По данной заявке испрашивается приоритет предварительной заявки США № 60/783746, поданной 17 марта 2006 года, описание которой включено в настоящий документ в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к анти-TAT226 антителам и их иммуноконъюгатам. Изобретение дополнительно относится к способам применения анти-TAT226 антител и их иммуноконъюгатов.
УРОВЕНЬ ТЕХНИКИ
Антитела, которые связываются с полипептидами, экспрессируемыми на поверхности раковых клеток, оказались эффективными в способах противораковой терапии. Такие антитела действуют посредством разнообразных механизмов, включающих, например, активацию антителозависимой клеточно-опосредованной цитотоксичности (ADCC), индукцию антителом комплементзависимой цитотоксичности (CDC), усиление высвобождения цитокинов и индукцию апоптоза. См., например, Cardarelli et al. (2002) Cancer Immunol. Immunother. 51:15-24. Например, ГЕРСЕПТИН® и РИТУКСАН® (оба от Genentech Inc., South San Francisco, California) представляют собой антитела, которые успешно применяют для лечения рака молочной железы и неходжкинской лимфомы соответственно. ГЕРСЕПТИН® представляет собой полученное на основе рекомбинантной ДНК гуманизированное моноклональное антитело, которое селективно связывается с внеклеточным доменом протоонкогена рецептора 2 (HER2) эпидермального фактора роста человека. Сверхэкспрессию белка HER2 наблюдают в 25-30% первичных форм рака молочной железы. РИТУКСАН® представляет собой генно-инженерное химерное моноклональное антитело мыши/человека, направленное против антигена CD20, обнаруженного на поверхности нормальных и злокачественных B-лимфоцитов. Оба этих антитела получают рекомбинантным способом в клетках CHO. Предполагают, что ГЕРСЕПТИН® действует, по меньшей мере, частично, путем ингибирования ангиогенеза (Izumi et al. (2002) Nature 416:279-280) и, как предполагают, РИТУКСАН® действует, по меньшей мере, частично, путем индуцирования апоптоза (Cardarelli et al. (2002) Cancer Immunol. Immunother. 51:15-24).
Иммуноконъюгаты, или «конъюгаты антитело-лекарственное средство», пригодны для местной доставки цитотоксических средств при лечении рака. См., например, Syrigos et al. (1999) Anticancer Research 19:605-614; Niculescu-Duvaz et al. (1997) Adv. Drug Deliv. Rev. 26:151-172; патент США № 4975278. Иммуноконъюгаты позволяют обеспечить направленную доставку компонента лекарственного средства в опухоль, в то время как системное введение неконъюгированных цитотоксических средств может привести к нежелательным уровням токсичности не только в опухолевых клетках, которые необходимо удалить, но и в нормальных клетках. См. Baldwin et al. (Mar. 15, 1986) Lancet p. 603-05; Thorpe (1985) «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review», в Monoclonal Antibodies '84: Biological and Clinical Applications (A. Pinchera et al., eds.) p. 475-506. Иммуноконъюгаты, которые направлены против полипептидов на поверхности клеток, разрабатывали и продолжают разрабатывать для лечения рака. Для обзора см., например, Hamann et al. (2005) Expert Opin. Ther. Patents (2005) 15:1087-1103.
Очевидно, что постоянно существует потребность для диагностических и/или терапевтических целей в средствах, которые направлены против полипептидов на поверхности клеток. Описанное здесь изобретение отвечает этой потребности и предоставляет другие преимущества.
Все цитированные в настоящем описании ссылки, включая патентные заявки и публикации, приведены в качестве ссылки в полном объеме.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к анти-TAT226 антителам и способам их применения.
Один из аспектов относится к антителу, которое связывается с TAT226, где антитело содержит, по меньшей мере, одну, две, три, четыре, пять или шесть HVR, выбранных из:
(1) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:4;
(2) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:5;
(3) HVR-H3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:11;
(4) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12;
(5) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и
(6) HVR-L3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19.
В другом аспекте антитело, которое связывается с TAT226, содержит (a) HVR-H3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:11, и (b), по меньшей мере, одну, две, три, четыре или пять HVR, выбранных из:
(1) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:4;
(2) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:5;
(3) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12;
(4) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и
(5) HVR-L3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19.
В одном из вариантов осуществления антитело содержит HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19. В одном из вариантов осуществления антитело дополнительно содержит HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5. В одном из вариантов осуществления антитело дополнительно содержит HVR-L1, содержащую аминокислотную последовательность SEQ NO:12, и HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13.
В одном из вариантов осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-10. В одном из вариантов осуществления антитело дополнительно содержит HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-18. В одном из вариантов осуществления HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9 и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17. В одном из вариантов осуществления HVR-H3 содержит аминокислотную последовательность SEQ ID NO:10 и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:18. В одном из вариантов осуществления антитело дополнительно содержит HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5. В одном из вариантов осуществления антитело дополнительно содержит HVR-L1, содержащую аминокислотную последовательность SEQ NO:12, и HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13.
Один аспект относится к антителу, которое связывается с TAT226, где антитело содержит, по меньшей мере, одну, две, три, четыре, пять или шесть HVR, выбранных из:
(1) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:1;
(2) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:2;
(3) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:3;
(4) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12;
(5) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и
(6) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:14.
В другом аспекте антитело, которое связывается с TAT226, содержит (a) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:3, и (b), по меньшей мере, одну, две, три, четыре или пять HVR, выбранных из:
(1) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:1;
(2) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:2;
(3) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12;
(4) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и
(5) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:14.
В одном из вариантов осуществления антитело содержит HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:14. В одном из вариантов осуществления антитело дополнительно содержит HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:1, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:2. В одном из вариантов осуществления антитело дополнительно содержит HVR-L1, содержащую аминокислотную последовательность SEQ NO:12, и HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13.
В некоторых вариантах осуществления любое из вышеописанных антител дополнительно содержит, по меньшей мере, одну каркасную область, выбранную из консенсусной каркасной области VH подгруппы III и консенсусной каркасной области VL подгруппы I.
Один из аспектов относится к антителу, которое связывается с TAT226, где антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:21-25. В одном из вариантов осуществления антитело дополнительно содержит вариабельный домен легкой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31. В одном из вариантов осуществления антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24. В одном из вариантов осуществления антитело дополнительно содержит вариабельный домен легкой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:29. В одном из вариантов осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:24 и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:29. В одном из вариантов осуществления антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:25. В одном из вариантов осуществления антитело дополнительно содержит вариабельный домен легкой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:30. В одном из вариантов осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:25 и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:30.
Один из аспектов относится к антителу, которое связывается с TAT226, где антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:20. В одном из вариантов осуществления антитело дополнительно содержит вариабельный домен легкой цепи, обладающий, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:26. В одном из вариантов осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:20 и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:26.
Некоторые варианты осуществления относятся к полинуклеотиду, кодирующему любое из вышеописанных антител. Один из вариантов осуществления относится к вектору, содержащему полинуклеотид. Один из вариантов осуществления относится к клетке-хозяину, содержащей вектор. В одном из вариантов осуществления клетка-хозяин является эукариотической. В одном из вариантов осуществления клетка-хозяин является клеткой CHO. Один из вариантов осуществления относится к способу получения анти-TAT226 антитела, где способ включает культивирование клетки-хозяина в условиях, подходящих для экспрессии полинуклеотида, кодирующего антитело, и выделение антитела.
Один из аспектов относится к антителу, которое связывается с TAT226, экспрессируемым на поверхности клетки. В одном из вариантов осуществления антитело связывается с эпитопом внутри области TAT226 из аминокислот 21-115 SEQ ID NO:75. В одном из вариантов осуществления клетка является раковой клеткой. В одном из вариантов осуществления раковая клетка является клеткой рака яичника, клеткой опухоли мозга или клеткой опухоли Вильмса.
В некоторых вариантах осуществления любое из вышеописанных антител представляет собой моноклональное антитело. В одном из вариантов осуществления антитело является фрагментом антитела, выбранным из фрагмента Fab, Fab'-SH, Fv, scFv или (Fab')2. В одном из вариантов осуществления антитело является гуманизированным. В одном из вариантов осуществления антитело является человеческим. В одном из вариантов осуществления антитело связывается с тем же эпитопом, что и антитело, выбранное из YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6.
Один из аспектов относится к способу детекции присутствия TAT226 в биологическом образце, где способ включает контактирование биологического образца с любым из вышеописанных антител в условиях, позволяющих антителу связываться с TAT226, и детекцию того, образован ли комплекс между антителом и TAT226. В одном из вариантов осуществления биологический образец содержит клетки опухоли яичника, клетки опухоли мозга или клетки опухоли Вильмса.
Один из аспектов относится к способу диагностики нарушения пролиферации клеток, связанного с повышенной экспрессией TAT226, где способ включает контактирование тестируемой клетки с любым из вышеописанных антител; определение уровня экспрессии TAT226 путем детекции связывания антитела с TAT226; и сравнение уровня экспрессии TAT226 тестируемой клеткой с уровнем экспрессии TAT226 контрольной клеткой, где более высокий уровень экспрессии TAT226 тестируемой клеткой по сравнению с контрольной клеткой указывает на наличие нарушения пролиферации клеток, связанного с повышенной экспрессией TAT226. В одном из вариантов осуществления тестируемая клетка представляет собой клетку пациента, у которого предполагают наличие нарушения пролиферации клеток. В одном из вариантов осуществления нарушение пролиферации клеток выбрано из рака яичника и опухоли Вильмса. В одном из вариантов осуществления способ включает определение уровня экспрессии TAT226 на поверхности тестируемой клетки и сравнение уровня экспрессии TAT226 на поверхности тестируемой клетки с уровнем экспрессии TAT226 на поверхности контрольной клетки.
Изобретение дополнительно относится к иммуноконъюгатам и способам их применения.
В одном из аспектов иммуноконъюгат содержит любое из вышеописанных анти-TAT226 антител, ковалентно присоединенное к цитотоксическому средству. В одном из вариантов осуществления цитотоксическое средство выбрано из токсина, химиотерапевтического средства, антибиотика, радиоактивного изотопа и нуклеолитического фермента.
Один из аспектов относится к иммуноконъюгату, имеющему формулу Ab-(L-D)p, где:
(a) Ab представляет собой любое из вышеописанных анти-TAT226 антител,
(b) L представляет собой линкер;
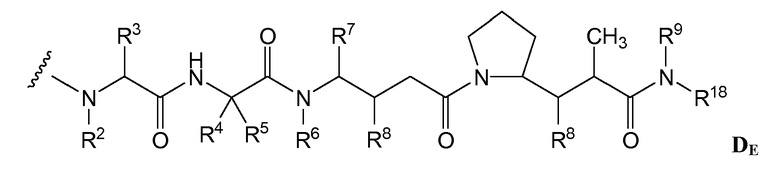
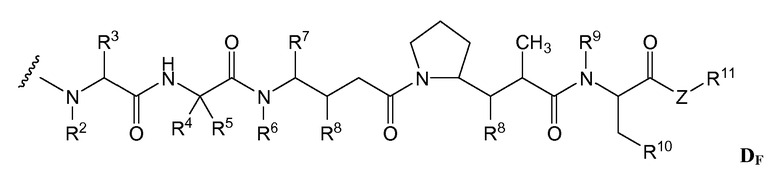
(c) D представляет собой лекарственное средство формулы DE или DF
 ,
,
и где каждый R2 и R6 представляет собой метил, каждый R3 и R4 представляет собой изопропил, R7 представляет собой втор-бутил, каждый R8 независимо выбран из CH3, O-CH3, OH и H; R9 представляет собой H; R10 представляет собой арил; Z представляет собой -O- или -NH-; R11 представляет собой H, C1-C8алкил или -(CH2)2-O-(CH2)2-O-(CH2)2-O-CH3; и R18 представляет собой -C(R8)2-C(R8)2-арил; и
(d) p варьирует приблизительно от 1 до 8.
В одном из вариантов осуществления антитело (Ab) содержит 1) HVR-H3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:11, и 2) по меньшей мере, одну, две, три, четыре или пять HVR, выбранных из:
(i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:4;
(ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:5;
(iii) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12;
(iv) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и
(v) HVR-L3, содержащей аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19.
В одном из вариантов осуществления антитело содержит HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19. В одном из вариантов осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:9, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:17. В одном из вариантов осуществления антитело содержит HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:10, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:18. В одном из вариантов осуществления антитело дополнительно содержит HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4, HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5, HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12, и HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13. В одном из вариантов осуществления антитело содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:21-25, и вариабельную область легкой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31. В одном из вариантов осуществления антитело содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24, и вариабельную область легкой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:29. В одном из вариантов осуществления антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:24, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:29. В одном из вариантов осуществления антитело содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:25, и вариабельную область легкой цепи, обладающую, по меньшей мере, 90% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:30. В одном из вариантов осуществления антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:25, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30.
Следующие варианты осуществления далее относятся к любому из вышеописанных иммуноконъюгатов. В одном из вариантов осуществления иммуноконъюгат обладает активностью в отношении уничтожения клеток in vitro или in vivo. В одном из вариантов осуществления линкер присоединен к антителу посредством тиольной группы на антителе. В одном из вариантов осуществления линкер является расщепляемым протеазой. В одном из вариантов осуществления линкер содержит дипептид val-cit. В одном из вариантов осуществления линкер содержит звено п-аминобензила. В одном из вариантов осуществления звено п-аминобензила расположено между лекарственным средством и расщепляемым протеазой участком линкера. В одном из вариантов осуществления звено п-аминобензила представляет собой п-аминобензилоксикарбонил (PAB). В одном из вариантов осуществления линкер содержит 6-малеимидокапроил. В одном из вариантов осуществления 6-малеимидокапроил расположен между антителом и расщепляемым протеазой участком линкера. Вышеописанные варианты осуществления можно осуществлять по отдельности или в любом сочетании друг с другом.
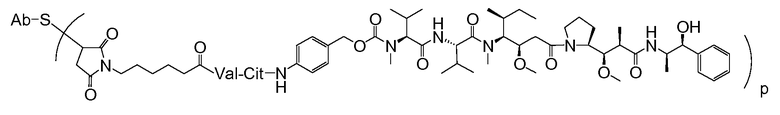
В одном из вариантов осуществления лекарственное средство выбрано из MMAE и MMAF. В одном из вариантов осуществления иммуноконъюгат имеет формулу
 ,
,
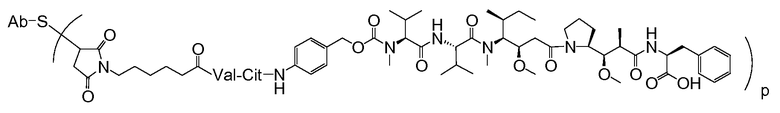
где Ab является любым из вышеописанных анти-TAT226 антител, S представляет собой атом серы и p варьирует от 2 до 5. В одном из вариантов осуществления иммуноконъюгат имеет формулу
 ,
,
где Ab является любым из вышеописанных анти-TAT226 антител, S представляет собой атом серы и p варьирует от 2 до 5.
Один из аспектов относится к фармацевтической композиции, содержащей любой из вышеописанных иммуноконъюгатов и фармацевтически приемлемый носитель. Один из аспектов относится к способу лечения нарушения пролиферации клеток, где способ включает введение индивидууму фармацевтической композиции. В одном из вариантов осуществления нарушение пролиферации клеток выбрано из рака яичника, рака тела матки, опухоли мозга и опухоли Вильмса. В одном из вариантов осуществления нарушение пролиферации клеток связано с повышенной экспрессией TAT226 на поверхности клетки.
Один из аспектов относится к способу ингибирования пролиферации клеток, где способ включает воздействие на клетку любым из вышеописанных иммуноконъюгатов в условиях, позволяющих иммуноконъюгату связываться с TAT226. В одном из вариантов осуществления клетка представляет собой опухолевую клетку. В одном из вариантов осуществления опухолевая клетка является клеткой опухоли яичников, клеткой опухоли тела матки, клеткой опухоли мозга или клеткой опухоли Вильмса. В одном из вариантов осуществления клетка представляет собой ксенотрансплантат. В одном из вариантов осуществления воздействие осуществляют in vitro. В одном из вариантов осуществления воздействие осуществляют in vivo.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1 представлено выравнивание TAT226 человека, макака-крабоеда («cyno»), мыши и крысы. Остатки, заключенные в рамки, идентичны у данных видов. Остальные остатки отличаются, по меньшей мере, у двух из четырех видов. Процентная доля аминокислотной идентичности среди последовательностей TAT226 человека, макака-крабоеда, мыши и крысы представлена в таблице под выравниванием. Процентную идентичность вычисляли с использованием программы ClustalW.
На фигуре 2 представлены последовательности H1, H2 и H3 гипервариабельной области (HVR) тяжелой цепи моноклональных анти-TAT226 антител, обозначенных как YWO.32 и YWO.49, как описано в примере B. Положения аминокислот пронумерованы согласно системе нумерации по Kabat, как описано ниже.
На фигуре 3 представлены последовательности L1, L2 и L3 легкой цепи HVR моноклональных анти-TAT226 антител, обозначенных как YWO.32 и YWO.49, как описано в примере B. Положения аминокислот пронумерованы согласно системе нумерации по Kabat, как описано ниже.
На фигуре 4 представлены последовательности HVR-H3 и HVR-L3 YWO.49 и YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6, которые получены аффинным созреванием YWO.49 с использованием мягко рандомизированных библиотек на основе HVR-H3 и HVR-L3, как описано в примере B. Также представлены консенсусные последовательности HVR-H3 и HVR-L3.
На фигурах 5A и 5B представлены иллюстративные акцепторные последовательности вариабельной тяжелой (VH) консенсусной каркасной области человека для использования на практике настоящего изобретения с идентификаторами последовательностей, как следует далее:
- консенсусная каркасная область «A» VH подгруппы I человека за вычетом CDR по Kabat (SEQ ID NO:32, 33, 34, 35);
- консенсусные каркасные области «B», «C» и «D» VH подгруппы I человека за вычетом расширенных гипервариабельных областей (SEQ ID NO:36, 37, 34, 35; SEQ ID NO:36, 37, 38, 35; и SEQ ID NO:36, 37, 39, 35);
- консенсусная каркасная область «A» VH подгруппы II человека за вычетом CDR по Kabat (SEQ ID NO:40, 41, 42, 35);
- консенсусные каркасные области «B», «C» и «D» VH подгруппы II человека за вычетом расширенных гипервариабельных областей (SEQ ID NO:43, 44, 42, 35; SEQ ID NO:43, 44, 45, 35; и SEQ ID NO:43, 44, 46 и 35);
- консенсусная каркасная область «A» VH подгруппы III человека за вычетом CDR по Kabat (SEQ ID NO:47, 48, 49, 35);
- консенсусные каркасные области «B», «C» и «D» VH подгруппы III человека за вычетом расширенных гипервариабельных областей (SEQ ID NO:50, 51, 49, 35; SEQ ID NO:50, 51, 52, 35; и SEQ ID NO:50, 51, 53, 35);
- акцепторная каркасная область «A» VH человека за вычетом CDR по Kabat (SEQ ID NO:54, 48, 55, 35);
- акцепторные каркасные области «B» и «C» VH человека за вычетом расширенных гипервариабельных областей (SEQ ID NO:50, 51, 55, 35; и SEQ ID NO:50, 51, 56, 35);
- акцепторная каркасная область 2 «A» VH человека за вычетом CDR по Kabat (SEQ ID NO:54, 48, 57, 35);
- акцепторные каркасные области 2 «B», «C» и «D» VH человека за вычетом расширенных гипервариабельных областей (SEQ ID NO:50, 51, 57, 35; SEQ ID NO:50, 51, 58, 35; и SEQ ID NO:50, 51, 59, 35).
На фигурах 6A и 6B представлены иллюстративные акцепторные последовательности вариабельной легкой (VL) консенсусной каркасной области человека для использования на практике настоящего изобретения с идентификаторами последовательностей, как следует далее:
- консенсусная каркасная область (κv1) подгруппы I VL каппа человека: SEQ ID NO:60, 61, 62, 63;
- консенсусная каркасная область (κv2) подгруппы II VL каппа человека: SEQ ID NO:64, 65, 66, 63;
- консенсусная каркасная область (κv3) подгруппы III VL каппа человека: SEQ ID NO:67, 68, 69, 63;
- консенсусная каркасная область (κv4) подгруппы IV VL каппа человека: SEQ ID NO:70, 71, 72, 63.
На фигуре 7 представлены последовательности каркасной области легких и тяжелых цепей huMAb4D5-8. Числа в надстрочных индексах/полужирным шрифтом указывают на положения аминокислот согласно Kabat.
На фигуре 8 представлены последовательности каркасной области легких и тяжелых цепей huMAb4D5-8 с указанными модификациями. Числа в надстрочных индексах/полужирным шрифтом указывают на положения аминокислот согласно Kabat.
На фигуре 9 представлены последовательности вариабельной области тяжелой цепи (VH) YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6. HVR подчеркнуты.
На фигуре 10 представлены последовательности вариабельной области легкой цепи (VL) YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6. VL-последовательности гуманизированного моноклонального антитела 4D5-8 («huMAb4D5-8») и «модифицированного» huMAb4D5-8 также представлены на SEQ ID NO:31 и SEQ ID NO:26 соответственно. YWO.32 и YWO.49 обладают такой же VL-последовательностью, как и «модифицированная» VL huMAb4D5-8 (SEQ ID NO:26), которая содержит следующие замещения относительно SEQ ID NO:31: N30S, R66G и H91S. HVR подчеркнуты.
На фигуре 11 представлено выравнивание последовательностей вариабельной области тяжелой цепи YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6. HVR заключены в рамки. Остатки HVR-H3 YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6, которые отличаются от соответствующих остатков HVR-H3 YWO.49, затенены.
На фигуре 12 представлено выравнивание последовательностей вариабельной области легкой цепи YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6. HVR заключены в рамки. Остатки HVR-L3 YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6, которые отличаются от соответствующих остатков HVR-L3 YWO.49, затенены.
На фигуре 13 показано графическое представление уровней экспрессии гена TAT226 человека в различных тканях, как описано в примере A.
На фигуре 14 показано графическое представление уровней экспрессии гена TAT226 человека в нормальном яичнике; нормальной фаллопиевой трубе; раке яичника светлоклеточного, муцинозного и серозного подтипов цистоаденокарциномы; метастатическом раке яичника и других типах рака яичника, как описано в примере A.
На фигуре 15 представлены результаты сортировки клеток с активацией флюоресценции (FACS) клеток OVCAR3 в отсутствие или присутствии указанных анти-TAT226 антител, как описано в примере D.
На фигуре 16 показана экспрессия мРНК и белка TAT226, как определено анализом с использованием 5'-нуклеазы (TaqMan®) и иммуногистохимией (IHC), выполненной на клетках OVCAR3 и панели образцов рака яичника, как описано в примере F.
На фигуре 17 представлена активность in vitro различных конъюгатов антитело-лекарственное средство (ADC) YWO.49.H2 и YWO.49.H6 в анализе уничтожения клеток OVCAR3, как описано в примере H.
На фигуре 18 представлена активность in vitro различных ADC YWO.49.H2 и YWO.49.H6 в анализе уничтожения клеток с использованием стабильных трансфектантов HCT116#9-4, как описано в примере H.
На фигуре 19 представлена активность in vivo ADC YWO.49.H6 с использованием ксенотрансплантатов у мышей, как описано в примере H.
На фигуре 20 представлена активность in vivo ADC YWO.49.H6 с использованием ксенотрансплантатов у мышей, полученных из опухолей пациентов-людей, как описано в примере H.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Предложены выделенные антитела, которые связываются с TAT226. Дополнительно предложены иммуноконъюгаты, содержащие анти-TAT226 антитела. Антитела и иммуноконъюгаты по изобретению пригодны, например, для диагностики или лечения нарушений, связанных с измененной экспрессией, например повышенной экспрессией, TAT226. В некоторых вариантах осуществления антитела или иммуноконъюгаты по изобретению пригодны для диагностики или лечения нарушения пролиферации клеток, такого как опухоль или рак. В некоторых вариантах осуществления антитела или иммуноконъюгаты по изобретению пригодны для детекции TAT226, например TAT226, экспрессируемого на клеточной поверхности.
Предложены полинуклеотиды, кодирующие анти-TAT226 антитела. Предложены векторы, содержащие полинуклеотиды, кодирующие анти-TAT226 антитела, и предложены клетки-хозяева, содержащие такие векторы. Также предложены композиции, включающие фармацевтические составы, содержащие что-либо из одного или нескольких полинуклеотидов, анти-TAT226 антител или иммуноконъюгатов по изобретению.
I. ОБЩИЕ СПОСОБЫ
Способы и процедуры, описанные или цитированные здесь, как правило, хорошо понятны и применяются в общепринятой практике с использованием общепринятых способов специалистами в данной области, такие как, например, широко используемые способы, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Current Protocols in Molecular Biology (F. M. Ausubel et al. eds., (2003)); серия Methods in Enzymology (Academic Press, Inc.): Pcr 2: A Practical Approach (M. J. MacPherson, B. D. Hames and G. R. Taylor eds. (1995)), Harlow and Lane eds. (1988) Antibodies, A Laboratory Manual, и Animal Cell Culture (R. I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M. J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J. P. Mather and P. E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths and D. G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D. M. Weir and C. C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J. E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C. A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999)); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (V. T. DeVita et al., eds., J.B. Lippincott Company, 1993).
II. ОПРЕДЕЛЕНИЯ И СОКРАЩЕНИЯ
A. Определения
«Выделенное» антитело представляет собой антитело, которое было идентифицировано и выделено и/или извлечено из компонента его природного окружения. Загрязняющие компоненты его природного окружения представляют собой вещества, которые могут препятствовать применению антитела для исследования, диагностики или лечения, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах осуществления антитело очищают (1) до более чем 95% масс. антитела, как определяют, например, способом Лоури, и в некоторых вариантах осуществления до более чем 99% масс., (2) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием, например, секвенатора с вращающимся стаканом, или (3) до гомогенности при SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием, например, кумасси голубого или окрашивания серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку в этом случае отсутствует, по меньшей мере, один компонент природного окружения антитела. Однако, как правило, выделенное антитело получают посредством, по меньшей мере, одной стадии очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которую отделяют, по меньшей мере, от одной другой молекулы нуклеиновой кислоты, с которой она обычно ассоциирована, например, в своем природном окружении. Выделенная молекула нуклеиновой кислоты дополнительно включает молекулу нуклеиновой кислоты, содержащуюся в клетках, в которых обычно экспрессируется молекула нуклеиновой кислоты, но расположение молекулы нуклеиновой кислоты вне хромосомы или на хромосоме отличается от ее природного расположения на хромосоме.
«Очищенный» означает, что молекула присутствует в образце в концентрации, по меньшей мере, 95% масс., или, по меньшей мере, 98% масс. образца, в котором она содержится.
Термин «по существу сходный» или «по существу одинаковый», как используют в настоящем документе, обозначает достаточно высокую степень сходства между двумя числовыми величинами (например, одной величиной, связанной с антителом по изобретению, и другой величиной, связанной со стандартным антителом/антителом для сравнения) так, что специалист в данной области может считать, что различие между двумя величинами является незначительным, или отсутствует биологическая и/или статистическая значимость в контексте биологической характеристики, измеренной при помощи указанных величин (например, величин Kd). Различие между указанными двумя величинами составляет, например, менее чем приблизительно 50%, менее чем приблизительно 40%, менее чем приблизительно 30%, менее чем приблизительно 20% и/или менее чем приблизительно 10% в виде функции от стандартной величины/величины для сравнения.
Фраза «по существу сниженный» или «по существу отличающийся», как используют в настоящем документе, обозначает достаточно высокую степень отличия между двумя числовыми величинами (как правило, одной величиной, связанной с молекулой, и другой величиной, связанной со стандартной молекулой/молекулой для сравнения) так, что специалист в данной области может считать, что различие между двумя величинами является статистически значимым в контексте биологической характеристики, измеренной при помощи указанных величин (например, величин Kd). Различие между указанными двумя величинами составляет, например, более чем приблизительно 10%, более чем приблизительно 20%, более чем приблизительно 30%, более чем приблизительно 40% и/или более чем приблизительно 50% в виде функции от величины для стандартной молекулы/молекулы для сравнения.
Термин «вектор», как используют в настоящем документе, предназначен для обозначения молекулы нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой она связана. Один тип вектора представляет собой «плазмиду», которая обозначает кольцевую двухцепочечную ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другой тип вектора представляет собой фаговый вектор. Другой тип вектора представляет собой вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, обладающие бактериальной точкой начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) можно интегрировать в геном клетки-хозяина при введении в клетку-хозяина, и, таким образом, они реплицируются вместе с геномом хозяина. Более того, определенные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы обозначают в настоящем документе как «рекомбинантные экспрессирующие векторы» или просто «экспрессирующие векторы». Как правило, экспрессирующие векторы, которые используют в способах рекомбинантных ДНК, часто находятся в форме плазмиды. В настоящем описании «плазмида» и «вектор» можно использовать взаимозаменяемо, так как плазмида является наиболее широко используемой формой вектора.
«Полинуклеотид» или «нуклеиновая кислота», как используют взаимозаменяемо в данном описании, относится к полимерам нуклеотидов любой длины и включает ДНК и РНК. Нуклеотиды могут являться дезоксирибонуклеотидами, рибонуклеотидами, модифицированными нуклеотидами или основаниями, и/или их аналогами, или любым веществом, которое можно встроить в полимер с помощью ДНК- или РНК-полимеразы, или с помощью синтетической реакции. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. При наличии модификацию структуры нуклеотида можно осуществить перед сборкой полимера или после нее. Последовательность нуклеотидов можно прерывать ненуклеотидными компонентами. Полинуклеотид может содержать модификацию(и), полученную(ые) после синтеза, как, например, конъюгация с меткой. Другие типы модификаций включают, например, «кэпы», замену одного или нескольких из природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, модификации незаряженными мостиками (например, метилфосфонатами, фосфотриэфирами, фосфоамидатами, карбаматами и т.п.) и заряженными мостиками (например, фосфоротиоатами, фосфородитиоатами и т.п.), модификации, содержащие подвешенные компоненты, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.п.), модификации интеркалирующими веществами (например, акридином, псораленом и т.п.), модификации, содержащие хелаторы (например, металлы, радиоактивные металлы, бор, металлы-окислители и т.п.), модификации, содержащие алкилирующие агенты, модификации модифицированными мостиками (например, альфа-аномерными нуклеиновыми кислотами и т.п.), а также немодифицированные формы полинуклеотида(ов). Далее, любые гидроксильные группы, обычно присутствующие в сахарах, можно заменить, например, фосфонатными группами, фосфатными группами, защищенными общепринятыми защитными группами или активированными для получения дополнительных связей с дополнительными нуклеотидами, или их можно конъюгировать с твердыми или полутвердыми носителями. 5'- и 3'-концевой OH можно фосфорилировать или заменить аминами или компонентами органической блокирующей группы из 1-20 атомов углерода. Можно также получить производные других гидроксилов с общепринятыми защитными группами. Полинуклеотиды могут также содержать аналогичные формы сахаров рибоз или дезоксирибоз, которые широко известны в данной области, включая, например, 2'-O-метил-, 2'-O-аллил, 2'-фтор- или 2'-азидорибозу, карбоциклические аналоги сахаров, α-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные формы сахаров, фуранозные формы сахаров, седогептулозы, ациклические аналоги и основные аналоги нуклеозидов, такие как метилрибозид. Одну или несколько фосфодиэфирных связей можно заменить альтернативными связующими группами. Эти альтернативные связующие группы включают в качестве неограничивающих примеров варианты осуществления, где фосфат замещен P(O)S («тиоатом»), P(S)S («дитиоатом»), «(O)NR2 («амидатом»), P(O)R, P(O)OR', CO или CH2 («формацеталем»), в которых каждый R или R' независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно содержащий эфирную (-O-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралкил. Не требуется, чтобы все связи в полинуклеотиде являлись идентичными. Предшествующее описание применимо ко всем упоминаемым здесь полинуклеотидам, включая РНК и ДНК.
«Олигонуклеотид», как используют в настоящем документе, как правило, относится к коротким, как правило, одноцепочечным, как правило, синтетическим полинуклеотидам, которые, как правило, но не обязательно, состоят из менее чем приблизительно 200 нуклеотидов в длину. Термины «олигонуклеотид» и «полинуклеотид» не являются взаимоисключающими. Вышеприведенное описание для полинуклеотидов в равной степени и полностью применимо для олигонуклеотидов.
«Процентную (%) идентичность аминокислотной последовательности» по отношению к стандартной полипептидной последовательности определяют как процентную долю аминокислотных остатков в рассматриваемой последовательности, которые идентичны аминокислотным остаткам в стандартной полипептидной последовательности после выравнивания последовательностей и, в случае необходимости, включения пропусков, для достижения максимальной процентной идентичности последовательности, и не рассматривая любые консервативные замещения в контексте идентичности последовательности. Выравнивание в целях определения процентной идентичности аминокислотной последовательности можно получить различными способами, которые доступны специалисту в данной области, например, с использованием общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, которые необходимы для достижения максимального выравнивания по всей полной длине сравниваемых последовательностей. Однако для целей данной работы величины % идентичности аминокислотной последовательности получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 разработана Genentech, Inc., и исходная программа с документацией для пользователя подана в U.S. Copyright Office, Washington D.C, 20559, где она зарегистрирована под U.S. Copyright Registration № TXU510087. Программа ALIGN-2 общедоступна от Genentech, Inc., South San Francisco, California, или ее можно компилировать с исходной программы. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, предпочтительно digital UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются.
В тех ситуациях, где используют ALIGN-2 для сравнений аминокислотных последовательностей, % идентичность аминокислотной последовательности для данной аминокислотной последовательности A по сравнению с данной аминокислотной последовательностью B (которую можно альтернативно перефразировать как данную аминокислотную последовательность A, которая обладает определенной % идентичностью аминокислотной последовательности или включает ее по сравнению с данной аминокислотной последовательностью B) вычисляют, как указано далее:
100 умножить на отношение X/Y,
где X представляет собой количество аминокислотных остатков, отмечаемых как идентичные совпадения программой для выравнивания последовательностей ALIGN-2 при выравнивании A и B этой программой, и где Y представляет собой общее количество аминокислотных остатков в B. Будет принято во внимание, что там, где длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичность аминокислотной последовательности A по сравнению с B не будет равна % идентичности аминокислотной последовательности B по сравнению с A. Если специально не указано иначе, все используемые здесь величины % идентичности аминокислотной последовательности получены, как описано в непосредственно предшествующем параграфе, с использованием компьютерной программы ALIGN-2.
Термин «TAT226», как используют в настоящем документе, относится к любому природному TAT226, полученному, если не указано иначе, из любого позвоночного, включая млекопитающих, таких как приматы (например, человек) и грызуны (например, мыши и крысы). Термин включает «полноразмерный», непроцессированный TAT226, а также любую форму TAT226, которая возникает при процессинге в клетке. Термин также включает природные варианты TAT226, например сплайсированные варианты или аллельные варианты. «Зрелая форма» TAT226 представляет собой форму TAT226, которая подверглась процессингу, например форму TAT226, которая подверглась N-концевому (например, отщепление сигнальной последовательности) и/или C-концевому расщеплению и/или модификации с присоединением GPI-якоря. В одном из вариантов осуществления «зрелая форма» TAT226 экспрессируется на клеточной поверхности.
«Антитела» (Ab) и «иммуноглобулины» (Ig) представляют собой гликопротеины, обладающие сходными структурными характеристиками. В то время как антитела обладают специфичностью связывания в отношении конкретного антигена, иммуноглобулины включают как антитела, так и другие антителоподобные молекулы, которые, вообще говоря, лишены антигенной специфичности. Полипептиды последнего типа, например, продуцируются в низких количествах лимфатической системой и в повышенных количествах миеломами.
Термины «антитело» и «иммуноглобулин» используют взаимозаменяемо в самом широком смысле, и они включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, моновалентные антитела, поливалентные антитела, полиспецифические антитела (например, биспецифические антитела при условии, что они обладают требуемой биологической активностью) и могут также включать определенные фрагменты антител (как здесь описано более подробно). Антитело может являться химерным, человеческим, гуманизированным и/или аффинно-зрелым.
Термин «анти-TAT226 антитело» или «антитело, которое связывается с TAT226» относится к антителу, которое способно связываться с TAT226 с достаточной аффинностью, такой что антитело пригодно в качестве диагностического и/или терапевтического средства при воздействии на TAT226. Предпочтительно степень связывания анти-TAT226 антитела с не относящимся к TAT226 белком составляет менее чем приблизительно 10% от связывания антитела с TAT226 при измерении, например, в радиоиммунном анализе (RIA). В некоторых вариантах осуществления антитело, которое связывается с TAT226, обладает константой диссоциации (Kd) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ или ≤ 0,1 нМ. В некоторых вариантах осуществления анти-TAT226 антитело связывается с эпитопом TAT226, который является консервативным для TAT226 из разных видов.
«Вариабельная область» или «вариабельный домен» антитела относится к аминоконцевым доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи можно обозначить как «VH». Вариабельный домен легкой цепи можно обозначить как «VL». Эти домены, как правило, являются наиболее вариабельными частями антитела и содержат антигенсвязывающие участки.
Термин «вариабельный» относится к тому факту, что последовательности определенных частей вариабельных доменов значительно различаются среди антител и используются в связывании и специфичности каждого конкретного антитела в отношении его конкретного антигена. Однако вариабельность не распределена равномерно на протяжении вариабельных доменов антител. Она сконцентрирована в трех сегментах, называемых областями, определяющими комплементарность, (CDR) или гипервариабельными областями (HVR), в вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высококонсервативные части вариабельных доменов называют каркасными областями (FR). Каждый из вариабельных доменов природных тяжелой и легкой цепей содержит четыре FR-области, в значительной степени принимающие конфигурацию бета-слоя, соединенную тремя CDR, которые формируют петли, соединяющие структуру бета-слоев и в некоторых случаях формирующие ее часть. CDR в каждой цепи расположены вместе в непосредственной близости с помощью областей FR и совместно с CDR другой цепи участвуют в формировании антигенсвязывающего участка антител (см. Kabat et al., Sequences of Protein of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, MD (1991)). Константные домены не вовлечены непосредственно в связывание антитела с антигеном, но они обладают различными эффекторными функциями, такими как участие антитела в антителозависимой клеточной токсичности.
«Легкие цепи» антител (иммуноглобулинов) из любых видов позвоночных на основе аминокислотных последовательностей их константных доменов можно отнести к одному из двух четко отличающихся типов, называемых каппа (κ) и лямбда (λ).
В зависимости от аминокислотных последовательностей константных доменов своих тяжелых цепей антитела (иммуноглобулины) можно отнести к разным классам. Существуют пять главных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно далее разделить на подклассы (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называют α, δ, ε, γ и µ соответственно. Структуры субъединиц и трехмерные конфигурации разных классов иммуноглобулинов хорошо известны и описаны в основном, например, в Abbas et al. Cellular and Mol. Immunology. 4th ed. (2000). Антитело может являться частью более крупной слитой молекулы, образованной ковалентным или нековалентным связыванием антитела с одним или несколькими другими белками или пептидами.
Термины «полноразмерное антитело», «интактное антитело» и «целое антитело» используют здесь взаимозаменяемо для обозначения антитела в его, по существу, интактной форме, не в виде фрагментов антитела, как определено ниже. Термины, в частности, относятся к антителу с тяжелыми цепями, которые содержат Fc-область.
«Фрагменты антитела» содержат только часть интактного антитела, где часть сохраняет, по меньшей мере, одну и их максимально возможное большинство или все функции, в норме связанные с этой частью, когда она присутствует в интактном антителе. В одном из вариантов осуществления фрагмент антитела содержит антигенсвязывающий участок интактного антитела и, таким образом, сохраняет способность связываться с антигеном. В другом варианте осуществления фрагмент антитела, например фрагмент, который содержит Fc-область, сохраняет, по меньшей мере, одну из биологических функций, в норме связанных с Fc-областью, когда она присутствует в интактном антителе, такую как связывание FcRn, модулирование времени полужизни антитела, функцию ADCC и связывание комплемента. В одном из вариантов осуществления фрагмент антитела является моновалентным антителом, которое обладает in vivo временем полужизни, по существу сходным с интактным антителом. Например, такой фрагмент антитела может содержать антигенсвязывающее плечо, связанное с Fc-последовательностью, способной придавать in vivo стабильность фрагменту.
Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых «Fab-фрагментами», каждый с единственным антигенсвязывающим участком, и остаточный «Fc-фрагмент», название которого отражает его способность легко кристаллизоваться. Обработка пепсином дает F(ab')2-фрагмент, который обладает двумя антигенсвязывающими участками, и он все еще способен образовывать поперечные сшивки с антигеном.
«Fv» представляет собой минимальный фрагмент антитела, который содержит полный антигенсвязывающий участок. В одном из вариантов осуществления двухцепочечная разновидность Fv состоит из димера вариабельного домена одной тяжелой и одной легкой цепи с прочной нековалентной связью. В одноцепочечной разновидности Fv (scFV) вариабельный домен одной тяжелой и одной легкой цепи можно ковалентно связать посредством гибкого пептидного линкера, так что легкую и тяжелую цепи можно соединить в «димерную» структуру, аналогичную структуре в двухцепочечной разновидности Fv. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего участка на поверхности димера VH-VL. В совокупности шесть CDR придают антигенсвязывающую специфичность антителу. Однако даже отдельный вариабельный домен (или половина Fv, содержащая только три CDR, специфичных в отношении антигена) обладает способностью распознавать и связывать антиген, хотя и с меньшей аффинностью, чем полный участок связывания.
Fab-фрагмент содержит вариабельные домены тяжелой и легкой цепей и также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на C-конце CH1-домена тяжелой цепи, включая один или несколько цистеинов из шарнирной области антитела. Fab'-SH служит здесь обозначением для Fab', в котором остаток(ки) цистеина константных доменов несет(ут) свободную тиоловую группу. Фрагменты антител F(ab')2 исходно получали в виде пар Fab'-фрагментов, которые имеют шарнирные цистеины между собой. Также известны другие химические сшивки фрагментов антител.
«Одноцепочечные Fv-» или «scFv-» фрагменты антитела содержат VH- и VL-домены антитела, где эти домены присутствуют в одной полипептидной цепи. Как правило, полипептид scFv дополнительно содержит полипептидный линкер между VH- и VL-доменами, который позволяет scFv сформировать требуемую структуру для связывания антигена. Для обзора по scFv см. Pluckthun, в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, p. 269-315 (1994).
Термин «димерные антитела» относится к малым фрагментам антител с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) на той же самой полипептидной цепи (VH-VL). При использовании линкера, который является слишком коротким, чтобы обеспечить спаривание между двумя доменами на одной и той же цепи, домены вынуждены спариваться с комплементарными доменами другой цепи и формировать два антигенсвязывающих участка. Димерные антитела могут являться бивалентными или биспецифическими. Димерные антитела более полно описаны, например, в EP 404097; WO 93/1161; Hudson et al. (2003) Nat. Med. 9:129-134; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993). Тримерные антитела и четырехмерные антитела также описаны в Hudson et al. (2003) Nat. Med. 9:129-134.
Термин «моноклональное антитело», как используют в настоящем документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными за исключением возможных мутаций, например естественно возникающих мутаций, которые могут присутствовать в минорных количествах. Таким образом, определение «моноклональные» указывает на характер антител, поскольку они не являются смесью отдельных антител. В некоторых вариантах осуществления такое моноклональное антитело, как правило, включает антитело, содержащее полипептидную последовательность, которая связывает мишень, где связывающую мишень полипептидную последовательность получали посредством процесса, который включает отбор одной связывающей мишень полипептидной последовательности из множества полипептидных последовательностей. Например, процесс отбора может представлять собой отбор определенного клона из множества клонов, таких как совокупность гибридомных клонов, фаговых клонов или клонов рекомбинантных ДНК. Следует понимать, что отобранную связывающую мишень последовательность можно дополнительно изменять, например, для повышения аффинности в отношении мишени, для гуманизации связывающей мишень последовательности, для повышения ее продукции в клеточной культуре, для снижения ее иммуногенности in vivo, для создания полиспецифического антитела и т.д. и что антитело, содержащее измененную связывающую мишень последовательность, также представляет собой моноклональное антитело по настоящему изобретению. В противоположность препаратам поликлонального антитела, которые, как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной детерминанты в антигене. В дополнение к их специфичности препараты моноклональных антител обладают преимуществом в том, что они, как правило, не содержат примесей других иммуноглобулинов.
Определение «моноклональный» указывает на тот признак антитела, что его получают по существу из гомогенной популяции антител, и не подразумевает того, что антитело должно быть получено каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, можно получать разнообразными способами, в том числе, например, способом гибридом (например, Kohler et al., Nature, 256:495 (1975); Harlow et al., Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N. Y., 1981)), способом рекомбинантных ДНК (см., например, патент США № 4816567), технологией фагового дисплея (см., например, Clackson et al., Nature, 352:624-628 (1991); Marks et al., J. Mol. Biol, 222:581-597 (1992); Sidhu et al., J. Mol. Biol. 338(2):299-310 (2004); Lee et al., J. Mol. Biol., 340(5):1073-1093 (2004); Fellouse, Proc. Nat. Acad. Sci. USA 101(34):12467-12472 (2004); и Lee et al. J. Immunol. Methods 284(1-2):119-132 (2004), и способами продукции антител человека или антител, подобных антителам человека, у животных, которые обладают частью или всеми локусами иммуноглобулина человека или генами, кодирующими последовательности иммуноглобулина человека (см., например, WO98/24893; WO96/34096; WO96/33735; WO91/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immunol. 7:33 (1993); патенты США № 5545807; 5545806; 5569825; 5625126; 5633425; 5661016; Marks et al., Bio. Technology, 10:779-783 (1992); Lonberg et al., Nature, 368:856-859 (1994); Morrison, Nature, 368:812-813 (1994); Fishwild et al., Nature Biotechnology, 14:845-851 (1996); Neuberger, Nature Biotechnology, 14:826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol., 13:65-93 (1995).
