Изобретение относится к области молекулярной биологии, молекулярной генетики, биотехнологии. Способ основан на использовании двух важнейших свойств ДНК молекул - (первое) комплементарности, которое позволяет одноцепочечным молекулам ДНК гибридизоваться, и (второе) полуконсервативном способе репликации. В результате общеизвестного процесса два отдельных фрагмента «сливаются» в один более крупный. Данный способ может найти широкое применение при поиске мутаций в генах, т.к. будет приводить к снижению трудозатрат и себестоимости молекулярно-генетических исследований. При определенной модификации данное изобретение может найти применение и в других областях молекулярной биологии.
УРОВЕНЬ ТЕХНИКИ
Получение сборных фрагментов ДНК широко используется в биотехнологии для получения рекомбинантных белков, для амплификации фрагментов ДНК в клеточной системе, а также с другими целями, при этом основным подходом является получение «липких» концов у фрагментов ДНК, которые предполагается срастить. Последнее достигается использованием рестрикции, которая формирует коплементарные концы у ДНК, представляющей интерес, и вектора (1, 2), в дальнейшем проводится сшивание вектора и ДНК-вставки с помощью лигаз (2, 3, 4). Благодаря развитию техники ПЦР наиболее популярным подходом для создания липких концов у ДНК мишени является использование праймеров с наличием «хвостов», содержащих необходимые для клонирования сайты рестрикции. Полученные ампликоны подвергаются обработке эндонуклеазами, затем очистке и лигированию с вектором.
Другой подход к получению «липких» концов у ДНК для ее клонирования - использование дезоксиурацила в «хвостах» используемых праймеров. После проведения ПЦР продукт реакции обрабатывается дезоксиурацил гликозилазой, что завершает формирование требуемых комплементарных выступов без использования рестрикции (5, 6). Была продемонстрирована попытка упростить процесс генерирования липких концов у ДНК-мишени, исключив из методики клонирования рестрикцию, а также обработку продуктов ПЦР дезоксиурацил гликозилазой (7). Принцип состоит в использовании 2 отдельных реакций ПЦР для одного продукта. В каждой из реакций используется только один праймер, содержащий «хвост» с последовательностью сайта рестрикции. Тогда как второй праймер полностью комплементарен матрице. Независимо получаются 2 продукта - у одного сайт рестрикции на левом конце, у другого - на правом. После окончания ПЦР продукты двух реакций соединяют, денатурируют нагреванием до 95°C. После охлаждения в смеси образуются гомодимеры, часть из которых имеет слева и справа «липкий» конец. В дальнейшем смесь используется для лигирования.
Рассматривались также принципы сращивания 2 фрагментов ПЦР с помощью частично перекрывающихся последовательностей, когда смешиваются 2 фрагмента каждый из которых комплементарен с одного конца другому, и с помощью 2 праймеров (прямого и обратного), фланкирующих общую последовательность, необходимую получить, амплифицируется один сборный фрагмент (8, 9). Этод подход позволяет реконструировать ДНК молекулы с высокой степенью деградации (10).
Была разработана технология получения сборных ДНК молекул из нескольких фрагментов с использованием химерических праймеров, имеющих на 5' концах последовательности комплементарные соответствующим концам других присоединяемых фрагментов ДНК для генерирования на изначально амплифицируемых фрагментах ДНК липких концов. Процесс состоит из нескольких стадий. На первом этапе получают первичные ПЦР фрагменты с липкими концами. Каждый фрагмент синтезируется отдельно от других в индивидуальной пробирке. Затем производится очистка фрагментов вырезанием их из агарозного геля и т.д.. Следующий этап заключается в соединении фрагментов методом ПЦР, но без использования праймеров. Фрагменты соединяются с помощью липких концов, элонгируются, формируя более крупные. На последнем этапе в реакционную смесь добавляют фланкирующие общий фрагмент праймеры и проводят его амплификацию. Данный этап предполагает использование nested ПЦР (11). Несколько ранее примерно аналогичный подход был использован H.Kuwayama и соавт., которые сумели с помощью ПЦР срастить в один 3 разных ПЦР фрагмента с липкими концами для чего, согласно разработанной авторами схеме, потребовалось получение «первичных» 3-х ПЦР продуктов, с использованием 3-х независимых реакций, их очистка. Затем 3 продукта смешивались и проводилась ПЦР с использованием фланкирующих общий фрагмент праймеров. Данный этап предполагал одновременную сборку требуемого сложного фрагмента ДНК и его амплификацию (12). Известен также подход для сращивания 2-х фрагментов ДНК или для получения делеции внутри гена, использующий 3 праймера - два из которых фланкируют общий фрагмент, а третий имеет последовательности двух сращиваемых фрагментов и обозначается как праймер сращивания ('linking oligo'). Процесс предполагал использование заранее клонированных 2-х фрагментов в определенной концентрации, состоял из 4 стадий (13).
