроматографии HPLC и rpHPLC и/или HPLC с гепарином.
Значительное уменьшение объема для процесса (5-кратное) при сохранении выхода выделенного димера VEGF наблюдали при рефолдинге в буфере с высоким pH, содержащим умеренные количества денатурирующих веществ и восстановителей. Ожидается, что этот способ применим при рефолдинге другого рекомбинантного белка, например других факторов роста.
ПРИМЕР II: Одностадийное растворение и рефолдинг рекомбинантного VEGF человека, экспрессированного в Escherichia coli
Растворение и рефолдинг . Осадок суспендируют в объемах 10-39 литров буфера для рефолдинга (в этом случае, обозначенном как «комбинированный буферный раствор») на каждый кг клеточного осадка, где комбинированный буферный раствор содержит 1 M мочевину, 15 мМ цистеин, 0,5 или 2 мМ DTT, 100 мМ аргинин, 10 мМ Трис или CHES, 5 мМ EDTA, pH 9,5-10,5, в конечной концентрации. Смотри фиг. 6 для эффекта добавления мочевины и аргинина в буферный раствор для рефолдинга. На фиг. 6 приведены результаты 1-стадийного рефолдинга осадка (комбинированный буферный раствор), как описано в этом примере, при pH 9,5 в течение 15 часов при комнатной температуре. Концентрации денатурирующего вещества варьируют, как следует далее: (1) 1 M мочевина и 100 мМ аргинин; (2) 1 M мочевина (и 0 мМ аргинин); (3) 2 M мочевина (и 0 мМ аргинин), в то время как все остальные компоненты буфера (например, Трис или CHES, DTT и т.д.) остаются в той же самой концентрации. Титр VEGF, экстрагированного из них, эквивалентен титру, определенному анализом с помощью катионообменной HPLC. На фиг. 6 показано, что профили rpHPLC сравнимы в присутствии или отсутствие аргинина.
Растворение и инкубацию для рефолдинга проводят при комнатной температуре в течение 3-24 часов и барботируют воздухом с коэффициентом массообмена kLa = от 0,004 до 0,1 мин-1 (например, для флакона на 2,5 л с импеллером морского типа, скорость барботирования воздухом может составлять от 0,3 до 10 куб. см/мин/л, или 0,3-1 куб. см/мин/л, или 1 куб. см/мин/л, или 25 куб. см/мин/л, в то время как перемешивание составляет 200-400 об/мин). Смотри фиг. 8 по поводу скоростей барботирования воздухом на условия рефолдинга. Например, осадок, содержащий VEGF, добавляли к комбинированному буферному раствору при pH 10 в соотношении 1:39 (кг осадка к л буфера). Готовили три реакционные емкости на 2,5 л и скорость перемешивания и скорость барботирования воздуха варьировали для каждой емкости до достижения kLa (a) 0,004, (b) 0,01, (c) 0,1 мин-1. Тестируемые скорости перемешивания в каждой емкости составляли 314 об/мин, в то время как диапазон для барботирования воздухом составлял от 1 куб. см/л/мин до 25 куб. см/л/мин. Реакции отслеживали с течением времени для выхода и качества продукта. Профили rpHPLC, представленные на фиг. 8, показывают сравнимое качество продукта для полученного свернутого VEGF после 12 час. Необязательно инкубацию можно проводить при комнатной температуре в течение вплоть до приблизительно 48 часов. Необязательно VEGF можно стабилизировать в буфере для рефолдинга добавлением азота вместо воздуха при той же самой скорости барботирования и перемешивания после 6 часов. Смотри фиг. 7, на которой представлены результаты по 2-стадийному рефолдингу осадка (как описано в примере I) в присутствии воздуха (например, kLa = 0,004 мин-1 или 0,3 куб. см/мин/л для емкости на 2,5 л), где на 6 часах пик мономера снижен (таким образом, указывая на то, что реакция рефолдинг по существу закончена). Барботирование воздухом заменяли на N2 (например, kLa = 0,004 мин-1) в течение вплоть до 48 часов. Как представлено на фиг. 7, материал остается стабильным, как показано на графиках rpHPLC. Фолдинг контролируют по SDS-PAGE, катионообменным хроматографическим колонкам HPLC и rpHPLC и/или HPLC с гепарином.
ПРИМЕР III: Безосадочный рефолдинг рекомбинантного белка
Массу целых клеток Escherichia coli, продуцирующих рекомбинантный белок, гомогенизировали в лабораторном гомогенизаторе Gaulin 15M модели 15 M (мелкомасштабный) или M3 (крупномасштабный) (Gaulin Corporation, Everett, MA) и разводили 1:4 (об/об) в буфере для рефолдинга на объем гомогената, барботировали воздухом с коэффициентом массообмена kLa = от 0,004 до 0,01 мин-1 (например, для флакона на 2,5 л с импеллером морского типа, скорость барботирования воздухом может составлять от 0,3 до 3 куб. см/мин/л, или 0,3-1 куб. см/мин/л, или 1 куб. см/мин/л, или 3 куб. см/мин/л, в то время как перемешивание составляет 200-400 об/мин). Буфер для рефолдинга содержит 1 M мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ Трис или CHES, 5 мМ EDTA, pH 9-10, в конечной концентрации. Инкубацию для рефолдинга проводили при комнатной температуре в течение 3-24 часов. Необязательно VEGF можно стабилизировать в буфере для рефолдинга путем добавления азота вместо воздуха после 3 часов инкубации для рефолдинга. Фолдинг контролировали по катионообменной хроматографии HPLC, rpHPLC и/или HPLC с гепарином.
