Изобретение относится к микробиологической промышленности и может быть использовано для увеличения продуктивности грибов Rhizopus oryzae по целевому продукту.
Мицелиальные грибы рода Rhizopus, относящиеся к семейству Мукоровых, являются важным биотехнологическим продуцентом ряда карбоновых кислот и ферментов, среди которых можно выделить ферменты: липазу и амилазу, а также кислоты: молочную и фумаровую [Biosci Biotechnol Biochem, 67 (8): 1725-1731, 2003; Can J Microbiol, 30: 126-128, 1984].
Одним из основных подходов к увеличению продуктивности штаммов являются методы генной инженерии, когда в микроорганизм в составе соответствующего экспрессионного вектора вводят дополнительные копии структурного гена, кодирующего целевой продукт. Однако необходимым условием для осуществления подобной генетической модификации является принципиальная возможность получения митотически стабильных трансформантов, что затруднительно как для грибов рода Rhizopus, так и для мукоровых грибов в целом (Mol Gen Genomics, 268: 397-406, 2002;).
Определения
1. Трансформант считается митотически стабильным (далее: стабильный трансформант), если при прохождении споруляции он не теряет введенный генетический материал. Величина митотической стабильности определяется отношением числа потомков трансформантов, сохранивших введенный генетический материал после споруляции, к общему числу полученных потомков и рассчитывается как отношение числа колоний, выросших на селективной среде, к числу колоний, выросших на неселективной среде, выраженное в процентах.
2. Ген pyrG - ген, кодирующий фермент оротидин-5′-фосфатдекарбоксилазу. Фермент принимает участие в синтезе урацила и его недостаточность ведет к ауксотрофности по урацилу. Ауксотрофность по урацилу может быть компенсирована добавлением в среду урацила или уридина.
3. Автономно реплицирующийся конкатамер - не встроенная в хромосому полимерная кольцевая или линейная структура, содержащая большое число копий плазмиды.
4. Геномные повторы (повторяющиеся геномные последовательности) - последовательности, представленные в геноме в некотором количестве копий, имеющих высокую гомологию между собой.
5. Тандемные повторы - повторы, расположенные друг за другом в прямой ориентации.
6. Гомокарион - организм, в котором все ядра несут одинаковую генетическую информацию.
7. Интегративный трансформант - трансформант, у которого введенный генетический материал интегрирован в состав генома.
8. Грибы вида Rhizopus oryzae - грибы рода Rhizopus, отнесенные к виду oryzae по классификации М. A.A.Schipper A revision of the genus Rhizopus Studies in Mycology, No.25 1 June 1984.
В качестве возможных причин низкой эффективности встраивания чужеродной ДНК в геном Rhizopus приводят (Mol Gen Genomics, 268: 397-406, 2002; Mol Gen Genomics, 271: 499-510, 2004), в частности, низкую частоту гомологичной рекомбинации или наличие модифицированных оснований в собственной ДНК, позволяющих отличать ее от гетерологичной.
Число работ, описывающих успешные эксперименты по стабильной трансформации грибов рода Rhizopus, довольно ограничено, при этом до настоящего времени не известна эффективная система трансформации и векторов, позволяющих воспроизводимо получать интегративные трансформанты.
В работе (Mol Gen Genomics, 271: 499-510, 2004) для введения ДНК в протопласты Rhizopus oryzae использована система агробактериальной трансформации, в результате которой у всех полученных стабильных трансформантов в хромосому интегрировала лишь последовательность маркера, но не вектора, то есть система оказалась не пригодной для клонирования рекомбинантной ДНК.
В работе (Curr Genet, 45: 302-310, 2004) приведена система трансформации, где штамм грибов Rhizopus oryzae, содержащий в качестве селективного маркера мутантный ген pyrG, трансформирован плазмидой, включающей тот же селективный маркер - инактивированный ген pyrG, но содержащий мутацию, локализованную в другой части гена. Предполагалось, что в результате рекомбинации в области ДНК, расположенной между этими мутациями, будет получен стабильный интегративный трансформант с одной активной и одной неактивной копией маркера. Однако большая часть полученных трансформантов содержала автономно реплицирующийся конкатамер с исправленной копией гена pyrG и, следовательно, была нестабильна.