Таким образом, получение сборных фрагментов - многостадийный процесс, включающий в себя получение первичных фрагментов ДНК, создание у сливаемых фрагментов «липких» (взаимно комплементарных) концов, слияние фрагментов в один и амплификацию общего фрагмента.
В наиболее близком аналоге, представленной нами ниже схеме - работе A.N.Warrens и др. для соединения 2-х молекул ДНК генерировались «липкие» концы в 2-х раздельных реакциях ПЦР с использованием в каждой реакции 2-х праймеров, один из которых содержал на 5' конце соответствующую другому фрагменту последовательность. Затем ПЦР продукты очищались от посторонних примесей, для чего кусочки геля, содержащие требуемые ДНК фрагменты, вырезались после электрофореза, ДНК элюировалась. Следующий этап использовал смесь очищенных фрагментов и 2 праймера, фланкирующих сборный продукт (14). Т.е. каждый этап работы был четко отделен от остальных. При амплификации конечного продукта в реакционной смеси не присутствовало ничего, что теоретически могло бы помешать его синтезу.
Мы разработали способ получения объединенных фрагментов ДНК, позволяющий получить конечный продукт из сложной смеси (геномной) ДНК при проведении 2-х раундов ПЦР, не требующих раздельного получения отдельных фрагментов и очистки полученных первичных фрагментов от праймеров и геномной ДНК. После второго раунда был получен конечный продукт в концентрации, достаточной для сиквенса и не требующий вырезаний из геля для очистки от балластной ДНК. Принцип заключается в получении нескольких фрагментов ПЦР в одной пробирке (мультиплексная ПЦР) с использованием для каждого фрагмента пары праймеров, каждый из которых с температурой плавления не выше 57°С. Во втором раунде используются праймеры, фланкирующие общий фрагмент, и праймеры для соединения фрагментов - см. схемы 1 и 2. 3' последовательность праймеров, предназначенных для соединения фрагментов, комплементарна концевой последовательности одного фрагмента, а 5' последовательность комплементарна концевой последовательности другого фрагмента, который планируется присоединить к первому. Т.е. на каждые 2 фрагмента предназначается 1 праймер для слияния. Температура плавления как 3' последовательности, так и 5' последовательности этих праймеров не ниже 70°С. Столь же высокую температуру плавления имеют праймеры (2 праймера), фланкирующие общий фрагмент, который предполагается получить, и предназначенные для его амплификации. Во втором раунде ПЦР объединены следующие стадии: получение липких концов у фрагментов ДНК, слияние отдельных ПЦР фрагментов в общую молекулу и ее амплификация. В первом раунде в качестве матрицы можно использовать полногеномную ДНК человека, во второй раунд предполагается брать неочищенную аликвоту из пробирки после проведения первого раунда. Использование в первом раунде праймеров с относительно низкой температурой плавления, а во втором с высокой, позволяет исключить помехи, которые могли бы оказывать во втором раунде ПЦР пары праймеров, предназначенные для генерации первичных ПЦР фрагментов. Т.к. они переносятся вместе с другими компонентами из пробирки «первого раунда» в пробирку для проведения второго раунда ПЦР. Такая избирательность достигается с помощью повышения температуры отжига праймеров до того уровня, что относительно короткие праймеры из первого раунда ПЦР теряют возможность гибридизоваться с ДНК фрагментами. Праймеры, используемые во втором раунде ПЦР для амплификации общего фрагмента, берутся в большей концентрации, нежели праймеры, предназначенные для слияния фрагментов, чтобы создать преимущество для конечной стадии процесса объединения фрагментов. Поскольку т.н. липкие концы, генерируемые у первичных ПЦР фрагментов праймерами для слияния, представляют собой последовательности, комплементарные концевым участкам соответствующих присоединяемых фрагментов, данный подход позволяет получать сборные фрагменты ДНК, не имеющие в своем составе последовательностей, не входящих в состав первичных ДНК фрагментов и соответственно геномной ДНК.
