ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение в основном относится к развитию нервной системы и к неврологическим расстройствам. Конкретно настоящее изобретение относится к идентификации новых модуляторов ассоциированной с миелином ингибирующей системы и к различным вариантам использования идентифицированных таким образом модуляторов.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Миелин и миелин-ассоциированные белки
Известно, что аксоны нейронов ЦНС взрослых млекопитающих обладают очень ограниченной способностью регенерировать после повреждения, тогда как аксоны в периферической нервной системе (ПНС) регенерируют быстро. Ограниченная способность нейронов ЦНС к регенерации является собственным свойством аксонов ЦНС, но, кроме того, определяется также особенностями среды, которая не способствует такой возможности. Миелин ЦНС, хотя он и не является единственным источником, определяющим ингибирующее воздействие на рост нейритов, содержит множество молекул ингибирующего действия, которые активно блокируют рост аксонов и, в этой связи, создают значительный барьер для регенерации. Были идентифицированы три таких миелин-ассоциированных белка (МАВ): Nogo (также известный как NogoA) является членом семейства белков Reticulon, имеющих два трансмембранных домена; миелин-ассоциированный гликопротеин (MAG) представляет собой трансмембранный белок из надсемейства Ig; и OMgp представляет собой белок с лейцин-обогащенными повторами (LRR) с гликозилфосфатидилинозитоловым (GPI) фрагментом (Chen et al., Nature 403:434-39 (2000); GrandPre et al., Nature 417:439-444 (2000); Prinjha et al., Nature 403:383-384 (2000); McKerracher et al, Neuron 13:805-11 (1994); Wang et al, Nature 417:941-4 (2002); Kottis et al J. Neurochem 82:1566-9 (2002)). Nogo66, представляющий собой часть NogoA, был описан как внеклеточный полипептид из 66 аминокислот, обнаруженный во всех трех изоформах Nogo.
Несмотря на структурные различия, все три ингибирующих белка (а также Nogo66), как было показано, связываются с одним и тем же рецептором с GPI-фрагментом, называемым Nogo-рецептором (NgR; также известным как Nogo-рецептор-1 или NgR1), и было высказано предположение, что NgR может быть необходим в качестве посредника ингибирующего действия Nogo, MAG и OMgp (Fournier et al., Nature 409:34'-346 (2001). Были также идентифицированы два NgR1 гомолога (NgR2 и NgR3) (патент США 2005/0048520 A1 (Strittmatter et al.), опубликованный 3 марта 2005 года). Ввиду того, что NgR представляет собой белок клеточной поверхности с GPI-фрагментом, маловероятно, что он может представлять собой непосредственный сигнальный трансдуктор (Zheng et al, Proc. Natl. Acad. Sci. USA 102:1205-1210 (2005)). Другие авторы высказали предположение, что рецептор нейротрофина p75NTR действует как ко-рецептор для NgR и обеспечивает сигнал-трансдуцирующий фрагмент в рецепторном комплексе (Wang et al., Nature 420:74-78 (2002); Wong et al., Nat. Neurosci. 5:1302-1308 (2002)).
Однако последние исследования рецепторного комплекса NgR/p75NTR подняли вопрос о роли NgR в миелин-ассоциированной ингибирующей системе. Zheng et al. показали, что генетическая делеция NgR не снижает ингибирование нейрита in vitro, как и не стимулирует регенерацию пирамидного пути (CST) in vivo. (Zheng et al. (2005), выше). Аналогично по результатам другого исследования также не удалось обнаружить какой-либо повышенной регенерации CST у NgR мутантных мышей. (Kim et al, Neuron 44:439-451 (2004)). Эти наблюдения противоречат гипотезе о том, что рецепторный комплекс NgR/p75NTR представляет собой ключевую точку в конвергенции множества ингибирующих сигналов. Неспособность к регенерации CST у NgR мутантных мышей не согласуется с данными о регенерации CST, наблюдаемой у животных дикого типа, которым вводили пептидный антагонист взаимодействия Nogo66/NgR (GrandPre et al. Nature 417:5470551 (2002) и Li и Strittmatter, Nature 23:4219-4227 (2002)). В другом исследовании было показано, что экспрессия доминант-отрицательного фрагмента NgR ведет к повышенной регенерации аксонов зрительного нерва в сочетании с соответствующим повреждением. Причем в обоих экспериментах не удалось определить непосредственно вовлечения NgR, поскольку оба антагонистических пептида обладают способностью препятствовать действию других ингибирующих лигандов/рецепторов.
Указанные несоответствия, имеющиеся в результатах проведенных экспериментов, представляют собой очень значимое указание на то, что NgR или рецепторный комплекс NgR/p75NTR может играть лишь ограниченную роль в миелин-ассоциированном ингибировании регенерации ЦНС и что другие компоненты, такие как дополнительные рецепторы, соответствующие связывающие молекулы и другие компоненты, могут принимать участие в передаче ингибирующего сигнала.
PirB и ортологи человека
Главная система тканевой совместимости (MHC) класса I была первоначально идентифицирована как область, кодирующая семейство молекул, важных для иммунной системы. В последнее время были получены данные, подтверждающие тот факт, что молекулы MHC класса I обладают дополнительными функциями в развитии и созревании ЦНС взрослого организма. (Boulanger и Shatz, Nature Rev Neurosci. 5:521-531 (2004); патент США 2003/0170690 (Shatz and Syken), опубликованный 11 сентября 2003 года). Было показано, что многие представители MHC класса I и соответствующие им связующие молекулы экспрессируются в нейронах ЦНС. Проведенные в последнее время генетические и молекулярные исследования были сконцентрированы на изучении физиологических функций молекул MHC класса I ЦНС и первоначальные результаты позволили предположить, что молекулы MHC класса I могут вовлекаться в синаптическую пластичность, зависимую от активности, процесс, в ходе которого сила существующих синаптических соединений повышается или понижается в ответ на нейрональную активность, с последующим созданием длительных структурных изменений в системе. Более того, область, кодирующая молекулы MHC класса I, также генетически связана с широким спектром расстройств, характеризующихся неврологическими симптомами, и полагают, что аномальные функции молекул MHC класса I вносят определенный вклад в нарушение нормального развития головного мозга и его пластичности.
Одним из известных рецепторов соединений MHC класса I в иммунном наборе является PirB, мышиный полипетид, который был впервые описан Кубагавой с соавт. (Kubagawa et al, Proc. Nat. Acad. Sci. USA 94:5261-6 (1997)). Мышиный PirB имеет несколько ортологов человеческого происхождения, которые являются членами надсемейства В лейкоцитарных иммоноглобулин-подобных рецепторов (LILRB), также обозначаемых как «иммуноглобулин-подобные транскрипты» (ILT). Человеческие ортологи демонстрируют выраженную гомологию с последовательностью мышиных молекул, в следующем порядке, начиная от высшей в направлении к низшей степени гомологии: LILRB3/ILT5, LILRB1/ILT2, LILRB5/ILT3, LILRB2/ILT4 и, как и PirB, все они являются ингибирующими рецепторами. LILRB3/ILT5 (NP_006855) и LILRB1/ILT2 (NP_006660) были впервые описаны Samaridis and Colonna, Eur. J. Immunol. 27(3):660-665 (1997). LILRB5/ILT3 (NP_006831) был идентифицирован Borges et al., J. Immunol. 159(11):5192-5196 (1997). LILRB2/ILT4 (также известный как MIR10) был идентифицирован Colonna et al, J. Exp. Med. 186:1809-18 (1997). PirB и его человеческие ортологи демонстрируют высокую степень структурной вариабельности. Последовательности различных форм с альтернативным сплайсингом доступны от EMBL/GenBank, включая, например, последовательности с указанными ниже номерами доступа для кДНК человека ILT4: ILT4-c11 AF009634; ILT4-c117 AF11566; ILT4-c126 AF11565. Как указывалось выше, полипептиды PirB/LILRB представляют собой ингибирующие рецепторы соединений MHC класса I (MHCI) и хорошо известны по той роли, которую они выполняют в регулировании активации иммунных клеток (Kubagawa et al., выше; Hayami et al., J. Biol. Chem. 272:7320 (1997); Takai et al., Immunology 115:433 (2005); Takai et al, Immunol. Rev. 181:215 (2001); Nakamura et al, Nat. Immunol. 5:623 (2004); Liang et al., Eur. J. Immunol. 32:2418 (2002)).
В недавнем исследовании, проведенном Сайкен с соавт. (Syken et al. (Science 313:1795-800 (2006)), было показано, что PirB экспрессируется в подмножестве нейронов по всему головному мозгу. В случае мутантных мышей, у которых отсутствует функциональный PirB, пластичность корковой зрительной доминантности (OD) значительно повышается во всех возрастных группах, указывая на возможную функцию PirB в ограничении зависимой от активности пластичности в зрительной коре.
Настоящее изобретение основано, по меньшей мере частично, на неожиданном выявлении того факта, что PirB/LILRB являются связывающими партнерами для Nogo (Nogo66) и MAG, и что PirB/LILRB являются антагонистами и сниженная активность PirB/LILRB эффективно разрушает миелин-ассоциированный ингибирующий путь, ускоряя тем самым нейрональную регенерацию.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном аспекте настоящее изобретение относится к способу идентификации антагониста PirB/LILRB, включающему контактирование исследуемого агента с рецепторным комплексом, который включает PirB/LILRB и миелин или миелин-ассоциированный белок или его фрагмент и детекцию способности исследуемого агента ингибировать взаимодействие между PirB/LILRB и миелин-ассоциированным белком или его фрагментом, где указанный исследуемый агент идентифицируют как антагонист, если взаимодействие ингибируется.
В одном варианте выявляемое взаимодействие представляет собой связывание.
В другом варианте выявляемое взаимодействие представляет собой клеточную передачу сигнала.
В еще одном варианте клеточная передача сигнала приводит к ингибированию роста аксона или нейрональной регенерации.
В еще одном варианте миелин-ассоциированный белок выбран из группы, состоящей из Nogo, MAG, OMgp и их фрагментов.
В другом варианте PirB/LILRB представляют собой белок человека LILRB, такой как LILRB1, LILRB2, LILRB3 или LILRB5.
В некоторых конкретных вариантах осуществления настоящего изобретения PirB/LILRB выбран из группы, состоящей из: LILRB2, варианта транскрипта 1 (SEQ ID NO: 2), LILRB2, варианта транскрипта 2 (SEQ ID NO: 14), LILRB1, варианта транскрипта 1 (SEQ ID NO: 10), LILRB1, варианта транскрипта 2 (SEQ ID NO: 11), LILRB1, варианта транскрипта 3 (SEQ ID NO: 12), LILRB1, варианта транскрипта 4 (SEQ ID NO: 13), L1LRB3, варианта транскрипта 1 (SEQ ID 30 NO: 15), LILRB3, варианта транскрипта 2 (SEQ ID NO: 16), LILRB5, варианта транскрипта 1 (SEQ ID NO: 17), LILRB5, варианта транскрипта 2 (SEQ ID NO: 18) и LILRB5, варианта транскрипта 3 (SEQ ID NO: 19).
В еще одном дополнительном варианте осуществления настоящего изобретения рецепторный комплекс дополнительно содержит NgR.
В других вариантах исследуемый агент выбран из группы, состоящей из антител, полипептидов, пептидов, нуклеиновых кислот, малых органических молекул, полисахаридов и полинуклеотидов и, предпочтительно, он представляет собой антитело или короткую интерферирующую РНК (киRNA). Указанное антитело предпочтительно специфически связывается с PirB/LILRB, таким как LIRB2, и включает, без ограничения, химерные, гуманизированные, антитела человека и фрагменты антител.
В конкретном варианте указанный фрагмент антитела выбран из группы, состоящей из Fv, Fab, Fab' и F(ab')2 фрагментов.
В еще одном варианте по меньшей мере один из PirB/LILRB и миелина или миелин-ассоциированного белка или его фрагмента иммобилизован.
В еще одном варианте используемый анализ представляет собой клеточный анализ.
В конкретном варианте клеточный анализ включает культивирование нейрональных клеток с миелином или миелин-ассоциированным белком, или его фрагментом, в присутствии или в отсутствие исследуемого агента, и определение наличия изменений в длине нейрита, где исследуемый агент идентифицирован как антагонист, когда длина нейрита становится больше в присутствии исследуемого агента.
В указанном выше клеточном анализе нейрональные клетки могут представлять собой первичные нейроны или, например, могут быть получены из клеток или клеточных линий, включающих стволовые клетки, например эмбриональные стволовые клетки (ES). В других вариантах указанные нейроны могут быть выбраны, например, из группы, состоящей из гранулярных нейронов мозжечка, нейронов спинномозговых узлов и корковых нейронов.
В одном варианте описанные выше способы дополнительно включают стадию использования идентифицированного антагониста с целью усиления роста нейрита и/или для стимуляции нейронального роста, восстановления и/или регенерации.
В другом варианте описанные выше способы дополнительно включают стадию введения идентифицированного антагониста субъекту, имеющему указанное заболевание или состояние, которое может быть ослаблено за счет усиления роста нейрита, стимуляции нейронального роста, восстановления или регенерации. Такое заболевание или состояние может, например, представлять собой неврологическое расстройство, которое может характеризоваться физически поврежденным нервом или может быть выбрано из группы, состоящей из повреждения периферического нерва, вызванного физическим повреждением, диабетом; физического повреждения центральной нервной системы; повреждения головного мозга, ассоциированного с инсультом, невралгии тройничного нерва, глоссофарингеальной невралгии, паралича Белла, миастении гравис, мышечной дистрофии, бокового амиотрофического склероза (ALS), прогрессирующей мышечной атрофии, прогрессирующей бульбарной наследственной мышечной атрофии, грыжи, разрыва и пролапса межпозвоночных дисков, шейного спондилеза, расстройств, затрагивающих сплетение, синдрома лестничной мышцы (сдавления плечевого сплетения и подключичной артерии), периферической нейропатии, порфирии, синдрома Гийена-Барре, болезни Альцгеймера, болезни Хантингтона и болезни Паркинсона.
В другом аспекте настоящее изобретение относится к агенту, идентифицированному по одному из приведенных в описании способу.
В одном варианте агент выбран из группы, состоящей из антител, полипептидов, пептидов, нуклеиновых кислот, малых органических молекул, полисахаридов и полинуклеотидов и, предпочтительно, он представляет собой антитело или короткую интерферирующую РНК (киRNA).
В другом аспекте настоящее изобретение относится к композиции, содержащей агент, идентифицированный согласно приведенным в описании способам, для стимуляции нейрональной регенерации.
В еще одном аспекте настоящее изобретение относится к набору, содержащему агент, идентифицированный согласно приведенным в описании способам, и инструкции по нейрональной регенерации.
В еще одном аспекте настоящее изобретение относится к применению комплекса PirB/LILRB и миелина или миелин-ассоциированного белка, или его фрагмента для идентификации антагониста PirB/LILRB.
В другом аспекте настоящее изобретение относится к применению антагониста PirB/LILRB при получении лекарственного средства для лечения заболевания или состояния, в случае которых достигается улучшение за счет ускорения роста нейрита, стимуляции нейронального роста, их восстановления или регенерации.
В другом аспекте настоящее изобретение относится к применению антагониста PirB/LILRB при получении лекарственного средства для лечения неврологического расстройства, где указанное неврологическое расстройство может характеризоваться физически поврежденным нервом или указанное расстройство может быть выбрано, например, из группы, состоящей из повреждения периферического нерва, вызванного физическим повреждением, диабетом; физического повреждения центральной нервной системы; повреждения головного мозга, ассоциированного с инсультом, невралгии тройничного нерва, глоссофарингеальной невралгии, паралича Белла, миастении гравис, мышечной дистрофии, бокового амиотрофического склероза (ALS), прогрессирующей мышечной атрофии, прогрессирующей бульбарной наследственной мышечной атрофии, грыжи, разрыва и пролапса межпозвоночных дисков, шейного спондилеза, расстройств, затрагивающих сплетение, синдрома лестничной мышцы (сдавления плечевого сплетения и подключичной артерии), периферической нейропатии, порфирии, синдрома Гийена-Барра, болезни Альцгеймера, болезни Хантингтона и болезни Паркинсона.
В другом аспекте настоящее изобретение относится к антагонисту PirB/LILRJB для применения при лечении заболевания или состояния, в случае которых достигается улучшение за счет ускорения роста нейрита, стимуляции нейронального роста, их восстановления или регенерации.
В еще одном аспекте настоящее изобретение относится к антагонистам PirB/LILRB для применения при лечении неврологического расстройства, где указанное неврологическое расстройство описано выше.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг. 1 продемонстрирована активность щелочной фосфатазы на трансфицированных клетках COS после инкубации с AP-Nogo66. AP-Nogo66 связывается с PirB и LILRB2.
На фиг. 2 продемонстрирована иммунореактивность на трансфицированных клетках COS после инкубации с MAG-Fc. MAG связывается с PirB и LILRB2.
На фиг. 3 показаны результаты анализа по методу ОТ-ПЦР, демонстрирующие наличие экспрессии SMAGP, PAN-PirA и PirB в разных частях нервной системы.
На фиг. 4A-4C приведены иллюстрации, подтверждающие в рамках проведенной гибридизации in situ экспрессию PirB в переднем мозге взрослого организма (A), в мозжечке взрослого организма (B) и в P10 спинномозговом узле (C).
На фиг. 5 показана аминокислотная последовательность мышиного PirB (SEQ ID NO: 1) и человеческого LILRB2, аминокислотная последовательность транскрипта варианта 1 (SEQ ID NO: 2).
На фиг. 6 проиллюстрировано ингибирование под действием Nogo66 роста аксона и реверсия такого ингибирования под действием PirB ECD (и PirBFc, и PirBHis). Для теста были использованы гранулярные нейроны мозжечка.
На фиг. 7 показаны результаты совместной иммунопреципитации PirB и NgR. NgR четко осаждается вместе с PirB (левая панель). На правой панели показан суммарный белок из лизатов целых клеток после проведения иммуноблоттинга с анти-NgR. Множественные полосы (показаны стрелками) отражают NgR, подвергнутый гликозилирующему процессингу в разной степени.
На фиг. 8 показано, что ингибирование роста аксона под действием Nogo66 частично снимается анти-PirB антителами в гранулярных нейронах мозжечка.
На фиг. 9 показана аминокислотная последовательность LILRB1, вариант транскрипта 1 (SEQ ID NO: 10).
На фиг. 10 показана аминокислотная последовательность LILRB1, вариант транскрипта 2 (SEQ ID NO: 11).
На фиг. 11 показана аминокислотная последовательность LILRB1, вариант транскрипта 3 (SEQ ID NO: 12).
На фиг. 12 показана аминокислотная последовательность LILRB1, вариант транскрипта 4 (SEQ ID NO: 13).
На фиг. 13 показана аминокислотная последовательность LILRB2, вариант транскрипта 2 (SEQ ID NO: 14).
На фиг. 14 показана аминокислотная последовательность LILRB3, вариант транскрипта 1 (SEQ ID NO: 15).
На фиг. 15 показана аминокислотная последовательность LILRB3, вариант транскрипта 2 (SEQ ID NO: 16).
На фиг. 16 показана аминокислотная последовательность LILRB5, вариант транскрипта 1 (SEQ ID NO: 17).
На фиг. 17 показана аминокислотная последовательность LILRB5, вариант транскрипта 2 (SEQ ID NO: 18).