Задача заявляемого изобретения состоит в создании генетической конструкции для получения стабильных трансформантов грибов Rhizopus oryzae.
Поставленная задача решена путем разработки генетической конструкции, содержащей селективный генетический маркер и нуклеотидную последовательность, содержащую последовательность, приведенную в списке последовательностей под номером SEQ ID NO:1 или SEQ ID NO:2, или последовательность, гомологичную одной из указанных не менее чем на 85%.
Кроме указанных элементов, конструкция, как правило, содержит генетические элементы, необходимые для ее поддержания и распространения в других организмах, в частности в Escherichia coli (E.coli).
Дополнительно, заявляемая генетическая конструкция может содержать генетические элементы, обеспечивающие экспрессию гетерологичных генов либо повышение или снижение уровня экспрессии гомологичных генов.
Известно, что последовательности, имеющие большое число гомологичных копий в геноме, используют в составе вектора для получения многокопийных интегративных трансформантов при работе с аскомицетовыми дрожжами и грибами. При этом как рибосомальные гены rDNA (US 5786212, US 609057), так и фрагменты транспозонов (US 5629203, US 6582951) используют с целью увеличения числа копий вектора в геноме, а следовательно, для увеличения уровня экспрессии встроенного в вектор целевого гена.
При решении поставленной задачи мы предположили, что использование при трансформации в составе генетической конструкции фрагмента ДНК с геномной последовательностью, имеющей в геноме большое число высокогомологичных копий, может повысить вероятность возникновения рекомбинационного комплекса и, как следствие, обеспечить встраивание конструкции в геном реципиента, а следовательно, возникновение стабильных интегративных трансформантов.
Принципиальным отличием заявляемого изобретения является то, что повторяющиеся генетические элементы используют не для увеличения числа копий вектора в геноме, а для получения стабильных трансформантов, в отличие от нестабильных трансформантов, как правило, образующихся в процессе трансформации грибов Rhizopus oryzae.
Для иллюстрации изобретения и получения митотически стабильных трансформантов в качестве генетического маркера использован ген pyrG, а в качестве повторяющейсяся геномной последовательности - последовательность SEQ1, указанная в перечне последовательностей под номером SEQ ID NO:1, и последовательность SEQ2, указанная в перечне последовательностей под номером SEQ ID NO:2.
Последовательность SEQ1 в геноме штамма Rhizopus oryzae RA 99-880 (http://www.broad.mit.edu/annotation/fungi/fgi/) имеет 26 гомологичных участков, степень ее гомологии к которым варьирует от 96% до 97%. Последовательность SEQ2 в этом геноме имеет более 100 гомологичных участков, степень гомологии с которыми варьирует от 94% до 96%.
Последовательность SEQ1 длиной 2053 п.н. представляет собой фрагмент открытой рамки считывания ретротранспозона, а последовательность SEQ2 имеет длину 575 п.н. и, возможно, также представляет собой фрагмент мобильного элемента, хотя соответствующей рамки считывания вблизи этого элемента в предварительном исследовании не обнаружено.
Изобретение проиллюстрировано следующими фигурами:
Фиг.1. Схема плазмиды pPyrG-SEQ1.
Фиг.2. Схема плазмиды pPyrG-SEQ2.
Фиг.3. Гибридизация по Саузерну геномной ДНК трансформантов плазмидами pPyrG-SEQ1 и pPyrG-SEQ2.
Фиг.4. Гибридизация по Саузерну геномной ДНК трансформантов плазмидой pPyrG-SEQ1 после десяти раундов споруляции на неселективной среде.
