ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
В настоящей заявке описаны RGM A-связывающие белки, в частности, моноклональные антитела, и, в частности, их CDR-привитые, гуманизированные варианты, которые способны связываться с RGM A и предотвращать связывание белков RGM с рецептором RGM A и другими RGM A-связывающими белками, и, следовательно, нейтрализовать функцию RGM A. Эти антитела могут использоваться для лечения некоторых состояний, включая, но ими не ограничиваясь, рассеянный склероз, травму головного мозга млекопитающих, повреждение спинного мозга, инсульт, нейродегенеративные заболевания и шизофрению.
УРОВЕНЬ ТЕХНИКИ
Регенерация аксонов после повреждения или после воспалительных поражений или после нейродегенеративных заболеваний центральной нервной системы млекопитающих (ЦНС) практически всегда является невозможной; результат зависит от баланса между врожденной способностью нервных волокон ЦНС к возобновлению роста и ингибирующими факторами ЦНС, расположенными в сайте поражения или повреждения, которые активно препятствуют возобновлению роста, и, следовательно, регенерации поврежденных путей нервных волокон.
Было установлено, что миелин ЦНС, вырабатываемый олигодендроцитами, и рубцовая ткань повреждения являются наиболее значимыми тормозящими структурами для роста аксонов на ранней фазе повреждения, вследствие коллапса конуса роста и ингибирования роста нейритов in vitro, а также in vivo, что приводит к непосредственному ингибированию роста аксонов. Были идентифицированы белки RGM, главные ингибирующие факторы миелина ЦНС и рубцовой ткани (Monnier et al., Nature 419: 392-395, 2002; Schwab et al., Arch. Neurol.62: 1561-8, 2005a; Schwab et al. Eur. J. Neurosci. 21: 1569-76, 2005 b; Hata et al. J. Cell Biol. 173:47-58, 2006; в отношении обзоров см.: Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361: 1513-29, 2006; Yamashita et al. Curr. Opin. Neurobiol. 17: 29-34, 2007). Белки RGM стимулированы в участках поражения или повреждения у людей, умерших от травмы головного мозга или ишемического инсульта (Schwab et al., Arch. Neurol. 62: 1561-8, 2005a) и стимулированы у крыс с повреждением спинного мозга (Schwab et al. Eur. J. Neurosci. 21: 1569-76, 2005 b; Hata et al. J. Cell Biol. 173:47-58, 2006, в отношении обзора см.: Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361: 1513-29, 2006; Yamashita et al. Curr. Opin. Neurobiol. 17: 29-34, 2007). Кроме того, первые результаты с использованием клинических образцов пациентов с рассеянным склерозом и здоровых лиц сделали возможность предположить, что RGM A человека стимулирован в цереброспинальной жидкости пациентов, страдающих MS (данные не показаны).
Для оценки стимулирующего регенерацию эффекта RGM A-специфического поликлонального антитела, эти антитела вводили в умеренную тяжелую модель повреждения спинного мозга, где приблизительно 60% спинного мозга на торакальном уровне 9/10 были рассечены. Гистологическое исследование выявило, что такое повреждение приводило к разрыву всех дорсальных и латеральных волокон кортикоспинального пути. RGM A-специфическое поликлональное антитело, вводимое местно посредством насоса в течение двух недель, индуцировало отдаленную регенерацию поврежденных нервных волокон (Hata et al., J. Cell Biol. 173:47-58, 2006).
Сотни нервных волокон, расположенных за пределами участка повреждения, и самые длинные волокна регенерировали более чем на 10 мм за пределами этого повреждения, тогда как регенерирующие волокна не были обнаружены дистально относительно этого повреждения в обработанных контрольным антителом животных. Функциональное восстановление анти-RGM A-обработанных крыс значительно улучшалось в сравнении с обработанными контрольным антителом крысами со спинномозговым повреждением, подтверждая тем самым, что RGM A является мощным ингибитором нейрорегенерации и ценной мишенью для стимуляции восстановления в показаниях, характеризующихся повреждением аксонов или повреждением нервных волокон. (Hata et al., J. Cell Biol. 173:47-58, 2006; Kyoto et al. Brain Res. 1186: 74-86, 2007). Кроме того, нейтрализация белка RGM A блокирующим функцию поликлональным антителом не только стимулировала рост поврежденных нервных волокон в крысах со спинномозговым повреждением, но и усиливала образование их синапса, тем самым делая возможным коррекцию или восстановление поврежденных нейронных цепей. (Kyoto et al. Brain Res. 1186: 74-86, 2007).
Семейство rgm-генов включает три разных гена, два из которых, rgm a и b, экспрессируются в ЦНС млекопитающих, давая начало белкам RGM A и RGM B, тогда как третий член, rgm c, экспрессируется на периферии (Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361: 1513-29, 2006), где RGM C играет важную роль в метаболизме железа. In vitro, RGM A ингибирует рост нейритов связыванием с Неогенином, который был идентифицирован как рецептор RGM (Rajagopalan et al. Nat Cell Biol.: 6(8), 756-62, 2004). Неогенин впервые был описан как нетринсвязывающий белок (Keino-Masu et al. Cell, 87(2):175-85, 1996). Это открытие является важным, так как сообщалось, что связывание Нетрина-1 и Неогенина или его близкородственного рецептора DCC (делетирован в колоректальном раке) скорее стимулирует, а не ингибирует рост нейритов (Braisted et al. J. Neurosci. 20: 5792-801, 2000). Таким образом, блокирование RGM A запускает RGM-опосредованное ингибирование роста, обеспечивая связывание неогенина с его стимулирующим рост нейритов лигандом Нетрином. На основании этих наблюдений, можно сделать предположение, что нейтрализация RGM A превосходит нейтрализацию неогенина на моделях повреждения спинного мозга человека. Кроме связывания RGM A с Неогенином и индукции ингибирования роста нейритов, связывание RGM A или B с морфогенетическими белками кости BMP-2 и BMP-4 может препятствовать успешной нейрорегенерации и функциональному восстановлению (Mueller et al., Philos. Trans. R. Soc. Lond. B. Biol. Sci. 361: 1513-29, 2006).
В данной области существует необходимость в улучшенных антителах, способных связывать RGM A, предпочтительно, в моноклональном антителе, которое блокирует RGM A и препятствует взаимодействию RGM A и его рецептора, и/или связывающих белков, т.е. Неогенина и BMP-2, BMP-4.
Настоящая заявка относится (a) к получению нейтрализующего моноклонального антитела против RGM A, которое селективно ингибирует связывание RGM A и рецептора, Неогенина и морфогенетических белков кости 2 и 4 (BMP-2, BMP-4), и (b) к получению нейтрализующего моноклонального антитела против RGM A, которое селективно ингибирует связывание RGM A с морфогенетическими белками кости 2 и 4 (BMP-2, BMP-4). Полагают, что эти нейтрализующие моноклональные антитела по настоящему изобретению стимулируют рост пораженных или поврежденных нервных волокон и формирование функциональных синапсов регенерирующих нервных волокон, так как одно из нейтрализующих моноклональных антител по настоящему изобретению, по-видимому, преобразует ингибиторную природу RGM A в состояние, при котором нервные клетки предпочитают мигрировать и расти на RGM А-субстрате, а не на пермиссивном субстрате, таком как Коллаген I. Кроме того, антитело способно индуцировать отдаленную регенерацию на модели in vivo повреждения зрительного нерва крыс, а также усиливает миелинизацию поврежденных и регенерирующих нервных волокон.
Таким образом, было сделано предложение, что нейтрализующие моноклональные антитела по настоящему изобретению стимулируют регенерацию нейронов и рост поврежденных или разрушенных нейронных соединений в поврежденной или воспаленной ЦНС человека, например, при рассеянном склерозе, после острого повреждения спинного мозга, травмы головного мозга или при нейродегенеративных заболеваниях, таких как, например, хорея Хантингтона, болезнь Паркинсона, болезнь Альцгеймера.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Согласно одному из аспектов настоящее изобретение относится к связывающему белку, который диссоциирует из RGM A человека (hRGM A) с значением KD, равным 1×10-7 M или менее, и константой скорости диссоциации koff, равной 1×10-2 с-1 или менее, причем обе константы определяют способом резонанса поверхностных плазмонов.
Согласно другому аспекту изобретение относится к связывающему белку, например, к связывающему белку, имеющему вышеуказанные кинетические свойства, который связывается с RGM A человека и нейтрализует активность, ингибирующую разрастание нейритов RGM A человека, как определено стандартным анализом in vitro, например, анализом нейронного роста Ntera, приведенным в качестве примера 3 ниже.
Настоящее изобретение также относится к связывающему белку, как определено выше, имеющему по меньшей мере одну из следующих дополнительных функциональных характеристик:
способности связывать RGM A крысы,
способности связывать RGM C человека и
способности связывать RGM C крысы.
В частности, описанный в настоящем документе связывающий белок модулирует способность RGM связываться по меньшей мере с одним из его рецепторов.
Такой связывающий белок, в частности, связывается с рецепторсвязывающим доменом RGM A человека. Были идентифицированы N- и C-концевые рецепторсвязывающие домены. Конкретные варианты связывающих белков по изобретению способны связываться с N-концевым рецепторсвязывающим доменом RGM A, как показано с помощью ингибирования связывания N-концевого фрагмента hRGM A, например, 47-168, и молекул рецептора, такими как Неогенин и BMP-4. Указанный N-концевой фрагмент hRGM A может иметь общую длину от приблизительно 30 до приблизительно 150 аминокислотных остатков или от приблизительно 30 до приблизительно 122 аминокислотных остатков. В качестве не ограничивающего примера, можно указать Фрагмент 0 (соответствующий N-концевым остаткам 47-168) hRGM A, как описано в настоящем документе, или любой более короткий рецепторсвязывающий фрагмент.
В частности, указанный связывающий белок модулирует, предпочтительно ингибирует, по меньшей мере одно из следующих взаимодействий:
связывание RGM A человека с BMP-4 человека,
связывание hRGM A с Неогенином человека,
связывание hRGM C с Неогенином человека,
связывание RGM A человека с BMP-2 человека.
Согласно одному из конкретных вариантов осуществления, связывающий белок, определенный в настоящем документе, является гуманизированным антителом.
Описанный выше связывающий белок может иметь антигенсвязывающий домен, причем указанный антигенсвязывающий домен способен связывать эпитоп молекулы RGM и указанный антигенсвязывающий домен содержит по меньшей мере один CDR, содержащий аминокислотную последовательность, выбранную из группы, состоящей из
модифицированных аминокислотных последовательностей CDR, последовательность которых по меньшей мере на 50% идентична одной из указанных последовательностей. В другом варианте осуществления изобретение относится к связывающему белку, содержащему антигенсвязывающий домен, причем указанный связывающий домен способен связывать эпитоп молекулы RGM, причем указанный антигенсвязывающий домен содержит по меньшей мере один CDR, содержащий аминокислотную последовательность, выбранную из группы, состоящей из:
модифицированных аминокислотных последовательностей CDR, последовательность которых по меньшей мере на 50% идентична, например, по меньшей мере на 55, 60, 65, 70, 75, 80, 85, 90, 95% идентична одной из указанных последовательностей.
Например, указанный связывающий белок может содержать два из указанных CDR, например, SEQ ID NO: 59 и 62; или SEQ ID NO: 65 и 68; где по меньшей мере один из указанных CDR может быть модифицированным и иметь последовательность, которая по меньшей мере на 50%, например, по меньшей мере на 55, 60, 65, 70, 75, 80, 85, 90, 95% идентична одной из указанных последовательностей.
Указанный связывающий белок может также содержать по меньшей мере один CDR, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 57, 58, 60, 61, 63, 64, 66, 67 и модифицированных аминокислотных последовательностей CDR, которые по меньшей мере на 50%, например, по меньшей мере на 55, 60, 65, 70, 75, 80, 85, 90, 95% идентичны одной из указанных последовательностей.
Другой вариант осуществления относится к связывающему белку, в котором указанный по меньшей мере один CDR содержит аминокислотную последовательность, выбранную из группы, состоящей из:
В одном из конкретных вариантов осуществления, указанный связывающий белок содержит по меньшей мере 3 CDR, которые выбраны из набора CDR вариабельных доменов, состоящего из:
или набора вариабельных доменов, в которых по меньшей мере один из указанных 3 CDR является модифицированной аминокислотной последовательностью CDR, которая по меньшей мере на 50%, например, по меньшей мере на 55, 60, 65, 70, 75, 80, 85, 90, 95% идентична исходной последовательности.
В частности, каждая из вышеуказанных модификаций может быть получена добавлением, делецией или, в частности, заменой одной аминокислоты или нескольких аминокислот, или их комбинациями.
В другом из вариантов осуществления, связывающий белок содержит по меньшей мере два набора CDR вариабельных доменов.
В частности, указанные по меньшей мере два набора CDR выбраны из группы, состоящей из:
VH 5F9-набора & VL 5F9-набора; и
VH 8D1-набора & VL 8D1-набора.
Связывающий белок по настоящему изобретению дополнительно содержит акцепторный каркасный участок человека.
Указанный акцепторный каркасный участок человека может содержать по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32 и 33.
Связывающий белок по изобретению может, в частности, содержать по меньшей мере один из наборов каркасных последовательностей, выбранных из группы, состоящей из наборов:
каждый из которых комбинирован с дополнительной каркасной последовательностью, выбранной из
JH3 (SEQ ID NO:18),
JH4 (SEQ ID NO:19),
JH6 (SEQ ID NO:20);
или
каждый из которых объединен с дополнительной каркасной последовательностью, выбранной из группы, состоящей из JK2 (SEQ ID NO:2).
В соответствии с конкретными вариантами осуществления, связывающий белок любого варианта, описанного выше, содержит по меньшей мере один CDR-привитый вариабельный домен тяжелой цепи, выбранный из SEQ ID NO: 35, 36, 37, 38, 39, 40, 41, 42 и 43; и/или по меньшей мере один CDR-привитый вариабельный домен легкой цепи, выбранный из SEQ ID NO: 44, 45 и 46.
Более конкретно, связывающий белок по изобретению содержит комбинацию двух вариабельных доменов, где указанные два вариабельные домена имеют аминокислотные последовательности, выбранные из:
SEQ ID NO: 35 & 44; 36 & 44; 37 & 44; 38 & 44; 39 & 44; 40 & 44; 41 & 44; 42 & 44; 43 & 44;
SEQ ID NO: 35 & 45; 36 & 45; 37 & 45; 38 & 45; 39 & 45; 40 & 45; 41 & 45; 42 & 45; 43 & 45;
SEQ ID NO: 35 & 46; 36 & 46; 37 & 46; 38 & 46; 39 & 46; 40 & 46; 41 & 46; 42 & 46; 43 & 46;
В другом варианте осуществления по изобретению, указанный акцепторный каркасный участок человека связывающего белка содержит по меньшей мере одну аминокислотную замену каркасного участка в ключевом остатке, причем указанный ключевой остаток выбран из группы, состоящей из:
остатка, смежного с CDR;
остатка сайта гликозилирования;
редкого остатка;
остатка, способного взаимодействовать с эпитопом RGM;
остатка, способного взаимодействовать с CDR;
канонического остатка;
контактного остатка между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи;
остатка в зоне Vernier;
N-концевого остатка, способного образовывать параглутамат, и
остатка в области перекрытия CDR1 вариабельной области тяжелой цепи по Хотиа и первым каркасным участком тяжелой цепи по Кабату.
В частности, указанные ключевые остатки выбраны из группы, состоящей из
(положения в последовательности тяжелой цепи) 1, 5, 37, 48, 49, 88, 98
(положения в последовательности легкой цепи): 2, 4, 41, 51.
В одном из конкретных вариантов осуществления связывающий белок по изобретению является консенсусным вариабельным доменом человека или содержит консенсусный вариабельный домен человека.
В соответствии с другим вариантом осуществления связывающий белок по изобретению, указанный акцепторный каркасный участок человека содержит по меньшей мере одну аминокислотную замену каркасной области, где аминокислотная последовательность каркасного участка по меньшей мере на 65%, например, по меньшей мере на 70, 75, 80, 85, 90, 95, 96, 97, 98 или 99%, идентична последовательности указанного акцепторного каркасного участка человека и содержит по меньшей мере 70 аминокислотных остатков, например, по меньшей мере 75, 80 или 85 остатков, идентичных указанному акцепторному каркасному участку человека.
В соответствии с конкретным вариантом осуществления, связывающий белок по изобретению содержит по меньшей мере один вариабельный домен с мутированным каркасным участком, имеющий аминокислотную последовательность, выбранную из группы, состоящей из:
SEQ ID NO: 47, 48, 49, 50; (VH-домен),
и/или выбранную из группы, состоящей из:
SEQ ID NO: 51, 52, 53 и 54 (VL-домен).
В частности, указанный связывающий белок содержит два необязательно мутированных в каркасном участке вариабельных доменов, где указанные два вариабельные домена имеют аминокислотные последовательности, выбранные из групп, состоящих из:
SEQ ID NO: 47 & 44; 47 & 45; 47 & 46; 47 & 51; 47 & 52; 47 & 53; 47 & 54;
SEQ ID NO: 48 & 44; 48 & 45; 48 & 46; 48 & 51; 48 & 52; 48 & 53; 48 & 54;
SEQ ID NO: 49 & 44; 49 & 45; 49 & 46; 49 & 51; 49 & 52; 49 & 53; 49 & 54;
SEQ ID NO: 50 & 44; 50 & 45; 50 & 46; 50 & 51; 50 & 52; 50 & 53; 50 & 54;
Связывающие белки по изобретению, описанные в настоящем описании, способны связывать по меньшей мере одну мишень, выбранную из молекул RGM.
В частности, белки способны связывать RGM A человека и, необязательно, одну дополнительную молекулу RGM человека или собакоподобных обезьян, крысы, курицы, лягушки и рыбы.
Например, белки могут дополнительно связываться с RGM A крысы, RGM C человека и/или RGM C крысы.
В частности, связывающий белок по изобретению способен модулировать, в частности, способен нейтрализовать или ингибировать биологическую функцию мишени, выбранной из молекул RGM, определенных выше.
В частности, связывающий белок по изобретению модулирует, в частности, ингибирует способность RGM связываться по меньшей мере с одним из его рецепторов, например, Неогенином и ВМР, таким как BMP-2 и BMP-4.
Например, указанный связывающий белок модулирует, в частности, уменьшает и, предпочтительно, ингибирует по меньшей мере одно из следующих взаимодействий:
связывание RGM A человека с BMP-4 человека,
связывание hRGM A с Неогенином человека,
связывание hRGM C с Неогенином человека,
связывание RGM A человека с BMP-2 человека.
Связывающие белки с различными комбинациями функциональных признаков и, следовательно, демонстрирующие разные функциональные характеристики, как описано в настоящем описании, входят в рамки настоящего изобретения. Неограничивающие примеры таких характеристик приведены в списке ниже.
Например, характеристика 1 встречается в случае антитела 5F9, описанного в настоящем изобретении, и его производных, описанных в настоящем описании.
Например, характеристика 2 встречается в случае антитела 8D1, описанного в настоящем изобретении, и его производных, описанных в настоящем описании.
В частности, связывающий белок по изобретению способен ингибировать по меньшей мере одну биологическую активность RGM, в частности, RGM A, где указанный RGM A выбран из RGM A человека, собакоподобных обезьян, крысы, курицы, лягушки и рыбы.
В соответствии с другим вариантом осуществления, связывающий белок по изобретению имеет один или несколько из следующих кинетических признаков:
(a) константу скорости ассоциации (Kon) для указанной мишени, выбранную из группы, состоящей из: по меньшей мере приблизительно 102 M-1c-1; по меньшей мере приблизительно 103 M-1c-1; по меньшей мере приблизительно 104 M-1c-1; по меньшей мере приблизительно 105 M-1c-1; по меньшей мере приблизительно 106 M-1c-1 и по меньшей мере приблизительно 107 M-1c-1, как измерено способом резонанса поверхностных плазмонов;
(b) константу скорости диссоциации (koff) для указанной мишени, выбранную из группы, состоящей из: самое большее приблизительно 10-2 с-1; самое большее 10-3 с-1; самое большее приблизительно 10-4 с-1; самое большее 10-5 с-1; и самое большее приблизительно 10-6 с-1, как измерено способом резонанса поверхностных плазмонов; или
(c) константу диссоциации (KD) для указанной мишени, выбранную выбрана из группы, состоящей из: самое большее приблизительно 10-7 M; самое большее приблизительно 10-8 M; самое большее приблизительно 10-9 M; самое большее приблизительно 10-10 M; самое большее приблизительно 10-11 M; самое большее приблизительно 10-12 M и самое большее 10-13 M.
В соответствии с дополнительным аспектом, изобретение относится к конструкции антитела, содержащей вышеописанный связывающий белок, причем указанная конструкция антитела содержит линкерный полипептид или константный домен иммуноглобулина.
Указанная конструкция антитела или связывающий белок по изобретению могут быть выбраны из группы, состоящей из: молекулы иммуноглобулина, моноклонального антитела, химерного антитела, CDR-привитого антитела, гуманизированного антитела, Fab, Fab', F(ab')2, Fv, дисульфид-связанного Fv, scFv, однодоменного антитела, диатела, мультиспецифического антитела, антитела с двойной специфичностью, иммуноглобулина с двумя вариабельными доменами и биспецифического антитела.
В конструкции антитела по изобретению указанный связывающий белок содержит константный домен тяжелой цепи иммуноглобулина, выбранный из группы, состоящей из:
константного домена IgM человека,
константного домена IgG1 человека,
константного домена IgG2 человека,
константного домена IgG3 человека,
константного домена IgG4 человека,
константного домена IgE человека,
константного домена IgD человека,
константного домена IgA1 человека,
константного домена IgA2 человека,
константного домена IgY человека и
соответствующих мутированных константных доменов.
В частности, конструкция антитела по изобретению содержит константный домен иммуноглобулина с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 11, 12, 13 и 14.
Согласно другому аспекту настоящее изобретение относится к конъюгату антитела, содержащему описанную в настоящем описании конструкцию антитела, причем указанное антитело дополнительно содержит средство, выбранное из группы, состоящей из: молекулы иммуноадгезии, средства визуализации, терапевтического средства и цитотоксического средства, причем каждое средство является конъюгированным, например, ковалентно связанным, с указанным связывающим белком.
Например, указанным средством является средство визуализации, выбранное из группы, состоящей из радиоактивной метки, фермента, флуоресцентной метки, люминесцентной метки, биолюминесцентной метки, магнитной метки и биотина. В частности, указанным средством визуализации является радиоактивная метка, выбранная из группы, состоящей из: 3H, 14C, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho и 153Sm.
Например, указанное средство является терапевтическим или цитотоксическим средством, выбранным из группы, состоящей из: антиметаболита, алкилирующего средства, антибиотика, фактора роста, цитокина, антиангиогенного средства, антимитотического средства, антрациклина, токсина и апоптотического средства.
В соответствии с другим вариантом осуществления, указанный связывающий белок по изобретению, как описано в настоящем описании, имеет характер гликозилирования, характерный для человека.
Кроме того, связывающие белки, конструкции антител и конъюгат антитела по изобретению могут находиться в виде кристалла (в кристаллической форме), предпочтительно, с сохранением биологической активности.
В частности, указанный кристалл представляет собой фармацевтический кристалл, не содержащий носителя с контролируемым высвобождением. Благодаря указанной кристаллической формы связывающий белок, конструкция антитела или конъюгат антитела может иметь большее время полужизни in vivo, чем соответствующая растворимая копия.
В другом аспекте изобретение относится к выделенной нуклеиновой кислоте, кодирующей аминокислотную последовательность связывающего белка, аминокислотную последовательность конструкции антитела и аминокислотную последовательность конъюгата антитела, как описано в настоящем описании.
Настоящее изобретение также относится к вектору, содержащему выделенную нуклеиновую кислоту, описанную в настоящем описании. В частности, вектор выбран из группы, состоящей из pcDNA, pTT, pTT3, pEFBOS, pBV, pJV и pBJ.
Изобретение также относится к клетке-хозяину, содержащей такой вектор. В частности, указанная клетка-хозяин является прокариотической клеткой, например, E.coli; или эукариотической клеткой и может быть выбрана из группы, состоящей из клетки одноклеточного организма, клетки животного, клетки растения и грибной клетки. В частности, указанной эукариотической клеткой является клетка животного, выбранная из группы, состоящей из: клетки млекопитающего, клетки птицы и клетки насекомого. Предпочтительно, указанная клетка-хозяин выбрана из клеток HEK, клеток CHO, клеток COS и клеток дрожжей. Клеткой дрожжей может быть Saccharomyces cerevisiae, а клеткой насекомого может быть клетка Sf9.
Изобретение также относится к способу получения белка, способного связывать RGM, предусматривающий культивирование клетки-хозяина, определенной в настоящем описании, в культуральной среде при условиях, достаточных для получения связывающего белка, способного связывать RGM.
Изобретение также относится к белку, получаемому в соответствии с указанным способом.
Изобретение также относится к композиции для высвобождения связывающего белка, причем указанная композиция содержит
(a) композицию, где указанная композиция содержит кристаллизованный продукт-белок, определенный в настоящем описании, и ингредиент; и
(b) по меньшей мере один полимерный носитель.
Указанным полимерным носителем может быть полимер, выбранный из группы, состоящей из: поли(акриловой кислоты), поли(цианоакрилатов), поли(аминокислот), поли(ангидридов), поли(депсипептида), поли(эфиров), поли(молочной кислоты), поли(сополимера молочной и гликолевой кислот) или PLGA, поли(b-гидроксибутирата), поли(капролактона), поли(диоксанона); поли(этиленгликоля), поли((гидроксипропил)метакрилата, поли[(органо)фосфазена], поли(ортоэфиров), поли(винилового спирта), поли(винилпирролидона), сополимеров малеинового ангидрида и алкилвинилового эфира, плюрониковых полиолов, альбумина, альгината, целлюлозы и производных целлюлозы, коллагена, фибрина, желатина, гиалуроновой кислоты, олигосахаридов, гликаминогликанов, сульфатированных полисахаридов, их смесей и сополимеров.
Указанный ингредиент может быть выбран из группы, состоящей из альбумина, сахарозы, трегалозы, лактита, желатина, гидроксипропил-β-циклодекстрина, метоксиполиэтиленгликоля и полиэтиленгликоля.
Согласно другому аспекту, изобретение относится к способу лечения млекопитающего, предусматривающий стадию введения млекопитающему эффективного количества композиции, определенной в настоящем описании.
Согласно другому аспекту, изобретение относится к фармацевтической композиции, содержащей продукт (в частности, связывающий белок, конструкцию или конъюгат, описанные выше) и фармацевтически приемлемый носитель.
Указанный фармацевтически приемлемый носитель может функционировать в качестве адъюванта, который можно использовать для увеличения абсорбции или дисперсии указанного связывающего белка.
Например, указанным адъювантом является гиалуронидаза.
В соответствии с другим вариантом осуществления, указанный фармацевтический препарат дополнительно содержит по меньшей мере одно дополнительное терапевтическое средство для лечения нарушения, в котором активность RGM оказывает негативное действие. Например, указанное средство выбрано из группы, состоящей из терапевтического средства, средства визуализации, цитотоксического средства, ингибиторов ангиогенеза; ингибиторов киназы; блокаторов молекул костимуляции; блокаторов молекул адгезии; антицитокин-антитела или его функционального фрагмента; метотрексата; циклоспорина; рапамицина; FK506; детектируемой метки или репортера; антагониста TNF; противоревматического средства; миорелаксанта, наркотического средства, нестероидного противовоспалительного лекарственного средства (NSAID), аналгезирующего средства, анестетического средства, седативного средства, местного анестетика, нейромышечного блокатора, противомикробного средства, антипсориатического средства, кортикостероида, анаболического стероида, эритропоэтина, иммунизирующего средства, иммуноглобулина, иммуносупрессивного средства, гормона роста, средства заместительной гормональной терапии, радиофармацевтического препарата, антидепрессанта, антипсихотического средства, стимулятора, противоастматического средства, бета-агониста, ингалируемого стероида, эпинефрина или его аналога, цитокина и антагониста цитокина.
Настоящее изобретение также относится к способу уменьшения активности RGM A человека, предусматривающему контактирование RGM A человека по меньшей мере с одним продуктом (в частности, связывающим белком, конструкцией или конъюгат, описанными в настоящем описании выше), так что уменьшается по меньшей мере одна активность RGM A человека.
Настоящее изобретение также относится к способу уменьшения связывания hRGM A с рецептором Неогенином у индивида, при необходимости, предусматривающему стадию введения индивиду продукта по изобретению (в частности, связывающего белка, конструкции или конъюгата, описанных в настоящем описании выше).
Настоящее изобретение также относится к способу уменьшения связывания hRGM A с морфогенетическим белком кости 2 и/или морфогенетическим белком кости 4 (BMP-2 и BMP-4) у индивида, при необходимости, предусматривающему стадию введения индивиду продукта по изобретению (в частности, связывающего белка, конструкции или конъюгата, описанных в настоящем описании выше).
Настоящее изобретение также относится к способу лечения нарушения, связанного с активностью RGM A у индивида, предусматривающему стадию введения одного или в комбинации с другими терапевтическими средствами продукта по изобретению (в частности, связывающего белка, конструкции или конъюгата, описанных в настоящем описании выше).
Настоящее изобретение также относится к способу уменьшения активности RGM A у индивида, страдающего нарушением, при котором активность RGM A оказывает негативное воздействие, предусматривающему введение индивиду продукта по изобретению (в частности, связывающего белка, конструкции или конъюгата, описанных в настоящем описании выше), одного или в комбинации с другими терапевтическими средствами.
Указанное нарушение предпочтительно включает неврологические заболевания, выбранные из группы, состоящей из бокового амиотрофического склероза, повреждения плечевого сплетения, повреждения головного мозга, в том числе травматического повреждения головного мозга, церебрального паралича, болезни Гийена-Барре, лейкодистрофий, рассеянного склероза, поствакцинального полиомиелита, spina bifida (расщелины позвоночника), повреждения спинного мозга, спинально-мышечной атрофии, спинальных опухолей, инсульта, поперечного миелита; деменции, сенильной деменции, легкого когнитивного нарушения, связанной с болезнью Альцгеймера деменции, хореи Хантингтона, поздней дискинезии, гиперкинезий, маний, болезни Паркинсона, синдрома Стила-Ричардсона, синдрома Дауна, тяжелой псевдопаралитической миастении, травмы нерва, сосудистого амилоидоза, кровоизлияния I в головной мозг с амилоидозом, воспаления головного мозга, острого нарушения со спутанностью сознания, бокового амиотрофического склероза, глаукомы и болезни Альцгеймера.
Другие конкретные варианты по изобретению определены ниже:
выделенный связывающий белок, который специфически взаимодействует по меньшей мере с одним эпитопом белка hRGM A;
указанный выделенный белок, являющийся моноклональным нейтрализующим антителом или его антигенсвязывающим фрагментом;
указанный антигенсвязывающий фрагмент, содержащий домен VH и VL;
указанное нейтрализующее антитело, уменьшающее способность hRGM A связываться с его рецептором;
указанное нейтрализующее антитело, способное ингибировать биологическую активность hRGM A;
указанное антитело, распознающее рецептор RGM A, выбранный из рецептора RGM A человека, собакоподобных обезьян, крысы, курицы, лягушки и рыбы;
указанное антитело, распознающее белок RGM A, аминокислотная последовательность которого на 90% гомологична аминокислотной последовательности SEQ ID NO:2;
указанное антитело, где белок RGM A кодирует нуклеиновая кислота, которая на 90% гомологична последовательности нуклеиновой кислоты SEQ ID NO:1;
указанное антитело, которое по меньшей мере на 90% идентично аминокислотной последовательности с последовательностью, содержащей вариабельную область тяжелой цепи (область VH), содержащий последовательность SEQ ID NO:9 или 34, или гуманизированную, необязательно, дополнительно мутированную версию указанной области VH;
указанное антитело, которое по меньшей мере на 90% идентично аминокислотной последовательности с последовательностью, содержащей вариабельную область легкой цепи (область VL), содержащий последовательность SEQ ID NO:10, или гуманизированную, необязательно, дополнительно мутированную версию указанной области VL,
указанное антитело, которое связывается с hRGM A, где это антитело является гликозилированным;
указанное антитело или антигенсвязывающий фрагмент, где указанное антитело или антигенсвязывающий фрагмент является мышиным антителом, гуманизированным антителом, полностью человеческим антителом, химерным антителом, антигенсвязывающим фрагментом гуманизированного антитела или антигенсвязывающим фрагментом химерного антитела;
указанное антитело или антигенсвязывающий фрагмент, где указанное антитело или антигенсвязывающий фрагмент является антигенсвязывающим фрагментом, выбранным из группы, состоящей из Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента и Fv-фрагмента;
указанное моноклональное антитело, которое специфически связывается по меньшей мере с одним эпитопом hRGM A, где указанное моноклональное антитело является моноклональным антителом, секретируемым гибридомной клеточной линией, описанной в настоящем описании;
указанное моноклональное антитело, связывание с которым приводит к инактивации взаимодействия hRGM А с его рецептором;
указанная гибридомная клеточная линия, которая продуцирует моноклональное антитело, которое специфически связывается по меньшей мере с одним эпитопом hRGM A;
указанная гибридомная клеточная линия, где гибридома выбрана из группы, состоящей из гибридомы человека, мыши, крысы, овцы, свиньи, крупного рогатого скота, козы и лошади;
указанное моноклональное антитело, связывание с которым приводит к инактивации hRGM A;
указанная гибридомная клеточная линия, которая продуцирует моноклональное антитело, которое специфически связывается по меньшей мере с одним эпитопом hRGM A;
указанная гибридомная клеточная линия, где гибридома выбрана из группы, состоящей из гибридомы человека, мыши, крысы, овцы, свиньи, крупного рогатого скота, козы и лошади;
указанное моноклональное нейтрализующее антитело или его антигенсвязывающий фрагмент, имеющий по меньшей мере одну характеристику, выбранную из группы, состоящей из:
a) связывания с RGM A млекопитающего с аффинностью в диапазоне нМ или менее;
b) функционального противодействия активности RGM A in vitro в анализе роста нейритов с эффективностью мкМ, нМ или меньшей;
c) индукции in vivo спрутинга (роста нервных волокон) в модели раздавливания зрительного нерва;
d) индукции in vivo спрутинга в модели повреждения спинного мозга;
e) облегчения in vivo экспериментального повреждения спинного мозга усилением регенеративного роста поврежденных нервных волокон или
f) облегчения in vivo экспериментального повреждения спинного мозга усилением образования синапсов;
выделенная нуклеиновая кислота, кодирующая указанное моноклональное нейтрализующее антитело или антигенсвязывающий фрагмент;
вектор, содержащий указанную выделенную нуклеиновую кислоту;
указанный вектор, выбранный из группы, состоящей из pcDNA; pTT; pTT3; pEFBOS; pBV; pJV; pHybE и pBJ;
клетка-хозяин, трансформированная указанным вектором, где клетка-хозяин выбрана из группы, состоящей из клетки одноклеточного организма, клетки животного, клетки растения и грибной клетки;
указанная клетка-хозяин, где клеткой животного является клетка млекопитающего, выбранная из группы, состоящей из HEK293, CHO и COS;
клетка-хозяин, трансформированная вектором, где клеткой-хозяином является эукариотическая клетка;
способ получения связывающего белка, который связывает hRGM A, предусматривающий культивирование клетки-хозяина в культуральной среде при условиях, достаточных для получения связывающего белка, который связывает hRGM A;
фармацевтическая композиция, содержащая указанное моноклональное антитело или антигенсвязывающую часть и фармацевтически приемлемый носитель;
способ уменьшения связывания hRGM A с рецептором Неогенином у индивида, при необходимости, предусматривающий стадию введения индивиду указанного антитела;
способ уменьшения связывания hRGM A с морфогенетическим белком кости 2 и морфогенетическим белком кости 4 (BMP-2 и BMP-4) у индивида, при необходимости, предусматривающий стадию введения индивиду указанного антитела;
способ лечения нарушения, связанного с активностью RGM A индивида, предусматривающий стадию введения указанного антитела, одного или в комбинации с другими терапевтическими средствами;
способ уменьшения активности RGM A у индивида, страдающего нарушением, при котором активность RGM A оказывает негативное влияние, предусматривающий введение индивиду указанного антитела, одного или в комбинации с другими терапевтическими средствами;
указанное антитело, содержащее по меньшей мере одну область VH, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 35, 36, 37, 38, 39, 40, 41, 42 и 43;
указанное антитело, содержащее по меньшей мере одну область VL, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 44, 45 и 46;
указанное антитело, дополнительно модифицированное 1-5 мутациями в последовательности VH или VL;
указанное антитело, где эти мутации выбраны из обратных мутаций каркасного участка и мутаций Vernier и VH/VL-смежных остатков.
Любое указание на SEQ ID NO: 34 или любая ссылка на SEQ ID NO: 34, описанная в настоящем документе, также относится к SEQ ID NO:9.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1A показано связывание моноклональных антител с hRGM A в анализе ELISA.
На фигуре 1В изображено связывание моноклональных антител с hRGM A, экспрессируемым в клетках HEK 293.
На фигуре 1C изображено связывание моноклональных антител с RGM A крысы, экспрессируемым в клетках HEK 293.
На фигуре 2 показано связывание полноразмерного RGM A с Неогенином. MAB 5F9 ингибирует связывание полноразмерного fc-связанного hRGM A с Неогенином.
На фигуре 3 изображено связывание полноразмерного RGM A с BMP-4. MAB 5F9 ингибирует связывание fc-связанного фрагмента (47-422) полноразмерного hRGM A с BMP-4.
На фигуре 4 изображено связывание фрагмента 0 RGM A с BMP-4. MAB 5F9 ингибирует связывание fc-связанного фрагмента 0 (47-168) с BMP-4.
На фигуре 5 показано связывание полноразмерного RGM A с BMP-2. MAB 5F9 ингибирует связывание fc-связанного фрагмента полноразмерного hRGM A (47-422) с BMP-2.
На фигуре 6 представлено объединение микрофотографий, показывающих mAb5F9-нейтрализацию фрагмента RGM A в анализе роста нейритов клеток NTera. MAB 5F9 нейтрализует ингибирующую рост активность fc-конъюгированного, сильного ингибирующего фрагмента hRGM A в анализах роста нейритов с агрегатами Ntera человека. A. Контрольная культура, рост нейронов Ntera на ламинине, B. на субстрате ламинин-фрагмент hRGM A (47-168), C - E. на субстрате ламинин-фрагмент hRGM A (47-168) в присутствии 0,1 мкг/мл MAB 5F9 (C), 1 мкг/мл MAB 5F9 (D.), 10 мкг/мл MAB 5F9 (E.).