На фиг. 18 показана аминокислотная последовательность LILRB5, вариант транскрипта 3 (SEQ ID NO: 19).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
А. Определения
Термины «спаренный иммуноглобулин-подобный рецептор B» и «PirB» используются в данном описании взаимозаменяемо и относятся к нативной последовательности мышиного ингибирующего белка из 841 аминокислоты с SEQ ID NO: 1 (фиг. 5) (NP_035225) и к его гомологам с нативной последовательностью у крыс и других млекопитающих, отличных от человека, включая все природные варианты, такие как варианты транскриптов с альтернативным сплайсингом и аллельные варианты и изоформы, а также их растворимые формы.
Термины «LILRB», «ILT» и «MIR» используются в настоящем описании взаимозаменяемо и относятся ко всем членам надсемейства В на основе «лейкоцитарного иммуноглобулин-подобного рецептора» человека, включая все природные варианты, такие как варианты транскриптов с альтернативным сплайсингом и аллельные варианты и изоформы, а также их растворимые формы. Индивидуальные представители данного семейства обозначаются номерами, следующими за соответствующим буквенным обозначением, например, LILRB3/ILT5, LILRB1/ILT2, LILRB5/ILT3 и LIRB2/ILT4, где ссылка на любого отдельного представителя, если особо не указано иное, также включает ссылку на все природные варианты, такие как варианты транскриптов с альтернативным сплайцсингом и аллельные варианты и изоформы, а также их растворимые формы. Таким образом, например, термин «LILRB1» в контексте настоящего описания специфически включает варианты транскриптов 1-4 (SEQ ID NoNo. 10, 11, 12 и 13, показанные на фиг. 9-12), а также все другие природные варианты, такие как варианты транскриптов с альтернативным сплайсингом и аллельные варианты и изоформы, а также их растворимые формы. Термин «"LILRB2» в контексте настоящего описания специфически включает LILRB2, вариант транскрипта 1 (SEQ ID NO: 2, показанный на фиг. 5) и вариант транскрипта 2 (SEQ ID NO: 14, показанный на фиг. 13), а также все другие природные варианты, такие как другие варианты транскриптов с альтернативным сплайсингом, аллельные варианты и изоформы, а также их растворимые формы. Термин «LILRB3» также специфически включает в контексте настоящего описания LILRB3 вариант транскрипта 1 (SEQ ID NO: 15, показанный на фиг. 14) и вариант транскрипта 2 (SEQ ID NO: 16, показанный на фиг. 15), а также все другие природные варианты, такие как варианты транскриптов с альтернативным сплайсингом, аллельные варианты и изоформы, а также их растворимые формы. Термин «LILRB5» специфически включает варианты транскриптов 1-3 (SEQ ID NoNo: 17-19, показанные на фиг. 16-18), а также все другие природные варианты, такие как другие варианты транскриптов с альтернативным сплайсингом, аллельные варианты и изоформы, а также их растворимые формы.
Термин «PirB/LILRB» в контексте настоящего описания обозначает сокращенное обозначение, относящееся к любому индивидуальному мышиному PirB и человеческим LILRB белкам, а также к их гомологам с нативной последовательностью у других млекопитающих, отличных от человека, включая все природные варианты, такие как варианты транскриптов с альтернативным сплайсингом и аллельные варианты и изоформы, а также их растворимые формы.
Термин «миелин-ассоциированный белок» используется в широком смысле и включает все белки, присутствующие в миелиновых волокнах ЦНС, которые ингибируют регенерацию нейронов, включая Nogo, MAG и OMgp.
Термин «выделенный» применительно к описанию различных рассматриваемых в настоящем изобретении белков обозначает белок, который был идентифицирован и отделен от и/или выделен из компонентов природного окружения. Контаминирующие компоненты природного окружения представляют собой материалы, которые в типичном случае мешают диагностическому использованию белка и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления настоящего изобретения указанный белок очищают: (1) до степени, достаточной для того, чтобы получить по меньшей мере 15 остатков на N-концевой или внутренней аминокислотной последовательности при использовании секвенатора с вращающимся стаканом и (2) до гомогенности, по данным оценки в ДСН-ПААГ в невосстанавливающих или восстанавливающих условиях, с использованием красителя кумасси-голубого или предпочтительно, серебряного красителя, или (3) до гомогенности, по данным масс-спектрохимического анализа или по результатам пептидного картирования. Выделенный белок включает белок in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природного окружения рассматриваемого белка не присутствует. Однако обычно выделенный белок получают по меньшей мере в ходе одной стадии очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой такую молекулу нуклеиновой кислоты, которая была идентифицирована и отделена по меньшей мере от одной контаминирующей молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в природном источнике данной нуклеиновой кислоты. Выделенная молекула нуклеиновой кислоты находится в иной форме или в составе иного набора, которые характерны для ее природного состояния. В этой связи выделенные молекулы нуклеиновой кислоты отличаются от молекул нуклеиновой кислоты, которые существуют в клетках в их естественном окружении. Однако термин «выделенная молекула нуклеиновой кислоты» охватывает также молекулы нуклеиновой кислоты, которые содержатся в клетках и которые обычно экспрессируют такую нуклеиновую кислоту, где, например, указанная молекула нуклеиновой кислоты локализована в хромосоме в месте, отличном от места, характерного для клеток в их естественном окружении.
В контексте настоящего описания термин «антагонист PirB/LILRB» используется для обозначения агента, способного блокировать, нейтрализовать, ингибировать, устранять, снижать или препятствовать активности PirB/LILRB. Конкретно антагонист PirB/LILRB мешает проявлению ингибирующей активности миелин-ассоциированных белков, что приводит к повышению роста нейрита и/или к стимуляции нейронального роста, восстановления и/или регенерации. В предпочтительном варианте осуществления настоящего изобретения указанный антагонист PirB/LILRB ингибирует связывание PirB/LILRB с Nogo66 и/или MAG, и/или OMgp, за счет связывания с PirB/LILRB. Антагонисты PirB/LILRB включают, например, антитела против PirB/LILRB и их антиген-связывающие фрагменты, усеченные или растворимые фрагменты PirB/LILRB, Nogo 66, MAG или OMgp, которые способны ослаблять связывание между PirB/LILRB и Nogo66 или между PirB/LILRB и MAG, или между PirB/LILRB и OMgp, и малыми молекулами ингибиторов на PirB/LILRB-связанном пути ингибирования. Антагонисты PirB/LILRB также включают короткие интерферирующие молекулы РНК (киРНК), способные ингибировать или снижать экспрессию мРНК PirB/LILRB.
Термин «антитело» в контексте настоящего описания используется в самом широком смысле и конкретно охватывает интактные антитела, моноклональные антитела, поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), образованные на основе по меньшей мере двух интактных антител, а также фрагменты антител, при условии, что они проявляют желаемую биологическую активность.
Термин «моноклональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции по существу гомогенных антител, например индивидуальных антител, включающих практически идентичную популяцию, за исключением возможных природных мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высоко специфическими молекулами, направленными против одного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, которые включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Характерные и полезные свойства моноклональных антител включают не только специфичность этих молекул, но также возможность синтезировать их в неконтаминированном другими антителами состоянии. Прилагательное «моноклональный», в данном сочетании, указывает на характер антител, которые получают из гомогенной, по существу, популяции антител, и этот термин не следует трактовать в контексте получения антитела по какому-либо одному конкретному методу. Так, например, моноклональные антитела, используемые согласно настоящему изобретению, могут быть получены по гибридомному методу, впервые описанному Колером с соавт. (Kohler et al, Nature, 256:495 (1975)), или могут быть получены согласно технологии рекомбинантных ДНК (см., например, патент США No. 4816567). «Моноклональные антитела» могут быть также выделены из библиотеки фаговых антител с использованием методик, описанных, например, Clackson et al, Nature, 352:624-628 (1991) и Marks et al, J. Mol Biol, 222:581-597 (1991).
В конкретных вариантах рассматриваемые антитела включают химерные антитела, где часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, тогда как оставшиеся одна или несколько цепей идентичны или гомологичны соответствующим последовательностям, полученных из другого вида или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют желаемую биологическую активность (патент США No. 4816567; и Morrison et al, Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). Химерные антитела, представляющие интерес в настоящем контексте, включают приматизированные антитела, содержащие антиген-связывающие последовательности вариабельного домена, полученные от приматов, отличных от человека (например, от обезьян Старого Света, обезьян Ape и т. п.), и последовательности константного участка молекул человека.
«Фрагменты антител» включают часть интактного антитела, предпочтительно содержащий его антиген-связывающий или вариабельный участок. Примеры фрагментов антител включают Fab, Fab1, F(ab')2 и Fv фрагменты; димерные антитела; линейные антитела; одноцепочечные молекулы антител; и полиспецифические антитела, образованные из фрагментов антител.
«Интактное антитело» представляет собой такое антитело, которое включает антиген-связывающий вариабельный участок, а также константный домен легкой цепи (CL) и константные домены тяжелой цепи CHI, CH2 и CH3. Константные домены могут представлять собой нативные последовательности константных доменов (например, константные домены нативной последовательности молекулы человека) или их варианты по аминокислотной последовательности. Предпочтительно интактное антитело характеризуется наличием одной или нескольких эффекторных функций.
«Гуманизированные» формы антител из источников, отличных от человека (например, грызунов), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, отличного от человеческого. В основном гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельного участка реципиентной молекулы замещены остатками из гипервариабельного участка молекулы из вида, отличного от человека (донорное антитело), такого как мышь, крыса, кролик или примат, отличный от человека, который имеет желательные значения специфичности, аффинности и емкости. В некоторых случаях остатки в рамке считывания (FR) в молекуле иммуноглобулина человека замещены соответствующими остатками из молекулы, отличной от человека. Кроме того, гуманизированные антитела могут включать остатки, которые не встречаются в реципиентном антителе или в донорном антителе. Такие модификации вводят с целью дополнительного улучшения характеристик антитела. В основном гуманизированные антитела включают по существу все или по меньшей мере один и, в типичном случае, два вариабельных домена (Fab, Fab', F(ab')2, Fabc, Fv), в которых все, или по существу все, вариабельные петли соответствуют петлям, имеющимся в иммуноглобулине из источника, отличного от человека, и все или по существу все FR соответствуют тем FR, которые имеются в последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также включает по меньшей мере часть константного участка иммуноглобулина (Fc), в типичном случае иммуноглобулина человека. Дополнительные детали приведены, например, в следующих работах: Jones et al, Nature 321:522-525 (1986); Riechmann et al, Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992).
Термин «гипервариабельная область» в контексте настоящего описания относится к участкам вариабельного домена антитела, которые в значительной мере варьируют по последовательности и/или формируют структурно определенные петли. Указанная гипервариабельная область включает аминокислотные остатки из «участка, определяющего комплементарность», или «CDR» (например, остатки 24-34, 50-56 и 89-97 в вариабельном домене легкой цепи и 31-35, 50-65 и 95-102 в вариабельном домене тяжелой цепи (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)), а также остатки из «гипервариабельной петли» (например, остатки 26-32, 50-52 и 91-96 в вариабельном домене легкой цепи и 26-32, 53-55 и 96-101 в вариабельном домене тяжелой цепи (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). В обоих случаях остатки вариабельного домена нумеруются в соответствии с системой, введенной Kabat et al., выше, как будет более подробно описано ниже. Остатки из «рамки считывания», или «FR», представляют собой такие остатки вариабельного домена, которые отличаются от остатков в гипервариабельных участках, определенных выше.
«Родительское антитело» или антитело «дикого типа» представляет собой антитело, включающее аминокислотную последовательность, в которой отсутствует одно или несколько аминокислотных изменений, в сравнении с вариантом антитела согласно настоящему описанию. Таким образом, родительское антитело в основном содержит по меньшей мере одну гипервариабельную область, которая отличается по аминокислотной последовательности от той аминокислотной последовательности, которая соответствует гипервариабельной области в варианте антитела согласно настоящему описанию. Родительский полипептид будет включать нативную последовательность (то есть природную последовательность) антитела (включая природный аллельный вариант) или антитела с уже имеющимися аминокислотными модификациями (такими как вставки, делеции и/или другие изменения) в природной последовательности. В тексте описания такие термины: антитело «дикого типа», «WT» и «родительские» или «исходные» антитела используются взаимозаменяемо.
В контексте настоящего описания термины «вариант антитела» или «вариантное антитело» относятся к антителу, которое имеет аминокислотную последовательность, отличающуюся от аминокислотной последовательности родительского антитела. Предпочтительно, вариант антитела включает вариабельный домен тяжелой цепи или вариабельный домен легкой цепи, содержащие аминокислотную последовательность, которая не встречается в природном состоянии. Такие варианты обязательно характеризуются наличием менее чем 100% идентичности или сходства по последовательности с родительским антителом. В предпочтительном варианте осуществления настоящего изобретения вариант антитела будет иметь аминокислотную последовательность с показателем идентичности или сходства по аминокислотной последовательности от примерно 75% до менее чем 100%, с аминокислотной последовательностью вариабельного домена тяжелой или легкой цепей родительского антитела, более предпочтительно от примерно 80% до менее чем 100%, более предпочтительно от примерно 85% до менее чем 100%, еще более предпочтительно от примерно 90% до менее чем 100%, и наиболее предпочтительно от примерно 95% до менее чем 100%. Вариант антитела в основном представляет собой такое антитело, которое включает одно или несколько аминокислотных изменений в одном или нескольких гипервариабельных участках или вблизи этих участков.
Термин «аминокислотное изменение» относится к изменению в аминокислотной последовательности относительно заданной аминокислотной последовательности. Примеры таких изменений включают вставки, замещения и делеции. «Аминокислотное замещение» относится к замене существующего аминокислотного остатка в заданной аминокислотной последовательности остатком другой аминокислоты.
Термин «замещение» применительно к аминокислотному остатку относится к аминокислотному остатку, который замещает или заменяет другой аминокислотный остаток в аминокислотной последовательности. Замещающий остаток может быть природным или неприродным аминокислотным остатком.
Термин «аминокислотная вставка (инсерция)» относится к встраиванию одного или нескольких аминокислотных остатков в заданную аминокислотную последовательность. Аминокислотная вставка может включать «пептидную вставку», где пептид, включающий два или более аминокислотных остатков, соединенных одной или несколькими пептидными связями, вводят в заданную аминокислотную последовательность. В том случае, когда аминокислотная вставка вовлекает встраивание пептида, указанный пептид со вставкой может быть получен путем случайного мутагенеза, так что он будет иметь аминокислотную последовательность, которая не существует в природе. Аминокислотное изменение «вблизи гипервариабельной области» относится к введению или замещению одного или нескольких аминокислотных остатков в N-концевой и/или С-концевой области гипервариабельной области, так что по меньшей мере один из встроенных или замещающих аминокислотных остатков формирует пептидную связь с N-концевым или С-концевым аминокислотным остатком в рассматриваемой гипервариабельной области.
«Природный аминокислотный остаток» представляет собой такой остаток, который кодируется соответствующим генетическим кодом, в основном выбираемым из группы, состоящей из: аланина (Ala); аргинина (Arg); аспарагина (Asn); аспарагиновой кислоты (Asp); цистеина (Cys); глютамина (Gln); глютаминовой кислоты (Glu); глицина (Gly); гистидина (His); изолейцина (Ile); лейцина (Leu); лизина (Lys); метионина (Met); фенилаланина (Phe); пролина (Pro); серина (Ser); треонина (Thr); типтофана (Trp); тирозина (Tyr) и валина (Val).
В контексте настоящего описания «неприродный аминокислотный остаток» представляет собой такой аминокислотный остаток, который отличается от природных аминокислотных остатков, перечисленных выше, и который способен ковалентно связываться с одним или несколькими соседними аминокислотными остатками в полипептидной цепи. Примеры неприродных аминокислотных остатков включают норлейцин, орнитин, норвалин, гомосерин и другие аналоги аминокислотных остатков, такие как, например, описанные в руководстве Эллмана с соавт. (Ellman et al. Meth. Enzym. 202:301-336 (1991)). Для получения таких неприродных аминокислотных остатков могут использоваться процедуры, описанные в литературе (Noren et al. Science 244:182 (1989) и Ellman et al., выше). В общих чертах эти процедуры включают химическую активацию супрессорной тРНК неприродным аминокислотным остатком, с последующим проведением in vitro транскрипции и трансляции РНК.
В тексте настоящего описания даются ссылки, в соответствии с системой нумерации, введенной Кабатом с соавт. (Kabat, Е. А., et al., Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987) и (1991)). В указанном руководстве Кабат (Kabat) перечисляет многие аминокислотные последовательности для антител каждого подкласса и перечисляет наиболее часто встречающиеся аминокислоты применительно к каждому из положений остатка в каждом подклассе. При этом Кабат использует способ, согласно которому каждому остатку в перечисленном перечне аминокислот присваивается соответствующий номер, и этот метод присваивания соответствующих номеров аминокислотным остаткам стал стандартным в данной области. Соответственно, далее в тексте описания используется схема нумерации Кабата. В случае отсутствия в схеме Кабата нужного элемента, в тексте настоящего описания, для присвоения соответствующих номеров остаткам в аминокислотной последовательности исследуемого антитела используют следующий подход. Так, в основном такая исследуемая последовательность сопоставляется с последовательностью иммуноглобулина или любой консенсусной последовательностью в схеме Кабата. Указанное сопоставление должно быть сделано вручную или с использованием компьютера в рамках соответствующих компьютерных программ, например, такой программы, как Align 2. При этом процесс сопоставления может быть облегчен за счет использования определенных аминокислотных остатков, которые являются общими для большинства Fab последовательностей. Так, например, легкая и тяжелая цепи, каждая, в типичном случае содержит два цистеина, которые имеют один и тот же номер; в VL домене два остатка цистеина представляют собой в типичном случае остатки с номерами 23 и 88, а в VH домене оба остатка цистеина в типичном случае нумеруются как 22 и 92. Остатки в рамке считывания в основном, хотя не всегда, имеют приблизительно одни и те же номера, однако CDR могут варьировать по размеру. Так, например, в случае CDR в исследуемой последовательности, которая длиннее, чем CDR в последовательности Кабат, с которой ее сопоставляют, в типичном случае добавляют суффиксы к номеру остатка, с тем чтобы обозначить наличие вставки из дополнительных остатков (см., например, остатки 100 abc на фиг. 1В). В случае исследуемых последовательностей, которые, например, сопоставляются с последовательностью Кабата по остаткам 34 и 36, но где между ними отсутствует соответствующий остаток для номера 35, никакому из остатков указанный номер 35 просто не присваивается.
В контексте настоящего описания антитело с «высокой аффинностью» представляет собой антитело, имеющее значение Kd, или константу диссоциации, в наномолярном (нМ) диапазоне значений или ниже. Показатель KD в «наномолярном диапазоне значений или ниже» может быть обозначен как X нМ, где Х обозначает число менее чем примерно 10.
Термин «нитевидный фаг» относится к вирусной частице, способной проявлять гетерогенный полипептид на своей поверхности, которая включает, без ограничения, fl, fd, Pf1 и M13. Нитевидный фаг может содержать селектируемый маркер, такой как тетрациклин (например, «fd-tet»). Различные системы проявления нитевидных фагов известны специалистам в данной области (см., например, Zacher et al. Gene 9: 127-140 (1980), Smith et al. Science 228: 1315-1317 (1985); и Parmley and Smith Gene 73: 305-318 (1988)).
Термин «пэннинг» в контексте настоящего описания относится к множественным циклам процесса скрининга для идентификации и выделения фагов, содержащих соединения, такие как антитела с высокой аффинностью и специфичностью к мишени.