Фиг.5. Гибридизация по Саузерну геномной ДНК ряда штаммов рода Rhizopus с последовательностью SEQ1 в качестве зонда.
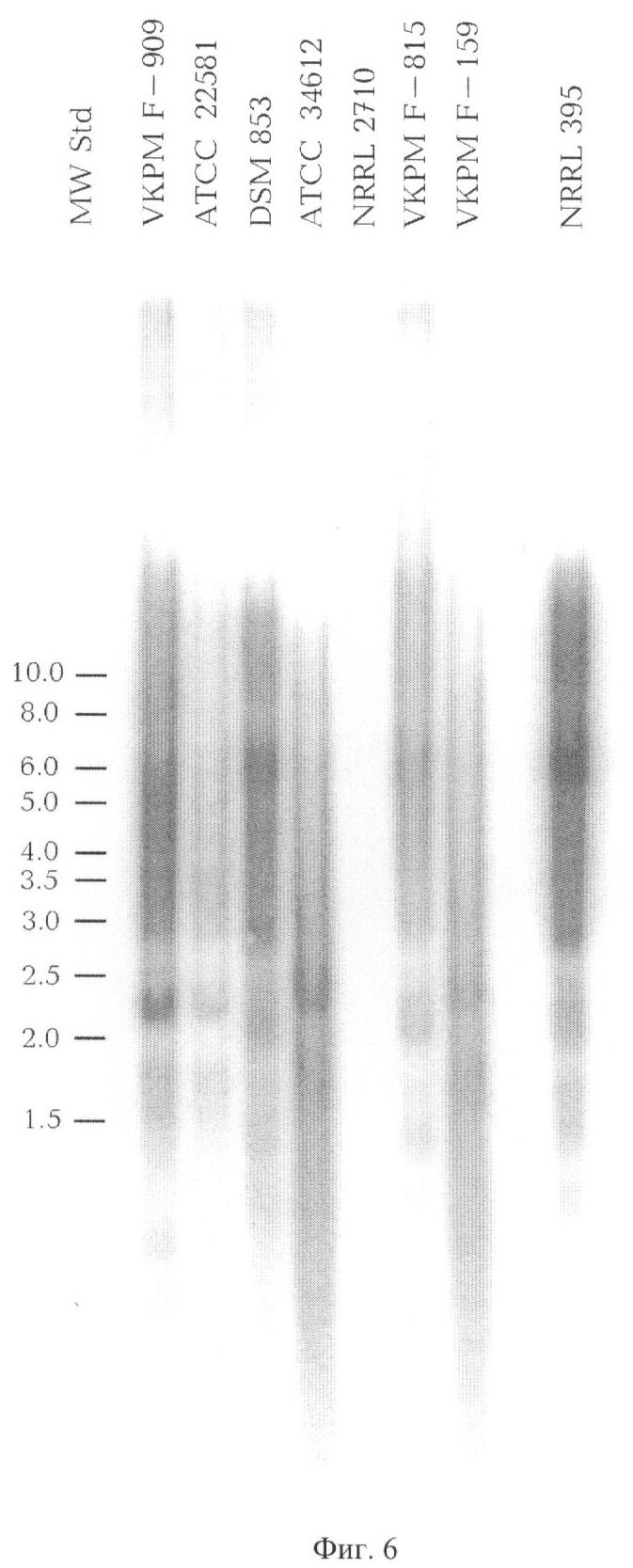
Фиг.6. Гибридизация по Саузерну геномной ДНК ряда штаммов рода Rhizopus с последовательностью SEQ2 в качестве зонда.
Пример 1: Получение плазмиды pPyrG
Фрагмент ДНК размером 2268 п.н., кодирующий аминокислотную последовательность, промоторную и терминаторную области гена pyrG Rhizopus oryzae, получают при помощи ПЦР с использованием Pfu-полимеразы (Fermentas Inc.) и праймеров: CCCGGGGAATTCTTCTTTTAGGTTAAAAACTC (PyrG225-F) и GGATCCGAATTCCAAAGCTTTTCATATATTG (PyrG225-R) (все используемые в работе праймеры синтезированы фирмой «Синтол», Москва).