На фигуре 7 показан количественный анализ результатов анализа NTera 2. MAB 5F9 нейтрализует зависимым от дозы образом ингибирующую рост активность fc-конъюгированного, сильного ингибирующего фрагмента hRGM A (фрагмента 0, 47-168) в анализах роста нейритов с агрегатами Ntera человека.
На фигуре 8 показан количественный анализ теста с полосами SH-SY5Y. MAB 5F9 нейтрализует отталкивание, индуцируемое полосами, состоящими из полноразмерного RGM A человека нейронных клеток SH-SY5Y человека в полосатых покрытиях мембран. В отсутствие MAB 5F9 (A) или в присутствии низких концентраций MAB нейроны SH-SY5Y предпочитают избегать RGM A-полос. Это поведение обращается увеличением концентраций MAB 5F9. (B-D). При наивысшей концентрации MAB (10 мкг/мл) (E), нейроны SH-SY5Y обнаруживают сильное предпочтение в отношении полос RGM А в сравнении с полосами Коллагена I.
На фигуре 9 суммирован количественный анализ характеристик связывания mABs 5F9 и 8D1. MAB 5F9 и 8D1 оценивали в анализах связывания hRGM A - неогенин, hRGM A - BMP-2 и hRGM A - BMP-4 при различных концентрациях.
На фигуре 10 показана нейтрализующая активность в отношении хемоотталкивающей активности гуманизированных антител 5F9 (h5F9.21, h5F9.23, h5F9.25) в анализе хемотаксиса SH-SY5Y.
На фигуре 11 показана in vivo нейрорегенеративная активность локального нанесения 5F9 на модели повреждения зрительного нерва животного. Локальное нанесение MAB 5F9 нейтрализует RGM A и стимулирует регенеративный рост аксонов поврежденного зрительного нерва в модели раздавливания зрительного нерва животного (крысы). В обработанных 5F9 животных (A), многие GAP-43-положительные волокна удлиняются за пределы участка раздавливания в отличие от контрольного MAB 8D1 (B), которое не связывается с RGM A крысы.
Фигуры 12A и 12B показан количественный анализ локального нанесения 5F9 в модели повреждения зрительного нерва животного. (A) 5F9, но не контрольное MAB 8D1, значительно увеличивало количество регенерирующихся GAP-43-положительных волокон. Значимо больше волокон (p<0,05) наблюдали у животных, обработанных 5F9, при расстояниях 200 мкм, 400 мкм и 600 мкм, а при 1200 мкм волокна обнаруживали только у 5F9-обработанных животных, но не у контрольных животных. (B) 5F9 значительно увеличивало GAP-43-положительную площадь в участке повреждения зрительного нерва по сравнению с контрольным антителом 8D1 и контролем-носителем ЗФР. Площадь регенеративного роста (GAP-43-положительную площадь) измеряли с использованием программного обеспечения Axiovision software (Zeiss).
На фигуре 13 показана in vivo нейрорегенеративная активность системного применения 5F9 на модели повреждения зрительного нерва животного. Животных обрабатывали 5F9 на день 0 и день 21 с использованием 2 мг/кг и 10 мг/кг, соответственно. Антитело или носитель вводили внутрибрюшинно или внутривенно. Показаны объединенные изображения зрительных нервов крыс. В обработанных 5F9 животных (A), многочисленные GAP-43-волокна удлинялись за пределы участка разрушения в отличие от контрольных животных, обработанных ЗФР (B). Участок раздавливания локализован в левом крае и регенерирующие волокна окрашены антителом против GAP-43. Многочисленные волокна наблюдаются в верхнем и нижнем крае зрительного нерва в обработанных 5F9 животных, но не в обработанных ЗФР животных.
На фигуре 14A и на фигуре 14B показан количественный анализ системного применения 5F9 в модели повреждения зрительного нерва животного.
На фигуре 15 показана активность повторной миелинизации (ремиелинизации) in vivo системного применения 5F9 на модели повреждения зрительного нерва животного. Животных обрабатывали 5F9 на день 0 и день 21 с использованием 2 мг/кг и 10 мг/кг, соответственно. Антитело или носитель вводили внутрибрюшинно или внутривенно. Показаны объединенные изображения зрительных нервов крыс. Миелинизацию визуализируют с использованием антитела, направленного против маркера миелина, базального белка миелина MBP. Участки раздавливания расположены в середине обработанных нервов и эта зона является свободной в обработанных носителем контрольных животных (A и B). В обработанных 5F9 животных (C и D), наблюдаются многочисленные MBP-положительные структуры в средней зоне (центре раздавливания) зрительных нервов.
На фигуре 16 показано количественное действие ремиелинизации системного применения 5F9 на модели повреждения зрительного нерва животного.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение относится к RGM A-связывающим белкам, более конкретно, к моноклональным RGM A-антителам, в частности, гуманизированным моноклональным RGM A-антителам, или их антигенсвязывающим частям, которые связывают RGM A. Различные аспекты этой заявки относятся к антителам и фрагментам антител и их фармацевтическим композициям, а также к нуклеиновым кислотам, рекомбинантным экспрессирующим векторам и клеткам-хозяевам для получения таких антител и фрагментов. Способы применения антител по изобретению для детектирования RGM A человека; для нейтрализации RGM человека и/или активности RGM A человека, in vivo или in vitro, и для регуляции экспрессии генов также включены в настоящее изобретение.
1. Общие определения
Если нет другого определения, научные и технические термины, используемые в настоящем изобретении, будут иметь значения, которые известны специалистам с обычной квалификацией в данной области. Значение и объем этих терминов ясны, однако, в случае любой неопределенности, определения, представленные в настоящем описании, должны учитываться в первую очередь по сравнению со словарным или другими определениями. Кроме того, если контекст не требует иного, термины в единственном числе должны включать и множественное число, а термины во множественном числе включают и единственное число. В этой заявке, использование слова "или" означает "и/или", если нет другого указания. Кроме того, использование термина "включающий", а также других форм, таких как "включает" и "включенные", не являются ограничивающим. Термины, такие как "элемент" или "компонент", включают как элементы и компоненты, содержащие одну единицу, так и элементы и компоненты, которые содержат более одной субъединицы, если нет другого указания.
Обычно, номенклатура и способы, используемые в отношении культуры клеток и ткани, молекулярной биологии, иммунологии, микробиологии, генетики и химии белков и нуклеиновых кислот и гибридизации, используемые в настоящем описании, являются хорошо известны и обычно используемыми в данной области. Методы и способы по изобретению обычно осуществляют в соответствии с общепринятыми способами, хорошо известными в данной области и описанными в различных общих и более конкретных ссылках, которые цитируются и описаны в описании, если нет других указаний. Ферментативные реакции и способы очистки проводят в соответствии с описаниями изготовителя, как обычно осуществляется в данной области или как описано в настоящем описании. Лабораторные процедуры и способы аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, используемые в настоящем описании, и номенклатуры, используемые в связи с ними, хорошо известны и обычно используются в данной области. Стандартные способы используются для химических синтезов, химических анализов, получения фармацевтических средств, получения композиций и их доставки и лечения пациентов.
Для большей ясности настоящего изобретения выбранные термины определены ниже.
Термин "полипептид" относится в контексте настоящего описания к полимерной цепи аминокислот. Термины "пептид" и "белок" используются взаимозаменяемо с термином полипептид и также относятся к полимерной цепи аминокислот. Термин "полипептид" включает природные или искусственные белки, фрагменты белков и полипептидные аналоги белковой последовательности. Полипептид может быть мономерным или полимерным.
Термин "выделенный белок" или "выделенный полипептид" обозначает белок или полипептид, которые вследствие его происхождения или источника получения не связан с встречающимися в природе компонентами, которые окружают его в природном состоянии; по существу свободен от других белков того же вида; экспрессируется клеткой другого вида или не встречается в природе. Таким образом, полипептид, который химически синтезирован или синтезирован в клеточной системе, отличающейся от клетки, из которой он природно происходит, будет "выделенным” из его природных компонентов. Белок может быть также получен по существу свободным от его природных компонентов посредством выделения с использованием способов очистки белков, хорошо известных в данной области.
Термин "выделение" относится в контексте настоящего описания к способу получения химической молекулы, такой как полипептид, по существу свободной от природных компонентов посредством выделения, например, с использованием способов очистки белков, хорошо известных в данной области.
Термин "RGM A человека" (сокращаемый в настоящем описании как hRGM A), относящийся к соединенному с гликозилфосфатидилинозитом (gpi) гликопротеину с 450 аминокислотами, был впервые описан как репеллент роста нейритов или ингибитор роста нейритов во время развития топографических выступов (выростов) (Stahl et al. Neuron 5: 735-43, 1990; Mueller, in Molecular Basis of Axon Growth and Nerve Pattern Formation, Edited by H. Fujisawa, Japan Scientific Societies Press, 215-229, 1997). Семейство генов rgm включает три разных гена, два из них, rgm a и b, экспрессируются в ЦНС млекопитающих, тогда как третий член, rgm c, экспрессируется в периферии (Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361:1513-29, 2006), где он играет важную роль в метаболизме железа. Белки RGM человека имеют идентичность последовательности 43% - 50%; гомология аминокислот RGM A человека и крысы равна 89%. Белки RGM человека не имеют значительной гомологии с последовательностью других известных белков. Они являются богатыми пролином белками, содержащими область RGD, и имеют структурную гомологию с доменом фактора Фон-Виллебранда и расщепляются при N-концевой аминокислоте 168 неизвестной протеазой с образованием функционально активного белка (Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361: 1513-29, 2006).
In vitro RGM A ингибирует рост нейрита (аксона) при пикомолярных концентрациях посредством связывания с Неогенином, который был идентифицирован как рецептор RGM (Rajagopalan et al. Nat Cell Biol.: 6(8), 756-62,2004). Неогенин был сначала описан как нетринсвязывающий белок (Keino-Masu et al. Cell, 87(2):175-85, 1996), но его аффинность в отношении Нетрина (KD 2 нМ) является на порядок величин более низкой, чем в отношении RGM (KD 0,2 нМ) (Rajagopalan et al. Nat Cell Biol.: 6(8), 756-62, 2004). Это является важным открытием, так как сообщалось, что связывание Нетрина-1 с Неогенином или с его близкородственным рецептором DCC (делетированным в колоректальном раке) стимулировало, а не ингибировало рост нейритов (Braisted et al. J. Neurosci. 20: 5792-801, 2000).
Наряду со связыванием RGM A с Неогенином и индукцией ингибирования роста нейритов, связывание RGM A или B с морфогенетическими белками кости BMP-2 и BMP-4 могло представлять другое препятствие успешной нейрорегенерации и функциональному восстановлению (Mueller et al., Philos. Trans. R. Soc. Lond. B Biol. Sci. 361:1513-29, 2006). Сообщалось, что оба класса белков (Неогенин и BMP) трансдуцируют ингибирующий рост нейритов сигнал RGM A через совершенно различные и независимые пути трансдукции сигналов. Обычно, экспрессия этих белков BMP является относительно низкой в большинстве областей ЦНС взрослых, но сообщалось, что она быстро увеличивается в экспрессии и накапливании некоторых BMP (например, BMP-2, BMP-6, BMP-7) в ответ на повреждение и инсульт (Lai et al., Neuroreport 8: 2691-94, 1997; Martinez et al. Brain Res. 894: 1-11, 2001; Hall and Miller, J. Neurosci. Res. 76: 1-8, 2004; Setoguchi et al., Exp. Neurol. 189: 33-44, 2004). Кроме того, на модели рассеянного склероза, модели экспериментального аутоиммунного энцефаломиелита (EAE), BMP-4, BMP-6 и BMP-7 повышающим образом регулировались в спинном мозгу мыши (Ara et al., J. Neurosci. Res. 86: 125-35, 2008). Сообщалось, что BMP-2 ингибировал рост нейритов связыванием с клеточной поверхностью RGM A, BMP-рецепторов I и II и прямой активацией LIM-киназы (Matsuura et al. Biochem Biophys Res Commun., 360: 868-73, 2007), и, следовательно, полагают, что блокирование взаимодействия RGM A-BMP-2 будет дополнительно увеличивать функциональное восстановление после повреждения ЦНС.
Как упоминалось выше, у крыс с повреждением спинного мозга и людей с повреждением головного мозга были обнаружены массивные накапливания клеточного RGM в участке повреждения, и характер окрашивания RGM A в крысах в участке спинномозгового повреждения очень сходен с окрашиванием pan RGM-антителом у людей, что предполагает, что большая часть pan RGM-окрашивания у людей связана с локализацией RGM A, а не с локализацией RGM B (Schwab et al., Arch. Neurol.62: 1561-8, 2005a; Schwab et al. Eur. J. Neurosci. 21:1569-76, 2005 b; Hata et al. J. Cell Biol. 173:47-58, 2006). В головном мозге здорового человека, pan RGM-окрашивание (иммунореактивность RGM A & B) детектировали на волокнах белого вещества, олигодендроцитах, перикарионах немногих нейронов, некоторых гладких мышцах сосудов и немногих эндотелиальных клетках. Окрашивания астроцитов не наблюдали. Характер окрашивания RGM в головном мозге взрослых людей очень сходен с характером окрашивания, наблюдаемым в спинном мозге взрослой крысы (Schwab et al. Eur. J. Neurosci. 21:1569-76, 2005 b; Hata et al. J. Cell Biol. 173:47-58, 2006).
На основании накапливания RGM A в участках повреждений в повреждении в головном мозге и спинном мозге вследствие ингибирующей рост клеточных нейритов активности, полагают, что этот белок проявляет ингибирующую рост нейритов активность, и нейтрализация ее антителами или их антигенсвязывающим фрагментом, который связывается по меньшей мере с одним эпитопом RGM А человека, может привести к улучшенному повторному росту поврежденных нервных волокон и к усилению функционального восстановления в показаниях, характеризуемых повреждением нервных волокон и накапливанием RGM.
Если нет других указаний, термин "RGM A" включает также молекулы RGM A, выделенные или полученные из других видов, например, грызунов, таких как мыши или крысы; конкретно, полученную из крысы молекулу называют в настоящем описании "RGM A крысы".
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ МОЛЕКУЛ, СХОДНЫХ С RGM A
"Биологическая активность" относится в контексте настоящего описания ко всем неотъемлемым биологическим свойствам RGM A, определенным в настоящем описании.
Термины "специфическое связывание" или "специфически связывающиеся", в контексте настоящего описания, при ссылке на взаимодействие антитела, белка или пептида со второй химической молекулой, означают, что это взаимодействие зависит от присутствия конкретной структуры (например, “антигенной детерминанты” или "эпитопа", определенных ниже) на этой химической молекуле; например, антитело распознает специфическую структуру белка и связывается со специфической структурой белка, а не с белками вообще. Если антитело является специфическим в отношении эпитопа "A", присутствие молекулы, содержащей эпитоп A (или свободного, немеченого A), в реакции, содержащей меченый "A" и это антитело, будет уменьшать количество меченого А, связанного с этим антителом.
Термин "антитело”, в контексте настоящего описания, в широком смысле относится к любой молекуле иммуноглобулина (Ig), состоящей из четырех полипептидных цепей, двух тяжелых (H) цепей и двух легких (L) цепей, или ее любому функциональному фрагменту, мутанту, варианту или производному, которые сохраняют основные признаки связывания эпитопа молекулы Ig. Такие мутантные, вариантные или производные варианты известны в данной области. Их не ограничивающие изобретение варианты описаны ниже. Говорят, что антитело "способно связывать" молекулу, если оно способно специфически реагировать с этой молекулой со связыванием посредством этого данной молекулы с этим антителом.
Термин "моноклональное антитело" обозначает в контексте настоящего описания препарат молекул антитела, которые имеют общую аминокислотную последовательность тяжелой цепи и общую аминокислотную последовательность легкой цепи, в отличие от препаратов “поликлонального” антитела, которые содержат смесь различных антител. Моноклональные антитела могут быть получены несколькими новыми технологиями, такими как фаговый, бактериальный, дрожжевой или рибосомный дисплей, а также классическими способами, примерами которых являются полученные из гибридом антитела (например, антитело, секретируемое гибридомой, полученной гибридомной технологией, такой как стандартная гибридомная технология Kohler and Milstein hybridoma methodology ((1975) Nature 256:495-497).
В полноразмерном антителе, каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращаемого в настоящем описании как HCVR или VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращаемого в настоящем описании как LCVR или VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Эти VH- и VL-области могут быть дополнительно подразделены на области гипервариабельности, называемые комплементарность определяемыми областями (CDR), чередующиеся с областями, которые являются более консервативными, названными каркасными участками (FR). Каждая из VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Молекулы иммуноглобулинов могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса.
Термин "антигенсвязывающая часть" или "антигенсвязывающий фрагмент" антитела (или просто "часть антитела" или "фрагмент антитела"), в контексте настоящего описания, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, hRGM A). Было показано, что антигенсвязывающую функцию антител могут выполнять фрагменты полноразмерного антитела. Такие варианты антител могут быть также биспецифичными, двояко специфичными или мультиспецифичными форматами; могут специфично связываться с двумя или более различными антигенами. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающая часть" антитела включают (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из VH- и CH1-доменов; (iv) Fv-фрагмент, состоящий из VL- и VH-доменов антитела с единственном плечом, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546, Winter et al., PCT-публикацию WO 90/05144 A1, включенные в настоящем описании посредством ссылки), который содержит единственный вариабельный домен; и (vi) выделенная определяющая комплементарность область (CDR). Кроме того, хотя эти два домена Fv-фрагмента, VL и VH, кодируются раздельными генами, они могут быть соединены, с использованием, рекомбинантных способов, синтетическим линкером, который позволяет им быть полученными в виде единой белковой цепи, в которой VL- и VH-области соединяются с образованием моновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426 и Huston et al. (1988) Proc. Natl. Acad. Sci USA 85:5879-5883). Предполагается, что такие одноцепочечные антитела также включены в термин "антигенсвязывающая часть" антитела. Другие формы одноцепочечных антител, такие как диатела, также входят в рамки этого термина. Диатела являются двухвалентными, биспецифическими антителами, в которых VH- и VL-домены экспрессируются на единой полипептидной цепи, но используют такой короткий линкер, который является слишком коротким для возможности спаривания между двумя этими доменами на одной и той же цепи, заставляя тем самым эти домены соединяться с комплементарными доменами на другой цепи и создавая два антигенсвязывающих сайта (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci USA 90:6444-6448; Poljak, RJ., et al. (1994) Structure 2:1121-1123). Такие антигенсвязывающие части известны в данной области (Kontermann and Dubel eds., Antibody Engineering (2001) Springer-Veriag. New York. 790 pp. (ISBN 3-540-41354-5).
Термин "конструкция антитела" относится в контексте настоящего описания к полипептиду, содержащему одну или несколько антигенсвязывающих частей по изобретению, связанных с линкерным полипептидом или константным доменом иммуноглобулина. Линкерные полипептиды содержат два аминокислотного остатка или более аминокислотных остатков, соединенных пептидными связями, и используются для связывания одной или нескольких антигенсвязывающих частей. Такие линкерные полипептиды хорошо известны в данной области (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci USA 90:6444-6448; Poljak, R.J., et al. (1994) Structure 2:1121-1123). Константным доменом иммуноглобулина называют константный домен тяжелой цепи или константный домен легкой цепи. Аминокислотные последовательности константного домена тяжелой цепи и константного домена легкой цепи IgG человека известны в данной области и представлены в таблице 2.
ПОСЛЕДОВАТЕЛЬНОСТЬ КОНСТАНТНОГО ДОМЕНА ТЯЖЕЛОЙ ЦЕПИ И КОНСТАНТНОГО ДОМЕНА ЛЕГКОЙ ЦЕПИ IgG ЧЕЛОВЕКА
DYFPEPVTVSWNSGALTSGVHTFPAVLQSS
GLYSLSSVVTVPSSSLGTQTYICNVNHKPS
NTKVDKKVEPKSCDKTHTCPPCPAPELLGG
PSVFLFPPKPKDTLMISRTPEVTCVVVDVS
HEDPEVKFNWYVDGVEVHNAKTKPREEQYN
STYRVVSVLTVLHQDWLNGKEYKCKVSNKA
LPAPIEKTISKAKGQPREPQVYTLPPSREE
MTKNQVSLTCLVKGFYPSDIAVEWESNGQP
ENNYKTTPPVLDSDGSFFLYSKLTVDKSRW
QQGNVFSCSVMHEALHNHYTQKSLSLSPGK
DYFPEPVTVSWNSGALTSGVHTFPAVLQSS
GLYSLSSVVTVPSSSLGTQTYICNVNHKPS
NTKVDKKVEPKSCDKTHTCPPCPAPEAAGG
PSVFLFPPKPKDTLMISRTPEVTCVVVDVS
HEDPEVKFNWYVDGVEVHNAKTKPREEQYN
STYRVVSVLTVLHQDWLNGKEYKCKVSNKA
LPAPIEKTISKAKGQPREPQVYTLPPSREE
MTKNQVSLTCLVKGFYPSDIAVEWESNGQP
ENNYKTTPPVLDSDGSFFLYSKLTVDKSRW
QQGNVFSCSVMHEALHNHYTQKSLSLSPGK
FYPREAKVQWKVDNALQSGNSQESVTEQDS
KDSTYSLSSTLTLSKADYEKHKVYACEVTH
QGLSSPVTKSFNRGEC
DFYPGAVTVAWKADSSPVKAGVETTTPSKQ
SNNKYAASSYLSLTPEQWKSHRSYSCQVTH
EGSTVEKTVAPTECS
Кроме того, антитело или его антигенсвязывающая часть могут быть частью более крупных молекул иммуноадгезии, образованных ковалентной или нековалентной связью этого антитела или части антитела с одним или несколько другими белками или пептидами. Примеры таких молекул иммуноадгезии включают использование коровой области стрептавидина для получения тетрамерной молекулы scFv (Kipriyanov, S. M., et al. (1995) Human Antibodies and Hybridomas 6:93-101) и использование остатка цистеина, маркерного пептида и С-концевой полигистидиновой метки для получения бивалентных и биотинилированных молекул scFv (Kipriyanov, S.M., et al. (1994) Mol. Immunol. 31:1047-1058). Части антител, такие как Fab- и F(ab')2-фрагменты, могут быть получены из полноразмерных антител с использованием общепринятых способов, таких как расщепление папаином или пепсином, соответственно, полноразмерных антител. Кроме того, антитела, части антител и молекулы иммуноадгезии могут быть получены с использованием стандартных способов рекомбинантных ДНК, описанных в настоящем описании.
"Выделенное антитело", в контексте настоящего описания, представляет собой антитело, которое по существу свободно от других антител, имеющих отличающиеся антигенные специфичности (например, выделенное антитело, которое специфически связывается с hRGM A, является по существу свободным от антител, которые специфически связывают антигены, другие, чем hRGM A). Однако, выделенное антитело, которое специфически связывается с hRGM A, может иметь перекрестную реактивность в отношении других антигенов, таких как молекулы RGM A других видов. Кроме того, выделенное антитело может быть по существу свободным от другого клеточного материала и/или от других химикалиев.
Термин "антитело человека", в контексте настоящего изобретения, включает антитела, имеющие вариабельные и константные области, полученные из последовательностей иммуноглобулина зародышевой линии человека. Эти антитела человека по изобретению могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro или соматической мутацией in vivo), например, в CDR и, в частности, CDR3. Однако, термин "антитело человека", в контексте настоящего описания, не включает в себя антитела, в которых CDR-последовательности, полученные из другого вида млекопитающего, такого как мышь, были привиты на каркасные последовательности человека.
Термин "рекомбинантное антитело человека", в контексте настоящего изобретения включает все антитела человека, которые получают, экспрессируют или выделяют рекомбинантными способами, таких как антитела, экспрессируемые с использованием рекомбинантного экспрессирующего вектора, трансфицированного в клетку-хозяина (описанного дополнительно ниже), антитела, выделенные из рекомбинантной, комбинаторной библиотеки антител человека (Hoogenboom H. R., (1997) TIB Tech. 15:62-70; Azzazy H. and Highsmith W.E., (2002) Clin. Biochem. 35:425-445; Gavilondo J.V., and Larrick J.W. (2002) BioTechniques 29:128-145; Hoogenboom H., and Chames P. (2000) Immunology Today 21: 371-378), антитела, выделенные из животного (например, мыши), которое является трансгенным в отношении генов иммуноглобулина человека (см., например, Taylor, L. D., et al. (1992) Nucl. Acids Res. 20:6287-6295; Kellermann S-A., and Green L.L. (2002) Current Opinion in Biotechnology 13:593-597; Little M. et al (2000) Immunology Today 21: 364-370), или антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, которые включают сплайсинг последовательностей гена иммуноглобулина человека в другие ДНК-последовательности. Такие рекомбинантные антитела человека имеют вариабельные и константные области, полученные из последовательностей иммуноглобулина зародышевой линии человека. Однако, в некоторых вариантах осуществления, такие рекомбинантные антитела человека подвергают мутагенезу in vitro (или, при использовании животного, трансгенного в отношении последовательностей Ig человека, соматическому мутагенезу in vivo), и, следовательно, аминокислотные последовательности VH- и VL-областей этих рекомбинантных антител являются последовательностями, которые, хотя и полученные из последовательностей VH- и VL-последовательностей зародышевой линии человека и родственные им, не могут существовать в природе в репертуаре (спектре) зародышевой линии антитела человека in vivo.
Термин "химерное антитело" относится к антителам, которые содержат последовательности вариабельной области тяжелой цепи и легкой цепи одного вида и последовательности константной области другого вида, такие как антитела, имеющие вариабельные области тяжелой цепи и легкой цепи мыши, связанные с константными областями человека. Это химерное антитело может быть получено рекомбинантными молекулярно-биологическими способами или может быть получено физической конъюгацией указанных областей.
Термин "CDR-привитое антитело" относится к антителам, которые содержат последовательности вариабельной области тяжелой цепи и легкой цепи одного вида, но в которых последовательности одного или нескольких из этих CDR-области VH и/или VL заменены CDR-последовательностями другого вида, таким как антитела, имеющие вариабельные области тяжелой цепи и легкой цепи мыши, в которых один или несколько CDR мыши (например, CDR3) были заменены последовательностями CDR человека.
Термины "нумерация Кабата", "определение по Кабату” и "мечение по Кабату" используются в настоящем описании взаимозаменяемо. Эти термины, которые являются хорошо известными в данной области, относятся к системе нумерации аминокислотных остатков, которые являются более вариабельными (т.е. гипервариабельными), чем другие аминокислотные остатки, в вариабельных областях тяжелой цепи и легкой цепи антитела или его антигенсвязывающей части (Kabat et al. (1971) Ann. NY Acad, Sci. 190: 382-391 и Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Для вариабельной области тяжелой цепи, эта гипервариабельная область находится в диапазоне положений аминокислот 31-35 для CDR1, положений аминокислот 50-65 для CDR2 и положений аминокислот 95-102 для CDR3. Для вариабельной области легкой цепи, эта гипервариабельная область находится в диапазоне положений аминокислот 24-34 для CDR1, положений аминокислот 50-56 для CDR2 и положений аминокислот 89-97 для CDR3.
В контексте настоящего описания, термины "акцептор" и "акцепторное антитело" относится к последовательности антитела или последовательности нуклеиновой кислоты, обеспечивающей или кодирующей по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или нескольких каркасных участков. В некоторых вариантах осуществления, термин "акцептор" относится к аминокислотной последовательности или последовательности нуклеиновой кислоты, обеспечивающей или кодирующей константную область (константные области). Еще в одном варианте осуществления, термин "акцептор" относится к аминокислотной последовательности или последовательности нуклеиновой кислоты, обеспечивающей или кодирующей одну или несколько каркасных участков и константной области (константных областей). В одном конкретном варианте осуществления, термин "акцептор" относится к аминокислотной последовательности или последовательности нуклеиновой кислоты антитела человека, которая обеспечивает или кодирует по меньшей мере 80%, предпочтительно, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100% аминокислотных последовательностей одной или нескольких каркасных областей. Согласно этому варианту осуществления, акцептор может содержать по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5 или по меньшей мере 10 аминокислотных остатков, который (которые) не встречаются в одном или нескольких конкретных положениях антитела человека. Акцепторная каркасная область и/или акцепторные константные области могут быть получены из гена антитела зародышевой линии, гена созревшего антитела, функционального антитела (например, антител, хорошо известных в данной области, антител в развитии или антител, коммерчески доступных).
В контексте настоящего описания, термин "CDR" относится к определяющей комплементарность области в вариабельных последовательностях антител. Имеются три CDR в каждой из вариабельных областей тяжелой цепи и легкой цепи, которые обозначают CDR1, CDR2 и CDR3, для каждой из этих вариабельных областей. Термин "набор CDR" относится в этом контексте к группе из трех CDR, которые находятся в единой вариабельной области, способной связывать антиген. Точные границы этих CDR определялись по-разному в соответствии с различными системами. Эта система, описанная Кабатом (Kabat et al., Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md. (1987) и (1991)), обеспечивает не только недвусмысленную систему нумерации остатков, которую можно использовать в любой вариабельной области антитела, но обеспечивает также точные границы остатков, определяющие эти три CDR. Эти CDR могут называться CDR по Кабату. В статье Chothia et al. (Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987) и Chothia et al., Nature 342:877-883 (1989)) обнаружили, что некоторые подчасти в CDR по Кабату принимают почти идентичные конформации пептидного скелета, несмотря на наличие большого разнообразия на уровне аминокислотной последовательности. Эти подчасти были названы L1, L2 и L3 или H1, H2 и H3, где "L" и "H" обозначают области легкой цепи и тяжелой цепи, соответственно. Эти области могут называться CDR Хотиа, которые имеют границы, которые перекрываются с CDR Кабата. Другие границы, определяющие CDR, были описаны Padlan (FASEB J. 9:133-139 (1995)) and MacCallum (J Mol Biol 262(5):732-45 (1996)). Другие определения областей CDR могут также не подчиняться строго одной из этих систем, но тем не менее будут перекрываться с CDR Кабата, хотя они могут быть укороченными или удлиненными в свете предсказания или экспериментальных открытий того, что конкретные остатки или группы остатков или даже все CDR не влияют значительно на связывание антигена. Используемые в настоящем описании способы могут использовать CDR, определенные в соответствии с любой из этих систем, хотя предпочтительные варианты осуществления используют CDR, определенные по Кабату или Хотиа.
В контексте настоящего описания, термин "канонический" остаток относится к остатку в CDR или каркасной области, которая определяет конкретную каноническую структуру CDR, определенную Chothia et al. (J. Mol. Biol. 196:901-907 (1987); Chothia et al., J. Mol. Biol. 227:799 (1992), обе статьи включены в настоящее описание посредством ссылки). Согласно Chothia et al., критические части CDR многих антител имеют почти идентичные конформации пептидного скелета этих CDR несмотря на большое разнообразие на уровне аминокислотной последовательности. Каждая каноническая структура точно определяет первично набор торсионных углов пептидного скелета для смежного сегмента аминокислотных остатков, образующего петлю.
В контексте настоящего описания, термины "донор" и "донорное антитело" относится к антителу, обеспечивающему один или несколько CDR. В предпочтительном варианте осуществления, донорным антителом является антитело вида, отличающегося от вида, из которого получено антитело, из которого получены или произведены каркасные области. В контексте гуманизированного антитела, термин "донорное антитело" относится к антителу не человека, обеспечивающему один или несколько CDR.
В контексте настоящего описания, термин "каркасный участок" или "каркасная последовательность" относится к оставшимся последовательностям вариабельной области минус эти CDR (т.е. после вычитания CDR). Поскольку точное определение CDR-последовательности может определяться различными системами, значение каркасной последовательности является объектом соответственно различных интерпретаций. Эти шесть CDR (CDR-L1, -L2 и -L3 легкой цепи и CDR-H1, -H2 и -H3 тяжелой цепи) также делят каркасные области на легкой цепи и тяжелой цепи на четыре субобласти (FR1, FR2, FR3 и FR4) на каждой цепи, в которой CDR1 расположен между FR1 и FR2, CDR2 между FR2 и FR3 и CDR3 между FR3 и FR4. Без точного определения конкретных субобластей как FR1, FR2, FR3 или FR4, каркасная область, называемая другими авторами, представляет объединенные FR в вариабельной области единой природно-встречающейся цепи иммуноглобулина. В контексте настоящего описания, один FR представляет один из четырех субобластей и FR во множественном числе представляют два или более из четырех субобластей, составляющих каркасную область.
В данной области известны акцепторные последовательности тяжелой цепи и легкой цепи человека. В одном варианте осуществления по изобретению акцепторные последовательности тяжелой цепи и легкой цепи человека выбраны из последовательностей, описанных в таблице 3 и таблице 4. В указанных таблицах описаны различные комбинации в отношении каркасных последовательностей FR1-FR4 человека.
АКЦЕПТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ТЯЖЕЛОЙ ЦЕПИ ЧЕЛОВЕКА
АКЦЕПТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ЛЕГКОЙ ЦЕПИ ЧЕЛОВЕКА
В контексте настоящего описания, термин "ген антитела зародышевой линии" или "фрагмент гена" относится к последовательности иммуноглобулина, кодируемой нелимфоидными клетками, которые не были подвергнуты процессу созревания, который приводит к генетической реаранжировке и мутации для экспрессии конкретного иммуноглобулина. (См., например, Shapiro et al., Crit. Rev. Immunol. 22(3): 183-200 (2002); Marchalonis et al., Adv Exp Med Biol. 484:13-30 (2001)). Одно из преимуществ, обеспечиваемых различными вариантами осуществления данного изобретения, возникает из признания того, что гены антител зародышевой линии с большей вероятностью, чем гены созревших антител, сохраняют незаменимые структуры аминокислотных последовательностей, характерные для индивидуумов в этих видах, следовательно, с меньшей вероятностью узнаются как гены из чужеродного источника при использовании терапевтически в этих видах.
В контексте настоящего описания, термин "ключевые" остатки относится к определенным остаткам в вариабельной области, которые имеют большее влияние на специфичность связывания и/или аффинность антитела, в частности, гуманизированного антитела. Ключевой остаток включает, но не ограничивается ими, один или несколько из следующих остатков: остаток, который является смежным с CDR, потенциальный сайт гликозилирования (может быть сайтом или N-, или O-гликозилирования), редкий остаток, остаток, способный взаимодействовать с антигеном, остаток, способный взаимодействовать с CDR, канонический остаток, контактный остаток между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, остаток в зоне Vernier и остаток в области перекрытия CDR1 вариабельной области тяжелой цепи по Хотия и первым каркасным участком тяжелой цепи по Кабату.
Термин "гуманизированное антитело" обычно относится к антителам, которые содержат последовательности вариабельной области тяжелой и легкой цепи из видов, не являющихся человеком (например, мыши), но в которых по меньшей мере часть VH- и/или VL-последовательности была изменена таким образом, что она стала более "человекоподобной", т.е. более сходной с вариабельными последовательностями зародышевой линии человека. Одним типом гуманизированного антитела является CDR-привитое антитело, в котором последовательности CDR человека введены в последовательности VH и VL для замены соответствующих CDR-последовательностей не человека.
В частности, термин "гуманизированное антитело" в контексте настоящего описания обозначает антитело или его вариант, производное, аналог или фрагмент, которые иммуноспецифически связываются с представляющим интерес антигеном и которые содержат каркасную (FR) область, имеющую по существу аминокислотную последовательность антитела человека, и определяющую комплементарность область (CDR), имеющую по существу аминокислотную последовательность антитела, не относящегося к человеку. В контексте настоящего описания, термин "по существу" в контексте с CDR относится к CDR, имеющему аминокислотную последовательность, по меньшей мере на 50, 55, 60, 65, 70, 75 или 80%, предпочтительно по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 98% или по меньшей мере на 99% идентичную аминокислотной последовательности CDR антитела не человека. Гуманизированное антитело содержит по существу все из по меньшей мере одного, обычно двух, вариабельных доменов (Fab, Fab', F(ab')2, FabC, Fv), в которых все или по существу все из областей CDR соответствуют областям CDR иммуноглобулина не человека (т.е. донорного антитела), и все или по существу все из каркасных областей являются каркасными областями консенсусной последовательности иммуноглобулина человека. Предпочтительно, гуманизированное антитело содержит по меньшей мере часть константной области иммуноглобулина (Fc), обычно часть константной области иммуноглобулина человека. В некоторых вариантах осуществления, гуманизированное антитело содержит как легкую цепь, так и по меньшей мере вариабельный домен тяжелой цепи. Это антитело может включать также области CH1, шарнирную область, области CH2, CH3 и CH4 тяжелой цепи. В некоторых вариантах осуществления, гуманизированное антитело содержит только гуманизированную легкую цепь. В некоторых вариантах осуществления, гуманизированное антитело содержит только гуманизированную тяжелую цепь. В конкретных вариантах осуществления, гуманизированное антитело содержит только гуманизированный вариабельный домен легкой цепи и/или гуманизированную тяжелую цепь.
Гуманизированное антитело может быть выбрано из любого класса иммуноглобулинов, в том числе IgY, IgM, IgG, IgD, IgA и IgE, и любого изотипа, в том числе, без ограничения, IgA1, IgA2, IgG1, IgG2, IgG3 и IgG4. Гуманизированное антитело может содержать последовательности из более чем одного класса или изотипа и конкретные константные домены могут быть выбраны для оптимизации желаемых эффекторных функций с использованием способов, хорошо известных в данной области.
Каркасные и CDR-области гуманизированного антитела не должны точно соответствовать исходным (родительским) последовательностям, например, CDR или консенсусный каркасный участок донорного антитела могут быть мутагенизированы заменой, инсерцией и/или делецией по меньшей мере одного аминокислотного остатка, так что остаток CDR или каркасный остаток в этом сайте не соответствует ни донорному антителу, ни консенсусному каркасному участку. Однако в предпочтительном варианте осуществления, такие мутации не будут экстенсивными. Обычно, по меньшей мере 50, 55, 60, 65, 70, 75 или 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% остатков гуманизированного антитела будут соответствовать остаткам в исходных последовательностях FR и CDR. В контексте настоящего описания, термин "консенсусный каркасный участок" относится к каркасной области в консенсусной последовательности иммуноглобулина. В контексте настоящего описания, термин "консенсусная последовательность иммуноглобулина" относится к последовательности, образованной из наиболее часто встречающихся аминокислот (или нуклеотидов) в семействе родственных последовательностей иммуноглобулинов (См., например, Winnaker, From Genes to Clones (Verlagsgesellschaft, Weinheim, Germany 1987). В семействе иммуноглобулинов, каждое положение в консенсусной последовательности занято аминокислотой, встречающейся наиболее часто в этом положении в этом семействе. Если две аминокислоты встречаются с одинаковой частотой, любая может быть включена в эту консенсусную последовательность.