Термин «короткая интерферирующая РНК (киРНК)» относится к малым двуцепочечным молекулам РНК, которые препятствуют генной экспрессии. Молекулы киРНК являются интермедиатами интерференции РНК, процесса, в ходе которого двуцепочечная РНК вызывает молчание гомологичных генов. Молекулы киРНК в типичном случае включают две одноцепочечных РНК из примерно 15-25 нуклеотидов в длину, формирующих дуплекс, который может включать, один или несколько, одноцепочечных выступающих фрагментов. Процессинг двуцепочечной РНК ферментным комплексом, например, полимеразным комплексом, приводит к расщеплению двуцепочечной РНК с образованием киРНК. Антисмысловую цепь киРНК используют в составе РНК-интерферирующего (РНКи) молчащего комплекса для проведения расщепления мРНК, что ускоряет деградацию мРНК. Для достижения молчания специфического гена с использованием киРНК, например в клетке млекопитающего, выбирают участок спаренных оснований, с тем чтобы избежать комплементарности к неродственной мРНК. РНКи молчащие комплексы были идентифицированы в данной области, и например, описаны в работах Fire et al., Nature 391:806-811 (1998) и McManus et al., Nat. Rev. Genet. 3(10):737-47 (2002).
Термин «интерферирующая РНК (РНКи)» в контексте настоящего описания относится к двуцепочечной РНК, которая приводит к каталитической деградации специфических мРНК и, таким образом, может использоваться для ингибирования/снижения экспрессии конкретного гена.
Термин «полиморфизм» в контексте настоящего описания относится к более чем одной форме гена или его части (например, с учетом аллельного варианта). Часть гена, для которого имеется по меньшей мере две разных формы, обозначается как «полиморфный участок» гена. Специфическая генная последовательность в полиморфном участке гена представляет собой «аллель». Полиморфный участок может представлять собой одиночный нуклеотид, который различен в разных аллелях или он может включать несколько нуклеотидов в длину.
В контексте настоящего описания термин «расстройство» в основном используют для описания любого состояния, в случае которого может быть применимо лечение соединениями согласно настоящему изобретению и которое включает любое заболевание или расстройство, способное поддаваться лечению эффективными количествами антагонистов PirB/LILRB. Неограничивающие примеры расстройств, подлежащих лечению соединениями согласно настоящему изобретению, включают, без ограничения, заболевания или состояния, для которых будет благоприятно усиление роста нейрита, стимуляция нейронального роста, восстановления или регенерации, включая неврологические расстройства, такие как физические повреждения нервов и нейродегенеративные заболевания. Такие расстройства конкретно включают повреждение периферического нерва, вызванное физическим повреждением или болезненными состояниями, такими как диабет; физическое повреждение центральной нервной системы (спинного мозга и головного мозга); повреждение головного мозга, ассоциированное с инсультом, и неврологические расстройства, связанные с нейродегенерацией, такие как, например, невралгия тройничного нерва, глоссофарингеальная невралгия, паралич Белла, миастения гравис, мышечная дистрофия, боковой амиотрофический склероз (ALS), прогрессирующая мышечная атрофия, прогрессирующая бульбарная наследственная мышечная атрофия, грыжа, разрыв и пролапс межпозвоночных дисков, шейный спондилез, расстройства, затрагивающие сплетение, синдром лестничной мышцы (сдавления плечевого сплетения и подключичной артерии), периферическая нейропатия, такие как расстройства, которые могут быть вызваны свинцом, даксоном, клещами, порфирия, синдром Гийена-Барра, болезнь Альцгеймера, болезнь Хантингтона или болезнь Паркинсона.
Термины «лечение», «терапия» и другие образованные от них формы относятся к излечивающей терапии, профилактической терапии и превентивной терапии. Последовательное лечение, или введение, относится к лечению, по меньшей мере, на ежедневной основе, без перерыва в курсе лечения на один или несколько дней. Перемежающееся лечение, или введение, или лечение, или введение в интермиттитрующем режиме, относится к лечению, которое не является последовательным, но, скорее, циклическим по своей природе.
Термин «предупреждение нейродегенерации» в контексте настоящего описания включает: (1) способность ингибировать или предупреждать нейродегенерацию у пациентов с только что поставленным диагнозом нейродегенеративного расстройства или в случае пациентов, у которых имеется риск развития нового для них нейродегенеративного заболевания; (2) способность ингибировать или предупреждать дальнейшую нейродегенерацию у пациентов, которые уже имеют нейродегенеративное заболевание или его симптомы.
Термин «млекопитающее» в контексте настоящего описания обозначает любое млекопитающее, относящееся к данному классу млекопитающих, включая людей, высших приматов, отличных от человека, грызунов, домашних и фермерских животных, таких как коровы, лошади, собаки и кошки. В предпочтительном варианте осуществления настоящего изобретения указанное млекопитающее представляет собой человека.
Введение «в сочетании с» одним или несколькими другими терапевтическими агентами включает одновременное (конкурентное) и последовательное введение в любом порядке.
Термин «эффективное количество» представляет собой количество, достаточное для проявления благоприятного или желаемого терапевтического (включая превентивный) результата. Эффективное количество может вводиться в ходе одного или нескольких введений.
В контексте настоящего описания выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо и такие обозначения включают также соответствующее потомство. Таким образом, слова «трансформанты» и «трансформируемые клетки» включают первичную клетку и культуры, полученные на ее основе, независимо от количества переносов. Следует также понимать, что все потомство необязательно будет идентичным по содержанию ДНК, в связи с наличием незначительных или невредных мутаций. Термин «потомство» относится к любым и всем последующим представителям каждой из генераций после исходно трансформированной клетки и клеточной линии. При этом включается также мутантное потомство, которое имеет ту же функцию или биологическую активность, как и исходная трансформированная клетка, по результатам скрининга. В том случае, когда следует ввести определенное различие применительно к описанию потомства, это будет видно из контекста.
«Процент (%) идентичности по аминокислотной последовательности», применительно к последовательностям согласно настоящему изобретению, определяется как процентное содержание аминокислотных остатков в исследуемой последовательности, которые идентичны аминокислотным остаткам в эталонной последовательности, после сопоставления последовательностей и встраивания пропусков, при необходимости, с достижением максимальной процентной идентичности по последовательности, где при этом не рассматриваются какие-либо консервативные замещения в качестве части такой идентичности. Сопоставление с целью определения процента идентичности аминокислот может быть достигнуто различными способами, которые известны специалистам в данной области, так что любой специалист может определить соответствующие параметры, необходимые для проведения такого сопоставления, включая выбор алгоритмов, необходимых для достижения максимального сопоставления по всей длине сравниваемых последовательностей. Для целей настоящего описания значение процентной идентичности по аминокислотному составу может быть получено с использованием компьютерной программы для сравнения последовательностей ALIGN-2 от компании Genentech, Inc., для которой кодирующие параметры были зарегистрированы пользователем документации в Управлении США по контролю за соблюдением авторских прав (US Copyright Office, Washington, DC, 20559), при регистрации в соответствия с законом об авторских правах США (US Copyright Registration No. TXU510087). Программа ALIGN-2 доступна для пользователей от компании Genentech, Inc., South San Francisco, CA. Все параметры для сравнения последовательностей установлены в программе ALIGN-2 и не варьируются.
«Жесткость» реакции гибридизации может быть легко определена любым специалистом в данной области и в основном определяется эмпирическими расчетами, учитывающими длину зонда, температуру на стадии промывки и соответствующую концентрацию соли на этой стадии. В основном более длинные зонды требуют более высоких температур для соответствующего отжига, тогда как более короткие зонды могут использоваться при сниженных температурах. Гибридизация в основном зависит от способности денатурированной ДНК к повторного отжигу при наличии комплементарных цепей в среде ниже их температуры плавления. Чем выше степень желательной идентичности между зондом и гибридизуемой последовательностью, тем выше относительная температура, которая может использоваться. Из этого следует, что более высокие относительные температуры будут делать реакционные условия более жесткими, тогда как снижение температур соответствующим образом уменьшает жесткость условий реакции. Дополнительные детали и пояснения принципа жесткости условий реакции гибридизации описаны, например, в работе Аусубеля с соавт. (Ausubel et al, Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995)).
Термин «условия высокой жесткости» в контексте настоящего определения означает такие условия, при которых: (1) для промывки используются условия низкой ионной силы и высокой температуры; 0015 М хлорид натрия/0,0015 М цитрат натрия/0,1 М додецилсульфат натрия при температуре 50°C; (2) в ходе гибридизации используют денатурирующие агенты; 50 объемн.% формамида с добавкой 0,1% бычьего сывороточного альбумина/0,1% Фиколл/ 0,1% поливинилпирролидон/ 50 мМ натрий-фосфатный буфер при pH 6,5 с содержанием 750 мМ хлорида натрия, 75 мМ цитрата натрия при температуре 42°C; или (3) используют 50% формамид, 5х SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5 × раствор Денхардта, озвученная ДНК спермы лосося (50 мкг/мл), 0,1% ДСН и 10% сульфат декстрана при температуре 42°C, в условиях промывок при температуре 42°C в 0,2 × SSC (хлорид натрия/цитрат натрия) и 50% формамида при 55°С, с последующей промывкой в условиях высокой жесткости, включающих использование 0,1 × SSC, содержащего ЭДТА, при температуре 55°C.
Термин «условия умеренной жесткости» может быть определен в соответствии с идентификацией, приведенной в руководстве Самбрука с соавт. (Sambrook et al, Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989), и включает инкубацию в течение ночи при температуре 37°C в растворе, содержащем: 20% формамида, 5 × SSC (150 мМ NaCl, 15 мМ цитрат тринатрия), 50 мМ фосфата натрия (pH 7,6), 5 х раствора Денхардта, 10% сульфат декстрана и 20 мг/мл денатурированной ДНК спермы лосося, с последующей промывкой фильтров в 1 х SSC при температуре примерно 37-50°C. Для специалистов в данной области очевидно, как можно корректировать температуру, ионные силы и другие параметры, при необходимости, в соответствии с таким факторами, как длина зонда и т.п.
Термин «регуляторные последовательности» относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Регуляторные последовательности, которые приемлемы для прокариот, включают, например, промоторы, необязательно последовательности оператора и сайт связывания рибосом. Эукариотические клетки, как известно, используют промоторы, сигналы полиаденилирования и энхансеры.
Считается, что нуклеиновая кислота «функционально связана», если она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Так, например, ДНК для предпоследовательности или секреторного лидера функционально связана с ДНК для полипептида, если она экспрессируется в виде предбелка, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. В основном термин «функционально связанный» означает, что последовательности ДНК, будучи связанными, являются непрерывными, а в случае секреторного лидера, непрерывными и в фазе считывания. Однако энхансеры необязательно должны быть непрерывными. Связывание сопровождается лигированием в соответствующих сайтах рестрикции. Если такие сайты отсутствуют, используют синтетические олигонуклеотидные адапторы или линкеры, в соответствии с традиционной практикой.
Термин «малая молекула» относится к молекулам с молекулярной массой ниже примерно 1000 Дальтон, предпочтительно, ниже примерно 500 Дальтон.
B. Анализы скрининга для идентификации стимуляторов нейрональной регенерации
Первичные анализы согласно настоящему изобретению основаны, по меньшей мере частично, на том факте, что PirB/LILRB является рецептором миелинового белка Nogo (Nogo66) и MAG и что антагонисты PirB/LILRB, которые препятствуют ассоциации PirB/LILRB с Nogo и/или MAG, способны усиливать рост нейрита и/или стимулировать нейрональный рост, восстановление и/или их регенерацию. В общих чертах такие агенты будут описаны ниже как стимуляторы нейрональной регенерации.
Анализы скрининга на выявление возможных средств-антагонистов могут быть разработаны таким образом, чтобы идентифицировать соединения, которые связываются или образуют комплекс с PirB/LILRB или иным образом препятствуют взаимодействию PirB/LILRB с Nogo, MAG или другими соединениями в составе миелин-ассоциированной ингибирующей системы. Анализы скрининга согласно настоящему описанию включают анализы, позволяющие проводить крупномасштабный скрининг химических библиотек, делая их пригодными для идентификации возможных лекарственных средств на основе малых молекул. В основном описываются анализы на связывание и анализы на определение активности.
Указанные анализы могут проводиться в разных форматах, которые включают, без ограничения, анализы на белок-белковое связывание, биохимические анализы скрининга, иммунологический анализ и клеточные анализы, которые хорошо известны в данной области.
Все анализы на выявление антагонистов характеризуются общими для них особенностями, связанными с тем, что для них необходим контакт рассматриваемого лекарственного средства с PirB/LILRB полипептидом в определенных условиях и в течение периодов времени, достаточных для того, чтобы эти два компонента вступили во взаимодействие.
В анализах на связывание взаимодействие представляет собой связывание и образованный комплекс может быть выделен или детектирован в реакционной смеси. В конкретном варианте осуществления настоящего изобретения проводят иммобилизацию либо PirB/LILRB полипептида, либо исследуемого лекарственного средства, на твердой фазе, например на микротитрационном планшете, за счет ковалентного или нековалентного присоединения. Нековалентное присоединение в основном достигается при нанесении на твердую поверхность покрытия из раствора PirB/LILRB полипептида с последующей сушкой. Альтернативно может использоваться иммобилизованное антитело, например моноклональное антитело, специфичное для PirB/LILRB полипептида, который подлежит иммобилизации, для прикрепления его к твердой поверхности. Указанный анализ проводят при добавлении неиммобилизованного компонента, который может быть помечен детектируемой меткой, к иммобилизованному компоненту так, чтобы он покрывал поверхность, содержащую прикрепленный компонент. После завершения реакции непрореагировавшие компоненты удаляют, например путем промывки, и выявляют комплексы, фиксированные на твердой поверхности. Когда исходно неиммобилизованный компонент содержит детектируемую метку, выявление метки, иммобилизованной на поверхности, указывает на наличие комплексообразования. Когда исходно неиммобилизованный компонент не несет метки, комплексообразование может быть выявлено, например, с использованием меченого антитела, которое специфически связывается с иммобилизованным комплексом.
Если исследуемое соединение представляет собой полипептид, который взаимодействует, но не связывается с PirB/LILRB, то указанное взаимодействие PirB/LILRB с соответствующим полипептидом может быть обнаружено по методам, известным как анализы для выявления белок-белковых взаимодействий. Соответствующие для этого случая анализы включают традиционные подходы, такие как, например, перекрестная сшивка, совместная иммунопреципитация и совместная очистка с использованием градиентов или хроматографических колонок. Кроме того, белок-белковые взаимодействия могут отслеживаться при использовании генетических систем, основанных на дрожжах, описанных Фильдсом с соавт. (Fields and Song, Nature (London), 340:245-246 (1989); Chien et al., Proc. Natl. Acad. Sci. USA. 88:9578-9582 (1991)), а также другими авторами (Chevray и Nathans, Proc. Natl. Acad. Sci. USA, 89: 5789-5793 (1991)). Множество активаторов транскрипции, таких как дрожжевой GAL4, состоят из двух физически дискретных модулярных доменов, один из которых действует как ДНК-связывающий домен, а другой функционирует как домен активации транскрипции. Дрожжевая система экспрессии, описанная в указанных выше публикациях (в основном известная как «двугибридная система»), имеет преимущество, определяемое этим ее свойством и использует два гибридных белка, в одном из которых целевой белок слит с ДНК-связывающим доменом GAL4, а в другом исследуемый активирующий белок слит с доменом активации. Экспрессия GALl-lacZ репортерного гена под контролем GAL4-активирующего промотора зависит от восстановления активности GAL4 в результате белок-белкового взаимодействия. Колонии, содержащие взаимодействующие полипептиды, выявляют с использованием хромогенного субстрата для β-галактозидазы. Полный набор (MATCHMAKER™) для идентификации белок-белковых взаимодействий между двумя специфическими белками с использованием двугибридной методики доступен в коммерческом варианте от компании Clontech. Сфера применения этой системы может быть расширена для маркирования белковых доменов, вовлекаемых в специфические белковые взаимодействия, а также для выявления аминокислотных остатков, которые являются решающими для таких взаимодействий.
Соединения, которые препятствуют взаимодействию PirB/LILRB и других внутри- и внеклеточных компонентов, в частности Nogo или MAG, могут быть проанализированы следующим образом. Обычно готовят реакционную смесь, которая содержит PirB/LILRB и внутри- или внеклеточный компонент, и выдерживают ее в определенных условиях и в течение периода времени, позволяющих достичь взаимодействия двух продуктов. Для тестирования способности исследуемого соединения ингибировать взаимодействие PirB/LILRB и Nogo или MAG указанную реакцию проводят в отсутствие и в присутствии исследуемого соединения. Кроме того, в исследование может быть включена третья реакционная смесь с плацебо, которая будет служить в качестве позитивного контроля.
Следует подчеркнуть, что анализы скрининга, которые конкретно и подробно обсуждаются в описании, являются лишь иллюстративными и приводятся здесь лишь для пояснения. Может быть выбрано множество других анализов в зависимости от типа скринируемых антагонистов (например, полипептидов, пептидов, непептидных малых органических молекул, нуклеиновых кислот и т. п.), которые хорошо известны специалистам в данной области и которые также будут приемлемы для целей настоящего изобретения.
Приведенные в настоящем описании анализы могут использоваться для скринирования библиотек соединений, включающих, без ограничения, библиотеки химических соединений, библиотеки природных продуктов (например, собрания природных продуктов из микроорганизмов, животных, растений и т.п.) и комбинаторные библиотеки, состоящие из рандомизированных пептидов, олигонуклеотидов или малых органических молекул. В конкретном варианте осуществления настоящего изобретения приведенные в описании анализы используют для скрининга библиотек антител, включающих, без ограничения, библиотеки «наивных» человеческих, рекомбинантных, синтетических и полусинтетических антител. Указанная библиотека антител может представлять собой, например, библиотеку проявления фагов, включающую моновалентные библиотеки, проявляющие, в среднем, одно одноцепочечное антитело или фрагмент антитела на фаговую частицу, поливалентные библиотеки, проявляющие, в среднем, два или три антитела или фрагмента антитела на вирусную частицу. Однако библиотеки антител, подлежащие скринингу, согласно настоящему изобретению не ограничены библиотеками проявления фагов. Другие методики индикации включают, например, процедуру индикации рибосомы или мРНК (Mattheakis et al„ Proc. Natl. Acad. Sci. USA 91:9022-9026 (1994); Hanes and Pluckthun, Proc. Natl. Acad. Sci. USA 94:4937-4942 (1997)), метод индикации микробных клеток, такой как бактериальная визуализация (Georgiou et al., Nature Biotech. 15:29-34 (1997)) или метод индикации на основе дрожжевых клеток (Kieke et al., Protein Ene. 10:1303-1310 (1997)), метод индикации на основе клеток млекопитающих, на основе спор, процедуру проявления вирусов, например проявления ретровирусов (Urban et. al., Nucleic Acids Res. 33:e35 (2005)), процедуру индикации, основанную на наличии связи белок-ДНК (Odegrip et al., Proc. Acad. Natl. Sci. USA 101:2806-2810 (2004); Reiersen et al., Nucleic Acids Res. 33:el0 (2005)), а также методики индикации, основанные на использовании микрошариков (Sepp et al., FEBS Lett. 532:455-458 (2002)).
Результаты, полученные при проведении первичных анализов на связывание/взаимодействие, согласно настоящему изобретению могут быть подтверждены в тестах in vitro и/или in vivo на регенерацию нейронов. Альтернативно тесты in vitro и/или in vivo на нейрональную регенерацию могут использоваться в качестве первичных анализов для идентификации наличия антагонистов PirB/LILRB.