ПРИМЕР IV: Очистка рекомбинантного VEGF человека (rhVEGF) после рефолдинга
Очистка: Пул для рефолдинга осветляют добавлением Тритона X-100 до конечной концентрации 1%, доведением pH до 9 и последующим центрифугированием (10000×g в течение 20 минут при 4°C. Супернатант затем фильтруют (объемный фильтр Cuno + 0,22 или 0,45 мкм мембранный фильтр) перед посадкой на смолу комбинированного типа (CaptoMMC™, GE Healthcare, Piscataway, NJ) при pH 9 и проводимости <10 мСм/см. Необязательно пул для рефолдинга разводят, по меньшей мере, 1:5 в равновесии с буфером для нанесения и затем фильтруют (объемный фильтр Cuno + 0,22 или 0,45 мкм мембранный фильтр) перед нанесением на смолу комбинированного типа (CaptoMMC™, GE Healthcare, Piscataway, NJ) при pH 9 и проводимости <10 мСм/см. Набитую колонку уравновешивают 25 мМ HEPES, pH 9, перед нанесением образца на колонку. VEGF элюируют с колонки MMC изократически 1 M аргинином/25 мМ HEPES, при pH 6-9 (например, pH 7,5). Смотри фиг. 2.
Пул с CaptoMMC™ доводят до pH 7,5 0,1 н. гидроксидом натрия и разводят WFI до проводимости 20 мСм/см перед нанесением на HP-колонку SP-Sepharose (уравновешивают 50 мМ HEPES, pH 7,5). VEGF элюируют с использованием линейного солевого градиента, образованного 50 мМ HEPES/0-1,2 M ацетата натрия, pH 7,5, 10-20 объемов колонки (например, 15 объемов колонки), и фракции собирают (1 объем колонки). Фракции с наибольшим поглощением A280 нм (макс. OD при ~42 мСм/см) в основном содержат >90% VEGF, и их объединяют для последующей обработки. Смотри фиг. 3.
Третья хроматографическая стадия включает гидрофобную смолу (например, Hi Propyl, J. T. Baker, Phenyl Sepharose Fast Flow (низкозамещенная), GE Healthcare, Piscataway, NJ). Пул с SP-Sepharose HP доводили до проводимости 50 мСм/см с использованием или ацетата натрия, или сульфата натрия перед нанесением на уравновешенную колонку (50 мМ HEPES, 1,2 M ацетат натрия, pH 7,5). Смотри фиг. 4. VEGF элюируется изократически в 50 мМ HEPES, pH 7,5, и пул анализируют на остающиеся примеси из клетки-хозяина и растворимые агрегаты. Фракции собирают и те, которые содержат правильно свернутый VEGF, как определено описанными здесь анализами, объединяют. Необязательно выполняют дополнительную хроматографическую стадию, например, с использованием второй гидрофобной смолы (например, Phenyl TSK) или ионообменной смолы.
Ультрафильтрация/диафильтрация. Объединенный VEGF можно ультрафильтровать на регенерированной целлюлозной мембране 5 кД на системе TFF лабораторного масштаба до концентрации 6 г/л (UF1). Образец диафильтруют с 7-14 DV (Diavolume) 5 мМ сукцинатом натрия через систему TFF до 10 г/л и получают композицию при 5 г/л для хранения при -80°C. Используемый буфер для композиции представляет собой 5 мМ сукцинат натрия/275 мМ дегидрат трегалозы/0,01% полисорбат 20/pH 5,0.
ПРИМЕР V: Очистка рекомбинантного VEGF человека (rhVEGF) после рефолдинга
Очистка: Пул рефолдинга осветляют добавлением Тритона X-100 до конечной концентрации 1%, доводят pH до 8,5-9,5 (например, pH 8,7) и оставляют при 25-30°C в течение от 1 до 10 часов перед центрифугированием. После обработки на центрифуге (10000×g в течение 20 минут при 4°C) для удаления больших плотных частиц, полученную жидкость (центрифугат) пропускают через ряд объемных фильтров и фильтры со стерильным барьером (мембрана 0,22 или 0,45 мкм) для удаления мелких частиц. rhVEGF затем собирали на смоле комбинированного типа (CaptoMMC™, GE Healthcare, Piscataway, NJ) при pH 8,7 и проводимостью <10 мСм/см. Набитую колонку уравновешивают 25 мМ CHES pH 8,7 перед нанесением образца на колонку. VEGF элюируют с колонки MMC изократически 0,9 M L-аргинином-HCl/25 мМ HEPES при pH 6-9 (например, pH 7,5).