Моноклональные антитела в данном описании, в частности, включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретных видов или принадлежащих конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) является(ются) идентичной(ыми) или гомологичной(ыми) соответствующим последовательностям в антителах, полученных из других видов или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител при условии, что они обладают требуемой биологической активностью (патент США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
«Гуманизированные» формы не являющихся человеческими (например, мышиных) антител представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. В одном из вариантов осуществления гуманизированное антитело представляет собой иммуноглобулин человека (реципиентное антитело), в котором остатки гипервариабельной области реципиента заменяют остатками гипервариабельной области видов, не относящихся к человеку (донорное антитело), таких как мышь, крыса, кролик или не являющийся человеком примат, который обладает требуемой специфичностью, аффинностью и/или активностью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими не являющимися человеческими остатками. Более того, гуманизированные антитела могут содержать остатки, которые не представлены в реципиентном антителе или в донорном антителе. Эти модификации можно осуществить для дополнительного улучшения свойств антител. Как правило, гуманизированное антитело содержит по существу все, по меньшей мере, из одного и, как правило, из двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют петлям иммуноглобулина, не являющегося человеческим, и все или по существу все FR представляют собой последовательности FR иммуноглобулина человека. Также гуманизированные антитела, не обязательно, содержат, по меньшей мере, часть константной области (Fc) иммуноглобулина, как правило, константной области иммуноглобулина человека. Для более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). См. также следующие цитированные здесь обзорные статьи и ссылки: Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994).
«Антитело человека» представляет собой антитело, которое обладает аминокислотной последовательностью, которая соответствует последовательности антитела, вырабатываемого человеком, и/или которое получают с использованием любых способов получения антител человека, как описано в данной работе. Это определение антитела человека, в частности, исключает гуманизированное антитело, содержащее не являющиеся человеческими антигенсвязывающие остатки.
Термин «гипервариабельная область», «HVR» или «HV» при использовании в данном описании относится к областям вариабельного домена антитела, которые являются гипервариабельными по последовательности и/или формируют структурно оформленные петли. Как правило, антитела содержат шесть гипервариабельных областей; три в VH (H1, H2, H3) и три в VL (L1, L2, L3). В природных антителах H3 и L3 обладают наибольшим разнообразием из шести гипервариабельных областей, и предполагают, что H3, в частности, играет исключительную роль в придании тонкой специфичности антителам. Xu et al. (2000) Immunity 13:37-45; Johnson and Wu (2003) in Methods in Molecular Biology 248:1-25 (Lo, ed., Humana Press, Totowa, NJ). Действительно, существующие в природе у верблюдовых антитела, состоящие только из тяжелой цепи, являются функциональными и стабильными в отсутствии легкой цепи. Hamers-Casterman et al. (1993) Nature 363:446-448; Sheriff et al. (1996) Nature Struct. Biol. 3:733-736.
Используют ряд определений гипервариабельной области, и они рассмотрены в данном описании. По Kabat, области, определяющие комплементарность (CDR), определяют на основе вариабельности последовательности и их используют чаще всего (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Вместо этого Chothia ссылается на местоположение структурных петель (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). Гипервариабельные области по AbM представляют собой компромисс между CDR по Kabat и структурными петлями по Chothia, и их используют в программном обеспечении для моделирования антител Oxford Molecular's AbM. «Контактные» гипервариабельные области определяют на основе анализа доступных кристаллических структур комплексов. Остатки каждой из этих гипервариабельных областей отмечены ниже.
Гипервариабельные области могут содержать «расширенные гипервариабельные области», как указано далее: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2), и 89-97 или 89-96 (L3) в VL, и 26-35 (H1), 50-65 или 49-65 (H2), и 93-102 или 95-102 (H3) в VH. Остатки вариабельного домена пронумерованы согласно Kabat et al. выше для каждого из этих определений.
«Каркасные» или «FR-» остатки являются такими остатками вариабельного домена, которые отличаются от остатков гипервариабельной области, как определено в данном описании.
Термин «нумерация остатка вариабельного домена, как у Kabat» или «нумерация положения аминокислоты, как у Kabat» и его вариации относится к системе нумерации, используемой для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи, из собрания данных по антителам в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). При использовании этой системы нумерации действительная линейная аминокислотная последовательность может содержать меньшее количество аминокислот или дополнительные аминокислоты, что соответствует укорочению FR или HVR вариабельного домена или вставке в них. Например, вариабельный домен тяжелой цепи может содержать единственную вставку аминокислоты (остаток 52a согласно Kabat) после остатка 52 H2 и вставленные остатки (например, остатки 82a, 82b и 82c и т.п. согласно Kabat) после остатка 82 тяжелой цепи FR. Нумерацию остатков по Kabat можно определить для данного антитела посредством выравнивания областей гомологии последовательности антитела со «стандартной» занумерованной по Kabat последовательностью.
«Аффинно-зрелое» антитело представляет собой антитело с одним или несколькими изменениями в одной или нескольких своих HVR, что приводит к повышению аффинности антитела в отношении антигена по сравнению с родительским антителом, которое не обладает этим(и) изменением(ями). В одном из вариантов осуществления аффинно-зрелое антитело обладает наномолярными или даже пикомолярными значениями аффинности в отношении антигена-мишени. Аффинно-зрелые антитела получают с помощью способов, известных в данной области. В Marks et al. Bio/Technology 10:779-783 (1992) описано аффинное созревание при перестановке VH- и VL-доменов. Случайный мутагенез остатков в HVR и/или каркасных остатков описан в Barbas et al. Proc Nat. Acad. Sci. USA 91:3809-3813 (1994); Schier et al. Gene 169: 147-155 (1995); Yelton et al. J. Immunol. 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-9 (1995); и Hawkins et al., J. Mol. Biol. 226:889-896 (1992).
«Блокирующее» антитело или «антитело-антагонист» представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается. Определенные блокирующие антитела или антитела-антагонисты по существу или полностью ингибируют биологическую активность антигена.
«Антитело-агонист», как используют в настоящем документе, представляет собой антитело, которое мимикрирует, по меньшей мере, одну из функциональных активностей представляющего интерес полипептида.
«Эффекторные функции» антитела относятся к таким видам биологической активности, которые свойственны Fc-области (природной последовательности Fc-области или варианту аминокислотной последовательности Fc-области) антитела, и варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточно-опосредуемую цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора); и активацию B-клеток.
Терминами «Fc-рецептор» или «FcR» описывают рецептор, который связывается с Fc-областью антитела. В некоторых вариантах осуществления FcR является природным FcR человека. В некоторых вариантах осуществления FcR является рецептором, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно-сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые обладают сходными аминокислотными последовательностями, которые отличаются преимущественно своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA в своем цитоплазматическом домене содержит иммунорецепторный активирующий мотив на основе тирозина (ITAM). Ингибирующий рецептор FcγRIIB в своем цитоплазматическом домене содержит иммунорецепторный ингибирующий мотив на основе тирозина (ITIM) (см. Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). FcR рассмотрены в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). К термину «FcR» в настоящем описании относятся другие FcR, в том числе FcR, которые будут выявлены в будущем.
Термин «Fc-рецептор» или «FcR» также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регуляцию гомеостаза иммуноглобулинов. Известны способы измерения связывания с FcRn (см., например, Ghetie 1997, Hinton 2004). Можно анализировать связывание с FcRn человека in vivo и время полужизни в сыворотке полипептидов с высокой аффинностью связывающихся с FcRn человека, например, у трансгенных мышей или в трансфицированных линиях клеток человека, экспрессирующих FcRn человека, или у приматов, которым вводят полипептиды вариантов Fc.
В WO00/42072 (Presta) описаны варианты антител с повышенным или сниженным связыванием с FcR. Содержание этой патентной публикации, в частности, приведено в настоящем документе в качестве ссылки. См. также Shields et al. J. Biol. Chem. 9(2):6591-6604 (2001).
«Эффекторные клетки человека» представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. В некоторых вариантах осуществления клетки экспрессируют, по меньшей мере, FcγRIII и выполняют эффекторную(ые) функцию(и) ADCC. Примеры лейкоцитов человека, которые осуществляют ADCC, включают мононуклеарные клетки периферической крови (PBMC), природные клетки-киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы. Эффекторные клетки можно выделять из нативного источника, например из крови.
«Антителозависимая клеточно-опосредуемая цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой секретируемый Ig, связанный с Fc-рецепторами (FcR), присутствующими на определенных цитотоксических клетках (например, природных клетках-киллерах (NK), нейтрофилах и макрофагах), вызывает специфичное для этих цитотоксических эффекторных клеток связывание с несущей антиген клеткой-мишенью, и они затем уничтожают клетку-мишень посредством цитотоксинов. Основные клетки для осуществления ADCC, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках суммирована в таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Для оценки активности представляющей интерес молекулы в отношении ADCC можно осуществить анализ ADCC in vitro, такой как анализ, описанный в патенте США № 5500362 или 5821337 или в патенте США № 6737056, Presta. Пригодные для таких анализов эффекторные клетки включают мононуклеарные клетки периферической крови (PBMC) и природные клетки-киллеры (NK). Альтернативно или дополнительно активность представляющей интерес молекулы в отношении ADCC можно оценивать in vivo, например, в модели на животных, такой как описана в Clynes et al. PNAS (USA) 95:652-656 (1998).
«Комплементзависимая цитотоксичность» или «CDC» относится к лизису клетки-мишени в присутствии комплемента. Активация классического каскада комплемента начинается со связывания первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), которые связаны с распознаваемым ими антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996).
Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или пониженной способностью связывать C1q описаны в патенте США № 6194551B1 и WO99/51642. Содержание этих патентных публикаций, в частности, приведено в настоящем документе в качестве ссылки. См. также Idusogie et al. J. Immunol. 164:4178-4184 (2000).
Термин «полипептид, содержащий Fc-область» относится к полипептиду, такому как антитело или иммуноадгезин, который содержит Fc-область. C-концевой лизин (остаток 447 согласно системе нумерации EU) Fc-области можно удалить, например, в ходе очистки полипептида или путем рекомбинантной инженерии нуклеиновой кислоты, кодирующей полипептид. Таким образом, композиция, содержащая полипептид, обладающий Fc-областью согласно этому изобретению, может содержать полипептиды с K447 со всеми удаленными K447 или смесь полипептидов с остатком K447 и без него.
«Акцепторная каркасная область человека» для целей данной работы представляет собой каркасную область, содержащую аминокислотную последовательность каркасной области VL или VH, происходящей из каркасной области иммуноглобулина человека или консенсусной каркасной области человека. Акцепторная каркасная область человека, «происходящая из» каркасной области иммуноглобулина человека или консенсусной каркасной области человека, может содержать такую же, как у них, аминокислотную последовательность, или она может содержать предсуществующие изменения в аминокислотной последовательности. В некоторых вариантах осуществления количество предсуществующих изменений аминокислот составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. Там, где предсуществующие изменения аминокислот присутствуют в VH, предпочтительно, чтобы эти изменения происходили только в трех, двух или одном из положений 71H, 73H и 78H; например, аминокислотные остатки в этих положениях могут представлять собой 71A, 73T и/или 78A. В одном из вариантов осуществления акцепторная каркасная область VL человека идентична по последовательности с последовательностью каркасной области VL иммуноглобулина человека или последовательностью консенсусной каркасной области человека.
«Консенсусная каркасная область человека» является каркасной областью, которая представляет собой наиболее часто встречающиеся аминокислотные остатки при селекции последовательностей каркасной области VL или VH иммуноглобулина человека. Как правило, селекцию последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельного домена. Как правило, подгруппа последовательностей является подгруппой, как у Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). В одном из вариантов осуществления для VL подгруппа является подгруппой каппа I, как у Kabat et al., выше. В одном из вариантов осуществления для VH подгруппа является подгруппой III, как у Kabat et al., выше.
«Консенсусная каркасная область VH подгруппы III» содержит консенсусную последовательность, полученную из аминокислотных последовательностей вариабельной подгруппы III тяжелой цепи по Kabat et al., выше. В одном из вариантов осуществления аминокислотная последовательность консенсусной каркасной области VH подгруппы III содержит, по меньшей мере, часть или все из любой из следующих последовательностей: EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:50)-H1-WVRQAPGKGLEWV (SEQ ID NO:51)-H2-RFTISADTSKNTAYLQMNSLRAEDTAVYYC (SEQ ID NO:59)-H3-WGQGTLVTVSS (SEQ ID NO:35).
«Консенсусная каркасная область VL подгруппы I» содержит консенсусную последовательность, полученную из аминокислотных последовательностей вариабельной подгруппы I легкой цепи каппа по Kabat et al., выше. В одном из вариантов осуществления аминокислотная последовательность консенсусной каркасной области VL подгруппы I содержит, по меньшей мере, часть или все из любой из следующих последовательностей: DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:60)-L1-WYQQKPGKAPKLLIY (SEQ ID NO:61)-L2-GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:62)-L3-FGQGTKVEIK (SEQ ID NO:63).
«Аффинность связывания», как правило, относится к силе общей суммы нековалентных взаимодействий между одиночным участком связывания молекулы (например, антитела) и связывающимся с ним партнером (например, антигеном). Если не указано иначе, как используют в настоящем документе, «аффинность связывания» относится к собственной аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антитела и антигена). Аффинность молекулы X по отношению к ее партнеру Y можно, как правило, представить посредством константы диссоциации (Kd). Аффинность можно измерять общепринятыми способами, известными в данной области, включая описанные в настоящем документе способы. Антитела с низкой аффинностью, как правило, связываются с антигеном медленно и обладают тенденцией к легкой диссоциации, в то время как антитела с высокой аффинностью, как правило, связываются с антигеном быстрее и обладают тенденцией дольше оставаться связанными. Множество способов измерения аффинности связывания известно в данной области, любые из которых можно использовать для целей настоящего изобретения. Конкретные наглядные варианты осуществления описаны далее.
В одном из вариантов осуществления «Kd» или «величину Kd» согласно этому изобретению измеряют с помощью анализа связывания радиоактивно меченного антигена (RIA), осуществляемого с Fab-вариантом представляющего интерес антитела и его антигеном, как описано с помощью следующего анализа. Аффинность связывания в растворе для Fab в отношении антигена измеряют посредством уравновешивания Fab минимальной концентрацией (125I)-меченного антигена в присутствии серийного разведения немеченого антигена с последующим улавливанием связанного антигена на планшете, покрытом антителами к Fab (Chen et al. (1999) J. Mol. Biol. 293:865-881). Для определения условий анализа планшеты для микротитрования (Dynex) покрывают в течение ночи 5 мкг/мл улавливающего антитела анти-Fab (Cappel Labs) в 50 мМ карбонате натрия (pH 9,6) и затем блокируют 2% (масс./об.) бычьего сывороточного альбумина в PBS в течение от двух до пяти часов при комнатной температуре (приблизительно 23°C). В неадсорбирующем планшете (Nunc #269620) 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями представляющего интерес Fab (например, в соответствии с оценкой антитела анти-VEGF, Fab-12, у Presta et al. (1997) Cancer Res. 57:4593-4599). Представляющий интерес Fab затем инкубируют в течение ночи; однако инкубацию можно продолжать в течение более длительного периода (например, 65 часов), чтобы гарантировать достижение равновесия. После этого смеси переносят в планшет для улавливания, чтобы инкубировать при комнатной температуре (например, в течение одного часа). Раствор затем удаляют и планшет промывают восемь раз 0,1% твином-20 в PBS. После того как планшеты высушивают, добавляют сцинтиллятор (MicroScint-20; Packard) из расчета 150 мкл/лунка и снимают показания с планшетов на гамма-счетчике Topcount (Packard) в течение десяти минут. Концентрации каждого Fab, которые дают менее чем 20% или равняются 20% от максимального связывания, выбирают для использования в анализах конкурентного связывания.
Согласно другому варианту осуществления Kd или величину Kd измеряют с использованием способов анализа на основе поверхностного плазмонного резонанса с использованием BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°C с чипами CM5 с иммобилизованным антигеном при ~10 единицах резонансного сигнала (RU). В кратком изложении, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) согласно инструкциям производителя. Антиген разводят 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл (~0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/минута для достижения приблизительно 10 единиц резонансного сигнала (RU) связанного белка. После инъекции антигена 1 М этаноламин инъецируют для блокирования непрореагировавших групп. Для измерений кинетики двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS с 0,05% твином-20 (PBST) инъецируют при 25°C при скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют с использованием простой модели связывания один к одному Лэнгмюра (программное обеспечение для вычисления BIAcore, версия 3.2) посредством одновременной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (Kd) вычисляют как отношение koff/kon. См., например, Chen, Y. et al. (1999) J. Mol. Biol. 293:865-881. Если скорость ассоциации превышает 106 M-1с-1 в вышеописанном анализе на основе поверхностного плазмонного резонанса, то скорость ассоциации можно определить с использованием способа гашения флюоресценции, которым измеряют повышение или понижение интенсивности испускания флюоресценции (возбуждение = 295 нМ; испускание = 340 нМ, полоса пропускания 16 нМ) при 25°C 20 нМ антитела к антигену (Fab-форма) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена при измерении в спектрометре, таком как спектрофотометр, оборудованный остановленной струей (Aviv Instruments), или спектрофотометре SLM-Aminco серии 8000 (ThermoSpectronic) с перемешиваемой кюветой.
«Скорость ассоциации», или «скорость связывания», или «kon», согласно изобретению можно также определить, как описано выше, с использованием системы BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ).
«Нарушение» представляет собой любое состояние или заболевание, на которое может оказывать благоприятное действие лечение веществом/молекулой или способом по изобретению. Оно включает хронические и острые нарушения, включающие такие патологические состояния, которые делают млекопитающее предрасположенным к рассматриваемому нарушению. Неограничивающие примеры нарушений, которые подлежат лечению по данному описанию, включают раковые состояния, такие как опухоли, например карциномы (эпителиальные опухоли) и бластомы (эмбриональные опухоли, возникающие из тканей), и в некоторых вариантах осуществления рак яичника, рак тела матки (в том числе рак эндометрия), опухоли мозга (например, астроцитомы и глиомы) и рак почки, в том числе нефробластомы (например, опухоль Вильмса).
Термины «нарушение пролиферации клеток» и «пролиферативное нарушение» относятся к нарушениям, которые связаны с некоторой степенью патологической пролиферации клеток. В одном из вариантов осуществления нарушение пролиферации клеток представляет собой рак.
«Опухоль», как используют в настоящем документе, относится к любому неопластическому росту и пролиферации клеток, или злокачественному, или доброкачественному, и любым предраковым и раковым клеткам и тканям. Как используют в данном описании, термины «рак», «раковый», «нарушение пролиферации клеток», «пролиферативное нарушение» и «опухоль» не являются взаимоисключающими.
Термины «рак» и «раковый» обозначают или описывают физиологическое состояние у млекопитающих, которое, как правило, отличается нерегулируемым клеточным ростом/пролиферацией. Примеры рака включают без ограничения карциному, лимфому (например, ходжкинскую и неходжкинскую лимфому), бластому, саркому и лейкоз. Более конкретные примеры таких форм рака включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, печеночно-клеточный рак, желудочно-кишечный рак, рак поджелудочной железы, глиому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак толстой кишки, рак ободочной и прямой кишки, карциному эндометрия или матки, карциному слюнных желез, рак почки, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, гепатокарциному, лейкоз и другие лимфопролиферативные нарушения и различные типы рака головы и шеи.
Как используют в настоящем описании, «лечение» (и вариации, такие как «лечить» или «проведение лечения») относится к клиническому вмешательству с целью изменения естественного течения заболевания индивидуума или клетки, подвергаемых лечению, и его можно осуществлять или для профилактики, или в ходе течения клинической патологии. Желательные эффекты лечения включают предотвращение возникновения или рецидивирования заболевания, смягчение симптомов, уменьшение любых непосредственных или опосредованных патологических последствий заболевания, предотвращение метастазов, снижение скорости прогрессирования заболевания, улучшение или временное облегчение состояния заболевания и ремиссию или улучшение прогноза. В некоторых вариантах осуществления антитела по изобретению применяют для задержки развития заболевания или нарушения или замедления прогрессирования заболевания или нарушения.
«Индивидуум» является позвоночным животным. В определенных вариантах осуществления позвоночное является млекопитающим. Млекопитающие включают в качестве неограничивающих примеров сельскохозяйственных животных (таких, как коровы), животных, используемых в спорте, домашних животных (таких, как кошки, собаки и лошади), приматов, мышей и крыс. В определенных вариантах осуществления млекопитающее является человеком.
«Эффективное количество» относится к количеству, эффективному, в дозировках и в течение необходимых периодов времени, для достижения требуемого терапевтического или профилактического результата.
«Терапевтически эффективное количество» вещества/молекулы по изобретению может варьировать благодаря таким факторам, как состояние заболевания, возраст, пол и масса индивидуума и способность вещества/молекулы вызывать требуемый ответ у индивидуума. Терапевтически эффективное количество включает количество, при котором любые токсические или вредные эффекты вещества/молекулы перевешиваются терапевтически полезными эффектами. «Профилактически эффективное количество» относится к количеству, эффективному, в дозировках и в течение необходимых периодов времени, для достижения требуемого профилактического результата. Как правило, но не обязательно, так как профилактическую дозу используют у субъектов перед заболеванием или на его ранней стадии, профилактически эффективное количество будет меньше терапевтически эффективного количества.
Термин «цитотоксическое средство», как используют в настоящем описании, относится к веществу, которое ингибирует функционирование клеток или препятствует ему и/или вызывает гибель или разрушение клеток. Предполагается, что термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu), химиотерапевтические средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин C, хлорамбуцил, даунорубицин или другие интеркалирующие средства, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, в том числе их фрагменты и/или варианты, и различные противоопухолевые или противораковые средства, описанные ниже). Другие цитотоксические средства описаны ниже. Тумороцидное средство вызывает разрушение опухолевых клеток.
«Токсин» представляет собой любое вещество, способное оказывать негативное воздействие на рост или пролиферацию клетки.
«Химиотерапевтическое средство» представляет собой химическое соединение, пригодное для лечения рака. Примеры химиотерапевтических средств включают алкилирующие средства, такие как тиотепа и циклофосфамид ЦИТОКСАН®; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, в том числе альтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогинины (особенно буллатацин и буллатацинон); дельта-9-тетрагидроканнабиол (дронабинол, МАРИНОЛ®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (в том числе синтетический аналог топотекан (ГИКАМТИН®), CPT-11 (иринотекан, КАМПТОСАР®), ацетилкамптотецин, скополетин и 9-аминокамптотецин); бриостатин; каллистатин; CC-1065 (в том числе его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофиллиновую кислоту; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (в том числе синтетические аналоги KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотные аналоги горчичного газа, такие как хлорамбуцил, хлорнафазин, холофосфамид, эстрамустин, ифосфамид, мехлорэтамин, мехлорэтамина оксида гидрохлорид, мелфалан, новэмбихин, фенестерин, преднемустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорзотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как эндииновые антибиотики (например, калихеамицин, особенно, калихеамицин гамма1I и калихеамицин омегаI1 (см., например, Agnew, Chem Intl. Ed. Engl., 33:183-186 (1994)); динемицин, в том числе динемицин A, эсперамицин; а также хромофор неокарциностатин и сходные хромофоры хромопротеиновых эндииновых антибиотиков, аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, декторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин АДРИАМИЦИН® (в том числе морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, порфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, циностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пуринов, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; аналоги пиримидинов, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолон пропионат, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие функцию коры надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрарубицил; бисантрен; эдатраксат; дефофамин; демеколцин; диазиквон; эфлорнитин; ацетат эллиптиния; эпотилон; этоглюцид; нитрат галлия; гидроксимочевину; лентинан; лонидамин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопидамол; нитракрин; пентостатин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2''-трихлортриэтиламин; трихотецены (особенно токсин T-2, верракурин A, роридин A и ангвидин); уретан; виндезин (ЭЛДИЗИН®, ФИЛДИЗИН®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («Ara-C»); тиотепу; таксоиды, например паклитаксел ТАКСОЛ® (Bristol-Myers Squibb Oncology, Princeton, NJ.), сконструированный на основе альбумина препарат из наночастиц паклитаксела, без кремофора, ABRAXANE™ (American Pharmaceutical Partners, Schaumberg, Illinois) и доцетаксел ТЕКСОТЕР® (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин (ГЕМЗАР®); 6-тиогуанин; меркаптопурин; метотрексат; аналоги платины, такие как цисплатин и карбоплатин; винбластин (ВЕЛБАН®); платину; этопзид (VP-16); ифосфамид; митоксантрон; винкристин (ОНКОВИН®), оксалиплатин; лейковорин; винорелбин (НАВЕЛБИН®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин (КСЕЛОДА®); фармацевтически приемлемые соли, кислоты или производные любого из вышеперечисленного; а также сочетания двух или более из вышеперечисленного, такие как CHOP, сокращение для комбинированной терапии циклофосфамидом, доксорубицином, винкристином и преднизолоном, и FOLFOX, сокращение для режима лечения оксалиплатином (ЭЛОКСАТИН™) в сочетании с 5-FU и лейковорином.
Также в это определение включены антигормональные средства, которые действуют посредством регуляции, снижения, блокирования или ингибирования воздействий гормонов, которые могут стимулировать рост раковых опухолей, и их часто используют в виде системного лечения или лечения всего организма. Они могут сами являться гормонами. Примеры включают антиэстрогены и селективные модуляторы рецепторов эстрогена (SERM), включая, например, тамоксифен (в том числе тамоксифен НОЛВАДЕКС®), ралоксифен ЭВИСТА®, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен ФАРЕСТОН®; антипрогестероны; отрицательные регуляторы рецепторов эстрогенов (ERD); средства, которые действуют, тормозя или прекращая функционирование яичников, например агонисты высвобождающего гормона лютеинизирующего гормона (LHRH), такие как ЛУПРОН® и леупролида ацетат ЭЛИГАРД®, гозерелина ацетат, бузерелина ацетат и триптерелин; другие антиандрогены, такие как флутамид, нилутамид и бикалутамид; и ингибиторы ароматазы, которые ингибируют фермент ароматазу, который регулирует выработку эстрогена в надпочечниках, такие как, например, 4(5)-имидазолы, аминоглутетимид, магестрола ацетат МЕГАС®, экземестан АРОМАЗИН®, форместан, фадрозол, ворозол РИВИЗОР®, летрозол ФЕМАРА® и анастрозол АРИМИДЕКС®. Кроме того, такое определение химиотерапевтических средств включает бифосфонаты, такие как клодронат (например, БОНЕФОС® или ОСТАК®), этидронат ДИДРОКАЛ®, NE-58095, золедроновая кислота/золедронат ЗОМЕТА®, алендронат ФОСАМАКС®, памидронат АРЕДИЯ®, тилудронат СКЕЛИД® или ризедронат АКТОНЕЛ®; а также троксацитабин (аналог нуклеозида цитозина на основе 1,3-диоксолана); антисмысловые олигонуклеотиды, в частности олигонуклеотиды, которые ингибируют экспрессию генов сигнальных путей, вовлеченных в нарушенную клеточную пролиферацию, таких как, например, PKC-альфа, Raf, H-Ras, и ген рецептора эпидермального фактора роста (EGF-R); вакцины, такие как вакцина ТЕРАТОП® и вакцины для генной терапии, например вакцина АЛЛОВЕКТИН®, вакцина ЛЕУВЕКТИН® и вакцина ВАКСИД®; ингибитор топоизомеразы 1 ЛУРТОТЕКАН®; АБАРЕЛИКС® rmRH; лапатиниб дитозилат (низкомолекулярный двойной ингибитор тирозинкиназ ErbB-2 и EGFR, также известный как GW572016); и фармацевтически приемлемые соли, кислоты или производные любого из вышеперечисленного.
«Средство ингибирования роста» при использовании в данном описании относится к соединению или композиции, которая ингибирует рост клетки (такой, как клетка, экспрессирующая TAT226) или in vitro, или in vivo. Таким образом, средство ингибирования роста может являться средством, которое значительно снижает процентную долю клеток (таких, как клетка, экспрессирующая TAT226) в S-фазе. Примеры средств ингибирования роста включают средства, которые блокируют прохождение клеточного цикла (на другой стадии по сравнению с S-фазой), такие как средства, которые индуцируют остановку в G1 и остановку в M-фазе. Классические блокаторы M-фазы включают алкалоиды барвинка (винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, эпирубицин, даунорубицин, этопозид и блеомицин. Те средства, которые вызывают остановку в G1, также в качестве побочного результата вызывают остановку в S-фазе, например средства, алкилирующие ДНК, такие как тамоксифен, преднизон, дакарбазин, мехлорэтамин, цисплатин, метотрексат, 5-фторурацил и ара-C. Дополнительную информацию можно найти в The Molecular Basis of Cancer, Mendelsohn and Israel, eds., Chapter 1, озаглавленном «Cell cycle regulation, oncogenes, and antineoplastic drugs», Murakami et al. (WB Saunders: Philadelphia, 1995), в особенности p. 13. Таксаны (паклитаксел и доцетаксел) являются противораковыми лекарственными средствами, которые, оба, получены из тисса. Доцетаксел (ТАКСОТЕР®, Rhone-Poulenc Rorer), полученный из европейского тиса, является полусинтетическим аналогом паклитаксела (ТАКСОЛ®, Bristol-Myers Squibb). Паклитаксел и доцетаксел стимулируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки путем предотвращения деполимеризации, что приводит к ингибированию митоза в клетках.
Термин «внутриклеточный метаболит» относится к соединению, образующемуся в метаболическом процессе или реакции внутри клетки из конъюгата антитело-лекарственное средство (ADC). Метаболический процесс или реакция может являться ферментативным процессом, таким как протеолитическое расщепление пептидного линкера ADC, или гидролиз функциональной группы, такой как гидразон, сложный эфир или амид. Внутриклеточные метаболиты включают в качестве неограничивающих примеров антитела и свободное лекарственное средство, которые подвергаются внутриклеточному расщеплению после поступления, диффузии, поглощения или транспорта в клетку.
Термины «внутриклеточно расщепленный» и «внутриклеточное расщепление» относятся к метаболическому процессу или реакции внутри клетки для конъюгата антитело-лекарственное средство (ADC), при которых ковалентный мостик, т.е. линкер, между компонентом лекарственного средства (D) и антителом (Ab) разрушается, что приводит к диссоциации свободного лекарственного средства и антитела внутри клетки. Расщепленные компоненты ADC, таким образом, являются внутриклеточными метаболитами.
Термин «биодоступность» относится к системной доступности (т.е. уровням кровь/плазма) данного количества лекарственного средства, введенного пациенту. Биодоступность является абсолютным термином, который указывает на измерение как времени (скорости) для лекарственного средства, так и его общего количества (степени), которое достигает общего кровотока из введенной лекарственной формы.
Термин «цитотоксическая активность» относится к эффекту уничтожения клеток, цитостатическому эффекту или эффекту ингибирования роста для конъюгата антитело-лекарственное средство или внутриклеточного метаболита конъюгата антитело-лекарственное средство. Цитотоксическую активность можно выразить в виде величины IC50, которая представляет собой концентрацию (молярную или массовую) на единицу объема, в котором остается в живых половина клеток.
«Алкил» представляет собой углеводород C1-C18, содержащий нормальные, вторичные, третичные или циклические углеродные атомы. Примерами являются метил (Me, -CH3), этил (Et, -CH2CH3), 1-пропил (n-Pr, н-пропил, -CH2CH2CH3), 2-пропил (i-Pr, изопропил, -CH(CH3)2), 1-бутил (n-Bu, н-бутил, -CH2CH2CH2CH3), 2-метил-1-пропил (i-Bu, изобутил, -CH2CH(CH3)2), 2-бутил (s-Bu, втор-бутил, -CH(CH3)CH2CH3), 2-метил-2-пропил (t-Bu, трет-бутил, -C(CH3)3), 1-пентил (н-пентил, -CH2CH2CH2CH2CH3), 2-пентил (-CH(CH3)CH2CH2CH3), 3-пентил (-CH(CH2CH3)2), 2-метил-2-бутил (-C(CH3)2CH2CH3), 3-метил-2-бутил (-CH(CH3)CH(CH3)2), 3-метил-1-бутил (-CH2CH2CH(CH3)2), 2-метил-1-бутил (-CH2CH(CH3)CH2CH3), 1-гексил (-CH2CH2CH2CH2CH2CH3), 2-гексил (-CH(CH3)CH2CH2CH2CH3), 3-гексил (-CH(CH2CH3)(CH2CH2CH3)), 2-метил-2-пентил (-C(CH3)2CH2CH2CH3), 3-метил-2-пентил (-CH(CH3)CH(CH3)CH2CH3), 4-метил-2-пентил (-CH(CH3)CH2CH(CH3)2), 3-метил-3-пентил (-C(CH3)(CH2CH3)2), 2-метил-3-пентил (-CH(CH2CH3)CH(CH3)2), 2,3-диметил-2-бутил (-C(CH3)2CH(CH3)2), 3,3-диметил-2-бутил (-CH(CH3)C(CH3)3.
Термин «C1-C8алкил», как используют в настоящем документе, относится к неразветвленному или разветвленному, насыщенному или ненасыщенному углеводороду, содержащему от 1 до 8 углеродных атомов. Репрезентативные «C1-C8алкильные группы» включают в качестве неограничивающих примеров -метил, -этил, -н-пропил, -н-бутил, -н-пентил, -н-гексил, -н-гептил, -н-октил, -н-нонил и -н-децил; в то время как разветвленные C1-C8алкилы включают в качестве неограничивающих примеров -изопропил, -втор-бутил, -изобутил, -трет-бутил, -изопентил, 2-метилбутил, ненасыщенные C1-C8алкилы включают в качестве неограничивающих примеров -винил, -аллил, -1-бутенил, -2-бутенил, -изобутиленил, -1-пентенил, -2-пентенил, -3-метил-1-бутенил, -2-метил-2-бутенил, -2,3-диметил-2-бутенил, 1-гексил, 2-гексил, 3-гексил, -ацетиленил, -пропинил, -1-бутинил, -2-бутинил, -1-пентинил, -2-пентинил, -3-метил-1-бутинил, метил, этил, пропил, изопропил, н-бутил, изобутил, втор-бутил, трет-бутил, н-пентил, изопентил, неопентил, н-гексил, изогексил, 2-метилпентил, 3-метилпентил, 2,2-диметилбутил, 2,3-диметилбутил, 2,2-диметилпентил, 2,3-диметилпентил, 3,3-диметилпентил, 2,3,4-триметилпентил, 3-метилгексил, 2,2-диметилгексил, 2,4-диметилгексил, 2,5-диметилгексил, 3,5-диметилгексил, 2,4-диметилпентил, 2-метилгептил, 3-метилгептил, н-гептил, изогептил, н-октил и изооктил. C1-C8алкильная группа может являться незамещенной или замещенной одним или несколькими группами, включая в качестве неограничивающих примеров -C1-C8алкил, -O- (C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2-NHC(O)R', -SO3R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«Алкенил» представляет собой C2-C18углеводород, содержащий нормальные, вторичные, третичные или циклические углеродные атомы, по меньшей мере, с одним ненасыщенным участком, т.е. углерод-углеродной, двойной sp 2-связью. Примеры включают без ограничения: этилен или винил (-CH=CH2), аллил (-CH2CH=CH2), циклопентенил (-C5H7) и 5-гексенил (-CH2CH2CH2CH2CH=CH2).
«Алкинил» представляет собой C2-C18углеводород, содержащий нормальные, вторичные, третичные или циклические углеродные атомы, по меньшей мере, с одним ненасыщенным участком, т.е. углерод-углеродной, тройной sp-связью. Примеры включают без ограничения: ацетиленил (-C≡CH) и пропаргил (-CH2C≡CH).
«Алкилен» относится к насыщенному, разветвленному или неразветвленному, или циклическому углеводородному радикалу из 1-18 углеродных атомов и обладающему двумя одновалентными радикальными центрами, которые получаются в результате удаления двух водородных атомов от одного и того же или двух разных углеродных атомов исходного алкана. Типичные алкиленовые радикалы включают в качестве неограничивающих примеров: метилен (-CH2-) 1,2-этил (-CH2CH2-), 1,3-пропил (-CH2CH2CH2-), 1,4-бутил (-CH2CH2CH2CH2-) и т.п.
«C1-C10алкилен» представляет собой неразветвленную, насыщенную углеводородную группу формулы -(CH2)1-10-. Примеры C1-C10алкиленов включают метилен, этилен, пропилен, бутилен, пентилен, гексилен, гептилен, октилен, нонилен и декален.
«Алкенилен» относится к ненасыщенному, разветвленному или неразветвленному, или циклическому углеводородному радикалу из 2-18 углеродных атомов и обладающему двумя одновалентными радикальными центрами, которые получаются в результате удаления двух водородных атомов от одного и того же или двух разных углеродных атомов исходного алкена. Типичные алкениленовые радикалы включают в качестве неограничивающих примеров: 1,2-этилен (-CH=CH-).
«Алкинилен» относится к ненасыщенному, разветвленному или неразветвленному, или циклическому углеводородному радикалу из 2-18 углеродных атомов и обладающему двумя одновалентными радикальными центрами, которые получаются в результате удаления двух водородных атомов от одного и того же или двух разных углеродных атомов исходного алкина. Типичные алкиниленовые радикалы включают в качестве неограничивающих примеров: ацетилен (-C≡C-), пропаргил (-CH2C≡C-) и 4-пентинил (-CH2CH2CH2C≡C-).
«Арил» относится к карбоциклической ароматической группе. Примеры арильных групп включают в качестве неограничивающих примеров фенил, нафтил и антраценил. Карбоциклическая ароматическая группа или гетероциклическая ароматическая группа может являться незамещенной или замещенной одной или несколькими группами, включая в качестве неограничивающих примеров -C1-C8алкил, -O-(C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, - C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), - N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«Арилен» представляет собой арильную группу, которая обладает двумя ковалентными связями и может находиться в орто-, мета- или параконфигурациях, как представлено на следующих структурах:
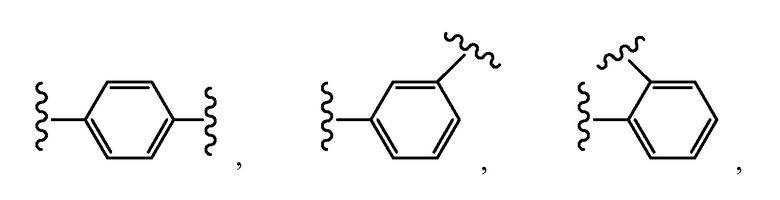
в которых фенильная группа может являться незамещенной или замещенной группами, вплоть до четырех, включающими в качестве неограничивающих примеров -C1-C8алкил, -O-(C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, - C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), - N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«Арилалкил» относится к ациклическому алкильному радикалу, в котором один из водородных атомов, связанный с углеродным атомом, как правило, концевым или sp 3-атомом углерода, замещен арильным радикалом. Типичные арилалкильные группы включают в качестве неограничивающих примеров бензил, 2-фенилэтан-1-ил, 2-фенилэтен-1-ил, нафтилметил, 2-нафтилэтан-1-ил, 2-нафтилэтен-1-ил, нафтобензил, 2-нафтофенилэтан-1-ил и т.п. Арилалкильная группа содержит от 6 до 20 углеродных атомов, например алкильная группа, включая алканильную, алкенильную или алкинильную группы, арилалкильной группы состоит из 1-6 углеродных атомов, и арильная группа состоит из 5-14 углеродных атомов.
«Гетероарилалкил» относится к ациклическому алкильному радикалу, в котором один из водородных атомов, связанный с углеродным атомом, как правило, концевым или sp 3-атомом углерода, замещен гетероарильным радикалом. Типичные гетероарилалкильные группы включают в качестве неограничивающих примеров 2-бензимидазолилметил, 2-фурилэтил и т.п. Гетероарилалкильная группа содержит от 6 до 20 углеродных атомов, например алкильная группа, включая алканильную, алкенильную или алкинильную группы, гетероарилалкильной группы состоит из 1-6 углеродных атомов, и гетероарильная группа состоит из 5-14 углеродных атомов, и от 1 до 3 гетероатомов выбраны из N, O, P и S. Гетероарильный компонент гетероарилалкильной группы может представлять собой моноцикл, содержащий от 3 до 7 элементов кольца (2-6 углеродных атомов) или бицикл, содержащий от 7 до 10 элементов кольца (4-9 углеродных атомов, и от 1 до 3 гетероатомов выбраны из N, O, P и S), например систему бицикло[4.5], [5.5], [5.6] или [6.6].
«Замещенный алкил», «замещенный арил» и «замещенный арилалкил» означает соответственно алкил, арил и арилалкил, в котором любой из одного или нескольких водородных атомов независимо замещен заместителем. Типичные заместители включают в качестве неограничивающих примеров -X, -R, -O-, -OR, -SR, -S-, -NR2, -NR3, =NR, -CX3, -CN, -OCN, -SCN, -N=C=O, -NCS, -NO, -NO2, =N2, -N3, NC(=O)R, -C(=O)R, -C(=O)NR2, -SO3 -, -SO3H, -S(=O)2R, -OS(=O)2OR, -S(=O)2NR, -S(=O)R, -OP(=O)(OR)2, -P(=O)(OR)2, -PO- 3, -PO3H2, -C(=O)R, -C(=O)X, -C(=S)R, -CO2R, -CO2 -, -C(=S)OR, -C(=O)SR, -C(=S)SR, -C(=O)NR2, -C(=S)NR2, -C(=NR)NR2, где каждый X независимо представляет собой галоген: F, Cl, Br или I; и каждый R независимо представляет собой -H, C2-C18алкил, C6-C20арил, C3-C14гетероцикл, защитную группу или компонент пролекарства. Алкиленовые, алкениленовые и алкиниленовые группы, как описано выше, можно заместить сходным образом.
«Гетероарил» и «гетероцикл» относится к кольцевой системе, в которой один или несколько атомов кольца являются гетероатомами, например азотом, кислородом и серой. Гетероциклический радикал содержит от 1 до 20 углеродных атомов, и от 1 до 3 гетероатомов выбраны из N, O, P и S. Гетероцикл может представлять собой моноцикл, содержащий от 3 до 7 элементов кольца (2-6 углеродных атомов, и от 1 до 3 гетероатомов выбраны из N, O, P и S) или бицикл, содержащий от 7 до 10 элементов кольца (4-9 углеродных атомов, и от 1 до 3 гетероатомов выбраны из N, O, P и S), например систему бицикло[4.5], [5.5], [5.6] или [6.6].
Гетероциклы описаны в Paquette, Leo A., «Principles of Modern Heterocyclic Chemistry» (W.A. Benjamin, New York, 1968), в частности главы 1, 3, 4, 6, 7 и 9; «The Chemistry of Heterocyclic Compounds, A series of Monographs» (John Wiley & Sons, New York, 1950 до настоящего времени), в частности тома 13, 14, 16, 19 и 28; и J. Am. Chem. Soc. (1960) 82:5566.
Примеры гетероциклов включают в качестве неограничивающих примеров пиридил, дигидропиридил, тетрагидропиридил (пиперидил), тиазолил, тетрагидротиофенил, тетрагидротиофенил с окисленной серой, пиримидинил, фуранил, тиенил, пирролил, пиразолил, имидазолил, тетразолил, бензофуранил, тианафталенил, индолил, индоленил, хинолинил, изохинолинил, бензимидазолил, пиперидинил, 4-пиперидонил, пирролидинил, 2-пирролидонил, пирролинил, тетрагидрофуранил, бис-тетрагидрофуранил, тетрагидропиранил, бис-тетрагидропиранил, тетрагидрохинолинил, тетрагидроизохинолинил, декагидрохинолинил, октагидроизохинолинил, азоцинил, триазинил, 6H-1,2,5-тиадиазинил, 2H,6H-1,5,2-дитиазинил, тиенил, тиантренил, пиранил, изобензофуранил, хроменил, ксантенил, феноксатинил, 2H-пирролил, изотиазолил, изоксазолил, пиразинил, пиридазинил, индолизинил, изоиндолил, 3H-индолил, 1H-индазолил, пуринил, 4H-хинолизинил, фталазинил, нафтиридинил, хиноксалинил, хиназолинил, циннолинил, птеридинил, 4aH-карбазолил, карбазолил, β-карболинил, фенантридинил, акридинил, пиримидинил, фенантролинил, феназинил, фенотиазинил, фуразанил, феноксазинил, изохроманил, хроманил, имидазолидинил, имидазолинил, пиразолидинил, пиразолинил, пиперазинил, индолинил, изоиндолинил, хинуклидинил, морфолинил, оксазолидинил, бензотриазолил, бензизоксазолил, оксиндолил, бензоксазолинил и изатиноил.