Тесты in vitro на рост нейритов известны специалистам в данной области и описаны, например, в ряде работ (Jin and Strittmatter, J Neurosci 17:6256-6263 (1997); Fournier et al., Methods Enzymol. 325:473-482 (2000); Zheng et al., Neuron 38:213-224 (2003); Wang et al, Nature 417:941-944 (2002) и Neumann et al., Neuron 34:885-893 (2002)). Соответственно коммерчески доступны наборы для определения и количественной оценки роста нейритов. Так, например, в наборе для тестирования роста нейрита CHEMICON Neurite Outgrowth Assay Kit (номер по каталогу NS200) используется технология микропористых фильтров для количественного тестирования соединений, которые влияют на образование нейрита и соответствующий процесс сокращения. При использовании данной системы можно провести скрининг в одновременном режиме биологических и фармакологических агентов, при непосредственной оценке адгезивной и направляющей рецепторной функций, ответственных за процессы расширения и сокращения нейрита, а также при анализе генной функции в трансфицированных клетках. Микропористый фильтр позволяет провести биохимическое разделение и очистку нейритов и клеточных тел для более точного молекулярного анализа экспрессии белка, процессов сигнальной трансдукции и идентификации лекарственных мишеней, которые регулируют процессы роста нейрита или его сокращения.
В формате типичной процедуры первичные нейроны, выделенные из невральной ткани грызунов (включающие гранулярные нейроны мозжечка, нейроны спинномозгового узла и корковые нейроны), культивируют в 96-ячеечных планшетах для культивирования ткани, на которые наслаивают иммобилизованный миелин или миелин-ассоциированные белки (например, Nogo66, MAG и/или OMgp). После культивирования в течение определенного периода времени, в типичном случае 24-48 часов, нейроны фиксируют 4% параформальдегидом и окрашивают маркером для нейронов (анти-b-тубулиновое антитело, класс III, Covance). Для получения снимков и последующего их анализа используют автоматизированную визуализирующую систему ImageXpress (Molecular Devices). Проводят анализ с целью выявления изменений в максимальной или общей длине нейрита в расчете на нейрон.
Тесты in vivo включают использование моделей животных с различными нейродегенеративными заболеваниями, таких как модели животных с повреждением спинного мозга, модели, используемые для оценки пластичности зрительной коры, и другие известные модели. Таким образом, регенерация и пластичность могут быть исследованы на моделях пластичности после проведения односторонней пирамидотомии и на моделях или в реальных ситуациях травматического повреждения головного мозга. Другие модели для исследования нейродегенерации включают мышей с рассеянным склерозом, такие как животные с экспериментальным аутоиммунным энцефалитом (ЭАЭ), модели животных с боковым амиотрофическим склерозом (ALS), такие как модели с использованием SODI мутантных мышей, модели на основе трансгенных животных с болезнью Альцгеймера и модели животных с болезнью Паркинсона.
С. Получение антител, действующих в качестве стимуляторов нейрональной регенерации
Антитела, идентифицированные в анализах на связывание и в анализах по оценке активности согласно настоящему изобретению, могут быть получены по методам, известным в данной области, которые включают процедуры, основанные на технологии рекомбинантных ДНК.
i) Получение антигена
Растворимые антигены или их фрагменты, необязательно конъюгированные с другими молекулами, могут использоваться в качестве иммуногенов для получения антител. Для получения трансмембранных молекул, таких как рецепторы, в качестве иммуногена могут использоваться их фрагменты (например, внеклеточный домен рецептора). Альтернативно в качестве иммуногена могут использоваться клетки, экспрессирующие трансмембранную молекулу. Такие клетки могут быть получены из природного источника (например, линий раковых клеток) или могут представлять собой клетки, которые были трансформированы в рамках рекомбинантных методик с целью экспрессии трансмембранной молекулы. Однако специалистам в данной области известны также другие антигены и их формы, подходящие для целей получения антител.
(ii) Поликлональные антитела
Поликлональные антитела предпочтительно образуются у животных при проведении множественных подкожных (п/к) или внутрибрюшинных (в/б) инъекций релевантного антигена и адъюванта. При этом полезно провести конъюгирование релевантного антигена с белком, который является иммуногенным для иммунизируемого вида животного, таким как, например, гемоцианин лимфы улитки, сывороточный альбумин, бычий тиреоглобулин или ингибитор соевого трипсина, с использованием бифункционального или дериватизирующего агента, например сложного малеимидобензоилсульфосукцинимидного эфира (конъюгированного через цистеиновые остатки), N-гидроксисукцинимида (за счет лизиновых остатков), глютаральдегида, янтарного альдегида, SOCl2 или R1N=C=NR1, где R и R1 представляют собой разные алкильные группы.
Животных иммунизируют путем введения антигена, иммуногенных конъюгатов или их производных при объединении, например, 100 мкг или 5 мкг белка или конъюгата (для кроликов или мышей, соответственно) с 3 объемами полного адъюванта Фрейнда, с последующей внутрикожной инъекцией раствора во множественные сайты. Через один месяц животных подвергают бустинг-инъекциям с использованием от 1/5 до 1/10 от исходного количества пептида или конъюгата в полном адъюванте Фрейнда, путем подкожных инъекций во множественные сайты. Через 7-14 дней у животных отбирают кровь для анализа титра антител. Животных подвергают бустингу до тех пор, пока титр антител не выйдет на плато. Предпочтительно животного подвергают бустингу конъюгатом того же антигена, но конъюгированного с другим белком и/или с использованием другого сшивающего реагента. Конъюгаты также могут быть получены в рекомбинантной клеточной культуре в качестве белков слияния. Кроме того, используют агрегирующие агенты, такие как квасцы, для повышения иммунного ответа.
(iii) Моноклональные антитела
Моноклональные антитела могут быть получены с использованием гибридомного метода, впервые описанного Колером с соавт. (Kohler et al., Nature, 256:495 (1975)), или могут быть получены по технологии рекомбинантных ДНК (патент США No. 4816567). В рамках гибдридомной процедуры проводят иммунизацию мыши или другого подходящего животного-хозяина, такого как хомяк или обезьяна-макака, по описанной выше процедуре для проявления лимфоцитов, которые продуцируют или способны продуцировать антитела, которые, в свою очередь, специфически связываются с белками, используемыми для иммунизации. Альтернативно лимфоциты могут быть иммунизированы in vitro. Далее лимфоциты сливают с миеломными клетками с использованием подходящего агента слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки (Goding, Monoclonal Antibodies; Principles and Practice, pp.59-103 (Academic Press, 1986)).
Полученные таким образом гибридомные клетки высевают и выращивают в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, ингибирующих рост или выживание неслитых исходных миеломных клеток. Так, например, если родительские миеломные клетки не содержали фермент гипоксантингуанинфосфорибозилтрансферазу (HGPRT или HPRT), культуральная среда для гибридом в типичном случае включает гипоксантин, аминоптерин и тимидин (HAT-среда), где указанные вещества будут препятствовать росту HGPRT-дефицитных клеток.
Предпочтительными миеломными клетками являются такие клетки, которые эффективно сливаются, стабильно и на высоком уровне поддерживают продукцию антител выбранными антителопродуцирующими клетками и которые чувствительны к определенной, выбранной для культивирования среде, такой как HAT-среда. Среди них предпочтительными линиями миеломных клеток являются мышиные миеломные линии, полученные, например, из MOPC-21 и MPC-11 мышиных опухолей, которые доступны от Центра клеточных культур в Сан Диего (Salk Institute Cell Distribution Center, San Diego, Calif. USA), а также SP-2 или X63-Ag8-653 клетки, доступные от Американской Коллекции типовых культур (American Type Culture Collection, Rockville, Md. USA). Человеческие миеломные клеточные линии и мышиные-человеческие гетеромиеломные клеточные линии были также описаны для целей продукции моноклональных антител человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al, Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой растят гибридомные клетки, тестируют с целью выявления продукции моноклональных антител против антигена. Предпочтительно определяют связывающую специфичность моноклональных антител, образованных гибридомными клетками, путем иммунопреципитации или в анализе на связывание in vitro, таком как радиоиммуноанализ (РИА) или иммуноферментный твердофазный анализ (ИФТФА).
После идентификации гибридомных клеток, которые продуцируют антитела желательной специфичности, аффинности и/или активности, указанные клоны могут быть подвергнуты субклонированию в процедурах ограниченного разведения с последующим выращиванием по стандартным методикам (Goding, Monoclonal Antibodies; Principles and Practice, pp.59-103 (Academic Press, 1986)). Соответствующие для данной цели культуральные среды включают, например, D-MEM или RPMI-1640 среду. Кроме того, гибридомные клетки могут выращиваться в виде опухолевых асцитов у животных.
Моноклональные антитела, секретируемые субклонами, отделяют соответствующим образом от культуральной среды, асцитной жидкости или сыворотки, с использованием стандартных методов очистки иммуноглобулинов, таких как, например, процедуры, включающие использование белком A-Сефарозы, хроматографию на гидроксилапатите, гель-электрофорез, диализ или аффинную хроматографию.
ДНК, кодирующая моноклональные антитела, может быть легко выделена и секвенирована в рамках стандартных процедур (например, при использовании олигонуклеотидных зондов, которые способны к специфическому связыванию с генами, кодирующими тяжелую и легкую цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. После выделения указанная ДНК может быть встроена в вектор экспрессии, который затем подвергают трансфекции в клетки-хозяева, такие как клетки E. coli, COS клетки обезьяны, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые до этого не продуцировали иммуноглобулиновый белок, с достижением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Рекомбинантная продукция антител ниже описана более подробно.
В еще одном варианте осуществления настоящего изобретения антитела или фрагменты антител могут быть отделены от антител из фаговых библиотек, созданных с использованием известных методик (McCafferty et al, Nature, 348:552-554 (1990)).
Клаксон с соавт. и Маркс с соавт. (Clackson et al, Nature, 352:624-628 (1991) и Marks et al., J. Mol. Biol., 222:581-597 (1991)) описывают процедуру выделения антител мыши и человека, соответственно, с использованием фаговых библиотек. В более поздних публикациях Маркс с соавт. описывают продукцию антител человека с высокой аффинностью (в нМ диапазоне) путем перестановки цепей (Marks et al., Bio/Technology, 10:779-783 (1992)), а также путем комбинаторной инфекции и последующей рекомбинации in vivo, где указанная стратегия используется для конструирования крупных фаговых библиотек (Waterhouse et al., Nuc. Acids. Res., 21:2265-2266 (1993)). Таким образом, указанные методики представляют собой жизнеспособную альтернативу относительно традиционных методик получения моноклональных антител с помощью гибридом с целью выделения моноклональных антител.
ДНК может быть также модифицирована, например, путем замещения с использованием кодирующей последовательности для константных доменов тяжелой и легкой цепей молекулы человека, а не гомологичных мышиных последовательностей (патент США No. 4816567; Morrison, et al., Proc. Natl Acad. Sci. USA, 81:6851 (1984)), или путем ковалентного присоединения к кодирующей последовательности для иммуноглобулина, полноразмерной или частичной, кодирующей последовательности для полипептида, отличного от иммуноглобулина.
В типичном случае такие полипептиды, отличные от иммуноглобулина, используют для замещения константных доменов антитела или они используются для замещения вариабельных доменов одного антиген-связывающего сайта антитела созданием химерного бивалентного антитела, включающего один антиген-связывающий сайт, обладающий специфичностью для одного антигена, и второй антиген-связывающий сайт, обладающий специфичностью для другого антигена.
(iv) Гуманизированные и антитела человека
Гуманизированные антитела включают один или несколько аминокислотных остатков, встроенных в молекулу из источника, который отличается от человеческого. Указанные аминокислотные остатки, отличные от остатков молекулы человека, часто обозначают как «импортные» остатки, которые в типичном случае отбирают из «импортного» вариабельного домена. Гуманизация может быть по существу проведена по методу, описанному Винтером с соавт. (Winter and co-workers) (Jones et al., Nature, 321:522-525 (1986); Riechmann et al., Nature, 332:323-327 (1988); Verhoeyen et al., Science, 239:1534-1536 (1988)), путем замещения одной или нескольких CDR последовательностей грызунов соответствующими последовательностями из антитела человека. Соответственно такие «гуманизированные» антитела представляют собой химерные антитела (патент США No. 4816567), в которых по существу меньшая часть, чем полный интактный вариабельный домен, молекулы человека был замещен соответствующей последовательностью из вида, отличного от человека. Практически гуманизированные антитела в типичном случае представляют собой антитела человека, в которых некоторые CDR остатки и, возможно, некоторые FR остатки, замещены остатками из аналогичных сайтов, имевшихся в антителах грызунов.
Выбор вариабельных доменов легкой и тяжелой цепей антитела человека, которые используются для создания гуманизированных антител, очень важен для снижения уровня антигенности. В соответствии с так называемой методикой «наибольшего соответствия», последовательность вариабельного домена из антитела грызуна подвергают скринингу с использованием полной библиотеки известных последовательностей вариабельного домена человека. И та последовательность молекулы человека, которая ближе всего соответствует таковой у грызуна, далее используется в качестве рамки считывания из молекулы человека (FR) в случае гуманизированного антитела (Sims et al., J. Immunol., 151:2296 (1993); Chothia et al., J. Mol. Biol,, 196:901 (1987)). В другом методе используется конкретная рамка считывания, полученная из консенсусной последовательности всех антител человека, оцениваемых применительно к определенной подгруппе легкой или тяжелой цепи. Одна и та же рамка считывания может использоваться для нескольких разных гуманизированных антител (Carter et al., Proc. Natl. Acad Sci. USA, 89:4285 (1992); Prestaetal., J. Immnol., 151:2623 (1993)).
Важно также, чтобы антитела подвергались гуманизации с сохранением высокой аффинности для антигена и с поддержанием других важных и подходящих биологических свойств. Для достижения этой цели согласно предпочтительному способу осуществления настоящего изобретения гуманизированные антитела получают по способу, включающему анализ родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Информация по трехмерным моделям иммуноглобулина является доступной и известна специалистам в данной области. Доступны также компьютерные программы, которые иллюстрируют и проявляют возможные трехмерные конформационные структуры выбранных последовательностей иммуноглобулина. Осмотр такого рода изображений позволяет проанализировать вероятную роль определенных остатков в функционировании рассматриваемых в качестве возможных кандидатов иммуноглобулиновых последовательностей, например провести анализ остатков, которые оказывают влияние на способность исследуемого иммуноглобулина связываться со своим антигеном. Таким образом, могут быть отобраны и объединены FR остатки из реципиентных и импортных источников, с достижением в результате желаемых характеристик антител, таких как повышенная аффинность для одного или нескольких целевых антигенов. В основном CDR остатки непосредственно и более значимо вовлекаются в связывание антигенов в качестве влияющих на этот процесс факторов.
Альтернативно в настоящее время можно создавать трансгенных животных (например, мышей), которые способны при иммунизации продуцировать полный репертуар антител человека, в отсутствие у них эндогенной продукции иммуноглобулина. Так, например, было описано, что гомозиготные делеции в гене, определяющем связующий участок тяжелой цепи антитела (JH), у химерных мышей и мутантных мышей зародышевой линии приводят к полному ингибированию эндогенной продукции антитела. Перенос совокупности генов зародышевой линии иммуноглобулинов человека у таких мутантных мышей зародышевой линии приводит к продукции антител человека при провокации антигеном (см., например, Jakobovits et al, Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggermann et al, Year in Immuno., 7:33 (1993); и Duchosal et al. Nature 355:258 (1992)). Антитела человека могут быть также получены из библиотеки проявления фагов (Hoogenboom et al, J. Mol. Biol., 227:381 (1991); Marks et al, J. MoL Biol., 222:581-597 (1991); Vaughan et al. Nature Biotech 14:309 (1996)). Получение антител человека из библиотек проявления фаговых антител описано более подробно ниже.
(v) Фрагменты антител
Были разработаны различные методики для создания фрагментов антител. Обычно эти фрагменты получали путем протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); и Brennan et al. Science, 229:81 (1985)). Однако указанные фрагменты в настоящее время могут быть получены непосредственно с использованием для этого рекомбинантных клеток-хозяев. Например, фрагменты антител могут быть выделены из библиотек фаговых антител, описанных выше. Альтернативно фрагменты Fab'-SH могут быть непосредственно восстановлены из E. coli и далее химически связаны с получением F(ab')2 фрагментов (Carter et al., Bio/Technology 10:163-167 (1992)). В другом варианте, описанном ниже в качестве примера, F(ab')2 получают с использованием лейцинового зиппера GCN4 для ускорения сборки F(ab')2 молекулы. Согласно другому подходу F(ab')2 фрагменты могут быть выделены непосредственно из культуры рекомбинантных клеток-хозяев. Другие методики получения фрагментов антител также известны специалистам в данной области. В других вариантах выбранное антитело представляет собой Fv-фрагмент одной цепи (scFv) (см., например, WO 93/16185).
(vi) Полиспецифические антитела
Полиспецифические антитела обладают связывающими специфичностями по меньшей мере для двух разных эпитопов, где указанные эпитопы обычно происходят из разных антигенов. Тогда как в норме такие молекулы будут связываться только с двумя разными эпитопами (например, биспецифические антитела, BsAb), антитела с дополнительными специфичностями, такие как триспецифические антитела, также охватываются данным выражением при его соответствующем использовании. Примеры BsAb включают такие антитела, в которых одно плечо направлено против PirB/LILRB, а другое плечо направлено против Nogo или MAG или OMgp. Другим примером BsAb являются антитела, где одно плечо направлено против PirB/LILRB, а второе плечо направлено против NgR.
Способы получения биспецифических антител известны в данной области. Традиционная продукция полноразмерных биспецифических антител основана на совместной экспрессии двух пар тяжелой цепи - легкой цепи иммуноглобулина, где обе цепи обладают разными специфичностями (Millstein et al., Nature, 305:537-539 (1983)). В связи со случайной пересортировкой легкой и тяжелой цепи иммуноглобулина указанные гибридомы (квадромы) продуцируют потенциальную смесь из 10 разных молекул антител, в числе которых только одна имеет корректную биспецифическую структуру. Очистка такой корректной молекулы, при использовании для этой процедуры нескольких стадий афинной хроматографии, является сложным процессом, выход продукта при этом достаточно бывает низким. Аналогичные процедуры описаны в WO 93/08829 и в работе Traunecker et al., EMBO J., 10:3655-3659 (1991). Согласно другому подходу вариабельные домены антитела с желательными связывающими специфичностями (антитело-антиген-связывающие сайты), сливают с последовательностями константного домена иммуноглобулина. Полученный гибрид представляет собой иммуноглобулин с константным доменом тяжелой цепи, включающий по меньшей мере часть шарнира, CH2 и CH3 участки. Предпочтительно, чтобы имелся константный участок первой тяжелой цепи (CH1), содержащий сайт, необходимый для связывания легкой цепи, присутствующий по меньшей мере в одной из сливаемых молекул. ДНК, кодирующие тяжелые цепи сливаемых иммуноглобулинов и, при желании, легкие цепи иммуноглобулинов встраивают в отдельный вектор экспрессии и подвергают совместной трансфекции в соответствующий организм-хозяин. Эта процедура обеспечивает большую гибкость при корректировке общих пропорций трех полипептидных фрагментов в тех случаях, когда используют неравные доли трех полипептидных цепей в конструкции, обеспечивая оптимальный выход. Однако можно встраивать кодирующие последовательности для двух или всех трех полипептидных цепей в один вектор экспрессии, так что экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к высокому выходу, или это возможно в тех случаях, когда указанные соотношения не имеют особого значения.
