Перекрестная ссылка на родственные заявки
В настоящей заявке испрашивается приоритет заявки на патент США рег. № 60/732243, поданной 1 ноября 2005 г.; заявки на патент США рег. № 60/748317, поданной 7 декабря 2005 г.; и заявки на патент США рег. № 60/799000, поданной 9 мая 2006 г. Содержание каждой из указанных предварительных заявок во всей своей полноте вводится в настоящее описание посредством ссылки.
Область, к которой относится изобретение
Настоящее изобретение относится к терапии гриппа и к композициям и способам, применяемым для модуляции репликации вируса, а более конкретно для негативной регуляции гена(ов) вируса гриппа с использованием олигонуклеотидов, которые действуют посредством интерференции РНК и которые вводят местно в легкие и в носовые ходы путем ингаляции/интраназально или системно, например путем внутривенной инъекции.
Предшествующий уровень техники
Термин интерференция РНК или “РНКи" был впервые введен в употребление Fire и сотрудниками для описания проводимых наблюдений, в процессе которых было обнаружено, что при введении двухцепочечной РНК (дцРНК) червям она может блокировать экспрессию гена (Fire et al., Nature 391:806-811, 1998). Короткоцепочечная дцРНК регулирует геноспецифический посттранскрипционный сайленсинг во многих организмах, включая позвоночных, и используется как новый инструмент для исследования функции генов. За последнее время эта технология была уже много раз описана в литературе, см., например, публикации Novina, C. D. & Sharp, P., Nature 2004, 430:161, и Sandy, P., et al., Biotechniques 2005, 39:215, которые вводятся в настоящее описание посредством ссылки.
Грипп является одним из наиболее широко распространенных инфекционных заболеваний во всем мире. Это может показаться невероятным, но, по оценкам специалистов, в 1918 году при пандемии гриппа, вызванного вирусом штамма А, умерло от 20 до 40 миллионов человек. В Соединенных Штатах ежегодно от гриппа, вызываемого вирусом штамма А, или от его осложнений умирает от 20 до 40 тысяч человек. Во время эпидемий только за одну зиму число случаев госпитализаций по поводу гриппа может достигать 300000.
Эпидемиологическому распространению вируса гриппа благоприятствуют некоторые свойства этого вируса. Во-первых, он легко передается от человека к человеку воздушно-капельным путем (воздушно-капельная инфекция). Во-вторых, в антигенах вируса гриппа часто возникают небольшие изменения (антигенный дрейф), в результате чего этот вирус легко “ускользает” от иммунного “надзора”, индуцированного предварительным воздействием другого варианта этого вируса. В-третьих, благодаря реассортации или смешиванию генетического материала различных штаммов (антигенный дрейф) могут легко продуцироваться новые штаммы вируса гриппа. В случае вируса гриппа А такое смешивание может происходить между подтипами или штаммами, которые поражают организмы различных видов. Считается, что пандемия 1918 года была вызвана возникновением гибридного штамма вируса, продуцированного в результате реассортации свиного и человеческого вируса гриппа А. В настоящее время существует большая опасность возможного распространения новых штаммов гриппа, являющихся инфекционными для человека, в частности, вариантов птичьего гриппа, а более конкретно штамма H5N1, которые образуются в результате смешивания человеческого и птичьего вирусов при одновременном их воздействии на человека. Эксперты уже не ставят под сомнение тот факт, что тесный контакт между домашней птицей и птицеводами, существующий во многих странах Азии, приведет к возникновению такого смешанного штамма, и они убеждены, что это лишь вопрос времени. При быстром развитии такой пандемии во всем мире ее последствия могут быть гораздо серьезнее, чем в 1918 году.
Несмотря на огромные усилия, предпринимаемые в настоящее время, пока еще не были разработаны эффективные способы лечения инфекций, вызываемых вирусом гриппа, а действие уже существующих вакцин ограничено отчасти их склонностью к антигенной изменчивости и антигенному дрейфу, как указывается выше. По этим причинам глобальные исследования вируса гриппа А продолжаются уже в течение многих лет, и Национальный институт здоровья США назвал эти исследования одним из самых приоритетных направлений в борьбе против вирусных патогенов. Хотя современные вакцины, которые были получены на основе инактивированных вирусов, способны предупреждать заболевание примерно у 70-80% здоровых индивидуумов в возрасте до 65 лет, однако у индивидуумов более старшего возраста или у индивидуумов с нарушенным иммунитетом этот процент гораздо ниже. Кроме того, высокая стоимость этих вакцин и возможные побочные эффекты, ассоциированные с введением этих вакцин, снижают эффективность таких вакцин до уровня ниже оптимального. Хотя современные противовирусные лекарственные средства, разрешенные для применения в Соединенных Штатах, являются эффективными для лечения и/или профилактики гриппа, однако они находят ограниченное применение из-за побочных эффектов, неудобства их употребления пациентом и возможного возникновения резистентных штаммов.
В заявке на патент США 20040242518 и в соответствующей заявке WO 04/028471, поданных 29 сентября 2003 г., предлагается небольшое число РНКи-агентов для лечения гриппа. Однако их эффективность для человека не описана.
Поэтому необходимость в разработке эффективной терапии, направленной на лечение и предупреждение инфекций, вызываемых вирусом гриппа у человека и животных, а в частности высокоэффективной терапии, направленной на уничтожение подтипов вируса гриппа широкого ряда, остается актуальной. Одним из необходимых условий высокой эффективности терапии является медленное разложение активного ингредиента в физиологических условиях.
Описание сущности изобретения
Настоящее изобретение основано на обнаружении in vitro и in vivo того факта, что инфекция, вызываемая вирусом гриппа, может быть ингибирована путем интраназального введения иРНК-агентов, а также путем парентерального введения таких агентов и идентификации эффективных иРНК-агентов, которые происходят от генов MP, NP, PB1, PB2 или PA вируса гриппа и которые могут снижать уровни РНК вируса гриппа некоторых подтипов. На основании этого было разработано настоящее изобретение, которое относится к конкретным композициям и к способам, применяемым для снижения уровней мРНК вируса гриппа, уровней белка вируса гриппа и титров вируса гриппа у индивидуума, например у млекопитающего, такого как человек.
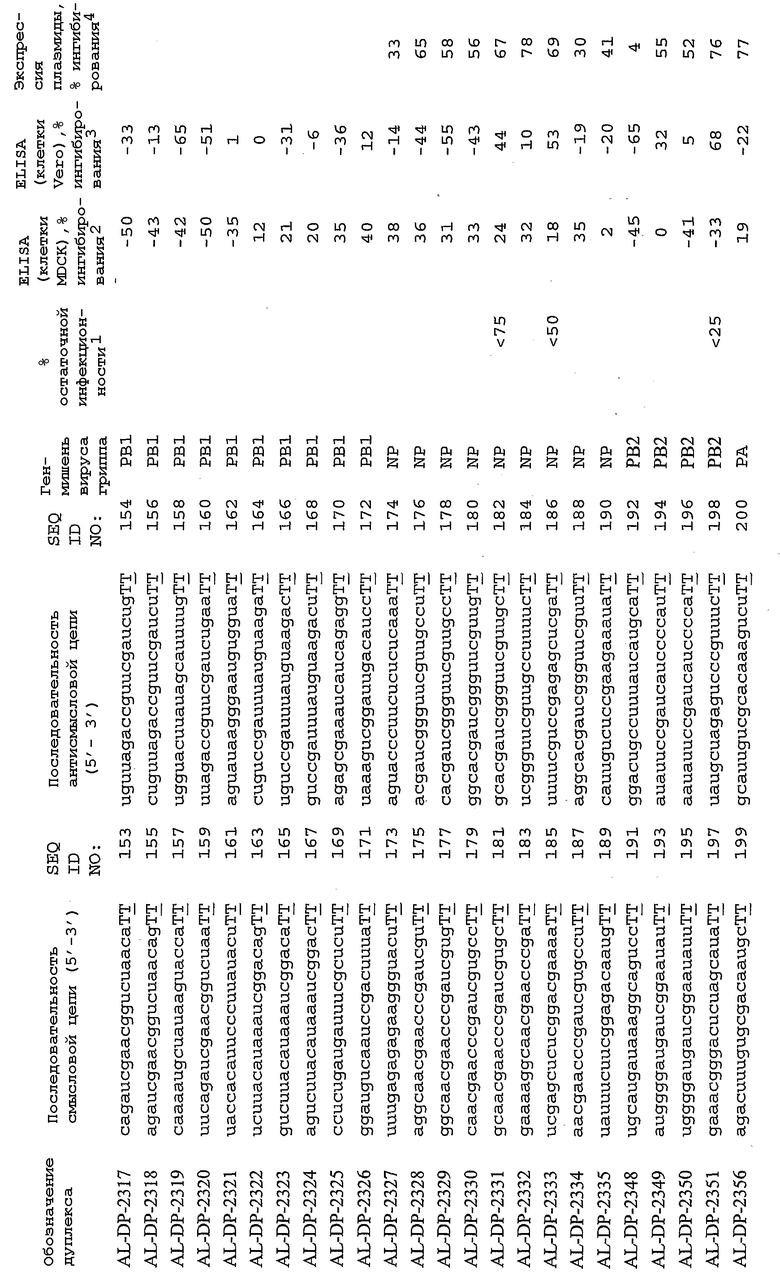
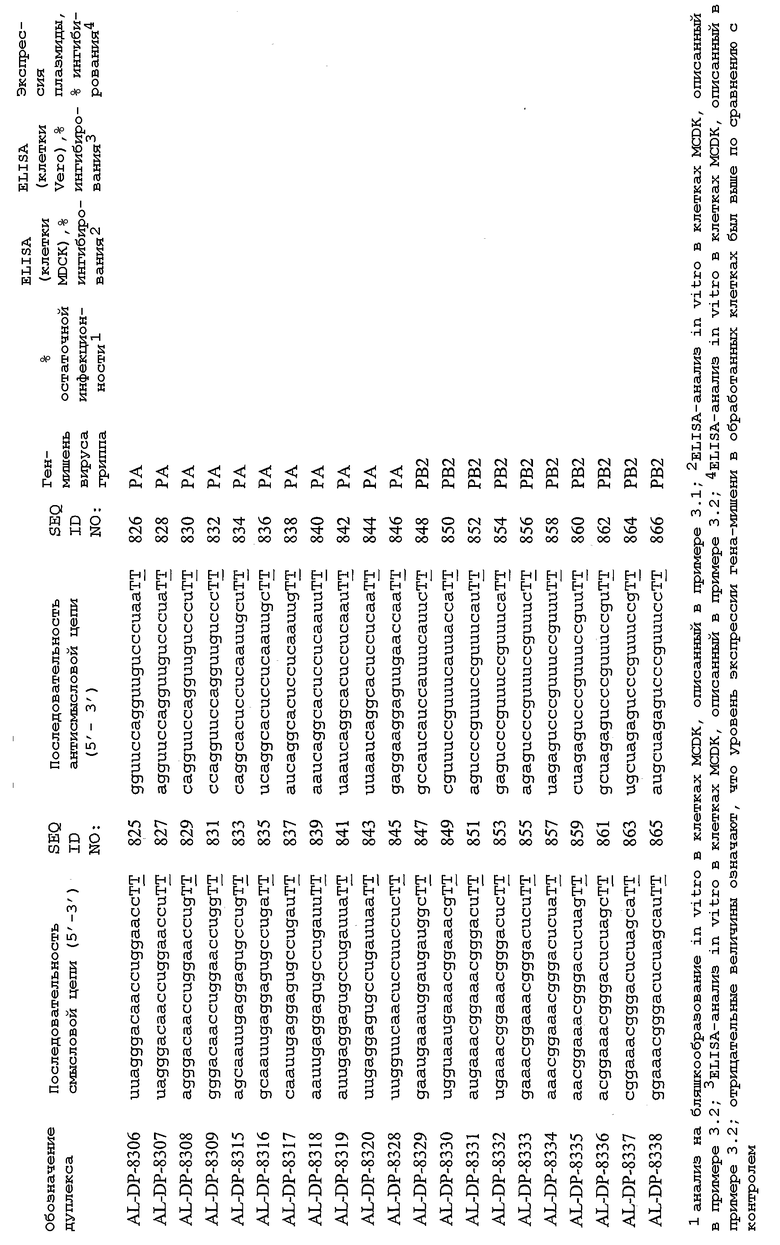
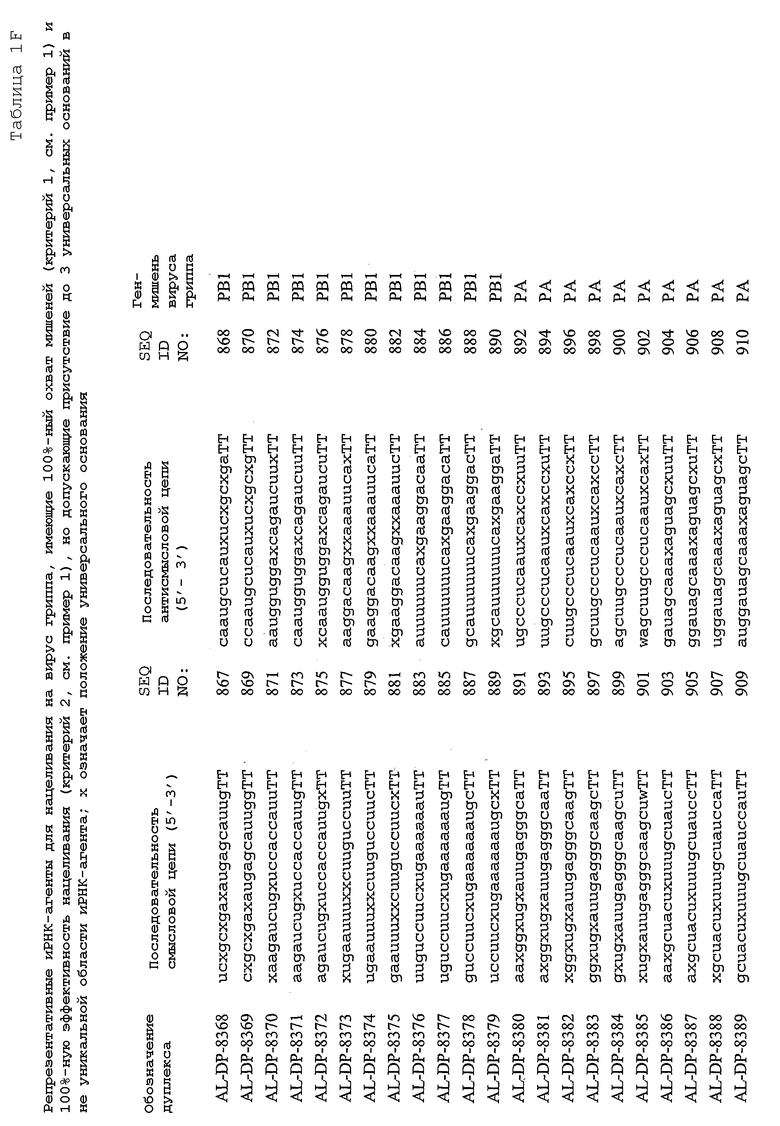
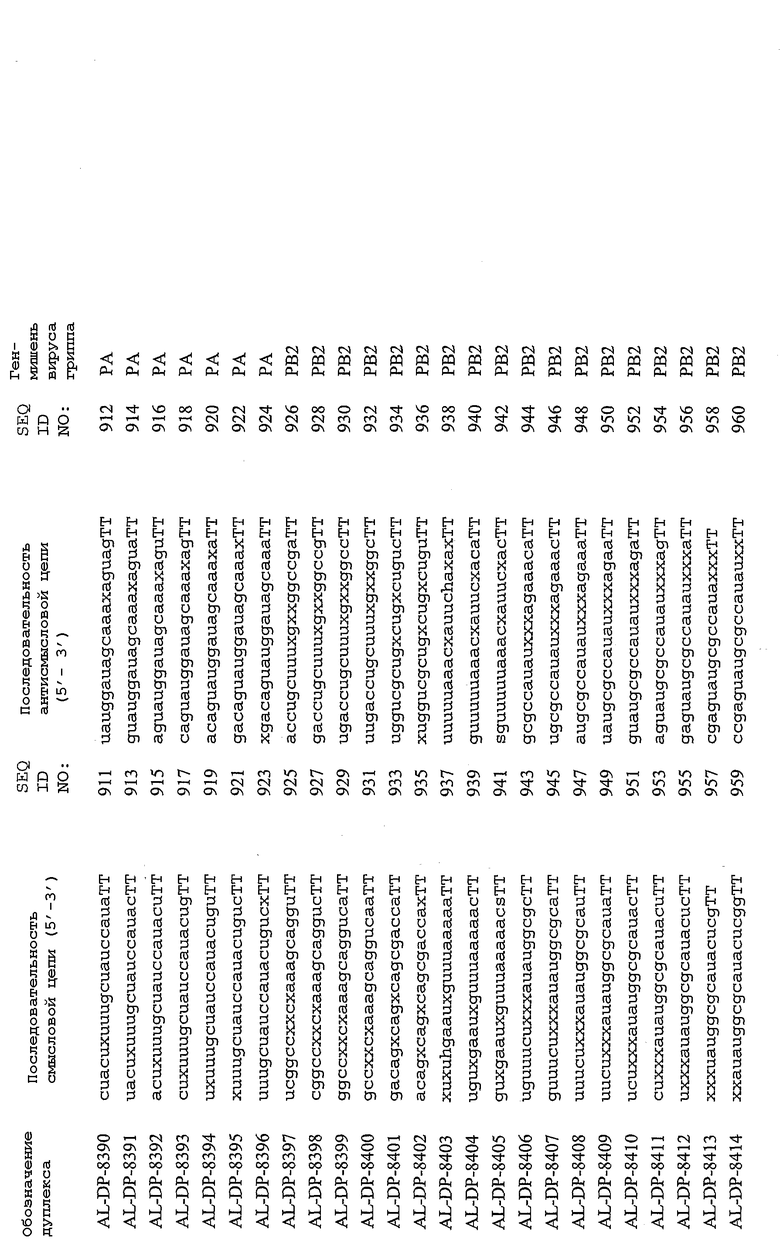
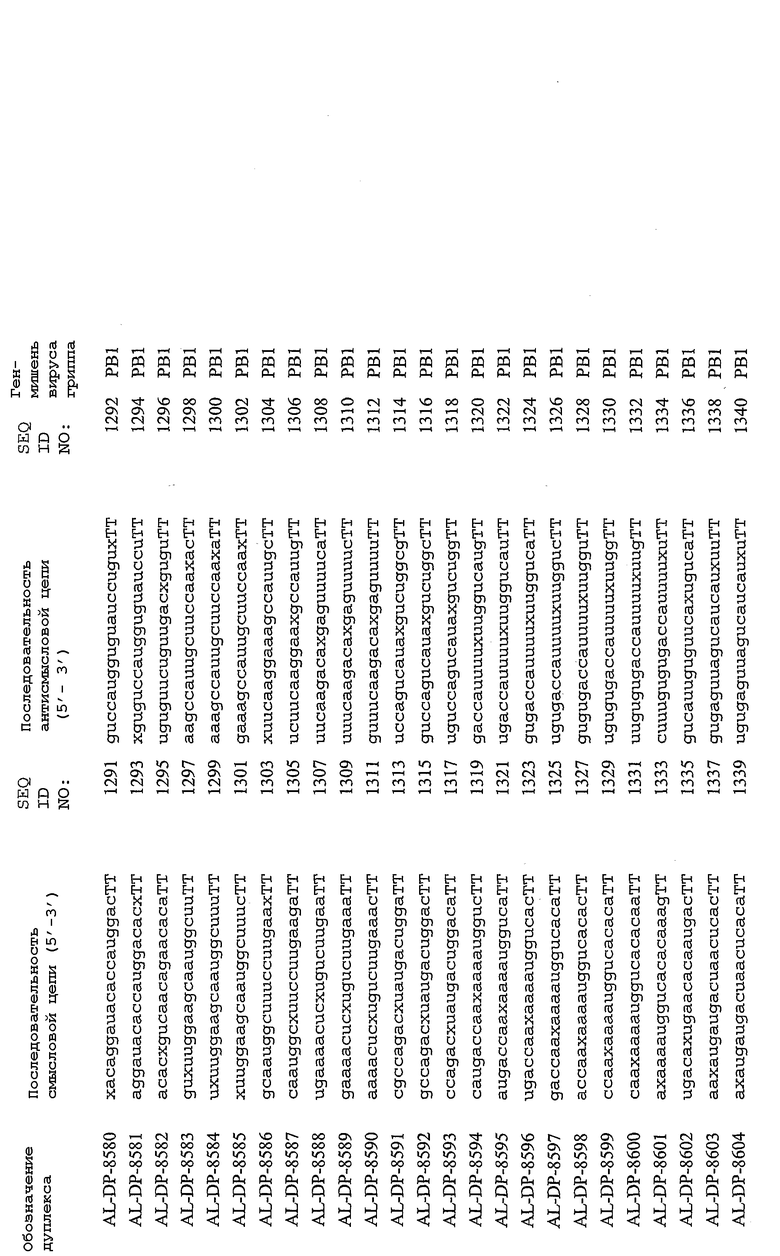
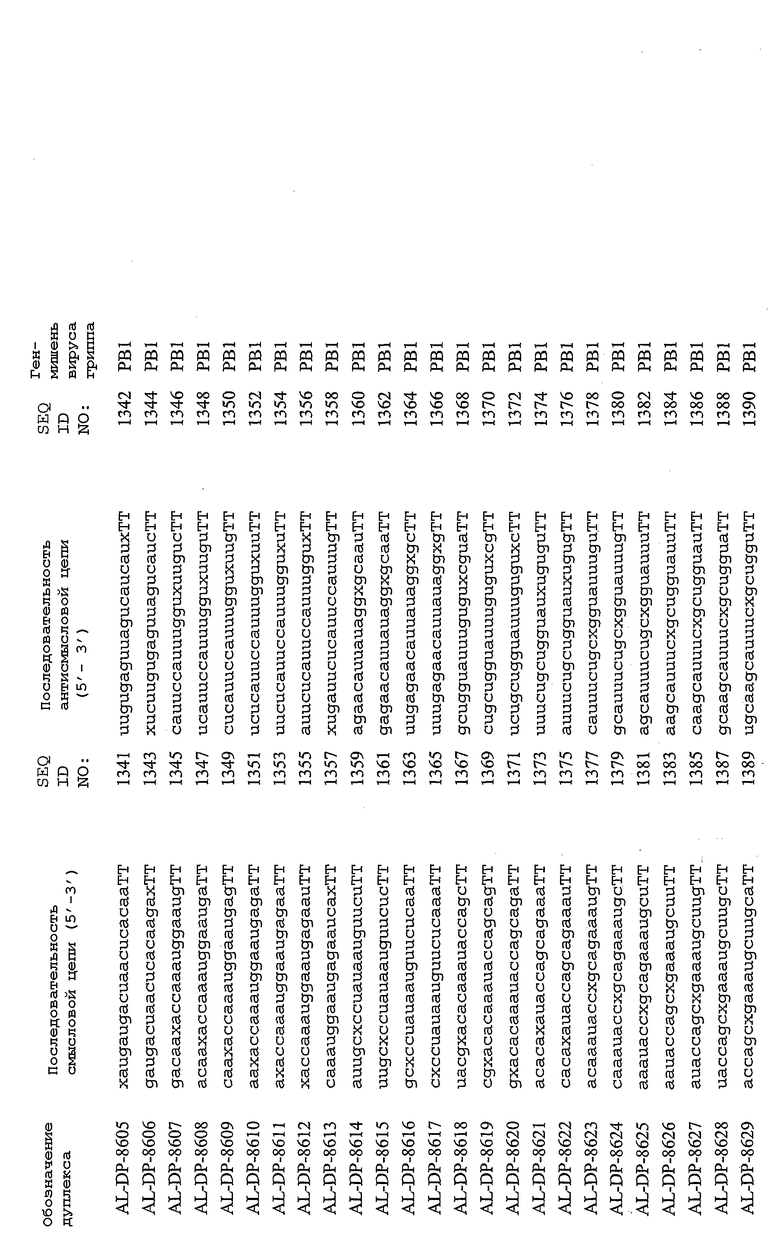
Настоящее изобретение, в частности, относится к иРНК-агентам, включающим либо состоящим или, в основном, состоящим из них, по меньшей мере, 15 или более смежных нуклеотидов одного из генов вируса гриппа, а в частности генов MP, NP, PB1, PB2 и PA вируса гриппа, а более конкретно к агентам, содержащим 15 или более смежных нуклеотидов одной из последовательностей, представленных в таблицах 1A-1H. иРНК-агент, предпочтительно, содержит менее чем 30 нуклеотидов на цепь, например 21-23 нуклеотида, таких как нуклеотиды, описанные в таблицах 1А-1Н. Двухцепочечный иРНК-агент либо может иметь тупые концы, либо он может, что предпочтительно, иметь выступающие концы, состоящие из 1-4 нуклеотидов, расположенных со стороны одного или обоих 3'-концов этого агента.
Кроме того, иРНК-агент может содержать либо только природные рибонуклеотидные субъединицы, либо он может быть синтезирован так, чтобы он содержал одну или несколько модификаций сахара или оснований в одной или в нескольких рибонуклеотидных субъединицах, включенных в этот агент. иРНК-агент может быть дополнительно модифицирован так, чтобы он был присоединен к лиганду, выбранному в целях повышения стабильности, распределения или поглощения вещества, например холестерина, в клетках. иРНК-агенты могут быть также получены в отдельной форме либо они могут быть частью фармацевтической композиции, используемой в описанных здесь способах, а в частности в фармацевтической композиции, предназначенной для доставки в легкие или в носовые ходы или для парентерального введения. Такие фармацевтические композиции могут содержать один или несколько иРНК-агентов, а в некоторых вариантах изобретения они могут содержать два или более иРНК-агентов, каждый из которых направлен на различные сегменты гена вируса гриппа или на различные гены вируса гриппа.
В одном из своих аспектов настоящее изобретение относится к двухцепочечному олигонуклеотиду, содержащему, по меньшей мере, одно неприродное нуклеотидное основание. В некоторых вариантах изобретения таким неприродным нуклеотидным основанием является дифтортолил, нитроиндолил, нитропирролил или нитроимидазолил. В предпочтительном варианте изобретения таким неприродным нуклеотидным основанием является дифтортолил. В некоторых вариантах изобретения только одна из двух олигонуклеотидных цепей, включающих двухцепочечный олигонуклеотид, содержит неприродное нуклеотидное основание. В некоторых вариантах изобретения обе олигонуклеотидные цепи, включающие двухцепочечный олигонуклеотид, независимо друг от друга содержат неприродное нуклеотидное основание.
Настоящее изобретение также относится к способам снижения уровня РНК вируса гриппа в клетке. Такие способы включают стадию введения одного из иРНК-агентов согласно изобретению индивидууму, подробно описанную ниже. В способах согласно изобретению используются клеточные механизмы, участвующие в интерференции РНК, приводящей к селективному разрушению вирусной РНК в клетке, и такие способы включают стадию контактирования клетки с одним из антивирусных иРНК-агентов согласно изобретению. Такие способы могут быть осуществлены непосредственно на клетках, либо они могут быть осуществлены путем введения индивидууму-млекопитающему одного из таких средств, как иРНК-агенты/фармацевтические композиции согласно изобретению. Снижение уровня вирусной РНК в клетке приводит к уменьшению количества продуцируемого вирусного белка, а в организме индивидуума - к снижению титра реплицирующегося вируса (как показано в примерах).
Способы и композиции согласно изобретению, например способы, в которых используется иРНК-агент, и композиции, содержащие иРНК-агент, могут быть применены в любой из описанных здесь лекарственных форм и/или препаратов, и такие формы и препараты могут быть также введены любым из описанных здесь способов. Особенно важным аспектом настоящего изобретения является интраназальное введение иРНК-агента и его способность ингибировать репликацию вируса в тканях дыхательных путей.
Подробное описание одного или нескольких вариантов осуществления настоящего изобретения приводится ниже в описании графического материала и в описании изобретения. Другие признаки, цели и преимущества изобретения будут более очевидны из описания изобретения, из графического материала и из формулы изобретения. Все цитируемые здесь работы, патенты и патентные заявки во всей своей полноте и во всех целях вводятся в настоящее описание посредством ссылки.
Краткое описание графического материала
Фиг. 1A-1I: Кривые “доза-ответ”, иллюстрирующие ингибирование экспрессии гена-мишени с использованием выбранных РНКи-агентов. Соответствующий ген-мишень рекомбинантно клонировали в плазмиде в клетки Cos-7, что приводило к экспрессии мРНК, кодирующей ген-мишень и люциферазу Renilla, а затем эти клетки обрабатывали РНКи-агентом и определяли количество люциферазы Renilla. Клетки обрабатывали РНКи-агентом при концентрациях 100 нМ, 25 нМ, 6,3 нМ, 1,6 нМ, 400 пМ, 100 пМ, 24 пМ, 6 пМ, 1,5 пМ и 380 фМ и величины IC50 определяли путем построения параметрической кривой с помощью программы XLfit.
Подробное описание изобретения
Используемый здесь термин “вирус гриппа” означает любой штамм вируса гриппа, который способен вызывать заболевание у животных или человека или который представляет интерес как кандидат для экспериментального анализа. Вирусы гриппа описаны в публикациях Fields, B., et al., Fields' Virology, 4th ed. 2001, Lippincott Williams and Wilkins; Philadelphia, ISBN: 0781718325. В частности, этот термин включает любой штамм вируса гриппа А, который способен вызывать заболевание у животного или человека или который представляет интерес как кандидат для экспериментального анализа. Было частично или полностью секвенировано большое число изолятов вируса гриппа А. В таблице 6 представлен лишь неполный список полноразмерных последовательностей сегментов генома вируса гриппа А, которые были депонированы в общедоступной базе данных (база данных последовательностей вируса гриппа (ISD), см. Macken, C, Lu, H., Goodman, J., & Boykin, L., "The value of a database in surveillance and vaccine selection." in Options for the Control of influenza IV. A. D. M. E. Osterhaus, N. Cox & A. W. Hampson (Eds.) 2001, Elsevier Science, Amsterdam, pp 103-106). Эта база данных также содержит полноразмерные последовательности сегментов генома вируса гриппа B и C. Указанная база данных имеется в Интернете и включает удобный механизм поиска, который позволяет пользователю осуществлять поиск по сегментам генома, по видам, инфицированным вирусом, и по году выделения штамма. Последовательности вируса гриппа также имеются в базе данных Genbank. Поэтому последовательности генов вируса гриппа являются легко доступными для среднего специалиста в данной области, либо они могут быть легко определены специалистом.
Для лучшего понимания используемых здесь терминов “нуклеотид" или “рибонуклеотид" следует отметить, что иногда эти термины применяются к одной или нескольким мономерным субъединицам РНК-агента. Кроме того, следует отметить, что используемый здесь термин “рибонуклеотид" или “нуклеотид", в том случае, если он относится к модифицированной РНК или нуклеотидному заменителю, также означает модифицированный нуклеотид или заменяющую его часть, подробно описанную ниже, в одном или нескольких положениях.
Используемый здесь термин “РНК-агент" означает немодифицированную РНК, модифицированную РНК или нуклеозидную замену, каждая из которых описана в настоящей заявке и хорошо известна специалистам в области синтеза РНК. Хотя в настоящей заявке описано множество модифицированных РНК и нуклеозидных замен, однако предпочтительными примерами являются молекулы, которые обладают большей резистентностью к расщеплению нуклеазой, чем немодифицированные РНК. Предпочтительными примерами являются молекулы, имеющие модификацию в 2'-положении сахара, модификацию в одноцепочечном выступающем конце, а в частности в одноцепочечном выступающем 3'-конце, или, в случае одноцепочечной молекулы, 5'-модификацию, которая включает одну или несколько фосфатных групп или один или несколько аналогов фосфатной группы.
Используемый здесь термин “иРНК-агент" (данное сокращение означает “интерферирующий РНК-агент") означает РНК-агент, который может ингибировать экспрессию гена-мишени, например, вируса гриппа. Не ограничиваясь какой-либо теорией, следует отметить, что иРНК-агент может действовать по одному или нескольким механизмам, включая посттранскрипционное расщепление мРНК-мишени, иногда называемое специалистами РНКи, или по претранскрипционным или претрансляционным механизмам. иРНК-агент может представлять собой двухцепочечный иРНК-агент.
Используемый здесь термин “дц-иРНК-агент" (данное сокращение означает “двухцепочечный иРНК-агент") представляет собой иРНК-агент, имеющий более чем одну, а предпочтительно две цепи, в которых межцепьевая гибридизация может приводить к образованию области с дуплексной структурой. Используемый здесь термин “цепь” означает последовательность из смежных нуклеотидов (включая неприродные или модифицированные нуклеотиды). Такие две цепи или несколько цепей могут представлять собой отдельные молекулы, либо каждый из них может быть частью отдельных молекул, либо они могут быть ковалентно связаны между собой, например, посредством линкера, такого как полиэтиленгликолевый линкер, с образованием одной молекулы. По меньшей мере, одна цепь может включать область, которая является в достаточной степени комплементарной РНК-мишени. Такая цепь называется “антисмысловой цепью”. Вторая цепь дцРНК-агента, содержащая область, комплементарную антисмысловой цепи, называется “смысловой цепью”. Однако дц-иРНК-агент может быть также образован из одной молекулы РНК, которая является по меньшей мере частично аутокомплементарной, образуя, например, шпилечную структуру или структуру, которая по своей форме напоминает “ручку сковородки” и включает область дуплекса. Последнюю из упомянутых выше структур называют короткой шпилечной РНК или кшРНК. В этом случае термин “цепь” означает одну из областей РНК-молекул, которая комплементарна другой области той же самой молекулы РНК.
В клетках млекопитающих длинные двухцепочечные иРНК-агенты могут индуцировать интерфероновый ответ, который часто является нежелательным, однако короткие двухцепочечные иРНК-агенты не индуцируют интерфероновый ответ, по меньшей мере, на таком уровне, который мог бы негативно воздействовать на клетки и/или на организм хозяина (Manche et al., Mol. Cell. Biol. 12:5238, 1992; Lee et al., Virology 199:491, 1994; Castelli et al., J. Exp. Med. 186:967, 1997; Zheng et al., RNA 10:1934, 2004; Heidel et al., "Lack of interferon response in animals to naked siRNAs" Nature Biotechn. advance online publication doi:10.1038/nbtl038, Nov. 21, 2004). иРНК-агенты согласно изобретению включают молекулы, которые являются достаточно короткими, то есть они не способны индуцировать нежелательный неспецифический интерфероновый ответ в нормальных клетках млекопитающих. Так, например, введение индивидууму композиции, включающей иРНК-агент (например, приготовленной, как описано в настоящей заявке), может быть осуществлено в целях снижения уровня экспрессии генов вируса гриппа в клетках, экспрессирующих вирус гриппа у индивидуума, что позволяет избежать вырабатывания интерферонового ответа. Молекулы, которые являются достаточно короткими и не способны индуцировать нежелательный интерфероновый ответ, называются здесь киРНК-агентами или киРНК. Используемый здесь термин “киРНК-агент" или “киРНК" означает иРНК-агент, например дц-иРНК-агент, который является достаточно коротким и не способен индуцировать нежелательный интерфероновый ответ в клетках млекопитающих, а в частности в клетках человека, например, он имеет дуплексную область, содержащую менее чем 60, а предпочтительно менее чем 50, 40 или 30 пар нуклеотидов.
Описанные здесь выделенные иРНК-агенты, включая дц-иРНК-агенты и киРНК-агенты, могут опосредовать снижение уровня экспрессии нуклеиновой кислоты вируса гриппа, например, посредством разрушения РНК. Для удобства такая РНК будет здесь также называться молчащей РНК. Такая нуклеиновая кислота также называется здесь геном-мишенью. Предпочтительно, молчащая РНК представляет собой генный продукт гена вируса гриппа, который является частью штамма вируса гриппа, патогенного для человека.
Используемый здесь термин “опосредует РНКи" означает способность данного агента к последовательность-специфической инактивации гена-мишени. Термин “сайленсинг гена-мишени” означает процесс, при котором клетка, содержащая и/или экспрессирующая определенный продукт гена-мишени, в том случае, когда она вступает в контакт с данным агентом, будет содержать и/или экспрессировать, по меньшей мере, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% или менее указанного генного продукта по сравнению с аналогичной клеткой, которая не контактировала с данным агентом. Указанный продукт гена-мишени может представлять собой, например, матричную РНК (мРНК), белок или регуляторный элемент.
Используемый здесь термин “комплементарный” указывает на достаточную степень комплементарности, то есть на осуществление стабильного и специфического связывания соединения согласно изобретению с РНК-молекулой-мишенью, например мРНК вируса гриппа. Для специфического связывания необходима достаточная степень комплементартности, что позволяет избежать неспецифического связывания олигомерного соединения с последовательностями, не являющимися мишенями, в условиях, при которых желательно специфическое связывание, а именно в физиологических условиях, во время проведения in vivo анализов или терапевтического лечения, или во время осуществления in vitro анализов в условиях, при которых проводят такие анализы. Не являющиеся мишенями последовательности обычно отличаются от последовательностей-мишеней, по меньшей мере, на 2, 3 или 4 нуклеотида.
Используемый здесь иРНК-агент является “достаточно комплементарным” РНК-мишени, например мРНК-мишени (например, мРНК-мишени вируса гриппа), если такой иРНК-агент способствует снижению уровня продуцирования белка, кодируемого РНК-мишенью в клетке. Такой иРНК-агент может быть также “абсолютно комплементарным” РНК-мишени, например РНК-мишени, и такой иРНК-агент гибридизуется, предпочтительно, с образованием гибрида, состоящего исключительно из пар оснований Уотсона-Крика в области абсолютной комплементарности. “Достаточно комплементарный” иРНК-агент может включать внутреннюю область (например, по меньшей мере, из 10 нуклеотидов), которая является абсолютно комплементарной РНК-мишени вируса гриппа. Кроме того, в некоторых вариантах изобретения иРНК-агент, в частности, отличается лишь одним нуклеотидом. В этом случае иРНК-агент опосредует РНКи, только если в области (например, из 7 нуклеотидов), отличающейся одним нуклеотидом, имеет место абсолютная комплементарность. Предпочтительные иРНК-агенты созданы на основе смысловых и антисмысловых последовательностей, представленных в таблицах 1A-1H, либо они состоят из этих последовательностей или включают эти последовательности.
Используемый здесь термин “по существу идентичный”, если он используется при сравнении первой нуклеотидной последовательности со второй нуклеотидной последовательностью, означает, что первая нуклеотидная последовательность идентична второй нуклеотидной последовательности, за исключением одной, двух или трех нуклеотидных замен (например, аденозин заменен урацилом). Используемое здесь выражение “по существу сохраняет способность ингибировать экспрессию вируса гриппа в культивированных клетках, экспрессирующих человеческий вирус гриппа”, если оно относится к иРНК-агенту, который не является идентичным одному из иРНК-агентов, указанных в таблицах 1A-1H, но происходит от одного из таких иРНК-агентов, указанных в таблицах 1A-1H, вследствие делеции, добавления или замены нуклеотидов, означает, что такой производный иРНК-агент обладает ингибирующей активностью, составляющей не менее чем 20% от ингибирующей активности ИРНК-агента, представленного в таблицах 1A-1H, от которого он происходит. Так, например, иРНК-агент, происходящий от иРНК-агента, представленного в таблицах 1А-1Н и на 70% снижающего количество мРНК вируса гриппа, присутствующего в культивированных человеческих клетках, инфицированных вирусом гриппа, сам может снижать количество мРНК вируса гриппа, присутствующей в культивированных человеческих клетках, инфицированных вирусом гриппа, по меньшей мере, на 50%, что означает, что он, по существу, сохраняет способность ингибировать репликацию вируса гриппа в культивированных человеческих клетках, инфицированных вирусом гриппа. иРНК-агент согласно изобретению может, но необязательно, снижать количество мРНК вируса гриппа, присутствующее в культивированных человеческих клетках, инфицированных вирусом гриппа, по меньшей мере, на 50%.
Используемый здесь термин “индивидуум” означает млекопитающее, подвергаемое лечению расстройства, опосредуемого инфицированием вирусом гриппа. Таким индивидуумом может быть любое млекопитающее, такое как корова, лошадь, мышь, крыса, собака, свинья, коза или примат. В предпочтительном варианте изобретения указанным индивидуумом является человек.
Свойства вируса гриппа
Вирусы гриппа представляют собой вирусы, имеющие оболочку и минус-цепь РНК и принадлежащие к семейству Ортомиксовирусов. Они подразделяются на подтипы A, B и C, из которых вирус гриппа А является наиболее патогенным и, вероятно, представляет собой один тип, способный подвергаться реассортации (перетасовке) со штаммами животных. Вирусы гриппа A, B и C могут отличаться друг от друга по их нуклеопротеинам и матриксным белкам. Как обсуждается ниже, подтипы вируса гриппа А определяются по отличию их генов гемаглютинина (HA) и нейраминидазы (NA) и обычно идентифицируются по антителам, которые связываются с соответствующими белками.
Геном вируса гриппа A состоит из десяти генов, расположенных в восьми РНК-сегментах. Эти гены кодируют 10 белков: гликопротеины оболочки: гемаглютинин (HA) и нейраминидазу (NA); матриксный белок (обозначаемый здесь M1 или MP); нуклеопротеин (NP); три полимеразы (PB1, PB2 и PA), которые являются компонентами РНК-зависимой РНК-транскриптазы, также называемой здесь полимеразой или полимеразным комплексом; белок ионных каналов (M2) и неструктурные белки (NS1 и NS2). Более подробное описание вируса гриппа А и его молекулярного патогенеза можно найти в публикации Julkunen, I., et al., Cytokine and Growth Factor Reviews, 12: 171-180, 2001. См. также Fields, B., et al., Fields' Virology, 4.sup.th. ed., Philadelphia: Lippincott Williams and Wilkins; ISBN: 0781718325, 2001. Организация генома вируса гриппа В очень похожа на организацию генома вируса гриппа А, а геном вируса гриппа С содержит семь РНК-сегментов и не содержит гена NA.
Классификацию вирусов гриппа A проводят на основе генов гемаглютинина (H1-H15) и нейраминидазы (N1-N9). В соответствии с номенклатурой, принятой Всемирной организацией здравоохранения (ВОЗ), каждый штамм вируса идентифицируется по животному-хозяину, которое он инфицирует (обычно человек, если это не оговорено особо); географическому региону, в котором он был обнаружен; числу штаммов; году выделения; и по антигенным свойствам HA и NA. Так, например, A/Пуэрто-Рико/8/34 (H1N1) означает штамм А, изолят 8, обнаруженный у человека в Пуэрто-Рико в 1934 году и имеющий антигены HA и NA подтипа 1. В другом примере A/куры/Гонконг/258/97 (H5N1) означает штамм А, изолят 258, обнаруженный у кур в Гонконге в 1997 г. и имеющий антиген HA подтипа 5 и антиген NA подтипа 1. Эпидемия у людей была вызвана вирусами, имеющими HA типов H1, H2 и H3 и NA типов N1 и N2.
Как упоминалось выше, генетические изменения в вирусе гриппа А происходят по двум основным механизмам. Антигенный дрейф происходит в результате точковых мутаций, которые часто возникают в антиген-специфических положениях вследствие давления отбора, вследствие иммунных ответов у хозяина и вследствие антигенной изменчивости (также называемой реассортацией), обусловленной заменой целого сегмента вирусного генома одного подтипа на сегмент другого подтипа. Многие различные типы животных, включая человека, свиней, птиц, лошадей, водных млекопитающих и т.п., могут быть инфицированы вирусами гриппа А. Некоторые вирусы гриппа A инфицируют животных конкретных видов и обычно не инфицируют животных других видов. Однако некоторые вирусы гриппа А могут инфицировать животных нескольких различных видов, главным образом птиц (в частности, перелетных водоплавающих птиц), свиней и человека. Считается, что такое свойство этого вируса ответственно за основную антигенную изменчивость вируса гриппа А. Так, например, было высказано предположение, что свинья инфицируется вирусом гриппа А, передаваемым от человека, и одновременно инфицируется другим вирусом гриппа А, передаваемым от уток. Если эти два различных вируса репродуцируются в клетках свиней, то гены человеческого и утиного штаммов могут “смешиваться”, что может приводить к образованию нового вируса с уникальной комбинацией РНК-сегментов. Этот процесс называется генетической реассортацией. (Следует отметить, что такой тип генетической реассортации отличается от изменения генетической информации, которое происходит между хромосомами в процессе мейоза).
Вирусы гриппа, подобно другим вирусам и некоторым бактериальным вирусам, реплицируются внутри клеток. Вирусы гриппа А реплицируются в эпителиальных клетках верхних дыхательных путей. Однако этим вирусом могут также инфицироваться моноциты/макрофаги и другие лейкоциты. Множество других клеток с поверхностными гликопротеинами, содержащими сиаловую кислоту, являются восприимчивыми к инфицированию in vitro, поскольку вирус использует эти молекулы в качестве рецептора.
Конструирование и отбор иРНК-агентов
Используемый здесь термин “расстройства, ассоциированные с экспрессией вируса гриппа” означает любое биологическое или патологическое состояние, (1) которое опосредуется, по меньшей мере, частично, присутствием вируса гриппа и (2) на исход которого влияет снижение уровня присутствующего вируса гриппа. Конкретные расстройства, ассоциированные с экспрессией вируса гриппа, приводятся ниже.
Настоящее изобретение основано на конструировании, синтезе и продуцировании иРНК-агентов, которые нацелены на гены вируса гриппа, и на выявление сайленсинга вирусного гена in vitro в культивированных клетках после их инкубирования с иРНК-агентом, а также на обеспечение защиты от инфекции, вызываемой вирусом гриппа.
иРНК-агент может быть соответствующим образом сконструирован, исходя из информации о последовательности и нужных свойствах этого агента. Так, например, иРНК-агент может быть сконструирован с учетом относительной температуры плавления дуплекса-кандидата. Обычно такой дуплекс должен иметь более низкую температуру плавления у 5'-конца, чем у 3'-конца антисмысловой цепи.
Настоящее изобретение относится к композициям, содержащим киРНК и/или кшРНК, направленные на один или несколько транскриптов вируса гриппа. Как продемонстрировано выше в описании репликативного цикла вируса гриппа, в клетках, инфицированных вирусом гриппа, присутствуют различные типы вирусных РНК-транскриптов (первичная и вторичная вирусная РНК (вРНК), первичная и вторичная вирусная мРНК и вирусная кРНК), и эти вирусные транскрипты играют важную роль в жизненном цикле вируса. Любые из этих транскриптов являются соответствующими мишенями для киРНК-опосредуемого ингибирования по прямому или опосредуемому механизму, описанному в настоящем изобретении. киРНК и кшРНК, которые нацелены на любой вирусный МРНК-транскрипт, будут специфически и непосредственно снижать уровень самого транскрипта, то есть посредством инициации деградации транскрипта. Кроме того, как обсуждается ниже, киРНК и кшРНК, которые направлены на некоторые вирусные транскрипты (например, MP, PA, PB1), будут вызывать опосредуемое снижение уровней вирусных транскриптов, по отношению к которым они не являются специфическими. В случаях, когда возможен альтернативный сплайсинг, как в случае мРНК, кодирующей MP и M2, и мРНК, кодирующей NS1 и NS2, в качестве транскрипта-мишени может служить несплайсированный или сплайсированный транскрипт.
Потенциальными вирусными транскриптами, которые могут служить в качестве мишени для терапии на основе РНКи согласно изобретению, являются, например, 1) любой геномный сегмент вируса гриппа; 2) транскрипты, кодирующие любые вирусные белки, включая транскрипты, кодирующие белки PB1, PB2, PA, NP, NS1, NS2, MP, M2, HA или NA. Следует отметить, что одна из киРНК или кшРНК может быть направлена на транскрипты в форме вРНК, кРНК, и/или мРНК. Однако, по мнению Ge et al., WO 04/028471, такой вирусной мРНК может быть единственная или первичная мишень РНКи.
Для любого конкретно выбранного гена-мишени конструирование киРНК или кшРНК в целях их использования в соответствии с настоящим изобретением, предпочтительно, осуществляют в соответствии с некоторыми критериями. Вообще говоря, желательными являются последовательности-мишени, которые специфичны для данного вируса (по сравнению с последовательностями, специфичными для хозяина) и которые, предпочтительно, играют важную или главную роль в функционировании вируса. Хотя некоторые вирусные гены, особенно гены, кодирующие HA и NA, характеризуются высоким уровнем мутации и толерантны к мутациям, однако некоторые их области и/или последовательности имеют тенденцию к сохранению консервативности. В соответствии с некоторыми вариантами изобретения, такие последовательности могут быть особенно подходящими мишенями. Как будет подробно описано ниже, такие консервативные области могут быть идентифицированы, например, путем поиска в литературе и/или путем сравнения с генными последовательностями вируса гриппа, имеющимися в базах данных, большое число которых является общедоступным. Кроме того, во многих случаях агент, доставляемый в клетку в соответствии с настоящим изобретением, может подвергаться одной или нескольким стадиям процессинга с образованием активного супрессорного агента (как подробно обсуждается ниже); причем в таких случаях для каждого среднего специалиста в данной области очевидно, что подходящий агент, может быть предпочтительно сконструирован так, чтобы он включал последовательности, которые могут оказаться необходимыми для его процессинга. Одним из аспектов настоящего изобретения является распознавание, которое, в случае присутствия множества штаммов, подтипов и т.п. (называемых общим термином варианты) существующих инфекционных агентов с варьирующимися последовательностями геномов, часто оказывается желательным для отбора и/или конструирования киРНК и кшРНК, направленных на области, которые являются в высокой степени консервативными у различных вариантов. В частности, путем сравнения достаточного числа последовательностей и отбора высококонсервативных областей можно получить множество вариантов с одной киРНК, дуплексная часть которых включает такую высококонсервативную область. Вообще говоря, длина каждой области должна быть такой, чтобы она включала всю дуплексную часть киРНК (например, 19 нуклеотидов) и необязательно один или несколько выступающих 3'-концов, хотя могут быть также использованы области, длина которых короче длины полноразмерного дуплекса (например, 15, 16, 17 или 18 нуклеотидов). В соответствии с некоторыми вариантами изобретения, область является высококонсервативной для множества вариантов, если она идентична для всех этих вариантов. В соответствии с некоторыми вариантами изобретения, область (независимо от длины дуплексной части киРНК, например, 15, 16, 17, 18 или, предпочтительно, 19 нуклеотидов) является высококонсервативной, если она отличается, самое большее, на один нуклеотид (то есть 0 или 1 нуклеотид) у различных вариантов. В соответствии с некоторыми вариантами изобретения, такая область является высококонсервативной для множества вариантов, если она отличается, самое большее, на два нуклеотида (то есть 0, 1 или 2 нуклеотида) у различных вариантов. В соответствии с некоторыми вариантами изобретения, область является высококонсервативной для множества вариантов, если она отличается, самое большее, на три нуклеотида (то есть 0, 1, 2 или 3 нуклеотида) для различных вариантов. В соответствии с некоторыми вариантами изобретения, киРНК включает дуплексную часть, которая направлена на область, являющуюся высококонсервативной, по меньшей мере, у 5 вариантов, по меньшей мере, 15 вариантов, по меньшей мере, 20 вариантов, по меньшей мере, 25 вариантов, по меньшей мере, 30 вариантов, по меньшей мере, 40 вариантов, по меньшей мере, 50 вариантов или более.
Для того чтобы определить, является ли область высококонсервативной для ряда вариантов, может быть проведена нижеследующая процедура. Один член ряда последовательностей отбирают как базовую последовательность, то есть последовательность, с которой сравнивают другие последовательности. Обычно длина базовой последовательности имеет длину, желательную для дуплексной части киРНК, например 15, 16, 17, 18 или, предпочтительно, 19 нуклеотидов. В соответствии с различными вариантами изобретения, базовой последовательностью может быть любая одна из последовательностей в наборе сравниваемых последовательностей, либо такой последовательностью может быть происходящая от нее консенсусная последовательность, например, выбранная путем определения каждого положения для каждого нуклеотида, где данный нуклеотид наиболее часто встречается в данном положении в наборе сравниваемых последовательностей.
После отбора базовой последовательности последовательность каждого члена набора из множества вариантов сравнивают с этой базовой последовательностью. Для того чтобы определить, является ли базовая последовательность высококонсервативной в конкретной представляющий интерес области и какой из ее членов является высококонсервативным, учитывается число отличий между базовой последовательностью и любым членом набора из множества вариантов по всей области данной последовательности. Как указывалось выше в различных вариантах согласно изобретению, если число различий в последовательностях между двумя областями составляет 0; 0 или 1; 0, 1 или 2; или 0, 1, 2 или 3, то считается, что такие области являются высококонсервативными. В положениях, в которых присутствуют такие отличия, последовательность киРНК может быть выбрана так, чтобы она была идентична базовой последовательности или одной из других последовательностей. Обычно выбирают нуклеотид, присутствующий в базовой последовательности. Однако в некоторых вариантах изобретения, а в частности в тех вариантах, когда нуклеотид, присутствующий в конкретном положении во второй последовательности сравниваемого набора последовательностей, находится во многих сравниваемых последовательностях, но отсутствует в базовой последовательности, последовательность киРНК может быть выбрана так, чтобы она была идентична этой второй последовательности. Кроме того, в соответствии с некоторыми вариантами изобретения, если консенсусный нуклеотид (наиболее часто встречающийся нуклеотид), присутствующий в положении, где обнаруживаются различия, отличается от нуклеотида базовой последовательности, то может быть использован консенсусный нуклеотид. При этом следует отметить, что в результате может быть получена последовательность, которая не является идентичной любой из сравниваемых последовательностей (поскольку консенсусная последовательность может быть использована в качестве базовой последовательности).
Авторами настоящего изобретения было установлено, что значительное число последовательностей, выбранных с использованием описанных ниже параметров конструирования (см. пример 1), являются эффективными для подавления репликации вируса, если они включены в киРНК или в кшРНК, как показал тест, описанный ниже.