Пул с CaptoMMC™ доводят до pH 7,5 0,1 н. гидроксидом натрия и разводят WFI до проводимости 20 мСм/см перед нанесением на колонку SP-Sepharose High Performance (уравновешивают 25 мМ HEPES pH 7,5). VEGF элюируют с использованием линейного солевого градиента, образованного 50 мМ HEPES/0-1,2 M ацетата натрия, pH 7,5, 10-20 объемов колонки (например, 15 объемов колонки), и фракции собирают (1 объем колонки). Фракции с наибольшим поглощением A280 нм (макс. OD при ~42 мСм/см) в основном содержат >90% VEGF, и их объединяют для последующей обработки. Третья хроматографическая стадия включает гидрофобную смолу (например, Hi Propyl, J. T. Baker, Phenyl Sepharose Fast Flow (низкозамещенная), GE Healthcare, Piscataway, NJ). Пул с SP-Sepharose HP доводили до проводимости 50 мСм/см с использованием или ацетата натрия или сульфата натрия перед нанесением на уравновешенную колонку (50 мМ HEPES, 1,2 M ацетат натрия, pH 7,5). Смотри фиг. 5. VEGF элюируется изократически в 50 мМ HEPES, pH 7,5, и пул анализируют на остающиеся примеси из клетки-хозяина и растворимые агрегаты. Фракции собирают и те, которые содержат правильно свернутый VEGF, как определено описанными здесь анализами, объединяют. Необязательно выполняют дополнительную хроматографическую стадию, например, с использованием второй гидрофобной смолы (например, Phenyl TSK) или ионообменной смолы.
Ультрафильтрация/диафильтрация. Объединенный VEGF можно ультрафильтровать на регенерированной целлюлозной мембране 5 кД в коммерческой системе TFF (2 кассеты Pellicon, Millipore, Billerica, MA) до концентрации 10 г/л, затем диафильтровать 7-14 диаобъемами (например, 10 DV) в буфер для композиции. Конечная обработка дает раствор, содержащий 5 г/л VEGF в 5 мМ сукцинате натрия/275 мМ дегидрате трегалозы/0,01% полисорбате 20/pH 5,0, который можно хранить при -80°C.
ПРИМЕР VI: Анализы для определения свернутого и/или очищенного рекомбинантного белка
В описанных здесь способах и процессах конечную чистоту и/или активность можно оценить пептидным картированием, дисульфидным картированием, SDS-PAGE (как восстановительным, так и невосстановительным), круговым дихроизмом, лизатом амебоцитов Limulus (LAL), катионообменной HPLC, HPLC с гепарином (например, HPLC с гепарином можно использовать для определения концентрации димера VEGF и уровня неправильно свернутых форм), обращенно-фазной (rp) хроматографией HPLC (например, rpHPLC востановленных образцов можно использовать для определения общей концентрации VEGF, в то время как rpHPLC нативных образцов может оценить качество повторно свернутого VEGF), связыванием рецептора (например, для VEGF, например, связыванием рецептора KDR-Bioanalytic R&D и/или связыванием Flt1-рецептора), анализом SEC, клеточными анализами, анализами активности HUVEC, с помощью ELISAs с антителами к VEGF, масс-спектрометрическим анализом и т.д.
Анализ определения общей экспрессии VEGF
(1) rpHPLC восстановленных образцов - Качество экспрессированного VEGF измеряют с использованием анализа посредством обращенно-фазной HPLC на колонке C18 (колонка Jupiter C18, 4,6×250 мм, 5 микрон, Phenomenex, Torrance, CA). Колонку уравновешивают 0,22% трифторуксусной кислотой и элюируют с использованием линейного градиента от 25% до 45% ацетонитрила, содержащего 0,2% трифторуксусной кислоты, за 30 мин со скоростью потока 1 мл/мин. Элюант контролируют при 280 нм. Образец обрабатывают и полностью восстанавливают гуанидином и DTT перед инжекцией. Восстановленный белок VEGF элюирует приблизительно 26 мин, и область пика используют для подсчета количества общего VEGF в образце по известной стандартной кривой.
Анализы повторно свернутого VEGF
(1) Анализ с помощью катионообменной HPLC - Количество правильно повторно свернутых димеров VEGF определяли с использованием аналитической катионообменной колонки, например колонки SP-5PW (гель TSK SP-5PW, 7,5×75 мм, 10 микрон, Tosoh Biosciences LLC, Japan). Колонку уравновешивают 50 мМ фосфатом натрия, pH 7,5. При скорости потока 1 мл/мин колонку элюируют с использованием линейного градиента от 0 до 2 M хлорида натрия в уравновешивающем буфере в течение 60 мин. Элюант контролируют при 280 нм или 214 нм. В основном большую часть белка элюируют в первые 30 мин и VEGF элюируют приблизительно 40 мин. Смотри фиг. 9.
