Область техники
Настоящее изобретение относится к областям иммунологии и вакцинологии. В частности, изобретение относится к улучшенным виросомам, композициям, включающим виросомы, и их применению.
Предпосылки создания изобретения
Одной из первостепенных задач медицины является разработка современных вакцин для профилактики и эффективной доставки терапевтических веществ для лечения заболеваний. В настоящее время виросомы известны как везикулы, которые можно использовать в качестве средств для доставки антигенов и/или в качестве носителей терапевтических веществ.
Виросомы представляют собой комплексы, состоящие из липидов и как минимум одного белка вирусной оболочки, которые получают in vitro. Липиды либо выделяют из яиц или растений, либо получают искусственно, а часть липидов получают из того же вируса, из которого получен белок оболочки. В сущности, виросомы представляют собой восстановленные (реконструированные) пустые вирусные оболочки без нуклеокапсида, содержащего генетический материал вируса(вирусов)-источника. Виросомы не способны к самовоспроизведению и являются всего лишь пузырьками, обладающими способностью к слиянию. Функциональность таких виросом заключается в том, что активность их мембран в отношении слияния весьма сходна с требующей низких значений рН способностью интактного вируса вызывать слияние мембран, которую обеспечивает исключительно вирусный белок слияния. Как и вирусы, виросомы быстро интернализуются в клетки по механизму опосредованного рецепторами эндоцитоза или путем слияния с клеточной мембраной.
В основном, используемые виросомы представляют собой так называемые иммуностимулирующие реконструированные виросомы гриппа (IRIV). IRIV - это сферические однослойные пузырьки со средним диаметром 150 нм, состоящие из липидного бислоя, состоящего из фосфолипидов, в основном, фосфатидилхолинов (PC) и фосфатидилэтаноламинов (РЕ). Виросомы IRIV содержат функциональные гликопротеины вирусной оболочки - гемагглютинин (НА) и нейраминидазу (NA) вируса гриппа, встроенные в мембрану, представляющую собой фосфолипидный бислой.
Биологически активный гемагглютинин не только придает структурную стабильность и гомогенность составам, содержащим виросомы, но и вносит значительный вклад в иммунологические характеристики, поскольку обеспечивает сохранение способности вируса вызывать слияние. При необходимости, IRIV содержат молекулы гемагглютинина более чем одного штамма вируса и таким образом представляют собой химерные IRIV.
IRIV были разработаны путем внедрения гемагглютинина (НА, ГА) штамма вируса гриппа А в липосомы, состоящие из фосфатидилхолина. Гликопротеин гемагглютинин оболочки вируса гриппа специфичным образом направляет виросомы к антигенпрезентирующим клеткам (АПС) и опосредует слияние с их мембраной их эидосом. Данный процесс обеспечивает оптимальное процессирование и презентацию антигенов иммунокомпетентным клеткам. Происходит активация продуцирования цитокинов Т-лимфоцитами, что, в свою очередь, стимулирует продуцирование больших количеств специфичных антител В-лимфоцитами. Более того, также происходит стимуляция В-лимфоцитов в результате прямого взаимодействия с комплексом антиген-виросома.
Виросомы являются высокоэффективными системами адъювант/переносчик в современной вакцинации/терапии, обладающими отличными характеристиками в качестве средств доставки антигена и высоким иммуногенным потенциалом, в то же время позволяющими минимизировать риск возникновения побочных эффектов. Кроме того, виросомы обладают эффектом адъюванта (WO 92/19267), эффектом транс-адъюванта (заявка на европейский патент ЕР 05027624) и неспецифичным иммуностимулирующим (Европейский патент на изобретение ЕР 06027120) эффектом.
Более 50 лет противогриппозные вакцины производили в клетках куриных эмбрионов. Тем не менее, традиционная стандартная методика чрезвычайно длительна и трудоемка. В настоящее время производство вакцин в эмбрионах занимает до 9 месяцев с момента выделения нового идентифицированного штамма вируса до получения конечного продукта. Это может затруднить решение неожиданных проблем, таких как выявление пандемичных штаммов, неудачи при производстве и сезонные изменения штаммов вируса гриппа. Более того, традиционная методика на основе эмбрионов требует огромных количеств яиц, переноса вирусного изолята в яйца и дополнительной очистки для уменьшения количества примесей эмбриональных белков и минимализации риска возникновения аллергии на альбумины яиц.
Способ, основанный на использовании клеточных линий, является более быстрым и гибким в отношении размножения вирусов и позволяет получать штаммы, которые не удается эффективно выращивать в куриных эмбрионах (например, Гонконгский грипп птиц в 1997). Кроме того, использование линий клеток при получении вирусов имеет некоторые преимущества с точки зрения безопасности получаемой вакцины: вакцина не содержит добавок антибиотиков, нет необходимости в токсичных консервантах (таких, как тиомерсал), уровень эндотоксинов понижен, исключено возникновение аллергии на яичные белки, рост вирусов происходит в среде, не содержащей белка и сыворотки (нет вспомогательных агентов/BSE), высока степень очистки вакцины.
В последнее время значительные усилия были направлены на разработку систем культур клеток для производства вакцин. Большая часть упомянутых систем основана на линиях клеток млекопитающих, таких как клетки Vero, MDCK, BHK и PerC6. Опубликовано несколько сообщений о разработке вакцин в культурах клеток млекопитающих. Тем не менее, существенным недостатком противовирусных вакцин, полученных в упомянутых культурах клеток, является риск возникновения аутоиммунных реакций к белкам клеток млекопитающих.
Процесс слияния виросом необходим для эффективной доставки антигенов/лекарственных средств (Schoen P и др., 1999). Соответственно, существует необходимость в разработке способа получения виросом с улучшенными характеристиками фузогенности (способности к слиянию) и иммуногенности.
Сущность изобретения
Настоящее изобретение решает указанную задачу путем создания новых виросом, содержащих гемагглютинин вируса гриппа, полученного в линиях клеток птиц. Такие новые виросомы характеризуются как повышенной способностью вызывать слияние и повышенной иммуногенностью по сравнению с виросомами, содержащими гемагглютинин вируса гриппа, полученными стандартным методом в куриных эмбрионах.
Таким образом, первый аспект настоящего изобретения относится к виросомам, содержащим гемагглютинин, причем гемагглютинин выделен из вируса гриппа, полученного в линии клеток птиц.
«Линия клеток птиц» в контексте настоящего изобретения - это культура клеток, выбранная по признаку однородности из популяции клеток, полученной из, как правило, гомогенной ткани птицы (такой, как орган). Данный термин не включает яйца птиц, такие как, например, куриные. Таким образом, «гемагглютинин, выделенный из вируса гриппа, полученного в линии клеток птиц», означает, что гемагглютинин выделен из вирусов, выращенных в культуре клеток птицы, а не из вирусов, выращенных в яйцах (куриных эмбрионах). Предпочтительные линии клеток птиц включают, но не ограничиваются перечисленными, первичные линии клеток, такие как фибробласты куриных эмбрионов (CEF); постоянные/иммортализованные линии клеток, например, DF-1 (US 5672485), PBS (US 5989805) и HD11.
Кроме того, настоящее изобретение относится к виросомам, содержащим гемагглютинин, причем способность к слиянию указанных виросом как минимум на 50% выше, чем способность к слиянию виросом, содержащих гемагглютинин, выделенный из вирусов гриппа, выращенных в куриных эмбрионах, и имеющий ту же первичную структуру, или пептидную последовательность. Согласно предпочтительному варианту осуществления виросомы согласно настоящему изобретению обладают значительно более высокой иммуногенностью, чем виросомы, содержащие гемагглютинин, полученный из вирусов гриппа, выращенных в куриных эмбрионах. В предпочтительном случае способность к слиянию виросом согласно настоящему изобретению как минимум на 30% выше способности к слиянию виросом, содержащих гемагглютинин, выделенный из вирусов гриппа, выращенных в клетках млекопитающих.
Неожиданно оказалось, что качество способности виросом к слиянию зависит от процесса выращивания вируса гриппа, из которого получают виросомы. Согласно предпочтительному варианту осуществления гемагглютинин, входящий в состав виросомы, выделяют из вирусов гриппа, полученных в клеточной линии. Предпочтительно гемагглютинин выделяют из вирусов гриппа, выращенных в линии клеток птиц.
Заявка на патент WO 2006/108846 (автор Vivalis) относится к использованию стволовых клеток эмбриона птицы, предпочтительно линии клеток ЕВх, для получения вирусных векторов и вирусов. Однако в WO 2006/108846 не содержится ни предпосылок для использования в виросомах гемагглютинина, полученного из вирусов, выращенных в линиях клеток.
Виросома может представлять собой химерную виросому, в которой когда гемагглютинин выделен из, по меньшей мере, двух разных штаммов вируса гриппа. Кроме того, виросома может быть лиофилизирована. В предпочтительном варианте осуществления изобретения виросома нагружена антигеном. В более предпочтительном варианте виросома согласно настоящему изобретению остается ненагруженной/пустой.
Согласно другому аспекту настоящее изобретение относится к композициям, включающим виросому согласно настоящему изобретению. В предпочтительном варианте осуществления, указанная композиция представляет собой вакцину. В другом предпочтительном варианте осуществления композиция является иммуногенной и дополнительно включает липосому и как минимум одну молекулу антигена. В предпочтительном случае указанная по меньшей мере одна молекула антигена заключена в липосому. внедрить по меньшей мере одну молекулу антигена в липосому.
Согласно следующему аспекту настоящее изобретение относится к применению виросом согласно настоящему изобретению в качестве средства доставки антигена в фармацевтической композиции для получения иммунного ответа против антигенов различного происхождения. Виросомы согласно настоящему изобретению также можно применять для получения фармацевтических композиций для вакцинации или иммунизации. Кроме того, настоящее изобретение относится к иммуностимулирующим виросомам, не нагруженным антигеном. Соответственно, настоящее изобретение относится к применению виросом согласно настоящему изобретению в качестве неспецифичных иммуностимулирующих агентов для получения фармацевтических композиций для стимуляции эффективного иммунного ответа против антигенов различного происхождения. Наконец, изобретение относится к применению виросом согласно настоящему изобретению в получении фармацевтических композиций для лечения или предотвращения заболевания или нарушения.
Согласно другому аспекту данное изобретение относится к наборам, включающим виросому или композицию согласно настоящему изобретению.
Следующий аспект включает способ вакцинации или иммунизации пациента виросомами или композициями согласно настоящему изобретению, включающий собой введение указанных виросом или композиций пациенту для стимуляции иммунного ответа. Также в настоящее изобретение включает способ лечения или предотвращения заболевания или нарушения (такого как инфекционные заболевания и/или злокачественные опухоли) у пациента с использованием виросом или композиций согласно настоящему изобретению, включающий введение указанных виросом или композиций указанному пациенту.
Согласно другому аспекту настоящее изобретение относится к способу получения виросом согласно настоящему изобретению, включающему этапы обработки целого вируса гриппа детергентом или короткоцепочечным фосфолипидом, отделение фракции, содержащей гемагглютинин, и удаление детергента, в результате чего происходит восстановление (формирование) виросомы. В качестве альтернативы, этап отделения может осуществлять путем добавления фосфолипидов. Настоящее изобретение также относится к виросомам, полученным упомянутым способом.
Краткие описания графических материалов
На фиг.1 показан Вестерн-блот анализ препаратов виросом с использованием гемагглютинина из линии инфицированных штаммом Новая Каледония гриппа А куриных клеток (полосы 1 и 4), утиных клеток (полосы 2 и 5) или вируса, полученного путем размножения в оплодотворенных яйцах (полосы 3 и 6). Блот А был получен с использованием специфичной в отношении вируса гриппа А поликлональной сыворотки кролика, блот В был получен с использованием моноклональных антител, распознающих определенный эпитоп на субъединице НА 1 гемагглютинина.
На фиг.2 показана способность к слиянию виросом гриппа. Верхняя часть: Графическое изображение способности к слиянию, результаты приведены в Таблице 2, Эксперимент 2, Пример 4.5. Нижняя часть: Соотношение способности к слиянию виросом гриппа, выделенных из клеток и из яиц. Столбики представляют средние соотношения активности образцов при различных разбавлениях, соответствующих концентрации гемагглютинина, от 1 до 6 мкг/ в общем объеме, равном 0,8 мл.