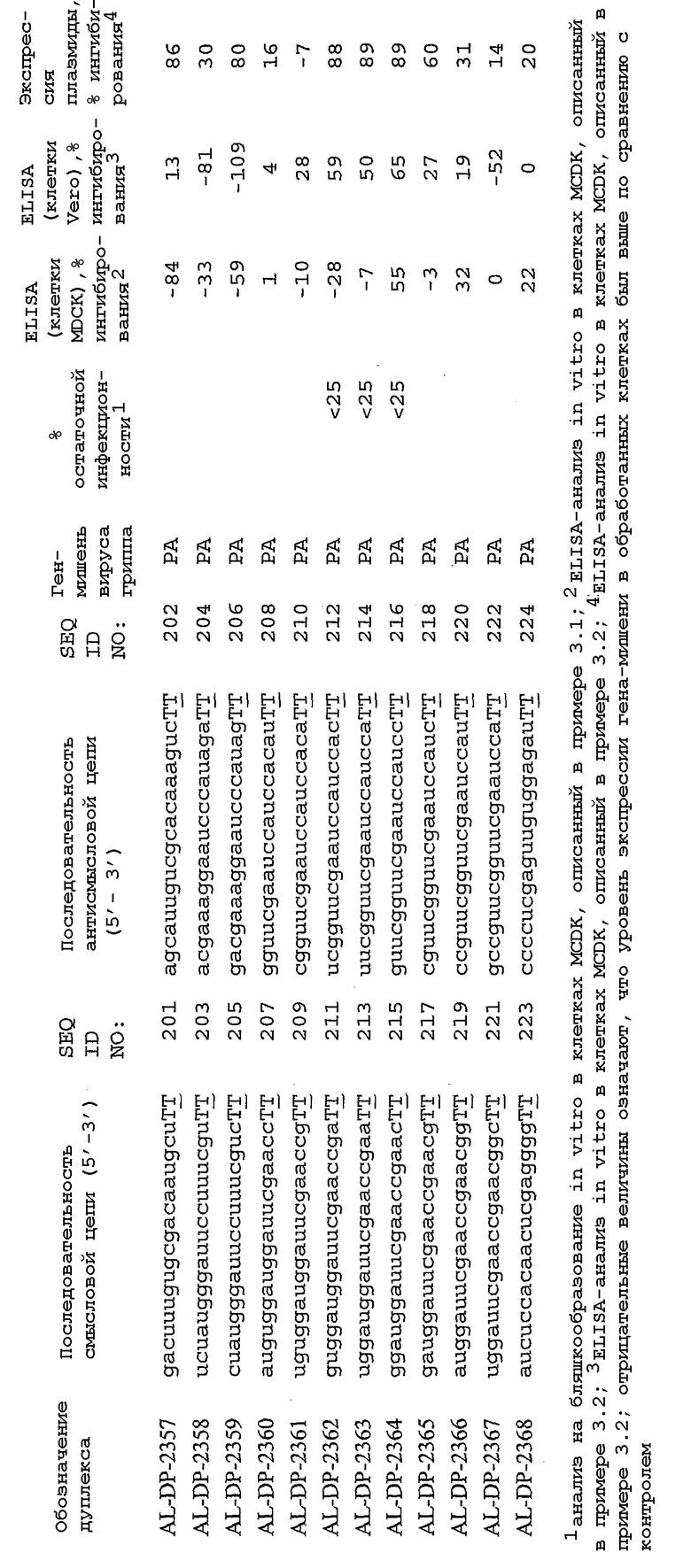
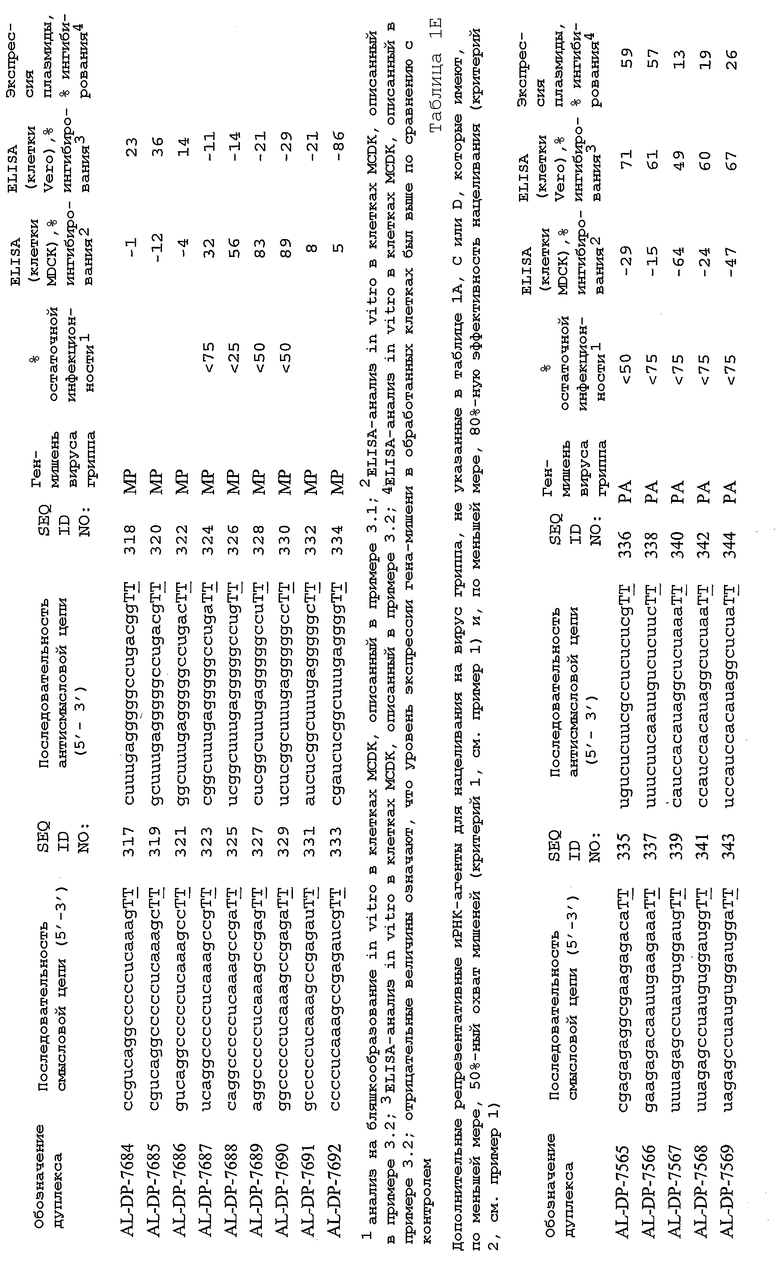
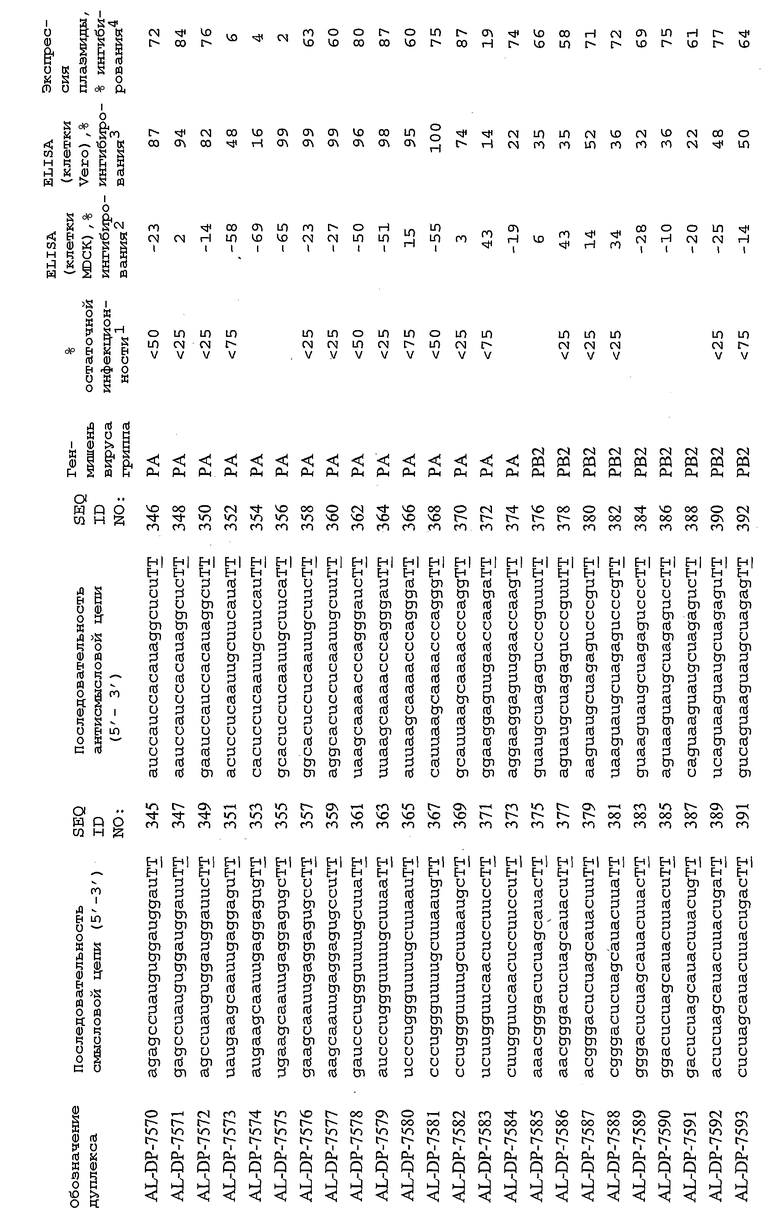
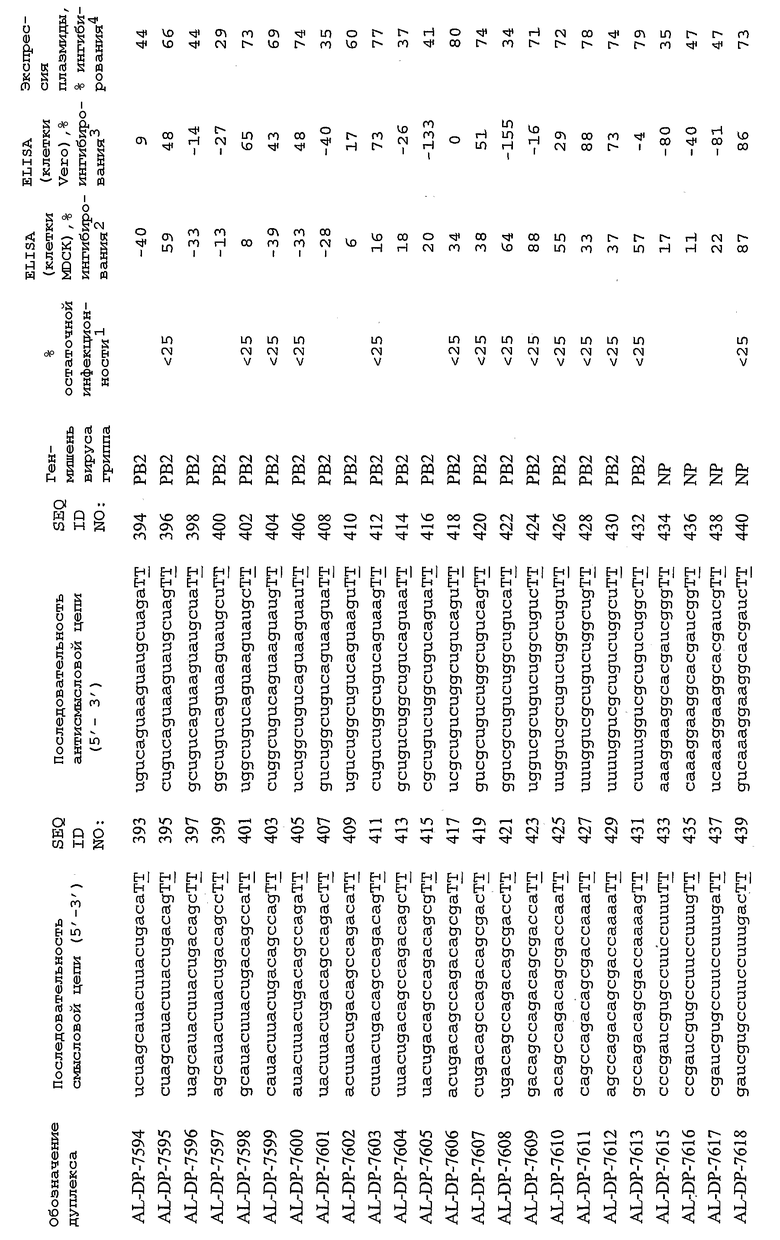
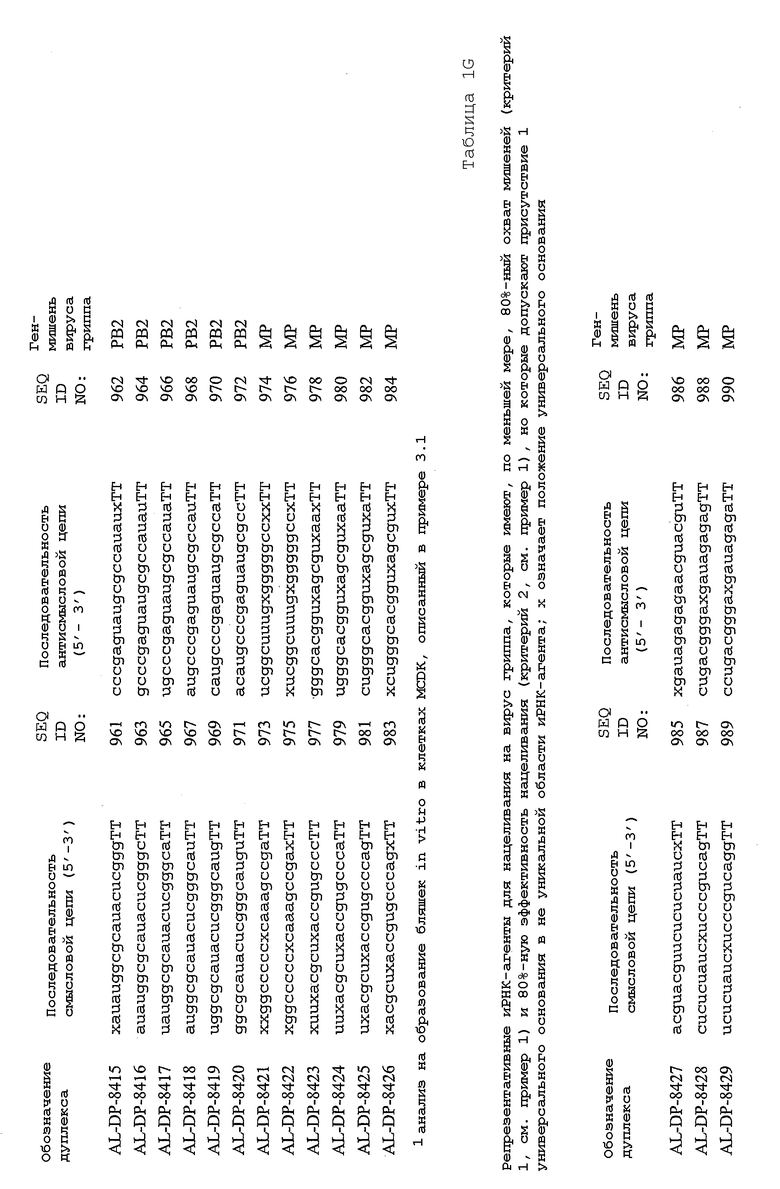
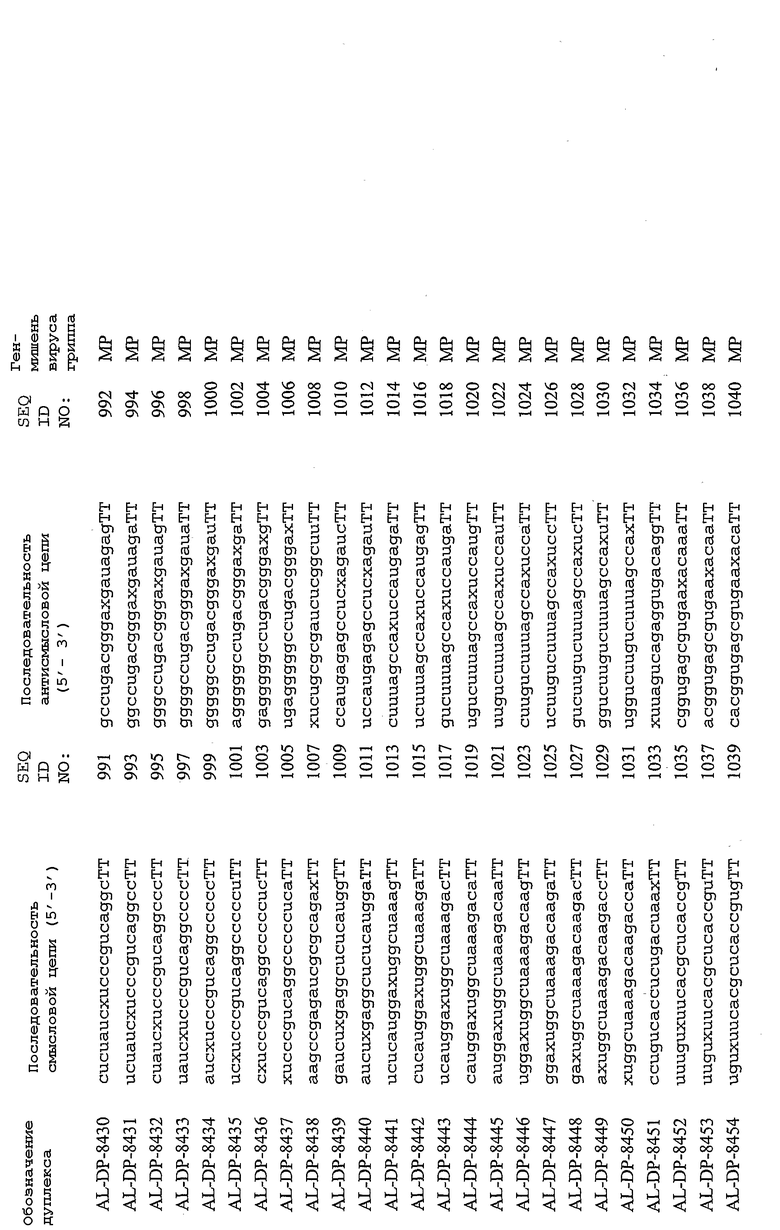
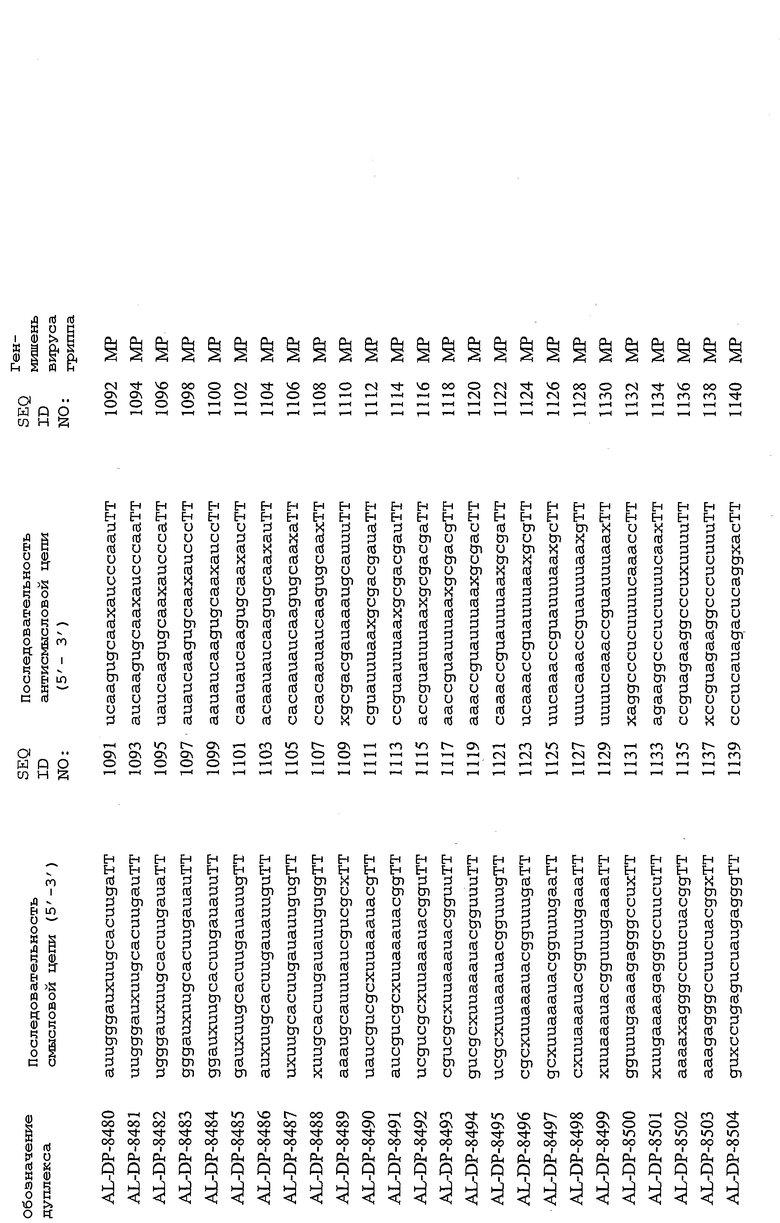
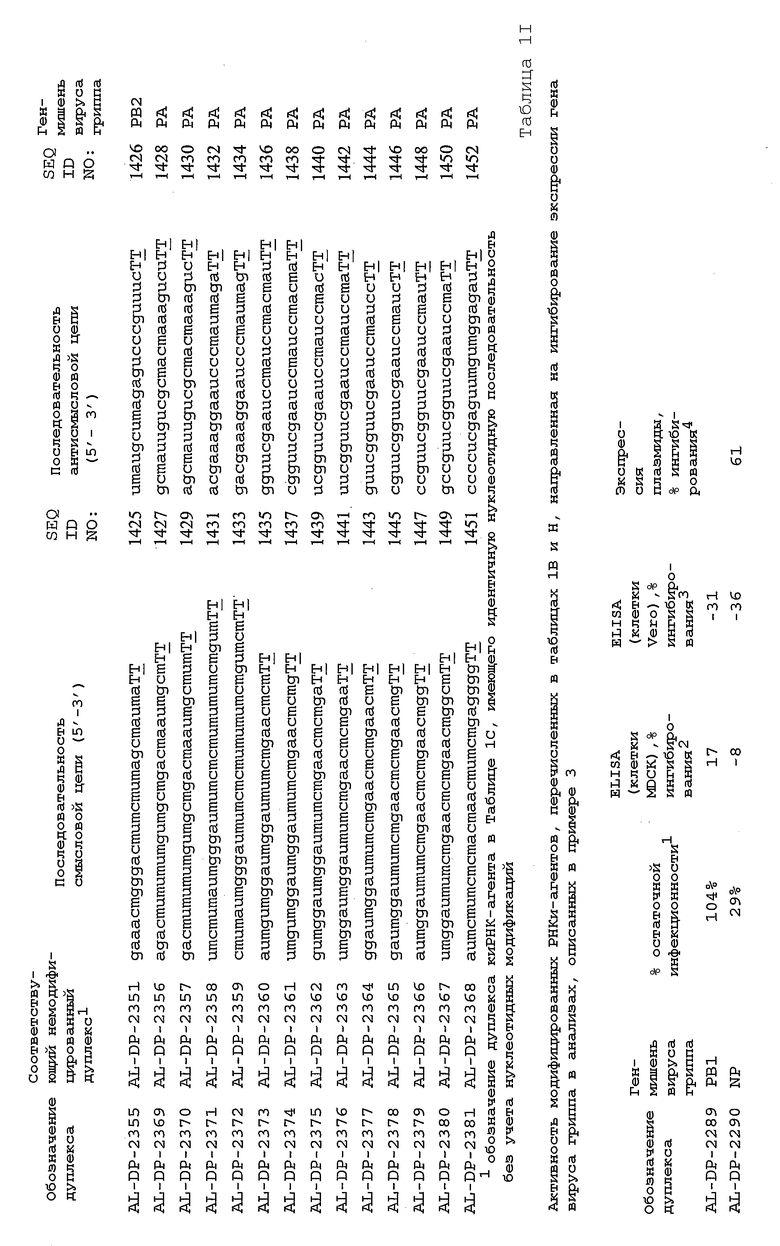
В соответствии с результатами, описанными в настоящей заявке, настоящее изобретение относится к иРНК-агентам, которые снижают уровень репликации вируса гриппа в культивированных клетках, инфицированных вирусом гриппа, и у индивидуума, например у млекопитающего, например у человека. В таблицах 1A-1H представлены репрезентативные иРНК-агенты, направленные на вирус гриппа. В таблицах 1A, C, D и E представлен список киРНК, не содержащих нуклеотидных модификаций, за исключением одной фосфортиоатной связи между 3'-концом и предпоследними тимидинами. В таблицах 1B и H представлен список киРНК, где все нуклеотиды, содержащие пиримидиновые основания, представляют собой 2'-О-метил-модифицированные нуклеотиды смысловой цепи, а все уридины в окружении последовательности 5'-ua-3', а также все цитидины в окружении последовательности 5'-ca-3' представляют собой 2'-О-метил-модифицированные нуклеотиды в антисмысловой цепи, за исключением иРНК-агентов с дуплексом, идентифицированным как AL-DP-2295, AL-DP-2301 и AL-DP-2302, где все уридины в окружении последовательности 5'-ug-3' представляют собой 2'-О-метил-модифицированные нуклеотиды в антисмысловой цепи. Эти последние киРНК не встречаются в мотивах последовательностей 5'-ua-3' или 5'-ca-3', а анализ фрагментов расщепления после инкубирования этих агентов в мышиной сыворотке показал, что мотив последовательности 5'-ug-3' представляет собой главный участок эндонуклеолитической атаки.
Исходя из этих результатов, было разработано настоящее изобретение, которое, в частности, относится к иРНК-агенту, включающему смысловую цепь, имеющую, по меньшей мере, 15 смежных нуклеотидов в последовательностях смысловой цепи агентов, представленных в таблицах 1A-1H, и антисмысловую цепь, имеющую, по меньшей мере, 15 смежных нуклеотидов в последовательностях антисмысловой цепи агентов, представленных в таблицах 1A-1H.
иРНК-агенты, представленные в таблицах 1A-1H, состоят из двух цепей длиной в 19 нуклеотидов, которые комплементарны или идентичны последовательности-мишени, и из выступающего 3'-TT-конца. Настоящее изобретение относится к агентам, содержащим, по меньшей мере, 15 или, по меньшей мере, 16, 17, 18 или 19 смежных нуклеотидов, происходящих от этих последовательностей. Однако, хотя такие длины могут быть оптимальными, тем не менее, это не означает, что иРНК-агенты должны иметь только такую длину. Специалисту в данной области хорошо известно, что более короткие или более длинные иРНК-агенты могут обладать аналогичной эффективностью, поскольку при определенных длинах эффективность нуклеотидной последовательности будет зависеть скорее от функции, чем от длины этой нуклеотидной последовательности. Так, например, в работе Yang et al., PNAS 99:9942-9947 (2002), было продемонстрировано, что иРНК-агенты обладают аналогичной эффективностью при длинах последовательностей в пределах от 21 до 30 пар оснований. Другими исследователями был обнаружен эффективный сайленсинг генов при меньшей длине иРНК-агентов, составляющей до 15 пар оснований (Byrom, et al., "Inducing RNAi with siRNA Cocktails Generated by RNase III" Tech Notes 10(1), Ambion, Inc., Austin, TX).
Поэтому для получения иРНК-агента, происходящего от одной из последовательностей, представленных в таблицах 1А-1Н, неполная последовательность согласно изобретению длиной в 15-19 нуклеотидов может быть выбрана из последовательностей, представленных в таблицах 1A-1H. Альтернативно, к одной из последовательностей, представленных в таблицах 1A-1H, или к агенту, содержащему 15 смежных нуклеотидов, происходящих от одного из этих агентов, предпочтительно, но необязательно, могут быть добавлены один или несколько нуклеотидов так, чтобы такие добавленные нуклеотиды были комплементарны соответствующей последовательности гена-мишени, например гена вируса гриппа. Так, например, первые 15 нуклеотидов, происходящих от одного из этих агентов, могут быть объединены с 8 нуклеотидами, присутствующими со стороны 5'-конца от этих последовательностей мРНК вируса гриппа, в результате чего может быть получен агент, имеющий 23 нуклеотида в смысловой и в антисмысловой цепях. Все указанные производные иРНК-агентов входят в определение иРНК-агентов согласно изобретению, при условии, что они, по существу, сохраняют способность ингибировать репликацию вируса гриппа в культивируемых человеческих клетках, инфицированных вирусом гриппа.







Антисмысловая цепь иРНК-агента должна иметь длину, равную или составляющую не менее чем 14, 15, 16, 17, 18, 19, 25, 29, 40 или 50 нуклеотидов. Указанная цепь должна иметь длину, равную 60, 50, 40 или 30 нуклеотидов или менее. Предпочтительная длина составляет 15-30, 17-25, 19-23 и 19-21 нуклеотидов.
Смысловая цепь иРНК-агента должна иметь длину, равную или составляющую не менее чем 14, 15, 16, 17, 18, 19, 25, 29, 40 или 50 нуклеотидов. Указанная цепь должна иметь длину, равную 60, 50, 40 или 30 нуклеотидов или менее. Предпочтительная длина составляет 15-30, 17-25, 19-23 и 19-21 нуклеотидов.
Двухцепочечная часть иРНК-агента должна иметь длину, равную или составляющую не менее чем 15, 16 17, 18, 19, 20, 21, 22, 23, 24, 25, 29, 40 или 50 пар нуклеотидов. Указанная цепь должна иметь длину, равную 60, 50, 40 или 30 пар нуклеотидов или менее. Предпочтительная длина составляет 15-30, 17-25, 19-23 и 19-21 пар нуклеотидов.
Вообще говоря, иРНК-агенты согласно изобретению включают область, достаточно комплементарную соответствующему гену вируса гриппа и имеющую длину нуклеотидной цепи, достаточную для того, чтобы иРНК-агент или его фрагмент могли опосредовать ингибирование гена вируса гриппа. При этом, необязательно, чтобы иРНК-агент и ген-мишень были абсолютно комплементарными, однако их соответствие должно быть таким, чтобы иРНК-агент или продукт его расщепления могли обеспечивать непосредственный последовательность-специфический сайленсинг, например, посредством РНКи-расщепления РНК вируса гриппа.
Поэтому иРНК-агентами согласно изобретению являются агенты, содержащие смысловую цепь и антисмысловую цепь, каждая из которых имеет последовательность, составляющую, по меньшей мере, 16, 17 или 18 нуклеотидов, которые, как определено ниже, в основном, идентичны нуклеотидам одной из последовательностей, представленных в таблицах 1А-1Н, за исключением не более чем 1, 2 или 3 нуклеотидов на цепь, соответственно, которые были заменены другими нуклеотидами (например, аденозин был заменен урацилом), где указанная замена, по существу, не влияет на способность этих агентов ингибировать репликацию вируса гриппа в культивированных человеческих клетках, инфицированных вирусом гриппа, соответственно. Следовательно, такие агенты имеют, по меньшей мере, 15 нуклеотидов, идентичных нуклеотидам одной из последовательностей, представленных в таблицах 1A-1H, но при этом они либо имеют 1, 2 или 3 несоответствия по сравнению с последовательностью РНК-мишени вируса гриппа, либо введены между смысловой и антисмысловой цепью. Несоответствия с последовательностью РНК-мишени вируса гриппа, а в частности в антисмысловой цепи, являются наиболее толерантными в концевых областях, и если они присутствуют, то предпочтительно, в концевой области или областях, например, в пределах 6, 5, 4 или 3 нуклеотидов у 5'- и/или 3'-конца, а наиболее предпочтительно, в пределах 6, 5, 4 или 3 нуклеотидов у 5'-конца смысловой цепи или у 3'-конца антисмысловой цепи. Для сохранения всей двухцепочечной структуры молекулы смысловая цепь должна быть в достаточной степени комплементарна лишь антисмысловой цепи.
При этом предпочтительно, чтобы смысловая и антисмысловая цепи были выбраны так, чтобы иРНК-агент включал одну цепь или неспаренную область у одного или обоих концов молекулы. Таким образом, иРНК-агент содержит смысловую и антисмысловую цепи, предпочтительно, спаренные так, чтобы они содержали выступающий конец, например один или два выступающих 5'- или 3'-конца, а предпочтительно, выступающий 3'-конец, содержащий 2-3 нуклеотида. Большинство вариантов имеют выступающий 3'-конец. Предпочтительные киРНК-агенты имеют одноцепочечные выступающие концы, а предпочтительно, выступающие 3'-концы, содержащие 1-4 или, предпочтительно, 2 или 3 нуклеотида, расположенные у одного или обоих концов указанного иРНК-агента. Образование выступающих концов обусловлено тем, что одна цепь длиннее другой, либо тем, что две цепи, имеющие одну и ту же длину, сдвинуты относительно друг друга. Неспаренными нуклеотидами, образующими выступающий конец, могут быть рибонуклеотиды, либо ими могут быть дезоксирибонуклеотиды, а предпочтительно, тимидин. 5'-концы являются, предпочтительно, фосфорилированными, либо они могут быть нефосфорилированными.
Длина дуплексной области, например, в киРНК-агенте, обсуждаемом выше, предпочтительно, составляет 15-30 нуклеотидов, а наиболее предпочтительно, 18, 19, 20, 21, 22 и 23 нуклеотида. Длина и структура киРНК-агентов может быть аналогична длине и структуре природных продуктов, процессированных ферментом Dicer из длинных дцРНК. Настоящее изобретение также включает варианты, в которых две цепи киРНК-агента связаны, например, ковалентной связью. Настоящее изобретение также относится к “шпилечным” или к другим одноцепочечным структурам, которые образуют необходимую двухцепочечную область, а предпочтительно, выступающий 3'-конец.
Оценка кандидатов на иРНК-агенты
Как было упомянуто выше, настоящее изобретение относится к системе идентификации киРНК, которые могут быть использованы в качестве ингибиторов инфекций, вызываемых вирусом гриппа, и/или репликации этого вируса. Поскольку, как упоминалось выше, кшРНК процессируются внутри клеток с образованием киРНК, имеющих дуплексные части с такой же последовательностью, как и стволовая структура кшРНК, то эта система в равной степени может быть использована для идентификации киРНК, которые могут служить в качестве ингибиторов инфекции, вызываемой вирусом гриппа. В этом разделе киРНК приводятся лишь в качестве примера, и данная система также включает соответствующие кшРНК. В частности, в настоящем изобретении продемонстрировано успешное получение киРНК, нацеленных на вирусные гены для блокирования или ингибирования вирусной инфекции и/или репликации вируса. Описанные здесь методы и реагенты могут быть легко применены для конструирования потенциально новых киРНК, нацеленных на другие гены или генные области, и протестированы на их активность в отношении ингибирования инфекции, вызываемой вирусом гриппа, и/или репликации вируса гриппа, обсуждаемых в настоящей заявке. При этом предполагается, что вирусы гриппа будут подвергаться мутации и реассортации, а поэтому необходимо продолжать разработку и тестирование новых киРНК, нацеленных на другие мишени.
В различных вариантах изобретения потенциальные ингибиторы вируса гриппа могут быть протестированы путем введения киРНК-кандидата(ов) в клетки (например, путем экзогенного введения или путем введения вектора или конструкции, которые направляют эндогенный синтез киРНК в клетку), или лабораторным животным до, во время или после трансфекции геномом вируса гриппа или его частью (например, в течение нескольких минут, часов или, максимум, нескольких дней) или до, во время или после инфицирования вирусом гриппа. Альтернативно, потенциальные ингибиторы вируса гриппа могут быть протестированы путем введения киРНК-кандидата(ов) в клетки или лабораторным животным, которые были стабильно инфицированы вирусом гриппа (то есть клетки, которые продуцируют вирусное потомство). Затем была оценена способность киРНК-кандидата(ов) снижать уровни транскрипта-мишени и/или ингибировать или подавлять один или несколько показателей или признаков жизненного цикла вируса, таких как репликация вируса, его патогенность и/или инфекционность. Так, например, продуцирование вирусных частиц и/или продуцирование вирусных белков и т.п. может быть оценено прямыми или непрямыми методами, хорошо известными специалистам.
Клетки или лабораторные животные, которым могут быть доставлены киРНК-композиции согласно изобретению (тестируемые клетки/животные), можно сравнить с аналогичными или со сравнимыми клетками или лабораторными животными, которым не была введена композиция согласно изобретению (контрольными клетками/животными, например клетками/животными, которым либо не вводили киРНК, либо вводили контрольную киРНК, такую как киРНК, нацеленную на невирусный транскрипт, такой как GFP). Восприимчивость тестируемых клеток/животных к инфекции, вызываемой вирусом гриппа, можно сравнить с восприимчивостью контрольных клеток/животных к таким инфекциям. Продуцирование вирусного(ых) белка(ов) и/или вирусного потомства в тестируемых клетках/животных можно сравнить с их продуцированием в контрольных клетках/животных. Аналогичным образом можно сравнивать и другие показатели, такие как инфекционность, репликация, патогенность вируса и т.п. Стандартные вирусологические тесты in vitro могут быть проведены путем анализа на ингибирование вирусных бляшек, на цитопатический эффект вируса (CPE), на наличие гемаглютинина или других вирусных белков, на ингибирование выхода вируса и т.п. CPE может быть определен визуально и путем поглощения цвета. См., например, Sidwell, R. W. and Smee, D. F, "In vitro and in vivo assay systems for study of influenza virus inhibitors" Antiviral Res 2000, 48:1. Вообще говоря, тестируемые клетки/животные и контрольные клетки/животные должны быть одного и того же вида, а тестируемые клетки должны быть аналогичного или идентичного типа. Так, например, могут быть подвергнуты сравнению клетки от одной и той же клеточной линии. Если тестируемой клеткой является первичная клетка, то обычно контрольная клетка также должна быть первичной клеткой. Для сравнения тестируемых клеток/животных с контрольными клетками/животными должны быть использованы одни и те же штаммы вируса гриппа.
Так, например, способность киРНК-кандидата ингибировать продуцирование вируса гриппа может быть легко определена путем (i) доставки киРНК-кандидата в клетки (до, во время или после инфицирования вирусом гриппа); (ii) оценки уровня продуцирования вирусного гемаглютинина с помощью анализа на гемаглютинин; и (iii) сравнения количества гемаглютинина, продуцируемого в присутствии киРНК, с количеством, продуцируемым в отсутствие киРНК. (Этот тест необязательно должен включать контроль, в котором отсутствует киРНК, но при этом можно использовать уже имеющуюся информацию о количестве гемаглютинина, продуцируемого в отсутствие ингибирования.) Снижение количества гемаглютинина со всей очевидностью свидетельствует о снижении уровня продуцирования вируса. Этот анализ может быть применен для тестирования киРНК, нацеленных на любой вирусный транскрипт, и не ограничивается киРНК, нацеленными на транскрипт, кодирующий вирусный гемаглютинин.
Способность киРНК-кандидата снижать уровень транскрипта-мишени может быть также оценена путем определения количества указанного транскрипта-мишени с помощью, например, Нозерн-блот-анализов, анализов на защиту от нуклеазы, ПЦР с обратной транскриптазой (ОТ)-ПЦР, ОТ-ПЦР в реальном времени, анализов с использованием микромассивов и т.п. Способность киРНК-кандидата ингибировать продуцирование полипептида, кодируемого транскриптом-мишенью (либо на транскрипционном, либо на посттранскрипционном уровне) может быть оценена с применением ряда методов на основе антител, включая, но не ограничиваясь ими, вестерн-блот-анализы, иммуноанализы, ELISA, анализы с помощью проточной цитометрии, анализы с использованием микромассивов белков и т.п. В общих чертах, может быть применен любой метод определения количества транскрипта-мишени или полипептида, кодируемого таким транскриптом-мишенью.
В общих чертах, некоторые предпочтительные ингибиторы вируса гриппа снижают уровень транскрипта-мишени, по меньшей мере, примерно в 2 раза, предпочтительно, по меньшей мере, примерно в 4 раза, более предпочтительно, по меньшей мере, примерно в 8 раз, по меньшей мере, примерно в 16 раз, по меньшей мере, примерно в 64 раза или даже более, по сравнению с уровнем, наблюдаемым в отсутствие ингибитора (например, в сравниваемых контрольных клетках, не содержащих ингибитора). В общих чертах, некоторые предпочтительные ингибиторы вируса гриппа осуществляют такое ингибирование репликации вируса, что уровень его репликации в клетках, содержащих указанный ингибитор, снижается, по меньшей мере, примерно в 2 раза, предпочтительно, по меньшей мере, примерно в 4 раза, более предпочтительно, по меньшей мере, примерно в 8 раз, по меньшей мере, примерно в 16 раз, по меньшей мере, примерно в 64 раза, по меньшей мере, примерно в 100 раз, по меньшей мере, примерно в 200 раз, или даже более, по сравнению с уровнем, наблюдаемым в контрольных клетках, не содержащих такого ингибитора.
Некоторые предпочтительные ингибиторы вируса гриппа ингибируют репликацию вируса до такого уровня, при котором продуцирование детектируемого титра вируса предотвращается, по меньшей мере, в течение 24 часов, по меньшей мере, 36 часов, по меньшей мере, 48 часов или, по меньшей мере, 60 часов после введения киРНК и инфицирования клеток. Некоторые предпочтительные ингибиторы вируса гриппа способны предупреждать (то есть снижать до недетектируемых уровней) или значительно снижать уровень репликации вируса, по меньшей мере, в течение 24 часов, по меньшей мере, 36 часов, по меньшей мере, 48 часов или, по меньшей мере, 60 часов после введения киРНК. В соответствии с различными вариантами изобретения, значительное снижение уровня репликации вируса означает снижение до уровня, составляющего приблизительно менее чем 90%, приблизительно менее чем 75%, приблизительно менее чем 50%, приблизительно менее чем 25% или приблизительно менее чем 10% от уровня, наблюдаемого в отсутствие киРНК. Снижение уровня репликации вируса может быть определено любым подходящим методом, включая, но не ограничиваясь ими, определение титра HA.
Тест на стабильность, модификация и повторное тестрование иРНК-агентов
иРНК-агент-кандидат может быть оценен на его стабильность, например на его чувствительность к расщеплению эндонуклеазой или экзонуклеазой, например, при введении иРНК-агента в организм индивидуума. Для идентификации участков, чувствительных к модификациям, а в частности, к расщеплению, например к расщеплению компонентом, присутствующим в организме индивидуума, могут быть применены различные методы. Такими методами могут быть выделение и идентификация наиболее часто встречающихся фрагментов, образованных в результате расщепления иРНК-агента-кандидата после его инкубирования с выделенной биологической средой in vitro, например с сывороткой, плазмой, мокротой, цереброспинальной жидкостью или с гомогенатами клеток или тканей, или после контактирования индивидуума с иРНК-агентом-кандидатом in vivo, и, тем самым, идентификация сайтов, подверженных расщеплению. Эти и другие методы описаны, например, в совместной международной патентной заявке № PCT/US2005/018931, поданной 27 мая 2005 г.
После идентификации сайтов, подверженных расщеплению, может быть сконструирован и/или синтезирован другой иРНК-агент, имеющий потенциальный сайт расщепления, который является резистентным к такому расщеплению, например, путем введения 2'-модификации в сайт расщепления, например, 2'-О-метильной группы. Такой дополнительный иРНК-агент может быть снова протестирован на стабильность, и эту процедуру можно повторять до тех пор, пока не будет установлено, что иРНК-агент обладает желаемой стабильностью.
Тест in vivo
иРНК-агент, идентифицированный как агент, способный ингибировать экспрессию гена вируса гриппа, может быть протестирован на его функциональность in vivo на животном-модели (например, млекопитающего, такого как мышь или крыса). Так, например, иРНК-агент может быть введен животному, а затем оценен на его биологическое распределение, стабильность и способность ингибировать репликацию вируса гриппа или снижать биологические или патологические процессы, опосредуемые, по меньшей мере, частично, вирусом гриппа.
иРНК-агент может быть введен непосредственно в нужную ткань, например, путем инъекции, либо такой иРНК-агент может быть введен животному-модели способом, аналогичным способу его введения человеку. иРНК-агент, предпочтительно, вводят в дыхательные пути индивидуума, например, интраназально.
иРНК-агент может быть также оценен на его внутриклеточное распределение. Такая оценка может включать определение наличия поглощения иРНК-агента клеткой. Такая оценка может также включать определение стабильности (например, времени полужизни) иРНК-агента. Оценку иРНК-агента in vivo можно облегчить путем использования иРНК-агента, конъюгированного с детектируемым маркером (например, флуоресцентным маркером, таким как флуоресцеин; радиоактивная метка, такая как 35S, 32Р, 33Р или 3Н; золотые частицы или антигенные частицы для иммуногистохимического анализа).
иРНК-агент может быть оценен на его способность ингибировать репликацию вируса гриппа. Уровни экспрессии гена вируса гриппа in vivo могут быть измерены, например, путем гибридизации in situ или путем выделения РНК из ткани до или после инфицирования иРНК-агентом. Если для сбора тканей животных необходимо умерщвлять, то в качестве контроля для сравнения служат необработанные животные. РНК вируса гриппа может быть детектирована любым подходящим методом, включая, но не ограничиваясь ими, ОТ-ПЦР, Нозерн-блот-анализ, анализ на разветвленную ДНК или анализ на защиту от РНКазы. Альтернативно или дополнительно, мониторинг экспрессии гена вируса гриппа может быть осуществлен путем проведения вестерн-блот-анализа или анализов на образование бляшек на тканевых экстрактах, обработанных иРНК-агентом.
Потенциальные ингибиторы вируса гриппа могут быть протестированы с использованием любых выведенных животных-моделей. Композиции, содержащие киРНК-кандидаты или включающие их конструкции или векторы, способные регулировать синтез таких киРНК в клетке-хозяине или в клетках, которые были сконструированы или в которые были введены мутации, так, чтобы они содержали киРНК-кандидаты, могут быть введены животному до, во время или после его инфицирования вирусом гриппа. Может быть оценена способность такой композиции предупреждать вирусную инфекцию, и/или замедлять ее развитие или предотвращать появление симптомов, ассоциированных с инфицированием вирусом гриппа, и/или ослаблять тяжесть состояния животных, инфицированных вирусом гриппа, по сравнению с животными, которым не вводили потенциальный ингибитор вируса гриппа. Такими животными, служащими в качестве моделей гриппа, являются, но не ограничиваются ими, мыши, куры, хорьки и приматы, не являющиеся человеком, известные специалистам и обычно используемые для анализа эффективности потенциальных терапевтических методов и вакцин против вируса гриппа. См., например, публикацию Sidwell, R. W. & Smee, D. F, указанные выше. Такие модели могут быть заражены природными штаммами вируса гриппа и/или штаммами, которые были модифицированы или адаптированы для их существования в организме конкретного хозяина (например, штаммы WSN или PR8, которые были адаптированы для репликации у мышей). Вышеуказанные животные-модели могут быть также использованы в целях определения концентрации, необходимой для достижения определенного желаемого эффекта (например, EC50).
Химическая структура иРНК
Описанные здесь агенты представляют собой выделенные иРНК-агенты, например дцРНК-агенты, которые опосредуют интерференцию РНК (РНКи), ингибирующую экспрессию гена вируса гриппа.
Обсуждаемыми здесь РНК-агентами являются либо немодифицированные РНК, либо также РНК, которые были модифицированы, например, для повышения эффективности; и полимеры нуклеозидных суррогатов. Немодифицированная РНК представляет собой молекулу, в которой компоненты нуклеиновой кислоты, а именно сахара, основания и фосфатные молекулы, являются такими же или, в основном, такими же, как и в природной молекуле, а предпочтительно такими же, как в организме человека. Специалисты в данной области указывают на то, что в качестве модифицированных РНК могут быть использованы редкие или необычные, но встречающиеся в природе РНК, см., например, Limbach et al. Nucleic Acids Res. 22: 2183-2196, 1994. Такие редкие или необычные РНК, часто называемые модифицированными РНК (очевидно потому, что они обычно образуются в результате посттранскрипционной модификации), охватываются используемым здесь термином “немодифицированная” РНК. Используемый здесь термин модифицированная РНК означает молекулу, в которой один или несколько компонентов нуклеиновой кислоты, а именно сахара, основания и фосфатные молекулы, отличаются от компонентов, присутствующих в природной молекуле, а предпочтительно, присутствующих в человеческом организме. Хотя такие молекулы называются модифицированными “РНК”, однако из-за имеющихся в них модификаций они включают молекулы, которые не являются РНК. Нуклеозидными суррогатами являются молекулы, в которых рибофосфатный остов заменен нерибофосфатной конструкцией, например незаряженными миметиками рибофосфатного остова, что позволяет получить основания в “правильной” пространственной взаимосвязи, так чтобы их спаривание было, в основном, аналогичным спариванию, наблюдаемому для рибофосфатного остова. Примеры вышеуказанных молекул обсуждаются в настоящей заявке.
Описанные здесь модификации могут быть введены в любую описанную здесь двухцепочечную РНК и РНК-подобную молекулу, например иРНК-агент. Может оказаться желательным модифицировать одну или обе антисмысловые и смысловые цепи иРНК-агента. Поскольку нуклеиновые кислоты представляют собой полимеры, состоящие из субъединиц или мономеров, то многие из описанных ниже модификаций присутствуют в соответствующем положении и повторяются в данной нуклеиновой кислоте, например модификация основания или фосфатной молекулы или О-несвязанной фосфатной молекулы. В некоторых случаях данная модификация встречается во всех рассматриваемых положениях нуклеиновой кислоты, но во многих случаях, а фактически в большинстве случаев, она отсутствует. Так, например, модификация может присутствовать только в 3'- или 5'-концевом положении, только в концевой области, например в положении концевого нуклеотида или в последних 2, 3, 4, 5 или 10 нуклеотидах цепи. Модификация может присутствовать в двухцепочечной области, в одноцепочечной области или в обеих областях. Так, например, фосфортиоатная модификация в О-несвязанном положении может присутствовать только у одного или обоих концов, только в концевых областях, например в положении концевого нуклеотида, или в последних 2, 3, 4, 5 или 10 нуклеотидах цепи, или в двухцепочечных и в одноцепочечных областях, а в частности у их концов. Аналогичным образом, модификация может присутствовать на смысловой цепи, антисмысловой цепи или в обеих цепях. В некоторых случаях смысловая и антисмысловая цепи имеют одни и те же модификации или модификации одного и того же класса, а в других случаях смысловая и антисмысловая цепи имеют различные модификации, например, в некоторых случаях может оказаться желательным модифицировать только одну цепь, например смысловую цепь.
Двумя главными целями введения модификаций в иРНК-агенты является их защита от деградации в биологических средах и улучшение их фармакологических свойств, например фармакологических свойств, которые подробно обсуждаются ниже. Другие подходящие модификации сахара, оснований или остова иРНК-агента описаны в совместной заявке РСТ № PCT/US2004/01193, поданной 16 января 2004 г. иРНК-агент может включать неприродные основания, такие как основания, описанные в совместной заявке РСТ № PCT/US2004/011822, поданной 16 апреля 2004 г. иРНК-агент может включать неприродный сахар, такой как молекула, несущая неуглеводную циклическую группу. Репрезентативные признаки неприродных сахаров, используемых в иРНК-агентах, описаны в совместной заявке РСТ № PCT/US2004/11829, поданной 16 апреля 2003 г.
иРНК-агент может включать межнуклеотидную связь (например, хиральную фосфортиоатную связь), используемую для повышения резистентности к нуклеазе. Дополнительно или альтернативно, иРНК-агент может включать рибозный миметик, используемый для повышения резистентности к нуклеазе. Репрезентативные межнуклеотидные связи и рибозные миметики, используемые для повышения резистентности к нуклеазе, описаны в совместной заявке PCT № PCT/US2004/07070, поданной 8 марта 2004 г.
иРНК-агент может включать конъюгированные с лигандом мономерные субъединицы и мономеры для синтеза олигонуклеотидов. Репрезентативные мономеры описаны в совместной заявке на патент США № 10/916185, поданной 10 августа 2004 г.
иРНК-агент может иметь структуру ZXY, например структуру, описанную в совместной заявке PCT № PCT/US2004/07070, поданной 8 марта 2004 г.
иРНК-агент может образовывать комплекс с амфипатической группой. Репрезентативные примеры амфипатических групп, используемых в иРНК-агентах, описаны в совместной заявке РСТ № PCT/US2004/07070, поданной 8 марта 2004 г.
В другом варианте изобретения иРНК-агент может образовывать комплекс с агентом для доставки, который имеет модульный комплекс. Такой комплекс может включать агент-носитель, связанный с одним или несколькими из нижеследующих компонентов (предпочтительно, с двумя или более, а более предпочтительно, с тремя компонентами), таких как (а) конденсирующий агент (например, агент, способный присоединять нуклеиновую кислоту, например связываться с нуклеиновой кислотой, например, посредством ионных или электростатических взаимодействий); (b) фузогенный агент (например, агент, способный связываться с клеточной мембраной и/или транспортироваться через клеточную мембрану); и (с) нацеливающая группа, например агент, доставляемый в клетку или ткань, например лектин, гликопротеин, липид или белок, например антитело, которое связывается с клеткой конкретного типа. иРНК-агенты, образующие комплексы с агентом для доставки, описаны в совместной заявке PCT № PCT/US2004/07070, поданной 8 марта 2004 г.
иРНК-агент может иметь неканонические пары, такие как пары между смысловыми и антисмысловыми последовательностями иРНК-дуплекса. Репрезентативные примеры неканонических иРНК-агентов описаны в совместной заявке PCT № PCT/US2004/07070, поданной 8 марта 2004 г.
Повышенный уровень резистентности к нуклеазе
иРНК-агент, например иРНК-агент, нацеленный на вирус гриппа, может иметь повышенный уровень резистентности к нуклеазам.
Одним из способов повышения уровня резистентности является идентификация сайтов расщепления и модификация таких сайтов для ингибирования расщепления, описанные в совместной заявке на патент США № 60/559917, поданной 4 мая 2004 г. Так, например, в качестве сайтов расщепления могут служить динуклеотиды 5'-ua-3', 5'-ca-3', 5'-ug-3', 5'-uu-3' или 5'-cc-3'. В некоторых вариантах изобретения все пиримидины иРНК-агента несут 2'-модификацию либо в смысловой цепи, либо в антисмысловой цепи, либо в обеих цепях, а поэтому иРНК-агент обладает повышенной резистентностью к эндонуклеазам. Повышенная резистентность в нуклеазе может быть также достигнута путем модификации 5'-нуклеотида, в результате чего образуется, например, по меньшей мере, один 5'-уридин-аденин-3'(5'-ua-3')-динуклеотид, где указанный уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере, один 5'-цитидин-аденин-3'(5'-са-3')-динуклеотид, где 5'-цитидин представляет собой 2'-модифицированный нуклеотид; по меньшей мере, один 5'-уридин-гуанин-3'(5'-ug-3')-динуклеотид, где 5'-уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере, один 5'-уридин-уридин-3'(5'-uu-3')-динуклеотид, где 5'-уридин представляет собой 2'-модифицированный нуклеотид; или, по меньшей мере, один 5'-цитидин-цитидин-3'(5'-сс-3')-динуклеотид, где 5'-цитидин представляет собой 2'-модифицированный нуклеотид, описанные в совместной международной заявке № PCT/US2005/018931, поданной 27 мая 2005 г. иРНК-агент может включать, по меньшей мере, 2, по меньшей мере, 3, по меньшей мере, 4 или, по меньшей мере, 5 таких динуклеотидов. В особенно предпочтительном варианте изобретения 5'-нуклеотид во всех мотивах последовательности 5'-ua-3' и 5'-ca-3' либо в смысловой цепи, либо в антисмысловой цепи, либо в обеих цепях представляет собой модифицированный нуклеотид. Предпочтительно, 5'-нуклеотид во всех мотивах последовательности 5'-ua-3', 5'-ca-3' и 5'-ug-3' либо в смысловой цепи, либо в антисмысловой цепи, либо в обеих цепях представляет собой модифицированный нуклеотид. Более предпочтительно, все пиримидиновые нуклеотиды в смысловой цепи представляют собой модифицированные нуклеотиды, и 5'-нуклеотиды во всех мотивах последовательности 5'-ua-3' и 5'-ca-3' в антисмысловой цепи представляют собой модифицированные нуклеотиды, либо антисмысловая цепь не содержит ни мотива 5'-ua-3', ни 5'-ca-3', но во всех случаях содержит мотив 5'-ug-3'.
Предпочтительно, 2'-модифицированные нуклеотиды включают, например, 2'-модифицированное рибозное звено, например, 2'-гидроксильная группа (OH) может быть модифицирована или заменена рядом различных “окси”- или “дезокси”-заместителей.
Примерами “окси"-2'-модификаций гидроксильной группы являются алкокси или арилокси (OR, например, R = H, алкил, циклоалкил, арил, аралкил, гетероарил или сахар); полиэтиленгликоли (ПЭГ), О(CH2CH2O)nCH2CH2OR; “блокированные” нуклеиновые кислоты (LNA), в которых 2'-гидроксил связан, например, метиленовым мостиком, с 4'-углеродом того же самого сахара рибозы; O-амин и аминоалкокси, О(СН2)nамин, (например, амин = NH2; алкиламино, диалкиламино, гетероциклиламино, ариламино, диариламино, гетероариламино или дигетероариламино, этилендиамин, полиамино). При этом следует отметить, что стабильностью к нуклеазе обладают олигонуклеотиды, содержащие только метоксиэтильную группу (MOE), (OCH2CH2OCH3, ПЭГ-производное), в отличие от олигонуклеотидов, модифицированных только фосфортиоатными группами.
“Дезокси"-модификациями являются водород (то есть дезоксирибозные сахара, которые являются особенно подходящими для выступающих участков неполной дцРНК); галоген (например, фтор); амино (например, NH2; алкиламино, диалкиламино, гетероциклил, ариламино, диариламино, гетероариламино, дигетероариламино или аминокислота); NH(CH2CH2NH)nCH2CH2-амин (амин = NH2; алкиламино, диалкиламино, гетероциклиламино, ариламино, диариламино, гетероариламино или дигетероариламино); -NHC(O)R (R = алкил, циклоалкил, арил, аралкил, гетероарил или сахар), циано; меркапто; алкилтиоалкил; тиоалкокси; и алкил, циклоалкил, арил, алкенил и алкинил, которые могут быть замещены, например, но необязательно, функциональной аминогруппой.
Предпочтительными заместителями являются 2'-метоксиэтил, 2'- OCH3, 2'-О-аллил, 2'-С-аллил и 2'-фтор.
Включение фуранозных сахаров в олигонуклеотидный остов может также приводить к снижению уровня эндонуклеотического расщепления. иРНК-агент может быть затем модифицирован путем включения 3'-катионной группы или путем инвертирования нуклеозида у 3'-конца 3'-3'-связью. В другом альтернативном варианте 3'-конец может быть блокирован аминоалкильной группой, например 3'-С5-аминоалкил-dT. Другие 3'-конъюгаты могут ингибировать 3'-5'-экзонуклеолитическое расщепление. Не ограничиваясь какой-либо теорией, можно лишь отметить, что 3'-конъюгат, такой как напроксен или ибупрофен, могут ингибировать экзонуклеолитическое расщепление путем стерического блокирования связывания экзонуклеазы с 3'-концом олигонуклеотида. 3'-5'-экзонуклеазы могут быть блокированы даже небольшими алкильными цепями, арильными группами или гетероциклическими конъюгатами или модифицированными сахарами (D-рибозой, дезоксирибозой, глюкозой и т.п.).
Нуклеолитическое расщепление может быть также ингибировано введением модификаций в фосфатный линкер, например фосфортиоатных связей. Таким образом, предпочтительными иРНК-агентами являются чистые нуклеотидные димеры или нуклеотидные димеры, обогащенные конкретной хиральной формой модифицированной фосфатной группы, содержащей гетероатом в немостиковом положении, обычно занимаемом кислородом. Таким гетероатомом может быть S, Se, Nr2 или Br3. Если гетероатомом является S, то предпочтительной связью является обогащенная или хирально чистая Sp-связь. Термин “обогащенный” относится к форме, которая составляет, по меньшей мере, 70, 80, 90, 95 или 99% от предпочтительной формы. Модифицированные фосфатные связи являются особенно эффективными для ингибирования экзонуклеолитического расщепления при их введении рядом с 5'- или 3'-концевыми положениями, а предпочтительно, 5'-концевыми положениями иРНК-агента.
5'-конъюгаты могут также ингибировать 5'-3'-экзонуклеолитическое расщепление. Не ограничиваясь какой-либо теорией, можно лишь отметить, что 5'-конъюгат, такой как напроксен или ибупрофен, могут ингибировать экзонуклеолитическое расщепление путем стерического блокирования связывания экзонуклеазы с 5'-концом олигонуклеотида. 3'-5'-экзонуклеазы могут быть блокированы даже небольшими алкильными цепями, арильными группами или гетероциклическими конъюгатами или модифицированными сахарами (D-рибозой, дезоксирибозой, глюкозой и т.п.).
иРНК-агент может обладать повышенной резистентностью к нуклеазам, если дуплексный иРНК-агент включает одноцепочечный нуклеотид, выступающий, по меньшей мере, с одного конца. В предпочтительных вариантах изобретения выступающие нуклеотиды включают 1-4, а предпочтительно, 2-3 неспаренных нуклеотида. В предпочтительном варианте изобретения такой неспаренный нуклеотид одноцепочечного выступающего конца, который является смежным с концевой нуклеотидной парой, содержит пуриновое основание, а концевой нуклеотидной парой является пара G-C, или, по меньшей мере, две из последних четырех комплементарных нуклеотидных пар представляют собой пары G-C. В других вариантах изобретения нуклеотидный выступающий конец может иметь 1 или 2 неспаренных нуклеотида, а в репрезентативном варианте изобретения таким нуклеотидным выступающим концом является 5'-gc-3'. В предпочтительных вариантах изобретения выступающий нуклеотид находится на 3'-конце антисмысловой цепи. В одном из вариантов изобретения иРНК-агент включает мотив 5'-cgc-3' на 3'-конце антисмысловой цепи, в результате чего образуется выступающий конец из двух нуклеотидов, 5'-gc-3'.
Таким образом, иРНК-агент может включать модификации, способствующие ингибированию расщепления, например, нуклеазами, например эндонуклеазами или экзонуклеазами, присутствующими в организме индивидуума. Такие мономеры называются здесь NRM или мономерами, стимулирующими резистентность к нуклеазе, а соответствующие модификации обозначаются NRM-модификациями. Во многих случаях такие модификации могут также модулировать другие свойства иРНК-агента, например способность взаимодействовать с белком, например с транспортным белком, например с сывороточным альбумином или с членом RISC, или способность первой и второй последовательности образовывать дуплекс друг с другом или с другой последовательностью, например с молекулой-мишенью.
Одна или несколько различных NRM-модификаций могут быть введены в иРНК-агент или в последовательность иРНК-агента. NRM-модификация может быть использована в такой последовательности иРНК-агента или в иРНК-агенте более чем один раз.