(2) Анализ rp-HPLC - Качество правильно повторно свернутого VEGF определяют с использованием колонки Zorbax 300SB-C8 (4,6×150 мм, 3,5 микрон, Agilent Technologies, Santa Clara, CA). Колонку уравновешивают 0,1% трифторуксусной кислотой и элюируют с использованием линейного градиента от 0 до 50% ацетонитрила, содержащего 0,08% трифторуксусной кислоты, в течение 50 мин со скоростью потока 1 мл/мин. Элюант контролируют при 214 нм. В основном VEGF элюирует приблизительно 35 мин, и профиль пика оценивают для процентного содержания переднего фронта гидрофобных форм относительно основного пика. Несвернутый мономер VEGF элюирует на 2-3 мин позднее.
(3) Гепаринсвязывающий анализ с помощью HPLC - Качество и количество правильно повторно свернутого VEGF определяют с использованием колонки, содержащей иммобилизованный гепарин. Колонку Heparin-5PW (7,5×75 мм, 10 микрон, гель TSK, Tosoh Biosciences LLC, Japan) уравновешивают 10 мМ фосфатом натрия, pH 7,4, содержащим 0,15 M хлорида натрия. Со скоростью потока 1 мл/мин колонку элюируют с использованием линейного градиента от 0,15 M до 1,6 M хлорида натрия в уравновешивающем буфере в течение 20 мин. В некоторых анализах элюцию осуществляют в течение 16 мин. Элюант контролируют при 280 нм. В основном большая часть белка элюирует в пустом объеме, и можно идентифицировать 3 класса VEGF. Наиболее высокоаффинные, позднее всего элюирующие формы идентифицируют как правильно свернутый VEGF и иногда идентифицируют как «пик 3 VEGF».