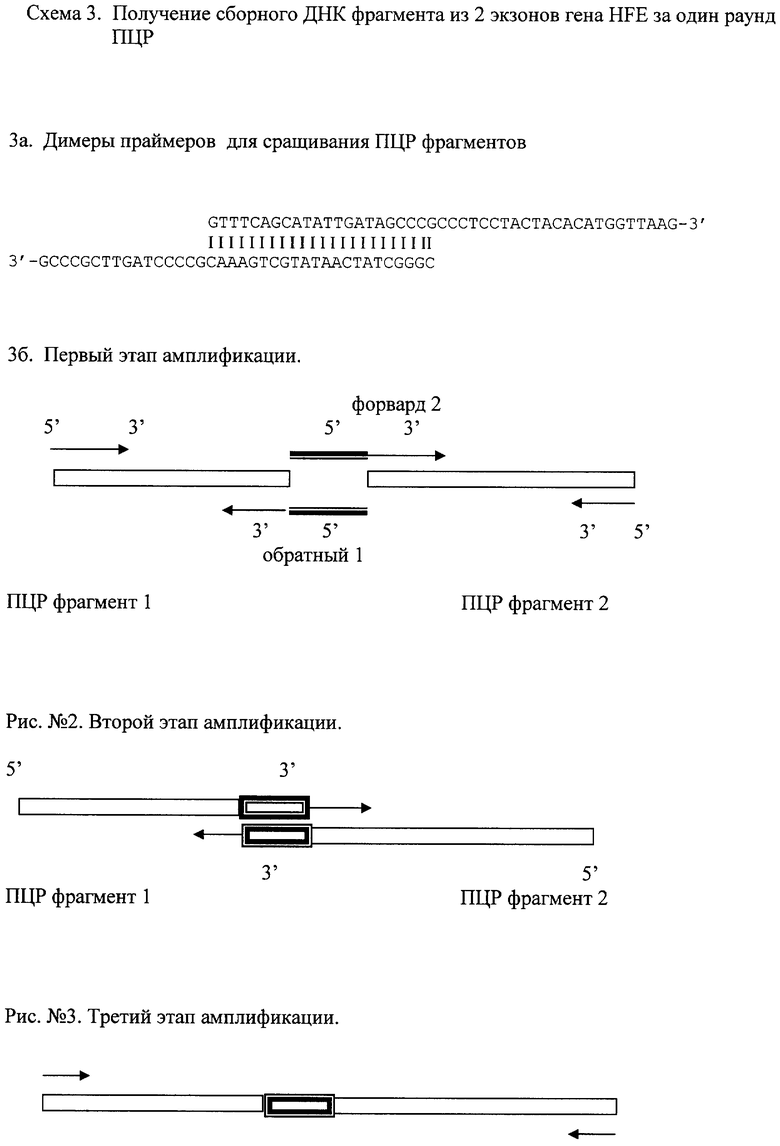
Когда необходимость получения точных (без вставок) фрагментов отсутствует, есть возможность объединить в одной реакции ПЦР 4 процесса: получение первичных фрагментов, генерирование у первичных фрагментов липких концов, слияние первичных фрагментов в один, амплификацию конечного продукта. Здесь в качестве матрицы также может быть использована смесь высокомолекулярной ДНК. Конечный продукт пригоден для сиквенса без электрофоретической очистки. Достаточно развести полученный продукт в необходимой пропорции деионизованной водой. В качестве первичного источника ДНК может быть использована смесь высокомолекулярной ДНК (полногеномной ДНК человека) см. примеры. В реакции используются 2 праймера, фланкирующих конечный сборный продукт (общие для первичных фрагментов и конечного), и 2 праймера (при сборке 2-х фрагментов), каждый из которых специфичен только одному из первичных фрагментов и имеет «липкий» 5' конец к 5' концу праймера другого фрагмента, предназначенного для соединения - см. схему. В отношении высокомолекулярной ДНК эти липкие концы должны быть уникальны. Фланкирующие праймеры присутствуют в более высокой концентрации, нежели праймеры с взаимно липкими 5' концами для обеспечения преимущества амплификации конечного сборного продукта. Поскольку праймеры находятся в реакционной смеси в молярной избыточности по отношению к геномной ДНК, праймеры с взаимно «липкими» 5' концами предположительно образуют гомодимеры с выступающими 3' концами (см. схему 3а). Это снижает вероятность получения избыточных артефактов реакции. Конечная ДНК из 2-х фрагментов будет содержать на стыке вставку в виде уникальной последовательности «липких» 5' концов праймеров для сборки.