В контексте настоящего описания, зоной "Vernier" называют субнабор каркасных остатков, которые могут корректировать структуру CDR и осуществлять тонкую настройку соответствия антигену, как описано в статье Foote and Winter (1992, J. Mol. Biol. 224:487-499), которая включена в настоящее описание посредством ссылки). Остатки зоны Vernier образуют слой, подстилающий CDR, и могут влиять на структуру CDR и аффинность этого антитела.
Термин "ингибирование связывания" RGM с одним из его рецепторов, используемых в настоящем описании, включает частичное (например, приблизительно на 20%, 40%, 60%, 80%, 85%, 90%, 95% или более) или полное уменьшение активности связывания указанного рецептора. Указанное "ингибирование связывания" может быть определено любым подходящим способом, доступным в данной области, предпочтительно любым способом, приведенным в настоящем описании в качестве примера, таким как, например, анализы связывания на основе ELISA.
В контексте настоящего описания, термин "нейтрализация" относится к нейтрализации биологической активности белка-мишени при специфическом связывании связывающим белком белка-мишени. Нейтрализация может быть результатом различных путей связывания указанного связывающего белка с мишенью. Например, нейтрализация может быть обусловлена связыванием связывающего белка в области мишени, которая не влияет на связывание рецептора с молекулой-мишенью. Альтернативно, связывание связывающего белка может приводить к блокаде связывания этого рецептора с мишенью, и эта блокада в конце концов нейтрализует активность белка-мишени. Согласно этому изобретению, может встречаться каждый из указанных различных механизмов. Предпочтительно нейтрализующий связывающий белок является нейтрализующим антителом, связывание которого с hRGM A приводит к нейтрализации биологической активности hRGM A. Предпочтительно этот нейтрализующий связывающий белок связывает hRGM A и уменьшает биологическую активность hRGM A по меньшей мере приблизительно на 20%, 40%, 60%, 80%, 85% или более. Нейтрализация биологической активности hRGM A нейтрализующим связывающим белком может оцениваться измерением одного или нескольких индикаторов биологической активности hRGM A, хорошо известных в данной области. Например, нейтрализация hRGM A обращает ингибирование в анализе нейронного роста в Ntera (см. пример 3 ниже). Анализ роста нейритов Ntera определяет ингибирование роста нейритов. В отсутствие ингибирующего белка или фрагмента RGM A и в присутствии стимулирующего рост нейритов субстрата ламинина нейронные агрегаты NTera обнаруживают экстенсивную и плотную сеть вырастающих нейритов. RGM A или фрагменты RGM A ингибируют рост нейритов, приводя к уменьшенной длине и уменьшенным количествам нейритов. Блокирующие функцию RGM A антагонисты или MAB, такие как mAb 5F9, нейтрализовали ингибирующую рост нейритов активность сильного фрагмента легкой цепи (аминокислоты 47-168) fc-конъюгированного hRGM A белка RGM A человека в анализах роста нейритов с агрегатами дифференцированных нейронов NTera человека, приводя к сильному увеличению длины и количества нейритов.
"Нейтрализующее моноклональное антитело” относится в контексте настоящего описания к препарату молекул антител, которые после связывания со специфическим антигеном способны конкурировать за связывание и ингибировать связывание природного лиганда для указанного антигена. В одном конкретном варианте данной заявки нейтрализующие антитела данного изобретения способны конкурировать с RGM A за связывание с Неогенином и/или с BMP-2 и/или BMP-4 и предотвращать биологическую активность или функцию RGM A. В частности, нейтрализующие антитела данного изобретения способны связываться с RGM A и предотвращать связывание с Неогенином и/или с BMP-2 и/или ВМР-4 и предотвращать биологическую активность или функцию RGM A. Термин "активность" включает такие активности, как специфичность/аффинность связывания антитела в отношении антигена, например, анти-hRGM A-антитела, которое связывается с антигеном RGM A, и/или нейтрализующую силу антитела, например, анти-hRGM A-антитела, связывание которого с hRGM A ингибирует биологическую активность hRGM A, например, как определено в анализе связывания hRGM A-Неогенина, анализе связывания hRGM A-BMP-2 или анализе связывания hRGM A-BMP-4, как описано ниже в экспериментальном разделе.
Биологическая активность RGM A может быть описана как регуляция клеточной миграции. Особым примером клеточной миграции является рост нейритов, который задерживается или ингибируется белками RGM A. Кроме того, было показано, что белки RGM модулируют активность BMP-белков. Опубликованные в настоящем описании примеры описывают синергическую, усиливающую активность белков RGM на BMP-пути, с одной стороны, и ингибирующую активность белков RGM на BMP-пути, который является важным для регуляции метаболизма железа, регенерации костей и хрящей и в ЦНС для ремиелинизации и регенерации.
Термин "эпитоп" или "антигенная детерминанта" включает любую полипептидную детерминанту, способную специфически связываться с иммуноглобулином или Т-клеточным рецептором. В некоторых вариантах осуществления, эпитопные детерминанты включают химически активные поверхностные группировки молекул, таких как аминокислоты, сахарные боковые цепи, фосфорил или сульфонил, и, в некоторых вариантах осуществления, могут иметь специфические трехмерные структурные характеристики и/или характеристики специфического заряда. Эпитоп является областью антигена, которая связывается антителом. В некоторых вариантах осуществления, говорят, что антитело специфически связывает антиген, когда оно преимущественно узнает его антигенмишень в комплексной смеси белков и/или макромолекул.
Термин "резонанс поверхностных плазмонов" относится в контексте настоящего описания к оптическому феномену, который позволяет анализировать биоспецифические взаимодействия реального времени детектированием изменений концентраций белка в биосенсорном матриксе, например, с использованием системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, NJ). В отношении дополнительных описаний, см. Jόnsson, U., et al. (1993) Ann. Biol. Clin. 51:19-26; Jόnsson, U., et al. (1991) Biotechniques 11:620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8:125-131; and Johnnson, B., et al. (1991) Anal. Biochem. 198:268-277.
Термин "kon" в контексте настоящего описания относится к константе скорости для ассоциации антитела с антигеном для образования комплекса антитело/антиген, как известно в данной области.
Термин "koff" относится в контексте настоящего описания к константе скорости для диссоциации антитела от комплекса антитело/антиген, как известно в данной области.
Термин "KD” относится в контексте настоящего описания к константе диссоциации конкретного взаимодействия антитело/антиген, как известно в данной области.
Термин "меченый связывающий белок" относится в контексте настоящего описания к белку с включенной меткой, которая обеспечивает идентификацию связывающего белка. Предпочтительно, меткой является детектируемый маркер, например, включение радиоактивно меченой аминокислоты или присоединение к полипептиду биотинилированных частей молекулы, которые могут быть детектированы меченым авидином (например, стрептавидином, содержащим флуоресцентный маркер или ферментативную активность, которые могут быть детектированы оптическим или колориметрическим способами). Примеры меток для полипептидов включают, но не ограничиваются ими, следующие метки: радиоизотопы или радионуклиды (например, 3Н, 14C, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho или 153Sm); флуоресцентные метки (например, FITC, родамин, лантанидные люминофоры), ферментные метки (например, пероксидаза хрена, люцифераза, щелочная фосфатаза); хемилюминесцентные маркеры; биотинилированные группы; предварительно определенные полипептидные эпитопы, узнаваемые вторичным репортером (например, пары последовательностей лейциновой молнии, сайты связывания для вторичных антител, металлсвязывающие домены, эпитопные метки (тэги)); и магнитные агенты, такие как хелаты гадолиния.
Термин "конъюгат антитела" относится к связывающему белку, такому как антитело, химически связанному со второй химической частью молекулы, такой как терапевтический или цитотоксический агент. Термин "агент" используется в настоящем описании для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, приготовленного из биологических материалов. Предпочтительно, эти терапевтические или цитотоксические средства включают, но не ограничиваются ими, токсин коклюша, таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропанолол и пуромицин и их аналоги или гомологи.
Термины "кристалл" и "кристаллизуемый" относятся в данном аспекте к антителу или его антигенсвязывающей части, которая существует в форме кристалла. Кристаллы являются одной формой твердого состояния вещества, которая отличается от других форм, таких как аморфное твердое состояние или жидкокристаллическое состояние. Кристаллы состоят из регулярных, повторяющихся, трехмерных упорядоченных рядов атомов, ионов, молекул (например, белков, таких как антитела) или молекулярных комплексов (например, комплексов антиген/антитело). Эти трехмерные упорядоченные ряды выстроены в соответствии с конкретными математическими взаимосвязями, которые хорошо понятны в данной области. Фундаментальное звено, или элемент структуры, который является повторяющимся в кристалле, называют асимметричным звеном. Повторение этого асимметричного звена в расположении, которое подчиняется конкретной, хорошо определенной кристаллографической симметрии, обеспечивает "элементарную ячейку" этого кристалла. Повторение этой элементарной ячейки посредством регулярных трансляций (переносов) во всех трех измерениях обеспечивает кристалл. См. Giege, R. and Ducruix, A. Barrett, Crystallization of Nucleic Acids and Proteins, a Practical Approach, 2nd ea., pp. 20 1-16, Oxford University Press, New York, New York, (1999).
Термин "полинуклеотид" в контексте настоящего описания обозначает полимерную форму двух или более нуклеотидов, либо рибонуклеотидов, либо дезоксинуклеотидов, или модифицированной формы любого типа нуклеотида. Этот термин включает одноцепочечные и двухцепочечные формы ДНК, но предпочтительно двухцепочечную ДНК.
Термин "выделенный полинуклеотид" в контексте настоящего описания будет обозначать полинуклеотид (например, геномный, кДНК или полинуклеотид синтетического происхождения или некоторые их комбинации), который, вследствие его происхождения в качестве "выделенного полинуклеотида": не связан со всем полинуклеотидом или частью полинуклеотида, с которыми этот "выделенный полинуклеотид” обнаруживается в природе; функционально связан с полинуклеотидом, который не является связанным с ним в природе; или не встречается в природе в качестве части большей последовательности.
Термин "вектор", в контексте настоящего описания, относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. Одним типом вектора является "плазмида", которая является кольцевой двухцепочечной петлей ДНК, в которую могут быть лигированы дополнительные ДНК-сегменты. Другим типом вектора является вирусный вектор, причем в этот вирусный геном могут быть лигированы дополнительные ДНК-сегменты. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации (ориджин репликации), и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина и посредством этого реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называют "рекомбинантными экспрессирующими векторами" (или просто "экспрессирующими векторами"). Обычно, экспрессирующие векторы, применимые в способах рекомбинантных ДНК, часто находятся в форме плазмид. В данном описании, термины "плазмида" и "вектор" могут использоваться взаимозаменяемо, так как плазмида является наиболее часто используемой формой вектора. Однако, предполагается, что это изобретение включает такие другие формы экспрессирующих векторов, как вирусные векторы (например, ретровирусы с дефектной репликацией, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Термин "функционально связанные" относится к непосредственному соседству, в котором описанные компоненты находятся в связи, позволяющей им функционировать предполагаемым для них образом. Регуляторную последовательность, “функционально связанную” с кодирующей последовательностью, лигируют таким образом, что экспрессия этой кодирующей последовательности достигается при условиях, совместимых с этими регуляторными последовательностями. "Функционально связанные" последовательности включают как регуляторные последовательности экспрессии, которые являются смежными с представляющим интерес геном, так и регуляторные последовательности экспрессии, которые действуют в транс-положении или на расстоянии для регуляции представляющего интерес гена. Термин "регуляторная последовательность экспрессии" относится в контексте настоящего описания к полинуклеотидной последовательности, которая является необходимой для осуществления экспрессии и процессинга кодирующих последовательностей, с которыми они лигированы. Регуляторные последовательности экспрессии включают подходящие последовательности инициации, терминации транскрипции, промоторные и энхансерные последовательности; эффективные сигналы процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, которые стабилизируют цитоплазматическую мРНК; последовательности, которые увеличивают эффективность трансляции (т.е. консенсусную последовательность Козака); последовательности, которые увеличивают стабильность белка; и, когда желательно, последовательности, которые увеличивают секрецию белка. Природа таких регуляторных последовательностей различается в зависимости от организма-хозяина; в прокариотах, такие регуляторные последовательности обычно включают промотор, сайт связывания рибосом и последовательность терминации транскрипции. Предполагается, что термин "регуляторные последовательности" включает компоненты, присутствие которых является важным для экспрессии и процессинга, и может также включать дополнительные компоненты, присутствие которых является выгодным, например, лидерные последовательности и последовательности партнера слияния.
"Трансформация", определенная в настоящем описании, относится к любому процессу, при помощи которого экзогенная ДНК входит в клетку-хозяина. Трансформация может происходить при природных или искусственных условиях с использованием различных способов, хорошо известных в данной области. Трансформация может основываться на любом известном способе для введения чужеродных последовательностей нуклеиновых кислот в прокариотическую или эукариотическую клетку-хозяина. Этот способ выбирают на основании подлежащей трансформации клетки-хозяина и может включать, без ограничения, вирусную инфекцию, электропорацию, липофекцию и бомбардировку частицами. Такие "трансформированные” клетки включают стабильно трансформированные клетки, в которых введена ДНК, способная к репликации либо в виде автономно реплицирующейся плазмиды, либо в виде части хромосомы хозяина. Они также включают клетки, которые транзиторно экспрессируют инсертированную ДНК или РНК в течение ограниченных периодов времени.
Термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин") относится в контексте настоящего описания к клетке, в которую была введена экзогенная ДНК. Должно быть понятно, что такие термины относятся не только к конкретной рассматриваемой клетке, но и к потомству такой клетки. Поскольку некоторые модификации могут встречаться в последующих генерациях вследствие мутации или влияний окружающей среды, такие потомки могут фактически не быть идентичными родительской клетке, но все еще включаются в объем термина "клетка-хозяин" в контексте настоящего описания. Предпочтительно клетки-хозяева включают прокариотические и эукариотические клетки, выбранные из любого из царств растений или животных. Предпочтительные эукариотические клетки включают клетки одноклеточных организмов (протистов), клетки грибов, растений и животных. Наиболее предпочтительно клетки-хозяева включают, но не ограничиваются ими, прокариотическую клеточную линию E. coli, линии клеток млекопитающих CHO, HEK 293 и COS; линию клеток насекомых Sf9 и клетки грибов Saccharomyces cerevisiae.
Для рекомбинантных ДНК, синтеза олигонуклеотидов и культуры ткани и трансформации (например, электропорации, липофекции) могут быть использованы стандартные способы. Ферментативные реакции и способы очистки могут выполняться в соответствии с описаниями изготовителя или в соответствии с тем, как они обычно выполняются в данной области или как описано в настоящем описании. Предшествующие способы и процедуры могут обычно выполняться в соответствии с общепринятыми способами, хорошо известными в данной области, описанными в различных общих и более конкретных ссылках, которые цитируются и обсуждаются в данном описании. См., например, руководство Sambrook et al. Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989)), которое включено в настоящее описании посредством ссылки для любой цели.
Термин "трансгенный организм", известный в данной области и используемый в настоящем описании, относится к организму, имеющему клетки, которые содержат трансген, где этот трансген, введенный в организм (или в организм предка этого организма), экспрессирует полипептид, не экспрессируемый в природе в этом организме. "Трансген" является ДНК-конструкцией, которая стабильно и функционально интегрирована в геном клетки, из которой развивается трансгенный организм, и которая управляет экспрессией продукта кодируемого гена в одном или нескольких типах клеток или тканей этого трансгенного организма.
Термины "регулируют" и "модулируют" используются взаимозаменяемо и, в контексте настоящего описания, относятся к изменению или смещению активности представляющей интерес молекулы (например, биологической активности hRGM A). Модуляция может быть увеличением или уменьшением величины определенной активности или функции представляющей интерес молекулы. Примеры активностей и функций молекулы включают, но не ограничиваются ими, характеристики связывания, ферментативную активность, активацию клеточных рецепторов и трансдукцию сигналов.
Соответственно, термин "модулятор" обозначает в контексте настоящего описания соединение, способное изменять или смещать активность или функцию представляющей интерес молекулы (например, биологическую активность hRGM A). Например, модулятор может вызывать увеличение или уменьшение величины определенной активности или функции молекулы в сравнении с величиной этой активности или функции, наблюдаемой в отсутствие модулятора. Термин "агонист" относится в контексте настоящего описания к модулятору, который, при контакте с представляющей интерес молекулой, вызывает увеличение величины этой активности или функции, наблюдаемой в отсутствие этого агониста. Конкретные представляющие интерес агонисты могут включать, но не ограничиваются ими, полипептиды hRGM A или полипептиды, нуклеиновые кислоты, углеводы или любые другие молекулы, которые связываются с hRGM A. Термин "антагонист" относится в контексте настоящего описания к модулятору, который, при контакте с представляющей интерес молекулой, вызывает уменьшение величины активности или функции молекулы, наблюдаемых в отсутствие этого антагониста. Примеры антагонистов включают, но не ограничиваются ими, белки, пептиды, антитела, пептитела, углеводы или малые органические молекулы. Пептитела описаны, например, в WO01/83525.
Конкретные представляющие интерес антагонисты включают антагонисты, которые блокируют или модулируют биологическую или иммунологическую активность hRGM A. Антагонисты hRGM A могут включать, но не ограничиваются ими, белки, нуклеиновые кислоты, углеводы или другие молекулы, которые связываются с hRGM A, такие как моноклональные антитела, которые взаимодействуют с молекулой RGM A. Следует отметить, что взаимодействие с RGM A может приводить к связыванию и нейтрализации других лигандов/компонентов клеточной мембраны и может быть полезным для аддитивного или синергического функционирования против многочисленных заболеваний.
В контексте настоящего описания, термин "эффективное количество" относится к количеству терапевтического средства, которое является достаточным для уменьшения или ослабления тяжести и/или продолжительности нарушения или одного или нескольких симптомов нарушения, предотвращения рецидива, развития, начала прогрессирования одного или нескольких симптомов, ассоциированных с нарушением, детектирования нарушения или увеличения или улучшения профилактических или терапевтических эффектов другого терапевтического средства (например, профилактического или терапевтического средства).
Термин "образец" используется в контексте настоящего описания в его самом широком смысле. "Биологический образец", в контексте настоящего описания, включает, но не ограничивается этим, любое количество вещества из живого существа или ранее живого существа. Такие живые существа включают, но не ограничиваются ими, людей, мышей, крыс, обезьян, собак, кроликов и других животных. Такие вещества включают, но не ограничиваются ими, кровь, сыворотку, мочу, синовиальную жидкость, клетки, органы, ткани, костный мозг, лимфатические узлы и селезенку.
2. Полипептиды, которые связывают hRGM A
Основной вариант данной заявки включает выделенные белки или полипептиды, которые специфически связываются по меньшей мере с одним эпитопом белка RGM A. Выделенные белки или полипептиды, которые специфически связываются по меньшей мере с одним эпитопом белка RGM A, способны ингибировать связывание RGM A и рецептора, Неогенина и/или морфогенетическими белками кости 2 и 4 (BMP-2, BMP-4).
Наиболее предпочтительный вариант данной заявки включает антитела, которые связываются с RGM A или его антигенсвязывающими частями или фрагментами.
Предпочтительно, анти-RGM A-антитела данного изобретения проявляют высокую способность уменьшения или нейтрализации активности RGM A, например, оцениваемой любым из нескольких анализов in vitro и in vivo, известных в данной области или описанных ниже.
Наиболее предпочтительно настоящая заявка включает нейтрализующие моноклональные антитела против RGM A, которые селективно предотвращают связывание RGM A и рецептора, Неогенина и морфогенетических белков кости 2 и 4 (BMP-2, BMP-4), и генерирование нейтрализующего моноклонального антитела против RGM A, которое селективно предотвращает связывание RGM A с его корецепторами морфогенетическими белками кости 2 и 4 (BMP-2, BMP-4).
Предпочтительно, этим моноклональным нейтрализующим антителом данной заявки является антитело человека или гуманизированное антитело. Термин "антитело человека" относится к антителам, имеющим вариабельные и константные области, соответствующие последовательностям иммуноглобулина зародышевой линии человека, или полученные из них, (например, см. Kabat et al. Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242, 1991). Однако, антитела человека данной заявки могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro, или соматические мутации in vivo), например, в CDR и, в частности, CDR3.
В различных вариантах осуществления, антитело является рекомбинантным антителом или моноклональным антителом. Наиболее предпочтительные нейтрализующие антитела данной заявки называют mAb5F9 и mAb8D1 и их функциональные фрагменты, и предполагается, что другие антитела и функциональные фрагменты антитела с эквивалентными свойствами относительно mAb5F9 и mAb8D1, такими как высокоаффинное связывание с RGM A с низкой кинетикой диссоциации и высокой нейтрализующей способностью, являются частью данного изобретения. Аффинность связывания и скорость диссоциации анти-RGM A-антитела данной заявки в отношении иммуногенного полипептида RGM A или его фрагмента могут быть определены любым способом, известным в данной области. Например, аффинность связывания может быть измерена при помощи конкурентных анализов ELISA, RIA, BIAcore или технологии KinExA. Скорость диссоциации может быть также измерена при помощи BIAcore или технологии KinExA. Аффинность связывания и скорость диссоциации измеряют по резонансу поверхностных плазмонов с использованием, например, BIAcore.
Одно из предпочтительных моноклональных антител данного изобретения, антитело mAb5F9, имеет по меньшей мере 90% идентичность аминокислотной последовательности с последовательностью, содержащей вариабельную область тяжелой цепи (VH-область), содержащую последовательность SEQ ID NO:9 или 34, и вариабельную область легкой цепи (VL-область), содержащую последовательность SEQ ID NO:10.
Предполагается также, что выделенные моноклональные антитела, которые взаимодействуют с RGM A данной заявки, могут быть гликозилированным связывающим белком, в котором антитело или его антигенсвязывающая часть содержит один или несколько углеводных остатков. Получение возникающих in vivo белков может подвергаться дополнительному процессингу, известному как посттрансляционная модификация. В частности, сахарные (гликозильные) остатки могут добавляться ферментативно в процессе, известном как гликозилирование. Полученные белки, несущие ковалентно связанные олигосахаридные боковые цепи, известны как гликозилированные белки или гликопротеины. Гликозилирование белков зависит от аминокислотной последовательности представляющего интерес белка, а также от клетки-хозяина, в которой экспрессируется этот белок. Различные организмы могут продуцировать различные ферменты гликозилирования (например, гликозилтрансферазы и гликозидазы) и имеют различные доступные субстраты (нуклеотидсахара). Вследствие таких факторов, характер гликозилирования белков и состав гликозильных остатков могут различаться в зависимости от системы-хозяина, в которой экспрессируется конкретный белок. Гликозильные остатки, применимые в этом изобретении, могут включать, но не ограничиваются ими, глюкозу, галактозу, маннозу, фукозу, н-ацетилглюкозамин и сиаловую кислоту. Предпочтительно, этот гликозилированный связывающий белок содержит такие гликозильные остатки, что характер гликозилирования является характером гликозилирования, свойственным человеку.
Антитела данной заявки содержат константную область тяжелой цепи, такую как константная область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM, IgY или IgD. Кроме того, это антитело может содержать константную область легкой цепи, либо константную область легкой цепи каппа, либо константную область легкой цепи лямбда. Альтернативно, частью-антителом может быть, например, Fab-фрагмент или одноцепочечный Fv-фрагмент. Замены аминокислотных остатков в Fc-части для изменения эффекторной функции антитела известны в данной области (Winter, et al. Патенты США № 5648260; 5624821). Fc-часть антитела опосредует несколько важных эффекторных функций, например, индукцию цитокинов, ADCC, фагоцитоз, комплемент-зависимую цитотоксичность (CDC) и время полужизни/скорость клиренса антитела и комплексов антигенантитело. В некоторых случаях эти эффекторные функции являются желательными для терапевтического антитела, но в других случаях могут быть необязательными или даже вредными, в зависимости от терапевтических целей. Некоторые изотипы IgG человека, в частности, IgG1 и IgG3, опосредуют ADCC и CDC посредством связывания с FcγR и комплементом C1q, соответственно. Неонатальные Fc-рецепторы (FcRn) являются критическими компонентами, определяющими время полужизни антител в кровотоке. Еще в одном варианте осуществления по меньшей мере один аминокислотный остаток заменен в константной области этого антитела, например, Fc-область антитела, так что эффекторные функции антитела являются измененными.
3. Генерирование анти-hRGM A-антител
3.1. Общая часть
Антитела этой заявки могут быть генерированы иммунизацией подходящего хозяина (например, позвоночных, в том числе людей, мышей, крыс, овец, коз, свиней, крупного рогатого скота, лошадей, пресмыкающихся, рыб, земноводных, и в яйцах птиц, земноводных и рыб). Для генерирования антител данной заявки хозяина иммунизируют иммуногенным полипептидом RGM A или его фрагментом по изобретению. Термин "иммунизация" относится в настоящем описании к способу предоставления антигена иммунному репертуару (спектру), который существует в природном не измененном генетически организме или трансгенном организме, в том числе трансгенном организме, модифицированном для представления искусственного иммунного репертуара человека. Подобным образом, "иммуногенный препарат" является композицией антигена, который содержит адъюванты или другие добавки, которые могли бы увеличивать иммуногенность антигена.
Иммунизация животных может выполняться любым способом, известным в данной области. См., например, Harlow and Lane, Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Press, 1990. Способы иммунизации животных (не человека), таких как мыши, крысы, овцы, козы, свиньи, крупный рогатый скот и лошади, хорошо известны в данной области. См., например, Harlow and Lane и Патент США № 5994619. В одном предпочтительном варианте осуществления, антиген RGM A вводят с адъювантом для стимуляции иммунной реакции. Такие адъюванты включают полный или неполный адъювант Фрейнда, RIBI (мурамилдипептиды) или ISCOM (иммуностимулирующие комплексы). Такие адъюванты могут защищать этот полипептид от быстрого рассеивания заключением его в локальное покрытие, или они могут содержать вещества, которые стимулируют секрецию хозяином факторов, которые являются хемотаксическими в отношении макрофагов и других компонентов иммунной системы. Предпочтительно, при введении полипептида, схема иммунизации будет включать два или более введений полипептида, распределяемых на протяжении нескольких недель.
Предполагается, что животное-хозяина иммунизируют антигеном, связанным с клеточной мембраной интактной или разрушенной клетки, и антитела данной заявки идентифицируют связыванием с иммуногенным полипептидом по изобретению. После иммунизации хозяина-животного антигеном, антитела могут быть получены из этого животного. Из этого животного посредством кровоизвлечения или умерщвления получают содержащую антитело сыворотку. Эта сыворотка может быть использована в том виде, в каком она получена, или эти антитела могут быть очищены из этой сыворотки. Сыворотка или иммуноглобулины, полученные таким образом, являются поликлональными, следовательно, имеющими гетерогенный ряд свойств.
3.2. Моноклональные анти-RGM A-антитела, полученные с использованием гибридомной технологии
Моноклональные антитела могут быть получены с использованием большого разнообразия способов, известных в данной области, включающих гибридомную, рекомбинантную технологии и технологию фагового дисплея или их комбинацию. Например, моноклональные антитела могут быть получены с использованием гибридомных способов, в том числе способов, известных в данной области и описанных, например, Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling, et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981) (указанные ссылки включены посредством ссылки в их полном объеме). Термин "моноклональное антитело" в контексте настоящего описания не ограничивается антителами, получаемыми посредством гибридомной технологии. Термин "моноклональное антитело" относится к антителу, которое получено из единственного клона, в том числе эукариотического, прокариотического или фагового клона, а не к способу, при помощи которого оно получено.
Способы получения и скрининга специфических антител с использованием гибридомной технологии являются рутинными и хорошо известными в данной области. В одном варианте осуществления, настоящее изобретение обеспечивает способы генерирования моноклональных антител, а также антитела, полученные этим способом, предусматривающие культивирование гибридомной клетки, секретирующей антитело по изобретению, где, предпочтительно, гибридому генерируют слиянием спленоцитов, выделенных из мыши, иммунизированной антигеном по изобретению, с миеломными клетками и затем скрининг гибридом, полученных из этого слияния, на гибридомные клоны, которые секретируют антитело, способное связывать полипептид по изобретению. Вкратце, мыши могут иммунизироваться антигеном RGM A. В предпочтительном варианте осуществления, антиген RGM A вводят с адъювантом для стимуляции иммунной реакции. Такие адъюванты включают полный или неполный адъювант Фрейнда, RIBI (мурамилдипептиды) или ISCOM (иммуностимулирующие комплексы). Такие адъюванты могут защищать полипептид от быстрого рассеивания заключением его в локальное покрытие, или они могут содержать вещества, которые стимулируют секрецию хозяином факторов, которые являются хемотаксическими в отношении макрофагов и других компонентов иммунной системы. Предпочтительно, при введении полипептида, схема иммунизации будет включать два или более введений этого полипептида, распределяемых на протяжении нескольких недель.
После детектирования иммунной реакции, например, детектирования антител, специфических в отношении антигена RGM A, в сыворотке мыши, извлекают селезенку мыши и выделяют спленоциты. Затем спленоциты сливают хорошо известными способами с любыми подходящими клетками миеломы, например, клетками из клеточной линии SP20, доступной из ATCC. Гибридомы отбирают и клонируют посредством лимитирующего разведения. Затем гибридомные клоны анализируют способами, известными в данной области, на клетки, которые секретируют антитела, способные связывать RGM A. Посредством иммунизации мышей положительными гибридомными клонами может быть генерирована асцитическая жидкость, которая обычно содержит высокие уровни антител.
В другом варианте осуществления, из этого иммунизированного животного могут быть получены продуцирующие антитело иммортализованные гибридомы. После иммунизации животное умерщвляют и В-клетки селезенки сливают с иммортализованными клетками миеломы, также хорошо известными в данной области. См., например, Harlow and Lane, supra. В предпочтительном варианте осуществления, эти клетки миеломы не секретируют полипептиды иммуноглобулина (несекретирующая клеточная линия). После слияния и отбора с использованием антибиотиков эти гибридомы подвергают скринингу с использованием RGM A или его части или клетки, экспрессирующей RGM A. В предпочтительном варианте осуществления, выполняют начальный скрининг с использованием твердофазного иммуноферментного анализа (ELISA) или радиоиммуноанализа (RIA), предпочтительно ELISA. Пример скрининга ELISA обеспечен в WO 00/37504, включенном в настоящее описание посредством ссылки.
Продуцирующие анти-RGM A-антитело гибридомы отбирают, клонируют и подвергают дополнительному скнринингу на желаемые характеристики, включающие сильный рост гибридом, высокое продуцирование антител и желаемые характеристики антител, обсуждаемые дополнительно ниже. Гибридомы могут культивироваться и размножаться in vivo в сингенных животных, в животных, которые лишены иммунной системы, например, “голых” мышах, или в культуре клеток in vitro. Способы отбора, клонирования и размножения гибридом хорошо известны квалифицированным в данной области специалистам.
В предпочтительном варианте осуществления, гибридомы являются гибридомами мыши, как описано выше. В другом предпочтительном варианте осуществления, гибридомы получают в видах, иных чем человек и мышь, таких как крысы, овцы, свиньи, козы, крупный рогатый скот или лошади. В другом варианте осуществления, гибридомы являются гибридомами человека, в которых несекретирующая миелома человека слита с клеткой человека, экспрессирующей анти-RGM A-антитело.
Фрагменты антител, которые узнают специфические эпитопы, могут быть генерированы известными способами. Например, фрагменты Fab и F(ab')2 по изобретению могут быть получены протеолитическим расщеплением молекул иммуноглобулина с использованием ферментов, таких как папаин (для получения Fab-фрагментов) или пепсин (для получения F(ab')2-фрагментов). F(ab')2-фрагменты содержат вариабельную область, константную область легкой цепи и СН1-домен тяжелой цепи.
3.3. Анти-RGM A-моноклональные антитела, полученные с использованием SLAM
В другом аспекте изобретения, рекомбинантные антитела генерируют из отдельных выделенных лимфоцитов с использованием процедуры, называемой в данной области способом с отобранными на антитела лимфоцитами (SLAM), описанным в Патенте США № 5627052, PCT Publication WO 92/02551 и Babcock, J.S. et al. (1996) Proc. Natl. Acad. Sci USA 93:7843-7848. В этом способе, отдельные клетки, секретирующие представляющие интерес антитела, например, лимфоциты, полученные из любого из описанных выше иммунизированных животных, подвергают скринингу с использованием антигенспецифическиого гемолитического анализа бляшек, где антиген RGM A, субъединицу RGM A или его фрагмент связывают с эритроцитами овцы при помощи линкера, такого как биотин, и используют для идентификации отдельных клеток, которые секретируют антитела, специфические в отношении RGM A. После идентификации представляющих интерес секретирующих антитело клеток кДНК вариабельной области тяжелой цепи и легкой цепи высвобождают из этих клеток при помощи ПЦР с обратной транскриптазой и эти вариабельные области могут быть затем экспрессированы, в контексте подходящих константных областей иммуноглобулина (например, константных областей иммуноглобулина человека), в клетках-хозяевах млекопитающего, таких как клетки COS или CHO. Клетки-хозяева, трансфицированные этими амплифицированными последовательностями иммуноглобулина, полученными из отобранных in vivo лимфоцитов, могут быть затем подвергнуты дополнительному анализу и отбору in vitro, например, посредством пэннинга трансфицированных клеток для выделения клеток, экспрессирующих антитела к RGM A. Эти амплифицированные последовательности иммуноглобулина могут быть затем подвергнуты манипуляциям in vitro, таким как способы созревания аффинности in vitro, такие как способы, описанные в PCT Publication WO 97/29131 и PCT Publication WO 00/56772.
3.4. Моноклональные анти-RGM A-антитела, полученные с использованием трансгенных животных
В другом варианте осуществления изобретения, антитела получают иммунизацией животного (не человека), содержащего несколько локусов или все локусы иммуноглобулина человека, антигеном RGM A. В предпочтительном варианте осуществления, этим животным (не человеком) является трансгенная мышь XENOMOUSE, полученный генной инженерией штамм мыши, который содержит большие фрагменты локусов иммуноглобулина человека и является дефектным в продуцировании антител мыши. См., например, Green et al. Nature Genetics 7:13-21 (1994) и Патенты США № 5916771, 5939598, 5985615, 5998209, 6075181, 6091001, 6114598 и 6130364. См., также WO 91/10741, опубликованный 25 июля 1991 года, WO 94/02602, опубликованный 3 февраля 1994 года, WO 96/34096 и WO 96/33735, оба опубликованные 31 октября 1996 года, WO 98/16654, опубликованный 23 апреля 1998 года, WO 98/24893, опубликованный 11 июня 1998 года, WO 98/50433, опубликованный 12 ноября 1998 года, WO 99/45031, опубликованный 10 сентября 1999 года, WO 99/53049, опубликованный 21 октября 1999 года, WO 00 09560, опубликованный 24 февраля 2000 года и WO 00/037504, опубликованный 29 июня 2000 года. Трансгенная мышь XENOMOUSE продуцирует подобный репертуару взрослого человека репертуар полностью человеческих антител и генерирует антигенспецифические mAb человека. Трансгенная мышь XENOMOUSE содержит приблизительно 80% репертуара антител человека посредством введения в размере мега п.н. YAC-фрагментов локусов тяжелой цепи человека х локусов легкой цепи конфигурации зародышевой линии. См. Mendez et al., Nature Genetics 15:146-156 (1997), Green and Jakobovits J. Exp. Med. 188:483-495 (1998), описания которых включены в настоящее описание посредством ссылки.
3.5. Моноклональные анти-RGM A-антитела, полученные с использованием библиотек рекомбинантных антител
Способы in vitro могут быть также использованы для получения антител по изобретению, в котором библиотеку антител подвергают скринингу для идентификации антитела, имеющего желаемую специфичность связывания. Способы для такого скрининга библиотек рекомбинантных антител хорошо известны в данной области и включают способы, описанные, например, в Ladner et al. Патенте США № 5223409; Kang et al. Публикации PCT № WO 92/18619; Dower et al. Публикации PCT № WO 91/17271; Winter et al. Публикации № WO 92/20791; Markland et al. Публикации PCT № WO 92/15679; Breitling et al. Публикации PCT № WO 93/01288; McCafferty et al. Публикации PCT № WO 92/01047; Garrard et al. Публикации PCT № WO 92/09690; Fuchs et al. (1991) Bio/Technology 9:1370-1372; Hay et al. (1992) Hum Antibod Hybridomas 3:81-85; Huse et al. (1989) Science 246:1275-1281; McCafferty et al., Nature (1990) 348:552-554; Griffiths et al. (1993) EMBO J 12:725-734; Hawkins et al. (1992) J Mol Biol 226:889-896; Clackson et al. (1991) Nature 352:624-628; Gram et al. (1992) PNAS 89:3576-3580; Garrad et al. (1991) Bio/Technology 9:1373-1377; Hoogenboom et al. (1991) Nuc Acid Res 19:4133-4137; и Barbas et al. (1991) PNAS 88:7978-7982, Публикации заявки на патент США 20030186374 и Публикации РСТ № WO 97/29131, содержание которых включено в настоящее описание посредством ссылки.
Библиотека рекомбинантных антител может быть получена у индивида, иммунизированного RGM A или частью RGM A. Альтернативно, библиотека рекомбинантных антител может происходить от необработанного индивида, т.е. индивида, который не был иммунизирован RGM A, например, библиотека антител человека из индивида-человека, который не был иммунизирован RGM A человека. Антитела по изобретению выбирают скринингом библиотеки рекомбинантных антител пептидом, содержащим RGM A человека, для отбора посредством этого антител, которые узнают RGM A. Способы проведения такого скрининга и отбора хорошо известны в данной области, такие как способы, описанные в ссылках в предыдущем абзаце. Для отбора антител по изобретению, имеющих конкретные аффинности связывания в отношении hRGM A, таких как антитела, которые диссоциируются от RGM A человека с конкретной константой скоростью koff, может быть использован известный в данной области способ резонанса поверхностных плазмонов для отбора антител, имеющих желаемую константу скорости koff. Для отбора антител по изобретению, имеющих конкретную нейтрализующую активность в отношении hRGM A, таких как антитела с конкретной IC50, могут быть использованы стандартные способы, известные в данной области, для оценки ингибирования активности hRGM A.
В одном аспекте, это изобретение относится к выделенному антителу, или его антигенсвязывающей части, которые связывают RGM A человека. Предпочтительно, антитело является нейтрализующим антителом. В различных вариантах осуществления, антитело является рекомбинантным антителом или моноклональным антителом.