NRM-модификации включают некоторые модификации, которые могут присутствовать только на концах, и другие модификации, которые могут присутствовать в любом положении. Некоторые NRM-модификации могут ингибировать гибридизацию, а поэтому их предпочтительно использовать только в концевых областях, а предпочтительно, не использовать в сайте расщепления или в области расщепления последовательности, которая направлена на рассматриваемую последовательность или ген, в частности на антисмысловую цепь. Эти модификации могут быть использованы в любом положении смысловой цепи, при условии, что между двумя цепями дц-иРНК-агента будет сохраняться достаточный уровень гибридизации. В некоторых вариантах изобретения желательно вводить NRM в сайт расщепления или в область расщепления смысловой цепи, поскольку это может минимизировать нежелательный сайленсинг.
В большинстве случаев NRM-модификации будут распределяться в зависимости от того, присутствуют ли они на смысловой или на антисмысловой цепи. Если модификации присутствуют на антисмысловой цепи, то модификации, подавляющие или ингибирующие эндонуклеазное расщепление, не должны быть встроены в область, подвергаемую RISC-опосредуемому расщеплению, например в сайт расщепления или в область расщепления (как описано в публикации Elbashir et al., 2001, Genes and Dev. 15: 188, которая вводится в настоящее описание посредством ссылки). Расщепление мишени происходит приблизительно в середине антисмысловой цепи, состоящей из 20 или 21 нуклеотида, или примерно в положениях 10 или 11 нуклеотидов, расположенных выше первого нуклеотида на мРНК-мишени, которая является комплементарной антисмысловой цепи. Используемый здесь термин “сайт расщепления” означает нуклеотиды, находящиеся на любой стороне сайта расщепления, на мишени или на цепи иРНК-агента, которая гибридизуется с этой мишенью. Термин “область расщепления” означает сайт расщепления, состоящий из 1, 2 или 3 нуклеотидов, расположенных в любом направлении.
Такие модификации могут быть введены в концевые области, например в концевое положение или в положения 2, 3, 4 или 5 от конца смысловой или антисмысловой цепи.
Связанные лиганды
Свойства иРНК-агента, включая его фармакологические свойства, могут быть изменены и модулированы, например, путем введения лигандов, например связанных лигандов. Кроме того, фармакологические свойства иРНК-агента могут быть улучшены путем введения лиганда в композицию иРНК-агента в том случае, если такой иРНК-агент содержит связанный лиганд.
Молекулы широкого ряда, например лиганды, могут быть связаны с иРНК-агентом, либо они могут быть использованы в качестве препарата-конъюгата или добавки, присоединенной, например, к носителю, то есть к конъюгированной с лигандом мономерной субъединице. Примеры таких молекул описаны ниже в обсуждении конъюгированной с лигандом мономерной субъединицы, однако эти молекулы являются лишь предпочтительными вариантами и могут быть связаны с иРНК-агентом в других участках.
Предпочтительными молекулами являются лиганды, которые связаны с носителем, предпочтительно ковалентно, либо прямо, либо опосредованно через внутреннюю связь. В предпочтительном варианте изобретения лиганд связан с носителем посредством внутренней связи. Лиганд или связанный лиганд могут присутствовать на конъюгированном с лигандом мономере в том случае, если такой конъюгированный с лигандом мономер включен в растущую цепь. В некоторых вариантах изобретения этот лиганд может быть введен в конъюгированную с лигандом мономерную субъединицу-“предшественник” после включения такой конъюгированной с лигандом мономерной субъединицы-“предшественника” в растущую цепь. Так, например, мономер, имеющий, например, амино-концевую связь, например TAP-(CH2)nNH2, может быть включен в растущую смысловую или антисмысловую цепь. В последующей операции, то есть после включения мономерной субъединицы-предшественника в цепь, лиганд, имеющий электрофильную группу, например группу пентафторфенилового эфира или альдегидную группу, может быть затем присоединен к конъюгированному с лигандом мономеру-предшественнику посредством взаимодействия электрофильной группы указанного лиганда с концевой нуклеофильной группой субъединичной связи конъюгированного с лигандом мономера-предшественника.
В предпочтительных вариантах изобретения лиганд после его включения в иРНК-агент влияет на характер распределения, нацеливающие свойства или время жизни этого иРНК-агента. В предпочтительных вариантах изобретения лиганд обеспечивает повышенную аффинность к выбранной мишени, например к молекуле, клетке или к типу клетки, к компартменту, например к компартменту клетки или органа, к ткани, к органу или к участку организма, по сравнению с вариантами, в которых этот лиганд отсутствует.
Предпочтительные лиганды могут усиливать транспорт, гибридизацию и специфичность, а также они могут повышать резистентность к нуклеазе у полученного природного или модифицированного олигорибонуклеотида или полимерной молекулы, содержащей любую комбинацию мономеров, описанных в настоящей заявке, и/или природных или модифицированных рибонуклеотидов.
Вообще говоря, лиганды могут включать терапевтические модификаторы, например, для повышения уровня поглощения; диагностические соединения или репортерные группы, например, для мониторинга характера распределения; перекрестносшивающие агенты; молекулы, сообщающие резистентность к нуклеазе, и природные или редко встречающиеся нуклеотидные основания. Основными примерами являются липофильные молекулы, липиды, лектины, стероиды (например, уваол, гецигенин, диосгенин), терпены (например, тритерпены, например сарсасапогенин, фриделин, литохолевая кислота, дериватизированная эпифриделанолом), витамины, углеводы (например, декстран, пуллулан, хитин, хитозан, инулин, циклодекстрин или гиалуроновая кислота), белки, белок-связывающие агенты, интегрин-нацеливающие молекулы, поликатионы, пептиды, полиамины и пептидомиметики.
Лиганд может представлять собой природную или рекомбинантную или синтетическую молекулу, такую как синтетический полимер, например синтетическую полиаминокислоту. Примерами полиаминокислот является полилизин (PLL), поли-L-аспарагиновая кислота, поли-L-глутаминовая кислота, сополимер стирола и ангидрида малеиновой кислоты, сополимер L-лактида и гликолида, сополимер дивинилового эфира и ангидрида малеиновой кислоты, сополимер N-(2-гидроксипропил)метакриламида (HMPA), полиэтиленгликоль (ПЭГ), поливиниловый спирт (ПВС), полиуретан, поли(2-этилакриловая кислота), полимеры N-изопропилакриламида или полифосфазин. Примерами полиаминов являются полиэтиленимин, полилизин (PLL), спермин, спермидин, полиамин, псевдопептид-полиамин, полиамин-пептидомиметик, полиамин-дендример, аргинин, амидин, протамин, катионогенные молекулы, например, катионогенный липид, катионогенный порфирин, четвертичная соль полиамина или альфа-спиральный пептид.
Лигандами могут быть также нацеливающие группы, например агент для доставки в клетку или в ткань, например тиротропин, меланотропин, поверхностно-активный белок А, углевод муцин, гликозилированная полиаминокислота, трансферрин, бисфосфонат, полиглутамат, полиаспартат или пептид RGD или пептидомиметик RGD.
Лигандами могут быть белки, например гликопротеины, липопротеины, например липопротеин низкой плотности (ЛНП) или альбумины, например альбумин человеческой сыворотки (HSA) или пептиды, например молекулы, обладающие специфичной аффинностью к ко-лиганду, или антитела, например антитело, которое связывается с клеткой конкретного типа, такой как раковая клетка, эндотелиальная клетка или остеоцит. Лигандами могут быть также гормоны и рецепторы гормонов. Они могут также включать непептидные части, такие как кофакторы, поливалентную лактозу, поливалентную галактозу, N-ацетил-галактозамин, N-ацетил-глюкозамин, поливалентную маннозу или поливалентную фукозу. Лигандом может быть, например, липополисахарид, активатор MAP-киназы p38 или активатор NF-kB.
Лигандом может быть вещество, например лекарственное средство, которое может стимулировать проникновение иРНК-агента в клетку, например, путем дизрупции клеточного цитоскелета, например путем дизрупции микротрубочек, микроволокон и/или промежуточных филаментов клетки. Лекарственным средством может быть, например, таксон, винкристин, винбластин, циитохалазин, нокодазол, яплакинолид, латрункулин A, фаллоидин, свинхолид A, инданоцин или миосервин.
В одном из аспектов изобретения лигандом является липид или липид-содержащая молекула. Такой липид или липид-содержащая молекула, предпочтительно, связывается с сывороточным белком, например альбумином человеческой сыворотки (HSA). HSA-связывающий лиганд обеспечивает распределение данного конъюгата в ткани-мишени, например в ткани печени, включая клетки паренхимы печени. В качестве лигандов могут быть также использованы и другие молекулы, которые связываются с HSA. Так, например, могут быть использованы непроксин или аспирин. Липид или липид-содержащий лиганд может (a) повышать резистентность к расщеплению конъюгата, (b) повышать способность к нацеливанию или к транспорту конъюгата в клетку-мишень или клеточную мембрану и/или (c) может быть использован для регуляции связывания с сывороточным белком, например с HSA.
Липидный лиганд может быть использован для модуляции, например для регуляции уровня связывания конъюгата с тканью-мишенью. Так, например, липид или липидный лиганд, который более сильно связывается с HSA, с меньшей вероятностью будет нацеливаться на почки, а поэтому менее вероятно, что он будет выводиться из организма. Липид или липидный лиганд, который менее сильно связывается с HSA, может быть использован для доставки конъюгата в почки.
В предпочтительном варианте изобретения указанный липидный лиганд связывается с HSA. При этом предпочтительно, чтобы он связывался с HSA с аффинностью, достаточной для того, чтобы такой конъюгат распределялся преимущественно в непочечной ткани. Однако предпочтительно, чтобы аффинность не была настолько сильной, чтобы такое связывание HSA с лигандом было необратимым.
В другом аспекте изобретения указанным лигандом является молекула, например витамин или микроэлемент, который поглощается клеткой-мишенью, например пролиферирующейся клеткой. Эти молекулы являются особенно подходящими для лечения расстройств, характеризующихся нежелательной пролиферацией клеток, например злокачественных или незлокачественных клеток, например раковых клеток. Репрезентативными витаминами являются витамин А, Е и К. Другими репрезентативными витаминами являются витамины В, например фолиевая кислота, В12, рибофлавин, биотин, пиридоксаль или другие витамины или микроэлементы, поглощаемые раковыми клетками.
В другом аспекте изобретения указанным лигандом является агент, способный проникать в клетки, а предпочтительно агент со спиральной структурой, проникающий в клетки. Предпочтительным является амфипатический агент. Репрезентативным агентом является пептид, такой как tat или пептид усиков насекомых (antennapedia). Если агентом является пептид, то он может быть модифицирован, и такие пептиды представляют собой пептиды, включая пептидилмиметики, инвертомеры, непептидные или псевдопептидные связи и могут содержать D-аминокислоты. Таким спиральным агентом предпочтительно является агент, который имеет альфа-спиральную структуру, и предпочтительно, содержит липофильную и липофобную фазу.
5'-фосфатные модификации
В предпочтительных вариантах изобретения иРНК-агентами являются 5'-фосфорилированные агенты или агенты, включающие фосфорильный аналог у 5'-конца. 5'-фосфатными модификациями антисмысловой цепи являются модификации, совместимые с RISC-опосредуемым генным сайленсингом. Подходящими модификациями являются 5'-монофосфат ((HO)2(O)P-O-5'); 5'-дифосфат ((HO)2(O)P-O-P(HO)(O)-O-5'); 5'-трифосфат ((HO)2(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-гуанозиновый кэп (7-метилированный или неметилированный) (7m-G-O-5'-(HO)(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-аденозиновый кэп (Appp), и любая модифицированная или немодифицированная структура нуклеотидного кэпа (N-O-5'-(HO)(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-монотиофосфат (фосфортиоат; (HO)2(S)P-О-5'); 5'-монодитиофосфат (фосфордитиоат; (HO)(HS)(S)P-O-5'), 5'-фосфортиолат ((HO)2(O)P-S-5'); любая дополнительная комбинация монофосфата, дифосфата и трифосфата, в которой атом кислорода заменен атомом серы (например, 5'-альфа-тиотрифосфат, 5'-гамма-тиотрифосфат и т.п.), 5'-фосфорамидаты ((HO)2(O)P-NH-5', (HO)(NH2)(O)P-O-5'), 5'-алкилфосфонаты (R=алкил=метил, этил, изопропил, пропил и т.п., например RP(OH)(O)-O-5'-, (OH)2(O)P-5'-CH2-), 5'-алкиловый эфир фосфорной кислоты (R = алкиловый эфир = метоксиметил (MeOCH2-), этоксиметил и т.п., например RP(OH)(O)-O-5'-).
Смысловая цепь может быть модифицирована для ее инактивации и для предотвращения образования активного RISC и, тем самым, возможного снижения нежелательных эффектов. Это может быть достигнуто посредством модификации, которая предотвращает 5'-фосфорилирование смысловой цепи, например посредством модификации 5'-O-метилрибонуклеотидом (см. Nykänen et al., (2001) ATP requirements and small interfering RNA structure in the RNA interference pathway. Cell 107, 309-321). Могут быть также использованы и другие модификации, предотвращающие фосфорилирование, например простое замещение 5'-OH атомом H, а не OMe. Альтернативно, к 5'-фосфату может быть присоединена большая объемная группа, в результате чего он превращается в фосфодиэфирную связь.
Неприродные нуклеотидные основания
Нитропирролил и нитроиндолил представляют собой неприродные нуклеотидные основания, которые являются членами класса соединений, известных как универсальные основания. Универсальными основаниями являются соединения, в которых любое из четырех природных оснований может быть заменено без какого-либо значительного воздействия на характер плавления или активность олигонуклеотидного дуплекса. Было сделано предположение, что в отличие от стабилизирующих водородных связей, ассоциированных с природными нуклеотидными основаниями, олигонуклеотидные дуплексы, содержащие 3-нитропирролильные нуклеотидные основания, стабилизируются только посредством “стэкинг”-взаимодействий. Отсутствие значительного взаимодействия водородных связей с нитропирролильными нуклеотидными основаниями позволяет устранять специфичность к конкретному комплементарному основанию. Кроме того, в различных работах было подтверждено, что 4-, 5- и 6-нитроиндолил обладает очень незначительной специфичностью к четырем природным основаниям. Интересно отметить, что олигонуклеотидный дуплекс, содержащий 5-нитроиндолил, является более стабильным, чем соответствующие олигонуклеотиды, содержащие 4-нитроиндоллил и 6-нитроиндолил. Процедуры получения 1-(2'-O-метил-β-D-рибофуранозил)-5-нитроиндола описаны в работе Gaubert, G.; Wengel, J. Tetrahedron Letters 2004, 45, 5629. Другими универсальными основаниями, подходящими для использования в настоящем изобретении, являются гипоксантинил, изоинозинил, 2-азаинозинил, 7-деазаинозинил, нитроимидазолил, нитропиразолил, нитробензимидазолил, нитроиндазолил, аминоиндолил, пирролпиримидинил и их структурные производные. Более подробное обсуждение, включая процедуры синтеза нитропирролила, нитроиндолила и других универсальных оснований, упомянутых выше, можно найти в публикациях Vallone et al., Nucleic Acids Research, 27(17):3589-3596 (1999); Loakes et al., J. Mol. Bio., 270:426-436 (1997); Loakes et al., Nucleic Acids Research, 22(20):4039-4043 (1994); Oliver et al., Organic Letters, Vol. 3(13):1977-1980 (2001); Amosova et al., Nucleic Acids Research, 25(10):1930-1934 (1997); Loakes et al., Nucleic Acids Research, 29(12):2437-2447 (2001); Bergstrom et al., J. Am. Chem. Soc., 117:1201-1209 (1995); Franchetti et al., Biorg. Med. Chem. Lett. 11 :67-69 (2001); and Nair et al., Nucleosides, Nucleotides & Nucleic Acids, 20(4-7):735-738 (2001).
Дифтортолил представляет собой неприродное нуклеотидное основание, которое действует как универсальное основание. Дифтортолил представляет собой изостер природного нуклеотидного основания тимина. Но дифтортолил в отличие от тимина не обнаруживает заметной селективности к любому из природных оснований. Другими ароматическими соединениями, которые действуют как универсальные основания и могут быть использованы в настоящем изобретении, являются 4-фтор-6-метилбензимидазол и 4-метилбензимидазол. Кроме того, универсальными основаниями, которые вызывают лишь небольшую дестабилизацию олигонуклеотидных дуплексов, по сравнению с олигонуклеотидной последовательностью, содержащей только природные основания, являются относительно гидрофобные изокарбостирилиловые производные, такие как 3-метилизокарбостирилил, 5-метилизокарбостирилил и 3-метил-7-пропинилизокарбостирилил. Другими неприродными нуклеотидными основаниями, рассматриваемыми в настоящем изобретении, являются 7-азаиндолил, 6-метил-7-азаиндолил, имидазопиридинил, 9-метилимидазопиридинил, пирролпиризинил, изокарбостирилил, 7-пропинилизокарбостирилил, пропинил-7-азаиндолил, 2,4,5-триметилфенил, 4-метилиндолил, 4,6-диметилиндолил, фенил, нафталенил, антраценил, фенантраценил, пиренил, стильбенил, тетраценил, пентаценил и их структурные производные. Более подробное обсуждение, включая процедуры синтеза дифтортолила, 4-фтор-6-метилбензимидазола, 4-метилбензимидазола и других неприродных оснований, упомянутых выше, можно найти в публикациях: Schweitzer et al., J. Org. Chem., 59:7238-7242 (1994); Berger et al., Nucleic Acids Research, 28(15):2911-2914 (2000); Moran et al., J. Am. Chem. Soc., 119:2056-2057 (1997); Morales et al., J. Am. Chem. Soc., 121:2323-2324 (1999); Guckian et al., J. Am. Chem. Soc, 118:8182-8183 (1996); Morales et al., J. Am. Chem. Soc., 122(6):1001-1007 (2000); McMinn et al., J. Am. Chem. Soc., 121:11585-11586 (1999); Guckian et al., J. Org. Chem., 63:9652-9656 (1998); Moran et al., Proc. Natl. Acad. Sci., 94:10506-10511 (1997); Das et al., J. Chem. Soc., Perkin Trans., 1:197-206 (2002); Shibata et al., J. Chem. Soc., Perkin Trans., 1:1605-1611 (2001); Wu et al., J. Am. Chem. Soc, 122(32):7621-7632 (2000); O'Neill et al., J. Org. Chem., 67:5869-5875 (2002); Chaudhuri et al., J. Am. Chem. Soc., 117:10434-10442 (1995); и в патенте США № № 6218108.
Более подробное описание синтеза и применения универсальных оснований приводится в совместной и одновременно рассматриваемой заявке PCT/US2005/025967, поданной 21 июля 2005, которая во всей своей полноте вводится в настоящее описание посредством ссылки.
Использование универсальных оснований может быть особенно полезным в тех случаях, когда ген-мишень находится в микрорганизме, обнаруживающим вариабельность между различными штаммами этого микроорганизма. Это часто относится даже к областям вирусного генома, которые считаются высококонсервативными. Введение универсального основания позволяет получать иРНК-агенты, которые нацелены на большое число различных штаммов вируса, даже если они отличаются в одном или в нескольких, например, в трех положениях нуклеотидов.
Наилучшими универсальными основаниями являются основания, включенные в область иРНК-агента, который является, по меньшей мере, чувствительным к несоответствиям нуклеотидов в отношении специфичности и активности иРНК-агента. Было показано, что положения 2-9 антисмысловой цепи иРНК-агента являются наиболее восприимчивыми к несоответствиям между антисмысловой цепью и мРНК-мишенью, и такая область называется “уникальной областью" (seed-область (область-“зерно”)) иРНК-агента. Следовательно, при введении одного или нескольких универсальных оснований в иРНК-агент это(и) основание(я) предпочтительно вводить в положения, находящиеся за пределами этой уникальной области.
В таблице 1F и в таблице 1G представлены иРНК-агенты, содержащие универсальные основания во взаимно комплементарных положениях в смысловой и антисмысловой цепи. Хотя этот вариант является одним из предпочтительных вариантов иРНК-агентов согласно изобретению, однако эффект универсального основания в иРНК-агенте является более выраженным в том случае, если это универсальное основание присутствует в антисмысловой цепи. Поэтому очевидно, что основание в смысловой цепи в положении, в котором оно образует пару с универсальным основанием в антисмысловой цепи, может представлять собой либо универсальное основание, либо любое другое подходящее основание, такое как a, u, c или g. При этом желательно определить, какое основание в данном положении смысловой цепи будет обладать наивысшей активностью и/или селективностью для данного иРНК-агента. Альтернативно, может быть выбрано основание, которое присутствует в этом конкретном положении у большинства вариантов генов-мишеней, экспрессию которых необходимо ингибировать посредством рассматриваемого иРНК-агента.
Транспорт иРНК-агентов в клетки
Не ограничиваясь какой-либо конкретной теорией, следует отметить, что химическое сходство между конъюгированными с холестерином иРНК-агентами и некоторыми компонентами липопротеинов (например, холестерином, холестериновыми эфирами, фосфолипидами) может приводить к связыванию иРНК-агентов с липопротеинами (например, ЛНП, ЛВП) в крови и/или к взаимодействию иРНК-агента с клеточными компонентами, обладающими аффинностью к холестерину, например к компонентам пути транспорта холестерина. Липопротеины, а также их компоненты поглощаются и процессируются клетками под действием различных механизмов активного и пассивного транспорта, например, таких как, но не ограничивающихся ими, эндоцитоз ЛНП-рецептора, связанного с ЛНП; эндоцитоз окисленных или как-либо иначе модифицированных ЛНП посредством взаимодействия с акцепторным рецептором А; поглощение ЛВП-холестерина печенью, опосредуемое акцепторным рецептором В1; пиноцитоз или транспорт холестерина через мембраны под действием белков-переносчиков ABC (ATP-связывающего кластера), например ABC-A1, ABC-G1 или ABC-G4. Следовательно, конъюгированные с холестерином иРНК-агенты могут легко поглощаться клетками, обладающими такими механизмами транспорта, например клетками печени. В целом, настоящее изобретение относится к конкретным и общим методам доставки иРНК-агентов в клетки, экспрессирующие некоторые компоненты клеточной поверхности, например рецепторы, путем конъюгирования природного лиганда для такого компонента (например, холестерина) с иРНК-агентом или путем конъюгирования химической молекулы (например, холестерина) с иРНК-агентом, который взаимодействует или связывается с природным лигандом для такого компонента (например, ЛНП, ЛВП).
Другие варианты осуществления изобретения
РНК, например иРНК-агент, может быть продуцирован в клетке in vivo, например, из экзогенных ДНК-матриц, доставляемых в эту клетку. Так, например, ДНК-матрицы могут быть встроены в векторы и использованы в качестве векторов для генотерапии. Векторы для генотерапии могут быть доставлены индивидууму, например, путем внутривенной инъекции, местного введения (патент США № 5328470) или путем стереотаксической инъекции (см., например, Chen et al. Proc. Natl. Acad. Sci. USA 91:3054-3057, 1994). Фармацевтический препарат вектора для генотерапии может включать вектор для генотерапии в подходящем разбавителе, либо он может включать матрицу для медленного высвобождения, в которую включают носитель для доставки генов. ДНК-матрицы, например, могут включать две транскрипционные единицы, одна из которых продуцирует транскрипт, включающий верхнюю цепь иРНК-агента, а другая продуцирует транскрипт, включающий нижнюю цепь иРНК-агента. После транскрипции матриц продуцируется иРНК-агент, который затем процессируется во фрагменты киРНК-агента, опосредующие сайленсинг генов.
Композиции
Настоящее изобретение также включает фармацевтические композиции и препараты, содержащие соединения дцРНК согласно изобретению. Фармацевтические композиции согласно изобретению могут быть введены различными способами в зависимости от нужного курса лечения, то есть местного или системного, и от участка организма, подвергаемого лечению. Введением может быть местное введение, внутрилегочное введение, например, путем ингаляции или инсуффляции порошков или аэрозолей, включая введение с помощью аэрозольного распылителя; интратрахеальное введение, интраназальное введение, эпидермальное введение и трансдермальное введение, а также пероральное или парентеральное введение. Парентеральное введение включает внутривенные, внутриартериальные, подкожные, внутрибрюшинные или внутримышечные инъекции или вливания; или интракраниальное, например интратекальное или интравентрикулярное введение.
Фармацевтическими композициями и препаратами для местного введения могут быть чрескожные пластыри, мази, лосьоны, кремы, гели, капли, суппозитории, спреи, жидкости и порошки. Необходимыми или желательными могут быть стандартные фармацевтические носители, водные, порошковые или масляные основы, загустители и т.п. Могут быть также использованы презервативы и перчатки с покрытиями и т.п. Предпочтительными препаратами для местного применения являются препараты, в которых дцРНК согласно изобретению представляют собой смесь с агентами для местной доставки, такими как липиды, липосомы, жирные кислоты, сложные эфиры жирных кислот, стероиды, хелатообразующие агенты и поверхностно-активные вещества. Предпочтительные липиды и липосомы являются нейтральными (например, диолеоилфосфатидилэтаноламин = DOPE, димиристоилфосфатидилхолин = DMPC, дистеароилфосфатидилхолин), отрицательно заряженными (например, димиристоилфосфатидилглицерин = DMPG) и катионогенными (например, диолеоилтетраметиламинопропил = DOTAP и диолеоилфосфатидилэтаноламин = DOTMA). дцРНК согласно изобретению могут быть инкапсулированы в липосомы или они могут образовывать комплексы с липосомами, а в частности с катионогенными липосомами. Альтернативно, дцРНК могут образовывать комплексы с липидами, а в частности с катионными липидами. Предпочтительными жирными кислотами и эфирами жирных кислот являются, но не ограничиваются ими, арахидоновая кислота, олеиновая кислота, эйкозановая кислота, лауриновая кислота, каприловая кислота, каприновая кислота, миристиновая кислота, пальмитиновая кислота, стеариновая кислота, линолевая кислота, линоленовая кислота, дикапрат, трикапрат, моноолеин, дилаурин, глицерил-1-монокапрат, 1-додецилазациклогептан-2-он, ацилкарнитин, ацилхолин или С1-10алкиловый эфир (например, изопропилмиристат, IPM), моноглицерид, диглицерид или их фармацевтически приемлемые соли. Препараты для местного введения подробно описаны в заявке на патент США, рег. № 09/315298, поданной 20 мая 1999 г., которая во всей своей полноте вводится в настоящее описание посредством ссылки.
Композициями и препаратами для перорального введения являются порошки или гранулы, микрочастицы, наночастицы, суспензии или растворы в воде или в безводных средах, капсулы, гелевые капсулы, саше, таблетки или мини-таблетки. Для их получения может оказаться желательным использовать загустители, ароматизаторы, разбавители, эмульгаторы, диспергирующие добавки или связующие агенты. Предпочтительными препаратами для перорального введения являются препараты, в которых дцРНК согласно изобретению вводят в сочетании с одним или несколькими усилителями проницаемости, поверхностно-активными веществами и хелатообразующими агентами. Предпочтительными поверхностно-активными веществами являются жирные кислоты и/или их сложные эфиры или соли, желчные кислоты и/или их соли. Предпочтительными желчными кислотами/солями являются хенодезоксихолевая кислота (CDCA) и урзодезоксихенодезоксихолевая кислота (UDCA), холевая кислота, дегидрохолевая кислота, дезоксихолевая кислота, глюкохолевая кислота, гликохолевая кислота, гликодезоксихолевая кислота, таурохолевая кислота, тауродезоксихолевая кислота, тауро-24,25-дигидрофузидат натрия и гликодигидрофузидат натрия. Предпочтительными жирными кислотами являются арахидоновая кислота, ундекановая кислота, олеиновая кислота, лауриновая кислота, каприловая кислота, каприновая кислота, миристиновая кислота, пальмитиновая кислота, стеариновая кислота, линолевая кислота, линоленовая кислота, дикапрат, трикапрат, моноолеин, дилаурин, глицерил-1-монокапрат, 1-додецилазациклогептан-2-он, ацилкарнитин, ацилхолин или моноглицерид, диглицерид или их фармацевтически приемлемые соли (например, соли натрия). Также предпочтительными являются комбинации усилителей проницаемости, например жирные кислоты/соли в сочетании с желчными кислотами/солями. Особенно предпочтительной комбинацией является натриевая соль лауриновой кислоты, каприновой кислоты и UDCA. Другими усилителями проницаемости являются полиоксиэтилен-9-лауриловый эфир, полиоксиэтилен-20-цетиловый эфир. дцРНК согласно изобретению могут быть введены перорально, в форме гранул, включающих тонкодисперсные сухие частицы, или в виде комплекса с микро- или наночастицами. Агентами, образующими комплексы с дцРНК, являются полиаминокислоты; полимимины; полиакрилаты; полиалкилакрилаты, полиоксэтаны, полиалкилцианоакрилаты; катионизированные желатины, альбумины, крахмалы, акрилаты, полиэтиленгликоли (ПЭГ) и крахмалы; полиалкилцианоакрилаты; DEAE-дериватизированные полиимины, пуллуланы, целлюлозы и крахмалы. Особенно предпочтительными комплексообразующими агентами являются хитозан, N-триметилхитозан, поли-L-лизин, полигистидин, полиорнитин, полиспермины, протамин, поливинилпиридин, политиодиэтиламинометилэтилен P(TDAE), полиаминостирол (например, п-амино), поли(метилцианоакрилат), поли(этилцианоакрилат), поли(бутилцианоакрилат), поли(изобутилцианоакрилат), поли(изогексилцианоакрилат), DEAE-метакрилат, DEAE-гексилакрилат, DEAE-акриламид, DEAE-альбумин и DEAE-декстран, полиметилакрилат, полигексилакрилат, поли(D,L-молочная кислота), сополимер DL-молочной и гликолевой кислоты (PLGA), альгинат и полиэтиленгликоль (ПЭГ). Препараты для перорального введения дцРНК и их получение подробно описаны в заявках на патент США рег. № 08/886829 (поданной 1 июля 1997 г.), рег. № 09/108673 (поданной 1 июля 1998 г.), рег. № 09/256515 (поданной 23 февраля 1999 г.), рег. № 09/082624 (поданной 21 мая 1998 г.) и рег. № 09/315298 (поданной 20 мая 1999 г.), каждая из которых во всей своей полноте вводится в настоящее описание посредством ссылки.
Композиции и препараты для парентерального, интратекального или интравентрикулярного введения могут включать стерильные водные растворы, которые могут также содержать буферы, разбавители и другие подходящие добавки, такие как, но не ограничивающиеся ими, усилители проницаемости, соединения-носители и другие фармацевтически приемлемые носители или наполнители.
Фармацевтические композиции согласно изобретению могут представлять собой, но не ограничиваются ими, растворы, эмульсии и препараты, содержащие липосомы. Эти композиции могут быть приготовлены из различных компонентов, которыми являются, но не ограничиваются ими, жидкие образцы, самоэмульгирующиеся твердые вещества и самоэмульгирующиеся полутвердые вещества.
Фармацевтические препараты согласно изобретению, которые обычно могут представлять собой унифицированную лекарственную форму, могут быть получены стандартными методами, хорошо известными фармацевтам. Такие методы включают стадию смешивания активных ингредиентов с фармацевтически приемлемым(и) носителем(ями) и наполнителем(ями). В общих чертах, такие препараты получают путем равномерного и тщательного смешивания активных ингредиентов с жидкими носителями и с тонко измельченными твердыми носителями либо с теми и другими, с последующим, если это необходимо, формованием продукта.
Композиции согласно изобретению могут быть приготовлены в любой из многих возможных лекарственных форм, таких как, но не ограничивающихся ими, таблетки, капсулы, гелевые капсулы, жидкие сиропы, мягкие гели, суппозитории и клизмы. Композиции согласно изобретению могут быть также приготовлены в виде суспензий в воде, в безводной среде или в смешанной среде. Водные суспензии могут также содержать вещества, которые повышают вязкость суспензии, включая, например, натрийсодержащую карбоксиметилцеллюлозу, сорбит и/или декстран. Такая суспензия может также содержать стабилизаторы.
В одном из вариантов изобретения фармацевтические композиции могут быть приготовлены и использованы в виде пены. Фармацевтическими препаратами в виде пен могут быть, но не ограничиваются ими, такие препараты, как эмульсии, микроэмульсии, кремы, желе и липосомы. Хотя эти композиции, в основном, аналогичны по своей природе, однако они могут отличаться по своим компонентам и по составу конечного продукта. Методы получения таких композиций и препаратов, по существу, известны специалистам-фармацевтам и могут быть применены для приготовления композиций согласно изобретению.
Эмульсии
Композиции согласно изобретению могут быть получены и приготовлены в виде эмульсий. Эмульсии обычно представляют собой гетерогенные системы, состоящие из одной жидкости, диспергированной в другой жидкости в виде капелек, диаметр которых обычно не превышает 0,1 мкм (Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199; Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., Volume 1, p. 245; Block in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 2, p. 335; Higuchi et al., in Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 1985, p. 301). Эмульсии часто представляют собой двухфазные системы, содержащие две несмешивающиеся жидкие фазы, однородно смешанные и диспергированные одна в другой. В общих чертах, эмульсии могут представлять собой либо эмульсии типа “вода в масле” (в/м), либо “масло в воде” (м/в). Если водная фаза является тонкодисперсной и диспергирована в виде небольших капелек в объемной масляной фазе, то такая композиция называется эмульсией типа “вода в масле” (в/м). Альтернативно, если масляная фаза является тонкодисперсной и диспергирована в виде небольших капелек в объемной водной фазе, то такая композиция называется эмульсией типа “масло в воде” (м/в). Помимо диспергированных фаз, эмульсии могут содержать дополнительные компоненты и активное лекарственное средство, которое может присутствовать в виде раствора в водной фазе, масляной фазе или в виде отдельной фазы. При необходимости, в эмульсии могут быть также добавлены фармацевтические наполнители, такие как эмульгаторы, стабилизаторы, красители и антиоксиданты. Фармацевтические эмульсии могут также представлять собой различные эмульсии, состоящие из более чем двух фаз, например, в случае эмульсий типа “масло - в воде- в масле” (м/в/м) и типа “вода - в масле - в воде” (в/м/в). В большинстве случаев такие комплексные препараты имеют определенные преимущества, которыми не обладают простые бинарные эмульсии. Многие эмульсии, в которых отдельные масляные капельки эмульсии типа м/в заключены в небольшие капельки воды, составляют эмульсию типа в/м/в. Аналогичным образом, система маслянных капелек, включенных в глобулы воды, стабилизированные в масляной непрерывной фазе, составляет эмульсию типа м/в/м.
Эмульсии характеризуются невысокой термодинамической стабильностью или ее отсутствием. В большинстве случаев дисперсная или прерывистая фаза эмульсии хорошо диспергируется во внешней или в непрерывной фазе и поддерживается в этой форме благодаря эмульгаторам или вязкости данного препарата. Любая из фаз данной эмульсии может быть полутвердой или твердой, как, например, в случае эмульсии на мазевой основе и кремов. Другим способом стабизизации эмульсии является использование эмульгаторов, которые могут быть включены в любую фазу данной эмульсии. В общих чертах, эмульгаторы могут быть подразделены на четыре категории: синтетические поверхностно-активные вещества, природные эмульгаторы, абсорбционные основы и тонкодисперсные твердые вещества (Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199).
Синтетические поверхностно-активные вещества, также известные как поверхностно-активные агенты, находят широкое применение в изготовлении эмульсионных препаратов и описаны в литературе (Rieger, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 285; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), Marcel Dekker, Inc., New York, N.Y., 1988, volume 1, p. 199). Поверхностно-активные вещества обычно являются амфифильными и содержат гидрофильную и гидрофобную часть. Отношение гидрофильности к гидрофобности поверхностно-активного вещества называется гидрофильно-липофильным балансом (ГЛБ) и является ценным инструментом при определении категорий и при отборе поверхностно-активных веществ для приготовления препаратов. Исходя из природы своей гидрофильной группы, поверхностно-активные вещества могут быть подразделены на различные классы: неионогенные, анионогенные, катионогенные и амфотерные (Rieger, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 285).
Природными эмульгаторами, используемыми в эмульсионных препаратах, являются ланолин, пчелиный воск, фосфатиды, лецитин и аравийская камедь. Абсорбционные основы обладают гидрофильными свойствами, а поэтому они могут впитывать воду и образовывать эмульсии типа в/м, которые еще сохраняют свою полутвердую консистенцию, и такими основами являются безводный ланолин и гидрофильная вазелиновая мазь. Хорошими эмульгаторами, особенно в комбинации с поверхностно-активными веществами и в вязких препаратах, могут быть также тонко измельченные твердые вещества. Указанными веществами являются полярные неорганические твердые вещества, такие как гидроксиды тяжелых металлов, ненабухающие глины, такие как бентонит, аттапульгит, гекторит, каолин, монтмориллонит, коллоидный силикат алюминия и коллоидный силикат магния-алюминия, пигменты и неполярные твердые вещества, такие как уголь или глицерилтристеарат.
В эмульсионные препараты могут быть также включены неэмульгирующиеся вещества широкого ряда, придающие таким эмульсиям определенные свойства. Такими веществами являются жиры, масла, воски, жирные кислоты, жирные спирты, сложные эфиры жирных кислот, увлажнители, гидрофильные коллоиды, консерванты и антиоксиданты (Block, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 335; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199).
Гидрофильными коллоидами или гидроколлоидами являются природные смолы и синтетические полимеры, такие как полисахариды (например, аравийская камедь, агар, альгиновая кислота, каррагенан, гуаровая камедь, камедь карайи и трагакантовая камедь), производные целлюлозы (например, карбоксиметилцеллюлоза и карбоксипропилцеллюлоза) и синтетические полимеры (например, карбомеры, эфиры целлюлозы и карбоксивиниловые полимеры). Эти вещества диспергируются или набухают в воде с образованием коллоидных растворов, которые стабилизируют эмульсию посредством образования прочных межфазных пленок вокруг капелек дисперсионной фазы и посредством повышения вязкости внешней фазы.
Поскольку эмульсии часто содержат ряд ингредиентов, таких как углеводы, белки, стиролы и фосфатиды, которые могут создавать благоприятную основу для роста микробов, то в большинстве случаев такие препараты включают консерванты. Консервантами, обычно используемыми в эмульсионных препаратах, являются метилпарабен, пропилпарабен, четвертичные аммониевые соли, хлорид бензалкония, сложные эфиры п-гидроксибензойной кислоты и борная кислота. В эмульсионные препараты, для предотвращения разложения эмульсии, обычно также добавляют антиоксиданты. Используемыми антиоксидантами могут быть акцепторы свободных радикалов, такие как токоферолы, алкилгаллаты, бутилированный гидроксианизол, бутилированный гидрокситолуол, или восстановители, такие как аскорбиновая кислота и метабисульфит натрия, и антиоксиданты-синергисты, такие как лимонная кислота, винная кислота и лецитин.
Применение эмульсионных препаратов посредством чрескожного, перорального и парентерального введения и методы их получения описаны в литературе (Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199). Эмульсионные препараты для перорального введения находят очень широкое применение благодаря легкости их приготовления, а также их эффективности с точки зрения абсорбции и биологической доступности (Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199). Слабительные на основе минерального масла, растворимые в масле витамины и питательные препараты с высоким содержание жира представляют собой вещества, которые обычно вводят перорально в виде эмульсий “м/в”.
В одном из вариантов изобретения композиции дцРНК и нуклеиновых кислот получают в виде микроэмульсий. Микроэмульсия может быть определена как система, состоящая из воды, масла и амфифильного вещества, которое представляет собой оптически чистый изотропный и термодинамически стабильный жидкий раствор (Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245). Обычно микроэмульсии представляют собой системы, которые получают сначала путем диспергирования масла в водном растворе поверхностно-активного вещества, с последующим добавлением достаточного количества четвертого компонента, в основном спирта со средней длиной цепи, и с образованием прозрачной системы. Поэтому микроэмульсии были также описаны как термодинамически стабильные изотропно чистые дисперсии двух несмешивающихся жидкостей, которые стабилизируются межфазными пленками, состоящими из поверхностно-активных молекул (Leung and Shah, in: Controlled Release of Drugs: Polymers and Aggregate Systems, Rosoff, M., Ed., 1989, VCH Publishers, New York, pages 185-215). Микроэмульсии обычно получают путем объединения 3-5 компонентнов, которыми являются масло, вода, поверхностно-активное вещество, вторичное поверхностно-активное вещество и электролит. Тип микроэмульсии, а именно “вода в масле” (в/м) или “масло в воде” (м/в), зависит от свойств масла и от используемого поверхностно-активного вещества, а также от структуры и геометрической упаковки полярных головок и углеводородных хвостов поверхностно-активных молекул (Schott, in Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 1985, p. 271).
Феноменологические подходы были глубоко исследованы с использованием фазовых диаграмм, в результате чего были получены исчерпывающие сведения, которые позволяют специалисту в данной области понять механизм образования микроэмульсий (Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245; Block, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 335). По сравнению со стандартными эмульсиями преимущество микроэмульсий заключается в способности к солюбилизации не растворимых в воде лекарственных средств в препарате, состоящем из термодинамически стабильных капелек, которые образуются спонтанно.
Поверхностно-активными веществами, используемыми при получении микроэмульсий, являются, но не ограничиваются ими, ионогенные поверхностно-активные вещества, неионогенные поверхностно-активные вещества, Brij 96, олеиловые эфиры полиоксиэтилена, сложные эфиры полиглицерина и жирных кислот, монолаурат тетраглицерина (ML310), моноолеат тетраглицерина (MO310), моноолеат гексаглицерина (PO310), пентаолеат гексаглицерина (PO500), монокапрат декаглицерина (MCA750), моноолеат декаглицерина (MO750), секвиолеат декаглицерина (SO750), декаолеат декаглицерина (DAO750), которые могут быть использованы отдельно или в комбинации со вторичными поверхностно-активными веществами. Вторичное поверхностно-активное вещество, обычно спирт с короткой цепью, такой как этанол, 1-пропанол и 1-бутанол, способствуют повышению текучести на границе раздела фаз посредством проникновения в поверхностно-активную пленку с последующим образованием неупорядоченной пленки в результате образования пустого пространства между молекулами поверхностно-активного вещества.
Однако микроэмульсии могут быть получены и без использования вторичных поверхностно-активных веществ, и специалистам известны самоэмульгирующиеся микроэмульсионные системы, не содержащие спирта. Водной фазой обычно могут служить, но не ограничиваются ими, вода, водный раствор лекарственного средства, глицерин, ПЭГ300, ПЭГ400, полиглицерины, пропиленгликоли и производные этиленгликоля. Масляной фазой могут служить, но не ограничиваются ими, такие вещества, как Captex 300, Captex 355, Capmul MCM, сложные эфиры жирных кислот, моно-, ди- и триглицериды со средней длиной цепи (C8-C12), полиоксиэтилированные глицериловые эфиры жирных кислот, жирные спирты, полигликолевые глицериды, насыщенные полигликолевые С8-С10-глицериды, растительные масла и силиконовое масло.
Микроэмульсии представляют особый интерес с точки зрения солюбилизации лекарственных средств и усиления абсорбции лекарственных средств. Было высказано предположение, что микроэмульсии на основе липидов (как м/в, так и в/м) повышают биологическую доступность лекарственных средств, включая пептиды, при их пероральном введении (Constantinides et al., Pharmaceutical Research, 1994, 11, 1385-1390; Ritschel, Meth. Find. Exp. Clin. Pharmacol., 1993, 13, 205). Преимущества микроэмульсий заключаются в улучшенной солюбилизации лекарственных средств, защите лекарственного средства от ферментативного гидролиза, возможном усилении абсорбции лекарственного средства, обусловленном индуцированным поверхностно-активным веществом изменением текучести и проницаемости мембраны, легкости приготовления, легкости перорального введения в виде твердых лекарственных форм, улучшенной клинической эффективности и низкой токсичности (Constantinides et al., Pharmaceutical Research, 1994, 11, 1385; Ho et al., J. Pharm. Sci., 1996, 85, 138-143). В большинстве случаев микроэмульсии могут образовываться спонтанно при взаимодействии их компонентов при температуре окружающей среды. Это может быть особенно предпочтительным при получении термолабильных лекарственных средств, пептидов или дцРНК. Микроэмульсии также являются эффективными при чрескожной доставке активных компонентов, содержащихся в косметических и в фармацевтических препаратах. Предполагается, что микроэмульсионные композиции и препараты согласно изобретению способствуют повышению уровня системной абсорбции дцРНК и нуклеиновых кислот из желудочно-кишечного тракта, а также повышению уровня локального поглощения дцРНК и нуклеиновых кислот клетками желудочно-кишечного тракта, влагалища, щечного кармана и других участков организма, в которые были введены такие композиции и препараты.
Микроэмульсии согласно изобретению могут также содержать дополнительные компоненты и добавки, такие как сорбитанмоностеарат (Grill 3), лабразол и усилители проницаемости, улучшающие свойства данного препарата и усиливающие абсорбцию дцРНК и нуклеиновых кислот согласно изобретению. Усилители проницаемости, используемые в микроэмульсиях согласно изобретению, могут быть классифицированы как вещества, принадлежащие к одной из пяти широких категорий соединений, таких как поверхностно-активные вещества, жирные кислоты, соли желчных кислот, хелатообразующие вещества и вещества, не являющиеся хелатообразующими и поверхностно-активными веществами (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p. 92). Каждое соединение этого класса обсуждалось выше.
Липосомы
Помимо микроэмульсий, которые были исследованы и использованы для получения лекарственных средств, существует множество поверхностно-активных веществ с упорядоченной структурой. Такими структурами являются монослои, мицеллы, бислои и везикулы. Везикулы, такие как липосомы, представляют большой интерес с точки зрения специфичности и продолжительного действия лекарственных средств, доставку которых они обеспечивают. Используемый в настоящем изобретении термин “липосома” означает везикулу, состоящую из амфифильных липидов, присутствующих в сферическом бислое или бислоях.
Липосомы представляют собой униламеллярные или мультиламеллярные везикулы, которые имеют мембрану, образованную из липофильного материала и водной внутренней части. Водная часть содержит композицию, предназначенную для доставки. Катионные липосомы имеют то преимущество, что они способны сливаться с клеточной стенкой. Некатионные липосомы, хотя они и неспособны подвергаться эффективному слиянию с клеточной стенкой, поглощаются макрофагами in vivo.
Для проникновения через неповрежденную кожу млекопитающего липидные везикулы должны проходить через серию тонких пор, каждая из которых имеет диаметр менее чем 50 нм, под воздействием подходящего градиента, создаваемого при прохождении через кожу. Поэтому желательно использовать липосомы, которые поддаются высокой степени деформации и способны проходить через тонкие поры.
Другие преимущества липосом заключаются в том, что липосомы, полученные из природных фосфолипидов, являются биологически совместимыми и биологически разлагаемыми; липосомы могут включать широкий ряд растворимых в воде и липидах лекарственных средств; и липосомы могут защищать инкапсулированные лекарственные средства в их внутренних компартментах от метаболизма и расщепления (Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245). При получении липосомных препаратов следует учитывать такие важные факторы, как поверхностный заряд липидов, размер везикулы и водный объем липосом.
Липосомы могут быть использованы для переноса и доставки активных ингредиентов в участок их действия. Поскольку липосомная мембрана по своей структуре аналогична биологическим мембранам, то при нанесении липосом на ткань эти липосомы начинают проникать в клеточные мембраны и по мере проникновения липосомы в клетку и ее продвижения по клетке, содержимое липосом высвобождается в клетку, в которой может действовать активное вещество.
Липосомные препараты, как способ доставки многих лекарственных средств, являются объектом интенсивных исследований. Появляется все больше данных о том, что липосомы для местного введения имеют несколько преимуществ по сравнению с другими препаратами. Такими преимуществами являются относительно низкий уровень побочных эффектов, ассоциированных с высокой системной абсорбцией вводимого лекарственного средства, повышенный уровень аккумуляции вводимого лекарственного средства в нужном участке-мишени и возможность введения широкого ряда лекарственных средств, как гидрофильных, так и гидрофобных, в кожу.
В нескольких работах подробно описана способность липосом доставлять агенты, включая высокомолекулярную ДНК, в кожу. В кожу были введены соединения, включая аналгетики, антитела, гормоны и высокомолекулярные ДНК. В большинстве случаев такие липосомы предназначены для доставки в верхний эпидермис.
Липосомы подразделяются на два широких класса. Катионными липосомами являются положительно заряженные липосомы, которые взаимодействуют с отрицательно заряженными молекулами ДНК с образованием стабильного комплекса. Комплекс “положительно заряженная ДНК/липосома” связывается с отрицательно заряженной клеточной поверхностью и интернализуется в эндосому. Из-за кислотного рН в эндосоме липосомы разрушаются, и их содержимое высвобождается в клеточную цитоплазму (Wang et al., Biochem. Biophys. Res. Commun., 1987, 147, 980-985).
Липосомы, которые являются рН-чувствительными или отрицательно заряженными, захватывают ДНК, но не образуют с ней комплекса. Поскольку ДНК и липид имеют одинаковый заряд, они будут отталкиваться друг от друга и не будут образовывать комплекс. Тем не менее, некоторые ДНК захватываются во внутреннем водном пространстве этих липосом. рН-чувствительные липосомы были использованы для доставки ДНК, содержащей ген тимидинкиназы, в монослои клеточной культуры. Экспрессия экзогенного гена была детектирована в клетках-мишенях (Zhou et al., Journal of Controlled Release, 1992, 19, 269-274).
Липосомная композиция одного из главных типов содержит фосфолипиды, не являющиеся природными фосфатидилхолинами. Нейтральные липосомные композиции, например, могут быть образованы из димиристоилфосфатидилхолина (DMPC) или дипальмитоилфосфатидилхолина (DPPC). Анионные липосомные композиции обычно образуются из димиристоилфосфатидилглицерина, а анионные фузогенные липосомы образуются, главным образом, из диолеоилфосфатидилэтаноламина (DOPE). Липосомную композицию другого типа получают из фосфатидилхолина (РС), такого как, например, соевый РС и яичный РС. Липосомную композицию другого типа получают из смесей фосфолипида и/или фосфатидилхолина и/или холестерина.
Было проведено несколько исследований по местной доставке липосомных лекарственных препаратов в кожу. Нанесение липосом, содержащих интерферон, на кожу морских свинок приводило к уменьшению степени поражения кожи герпесом, а доставка интерферона другими средствами (например, в виде раствора или эмульсии) оказалась неэффективной (Weiner et al., Journal of Drug Targeting, 1992, 2, 405-410). Кроме того, дополнительные исследования продемонстрировали эффективность интерферона, вводимого как часть липосомного препарата, предназначенного для введения интерферона с использованием водной системы, и был сделан вывод, что такой липосомный препарат был более эффективным, чем препарат для введения с использованием водной системы (du Plessis et al., Antiviral Research, 1992, 18, 259-265).
Неионные липосомные системы, а в частности системы, содержащие неионогенное поверхностно-активное вещество и холестерин, были также проанализированы на их способность доставлять лекарственные средства в кожу. Неионные липосомные препараты, содержащие Novasome.TM. I (глицерилдилаурат/холестерин/стеариловый эфир полиоксиэтилена- 10) и Novasome.TM. II (глицерилдистеарат/холестерин/стеариловый эфир полиоксиэтилена-10), были использованы для доставки циклоспорина А в дерму мышиной кожи. Полученные результаты показали, что такие неионогенные липосомные системы эффективно способствуют отложению циклоспорина А в различных слоях кожи (Hu et al. S.T.P.Pharma. Sci, 1994, 4, 6, 466).
Липосомами также являются “стерически стабилизированные” липосомы, и этот используемый здесь термин означает липосомы, содержащие один или несколько специализированных липидов, которые при их включении в липосомы повышают время их жизни в кровотоке по сравнению с липосомами, не содержащими таких специализированных липидов. Примерами стерически стабилизированных липосом являются липосомы, в которых часть везикулообразующей липидной молекулы липосомы (А) содержит один или несколько гликолипидов, таких как моносиалоганглиозид Gm1, или (B) является дериватизированной одним или несколькими гидрофильными полимерами, такими как полиэтиленгликоль (ПЭГ). Не ограничиваясь какой-либо конкретной теорией, следует отметить, что по меньшей мере для стерически стабилизированных липосом, содержащих ганглиозиды, сфингомиелин или ПЭГ-дериватизированные липиды, повышенное время полужизни этих стерически стабилизированных липосом в кровотоке обусловлено снижением уровня их поглощения клетками ретикулоэндотелиальной системы (RES) (Allen et al., FEBS Letters, 1987, 223, 42; Wu et al., Cancer Research, 1993, 53, 3765).
Различные липосомы, содержащие один или несколько гликолипидов, известны специалистам. В публикации Papahadjopoulos et al. (Ann. N.Y. Acad. Sci., 1987, 507, 64) описана способность моносиалоганглиозида Gm1, сульфата галактоцереброзида и фосфатидилинозита увеличивать время полужизни липосом в кровотоке. Эти данные были интерпретированы Gabizon et al. (Proc. Natl. Acad. Sci. U.S.A., 1988, 85, 6949). В патенте США № 4837028 и в заявке WO 88/04924, Allen et al., описаны липосомы, содержащие (1) сфингомиелин и (2) ганглиозид Gm1 или сложный эфир сульфата галактоцереброзида. В патенте США № 5543152 (Webb et al.) описаны липосомы, содержащие сфингомиелин. Липосомы, содержащие 1,2-sn-димиристоилфосфатидилхолин, описаны в WO 97/13499 (Lim et al.).
Многие липосомы, содержащие липиды, дериватизированные одним или несколькими гидрофильными полимерами, и методы их получения известны специалистам. В публикации Sunamoto et al. (Bull. Chem. Soc. Jpn., 1980, 53, 2778) описаны липосомы, содержащие неионогенный детергент, 2C1215G, который включает молекулу ПЭГ. В публикации Illum et al. (FEBS Lett., 1984, 167, 79) указывается, что гидрофильное покрытие полистироловых частиц полимерными гликолями значительно увеличивает время полужизни липосом в крови. Синтетические фосфолипиды, модифицированные путем присоединения карбоксильных групп полиалкиленгликолей (например, ПЭГ), описаны Sears (патенты США №№ 4426330 и 4534899). В публикации Klibanov et al. (FEBS Lett., 1990, 268, 235) описаны эксперименты, продемонстрировавшие, что липосомы, содержащие фосфатидилэтаноламин (ФЭ), дериватизированный ПЭГ или стеаратом ПЭГ, имеют значительно более длительное время полужизни в кровотоке. В публикации Blume et al. (Biochimica et Biophysica Acta, 1990, 1029, 91) такие наблюдения были направлены на другие ПЭГ-дериватизированные фосфолипиды, например DSPE-ПЭГ, образованные из комбинации дистеароилфосфатидилэтаноламина (DSPE) и ПЭГ. Липосомы, имеющие ковалентно связанные молекулы ПЭГ на своей внешней поверхности, описаны в европейском патенте № EP 0445131 B1 и WO 90/04384, Fisher. Липосомные композиции, содержащие 1-20% моль ПЭГ-дериватизированного ФЭ, и методы их применения описаны Woodle et al. (патенты США №№ 5013556 и 5356633) и Martin et al. (патент США № 5213804 и европейский патент № EP 0496813 B1). Липосомы, содержащие ряд других конъюгатов “липид-полимер”, описаны в заявке WO 91/05545 и в патенте США № 5225212 (оба Martin et al.), а также в заявке WO 94/20073 (Zalipsky et al.). Липосомы, содержащие ПЭГ-модифицированные церамидные липиды, описаны в WO 96/10391 (Choi et al.). В патенте США № 5540935 (Miyazaki et al.) и в патенте США № 5556948 (Tagawa et al.) описаны ПЭГ-содержащие липосомы, которые на своих поверхностях могут быть также дериватизированы функциональными молекулами.
Специалистам известно ограниченное число липосом, содержащих нуклеиновые кислоты. В WO 96/40062, Thierry et al., описаны методы инкапсуляции высокомолекулярных нуклеиновых кислот в липосомы. В патенте США № 5264221, Tagawa et al., описаны связанные с белком липосомы, и утверждается, что такие липосомы могут включать дцРНК. В патенте США № 5665710, Rahman et al., описаны некоторые методы инкапсуляции олигодезоксинуклеотидов в липосомы. В WO 97/04787, Love et al., описаны липосомы, содержащие дцРНК, нацеленные на ген raf.
Трансферсомы представляют собой еще один тип липосом, являющихся в высокой степени деформируемыми липидными агрегатами, которые могут быть привлекательными кандидатами на их использование в качестве везикул для доставки лекарственных средств. Трансферсомы могут быть описаны как липидные капельки, степень деформируемости которых настолько высока, что они могут легко проникать через поры, размеры которых меньше, чем сами капли. Трансферсомы могут быть адаптированы к условиям окружающей среды, в которых они используются, например, они подвергаются самооптимизации (адаптируются к форме пор в коже), саморепарации, часто достигают своих мишеней без фрагментации и подвергаются самозагрузке. Для получения трансферсом в стандартную липосомную композицию могут быть добавлены активаторы поверхности раздела фаз, обычно поверхностно-активные вещества. Трансферсомы были использованы для доставки сывороточного альбумина в кожу. Было показано, что опосредуемая трансферсомами доставка сывороточного альбумина является эффективной при подкожной инъекции раствора, содержащего сывороточный альбумин.
Поверхностно-активные вещества находят широкое применение в таких препаратах, как эмульсии (включая микроэмульсии) и липосомы. Наиболее распространенным путем классификации и оценки свойств многих поверхностно-активных веществ различных типов, как природных, так и синтетических, является определение гидрофильно-липофильного баланса (ГЛБ). При определении категории различных поверхностно-активных веществ, используемых в препаратах, наиболее часто применяемым критерием является природа гидрофильной группы (также известной как “головная” группа) (Rieger, in Pharmaceutical Dosage Forms, Marcel Dekker, Inc., New York, N.Y., 1988, p. 285).
Если молекула поверхностно-активного вещества не ионизована, то такая молекула принадлежит к классу неионогенных поверхностно-активных веществ. Неионогенные поверхностно-активные вещества широко применяются в фармацевтической и косметической промышленности и используются в широком интервале величин рН. Обычно их величины ГЛБ составляют в пределах от 2 до примерно 18, в зависимости от их структуры. Неионогенными поверхностно-активными веществами являются неионогенные сложные эфиры, такие как сложные эфиры этиленгликоля, сложные эфиры пропиленгликоля, глицериловые сложные эфиры, полиглицериловые сложные эфиры, сложные эфиры сорбитана, сложные эфиры сахарозы и этоксилированные сложные эфиры. В класс таких соединений входят также неионогенные алканоламиды и простые эфиры, такие как этоксилаты жирного спирта, пропоксилированные спирты и этоксилированные/пропоксилированные блоксополимеры. Наиболее известными членами класса неионогенных поверхностно-активных веществ являются полиоксиэтиленовые поверхностно-активные вещества.