В предпочтительном варианте осуществления данного подхода биспецифические антитела состоят из гибрида тяжелой цепи иммуноглобулина с первой связывающей специфичностью в одном плече, а парный элемент гибридной тяжелой цепи - легкой цепи иммуноглобулина (обеспечивающий вторую связывающую специфичность) находится во втором плече. Было обнаружено, что такая ассиметричная структура облегчает отделение соединения с желательной специфической активностью от нежелательных сочетаний цепей иммуноглобулина, поскольку присутствие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает более легкий способ отделения. Такого рода подход описан в WO 94/04690. Дополнительные детали, относящиеся к получению биспецифических антител, описаны например, в работе Suresh et al., Methods in Enzymology, 121:210(1986).
Согласно другому подходу, описанному в WO 96/27011, интерфейс между парой молекул антител может быть сконструирован таким образом, чтобы максимизировать процент гетеродимеров, восстанавливаемых из культуры рекомбинантных клеток. Предпочтительный интерфейс включает по меньшей мере часть CH3 домена константного домена антитела. В рамках данного метода одну или несколько аминокислот с малыми боковыми цепями в интерфейсе первой молекулы антитела замещают молекулами с более крупными боковыми цепями (например, тирозином или триптофаном). «Компенсаторную» полость идентичного или близкого размера, относительно одной или нескольких крупных боковых цепей, создают на интерфейсе второй молекулы антитела путем замещения аминокислот с крупными боковыми цепями более мелкими (например, аланином или треонином). И это обеспечивает механизм повышения выхода гетеродимеров относительно нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела включают сшитые антитела или так называемые «гетероконъюгатные» антитела. Так, например, одно из антител в указанном гетероконъюгате может быть связано с авидином, тогда как второе - с биотином. Такие антитела были, в частности, предложены для того, чтобы клетки иммунной системы могли воздействовать на нежелательные клетки (патент США No. 4676980), а также для лечения ВИЧ-инфекции (WO 91/00360, WO 92/200373). Гетероконъюгатные антитела могут быть созданы с использованием удобных методов сшивки. Подходящие сшивающие агенты известны в данной области и описаны, например, в патенте США No. 4676980, где приведено также множество методик сшивки.
Способы создания биспецифических антител из фрагментов антител также описаны в литературе. Так, например, биспецифические антитела могут быть получены с использованием химического связывания. Бреннан с соавт. (Brennan et al., Science 229: 81 (1985)) описывают процедуру, в рамках которой интактные антитела подвергают протеолитическому расщеплению с созданием фрагментов F(ab')2. Эти фрагменты восстанавливают в присутствии арсенита натрия, образующего комплекс с дитиолами, для стабилизации соседних дитиоловых групп и предупреждения образования межмолекулярных дисульфидных связей. Образованные при этом Fab' фрагменты далее превращают в тионитробензоатные (TNB) производные. Одно из таких производных, Fab'-TNB, затем превращают в Fab'-тиол путем восстановления меркаптоэтиламином и смешивают с эквимолярным количеством другого Fab'-TNB-производного с образованием биспецифического антитела. Полученные при этом биспецифические антитела могут далее использоваться как агенты для селективной иммобилизации ферментов.
Fab'-SH фрагменты также могут быть непосредственно восстановлены из E. coli и далее химически связаны с формированием биспецифических антител. Шалаби с соавт. (Shalaby et al., J. Exp. Med.. 175: 217-225 (1992)) описывают получение полностью гуманизированной биспецифической молекулы антитела F(ab')2. Каждый Fab' фрагмент отдельно секретируется из E. coli и подвергается направленному химическому связыванию in vitro с образованием биспецифического антитела.
В литературе описаны различные методики получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Так, например, биспецифические антитела получают с использованием лейциновых зипперов (Kostelny et al., J. Immunol., 148(5):1547-1553 (1992)). Лейциновые зипперы из Fos и Jun белков связывают с Fab' частями двух разных антител путем слияния генов. Гомодимеры антител восстанавливают в шарнирной области с образованием мономеров и затем повторно окисляют с получением гетеродимеров антител. Этот метод может также использоваться для получения гомодимеров антител. Стратегия на основе создания «димерных антител», описанная Холлингером в соавт. (Hollinger et al., Proc. Natl Acad. Sci. USA, 90:6444-6448 (1993)), представляет альтернативный механизм создания фрагментов биспецифических антител. Указанные фрагменты включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) через линкер, который слишком короткий для того, чтобы происходило спаривание между двумя доменами на одной цепи. Соответственно VH и VL домены одного фрагмента будут спариваться с комплементарными VL и VH доменами другого фрагмента, образуя два антиген-связывающего сайта. Была также описана другая стратегия создания фрагментов биспецифических антител за счет использования димеров одноцепочечных Fv (sFv) (см. Gruber et al, J. Immunol, 152:5368 (1994)).
Рассматриваются также антитела, обладающие более чем двумя валентностями. Так, например, могут быть получены триспецифические антитела (Tuft et al. J. Immunol. 147: 60 (1991)).
(vii) Разработка эффекторных функций
Может быть желательно модифицировать антитело согласно настоящему изобретению применительно к его эффекторной функции, с тем чтобы повысить эффективность функционирования полученного антитела. Так, например, один или несколько цистеиновых остатков могут быть введены в Fc-участок, что будет способствовать образованию в этом месте межцепьевой дисульфидной связи. Образованное таким образом гомодимерное антитело может обладать повышенной способностью к интернализации и/или повышенной способностью к лизису клеток, опосредованному комплементом, а также повышенной антитело-зависимой клеточной цитотоксичностью (ADCC) (см. Caron et al., J. Exp Med. 176:1191-1195 (1992); и Shopes, B. J. Immunol. 148:2918-2922 (1992)). Могут быть также получены гомодимерные антитела с повышенной противоопухолевой активностью при использовании гетеробифункциональных сшивающих линкеров, описанных в работе Вольфа с соавт. (Wolff et al. Cancer Research 53:2560-2565 (1993)). Альтернативно антитело может быть сконструировано таким образом, что оно будет иметь двойные Fc участки и, в этой связи, будет обладать повышенными способностями к комплемент-зависимому лизису и ADCC (см. Stevenson et al Anti-Cancer Drug Design 3:219-230 (1989)).
(viii) Слияние на основе эпитопа, связывающего Salvage рецептор антитела
В некоторых вариантах осуществления настоящего изобретения может быть желательно использовать фрагменты антител, а не интактные антитела, с тем чтобы, в частности, повысить проникновение в опухоль. В этом случае может быть желательно модифицировать фрагмент антитела таким образом, чтобы повысить период его полувыведения из сыворотки. Эта цель может быть достигнута, например, при включении во фрагмент антитела эпитопа, связывающего salvage-рецептор (например, за счет мутации соответствующего участка во фрагменте антитела или путем встраивания эпитопа в пептидный фрагмент, который затем сливают с фрагментом антитела на любом его конце или в середине, например, путем синтеза ДНК или пептидного синтеза).
Эпитоп, связывающийся с salvage-рецептором, предпочтительно состоит из участка, в котором один или несколько аминокислотных остатков из одной или двух петлевых структур Fc домена переносят в аналогичное положение во фрагменте антитела. Еще более предпочтительно, переносят три или более остатков из одной или двух петлевых структур Fc домена. Еще более предпочтительно, указанный эпитоп отбирают из CH2 домена Fc участка (например, из IgG) и переносят в CH1, CH3 и VsubH участок или в более, чем один такой участок антитела. Альтернативно указанный эпитоп отбирают из CH2 домена Fc участка и переносят в CL участок или VL участок, или на оба участка в указанном фрагменте антитела.
(ix) Другие ковалентные модификации антител
В область настоящего изобретения включаются также ковалентные модификации антител. Они могут быть выполнены путем химического синтеза или путем ферментативного или химического расщепления антитела, если это возможно. Ковалентные модификации антител другого типа могут быть встроены в молекулы путем проведения реакции целевых аминокислотных остатков антитела с органическим дериватизирующим агентом, который способен взаимодействовать с выбранными боковыми цепями или с N-, или C-концевыми остатками. Примеры ковалентных модификаций описаны в патенте США No. 5534615 и включены в настоящее описание в качестве ссылки. Предпочтительный тип ковалентной модификации антитела включает связывание антитела с одним или множеством небелковых полимеров, например с полиэтиленгликолем, полипропиленгликолем или полиоксиалкиленами, по процедуре, описанной в ряде патентов США: NoNo. 4640835; 4496689; 4301144; 4670417; 4791192 или 4179337.
(x) Создание антител с использованием библиотеки синтезированных фаговых антител
В предпочтительном варианте настоящее изобретение относится к способу создания и отбора новых антител с использованием уникальной стратегии фагового дисплея. Указанная стратегия включает создание библиотеки синтетических фаговых антител, основанных на одной рамочной матрице, с включением достаточного уровня различий внутри вариабельных доменов, детекцию полипептидов, обладающих различающимися вариабельными доменами, отбор антител-кандидатов с высокой аффинностью к целевому антигену и выделение выбранных антител.
Детали указанных методов фагового дисплея можно найти, например, в патенте WO 03/102157, опубликованном 11 декабря 2003 г., полное писание которого включено в настоящее изобретение в качестве ссылки.
В одном аспекте используемые в рамках настоящего изобретения библиотеки антител могут быть получены путем мутации доступных для растворителя и/или отличающихся высоким разнообразием положений по меньшей мере в одном CDR в вариабельном домене антитела. Некоторые или все CDR могут быть мутированы с использованием приведенных в настоящем описании способов. В некоторых вариантах может быть предпочтительно создать библиотеки дивергентных антител путем мутации в положениях CDRH1, CDRH2 и CDRH3 с образованием одной библиотеки или путем мутации в положениях CDRL3 и CDRH3 с образованием одной библиотеки или путем мутации в положениях CDRL3 и CDRH1, CDRH2 и CDRH3 с образованием одной библиотеки.
Может быть получена библиотека вариабельных доменов антител, которая содержит, например, мутации в доступных для растворителя и/или отличающихся высоким разнообразием положениях CDRH1, CDRH2 и CDRH3. Может быть создана другая библиотека, содержащая мутации в CDRL1, CDRL2 и CDRL3. Эти библиотеки могут также использоваться в сочетании друг с другом для создания связующих участков с желательной аффинностью. Так, например, после одного или нескольких раундов отбора тяжелых цепей для библиотек, подходящих для связывания с целевым антигеном, библиотека легких цепей может быть замещена с получением популяции связующих элементов тяжелой цепи, для проведения дальнейших раундов отбора с целью повышения аффинности связок.
Предпочтительно библиотеку создают путем замещения исходных аминокислот вариантными аминокислотами в CDRH3 участке последовательности вариабельного участка тяжелой цепи. Полученная библиотека может содержать множество последовательностей антител, где разнообразие последовательностей связано, с первую очередь, с CDRH3 участком в последовательности тяжелой цепи.
В одном аспекте библиотеку создают в контексте 4D5 последовательности гуманизированного антитела или последовательности аминокислот на рамочном участке последовательности гуманизированного антитела 4D5. Предпочтительно библиотеку создают путем замещения по меньшей мере остатков 95-100а в тяжелой цепи аминокислотами, кодируемыми кодоновым набором DVK, где кодоновый набор DVK используют для кодирования множества вариантных аминокислот для каждого из указанных положений. Пример олигонуклеотидного набора, который может использоваться для создания таких замещений, включает последовательность (DVK) 7. В некоторых вариантах осуществления настоящего изобретения указанную библиотеку создают путем замещения остатков 95-100а аминокислотами, кодируемыми кодоновыми наборами DVK и NNK. Примером олигонуклеотидного набора, который может использоваться для создания таких замещений, является набор, который включает последовательность (DVK) 6 (NNK). В другом варианте осуществления настоящего изобретения указанную библиотеку создают путем замещения остатков 95-100а аминокислотами, кодируемыми кодоновыми наборами и DVK, и NNK, обоими наборами. Примером олигонуклеотидного набора, который может использоваться для создания таких замещений, является набор, который содержит последовательность (DVK) 5 (NNK). Другой пример олигонуклеотидного набора, который используется для создания таких замещений, включает последовательность (NNK) 6. Другие примеры подходящих олигонуклеотидных последовательностей могут быть определены любым специалистом в данной области согласно критериям, приведенным в настоящем описании.
В другом варианте осуществления настоящего изобретения используют разные модели CDRH3 для выделения связующих компонентов с высокой активностью и для выделения указанных связок для множества эпитопов. Диапазон длин CDRH3, получаемых в этой библиотеке, составляет от 11 до 13 аминокислот, хотя могут быть получены также фрагменты с длиной, отличающейся от указанной выше. Разнообразие по H3 может быть расширено за счет использования NNK, DVK и NVK кодоновых наборов, а также в случае более ограниченного разнообразия на N- и/или C-концах.
Разнообразие может быть создано в рамках CDRH1 и CDRH2. Варианты с CDRHl и H2 различиями создают согласно стратегиям, направленным на достижение репертуара имитирующих природных антител, как было описано, с встраиванием модификаций, направленных на указанную дивергентность, с тем чтобы более точно соответствовать природному разнообразию, чем это было в более ранних конструкциях.
Для достижения разнообразия по CDRH3 могут быть сконструированы, по отдельности, различные библиотеки с разными длинами H3 и затем они могут быть объединены для выбора соответствующих связок, с тем чтобы воздействовать на антигены. Указанные различные библиотеки могут быть объединены и далее сортированы с использованием методов отбора на твердых носителях и методов сортировки в растворе, как было описано ранее и как приведено ниже в описании. При этом могут использоваться различные стратегии сортировки. Так, например, в одном варианте используют сортировку на мишени, связанной с твердым носителем, с последующей сортировкой по фрагменту, который может присутствовать в гибридном полипептиде (например, по анти-gD фрагменту), с последующим проведением другого рода сортировки на мишени, соединенной с твердым носителем. Альтернативно библиотеки могут быть отсортированы вначале по мишени, связанной с твердой поверхностью, с последующей их сортировкой, после элюции связующих веществ, с использованием процедуры связывания в фазе раствора при снижении концентраций целевого антигена. Использование сочетаний разных методов сортировки обеспечивает минимизацию выборки, содержащей только высоко экспрессируемые последовательности, и обеспечивает отбор множества различных клонов с высокой аффинностью.
Высокоаффинные связки для целевого антигена могут быть выделены из соответствующих библиотек. Ограниченное разнообразие в участке H1/H2 снижает вырожденность кода от 104 до 105 раз и позволяет достичь большего разнообразия по H3, что обеспечивает получение связующих веществ с более высокой аффинностью. Использование библиотек с разными типами дивергентности по CDRH3 (например, при использовании DVK или NVT) обеспечивает выделение связок, которые могут связываться с разными эпитопами целевого антигена.
Для связующих компонентов, выделенных из объединенных библиотек, указанных выше, было показано, что аффинность может быть далее повышена за счет обеспечения ограниченного разнообразия по легкой цепи. Разнообразие по легкой цепи создается согласно данному варианту следующим образом: в CDRL1 - аминокислота в положении 28 кодируется RDT; аминокислота в положении 29 кодируется RKT; аминокислота в положении 30 кодируется RVW; аминокислота в положении 31 кодируется ANW; аминокислота в положении 32 кодируется THT; необязательно, аминокислота в положении 33 кодируется CTG ; в CDRL2 - аминокислота в положении 50 кодируется KBG; аминокислота в положении 53 кодируется AVC; и необязательно, аминокислота в положении 55 кодируется GMA; в CDRL3 - аминокислота в положении 91 кодируется TMT или SRT или ими обоими; аминокислота в положении 92 кодируется DMC; аминокислота в положении 93 кодируется RVT; аминокислота в положении 94 кодируется NHT; и аминокислота в положении 96 кодируется TWT или YKG, или ими обоими.
В другом варианте создают библиотеку, одну или несколько, с разнообразием по CDRH1, CDRH2 и CDRH3 участкам. В рамках данного варианта разнообразие по CDRH3 создают при использовании Н3 участков различной длины и при использовании, главным образом, кодоновых наборов XYZ и NNK или NNS. Библиотеки могут быть созданы с использованием отдельных олигонуклеотидов или их объединений либо олигонуклеотиды могут быть объединены с образованием подмножества библиотек. Согласно этому варианту библиотеки могут быть сортированы относительно мишеней, связанных с твердым носителем. Клоны, выделенные в результате множества сортировок, могут быть далее скринированы на специфичность и аффинность с использованием тестов по процедуре ИФТФА. Клоны, для оценки их специфичности, могут быть скринированы относительно целевых антигенов, а также других нецелевых антигенов. Указанные связующие компоненты для целевого антигена могут быть далее скринированы на аффинность в конкурентном анализе по процедуре ИФТФА на связывание в фазе раствора или в конкурентном анализе с использованием спот-процедуры. Высокоаффинные связующие компоненты могут быть выделены из библиотеки, включающей кодоновые наборы XYZ, согласно описанным выше методам. Указанные связующие компоненты могут быть легко получены в клеточной культуре в виде антител или антиген-связывающих фрагментов с высоким выходом.
В некоторых вариантах осуществления настоящего изобретения может быть желательно создать библиотеки с большим разнообразием по длинам CDRH3 участка. Так, например, может быть желательно создать библиотеки с участками CDRH3, варьирующими по длине примерно от 7 до 19 аминокислот.
Высокоаффинные связки, выделяемые из библиотек согласно указанным вариантам, могут быть легко получены в культуре бактериальных и эукариотических клеток, с высоким выходом. При этом могут быть созданы векторы для удаления последовательностей, таких как gD фрагменты, последовательность белка вирусной оболочки, и/или указанные векторы могут быть использованы для добавления к последовательностям константного участка, с тем чтобы обеспечить продукцию полноразмерных антител или антиген-связывающих фрагментов, с высоким выходом.
Библиотека с мутациями по CDRH3 может быть объединена с библиотекой, содержащей вариантные версии других CDR, например CDRL1, CDRL2, CDRL3, CDRH1 и/или CDRH2. Так, например, в одном варианте библиотеку CDRH3 объединяют с библиотекой CDRL3, созданной согласно процедуре получения последовательности гуманизированной последовательности 4D5 антитела, с вариантными аминокислотами в положениях 28, 29, 30, 31 и/или 32 с использованием заданных кодоновых наборов. В другом варианте библиотека с мутациями в CDRH3 может быть объединена с библиотекой, включающей вариантный CDRH1 и/или CDRH2 вариабельного домена тяжелой цепи. В одном варианте библиотеку CDRH1 создают с включением в нее последовательности гуманизированного антитела 4D5 с вариантными аминокислотами в положениях 28, 30, 31, 32 и 33. Библиотека CDRH2 может быть создана с включением в нее последовательности гуманизированного антитела 4D5 с вариантными аминокислотами в положениях 50, 52, 53, 54, 56 и 58 с использованием заданных кодоновых наборов.