Понятно, что депонируемые материалы, примеры и варианты осуществления, описанные здесь, приведены только с иллюстративными целями и что их разнообразные модификации или небольшие изменения будут предложены специалистам в данной области и их следует относить к сущности и сфере действия этой заявки и объему приложенной формулы изобретения. Все цитированные здесь публикации, цитирования, патенты и патентные заявки приведены здесь в качестве ссылки в полном объеме для всех целей.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СВЯЗЫВАЮЩИХ ГЕПАРИН БЕЛКОВ | 2006 |
|
RU2385877C1 |
| СПОСОБ ПРОМЫШЛЕННОЙ ОЧИСТКИ РОМИПЛОСТИМА | 2021 |
|
RU2804622C2 |
| СПОСОБЫ ОЧИСТКИ БЕЛКОВ, СОДЕРЖАЩИХ ОБЛАСТЬ Fc | 2006 |
|
RU2415865C2 |
| НОВЫЙ СПОСОБ КЛОНИРОВАНИЯ, ЭКСПРЕССИИ И ОЧИСТКИ ДЛЯ ПОЛУЧЕНИЯ РАНИБИЗУМАБА | 2014 |
|
RU2646098C2 |
| ОПТИМИЗИРОВАННЫЙ СПОСОБ ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ФАКТОРА РОСТА | 2008 |
|
RU2491290C2 |
| АГЕНТ ДЛЯ ИСПОЛЬЗОВАНИЯ ПРИ ЛЕЧЕНИИ ИЛИ ПРОФИЛАКТИКЕ ОФТАЛЬМОЛОГИЧЕСКИХ РАССТРОЙСТВ | 2020 |
|
RU2812055C1 |
| СПОСОБЫ ОЧИСТКИ АНТИТЕЛ | 2017 |
|
RU2759909C2 |
| СПОСОБ ОЧИСТКИ АРО-2 LIGAND/TRAIL ПОСРЕДСТВОМ КРИСТАЛЛИЗАЦИИ НА ХОЛОДЕ | 2006 |
|
RU2435781C2 |
| ОЧИСТКА АНТИТЕЛ С ПОМОЩЬЮ КАТИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2008 |
|
RU2498991C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2824045C2 |