Если молекула поверхностно-активного вещества несет отрицательный заряд, в том случае, если она растворена или диспергирована в воде, то такое поверхностно-активное вещество классифицируется как анионогенное. Анионогенными поверхностно-активными веществами являются карбоксилаты, такие как мыла, ациллактилаты, ациламиды аминокислот, сложные эфиры серной кислоты, такие как алкилсульфаты и этоксилированные алкилсульфаты, сульфонаты, такие как алкилбензолсульфонаты, ацилизетионаты, ацилтаураты и сульфосукцинаты, и фосфаты. Наиболее важными членами класса анионогенных поверхностно-активных веществ являются алкилсульфаты и мыла.
Если молекула поверхностно-активного вещества несет положительный заряд, в том случае, если она растворена или диспергирована в воде, то такое поверхностно-активное вещество классифицируется как катионогенное. Катионогенными поверхностно-активными веществами являются четвертичные аммониевые соли и этоксилированные амины. Наиболее часто используемыми членами соединений этого класса являются четвертичные аммониевые соли.
Если молекула поверхностно-активного вещества несет либо положительный, либо отрицательный заряд, то такое поверхностно-активное вещество классифицируется как амфотерное. Амфотерными поверхностно-активными веществами являются производные акриловой кислоты, замещенные алкиламиды, N-алкилбетаины и фосфатиды.
Применение поверхностно-активных веществ для получения лекарственных средств, фармацевтических препаратов и эмульсий описано в литературе (Rieger, in Pharmaceutical Dosage Forms, Marcel Dekker, Inc., New York, N.Y., 1988, p. 285).
Усилители проницаемости
В одном из вариантов изобретения предусматривается использование различных усилителей проницаемости в целях обеспечения эффективной доставки нуклеиновых кислот, а в частности дцРНК, в кожу животных. Большинство лекарственных средств присутствует в растворе в ионизованной и в неионизованной формах. Однако через клеточные мембраны легко проходят только растворимые в липидах или липофильные лекарственные средства. Было установлено, что через клеточную мембрану могут проходить даже нелипофильные лекарственные средства, но только в том случае, если такая пересекаемая мембрана обработана усилителем проницаемости. Усилители проницаемости, помимо стимуляции диффузии нелипофильных лекарственных средств через клеточные мембраны, также повышают способность к прохождению липофильных лекарственных средств.
Усилители проницаемости могут быть классифицированы как соединения, принадлежащие к одной из пяти широких категорий, то есть поверхностно-активные вещества, жирные кислоты, соли желчных кислот, хелатообразующие агенты и соединения, не образующие хелатные комплексы и не являющиеся поверхностно-активными веществами (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p.92). Каждый из вышеупомянутых классов усилителей проницаемости более подробно описан ниже.
Поверхностно-активные вещества: В соответствии с настоящим изобретением, поверхностно-активные вещества (или “поверхностно-активные агенты”) представляют собой химические молекулы, которые при их растворении в водном растворе снижают поверхностное натяжение раствора или межфазное поверхностное натяжение между водным раствором и другой жидкостью, в результате чего повышается уровень абсорбции дцРНК через слизистую. Помимо солей желчных кислот и жирных кислот, такими усилителями проницаемости являются, например, лаурилсульфат натрия, лауриловый эфир полиоксиэтилена-9 и цетиловый эфир полиоксиэтилена-20 (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p.92); и перфторированные эмульсии, такие как FC-43 (Takahashi et al., J. Pharm. Pharmacol., 1988, 40, 252).
Жирные кислоты: Различными жирными кислотами и их производными, которые действуют как усилители проницаемости, являются, например, олеиновая кислота, лауриловая кислота, каприновая кислота (н-декановая кислота), миристиновая кислота, пальмитиновая кислота, стеариновая кислота, линолевая кислота, линоленовая кислота, дикапрат, трикапрат, моноолеин (1-моноолеоил-рац-глицерин), дилаурин, каприловая кислота, арахидоновая кислота, 1-монокапрат глицерина, 1-додецилазациклогептан-2-он, ацилкарнитины, ацилхолины или их С1-10 алкиловые эфиры (например, метиловый, изопропиловый и трет-бутиловый эфиры) и моно- и диглицериды (то есть олеат, лаурат, капрат, миристат, пальмитат, стеарат, линолеат и т.п.) (Lee et al., Critical Reviews in Therapeutic Drug Carryier Systems, 1991, p.92; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; El Hariri et al., J. Pharm. Pharmacol., 1992, 44, 651-654).
Соли желчных кислот: Физиологическая роль желчных кислот состоит в облегчении диспергирования и абсорбции липидов и жирорастворимых витаминов (Brunton, Chapter 38 in: Goodman & Gilman's The Pharmacological Basis of Therapeutics, 9th Ed., Hardman et al. Eds., McGraw-Hill, New York, 1996, pp. 934-935). Различные природные соли желчных кислот и их синтетические производные действуют как усилители проницаемости. Таким образом, термин “соли желчных кислот” включает любой из природных компонентов желчных кислот, а также любое из их синтетических производных. Солями желчных кислот согласно изобретению являются, например, холевая кислота (или ее фармацевтически приемлемая натриевая соль, холат натрия), дегидрохолевая кислота (дегидрохолат натрия), дезоксихолевая кислота (дезоксихолат натрия), глюкохолевая кислота (глюкохолат натрия), гликохолевая кислота (гликохолат натрия), гликодезоксихолевая кислота (гликодезоксихолат натрия), таурохолевая кислота (таурохолат натрия), тауродезоксихолевая кислота (тауродезоксихолат натрия), хенодезоксихолевая кислота (хенодезоксихолат натрия), урсодезоксихолевая кислота (UDCA), тауро-24,25-дигидрофузидат натрия (STDHF), гликодигидрофузидат натрия и лауриловый эфир полиоксиэтилена-9 (POE) (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92; Swinyard, Chapter 39 In: Remington's Pharmaceutical Sciences, 18th Ed., Gennaro, ed., Mack Publishing Co., Easton, Pa., 1990, pages 782-783; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; Yamamoto et al., J. Pharm. Exp. Ther., 1992, 263, 25; Yamashita et al., J. Pharm. Sci., 1990, 79, 579-583).
Хелатообразующие агенты: Хелатообразующие агенты, используемые в настоящем изобретении, могут быть определены как соединения, которые удаляют ионы металла из раствора посредством образования комплексов с этими ионами, что приводит к повышению уровня абсорбции дцРНК через слизистую. Что касается использования таких агентов в качестве усилителей проницаемости согласно изобретению, то следует отметить, что такие хелатообразующие агенты имеют дополнительное преимущество, которое состоит в том, что они также являются ингибиторами ДНКазы, поскольку наиболее известным ДНК-нуклеазам для их катализа необходим ион двухвалентного металла, и такие нуклеазы ингибируются указанными хелатообразующими агентами (Jarrett, J. Chromatogr., 1993, 618, 315-339). Хелатообразующими агентами согласно изобретению являются, но не ограничиваются ими, динатрийэтилендиаминотетраацетат (EDTA), лимонная кислота, салицилаты (например, салицилат натрия, 5-метоксисалицилат и гомованилат), N-ацильные производные коллагена, лаурет-9 и N-аминоацильные производные бета-дикетонов (енаминов) (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; Buur et al., J. Control Rel., 1990, 14, 43-51).
Соединения, не образующие хелатные комплексы и не являющиеся поверхностно-активными веществами: Используемые здесь усилители проницаемости, не образующие хелатные комплексы и не являющиеся поверхностно-активными веществами, могут быть определены как соединения, которые обладают незначительной активностью как хелатообразующие агенты или как поверхностно-активные вещества, но, тем не менее, повышают уровень абсорбции дцРНК через слизистую пищеварительного тракта (Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33). Такой класс усилителей проницаемости включает, например, ненасыщенные циклические мочевины, производные 1-алкил- и 1-алкенилазациклоалканонов (Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92); и нестероидные противовоспалительные средства, такие как диклофенак-натрий, индометацин и фенилбутазон (Yamashita et al., J. Pharm. Pharmacol., 1987, 39, 621-626).
В фармацевтические и другие композиции согласно изобретению могут быть также добавлены агенты, повышающие уровень поглощения дцРНК клетками. Так, например, известно, что катионогенные липиды, такие как липофектин (Junichi et al., патент США № 5705188), катионогенные производные глицерина и поликатионогенные молекулы, такие как полилизин (Lollo et al., заявка PCT WO 97/30731), усиливают поглощение дцРНК клетками.
Другими агентами, которые могут быть использованы для усиления проницаемости вводимых нуклеиновых кислот, являются гликоли, такие как этиленгликоль и пропиленгликоль, пирролы, такие как 2-пиррол, азоны и терпены, такие как лимонен и ментон.
Носители
В состав некоторых композиций согласно изобретению также входят соединения-носители. Используемый здесь термин “соединение-носитель” или “носитель” может означать нуклеиновую кислоту или ее аналог, которые являются инертными (то есть не обладают биологической активностью per se), но распознаются как нуклеиновая кислота в процессах in vivo, которые снижают биологическую доступность нуклеиновой кислоты, обладающей биологической активностью, посредством, например, расщепления биологически активной нуклеиновой кислоты или стимуляции ее удаления из кровотока. Совместное введение нуклеиновой кислоты и соединения-носителя, обычно с избытком носителя, может приводить к значительному снижению количества нуклеиновой кислоты, высвобождаемой в печень, почки или в другие системы, не относящиеся к органам кровообращения, что, вероятно, обусловлено конкуренцией между соединением-носителем и нуклеиновой кислотой за связывание с их общим рецептором. Так, например, высвобождение частично фосфортиоатной дцРНК в ткань печени может быть уменьшено при совместном введении такой дцРНК с полиинозиновой кислотой, сульфатом декстрана, полицитидиновой кислотой или 4-ацетамидо-4'-изотиоциано-стильбен-2,2'-дисульфоновой кислотой (Miyao et al., Antisense Res. Dev., 1995, 5, 115-121; Takakura et al., Antisense & Nucl. Acid Drug Dev., 1996, 6, 177-183.
Наполнители
В отличие от соединений-носителей “фармацевтический носитель” или “наполнитель” представляет собой фармацевтически приемлемый растворитель, суспендирующий агент или любой другой фармакологически инертный носитель для доставки одной или нескольких нуклеиновых кислот в организм животного. Такой наполнитель может быть жидким или твердым, и он может быть выбран в соответствии с нужным способом введения, так чтобы при объединении с нуклеиновой кислотой и с другими компонентами данной фармацевтической композиции он обеспечивал желаемые объем, консистенцию и т.п. этой композиции. Типичными фармацевтическими носителями являются, но не ограничиваются ими, связующие вещества (например, предварительно желатинизированный кукурузный крахмал, поливинилпирролидон или гидроксипропилметилцеллюлоза и т.п.); наполнители (например, лактоза и другие сахара, микрокристаллическая целлюлоза, пектин, желатин, сульфат кальция, этилцеллюлоза, полиакрилаты или бифосфат кальция и т.п.); замасливатели (например, стеарат магния, тальк, двуокись кремния, коллоидная двуокись кремния, стеариновая кислота, стеараты металлов, гидрогенизированные растительные масла, кукурузный крахмал, полиэтиленгликоли, бензоат натрия, ацетат натрия и т.п.); дезинтегрирующие агенты (например, крахмал, гликолят натрия-крахмала и т.п.); и смачивающие агенты (например, лаурилсульфат натрия и т.п.).
Для получения композиций согласно изобретению могут быть также использованы фармацевтически приемлемые органические или неорганические наполнители, подходящие для непарентерального введения и не оказывающие какого-либо нежелательного взаимодействия с нуклеиновыми кислотами. Подходящими фармацевтически приемлемыми носителями являются, но не ограничиваются ими, вода, солевые растворы, спирты, полиэтиленгликоли, желатин, лактоза, амилоза, стеарат магния, тальк, кремниевая кислота, вязкое вазелиновое масло, гидроксиметилцеллюлоза, поливинилпирролидон и т.п.
Препараты для местного введения нуклеиновых кислот могут включать стерильные и нестерильные водные растворы, безводные растворы в стандартных растворителях, таких как спирты, или растворы нуклеиновых кислот в жидких или твердых масляных основах. Эти растворы могут также содержать буферы, разбавители и другие подходящие добавки. Могут быть также использованы фармацевтически приемлемые органические или неорганические наполнители, подходящие для непарентерального введения, которые не оказывают какого-либо нежелательного взаимодействия с нуклеиновыми кислотами.
Подходящими фармацевтически приемлемыми наполнителями являются, но не ограничиваются ими, вода, солевые растворы, спирты, полиэтиленгликоли, желатин, лактоза, амилоза, стеарат магния, тальк, кремниевая кислота, вязкое вазелиновое масло, гидроксиметилцеллюлоза, поливинилпирролидон и т.п.
Фармацевтические композиции для доставки в дыхательные пути
В другом своем аспекте настоящее изобретение относится к доставке иРНК-агентов в дыхательные пути, а в частности для лечения кистозного фиброза. Дыхательные пути состоят из верхних дыхательных путей, включая ротовую часть глотки и гортань, и нижних дыхательных путей, которые включают трахею с последующим ее разветвлением на бронхи и бронхиолы. Верхние и нижние дыхательные пути называются воздухопроводящими путями. Терминальные бронхиолы, в свою очередь, делятся на дыхательные бронхиолы, которые затем ведут к конечной респираторной зоне, альвеолам или глубокому легкому. Глубокое легкое или альвеолы являются основными мишенями для вдыхаемых терапевтических аэрозолей, предназначенных для системной доставки иРНК-агентов.
Композиции для доставки в легкие могут быть введены путем вдыхания пациентом дисперсии, так чтобы композиция, а предпочтительно иРНК-агент, в данной дисперсии могли достигать легких, в которых они могут, например, легко абсорбироваться через альвеолярную область непосредственно в кровоток. Доставка в легкие может быть эффективной при системном введении и при местном введении для лечения легочных заболеваний.
Доставка в легкие может быть достигнута различными методами, включая использование препаратов для ингаляций, аэрозольных препаратов, мицеллярных препаратов и препаратов для инсуффляции; при этом введение путем ингаляции может быть осуществлено перорально и/или интраназально. Доставка может быть осуществлена с использованием жидких аэрозолей, аэрозольных ингаляторов и устройств для диспергирования сухих порошков. Предпочтительными являются устройства с дозирующим клапаном. Одно из преимуществ использования распылителей или ингаляторов состоит в том, что в данном случае возможность загрязнения сведена к минимуму, поскольку такие устройства являются автономными. Устройства для диспергирования сухих порошков, например, обеспечивают доставку лекарственных средств, которые могут быть легко приготовлены в виде сухих порошков. Композиция иРНК может стабильно храниться в виде лиофилизованных или осушенных распылением порошков отдельно или в комбинации с подходящими порошками-носителями. Доставка композиции путем ингаляции может быть осуществлена с помощью элемента-таймера доз, который может включать таймер, дозиметр, счетчик времени или индикатор времени, которые при их монтировании в данное устройство позволяют прослеживать поступление дозы, проводить мониторинг соблюдения пациентом режима введения доз и/или инициировать запуск дозы пациенту во время введения аэрозольного препарата.
Примерами фармацевтических устройств для доставки аэрозолей являются ингаляторы с дозирующим клапаном (MDI), инсуффляторы (DPI) и струйные ингаляторы. Примеры систем для доставки посредством ингаляции, которые могут быть легко адаптированы для введения рассматриваемых иРНК-агентов, описаны, например, в патентах США №№ 5756353; 5858784 и в заявках PCT WO98/31346; WO98/10796; WO00/27359; WO01/54664; WO02/060412. Другие аэрозольные препараты, которые могут быть использованы для доставки иРНК-агентов, описаны в патентах США №№ 6294153; 6344194; 6071497 и в заявках PCT WO02/066078; WO02/053190; WO01/60420; WO00/66206. Кроме того, способы доставки иРНК-агентов могут быть разработаны на основе способов, применяемых для доставки других олигонуклеотидов (например, антисмыслового олигонуклеотида) путем ингаляции, таких как способы, описанные в публикации Templin et al., Antisense Nucleic Acid Drug Dev, 2000, 10:359-68; Sandrasagra et al., Expert Opin Biol Ther, 2001, 1:979-83; Sandrasagra et al., Antisense Nucleic Acid Drug Dev, 2002, 12:177-81.
Доставка агентов согласно изобретению может также включать введение так называемых “пролекарств”, то есть препаратов или химических модификаций терапевтических средств, которым для высвобождения терапевтического вещества в данный организм, а предпочтительно в участок данного организма, в котором желательно такое высвобождение, необходима определенная форма процессинга или транспорта в соответствии с природным механизмом; при этом доставка в нужный участок организма может быть осуществлена в сочетании с доставкой в дыхательные пути, а также в сочетании с другими вариантами настоящего изобретения. Так, например, легкие человека могут удалять или быстро разлагать гидролитически расщепляемые осажденные аэрозоли в течение периода времени от нескольких минут до нескольких часов. В верхних дыхательных путях реснитчатый эпителий служит “эскалатором”, по которому частицы удаляются из дыхательных путей по направлению к ротовой полости. Pavia, D., "Lung Mucociliary Clearance," in Aerosols and the Lung: Clinical and Experimental Aspects, Clarke, S. W. and Pavia, D., Eds., Butterworths, London, 1984. В глубоких легких альвеолярные макрофаги обладают способностью к фагоцитозу частиц вскоре после их осаждения. Warheit et al. Microscopy Res. Tech., 26: 412-422 (1993); and Brain, J. D., "Physiology and Pathophysiology of Pulmonary Macrophages," in The Reticuloendothelial System, S. M. Reichard and J. Filkins, Eds., Plenum, New. York., pp. 315-327, 1985.
В предпочтительных вариантах изобретения, а в частности в тех случаях, когда желательно системное введение иРНК-агента, аэрозольные иРНК-агенты приготавливают в виде микрочастиц. Микрочастицы, имеющие диаметр от 0,5 до 10 микрон, могут проникать через легкие, проходя через большинство природных барьеров. Для прохождения не через горло, а в его обход частицы должны иметь диаметр менее чем 10 микрон, а для предотвращения выдыхания частицы должны иметь диаметр 0,5 микрон или более.
Другие компоненты
Композиции согласно изобретению могут дополнительно содержать и другие вспомогательные компоненты, обычно присутствующие в фармацевтических композициях на уровнях, устанавливаемых специалистом. Так, например, такие композиции могут содержать дополнительные совместимые фармацевтически активные вещества, такие как, например, противозудное средство, вяжущие средства, местные анестезирующие средства или противовоспалительные средства, либо они могут содержать дополнительные вещества, которые могут быть использованы при изготовлении композиций согласно изобретению в виде различных физически дискретных лекарственных форм, а именно, такие как красители, ароматизаторы, консерванты, антиоксиданты, замутнители, загустители и стабилизаторы. Однако такие вещества при их добавлении не должны оказывать чрезмерного влияния на биологическую активность компонентов композиций согласно изобретению. Эти препараты могут быть стабилизированы и при необходимости смешаны со вспомогательными агентами, например замасливателями, консервантами, стабилизаторами, смачивающими агентами, эмульгаторами, солями для изменения осмотического давления, буферами, красителями, отдушками и/или ароматическими веществами и т.п., которые не участвуют в каком-либо нежелательном взаимодействии с нуклеиновой(ыми) кислотой(ами) данного препарата.
Водные суспензии могут содержать вещества, которые повышают вязкость суспензии, включая, например, натрий-содержащую карбоксиметилцеллюлозу, сорбит и/или декстран. Такая суспензия может также содержать стабилизаторы.
Некоторые варианты согласно изобретению относятся к фармацевтическим композициям, содержащим (a) один или несколько дцРНК-агентов и (b) один или несколько других химиотерапевтических средств, которые действуют не по механизму интерференции РНК, а по какому-либо другому механизму. Примерами таких химиотерапевтических агентов являются, но не ограничиваются ими, даунорубицин, дауномицин, дактиномицин, доксорубицин, эпирубицин, идарубицин, эзорубицин, блеомицин, мафосфамид, ифосфамид, цитозинарабинозид, бис-хлорэтилнитрозомочевина, бисульфан, митомицин C, актиномицин D, митрамицин, преднизон, гидроксипрогестерон, тестостерон, тамоксифен, дакарбазин, прокарбазин, гексаметилмеламин, пентаметилмеламин, митоксантрон, амсакрин, хлорамбуцил, метилциклогексилнитрозомочевина, азотные аналоги горчичного газа, мелфалан, циклофосфамид, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-азацитидин, гидроксимочевина, дезоксикоформицин, 4-гидроксипероксициклофосфорамид, 5-фторурацил (5-FU), 5- фтордезоксиуридин (5-FUdR), метотрексат (MTX), колхицин, таксол, винкристин, винбластин, этопозид (VP-16), триметрексат, иринотекан, топотекан, гемцитабин, тенипозид, цисплатин и диэтилстилбестрол (DES). В общих чертах, см. The Merck Manual of Diagnosis and Therapy, 15th Ed. 1987, pp. 1206-1228, Berkow et al., eds., Rahway, NJ. При применении соединений согласно изобретению в сочетании с химиотерапевтическими агентами такие химиотерапевтические агенты могут быть использованы отдельно (например, 5-FU и олигонуклеотид), последовательно (например, в течение некоторого периода времени могут быть введены 5-FU и олигонуклеотид, а затем MTX и олигонуклеотид) или в комбинации с одним или несколькими другими химиотерапевтическими агентами (например, 5-FU, MTX и олигонуклеотид или 5-FU, лучевая терапия и олигонуклеотид). В композициях согласно изобретению соединения согласно изобретению могут быть также использованы в комбинации с противовоспалительными лекарственными средствами, включая, но не ограничиваясь ими, нестероидные противовоспалительные средства, кортикостероиды, и противовирусные лекарственные средства, включая, но не ограничиваясь ими, рибивирин, видарабин, ацикловир и ганцикловир. В общих чертах, см. The Merck Manual of Diagnosis and Therapy, 15th Ed., Berkow et al., eds., 1987, Rahway, NJ., pages 2499-2506 and 46-49 соответственно. В объем настоящего изобретения также входят и другие химиотерапевтические агенты, не содержащие дцРНК. Два или несколько совместно используемых соединений могут быть введены вместе или последовательно.
Токсичность и терапевтическая эффективность таких соединений может быть определена с помощью стандартных фармацевтических процедур с использованием клеточных культур или экспериментальных животных, например для определения LD50 (средняя летальная доза, при которой погибает 50% особей) и ED50 (средняя эффективная доза, индуцирующая терапевтический эффект у 50% особей). Отношение дозы, оказывающей токсическое действие, к дозе, оказывающей терапевтическое действие, называется терапевтическим индексом и может быть выражено как отношение LD50/ED50. Предпочтительными являются соединения, которые имеют высокие терапевтические индексы.
Данные, полученные в результате анализов клеточной культуры или в исследованиях, проводимых на животных, могут быть использованы в целях приготовления ряда лекарственных средств для введения человеку. Дозы композиций согласно изобретению обычно составляют в пределах величин, которые, по расчетам, дают нужные концентрации в кровотоке, включая ED50, и имеют незначительный уровень токсичности или вообще не обладают токсичностью. Эти дозы могут варьироваться в указанных пределах в зависимости от используемой лекарственной формы и от способа введения. Для любого соединения, используемого в способе согласно изобретению, терапевтически эффективная доза может быть предварительно оценена с помощью анализов клеточных культур. У животных-моделей доза может быть расчитана так, чтобы концентрация соединения или, если это необходимо, полипептидного продукта нацеливаемой последовательности в плазме достигала определенной концентрации (например, с достижением пониженной концентрации полипептида), например IC50 (то есть концентрации тестируемого соединения, которая обеспечивает полумаксимальное подавление симптомов), как может быть определено в анализе клеточной культуры. Такая информация может быть использована в целях более точного определения доз, подходящих для введения человеку. Уровни в плазме могут быть измерены, например, с помощью высокоэффективной жидкостной хроматографии.
дцРНК согласно изобретению, помимо их введения отдельно или одновременно, как обсуждалось выше, могут быть введены в комбинации с другими известными агентами, которые являются эффективными для лечения инфекции, вызываемой вирусом гриппа. В любом случае, практический врач может скорректировать количество и время введения дцРНК, исходя из результатов, полученных с использованием стандартных критериев эффективности, известных специалистам или описанных в настоящей заявке.
Методы лечения и способы доставки
Композиция, включающая иРНК-агент, например иРНК-агент, нацеленный на вирус гриппа, может быть введена индивидууму различными способами в целях ее локальной доставки в нужный участок, подвергаемый воздействию, или системной доставки индивидууму. Репрезентативные способы введения включают прямое местное введение в участок, подвергаемый лечению, такой как легкие и носовые ходы, а также внутривенное, интраназальное, пероральное и офтальмическое введение. Предпочтительным способом введения иРНК-агентов согласно изобретению является прямое введение в легкие и носовые ходы в виде жидкости, аэрозоля или аэрозольного раствора.
В общих чертах, иРНК-агенты согласно изобретению вводят для осуществления их доставки индивидууму в область поражения инфекцией. Предпочтительным способом достижения такой доставки является местное введение в легкие или в носовые ходы, например в ткани дыхательных путей, посредством ингаляции или интраназального введения, или путем системного введения, например парентерального введения.
Препараты для ингаляции или парентерального введения хорошо известны специалистам. Такие препараты могут включать стерильные водные растворы, которые могут также содержать буферы, разбавители и другие подходящие добавки. Для внутривенного введения общая концентрация растворенных веществ должна быть скорректирована так, чтобы такой препарат обладал изотоничностью.
Описанные здесь активные соединения, предпочтительно, вводят в легкое(ие) или носовые ходы индивидуума любым подходящим способом. Активные соединения могут быть доставлены путем введения аэрозольной суспензии частиц, состоящих из активного соединения или активных соединений и вдыхаемых данным индивидуумом. Активное соединение может быть приготовлено в виде аэрозоля в различных формах, таких как, но не ограничивающихся ими, инсуффляторы, ингаляторы с дозирующим клапаном или суспензии типа “жидкость/жидкость”. Вдыхаемые частицы могут быть жидкими или твердыми. Такие частицы могут, но необязательно, содержать другие терапевтические ингредиенты, такие как амилорид, бензамил или фенамил, при этом выбранное соединение содержится в количестве, эффективном для ингибирования реабсорбции воды из секрета слизистой дыхательных путей, как описано в патенте США № 4501729.
Для облегчения диспергирования или транспорта фармацевтическая композиция, состоящая из частиц, может быть, но необязательно, объединена с носителем. Подходящий носитель, такой как сахар (то есть лактоза, сахароза, трегалоза, маннит), может быть смешан с активным соединением или с соединениями в любом подходящем соотношении (например, в отношении 1:1 по массе).
Частицы, состоящие из активного соединения, подходящего для осуществления настоящего изобретения, должны иметь соответствующий размер, обеспечивающий их вдыхание, то есть они должны иметь достаточно малый размер, такой, чтобы после их вдыхания они могли проходить через рот, нос и гортань и поступать в бронхи и альвеолы легких. В общих чертах, вдыхаемые частицы должны иметь размер примерно от 1 до 10 микрон (более конкретно, менее чем примерно 5 микрон). Частицы аэрозоля, имеющие размер, затрудняющий их вдыхание, обычно осаждаются в горле и проглатываются, а поэтому предпочтительно минимизировать количество частиц аэрозоля, не пригодных для вдыхания. При интраназальном введении для гарантии удерживании частиц в носовой полости их размер предпочтительно должен составлять в пределах 10-500 мкМ.
Жидкие фармацевтические композиции активного соединения, используемые для приготовления аэрозоля, могут быть получены путем смешивания активного соединения с подходящим носителем, таким как стерильная апирогенная вода. Гипертоническими физиологическими растворами, используемыми для осуществления настоящего изобретения, предпочтительно, являются стерильные апирогенные растворы, содержащие от одного до пятнадцати процентов (по массе) физиологически приемлемой соли, а более предпочтительно, от трех до семи процентов (по массе) физиологически приемлемой соли.
Аэрозоли, состоящие из жидких частиц, содержащих активное соединение, могут быть получены любым подходящим способом, например с использованием струйного распылителя под давлением или ультразвукового ингалятора. См., например, патент США № 4501729. Ингаляторы представляют собой коммерчески доступные устройства, которые превращают растворы или суспензии активного ингредиента в терапевтический аэрозольный туман посредством ускорения подачи сжатого газа, обычно воздуха или кислорода, через узкое отверстие трубки Вентури или посредством ультразвукового перемешивания.
Композиции, подходящие для использования в ингаляторах, состоят из активного ингредиента в жидком носителе, где указанный активный ингредиент составляет до 40%, а предпочтительно, менее чем 20% по массе данной композиции. Носителем обычно является вода (а наиболее предпочтительно, стерильная апирогенная вода) или разбавленный водный спиртовой раствор, предпочтительно изотонический, но он может быть также сделан гипертоническим по отношению к физиологической жидкости организма путем добавления, например, хлорида натрия. Необязательными вспомогательными добавками являются консерванты, если данная композиция не является стерильной, например метилгидроксибензоат, антиоксиданты, отдушки, летучие масла, забуферивающие агенты и поверхностно-активные вещества.
Аэрозоли твердых частиц, содержащих активное соединение, могут быть аналогичным образом получены с помощью любого генератора терапевтических аэрозолей, содержащих твердые частицы. Аэрозольные генераторы, предназначенные для введения индивидууму терапевтических средств, состоящих из твердых частиц, продуцируют частицы, пригодные для вдыхания, и создают объем аэрозоля, содержащего предварительно определенную дозу терапевтического средства, подаваемую со скоростью, подходящей для введения человеку. Одним из репрезентативных типов аэрозольных генераторов, состоящих из твердых частиц, является инсуффлятор. Подходящими препаратами для введения путем инсуффляции являются тонкоизмельченные порошки, которые могут быть доставлены с помощью инсуффлятора или которые могут проникать в носовую полость при вдыхании. В таком инсуффляторе порошок (например, в его отмеренной дозе, эффективной для лечения описанных здесь заболеваний) содержится в капсулах или в картриджах, обычно изготовленных из желатина или пластика, которые либо разрушаются, либо раскрываются in situ, и порошок поступает с потоком воздуха, подаваемым с помощью такого устройства после ингаляции или с помощью ручного насоса. Порошок, используемый в инсуффляторе, состоит либо только из активного ингредиента, либо из порошкообразной смеси, содержащей активный ингредиент, подходящий порошкообразный разбавитель, такой как лактоза, и, необязательно, поверхностно-активное вещество. Активный ингредиент обычно составляет от 0,1 до 100% по массе данного препарата.
Репрезентативный аэрозольный генератор второго типа содержит ингалятор с дозирующим клапаном. Ингаляторы с дозирующим клапаном представляют собой герметичные аэрозольные ингаляторы, обычно содержащие препарат, включающий активный ингредиент в виде суспензии или раствора в ожиженном пропелленте. В процессе применения этого устройства указанный препарат высвобождается через клапан, приспособленный для подачи определенного объема, обычно от 10 до 200 мкл, с продуцированием тонкодисперсного спрея, содержащего активный ингредиент. Подходящими пропеллентами являются некоторые хлорфторуглеродные соединения, например дихлордифторметан, трихлорфторметан, дихлортетрафторэтан и их смеси. Такой препарат может дополнительно содержать один или несколько сорастворителей, например этанол, поверхностно-активные вещества, такие как олеиновая кислота или триолеат сорбитана, антиоксидант и подходящие ароматизаторы.
В фармацевтические композиции, подходящие для введения, может быть включен иРНК-агент. Так, например, композиции могут включать иРНК-агенты одного или нескольких видов и фармацевтически приемлемый носитель. Используемый здесь термин “фармацевтически приемлемый носитель” включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, агенты, придающие изотоничность и замедляющие абсорбцию, и т.п., совместимые со способом введения фармацевтического средства. Применение таких сред и агентов для введения фармацевтически активных веществ хорошо известно специалистам. Рассматривается также возможность применения в данных композициях любых стандартных сред или агентов, за исключением тех сред или агентов, которые являются несовместимыми с активным соединением. В указанные композиции могут быть также введены вспомогательные активные соединения.
Введение может быть осуществлено самим индивидуумом или другим человеком, например лицом, осуществляющим уход за данным индивидуумом или его лечение. Таким лицом может быть любое физическое или юридическое лицо, осуществляющее уход за данным индивидуумом или его лечение, например больница, хоспис, врачебный кабинет, амбулатория, медицинский работник, такой как врач, медсестра или другой специалист-медик, либо супруги или опекуны, такие как родители. Медикаментозное лечение может быть осуществлено путем введения предварительно рассчитанных доз или с помощью ингалятора, доставляющего такую отмеренную дозу.
Термин “терапевтически эффективное количество” означает количество, присутствующее в композиции и необходимое для достижения нужного уровня лекарственного средства у индивидуума, подвергаемого лечению, в целях вырабатывания у него ожидаемого физиологического ответа.
Термин “физиологически эффективное количество” означает количество, доставляемое индивидууму в целях продуцирования у него желаемого паллиативного или лечебного эффекта.
Термин “фармацевтически приемлемый носитель” означает носитель, который может проникать в легкие, не вызывая каких-либо значительных нежелательных токсических эффектов.
Термин “совместное введение” означает введение индивидууму двух или более средств, а в частности двух или более иРНК-агентов. Такие агенты могут присутствовать в одной фармацевтической композиции и могут быть введены одновременно, либо такие агенты могут присутствовать в отдельных препаратах и могут быть введены индивидууму последовательно. Если эти два агента могут быть детектированы у индивидуума одновременно, то считается, что эти агенты вводятся одновременно.
Фармацевтическими наполнителями, которые могут быть использованы в качестве носителя, являются стабилизаторы, такие как альбумин человеческой сыворотки (HSA); агенты, создающие объем, такие как углеводы, аминокислоты и полипептиды; агенты или буферы, корректирующие рН; соли, такие как хлорид натрия, и т.п. Такие носители могут присутствовать в кристаллической или аморфной форме, либо они могут представлять собой смесь из двух этих форм.
Агенты, придающие объем, которые являются особенно ценными, включают совместимые углеводы, полипептиды, аминокислоты или их комбинации. Подходящими углеводами являются моносахариды, такие как галактоза, D-манноза, сорбоза и т.п.; дисахариды, такие как лактоза, трегалоза и т.п.; циклодекстрины, такие как 2-гидроксипропил-бета-циклодекстрин; и полисахариды, такие как раффиноза, мальтодекстрины, декстраны и т.п.; альдиты, такие как маннит, ксилит и т.п. Предпочтительными углеводами являются лактоза, трегалоза, раффиноза, мальтодекстрины и маннит. Подходящим полипептидом является аспартам. Подходящими аминокислотами являются аланин и глицин, при этом предпочтительным является глицин.
Подходящими pH-корректирующими агентами или буферами являются органические соли, полученные из органических кислот и оснований, такие как цитрат натрия, аскорбат натрия и т.п., при этом предпочтительным является цитрат натрия.
Дозы
иРНК-агент может быть введен в унифицированной дозе, составляющей примерно менее чем 75 мг на кг массы тела или примерно менее чем 70, 60, 50, 40, 30, 20, 10, 5, 2, 1, 0,5, 0,1, 0,05, 0,01, 0,005, 0,001 или 0,0005 мг на кг массы тела и менее чем 200 нмоль (например, примерно 4,4×1016 копий) на кг массы тела или менее чем 1500, 750, 300, 150, 75, 15, 7,5, 1,5, 0,75, 0,15, 0,075, 0,015, 0,0075, 0,0015, 0,00075, 0,00015 нмоль на кг массы тела. Так, например, эта унифицированная доза может быть введена путем инъекции (например, внутривенно или внутримышечно, интратекально или непосредственно в орган), путем ингаляции или путем местного применения.
Доставка иРНК-агента непосредственно в орган (например, в легкие) может быть осуществлена в дозе, составляющей примерно от 0,00001 мг до 3 мг на орган или предпочтительно примерно 0,0001-0,001 мг на орган, примерно 0,03-3,0 мг на орган, примерно 0,1-3,0 мг на глаз или примерно 0,3-3,0 мг на орган.
Эта доза может представлять собой количество, эффективное для лечения или предупреждения заболевания или расстройства. Она может быть введена в профилактических целях или в качестве главной или частичной составляющей схемы лечения.
В одном из вариантов изобретения такую унифицированную дозу вводят не чаще, чем один раз в день, например не чаще, чем через 2, 4, 8 или 30 дней. В другом варианте изобретения унифицированную дозу не вводят с какой-либо определенной частотой (например, вводят нерегулярно). Так, например, унифицированная доза может быть введена один раз. Поскольку опосредуемый иРНК-агентом сайленсинг может поддерживаться в течение нескольких дней после введения композиции иРНК-агента, то во многих случаях можно вводить композицию не чаще, чем один раз в день, или в некоторых случаях только один раз за весь курс терапии.
В одном из вариантов изобретения индивидууму вводят начальную дозу и одну или несколько поддерживающих доз иРНК-агента, например двухцепочечного иРНК-агента или киРНК-агента (например, предшественника, то есть более крупного иРНК-агента, который может быть процессирован в киРНК-агент, или ДНК, которая кодирует иРНК-агент, например двухцепочечный иРНК-агент, или киРНК-агента или его предшественника. Поддерживающая доза или дозы обычно ниже первоначальной дозы, например, они составляют меньше половины от первоначальной дозы. Поддерживающий режим может включать введение индивидууму дозы или доз в пределах от 0,01 до 75 мг/кг массы тела в день, например 70, 60, 50, 40, 30, 20, 10, 5, 2, 1, 0,5, 0,1, 0,05, 0,01, 0,005, 0,001 или 0,0005 мг на кг массы тела в день. Поддерживающие дозы предпочтительно вводят не более чем один раз в 5, 10 или 30 дней. Кроме того, схема лечения может быть продлена на определенное время, зависящее от природы конкретного заболевания, его тяжести и общего состояния здоровья пациента. В предпочтительных вариантах изобретения доза может быть введена не чаще, чем один раз в день, например не чаще, чем один раз в 24, 36, 48 или более часов, например не чаще, чем один раз в 5 или 8 дней. После лечения пациент может быть обследован на изменение состояния его здоровья и на ослабление симптомов указанного патологического состояния. Доза соединения может быть либо увеличена, в случае, если у данного пациента не наблюдается значимого ответа на вводимые уровни доз, либо эта доза может быть снижена, в случае, если у пациента наблюдается ослабление симптомов патологического состояния или ремиссия данного патологического состояния или если наблюдаются нежелательные побочные эффекты.
Эффективная доза может быть введена в виде разовой дозы или в виде двух или более доз, по желанию или в зависимости от конкретной ситуации. Если необходимо проводить повторные или частые вливания, то может оказаться желательной имплантация устройства для доставки, например насос, стент для полуперманентного введения (например, внутривенного, внутрибрюшинного, интрацистернального или интракапсулярного) или резервуар.
После успешного лечения может оказаться желательным проведение данному пациенту поддерживающей терапии для предупреждения рецидива патологического состояния, где указанное соединение согласно изобретению вводят в поддерживающих дозах, составляющих от 0,001 г до 100 г на кг массы тела (см. патент США 6107094).
Концентрация композиции иРНК-агента представляет собой количество, достаточное для эффективного лечения или предупреждения состояния или расстройства или для регуляции физиологического состояния у человека. Концентрация вводимого количества иРНК-агента зависит от параметров, определенных для данного агента, и способа введения, например интраназального, трансбуккального или внутрилегочного введения. Так, например, во избежание раздражения или ожогов носовых ходов в интраназальных композициях должны быть использованы гораздо меньшие концентрации некоторых ингредиентов. Для приготовления подходящей интраназальной композиции иногда может оказаться желательным разведение, которое примерно в 10-100 раз меньше, чем это требуется для пероральной композиции.
Доза, необходимая для эффективного лечения индивидуума, может зависеть от некоторых факторов, включая, но не ограничиваясь ими, тяжесть заболевания или расстройства, проводимое ранее лечение, общее состояние здоровья, и/или возраст данного индивидуума и наличие других заболеваний. Следует также отметить, что эффективная доза иРНК-агента, такого как киРНК, используемого для лечения, может быть увеличена или снижена во время проведения курса лечения. Изменение доз может быть осуществлено, исходя из результатов диагностических анализов. Так, например, индивидуум может быть обследован после введения композиции иРНК-агента. Исходя из результатов такого обследования, может быть введено дополнительное количество композиции иРНК-агента.
Доза зависит от тяжести патологического состояния, подвергаемого лечению, и от его восприимчивости к данному лечению, при этом курс лечения может продолжаться от нескольких дней до нескольких месяцев или до тех пор, пока не будет достигнут нужный эффект или не будет наблюдаться ослабление патологического состояния. Оптимальная схема введения доз может быть рассчитана, исходя из оценки аккумуляции лекарственного средства в организме пациента. Средний специалист в данной области может легко определить оптимальные дозы и выбрать методику введения доз и частоту их повторного введения. Оптимальные дозы могут варьироваться в зависимости от относительной эффективности отдельных соединений и могут быть, в основном, рассчитаны, исходя из концентраций EC50, которые были определены как эффективные в анализах, проводимых in vitro и in vivo на животных-моделях, описанных выше.
Настоящее изобретение подробно проиллюстрировано на нижеследующих примерах, которые, однако, не должны рассматриваться как ограничение настоящего изобретения.
ПРИМЕРЫ
Последовательности нуклеиновых кислот представлены ниже в таблице 3, где конкретные сокращения даны в соответствии со стандартной номенклатурой.
Источник реагентов
Если источник реагента конкретно не указан, то такой реагент может быть получен от любого поставщика реагентов для исследований в области молекулярной биологии при стандартном отношении качество/чистота, обычно применяемом в молекулярной биологии.
Пример 1 : Выбор последовательностей
Конструирование киРНК осуществляют в целях идентификации киРНК, нацеленных на мРНК белков MP, NP, PA, PB1 и PB2 вируса гриппа. В первом раунде отбор киРНК in silico привел к получению 44 последовательностей, нацеливающих на MP; 3 последовательностей, нацеливающих на NP; и 1 последовательности, нацеливающей на PB1. Ни одна из киРНК, специфичная к генам PA или PB2 вируса гриппа, не прошла первый раунд отбора, который требовал 80% охвата мишеней и 80% эффективности нацеливания (см. ниже).
Для установления операционной программы в целях анализа последовательности загружали пакет программ fastA (Pearson, W.R., & Lipman, D.J., PNAS 1988, 85:2444) из сайта ftp://ftp.virginia.edu/pub/ и инсталлировали на рабочей станции под операционной системой Suse Linux® 9.3 в соответствии со стандартными параметрами инсталляции. Для запуска perl-скриптов необходимо убедиться, что интерпретатор perl, версии 5.8.6 (copyright 1987-2004, Larry Wall) вместе с инсталлированной стандартной Suse Linux 9.3 является функциональным. Редактор выравнивания последовательностей BioEdit (Hall, T.A., Nucl. Acids. Symp. Ser. 1999, 41:95) загружали из сервера Brown Lab Web Server на веб-сайте Университета штата Северная Каролина и инсталлировали на компьютере с опреационной системой Microsoft Windows 2000®.
Операцию для отбора in silico проводили в следующем порядке: сначала загружали представляющие интерес последовательности вируса гриппа A, затем их выравнивали и проводили статистический анализ для получения данных о распределении оснований в каждом положении по сравнению с вычисленной консенсусной последовательностью. Perl-скрипт использовали для идентификации кандидатов на области-мишени, удовлетворяющих определенным критериям отсечки. Последовательности киРНК для кандидатов на области-мишени анализировали на специфичность с помощью алгоритма fastA и с использованием базы данных человеческих RefSeq. Другой perl-скрипт использовали для оценки киРНК в соответствии с предсказанной специфичностью. И, наконец, вручную отбирали те киРНК, которые удовлетворяли критериям специфичности.
Представляющие интерес последовательности вируса гриппа А, имеющиеся в базе данных на 24 июня 2005 г., загружали из базы данных последовательностей вируса гриппа NCBI, имеющейся на веб-сайте Национального центра биотехнологической информации. Число последовательностей на ген для белков MP, NP, PB1, PB2 и PA представлено в таблице 4, и в этой таблице представлены их соответствующие регистрационные номера.
Функцию множественного выравнивания ClustalW (Thompson, J.D., et al., Nucleic Acids Res. 1994; 22:4673) программы редактора выравнивания последовательностей BioEdit использовали для осуществления глобального выравнивания всех последовательностей с применением параметров по умолчанию для каждой мишени соответственно. Конечный результат вычислений нуклеотидов в данном положении получали, исходя из информации о распределении нуклеотидов в каждом положении по отношению к вычисленной консенсусной последовательности для каждой мишени.
Для идентификации кандидатов областей-мишеней длиной в 19 нуклеотидов использовали следующие критерии отсечки:
Критерий 1, охват мишеней: по меньшей мере, 80% всех последовательностей, имеющихся для соответствующего гена вируса гриппа А, представляемого в области-кандидате.
Критерий 2, эффективность нацеливания: по меньшей мере, 80% от всех последовательностей, в которых области-кандидаты должны быть идентичными.
Критерий 1 был установлен для исключения областей, о последовательности которых имеется мало информации, а критерий 2 обеспечивает нацеливание на большое число подтипов.
Perl-скрипт используется для скрининга файла суммарных численных данных о положениях нуклеотидов в целях идентификации кандидатов на области-мишени длиной в 19 оснований, соответствующих критериям отсечки, и для создания файла, используемого в качестве входных данных fastA в следующей стадии анализа. Для входных данных скрипта вводили общее число последовательностей для каждой мишени и величину 80 для установления процента. Все кандидаты на смысловые последовательности киРНК, соответствующие наиболее часто встречающимся последовательностям в подходящих областях-мишенях, отбирали и сохраняли в fastA-отформатированном файле. Для учета возможных взаимодействий выступающих dTdT-концов киРНК с последовательностью-мишенью все последовательности удлиняли у 5'-конца путем добавления 'AA' с получением 21-мерных исходных последовательностей. Для каждого кандидата на область-мишень создавали дополнительный файл, исходя из информации о свойствах этой области: зона охвата (присутствующие последовательности), эффективности нацеливания, общего показателя несоответствий, числа консервативных последовательностей и числа присутствующих последовательностей.
Для дополнительного отбора киРНК-кандидаты распределяли по их предсказанной способности к взаимодействию с генами хозяина (в данном случае, но не ограничиваясь им, человека) (по потенциалу взаимодействия с нерелевантными мишенями (off-target potential)). При этом высказывается предположение, что киРНК с низким потенциалом нерелевантного взаимодействия является наиболее специфической in vivo.
Для предсказания киРНК-специфического потенциала нерелевантного взаимодействия были сделаны следующие предположения, что:
1) положения 2-9 (в направлении 5'→3') цепи (уникальная область) вносят более важный вклад в потенциал нерелевантного взаимодействия, чем остальная последовательность (область за пределами указанной уникальной области (неосновная область)) (Haley, B. & Zamore, P.D., Nat Struct Mol Biol. 2004, 11:599),
2) показатель нерелевантных взаимодействий может быть вычислен для каждого совпадения, исходя из идентичности по отношению к последовательности киРНК и положения несоответствий,
3) путем введения соответствующих нуклеотидных модификаций в смысловую цепь (например, модификаций, при которых все нуклеотиды, содержащие пиримидиновое основание, представляют собой 2'-O-метил-модифицированные нуклеотиды) может быть получена смысловая цепь, не обладающая интерферирующей активностью по отношению к РНК; следовательно, необходимо учитывать только потенциал нерелевантных взаимодействий антисмысловой цепи.
Для идентификации потенциальных нерелевантных генов 21-мерные последовательности, соответствующие подходящим областям-мишеням, плюс 3'-концевой хвост AA (с учетом выступающих концов ТТ), были подвергнуты скринингу на гомологию с известными человеческими последовательностями мРНК, имеющимися в общедоступных базах данных. Для этой цели с помощью fastA (version 3.4) проводили поиск для всех 21-мерных исходных последовательностей в базе данных человеческих последовательностей RefSeq (загруженной из версии, имеющейся на ftp://ftp.ncbi.nih.gov/refseq/, 2005-07-25). Для определения гомологии по всей длине 21-меров fastA-поиск проводили по параметрам-величинам-парам - b 30 - g 30. В результате этого поиска был получен список потенциальных нерелевантных мишеней для киРНК-кандидатов.
Для сортировки полученного списка потенциальных нерелевантных мишеней выходные файлы fastA анализировали на идентичность гена хозяина с наиболее высоким показателем несоответствий. Для вычисления показателя несоответствий для каждой потенциальной нерелевантной мишени были установлены нерелевантные свойства для каждой 21-мерной исходной последовательности.
1. Число идентичных нуклеотидов для 21-мерной последовательности (идентичность).
2. Показатель несоответствий в уникальной области.
Показатель несоответствий вычисляли, исходя из предположения, что 1 и 2 имеют следующую зависимость:
Идентичность - 0,2 × число несоответствий в уникальной области.
Все киРНК были отсортированы по их наиболее высоким оценкам несоответствий (по возрастанию). Показатель несоответствий 16,8 был использован как величина отсечки для отбора киРНК. 42 киРНК, специфичные к матриксному белку (MP) вируса гриппа А; 3 киРНК, специфичные к нуклеокапсидному белку (NP) вируса гриппа А; и 1 киРНК, специфичная к основному полимеразному белку вируса гриппа А (PB1), имели показатель несоответствий, равный этому пороговому значению или ниже.
Принимая во внимание сравнительно низкое число киРНК-кандидатов, полученных после вышеописанной процедуры отбора, численная суммарная оценка нуклеотидов в каждом положении была снова проведена с использованием критерия отсечки 1 (охват мишеней), установленной 70%, и критерия 2 (специфичность нацеливания) при ранее установленном 80%, после чего снова проводили распределение по оценкам несоответствий. В результате были дополнительно отобраны 2 киРНК, специфичные к мРНК белка MP вируса гриппа A с эффективностью нацеливания 79,9%, а всего было отобрано 48 киРНК-кандидатов. Последовательности этих 48 киРНК-кандидатов показаны в таблице 1А.
Поскольку описанный выше процесс отбора приводил только к ограниченному числу агентов-кандидатов, то критерий отбора был до некоторой степени ослаблен с получением дополнительных агентов-кандидатов. В частности, вышеуказанный критерий 1 был ослаблен до 50% охвата мишеней, критерий 2, эффективность нацеливания сохраняли на прежнем уровне 80%, после чего снова повторяли вышеописанный процесс отбора. В результате этой процедуры получали дополнительные агенты AL-DP-8001 - AL-DP-8040, перечисленные в таблице 1C.
В процессе осуществления этой процедуры было установлено, что стадия определения показателя несоответствий приводит к более высокому уровню отсева потенциальных агентов. Поэтому для получения большего числа агентов-кандидатов процедуру отбора повторяли еще раз с использованием критерия 1 с охватом мишеней 80%, критерия 2 с эффективностью нацеливания 80%, а определение показателя несоответствий не проводили. В результате этой процедуры были получены дополнительные агенты, перечисленные в таблице 1D. Дополнительные агенты-кандидаты были получены путем повторения процедуры отбора еще раз с использованием критерия 1 с охватом мишеней 50% и критерия 2 с эффективностью нацеливания 80%, без определения показателя несоответствий.
Дополнительные иРНК-агенты-кандидаты были идентифицированы по включению универсальных оснований. Perl-скрипт использовали для предварительной идентификации последовательностей-кандидатов, имеющих охват мишеней и эффективность нацеливания 100% в случае включения до 3 универсальных оснований на цепь в неосновной области (соответствующей положениям 2-9 антисмысловой цепи) иРНК-агента на цепь. В таблице 1F представлены агенты, идентифицированные таким способом. Во втором раунде были идентифицированы дополнительные иРНК-агенты, которые имеют охват мишеней и эффективность нацеливания 80% при включении универсального основания. Эти иРНК-агенты представлены в таблице 1G.
Пример 2: Синтез киРНК
Синтез нуклеотидов, содержащих природные основания
Одноцепочечные RNA получали методом твердофазного синтеза в масштабе 1 мкмоль с использованием синтезатора Expedite 8909 (Applied Biosystems, Applera Deutschland GmbH, Darmstadt, Germany) и стекла с регулируемым размером пор (CPG, 500Å, Glen Research, Sterling VA) в качестве твердого носителя. РНК и РНК, содержащую 2'-O-метил-нуклеотиды, получали методом твердофазного синтеза с использованием соответствующих фосфорамидитов и 2'-O-метил-фосфорамидитов соответственно (Proligo Biochemie GmbH, Hamburg, Germany). Эти структурные блоки включали в отобранные сайты в последовательности олигорибонуклеотидной цепи с применением стандартного нуклеозид-фосфорамидитного метода, такого как метод, описанный в руководствах “Current protocols in nucleic acid chemistry”, Beaucage, S.L. et al. (Edrs.), John Wiley & Sons, Inc., New York, NY, USA. Фосфортиоатные связи были введены путем замены раствора иодного окислителя на раствор реагента Бокажа (Chruachem Ltd, Glasgow, UK) в ацетонитриле (1%). Дополнительные вспомогательные реагенты были получены от компании Mallinckrodt Baker (Griesheim, Germany).
Снятие защиты и очистку с помощью анионообменной ВЭЖХ неочищенных олигорибонуклеотидов проводили в соответствии с установленными процедурами. Выходы и концентрации определяли по УФ-поглощению раствора соответствующей РНК на длине волны 260 нМ с использованием спектрального фотометра (DU 640B, Beckman Coulter GmbH, Unterschleißheim, Germany). Двухцепочечную РНК получали путем смешивания эквимолярного раствора комплементарных цепей в буфере для отжига (20 мМ фосфат натрия, pH 6,8; 100 мМ хлорид натрия), с последующим нагреванием на водяной бане при 85-90°С в течение 3 минут и охлаждением до комнатной температуры в течение 3-4 часов. Очищенный раствор РНК хранили при -20°С до его использования.
Все олигонуклеотиды, полученные описанным выше методом синтеза, не содержали фосфатной группы на своих самых крайних 5'-концевых нуклеотидах.
Синтез нуклеотидов, содержащих универсальные основания
Синтез фосфорамидита и 5'-О-(4,4'-диметокситритил)-2'-O-(трет-бутилдиметилсилил)-1'-(5-нитроиндол)-D-рибозида на стекле с регулируемым размером пор, используемым в качестве носителя
Стадия A: 1-O-метил-D-рибозид (102)
К раствору D-рибозы (25 г) в сухом метаноле (300 мл) добавляли концентрированную серную кислоту (1,88 мл) и перемешивали при комнатной температуре в течение 3 дней. Затем реакционную смесь нейтрализовали 1н. раствором гидроксида натрия и концентрировали с получением неочищенного остатка. Полученный неочищенный остаток растворяли в метаноле (200 мл) и твердые вещества отфильтровывали. Фильтрат концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью дихлорметан-метанол (5:1), в результате чего получали чистое соединение (23,0 г, 82%) в виде сиропа.
Стадия B: 1-О-метил-2,3,5-три-О-(2,4-дихлорбензил)-D-рибозид (103)
К раствору 1-О-метил-D-рибозида (13,43 г, 81,83 ммоль), 18-краун-6 (1,34 г) в сухом ТГФ (100 мл) добавляли порошкообразный гидроксид калия (69 г, 1,23 моль) и перемешивали при комнатной температуре в течение 40-60 минут. Затем по каплям добавляли 2,4-дихлорбензилхлорид (51 мл, 368,2 ммоль) и реакционную смесь перемешивали при той же температуре в течение ночи. Твердые вещества отфильтровывали и фильтрат концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью гексан-этилацетат (4:1), в результате чего получали чистое соединение (48 г, 92%) в виде белого твердого вещества.
1H-ЯМР (CDCl3, 400 МГц): δ 7,46-7,34 (м, 5H, ArH), 7,24-7,16 (м, 4H, ArH), 4,99 (с, 1H, H-1), 4,71 (дд, 2H, Jgem=12,8 Гц, OCH2Ar), 4,63-4,61 (м, 4Н, 2 OCH2Ar), 4,38-4,36 (м, 1H), 4,19-4,16 (дд, 1H), 3,98 (д, 1H, J=4,4 Гц), 3,75 (дд, 1H, J=3,6, 7=10,2 Гц, H-5a), 3,66 (дд, 1H, J=3,6, J=10,4 Гц, H-5b), 3,37 (с, 3H, OCH3).
Стадия B: 1-бром-2,3,5-три-О-(2,4-дихлорбензил)-D-рибоза (104).
К холодному раствору 1-O-метил-2,3,5-три-O-(2,4-дихлорбензил)-D-рибозида (3,22 г, 5,02 ммоль) в сухом дихлорметане (50 мл), охлажденном на ледяной бане, добавляли HOAc-HBr (5,3 мл, 30%) и перемешивали при 0-25°С в течение 3 часов. Реакционную смесь концентрировали с получением неочищенного остатка, который выпаривали с толуолом (3×30 мл) и получали неочищенный остаток в виде сиропа, который сушили в условиях высокого вакуума и использовали в следующей реакции без очистки и идентификации.
Стадия D: 1-(5-нитроиндол)-2,3,5-три-O-(2,4-дихлорбензил)-D-рибозид (105)
К раствору 5-нитроиндола (2,44 г, 15,06 ммоль) в сухом CH3CN (30 мл) добавляли гидрид натрия (602 мг, 15,06 ммоль, 60%) и перемешивали при комнатной температуре в течение 3-4 часов в атмосфере аргона. Затем добавляли сахар, полученный, как описано выше, и используемый в качестве донора (104), в сухом CH3CN (10 мл), и смесь перемешивали при той же температуре в атмосфере аргона в течение ночи. Твердые вещества отфильтровывали и фильтрат концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью гексан-этилацетат (3:1), в результате чего получали чистое соединение 105 (2,16 г, 60%) в виде α- и β-смеси (1:1).
Стадии E, F: 5'-O-(4,4'-диметокситритил)-1'-(5-нитроиндол)-D-рибозид (106) и (107)
К холодному раствору 1-(5-нитроиндол)-2,3,5-три-О-(2,4-дихлорбензил)-D-рибозида 105 (1,16 г, 1,51 ммоль) в сухом дихлорметане (100 мл) при -78°С добавляли BCl3 в дихлорметане (23 мл, 1,0M) и перемешивали при той же температуре в течение 2 часов в атмосфере аргона и при -40°С в течение 2 часов. Реакцию гасили добавлением метанола-дихлорметана (1:1, 50 мл) и смесь нейтрализовали раствором аммиака-метанола. Твердые вещества отфильтровывали и фильтрат концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью дихлорметан-метанол (10:1), в результате чего получали чистое соединение (300 мг, 68%) в виде α- и β-смеси (1:1). К раствору соединения, полученного, как описано выше (840 мг, 2,86 ммоль), в сухом пиридине (3-4 мл) и DMAP (90 мг) добавляли DMTrCl (1,06 г) и смесь перемешивали при комнатной температуре в атмосфере аргона в течение ночи. Реакционную смесь концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью гексан-этилацетат (1:1), в результате чего получали чистое соединение 106 (550 мг) и соединение 107 (190 мг), а также смесь соединений 106 и 107 (360 мг).