ОПИСАНИЕ ПОСТАНОВКИ ЭКСПЕРИМЕНТА
ПЦР проводится в термоциклере.
Вариант первый - получение сборных фрагментов без вставок. 2 раунда ПЦР.
Необходимое условие - разница в температуре плавления между праймерами для первого и второго раунда ПЦР должна быть такой, чтобы исключить отжиг праймеров, предназначенных для первого раунда ПЦР, во втором раунде ПЦР. Для проведения первого раунда используется столько пар праймеров, сколько фрагментов ДНК предполагается срастить (мультиплексная ПЦР). Во втором раунде используются фланкирующие общий фрагмент праймеры и праймеры, имеющие «липкие» концы к присоединяемым фрагментам, праймеры с липкими концами берутся в меньшей концентрации, чем фланкирующие праймеры. В качестве источника ДНК могут использоваться продукты, полученные в результате первого раунда ПЦР без очистки. Время элонгации выбирается соответственно размеру получаемого фрагмента. И в первом и во втором раунде используется «горячий старт».
Вариант второй - получение сборных фрагментов ДНК, имеющих в составе «постороннюю» последовательность ДНК. Мультиплексная ПЦР. Используется столько пар праймеров, сколько фрагментов планируется срастить. Фланкирующие общий фрагмент праймеры не имеют существенных отличий от ДНК матрицы. Все другие праймеры имеют взаимные липкие 5' концы для создания таковых у фрагментов. В реакцию праймеры с липкими концами берутся в меньшей концентрации, чем фланкирующие праймеры. Время элонгации устанавливается соответственно размеру получаемого фрагмента. Желательно, чтобы праймеры имели примерно равную температуру плавления. Используется «горячий старт». В остальном никаких особенностей. Перед сиквенсом полученный фрагмент очищается вырезанием из агарозного геля, либо с помощью хроматографических колонок (необязательная стадия).
ПРИМЕРЫ ПРИМЕНЕНИЯ
Вариант первый
Пример №1. Соединение 3 экзонов (2, 3 и 4) гена CFTR (cystic fibrosis conductance regulator gene) (сплайсинг)
ДНК выделена из лейкоцитов крови с помощью набора для выделения ДНК «ДНК-ЭКСПРЕСС» производства «Литех» согласно инструкции, прилагаемой к набору. В первом раунде ПЦР были использованы следующие праймеры:
TTGGGGGCAGTTATTTTTT-2F
CTTTCCAATTTTTCAGATAGA-2R - для синтеза 2 экзона
AGAATGGGATAGAGAGCTG-3F
CCCCTAAATATAAAAAGATTC-3R - для синтеза 3 экзона
AAGTCACCAAAGCAGTACAG-4F
GCATTTATCCCTTACTTGT-4R - для синтеза 4 экзона
В реакционную смесь, содержащую буфер, смесь 4-х дезоксинуклеотид-3'фосфатов (5 mMol каждого), добавляли указанные выше 3 пары праймеров (10 pMol каждого праймера), 2 mkl геномной человеческой ДНК (концентрация не измерялась) из полученного препарата ДНК. Taq ДНК полимераза (5 ед) была добавлена в раствор после установления в блоке термоциклера температуры 85°С («горячий» старт).
Амплификация проводилась по следующей схеме:
95°С - 5 мин,
85°С - 5 мин (добавление Taq ДНК полимеразы),
55°С - 30 сек,
72°С - 45 сек.
После чего 35 циклов по программе: 95 - 30 сек, 55 - 30 сек, 72 - 45 сек
Во втором раунде были использованы следующие праймеры:
CGCCAGGGTTTCCCAGTCACGACGGTGTAGCCTGTAAGAGATGAAG-2fCM13F
TCACACAGGAAACAGCTATGACAGCATTTATCCCTTACTTGTACCAG-4fCM13R
- фланкирующие общий сборный фрагмент праймеры с последовательностями на 5' концах праймеров для сиквенса (вставки праймеров для сиквенса выделены курсивом и подчеркнуты - используются только для удобства последующего секвенирования, можно последовательности праймеров для сиквенса не использовать)
TTCTTTGAAGCCAGCTCTCTATCCCATTCTCTTTCCAATTTTTCAGATAGATTGTCAGCA-2FCR3F
- обратный праймер для второго экзона содержит на 5' конце участок, обратно комплементарный 5' концу 3-го экзона данного гена.
AAGAGAGGCTGTACTGCTTTGGTGACTTCCCCTAAATATAAAAAGATTCCATAGAACATAA-3CFR4f
- обратный праймер для третьего экзона содержит на 5' конце участок, обратно комплементарный 5' концу 4-го экзона данного гена.
В реакционную смесь, содержащую буфер, смесь 4-х дезоксинуклеотид-3' фосфатов (5 mMol каждого), добавляли указанные выше праймеры 2fCM13F и 4fCM13R.
(10 pMol каждого праймера), а также праймеры 2FCR3F и 3CFR4f (1 pMol каждого праймера), 2 mkl ДНК аликвота из пробирки после первого раунда амплификации (концентрация не измерялась). Taq ДНК полимераза (5 ед) была добавлена в раствор после установления в блоке термоциклера температуры 85°С («горячий» старт).
Амплификация проводилась по следующей схеме:
95°С - 5 мин,
85°С - 5 мин (добавление Taq ДНК полимеразы),
70°С - 1 мин 30 сек.
После чего 35 циклов по программе: 95 - 30 сек, 70°С - 1 мин 30 сек.
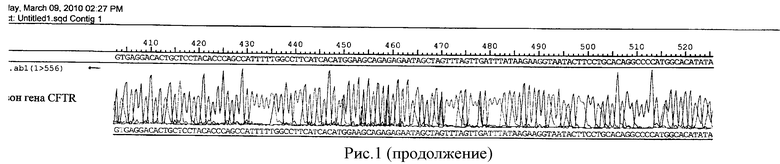
После проведения второго раунда ПЦР, продукт реакции был разбавлен в 15 раз и без очистки секвенирован - см. рис.1.
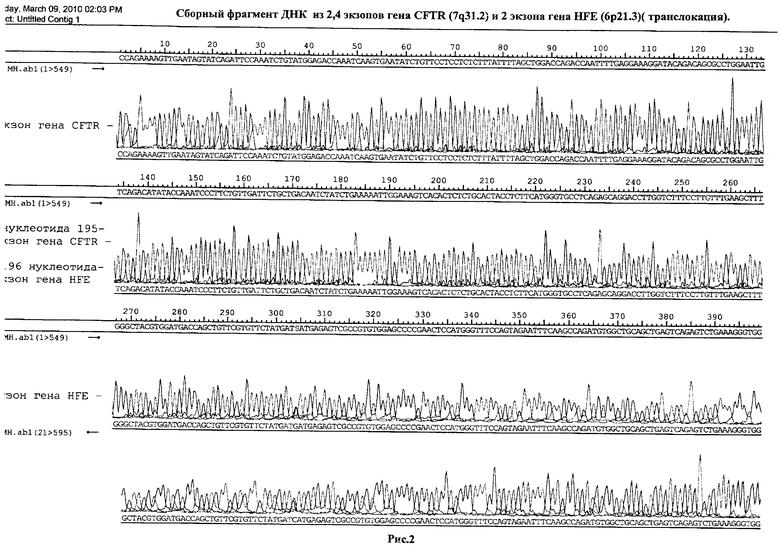
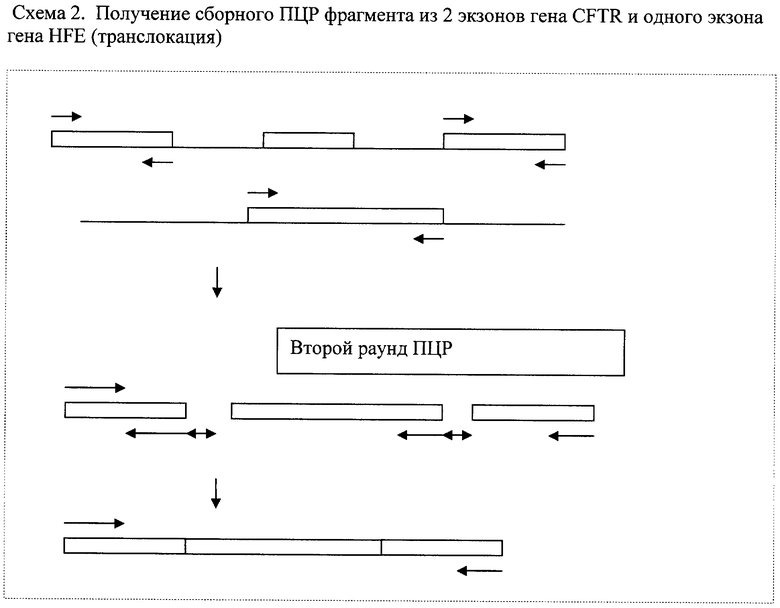
Пример №2. Получение сборного фрагмента ДНК из 2,4 экзонов гена CFTR, находящегося на длинном плече 7 хромосомы (7q31.2) и 2 экзона гена HFE, расположенного на коротком плече 6 хромосомы (6р21.3) (транслокация).