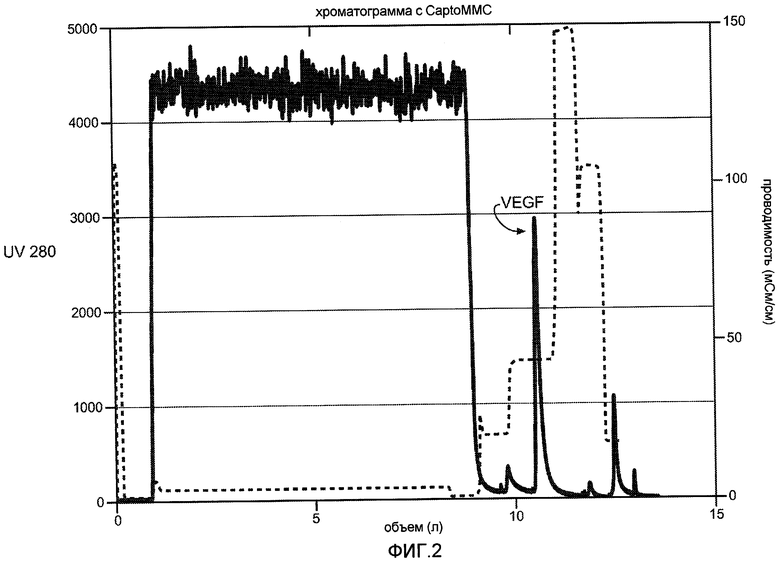
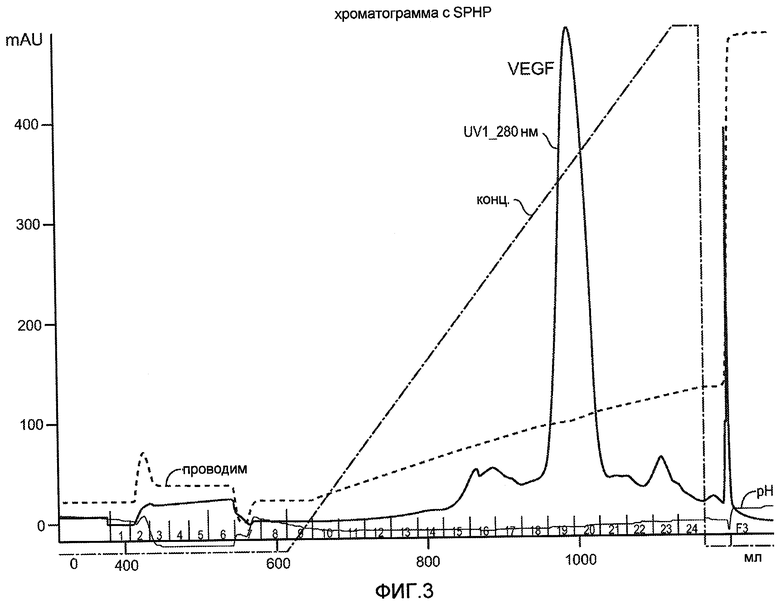
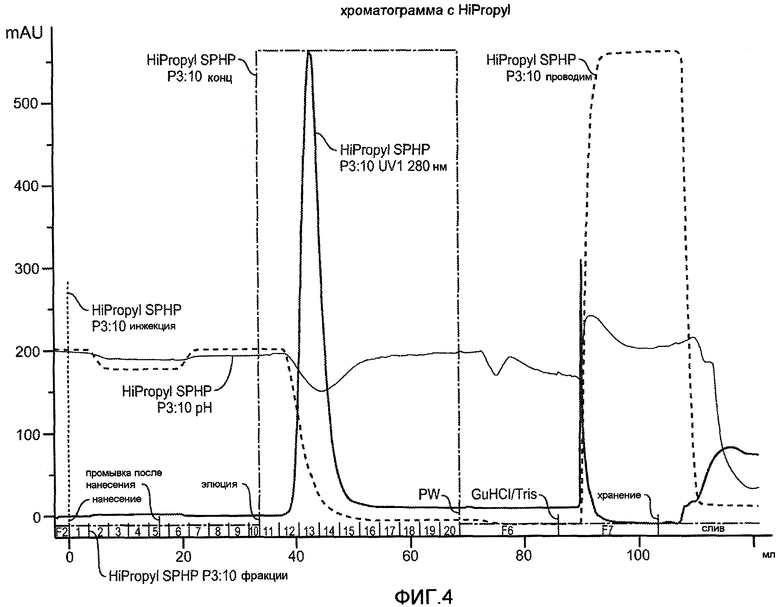
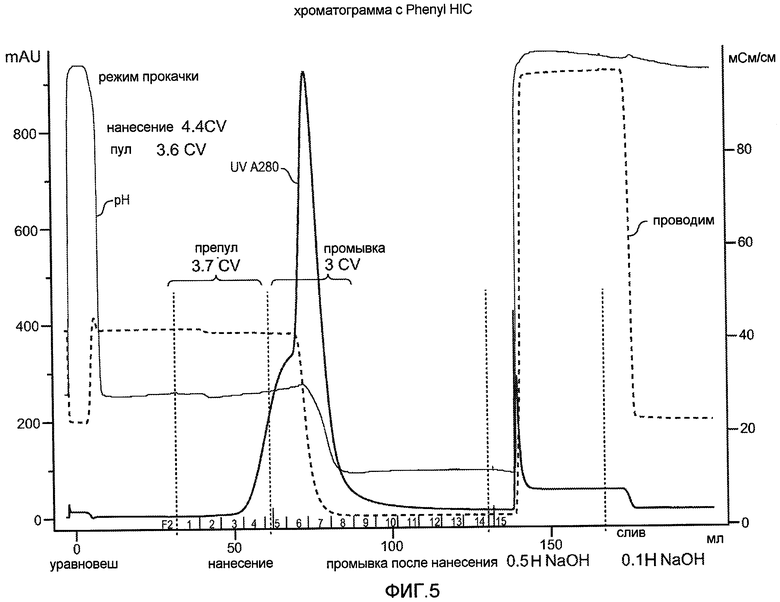
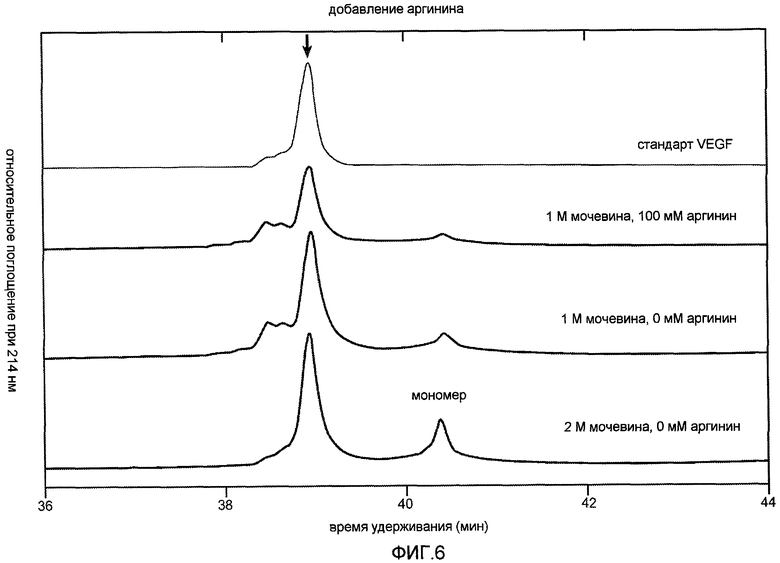
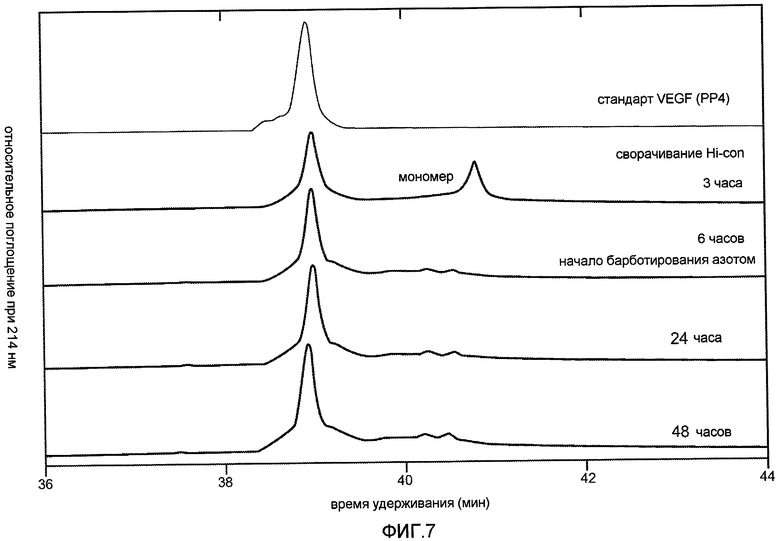
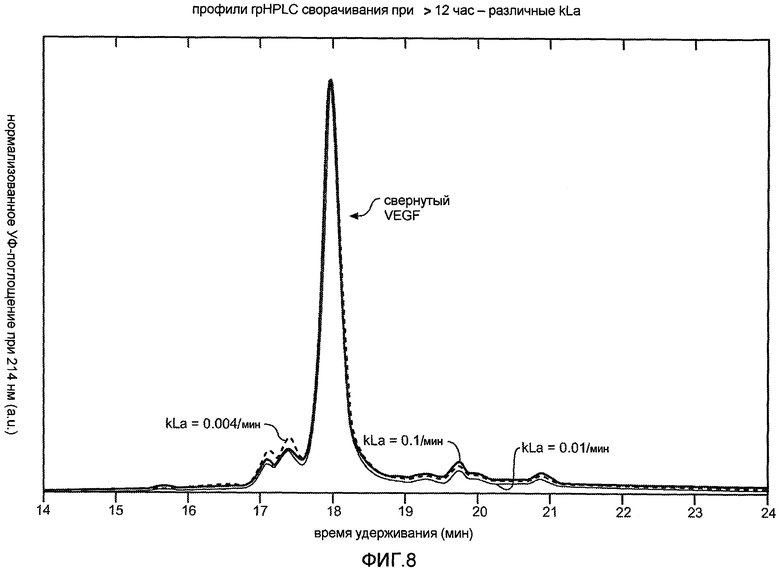
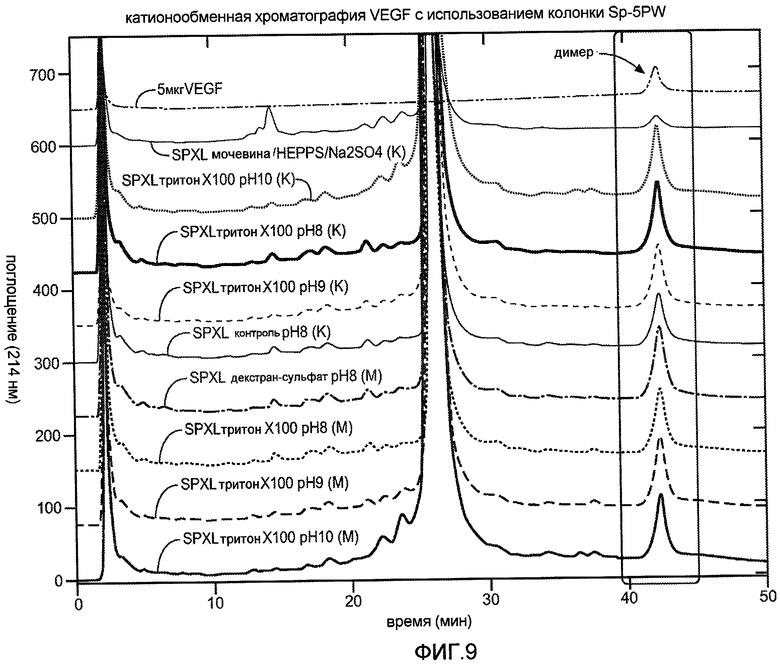
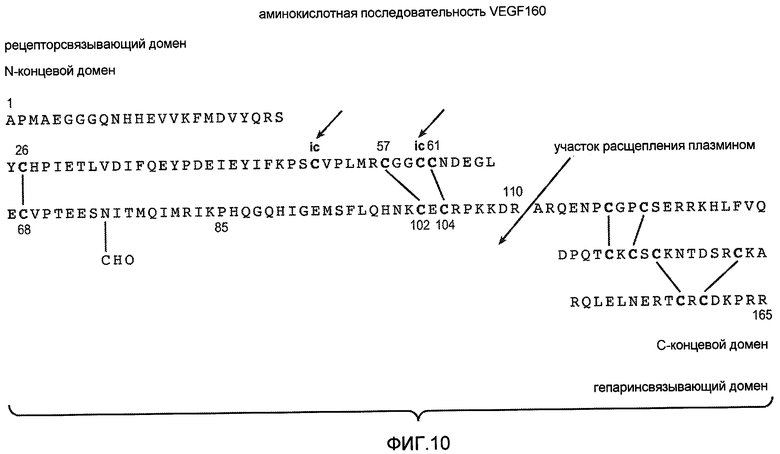
Изобретение относится к области биотехнологии и может быть использовано для выделения рекомбинантного фактора роста эндотелия сосудов (VEGF) из культуры прокариотических клеток. Проводят стадии выделения рекомбинантного белка VEGF из культуры клеток и растворения в первом буферном растворе, рН более 9, который содержит в конечной концентрации: 1 М мочевину, 300 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 11, или 1 М мочевину, 300 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, рН 11. Затем осуществляют рефолдинг растворенного белка во втором буферном растворе с добавлением воздуха, рН>9, но ≤11, который содержит в конечной концентрации: 1 М мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 10, или 1 М мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, рН 10, и выделяют повторно свернутый рекомбинантный белок. Как вариант возможно проведение растворения и рефолдинга указанного белка в комбинированном буферном растворе, рН>9, но ≤11, с добавлением воздуха или кислорода, где комбинированный буферный раствор содержит в конечной концентрации: 1 М мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 10, или 1 М мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, рН 10. Изобретение позволяет выделять повторно свернутый рекомбинантный белок VEGF из культуры прокариотических клеток. 2 н. и 14 з. п. ф-лы, 10 ил.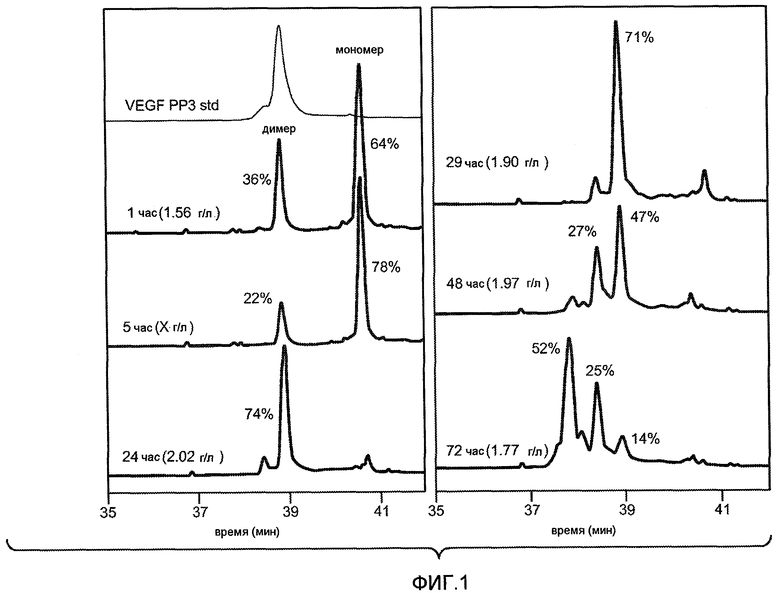
1. Способ выделения повторно свернутого рекомбинантного белка из культуры прокариотических клеток, причем способ включает стадии:
(a) выделения рекомбинантного белка из культуры прокариотических клеток, где рекомбинантный белок представляет собой фактор роста эндотелия сосудов (VEGF);
(b) растворения указанного белка в первом буферном растворе, рН более 9;
где первый буферный раствор содержит
в конечной концентрации 1 М мочевину, 300 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 11, или
в конечной концентрации 1 М мочевину, 300 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, pH 11;
(c) рефолдинга указанного растворенного белка во втором буферном растворе, рН>9, но<11, где второй буферный раствор содержит в конечной концентрации 1 М мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 10, или
в конечной концентрации 1 М мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, рН 10, и добавления воздуха или кислорода в течение такого времени и при таких условиях, при которых происходит рефолдинг рекомбинантного белка; и
(d) выделения указанного повторно свернутого рекомбинантного белка.