Например, антитела по данному изобретению могут быть также генерированы с использованием различных способов фагового дисплея, известных в данной области. В способах фагового дисплея, функциональные домены антитела представлены на поверхности фаговых частиц, которые несут кодирующие их полинуклеотидные последовательности. В частности, такой фаг может быть использован для представления антигенсвязывающих доменов, экспрессируемых из репертуара или комбинированной библиотеки антител (например, человека или мыши). Фаг, экспрессирующий антигенсвязывающий домен, который связывает представляющий интерес антиген, может быть отобран или идентифицирован с использованием антигена, например, с использованием меченого антигена или антигена, связанного с твердой поверхностью или гранулой или прикрепленного к твердой поверхности или грануле. Фаг, используемый в этих способах, является обычно нитчатым фагом, включающим связывающие домены fd и M13, экспрессируемые из фага, с доменами Fab, Fv или стабилизированного дисульфидом Fv антитела, слитыми с белком либо гена III, либо гена VIII фага. Примеры способов фагового дисплея, которые могут быть использованы для получения антител данного изобретения, включают примеры способов, описанные в Brinkman et al., J. Immunol. Methods 182:41-50 (1995); Ames et al., J. Immunol. Methods 184:177-186 (1995); Kettleborough et al., Eur. J. Immunol. 24:952-958 (1994); Persic et al., Gene 187 9-18 (1997); Burton et al., Advances in Immunology 57:191-280 (1994); Публикации PCT № PCT/GB91/01134; Публикациях PCT № WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и Патентах США № 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743 и 5969108; указанные ссылки включены в настоящее описание посредством ссылки в их полном объеме.
Как описано в приведенных выше ссылках, после отбора фага, кодирующие области антитела из фага могут быть выделены и использованы для генерирования полноразмерных антител, в том числе антител человека или любого другого желаемого антигенсвязывающего фрагмента, и экспрессированы в любом желаемом хозяине, в том числе клетках млекопитающего, клетках насекомого, клетках растения, дрожжах и бактериях, например, как описано более подробно ниже. Например, могут быть также использованы способы рекомбинантого получения фрагментов Fab, Fab' и F(ab')2, с использованием способов, известных в данной области, таких как способы, описанные в Публикации PCT WO 92/22324; Mullinax et al., BioTechniques 12(6):864-869 (1992); и Sawai et al., AJRI 34:26-34 (1995); и Better et al., Science 240:1041-1043 (1988) (указанные ссылки включены в настоящее описание посредством ссылки в их полном объеме). Примеры способов, которые могут быть использованы для получения одноцепочечных Fv и антител, включают способы, описанные в Патентах США 4946778 и 5258498; Huston et al., Methods in Enzymology 203:46-88 (1991); Shu et al., PNAS 90:7995-7999 (1993) и Skerra et al., Science 240:1038-1040 (1988).
Альтернативой скринингу библиотек рекомбинантных антител посредством фагового дисплея, другие методологии, известные в данной области для скрининга больших комбинаторных библиотек, могут быть применены к идентификации антител по изобретению с двойной специфичностью. Одним типом альтернативной системы экспрессии является тип, в котором библиотеку рекомбинантных антител экспрессируют в виде слияний РНК-белок, как описано в Публикации РСТ № WO 98/31700 (Szostak and Roberts) и в Roberts, R.W. and Szostak, J.W. (1997) Proc. Natl. Acad. Sci. USA 94:12297-12302. В этой системе, создается ковалентное слияние между мРНК и пептидом или белком, который она кодирует, посредством in vitro трансляции синтетических мРНК, которые несут пуромицин, пептидил-акцепторный антибиотик, на их 3'-конце. Таким образом, специфическая мРНК может быть обогащена из комплексной смеси мРНК (например, комбинаторной библиотеки) на основе свойств кодируемого пептида или белка, например, антитела или его части, таких как связывание этого антитела или его части с антигеном двойной специфичности. Последовательности нуклеиновых кислот, кодирующие антитела или их части, извлеченные из скрининга таких библиотек, могут быть экспрессированы рекомбинантными способами, как описано выше (например, в клетках млекопитающего-хозяина), и, кроме того, могут быть подвергнуты дополнительному созреванию аффинности либо дополнительными раундами скрининга слияний мРНК-пептид, который были введены мутации в исходно выбранную последовательность (исходно выбранные последовательности), либо другими способами созревания аффинности in vitro рекомбинантных антител, описанными выше.
В другом подходе антитела данного изобретения могут быть также генерированы с использованием способов дрожжевого дисплея, известных в данной области. В способах дрожжевого дисплея, используют генетические способы для прикрепления доменов антител к клеточной стенке дрожжей и представления их на поверхности дрожжей. В частности, такие дрожжи могут быть использованы для представления антигенсвязывающих доменов, экспрессируемых из репертуара или комбинаторной библиотеки антител (например, человека или мыши). Примеры способов дрожжевого дисплея, которые могут быть использованы для получения антител данного изобретения, включают способы, описанные Wittrup, et al. Патент США № 6699658, включенный в настоящее описание посредством ссылки.
4. Получение конкретных рекомбинантных RGM A-антител по изобретению
Антитела данного изобретения могут быть получены любым из ряда способов, известных в данной области. Например, может быть использована экспрессия из клеток-хозяев, где экспрессирующий вектор (экспрессирующие векторы), кодирующий тяжелую и легкую цепи, трансфицируют в клетку-хозяина стандартными способами. Предполагается, что различные формы термина "трансфекция" включают большое разнообразие способов, обычно используемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, осаждения фосфатом кальция, трансфекцию с использованием ДЭАЭ-декстрана и т.п. Хотя можно экспрессировать антитела по изобретению в любых прокариотических или эукариотических клетках, предпочтительной является экспрессия антител в эукариотических клетках-хозяевах и наиболее предпочтительной является экспрессия в клетках-хозяевах млекопитающих, так как такие эукариотические клетки (и, в частности, клетки млекопитающих) с большей вероятностью, чем прокариотические клетки, собирают и секретируют правильно уложенное и иммунологически активное антитело.
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по изобретению включают клетки яичника китайского хомячка (клетки СНО) (в том числе клетки dhfr-CHO, описанные в Urlaub and Chasin, (1980) Proc. Natl. Acad. Sci USA 77:4216-4220, используемые с DHFR-селектируемым маркером, например, как описано в R.J. Kaufman and P.A. Sharp (1982) Mol. Biol. 159:601-621), клетки миеломы NSO, клетки COS и клетки SP2. При введении рекомбинантных экспрессирующих векторов, кодирующих гены антител, в клетки-хозяева млекопитающих, антитела получают культивированием клеток-хозяев в течение периода времени, достаточного для возможности экспрессии антитела в клетках-хозяевах, или, более предпочтительно, секреции антитела в культуральную среду, в которой выращиваются клетки-хозяева. Антитела могут быть извлечены из культуральной среды с использованием стандартных способов очистки белка.
Клетки-хозяева могут быть также использованы для получения функциональных фрагментов антител, таких как Fab-фрагменты или молекулы scFv. Будет понятно, что вариации вышеописанной процедуры находятся в объеме данного изобретения. Например, может быть желательной трансфекция клетки-хозяина ДНК, кодирующей функциональные фрагменты или легкой цепи, и/или тяжелой цепи антитела по изобретению. Технология рекомбинантных ДНК может быть также использована для удаления некоторой части или всей ДНК, кодирующей любую из легкой цепи и тяжелой цепи или обе эти цепи, которая не является обязательной для связывания представляющих интерес антигенов. Молекулы, экспрессируемые из таких укороченных молекул ДНК, также включены в антитела по изобретению. Кроме того, могут быть получены бифункциональные антитела, в которых одна тяжелая цепь и одна легкая цепь являются цепями антитела по изобретению, а другая тяжелая цепь и другая легкая цепь являются специфическими в отношении антигена, другого, чем представляющие интерес антигены, в результате сшивания антитела по изобретению со вторым антителом стандартными химическими способами сшивания.
В одной предпочтительной системе для рекомбинантной экспрессии антитела или его антигенсвязывающей части по изобретению, рекомбинантный экспрессирующий вектор, кодирующий как тяжелую цепь антитела, так и легкую цепь антитела, вводят в клетки dhfr-CHO опосредуемой фосфатом кальция трансфекцией. В этом рекомбинантном экспрессирующем векторе каждый из генов тяжелой и легкой цепи антитела функционально связан с регуляторными элементами энхансер CMV/промотор AdMLP для запуска высоких уровней транскрипции этих генов. Этот рекомбинантный экспрессирующий вектор несет также ген DHFR, который позволяет производить отбор клеток CHO, которые были трансфицированы этим вектором, с использованием метотрексатного отбора/амплификации. Отобранные трансформированные клетки-хозяева культивируют для возможности экспрессии тяжелой и легкой цепей этого антитела и из культуральной среды извлекают интактное антитело. Для получения рекомбинантного экспрессирующего вектора, трансфекции клеток-хозяев, отбора на трансформанты, культивирования клеток-хозяев и извлечения этого антитела из культуральной среды используют стандартные способы молекулярной биологии. Кроме того, это изобретение обеспечивает способ синтеза рекомбинантного антитела по изобретению культивированием клетки-хозяина по изобретению в подходящей культуральной среде, пока не будет синтезировано рекомбинантное антитело по изобретению. Способ может дополнительно предусматривать выделение этого рекомбинантного антитела из культуральной среды.
4.1. Анти-RGM A-антитела
Таблица 5 является списком аминокислотных последовательностей областей VH и VL предпочтительных анти-hRGM A-антител по изобретению.
СПИСОК АМИНОКИСЛОТНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ОБЛАСТЕЙ VH И VL АНТИ-hRGM A-АНТИТЕЛ 5F9 И 8D1
31-35
SEQ ID NO:34
50-66
SEQ ID NO:34
99-104
SEQ ID NO:34
24-39
SEQ ID NO:10
55-61
SEQ ID NO:10
94-102
SEQ ID NO:10
31-35
SEQ ID NO:55
50-66
SEQ ID NO:55
97-110
SEQ ID NO:55
24-34
SEQ ID NO:56
50-56
SEQ ID NO:56
89-97
SEQ ID NO:56
Приведенные выше выделенные CDR-последовательности анти-RGM A-антитела образуют новое семейство RGM A-связывающих белков, выделенных в соответствии с этим изобретением. Для генерирования и для отбора CDR по изобретению, имеющих предпочтительную RGM A-связывающую и/или нейтрализующую активность в отношении hRGM A, могут быть использованы стандартные способы, известные в данной области для генерирования связывающих белков данного изобретения и оценки связывания RGM A и/или нейтрализующих характеристик этого связывающего белка, в том числе, но не только, способы, конкретно описанные в настоящем описании.
4.2. Химерные анти-RGM A-антитела
Химерное антитело является молекулой, в которой различные части антитела получены из разных видов животных, такой как антитела, имеющие вариабельную область, полученную из моноклонального антитела мыши, и константная область иммуноглобулина человека. Способы получения химерных антител известны в данной области. См., например, Morrison, Science 229:1202 (1985); Oi et al., BioTechniques 4:214 (1986); Gillies et al., (1989) J. Immunol. Methods 125:191-202; Патенты США № 5807715; 4816567 и 4816397, которые включены в настоящее описание посредством ссылки в их полном объеме. Кроме того, могут быть использованы способы, разработанные для получения "химерных антител" (Morrison et al., 1984, Proc. Natl. Acad. Sci. 81:851-855; Neuberger et al., 1984, Nature 312:604-608; Takeda et al., 1985, Nature 314:452-454, которые включены в настоящее описание посредством ссылки в их полном объеме) сплайсингом генов из молекулы антитела мыши подходящей антигенной специфичности вместе с генами из молекулы антитела человека подходящей биологической активности.
В одном варианте осуществления, химерные антитела по изобретению получают заменой константной области тяжелой цепи моноклональных антител мыши против RGM A антител человека, описанных в настоящем описании, константной областью IgG1 человека.
4.3. CDR-привитые анти-RGM A-антитела
CDR-привитые антитела по изобретению содержат последовательности вариабельной области тяжелой и легкой цепи антитела человека, в которых одна или несколько CDR-областей VH и/или VL заменены CDR-последовательностями антител не человека, например, антител мыши по изобретению. Каркасная последовательность из любого антитела человека может служить в качестве шаблона для трансплантации CDR. Однако, прямая замена цепи на таком каркасном участке часто приводит к некоторой потере связывающей аффинности в отношении этого антигена. Чем более гомологичным антитело человека является в отношении исходного антитела мыши, тем менее вероятной является возможность того, что объединение CDR мыши с каркасным участком человека будет вводить нарушения в эти CDR, которые могут уменьшать аффинность. Таким образом, предпочтительно, вариабельный каркасный участок человека, который выбран для замены вариабельного каркасного участка мыши, имеет по меньшей мере 65% идентичность последовательности с вариабельным каркасным участком антитела мыши. Более предпочтительно, вариабельные области человека и мыши, отдельно от этих CDR, имеют по меньшей мере 70% идентичность последовательности. Даже еще более предпочтительно, вариабельные области человека и мыши, отдельно от этих CDR, имеют по меньшей мере 75% идентичность последовательности. Наиболее предпочтительно, вариабельные области человека и мыши, отдельно от этих CDR, имеют по меньшей мере 80% идентичность последовательности. Способы получения CDR-привитых антител известны в данной области (Jones et al., Nature 321:522-525 (1986); Патент США № 5225539). В одном конкретном варианте осуществления это изобретение обеспечивает CDR-привитые антитела с цепями VH и/или VL, описанными в таблице 6.
CDR-ПРИВИТЫЕ АНТИТЕЛА
(15)
(16)
(17)
(18)
(VH3-48/JH3 FR1)
(VH3-48/JH3 FR2)
(VH3-48/JH3 FR3)
(VH3-48/JH3 FR4)
(15)
(16)
(17)
(19)
(VH3-48/JH4 FR1)
(VH3-48/JH4 FR2)
(VH3-48/JH4 FR3)
(VH3-48/JH4 FR4)
(15)
(16)
(17)
(20)
(VH3-48/JH6 FR1)
(VH3-48/JH6 FR2)
(VH3-48/JH6 FR3)
(VH3-48/JH6 FR4)
(21)
(22)
(23)
(18)
(VH3-33/JH3 FR1)
(VH3-33/JH3 FR2)
(VH3-33/JH3 FR3)
(VH3-33/JH3 FR4)
(21)
(22)
(23) (19)
(VH3-33/JH4 FR1)
(VH3-33/JH4 FR2)
(VH3-33/JH4 FR3)
(VH3-33/JH4 FR4)
(21)
(22)
(23) (20)
(VH3-33/JH6 FR1)
(VH3-33/JH6 FR2)
(VH3-33/JH6 FR3)
(VH3-33/JH6 FR4)
(24)
(25)
(26)
(18)
(VH3-23/JH3 FR1)
(VH3-23/JH3 FR2)
(VH3-23/JH3 FR3)
(VH3-23/JH3 FR4)
(24)
(25)
(26) (19)
(VH3-23/JH4 FR1)
(VH3-23/JH4 FR2)
(VH3-23/JH4 FR3)
(VH3-23/JH4 FR4)
(24)
(25)
(26) (20)
(VH3-23/JH6 FR1)
(VH3-23/JH6 FR2)
(VH3-23/JH6 FR3)
(VH3-23/JH6 FR4)
(27)
(28)
(29)
(30)
(A18/JK2 FR1)
(A18/JK2 FR2)
(A18/JK2 FR3)
(A18/JK2 FR4)
(31)
(32)
(33)
(30)
(A17/JK2 FR1)
(A17/JK2 FR2)
(A17/JK2 FR3)
(A17/JK2 FR4)
(31)
(28)
(29)
(30)
(A17/JK2 FR1)
(A18/JK2 FR2)
(A18/JK2 FR3)
(A18/JK2 FR4)
CDR-последовательности, полученные из mAb 5F9 указаны жирным шрифтом, делается также ссылка на конкретные каркасные последовательности (FR1-FR4) посредством указания соответствующих SEQ ID NO: (см. также таблицы 3 и 4)
4.4. Гуманизированные анти-RGM A-антитела
Гуманизированные антитела являются молекулами антител из антитела вида не человека, которое связывает желаемый антиген, имеющий одну или несколько определяющих комплементарность областей (CDR) из вида не человека и каркасные области из молекулы иммуноглобулина человека. Известные последовательности Ig человека описаны, например, в www.ncbi.nlm.nih.gov/entrez-/query.fcgi; www.atcc.org/phage/hdb.html; www.sciquest.com/; www.abcam.com/; www.antibodyresource.com/onlinecomp.html; www.public.iastate.edu/.about.pedro/research_tools.html; www.mgen.uni-heidelberg.de/SD/IT/IT.html; www.whfreeman.com/immunology/CH-05/kuby05.htm; www.library.thinkquest.org/12429/lmmune/Antibody.html; www.hhmi.org/grants/lectures/1996/vlab/; www.path.cam.ac.uk/.about.mrcZ/m-ikeimages.html; www.antibodyresource.com/; mcb.harvard.edu/BioLinks/lmmuno- logy.html.www.immunologylink.com/; pathbox.wustl.edu/.about.hcenter/index.-html; www.biotech.ufl.edu/.about.hcl/; www.pebio.com/pa/340913/340913.html-; www.nal.usda.gov/awic/pubs/antibody/; www.m.ehime-u.acjp/.about.yasuhito-/Elisa.html; www.biodesign.com/table.asp; www.icnet.uk/axp/facs/davies/lin-ks.html; www.biotech.ufl.edu/.about.fccl/protocol.html; www.isac-net.org/sites_geo.html; aximtl.imt.uni-marburg.de/.about.rek/AEP-Start.html; baserv.uci.kun.nl/.about.jraats/linksl.html; www.recab.uni- hd.de/immuno.bme.nwu.edu/; www.mrc-cpe.cam.ac.uk/imt-doc/pu- blic/INTRO.html; www.ibt.unam.mx/vir/V_mice.html; imgt.cnusc.fr:8104/; www.biochem.ucl.ac.uk/.about. martin/abs/index.html; antibody.bath.ac.uk/; abgen.cvm.tamu.edu/lab/wwwabgen.html; www.unizh.ch/.about.honegger/AHOsem-inar/Slide01.html; www.cryst.bbk.ac.uk/.about.ubcg07s/; www.nimr.mrc.ac.uk/CC/ccaewg/ccaewg.htm; www.path.cam.ac.uk/.about.mrc7/h-umanisation/TAHHP.html; www.ibt.unam.mx/vir/structure/stat_aim.html; www.biosci.missouri.edu/smithgp/index.html; www.cryst.bioc.cam. ac.uk/.about.fmolina/Web-pages/Pept/spottech.html; www.jerini.de/fr roducts.htm; www.patents.ibm.com/ibm.html. Kabat et al., Sequences of Proteins of Immunological Interest, U.S. Dept. Health (1983), каждая из которых включена полностью посредством ссылки. Такие вводимые последовательности могут быть использованы для уменьшения иммуногенности или уменьшения, увеличения или модификации связывания, аффинности, скорости ассоциации, скорости диссоциации, авидности, специфичности, времени полужизни или любой другой подходящей характеристики, как известно в данной области.
Каркасные остатки в каркасных областях человека могут быть заменены соответствующим остатком из антитела-донора CDR для изменения, предпочтительно улучшения, связывания антигена. Эти каркасные замены идентифицируют способами, хорошо известными в данной области, например, моделированием взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания антигена, и идентификацией необычных каркасных остатков в конкретных положениях. (См., например, Queen et al., Патент США № 5585089; Riechmann et al., Nature 332:323 (1988), которые включены в настоящее описание посредством ссылки в их полном объеме). Трехмерные модели иммуноглобулина обычно являются доступными и известны квалифицированным в данной области специалистам. Доступны компьютерные программы, которые иллюстрирут и показывают возможные трехмерные конформационные структуры отобранных кандидатных последовательностей иммуноглобулина. Исследование этих дисплеев позволяет анализировать вероятную роль этих остатков в функционировании такой кандидатной последовательности иммуноглобулина, т.е. анализировать остатки, которые влияют на способность этого кандидатного иммуноглобулина связывать его антиген. Таким путем, остатки FR могут быть отобраны из консенсусной и вводимой (импортируемой) последовательностей и объединены, так что получают желаемую характеристику антитела, такую как увеличенная аффинность в отношении антигена-мишени (антигенов-мишеней). Обычно, эти остатки CDR прямо и наиболее существенно участвуют во влиянии на связывание антигена. Антитела могут быть гуманизированы с использованием способов, известных в данной области, таких как, но ими не ограничиваясь, способы, описанные в Jones et al., Nature 321:522 (1986); Verhoeyen et al., Science 239:1534 (1988)), Sims et al., J. Immunol. 151: 2296 (1993); Chothia and Lesk, J. Mol. Biol. 196:901 (1987), Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285 (1992); Presta et al., J. Immunol. 151:2623 (1993), Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka et al., Protein Engineering 7(6):805-814 (1994); Roguska. et al., PNAS 91:969-973 (1994); Публикации PCT WO 91/09967, PCT/: US98/16280, US96/18978, US91/09630, US91/05939, US94/01234, GB89/01334, GB91/01134, GB92/01755; WO90/14443, WO90/14424, WO90/14430, EP 229246, EP 592106; EP 519596, EP 239400, Патенты США № 5565332, 5723323, 5976862, 5824514, 5817483, 5814476, 5763192, 5723323, 5766886, 5714352, 6204023, 6180370, 5693762, 5530101, 5585089, 5225539; 4816567, каждая из этих ссылок включена в настоящее описание посредством ссылки в полном объеме, включающем цитированные в них ссылки.
5. Дополнительные варианты антител по изобретению
5.1. Слитые антитела и иммуноадгезины
Данная заявка описывает также слитое антитело или иммуноадгезин, которое может быть получено и которое содержит все RGM A-антитело или часть RGM A-антитела данного изобретения, связанные с другим полипептидом. В некоторых вариантах осуществления, только вариабельная область RGM A-антитела связана с этим полипептидом. В других вариантах осуществления, VH-домен RGM A-антитела этой заявки связан с первым полипептидом, тогда как VL-домен антитела связан со вторым полипептидом, который ассоциируется с первым полипептидом таким образом, что это позволяет VH- и VL-доменам взаимодействовать друг с другом с образованием антигенсвязывающего сайта. В других вариантах осуществления, VH-домен отделен от VL-домена линкером, который позволяет VH- и VL-доменам взаимодействовать друг с другом (см. ниже в разделе под заголовком Одноцепочечные Антитела). Затем антитело VH-линкер-VL связывают с представляющим интерес полипептидом. Это слитое антитело применимо в направлении полипептида в клетку или ткань, которая экспрессирует RGM A. Представляющим интерес полипептидом может быть терапевтическое средство, такое как токсин, или может быть диагностическое средство, такое как фермент; который может быть легко визуализирован, например, такой как пероксидаза хрена. Кроме того, могут быть созданы слитые антитела, в которых два (или более) одноцепочечных антитела связаны друг с другом. Это применимо, если желательным является создание двухвалентного или поливалентного антитела на единой полипептидной цепи или если желательным является создание биспецифического антитела.
Один вариант обеспечивает меченый связывающий белок, в котором антитело или часть антитела данной заявки дериватизовано или связано с другой функциональной молекулой (например, другим пептидом или белком). Например, меченый связывающий белок данной заявки может быть получен функциональным связыванием антитела или части антитела данной заявки (химическим связыванием, генетическим слиянием, нековалентной связью или другим образом) с одной или несколькими другими молекулярными частицами, такими как нуклеиновая кислота, другое антитело (например, биспецифическое антитело или диатело), детектируемый агент, цитотоксический агент, фармацевтический агент и/или белок или пептид, который может опосредовать ассоциацию этого антитела или части антитела с другой молекулой (такой как область коры (сердцевины) стрептавидина или полигистидиновая метка)).
Применимые детектируемые агенты, которыми могут быть дериватизованы антитело или часть антитела данной заявки, включают флуоресцентные соединения. Примеры флуоресцентных детектируемых агентов включают флуоресцеин, флуоресцеинизотиоцианат, родамин, 5-диметиламин-1-нафталинсульфанилхлорид, фикоэритрин и т.п. Антитело может быть также дериватизовано детектируемыми ферментами, такими как щелочная фосфатаза, пероксидаза хрена, глюкозооксидаза и т.п. При дериватизации антитела детектируемым ферментом, его детектируют добавлением дополнительных реагентов, которые этот фермент использует для образования детектируемого продукта реакции. Например, когда присутствует пероксидаза хрена в качестве детектируемого агента, добавление пероксида водорода и диаминобензидина приводит к окрашенному продукту реакции, который является детектируемым. Антитело может быть также дериватизовано нуклеиновой кислотой, биотином и детектировано посредством опосредованного измерения связывания авидина или стрептавидина.
5.2. Одноцепочечные антитела
Данная заявка включает одноцепочечное антитело (scFv), которое связывает иммуногенный RGM A по изобретению. Для получения scFv VH- и V-кодирующую ДНК функционально связывают с ДНК, кодирующей гибкий линкер, например, кодирующую аминокислотную последовательность (Gly4-Ser), так что эти VH- и VL-последовательности могут экспрессироваться в виде смежного одноцепочечного белка, с VL- и VH-областями, соединенными этим гибким линкером (см., например, Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., 30 Nature (1990) 34 8:552-554). Это одноцепочечное антитело может быть моновалентным, если используют только единственные VH и VL, бивалентным, если используют два VH и VL, или поливалентным, если используют более двух VH и VL. Два из указанных scFv-фрагментов, связанные через линкер, называют "диателом", причем эта форма также включена в данное изобретение.
5.3. Биспецифические антитела
Данная заявка включает далее биспецифическое антитело или его антигенсвязывающий фрагмент, в котором одна специфичность является специфичностью в отношении иммуногенного RGM A-полипептида данной заявки. Например, может быть генерировано биспецифическое антитело, которое специфически связывается с иммуногенным полипептидом RGM A по изобретению через один связывающий домен и со второй молекулой через второй связывающий домен. Кроме того, может быть генерировано одноцепочечное антитело, содержащее более одного VH и VL, которое связывается специфически с иммуногенным полипептидом по изобретению и с другой молекулой, которая связана с ослаблением опосредованного миелином коллапса конуса роста и ингибированием роста нейритов (аксонов) и спрутинга (роста нервных волокон). Такие биспецифические антитела могут быть генерированы с использованием способов, которые хорошо известны, например, Fanger et al. Immunol Methods 4:72-81 (1994) и Wright and Harris, 20 (supra).
В некоторых вариантах осуществления, биспецифические антитела получают с использованием одного или нескольких вариабельных областей из антитела по изобретению. В другом варианте осуществления, это биспецифическое антитело получают с использованием одной или нескольких областей CDR из указанного антитела.
5.4. Дериватизованные и меченые антитела
Антитело или антигенсвязывающий фрагмент данной заявки могут быть дериватизованы или связаны с другой молекулой (например, другим пептидом или белком). Обычно, антитело или антигенсвязывающий фрагмент дериватизуют таким образом, что эта дериватизация или это мечение не оказывает негативное действие на связывание с иммуногенным полипептидом по изобретению.
Например, антитело или часть антитела данной заявки могут быть функционально связаны (химическим связыванием, генетическим слиянием, нековалентной связью или другим образом) с одной или несколькими другими молекулярными частицами, такими как нуклеиновая кислота, другое антитело (например, биспецифическое антитело или диатело), детектируемый агент, цитотоксический агент, фармацевтический агент и/или белок или пептид, который может опосредовать ассоциацию антитела или части антитела с другой молекулой (такой как область кора (сердцевины) стрептавидина или полигистидиновая метка)). Кроме того, антитело или его антигенсвязывающая часть могут быть частью более крупной молекулы иммуноадгезии, образованной ковалентной или нековалентной связью этого антитела или части антитела с одним или несколькими другими белками или пептидами. Примеры таких молекул иммуноадгезии включают использование сердцевинной области (кора) стрептавидина для получения тетрамерной молекулы scFv (Kipriyanov, S. M., et al. (1995) Human Antibodies and Hybridomas 6:93-101) и использование остатка цистеина, маркерного пептида и С-концевой полигистидиновой метки для получения бивалентных и биотинилированных молекул scFv (Kipriyanov, S.M., et al. (1994) Mol. Immunol. 31:1047-1058). Части антител, такие как Fab- и F(ab')2-фрагменты, могут быть получены из полноразмерных антител с использованием общепринятых способов, таких как расщепление папаином или пепсином, соответственно, полноразмерных антител. Кроме того, антитела, части антител и молекулы иммуноадгезии могут быть получены с использованием стандартных способов рекомбинантных ДНК.
Дериватизованное антитело может быть получено сшиванием двух или более антител (одного и того же типа или различных типов, например, для создания биспецифических антител). Подходящие сшивающие линкеры включают линкеры, которые являются гетеробифункциональными, имеющими две по-разному реакционноспособные группы, разделенные подходящим спейсером (например, сложным эфиром м-малеидобензоил-N-гидроксисукцинимида), или гомофункциональные линкеры (например, дисукцинимидилсуберат). Такие линкеры доступны из Pierce Chemical Company, Rockford, III.
Дериватизованное антитело может также быть меченым антителом. Например, агентами детектирования, которыми могут быть дериватизованы антитело или часть антитела по изобретению, являются флуоресцентные соединения, включающие флуоресцеин, флуоресцеинизотиоцианат, родамин, 5-диметиламин-1-нафталинсульфонилхлорид, фикоэритрин, лантанидные люминофоры и т.п. Антитело может быть также помечено ферментами, которые применимы для детектирования, такими пероксидаза хрена, галактозидаза, люцифераза, щелочная фосфатаза, глюкозооксидаза и т.п. В вариантах, которые метятся детектируемым ферментом, антитело детектируют добавлением дополнительных реагентов, которые этот фермент использует для образования детектируемого продукта реакции. Например, когда присутствует пероксидаза хрена в качестве детектируемого агента, добавляют пероксид водорода и диаминобензидин. Антитело может быть также помечено биотином и детектировано посредством опосредованного измерения связывания авидина или стрептавидина. Антитело может быть также помечено предварительно определенным полипептидным эпитопом, узнаваемым вторичным репортером (например, парой последовательностей лейциновой молнии, сайтами связывания для вторичных антител, металлсвязывающими доменами, эпитопными метками (тэгами)). RGM A-антитело или его антигенсвязывающий фрагмент могут быть также помечены радиоактивно меченой аминокислотой. Эта радиоактивная метка может быть использована как для диагностических, так и для терапевтических целей. Радиоактивно меченое RGM A-антитело может использоваться диагностически, например, для определения уровней рецептора RGM A у индивида. Кроме того, радиоактивно меченое RGM A-антитело может использоваться терапевтически для лечения повреждения спинного мозга.
Примеры меток для полипептидов включают, но не ограничиваются ими, следующие радиоизотопы или радионуклиды 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho, 153Sm. RGM A-антитело или его антигенсвязывающий фрагмент могут быть также дериватизованы химической группой, такой как полиэтиленгликоль (PEG), метильная или этильная группа или углеводная группа. Эти группы могут быть применимы для улучшения биологических характеристик антитела, например, для увеличения времени полужизни в сыворотке или для увеличения связывания ткани. Кроме того, метка для полипептидов может включать нуклеиновую кислоту, например, ДНК для детектирования при помощи ПЦР или увеличения экспрессии генов или siRNA для супрессии экспрессии генов в RGM A-несущих клетках или тканях.
Класс и подкласс RGM A-антител может быть определен любым способом, известным в данной области. Обычно, класс и подкласс антитела могут быть определены с использованием антител, которые являются специфическими в отношении конкретного класса и подкласса антитела. Такие антитела коммерчески доступны. Класс и подкласс антител могут быть определены при помощи ELISA, Вестерн-блоттинга, а также других способов. Альтернативно, класс и подкласс могут быть определены секвенированием всех или части константных доменов тяжелой и/или легкой цепей этих антител, сравнением их аминокислотных последовательносткй с известными аминокислотными последовательностями различных классов и подклассов иммуноглобулинов и определением класса и подкласса этих антител.
5.5. Иммуноглобулины с двойным вариабельным доменом
Связывающие белки или иммуноглобулины с двойным вариабельным доменом (DVD), используемые в настоящем описании, являются связывающими белками, которые содержат два или более антигенсвязывающих сайта и являются мультивалентными белками, например, мультивалентными связывающими белками, например, двухвалентными и тетравалентными. Термин "мультивалентный связывающий белок” используется в данной заявке для обозначения связывающего белка, содержащего два или более антигенсвязывающих сайтов. Этот мультивалентный связывающий белок предпочтительно конструируют таким образом, что он имеет два или более антигенсвязывающих сайтов и обычно является не встречающимся в природе антителом. Термин "мультиспецифический связывающий белок" относится к связывающему белку, способному связывать две или более мишеней, или не связывать. Такие DVD могут быть моноспецифическими, т.е. способными связывать один антиген, или мультиспецифическими, т.е. способными связывать два или более антигенов. Связывающие DVD-белки, содержащие два полипептида тяжелой цепи DVD и два полипептида легкой цепи DVD, называют DVD-Ig. Каждая половина DVD-Ig содержит DVD-полипептид тяжелой цепи и DVD-полипептид легкой цепи и два антигенсвязывающих сайта. Каждый связывающий сайт содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, т.е. имеет в целом 6 CDR, участвующих в связывании антигена, на один антигенсвязывающий сайт. Связывающие DVD-белки и способы получения связывающих DVD-белков описаны в Заявке на патент США № 11/507050 и включены в настоящее описание посредством ссылки. Предполагается, что настоящее изобретение включает связывающий DVD-белок, содержащий белки, способные связывать RGM A. Предпочтительно, этот связывающий DVD-белок способен связывать RGM A и вторую мишень. Эта вторая мишень выбрана из группы, состоящей из противовоспалительных MAB-активностей (IL-1, IL-6, IL-8, IL-11, IL-12, IL-17, IL-18, IL-23, TNF альфа/бета, IFN-бета, гамма, LIF, OSM, CNTF, PF-4, щелочного белка тромбоцитов (PBP), NAP-2, бета-TG, MIP-1, MCP2/3, RANTES, лимфотактина), опосредующих транспорт белков (рецептора инсулина, рецептора траснферрина, рецептора тромбина, рецептора лептина, рецептора LDL), других нейрорегенеративных MAB (NgR, Lingo, p75, CSPG (например, NG-2, нейрокана, бревикана, версикана, агрекана) гиалуроновой кислоты, mAG, тенасцина, NI-35, NI-250, IMP, перлекана, нейрокана, фосфакана, nogo-A, OMGP, Sema4D, Sema 3A, эфрина B3, эфрина A2, эфрина A5, MAG, EphA4, плексина B1, TROY, wnts, ryk rec, BMP-2, BMP-4, BMP-7), нейропротективных активностей MAB (EGF, EGFR, Sema 3), антиамилоидных бета-MAB (например, m266, 3D6 (бапинейцумаб), анти-глобулярных MAB 7C6), локализованных в ЦНС рецепторов и транспортеров (рецепторов серотонина, рецепторов допамина, DAT, Asc-1, GlyT1).
5.6. Антитела с двойной специфичностью
Данная заявка описывает также технологию "антител с двойной специфичностью". Антитела с двойной специфичностью могут служить в качестве агонистов, антагонистов или обоих в различных комбинациях. Антитела с двойной специфичностью являются антителами, в которых VH-цепь связывается с первым антигеном и VL-цепь связывается со вторым антигеном, как приведено в примерах WO2008082651.
5.7. Кристаллизуемые антитела
Другой вариант данной заявки обеспечивает кристаллизуемый связывающий белок. Термин "кристаллизуемые" относится в данном констексте к антителу или его антигенсвязывающей части, которые существуют в виде кристалла. Кристаллы являются одной формой твердого состояния вещества, которая отличается от других форм, таких как аморфное твердое состояние или жидкокристаллическое состояние. Кристаллы состоят из регулярных, повторяющихся трехмерных упорядоченных рядов атомов, ионов, молекул (например, белков, таких как антитела) или молекулярных комплексов (например, комплексов антиген/антитело). Эти трехмерные упорядоченные ряды выстроены в соответствии со специфическими математическими уравнениями, которые хорошо известны в данной области. Фундаментальная единица, или элементарное звено молекулы, которые повторяются в кристалле, называют асимметрическим звеном. Повторение асимметрического звена в кристалле в расположении, которое соответствует конкретной, хорошо определенной кристаллографической симметрии, обеспечивает “элементарную ячейку” кристалла. Повторение этой ячейки кристалла посредством регулярных трансляций (переносов) во всех трех направлениях обеспечивает кристалл. См. Giege, R. and Ducruix, A. Barrett, Crystallization of Nucleic Acids and Proteins, a Practical Approach, 2nd ed., pp. 201-16, Oxford University Press, New York, New York, (1999).
Предпочтительно настоящая заявка описывает кристаллы полноразмерных RGM A-антител и их фрагментов, описанных в настоящем описании, и готовые составы и композиции, содержащие такие кристаллы. В одном варианте осуществления, кристаллизованный связывающий белок имеет более высокое время полужизни in vivo, чем растворимая копия этого связывающего белка. В другом варианте осуществления этот связывающий белок сохраняет биологическую активность после кристаллизации.
Кристаллизуемый связывающий белок по изобретению может быть получен в соответствии со способами, известными в данной области, такими как способы, описанные в WO 02072636, включенном в настоящее описание посредством ссылки.
5.8. Гликозилированные антитела
Другой вариант осуществления по изобретению обеспечивает гликозилированный связывающий белок, где антитело или его антигенсвязывающая часть содержит один или несколько углеводных остатков. Продукция формирующегося in vivo белка может подвергаться дополнительному процессингу, известному как посттрансляционная модификация. В частности, сахарные (гликозильные) остатки могут добавляться ферментативно в процессе, известном как гликозилирование. Полученные белки, несущие ковалентно связанные олигосахаридные боковые цепи, известны как гликозилированные белки или гликопротеины. Антитела являются гликопротеинами с одним или несколькими углеводными остатками в Fc-домене, а также вариабельном домене. Углеводные остатки в Fc-домене оказывают важное действие на эффекторную функцию Fc-домена с минимальным действием на связывание антигена или время полужизни этого антитела (R. Jefferis, Biotechnol. Prog. 21 (2005), pp. 11-16). В противоположность этому, гликозилирование вариабельного домена может оказывать действие на антигенсвязывающую активность этого антитела. Гликозилирование в вариабельном домене может оказывать негативное действие на аффинность связывания антитела, вероятно, вследствие пространственного затруднения (стерического препятствия) (Co, M.S., et al., Mol. Immunol. (1993) 30:1361-1367), или приводить к увеличенной аффинности в отношении этого антигена (Wallick, S.C., et al., Exp. Med. (1988) 168:1099-1109; Wright, A., et al., EMBO J. (1991) 10:2717 2723).
Один аспект данного изобретения направлен на получение мутантов сайта гликозилирования, в которых был мутирован O- или N-связанный сайт гликозилирования связывающего белка. Квалифицированный в данной области специалист может генерировать такие мутанты с использованием стандартных хорошо известных технологий. Мутанты сайтов гликозилирования, которые сохраняют биологическую активность, но имеют увеличенную или уменьшенную связывающую активность, являются следующим объектом данного изобретения.
Еще в одном варианте осуществления, гликозилирование антитела или его антигенсвязывающей части по изобретению является модифицированным. Например, может быть получено агликозилированное антитело (т.е. антитело, лишенное гликозилирования). Гликозилирование может быть изменено, например, для увеличения аффинности этого антитела в отношении антигена. Такие углеводные модификации могут выполняться, например, изменением одного или нескольких сайтов гликозилирования в последовательности этого антитела. Например, могут быть получены одна или несколько аминокислотных замен, которые приводят к элиминации одного или нескольких сайтов гликозилирования вариабельной области для элиминации посредством этого гликозилирования в этом сайте. Такое агликозилирование может увеличивать аффинность этого антитела в отношении антигена. Такой подход описан более подробно в Публикации РСТ WO2003016466A2 и Патентах США № 5714350 и 6350861, все ссылки включены в настоящее описание посредством ссылки в полном объеме.
Дополнительно или альтернативно, может быть получено модифицированное антитело по изобретению, которое имеет измененный тип гликозилирования, которое является гипофукозилированным антителом, имеющим уменьшенное количество фукозильных остатков, или антителом, имеющим увеличенное количество делящих пополам структур GlcNAc. Было показано, что такие измененные распределения (паттерны) гликозилирования увеличивают ADCC-способность антител. Такие углеводные модификации могут осуществляться, например, экспрессией этого антитела в клетке-хозяине с измененным аппаратом гликозилирования. Клетки с измененным аппаратом гликозилирования были описаны в данной области и могут быть использованы в качестве клеток-хозяев для экспрессии в них рекомбинантных антител по изобретению для получения посредством этого антитела с измененным гликозилированием. См., например, Shields, R. L. et al. (2002) J. Biol. Chem. 277:26733-26740; Umana et al. (1999) Nat. Biotech. 17:176-1, а также European Patent No: EP 1176195; Публикации PCT WO 03/035835; WO 99/5434280, все эти ссылки включены в настоящее описание посредством ссылки в их полном объеме.
Гликозилирование белков зависит от аминокислотной последовательности представляющего интерес белка, а также от клетки-хозяина, в которой экспрессируется этот белок. Различные организмы могут продуцировать различные ферменты гликозилирования (например, гликозилтрансферазы и гликозидазы) и имеют различные доступные субстраты (нуклеотидсахара). Вследствие таких факторов, характер гликозилирования белков и состав гликозильных остатков могут различаться в зависимости от системы-хозяина, в которой экспрессируется конкретный белок. Гликозильные остатки, применимые в изобретении, могут включать, но не ограничиваются ими, глюкозу, галактозу, маннозу, фукозу, н-ацетилглюкозамин и сиаловую кислоту. Предпочтительно, этот гликозилированный связывающий белок содержит такие гликозильные остатки, что характер гликозилирования является характером гликозилирования, свойственным человеку.
Квалифицированному в данной области специалисту известно, что различающееся гликозилирование белков может приводить к различающимся характеристикам белков. Например, эффективность терапевтического белка, полученного из микроорганизма-хозяина, такого как дрожжи, и гликозилированного с использованием эндогенного пути дрожжей, может быть уменьшенным в сравнении с эффективностью того же самого белка, экспрессируемого в клетке млекопитающего, такой как клеточная линия СНО. Такие гликопротеины могут быть также иммуногенными в людях и обнаруживать уменьшенное время жизни in vivo после введения. Специфические рецепторы в людях и других животных могут узнавать специфические гликозильные остатки и стимулировать быстрый клиренс этого белка из кровотока. Другие негативные эффекты могут включать изменения в фолдинге белков, растворимости, чувствительности к протеазам, направленной миграции, транспорте, компартментализации, секреции, узнавании других белков или факторов, антигенности или аллергенности. Таким образом, практикующий врач может предпочесть терапевтический белок с конкретным составом и характером гликозилирования, например, составом и распределением гликозилирования, идентичным, или по меньшей мере сходным, с составом и распределением гликозилирования, получаемым в клетках человека или в видоспецифических клетках предполагаемого индивида-животного.
Экспрессия гликозилированных белков, отличных от гликозилированных белков клетки-хозяина, может быть достигнута генетической модификацией клетки-хозяина для экспрессии гетерологичных ферментов гликозилирования. При помощи способов, известных в данной области, практикующий врач может генерировать антитела или их антигенсвязывающие части, проявляющие гликозилирование белков человека. Например, штаммы дрожжей были генетически модифицированы для экспрессии не встречающихся в природе ферментов гликозилирования, так что гликозилированные белки (гликопротеины), продуцируемые этими штаммами дрожжей, обнаруживают гликозилирование белков, идентичное с гликозилированием белков клеток животных, в частности, клеток человека (Заявки на патент США 20040018590 и 20020137134 и Публикация PCT WO2005100584 A2).
Кроме того, квалифицированному в данной области специалисту будет понятно, что представляющий интерес белок может быть экспрессирован с использованием библиотеки клеток-хозяев, генетически сконструированной для экспрессии различных ферментов гликозилирования, так что клетки-хозяева, являющиеся членами этой библиотеки, продуцируют представляющий интерес белок с конкретными новыми распределениями гликозилирования. Предпочтительно, белок, имеющий конкретно отобранное новое распределение гликозилирования, проявляет улучшенные или измененные биологические свойства.
5.9. Антиидиотипические антитела
Кроме этих связывающих белков, настоящее изобретение также относится к антиидиотипическому (анти-Id) антителу, специфическому в отношении таких связывающих белков по изобретению. Анти-Id-антитело является антителом, которое узнает уникальные детерминанты, обычно связанные с антигенсвязывающей областью другого антитела. Это анти-Id может быть получено иммунизацией животного этим связывающим белком или его CDR-содержащей областью. Иммунизированное животное будет узнавать идиотипические детерминанты иммунизирующего антитела и отвечать на них и продуцировать анти-Id-антитело. Анти-Id-антитело может быть также использовано в качестве "иммуногена" для индукции иммунной реакции еще в одном животном с получением так называемого анти-анти-Id-антитела.
6. Применения антител
Вследствие их способности связываться с RGM A человека, нейтрализующие антитела данного изобретения, или их части, могут быть использованы для детектирования RGM A человека (например, в биологическом образце, таком как сыворотка или плазма), с использованием общепринятого иммуноанализа, такого как твердофазные иммуноферментные анализы (ELISA), радиоиммуноанализ (RIA) или иммуногистохимия ткани. Данная заявка обеспечивает способ детектирования RGM A человека в биологическом образце, предусматривающий контактирование биологического образца с антителом или частью антитела по изобретению и детектирование либо антитела (или части антитела), связанного с RGM A человека, либо несвязанного антитела (или части антитела) для детектирования посредством этого RGM A человека в биологическом образце. Это антитело прямо или опосредованно метят детектируемым веществом для облегчения детектирования связанного или несвязанного антитела. Подходящие детектируемые вещества включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества и радиоактивные вещества. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных веществ включают умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, дансилхлорид или фикоэритрин; примеры люминесцентного вещества включают люминол и примеры подходящего радиоактивного материала включают 3H, 14C, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho, 153Sm.
Антитела и части антител данной заявки предпочтительно способны нейтрализовать активность RGM A человека как in vitro, так и in vivo. Таким образом, такие антитела и части антител по изобретению могут быть использованы для ингибирования связывания RGM A с его рецептором Неогенином, с BMP-2 или BM-4, и, следовательно, ингибировать полученную активность.
В другом варианте осуществления, настоящая заявка обеспечивает способ уменьшения активности RGM A у индивида, преимущественно у индивида, страдающего от заболевания или нарушения, в котором полученная активность RGM A оказывает негативное воздействие. Данная заявка обеспечивает способы уменьшения активности RGM A у индивида, страдающего от такого заболевания или нарушения, предотвращением связывания RGM A и рецептора, Неогенина, и/или с BMP-2 и/или BM-4, посредством применения моноклональных антител данной заявки. Антитела данного изобретения, в частности, гуманизированные антитела, описанные в настоящем описании, могут быть введены индивиду-человеку для терапевтических целей. Кроме того, антитела данной заявки могут быть введены млекопитающему не человеку, экспрессирующему RGM A, с которым это антитело способно связываться, для ветеринарных целей или животному, используемому в качестве модели заболевания человека. Что касается последнего применения, такие животные-модели могут быть применимы для оценки терапевтической эффективности антител по изобретению (например, испытания доз и временных схем введения).
В контексте настоящего описания, термин "нарушение, в котором активность RGM A оказывает негативное воздействие", включает заболевания и другие нарушения, в которых, как было показано, присутствие RGM A или его полученная активность у индивида, страдающего от этого нарушения, является или предположительно является ответственной за патофизиологию этого нарушения, или является фактором, который способствует ухудшению этого нарушения. Таким образом, полагают, что нарушение, в котором активность RGM A оказывает негативное воздействие, является нарушением, в котором уменьшение активности RGM A ослабляет симптомы и/или прогрессирование этого нарушения. Не ограничивающие изобретение примеры нарушений, которые могут лечиться антителами по изобретению, включают нарушения, обсуждаемые в разделе ниже, относящемся к фармацевтическим композициям антител по изобретению.
Признается, что RGM A играет важную роль в патологии, ассоциированной с различными заболеваниями, в том числе неврологическими заболеваниями, ассоциированными с нейродегенерацией или ингибированием нейрорегенеративных процессов, приводящим к параличу. Она включает деменцию, сенильную деменцию, умеренное когнитивное расстройство, деменцию, связанную с болезнью Альцгеймера, хорею Хангтингтона, позднюю дискинезию, гиперкинезии, мании, болезнь Паркинсона, синдром Стила-Ричардсона, синдром Дауна, тяжелую псевдопаралитическую миастению, повреждение нерва, васкулярный амилоидоз, внутримозговое кровоизлияние I с амилоидозом, воспаление головного мозга, наследственную атаксию Фридрейха, острое нарушение с дезориентацией во времени и пространстве, глаукому, болезнь Альцгеймера, боковой амиотрофический склероз, повреждение плечевого сплетения, повреждение головного мозга, в том числе травматическое повреждение головного мозга, детский церебральный паралич, болезнь Гийена-Барре, лейкодистрофии, рассеянный склероз, поствакцинальный полиомиелит, Spina Bifida (расщелину позвоночника), повреждение спинного мозга, атрофию мыщц позвоночника, опухоли позвоночника, инсульт и поперечный миелит.
Кроме того, как обсуждалось ранее, между любыми из описанных выше партнеров могут использоваться DVD-иммуноглобулины, или антитела с двойной специфичностью. Такие препараты антител, как описанные выше, могут быть применимы для лечения болезни Альцгеймера, болезни Паркинсона, повреждения спинного мозга, травматического повреждения головного мозга, рассеянного склероза, повреждения периферических нервов, шизофрении, депрессии, тревоги, а также любой пластичности и роста нейритов и связанного с нейротоксичностью заболевания, цитируемого выше.
Антитела данной заявки могут также объединяться с пептидами, делая возможным трансмембранный перенос для включения нацеливания на внутриклеточные белки-мишени. Такие пептидные последовательности могут включать, но не ограничиваются ими, tat, antennapedia, полиаргинины, некоторые антимикробные пептиды. Такие пептиды могут делать возможным перенос через мембраны, в том числе клеточные плазматические мембраны, но также эпителиальные и эндотелиальные мембраны, в том числе гематоэнцефалический барьер, слизистую оболочку кишечника, мозговую оболочку и другие.
Антитело, или часть антитела данной заявки может также вводиться с одним или несколькими дополнительными агентами с малыми молекулами, применимыми в лечении нарушений, в которых участвует активность RGM A, как обсуждалось в предыдущих абзацах. Должно быть понятно, что антитела данной заявки или их антигенсвязывающие части могут быть использованы отдельно или в комбинации с дополнительным средством, например, терапевтическим средством, причем указанное дополнительное средство выбирается квалифицированным специалистом для предполагаемой для него цели. Например, этим дополнительным средством может быть терапевтическое средство, признаваемое в данной области в качестве средства, которое применимо для лечения заболевания или состояния, которое подлежит лечению антителом данного изобретения. Это дополнительное средство может быть также средством, которое придает выгодный признак этой терапевтической композиции, например, средством, которое влияет на вязкость этой композиции.
7. Фармацевтические композиции
Это изобретение обеспечивает также фармацевтические композиции, содержащие антитело или его антигенсвязывающую часть по изобретению и фармацевтически приемлемый носитель. Фармацевтические композиции, содержащие антитела по изобретению, применимы, но не ограничиваются ими, в диагностике, детектировании или мониторинге нарушения, в предотвращении, лечении, контролировании или уменьшении интенсивности нарушения или одного или нескольких его симптомов и/или в исследовании. В конкретном варианте осуществления, композиция содержит один или несколько антител по изобретению. В другом варианте осуществления, эта фармацевтическая композиция содержит одно или несколько антител по изобретению и одно или несколько профилактических или терапевтических средств, других, чем антитела по изобретению, для лечения нарушения, в котором активность RGM A оказывает негативное воздействие. Предпочтительно, известно, что эти профилактические или терапевтические средства являются применимыми или использовались или используются в настоящее время в предупреждении, лечении, контролировании или уменьшении интенсивности нарушения или одного или нескольких его симптомов. В соответствии с этими вариантами осуществления, эта композиция может дополнительно содержать носитель, разбавитель или эксципиент.
Антитела и части антител по изобретению могут быть включены в фармацевтические композиции, подходящие для введения индивиду. Обычно, эта фармацевтическая композиция содержит антитело или часть антитела по изобретению и фармацевтически приемлемый носитель. В контексте настоящего описания, термин "фармацевтически приемлемый носитель" включает любой или все из растворителей, диспергирующих сред, покрытий, антибактериальных и противогрибковых агентов, изотонических и задерживающих абсорбцию агентов и т.п., которые являются физиологически совместимыми. Примеры фармацевтически приемлемых носителей включают один или несколько компонентов из воды, солевого раствора, забуференного фосфатом солевого раствора, декстрозы, глицерина, этанола и т.п., а также их комбинаций. Во многих случаях будет предпочтительным включение изотонических агентов, например, сахаров, полиспиртов, таких как маннит, сорбит, или хлорид натрия, в этой композиции. Фармацевтически приемлемые носители могут дополнительно содержать минорные количества вспомогательных веществ, таких как увлажняющие или эмульгирующие агенты, консерванты или буферы, которые увеличивают срок хранения или эффективность этого антитела или части антитела.
Известны различные системы доставки, и они могут быть использованы для введения одного или нескольких антител по изобретению или комбинации одного или нескольких антител по изобретению и профилактического средства или терапевтического средства, применимого для предотвращения, контролирования, лечения или уменьшения интенсивности нарушения или одного или нескольких его симптомов, например, инкапсулирование в липосомах, микрочастицах, микрокапсулах, рекомбинантные клетки, способные экспрессировать антитело или фрагмент антитела, опосредованный рецептором эндоцитоз (см., например, Wu and Wu, J. Biol. Chem. 262:4429-4432 (1987)), конструирование нуклеиновой кислоты как части ретровирусного или другого вектора, и т.д. Способы введения профилактического или терапевтического агента по изобретению включают, но не ограничиваются ими, парентеральное введение (например, внутрикожное, внутримышечное, интраперитонеальное, внутривенное и подкожное), эпидуральное введение, внутриопухолевое введение, и введение в слизистую оболочку (например, введение в нос и в полость рта). Кроме того, может быть использовано легочное введение, например, с использованием ингалятора или распылителя и композиции с аэрозольным агентом. См, например, Патенты США № 6019968, 5985320, 5985309, 5934272, 5874064, 5855913, 5290540 и 4880078; и Публикации РСТ № WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903, все из которых включены в настоящее описание посредством ссылки в их полном объеме. В одном варианте осуществления, антитело по изобретению, комбинированное терапевтическое средство или композицию по изобретению вводят с использованием технологии легочной доставки лекарственного средства Alkermes AIR® (Alkermes, Inc., Cambridge, Mass.). В одном конкретном варианте осуществления, профилактические или терапевтические средства по изобретению вводят внутримышечно, внутривенно, внутрь опухоли, перорально, интраназально, легочным путем или подкожно. Эти профилактические или терапевтические средства могут вводиться любым удобным способом, например, инфузией или болюсной инъекцией, абсорбцией через эпителиальные или кожно-слизистые выстилки (например, через слизистую оболочку полости рта, ректально или через кишечную слизистую оболочку и т.д.) и могут вводиться вместе с другими биологически активными средствами. Введение может быть системным или местным.
В конкретном варианте осуществления, может быть желательным введение профилактических или терапевтических агентов по изобретению локально в зону, нуждающуюся в лечении; это может быть достигнуто, например, без ограничения, локальной инфузией, инъекцией или посредством имплантата, причем указанный имплантат является пористым или непористым материалом, включающим мембраны и матриксы, такие как мембраны из силастика, полимеры, волокнистые матриксы (например, Tisseel®) или коллагеновые матриксы. В одном варианте осуществления, эффективное количество одного или нескольких антител антагонистов по изобретению вводят локально в пораженную зону индивиду для предупреждения, лечения, контролирования и/или уменьшения интенсивности нарушения или его симптома. В другом варианте осуществления, эффективное количество одного или нескольких антител антагонистов по изобретению вводят локально в пораженную зону в комбинации с эффективным количеством одного или нескольких терапевтических средств (например, одного или нескольких профилактических или терапевтических средств), других, чем антитело по изобретению, для предупреждения, лечения, контролирования и/или уменьшения интенсивности нарушения или одного или нескольких его симптомов.
В другом варианте осуществления, это профилактическое или терапевтическое средство может быть доставлено в системе регулируемого или пролонгированного высвобождения. В одном варианте осуществления, для достижения регулируемого или пролонгированного высвобождения терапевтических средств по изобретению может быть использован насос (см. Langer, supra; Sefton, 1987, CRC Crit. Ref. Biomed. Eng. 14:20; Buchwald et al., 1980, Surgery 88:507; Saudek et al., 1989, N. Engl. J. Med. 321:574). В другом варианте осуществления, для достижения регулируемого или пролонгированного высвобождения терапевтических средств по изобретению могут быть использованы полимерные материалы (см., например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, FIa. (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, 1983, J. Macromol. Sci. Rev. Macromol. Chem. 23:61; см. также Levy et al., 1985, Science 228:190; During et al., 1989, Ann. Neurol. 25:351; Howard et al., 1989, J. Neurosurg. 71:105); Патенты США № 5679377; Патент США № 5916597; Патент США № 5912015; Патент США № 5989463; Патент США № 5128326; Публикацию PCT № WO 99/15154 и Публикацию PCT № WO 99/20253. Примеры полимеров, используемых в формах пролонгированного высвобождения, включают, но не ограничиваются ими, поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), поли(акриловую кислоту), сополимер(этилен-винилацетат), поли(метакриловую кислоту), полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поли(виниловый спирт), полиакриламид, поли(этиленгликоль), полилактиды (PLA), сополимеры (лактидов и гликолидов) (PLGA) и полиортоэфиры. В одном предпочтительном варианте осуществления, полимер, используемый в форме пролонгированного действия, является инертным, свободным от выщелачивающихся примесей, стабильным при хранении, стерильным и биодеградируемым. В другом варианте осуществления, система регулируемого или пролонгированного высвобождения может быть помещена вблизи профилактической или терапевтической мишени, что позволяет вводить только малую часть системной дозы (см., например, Goodson, in Medical Applications of Controlled Release, supra, vol. 2, pp. 115-138 (1984)).
Системы регулируемого высвобождения описаны в обзоре Langer (1990, Science 249:1527-1533). Любой способ, известный квалифицированному в данной области специалисту, может быть использован для получения композиции пролонгированного высвобождения, содержащий один или более терапевтических средств по изобретению. См. например, Патент США № 4526938, Публикацию PCT WO 91/05548, Публикацию PCT WO 96/20698, Ning et al., 1996, "Intratumoral Radioimmunotheraphy of a Human Colon Cancer Xenograft Using a Sustained-Release Gel", Radiotherapy & Oncology 39:179-189, Song et al., 1995, "Antibody Mediated Lung Targeting of Long-Circulating Emulsions", PDA Journal of Pharmaceutical Science & Technology 50:372-397, Cleek et al., 1997, "Biodegradable Polymeric Carriers for a bFGF Antibody for Cardiovascular Application", Pro. Int'l. Symp. Control. Rel. Bioact. Mater. 24:853-854 и Lam et al., 1997, "Microencapsulation of Recombinant Humanized Monoclonal Antibody for Local Delivery", Proc. Int'l. Symp. Control Rel. Bioact. Mater. 24:759-760, каждая из этих ссылок включена в настоящее описание посредством ссылки в ее полном объеме.
В одном конкретном варианте осуществления, где композиция по изобретению является нуклеиновой кислотой, кодирующей профилактическое или терапевтическое средство, нуклеиновая кислота может вводиться in vivo для стимуляции экспрессии кодируемого ею профилактического или терапевтического средства конструированием ее в виде части подходящего экспрессирующего вектора нуклеиновой кислоты и введением ее таким образом, что она становится внутриклеточной, например, с использованием ретровирусного вектора (см. Патент США №4980286), или прямой инъекцией или с использованием бомбардировки микрочастицами (например, генного пистолета; Biolistic, Dupont) или покрытия липидами или рецепторами поверхности клеток или трансфицирующими агентами или введением ее в связи с гомеобокс-подобным пептидом, который, как известно, входит в ядро (см. Joliot et al., 1991, Proc. Natl. Acad. Sci. USA 88:1864-1868). Альтернативно, нуклеиновая кислота может быть введена внутриклеточно и включена в ДНК клетки-хозяина для экспрессии гомологичной рекомбинацией.
Фармацевтическую композицию по изобретению готовят таким образом, что она совместима с предполагаемым способом введения. Примеры способов введения включают, но не ограничиваются ими, парентеральное, например, внутривенное, внутрикожное, подкожное, пероральное, интраназальное (например, ингаляцией), чрезкожное (например, местное), трансмукозное и ректальное введение. В одном конкретном варианте осуществления, эту композицию готовят в соответствии с рутинными процедурами в виде фармацевтической композиции, адаптированной для внутривенного, подкожного, внутримышечного, перорального, интраназального или местного введения людям. Обычно, композиции для внутривенного введения являются растворами в стерильном изотоническом водном буфере. В случае необходимости, эта композиция может также включать солюбилизирующий агент и местный анестетик, такой как лидокаин, для облегчения боли в месте инъекции.
Если композиции по изобретению должны вводиться местно, эти композиции могут быть приготовлены в форме мази, крема, трансдермального пластыря, лосьона, геля, шампуня, спрея, аэрозоля, раствора, эмульсии или другой формы, хорошо известной квалифицированному в данной области специалисту. См., например, Remington's Pharmaceutical Sciences and Introduction to Pharmaceutical Dosage Forms, 19th ed., Mack Pub. Co., Easton, Pa. (1995). Для непригодных для аэрозолей местных лекарственных форм, обычно используют вязкие - полутвердые или твердые формы, содержащие носитель или один или несколько эксципиентов, совместимых с местным применением, и имеющие динамическую вязкость, предпочтительно более высокую, чем вязкость воды. Подходящие готовые формы включают, без ограничения, растворы, суспензии, эмульсии, кремы, мази, порошки, линименты (жидкие мази), целебные мази и т.п., которые, если желательно, стерилизуют или смешивают с вспомогательными агентами (например, консервантами, стабилизаторами, увлажняющими агентами, буферами или солями) для влияния на различные свойства, такие как, например, осмотическое давление. Другие подходящие лекарственные формы для местного применения включают разбрызгиваемые аэрозольные препараты, в которых активный ингредиент, предпочтительно в комбинации с твердым или жидким инертным носителем, упакован в смеси с находящимся под давлением летучим (например, газообразным) пропеллентом, таким как фреон, или в сжимаемом флаконе. Если желательно, к фармацевтическим композициям и лекарственным формам могут быть также добавлены увлажнители или смачивающие средства. Примеры таких дополнительных ингредиентов хорошо известны в данной области.
Если способ по изобретению предусматривает интраназальное введение композиции, эта композиция может быть приготовлена в аэрозольной форме, в виде спрея, тумана или в форме капель. В частности, профилактические или терапевтические средства для применения в соответствии с данным изобретением могут удобным образом доставляться в форме предоставления аэрозольного спрея из находящихся под давлением упаковок или распылителя, с использованием подходящего пропеллента (например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа). В случае находящегося под давлением аэрозоля, единица дозы может быть определена обеспечением клапана для доставки отмеренного количества. Для применения в ингаляторе или инсуффляторе могут быть приготовлены капсулы и картриджи (кассеты) (состоящие, например, из желатина)), содержащие порошкообразную смесь рассматриваемого соединения и подходящей порошкообразной основы, такой как лактоза или крахмал.
Если способ по изобретению предусматривает пероральное введение, композиции могут быть приготовлены для перорального введения в форме таблеток, капсул, крахмальных облаток (капсул), гелевых желатиновых капсул, растворов, суспензий и т.п. Таблетки или капсулы могут быть приготовлены общепринятыми способами с фармацевтически приемлемыми эксципиентами, такими как связывающие агенты (например, предварительно клейстеризованный кукурузный крахмал, поливинилпирролидон или гидроксипропилметилцеллюлоза); наполнители (например, лактоза, микрокристаллическая целлюлоза или гидрофосфат кальция); смазывающие агенты (например, стеарат магния, тальк или диоксид кремния); дезинтегрирующие агенты (например, картофельный крахмал или гликолат натрий-крахмала) или увлажняющие агенты (например, лаурилсульфат натрия). Эти таблетки могут иметь покрытие, нанесенное способами, хорошо известными в данной области. Жидкие препараты для перорального введения могут иметь (но не ограничиваются ими) форму растворов, сиропов или суспензий, или они могут быть представлены в виде сухого продукта для восстановления водой или другим подходящим носителем перед использованием. Такие жидкие препараты могут быть приготовлены общепринятыми способами с фармацевтически приемлемыми добавками, такими как суспендирующие агенты (например, сироп из сорбита, производные целлюлозы или гидрогенизированные годные в пищу жиры); эмульгирующие агенты (например, лецитин или аравийская камедь); неводные носители (например, миндальное масло, масляные эфиры, этиловый спирт или фракционированные растительные масла) и консерванты (например, метил- или пропил-п-гидроксибензоаты или сорбиновая кислота). Эти препараты могут также содержать необязательно буферные соли, ароматизирующие, красящие и подслащивающие агенты. Препараты для перорального введения могут быть удобным образом приготовлены для медленного высвобождения, регулируемого высвобождения или пролонгированного высвобождения профилактического или терапевтического средства (профилактических или терапевтических средств).
Способ по изобретению может предусматривать легочное введение, например, с использованием ингалятора или распылителя, композиции, приготовленной с образующим аэрозоль агентом. См., например, Патенты США № 6019968, 5985320, 5985309, 5934272, 5874064, 5855913, 5290540 и 4880078; и Публикации PCT № WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903, каждая из этих ссылок включена в настоящее описание посредством ссылки в ее полном объеме. В одном конкретном варианте осуществления, антитело по изобретению, средство комбинированной терапии и/или композицию по изобретению вводят с использованием технологии легочной доставки лекарственного средства Alkermes AIR® (Alkermes, Inc., Cambridge, Mass.).
Способ по изобретению может предусматривать введение композиции, приготовленной для парентерального введения инъекцией (например, болюсной инъекцией или непрерывной инфузией). Композиции для инъекции могут быть приготовлены в форме унифицированной (стандартной) дозы (например, в ампулах или многодозовых контейнерах) с добавленным консервантом. Эти композиции могут иметь такие формы, как суспензии, растворы или эмульсии в масляных или водных носителях и могут содержать способствующие приготовлению агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, активный ингредиент может находиться в форме порошка для восстановления подходящим носителем (например, стерильной апирогенной водой) перед использованием. Способы по изобретению могут дополнительно предусматривать введение композиций, приготовленных в виде депо-препаратов. Такие формы продолжительного действия могут вводиться имплантацией (например, подкожно или внутримышечно) или внутримышечной инъекцией. Таким образом, эти композиции могут быть, например, приготовлены с подходящими полимерными или гидрофобными веществами (например, в виде эмульсии в приемлемом масле) или с ионообменными смолами или с умеренно растворимыми производными (например, с умеренно растворимой солью).
Способы по изобретению предусматривают введение композиций, приготовленных в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают соли, образованные с анионами, такими как анионы, полученные из хлористоводородной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образованные с катионами, такими как катионы, полученные из натрия, калия, аммония, кальция, гидроксидов трехвалентного железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и т.д.
Обычно, ингредиенты композиций поставляются либо раздельно, либо смешанными вместе в виде унифицированной (стандартной) лекарственной формы, например, в виде сухого лиофилизированного порошка, или в безводном коцентрате в герметичном контейнере, таком как ампула или пакетик, с указанием количества активного агента. Когда способом введения является инфузия, может быть обеспечена ампула стерильной воды для инъекции, так чтобы эти ингредиенты могли быть смешаны перед введением.
В частности, это изобретение обеспечивает также упаковку одного или нескольких профилактических или терапевтических средств или фармацевтических композиций по изобретению в герметичном контейнере, таком как ампула или пакетик, с указанием количества этого средства. В одном варианте осуществления, один или несколько профилактических или терапевтических средств или фармацевтических композиций по изобретению поставляются в виде сухого стерилизованного лиофилизированного порошка или безводного концентрата в герметичном контейнере и могут быть восстановлены (например, водой или солевым раствором) до подходящей концентрации для введения индивиду. Предпочтительно, одно или несколько профилактических или терапевтических средств или фармацевтических композиций по изобретению поставляются в виде сухого стерилизованного лиофилизированного порошка в герметичном контейнере при унифицированной дозе по меньшей мере 5 мг, более предпочтительно по меньшей мере 10 мг, по меньшей мере 15 мг, по меньшей мере 25 мг, по меньшей мере 35 мг, по меньшей мере 45 мг, по меньшей мере 50 мг, по меньшей мере 75 мг или по меньшей мере 100 мг. Эти лиофилизированные профилактические или терапевтические средства или фармацевтические композиции по изобретению должны храниться при 2°C - 8°C в их первоначальном контейнере и эти профилактические или терапевтические средства или фармацевтические композиции по изобретению должны вводиться в пределах 1 недели, предпочтительно в пределах 5 дней, в пределах 72 часов, в пределах 48 часов, в пределах 24 часов, в пределах 12 часов, в пределах 6 часов, в пределах 5 часов, в пределах 3 часов или в пределах 1 часа после их восстановления. В альтернативном варианте осуществления, одно или несколько профилактических или терапевтических средств или фармацевтических композиций по изобретению поставляются в жидкой форме в герметичном контейнере с указанием количества и концентрации этого средства. Предпочтительно, жидкая форма вводимой композиции поставляется в герметичном контейнере при по меньшей мере 0,25 мг/мл, более предпочтительно по меньшей мере 0,5 мг/мл, по меньшей мере 1 мг/мл, по меньшей мере 2,5 мг/мл, по меньшей мере 5 мг/мл, по меньшей мере 8 мг/мл, по меньшей мере 10 мг/мл, по меньшей мере 15 мг/мл, по меньшей мере 25 мг/мл, по меньшей мере 50 мг/мл, по меньшей мере 75 мг/мл или по меньшей мере 100 мг/мл. Эта жидкая форма должна храниться при 2°C - 8°C в ее первоначальном контейнере.
Антитела и части антител по изобретению могут быть включены в фармацевтическую композицию, подходящую для парентерального введения. Предпочтительно, эти антитела или части антител готовят в виде инъекционного раствора, содержащего 0,1-250 мг/мл антитела. Этот инъекционный раствор может состоять из жидкой или лиофилизированной дозированной лекарственной формы во флаконе из кварца или янтаря, ампуле или заранее наполненном шприце. Буфером может быть L-гистидин (1-50 мМ), оптимально 5-10 мМ, при рН 5,0-7,0 (оптимально рН 6,0). Другие подходящие буферы включают, но не ограничиваются ими, сукцинат натрия, цитрат натрия, фосфат натрия или фосфат калия. Для модификации токсичности этого раствора может быть использован хлорид натрия в концентрации 0-300 мМ (оптимально 150 мМ для жидкой лекарственной формы). Для лиофилизированной лекарственной формы могут быть включены криопротекторы, преимущественно 0-10% сахароза (оптимально 0,5-1,0%). Другие подходящие криопротекторы включают трегалозу и лактозу. Для лиофилизированной лекарственной формы могут быть включены наполнители, преимущественно 1-10% маннит (оптимально 2-4%). Как для жидкой, так и для лиофилизированной лекарственной формы могут быть использованы стабилизаторы, преимущественно 1-50 мМ L-метионин (оптимально 5-10 мМ). Другие подходящие наполнители включают глицин, аргинин, и могут быть включены в виде 0-0,05% полисорбата-80 (оптимально 0,005-0,01%). Дополнительные поверхностно-активные вещества включают, но не ограничиваются ими, полисорбат 20 и поверхностно-активные вещества BRIJ. Фармацевтическая композиция, содержащая антитела и части антител по изобретению, приготовленная в виде инъекционного раствора для парентерального введения, может дополнительно содержать агент, применимый в качестве адъюванта, такой как агенты, используемые для увеличения абсорбции или диспергирования терапевтического белка (например, антитела). Особенно применимым адъювантом является гиалуронидаза, такая как Hylenex® (рекомбинантная гиалуронидаза человека). Добавление гиалуронидазы в инъекционный раствор улучшает биодоступность для человека после парентерального введения, в частности, подкожного введения. Оно позволяет также использовать более высокие объемы для участков инъекции (т.е. превышающие 1 мл) с меньшей болью и меньшим дискомфортом и получать минимальное количество реакций в участке инъекции (см. WO2004078140, US2006104968, включенные в настоящее описание посредством ссылки).
Композиции по изобретению могут находиться в различных формах. Они включают, например, жидкие, полутвердые и твердые лекарственные формы, такие как жидкие растворы (например, инъекционные и инфузионные растворы), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения. Типичные предпочтительные композиции находятся в форме инъекционных или инфузионных растворов, таких как композиции, сходные с композициями, используемыми для пассивной иммунизации людей другими антителами. Предпочтительным способом введения является парентеральный способ (например, внутривенный, подкожный, внутрибрюшинный, внутримышечный). В одном предпочтительном варианте осуществления, это антитело вводят внутривенной инфузией или инъекцией. В другом предпочтительном варианте осуществления, это антитело вводят внутримышечным или подкожным введением.
Обычно терапевтические композиции должны быть стерильными и стабильными в условиях приготовления и хранения. Эта композиция может быть приготовлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Стерильные инъекционные растворы могут быть приготовлены включением этого активного соединения (т.е. антитела или части антитела) в требуемом количестве в подходящем растворителе с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, если требуется, с последующей стерилизацией при помощи стерилизационного фильтра. Обычно, дисперсии готовят включением этого активного соединения в стерильный носитель, который содержит базовую дисперсионную среду и требующиеся другие ингредиенты из ингредиентов, перечисленных выше. В случае стерильных, лиофилизированных порошков для приготовления стерильных инъекционных растворов, предпочтительными способами приготовления являются вакуумная сушка и распылительная сушка, которые дают порошок активного ингредиента плюс любой дополнительный желаемый ингредиент из его предварительно профильтрованного через стерилизационный фильтр раствора. Должная текучесть раствора может поддерживаться, например, при помощи покрытия, такого как лецитин, поддержанием требуемого размера частиц в случае дисперсии и при помощи поверхностно-активных веществ. Пролонгированная абсорбция инъецируемых композиций может осуществляться включением в эту композицию агента, который задерживает абсорбцию, например, моностеаратных солей и желатина.
Антитела и части антител данного изобретения могут вводиться различными способами, известными в данной области, хотя для многих терапевтических применений предпочтительным способом/методом введения является подкожная инъекция, внутривенная инъекция или инфузия. Как будет понятно квалифицированному в данной области специалисту, этот способ и/или метод введения будет варьироваться в зависимости от желаемых результатов. В некоторых вариантах осуществления, активное соединение может быть приготовлено с носителем, который будет защищать это соединение от быстрого высвобождения, например, в виде формы регулируемого высвобождения, включающей имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биодеградируемые, биосовместимые полимеры, такие как этилен-винилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие способы приготовления таких готовых форм запатентованы или обычно известны квалифицированным в данной области специалистам. См., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
В некоторых вариантах осуществления, антитело или часть антитела по изобретению может вводиться перорально, например, с инертным разбавителем или ассимилируемым съедобным носителем. Это соединение (и другие ингредиенты, если желательно) могут быть также заключено в желатиновую капсулу с твердой или мягкой оболочкой, спрессовано в таблетки или включено непосредственно в рацион индивида. Для перорального терапевтического введения, эти соединения могут быть включены с эксципиентами и использованы в форме пригодных для проглатывания таблеток, трансбуккальных таблеток, пастилок, капсул, эликсиров, суспензий, сиропов, облаток и т.п. Для введения соединения по изобретению другим способом, чем парентеральное введение, может быть необходимым покрытие этого соединения материалом для предотвращения его инактивации или совместное введение этого соединения с таким материалом.
В эти композиции могут быть также включены дополнительные активные соединения. В некоторых вариантах осуществления, антитело или часть антитела по изобретению готовят совместно и/или вводят совместно с одним или несколькими дополнительными терапевтическими средствами, которые применимы для лечения нарушений, в которых активность RGM A оказывает негативное воздействие. Например, анти-RGM A-антитело или часть антитела по изобретению может быть совместно приготовлено и/или совместно введено с одним или несколькими дополнительными антителами, которые связывают другие мишени (например, антителами, которые связывают цитокины или которые связывают молекулы клеточной поверхности). Кроме того, одно или несколько антител по изобретению могут быть использованы в комбинации с двумя или более из предыдущих терапевтических средств. Такие комбинированные терапии могут выгодным образом использовать более низкие дозы вводимых терапевтических средств, что позволяет избежать возможных токсичностей или осложнений, связанных с различными монотерапиями.
В некоторых вариантах осуществления, антитело к RGM A или его фрагмент связан с продлевающим время полужизни носителем, известным в данной области. Такие носители включают, но не ограничиваются ими, Fc-домен, полиэтиленгликоль и декстран. Такие носители описаны, например, в Заявке США № 09/428082 и опубликованной Заявке РСТ № WO 99/25044, которые включены в настоящее описание посредством ссылки для любой цели.
В одном конкретном варианте осуществления, вводят последовательности нуклеиновых кислот, содержащие нуклеотидные последовательности, кодирующие антитело по изобретению или другое профилактическое или терапевтическое средство по изобретению для лечения, предупреждения, контролирования или уменьшения интенсивности нарушения или его одного или нескольких симптомов посредством генной терапии. Генной терапией называют терапию, выполняемую введением индивиду экспрессируемой или способной экспрессироваться нуклеиновой кислоты. В этом варианте осуществления данного изобретения, эти нуклеиновые кислоты продуцируют кодируемое ими антитело или профилактическое или терапевтическое средство по изобретению, которое опосредует профилактическое или терапевтическое действие.
Любой из способов генной терапии, доступных в данной области, может быть использован в соответствии с данным изобретением. В отношении обзоров способов генной терапии, см. Goldspiel et al., 1993, Clinical Pharmacy 12:488-505; Wu and Wu, 1991, Biotherapy 3:87-95; Tolstoshev, 1993, Ann. Rev. Pharmacol. Toxicol. 32:573-596; Mulligan, Science 260:926-932 (1993); и Morgan and Anderson, 1993, Ann. Rev. Biochem. 62:191-217; May, 1993, TIBTECH 11(5):155-215. Способы, обычно известные в области технологии рекомбинантных ДНК, которые могут быть использованы, описаны в Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley &Sons, NY (1993) и Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY (1990). Подробное описание различных способов генной терапии описано в US20050042664 A1, который включен в настоящее описание посредством ссылки.
RGM A играет критическую роль в патологии, связанной с различными определенными выше заболеваниями. Было описано, что RGM A и белки RGM являются повышающим образом регулируемыми в участках повреждений у людей, страдающих от травматического повреждения головного мозга (Schwab et al., Arch. Neurol. 62:1561-8, 2005a), в участках полутени и кора поврежденного инсультом головного мозга человека (Schwab et al., Arch. Neurol. 62:1561-8, 2005a), в субстанции негра пациентов, страдающих от болезни Паркинсона (Bossers et al. Brain Pathology vol. 19:91-107, 2008). Таким образом, RGM A-антитела являются подходящими агентами для комбинированной терапии внутримозгового инсульта, травматического повреждения головного мозга, болезни Альцгеймера и других нейродегенеративных нарушений нервной системы человека. В случае пациентов с инсультом лечение в пределах первых трех часов состоит из доставки тканевого активатора плазминогена для лизиса сгустков крови (Liang et al. Arch. Neurol. 65:1429-33, 2008), и такое лечение могло бы в принципе комбинироваться с доставкой RGM A-антитела, которая предоставляет подход с использованием отличающегося лечения и гораздо более расширенного окна терапии. В случае болезни Альцгеймера, комбинирование лекарственных средств с RGM A-антителами возможно с одобренными усилителями познавательной способности, Донепезилом, Мемантином, и такой подход может существенным образом замедлять прогрессирующую невропатологию. Интраназальная доставка инсулина оказывает положительные действия на внимание и память (Hanson and Frey, BMC Neurosci. 9:S5, 2008) и является возможным способом введения для RGM A-антител, позволяющим обходить гематоэнцефалический барьер. У пациентов с болезнью Паркинсона (PD), существующее в настоящее время лечение основано главным образом на допаминергических агентах, подобных леводопа, пролекарству допамина (Khor and Hsu, Curr. Clin. Pharmacol. 2:234-43, 2007), ропинролу, неэрголиническому агонисту допамина (Jost et al. J. Neurol. 255 Suppl. 5:60-63, 2008), ингибиторам моноаминоксидазы В Расагилину и Селегилину (Elmer and Bertoni, Expert Opin. Pharmacother. 9:2759-72, 2008). Несмотря на их благоприятные действия в случае ранней и умеренной болезни Паркинсона (PD), ни одно из этих лекарственных средств не способно предупреждать прогрессирующую дегенерацию субстанции негра и ассоциированных субкортикальных и кортикальных участков головного мозга, и, следовательно, комбинированная терапия со стимулирующими регенерацию RGM A-антителами могла бы замедлять процесс этого заболевания.
Любое нейропротективное средство, будь то антиоксидант, акцепторы радикалов, противосудорожное лекарственное средство, такое как Фенитоин, или лекарственное средство Эритропоэтин, является подходящим для комбинированной терапии с про-регенеративными RGM A-антителами, расширяя посредством этого обычно очень короткое окно терапевтического лечения нейропротекторов.
Антитела и части антител по изобретению могут быть использованы для лечения людей, страдающих от таких заболеваний.
Должно быть понятно, что антитела по изобретению или их антигенсвязывающие части могут быть использованы отдельно или в комбинации с дополнительным средством, например, терапевтическим средством, причем указанное дополнительное средство выбирается квалифицированным специалистом для его предполагаемой цели. Например, дополнительным средством может быть терапевтическое средством, которое признано в данной области в качестве применимого средства для лечения заболевания или состояния, подлежащего лечению антителом данного изобретения. Этим дополнительным средством может быть также средство, которое придает предпочтительный признак этой терапевтической композиции, например, средство, которое действует на вязкость этой композиции.
Кроме того, должно быть понятно, что комбинациями, которые должны быть включены в это изобретение, является комбинации, применимые для предполагаемой для них цели. Агенты, представленные ниже, являются иллюстративными для такой цели, но не предназначены для ограничения. Комбинациями, которые являются частью по изобретению, могут быть антитела данного изобретения и по меньшей мере одно дополнительное средство, выбранное из списков, приведенных ниже. Комбинация может также включать более одного дополнительного средства, например, два или три дополнительных средств, если эта комбинация является такой, что может выполняться ее предполагаемая функция.
Неограничивающие примеры терапевтических средств для рассеянного склероза, с которыми антитело или часть антитела по изобретению могут быть объединены, включают следующее: кортикостероиды; преднизолон; метилпреднизолон; азатиоприн; циклофосфамид; циклоспорин; метотрексат; 4-аминопиридин; тизанидин; интерферон-β1a (AVONEX; Biogen); интерферон-β1b (BETASERON; Chiron/Berlex); интерферон α-n3) (Interferon Sciences/Fujimoto), интерферон-α (Alfa Wassermann/J&J), интерферон β1A-IF (Serono/lnhale Therapeutics), Пегинтерферон α2b (Enzon/Schering-Plough), Сополимер 1 (Cop-1; COPAXONE; Teva Pharmaceutical Industries, Inc.); гипербарический кислород; внутривенный иммуноглобулин; кладрибин; антитела к другим цитокинам человека, факторам роста и их рецепторам или антагонисты других цитокинов человека, факторов роста и их рецепторов, например, TNF, LT, IL-1, IL-2, IL-6, IL-7, IL-8, IL-23, IL-15, IL-16, IL-18, EMAP-II, GM-CSF, FGF и PDGF. Антитела по изобретению или их антигенсвязывающие части могут комбинироваться с антителами к молекулам клеточной поверхности, таким как CD2, CD3, CD4, CD8, CD19, CD20, CD25, CD28, CD30, CD40, CD45, CD69, CD80, CD86, CD90, или их лигандам. Антитела по изобретению или их антигенсвязывающие части могут также комбинироваться со средствами, такими как метотрексат, циклоспорин, FK506, рапамицин, микофенолат-мофетил, лефлуномид, NSAID, например, ибупрофен, кортикостероиды, такие как преднизолон, ингибиторы фосфодиэстеразы, агонисты аденозина, антитромботические агенты, ингибиторы комплемента, адренергические агенты, агенты, которые препятствуют передаче сигнала провоспалительных цитокинов, таких как TNFα или IL-1 (например, IRAK, NIK, IKK, p38 или ингибиторы MAP-киназы), ингибиторы IL-1β-превращающего фермента, ингибиторы TACE, ингибиторы передачи сигнала Т-клеток, такие как ингибиторы киназы, ингибиторы металлопротеиназы, сульфасалазин, азатиоприн, 6-меркаптопурины, ингибиторы превращающего ангиотензин фермента, растворимые рецепторы цитокинов и их производные (например, растворимые рецепторы p55 или p75 TNF, sIL-1RI, SIL-1RII, sIL-6R) и противовоспалительные цитокины (например, IL-4, IL-10, IL-13 и TGFβ).
Предпочтительные примеры терапевтических средств для рассеянного склероза, с которыми может комбинироваться это антитело или его антигенсвязывающая часть, включают интерферон-β, например, IFNβ1а и IFNβ1b; копаксон, кортикостероиды, ингибиторы каспазы, например, ингибиторы каспазы-1, ингибиторы IL-1, ингибиторы TNF и антитела к лиганду CD40 и CD80.
Антитела по изобретению или их антигенсвязывающие части могут также комбинироваться со средствами, такими как алемтуцумаб, дронабинол, Унимед, даклицумаб, митоксантрон, ксалипродена гидрохлорид, фампридин, глатирамера ацетат, наталицумаб, синнабидол, а-иммунокин NNSO3, ABR-215062, AnergiX.MS, антагонисты рецепторов хемокинов, BBR-2778, калаквалин, CPI-1189, LEM (инкапсулированный в липосоме митоксантрон), THCCBD (агонист каннабиноида) MBP-8298, мезопрам (ингибитор PDE4), MNA-715, анти-IL-6-рецептор-антитело, нейровакс, пирфенидон-аллотрап 1258 (RDP-1258), STNF-R1, талампанел, терифлуномид, TGF-бета2, типлимотид, антагонисты VLA-4 (например, TR-14035, VLA4 Ultrahaler, Antegran-ELAN/Biogen), антагонисты интерферона гамма, агонисты IL-4.
Фармацевтические композиции по изобретению могут включать "терапевтически эффективное количество" или "профилактически эффективное количество" антитела или части антитела по изобретению. "Терапевтически эффективным количеством" называют количество, эффективное, при необходимых дозах и в течение необходимых периодов времени, для достижения желаемого терапевтического результата. Терапевтически эффективное количество антитела или части антитела может быть определено квалифицированным в данной области специалистом и может варьироваться в соответствии с такими факторами, как состояние заболевания, возраст, пол и масса индивидуума и способность этого антитела или этой части антитела индуцировать желаемую реакцию в этом индивидууме. Терапевтически эффективным количеством является также количество, в котором терапевтически благоприятные действия превышают любые токсичные или вредные для здоровья действия этого антитела или части антитела. "Профилактически эффективным количеством" называют количество, эффективное, при необходимых дозах и в течение необходимых периодов времени, для достижения желаемого профилактического результата. Обычно, поскольку профилактическую дозу используют в индивидах до или на ранних стадиях заболевания, профилактически эффективное количество будет меньшим, чем терапевтически эффективное количество.
Схемы введения доз могут корректироваться для обеспечения оптимальной желаемой реакции (например, терапевтической или профилактической реакции). Например, может вводиться единственный болюс, могут вводиться несколько разделенных доз на протяжении времени или эта доза может быть пропорционально уменьшена или увеличена в случае острой необходимости терапевтической ситуации. Особенно предпочтительным является приготовление парентеральных композиций в форме дозы на один прием для легкости введения и однородности дозы. Формой унифицированной (стандартной) дозы (дозы на один прием) называют физически дискретные единицы, подходящие в качестве унифицированных (стандартных) доз для подлежащих лечению индивидов-млекопитающих; каждая унифицированная (стандартная) доза содержит заранее определенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, в учетом требуемого фармацевтического носителя. Спецификация в отношении унифицированных (стандартных) форм по изобретению диктуется (a) уникальными характеристиками активного соединения и конкретным терапевтическим или профилактическим эффектом, который должен быть достигнут, и (b) недостатками, присущими в данной области компаундированию такого активного соединения, в отношении влияния на чувствительность индивидуума, и непосредственно зависит от них.
Примерным, неограничивающим диапазоном для терапевтически или профилактически эффективного количества антитела или части антитела по изобретению является 0,1-20 мг/кг, более предпочтительно 1-10 мг/кг. Следует отметить, что величины доз могут варьироваться в зависимости от типа и тяжести состояния, симптомы которого должны быть уменьшены. Кроме того, должно быть понятно, что для любого конкретного индивида, конкретные схемы введения доз должны корректироваться на протяжении времени в соответствии с потребностью индивидуума и профессиональным мнением лица, вводящего эти композиции или контролирующего введение этих композиций, и что диапазоны доз, установленные в настоящем описании, являются только примерными и не предназначены для ограничения объема или практики заявленной композиции.
Для квалифицированных в данной области специалистов будет легко понятным, что другие подходящие модификации и адаптации способов описанного в настоящем описании изобретения являются очевидными и могут быть получены с использованием подходящих эквивалентов без отклонения от объема по изобретению или описанных в настоящем описании вариантов. После подробного описания данного изобретения, это изобретение будет более понятным при ссылке на следующие примеры, которые включены только для целей иллюстрации и не предназначены для ограничения изобретения.
ПРИМЕРЫ
Способы
Следующие способы описывают подробно экспериментальные процедуры, используемые в разделе Примеры.
(i) ELISA-планшеты для прямого связывания покрывали hRGM A (R&D) в концентрации 2 мкг/мл в карбонатном буфере. Затем лунки блокировали 2% блокирующим раствором (Bio-Rad) в течение 1 часа при комнатной температуре. Биотинилированные антитела серийно разводили с фактором разведения 1:5 в 0,1% БСА/ЗФР вдоль по планшету и инкубировали в течение 1 часа при комнатной температуре. Детектирующим реагентом было разведение 1:10000 страптавидина-HRP в 0,1% БСА/ЗФР. Детектирование выполняли с реагентом TMB, которое останавливали 2 н H2SO4 и считывали OD при 450 нм.
(ii) FACS-анализ. Стабильные трансфектанты клеток HEK293, сверхэкспрессирующие hRGM A, или клеток BAF3, сверхэкспрессирующие ratRGM A, подвергали окрашиванию немечеными МАВ 5F9 или 8D1 в течение более 15 минут при 4°С в 0,1% буфере БСА/ЗФР. Детектирование проводили с антителом РЕ мыши против IgG крысы.
(iii) Твердофазные анализы ELISA для оценки MAB 5F9 в анализах связывания hRGM A - неогенин.
Планшеты ELISA (Immuno Plate Cert. Maxi Sorb. F96 NUNC, 439454) покрывали в течение 1 часа при 37°C концентрацией 2,5 мкг/мл внеклеточного домена His-меченого белка Неогенина человека (концентрация исходного раствора: 30 мкг/мл). После ингибирования, несвязанный Неогенин удаляли в 3 отдельных стадиях промывок с использованием ЗФР, содержащего 0,02% Твин 20. Блокирование покрытых Неогенином планшетов выполняли добавлением 200 мкл на лунку блокирующего раствора 3% бычьего сывороточного альбумина (БСА), ЗФР, Твин 20 (0,02%). После ингибирования в течение 1 часа при 37°C, этот блокирующий раствор удаляли и добавляли фрагменты RGM A или полноразмерный белок, конъюгированный с fc-меткой (тэгом) человека, с антителом или без антитела. В некоторых вариантах осуществления антитела предварительно инкубировали с fc-конъюгированными белками hRGM A в течение 1 часа при комнатной температуре. Покрытые Неогенином планшеты инкубировали с hRGM A с антителами или без антител в течение 1 часа при 37°С. После 3 стадий промывки ЗФР-Твин 20 (0,02%), планшеты инкубировали с меченым биотином антителом против fc человека (1 мг/мл, разведенный 1:200 в ЗФР, содержащем 0,6% БСА, 0,02% Твин 20), номера по каталогу Jackson ImmunoResearch: 709-065-149, в течение 1 часа при 37°С. Несвязанное антитело удаляли 3 стадиями промывок с использованием ЗФР/Твин 20 (0,02%). Для визуализации меченого биотином fc-антитела, добавляли комплекс, состоящий из Стрептавидина-пероксидазы (Roche, cat.# 11089153001), разведенный 1:5000 ЗФР, содержащим 0,6% BSA, 0,02% Твин 20, с последующим ингибированием при 37°С в течение 1 часа. Несвязанный комплекс пероксидазы удаляли в 3 последовательных стадиях промывок (ЗФР-Твин 20 (0,02%) перед добавлением субстрата пероксидазы (Immuno Pure TMB, Pierce # 34021). Реакцию субстрата останавливали спустя 1-30 минут после его добавления к лункам с использованием 2,5 M H2SO4. Планшеты анализировали (определением OD) при длине волны 450 нм с использованием фотометра Anthos.
(iv) Твердофазные анализы ELISA для оценки MAB 5F9 в анализах связывания hRGM A - BMP-4.
Планшеты ELISA (Immuno Plate Cert. Maxi Sorb. F96 NUNC, 439454) покрывали в течение 1 часа при 37°C раствором, содержащим концентрацию 2,5 мкг/мл рекомбинантного белка BMP-4 человека (R&D Systems, # 314-BP, Lot # BEM316061). После инкубирования, несвязанный BMP-4 удаляли в 3 отдельных стадиях промывок с использованием ЗФР, содержащего 0,02% Твин 20. Блокирование покрытых BMP-4 планшетов выполняли добавлением 200 мкл на лунку блокирующего раствора 3% бычьего сывороточного альбумина (БСА), ЗФР, Твина 20 (0,02%). После инкубирования в течение 1 часа при 37°С, этот блокирующий раствор удаляли и добавляли фрагменты RGM A или полноразмерный белок, конъюгированный с fc-меткой (тэгом) человека, с антителом или без антитела. В некоторых вариантах осуществления антитела предварительно инкубировали с fc-конъюгированными белками hRGM A в течение 1 часа при комнатной температуре. Покрытые BMP-4 планшеты инкубировали с hRGM A с антителами или без антител в течение 1 часа при 37°С. После 3 стадий промывки ЗФР-Твин 20 (0,02%), планшеты инкубировали с меченым биотином антителом против fc человека (1 мг/мл, разведенный 1:200 в ЗФР, содержащем 0,6% БСА, 0,02% Твин 20), номера по каталогу Jackson ImmunoResearch: 709-065-149, в течение 1 часа при 37°С. Несвязанное антитело удаляли 3 стадиями промывок с использованием ЗФР/Твин 20 (0,02%). Для визуализации меченого биотином fc-антитела, добавляли комплекс, состоящий из Стрептавидина-пероксидазы (Roche, cat.# 11089153001), разведенный 1:5000 ЗФР, содержащим 0,6% BSA, 0,02% Твин 20, с последующим ингибированием при 37°С в течение 1 часа. Несвязанный комплекс пероксидазы удаляли в 3 последовательных стадиях промывок (ЗФР-Твин 20 (0,02%) перед добавлением субстрата пероксидазы (Immuno Pure TMB, Pierce # 34021). Реакцию субстрата останавливали спустя 1-30 минут после его добавления к лункам с использованием 2,5 M H2SO4. Планшеты анализировали (определением OD) при длине волны 450 нм с использованием фотометра Anthos.
(v) Твердофазные анализы ELISA для оценки MAB 5F9 в анализах связывания hRGM A - BMP-2.
Планшеты ELISA (Immuno Plate Cert. Maxi Sorb. F96 NUNC, 439454) покрывали в течение 1 часа при 37°C раствором, содержащим концентрацию 2,5 мкг/мл рекомбинантного белка BMP-2 человека (R&D Systems, # 355-BM, Lot # MSA04). После инкубирования, несвязанный BMP-2 удаляли в 3 отдельных стадиях промывок с использованием ЗФР, содержащего 0,02% Твин 20. Блокирование покрытых BMP-4 планшетов выполняли добавлением 200 мкл на лунку блокирующего раствора 3% бычьего сывороточного альбумина (БСА), ЗФР, Твина 20 (0,02%). После инкубирования в течение 1 часа при 37°С, этот блокирующий раствор удаляли и добавляли фрагменты RGM A или полноразмерный белок, конъюгированный с fc-меткой (тэгом) человека, с антителом или без антитела. В некоторых вариантах осуществления антитела предварительно инкубировали с fc-конъюгированными белками hRGM A в течение 1 часа при комнатной температуре. Покрытые BMP-2 планшеты инкубировали с hRGM A с антителами или без антител в течение 1 часа при 37°С. После 3 стадий промывки ЗФР-Твин 20 (0,02%), планшеты инкубировали с меченым биотином антителом против fc человека (1 мг/мл, разведенный 1:200 в ЗФР, содержащем 0,6% БСА, 0,02% Твин 20), номера по каталогу Jackson ImmunoResearch: 709-065-149, в течение 1 часа при 37°С. Несвязанное антитело удаляли 3 стадиями промывок с использованием ЗФР/Твин 20 (0,02%). Для визуализации меченого биотином fc-антитела, добавляли комплекс, состоящий из Стрептавидина-пероксидазы (Roche, cat.# 11089153001), разведенный 1:5000 ЗФР, содержащим 0,6% BSA, 0,02% Твин 20, с последующим ингибированием при 37°С в течение 1 часа. Несвязанный комплекс пероксидазы удаляли в 3 последовательных стадиях промывок (ЗФР-Твин 20 (0,02%) перед добавлением субстрата пероксидазы (Immuno Pure TMB, Pierce # 34021). Реакцию субстрата останавливали спустя 1-30 минут после его добавления к лункам с использованием 2,5 M H2SO4. Планшеты анализировали (определением OD) при длине волны 450 нм с использованием фотометра Anthos.
(vi) Культура клеток Ntera-2
Клетки человека Ntera-2 получали из German Collection of Microorganisms and Cell Cultures (DMSZ, Braunschweig). Замороженные исходные растворы недифференцированных клеток Ntera-2 оттаивали в среде DMEM, содержащей 10% фетальную телячью сыворотку (ФТС; JRH Bioscience, Kansas, USA) и 5% лошадиную сыворотку (HS; Sigma, Germany). Клетки выращивали в колбах для культивирования (Greiner, Germany), пока они не достигали конфлюэнтности 80%.
Для дифференцировки нейронов, клетки Ntera-2 высевали при плотности 2,5×106 клеток/175 см2 в среде для дифференцировки (среде DMEM, содержащей 10% ФТС, 5% HS, 1% пенициллин-стрептомицин, ретиноевую кислоту 10 мкМ). Клетки дифференцировались в течение 3 недель и эту среду заменяли два раза в неделю.
После дифференцировки клетки отделяли с использованием трипсина-ЭДТА и разделяли при соотношении 1:6. Спустя 48 часов нейронные клетки отделяли постукиванием от подстилающих клеток. Смещенные клетки переносили для агрегации в новой среде в новые встряхиваемые колбы для культивирования (Corning, USA). Дифференцированным клеткам Ntera-2 давали агрегироваться в условиях однородного горизонтального встряхивания при 37°С в течение 24 часов в базальной среде для нейронов (Neurobasal) (Gibco), дополненной B27 (Gibco), глутамином (Gibco) и пенициллином-стрептомицином. Агрегаты Ntera-2 высевали при плотности приблизительно 20-30 агрегатов на покровное стекло в 24-луночных чашках. Покрытые предварительно полилизином покровные стекла покрывали ламинином (20 мкг/мл, Sigma) и рекомбинантным fc-связанным фрагментом №786 (аминокислоты 47-168) RGM A человека в концентрации 10 мкг/мл. После посева культуры обрабатывали MAB 5F9, добавляемым при трех различных концентрациях (0,1 мкг/мл; 1 мкг/мл; 10 мкг/мл) к культуральной среде, и дополнительно инкубировали в течение 24 часов при 37°С в базальной среде для нейронов (Neurobasal). Затем агрегаты фиксировали в 4% параформальдегиде (2 часа, комнатная температура) и повышали проницаемость клеток добавлением 0,1% Тритона Х-100 в ЗФР (20 минут, комнатная температура). Для флуоресцентного окрашивания культуры блокировали ЗФР, содержащим 1% БСА, в течение 1 часа при комнатной температуре. После блокирования клетки Ntera инкубировали с моноклональным антителом мыши против изотипа 3 β-тубулина (клона SDL3D10, Sigma # T8660) в течение 2 часов при комнатной температуре. Несвязанное антитело удаляли 3 разными стадиями промывок (5-15 мин каждая) и клетки Ntera инкубировали с Cy-3-конъюгированным ослиным антителом против мышиного Ig. (Jackson ImmunoResearch Lot 62597), разведенным 1:350-кратно в ЗФР/0,5% БСА и 0,5 мкг/мл бисбензимида. После инкубирования в течение 1 часа культуры промывали 3 раза для удаления несвязанного вторичного антитела. Для флуоресцентной микроскопии, покровные стекла заливали в Fluoromount G (Southern Biotech, Eching).
Изображения агрегатов Ntera-2 получали с использованием флуоресцентного микроскопа Zeiss Axiovert 200 и рост нейритов этих культур автоматически анализировали с использованием собственной системы получения изображения и системы анализа. Автоматический анализ роста нейритов выполняли с использованием Image Pro Plus 4.5 и статистический анализ этих данных выполняли с использованием Graph Pad Prism 4. Вырастание нейритов нормализовали относительно контрольных культур, выращиваемых в отсутствие фрагмента #786 RGM A человека.
(vii) Культура SH-SY5Y
Клетки SH-SY5Y (ATCC, CRL-2266) являются клетками нейробластомы человека, полученными из метастатической опухоли головного мозга. Эти клетки выращивали в среде, состоящей из 50% сбалансированного солевого раствора Эрла (Invitrogen Life Technologies, Cat. # 24010-043) и 50% F12 (Ham) питательная смесь + GlutaMAX-1 (Invitrogen Life Technologies, Cat. # 31765-027). Эта среда дополнительно дополнена инактивированной нагреванием 10% фетальной телячьей сывороткой (ФТС, JRH Biosciences, Kansas Cat. # 12107-1000M), 1% NEAA (раствор заменимых аминокислот MEM (Sigma-Aldrich Cat.# M 1745) и 1% пенициллином (10000 Е/мл)/стрептомицином (10000 мкг/мл) (Invitrogen Life Technologies, Cat. # 15140-122). Для стимуляции дифференцировки нейронов и роста нейронных выступов, клетки SH-SY5Y культивировали в среде, дополненной 10 мкМ ретиноевой кислотой (RA, Sigma-Aldrich Cat. # R2625-050MG)), в течение нескольких дней. Дифференцированные клетки SH-SY5Y выращивали в колбах для культуры ткани и удаляли осторожной трипсинизацией и высевали на стеклянные покровные стекла, покрытые полосатым рисунком (паттерном) белка RGM A или его фрагмента и Коллагеном I.
(viii) Получение полосатых покровных стекол
Модифицированную версию анализа полос на покровных стеклах выполняли слегка отличающимся путем, как описано ранее (Knoell et al. Nature Protocols 2:1216 - 1224, 2007) и суммировано ниже.
Стерильные кремниевые матриксы для получения полосок очищенных белков спрессовывали на поверхности чашек Петри с шершавой стороной, обращенной кверху. Промытые этанолом, чистые покровные стекла укладывали на этот матрикс и углы этого матрикса помечали шариковой ручкой с чернилами на обратной стороне покровного стекла. Матрикс, несущий это покровное стекло, осторожно переворачивали вверх дном с покровным стеклом, обращенным к дну этой чашки Петри. Fc-конъюгированное полноразмерное ингибирующее антитело RGM A или его fc-фрагменты или рекомбинантный RGM A человека (R&D Systems Cat. # 2459 RM) смешивали с 10 мкл антимышиного FITC-меченого антитела (Fab-специфического козьего антитела против мышиного IgG, Sigma-Aldrich Cat. # F-4018) для визуализации полосок RGM A. С использованием шприца Гамильтона, 50 мкл RGM A - FITC-антитела осторожно инъецировали через входной канал. Избыточная жидкость оставляла этот матрикс через выходной канал и удалялась тканью Kleenex. После инкубирования этого матрикса-покровного стекла при 37°С в течение 2 часов, первый покрывающий раствор (содержащий RGM A), вымывали 200 мкл ЗФР. На следующей стадии, покровное стекло с полосками RGM A переносили в 24-луночную чашку, покрытую 500 мкл Коллагена I (Коллаген хвоста крысы, Becton Dickinson Biosciences Cat. # 354236), для заполнения пустых пространств между полосами RGM A и инкубировали при 37°С в течение 2 часов. В конце концов получали рисунок чередующихся полос RGM A и Коллагена на этом покровном стекле. После инкубирования, несвязанный Коллаген I вымывали тремя отдельными стадиями промывок с использованием ЗФР и дифференцированные клетки SH-SY5Y высевали на эти покровные стекла. Инкубирование клеток SH-SY5Y на этом имеющем рисунок (паттерн) субстрате продолжали при 37°С в течение 20-24 часов в присутствии или в отсутствие моноклональных антител, направленных против RGM A человека.
Для анализа иммунофлуоресценции клетки фиксировали в 4% параформальдегиде в течение 2 часов при комнатной температуре или в течение ночи при 4°С и проницаемость клеток увеличивали инкубированием с ЗФР, содержащим 0,1% Тритон X-100, в течение 10-20 минут при комнатной температуре. После блокирования 3% БСА в течение 60 минут клетки инкубировали с первичным антителом (моноклональным антителом против клона изотипа 3 β-тубулина SDL 3D10, Sigma-Aldrich Cat. # T8660) в течение 2 часов при комнатной температуре и после нескольких стадий промывок со вторичным антителом (ослиным антимышиным антителом Cy-3 Jacksonlmmuno Research Lot:62597), разведенным в ЗФР с 0,1% БСА, в течение 1 часа. Ядра контрастировали с использованием бисбензимида H33258 (Riedel-De-Haen, Cat. # A-0207). Наконец, клетки заливали в Fluoromount G (Southern Biotechnology Associates Inc.: Cat. # 010001). Клетки анализировали с использованием флуоресцентного микроскопа Axioplan2 (Zeiss).
(ix) Конструирование и экспрессия рекомбинантных анти-RGM A-антител
ДНК, кодирующую кДНК-фрагменты вариабельной области тяжелой цепи антител крысы против моноклональных антител против RGM A человека 5F9 и 8D1, клонировали в экспрессирующий вектор pHybE, содержащий константную область IgG1 человека, который содержит 2 аминокислотные мутации шарнирной области, гомологичной рекомбинацией в бактериях. Этими мутациями являются замена лейцина на аланин в положениях 234 и 235 (EU numbering, Lund et al., 1991, J. Immunol., 147:2657). Вариабельную область легкой цепи моноклональных антител 5F9 и 8D1 клонировали в вектор pHybE, содержащий константную область каппа человека. Примеры векторов pHyb-E включают pHybE-hCk и pHybE-hCg1,z,non-a (см. Заявку на патент США № 61/021282). Полноразмерные антитела транзиторно экспрессировали в клетках 293E котрансфекцией химерных кДНК тяжелой цепи и легкой цепи, лигированных в экспрессионную плазмиду pHybE. Супернатанты клеток, содержащие рекомбинантное антитело, очищали хроматографией на Протеин А-Сефарозе и связанное антитело элюировали добавлением кислого буфера. Антитела нейтрализовали и диализовали в ЗФР. Затем эти очищенные моноклональные антитела против RGM A человека испытывали на их способность связывать RGM A при помощи ELISA, описанного в Примере 1, и конкурентного ELISA, описанного в Примере 7.
Пример 1: Генерирование моноклональных антител против RGMA человека
Моноклональные антитела крысы против RGMA человека получали следующим образом:
Пример 1А: Иммунизация крыс антигеном RGMA человека
Двадцать пять микрограммов рекомбинантного очищенного RGMA человека (R&D Systems Cat#2459-RM lot MRH02511A), смешанного с полным адъювантом Фрейнда (Sigma), инъецировали подкожно в четыре 6-8-недельные крысы Harlan Sprague Dawley в день 1. В дни 21, 42 и 63, двадцать пять микрограммов рекомбинантного очищенного RGMA человека, смешанного с неполным адъювантом Фрейнда (Sigma), инъецировали подкожно в тех же самых 4 крыс Harlan Sprague Dawley. В день 144 или день 165 крыс инъецировали внутривенно 10 мкг рекомбинантного очищенного RGMA человека.
Пример 1В: Генерирование гибридомы
Спленоциты, полученные из иммунизированных крыс, описанные в примере 1.2.А, сливали с SP2/O-клетками при соотношении 2:1 в соответствии с установленным способом, описанным в Kohler, G. and Milstein 1975, Nature, 256:495, для генерирования гибридом. Продукты слияния высевали в селективные среды, содержащие азасерин и гипоксантин, в 96-луночных планшетах при плотности 1,5×105 клеток селезенки на лунку. Спустя семь - десять дней после слияния, наблюдали макроскопические гибридомные колонии. Супернатант из каждой лунки, содержащий колонии гибридом, испытывали прямым ELISA (см. пример 2) на присутствие антитело к RGMA человека. ELISA-положительные клеточные линии тестировали в FACS против стабильно трансфицированных клеток HEK293, экспрессирующих RGMA человека или крысы. Линии гибридомных клеток крыс затем тестировали в прямом ELISA на перекрестную реактивность с крысиным RGMA и ELISA-связывание со слитым белком HuRGMA 47-168.
Связывание крысиных моноклональных антител против RGMA
Пример 2. Прямое ELISA-связывание mAb 5F9 and 8D1.
Как показано на фигуре 1А, mAb 5F9 и 8D1 связываются с hRGM A со сходными титрами, как описано выше в разделе (i). Было также показано, что mAb 5F9 связывается с крысиным RGM A в ELISA, тогда как 8D1 не способно связываться с крысиным RGM A (данные не показаны). Фигура 1В показывает, что mAb 5F9 и 8D1 связываются с клетками HEK293, сверэкспрессирующими hRGM A, в FACS. Фигура 1С показывает, что 5F9, но не 8D1, было способно связывать клетки BAF3, сверхэкспрессирующие крысиный RGM A, в FACS. FACS проводили, как описано в разделе (ii).
Для оценки связывания mAb 5F9 в конкурентных анализах связывания hRGM A-неогенин, использовали твердофазные анализы ELISA. Готовили планшеты ELISA и использовали их, как описано в разделе (iii) данной заявки. hRGM A добавляли в концентрации 0,5 мкг/мл с антителами 5F9 в течение 1 часа при 37°С. mAb 5F9 использовали в следующих концентрациях: 1,25 мкг/мл; 0,63 мкг/мл; 0,32 мкг/мл; 0,16 мкг/мл; 0,08 мкг/мл; 0,04 мкг/мл; 0,02 мкг/мл; 0,01 мкг/мл. Связывание hRGM A визуализировали с использованием меченого биотином анти-fc-антитела и комплекса стрептавидин-пероксидаза. Планшеты анализировали (определение OD) при длине волны 450 нм с использованием фотометра Anthos. Как показано на фигуре 2, три наивысшие концентрации антител ингибировали зависимым от дозы образом связывание полноразмерного RGM A человека с неогенином.
Твердофазные анализы ELISA использовали также для оценки mAb 5F9 в конкурентных анализах связывания hRGM A - BMP-4. Готовили планшеты ELISA и использовали их, как описано в разделе (iv) данной заявки. hRGM A добавляли в концентрации 0,5 мкг/мл с антителами 5F9 в течение 1 часа при 37°С. mAb 5F9 использовали в следующих концентрациях: 1,25 мкг/мл; 0,63 мкг/мл; 0,32 мкг/мл; 0,16 мкг/мл; 0,08 мкг/мл; 0,04 мкг/мл; 0,02 мкг/мл; 0,01 мкг/мл. Связывание hRGM A визуализировали с использованием меченого биотином анти-fc-антитела и комплекса стрептавидин-пероксидаза. Планшеты анализировали (определение OD) при длине волны 450 нм с использованием фотометра Anthos. Как показано на фигуре 3, четыре наивысшие концентрации антител ингибировали зависимым от дозы образом связывание полноразмерного RGM A человека с BMP-4.
Твердофазные анализы ELISA использовали также для оценки ингибирования связывания mAb 5F9 фрагмента 0 (47-168) hRGM A с BMP-4. Планшеты ELISA покрывали в течение 1 часа при 37°С концентрацией 2,5 мкг/мл рекомбинантного белка BMP-4 человека. Легкую цепь hRGM A (фрагмент 0, 47-168) добавляли в концентрации 0,5 мкг/мл с антителами 5F9 в течение 1 часа при 37°С. mAb 5F9 использовали в следующих концентрациях: 1,25 мкг/мл; 0,63 мкг/мл; 0,32 мкг/мл; 0,16 мкг/мл; 0,08 мкг/мл; 0,04 мкг/мл; 0,02 мкг/мл; 0,01 мкг/мл. Связывание hRGM A визуализировали с использованием меченого биотином анти-fc-антитела и комплекса стрептавидин-пероксидаза. Планшеты анализировали (определение OD) при длине волны 450 нм с использованием фотометра Anthos. Фигура 4 изображает концентрации антитела 1,25 мкл/мл, 0,63 мкл/мл и 0,32 мкг/мл, ингибирующие зависимым от дозы образом связывание легкой цепи RGM A человека с BMP-4.
Твердофазные анализы ELISA использовали также для оценки mAb 5F9 в конкурентных анализах связывания hRGM A - BMP-2. Готовили планшеты ELISA и использовали их, как описано в разделе (v) данной заявки. Полноразмерный hRGM A добавляли в концентрации 0,5 мкг/мл с антителами 5F9 в течение 1 часа при 37°С. mAb 5F9 использовали в следующих концентрациях: 5 мкг/мл; 2,5 мкг/мл; 1,25 мкг/мл; 0,63 мкг/мл; 0,32 мкг/мл; 0,16 мкг/мл. Связывание hRGM A визуализировали с использованием меченого биотином анти-fc-антитела и комплекса стрептавидин-пероксидаза. Планшеты анализировали (определение OD) при длине волны 450 нм с использованием фотометра Anthos. Фигура 5 изображает концентрации антитела 5 мкг/мл, 2,5 мкг/мл, 1,25 мкг/мл, 0,63 мкг/мл, ингибирующие связывание полноразмерного RGM A человека с BMP-2.
Твердофазные анализы ELISA использовали также для оценки mAb 5F9 и 8D1 в анализах связывания hRGM A - неогенин, hRGM A - BMP-2 и hRGM A - BMP-4 (фигура 9). Как описано, планшеты ELISA покрывали в течение 1 часа при 37°С концентрацией 2,5 мкг/мл внеклеточного домена His-меченого белка Неогенина человека или 2,5 мкг/мл BMP-2 или BMP-4. Добавляли полноразмерный fc-конъюгированный hRGM A в концентрации 0,5 мкг/мл с антителами в течение 1 часа при 37°С. mAb 5F9 и 8D1 использовали при следующих концентрациях: 5 мкг/мл; 2,5 мкг/мл; 1,25 мкг/мл; 0,63 мкг/мл; 0,32 мкг/мл; 0,16 мкг/мл; 0,08 мкг/мл. Связывание hRGM A визуализировали с использованием меченого биотином анти-fc-антитела и комплекса стрептавидин-пероксидаза. Планшеты анализировали (определение OD) при длине волны 450 нм с использованием фотометра Anthos. Как показано на фигуре 9, крысиное моноклональное антитело 8D1 ингибирует или уменьшает связывание RGM A человека с BMP-2 и BMP-4, но не способно ингибировать его связывание с Неогенином.
Пример 3. Активность mAb 5F9 в анализах роста нейритов с агрегатами дифференцированных нейронов Ntera человека
Клетки Ntera получали и культивировали, как описано в разделе (vi) Способов данной заявки. mAb 5F9 нейтрализовал ингибирующую рост нейритов активность сильной fc-конъюгированной легкой цепи (аминокислоты 47-168) белка RGM A человека в анализах роста нейритов с агрегатами дифференцированных нейронов Ntera человека. Как показано на фигуре 6, в отсутствие ингибирующего RGM A белка или фрагмента и в присутствии стимулирующего врост нейритов субстрата ламинина, нейронные агрегаты Ntera обнаруживают экстенсивную и плотную сеть вырастающих нейритов (A). На фигуре 6 также показано, что присутствие легкой цепи hRGM А разительно уменьшает количество, плотность и длину нейритов NTera, подтверждая сильную ингибирующую активность этого фрагмента hRGM A. Немногие нейриты, покидающие агрегат, являются короткими и прочно связанными (В). Части С-Е фигуры 6 показывают, что увеличивающиеся концентрации mAb 5F9, добавляемые к культурам, нейтрализовали или дерепрессировали зависимым от дозы образом ингибирующую рост нейритов активность фрагмента легкой цепи hRGM A. С увеличением концентраций mAb, рост нейронных агрегатов Ntera полностью разрушается, несмотря на присутствие ингибитора RGM A (C: 0,1 мкг/мл mAb 5F9; D: 1 мкг/мл mAb 5F9; E: 10 мкг/мл mAb 5F9).
Количественный анализ нейтрализующей активности mAb 5F9 в анализах роста нейритов с агрегатами Ntera человека выполняли для тестирования сильного fc-конъюгированного ингибирующего фрагмента легкой цепи (аминокислоты 47-168) белка RGM A человека. Вырастание этих культур автоматически анализировали по наличию агрегатов, окрашенных бисбензимидом, и затем фотографировали. Это окрашивание метило только агрегаты, но не вырастающие нейриты. Однако они окрашивались антителом к β3-тубулину и меченым флуорофором вторичным антителом. Вырастание нейритов автоматически определяли расчетом индекса роста нейритов, индекса, определяемого вычитанием площади клеточных тел (протопластов) из окрашенной β3-тубулином площади этого агрегата и его выступов. Затем этот фактор делят на площадь клеточных тел, как описано в Lingor et al. J. Neurochem. 103:181-189, 2007. Фигура 7 показывает, что mAb 5F9 зависимым от дозы образом (0,1-10,0 мкг) нейтрализовал ингибирующую рост нейритов активность fc-конъюгированного, сильного ингибирующего фрагмента hRGM A (фрагмента 0, 47-168; 10 мкг) в анализах роста нейритов с агрегатами Ntera человека.
Пример 4. Активность mAb 5F9 в полосах hRGM A/Коллаген I
Клетки SH-SY5Y культивировали и использовали, как описано в разделе (vii) данной заявки. Покровные стекла с полосами RGM A и Коллагена I на стекле готовили, как описано в разделе (viii) данной заявки. Покрытие с чередующимися полосами hRGM A/Коллаген I и Коллагена I получали для этих экспериментов в соответствии с протоколом, описанным в литературе (Knoell et al. Nature Protocols 2: 1216-1224, 2007). В отсутствие mAb 5F9 (A), нейронные клетки SH-SY5Y обнаруживают явное предпочтение в отношении полосы Коллагена I с более 90% этих клеток, предпочитающих полосы Коллагена I полосам hRGM A. С увеличивающимися концентрациями mAb 5F9 нейронные клетки SH-SY5Y предпочитают полосы hRGM A полосам Коллагена I (B-E). При наивысшей используемой концентрации mAb (E)), нейроны SH-SY5Y обнаруживают сильное предпочтение в отношении полос hRGM A в сравнении с полосами Коллагена I (см. фигуру 8). Это может интерпретироваться как уникальная характеристика mAb 5F9, так как она превратила ингибирующую природу RGM A в привлекательную активность. В присутствии увеличивающихся концентраций 5F9, нейронные клетки предпочитают мигировать и расти на RGM A-субстрате, а не на пермиссивном субстрате, таком как Коллаген I. Такой уникальнрый признак никогда не описывался прежде для моноклонального антитела.
Пример 5: Конструирование CDR-привитых антител
С применением стандартных способов, хорошо известных в данной области, CDR-последовательности цепей VH и VL моноклонального антитела 5F9 (см. таблицу 5 выше) трансплантировали в различные акцепторные последовательности тяжелой и легкой цепи человека. На основе сопоставлений последовательностей VH и VL с последовательностями VH и VL моноклонального антитела 5F9 данного изобретения отобраны следующие известные последовательности человека:
a) VH3-48, VH3-33 и VH3-23, а также соединительные последовательности hJH3, hJH4 и hJH6 для конструирования акцепторных последовательностей тяжелой цепи (в соответствии с приведенной выше таблицей 3);
b) A17 и A18, а также hJK2 для конструирования акцепторных последовательностей легкой цепи (в соответствии с приведенной выше таблицей 4).
Трансплантацией соответствующих CDR VH и VL 5F9 в указанные акцепторные последовательности были получены следующие CDR-привитые, гуманизированные, модифицированные последовательности VH и VL (см. также таблицу 6 выше): VH 5F9.1-GL, VH 5F9.2-GL, VH 5F9.3-GL, VH 5F9.4-GL, VH 5F9.5-GL, VH 5F9.6-GL, VH 5F9.7-GL и VH 5F9.8-GL; VL 5F9.1-GL, VL 5F9.2-GL, and VL 5F9.3-GL.
Пример 6: Конструирование каркасных обратных мутаций в CDR-привитых антителах
Для генерирования каркасных обратных мутаций гуманизированного антитела, мутации вводят в последовательности CDR-привитых антител, полученных в соответствии с примером 5, посредством синтеза de novo этого вариабельного домена и/или с использованием мутагенных праймеров и ПЦР, и способами, хорошо известными в данной области. Различные комбинации обратных мутаций и других мутаций конструируют для каждого из CDR-трансплантатов следующим образом.
Для тяжелых цепей VH 5F9.1-GL, VH 5F9.2-GL и VH 5F9.3-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: V37→I, V48→I, S49→G и/или R98→K.
Для тяжелых цепей VH 5F9.4-GL, VH 5F9.5-GL и VH 5F9.6-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: V37→I, V48→I, A49→G, R98→K.
Для тяжелых цепей VH 5F9.7-GL, VH 5F9.8-GL и VH 5F9.9-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: V37→I, V48→I, S49→G.
Дополнительные мутации включают следующие:
для тяжелых цепей VH 5F9.1-GL, VH 5F9.2-GL и VH 5F9.3-GL: D88→A,
для тяжелых цепей VH 5F9.4-GL, VH 5F9.5-GL и VH 5F9.6-GL: Q1→E и
для тяжелых цепей VH 5F9.7-GL, VH 5F9.8-GL и VH 5F9.9-GL: L5→V.
Для легкой цепи VL 5F9.1-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: I2→V, M4→L, Y41→F.
Для легкой цепи VL 5F9.2-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: M4→L, R51→L.
Для легкой цепи VL 5F9.3-GL один или несколько следующих смежных с зоной Vernier и VH/VL остатков обратно мутируют следующим образом: M4→L, Y41→F.
Пример 7: Конструирование и экспрессия рекомбинантных гуманизированных анти-RGMA-антител
Экспрессирующие векторы pHybE, несущие тяжелые и легкие цепи, содержащие каркасные обратные мутации, котрансфицировали в клетки 293-6E для транзиторного получения полноразмерных гуманизированных антител, как описано в разделе (ix) выше. Мутации вводили в последовательности CDR-привитых антител, полученных в соответствии с примером 5, посредством синтеза de novo этого вариабельного домена и/или с использованием мутагенных праймеров и ПЦР и способами, хорошо известными в данной области. Аминокислотные последовательности областей VH и VL гуманизированных антител описаны в таблице 8.
Конкретно, для тяжелых цепей:
VH 5F9.1, VH 5F9.5 и VH 5F9.9 содержат VH 5F9.4-GL с мутацией Q1→E.
VH 5F9.2, VH 5F9.6, VH 5F9.10, VH 5F9.19, VH 5F9.20, VH 5F9.21 и VH 5F9.22 содержат VH 5F9.4-GL с мутацией Q1→E и следующими обратными мутациями смежных с зоной Vernier и VH/VL остатков: V37→I, V48→I, A49→G, R98→K.
VH 5F9.3, VH 5F9.7 и VH 5F9.11 содержат VH 5F9.7-GL с мутацией L5→V. VH 5F9.4, VH 5F9.8, VH 5F9.12, VH 5F9.23, VH 5F9.24, VH 5F9.25 и VH 5F9.26 содержат VH 5F9.7-GL с мутацией L5→V и следующими обратными мутациями смежных с зоной Vernier и VH/VL остатков: V37→I, V48→I, S49→G.
Для легких цепей:
VL 5F9.1, VL 5F9.2, VL 5F9.3 и VL 5F9.4 являются идентичными VL 5F9.1-GL.
VL 5F9.5, VL 5F9.6, VL 5F9.7 и VL 5F9.8 являются идентичными VL 5F9.2-GL.
VL 5F9.9, VL 5F9.10, VL 5F9.11 и VL 5F9.12 являются идентичными VL 5F9.3-GL.
VL 5F9.19 и VL 5F9.23 содержат VL 5F9.2-GL со следующими обратными мутациями смежных с зоной Vernier и VH/VL остатков: M4→L, R51→L.
VL 5F9.20 и VL 5F9.24 содержат VL 5F9.2-GL со следующей обратной мутацией смежного с зоной Vernier и VH/VL остатка: M4→L.
VL 5F9.21 И VL 5F9.25 содержат VL 5F9.3-GL со следующими обратными мутациями смежных с зоной Vernier и VH/VL остатков: M4→L, Y41→F.
VL 5F9.22 и VL 5F9.26 содержат VL 5F9.3-GL со следующей обратной мутацией смежного с зоной Vernier и VH/VL остатка: M4→L.
ЭКСПРЕССИЯ ГУМАНИЗИРОВАННЫХ АНТИТЕЛ
Пример 8: Характеристика гуманизированных антител 5F9 с использованием конкурентного ELISA
Планшеты ELISA (Costar 3369) покрывали в течение ночи при 4°C 50 мкл на лунку 0,25 мкг/мл hRGMA в 0,2 M натрий-карбонатном-бикарбонатном буфере, рН 9,4, промывали промывочным буфером (ЗФР, содержащим 0,1% Твин 20) и блокировали в течение 1 часа при комнатной температуре 200 мкл на лунку 2% обезжиренным сухим молоком в ЗФР. После промывания промывочным буфером добавляли в двух повторностях смесь биотинилированного химерного антитела 5F9 (конечная концентрация 0,1 мкг/мл) и немеченого конкурентного тест-антитела (начиная при конечной концентрации 50 мг/мл и серийно разведенного в 5-раз) в 50 мкл на лунку буфера ELISA. После инкубирования этих планшетов в течение 1 часа при комнатной температуре и промывания промывочным буфером, связанные антитела детектировали с использованием 100 мкл на лунку разведения 1:10000 конъюгированного с HRP стрептавидина (Fitzgerald) в буфере ELISA. После инкубирования в течение 1 часа при комнатной температуре и промывания промывочным буфером развитие окраски выполняли добавлением 100 мкл на лунку буфера TMB (Zymed). После инкубирования в течение 15 минут при комнатной температуре развитие окраски останавливали добавлением 50 мкл на лунку 1 н хлористоводородной кислоты. Оптическую плотность считывали при 490 нм.
Таблица 9 показывает величины IC50 гуманизированных антител 5F9, полученные с использованием компьютерной программы GraphPad Prism (GraphPad Software Inc., San Diego, CA).
Величины IC50 гуманизированных антител 5F9 в конкурентных анализах ELISA
Пример 9: Определения аффинности химерных и гуманизированных антител с использованием технологии BIACORE
Анализ BIACORE (Biacore, Inc, Piscataway, NJ) определяет аффинность антител кинетическими измерениями констант скорости ассоциации, скорости диссоциации. Связывание антител с рекомбинантным очищенным RGM A человека определяли измерениями, основанными на резонансе поверхностных плазмонов, прибором Biacore® 3000 (Biacore® AB, Uppsala, Sweden) с использованием пропускания HBS-EP (10 мМ HEPES [pH 7,4], 150 мМ NaCl, 3 мМ ЭДТА и 0,005% поверхностно-активное вещество P20) при 25°C. Все химикалии получали из Biacore® AB (Uppsala, Sweden). Приблизительно 5000 единиц RU козьего антитела против IgG человека, (Fcγ), фрагмента специфического поликлонального антитела (Pierce Biotechnology Inc, Rockford, IL), разведенного в 10 мМ ацетате натрия (pH 4,5) иммобилизовали непосредственно через биосенсорный чип СМ5 с категорией, достаточной для исследований, с использованием стандартного набора для связывания амина в соответствии с инструкциями и процедурами изготовителя при 25 мкг/мл. Непрореагировавшие части молекул на биосенсорной поверхности блокировали этаноламином. В качестве поверхности реакции использовали поверхность модифицированного карбоксиметилдекстрана в проточной ячейке 2 и 4. Немодифицированный карбоксиметилдекстран без козьего антитела против IgG человека использовали в проточной ячейке 1 и 3 в качестве ссылочной поверхности. Очищенные антитела разводили в забуференном HEPES солевом растворе для захвата при прохождении через поверхности реакции, специфические в отношении козьих антител против IgG человека. Антитела человека, подлежащие захвату в качестве лиганда (25 мкг/мл), инъецировали на матриксы реакции при скорости тока 5 мкл/мин. Константы скорости ассоциации и диссоциации, Kon (единицы M-1с-1) и Koff (единицы с-1), определяли при непрерывной скорости тока 25 мкл/миг. Константы скорости получали выполнением кинетических измерений при десяти различных концентрациях антигена в диапазоне от 0,30 до 50 нМ. Для кинетического анализа, уравнения скорости, полученные из модели связывания 1:1 Langmuir подгоняли одновременно к фазам ассоциации и диссоциации всех восьми инъекций (с использованием анализа глобальной подгонки) с использованием программы Biaevaluation 4.0.1. Затем равновесную константу диссоциации (unit M) реакции между гуманизированными антителами и рекомбинантным очищенным RGM A человека рассчитывали из кинетических констант скорости с использованием формулы: KD = koff/kon.
Аффинность химерных и гуманизированных моноклональных антител против RGM A
Пример 10: Гуманизированные антитела 5F9 нейтрализуют хемоотталкивающую активность RGM A человека в анализе хемотаксиса нейронных клеток SH-SY5Y
Анализ хемотаксиса измеряет поведение клеток в реакции на диффундируемые факторы, которые могут проявлять хемоаттрагирующую или хемоотталкивающую активности. RGM A был описан как белок, действующий как в мембраносвязанной (в зависимом от контакта отталкивании), так и в растворимой диффундируемой форме (хемоотталкивающей) и, следовательно, оценивался в анализе хемотаксиса hRGM A. Для этой цели использовали RGM A-чувствительные клетки SH-SY5Y нейробластомы человека, несущие RGM-рецептор Неогенин (Schaffar et al. J. Neurochemistry: 107:418-431, 2008). Клетки SH-SY5Y выращивали в среде сбалансированного солевого раствора Эрла/F12 (EBSS/F12), дополненной 10% фетальной телячьей сывороткой и 1% заменимыми аминокислотами (MEM-NEAA). Для индукции роста нейритов клетки культивировали в среде, дополненной 10 мкМ ретиноевой кислотой (RA). Спустя 5-6 часов, клетки трипсинизировали и считали для посева в 24-луночные камеры Бойдена (BD Falcon 351185, HTS Multiwell System). 500 мкл суспензии клеток (соответствующих 1×105 клеток) добавляли во внутренний кружок каждой лунки. Этот внутренний кружок отделен от большего наружного кружка каждой лунки PET-мембраной с диаметром пор 8 мкм. 600 мкл среды +/- RGM A +/- антитела пипетировали в наружный кружок и эти клетки культивировали в мультилуночных камерах Бойдена в течение ночи при 37°С. После инкубирования среду аспирировали и заменяли фиксатором (2% параформальдегидом). Фиксацию продолжали в течение 2 часов при комнатной температуре и после нескольких стадий промывки ЗФР выполняли увеличение проницаемости с использованием ЗФР, содержащего 0,1% Тритон X-100 (15 мин, кт). Окрашивание клеток выполняли инкубированием их в течение 1 часа в темноте в растворе Alexa Fluor 488 Phalloidin 1:100 (Invitrogen A12379) и бисбензимида (H332456) 1:100. После 2 стадий промывки ЗФР, культуры заливали ЗФР, заделывали парафилмом и хранили в темноте для анализа с использованием флуоресцентного микроскопа (Zeiss Axiovert).
В отсутствие hRGM A клетки мигрируют через поры мембран и могут быть подсчитаны после фиксации и окрашивания. Считают только те клетки, которые прикреплены к нижней части мембраны, так как эти клетки мигрировали через мембрану РЕТ. Клетки на верхней стороне этой мембраны могут быть осторожно удалены перед процедурой фиксации. Этот анализ хемотаксиса доказал, что присутствие hRGM A значительно уменьшает количество клеток SH-SY5Y, мигрирующих через мембрану, более чем на 80%. Крысиное моноклональное антитело 5F9, химерное “человек-крыса” антитело 5F9, но не изотипически сходное контрольное крысиное моноклональное антитело (p21), частично или полностью нейтрализовали хемоотталкивающую активность при 10 мкг/мл, что проявлялось в виде больших количеств клеток, обнаруживаемых на дне этой мембраны (фигура 10).
Пример 11: 5F9 индуцирует регенерацию раздавленных, поврежденных аксонов зрительного нерва в крысиной модели повреждения зрительного нерва
Модель раздавливания зрительного нерва (или повреждения зрительного нерва) обеспечивает модель животного для тестирования различных веществ, которые стимулируют регенерацию волокон зрительного нерва и уменьшают массивную гибель клеток ретинального ганглия.
Эксперименты проводили во взрослых самцах крыс Sprague Dawley и самцах крыс Wistar, полученных из Charles River (D) Laboratories (Germany). Животных содержат в отдельных клетках при цикле свет/темнота 12:12 часов с доступом к корму и воде ad libitum. Раздавливание зрительного нерва выполняют только в левом глазе при помощи минимальной хирургии передней камеры глазного яблока. Этот способ является минимально инвазивным способом повреждения зрительного нерва и был разработан авторами по изобретению в соответствии с хирургическими способами для передней части зрительного нерва человека. Перед процедурой операции и во время процедуры операции животных анестезируют ингаляционной анестезией с использованием Севофлурана (Abbott GmbH Co. & KG, Delkenheim, Germany) и фиксируют на операционном столе при помощи челюстного зажима и лейкопластыря для конечностей. Падение температуры тела предотвращают помещением животных на грелку-подушку. Для раздавливающей хирургии зрительного нерва крысы в передней камере глазного яблока, левый глаз осторожно освобождают от связки и соединительной ткани. В качестве первой стадии, выполняют микрохирургический разрез (2-3 мм) смежной ткани в наружном углу этого глаза. Затем зрительный нерв обнажают при помощи пары пинцетов для смещения вбок мышц глаза и слезной железы для их сохранения. В следующей стадии, мозговые оболочки раскрывают продольно с использованием микроножниц для экспонирования зрительного нерва.
Это приводит к большей подвижности глаза и делает возможной латеральное вращение глаза и доступ к его левому зрительному нерву. Зрительный нерв повреждают на расстоянии приблизительно 1-3 мм за глазом с использованием пары зажимов, устанавливаемых для обеспечения фиксированного максимального давления в течение 20-30 секунд. Соблюдается особая осторожность во избежание повреждения васкулярного снабжения этого глаза.
Локальное введение антител и буферного раствора
После повреждения раздавливанием зрительного нерва самцов крыс Sprague Dawley обрабатывали локально антителом 5F9 (n = 10 животных), контрольным антителом 8D1 (n = 10 животных) или контролем-носителем ЗФР (n = 10 животных). Экспериментаторам не были известны группы разной обработки (слепой эксперимент). Для локального применения антител небольшие кусочки рассасывающейся желатиновой губки (длина: 2,5 мм, ширина: 2,5 мм, высота: 2,5 мм) пропитывали 20 мкл раствора антитела 10 мг/мл или 20 мкл ЗФР и помещали непосредственно рядом с участком повреждения зрительного нерва. После минимальной инвазивной хирургии и нанесения антител животных помещали на бумажные полотенца в чистой клетке, установленной на грелку (нагревательный прибор) для контроля температуры тела, пока они не начинали двигаться. На этот глаз наносили мазь, которая содержит антибиотик (Gentamytrex, Dr. Mann Pharma) во избежание бактериальной инфекции и высыхания склеры. Внутрибрюшинно вводили карпрофен (Римадил, 5 мг/кг, Pfizer GmbH, Karlsruhe) для терапии послеоперационной боли непосредственно после операции и затем дважды в день в течение 3-дневного периода. Животных наблюдали и контролировали регулярно несколько часов непосредственно после операции и в последующие дни, чтобы быть уверенными, что все эти животные выживали и восстанавливались от анестезии и хирургии. Спустя 5 недель после хирургии и применения антитела/носителя, животных анестезировали избыточной дозой Наркорена (40-60 мг/кг) и перфузировали инъекцией 4% раствора параформальдегида в сердце. Зрительные нервы выделяли и переносили в 4% раствор параформальдегида и оставляли в течение 1 часа при комнатной температуре для гарантии должной фиксации этой ткани. После постфиксации, зрительные нервы выдерживали в течение ночи в 30% растворе сахарозы (4°С). На следующий день зрительные нервы заливали в Tissue Tek, замораживали и делали продольные срезы с толщиной 16 мкм при помощи Криостата.
Для иммуноокрашиваний (с иммунной меткой) срезы зрительных нервов фиксировали холодным ацетоном (10 минут), промывали 3х (5 минут) забуференным Трисом раствором (TBS, Fluka 93312) и блокировали и их проницаемость увеличивали с использованием TBS, содержащего 5% бычий сывороточный альбумин и 1% Тритон Х-100 (30 минут) при комнатной температуре). Оставшиеся БСА и детергент удаляли 2 отдельными стадиями промывки (5 минут каждая) с использованием TBS. Срезы инкубировали в течение 1 часа при комнатной температуре с поликлональным кроличьим антителом против анти-GAP-43-антитела (Abeam, ab 7562), разведенным 1:100 в 5% растворе БСА/TBS. После 3 стадий промывки TBS, 0,1% Твин, срезы инкубировали в течение 1 часа при комнатной температуре с Alexa Fluor 488-конъюгированным козьим вторичным антителом против IgG кролика (Molecular Probes A11034), разведенным 1:1000 в 5% BSA/TBS, содержащем разведение 1:100 бисбензамида (H33258, 50 мкг/мл), для визуализации клеточных ядер. Перед заливкой, окрашенные срезы промывали 3 раза TBS, 0,1% Твином (каждая стадия 5 минут) и дистиллированной водой. Срезы заливали в Fluoromount G, покрывали покровным стеклом и хранили в темноте для микроскопической документации.
Полученные с использованием флуоресцентного микроскопа Zeiss изображения (фигура 11) окрашенных продольных срезов хранили при помощи программного обеспечения Zeiss Axiovison. Отдельные снимки каждого нерва устанавливали для анализа с использованием программы Photoshop Image Analysis (Adobe). Количественный анализ выполняли двумя различными способами с использованием комбинированных изображений этих зрительных нервов. GAP-43-положительную площадь в каждом участке повреждения измеряли с использованием программы Axiovision (фигура 12B). Независимо от этого первого количественного анализа отдельные регенерирующие волокна (GAP-43-положительные) считали в 4 различных зонах: 0-200 мкм, 200-400 мкм, 400-600 мкм и 600-1200 мкм за пределами участка раздавливания. Анализ данных и статистическую оценку данных выполняли с использованием программного обеспечения Graphpad Prism (фигура 12A).
Системное введение антител и буферного раствора
Для системной доставки антител самцов крыс Wistar обрабатывали системно (внутрибрюшинно, ip) или внутривенно iv) антителом 5F9 (n = 10 животных) или контролем-носителем ЗФР (n = 10 животных). Животных инъецировали два раза и инъекции выполняли в день 0, вскоре после раздавливания нерва и в день 21 после раздавливания. Дозы предоставляемого антитела были равны 2 мг/кг в день 0 и 10 мг/кг в день 21. Животных убивали спустя пять недель после повреждения раздавливанием и выделение ткани, приготовление срезов, окрашивания и количественный анализ выполняли, как описано выше. Как и прежде, экспериментаторам не были известны эти две различные группы обработки. Комбинированные изображения зрительных нервов крыс показаны на фигуре 13. В обработанных 5F9 животных (A), многие GAP-43-положительные волокна простирались за пределы участка раздавливания в отличие от контрольных животных, обработанных ЗФР (В). Участок раздавливания расположен на левой границе и регенерирующиеся волокна окрашены антителом к GAP-43. Многочисленные волокна наблюдаются на верхнем и нижнем крае зрительного нерва в 5F9-обработанных животных, но не в получавших ЗФР животных.
5F9, но не контроль-носитель ЗФР значительно увеличивал количество регенерирующихся GAP-43-положительных волокон. Значимо больше волокон (p < 0,001) были обнаружены в животных, обработанных 5F9, при расстояниях 300 мкм - 1800 мкм, чем в обработанных носителем животных. Животных обрабатывали 5F9 в день 0 и день 21 с использованием 2 мг/кг и 10 мг/кг, соответственно. Антитело или носитель вводили внутрибрюшинно или внутривенно. Данные приведены из анализа 9 животных на группу. Анализировали 3 серии криостатных срезов на одно животное (фигура 14А).
Во втором варианте осуществления, самцов крыс Wistar обрабатывали после повреждения зрительного нерва системно (iv) антителом 5F9 (n = 10 животных), контрольным антителом 8D1(n = 10 животных) или контролем-носителем ЗФР (n = 10 животных). Крыс инъецировали один раз в неделю 2 мг/кг антитела, вводимого iv, и инъекции начинали немедленно после раздавливания зрительного нерва. Все крысы получали 4 инъекции и животных эвтаназировали спустя 5 недель после раздавливания нерва. Экспериментаторам не были известны варианты (слепой эксперимент), и обработку ткани и количественный анализ выполняли, как описано ранее. 5F9, но не контроль-носитель ЗФР, значительно увеличивал количество регенерирующихся GAP-43-положительных волокон. Значимо больше волокон (p < 0,001) были обнаружены в животных, обработанных 5F9, при расстояниях 200 мкм - 1400 мкм, чем в обработанных носителем или контрольных животных. Животных обрабатывали iv один раз в неделю в течение 4 недель, начиная со дня 0, антителом 5F9 (2 мг/кг на дозу), контрольным антителом 8D1 (2 мг/кг на дозу) или ЗФР (фигура 14В).
Пример 11: 5F9 индуцирует ремиелинизацию аксонов раздавленных зрительных нервов в крысиной модели повреждения зрительного нерва
Маркером для олигодендроцитов и миелина является базальный белок миелина (MBP). Для ответа на вопрос, имеют ли место различия в ремиелинизации в различных группах обработки, использовали антитело, направленное против МВР. Для визуализации процуесса ремиелинизации, срезы зрительного нерва животных, обработанных системно, фиксировали охлаждением (-20°C) ацетоном (10 минут), промывали 3х (5 минут) забуференным Трисом солевым раствором (TBS, Fluka 93312) и блокировали и увеличивали проницаемость TBS, содержащим 5% бычий сывороточный альбумин и 1% Тритон Х-100 (30 минут), при комнатной температуре). Оставшиеся БСА и детергент удаляли 2 отдельными стадиями промывки (5 минут каждая) с использованием TBS. Срезы инкубировали в течение 3 часов или в течение ночи при 4°С с поликлональным кроличьим антителом против анти-МВР-антитела (Abeam, ab 2404), разведенным 1:100 в 5% растворе БСА/TBS. После 3 стадий промывки TBS, 0,1% Твин, срезы инкубировали в течение 1 часа при комнатной температуре с Alexa Fluor 488-конъюгированным козьим вторичным антителом против IgG кролика (Molecular Probes A11034), разведенным 1:1000 в 5% BSA/TBS, содержащем разведение 1:100 бисбензимида (H33258, 50 мкг/мл), для визуализации клеточных ядер. Перед заливкой, окрашенные срезы промывали 3 раза TBS, 0,1% Твином (каждая стадия 5 минут) и дистиллированной водой. Срезы заливали в Fluoromount G, покрывали покровным стеклом и хранили в темноте для микроскопической документации.
Полученные с использованием флуоресцентного микроскопа Zeiss изображения окрашенных продольных срезов хранили при помощи программного обеспечения Zeiss Axiovison. Отдельные снимки каждого нерва устанавливали для анализа с использованием программы Photoshop Image Analysis (Adobe). Количественный анализ выполняли двумя различными способами с использованием объединенных изображений этих зрительных нервов. МВР-положительную площадь в каждом участке повреждения измеряли с использованием программы Axiovision. Анализ данных и статистическую оценку данных выполняли с использованием программного обеспечения Graphpad Prism.
Животных обрабатывали 5F9 в день 0 и день 21 с использованием 2 мг/кг и 10 мг/кг, соответственно. Антитело или носитель вводили внутрибрюшинно или внутривенно. Получали объединенные изображения зрительных нервов крыс.
Миелинизацию визуализировали с использованием антитела, направленного против маркера миелина, базального белка миелина МВР. Участки раздавливания расположены в середине этих комбинированных нервов, и эта зона является свободной в обработанных носителем контрольных животных (А и В). В обработанных антителом 5F9 животных (C и D), наблюдали многочисленные MBP-положительные структуры в этой средней зоне (центре раздавливания) зрительных нервов (фигура 15).
Миелинизацию визуализировали с использованием антитела, направленного против маркера миелина, базального белка миелина МВР. Площадь MBP измеряли с использованием программы Zeiss Axiovison. M1 и M2 являются двумя независимыми измерениями и М является средней измеренной MPB-положительной площадью. 5F9 увеличивает значительно (p<0,001 в сравнении с контролем-носителем) MBP-площадь участка раздавливания зрительного нерва (в 3,5 раза) (фигура 16).