Соединение 106: 1H-ЯМР (CDCl3, 2D g-COSY и 2D NOESY, 400 МГц): δ 8,49 (д, 1Н, J=1,6 Гц), 8,35 (д, 1Н), 8,03 (дд, 1Н, J=2,0, J=9,0 Гц), 7,70-7,69 (м, 2H), 7,47-7,14 (м, 8H, ArH), 6,86-6,81 (м, 5Н, ArH), 6,71 (д, 1Н, J=3,6 Гц), 6,41 (д, J=5,2 Гц, H'-1), 4,73 (т, 1Н, J=4,8Гц, H'-2), 4,46-4,42 (м, 3H, H'-3, H'-4, H'-5), 3,79 (с, 6H, 2OCH3), 3,51 (дд, 1Н, J=3,2, J=10,4 Гц, H'-5a), 3,26 (дд, 1Н, J=3,2, J=10,6 Гц, H'-5b).
Соединение 107: 1H-ЯМР (CDCl3, 2D g-COSY и 2D NOESY, 400 МГц): δ 8,55 (д, 1Н, J=2,0 Гц), 7,98 (дд, 1Н, J=2,4, J=9,2 Гц), 7,60 (д, 1Н, J=9,2 Гц), 7,53 (д, 1Н, J=3,2 Гц), 7,44-7,42 (м, 2H), 7,34-7,24 (м, 7H, ArH), 6,84-6,81 (м, 4Н, ArH), 6,68 (д, 1Н, J=3,2 Гц), 6,00 (д, 1Н, J=5,2 Гц, H'-1), 4,53 (т, 1Н, J=7,6 Гц), 4,46-4,44 (м, 1Н), 4,23-4,20 (м, 1Н), 3,80-3,76 (м, 7H, 2OCH3, H'-5), 3,55 (дд, 1Н, H'-5a), 3,43 (дд, 1Н, H'-5b).
Стадия G: 5'-O-(4,4'-диметокситритил)-2'-O-(трет-бутилдиметилсилил)-1'-(5-нитроиндол)-D-рибозид (108) и 5'-О-(4,4'-диметокситритил)-3'-О-(трет-бутилдиметилсилил)-1'-(5-нитроиндол)-D-рибозид (109)
К раствору 5'-O-(4,4'-диметокситритил)-1'-(5-нитроиндол)-D-рибозида (106) (550 мг, 0,92 ммоль), AgNO3 (188 мг, 1,104 ммоль) и пиридина (0,74 мл, 9,2 ммоль) в сухом ТГФ (9,2 мл) добавляли TBDMSCl (188 мг, 1,196 ммоль) и перемешивали при комнатной температуре в атмосфере аргона в течение ночи. Твердые вещества отфильтровывали и фильтрат концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, элюируя смесью гексан-этилацетат (4:1), в результате чего получали чистое соединение 108 (230 мг, 35%), соединение 109 (150 мг, 23%) и смесь соединений 108 и 109 (110 мг, 17%) с общим выходом 75%.
Соединение 108: 1H-ЯМР (CDCl3, 2D g-COSY, 2D NOESY, 400 МГц): δ 8,56 (д, 1Н, J=2,4 Гц), 7,88 (дд, 1Н, J=2,4, J=8,8 Гц), 7,62 (д, 1Н, J=9,2 Гц), 7,54 (д, 1Н, J=3,6 Гц), 7,46-7,44 (м, 2H), 7,36-7,25 (м, 6H, ArH), 6,85-6,83 (д, 5Н, ArH), 6,69 (д, 1Н, J=3,6 Гц), 5,94 (д, 1Н, J=7,2 Гц, H'-1), 4,69 (дд, 1Н, H'-2), 4,31-4,29 (м, 2H, H'-3, H'-4), 3,80 (с, 6H, 2OCH3), 3,58 (дд, 1Н, J=2,0, J=10,6 Гц, H'-5a), 3,40 (дд, 1Н, J=2,0, J=10,4 Гц, H'-5b), 2,85 (д, 1Н, J=0,8 Гц, 3'-OH), 0,78 (с, 9H, t-Bu), -0,016 (с, 3Н, SiCH3), -0,43 (с, 3Н, SiCH3).
Соединение 109: 1H-ЯМР (CDCl3, 2D g-COSY, 2D NOESY, 400 МГц): δ 8,61 (д, 1Н, J=2,4 Гц), 8,05 (дд, 1Н, J=2,0, J=8,8 Гц), 7,69-7,65 (м, 2H), 7,47-7,45 (м, 2H, ArH), 7,36-7,27 (м, 5Н, ArH), 6,86-6,83 (м, 3Н, ArH), 6,71 (д, 1Н, J=3,2 Гц), 5,99 (д, 1Н, J=4,8 Гц, H'-1), 4,51 (т, 1Н, J=4,8 Гц, J=5,6 Гц, H'-3), 4,40-4,36 (м, 1Н, H'-2), 4,17-4,15 (м, 2H, H'-4, H'-5), 3,82 (с, 3Н, OCH3), 3,81 (с, 3Н, OCH3), 3,63 (дд, 1Н, J=2,4, J=11,0 Гц, H'-5a), 3,31 (дд, 1Н, J=2,8, J=11,0 Гц, H'-5b), 2,95 (д, 1Н, J=6,0 Гц, 2'-OH), 0,91 (с, 9H, t-Bu), 0,05 (с, 3Н, SiCH3), 0,00 (с, 3Н, SiCH3).
Стадия H: 5'-O-(4,4'-диметокситритил)-2'-O-(трет-бутилдиметилсилил)-1'-(5'-нитроиндол)-D-рибозид-3'-0-цианоэтил-N,N-диизопропилфосфорамидат (110)
2-цианоэтил-N,N-диизопропилхлорфосфорамидит (153 мг, 0,646 ммоль) добавляли к раствору 5'-O-(4,4'-диметокситритил)-3'-O-(трет-бутилдиметилсилил)-1-(5-нитроиндол)-D-β-рибозида 108 (230 мг, 0,323 ммоль), диизопропилэтиламина (306 мкл, 1,78 ммоль) и DMAP (10 мг) в сухом дихлорметане (3 мл) и перемешивали при комнатной температуре в течение 4-6 часов в атмосфере аргона. Реакционную смесь концентрировали с получением неочищенного остатка, который наносили на колонку с силикагелем, которую насыщали 2% триэтиламином в гексане и элюировали смесью гексан-этилацетат (2:1), в результате чего получали чистое указанное в заголовке соединение 110 (250 мг, 85%) в виде аморфного твердого вещества.
31P-ЯМР (CDCl3, 400 МГц): δ 149,54 (с), 146,57 (с). Анализ для C50H65N4O9PSi. Вычислено: 924,43, найдено: 947,43 [М+Na]+.
Стадия I: 2'-гидрокси- или 3'-гидрокси-5'-О-(4,4'-диметокситритил)-1-(5-нитроиндол)-D-рибозид (111) на твердых носителях
К смеси 2'-OTBDMS (108) или 3'-О-TBDMS 5'-O-(4,4'-диметокситритил)-1-(5-нитроиндол)-Z)-D-β-рибозида (109) и DMAP в сухом дихлорметане добавляли ангидрид янтарной кислоты. Реакционную смесь перемешивали при комнатной температуре в течение 6 часов в атмосфере аргона. Затем добавляли еще одну порцию ангидрида янтарной кислоты и DMAP и смесь перемешивали всего 16 часов. Смесь концентрировали с получением неочищенного остатка, который растворяли в этилацетате (50 мл), промывали лимонной кислотой (400 мг/20 мл) и насыщенным раствором соли, а затем сушили (Na2SO4). Органический слой концентрировали с получением неочищенного нуклеозид-сукцината, который непосредственно использовали в последующей реакции без дополнительной очистки.
Нуклеозид-сукцинат, DMAP, DTNP и Ph3P перемешивали при комнатной температуре в течение 20 минут [Nucleoside and nucleotides, 1996, 15(4), 879-888]. Затем добавляли lcaa-CPG и перемешивали при той же температуре в течение 45 минут. Твердые вещества отфильтровывали и промывали CH3CN, дихлорметаном и эфиром. Твердые носители сушили, кэпировали в соответствии со стандартной процедурой и промывали с получением твердого носителя.
Полученный таким образом нитроиндол-содержащий стеклянный носитель с регулируемым размером пор и фосфорамидат использовали в стандартном методе синтеза олигонуклеотидов, описанном выше, для получения олигонуклеотидов, содержащих природные основания.
Пример 3 : Анализ киРНК in vitro
Способность иРНК-агентов ингибировать репликацию вируса гриппа оценивали в человеческих клеточных линиях in vitro или у мышей in vivo. иРНК-агент вводили в клетки, например, путем трансфекции или электропорации для определения его действия на клетки в течение определенного периода времени, например 24 часов, и уровни инфекционности определяли с помощью анализа на бляшкообразование или ELISA-анализа. В дополнение к таким прямым анализам, авторами настоящего изобретения было протестировано ингибирование экспрессии гена-мишени под действием РНКи-агентов для некоторых генов вирусов гриппа, рекомбинантно экспрессируемых в клетках-хозяевах млекопитающих.
Вирусы и клеточные линии
Вирус гриппа A/PR/8/34 (PR8), подтипа H1N1, получали из лаборатории Charles River Laboratories (ATCC # VR-1469). Штамм A/WSN/33 (WSN), подтипа H1N1, может быть получен от Thomas Chambers, University of Kentucky, Lexington, KY. USA (см. Castrucci, М.R., et al., J Virol. 1992, 66:4647) или от Dr. Peter Palese, Mount Sinai School of Medicine New York City, NY, USA (см. WO 04/028471). Вирусные штаммы размножали в аллантоисном мешочке оплодотворенных куриных яиц при 34°С в течение 48-72 часов (PR8) или 37°С в течение 24 часов (WSN) (Tompkins, S.М., et al. Proc. Natl. Acad. Sci. 2004, 101:8682).
Клетки MDCK брали из Американской коллекции типовых культур (ATCC, Rockville MD, USA; ATCC # CCL-34) и культивировали в среде МЕМ, содержащей 8%-ную термоинактивированную фетальную бычью сыворотку (FBS), 2 мМ L-глутамин, 1 мМ пируват натрия, 1,5 г/л бикарбоната натрия и заменимые аминокислоты при 37°С в атмосфере 5% CO2 c влажностью 95%.
Эпителиальные клетки почек африканской зеленой мартышки Vero E6 брали из ATCC (Rockville MD, USA, ATCC # CRL-1586) и культивировали в среде DMEM, в которую были добавлены 4,5 г/л D-глюкозы, 2 мМ L-глутамина, 110 мг/л пирувата натрия, 10% фетальная бычья сыворотка (Hyclone, Cat # 30070.03) и 0,1% пенициллина/стрептомицина при 37°С в 5% СО2 с влажностью 95%.
Клетки почек африканской зеленой мартышки Cos-7 брали из Немецкой коллекции микроорганизмов и клеточных культур (DSMZ, Braunschweig, Germany, DSMZ # ACC 60) и культивировали в модифицированной по способу Дульбекко среде MEM, содержащей 10% фетальную телячью сыворотку, 2 мМ L-глутамин, 1,2 мкг/мл бикарбоната натрия, 100 ед. пенициллина/ 100 мкг/мл стрептомицина (Biochrom AG, Berlin, Germany).
Пример 3.1: Анализ на бляшкообразование
Культивирование клеток, киРНК-трансфекция и инфицирование вирусом
За один день до трансфекции клетки MDCK засевали в 24-луночные планшеты при плотности 7,5×104 клеток на лунку в 0,5 мл среды для культивирования. В день трансфекции клетки MDCK достигали 80%-ной конфлюентности. Перед трансфекцией клетки подпитывали 0,25 мл культуральной среды.
Перед добавлением в клетки 1,5 мл (50 мкл на лунку) реагента Optimem I (Invitrogen) и 90 мкл (3 мкл на лунку) липофектамина 2000 (Invitrogen) в количестве, достаточном для трансфекции 24-луночного планшета, объединяли в 2-миллилитровой пробирке Сарстедта и инкубировали в течение 10-15 минут при комнатной температуре. К смеси реагента Optimem/липофектамина 2000 добавляли соответствующее количество киРНК, растворенной в буфере для отжига, до получения нужной конечной концентрации, а затем смешивали и инкубировали еще 15-25 минут при комнатной температуре. После этого в каждую лунку по каплям добавляли 50 мкл комплекса киРНК/реагент в соответствии с протоколом эксперимента. Затем планшеты слегка встряхивали для лучшего смешивания и инкубировали при 37ºС при 5% СО2/влажности 95%, в течение 14 часов.
После этого среду для трансфекции осторожно отсасывали, клетки один раз промывали 0,25-0,5 мл PBS и в каждую лунку добавляли 100 мкл различных концентраций PR8 в среде MEM. После инкубирования при 37ºС в течение 1-2 часов добавляли 0,5 мл среды для покрытия (MEM, 20 мМ HEPES, 0,075% NaHCO3, 2 мМ глутамин, 0,6% агароза, 0,5 мкг/мл TPCK-трипсин) и планшеты инкубировали в течение 48 часов при 37ºС в инкубаторе при 5% СО2/влажности 95%. Затем планшеты фиксировали и подвергали иммунологическому окрашиванию на вирусные бляшки, как описано ниже.
Иммунологическое окрашивание и количественная оценка вируса
Через 48 часов после инфицирования клетки фиксировали в нейтрально забуференном 10% формалине в течение 45 минут и лунки промывали PBS. Затем лунки блокировали буфером для повышения проницаемости (1×PBS, 2% FBS, 0,5% сапонин, 0,1% азид натрия) в течение 15 минут при комнатной температуре и добавляли 125 мкл раствора, содержащего 0,5 мкг/мл биотинилированного мышиного антитела против вируса гриппа А, MAB8258B (Chemicon). После инкубирования в течение 1 часа при комнатной температуре лунки два раза промывали PBS для удаления несвязанного антитела, а затем добавляли 125 мкл раствора 1 мкг/мл конъюгированного с пероксидазой хрена (ПХ) стрептавидина (Vector Laboratories) в PBS на лунку и планшеты инкубировали в течение 45 минут, после чего их три раза промывали PBS. Затем добавляли 200 мкл субстрата TMB (Vector Laboratories #SK-4400) на лунку. После инкубирования в темноте в течение 5-10 минут при комнатной температуре колориметрическую реакцию прекращали добавлением дистиллированной воды, а затем воду сливали и планшеты сушили на воздухе. Окрашенные бляшки вируса гриппа подсчитывали на инвертированном оптическом микроскопе при увеличении 4X. Бляшкообразующую активность клеток сравнивали с активностью клеток, трансфицированных только липофектамином (ложнообработанных), и вычисляли по формуле: [(бляшкообразующая активность в обработанных клетках)/(бляшкообразующая активность в ложнообработанных клетках)] × 100 = % остаточная инфекционность.
Пример 3.2: ELISA-анализ
Клетки MDCK или Vero высевали в 96-луночные планшеты при плотности 104 клеток на лунку в 0,1 мл культуральной среды за день до трансфекции. На день киРНК-трансфекции клетки достигали 80%-ной конфлюэнтности. Перед трансфекцией клетки подпитывали 44 мкл культуральной среды.
1,08 мл (9 мкл на лунку) реагента Optimem I (Invitrogen) и 42 мкл (0,35 мкл на лунку) липофектамина 2000 (Invitrogen) в количестве, достаточном для трансфекции одного 96-луночного планшета, объединяли в 2-миллилитровой пробирке Сарстедта и инкубировали в течение 10-15 минут при комнатной температуре. К смеси реагента Optimem/липофектамина 2000 добавляли соответствующее количество киРНК, растворенной в буфере для отжига, до получения нужной конечной концентрации, а затем смешивали и инкубировали еще 15-25 минут при комнатной температуре. После этого в каждую лунку по каплям добавляли 10 мкл комплекса киРНК/реагент в соответствии с протоколом эксперимента. Затем планшеты слегка встряхивали для лучшего смешивания и инкубировали при 37ºС при 5% СО2/влажности 95% в течение 14 часов.
После этого клетки один раз промывали PBS, инфицировали вирусом гриппа PR8 в 50 мкл MEM на лунку и инкубировали в течение 1-2 часов. Затем планшеты один раз промывали PBS и добавляли 200 мкл среды MEM с 0,25/0,5 мкг/мл (MDCK/VERO соответственно) трипсина. Через два дня после инфицирования планшеты фиксировали в забуференном 10% формалине в течение 15 минут. Клетки промывали PBS, блокировали блокирующим буфером в течение 15 минут при комнатной температуре и добавляли 50 мкл раствора, содержащего 0,5 мкг/мл биотинилированного моноклонального антитела против вируса гриппа А, MAB8258B (Chemicon) на лунку. Планшеты инкубировали при комнатной температуре в течение 1 часа, два раза промывали PBS и добавляли 50 мкл на лунку раствора, содержащего 1 мкг/мл конъюгированного со щелочной фосфатазой (ЩФ) стрептавидина (Vector Laboratories) в блокирующем буфере. После инкубирования в течение 45 минут и 3-кратной промывки PBS добавляли 100 мкл раствора субстрата pNPP. Планшеты проявляли в темноте при комнатной температуре и считывали при 405 нм.
Пример 3.3: Ингибирование рекомбинантно экспрессируемых генов-мишеней вируса гриппа под действием киРНК
Консенсусные последовательности MP (SEQ ID NO: 1453), NP (SEQ ID NO: 1454), PA (SEQ ID NO: 1455), PB1 (SEQ ID NO: 1456) и PB2 (SEQ ID NO: 1457) (см. таблицу 5) синтезировали с помощью GENEART (Regensburg, Germany) и клонировали в стандартные векторы GENEART. MP и PA субклонировали в вектор psiCheck-2 (Promega, Mannheim, Germany) посредством AsiSI- и NotI-сайтов (оба от NEBn, Frankfurt, Germany), а NP (PB1) и PB2 - посредством XhoI- и NotI-сайтов, в результате чего получали конструкцию, в которой ген вируса гриппа был расположен между стоп-кодоном и polyA-сигналом люциферазы Renilla. “Правильность” клонирования подтверждали путем секвенирования концов, осуществляемого с помощью GATC Biotech (Konstanz, Germany).
Трансфекции
Клетки Cos-7 высевали при плотности 1,5×104 клеток/лунку на белые 96-луночные планшеты с прозрачным дном (Greiner Bio-One GmbH, Frickenhausen, Germany) в 75 мкл культуральной среды. Сразу после посева клеток 50 нг плазмиды/лунку трансфицировали липофектамином 2000 (Invitrogen), как описано ниже для киРНК, при этом эту плазмиду разводили в среде Opti-MEM до конечного объема 12,5 мкл/лунку, в результате чего получали маточную смесь для всего планшета.
киРНК-трансфекции осуществляли с четырьмя повторностями через 4 часа после трансфекции плазмиды. Для каждой лунки 0,5 мкл липофектамина 2000 (Invitrogen GmbH, Karlsruhe, Germany) смешивали с 12 мкл среды Opti-MEM (Invitrogen) и инкубировали в течение 15 минут при комнатной температуре. Для получения концентрации 50 нМ киРНК в 100 мкл объема смеси для трансфекции, 1 мкл 5 мкМ киРНК смешивали с 11,5 мкл Opti-MEM на лунку, а затем объединяли со смесью липофектамин 2000 - Opti-MEM и снова инкубировали в течение 15 минут при комнатной температуре. Во время инкубирования культуральную среду удаляли из лунок и заменяли 75 мкл/лунку свежей среды. На клетки полностью наносили комплексы киРНК-липофектамин 2000 (25 мкл каждого на лунку) и клетки инкубировали в течение 24 часов при 37ºС и 5% СО2 в инкубаторе с повышенной влажностью (Heraeus GmbH, Hanau, Germany).
Клетки собирали путем удаления культуральной среды и нанесения 150 мкл смеси 1:1, состоящей из среды и субстрата для люциферазы Dual-Glo, взятого из наборов для люциферазного анализа Dual-Glo (Promega, Mannheim, Germany). Люциферазный анализ проводили с помощью наборов для люциферазного анализа Dual-Glo в соответствии с инструкциями производителя и интенсивность люминесценции измеряли на люминесцентном счетчике Victor-Light 1420 (Perkin Elmer, Rodgau-Jügesheim, Germany). Величины, полученные для люциферазы Renilla, нормализовали на величины, полученные для люциферазы светляка. Величины, полученные для киРНК, направленных против гена вируса гриппа, нормализовали на величину, полученную для неспецифической киРНК (направленной против гена резистентности к неомицину) и принятую за 100%.
Эффективные киРНК, выявленные в результате скрининга, затем характеризовали с использованием дозозависимых кривых. Трансфекции, представленные на дозозависимых кривых, осуществляли при следующих концентрациях киРНК: 100 нМ, 25 нМ, 6,3 нМ, 1,6 нМ, 400 пМ, 100 пМ, 24 пМ, 6 пМ, 1,5 пМ, 380 фМ в соответствии с вышеописанным протоколом. Величины IC50 определяли путем построения параметрической кривой с помощью программы XLfit.
Таблица 5
Виртуальные консенсусные последовательности генов белков MP (SEQ ID NO: 1453), NP (SEQ ID NO: 1454), PA (SEQ ID NO: 1455), PB1 (SEQ ID NO: 1456) и PB2 (SEQ ID NO: 1457) вируса гриппа для клонирования в клетки Cos-7







В таблицах 1A, C, D и E указаны: идентификатор дуплекса, последовательности смысловой и антисмысловой цепей, гены-мишени для агентов и результаты вышеописанных анализов, проводимых для отбора репрезентативных агентов согласно изобретению. В таблицах 1B и H указаны: идентификатор дуплекса, идентификатор дуплекса соответствующей немодифицированной последовательности, последовательности смысловой и антисмысловой цепей, гены-мишени для агентов, а именно для отобранных репрезентативных агентов, несущих модифицированные группы нуклеиновых кислот, введенные для защиты этих агентов от расщепления, в которых все нуклеотиды, содержащие пиримидиновые основания, представляют собой 2'-О-метильную группу смысловой цепи и все нуклеотиды, содержащие пиримидиновые основания в окружении последовательности 5'-ca-3' или 5'-ua-3', содержат 2'-О-метильную группу в антисмысловой цепи, за исключением агентов, в которых антисмысловая цепь не содержит нуклеотидов в окружении последовательности 5'-ca-3' или 5'-ua-3', где все уридины в окружении последовательности 5'-ug-3' представляют собой 2'-О-метил-модифицированные нуклеотиды в антисмысловой цепи (например, AL-DP-2295, AL-DP-2301 и AL-DP-2302). В таблице 2 указаны концентрации для некоторых особенно предпочтительных РНКи-агентов, где указанные концентрации соответствуют 50% максимальному ингибированию и вычислены исходя из определений зависимости “доза-ответ” в клетках Cos-7, сконструированных так, чтобы они экспрессировали гены вируса гриппа.
Таблица 6
Последовательности, используемые в анализах матриксного белка (МР) вируса гриппа А
AY180470, ген матриксного белка (М) штамма (H1N1) вируса гриппа штамма A/перепел/Наньхань/12-340/2000, неполная кодирующая последовательность.
AY633213, ген матриксного белка (М) вируса гриппа А (A/кряква/Альберта/211/98(H1N1)), полноразмерная кодирующая последовательность.
AY664487, мРНК нефункционального матриксного белка вируса гриппа А (A/кряква/Альберта/119/98(H1N1)), неполная последовательность.
M55476, ген матриксного белка (M1), полноразмерная кодирующая последовательность, ген белка M2 (M2), полноразмерная кодирующая последовательность, вируса гриппа типа А.
M55479, ген матриксного белка (M1), полноразмерная кодирующая последовательность, ген белка M2 (M2), полноразмерная кодирующая последовательность, вируса гриппа типа А.
M55480, ген матриксного белка (M1), полноразмерная кодирующая последовательность, ген белка M2 (M2), полноразмерная кодирующая последовательность, вируса гриппа типа А.
M63528, гены мембранного белка M1 и мембранного белка M2 вируса гриппа A (A/индейка/Миннесота/166/81(H1N1)), полноразмерная кодирующая последовательность
U49119, гены матриксных белков М1 и М2 (М) вируса гриппа А, полноразмерная кодирующая последовательность.
Z26859, гены матриксных белков М и M2 вируса гриппа типа А
Z26860, гены матриксных белков М и M2 вируса гриппа типа А
AY422021, ген матриксного белка 1 (М) вируса гриппа А (A/утка/Хоккайдо/95/01(H2N2)), неполная кодирующая последовательность.
M12699, РНК-сегмент 7, кодирующий белки М1 и М2 вируса птичьего гриппа А, A/кряква/NY/6750/78, полноразмерная кодирующая последовательность.
AF213915, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/курица/Италия/5945/95 (H3N2)), неполная кодирующая последовательность.
AY180498, ген матриксного белка (М) штамма вируса гриппа А (A/курица/Наньхань/3-120/2001(H3N2)), неполная кодирующая последовательность.
AY664458, мРНК нефункционального матриксного белка вируса гриппа А (A/обыкновенная камнешарка/Делавер/142/99(H3N2)), неполная последовательность.
AY769614, ген матриксного белка вируса гриппа А (A/индейка/Огайо/313053/04 (H3N2)), неполная кодирующая последовательность.
AY779257, ген матриксного белка 2 (М), неполная кодирующая последовательность, и ген матриксного белка 1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/индейка/Северная Каролина/12344/03 (H3N2)).
AY779258, ген матриксного белка 2 (М), неполная кодирующая последовательность, и ген матриксного белка 1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/индейка/Миннесота/764-2/03 (H3N2)).
AY862623, ген матриксного белка (М) вируса гриппа А (A/Ck/Корея/S6/03(H3N2)), полноразмерная кодирующая последовательность.
AY862624, ген матриксного белка (М) вируса гриппа А (A/утка/Корея/S7/03 (H3N2)), полноразмерная кодирующая последовательность.
AY862625, ген матриксного белка (М) вируса гриппа А (А/Dk/Корея/S8/03 (H3N2)), полноразмерная кодирующая последовательность.
AY862626, ген матриксного белка (М) вируса гриппа А (A/утка/Корея/S9/03 (H3N2)), полноразмерная кодирующая последовательность.
AY862627, ген матриксного белка (М) вируса гриппа А (A/утка/Корея/S10/03 (H3N2)), полноразмерная кодирующая последовательность.
AY862628, ген матриксного белка (М) вируса гриппа А (A/голубь/Корея/S11/03(H3N2)), полноразмерная кодирующая последовательность.
Z26858, гены матриксных белков М и М2 вируса гриппа типа А.
AB166865, гены мембранного ионного канала матриксного белка М1 и М2 вируса гриппа А (A/курица/Ямагучи/7/2004(H5N1)), полноразмерная кодирующая последовательность.
AB188819, гены мембранного ионного канала 2, матриксного белка 1, М2, М1, вируса гриппа А (A/курица/Оита/8/2004 (H5N1)), полноразмерная кодирующая последовательность.
AF509043, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509044, ген белка М1 (М1) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509045, ген белка М1 (М1) вируса гриппа А (A/шелковая курица/Гонконг/SF189/01(H5N1)), полноразмерная кодирующая последовательность.
AF509046, ген белка М1 (М1) вируса гриппа А (A/перепел/Гонконг/SF203/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509047, ген белка М1 (М1) вируса гриппа А (A/голубь/Гонконг/SF215/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509048, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/SF219/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509049, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/715.5/01(H5N1)), полноразмерная кодирующая последовательность.
AF509050, ген белка М1 (М1) вируса гриппа А (А/курица/Гонконг/751.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509051, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/822.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509052, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/829.2/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509053, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/830.2/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509054, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/858.3/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509055, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/866.3/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509056, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/867.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509057, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/879.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509058, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/873.3/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509059, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/876.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509060, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/891.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509061, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/893.2/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509062, ген белка М1 (М1) вируса гриппа А (A/гусь/Гонконг/76.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509063, ген белка М1 (М1) вируса гриппа А (A/гусь/Гонконг/ww100/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509064, ген белка М1 (М1) вируса гриппа А (A/утка/Гонконг/573.4/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509065, ген белка М1 (М1) вируса гриппа А (A/утка/Гонконг/646.3/01 (H5N1)), полноразмерная кодирующая последовательность.
AY059506, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/гусь/Гонконг/ww26/2000 (H5N1)), неполная кодирующая последовательность.
AY059507 ген сегмента 7 матриксного белка (М) вируса гриппа А (A/гусь/Гонконг/ww28/2000 (H5N1)), неполная кодирующая последовательность.
AY059508, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/утка/Гонконг/ww381/2000 (H5N1)), неполная кодирующая последовательность.
AY059509, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/утка/Гонконг/ww461/2000 (H5N1)), неполная кодирующая последовательность.
AY059510, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/гусь/Гонконг/ww491/2000 (H5N1)), неполная кодирующая последовательность.
AY059511, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/утка/Гонконг/2986.1/2000 (H5N1)), неполная кодирующая последовательность.
AY059512, ген сегмента 7 матриксного белка (М) вируса гриппа А (A/гусь/Гонконг/3014.8/2000 (H5N1)), неполная кодирующая последовательность.
AY075029, ген матриксного белка 1 и матриксного белка 2 (М) вируса гриппа А (A/курица/Гонконг/317.5/2001(H5N1)), полноразмерная кодирующая последовательность.
AY075035, ген матриксного белка 1 и матриксного белка 2 (М) вируса гриппа А (A/утка/Гонконг/380.5/2001(H5N1)), полноразмерная кодирующая последовательность.
AY221530, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/NT873.3/01-MB(H5N1)), полноразмерная кодирующая последовательность.
AY221531, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/NT873.3/01 (H5N1)), полноразмерная кодирующая последовательность.
AY221532, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/FY150/01-MB (H5N1)), полноразмерная кодирующая последовательность.
AY221533, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), полноразмерная кодирующая последовательность.
AY221534, ген матриксного белка (М) вируса гриппа А (A/фазан/Гонконг/FY155/01-MB(H5N1)), полноразмерная кодирующая последовательность.
AY221535, ген матриксного белка (М) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), полноразмерная кодирующая последовательность.
AY221536, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/YU822.2/01-MB(H5N1)), полноразмерная кодирующая последовательность.
AY221537, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/YU822.2/01(H5N1)), полноразмерная кодирующая последовательность.
AY221538, ген матриксного белка (М) вируса гриппа А (A/курица/Гонконг/YU562/01(H5N1)), полноразмерная кодирующая последовательность.
AY518361, ген белка М2 мембранного ионного канала и матриксного белка М1 (М) вируса гриппа А (A/утка/Китай/E319-2/03 (H5N1)), полноразмерная кодирующая последовательность.
AY575895, ген матриксного белка (М) вируса гриппа А (A/Gs/HK/739.2/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575896, ген матриксного белка (М) вируса гриппа А (A/Eg/HK/757.3/02(H5N1)), неполная кодирующая последовательность.
AY575897, ген матриксного белка (М) вируса гриппа А (A/G.H/HK/793.1/02 (H5N1)), неполная кодирующая последовательность.
AY575898, ген матриксного белка (М) вируса гриппа А (A/Dk/HK/821/02 (H5N1)), неполная кодирующая последовательность.
AY575899, ген матриксного белка (М) вируса гриппа А (A/Ck/HK/31.4/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575900, ген матриксного белка (М) вируса гриппа А (A/Ck/HK/61.9/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575901, ген матриксного белка (М) вируса гриппа А (A/Ck/HK/YU777/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575902, ген матриксного белка (М) вируса гриппа А (A/Ck/HK/96.1/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575903, ген матриксного белка (М) вируса гриппа А (A/Ck/HK/409.1/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575904, ген матриксного белка (М) вируса гриппа А (A/Ph/HK/sv674.15/02 (H5N1)), полноразмерная кодирующая последовательность.
AY585378, мРНК матриксного белка вируса гриппа А (A/утка/Фуджиянь/01/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585379, мРНК матриксного белка вируса гриппа А (A/утка/Фуджиянь/13/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585380, мРНК матриксного белка вируса гриппа А (A/утка/Фуджиянь/17/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585381, мРНК матриксного белка вируса гриппа А (A/утка/Фуджиянь/19/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585382, мРНК матриксного белка вируса гриппа А (A/утка/Гуандун/01/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585383, мРНК матриксного белка вируса гриппа А (A/утка/Гуандун/07/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585384, мРНК матриксного белка вируса гриппа А (A/утка/Гуандун/12/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585385, мРНК матриксного белка вируса гриппа А (A/утка/Гуандун/22/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585386, мРНК матриксного белка вируса гриппа А (A/утка/Гуандун/40/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585387, мРНК матриксного белка вируса гриппа А (A/утка/Гуангкси/07/1999 (H5N1)), полноразмерная кодирующая последовательность.
AY585388, мРНК матриксного белка вируса гриппа А (А/утка/Гуангкси/22/2001 (H5N1)), неполная кодирующая последовательность.
AY585389, мРНК матриксного белка вируса гриппа А (A/утка/Гуангкси/35/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585390, мРНК матриксного белка вируса гриппа А (А/утка/Гуангкси/53/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585391, мРНК матриксного белка вируса гриппа А (А/утка/Шанхай/08/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585392, мРНК матриксного белка вируса гриппа А (A/утка/Шанхай/13/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585393, мРНК матриксного белка вируса гриппа А (А/утка/Шанхай/35/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585394, мРНК матриксного белка вируса гриппа А (A/утка/Шанхай/37/2002(H5N1)), полноразмерная кодирующая последовательность.
AY585395, мРНК матриксного белка вируса гриппа А (А/утка/Шанхай/38/2001(H5N1)), полноразмерная кодирующая последовательность.
AY585396, мРНК матриксного белка вируса гриппа А (A/утка/Чжеянь/11/2000(H5N1)), полноразмерная кодирующая последовательность.
AY585397, мРНК матриксного белка вируса гриппа А (A/утка/Чжеянь/52/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585398, мРНК матриксного белка вируса гриппа А (A/утка/Гуангкси/50/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY590578, ген матриксного белка М2 и матриксного белка М1 (М) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU- K2/2004 (H5N1)), неполная и полноразмерная кодирующая последовательность.
AY609315, сегмент 7 вируса гриппа А (A/курица/Гуандун/174/04 (H5N1)), полноразмерная последовательность.
AY651374, ген мембранного ионного канала 2 (М) вируса гриппа А, неполная кодирующая последовательность; и ген матриксного белка 1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/Ck/Индонезия/BL/2003 (H5N1)).
AY651375, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Dk/Индонезия/MS/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651376, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Индонезия/PA/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651377, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Индонезия/2A/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651378, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Индонезия/4/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651379, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Индонезия/5/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651380, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Таиланд/1/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651381, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/Таиланд/73/2004 (H5N1)).
AY651382, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/Таиланд/9.1/2004 (H5N1)).
AY651383, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Qa/Таиланд/57/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651384, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/птичий/Таиланд/3.1/2004 (H5N1)).
AY651385, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/Таиланд/71.1/2004 (H5N1)).
AY651386, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Gs/Таиланд/79/2004 (H5N1)).
AY651391, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Вьетнам/33/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651392, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Вьетнам/35/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651393, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/Вьетнам/36/2004 (H5N1)).
AY651394, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Вьетнам/37/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651395, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/Вьетнам/38/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651396, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/Вьетнам/39/2004 (H5N1)).
AY651397, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/Вьетнам/C57/2004 (H5N1)).
AY651398, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Dk/Вьетнам/11/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651399, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Gf/HK/38/2002 (H5N1)), неполная кодирующая последовательность.
AY651400, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/31.2/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651401, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/37.4/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651402, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/SCk/HK/YU100/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651403, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/YU22/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651404, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/3176.3/2002 (H5N1)), неполная кодирующая последовательность.
AY651405, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/3169.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651406, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/FY157/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651407, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/YU324/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651408, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/2133.1/2003 (H5N1)), неполная кодирующая последовательность.
AY651409, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (А/Ck/HK/NT93/2003 (H5N1)).
AY651410, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/HK/SSP141/2003 (H5N1)).
AY651411, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/Ck/HK/WF157/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651412, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/сапсан/HK/D0028/2004 (H5N1)).
AY651413, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/черноголовая чайка/HK/12.1/2003 (H5N1)), полноразмерная кодирующая последовательность.
AY651414, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/серая цапля/HK/861.1/2002 (H5N1)).
AY651415, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/одичавший голубь/HK/862.7/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651416, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/древесный воробей/HK/864/2002 (H5N1)).
AY651417, ген мембранного белка ионного канала 2 и матриксного белка 1 (М) вируса гриппа А (A/чирок/Китай/2978.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651418, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/HN/5806/2003 (H5N1)).
AY651419, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/ST/4003/2003 (H5N1)).
AY651420, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/ST/4231/2003 (H5N1)).
AY651421, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/YN/6255/2003 (H5N1)).
AY651422, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (ADk/YN/6445/2003 (H5N1)).
AY651423, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ck/YN/374/2004 (H5N1)).
AY651424, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/HN/101/2004 (H5N1)).
AY651425, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Dk/HN/303/2004 (H5N1)).
AY651426, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/Ph/ST/44/2004 (H5N1)).
AY651427, ген мембранного белка ионного канала 2 (М) (неполная кодирующая последовательность); и ген матриксного белка 1 (М), (полноразмерная кодирующая последовательность) вируса гриппа А (A/курица/YN/115/2004 (H5N1)).
AY653194, сегмент 7 вируса гриппа А (A/курица/Jilin/9/2004 (H5N1)), полноразмерная последовательность.
AY676045, ген мембранного белка (М) штамма вируса гриппа А (A/утка/Гонконг/821/02 (H5N1)), полноразмерная кодирующая последовательность.
AY676046, ген мембранного белка (М) штамма вируса гриппа А (A/белая цапля/Гонконг/757.2/03 (H5N1)) полноразмерная кодирующая последовательность.
AY676047, ген мембранного белка (М) штамма вируса гриппа А (A/курица/Корея/ES/03 (H5N1)), полноразмерная кодирующая последовательность.
AY676048, ген мембранного белка (М) штамма вируса гриппа А (A/утка/Корея/ESD1/03 (H5N1)), полноразмерная кодирующая последовательность.
AY684709, гены матриксного белка 2 (М2) и матриксного белка 1 (М1) вируса гриппа А (A/курица/Хубей/327/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY737292, сегмент 7 вируса гриппа А (A/курица/Гуандун/191/04 (H5N1), полноразмерная последовательность.
AY737298, сегмент 7 вируса гриппа А (A/курица/Гуандун/178/04 (H5N1)), полноразмерная последовательность.
AY737306, сегмент 7 вируса гриппа А (A/утка/Гуандун/173/04 (H5N1)), полноразмерная последовательность.
AY770077, гены матриксного белка 2 (М2) и матриксного белка 1 (М1) вируса гриппа А (A/курица/Хубей/489/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY770998, ген матриксного белка вируса гриппа А (A/курица/Аюттхая/Таиланд/CU-23/04 (H5N1)), полноразмерная кодирующая последовательность.
AY818145, ген матриксного белка М1 вируса гриппа А (A/курица/Вьетнам/C58/04 (H5N1)), полноразмерная кодирующая последовательность.
AY818146, ген матриксного белка М1 вируса гриппа А (A/перепел/Вьетнам/36/04 (H5N1)), полноразмерная кодирующая последовательность.
AY856865, сегмент 7 вируса гриппа А (A/утка/Шанду/093/2004 (H5N1)), полноразмерная последовательность.
DQ055851, ген матриксного белка М1 вируса гриппа А (A/курица/Юнань/K001/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189048, гены мембранного белка ионного канала М2, М1; М2 и матриксного белка 1 вируса гриппа А (A/курица/Киото/3/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189056, гены мембранного белка ионного канала М2, М1; М2 и матриксного белка 1 вируса гриппа А (А/ворон/Киото/53/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189064, гены мембранного белка ионного канала М2, М1; М2 и матриксного белка 1 вируса гриппа А (A/ворон/Осака/102/2004 (H5N1)), полноразмерная кодирующая последовательность.
AF046082, гены матриксного белка 2 (М2) и матриксного белка 1 (М1) вируса гриппа А (A/курица/Гонконг/220/97(H5N1)), полноразмерная кодирующая последовательность.
AF098560, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (А/курица/Гонконг/258/97 (H5N1)), неполная кодирующая последовательность.
AF098561, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (А/курица/Гонконг/y388/97 (H5N1)), неполная кодирующая последовательность.
AF098562, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (A/курица/Гонконг/728/97 (H5N1)), неполная кодирующая последовательность.
AF098563, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (A/курица/Гонконг/786/97 (H5N1)), неполная кодирующая последовательность.
AF098564, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (А/курица/Гонконг/915/97 (H5N1)), неполная кодирующая последовательность.
AF098566, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (A/утка/Гонконг/p46/97 (H5N1)), неполная кодирующая последовательность.
AF098567, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (A/утка/Гонконг/y283/97 (H5N1)), неполная кодирующая последовательность.
AF098568, гены матриксного белка М2 (М) и матриксного белка М1 (М) вируса гриппа А (A/гусь/Гонконг/w355/97 (H5N1)), неполная кодирующая последовательность.
AF144306, гены матриксных белков М1 и М2 (М) вируса гриппа А (A/гусь/Гуандун/1/96 (H5N1)), продукты альтернативного сплайсинга, полноразмерная кодирующая последовательность.
AF216711, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-4/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216719, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-6/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216727, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-8/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216735, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-10/99 (H5N1)), полноразмерная кодирующая последовательность.
AF359560, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/гусь/Гуандун/3/97 (H5N1)), полноразмерная кодирующая последовательность.
AF398429, ген матриксного белка 1 (М) вируса гриппа А (A/гусь/Гонконг/385.3/2000 (H5N1)), неполная кодирующая последовательность.
AF398430, ген матриксного белка 1 (М) вируса гриппа А (A/гусь/Гонконг/385.5/2000 (H5N1)), неполная кодирующая последовательность.
AF468843, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/утка/Аньян/AVL-1/2001 (H5N1)), полноразмерная кодирующая последовательность.
AF509040, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/FY77/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509041, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509042, ген белка М1 (М1) вируса гриппа А (A/курица/Гонконг/YU563/01 (H5N1)), полноразмерная кодирующая последовательность.
AF073180, гены матриксного белка 1 (М1) и матриксного белка 2 (М2) вируса гриппа А (A/курица/Нью-Джерси/15086-3/94 (H7N3NSA)), полноразмерная кодирующая последовательность.
AF073197, гены матриксного белка 1 (М1) и матриксного белка 2 (М2) вируса гриппа А (A/индейка/Орегон/71 (H7N3NSB)), полноразмерная кодирующая последовательность.
AY664433, мРНК нефункционального матриксного белка вируса гриппа А (A/обыкновенная камнешарка/Нью-Джерси/65/85 (H7N3)), неполная последовательность.
AY677732, ген матриксного белка 1 вируса гриппа А (A/курица/Британская Колумбия/CN7-3/04 (H7N3)), полноразмерная кодирующая последовательность.
AF073198, гены матриксного белка 1 (М1) и матриксного белка 2 (М2) вируса гриппа А (A/индейка/Колорадо/13356/91 (H7N3NSA)), полноразмерная кодирующая последовательность.
AF073200, гены матриксного белка 1 (М1) и матриксного белка 2 (М2) вируса гриппа А (A/перепел/Арканзас/16309-7/94 (H7N3NSA)), полноразмерная кодирующая последовательность.
AF073201, гены матриксного белка 1 (М1) и матриксного белка 2 (М2) вируса гриппа А (A/индейка/Юта/24721-10/95 (H7N3NSA)), полноразмерная кодирующая последовательность.
AJ627492, ген мембранного белка 1 и ген мембранного белка 2 вируса гриппа А (A/индейка/Италия/214845/2002 (H7N3)), геномная РНК.
AJ627497, ген мембранного белка 1 и ген мембранного белка 2 вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), геномная РНК.
AY241600, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/Нью-Йорк/12273-11/99 (H7N3)), полноразмерная кодирующая последовательность.
AY241602, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/NY/14714-9/99 (H7N3)), полноразмерная кодирующая последовательность.
AY241615, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/утка/NJ/117228-7/01 (H7N3)), полноразмерная кодирующая последовательность.
AY241616, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/утка/PA/143585/01 (H7N3)), полноразмерная кодирующая последовательность.
AY300975, ген мембранного белка (М) вируса гриппа А (A/синекрылый чирок/TX/2/01 (H7N3), полноразмерная кодирующая последовательность.
AY303652, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/Чили/176822/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303653, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/Чили/4322/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303654, ген матриксного белка 1, полноразмерная кодирующая последовательность; и ген матриксного белка 2, неполная кодирующая последовательность, вируса гриппа А (A/курица/Чили/4957/02 (H7N3)).
AY303655, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/Чили/4968/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303656, гены матриксного белка 1 и матриксного белка 2 вируса гриппа А (A/курица/Чили/4977/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303657, ген матриксного белка 1, полноразмерная кодирующая последовательность; и ген матриксного белка 2, неполная кодирующая последовательность, вируса гриппа А (А/индейка/Чили/4418/02 (H7N3)).
AY586427, ген матриксного белка вируса гриппа А (А/индейка/Италия/214845/02 (H7N3)), неполная кодирующая последовательность.
AY586428, ген матриксного белка вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), неполная кодирующая последовательность.
AY586429, ген матриксного белка вируса гриппа А (A/кряква/Италия/43/01 (H7N3)), неполная кодирующая последовательность.
AY586430, ген матриксного белка вируса гриппа А (A/кряква/Италия/33/01 (H7N3)), неполная кодирующая последовательность.
AY611525, гены матриксного белка 2 (М) и матриксного белка 1 (М) вируса гриппа А (A/курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY646079, гены матриксного белка 2 и матриксного белка 1 (М) вируса гриппа А (А/курица/Британская Колумбия/GSC_человек_B/04 (H7N3)), полноразмерная кодирующая последовательность.
AY648288, гены матриксного белка 2 (М) и матриксного белка 1 (М) вируса гриппа А (A/GSC_курица_B/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY650271, гены матриксного белка 2 (М) и матриксного белка 1 (М) вируса гриппа А (A/GSC_курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AJ619676, ген М1 мембранного белка 1 вируса гриппа А (A/курица/Германия/R28/03 (H7N7)), геномная РНК.
AY340086, ген матриксного белка вируса гриппа А (A/Нидерланды/124/03 (H7N7)), неполная кодирующая последовательность.
AY340087, ген матриксного белка вируса гриппа А (A/Нидерланды/126/03 (H7N7)), неполная кодирующая последовательность.
AY340088, ген матриксного белка вируса гриппа А (A/Нидерланды/127/03 (H7N7)), неполная кодирующая последовательность.
AY340089, ген матриксного белка вируса гриппа А (A/Нидерланды/219/03 (H7N7)), полноразмерная кодирующая последовательность.
AY340090, ген матриксного белка вируса гриппа А (A/Нидерланды/33/03 (H7N7)), полноразмерная кодирующая последовательность.
AY340091, ген матриксного белка вируса гриппа А (A/курица/Нидерланды/1/03 (H7N7)), полноразмерная кодирующая последовательность.
AY664468, мРНК нефункционального матриксного белка вируса гриппа А (A/обыкновенная камнешарка/Делавер/134/99 (H7N7)), неполная последовательность.
L37795, ген матриксного белка (М1) и ген трансмембранного белка (М2) вируса гриппа А (A/курица/Бресция/1902 (H7N7), полноразмерная кодирующая последовательность.
L37796, ген матриксного белка (М1) и ген трансмембранного белка (М2) вируса гриппа А (A/FPV/Добсон (H7N7)), полноразмерная кодирующая последовательность.
L37797, ген матриксного белка (М1) и ген трансмембранного белка (М2) вируса гриппа А (A/FPV/Вейбридж (H7N7)), полноразмерная кодирующая последовательность.
M23917, ген матриксного белка М1 вируса гриппа А (A/курица/FPV/Вейбридж (H7N7)), полноразмерная кодирующая последовательность.
M23921, ген матриксного белка М2 вируса гриппа А (A/курица/FPV/Вейбридж (H7N7)), полноразмерная кодирующая последовательность.
M38299, ген (сегмент 7) матриксного белка (М) вируса гриппа А (A/FPV/Вейбридж (H7N7)), полноразмерная кодирующая последовательность.
M63523, гены мембранного белка М1 и мембранного белка М2 вируса гриппа А (A/курица/Виктория/1/85 (H7N7)), полноразмерная кодирующая последовательность.
M63526, гены мембранного белка М1 и мембранного белка М2 вируса гриппа А (штамм A/FPV/Добсон/27 (H7N7)), полноразмерная кодирующая последовательность.
AB049165, гены мембранного белка ионного канала М1, М2, матриксного белка вируса гриппа А (A/попугай/Чиба/1/97 (H9N2)), полноразмерная кодирующая последовательность.
AB049166, гены мембранного белка ионного канала М1, М2, матриксного белка вируса гриппа А (A/попугай/Нарита/92A/98 (H9N2)), полноразмерная кодирующая последовательность.
AF222671, ген сегмента 7 М1 (М1) вируса гриппа А (A/шелковая курица/Гонконг/SF44/99(H9N2)), неполная кодирующая последовательность.
AF508684, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/страус/Южная Африка/9508103/95 (H9N2)), полноразмерная кодирующая последовательность.
AF508685, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Пакистан/4/99 (H9N2)), неполная кодирующая последовательность.
AF508686, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Пакистан/5/99 (H9N2)), неполная кодирующая последовательность.
AF508687, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Германия/R45/98 (H9N2)), полноразмерная кодирующая последовательность.
AF508688, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/утка/Германия/113/95 (H9N2)), полноразмерная кодирующая последовательность.
AF508689, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Иран/11T/99 (H9N2)), неполная кодирующая последовательность.
AF508690, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Саудовская Аравия/532/99 (H9N2)), неполная кодирующая последовательность.
AF508691, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/фазан/Ирландия/PV18/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508692, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Корея/99029/99 (H9N2)), полноразмерная кодирующая последовательность.
AF508693, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Бейжин/8/98 (H9N2)), полноразмерная кодирующая последовательность.
AF508694, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Гуандун/10/00 (H9N2)), полноразмерная кодирующая последовательность.
AF508695, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Гуандун/11/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508696, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (А/курица/Хейлоньян/10/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508697, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Ханан/62/00 (H9N2)), полноразмерная кодирующая последовательность.
AF508698, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Нингсия/5/99 (H9N2)), полноразмерная кодирующая последовательность.
AF508699, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Сихуа/5/97 (H9N2)), неполная кодирующая последовательность.
AF508700, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Шанду/6/96 (H9N2)), полноразмерная кодирующая последовательность.
AF508701, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (А/курица/Шиячжуан/2/99 (H9N2)), неполная кодирующая последовательность.
AF508702, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/курица/Шенчжен/9/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508703, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/утка/Наньян/1/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508704, ген сегмента 7 матриксного белка М1 (М) вируса гриппа А (A/перепел/Шанхай/8/96 (H9N2)), полноразмерная кодирующая последовательность.
AF523482, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/1043/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523483, ген матриксного белка (М) вируса гриппа А (А/утка/Шанту/2134/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523484, ген матриксного белка (М) вируса гриппа А (А/дикая утка/Шанту/4808/01(H9N2)), полноразмерная кодирующая последовательность.
AF523485, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/1042/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523486, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/2143/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523487, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/2144/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523488, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/1881/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523489, ген матриксного белка (М) вируса гриппа А (А/утка/Шанту/1796/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523490, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/2102/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523491, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/830/00 (H9N2)), полноразмерная кодирующая последовательность.
AF523492, ген матриксного белка (М) вируса гриппа А (A/утка/Шанту/2088/01 (H9N2)), полноразмерная кодирующая последовательность.
AF523493, ген матриксного белка (М) вируса гриппа А (А/утка/Шанту/1605/01 (H9N2)), полноразмерная кодирующая последовательность.
AF523494, ген матриксного белка (М) вируса гриппа А (A/утка/Гонконг/610/79 (H9N2)), полноразмерная кодирующая последовательность.
AF523495, ген матриксного белка (М) вируса гриппа А (A/утка/Гонконг/552/79 (H9N2)), полноразмерная кодирующая последовательность.
AF523496, ген матриксного белка (М) вируса гриппа А (А/утка/Гонконг/289/78 (H9N2)), полноразмерная кодирующая последовательность.
AF523497, ген матриксного белка (М) вируса гриппа А (A/утка/Гонконг/86/76 (H9N2)), полноразмерная кодирующая последовательность.
AF523498, ген матриксного белка (М) вируса гриппа А (A/утка/Гонконг/366/78 (H9N2)), неполная кодирующая последовательность.
AF536719, ген нефункционального матриксного белка вируса гриппа А (A/курица/Бейжин/1/95 (H9N2)), неполная последовательность.
AF536720, ген нефункционального матриксного белка вируса гриппа А (А/курица/Бейжин/2/97 (H9N2)), неполная последовательность.
AF536721, ген нефункционального матриксного белка вируса гриппа А (A/курица/Бейжин/3/99 (H9N2)), неполная последовательность.
AF536722, ген нефункционального матриксного белка вируса гриппа А (A/курица/Гуандун/97 (H9N2)), неполная последовательность.
AF536723, ген нефункционального матриксного белка вируса гриппа А (A/курица/Хэбей/1/96 (H9N2)), неполная последовательность.
AF536724, ген нефункционального матриксного белка вируса гриппа А (A/курица/Хэбей/2/98 (H9N2)), неполная последовательность.
AF536725, ген нефункционального матриксного белка вируса гриппа А (A/курица/Хэбей/3/98 (H9N2)), неполная последовательность.
AF536726, ген нефункционального матриксного белка вируса гриппа А (A/курица/Хэнань/98 (H9N2)), неполная последовательность.
AF536727, ген нефункционального матриксного белка вируса гриппа А (A/курица/Ляонин/99 (H9N2)), неполная последовательность.
AF536728, ген нефункционального матриксного белка вируса гриппа А (A/курица/Шанду/98 (H9N2)), неполная последовательность.
AJ291398, ген матриксного белка 1 (М1) (экзон 1) и ген матриксного белка 2 (М2) (экзоны 1 и 2) вируса гриппа А (A/курица/Пакистан/2/99 (H9N2)), геномная РНК.
AJ427865, неполный ген матриксного белка (М) вируса гриппа А (A/перепел/Гонконг/FY298/00 (H9N2)), геномная РНК
AY180461, ген матриксного белка (М) вируса гриппа А (штамм A/голубь/Наньхань/2-0461/2000 (H9N2), неполная кодирующая последовательность.
AY180462, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/11-290/2000 (H9N2)), неполная кодирующая последовательность.
AY180463, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/11-197/2000 (H9N2)), неполная кодирующая последовательность.
AY180464, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/11-392/2000 (H9N2)), неполная кодирующая последовательность.
AY180477, ген матриксного белка (М) вируса гриппа А (штамм A/курица/Наньхань/4-361/2001 (H9N2)), неполная кодирующая последовательность.
AY180485, ген матриксного белка (М) вируса гриппа А (штамм A/голубь/Наньхань/11-145/2000 (H9N2)), неполная кодирующая последовательность.
AY180486, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/1-0070/2000 (H9N2)), неполная кодирующая последовательность.
AY180489, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/10-389/2000 (H9N2)), неполная кодирующая последовательность.
AY180490, ген матриксного белка (М) вируса гриппа А (штамм A/курица/Наньхань/1-0016/2000 (H9N2), неполная кодирующая последовательность.
AY180492, ген матриксного белка (М) вируса гриппа А (штамм A/голубь/Наньхань/7-058/2000 (H9N2)), неполная кодирующая последовательность.
AY180495, ген матриксного белка (М) вируса гриппа А (штамм A/перепел/Наньхань/2-0460/2000 (H9N2)), неполная кодирующая последовательность.
AY180502, ген матриксного белка (М) вируса гриппа А (штамм A/курица/Наньхань/4-010/2000 (H9N2)), неполная кодирующая последовательность.
AY180504, ген матриксного белка (М) вируса гриппа А (штамм A/перепел/Наньхань/4-040/2000 (H9N2)), неполная кодирующая последовательность.
AY180506, ген матриксного белка (М) вируса гриппа А (штамм A/курица/Наньхань/4-301/2001 (H9N2)), неполная кодирующая последовательность.
AY180516, ген матриксного белка (М) вируса гриппа А (штамм A/утка/Наньхань/7-092/2000 (H9N2)), неполная кодирующая последовательность.
AY180519, ген матриксного белка (М) вируса гриппа А (штамм A/дикая утка/Наньхань/2-0480/2000 (H9N2)), неполная кодирующая последовательность.
AY253755, гены матриксного белка М1 и мембранного белка ионного канала М2 вируса гриппа А (A/курица/Шанхай/F/98 (H9N2)), полноразмерная кодирующая последовательность.
AY496852, мРНК матриксного белка (М1) вируса гриппа А (A/курица/Муданьцзян/0823/2000 (H9N2)), полноразмерная кодирующая последовательность.
AY633165, ген матриксного белка (М) вируса гриппа А (A/кряква/Альберта/17/91 (H9N2)), полноразмерная кодирующая последовательность.
AY633277, ген матриксного белка (М) вируса гриппа А (A/кряква/Альберта/321/88 (H9N2)), полноразмерная кодирующая последовательность.
AY633293, ген матриксного белка (М) вируса гриппа А (А/кряква/Альберта/11/91 (H9N2)), полноразмерная кодирующая последовательность.
AY664464, мРНК нефункционального матриксного белка (A/ржанка/Делавер/276/99 (H9N2)), неполная последовательность.
AY664679, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/CSW153/03 (H9N2)).
AY664680, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/AP45/03 (H9N2)).
AY664681, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/BD90/03 (H9N2)).
AY664682, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/CSW291/03 (H9N2)).
AY664683, гены мембранного белка М2 (М) и мембранного белка М1 (М) вируса гриппа А (A/курица/Гонконг/CSW304/03 (H9N2)), неполная кодирующая последовательность.
AY664684, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/FY23/03 (H9N2)).
AY664685, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/цесарка/Гонконг/NT101/03 (H9N2)).
AY664686, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/NT142/03 (H9N2)).
AY664687, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (А/курица/Гонконг/SF1/03 (H9N2)).
AY664688, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/SSP101/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664689, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/TP38/03 (H9N2)).
AY664690, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/WF126/03 (H9N2)).
AY664691, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/голубь/Гонконг/WF53/03 (H9N2)).
AY664692, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/фазан/Гонконг/WF54/03 (H9N2)).
AY664693, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (А/цесарка/Гонконг/NT184/03 (H9N2)).
AY664694, ген мембранного белка М2 (М) и ген мембранного белка М1 (М), вируса гриппа А (A/курица/Гонконг/WF120/03 (H9N2)), неполная кодирующая последовательность.