2. Способ по п.1, где VEGF представляет собой VEGF165.
3. Способ по п.1, где рекомбинантный белок инкубируют в первом буферном растворе в течение, по меньшей мере, 1 ч.
4. Способ по п.3, где инкубацию выполняют при 2-40°С.
5. Способ по п.1, где растворенный белок инкубируют во втором буферном растворе в течение приблизительно 3-24 ч.
6. Способ по п.5, где инкубацию выполняют при 2-40°С.
7. Способ по п.1, где добавление воздуха или кислорода обеспечивают при kLa=от 0,001 до 0,1 мин-1.
8. Способ по п.1, дополнительно включающий стабилизацию повторно свернутого рекомбинантного белка добавлением азота.
9. Способ по п.1, где стадия выделения включает осветление второго буферного раствора с рекомбинантным белком и последовательное приведение указанного повторно свернутого рекомбинантного белка в контакт с хроматографическим носителем комбинированного типа, катионным хроматографическим носителем и первым гидрофобным хроматографическим носителем, и селективное элюирование повторно свернутого рекомбинантного белка с каждого носителя.
10. Способ выделения повторно свернутого рекомбинантного белка из культуры прокариотических клеток, причем способ включает стадии:
(a) выделения рекомбинантного белка из культуры прокариотических клеток, где рекомбинантный белок представляет собой фактор роста эндотелия сосудов (VEGF);
(b) растворения и рефолдинга указанного белка в комбинированном буферном растворе, рН>9, но<11, с добавлением воздуха или кислорода, где комбинированный буферный раствор содержит
в конечной концентрации 1 М мочевину, 15 мМ цистеин, 2 мМ DTT, 100 мМ аргинин, 10 мМ CHES, 5 мМ EDTA, рН 10, или
в конечной концентрации 1 М мочевину, 15 мМ цистеин, 0,5-2 мМ DTT, 100 мМ аргинин, 10 мМ ТРИС, 5 мМ EDTA, рН 10; и
(c) выделения указанного повторно свернутого рекомбинантного белка.
11. Способ по п.10, где стадия выделения включает осветление комбинированного раствора с рекомбинантным белком и последовательное приведение указанного повторно свернутого рекомбинантного белка в контакт с хроматографическим носителем комбинированного типа, катионным хроматографическим носителем и первым гидрофобным хроматографическим носителем и селективное элюирование повторно свернутого рекомбинантного белка с каждого носителя.
12. Способ по п.11, где стадия осветления включает: добавление детергента до конечной концентрации 1%, доведение рН до приблизительно 8,5-9,5, инкубирование раствора в течение 1-10 ч при 25-30°С, центрифугирование раствора и фильтрование жидкости, полученной со стадии центрифугирования.
13. Способ по п.10, где рекомбинантный белок инкубируют в комбинированном буферном растворе в течение приблизительно 3-24 ч.
14. Способ по п.13, где инкубацию выполняют при 2-40°С.
15. Способ по п.11, дополнительно включающий приведение указанного повторно свернутого рекомбинантного белка в контакт со вторым гидрофобным хроматографическим носителем или ионообменным носителем и селективное элюирование повторно свернутого рекомбинантного белка с носителя.
16. Способ по п.10 или 15, где указанные первый и второй хроматографические носители для гидрофобного взаимодействия выбраны из группы, состоящей из бутил-, пропил-, октил-, фенил- и арилагарозных смол.
| Многоканальный спектрометр | 1987 |
|
SU1449848A1 |
| СПОСОБ МИНИМИЗАЦИИ ДЕГРАДАЦИИ АКТИВИРОВАННОГО ПРОТЕИНА С (ВАРИАНТЫ), СТАБИЛИЗИРОВАННАЯ КОМПОЗИЦИЯ АКТИВИРОВАННОГО ПРОТЕИНА С | 1995 |
|
RU2167936C2 |
| 0 |
|
SU155174A1 | |
| US 5410026 A, 25.04.1995 | |||
| ®С?СОЮЗНА'1 | 0 |
|
SU373325A1 |
| Литьевая форма с двухступенчатой выталкивающей системой | 1976 |
|
SU596459A1 |
| WO 9942486 A1, 26.08.1999 | |||
| US 4929700 A, 29.05.1990. | |||