(xi) Мутанты антител
Новые антитела, полученные из фаговых библиотек, могут быть затем модифицированы для создания мутантных антител с улучшенными физическими, химическими и/или биологическими свойствами относительно родительского антитела. В случае, когда используемый анализ основан на биологической активности, мутантное антитело предпочтительно обладает биологической активностью в рамках данного выбранного анализа, которая по меньшей мере в 10 раз выше, предпочтительно по меньшей мере в 20 раз выше, более предпочтительно по меньшей мере в 50 раз выше и иногда по меньшей мере в 100 раз или 200 раз выше, чем биологическая активность исходного антитела в данном анализе. Например, анти-PirB/LILRB мутантное антитело предпочтительно обладает связывающей аффинностью для PirB/LILRB, которая по меньшей мере примерно в 10 раз выше, предпочтительно, по меньшей мере в 20 раз выше, более предпочтительно, по меньшей мере примерно в 50 раз выше, и иногда по меньшей мере примерно в 100 раз или 2000 раз выше, чем связывающая аффинность исходного родительского антитела.
Для создания мутантного антитела одно или несколько аминокислотных изменений (например, замещения) встраивают в один или несколько гипервариабельных участков родительского антитела. Альтернативно или дополнительно одно или несколько изменений (например, замещений) остатков в рамке считывания могут встраиваться в исходное родительское антитело, что приводит к повышению связывающей аффинности получаемого мутантного антитела для антигена, взятого из второго вида млекопитающего. Примеры остатков из рамки считывания, подлежащих модификации, включают такие остатки, которые нековалентно связываются непосредственно с антигеном (Amit et al. (1986) Science 233:747-753); взаимодействуют/влияют с/на CDR (Chothia et al. (1987) J. Mol. Biol. 196:901-917); и/или участвуют в интерфейсе VL - VH (EP 239400B1). В некоторых вариантах модификация одного или нескольких таких остатков в рамочном участке приводит к повышению связывающей аффинности антитела для антигена, взятого из второго вида млекопитающего. Так, например, согласно данному варианту осуществления настоящего изобретения могут быть изменены от примерно одного до примерно пяти остатков в рамке считывания. Иногда этого может быть достаточно для достижения выхода мутантного антитела, приемлемого для его использования в доклинических испытаниях, даже в тех случаях, когда ни один из остатков гипервариабельного участка не был изменен. Однако обычно мутантное антитело включают дополнительное одно или несколько изменений в гипервариабельном участке.
Остатки гипервариабельного участка, которые подлежат изменению, могут меняться в рандомизированном варианте, особенно в тех случаях, когда исходная связывающая аффинность родительского антитела такова, что указанный рандомизированный подход позволяет получить мутантные антитела, которые легко поддаются скринингу.
Одна из используемых процедур для создания таких мутантных антител носит название «аланин-сканирующий мутагенез» (Cunningham and Wells (1989) Science 244:1081-1085). В этом случае один или несколько остатков в гипервариабельном участке замещают аланином или одним, или несколькими остатками полиаланина с тем, чтобы оказать эффект на взаимодействие аминокислот с антигеном, взятым из второго вида млекопитающего. Указанные один или несколько остатков гипервариабельного участка, демонстрирующие функциональную чувствительность к таким замещениям, далее корректируют путем введения дополнительных или других мутаций в сайтах замещения. Таким образом, несмотря на то, что сайт введения вариаций по аминокислотной последовательности заранее определен, природа мутации per se не определена. Получаемые таким образом ala-мутанты скринируют на их биологическую активность согласно приведенному описанию.
В норме обычно начинают с введения консервативного замещения, например согласно графе «Предпочтительные замещения» в приведенной ниже таблице. Если такие замещения приводят к изменению биологической активности (например, в связывающей аффинности), вводят более значимые изменения, обозначенные в таблице как «иллюстративные замещения», как это будет описано ниже, применительно к различным классам аминокислот, и полученные продукты подвергают скринингу.
Еще более существенные модификации биологических свойств антител могут быть достигнуты при выборе замещений, которые значительно отличаются по их эффекту на поддержание (a) структуры полипептидного скелета в зоне замещения, например, в виде складчатой или спиральной конформации, (b) заряженности или гидрофобности молекулы в сайте-мишени или (c) объемности боковой цепи. Природные остатки делятся на группы на основании общих свойств боковой цепи:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr, asn, gln;
(3) кислотные: asp, glu;
(4) основные: his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи в пространстве: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замещения относятся к обмену члена одного из этих классов на члена другого класса.
В другом варианте сайты, отобранные для модификации, характеризуются зрелостью по аффинности, при оценке по проявлению фагов (см. приведенное выше описание).
Молекулы нуклеиновой кислоты, кодирующие последовательность мутантных аминокислот, получают с использованием известных в данной области методов. Эти методы включают, без ограничения, олигонуклеотидный (или сайт-направленный) мутагенез, мутагенез с использованием ПЦР и кассетный мутагенез ранее полученной мутантной или немутантной версии родительского антитела. Предпочтительным способом получения мутантов является сайт-направленный мутегенез (см., например, Kunkel (1985) Proc. Natl. Acad. Sci. USA 82:488).
В некоторых вариантах осуществления настоящего изобретения мутантное антитело содержит только один замещенный остаток в гипервариабельной области. В других вариантах два или более остатков в гипервариабельной области родительского антитела замещаются, например вводятся от примерно двух до примерно десяти остатков в гипервариабельной области с целью замещения.
Обычно мутантное антитело с улучшенными биологическими свойствами имеет аминокислотную последовательность, характеризующуюся наличием по меньшей мере 75% идентичности по аминокислотной последовательности или сходства по аминокислотным последовательности с аминокислотной последовательностью вариабельного домена тяжелой или легкой цепи родительского антитела, более предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, более предпочтительно, по меньшей мере 90%, и наиболее предпочтительно, по меньшей мере 95%, идентичностью или сходством. Указанные идентичность или сходство относительно данной последовательности определяются в контексте настоящего описания как процент аминокислотных остатков в исследуемой последовательности, которая идентична (то есть, имеются одинаковые остатки) или близка (то есть, имеются аминокислотные остатки из одной и той же группы, определяемой по свойствам боковой цепи, см. выше) к остаткам родительского антитела, по результатам сопоставления последовательностей и встраивания пропусков, при необходимости, для достижения максимальной процентной идентичности по последовательности. При этом ни N-концевые, ни С-концевые или внутренние расширения, делеции или вставки в аминокислотной последовательности за пределами вариабельного участка не должны влиять на указанные идентичность или сходство по последовательности.
После получения мутантного антитела определяют биологическую активность такой молекулы в сравнении с родительским антителом. Как указывалось выше, эта процедура может включать определение связывающей аффинности и/или других видов биологической активности антитела. В предпочтительном варианте осуществления настоящего изобретения получают панель мутантных антител и подвергают их скринингу для оценки связывающей аффинности относительно антигена или его фрагмента. Одно или несколько мутантных антител, отобранных в рамках исходного скрининга, необязательно подвергают одному или нескольким тестам для определения их биологической активности, с тем чтобы подтвердить, что указанные одно или несколько антител с повышенной связывающей аффинностью действительно будут применимы, например, в доклинических испытаниях.
Выбранные таким образом одно или несколько мутантных антител могут быть далее подвергнуты процедурам встраивания в них других модификаций, которые зачастую зависят от предполагаемого варианта использования такого антитела. Указанные модификации могут вовлекать дополнительные изменения в аминокислотной последовательности, слияние с одним или несколькими гетерологичными полипептидами и/или введение ковалентных модификаций, например, описанных ниже. Что касается изменения аминокислотной последовательности, то репрезентативные модификации были описаны ранее. Так, например, любой цистеиновый остаток, не участвующий в поддержании соответствующей конформации мутантного антитела, также может быть замещен, в основном серином, для повышения стабильности молекулы к окислению и предупреждения аберрантной сшивки. И наоборот, одна или несколько цистеиновых связей могут быть добавлены к антителу для повышения стабильности (в особенности в том случае, когда антитело представлено фрагментом антитела, таким как Fv фрагмент). Другой тип аминокислотного мутанта характеризуется измененным характером гликозилирования. Указанные изменения могут быть достигнуты путем делеции одного или нескольких углеводных фрагментов, имеющихся в антителе, и/или путем добавления одного или нескольких сайтов гликозилирования, которые не присутствуют в антителе. Гликозилирование антител в типичном случае является N-связанным или O-связанным. N-связанный характер гликозилирования относится к присоединению углеводного фрагмента к боковой цепи аспарагинового остатка. Трипептидные последовательности: аспарагин-X-серин и аспарагин-Х-треонин, где Х может быть любой аминокислотой, кроме пролина, представляют собой сайты распознавания для ферментативного присоединения углеводного фрагмента к аспарагину боковой цепи. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. О-связанный характер гликозилирования относится к присоединению одного из сахаров: N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиламинокислоте, чаще всего, это серин или треонин, хотя могут также использоваться 5-гидроксипролин или 5-гидроксилизин. Добавление сайтов гликозилирования к антителу обычно осуществляют путем изменения аминокислотной последовательности, так чтобы она содержала одну или несколько описанных выше трипептидных последовательностей (в случае N-связанных сайтов гликозилирования). Указанное изменение может быть также внесено путем добавления или замещения одного или нескольких остатков серина или треонина к последовательности исходного антитела (в случае О-связанных сайтов гликозилирования).
(xii) Рекомбинантное получение антител
Для рекомбинантной продукции антитела выделяют нуклеиновую кислоту, кодирующую его, и встраивают в реплицируемый вектор для дальнейшего клонирования (амплификация ДНК) или для экспрессии. ДНК, кодирующую моноклональное антитело, можно легко выделить и затем секвенировать с использованием стандартных процедур (например, при использовании олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими легкую и тяжелую цепи антитела). Для этой цели доступно множество векторов. Векторы в основном включают, без ограничения, один или несколько следующих компонентов: сигнальную последовательность, ориджин репликации, один или несколько маркерных генов, энхансер, промотор и последовательность терминации транскрипции (например, как описано в патенте США No. 5534615, который включен в настоящее описание в качестве ссылки).
Клетки-хозяева, подходящие для клонирования или экспрессии ДНК в векторах, согласно настоящему описанию включают прокариоты, дрожжи или клетки высших эукариотов, описанные ранее. Подходящие для этой цели прокариотические клетки включают эубактерии, такие как грамотрицательные, грамположительные организмы, например Enterobacteriaceae, такие как Escherichia, например E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, а также виды Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанный в DD 266710, опубл. 12 апреля 1989 г.), виды Pseudomonas, такие как P. aeruginosa, и Streptomyces. Предпочтительными для клонирования клетками-хозяивами E. coli являются E. coli 294 (ATCC 31,446), хотя могут быть приемлемы также другие штаммы, такие как E. coli B, E. coli X 1776 (ATCC 31,537) и Е. coli W3110 (ATCC 27325). Однако, приведенные примеры являются лишь иллюстративными, которые не следует трактовать в ограничительном смысле.
Кроме прокариотов, эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, также представляют собой подходящие организмы-хозяева для клонирования или экспрессии векторов, кодирующих антитела. При этом чаще всего среди низших эукариотических микроорганизмов-хозяев используют Saccharomyces cerevisiae, или обычные пекарские дрожжи. Однако доступно и может использоваться, в контексте настоящего описания, множество представителей других родов, видов и штаммов в качестве организмов-хозяев, такие как Schizosaccharomyces pombe; виды Kluyveromyces, такие как K. lactis, K. fragilis (ATCC 12,424), K. bulgaricus (ATCC 16,045), K. wickeramii (ATCC 24,178), K. waltii (ATCC 56,500), K. drosophilarum (ATCC 36,906), K. thermotolerans и K.marxianus; Yarrowia (EP 402,226); Pichia pastoris (EP 183,070); Candida; Trichoderma reesia (EP 244,234); Neurospora crassa; виды Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, такие как Neurospora, Penicillium, Tolypocladium и Aspergillus, в качестве хозяев, в частности, A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированных антител получают из многоклеточных организмов. Примеры клеток беспозвоночных организмов включают клетки растений и насекомых. Было идентифицировано множество штаммов бакуловируса и их варианты, а также соответствующие пермиссивные клетки насекомого-хозяина из таких организмов-хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. В настоящее время доступно для использования множество штаммов вирусов, применимых для трансфекции, например L-1 вариант Autographa califomica NPV и Bm-5 штамм Bombyx mori NPV, и такие вирусы могут использоваться в качестве вирусов согласно настоящему изобретению, в особенности для трансфекции клеток Spodoptera frugiperda. Культуры клеток растений, таких как хлопок, кукуруза, картофель, соевые бобы, петунья, томаты и табак, также могут использоваться в качестве организмов-хозяев.
Однако особый интерес вызвали клетки позвоночных животных, и процедура размножения клеток позвоночных в культуре (тканевая культура) стала в настоящее время достаточно рутинной. Примеры используемых клеточных линий клеток млекопитающих-хозяев включают: клеточную линию клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); клеточную линию клеток почки эмбриона человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре, Graham et al, J, Gen Virol. 36:59 (1977)); клетки почки детеныша хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad, Sci, USA 77:4216 (1980)); клетки Сертоли мышей (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CVl ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки цервикальной карциномы человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); TRI клетки (Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); MRC 5 клетки; FS4 клетки и клетки линии гепатомы человека (Hep G2).
Клетки-хозяева подвергают трансформации описанными выше векторами экспрессии или клонирования для продукции антител и культивируют в традиционных питательных средах, модифицированных соответствующим образом для индукции промоторов, выбора трансформантов или амплификации генов, кодирующих желательные последовательности.
Клетки-хозяева, используемые для продукции антител согласно настоящему изобретению, могут культивироваться в различных средах. Для культивированная клеток-хозяев приемлемы коммерчески доступные среды, такие как среда Хэмса F10 (Sigma), минимальная эссенциальная среда (MEM), (Sigma), среда RPMI-1640 (Sigma) и среда Игла в модификации Дульбекко ((DMEM), Sigma). Кроме того, любая из сред, описанных в работах Ham et al, Meth. Enz. 58:44 (1979), Barnes et al, Anal. Biochem, 102:255 (1980), патентах США NoNo. 4767704; 4657866; 4927762; 4560655; или 5122469; WO 90/03430; WO 87/00195; или патенте США No. Re. 30985 может использоваться в качестве среды для культивирования клеток-хозяев. Любая из указанных сред может содержать добавку, при необходимости, в виде гормонов и/или других факторов роста (таких как инсулин, трансферрин или эпидермальный фактор роста), соли (такой как хлорид натрия, кальция, магния и фосфат), буферы (такой как HEPES), нуклеотидов (таких как аденозин и тимидин), антибиотиков (таких как гентамицин (GENTAMYCIN™)), микроэлементов (определяемых как неорганические соединения, обычно присутствующие в конечной концентрации в микромолярном диапазоне) и глюкозы или эквивалентного источника энергии. Любые другие необходимые добавки также могут быть включены в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т. п., представляют собой такие условия, которые обычно используют для экспрессии выбранных клеток-хозяев и такие условия известны специалистам в данной области.
В случае использования рекомбинантных методик антитело может быть получено внутри клетки, в периплазматическом пространстве, или оно может быть непосредственно секретировано в среду. Если антитело получают внутри клетки в качестве первой стадии, то осколки частиц, клеток-хозяев или лизированных клеток, удаляют, например, путем центрифугирования или ультрафильтрации. В том случае, когда антитело секретируется в среду, супернатанты в таких системах экспрессии в основном вначале концентрируют с использованием коммерчески доступных фильтров для концентрирования белка, таких как фильтры в системах ультрафильтрации Amicon или Millipore Pellicon. На любой из указанных выше стадий в среду может включаться ингибитор протеазы, такой как PMSF, для ингибирования протеолиза, а также антибиотики для предотвращения роста вредных контаминантов.
Композиция антитела, полученная на основе указанных клеток, может быть очищена с использованием, например, хроматографии на гидроксилапатите, гель-электрофореза, диализа и аффинной хроматографии, где аффинная хроматография представляет собой предпочтительную методику для очистки. Применимость белка А, в качестве аффинного лиганда, зависит от вида и изотипа любого Fc домена иммуноглобулина, который присутствует в антителе. Белок А может использоваться для очистки антител, которые основаны на тяжелых цепях антител человека гамма 1, гамма 2 или гамма 4 (Lindmark et al, J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендуется для очистки всех мышиных изотипов и для очистки антитела человека гамма 3 (Guss et al., EMBO J. 5:1567-1575 (1986)). Матрица, к которой присоединяется аффинный лиганд, чаще всего представляет собой агарозу, но могут использоваться и доступные другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, позволяют ускорить протекание и сократить время обработки, в сравнении с вариантом использования агарозы. В том случае, когда антитело включает CH3 домен, для очистки используют смолу Bakerbond ABX™ (J. T. Baker, Phillipsburg, N.J.). Применимы также другие методики, используемые для очистки белка, в зависимости от восстанавливаемого антитела, такие как фракционирование на ионообменной колонке, осаждение этанолом, ВЭЖХ с обращением фазы, хроматография на силикагеле, хроматография на гепарин-Сефарозе (SEPHAROSE™), хроматография на анионообменной или катионообменной смоле (например, с использованием колонки с полиаспарагиновой кислотой), хроматофокусирование, электрофорез в ДСН-ПААГ и осаждение сульфатом аммония.
D. Использование стимуляторов нейрональной регенерации
Молекулы, идентифицированные в анализах скрининга согласно настоящему изобретению, как предполагается, могут использоваться в качестве агентов для повышения выживаемости нервных клеток или для индукции образования отростков нервных клеток. В этой связи они могут использоваться в терапии дегенеративных расстройств нервной системы («нейродегенеративные заболевания»), включающих, например, повреждение периферических нервов, вызванное физическим повреждением (например, ожоги, раны), или заболевания, такие как диабет, дисфункция почки, или токсическое воздействие химиотерапевтических средств, используемых для лечения рака и СПИД; физическое повреждение центральной нервной системы (спинного мозга и головного мозга); повреждение головного мозга, ассоциированное с инсультом; и неврологические расстройства, связанные с нейродегенерацией, такие как, например, невралгия тройничного нерва, глоссофарингеальная невралгия, паралич Белла, миастения гравис, мышечная дистрофия, боковой амиотрофический склероз (ALS), прогрессирующая мышечная атрофия, прогрессирующая бульбарная наследственная мышечная атрофия, грыжа, разрыв или пролапс межпозвоночных дисков, шейный спондилез, расстройства, затрагивающие сплетение, синдром лестничной мышцы (сдавления плечевого сплетения и подключичной артерии), периферические нейропатии, например, вызванные свинцом, даксоном, клещами, порфирия, синдром Гийена-Барра, болезнь Альцгеймера, болезнь Хантингтона или болезнь Паркинсона.
Соединения, идентифицированные согласно настоящему изобретению, также используются в качестве компонентов культуральных сред для культивирования нервных клеток in vitro.
И наконец, препараты, включающие соединения, идентифицированные в рамках описанных тестов, применимы в качестве стандартов в конкурентных анализах на связывание, при использовании для мечения радиоактивного йода, ферментов, флуорофоров, спиновых меток и т.п.
Терапевтические композиции соединения согласно настоящему изобретению получают таким образом, что они могут храниться при смешивании идентифицированного соединения (такого как антитело), характеризующегося желательной степенью чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences, выше), в форме лиофилизированной лепешки или в составе водных растворов. Подходящие носители, эксципиенты или стабилизаторы, которые должны быть нетоксичными для реципиентов в используемых дозировках и концентрациях, включают буферы, такие как фосфатный, цитратный и на основании других органических кислот; антиоксиданты, включающие аскорбиновую кислоту; низкомолекулярные полипептиды (содержащие менее, чем примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон, аминокислоты, такие как глицин, глютамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как Твин, Плуроники или ПЭГ.