ДНК выделена из лейкоцитов крови с помощью набора для выделения ДНК «ДНК-ЭКСПРЕСС» производства «Литех» согласно инструкции, прилагаемой к набору. В первом раунде ПЦР были использованы следующие праймеры:
TTGGGGGCAGTTATTTTTT-2F
CTTTCCAATTTTTCAGATAGA-2R - для синтеза 2 экзона гена CFTR
TCACACTCTCTGCACTAC-2FH
CCTTGCTGTGGTTGTGAT-2RH - для синтеза 2 экзона гена HFE
AAGTCACCAAAGCAGTACAG-4F
GCATTTATCCCTTACTTGT-4R - для синтеза 4 экзона гена CFTR
В реакционную смесь, содержащую буфер, смесь 4-х дезоксинуклеотид-3' фосфатов (5 mMol каждого), добавляли указанные выше 3 пары праймеров (10 pMol каждого праймера), 2 mkl геномной человеческой ДНК (концентрация не измерялась) из полученного препарата ДНК. Taq ДНК полимераза (5 ед) была добавлена в раствор после установления в блоке термоциклера температуры 85°С («горячий» старт).
Амплификация проводилась по следующей схеме:
95°С - 5 мин,
85°С - 5 мин (добавление Taq ДНК полимеразы),
55°С - 30 сек,
72°С - 45 сек.
После чего 35 циклов по программе: 95 - 30 сек, 55 - 30 сек, 72 - 45 сек.
Во втором раунде были использованы следующие праймеры:
CGCCAGGGTTTCCCAGTCACGACGGTGTAGCCTGTAAGAGATGAAG-2fCM13F
TCACACAGGAAACAGCTATGACAGCATTTATCCCTTACTTGTACCAG-4fCM13R
- фланкирующие общий сборный фрагмент праймеры с последовательностями на 5' концах праймеров для сиквенса (вставки праймеров для сиквенса выделены курсивом и подчеркнуты - используются только для удобства последующего секвенирования, можно последовательности праймеров для сиквенса не использовать)
CCATGAAGAGGTAGTGCAGAGAGTGTGACTTTCCAATTTTTCAGATAGATTGTCAGCA-2FCR2H
- обратный праймер для второго экзона гена CFTR содержит на 5' конце участок, обратно комплементарный 5' концу 2-го экзона гена HFE
AAGAGAGGCTGTACTGCTTTGGTGACTTCCTTGCTGTGGTTGTGATTTTCCAT-2H4FCR
- обратный праймер для второго экзона гена HFE
CFTR содержит на 5' конце участок, обратно комплементарный 5' концу 4-го экзона гена CFTR
В реакционную смесь, содержащую буфер, смесь 4-х дезоксинуклеотид-3' фосфатов (5 mMol каждого), добавляли указанные выше праймеры 2fCM13F и 4 fCM13R
(10 pMol каждого праймера), а также праймеры 2FCR2H и 2H4FCR (1 pMol каждого праймера), 2 mkl ДНК аликвота из пробирки после первого раунда амплификации (концентрация не измерялась). Taq ДНК полимераза (5 ед) была добавлена в раствор после установления в блоке термоциклера температуры 85°С («горячий» старт).
Амплификация проводилась по следующей схеме:
95°С - 5 мин,
85°С - 5 мин (добавление Taq ДНК полимеразы),
70°С - 1 мин 30 сек.
После чего 35 циклов по программе: 95 - 30 сек, 70°С - 1 мин 30 сек.
После проведения второго раунда ПЦР, продукт реакции был разбавлен в 15 раз и без очистки секвенирован - см. рис.2.