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ ОСТЕОАРТРИТА И БОЛИ | 2012 |
|
RU2563830C2 |
| СВЯЗЫВАЮЩИЙ RGMA БЕЛОК И ЕГО ИСПОЛЬЗОВАНИЕ | 2016 |
|
RU2705304C2 |
| ИММУНОГЛОБУЛИН С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ДОМЕНАМИ И ЕГО ПРИМЕНЕНИЯ | 2006 |
|
RU2515108C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ КОНЕЧНЫХ ПРОДУКТОВ ГЛУБОКОГО ГЛИКИРОВАНИЯ (RAGE) И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2518351C2 |
| ТЕРАПЕВТИЧЕСКИЕ DLL4-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2011 |
|
RU2605928C2 |
| ТЕРАПЕВТИЧЕСКИЕ DLL4-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2570639C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С ДЕГЕНЕРАЦИЕЙ НЕЙРИТОВ | 2013 |
|
RU2644337C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ВЕКТОРЫ ЭКСПРЕССИИ МЛЕКОПИТАЮЩИХ И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2502800C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD137 И ОПУХОЛЕВЫЕ АНТИГЕНЫ, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2805648C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2605327C2 |





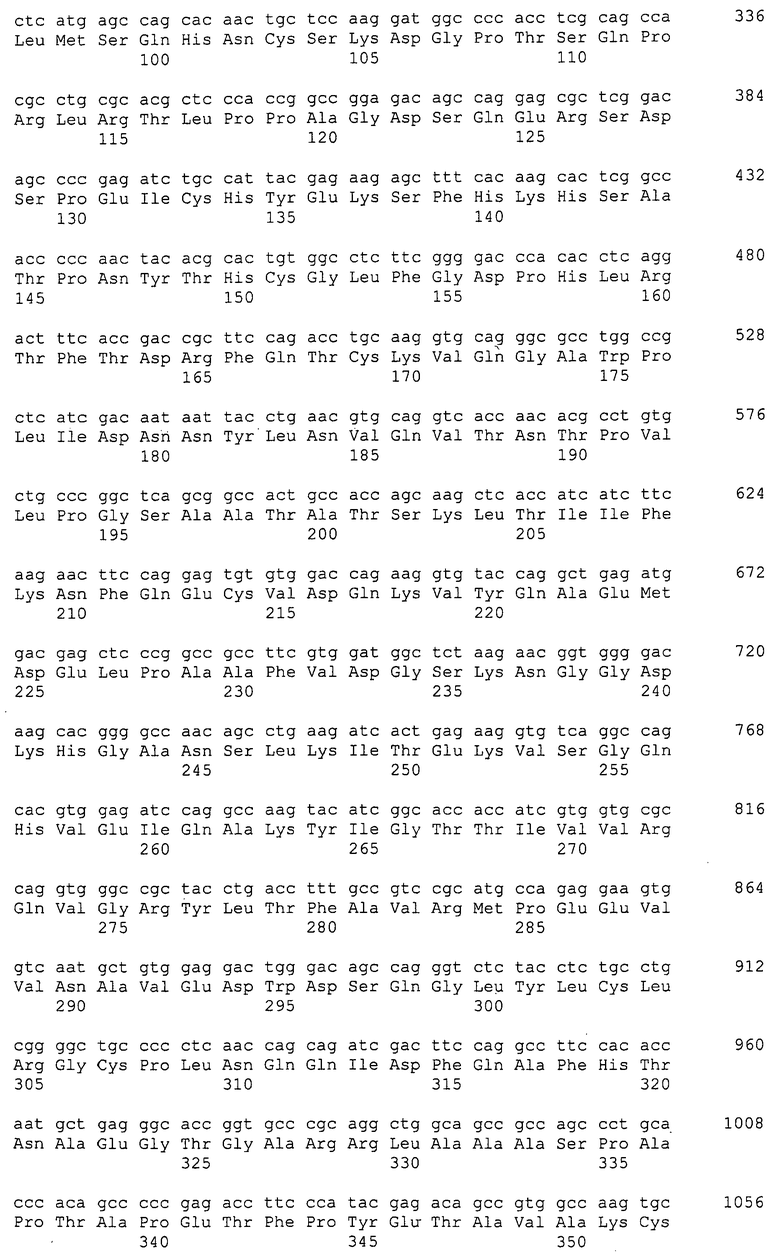









































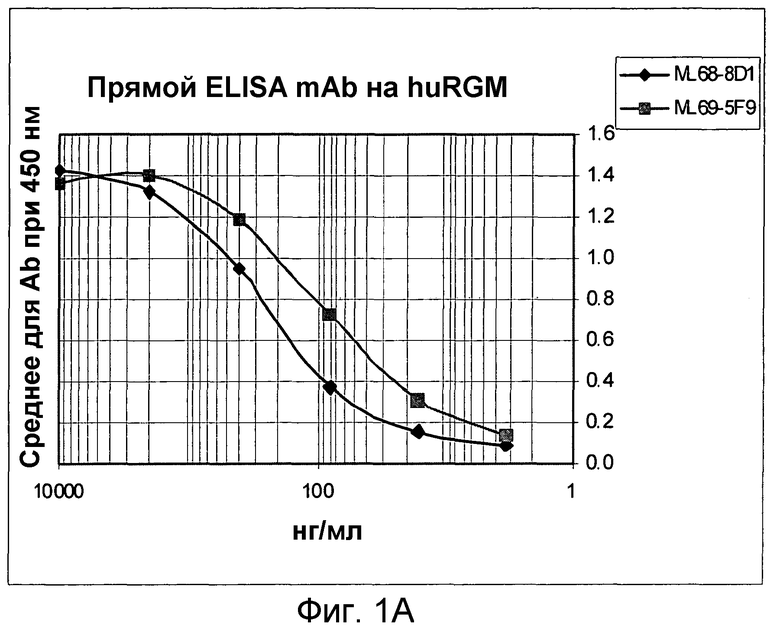
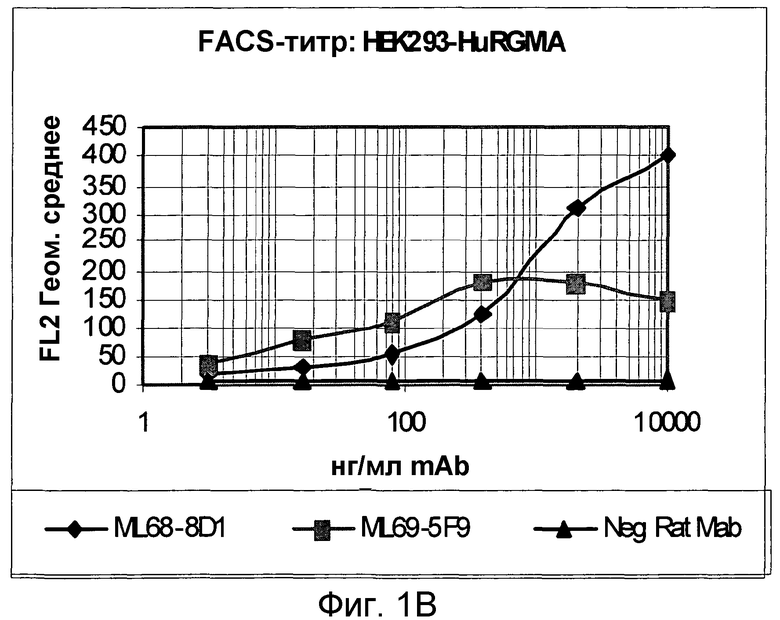
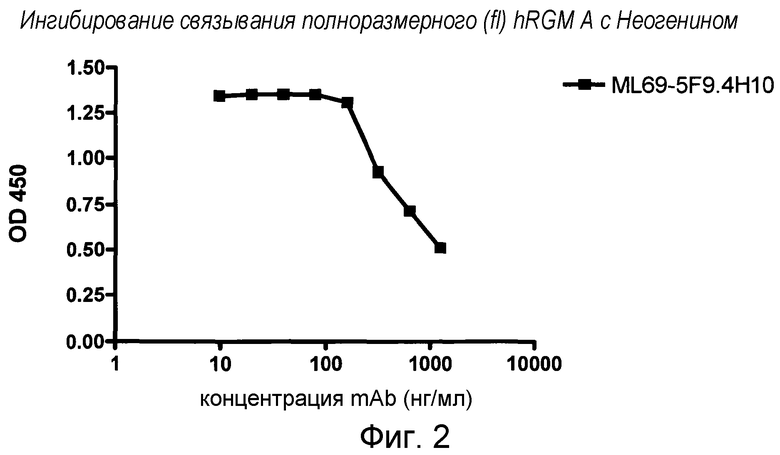
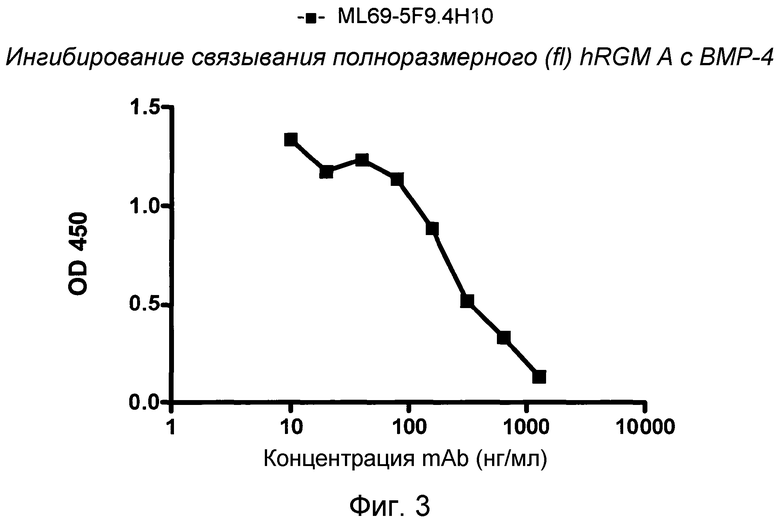
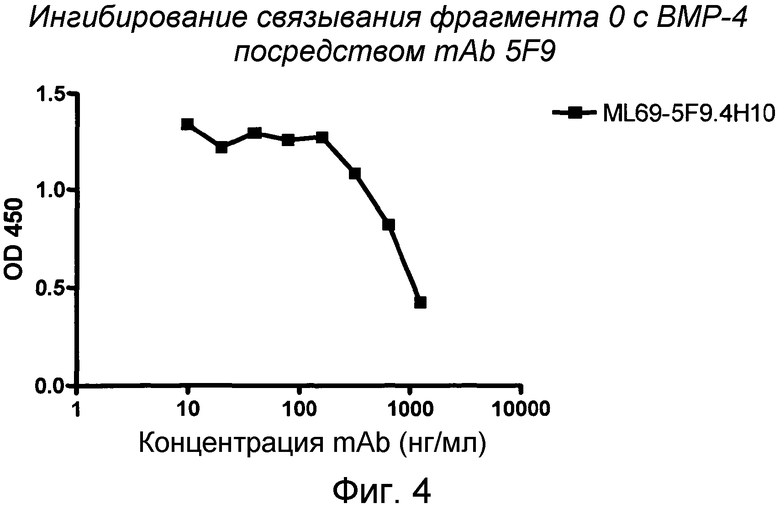
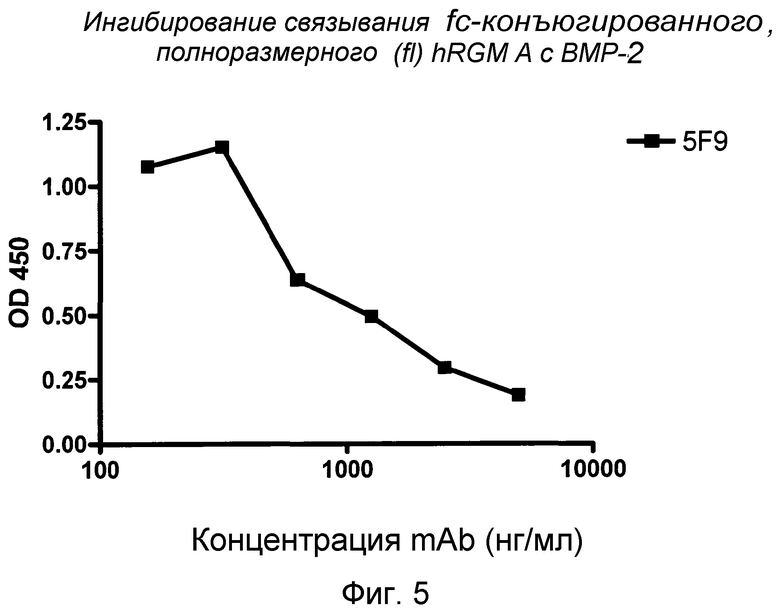
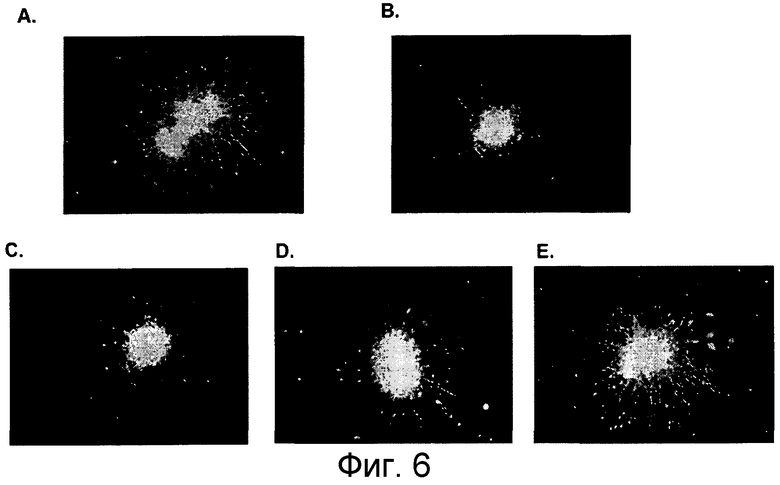
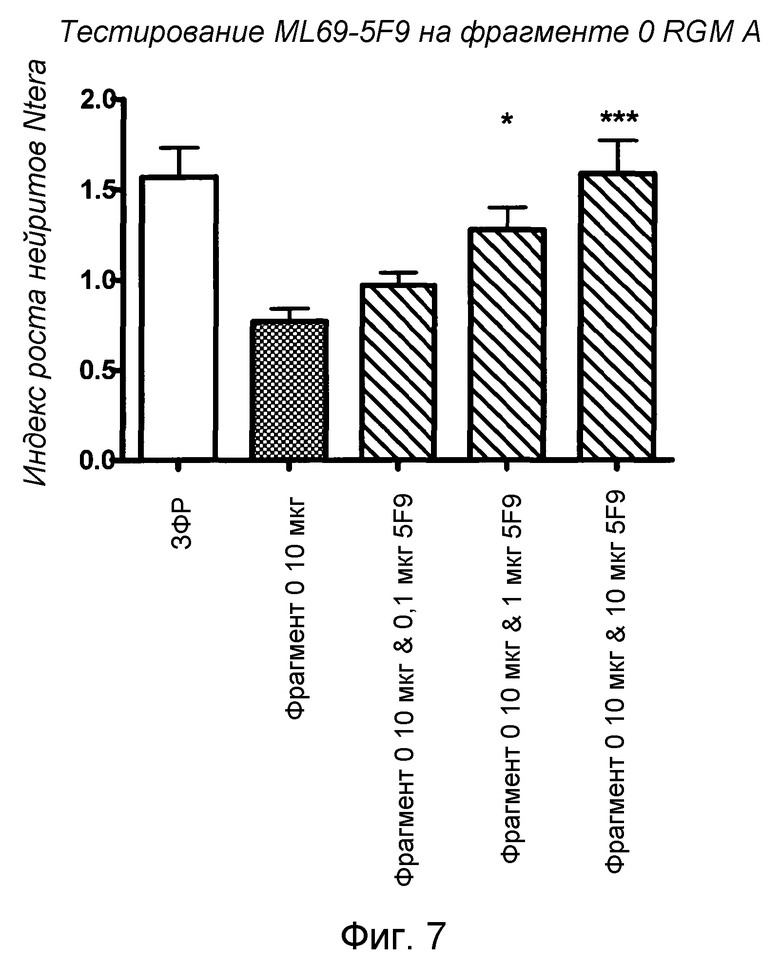
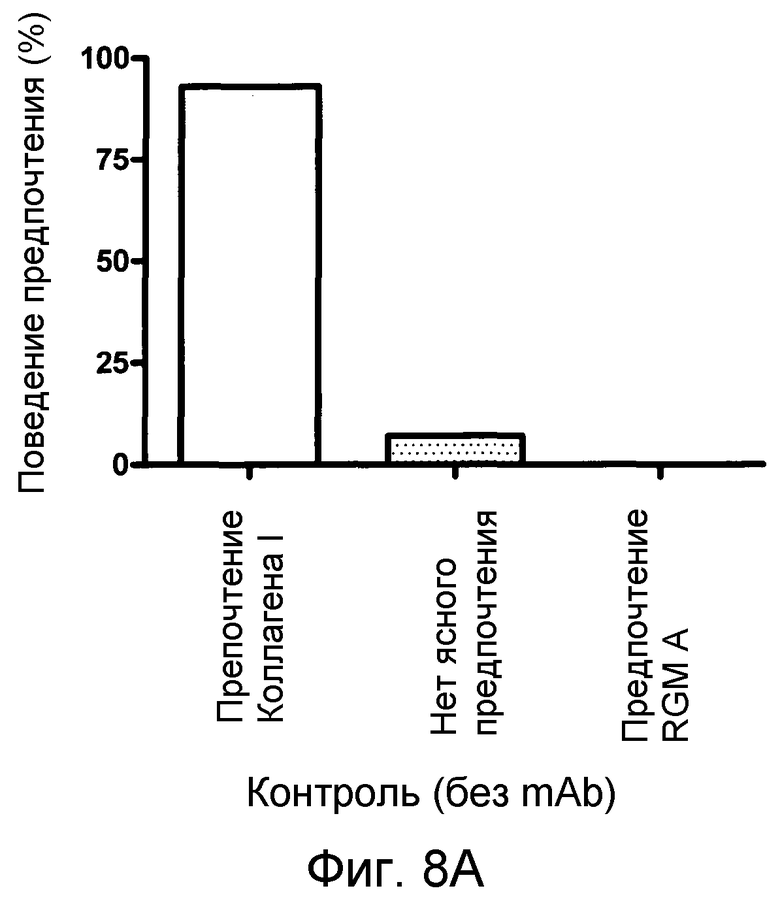
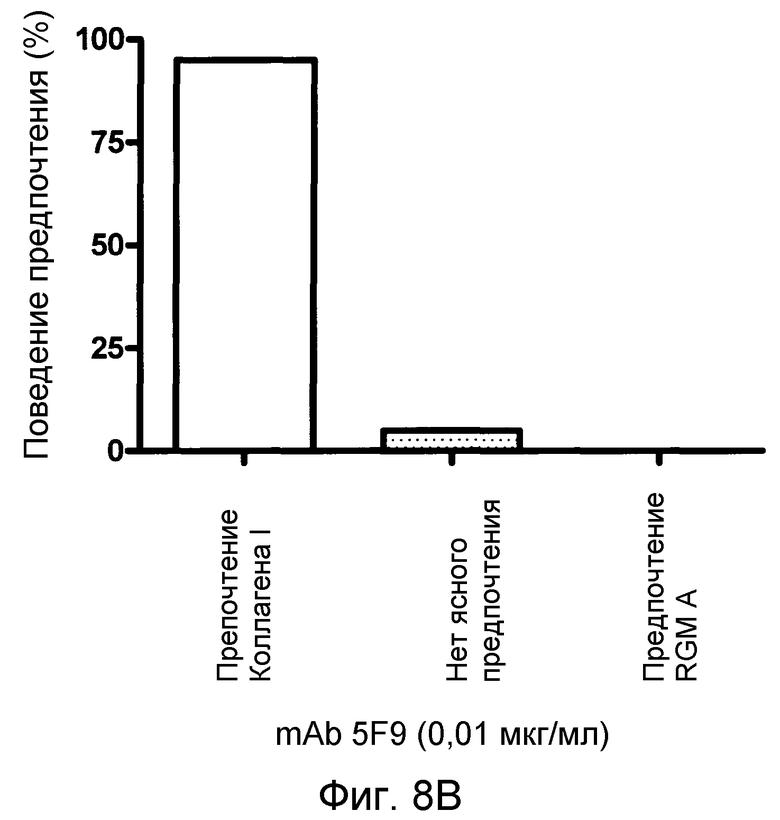
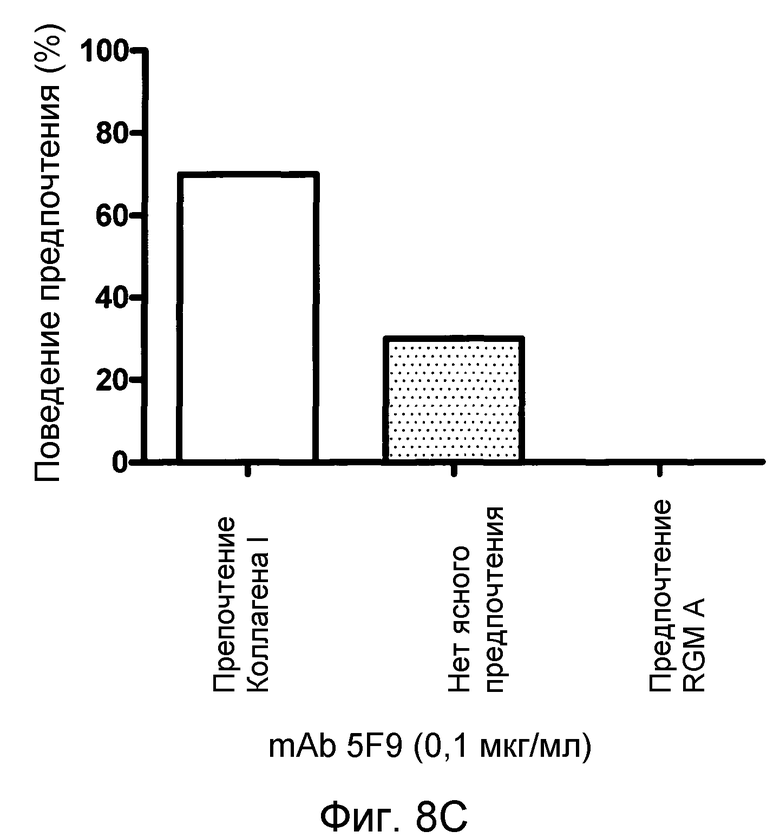
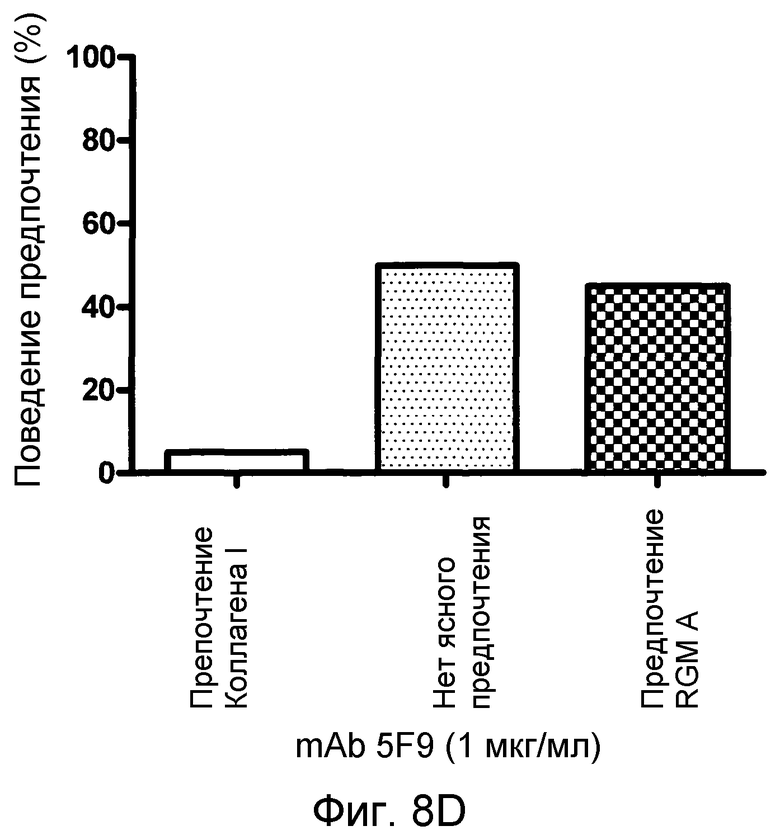
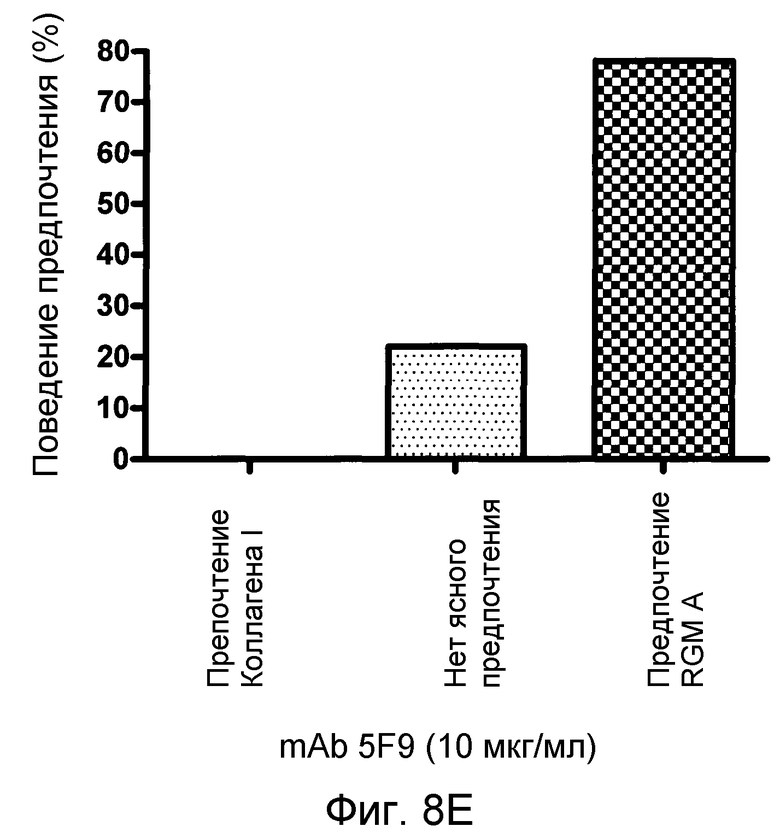
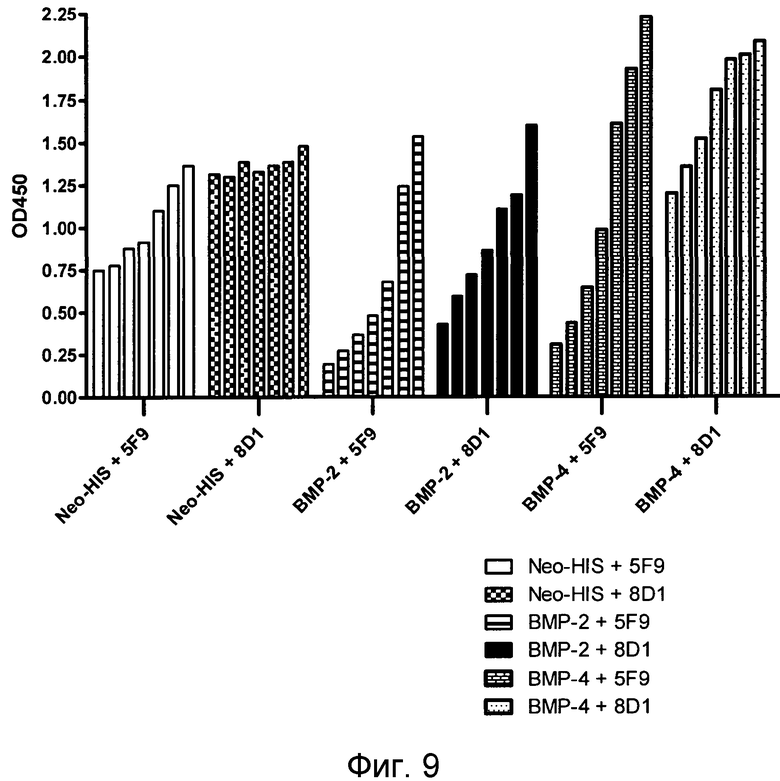
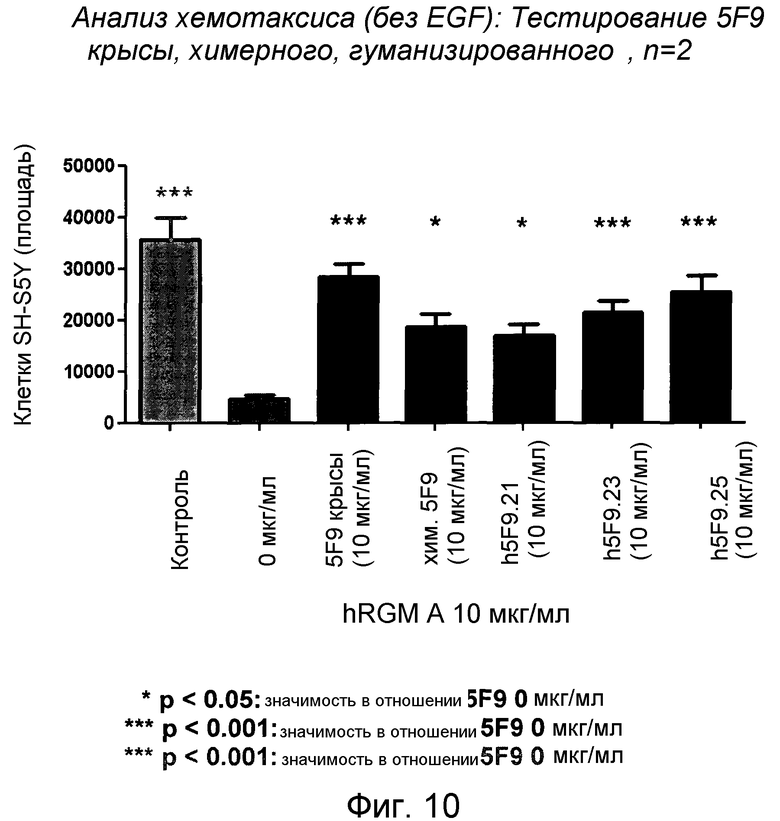
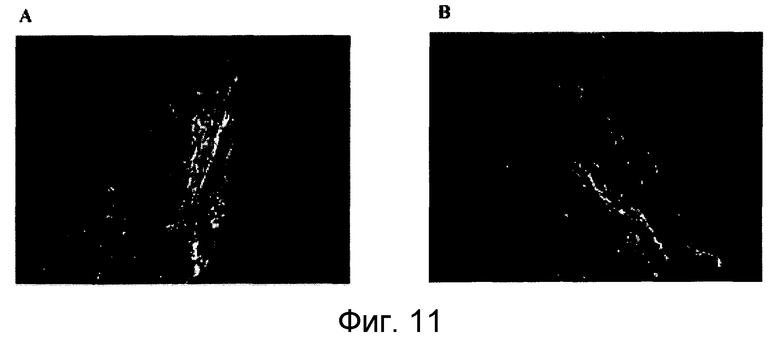
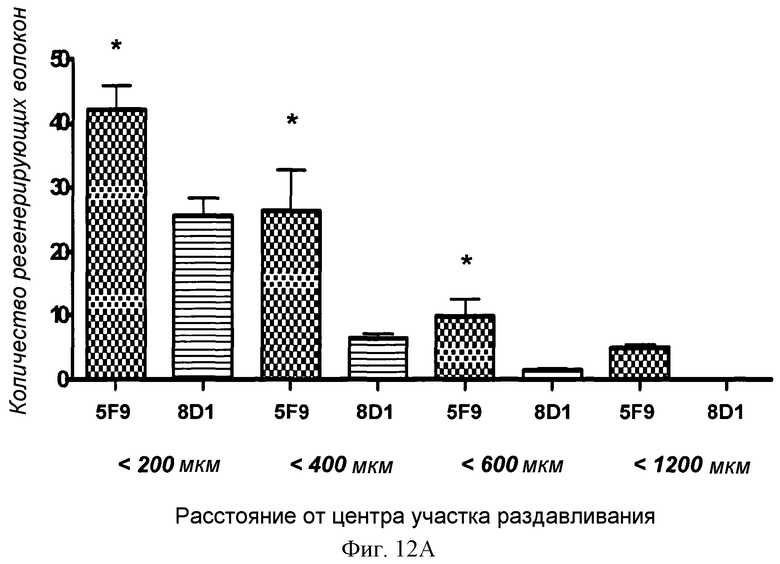
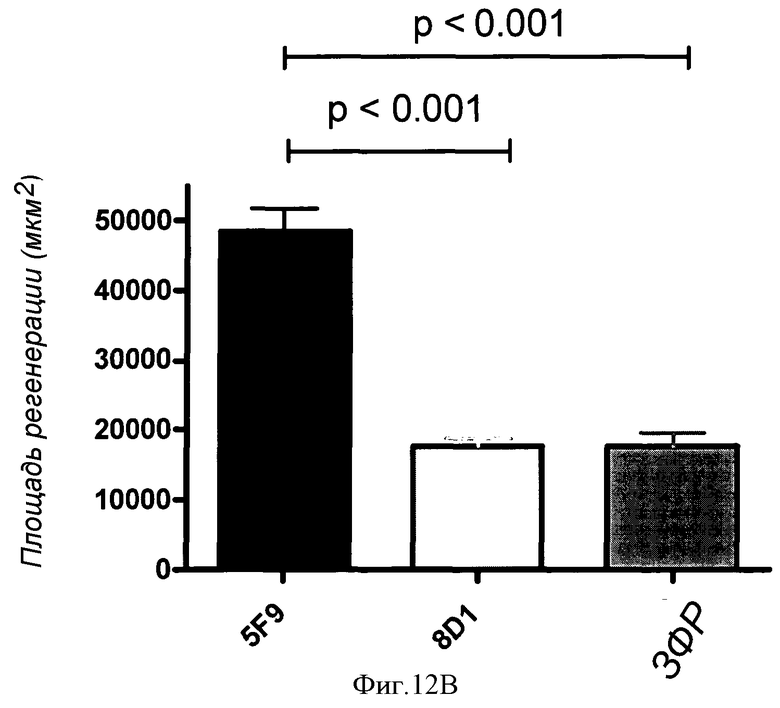

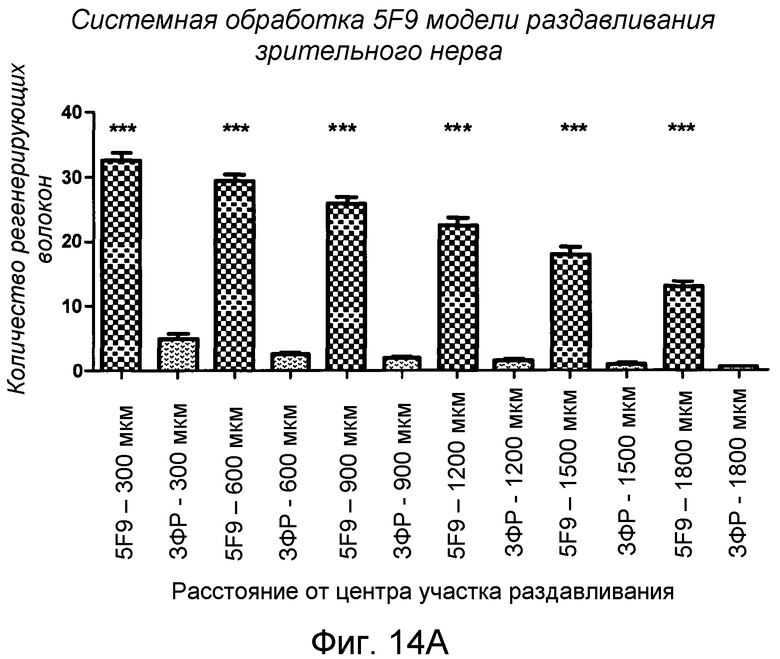
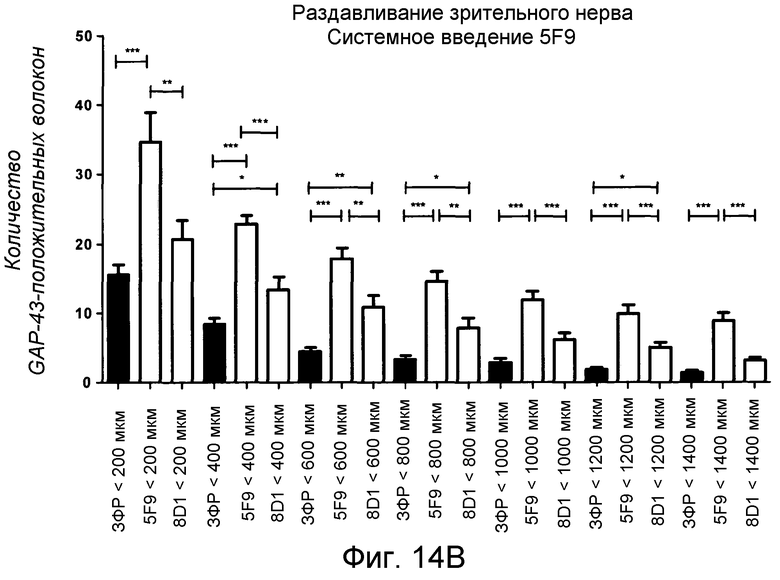
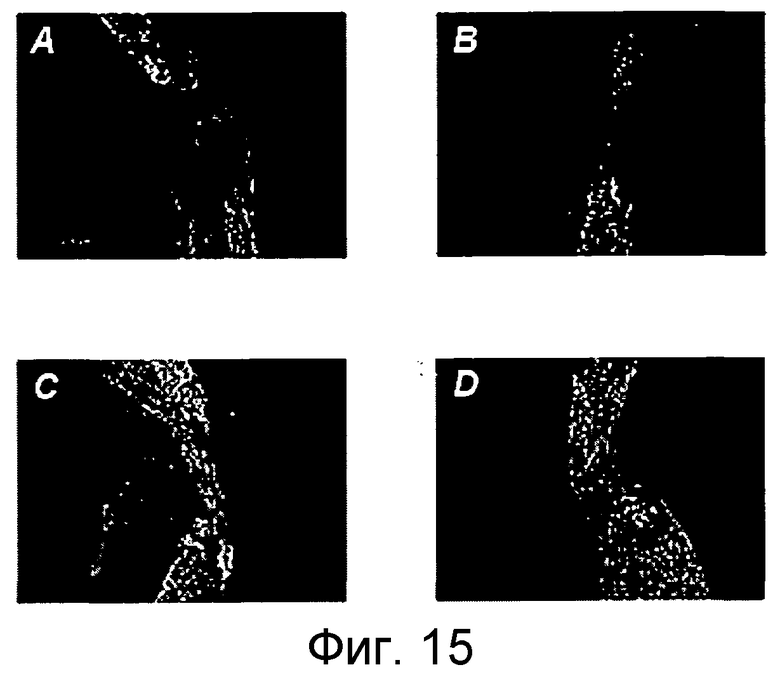
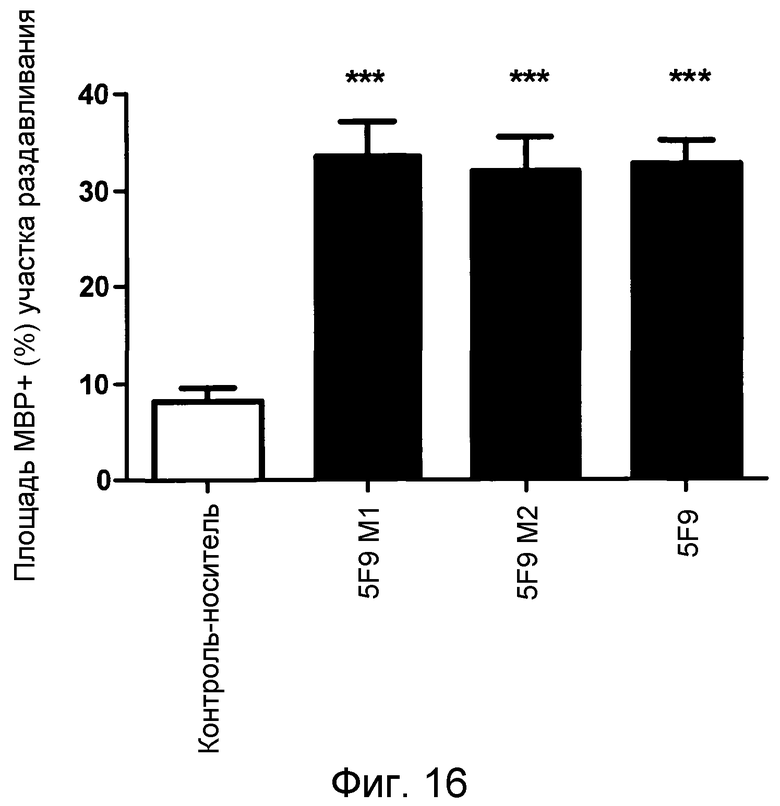
Изобретение относится к области биотехнологии и иммунологии. Описаны варианты антител, связывающих молекулу GRM, а также их антигенсвязывающие фрагменты, аминокислотные последовательности вариабельных областей которых представлены в материалах заявки. Представлена нуклеиновая кислота, кодирующая указанные антитела. Предложен способ получения белка, связывающего RGM, включающий культивирование клетки-хозяина в культуральной среде в условиях, подходящих для получения связывающего белка, способного связываться с RGM, где клетка-хозяин содержит вектор экспрессии, содержащий выделенную нуклеиновую кислоту, кодирующую указанное антитело. Описана фармацевтическая композиция для лечения заболевания, в котором активность RGM А оказывает негативное воздействие, содержащая терапевтически эффективное количество указанного антитела и фармацевтически приемлемый носитель. Предложено применение указанного антитела для получения лекарственного средства, используемого для a) снижения связывания hRGM А с рецептором Neogenin больного; или b) для снижения связывания hRGM А с ВМР-2 и ВМР-4 у больного. Изобретение позволяет получить антитела против GRM, которые используются для лечения заболеваний, связанных с избыточным взаимодействием RGM с рецептором Neogenin, ВМР-2 и ВМР-4. 9 н. и 4 з.п. ф-лы, 16 ил., 10 табл., 11 пр.
1. Выделенное моноклональное антитело, содержащее антигенсвязывающий домен, где указанное антитело способно связывать эпитоп молекулы RGM, указанный антигенсвязывающий домен содержит наборы CDR вариабельного домена, состоящие из:
a) VH 5F9 CDR-H1 с последовательностью SEQ ID NO:57, VH5F9 CDR-H2 с последовательностью SEQ ID NO:58 и VH5F9 CDR-Н3 с последовательностью SEQ ID NO:59; и
b) VL5F9 CDR-L1 с последовательностью SEQ ID NO:60, VL5F9 CDR-L2 с последовательностью SEQ ID NO:61 и VL5F9 CDR-L3 с последовательностью SEQ ID NO:62.
2. Антитело по п.1, дополнительно содержащее акцепторную каркасную область человека, где указанная акцепторная каркасная область человека содержит по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32 и 33.
3. Антитело по п.1, содержащее вариабельный домен тяжелой цепи, выбранный из SEQ ID NO:35, 36, 37, 38, 39, 40, 41, 42 и 43; и/или вариабельный домен легкой цепи, выбранный из SEQ ID NO:44, 45 и 46.
4. Антитело по п.1, содержащее вариабельный домен тяжелой цепи с аминокислотной последовательностью SEQ ID NO:34 и легкой цепи с аминокислотной последовательностью SEQ ID NO:10.
5. Выделенное моноклональное антитело, которое способно связывать эпитоп молекулы RGM, содержащее:
(a) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:44;
(b) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:44;
(c) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:44;
(d) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:44;
(e) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:45;
(f) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:45;
(g) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:45;
(h) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:45;
(i) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:46;
(j) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:46;
(k) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:46;
(l) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:46;
(m) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:51;
(n) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:52;
(o) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:53;
(p) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:48 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:54;
(q) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:51;
(r) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:52;
(s) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:53; или
(t) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:50 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:54.
6. Выделенное моноклональное антитело, которое способно связывать эпитоп молекулы RGM, содержащее:
(a) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:51;
(b) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:52;
(c) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:53;
(d) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:47 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:54;
(e) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:51;
(f) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:52;
(g) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:53; или
(h) вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:49 и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:54.
7. Антигенсвязывающий фрагмент выделенного моноклонального антитела по любому из пп.1-6.
8. Выделенная нуклеиновая кислота, кодирующая молекулу антитела по любому из пп.1-6.
9. Способ получения белка, способного связываться с RGM, включающий культивирование клетки-хозяина в культуральной среде в условиях, подходящих для получения связывающего белка, способного связываться с RGM, где клетка-хозяин содержит вектор экспрессии, содержащий выделенную нуклеиновую кислоту, кодирующую молекулу антитела по любому из пп.1-6.
10. Фармацевтическая композиция для лечения заболевания, в котором активность RGM А оказывает негативное воздействие, содержащая терапевтически эффективное количество антитела по любому из пп.1-6, и фармацевтически приемлемый носитель.
11. Применение антитела по любому из пп.1-6 для получения лекарственного средства, используемого для a) снижения связывания hRGM А с рецептором Neogenin больного; или b) для снижения связывания hRGM А с костным морфогенетическим белком-2 и костным морфогенетическим белком-4 (ВМР-2 и ВМР-4) у больного.
12. Применение антитела по любому из пп.1-6 для получения лекарственного средства для снижения активности RGM А у больного, страдающего заболеванием, при котором активность RGM А является патологической.
13. Применение по п.12, где заболевание включает неврологические заболевания, выбранные из группы, включающей боковой амиотрофический склероз, повреждение плечевого сплетения, повреждение головного мозга, в том числе травматическое повреждение головного мозга, церебральный паралич, болезнь Гийена-Барре, лейкодистрофию, рассеянный склероз, поствакцинальный полиомиелит, расщелину позвоночника, повреждение спинного мозга, спинально-мышечную атрофию, спинальные опухоли, инсульт, поперечный миелит; деменцию, сенильную деменцию, когнитивные нарушения умеренной степени, деменцию, связанную с болезнью Альцгеймера, хорею Хантингтона, позднюю дискинезию, гиперкинезию, мании, болезнь Паркинсона, синдром Стила-Ричардсона, синдром Дауна, тяжелую псевдопаралитическую миастению, травму нерва, сосудистый амилоидоз, кровоизлияние в головной мозг I степени с амилоидозом, воспаление головного мозга, острое нарушение со спутанностью сознания, боковой амиотрофический склероз, глаукому и болезнь Альцгеймера.
| KATSUHIKO HATA, et al "RGMa inhibition promotes axonal growth and recovery after spinal cord injury", The Journal of Cell Biology, 10 apr | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Джино-прядильная машина | 1922 |
|
SU173A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| US 7265212 B2, 04.09.2007 | |||
| HERMANSON GREG T | |||
| "Antibody modification and conjugation", Bioconjugate techniques, copyright 1996, Academic press, | |||