Соединения, используемые для введения in vivo, должны быть стерильными. Указанная стерильность может быть легко достигнута путем фильтрации через стерильные мембраны для фильтрования, перед или после лиофилизации и восстановления.
Терапевтические композиции могут быть внесены в контейнеры, включающие стерильный порт доступа, например пакет с раствором для внутривенного введения или флакон, который содержит пробку, прокалываемую гиподермной иглой для инъекций.
Соединения, идентифицированные с использованием тестов согласно настоящему изобретению, могут быть необязательно объединены с конкретными нейротропными факторами или введены в сочетании с ними, где указанные нейротропные факторы включают NGF, NT-3, и/или BDNF, которые могут использоваться в сочетании с другими стандартными видами терапии, применяемыми при дегенеративных нервных расстройствах.
Способ введения соответствует известным способам, то есть это может быть инъекция или инфузия путем внутривенного, внутрибрюшинного, интрацеребрального, внутримышечного, внутриглазного, интраартериального введения или путем введения внутрь повреждения, это может быть местное введение или введение с использованием систем пролонгированного высвобождения, описанных ниже.
В случае интрацеребрального использования соединения могут вводиться непрерывно путем инфузии во внутренние полости ЦНС, хотя приемлемы также инъекции болюсом. Указанное соединение предпочтительно вводят в желудочки головного мозга или иным способом вводят в ЦНС или в спинномозговую жидкость. Введение может осуществляться через катетер в режиме непрерывного введения с использованием таких устройств, как насос, или указанное введение может быть достигнуто путем имплантации, например, интрацеребральной имплантации, или с использованием носителя с пролонгированным высвобождением. Более конкретно указанные соединения могут инъецироваться в хроническом режиме с использованием имплантируемых канюль или в режиме хронической инфузии с помощью осмотических мини-насосов. Для этих целей доступны подкожные насосы, которые доставляют белки через маленькую трубку, подсоединенную к церебральным желудочкам. Очень сложно выполненный насос может наполняться через кожу, и скорость доставки может устанавливаться без хирургической интервенции. Примеры подходящих протоколов введения и систем доставки, в которых используется подкожное насосное устройство или проводится непрерывная интрацеребровентрикулярная инфузия, за счет полностью имплантируемой системы доставки лекарственного средства, включают такие протоколы, которые применимы для введения допамина, агонистов допамина и холинергических агонистов пациентам с болезнью Альцгеймера и животным, в рамках моделей болезни Паркинсона, как описанным в работах Harbaugh, J. Neural Transm. Sxippl., 24:271 (1987); и DeYebenes, et al, Mov. Disord. 2:143 (1987).
Соответствующие примеры препаратов с пролонгированным высвобождением включают полупроницаемые полимерные матрицы в форме определенных структурированных продуктов, например в виде пленок или микрокапсул. Матрицы, применяемые в системах пролонгированного высвобождения, включают полиэфиры, гидрогели, полилактиды (патент США No. 3773919, EP 58481), сополимеры L-глютаминовой кислоты и гамма-этил-L-глютамата (Sidman, et al., 1983, Biopolymers 22:547), поли-(2-гидроксиэтилметакрилат) (Langer, et al., 1981, J. Biomed. Mater. Res. 15:167; Langer, 1982, Chem. Tech. 12:98), этиленвинилацетат (Langer, et al., Id.) или poly-D-(-)-3-гидроксимасляную кислоту (EP 133,988A). Композиции с пролонгированным высвобождением также включают соединения, введенные в состав липосом, которые могут быть получены способом, известным per se (Epstein, et al., Proc. Natl. Acad. Sci. 82:3688 (1985); Hwang, et al., Proc. Natl. Acad. Sci. USA 77:4030 (1980); патенты США NoNo. 4485045 и 4544545; и EP 102324A). Обычно липосомы представляют собой небольшие (примерно 200-800 ангстрем) однослойные тела, в которых липидное содержание превышает примерно 30 моль.% по холестерину, где выбранная пропорция была откорректирована с целью достижения оптимальных результатов терапии.
Эффективное количество активного соединения, используемого в ходе терапии, будет зависеть, например, от целей лечения, способа введения и состояния пациента. Соответственно может быть необходимо, чтобы лечащий врач провел титрование дозы и модифицировал способ введения в соответствии с потребностями данного случая для достижения оптимального терапевтического эффекта. Типичная дневная доза варьирует от примерно 1 мкг/кг до 1000 мг/кг или более, в зависимости от указанных выше факторов. В типичном случае врач-клиницист будет вводить активное соединение до достижения дозировки, которая приводит к восстановлению, подержанию и, в оптимальном варианте, восстановлению функции нейрона. Прогресс такой терапии легко отслеживать с использованием традиционных тестов.
Дополнительные детали настоящего изобретения иллюстрируются приведенными ниже неограничивающими примерами.
Пример 1
Экспрессия вектора клонирования LILRB2
С целью идентификации новых рецепторов для ингибирующих миелиновых белков используют стратегию на основе клонирования вектора экспрессии. В качестве имитирующих агентов создают слитый гибридный белок на основе щелочной фосфатазы (AP) с N- и/или C-концом следующих хорошо известных миелиновых ингибиторов (используют кДНК человека): Nogo66, два дополнительных ингибирующих домена NogoA (NiR<delta>D2 и NiG<delta>20) (Oertle T, J Neurosci. 2003, 23(13): 5393-406), MAG и OMgp. Эти конструкции переносят в клетки 293 для продукции в кондиционированной среде (в DMEM/2% ФБР), содержащей имитирующие белки. Библиотека кДНК, используемая для скрининга, состоит из полноразмерных клонов кДНК человека в готовых для экспрессии векторах, от компании Origene. Указанные кДНК объединяют, создают на их основе чипы и объединяют в пулы. Пулы, содержащие примерно 100 кДНК, подвергают временной трансфекции в COS7 клетки.
В частности, в первый день COS7 клетки высевают с плотностью 85000 клеток на ячейку в 12-ячеечные планшеты. На 2-й день проводят трансфекцию, используя по 1 мг объединенной кДНК на ячейку, с включением липидного реагента трансфекции FuGENE 6 (Roche). На 4-й день проводят скрининг. В общих чертах процедура состоит в том, что культуральную среду отбирают от клеток и замещают 0,5 мл кондиционированной среды для клеток 293, содержащей затравочные белки слияния на основе AP (20-50 нМ). Клетки инкубируют при комнатной температуре в течение 90 минут. Затем клетки промывают три раза фосфатно-буферным раствором (ФБР), фиксируют в течение 7 минут с использованием 4% параформальдегида, промывают 3 раза в HEPES-буферном солевом растворе (HBS) и инактивируют нагреванием при температуре 65°C в течение 90 минут для разрушения активности эндогенной AP. Далее клетки промывают один раз в AP буфере (100 мМ NaCl, 5 мМ MgCl2, 100 мМ Трис, pH 9,5), инкубируют в хромогенном субстрате (Western Blue, Promega) и далее анализируют для выявления наличия продукта реакции через час после инкубации и после ночной инкубации. Положительные клетки идентифицируют по наличию темно-синего осадка на поверхности мембраны. Далее положительные пулы разрушают для идентификации отдельных положительных клонов с последующими раундами скрининга.
В результате скрининга были идентифицированы следующие положительные пики:
затравочный компонент MAG-AP дает 4 положительных пика. Один был ранее охарактеризован как Nogo-рецептор (Fournier et al., Nature 409, 342-346 (2001)). Два из указанных пиков представляли собой ферменты гликолитического процессинга и, по всей видимости, не были релевантными. Четвертый был обозначен как «гипотетический белок Homo sapiens из клона 643 (LOC57228), мРНК». Более точный анализ кДНК выявил альтернативную открытую рамку считывания (ORF), которая была гомологична таковой для ранее описанного белка SMAG.
Затравочный компонент AP-Nogo66 привел к получению двух положительных пиков. Один был описан ранее как Nogo-рецептор. Другой был обозначен как «лейкоцитарный иммуноглобулин-подобный рецептор Homo sapiens, подсемейство В (с доменами TM и ITIM), представитель 2 (LILRB2), мРНК» (SEQ ID NO: 2). Этот ген также известен и описан в различных альтернативных номенклатурах, включающих такие наименования, как MIG10, ILT4 и LIR2 (Kubagawa et al., Proc. Natl. Acad. Sci. USA 94:5261-6 (1997): Colonna et al., J. Exp. Med. 186:1809-18 (1997)).
Пример 2
AP-Nogo66 связывается с PirB и LILRB2
Для подтверждения наличия связывания Nogo66 с LILRB2 (SEQ ID NO: 2) и для тестирования факта связывания Nogo66 с мышиным ортологом PirB (SEQ ID NO: 1), проводят анализы на связывание, аналогичные анализам, описанным в примере 1. В общих чертах процедуру проводят следующим образом: COS7 клетки трансфицируют кДНК, кодирующей PirB, LILRB2 или NgR, в качестве положительного контроля. Через 48 часов после трансфекции клетки инкубируют с кондиционированной средой для клеток 293, содержащей AP-Nogo66, в течение 90 минут при комнатной температуре. Далее клетки тщательно промывают, фиксируют и нейтрализуют эндогенную AP активность тепловой инактивацией. После чего указанные клетки подвергают реакции с хромогенным субстратом (Western Blue, Promega).
Как показано на фиг. 1, выявляется мощный положительный сигнал как на PirB-, так и на LILRB2-экспрессирующих клетках.
Пример 3
MAG связывается с PirB и LILRB2
Для тестирования, связывается или нет MAG с PirB и LILRB2, проводят анализы на связывание с MAG-Fc, который, по предположениям, должен быть более активным, чем MAG-AP. Клетки COS7 трансфицируют кДНК, кодирующей PirB, LILRB2 или mSMAGP, в качестве положительного контроля. Через 48 часов после трансфекции клетки инкубируют с кондиционированной средой для клеток 293, содержащей MAG-Fc, в течение 90 минут при комнатной температуре. Далее клетки промывают четыре раза забуференным солевым раствором Хэнкса (HBSS), фиксируют в течение пяти минут 2% параформальдегидом, промывают четыре раза HBSS и блокируют в течение 15 минут инкубированием с инактивированной нагреванием 10% козьей сывороткой (HIGS) в HBSS. После этого клетки инкубируют в течение одного часа с антителом против человеческого Fc (1:500, Jackson Immunochemicals), промывают 10% HIGS в ФБР и инкубируют со вторичным антителом (AlexaFluor 568-конъюгированное козье противомышиное антитело, Molecular Probes). Клетки промывают ФБР, накрывают покровным стеклом и выявляют наличие иммунофлуоресценции с использованием инвертированного флуоресцентного микроскопа.
Полученные результаты показаны на фиг. 2. В сравнении с контрольными клетками PirB-, и LILRB2-экспрессирующие клетки демонстрируют связывание с MAG-Fc. В приведенном примере mSMAGP-экспрессирующие клетки служат в качестве положительного контроля на связывание.
Пример 4
PirB экспрессируется в нервной системе
Для проверки, экспрессируется ли PirB в нервной системе, проводят ОТ-ПЦР анализ на мРНК, выделенной из разных невральных тканей. P7 мозжечок, P10 спинномозговые узлы (DRG), головной мозг взрослого животного и селезенку взрослого животного (в качестве положительного контроля) вырезают из CD1 мышей и сразу же помещают в РНКlater (Ambion). мРНК экстрагируют с использованием набора для выделения РНEasy (Qiagen). кДНК получают на основе мРНК с использованием системы синтеза первой цепи Superscript III (Invitrogen). Проводят От-ПЦР с использованием праймеров, специфичных для PirB 5'TGAAGGCTCTCATTGGAGTGTCTG3' (SEQ ID NO:20) и 5'GGCATAGGTCACATCCTGGGAC3' (SEQ ID NO: 3), праймеров, реактивных в перекрестном режиме с различными представителями подсемейства PirA (5'GTCTCAGAAACCATTGAATCC3' (SEQ ID NO: 4) и 5'GACAGAAAACTTTGGGTCATCAG3' (SEQ ID NO: 5)), или праймеров, специфичных для мышиного SMAGP (5'CCCTCAGCAACGATGAACAACC3' (SEQ ID NO: 6) и 5'TGGACCCTGGAGTCAGTGATTC3' (SEQ ID NO: 7)).
Как показано на фиг. 3, анализ по методу ОТ-ПЦР показывает, что и PirB, и PirA изоформы могут быть обнаружены в мозжечке, DRG и спинном мозге.
Пример 5
Результаты гибридизации in situ
Для целей более детального анализа нейрональной экспрессии PirB проводят гибридизационные анализы in situ. Проводят перфузию взрослых или постнатальных мышей, выделяют и нарезают головной мозг и DRG и фиксируют все в течение ночи. Далее ткани подвергают криообработке в 30% сахарозе и погружают и замораживают в O.C.T. соединение (Tissue-Tek). Замороженные ткани нарезают на образцы толщиной 12 микрон. Получают радиоактивные зонды с использованием набора для транскрипции in vitro MAXIscript (Ambion) по соответствующим протоколам от производителя. Гибридизацию in situ проводят с использованием набора мРНКlocator ISH (Ambion) в соответствии с протоколами производителя.
PirB зонд был разработан таким образом, чтобы его можно было использовать для гибридизации с нуклеотидами No.1922-2385, которые находятся в трансмембранном и внутриклеточном домене и уникальны для PirB. Праймеры, используемые для амплификации данного участка, представляют собой: 5TGAAGGCTCTCATTGGAGTGTCTG3' (SEQ ID NO: 8) и 5'GGCATAGGTCACATCCTGGGAC3' (SEQ ID NO: 9). Фрагмент размером 464 п.н. клонируют в pCRII-TOPO (Invitrogen).
Результаты гибридизации in situ показаны на фиг. 4A-4C. Положительный сигнал гибридизации отмечается в коре головного мозга, в гиппокампе и в мозжечке, а также в клетках DRG.
Пример 6
Ингибирование аксонального роста под действием Nogo66 и «спасение» от ингибирования с помощью PirB ECD в гранулярных нейронах мозжечка
В данном эксперименте была подтверждена способность Nogo66 ингибировать аксональный рост. Кроме того, были исследованы конструкции PirB внеклеточного домена на их способность препятствовать ингибирующей активности Nogo66.
AP-Nogo66 получают при клонировании ингибирующей петли из 66 аминокислот в NogoA по направлению считывания информации для гена человека плацентарной щелочной фосфатазы (AP). Указанную конструкцию, содержащую FLAG-фрагмент на N-конце для целей более легкой очистки, встраивают в скелет pRK вектора (Genentech). Для создания белков PirB внеклеточного домена (ECD), аминокислоты No. 1-638 в PirB клонируют в векторах экспрессии pRK против направления считывания информации для 8-His фрагмента (PirBHis) или человеческого Fc (PirBFc). Указанные конструкции экспрессии подвергают временной трансфекции в CHO клетки, секретированные белки выделяют из кондиционированной среды и очищают с помощью аффинной хроматографии.
Гранулярные нейроны мозжечка (CGN) выделяют из P7 CD1 мышей и культивируют на иммобилизованном AP-Nogo66 белке для проведения тестов на ингибирование. В общих чертах процедура состоит в том, что на ячейки 96-ячеечного планшета, содержащие покрытие из поли-D-lysine (Biocoat, Becton Dickinson), наносят в спот-режиме рекомбинантный AP-Nogo66 (180 или 300 нг/пятно в 3 мкл). AP-Nogo66 наносят либо один, либо в смеси с избытком PirBFc (850 нг/пятно в 3 мкл) или PirBHis (1000 нг/пятно в 3 мкл). Это приводит к получению пятен, содержащих 2-4-кратный молярный избыток PirB ECD. Нанесенные в спот-режиме белки оставляют для адгезии в течение 2 часов и затем планшет обрабатывают с использованием 10 мкг/мл ламинина (Invitrogen) в течение 2 часов. Клетки мышиного мозжечка P7 подготавливают по описанной ранее процедуре (Zheng et al., 2005) и высевают с плотностью 2×104 клеток/ячейку. Культуры инкубируют при температуре 37°C в течение 22 часов, фиксируют с использованием 4% параформальдегида/4% сахарозы и окрашивают антитубулиновым антителом (TuJl, Covance). Снимки получают с использованием визуализирующей системы ImageXpress (Molecular Devices).
Как показано на фиг. 6, AP-Nogo66 в значительной мере ингибирует рост аксона в нейронах мозжечка P7. Наличие избытка PirB ECD либо в виде PirBFc, либо PirBHis, существенно снижает это ингибирование. Включение других контрольных белков (Fc или Robo4Fc) вместе с AP-Nogo66 не снижает ингибирования.
Пример 7
Совместная иммунопреципитация PirB и NgR
Данный эксперимент был разработан таким образом, чтобы исследовать взаимосвязь и потенциальное взаимодействие PirB и NgR при их совместной экспрессии в клетках-хозяевах in vitro.
Клетки COS7 подвергают временной трансфекции контрольным вектором, полноразмерным PirB или смесью полноразмерного PirB и полноразмерного NgR. Через 48 часов после трансфекции клетки лизируют с использованием буфера для лизиса клеток (Cell Signaling Technology) и лизаты подвергают иммунопреципитации с использованием анти-PirA/B (6C1, Pharmingen). Образцы разделяют путем электрофореза в ДСН-ПААГ, переносят на нитроцеллюлозу и зондируют с использованием анти-NgR (Alpha Diagnostics International).
Как показано на фиг. 7, NgR подвергается жесткой совместной преципитации с PirB (левая панель). На правой панели проиллюстрирован результат анализа суммарного белка из лизатов целых клеток после иммуноблоттинга анти-NgR. Множественные полосы (показаны стрелками) отражают NgR, в разной степени процессированный в ходе гликозилирования.
Пример 8
PirB антитела и их использование для того, чтобы препятствовать ингибирующей активности Nogo66 на аксональный рост
В данном эксперименте были проанализированы антитела против PirB на их способность препятствовать Nogo66-индуцированному ингибированию аксонального роста.
Гранулярные нейроны мозжечка (CGN) выделяют из P7 CD1 мышей и культивируют на иммобилизованном белке AP-Nogo66 для проведения тестов на ингибирование, как было описано в приведенных выше примерах. Антитела, блокирующие указанную функцию, получают при скрининге PirB ECD с использованием фаговой библиотеки с синтетическими антителами человека, по существу, как описано в работе Лианга с соавт. (Liang et al., J. Mol. Biol. 366(3):815-819 (2006)), которая включена в настоящее описание полностью в качестве ссылки. Клоны отбирают по их способности конкурировать с AP-Nogo66 за связывание с PirB. В данном эксперименте анти-PirB или контрольные антитела инкубируют с нейронами, растущими на AP-Nogo66. Как видно из фиг. 8, анти-PirB антитела приводят к значительному снижению ингибирования под действием AP-Nogo66.
Все ссылки, приведенные в данном описании, включены в него в полном объеме в качестве ссылки.