Вариант второй
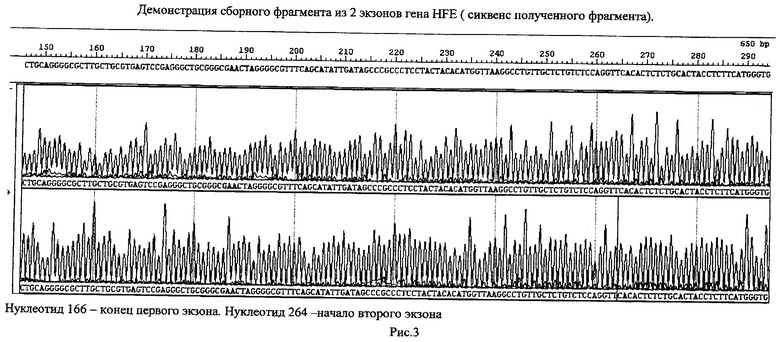
Пример - получение сборного фрагмента ДНК из смеси высокомолекулярной (геномная человеческая ДНК), состоящего из 2-х фрагментов гена HFE в процессе проведении одной реакции ПЦР.
ДНК выделена из лейкоцитов крови с помощью набора для выделения ДНК «ДНК-ЭКСПРЕСС» производства «Литех» согласно инструкции, прилагаемой к набору. В первом раунде ПЦР были использованы следующие праймеры:
GGTGGCGACGACTCCTGGAGCCCGCTGGAGCCATCCCCGTTT-1HF - прямой праймер для амплификации первого экзона гена HFE, содержит последовательность праймера для сиквенса (выделено курсивом и подчеркнуто)
CGGGCTATCAATATGCTGAAACgcccctagttcgcccg-1HRrevJ -
Обратный праймер для первого экзона содержит на 5' конце последовательность, обратно комплементарную последовательности на 5' конце прямого праймера для второго экзона гена HFE (выделена прописными буквами)
GTTTCAGCATATTGATAGCCCGccctcctactacacatggttaag-2HFforJ
Прямой праймер для второго экзона содержит на 5' конце последовательность, обратно комплементарную последовательности на 5' конце обратного праймера для первого экзона гена HFE (выделена прописными буквами), эта часть праймера является фрагментом генома вируса японского энцефалита
TGACACGAGACGCAACGCTGCTCAGCAACCTGCTCCCACAAGACCTCAG-2HR - обратный праймер для амплификации второго экзона гена HFE, содержит последовательность праймера для сиквенса (выделено курсивом и подчеркнуто)
Наличие в праймерах, фланкирующих общий ПЦР фрагмент, не является обязательным условием и используется для удобства секвенирования.
В реакционную смесь, содержащую буфер, смесь 4-х дезоксинуклеотид-3' фосфатов (5 mMol каждого), добавляли указанные выше праймеры 1HF и 2HR
(10 pMol каждого праймера), а также праймеры 1HRrevJ и 2HFforJ (1 pMol каждого праймера), 2 mkl ДНК аликвота из пробирки после первого раунда амплификации (концентрация не измерялась). Taq ДНК полимераза (5 ед) была добавлена в раствор после установления в блоке термоциклера температуры 85°С («горячий» старт).
Амплификация проводилась по следующей схеме:
95°С - 5 мин,
85°С - 5 мин (добавление Taq ДНК полимеразы),
60°С - 30 сек,
72°С - 1 мин 30 сек.
После чего 35 циклов по программе: 95 - 30 сек, 60 - 30 сек, 72 - 1 мин 30 сек.
После проведения амплификации продукт реакции разводили в 5 раз деионизованной водой и использовали для сиквенса - см. рис.3.
Источники информации
1. Proc. Nat. Acad. Sci. USA, 1973, 70, 3240-3244.
2. Current Opinion in Biotechnology 1995, 6: 37-43.
3. Nucleic Acids Research 2002, 30, 2e8. (A genetic system for direct selection of gene-positive clones during recombinational cloning in yeast).
4. Science 2002 296: 550-553.
5. PCR Methods Appl 1991, 1: 121-123.
6. Anal Biochem 1992 286: 91-97.
7. Plasmid 2008 59, 155-162.
8. Nucl. Acids Res. 1988 16, 7351-7367.
9. Gene 1989 77, 61-68.
10. Nucleic Acids Research 1996 24, 24, 5026-5033.
11. Nucleic Acids Research, 2004, Vol.32, No. 2 e19 (Construction of long DNA molecules using long PCR-based fusion of several fragments Simultaneously).
12. Nucleic Acids Research 2002 30, 2e2. (PCR-mediated generation of a gene disruption construct without the use of DNA ligase and plasmid vectors).