В качестве матрицы для ПЦР используют геномную ДНК штамма Rhizopus oryzae NRRL 395, полученную по методу, описанному в (Molecular cloning: a laboratory manual (2nd edn). Cold Spring Harbor Laboratory Press, 1989).
Полученный фрагмент амплифицированной ДНК очищают после электрофореза в 1% агарозном геле методом экстракции ДНК (Kit #K0513, Fermentas Inc.).
0,5 мкг полученной ДНК лигируют с 0,2 мкг ДНК вектора pUC19 (Fermentas Inc.), обработанного эндонуклеазой рестрикции SmaI, и трансформируют в Е.coli XL 1-Blue. Клоны, содержащие необходимую вставку амплифицированной ДНК размером 2268 п.н., отбирают на чашках по устойчивости к ампициллину и стандартному тесту на отсутствие активности β-галактозидазы. Плазмидную ДНК, выделенную из полученных клонов, проверяют методом рестрикционного анализа.
Полученная плазмида размером 4954 п.н. названа pPyrG.
Пример 2: Получение плазмиды pPyrG-SEQ1
Фрагмент ДНК размером 2065 п.н., содержащий последовательность SEQ1, получают при помощи ПЦР с использованием Pfu-полимеразы и праймеров: GCTAGCGTTGAAACGTCTTACTGCTGC (HF-1) и GCTAGCTGCTTGAATCTCATAATGTTGG (HR-1). В качестве матрицы для ПЦР используют геномную ДНК штамма Rhizopus oryzae NRRL 395.
Полученный фрагмент амплифицированной ДНК очищают после электрофореза в 1% агарозном геле методом экстракции ДНК.
0,5 мкг полученной ДНК лигируют с 0,2 мкг ДНК вектора pUC19, обработанного эндонуклеазой рестрикции Ecl136II и трансформируют в Е.coli XL1-Blue. Клоны, содержащие необходимую вставку амплифицированной ДНК размером 2065 п.н., отбирают на чашках по устойчивости к ампициллину и стандартному тесту на отсутствие активности β-галактозидазы. Плазмидную ДНК, выделенную из полученных клонов, анализируют методом рестрикции. Последовательность SEQ1, входящую в состав плазмиды, секвенируют.
Полученная плазмида размером 4751 п.н. названа pUC-SEQ1.
0,4 мкг фрагмента ДНК SEQ1 размером 2059 п.н., полученного после обработки плазмиды pUC-SEQ1 эндонуклеазой NheI и очистки после электрофореза в 1% агарозном геле, лигируют с 0,2 мкг ДНК плазмиды pPyrG, обработанной эндонуклеазой рестрикции XbaI и щелочной фосфатазой CIAP (Fermentas Inc.). Лигазную смесь далее трансформируют в Е.coli XL1-Blue. Плазмидную ДНК, выделенную из полученных клонов, проверяют рестрикционным анализом.
Полученная плазмида содержит ген pyrG Rhizopus oryzae и последовательность SEQ1, имеет размер 7013 п.н. и названа pPyrG-SEQ1 (фиг.1).
Пример 3: Получение плазмиды pPyrG-SEQ2
Фрагмент ДНК размером 589 п.н., содержащий последовательность SEQ2, получают при помощи ПЦР с использованием Pfu-полимеразы и праймеров: TGAGCTCTGTTGTAGAACAGAGTCTGAT (ZF-1) и TGAGCTCTGTTATGTTCTGGTAAAGTAAAAG (ZR-1). В качестве матрицы для ПЦР используют геномную ДНК штамма Rhizopus oryzae NRRL 395.
Полученный фрагмент амплифицированной ДНК очищают после электрофореза в 1% агарозном геле методом экстракции ДНК и обрабатывают эндонуклеазой рестрикции SacI.