Несмотря на то, что настоящее описание было описано со ссылкой на конкретные варианты его осуществления, следует понимать, что настоящее изобретение не ограничивается такими вариантами. И в этой связи настоящее изобретение будет охватывать различные модификации и эквиваленты, соответствующие принципам и области, описанным в прилагаемой формуле изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОЧИЩЕННЫЙ ИММУНОГЕННЫЙ БЕЛОК, ЕГО ФРАГМЕНТЫ И ПРОИЗВОДНЫЕ | 1999 |
|
RU2304585C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПОЛИКЛОНАЛЬНЫХ АНТИТЕЛ, АНТИСЫВОРОТКА, СОДЕРЖАЩАЯ ПОЛИКЛОНАЛЬНЫЕ АНТИТЕЛА, И СПОСОБЫ ИММУНИЗАЦИИ | 1999 |
|
RU2358982C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ БЕЛКА RGM А И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2524136C2 |
| ГЕН НОВОЙ СЕРИНОВОЙ ПРОТЕАЗЫ, РОДСТВЕННОЙ DPPIV | 2001 |
|
RU2305133C2 |
| ЛЕЧЕНИЕ РАССЕЯННОГО СКЛЕРОЗА | 2010 |
|
RU2539034C2 |
| АНТИТЕЛА ПРОТИВ МИЕЛИН-АССОЦИИРОВАННОГО ГЛИКОПРОТЕИНА (MAG) | 2003 |
|
RU2303461C9 |
| АНТИТЕЛА К РЕЦЕПТОРУ КОНЕЧНЫХ ПРОДУКТОВ ГЛУБОКОГО ГЛИКИРОВАНИЯ (RAGE) И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2518351C2 |
| ЛЕЧЕНИЕ ИНТЕРСТИЦИАЛЬНОГО ЦИСТИТА | 2008 |
|
RU2570559C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С ДЕГЕНЕРАЦИЕЙ НЕЙРИТОВ | 2013 |
|
RU2644337C2 |
| АНТИ-Nogo АНТИТЕЛА ("11С7") И ИХ ФАРМАЦЕВТИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ | 2003 |
|
RU2350623C2 |



















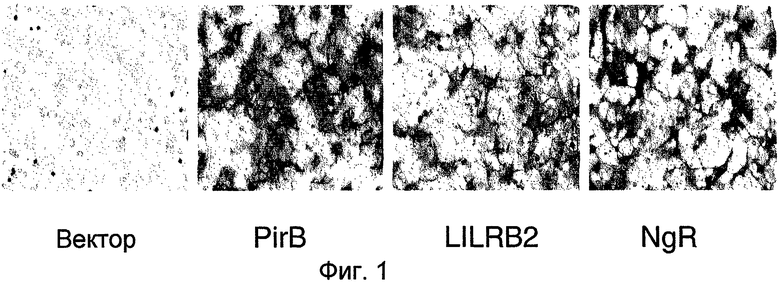
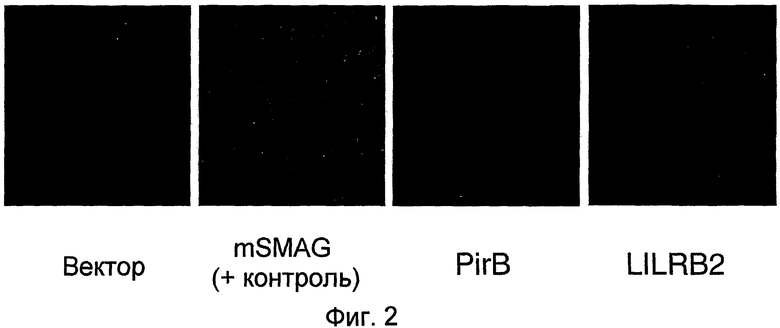

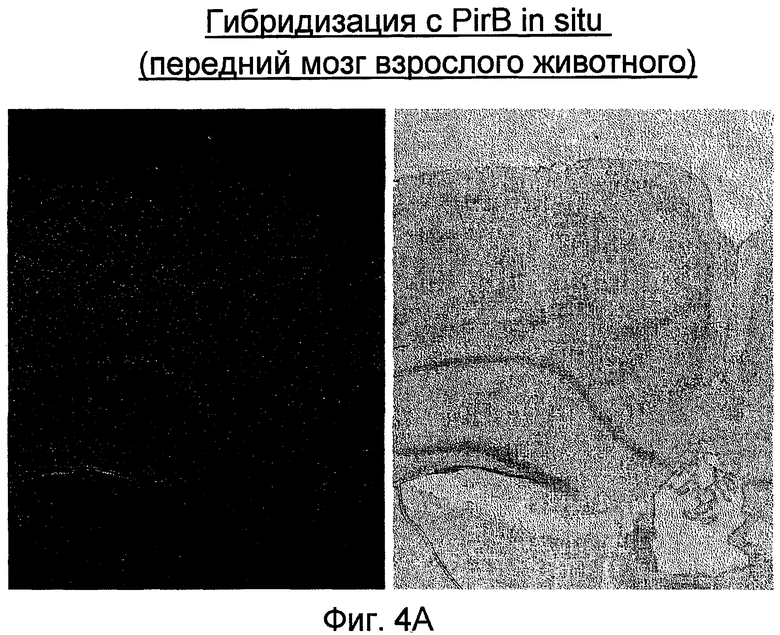
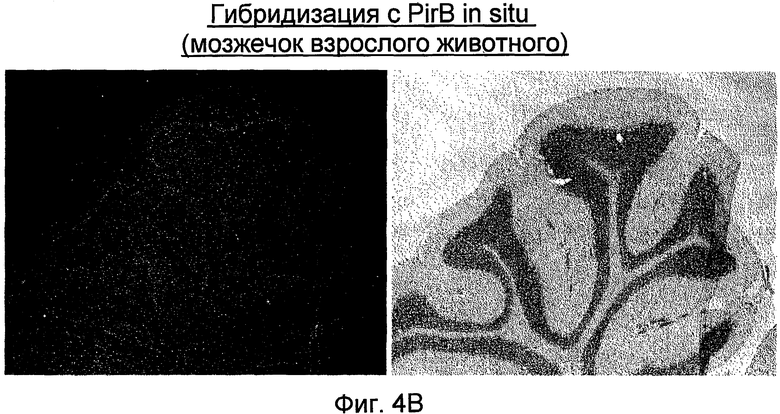
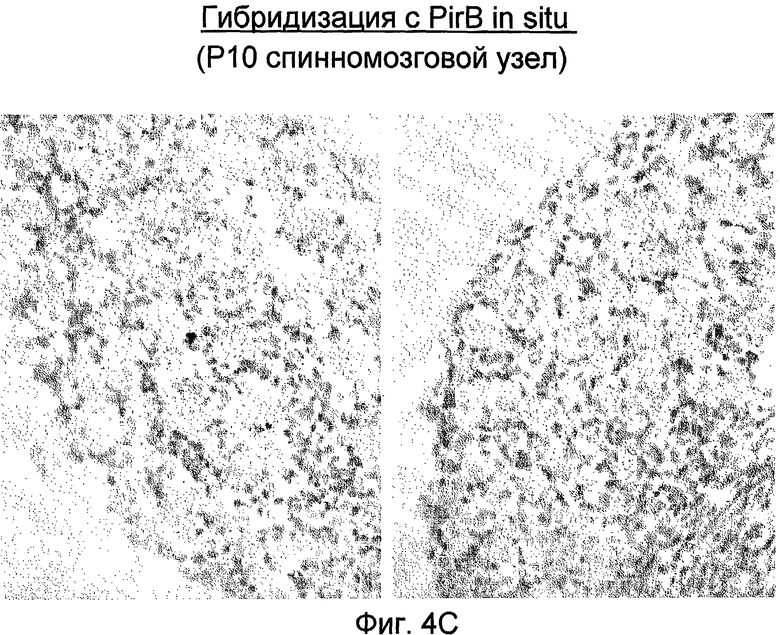

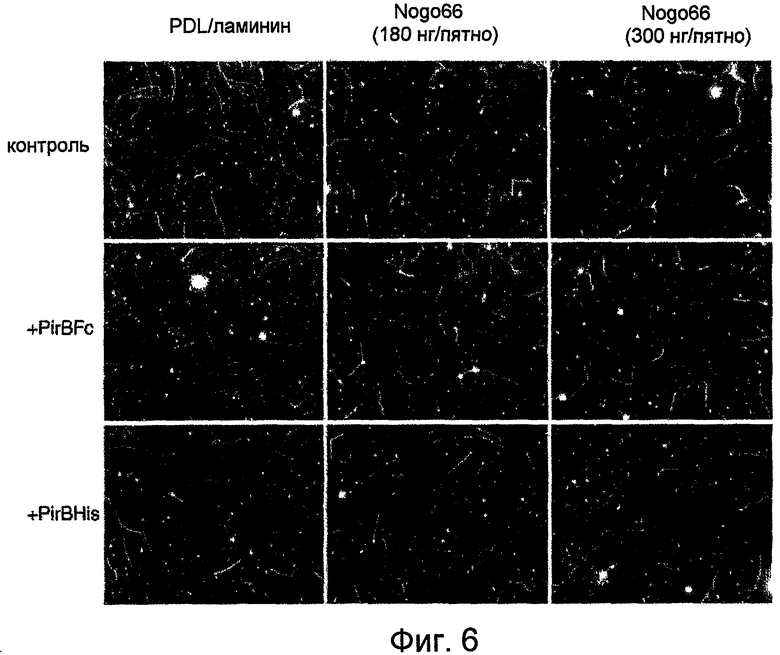
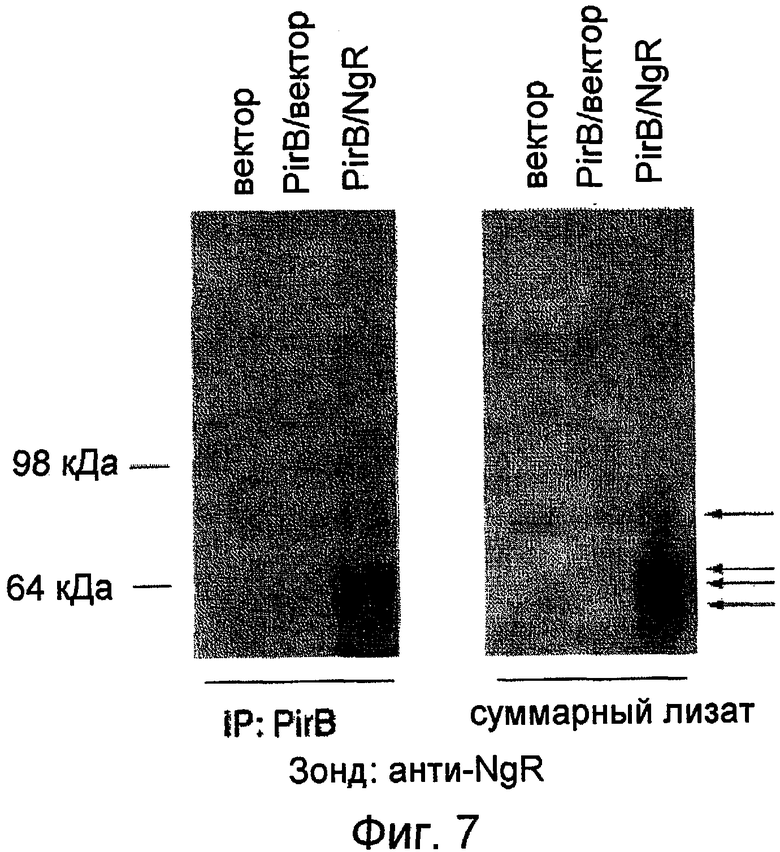











Изобретение относится к способам, связанным с функцией ЦНС и заболеваниями ЦНС, и касается способа идентификации антагониста PirB/LILRB, включающего контактирование исследуемого агента с рецепторным комплексом, который включает PirB/LILRB и миелин или миелин-ассоциированный белок, или его фрагмент, и детекцию способности исследуемого агента ингибировать взаимодействие между PirB/LILRB и миелин-ассоциированным белком или его фрагментом. Данный способ приводит к снижению ингибирования и стимуляции роста аксона в нейроне ЦНС. Изобретение также относится к способу лечения неврального повреждения у субъекта и способу поддержания жизнеспособности нейрона в ЦНС, включающему контактирование указанного нейрона с антагонистом PirB/LILRB. Сниженная активность PirB/LILRB эффективно разрушает миелин-ассоциированный ингибирующий путь, ускоряя нейрональную регенерацию. 3 н. и 32 з.п. ф-лы, 1 табл., 8 пр., 18 ил.
1. Способ идентификации антагониста PirB/LILRB, включающий контактирование исследуемого агента с комплексом, включающим PirB/LILRB и миелин или миелин-ассоциированный белок, или его фрагмент, и обнаружение способности указанного исследуемого агента ингибировать взаимодействие между PirB/LILRB и указанным миелином или миелин-ассоциированным белком, или его фрагментом, где указанный исследуемый агент идентифицируют как антагонист, если такое взаимодействие ингибируется.
2. Способ по п.1, отличающийся тем, что указанное взаимодействие представляет собой связывание.
3. Способ по п.1, отличающийся тем, что указанное взаимодействие представляет собой клеточную передачу сигнала.
4. Способ по п.3, отличающийся тем, что указанная клеточная передача сигнала приводит к ингибированию аксонального роста или нейрональной регенерации.
5. Способ по п.1, отличающийся тем, что миелин-ассоциированный белок выбран из группы, состоящей из Nogo, MAG и OMgp.
6. Способ по п.5, отличающийся тем, что указанный PirB/LILRB выбран из группы, состоящей из LILRB1, LILRB2, LILRB3 и LILRB5.
7. Способ по п.6, отличающийся тем, что указанный PirB/LILRB выбран из группы, состоящей из LILRB2, вариант транскрипта 1 (SEQ ID NO:2), LILRB2, вариант транскрипта 2 (SEQ ID NO:14), LILRB1, вариант транскрипта 1 (SEQ ID NO:10), LILRB1, вариант транскрипта 2 (SEQ ID NO:11), LILRB1, вариант транскрипта 3 (SEQ ID NO:12), LILRB1, вариант транскрипта 4 (SEQ ID NO:13), LILRB3, вариант транскрипта 1 (SEQ ID NO:15), LILRB3, вариант транскрипта 2 (SEQ ID NO:16), LILRB5, вариант транскрипта 1 (SEQ ID NO:17), LILRB5, вариант транскрипта 2 (SEQ ID NO:18) и LILRB5, вариант транскрипта 3 (SEQ ID NO:19).
8. Способ по п.7, отличающийся тем, что PirB представляет собой LILRB2, вариант транскрипта 1 (SEQ ID NO:2) или LILRB2, вариант транскрипта 2 (SEQ ID NO:14).
9. Способ по п.5, отличающийся тем, что комплекс дополнительно содержит NgR.
10. Способ по п.1, отличающийся тем, что исследуемый агент выбран из группы, состоящей из антител, полипептидов, пептидов, нуклеиновых кислот, коротких интерферирующих молекул РНК (киРНК), малых органических молекул, полисахаридов и полинуклеотидов.
11. Способ по п.10, отличающийся тем, что исследуемый агент представляет собой антитело.
12. Способ по п.11, отличающийся тем, что указанное антитело специфически связывается с PirB/LILRB.
13. Способ по п.12, отличающийся тем, что указанное антитело специфически связывается с LILRB2.
14. Способ по п.12, отличающийся тем, что указанное антитело представляет собой моноклональное антитело.
15. Способ по п.12, отличающийся тем, что указанное антитело представляет собой химерное антитело.
16. Способ по п.12, отличающийся тем, что указанное антитело представляет собой гуманизированное антитело.
17. Способ по п.12, отличающийся тем, что указанное антитело представляет собой антитело человека.
18. Способ по п.12, отличающийся тем, что указанное антитело представляет собой антиген-связывающий фрагмент.
19. Способ по п.18, отличающийся тем, что указанный фрагмент антитела выбран из группы, состоящей из Fv, Fab, Fab' и F(ab')2 фрагментов.
20. Способ по п.10, отличающийся тем, что исследуемый агент представляет собой короткую интерферирующую РНК (киРНК).
21. Способ по п.1, отличающийся тем, что по меньшей мере один из указанного PirB/LILRB и указанного миелина или миелинассоциированного белка, или его фрагмента иммобилизован.
22. Способ по п.1, который представляет собой клеточный анализ.
23. Способ по п.22, отличающийся тем, что указанный клеточный анализ включает культивирование нейрональных клеток с указанным миелином или миелинассоциированным белком, или его фрагментом в присутствии и в отсутствие указанного исследуемого агента, и определение изменения длины нейрита, где указанный исследуемый агент идентифицируют как антагонист, если длина нейрита будет больше в присутствии указанного исследуемого агента.
24. Способ по п.23, отличающийся тем, что указанные нейрональные клетки представляют собой первичные нейроны.
25. Способ по п.23, отличающийся тем, что указанные нейрональные клетки получают из эмбриональных стволовых клеток (ES) или клеточных линий.
26. Способ по п.25, отличающийся тем, что указанные нейрональные клетки получают из нейробластомы.
27. Способ по п.23, отличающийся тем, что указанные нейрональные клетки выбраны из группы, состоящей из гранулярных нейронов мозжечка, нейронов спинномозгового узла и кортикальных нейронов.
28. Способ по любому из пп.1-27, дополнительно включающий стадию введения идентифицированного антагониста субъекту с заболеванием или состоянием, в случае которых наблюдается улучшение за счет усиления роста нейрита, стимуляции роста нейронов, их восстановления или регенерации, где антагонист представляет собой антитело, которое специфически связывается с PirB/LILRB.
29. Способ по п.28, отличающийся тем, что указанное заболевание или состояние представляет собой неврологическое расстройство.
30. Способ по п.29, отличающийся тем, что указанное неврологическое расстройство характеризуется физически поврежденным нервом.
31. Способ по п.29, отличающийся тем, что указанное неврологическое расстройство выбрано из группы, состоящей из повреждения периферических нервов, вызванного физическим повреждением, диабетом; физического повреждения центральной нервной системы; повреждения головного мозга, ассоциированного с инсультом; невралгии тройничного нерва, глоссофарингеальной невралгии, паралича Белла, миастении гравис, мышечной дистрофии, бокового амиотрофического склероза (ALS), прогрессирующей мышечной атрофии, прогрессирующей бульбарной наследственной мышечной атрофии, грыжи, разрыва или пролапса межпозвоночных дисков, шейного спондилеза, расстройств, затрагивающих сплетение, синдрома лестничной мышцы (сдавление плечевого сплетения и подключичной артерии), периферических нейропатий, порфирии, синдрома Гийена-Барре, болезни Альцгеймера, болезни Хантингтона и болезни Паркинсона.
32. Способ снижения ингибирования аксонального роста в нейроне ЦНС, включающий контактирование указанного нейрона с антагонистом PirB/LILRB, идентифицированным согласно пп.1-29, где антагонист представляет собой антитело, которое специфически связывается с PirB/LILRB.
33. Способ стимуляции роста аксона в нейроне ЦНС, включающий контактирование указанного нейрона с антагонистом PirB/LILRB, идентифицированным согласно пп.1-29, где антагонист представляет собой антитело, которое специфически связывается с PirB/LILRB.
34. Способ лечения неврального повреждения у субъекта, включающий введение указанному субъекту антагониста PirB/LILRB, идентифицированного согласно пп.1-29, где антагонист представляет собой антитело, которое специфически связывается с PirB/LILRB.
35. Способ поддержания жизнеспособности нейрона в ЦНС, включающий контактирование указанного нейрона с антагонистом PirB/LILRB, идентифицированным согласно пп.1-29, где антагонист представляет собой антитело, которое специфически связывается с PirB/LILRB.
| WO 03051834 A2, 2003.06.26 | |||
| SYKEN J | |||
| et all | |||
| PirB restricts ocular-dominance plasticity in visual cortex | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| HIRAYASU K | |||
| et all | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Toshio Yabe | |||
| Hum Genet | |||