13. Nucleic Acids Research 1989 17, 12, 4895.
14. Gene 1997 186, 29-35.


Изобретение относится к области биотехнологии. Описаны способы слияния ПЦР фрагментов, которые не требуют проведения промежуточных очисток, предварительного клонирования первичных ДНК фрагментов, а также раздельной амплификации первичных фрагментов. Проведение двух раундов ПЦР достаточно для получения конечного продукта. В первом раунде с помощью мультиплексной ПЦР получают первичные фрагменты ДНК, во втором раунде у полученных ДНК фрагментов появляются «липкие» концы, фрагменты сливаются и результирующий продукт амплифицируется. В качестве первичного материала в первом раунде можно использовать смесь высокомолекулярной ДНК или геномную ДНК человека, во втором раунде используются ПНР продукты из первого раунда. Способ позволяет получать очень точные фрагменты, состоящие из одних экзонов. Если необходимость получения сборных фрагментов без вставок отсутствует, предлагается получать сборные фрагменты из смеси высокомолекулярной ДНК в одной реакции ПЦР. Мультиплексная ПЦР с использованием особых праймеров, которые содержат уникальные для геномной ДНК 5'-концы, которые взаимно комлементарны, чем достигается формирование липких концов у синтезируемых первичных фрагментов. При этом используется также пара фланкирующих общий составной фрагмент праймеров, которые полностью комплементарны внешним краям желаемого составного ДНК фрагмента и предназначенные для его амплификации. 2 н.п. ф-лы, 7 ил.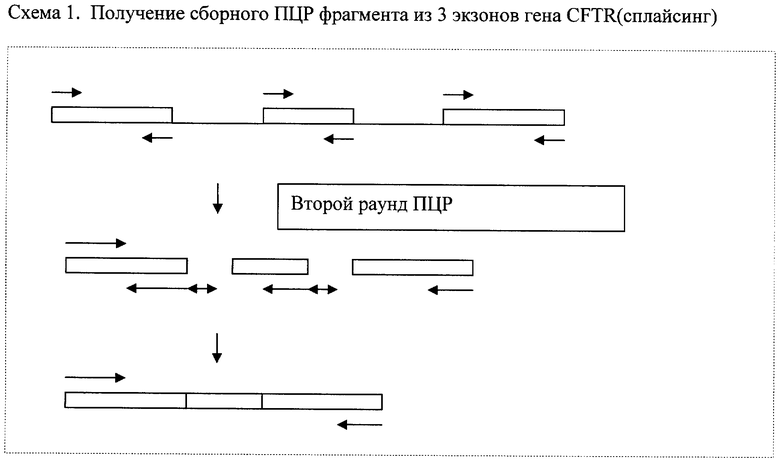
1. Способ получения объединенных фрагментов ДНК в ПЦР с использованием для их слияния генерируемых липких концов, включающий синтез первичных фрагментов, генерирование у сливаемых фрагментов липких концов, слияние фрагментов в один и амплификацию общего фрагмента, отличающийся тем, что все стадии осуществляются в пределах двух раундов мультиплексной ПЦР, использованием двух групп специфических праймеров с разной температурой отжига: не выше 57°С - для первого раунда, в котором происходит синтез первичных ПЦР фрагментов, не ниже 70°С - для второго раунда, в котором объединены все остальные стадии процесса, которые реализуются благодаря комплементарности 5' концов, используемых во втором раунде ПЦР праймеров, концами тех ДНК фрагментов, которые надо присоединить, что позволяет генерировать у ДНК фрагментов взаимно комплементарные концы, общий сборный фрагмент амплифицируется с помощью двух праймеров, также имеющих температуру отжига не ниже 70°С, при этом в качестве источника материала для получения начальных фрагментов используется смесь высокомолекулярной ДНК/геномной ДНК, а вторая стадия использует в качестве матриц продукты первого раунда ПЦР без очистки.
2. Способ получения объединенных фрагментов ДНК в ПЦР, предусматривающий амплификацию и сращивание фрагментов ДНК в течение одного раунда мультиплексной ПЦР с использованием праймеров, которые одновременно с синтезом первичных фрагментов формируют у них комплементарные концы, что достигается через взаимную комплементарность 5' концов праймеров сращиваемых фрагментов, при этом последовательность 5' концов данных праймеров в отношении изначальной смеси ДНК, а также остальных праймеров, которые используются в данном процессе, должна быть уникальной.
| KUWAYAMA H | |||
| et al., "PCR-mediated generation of a gene disruption construct without the use of DNA ligase and plasmid vectors", Nucleic Acids Res | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| SHEVCHUK N.A | |||
| et al., "Construction of long DNA molecules using long PCR-based fusion of several fragments simultaneously", Nucleic Acids Res | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| NOSKOV V. | |||