В качестве неограничивающих примеров, связанные с углеродом гетероциклы связаны в положении 2, 3, 4, 5 или 6 пиридина, положении 3, 4, 5 или 6 пиридазина, положении 2, 4, 5 или 6 пиримидина, положении 2, 3, 5 или 6 пиразина, положении 2, 3, 4 или 5 фурана, тетрагидрофурана, тиофурана, тиофена, пиррола или тетрагидропиррола, положении 2, 4 или 5 оксазола, имидазола или тиазола, положении 3, 4 или 5 изоксазола, пиразола или изотиазола, положении 2 или 3 азиридина, положении 2, 3 или 4 азетидина, положении 2, 3, 4, 5, 6, 7 или 8 хинолина или положении 1, 3, 4, 5, 6, 7 или 8 изохинолина. Более конкретно, связанные с углеродом гетероциклы включают 2-пиридил, 3-пиридил, 4-пиридил, 5-пиридил, 6-пиридил, 3-пиридазинил, 4-пиридазинил, 5-пиридазинил, 6-пиридазинил, 2-пиримидинил, 4-пиримидинил, 5-пиримидинил, 6-пиримидинил, 2-пиразинил, 3-пиразинил, 5-пиразинил, 6-пиразинил, 2-тиазолил, 4-тиазолил или 5-тиазолил.
В качестве неограничивающих примеров, связанные с азотом гетероциклы связаны в положении 1 азиридина, азетидина, пиррола, пирролидина, 2-пирролина, 3-пирролина, имидазола, имидазолидина, 2-имидазолина, 3-имидазолина, пиразола, пиразолина, 2-пиразолина, 3-пиразолина, пиперидина, пиперазина, индола, индолина, 1H-индазола, положении 2 изоиндола или изоиндолина, положении 4 морфолина и положении 9 карбазола или β-карболина. Более конкретно, связанные с азотом гетероциклы включают 1-азиридил, 1-азетедил, 1-пирролил, 1-имидазолил, 1-пиразолил и 1-пиперидинил.
«C3-C8гетероцикл» относится к ароматическому или неароматическому C3-C8 карбоциклу, в котором от одного до четырех углеродных атомов кольца независимо замещены гетероатомом из группы, состоящей из O, S и N. Репрезентативные примеры C3-C8гетероциклов включают без ограничения бензофуранил, бензотиофен, индолил, бензопиразолил, кумаринил, изохинолинил, пирролил, тиофенил, фуранил, тиазолил, имидазолил, пиразолил, триазолил, хинолинил, пиримидинил, пиридинил, пиридонил, пиразинил, пиридазинил, изотиазолил, изоксазолил и тетразолил. C3-C8гетероцикл может являться незамещенным или замещенным группами, вплоть до семи, включающими в качестве неограничивающих примеров -C1-C8алкил, -O-(C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«C3-C8гетероцикло» относится к определенной выше C3-C8гетероциклической группе, где один из водородных атомов гетероциклической группы замещен связью. C3-C8гетероцикло может являться незамещенным или замещенным группами, вплоть до шести, включающими в качестве неограничивающих примеров -C1-C8алкил, -O-(C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, -C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), - N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«Карбоцикл» означает насыщенное или ненасыщенное кольцо, содержащее от 3 до 7 углеродных атомов в виде моноцикла или от 7 до 12 углеродных атомов в виде бицикла. Моноциклические карбоциклы содержат от 3 до 6 атомов в кольце, более конкретно, 5 или 6 атомов в кольце. Бициклические карбоциклы содержат от 7 до 12 атомов в кольце, например, расположенных в виде системы бицикло [4,5], [5,5], [5,6] или [6,6], или 9 или 10 атомов в кольце, расположенных в виде системы бицикло [5,6] или [6,6]. Примеры моноциклических карбоциклов включают циклопропил, циклобутил, циклопентил, 1-циклопент-1-енил, 1-циклопент-2-енил, 1-циклопент-3-енил, циклогексил, 1-циклогекс-1-енил, 1-циклогекс-2-енил, 1-циклогекс-3-енил, циклогептил и циклооктил.
«C3-C8карбоцикл» представляет собой 3-, 4-, 5-, 6-, 7- или 8-членное насыщенное или ненасыщенное неароматическое карбоциклическое кольцо. Репрезентативные C3-C8карбоциклы включают в качестве неограничивающих примеров -циклопропил, -циклобутил, -циклопентил, -циклопентадиенил, -циклогексил, -циклогексенил, -1,3-циклогексадиенил, -1,4-циклогексадиенил, -циклогептил, -1,3-циклогептадиенил, -1,3,5-циклогептатриенил, -циклооктил и -циклооктадиенил. C3-C8карбоциклическая группа может являться незамещенной или замещенной одной или несколькими группами, включающими в качестве неограничивающих примеров -C1-C8алкил, -O-(-C1-C8алкил), -арил, -C(O)R', -OC(O)R', -C(O)OR', -C(O)NH2, - C(O)NHR', -C(O)N(R')2, -NHC(O)R', -S(O)2R', -S(O)R', -OH, -галоген, -N3, -NH2, -NH(R'), -N(R')2 и -CN; где каждый R' независимо выбран из H, -C1-C8алкила и арила.
«C3-C8карбоцикло» относится к определенной выше C3-C8карбоциклической группе, где один из водородных атомов карбоциклической группы замещен связью.
«Линкер» относится к химическому компоненту, содержащему ковалентную связь или цепь атомов, которые ковалентно присоединяют антитело к компоненту лекарственного средства. В различных вариантах осуществления линкеры включают двухвалентные радикалы, такие как алкилдиил, арилдиил, гетероарилдиил, компоненты, такие как: -(CR2)nO(CR2)n-, повторяющиеся звенья алкилокси (например, полиэтиленокси, PEG, полиметиленокси) и алкиламино (например, полиэтиленамино, Jeffamine™); и сложные эфиры и амиды двухосновных кислот, включая сукцинат, сукцинамид, дигликолят, малонат и капроамид.
Термин «хиральный» относится к молекулам, которые обладают свойством невозможности совмещения со своим зеркальным отображением, в то время как термин «ахиральный» относится к молекулам, которые можно совместить со своим зеркальным отображением.
Термин «стереоизомеры» относится к соединениям, которые обладают идентичным химическим строением, но отличаются в отношении расположения атомов или групп в пространстве.
«Диастереомер» относится к стереоизомеру с двумя или несколькими центрами хиральности и тем молекулам, которые не являются зеркальными отображениями друг друга. Диастереомеры обладают разными физическими свойствами, например температурами плавления, температурами кипения, спектральными характеристиками и реакционными способностями. Смеси диастереомеров можно разделить аналитическими способами высокого разрешения, такими как электрофорез и хроматография.
«Энантиомеры» относятся к двум стереоизомерам соединения, которые являются несовместимыми зеркальными отображениями друг друга.
Используемые здесь стереохимические определения и соглашения, как правило, следуют S. P. Parker, Ed., McGraw-Hill Dictionary of Chemical Terms (1984) McGraw-Hill Book Company, New York; и Eliel, E. and Wilen, S., Stereochemistry of Organic Compounds (1994) John Wiley & Sons, Inc., New York. Многие органические соединения существуют в оптически активных формах, т.е. они обладают способностью вращать плоскость плоскополяризованного света. В описании оптически активного соединения приставки D и L, или R и S, используют для обозначения абсолютной конфигурации молекулы в районе ее хирального(ых) центра(ов). Приставки d и l, или (+) и (-), используют для обозначения знака вращения плоскополяризованного света соединением, где (-) или l означает, что соединение является левовращающим. Соединение с приставкой (+) или d является правовращающим. Для данной химической структуры эти стереоизомеры идентичны за исключением того, что они являются зеркальными отображениями друг друга. Конкретный стереоизомер можно также обозначить как энантиомер, и смесь таких изомеров часто называют энантиомерной смесью. Смесь энантиомеров 50:50 обозначают как рацемическую смесь или рацемат, которая может образоваться там, где отсутствует стереоселективность или стереоспецифичность химической реакции или процесса. Термины «рацемическая смесь» и «рацемат» относятся к эквимолярной смеси двух форм энантиомеров, которая лишена оптической активности.
«Уходящая группа» относится к функциональной группе, которую можно заместить другой функциональной группой. Определенные уходящие группы хорошо известны в данной области, и примеры включают без ограничения галогенид (например, хлорид, бромид, йодид), метансульфонил (мезил), п-толуолсульфонил (тозил), трифторметилсульфонил (трифлат) и трифторметилсульфонат.
B. Сокращения
КОМПОНЕНТЫ ЛИНКЕРА:
MC = 6-малеимидокапроил
Val-Cit или «vc» = валин-цитруллин (иллюстративный дипептид в расщепляемом протеазой линкере)
Цитруллин = 2-амино-5-уреидопентановая кислота
PAB = п-аминобензилоксикарбонил (пример «жертвующего собой» компонента линкера)
Me-Val-Cit = N-метил-валин-цитруллин (где линкерная пептидная связь модифицирована для предотвращения ее расщепления катепсином B)
MC(PEG)6-OH = малеимидокапроилполиэтиленгликоль (можно присоединять к цистеинам антитела)
ЦИТОТОКСИЧЕСКИЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА:
MMAE = монометилауристатин E (MW 718)
MMAF = вариант ауристатина E (MMAE) с фенилаланином на C-конце лекарственного средства (MW 731.5)
MMAF-DMAEA = MMAF с DMAEA (диметиламиноэтиламином) в амидной связи с C-концевым фенилаланином (MW 801.5)
MMAF-TEG = MMAF с тетраэтиленгликолем, этерифицированым фенилаланином
MMAF-NtBu = N-трет-бутил, присоединенный в виде амида к C-концу MMAF
Дополнительные сокращения представляют собой, как указано далее: AE представляет собой ауристатин E, Boc представляет собой N-(трет-бутоксикарбонил), cit представляет собой цитруллин, dap представляет собой долапроин, DCC представляет собой 1,3-дициклогексилкарбодиимид, DCM представляет собой дихлорметан, DEA представляет собой диэтиламин, DEAD представляет собой диэтилазодикарбоксилат, DEPC представляет собой диэтилфосфорилцианидат, DIAD представляет собой диизопропилазодикарбоксилат, DIEA представляет собой N,N-диизопропилэтиламин, dil представляет собой долаизолейцин, DMA представляет собой диметилацетамид, DMAP представляет собой 4-диметиламинопиридин, DME представляет собой диметиловый эфир этиленгликоля (или 1,2-диметоксиэтан), DMF представляет собой N,N-диметилформамид, DMSO представляет собой диметилсульфоксид, doe представляет собой долафенин, dov представляет собой N,N-диметилвалин, DTNB представляет собой 5,5'-дитиобис(2-нитробензойную кислоту), DTPA представляет собой диэтилентриаминопентауксусную кислоту, DTT представляет собой дитиотреитол, EDCI представляет собой 1-(3-диметиламинопропил)-3-этилкарбодиимид-гидрохлорид, EEDQ представляет собой 2-этокси-1-этоксикарбонил-1,2-дигидрохинолин, ES-MS представляет собой масс-спектрометрию с электрораспылением, EtOAc представляет собой этилацетат, Fmoc представляет собой N-(9-флюоренилметоксикарбонил), gly представляет собой глицин, HATU представляет собой O-(7-азабензотриазол-1-ил)-N,N,N',N'-тетраметилуроний-гексафторфосфат, HOBt представляет собой 1-гидроксибензотриазол, HPLC представляет собой жидкостную хроматографию высокого давления, ile представляет собой изолейцин, lys представляет собой лизин, MeCN (CH3CN) представляет собой ацетонитрил, MeOH представляет собой метанол, Mtr представляет собой 4-анизилдифенилметил (или 4-метокситритил), nor представляет собой (1S,2R)-(+)-норэфедрин, PBS представляет собой фосфатно-буферный солевой раствор (pH 7,4), PEG представляет собой полиэтиленгликоль, Ph представляет собой фенил, Pnp представляет собой п-нитрофенил, MC представляет собой 6-малеимидокапроил, phe представляет собой L-фенилаланин, PyBrop представляет собой бром-трис-пирролидинoфосфоний-гексафторфосфат, SEC представляет собой эксклюзионную хроматографию, Su представляет собой сукцинимид, TFA представляет собой трифторуксусную кислоту, TLC представляет собой тонкослойную хроматографию, UV представляет собой ультрафиолет, и val представляет собой валин.
III. КОМПОЗИЦИИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ
Представлены антитела, которые связываются с TAT226. Представлены иммуноконъюгаты, содержащие анти-TAT226 антитела. Антитела и иммуноконъюгаты по изобретению пригодны, например, для диагностики или лечения нарушений, связанных с измененной экспрессией, например повышенной экспрессией, TAT226. В некоторых вариантах осуществления антитело или иммуноконъюгаты по изобретению пригодны для диагностики или лечения нарушения пролиферации клеток, такого как рак.
A. Анти-TAT226 антитела
TAT226 («опухоль-ассоциированная антигенная мишень № 226») представляет собой белок, который процессируется и экспрессируется на поверхности определенных типов клеток, включая опухолевые клетки. В частности, ранее сообщалось, что TAT226 человека сверхэкспрессируется в определенных типах опухолей, включая опухоли яичников, матки, эндометрия, почки, легкого, поджелудочной железы, надпочечников и клеток печени. См., например, публикации патентных заявок США № US 2003/0148408 A1, US 2004/0229277 A1 и US 2003/0100712 A1 (где TAT226 обозначен как «PRO9917»); и патент США № 6710170 B2 (SEQ ID NO:215). Другие компоненты и описания в базах данных, связанные с TAT226, представляют собой, как указано далее: депозитарный № NCBI AY358628_1 (где TAT226 человека обозначен как «PSCA Hlog»); депозитарный № NCBI AAQ88991.1 и «gi» № 37182378 (где TAT226 человека обозначен как «PSCA Hlog»); RIKEN cDNA 2700050C12; US 2003/0096961 A1 (SEQ ID NO:16); US 2003/0129192 A1 (SEQ ID NO:215); US 2003/0206918 A1 (пример 5; SEQ ID NO:215); US 2003/0232056 A1 (SEQ ID NO:215); US 2004/0044179 A1 (SEQ ID NO:16); US 2004/0044180 A1 (SEQ ID NO:16); US 2005/0238649 A1 (где TAT226 человека обозначен как «PSCA Hlog»); WO 2003/025148 (SEQ ID NO:292); WO 2003/105758 (SEQ ID NO:14) и EP 1347046 (SEQ ID NO:2640).
Полноразмерный TAT226 подвергается процессингу в клетке с образованием зрелой формы белка, которая экспрессируется на клеточной поверхности. Например, полноразмерный TAT226 человека, как представлено в SEQ ID NO:75, содержит предполагаемую последовательность сигнального пептида из аминокислот 1-20 или 1-22, которая, как предполагают, отщепляется от белка. C-конец из аминокислот 116-141, как предполагают, отщепляется от белка, и компонент GPI присоединяется к аминокислоте 115 белка. Как предполагают, зрелая форма TAT226 человека из аминокислот 21-115 или 23-115 SEQ ID NO:75 заякорена на клеточной поверхности с помощью компонента GPI. TAT226 обезьян и грызунов (см., например, SEQ ID NO:76-78) в высокой степени сходен с TAT226 человека и, таким образом, как предполагают, отщепляется и модифицируется в эквивалентных положениях аминокислот. См. фигуру 1. Получающиеся зрелые формы TAT226 человека, обезьян и грызунов из аминокислот 21-115 или 23-115 (как представлено на фигуре 1) идентичны на 100%.
Другие характеристики TAT226 человека включают предполагаемый участок N-гликозилирования в положении аминокислоты 45, который подтвержден экспериментально, и предполагаемый домен Ly6/u-PAR из аминокислот 94-107 SEQ ID NO:75. TAT226 человека обладает приблизительно 32% аминокислотной гомологией с антигеном стволовых клеток предстательной железы (PSCA), специфичным для рака предстательной железы опухолевым антигеном, который экспрессируется на клеточной поверхности с помощью GPI-якоря. См. Reiter et al. (1998) Proc. Natl. Acad. Sci. USA 95:1735-1740. PSCA сверхэкспрессируется в свыше 80% случаев рака предстательной железы. Id. Подобно TAT226, он содержит предполагаемый домен Ly6/u-PAR, который вовлечен в клеточные функции, такие как передача сигнала и адгезия клеток. Id.
В одном из аспектов изобретение относится к антителам, которые связываются с TAT226. В некоторых вариантах осуществления представлены антитела, которые связываются со зрелой формой TAT226. В одном из таких вариантов осуществления зрелая форма TAT226 обладает аминокислотной последовательностью из аминокислот 21-115 или 23-115 SEQ ID NO:75. В некоторых вариантах осуществления антитело к TAT226 связывается со зрелой формой TAT226, экспрессирующейся на клеточной поверхности. В некоторых вариантах осуществления антитело, которое связывается со зрелой формой TAT226, экспрессирующейся на клеточной поверхности, ингибирует рост клетки. В некоторых вариантах осуществления анти-TAT226 антитело связывается со зрелой формой TAT226, экспрессирующейся на клеточной поверхности, и ингибирует пролиферацию клеток. В некоторых вариантах осуществления анти-TAT226 антитело связывается со зрелой формой TAT226, экспрессирующейся на клеточной поверхности, и вызывает гибель клеток. В некоторых вариантах осуществления анти-TAT226 антитело связывается со зрелой формой TAT226, экспрессирующейся на поверхности раковых клеток. В некоторых вариантах осуществления анти-TAT226 антитело связывается со зрелой формой TAT226, которая сверхэкспрессируется на поверхности раковых клеток по сравнению с нормальными клетками, происходящими из той же ткани.
В одном из аспектов анти-TAT226 антитело является моноклональным антителом. В одном из аспектов анти-TAT226 антитело является фрагментом антитела, например фрагментом Fab, Fab'-SH, Fv, scFv или (Fab')2. В одном из аспектов анти-TAT226 антитело является химерным, гуманизированным или человеческим антителом. В одном из аспектов любое из описанных здесь анти-TAT226 антител является очищенным.
Иллюстративные моноклональные антитела, полученные из фаговой библиотеки, представлены здесь, как описано в примере B. Антиген, используемый для скрининга библиотеки, представлял собой полипептид, обладающий последовательностью из аминокислот 1-115 SEQ ID NO:75, что соответствует форме TAT226, лишенной аминокислот, которые являются C-концевыми для предполагаемого участка прикрепления GPI. Антитела, полученные из скрининга библиотеки, обозначены YWO.32 и YWO.49. YWO.49 подвергли аффинному созреванию с образованием YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6. Выравнивания последовательностей вариабельных доменов тяжелой и легкой цепей YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6 представлены на фигурах 11 и 12 соответственно.
В одном из аспектов представлены моноклональные антитела, которые конкурируют с YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6 за связывание с TAT226. Также представлены моноклональные антитела, которые связываются с тем же эпитопом, что и YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6.
В одном из аспектов изобретения представлены полинуклеотиды, кодирующие анти-TAT226 антитело. В некоторых вариантах осуществления представлены векторы, содержащие полинуклеотиды, кодирующие анти-TAT226 антитела. В некоторых вариантах осуществления представлены клетки-хозяева, содержащие такие векторы. В другом аспекте изобретения представлены композиции, содержащие анти-TAT226 антитела или полинуклеотиды, кодирующие анти-TAT226 антитела. В некоторых вариантах осуществления композиция по изобретению является фармацевтической композицией для лечения нарушения пролиферации клеток, такого как перечисленные здесь нарушения.
Подробное описание иллюстративных анти-TAT226 антител представляет собой, как указано далее:
1. Конкретные варианты осуществления анти-TAT226 антител
В одном из аспектов изобретение относится к антителу, содержащему, по меньшей мере, одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:4; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:5; (c) HVR-H3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11; (d) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO:14-19.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему, по меньшей мере, одну, по меньшей мере, две или все три последовательности VH HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:4; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:5; и (c) HVR-H3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:4. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:5. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:9 или 10.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11, и HVR-H1, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:1 и SEQ ID NO:4. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-11, и HVR-H1, содержащую SEQ ID NO:4. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую SEQ ID NO:9 или 10, и HVR-H1, содержащую SEQ ID NO:4. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую SEQ ID NO:3, и HVR-H1, содержащую SEQ ID NO:1.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11, и HVR-H2, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:2 и SEQ ID NO:5. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-11, и HVR-H2, содержащую SEQ ID NO:5. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую SEQ ID NO:9 или 10, и HVR-H2, содержащую SEQ ID NO:5. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H3, содержащую SEQ ID NO:3, и HVR-H2, содержащую SEQ ID NO:2.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:4; HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:5; и HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11. В одном из вариантов осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO:4; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:5; и HVR-H3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:6-11. В одном из вариантов осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO:4; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:5; и HVR-H3 содержит SEQ ID NO:9 или 10. В одном из вариантов осуществления HVR-H1 содержит аминокислотную последовательность SEQ ID NO:1; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:2; и HVR-H3 содержит аминокислотную последовательность SEQ ID NO:3.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему, по меньшей мере, одну, по меньшей мере, две, или все три последовательности VL HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:12; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:13; и (c) HVR-H3, содержащей аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:17 или 18.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11, и (b) HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:3, и (b) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:14. В некоторых вариантах осуществления антитело к TAT226 дополнительно содержит (a) HVR-H1, содержащую SEQ ID NO:1, и HVR-H2, содержащую SEQ ID NO:2.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-11, и (b) HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В некоторых вариантах осуществления HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17. В некоторых вариантах осуществления HVR-H3 содержит аминокислотную последовательность SEQ ID NO:10, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:18. В некоторых вариантах осуществления антитело к TAT226 дополнительно содержит HVR-H1, содержащую SEQ ID NO:4, и HVR-H2, содержащую SEQ ID NO:5.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:1 или SEQ ID NO:4; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:5; (c) HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:3 и 6-11; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-19.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:1; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:2; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:3; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:14.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5; (c) HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-11; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-19.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:9; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:17.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:10; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:18.
В некоторых вариантах осуществления анти-TAT226 антитело подвергают аффинному созреванию для получения требуемой аффинности связывания с мишенью. В некоторых вариантах осуществления любую из одной или нескольких аминокислот антитела замещают в следующих положениях HVR (нумерация по Kabat): H98, H99, H100, H100B, L90, L92, L93, L96 и L97. Например, в некоторых вариантах осуществления любое из одного или нескольких из следующих замещений можно получить в любом сочетании:
- в HVR-H3 (SEQ ID NO:6): V98I; S99T; R100L или I; G100bA, S или P;
- в HVR-L3 (SEQ ID NO:14): Q90R, K, H или N; Y92V; T93F, N, G или A; P96F; T97I или A.
Все возможные сочетания вышеупомянутых замещений охватываются консенсусными последовательностями SEQ ID NO:11 (HVR-H3) и SEQ ID NO:19 (HVR-L3).
Анти-TAT226 антитело может содержать любую подходящую последовательность каркасной области вариабельного домена при условии, что антитело сохраняет способность связываться с TAT226. Например, в некоторых вариантах осуществления анти-TAT226 антитела по изобретению содержат консенсусную последовательность каркасной области тяжелой цепи подгруппы III человека. В одном из вариантов осуществления этих антител консенсусная последовательность каркасной области тяжелой цепи содержит замещение(я) в положении 71, 73 и/или 78. В одном из вариантов осуществления этих антител положение 71 представляет собой A, положение 73 представляет собой T и/или положение 78 представляет собой A. В одном из вариантов осуществления эти антитела содержат последовательность каркасной области вариабельного домена тяжелой цепи huMAb4D5-8, например SEQ ID NO:50, 51, 59, 35 (FR1, 2, 3, 4 соответственно). huMAb4D5-8 обладает коммерческим названием ГЕРСЕПТИН®, Genentech, Inc., South San Francisco, CA, USA; также оно рассмотрено в патентах США № 6407213 & 5821337, и Lee et al., J. Mol. Biol. (2004), 340(5):1073-93. В одном из таких вариантов осуществления эти антитела дополнительно содержат консенсусную последовательность каркасной области легкой цепи κI человека. В одном из таких вариантов осуществления эти антитела содержат последовательность каркасной области вариабельного домена легкой цепи huMAb4D5-8.
В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен тяжелой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4, выбранные из последовательностей, представленных на фигурах 5A и 5B; HVR-H1 содержит аминокислотную последовательность SEQ ID NO:4; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:5; и HVR-H3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:6-11. В одном из вариантов осуществления этих антител HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9 или 10. В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен легкой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4, выбранные из последовательностей, представленных на фигурах 6A и 6B; HVR-L1 содержит аминокислотную последовательность SEQ ID NO:12; HVR-L2 содержит аминокислотную последовательность SEQ ID NO:13; и HVR-L3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из вариантов осуществления этих антител HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17 или 18.
В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен тяжелой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4 SEQ ID NO:50, 51, 59 и 35, как представлено на фигуре 7; HVR-H1 содержит аминокислотную последовательность SEQ ID NO:4; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:5; и HVR-H3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:6-11. В одном из вариантов осуществления этих антител HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9 или 10. В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен легкой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4 SEQ ID NO:60, 61, 62 и 63, как представлено на фигурах 6A и 6B; HVR-L1 содержит аминокислотную последовательность SEQ ID NO:12; HVR-L2 содержит аминокислотную последовательность SEQ ID NO:13; и HVR-L3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из вариантов осуществления этих антител HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17 или 18.
В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен тяжелой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4 SEQ ID NO:50, 51, 53 и 35, как представлено на фигуре 8; HVR-H1 содержит аминокислотную последовательность SEQ ID NO:4; HVR-H2 содержит аминокислотную последовательность SEQ ID NO:5; и HVR-H3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:6-11. В одном из вариантов осуществления этих антител HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9 или 10. В одном из вариантов осуществления анти-TAT226 антитело содержит вариабельный домен легкой цепи, содержащий последовательность каркасной области и гипервариабельные области, где последовательность каркасной области содержит последовательности FR1-FR4 SEQ ID NO:60, 61, 62 и 74, как представлено на фигуре 8; HVR-L1 содержит аминокислотную последовательность SEQ ID NO:12; HVR-L2 содержит аминокислотную последовательность SEQ ID NO:13; и HVR-L3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:14-19. В одном из вариантов осуществления этих антител HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17 или 18.
В некоторых вариантах осуществления изобретение относится к анти-TAT226 антителу, содержащему вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, обладающую, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:20-25. В некоторых вариантах осуществления аминокислотная последовательность, обладающая, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности, содержит замещения, вставки или делеции относительно контрольной последовательности, но антитело, содержащее эту аминокислотную последовательность, сохраняет способность связываться с TAT226. В некоторых вариантах осуществления суммарно от 1 до 10 аминокислот замещено, вставлено или удалено в последовательности, выбранной из SEQ ID NO:20-25. В некоторых вариантах осуществления замещения вставки или делеции возникают в областях за пределами HVR (т.е. в FR). В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из SEQ ID NO:20-25.
В некоторых вариантах осуществления изобретение относится к анти-TAT226 антителу, содержащему вариабельный домен легкой цепи гуманизированного антитела 4D5 (huMAb4D5-8) (ГЕРСЕПТИН®, Genentech, Inc., South San Francisco, CA, USA) (также рассмотренному в патенте США № 6407213 и Lee et al. J. Mol. Biol. (2004), 340(5):1073-93), как представлено на SEQ ID NO:31 ниже.
1 Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg 108 (SEQ ID NO:31) (остатки HVR подчеркнуты).
В некоторых вариантах осуществления последовательность вариабельного домена легкой цепи huMAb4D5-8 модифицирована в одном или нескольких положениях 30, 66 и 91 (Asn, Arg и His соответственно, как указано выше полужирным шрифтом/курсивом). В одном из вариантов осуществления модифицированная последовательность huMAb4D5-8 содержит Ser в положении 30, Gly в положении 66 и/или Ser в положении 91. Соответственно, в одном из вариантов осуществления антитело по изобретению содержит вариабельный домен легкой цепи, содержащий последовательность, представленную на SEQ ID NO:26 ниже:
1 Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Thr Thr Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg 108 (SEQ ID NO:26) (остатки HVR подчеркнуты).
Замещенные остатки относительно huMAb4D5-8 указаны выше полужирным шрифтом/курсивом.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему вариабельный домен легкой цепи, содержащий аминокислотную последовательность, обладающую, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31. В некоторых вариантах осуществления аминокислотная последовательность, обладающая, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности, содержит замещения, добавления или делеции относительно контрольной последовательности, но антитело, содержащее такую аминокислотную последовательность, сохраняет способность связываться с TAT226. В некоторых вариантах осуществления суммарно от 1 до 10 аминокислот замещено, вставлено или удалено в последовательности, выбранной из SEQ ID NO:26-31. В некоторых вариантах осуществления замещения, вставки или делеции возникают в областях за пределами HVR (т.е. в FR). В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из SEQ ID NO:26-31.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, обладающую, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:20-25; и (b) вариабельный домен легкой цепи, содержащий аминокислотную последовательность, обладающую, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31. В некоторых вариантах осуществления аминокислотная последовательность, обладающая, по меньшей мере, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности, содержит замещения, добавления или делеции относительно контрольной последовательность, но антитело, содержащее такую аминокислотную последовательность, сохраняет способность связываться с TAT226. В некоторых вариантах осуществления суммарно от 1 до 10 аминокислот замещено, вставлено или удалено в контрольной последовательности. В некоторых вариантах осуществления замещения, вставки или делеции возникают в областях за пределами HVR (т.е. в FR). В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из SEQ ID NO:20-25, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из SEQ ID NO:26-31.
В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:20, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:26. В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:21, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:26. В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:22, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:27. В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:23, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:28. В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:24, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:29. В некоторых вариантах осуществления анти-TAT226 антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:25, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30.
В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему (a) одну, две или три VH HVR, выбранные из VH HVR, представленных на фигурах 2 и 4, и/или (b) одну, две или три VL HVR, выбранные из VL HVR, представленных на фигурах 3 и 4. В одном из аспектов изобретение относится к анти-TAT226 антителу, содержащему вариабельный домен тяжелой цепи, выбранный из вариабельных доменов, представленных на фигурах 9 и 11, и вариабельный домен легкой цепи, выбранный из вариабельных доменов, представленных на фигурах 10 и 12.
2. Фрагменты антител
Настоящее изобретение относится к фрагментам антител. Фрагменты антител можно получать традиционными способами, такими как ферментативное расщепление или рекомбинантные способы. В определенных условиях существуют преимущества использования фрагментов антител вместо целых антител. Меньший размер фрагментов обеспечивает быстрый клиренс и может вести к улучшению доступа к солидным опухолям. Для обзора некоторых фрагментов антител см. Hudson et al. (2003) Nat. Med. 9:129-134.
Для получения фрагментов антител были разработаны различные способы. Традиционно эти фрагменты получали посредством протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако эти фрагменты в настоящее время можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Все Fab-, Fv- и ScFv-фрагменты антител можно экспрессировать в E. coli, и они могут секретироваться из E. coli, обеспечивая, таким образом, легкое получение больших количеств этих фрагментов. Фрагменты антител можно выделять из фаговых библиотек антител, рассмотренных выше. Альтернативно фрагменты Fab'-SH можно выделять непосредственно из E. coli и химически соединять с образованием фрагментов F(ab')2 (Carter et al., Bio/Technology 10:163-167 (1992)). В соответствии с другим подходом фрагменты F(ab')2 можно выделять непосредственно из рекомбинантной культуры клеток-хозяев. Фрагмент Fab и F(ab')2 с повышенным временем полужизни in vivo, содержащий остатки эпитопа, связывающего рецептор спасения, описан в патенте США № 5869046. Другие способы получения фрагментов антител будут очевидны квалифицированному специалисту. В некоторых вариантах осуществления антитело представляет собой одноцепочечный Fv-фрагмент (scFv). См. WO 93/16185, патенты США № 5571894 и 5587458. Fv и sFv представляют собой единственные типы с интактными активными центрами, которые лишены константных областей; таким образом, они могут быть пригодны для снижения неспецифического связывания при применении in vivo. Слитые белки sFv можно сконструировать с получением эффекторного белка, слитого или с N-концом, или с C-концом sFv. См. Antibody Engineering, ed. Borrebaeck, выше. Также фрагмент антитела может представлять собой «линейное антитело», например, как описано в патенте США № 5641870. Такие линейные антитела могут быть моноспецифичными или биспецифичными.
3. Гуманизированные антитела
Настоящее изобретение относится к гуманизированным антителам. Разнообразные способы гуманизации антител, не являющихся человеческими, известны в данной области. Например, гуманизированное антитело может обладать одним или несколькими аминокислотными остатками, встроенными в него из источника, который не является человеческим. Такие не являющиеся человеческими аминокислотные остатки часто называют «импортными» остатками, которые, как правило, взяты из «импортного» вариабельного домена. Гуманизацию, главным образом, можно проводить в соответствии со способом Winter и коллег (Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-327; Verhoeyen et al. (1988) Science 239:1534-1536) посредством замены соответствующих последовательностей антитела человека последовательностями гипервариабельных областей. Таким образом, такие «гуманизированные» антитела представляют собой химерные антитела (патент США № 4816567), где, по существу, менее интактного вариабельного домена человека замещено соответствующей последовательностью вида, не относящегося к человеку. На практике гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки гипервариабельной области и, возможно, некоторые остатки FR замещены остатками из аналогичных участков антител грызунов.
Выбор вариабельных доменов человека, как легких, так и тяжелых цепей, для использования при получении гуманизированных антител является очень важным для снижения антигенности. В соответствии с так называемым способом «наилучшего соответствия» последовательность вариабельного домена антитела грызуна анализируют относительно целой библиотеки известных последовательностей вариабельных доменов человека. Затем последовательность человека, которая является наиболее сходной с последовательностью грызуна, считают приемлемой в качестве каркасной области человека для гуманизированного антитела (Sims et al., J. Immunol., 151:2296 (1993); Chothia et al., J. Mol. Biol., 196:901 (1987)). В другом способе используют конкретную каркасную область, полученную из консенсусной последовательности всех антител человека конкретной подгруппы легких или тяжелых цепей. Одну и ту же каркасную область можно использовать для нескольких различных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA, 89:4285 (1992); Presta et al., J. Immunol., 151:2623 (1993)).
Более того, как правило, требуется, чтобы антитела были гуманизированными с сохранением высокой аффинности в отношении антигена и других положительных биологических свойств. Для достижения этой цели согласно одному способу гуманизированные антитела получают с помощью процесса анализа родительских последовательностей и различных предполагаемых гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются широкодоступными и известны специалистам в данной области. Доступными являются компьютерные программы, которые иллюстрируют и выводят на экран возможные трехмерные конформационные структуры выбранных предполагаемых последовательностей иммуноглобулинов. Исследование этих выведенных на экран данных позволяет проводить анализ возможной роли остатков в функционировании предполагаемых последовательностей иммуноглобулинов, т.е. анализ остатков, которые влияют на способность предполагаемого иммуноглобулина связывать свой антиген. Таким образом, остатки FR можно отбирать и комбинировать из реципиентной и импортированной последовательностей таким образом, чтобы добиться требуемых свойств антитела, таких как повышенная аффинность к антигену(ам)-мишени(ям). Как правило, непосредственное и наиболее значительное влияние на связывание антигена оказывают остатки гипервариабельной области.
4. Человеческие антитела
Анти-TAT226 антитела человека по изобретению можно получать сочетанием последовательности(ей) вариабельного домена Fv-клона, отобранной(ых) из библиотек фагового дисплея, происходящих из человека, с известной(ыми) последовательностью(ями) константного домена человека, как описано выше. Альтернативно человеческие моноклональные анти-TAT226 антитела по изобретению можно получать гибридомным способом. Линии клеток миеломы человека и гетеромиеломы мыши-человека для получения человеческих моноклональных антител описаны, например, в Kozbor J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, p.51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147:86 (1991).
В настоящее время возможно получение трансгенных животных (например, мышей), которые при иммунизации способны продуцировать полный репертуар антител человека в отсутствии эндогенной продукции иммуноглобулинов. Например, было описано, что гомозиготная делеция гена области соединения тяжелых цепей антитела (JH) у химерных и несущих мутации в зародышевой линии мышей приводит к полному ингибированию эндогенной продукции антител. Перенос системы генов иммуноглобулинов зародышевой линии человека таким несущим мутации в зародышевой линии мышам при антигенной стимуляции будет приводить к продукции антител человека. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255 (1993); Bruggermann et al., Year in Immunol., 7:33 (1993).
Для получения антител человека из не принадлежащих человеку антител, например антител грызунов, где антитело человека обладает сходными аффинностями и специфичностями по отношению к исходному, не принадлежащему человеку антителу, также можно использовать перестановку генов. Согласно этому способу, который также называют «эпитопным импринтингом», вариабельную область или тяжелой, или легкой цепи не принадлежащего человеку фрагмента антитела, который получают способами фагового дисплея, как описано в данной работе, замещают репертуаром генов V-домена человека, получая популяцию химерных молекул scFv или Fab, состоящих из не принадлежащих человеку/принадлежащих человеку цепей. Отбор по отношению к антигену приводит к выделению химерных молекул scFv или Fab, состоящих из не принадлежащих человеку/принадлежащих человеку цепей, где в цепи человека восстанавливается антигенсвязывающий участок, нарушенный при удалении соответствующей не принадлежащей человеку цепи в первичном клоне фагового дисплея, т.е. эпитоп обуславливает (фиксирует) выбор партнера цепи человека. Антитела человека получают при повторении процесса замещения остающейся, не принадлежащей человеку цепи (см. PCT WO 93/06213, опубликованный 1 апреля 1993). В отличие от традиционной гуманизации не принадлежащих человеку антител привитием CDR этот способ приводит к получению полностью человеческих антител, которые не содержат остатки FR или CDR не связанного с человеком происхождения.
5. Биспецифические антитела
Биспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностями связывания в отношении, по меньшей мере, двух разных антигенов. В некоторых вариантах осуществления биспецифические антитела представляют собой человеческие или гуманизированные антитела. В некоторых вариантах осуществления одна из специфичностей связывания относится к TAT226 и другая относится к любому другому антигену. В некоторых вариантах осуществления биспецифические антитела могут связываться с двумя разными эпитопами TAT226. Биспецифические антитела можно также использовать для направленной доставки цитотоксических средств к клеткам, которые экспрессируют TAT226. Эти антитела обладают частью для связывания с TAT226 и частью, которая связывается с цитотоксическим средством, таким как, например, сапорин, анти-интерферон-α, алкалоид барвинка, A-цепь рицина, метотрексат или гаптен с радиоактивным изотопом. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител (например, биспецифических антител F(ab')2).
Способы получения биспецифических антител известны в данной области. Традиционно рекомбинантное получение биспецифических антител основано на совместной экспрессии двух пар тяжелая цепь-легкая цепь иммуноглобулинов, где две тяжелые цепи обладают различными специфичностями (Milstein and Cuello, Nature, 305:537 (1983)). Вследствие случайной сборки тяжелых и легких цепей иммуноглобулинов такие гибридомы (квадромы) продуцируют смесь потенциально из 10 разных молекул антител, из которых только одна обладает правильной биспецифической структурой. Очистка правильных молекул, которую обычно осуществляют посредством стадий аффинной хроматографии, является довольно трудоемкой, и выход продукта является низким. Аналогичные способы описаны в WO 93/08829 и в Traunecker et al., EMBO J., 10:3655 (1991).
В соответствии с другим подходом проводят слияние вариабельных доменов антител с требуемыми специфичностями связывания (участками связывания антитело-антиген) с последовательностями константных доменов иммуноглобулинов. Слияние, например, проводят с константным доменом тяжелой цепи иммуноглобулина, содержащим, по меньшей мере, часть шарнирных областей CH2 и CH3. В некоторых вариантах осуществления первая константная область тяжелой цепи (CH1), содержащая участок, необходимый для связывания легкой цепи, присутствует, по меньшей мере, в одном из компонентов, предназначенных для слияния. Молекулы ДНК, кодирующие предназначенные для слияния компоненты тяжелых цепей иммуноглобулинов и, если требуется, легкую цепь иммуноглобулина, встраивают в отдельные экспрессирующие векторы и осуществляют совместную трансфекцию ими пригодного организма-хозяина. Это встраивание обеспечивает значительную гибкость регулирования взаимных соотношений трех полипептидных фрагментов в вариантах осуществления, где неравные соотношения трех полипептидных цепей, используемых в конструкции, обеспечивают оптимальный выход продукта. Однако возможно встраивание кодирующих последовательностей двух или всех трех полипептидных цепей в один экспрессирующий вектор, если экспрессия, по меньшей мере, двух полипептидных цепей в равных соотношениях приводит к высокому выходу продукта или если соотношения не имеют особого значения.
В одном из вариантов осуществления этого подхода биспецифические антитела состоят из тяжелой цепи гибридного иммуноглобулина с первой специфичностью связывания в одной части и пары тяжелая цепь-легкая цепь гибридного иммуноглобулина (обладающей второй специфичностью связывания) в другой части. Обнаружено, что такая асимметричная структура облегчает разделение требуемого биспецифического соединения и нежелательных сочетаний цепей иммуноглобулинов, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ разделения. Этот способ описан в WO 94/04690. Для более подробного описания получения биспецифических антител см., например, Suresh et al., Methods in Enzymology, 121:210 (1986).
Согласно другому способу можно сконструировать область контакта между парой молекул антител для максимального увеличения процентной доли гетеродимеров, которые выделяют из рекомбинантной клеточной культуры. Область контакта содержит, по меньшей мере, часть CH3-домена константного домена антител. В этом способе одну или несколько небольших аминокислотных боковых цепей из области контакта первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозином или триптофаном). В области контакта второй молекулы антитела создают компенсирующие «полости» идентичного или сходного с крупной(ыми) боковой(ыми) цепью(ями) размера посредством замены крупных аминокислотных боковых цепей цепями меньших размеров (например, аланином или треонином). Эта замена обеспечивает механизм для повышения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают поперечно сшитые антитела или «гетероконъюгаты» антител. Например, одно из антител в гетероконъюгате можно соединить с авидином, а другое с биотином. Такие антитела, например, предложены для нацеливания клеток иммунной системы против нежелательных клеток (патент США № 4676980) и для лечения ВИЧ-инфекции (WO 91/00360, WO 92/00373 и EP 03089). Гетероконъюгаты антител можно получать с использованием любых пригодных способов поперечного сшивания. Пригодные вещества для поперечного сшивания хорошо известны в данной области и описаны в патенте США № 4676980 совместно с рядом способов поперечного сшивания.
Способы получения биспецифических антител из фрагментов антител также описаны в литературе. Например, биспецифические антитела можно получать с использованием образования химических связей. В Brennan et al., Science, 229:81 (1985) описан способ, в котором интактные антитела протеолитически расщепляют с получением F(ab')2-фрагментов. Эти фрагменты восстанавливают в присутствии арсенита натрия, вещества, образующего комплексы с дитиолами, для стабилизации соседних дитиолов и предотвращения образования межмолекулярного дисульфида. Затем полученные Fab'-фрагменты превращают в производные тионитробензоата (TNB). Затем одно из производных Fab'-TNB повторно превращают в Fab'-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB с получением биспецифического антитела. Полученные биспецифические антитела можно использовать в качестве средств для селективной иммобилизации ферментов.
Последние достижения упростили прямое выделение из E. coli фрагментов Fab'-SH, которые можно химически соединять с образованием биспецифических антител. В Shalaby et al., J. Exp. Med., 175:217-225 (1992) описано получение полностью гуманизированной молекулы F(ab')2 биспецифического антитела. Осуществляли секрецию из E. coli каждого Fab'-фрагмента по отдельности и их подвергали прямому химическому связыванию in vitro с образованием биспецифического антитела. Биспецифическое антитело, полученное таким образом, обладало способностью связываться с клетками, сверхэкспрессирующими рецептор HER2, и нормальными T-клетками человека, а также запускать литическую активность цитотоксических лимфоцитов человека против мишеней опухоли молочной железы человека.
Также описаны различные способы получения и выделения фрагментов биспецифических антител непосредственно из рекомбинантной клеточной культуры. Например, получены биспецифические антитела с использованием «лейциновых молний», Kostelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды «лейциновой молнии» из белков Fos и Jun соединяли с Fab'-частями двух различных антител посредством слияния генов. Гомодимеры антител восстанавливали в шарнирной области с образованием мономеров и затем повторно окисляли с образованием гетеродимеров антител. Этот способ также можно использовать для получения гомодимеров антител. Способ «димерных антител», описанный в Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993), предусматривает альтернативный механизм получения биспецифических фрагментов антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким для возможности образования пары между двумя доменами на одной и той же цепи. Таким образом, домены VH и VL одного фрагмента вынуждены образовывать пару с комплементарными доменами VL и VH другого фрагмента, таким образом формируя два антигенсвязывающих участка. Также описан другой способ получения фрагментов биспецифических антител с использованием димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol., 152:5368 (1994).
Также предусмотрены антитела более чем с двумя валентностями. Например, можно получать триспецифические антитела, Tutt et al. J. Immunol. 147:60 (1991).
6. Поливалентные антитела
Поливалентное антитело может быть быстрее, чем бивалентное антитело, интернализовано (и/или катаболизировано) клеткой, экспрессирующей антиген, с которой связываются антитела. Антитела по настоящему изобретению могут представлять собой поливалентные антитела (которые являются отличными от класса IgM) с тремя антигенсвязывающими участками или более (например, тетравалентные антитела), которые можно легко получать посредством рекомбинантной экспрессии нуклеиновой кислоты, кодирующей полипептидные цепи антитела. Поливалентное антитело может содержать димеризационный домен и три или более антигенсвязывающих участка. В некоторых вариантах осуществления димеризационный домен содержит (или состоит из) Fc-область или шарнирную область. В этом случае антитело содержит Fc-область и три или более антигенсвязывающих участка со стороны N-конца Fc-области. В некоторых вариантах осуществления поливалентное антитело содержит (или состоит из) от трех до приблизительно восьми антигенсвязывающих участков. В одном из таких вариантов осуществления поливалентное антитело содержит (или состоит из) четыре антигенсвязывающих участка. Поливалентное антитело содержит, по меньшей мере, одну полипептидную цепь (например, две полипептидные цепи), где полипептидная(ые) цепь(и) содержит(ат) два или более вариабельных домена. Например, полипептидная(ые) цепь(и) может(могут) содержать VD1-(X1)n-VD2-(X2)n-Fc, где VD1 представляет собой первый вариабельный домен, VD2 представляет собой второй вариабельный домен, Fc представляет собой одну полипептидную цепь Fc-области, X1 и X2 представляют собой аминокислоту или полипептид и n представляет собой 0 или 1. Например, полипептидная(ые) цепь(и) может(могут) содержать: цепь VH-CH1-гибкий линкер-VH-CH1-Fc-область; или цепь VH-CH1-VH-CH1-Fc-область. В данной работе поливалентное антитело может дополнительно содержать, по меньшей мере, два (например, четыре) полипептида вариабельных доменов легкой цепи. В данной работе поливалентное антитело, например, может содержать от приблизительно двух до приблизительно восьми полипептидов вариабельного домена легкой цепи. Полипептиды вариабельного домена легкой цепи, предусмотренные в данной работе, содержат вариабельный домен легкой цепи и, необязательно, дополнительно содержат домен CL.
7. Однодоменные антитела
В некоторых вариантах осуществления антитело по изобретению представляет собой однодоменное антитело. Однодоменное антитело представляет собой одиночную полипептидную цепь, содержащую весь вариабельный домен тяжелой цепи антитела или его часть или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления однодоменное антитело является однодоменным антителом человека (Domantis, Inc., Waltham, MA; см., например, патент США № 6248516 B1). В одном из вариантов осуществления однодоменное антитело состоит из всего вариабельного домена тяжелой цепи антитела или его части.
8. Варианты антител
В некоторых вариантах осуществления предусмотрена(ы) модификация(и) аминокислотных последовательностей описанных в данной работе антител. Например, может быть желательным улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотных последовательностей антитела можно получать посредством внесения соответствующих замен в нуклеотидную последовательность, кодирующую антитело, или посредством пептидного синтеза. Такие модификации включают, например, делеции остатков, и/или вставки остатков, и/или замены остатков аминокислотных последовательностей антитела. Для получения конечной конструкции можно осуществлять любое сочетание из делеции, вставки и замены при условии, что конечная конструкция обладает требуемыми свойствами. Модификации аминокислот можно вносить в рассматриваемую аминокислотную последовательность антитела во время получения последовательности.
Пригодный способ выявления определенных остатков или областей антитела, которые являются предпочтительными участками для мутагенеза, называют «сканирующим аланином мутагенезом», как описано в Cunningham and Wells (1989) Science, 244:1081-1085. В данной работе выявляют остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), чтобы влиять на взаимодействие аминокислот с антигеном. Те положения аминокислот, которые проявляют функциональную чувствительность к замещениям, затем корректируют посредством введения дополнительных или других вариантов в участки замещения или вместо них. Таким образом, несмотря на то что участок для внесения изменения в аминокислотную последовательность является предопределенным, нет необходимости в непосредственной предопределенности характера мутации. Например, для анализа действия мутации в данном участке, в кодоне-мишени или области-мишени проводят сканирование аланином или случайный мутагенез и для экспрессированных иммуноглобулинов проводят скрининг на наличие требуемой активности.
Вставки аминокислотных последовательностей включают предназначенные для слияния на N- и/или C-конце последовательности, обладающие длиной, находящейся в диапазоне от одного остатка до полипептидов, содержащих сотню или более остатков, а также вставки внутрь последовательности одного или нескольких аминокислотных остатков. Примеры концевых вставок включают антитело с N-концевым остатком метионила. Другие варианты вставок в молекулу антитела включают слияние N- или C-конца антитела с ферментом (например, для ADEPT) или полипептидом, который повышает время полужизни антитела в сыворотке.
В некоторых вариантах осуществления антитело по изобретению изменяют для повышения или снижения степени гликозилирования антитела. Гликозилирование полипептидов, как правило, представляет собой или N-гликозилирование, или O-гликозилирование. N-гликозилирование относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту за исключением пролина, представляют собой последовательности, распознаваемые для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. O-гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, как правило, к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление или удаление участков гликозилирования антитела удобно осуществлять посредством изменения аминокислотной последовательности таким образом, чтобы происходило создание или удаление одной или нескольких из вышеописанных трипептидных последовательностей (в случае участков N-гликозилирования). Также изменение можно осуществлять добавлением или удалением одного или нескольких остатков серина или треонина или заменой на один или несколько остатков серина или треонина в последовательности исходного антитела (в случае участков O-гликозилирования).
В том случае, если антитела содержат Fc-область, можно изменить присоединенный к ней углевод. Например, антитела со зрелой углеводной структурой, в которой отсутствует фукоза, присоединенная к Fc-области антитела, описаны в патентной заявке США № US 2003/0157108 (Presta, L.). См. также US 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Антитела, содержащие разделяющий N-ацетилглюкозамин (GlcNAc) в углеводе, присоединенном к Fc-области антитела, рассмотрены в WO 2003/011878, Jean-Mairet et al., и патенте США № 6602684, Umana et al. Антитела, обладающие, по меньшей мере, одним остатком галактозы в олигосахариде, присоединенном к Fc-области антител, рассмотрены в WO 1997/30087, Patel et al. См. также WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.) относительно антител с измененным углеводом, присоединенным к Fc-области антитела. См. также US 2005/0123546 (Umana et al.) относительно антигенсвязывающих молекул с модифицированным гликозилированием.
В некоторых вариантах осуществления вариант гликозилирования содержит Fc-область, где в углеводной структуре, присоединенной к Fc-области, отсутствует фукоза. Такие варианты обладают улучшенной функцией в отношении ADCC. Необязательно Fc-область дополнительно содержит одну или несколько аминокислотных замен, которые еще более улучшают ADCC, например замены в положениях 298, 333 и/или 334 Fc-области (нумерация остатков Eu). Примеры публикаций, относящихся к «дефукозилированным» или «лишенным фукозы» антителам, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87:614 (2004). Примеры линий клеток, продуцирующих дефукозилированные антитела, включают клетки Lec13 CHO, у которых нарушено фукозилирование белка (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США № US 2003/0157108 A1, Presta, L; и WO 2004/056312 A1, Adams et al., в особенности в примере 11), и нокаутные линии клеток, такие как нокаутные клетки CHO по гену альфа-1,6-фукозилтрансферазы, FUT8 (Yamane-Ohnuki et al. Biotech. Bioeng. 87:614 (2004)).
Другим типом варианта является вариант с заменой аминокислоты. Такие варианты обладают, по меньшей мере, одним остатком аминокислоты в молекуле антитела, который заменен другим остатком. Участки, представляющие интерес для проведения мутагенеза с заменой, включают гипервариабельные участки, но также предполагаются изменения FR. Консервативные замены представлены в таблице 1 под заголовком «предпочтительные замены». Если результатом таких замен является желаемое изменение биологической активности, тогда можно вносить более значительные изменения, обозначенные в таблице 1 как «иллюстративные замены», или как описано далее относительно классов аминокислот, и проводить скрининг продуктов.
Phe; норлейцин
Met; Ala; Phe
Ala; норлейцин
Существенные модификации биологических свойств антитела проводят, выбирая замены, которые значительно различаются по их эффекту на поддержание (a) структуры полипептидного остова в области замены, например, такого как конформация в виде слоя или спирали, (b) заряда или гидрофобности молекулы в участке-мишени или (c) размера боковой цепи. Аминокислоты можно объединить в группы согласно сходству свойств их боковых цепей (в A. L. Lehninger, в Biochemistry, second ed., p.73-75, Worth Publishers, New York (1975)):
(1) неполярные: Ala (A), Val (V), Leu (L), Ile (I), Pro (P), Phe (F), Trp (W), Met (M;)
(2) незаряженные полярные: Gly (G), Ser (S), Thr (T), Cys (C), Tyr (Y), Asn (N), Gln (Q);
(3) кислые: Asp (D), Glu (E);
(4) основные: Lys (K), Arg (R), His(H).
Альтернативно встречающиеся в природе остатки можно разделить на группы на основе типичных свойств боковых цепей:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro; и
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены будут приводить к замене члена одного из этих классов на член из другого класса. Такие замещенные остатки также можно вводить в участки консервативного замещения или в остающиеся (неконсервативные) участки.
Один из типов варианта с заменой включает замену одного или нескольких остатков гипервариабельной области родительского антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный(ые) вариант(ы), отобранный(ые) для дальнейшей разработки, обладает(ют) модифицированными (улучшенными) биологическими свойствами относительно родительского антитела, из которого он(они) получен(ы). Удобный способ получения таких вариантов с заменами включает аффинное созревание с использованием фагового дисплея. В кратком изложении, в несколько участков гипервариабельной области (например, в 6-7 участков) вносят мутации для получения всех возможных аминокислотных замен в каждом участке. Антитела, полученные таким образом, продуцируют в частицах нитевидных фагов в качестве антител, слитых, по меньшей мере, с частью белка оболочки фага (например, продукта гена III M13), упакованного в каждой частице. Затем проводят скрининг продуцированных в фагах вариантов на их биологическую активность (например, аффинность связывания). В целях выявления участков гипервариабельной области, являющихся кандидатами для модификации, можно проводить сканирующий мутагенез (например, сканирующий аланином мутагенез) для выявления остатков гипервариабельной области, в значительной степени участвующих в связывании антигена. Альтернативно или дополнительно может быть целесообразным проведение анализа кристаллической структуры комплекса антиген-антитело для определения участков контакта между антителом и антигеном. Такие формирующие контакт остатки и соседние остатки являются кандидатами для замены в соответствии с известными в данной области способами, включая способы, разработанные в данной работе. После получения таких вариантов панель вариантов подвергают скринингу, используя способы, известные в данной области, включая способы, описанные в данной работе, и для дальнейшей разработки можно отбирать антитела с наилучшими свойствами посредством одного или нескольких пригодных способов анализа.
Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности антитела, получают множеством способов, известных в данной области. Эти способы включают в качестве неограничивающих примеров выделение из природного источника (в случае вариантов встречающейся в природе аминокислотной последовательности) или получение с помощью олигонуклеотид-опосредованного (или сайт-специфического) мутагенеза, мутагенеза с помощью PCR и кассетного мутагенеза ранее полученного вариантного или невариантного вида антител.
Может являться желательным внесение одной или нескольких модификаций аминокислот в Fc-область антител по изобретению, таким образом получая вариант Fc-области. Вариант Fc-области может содержать последовательность Fc-области человека (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) в одном или нескольких положениях аминокислот, включая положения шарнирного цистеина.
В соответствии с данным описанием и представлениями в данной области предполагают, что в некоторых вариантах осуществления антитело по изобретению может содержать одну или несколько модификаций по сравнению с аналогичными антителами дикого типа, например, в Fc-области. Эти антитела могут, тем не менее, сохранять по существу те же характеристики, необходимые для терапевтического применения по сравнению с их аналогом дикого типа. Например, предполагают, что в Fc-области можно осуществить определенные модификации, которые могут приводить к изменению (т.е. или улучшению, или ухудшению) связывания с C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в WO 99/51642. См. также Duncan & Winter Nature 322:738-40 (1988); патент США № 5648260; патент США № 5624821; и WO 94/29351 относительно других примеров вариантов Fc-области. В WO 00/42072 (Presta) и WO 2004/056312 (Lowman) описаны варианты антител с улучшенным или ухудшенным связыванием с FcR. Содержание этих патентных публикаций, в частности, приведено в настоящем документе в качестве ссылки. См. также Shields et al. J. Biol. Chem. 9(2):6591-6604 (2001). Антитела с увеличенным временем полужизни и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Эти антитела содержат Fc-область с одной или несколькими заменами, которые улучшают связывание Fc-области с FcRn. Варианты полипептидов с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной способностью связываться с C1q описаны в патенте США № 6194551 B1, WO 99/51642. Содержание этих патентных публикаций, в частности, приведено в настоящем документе в качестве ссылки. См. также Idusogie et al. J. Immunol. 164:4178-4184 (2000).
В одном из аспектов изобретение относится к антителам, содержащим модификации в области контакта Fc-полипептидов, содержащих Fc-область, где модификации облегчают гетеродимеризацию и/или способствуют ей. Эти модификации включают введение выступающей части в первый Fc-полипептид и полости во второй Fc-полипептид, где выступающая часть располагается в полости так, чтобы способствовать образованию комплекса из первого и второго Fc-полипептидов. Способы получения антител с такими модификациями известны в данной области, например, как описано в патенте США № 5731168.
9. Производные антител
Антитела по настоящему изобретению можно дополнительно модифицировать так, чтобы они содержали дополнительные небелковые группы, которые известны в данной области и легко доступны. Предпочтительно группы, подходящие для получения производных антитела, представляют собой водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают без ограничения полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (или гомополимеры или статистические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры пролипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликольпропиональдегид может обладать преимуществами в производстве благодаря своей стабильности в воде. Полимер может обладать любой молекулярной массой и может являться разветвленным или неразветвленным. Количество присоединенных к антителам полимеров может варьировать, и если присоединено более одного полимера, то они могут представлять собой одинаковые или разные молекулы. Как правило, количество и/или тип полимеров, используемых для получения производных, можно определить на основе факторов, которые включают в качестве неограничивающих примеров конкретные свойства или функции антитела, которое нуждается в улучшении, тот факт, будут ли использовать производное антитела в лечении при определенных условиях, и т.п.
В другом варианте осуществления представлены конъюгаты антитела и небелкового компонента, который можно избирательно нагревать посредством воздействия облучения. В одном из вариантов осуществления небелковый компонент представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. 102:11600-11605 (2005)). Облучение может обладать любой длиной волны и включает в качестве неограничивающих примеров длины волн, которые не повреждают обычные клетки, но которые нагревают небелковый компонент до температуры, при которой погибают клетки, расположенные в непосредственной близости к конъюгату антитело-небелковый компонент.
B. Некоторые способы получения антител
1. Некоторые способы на основе гибридом
Моноклональные анти-TAT226 антитела по изобретению можно получать с использованием гибридомного способа, впервые описанного в Kohler et al., Nature, 256:495 (1975), или их можно получать способами рекомбинантных ДНК (патент США № 4816567).
В гибридомном способе мышь или другое соответствующее животное-хозяин, такое как хомяк, иммунизируют для получения лимфоцитов, которые вырабатывают или которые способны вырабатывать антитела, которые будут специфически связываться с белком, используемым для иммунизации. Индуцирование анти-TAT226 антител, как правило, осуществляют у животных множественными подкожными (sc) или внутрибрюшинными (ip) инъекциями TAT226 и адъюванта. TAT226 можно получать с использованием способов, хорошо известных в данной области, некоторые из которых далее здесь описаны. Например, TAT226 можно получать рекомбинантным способом. В одном из вариантов осуществления животных иммунизируют производным TAT226, которое содержит внеклеточную часть TAT226, слитую с Fc-частью тяжелой цепи иммуноглобулина. В одном из вариантов осуществления животных иммунизируют слитым белком TAT226-IgG1. В одном из вариантов осуществления животных иммунизируют иммуногенными производными TAT226 в растворе с монофосфорил-липидом A (MPL)/дикриномиколатом трегалозы (TDM) (Ribi Immunochem. Research, Inc., Hamilton, MT) и раствор инъецируют интрадермально во многих участках. Две недели спустя животным проводят повторную иммунизацию. От семи до четырнадцати суток спустя у животных отбирают кровь и сыворотку анализируют на титр анти-TAT226. Животным проводят повторную иммунизацию до тех пор, пока титр не стабилизируется.
Альтернативно лимфоциты можно иммунизировать in vitro. Затем производят слияние лимфоцитов с миеломными клетками с использованием подходящего вещества, вызывающего слияние, такого как полиэтиленгликоль, для получения гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, p.59-103 (Academic Press, 1986)).
Таким образом полученные гибридомные клетки высевают в подходящую культуральную среду и выращивают в ней, например в среде, которая содержит одно или несколько веществ, которые ингибируют рост или выживаемость не подвергшихся слиянию родительских миеломных клеток. Например, если у родительских миеломных клеток отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), то культуральная среда для гибридом, как правило, будет включать гипоксантин, аминоптерин и тимидин (среда HAT), вещества, которые предотвращают рост клеток с дефектом HGPRT.
В некоторых вариантах осуществления миеломные клетки представляют собой клетки, которые эффективно подвергаются слиянию, поддерживают стабильную выработку антител на высоком уровне селектированными продуцирующими антитела клетками и являются чувствительными к среде, такой как среда HAT. Иллюстративные миеломные клетки включают в качестве неограничивающих примеров мышиные миеломные линии, такие как линии, происходящие из опухолей мышей MOPC-21 и MPC-11, доступных в Salk Institute Cell Distribution Center, San Diego, California USA, и клеток SP-2 или X63-Ag8-653, доступных в American Type Culture Collection, Rockville, Maryland USA. Линии клеток миеломы человека и гетеромиеломы мыши-человека также описаны относительно получения человеческих моноклональных антител (Kozbor, J. Immunol., 133:3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, p.51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой выращивают гибридомные клетки, анализируют относительно выработки моноклональных антител, которые связываются с TAT226. Предпочтительно специфичность связывания моноклональных антител, вырабатываемых гибридомными клетками, определяют иммунопреципитацией или анализом связывания in vitro, таким как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Аффинность связывания моноклонального антитела, например, можно определить с помощью анализа Скэтчарда по Munson et al., Anal. Biochem., 107:220 (1980).
После идентификации тех гибридомных клеток, которые вырабатывают антитела требуемой специфичности, аффинности и/или активности, клоны можно субклонировать посредством способов серийных разведений и выращивать с помощью общепринятых способов (Goding, Monoclonal Antibodies: Principles and Practice, p.59-103 (Academic Press, 1986)). Подходящие культуральные среды для этой цели включают, например, среду D-MEM или RPMI-1640. Кроме того, гибридомные клетки можно выращивать in vivo в виде асцитных опухолей в животном. Моноклональные антитела, секретируемые субклонами, соответствующим образом выделяют из культуральной среды, асцитической жидкости или сыворотки с помощью общепринятых способов очистки иммуноглобулинов, таких как, например, протеин-A-сефароза, хроматография с гидроксилапатитом, гель-электрофорез, диализ или аффинная хроматография.
2. Некоторые способы скрининга библиотек
Анти-TAT226 антитела по изобретению можно получать с использованием комбинаторных библиотек для скрининга на антитела с требуемой активностью или активностями. Например, множество способов известно в данной области для получения библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие требуемыми характеристиками связывания. Такие способы описаны в общих чертах в Hoogenboom et al. (2001) в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ) и в некоторых вариантах осуществления в Lee et al. (2004) J. Mol. Biol. 340:1073-1093.
По существу клоны синтетических антител отбирают с помощью скрининга фаговых библиотек, содержащих фаг, на котором экспонированы различные фрагменты вариабельной области (Fv) антитела, слитой с белком оболочки фага. Такие фаговые библиотеки отбирают аффинной хроматографией относительно требуемого антигена. Клоны, экспрессирующие Fv-фрагменты, способные связываться с требуемым антигеном, адсорбируются на антигене и, таким образом, отделяются от несвязавшихся клонов в библиотеке. Связавшиеся клоны затем элюируют с антигена, и их можно далее обогащать с помощью дополнительных циклов адсорбции на антигене/элюции с него. Любые из анти-TAT226 антител по изобретению можно получать путем разработки подходящего способа скрининга с использованием антигена для селекции представляющего интерес фагового клона с последующим получением клона полноразмерного анти-TAT226 антитела с использованием Fv-последовательностей из представляющего интерес фагового клона и последовательностей подходящей константной области (Fc), описанных у Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), vols. 1-3.
В некоторых вариантах осуществления антигенсвязывающий домен антитела образован двумя вариабельными (V) областями приблизительно из 110 аминокислот, по одной области из легкой (VL) и тяжелой (VH) цепей, которые, обе, дают три гипервариабельные петли (HVR) или области, определяющие комплементарность (CDR). Вариабельные домены можно функционально экспонировать на фаге, или в виде одноцепочечных Fv-фрагментов (scFv), в которых VH и VL ковалентно связаны посредством короткого, гибкого пептида, или в виде Fab-фрагментов, в которых каждый из них слит с константным доменом, и они взаимодействуют нековалентно, как описано в Winter et al., Ann. Rev. Immunol., 12:433-455 (1994). Как используют в данной работе, фаговые клоны, кодирующие scFv, и фаговые клоны, кодирующие Fab, собирательно обозначают как «фаговые Fv-клоны» или «Fv-клоны».
Репертуары генов VH и VL можно по отдельности клонировать с помощью полимеразной цепной реакции (PCR) и случайным образом рекомбинировать в фаговых библиотеках, в которых затем можно осуществлять поиск антигенсвязывающих клонов, как описано в Winter et al., Ann. Rev. Immunol., 12:433-455 (1994). Библиотеки из иммунизированных источников дают высокоаффинные антитела к иммуногену без необходимости создания гибридом. Альтернативно «наивный» репертуар можно клонировать для создания единого источника антител человека к широкому диапазону неауто-, а также аутоантигенов без какой-либо иммунизации, как описано в Griffiths et al., EMBO J, 12:725-734 (1993). В заключение, «наивные» библиотеки можно также получать синтетически клонированием неперестроенных сегментов V-генов из стволовых клеток и с использованием праймеров для PCR, содержащих случайную последовательность для кодирования высоковариабельных областей CDR3 и для осуществления перестройки in vitro, как описано в Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992).
В некоторых вариантах осуществления нитевидный фаг применяют для экспонирования фрагментов антител посредством слияния с минорным белком оболочки pIII. Фрагменты антител можно экспонировать в виде одноцепочечных Fv-фрагментов, в которых VH- и VL-домены связаны на одной полипептидной цепи гибким полипептидным спейсером, например, как описано в Marks et al., J. Mol. Biol., 222:581-597 (1991), или в виде Fab-фрагментов, в которых одну цепь подвергают слиянию с pIII и другая секретируется в периплазму бактериальной клетки-хозяина, где происходит сборка структуры Fab-белок оболочки, которая начинает экспонироваться на поверхности фага путем замещения некоторых белков оболочки дикого типа, например, как описано в Hoogenboom et al., Nucl. Acids Res., 19:4133-4137 (1991).
Как правило, нуклеиновые кислоты, кодирующие фрагменты генов антител, получают из иммунных клеток, выделенных из человека или животных. Если требуется библиотека, обогащенная клонами против TAT226, то субъект иммунизируют посредством TAT226 для получения гуморального иммунного ответа, и клетки селезенки и/или циркулирующие B-клетки из других периферических лимфоцитов крови (PBL) выделяют для создания библиотеки. В предпочтительном варианте осуществления библиотеку фрагментов гена антител человека, обогащенную клонами против TAT226, получают вызыванием гуморального иммунного ответа по отношению TAT226 у трансгенных мышей, несущих функциональный набор генов иммуноглобулинов человека (и лишенных функциональной системы выработки эндогенных антител) таким образом, что иммунизация посредством TAT226 приводит к выработке B-клетками человеческих анти-TAT226 антител. Получение трансгенных мышей, вырабатывающих человеческие антитела, описано ниже.
Дополнительное обогащение популяциями активированных клеток против TAT226 можно получать с использованием подходящего способа скрининга для выделения B-клеток, экспрессирующих специфичное по отношению к TAT226 мембраносвязанное антитело, например, путем отделения клеток аффинной хроматографией на основе TAT226 или адсорбцией клеток к меченному флюорохромом TAT226 с последующим флюоресцентно-активированным клеточным сортингом (FACS).
Альтернативно использование клеток селезенки и/или B-клеток или других PBL из неиммунизированного донора дает лучшее представление возможного репертуара антител, а также позволяет создание библиотеки антител с использованием любого вида животного (человека или не являющегося человеком), в котором TAT226 не является антигенным. Для библиотек, включающих конструирование генов антител in vitro, стволовые клетки выделяют из субъекта для получения нуклеиновых кислот, кодирующих неперестроенные сегменты генов антител. Представляющие интерес иммунные клетки можно получать из множества видов животных, таких видов, как человек, мышь, крыса, зайцеобразные, кроличьи, собачьи, кошачьи, свиньи, бычьи, лошадиные и птицы и т.п.
Нуклеиновую кислоту, кодирующую сегменты вариабельного гена антител (включая VH- и VL-сегменты), выделяют из представляющих интерес клеток и амплифицируют. В случае библиотек перестроенных VH- и VL-генов требуемую ДНК можно получать выделением геномной ДНК или мРНК из лимфоцитов с последующей полимеразной цепной реакцией (PCR) с праймеров, комплементарных 5'- и 3'-концам перестроенных VH- и VL-генов, как описано в Orlandi et al., Proc. Natl. Acad. Sci. (USA), 86:3833-3837 (1989), таким образом создавая разнообразные репертуары V-генов для экспрессии. V-гены можно амплифицировать исходя из кДНК и геномной ДНК с обратными праймерами на 5'-конце экзона, кодирующего зрелый V-домен, и прямыми праймерами, расположенными в пределах J-сегмента, как описано в Orlandi et al. (1989) и в Ward et al., Nature, 341:544-546 (1989). Однако для амплификации с кДНК обратные праймеры можно также располагать в лидерном экзоне, как описано в Jones et al., Biotechnol., 9:88-89 (1991), и прямые праймеры в пределах константной области, как описано в Sastry et al., Proc. Natl. Acad. Sci. (USA), 86:5728-5732 (1989). Для максимизации комплементарности в праймеры можно включить вырожденность, как описано в Orlandi et al. (1989) или Sastry et al. (1989). В некоторых вариантах осуществления разнообразие библиотеки максимизируют с использованием праймеров для PCR, нацеленных на каждое семейство V-генов, для того, чтобы амплифицировать все доступные VH- и VL-конфигурации, присутствующие в образце нуклеиновой кислоты из иммунных клеток, например, как описано в способе по Marks et al., J. Mol. Biol., 222:581-597 (1991), или как описано в способе по Orum et al., Nucleic Acids Res., 21:4491-4498 (1993). Для клонирования амплифицированной ДНК в экспрессирующие векторы можно вводить редкие участки рестрикции в праймер для PCR в виде метки на один конец, как описано в Orlandi et al. (1989), или дополнительной амплификацией посредством PCR с меченого праймера, как описано в Clackson et al., Nature, 352:624-628 (1991).
Репертуары синтетически перестроенных V-генов можно получать in vitro исходя из сегментов V-генов. Большинство сегментов VH-генов человека клонировано и секвенировано (сообщено в Tomlinson et al., J. Mol. Biol., 227:776-798 (1992)), и картировано (сообщено в Matsuda et al., Nature Genet., 3:88-94 (1993); эти клонированные сегменты (включая все мажорные конформации H1- и H2-петли) можно использовать для получения различных репертуаров VH-генов с праймеров для PCR, кодирующих H3-петли разной последовательности и длины, как описано в Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992). VH-репертуары можно также получать во всем разнообразии последовательности, сосредоточенном в длинной H3-петле одной длины, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 89:4457-4461 (1992). Сегменты Vκ и Vλ человека клонированы и секвенированы (сообщено в Williams and Winter, Eur. J. Immunol., 23:1456-1461 (1993)), и их можно использовать для получения репертуаров синтетической легкой цепи. Репертуары синтетического V-гена на основе диапазона VH- и VL-изгибов и L3- и H3-длин кодируют антитела со значительным структурным разнообразием. После амплификации ДНК, кодирующих V-ген, сегменты V-генов зародышевой линии можно перестраивать in vitro согласно способам по Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992).
Репертуары фрагментов антител можно конструировать сочетанием вместе репертуаров VH- и VL-генов несколькими способами. Каждый репертуар можно получать в разных векторах, и векторы рекомбинируют in vitro, например, как описано в Hogrefe et al., Gene, 128:119-126 (1993), или in vivo сочетанной инфекцией, например, посредством системы loxP, описанной в Waterhouse et al., Nucl. Acids Res., 21:2265-2266 (1993). В способе рекомбинации in vivo используют двухцепочечную природу Fab-фрагментов для преодоления ограничения размера библиотеки, на который накладывает ограничение эффективность трансформации E. coli. «Наивные» VH- и VL-репертуары клонируют по отдельности, один в фагмидный и другой в фаговый вектор. Две библиотеки затем комбинируют путем фаговой инфекции содержащих фагмиду бактерий таким образом, что каждая клетка содержит разное сочетание и размер библиотеки ограничен только количеством присутствующих клеток (приблизительно 1012 клонов). Оба вектора содержат сигналы рекомбинации in vivo таким образом, что VH- и VL-гены рекомбинируют в единый репликон и совместно упаковываются в фаговые вирионы. Эти гигантские библиотеки дают большие количества различных антител с хорошей аффинностью (Kd-1 приблизительно 10-8 М).
Альтернативно репертуары можно клонировать последовательно в тот же самый вектор, например, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или собирать вместе с помощью PCR и затем клонировать, например, как описано в Clackson et al., Nature, 352:624-628 (1991). Сборку с помощью PCR можно также использовать для соединения ДНК VH и VL с ДНК, кодирующей гибкий пептидный спейсер, для образования репертуаров одноцепочечных Fv (scFv). В другом способе «сборку с помощью PCR в клетке» используют для объединения VH- и VL-генов в лимфоцитах с помощью PCR и затем клонирования репертуаров соединенных генов, как описано в Embleton et al., Nucl. Acids Res., 20:3831-3837 (1992).
Антитела, образуемые «наивными» библиотеками (или природными, или синтетическими), могут обладать умеренной аффинностью (Kd-1 приблизительно от 106 до 107 M-1), но аффинное созревание можно также имитировать in vitro конструированием и повторной селекцией исходя из вторичных библиотек, как описано в Winter et al. (1994) выше. Например, мутацию можно случайно вводить in vitro с использованием неточной полимеразы (сообщено в Leung et al., Technique, 1:11-15 (1989)) в способе по Hawkins et al., J. Mol. Biol., 226:889-896 (1992) или в способе по Gram et al., Proc. Natl. Acad. Sci. USA, 89:3576-3580 (1992). Кроме того, аффинное созревание можно осуществлять случайным мутагенезом одной или нескольких CDR, например, с использованием PCR с праймеров, несущих случайную последовательность, охватывающую представляющую интерес CDR, в отобранных индивидуальных Fv-клонах, и скринингом на клоны с более высокой аффинностью. В WO 9607754 (опубликован 14 марта 1996) описан способ индуцирования мутагенеза в определяющей комплементарность области легкой цепи иммуноглобулина для получения библиотеки генов легких цепей. Другой эффективный способ представляет собой рекомбинацию VH- или VL-доменов, отобранных при помощи фагового дисплея, с репертуарами встречающихся в природе вариантов V-домена, полученных от неиммунизированных доноров, и скрининг на более высокую аффинность в нескольких циклах перестройки цепи, как описано в Marks et al., Biotechnol., 10:779-783 (1992). Этот способ позволяет получать антитела и фрагменты антител с аффинностями приблизительно 10-9 М или менее.
Скрининг библиотек можно осуществлять различными способами, известными в данной области. Например, TAT226 можно использовать для покрытия лунок адсорбционных планшетов, экспрессировать на клетках-хозяевах, прикрепленных к адсорбционным планшетам, или использовать в клеточном сортинге, или конъюгировать с биотином для улавливания покрытыми стрептавидином бусами, или использовать в любом другом способе для пэннинга библиотек фагового дисплея.
Образцы фаговой библиотеки контактируют с иммобилизованным TAT226 в условиях, подходящих для связывания, по меньшей мере, части частиц фага с адсорбентом. Обычно условия, включая pH, ионную силу, температуру и т.п., подбирают, чтобы имитировать физиологические условия. Фаги, связанные с твердой фазой, промывают и затем элюируют кислотой, например, как описано в Barbas et al., Proc. Natl. Acad. Sci. USA, 88:7978-7982 (1991), или щелочью, например, как описано в Marks et al., J. Mol. Biol., 222:581-597 (1991), или конкуренцией с антигеном TAT226, например, в способе, сходном со способом конкуренции с антигеном в Clackson et al., Nature, 352:624-628 (1991). Фаги можно обогащать в 20-1000 раз в одном цикле селекции. Более того, обогащенные фаги можно выращивать в бактериальной культуре и подвергать дальнейшим циклам селекции.
Эффективность селекции зависит от многих факторов, включая кинетику диссоциации в ходе отмывки, и от того, могут ли множественные фрагменты антител на одном фаге одновременно контактировать с антигеном. Антитела с быстрой кинетикой диссоциации (и низкими аффинностями связывания) можно удерживать при использовании коротких отмывок, поливалентного фагового дисплея и высокой плотности покрытия антигеном твердой фазы. Высокая плотность не только стабилизирует фаг посредством поливалентных взаимодействий, но способствует повторному связыванию диссоциировавшего фага. Селекцию антител с медленной кинетикой диссоциации (и хорошими аффинностями связывания) можно облегчать использованием длительных отмывок и моновалентного фагового дисплея, как описано в Bass et al., Proteins, 8:309-314 (1990) и в WO 92/09690, и низкой плотности покрытия антигеном, как описано в Marks et al., Biotechnol., 10:779-783 (1992).
Возможно осуществление селекции на TAT226 между фаговыми антителами разных аффинностей, даже аффинностей, которые незначительно различаются. Однако случайный мутагенез селектируемого антитела (например, в ходе осуществления некоторых способов аффинного созревания), вероятно, приведет к образованию многих мутантов, большинство из которых связываются с антигеном, и немногих с более высокой аффинностью. При ограниченном количестве TAT226 немногочисленные фаги с высокой аффинностью могут отсеиваться. Для сохранения всех мутантов с более высокой аффинностью фаги можно инкубировать с избытком биотинилированного TAT226, но с биотинилированным TAT226 в концентрации более низкой молярности, чем планируемое значение молярной константы сродства для TAT226. Фаги с высокой аффинностью связывания можно затем улавливать на покрытых стрептавидином парамагнитных бусах. Такое «равновесное улавливание» дает возможность селектировать антитела согласно их аффинности связывания с чувствительностью, которая позволяет выделять мутантные клоны с аффинностью, до двух раз более высокой, из большого избытка фагов с более низкой аффинностью. Используемыми условиями отмывки фагов, связанных с твердой фазой, также можно манипулировать для разделения на основе кинетики диссоциации.
Клоны анти-TAT226 можно селектировать по активности. В некоторых вариантах осуществления изобретение относится к анти-TAT226 антителам, которые связываются с живыми клетками, которые экспрессируют TAT226 в естественных условиях. В одном из вариантов осуществления изобретение относится к анти-TAT226 антителам, которые блокируют связывание между лигандом TAT226 и TAT226, но не блокируют связывание между лигандом TAT226 и вторым белком. Fv-клоны, соответствующие таким анти-TAT226 антителам, можно селектировать (1) выделением клонов анти-TAT226 из фаговой библиотеки, как описано выше, и необязательно амплификацией выделенной популяции фаговых клонов путем выращивания популяции в подходящем бактериальном хозяине; (2) отбором TAT226 и второго белка, по отношению к которым соответственно требуются блокирующая и неблокирующая активности; (3) адсорбцией фаговых клонов анти-TAT226 на иммобилизованном TAT226; (4) использованием избытка второго белка для элюции любых нежелательных клонов, которые распознают детерминанты связывания TAT226, которые перекрываются с детерминантами связывания второго белка или совпадают с ними; и (5) элюцией клонов, которые остаются адсорбированными в ходе стадии (4). Необязательно клоны с требуемыми свойствами блокирования/неблокирования можно далее обогатить повторением описанных здесь процедур селекции один или несколько раз.
ДНК, кодирующую полученные из гибридом моноклональные антитела или Fv-клоны фагового дисплея по изобретению, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных праймеров, сконструированных для специфической амплификации представляющих интерес областей, кодирующих тяжелую и легкую цепи, с гибридомной или фаговой матричной ДНК). После выделения ДНК можно встроить в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, клетки COS обезьян, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые в других случаях не вырабатывают белок-иммуноглобулин, для осуществления синтеза требуемых моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитела, включают Skerra et al., Curr. Opinion in Immunol., 5:256 (1993) и Pluckthun, Immunol. Revs, 130:151 (1992).
ДНК, кодирующую Fv-клоны по изобретению, можно объединять с известными последовательностями ДНК, кодирующими константные области тяжелой цепи и/или легкой цепи (например, подходящие последовательности ДНК можно получать от Kabat et al., выше), для получения клонов, кодирующих тяжелые и/или легкие цепи полной или частичной длины. Будет принято во внимание, что можно использовать для этой цели константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получать из любого вида человека или животного. Как используют в настоящем описании, Fv-клон, происходящий из ДНК вариабельного домена одного вида животного (такого, как человек) и затем слитый с ДНК константной области другого вида животного с образованием последовательности(ей), кодирующей(их) «гибридную», полноразмерную тяжелую цепь и/или легкую цепь, включают в определение «химерных» и «гибридных» антител. В некоторых вариантах осуществления Fv-клон, происходящий из вариабельной ДНК человека, подвергают слиянию с ДНК константной области человека с образованием последовательности(ей), кодирующей(их) тяжелые и/или легкие цепи человека полной или частичной длины.
ДНК, кодирующую анти-TAT226 антитела, происходящую из гибридомы по изобретению, можно также модифицировать, например, замещением последовательности, кодирующей константные домены тяжелой и легкой цепей человека, вместо гомологичных мышиных последовательностей, происходящих из клона гибридомы (например, как в способе Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). ДНК, кодирующую антитело или фрагмент, происходящие из гибридомы или Fv-клона, можно далее модифицировать ковалентным присоединением к последовательности, кодирующей иммуноглобулин, всей или части последовательности, кодирующей полипептид, не являющийся иммуноглобулином. Таким образом, получают «химерные» или «гибридные» антитела, которые обладают специфичностью связывания антител по изобретению, происходящих из Fv-клона или клона гибридомы.
3. Векторы, клетки-хозяева и рекомбинантные способы
Для рекомбинантного получения антитела по изобретению кодирующую его нуклеиновую кислоту выделяют и вставляют в реплицирующийся вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Доступны многие векторы. Выбор вектора зависит частично от используемой клетки-хозяина. Как правило, клетки-хозяева обладают или прокариотическим, или эукариотическим (как правило, из млекопитающих) происхождением. Принимают во внимание, что для этой цели можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE, и что такие константные области можно получать из любого вида человека или животных.
a) Получение антител с использованием прокариотических клеток-хозяев:
(1) Конструирование векторов
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела по изобретению, можно получать с использованием общепринятых рекомбинантных способов. Требуемые полинуклеотидные последовательности можно выделять и секвенировать из продуцирующих антитело клеток, таких как клетки гибридом. Альтернативно полинуклеотиды можно синтезировать с использованием нуклеотидного синтезатора или способов на основе PCR. После получения последовательности, кодирующие полипептиды, вставляют в рекомбинантный вектор, способный к репликации и экспрессии гетерологичных полинуклеотидов в прокариотических хозяевах. Многие векторы, которые доступны и известны в данной области, можно использовать для цели настоящего изобретения. Выбор подходящего вектора зависит в основном от размера нуклеиновых кислот, которые вставляют в вектор, и конкретной клетки-хозяина, которую трансформируют вектором. Каждый вектор содержит различные компоненты в зависимости от своей функции (амплификация или экспрессия гетерологичного полинуклеотида или и то, и другое) и его совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора, как правило, включают в качестве неограничивающих примеров: ориджин (начало) репликации, маркерный ген для селекции, промотор, участок связывания с рибосомой (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
Как правило, плазмидные векторы, содержащие репликон и контрольные последовательности, которые происходят из видов, совместимых с клеткой-хозяином, используют применительно к этим хозяевам. Вектор, как правило, несет участок репликации, а также маркерные последовательности, которые способны обеспечить селекцию трансформированных клеток по фенотипу. Например, E. coli, как правило, трансформируют с использованием pBR322, плазмиды, полученной из вида E. coli. pBR322 содержит гены, кодирующие устойчивость к ампициллину (Amp) и тетрациклину (Tet) и, таким образом, дает легкий способ выявления трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаг могут также содержать - или их можно модифицировать так, чтобы они содержали - промоторы, которые можно использовать в микробном организме для экспрессии эндогенных белков. Примеры производных pBR322, используемых для экспрессии конкретных антител, подробно описаны в Carter et al., патент США № 5648237.
Кроме того, фаговые векторы, содержащие репликон и контрольные последовательности, которые совместимы с микроорганизмом-хозяином, можно использовать в качестве трансформирующих векторов применительно к этим хозяевам. Например, бактериофаг, такой как λGEM.TM.-11, можно использовать для получения рекомбинантного вектора, который можно использовать для трансформации чувствительных клеток-хозяев, таких как E. coli LE392.
Экспрессирующий вектор по изобретению может содержать две или более пар промотор-цистрон, которые кодируют каждый из полипептидных компонентов. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5') цистрона, которая модулирует его экспрессию. Прокариотические промоторы, как правило, относятся к двум классам, индуцибельные и конститутивные. Индуцибельный промотор представляет собой промотор, который стимулирует повышение уровней транскрипции цистрона, находящегося под его управлением, в ответ на изменения в условиях культивирования, например присутствие или отсутствие питательного вещества или изменение температуры.
Хорошо известно большое количество промоторов, которые распознаются множеством потенциальных клеток-хозяев. Выбранный промотор можно функционально связать с цистронной ДНК, кодирующей легкую или тяжелую цепь, выделяя промотор из исходной ДНК посредством расщепления ферментами рестрикции и вставляя выделенную промоторную последовательность в вектор по изобретению. Как природную промоторную последовательность, так и множество гетерологичных промоторов можно использовать для управления амплификацией и/или экспрессией целевых генов. В некоторых вариантах осуществления используют гетерологичные промоторы, так как они, как правило, обеспечивают более высокий уровень транскрипции и более высокий выход экспрессируемого целевого гена по сравнению с природным промотором целевого полипептида.
Промоторы, подходящие для использования в прокариотических хозяевах, включают промотор PhoA, промоторные системы β-галактамазы и лактозы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac или trc. Однако также пригодны другие промоторы, которые функциональны в бактериях (такие, как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, таким образом давая возможность квалифицированному специалисту функционально лигировать их с цистронами, кодирующими целевые легкие и тяжелые цепи (Siebenlist et al. (1980) Cell 20:269) с использованием линкеров или адаптеров для обеспечения любыми необходимыми участками рестрикции.
В одном из аспектов изобретения каждый цистрон в рекомбинантном векторе содержит компонент секреторной сигнальной последовательности, который направляет перемещение экспрессируемых полипептидов через мембрану. Как правило, сигнальная последовательность может являться компонентом вектора, или она может являться частью кодирующей целевой полипептид ДНК, которая вставлена в вектор. Сигнальная последовательность, выбранная для целей данного изобретения, должна распознаваться клеткой-хозяином и процессироваться (т.е. расщепляться сигнальной пептидазой) в ней. Для прокариотических клеток-хозяев, которые не распознают и не процессируют сигнальные последовательности, свойственные гетерологичным полипептидам, сигнальную последовательность замещают прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Ipp или термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и MBP. В одном из вариантов осуществления изобретения сигнальные последовательности, используемые для обоих цистронов экспрессионной системы, представляют собой сигнальные последовательности STII или их варианты.
В другом аспекте продукция иммуноглобулинов по изобретению может происходить в цитоплазме клетки-хозяина и, таким образом, не требует наличия секреторных сигнальных последовательностей в каждом цистроне. В этом случае легкие и тяжелые цепи иммуноглобулинов экспрессируются, сворачиваются и собираются с образованием функциональных иммуноглобулинов в цитоплазме. Некоторые штаммы-хозяева (например, штаммы E. coli trxB-) обеспечивают условия в цитоплазме, которые благоприятны для образования дисульфидной связи, таким образом обеспечивая правильное сворачивание и сборку экспрессируемых белковых субъединиц, Proba and Pluckthun, Gene, 159:203 (1995).
Антитела по изобретению можно также продуцировать с использованием экспрессионной системы, в которой количественное соотношение экспрессируемых полипептидных компонентов можно модулировать для достижения максимального выхода секретируемых и правильно собранных антител по изобретению. Такую модуляцию осуществляют, по меньшей мере, частично одновременным модулированием эффективностей трансляции полипептидных компонентов.
Один из способов модулирования эффективности трансляции описан в Simmons et al., патент США № 5840523. В нем используют варианты области инициации трансляции (TIR) в цистроне. Для данной TIR можно получить ряд вариантов аминокислотной последовательности или последовательности нуклеиновой кислоты с диапазоном эффективностей трансляции, таким образом давая удобные средства для приспособления этого фактора к требуемому уровню экспрессии конкретной цепи. Варианты TIR можно получать общепринятыми способами мутагенеза, которые приводят к заменам кодонов, которые могут изменять аминокислотную последовательность. В некоторых вариантах осуществления замены в нуклеотидной последовательности являются молчащими. Изменения в TIR могут включать, например, изменения количества или расположения последовательностей Шайна-Дальгарно, вместе с изменениями в сигнальной последовательности. Один из способов получения мутантных сигнальных последовательностей состоит в получении «банка кодонов» в начале кодирующей последовательности, который не изменяет аминокислотную последовательность сигнальной последовательности (т.е. замены являются молчащими). Это можно осуществить изменением третьего нуклеотидного положения каждого кодона; кроме того, некоторые аминокислоты, такие как лейцин, серин и аргинин, обладают несколькими первыми и вторыми положениями, что может усложнять получение банка. Этот способ мутагенеза подробно описан в Yansura et al. (1992) METHODS: A Companion to Methods in Enzymol. 4:151-158.
В одном из вариантов осуществления набор векторов получают с диапазоном эффективностей TIR для каждого цистрона в этих векторах. Этот ограниченный набор обеспечивает сравнение уровней экспрессии каждой цепи, а также выхода требуемых продуктов антител при различных сочетаниях эффективностей TIR. Эффективности TIR можно количественно определить по уровню экспрессии репортерного гена, как подробно описано в Simmons et al., патент США № 5840523. На основе сравнения эффективности трансляции отбирают желательные конкретные TIR для сочетания в конструкциях экспрессирующих векторов по изобретению.
Прокариотические клетки-хозяева, подходящие для экспрессии антител по изобретению, включают архебактерии и эубактерии, такие как грамотрицательные или грамположительные организмы. Примеры пригодных бактерий включают Escherichia (например, E. coli), Bacilli (например, B. subtilis), энтеробактерии, виды Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления используют грамотрицательные клетки. В одном из вариантов осуществления используют клетки E. coli в качестве хозяев для конструкций по изобретению. Примеры штаммов E. coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), p.1190-1219; депонирование в ATCC № 27325) и его производные, включая штамм 33D3, обладающий генотипом W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTΔ(nmpc-fepE) degP41 kanR (патент США № 5639635). Также пригодны другие штаммы и их производные, такие как E. coli 294 (ATCC 31446), E. coli B, E. coli λ 1776 (ATCC 31537) и E. coli RV308 (ATCC 31608). Эти примеры являются иллюстративными, а не ограничивающими. Способы конструирования производных любой из вышеупомянутых бактерий, обладающих определенными генотипами, известны в данной области и описаны, например, в Bass et al., Proteins, 8:309-314 (1990). Как правило, необходимо выбирать соответствующие бактерии, принимая во внимание реплицируемость репликона в клетках бактерии. Например, виды E. coli, Serratia или Salmonella можно подходящим образом использовать в качестве хозяина, если для обеспечения репликации используют хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Как правило, клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и по желанию в культуру клеток можно вносить дополнительные ингибиторы протеаз.
(2) Продукция антител
Клетки-хозяева трансформируют вышеописанными экспрессирующими векторами и культивируют в общепринятых питательных средах, модифицированных соответствующим образом для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности.
Трансформация означает введение ДНК в прокариотического хозяина таким образом, что ДНК реплицируется или в качестве внехромосомного элемента, или в качестве хромосомного интегранта. В зависимости от используемой клетки-хозяина трансформацию осуществляют с использованием общепринятых способов, пригодных для таких клеток. Обработку кальцием, используя хлорид кальция, как правило, используют для бактериальных клеток, которые обладают значительными барьерами в виде клеточной стенки. В другом способе трансформации используют полиэтиленгликоль/DMSO. Еще одним используемым способом является электропорация.
Прокариотические клетки, используемые для продукции полипептидов по изобретению, выращивают в средах, известных в данной области и подходящих для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают среду Лурия (LB) плюс необходимые питательные вещества. В некоторых вариантах осуществления среды также содержат вещество для осуществления отбора, выбираемое на основе конструкции экспрессирующего вектора, чтобы обеспечить избирательный рост прокариотических клеток, содержащих экспрессирующий вектор. Например, ампициллин добавляют к средам для роста клеток, экспрессирующих ген устойчивости к ампициллину.
Любые необходимые добавки помимо источников углерода, азота и неорганического фосфата также можно включать в соответствующих концентрациях, внося их по отдельности или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно культуральная среда может содержать одно или несколько восстанавливающих средств, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при подходящих температурах. В некоторых вариантах осуществления для выращивания E. coli температуры для выращивания варьируют от приблизительно 20°C до приблизительно 39°C, от приблизительно 25°C до приблизительно 37°C или приблизительно 30°C. pH среды может представлять собой любой pH, варьирующий от приблизительно 5 до приблизительно 9, в основном в зависимости от организма-хозяина. В некоторых вариантах осуществления для E. coli pH составляет от приблизительно 6,8 до приблизительно 7,4 или приблизительно 7,0.
Если в экспрессирующем векторе по изобретению используют индуцируемый промотор, то экспрессию белка индуцируют в условиях, подходящих для активации промотора. В одном из аспектов изобретения используют промоторы PhoA для регуляции транскрипции полипептидов. Таким образом, трансформированные клетки-хозяева культивируют в среде для индукции, содержащей ограниченное количество фосфата. В некоторых вариантах осуществления среда, содержащая ограниченное количество фосфата, представляет собой среду CRAP (см., например, Simmons et al., J. Immunol. Methods (2002), 263:133-147). Можно использовать множество других индукторов в соответствии с используемой конструкцией вектора, как известно в данной области.
В одном из вариантов осуществления экспрессируемые полипептиды по настоящему изобретению секретируются в периплазму клеток-хозяев, и их выделяют из нее. Выделение белка, как правило, включает разрушение микроорганизма, как правило, такими способами, как осмотический шок, обработка ультразвуком или лизис. После разрушения клеток остатки клеток или целые клетки можно удалить центрифугированием или фильтрацией. Белки можно дополнительно очищать, например, хроматографией на аффинной смоле. Альтернативно белки можно переносить в культуральные среды и выделять из них. Клетки можно удалять из культуры и культуральный супернатант фильтровать и концентрировать для дальнейшей очистки продуцированных белков. Экспрессируемые полипептиды можно далее выделять и идентифицировать с использованием общепринятых способов, таких как электрофорез в полиакриламидном геле (PAGE) и анализ с помощью вестерн-блоттинга.
В одном из аспектов изобретения продукцию антитела осуществляют в больших количествах при помощи процесса ферментации. Различные крупномасштабные процедуры ферментации при культивировании с подпиткой доступны для продукции рекомбинантных белков. Крупномасштабные способы ферментации предусматривают, по меньшей мере, емкость в 1000 литров и в некоторых вариантах осуществления емкость приблизительно от 1000 до 100000 литров. В этих ферментерах используют лопастные перемешиватели для распределения кислорода и питательных веществ, в особенности глюкозы (предпочтительный источник углерода/энергии). Мелкомасштабная ферментация означает, как правило, ферментацию в ферментере, объемная емкость которого не превышает приблизительно 100 литров и может варьировать от приблизительно 1 литра до приблизительно 100 литров.
В процессе ферментации индукцию экспрессии белка, как правило, начинают после того, как клетки вырастают при подходящих условиях до требуемой плотности, например OD550 приблизительно 180-220, на этой стадии клетки находятся в ранней стационарной фазе. Можно использовать множество индукторов в соответствии с используемой конструкцией вектора, как известно в данной области и описано выше. Клетки можно выращивать в течение более коротких периодов перед индукцией. Клетки, как правило, индуцируют приблизительно 12-50 часов, хотя можно использовать более длительное или более короткое время индукции.
Для улучшения выхода продукции и качества полипептидов по изобретению можно изменять различные условия ферментации. Например, для улучшения правильной сборки и сворачивания секретируемых полипептидов антител при совместной трансформации прокариотических клеток-хозяев можно использовать дополнительные векторы, сверхэкспрессирующие белки-шапероны, такие как белки Dsb (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидилпролил-цис,транс-изомераза с шаперонной активностью). Показано, что белки-шапероны способствуют правильному сворачиванию и растворимости гетерологичных белков, продуцируемых бактериальными клетками-хозяевами, Chen et al. (1999) J. Biol. Chem. 274:19601-19605; Georgiou et al., патент США № 6083715; Georgiou et al., патент США № 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275:17100-17105; Ramm and Pluckthun (2000) J. Biol. Chem. 275:17106-17113; Arie et al. (2001) Mol. Microbiol. 39:199-210.
Для снижения до минимума протеолиза экспрессируемых гетерологичных белков (в особенности белков, которые чувствительны к протеолизу) можно использовать для настоящего изобретения определенные штаммы-хозяева с нарушенными протеолитическими ферментами. Например, штаммы клеток-хозяев можно модифицировать для получения генетической(их) мутации(й) в генах, кодирующих известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их сочетания. Некоторые штаммы E. coli с нарушенными протеазами доступны и описаны, например, в Joly et al. (1998), выше; Georgiou et al., патент США № 5264365; Georgiou et al., патент США № 5508192; Hara et al., Microbial Drug Resistance, 2:63-72 (1996).
В одном из вариантов осуществления штаммы E. coli с нарушенными протеолитическими ферментами и трансформированные плазмидами, сверхэкспрессирующими один или несколько белков-шаперонов, используют в качестве клеток-хозяев в экспрессионной системе по изобретению.
(3) Очистка антител
В одном из вариантов осуществления полученный в данной работе белок антитела дополнительно очищают до получения препаратов, которые являются по существу гомогенными для дальнейших анализов и применений. Можно использовать общепринятые способы очистки белка, известные в данной области. Следующие способы приведены из подходящих способов очистки в качестве примера: фракционирование на иммуноаффинных или ионообменных колонках, осаждение этанолом, HPLC с обращенной фазой, хроматография на кремнеземе или катионообменной смоле, такой как DEAE, хроматофокусирование, SDS-PAGE, осаждение сульфатом аммония и гель-фильтрация с использованием, например, сефадекса G-75.
В одном из аспектов протеин A, иммобилизованный на твердой фазе, используют для иммуноаффинной очистки продуктов антител по изобретению. Протеин A представляет собой белок клеточной стенки массой 41 кДа из Staphylococcus aureas, который связывается с высокой аффинностью с Fc-областью антител, Lindmark et al. (1983) J. Immunol. Meth. 62:1-13. Твердая фаза, на которой иммобилизован протеин A, может являться колонкой, содержащей стеклянную или кремнеземную поверхность, или стеклянной колонкой с контролируемым размером пор, или колонкой на основе кремниевой кислоты. В некоторых вариантах осуществления колонку покрывают реагентом, таким как глицерин, с целью возможного предотвращения неспецифической сорбции примесей.
В качестве первой стадии очистки препарат, полученный из культуры клеток, как описано выше, помещают на твердую фазу с иммобилизованным протеином A для обеспечения специфического связывания представляющего интерес антитела с протеином A. Твердую фазу можно затем промывать для удаления примесей, неспецифически связавшихся с твердой фазой. В заключение, представляющее интерес антитело удаляют с твердой фазы элюцией.
b) Получение антител с использованием эукариотических клеток-хозяев:
Вектор для использования в эукариотической клетке-хозяине, как правило, содержит в качестве неограничивающих примеров одно или несколько из следующего: сигнальная последовательность, ориджин (начало) репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
(1) Компонент сигнальной последовательности
Вектор для использования в эукариотической клетке-хозяине может также содержать сигнальную последовательность или другой полипептид, обладающий специфическим участком расщепления на N-конце представляющего интерес зрелого белка или полипептида. Выбранная гетерологичная сигнальная последовательность может являться последовательностью, которая распознается клеткой-хозяином и процессируется (т.е. расщепляется сигнальной пептидазой) в ней. Для экспрессии в клетках млекопитающих доступны сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, gD-сигнал вируса простого герпеса. ДНК для такой области-предшественника лигируют в рамке считывания с кодирующей антитело ДНК.
(2) Ориджин (начало) репликации
Как правило, компонент ориджина (начала) репликации не является необходимым для экспрессирующих векторов млекопитающих. Например, ориджин SV40 можно, как правило, использовать только потому, что он содержит ранний промотор.
(3) Компонент гена, обеспечивающего селекцию
Экспрессирующие и клонирующие векторы могут содержать ген, обеспечивающий селекцию, также называемый селектируемым маркером. Типичные гены, обеспечивающие селекцию, кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют ауксотрофную недостаточность в соответствующих условиях или (c) обеспечивают жизненно важными питательными веществами, которые недоступны в комплексных средах.
В одном из примеров процедуры селекции используют лекарственное средство для остановки роста клетки-хозяина. Те клетки, которые успешно трансформированы гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному средству, и, таким образом, выживают в условиях селекции. Примеры такой доминантной селекции включают лекарственные средства неомицин, микофеноловую кислоту и гигромицин.
Другим примером подходящих селектируемых маркеров для клеток млекопитающих являются маркеры, такие как DHFR, тимидинкиназа, металлотионеин-I и -II, предпочтительно гены металлотионеина приматов, аденозиндезаминаза, орнитиндекарбоксилаза и т.п., которые позволяют идентифицировать клетки, компетентные в отношении поглощения нуклеиновой кислоты, кодирующей антитело.
Например, в некоторых вариантах осуществления клетки, трансформированные обеспечивающим селекцию геном DHFR, сначала выявляют при культивировании всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. В некоторых вариантах осуществления при использовании DHFR дикого типа соответствующей клеткой-хозяином является линия клеток яичника китайского хомяка (CHO) с нарушенной активностью DHFR (например, ATCC CRL-9096).
Альтернативно клетки-хозяева (в частности, хозяева дикого типа, которые содержат эндогенный DHFR), трансформированные или совместно трансформированные последовательностями ДНК, кодирующими антитело, белок DHFR дикого типа и другой селектируемый маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), можно отбирать по росту клеток в среде, содержащей обеспечивающее селекцию вещество для селектируемого маркера, такое как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США № 4965199.
(4) Компонент промотора
Экспрессирующие и клонирующие векторы, как правило, содержат промотор, который распознается организмом-хозяином и который функционально связан с нуклеиновой кислотой, кодирующей представляющий интерес полипептид (например, антитело). Промоторные последовательности известны для эукариот. Например, практически все эукариотические гены обладают AT-богатой областью, расположенной приблизительно на 25-30 оснований выше участка инициации транскрипции. Другая последовательность, расположенная на 70-80 оснований выше от начала транскрипции во многих генах, представляет собой область CNCAAT, где N может являться любым нуклеотидом. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может представлять собой сигнал для добавления поли-A-хвоста на 3'-конец кодирующей последовательности. В некоторых вариантах осуществления любую или все из этих последовательностей можно надлежащим образом вставить в эукариотические экспрессирующие векторы.
Транскрипцией векторов в клетках-хозяевах млекопитающих управляют, например, промоторы, получаемые из геномов вирусов, таких как вирус полиомы, вирус чумы птиц, аденовирус (такой, как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B и вирус обезьян 40 (SV40), из гетерологичных промоторов млекопитающих, например промотора актина или промотора иммуноглобулина, из промоторов теплового шока при условии, что такие промоторы совместимы с системами клетки-хозяина.
Ранние и поздние промоторы вируса SV40 удобно получать в виде рестрикционного фрагмента SV40, который также содержит вирусный ориджин (начало) репликации SV40. Предранний промотор цитомегаловируса человека удобно получать в виде рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в млекопитающих-хозяевах с использованием вируса бычьей папилломы в качестве вектора описана в патенте США № 4419446. Модификация этой системы описана в патенте США № 4601978. См. также Reyes et al., Nature 297:598-601 (1982) в отношении экспрессии кДНК β-интерферона человека в клетках мышей под контролем промотора тимидинкиназы из вируса простого герпеса. Альтернативно длинный концевой повтор вируса саркомы Рауса можно использовать в качестве промотора.
(5) Компонент энхансерного элемента
Транскрипция ДНК, кодирующей антитело по данному изобретению, у высших эукариот часто усиливается посредством вставки энхансерной последовательности в вектор. В настоящее время известны многие энхансерные последовательности из генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Как правило, однако, используют энхансер из вируса эукариотических клеток. Примеры включают энхансер SV40 на поздней стороне от ориджина репликации (100-270 п.о.), энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы на поздней стороне от ориджина репликации и энхансеры аденовируса. См. также Yaniv, Nature 297:17-18 (1982) относительно энхансерных элементов для активации эукариотических промоторов. Энхансер можно встроить в вектор в 5'- или 3'-положении по отношению к последовательности, кодирующей полипептид антитела, но, как правило, он расположен на 5'-участке от промотора.
(6) Компонент терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах, могут также содержать последовательности, необходимые для терминации транскрипции и стабилизации мРНК. Такие последовательности обычно расположены в 5'- и иногда в 3'-нетранслируемых областях эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибируемые как полиаденилированные фрагменты в нетранслируемой части мРНК, кодирующей антитело. Один из пригодных компонентов терминации транскрипции представляет собой область полиаденилирования гормона роста быка. См. WO94/11026 и описанный в этой работе экспрессирующий вектор.
(7) Селекция и трансформация клеток-хозяев
В настоящей работе подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах включают клетки высших эукариот, описанные в настоящей работе, включая клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (культуре тканей) стало обычной процедурой. Примеры пригодных линий клеток-хозяев млекопитающих представляют собой линию почки мартышки CV1, трансформированную SV40 (COS-7, ATCC CRL 1651); линию эмбриональной почки человека (293 или клетки 293, субклонированные для выращивания в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почки новорожденного хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мышей (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки мартышки (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крыс линии Буффало (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мышей (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N. Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4 и линию гепатомы человека (Hep G2).
Клетки-хозяева трансформируют вышеописанными экспрессирующими или клонирующими векторами для продукции антител и культивируют в общепринятых питательных средах, модифицированных соответствующим образом для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности.
(8) Культивирование клеток-хозяев
Клетки-хозяева, используемые для получения антител по данному изобретению, можно культивировать во множестве сред. Для культивирования клеток-хозяев подходят коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная питательная среда (MEM, Sigma), RPMI-1640 (Sigma) и среда Игла в модификации Дульбекко (DMEM, Sigma). Кроме того, в качестве культуральных сред для клеток-хозяев можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), патентах США № 4767704, 4657866, 4927762, 4560655 или 5122469; WO 90/03430; WO 87/00195; или патенте США Re. 30985. В любую из этих сред можно добавить в случае необходимости гормоны и/или другие факторы роста (такие, как инсулин, трансферрин или фактор роста эпидермиса), соли (такие, как хлорид натрия, кальций, магний и фосфат), буферы (такие, как HEPES), нуклеотиды (такие, как аденозин и тимидин), антибиотики (такие, как лекарственное средство гентамицин™), микроэлементы (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях порядка микромолей) и глюкозу или эквивалентный источник энергии. Любые другие добавки можно также вносить в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, использованные ранее для клетки-хозяина, выбранной для экспрессии, и будут очевидны для обычного специалиста.
(9) Очистка антител
С использованием рекомбинантных способов антитело можно продуцировать внутриклеточно или непосредственно секретировать в среду. Если антитело продуцируют внутриклеточно, то в качестве первой стадии дебрис в виде частиц или клетки-хозяева, или лизированные фрагменты можно удалить, например, центрифугированием или ультрафильтрацией. Если антитело секретируется в среду, то супернатанты из таких экспрессионных систем можно сначала сконцентрировать с использованием коммерчески доступного фильтра для концентрирования белка, например, комплекта для ультрафильтрации Amicon или Millipore Pellicon. Можно вносить ингибитор протеаз, такой как PMSF, на любой из вышеописанных стадий для ингибирования протеолиза и можно вносить антибиотики для предотвращения роста случайных контаминантов.
Композицию антител, полученную из клеток, можно очищать с использованием, например, хроматографии с гидроксилапатитом, гель-электрофореза, диализа и аффинной хроматографии, где аффинная хроматография является подходящим способом очистки. Пригодность протеина A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который находится в антителе. Протеин A можно использовать для очистки антител, в основе которых лежат тяжелые цепи γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Methods 62:1-13 (1983)). Протеин G рекомендован для всех изотипов мыши и для γ3 человека (Guss et al., EMBO J. 5:1567-1575 (1986)). Матрица, к которой присоединен аффинный лиганд, может представлять собой агарозу, однако доступны и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и более короткое время обработки, чем те, которые можно достичь с помощью агарозы. В случае когда антитело содержит домен CH3, для очистки пригодна смола Bakerbond ABX™ (J. T. Baker, Phillipsburg, NJ). Также в зависимости от антитела, подлежащего выделению, доступны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, HPLC с обращенной фазой, хроматография на кремнеземе, хроматография на сефарозе™ с гепарином, хроматография на анионной или катионной обменной смоле (такой, как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
После любой(ых) предварительной(ых) стадии(й) очистки смесь, содержащую представляющее интерес антитело и примеси, можно подвергать дополнительной очистке, например, хроматографией гидрофобного взаимодействия с низким значением pH с использованием буфера для элюции со значением pH приблизительно между 2,5-4,5, которую предпочтительно проводят при низких концентрациях соли (например, приблизительно 0-0,25 М соль).
Как правило, различные способы получения антител для использования в исследовании, тестировании и клиническом применении хорошо известны в данной области, согласуются с вышеописанными способами и/или предполагаются подходящими специалистом в данной области для конкретного представляющего интерес антитела.
C. Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам (взаимозаменяемо обозначаемым как «конъюгаты антитело-лекарственное средство» или «ADC»), содержащим любые анти-TAT226 антитела по изобретению, конъюгированные с одним или несколькими цитотоксическими средствами, такими как химиотерапевтическое средство, лекарственное средство, средство ингибирования роста, токсин (например, ферментативно активный токсин бактериального, грибного, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (т.е. радиоконъюгат).
Иммуноконъюгаты можно использовать при лечении рака для местной доставки цитотоксических средств, т.е. лекарственных средств, которые уничтожают или ингибируют рост или пролиферацию опухолевых клеток (Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drug Del. Rev. 26:151-172; патент США № 4975278). Иммуноконъюгаты обеспечивают целенаправленную доставку компонента лекарственного средства к опухоли и его внутриклеточное накопление в ней, если системное введение неконъюгированных лекарственных средств может приводить к неприемлемым уровням токсичности для нормальных клеток, а также когда стремятся элиминировать опухолевые клетки (Baldwin et al. (1986) Lancet (Mar. 15, 1986) p.603-05; Thorpe (1985) «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review» в Monoclonal Antibodies'84: Biological And Clinical Applications (A. Pinchera et al., eds) p.475-506). Сообщалось, что как поликлональные антитела, так и моноклональные антитела пригодны для этих способов (Rowland et al. (1986) Cancer Immunol. Immunother., 21:183-87). Лекарственные средства, используемые в этих способах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al. (1986), выше). Токсины, используемые в конъюгатах антитело-токсин, включают бактериальные токсины, такие как дифтерийный токсин, растительные токсины, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al. (2000) Jour. of the Nat. Cancer Inst. 92(19):1573-1581; Mandler et al. (2000) Bioorganic & Med. Chem. Letters 10:1025-1028; Mandler et al. (2002) Bioconjugate Chem. 13:786-791), майтанзиноиды (EP 1391213; Liu et al. (1996) Proc. Natl. Acad. Sci. USA 93:8618-8623) и калихеамицин (Lode et al. (1998) Cancer Res. 58:2928; Hinman et al. (1993) Cancer Res. 53:3336-3342). Токсины могут вызывать цитотоксические и цитостатические эффекты посредством механизмов, включая связывание тубулина, связывание ДНК или ингибирование топоизомеразы. Некоторые цитотоксические лекарственные средства обладают тенденцией инактивироваться или становиться менее активными при конъюгировании с крупными антителами или белковыми лигандами рецепторов.
ЗЕВАЛИН® (ибритумомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело-радиоизотоп, состоящий из мышиного моноклонального антитела IgG1 каппа, направленного против антигена CD20, представленного на поверхности нормальных и малигнизированных B-лимфоцитов, и радиоизотопа 111In или 90Y, связанных тиомочевинным хелатным линкером (Wiseman et al. (2000) Eur. Jour. Nucl. Med. 27(7):766-77; Wiseman et al. (2002) Blood 99(12):4336-42; Witzig et al. (2002) J. Clin. Oncol. 20(10):2453-63; Witzig et al. (2002) J. Clin. Oncol. 20(15):3262-69). Хотя ЗЕВАЛИН обладает активностью против B-клеточной неходжкинской лимфомы (NHL), его введение вызывает тяжелые и длительные цитопении у большинства пациентов. МИЛОТАРГ™ (гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитела с лекарственным средством, состоящий из антитела к huCD33, связанного с калихеамицином, получил одобрение с 2000 для лечения острого миелоидного лейкоза при помощи инъекции (Drugs of the Future (2000) 25(7):686; патенты США № 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Кантузумаб мертанзин (Immunogen, Inc.), конъюгат антитела с лекарственным средством, состоящий из антитела huC242, связанного посредством дисульфидного линкера SPP с компонентом лекарственного средства на основе майтанзиноидов, DM1, проходит фазу II исследований для лечения видов рака, при которых экспрессируется CanAg, таких как рак толстой кишки, поджелудочной железы, желудка и др. MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen, Inc.), конъюгат антитела с лекарственным средством, состоящий из моноклонального антитела против специфичного для предстательной железы мембранного антигена (PSMA), связанного с компонентом лекарственного средства на основе майтанзиноидов, DM1, находится в состоянии разработки для возможного лечения опухолей предстательной железы. Ауристатиновые пептиды, ауристатин E (AE) и монометилауристатин (MMAE), синтетические аналоги доластатина, конъюгировали с химерными моноклональными антителами cBR96 (специфичными к Lewis Y на карциномах) и cAC10 (специфичными к CD30 на гематологических опухолях) (Doronina et al. (2003) Nature Biotechnology 21(7):778-784), и они находятся в состоянии терапевтической разработки.
В некоторых вариантах осуществления иммуноконъюгат содержит анти-TAT226 антитело и химиотерапевтическое средство или другой токсин. Химиотерапевтические средства, пригодные для получения иммуноконъюгатов, описаны в данной работе (например, выше). Можно также использовать ферментативно активные токсины и их фрагменты, и они описаны в данной работе.
В некоторых вариантах осуществления иммуноконъюгат содержит анти-TAT226 антитело и один или несколько низкомолекулярных токсинов, включая в качестве неограничивающих примеров низкомолекулярные лекарственные средства, такие как калихеамицин, майтанзиноид, доластатин, ауристатин, трихотецен и CC1065, и производные этих лекарственных средств, которые обладают цитотоксической активностью. Примеры таких иммуноконъюгатов далее подробно рассмотрены ниже.
1. Иллюстративные иммуноконъюгаты
Иммуноконъюгат (или «конъюгат антитело-лекарственное средство» («ADC»)) по изобретению может обладать формулой I, ниже, где анти-TAT226 антитело конъюгировано (т.е. ковалентно присоединено) с одним или несколькими компонентами лекарственного средства (D) посредством подходящего линкера (L).
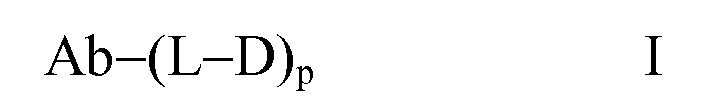
Соответственно, анти-TAT226 антитело можно конъюгировать с лекарственным средством или непосредственно, или посредством линкера. В формуле I p представляет собой среднее количество компонентов лекарственного средства на антитело, которое может варьировать, например, от приблизительно 1 до приблизительно 20 компонентов лекарственного средства на антитело и в некоторых вариантах осуществления от 1 до приблизительно 8 компонентов лекарственного средства на антитело.
a) Иллюстративные линкеры
Линкер может состоять из одного или нескольких компонентов линкера. Иллюстративные компоненты линкеров включают 6-малеимидокапроил («MC»), малеимидопропаноил («MP»), валин-цитруллин («val-cit» или «vc»), аланин-фенилаланин («ala-phe»), п-аминобензилоксикарбонил («PAB»), N-сукцинимидил-4-(2-пиридилтио)пентаноат («SPP»), N-сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат («SMCC) и N-сукцинимидил-(4-йодацетил)аминобензоат («SIAB»). Различные компоненты линкеров известны в данной области, некоторые из которых описаны ниже.
Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение лекарственного средства в клетке. Например, можно использовать неустойчивый к кислотам линкер (например, гидразон), чувствительный к протеазам (например, чувствительный к пептидазам) линкер, фотолабильный линкер, диметиловый линкер или дисульфидсодержащий линкер (Chari et al. Cancer Research 52:127-131 (1992); патент США № 5208020).
В некоторых вариантах осуществления компонент линкера может содержать «растягивающее звено», которое связывает антитело с другим компонентом линкера или с компонентом лекарственного средства. Иллюстративные растягивающие звенья представлены ниже (где волнистая линия указывает на участки ковалентного прикрепления к антителу):
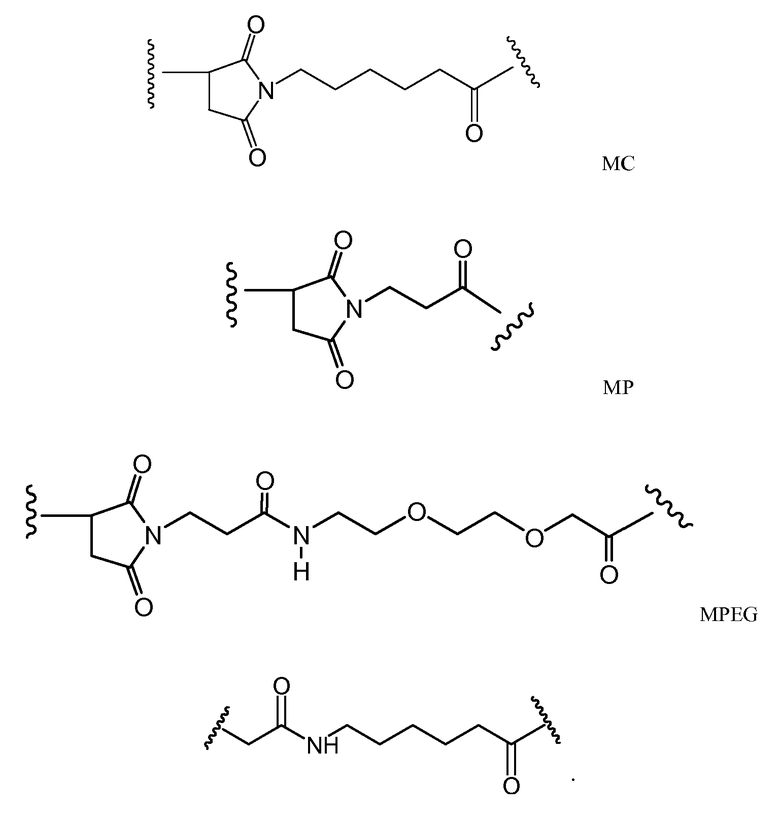
В некоторых вариантах осуществления компонент линкера может содержать аминокислотное звено. В одном из таких вариантов осуществления аминокислотное звено обеспечивает расщепление линкера протеазой, таким образом облегчая высвобождение лекарственного средства из иммуноконъюгата при воздействии внутриклеточных протеаз, таких как лизосомальные ферменты. См., например, Doronina et al. (2003) Nat. Biotechnol. 21:778-784. Иллюстративные аминокислотные звенья включают в качестве неограничивающих примеров дипептид, трипептид, тетрапептид и пентапептид. Иллюстративные дипептиды включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe), фенилаланин-лизин (fk или phe-lys) или N-метил-валин-цитруллин (Me-val-cit). Иллюстративные трипептиды включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотное звено может содержать аминокислотные остатки, которые существуют в природе, а также минорные аминокислоты и не существующие в природе аналоги аминокислот, такие как цитруллин. Аминокислотные звенья можно конструировать и оптимизировать по их избирательности по отношению к ферментативному расщеплению конкретным ферментом, например связанной с опухолью протеазой, катепсином B, C и D или протеазой плазмина.
В некоторых вариантах осуществления компонент линкера может содержать «спейсерное» звено, которое связывает антитело с компонентом лекарственного средства, или непосредственно, или посредством растягивающего звена и/или аминокислотного звена. Спейсерное звено может являться «жертвующим собой» или «нежертвующим собой». «Нежертвующее собой» спейсерное звено может представлять собой звено, в котором часть спейсерных звеньев или все из них остаются связанными с компонентом лекарственного средства при ферментативном (например, протеолитическом) расщеплении ADC. Примеры нежертвующих собой спейсерных звеньев включают в качестве неограничивающих примеров глициновое спейсерное звено и глицин-глициновое спейсерное звено. Также предусмотрены другие сочетания пептидных спейсеров, подверженных специфичному по отношению к последовательности ферментативному расщеплению. Например, ферментативное расщепление ADC, содержащего глицин-глициновое спейсерное звено, связанной с опухолевыми клетками протеазой может приводить к высвобождению глицин-глицинового компонента лекарственного средства из остатка ADC. В одном из таких вариантов осуществления глицин-глициновый компонент лекарственного средства затем подвергается отдельной стадии гидролиза в опухолевой клетке, таким образом отщепляя глицин-глициновое спейсерное звено от компонента лекарственного средства.
«Жертвующее собой» спейсерное звено обеспечивает высвобождение компонента лекарственного средства без отдельной стадии гидролиза. В некоторых вариантах осуществления спейсерное звено линкера содержит звено п-аминобензила. В одном из таких вариантов осуществления п-аминобензиловый спирт присоединен к аминокислотному звену посредством амидной связи и между бензиловым спиртом и цитотоксическим средством образуется карбамат, метилкарбамат или карбонат. См., например, Hamann et al. (2005) Expert Opin. Ther. Patents (2005) 15:1087-1103. В одном из вариантов осуществления спейсерное звено представляет собой п-аминобензилоксикарбонил (PAB). В некоторых вариантах осуществления фениленовая часть звена п-аминобензила замещена Qm, где Q представляет собой -C1-C8алкил, -O-(C1-C8алкил), -галоген, -нитро или -циано и m представляет собой целое число в диапазоне 0-4. Примеры жертвующих собой спейсерных звеньев дополнительно включают в качестве неограничивающих примеров ароматические соединения, которые электронно сходны с п-аминобензиловым спиртом (см., например, US 2005/0256030 A1), такие как производные 2-аминоимидазол-5-метанола (Hay et al. (1999) Bioorg. Med. Chem. Lett. 9:2237) и орто- или пара-аминобензилацетали. Можно использовать спейсеры, которые подвергаются циклизации при гидролизе амидной связи, такие как замещенные и незамещенные амиды 4-аминомасляной кислоты (Rodrigues et al., Chemistry Biology, 1995, 2, 223); соответствующим образом замещенные циклические системы бицикло[2.2.1] и бицикло[2.2.2] (Storm, et al., J. Amer. Chem. Soc., 1972, 94, 5815); и амиды 2-аминофенилпропионовой кислоты (Amsberry, et al., J. Org. Chem., 1990, 55, 5867). Удаление аминосодержащих лекарственных средств, которые замещены в α-положении глицина (Kingsbury et al., J. Med. Chem., 1984, 27, 1447), также представляет собой примеры жертвующих собой спейсеров, пригодных для ADC.
В одном из вариантов осуществления спейсерное звено представляет собой разветвленное звено бис(гидроксиметил)стирола (BHMS), как представлено ниже, которое можно использовать для включения и высвобождения нескольких лекарственных средств.
где Q представляет собой -C1-C8алкил, -O-(C1-C8алкил), -галоген, -нитро или -циано; m представляет собой целое число в диапазоне 0-4; n представляет собой 0 или 1; и p варьирует от 1 до приблизительно 20.
Линкер может содержать любой из одного или нескольких вышеописанных компонентов линкера. В некоторых вариантах осуществления линкер является таким, как представлено в скобках в следующей формуле II ADC:

где A представляет собой растягивающее звено и a представляет собой целое число от 0 до 1; W представляет собой аминокислотное звено и w представляет собой целое число от 0 до 12; Y представляет собой спейсерное звено и y представляет собой 0, 1 или 2; и Ab, D и p определены, как и выше для формулы I. Иллюстративные варианты осуществления таких линкеров описаны в US 2005-0238649 A1, который в явной форме приведен здесь в качестве ссылки.
Иллюстративные компоненты линкера и их сочетания представлены ниже в контексте ADC формулы II:
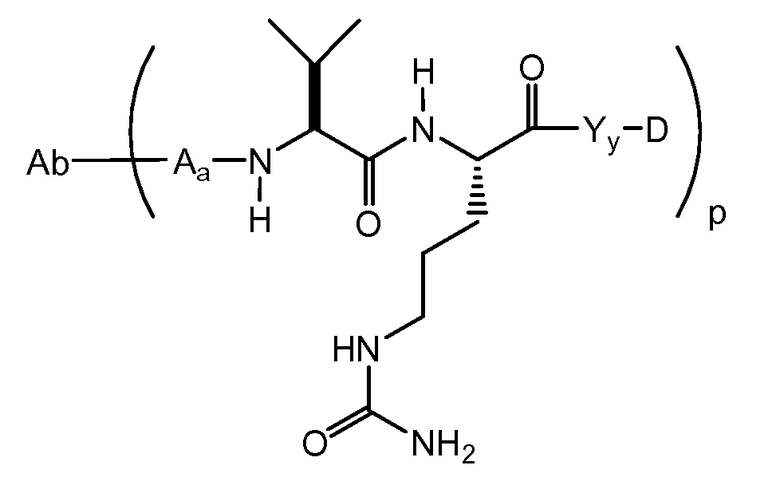 Val-Cit или VC
Val-Cit или VC
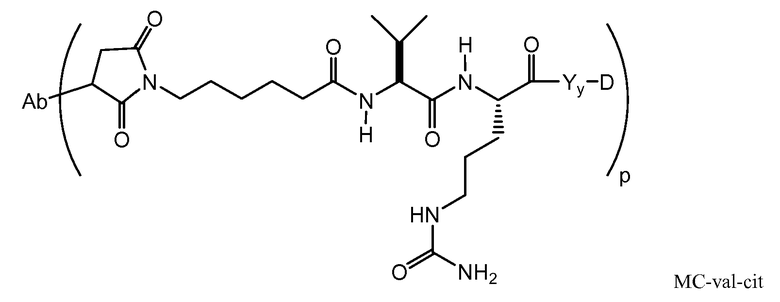
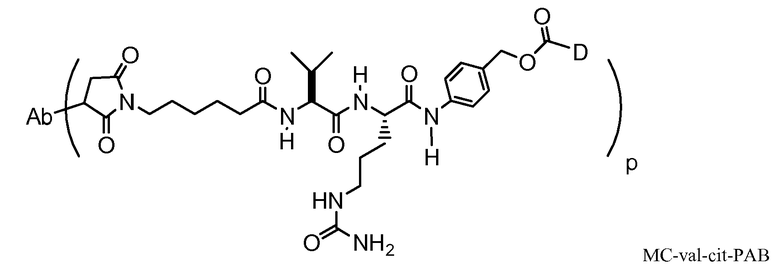
Компоненты линкеров, включая растягивающие, спейсерные и аминокислотные звенья, можно синтезировать способами, известными в данной области, такими как способы, описанные в US 2005-0238649 A1.
b) Иллюстративные компоненты лекарственного средства
(1) Майтанзин и майтанзиноиды
В некоторых вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с одной или несколькими молекулами майтанзиноидов. Майтанзиноиды представляют собой ингибиторы митоза, которые действуют посредством ингибирования полимеризации тубулина. Впервые майтанзин был выделен из восточноафриканского кустарника Maytenus serrata (патент США № 3896111). Затем было обнаружено, что определенные микробы также продуцируют майтанзиноиды, такие как майтанзинол и сложные эфиры майтанзинола C-3 (патент США № 4151042). Синтетический майтанзинол и его производные и аналоги описаны, например, в патентах США № 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663 и 4371533.
Компоненты лекарственного средства на основе майтанзиноидов являются привлекательными компонентами лекарственных средств в конъюгатах антител с лекарственным средством вследствие того, что они: (i) относительно доступны для получения ферментацией или химической модификацией при получении производных продуктов ферментации, (ii) пригодны для получения производных с помощью функциональных групп, подходящих для конъюгации посредством недисульфидных линкеров с антителами, (iii) стабильны в плазме и (iv) эффективны против множества линий опухолевых клеток.
Соединения на основе майтанзина, пригодные для использования в качестве компонентов лекарственного средства на основе майтанзиноидов, хорошо известны в данной области, и их можно выделять из природных источников согласно известным способам или получать с использованием способов генетической инженерии (см. Yu et al. (2002) PNAS 99:7968-7973). Майтанзинол и аналоги майтанзинола можно также получать синтетически согласно известным способам.
Иллюстративные компоненты лекарственного средства на основе майтанзиноидов включают компоненты, обладающие модифицированным ароматическим кольцом, таким как: C-19-дехлоро (патент США № 4256746) (получено восстановлением гидридом лития-алюминия анзамитоцина P2); C-20-гидрокси (или C-20-деметил) +/-C-19-дехлоро (патенты США № 4361650 и 4307016) (получено деметилированием с использованием Streptomyces или Actinomyces или отщеплением хлора с использованием LAH); и C-20-деметокси, C-20-ацилокси (-OCOR), +/-дехлоро (патент США № 4294757) (получено ацилированием с использованием ацилхлоридов), и компоненты, обладающие модификациями в других положениях.
Иллюстративные компоненты лекарственного средства на основе майтанзиноидов также включают компоненты, обладающие модификациями, такими как: C-9-SH (патент США № 4424219) (получено взаимодействием майтанзинола с H2S или P2S5); C-14-алкоксиметил(деметокси/CH2OR)(US 4331598); C-14-гидроксиметил или ацилоксиметил (CH2OH или CH2OAc) (патент США № 4450254) (получено из Nocardia); C-15-гидрокси/ацилокси (US 4364866) (получено превращением майтанзинола с помощью Streptomyces); C-15-метокси (патенты США № 4313946 и 4315929) (выделено из Trewia nudlflora); C-18-N-деметил (патент США № 4362663 и 4322348) (получено деметилированием майтанзинола с помощью Streptomyces); и 4,5-дезокси (US 4371533) (получено восстановлением трихлоридом титана/LAH майтанзинола).
Иллюстративные варианты осуществления компонентов лекарственного средства на основе майтанзиноидов включают: DM1, DM3 и DM4, обладающие структурами:

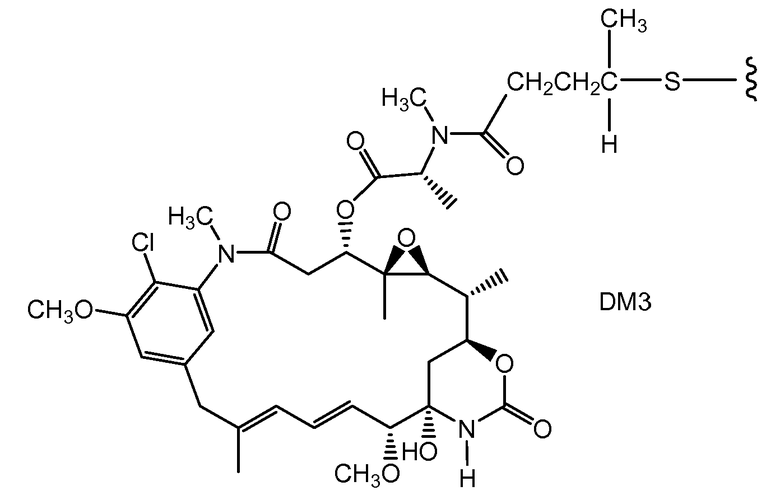
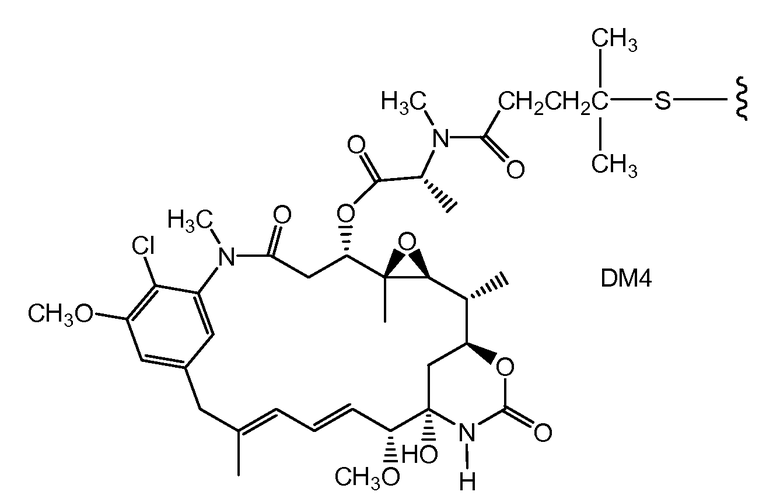
где волнистая линия указывает на ковалентное присоединение атома серы лекарственного средства к линкеру (L) конъюгата антитело-лекарственное средство. Сообщалось про ГЕРСЕПТИН® (трастузумаб), связанный посредством SMCC с DM1 (WO 2005/037992; US 2005/0276812 A1).
Другие иллюстративные конъюгаты антитело-лекарственное средство на основе майтанзиноидов обладают следующими структурами и сокращениями (где Ab представляет собой антитело и p составляет от 1 до приблизительно 8):
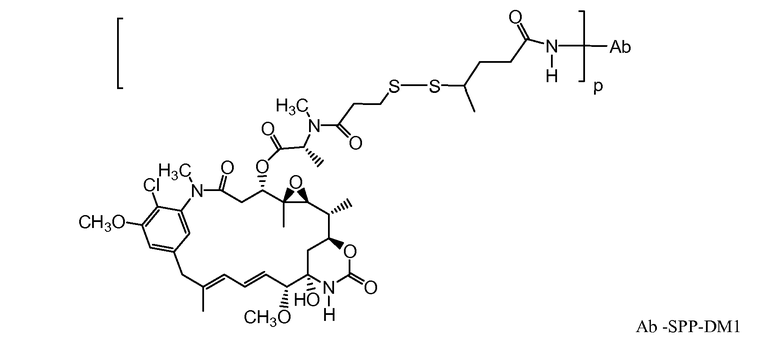
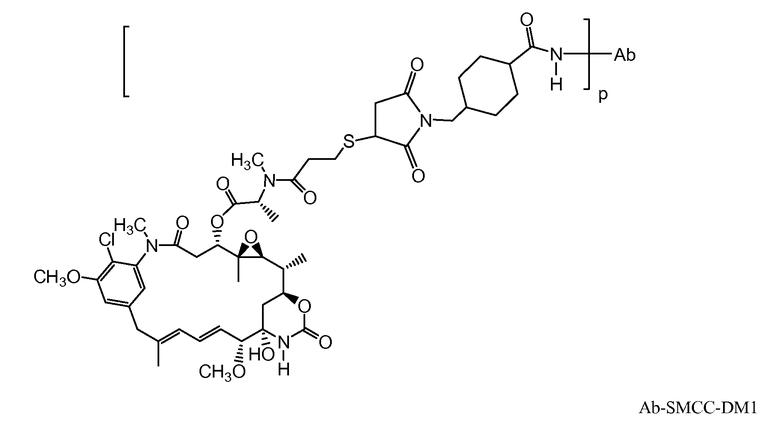
Иллюстративные конъюгаты антитело-лекарственное средство, где DM1 связан посредством линкера BMPEO с тиольной группой антитела, обладают структурой и сокращением:
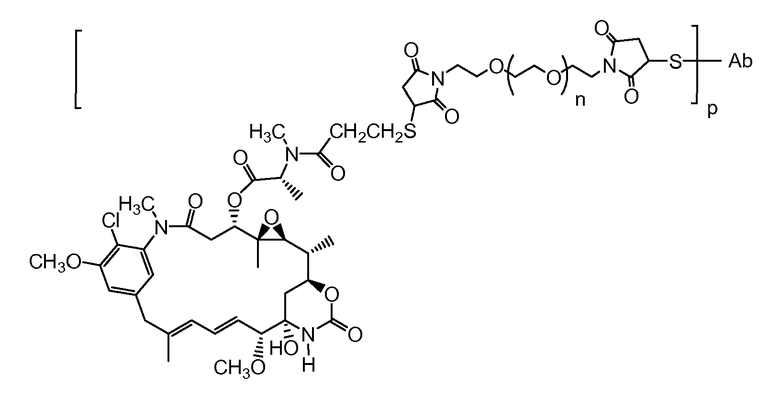
где Ab представляет собой антитело; n представляет собой 0, 1 или 2; и p представляет собой 1, 2, 3 или 4.
Иммуноконъюгаты, содержащие майтанзиноиды, способы их получения и их терапевтическое применение описаны, например, в патентах США № 5208020, 5416064, US 2005/0276812 A1 и в европейском патенте EP 0425235 B1, описание которых приведено в настоящем документе явно в качестве ссылки. В Liu et al., Proc. Natl. Acad. Sci. USA 93:8618-8623 (1996) описаны иммуноконъюгаты, содержащие майтанзиноид, обозначаемый как DM1, связанный с моноклональными антителами C242, направленными против колоректального рака человека. Обнаружено, что конъюгат является высокоцитотоксичным в отношении культивируемых клеток рака толстого кишечника, и он проявлял противоопухолевую активность в анализе роста опухоли in vivo. В Chari et al. Cancer Research 52:127-131 (1992) описаны иммуноконъюгаты, в которых майтанзиноид конъюгировали посредством дисульфидного линкера с антителом мыши A7, связывающимся с антигеном на линиях клеток рака толстого кишечника человека, или с другим моноклональным антителом мыши TA.1, которое связывается с онкогеном HER-2/neu. Цитотоксичность конъюгата TA.1-майтанзиноид тестировали in vitro на линии клеток рака молочной железы человека SK-BR-3, которая экспрессирует 3×105 поверхностных антигенов HER-2 на клетку. Конъюгат с лекарственным средством достигал степени цитотоксичности, сходной со свободным лекарственным средством на основе майтанзиноидов, которую можно повышать путем увеличения количества молекул майтанзиноида на молекулу антитела. Для конъюгата A7-майтанзиноид на мышах показана низкая системная цитотоксичность.
Конъюгаты антитело-майтанзиноид получают, химически связывая антитело с молекулой майтанзиноида без значительного снижения биологической активности или антитела, или молекулы майтанзиноида. См., например, патент США № 5208020 (описание которого приведено в настоящем документе явно в качестве ссылки). В среднем 3-4 молекулы майтанзиноида, конъюгированные с молекулой антитела, обладают эффективностью в усилении цитотоксичности по отношению к клеткам-мишеням без отрицательного воздействия на функцию или растворимость антитела, хотя даже одна молекула токсина на антитело, как можно ожидать, усиливает цитотоксичность по сравнению с использованием неконъюгированного антитела. Майтанзиноиды хорошо известны в данной области, и их можно синтезировать известными способами или выделять из природных источников. Подходящие майтанзиноиды описаны, например, в патенте США № 5208020 и в других патентах и не являющихся патентами публикациях, ссылки на которые приведены выше в данном описании. Предпочтительные майтанзиноиды представляют собой майтанзинол и аналоги майтанзинола, модифицированные по ароматическому кольцу или по другим положениям молекулы майтанзинола, такие как различные сложные эфиры майтанзинола.
В данной области известно множество связующих групп для получения конъюгатов антитело-майтанзиноид, включая, например, группы, описанные в патенте США № 5208020 или в европейском патенте 0425235 B1, Chari et al. Cancer Research 52:127-131 (1992), и US 2005/016993 A1, описания которых приведены здесь явно в качестве ссылки. Конъюгаты антитело-майтанзиноид, содержащие линкерный компонент SMCC, можно получать, как описано в US 2005/0276812 A1 «Antibody-drug conjugates and Methods». Линкеры включают дисульфидные группы, тиоэфирные группы, неустойчивые к кислотам группы, фотолабильные группы, неустойчивые к действию пептидаз группы или неустойчивые к действию эстераз группы, как описано в указанных выше патентах. Дополнительные линкеры описаны и приведены в качестве примеров в данной работе.
Конъюгаты антитела и майтанзиноида можно получать с использованием множества бифункциональных веществ для присоединения белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие, как диметиладипимидат-HCl), активные сложные эфиры (такие, как дисукцинимидилсуберат), альдегиды (такие, как глутаральдегид), бис-азидосоединения (такие, как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие, как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие, как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). В некоторых вариантах осуществления вещество для присоединения представляет собой N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) (Carlsson et al., Biochem. J. 173:723-737 (1978)) или N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP) для обеспечения образования дисульфидной связи.
Линкер можно присоединять к молекуле майтанзиноида в различных положениях в зависимости от типа связи. Например, сложноэфирная связь может образоваться посредством реакции с гидроксильной группой с использованием общепринятых способов присоединения. Реакция может протекать в положении C-3, обладающем гидроксильной группой, в положении C-14, модифицированном гидроксиметилом, в положении C-15, модифицированном гидроксильной группой, и в положении C-20, обладающем гидроксильной группой. В одном из вариантов осуществления связь образуется в положении C-3 майтанзинола или аналога майтанзинола.
(2) Ауристатины и доластатины
В некоторых вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с доластатином или пептидным аналогом и производным доластатина, например ауристатином (патенты США № 5635483; 5780588). Показано, что доластатины и ауристатины нарушают динамику микротрубочек, гидролиз GTP и ядерное и клеточное деление (Woyke et al. (2001) Antimicrob. Agents and Chemother. 45(12):3580-3584) и обладают противораковой (патент США № 5663149) и противогрибковой активностью (Pettit et al. (1998) Antimicrob. Agents Chemother. 42:2961-2965). Компонент лекарственного средства на основе доластатина или ауристатина можно присоединять к антителу посредством N-конца (амино) или C-конца (карбоксил) пептидного компонента лекарственного средства (WO 02/088172).
Иллюстративные варианты осуществления с применением ауристатина включают компоненты лекарственного средства на основе связанного с N-концом монометилауристатина DE и DF, описанные в Senter et al., Proceedings of the American Association for Cancer Research, Volume 45, Abstract Number 623, представленной 28 марта, 2004, описание которой явно приведено в качестве ссылки в полном объеме.
Пептидный компонент лекарственного средства можно выбрать из формул DE и DF ниже:
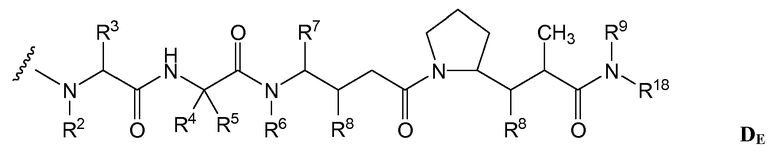
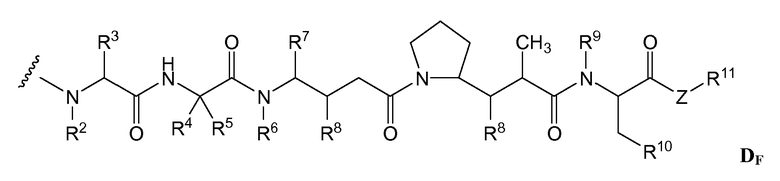 ,
,
где волнистая линия для DE и DF указывает на участок ковалентного присоединения к антителу или антителу-компоненту линкера и независимо в каждом положении:
R2 выбран из H и алкила C1-C8;
R3 выбран из H, C1-C8алкила, C3-C8карбоцикла, арила, C1-C8алкиларила, C1-C8алкила-(C3-C8карбоцикл), C3-C8гетероцикла и C1-C8алкила-(C3-C8гетероцикл);
R4 выбран из H, C1-C8алкила, карбоцикла C3-C8, арила, C1-C8алкиларила, C1-C8алкила-(C3-C8карбоцикл), C3-C8гетероцикла и C1-C8алкила-(C3-C8гетероцикл);
R5 выбран из H и метила;
или R4 и R5 совместно образуют карбоциклическое кольцо и обладают формулой -(CRaRb)n-, где Ra и Rb независимо выбраны из H, C1-C8алкила и C3-C8карбоцикла и n выбрано из 2, 3, 4, 5 и 6;
R6 выбран из H и C1-C8алкила;
R7 выбран из H, C1-C8алкила, C3-C8карбоцикла, арила, C1-C8алкиларила, C1-C8алкила-(C3-C8карбоцикл), C3-C8гетероцикла и C1-C8алкила-(C3-C8гетероцикл);
каждый R8 независимо выбран из H, OH, C1-C8алкила, C3-C8карбоцикла и O-(C1-C8алкил);
R9 выбран из H и C1-C8алкила;
R10 выбран из арила или C3-C8гетероцикла;
Z представляет собой O, S, NH или NR12, где R12 представляет собой C1-C8алкил;
R11 выбран из H, C1-C20алкила, арила, C3-C8гетероцикла, -(R13O)m-R14 или -(R13O)m-CH(R15)2;
m представляет собой целое число в диапазоне 1-1000;
R13 представляет собой C2-C8алкил;
R14 представляет собой H или C1-C8алкил;
каждое вхождение R15 независимо представляет собой H, COOH, -(CH2)n-N(R16)2, -(CH2)n-SO3H или -(CH2)n-SO3-C1-C8алкил;
каждое вхождение R16 независимо представляет собой H, C1-C8алкил или -(CH2)n-COOH;
R18 выбран из -C(R8)2-C(R8)2-арила, -C(R8)2-C(R8)2-(C3-C8гетероцикл) и -C(R8)2-C(R8)2-(C3-C8карбоцикл); и
n представляет собой целое число в диапазоне 0-6.
В одном из вариантов осуществления R3, R4 и R7 независимо представляют собой изопропил или втор-бутил и R5 представляет собой -H или метил. В иллюстративном варианте осуществления каждый R3 и R4 представляет собой изопропил, R5 представляет собой -H и R7 представляет собой втор-бутил.
В другом варианте осуществления каждый R2 и R6 представляет собой метил и R9 представляет собой -H.
В другом варианте осуществления каждое вхождение R8 представляет собой -OCH3.
В иллюстративном варианте осуществления каждый R3 и R4 представляет собой изопропил, каждый R2 и R6 представляет собой метил, R5 представляет собой -H, R7 представляет собой втор-бутил, каждое вхождение R8 представляет собой -OCH3 и R9 представляет собой -H.
В одном из вариантов осуществления Z представляет собой -O- или -NH-.
В одном из вариантов осуществления R10 представляет собой арил.
В иллюстративном варианте осуществления R10 представляет собой -фенил.
В иллюстративном варианте осуществления, где Z представляет собой -O-, R11 представляет собой -H, метил или трет-бутил.
В одном из вариантов осуществления, где Z представляет собой -NH, R11 представляет собой -CH(R15)2, где R15 представляет собой -(CH2)n-N(R16)2 и R16 представляет собой -C1-C8алкил или -(CH2)n-COOH.
В другом варианте осуществления, где Z представляет собой -NH, R11 представляет собой -CH(R15)2, где R15 представляет собой -(CH2)n-SO3H.
Иллюстративный вариант осуществления с применением ауристатина формулы DE представляет собой MMAE, где волнистая линия указывает на ковалентное присоединение к линкеру (L) конъюгата антитело-лекарственное средство:
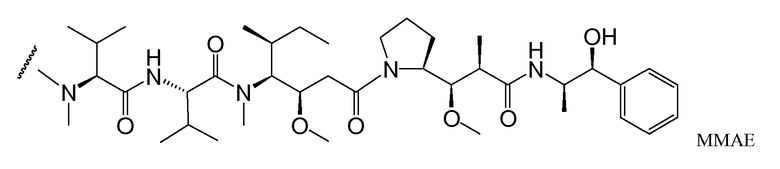
Иллюстративный вариант осуществления с применением ауристатина формулы DF представляет собой MMAF, где волнистая линия указывает на ковалентное присоединение к линкеру (L) конъюгата антитело-лекарственное средство (см. US 2005/0238649 и Doronina et al. (2006) Bioconjugate Chem. 17:114-124):
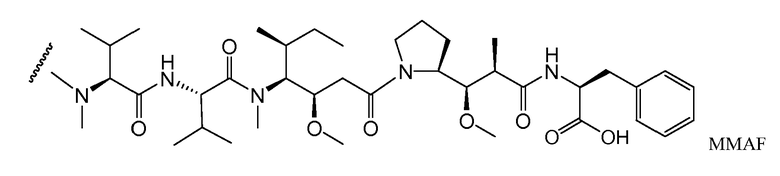
Другие компоненты лекарственного средства включают следующие производные MMAF, где волнистая линия указывает на ковалентное присоединение к линкеру (L) конъюгата антитело-лекарственное средство:
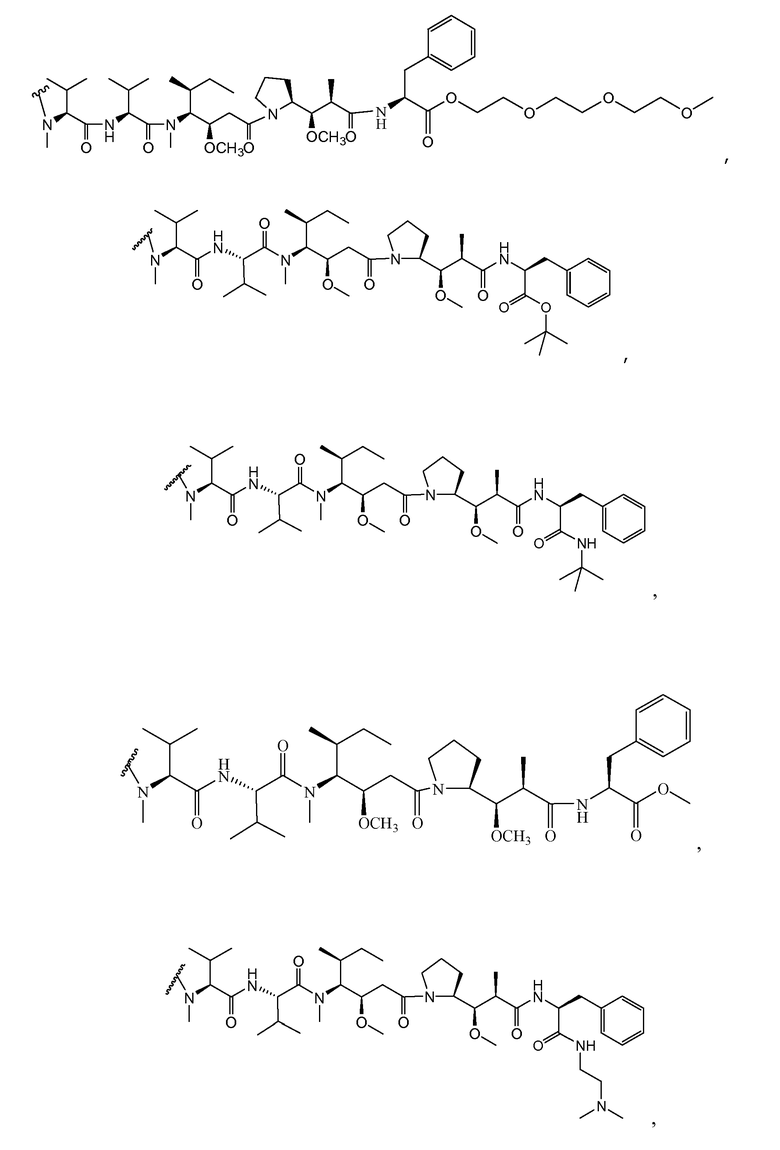
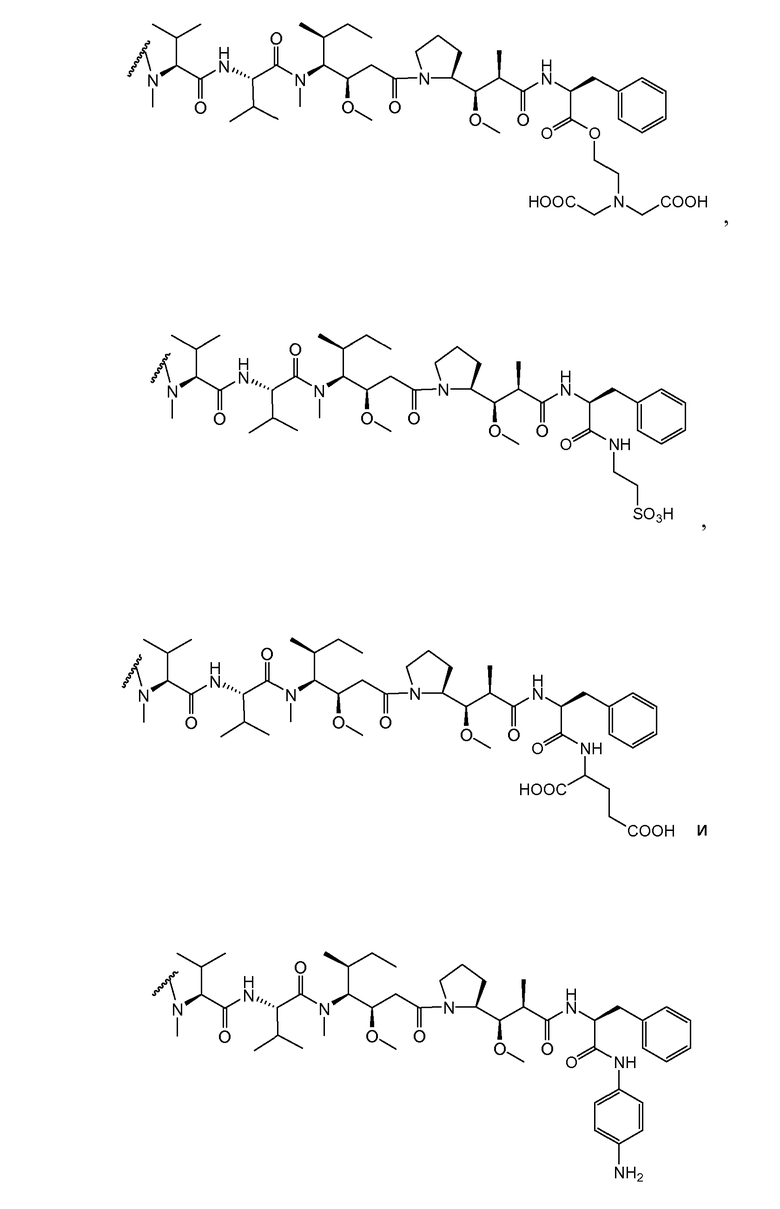
В одном из аспектов гидрофильные группы, включающие в качестве неограничивающих примеров сложные эфиры триэтиленгликоля (TEG), как показано выше, можно присоединять к компоненту лекарственного средства в положении R11. Не вдаваясь в конкретные теоретические подробности, гидрофильные группы содействуют интернализации и отсутствию агрегации компонента лекарственного средства.
Иллюстративные варианты осуществления с применением ADC формулы I, содержащего ауристатин/доластатин или их производное, описаны в US 2005-0238649 A1 и Doronina et al. (2006) Bioconjugate Chem. 17:114-124, которое явно приведено в настоящем документе в качестве ссылки. В иллюстративных вариантах осуществления ADC формулы I, содержащие MMAE или MMAF и различные компоненты линкера, обладают следующими структурами и сокращениями (где «Ab» представляет собой антитело; p составляет от 1 до приблизительно 8, «Val-Cit» представляет собой дипептид валин-цитруллин; и «S» представляет собой атом серы):
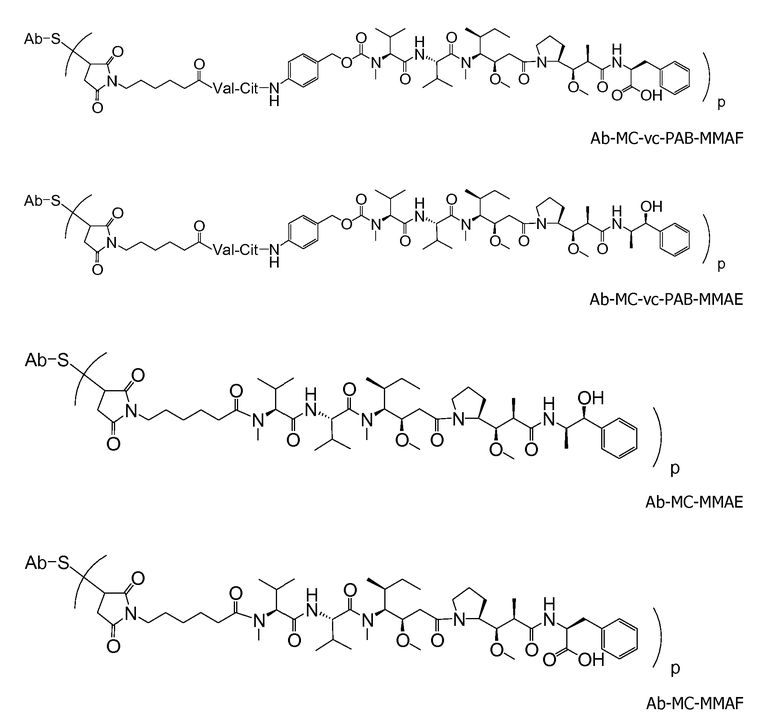
Иллюстративные варианты осуществления с применением ADC формулы I, содержащих MMAF и различные компоненты линкера, дополнительно включают Ab-MC-PAB-MMAF и Ab-PAB-MMAF. Интересно отметить, что иммуноконъюгаты, содержащие MMAF, присоединенный к антителу при помощи линкера, который не является протеолитически расщепляемым, как показано, обладают активностью, сравнимой с иммуноконъюгатами, содержащими MMAF, присоединенный к антителу при помощи протеолитически расщепляемого линкера. См., Doronina et al. (2006) Bioconjugate Chem. 17:114-124. В таких случаях, как предполагают, высвобождение лекарственного средства происходит при деградации антитела в клетке. Id.
Как правило, компоненты лекарственного средства на основе пептидов можно получать путем образования пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получать, например, согласно способу жидкофазного синтеза (см. E. Schroder and K. Lubke. «The Peptides», volume 1, p.76-136, 1965, Academic Press), который хорошо известен в области химии пептидов. Компоненты лекарственного средства на основе ауристатина/доластатина можно получать согласно способам: US 2005-0238649 A1; патент США № 5635483; патент США № 5780588; Pettit et al. (1989) J. Am. Chem. Soc. 111:5463-5465; Pettit et al. (1998) Anti-Cancer Drug Design 13:243-277; Pettit G.R. et al. Synthesis, 1996, 719-725; Pettit et al. (1996) J. Chem. Soc. Perkin Trans. 1 5:859-863; и Doronina (2003) Nat. Biotechnol. 21(7):778-784.
В частности, компоненты формулы DF лекарственного средства на основе ауристатина/доластатина, такие как MMAF и его производные, можно получать с использованием способов, описанных в US 2005-0238649 A1 и Doronina et al. (2006) Bioconjugate Chem. 17:114-124. Компоненты формулы DE лекарственного средства на основе ауристатина/доластатина, такие как MMAE и его производные, можно получать с использованием способов, описанных в Doronina et al. (2003) Nat. Biotech. 21:778-784. Компоненты лекарственного средства-линкера MC-MMAF, MC-MMAE, MC-vc-PAB-MMAF и MC-vc-PAB-MMAE можно подходящим образом синтезировать общепринятыми способами, например, как описано в Doronina et al. (2003) Nat. Biotech. 21:778-784 и публикации патентной заявки № US 2005/0238649 A1, и затем конъюгировать с представляющим интерес антителом.
(3) Калихеамицин
В других вариантах осуществления иммуноконъюгат содержит антитело по изобретению, конъюгированное с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицина способны создавать двухцепочечные разрывы ДНК в субпикомолярных концентрациях. Для получения конъюгатов семейства калихеамицина см. патенты США 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001, 5877296 (все выданы American Cyanamid Company). Структурные аналоги калихеамицина, которые можно использовать, включают в качестве неограничивающих примеров γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θI 1 (Hinman et al. Cancer Research 53:3336-3342 (1993), Lode et al. Cancer Research 58:2925-2928 (1998) и вышеупомянутые патенты США, выданные American Cyanamid). Другим противоопухолевым лекарственным средством, с которым можно конъюгировать антитело, является QFA, который представляет собой антифолат. Как калихеамицин, так и QFA обладают внутриклеточным действием, и они нелегко проходят через плазматическую мембрану. Таким образом, клеточный захват этих веществ посредством опосредуемой антителами интернализации значительно повышает их цитотоксические эффекты.
c) Другие цитотоксические средства
Другие противоопухолевые средства, которые можно конъюгировать с антителами по изобретению, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство средств, известное под собирательным названием комплекс LL-E33288, описанный в патентах США № 5053394, 5770710, а также эсперамицины (патент США № 5877296).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают A-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. См., например, WO 93/21232, опубликованный 28 октября 1993.
Настоящее изобретение, кроме того, относится к иммуноконъюгату, образованному антителом и соединением с нуклеолитической активностью (например, рибонуклеаза или ДНК-эндонуклеаза, такая как дезоксирибонуклеаза; ДНКаза).
В некоторых вариантах осуществления иммуноконъюгат может содержать высокорадиоактивный атом. Для получения радиоактивных конъюгированных антител доступны различные радиоактивные изотопы. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. В случае применения конъюгата для детекции он может содержать радиоактивный атом для сцинтиграфических исследований, например Tc99m или I123, или спиновую метку для получения изображения ядерного магнитного резонанса (NMR) (также известного как магнитно-резонансная томография, MRI), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивные или другие метки можно встраивать в конъюгат известными способами. Например, пептид можно биологически синтезировать или можно синтезировать посредством химического синтеза аминокислот с использованием пригодных предшественников аминокислот, содержащих, например, фтор-19 вместо водорода. Метки, такие как Tc99m или I123, Re186, Re188 и In111, можно присоединять через остаток цистеина в пептиде. Иттрий-90 можно присоединять через остаток лизина. Способ с ЙОДОГЕНОМ (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80:49-57) можно использовать для встраивания йода-123. В «Monoclonal Antibodies in Immunoscintigraphy» (Chatal, CRC Press 1989) подробно описаны другие способы.
В некоторых вариантах осуществления иммуноконъюгат может содержать анти-TAT226 антитело по изобретению, конъюгированное с активирующим пролекарство ферментом, который превращает пролекарство (например, пептидное химиотерапевтическое средство, см. WO 81/01145) в активное лекарственное средство, такое как противораковое лекарственное средство. Такие иммуноконъюгаты пригодны в антителозависимой фермент-опосредованной пролекарственной терапии («ADEPT»). Ферменты, которые можно конъюгировать с анти-TAT226 антителом по изобретению, включают в качестве неограничивающих примеров щелочные фосфатазы, которые пригодны для превращения фосфатсодержащих пролекарств в свободные лекарственные средства; арилсульфатазы, которые пригодны для превращения сульфатсодержащих пролекарств в свободные лекарственные средства; цитозиндезаминазу, которая пригодна для превращения нетоксичного 5-фторцитозина в противораковое лекарственное средство, 5-фторурацил; протеазы, такие как протеаза Serratia, термолизин, субтилизин, карбоксипептидазы и катепсины (такие, как катепсины B и L), которые пригодны для превращения пептидсодержащих пролекарств в свободные лекарственные средства; D-аланилкарбоксипептидазы, которые пригодны для превращения пролекарств, которые содержат D-аминокислотные заместители; углевод-расщепляющие ферменты, такие как β-галактозидаза и нейраминидаза, которые пригодны для превращения гликозилированных пролекарств в свободные лекарственные средства; β-лактамазу, которая пригодна для превращения лекарственных средств, модифицированных с помощью β-лактамов, в свободные лекарственные средства; и пенициллинамидазы, такие как пенициллин-V-амидаза и пеницилин-G-амидаза, которые пригодны для превращения лекарственных средств, модифицированных по атомам азота аминогруппы с помощью феноксиацетильных или фенилацетильных групп соответственно, в свободные лекарственные средства. Ферменты можно ковалентно связывать с анти-TAT226 антителами по изобретению при помощи способов рекомбинантных ДНК, хорошо известных в данной области. См., например, Neuberger et al., Nature 312:604-608 (1984).
d) Нагрузка лекарственного средства
Нагрузка лекарственного средства представляет собой p, среднее количество компонентов лекарственного средства на антитело в молекуле формулы I. Нагрузка лекарственного средства может варьировать от 1 до 20 компонентов лекарственного средства (D) на антитело. ADC формулы I включают собрание антител, конъюгированных с компонентами лекарственного средства в диапазоне от 1 до 20. Среднее количество компонентов лекарственного средства на антитело в препаратах ADC из реакций конъюгации можно охарактеризовать общепринятыми способами, такими как масс-спектроскопия, анализ ELISA и HPLC. Можно также определить количественное распределение ADC в терминах p. В некоторых случаях разделение, очистку и характеристику гомогенного ADC, где p представляет собой определенную величину из ADC с другими нагрузками лекарственного средства, можно достичь способами, такими как HPLC с обращенной фазой или электрофорез.
Для некоторых конъюгатов антитело-лекарственное средство, p может ограничиваться количеством участков присоединения на антителе. Например, там, где присоединение осуществляют по тиолу цистеина, как в иллюстративных вариантах осуществления выше, антитело может содержать только одну или несколько тиольных групп цистеина или может содержать только одну или несколько достаточно активных тиольных групп, по которым можно присоединять линкер. В некоторых вариантах осуществления более высокая нагрузка лекарственного средства, например p>5, может вызывать агрегацию, нерастворимость, токсичность или утрату способности проникать в клетки для определенных конъюгатов антитело-лекарственное средство. В некоторых вариантах осуществления нагрузка лекарственного средства для ADC по изобретению варьирует от 1 до приблизительно 8, от приблизительно 2 до приблизительно 6 или от приблизительно 3 до приблизительно 5. Действительно, показано, что для некоторых ADC оптимальное отношение компонентов лекарственного средства на антитело может составлять менее чем 8 и может составлять от приблизительно 2 до приблизительно 5. См. US 2005-0238649 A1.
В некоторых вариантах осуществления меньшее количество компонентов лекарственного средства по сравнению с теоретическим максимумом конъюгируют с антителом в ходе реакции конъюгации. Антитело может содержать, например, остатки лизина, которые не взаимодействуют с промежуточным соединением лекарственного средства с линкером или линкерным реагентом, как рассмотрено ниже. Как правило, антитела не содержат много свободных и активных тиольных групп цистеина, которые можно связать с компонентом лекарственного средства; на практике большинство тиольных остатков цистеина в антителах существует в виде дисульфидных мостиков. В некоторых вариантах осуществления для получения активных тиольных групп цистеина антитело можно восстанавливать восстанавливающим средством, таким как дитиотреитол (DTT) или трикарбонилэтилфосфин (TCEP) в частично или полностью восстанавливающих условиях. В некоторых вариантах осуществления антитело подвергают воздействию денатурирующих условий для выявления активных нуклеофильных групп, таких как лизин или цистеин.
Нагрузку (отношение лекарственное средство/антитело) ADC можно контролировать разными способами, например посредством: (i) ограничения молярного избытка промежуточного соединения лекарственного средства с линкером или линкерного реагента относительно антитела, (ii) ограничение времени или температуры реакции конъюгации и (iii) частичные или ограничивающие восстановительные условия для модификации тиола цистеина.
Следует понимать, что если с промежуточным соединением лекарственного средства с линкером или линкерным реагентом реагирует более одной нуклеофильной группы с последующим взаимодействием с реагентом компонента лекарственного средства, то полученный продукт представляет собой смесь соединений ADC с распределением одного или нескольких компонентов лекарственного средства, присоединенных к антителу. Среднее количество лекарственных средств на антитело можно вычислить из смеси с помощью двойного анализа ELISA для антител, который специфичен для антитела и специфичен для лекарственного средства. Индивидуальные молекулы ADC можно идентифицировать в смеси путем масс-спектроскопии и разделить путем HPLC, например, хроматографией на основе гидрофобного взаимодействия (см., например, Hamblett K. J. et al. «Effect of drug loading on the pharmacology, pharmacokinetics, and toxicity of an anti-CD30 antibody-drug conjugate», Abstract № 624, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of AACR, Volume 45, March 2004; Alley S.C. et al. «Controlling the location of drug attachment in antibody-drug conjugates», Abstract № 627, American Association for Cancer Research, 2004 Annual Meeting, March 27-31, 2004, Proceedings of AACR, Volume 45, March 2004). В некоторых вариантах осуществления гомогенный ADC с единой величиной нагрузки можно выделить из конъюгационной смеси электрофорезом или хроматографией.
e) Некоторые способы получения иммуноконъюгатов
ADC формулы I можно получать несколькими способами с использованием реакций органической химии, условий и реагентов, известных специалистам в данной области, включая: (1) взаимодействие нуклеофильной группы антитела с бивалентным линкерным реагентом с образованием Ab-L посредством ковалентной связи с последующим взаимодействием с компонентом лекарственного средства D; и (2) взаимодействие нуклеофильной группы компонента лекарственного средства с бивалентным линкерным реагентом с образованием D-L посредством ковалентной связи с последующим взаимодействием с нуклеофильной группой антитела. Иллюстративные способы получения ADC формулы I посредством последнего способа описаны в US 2005-0238649 A1, который явно приведен здесь в качестве ссылки.
Нуклеофильные группы на антителах включают в качестве неограничивающих примеров: (i) N-концевые аминогруппы, (ii) аминогруппы боковой цепи, например лизин, (iii) тиоловые группы боковой цепи, например цистеин, и (iv) гидроксильные группы или аминогруппы сахаров, если антитело гликозилировано. Аминогруппы, тиоловые и гидроксильные группы являются нуклеофилами и способны взаимодействовать с образованием ковалентных связей с электрофильными группами на компонентах линкера, где линкерные реагенты включают: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, трехгалоидные производные и галогениды кислот; (ii) алкил- и бензилгалогениды, такие как галоацетамиды; (iii) альдегидные, кетоновые, карбоксильные и малеинимидные группы. Некоторые антитела обладают восстанавливаемыми межцепочечными дисульфидами, т.е. цистеиновыми мостиками. Антитела можно получать в активной форме для конъюгации с линкерными реагентами обработкой восстанавливающим реагентом, таким как DTT (дитиотреитол) или трикарбонилэтилфосфин (TCEP), таким образом, что антитело полностью или частично восстановлено. Каждый цистеиновый мостик, таким образом, образует теоретически два активных тиоловых нуклеофила. Дополнительные нуклеофильные группы можно вводить в антитела с помощью модификации остатков лизина, например взаимодействия лизиновых остатков с 2-иминотиоланом (реагент Траута), что приводит к превращению амина в тиол. Активные тиоловые группы можно вводить в антитело введением одного, двух, трех, четырех или более остатков цистеина (например, получением вариантов антител, содержащих один или несколько неприродных аминокислотных остатков цистеина).
Конъюгаты антител с лекарственными средствами по изобретению можно также получать взаимодействием между электрофильной группой на антителе, такой как альдегидная или кетоновая карбонильная группа, и нуклеофильной группой на линкерном реагенте или лекарственном средстве. Пригодные нуклеофильные группы на линкерном реагенте включают в качестве неограничивающих примеров гидразид, оксим, амино, гидразин, тиосемикарбазон, гидразинкарбоксилат и арилгидразид. В одном из вариантов осуществления антитело модифицируют для введения электрофильных компонентов, которые способны взаимодействовать с нуклеофильными заместителями на линкерном реагенте или лекарственном средстве. В другом варианте осуществления сахара гликозилированных антител можно окислять, например, окисляющими реагентами на основе периодата с образованием альдегидных или кетоновых групп, которые могут реагировать с аминогруппой линкерных реагентов или компонентов лекарственного средства. Получаемые группы иминового основания Шиффа могут образовывать стабильную связь, или их можно восстанавливать, например, с помощью борогидридных реагентов с образованием стабильных аминных связей. В одном из вариантов осуществления взаимодействие углеводной части гликозилированного антитела с либо галактозооксидазой, либо метапериодатом натрия может приводить к образованию карбонильных (альдегидных и кетоновых) групп в антителе, которые могут реагировать с соответствующими группами на лекарственном средстве (Hermanson, Bioconjugate Techniques). В другом варианте осуществления антитела, содержащие N-концевые остатки серина или треонина, могут взаимодействовать с метапериодатом натрия, приводя к образованию альдегида на месте первой аминокислоты (Geoghegan & Stroh (1992) Bioconjugate Chem. 3:138-146; US 5362852). Такой альдегид может реагировать с компонентом лекарственного средства или нуклеофилом линкера.
Нуклеофильные группы на компоненте лекарственного средства включают в качестве неограничивающих примеров: аминогруппы, тиоловые, гидроксильные, гидразидные, оксимные, гидразиновые, тиосемикарбазоновые, гидразинкарбоксилатные и арилгидразидные группы, способные реагировать с образованием ковалентных связей с электрофильными группами на компонентах линкера, где линкерные реагенты включают: (i) активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, трехгалоидные производные и галогениды кислот; (ii) алкил- и бензилгалогениды, такие как галоацетамиды; (iii) альдегидные, кетоновые, карбоксильные и малеинимидные группы.
Соединения по изобретению явным образом включают в качестве неограничивающих примеров ADC, полученные с помощью следующих перекрестносшивающих реагентов: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые коммерчески доступны (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A); см. стр. 467-498, 2003-2004 Applications Handbook and Catalog.
Иммуноконъюгаты, содержащие антитело и цитотоксическое средство, можно также получать с использованием множества бифункциональных веществ для присоединения белка, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие, как диметиладипимидат-HCl), активные сложные эфиры (такие, как дисукцинимидилсуберат), альдегиды (такие, как глутаральдегид), бис-азидосоединения (такие, как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие, как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие, как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). Например, рициновый иммунотоксин можно получить, как описано в Vitetta et al., Science, 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминопентауксусная кислота (MX-DTPA) представляет собой иллюстративное хелатирующее средство для конъюгации радионуклеотида с антителом. См. WO94/11026.
Альтернативно слитый белок, содержащий антитело и цитотоксическое средство, можно получать, например, рекомбинантными способами или пептидным синтезом. Рекомбинантная молекула ДНК может содержать области, кодирующие антительную и цитотоксическую части конъюгата, или примыкающие друг к другу, или разделенные областью, кодирующей линкерный пептид, который не нарушает требуемые свойства конъюгата.
В другом варианте осуществления антитело можно конъюгировать с «рецептором» (таким, как стрептавидин) для использования в предварительном «превращении опухоли в мишень», где конъюгат антитело-рецептор вводят пациенту с последующим удалением несвязавшегося конъюгата из кровотока с использованием средства, обеспечивающего клиренс, и затем введением «лиганда» (например, авидина), который конъюгирован с цитотоксическим средством (например, радиоактивно меченным нуклеотидом).
D. Фармацевтические композиции
В одном из аспектов изобретение дополнительно относится к фармацевтическим композициям, содержащим, по меньшей мере, одно анти-TAT226 антитело по изобретению и/или, по меньшей мере, один его иммуноконъюгат. В некоторых вариантах осуществления фармацевтическая композиция содержит: 1) анти-TAT226 антитело и/или его иммуноконъюгат и 2) фармацевтически приемлемый носитель. В некоторых вариантах осуществления фармацевтическая композиция содержит: 1) анти-TAT226 антитело и/или его иммуноконъюгат и необязательно 2), по меньшей мере, одно дополнительное терапевтическое средство. Дополнительные терапевтические средства включают в качестве неограничивающих примеров средства, описанные ниже в разделе E.2.
Фармацевтические композиции, содержащие антитело или иммуноконъюгат по изобретению, получают в целях хранения посредством смешивания антитела или иммуноконъюгата, обладающего требуемой степенью чистоты, необязательно с физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в форме водных растворов или лиофилизированных или другим способом обезвоженных композиций. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях, и они включают буферы, такие как фосфатный, цитратный, гистидиновый и на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие, как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензэтония); фенол, бутиловый или бензиловый спирт; алкилпарабены (такие, как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (приблизительно менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как ТВИН™, ПЛЮРОНИК™ или полиэтиленгликоль (PEG). Фармацевтические композиции для применения при введении in vivo, как правило, являются стерильными. Этого легко достигают посредством фильтрации через стерильные фильтрационные мембраны.
Активные ингредиенты также можно включать в микрокапсулы, полученные, например, способами коацервации или посредством межфазной полимеризации, например микрокапсулы на основе гидроксиметилцеллюлозы или желатина и микрокапсулы на основе полиметилметакрилата, соответственно в коллоидных системах для доставки лекарственного средства (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Примеры пригодных препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело или иммуноконъюгат по изобретению, где матрицы находятся в форме обладающих определенной формой изделий, например пленок или микрокапсул. Примеры матриц для замедленного высвобождения включают сложные полиэфиры, гидрогели (например, поли-2-гидроксиэтилметакрилат или поливиниловый спирт), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как полимеры, такие как на основе этиленвинилацетата и молочной кислоты-гликолевой кислоты, способны высвобождать молекулы в течение 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Если инкапсулированные антитела или иммуноконъюгаты остаются в организме в течение длительного времени, то они могут денатурировать или агрегировать под действием влажности при 37°C, что приводит к утрате биологической активности и возможным изменениям иммуногенности. Можно разработать рациональные стратегии для стабилизации в зависимости от вовлеченного механизма. Например, если обнаружено, что механизм агрегации состоит в образовании межмолекулярных S-S связей посредством превращения тиола в дисульфид, то стабилизации можно достичь модификацией сульфгидрильных остатков, лиофилизацией из кислотных растворов, контролированием содержания воды с использованием подходящих добавок и разрабатывая специфические композиции на основе полимерных матриц.
E. Способы применения анти-TAT226 антител и иммуноконъюгатов
1. Диагностические способы и способы детекции
В одном из аспектов анти-TAT226 антитела и иммуноконъюгаты по изобретению пригодны для детекции присутствия TAT226 в биологическом образце. Термин «детекция», как используют в настоящем документе, относится к количественной или качественной детекции. В некоторых вариантах осуществления биологический образец содержит клетку или ткань, такую как ткани, перечисленные на фигуре 13. В некоторых вариантах осуществления такие ткани включают нормальные и/или раковые ткани, которые экспрессируют TAT226 на более высоких уровнях относительно других тканей, например тканей яичников, почек, мозга, эндометрия, надпочечников, костей, легких, кожи и мягких тканей.
В одном из аспектов изобретение относится к способу детекции присутствия TAT226 в биологическом образце. В некоторых вариантах осуществления способ включает контактирование биологического образца с анти-TAT226 антителом в условиях, обеспечивающих связывание анти-TAT226 антитела с TAT226 и детекцию того, произошло ли образование комплекса между анти-TAT226 антителом и TAT226.
В одном из аспектов изобретение относится к способу диагностики нарушения, связанного с повышенной экспрессией TAT226. В некоторых вариантах осуществления способ включает контактирование тестируемой клетки с анти-TAT226 антителом; определение уровня экспрессии (или количественно, или качественно) TAT226 тестируемой клеткой путем детекции связывания анти-TAT226 антитела с TAT226; и сравнение уровня экспрессии TAT226 тестируемой клеткой с уровнем экспрессии TAT226 контрольной клеткой (например, нормальной клеткой того же тканевого происхождения, что и тестируемая клетка, или клеткой, которая экспрессирует TAT226 на уровнях, сравнимых с уровнями нормальной клетки), где более высокий уровень экспрессии TAT226 тестируемой клеткой по сравнению с контрольной клеткой указывает на наличие нарушения, связанного с повышенной экспрессией TAT226. В некоторых вариантах осуществления тестируемую клетку получают из индивидуума, у которого предполагают наличие нарушения, связанного с повышенной экспрессией TAT226. В некоторых вариантах осуществления нарушение представляет собой нарушение пролиферации клеток, такое как рак или опухоль.
Иллюстративные нарушения клеточной пролиферации, которые можно диагностировать с использованием антитела по изобретению, включают раковые состояния, такие как опухоли, например карциномы (эпителиальные опухоли) и бластомы (опухоли, возникающие из эмбриональных тканей) и в некоторых вариантах осуществления рак яичника, рак тела матки (в том числе рак эндометрия) и рак почки, включая нефробластомы (например, опухоль Вильмса). К раку яичника, в частности, относится гетерогенная группа злокачественных опухолей, возникающих из яичника. Приблизительно 90% злокачественных опухолей яичника обладают эпителиальным происхождением; остальные представляют собой опухоли из зародышевых клеток и стромальные опухоли. Эпителиальные опухоли яичника классифицируют по следующим гистологическим подтипам: серозные аденокарциномы (составляющие приблизительно 50% эпителиальных опухолей яичника); эндометриоидные аденокарциномы (~20%); муцинозные аденокарциномы (~10%); светлоклеточные карциномы (~5-10%); опухоли Бреннера (переходно-клеточные) (относительно редкие). Прогноз для рака яичника, который стоит на шестом месте из наиболее распространенных видов рака у женщин, является обычно неблагоприятным, где уровни пятилетней выживаемости находятся в диапазоне 5-30%. Для обзоров по раку яичника см. Fox et al. (2002) «Pathology of epithelial ovarian cancer», в Ovarian Cancer ch. 9 (Jacobs et al., eds., Oxford University Press, New York); Morin et al. (2001) «Ovarian Cancer», в Encyclopedic Reference of Cancer, p.654-656 (Schwab, ed., Springer-Verlag, New York). По настоящему изобретению предусмотрены способы диагностики или лечения любого из вышеописанных подтипов эпителиальной опухоли яичника и, в частности, подтипа серозной аденокарциномы.
В некоторых вариантах осуществления способ диагностики или детекции, такой как вышеописанные способы, включает детекцию связывания анти-TAT226 антитела с TAT226, экспрессирующимся на поверхности клетки или в мембранном препарате, полученном из клетки, экспрессирующей TAT226 на своей поверхности. В некоторых вариантах осуществления способ включает контактирование клетки с анти-TAT226 антителом в условиях, обеспечивающих связывание анти-TAT226 антитела с TAT226 и детекцию того, произошло ли образование комплекса между анти-TAT226 антителом и TAT226 на клеточной поверхности. Иллюстративный анализ для детекции связывания анти-TAT226 антитела с TAT226, экспрессирующимся на поверхности клетки, представляет собой анализ «FACS», такой как анализ, описанный в примере D, ниже.
Можно использовать некоторые другие способы детекции связывания анти-TAT226 антител с TAT226. Такие способы включают в качестве неограничивающих примеров анализы связывания антигена, которые хорошо известны в данной области, такие как виды вестерн-блоттинга, виды радиоиммунного анализа, ELISA (твердофазный иммуноферментный анализ), «двухвалентные» иммуноанализы, анализы посредством иммунопреципитации, флюоресцентные иммуноанализы, иммуноанализы с протеином A и иммуногистохимия (IHC).
В некоторых вариантах осуществления анти-TAT226 антитела являются мечеными. Метки включают в качестве неограничивающих примеров метки или компоненты, которые детектируют непосредственно (такие, как флюоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также компоненты, такие как ферменты или лиганды, которые детектируют опосредованно, например, по ферментативной реакции или молекулярному взаимодействию. Иллюстративные метки включают в качестве неограничивающих примеров радиоизотопы 32P, 14C, 125I, 3H и 131I, флюорофоры, такие как редкоземельные хелаты или флюоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например люциферазу светляка и бактериальную люциферазу (патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахаридооксидазы, например глюкозооксидазу, галактозооксидазу, и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, связанные с ферментом, который использует пероксид водорода для окисления предшественника красителя, таким как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, метки на основе бактериофагов, стабильные свободные радикалы и т.п.
В некоторых вариантах осуществления анти-TAT226 антитела иммобилизованы на нерастворимой матрице. Иммобилизация приводит к отделению анти-TAT226 антител от любого TAT226, который остается свободным в растворе. Иммобилизацию общепринятым способом осуществляют или переводом в нерастворимую форму анти-TAT226 антитела перед процедурой анализа, как, например, адсорбцией на водонерастворимой матрице или поверхности (Bennich et al., US 3720760), или ковалентным связыванием (например, с использованием поперечной сшивки глутаральдегидом), или переводом в нерастворимую форму анти-TAT226 антитела после образования комплекса между анти-TAT226 антителом и TAT226, например иммунопреципитацией.
Любой из вышеописанных вариантов осуществления диагностики или детекции можно осуществлять с использованием иммуноконъюгата по изобретению вместо анти-TAT226 антитела или в дополнение к нему.
2. Терапевтические способы
Антитело или иммуноконъюгат по изобретению можно применять, например, в терапевтических способах in vitro, ex vivo и in vivo. В одном из аспектов изобретение относится к способам ингибирования клеточного роста или пролиферации или in vivo, или in vitro, где способ включает воздействие на клетку с помощью анти-TAT226 антитела или его иммуноконъюгата в условиях, обеспечивающих связывание иммуноконъюгата с TAT226. «Ингибирование клеточного роста или пролиферации» означает замедление клеточного роста или пролиферации, по меньшей мере, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 100% и включает индукцию гибели клеток. В некоторых вариантах осуществления клетка представляет собой опухолевую клетку. В некоторых вариантах осуществления клетка является клеткой опухоли яичников, клеткой опухоли тела матки, клеткой опухоли мозга или клеткой опухоли почек. В некоторых вариантах осуществления клетка представляет собой ксенотрансплантат, например, как приведено в качестве примера в данной работе.
В одном из аспектов антитело или иммуноконъюгат по изобретению применяют для лечения или профилактики нарушения пролиферации клеток. В некоторых вариантах осуществления нарушение пролиферации клеток связано с повышенной экспрессией и/или активностью TAT226. Например, в некоторых вариантах осуществления нарушение пролиферации клеток связано с повышенной экспрессией TAT226 на поверхности клетки. В некоторых вариантах осуществления нарушение пролиферации клеток представляет собой опухоль или рак. Примеры нарушений пролиферации клеток, которые подлежат лечению антителами или иммуноконъюгатами по изобретению, включают в качестве неограничивающих примеров раковые состояния, такие как опухоли, например карциномы (эпителиальные опухоли) и бластомы (опухоли, возникающие из эмбриональных тканей) и в некоторых вариантах осуществления рак эндометрия; опухоли мозга (например, астроцитомы, в том числе запущенные стадии глиом, также обозначаемые как мультиформная глиобластома); и рак яичника; рак тела матки, в том числе рак почки, в том числе нефробластомы (например, опухоль Вильмса).
В одном из аспектов изобретение относится к способам лечения нарушения пролиферации клеток, которые включают введение индивидууму эффективного количества анти-TAT226 антитела или его иммуноконъюгата. В некоторых вариантах осуществления способ лечения нарушения пролиферации клеток включает введение индивидууму эффективного количества фармацевтической композиции, содержащей анти-TAT226 антитело и необязательно, по меньшей мере, одно дополнительное терапевтическое средство, такое как представленные ниже средства. В некоторых вариантах осуществления способ лечения нарушения пролиферации клеток включает введение индивидууму эффективного количества фармацевтической композиции, содержащей: 1) иммуноконъюгат, содержащий анти-TAT226 антитело и цитотоксическое средство; и необязательно 2), по меньшей мере, одно дополнительное терапевтическое средство, такое как представленные ниже средства.
В одном из аспектов, по меньшей мере, некоторые антитела или иммуноконъюгаты по изобретению могут связываются с TAT226 из не являющихся человеком видов. Таким образом, антитела или иммуноконъюгаты по изобретению можно применять для связывания с TAT226, например, в культуре клеток, содержащей TAT226, в организме человека или в организме других млекопитающих, содержащих TAT226, с которым антитело или иммуноконъюгат по изобретению перекрестно реагирует (например, шимпанзе, бабуин, игрунка, Cynomolgus и макак-резус, свинья или мышь). В одном из вариантов осуществления анти-TAT226 антитело или иммуноконъюгат можно применять для ингибирования активности TAT226 путем контактирования антитела или иммуноконъюгата с TAT226 таким образом, что происходит ингибирование активности TAT226. В одном из вариантов осуществления TAT226 представляет собой TAT226 человека.
В одном из вариантов осуществления анти-TAT226 антитело или иммуноконъюгат можно применять в способе связывания TAT226 у субъекта, страдающего нарушением, связанным с повышенной экспрессией и/или активностью TAT226, где способ включает введение субъекту антитела или иммуноконъюгата таким образом, что происходит связывание TAT226 у субъекта. В одном из вариантов осуществления TAT226 представляет собой TAT226 человека и субъект является человеком. Альтернативно субъект может являться млекопитающим, экспрессирующим TAT226, с которым связывается анти-TAT226 антитело. Кроме того, субъект может являться млекопитающим, которому ввели TAT226 (например, при введении TAT226 или экспрессии трансгена, кодирующего TAT226).
Анти-TAT226 антитело или иммуноконъюгат можно вводить человеку с терапевтическими целями. Более того, анти-TAT226 антитело можно вводить не являющемуся человеком млекопитающему, экспрессирующему TAT226, с которым перекрестно реагирует антитело (например, примату, свинье, крысе или мыши), с ветеринарными целями или используя его в качестве модельного животного для заболевания человека. Относительно вышесказанного, такие модельные животные могут являться пригодными для оценки терапевтической эффективности антител или иммуноконъюгатов по изобретению (например, для тестирования дозировок и продолжительности курсов введения).
Антитело или иммуноконъюгаты по изобретению можно применять при лечении или по отдельности, или в сочетании с другими композициями. Например, антитело или иммуноконъюгат по изобретению можно совместно вводить, по меньшей мере, с одним дополнительным терапевтическим средством и/или адъювантом. В некоторых вариантах осуществления дополнительное терапевтическое средство является цитотоксическим средством, химиотерапевтическим средством или средством ингибирования роста. В одном из таких вариантов осуществления химиотерапевтическое средство представляет собой средство или сочетание средств, используемых при лечении рака яичника, такое как соединение платины (например, цисплатин или карбоплатин); таксан (например, паклитаксел или доцетаксел); топотекан; антрациклин (например, доксорубицин (АДРИАМИЦИН®) или липосомальный доксорубицин (ДОКСИЛ®)); гемцитабин; циклофосфамид; винорелбин (НАВЕЛБИН®); гексаметилмеламин; ифосфамид и этопозид. В других таких вариантах осуществления химиотерапевтическое средство представляет собой средство или сочетание средств, используемых при лечении рака матки или эндометрия, такое как цисплатин, карбоплатин, доксорубицин, паклитаксел, метотрексат, фторурацил и медроксипрогестерон. В других таких вариантах осуществления химиотерапевтическое средство представляет собой средство или сочетание средств, используемых при лечении опухолей мозга, такое как нитрозомочевина (например, кармустин или ломустин); цитотоксическое средство (например, иринотекан или темозоламид); антиангиогенное средство (например, талидомид, TNP-470, тромбоцитарный фактор 4, интерферон и эндостатин); средство дифференцировки (например, ретиноиды, фенилбутират, фенилацетат и антинеопластоны); антиинвазивное средство (например, ингибиторы матричных металлопротеаз, такие как маримастат); модулятор передачи сигнала (например, тамоксифен, бриостатин и O-6-бензогуанин); ингибитор топоизомеразы (например, иринотекан или топотекан); и ингибитор факторов роста (например, ингибитор тирозинкиназы). В других таких вариантах осуществления химиотерапевтическое средство представляет собой средство или сочетание средств, используемых при лечении рака почки (например, опухоли Вильмса), такое как винкристин, актиномицин D, адриамицин, доксорубицин, циклофосфамид, ифосфамид, этопозид и карбоплатин. В некоторых вариантах осуществления антитело по изобретению можно сочетать с противовоспалительным и/или антисептическим средством.
Такие указанные выше виды комбинированной терапии включают сочетанное введение (где два или более терапевтических средств входят в состав одинаковых или разных композиций) и раздельное введение, при котором введение антитела или иммуноконъюгата по изобретению можно осуществлять до введения дополнительного терапевтического средства и/или адъюванта, одновременно с ним и/или после него. Антитела или иммуноконъюгаты по изобретению можно также применять в сочетании с радиотерапией.
Антитело по изобретению (и любое дополнительное терапевтическое средство или адъювант) вводят любыми подходящими способами, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и интраназальное и, если требуется для местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, антитело или иммуноконъюгат соответствующим образом вводят пульсовой инфузией, в частности, со снижением доз антитела или иммуноконъюгата. Введение доз можно осуществлять любым подходящим способом, например, инъекциями такими как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение краткосрочным или хроническим.
Если мишень для связывания находится в мозге, то в определенных вариантах осуществления изобретения предусмотрено, что антитело или иммуноконъюгат проходит через гематоэнцефалический барьер. Существует несколько известных в данной области способов для транспорта молекул через гематоэнцефалический барьер, включая в качестве неограничивающих примеров физические способы, способы с применением липидов, способы с применением стволовых клеток и способы с применением рецепторов и каналов.
Физические способы транспорта антитела или иммуноконъюгата через гематоэнцефалический барьер включают в качестве неограничивающих примеров полный обход гематоэнцефалического барьера или создание отверстий в гематоэнцефалическом барьере. Способы обхода включают в качестве неограничивающих примеров прямую инъекцию в мозг (см., например, Papanastassiou et al., Gene Therapy 9:398-406 (2002)), интерстициальную инфузию/конвекционную доставку (см., например, Bobo et al., Proc. Natl. Acad. Sci. USA 91:2076-2080 (1994)) и имплантацию средства доставки в мозг (см., например, Gill et al., Nature Med. 9:589-595 (2003); и Gliadel Wafers™, Guildford Pharmaceutical). Способы создания отверстий в барьере включают в качестве неограничивающих примеров ультразвук (см., например, публикацию патента США № 2002/0038086), осмотическое давление (например, при введении гипертонического маннита (Neuwelt E. A., Implication of the Blood-Brain Barrier and its Manipulation, Vols 1 & 2, Plenum Press, N. Y. (1989)), пермеабилизацию, например, брадикинином или пермеабилизатором A-7 (см., например, патенты США № 5112596, 5268164, 5506206 и 5686416) и трансфекцию нейронов, которые окружают гематоэнцефалический барьер, векторами, содержащими гены, кодирующими антитело (см., например, публикацию патента США № 2003/0083299).
Способы с применением липидов для транспорта антитела или иммуноконъюгата через гематоэнцефалический барьер включают в качестве неограничивающих примеров инкапсулирование антитела или иммуноконъюгата в липосомы, которые связаны с антителосвязывающими фрагментами, которые связываются с рецепторами на сосудистом эндотелии гематоэнцефалического барьера (см., например, публикацию патентной заявки США № 20020025313), и заключение антитела или иммуноконъюгата в частицы липопротеинов низкой плотности (см., например, публикацию патентной заявки США № 20040204354) или аполипопротеина E (см., например, публикацию патентной заявки США № 20040131692).
Способы с применением стволовых клеток для транспорта антитела или иммуноконъюгата через гематоэнцефалический барьер включают генетическое конструирование нервных клеток-предшественников (NPC) для экспрессии представляющего интерес антитела или иммуноконъюгата и затем имплантирование стволовых клеток в мозг подвергаемого лечению субъекта. См. Behrstock et al. (2005) Gene Ther. 15 Dec. 2005, опережающая интерактивная публикация (в ней сообщается, что NPC, генетически сконструированные для экспрессии нейротрофического фактора GDNF, снижали выраженность симптомов при болезни Паркинсона, если их имплантировали в мозг в моделях на грызунах и приматах).
Способы с применением рецепторов и каналов для транспорта антитела или иммуноконъюгата через гематоэнцефалический барьер включают в качестве неограничивающих примеров использование глюкокортикоидных блокаторов для усиления проницаемости гематоэнцефалического барьера (см., например, публикации патентных заявок США № 2002/0065259, 2003/0162695 и 2005/0124533); активацию калиевых каналов (см., например, публикацию патентной заявки США № 2005/0089473); ингибирование ABC-переносчиков лекарственных средств (см., например, публикацию патентной заявки США № 2003/0073713); покрытие антител или иммуноконъюгатов трансферрином и модулирование активности одного или нескольких рецепторов трансферрина (см., например, публикацию патентной заявки США № 2003/0129186); и катионизацию антител или иммуноконъюгатов (см., например, патент США № 5004697).
Композицию антител или иммуноконъюгатов по изобретению составляют, дозируют и вводят в соответствии с надлежащей медицинской практикой. Факторы для рассмотрения в данном контексте включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние конкретного пациента, причину нарушения, участок доставки средства, способ введения, схему введения и другие факторы, известные практикующим врачам. Антитело или иммуноконъюгат необязательно составляют в композицию с одним или несколькими средствами, в настоящее время используемыми для профилактики или лечения рассматриваемого нарушения. Эффективное количество таких других средств зависит от количества антитела или иммуноконъюгата, присутствующего в композиции, типа нарушения или лечения и других факторов, рассмотренных выше. Их, как правило, используют в тех же дозировках и способах введения, как описано в данной работе, или приблизительно от 1 до 99% от описанных в данной работе дозировок, или в любой дозировке и любым способом, который эмпирически/клинически определен в качестве подходящего.
Для профилактики или лечения заболевания соответствующая дозировка антитела и иммуноконъюгата по изобретению (при использовании в виде монотерапии или в сочетании с одним или несколькими другими дополнительными терапевтическими средствами, такими как химиотерапевтические средства) зависит от типа подлежащего лечению заболевания, типа антитела и иммуноконъюгата, тяжести и течения заболевания, от того, вводят ли антитело и иммуноконъюгат с профилактическими или терапевтическими целями, предыдущего лечения, истории болезни пациента и ответа на антитело и иммуноконъюгат и усмотрения лечащего врача. Антитело и иммуноконъюгат подходящим образом вводят пациенту за один раз или в ходе курсов лечения. В зависимости от типа и тяжести заболевания приблизительно от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг-10 мг/кг) антитела и иммуноконъюгата может составлять начальную возможную дозировку для введения пациенту, или, например, в виде одного или нескольких отдельных введений, или в виде непрерывной инфузии. Одна типичная суточная дозировка может варьировать приблизительно от 1 мкг/кг до 100 мг/кг или более в зависимости от вышеупомянутых факторов. При повторных введениях в течение нескольких суток или более в зависимости от состояния лечение продолжают до тех пор, пока не наступит требуемое ослабление симптомов заболевания. Одна из иллюстративных дозировок антитела и иммуноконъюгата находится в диапазоне приблизительно от 0,05 мг/кг до приблизительно 10 мг/кг. Таким образом, пациенту можно вводить одну или несколько доз из расчета приблизительно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любые их сочетания). Такие дозы можно вводить периодически, например каждую неделю или каждые три недели (например, таким образом, что пациент получает от приблизительно двух до приблизительно двадцати, например приблизительно шесть доз антитела и иммуноконъюгата). Можно вводить начальную более высокую ударную дозу с последующими одной или несколькими более низкими дозами. Иллюстративный режим дозирования включает введение начальной ударной дозы из расчета приблизительно 4 мг/кг с последующей поддерживающей дозой в течение недели на уровне приблизительно 2 мг/кг антитела. Однако можно использовать другие режимы дозирования. Ход такого лечения легко отслеживать посредством общепринятых способов и анализов.
3. Анализы
Анти-TAT226 антитела и иммуноконъюгаты по изобретению можно характеризовать по их физическим/химическим свойствам и/или видам биологической активности с помощью различных анализов, известных в данной области.
a) Анализы активности
Один из аспектов относится к анализам для идентификации анти-TAT226 антител или их иммуноконъюгатов, обладающих биологической активностью. Биологическая активность может включать, например, способность ингибировать клеточный рост или пролиферацию (например, активность в «уничтожении клеток») или способность индуцировать гибель клеток, включая программированную гибель клеток (апоптоз). Также представлены антитела или иммуноконъюгаты, обладающие такой биологической активностью in vivo и/или in vitro.
В некоторых вариантах осуществления анти-TAT226 антитело или его иммуноконъюгат тестируют по его способности ингибировать клеточный рост или пролиферацию in vitro. Анализы ингибирования клеточного роста или пролиферации хорошо известны в данной области. В определенных анализах пролиферации клеток, иллюстрированных описанными здесь анализами «уничтожения клеток», измеряют жизнеспособность клеток. Один из таких анализов представляет собой люминесцентный анализ жизнеспособности клеток CellTiter-Glo™, который коммерчески доступен от Promega (Madison, WI). В этом анализе определяют количество живых клеток в культуре на основе определения количества присутствующей АТФ, которое является индикатором метаболически активных клеток. См. Crouch et al. (1993) J. Immunol. Meth. 160:81-88, патент США № 6602677. Анализ можно осуществлять в 96- или 384-луночном формате, делая его пригодным для автоматизированного высокопроизводительного скрининга (HTS). См. Cree et al. (1995) Anticancer Drugs 6:398-404. Процедура анализа включает добавление одного реагента (реагент CellTiter-Glo®) непосредственно к культивируемым клеткам. Это приводит к лизису клеток и образованию люминесцентного сигнала, продуцируемого люциферазной реакцией. Люминесцентный сигнал пропорционален количеству присутствующей АТФ, которое прямо пропорционально количеству живых клеток, присутствующих в культуре. Данные можно регистрировать люменометром или устройством для получения изображений с помощью CCD-камеры. Выход люминесценции выражают в относительных единицах света (RLU).
Другой анализ пролиферации клеток представляет собой анализ «MTT», колориметрический анализ, с помощью которого измеряют окисление бромида 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия в формазан при помощи митохондриальной редуктазы. Подобно анализу CellTiter-Glo™, этот анализ указывает на количество метаболически активных клеток, присутствующих в клеточной культуре. См., например, Mosmann (1983) J. Immunol. Meth. 65:55-63 и Zhang et al. (2005) Cancer Res. 65:3877-3882.
В одном из аспектов анти-TAT226 антитело тестируют по его способности индуцировать гибель клеток in vitro. Анализы индукции гибели клеток хорошо известны в данной области. В некоторых вариантах осуществления в таких анализах измеряют, например, утрату целостности мембран, на что указывает поглощение йодида пропидия (PI), трипанового синего (см. Moore et al. (1995) Cytotechnology, 17:1-11) или 7AAD. В иллюстративном анализе поглощения PI клетки культивируют в среде Игла в модификации Дульбекко (D-MEM):среде Хэма F-12 (50:50) с добавлением 10% инактивированной нагреванием FBS (Hyclone) и 2 мМ L-глутамина. Таким образом, анализ осуществляют в отсутствие комплемента и иммунных эффекторных клеток. Клетки высевают исходя из плотности 3×106 на чашку на 100×20 мм чашки и позволяют им прикрепиться в течение ночи. Среду удаляют и замещают чистой свежей средой или средой, содержащей различные концентрации антитела или иммуноконъюгата. Клетки инкубируют в течение 3 суток. После обработки монослой промывают PBS и отделяют обработкой трипсином. Клетки затем центрифугируют при 1200 об/мин в течение 5 минут при 4°C, осадок ресуспендируют в 3 мл холодного буфера для связывания Ca2+ (10 мМ Hepes, pH 7,4, 140 мМ NaCl, 2,5 мМ CaCl2) и разделяют на аликвоты по 12×75 мм пробиркам с надетым 35-мм фильтром (1 мл на пробирку, 3 пробирки на группу обработки) для удаления комков клеток. В пробирки затем добавляют PI (10 мкг/мл). Образцы анализируют с использованием проточного цитометра FACSCAN™ и программного обеспечения FACSCONVERT™ CellQuest (Becton Dickinson). Таким образом, идентифицируют антитела или иммуноконъюгаты, которые индуцируют статистически значимые уровни гибели клеток, как определяют по поглощению PI.
В одном из аспектов анти-TAT226 антитело или иммуноконъюгат тестируют по его способности индуцировать апоптоз (программируемую гибель клеток) in vitro. Иллюстративный анализ антител или иммуноконъюгатов, которые индуцируют апоптоз, представляет собой анализ связывания аннексина. В иллюстративном анализе связывания аннексина клетки культивируют и высевают на чашки, как рассмотрено в предыдущем абзаце. Среду удаляют и замещают чистой свежей средой или средой, содержащей от 0,001 до 10 мкг/мл антитела или иммуноконъюгата. После инкубации в течение трех суток монослой промывают PBS и отделяют обработкой трипсином. Клетки затем центрифугируют, ресуспендируют в буфере для связывания Ca2+ и разделяют на аликвоты по пробиркам, как рассмотрено в предыдущем параграфе. В пробирки затем добавляют меченый аннексин (например, аннексин-V-FITC) (1 мкг/мл). Образцы анализируют с использованием проточного цитометра FACSCAN™ и программного обеспечения FACSCONVERT™ CellQuest (BD Biosciences). Таким образом, идентифицируют антитела или иммуноконъюгаты, которые индуцируют статистически значимые уровни связывания аннексина относительно контроля. Другой иллюстративный анализ антител или иммуноконъюгатов, которые индуцируют апоптоз, представляет собой колориметрический анализ ELISA гистоновой ДНК для детекции межнуклеосомной деградации геномной ДНК. Такой анализ можно осуществлять с использованием, например, набора Cell Death Detection ELISA (Roche, Palo Alto, CA).
Клетки для использования в любом из вышеописанных анализов in vitro включают клетки или линии клеток, которые естественным образом экспрессируют TAT226 или которые сконструированы для экспрессии TAT226. Такие клетки включают опухолевые клетки, которые сверхэкспрессируют TAT226 относительно нормальных клеток, которые происходят из той же ткани. Такие клетки также включают линии клеток (в том числе опухолевые линии клеток), которые экспрессируют TAT226, и линии клеток, которые в норме не экспрессируют TAT226, но которые трансфицированы нуклеиновой кислотой, кодирующей TAT226. Иллюстративные линии клеток, представленные здесь для использования в любом из вышеописанных анализов in vitro, включают линию клеток рака яичника человека OVCAR3, которая экспрессирует TAT226, и линию клеток рака толстой кишки человека HCT116, трансфицированную нуклеиновой кислотой, кодирующей TAT226.
В одном из аспектов анти-TAT226 антитело или его иммуноконъюгат тестируют по его способности ингибировать клеточный рост или пролиферацию in vivo. В некоторых вариантах осуществления анти-TAT226 антитело или его иммуноконъюгат тестируют по его способности ингибировать опухолевый рост in vivo. Для такого тестирования можно использовать модельные системы in vivo, такие как модели с использованием ксенотрансплантата. В иллюстративной системе с использованием ксенотрансплантата опухолевые клетки человека вводят подходящему не являющемуся человеком животному с ослабленным иммунитетом, например бестимусной «голой» мыши. Антитело или иммуноконъюгат по изобретению вводят животному. Измеряют способность антитела или иммуноконъюгата ингибировать или замедлять опухолевый рост. В определенных вариантах осуществления вышеописанной системы с использованием ксенотрансплантата опухолевые клетки человека являются опухолевыми клетками пациента. Такие модели с использованием ксенотрансплантата коммерчески доступны от Oncotest GmbH (Frieberg, Germany). В некоторых вариантах осуществления опухолевые клетки человека являются клетками опухолевой линии клеток человека, такой как клетки OVCAR3, как представлено здесь. В некоторых вариантах осуществления опухолевые клетки человека вводят подходящему, не являющемуся человеком животному с ослабленным иммунитетом подкожной инъекцией или трансплантацией в подходящий участок, такой как скопление жировой ткани молочной железы.
b) Анализы связывания и другие анализы
В одном из аспектов анти-TAT226 антитело тестируют по его антигенсвязывающей активности. Например, в некоторых вариантах осуществления анти-TAT226 антитело тестируют по его способности связываться с TAT226, экспрессирующимся на поверхности клетки. Для такого тестирования можно использовать анализ FACS, такой, как описано в примере D.
В одном из аспектов можно использовать конкурентные анализы для идентификации моноклонального антитела, которое конкурирует с YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6 за связывание с TAT226. В некоторых вариантах осуществления такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), с которым связывается YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6. Иллюстративные конкурентные анализы включают в качестве неограничивающих примеров общепринятые анализы, такие как анализы, представленные в Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). Подробные иллюстративные способы картирования эпитопа, с которым связывается антитело, представлены в Morris (1996) «Epitope Mapping Protocols», в Methods in Molecular Biology, vol. 66 (Humana Press, Totowa, NJ). Говорят, что два антитела связываются с одинаковым эпитопом, если каждое блокирует связывание другого на 50% или более.
В иллюстративном конкурентном анализе иммобилизованный TAT226 инкубируют в растворе, содержащем первое меченое антитело, которое связывается с TAT226 (например, YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6), и второе немеченое антитело, которое тестируют по его способности конкурировать с первым антителом за связывание с TAT226. Второе антитело может присутствовать в гибридомном супернатанте. В качестве контроля иммобилизованный TAT226 инкубируют в растворе, содержащем первое меченое антитело, но без второго немеченого антитела. После инкубации в условиях, обеспечивающих связывание первого антитела с TAT226, удаляют избыток несвязанного антитела и измеряют количество метки, связанной с иммобилизованным TAT226. Если количество метки, связанной с иммобилизованным TAT226, существенно снижено в тестируемом образце относительно контрольного образца, то это указывает на то, что второе антитело конкурирует с первым антителом за связывание с TAT226. В некоторых вариантах осуществления иммобилизованный TAT226 присутствует на поверхности клетки или в мембранном препарате, полученном из клетки, экспрессирующей TAT226 на своей поверхности.
В одном из аспектов очищенные анти-TAT226 антитела можно дополнительно характеризовать в ряде анализов, включающих в качестве неограничивающих примеров N-концевое секвенирование, анализ аминокислот, эксклюзионную жидкостную хроматографию высокого давления (HPLC) в неденатурирующих условиях, масс-спектрометрию, ионообменную хроматографию и расщепление папаином.
В одном из вариантов осуществления изобретение относится к измененному антителу, которое обладает некоторыми, но не всеми эффекторными функциями, которые делают его требуемым кандидатом для многих применений, в которых важно время полужизни антитела in vivo, однако некоторые эффекторные функции (такие, как связывание комплемента и ADCC) являются необязательными или вредными. В конкретных вариантах осуществления измеряют активности Fc антитела для уверенности в том, что сохраняются только требуемые свойства. Анализы цитотоксичности in vitro и/или in vivo можно проводить для подтверждения снижения/ослабления активностей в отношении CDC и/или ADCC. Например, можно проводить анализы связывания Fc-рецептора (FcR) для уверенности в том, что у антитела отсутствует связывание с FcγR (таким образом, вероятное отсутствие активности в отношении ADCC), но сохраняется способность связываться с FcRn. Основные клетки для осуществления ADCC, NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на кроветворных клетках суммирована в таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991). Пример анализа in vitro для оценки активности представляющей интерес молекулы в отношении ADCC описан в патенте США № 5500362 или 5821337. Пригодные для таких анализов эффекторные клетки включают мононуклеарные клетки периферической крови (PBMC) и природные клетки-киллеры (NK). Альтернативно или дополнительно активность представляющей интерес молекулы в отношении ADCC можно оценивать in vivo, например, в модели на животных, такой как описана в Clynes et al. PNAS (USA) 95:652-656 (1998). Можно также осуществлять анализы связывания C1q для подтверждения того, что антитело неспособно связываться с C1q и, таким образом, утратило активность в отношении CDC. Для оценки активации комплемента можно осуществлять анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996). Связывание FcRn и определения клиренса/времени полужизни in vivo можно также осуществлять с использованием способов, известных в данной области.
F. Изделия
В другом варианте осуществления изобретение относится к изделию, содержащему вещества, пригодные для лечения, профилактики и/или диагностики описанных выше нарушений. Изделие содержит контейнер и этикетку или вкладыш в упаковку на контейнере или в связи с ним. Пригодные контейнеры включают, например, бутыли, флаконы, шприцы и т.д. Контейнеры могут быть образованы из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая сама по себе или при сочетании с другой композицией является эффективной для лечения, профилактики и/или диагностики состояния и может обладать отверстием для стерильного доступа (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, обладающий пробкой, проницаемой для иглы для подкожных инъекций). По меньшей мере, одно активное средство в композиции представляет собой антитело или иммуноконъюгат по изобретению. На этикетке или вкладыше в упаковку указано, что композицию используют для лечения выбранного состояния. Более того, изделие может содержать (a) первый контейнер с содержащейся в нем композицией, где композиция содержит антитело и иммуноконъюгат по изобретению; и (b) второй контейнер с содержащейся в нем композицией, где композиция содержит дополнительное цитотоксическое или другое терапевтическое средство. Изделие в этом варианте осуществления изобретения может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного состояния. Альтернативно или дополнительно изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Кроме того, оно может дополнительно содержать другие необходимые с коммерческой и потребительской точки зрения материалы, включая другие буферы, разбавители, фильтры, иглы и шприцы.
IV. ПРИМЕРЫ
Далее следуют примеры способов и композиций по изобретению. Следует понимать, что можно применять различные другие варианты осуществления на основе представленного выше общего описания.
A. Анализ экспрессии гена TAT226
Экспрессию гена TAT226 человека анализировали с использованием частной базы данных, содержащей информацию по экспрессии генов (GeneExpress®, Gene Logic Inc., Gaithersburg, MD). Графический анализ базы данных GeneExpress® проводили с использованием просмотрщика профилей микрочипов. Фигура 13 представляет собой графическое представление экспрессии гена TAT226 в различных тканях, которые перечислены слева. Шкала в верхней части графика указывает на уровни экспрессии гена на основе интенсивности сигнала гибридизации. Точки расположены как сверху, так и снизу линии, соответствующей каждой из перечисленных тканей. Точки, расположенные выше линии, представляют собой экспрессию гена в нормальной ткани, и точки, расположенные ниже линии, представляют собой экспрессию гена в опухолевой или больной ткани. На фигуре 13 представлен тренд в сторону повышения экспрессии гена TAT226 в опухолевых или больных тканях относительно их нормальных вариантов. В частности, для TAT226 показана существенная сверхэкспрессия в опухолевом и больном яичнике относительно нормального яичника и в опухоли Вильмса относительно нормальной почки. Другие ткани, для которых показана сверхэкспрессия в опухолевой или больной ткани относительно нормальной ткани, включают ткань эндометрия, надпочечников, кости, легкого, кожи и мягкую ткань. Кроме того, TAT226 в значительной степени экспрессируется в нормальной ткани мозга (такой, как обонятельный мозг, гиппокамп и базальные ганглии) и в опухолевой или больной ткани мозга, такой как глиомы.
Базу данных GeneExpress® также использовали для анализа экспрессии гена TAT226 человека в нормальном яичнике; нормальной фаллопиевой трубе; раке яичника подтипов светлоклеточной, муцинозной и серозной цистоаденокарциномы; метастатическом раке яичника и других типах рака яичника. Результаты представлены графически на фигуре 14, где конкретные типы тканей указаны под графиком. Шкала по оси y графика указывает на уровни экспрессии гена на основе интенсивности сигнала гибридизации. Для серозной цистоаденокарциномы и метастатического рака яичника показана значительная сверхэкспрессия TAT226 относительно нормального яичника. Для светлоклеточного и муцинозного подтипов показана экспрессия, сравнимая с нормальным яичником. Для нормальной фаллопиевой трубы также показана существенная экспрессия TAT226. Примечательно то, что серозный подтип рака яичника гистологически в значительной степени похож на эпителий фаллопиевой трубы, и как яичник, так и фаллопиевы трубы происходят из одинаковой эмбриональной ткани. См. Fox et al. (2002) «Pathology of epithelial ovarian cancer», в Ovarian Cancer ch. 9 (Jacobs et al., eds., Oxford University Press, New York).
B. Получение анти-TAT226 антител
Антитела к TAT226 получали скринингом библиотеки фагового дисплея с помощью рекомбинантного слитого белка «TAT226-His», содержащего аминокислоты 1-115 SEQ ID NO:75 и C-концевую полигистидиновую метку. Библиотека фагового дисплея представляла собой синтетическую библиотеку (Fab')2, полученную с использованием системы Fab'-zip-phage. См. Lee et al. (2004) J. Immunol. Methods 284:119-132. Библиотека содержала библиотеку тяжелой цепи HVR в каркасной области вариабельной области тяжелой цепи huMAb4D5-8 (см. фигуры 5A и 5B, второй акцептор «B», SEQ ID NO:50, 51, 57, 35) и фиксированную вариабельную область легкой цепи huMAb4D5-8, как представлено на SEQ ID NO:26. Клоны, отобранные с использованием фагового дисплея, подвергали скринингу против TAT226-His с использованием фагового ELISA (см., например, Sidhu et al. (2004) J. Mol. Biol. 338:299-310). Клоны YWO.32 и YWO.49 отобрали для дальнейшего анализа.
Для улучшения аффинности YWO.49 библиотеки фагового дисплея получали на основе YWO.49, где HVR-H3 и HVR-L3 служили мишенями для мягкой рандомизации, в которой отобранные аминокислотные остатки в обозначенных HVR сохраняли константными, в то время как другие подвергали мутагенезу. Отобранные клоны подвергали скринингу с помощью фагового ELISA. Аффинно зрелые антитела, обозначенные YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6, отбирали для дальнейшего анализа. Нуклеотидные последовательности и последовательности кодируемого полипептида областей VH и VL YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6 определяли, как представлено на фигурах 9 и 10. Последовательности тяжелой и легкой цепей HVR YWO.32, YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6 представлены на фигурах 2-4. Консенсусные последовательности HVR-H3 и HVR-L3, полученные из YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6, также представлены на фигуре 4. YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6 «преобразовывали» в виде полноразмерных IgG привитием фрагментов Fab' на соответствующие константные области с использованием рекомбинантных способов. Описанные далее эксперименты осуществляли с использованием преобразованных антител.
C. Характеристика аффинности связывания с рекомбинантным антигеном
Аффинность связывания YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 и YWO.49.H6 с рекомбинантным антигеном определяли измерением при помощи поверхностного плазмонного резонанса с использованием системы BIACORE® 3000 (Biacore, Inc., Piscataway, NJ). В кратком изложении, биосенсорные чипы с карбоксиметилированным декстраном (CM5, BIAcore Inc.) активировали гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) согласно инструкциям поставщика. Анти-TAT226 антитело разводили 10 мМ ацетатом натрия, pH 4,8, до 5 мкг/мл перед инъекцией со скоростью течения 5 мкл/минута для достижения приблизительно 500 единиц резонансного сигнала (RU) связанного антитела. Далее 1 М этаноламин инъецировали для блокирования непрореагировавших групп. Для измерений кинетики двукратные серийные разведения TAT226-His (от 0,7 нМ до 500 нМ) инъецировали в PBS с 0,05% твином-20 (PBST) при 25°C при скорости потока приблизительно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляли с использованием простой модели связывания один к одному Лэнгмюра (программное обеспечение BIAevaluation, версия 3.2). Равновесную константу диссоциации (Kd) вычисляли как отношение koff/kon. Результаты этого эксперимента представлены в таблице 2 ниже.
D. Характеристика связывания антитела с TAT226 на клеточной поверхности
Исследовали способность анти-TAT226 антител связываться с TAT226, экспрессирующимся на поверхности OVCAR3, линии клеток рака яичника человека. Осуществляли флюоресцентно-активированный клеточный сортинг (FACS) на клетках OVCAR3 в отсутствие и в присутствии YWO.49, YWO.49.B7, YWO.49.C9, YWO.49.H2 или YWO.49.H6. В кратком изложении, открепленные клетки инкубировали с 5 мкг/мл первичного антитела в течение 1 часа на льду, промывали и инкубировали с вторичным антителом (IgG против человека, конъюгированный с фикоэритрином) в течение 30 минут на льду. FACS осуществляли с использованием проточного цитометра FACScan™ (BD Biosciences, San Jose, CA).
Результаты анализа FACS для YWO.49, YWO.49.H2 и YWO.H6 представлены на фигуре 15. Пики с левой стороны каждого графика представляют собой «фоновое» связывание, т.е. связывание только с вторичным антителом. Пики с правой стороны каждого графика представляют собой связывание с указанным анти-TAT226 антителом. YWO.49.B7 не связывается в значительной степени с клетками OVCAR3 по данным FACS, хотя оно связывается с рекомбинантным антигеном с Kd в диапазоне Kd, найденных для YWO.49 и других аффинно-зрелых антител (см. результаты анализа BIACORE®, таблица 2 выше). Связывание YWO.49.C9 оказалось сравнимо с найденным для YWO.49.H2 и YWO.H6.
E. Характеристика аффинности связывания с антигеном на клеточной поверхности
Аффинность связывания YWO.49.H2 и YWO.49.H6 с TAT226, экспрессирующимся на поверхности клеток OVCAR3, исследовали с использованием конкурентного анализа. В кратком изложении, меченому (йодированному) YWO.49.H2 или YWO.49.H6 позволяли связываться с клетками OVCAR3 в присутствии немеченого антитела. Аффинность связывания антител определяли в соответствии со способом анализа по Скэтчарду, первоначально описанному в Munson et al., Anal. Biochem. 107:220 (1980). Результаты этого эксперимента представлены в таблице 3 ниже.
Kd YWO.49.H2 и YWO.49.H6 выше для TAT226, экспрессирующегося на поверхности клеток OVCAR3, по сравнению с рекомбинантным TAT226-His (ср. Kd для YWO.49.H2 и YWO.49.H6 в таблицах 2 и 3), указывая на то, что YWO.49.H2 и YWO.49.H6 связываются с незначительно более высокой аффинностью с рекомбинантным TAT226-His, чем с TAT226, экспрессирующимся на поверхности клеток OVCAR3.
F. Экспрессия мРНК и белка TAT226
Экспрессию мРНК TAT226 в клетках OVCAR3 и в панели образцов рака яичника анализировали с использованием анализа 5'-нуклеазой (TaqMan®) и количественной PCR в реальном времени. Образцы рака яичника, обозначенные «HF ####» на фигуре 16, представляли собой замороженные срезы тканей. РНК выделяли из срезов тканей, амплифицировали с использованием набора Ambion's Message Amp II (Ambion, Austin, TX) и подвергали обратной транскрипции в кДНК. РНК выделяли из клеток OVCAR3 и подвергали обратной транскрипции в кДНК. кДНК TAT226 амплифицировали с помощью PCR в реальном времени в присутствии недостраиваемого репортерного зонда, специфичного в отношении продукта амплификации. Пороговый цикл или «Ct» (цикл, в котором сигнал, полученный при расщеплении репортерного зонда, превышал фон) определяли и использовали для вычисления начальных уровней мРНК TAT226. Уровень мРНК TAT226 в панели образцов рака яичника выражали относительно уровня мРНК TAT226 в клетках OVCAR3, как представлено на столбчатой диаграмме на фигуре 16.
Экспрессию белка TAT226 в клетках OVCAR3 и вышеописанной панели образцов рака яичника анализировали с использованием иммуногистохимии (IHC), как указано далее. Срезы тканей (замороженных или заключенных в парафин) образцов рака яичника фиксировали в течение 5 минут в ацетоне/этаноле. Срезы промывали в PBS, блокировали авидином и биотином (Vector Laboratories, Inc., Burlingame, CA) в течение 10 минут для каждого и снова промывали в PBS. Срезы затем блокировали 10% сывороткой в течение 20 минут и подвергали блоттингу для удаления избытка сыворотки. Первичное антитело (YWO.49.H2 или YWO.49.H6) затем добавляли к срезам в концентрации 10 мкг/мл на 1 час. Срезы затем промывали в PBS. Биотинилированное вторичное антитело против человека добавляли к образцам на 30 минут и затем срезы промывали PBS. Срезы затем обрабатывали реагентами набора Vector ABC (Vector Laboratories, Inc., Burlingame, CA) в течение 30 минут и затем промывали в PBS. Образцы затем обрабатывали диаминобензидином (Pierce) в течение 5 минут и промывали в PBS. Образцы затем контрастировали гематоксилином Майера, покрывали покровным стеклом и визуализировали. IHC осуществляли на клетках OVCAR3 с использованием того же протокола за исключением того, что клетки сначала осаждали, замораживали и затем делали из них срезы. Срезы затем обрабатывали согласно вышеописанному протоколу.
Результаты качественно представлены на фигуре 16, где уровни экспрессии ранжированы как «-», «±» или «+». Как правило, в целом существовала корреляция между уровнем экспрессии мРНК TAT226 и экспрессии белка TAT226 на поверхности клеток OVCAR3. Эксперименты с IHC также подтвердили, что антитела распознают TAT226 на клеточной поверхности. Гистологический анализ каждой клетки в панели клеток рака яичника также представлен на фигуре 16 с сокращением «adenoca», обозначающим «аденокарциному».
G. Получение ADC анти-TAT226
ADC анти-TAT226 получали конъюгированием YWO.49.H2 и YWO.49.H6 со следующими компонентами лекарственного средства-линкера: MC-vc-PAB-MMAE, MC-vc-PAB-MMAF и MC-MMAF, которые представлены выше в разделе III.C.1.b.2. Перед конъюгацией антитела частично восстанавливали с помощью TCEP с использованием общепринятых способов согласно способу, описанному в WO 2004/010957 A2. Частично восстановленные антитела конъюгировали с отмеченными выше компонентами лекарственного средства-линкера с использованием общепринятых способов согласно способу, описанному в Doronina et al. (2003) Nat. Biotechnol. 21:778-784 и US 2005/0238649 A1. В кратком изложении, частично восстановленные антитела сочетали с компонентами лекарственного средства-линкера для обеспечения конъюгации компонентов с цистеиновыми остатками. Останавливали реакции конъюгации и очищали ADC. Нагрузку лекарственного средства (среднее количество компонентов лекарственного средства на антитело) для каждого ADC определяли путем HPLC, как указано далее:
H. Анализы уничтожения клеток
Конъюгаты антитело-лекарственное средство (ADC) тестировали по способности ингибировать пролиферацию клеток, экспрессирующих TAT226, в следующих анализах уничтожения клеток in vitro и in vivo:
1. Анализ уничтожения клеток OVCAR3 in vitro
ADC YWO.49.H2 и YWO.49.H6 тестировали по способности ингибировать пролиферацию клеток OVCAR3. Клетки OVCAR3 высевали в 96-луночные планшеты в RPMI с 20% FBS. Клетки OVCAR3 в плотности 3000 клеток на лунку инкубировали с различными концентрациями ADC, как представлено на фигуре 17. Анти-MUC16/CA125 антитело, конъюгированное с MC-vc-PAB-MMAE, использовали в качестве положительного контроля. MUC16/CA125 представляет собой известный антиген рака яичника. См., например, Yin et al. (2001) J. Biol. Chem. 276:27371-27375. Анти-IL-8 антитело, конъюгированное с MC-vc-PAB-MMAE, использовали в качестве отрицательного контроля. После 5 суток инкубации жизнеспособность клеток измеряли с использованием люминесцентного анализа жизнеспособности клеток CellTiter-Glo™ (Promega, Madison, WI) согласно инструкциям производителя. Шкала по оси y фигуры 17 указывает на относительные единицы света или «RLU» при люциферазной люминесценции, которая является мерой жизнеспособности клеток.
На фигуре 17 показано, что аналогично положительному контролю YWO.49.H2-MC-vc-PAB-MMAF и YWO.49.H6-MC-vc-PAB-MMAF обладали заметной активностью в отношении уничтожения клеток, в частности при концентрации из расчета и приблизительно 0,01 и 0,1 мкг/мл. YWO.49.H2-MC-vc-PAB-MMAE и YWO.49.H6-MC-vc-PAB-MMAE также обладали активностью в отношении уничтожения клеток, но в меньшей степени, чем активность YWO.49.H2-MC-vc-PAB-MMAF и YWO.49.H6-MC-vc-PAB-MMAF. IC50 для YWO.49.H2-MC-vc-PAB-MMAF и YWO.49.H6-MC-vc-PAB-MMAF составила приблизительно 0,005 нМ, и IC50 для YWO.49.H2-MC-vc-PAB-MMAE и YWO.49.H6-MC-vc-PAB-MMAE составила приблизительно 0,2 нМ. IC50 для свободного MMAE составила приблизительно 0,1 нМ. YWO.49.H2-MC-MMAF и YWO.49.H6-MC-MMAF не обладали значительной активностью в отношении уничтожения клеток в этом анализе. Для этой конкретной системы анализа следует отметить, что при высоких концентрациях ADC, включая ADC в качестве отрицательного контроля, жизнеспособность клеток снижается существенно вследствие общей высокой концентрации MMAE и MMAF.
2. Анализ уничтожения клеток in vitro с использованием трансфицированных клеток HCT116
ADC YWO.49.H2 и YWO.49.H6 тестировали по способности ингибировать пролиферацию клеток HCT116, линия клеток рака толстой кишки, которых стабильно трансфицировали нуклеиновой кислотой, кодирующей TAT226 человека. Нетрансфицированные клетки HCT116 в норме приблизительно в 5-6 раз менее чувствительны к свободному (неконъюгированному) MMAE, чем клетки OVCAR3.
В кратком изложении, клетки HCT116 трансфицировали, как указано далее. Нуклеиновую кислоту, кодирующую связанный с эпитопом TAT226 человека, конструировали в экспрессирующем векторе млекопитающих pcDNA3.1 (Invitrogen, Carlsbad, CA). Эпитопная метка состояла из аминокислот 1-53 гликопротеина D вируса простого герпеса 1 типа (метка «gD»), которая заменила сигнальную последовательность из аминокислот 1-22 на N-конце TAT226 человека. Рекомбинантный вектор трансфицировали в клетки HCT116 с использованием Lipofectamine200 (Invitrogen) согласно протоколу производителя. Трансфицированные клетки HCT116 культивировали в среде МакКоя 5a с 10% FBS и 0,4 мг/мл G418. Клетки окрашивали с использованием антител анти-gD и сортировали путем FACS для отбора индивидуальных клонов, экспрессирующих рекомбинантный слитый белок gD:TAT226 человека. Один из клонов, обозначенный HCT116#9-4, отобрали для дальнейшего анализа.
Для осуществления анализа уничтожения клеток клетки HCT116#9-4 высевали в 96-луночные планшеты. Клетки HCT116#9-4 в плотности 1000 клеток на лунку инкубировали с различными концентрациями ADC, как представлено на фигуре 18. Анти-gp120 антитело, конъюгированное с MC-vc-PAB-MMAE, использовали в качестве отрицательного контроля. После 3 суток инкубации жизнеспособность клеток измеряли с использованием люминесцентного анализа жизнеспособности клеток CellTiter-Glo™ (Promega, Madison, WI) согласно инструкциям производителя.
На фигуре 18 показано, что YWO.49.H2-MC-vc-PAB-MMAF и YWO.49.H6-MC-vc-PAB-MMAF обладали заметной активностью в отношении уничтожения клеток, в частности, при приблизительно 0,01 мкг/мл и вплоть до самых высоких из тестируемых концентраций. IC50 для YWO.49.H2-MC-vc-PAB-MMAF и YWO.49.H6-MC-vc-PAB-MMAF составила приблизительно 0,05 нМ, и IC50 для свободного MMAE составила приблизительно 0,9 нМ. YWO.49.H2-MC-vc-PAB-MMAE и YWO.49.H6-MC-vc-PAB-MMAE не обладали существенной активностью в отношении уничтожения клеток относительно отрицательного контроля в этом конкретном анализе. Отличие между активностью в отношении уничтожения клеток для YWO.49.H2-MC-vc-PAB-MMAE и YWO.49.H6-MC-vc-PAB-MMAE в этом анализе по сравнению с анализом уничтожения клеток OVCAR3 (выше) можно отнести за счет разнообразных факторов, например плотности клеток и/или различий в чувствительности к лекарственному средству.
Следует отметить, что для некоторых других тестируемых линий клеток, которые в норме экспрессируют мРНК или белок TAT226, ADC YWO.49.H2 и YWO.49.H6 не обладали значительной активностью в отношении уничтожения клеток. Это можно отнести за счет разнообразных факторов, например эффектов, специфичных для типов клеток, уровней экспрессии TAT226 на клеточной поверхности и/или различиях в чувствительности к лекарственному средству.
3. Анализ in vivo с использованием ксенотрансплантата HCT116#9-4
Модель in vivo с применением ксенотрансплантата использовали для тестирования неконъюгированного и конъюгированного YWO.49.H6 по способности ингибировать пролиферацию экспрессирующих TAT226 опухолевых клеток in vivo. Образование опухолей индуцировали у бестимусных голых мышей «nu-nu» подкожной инъекцией приблизительно 5×106 клеток HCT116#9-4 в дорсальные паховые области мышей. Опухолям давали возможность расти до тех пор, пока они не достигали среднего объема опухоли 200 мм3. Этот момент времени обозначали как «0 сутки». Как представлено на фигуре 19, мыши получали внутривенные инъекции из расчета 3 мг/кг указанных неконъюгированных антител или ADC на 0, 7 и 16 сутки. Неконъюгированные и конъюгированные антитела к агглютинину амброзии (Ab) служили в качестве отрицательных контролей. Средний объем опухоли измеряли на 3, 7, 10, 16 и 21 сутки. Как представлено на фигуре 19, YWO.49.H6-MC-vc-PAB-MMAF обладал значительной активностью в отношении уничтожения опухолевых клеток при измерении среднего объема опухоли по сравнению с анти-агглютинином амброзии Ab-MC-vc-PAB-MMAF в этой конкретной модели с применением ксенотрансплантата. YWO.49.H6-MC-vc-PAB-MMAE не обладал значительной активностью в отношении уничтожения опухолевых клеток относительно анти-агглютинина амброзии Ab-MC-vc-PAB-MMAE в этой модели с применением ксенотрансплантата. Однако эта модель с применением ксенотрансплантата может не отражать активность в отношении уничтожения клеток YWO.49.H6-MC-vc-PAB-MMAE, наблюдаемую in vitro, вследствие множества факторов, например вследствие различий в чувствительности к лекарственному средству или уровней экспрессии TAT226 на клеточной поверхности в микроокружении ксенотрансплантата опухоли.
4. Другие модели с применением ксенотрансплантата
Другие модели с применением ксенотрансплантата можно использовать для тестирования неконъюгированных и конъюгированных анти-TAT226 антител по способности ингибировать пролиферацию экспрессирующих TAT226 опухолевых клеток in vivo. Например, модели с применением ксенотрансплантата для опухолей яичника и мозга можно получить из общедоступных источников, таких как Oncotest GmbH (Frieberg, Germany) и Southern Research Institute (Birmingham, AL). Модели Oncotest, в частности, разработаны путем выращивания опухолей пациентов в голых мышах с ослабленным иммунитетом. Ксенотрансплантаты, которые экспрессируют мРНК и/или белок TAT226, можно использовать для демонстрации активности в отношении уничтожения клеток анти-TAT226 антителами in vivo.
Конъюгированное YWO.49.H6 тестировали в модели Oncotest OVXF1023 по способности ингибировать пролиферацию клеток опухоли яичника in vivo. Модель Oncotest OVXF1023 происходит из метастатической слабо дифференцированной папиллярной серозной аденоматозной карциномы яичников, стадия М1. Мышей OVXF1023 обрабатывали ADC, указанными на фигуре 20. На фигуре 20 YWO.49.H6 обозначено как «H6»; антитело к агглютинину амброзии (контроль) обозначено как «RW»; и линкер -MC-vc-PAB- сокращенно обозначен как «vc». ADC вводили на сутки и в концентрациях, которые указаны на фигуре 20. Результаты, представленные на фигуре 20, указывают на то, что YWO.49.H6-MC-vc-PAB-MMAF и YWO.49.H6-MC-MMAF значительно уменьшают объем опухоли относительно других ADC.
Вышеописанный эксперимент повторяли с использованием OVXF1023 в сходных условиях, но с изменениями в дозировке и в контрольных ADC. В повторном эксперименте мышей обрабатывали более высокой дозой (5 мг/кг) YWO.49.H6-MC-vc-PAB-MMAE и более низкими дозами (5 мг/кг) YWO.49.H6-MC-vc-PAB-MMAF и YWO.49.H6-MC-MMAF. Результаты показали, что ADC H6 (и, в частности, YWO.49.H6-MC-vc-PAB-MMAE) снижали объем опухоли относительно соответствующих им контрольных ADC (анти-gp120-MC-vc-PAB-MMAE исходя из 6,4 мг/кг; анти-gp120-MC-vc-PAB-MMAF исходя из 7,2 мг/кг; и анти-gp120-MC-MMAF исходя из 5,4 мг/кг), хотя различие в эффективности между ADC H6 и соответствующими им контрольными ADC не являлось статистически значимым для некоторых значений данных (данные не представлены). Различие между этими результатами и результатами, полученными в первом эксперименте с OVXF1023, можно отнести за счет различий в дозировках ADC H6 и выявлению того факта, что контрольный ADC анти-gp120 обладал непредвиденной активностью в снижении объема опухоли.
Кроме того, ADC H6 также тестировали в другой модели Oncotest, OVXF899, которая происходит из умеренно дифференцированной папиллярной серозной карциномы яичников (первичная опухоль). ADC H6 не снижали объем опухоли в этой модели (данные не представлены). Однако эта конкретная модель Oncotest обладала низкой экспрессией TAT226, что может отвечать за наблюдаемые результаты.
I. ТиоMAb
Получены антитела с конструированным цистеином или «тиоMAb», в которых выбранный остаток YWO.49.H6 замещен на цистеин для обеспечения дополнительного участка для конъюгации с компонентом линкера-лекарственного средства. В частности, замещение A118C (нумерация EU) получено в тяжелой цепи YWO.49.H6, или замещение V205C (нумерация по Kabat) получено в легкой цепи YWO.49.H6. Полученное тиоMAb A118C затем конъюгировали с MC-MMAF и полученное тиоMAb V205C затем конъюгировали или с MC-MMAF, или с MC-vc-PAB-MMAE. Все тиоMAb оказались способными связываться с клетками OVCAR3 по данным анализа FACS (данные не представлены).
Хотя вышеописанное изобретение довольно подробно описано посредством иллюстрации и примера с целями ясности понимания, описания и примеры не следует толковать как ограничивающие объем изобретения. Описания всех патентных и научных публикаций, цитированных в настоящем документе, явно приведены в качестве ссылки в полном объеме.
























| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2436796C9 |
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2639543C9 |
| АНТИ-LRP6 АНТИТЕЛА | 2011 |
|
RU2587625C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2511410C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2553566C2 |
| ЛЕЧЕНИЕ ВИРУСА ГРИППА А H7N9 | 2015 |
|
RU2682049C2 |
| АНТИТЕЛА ПРОТИВ 5-БРОМ-2'-ДЕЗОКСИУРИДИНА И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2705299C2 |
| ПИРРОЛОБЕНЗОДИАЗЕПИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2736725C1 |
| АНТИ-FcRH5 АНТИТЕЛА | 2014 |
|
RU2687132C2 |
| СТАБИЛЬНЫЕ КОМПОЗИЦИИ ИММУНОКОНЪЮГАТОВ, СОДЕРЖАЩИХ АНТИТЕЛО К CD79b | 2019 |
|
RU2800803C2 |
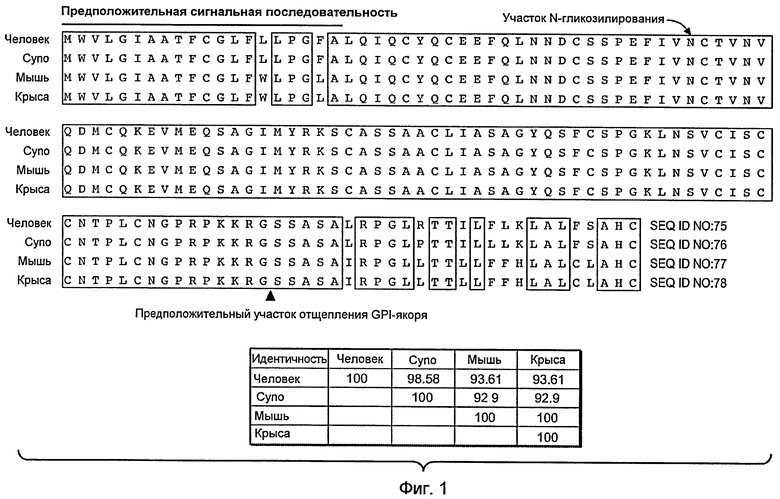
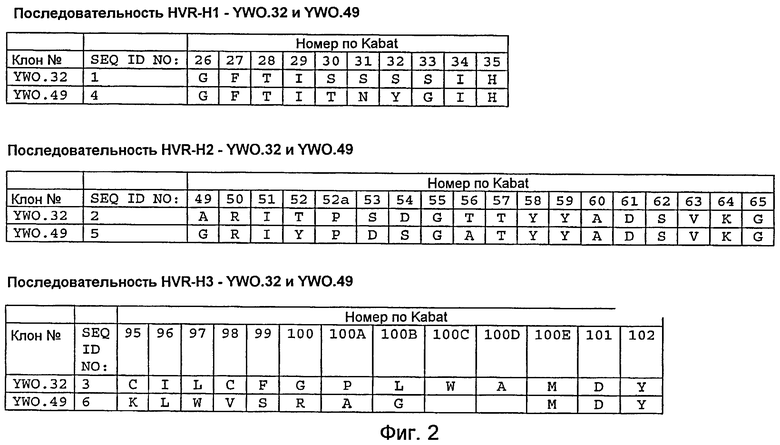
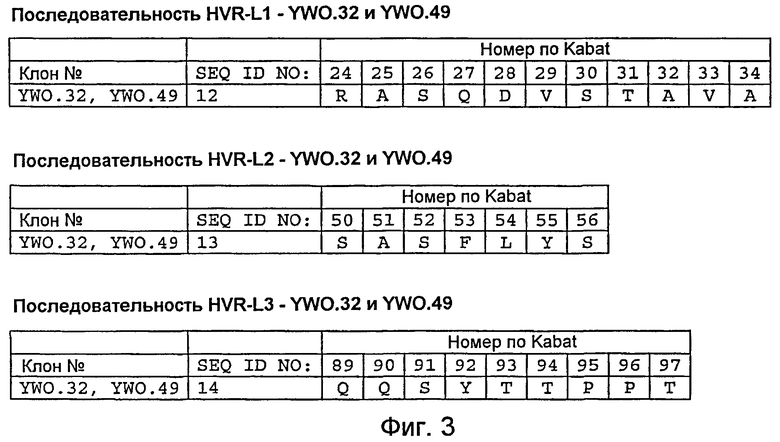
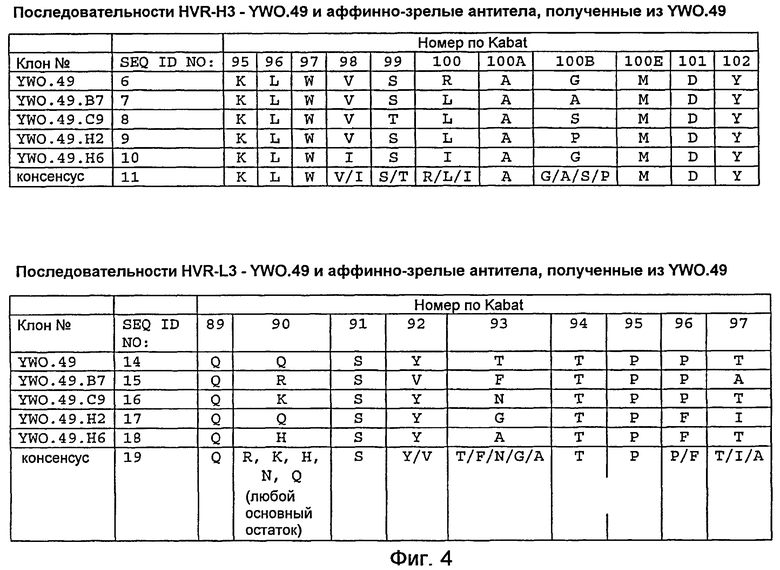
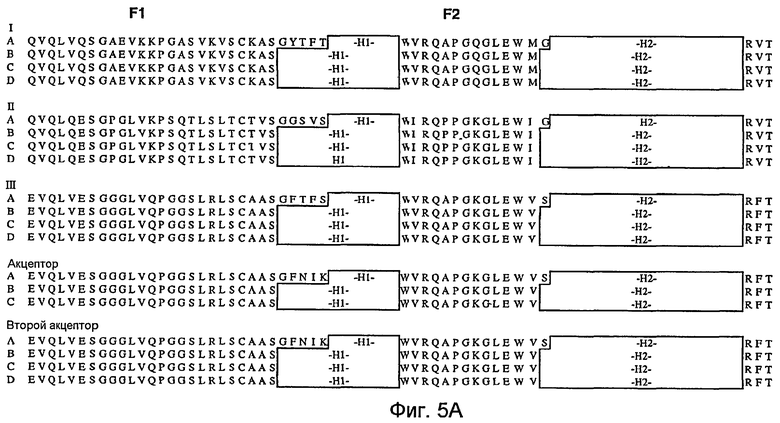
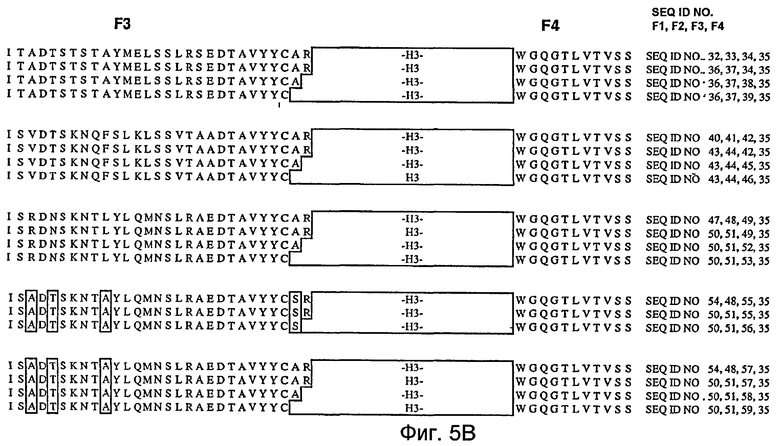
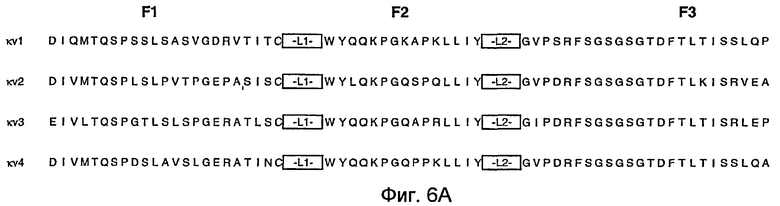
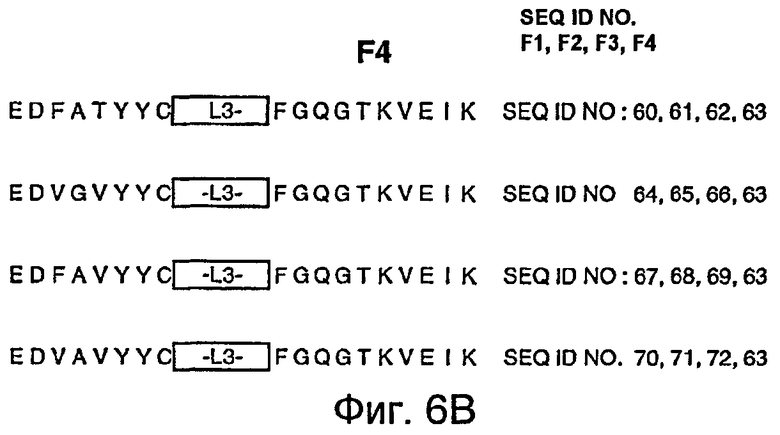





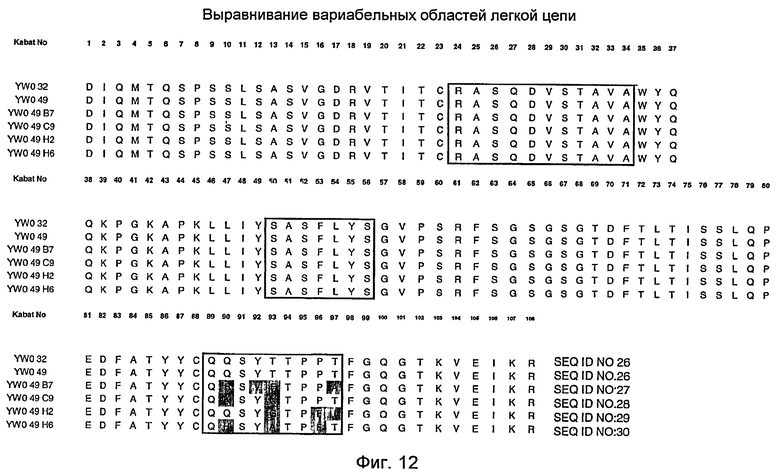

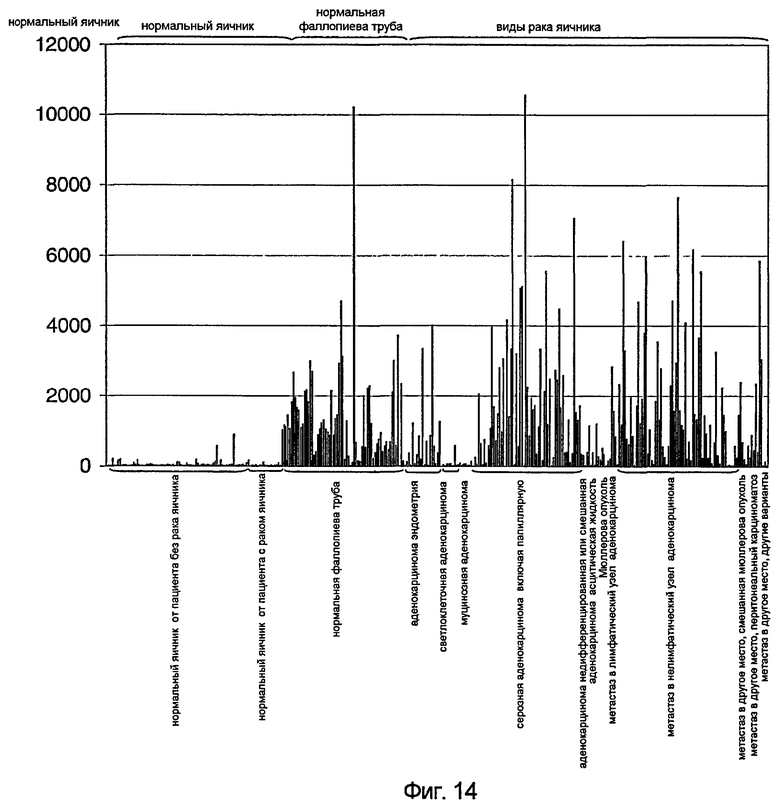
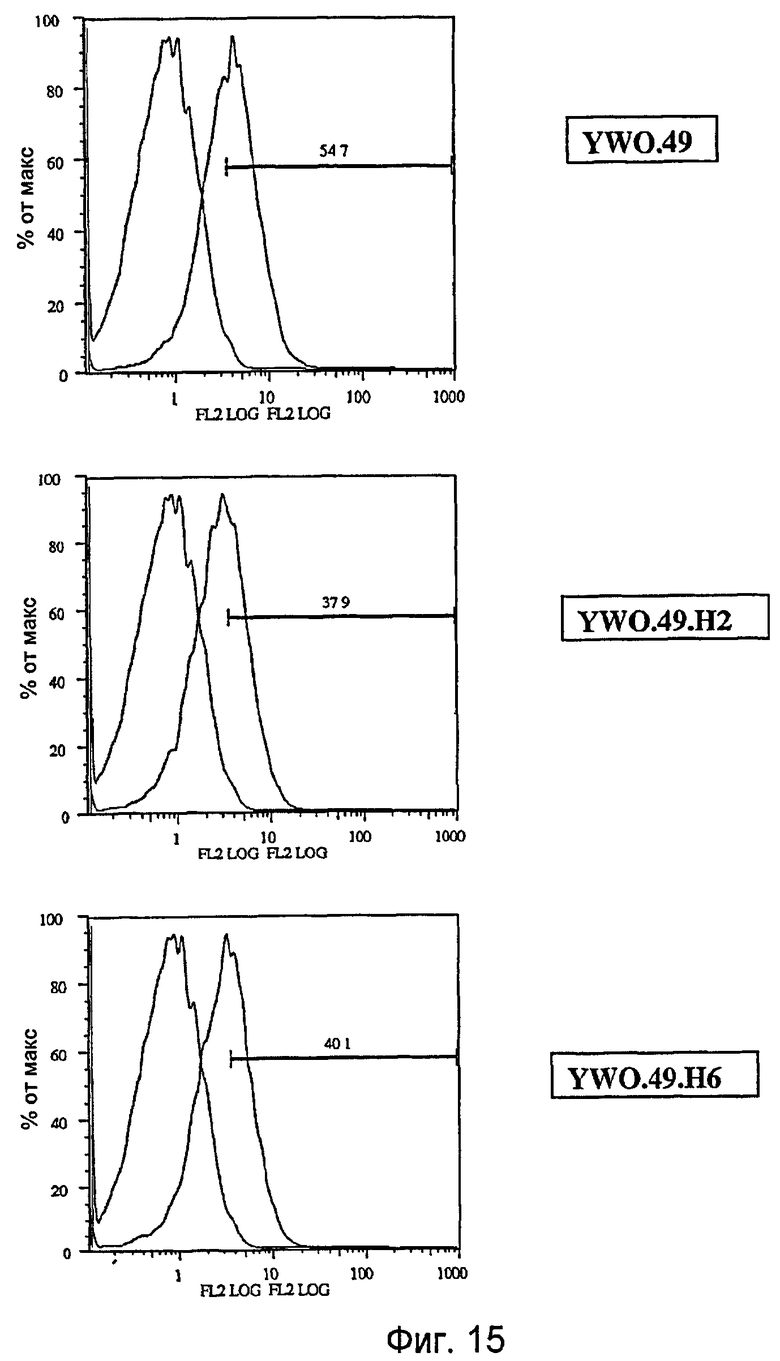
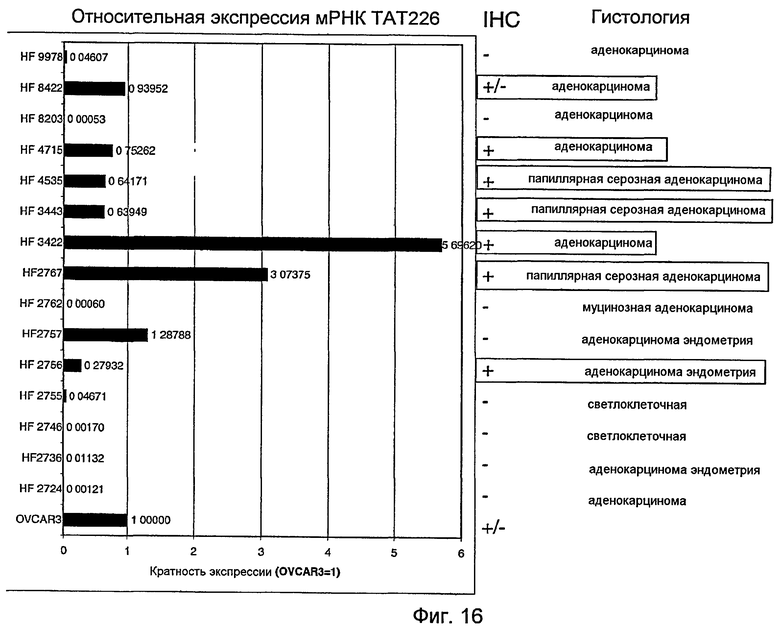
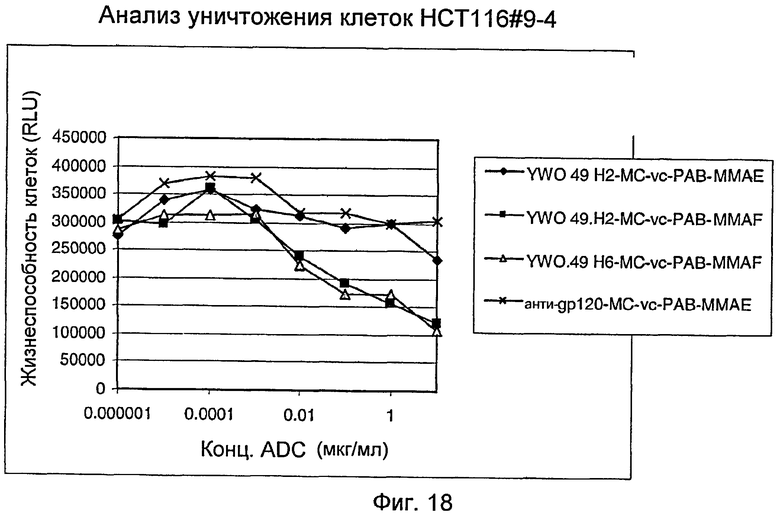
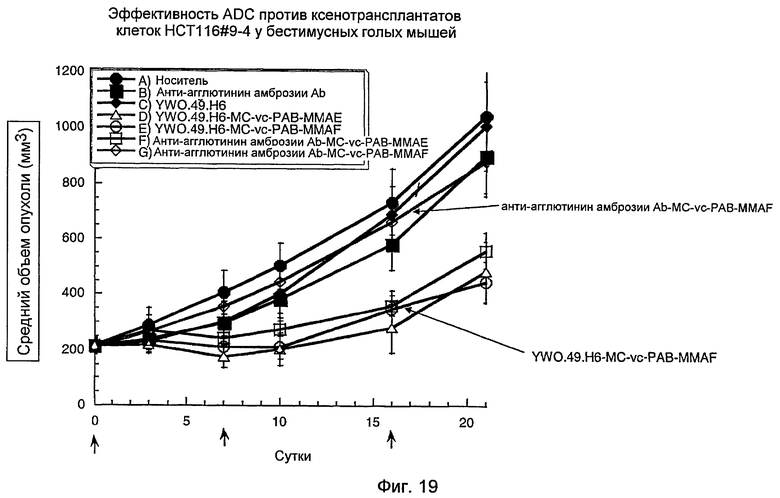
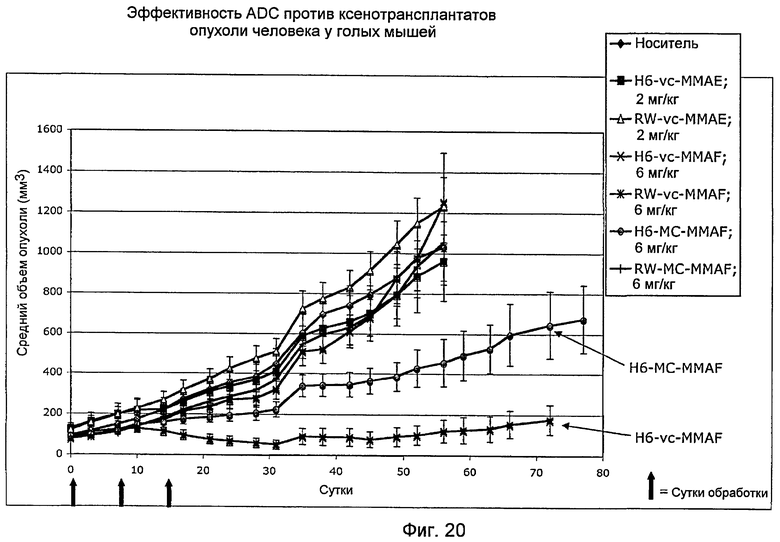
Изобретение относится к биохимии и представляет собой варианты моноклональных анти-ТАТ226 антител, их иммуноконъюгаты и фармацевтические композиции. Антитела по изобретению могут эффективно применяться для ингибирования пролиферации опухолевых клеток. 6 н. и 35 з.п. ф-лы, 22 ил., 3 табл., 4 пр.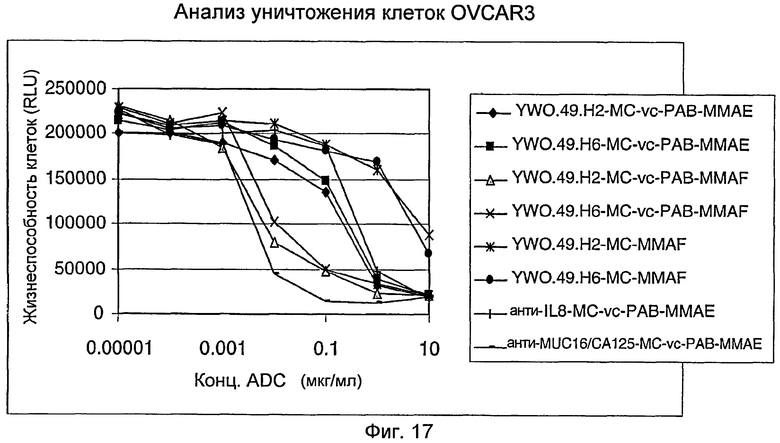
1. Моноклональное антитело, которое связывается с ТАТ226, где антитело содержит
(1) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4;
(2) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5;
(3) HVR-H3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:11;
(4) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12;
(5) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и
(6) HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19.
2. Антитело по п.1, где антитело содержит HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-10, и HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-18.
3. Антитело по п.2, где HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17.
4. Антитело по п.2, где HVR-H3 содержит аминокислотную последовательность SEQ ID NO:10, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:18.
5. Антитело по п.1, дополнительно содержащее, по меньшей мере, одну каркасную область, выбранную из консенсусной каркасной области VH подгруппы III и консенсусной каркасной области VL подгруппы I.
6. Моноклональное антитело, которое связывается с ТАТ226, где антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:21-25, и вариабельный домен легкой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31, и где указанное антитело, которое связывается с ТАТ226, обладает константой диссоциации (КD)≤10 нМ.
7. Антитело по п.6, где антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24, и вариабельный домен легкой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:29.
8. Антитело по п.7, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:24, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:29.
9. Антитело по п.6, где антитело содержит вариабельный домен тяжелой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:25, и вариабельный домен легкой цепи, обладающий, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:30.
10. Антитело по п.9, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:25, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:30.
11. Антитело по п.1 или 6, где антитело является фрагментом антитела, выбранным из фрагмента Fab, Fab'-SH, Fv, scFv или (Fab')2.
12. Антитело по п.1 или 6, где антитело является гуманизированным.
13. Антитело по п.1 или 6, где антитело является человеческим.
14. Иммуноконъюгат, содержащий антитело, которое связывается с ТАТ226 по п.1 или 6, ковалентно присоединенное к цитотоксическому средству.
15. Иммуноконъюгат по п.14, где цитотоксическое средство выбрано из токсина, химиотерапевтического средства, антибиотика, радиоактивного изотопа и нуклеолитического фермента.
16. Иммуноконъюгат, имеющий формулу Ab-(L-D)p, где:
(a) Ab представляет собой антитело, которое связывается с ТАТ226, и которое содержит
(i) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:4;
(ii) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:5;
(iii) HVR-H3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:11;
(iv) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:12;
(v) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:13; и
(vi) HVR-L3, содержащую аминокислотную последовательность, которая соответствует консенсусной последовательности SEQ ID NO:19;
(b) L представляет собой линкер;
(с) D представляет собой лекарственное средство формулы DE или DF
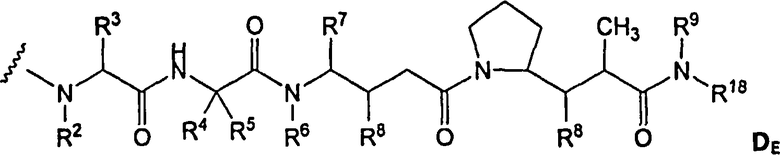
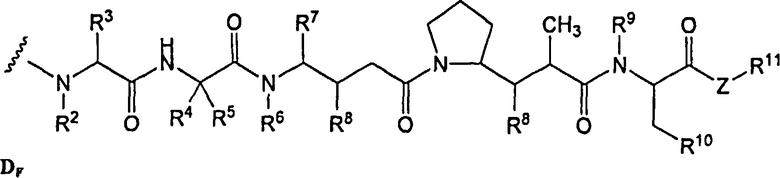 ,
,
и где каждый R2 и R6 представляет собой метил, каждый R3 и R4 представляет собой изопропил, R7 представляет собой втор-бутил, каждый R8 независимо выбран из СН3, O-СН3, ОН и Н; R9 представляет собой H; R10 представляет собой арил; Z представляет собой -О- или -NH-; R11 представляет собой Н, С1-С8алкил или -(СН2)2-O-(СН2)2-O-(СН2)2-O-СН3; и R18 представляет собой -С(R8)2-С(R8)2-арил; и
(d) p варьирует приблизительно от 1 до 8.
17. Иммуноконъюгат по п.16, где антитело содержит HVR-H3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:6-10, и HVR-L3, содержащую аминокислотную последовательность, выбранную из SEQ ID NO:14-18.
18. Иммуноконъюгат по п.17, где HVR-H3 содержит аминокислотную последовательность SEQ ID NO:9, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:17.
19. Иммуноконъюгат по п.17, где HVR-H3 содержит аминокислотную последовательность SEQ ID NO:10, и HVR-L3 содержит аминокислотную последовательность SEQ ID NO:18.
20. Иммуноконъюгат, имеющий формулу Ab-(L-D)p, где:
(а) Ab представляет собой антитело, которое связывается с ТАТ226 и которое содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:21-25, и вариабельную область легкой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью, выбранной из SEQ ID NO:26-31;
(b) L представляет собой линкер;
(c) D представляет собой лекарственное средство формулы DE или DF
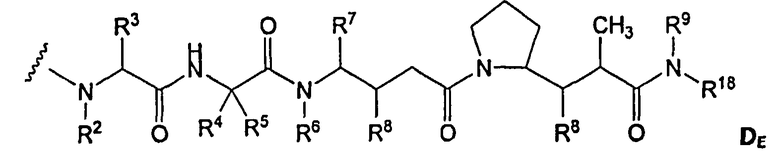
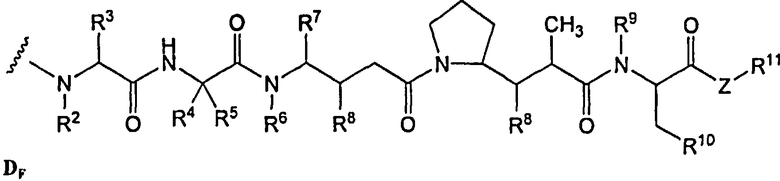 ,
,
и где каждый R2 и R6 представляет собой метил, каждый R3 и R4 представляет собой изопропил, R7 представляет собой втор-бутил, каждый R8 независимо выбран из СН3, O-СН3, ОН и Н; R9 представляет собой Н; R10 представляет собой арил; Z представляет собой -O- или -NH-; R11 представляет собой Н, С1-С8алкил или -(СН2)2-O-(СН2)2-O-(СН2)2-O-СН3; и R18 представляет собой -C(R8)2-С(R8)2-арил; и
(d) p варьирует приблизительно от 1 до 8,
и где указанное антитело, которое связывается с ТАТ226, обладает константой диссоциации (КD)≤10 нМ.
21. Иммуноконъюгат по п.20, где антитело содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:24, и вариабельную область легкой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:29.
22. Иммуноконъюгат по п.21, где антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:24, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:29.
23. Иммуноконъюгат по п.20, где антитело содержит вариабельную область тяжелой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:25, и вариабельную область легкой цепи, обладающую, по меньшей мере, 95% идентичностью последовательности с аминокислотной последовательностью SEQ ID NO:30.
24. Иммуноконъюгат по п.23, где антитело содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:25, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30.
25. Иммуноконъюгат по п.16 или 20, где линкер присоединен к антителу посредством тиольной группы на антителе.
26. Иммуноконъюгат по п.16 или 20, где лекарственное средство выбрано из ММАЕ и MMAF.
27. Иммуноконъюгат по п.26, где лекарственное средство представляет собой ММАЕ.
28. Иммуноконъюгат по п.26, где лекарственное средство представляет собой MMAF.
29. Иммуноконъюгат по п.26, где линкер является расщепляемым протеазой.
30. Иммуноконъюгат по п.29, где линкер содержит дипептид val-cit.
31. Иммуноконъюгат по п.29, где линкер содержит звено п-аминобензила.
32. Иммуноконъюгат по п.29, где линкер содержит 6-малеимидокапроил.
33. Иммуноконъюгат по п.21, где иммуноконъюгат имеет формулу
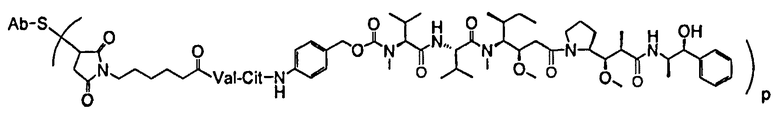 ,
,
где S представляет собой атом серы, и р варьирует от 2 до 5.
34. Иммуноконъюгат по п.33, где Ab представляет собой моноклональное антитело по п.3 или 8.
35. Иммуноконъюгат по п.33, где Ab представляет собой моноклональное антитело по п.4 или 10.
36. Иммуноконъюгат по п.28, где иммуноконъюгат имеет формулу
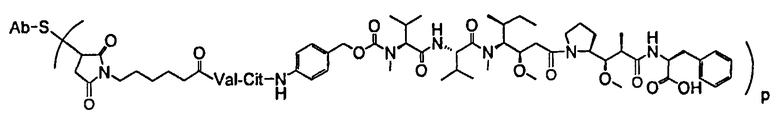 ,
,
где S представляет собой атом серы, и р варьирует от 2 до 5.
37. Иммуноконъюгат по п.36, где Ab представляет собой моноклональное антитело по п.3 или 8.
38. Иммуноконъюгат по п.36, где Ab представляет собой моноклональное антитело по п.4 или 10.
39. Фармацевтическая композиция, содержащая эффективное количество иммуноконъюгата по любому из пп.14-38, для применения при лечении нарушения пролиферации клеток.
40. Фармацевтическая композиция по п.39, где нарушение пролиферации клеток выбрано из рака яичника, рака тела матки, опухоли мозга и опухоли Вильмса.
41. Фармацевтическая композиция по п.39, где иммуноконъюгат представляет собой иммуноконъюгат по п.16 или 20.
| US 2006057141, 16.03.2006 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ ОСНОВНОГО ОБМЕНА ОРГАНИЗМА ЧЕЛОВЕКА | 1992 |
|
RU2057480C1 |
| РОЙТ А | |||
| и др | |||
| Иммунология | |||
| - М.: Мир, 2000, с.537-539. | |||