0,5 мкг полученной ДНК лигируют с 0,2 мкг ДНК вектора pUC19, также обработанного эндонуклеазой рестрикции SacI, и трансформируют в Е.coli XL1-Blue. Клоны, содержащие необходимую вставку амплифицированной ДНК размером 581 п.н., отбирают на чашках по устойчивости к ампициллину и стандартному тесту на отсутствие активности β-галактозидазы. Плазмидную ДНК, выделенную из полученных клонов, анализируют методом рестрикции. Последовательность SEQ2, входящую в состав плазмиды, секвенируют.
Полученная плазмида размером 3267 п.н. названа pUC-SEQ2.
0,4 мкг фрагмента ДНК SEQ2 размером 581 п.н., полученного после обработки плазмиды pUC-SEQ2 эндонуклеазой SacI и очистки после электрофореза в 1% агарозном геле, лигируют с 0,2 мкг ДНК плазмиды pPyrG, обработанной эндонуклеазой рестрикции SacI и щелочной фосфатазой CIAP. Лигазную смесь далее трансформируют в Е.coli XL1-Blue. Плазмидную ДНК, выделенную из полученных клонов, анализируют методом рестрикции.
Полученная плазмида содержит ген pyrG гриба Rhizopus oryzae и последовательность SEQ2, имеет размер 5535 п.н. и названа pPyrG-SEQ2 (фиг.2).
Пример 4. Трансформация штамма Rhizopus oryzae PYR-17 плазмидой pPyrG-SEQ1, отбор стабильных клонов
Для трансформации используют штамм Rhizopus oryzae PYR-17, полученный из штамма Rhizopus oryzae NRRL 395 путем химического мутагенеза и последующей селекци на среде с 5-фтороротовой кислотой, и содержащий мутацию в гене pyrG (Mol Gen Genomics, 268: 397-406, 2002).
Трансформацию спор штамма Rhizopus oryzae PYR-17 проводят методом бомбардировки микрочастицами (Mol Genet Genomics, 268: 397-406, 2002). В качестве трансформирующей ДНК используют кольцевую ДНК плазмид pPyrG-SEQ1. В качестве контроля используют кольцевую ДНК плазмиды pPyrG. Здесь и далее культивирование штаммов рода Rhizopus осуществляют при 30°С.
Селекцию трансформантов ведут по комплементации ауксотрофности по урацилу на агаризованной минимальной среде RZ (Appl Environ Microbiol, 66 (6): 2343-2348, 2000) с добавлением глюкозы (2 мас.%).
Трансформанты инкубируют 5-7 дней до полной споруляции.
Смесь спор трансформантов с каждой чашки в различных разведениях повторно высевают на среду RZ с добавлением глюкозы (2 мас.%) и Triton-Х100 (0,01 мас.%). Процедуру споруляции и последующего высева повторяют еще 2 раза с целью получения гомокариотичной культуры. На заключительном этапе селекции суспензию спор высевают в большом разведении. Отбор стабильных трансформантов осуществляют по более крупному размеру колоний (т.к. нестабильные трансформанты обладают меньшей скоростью роста). Выбранные крупные клоны, значительно превосходящие остальные в размере, пересевают на неселективную среду PDA (Himedia) с добавлением уридина (0,05 мас.%). Для получения независимых трансформантов споры, полученные с одной чашки после трансформации, используют для получения только одного трансформанта.
После споруляции на неселективной среде подтверждают митотическую стабильность трансформантов.
Для подтверждения митотической стабильности полученного трансформанта сравнивают количество колоний, выросших при высеве образованных им спор на минимальную среду RZ с добавлением Triton-X100 (0,01 мас.%) и глюкозы (2 мас.%), с количеством колоний, выросших на той же среде, но дополнительно содержащей уридин (0,05 мас.%).