AY664695, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/NT366/03 (H9N2)).
AY664696, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/SSP418/03 (H9N2)).
AY664697, ген мембранного белка М2 (М), неполная кодирующая последовательность; и ген мембранного белка М1 (М), полноразмерная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/YU427/03 (H9N2)).
AY800234, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S1/2003 (H9N2)), полноразмерная кодирующая последовательность.
AY862614, ген матриксного белка (М) вируса гриппа А (A/шелковая курица/Корея/S3/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862615, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S4/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862616, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S5/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862617, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S12/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862618, ген матриксного белка (М) вируса гриппа А (A/утка/Корея/S13/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862619, ген матриксного белка (М) вируса гриппа А (A/голубь/Корея/S14/03 (H9N2)), неполная кодирующая последовательность.
AY862620, ген матриксного белка (М) вируса гриппа А (А/курица/Корея/S15/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862621, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S16/03 (H9N2)), полноразмерная кодирующая последовательность.
AY862622, ген матриксного белка (М) вируса гриппа А (A/курица/Корея/S18/03 (H9N2)), полноразмерная кодирующая последовательность.
AF156458, гены сегмента 7 матриксного белка М1 (М1) и матриксного белка М2 (М2) вируса гриппа А (A/курица/Гонконг/G9/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156459, гены сегмента 7 матриксного белка М1 (М1) и матриксного белка М2 (М2) вируса гриппа А (A/курица/Гонконг/G23/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156460, гены сегмента 7 матриксного белка М1 (М1) и матриксного белка М2 (М2) вируса гриппа А (А/голубь/Гонконг/Y233/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156461, гены сегмента 7 матриксного белка М1 (М1) и матриксного белка М2 (М2) вируса гриппа А (A/утка/Гонконг/Y280/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156462, гены сегмента 7 матриксного белка М1 (М1) и матриксного белка М2 (М2) вируса гриппа А (A/утка/Гонконг/Y439/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156463, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/перепел/Гонконг/G1/97 (H9N2)).
AF156464, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/курица/Гонконг/739/94 (H9N2)).
AF156465, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/перепел/Гонконг/AF157/92 (H9N2)), неполная кодирующая последовательность.
AF156466, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/курица/Бейжин/1/94 (H9N2)).
AF156467, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/курица/Корея/38349-p96323/96 (H9N2)).
AF156468, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/курица/Корея/25232-96006/96 (H9N2)).
AF156469, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/Ржанка/Делавер/9/96 (H9N2)).
AF156470, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/перепел/Арканзас/29209-1/93 (H9N2)).
AF156471, гены сегмента 7 матриксного белка М1 (М1), полноразмерная кодирующая последовательность; и матриксного белка М2 (М2), неполная кодирующая последовательность, вируса гриппа А (A/индейка/Калифорния/189/66 (H9N2)).
AF203788, мРНК матриксного белка 1, полноразмерная кодирующая последовательность; и мРНК матриксного белка 2, неполная кодирующая последовательность, вируса гриппа А (A/курица/Корея/MS96/96 (H9N2)).
AF222662, ген сегмента 7 М1 (М1) вируса гриппа А (A/перепел/Гонконг/A17/99 (H9N2)), неполная кодирующая последовательность.
AF222663, ген сегмента 7 М1 (М1) вируса гриппа А (A/голубь/Гонконг/FY6/99 (H9N2)), неполная кодирующая последовательность.
AF222664, ген сегмента 7 М1 (М1) вируса гриппа А (A/курица/Гонконг/NT16/99 (H9N2)), неполная кодирующая последовательность.
AF222665, ген сегмента 7 М1 (М1) вируса гриппа А (A/перепел/Гонконг/SSP10/99 (H9N2)), неполная кодирующая последовательность.
AF222666, ген сегмента 7 М1 (М1) вируса гриппа А A/фазан/Гонконг/SSP11/99 (H9N2)), неполная кодирующая последовательность.
AF222667, ген сегмента 7 М1 (М1) вируса гриппа А (A/курица/Гонконг/FY20/99 (H9N2)), неполная кодирующая последовательность.
AF222668, ген сегмента 7 М1 (М1) вируса гриппа А (A/курица/Гонконг/KC12/99 (H9N2)), неполная кодирующая последовательность.
AF222669, ген сегмента 7 М1 (М1) вируса гриппа А (A/перепел/Гонконг/NT28/99 (H9N2)), неполная кодирующая последовательность.
AF222670, ген сегмента 7 М1 (М1) вируса гриппа А (A/курица/Гонконг/SF2/99 (H9N2)), неполная кодирующая последовательность.
Последовательности, используемые в анализе нуклеокапсидного белка (NP) вируса гриппа А
AF156415, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/индейка/Калифорния/189/66(H9N2)), неполная кодирующая последовательность.
AF523423, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/86/76 (H9N2)), полноразмерная кодирующая последовательность.
AF523424, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/366/78 (H9N2)), неполная кодирующая последовательность.
AF523421, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/289/78 (H9N2)), полноразмерная кодирующая последовательность.
AF523422, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/552/79 (H9N2)), неполная кодирующая последовательность.
AY633279, ген нуклеокапсидного белка (NP) вируса гриппа А (A/кряква/Альберта/321/88 (H9N2)), полноразмерная кодирующая последовательность.
AY633295, ген нуклеокапсидного белка (NP) вируса гриппа А (A/кряква/Альберта/11/91 (H9N2)), неполная кодирующая последовательность.
AY633167, ген нуклеокапсидного белка (NP) вируса гриппа А (A/кряква/Альберта/17/91 (H9N2)), полноразмерная кодирующая последовательность.
AF156410, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Гонконг/AF157/92 (H9N2)), полноразмерная кодирующая последовательность.
AF156414, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/перепел/Арканзас/29209-1/93 (H9N2)), неполная кодирующая последовательность.
AF156408, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/739/94 (H9N2)), полноразмерная кодирующая последовательность.
AF156409, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Бейжин/1/94 (H9N2)), полноразмерная кодирующая последовательность.
AF536699, ген нуклеопротеина (NP) вируса гриппа А (А/курица/Бейжин/1/95 (H9N2)), неполная кодирующая последовательность.
AF508596, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/страус/Южная Африка/9508103/95 (H9N2)), неполная кодирующая последовательность.
AF508600, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/утка/Германия/113/95 (H9N2)), неполная кодирующая последовательность.
AB020778, ген нуклеопротеина вируса гриппа А, полноразмерная кодирующая последовательность.
AF508613, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Шанду/6/96 (H9N2)), неполная кодирующая последовательность.
AF508617, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Шанхай/8/96 (H9N2)), неполная кодирующая последовательность.
AF536703, ген нуклеопротеина (NP) вируса гриппа А (А/курица/Хэбей/1/96 (H9N2)), неполная кодирующая последовательность.
AF156411, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Корея/38349-96323/96 (H9N2)), полноразмерная кодирующая последовательность.
AF156412, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Корея/25232-96006/96 (H9N2)), неполная кодирующая последовательность.
M63779, мРНК нуклеопротеина вируса гриппа А/FPV/Добсон/”голландский”/27 (H7N7), полноразмерная кодирующая последовательность.
M63784, мРНК нуклеопротеина вируса гриппа A/чирок/Исландия/29/80 (H7N7), полноразмерная кодирующая последовательность.
AJ620352, ген нуклеопротеина NP вируса гриппа А (A/курица/Германия/R28/03 (H7N7)), геномная РНК.
AY342425, ген нуклеокапсидного белка вируса гриппа А (A/Нидерланды/219/03 (H7N7)), полноразмерная кодирующая последовательность.
AY342426, ген нуклеокапсидного белка вируса гриппа А (A/Нидерланды/033/03 (H7N7)), полноразмерная кодирующая последовательность.
AY342427, ген нуклеокапсидного белка вируса гриппа А (A/курица/Нидерланды/1/03 (H7N7)), полноразмерная кодирующая последовательность.
AF156413, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/ржанка/Делавер/9/96 (H9N2)), неполная кодирующая последовательность.
AF203787, мРНК нуклеопротеина вируса гриппа А (A/курица/Корея/MS96/96 (H9N2)), полноразмерная кодирующая последовательность.
AF156402, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Гонконг/G9/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156403, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Гонконг/G23/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156404, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/голубь/Гонконг/Y233/97 (H9N2)), неполная кодирующая последовательность.
AF156405, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/утка/Гонконг/Y280/97 (H9N2)), неполная кодирующая последовательность.
AF156406, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/утка/Гонконг/Y439/97 (H9N2)), полноразмерная кодирующая последовательность.
AF156407, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Гонконг/G1/97 (H9N2)), неполная кодирующая последовательность.
AF508612, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Сихуань/5/97 (H9N2)), неполная кодирующая последовательность.
AF536702, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гуандун/97 (H9N2)), неполная кодирующая последовательность.
AF536700, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Бейжин/2/97 (H9N2)), неполная кодирующая последовательность.
AF508615, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Шензен/9/97 (H9N2)), неполная кодирующая последовательность.
AF508616, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/утка/Наньян/1/97 (H9N2)), неполная кодирующая последовательность.
AB049161, ген нуклеопротеина (NP) вируса гриппа А (A/попугай/Чиба/1/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508603, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/фазан/Ирландия/PV18/97 (H9N2)), неполная кодирующая последовательность.
AF508607, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Гуандун/11/97 (H9N2)), неполная кодирующая последовательность.
AF508609, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Хейлоньян/10/97 (H9N2)), неполная кодирующая последовательность.
AF508608, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Хэбей/4/98 (H9N2)), неполная кодирующая последовательность.
AF508605, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Бейжин/8/98 (H9N2)), неполная кодирующая последовательность.
AF508599, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Германия/R45/98 (H9N2)), неполная кодирующая последовательность.
AF536708, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Шанду/98 (H9N2)), неполная кодирующая последовательность.
AY253753, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Шанхай/F/98 (H9N2)), полноразмерная кодирующая последовательность.
AF536704, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Хэбей/2/98 (H9N2)), неполная кодирующая последовательность.
AF536705, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Хэбей/3/98 (H9N2)), неполная кодирующая последовательность.
AF536706, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Хэнань/98 (H9N2)), неполная кодирующая последовательность.
AB049162, ген нуклеопротеина (NP) вируса гриппа А (A/попугай/Нарита/92A/98 (H9N2)), полноразмерная кодирующая последовательность.
AF186270, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Гонконг/NT28/99 (H9N2)), неполная кодирующая последовательность.
AF186271, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/шелковая курица/Гонконг/SF43/99 (H9N2)), неполная кодирующая последовательность.
AF186272, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/SF2/99 (H9N2)), неполная кодирующая последовательность.
AF222614, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Гонконг/A17/99 (H9N2)), неполная кодирующая последовательность.
AF222615, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/голубь/Гонконг/FY6/99 (H9N2)), неполная кодирующая последовательность.
AF222616, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/NT16/99 (H9N2)), неполная кодирующая последовательность.
AF222617, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/перепел/Гонконг/SSP10/99 (H9N2)), неполная кодирующая последовательность.
AF222618, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/фазан/Гонконг/SSP11/99 (H9N2)), неполная кодирующая последовательность.
AF536707, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Ляонин/99 (H9N2)), неполная кодирующая последовательность.
AJ291394, ген нуклеопротеина NP вируса гриппа А (A/курица/Пакистан/2/99 (H9N2)), геномная РНК.
AF536701, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Бейжин/3/99 (H9N2)), неполная кодирующая последовательность.
AF508611, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Нингсия/5/99 (H9N2)), неполная кодирующая последовательность.
AF508614, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Шиячжуан/2/99 (H9N2)), неполная кодирующая последовательность.
AF508604, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Корея/99029/99 (H9N2)), неполная кодирующая последовательность.
AF222619, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/FY20/99 (H9N2)), неполная кодирующая последовательность.
AF222620, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (А/курица/Гонконг/KC12/99 (H9N2)), неполная кодирующая последовательность.
AF222621, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/шелковая курица/Гонконг/SF44/99 (H9N2)), неполная кодирующая последовательность.
AF508601, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Иран/11T/99 (H9N2)), неполная кодирующая последовательность.
AF508602, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Саудовская Аравия/532/99 (H9N2)), неполная кодирующая последовательность.
AF508597, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Пакистан/4/99 (H9N2)), неполная кодирующая последовательность.
AF508598, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Пакистан/5/99 (H9N2)), неполная кодирующая последовательность.
AF508606, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Гуандун/10/00 (H9N2)), неполная кодирующая последовательность.
AF508610, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Henan/62/00 (H9N2)), неполная кодирующая последовательность.
AF523410, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/1043/00 (H9N2)), неполная кодирующая последовательность.
AF523411, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/2134/00 (H9N2)), неполная кодирующая последовательность.
AF523413, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/1042/00 (H9N2)), неполная кодирующая последовательность.
AF523415, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/2102/00 (H9N2)), неполная кодирующая последовательность.
AF523416, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/830/00 (H9N2)), неполная кодирующая последовательность.
AF523417, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/2144/00 (H9N2)), неполная кодирующая последовательность.
AF523419, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/2143/00 (H9N2)), неполная кодирующая последовательность.
AF523420, ген нуклеокапсидного белка (NP) вируса гриппа А (А/утка/Шанту/1881/00 (H9N2)), неполная кодирующая последовательность.
AJ427864, неполный ген нуклеопротеина NP вируса гриппа А (A/перепел/Гонконг/FY298/00 (H9N2)), геномная РНК.
AY180525, ген нуклеопротеина (NP) вируса гриппа А (A/голубь/Наньхань/2-0461/2000 (H9N2)), неполная кодирующая последовательность.
AY180534, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/7-092/2000 (H9N2), неполная кодирующая последовательность.
AY180537, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/11-392/2000 (H9N2), неполная кодирующая последовательность.
AY180538, ген нуклеопротеина (NP) вируса гриппа А (A/голубь/Наньхань/11-145/2000 (H9N2)), неполная кодирующая последовательность.
AY180542, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/11-197/2000 (H9N2), неполная кодирующая последовательность.
AY180544, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/11-290/2000 (H9N2), неполная кодирующая последовательность.
AY180560, ген нуклеопротеина (NP) вируса гриппа А (A/голубь/Наньхань/7-058/2000 (H9N2)), неполная кодирующая последовательность.
AY180562, ген нуклеопротеина (NP) вируса гриппа А штамма A/курица/Наньхань/4-010/2000 (H9N2), неполная кодирующая последовательность.
AY180563, ген нуклеопротеина (NP) вируса гриппа А штамма A/перепел/Наньхань/4-040/2000 (H9N2), неполная кодирующая последовательность.
AY180564, ген нуклеопротеина (NP) вируса гриппа А (A/дикая утка/Наньхань/2-0480/2000 (H9N2)), неполная кодирующая последовательность.
AY180575, ген нуклеопротеина (NP) вируса гриппа А (A/перепел/Наньхань/2-0460/2000 (H9N2)), неполная кодирующая последовательность.
AY496851, мРНК нуклеопротеина (NP) вируса гриппа А (A/курица/Муданьцзян/0823/2000 (H9N2)), полноразмерная кодирующая последовательность.
AY180581, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Наньхань/1-0016/2000 (H9N2)), неполная кодирующая последовательность.
AY180583, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/10-389/2000 (H9N2), неполная кодирующая последовательность.
AY180584, ген нуклеопротеина (NP) вируса гриппа А штамма A/утка/Наньхань/1-0070/2000 (H9N2), неполная кодирующая последовательность.
AY768567, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0028/00 (H9N2)), неполная кодирующая последовательность.
AY768568, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0037/00 (H9N2)), неполная кодирующая последовательность.
AY768569, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0057/00 (H9N2)), неполная кодирующая последовательность.
AY768570, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0073/00 (H9N2)), неполная кодирующая последовательность.
AY768571, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0091/00 (H9N2)), неполная кодирующая последовательность.
AY768572, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0140/00 (H9N2)), неполная кодирующая последовательность.
AY768573, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU0146/00 (H9N2)), неполная кодирующая последовательность.
AY768574, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/SNU1035C/00 (H9N2)), неполная кодирующая последовательность.
AY268949, мРНК нуклеопротеина вируса гриппа А (A/курица/Наньхань/4/2001 (H9N2)), полноразмерная кодирующая последовательность.
AY180578, ген нуклеопротеина (NP) вируса гриппа А штамма A/курица/Наньхань/4-301/2001 (H9N2), неполная кодирующая последовательность.
AY180551, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Наньхань/4-361/2001 (H9N2)), неполная кодирующая последовательность.
AF523418, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/2088/01 (H9N2)), неполная кодирующая последовательность.
AF523414, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Шанту/1605/01 (H9N2)), неполная кодирующая последовательность.
AF523412, ген нуклеокапсидного белка (NP) вируса гриппа А (A/дикая утка/Шанту/4808/01 (H9N2)), неполная кодирующая последовательность.
AY800236, ген нуклеопротеина (NP) (A/курица/Корея/S1/2003 (H9N2)), неполная кодирующая последовательность.
AY862646, ген нуклеокапсидного белка (NP) вируса гриппа А (A/шелковая курица/Корея/S3/03 (H9N2)), неполная кодирующая последовательность.
AY862647, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S4/03 (H9N2)), неполная кодирующая последовательность.
AY862648, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S5/03 (H9N2)), неполная кодирующая последовательность.
AY862649, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S12/03 (H9N2)), неполная кодирующая последовательность.
AY862650, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Корея/S13/03 (H9N2)), неполная кодирующая последовательность.
AY862651, ген нуклеокапсидного белка (NP) вируса гриппа А (A/голубь/Корея/S14/03 (H9N2)), неполная кодирующая последовательность.
AY862652, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S15/03 (H9N2)), неполная кодирующая последовательность.
AY862653, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S16/03 (H9N2)), неполная кодирующая последовательность.
AY862654, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S18/03 (H9N2)), неполная кодирующая последовательность.
AY664717, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/CSW153/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664718, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/AP45/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664719, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/BD90/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664720, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/CSW291/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664721, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/CSW304/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664722, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/FY23/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664723, ген нуклеопротеина (NP) вируса гриппа А (A/цесарка/Гонконг/NT101/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664724, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/NT142/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664725, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/SF1/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664726, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/SSP101/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664727, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/TP38/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664728, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/WF126/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664729, ген нуклеопротеина (NP) вируса гриппа А (A/голубь/Гонконг/WF53/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664730, ген нуклеопротеина (NP) вируса гриппа А (A/фазан/Гонконг/WF54/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664731, ген нуклеопротеина (NP) вируса гриппа А (A/цесарка/Гонконг/NT184/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664732, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/WF120/03 (H9N2)), полноразмерная кодирующая последовательность.
AY664733, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/NT366/03 (H9N2)), неполная кодирующая последовательность.
AY664734, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/SSP418/03 (H9N2)), неполная кодирующая последовательность.
AY664735, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/YU427/03 (H9N2)), неполная кодирующая последовательность.
AY788915, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Китай/HSS2004 (H9N2)), полноразмерная кодирующая последовательность.
AY586423, ген нуклеопротеина вируса гриппа А (A/кряква/Италия/33/01 (H7N3)), неполная кодирующая последовательность.
AY586424, ген нуклеопротеина вируса гриппа А (A/кряква/Италия/43/01 (H7N3)), неполная кодирующая последовательность.
AY586425, ген нуклеопротеина вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), неполная кодирующая последовательность.
AY586426, ген нуклеопротеина вируса гриппа А (A/индейка/Италия/214845/02 (H7N3)), неполная кодирующая последовательность.
AJ627486, ген нуклеопротеина NP вируса гриппа А (A/индейка/Италия/214845/2002 (H7N3)), геномная РНК.
AJ627495, ген нуклеопротеина NP вируса гриппа А (А/индейка/Италия/220158/2002 (H7N3)), геномная РНК.
AY303658, ген нуклеопротеина NP вируса гриппа А (A/курица/Чили/176822/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303659, ген нуклеопротеина NP вируса гриппа А (A/курица/Чили/4957/02 (H7N3)), полноразмерная кодирующая последовательность.
AY611527, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY646081, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Британская Колумбия/GSC_человек_B/04 (H7N3)), полноразмерная кодирующая последовательность.
AY648290, ген нуклеопротеина (NP) вируса гриппа А (A/GSC_курица_B/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY650273, ген нуклеопротеина (NP) вируса гриппа А (A/GSC_курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AF144303, ген нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гуандун/1/96 (H5N1)), полноразмерная кодирующая последовательность.
AF046084, ген нуклеопротеина вируса гриппа А (A/курица/Гонконг/220/97 (H5N1)), полноразмерная кодирующая последовательность.
AF057293, мРНК нуклеопротеина вируса гриппа А (A/курица/Гонконг/258/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098617, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/y388/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098618, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/728/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098619, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/786/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098620, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Гонконг/915/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098621, ген нуклеопротеина (NP) вируса гриппа А (A/утка/Гонконг/p46/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098622, ген нуклеопротеина (NP) вируса гриппа А (A/утка/Гонконг/y283/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098623, ген нуклеопротеина (NP) вируса гриппа А (A/гусь/Гонконг/w355/97 (H5N1)), полноразмерная кодирующая последовательность.
AF370122, ген нуклеопротеина вируса гриппа А (A/гусь/Гуандун/3/97 (H5N1)), полноразмерная кодирующая последовательность.
AF216712, ген нуклеопротеина вируса гриппа А (A/окружающая среда/Гонконг/437-4/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216720, ген нуклеопротеина вируса гриппа А (A/окружающая среда/Гонконг/437-6/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216728, ген нуклеопротеина вируса гриппа А (A/окружающая среда/Гонконг/437-8/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216736, ген нуклеопротеина вируса гриппа А (A/окружающая среда/Гонконг/437-10/99 (H5N1)), полноразмерная кодирующая последовательность.
AY585429, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуангкси/07/1999 (H5N1)), полноразмерная кодирующая последовательность.
AY585439, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Цайянь/11/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585440, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Цайянь/52/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585428, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуандун/40/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585423, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Фуджиянь/19/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585425, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуандун/07/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585426, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуандун/12/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY059492, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/ww26/2000 (H5N1)), неполная кодирующая последовательность.
AY059493, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/ww28/2000 (H5N1)), неполная кодирующая последовательность.
AY059494, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/ww381/2000 (H5N1)), неполная кодирующая последовательность.
AY059495, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/ww461/2000 (H5N1)), неполная кодирующая последовательность.
AY059496, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/ww491/2000 (H5N1)), неполная кодирующая последовательность.
AY059497, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/2986.1/2000 (H5N1)), неполная кодирующая последовательность.
AY059498, ген сегмента 5 нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/3014.8/2000 (H5N1)), неполная кодирующая последовательность.
AF398419, ген нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/385.3/2000 (H5N1)), неполная кодирующая последовательность.
AF398420, ген нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/385.5/2000 (H5N1)), неполная кодирующая последовательность.
AF468842, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Аньян/AVL-1/2001 (H5N1)), полноразмерная кодирующая последовательность.
AF509117, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/FY77/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509118, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509119, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/YU563/01 (H5N1)), полноразмерная кодирующая последовательность.
AY585438, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Шанхай/38/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY221548, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/NT873.3/01-MB(H5N1)), неполная кодирующая последовательность.
AY221549, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/NT873.3/01 (H5N1)), неполная кодирующая последовательность.
AY221550, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/FY150/01-MB (H5N1)), неполная кодирующая последовательность.
AY221551, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
AY221552, ген нуклеокапсидного белка (NP) вируса гриппа А (A/фазан/Гонконг/FY155/01-MB (HSN1)), неполная кодирующая последовательность.
AY221553, ген нуклеокапсидного белка (NP) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
AY221554, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/YU822.2/01-MB (H5N1)), неполная кодирующая последовательность.
AY221555, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/YU822.2/01 (H5N1)), неполная кодирующая последовательность.
AY221556, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/YU552/01 (H5N1)), неполная кодирующая последовательность.
AF509120, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509121, ген нуклеокапсидного белка (NP) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509122, ген нуклеокапсидного белка (NP) вируса гриппа А (A/шелковая курица/Гонконг/SF189/01 (H5N1)), неполная кодирующая последовательность.
AF509123, ген нуклеокапсидного белка (NP) вируса гриппа А (A/перепел/Гонконг/SF203/01 (H5N1)), неполная кодирующая последовательность.
AF509124, ген нуклеокапсидного белка (NP) вируса гриппа А (A/голубь/Гонконг/SF215/01 (H5N1)), неполная кодирующая последовательность.
AF509125, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/SF219/01 (H5N1)), неполная кодирующая последовательность.
AF509126, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/715.5/01 (H5N1)), неполная кодирующая последовательность.
AF509127, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/751.1/01 (H5N1)), неполная кодирующая последовательность.
AF509128, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/822.1/01 (H5N1)), неполная кодирующая последовательность.
AF509129, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/829.2/01 (H5N1)), неполная кодирующая последовательность.
AF509130, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/830.2/01 (H5N1)), неполная кодирующая последовательность.
AF509131, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/858.3/01 (H5N1)), неполная кодирующая последовательность.
AF509132, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/866.3/01 (H5N1)), неполная кодирующая последовательность.
AF509133, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/867.1/01 (H5N1)), неполная кодирующая последовательность.
AF509134, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/879.1/01 (H5N1)), неполная кодирующая последовательность.
AF509135, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/873.3/01 (H5N1)), неполная кодирующая последовательность.
AF509136, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/876.1/01 (H5N1)), неполная кодирующая последовательность.
AF509137, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/891.1/01 (H5N1)), неполная кодирующая последовательность.
AF509138, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Гонконг/893.2/01 (H5N1)), неполная кодирующая последовательность.
AF509139, ген нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/76.1/01 (H5N1)), полноразмерная кодирующая последовательность.
AF509140, ген нуклеокапсидного белка (NP) вируса гриппа А (A/гусь/Гонконг/ww100/01 (H5N1)), неполная кодирующая последовательность.
AF509141, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/573.4/01 (H5N1)), неполная кодирующая последовательность.
AF509142, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Гонконг/646.3/01 (H5N1)), неполная кодирующая последовательность.
AY585424, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуандун/O1/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585422, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Фуджиянь/17/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585430, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуангкси/22/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585431, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуангкси/35/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585432, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуангкси/50/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585434, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Шанхай/08/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585435, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Шанхай/13/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585436, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Шанхай/35/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585437, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Шанхай/37/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585433, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуангкси/53/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585427, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Гуандун/22/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585420, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Фуджиянь/01/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585421, мРНК нуклеопротеина (NP) вируса гриппа А (A/утка/Фуджиянь/13/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY575907, ген нуклеокапсидного белка (NP) вируса гриппа А (А/Gs/HK/739.2/02 (H5N1)), неполная кодирующая последовательность.
AY575908, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Eg/HK/757.3/02 (H5N1)), неполная кодирующая последовательность.
AY575909, ген нуклеокапсидного белка (NP) вируса гриппа А (A/G.H/HK/793.1/02 (H5N1)), неполная кодирующая последовательность.
AY575910, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/HK/821/02 (H5N1)), неполная кодирующая последовательность.
AY575911, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/31.4/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575912, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/61.9/02 (H5N1)), полноразмерная кодирующая последовательность.
AY575913, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/YU777/02 (H5N1)), неполная кодирующая последовательность.
AY575914, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/96.1/02 (H5N1)), неполная кодирующая последовательность.
AY575915, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/409.1/02 (H5N1)), неполная кодирующая последовательность.
AY575916, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ph/HK/sv674.15/02 (H5N1)), неполная кодирующая последовательность.
DQ023146, мРНК нуклеопротеина (NP) вируса гриппа А A/курица/sd/1/02 (H5N1)), полноразмерная кодирующая последовательность.
AY676037, ген нуклеопротеина (NP) вируса гриппа А (A/утка/Гонконг/821/02 (H5N1)), полноразмерная кодирующая последовательность.
AY651510, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Gf/HK/38/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651511, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/31.2/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651512, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/37.4/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651513, ген нуклеокапсидного белка (NP) вируса гриппа А (A/SCk/HK/YU100/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY651514, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/YU22/2002 (H5N1)), неполная кодирующая последовательность.
AY651521, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/3176.3/2002 (H5N1)), неполная кодирующая последовательность.
AY651522, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/3169.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651524, ген нуклеокапсидного белка (NP) вируса гриппа А (A/одичавший голубь/HK/862.7/2002 (H5N1)), неполная кодирующая последовательность.
AY651525, ген нуклеокапсидного белка (NP) вируса гриппа А (A/древесный воробей/HK/864/2002 (H5N1)), неполная кодирующая последовательность.
AY651526, ген нуклеокапсидного белка (NP) вируса гриппа А (A/серая цапля/HK/861.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651527, ген нуклеокапсидного белка (NP) вируса гриппа А (A/чирок/Китай/2978.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651523, ген нуклеокапсидного белка (NP) вируса гриппа А (A/черноголовая чайка/HK/12.1/2003 (H5N1)), неполная кодирующая последовательность.
AY651487, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Индонезия/PA/2003 (H5N1)), неполная кодирующая последовательность.
AY651515, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/2133.1/2003 (H5N1)), неполная кодирующая последовательность.
AY651516, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/NT93/2003 (H5N1)), неполная кодирующая последовательность.
AY651517, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/SSP141/2003 (H5N1)), неполная кодирующая последовательность.
AY651518, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/WF157/2003 (H5N1)), неполная кодирующая последовательность.
AY651519, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/FY157/2003 (H5N1)), неполная кодирующая последовательность.
AY651520, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/HK/YU324/2003 (H5N1)), неполная кодирующая последовательность.
AY651490, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Индонезия/2A/2003 (H5N1)), неполная кодирующая последовательность.
AY676038, ген нуклеопротеина (NP) вируса гриппа А (A/белая цапля/Гонконг/757.2/03 (H5N1)), полноразмерная кодирующая последовательность.
AY676039, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Корея/ES/03 (H5N1)), полноразмерная кодирующая последовательность.
AY676040, ген нуклеопротеина (NP) вируса гриппа А (A/утка/Корея/ESD1/03 (H5N1)), полноразмерная кодирующая последовательность.
AY651529, ген нуклеопротеина (NP) вируса гриппа А (А/Dk/HN/5806/2003 (H5N1)), неполная кодирующая последовательность.
AY651532, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/ST/4231/2003 (H5N1)), неполная кодирующая последовательность.
AY651534, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/ST/4003/2003 (H5N1)), неполная кодирующая последовательность.
AY651535, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/YN/6255/2003 (H5N1)), неполная кодирующая последовательность.
AY651536, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/YN/6445/2003 (H5N1)), неполная кодирующая последовательность.
AY651485, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Индонезия/BL/2003 (H5N1)), неполная кодирующая последовательность.
AY518364, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Китай/E319-2/03 (H5N1)), полноразмерная кодирующая последовательность.
AY574189, ген нуклеопротеина вируса гриппа А (A/курица/Вьетнам/HD1/2004 (H5N1)), неполная кодирующая последовательность.
AY574192, ген нуклеопротеина вируса гриппа А (A/курица/Вьетнам/HD2/2004 (H5N1)), неполная кодирующая последовательность.
AJ867076, ген нуклеопротеина NP вируса гриппа А (A/Хатэй/2004/(H5N1)), геномная РНК.
AY651486, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/Индонезия/MS/2004 (H5N1)), неполная кодирующая последовательность.
AY590579, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU-K2/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY609313, сегмент 5 вируса гриппа А (A/курица/Гуандун/174/04 (H5N1)), полноразмерная последовательность.
AY576929, ген сегмента 5 нуклеопротеина вируса гриппа А (A/курица/Вьетнам/CM/2004 (H5N1)), неполная кодирующая последовательность.
AY576931, ген сегмента 5 нуклеопротеина вируса гриппа А (A/мускусная утка/Вьетнам/MdGL/2004 (H5N1)), неполная кодирующая последовательность.
AB166863, ген нуклеопротеина NP вируса гриппа А (A/курица/Ямагучи/7/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB188817, ген нуклеопротеина NP вируса гриппа А (A/курица/Оита/8/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651537, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/YN/374/2004 (H5N1)), неполная кодирующая последовательность.
AY651538, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/YN/115/2004 (H5N1)), неполная кодирующая последовательность.
AY653196, сегмент 5 вируса гриппа А (A/курица/Жилинь/9/2004 (H5N1)), полноразмерная последовательность.
AY651533, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ph/ST/44/2004 (H5N1)), неполная кодирующая последовательность.
AY651530, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/HN/303/2004 (H5N1)), неполная кодирующая последовательность.
AY651531, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/HN/101/2004 (H5N1)), неполная кодирующая последовательность.
AY684707, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Хубей/327/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY737290, сегмент 5 вируса гриппа А (A/курица/Гуандун/191/04 (H5N1)), полноразмерная последовательность.
AY737297, сегмент 5 вируса гриппа А (A/курица/Гуандун/178/04 (H5N1)), полноразмерная последовательность.
AY737305, сегмент 5 вируса гриппа А (A/утка/Гуандун/173/04 (H5N1)), полноразмерная последовательность.
AY770081, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Хубей/489/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY770996, ген нуклеопротеина вируса гриппа А (A/курица/Аюттхая/Таиланд/CU-23/04 (H5N1)), неполная кодирующая последовательность.
AY818139, ген нуклеопротеина NP вируса гриппа А (A/курица/Вьетнам/C58/04 (H5N1)), полноразмерная кодирующая последовательность.
AY818140, ген нуклеопротеина NP вируса гриппа А (A/перепел/Вьетнам/36/04 (H5N1)), полноразмерная кодирующая последовательность.
AY856864, сегмент 5 вируса гриппа А (A/утка/Шанду/093/2004 (H5N1)), полноразмерная последовательность.
AB189046, ген нуклеопротеина NP вируса гриппа А (A/курица/Киото/3/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189054, ген нуклеопротеина NP вируса гриппа А (A/ворон/Киото/53/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189062, ген нуклеопротеина NP вируса гриппа А (A/ворон/Осака/102/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY651491, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Таиланд/1/2004 (H5N1)), неполная кодирующая последовательность.
AY651492, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Таиланд/73/2004 (H5N1)), неполная кодирующая последовательность.
AY651493, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Таиланд/9.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651494, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Qa/Таиланд/57/2004 (H5N1)), неполная кодирующая последовательность.
AY651495, ген нуклеокапсидного белка (NP) вируса гриппа А (A/птичий/Таиланд/3.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651496, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/Таиланд/71.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651497, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Gs/Таиланд/79/2004 (H5N1)), неполная кодирующая последовательность.
AY651502, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/33/2004 (H5N1)), неполная кодирующая последовательность.
AY651503, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/35/2004 (H5N1)), неполная кодирующая последовательность.
AY651504, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/36/2004 (H5N1)), неполная кодирующая последовательность.
AY651505, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/37/2004 (H5N1)), неполная кодирующая последовательность.
AY651506, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/38/2004 (H5N1)), неполная кодирующая последовательность.
AY651507, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/39/2004 (H5N1)), неполная кодирующая последовательность.
AY651508, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Вьетнам/C57/2004 (H5N1)), неполная кодирующая последовательность.
AY651509, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Dk/Вьетнам/11/2004 (H5N1)), неполная кодирующая последовательность.
AY651488, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Индонезия/4/2004 (H5N1)), неполная кодирующая последовательность.
AY651489, ген нуклеокапсидного белка (NP) вируса гриппа А (A/Ck/Индонезия/5/2004 (H5N1)), неполная кодирующая последовательность.
AY651528, ген нуклеокапсидного белка (NP) вируса гриппа А (A/сапсан/HK/D0028/2004 (H5N1)), неполная кодирующая последовательность.
M22573, РНК нуклеопротеина (сегмент 5) вируса гриппа A/утка/Гонконг/7/75 (H3N2).
AY180555, ген нуклеопротеина (NP) вируса гриппа А (A/курица/Наньхань/3-120/2001 (H3N2)), неполная кодирующая последовательность.
AY779261, ген нуклеопротеина (NP) вируса гриппа А (A/индейка/Северная Каролина/12344/03 (H3N2)), неполная кодирующая последовательность.
AY779262, ген нуклеопротеина (NP) вируса гриппа А (A/индейка/Миннесота/764-2/03 (H3N2)), неполная кодирующая последовательность.
AY862655, ген нуклеокапсидного белка (NP) вируса гриппа А (A/курица/Корея/S6/03 (H3N2)), неполная кодирующая последовательность.
AY862656, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Корея/S7/03 (H3N2)), неполная кодирующая последовательность.
AY862657, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Корея/S8/03 (H3N2)), неполная кодирующая последовательность.
AY862658, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Корея/S9/03 (H3N2)), неполная кодирующая последовательность.
AY862659, ген нуклеокапсидного белка (NP) вируса гриппа А (A/утка/Корея/S10/03 (H3N2)), неполная кодирующая последовательность.
AY862660, ген нуклеокапсидного белка (NP) вируса гриппа А (A/голубь/Корея/S11/03 (H3N2)), неполная кодирующая последовательность.
D00050, ген нуклеопротеина вируса гриппа А, полноразмерная кодирующая последовательность.
M14921, РНК нуклеопротеина (сегмент 5) вируса гриппа А/кряква/NY/6750/78 (H2N2), полноразмерная кодирующая последовательность.
AY422026, ген нуклеопротеина вируса гриппа А, (A/утка/Хоккайдо/95/01 (H2N2)), неполная кодирующая последовательность.
U49093, мРНК нуклеопротеина (NP) вируса гриппа А, неполная кодирующая последовательность.
M22574, РНК нуклеопротеина (сегмент 5) вируса гриппа А/утка/Бавария/2/77 (H1N1).
M76603, мРНК вируса гриппа A/индейка/Англия/647/77 (H1N1), полноразмерная кодирующая последовательность.
M63783, мРНК нуклеопротеина вируса гриппа A/утка/Австралия/749/80 (H1N1), полноразмерная кодирующая последовательность.
M63778, мРНК нуклеопротеина вируса гриппа A/Индейка/Миннесота/1661/81 (H1N1), полноразмерная кодирующая последовательность.
Z26855, ген нуклеопротеина NP вируса гриппа типа А.
M76609, мРНК вируса гриппа A/индейка/Северная Каролина/1790/88 (H1N1), полноразмерная кодирующая последовательность.
Z26857, ген нуклеопротеина NP вируса гриппа типа А.
AF213905, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/кряква/Италия/24/95 (H1N1)), неполная кодирующая последовательность.
AF213906, ген сегмента 5 нуклеопротеина (NP) вируса гриппа А (A/курица/Италия/24/95 (H1N1)), неполная кодирующая последовательность.
AY633215, ген нуклеопротеина (NP) вируса гриппа А (A/кряква/Альберта/211/98 (H1N1)), полноразмерная кодирующая последовательность.
AY180543, ген нуклеопротеина (NP) вируса гриппа А (A/перепел/Наньхань/12-340/2000 (H1N1)), неполная кодирующая последовательность.
Последовательности, используемые в анализе основного полимеразного белка 1 (PB1) вируса гриппа А
AY633218, ген РНК-зависимой РНК-полимеразной субъединицы Р1 (PB1) вируса гриппа А (A/кряква/Альберта/211/98 (H1N1)), неполная кодирующая последовательность.
U48284, мРНК полимеразы (PB1) вируса гриппа А, неполная кодирующая последовательность.
AY180855, ген полимеразной субъединицы PB1 (PB1) штамма вируса гриппа A/перепел/Наньхань/12-340/2000 (H1N1), неполная кодирующая последовательность.
AY422038, ген полимеразной субъединицы PB1 (PB1) штамма вируса гриппа А (A/утка/Хоккайдо/95/01 (H2N2)), неполная кодирующая последовательность.
M25926, ген PB1 вируса гриппа А/кряква/Нью-Йорк/6750/78 (H2N2), полноразмерная кодирующая последовательность.
AY180871, ген полимеразной субъединицы PB1 (PB1) штамма вируса гриппа A/курица/Наньхань/3-120/2001 (H3N2), неполная кодирующая последовательность.
AY779265, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/индейка/Северная Каролина/12344/03 (H3N2)), неполная кодирующая последовательность.
AY779266, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/индейка/Миннесота/764-2/03 (H3N2)), неполная кодирующая последовательность.
AY862703, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S6/03 (H3N2)), неполная кодирующая последовательность.
AY862704, ген PB1 (PB1) вируса гриппа А (A/утка/Корея/S7/03 (H3N2)), неполная кодирующая последовательность.
AY862705, ген PB1 (PB1) вируса гриппа А (A/утка/Корея/S8/03 (H3N2)), неполная кодирующая последовательность.
AY862706, ген PB1 (PB1) вируса гриппа А (A/утка/Корея/S9/03 (H3N2)), неполная кодирующая последовательность.
AY862707, ген PB1 (PB1) вируса гриппа А (A/утка/Корея/S10/03 (H3N2)), неполная кодирующая последовательность.
AY862708, ген PB1 (PB1) вируса гриппа А (A/голубь/Корея/S11/03 (H3N2)), неполная кодирующая последовательность.
AF213911, ген сегмента 8 полимеразного белка PB1 вируса гриппа А (A/курица/Италия/5945/95 (H3N2)), неполная кодирующая последовательность.
AB166860, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Ямагучи/7/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB188814, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Оита/8/2004 (H5N1)), полноразмерная кодирующая последовательность.
AF398423, ген полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/385.3/2000 (H5N1)), неполная кодирующая последовательность.
AF398424, ген полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/385.5/2000 (H5N1)), неполная кодирующая последовательность.
AF468839, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Аньян/AVL-1/2001 (H5N1)), полноразмерная кодирующая последовательность.
AF509169, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/FY77/01 (H5N1)), неполная кодирующая последовательность.
AF509170, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
AF509171, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/YU563/01 (H5N1)), неполная кодирующая последовательность.
AF509172, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
AF509173, ген полимеразы (PB1) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
AF509174, ген полимеразы (PB1) вируса гриппа А (A/шелковая курица/Гонконг/SF189/01 (H5N1)), неполная кодирующая последовательность.
AF509175, ген полимеразы (PB1) вируса гриппа А (A/перепел/Гонконг/SF203/01 (H5N1)), неполная кодирующая последовательность.
AF509176, ген полимеразы (PB1) вируса гриппа А (A/голубь/Гонконг/SF215/01 (H5N1)), неполная кодирующая последовательность.
AF509177, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/SF219/01 (H5N1)), неполная кодирующая последовательность.
AF509178, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/715.5/01 (H5N1)), неполная кодирующая последовательность.
AF509179, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/751.1/01 (H5N1)), неполная кодирующая последовательность.
AF509180, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/822.1/01 (H5N1)), неполная кодирующая последовательность.
AF509181, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/829.2/01 (H5N1)), неполная кодирующая последовательность.
AF509182, ген полимеразы (PB1) вируса гриппа А (А/курица/Гонконг/830.2/01 (H5N1)), неполная кодирующая последовательность.
AF509183, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/858.3/01 (H5N1)), неполная кодирующая последовательность.
AF509184, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/866.3/01 (H5N1)), неполная кодирующая последовательность.
AF509185, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/867.1/01 (H5N1)), неполная кодирующая последовательность.
AF509186, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/879.1/01 (H5N1)), неполная кодирующая последовательность.
AF509187, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/873.3/01 (H5N1)), неполная кодирующая последовательность.
AF509188, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/876.1/01 (H5N1)), неполная кодирующая последовательность.
AF509189, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/891.1/01 (H5N1)), неполная кодирующая последовательность.
AF509190, ген полимеразы (PB1) вируса гриппа А (A/курица/Гонконг/893.2/01 (H5N1)), неполная кодирующая последовательность.
AF509191, ген полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/76.1/01 (H5N1)), неполная кодирующая последовательность.
AF509192, ген полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/ww100/01 (H5N1)), неполная кодирующая последовательность.
AF509193, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/573.4/01 (H5N1)), неполная кодирующая последовательность.
AF509194, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/646.3/01 (H5N1)), неполная кодирующая последовательность.
AY035888, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/гусь/Гуандун/3/97 (H5N1)), полноразмерная кодирующая последовательность.
AY059513, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/ww26/2000 (H5N1)), неполная кодирующая последовательность.
AY059514, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/ww28/2000 (H5N1)), неполная кодирующая последовательность.
AY059515, ген сегмента 2 полимеразы (PB1) вируса гриппа А (А/утка/Гонконг/ww381/2000 (H5N1)), неполная кодирующая последовательность.
AY059516, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/ww461/2000 (H5N1)), неполная кодирующая последовательность.
AY059517, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/ww491/2000 (H5N1)), неполная кодирующая последовательность.
AY059518, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/2986.1/2000 (H5N1)), неполная кодирующая последовательность.
AY059519, ген сегмента 2 полимеразы (PB1) вируса гриппа А (A/гусь/Гонконг/3014.8/2000 (H5N1)), неполная кодирующая последовательность.
AY221575, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/NT873.3/01-MB (H5N1)), неполная кодирующая последовательность.
AY221576, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/NT873.3/01 (H5N1)), неполная кодирующая последовательность.
AY221577, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/FY150/01-MB (H5N1)), неполная кодирующая последовательность.
AY221578, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
AY221579, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/фазан/Гонконг/FY155/01-MB (H5N1)), неполная кодирующая последовательность.
AY221580, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
AY221581, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/YU822.2/01-MB (H5N1)), неполная кодирующая последовательность.
AY221582, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/YU822.2/01 (H5N1)), неполная кодирующая последовательность.
AY221583, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
AY518366, ген полимеразной субъединицы РВ1 (PB1) вируса гриппа А (A/утка/Китай/E319-2/03 (H5N1)), полноразмерная кодирующая последовательность.
AY576394, ген полимеразы (PB1) вируса гриппа А (A/Gs/HK/739.2/02 (H5N1)), неполная кодирующая последовательность.
AY576395, ген полимеразы (PB1) вируса гриппа А (A/Eg/HK/757.3/02 (H5N1)), неполная кодирующая последовательность.
AY576396, ген полимеразы (PB1) вируса гриппа А (A/G.H/HK/793.1/02 (H5N1)), неполная кодирующая последовательность.
AY576397, ген полимеразы (PB1) вируса гриппа А (A/Dk/HK/821/02 (H5N1)), полноразмерная кодирующая последовательность.
AY576398, ген полимеразы (PB1) вируса гриппа А (A/Ck/HK/31.4/02 (H5N1)), неполная кодирующая последовательность.
AY576399, ген полимеразы (PB1) вируса гриппа А (A/Ck/HK/61.9/02 (H5N1)), неполная кодирующая последовательность.
AY576400, ген полимеразы (PB1) вируса гриппа А (A/Ck/HK/YU777/02 (H5N1)), неполная кодирующая последовательность.
AY576401, ген полимеразы (PB1) вируса гриппа А (A/Ck/HK/96.1/02 (H5N1)), неполная кодирующая последовательность.
AY576402, ген полимеразы (PB1) вируса гриппа А (A/Ck/HK/409.1/02 (H5N1)), неполная кодирующая последовательность.
AY576403, ген полимеразы (PB1) вируса гриппа А (A/Ph/HK/674.15/02 (H5N1)), неполная кодирующая последовательность.
AY585483, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Фуйян/01/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585484, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Фуйян/13/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585485, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Фуйян/17/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585486, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Фуйян/19/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585487, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуандун/01/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585488, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуандун/07/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585489, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуандун/12/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585490, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуандун/22/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585491, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуандун/40/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585492, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуангкси/07/1999 (H5N1)), полноразмерная кодирующая последовательность.
AY585493, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуангкси/22/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585494, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуангкси/35/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585495, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуангкси/50/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585496, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Гуангкси/53/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585497, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Шанхай/08/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585498, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Шанхай/13/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585499, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Шанхай/35/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585500, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Шанхай/37/2002 (H5N1)), полноразмерная кодирующая последовательность.
AY585501, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Шанхай/38/2001 (H5N1)), полноразмерная кодирующая последовательность.
AY585502, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Чжеянь/11/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY585503, мРНК полимеразного основного белка 1 (PB1) вируса гриппа А (A/утка/Чжеянь/52/2000 (H5N1)), полноразмерная кодирующая последовательность.
AY590582, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU-K2/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY609310, сегмент 2 вируса гриппа А (A/курица/Гуандун/174/04 (H5N1)), полноразмерная последовательность.
AY651651, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Индонезия/BL/2003 (H5N1)), неполная кодирующая последовательность.
AY651652, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/Индонезия/MS/2004 (H5N1)), неполная кодирующая последовательность.
AY651653, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Индонезия/PA/2003 (H5N1)), неполная кодирующая последовательность.
AY651654, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Индонезия/4/2004 (H5N1)), неполная кодирующая последовательность.
AY651655, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/курица/Индонезия/2A/2003 (H5N1)), неполная кодирующая последовательность.
AY651656, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Индонезия/5/2004 (H5N1)), неполная кодирующая последовательность.
AY651657, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Таиланд/1/2004 (H5N1)), неполная кодирующая последовательность.
AY651658, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Таиланд/73/2004 (H5N1)), неполная кодирующая последовательность.
AY651659, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Таиланд/9.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651660, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Qa/Таиланд/57/2004 (H5N1)), неполная кодирующая последовательность.
AY651661, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/птичий/Таиланд/3.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651662, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/Таиланд/71.1/2004 (H5N1)), неполная кодирующая последовательность.
AY651663, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Gs/Таиланд/79/2004 (H5N1)), неполная кодирующая последовательность.
AY651668, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/33/2004 (H5N1)), неполная кодирующая последовательность.
AY651669, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/35/2004 (H5N1)), неполная кодирующая последовательность.
AY651670, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/36/2004 (H5N1)), неполная кодирующая последовательность.
AY651671, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/37/2004 (H5N1)), неполная кодирующая последовательность.
AY651672, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/38/2004 (H5N1)), неполная кодирующая последовательность.
AY651673, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/39/2004 (H5N1)), неполная кодирующая последовательность.
AY651674, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/Вьетнам/C57/2004 (H5N1)), неполная кодирующая последовательность.
AY651675, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/Вьетнам/11/2004 (H5N1)), неполная кодирующая последовательность.
AY651676, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Gf/HK/38/2002 (H5N1)), неполная кодирующая последовательность.
AY651677, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/31.2/2002 (H5N1)), неполная кодирующая последовательность.
AY651678, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/37.4/2002 (H5N1)), неполная кодирующая последовательность.
AY651679, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/SCk/HK/YU100/2002 (H5N1)), неполная кодирующая последовательность.
AY651680, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/YU22/2002 (H5N1)), неполная кодирующая последовательность.
AY651681, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/3176.3/2002 (H5N1)), неполная кодирующая последовательность.
AY651682, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/3169.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651683, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/FY157/2003 (H5N1)), неполная кодирующая последовательность.
AY651684, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/YU324/2003 (H5N1)), неполная кодирующая последовательность.
AY651685, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/2133.1/2003 (H5N1)), неполная кодирующая последовательность.
AY651686, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/NT93/2003 (H5N1)), неполная кодирующая последовательность.
AY651687, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/SSP141/2003 (H5N1)), неполная кодирующая последовательность.
AY651688, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/HK/WF157/2003 (H5N1)), неполная кодирующая последовательность.
AY651689, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/черноголовая чайка/HK/12.1/2003 (H5N1)), неполная кодирующая последовательность.
AY651690, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/одичавший голубь/HK/862.7/2002 (H5N1)), неполная кодирующая последовательность.
AY651691, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/серая цапля/HK/861.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651692, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/древесный воробей/HK/864/2002 (H5N1)), неполная кодирующая последовательность.
AY651693, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/чирок/Китай/2978.1/2002 (H5N1)), неполная кодирующая последовательность.
AY651694, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/сапсан/HK/D0028/2004 (H5N1)), неполная кодирующая последовательность.
AY651695, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/HN/5806/2003 (H5N1)), неполная кодирующая последовательность.
AY651696, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/ST/4003/2003 (H5N1)), неполная кодирующая последовательность.
AY651697, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/ST/4231/2003 (H5N1)), неполная кодирующая последовательность.
AY651698, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/YN/6255/2003 (H5N1)), неполная кодирующая последовательность.
AY651699, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/YN/6445/2003 (H5N1)), неполная кодирующая последовательность.
AY651700, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ph/ST/44/2004(H5N1)), неполная кодирующая последовательность.
AY651701, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/HN/303/2004 (H5N1)), неполная кодирующая последовательность.
AY651702, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Dk/HN/101/2004 (H5N1)), неполная кодирующая последовательность.
AY651703, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/YN/374/2004 (H5N1)), неполная кодирующая последовательность.
AY651704, ген полимеразной основной субъединицы 1 (PB1) вируса гриппа А (A/Ck/YN/115/2004 (H5N1)), неполная кодирующая последовательность.
AY653199, сегмент 2 вируса гриппа А (A/курица/Жилинь/9/2004 (H5N1)), полноразмерная последовательность.
AY676025, ген полимеразного основного белка 1 (PB1) вируса гриппа штамма А (A/утка/Гонконг/821/02 (H5N1)), полноразмерная кодирующая последовательность.
AY676026, ген полимеразного основного белка 1 (PB1) вируса гриппа штамма А (A/белая цапля/Гонконг/757.2/03 (H5N1)), полноразмерная кодирующая последовательность.
AY676027, ген полимеразного основного белка 1 (PB1) вируса гриппа штамма А (A/курица/Корея/ES/03 (H5N1)), полноразмерная кодирующая последовательность.
AY676028, ген полимеразного основного белка 1 (PB1) вируса гриппа штамма А (A/утка/Корея/ESD1/03 (H5N1)), полноразмерная кодирующая последовательность.
AY684704, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Хубей/327/2004 (H5N1)), полноразмерная кодирующая последовательность.
AY737287, сегмент 2 вируса гриппа А (A/курица/Гуандун/191/04 (H5N1)), полноразмерная последовательность.
AY737294, сегмент 2 вируса гриппа А (A/курица/Гуандун/178/04 (H5N1)), полноразмерная последовательность.
AY737302, сегмент 2 вируса гриппа А (A/утка/Гуандун/173/04 (H5N1)), полноразмерная последовательность.
AY770083, ген нефункционального полимеразного основного белка 1 вируса гриппа А (A/курица/Хубей/489/2004 (H5N1)), полноразмерная последовательность.
AY770994, ген полимеразного основного белка 1 вируса гриппа А (A/курица/Аюттхая/Таиланд/CU-23/04 (H5N1)), неполная кодирующая последовательность.
AY818130, ген полимеразного белка РВ1 вируса гриппа А (A/курица/Вьетнам/C58/04 (H5N1)), полноразмерная кодирующая последовательность.
AY818131, ген полимеразного белка PB1 вируса гриппа А (A/перепел/Вьетнам/36/04 (H5N1)), полноразмерная кодирующая последовательность.
AY856862, сегмент 2 вируса гриппа А (A/утка/Шанду/093/2004 (H5N1)), полноразмерная последовательность.
AB188822, ген полимеразного основного белка 1 вируса гриппа А (A/курица/Киото/3/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189051, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/ворон/Киото/53/2004 (H5N1)), полноразмерная кодирующая последовательность.
AB189060, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/ворон/Осака/102/2004 (H5N1)), полноразмерная кодирующая последовательность.
AF046085, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/220/97 (H5N1)), полноразмерная кодирующая последовательность.
AF098590, ген белка PB1 (PB1) вируса гриппа А (A/курица/Гонконг/258/97 (H5N1)), неполная кодирующая последовательность.
AF098591, ген белка PB1 (PB1) вируса гриппа А (A/курица/Гонконг/y388/97 (H5N1)), неполная кодирующая последовательность.
AF098592, ген белка PB1 (PB1) вируса гриппа А (A/курица/Гонконг/728/97 (H5N1)), неполная кодирующая последовательность.
AF098593, ген белка PB1 (PB1) вируса гриппа А (A/курица/Гонконг/786/97 (H5N1)), неполная кодирующая последовательность.
AF098594, ген белка PB1 (PB1) вируса гриппа А (A/курица/Гонконг/915/97 (H5N1)), неполная кодирующая последовательность.
AF098595, ген белка PB1 (PB1) вируса гриппа А (A/утка/Гонконг/p46/97 (H5N1)), неполная кодирующая последовательность.
AF098596, ген белка PB1 (PB1) вируса гриппа А (A/утка/Гонконг/y283/97 (H5N1)), неполная кодирующая последовательность.
AF098598, ген белка PB1 (PB1) вируса гриппа А (A/гусь/Гонконг/w355/97 (H5N1)), неполная кодирующая последовательность.
AF144301, ген полимеразы (PB1) вируса гриппа А (A/гусь/Гуандун/1/96 (H5N1)), полноразмерная кодирующая последовательность.
AF216716, ген полимеразного основного белка 1 вируса гриппа А (A/окружающая среда/Гонконг/437-4/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216724, ген полимеразного основного белка 1 вируса гриппа А (A/окружающая среда/Гонконг/437-6/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216732, ген полимеразного основного белка 1 вируса гриппа А (A/окружающая среда/Гонконг/437-8/99 (H5N1)), полноразмерная кодирующая последовательность.
AF216740, ген полимеразного основного белка 1 вируса гриппа А (A/окружающая среда/Гонконг/437-10/99 (H5N1)), полноразмерная кодирующая последовательность.
AY303663, ген полимеразного основного белка 1 вируса гриппа А (A/курица/Чили/176822/02 (H7N3)), полноразмерная кодирующая последовательность.
AY303664, ген полимеразного основного белка 1 вируса гриппа А (A/курица/Чили/4957/02 (H7N3)), неполная кодирующая последовательность.
AY586435, ген PB1 вируса гриппа А (A/индейка/Италия/214845/02 (H7N3)), неполная кодирующая последовательность.
AY586436, ген PB1 вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), неполная кодирующая последовательность.
AY586437, ген PB1 вируса гриппа А (A/кряква/Италия/33/01 (H7N3)), неполная кодирующая последовательность.
AY586438, ген PB1 вируса гриппа А (A/кряква/Италия/43/01 (H7N3)), неполная кодирующая последовательность.
AY616765, ген полимеразной субъединицы РВ1 (PB1) вируса гриппа А (A/курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY646084, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Британская Колумбия/GSC_человек_B/04 (H7N3)), полноразмерная кодирующая последовательность.
AY648293, ген полимеразной субъединицы РВ1 (PB1) вируса гриппа А (A/GSC_курица_B/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AY653039, ген полимеразной субъединицы РВ1 (PB1) вируса гриппа А (A/GSC_курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
AJ620348, ген РНК-полимеразы PB1 вируса гриппа А (A/курица/Германия/R28/03 (H7N7)), геномная РНК.
AY340080, ген полимеразы (PB1) вируса гриппа А (A/Нидерланды/124/03 (H7N7)), неполная кодирующая последовательность.
AY340081, ген полимеразы (PB1) вируса гриппа А (A/Нидерланды/126/03 (H7N7)), неполная кодирующая последовательность.
AY340082, ген полимеразы (PB1) вируса гриппа А (A/Нидерланды/127/03 (H7N7)), неполная кодирующая последовательность.