Фигуры 3 и 4 иллюстрируют результаты исследования иммуногенности на мышах. Согласно фиг.4 видна повышенная иммуногенность виросом, содержащих гемагглютинин, выделенный из вируса гриппа, полученного в линии клеток птиц, и содержащих гетерологичный антиген (UK39). Фиг.3А показывает, что источник вируса (линия клеток/культура клеток или яйцо), использованный для получения виросом согласно изобретению, не оказывает значительного влияния на титр антител к гемагглютинину, выделенному из яиц, после одной иммунизации. Фиг.3В демонстрирует повышенную иммуногенность гемагглютинина: более высокий титр антител к гемагглютинину, выделенному из ЕВх, после первой иммунизации виросомами, содержащими гемагглютинин, полученный из вирусов, полученных в клетках Евх. На фиг.4 показаны индивидуальные титры антител, специфичных к гетерологичному антигену UK39. Результаты получены путем вычисления степени разбавления, соответствующей величине оптической плотности, равной 20% максимальной оптической плотности - значение для контрольной сыворотки, присутствующей в каждом планшете. В показанном примере разницу, которую наблюдали между виросомами, содержащими гемагглютинин, выделенный из вирусов, полученных в яйцах, и виросомами, содержащими гемагглютинин, выделенный из вирусов, полученных в линиях клеток, в отношении иммуногенности гетерологичного антигена UK39, была значительной: р=0.002 для линии куриных клеток в сравнении с яйцами и р=0.009 для культуры клеток утки в сравнении с яйцами, с использованием критерия Вилкоксона.
На фиг.5 показана повышенная индукция CD8+ Т-лимфоциток, специфичных к гетерологичному антигену (не гемагглютинину) виросомами, содержащими гемагглютинин, выделенный из вирусов, полученных в линии клеток птиц и нагруженных гетерологичным антигеном, в сравнении с виросомами, содержащими гемагглютинин из вирусов, выделенных из яйца.
Подробное описание изобретения
Определения
В настоящем описании термин «виросома» относится к пузырьку (везикуле), полученному в результате процедуры in vitro и состоящему из липидов и, по меньшей мере, одного белка оболочки вируса. Липиды либо выделены из биологического источника (например, яиц, растений, животных, культур клеток, бактерий, вирусов и т.д.), либо получены синтетически (химический синтез). Виросома может представлять собой восстановленную оболочку вируса, которая может быть получена из различных вирусов и которая не содержит инфекционные нуклеокапсиды и генетический материал вируса-источника, например, иммуностимулирующая реконструированная виросома гриппа (IRIV). Таким образом, виросома представляет собой особый тип липидных везикул, состоящих из липидной мембраны и, по меньшей мере, одного белка оболочки вируса. Термин «белок оболочки вируса» в настоящем описании относится к любому белку, кодируемому оболочечным вирусом, из которого полностью или частично получены виросома согласно настоящему изобретению и который присутствует в липидной мембране виросомы. Белки оболочки вируса иногда выполняют функцию «белков слияния вируса» (фузионные белки), в случае когда они участвуют в слиянии вирусов или виросом с мембранами клеток-мишеней.
Виросома согласно настоящему изобретению может содержать больше одного типа белка оболочки. Источником указанных дополнительных белков, содержащихся в мембране виросомы, необязательно происходят от оболочечных вирусов, может быть любой живой организм (включая микроорганизмы, такие, как бактерии, грибы или паразиты).
Белки оболочки могут быть рекомбинантными белками, при условии, что их биохимические свойства допускают физическое присоединение к липидной мембране. Такие белки оболочки ответственны за функциональность виросомы.
В отличие от вирусных систем виросомы безопасны, поскольку инфекционный нуклеокапсид вируса удален. В настоящее время виросомы, в основном, применяют в качестве вакцин после внедрения антигена в поверхность или в полость виросом. В отличие от вирусоподобных частиц виросомы не формируются самопроизвольно в процессе рекомбинантной экспрессии белка в подходящей экспрессионной системе, а являются продуктом контролируемого процесса in vitro, обеспечивающего крупномасштабное промышленное производство виросом.
В настоящем описании термин «средство доставки антигена» относится к виросоме, содержащей один специфичный для определенного заболевания антиген, помещенный в полость виросомы или встроенный в ее поверхность.
В настоящем описании термин «способность к слиянию» (фузионная активность) относится к способности виросомы к слиянию с клеточной и/или синтетической мембраной. В то время как in vivo виросомы сливаются с наружной мембраной клетки или с мембраной эндосомы, слияние с липосомами является признанной моделью системы для определения способности к слиянию виросом in vitro (Smith, J M и др., 2003). Показано, что слияние вируса гриппа и виросом с липосомами обладает характеристиками, близкими к слиянию с биологическими оболочками мишеней (Stagmann, T. и др., 1989).
Термин «мембрана клетки» (клеточная мембрана) в настоящем описании относится к биологической мембране, присутствующей в клетках в естественных условиях, такой как наружная клеточная мембрана или мембрана эндосом внутри клеток. Термин «синтетическая мембрана», напротив, относится к искусственным мембранам, таким как липидные мембраны липосом. Примером синтетических мембран являются мембраны липосом, состоящие только из фосфатидилхолина (ФХ) и ДПФГ (дипальмитилфосфатидилглицерина) и не содержащие белков, обычно присутствующих в клеточных мембранах.
Способность к слиянию вирусов и виросом обычно измеряют методом флуоресцентного резонансного переноса энергии (FRET) (Struck DK и др., 1981). В данном методе используется фотофизический процесс, вызывающий уменьшение флуоресценции одного элемента (донора) путем передачи энергии возбуждения другому элементу (акцептору) без излучения. Необходимо, чтобы спектр испускания донора перекрывал спектр поглощения акцептора. Эффект уменьшения флуоресценции напрямую зависит от расстояния между двумя молекулами: каждое событие, вызывающее некоторое изменение расстояния между молекулами, подавляет эффект гашения (уменьшения флуоресценции), в результате чего происходит высвобождение энергии, которое может быть измерено. Таким образом, данный метод представляет собой ценный in vitro тест для изучения многих биологических явлений, таких как слияние вирусных частиц и биологических клеточных мембран. Были разработаны различные методы изучения слияния, основанные на флуоресцентном резонансном переносе энергии (FRET) для демонстрации способности к слиянию вирусных мембран (вирусов или виросом) с липосомами или «тенями» эритроцитов (пустыми эритроцитами) in vitro (Smit JM и др., 2003). Некоторые из этих методов включают мечение мембран мишеней (липосом), другие - мечение исследуемых образцов, а именно вирусов или виросом. Тем не менее, необходимость мечения исследуемого образца несовместимо с контролем качества фармацевтической продукции по цГМФ. Более чувствительный тест для определения слияния, основанный на флуоресцентном резонансном переносе энергии без мечения исследуемых образцов, был разработан компанией Pevion Biotech (Amacker M. и др., 2005).
Способность к слиянию виросом согласно настоящему изобретению может быть измерена методом флюоресцентного резонансного переноса энергии (FRET), как описано в примерах ниже. Чтобы определить, повышена ли способность к слиянию виросом согласно настоящему изобретению по сравнению с другими виросомами, выполняют следующие этапы: (а) измерение способности к слиянию виросом, содержащих различные количества гемагглютинина, выделенного из вирусов, полученных в линии клеток, и соответствующей виросомы, содержащей такое же количество гемагглютинина, выделенного из вирусов, полученных в яйцах; (b) определение отношения значений способности к слиянию, определенных на этапе (а) (т.е. виросомы, содержащей гемагглютинин из линии клеток, и виросомы, содержащей гемагглютинин из яйца); и (с) усреднение полученных отношений. Таким образом, для сравнения значений способности к слиянию необходимо сделать несколько измерений с разным количеством гемагглютинина для каждого типа виросом. Согласно предпочтительному варианту осуществления способность к слиянию измеряют для виросом, содержащих 3-6 мкг в общем объеме 0,8 мл. Пример вычислений приведен в подразделе 4.5 раздела Примеры ниже. Способность к слиянию рассматриваемой виросомы «на 50% выше», если упомянутое среднее отношение имеет значение ≥1.5.
В настоящем описании термин «иммуногенность» относится к способности определенного вещества (антигена) вызывать иммунный ответ. Чтобы определить, обладает ли виросома согласно настоящему изобретению повышенной (т.е. улучшенной), субъекта иммунизируют виросомой или композицией согласно настоящему изобретению, содержащей гемагглютинин или гемагглютинина в комбинации с дополнительным специфичным (гетерологичным) антигеном, и измеряют титр антител к гемагглютинину или указанному антигену в сыворотке указанного субъекта. Для сравнения, другого субъекта иммунизируют аналогичной виросомой, содержащей гемагглютинин, выделенный из вирусов, полученных в яйце. Виросома обладает «значительно улучшенной» или «значительно повышенной» иммуногенностью, если в применение критерия Вилкоксона к титрам антител к виросомам согласно настоящему изобретению (с гемагглютинином, выделенным из вирусов, полученных в линии клеток) и виросом с гемагглютинином, выделенным из вирусов, полученных в яйце, дает р-значение ниже 0,05. Пример расчетов приведен в подразделе 5.1 раздела Примеры ниже.
Термины «выделенный из линии клеток», «полученный в линии клеток», «продуцированный в линии клеток» взаимозаменяемы и означают, что что-либо получено или произведено в линии клеток или культуре клеток.
Термин «нагруженный антигеном» в настоящем описании относится к виросомам, содержащим дополнительный антиген, отличный от гемагглютинина (то есть «гетерологичный антиген» или «антиген, отличный от гемагглютинина»). Антиген может быть внедрен в виросому (например, внутрь полости), адсорбирован на/связан с поверхностью виросомы, интегрирован в липидную мембрану виросомы и т.п. Виросома с внедренным антигеном может быть использована как средство доставки антигена.
В настоящем описании термин «химерная виросома» относится к виросоме, содержащей гемагглютинин, полученный из по меньшей мере двух различных штаммов вируса гриппа.
В настоящем описании термины «ненагруженный» или «пустой» применительно к виросомам взаимозаменяемы и обозначают тот факт, что характеризуемые таким образом хромосомы не содержат специфичных для какого-либо заболевания антигенов полости и не несут таких антигенов в липидном бислое. Таким образом, «ненагруженные» или «пустые» виросомы содержат только окружающий раствор внутри полости и не содержит белка, за исключением белка гемагглютинина вирусной оболочки и возможных следов нейраминазы, в мембране.
В настоящем описании термины «терапевтический», «терапия», «лечение» и т.п. относятся к действиям, направленным против уже присутствующего заболевания или нарушения или заболевания или нарушения, в отношении которого предполагают, что оно уже присутствует, независимо от того, проявились ли соответствующие симптомы. В этом случае «лечение» или «терапия» относятся к устранению заболевания или нарушения или, по меньшей мере, снижению выраженности симптомов заболевания или нарушения; при этом, если симптомы уже присутствуют, их смягчают, а если симптомы отсутствуют, снижают тяжесть возникающих симптомов или полностью их устраняют. Термины «профилактический», «профилактика», «предотвратить», «предотвращение» в настоящем описании относятся к действиям, предпринимаемым для того, чтобы предотвратить возникновение заболевания у субъекта в случае, если нет подозрения, что в прошлом у данного субъекта уже было указанное заболевание, но предполагают, что субъекту угрожает или будет угрожать опасность возникновения указанного заболевания. Кроме того, указанные термины относятся к предотвращению заболевания, в случае, если пациент уже подвергся соответствующей вакцинации/иммунизации, эффект от которой, тем не менее, недолгосрочен.
В настоящем описании термин «фармацевтический» относится к характеристикам веществ и/или лекарственных препаратов, которые обеспечивают возможность из введения живому животному, предпочтительно человеку.
Термины «потенцирующий», «иммунопотенцирующий», «стимулирующий», «иммуностимулирующий» и т.п. в настоящем описании относятся к веществу или усиливающему влиянию на иммунные функции, которые могут обеспечить разрушение или выведение антигенсодержащих патогенов или злокачетственных новообразований и/или к возникновению иммунитета к ним.
В настоящем описании термин «неспецифический» или «неспецифичный» относится к общей иммуностимулирующей активности виросомы согласно настоящему изобретению и означает, что происходит стимуляция способности иммунной системы предотвращать, противостоять и/или устранять любое из многих заболеваний или нарушений, а не какое-то одно конкретное. Специфическая иммуностимулирующая активность, напротив, относится к стимуляции способности иммунной системы предотвращать, противостоять и/или устранять одно конкретное заболевание. Так, вакцинация против какого-либо конкретного заболевания является примером достижения специфической иммуностимулирующей активности.
Термин «заболевание» или «нарушение» в данном описании относится к аномальному состоянию тела или психики, вызывающему неудобство. Заболевания и нарушения делятся на инфекционные, неинфекционные, неопластические, иммунные или метаболические.