Полученные результаты показали, что для всех исследованных трансформантов число клонов, выросших в течение 2-х суток на среде с уридином (неселективной среде) и на среде без уридина (селективной среде), отличалось незначительно, что свидетельствует о стабильности проанализированных трансформантов.
В качестве контроля для трансформации использована плазмида pPyrG, не содержащая повторяющихся генетических элементов.
В серии контрольных экспериментов крупных колоний не обнаружено, а типичные, медленно растущие трансформанты образуют после споруляции в селективных условиях незначительное количество прототрофных колоний, что свидетельствует об их нестабильности.
Пример 5: Определение стабильности трансформантов с плазмидой pPyrG-SEQ1 методом гибридизации
Для выделения геномной ДНК выращивают биомассу трех независимых трансформантов Т1-1, Т1-2 и Т1-3, полученных при введении плазмиды pPyrG-SEQ1 в штамм реципиента Rhizopus oryzae PYR-17, засевая спорами жидкую среду YPD следующего состава (мас.%): пептон - 0,5; дрожжевой экстракт - 0,3; глюкоза - 2; вода - остальное, с добавлением Tween-80 - 0,1 мас.%.
Геномную ДНК трансформантов и штамма-реципиента обрабатывают эндонуклеазой рестрикции Eco88I, сайт узнавания которой является уникальным для используемой плазмиды (фиг.1). После электрофоретического разделения в агарозном геле образцы используют для гибридизации по Саузерну (Molecular cloning: a laboratory manual (2nd edn). Cold Spring Harbor Laboratory Press, 1989).
В качестве зонда используют фрагмент ДНК длиной 718 п.н., представляющий собой часть промоторной области гена pyrG. Для его получения 0,5 мкг ДНК плазмиды pPyrG обрабатывают эндонуклеазой рестрикции EcoRI. Необходимый фрагмент выделяют после электрофореза в 1% агарозном геле методом экстракции ДНК.
На фиг.3 отображены результаты гибридизации геномной ДНК трансформантов T1-1, T1-2 и Т1-3, а также штамма реципиента (PYR-17). Для контроля величины фрагментов ДНК при электрофорезе использован молекулярный маркер GeneRuler 1 kb DNA ladder (Fermentas) (обозначен MW Std, размеры фрагментов указаны слева) и λ/EcoRI маркер (Fermentas) (обозначен λ/EcoRI, размер фрагмента указан справа). Из результатов гибридизации следует, что помимо хромосомальной копии гена pyrG (полоса около 4 т.п.н.), присутствующей у реципиента, у трансформантов наблюдается большое число дополнительных копий, основная масса которых представлена полосой в районе 7 т.п.н., что соответствует плазмиде pPyrG-SEQ1.
Возможным объяснением наличия этих копий может быть как образование тандемных повторов при интеграции, так и наличие дополнительных копий вектора вне хромосомы в виде конкатамера.
Таким образом, исходя из полученных результатов нельзя однозначно сделать вывод о том, встроилась ли плазмида в состав хромосомы в виде тандемных копий или часть копий присутствует в виде конкатамера.
Так как в случае автономной репликации конкатамер должен быстро теряться в ходе роста при интенсивном делении ядер, поведение ДНК плазмиды pPyrG-SEQ1 у трансформантов Т1-1 и Т1-2 изучено дополнительно через десять поколений споруляции в неселективных условиях.
Споры трансформантов Т1-1 и Т1-2 высевают на неселективную среду PDA с добавлением уридина и Triton-X100. Отбирают по три отдельных клона каждого из трансформантов и наращивают их до стадии споруляции на чашках со средой PDA с добавлением уридина. Полученные в шести независимых экспериментах споры снова рассевают на PDA с добавлением уридина и Triton-X100. Далее в каждом эксперименте отбирают по одному клону и операцию повторяют еще девять раз. В последнем эксперименте полученные споры используют для выделения геномной ДНК и гибридизации по Саузерну аналогично тому, как описано выше. Результаты гибридизации отображены на фиг.4.
Из результатов видно, что потомки трансформантов Т1-1 и Т1-2, названые Т1-1-1, T1-1-2, T1-1-3 и T1-2-1, T1-2-2, T1-2-3, соответственно, содержат характерный набор фрагментов ДНК, схожий с таковым у исходных трансформантов, при этом основная доля фрагментов имеет размер около 7 т.п.н., что соответствует немодифицированной плазмиде. Таким образом, полученные результаты говорят в пользу многокопийного тандемного типа встраивания плазмиды pPyrG-SEQ1 и подтверждают стабильность трансформантов.
Пример 6. Трансформация штамма Rhizopus oryzae PYR-17 плазмидой pPyrG-SEQ2, отбор стабильных клонов
Трансформацию штамма Rhizopus oryzae PYR-17 плазмидой pPyrG-SEQ2 и отбор стабильных клонов осуществляют по примеру 4. В качестве трансформирующей ДНК используют кольцевую ДНК плазмиды pPyrG-SEQ2.
Гибридизацию полученных трансформантов Т2-1, Т2-2 и Т2-3 осуществляют по примеру 5. Геномную ДНК обрабатывают эндонуклеазой рестрикции Eco88I, сайт узнавания которой является уникальным для данной плазмиды (фиг.2).
Из результатов, приведенных на фиг.3, видно, что для трансформантов Т2-1, Т2-2 и Т2-3 полоса, соответствующая по размеру геномной копии гена pyrG (4 т.п.н.), отсутствует, и можно заключить, что интеграции произошла в геномный локус pyrG. При этом появление полос, соответствующих размерам 2,5; 5,5 и 7,5 т.п.н., и их интенсивность говорят о наличии 2; 4 и 1 копий плазмиды, интегрировавшихся в геном трансформантов Т2-1, Т2-2 и Т2-3 соответственно. Факт интегративного встраивания плазмиды свидетельствует о стабильности трансформантов Т2-1, Т2-2 и Т2-3.
Пример 7. Выявление повторяющихся геномных последовательностей SEQ1 и SEQ2 у штаммов рода Rhizopus
Геномную ДНК семи штаммов Rhizopus oryzae и одного штамма Rhizopus oligosporus (таблица), полученную по примеру 5, обрабатывают эндонуклеазой рестрикции ScaI, сайт узнавания которой отсутствует в повторяющихся геномных последовательностях SEQ1 и SEQ2.
Полученные образцы ДНК далее используют для гибридизации по Саузерну. Используемые в качестве зондов фрагменты ДНК, содержащие последовательности SEQ1 и SEQ2, получают методом ПЦР по примерам 2 и 3 соответственно.
На фиг.5 и фиг.6 приведены результаты гибридизации геномной ДНК восьми штаммов (таблица) с последовательностями SEQ1 и SEQ2 в качестве зондов соответственно (при этом каждая копия повторяющейся в геноме последовательности проявляется в качестве отдельной полосы).
Наблюдающиеся для всех штаммов Rhizopus oryzae многочисленные полосы показывют, что последовательности SEQ1 и SEQ2 имеют большое число повторов во всех исследованных штаммах Rhizopus oryzae. В штамме Rhizopus oligosporus NRRL 2710 гомология к последовательности SEQ2 отсутствует и наблюдается лишь ограниченная гомология к последовательности SEQ1.
Наличие описанных повторяющихся в геноме последовательностей во всех исследованных штаммах Rhizopus oryzae позволяет использовать генетическую конструкцию, полученную на основе последовательности, повторяющейся в геноме одного штамма, для стабильной интегративной трансформации широкого круга реципиентов этого вида.
Таким образом, заявленная генетическая конструкция позволяет получить митотически стабильные трансформанты грибов вида Rhizopus oryzae, причем генетическая конструкция, полученная на основе последовательности, повторяющейся в геноме одного штамма, может быть использована также для стабильной интегративной трансформации широкого круга реципиентов этого вида.
Важно также отметить, что заявленная конструкция может быть использована, в том числе, для получения штаммов с более высоким уровнем продукции того или иного целевого продукта. Для этого в описанную генетическую конструкцию должны быть добавлены элементы, обеспечивающие повышение или снижение уровня экспрессии выбранных генов или экспрессию гетерологичных генов. Например, для повышения продукции молочной кислоты в конструкцию pPyrG-SEQ1 может быть добавлен ген Rhizopus oryzae ldhA под гомологичным промотором pgk2.
| название | год | авторы | номер документа |
|---|---|---|---|
| ДРОЖЖИ РОДА YARROWIA, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ ВНУТРИКЛЕТОЧНО НАКАПЛИВАТЬ СЛОЖНЫЕ ЭФИРЫ ЖИРНЫХ КИСЛОТ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ТАКИХ ЭФИРОВ | 2013 |
|
RU2539744C1 |
| ИНТЕГРАТИВНЫЙ ВЕКТОР Random-URA3-RPT ДЛЯ ПОСЛЕДОВАТЕЛЬНОГО ВВЕДЕНИЯ МНОЖЕСТВЕННЫХ КОПИЙ ГЕНЕТИЧЕСКИХ ЭЛЕМЕНТОВ В ДРОЖЖИ Yarrowia lipolytica | 2006 |
|
RU2376376C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА МОЛОЧНОЙ КИСЛОТЫ И РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ SCHIZOSACCHAROMYCES POMBE ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2004 |
|
RU2268304C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ КЛЕТОЧНО-СВЯЗАННОЙ ЛИПАЗЫ | 2012 |
|
RU2475532C1 |
| ПРЕПАРАТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО СЫВОРОТОЧНОГО АЛЬБУМИНА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2337966C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ YARROWIA-LIPOLYTICA - ПРОДУЦЕНТ ЛИПАЗЫ | 2011 |
|
RU2451075C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ЦЕЛЕВЫХ ГОМОЛОГИЧНЫХ И ГЕТЕРОЛОГИЧНЫХ ГЕНОВ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ ШТАММА ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА | 2008 |
|
RU2378372C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI-ПРОДУЦЕНТ L-ТРЕОНИНА | 2013 |
|
RU2546237C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ ЛИПАЗЫ | 2007 |
|
RU2355754C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ЭКСПОЗИЦИИ БЕЛКА НА ПОВЕРХНОСТИ КЛЕТОЧНОЙ СТЕНКИ ДРОЖЖЕЙ Yarrowia lipolytica | 2010 |
|
RU2451749C1 |





Изобретение относится к микробиологической промышленности и касается вектора, предназначенного для получения стабильно трансформированных штаммов грибов Rhizopus oryzae. Охарактеризованный вектор содержит селективный генетический маркер и нуклеотидную последовательность, представленную в SEQ ID NO:1 или SEQ ID NO:2. Представленная генетическая конструкция может быть использована для получения высокостабильных штаммов грибов рода Rhizopus с высоким уровнем продукции целевого продукта. 1 з.п. ф-лы, 6 ил., 1 табл.
1. Вектор для получения стабильных трансформированных штаммов грибов Rhizopus oryzae, содержащий селективный генетический маркер и нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:2 или последовательность, гомологичную одной из указанных не менее чем на 85%.
2. Вектор по п.1, отличающийся тем, что дополнительно содержит генетические элементы, обеспечивающие экспрессию гетерологичных генов, либо повышение, или снижение уровня экспрессии гомологичных генов.
| US 6090574, 18.06.2000 | |||
| US 6582951, 24.06.2003 | |||
| MICHIELSE C.B | |||
| et al | |||
| Development of a system for integrative and stable transformation of the zygomycete Rhizopus oryzae by Agrobacterium-mediated DNA transfer, Mol Gen Genomics, 2004, Vol.271, p.p.499-510. |