AY340083, ген полимеразы (PB1) вируса гриппа А (A/Нидерланды/219/03 (H7N7)), полноразмерная кодирующая последовательность.
AY340084, ген полимеразы (PB1) вируса гриппа А (A/Нидерланды/033/03 (H7N7)), полноразмерная кодирующая последовательность.
AY340085, ген полимеразы (PB1) вируса гриппа А (A/курица/Нидерланды/1/03 (H7N7)), полноразмерная кодирующая последовательность.
AB049155, ген PB1 полимеразного основного белка 1 вируса гриппа А (A/попугай/Чиба/1/97 (H9N2)), полноразмерная кодирующая последовательность.
AB049156, ген PB1 полимеразного основного белка 1 вируса гриппа А (A/попугай/Нарита/92A/98 (H9N2)), полноразмерная кодирующая последовательность.
AF508618, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/страус/Южная Африка/9508103/95 (H9N2)), неполная кодирующая последовательность.
AF508619, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Пакистан/4/99 (H9N2)), неполная кодирующая последовательность.
AF508620, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Пакистан/5/99 (H9N2)), неполная кодирующая последовательность.
AF508621, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Германия/R45/98 (H9N2)), неполная кодирующая последовательность.
AF508622, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/утка/Германия/113/95 (H9N2)), неполная кодирующая последовательность.
AF508623, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Иран/11T/99 (H9N2)), неполная кодирующая последовательность.
AF508624, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Саудовская Аравия/532/99 (H9N2)), неполная кодирующая последовательность.
AF508625, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/фазан/Ирландия/PV18/97 (H9N2)), неполная кодирующая последовательность.
AF508626, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Корея/99029/99 (H9N2)), неполная кодирующая последовательность.
AF508627, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Бейжин/8/98 (H9N2)), полноразмерная кодирующая последовательность.
AF508628, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Гуандун/10/00 (H9N2)), неполная кодирующая последовательность.
AF508629, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Гуандун/11/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508630, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Хэбей/4/98 (H9N2)), полноразмерная кодирующая последовательность.
AF508631, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Хейлоньян/10/97 (H9N2)), неполная кодирующая последовательность.
AF508632, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Хэнань/62/00 (H9N2)), полноразмерная кодирующая последовательность.
AF508633, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Ningxia/5/99 (H9N2)), полноразмерная кодирующая последовательность.
AF508634, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Сихуань/5/97 (H9N2)), неполная кодирующая последовательность.
AF508635, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Шанду/6/96 (H9N2)), неполная кодирующая последовательность.
AF508636, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Шиячжуан/2/99 (H9N2)), полноразмерная кодирующая последовательность.
AF508637, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/курица/Шензен/9/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508638, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/утка/Наньян/1/97 (H9N2)), полноразмерная кодирующая последовательность.
AF508639, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/перепел/Шанхай/8/96 (H9N2)), полноразмерная кодирующая последовательность.
AF523427, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/830/00 (H9N2)), неполная кодирующая последовательность.
AF523428, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/2102/00 (H9N2)), неполная кодирующая последовательность.
AF523429, ген сегмента 2 полимеразы PB1 (PB1) вируса гриппа А (A/утка/Шанту/1043/00 (H9N2)), неполная кодирующая последовательность.
AF523430, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/2134/00 (H9N2)), неполная кодирующая последовательность.
AF523431, ген полимеразы (PB1) вируса гриппа А (A/дикая утка/Шанту/4808/01 (H9N2)), неполная кодирующая последовательность.
AF523432, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/2144/00 (H9N2)), неполная кодирующая последовательность.
AF523433, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/2143/00 (H9N2)), неполная кодирующая последовательность.
AF523434, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/1796/00 (H9N2)), неполная кодирующая последовательность.
AF523435, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/2088/01 (H9N2)), неполная кодирующая последовательность.
AF523436, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/1881/00 (H9N2)), неполная кодирующая последовательность.
AF523437, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/366/78 (H9N2)), неполная кодирующая последовательность.
AF523438, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/552/79 (H9N2)), неполная кодирующая последовательность.
AF523439, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/86/76 (H9N2)), неполная кодирующая последовательность.
AF523440, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/289/78 (H9N2)), неполная кодирующая последовательность.
AF523441, ген полимеразы (PB1) вируса гриппа А (A/утка/Гонконг/610/79 (H9N2)), неполная кодирующая последовательность.
AF523442, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/1605/01 (H9N2)), неполная кодирующая последовательность.
AF523443, ген полимеразы (PB1) вируса гриппа А (A/утка/Шанту/1042/00 (H9N2)), неполная кодирующая последовательность.
AF536659, ген PB1 вируса гриппа А (A/курица/Бейжин/1/95 (H9N2)), неполная кодирующая последовательность.
AF536660, ген PB1 вируса гриппа А (A/курица/Бейжин/2/97 (H9N2)), неполная кодирующая последовательность.
AF536661, ген PB1 вируса гриппа А (A/курица/Бейжин/3/99 (H9N2)), неполная кодирующая последовательность.
AF536662, ген PB1 вируса гриппа А (A/курица/Гуандун/97 (H9N2)), неполная кодирующая последовательность.
AF536663, ген PB1 вируса гриппа А (A/курица/Хэбей/1/96 (H9N2)), неполная кодирующая последовательность.
AF536664, ген PB1 вируса гриппа А (A/курица/Хэбей/2/98 (H9N2)), неполная кодирующая последовательность.
AF536665, ген PB1 вируса гриппа А (A/курица/Хэбей/3/98 (H9N2)), неполная кодирующая последовательность.
AF536666, ген PB1 вируса гриппа А (A/курица/Хэнань/98 (H9N2)), неполная кодирующая последовательность.
AF536667, ген PB1 вируса гриппа А (A/курица/Лиаонь/99 (H9N2)), неполная кодирующая последовательность.
AF536668, ген PB1 вируса гриппа А (A/курица/Шанду/98 (H9N2)), неполная кодирующая последовательность.
AJ291396, ген PB1 полимеразы PB1 вируса гриппа А (A/курица/Пакистан/2/99 (H9N2)), геномная РНК.
AJ427862, неполный ген pb1 полимеразного белка PB1 вируса гриппа А (A/перепел/Гонконг/FY298/00 (H9N2)), геномная РНК
AY180840, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/голубь/Наньхань/7-058/2000 (H9N2), неполная кодирующая последовательность.
AY180843, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/перепел/Наньхань/2-0460/2000 (H9N2), неполная кодирующая последовательность.
AY180844, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/голубь/Наньхань/2-0461/2000 (H9N2), неполная кодирующая последовательность.
AY180851, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/голубь/Наньхань/11-145/2000 (H9N2), неполная кодирующая последовательность.
AY180852, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/11-197/2000 (H9N2), неполная кодирующая последовательность.
AY180854, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/11-290/2000 (H9N2), неполная кодирующая последовательность.
AY180856, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/1-0070/2000 (H9N2), неполная кодирующая последовательность.
AY180866, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/7-092/2000 (H9N2), неполная кодирующая последовательность.
AY180867, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/курица/Наньхань/1-0016/2000 (H9N2), неполная кодирующая последовательность.
AY180873, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/курица/Наньхань/4-010/2000 (H9N2), неполная кодирующая последовательность.
AY180874, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/курица/Наньхань/4-301/2001 (H9N2), неполная кодирующая последовательность.
AY180875, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/курица/Наньхань/4-361/2001 (H9N2), неполная кодирующая последовательность.
AY180892, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/перепел/Наньхань/4-040/2000 (H9N2), неполная кодирующая последовательность.
AY180897, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/дикая утка/Наньхань/2-0480/2000 (H9N2), неполная кодирующая последовательность.
AY180900, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/10-389/2000 (H9N2), неполная кодирующая последовательность.
AY180901, ген полимеразной субъединицы PB1 (PB1) вируса гриппа штамма А/утка/Наньхань/11-392/2000 (H9N2), неполная кодирующая последовательность.
AY253751, ген полимеразного основного белка 1 (PB1) вируса гриппа штамма А (A/курица/Шанхай/F/98 (H9N2)), полноразмерная кодирующая последовательность.
AY307947, ген полимеразной субъединицы (PB1) вируса гриппа А (A/курица/Бейжин/1/00 (H9N2)), неполная кодирующая последовательность.
AY307948, ген полимеразной субъединицы (PB1) вируса гриппа А (A/курица/Хэбей/1/01 (H9N2)), неполная кодирующая последовательность.
AY633170, ген РНК-зависимой РНК-полимеразной субъединицы Р1 (PB1) вируса гриппа А (A/кряква/Альберта/17/91 (H9N2)), неполная кодирующая последовательность.
AY633282, ген РНК-зависимой РНК-полимеразной субъединицы Р1 (PB1) вируса гриппа А (A/кряква/Альберта/321/88 (H9N2)), неполная кодирующая последовательность.
AY633298, ген РНК-зависимой РНК-полимеразы Р1 (PB1) вируса гриппа А (A/кряква/Альберта/11/91 (H9N2)), неполная кодирующая последовательность.
AY664774, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/CSW153/03 (H9N2)), неполная кодирующая последовательность.
AY664775, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/AP45/03 (H9N2)), неполная кодирующая последовательность.
AY664776, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/BD90/03 (H9N2)), неполная кодирующая последовательность.
AY664777, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/CSW291/03 (H9N2)), неполная кодирующая последовательность.
AY664778, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/CSW304/03 (H9N2)), неполная кодирующая последовательность.
AY664779, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/FY23/03 (H9N2)), неполная кодирующая последовательность.
AY664780, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/цесарка/Гонконг/NT101/03 (H9N2)), неполная кодирующая последовательность.
AY664781, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/NT142/03 (H9N2)), неполная кодирующая последовательность.
AY664782, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/SF1/03 (H9N2)), неполная кодирующая последовательность.
AY664783, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/SSP101/03 (H9N2)), неполная кодирующая последовательность.
AY664784, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/TP38/03 (H9N2)), неполная кодирующая последовательность.
AY664785, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/WF126/03 (H9N2)), неполная кодирующая последовательность.
AY664786, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/голубь/Гонконг/WF53/03 (H9N2)), неполная кодирующая последовательность.
AY664787, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/фазан/Гонконг/WF54/03 (H9N2)), неполная кодирующая последовательность.
AY664788, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/цесарка/Гонконг/NT184/03 (H9N2), неполная кодирующая последовательность.
AY664789, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/WF120/03 (H9N2)), неполная кодирующая последовательность.
AY664790, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/NT366/03 (H9N2)), неполная кодирующая последовательность.
AY664791, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Гонконг/YU427/03 (H9N2)), неполная кодирующая последовательность.
AY800239, ген полимеразного основного белка 1 (PB1) вируса гриппа А (A/курица/Корея/S1/2003 (H9N2)), неполная кодирующая последовательность.
AY862694, ген PB1 (PB1) вируса гриппа А (A/шелковая курица/Корея/S3/03 (H9N2)), неполная кодирующая последовательность.
AY862695, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S4/03 (H9N2)), неполная кодирующая последовательность.
AY862696, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S5/03 (H9N2)), неполная кодирующая последовательность.
AY862697, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S12/03 (H9N2)), неполная кодирующая последовательность.
AY862698, ген PB1 (PB1) вируса гриппа А (A/утка/Корея/S13/03 (H9N2)), неполная кодирующая последовательность.
AY862699, ген PB1 (PB1) вируса гриппа А (A/голубь/Корея/S14/03 (H9N2)), неполная кодирующая последовательность.
AY862700, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S15/03 (H9N2)), неполная кодирующая последовательность.
AY862701, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S16/03 (H9N2)), неполная кодирующая последовательность.
AY862702, ген PB1 (PB1) вируса гриппа А (A/курица/Корея/S18/03 (H9N2)), неполная кодирующая последовательность.
AF156416, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Гонконг/G9/97 (H9N2)), неполная кодирующая последовательность.
AF156417, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Гонконг/G23/99 (H9N2)), неполная кодирующая последовательность.
AF156418, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/голубь/Гонконг/Y233/97 (H9N2)), неполная кодирующая последовательность.
AF156419, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/утка/Гонконг/Y280/97 (H9N2)), неполная кодирующая последовательность.
AF156420, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/утка/Гонконг/Y439/97 (H9N2)), неполная кодирующая последовательность.
AF156421, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/перепел/Гонконг/G1/97 (H9N2)), неполная кодирующая последовательность.
AF156422, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Гонконг/739/94 (H9N2)), неполная кодирующая последовательность.
AF156423, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Бейжин/1/94 (H9N2)), неполная кодирующая последовательность.
AF156424, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/перепел/Гонконг/AF157/92 (H9N2)), неполная кодирующая последовательность.
AF156425, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Корея/38349-p96323/96 (H9N2)), неполная кодирующая последовательность.
AF156426, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/курица/Корея/25232-96006/96 (H9N2)), неполная кодирующая последовательность.
AF156427, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/ржанка/Делавер/9/96 (H9N2)), неполная кодирующая последовательность.
AF156428, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/перепел/Арканзас/29209-1/93 (H9N2)), неполная кодирующая последовательность.
AF156429, ген сегмента 2 полимеразной субъединицы PB1 (PB1) вируса гриппа А (A/Индейка/Калифорния/189/66 (H9N2)), неполная кодирующая последовательность.
AF222632, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/перепел/Гонконг/A17/99 (H9N2)), неполная кодирующая последовательность.
AF222633, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/голубь/Гонконг/FY6/99 (H9N2)), неполная кодирующая последовательность.
AF222634, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/курица/Гонконг/NT16/99 (H9N2)), неполная кодирующая последовательность.
AF222635, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/перепел/Гонконг/SSP10/99 (H9N2)), неполная кодирующая последовательность.
AF222636, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/фазан/Гонконг/SSP11/99 (H9N2)), неполная кодирующая последовательность.
AF222637, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/курица/Гонконг/FY20/99 (H9N2)), неполная кодирующая последовательность.
AF222638, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/курица/Гонконг/KC12/99 (H9N2)), неполная кодирующая последовательность.
AF222639, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/перепел/Гонконг/NT2899 (H9N2)), неполная кодирующая последовательность.
AF222640, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/курица/Гонконг/SF2/99 (H9N2)), неполная кодирующая последовательность.
AF222641, ген сегмента 2 полимеразы 1 (PB1) вируса гриппа А (A/шелковая курица/Гонконг/SF44/99 (H9N2)), неполная кодирующая последовательность.
Последовательности, используемые в анализах основного полимеразного белка 2 (PB2) вируса гриппа А
gi|49356919|AY633219| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/кряква/Альберта/211/98 (H1N1)), неполная кодирующая последовательность.
gi|27466107|AY180748| ген полимеразной субъедницы PB2 (PB2) вируса гриппа штамма A/перепел/Наньхань/12-340/2000 (H1N1), неполная кодирующая последовательность.
gi|45272173|AY422042| ген полимеразной субъединицы (PB2) вируса гриппа А (A/утка/Хоккайдо/95/01 (H2N2)), неполная кодирующая последовательность.
gi|18091825|AF213910| ген сегмента 1 полимеразного белка PB2 вируса гриппа А (A/курица/Италия/5945/95 (H3N2)), неполная кодирующая последовательность.
gi|27466133|AY180761| ген полимеразной субъединицы PB2 (PB2) вируса гриппа штамма А/курица/Наньхань/3-120/2001 (H3N2), неполная кодирующая последовательность.
gi|56160002|AY779267| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/индейка/Северная Каролина/12344/03 (H3N2)), неполная кодирующая последовательность.
gi|56160004|AY779268| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/индейка/Миннесота/764-2/03 (H3N2)), неполная кодирующая последовательность.
gi|58429704|AY862719| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S6/03 (H3N2)), неполная кодирующая последовательность.
gi|58429706|AY862720| ген PB2 (PB2) вируса гриппа А (А/утка/Корея/S7/03 (H3N2)), неполная кодирующая последовательность.
gi|58429708|AY862721| ген PB2 (PB2) вируса гриппа А (А/утка/Корея/S8/03 (H3N2)), неполная кодирующая последовательность.
gi|58429710|AY862722| ген PB2 (PB2) вируса гриппа А (A/утка/Корея/S9/03 (H3N2)), неполная кодирующая последовательность.
gi|58429712|AY862723| ген PB2 (PB2) вируса гриппа А (A/утка/Корея/S10/03 (H3N2)), неполная кодирующая последовательность.
gi|58429714|AY862724| ген PB2 (PB2) вируса гриппа А (A/голубь/Корея/S11/03 (H3N2)), неполная кодирующая последовательность.
gi|5805276|AF144300| ген полимеразы (PB2) вируса гриппа А (A/гусь/Гуандун/1/96 (H5N1)), полноразмерная кодирующая последовательность.
gi|3335416|AF046086| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/220/97 (H5N1)), неполная кодирующая последовательность.
gi|60488411|AF098577| ген белка PB2 (PB2) вируса гриппа А (А/курица/Гонконг/258/97 (H5N1)), неполная кодирующая последовательность.
gi|6048843|AF098578| ген белка PB2 (PB2) вируса гриппа А (A/курица/Гонконг/y388/97 (H5N1)), неполная кодирующая последовательность.
gi|6048845|AF098579| ген белка PB2 (PB2) вируса гриппа А (А/курица/Гонконг/728/97 (H5N1)), неполная кодирующая последовательность.
gi|6048847|AF098580| ген белка PB2 (PB2) вируса гриппа А (A/курица/Гонконг/786/97 (H5N1)), неполная кодирующая последовательность.
gi|6048849|AF098581| ген белка PB2 (PB2) вируса гриппа А (A/курица/Гонконг/915/97 (H5N1)), неполная кодирующая последовательность.
gi|6048851|AF098582| ген белка PB2 (PB2) вируса гриппа А (A/утка/Гонконг/p46/97 (H5N1)), неполная кодирующая последовательность.
gi|6048853|AF098583| ген белка PB2 (PB2) вируса гриппа А (A/утка/Гонконг/y283/97 (H5N1)), неполная кодирующая последовательность.
gi|6048855|AF098584| ген белка PB2 (PB2) вируса гриппа А (A/гусь/Гонконг/w355/97 (H5N1)), неполная кодирующая последовательность.
gi|14860983|AY038798| ген белка PB2 (PB2) вируса гриппа А (A/гусь/Гуандун/3/1997 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156244|AY585513| мРНК полимеразного основного белка 2 (PB2) вируса гриппа А (A/утка/Гуангкси/07/1999 (H5N1)), полноразмерная кодирующая последовательность.
gi|9863884|AF216717| ген полимеразного основного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-4/99 (H5N1)), неполная кодирующая последовательность.
gi|9863903|AF216725| ген полимеразного основного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-6/99 (H5N1)), неполная кодирующая последовательность.
gi|9863921|AF216733| ген полимеразного основного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-8/99 (H5N1)), неполная кодирующая последовательность.
gi|9863939|AF216741| ген полимеразного основного белка 2 вируса гриппа А (A/окружающая среда/Гонконг/437-10/99 (H5N1)), неполная кодирующая последовательность.
gi|47156264|AY585523| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Чжеянь/11/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156266|AY585524| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Чжеянь/52/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156232|AY585507| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Фуджиянь/19/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156236|AY585509| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуандун/07/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156238|AY585510| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуандун/12/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156242|AY585512| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуандун/40/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|19697849|AY059520| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/ww26/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697851|AY059521| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/ww28/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697853|AY059522| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/ww381/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697855|AY059523| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/ww461/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697857|AY059524| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/ww491/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697859|AY059525| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/2986.1/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697867|AY059529| ген сегмента 1 полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/3014.8/2000 (H5N1)), неполная кодирующая последовательность.
gi|18092181|AF398425| ген полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/385.3/2000 (H5N1)), неполная кодирующая последовательность.
gi|18092183|AF398426| ген полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/385.5/2000 (H5N1)), неполная кодирующая последовательность.
gi|21359665|AF468840| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Аньян/AVL-1/2001 (H5N1)), неполная кодирующая последовательность.
gi|28849606|AF509143| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/FY77/01 (H5N1)), неполная кодирующая последовательность.
gi|28849608|AF509144| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
gi|28849610|AF509145| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/YU563/01 (H5N1)), неполная кодирующая последовательность.
gi|28849612|AF509146| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
gi|28849614|AF509147| ген полимеразы (РВ2) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
gi|28849616|AF509148| ген полимеразы (РВ2) вируса гриппа А (A/шелковая курица/Гонконг/SF189/01 (H5N1)), неполная кодирующая последовательность.
gi|28849618|AF509149| ген полимеразы (РВ2) вируса гриппа А (A/перепел/Гонконг/SF203/01 (H5N1)), неполная кодирующая последовательность.
gi|28849620|AF509150| ген полимеразы (РВ2) вируса гриппа А (A/голубь/Гонконг/SF215/01 (H5N1)), неполная кодирующая последовательность.
gi|28849622|AF509151| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/SF219/01 (H5N1)), неполная кодирующая последовательность.
gi|28849624|AF509152| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/715.5/01 (H5N1)), неполная кодирующая последовательность.
gi|28849626|AF509153| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/751.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849628|AF509154| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/822.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849630|AF509155| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/829.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28849632|AF509156| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/830.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28849634|AF509157| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/858.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849636|AF509158| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/866.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849638|AF509159| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/867.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849640|AF509160| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/879.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849642|AF509161| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/873.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849644|AF509162| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/876.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849646|AF509163| ген полимеразы (РВ2) вируса гриппа А (A/курица/Гонконг/891.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849648|AF509164| ген полимеразы (РВ2) вируса гриппа А (А/курица/Гонконг/893.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28849650|AF509165| ген полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/76.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849652|AF509166| ген полимеразы (РВ2) вируса гриппа А (A/гусь/Гонконг/ww100/01 (H5N1)), неполная кодирующая последовательность.
gi|28849654|AF509167| ген полимеразы (РВ2) вируса гриппа А (А/утка/Гонконг/573.4/01 (H5N1)), неполная кодирующая последовательность.
gi|28849656|AF509168| ген полимеразы (РВ2) вируса гриппа А (А/утка/Гонконг/646.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28823262|AY221584| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/NT873.3/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28823443|AY221585| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/NT873.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28823612|AY221586| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/FY150/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28823783|AY221587| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
gi|28823961|AY221588| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/фазан/Гонконг/FY155/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28824143|AY221589| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
gi|28824334|AY221590| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/YU822.2/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28824502|AY221591| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/YU822.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28824684|AY221592| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
gi|47156230|AY585506| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Фуйян/17/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156234|AY585508| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуандун/01/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156246|AY585514| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуангкси/22/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156248|AY585515| мРНК полимеразы 2 (РВ2) вируса гриппа А (A/утка/Гуангкси/35/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156250|AY585516| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуангкси/50/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156254|AY585518| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Шанхай/08/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156256|AY585519| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Шанхай/13/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156262|AY585522| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Шанхай/38/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156258|AY585520| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Шанхай/35/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156260|AY585521| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Шанхай/37/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156252|AY585517| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуангкси/53/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156240|AY585511| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Гуандун/22/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47834791|AY576382| ген полимеразы (PB2) вируса гриппа А (A/Gs/HK/739.2/02 (H5N1)), неполная кодирующая последовательность.
gi|47834793|AY576383| ген полимеразы (PB2) вируса гриппа А (A/Eg/HK/757.3/02 (H5N1)), неполная кодирующая последовательность.
gi|47834795|AY576384| ген полимеразы (PB2) вируса гриппа А (A/G.H/HK/793.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834797|AY576385| ген полимеразы (PB2) вируса гриппа А (A/утка/HK/821/02 (H5N1)), неполная кодирующая последовательность.
gi|47834799|AY576386| ген полимеразы (PB2) вируса гриппа А (A/курица/HK/31.4/02 (H5N1)), неполная кодирующая последовательность.
gi|47834801|AY576387| ген полимеразы (PB2) вируса гриппа А (A/Ck/HK/61.9/02 (H5N1)), неполная кодирующая последовательность.
gi|47834803|AY576388| ген полимеразы (PB2) вируса гриппа А (A/Ck/HK/YU777/02 (H5N1)), неполная кодирующая последовательность.
gi|47834805|AY576389| ген полимеразы (PB2) вируса гриппа А (A/Ck/HK/96.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834807|AY576390| ген полимеразы (PB2) вируса гриппа А (A/Ck/HK/409.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834809|AY576391| ген полимеразы (PB2) вируса гриппа А (A/Ph/HK/sv674.15/02 (H5N1)), неполная кодирующая последовательность.
gi|47156226|AY585504| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Фуйян/01/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156228|AY585505| мРНК полимеразного основного белка 2 (РВ2) вируса гриппа А (A/утка/Фуйян/13/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296597|AY651744| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/серая цапля/HK/861.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296599|AY651745| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/одичавший голубь/HK/862.7/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296601|AY651746| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/древесный воробей/HK/864/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296603|AY651747| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/чирок/Китай/2978.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|56548879|AY676021| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/утка/Гонконг/821/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296569|AY651730| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Gf/HK/38/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296571|AY651731| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/31.2/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296573|AY651732| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/37.4/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296575|AY651733| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/SCk/HK/YU100/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296577|AY651734| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/YU22/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296579|AY651735| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/3176.3/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296581|AY651736| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/3169.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|50296583|AY651737| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/FY157/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296585|AY651738| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/YU324/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296587|AY651739| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/2133.1/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296589|AY651740| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/NT93/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296591|AY651741| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/SSP141/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296593|AY651742| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/HK/WF157/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296595|AY651743| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/черноголовая чайка/HK/12.1/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296607|AY651749| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/HN/5806/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296609|AY651750| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/ST/4003/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296611|AY651751| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/ST/4231/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296613|AY651752| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/YN/6255/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296615|AY651753| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/YN/6445/2003 (H5N1)), неполная кодирующая последовательность.
gi|50296529|AY651710| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Индонезия/2A/2003 (H5N1)), неполная кодирующая последовательность.
gi|56548881|AY676022| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/белая цапля/Гонконг/757.2/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|56548883|AY676023| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/курица/Корея/ES/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|56548885|AY676024| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/утка/Корея/ESD1/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296519|AY651705| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Индонезия/PA/2003 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296523|AY651707| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (А/Ck/Индонезия/BL/2003 (H5N1)), полноразмерная кодирующая последовательность.
gi|41207501|AY518367| ген полимеразной субъединицы PB2 (РВ2) вируса гриппа А (A/утка/Китай/E319-2/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|45359369|AY550147| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU-K2/04 (H5N1)), неполная кодирующая последовательность.
gi|50296525|AY651708| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Индонезия/5/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296527|AY651709| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Индонезия/4/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296521|AY651706| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/Индонезия/MS/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|51094103|AY590581| ген полимеразной основного белка 2 (РВР2) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU-K2/2004 (H5N1)), неполная кодирующая последовательность.
gi|47716766|AY609309| сегмент 1 вируса гриппа А (A/курица/Гуандун/174/04 (H5N1)), полноразмерная последовательность.
gi|58531082|AB166859| ген полимеразного основного белка 2 РВ2 вируса гриппа А (A/курица/Ямагучи/7/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531114|AB188813| ген полимеразного основного белка 2 РВ2 вируса гриппа А (A/курица/Оита/8/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50956621|AY684703| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Хубей/327/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|57915957|AY737286| сегмент 1 вируса гриппа А (A/курица/Гуандун/191/04 (H5N1)), полноразмерная последовательность.
gi|57916006|AY737293| сегмент 1 вируса гриппа А (A/курица/Гуандун/178/04 (H5N1)), полноразмерная последовательность.
gi|57916060|AY737301| сегмент 1 вируса гриппа А (A/утка/Гуандун/173/04 (H5N1)), полноразмерная последовательность.
gi|55233237|AY770084| ген полимеразного основного белка 2 (РВ2) вируса гриппа А (A/курица/Хубей/489/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|54873461|AY770993| ген полимеразного основного белка 2 вируса гриппа А (A/курица/Ауттхая/Таиланд/CU-23/04 (H5N1)), неполная кодирующая последовательность.
gi|58618421|AY818127| ген полимеразного белка РВ2 вируса гриппа А (A/курица/Вьетнам/C58/04 (H5N1)), полноразмерная кодирующая последовательность.
gi|58618423|AY818128| ген полимеразного белка РВ2 вируса гриппа А (A/перепел/Вьетнам/36/04 (H5N1)), полноразмерная кодирующая последовательность.
gi|58374183|AY856861| сегмент 1 вируса гриппа А (A/утка/Шанду/093/2004 (H5N1)), полноразмерная последовательность.
gi|58531132|AB188821| ген полимеразного основного белка 2 РВ2 вируса гриппа А (A/курица/Киото/3/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531150|AB189050| ген полимеразного основного белка 2 РВ2 вируса гриппа А (A/ворон/Киото/53/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531168|AB189058| ген полимеразного основного белка 2 РВ2 вируса гриппа А (A/ворон/Осака/102/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296605|AY651748| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/сапсан/HK/D0028/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296531|AY651711| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Таиланд/1/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296533|AY651712| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Таиланд/73/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296535|AY651713| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Таиланд/9.1/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296537|AY651714| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Qa/Таиланд/57/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296539|AY651715| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/птичий/Таиланд/3.1/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296541|AY651716| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/Таиланд/71.1/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296543|AY651717| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Gs/Таиланд/79/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296553|AY651722| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/33/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296555|AY651723| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/35/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296557|AY651724| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/36/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296559|AY651725| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/37/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296561|AY651726| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/38/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296563|AY651727| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/39/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296565|AY651728| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/Вьетнам/C57/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296567|AY651729| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/Вьетнам/11/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|50296617|AY651754| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/YN/374/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296619|AY651755| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ck/YN/115/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296621|AY651756| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Ph/ST/44/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296623|AY651757| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/HN/303/2004 (H5N1)), неполная кодирующая последовательность.
gi|50296625|AY651758| ген полимеразной основной субъединицы 2 (РВ2) вируса гриппа А (A/Dk/HN/101/2004 (H5N1)), неполная кодирующая последовательность.
gi|50365712|AY653193| сегмент 1 вируса гриппа А (A/курица/Jilin/9/2004 (H5N1)), полноразмерная последовательность.
gi|47680940|AY586445| ген PB2 вируса гриппа А (A/кряква/Италия/43/01 (H7N3)), неполная кодирующая последовательность.
gi|47680930|AY586440| ген PB2 вируса гриппа А (A/кряква/Италия/33/01 (H7N3)), неполная кодирующая последовательность.
gi|47680932|AY586441| ген PB2 вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), неполная кодирующая последовательность.
gi|45124743|AJ627485| ген РНК-полимеразы PB2 вируса гриппа А (A/индейка/Италия/214845/2002 (H7N3)), геномная РНК.
gi|45124767|AJ627496| ген РНК-полимеразы PB2 вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), геномная РНК.
gi|34597782|AY303665| ген полимеразного основного белка 2 вируса гриппа А (A/курица/Чили/176822/02 (H7N3)), неполная кодирующая последовательность.
gi|34597784|AY303666| ген полимеразного основного белка 2 вируса гриппа А (A/курица/Чили/4957/02 (H7N3)), неполная кодирующая последовательность.
gi|47680928|AY586439| ген PB2 вируса гриппа А (A/индейка/Италия/214845/02 (H7N3)), неполная кодирующая последовательность.
gi|47834374|AYS16766| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50542651|AY646085| ген полимеразного основного белка 2 вируса гриппа А (A/курица/Британская Колумбия/GSC_человек_B/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50083053|AY648294| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/GSC_курица_B/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50059194|AY650276| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/GSC_курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|60700|Х58691| ген кэп-связывающего белка PB2 вируса гриппа А (A/FPV/Добсон/27 (H7N7)), геномная РНК.
gi|325001|M38291| ген (сегмент 3) полимеразного основного белка 2 (PB2) вируса гриппа A/FPV/Вейбридж, полноразмерная кодирующая последовательность.
gi|9988661|AF268120| ген полимеразного белка РВ2 вируса гриппа А (A/исландский песочник/Делавер/259/94 (H7N7)), неполная кодирующая последовательность.
gi|40732893|AJ620347| ген РНК-полимеразного белка PB2 вируса гриппа А (A/курица/Германия/R28/03 (H7N7)) A/курица/Германия/R28/03 (H7N7)), геномная РНК.
gi|37813157|AY342410| ген полимеразного белка 2 (A/Нидерланды/124/03 (H7N7)), неполная кодирующая последовательность.
gi|37813159|AY342411| ген полимеразного белка 2 вируса гриппа А (A/Нидерланды/126/03 (H7N7)), неполная кодирующая последовательность.
gi|37813161|AY342412| ген полимеразного белка 2 вируса гриппа А (A/Нидерланды/127/03 (H7N7)), неполная кодирующая последовательность.
gi|37813163|AY342413| ген полимеразного белка 2 вируса гриппа А (A/Нидерланды/219/03 (H7N7)), неполная кодирующая последовательность.
gi|37813165|AY342414| ген полимеразного белка 2 вируса гриппа А (A/курица/Нидерланды/1/03 (H7N7)), неполная кодирующая последовательность.
gi|5732354|AF156443| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/индейка/Калифорния/189/66 (H9N2)), неполная кодирующая последовательность.
gi|31339587|AF523469| ген полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/86/76 (H9N2)), неполная кодирующая последовательность.
gi|31339583|AF523467| ген полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/366/78 (H9N2)), неполная кодирующая последовательность.
gi|31339605|AF523478| ген полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/289/78 (H9N2)), неполная кодирующая последовательность.
gi|31339585|AF523468| ген полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/552/79 (H9N2)), неполная кодирующая последовательность.
gi|31339589|AP523470| ген полимеразы (РВ2) вируса гриппа А (A/утка/Гонконг/610/79 (H9N2)), неполная кодирующая последовательность.
gi|49356935|AY633283| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/кряква/Альберта/321/88 (H9N2)), неполная кодирующая последовательность.
gi|49356939|AY633299| ген полимеразного основного белка 2 (PB2) (PB2) (PB2)вируса гриппа А (A/кряква/Альберта/11/91 (H9N2)), неполная кодирующая последовательность.
gi|49356907|AY633171| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/кряква/Альберта/17/91 (H9N2)), неполная кодирующая последовательность.
gi|5732342|AF156437| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/перепел/Гонконг/|AF157/92 (H9N2)), неполная кодирующая последовательность.
gi|5732352|AF156442| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/перепел/Арканзас/29209-1/93 (H9N2)), неполная кодирующая последовательность.
gi|5732340|AF156436| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Гонконг/739/94 (H9N2)), неполная кодирующая последовательность.
gi|5732344|AF156438| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Бейжин/1/94 (H9N2)), неполная кодирующая последовательность.
gi|22759060|AF536679| ген PB2 вируса гриппа А (А/курица/Бейжин/1/95 (H9N2)), неполная кодирующая последовательность.
gi|33318110|AF508640| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/страус/Южная Африка/9508103/95 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318118|AF508644| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/утка/Германия/113/95 (H9N2)), неполная кодирующая последовательность.
gi|33318144|AF508657| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Шанду/6/96 (H9N2)), неполная кодирующая последовательность.
gi|33318152|AF508661| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/перепел/Шанхай/8/96 (H9N2)), неполная кодирующая последовательность.
gi|5732346|AF156439| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Корея/38349-p96323/96 (H9N2)), неполная кодирующая последовательность.
gi|5732348|AF156440| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Корея/25232- 96006/96 (H9N2)), неполная кодирующая последовательность.
gi|5732350|AF156441| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/ржанка/Делавер/9/96 (H9N2)), неполная кодирующая последовательность.
gi|22759068|AF536683| ген PB2 вируса гриппа А (A/курица/Хэбей/1/96 (H9N2)), неполная кодирующая последовательность.
gi|5732328|AF156430| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Гонконг/G9/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|5732330|AF156431| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/курица/Гонконг/G23/97 (H9N2)), неполная кодирующая последовательность.
gi|5732332|AF156432| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/голубь/Гонконг/Y233/97 (H9N2)), неполная кодирующая последовательность.
gi|5732334|AF156433| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/утка/Гонконг/Y280/97 (H9N2)), неполная кодирующая последовательность.
gi|5732336|AF156434| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/утка/Гонконг/Y439/97 (H9N2)), неполная кодирующая последовательность.
gi|5732338|AF156435| ген сегмента 1 полимеразной субъединицы PB2 (PB2) вируса гриппа А (A/перепел/Гонконг/G1/97 (H9N2)), неполная кодирующая последовательность.
gi|33318148|AF508659| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Шенчжен/9/97 (H9N2)), неполная кодирующая последовательность.
gi|33318150|AF508660| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/утка/Наньян/1/97 (H9N2)), неполная кодирующая последовательность.
gi|33318124|AF508647| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/фазан/Ирландия/PV18/97 (H9N2)), неполная кодирующая последовательность.
gi|13383266|AB049153| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/попугай/Чиба/1/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318132|AF508651| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Гуандун/11/97 (H9N2)), неполная кодирующая последовательность.
gi|33318136|AF508653| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Хейлоньян/10/97 (H9N2)), неполная кодирующая последовательность.
gi|22759062|AF536680| ген PB2 вируса гриппа А (A/курица/Бейжин/2/97 (H9N2)), неполная кодирующая последовательность.
gi|33318142|AF508656| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Сихуань/5/97 (H9N2)), неполная кодирующая последовательность.
gi|22759066|AF536682| ген PB2 вируса гриппа А (A/курица/Гуандун/97 (H9N2)), неполная кодирующая последовательность.
gi|22759078|AF536688| ген PB2 вируса гриппа А (A/курица/Шанду/98 (H9N2)), неполная кодирующая последовательность.
gi|33318116|AF508643| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Германия/R45/98 (H9N2)), неполная кодирующая последовательность.
gi|33318134|AF508652| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Хэбей/4/98 (H9N2)), неполная кодирующая последовательность.
gi|33318128|AF508649| ген сегмента 1 полимеразы PB2 (PB2) вируса гриппа А (A/курица/Бейжин/8/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|13383268|AB049154| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/попугай/Нарита/92A/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|22759070|AF536684| ген PB2 вируса гриппа А (A/курица/Хэбей/2/98 (H9N2)), неполная кодирующая последовательность.
gi|22759072|AF536685| ген PB2 вируса гриппа А (A/курица/Хэбей/3/98 (H9N2)), неполная кодирующая последовательность.
gi|22759074|AF536686| ген PB2 вируса гриппа А (A/курица/Хэнань/98 (H9N2)), неполная кодирующая последовательность.
gi|30025722|AY253750| ген РНК-полимеразы (PB2) вируса гриппа А (A/курица/Шанхай/F/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|12060631|AF222622| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/перепел/Гонконг/A17/99 (H9N2)), неполная кодирующая последовательность.
gi|12060633|AF222623| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/голубь/Гонконг/FY6/99 (H9N2)), неполная кодирующая последовательность.
gi|12060635|AF222624| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Гонконг/NT16/99 (H9N2)), неполная кодирующая последовательность.
gi|12060637|AF222625| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/перепел/Гонконг/SSP10/99 (H9N2)), неполная кодирующая последовательность.
gi|12060639|AF222626| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/фазан/Гонконг/SSP11/99 (H9N2)), неполная кодирующая последовательность.
gi|12060641|AF222627| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Гонконг/FY20/99 (H9N2)), неполная кодирующая последовательность.
gi|12060643|AF222628| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Гонконг/KC12/99 (H9N2)), неполная кодирующая последовательность.
gi|12060645|AF222629| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/перепел/Гонконг/NT28/99 (H9N2)), неполная кодирующая последовательность.
gi|12060647|AF222630| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Гонконг/SF2/99 (H9N2)), неполная кодирующая последовательность.
gi|12060649|AF222631| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/шелковая курица/Гонконг/SF44/99 (H9N2)), неполная кодирующая последовательность.
gi|22759076|AF536687| ген PB2 вируса гриппа А (A/курица/Ляонин/99 (H9N2)), неполная кодирующая последовательность.
gi|33318112|AF508641| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Пакистан/4/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318114|AF508642| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Пакистан/5/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318126|AF508648| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Корея/99029/99 (H9N2)), неполная кодирующая последовательность.
gi|33318120|AF508645| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Иран/11T/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318122|AF508646| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Саудовская Аравия/532/99 (H9N2)), неполная кодирующая последовательность.
gi|33318140|AF508655| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Нингсия/5/99 (H9N2)), неполная кодирующая последовательность.
gi|33318146|AF508658| ген сегмента 1 полимеразы 2 (PB2) вируса гриппа А (A/курица/Шиячжуан/2/99 (H9N2)), неполная кодирующая последовательность.
gi|12038893|AJ291395| ген полимеразы РВ2 вируса гриппа А (A/курица/Пакистан/2/99 (H9N2)), геномная РНК.
gi|22759064|AF536681| ген PB2 вируса гриппа А (A/курица/Бейжин/3/99(H9N2)), неполная кодирующая последовательность.
gi|31339607|AF523479| ген полимеразы (РВ2) вируса гриппа А (A/утка/Шанту/1881/00 (H9N2)), неполная кодирующая последовательность.
gi|31339601|AF523476| ген полимеразы (РВ2) вируса гриппа А (A/утка/Шанту/830/00 (H9N2)), неполная кодирующая последовательность.
gi|31339603|AF523477| ген полимеразы (РВ2) вируса гриппа А (A/утка/Шанту/1796/00 (H9N2)), неполная кодирующая последовательность.
gi|18496117 AJ427861| неполный ген pb2 полимеразного белка PB2 вируса гриппа А (A/перепел/Гонконг/FY298/00 (H9N2)), геномная РНК.
gi|27466041|AY180715| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/дикая утка/Наньхань/2-0480/2000 (H9N2), неполная кодирующая последовательность.
gi|27466043|AY180716| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/голубь/Наньхань/2-0461/2000 (H9N2), неполная кодирующая последовательность.
gi|27466045|AY180717| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/утка/Наньхань/1-0070/2000 (H9N2), неполная кодирующая последовательность.
gi|27466055|AY180722| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/утка/Наньхань/10-389/2000 (H9N2), неполная кодирующая последовательность.
gi|27466057|AY180723| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/голубь/Наньхань/7-058/2000 (H9N2), неполная кодирующая последовательность.
gi|27466067|AY180728| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/перепел/Наньхань/2-0460/2000 (H9N2), неполная кодирующая последовательность.
gi|27466085|AY180737| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/голубь/Наньхань/11-145/2000 (H9N2), неполная кодирующая последовательность.
gi|27466087|AY180738| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/утка/Наньхань/11-197/2000 (H9N2), неполная кодирующая последовательность.
gi|27466091|AY180740| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/утка/Наньхань/11-290/2000 (H9N2), неполная кодирующая последовательность.
gi|27466093|AY180741| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/утка/Наньхань/11-392/2000 (H9N2), неполная кодирующая последовательность.
gi|31339575|AF523463| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/2134/00 (H9N2)), неполная кодирующая последовательность.
gi|31339579|AF523465| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/1043/00 (H9N2)), неполная кодирующая последовательность.
gi|31339581|AF523466| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/1042/00 (H9N2)), неполная кодирующая последовательность.
gi|31339593|AF523472| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/2102/00 (H9N2)), неполная кодирующая последовательность.
gi|31339595|AF523473| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/2144/00 (H9N2)), неполная кодирующая последовательность.
gi|31339597|AF523474| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/2143/00 (H9N2)), неполная кодирующая последовательность.
gi|33318138|AF508654| ген сегмента 1 полимеразы (PB2) вируса гриппа А (A/курица/Хэнань/62/00 (H9N2)), неполная кодирующая последовательность.
gi|33318130|AF508650| ген сегмента 1 полимеразы (PB2) вируса гриппа А (A/курица/Гуандун/10/00 (H9N2)), неполная кодирующая последовательность.
gi|27466121|AY180755| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А штамма A/утка/Наньхань/7-092/2000 (H9N2), неполная кодирующая последовательность.
gi|27466141|AY180765| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А штамма A/курица/Наньхань/4-010/2000 (H9N2), неполная кодирующая последовательность.
gi|27466157|AY180773| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А штамма A/перепел/Наньхань/4-040/2000 (H9N2), неполная кодирующая последовательность.
gi|27466159|AY180774| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А штамма A/курица/Наньхань/1-0016/2000 (H9N2), неполная кодирующая последовательность.
gi|55469788|AY768575| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0028/00 (H9N2)), неполная кодирующая последовательность.
gi|55469790|AY768576| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0037/00 (H9N2)), неполная кодирующая последовательность.
gi|55469792|AY768577| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0073/00 (H9N2)), неполная кодирующая последовательность.
gi|55469794|AY768578| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0091/00 (H9N2)), неполная кодирующая последовательность.
gi|55469796|AY768579| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0140/00 (H9N2)), неполная кодирующая последовательность.
gi|55469798|AY768580| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU0146/00 (H9N2)), неполная кодирующая последовательность.
gi|55469800|AY768581| ген полимеразной основной субъединицы 2 (PB2) вируса гриппа А (A/курица/Корея/SNU1035C/00 (H9N2)), неполная кодирующая последовательность.
gi|27466143|AY180766| ген полимеразной субъединицы РВ2 (PB2) вируса гриппа А штамма A/курица/Наньхань/4-301/2001 (H9N2), неполная кодирующая последовательность.
gi|31339599|AF523475| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/2088/01 (H9N2)), неполная кодирующая последовательность.
gi|31339591|AF523471| ген полимеразы (PB2) вируса гриппа А (A/утка/Шанту/1605/01 (H9N2)), неполная кодирующая последовательность.
gi|31339577|AF523464| ген полимеразы (PB2) вируса гриппа А (A/дикая утка/Шанту/4808/01 (H9N2)), неполная кодирующая последовательность.
gi|27466097|AY180743| ген полимеразной субъединицы PB2 (PB2) вируса гриппа А штамма A/курица/Наньхань/4-361/2001 (H9N2), неполная кодирующая последовательность.
gi|54398631|AY664792| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/CSW153/03 (H9N2)), неполная кодирующая последовательность.
gi|54398633|AY664793| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/AP45/03 (H9N2)), неполная кодирующая последовательность.
gi|54398635|AY664794| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/BD90/03 (H9N2)), неполная кодирующая последовательность.
gi|54398637|AY664795| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/CSW291/03 (H9N2)), неполная последовательность.
gi|54398638|AY664796| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/CSW304/03 (H9N2)), неполная последовательность.
gi|54398639|AY664797| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/FY23/03 (H9N2)), неполная последовательность.
gi|54398640|AY664798| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/цесарка/Гонконг/NT101/03 (H9N2)), неполная кодирующая последовательность.
gi|54398642|AY664799| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/NT142/03 (H9N2), неполная кодирующая последовательность.
gi|54398644|AY664800| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/SF1/03 (H9N2)), неполная последовательность.
gi|54398645|AY664801| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/SSP101/03 (H9N2)), неполная последовательность.
gi|54398646|AY664802| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/TP38/03 (H9N2)), неполная кодирующая последовательность.
gi|54398648|AY664803| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/WF126/03 (H9N2)), неполная последовательность.
gi|54398649|AY664804| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/голубь/Гонконг/WF53/03 (H9N2)), неполная последовательность.
gi|54398650|AY664805| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/фазан/Гонконг/WF54/03 (H9N2)), неполная последовательность.
gi|54398651|AY664806| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/цесарка/Гонконг/NT184/03 (H9N2)), неполная кодирующая последовательность.
gi|54398653|AY664807| ген нефункционального полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/WF120/03 (H9N2)), неполная последовательность.
gi|54398654|AY664808| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/NT366/03 (H9N2)), неполная кодирующая последовательность.
gi|54398656|AY664809| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/SSP418/03 (H9N2)), неполная кодирующая последовательность.
gi|54398658|AY664810| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Гонконг/YU427/03 (H9N2)), неполная кодирующая последовательность.
gi|55793686|AY800240| ген полимеразного основного белка 2 (PB2) вируса гриппа А (A/курица/Корея/S1/2003 (H9N2)), неполная кодирующая последовательность.
gi|58429686|AY862710| ген PB2 (PB2) вируса гриппа А (A/шелковая курица/Корея/S3/03 (H9N2)), неполная кодирующая последовательность.
gi|58429688|AY862711| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S4/03 (H9N2)), неполная кодирующая последовательность.
gi|58429690|AY862712| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S5/03 (H9N2)), неполная кодирующая последовательность.
gi|58429692|AY862713| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S12/03 (H9N2)), неполная кодирующая последовательность.
gi|58429694|AY862714| ген PB2 (PB2) вируса гриппа А (A/утка/Корея/S13/03 (H9N2)), неполная кодирующая последовательность.
gi|58429696|AY862715| ген PB2 (PB2) вируса гриппа А (A/голубь/Корея/S14/03 (H9N2)), неполная кодирующая последовательность.
gi|58429698|AY862716| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S15/03 (H9N2)), неполная кодирующая последовательность.
gi|58429700|AY862717| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S16/03 (H9N2)), неполная кодирующая последовательность.
gi|58429702|AY862718| ген PB2 (PB2) вируса гриппа А (A/курица/Корея/S18/03 (H9N2)), неполная кодирующая последовательность.
Последовательности, используемые в анализе полимеразного кислотного белка вируса гриппа А (PA)
gi|27465935|AY180662| ген полимеразной субъединицы PA (PA) вируса гриппа А штамма A/перепел/Наньхань/12-340/2000 (H1N1), неполная кодирующая последовательность.
gi|49357063|AY633217| ген полимеразного белка А (PA) вируса гриппа А (A/кряква/Альберта/211/98 (H1N1)), неполная кодирующая последовательность.
gi|5918195|AJ243994| неполная мРНК белка РА вируса гриппа А (штамм A/кряква/Нью-Йорк/6750/78).
gi|45272157|AY422034| ген белка PA (PA) вируса гриппа А (A/утка/Хоккайдо/95/01 (H2N2)), неполная кодирующая последовательность.
gi|27465965|AY180677| ген полимеразной субъединицы PA (PA) вируса гриппа А штамма A/курица/Наньхань/3-120/2001 (H3N2), неполная кодирующая последовательность.
gi|56159994|AY779263| ген полимеразного кислотного белка PA (PA) вируса гриппа А (A/индейка/Северная Каролина/12344/03 (H3N2)), неполная кодирующая последовательность.
gi|56159996|AY779264| ген полимеразного кислотного белка PA (PA) вируса гриппа А (A/индейка/Миннесота/764-2/03 (H3N2)), неполная кодирующая последовательность.
gi|58429736|AY862687| ген PA (PA) вируса гриппа А (A/курица/Корея/S6/03 (H3N2)), неполная кодирующая последовательность.
gi|58429738|AY862688| ген PA (PA) вируса гриппа А (A/утка/Корея/S7/03 (H3N2)), неполная кодирующая последовательность.
gi|58429740|AY862689| ген PA (PA) вируса гриппа А (A/утка/Корея/S8/03 (H3N2)), неполная кодирующая последовательность.
gi|58429742|AY862690| ген PA (PA) вируса гриппа А (A/утка/Корея/S9/03 (H3N2)), неполная кодирующая последовательность.
gi|58429744|AY862691| ген PA (PA) вируса гриппа А (A/утка/Корея/S10/03 (H3N2)), неполная кодирующая последовательность.
gi|58429746|AY862692| ген PA (PA) вируса гриппа А (A/голубь/Корея/S11/03 (H3N2)), неполная кодирующая последовательность.
gi|18091833|AF213914| ген сегмента 3 полимеразного белка РА вируса гриппа А (A/курица/Италия/5945/95 (H3N2)), неполная кодирующая последовательность.
gi|58531086|AB166861| ген полимеразного кислотного белка РА вируса гриппа А (A/курица/Ямагучи/7/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531118|AB188815| ген полимеразного кислотного белка РА вируса гриппа А (A/курица/Оита/8/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|9863935|AF216739| ген полимеразного кислотного белка вируса гриппа А (A/окружающая среда/Гонконг/437-10/99 (H5N1)), полноразмерная кодирующая последовательность.
gi|14165201|AF380163| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/гусь/Гуандун/3/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|18092185|AF398427| ген полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/385.3/2000 (H5N1)), неполная кодирующая последовательность.
gi|18092187|AF398428| ген полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/385.5/2000 (H5N1)), неполная кодирующая последовательность.
gi|21359667|AF468841| ген полимеразного кислотного белка (PA) вируса гриппа А (A/утка/Аньян/AVL-1/2001 (H5N1)), неполная кодирующая последовательность.
gi|28849710|AF509195| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/FY77/01 (H5N1)), неполная кодирующая последовательность.
gi|28849712|AF509196| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
gi|28849714|AF509197| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/YU563/01 (H5N1)), полноразмерная кодирующая последовательность.
gi|28849716|AF509198| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
gi|28849718|AF509199| ген полимеразы (РА) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
gi|28849720|AF509200| ген полимеразы (РА) вируса гриппа А (A/шелковая курица/Гонконг/SF189/01 (H5N1)), неполная кодирующая последовательность.
gi|28849722|AF509201| ген полимеразы (РА) вируса гриппа А (A/перепел/Гонконг/SF203/01 (H5N1)), неполная кодирующая последовательность.
gi|28849724|AF509202| ген полимеразы (РА) вируса гриппа А (A/голубь/Гонконг/SF215/01 (H5N1)), неполная кодирующая последовательность.
gi|28849726|AF509203| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/SF219/01 (H5N1)), неполная кодирующая последовательность.
gi|28849728|AF509204| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/715.5/01 (H5N1)), неполная кодирующая последовательность.
gi|28849730|AF509205| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/751.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849732|AF509206| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/822.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849734|AF509207| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/829.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28849736|AF509208| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/830.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28849738|AF509209| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/858.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849740|AF509210| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/866.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849742|AF509211| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/867.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849744|AF509212| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/879.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849746|AF509213| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/873.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28849748|AF509214| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/876.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849750|AF509215| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/891.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849752]|AF509216| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/893.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849754|AF509217| ген полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/76.1/01 (H5N1)), неполная кодирующая последовательность.
gi|28849756|AF509218| ген полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/ww100/01 (H5N1)), неполная кодирующая последовательность.
gi|28849758|AF509219)| ген полимеразы (РА) вируса гриппа А (A/утка/Гонконг/573.4/01 (H5N1)), неполная кодирующая последовательность.
gi|28849760|AF509220| ген полимеразы (РА) вируса гриппа А (A/утка/Гонконг/646.3/01 (H5N1)), неполная кодирующая последовательность.
gi|19697861|AY059526| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/ww26/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697863|AY059527| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/ww28/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697865|AY059528| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/утка/Гонконг/ww381/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697869|AY059530| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/утка/Гонконг/ww461/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697871|AY059531| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/ww491/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697873|AY059532| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/утка/Гонконг/2986.1/2000 (H5N1)), неполная кодирующая последовательность.
gi|19697875|AY059533| ген сегмента 3 полимеразы (РА) вируса гриппа А (A/гусь/Гонконг/3014.8/2000 (H5N1)), неполная кодирующая последовательность.
gi|28821204|AY221566| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/NT873.3/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28821206|AY221567| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/NT873.3/01 (H5N1)), неполная кодирующая последовательность.
gi|28821208|AY221568| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/FY150/01-MB (HSN1)), неполная кодирующая последовательность.
gi|28821210|AY221569| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/FY150/01 (H5N1)), неполная кодирующая последовательность.
gi|28821212|AY221570| ген полимеразы (РА) вируса гриппа А (A/фазан/Гонконг/FY155/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28821214|AY221571| ген полимеразы (РА) вируса гриппа А (A/фазан/Гонконг/FY155/01 (H5N1)), неполная кодирующая последовательность.
gi|28821216|AY221572| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/YU822.2/01-MB (H5N1)), неполная кодирующая последовательность.
gi|28821218|AY221573| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/YU822.2/01 (H5N1)), неполная кодирующая последовательность.
gi|28821220|AY221574| ген полимеразы (РА) вируса гриппа А (A/курица/Гонконг/YU562/01 (H5N1)), неполная кодирующая последовательность.
gi|41207483|AY518365| ген полимеразы (РА) вируса гриппа А (A/утка/Китай/E319-2/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|51094114|AY551934| ген полимеразы (РА) вируса гриппа А (A/курица/Патомское нагорье/Таиланд/CU-K2/04 (H5N1)), полноразмерная кодирующая последовательность.
gi|47834839|AY576406| ген полимеразы (РА) вируса гриппа А (A/GS/HK/739.2/02 (H5N1)), неполная кодирующая последовательность.
gi|47834841|AY576407| ген полимеразы (РА) вируса гриппа А (А/Eg/HK/757.3/02 (H5N1)), неполная кодирующая последовательность.
gi|47834843|AY576408| ген полимеразы (РА) вируса гриппа А (А/G.H/HK/793.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834845|AY576409| ген полимеразы (РА) вируса гриппа А (А/Dk/HK/821/02 (H5N1)), неполная кодирующая последовательность.
gi|47834847|AY576410| ген полимеразы (РА) вируса гриппа А (A/Ck/HK/31.4/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|47834849|AY576411| ген полимеразы (РА) вируса гриппа А (A/Ck/HK/61.9/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|47834851|AY576412| ген полимеразы (РА) вируса гриппа А (A/Ck/HK/YU777/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|47834853|AY576413| ген полимеразы (РА) вируса гриппа А (А/Ck/HK/96.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834855|AY576414| ген полимеразы (РА) вируса гриппа А (A/курица/HK/409.1/02 (H5N1)), неполная кодирующая последовательность.
gi|47834857|AY576415| ген полимеразы (РА) вируса гриппа А (A/Ph/HK/sv674.15/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156478|AY585462| мРНК полимеразы (РА) вируса гриппа А (А/утка/Фуджиянь/01/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156480|AY585463| мРНК полимеразы (РА) вируса гриппа А (A/утка/Фуджиянь/13/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156482|AY585464| мРНК полимеразы (РА) вируса гриппа А (A/утка/Фуджиянь/17/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156484|AY585465| мРНК полимеразы (РА) вируса гриппа А (A/утка/Фуджиянь/19/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156486|AY585466| мРНК полимеразы (РА) вируса гриппа А (А/утка/Гуандун/01/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156488|AY585467| мРНК полимеразы (РА) вируса гриппа А (А/утка/Гуандун/07/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156490|AY585468| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуандун/12/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156492|AY585469| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуандун/22/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156494|AY585470| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуандун/40/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156496|AY585471| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуангкси/07/1999 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156498|AY585472| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуангкси/22/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156500|AY585473| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуангкси/35/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156502|AY585474| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуангкси/50/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156504|AY585475| мРНК полимеразы (РА) вируса гриппа А (A/утка/Гуангкси/53/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156506|AY585476| мРНК полимеразы (РА) вируса гриппа А (A/утка/Шанхай/08/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156508|AY585477| мРНК полимеразы (РА) вируса гриппа А (A/утка/Шанхай/13/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156510|AY585478| мРНК полимеразы (РА) вируса гриппа А (A/утка/Шанхай/35/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156512|AY585479| мРНК полимеразы (РА) вируса гриппа {A/утка/Шанхай/37/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156514|AY585480| мРНК полимеразы (РА) вируса гриппа А (A/утка/Шанхай/38/2001 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156516|AY585481| мРНК полимеразы (РА) вируса гриппа А (A/утка/Чжеянь/11/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47156518|AY585482| мРНК полимеразы (РА) вируса гриппа А (A/утка/Чжеянь/52/2000 (H5N1)), полноразмерная кодирующая последовательность.