Вирусы гриппа
Вирусы гриппа (Orthomyxoviridae) - это оболочечные РНК-вирусы с РНК, представленной отрицательной цепью, и сегментированным геномом. Вирусы гриппа делят на два рода: первый включает вирусы гриппа А и В, второй включает вирус гриппа С. Такое деление основано на значительных антигенных различиях в их нуклеопротеинах и белках мутрикса. Также указанные три типа вирусов различаются по патогенности и организации генома. Тип А встречается у многих теплокровных животных, а типы В и С являются преимущественно патогенами человека. Вирусы гриппа А далее разделяют по антигенным характеристикам гликопротеинов оболочки гемагглютинина и нейраминидазы, выступающих над поверхность вириона. В настоящее время выделяют 15 подтипов гемагглютинина и 9 подтипов нейраминидазы. Вирусы гриппа А поражают много видов животных, включая птиц, свиней, лошадей, людей и других млекопитающих. Водоплавающие птицы являются естественным резервуаром для всех известных подтипов гриппа А и, возможно, источником генетического материала для появления штаммов, вызывающих пандемии гриппа у людей.
Вирусы гриппа накапливают точечные мутации во время репликации, поскольку в их комплексе РНК-полимеры отсутствует механизм устранения ошибок. Мутации, при которых происходят изменения аминокислот в антигенных участках оболочечных гликопротеинов, могут обеспечить селективные преимущества штамму вируса, заключающееся в избегании существующего иммунитета. Гемагглютинин является основной антигенной детерминантой вируса гриппа, которую распознают и связывают нейтрализующие антитела. Молекула гемагглютинина инициирует инфекцию путем связывания с рецепторами (остатками сиаловой кислоты) на определенных (респираторных) клетках хозяина.
Молекула гемагглютинина состоит из двух различных доменов: корневая структура, выступающая на поверхности вириона и состоящая из НА2 и части НА1 полипептида НА, и глобулярная головка, полностью состоящая из НА1.
Антитела к белку НА ингибируют связывание с рецептором и являются очень эффективными в предотвращении повторного заражения тем же штаммом. Гемагглютинин может ускользнуть от существующего иммунитета благодаря дрейфу антигенов, при котором мутации уже имеющегося гена гемагглютинина препятствуют связыванию антителами, и антигенному сдвигу, при котором вирус приобретает гемагглютинин нового подтипа. Такого рода изменения в большей степени характерны для гемагглютинина, чем для нейраминидазы. Изменения в других белках вируса гриппа происходят гораздо реже. Влияние антигенного дрейфа наиболее выражено у штаммов гриппа, поражающих человека, менее характерно для штаммов, инфицирующих свиней и лошадей, и и в наименьшей степени присуще штаммам, поражающим птиц.
Штаммы гриппа могут быть охарактеризованы генетически путем сравнения отдельных сегментов генов.
В то время как разработка вакцин против штаммов гриппа, вызывающих ежегодные эпидемии, продолжается, мир озабочен угрозой пандемий гриппа. Власти и медицинские службы во всем мире сегодня заняты разработкой стратегий по подготовке к борьбе с пандемической формой гриппа.
Виросомы
Виросомы согласно настоящему изобретению могут быть использованы для доставки соединения (например, иммуногенной молекулы, лекарственного средства и/или гена) в клетку-мишень. В сравнении с липосомами виросомы обладают преимуществом возможности эффективно проникать в клетку, которую обеспечивает белок оболочки вируса, и с последующим высвобождением содержимого виросомы в клетку. Более того, если в мембрану виросомы встроены некоторые активные белки оболочки вируса, такие виросомы могут высвобождать содержимое в цитоплазму сразу после слияния с клеткой, что позволяет избежать разрушения лекарственного средства в кислой среде эндосомы.
Виросомы согласно настоящему изобретению особенно полезны в области вакцинации, когда задача состоит в стимуляции иммунного ответа на антиген, ассоциированный с конкретным заболеванием или нарушением. В таких случаях антиген обычно инкапсулирован в или связан с виросомой, которая доставляет этот антиген иммунной системе хозяина, которого подвергли вакцинации. Благодаря данному конкретному антигену достигнутое профилактическое и/или терапевтическое действие обязательно будет специфичным в отношении заболевания или нарушения, с которым ассоциирован которому данный антиген.
Кроме того, виросомы могут быть одновременно нагружены несколькими различными эпитопами В- и Т-лимфоцитов (Pδltl-Frank и др. (1999)), включая универсальные эпитопы Т-хелперов (Kumar и др. (1992)) и другие, известные специалистам в данной области. Таким образом, виросомы являются высокоэффективными адъювантами в современной вакцинации, обладающими прекрасными характеристиками в качестве средства доставки антигенов и высоким иммуногенным потенциалом, в то же время минимизирующими риск побочных эффектов.
Функциональность иммуностимулирующих реконструированных виросом гриппа (IRIV) проявляется в том, что их способность к слиянию мембраны в большой степени сходна с хорошо известной, зависимой от низкого рН способности к слиянию мембран интактного вируса, обеспечиваемой исключительно белком оболочки вируса. Как и вирусы, виросомы гриппа быстро интернализуются по механизму рецепторопосредуемого эндоцитоза или опсонизации. В отличие от вирусных систем виросомы безопасны, поскольку из них удален инфекционный нуклеокапсид вируса. Таким образом, виросомы согласно настоящему изобретению являются многообещающими транспортными системами для доставки широкого спектра различных соединений, заключенных во внутренней среде внутри виросом, в водной среде внутри виросомы или встроенных в ее мембрану. Совместное внедрение различных рецепторов в мембрану виросом обеспечивает нацеливание виросом в различные клетки и ткани. В основном, виросомы используют в качестве вакцин после связывания антигена с их поверхностью, или после инкапсуляции антигена в полость виросомы, или используют их адъювантный эффект в случае введения в комбинации с липосомами, связанными с антигеном.
IRIV реконструируют из оболочки вируса гриппа и используют рецептор-опосредуемый эндоцитоз, как и их вирусные аналоги. Медиатором связывания вируса гриппа с рецепторами и его слияния с мембраной эндосомами считается основной гликопротеин оболочки вируса - гемагглютинин (Bungener и др. (2002)). Как и в случае с вирусными векторами, слабокислый рН полости эндосомы запускает механизм слияния мембран виросомы и эндосомы и высвобождение инкапсулированного материала, такого как ДНК, РНК или белки, в цитоплазму клетки. Соответственно, экзогенные антигены, инкапсулированные в виросоме, достигают пути главного комплекса гистосовместимости I (ГКГС I) без необходимости синтеза белка de novo. В процессе слияния белки, присутствующие на поверхности виросом, остаются в полости эндосомы и поэтому, по всей видимости, становятся доступными для пути ГКГС I.
Показано, что коммерчески доступные вакцины на основе виросом (INFLEXAL® V, EPAXAL®) весьма эффективны и безопасны (Gluck R. и др. (2000)). Потенциал виросом в качестве систем доставки был продемонстрирован для вакцин на основе нуклеиновых кислот и белков, например, в случае малярии (Pőltl-Frank и др. (1999)). По результатам недавних исследований также можно заключить, что вакцины, содержащие синтетические пептиды и вводимые подкожно с использованием виросомам, способны индуцировать эффективный цитотоксический иммунный ответ (Amacker и др. (2005)).
Получение виросом
Получение виросом - процедура, хорошо знакомая специалистам в данной области. Применяемые методики получения виросом описаны, например, в ЕР 538437 и в Mischler and Metcalfe (2002).
Виросомы согласно настоящему изобретению могут быть реконструированы из липидов вирусной мембраны и белков после солюбилизации вируса гриппа октаэтиленгликоль-монододециловым эфиром, седиментации нуклеокапсида (вирусные гликопротеины и липиды остаются в верхней фракции) и удаления детергента из верхней фракции гидрофобной смолой (Bio-Beads SM2). Способы получения гриппозных виросом описаны в WO 92/19267, виросом других представителей семейства - в WO 04/071492.
Получение виросом, содержащих гемагглютинин, выделенный из разных штаммов вируса гриппа, может быть осуществлено путем использования равных количеств белков этих вирусов, солюбилизированных неионным детергентом октаэтиленгликоль-монододециловым эфиром. После удаления детергента смолой Bio-Beads SM2, становится возможным образование виросом, содержащих разные типы белков оболочки.
Протоколы получения виросом, содержащих материал, полученный из куриных яиц и из клеточных линий, идентичны.
Подтипы вируса гриппа, из которых могут быть получены виросомы согласно настоящему изобретению, включают грипп H1N1, грипп H1N2, грипп H2N2, грипп H3N2, грипп H3N8, грипп H5N1, грипп H5N2, грипп H5N3, грипп H5N8, грипп H5N9, грипп H7N1, грипп H7N2, грипп H7N3, грипп H7N4, грипп H7N7, грипп H9N2 и/или грипп H10N7. Кроме того, по меньшей мере, один белок оболочки вируса может быть получен из следующих штаммов: А/Бангкокский/1/79, грипп А/Пекинский/32/92, грипп А/Бразильский/11/78, грипп А/Калифорнийский/7/2004 (H3N2), грипп А/Чилийский/1/83, грипп А/Крайстчерчский/4/85, грипп А/Английский/42/72, грипп А/Фуцзяньский/411/2002 (H3N2), грипп А/Гуичжоуский/54/89, грипп А/Гонконгский/1 /68, грипп А/Йоханнесбургский/33/94, грипп А/Ленинградский/360/86, грипп А/Миссисипский/1/85, грипп А/Московский/10/99 (H3N2), грипп А/Новокаледонский/20/99 (H1N1), грипп А/Панамский/2007/99 -RESVIR- 17), грипп А/Филиппинский/2/82, грипп А/порт-Чалмерский/1/73, грипп А/Шотландский/840/74, грипп А/Шаньдунский/9/93, грипп А/Шанхайский/11/87, грипп А/Сичуаньский/2/87, грипп А/Сингапурский/6/86, грипп А/Сиднейский/5/97, грипп А/Техасский/1/77, грипп А/Советский/90/77, грипп А/Викторианский/3/75, грипп А/Висконсинский/67/2005 (H3N2), грипп А/Уханьский/359/95, грипп А/Вайомингский/3/2003 Х- 147), грипп В/Гонконгский/330/2001, грипп В/Цзилиньский/20/2003, грипп В/Малайзийский/2506/2004, грипп В/Шанхайский/361/2002, грипп А/Пекинский/262/95, грипп В/Викторианский/98926/70, грипп В/Сингапурский/222/79, грипп В/Советский/100/83, грипп В/Ямагатский/16/88, грипп В/Панамский/45/90, грипп В/Гонконгский/5/72, грипп В/Энн-Арборский/1/86, грипп А/Баварский/7/95, грипп В/Шаньдунский/7/97) и/или В/Цзянсусский/10/2003.
Иммуностимулирующие реконструированные виросомы гриппа (IRIV) содержат двойную липидную мембрану, состоящую по существу из фосфолипидов, преимущественно фосфатидилхолинов (НС) и фосфатидилэтаноламинов (РЕ). В отличие от липосом IRIV содержат дополнительно функциональные вирусные гликопротеины оболочки гемагглютинин и нейраминидазу, встроенные в фосфолипидный бислой мембраны. Биологически активный гемагглютинин вносит существенный вклад в иммунологические свойства, поскольку обеспечивает сохранение способности вирусов к слиянию.
IRIV являются весьма эффективными средствами усиления неспецифического иммунитета. Кроме того, они обладают превосходными характеристиками безопасности (Glück и др. (2000)) и, таким образом, оказываются подходящими для использования в лекарственных средствах для неспецифической иммуностимуляции, предназначенных для людей.
Виросома согласно настоящему изобретению также может представлять собой химерную виросому. Это значит, что она содержит белки оболочки как минимум двух штаммов вируса гриппа, например, штаммов Х-31 и А/Синг или любых других, перечисленных выше. Кроме того, для создания химерных виросом, способных к последовательному или раздельному слиянию, могут быть использованы белки оболочки других известных вирусов, как, например, белок G вируса везикулярного стоматита, белок Е1 вируса Семлики, белок F вируса Сендаи, белок G или F респираторно-синцитиального вируса или белок Е вируса гепатита С, а также многие другие.
Как было продемонстрировано ранее (Tsurudome et al. 1992), белки слияния из разных штаммов вирусов заметно различаются по температурным характеристикам слияния и инактивации. Например, гемагглютинин Х-31 эффективно инициирует слияние при рН 5.0 и низких температурах, в то время как штаммы PR8/34 или А/Сингапурский при той же кислотности требуют повышенной температуры (>25°С). Таким образом, мембраны химерных виросом могут содержать белки, инициирующие слияние при двух разных значениях температуры. Различная чувствительность к температуре является особенно полезной характеристикой белков слияния, позволяющей легко и удобно осуществлять контроль реакций слияния. Например, виросомы, содержащие гемагглютинин вирионов Х-31 и PR8/34, могут катализировать две различных реакции слияния при значениях рН 5: при низких температурах (4-10°С) и при повышенных (>25°С). Однако также известны другие белки слияния, имеющие различные характеристики слияния, такие как чувствительность к температуре, концентрации ионов, кислотности, специфичности к типу клеток и тканей.
Белки слияния с разными характеристиками слияния могут быть выделены из различных штаммов вируса гриппа, таких как MRC-11, Х-97, NIB24, NIB26, Х-47, А/Йоханнесбургский/33 и А/Сингапурский.
В предпочтительном случае виросома согласно настоящему изобретению преимущественно содержит липиды, выбранные из группы, включающей катионные липиды, синтетические липиды, гликолипиды, фосфолипиды, холестерин и их производные. Фосфолипиды преимущественно включают фосфатидилхолин, сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилглицерол, фосфатидная кислота, кардиолипин и фосфатидилинозитол с вариабельными остатками жирных кислот. Катионные липиды преимущественно выбраны из группы, включающей DOTMA (N-[(1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмония хлорид), DOTAP (N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмония хлорид), DODAC (N,N-диолеил-N,N-диметиламмония хлорид), DDAB (дидодецилдиметиламмония бромид), TC-Chol (холестерил N-(триметиламмониоэтил)карбамата хлорид), DC-Choi (холестерил N-(диметиламмониоэтил)карбамата хлорид) или другие катионные производные холестерина, стеариламина или других алифатических аминов, DPPE (дипальмитоилфосфатидилэтаноламинов), DOGS (диолеил-глицеро-сукцинат), DOSPA (2,3-диолеоилокси-N-[2(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминтрифторацетат), DOSPER (1,3-диолеилокси-2-(6-карбоксиспермил)пропиламид), THDOB (N,N,N',N'-тетраметил-N,N'-бис (2-гидроксиэтил)-2,3-диолеоилокси-1,4-бутандиаммония иодид), DOPA (диолеил-sn-глицерофосфат), DOTP (диоктил терефталат), DOSC (диолеил-сукцинил-глицерин), DOTB (диолеил-е-(4'-триметиламмонио)-бутаноил-sn-глицерин), DOPC (диолеил-sn-глицеро-фосфохолин) и т.п. Наиболее предпочитаемыми вариантами катионного липида являются катионные производные холестерина, как TC-Chol (холестерил N-(триметиламмониоэтил) карбамат) или DC-Chol (холестерил N-(диметиламмониоэтил) карбамат). Таковые могут быть обобщены как малые монослойные липосомы, смешанные с фосфатидилхолином. Виросомы согласно настоящему изобретению могут, предпочтительно, содержать фосфатидилхолин, полученный из яиц или, еще более предпочтительно 1-олеил-3-пальмитоил-гас-глицеро-фосфатидилэтаноламин.
Мембрана виросомы согласно настоящему изобретению, предпочтительно, содержит 1.9-37 моль% DC-Chol или TC-Chol от общего количества липидов в мембране. Согласно особенно предпочтительному варианту осуществления содержание DC-Chol или TC-Chol в мембране составляет 1.9-16 моль % от общего количества липидов в мембране. Оставшиеся липиды мембраны, в предпочтительном случае, являются фосфолипидами, в частности, фосфатидилхолином и фосфатидилэтаноламином в соотношении 4:1.
Коэмульгирующий агент может также быть использован для повышения жесткости и способности виросомы к замыканию. Примерами коэмульсифирующих агентов могут служить сложные эфиры холестерина, заряженные или нейтральные, такие как сульфат холестерина, а также производные на основе стеролов, такие как производные растительного происхождения, например, ситостерол, сигмастерол и их смеси.
Виросома согласно настоящему изобретению может, к примеру, быть получена способом, аналогичном любому из способов синтеза DOTAP-содержащих виросом, описанных в примерах 1-3 и 6 в WO 97/41834, за тем исключением, что DOTAP заменяют на DOSPER и что концентрацию DOSPER в мембране конечной виросомы подбирают соответствующим образом, как описано в WO 97/41834, и, в частности, она не должна составлять более 30% от общей массы всех липидов мембраны виросомы. В сущности, способ получения виросом согласно настоящему изобретению может состоять из следующих этапов:
a) приготовление буферного раствора, содержащего неионный детергент, DOSPER и другие лип иды и, по меньшей мере, один белок оболочки вируса;
b) доведение концентраций липидов до значений (в расчете на общую массу липидов в мембране) 5-30% DOSPER и, соответственно, 95-70% других упомянутых липидов, включая фосфатидилхолин и его производные или, в качестве альтернативы, фосфатидилэтаноламин и/или катионные липиды, исключая DOSPER; и
(с) удаление детергента диализом или путем обработки раствора гранулами-микропереносчиками, в результате чего получают указанные виросомы.
Применение виросом согласно настоящему изобретению
Виросомы согласно настоящему изобретению можно применять в приготовлении лекарственных средств для лечения и/или предотвращения, по меньшей мере, одного заболевания или нарушения. Указанное, по меньшей мере, одно заболевание или нарушение может быть инфекционным, неинфекционным, неопластическим, иммунным или метаболическим заболеванием или нарушением. Согласно одному варианту осуществления применение включает введение виросомы согласно настоящему изобретению здоровым субъектам, которые временно подвергаются повышенному риску приобрести одно или несколько инфекционных заболеваний или нарушений, или (пока еще) здоровым субъектам немедленно до появления подозрения на риск приобрести одно или несколько заболеваний или нарушений, до появления симптомов или подтверждения диагноза. Классификация воздействия на индивидуума как лечебного или профилактического описана выше.
Применение может также относиться к лечению одного или нескольких уже имеющихся заболеваний или нарушений, возможно, в качестве вспомогательных лечебных мер по борьбе с такими заболеваниями или нарушениями.
Согласно другому варианту осуществления по меньшей мере одно инфекционное заболевание или нарушение может представлять собой вирусное заболевание или нарушение, бактериальное заболевание или нарушение, грибковое заболевание или нарушение, паразитарное заболевание или нарушение или прионное заболевание или нарушение.
Согласно следующему варианту осуществления животное является млекопитающим. Млекопитающее предпочтительно представляет собой: человека, шимпанзе, яванского макака, гиббона, не человекоподобную обезьяну, макаку резус, мышь, крысу, кошку, собаку, лошадь, кролика, верблюда, ламу, жвачное или свинью. В предпочтительном случае жвачное представляет собой корову, быка, козу, овцу, буйвола, оленя или вола.
Согласно следующему варианту осуществления лекарственное средство пригодно для внутримышечного, внутрикожного, внутривенного (например, инъекцией), подкожного, внутриперитонального, парентерального введения, топического применения, введения в ухо, в сустав, в глаз, локального введения, введения путем полоскания, введения с помощью пластыря (например, кожного пластыря), в виде спрея (например, назо-фарингеального спрея), сублингвального, орального (например, таблетки, капсулы, драже), при помощи суппозитория (например, ректальные или вагинальные суппозитории) и в виде капель (например, глазных капель). Препарат можно вводить однократно или, в случае необходимости, несколькими дозами с подходящими интервалами, определенными лечащим врачом.
Возможно необднократное применение виросом согласно настоящему изобретению. Комбинирование виросом согласно настоящему изобретению с другими препаратами, например, адъювантами или иммуностимуляторами, может обеспечить синергетическое усиление общего эффекта. Общий эффект зависит от количества и типа вводимой виросомы, участка стимуляции и ко-стимулирующих агентов (инфекции, воздействия аллергенов и т.п.). Эффект временный, продолжительностью от нескольких часов до нескольких недель. Продолжительность эффекта зависит от дозы, периодичности введения, места введения и состава вводимого препарата.
Полученное согласно настоящему изобретению лекарственное средство вводят в фармацевтически приемлемых формах. Такие формы могут, как обычно, содержать фармацевтически приемлемые концентрации солей, буферных агентов, консервантов, совместимых переносчиков, дополнительных иммуностимулирующих агентов, таких как адъюванты и цитокины, и, при необходимости, других лекарственных веществ. Предпочтительное количество вводимых виросом зависит от заболевания или нарушения, которое нужно предотвратить или вылечить. Как правило, эффективными считаются количества в диапазоне от 1 нг/кг до 100 мкг/кг, где в килограммах указан вес животного. Считается, что предпочтительный диапазон составляет от 10 нг/кг до 10 мкг/кг. Абсолютное количество зависит от множества факторов, включая композицию, выбранную для введения, количество вводимых доз (одна или несколько доз) и индивидуальные характеристики пациента, такие как возраст, физическое состояние, телосложение, вес и стадия заболевания.
Путь и режим введения будут различаться в зависимости от стадии и тяжести заболевания, которое лечат, и должны быть определены опытным практикующим врачом. Лекарственное средство, приготовленное согласно настоящему изобретению, подходит для парентерального введения. Лекарственное средство содержит виросому, растворенную или суспендированную в подходящем переносчике, предпочтительно в водном. Может быть использовано множество водных переносчиков, например, вода, буферные водные растворы, 0,4% соляной раствор, 0,3% раствор глицина, гиалуроновой кислоты и т.п. Такие композиции могут быть стерилизованы обычными, общеизвестными методами или могут быть подвергнуты стерильной фильтрации. Полученные водные растворы могут быть упакованы для применения, как есть, или лиофилизированы, при перед введением лиофилизированный препарат смешивают со стерильным раствором.
Лекарственное средство, приготовленное согласно настоящему изобретению, может также содержать фармацевтически приемлемые вспомогательные вещества, необходимые для приближения к физиологическим условиям, такие как регулирующие рН и буферные вещества, агенты, влияющие на тоничность, увлажняющие агенты и т.п., например, ацетат натрия, лактат натрия, хлорид натрия, хлорид калия, хлорид кальция, монолаурат сорбитана, олеат триэтаноламина и многие другие. Собственно, способы приготовления составов, пригодных для парентерального введения, известны и очевидны для специалистов в данной области и подробно описаны, например, в Remington: The Sciene an Practice of Pharmacy ("Remington's Pharmaceutical Sciene") издательство Gennaro AR, 20-е издание, 2000: Williams & Wilkins PA, USA, включенной в настоящее описание путем ссылки.
Лекарственное средство, приготовленное согласно настоящему изобретению, можно также вводить перорально, например, в виде таблеток, капсул (включая составы с 5 высвобождением в течение заданного времени и отложенным высвобождением), пилюль, порошков, гранул, эликсиров, настоек, растворов, суспензий, сиропов и эмульсий, а также в форме инъекций. К примеру, при оральном введении в форме таблеток или капсул активный лекарственный компонент может быть введен в комбинации с подходящим для перорального введения фармацевтически неактивным инертным переносчиком, таким как 10 этанол, глицерин, вода и т.п.
Аналогично, лекарственное средство, приготовленное согласно настоящему изобретению, может быть введено внутривенно (путем болюсной инъекции или инфузии), интраперитонеально, подкожно, топически (с окклюзией или без) или внутримышечно. В предпочтительном варианте осуществления приготовленное лекарственное средство, приготовленное согласно настоящему изобретению, вводят внутримышечно, подкожно, внутрикожно, через слизистые или чрескожным путем. Все указанные формы хорошо известны специалистам в фармацевтике.
Режим дозирования, согласно которому вводят лекарственное средство, приготовленное согласно настоящему изобретению, выбирают в соответствии с множеством факторов, включая, например, вид, возраст, вес, пол и состояние здоровья пациента, стадию и тяжесть заболевания или нарушения, а также конкретный тип применяемой виросомы. Обычный квалифицированный врач может без труда определить и назначить эффективное количество лекарственного средства, необходимого для профилактики, борьбы с или остановки прогрессирования инфекционного заболевания или нарушения. Оптимальная точность при определении концентрации препарата с соблюдением эффективности и без токсичности или с приемлемым уровнем токсичности подразумевает режим, основанный на кинетике доставки виросомы (доступности) в целевые участки. Этот процесс, включающий учет распределения, равновесия и выведения виросомы, находится в пределах компетенции практикующего врача и может быть определен в ходе рутинных экспериментов.
Согласно одному варианту осуществления лекарственное средство, приготовленное согласно настоящему изобретению, можно вводить ежедневно в виде однократной дозы, либо общая суточная доза может быть определена и разделена на несколько частей, например, при приеме 2, 3 или 4 раза в день. Согласно другому варианту осуществления предусмотрено еженедельное или ежемесячное введение.
Суточная доза лекарственного средства, приготовленного согласно настоящему изобретению, лежит в пределах от 10 нг/кг до 10 мкг/кг виросом для взрослого пациента в день. В случае перорального введения лекарственного средства, приготовленного согласно настоящему изобретению, его предпочтительно изготавливают в форме таблеток, содержащих 0.001-1000 мг, предпочтительно 0.01-100 мг, еще более предпочтительно 0.05-50 мг и наиболее предпочтительно 0.1-20 мг виросом для симптоматического контроля дозы в зависимости от признаков и симптомов заболевания пациента в процессе лечения. Так, таблетки могут содержать 0.001, 0.01, 0.05, 0.1, 0.5, 1, 2.5, 10, 20, 50 или 100 мг виросом. Эффективное количество виросом в лекарственном средстве, приготовленном согласно настоящему изобретения, обычно достигается при уровне доз от 0.0001 мг/кг до 50 мг/кг веса тела в день. Более конкретно, уровень доз лежит в пределах от приблизительно 0.0001 мг/кг до 7 мг/кг веса тела в день. При введении детям доза может быть соответственно понижена соответствующим образом.
Кроме того, лекарственное средство, приготовленное согласно настоящему изобретению, может быть введен интраназально или чрескожным путем, известным специалистам в данной области. При чрескожном введении в форме системы для чрескожной доставки режим введения, безусловно, будет непрерывным, а не периодическим.
Лекарственное средство, приготовленное согласно настоящему изобретению, можно комбинировать с биоразлагаемыми полимерами, используемыми для контролируемого высвобождения лекарственного средства в организме, например, с полимолочной кислотой, полипепсилон капролактоном, полигидроксимасляной кислотой, полиортоэстерами, полиацеталями, полидигидропиранами, полицианоакрилатами и сшитыми или амфипатическими блок-сополимерами гидрогелей.
Подходящая форма лекарственного средства, приготовленного согласно настоящему изобретению, для наружного введения может быть, например, в виде раствора, крема, мази, геля, лосьона, шампуня или аэрозоля, подходящих для нанесения на кожу. Такие составы для топического применения, содержащие лекарственное средство согласно настоящему изобретению, обычно включают около 0.005-5 мас.% активного вещества, т.е. виросомы, в смеси с фармацевтически приемлемым носителем.
Независимо от способа и пути введения лекарственного средства, приготовленного согласно настоящему изобретению, его вводят в эффективном количестве. Эффективное количество - это такое количество фармацевтического средства, которое после однократного введения или вместе с последующими дозами стимулирует желаемый неспецифический иммунный ответ.
Более того, в лекарственное средство, приготовленное согласно настоящему изобретению, при необходимости или желании могут быть добавлены приемлемые агенты, лубриканты, дезинтегрирующие агенты или красители. Подходящие связывающие агенты включают, без ограничения, крахмал, желатин, натуральные сахара, такие как глюкоза или бета-лактоза, кукурузные подсластители, натуральные или синтетические смолы, такие как гуммиарабик, трагакантовая камедь или альгинат натрия, карбоксиметилцеллюлозу, полиэтиленгликоль, воски и т.п. Лубриканты, применяемые в таких формах лекарственного средства, включают, без ограничений, олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия и т.п. Дазинтегрирующие агенты включают, без ограничения, крахмал, метилцеллюлозу, агар, бентонит, ксантановую смолу и т.п.
Жидкие формы лекарственного средства, приготовленного согласно настоящему изобретению, могут быть соответствующим образом дополнены суспендирующими или диспергирующими агентами, такими как синтетические и натуральные смолы, например, трагакаитовая камедьгуммиарабик, метилцеллюлоза и т.п. Другие диспергирующие агенты, пригодные для использования, представляют собой глицерин и т.п. Для парентерального введения желательно использовать стерильные суспензии и растворы. Для внутривенного введения используют изотоничные составы, обычно содержащие соответствующие консерванты. Составы для наружного применения, содержащие активный лекарственный компонент, могут быть смешаны с различными материалами-носителями, широко известными в данной области, такими как, например, спирты, гель алоэ вера, аллантоин, глицерин, масла витаминов А и Е, минеральное масло, PPG2 миристоил пропионат и т.п., для получения, например, спиртовых растворов, наружных очищающих средств, очищающих кремов, гелей для кожи, лосьонов для кожи и шампуней в форме крема или геля.
В одном варианте осуществления лекарственное средство, приготовленное согласно настоящему изобретению, может дополнительно содержать, по меньшей мере, один адъювант, усиливающий или опосредующий иммунный ответ, например, врожденный иммунный ответ, ответ Th1 или Th2. Подходящие адъюванты могут усиливать иммунный ответ путем активации макрофагов и/или стимуляции определенных групп лимфоцитов. Приемлемый адъювант может представлять собой любой лиганд, способный активировать рецептор распознавания патогена. Соединения, активирующие иммунный ответ, классифицируют как адъюванты или цитокины. Адъюванты могут усиливать иммунный ответ путем обеспечения резервуара для антигена (внеклеточный или внутри макрофагов), активируя макрофаги и стимуляции определенных групп лимфоцитов.
В данной области известно много видов адъювантов; конкретные примеры включают адъювант Фрейнда (полный и неполный), микобактерии, такие как БЦЖ, М. vaccae или Corynebacterium parvum, токсины холеры или столбняка, теплочувствительный токсин кишечной палочки, смеси хил-сапонина, как QS-21 (SmithKline Beecham), MF59 (Chiron), и различные водно-масляные эмульсии (например, IDEC- AF). Другие адъюванты, пригодные для использования, включают (но не ограничиваются перечисленными): минеральные соли или минеральные гели, такие как гидроксид алюминия, фосфат алюминия и фосфат кальция; ПАВ, такие как лизолецитин, полиолы - плюроники, полианионы, пептиды, гемоцианин моллюска фиссуррелии, динитрефенол, иммуностимулирующие агенты, такие как производные сапонина, мурамиловых дипептидов и трипептидов, короткоцепочечные молекулы нуклеиновых кислот, такие как динуклеотиды ЦфГ, олигонуклеотиды ЦфГ, монофосфорил-липид А и полифосфазины, адъювантные частицы и микрочастицы, такие как эмульсии, липосомы, виросомы, со-хелаты или комплексные иммуностимулирующие адъюванты.
Цитокины также можно использовать благодаря их способности стимулировать лимфоциты. Специалистам в данной области знакомы многие цитокины, которые можно использовать для этих целей, такие, как интерлейкин-2 (IL-2), IL-12, колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSF) и многие другие. Кроме того, лиганды из семейства хемокинов, такие как RANTES (цитокин А5), липопротеин грам-положительных бактерий, компонент клеточной стенки дрожжей, двухцепочечная РНК, ЛПС грам-отрицательных бактерий, флагеллин, богатая урацилом одноцепочечная вирусная РНК, супрессорная малая интерферирующая РНК цитокиновых сигналов (SOCS siRNA), эпитоп Pan DR (PADRE) и их смеси также могут быть использованы.
Лекарственное средство, приготовленное согласно настоящему изобретению, можно вводить в комбинации с фармацевтически приемлемым переносчиком, подходящим для топического применения. Также лекарственное средство, приготовленное согласно настоящему изобретению, можно использовать для лечения и предотвращения злокачественных опухолей, новообразований и/или метастазов или вирусных инфекций в комбинации с другими агентами, о которых известно, что они подходят для лечения указанных заболеваний. При комбинированном лечении более чем одним активным агентом активные вещества можно вводить одновременно или раздельно, в разное время.
Следующий аспект настоящего изобретения относится к неспецифической стимуляции иммунной системы животного путем введения виросомы согласно настоящему изобретению. Желательно усилить общую сопротивляемость к заболеваниям, в частности, инфекционным заболеваниям и новообразованиям, посредством неспецифической стимуляции (пробуждения) иммунной системы организма. Такой неспецифический стимул можно обеспечить путем введения виросом. Такое введение, однократное или периодическое, может быть осуществлено до, во время и после воздействия инфицирующего агента или диагностирования заболевания, в качестве профилактического, метафилактического, терапевтического или вспомагательного средства.
Конкретные варианты осуществления изобретения и приведенные ниже примеры демонстрируют эффективность настоящего изобретения, однако их не следует интерпретировать как ограничивающие объем изобретения. В тех случаях, когда указаны конкретные материалы, такое указание приведено исключительно в иллюстративных целях и не предполагает ограничение объема изобретения. Если не указано другое, биохимические и относящиеся к молекулярной биологии процедуры проводят согласно описанию в следующих источниках: Voet, Biochemistry, Wiley, 1990; Stryer, Biochemistry, W.H. Freeman, 1995; Bodanszky, Peptide chemistry. A Practical Textbook, 2nd ed., Springer-Verlag, Berlin, 1993; Sambrook et al., Molecular cloning, Cold Spring Harbor Laboratory, 2001; Ausubel et al. (Eds.), Current Protocols in Molecular Biology, John Wiley & Sons, 2000. Специалисты в данной области могут разработать равноценные средства и реагенты, не привлекая дополнительной изобретательской активности и не выходя за пределы объема настоящего изобретения.
Очевидным является множество вариантов описанных в настоящем тексте вариантов композиций и процедур, которые не выходят за рамки объема настоящего изобретения. По замыслу авторов изобретения такие варианты включены в объем изобретения.
Примеры
Содержание
1. Вирусы
2. Размножение вирусов
3. Получение виросом
3.1 Реагенты
3.2 Получение стандартных виросом (иммуностимулирующих реконструированных виросом гриппа, IRIV)
3.3 Получение стандартных виросом с интегрированным гетерологичным антигеном (IRIV)
3.4 Получение стандартных виросом, содержащих TC-chol-холестерил-N-(триметиламмониоэтил)-карбамат-хлорид (TIRIV)
3.5 Получения TIRIV, содержащих гетерологичный антиген
3.6 Гетерологичные антигены, используемые для комплектации виросом
4. Аналитические методы
4.1 Электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE)
4.2 Определение размера частиц: средний диаметр/полидисперсность (Таблица 1)
4.3 Простая радиальная диффузия (SRD, концентрация гемагглютитнина)
4.4 Вестерн-Блот
4.5 Анализ повышенной способности к слиянию методом флуоресцентного резонансного переноса энергии (FRET)
5. Анализ иммуногенности
5.1 Улучшенные антигенные характеристики
5.2 Определение повышенной иммуногенности по окрашиванию, окрашивание интерферона-гамма (IFN-y) и повышенная иммуногенность
5.3 Сравнение гемагглютинина, полученного из вирусов, полученных в яйцах, и гемагглютинина, полученного из вирусов, полученных в клеточной культуре
1. Вирусы
Использованы штаммы вируса гриппа А/Новокаледонский/20/99 (H1 N1) и А/ Сингапурский/6/86 (H1N1).
2. Размножение вирусов
Вирусы были размножены либо в аллантоисной полости оплодотворенных яиц (Gerhard, 1996), либо в линии эмбриональных стволовых клеток птиц (WO 2006/108846). Вирусы, которые размножали в аллантоисной полости оплодотворенных яиц, получены от Berna Biotech AG (Bern, Швейцария); вирусы, которые размножали в линии эмбриональных стволовых клеток птиц, получены от Vivalis (Roussay, Франция).
Полученный из яиц вирус очищали и концентрировали с помощью ультрацентрифугирования в градиенте сахарозы и инактивировали бета-пропиолактоном (BPL). Полученный из линий клеток птиц вирус (ЕВх, курицы (ЕВ14) или утки) присутствовал в надосадочной жидкости культуры клеток Ebx, инфицированных гриппом А. Перед проведением анализа надосадочную жидкость концентрировали и осаждали ультрацентрифугированием. Количество белков вируса определяли методом простой радиальной диффузии (SRD) (Wood и др., 1977).
Отношение гемагглютинин/фосфолипиды определяли методом по Böttcher (Böttcher и др., 1961), путем определения содержания гемагтлютинина с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и экстракции Кумасси, как описано у Ball, 1986.
3. Получение виросом 3.1. Реагенты
Октаэтиленгликоль-моно-(п-додецил)эфир (OEG, Ci2E8), диметилсульфоксид (DMSO), гидроксиламин гидрохлорид, 1,2-дипальмитоил-sn-глицеро-3-фосфо-гас-(1-глицерол) (PG), ацетонитрил, раствор фосфата триэтиламмония (ТЕАР), сахароза, стрептомицин, HEPES, пенициллин и среда RPMI были приобретены в Fluka Chemie GmbH и Signia (Buchs, Швейцария) соответственно. Сахароза (Eur. Phar.) была приобретена в Merck (Dietikon, Швейцария). Флуоресцентный корреляционный спектроскоп (FCS) был приобретен в Gibco BRL (Basel, Швейцария). Яичный фосфатидил холин был приобретен в Lipoid (Cham, Швейцария). 1-олеил-3-пальмитоил-гас-глицеро-2-фосфоэтаноламин был приобретен в Bachem (Bubendorf, Швейцария). Био-частицы Bio-Beads SM2 были приобретены в Bio-Rad Laboratories (Glattbrugg, Швейцария). 1,2-дипальмитоил-sn-глицеро-3-фосфоэтаноламин-N-[4-(р-малеимидометил)циклогексан-карбоксамид] (N-МСС-РЕ) был приобретен в Avanti Polar Lipids (Alabaster, США). N-(4,4-дифторо-5,7-дифенил-бора-3а,4а-диаза-8-индацен-8-пропионил-1,2-дигексадеканоил-sn-глицеро-3-фосфоэтаноламин (Bodipy 530/550-DHPE), лиссамин-родамин В 1,2-дигексадеканоил-5п-глицеро-3-фосфаэтаноламин триметиламмония соль (N-Rh-DHPE) и биотин-DHPE (N-(биотиноил)-1,2-дигексадеканоил-sn-глицеро-3-фосфоэтаноламин, триэтиламмония соль) были получены в Molecular Probes Europe (Leiden, Нидерланды). Сефадекс (Sephadex) G-50, грубый, был приобретен в Amersham Biosciences (Otelfingen, Швейцария). Интерлейкин-2. (IL-2) был приобретен в EuroCetus B.V. (Amsterdam, Нидерланды). N- сукцинимидил-S-ацетилтиоацетат (SATA) is был получен в Pierce Biotechnology (Rockford, США). Холестерил-N-(триметиламмониоэтил)карбамата хлорид (TC-chol) был получен из Merck Eprova (Schaffhausen, Швейцария).
3.2 Получение стандартных виросом (иммуностимулирующих реконструированных виросом гриппа. IRIVы)
В общем объеме 4 мл растворяли 32 мг фосфатидилхолина из яиц, 8 мг фосфоэтаноламина растворяли в 3 мл фосфатно-солевого буфера, содержащего 100 мМ олигоэтиленгликоля (PBS/OEG). Инактивированный вирус гриппа, содержащий 2 мг гемагглютинина, центрифугировали при 100,000 × g в течение 1 ч при температуре 4°С, после чего осадок был растворен в 1 мл PBS/OEG. Фосфолипиды и вирусы, солюбилизированные в детергенте, смешивали в общем объеме 4 мл и диспергировали под действием ультразвука в течение 1 мин. Полученную смесь центрифугировали при 100,000 × g в течение 1 ч при 18°С. Затем виросомы путем удаления детергента, с использованием 1.5 г влажных гранул SM2 Bio-Beads обеспечивали образование виросом (BioRad, Glattbrugg, Швейцария) дважды в течение 1 часа при комнатной температуре, при встряхивании. Затем виросомы подвергали стерильной фильтрации (0.22 мкм) и хранили при 4°С.
3.3 Получение стандартных виросом с интегрированным гетерологичным антигеном (I I RIVы).
В общем объеме 4 мл 32 мг фосфатидилхолина из яиц, 8 мг фосфоэтаноламина и желаемое количество соединения гетерологичного антигена и фосфоэтаноламина растворяли в 3 мл фосфатно-солевого буфера, содержащего 100 мМ олигоэтиленгликоля (PBS/OEG). Инактивированный штамм А/Сингапурский/6/86 вируса гриппа, содержащий 2 мг гемагглютинина, центрифугировали при 100,000 × g в течение 1 ч при 4°С, после чего осадок растворяли в 1 мл PBS/OEG. Фосфолипиды и вирусы, солюбилизированные в детергенте, смешивали и диспергировали под действием ультразвука в течение 1 мин. Эта смесь была центрифугирована при 100,000 × g в течение 1 ч при 18°С. Затем путем удаления детергента, с использованием 1.5 г влажных гранул SM2 Bio-Beads (BioRad, Glattbrugg, Швейцария) дважды в течение 1 часа при комнатной температуре при встряхивании обеспечивали образование виросом. Затем виросомы подвергали стерильной фильтрации (0.22 мкм) и хранили при 4°С.
3.4 Получение стандартных виросом, содержащих TC-chol - холестерил-N-(триметиламмониоэтил)-карбамат-хлорид (TIRIV)
TIRIV получали методом путем детергента. В общем объеме 4 мл 32 мг фосфатидилхолина из яиц, 8 мг фосфоэтаноламина, 5 мг холестерил-Т-(триметиламмониоэтил)-карбамат-хлорида и 200 мг сахарозы растворяли в 3 мл фосфатно-солевого буфера, содержащего 100 мМ олигоэтиленгликоля (PBS/OEG). 1-2 мг гемагглютинина инактивированного вируса гриппа были центрифугированы при 100,000 × g в течение 1 ч при 4°С, после чего осадок растворяли в 1 мл PBS/OEG. Фосфолипиды и вирусы, солюбилизированные в детергенте, смешивали и диспергировали под действием ультразвука в течение 1 мин. Эту смесь центрифугировали при 100,000 × g в течение 1 ч при 18°С. Затем путем удаления детергента, с использованием 1.5 г влажных гранул SM2 Bio-Beads (BioRad, Glattbrugg, Швейцария) дважды в течение 1 часа при комнатной температуре при встряхивании обеспечивали получение виросом. Затем виросомы подвергали стерильной фильтрации (0.22 мкм) и разливали в стерильные стеклянные пробирки. Закрытые пробирки замораживали при -70°С, затем лиофилизированы при -40°С в течение 20 ч и при 10°С в течение 2 ч. Закрытые пробирки хранились замороженными до момента использования.
3.5 Получение TIRIV, содержащих гетерологичный антиген
Для получения TIRIV, содержащих выбранный гетерологичный антиген, указанный антиген растворили в воде при желаемой концентрации. Замороженные, лиофилизированные TIRIV извлекали из морозильной камеры и уравновешивали при комнатной температуре 2-5 мин, после чего в лиофилизат добавляли равное количество количества растворенного гетерологичного антигена (4°С). Затем пробирку ненадолго (около 10 сек) помещали в вихревую мешалку при средней скорости и хранили при 4°С до использования.
В качестве альтернативы, пептиды, связанные с фосфатидилэтаноламином, можно добавить к TIRIV во время процесса получения, описанного в примере 5. Пептид в желаемой концентрации добавляли перед диспергированием под действием ультразвука и стерильной фильтрацией раствора. Остальные стадии приготовления не меняли. Восстановление диофилизированных TIRIV осуществляли с равным количеством воды.
3.6 Гетерологичные антигены, используемые для получения виросом
Использованными гетерологичными антигенами являлись малярийный UK 39 (WO 2004/106366), полученный из Plasmodium falciparum (UK 39); и оболочечный антиген вируса гепатита С (HCV core 132).
4. Методы анализа
4.1 Электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE)
Исследуемые образцы смешивали с соответствующим буфером для анализа производства в Invitrogen (Basel, Швейцария) с восстанавливающим агентом или без него (Invitrogen) и инкубировали при 85°С 2 мин. 5-10 мкл образца нанесли на матрицу из полиакриламидного геля (Invitrogen, Basel, Швейцария) и провели анализ согласно инструкции производителя. Затем гели анализировали методом Вестерн-Блотт и/или окрашивалм серебром с использованием набора SilverQuest Kit (Invitrogen, Basle, Швейцария), следуя протоколу «быстрое окрашивание» ("fast staining"), предоставленного производителем.
4.2 Определение размера частиц: средний диаметр/полидисперсность
В таблице ниже показана концентрация гемагглютинина в мг/мл и средний диаметр частиц виросом. Полидисперсность, приведенная в последнем ряду, является показателем гомогенности размера частиц в растворе. Раствор частиц со значением полидисперсности ниже 0.3 приемлем для виросом, применяемых в качестве вакцин, значение ниже 0.1 указывает на значительную гомогенность (гомогенность определяли методом динамического светорассеивания на приборе Zetasizer 1000HS).
Размер определяли методом динамического светорассеивания на приборе Zetasizer 1000HS (Malvern Instruments), оснащенного стандартным 10 мВт гелий-неоновым лазером (λ=633 nm) и лавинным фотодиодом (APD). 5-20 мкл образца добавили в фильтрованный фосфатно-солевой буфер в кювете с конечным объемом 1 мл. Измерения осуществляли при температуре 25°С и фиксированном угле рассеивания 90°. Распределения размеров оценивали путем подбора соответствующей аппроксимации.
4.3 Простая радиальная диффузия (SRD, концентрация гемагглютитнина)
Анализ методом простой радиальной иммунодиффузии для определения гемагглютинина в описанных выше образцах вируса гриппа, полученного из яиц и клеточных линий, осуществляли согласно процедуре, описанной Вудом и др. (Wood et al, 1977). Вирионы разрушали путем инкубации в 1% Zwittergent (Calbiochem) в течение 30 мин при комнатной температуре и подвергали иммунодиффузии в течение 72 ч при комнатной температуре в агарозном геле с антителами. Измеряли диаметры зон осаждения комплексов антиген-антитело и определяли содержание антигена в препаратах вируса с использованием калибровочной кривой контрольного образца цельного вируса (NIBSC, London) с известным содержанием гемагглютинина, предоставленным производителем NIBSC. В качестве контрольных образцов цельного вируса, используемых для определения количества гемагглютинина в образцах вируса гриппа, полученного в линиях клеток, использовали полученные из яиц стандартные аналоги, полученные в NIBSC, а также используемые антисыворотки.
Среднее (нм)
4.4 Анализ методом Вестерн-Блот
Сравнительный анализ вируса гриппа, полученного в оплодотворенных яйцах и линиях клеток птиц, включал SDS-PAGE и анализ методом Вестерн-Блот препаратов вирусов, которые позволяли получить информацию о синтезе и процессинге гемагглютинина вируса в обоих типах клеток: анализ SDS-PAGE позволял определить чистоту и содержание белка в суспензии вирусов, а также идентифицировать белки/размер белков гемагглютинина и нейраминидазы.
Осуществляли анализ исследуемых образцов на геле SDS-PAGE, как описано выше. Гели помещали в соответствующий буфер для переноса, предоставленный производителем (Invitrogen, Basel, Швейцария). Параллельно поливинилилендифторидную (PVDF) мембрану (Invitrogen, Basel, Швейцария) инкубировали в метаноле и также помещали в буфер для переноса. По 4-5 фильтров для блоттинга и по 2 листа ватмана (Biorad, Reinach, Швейцария) для каждого геля пропитывали в буфере для переноса и получают иммуноблот. Перенос осуществляли приложением 25В, 125 мА, 17 Вт к каждому гелю в течение 1 ч 30 мин. Мембраны быстро промывали в фосфатно-солевом буфере, содержащем 0.2% Tween 20, неспецифическое связывание антител или сыворотки блокировали путем инкубации в 5% молоке в фосфатно-солевом буфере в течение 2 ч. После повторного промывания мембран в фосфатно-солевом буфере/0.2% Tween 20, иммуноблоты инкубировали со смесью первичных антител и сыворотки, растворенной в 0,5% молоке в фосфатно-солевом буфере/0.2% Tween 20, в соотношении от 1:100 до 1:1000, в зависимости от антител, при комнатной температуре в течение 1-2 ч. Мембраны трижды промыли в фосфатно-солевом буфере/0.2% Tween 20 по 5 минут и инкубировали в соответствующем растворе вторичных антител, меченных пероксидазой хрена (HRP), разведенных в пропорции от 1:1000 до 1:20000 в 0,5% молоке в фосфатно-солевом буфере/0.2% Tween 20. После 5-кратного промывания мембран в фосфатно-солевом буфере/0.2% Tween 20 осуществляли визуализацию с помощью набора SuperSignal West Dura (Pierce, Lausanne, Швейцария) в соответствии с инструкцией производителя.
4.5 Анализ повышенной способности к слиянию методом флуоресцентного резонансного переноса энергии (FRET)
Для измерения способности к слиянию in vitro методом флуоресцентного резонансного переноса энергии (FRET) была разработана следующая процедура: 0.75 мол.% N-(4,4-дифторо-5,7-дифенил-бора-3а,4а-диаза-s-индацен-S-пропионил-1,2-дигексадеканоил-sn-глицеро-S-фосфоэтаноламина (Bodipy 530/550-DHPE) и 0.25 мол.% 1,2-дигексадеканоил-sn-глицеро-3-фосфоэтаноламин триметиламмония соли (N-Rh-DHPE) внедряли в липосомы, содержащие фосфохолин/1,2-дипальмитоил-sn-глицеро-3-фосфо-rac-(1-глицерол) (PC/PG) (70:30). Измерение флуоресценции осуществляли в 5 мМ натрий-фосфатном буфере рН 7.5, 100 мМ NaCl, в конечном объеме 0.8 мл в 2.5 мл микрокюветах из полиметилметакрилата (VWR, Dietikon, Швейцария) с постоянным перемешиванием. Обычно 1 мкл меченых липосом (0.3 нмоль фосфолипидов) смешивали с 5-20 мкл виросом и стимулировали слияние при помощи 3.75-7 мкл 1 М HCl с результирующим рН 4.5. Усиление флуоресценции регистрировали каждые 5 сек при длинах волн возбуждения и испускания, равных соответственно 538 нм и 558 нм, с сечением возбуждения 2.5 нм и сечением испускания 15.0 нм. Измерения осуществляли на люминесцентном спектрометре LS 55 (Perkin Elmer Instruments, Schwerzenbach, Швейцария), оборудованном держателем кюветы и магнитной мешалкой. Значение температуры прибора было установлено на 42°С, при этом конечная температура образца составляла 35-37°С. Максимальную флуоресценцию при бесконечном разведении пробы получали путем добавления буфера Triton X-100 (конечная концентрация 0.5% об./об.). Для калибровки шкалы флуоресценции начальную остаточную флуоресценцию липосом приравнивали нулю, а флуоресценцию пробы с бесконечным разведением принимали за 100% (максимальная флуоресценция).
Общее содержание гемагглютинина в образцах для анализа методом FRET должно составлять 0.5-10 мкг. Для анализа составов виросом оптимальным было признано содержание 2-6 мкг гемагглютинина. Концентрацию гемагглютинина в образцах предварительно определяли методом простой радиальной диффузии (SRD). В зависимости от концентрации гемагглютинина в каждом отдельном образце необходимый для FRET объем образца виросом лежит в пределах от 3 до 40 мкл (соответственно, 2-6 мкг гемагглютинина). Если объем образца виросом был ниже 40 мкл, разница компенсировалась добавлением фосфатно-солевого буфера.
Важно, что соотношение гемагглютинина и липидов виросом оставалось неизменным при использовании различных объемов виросом в тесте FRET, например, в последовательных измерениях, описанных в Таблице 2.
Интерпретация результатов FRET
Поскольку процентные значения, полученные методом FRET, варьируются, абсолютное значение диапазона/порога было сложно установить. Напротив, соотношение между разными образцами было сравнительно устойчивым. Количество гемагглютинина, использованного в анализе, следует выбирать в диапазоне 3-6 мкг в общем объеме 0.8 мл. Множественные измерения с различными количествами в пределах указанного интервала (например, 3, 4, 5, 6 мкг) позволили построить кривую зависимости от дозы, предоставляющую дополнительную информацию (например, о насыщении системы).
Для каждого значения концентрации гемагглютинина, исследованной методом FRET, сравнивали результаты (выраженные в % способности к слиянию), полученные для IRIVa содержащих гемагглютинин, выделенный из вирусов, полученных в яйцах, и из вирусов, полученных в линиях клеток, сравнение осуществляли путем вычисления отношения значений для двух соответствующих образцов для каждого значения концентрации. Затем вычисляли среднее значение отношений для различных концентраций гемагглютинина. Необработанные показания (% способности к слиянию) в значительной степени варьируют для разных концентраций гемагглютинина (зависимость от дозы), а также в разных повторах теста (вариабельность теста) для одного и того же образца. Среднее отношение, напротив, показало значительно более низкую вариабельность и, таким образом, представляет собой надежный показатель, позволяющий воспроизводимо сравнивать образцы.
5 мкг: 39%:14%=2,78
3 мкг: 23:8=2,87
2 мкг: 26:13=2,0
1 мкг: 18:5=3.6
В результате получили среднее значение 2,81 (фиг.2, нижняя часть)
5. Иммуногенные методы. Исследование иммуногенности на мышах
Реакция антител: Если не указано другое, для исследования иммунного ответа группы из, по меньшей мере, 10 мышей линии BALB/c иммунизировали внутримышечно 0,1 мл пустых IRIV или IRIV, нагруженных гетерологичным антигеном (MIRIV). В этом эксперименте в качестве гетерологичного антигена использовали малярийный антиген UK39. Осуществляли две вакцинации с интервалом в три недели, пробы сыворотки брали через две недели после второй вакцинации.
Ответ CD8+ Т-клеток: Если не указано иное, для исследования иммунного ответа трансгенных мышей линии HLA-A2 6 иммунизировали внутримышечно 0,1 мл пустых IRIV или IRIV, нагруженных гетерологичным антигеном (MIRIV). Осуществляли две вакцинации с интервалом в три недели, пробы клеток селезенки брали через две недели после второй вакцинации.
5.1 Иммунологический анализ методом ИФА (твердофазный иммуноферментный анализ, реакция В-клеток)
Группы мышей из, по меньшей мере, 10 особей, линии BALB/c иммунизировали внутримышечно дважды с интервалом в три недели, после чего брали пробы сыворотки. График, дозировка и количество процедур иммунизации могут различаться при условии, что для сравниваемых групп применяют одну и ту же процедуру. Сыворотки были исследованы методом ИФА для определения реакции антител к гемагглютинину вируса гриппа (фиг.3) и к гетерологичному малярийному антигену UK39, полученному из Plasmodium falciparum (фиг.4). Для каждого образца сыворотки определяли титр антител к указанному антигену. С этой целью рассчитывали разведение, соответствующее значению оптической плотности (OD) 20% от максимального значения OD контрольной пробы сыворотки, присутствовавшей на каждом планшете.
Для каждой группы фиксировали все индивидуальные титры . Группы сравнивали с применением критерия Вилкоксона к данным (титры сыворотки). Если полученное значение р ниже 0.05, это значит, с 95% вероятностью две группы были различны. В данном случае тест Вилкоксона применяли для того, чтобы показать, что улучшенные виросомы являются «значительно более иммуногенными», т.е. согласно критерию Вилкоксона для титров сыворотки полученное р-значение ниже 0.05 при сравнении виросом, полученных с использованием яиц, и виросом, полученных с использованием линий клеток.
Для обнаружения антител к гетерологичным антигенам или гемагглютинину осуществляли иммуноферментный анализ (ИФА). Вкратце, микротитрационные планшеты с 96 ячейками (Nunc, Fisher Scientific, Wohlen, Швейцария) в течение ночи при температуре 4°С покрывали исследуемым антигеном в подходящей буферной системе, в объеме 100 мкл/лунка. Например, комплексы малярийного антигена UK39 с фосфатидилэтаноламином были нанесены в концентрации 10 мкг/мл раствора в фосфатно-солевом буфере (рН 7.4) на микротитрационные планшеты Polysorb, а белки - гемагглютинные белки (инактивированный целый вирус или образцы виросом) наносили в концентрации 1 мкг/мл раствора в 0.05М карбонатном буфере (рН 9.4) на микротитрационные планшеты Maxisorb.
После нанесения планшеты блокировали 5% молочным порошком в фосфатно-солевом буфере минимум в течение 2 ч при комнатной температуре, затем трижды промывали фосфатно-солевым буфером/0.2% Tween 20. Затем планшеты инкубировали с последовательными разведениям (начиная с 1:50) сыворотки мыши в фосфатно-солевом буфере, содержащем 0.05% Tween 20 и 0.5% молочного порошка, в течение 2 ч при 37°С. На каждом планшете присутствовала сыворотка в качестве положительного контроля. После цикла дополнительной промывки, планшеты инкубировали со связанными с пероксидазой хрена антителами козы к Ig мыши (BD Bioscience, Basel, Швейцария) в течение 1 ч при 37°С. После последнего цикла промывки добавили субстрат OPD (таблетки O-фенилендиамина Fluka, Buchs, Швейцария, 1 таблетка в 50 мл нитратного буфера +20 мкл H2O2), и планшеты инкубированы в темноте при комнатной температуре до получения достаточной окраски в результате колориметрической реакции, которую останавливали добавлением 100 мкл 1 М H2SO4, после чего измерили оптическую плотность на 492 нм с помощью Spectra Max Plus (Molecular Devices, Bucher Biotech, Basel, Швейцария).
Фиг.4 демонстрирует существенные различия в иммуногенности между IIRIV (иммуностимулирующими реконструированными виросомами гриппа, нагруженными UK39), содержащими гемагглютинин из вируса, полученного в яйцах, и IIRIV, содержащими гемагглютинин из вируса, полученного в линиях клеток: р=0.002 для культуры в клетках кур в сравнении с культурой в яйцах, и р=0.009 для культуры клеток утки при сравнении с культурами в яйцах. Показана улучшенная иммуногенность UK39, связанного с виросомами, содержащими гемагглютинин из вируса, полученного в Ebx, по сравнению с UK39, связанными с виросомами, содержащими гемагглютинин из вируса, полученного в яйцах. Серая пунктирная линия показывает значение OD, позволяющее вычислить титр антител к UK39, определяемый как 20% от максимального значения OD492 контроля, присутствовавшего на каждом планшете.
5.2 Иммунологический анализ путем окрашивания внутриклеточного интерферона-гамма IFN-v (ответ Т-клеток)
Окрашивание внутриклеточного IFNγ: Клетки селезенки (12×106) инкубировали с 10 мкг/мл конкретного пептида или пептида, не имеющего отношения к настоящему изобретению (отрицательный контроль), в полной среде RPMI, содержащей 2 мМ L-глутамина, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина, 5 мМ HEPES, 5% сыворотки фетальной бычьей сыворотки (FCS) и 5×10'5 М 2-меркаптоэтанола при 37°С и 5% CO2, в присутствии 5 мкг/мл Брефелдина А в течение 4 ч. Клетки окрашивали антителами к CD8, связанными с изотиоцианатом флуоресцеина (FITC), пермеабилизовали, окрашивали связанными с фосфатидилэтаноламином антителами к IFNy, с помощью набора Cytofix/Cytoperm, следуя инструкции производителя (BD Pharmingen, San Diego, США). Результаты считывали на поточном цитометре BD™ LSR II и анализировали с использованием программного обеспечения FlowJo. Количество синтезирующих клеток, синтезирующих IFNy, рассчитывали как процентную долю IFNy-положительных и CD8-положительных клеток от всех CD8-положительных клеток. Процент пептид-специфичных клеток получали путем вычитания процента в образцах, стимулированных пептидом, используемым в качестве отрицательного контроля, из процента в образцах, стимулированных специфичным пептидом.
Трансгенные мыши линии HLA-A2 были иммунизированы дважды подкожно с интервалом в три недели иммуностимулирующими реконструированными виросомами гриппа, содержащими холестерил-N-(триметиламмониоэтил)-карбамат-хлорид (ТС-Chol-IRIV, TIRIV), полученными из вируса гриппа, культивированного в клеточной культуре (TIRIV ЕВх утки) и TIRIV, полученными из полученных в яйцах вирусов (TIRIV яиц), в обоих вариантах снабженными гетерологичным антигеном оболочки гепатита С (HCV core 132). Количество гемагглютинина в обоих случаях было одинаковым. Контрольных мышей иммунизировали TIRIV без гетерологичного антигена. Через две недели после последней иммунизации определяли количество
CD8+Т-лимфоцитов, специфичных к гетерологичному антигену, методом окрашивания внутриклеточного иитерферона-гамма. Показаны средние значения ± стандартное отклонение.
5.3 Сравнение гемагглютинина, выделенного из вирусов, полученных в яйцах, и гемагглютинина, выделенного из вирусов, полученных в культуре клеток.
Как показано на фиг.1, поликлональные и моноклональные антитела реагируют с материалом, полученным из яиц и полученным из клеток линий птиц, различным образом. В то время как поликлональная сыворотка реагирует как с гемагглютинином, выделенным из полученного в линиях клеток птиц вируса, так и с гемагглютинином, выделенным из полученного в яйцах вируса, моноклональные антитела реагируют только с гемагглютинином, выделенным из полученного в яйцах вируса.
Дегликозилирование гемагглютинина
Разные штаммы гриппа А (А/Сингапурский (H1/N1), А/Новокаледонский (H1/N1), А/Панамский (H3/N2)) амплифицировали в оплодотворенных яйцах; гемагглютинин, полученный из соответствующих препаратов, распознается MAT, что указывает на отсутствие специфичности MAT к штаммам. Для дегликозилирования использовали набор "Enzymatic protein deglycosylation Kit", полученный в Sigma-Aldrich (Buchs, Швейцария). Все процедуры выполняли согласно инструкции производителя. Дегликозилирование препаратов вирусов указанных трех штаммов во всех случаях приводило к полной потере сигнала, в то время как поликлональная сыворотка, специфичная к гемагглютинину, распознала полосы меньшего размера, представляющие дегликозилированный белок (Таблица 3).
Пассирование вируса гриппа в линии клеток млекопитающих и анализ методом Вестерн-Блот.
Подготовили два планшета с 6 лунками и культурами клеток линии MDCK или линии Vera: в каждую лунку высевали 5×105 клеток в среде Episerf. На следующий день клетки инфицировали вирусом, полученным в оплодотворенных яйцах или в клетках ЕВх, с добавлением трипсина или без него, что обеспечивало наработку инфекционных вирионов (в результате расщепления гемагглютинина, в результате которого белок становится активным) или ограничение репликации до одного пассажа (в результате поддержания гемагглютинина в неактивной конформации НА0 в отсутствие трипсина), соответственно. Соскобы клеток брали через 1 день (для MDCK) или через 3 дня (для Vera), как только вирусная инфекция вызывала лизис клеток или выраженный цитопатический эффект соответственно. После этого инфицированные клетки были исследованы методом Вестерн-Блот с использованием поликлональной сывороткой кролика, специфичной к гемагглютинину, или с использованием специфичных к гемагглютинину МАТ.
Однако окрашивание моноклональными антителами показало положительный результат только в случае с вирусом, выделенным из инфицированных яиц (контроль), но оказалось отрицательным в случае с образцами гемагглютинина, полученного из вирусов, прошедших пассаж в клетках млекопитающих (клетках MDCK или Vera: эксперимент без трипсина, в котором образуются только вирусы, неспособные повторно инфицировать клетки). Таким образом, одного пассажа было достаточно для устранения сигналов от МАТ.
Один пассаж исключает возможность замены аминокислотам, и поэтому модификация эпитопа на уровне аминокислот как причина потери способности к связыванию может быть исключена. В отсутствие трипсина не происходит расщепления полипептида HA0 на НА1 и НА2. Разрушение эпитопа в результате расщепления гемагглютинина трипсином как причина невозможности связывания антителами также можно исключить.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
| УКОРОЧЕННАЯ СЕКРЕТИРУЕМАЯ АСПАРТИЛ-ПРОТЕИНАЗА 2 | 2008 |
|
RU2493255C2 |
| ФУНКЦИОНАЛЬНО РЕКОНСТРУИРОВАННЫЕ ВИРУСНЫЕ МЕМБРАНЫ, СОДЕРЖАЩИЕ АДЪЮВАНТ | 2004 |
|
RU2348428C2 |
| УЛУЧШЕННАЯ ВАКЦИНАЦИЯ ПРОТИВ ГРИППА | 2013 |
|
RU2599496C2 |
| ПЕРОРАЛЬНО ДИСПЕРГИРУЕМАЯ ВАКЦИНА, СОДЕРЖАЩАЯ ВИРОСОМЫ | 2019 |
|
RU2808276C2 |
| ВАКЦИНЫ И СПОСОБЫ ЛЕЧЕНИЯ СОБАЧЬЕГО ГРИППА | 2006 |
|
RU2396976C2 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ, СОДЕРЖАЩИЕ БЕЛКИ-МИШЕНИ, СЛИТЫЕ С БЕЛКАМИ ОБОЛОЧКИ РАСТИТЕЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2571223C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ОТ ПТИЧЬЕГО ГРИППА И ЕЁ ПРИМЕНЕНИЕ | 2009 |
|
RU2559527C2 |
| Пентавалентная субъединичная вакцина против респираторных инфекций и способ ее получения | 2022 |
|
RU2804948C2 |
| ОПТИМИЗИРОВАННЫЕ С ПОМОЩЬЮ ВЫЧИСЛИТЕЛЬНЫХ СРЕДСТВ АНТИГЕНЫ С ШИРОКИМ СПЕКТРОМ РЕАКТИВНОСТИ ДЛЯ ВИРУСОВ ГРИППА H5N1 И H1N1 | 2013 |
|
RU2639551C2 |
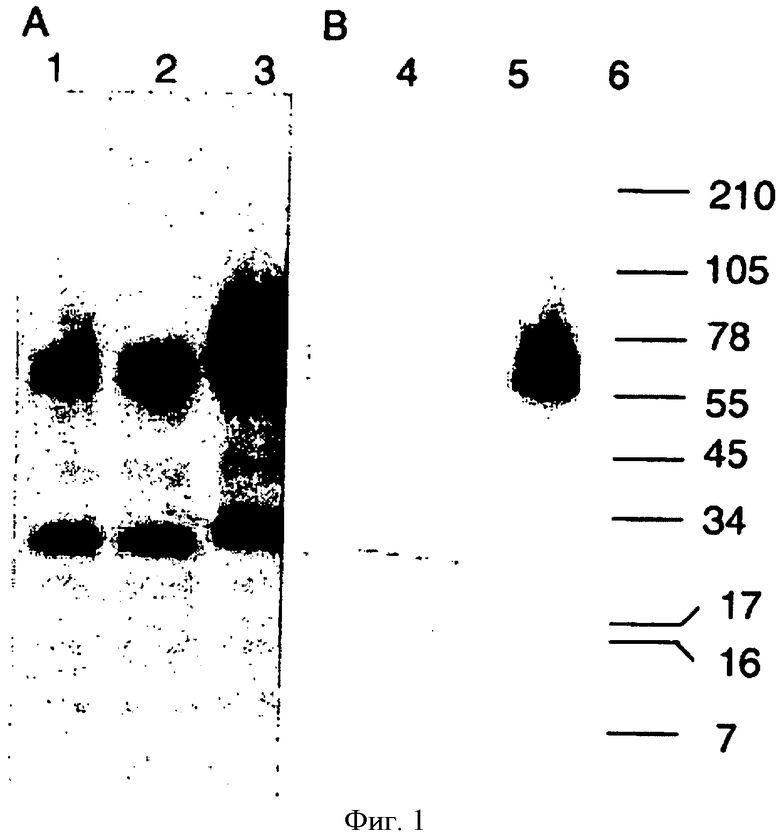

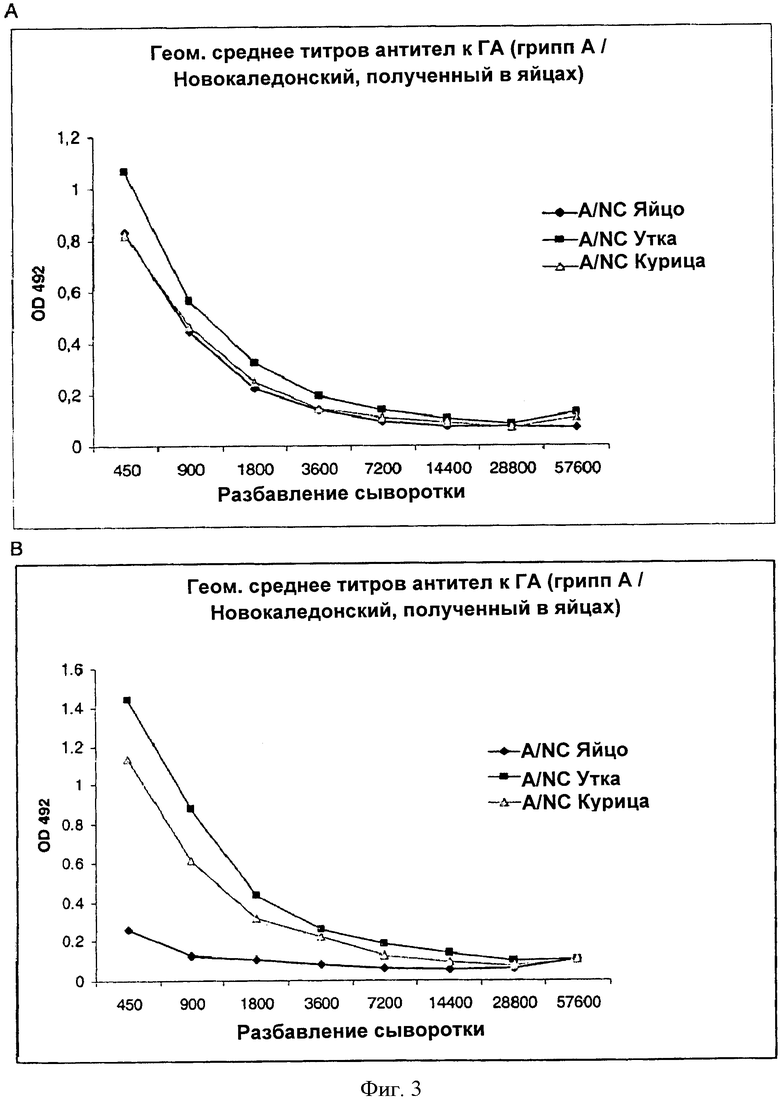

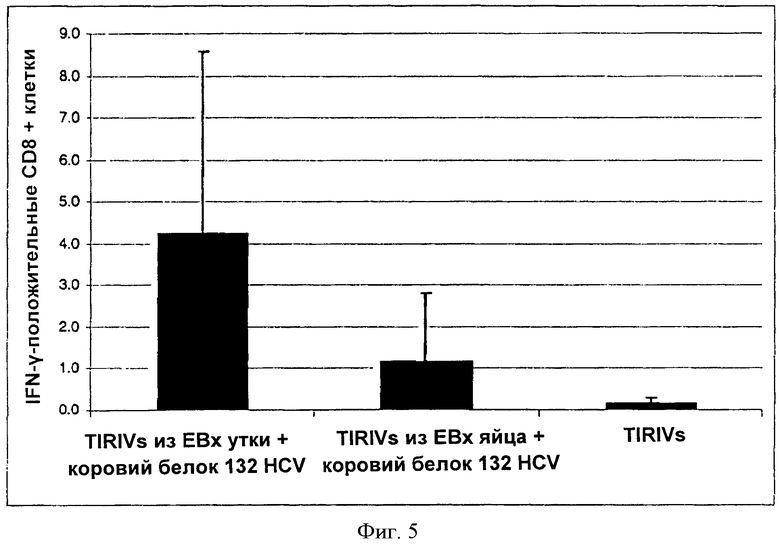
Группа изобретений относится к области медицины, в частности к области вирусологии, и касается виросом, включающим гемагглютинин, выделенный из вируса гриппа, полученного в линии клеток, композиций, содержащих указанные виросомы, способам изготовления и применения. Сущность изобретения включает виросомы, содержащие гемагглютинин, выделенный из вируса гриппа, полученного в линии клеток птицы, композицию, содержащую виросому, применение виросомы в качестве средства доставки, набор, способы вакцинации, способы лечения и способ получения виросомы. Преимущество группы изобретений заключается в получении виросом, обладающих улучшенной способностью к слиянию и повышенной иммуногенностью. 11 н. и 10 з.п. ф-лы, 3 табл., 5 ил.
1. Виросома, содержащая гемагглютинин, выделенный из вируса гриппа, полученного в линии клеток птицы.
2. Виросома по п.1, отличающаяся тем, что способность к слиянию указанной виросомы по меньшей мере на 50% выше средней способности к слиянию виросом, содержащих гемагглютинин, выделенный из вируса гриппа, полученного в яйцах кур.
3. Виросома по любому из пп.1 или 2, отличающаяся тем, что иммуногенность указанной виросомы значительно выше иммуногенности виросом, содержащих гемагглютинин, выделенный из вируса гриппа, полученного в яйцах кур.
4. Виросома по любому из пп.1 или 2, отличающаяся тем, что гемагглютинин получен из по меньшей мере двух различных штаммов вируса гриппа.
5. Виросома по любому из пп.1 или 2, отличающаяся тем, что виросома лиофилизирована.
6. Виросома по любому из пп.1 или 2, отличающаяся тем, что виросома нагружена антигеном.
7. Виросома по любому из пп.1 или 2, отличающаяся тем, что виросома пустая.
8. Композиция, содержащая виросому по любому из пп.1 или 2.
9. Композиция по п.8, представляющая собой вакцину.
10. Композиция по п.8, отличающаяся тем, что является иммуногенной и дополнительно содержит липосому и по меньшей мере одну молекулу антигена.
11. Композиция по п.10, в которой в указанную липосому внедрена по меньшей мере одна молекула антигена.
12. Применение виросомы по любому из пп.1 или 2 в качестве средства доставки антигена в фармацевтической композиции для обеспечения иммунного ответа на антиген.
13. Применение виросомы по п.7 в качестве неспецифического иммуностимулирующего агента для получения фармацевтических композиций для обеспечения иммунных ответов на антигены различного происхождения.
14. Применение виросомы по любому из пп.1 или 2 для получения фармацевтической композиции для вакцинации или иммунизации.
15. Применение виросомы по любому из пп.1 или 2 для получения фармацевтической композиции для лечения или предотвращения заболевания или нарушения.
16. Набор, включающий виросому по любому из пп.1 или 2 или композицию по любому из пп.9-11.
17. Способ вакцинации или иммунизации субъекта виросомой по любому из пп.1-7 или композицией по любому из пп.9-11, включающий введение указанной виросомы или указанной композиции субъекту.
18. Способ лечения или предотвращения заболевания или нарушения у нуждающегося в этом субъекта с применением виросомы по любому из пп.1 или 2 или композиции по любому из пп.9-11, включающий введение указанной виросомы или композиции субъекту.
19. Способ по п.18, в котором заболевание или нарушение представляет собой инфекционное заболевание или раковое заболевание.
20. Способ получения виросомы по любому из пп.1 или 2, включающий следующие этапы:
a) обработка целого вируса гриппа детергентом или короткоцепочечным фосфолипидом,
b) отделение фракции, содержащей гемагглютинин, возможно с добавлением фосфолипидов,
c) удаление детергента, что приводит к формированию виросомы.
21. Виросома, полученная способом по п.20.
| AMACKER M | |||
| et al | |||
| Peptide-loaded chimeric influenza virosomes for efficient in vivo induction of cytotoxic cells, Oxford Journal Life sciences & Medicine International Immunolog., 2005, v.17, №6, pp.695-704 | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| GLUCK R., WALTI E | |||
| Biophysical validation of EPAXAL Berna, a hepatitis A vaccine adjuvanted with | |||