gi|47716770|AY609311| сегмент 3 вируса гриппа А (A/курица/Гуандун/174/04 (H5N1)), полноразмерная последовательность.
gi|50313026|AY651597| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Индонезия/4/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313028|AY651598| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Индонезия/5/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313030|AY651599| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Индонезия/2A/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313032|AY651600| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/Индонезия/MS/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313034|AY651601| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Индонезия/BL/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313036|AY651602| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Индонезия/PA/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313038|AY651603| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Таиланд/1/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313040|AY651604| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Таиланд/73/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313042|AY651605| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Таиланд/9.1/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313044|AY651606| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Qa/Таиланд/57/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313046|AY651607| ген полимеразного кислотного белка (РА) вируса гриппа А (A/птичий/Таиланд/3.1/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313048|AY651608| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/Таиланд/71.1/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313050|AY651609| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Gs/Таиланд/79/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313060|AY651614| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/33/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313062|AY651615| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/35/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313064|AY651616| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/36/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313066|AY651617| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/37/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313068|AY651618| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/38/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313070|AY651619| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/39/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313072|AY651620| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/Вьетнам/C57/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313074|AY651621| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/Вьетнам/11/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313076|AY651622| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Gf/HK/38/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|50313078|AY651623| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/31.2/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|50313080|AY651624| ген полимеразного кислотного белка (РА) вируса гриппа А (A/курица/HK/37.4/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313082|AY651625| ген полимеразного кислотного белка (РА) вируса гриппа А (A/SCk/HK/YU100/2002 (H5N1)), полноразмерная кодирующая последовательность.
gi|50313084|AY651626| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/YU22/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313086|AY651627| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/3176.3/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313088|AY651628| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/3169.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313090|AY651629| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/FY157/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313092|AY651630| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/YU324/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313094|AY651631| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/2133.1/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313096|AY651632| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/NT93/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313098|AY651633| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/SSP141/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313100|AY651634| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/HK/WF157/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313102|AY651635| ген полимеразного кислотного белка (РА) вируса гриппа А (A/черноголовая чайка/HK/12.1/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313104|AY651636| ген полимеразного кислотного белка (РА) вируса гриппа А (A/серая цапля/HK/861.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313106|AY651637| ген полимеразного кислотного белка (РА) вируса гриппа А (A/одичавший голубь/HK/862.7/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313108|AY651638| ген полимеразного кислотного белка (РА) вируса гриппа А (A/древесный воробей/HK/864/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313110|AY651639| ген полимеразного кислотного белка (РА) вируса гриппа А (A/чирок/Китай/2978.1/2002 (H5N1)), неполная кодирующая последовательность.
gi|50313112|AY651640| ген полимеразного кислотного белка (РА) вируса гриппа А (A/сапсан/HK/D0028/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313114|AY651541| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/HN/5806/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313116|AY651642| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/HN/303/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313118|AY651543| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/HN/101/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313120|AY651644| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/ST/4003/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313122|AY651645| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ph/ST/44/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313124|AY651646| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/ST/4231/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313126|AY651647| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/YN/6255/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313128|AY651648| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Dk/YN/6445/2003 (H5N1)), неполная кодирующая последовательность.
gi|50313130|AY651649| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/YN/115/2004 (H5N1)), неполная кодирующая последовательность.
gi|50313132|AY651650| ген полимеразного кислотного белка (РА) вируса гриппа А (A/Ck/YN/374/2004 (H5N1)), неполная кодирующая последовательность.
gi|50365724|AY653198| сегмент 3 вируса гриппа А (A/курица/Жилинь/9/2004 (H5N1)), полноразмерная последовательность.
gi|56548923|AY676029| ген полимеразы (РА) вируса гриппа А (A/утка/Гонконг/821/02 (H5N1)), полноразмерная кодирующая последовательность.
gi|56548925|AY676030| ген полимеразы (РА) вируса гриппа А (A/белая цапля/Гонконг/757.2/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|56548927|AY676031| ген полимеразы (РА) вируса гриппа А (A/курица/Корея/ES/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|56548929|AY676032| ген полимеразы (РА) вируса гриппа А (A/утка/Корея/ESD1/03 (H5N1)), полноразмерная кодирующая последовательность.
gi|50956625|AY684705| ген полимеразы (РА) вируса гриппа А (A/курица/Хубей/327/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|56119221|AY720944| ген полимеразного кислотного белка (РА) вируса гриппа А (A/курица/Вьетнам/DT-171/2004 (H5N1)), неполная кодирующая последовательность.
gi|56119227|AY720947| ген полимеразного кислотного белка (РА) вируса гриппа А (A/утка/Вьетнам/TG-007A/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924419|AY724784| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/HCM-022/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924480|AY724786| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/DN-045/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924569]|AY724788| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/VL-008/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924680|AY724790| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/AG-010/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924765|AY724792| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/DT-015/2004 (H5N1)), неполная кодирующая последовательность.
gi|57924882|AY724796| ген полимеразы (РА) вируса гриппа А (A/курица/Вьетнам/LA-024/2004 (H5N1)), неполная кодирующая последовательность.
gi|57915971|AY737288| сегмент 3 вируса гриппа А (A/курица/Гуандун/191/04 (H5N1)), полноразмерная последовательность.
gi|57916018|AY737295| сегмент 3 вируса гриппа А (A/курица/Гуандун/178/04 (H5N1)), полноразмерная последовательность.
gi|57916074|AY737303| сегмент 3 вируса гриппа А (A/утка/Гуандун/173/04 (H5N1)), полноразмерная последовательность.
gi|55233234|AY770082| ген полимеразы (РА) вируса гриппа А (A/курица/Хубей/489/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|54873465|AY770995| ген полимеразы вируса гриппа А (A/курица/Аттайя/Таиланд/CU-23/04 (H5N1)), неполная кодирующая последовательность.
gi|58618433|AY818133| ген полимеразного белка (РА) вируса гриппа А (A/курица/Вьетнам/C58/04 (H5N1)), полноразмерная кодирующая последовательность.
gi|58618435|AY818134| ген полимеразы (РА) вируса гриппа А (A/перепел/Вьетнам/36/04 (H5N1)), полноразмерная кодирующая последовательность.
gi|58374187|AY856863| сегмент 3 вируса гриппа А (A/утка/Шанду/093/2004 (H5N1)), полноразмерная последовательность.
gi|58531136|AB188823| ген полимеразного кислотного белка РА вируса гриппа А (A/курица/Киото/3/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531154|AB189052| ген полимеразного кислотного белка РА вируса гриппа А (A/ворон/Киото/53/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|58531170|AB189059| ген полимеразного кислотного белка РА вируса гриппа А (A/ворон/Осака/102/2004 (H5N1)), полноразмерная кодирующая последовательность.
gi|3335418|AF046087| ген полимеразного кислотного белка (РА) вируса гриппа А (A/курица/Гонконг/220/97 (H5N1)), неполная кодирующая последовательность.
gi|6048895|AF098604| ген белка РА (РА) вируса гриппа А (A/курица/Гонконг/258/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048897|AF098605| ген белка РА (РА) вируса гриппа А (A/курица/Гонконг/y388/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048899|AF098606| ген белка РА (РА) вируса гриппа А (A/курица/Гонконг/728/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048901|AF098607| ген белка РА (РА) вируса гриппа А (A/курица/Гонконг/786/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048903|AF098608| ген белка РА (РА) вируса гриппа А (A/курица/Гонконг/915/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048905|AF098609| ген белка РА (РА) вируса гриппа А (A/утка/Гонконг/p46/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048907|AF098610| ген белка РА (РА) вируса гриппа А (A/утка/Гонконг/y283/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|6048909|AF098611| ген белка РА (РА) вируса гриппа А (A/гусь/Гонконг/w355/97 (H5N1)), полноразмерная кодирующая последовательность.
gi|5805280|AF144302| ген полимеразы PA (PA) вируса гриппа А (A/гусь/Гуандун/1/96 (H5N1)), полноразмерная кодирующая последовательность.
gi|9863880|AF216715| ген полимеразного кислотного белка вируса гриппа А (A/окружающая среда/Гонконг/437-4/99 (H5N1)), полноразмерная кодирующая последовательность.
gi|9863899|AF216723| ген полимеразного кислотного белка вируса гриппа А (A/окружающая среда/Гонконг/437-6/99 (H5N1)), полноразмерная кодирующая последовательность.
gi|9863917|AF216731| ген полимеразного кислотного белка вируса гриппа А (A/окружающая среда/Гонконг/437-8/99 (H5N1)), полноразмерная кодирующая последовательность.
gi|34597776|AY303660| ген полимеразного кислотного белка вируса гриппа А (A/курица/Чили/176822/02 (H7N3)), полноразмерная кодирующая последовательность.
gi|34597778|AY303661| ген полимеразного кислотного белка вируса гриппа А (A/курица/Чили/4957/02 (H7N3)), полноразмерная кодирующая последовательность.
gi|34597780|AY303662| ген полимеразного кислотного белка вируса гриппа А (A/курица/Чили/4322/02 (H7N3)), неполная кодирующая последовательность.
gi|47680912|AY586431| ген PA вируса гриппа А (A/кряква/Италия/43/01 (H7N3)), неполная кодирующая последовательность.
gi|47680914|AY586432| ген PA вируса гриппа А (A/кряква/Италия/33/01 (H7N3)), неполная кодирующая последовательность.
gi|47680916|AY586433| ген PA вируса гриппа А (A/индейка/Италия/220158/2002 (H7N3)), неполная кодирующая последовательность.
gi|47680918|AY586434| ген PA вируса гриппа А (A/индейка/Италия/214845/02 (H7N3)), неполная кодирующая последовательность.
gi|47834370|AY616764| ген полимеразного кислотного белка 2 (PA) вируса гриппа А (A/курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50542647|AY646083| ген полимеразного кислотного белка 2 (PA) вируса гриппа А (A/курица/Британская Колумбия/GSC_человек_B/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50083049|AY648292| ген полимеразного кислотного белка 2 (PA) вируса гриппа А (A/GSC_курица_B/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|50059192|AY650275| ген полимеразного кислотного белка 2 (PA) вируса гриппа А (A/GSC_курица/Британская Колумбия/04 (H7N3)), полноразмерная кодирующая последовательность.
gi|9988639|AF268109| ген полимеразного белка PA вируса гриппа А (A/исландский песочник/Делавер/259/94 (H7N7)), неполная кодирующая последовательность.
gi|40353080|AJ619677| ген субъединицы полимеразного комплекса PA вируса гриппа А (A/курица/Германия/R28/03 (H7N7)), геномная РНК.
gi|37813167|AY342415| ген полимеразного белка А вируса гриппа А (A/Нидерланды/124/03 (H7N7)), неполная кодирующая последовательность.
gi|37813169|AY342416| ген полимеразного белка А вируса гриппа А (A/Нидерланды/126/03 (H7N7)), неполная кодирующая последовательность.
gi|37813171|AY342417| ген полимеразного белка А вируса гриппа А (A/Нидерланды/127/03 (H7N7)), неполная кодирующая последовательность.
gi|37813173|AY342418| ген полимеразного белка А вируса гриппа А (A/Нидерланды/219/03 (H7N7)), полноразмерная кодирующая последовательность.
gi|37813175|AY342419| ген полимеразного белка А вируса гриппа А (A/Нидерланды/033/03 (H7N7)), полноразмерная кодирующая последовательность.
gi|37813177|AY342420| ген полимеразного белка А вируса гриппа А (A/курица/Нидерланды/1/03 (H7N7)), полноразмерная кодирующая последовательность.
gi|13383274|AB049157| ген полимеразного кислотного белка PA вируса гриппа А (A/попугай/Чиба/1/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|13383276|AB049158| ген полимеразного кислотного белка А вируса гриппа А (A/попугай/Нарита/92A/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318154|AF508662| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/страус/Южная Африка/9508103/95 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318156|AF508663| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Пакистан/4/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318158|AF508664| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Пакистан/5/99 (H9N2)), неполная кодирующая последовательность.
gi|33318160|AF508665| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Германия/R45/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318162|AF508666| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/утка/Германия/113/95 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318164|AF508667| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Иран/11T/99 (H9N2)), неполная кодирующая последовательность.
gi|33318166|AF508668| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Саудовская Аравия/532/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318168|AF508669| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/фазан/Ирландия/PV18/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318170|AF508670| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Корея/99029/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318172|AF508671| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Бейжин/8/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318174|AF508672| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Гуандун/10/00 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318176|AF508673| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Гуандун/11/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318178|AF508674| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Хэбей/4/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318180|AF508675| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Хейлоньян/10/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318182|AF508676| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Хэнань/62/00 (H9N2)), неполная кодирующая последовательность.
gi|33318184|AF508677| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Нингсия/5/99 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318186|AF508678| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Сихуань/5/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318188|AF508679| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Шанду/6/96 (H9N2)), неполная кодирующая последовательность.
gi|33318190|AF508680| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Шиячжуан/2/99 (H9N2)), неполная кодирующая последовательность.
gi|33318192|AF508681| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/курица/Шенчжен/9/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318194|AF508682| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/утка/Наньян/1/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|33318196|AF508683| ген сегмента 3 полимеразы PA (PA) вируса гриппа А (A/перепел/Шанхай/8/96 (H9N2)), неполная кодирующая последовательность.
gi|31339541|AF523446| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/1043/00 (H9N2)), неполная кодирующая последовательность.
gi|31339543|AF523447| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/1042/00 (H9N2)), неполная кодирующая последовательность.
gi|31339545|AF523448| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/2088/01 (H9N2)), неполная кодирующая последовательность.
gi|31339547|AF523449| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/830/00 (H9N2)), неполная кодирующая последовательность.
gi|31339549|AF523450| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/1796/00 (H9N2)), неполная кодирующая последовательность.
gi|31339551|AF523451| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/2143/00 (H9N2)), неполная кодирующая последовательность.
gi|31339553|AF523452| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/2134/00 (H9N2)), неполная кодирующая последовательность.
gi|31339555|AF523453| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/2144/00 (H9N2)), неполная кодирующая последовательность.
gi|31339557|AF523454| ген полимеразы (PA) вируса гриппа А (A/дикая утка/Шанту/4808/01 (H9N2)), неполная кодирующая последовательность.
gi|31339559|AF523455| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/1881/00 (H9N2)), неполная кодирующая последовательность.
gi|31339561|AF523456| ген полимеразы (PA) вируса гриппа А (A/утка/Шанту/2102/00 (H9N2)), неполная кодирующая последовательность.
gi|31339563|AF523457| ген полимеразы (PA) вируса гриппа А (A/утка/Гонконг/289/78 (H9N2)), неполная кодирующая последовательность.
gi|31339565|AF523458| ген полимеразы (PA) вируса гриппа А (A/утка/Гонконг/610/79 (H9N2)), неполная кодирующая последовательность.
gi|31339567|AF523459| ген полимеразы (PA) вируса гриппа А (A/утка/Гонконг/86/76 (H9N2)), неполная кодирующая последовательность.
gi|31339569|AF523460| ген полимеразы (PA) вируса гриппа А (A/утка/Гонконг/366/78 (H9N2)), неполная кодирующая последовательность.
gi|22759040|AF536669| ген PA вируса гриппа А (A/курица/Бейжин/1/95 (H9N2)), неполная кодирующая последовательность.
gi|22759042|AF536670| ген PA вируса гриппа А (A/курица/Бейжин/2/97 (H9N2)), неполная кодирующая последовательность.
gi|22759044|AF536671| ген PA вируса гриппа А (A/курица/Бейжин/3/99 (H9N2)), неполная кодирующая последовательность.
gi|22759046|AF536672| ген PA вируса гриппа А (A/курица/Гуандун/97 (H9N2)), неполная кодирующая последовательность.
gi|22759048|AF536673| ген PA вируса гриппа А (A/курица/Хэбей/1/96 (H9N2)), неполная кодирующая последовательность.
gi|22759050|AF536674| ген PA вируса гриппа А (A/курица/Хэбей/2/98 (H9N2)), неполная кодирующая последовательность.
gi|22759052|AF536675| ген PA вируса гриппа А (A/курица/Хэбей/3/98 (H9N2)), неполная кодирующая последовательность.
gi|22759054|AF536676| ген PA вируса гриппа А (A/курица/Хэнань/98 (H9N2)), неполная кодирующая последовательность.
gi|22759056|AF536677| ген PA вируса гриппа А (A/курица/Ляонин/99 (H9N2)), неполная кодирующая последовательность.
gi|22759058|AF536678| ген PA вируса гриппа А (A/курица/Шанду/98 (H9N2)), неполная кодирующая последовательность.
gi|12038897|AJ291397| ген полимеразы PA вируса гриппа А (A/курица/Пакистан/2/99 (H9N2)), геномная РНК.
gi|18496121|AJ427863| неполный ген ра полимеразного белка PA вируса гриппа А (A/перепел/Гонконг/FY298/00 (H9N2)), геномная РНК.
gi|27465911|AY180650| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/11-392/2000 (H9N2), неполная кодирующая последовательность.
gi|27465913|AY180651| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/11-290/2000 (H9N2), неполная кодирующая последовательность.
gi|27465915|AY180652| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/курица/Наньхань/1-0016/2000 (H9N2), неполная кодирующая последовательность.
gi|27465917|AY180653| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/11-197/2000 (H9N2), неполная кодирующая последовательность.
gi|27465937|AY180663| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/голубь/Наньхань/2-0461/2000 (H9N2), неполная кодирующая последовательность.
gi|27465941|AY180665| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/курица/Наньхань/4-301/2001 (H9N2), неполная кодирующая последовательность.
gi|27465943|AY180666| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/курица/Наньхань/4-361/2001 (H9N2), неполная кодирующая последовательность.
gi|27465961|AY180675| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/дикая утка/Наньхань/2-0480/2000 (H9N2), неполная кодирующая последовательность.
gi|27465983|AY180686| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/1-0070/2000 (H9N2), неполная кодирующая последовательность.
gi|27465989|AY180689| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/10-389/2000 (H9N2), неполная кодирующая последовательность.
gi|27465993|AY180691| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/голубь/Наньхань/11-145/2000 (H9N2), неполная кодирующая последовательность.
gi|27466001|AY180695| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/перепел/Наньхань/2-0460/2000 (H9N2), неполная кодирующая последовательность.
gi|27466003|AY180696| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/перепел/Наньхань/4-040/2000 (H9N2), неполная кодирующая последовательность.
gi|27466009|AY180699| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/курица/Наньхань/4-010/2000 (H9N2), неполная кодирующая последовательность.
gi|27466015|AY180702| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/утка/Наньхань/7-092/2000 (H9N2), неполная кодирующая последовательность.
gi|27466019|AY180704| ген полимеразной субъединицы PA (PA) вируса гриппа штамма А/голубь/Наньхань/7-058/2000 (H9N2), неполная кодирующая последовательность.
gi|30025973|AY253752| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Шанхай/F/98 (H9N2)), полноразмерная кодирующая последовательность.
gi|49357051|AY633169| ген полимеразного белка А (PA) вируса гриппа А (A/кряква/Альберта/17/91 (H9N2)), неполная кодирующая последовательность.
gi|49357079|AY633281| ген полимеразного белка А (PA) вируса гриппа А (A/кряква/Альберта/321/88 (H9N2)), неполная кодирующая последовательность.
gi|49357083|AY633297| ген полимеразного белка А (PA) вируса гриппа А (A/кряква/Альберта/11/91 (H9N2)), неполная кодирующая последовательность.
gi|54301528|AY664755| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/CSW153/03 (H9N2)), неполная кодирующая последовательность.
gi|54301530|AY664756| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/AP45/03 (H9N2)), неполная кодирующая последовательность.
gi|54301532|AY664757| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/BD90/03 (H9N2)), неполная кодирующая последовательность.
gi|54301534|AY664758| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/CSW291/03 (H9N2)), неполная кодирующая последовательность.
gi|54301536|AY664759| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/CSW304/03 (H9N2)), неполная кодирующая последовательность.
gi|54301538|AY664760| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/FY23/03 (H9N2)), неполная кодирующая последовательность.
gi|54301540|AY664761| ген полимеразного кислотного белка (PA) вируса гриппа А {A/цесарка/Гонконг/NT101/03 (H9N2)), неполная кодирующая последовательность.
gi|54301542|AY664762| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/NT142/03 (H9N2)), неполная кодирующая последовательность.
gi|54301544|AY664763| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/SF1/03 (H9N2)), неполная кодирующая последовательность.
gi|54301546|AY664764| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/SSP101/03 (H9N2)), неполная кодирующая последовательность.
gi|54301548|AY664765| ген, подобный гену полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/TP38/03 (H9N2)), полноразмерная последовательность.
gi|54301549|AY664766| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/WF126/03 (H9N2)), неполная кодирующая последовательность.
gi|54301551|AY664767| ген полимеразного кислотного белка (PA) вируса гриппа А (A/голубь/Гонконг/WF53/03 (H9N2)), неполная кодирующая последовательность.
gi|54301553|AY664768| ген полимеразного кислотного белка (PA) вируса гриппа А (A/фазан/Гонконг/WF54/03 (H9N2)), неполная кодирующая последовательность.
gi|54301555|AY664769| ген полимеразного кислотного белка (PA) вируса гриппа А (A/цесарка/Гонконг/NT184/03 (H9N2)), неполная кодирующая последовательность.
gi|54301557|AY664770| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/WF120/03 (H9N2)), неполная кодирующая последовательность.
gi|54301559|AY664771| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/NT366/03 (H9N2)), неполная кодирующая последовательность.
gi|54301561|AY664772| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/SSP418/03 (H9N2)), неполная кодирующая последовательность.
gi|54301563|AY664773| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Гонконг/YU427/03 (H9N2)), неполная кодирующая последовательность.
gi|55793682|AY800238| ген полимеразного кислотного белка (PA) вируса гриппа А (A/курица/Корея/S1/2003 (H9N2)), полноразмерная кодирующая последовательность.
gi|58429718|AY862678| ген PA (PA) вируса гриппа А (A/шелковая курица/Корея/S3/03 (H9N2)), неполная кодирующая последовательность.
gi|58429720|AY862679| ген PA (PA) вируса гриппа А (A/курица/Корея/S4/03 (H9N2)), неполная кодирующая последовательность.
gi|58429722|AY862680| ген PA (PA) вируса гриппа А (A/курица/Корея/S5/03 (H9N2)), полноразмерная кодирующая последовательность.
gi|58429724|AY862681| ген PA (PA) вируса гриппа А (A/курица/Корея/S12/03 (H9N2)), неполная кодирующая последовательность.
gi|58429726|AY862682| ген PA (PA) вируса гриппа А (A/утка/Корея/S13/03 (H9N2)), неполная кодирующая последовательность.
gi|58429728|AY862683| ген PA (PA) вируса гриппа А (A/голубь/Корея/S14/03 (H9N2)), неполная кодирующая последовательность.
gi|58429730|AY862684| ген PA (PA) вируса гриппа А (A/курица/Корея/S15/03 (H9N2)), неполная кодирующая последовательность.
gi|58429732|AY862685| ген PA (PA) вируса гриппа А (A/курица/Корея/S16/03 (H9N2)), неполная кодирующая последовательность.
gi|58429734|AY862686| ген PA (PA) вируса гриппа А (A/курица/Корея/S18/03 (H9N2)), неполная кодирующая последовательность.
gi|5732356|AF156444| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Гонконг/G9/97 (H9N2)), неполная кодирующая последовательность.
gi|5732358|AF156445| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Гонконг/G23/97 (H9N2)), полноразмерная кодирующая последовательность.
gi|5732360|AF156446| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/голубь/Гонконг/Y233/97 (H9N2)), неполная кодирующая последовательность.
gi|5732362|AF156447| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/утка/Гонконг/Y280/97 (H9N2)), неполная кодирующая последовательность.
gi|5732364|AF156448| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/утка/Гонконг/Y439/97 (H9N2)), неполная кодирующая последовательность.
gi|5732366|AF156449| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/перепел/Гонконг/G1/97 (H9N2)), неполная кодирующая последовательность.
gi|5732368|AF156450| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Гонконг/739/94 (H9N2)), неполная кодирующая последовательность.
gi|5732370|AF156451| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/перепел/Гонконг/|AF157/92 (H9N2)), неполная кодирующая последовательность.
gi|5732372|AF156452| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Бейжин/1/94 (H9N2)), неполная кодирующая последовательность.
gi|5732374|AF156453| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Корея/38349-р96323/96 (H9N2)), неполная кодирующая последовательность.
gi|5732376|AF156454| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/курица/Корея/25232-96006/96 (H9N2)), неполная кодирующая последовательность.
gi|5732378|AF156455| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/ржанка/Делавер/9/96 (H9N2)), неполная кодирующая последовательность.
gi|5732380|AF156456| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/перепел/Арканзас/29209-1/93 (H9N2)), неполная кодирующая последовательность.
gi|5732382|AF156457| ген сегмента 3 полимеразы (PA) вируса гриппа А (A/индейка/Калифорния/189/66 (H9N2)), неполная кодирующая последовательность.
gi|12060671|AF222642| ген сегмента 3 PA (PA) вируса гриппа А (A/перепел/Гонконг/A17/99 (H9N2)), неполная кодирующая последовательность.
gi|12060673|AF222643| ген сегмента 3 PA (PA) вируса гриппа А (A/голубь/Гонконг/FY6/99 (H9N2)), неполная кодирующая последовательность.
gi|12060675|AF222644| ген сегмента 3 PA (PA) вируса гриппа А (A/курица/Гонконг/NT16/99 (H9N2)), неполная кодирующая последовательность.
gi|12060677|AF222645| ген сегмента 3 PA (PA) вируса гриппа А (A/перепел/Гонконг/SSP10/99 (H9N2)), неполная кодирующая последовательность.
gi|12060679|AF222646| ген сегмента 3 PA (PA) вируса гриппа А (A/фазан/Гонконг/SSP11/99 (H9N2)), неполная кодирующая последовательность.
gi|120606811|AF222647| ген сегмента 3 PA (PA) вируса гриппа А (A/курица/Гонконг/FY20/99 (H9N2)), неполная кодирующая последовательность.
gi|12060683|AF222648| ген сегмента 3 PA (PA) вируса гриппа А (A/курица/Гонконг/KC12/99 (H9N2)), неполная кодирующая последовательность.
gi|12060685|AF222649| ген сегмента 3 PA (PA) вируса гриппа А (A/перепел/Гонконг/NT28/99 (H9N2)), неполная кодирующая последовательность.
gi|12060687|AF222650| ген сегмента 3 PA (PA) вируса гриппа А (A/курица/Гонконг/SF2/99 (H9N2)), неполная кодирующая последовательность.
gi|12060689|AF222651| ген сегмента 3 PA (PA) вируса гриппа А (A/шелковая курица/Гонконг/SF44/99 (H9N2)), неполная кодирующая последовательность.
| название | год | авторы | номер документа |
|---|---|---|---|
| УСЛОВНО ДЕФЕКТНАЯ ЧАСТИЦА ВИРУСА ГРИППА И СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ (ВАРИАНТЫ) | 2005 |
|
RU2420577C2 |
| ШТАММ ВИРУСА ГРИППА ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ И ИНАКТИВИРОВАННОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2011 |
|
RU2464309C1 |
| ВАКЦИНЫ НА ОСНОВЕ НУКЛЕИНОВЫХ КИСЛОТ | 2015 |
|
RU2746406C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ РЕПЛИКАТИВНОЙ ЧАСТИЦЫ ВИРУСА ГРИППА, КОМПОЗИЦИЯ КЛЕТОК (ВАРИАНТЫ), КОМПОЗИЦИЯ КЛЕТОЧНОЙ КУЛЬТУРЫ И ЕЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2435855C2 |
| ЭКСПРЕССИОННЫЕ СИСТЕМЫ | 2011 |
|
RU2609645C2 |
| ШТАММ ВИРУСА ГРИППА ГКВ 2389 ДЛЯ ПОЛУЧЕНИЯ ЖИВОЙ ИНТРАНАЗАЛЬНОЙ И ИНАКТИВИРОВАННОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2006 |
|
RU2318871C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ВИРУЛЕНТНЫХ И ПАТОГЕННЫХ ФОРМ ВИРУСОВ ГРИППА | 2011 |
|
RU2489488C1 |
| Штамм A/goose/Kalmykia/813/16 H5N8 вируса гриппа птиц Influenza virus avicum типа А подтипа H5 для контроля антигенной и иммуногенной активности вакцин против гриппа птиц и для изготовления биопрепаратов для диагностики и специфической профилактики гриппа птиц типа А подтипа Н5 | 2017 |
|
RU2647566C1 |
| ОПТИМИЗИРОВАННЫЕ С ПОМОЩЬЮ ВЫЧИСЛИТЕЛЬНЫХ СРЕДСТВ АНТИГЕНЫ С ШИРОКИМ СПЕКТРОМ РЕАКТИВНОСТИ ДЛЯ ВИРУСОВ ГРИППА H5N1 И H1N1 | 2013 |
|
RU2639551C2 |
| ШТАММ ВИРУСА ГРИППА А/Гонконг/1/68/162/35 (H3N2)-УНИВЕРСАЛЬНЫЙ ДОНОР ВНУТРЕННИХ ГЕНОВ ДЛЯ РЕАССОРТАНТОВ И РЕАССОРТАНТНЫЕ ШТАММЫ А/СПБ/ГК/09 (H1N1) И А/НК/Astana/6:2/2010 (H5N1), ПОЛУЧЕННЫЕ НА ЕГО ОСНОВЕ | 2011 |
|
RU2511431C2 |
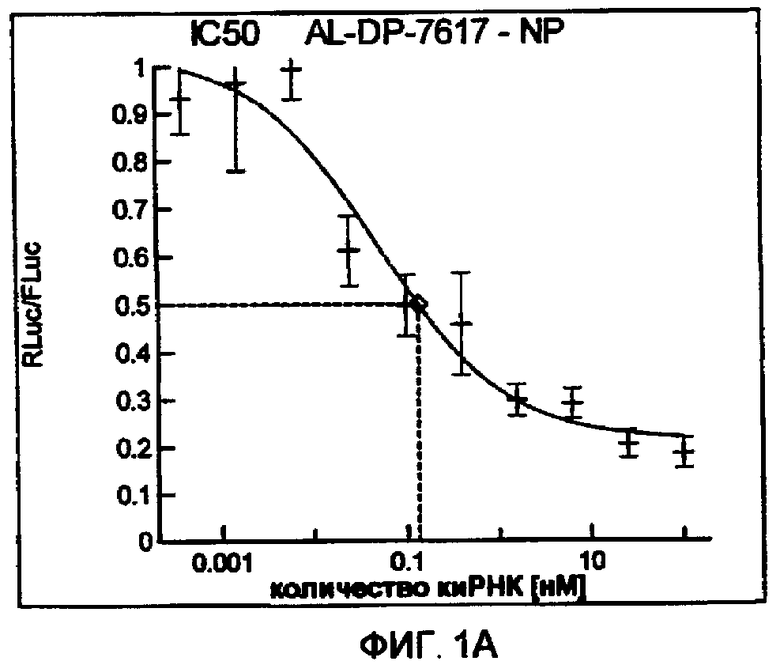
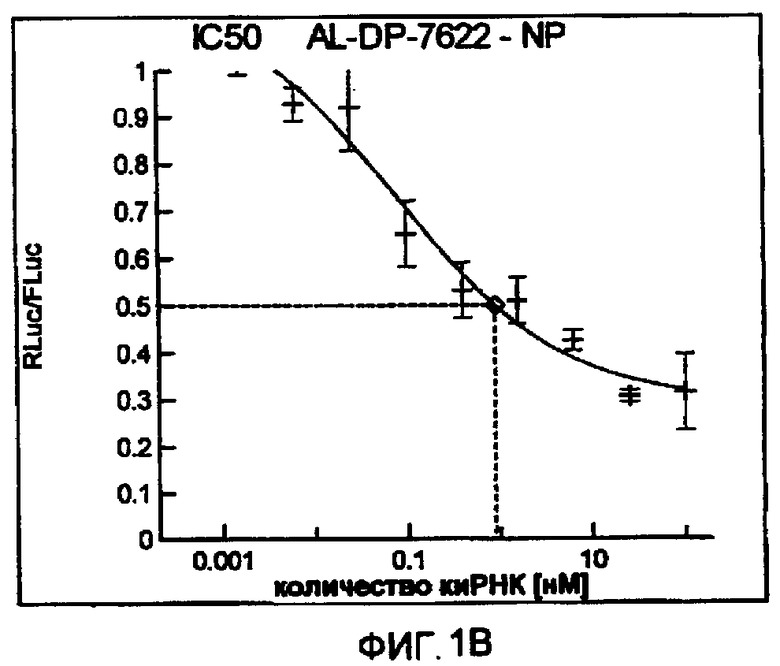
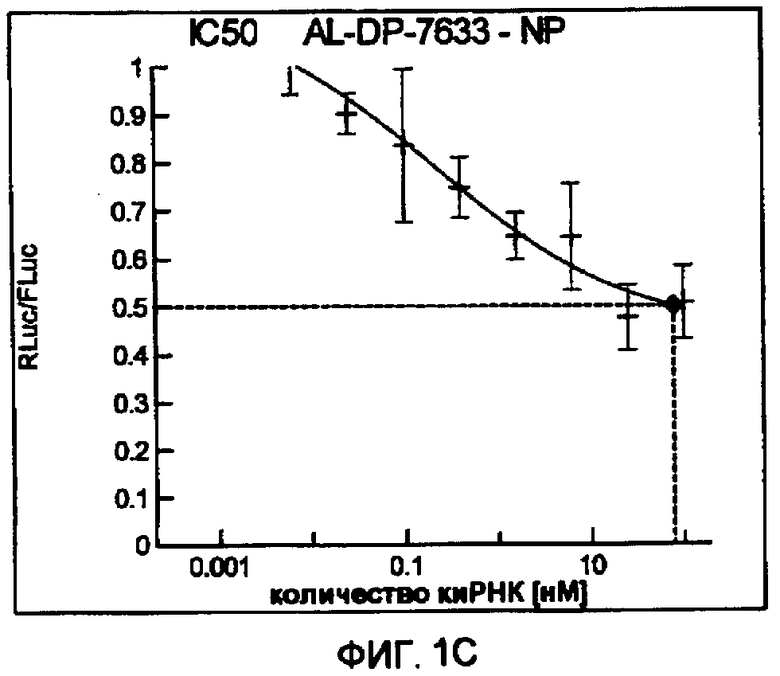
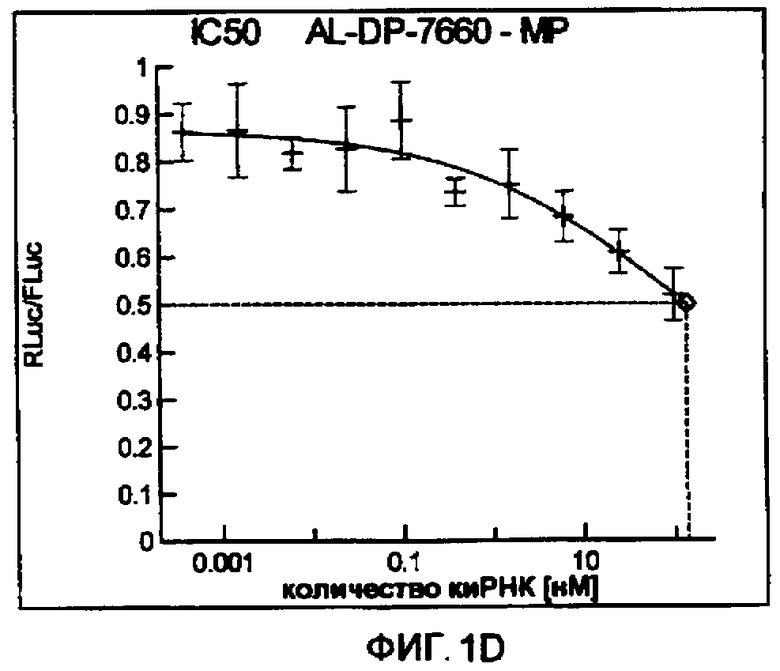
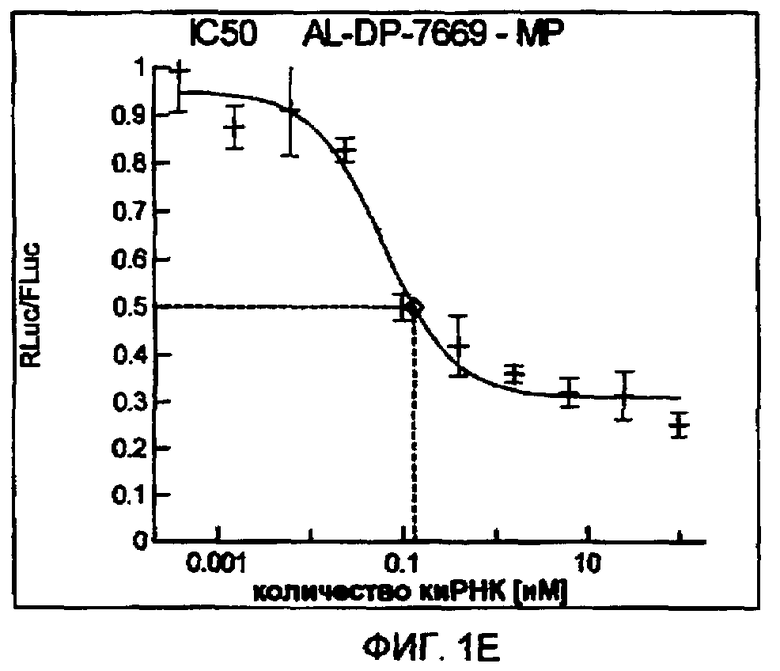
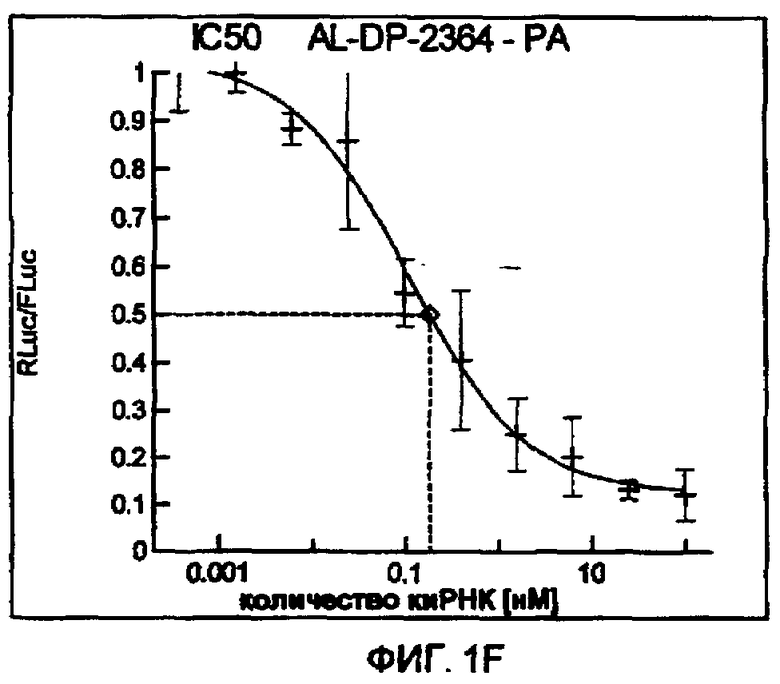
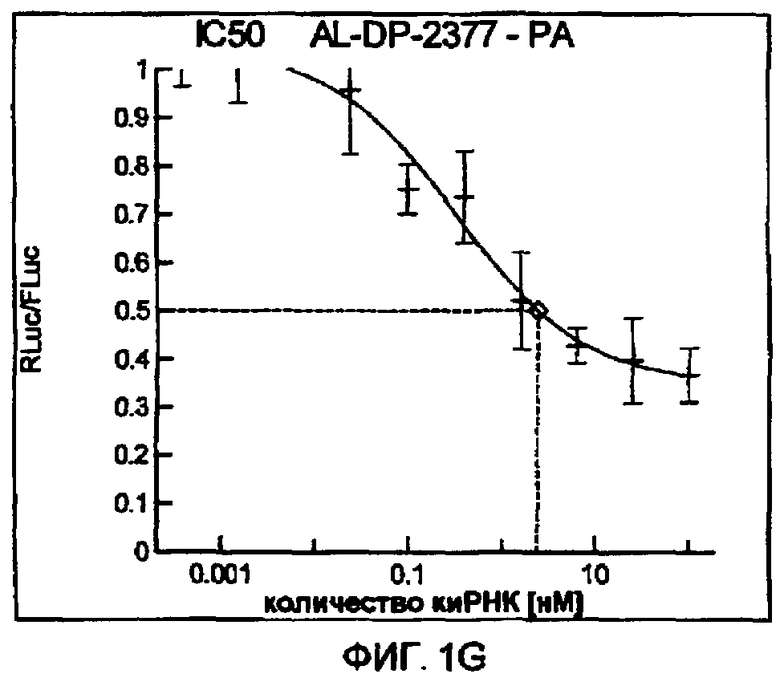
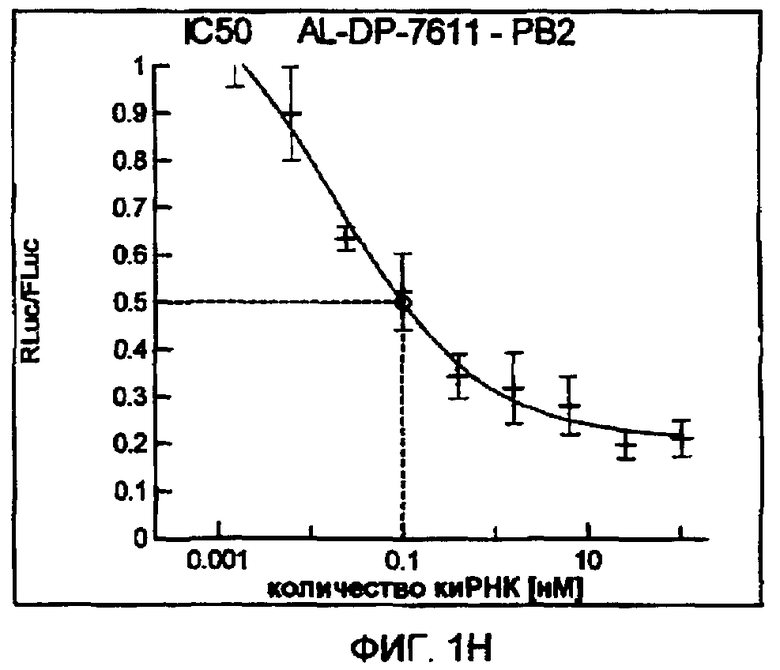
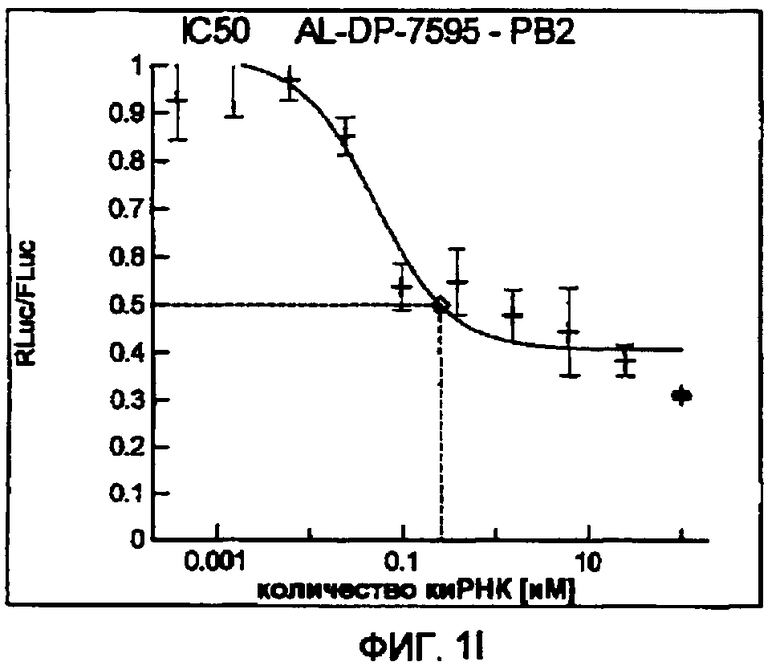
Группа изобретений относится к области биотехнологии. iPHK-агенты содержат смысловую цепь, где указанная смысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид. iPHK-агенты могут содержать модификации, которые повышают их стабильность в биологическом образце, молекулу холестерина, неприродное нуклеотидное основание. iPHK-агенты применяют для ингибирования экспрессии гена РВ2 вируса гриппа А. 11 н. и 62 з.п. ф-лы, 9 ил., 6 табл., 3 пр.
1. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А, где iPHK-агент содержит смысловую цепь, где указанная смысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид.
2. iPHK-агент по п.1, в котором длина указанной дуплексной области iPHK-агента составляет 19 нуклеотидных пар.
3. iPHK-агент по п.1, содержащий модификацию, которая повышает стабильность iPHK-агента в биологическом образце.
4. iPHK-агент по п.1, содержащий фосфоротиоат или 2'-модифицированный нуклеотид.
5. iPHK-агент по п.1, содержащий по меньшей мере один 5'-уридин-аденин-3'(5'-ua-3')-динуклеотид, в котором уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-уридин-гуанин-3'(5'-ug-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-цитидин-аденин-3'(5'-са-3')-динуклеотид, в котором 5'-цитидин представляет собой 2'-модифицированный нуклеотид; или по меньшей мере один 5'-уридин-уридин-3'(5'-uu-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид.
6. iPHK-агент по п.4, в котором указанная 2'-модификация выбрана из группы, состоящей из: 2'-дезокси, 2'-дезокси-2'-фтора, 2'-O-метила, 2'-O-метоксиэтила (2'-O-МОЕ), 2'-O-аминопропила (2'-О-АР), 2'-O-диметиламиноэтила (2'-O-DMAOE), 2'-O-диметиламинопропила (2'-O-DMAP), 2'-O-диметиламиноэтилоксиэтила (2'-O-DMAEOE) и 2'-O-N-метилацетамида (2'-O-NMA).
7. iPHK-агент по п.1, содержащий выступающий нуклеотидный конец, имеющий 1-4 неспаренных нуклеотида.
8. iPHK-агент по п.7, в котором нуклеотидный выступающий конец имеет 2 или 3 неспаренных нуклеотида.
9. iPHK-агент по п.7, в котором выступающий нуклеотид находится у 3'-конца антисмысловой цепи iPHK-агента.
10. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А, где iPHK-агент содержит смысловую цепь, где указанная смысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид, содержащий молекулу холестерина.
11. iPHK-агент по п.10, в котором молекула холестерина конъюгирована с 3'-концом смысловой цепи iPHK-агента.
12. iPHK-агент по п.1, в котором iPHK-агент нацеливают для захвата клетками легких.
13. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А, где iPHK-агент содержит смысловую цепь, где указанная смысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит во внутренней области 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид, содержащий по меньшей мере одно неприродное нуклеотидное основание.
14. iPHK-агент по п.13, в котором неприродным нуклеотидным основанием является дифтортолил, нитроиндолил, нитропирролил или нитроимидазолил.
15. iPHK-агент по п.13, в котором неприродным нуклеотидным основанием является дифтортолил.
16. iPHK-агент по п.13, в котором только одна из двух олигонуклеотидных цепей, составляющих двухцепочечный олигонуклеотид, содержит неприродное нуклеотидное основание.
17. iPHK-агент по п.13, в котором обе олигонуклеотидных цепи, составляющие двухцепочечный олигонуклеотид, независимо содержат неприродное нуклеотидное основание.
18. iPHK-агент по п.1, который снижает экспрессию гена вируса гриппа более чем на 20%, 30%, 40%, 50%, 60%, 70% или 80% в клетках, контактирующих с iPHK-агентом по сравнению с клетками, которые не контактируют с iPHK-агентом.
19. iPHK-агент по п.1, который, по существу, сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования.
20. iPHK-агент по п.1, в котором смысловая цепь состоит из нуклеотидной последовательности SEQ ID NO:427, а антисмысловая последовательность состоит из нуклеотидной последовательности SEQ ID NO:428.
21. iPHK-агент по п.1, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных нуклеотида.
22. iPHK-агент по п.1, содержащий выступающий нуклеотидный конец на каждом конце дцРНК, причем выступающий нуклеотидный конец содержит два неспаренных нуклеотида.
23. iPHK-агент по п.1, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных тимидина.
24. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь, где указанная смысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид, и где указанный iPHK-агент снижает уровень экспрессии гена в клетках Cos7, сконструированных для экспрессии указанного гена, на более чем на 20% по сравнению с экспрессией, наблюдаемой в сконструированных Cos7 клетках, которые не контактировали с указанным iPHK-агентом.
25. iPHK-агент по п.24, в котором длина указанной дуплексной области iPHK-агента составляет 19 пар.
26. iPHK-агент по п.24, содержащий модификацию, которая повышает стабильность iPHK-агента в биологическом образце.
27. iPHK-агент по п.24, содержащий фосфортиоат или 2'-модифицированный нуклеотид.
28. iPHK-агент по п.24, содержащий по меньшей мере один 5'-уридин-аденин-3'(5'-ua-3')-динуклеотид, в котором уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-уридин-гуанин-3'(5'-ug-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-цитидин-аденин-3'(5'-са-3')-динуклеотид, в котором 5'-цитидин представляет собой 2'-модифицированный нуклеотид; или по меньшей мере один 5'-уридин-уридин-3'(5'-uu-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид.
29. iPHK-агент по п.27, в котором указанная 2'-модификация выбрана из группы, состоящей из 2'-дезокси, 2'-дезокси-2'-фтора, 2'-O-метила, 2'-O-метоксиэтила (2'-O-МОЕ), 2'-O-аминопропила (2'-О-АР), 2'-O-диметиламиноэтила (2'-O-DMAOE), 2'-O-диметиламинопропила (2'-O-DMAP), 2'-O-диметиламиноэтилоксиэтила (2'-O-DMAEOE) и 2'-O-N-метилацетамида (2'-O-NMA).
30. iPHK-агент по п.24, содержащий выступающий нуклеотидный конец, имеющий 1-4 неспаренных нуклеотида.
31. iPHK-агент по п.30, в котором нуклеотидный выступающий конец имеет 2 или 3 неспаренных нуклеотида.
32. iPHK-агент по п.30, в котором выступающий нуклеотид находится у 3'-конца антисмысловой цепи iPHK-агента.
33. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь, где указанная смысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид, и где указанный iPHK-агент снижает уровень экспрессии гена в клетках Cos7, сконструированных для экспрессии указанного гена, на более чем на 20% по сравнению с экспрессией, наблюдаемой в сконструированных Cos7 клетках, которые не контактировали с указанным iPHK-агентом, содержащим молекулу холестерина.
34. iPHK-агент по п.33, в котором молекула холестерина конъюгирована с 3'-концом смысловой цепи iPHK-агента.
35. iPHK-агент по п.24, в котором iPHK-агент нацеливают для захвата клетками легких.
36. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь, где указанная смысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид, и где указанный iPHK-агент снижает уровень экспрессии гена в клетках Cos7, сконструированных для экспрессии указанного гена, на более чем на 20% по сравнению с экспрессией, наблюдаемой в сконструированных Cos7 клетках, которые не контактировали с указанным iPHK-агентом, содержащим по меньшей мере одно неприродное нуклеотидное основание.
37. iPHK-агент по п.36, в котором неприродным нуклеотидным основанием является дифтортолил, нитроиндолил, нитропирролил или нитроимидазолил.
38. iPHK-агент по п.36, в котором неприродным нуклеотидным основанием является дифтортолил.
39. iPHK-агент по п.36, в котором только одна из двух олигонуклеотидных цепей, составляющих двухцепочечный олигонуклеотид, содержит неприродное нуклеотидное основание.
40. iPHK-агент по п.36, в котором обе олигонуклеотидные цепи, составляющие двухцепочечный олигонуклеотид, независимо содержат неприродное нуклеотидное основание.
41. iPHK-агент по п.24, который снижает экспрессию гена вируса инфлюенции более чем на 20%, 30%, 40%, 50%, 60%, 70% или 80% в клетках, контактирующих с iPHK-агентом по сравнению с клетками, которые не контактируют с iPHK-агентом.
42. iPHK-агент по п.24, который, по существу, сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования.
43. iPHK-агент по п.24, в котором смысловая цепь состоит из нуклеотидной последовательности SEQ ID NO:427, а антисмысловая последовательность состоит из нуклеотидной последовательности SEQ ID NO:428.
44. iPHK-агент по п.24, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных нуклеотида.
45. iPHK-агент по п.24, содержащий выступающий нуклеотидный конец на каждом конце дцРНК, причем выступающий нуклеотидный конец содержит два неспаренных нуклеотида.
46. iPHK-агент по п.24, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных тимидина.
47. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь и антисмысловую цепь, причем каждая цепь содержит последовательность, которая состоит из 15-30 нуклеотидов, которые, по существу, идентичны SEQ ID NO:427 и SEQ ID NO:428 соответственно, за исключением того, что не более чем один нуклеотид в каждой цепи заменен другим нуклеотидом, при этом iPHK-агент сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования.
48. iPHK-агент по п.47, в котором длина указанной дуплексной области iPHK-агента составляет 19 пар.
49. iPHK-агент по п.47, содержащий модификацию, которая повышает стабильность iPHK-агента в биологическом образце.
50. iPHK-агент по п.47, содержащий фосфортиоат или 2'-модифицированный нуклеотид.
51. iPHK-агент по п.47, содержащий по меньшей мере один 5'-уридин-аденин-3'(5'-ua-3')-динуклеотид, в котором уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-уридин-гуанин-3'(5'-ug-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид; по меньшей мере один 5'-цитидин-аденин-3'(5'-са-3')-динуклеотид, в котором 5'-цитидин представляет собой 2'-модифицированный нуклеотид; или по меньшей мере один 5'-уридин-уридин-3'(5'-uu-3')-динуклеотид, в котором 5'-уридин представляет собой 2'-модифицированный нуклеотид.
52. iPHK-агент по п.50, в котором указанная 2'-модификация выбрана из группы, состоящей из 2'-дезокси, 2'-дезокси-2'-фтора, 2'-O-метила, 2'-O-метоксиэтила (2'-O-МОЕ), 2'-O-аминопропила (2'-О-АР), 2'-O-диметиламиноэтила (2'-O-DMAOE), 2'-O-диметиламинопропила (2'-O-DMAP), 2'-O-диметиламиноэтилоксиэтила (2'-O-DMAEOE) и 2'-O-N-метилацетамида (2'-O-NMA).
53. iPHK-агент по п.47, содержащий выступающий нуклеотидный конец, имеющий 1-4 неспаренных нуклеотида.
54. iPHK-агент по п.53, в котором нуклеотидный выступающий конец имеет 2 или 3 неспаренных нуклеотида.
55. iPHK-агент по п.53, в котором выступающий нуклеотид находится у 3'-конца антисмысловой цепи iPHK-агента.
56. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь и антисмысловую цепь, причем каждая цепь содержит последовательность, которая состоит из 15-30 нуклеотидов, которые, по существу, идентичны SEQ ID NO:427 и SEQ ID NO:428 соответственно, за исключением того, что не более чем один нуклеотид в каждой цепи заменен другим нуклеотидом, при этом iPHK-агент сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования, содержащий молекулу холестерина.
57. iPHK-агент по п.56, в котором молекула холестерина конъюгирована с 3'-концом смысловой цепи iPHK-агента.
58. iPHK-агент по п.47, в котором iPHK-агент нацеливают для захвата клетками легких.
59. iPHK-агент, ингибирующий экспрессию гена РВ2 вируса гриппа А и содержащий смысловую цепь и антисмысловую цепь, причем каждая цепь содержит последовательность, которая состоит из 15-30 нуклеотидов, которые, по существу, идентичны SEQ ID NO:427 и SEQ ID NO:428 соответственно, за исключением того, что не более чем один нуклеотид в каждой цепи заменен другим нуклеотидом, при этом iPHK-агент сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования, содержащий по меньшей мере одно неприродное нуклеотидное основание.
60. iPHK-агент по п.59, в котором неприродным нуклеотидным основанием является дифтортолил, нитроиндолил, нитропирролил или нитроимидазолил.
61. iPHK-агент по п.59, в котором неприродным нуклеотидным основанием является дифтортолил.
62. iPHK-агент по п.59, в котором только одна из двух олигонуклеотидных цепей, составляющих двухцепочечный олигонуклеотид, содержит неприродное нуклеотидное основание.
63. iPHK-агент по п.59, в котором обе олигонуклеотидные цепи, составляющие двухцепочечный олигонуклеотид, независимо содержат неприродное нуклеотидное основание.
64. iPHK-агент по п.47, который снижает экспрессию гена вируса инфлюенции более чем на 20%, 30%, 40%, 50%, 60%, 70% или 80% в клетках, контактирующих с iPHK-агентом по сравнению с клетками, которые не контактируют с iPHK-агентом.
65. iPHK-агент по п.47, который, по существу, сохраняет способность снижать количество образующихся в клетках бляшек вируса гриппа А, обнаруживаемых в анализе бляшкообразования.
66. iPHK-агент по п.47, в котором смысловая цепь состоит из нуклеотидной последовательности SEQ ID NO:427, а антисмысловая последовательность состоит из нуклеотидной последовательности SEQ ID NO:428.
67. iPHK-агент по п.47, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных нуклеотида.
68. iPHK-агент по п.47, содержащий выступающий нуклеотидный конец на каждом конце дцРНК, причем выступающий нуклеотидный конец содержит два неспаренных нуклеотида.
69. iPHK-агент по п.47, содержащий по меньшей мере один выступающий нуклеотидный конец, содержащий два неспаренных тимидина.
70. Применение iPHK-агента для ингибирования экспрессии гена РВ2 вируса группа А у индивида, где указанная смысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:427 не более чем на один нуклеотид, и антисмысловую цепь, где указанная антисмысловая цепь содержит 15-30 последовательных нуклеотидов, которые отличаются от нуклеотидов последовательности SEQ ID NO:428 не более чем на один нуклеотид.
71. Применение по п.70, где указанный iPHK-агент вводят индивиду в количестве, достаточном для снижения уровня репликации вируса гриппа в клетках или тканях указанного индивидуума.
72. Применение по п.70, где индивидуумом является человек.
73. Фармацевтическая композиция для ингибирования экспрессии гена РВ2 вируса гриппа А, содержащая:
a) iPHK-агент по любому из пп.1, 24 и 47; и
b) фармацевтически приемлемый носитель.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| US 5580767 03.12.1996 | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |

