Перекрестная ссылка на родственные заявки
По настоящей заявке испрашивается приоритет предварительной патентной заявки США № 61/317137, поданной 24 марта 2010 г. и предварительной патентной заявки США № 61/394836, поданной 20 октября 2010 г, описание которых полностью включено в настоящее описание путем ссылки.
Область техники
Настоящее изобретение относится к анти-LRP6 антителам и способам их применения для лечения раковых заболеваний или нарушений скелета.
Предпосылки создания изобретения
Аналогично путям передачи сигнала от большинства других морфогенов и ростовых факторов, в процессе развития и гомеостаза тканей у млекопитающих многократно запускается передача сигнала от Wnt, в которой задействовано 19 различных лигандов, 10 рецепторов и множество корецепторов, включая LRP5/6, Ror1/2 и Ryk (van Amerongen and Nusse, 2009). Кроме того, различные секретируемые антагонисты, которые связывают либо белки Wnt, такие как SFRP1/2/3/4/5 и WIF1, либо LRP5/6, включая DKK1/2/4 и SOST, модулируют взаимодействие между лигандами и рецепторами. Эти мембранные и внеклеточные белки, а также их многочисленные изоформы, обеспечивают специфичную регуляцию на уровне экспрессии и комбинаторных белковых взаимодействий. Большинство изоформ Wnt, по-видимому, способно связывать корецептор LRP5/6, и активация LRP5/6 определяет каноническую или β-катенин-зависимую передачу сигнала от Wnt. Wnt образует гетеродимеры с LRP5/6 и FZD, опосредуя фосфорилирование внутриклеточного домена LRP5/6 и связывание с Axin (Tamai et al., 2000; Semenov et al., 2001; Tamai et al., 2004). DVL входит в комплекс, напрямую связываясь как с Axin, так и с FZD, и олигомеризация DVL вероятно увеличивает эти белковые комплексы на цитоплазматической стороне мембраны, которые связывают GSK3 и ингибируют обусловленное этой киназой фосфорилирование и дестабилизацию β-катенина (Mi et al., 2006; Bilic et al., 2007; Schwarz-Romond et al., 2007; Cselenyi et al., 2008; Piao et al., 2008; Zeng et al., 2008; Wu et al., 2009).
Уникально большое число изоформ лиганда, имеющих значительные расхождения по первичной последовательности, которые опосредуют каноническую передачу сигнала от Wnt у млекопитающих, составляет контраст с парой высокогомологичных корецепторов. Внеклеточные домены LRP6 и LRP5 состоят в основном из четырех гомологичных областей, называемых Е1-Е4 в направлении от N- к С-концу, каждая из которых содержит β-пропеллер YWTD-типа и EGF-подобный домен (Jeon et al., 2001). Каждый повтор в аналогичном положении в LRP6 и LRP5 является высоко консервативным, в то время как разные повторы в одном белке отличаются в значительно большей степени. Примечательно, что группой Bourhis et al. (2010) было продемонстрировано, что Wnt9b связывается исключительно с областью Е1-Е2 in vitro, в то время как Wnt3a связывается только с фрагментом, содержащим E3-E4, позволяя предположить, что каждый повтор или комбинация двух соседних повторов связывает различные субпопуляции изоформ Wnt. Эта схема может соответствовать разнообразию Wnt-белков и, возможно, также позволяет их специфичную регуляцию с помощью лигандов-антагонистов LRP5/6. У Notch и VEGF-рецепторов, чьи внеклеточные области содержат повторы EGF-подобных и Ig-доменов, соответственно, связывание многочисленных изоформ лиганда локализовано в одной области одного или двух повторов, хотя присутствие других повторов может усиливать связывание. (Rebay et al., 1991; Davis-Smyth et al., 1996; Cunningham et al., 1997).
В случае рецепторных тирозиновых киназ, индуцируемая лигандами димеризация стимулирует киназную активность и сигнальные пути. Несмотря на то, что индуцируемая лигандами гетеродимеризация рецептора и корецептора необходима для канонической передачи сигнала от Wnt, точная роль гомодимеризации LRP5/6 или FZD не определена. Направленная димеризация различных рекомбинантных белков LRP6 может либо активировать, либо ингибировать передачу сигнала от Wnt.
β-катенин-зависимая передача сигнала от Wnt активируется связыванием изоформы Wnt как с рецептором FZD, так и с корецептором LRP5/6, которые затем собираются в мультимерный комплекс на цитоплазматической поверхности мембраны, связывая и инактивируя киназу GSK3. Остается необходимым определить, могут ли и каким образом могут механически различные взаимодействия между изоформами Wnt и рецепторами модулировать этот процесс.
Краткое изложение сущности изобретения
Один аспект изобретения относится к выделенному антителу, которое связывает LRP6, где антитело ингибирует передачу сигнала, индуцированную первой изоформой Wnt, и усиливает передачу сигнала, индуцированную второй изоформой Wnt. В одном варианте осуществления изобретения первая изоформа Wnt выбрана из группы, состоящей из Wnt3 и Wnt3a. В одном варианте осуществления изобретения вторая изоформа Wnt выбрана из группы, состоящей из Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и 10b. В другом варианте осуществления изобретения первая изоформа Wnt выбрана из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b, и вторая изоформа Wnt выбрана из группы, состоящей из Wnt3 и Wnt3a.
Один аспект изобретения относится к антителу, которое связывает область Е3-E4 белка LRP6. Другой аспект изобретения относится к антителу, которое связывает область Е1-E2 белка LRP6. Еще один аспект относится к антителу, которое связывает две различных области на LRP6, такие как область E1-E2 белка LRP6 и область E3-E4 белка LRP6. В одном аспекте эти антитела ингибируют передачу сигнала от Wnt, индуцированную комбинацией Wnt1 и Wnt3a. В одном аспекте эти антитела ингибируют аутокринную передачу сигнала от Wnt.
Один аспект изобретения относится к способу лечения индивидуума, страдающего раком, включающему введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt1 и Wnt3a, и выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b.
Другой аспект изобретения относится к способу лечения индивидуума, страдающего раком, который включает введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt3 и Wnt3a, и выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b.
Другой аспект изобретения относится к способу лечения индивидуума, страдающего раком, который включает введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt3 и Wnt3a, и выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и 10b.
Один аспект изобретения относится к способу лечения индивидуума, имеющего нарушение скелета, такое как остеопороз, остеоартрит, переломы костей и поражения костной ткани, включающему введение индивидууму эффективного количества анти-LRP6 антитела, описанного в настоящем описании.
Другой аспект изобретения относится к способу усиления передачи сигнала от Wnt, индуцированной изоформой Wnt, у индивидуума, включающему введение индивидууму эффективного количества анти-LRP6 антитела, описанного в настоящем описании, и изоформы Wnt для усиления передачи сигнала от Wnt, индуцированной изоформой Wnt.
Также изобретение относится к специфичным анти-LRP6 антителам, включая биспецифичные анти-LRP6 антитела. В одном варианте осуществления изобретения выделенное антитело, которое связывает LRP6, включает в себя VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 и SEQ ID NO:15. В одном варианте осуществления изобретения антитело дополнительно включает в себя VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:10 и SEQ ID NO:12. В одном варианте осуществления изобретения выделенное антитело, которое связывает LRP6, включает в себя VH, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 и SEQ ID NO:15. В одном варианте осуществления изобретения выделенное антитело, которое связывает LRP6, дополнительно включает в себя VL, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:10 и SEQ ID NO:12.
В одном варианте осуществления изобретения антителом является выделенное биспецифичное антитело, которое связывает две различные области LRP6, где антитело включает в себя VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 и SEQ ID NO:15. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:15, и вторую VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:9, SEQ ID NO:11 и SEQ ID NO:13. В одном варианте осуществления изобретения биспецифичное антитело дополнительно включает в себя VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:10 и SEQ ID NO:12.
В одном варианте осуществления изобретения биспецифичное антитело, которое связывает две различные области LRP6, включает в себя VH, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15. В одном варианте осуществления изобретения биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первую VH, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью SEQ ID NO:15, и вторую VH, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:9, SEQ ID NO:11 и SEQ ID NO:13. В одном варианте осуществления изобретения биспецифичное антитело дополнительно включает в себя VL, содержащую аминокислотную последовательность, имеющую по меньшей мере 90% гомологии с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:10 и SEQ ID NO:12.
В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, и (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24. В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24. В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24. В одном варианте осуществления изобретения выделенное биспецифичное антитело, которое связывает две различные области LRP6, включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20, (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения биспецифичное антитело по указанным выше вариантам осуществления дополнительно включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26; (c) HVR-L3 SEQ ID NO:27.
В одном варианте осуществления изобретения биспецифичное антитело по указанным выше вариантам осуществления дополнительно включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26; (c) HVR-L3 SEQ ID NO:28.
Один вариант осуществления изобретения относится к выделенному биспецифичному антителу, которое связывает две различные области LRP6, где антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:15, и вторую VH, выбранную из группы, состоящей из VH, содержащей аминокислотную последовательность SEQ ID NO:9, SEQ ID NO:11 и SEQ ID NO:13. В одном варианте осуществления изобретения это антитело дополнительно включает в себя VL, содержащую аминокислотную последовательность SEQ ID NO:10 или SEQ ID NO:12. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:15, и вторую VH, содержащую аминокислотную последовательность SEQ ID NO:9, и VL, содержащую аминокислотную последовательность SEQ ID NO:10.
В одном варианте осуществления изобретения биспецифичное антитело ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b. В одном варианте осуществления изобретения биспецифичное антитело дополнительно ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 4, 7a, 7b и 10a. В одном варианте осуществления изобретения биспецифичное антитело ингибирует аутокринную передачу сигнала от Wnt.
Один аспект изобретения относится к биспецифичному антителу, которое ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из 1, 2, 2b, 6, 8a, 9a, 9b и 10b. В одном варианте осуществления изобретения биспецифичное антитело дополнительно ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 4, 7a, 7b и 10a.
Один аспект изобретения относится к антителу, которое конкурирует за связывание с LRP6 с любым из анти-LRP6 антител, включая биспецифичные антитела, описанные в настоящем описании.
Другой аспект изобретения относится к антителу, которое связывает такие же два эпитопа, как биспецифичное антитело, описанное в настоящем описании. В одном варианте осуществления изобретения один из двух эпитопов содержит аминокислотные остатки R28, E51, D52, V70, S71, E73, L95, S96, D98, E115, R141 и N185 из LRP6. В одном варианте осуществления изобретения один из двух эпитопов содержит аминокислотные остатки R28, E51, D52, V70, S71, E73, L95, S96, D98, E115, R141, N185, R29, W188, K202, P225, H226, S243 и F266 из LRP6.
Другой аспект изобретения относится к выделенной нуклеиновой кислоте, кодирующей анти-LRP6 антитело, описанное в настоящем описании. Другой аспект относится к клетке-хозяину, содержащей такую нуклеиновую кислоту.
Один аспект изобретения относится к иммуноконъюгату, содержащему анти-LRP6 антитело, описанное в настоящем описании, и цитоплазматический агент. Другой аспект относится к фармацевтическому составу, содержащему анти-LRP6 антитело, описанное в настоящем описании, и фармацевтически приемлемый носитель.
Один аспект изобретения относится к анти-LRP6 антителу, описанному в настоящем описании, для применения в качестве лекарственного препарата. Один аспект относится к анти-LRP6 антителу, описанному в настоящем описании, для использования при лечении ракового заболевания или нарушения скелета. Один аспект относится к анти-LRP6 антителу, описанному в настоящем описании, для применения при ингибировании передачи сигнала, индуцированной первой изоформой Wnt, и усиления передачи сигнала, индуцированной второй изоформой Wnt. Один аспект относится к применению анти-LRP6 антитела, описанного в настоящем описании, при получении лекарственного препарата, пригодного для лечения, например, ракового заболевания или нарушения скелета.
Один аспект изобретения относится к способу лечения индивидуума, страдающего раковым заболеванием, таким как немелкоклеточный рак легких, рак молочных желез, рак поджелудочной железы, рак яичников, рак почек и рак предстательной железы, включающему введение индивидууму эффективного количества анти-LRP6 антитела, описанного в настоящем описании.
Краткое описание фигур
Фиг.1А. На графике показано ингибирование антителами к белку LRP6.E3-E4 активности Wnt-люциферазного репортера, индуцированной 0,1 мг/мл очищенного Wnt3a, в клетках HEK293.
Фиг.1В. Вестерн-блот анализ клеток НЕК293 либо нестимулированных, либо индуцированных Wnt3a, и получавших воздействие указанного антитела к LRP6 или очищенного белка.
Фиг.1С. На графике показано, что антитело YW210.09 усиливает активность Wnt-репортерного гена пропорционально концентрации Wnt3a в клетках HEK293.
Фиг.2А. На графике показано зависимое от концентрации ингибирование и усиление аутокринной передачи сигнала от Wnt в клетках тератокарциномы РА-1, трансфицированных люциферазными репортерами, и к которым были добавлены антитела к LRP6 либо отдельно, либо в комбинации, или белок Fzd8CRD-Fc.
Фиг.2В. На графике показан результат анализа экспрессии с помощью количественной ПЦР Wnt-индуцированных генов SAX1 и GAD1 и Wnt-репрессированного гена, LEFTY2, в клетках PA-1, получавших или не получавших воздействие 0,3 мг/мл белка Wnt3a, и получавших воздействие 10 мг/мл антитела YW211.31, анти-gD моноклонального антитела (отрицательный контроль) или белка Fzd8CRD-Fc (положительный контроль). Данные нормализованы по образцам из клеток без добавления (NA) белка Wnt3a.
Фиг.3. Итоговая таблица, в которой показаны эффекты антител к LRP6 и белка Fzd8CRD-Fc на аутокринную передачу сигнала в клеточных линиях.
Фиг.4А. На графике приведен результат анализа экспрессии с помощью количественной ПЦР мРНК AXIN2 в четырех клеточных линиях, получавших воздействие 25 мкг/мл антитела YW211.31.57 или белка Fzd8CRD-Fc, в присутствии или без (NA) 0,2 мкг/мл белка Wnt3a.
Фиг.4В. На графике изображено, что экспрессия Wnt-индуцируемых генов в клетках NCI-H23 усиливается под действием антитела YW211.31.57 и подавляется антителом YW210.09 (30 мкг/мл). Белок CD4-Fc (30 мкг/мл) служит в качестве отрицательного контроля.
Фиг.4С. На графике изображено, что экспрессия Wnt-индуцируемых генов в клетках M14 усиливается под действием антитела YW211.31.57 и подавляется антителом YW210.09 (30 мкг/мл).
Фиг.4D. На графике показано зависимое от концентрации ингибирование стимулированной Wnt3a передачи сигнала в результате действия антитела YW211.31.57 в клетках Hs578T со стабильно встроенным Wnt-люциферазным репортером.
Фиг.4Е. На графике показано зависимое от концентрации усиление аутокринной передачи сигнала от Wnt антителом YW211.31.57 в клетках Hs578T со стабильно встроенным Wnt-люциферазным репортером.
Фиг.4F. На графике показано, что клетки EKVX, трансфицированные Wnt-люциферазным репортером, демонстрируют усиление аутокринной передачи сигнала от Wnt (NA) и антагонизм в отношении Wnt3a-индуцированной передачи сигнала в результате действия антитела YW211.31.57.
Фиг.4G. На графике показано, что опосредованное антителами усиление аутокринной передачи сигнала от Wnt ингибируется 5 мкг/мл белка Fzd8CRD-Fc.
Фиг.5. Итоговая таблица эффектов 10 мг/мл антител к LRP6 или белка Fzd8CRD-Fc на сигнальный путь, индуцированный трансфекцией конструкций для экспрессии изоформ Wnt в клеточные линии HEK293 или Hs578T со стабильно встроенными Wnt-люциферазными репортерами. Экспрессию Wnt-люциферазного репортера нормализовали на число клеток и дополнительно нормализовали на уровень в клетках, трансфицированных такой же экспрессирующей конструкцией, но к которым не добавляли белки.
Фиг.6. Итоговая таблица эффектов 10 мг/мл антител к LRP6 или белка Fzd8CRD-Fc на передачу сигнала в клеточной лини HEK293 со стабильно встроенными Wnt-люциферазными репортерами. Передачу сигнала индуцировали трансфекцией конструкций для экспрессии химерных белков, состоящих из изоформ Wnt, слитых с изоформами FZD или LRP6. Экспрессию Wnt-люциферазного репортера нормализовали на число клеток и дополнительно нормализовали на уровень в клетках, трансфицированных такой же экспрессирующей конструкцией, но к которым не добавляли белки.
Фиг.7. Итоговая таблица эффектов 10 мг/мл антител к LRP6 или комбинаций антител на передачу сигнала, индуцированную трансфекцией конструкций для экспрессии изоформ Wnt, в клеточных линиях со стабильно встроенными Wnt-люциферазными репортерами. Экспрессию Wnt-люциферазного репортера нормализовали на число клеток и дополнительно нормализовали на уровень в клетках, трансфицированных такой же экспрессирующей конструкцией, но к которым не добавляли белки.
Фиг.8А. На графике показано, что комбинация антител YW211.31.57 и YW210.09 ингибирует передачу сигнала в клетках HEK293 со стабильно встроенным Wnt-люциферазным репортером, которые были трансфицированы для экспрессии либо Wnt3a, Wnt1, либо совместно Wnt3a и Wnt1. Анти-gD антитело и белок Fzd8CRD-Fc показаны в качестве отрицательного и положительного контролей, соответственно, для ингибирования передачи сигнала Wnt.
Фиг.8В. На графике показано, что комбинация антител YW211.31.57 и YW210.09 усиливает аутокринную передачу сигнала от Wnt в клетках Hs578T.
Фиг.8С. На графике показано, что комбинация антител YW211.31.57 и YW210.09 усиливает аутокринную передачу сигнала от Wnt в клетках EKVX.
Фиг.9А и В. Анализ взаимодействия между биослоями (интерферометрия биослоев) с биотинилированным белком LRP6 E1-E4, иммобилизованным на стрептавидиновых биосенсорах, указывает на то, что антитело YW211.31.57 ингибирует связывание Wnt3a и Wnt9b с LRP6, а антитело YW210.09 ингибирует только связывание Wnt9b.
Фиг.9С. Метод интерферометрии биослоев с меньшим, неперекрывающимся фрагментом LRP6 показал, что Wnt3a связывает область Е3-Е4, и это взаимодействие блокируется либо интактным, либо одноплечевым антителом YW211.31.
Фиг.9D. Метод интерферометрии биослоев с меньшим, неперекрывающимся фрагментом LRP6 показал, что антитело YW210.09 связывает фрагмент E1-E2 белка LRP6 и конкурирует за данное связывание с Wnt9b.
Фиг.9Е. Метод интерферометрии биослоев показал, что антитела YW211.31.57 и YW210.09 могут совместно связывать иммобилизованный белок LRP6.E1-E4 при последовательном добавлении в любом порядке, что подтверждает различные эпитопы для этих антител.
Фиг.10А. График, на котором показана регрессия роста аллотрансплантированных опухолей MMTV-Wnt1 при введении мышам антитела YW210.09, аналогично действию, наблюдаемому с использованием белка Fzd8CRD-Fc.
Фиг.10В. На графике с помощью количественного ПЦР-анализа показано, что ксенотрансплантированные опухоли Ntera-2 демонстрируют снижение экспрессии мРНК SP5 у мышей, получавших интактное или одноплечевое антитело YW211.31, но не антитело YW210.09.
Фиг.10С. На графике показано, что воздействие антитела YW210.09, но не антитела YW211.31.62, на эксплантаты свода черепа мышей в культуре значительно повышает плотность минерализации костной ткани (BMD) кальцифицированной теменной кости, аналогично воздействию белка RANK-Fc.
Фиг.11А. На графике показано, что биспецифичные анти-LRP6 антитела, полученные в E. coli или клетках HEK293, аналогично ингибируют зависимым от концентрации образом активность Wnt-люциферазного репортера в клетках HEK293, индуцированных 0,1 мг/мл очищенного Wnt3a. Величины IC50 составляют 0,032 и 0,014 мкг/мл, соответственно.
Фиг.11В. На графике показан эффект воздействия указанных контрольного буфера (PBS), антитела, комбинации антител или белка Fzd8CRD-Fc (концентрация каждого составляла 10 мкг/мл) на аутокринную передачу сигнала от Wnt в клетках PA-1 и M14 со стабильно встроенным Wnt-люциферазным репортером и клетках CAL-51, трансфицированных репортером, при наличии (С) или отсутствии (В) стимуляции 0,1 мкг/мл Wnt3a.
Фиг.11С. На графике показан эффект воздействия указанных контрольного буфера (PBS), антитела, комбинации антител или белка Fzd8CRD-Fc (концентрация каждого составляла 10 мкг/мл) на аутокринную передачу сигнала от Wnt в клетках PA-1 и M14 со стабильно встроенным Wnt-люциферазным репортером и клетках CAL-51, трансфицированных репортером, которые были стимулированы 0,1 мкг/мл Wnt3a.
Фиг.12. Итоговая таблица эффектов антител или белка Fzd8CRD (10 мкг/мл) на передачу сигнала, индуцированную трансфекцией конструкций для экспрессии изоформ Wnt в клеточных линиях HEK293 или Hs578T со стабильно встроенным Wnt-люциферазным репортером.
Фиг.13А. Вестерн-анализ клеток HEK293, трансфицированных или не трансфицированных Wnt3a, и получавших воздействие указанного антитела или белка Fzd8CRD-Fc (5 мкг/мл) в течение 18 часов. Уровни белков β-актина и GAPDH показаны в качестве контроля наноса образца для верхнего и нижнего гелей, соответственно.
Фиг.13В. На графике с помощью количественного ПЦР-анализа показано, что ксенотрансплантированные опухоли М14 у мышей SCID-bg, получавших в течение 16 часов 30 мг/кг биспецифичного антитела к LRP6 или белок Fzd8CRD, но не контрольное антитело анти-gD, демонстрируют снижение экспрессии мРНК AXIN2 и APCDD1.
Фиг.14. Подробное изображение взаимодействия CDR H3 с остатками в бороздке LRP6, которое показывает важную систему взаимодействий, создаваемую NAVK-мотивом.
Фиг.15. Подробное изображение взаимодействий, образуемых CDR H1,2, L1,2 и 3.
Фиг.16. Вариабельная область тяжелой цепи (VH) типовых анти-LRP6 антител, в которой показаны CDR по Кабату.
Фиг.17. Вариабельная область легкой цепи (VL) типовых анти-LRP6 антител, в которой показаны CDR по Кабату.
Подробное описание вариантов осуществления изобретения
I. ОПРЕДЕЛЕНИЯ
В контексте настоящего описания «акцепторная каркасная область человека» является каркасной областью, содержащей аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученные из каркасной области иммуноглобулина человека или из консенсусной каркасной области иммуноглобулина человека. Акцепторная каркасная область человека, «полученная из» каркасной области иммуноглобулина человека или консенсусной каркасной области иммуноглобулина человека, может содержать аминокислотную последовательность одинаковую с ними, или она может содержать аминокислотные замены. В некоторых вариантах осуществления изобретения число аминокислотных замен составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления изобретения акцепторная каркасная область VL человека идентична последовательности каркасной области VL иммуноглобулина человека или консенсусной каркасной области человека.
«Аффинность» относится к силе общей суммы нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иное, то используемый в данном описании термин «аффинность связывания» относится к подлинной аффинности связывания, которая отражает взаимодействие 1:1 между членами пары связывания (например, антитела и антигена). Аффинность молекулы Х к его партнеру Y, как правило, можно представить константой диссоциации (Kd). Аффинность можно измерить стандартными способами, известными в данной области, включая способы, описанные в настоящем описании. Конкретные иллюстративные и примеры вариантов осуществления измерения аффинности связывания изобретения описаны ниже.
«Аффинно зрелое» антитело относится к антителу с одним или несколькими изменениями в одной или нескольких гипервариабельных областях (HVR) относительно родительского антитела, которое не несет таких изменений, причем данные изменения приводят к увеличению аффинности антитела к антигену.
Термин «анти-LRP6 антитело» и «антитело, которое связывает LRP6» относятся к антителу, способному связывать LRP6 с достаточной аффинностью, при которой антитело пригодно в качестве диагностического и/или терапевтического агента для обнаружения LRP6. В одном варианте осуществления изобретения связывание анти-LRP6 антитела с неродственным белком (не LRP6) составляет менее примерно 10% от связывания антитела с LRP6, измеренного, например, радиоиммуноанализом (RIA). В некоторых вариантах осуществления изобретения антитело, которое связывает LRP6, имеет константу диссоциации (Kd) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 M или менее, например, от 10-8 M до 10-13 M, например, от 10-9 M до 10-13 M). В некоторых вариантах осуществления изобретения анти-LRP6 антитело связывает эпитоп LRP6, консервативный среди LRP6 из различных биологических видов.
Термин «антитело» в настоящем описании используется в максимально широком смысле и охватывает различные структуры антител, включая, но без ограничений, моноклональные антитела, поликлональные антитела, полиспецифичные антитела (например, биспецифичные антитела) и фрагменты антител, при условии, что они проявляют желаемую антиген-связывающую активность.
«Фрагмент антитела» относится к молекуле антитела, но не интактному антителу, которая содержит участок интактного антитела, который связывает антиген, с которым связывается интактное антитело. Примеры фрагментов антител включают, но без ограничений, Fv, Fab, Fab', Fab'-SH, F(ab')2; диантитела; линейные антитела; молекулы одноцепочечных антител (например, scFv); и полиспецифичные антитела, образуемые фрагментами антител.
«Антитело, которое связывается с тем же эпитопом», что и референсное антитело, относится к антителу, которое блокирует связывание референсного антитела с его антигеном в конкурентном анализе на 50% или более, и наоборот, референсное антитело блокирует связывание антитела с его антигеном в конкурентном анализе на 50% или более. Пример конкурентного анализа приведен в настоящем описании.
Термины «рак (раковое заболевание)» или «раковый» относятся или описывают физиологическое состояние у млекопитающих, которое обычно отличается неконтролируемым ростом/пролиферацией клеток. Примеры рака включают, но без ограничения, карциному, лимфому (например, Ходжскинскую и неходжскинскую лимфому), бластому, саркому и лейкемию. Более конкретные примеры таких раковых заболеваний включают сквамозноклеточную карциному, мелкоклеточный рак легких, немелкоклеточный рак легких, аденокарциному легких, сквамозную карциному легких, рак брюшины, гепатоцеллюлярный рак, гастроинтестинальный рак, рак поджелудочной железы, глиому, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак молочных желез, рак толстой кишки, рак толстой и прямой кишки, эндометриальную или маточную карциному, рак слюнных желез, рак почки, рак печени, рак простаты, рак вульвы, рак щитовидной железы, печеночную карциному, лейкемию и другие лимфопролиферативные заболевания, а также различные типы рака головы и шеи.
«Химиотерапевтический агент» относится к химическому соединению, пригодному для лечения рака. Примеры химиотерапевтических агентов включают химическое соединение, пригодное для лечения рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа и CYTOXAN® (циклофосфамид); алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метиламеламины, включая алтретамин, триэтиленмеламин, триэтиленфосфорамид, триэтилентиофосфорамид и триметиломеламин; ацетогенины (особенно буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновую кислоту; камптотецин (включая синтетический аналог топотекан (HYCAMITIN®), CPT-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; CC-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); криптофицины (особенно криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги, KW-2189 и CB1-TM1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорофосфамид, эстрамустин, ифосфамид, мехлоретамин, мехлоретамина оксида гидрохлорид, мелфалан, новембихин, фенестерин, преднимустин, трофосфамид, урамустин; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимнустин; антибиотики, такие как энедииновые антибиотики (например, калихимицин, особенно калихимицин-гамма1I и калихимицин-омегаI1 (см., например, Nicolaou et al., Angew. Chem Intl. Ed. Engl., 33: 183-186 (1994)); CDP323, пероральный ингибитор интегрина альфа-4; динемицин, включая динемицин A; эсперамицин; а также неокарциностатиновый хромофор и родственные ему хромофоры хромопротеиновых энедииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карминомицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, доксорубицин (включая ADRIAMYCIN®, морфолинодоксорубицин, цианоморфолинодоксорубицин, 2-пирролинодоксорубицин, липосомальную форму для инъекций доксорубицина-HCl (DOXIL®), липосомальный доксорубицин TCL D-99 (MYOCET®), пеглилированый липосомальный доксорубицин (CAELYX®) и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин C, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозоцин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат, гемцитабин (GEMZAR®), тегафур (UFTORAL®), капецитабин (XELODA®), эпотилон и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; пуриновые аналоги, такие как флударабин, 6-меркаптопурин, тиамиприн, тиогуанин; пиримидиновые аналоги, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, доксифлуридин, эноцитабин, флоксуридин; андрогены, такие как калустерон, дромостанолона пропионат, эпитиостанол, мепитиостан, тестолактон; средства, угнетающие деятельность коры надпочечников, такие как аминоглутетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; гликозид альдофосфамида; аминолевулиновую кислоту; энилурацил; амсакрин; бестрабуцил; бисантрен; эдатрексат; дефофамин; демеколцин; диазиквон; элфорнитин; эллиптиния ацетат; эпотилон; этоглуцид; нитрат галлия; гидроксимочевину; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; пентостатин; фенамет; пирарубицин; лозоксантрон; подофиллиновую кислоту; 2-этилгидразид; прокарбазин; полисахаридный комплекс PSK® (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновую кислоту; триазиквон; 2,2',2'-трихлортриэтиламин; трихотецены (особенно токсин T-2, верракурин A, роридин A и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид («ара-С»); тиотепу; таксоиды, например, паклитаксел (TAXOL®) препарат наночастиц паклитаксела, связанного с альбумином (ABRAXANE™) и доксетаксел (TAXOTERE®); хлоранбуцил; 6-тиогуанин; меркаптопурин; метотрексат; содержащие платину агенты, такие как цисплатин, оксалиплатин (ELOXATIN®) и карбоплатин; препараты барвинка, которые препятствуют полимеризации тубулина из образующихся микротрубочек, включая винбластин (VELBAN®), винкристин (ONCOVIN®), виндезин (ELDISINE®, FILDESIN®) и винорелбин (NAVELBINE®); этопозид (VP-16); ифосфамид; митоксантрон; лейковорин; новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота включая бексаротен (TARGRETIN®); бисфосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), этидронат (DIDRICAL®), NE-58095, золедроновая кислота/золедронат (ZOMETA®), алендронат (FOSAMAX®), памидронат (AREDIA®), тилудронат (SKELID®), или резидронат (ACTONEL®); троксацитабин (1,3-диоксолановый нуклеозидный аналог цитозина); антисмысловые олигонуклеотиды, в особенности такие, которые ингибируют экспрессию генов В сигнальных путях, вовлеченных в аберрантную пролиферацию клеток, такие как, например, PKC-альфа, Raf, H-Ras и рецептор фактора роста эпидермальных клеток (EGF-R); вакцины, такие как вакцина THERATOPE® и вакцины для генной терапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; ингибитор топоизомеразы I (например, LURTOTECAN®); rmRH (например, ABARELIX®); BAY439006 (сорафениб; Bayer); SU-11248 (сунитиниб, SUTENT®, Pfizer); перифозин, ингибитор COX-2 (например, целекоксиб или эторикоксиб); протеосомный ингибитор (например, PS341); бортезомиб (VELCADE®); CCI-779; типифарниб (R11577); орафениб, ABT510; ингибитор Bcl-2, такой как облимерсен-натрий (GENASENSE®); пиксантрон; ингибиторы EGFR (см. определение ниже); ингибиторы тирозинкиназ (см. определение ниже); ингбиторы серин-треониновых киназ, такие как рапамицин (сиролимус, RAPAMUNE®); ингибиторы фарнезилтрансферазы, такие как лонафарниб (SCH 6636, SARAZAR™) и фармацевтически приемлемые соли, кислоты или производные любых указанных выше соединений; а также комбинации двух или более указанных выше соединений, такие как CHOP, сокращенное название комбинированной терапии с использованием циклофосфамида, доксорубицина, винкристина и преднизолона, и FOLFOX, сокращенное название лечения, включающего оксиплатин (ELOXATIN™), объединенный с 5-FU и лейковорином.
Химиотерапевтические агенты в контексте настоящего документа включают «противогормональные агенты» или «эндокринные терапевтические средства», которые действуют, регулируя, снижая, блокируя или ингибируя эффекты гормонов, которые могут ускорять рост раковых опухолей. Эти средства могут представлять собой гормоны, включая, но без ограничения: антиэстрогены со смешанным агонистическим/антагонистическим профилем, включающие тамоксифен (NOLVADEX®), 4-гидрокситамоксифен, торемифен (FARESTON®), идоксифен, дролоксифен, ралоксифен (EVISTA®), триоксифен, кеоксифен и селективные модуляторы рецептора эстрогена (SERM), такие как SERM3; чистые антиэстрогены без агонистических свойств, такие как фулвестрант (FASLODEX®) и EM800 (такие агенты могут блокировать димеризацию рецептора эстрогена (ER), ингибировать связывание ДНК, повышать метаболизм ER и/или снижать уровень ER), ингибиторы ароматазы, включающие стероидные ингибиторы ароматазы, такие как форместан и экземестан (AROMASIN®), и нестероидные ингибиторы ароматазы, такие как анастразол (ARIMIDEX®), летрозол (FEMARA®) и аминоглютетимид, и другие ингибиторы ароматазы, включающие ворозол (RIVISOR®), ацетат мегестрола (MEGASE®), фадрозол и 4(5)-имидазолы; агонисты гормона, высвобождающего лютеинизирующий гормон, включая леупролид (LUPRON® и ELIGARD®), гозерелин, бузерелин и триптерелин; половые гормоны, включающие прогестины, такие как ацетат мегестрола и ацетат медроксипрогестерона, эстрогены, такие как диэтилстилбестрол и премарин, и андрогены/ретиноиды, такие как флуоксиместерон, все трансретиноевые кислоты и фенретинид; онапристон; антипрогестероны; отрицательные регуляторы рецептора эстрогена (ERD), антиандрогены, такие как флутамид, нилутамид и бикалутамид; тестолактон; и фармацевтически приемлемые соли, кислоты или производные любого из указанных выше соединений, а также комбинации двух или более указанных соединений.
Термин «химерное» антитело относится к такому антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или биологического вида, тогда как остаток тяжелой и/или легкой цепи получен из другого источника или биологического вида.
«Класс» антитела относится к типу константного домена или константной области в его тяжелой цепи. Существует пять основных классов природных антител: IgA, IgD, IgE, IgG и IgM, и несколько из них можно дополнительно разделить на подклассы (изотипы), например, IgG1, IgG2, IgG3 и IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам антител, называются соответственно α, δ, ε, γ и μ.
Используемый в настоящем описании термин «цитотоксический агент» относится к веществу, ингибирующему или предупреждающему функционирование клеток и/или вызывающему смерть или разрушение клеток. Цитотоксические агенты включают, но без ограничения, радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, Р32, Pb212 и радиоактивные изотопы Lu), химиотерапевтические агенты или лекарственные соединения (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин C, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); агенты, ингибирующие рост клеток; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, описанные ниже.
«Эффекторные функции» относятся к таким видам биологической активности, свойственным Fc-области антитела, которые варьируют в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплемент-зависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточно-опосредованную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора); и активацию B-клеток.
«Эффективное количество» агента, например, фармацевтического состава, относится к количеству, эффективному в дозировках и в течение необходимых периодов времени, для достижения желаемого терапевтического или профилактического результата.
Термин «Fc-область» в настоящем описании используют для определения C-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере участок константной области. Термин включает Fc-область с нативной последовательностью или вариант Fc-области. В одном варианте осуществления изобретения Fc-область тяжелой цепи IgG человека расположена от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc-области может присутствовать или нет. Если специально не указано обратное, то нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой EU-индексом, которая описана в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, но не остаткам гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно расположены в следующем порядке в VH (или в VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины «полноразмерное антитело», «интактное антитело» и «целое антитело» используются в настоящем описании взаимозаменяемо и относятся к антителу, имеющему структуру по существу аналогичную структуре нативного антитела или имеющему тяжелые цепи, которые содержат Fc-область, указанную в настоящем описании.
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была внесена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформанты» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство независимо от числа пассажей. Потомство может быть неполностью идентично родительской клетке по содержанию нуклеиновых кислот и может содержать мутации. В настоящее изобретение включено мутантное потомство, которое имеет такую же функцию или биологическую активность, по которой скринируют или отбирают исходно трансформированные клетки.
«Антитело человека» представляет собой антитело, которое несет аминокислотную последовательность, соответствующую последовательности антитела, вырабатываемого человеком или клеткой человека, либо полученную из источника кроме человека, который использует репертуар антител человека или другие последовательности, кодирующие антитела человека. Это определение антитела человека специально исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, не принадлежащие человеку.
«Консенсусная каркасная область человека» является каркасной областью, которая представляет собой наиболее часто встречающиеся аминокислотные остатки при селекции последовательностей каркасной области VL или VH иммуноглобулинов человека. Как правило, селекцию последовательностей VL или VH иммуноглобулинов человека осуществляют из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей является подгруппой, как у Kabat et al., Sequences of Proteins of Immunological Interest, 5 th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). В одном варианте осуществления изобретения для VL подгруппой является подгруппа каппа I, как у Kabat et al., supra. В одном варианте осуществления изобретения для VH подгруппой является подгруппа III, как у Kabat et al., supra.
«Гуманизированное антитело» относится к химерному антителу, содержащему аминокислотные остатки из HVR, не принадлежащие человеку, и аминокислотные остатки из FR человека. В некоторых вариантах осуществления изобретения гуманизированное антитело будет содержать по существу все по меньшей мере из одного и, обычно, двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют этим областям антитела, не принадлежащего человеку, и все или по существу все FR соответствуют этим областям антитела человека. Гуманизированное антитело необязательно может содержать по меньшей мере участок константной области антитела, полученной из антитела человека. «Гуманизированная форма» антитела, например, антитела, не принадлежащего человеку, относится к антителу, которое прошло гуманизацию.
Термин «гипервариабельная область» или «HVR» в контексте данного описания относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности и/или образуют структурно оформленные петли («гипервариабельные петли»). Как правило, нативные антитела из четырех цепей содержат шесть HVR; три в VH (H1, H2, H3) и три в VL (L1, L2, L3). HVR обычно содержат аминокислотные остатки из гипервариабельных петель и/или из «определяющих комплементарность областей» (CDR), причем последние имеют наиболее высокую вариабельность по последовательности и/или вовлечены в распознавание антигена. Иллюстративные гипервариабельные петли расположены между остатками 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2) и 96-101 (H3). (Chothia and Lesk, J. Mol. Biol., 196: 901-917 (1987)). Иллюстративные CDR (CDR-L1, CDR-L2, CDR-L3, CDR-H1, CDR-H2 и CDR-H3) расположены между аминокислотными остатками 24-34 для L1, 50-56 для L2, 89-97 для L3, 31-35 для H1, 50-65 для H2 и 95-102 для H3. (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). За исключением CDR1 в VH, CDR, как правило, содержат аминокислотные остатки, которые образуют гипервариабельные петли. CDR также содержат «определяющие специфичность остатки» или «SDR», которые представляют собой остатки, контактирующие с антигеном. SDR содержатся внутри областей CDR и называются сокращенными CDR или a-CDR. Иллюстративные a-CDR (a-CDR-L1, a-CDR-L2, a-CDR-L3, a-CDR-H1, a-CDR-H2 и a-CDR-H3) расположены между аминокислотными остатками 31-34 для L1, 50-55 для L2, 89-96 для L3, 31-35B для H1, 50-58 для H2 и 95-102 для H3. (См. Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008)). Если не указано иное, то остатки HVR и другие остатки в вариабельном домене (например, остатки FR) пронумерованы по Kabat et al., supra.
«Иммуноконъюгат» является антителом, конъюгированным с одной или несколькими гетерологичными молекулами, включая, но без ограничения, цитоплазматический агент.
«Индивидуум» или «субъект» является млекопитающим. Млекопитающие включают, но без ограничения, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и приматов, кроме человека, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых вариантах осуществления изобретения индивидуумом или субъектом является человек.
«Выделенное» антитело является антителом, которое отделено от компонента своего природного окружения. В некоторых вариантах осуществления изобретения антитело очищено более чем до 95%-й или 99%-й чистоты, определенной, например, электрофоретически (например, электрофорезом в ДСН-ПААГ, изоэлектрическим фокусированием (IEF), капиллярным электрофорезом) или хроматографически (например, ионообменной ВЭЖХ или с обращенной фазовой). Обзор способов оценки чистоты антител, см., например, в Flatman et al., J. Chromatogr. B 848:79-87 (2007).
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая отделена от компонента своего природного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулы нуклеиновых кислот, но молекула нуклеиновой кислоты присутствует в качестве внехромосомного элемента, или ее местоположение на хромосоме отличается от природного местоположения на хромосоме.
«Выделенная нуклеиновая кислота, кодирующая анти-LRP6 антитело» относится к одной или нескольким молекулам нуклеиновых кислот, кодирующим тяжелую и легкую цепи антитела (или их фрагменты), включая такие молекулы нуклеиновых кислот в одном векторе или в отдельных векторах, и такие молекулы нуклеиновых кислот присутствуют в одном или нескольких местах в клетке-хозяине.
Термин «моноклональное антитело», используемый в настоящем описании, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантов, например, содержащих природные мутации или возникающих в ходе получения препарата моноклонального антитела, причем такие варианты обычно присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело из препарата моноклонального антитела направлено против единственной детерминанты на антигене. Таким образом, определение «моноклональный» указывает на характер антитела, как полученного по существу из гомогенной популяции антител, и его не следует истолковывать как требование получения антитела каким-либо конкретным способом. Например, моноклональные антитела для применения по настоящему изобретению, можно получить с помощью ряда методик, включая, но без ограничения, гибридомный способ, способы рекомбинантных ДНК, способы фагового дисплея и способы с использованием трансгенных животных, содержащих все или часть иммуноглобулиновых локусов человека, причем такие способы и другие стандартные способы получения моноклональных антител описаны в настоящем описании.
«Голое антитело» относится к антителу, которое не конъюгировано с гетерологичной молекулой (например, цитотоксической молекулой) или радиоактивной меткой. Голое антитело может присутствовать в фармацевтическом составе.
«Нативные антитела» относятся к природным иммуноглобулиновым молекулам с различной структурой. Например, природные антитела IgG являются гетеродимерными гликопротеинами с массой около 150000 дальтон, состоящими из двух идентичных легких цепей и двух идентичных тяжелых цепей, соединенных дисульфидными связями. В направлении от N- до C-конца, каждая тяжелая цепь содержит вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которым следуют три константных домена (СН1, СН2 и СН3). Аналогичным образом, в направлении от N- до C-конца, каждая легкая цепь содержит вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которым следуют константный легкий домен (CL). «Легкую цепь» антитела можно отнести к одному из двух типов, называемых каппа (κ) и лямбда (λ), исходя из аминокислотных последовательностей ее константного домена.
Термин "вкладыш в упаковку" используют применительно к инструкциям, которые обычно вкладывают в коммерческие упаковки терапевтических продуктов, и которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предостережениях, касающихся использования таких терапевтических продуктов.
«Процент (%) идентичности аминокислотных последовательностей» относительно референсной полипептидной последовательности определяется как процентное содержание аминокислотных остатков в исследуемой последовательности, которые идентичны аминокислотным остаткам в референсной последовательности после выравнивания последовательностей и, при необходимости, введения разрывов для достижения максимального процента идентичности последовательностей, при этом не рассматривая какие-либо консервативные замены как часть идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотных последовательностей можно достичь различными путями, которые входят в компетенцию специалистов в данной области, например, с использованием общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Опытные специалисты в данной области смогут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Для целей настоящего изобретения, величину процента идентичности аминокислотных последовательностей можно вычислить, используя компьютерную программу для сравнения последовательностей, ALIGN-2. Компьютерная программа для сравнения последовательностей, ALIGN-2, принадлежит Genentech, Inc., и исходный код зарегистрирован с пользовательской документацией в Ведомстве по охране авторского права США, Washington D.C., 20559, под регистрационным номером авторского права США, TXU510087. Программа ALIGN-2 находится в общем доступе через компанию Genentech, Inc., South San Francisco, California, или ее можно скомпилировать из исходного кода. Программу ALIGN-2 следует компилировать для использования под операционной системой UNIX, включая Digital UNIX V4.0D. Все параметры сравнения последовательностей установлены программой ALIGN-2 и не изменяются.
В тех ситуациях, где используют ALIGN-2 для сравнений аминокислотных последовательностей, процент идентичности аминокислотных последовательностей для данной аминокислотной последовательности A по отношению к, по сравнению с или против данной аминокислотной последовательности B (что можно альтернативно перефразировать, как данная аминокислотная последовательность A, которая имеет или включает некоторый процент идентичности аминокислотных последовательностей по отношению к, по сравнению с или против данной аминокислотной последовательности B) вычисляют, как указано ниже:
100 умножить на долю X/Y,
где X представляет собой количество аминокислотных остатков, отмечаемых как идентичные совпадения программой для выравнивания последовательностей ALIGN-2 при выравнивании A и B этой программой, и где Y представляет собой общее количество аминокислотных остатков в B. Следует учитывать, что в случае, когда длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, процент идентичности аминокислотных последовательностей A по сравнению с B не будет равен проценту идентичности аминокислотных последовательностей B по сравнению с A. Если специально не указано иное, то все используемые в настоящем описании величины процентов идентичности аминокислотных последовательностей получены, как описано в непосредственно предшествующем параграфе, с использованием компьютерной программы ALIGN-2.
Термин «фармацевтический состав» относится к препарату в форме, которая позволяет проявление биологической активности действующего ингредиента, содержащегося в нем, и который не содержит дополнительных компонентов, неприемлемо токсичных для субъекта, которому водят состав.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтическом составе, но не действующему ингредиенту, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но без ограничения, буфер, эксципиент, стабилизатор или консервант.
Термин «LRP6» в контексте настоящего документа относится к любому нативному LRP6 любых позвоночных, включая млекопитающих, таких как приматы (например, человек) и грызуны (например, мыши и крысы), если не указано иное. Термин охватывает «полноразмерный» непроцессированный LRP6, а также любую форму LRP6, которая образуется в результате процессинга в клетке. Термин также охватывает природные варианты LRP6, например, сплайсинговые варианты или аллельные варианты. Аминокислотная последовательность иллюстративного LRP6 человека показана в SEQ ID NO:29. См. также номер доступа в NCBI - AAI43726, Strausberg, R.L., et al., Proc. Natl. Acad. Sci. U.S.A. 99: 16899-16903 (2002); He, X, et al., Development, 131: 1663-1677 (2004); Chen, M., et al., J. Biol. Chem., 284:35040-35048 (2009).
В контексте настоящего описания «воздействие» (и грамматические варианты «воздействовать» или «воздействующий») относится к клиническому вмешательству в попытке изменить естественное течение заболевания у индивидуума, получавшего воздействие, и его можно выполнять либо в профилактических целях, либо в при проявлениях клинической патологии. Желаемые эффекты воздействия включают, но без ограничения, предупреждение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любых прямых или непрямых патологических последствий заболевания, предупреждения метастазирования, снижение скорости развития заболевания, ослабление или облегчение болезненного состояния и ремиссию или улучшенный прогноз. В некоторых вариантах осуществления изобретения антитела настоящего изобретения используют для задержки развития заболевания или замедления прогресса заболевания.
Термин «вариабельная область» или «вариабельный домен» относится к домену легкой или тяжелой цепи антитела, который вовлечен в связывание антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (соответственно, VH и VL) нативного антитела обычно имеют аналогичные структуры, причем каждый домен содержит четыре консервативных каркасных области (FR) и три гипервариабельных области (HVR). (См., например, Kindt et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007)). Одного VH- или VL-домена может быть достаточно, чтобы придать антиген-связывающую специфичность. Кроме того, антитела, которые связывают конкретный антиген, можно выделить с использованием VH- или VL-домена антитела, который связывает антиген, для скрининга библиотеки комплементарных VL- или VH-доменов, соответственно. См., например, Portolano et al., J. Immunol. 150:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991).
Термин «вектор» в контексте настоящего описания относится к молекуле нуклеиновой кислоты, способной к размножению другой нуклеиновой кислоты, с которой она соединена. Термин включает вектор в качестве самореплицирующейся нуклеотидной структуры, а также вектор, включенный в геном клетки-хозяина, в которую он был внесен. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально соединены. Такие векторы называются в настоящем описании «экспрессионными векторами».
II. КОМПОЗИЦИИ И СПОСОБЫ
Настоящее изобретение относится к анти-LRP6 антителам с неожиданной способностью ингибировать передачу сигнала от некоторых изоформ Wnt и усиливать передачу сигнала от других изоформ. Как описано в разделе «Примеры», два описанных ниже анти-LRP6 антитела оказывают противоположное действие на большинство Wnt, причем одно антитело является антагонистическим, а другое усиливающим. Эти два антитела связываются с различными областями LRP6 (также как и различные изоформы Wnt), и ингибирование передачи сигнала является результатом блокирования связывания с Wnt.
Исходя из их функционального взаимодействия с анти-LRP6 антителами, 14 протестированных изоформ Wnt можно разделить на три класса: действие Wnt3 и Wnt3a ингибируется анти-LRP6 антителом YW211.31 и усиливается анти-LRP6 антителом YW210.09; действие Wnt 1, 2, 2B, 6, 8A, 9A, 9B и 10B усиливается анти-LRP6 антителом YW211.31 и ингибируется анти-LRP6 антителом YW210.09; и действие Wnt 4, 7A, 7B и 10A усиливается анти-LRP6 антителом YW211.31 и не ингибируется анти-LRP6 антителом YW210.09 (фиг.3C). Данное разделение по классам явно не соответствует предложенному филогенезу генов Wnt, хотя подсемейство Wnt3/3a является наиболее эволюционно разрозненным (Cho et al., 2010). Комбинации анти-LRP6 антител, которые ингибируют различные классы изоформ Wnt, можно использовать для создания эффективных терапевтических средств для лечения заболеваний, ассоциированных с передачей сигнала от Wnt.
Опосредованная антителами димеризация LRP6 может усиливать передачу сигнала, только когда изоформа Wnt также способна связывать комплекс, предположительно, рекрутируя FZD. Антитела к LRP6 могут либо противодействовать, либо усиливать эндогенную аутокринную передачу сигнала от Wnt в различных опухолевых клеточных линиях. Данная комплексность взаимодействий корецептора с лигандом может позволить дифференциальную регуляцию передачи сигнала от изоформ Wnt, и с использованием антител ее можно использовать для дифференциального воздействия на передачу сигнала от Wnt в определенных тканях или при определенных болезненных состояниях.
В некоторых вариантах осуществления изобретения анти-LRP6 антитела могут ингибировать аутокринную или эндогенную передачу сигнала от Wnt в некоторых типах клеток и усиливать аутокринную передачу сигнала в других типах клеток. В некоторых вариантах осуществления изобретения анти-LRP6 антитела опосредуют димеризацию LRP6 и увеличивают или усиливают передачу сигнала в присутствии изоформы Wnt, которая одновременно связывается с LRP6. В некоторых вариантах осуществления изобретения анти-LRP6 антитела усиливают передачу сигнала от Wnt, ингибируя связывание антагонистов Wnt, таких как изоформы DKK и SOST.
Анти-LRP6 антитела можно использовать для избирательного модулирования процессов, активируемых или ингибируемых передачей сигнала, индуцированной изоформой Wnt. Такие процессы включают, например, клеточную пролиферацию, направление клеточной дифференцировки и самообновление стволовых клеток при различных типах раковых заболеваний, и процессы развития. Анти-LRP6 антитела пригодны, например, для лечения Wnt-опосредованных заболеваний, таких как рак и нарушения костной ткани или скелета, и сосудистые расстройства. Примеры раковых заболеваний, которые можно лечить с использованием анти-LRP6 антител, включают мелкоклеточный рак легких, немелкоклеточный рак легких, гепатоцеллюлярный рак, гастроинтестинальный рак, рак поджелудочной железы, рак яичников, рак печени, рак мочевого пузыря, гепатому, рак молочных желез, рак толстой кишки, колоректальный рак, эндометриальную или маточную карциному, рак слюнных желез, рак почки (включая почечноклеточную карциному, рак печени, рак простаты. Примеры нарушений скелета или костной ткани, которые можно лечить с использованием анти-LRP6 антител, включают остеопороз, остеоартрит, переломы костей и поражения костной ткани. Примеры сосудистых расстройств, которые можно лечить с использованием анти-LRP6 антител, включают сосудистые заболевания сетчатки, такие как болезнь Норри, синдром остеопороза-псевдоглиомы (OPPG), семейную экссудативную витреоретинопатию (FEVR), ретинопатию недоношенных (ROP), диабетическую ретинопатию, возрастную макулярную дегенерацию, ретинопатию недоношенных, болезнь Коатса и реакцию, подобную болезни Коатса, и окклюзию вены или артерии сетчатки, а также связанные с миокардом состояния, такие как инфаркт миокарда и ишемическая болезнь сердца.
Соответственно, один аспект изобретения относится к антителу, которое связывает LRP6, где антитело ингибирует передачу сигнала, индуцированную изоформой Wnt, и усиливает передачу сигнала, индуцированную другой изоформой Wnt. В одном варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt3 и/или Wnt3a. В одном варианте осуществления изобретения антитело усиливает передачу сигнала от Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и/или 10b. В одном варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt3 и/или Wnt3a и усиливает передачу сигнала от Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и/или 10b. В одном варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt3 и Wnt3a и усиливает передачу сигнала от Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и 10b. В одном варианте осуществления изобретения анти-LRP6 антитело связывается с областью Е3-Е4 (первый и второй бета-пропеллеры) LRP6.
В другом варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt 1, 2, 2b, 6, 8a, 9a, 9b, и/или 10b. В одном варианте осуществления изобретения антитело усиливает передачу сигнала от Wnt3 и/или Wnt3a. В одном варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt 1, 2, 2b, 6, 8a, 9a, 9b и/или 10b и усиливает передачу сигнала от Wnt3 и/или Wnt3a. В одном варианте осуществления изобретения антитело ингибирует передачу сигнала от Wnt 1, 2, 2b, 6, 8a, 9a, 9b и/или 10b и усиливает передачу сигнала от Wnt3 и/или Wnt3a. В одном варианте осуществления изобретения анти-LRP6 антитело связывается с областью Е1-Е2 (третий и четвертый бета-пропеллеры) LRP6.
Другой аспект изобретения относится к полиспецифичным анти-LRP6 антителам. Как показано в примерах, полиспецифичные антитела в некоторых вариантах осуществления изобретения обладают преимуществом ингибирования всех трех классов изоформ Wnt. В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным антителом, способным связывать две или несколько областей или эпитопов LRP6. В одном варианте осуществления изобретения полиспецифичным антителом является биспецифичное антитело, способное специфично связывать две различные области LRP6. В одном варианте осуществления изобретения биспецифичное антитело связывает область Е1-Е2 LRP6. В одном варианте осуществления изобретения полиспецифичное антитело ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b. В одном варианте осуществления изобретения полиспецифичное антитело ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b, и дополнительно ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 4, 7a, 7b и 10a. В одном варианте осуществления изобретения полиспецифичное антитело ингибирует передачу сигнала, индуцированную комбинацией Wnt 1 и Wnt3a. В одном варианте осуществления изобретения полиспецифичное антитело ингибирует аутокринную передачу сигнала от Wnt.
В некоторых вариантах осуществления изобретения полиспецифичным антителом является биспецифичное антитело, которое ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b. В некоторых вариантах осуществления изобретения полиспецифичным антителом является биспецифичное антитело, которое ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b, и дополнительно ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 4, 7a, 7b и 10a. В некоторых вариантах осуществления изобретения полиспецифичным антителом является биспецифичное антитело, которое ингибирует передачу сигнала, индуцированную комбинацией Wnt 1 и Wnt3a. В одном варианте осуществления изобретения полиспецифичным антителом является биспецифичное антитело, которое ингибирует передачу сигнала, индуцированную комбинацией Wnt 1 и Wnt3a, более эффективно, чем комбинация моноспецифичных антител, имеющих такие же специфичности как биспецифичное антитело.
В некоторых вариантах осуществления изобретения полиспецифичным антителом является биспецифичное антитело, которое ингибирует аутокринную передачу сигнала от Wnt, более эффективно, чем комбинация моноспецифичных антител, имеющих такие же специфичности как биспецифичное антитело.
В некоторых вариантах осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело ингибирует передачу сигнала от Wnt по меньшей мере примерно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более. Ингибирование передачи сигнала от Wnt можно определить с использованием методов анализа, известных в данной области и описанных в настоящем описании. Например, ингибирование передачи сигнала от Wnt можно определить, используя Wnt-репортерный анализ, такой как Wnt-люциферазный репортерный анализ, описанный в примерах.
Ингибирование передачи сигнала от Wnt также можно определить мониторингом экспрессии генов-мишеней Wnt, таких как APCDD1, AXIN2, GAD1, LEFTY2 и SAX1, как описано в разделе «Примеры».
В некоторых вариантах осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело ингибирует экспрессию генов-мишеней Wnt, таких как APCDD1, AXIN2, GAD1, LEFTY2 и SAX1, по меньшей мере примерно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%. В одном варианте осуществления изобретения экспрессию генов-мишеней Wnt определяют, используя анализ экспрессии генов такой как ПЦР, в том числе кПЦР.
Другой аспект изобретения относится к антителам, которые связывают LRP6 и конкурируют за связывание с любыми анти-LRP6 антителами, описанными в настоящем описании. Другой аспект изобретения относится к антителам, которые связывают такой же эпитоп на LRP6, как любое из анти-LRP6 антител, описанных в настоящем описании.
А. Примеры анти-LRP6 антител
Один аспект изобретения относится к анти-LRP6 антителу, которое является моноклональным антителом, включая химерное антитело, гуманизированное антитело или антитело человека. В одном варианте осуществления изобретения анти-LRP6 создают, используя фаговые библиотеки. В одном варианте осуществления изобретения анти-LRP6 антителом является фрагмент антитела, например, фрагмент Fv, Fab, Fab', scFv, диантитела или F(ab')2. В другом варианте осуществления изобретения антителом является полноразмерное антитело, например, интактное антитело IgG1, или антитело другого класса или изотипа, описанное в настоящем описании.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность из таблицы 2. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность легкой цепи, содержащую аминокислотную последовательность из таблицы 2. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи и последовательность легкой цепи, содержащие аминокислотную последовательность из таблицы 2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:1. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:2. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:1, и последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:3. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:4. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:3, и последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:5. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:6. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:5, и последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:6.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7, и последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH) из аминокислотных последовательностей таблицы 3. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL) из аминокислотных последовательностей таблицы 3. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH и VL из аминокислотных последовательностей таблицы 3.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH) из тяжелой цепи аминокислотной последовательности SEQ ID NO:1. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL) из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:2. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH из тяжелой цепи аминокислотной последовательности SEQ ID NO:1 и VL из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO:9. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность SEQ ID NO:10. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность SEQ ID NO:9, и VL, содержащую аминокислотную последовательность SEQ ID NO:10.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH) из тяжелой цепи аминокислотной последовательности SEQ ID NO:3. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL) из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:4. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH из тяжелой цепи аминокислотной последовательности SEQ ID NO:3 и VL из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO:11. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность SEQ ID NO:12. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность SEQ ID NO:11, и VL, содержащую аминокислотную последовательность SEQ ID NO:12.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH) из тяжелой цепи аминокислотной последовательности SEQ ID NO:5. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL) из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:6. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH из тяжелой цепи аминокислотной последовательности SEQ ID NO:5 и VL из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:6.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO:13. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность SEQ ID NO:14. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность SEQ ID NO:13, и VL, содержащую аминокислотную последовательность SEQ ID NO:14.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH) из тяжелой цепи аминокислотной последовательности SEQ ID NO:7. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL) из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:8. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH из тяжелой цепи аминокислотной последовательности SEQ ID NO:7 и VL из последовательности легкой цепи аминокислотной последовательности SEQ ID NO:8.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен тяжелой цепи (VH), содержащий аминокислотную последовательность SEQ ID NO:15. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя вариабельный домен легкой цепи (VL), содержащий аминокислотную последовательность SEQ ID NO:16. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность SEQ ID NO:15, и VL, содержащую аминокислотную последовательность SEQ ID NO:16.
Другой аспект изобретения относится к полиспецифичному анти-LRP6 антителу. В одном варианте осуществления изобретения полиспецифичное антитело включает в себя тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения полиспецифичное антитело включает в себя тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере двух из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения полиспецифичное антитело является биспецифичным антителом, которое включает в себя тяжелую цепь, содержащую аминокислотную последовательность по меньшей мере одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:1, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:3, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:5, и вторую тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:7.
В одном варианте осуществления изобретения полиспецифичное антитело включает в себя VH из тяжелой цепи аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения полиспецифичное антитело включает в себя VH из тяжелой цепи аминокислотной последовательности по меньшей мере двух из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения полиспецифичное антитело является биспецифичным антителом, которое включает в себя VH из тяжелой цепи аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:1 и включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:3 и включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:7. В одном варианте осуществления изобретения биспецифичное антитело включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:5 и включает в себя VH тяжелой цепи аминокислотной последовательности SEQ ID NO:7.
В одном варианте осуществления изобретения полиспецифичное анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность по меньшей мере одной из SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15. В одном варианте осуществления изобретения полиспецифичное анти-LRP6 антитело включает в себя VH, содержащую аминокислотную последовательность по меньшей мере двух из SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15. В одном варианте осуществления изобретения полиспецифичное антитело является биспецифичным антителом, которое включает в себя VH, содержащую аминокислотную последовательность SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:9, и вторую VH, содержащую аминокислотную последовательность SEQ ID NO:15. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:11, и вторую VH, содержащую аминокислотную последовательность SEQ ID NO:15. В одном варианте осуществления изобретения биспецифичное антитело включает в себя первую VH, содержащую аминокислотную последовательность SEQ ID NO:13, и вторую VH, содержащую аминокислотную последовательность SEQ ID NO:15.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1 из аминокислотных последовательностей HVR-H1 в таблице 4; (b) HVR-H2 из аминокислотных последовательностей HVR-H2 в таблице 4; (c) HVR-H3 из аминокислотных последовательностей HVR-H3 в таблице 4; (d) HVR-L1 из аминокислотных последовательностей HVR-L1 в таблице 4; (e) HVR-L2 из аминокислотных последовательностей HVR-L2 в таблице 4; и (f) HVR-L3 из аминокислотных последовательностей HVR-L3 в таблице 4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; (c) HVR-H3 тяжелой цепи SEQ ID NO:1; (d) HVR-L1 легкой цепи SEQ ID NO:2; (e) HVR-L2 легкой цепи SEQ ID NO:2; и (f) HVR-L3 легкой цепи SEQ ID NO:2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; (c) HVR-H3 тяжелой цепи SEQ ID NO:1. В одном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:1. В другом варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:1 и HVR-L3 легкой цепи SEQ ID NO:2. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:1, HVR-L3 легкой цепи SEQ ID NO:2 и HVR-H2 тяжелой цепи SEQ ID NO:1. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; и (c) HVR-H3 тяжелой цепи SEQ ID NO:1.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19. В одном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19. В другом варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; и (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1 легкой цепи SEQ ID NO:2; (b) HVR-L2 легкой цепи SEQ ID NO:2; и (c) HVR-L3 легкой цепи SEQ ID NO:2. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:2; (b) HVR-L2 легкой цепи SEQ ID NO:2; и (c) HVR-L3 легкой цепи SEQ ID NO:2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1 тяжелой цепи SEQ ID NO:1, (ii) HVR-H2 тяжелой цепи SEQ ID NO:1 и (iii) HVR-H3 тяжелой цепи SEQ ID NO:1; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1 легкой цепи SEQ ID NO:2, (ii) HVR-L2 легкой цепи SEQ ID NO:2, и (c) HVR-L3 легкой цепи SEQ ID NO:2.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26, и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:27.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; (c) HVR-H3 тяжелой цепи SEQ ID NO:1; (d) HVR-L1 легкой цепи SEQ ID NO:2; (e) HVR-L2 легкой цепи SEQ ID NO:2; и (f) HVR-L3 легкой цепи SEQ ID NO:2.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:25; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:26; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя VH, содержащую по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; (c) HVR-H3 тяжелой цепи SEQ ID NO:3; (d) HVR-L1 легкой цепи SEQ ID NO:4; (e) HVR-L2 легкой цепи SEQ ID NO:4; и (f) HVR-L3 легкой цепи SEQ ID NO:4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; (c) HVR-H3 тяжелой цепи SEQ ID NO:3. В одном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:3. В другом варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:3 и HVR-L3 легкой цепи SEQ ID NO:4. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:3, HVR-L3 легкой цепи SEQ ID NO:4 и HVR-H2 тяжелой цепи SEQ ID NO:3. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; и (c) HVR-H3 тяжелой цепи SEQ ID NO:3.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21. В одном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:21. В другом варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:21 и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:28. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:21, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:28, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; и (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:21.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1 легкой цепи SEQ ID NO:4; (b) HVR-L2 легкой цепи SEQ ID NO:4; и (c) HVR-L3 легкой цепи SEQ ID NO:4. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:4; (b) HVR-L2 легкой цепи SEQ ID NO:4; и (c) HVR-L3 легкой цепи SEQ ID NO:4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25; (b) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26; и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:28. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:25; (b) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:26; и (c) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:28.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (a) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1 тяжелой цепи SEQ ID NO:3, (ii) HVR-H2 тяжелой цепи SEQ ID NO:3 и (iii) HVR-H3 тяжелой цепи SEQ ID NO:3; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1 легкой цепи SEQ ID NO:4, (ii) HVR-L2 легкой цепи SEQ ID NO:4 и (c) HVR-L3 легкой цепи SEQ ID NO:4.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26, и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:28.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; (c) HVR-H3 тяжелой цепи SEQ ID NO:3; (d) HVR-L1 легкой цепи SEQ ID NO:4; (e) HVR-L2 легкой цепи SEQ ID NO:4; и (f) HVR-L3 легкой цепи SEQ ID NO:4.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:21; (d) HVR-L1 содержащую аминокислотную последовательность SEQ ID NO:25; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:26; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:28.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; (c) HVR-H3 тяжелой цепи SEQ ID NO:5; (d) HVR-L1 легкой цепи SEQ ID NO:6; (e) HVR-L2 легкой цепи SEQ ID NO:6; и (f) HVR-L3 легкой цепи SEQ ID NO:6.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; (c) HVR-H3 тяжелой цепи SEQ ID NO:5. В одном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:5. В другом варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:5 и HVR-L3 легкой цепи SEQ ID NO:6. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:5, HVR-L3 легкой цепи SEQ ID NO:6, и HVR-H2 тяжелой цепи SEQ ID NO:5. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; и (c) HVR-H3 тяжелой цепи SEQ ID NO:5.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19. В одном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19. В другом варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19, и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:20; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; и (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1 легкой цепи SEQ ID NO:6; (b) HVR-L2 легкой цепи SEQ ID NO:6; и (c) HVR-L3 легкой цепи SEQ ID NO:6. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:6; (b) HVR-L2 легкой цепи SEQ ID NO:6; и (c) HVR-L3 легкой цепи SEQ ID NO:6.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (a) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1 тяжелой цепи SEQ ID NO:5, (ii) HVR-H2 тяжелой цепи SEQ ID NO:5 и (iii) HVR-H3 тяжелой цепи SEQ ID NO:5; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1 легкой цепи SEQ ID NO:6, (ii) HVR-L2 легкой цепи SEQ ID NO:6 и (c) HVR-L3 легкой цепи SEQ ID NO:6.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (a) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26, и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:27.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; (c) HVR-H3 тяжелой цепи SEQ ID NO:5; (d) HVR-L1 легкой цепи SEQ ID NO:6; (e) HVR-L2 легкой цепи SEQ ID NO:6; и (f) HVR-L3 легкой цепи SEQ ID NO:6.
В другом аспекте изобретение относится к антителу, включающему в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:20; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:19; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:25; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:26; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, две, три, четыре, пять или шесть HVR, выбранных из (a) HVR-H1 тяжелой цепи SEQ ID NO:7; (b) HVR-H2 тяжелой цепи SEQ ID NO:7; (c) HVR-H3 тяжелой цепи SEQ ID NO:7; (d) HVR-L1 легкой цепи SEQ ID NO:8; (e) HVR-L2 легкой цепи SEQ ID NO:8; и (f) HVR-L3 легкой цепи SEQ ID NO:8.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:7; (b) HVR-H2 тяжелой цепи SEQ ID NO:7; (c) HVR-H3 тяжелой цепи SEQ ID NO:7. В одном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:7. В другом варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:7 и HVR-L3 легкой цепи SEQ ID NO:8. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3 тяжелой цепи SEQ ID NO:7, HVR-L3 легкой цепи SEQ ID NO:8 и HVR-H2 тяжелой цепи SEQ ID NO:7. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:7; (b) HVR-H2 тяжелой цепи SEQ ID NO:7; и (c) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24. В одном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:24. В другом варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:24 и HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27. В дополнительном варианте осуществления изобретения антитело включает в себя HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:24, HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27, и HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:23. В дополнительном варианте осуществления изобретения антитело включает в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:22; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:23; и (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1 легкой цепи SEQ ID NO:8; (b) HVR-L2 легкой цепи SEQ ID NO:8; и (c) HVR-L3 легкой цепи SEQ ID NO:8. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:8; (b) HVR-L2 легкой цепи SEQ ID NO:8; и (c) HVR-L3 легкой цепи SEQ ID NO:8.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (a) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1 тяжелой цепи SEQ ID NO:7, (ii) HVR-H2 тяжелой цепи SEQ ID NO:7 и (iii) HVR-H3 тяжелой цепи SEQ ID NO:7; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1 легкой цепи SEQ ID NO:8, (ii) HVR-L2 легкой цепи SEQ ID NO:8 и (c) HVR-L3 легкой цепи SEQ ID NO:8.
В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя (a) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (i) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22, (ii) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23, и (iii) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24; и (b) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (i) HVR-L1, содержащей аминокислотную последовательность SEQ ID NO:25, (ii) HVR-L2, содержащей аминокислотную последовательность SEQ ID NO:26, и (c) HVR-L3, содержащей аминокислотную последовательность SEQ ID NO:27.
В другом варианте осуществления изобретение относится к антителу, включающему в себя (a) HVR-H1 тяжелой цепи SEQ ID NO:7; (b) HVR-H2 тяжелой цепи SEQ ID NO:7; (c) HVR-H3 тяжелой цепи SEQ ID NO:7; (d) HVR-L1 легкой цепи SEQ ID NO:8; (e) HVR-L2 легкой цепи SEQ ID NO:8; и (f) HVR-L3 легкой цепи SEQ ID NO:8.
В другом варианте осуществления изобретение относится к антителу, включающему в себя (a) HVR-H1, содержащую аминокислотную последовательность SEQ ID NO:22; (b) HVR-H2, содержащую аминокислотную последовательность SEQ ID NO:23; (c) HVR-H3, содержащую аминокислотную последовательность SEQ ID NO:24; (d) HVR-L1, содержащую аминокислотную последовательность SEQ ID NO:25; (e) HVR-L2, содержащую аминокислотную последовательность SEQ ID NO:26; и (f) HVR-L3, содержащую аминокислотную последовательность SEQ ID NO:27.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; (c) HVR-H3 тяжелой цепи SEQ ID NO:1, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1 тяжелой цепи SEQ ID NO:1; (b) HVR-H2 тяжелой цепи SEQ ID NO:1; (c) HVR-H3 тяжелой цепи SEQ ID NO:1, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; (c) HVR-H3 тяжелой цепи SEQ ID NO:3, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1 тяжелой цепи SEQ ID NO:3; (b) HVR-H2 тяжелой цепи SEQ ID NO:3; (c) HVR-H3 тяжелой цепи SEQ ID NO:3, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:17; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:21, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; (c) HVR-H3 тяжелой цепи SEQ ID NO:5, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности VH-HVR, выбранные из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2 содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1 тяжелой цепи SEQ ID NO:5; (b) HVR-H2 тяжелой цепи SEQ ID NO:5; (c) HVR-H3 тяжелой цепи SEQ ID NO:5, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1 тяжелой цепи SEQ ID NO:7; (e) HVR-H2 тяжелой цепи SEQ ID NO:7; (f) HVR-H3 тяжелой цепи SEQ ID NO:7.
В одном варианте осуществления изобретения анти-LRP6 антитело является полиспецифичным анти-LRP6 антителом, которое включает в себя первый VH-домен, содержащий все три последовательности VH-HVR из (a) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:20; (b) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:18; (c) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:19, и включает в себя второй VH-домен, содержащий все три последовательности VH-HVR из (d) HVR-H1, содержащей аминокислотную последовательность SEQ ID NO:22; (e) HVR-H2, содержащей аминокислотную последовательность SEQ ID NO:23; (f) HVR-H3, содержащей аминокислотную последовательность SEQ ID NO:24.
В любом из вышеуказанных вариантов полиспецифичных анти-LRP6 антител они дополнительно содержит по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1 легкой цепи SEQ ID NO:2; (b) HVR-L2 легкой цепи SEQ ID NO:2; и (c) HVR-L3 легкой цепи SEQ ID NO:2 или SEQ ID NO:4. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:2; (b) HVR-L2 легкой цепи SEQ ID NO:2; и (c) HVR-L3 легкой цепи SEQ ID NO:2. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1 легкой цепи SEQ ID NO:2; (b) HVR-L2 легкой цепи SEQ ID NO:2; и (c) HVR-L3 легкой цепи SEQ ID NO:4.
В любом из вышеуказанных вариантов полиспецифичных анти-LRP6 антител антитела дополнительно содержит по меньшей мере одну, по меньшей мере две или все три последовательности VL-HVR, выбранные из (a) HVR-L1, содержащей аминокислоты SEQ ID NO:25; (b) HVR-L2, содержащей аминокислоты SEQ ID NO:26; и (c) HVR-L3, содержащей аминокислоты SEQ ID NO:27 или SEQ ID NO:28. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1, содержащую аминокислоты SEQ ID NO:25; (b) HVR-L2, содержащую аминокислоты SEQ ID NO:26; и (c) HVR-L3, содержащую аминокислоты SEQ ID NO:27. В одном варианте осуществления изобретения антитело включает в себя (a) HVR-L1, содержащую аминокислоты SEQ ID NO:25; (b) HVR-L2, содержащую аминокислоты SEQ ID NO:26; и (c) HVR-L3, содержащую аминокислоты SEQ ID NO:28.
В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1X2K (SEQ ID NO:41). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1X2KN (SEQ ID NO:42). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1VK (SEQ ID NO:43). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1IK (SEQ ID NO:44). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1VKN (SEQ ID NO:45). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NX1IKN (SEQ ID NO:46). В вышеуказанных вариантах осуществления Х1 является аминокислотой, и Х2 представляет собой I или V; или Х1 является A, S, F, T, Y или L, и Х2 представляет собой I или V; или Х1 является A, S, F, T, Y или L, и Х2 представляет собой I; или Х1 является A, S, F, T, Y или L, и Х2 представляет собой V.
В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NAVK (SEQ ID NO:47). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NAIK (SEQ ID NO:48). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NAVKN (SEQ ID NO:49). В одном варианте осуществления изобретения анти-LRP6 антитело или полиспецифичное анти-LRP6 антитело включает в себя HVR-H3, содержащую аминокислотную последовательность NAIKN (SEQ ID NO:50).
В любом из вышеуказанных вариантов осуществления анти-LRP6 антитело является гуманизированным. В одном варианте осуществления изобретения анти-LRP6 антитело включает в себя HVR, как в любом из вышеуказанных вариантов осуществления, и дополнительно включает в себя акцепторную каркасную область человека, например, каркасную область иммуноглобулина человека или консенсусную каркасную область человека.
В другом аспекте анти-LRP6 антитело включает в себя последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с VH тяжелой цепи аминокислотной последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В другом аспекте анти-LRP6 антитело включает в себя последовательность вариабельного домена тяжелой цепи (VH), имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с аминокислотной последовательностью SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15. В некоторых вариантах осуществления изобретения последовательность VH, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность, содержит замены (например, консервативные замены), вставки или делеции относительно референсной последовательности, но анти-LRP6 антитело, содержащее эту последовательность, сохраняет способность связывать LRP6. В некоторых вариантах осуществления изобретения всего 1-10 аминокислот заменено, вставлено и/или удалено в VH из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, или в SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13 или SEQ ID NO:15.
В другом аспекте анти-LRP6 антитело включает в себя последовательность тяжелой цепи, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с аминокислотной последовательностью SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. В некоторых вариантах осуществления изобретения последовательность тяжелой цепи, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность, содержит замены (например, консервативные замены), вставки или делеции относительно референсной последовательности, но анти-LRP6 антитело, содержащее эту последовательность, сохраняет способность связывать LRP6. В некоторых вариантах осуществления изобретения всего 1-10 аминокислот заменено, вставлено и/или удалено в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
В некоторых вариантах осуществления изобретения замены, вставки или делеции встречаются в областях вне HVR (то есть, в FR). Необязательно, анти-LRP6 антитело содержит тяжелую цепь и/или VH тяжелой цепи SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, включающие пост-трансляционные модификации этой последовательности.
В другом аспекте изобретение относится к анти-LRP6 антителу, где антитело включает в себя вариабельный домен легкой цепи (VL), имеющий по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с аминокислотной последовательностью VL легкой цепи SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8. В другом аспекте изобретение относится к анти-LRP6 антителу, где антитело включает в себя вариабельный домен легкой цепи (VL), имеющий по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с аминокислотной последовательностью SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14 или SEQ ID NO:16. В некоторых вариантах осуществления изобретения последовательность VL, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции относительно референсной последовательности, но анти-LRP6 антитело, содержащее эту последовательность, сохраняет способность связывать LRP6. В некоторых вариантах осуществления изобретения всего 1-10 аминокислот заменено, вставлено и/или удалено в VL SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8, или в SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14 или SEQ ID NO:16.
В другом аспекте изобретение относится к анти-LRP6 антителу, причем антитело включает в себя легкую цепь, имеющую по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательностей с аминокислотной последовательностью SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8. В некоторых вариантах осуществления изобретения последовательность легкой цепи, имеющая по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность, содержит замены (например, консервативные замены), вставки или делеции относительно референсной последовательности, но анти-LRP6 антитело, содержащее эту последовательность, сохраняет способность связывать LRP6. В некоторых вариантах осуществления изобретения всего 1-10 аминокислот заменено, вставлено и/или удалено в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8.
В некоторых вариантах осуществления изобретения замены, вставки или делеции встречаются в областях вне HVR (то есть в FR). Необязательно, анти-LRP6 антитело содержит легкую цепь и/или последовательность VL в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:8, включающие пост-трансляционные модификации этой последовательности.
В другом аспекте изобретение относится к анти-LRP6 антителу, где антитело содержит VH, как в любом из приведенных выше вариантов, и VL, как в любом из приведенных выше вариантов.
В дополнительном аспекте изобретение относится к антителу, которое связывает такой же эпитоп, как анти-LRP6 антитело, приведенное в настоящем описании. Например, в некоторых вариантах осуществления изобретение относится к антителу, которое связывает такой же эпитоп, как анти-LRP6 антитело, выбранное из анти-LRP6 антитела, содержащего последовательность VH SEQ ID NO:9 и последовательность VL SEQ ID NO:10, или последовательность VH SEQ ID NO:11 и последовательность VL SEQ ID NO:12, или последовательность VH SEQ ID NO:13 и последовательность VL SEQ ID NO:14, или последовательность VH SEQ ID NO:15 и последовательность VL SEQ ID NO:16. В одном варианте осуществления изобретения анти-LRP6 антитело связывает эпитоп, который содержится в аминокислотных остатках в области Е1-Е2 LRP6. В одном варианте осуществления изобретения анти-LRP6 антитело связывает эпитоп, который содержится в аминокислотных остатках в области Е3-Е4 LRP6. В одном варианте осуществления изобретения анти-LRP6 антитело является биспецифичным антителом, которое связывает эпитоп, который состоит из аминокислотных остатков, присутствующих в области Е1-Е2 LRP6, и связывает эпитоп, который состоит из аминокислотных остатков, присутствующих в области Е3-Е4 LRP6.
В одном варианте осуществления изобретения анти-LRP6 антитело связывает конформационный эпитоп, который включает остатки R28, E51, D52, V70, S71, E73, L95, S96, D98, E115, R141 и N185 в LRP6. В одном варианте осуществления изобретения анти-LRP6 антитело связывает конформационный эпитоп, который включает остатки R28, E51, D52, V70, S71, E73, L95, S96, D98, E115, R141, N185, R29, W188, K202, P225, H226, S243 и F266 в LRP6.
В одном варианте осуществления изобретения анти-LRP6 антитело взаимодействует по меньшей мере с одним, по меньшей мере с двумя, по меньшей мере с тремя, по меньшей мере с четырьмя, по меньшей мере с пятью, по меньшей мере с шестью, по меньшей мере с семью, по меньшей мере с восемью, по меньшей мере с девятью, по меньшей мере с десятью, по меньшей мере с одиннадцатью или со всеми аминокислотными остатками R28, E51, D52, V70, S71, E73, L95, S96, D98, E115, R141 и N185 β-пропеллера E1 в LRP6. В дополнительном варианте осуществления изобретения анти-LRP6 антитело дополнительно взаимодействует по меньшей мере с одним, по меньшей мере с двумя, по меньшей мере с тремя, по меньшей мере с четырьмя, по меньшей мере с пятью, по меньшей мере с шестью, по меньшей мере с семью остатками R29, W188, K202, P225, H226, S243 и F266 в LRP6.
В дополнительном аспекте анти-LRP6 антитело по любому из вышеуказанных вариантов осуществления может включать любой из признаков (по отдельности или в комбинации), описанных в разделах 1-7 ниже:
1. Аффинность антител
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению имеет константу диссоциации (Kd) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 M или менее, например, от 10-8 M до 10-13 M, например, от 10-9 M до 10-13 M).
В одном варианте осуществления изобретения Kd измеряют методом анализа связывания с радиоактивно меченным антигеном (RIA), осуществляемого с использованием Fab-варианта исследуемого антитела и его антигена, как описано в нижеследующем методе анализа. Аффинность связывания в растворе Fab с антигеном измеряют уравновешиванием Fab с минимальной концентрацией (125I)-меченного антигена в присутствии титрационных разведений немеченого антигена, с последующей иммобилизацией связанного антигена на планшете, покрытом антителами к Fab-фрагменту (см., например, Chen et al., J. Mol. Biol. 293:865-881 (1999)). Для подбора условий проведения анализа многолуночные планшеты MICROTITER® (Thermo Scientific) покрывают в течение ночи 5 мкг/мл иммобилизующего антитела к Fab (Cappel Labs) в 50 мМ карбонате натрия (рН 9,6), и затем блокируют 2%-ным (масс./об.) бычьим сывороточным альбумином в PBS в течение 2-5 часов при комнатной температуре (приблизительно при 23°С). В неадсорбирующем планшете (Nunc #269620) 100 пМ или 26 пМ [125I]-антигена смешивают с серийными разведениями исследуемого Fab (например, при оценке антитела к VEGF - с Fab-12, в Presta et al., Cancer Res. 57:4593-4599 (1997)). Затем исследуемый Fab инкубируют в течение ночи, однако, для гарантии достижения равновесия инкубирование можно проводить в течение более длительного периода времени (например, примерно 65 часов). После этого, смеси переносят в планшет для иммобилизации и инкубируют при комнатной температуре (например, в течение одного часа). Затем раствор удаляют, и планшет восемь раз промывают 0,1% полисорбатом 20 (TWEEN-20®) в PBS. После сушки планшетов добавляют сцинтилляционную жидкость (MICROSCINT-20™; Packard) по 150 мкл/лунка, и измеряют радиоактивность планшетов на гамма-счетчике TOPCOUNT™ (Packard) в течение десяти минут. Концентрации каждого Fab, которые составляют 20% или меньше от максимального связывания, отбирают для использования в методах анализа конкурентного связывания.
В соответствии с другим вариантом изобретения Kd измеряют методами анализа поверхностного плазмонного резонанса с использованием BIACORE®-2000 или BIACORE ®-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С, используя чипы СМ5 с иммобилизованным на них антигеном при ~10 единицах отклика (RU). Кратко, биосенсорные чипы, покрытые карбоксиметилированным декстраном (CM5, BIACORE, Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разводят 10 мМ ацетатом натрия, рН 4,8, до 5 мкг/мл (~0,2 мкМ) и впрыскивают при скорости потока 5 мкл/минута до достижения ~10 единиц отклика (RU) связанного белка. После впрыскивания антигена вводят 1М этаноламин, чтобы блокировать непрореагировавшие группы. Для измерения кинетики связывания впрыскивают двукратные серийные разведения Fab (0,78 нМ-500 нМ) в PBS, содержащем 0,05% поверхностно-активного вещества, полисорбата 20 (TWEEN-20™) (PBST), при 25°С и скорости потока приблизительно 25 мкл/мин. Скорость ассоциации (kon) и скорость диссоциации (koff) вычисляют с использованием простой лангмуровской модели связывания 1:1 (BIACORE® Evaluation Software version 3.2) одновременной аппроксимацией сенсорограмм ассоциации и диссоциации. Константу равновесной диссоциации (Kd) вычисляют как отношение koff/kon (см., например, Chen et al., J. Mol. Biol. 293:865-881 (1999)). Если в вышеописанном методе анализа поверхностного плазмонного резонанса скорость ассоциации превышает 106 M-1с-1, то скорость ассоциации можно определить с использованием метода гашения флюоресценции, которым измеряют увеличение или снижение интенсивности испускания флюоресценции (возбуждение=295 нМ; испускание=340 нМ, полоса пропускания 16 нМ) при 25°C 20 нМ антитела к антигену (Fab-формы) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена, измеряемой в спектрометре, таком как спектрофотометр, оборудованный «остановленной струей» (Aviv Instruments), или спектрофотометре серии 8000 SLM-AMINCO™ (ThermoSpectronic) с кюветой с перемешиванием.
2. Фрагменты антител
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению является фрагментом антитела. Фрагменты антител включают, но без ограничения, фрагменты Fab, Fab', Fab'-SH, F(ab')2, Fv и scFv, и другие фрагменты, описанные ниже. Обзор по некоторым фрагментам антител см. в Hudson et al. Nat. Med. 9: 129-134 (2003). Для обзора по scFv-фрагментам см., например, Pluckthun, в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), pp. 269-315 (1994); см. также WO 93/16185; и патенты США 5571894 и 5587458. Обсуждение фрагментов Fab и F(ab')2, содержащих остатки эпитопа, связывающего «рецептор спасения» и имеющих увеличенное время полужизни in vivo, см. в патенте США 5869046.
Диантитела являются фрагментами антител с двумя антиген-связывающими сайтами, которые могут быть бивалентными или биспецифичными. См., например, в ЕР 404097; WO 93/11161; Hudson et al., Nat. Med. 9: 129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993). Триантитела и тетраантитела также описаны в Hudson et al., Nat. Med. 9: 129-134 (2003).
Однодоменные антитела являются фрагментами антител, содержащими весь вариабельный домен тяжелой цепи антитела или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления однодоменное антитело является однодоменным антителом человека (Domantis, Inc., Waltham, MA; см., например, патент США 6248516 B1).
Фрагменты антител можно получить различными методиками, включая, но без ограничения, протеолитическое расщепление интактного антитела, а также продукцию рекомбинантными клетками-хозяевами (например, E. coli или фагами), описанными в настоящем описании.
3. Химерные и гуманизированные антитела
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению является химерным антителом. Некоторые химерные антитела описаны, например, в патенте США 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)). В одном примере химерное антитело включает в себя не принадлежащую человеку вариабельную область (например, вариабельную область, полученную из мыши, крысы, хомячка, кролика или приматов кроме человека, таких как мартышка) и константную область человека. В дополнительном примере химерным антителом является антитела «переключенного класса», у которых класс или подкласс был изменен относительно родительского антитела. Химерные антитела включают их антиген-связывающие фрагменты.
В некоторых вариантах осуществления изобретения химерным антителом является гуманизированное антитело. Обычно антитело, не принадлежащее человеку, «гуманизируют», чтобы снизить иммуногенность у человека, одновременно сохраняя специфичность и аффинность родительского антитела, не принадлежащего человеку. В общем, гуманизированное антитело включает в себя один или несколько вариабельных доменов, в которых HVR, например, CDR, (или их участки) получены из антитела, не принадлежащего человеку, и FR (или их участи) получены из последовательностей антитела человека. Гуманизированное антитело необязательно будет также содержать по меньшей мере часть константной области антитела человека. В некоторых вариантах осуществления изобретения некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками из антитела, не принадлежащего человеку (например, антитела, из которого получены остатки HVR), например, чтобы восстановить или улучшить специфичность или аффинность антитела.
Обзоры гуманизированных антител и способы их получения приведены, например, в Almagro and Fransson, Front. Biosci. 13: 1619-1633 (2008), и дополнительно описаны, например, в Riechmann et al., Nature 332:323-329 (1988); Queen et al., Proc. Natl. Acad. Sci. USA 86: 10029-10033 (1989); патентах США 5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36:25-34 (2005) (описывающей перенос SDR (a-CDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (описывающей изменение поверхностной структуры антитела); DaU'Acqua et al., Methods 36:43-60 (2005) (описывающей «перетасовку» остатков в FR); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (описывающей подход «направленного отбора» при перетасовке остатков в FR).
Каркасные области антитела человека, которые можно использовать для гуманизация, включают, но без ограничения: каркасные области, отобранные с использованием способа «оптимальной подгонки» (см., например, Sims et al., J. Immunol. 151:2296 (1993)); каркасные области, полученные из консенсусной последовательности антител человека конкретной подгруппы вариабельных областей легкой или тяжелой цепей (см., например, Carter et al., Proc. Natl. Acad. Sci. USA, 89:4285 (1992); и Presta et al., J. Immunol., 151:2623 (1993)); зрелых (соматически мутированных) каркасных областей человека или каркасных областей зародышевой линии человека (см., например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)); и каркасных областей, полученных при скрининге библиотек FR (см., например, Baca et al., J. Biol. Chem. 272:10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
4. Антитела человека
В некоторых вариантах осуществления изобретения антителом, приведенным в настоящем описании, является антитело человека. Антитела человека получают, используя различные методики, известные в данной области. Антитела человека описаны в общем в van Dijk and van de Winkel, Curr. Opin. Pharmacol. 5: 368-74 (2001) и в Lonberg, Curr. Opin. Immunol. 20: 450-459 (2008).
Антитела человека можно получить введением иммуногена трансгенному животному, модифицированному, чтобы продуцировать интактные антитела человека или интактные антитела с вариабельными областями антител человека в ответ на стимуляцию антигеном. Такие животные обычно содержат все или участок иммуноглобулиновых локусов человека, которые замещают эндогенные иммуноглобулиновые локусы, или которые являются внехромосомными или встроенными случайным образом в хромосомы животного. У таких трансгенных мышей эндогенные иммуноглобулиновые локусы обычно инактивированы. Для обзора по способам получения антител человека в трансгенных животных см. Lonberg, Nat. Biotech. 23: 1117-1125 (2005). См. также, например, патенты США 6075181 и 6150584, описывающие технологию XENOMOUSE™; патент США 5770429, описывающий технологию HUMAB®; патент США 7041870, описывающий технологию K-M MOUSE®, и публикацию патентной заявки США № 2007/0061900, описывающую технологию VELOCIMOUSE®). Вариабельные области человека из интактных антител, синтезируемых такими животными, можно дополнительно модифицировать, например, объединяя с другой константной областью человека.
Антитела человека могут быть получены способами на основе гибридом. Описаны клеточные линии миеломы человека и гетеромиеломы человека и мыши для получения моноклональных антител человека. (См., например, Kozbor J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987); и Boerner et al., J. Immunol., 147: 86 (1991)). Антитела человека, полученные посредством В-клеточный гибридомной методики, также описаны в Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006). Дополнительные способы включают описанные, например, в патенте США 7189826 (описывающем получение моноклональных IgM-антител человека из гибридомных клеточных линий) и в Ni, Xiandai Mianyixue, 26(4):265-268 (2006) (описывающей гибридомы других биологических видов кроме человека). Технология гибридом человека (технология Trioma) также описана в Vollmers and Brandlein, Histology and Histopathology, 20(3): 927-937 (2005) и в Vollmers and Brandlein, Methods and Findings in Experimental and Clinical Pharmacology, 27(3): 185-91 (2005).
Антитела человека также можно создать путем выделения последовательностей Fv-клонов вариабельных доменов, отобранных из библиотек фагового дисплея на основе последовательностей антител человека. Такие последовательности вариабельных доменов можно затем объединить с желаемым константным доменом антитела человека. Методики отбора антител человека из библиотек антител описаны ниже.
5. Антитела, полученные из библиотек
Антитела настоящего изобретения могут быть выделены скринингом комбинаторных библиотек на антитела с требуемой активностью или активностями. Например, в данной области известно множество способов создания библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие требуемыми характеристиками связывания. Такие способы описаны в общих чертах в Hoogenboom et al. (2001) в Methods in Molecular Biology 178:1-37 (O'Brien et al., ed., Human Press, Totowa, NJ) и дополнительно описаны, например, в the McCafferty et al., Nature 348:552-554; Clackson et al., Nature 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Marks and Bradbury, в Methods in Molecular Biology 248: 161-175 (Lo, ed., Human Press, Totowa, NJ, 2003); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2): 119-132(2004).
В некоторых методах фагового дисплея репертуары VH и VL-генов отдельно клонируют полимеразной цепной реакцией (ПЦР) и рекомбинируют случайным образом в фаговых библиотеках, которые затем можно скринировать на антиген-связывающий фаг, как описано в Winter et al., Ann. Rev. Immunol., 12: 433-455 (1994). В фагах обычно представляют фрагменты антител, либо в виде одноцепочечных фрагментов Fv (scFv) или в виде фрагментов Fab. Библиотеки из иммунизированных источников обеспечивают высокоаффинные антитела к иммуногену, не требуя конструирования гибридом. В альтернативном варианте можно клонировать неиммунизированный репертуар (например, человека), чтобы обеспечить единый источник антител к широкому диапазону чужеродных и аутоантигенов без какой-либо иммунизации, как описано Griffiths et al., EMBO J, 12: 725-734 (1993). В заключение, неиммунизированные библиотеки также можно получать синтетическим способом, путем клонирования неперестроенных сегментов V-генов из стволовых клеток и используя ПЦР-праймеры, содержащие случайные последовательности, для кодирования высоковариабельных CDR3-областей и для осуществления перестройки in vitro, как описано в Hoogenboom and Winter, J. Mol. Biol., 227:381-388 (1992). Патентные публикации, описывающие фаговые библиотеки антител человека, включают, например: патент США 5750373 и патентные публикации США №№ 2005/0079574, 2005/0119455, 2005/0266000, 2007/0117126, 2007/0160598, 2007/0237764, 2007/0292936 и 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек антител человека, считаются в настоящем описании антителами человека или фрагментами антител человека.
6. Полиспецифичные антитела
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению является полиспецифичным антителом, например, биспецифичным антителом. Полиспецифичные антитела являются моноклональными антителами, которые имеют специфичность связывания по меньшей мере к двум различным сайтам. В некоторых вариантах осуществления изобретения первая специфичность связывания относится к LRP6, и вторая относится к какому-либо другому антигену. В некоторых вариантах осуществления изобретения биспецифичное антитело связывает два различных эпитопа на LRP6. Биспецифичные антитела также можно использовать для определения места действия цитотоксичных агентов на клетки, которые экспрессируют LRP6. Биспецифичные антитела можно получить в виде полноразмерных антител или фрагментов антител.
Методики получения полиспецифичных антител включают, но без ограничения, рекомбинантную совместную экспрессию двух пар тяжелая цепь-легкая цепь иммуноглобулина, где две тяжелые цепи обладают различной специфичностью (Milstein and Cuello, Nature, 305:537 (1983)), WO 93/08829 и Traunecker et al., EMBO J. 10: 3655 (1991)), и принцип конструирования «узел в полости» (см., например, патент США 5731168). Полиспецифичные антитела также могут быть получены конструированием эффектов электростатического управления для изготовления Fc-гетеродимерных молекул (WO 2009/089004A1); сшиванием двух или нескольких антител или фрагментов (см., например, патент США 4676980 и Brennan et al., Science, 229: 81 (1985)); используя лейциновые молнии для получения биспецифичных антител (см., например, Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992)); используя технологию «диантител» для создания биспецифичных фрагментов антител (см., например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993)); и используя димеры одноцепочечных Fv (sFv) (см., например, Gruber et al., J. Immunol., 152:5368 (1994)); и получая триспецифичные антитела, как описано, например, в Tutt et al., J. Immunol. 147: 60 (1991).
В настоящее описание также включены сконструированные антитела с тремя или несколькими функциональными сайтами связывания антигена, включая «антитела осьминога» (см., например, US 2006/0025576A1).
Антитело или фрагмент в настоящем описании также включает «Fab двойного действия» (или «DAF»), включающий антиген-связывающий сайт, который связывает LRP6, а также другой, отличный от него антиген (см., например, US 2008/0069820).
7. Варианты антител
В некоторых вариантах осуществления предусмотрены варианты антител, описанные в настоящем описании. Например, может быть желательным улучшить аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотных последовательностей антитела можно получить внесением соответствующих замен в нуклеотидную последовательность, кодирующую антитело, или пептидным синтезом. Такие модификации включают, например, делеции и/или вставки остатков, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно осуществлять любую комбинацию делеций, вставок и замен, при условии, что конечная конструкция обладает требуемыми характеристиками, например, антиген-связыванием.
а) Варианты замен, вставок и делециий
В некоторых вариантах осуществления изобретение относится к вариантам антител, имеющим одну или несколько аминокислотных замен. Участки, представляющие интерес для проведения замещающего мутагенеза, включают HVR и FR. Консервативные замены представлены в таблице 1 под заголовком «предпочтительные замены». Более значимые замены представлены в таблице 1 под заголовком «примеры замен» и дополнительно описаны ниже относительно классов боковых цепей аминокислот. Аминокислотные замены можно вводить в представляющее интерес антитело, и продукты скринировать на целевую активность, например, сохраненное/улучшенное связывание антигена, сниженную иммуногенность или повышенную ADCC или CDC.
Аминокислоты можно сгруппировать в соответствии с общими свойствами боковых цепей:
(1) гидрофобные: Норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro; и
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены будут вызывать замену члена одного из этих классов на другой класс.
Один тип вариантов замен включает замены одного или нескольких остатков гипервариабельной области родительского антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный(е) вариант(ы), отобранный(е) для дальнейшего исследования, должен(ны) будет(ут) иметь модификации (например, улучшение) некоторых биологических свойств (например, повышенную аффинность, сниженную иммуногенность) относительно родительского антитела и/или должен(ны) будет(ут) иметь по существу сохраненные некоторые биологические свойства родительского антитела. Примером варианта замены является аффинно зрелое антитело, которое обычно можно создать, например, используя методики созревания аффинности на основе фагового дисплея, такие, как описано в настоящем описании. Кратко, один или несколько остатков HVR мутируют и варианты антитела представляют на фагах и скринируют по конкретной биологической активности (например, аффинности связывания).
Изменения (например, замены) можно вносить в HVR, например, чтобы улучшить аффинность антитела. Такие изменения можно вносить в «горячие точки» HVR, то есть остатки, кодируемые кодонами, подверженными мутациям с высокой частотой в ходе процесса соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207: 179-196 (2008)), и/или в SDR (a-CDR), тестируя полученный вариант VH или VL на аффинность связывания. Созревание аффинности путем конструирования и повторного отбора из вторичных библиотек было описано, например, в Hoogenboom et al. в Methods in Molecular Biology 178: 1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, (2001)). В некоторых вариантах осуществления созревания аффинности вводят разнообразие в гены вариабельных доменов, выбранных для созревания с помощью любого из ряда способов (например, вносящей ошибки ПЦР, перетасовки цепей или олигонуклеотид-направленного мутагенеза). Затем создают вторичную библиотеку. Затем библиотеку скринируют, чтобы идентифицировать любые варианты антител с желаемой аффинностью. Другой способ для внесения разнообразия включает HVR-направленные подходы, в которых несколько остатков в HVR (например, 4-6 остатков одновременно) изменяют случайным образом. Остатки в HVR, участвующие в связывании антигена, можно специально идентифицировать, например, используя аланиновый сканирующий мутагенез или моделирование. Особенно часто мишенями являются CDR-H3 и CDR-L3.
В некоторых вариантах осуществления изобретения замены, вставки или делеции могут встречаться в одной или нескольких HVR, при условии, что такие изменения по существу не снижают способность антитела связывать антиген. Например, в HRV можно осуществить консервативные изменения (например, консервативные замены, описанные в настоящем описании), которые по существу не снижают аффинность связывания. Такие изменения могут находиться вне «горячих точек» HVR или SDR. В некоторых вариантах осуществления вариантов последовательностей VH и VL, описанных в настоящем описании, каждая HVR является либо неизмененной, либо содержит не более одной, двух или трех аминокислотных замен.
Способ, пригодный для идентификации остатков или областей антитела, которые могут быть мишенями при мутагенезе, носит название «аланиновый сканирующий мутагенез», описанный Cunningham and Wells (1989) Science, 244:1081-1085. В этом способе идентифицируют один или группу остатков-мишеней (например, заряженных остатков, таких как arg, asp, his, lys и glu) и замещают нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), чтобы определить, будет ли затронуто взаимодействие антитела с антигеном. Дополнительные замены можно ввести в положения аминокислот, демонстрирующих функциональную чувствительность к первоначальным заменам. Альтернативно или дополнительно, можно проанализировать кристаллическую структуру комплекса антиген-антитело, чтобы идентифицировать точки контакта между антителом и антигеном. Такие контактирующие остатки и соседние остатки могут являться мишенями или их можно исключить в качестве потенциальных остатков для замены. Варианты можно скринировать, с тем чтобы определить, имеют ли они искомые свойства.
Вставки в аминокислотную последовательность включают амино- и/или карбокси-концевые слияния в диапазоне длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или множества аминокислотных остатков внутри последовательности. Примеры концевых вставок включают антитело с N-концевым остатком метионина. Другие варианты молекулы антитела с вставками включают слияние N- или C-конца антитела с ферментом (например, для ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
b) Варианты гликозилирования
В некоторых вариантах осуществления изобретения антитело, описанное в настоящем описании, изменяют для повышения или снижения степени гликозилирования антитела. Добавление или удаление участков гликозилирования в антителе можно удобно осуществить, изменяя аминокислотную последовательность таким образом, чтобы создать или удалить один или несколько сайтов гликозилирования.
Если антитело содержит Fc-область, можно изменить присоединенный к ней углевод. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный биантеннальный олигосахарид, который обычно присоединен через азот к Asn297 CH2-домену Fc-области. См., например, Wright et al. TIBTECH 15:26-32 (1997). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc на «стебле» биантеннальной олигосахаридной структуры. В некоторых вариантах осуществления изобретения модификации олигосахарида в антителе настоящего изобретения могут быть выполнены для того, чтобы создать варианты антител с некоторыми улучшенными свойствами.
В одном варианте осуществления варианты антител по настоящему изобретению имеют углеводную структуру, в которой отсутствует фукоза, присоединенная (прямо или косвенно) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяют, вычисляя среднее количество фукозы в сахарной цепи по Asn 297 относительно суммарного количества всех гликоструктур, присоединенных к Asn 297 (например, комплексных, гибридных или с высоким содержанием маннозы), измеряемого с помощью MALDI-TOF масс-спектрометрии, описанной, например, в WO 2008/077546. Asn 297 относится к остатку аспарагина, находящемуся в положении 297 в Fc-области (Eu-нумерация остатков Fc-области); однако, вследствие небольших вариаций последовательностей в антителах Asn 297 также может находиться в положении ±3 аминокислоты выше или ниже от положения 297, то есть, между положениями 294 и 300. Такие варианты фукозилирования могут иметь улучшенную функцию ADCC. См., например, патентные публикации США №№ 2003/0157108 (Presta, L.); 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций относительно «дефукозилированных» или «лишенных фукозы» вариантов антител включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO2002/031140; Okazaki et al. J. Mol. Biol. 336:1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры клеточных линий, продуцирующих дефукозилированные антитела, включают клетки Lec13 CHO с нарушенным фукозилированием белков (Ripka et al., Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США 2003/0157108 A1, Presta L.; и WO 2004/056312 A1, Adams et al., в частности пример 11), и нокаутные линии клеток, такие как клетки СНО, нокаутные по гену альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki et al., Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94(4):680-688 (2006); и WO 2003/085107).
Дополнительно изобретение относится к вариантам антител с «разделенными» олигосахаридами, например, в которых биантеннальный олигосахарид, присоединенный к Fc-обалсти антитела, разделен на две части посредством GlcNAc. Такие варианты антител могут иметь меньшую степень фукозилирования и/или улучшенную функцию ADCC. Примеры таких вариантов антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США 6602684 (Umana et al.); и US 2005/0123546 (Umana et al.). Также изобретение относится к вариантам антител по меньшей мере с одним остатком галактозы, присоединенным к Fc-области. Такие варианты антител могут обладать улучшенной функцией CDC. Такие варианты антител описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.); и WO 1999/22764 (Raju, S.).
с) Варианты Fc-области
В некоторых вариантах осуществления изобретения одну или несколько аминокислотных модификаций можно ввести в Fc-область антитела по настоящему изобретению, таким образом, создавая вариант Fc-области. Вариант Fc-области может содержать последовательность Fc-области человека (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену) по одной или нескольким аминокислотным положениям.
В некоторых вариантах осуществления изобретение предусматривает вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его искомым кандидатом для вариантов применения, в которых важно время полужизни антитела in vivo, а некоторые эффекторные функции (такие, как активация системы комплемента и ADCC) являются необязательными или вредными. Для подтверждения снижения/ослабления CDC- и/или ADCC-активности можно провести анализ цитотоксичности in vitro и/или in vivo. Например, можно провести анализ связывания с Fc-рецепторами (FcR) для уверенности в том, что антитело не способно связываться с FcyR (таким образом, вероятно отсутствие ADCC-активности), но сохраняет способность связывать FcRn. Основные клетки, осуществляющие ADCC, NK-клетки, экспрессируют только FcyRIII, в то время как моноциты экспрессируют FcyRI, FcyRII и FcyRIII. Экспрессия FcR на гемопоэтических клетках суммирована в таблице 3 на странице 464 в Ravetch and Kinet, Annu. Rev. Immunol. 9:457-92 (1991). Неограничивающие примеры методов анализа in vitro для оценки ADCC-активности представляющей интерес молекулы описан в патенте США 5500362 (см. Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 83:7059-7063 (1986)) и Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 82: 1499-1502 (1985); 5821337 (см. Bruggemann, M. et al., J. Exp. Med. 166: 1351-1361 (1987)). Альтернативно, можно использовать нерадиоактивные методы анализа (см., например, нерадиоактивный метод анализа цитотоксичности, ACTI™, для проточной цитометрии (CellTechnology, Inc. Mountain View, CA; и нерадиоактивный метод анализа цитотоксичности CytoTox 96® (Promega, Madison, WI). Пригодные для таких анализов эффекторные клетки включают мононуклеарные клетки периферической крови (PBMC) и природные клетки-киллеры (NK). Альтернативно или дополнительно, ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, такой, как описано в Clynes et al., Proc. Natl. Acad. Sci. USA 95:652-656 (1998). Для того чтобы подтвердить, что антитело не способно связывать C1q и, соответственно, не имеет CDC-активности, можно провести анализ связывания C1q. См., например, ELISA на связывание с C1q и C3c в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно проводить анализ CDC (см., например, Gazzano-Santoro et al., J. Immunol. Methods 202:163 (1996); Cragg, M.S. et al., Blood 101: 1045-1052 (2003); и Cragg, M.S. and M.J. Glennie, Blood 103:2738-2743 (2004)). Определение связывания с FcRn и определение времени выведения/полужизни in vivo можно также провести, используя способы, известные в данной области (см., например, Petkova, S.B. et al., Int'l. Immunol. 18(12): 1759-1769 (2006)).
Антитела со сниженной эффекторной функцией включают антитела с заменой одного или нескольких остатков в Fc-области: 238, 265, 269, 270, 297, 327 и 329 (патент США 6737056). Такие мутанты Fc включают мутанты Fc с заменами по двум или нескольким аминокислотным положениям 265, 269, 270, 297 и 327, включая так называемый Fc-мутант «DANA» с заменой остатков 265 и 297 на аланин (патент США 7332581).
Описаны некоторые варианты антител с улучшенным или ухудшенным связыванием с FcR. (См., например, патент США 6737056; WO 2004/056312, и Shields et al., J. Biol. Chem. 9(2): 6591-6604 (2001)).
В некоторых вариантах осуществления изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые улучшают ADCC, например, заменами в положениях 298, 333 и/или 334 Fc-области (Eu-нумерация остатков).
В некоторых вариантах осуществления изобретения модификации выполнены в Fc-области, которые приводят к изменению (например, либо улучшению, либо снижению) связывания с C1q и/или комлемент-зависимой цитотоксичности (CDC), например, как описано в патенте США 6194551, WO 99/51642 и Idusogie et al. J. Immunol. 164: 4178-4184 (2000).
Антитела с увеличенным временем полужизни и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG плоду (Guyer et al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)), описаны в US 2005/0014934 A1 (Hinton et al.). Эти антитела содержат Fc-область с одной или несколькими заменами в ней, которые улучшают связывание Fc-области с FcRn. Такие варианты Fc включают варианты с заменами по одному или нескольким остаткам в Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, с заменой остатка 434 в Fc-области (патент США 7371826).
Относительно других примеров вариантов Fc-области, см. также Duncan & Winter, Nature 322:738-40 (1988); патент США 5648260; патент США 5624821; и WO 94/29351.
d) Варианты антител с введенными остатками цистеина
В некоторых вариантах осуществления изобретения может быть желательным создание антител с введенными цистеинами, например, «тиоMAb», в которых один или несколько остатков в антителе заменены цистеиновыми остатками. В конкретных вариантах осуществления изобретения замещенные остатки находятся в доступных сайтах антитела. В результате замещения этих остатков цистеинами реакционноспособные тиольные группы, таким образом, помещают в доступные сайты антитела, которые можно использовать для конъюгации антитела с другими молекулами, такими как молекулы лекарственных соединений или молекулы лекарственного соединения и линкера, для того чтобы создать иммуноконъюгат, как описано ниже в настоящем описании. В некоторых вариантах осуществления изобретения любой один или несколько из следующих остатков может быть замещен цистеином: V205 (нумерация по Кабату) легкой цепи; A118 (EU-нумерация) тяжелой цепи; и S400 (EU-нумерация) Fc-области тяжелой цепи. Антитела с введенными цистеинами можно создать, как описано, например, в патенте США 7521541.
е) Производные антител
В некоторых вариантах осуществления антитела по настоящему изобретению можно дополнительно модифицировать таким образом, чтобы они содержали дополнительные небелковые молекулы, которые известны в данной области и легкодоступны. Молекулы, подходящие для получения производных антител, включают, но без ограничения, полиэтиленгликоль (PEG), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (или гомополимеры или сополимеры со случайной последовательностью) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные многоатомные спирты (например, глицерин), поливиниловый спирт и их смеси. Пропионовый альдегид полиэтиленгликоля может обладать преимуществами при получении вследствие его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число присоединенных к антителу полимеров может варьировать, и если присоединено более одного полимера, они могут представлять собой одинаковые или разные молекулы. Как правило, количество и/или тип полимеров, используемых для получения производных, можно определить, исходя из факторов, включающих, но без ограничения: конкретные свойства и функции антитела, планируемого для улучшения, будут ли производное антитело использовать для терапии в определенных условиях и т.д.
Другой вариант осуществления относится к конъюгатам антитела и небелковой молекулы, которые можно избирательно нагревать в результате воздействия облучения. В одном из вариантов осуществления небелковая молекула представляет собой углеродную нанотрубку (Kam et al., Proc. Natl. Acad. Sci. USA 102: 11600-11605 (2005)). Облучение может быть любой волны и включает, но без ограничения, длины волн, которые не приносят вреда обычным клеткам, но которые нагревают небелковую молекулу до температуры, при которой клетки рядом с антителом-небелковой молекулой погибают.
В. Рекомбинантные способы и композиции
Антитела можно получить, используя рекомбинантные способы и композиции, например, описанные в патенте США 4816567. Один вариант осуществления изобретения относится к выделенной нуклеиновой кислоте, кодирующей анти-LRP6 антитело, описанное в настоящем описании. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). Дополнительный вариант осуществления относится к одному или нескольким векторам (например, экспрессионным векторам), содержащим такую нуклеиновую кислоту. Дополнительный вариант осуществления относится к клетке-хозяину, содержащей такую нуклеиновую кислоту. В одном таком варианте осуществления изобретения клетка-хозяин содержит (например, трансформирована): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном варианте осуществления изобретения клетка-хозяин является эукариотической, например, клеткой яичников китайского хомячка (СНО) или лимфоидной клеткой (например, клеткой Y0, NS0, Sp20). Один вариант осуществления относится к способу получения анти-LRP6 антитела, включающему культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, описанное выше, в условиях, подходящих для экспрессии антитела, и необязательно, выделение антитела из клетки-хозяина (или культуральной среды клеток-хозяев).
Чтобы получить рекомбинантное анти-LRP6 антитело, нуклеиновую кислоту, кодирующую антитело, например, описанную выше, выделяют и встраивают в один или несколько векторов для последующего клонирования и/или экспрессии в клетке-хозяине. Такую нуклеиновую кислоту можно легко выделить и секвенировать с использованием стандартных методик (например, используя олигонуклеотидные зонды, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии кодирующих антитела векторов включают прокариотические или эукариотические клетки, описанные в настоящем описании. Например, антитела можно продуцировать в бактериях, в частности, если не требуются гликозилирование и Fc-эффекторная функция. Для экспрессии фрагментов и полипептидов антител в бактериях, см., например, патенты США 5648237, 5789199 и 5840523. (См., также Charlton, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ, 2003), pp. 245-254, где описана экспрессия фрагментов антител в E. coli.) После экспрессии антитело можно выделить из массы бактериальных клеток в растворимой фракции и дополнительно очистить.
В дополнение к прокариотам, для клонирования или экспрессии кодирующих антитела векторов подходящими хозяевами являются эукариотические микроорганизмы, такие как филаментные грибы или дрожжи, включая штаммы грибов и дрожжей, чей процесс гликозилирования был «гуманизирован», что приводит к продукции антител, гликозилирование которых частично или полностью совпадает с гликозилированием антител человека. См. Gerngross, Nat. Biotech. 22: 1409-1414 (2004) и Li et al., Nat. Biotech. 24:210-215 (2006).
Подходящие клетки-хозяева для экспрессии гликозилированного антитела также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать совместно с клетками насекомых, в частности для трансфекции клеток Spodoptera frugiperda.
Культуры растительных клеток также можно использовать в качестве хозяев. См., например, патенты США 5959177, 6040498, 6420548, 7125978 и 6417429 (описывающие технологию PLANTIBODIES™ для получения антител в трансгенных растениях).
Клетки позвоночных также можно использовать в качестве хозяев. Например, можно использовать линии клеток млекопитающих, адаптированные для роста в суспензии. Другие примеры пригодных линий клеток-хозяев млекопитающих представляют собой линию клеток почки мартышки CV1, трансформированную SV40 (COS-7); линию эмбриональной почки человека (293 или клетки 293, описанные Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почки новорожденного хомячка (BHK); клетки Сертоли мышей (клетки TM4, описанные, например в Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки мартышки (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени серой крысы (BRL 3A); клетки легкого человека (W138); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мышей (MMT 060562); клетки TRI, описанные, например, в Mather et al., Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; и клетки FS4. Другие пригодные линии клеток-хозяев млекопитающих включают клетки яичников китайского хомячка (СНО), включая DHFR клетки СНО (Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); и миеломные клеточные линии, такие как Y0, NS0 и Sp2/0. Для обзора некоторых линий клеток-хозяев млекопитающих, подходящих для продукции антител, см., например, Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp. 255-268 (2003).
С. Методы анализа
Анти-LRP6 антитела по настоящему изобретению можно идентифицировать, скринировать или характеризовать по их физико-химическим свойствам и/или видам биологической активности с помощью различных методов анализа, известных в данной области.
1. Методы анализа связывания и другие методы анализа
В одном аспекте антитело настоящего изобретения тестируют по его антиген-связывающей активности, например, известными способами, такими как ELISA, Вестерн-блот и т.п.
В другом аспекте для идентификации антитела, которое конкурирует с анти-LRP6 антителом настоящего изобретения за связывание с LRP6, можно использовать конкурентные методы анализа. В некоторых вариантах осуществления изобретения такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), который связывает анти-LRP6 антитело настоящего изобретения. Подробные примеры способов картирования эпитопов, с которыми связывается антитело, приведены в Morris (1996) "Epitope Mapping Protocols" в Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ).
В иллюстративном конкурентном анализе иммобилизованный LRP6 инкубируют в растворе, содержащем первое меченое антитело, связывающееся с LRP6, и второе немеченое антитело, которое тестируют по его способности конкурировать с первым антителом за связывание с LRP6. Второе антитело может находиться в супернатанте гибридомы. В качестве контроля иммобилизованный LRP6 инкубируют в растворе, содержащем первое меченое антитело, но не второе немеченое антитело. После инкубации в условиях, позволяющих связывание первого антитела с LRP6, избыток несвязавшегося антитела удаляют и измеряют количество метки, связанной с иммобилизованным LRP6. Если количество метки, связанной с иммобилизованным LRP6 существенно снижается в тестируемом образце относительно контрольного образца, то это указывает на то, что второе антитело конкурирует с первым антителом за связывание с LRP6. См. Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
2. Методы анализа активности
Один аспект относится к методам анализа для идентификации анти-LRP6 антител, обладающих биологической активностью. Биологическая активность может включать, например, ингибирование или усиление передачи сигнала от изоформ Wnt, модуляцию массы/содержания костной ткани, ингибирование клеточной пролиферации, увеличение клеточной пролиферации. Также изобретение относится к антителам, проявляющим такую биологическую активность in vivo и/или in vitro.
В некоторых вариантах осуществления изобретения антитело настоящего изобретения тестируют на такую биологическую активность. Конкретные методы анализа, используемые для тестирования видов биологической активности, приведены в примерах.
D. Иммуноконъюгаты
Изобретение также относится к иммуноконъюгатам, содержащим анти-LRP6 антитело настоящего изобретения, конъюгированное с одним или несколькими цитотоксическими агентами, такими как химиотерапевтические агенты или лекарственные соединения, агенты, ингибирующие рост клеток, токсины (например, белковые токсины, ферментативно-активные токсины бактериального, грибкового, растительного или животного происхождения, или их фрагменты) или радиоактивные изотопы.
В одном варианте осуществления изобретения иммуноконъюгатом является конъюгат антитела с лекарственным средством (ADC), в котором антитело конъюгировано с одним или несколькими лекарственными средствами, включая, но без ограничения, майтанзиноиды (см. патенты США 5208020, 5416064 и европейский патент EP 0425235 B1); ауристатин, такой как лекарственные средства монометилауристатина DE и DF (MMAE и MMAF) (см. патенты США 5635483 и 5780588, а также 7498298); доластатин; калихеамицин или его производные (см. патенты США 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296; Hinman et al., Cancer Res. 53:3336-3342 (1993); и Lode et al., Cancer Res. 58:2925-2928 (1998)); антрациклин, такой как дауномицин или доксорубицин (см. Kratz et al., Current Med. Chem. 13:477-523 (2006); Jeffrey et al., Bioorganic & Med. Chem. Letters 16:358-362 (2006); Torgov et al., Bioconj. Chem. 16:717-721 (2005); Nagy et al., Proc. Natl. Acad. Sci. USA 97:829-834 (2000); Dubowchik et al., Bioorg. & Med. Chem. Letters 12: 1529-1532 (2002); King et al., J. Med. Chem. 45:4336-4343 (2002); и патент США 6630579); метотерксат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен; и CC1065.
В другом варианте осуществления изобретения иммуноконъюгат включает антитело, описанное в настоящем описании, конъюгированное с ферментативно-активным токсином и его фрагментом, включая, но без ограничения, цепь A дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь A экзотоксина (из Pseudomonas aeruginosa), цепь A рицина, цепь A абрина, цепь A модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из Momordica charantia, курцин, кротин, ингибитор из Saponaria officinalis, гелонин, митогелин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом варианте осуществления изобретения иммуноконъюгат включает антитело, описанное в настоящем описании, конъюгированное с радиоактивным атомом, образуя радиоконъюгат. Для получения радиоактивных конъюгированных антител доступны различные радиоактивные изотопы. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. В случае применения радиоконъюгата для детекции, он может содержать радиоактивный атом для сцинтиграфических исследований, например, Tc99m или I123, или спиновую метку для получения изображения с помощью ядерного магнитного резонанса (ЯМР) (также известного как магнитно-резонансная томография, МРТ), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Иммуноконъюгаты антитела и цитотоксического агента можно также получить, используя разнообразные бифункциональные агенты для присоединения белка, такие как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат-HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-реакционноспособные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Например, рициновый иммунотоксин можно получить, как описано в Vitetta et al., Science, 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатбензил-3-метилдиэтилентриаминопентауксусная кислота (MX-DTPA) представляет собой иллюстративное хелатирующее средство для конъюгации радионуклеотида с антителом. См. WO94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать неустойчивый к кислотам линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); патент США 5208020).
Иммуноконъюгаты или ADC в изобретении явным образом предусматривают, но без ограничения, конъюгаты, полученные с помощью сшивающих реагентов, включая, но без ограничения: BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A).
Е. Способы и композиции для диагностики и детекции
В некоторых вариантах осуществления изобретения любое из анти-LRP6 антител настоящего изобретения пригодно для детекции присутствия LRP6 в биологическом образце. Термин "детекция", используемый в настоящем описании, охватывает количественную или качественную детекцию. В некоторых вариантах осуществления биологический образец содержит клетку или ткань.
В одном варианте осуществления изобретение относится к анти-LRP6 антителу для применения в способе диагностики или детекции. В дополнительном аспекте изобретение относится к способу детекции наличия LRP6 в биологическом образце. В некоторых вариантах осуществления способ включает контактирование биологического образца с анти-LRP6 антителом в условиях, позволяющих связывание анти-LRP6 антитела с LRP6, и детекцию того, происходит ли образование комплекса между анти-LRP6 антителом и LRP6. Такой способ может быть in vitro или in vivo способом. В одном варианте осуществления анти-LRP6 антитело используют для отбора субъектов, подходящих для терапии с использованием анти-LRP6 антитела, например, когда LRP6 является биомаркером для отбора пациентов.
Примеры заболеваний, которые можно диагностировать с использованием антитела настоящего изобретения, включают рак и нарушения скелетной системы.
В некоторых вариантах осуществления анти-LRP6 антитела являются мечеными. Метки включают, но без ограничения, метки или молекулы, которые детектируются напрямую (такие как флуоресцентные, хромофорные, электроноплотные, хемилюминесцентные и радиоактивные метки), а также группы, такие как ферменты или лиганды, которые детектируются непрямым образом, например, посредством ферментативной реакции или молекулярного взаимодействия. Иллюстративные метки включают, но без ограничения, радиоактивные изотопы 32P, 14C, 125I, 3H и 131I, флуорофоры, такие как хелаты редкоземельных элементов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светлячка и бактериальную люциферазу (патент США 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, сахарид-оксидазы, например, глюкозо-оксидазу, галактозо-оксидазу и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, связанные с ферментом, который использует перекись водорода для окисления предшественника красителя, таким как HRP, лактопероксидаза или микропероксидаза, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
F. Фармацевтические композиции
Фармацевтические композиции анти-LRP6 антитела, описанного в настоящем описании, получают смешиванием такого антитела, имеющего желаемую степень чистоты, с одним или несколькими необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в форме лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители обычно являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают, но без ограничения: буферы, такие как фосфатный, цитратный и на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмонийхлорид; гексаметонийхлорид; бензалконийхлорид, бензэтонийхлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены (такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (приблизительно менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Иллюстративные фармацевтически приемлемые носители в настоящем описании дополнительно включают агенты для интерстициальной дисперсии лекарственных соединений, такие как растворимые нейтрально-активные гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы РН-20 человека, такие как rHuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые иллюстративные sHASEGP и способы применения, включая rHuPH20, описаны в патентных публикациях США №№ 2005/0260186 и 2006/0104968. В одном аспекте sHASEGP объединен с одной или несколькими дополнительными гликозаминогликаназами, такими как хондроитиназы.
Иллюстративные лиофилизированные составы антител описаны в патенте США 6267958. Водные составы антител включают описанные в патенте США 6171586 и WO2006/044908, причем последние составы включают гистидин-ацетатный буфер.
Составы настоящего изобретения также могут содержать более одного действующего ингредиента при необходимости для конкретного заболевания, в отношении которого проводится лечение, предпочтительно, ингредиенты с дополняющим друг друга действием, которые не влияют отрицательно друг на друга. Такие действующие ингредиенты обычно присутствуют в комбинации в количестве, которое эффективно для предполагаемой цели.
Действующие ингредиенты также можно включать в микрокапсулы, полученные, например, методиками коацервации или посредством межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методики описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Можно получать препараты с замедленным высвобождением. Примеры пригодных препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где матрицы находятся в форме формованных изделий, например, пленок или микрокапсул.
Фармацевтические композиции для применения при введении in vivo, как правило, являются стерильными. Стерильность легко достигается фильтрацией через стерильные фильтрационные мембраны.
G. Терапевтические способы и композиции
Любое из анти-LRP6 антител по настоящему изобретению можно использовать в терапевтических способах.
Один аспект изобретения относится к анти-LRP6 антителу настоящего изобретения для применения в качестве лекарственного препарата. Дополнительные аспекты относятся к анти-LRP6 антителу настоящего изобретения для использования при лечении Wnt-опосредованного заболевания, такого как рак или нарушение скелета или костной ткани. Некоторые варианты осуществления изобретения относятся к анти-LRP6 антителу для применения в способе лечения. В некоторых вариантах осуществления изобретение относится к анти-LRP6 антителу для применения в способе лечения индивидуума, страдающего раком или нарушением скелета или костной ткани, включающему введение индивидууму эффективного количества анти-LRP6 антитела. В одном таком варианте осуществления способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного терапевтического агента, например, описанного ниже. В дополнительных вариантах осуществления изобретение относится к анти-LRP6 антителу для использования при ингибировании передачи сигнала, индуцированной первой изоформой Wnt, и усиления передачи сигнала, индуцированной второй изоформой Wnt. В некоторых вариантах осуществления изобретение относится к анти-LRP6 антителу для использования в способе ингибирования передачи сигнала, индуцированной первой изоформой Wnt, и усиления передачи сигнала, индуцированной второй изоформой Wnt, у индивидуума, включающем введение индивидууму эффективного количества анти-LRP6 для ингибирования передачи сигнала, индуцированной первой изоформой Wnt, и усиления передачи сигнала, индуцированной второй изоформой Wnt. «Индивидуумом» по любому из вышеуказанных вариантов осуществления предпочтительно является человек.
В дополнительном аспекте изобретение относится к применению анти-LRP6 антитела для использования при получении или изготовлении лекарственного препарата. В одном варианте осуществления изобретения лекарственный препарат предназначен для лечения Wnt-опосредованного заболевания, такого как рак или нарушение скелета или костной ткани. В дополнительном варианте осуществления изобретения лекарственный препарат предназначен для применения в способе лечения Wnt-опосредованного заболевания, такого как рак или заболевание скелета или костной ткани, включающем введение индивидууму, страдающему Wnt-опосредованным заболеванием, эффективного количества лекарственного препарата. В одном таком варианте осуществления изобретения способ дополнительно включает введение индивидууму эффективного количества дополнительного терапевтического агента, например, описанного ниже. «Индивидуумом» по любому из вышеуказанных вариантов осуществления может быть человек.
В дополнительном аспекте изобретение относится к способу лечения Wnt-опосредованного заболевания, такого как рак или нарушение скелета или костной ткани. В одном варианте осуществления изобретения способ включает введение индивидууму, страдающему таким Wnt-опосредованным заболеванием, эффективного количества анти-LRP6 антитела. В одном таком варианте осуществления изобретения способ дополнительно включает введение индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, описанного ниже. «Индивидуумом» по любому из вышеуказанных вариантов осуществления может быть человек.
В одном варианте осуществления изобретения Wnt-опосредованным заболеванием является рак, такой как немелкоклеточный рак легких, рак молочных желез, рак поджелудочной железы, рак яичников, рак почек или рак предстательной железы. В другом варианте осуществления изобретения Wnt-опосредованным заболеванием является нарушение скелета или костной ткани, такое как остеопороз, остеоартрит, переломы костей или поражения костной ткани.
Один вариант осуществления изобретения относится к способу лечения индивидуума, страдающего раком, включающему введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b. Другой вариант осуществления изобретения относится к способу лечения индивидуума, страдающего раком, включающему введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt3 и Wnt3a, и антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b. Другой вариант осуществления изобретения относится к способу лечения индивидуума, страдающего раком, включающему введение индивидууму эффективного количества выделенного антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную изоформой Wnt3 и Wnt3a, и антитела, которое связывает LRP6 и ингибирует передачу сигнала, индуцированную Wnt 1, 2, 2b, 4, 6, 7a, 7b, 8a, 9a, 9b, 10a и 10b.
В дополнительном аспекте изобретение относится к способу усиления передачи сигнала от Wnt, индуцированной изоформой Wnt, у индивидуума, включающему введение индивидууму эффективного количества анти-LRP6 антитела, которое усиливает передачу сигнала от изоформы Wnt, и изоформы Wnt для усиления передачи сигнала от Wnt, индуцированной изоформой Wnt.
В дополнительном аспекте изобретение относится к фармацевтическим составам, содержащим любое из анти-LRP6 антител настоящего изобретения, например, для применения в любом из вышеуказанных терапевтических способах. В одном варианте осуществления изобретения фармацевтический состав содержит любое из анти-LRP6 антител настоящего изобретения и фармацевтически приемлемый носитель. В другом варианте осуществления изобретения фармацевтический состав включает любое из анти-LRP6 антител настоящего изобретения и по меньшей мере один дополнительный терапевтический агент, например, описанный ниже.
Антитела настоящего изобретения можно применять в терапии либо по отдельности, либо в сочетании с другими агентами. Например, антитело настоящего изобретения можно совместно вводить по меньшей мере с одним дополнительным терапевтическим агентом. В некоторых вариантах осуществления изобретения дополнительным терапевтическим агентом является химиотерапевтический агент. В другом варианте осуществления изобретения агентом является антитело, которое эффективно для лечения рака или для лечения нарушений скелета или костной ткани. Такие комбинированные методы лечения, указанные выше, охватывают совместное введение (в котором два или несколько терапевтических агентов включены в состав одного или отдельных составов) и раздельное введение, в случае которого введение антитела настоящего изобретения можно осуществлять до введения дополнительного терапевтического средства и/или адъюванта, одновременно с ним, и/или после него. Антитела настоящего изобретения можно также применять в сочетании с радиотерапией.
Антитело настоящего изобретения (и любой другой дополнительный терапевтический агент) можно вводить любым подходящим путем, включая парентеральное, внутрилегочное и интраназальное и, если требуется для местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Введение доз можно осуществлять любым подходящим способом, например, с помощью инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение краткосрочным или хроническим. В настоящем описании предусмотрены различные схемы введения, включая, но без ограничения, однократное или многократное ведение через различные интервалы времени, болюсное введение и пульсовую инфузию.
Антитела настоящего изобретения составляют, дозируют и вводят в соответствии с надлежащей медицинской практикой. Факторы, которые следует учитывать в данном контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние конкретного пациента, причину нарушения, участок доставки средства, способ введения, схему введения и другие факторы, известные практикующим врачам. Антитело необязательно, но по возможности, вводят в состав с одним или несколькими агентами, в настоящее время используемыми для профилактики или лечения рассматриваемого нарушения. Эффективное количество таких других агентов зависит от количества антитела, присутствующего в составе, типа нарушения или лечения и других факторов, рассмотренных выше. Их, как правило, используют в тех же дозировках и способах введения, как описано в данном описании, или в количестве приблизительно от 1 до 99% от описанных в данном описании дозировок, или в любой дозировке и любым способом, который эмпирически/клинически определен в качестве подходящего.
Для профилактики или лечения заболевания соответствующая дозировка антитела настоящего изобретения (при использовании индивидуально или в сочетании с одним или несколькими другими дополнительными терапевтическими агентами) зависит от типа подлежащего лечению заболевания, типа антитела, тяжести и течения заболевания, от того, вводят ли антитело с профилактическими или терапевтическими целями, предшествующего лечения, истории болезни пациента и ответа на антитело, а также от решения лечащего врача. Антитело вводят пациенту соответствующим образом за один раз или лечебными курсами. В зависимости от типа и тяжести заболевания примерно от 1 мкг/кг до 15 мг/кг (например, 0,1-10 мг/кг) антитела может представлять собой начальную возможную дозировку для введения пациенту, либо, например, посредством однократного или многократного введения, либо посредством непрерывной инфузии. Одна конкретная суточная дозировка может варьировать примерно от 1 мкг/кг до 100 мг/кг или более в зависимости от вышеуказанных факторов. При повторном введении в течение нескольких суток или более в зависимости от состояния лечение, как правило, продолжают до тех пор, пока не наступит требуемое подавление симптомов заболевания. Одна из иллюстративных дозировок антитела находится в диапазоне примерно от 0,05 мг/кг до примерно 10 мг/кг. Таким образом, пациенту можно вводить одну или несколько доз примерно по 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любые их комбинации). Такие дозы можно вводить периодически, например каждую неделю или каждые три недели (например, чтобы пациент получал приблизительно от двух до приблизительно двадцати, например, около шести доз антитела). Можно вводить начальную более высокую ударную дозу с последующей одной или несколькими более низкими дозами. Прогресс данного метода лечения легко отслеживать посредством общепринятых методик и методов анализа.
Следует понимать, что любые из вышеуказанных составов и методов лечения можно проводить, используя иммуноконъюгат настоящего изобретения вместо или добавочно к анти-LRP6 антителу.
Н. Готовые изделия
В другом варианте осуществления изобретение относится к готовому изделию, содержащему материалы, пригодные для лечения, профилактики и/или диагностики описанных выше нарушений. Готовое изделие включает контейнер и этикетку (или вкладыш в упаковку) на контейнере или ассоциированную с ним. Пригодные контейнеры включают, например, бутыли, флаконы, шприцы, мешки для внутривенных инъекций и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая сама по себе или при сочетании с другой композицией является эффективной для лечения, профилактики и/или диагностики состояния, и может иметь отверстие для стерильного доступа (например, контейнер может представлять собой мешок для раствора для внутривенных инъекций или флакон с пробкой, проницаемой для игл для подкожных инъекций). По меньшей мере один действующий агент в композиции является антителом настоящего изобретения. На этикетке или вкладыше в упаковку указано, что композицию используют для лечения выбранного состояния. Более того, готовое изделие может содержать (a) первый контейнер с содержащейся в нем композицией, где композиция содержит антитело настоящего изобретения; и (b) второй контейнер с содержащейся в нем композицией, где композиция содержит дополнительный цитотоксический агент или другой терапевтический агент. Готовое изделие в этом варианте осуществления изобретения может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного состояния. Альтернативно или дополнительно, готовое изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), забуференный фосфатом солевой буфер, раствор Рингера и раствор декстрозы. Оно может дополнительно содержать другие необходимые с коммерческой и потребительской точки зрения материалы, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Следует понимать, что любые из вышеуказанных готовых изделий могут включать иммуноконъюгат настоящего изобретения, вместо или добавочно к анти-LRP6 антителу.
III. ПРИМЕРЫ
Далее следуют примеры способов и композиций настоящего изобретения. Следует понимать, что можно использовать различные другие варианты осуществления на основе приведенного выше общего описания.
Пример 1
Экспериментальные методики
Культивирование клеток и клеточные методы анализа
Клеточные линии EKVX и M14 выращивали в среде RPMI-1640 с 10% фетальной бычьей сывороткой и 2 мМ глутамином; клетки JHH-1 выращивали в среде Williams' Medium E с такими же добавками. Все другие клеточные линии получали от американской коллекции типовых культур (АТСС) и культивировали в соответствии с рекомендациями.
Клетки трансфицировали с использованием трансфекционного реагента FuGENE 6 (Roche) в 24-луночных планшетах в соответствии с рекомендациями изготовителя. Для люциферазных репортерных методов анализа трансфицировали смесь ДНК экспрессионных плазмид: 7,5 нг TOPglow (Upstate) или TOPbrite (Zhang et al., 2009) (Wnt-репортера с люциферазой светлячка), 0,5 нг pRL-SV40 (люцифераза из Renilla, Promega) и 1 нг LEF1. Клетки обрабатывали антителами в течение 16-20 ч через 24 часа после трансфекции. К клеткам добавляли белок Wnt3a (очищенный в соответствии с X или приобретенный от R&D Systems), через 1 час после начала воздействия антител. Клетки собирали в 150 мкл буфера для лизиса (DeAlmeida et al., 2007), и люминесценцию измеряли для 30-50 мкл лизата, используя систему Dual-Glo Luciferase (Promega) и считывающее устройство Envision Multilabel (PerkinElmer). Уровень экспрессии люциферазы светлячка нормализовали по эффективности трансфекции на уровень экспрессии люциферазы из Renilla, и относительные люциферазные единицы (RLU) дополнительно нормализовали на уровень свечения в клетках, не стимулированных Wnt3a.
Клеточные линии HEK293 и Hs578T со стабильно интегрированным репортером TOPbrite отбирали по устойчивости к гигромицину. Экспрессию Wnt-люциферазного репортера нормализовали на число клеток, исходя из стабильно интегрированной люциферазы Renilla под контролем SV40-промотора в клетках HEK293 или исходя из анализа жизнеспособности клеток MultiTox-Fluor (Promega) в клетках Hs578T.
Конструкции химер Wnt создавали, клонируя полноразмерный Wnt1 или Wnt3a выше полноразмерного FZD4, FZD5 или LRP6 в экспрессионном векторе pRK5. 24-аминокислотный линкер (GGGSGGGT)3 встраивали между последовательностями Wnt и FZD или LRP6 (Cong et al., 2004).
Одноплечевой вариант антитела YW211.31 получали в E. coli совместной экспрессией тяжелой и легкой цепей YW211.31.62 с укороченным Fc-доменом, используя методику конструирования «узел в полости» (Ridgway, J.B.B. et al., Protein Engineering 9:617-621 (1996)). Для сшивания антител, козьи антитело или фрагмент F(ab')2, специфичные к Fc области IgG человека (Sigma-Aldrich), инкубировали с одноплечевым антителом YW211.31 в течение 1 часа перед добавлением смеси к клеткам.
Для Вестерн-анализа высевали 1,2×106 клеток HEK293 на 10-см чашки и через 3 дня обрабатывали 10 мкг/мл антитела или Х мкг/мл белка DKK1 (R&D Systems) или Fzd8CRD-Fc (DeAlmeida et al., 2007) в течение 1 часа перед добавлением 0,2 мкг/мл белка Wnt3a еще на 1 час. Клетки промывали дважды холодным PBS и лизировали в 0,5 мл буфера для лизиса на льду. 20 мкг белка разделяли электрофоретически в денатурирующем ДСН-полиакриламидном геле (4-12%), переносили на нитроцеллюлозную мембрану и блотировали с антителами к фосфо- и общему LRP6 (Cell Signaling Technology), β-катенину (BD Transduction Laboratories), β-актину и GAPDH. Белки визуализировали, используя инфракрасно-меченные вторичные антитела (Rockland Immunochemicals) и устройство для визуализации Odyssey imager (LI-COR).
Для анализа экспрессии с помощью количественной ПЦР в реальном времени (кПЦР) РНК выделяли из клеток, используя набор RNeasy (QIAGEN), и реакции проводили с набором TaqMan One-Step RT-PCR Master Mix Reagents (Applied Biosystems) на системе 7900 HT Fast Real-Time PCR (Applied Biosystems). Относительные уровни РНК вычисляли, используя метод ΔΔCt и нормализовали на уровни РНК GAPDH человека или Rpl19 мыши в одном образце, и дополнительно нормализовали на образцы из клеток без добавления (NA) Wnt3a, антитела или других белков. Наборы праймеров и зондов, указанных в 5'-3' направлении для последовательностей прямого праймера, обратного праймера и зонда, соответственно, являются:
SP5: AATGCTGCTGAACTGAATAGAAA (SEQ ID NO:32),
AACCGGTCCTAGCGAAAA (SEQ ID NO:33),
CCGAGCACTGTTTCAAATCTCCCA (SEQ ID NO:34);
ZNRF3: TGAGAGTGTGACATTGTTGGAA (SEQ ID NO:35),
GTAAAATCTGTGTGCAATTATCATGT (SEQ ID NO:36),
AATCATTGAAAATGACTAACACAAGACCCTGTAAAT (SEQ ID NO:37);
мышиная Mmp7: TGAGGACGCAGGAGTGAA (SEQ ID NO:38),
CCCAGAGAGTGGCCAAAT (SEQ ID NO:39),
CCTGTTTGCTGCCACCCATGA (SEQ ID NO:40).
Праймеры и зонды, используемые для APCDD1, AXIN2, GAD1, LEFTY2 и SAX1 человека, и для Rpl19 и Axin2 мыши, были описаны ранее (DeAlmeida, et al. (2007); Liu et al., (2010)).
Праймеры и зонд к GAPDH были приобретены от Applied Biosystems. Для методов анализа с использованием репортерного гена и кПЦР все фигуры представляют среднее и среднеквадратичное отклонение для трех или четырех повторов в эксперименте.
Скрининг антител к LRP6 и созревание аффинности
Фрагменты кДНК LRP6 человека, кодирующие области E1-E2 (аминокислоты A19-R644 в SEQ ID NO:29) и E3-E4 (аминокислоты V629-G1244 в SEQ ID NO:29) по отдельности клонировали в вектор для экспрессии в клетках млекопитающих, содержащий HSV-сигнальную последовательность и Fc-область IgG человека в качестве белкового тэга (SEQ ID NO:30 (E1-E2-fc); SEQ ID NO:31 (E3-E4-fc)). Белки LRP6.E1-E2-Fc и LRP6.E3-E4-Fc экспрессировали в клетках СНО с помощью транзиентной трансфекции и очищали, используя аффинную хроматографию на белках A/G. Белки LRP6.E1-E2-Fc и LRP6.E3-E4-Fc также использовали по отдельности для скрининга библиотеки фагового дисплея, содержащей синтезированные Fab человека. После отбора на иммобилизованном белке LRP6 клоны фагов выделяли, и связывание подтверждали с помощью фагового ELISA на связывание с фрагментом слитого белка LRP6-Fc, но не с белком Fc. Фаговые Fab-клоны затем реформатировали для экспрессии в качестве моноклональных антител IgG1 человека. 24 уникальных клона тяжелых цепей антител к LRP6.E1-E2-Fc и 22 клона к LRP6.E3-E4-Fc трансфицировали и транзиентно экспрессировали в клетках HEK293 с общей легкой каппа-цепью человека, полученной из антитела герцептин, и белок IgG очищали аффинной хроматографией. Впоследствии, получение препаратов антител в большом масштабе осуществляли транзиентной трансфекцией в клетках СНО.
Аффинное созревание антитела YW211.31 проводили, используя His-тэгированный белок LRP6.E3-E4. Три различные комбинации петель CDR (H1/L3, H2/L3 и H3/L3) являлись мишенями для рандомизации в отдельных библиотеках с использованием «мягкой» рандомизации выбранных остатков. Кроме того, для интенсивной («тяжелой») рандомизации мишенью являлась комбинация CDR-L1/L2/L3. В первом раунде отбора фаги из рандомизированных библиотек отбирали на иммобилизованный белок LRP6.E3-4-His с последующими пятью раундами жидкофазного сортинга, в котором концентрацию LRP6.E3-4-His постепенно снижали с 300 нМ до 0,5 нМ и добавляли 100-кратный избыток белка LRP6.E3-4-Fc для истощения антител с более быстрой скоростью диссоциации. Было очищено одиннадцать фаговых клонов, и все они показали увеличенную аффинность к LRP3.E3-E4, определенную с помощью фагового конкурентного ELISA. Последовательности этих клонов продемонстрировали 1-6 аминокислотных замен в CDR-H1, CDR-H3 и CDR-L3. Константы скорости диссоциации очищенных антител оценивали с помощью анализа поверхностного плазмонного резонанса, используя прибор BIAcore.
Анализ связывания с белком LRP6 с помощью интерферометрии биослоев
Интерферометрию биослоев проводили, как описано ранее (Bourhis et al., 2010). Кратко, биотинилированные, His-тэгированные белки LRP6 очищали из инфицированных бакуловирусами клеток насекомых, используя систему AviTag (GeneCopoeia). Кинетику связывания измеряли на системе Octet RED (ForteBio), используя Streptavidin High Binding FA Biosensors, нагруженные 20 мкг/мл белка LRP6. Свободный от носителя очищенный Wnt3a человека и Wnt9b мыши получали из R&D Systems, и очищенный белок DKK1 получали, как описано ранее (Bourhis et al., 2010).
Исследования опухолей и костной ткани
Опухоли из трансгенных мышей MMTV-Wnt1 пассировали в скоплениях жировой ткани в молочных железах мышей C57BL/6, механически и ферментативно разделяли, ресуспендировали в матригеле и сбалансированном солевом растворе Хенкса (HBSS), и вводили с помощью инъекций в скопления жировой ткани в молочных железах бестимусных голых мышей NCr (Taconic). Воздействие начинали после того, как объем опухоли достигал 250-800 мм3. Для каждой группы воздействия десяти мышам вводили по 30 мг/кг антитела или белка внутрибрюшинно (IP) каждые два дня. Объем опухоли анализировали, используя измерение штангенциркулем.
Выращивание ксенотрансплантированной опухоли Ntera-2 и исследования in vivo проводили, как описано ранее (DeAlmeida et al., 2007). Кратко, бестимусным голым мышам NU/NU (Charles River) вводили с помощью подкожных инъекций по 10 миллионов клеток Ntera-2 (в 50%-ом матригеле в HBSS) на мышь, которых разделяли на группы по 4-5 животных после того, как средний объем опухоли достигал 535-595 мм3, и вводили с помощью однократной внутрибрюшинной инъекции дозу антитела 100 мг/кг или 30 мкг/кг белка Fzd8CRD-Fc. Образцы опухолей и сыворотки крови собирали через 16 часов после воздействия. Опухоли гомогенизировали, используя систему TissueLyser (QIAGEN), и выделяли РНК, используя набор RNeasy (QIAGEN).
Своды черепа собирали и культивировали, как описано Mohammad et al., 2008. Кратко, своды черепа отсекали от 2-дневных детенышей мышей, разрезали пополам и отделяли от твердой оболочки, сосудов и кожи черепа. Своды черепа культивировали в планшетах для культуры тканей в среде BGJb с 0,1%-м бычьим сывороточным альбумином и 100 Ед/мл каждого из пенициллина и стрептомицина в течение 1 дня перед воздействием 10 мкг/мл антитела или белка в течение 7 дней. Костную ткань культивировали во влажной атмосфере с 5% CO2 при 37°C. Изображения сводов черепа мышей получали с использованием микросистемы для рентгеновской компьютерной томографии (КТ), μCT 40 (SCANCO Medical, Basserdorf, Switzerland). Данные микрокомпьютерной томографии получали, используя следующие параметры: уровень энергии рентгеновской трубки, равный 45 кВ, ток составлял 177 мкΑ, время накопления составляло 300 мс, 2000 проекций. Аксиальные изображения получали с изотропным разрешением 6 мкм. Для калибровки использовали гидроксиапатитный (НА) фантом. Изображения, полученные с помощью микро-КТ анализировали, используя Analyze (AnalyzeDirect Inc., Lenexa, KS, USA). Для каждого образца создавали проекции с максимальной интенсивностью и трехмерное изображение поверхности в поперечной плоскости. Границы теменной кости рисовали вручную, используя Trace tool для того, чтобы сегментировать теменную область. В этой области вычисляли объем образца и среднюю минеральную плотность костной ткани (BMD). Использовали пороговое значение 0,3 мг/см3 для этой области, чтобы вычислить среднюю BMD только кальцифицированной ткани внутри области. Пороговое значение также использовали для вычисления процента кальцифицированного объема теменной области путем деления числа кальцифицированных элементов объема на общее число элементов объема для теменной области. Следующие параметры анализировали для каждого образца: объем теменной области, BMD кальцифицированных элементов объема теменной области и процент кальцификации теменной области. Разницу между группами считали достоверной, если значения р были ниже 0,05 в тесте Дуннетта.
Все эксперименты с использованием мышей проводили в соответствии с инструкциями ведомственного комитета по содержанию и использованию животных компании Дженентек.
Пример 2
Выделение анти-LRP6 моноклональных антител, обладающих антагонистическим и усиливающим действием в отношении передачи сигнала от Wnt
Для разработки потенциальных терапевтических молекул для манипулирования передачи сигнала от Wnt, создавали антитела, которые могут либо ингибировать, либо усиливать передачу сигнала, индуцированную белком Wnt3a. Рекомбинантные белки LRP6.E1-E2-Fc (SEQ ID NO:30) и LRP6.E3-E4-Fc (SEQ ID NO:31) использовали для скрининга библиотеки фагового дисплея, содержащей синтезированные Fab человека, и связывание выделенных фаговых клонов с LRP6 подтверждали с помощью ELISA. 24 уникальных клона тяжелых цепей антител к LRP6.E1-E2 и 22 клона к LRP6.E3-E4 выделяли, реформатировали и экспрессировали в качестве антител IgG1 человека. Шесть анти-LRP6.E3-E4 антител ингибировали зависимым от концентрации образом активность Wnt-люциферазного репортера в клетках НЕК293, индуцированную 0,1 мг/мл очищенного Wnt3a (фиг.1A. Планки погрешностей на данном и всех других графиках, если не указано иное, представляют среднеквадратичное отклонение по меньшей мере для 3 повторных выборок). Эти антитела были названы YW211.03, YW211.08, YW211.11, YW211.12, YW211.31 и YW211.33. Ни одно из антител к LRP6.E1-E2 не проявляло ингибирующих свойств. Антитело YW211.31, распознающее домен LRP6.E3-E4, наиболее сильно ингибировало передачу сигнала в стимулированных Wnt3a клетках HEK293 с IC50 приблизительно 1 мкг/мл (или 6 нМ). Антитело YW211.31 ингибировало индуцированное Wnt3a фосфорилирование LRP6 и стабилизацию белка β-катенина, не влияя на уровень белка LRP6, аналогично очищенным белкам Fzd8CRD и DKK1 (фиг.1B, на которой показан Вестерн-анализ клеток, либо нестимулированных, либо активированных Wnt3a, и получавших воздействие указанного антитела к LRP6 или очищенного белка (уровни белков β-актина и GAPDH показаны в качестве контроля наноса образцов)). Эксперименты по РНКи продемонстрировали, что только полоса меньшей молекулярной массы, узнаваемая поликлональным антителом к β-катенину, представляет собой белок β-катенин. Антитело YW211.31 также может противодействовать функции мышиного LRP6, так как оно частично ингибирует индуцированную Wnt3a активность репортера в мышиных клетках NIH/3T3 и стабилизацию белка β-катенина в мышиных клетках L.
Антитело YW211.31 имеет аффинность связывания примерно 2 нМ по поверхностному плазмонному резонансу (SPR) и 0,6 нМ по анализу Скетчарда. Чтобы улучшить аффинность и возможную эффективность действия антитела YW211.31, проводили созревание аффинности клона, используя His-тэгированный белок LRP6 E3-E4 и комбинаторные библиотеки CDR, в которых отдельные остатки в CDR являлись мишенью для рандомизации. Четыре фаговых клона, показывающих наибольшее увеличение аффинности в фаговом конкурентном ELISA - YW211.31.11, YW211.31.11, 35, YW211.31.57 и YW211.31.62 - были реформатированы и экспрессированы в качестве полноразмерных IgG человека. Константы скорости диссоциации всех четырех аффинно-зрелых IgG были снижены, приводя к увеличению аффинности для наилучших двух антител, YW211.31.57 и YW211.31.62, с KD 0,27 и 0,17 нМ, соответственно. YW211.31.57 и YW211.31.62 также показывали увеличение эффективности ингибирования передачи сигнала в стимулированных Wnt3a клетках HEK293 с величинами IC50 приблизительно 0,1 мкг/мл (0,6 нМ).
Ни одно из антител, выделенных при скрининге, не активировало передачу сигнала в клетках НЕК293 при отсутствии стимуляции экзогенным белком Wnt3a, однако пять антител к LRP6 E1-E2 и два антитела к E3-E4, усиливали индуцированную Wnt3a передачу сигнала по меньшей мере в 2 раза. В мышиных клетках NIH/3T3 YW210.09, антитело к Е1-Е2, также усиливало индуцированную Wnt3a передачу сигнала по меньшей мере в 1,5 раза, что указывает на то, что оно также распознает мышиный LRP6. В клетках HEK293 величина усиления индуцированной Wnt3a передачи сигнала в результате действия антитела YW210.09 пропорциональна концентрации Wnt3a (фиг.1C). Антитело YW210.09 взаимодействует с белком LRP6 E1-E2 человека с KD 5 нМ, измеренной SPR-анализом. Тестирование методом ELISA показывает, что все антагонистические и усиливающие антитела специфично связывают только фрагмент белка LRP6, использованный при их выделении, и ни одно не распознает оба фрагмента E1-E2 и E3-E4. Анализ методом FACS указывает на то, что растворимый белок LRP6 E1-E4 эффективно и полностью блокирует связывание YW211.31.57 и YW210.09 с клетками HEK293, что указывает на то, что эти антитела не распознают другие белки клеточной поверхности.
Пример 3
Эффекты моноклональных антител к LRP6 на аутокринную передачу сигнала от Wnt
Способность антител к LRP6 противодействовать или усиливать эндогенную или аутокринную передачу сигнала от Wnt определяли с использованием ряда опухолевых клеточных линий (Bafico et al., 2004; DeAlmeida et al., 2007; Akiri et al., 2009). В клеточных линиях тератокарциномы PA-1 и NTERA-2 антитело YW211.31 ингибирует активность репортера, индуцированную аутокринной передачей сигнала от Wnt, с той же интенсивностью, которую наблюдали с экзогенным Wnt3a (на фиг.2A показано зависимое от концентрации ингибирование и усиление аутокринной передачи сигнала от Wnt в клетках тератокарционмы PA-1, трансфицированных люциферазными репортерами и получавших воздействие антител к LRP6, либо по отдельности, либо в комбинации, либо белка Fzd8CRD-Fc (положительный контроль)).
В клетках РА-1 ингибирование передачи сигнала от Wnt антителом YW211.31 также наблюдали в отношении экспрессии эндогенных генов-мишеней Wnt (фиг.2B). На фиг.2B показаны результаты анализа экспрессии с помощью кПЦР индуцированных Wnt генов SAX1 и GAD1 и подавляемого Wnt гена, LEFTY2, в клетках PA-1, не получавших или получавших воздействие 0,3 мг/мл белка Wnt3a, и получавших воздействие 10 мг/мл антитела YW211.31, анти-gD моноклонального антитела в качестве отрицательного контроля или белка Fzd8CRD-Fc в качестве образца положительного контроля, и дополнительно нормализованных на образцы из клеток без добавления Wnt3a (NA).
Антитело частично ингибирует экспрессию SAX1, GAD1 и APCDD1, которая либо индуцирована экзогенным белком Wnt3a, либо поддерживается эндогенной аутокринной передачей сигнала от Wnt. И наоборот, подавление экспрессии LEFTY2 либо белком Wnt3a, либо аутокринной передачей сигнала от Wnt, освобождается в результате действия антитела YW211.31. В отличие от YW211.31, антитело YW210.09 усиливает как индуцированную Wnt3a аутокринную передачу сигнала от Wnt в клеточных линиях PA-1 и NTERA-2 по анализу экспрессии репортерного гена (фиг.2A). Хотя ингибирование передачи сигнала от Wnt антителом YW211.31.57 увеличивается постепенно с увеличением концентрации антитела, усиление сигнала под действием YW210.09 и других антител может снижаться при высоких концентрациях антител в некоторых типах клеток, таких как клетки РА-1. Это позволяет предположить, что для усиления сигнала необходима димеризация рецептора LRP6, так как высокие концентрации антител могут благоприятствовать моновалентным взаимодействиям, и это ограничивает сшивку молекул LRP6. Воздействие на клетки PA-1 или NTERA-2 комбинации двух антител YW211.31.57 и YW210.09 противодействует как индуцированной Wnt3a, так и аутокринной передаче сигнала от Wnt, аналогично эффекту одного YW211.31.57.
Для идентификации дополнительных клеточных линий, которые имеют аутокринную передачу сигнала от Wnt, клеточные линии с относительно высокой экспрессией мРНК Axin2 или фосфо-LRP5/6 тестировали на ингибирование передачи сигнала от Wnt белком Fzd8CRD-Fc в анализах Wnt-люциферазного репортерного гена. Девять клеточных линий имели аутокринную передачу сигнала от Wnt, которую ингибировал белок Fzd8CRD-Fc, включая клетки немелкоклеточной легочной карциномы, NCI-H23 и NCI-H2030 и клетки саркомы мягких тканей, SW872 и HT-1080, которые, как ранее было опубликовано, имеют эндогенную передачу сигнала от Wnt, исходя из анализов с другими антагонистами Wnt (Guo et al., 2008; Akiri et al., 2009; Nguyen et al., 2009). На фиг.3 показан итоговый результат анализа данных с помощью однонаправленного дисперсионного анализа (ANOVA) (со значением p<0,01). Анализы проводили, используя 10 мг/мл антител, за исключением клеток NCI-H358 и HT-1080, которые обрабатывали 1 мг/мл антител YW211.31.57 или YW210.09, соответственно, для увеличения эффектов усиления передачи сигнала.
Передачу сигнала от Wnt дополнительно индуцировали во всех девяти клеточных линиях с помощью экзогенного белка Wnt3a, и антитело YW211.31.57 ингибировало этот ответ на Wnt3a (фиг.3, 4A, 4D и 4F). К удивлению, антитело YW211.31.57 усиливало аутокринную передачу сигнала от Wnt во всех этих девяти линиях, в то время как YW210.09 усиливало аутокринную передачу сигнала от Wnt в пяти линиях и ингибировало в трех линиях (фиг.3, 4A- 4C, 4E и 4F). Это противоположное действие антитела YW211.31.57 на аутокринную и индуцированную Wnt3a передачу сигнала наблюдали не только с использованием люциферазного репортера, но также и для экспрессии эндогенных генов-мишеней Wnt, таких как Axin2 в шести протестированных клеточных линиях (фиг.4A, 4B и 4C). В клеточных линиях EKVX и карциномы молочных желез, Hs578T, было подтверждено, что увеличение передачи сигнала от Wnt антителом YW211.31 зависит от аутокринного(ых) Wnt, демонстрацией того, что это увеличение блокировалось белком Fzd8CRD-Fc (фиг.4G). Усиление аутокринной передачи сигнала от Wnt в клетках EKVX и Hs578T также наблюдалось для других пяти антител-антагонистов индуцированной Wnt3a передачи сигнала, которые были идентифицированы при скрининге.
На фиг.4А анализ экспрессии мРНК AXIN2 с помощью кПЦР в HT-1080, EKVX, NCI-H358 и Hs578T указывает на то, что антитело YW211.31.57 (25 мг/мл) усиливает аутокринную (NA) передачу сигнала от Wnt и ингибирует передачу сигнала, индуцированную Wnt3a (0,2 мг/мл), в то время как Fzd8CRD-Fc (25 мг/мл) противодействует как аутокринной (NA), так и индуцированной Wn3a передаче сигнала. На фиг.4В и 4В экспрессия индуцированного Wnt гена в клетках NCI-H23 (B) и M14 (C) усиливалась антителом YW211.31.57 и ингибировалась антителом YW210.09 (30 мг/мл). Воздействие Wnt3a (0,2 мг/мл) и Fzd8CRD-Fc (30 мг/мл) показано в качестве положительных контролей для усиления и ингибирования, соответственно, аутокринной передачи сигнала от Wnt, и белок CD4-Fc (B) или анти-gD антитело (C) служат в качестве отрицательных контролей (30 мг/мл). Для клеток M14 (C), белок Wnt3a и антитело YW211.31.57 сильнее увеличивали экспрессию AXIN2 и SP5, чем экспрессию APCDD1 и ZNRF3. На фиг.4D и 4E показано, что в клетках Hs578T со стабильно встроенным люциферазным Wnt-репортером антитело YW211.31.57 показывает зависимое от концентрации ингибирование стимулированной Wnt3a передачи сигнала (E), в то время как белок Fzd8CRD-Fc ингибирует, а антитело YW210.09 усиливает передачу сигнала в присутствии или без (NA) стимуляции 0,1 мг/мл Wnt3a. Эксперименты РНКи указывают, что по меньшей мере 41% индуцированной Wnt3a передачи сигнала в клетках Hs578T зависит от экспрессии LRP5 и, предполагается, ингибируется белком Fzd8CRD-Fc, но не антителом YW211.31.57. В этом эксперименте люциферазу под контролем SV40 не трансфицировали для нормализации, а, наоборот, было независимо подтверждено, что воздействие антител и белка не имеют значительного влияния на выживаемость данной клеточной линии. На фиг.4F и 4G показано, что под действием антитела YW211.31.57 клетки EKVX, трансфицированные Wnt-люциферазным репортером, также показывают усиление аутокринной передачи сигнала от Wnt и блокирование индуцированной Wnt3a передачи сигнала. Опосредованное антителом усиление аутокринной передачи сигнала от Wnt ингибируется 5 мг/мл белка Fzd8CRD-Fc.
Пример 4
Противоположное действие анти-LRP6 антител на различные изоформы Wnt
Антитело YW211.31 ингибирует передачу сигнала, индуцированную экзогенным белком Wnt3a во всех клеточных линиях, но может либо ингибировать, либо усиливать аутокринную передачу сигнала от Wnt в зависимости от клеточной линии, что позволяет предположить, что конкретная изоформа Wnt, управляющая аутокринным сигналом, определяет активность антитела. Поэтому, определяли активность антитела на передачу сигнала, индуцированную экзогенной экспрессией Wnt3a и других изоформ Wnt. Антитело YW211.31.57 ингибирует передачу сигнала от Wnt, индуцированную трансфекцией Wnt3a, либо в клетках HEK293, либо в клетках Hs578T с эффективностью, аналогичной ингибированию передачи сигнала, индуцированной воздействием белка Wnt3a. К удивлению, антитело YW211.31.57 усиливало передачу сигнала, индуцированную экспрессией Wnt1 в обеих клеточных линиях. Оба сигнала, от Wnt1 и Wnt3a, как и ожидалось, ингибировались белком Fzd8CRD-Fc. Усиление передачи сигнала от Wnt1 также наблюдалось с другими Wnt3a-антагонистическими антителами, идентифицированными в скрининге. Антитело YW210.09 также проявляло противоположное действие в отношении передачи сигнала, индуцированной Wnt3a и Wnt1, которое было прямо противоположным действию YW211.31.57; то есть усиление сигнала от Wnt3a и ингибирование сигнала от Wnt1. Антитело YW210.09 также ингибировало передачу сигнала от Wnt1 в растущих в культуре опухолевых клетках из мышиных опухолей MMTV-Wnt1, что наблюдалось по снижению экспрессии генов-мишеней Wnt, Axin2 и Mmp7, в той же степени, что и при воздействии белка Fzd8CRD-Fc. В клетках MMTV-Wnt1 антитело YW211.31.57 не было способно усиливать передачу сигнала от Wnt1, возможно, вследствие того, что передача сигнала от Wnt1 уже максимальна в этих клетках.
Показав, что антитела YW211.31.57 и YW210.09 имеют противоположную активность в отношении передачи сигнала от Wnt, вызываемой Wnt3a и Wnt1, этот анализ также был проведен для дополнительных 11 из 19 генов, которые вызывают более чем двукратную активацию люциферазного репортера в клетках НЕК293. На фиг.5 представлены итоги данных анализов, проведенных с использованием 10 мкг/мл антител, 10 мкг/мл Fzd8CRD. Антитело W211.31.57 ингибировало только активность Wnt3a и Wnt3, и оба сигнала усиливало антитело YW210.09. Сигнал от семи изоформ Wnt в дополнение к Wnt1 усиливало антитело YW211.31.57 и ингибировало антитело YW210.09. Третий класс изоформ Wnt (Wnt7a, 7b и 10a) проявлял сигнальную активность, которую не ингибировало ни одно антитело, и усиливало по меньшей мере антитело YW211.31.57. В клетках Hs578T, трансфицированных различными изоформами Wnt, антитела проявляли большинство этих же самых активностей. В частности, YW211.31.57 ингибирует Wnt3 и Wnt3a и усиливает все 11 других изоформ Wnt, которые индуцируют люциферазный репортер по меньшей мере в два раза. YW210.09 также усиливает Wnt3 и Wnt3a в клетках Hs578T, а также ингибирует 5 из 7 изоформ Wnt, которые оно ингибировало в клетках HEK293, и которые подходили для тестирования в клетках Hs578T. На другие две изоформы Wnt в этом классе, Wnt8a и Wnt9b, антитело YW210.09 не оказывало влияние в клетках Hs578T. Поскольку эксперименты РНКи указывают на то, что передача сигнала от Wnt3a в клетках Hs578T, но не в НЕК293, передается как через LRP6, так и через LRP5, то Wnt8a и Wnt9b могут передавать сигнал преимущественно через LRP5 в клетках Hs578T. Как в клетках НЕК293, изоформы Wnt в третьем классе не ингибировало ни одно антитело в клетках Hs587T, и авторы могут добавить Wnt4 к этому классу, который не вызывает сигнал в клетках HEK293. В этом классе активность только Wnt7b отличается от других в том, что антитело YW210.09 усиливает передачу сигнала от него в клетках Hs578T, но не в клетках HEK293. В отличие от специфичной к изоформам Wnt активности антител YW211.31.57 и YW210.09, белок Fzd8CRD-Fc может эффективно ингибировать активность всех Wnt за исключением Wnt6 и Wnt9b в клетках HEK293. В клетках Hs578T аутокринную передачу сигнала от Wnt усиливают оба антитела, YW211.31.57 и YW210.09, также как передача сигнала индуцируется экспрессией Wnt4, Wnt7a и Wnt7b. Таким образом, эти три изоформы Wnt могут представлять собой изоформы Wnt, которые управляют аутокринной передачей сигнала в клетках Hs578T.
Многочисленные киРНК к Wnt7b, но не к другим изоформам Wnt, ингибируют аутокринную передачу сигнала в клетках Hs578T, что позволяет идентифицировать конкретный белок Wnt, обуславливающий передачу сигнала. Поскольку аутокринную передачу сигнала от Wnt в клетках РА-1 ингибировало антитело YW211.31.57 и усиливало антитело YW210.09, Wnt3 или Wnt3a, вероятно, активируют эндогенную передачу сигнала в этих клетках. Действительно, киРНК к Wnt3, но не к Wnt3a, ингибируют аутокринную передачу сигнала в клетках PA-1. В клетках немелкоклеточной легочной карциномы, NCI-H23, в клетках меланомы, M14, усиление аутокринной передачи сигнала от Wnt антителом YW211.31 и антагонизм антитела YW210.09 согласуются с тем, что РНКи к Wnt2 ингибирует передачу сигнала в клетках NCI-H23, и с эндогенной экспрессией Wnt1 в клетках M14. Используя многочисленные киРНК, авторы подтвердили, что экспрессия Wnt2 в клетках NCI-H23 и Wnt1 в клетках M14 необходима для аутокринной передачи сигнала от Wnt.
Поскольку все антитела, выделенные при скрининге в примере 2, которые противодействуют передаче сигнала в стимулированных Wnt3a клетках HEK293, также ингибируют стимуляцию Wnt3a во всех других протестированных клеточных линиях, а также ингибируют аутокринную передачу сигнала от Wnt в клеточных линиях тератокарциномы, было неожиданно, что эти антитела усиливают аутокринную передачу сигнала в других 9 протестированных клеточных линиях. В дополнение, антитело YW210.09 усиливает передачу сигнала от Wnt3a во всех протестированных клеточных линиях и усиливает аутокринную передачу сигнала от Wnt в 7 клеточных линиях, но оно ингибирует эндогенную передачу сигнала в 3 других линиях. Эти исследования показывают, что различные изоформы Wnt (экспрессированные в одной клеточной линии) определяют активность антител к LRP6, и что антитела, противодействующие и усиливающие Wnt3a, также могут обладать противоположными эффектами на большинство других белков Wnt. Исследования также показывают, что введение различных изоформ Wnt в одну клеточную линию определяет активность антител к LRP6, и что антитела, противодействующие и усиливающие Wnt3a, также обладают противоположными эффектами на большинство других белков Wnt. Исходя из их функционального взаимодействия с двумя антителами к LRP6, 14 изоформ Wnt можно сгруппировать по трем классам: сигнал от Wnt3 и Wnt3a ингибируется антителом YW211.31 и усиливается антителом YW210.09; сигнал от Wnt 1, 2, 2b, 6, 8a, 9a, 9b и 10b усиливается антителом YW211.31, и антагонизируется YW210.09; и сигнал от Wnt 4, 7a, 7b и 10a усиливается антителом YW211.31 и не ингибируется антителом YW210.09 (фиг.5). Данное разделение по классам явно не соответствует предложенному филогенезу генов Wnt, хотя подсемейство Wnt3/3a является наиболее эволюционно разрозненным (Cho et al., 2010).
Пример 5
Изоформы Wnt определяют отличающуюся активность антител к LRP6
Различные изоформы Wnt могут преимущественно связываться с различными изоформами FZD, экспрессирующимися эндогенно в различных клеточных линиях, и могут, соответственно, отвечать за отличающуюся активность антител к LRP6 настоящего изобретения. Для проверки этой возможности были созданы химерные белки, ковалентно соединяющие различные пары Wnt-FZD, чтобы проверить, могут ли определенные изоформы Wnt или FZD определять активность антитела к LRP6. Wnt3a или Wnt1, слитые с любым из FZD4 или FZD5, сильно активировали передачу сигнала от Wnt в клетках HEK293 в условиях предполагаемого отсутствия эндогенной экспрессии Wnt, в то время как сверхэкспрессия FZD4 или FZD5 не индуцировала передачу сигнала от Wnt. Антитело YW211.31.57 ингибирует сигнальную активность Wnt3a, слитого с любым из FZD4 или FZD5, и усиливает активность Wnt1, слитого с любым из FZD4 или FZD5 (на фиг.6 показаны итоговые данные, анализы, проведенные с использованием 10 мкг/мл антител, 10 мкг/мл Fzd8CRD). Антитело YW210.09 показывает противоположную активность в отношении химер Wnt1, ингибируя обе эти химеры. Поэтому, активность антитела коррелирует с изоформой Wnt, а не с изоформой FZD. Белок Fzd8CRD-Fc не обладал каким-либо эффектом на передачу сигнала, индуцированную любой из четырех химер Wnt-FZD, что согласуется с функционированием химер независимо от сайта связывания FZD на Wnt.
Экспрессия химер, в которых Wnt1 или Wnt3a слиты с LRP6, индуцирует передачу сигнала от Wnt гораздо сильнее, чем сверхэкспрессия LRP6. Антитела YW211.31.57 и YW210.09 не способны ингибировать эту индукцию, что согласуется с гипотезой о том, что ингибирующая функция антител зависит от блокирования связывания Wnt с LRP6 (фиг.6).
Это исследование подтверждает, что изоформа Wnt, а не FZD, определяет активность антитела. Химерные белки изоформ Wnt, слитые с LRP6, но не FZD, не чувствительны к ингибированию антителами к LRP6, позволяя предположить, что антагонистическое действие может быть обусловлено блокированием взаимодействий между лигандом и корецептором. Это подтверждается исследованиями связывания in vitro для Wnt3a и антитела YW210.09, которые оба связывают конкурентным образом участок в области E3-E4 белка LRP6, и для Wnt9b и антитела YW211.31, которые конкурируют за связывание в пределах области E1-E2. Каждый из эпитопов двух анти-LRP6 антител определяют сайт связывания для различного класса изоформ Wnt, один, расположенный в домене E1-E2, а другой - в домене E3-E4. По меньшей мере третий сайт связывания с Wnt предложен для изоформ, которые не ингибируют ни антитела, ни их комбинация, и, по-видимому, вероятно, что каждый из четырех повторяющихся доменов связывает различный подтип изоформ Wnt. Эта модульная организация может дать возможность структурному разнообразию различных Wnt и их сайтов связывания обеспечить дифференциальную регуляцию посредством антагонистов, связывающих Wnt и корецептор, таких как изоформы белков SFRP и DKK, соответственно.
Пример 6
Опосредованное антителами усиление передачи сигнала от Wnt включает димеризацию LRP6
Усиление аутокринной передачи сигнала от Wnt антителом YW211.31 требует эффектов авидности, вероятно, через димеризацию LRP6. Моновалентный Fab-фрагмент YW211.31 и рекомбинантное одноплечевое антитело YW211.31 не усиливают аутокринную передачу сигнала от Wnt в клетках EKVX и Hs578T при концентрациях, которые ингибируют индуцированную Wnt3a передачу сигнала в этих клеточных линиях. В отличие от этого, Fab-фрагмент YW211.31 и одноплечевое mAb, оба ингибируют как аутокринную, так и индуцированную Wnt3a передачу сигнала от Wnt в линии клеток тератокарциномы PA-1 с силой, аналогичной целому IgG. Для того чтобы проверить, будет ли сшивание одноплечевого антитела YW211.31 восстанавливать функцию усиления Wnt целой молекулы IgG, использовали линию клеток саркомы мягких тканей, HT-1080, которая имеет как аутокринную, так и индуцированную Wnt2 передачу сигнала от Wnt, усиливаемую антителом YW211.31.57, а также индуцируемый антителом Apomab апоптоз, который усиливается сшиванием Fc (Adams et al., 2008). Одноплечевое антитело YW211.31 не имело эффекта ни на аутокринную передачу сигнала от Wnt, ни на передачу сигнала, индуцированную трансфекцией Wnt2. В условиях сшивки анти-Fc антителами, которые вызывают опосредованный Apomab апоптоз, было установлено, что сшивка одноплечевого антитела частично восстанавливает усиление как аутокринной, так и Wnt2-индуцированной передачи сигнала от Wnt, наблюдаемые с целым антителом YW211.31.57.
Опосредованное антителами усиление сигнала от Wnt требует димеризации корецептора, поскольку одноплечевой и Fab-форматы антител не способны усиливать передачу сигнала от Wnt, если они не сшиты. Кроме того, клеточные и биохимические данные, представленные в настоящем описании, указывают на то, что связывание Wnt со сшитым LRP6 абсолютно необходимо для усиления сигнала, вероятно, отражая необходимость в опосредованном Wnt привлечении FZD в комплекс. Небольшую долю сверхэкспрессированного LRP6 можно идентифицировать в качестве гомодимера на клеточной поверхности, и для димеризации необходим внеклеточный домен, однако не ясно, может ли это вносить вклад в Wnt-независимую передачу сигнала через β-катенин, индуцированную сверхэкспрессией LRP6 (Liu et al., 2003). Делеция внеклеточного домена LRP6 также активирует передачу сигнала Wnt-независимым образом, и направленная внеклеточная димеризация этого рекомбинантного белка, выполненная различными способами, может либо усиливать, либо ингибировать эту активность (Liu et al., 2003; Cong et al., 2004). Wnt индуцирует агрегацию и фосфорилирование LRP6 на плазматической мембране, причем для обоих процессов необходима функция гомоолигомеризации внутриклеточного белка DVL (Bilic et al., 2007). Эти крупные агрегаты также содержат Axin и GSK3, и, вероятно, ингибируют деградацию β-катенина.
Пример 7
Опосредованное антителами усиление передачи сигнала от Wnt путем ингибирования связывания с антагонистами Wnt
Ингибирование активности внеклеточных антагонистов LRP6, таких как изоформы DKK1 и SOST, также может усиливать передачу сигнала от Wnt (Niida et al., 2004). Экзогенный белок DKK1 ингибирует индуцированную Wnt1 передачу сигнала в клетках HEK293, а антитело YW211.31.57 может блокировать этот антагонизм и даже усиливать передачу сигнала при достаточно высоких концентрациях в присутствии белка DKK1 (фиг.4G). В отличие от этого одноплечевое антитело YW211.31 только очень слабо ингибирует антагонизм DKK1 в отношении передачи сигнала от Wnt1 при таких же концентрациях. Целое антитело YW211.31.57 эффективно противодействует активности DKK1 при всех испытанных концентрациях DKK1, тогда как одноплечевое антитело имеет минимальный эффект или не оказывает эффекта даже при низкой концентрации DKK1. Сильный антагонизм в отношении активности экзогенного DKK1, наблюдаемый для целого, но не для одноплечевого антитела YW211.31, может вносить вклад в Wnt-усиливающую активность, специфичную для целого антитела. Альтернативно, поскольку белок DKK1 не был способен полностью ингибировать индуцированную Wnt1 передачу сигнала в этом анализе, также возможно, что целое антитело YW211.31 просто усиливает оставшийся сигнал посредством димеризации LRP6.
Поскольку ингибирование антителом взаимодействия DKK1 с LRP6 необязательно придает активность, усиливающую Wnt, и антагонизм в отношении DKK1, по-видимому, требует димеризации LRP6, ингибирование активности DKK1, вероятно, опосредовано преимущественно усилением остаточного сигнала от Wnt, связанного с корецептором.
Пример 8
Антагонизм в отношении передачи сигнала от Wnt превалирует для комбинаций антител к LRP6
Приведенные выше анализы указывают на то, что Wnt3a и Wnt связывают участок в области E1-E2 белка LRP6, и это связывание ингибировано антителом YW211.31.57. Изоформы Wnt в классе Wnt1 предположительно связывают участок в области E3-E4, и это связывание блокировано антителом YW210.09. Не придерживаясь какой-либо конкретной теории, усиление передачи сигнала от Wnt может возникнуть, когда и изоформа Wnt, и антитело способны связывать одну и ту же молекулу LRP6, что, вероятно, требует привлечения FZD белком Wnt и димеризации LRP6 антителом. Эта модель предполагает, что комбинация двух антител будет ингибировать передачу сигнала, индуцированную любым классом изоформ Wnt, поскольку, несмотря на то, что димеризация LRP6, вероятно, все еще возникает, связывание с Wnt будет блокировано одним или другим антителом. Как и предполагалось, воздействие на клетки НЕК293 одновременно антител YW211.31.57 и YW210.09 ингибирует передачу сигнала, инициируемую экспрессией либо Wnt3a, либо Wnt1 (фиг.7 и 8A). Анализ, показанный на фиг.8А проводили в клетках НЕК293 со стабильно встроенным Wnt-люциферазным репортером, которые были трансфицированы конструкциями для экспрессии или любого из Wnt3a, Wnt1, или совместно Wnt3a и Wnt1. Все антитела и белки использовали в концентрации 10 мкг/мл каждый. Этот анализ проводили еще с тремя другими изоформами Wnt в классе Wnt1, и было обнаружено, что комбинация антител YW211.31.57 и YW210.09 ингибирует передачу сигнала от Wnt в той же степени, что и одно YW210.09 (фиг.7). При одновременной экспрессии Wnt3a и Wnt1 ни одно из антител не противодействовало передаче сигнала от Wnt, но комбинация двух этих антител ингибирует передачу сигнала (фиг.8А). Одним возможным объяснением этого результата является то, что каждое антитело ингибирует связывание только одной изоформы Wnt, но оба антитела способны связывать молекулы LRP6 одновременно, блокируя оба сайта связывания Wnt.
Четыре изоформы Wnt в третьем классе, которым не противодействуют антитела YW211.31.57 или YW210.09, могут связывать сайт на LRP6, не блокируемый ни одним антителом, или, альтернативно, они могут иметь способность связываться с любым из сайтов связывания Wnt, определяемых антителами. Для каждой из этих изоформ Wnt комбинация двух антител также не ингибировала передачу сигнала от них, а только усиливала или не влияла на их активность, позволяя предположить, что эти изоформы Wnt могут связывать сайт, отличающийся от эпитопов YW211.31.57 и YW210.09 (фиг.7).
Наблюдаемое действие комбинации антител YW211.31.57 и YW210.09 на передачу сигнала от Wnt, индуцированную экзогенными изоформами Wnt, также распространяется на эндогенную аутокринную передачу сигнала от Wnt. В линиях клеток тератокарциномы, PA-1 и Ntera-2, в которых антитело YW211.31.57 ингибирует, а антитело YW210.09 усиливает аутокринную передачу сигнала от Wnt, комбинация антител ингибирует передачу сигнала (фиг.2А). В клетках Hs578T и EKVX, где оба антитела усиливают аутокринную передачу сигнала от Wnt, комбинация антител также ее усиливает (фиг.8В и 8С).
Пример 9
Антитела к LRP6 дифференциально ингибируют связывание Wnt с несколькими сайтами
Противоположное действие антител к LRP6 позволяет предположить, что YW211.31.57 и YW210.09 взаимодействуют с различными сайтами связывания изоформ Wnt на LRP6, и что антагонистическое антитело конкурирует со связыванием Wnt, а усиливающее антитело дает возможность связыванию. В анализе интерферометрии биослоев, которые измеряет связывание очищенных белков Wnt с иммобилизованными очищенными белковыми фрагментами внеклеточного домена LRP6 (см. пример 1), было продемонстрировано, что Wnt3a связывает область E3-E4 в LRP6, где находится эпитоп для антитела YW211.31.57, а Wnt9b (в том же классе, что и Wnt1, по взаимодействию с антителами) связывается только с областью Е1-Е2, в которой также связывается антитело YW210.09 (Bourhis et al. (2010)). Антитело YW211.31.57, но не YW210.09, ингибирует связывание Wnt3a с белковым фрагментом LRP6.E1-E4 (фиг.9A). В противоположность этому, YW210.09, но не YW211.31.57, ингибирует связывание Wnt9b с LRP6.E1-E4 (фиг.9B). В этих анализах связывание антител с белком LRP6 оставляли до достижения равновесия, и последующий сдвиг длины волны в картине интерференции показан для фаз ассоциации и диссоциации связывания белка Wnt.
Опосредованное антителами ингибирование связывания Wnt также можно детектировать, используя меньшие по размеру связывающие Wnt фрагменты - E3-E4 для Wnt3a и E1-E2 для Wnt9b (фиг.9C и 9D). Одноплечевое антитело YW211.31 также может ингибировать связывание Wnt3a с фрагментом E3-E4. Кроме того, YW211.31.57 и YW210.09 могут связываться последовательно с белком LRP6.E3-E4 без конкуренции и при добавлении в любом порядке (фиг.9Е). Конкуренция за связывание только между Wnt3a и антителом YW211.31.57, и только между Wnt9b и YW210.09, с разными сайтами на белке LRP6 коррелирует с ингибирующей активностью каждого белка в отношении передачи сигнала от определенной изоформы Wnt.
Анализ интерферометрии биослоев ранее показал, что очищенный белок DKK1 может связывать оба фрагмента, E3-E4 и E1-E2, белка LRP6, и что это связывание может ингибировать связывание Wnt3a и Wnt9b с этими соответствующими областями белка. (Bourhis et al. (2010)). Этот анализ использовали, чтобы показать, что каждое из антител YW211.31.57 и YW210.09 может ингибировать связывание DKK1 с белком LRP6.E1-E4. Антитело YW211.31.57 также ингибирует связывание DKK1 с белком LRP6.E3-E4, и антитело YW210.09 блокирует связывание DKK1 с фрагментом LRP6.E1-E2. Одноплечевое антитело YW211.31 полностью сохраняет эту ингибирующую активность, даже, несмотря на то, что оно не может ни усиливать передачу сигнала от Wnt, ни значительно противодействовать активности экзогенного DKK1 на передачу сигнала от Wnt в клетках. Этот результат позволяет предположить, что антагонизм с DKK1 вероятно не вносит значительного вклада в опосредованное антителами усиление передачи сигнала от Wnt.
Пример 10
Антитела к LRP6 активны в отношении Wnt-стимулируемых опухолей и формирования костной ткани
Для начала исследований противоопухолевой терапевтической эффективности антител к LRP6 обрабатывали две модели стимулируемых Wnt-лигандом опухолей. Аллотрансплантанты трансгенной опухоли молочных желез MTV-Wntl, зависимой от экспрессии Wnt1, и ксенотрансплантанты тератокарциномы человека Ntera-2, стимулируемые аутокринным Wnt-сигналом от неизвестной изоформы Wnt (DeAlmeida et al., 2007). Клетки, выделенные из трансгенных опухолей молочных желез MMTV-Wnt1, использовали для формирования опухолей у бестимусных голых мышей, которые получали антитело каждые два дня. Аналогично белку Fzd8CRD-Fc, при воздействии антитела YW210.09 наблюдалась быстрая и устойчивая регрессия опухоли (фиг.10А). Антитело YW211.31.57 не изменяло рост опухоли в этих условиях по сравнению с контрольным буфером (PBS) и воздействием анти-gD антитела. Мышам вводили по 30 мг/кг антитела или белка каждые два дня (указано стрелками) (фиг.10А). Эти результаты согласуются с описанными выше эффектами антител на экспрессию генов-мишеней Wnt для опухолевых клеток MMTV-Wnt1, получавших воздействие антител в клеточной культуре.
Также для образования ксенотрансплантированных опухолей у бестимусных голых мышей использовали клетки тератокарциномы Ntera-2, которые обрабатывали либо антителом, либо белком Fzd8CRD-Fc. В РНК, выделенной из опухолей, получавших воздействие антител YW211.31.57, одноплечевого YW211.31 или комбинации YW211.31.57 и YW210.09, было выявлено снижение экспрессии гена-мишени Wnt, SP5, до 41-57% от уровня в опухолях контрольных мышей, получавших инъекции буфера, тогда как воздействие белка Fzd8CRD-Fc снижало экспрессию SP5 до 8,0%. Уровень мРНК SP5 нормализовали на уровень мРНК GAPDH в той же опухоли и дополнительно нормализовали на уровень в опухолях, на которые воздействовали PBS. Все типы воздействия, за исключением YW210.09, имели значение p<0,005 по ANOVA по сравнению с контролем PBS. (фиг.10B). Экспрессия Axin2 была снижена только до 56,2% в результате действия Fzd8CRD-Fc, и каких-либо значительных изменений в экспрессии Axin2 не детектировалось ни с одним из антител. Воздействие антитела YW210.09 значительно не влияло на экспрессию ни SP5, ни Axin2. Образцы сыворотки, проанализированные на ингибирование или усиление индуцированной Wnt3a передачи сигнала в клетках HEK293, подтверждают, что введенные с помощью инъекций антитела и белок сохраняли по меньшей мере некоторую активность in vivo на протяжении 16-часового воздействия.
Поскольку активация или усиление передачи сигнала от Wnt может увеличивать массу костной ткани в результате усиления дифференцировки и функций остеобластов и, непрямым образом, в результате ингибирования дифференцировки остеокластов (Glass et al., 2005), тестировали активность антител к LRP6 в отношении костей свода черепа мышей в органотипической культуре. Эксплантаты микросрезов основания черепа культивировали с антителом или RANK-Fc, и затем анализировали объем и плотность теменной кости с помощью микрокомпьютерной томографии. С использованием гистограммного анализа контрольных образцов определяли границы ослабления рентгеновского облучения кальцифицированных (кость) и некальцифицированных (хрящ) тканей. Воздействие антитела YW210.09 значительно усиливало среднюю минеральную плотность костной ткани (BMD) кальцифицированной теменной кости на 7,4%, аналогично 6,8%-му увеличению, наблюдаемому при воздействии RANK-Fc для ингибирования дифференцировки остеокластов (фиг.10С; Hsu et al., 1999). Воздействие антитела YW211.31.62 по существу не изменяло BMD кальцифицированной теменной кости. Все воздействия проводили 10 мкг/мл антитела или белка в течение 7 дней. На фиг.10С точки данных представляют восемь половинок свода черепа от четырех мышей для каждой группы воздействия; среднее и стандартная ошибка среднего показаны горизонтальными и вертикальными линиями, соответственно. Только воздействие YW210.09 и RANK-Fc достоверно отличалось от не получавших воздействие образцов со значениями p меньше 0,01 и 0,05, соответственно, по тесту Дуннетта (<0,05 для обоих по t-тесту).
Объем общей области теменной кости (кальцифицированной и некальцифицированной) и доля кальцифицированной костной ткани в этой области по существу не менялись в результате воздействия антитела или RANK-Fc, позволяя предположить, что антитело YW210.09 может усиливать минерализацию без существенных изменений в пролиферации клеток.
Пример 11
Биспецифичное антитело к LRP6 действует как общий (пан) ингибитор Wnt
Для конструирования биспецифичного IgG-гибрида с гетеродимерами тяжелых цепей YW211.31.62 и YW210.09 использовали методику конструирования «узлы в полостях» (Atwell et al., 1997). Это биспецифичное антитело к LRP6, полученное в клетках либо E. coli, либо HEK293, противодействовало индуцированному Wnt3a (0,1 мкг/мл) сигналу в клетках HEK293 (фиг.11A) и опухолевых клеточных линиях PA-1, M14 и CAL-51 (фиг.11C). Следует отметить, что биспецифичное антитело ингибирует сигнал по меньшей мере также эффективно, как YW211.31, и не сохраняет активность YW210.09, усиливающую сигнал от Wnt3a. Биспецифичное антитело также ингибирует аутокринную передачу сигнала от Wnt во всех трех протестированных опухолевых клеточных линиях (фиг.11В), сохраняя ингибирующую активность антитела YW211.31 в клетках PA-1 и антитела YW210.09 в клетках M14. Интересно отметить, что хотя YW211.31 усиливает, а YW210.09 не обладает эффектом на аутокринную передачу сигнала от Wnt в клетках карциномы молочных желез, CAL-51, биспецифичное антитело ингибирует сигнал. Эта новая антагонистическая активность не наблюдалась в случае комбинации антител YW211.31 и YW210.09. В приведенных выше методах анализа, на клетки PA-1 и M14 со стабильно встроенным Wnt-люциферазным репортером и клетки CAL-51, трансфицированные репортером, воздействовали указанными контрольным буфером (PBS), антителом, комбинацией антител или белком Fzd8CRD-Fc (по 10 мкг/мл каждый) с (11С) или без (11B) стимуляции 0,1 мкг/мл Wnt3a.
При тестировании на передачу сигнала, индуцированную трансфекцией 13 изоформ Wnt в клетках HEK293, биспецифичное антитело сильно ингибировало действие всех Wnt, которые блокировались либо YW211.31, либо YW210.09 (фиг.12). Итоги анализов, приведенные в таблице 12, определяют эффекты антител или белка (10 мкг/мл) на передачу сигнала, индуцированную трансфекцией конструкций для экспрессии изоформ Wnt в клеточных линиях HEK293 или Hs578T со стабильно встроенным Wnt-люциферазным репортером. Репортерную активность нормализовали на количество клеток и дополнительно нормализовали на уровень в клетках, трансфицированных такой же экспрессионной конструкцией, но не получавших воздействие антитела или белка. В качестве контроля использовали анти-gD. Величины кратности изменения считались значимыми, когда они выходили за рамки диапазона, наблюдаемого при воздействии контрольного анти-gD антитела: меньше 0,80 для ингибирования и выше 1,30 для усиления в клетках HEK293, и меньше 0,65 для ингибирования и выше 1,30 для усиления в клетках Hs578T.
Аналогично, комбинации антител YW211.31 и YW210.09, и в отличие от любого антитела по отдельности, биспецифичное антитело блокирует сигнал, индуцируемый комбинацией Wnt1 и Wnt3a (фиг.12). Неожиданно, биспецифичное антитело также снижает сигнал от трех Wnt, которые не ингибировали гомодимерные антитела по отдельности или в комбинации. Эта антагонистическая активность биспецифичного антитела также наблюдалась в клетках Hs578T с возможным исключением в виде отсутствия эффекта на индуцированную Wnt7a передачу сигнала.
Исследовали способность биспецифичного антитела ингибировать индуцированную Wnt3a стабилизацию белка β-катенина. Клетки НЕК293 с трансфекцией Wnt3a или без нее обрабатывали YW211.31, YW210.09 или биспецифичным антителом, или контролями - белком Fzd8CRD-Fc или анти-gD, в концентрации 5 мкг/мл в течение 18 ч., и уровень белка β-катенина и уровень фосфорилированных LRP5/6 определяли Вестрен-блот-анализом. Фиг.13А. В клетках НЕК293 биспецифичное антитело ингибирует индуцированную Wnt3a стабилизацию белка β-катенина аналогично YW211.31, и в отличие от YW210.09, которое повышает уровень β-катенина (фиг.13A). Оба антитела, биспецифичное и YW211.31, но не YW210.09, блокируют индуцированное Wnt3a появление высокомолекулярных вариантов фосфорилированных LRP5/6. К удивлению, тогда как YW211.31 и YW210.09 не влияют на постоянный уровень общего белка LRP6, биспецифичное антитело увеличивает уровень белка LRP6 в присутствии или без индукции белком Wnt3a. В отсутствие Wnt-стимуляции этот стабилизированный LRP6 может иметь немного повышенное фосфорилирование Serl490, хотя биспецифичное антитело не влияет на активность Wnt-репортера в клетках HEK293 в отсутствие Wnt.
Также была определена способность биспецифичного антитела ингибировать передачу сигнала от Wnt in vivo. Мышам SCID-bg с ксенотрансплантированными опухолями меланомы М14 вводили с помощью инъекций по 30 мг/кг биспецифичного антитела к LRP6, белок Fzd8CRD (положительный контроль) или анти-gD антитело (отрицательный контроль). Через 16 часов после инъекции выделяли РНК и анализировали с помощью кПЦР на экспрессию генов-мишеней Wnt. Уровни мРНК нормализовали на уровни мРНК GAPDH в той же опухоли и дополнительно нормализовали на уровень мРНК в опухолях, получавших воздействие анти-gD. Для всех воздействий биспецифичного антитела и Fzd8CRD значения р были <0,001 по ANOVA по сравнению с контролем анти-gD. Как показано на фиг.13В, биспецифичное антитело к LRP6 ингибировало передачу сигнала от Wnt в клетках меланомы М14, растущих в виде ксенотрансплантированных опухолей. В РНК, выделенной из опухолей, получавших воздействие антитела, было показано снижение экспрессии генов-мишеней Wnt, AXIN2 и APCDD1 до 46-57% и 35-38%, соответственно, от уровня в опухолях, получавших воздействие контрольного анти-gD антитела. Это снижение уровня экспрессии аналогично наблюдаемому при инъекциях белка Fzd8CRD и указывает на то, что биспецифичное антитело стабильно и активно in vivo.
Пример 12
Структура комплекса LRP6 E1-YW210.09 Fab
Кристаллическую структуру первого β-пропеллера и EGF-домена LRP6 (также называемого E1) в комплексе с YW210.09 Fab (Fab-фрагментом антитела YW210.09) определяли молекулярным замещением и уточняли с разрешением 1,9 Å с R и Rfree, составляющими 0,175 и 0,220, соответственно. Кристаллографическая асимметричная единица состояла из одного домена LRP6 E1 и одного YW210.09 Fab. Интерпретируемая электронная плотность позволила отследить остатки от Ala 20 по Lys 324 для домена E1 и остатки от Asp 1 по Glu 213 и от Glu 1 по Lys 214, соответственно, для легкой и тяжелой цепей Fab, за исключением остатков тяжелой цепи Fab, Ser 127-Thr 131 (нумерацию по Кабату использовали по всей длине).
Домен LRP6 E1 собирали в модульной архитектуре, которая включает модуль β-пропеллера и модуль, подобный фактору роста эпидермиса (EGF). β-пропеллер состоит из шести лопастей, образованных четырехцепочечным антипараллельным β-листом, расположенным радиально с N-концевым краем, смотрящим на центральный канал, и YWTD-мотивами, расположенными во второй цепи каждой лопасти. Структура β-пропеллера домена LRP6 E1 близко напоминает структуру LDLr (Jeon, H., et al, 2001) с rmsd, составляющим 0,83 Å при наложении на 245 атомов С-α, несмотря на то, что идентичность последовательностей составляет только 36%. Большинство консервативных остатков сконцентрировано около расположенных в глубине YWTD-мотивов, образующих β-листы, необходимые для целостности структуры β-пропеллера, в то время как остатки на поверхности сильно варьируют, внося вклад в функциональное разнообразие этих рецепторов. LRP6 использует свой EGF-подобный домен для фиксации первой и шестой лопасти пропеллера и поддержания его механической прочности. EGF-подобный модуль выступает своим С-концом из β-пропеллера посредством линкера из десяти аминокислотных остатков и складывается обратно на нижней стороне β-пропеллера, фиксируя поверхность между третьей и четвертой лопастью. Взаимодействие между EGF и β-пропеллером является обширным, на что указывает большая общая площадь скрытой поверхности 1226 Å2, и комплементарность формы 0,74. Три остатка, Leu 296, Leu 298 и Met 299, в первой β-цепи EGF-модуля составляют гидрофобное ядро, которое пакуется в комплементарную полость β-пропеллера, окруженную некоторыми прямыми или опосредованными через воду полярными взаимодействиями. Эти признаки также наблюдаются в структурах LDLR (Jeon, H., et al., 2001; Rudenko, G., et al., 2002).
YW210.09 Fab распознает область в центре верха β-пропеллера, область, которую часто находят участвующей в белок-белковых взаимодействиях (Springer, T. A., 1998). Паратоп состоит из остатков из пяти CDR, включая три CDR тяжелой цепи (H1, H2, H3) и две CDR легкой цепи (L1 и L3). Связывание антител с β-пропеллером скрывает общую площадь 1691 Å2 с оценкой комплементарности формы 0,76. Кислый участок на верхней лицевой стороне β-пропеллера занимает примерно треть от общей площади, но едва перекрывается с эпитопом YW210. В противоположность этому, тяжелая цепь и легкая цепь распознают разные области. Прямые контакты, образованные CDR тяжелой цепи представляют 80% от скрытой площади с CDR H3, в одиночку отвечающей более чем за 50%. Этот сегмент состоит из 17 остатков, среди которых остатки от His 98 до Lys 100c образуют прямые контакты с β-пропеллером. Важно отметить, что Asn 100 антитела образует пару водородных связей с Asn 185 в LRP6, формируя взаимодействие по типу «рукопожатия» (фиг.16). Кроме того, необычное конформационное положение карбонильной группы основных цепей Val 100b и Lys 100c, которая взаимодействует с Arg 28 в LRP6 сзади, и две NH-группы, которые взаимодействуют с кислым участком через две молекулы воды (Wat1 и Wat2) спереди (фиг.14). Боковая цепь Lys 100c также нейтрализует кислый участок, образуя водородные связи с карбонилами главной цепи Val 70 и Ser 96 в LRP6. Arg 141 в LRP6 удерживается в центре и взаимодействует с образующей мостик водой Wat2, Asn 185 в LRP6, и Ala 100a в YW210.09. Arg 141, по-видимому, объединяет вместе две сети водородных связей. Кроме того, боковая цепь Val 100b фиксируется в гидрофобной полости в центральном канале β-пропеллера. Поэтому, последовательность NAVK в YW210.09 H3 проявляет необычную картину связывания с β-пропеллером E1 в LRP6. Другие CDR взаимодействуют с остатками в параметрах на верхушке β-пропеллера. Другие остатки, вовлеченные в связывание Н3 с LRP6, включают E51, D52, V70, S71, E73, L95, S96, D98 и E115. H1 и H2 соприкасаются с пятой и шестой лопастями, в то время как L1 и L3 соприкасаются с шестой, первой и второй лопастями (фиг.15). Дополнительные остатки в LRP6, вовлеченные в связывание YW210.09 с LRP6, включают R29, W188, K202, P225, H226, S243 и F266. Взаимодействия в кристаллической упаковке напрямую не вовлечены в области контакта YW210.09 с эпитопом LRP6, указывая на то, что кристаллическая структура должна отражать, как две молекулы взаимодействуют в растворе. Взаимодействие между отдельным NAVKN-мотивом (SEQ ID NO:49) из CDR H3 и β-пропеллером из LRP6 E1 очень похоже на взаимодействие, опубликованное для Laminin и Nidogen (Takagi, J., et al., 2003). В обоих случаях, значительные контакты осуществляются через «Asn-рукопожатие», описанное выше и разветвленные гидрофобные остатки, входящие в гидрофобную полость, образованную вершиной центрального канала β-пропеллера, который закрыт в обоих пропеллерах Phe-заслонкой. Dkk1 человека представляет мотив (аминокислоты 40-44: NAIKN (SEQ ID NO:50)), который помимо своего Ile 42 является идентичным мотиву, найденному в петле CDR H3 из YW210.09. Этот мотив является жестко консервативным среди биологических видов и членов семейства за исключением Dkk3, указывая на определенную функцию этого мотива в биологической роли белков Dkk. Консервативный мотив был обнаружен на N-конце Dkk1, области которую ранее не учитывали (Brott, B. K., and Sokol, S. Y., 2002), и которая, предположительно, не имела структуры. Дополнительно, этот конкретный мотив также консервативен в двух других белках, регулирующих передачу сигнала от Wnt через взаимодействие с LRP5/6, а именно Sclerostin (Semenov, M., et al., 2005) и Wise (Itasaki, N., et al., 2003). Эти два белка принадлежат тому же суперсемейству белков с «цистеиновыми узлами» (McDonald, N. Q., and Hendrickson, W. A., 1993) и представляют идентифицированный мотив в их петле номер 2, также называемой «каблуком» этой консервативной укладки (Lintern, K. B., et al., 2009; Veverka, V., et al., 2009).
Пример 13
Примеры анти-LRP6 антител
Аминокислотные последовательности некоторых анти-LRP6 антител приведены в списке последовательностей. В таблицах 2-4 приведено описание последовательностей. Выравнивания аминокислотных последовательностей VH- и VL-доменов конкретных анти-LRP6 антител приведены на фиг.16 и 17.
Тяжелые и легкие цепи
Вариабельные области тяжелых и легких цепей
HVR тяжелых и легких цепей
Хотя изложенное выше изобретение было описано подробно с помощью иллюстраций и примеров с целью ясности понимания, описание и примеры не следует истолковывать, как ограничивающие объем изобретения. Описание всей патентной и научной литературы, цитируемой в настоящем описании, определенно включено посредством ссылки во всей своей полноте.
Ссылки
Adams C, Totpal K, Lawrence D, Marsters S, Pitti R, Yee S, Ross S, Deforge L, Koeppen H, Sagolla M, Compaan D, Lowman H, Hymowitz S, Ashkenazi A. Structural and functional analysis of the interaction between the agonistic monoclonal antibody Apomab and the proapoptotic receptor DR5. Cell Death Differ. 2008 Apr; 15(4):751-61. Epub 2008 Jan 25.
Akiri G, Cherian MM, Vijayakumar S, Liu G, Bafico A, Aaronson SA. Wnt pathway aberrations including autocrine Wnt activation occur at high frequency in human non-small-cell lung carcinoma. Oncogene. 2009 May 28; 28(21):2163-72. Epub 2009 Apr 20.
Bafico A, Liu G, Goldin L, Harris V, Aaronson SA. An autocrine mechanism for constitutive Wnt pathway activation in human cancer cells. Cancer Cell. 2004 Nov; 6(5):497-506.
Bilic J, Huang YL, Davidson G, Zimmermann T, Cruciat CM, Bienz M, Niehrs C. Wnt induces LRP6 signalosomes and promotes dishevelled-dependent LRP6 phosphorylation. Science. 2007 Jun 15; 316(5831):1619-22.
Binnerts ME, Tomasevic N, Bright JM, Leung J, Ahn VE, Kim KA, Zhan X, Liu S, Yonkovich S, Williams J, Zhou M, Gros D, Dixon M, Korver W, Weis WI, Abo A. The first propeller domain of LRP6 regulates sensitivity to DKK1. Mol Biol Cell. 2009 Aug; 20(15):3552-60. Epub 2009 May 28.
Bourhis E, Tam C, Franke Y, Bazan JF, Ernst J, Hwang J, Costa M, Cochran AG, Hannoush RN. Reconstitution of a Frizzled8-Wnt3a-LRP6 Signaling Complex Reveals Multiple Wnt and Dkkl Binding Sites on LRP6. J Biol Chem. 2010 Mar 19; 285(12):9172-9. Epub 2010 Jan 21.
Brott, B.K, and Sokol, S.Y. (2002) Mol Cell Biol 22, 6100-6110.
Cho SJ, Vallès Y, Giani VC Jr, Seaver EC, Weisblat DA. Evolutionary dynamics of the Wnt gene family: a lophotrochozoan perspective. Mol Biol Evol. 2010 Feb 22. [Epub ahead of print].
Cong F, Schweizer L, Varmus H. Wnt signals across the plasma membrane to activate the beta-catenin pathway by forming oligomers containing its receptors, Frizzled and LRP. Development. 2004 Oct; 131 (20):5103-15.
Cselenyi CS, Jernigan KK, Tahinci E, Thorne CA, Lee LA, Lee E. LRP6 transducesa canonical Wnt signal independently of Axin degradation by inhibiting GSK3's phosphorylation of beta-catenin. Proc Natl Acad Sei USA. 2008 Jun 10; 105(23):8032-7. Epub 2008 May 28.
Cunningham SA, Stephan CC, Arrate MP, Ayer KG, Brock TA. Identification of the extracellular domains of Fit-1 that mediate ligand interactions. Biochem Biophys Res Commun. 1997 Feb 24; 231(3):596-9.
Davis-Smyth T, Chen H, Park J, Presta LG, Ferrara N. The second immunoglobulin-like domain of the VEGF tyrosine kinase receptor Flt-1 determines ligand binding and may initiate a signal transduction cascade. EMBO J. 1996 Sep 16; 15(18):4919-27.
DeAlmeida VI, Miao L, Ernst JA, Koeppen H, Polakis P, Rubinfeld В. The soluble wnt receptor Frizzled8CRD-hFc inhibits the growth of teratocarcinomas in vivo. Cancer Res. 2007 Jun 1; 67(11):5371-9.
Glass DA 2nd, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, Taketo MM, Long F, McMahon AP, Lang RA, Karsenty G. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell. 2005 May; 8(5):751-64.
Guo Y, Xie J, Rubin E, Tang YX, Lin F, Zi X, Hoang ВН. Frzb, a secreted Wntantagonist, decreases growth and invasiveness of fibrosarcoma cells associated with inhibition of Met signaling. Cancer Res. 2008 May 1; 68(9):3350-60.
Hsu H, Lacey DL, Dunstan CR, Solovyev I, Colombero A, Timms E, Tan HL, Elliott G, Kelley MJ, Sarosi I, Wang L, Xia XZ, Elliott R, Chiu L, Black T, Scully S, Capparelli C, Morony S, Shimamoto G, Bass MB, Boyle WJ. Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc Natl Acad Sei USA. 1999 Mar 30; 96(7):3540-5.
Itasaki, N., Jones, C.M., Mercurio, S., Rowe, A., Domingos, P. M., Smith, J. C., and Krumlauf, R. (2003) Development 130, 4295-4305.
Jeon H, Meng W, Takagi J, Eck MJ, Springer TA, Blacklow SC. Implications for familial hypercholesterolemia from the structure of the LDL receptor YWTD-EGF domain pair. Nat Struct Biol. 2001 Jun; 8(6):499-504.
Lintern, К.В., Guidato, S., Rowe, A., Saldanha, J.W., and Itasaki, N. (2009) J Biol Chem 284, 23159-23168.
Liu G, Bafico A, Harris VK, Aaronson SA. A novel mechanism for Wnt activation of canonical signaling through the LRP6 receptor. Mol Cell Biol.(2003) 16:5825-35.
Liu BY, Soloviev I, Chang P, Lee J, Huang X, et al. (2010) Stromal cell-derived factor-1/CXCL12 contributes to MMTV-Wntl tumor growth involving Gr1+CD11b+cells. PLoS One (2010) 5 (1):1-13; e8611.
McDonald, N.Q., and Hendrickson, W.A. (1993) Cell 73, 421-424.
Mi К, Dolan PJ, Johnson GV. The low density lipoprotein receptor-related protein 6 interacts with glycogen synthase kinase 3 and attenuates activity. J Biol Chem. 2006 Feb 24; 281(8):4787-94. Epub 2005 Dec 19.
Mohammad KS, Chirgwin JM, Guise ТА. Assessing new bone formation in neonatal calvarial organ cultures. Methods Mol Biol. 2008; 455:37-50.
Niida A, Hiroko T, Kasai M, Furukawa Y, Nakamura Y, Suzuki Y, Sugano S, Akiyama T. DKKl, a negative regulator of Wnt signaling, is a target of the beta-catenin/TCF pathway. Oncogene. 2004 Nov 4; 23(52):8520-6.
Nguyen DX, Chiang AC, Zhang XH, Kim JY, Kris MG, Ladanyi M, Gerald WL, Massagué J. WNT/TCF signaling through LEF1 and HOXB9 mediates lung adenocarcinoma metastasis. Cell. 2009 Jul 10; 138(1):51-62. Epub 2009 Jul 2.
Piao S, Lee SH, Kim H, Yum S, Stamos JL, Xu Y, Lee SJ, Lee J, Oh S, Han JK, Park BJ, Weis WI, Ha NC. Direct inhibition of GSK3beta by the phosphorylated cytoplasmic domain of LRP6 in Wnt/beta-catenin signaling. PLoS One. 2008; 3(12): e4046. Epub 2008 Dec 24.
Quarto N, Wan DC, Kwan MD, Panetta NJ, Li S, Longaker MT. Origin Matters: Differences in Embryonic Tissue Origin and Wnt Signaling Determine the Osteogenic Potential and Healing Capacity of Frontal and Parietal Calvarial Bones. J Bone Miner Res. 2009 Nov 23. [Epub ahead of print].
Rebay I, Fleming RJ, Fehon RG, Cherbas L, Cherbas P, Artavanis-Tsakonas S.
Specific EGF repeats of Notch mediate interactions with Delta and Serrate: implications for Notch as a multifunctional receptor. Cell. 1991 Nov 15; 67(4):687-99.
Rudenko, G., Henry, L., Henderson, K., Ichtchenko, K., Brown, M.S., Goldstein, J.L., and Deisenhofer, J. (2002) Science 298, 2353-2358.
Schwarz-Romond T, Metcalfe C, Bienz M. Dynamic recruitment of axin by Dishevelled protein assemblies. J Cell Sei. 2007 Jul 15; 120(Pt 14):2402-12.
Semenov MV, Tamai К, Brott BK, Kühl M, Sokol S, He X. Head inducer Dickkopf-1 is a ligand for Wnt coreceptor LRP6. Curr Biol. 2001 Jun 26; 11(12):951-61.
Semenov, M., Tamai, K., and He, X. (2005) J Biol Chem 280, 26770-26775.
Springer, T.A. (1998) J Mol Biol 283, 837-862.
Takagi, J., Yang, Y., Liu, J.H., Wang, J.H., and Springer, T.A. (2003) Nature 424, 969-974.
Tamai K, Semenov M, Kato Y, Spokony R, Liu C, Katsuyama Y, Hess F, Saint-Jeannet JP, He X. LDL-receptor-related proteins in Wnt signal transduction. Nature. 2000 Sep 28; 407(6803):530-5.
Tamai K, Zeng X, Liu C, Zhang X, Harada Y, Chang Z, He X. A mechanism for Wnt coreceptor activation. Mol Cell. 2004 Jan 16; 13(1):149-56.
van Amerongen R, Nüsse R. Towards an integrated view of Wnt signaling in development. Development. 2009 Oct; 136(19):3205-14. Veverka, V., Henry, A.J., Slocombe, P.M., Ventom, A., Mulloy, В., Muskett, F.W., Muzylak, M., Greenslade, K., Moore, A., Zhang, L., Gong, J., Qian, X., Paszty, C., Taylor, R.J., Robinson, M.K., and Carr, M.D. (2009) J Biol Chem 284, 10890-10900.
Veverka, V., Henry, A.J., Slocombe, P.M., Ventom, A., Mulloy, В., Muskett, F.W., Muzylak, M., Greenslade, K., Moore, A., Zhang, L., Gong, J., Qian, X., Paszty, C., Taylor, R.J., Robinson, M.K., and Carr, M.D. (2009) J Biol Chem 284, 10890-10900
Wu G, Huang H, Garcia Abreu J, He X. Inhibition of GSK3 phosphorylation of beta-catenin via phosphorylated PPPSPXS motifs of Wnt coreceptor LRP6. PLoS One. 2009; 4(3): e4926. Epub 2009 Mar 18.
Yasui N, Mihara E, Nampo M, Tamura-Kawakami K, Unno H, Matsumoto K, Takagi J. Detection of endogenous LRP6 expressed on human cells by monoclonal antibodies specific for the native conformation. J Immunol Methods. (2010) Jan 31; 352(1-2):153-60. Epub 2009 Nov 26.
Ye, X., et al, The Norrin/Frz4 signaling pathway in retinal vascular development and disease. (2010) Trends Mol Med. 16, 417-425.
Zhang Y, Appleton BA, Wiesmann С, Lau T, Costa M, Hannoush RN, Sidhu SS. Inhibition of Wnt signaling by Dishevelled PDZ peptides. Nat Chem Biol. 2009 Apr; 5(4):217-9. Epub 2009 Mar 1.
Zeng X, Huang H, Tamai K, Zhang X, Harada Y, Yokota C, Almeida K, Wang J, Doble B, Woodgett J, Wynshaw-Boris A, Hsieh JC, He X. Initiation of Wnt signaling: control of Wnt coreceptor Lrp6 phosphorylation/activation via frizzled, dishevelled and axin functions. Development. 2008 Jan; 135(2):367-75. Epub 2007 Dec 12.
Zhou H, Мак W, Kalak R, Street J, Fong-Yee C, Zheng Y, Dunstan CR, Seibel MJ. Glucocorticoid-dependent Wnt signaling by mature osteoblasts is a key regulator of cranial skeletal development in mice. Development. 2009 Feb; 136(3):427-36.
Zoltewicz JS, Ashique AM, Choe Y, Lee G, Taylor S, Phamluong K, Solloway M, Peterson AS. Wnt signaling is regulated by endoplasmic reticulum retention. PLoS One. 2009 Jul 10; 4(7): e6191.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К JAGGED И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2666990C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С БЕЛКАМИ LRP5, И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2832155C2 |
| АНТИТЕЛА ПРОТИВ NRR Notch1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2476443C2 |
| АНТИТЕЛА ПРОТИВ AXL И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2577986C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ АКСИАЛЬНОГО СВЯЗЫВАНИЯ PD-1 И VEGF АНТАГОНИСТОВ | 2013 |
|
RU2689760C2 |
| АНТИТЕЛА ПРОТИВ DLL4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2415869C2 |
| АНТИТЕЛА К PD-L1 И ИХ ПРИМЕНЕНИЕ ДЛЯ УСИЛЕНИЯ ФУНКЦИИ Т-КЛЕТОК | 2009 |
|
RU2636023C2 |
| СЕЛЕКТИВНЫЕ В ОТНОШЕНИИ ИЗОФОРМЫ АНТИТЕЛА К TGF-БЕТА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2839151C1 |
| АНТИТЕЛА ПРОТИВ FGFR3 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2568066C2 |
| АНТИТЕЛА ПРОТИВ NOTCH2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2580029C2 |
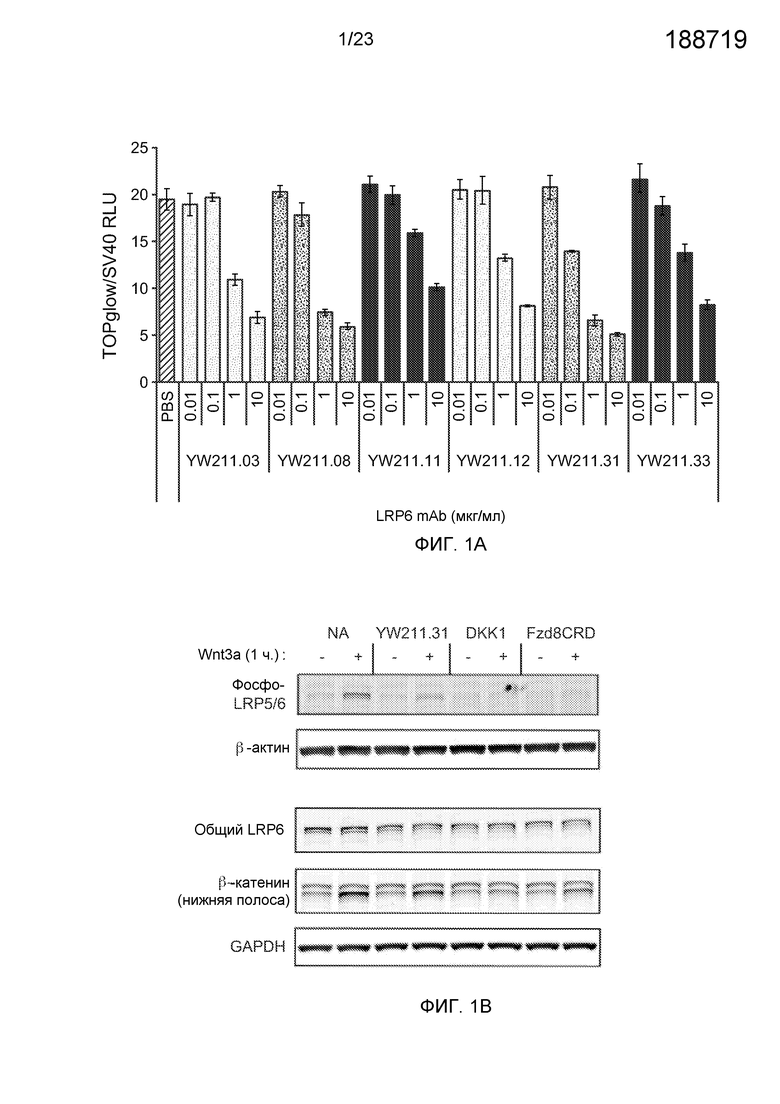
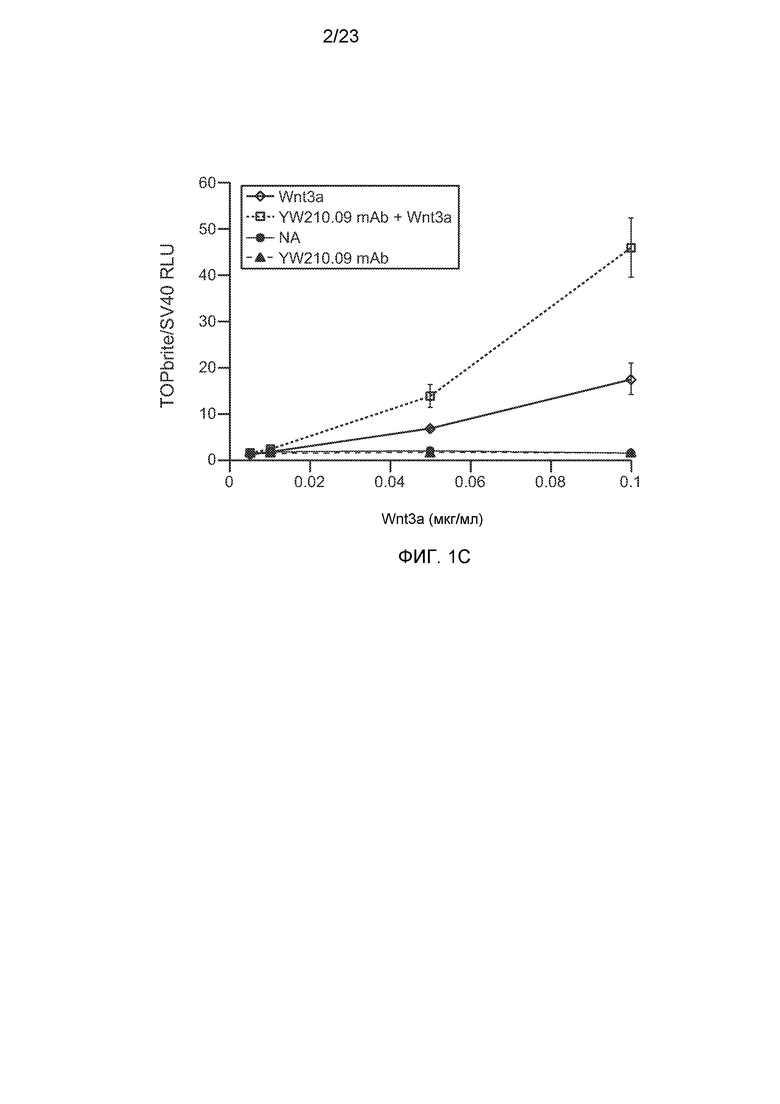
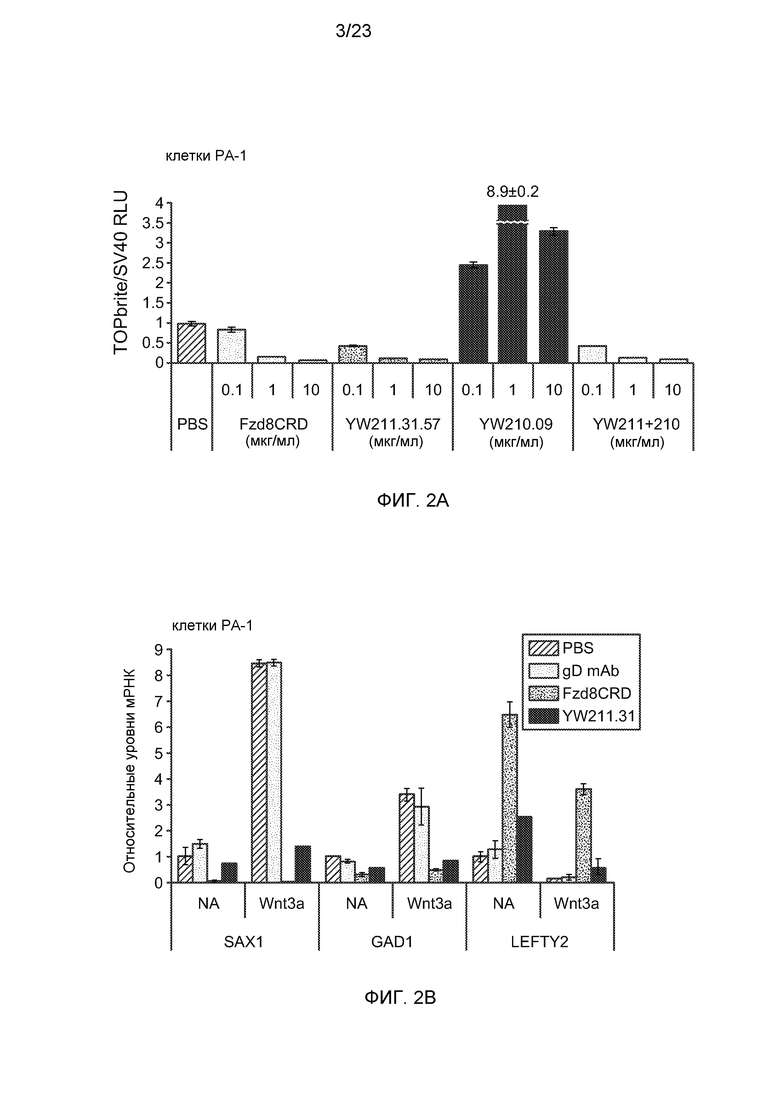
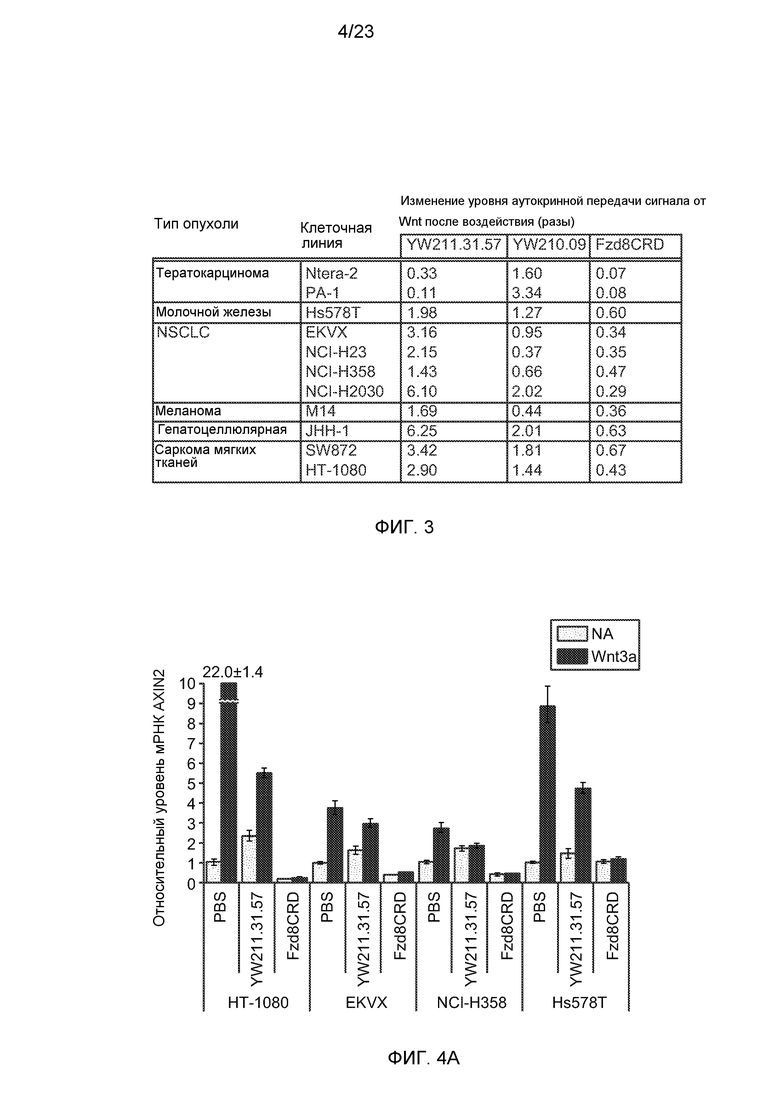
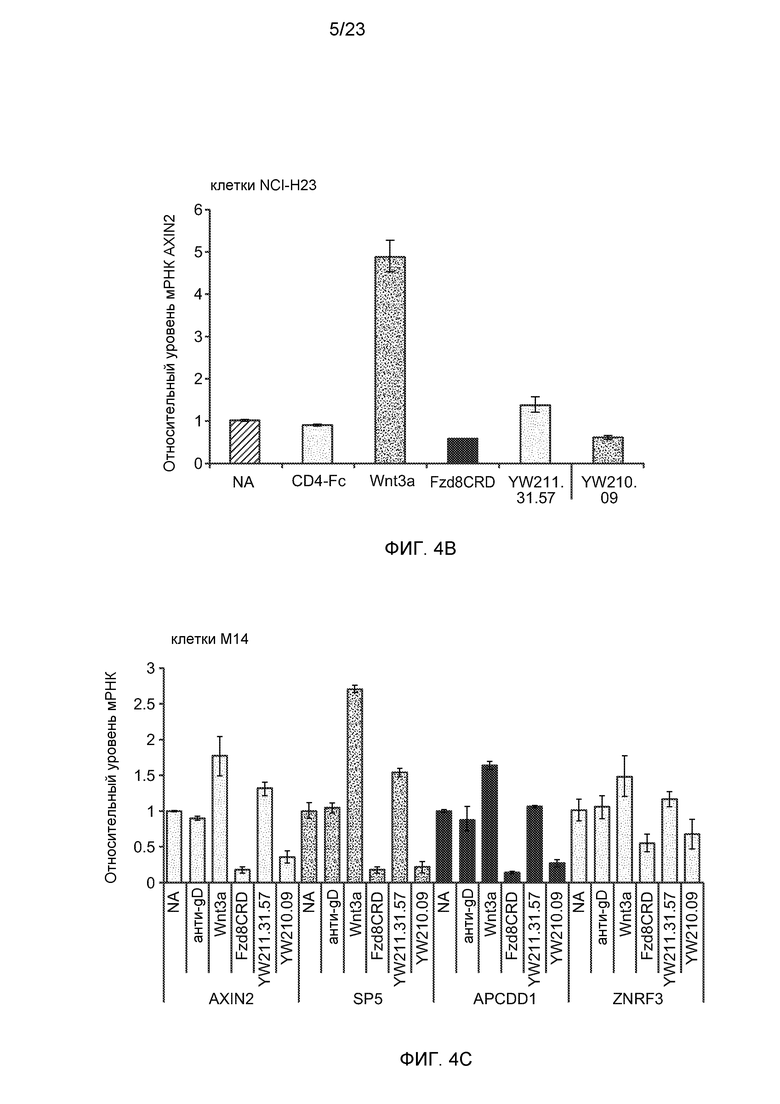
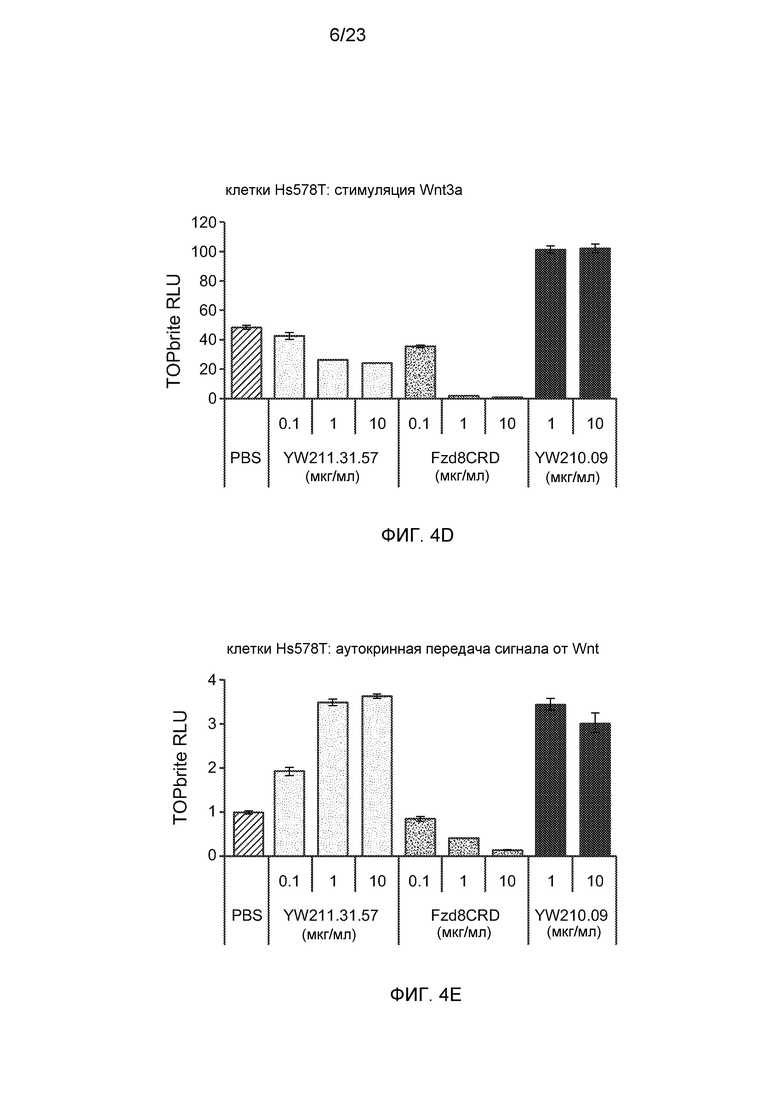
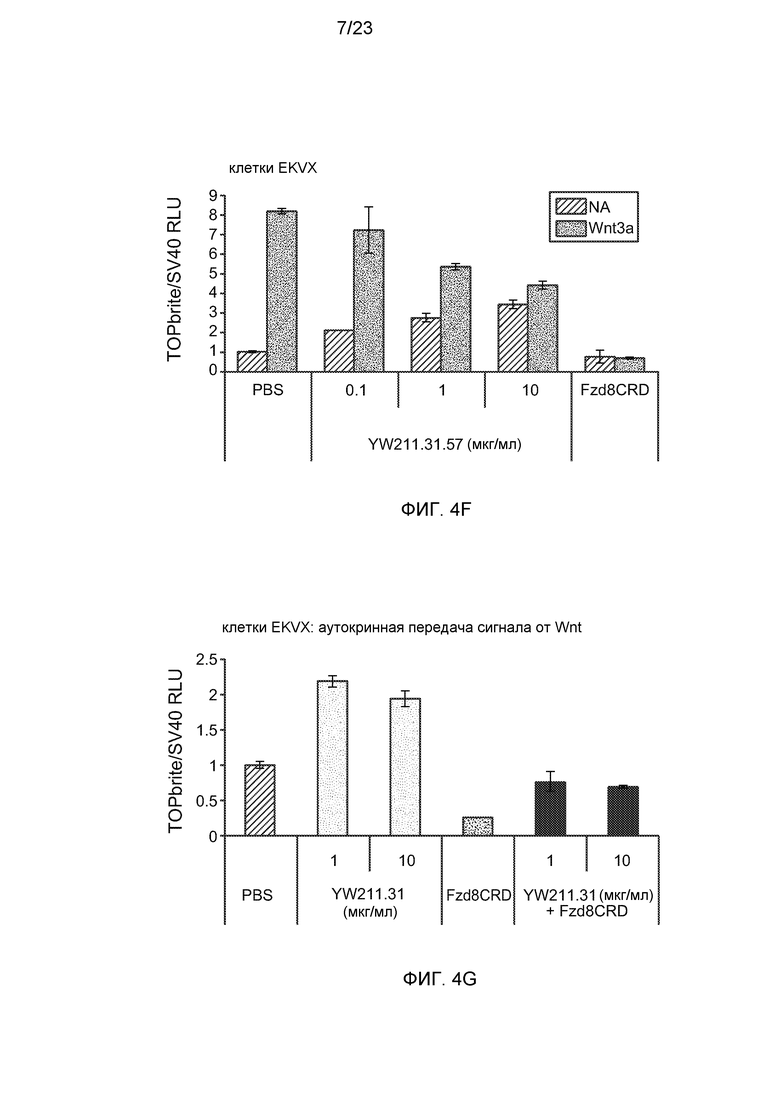
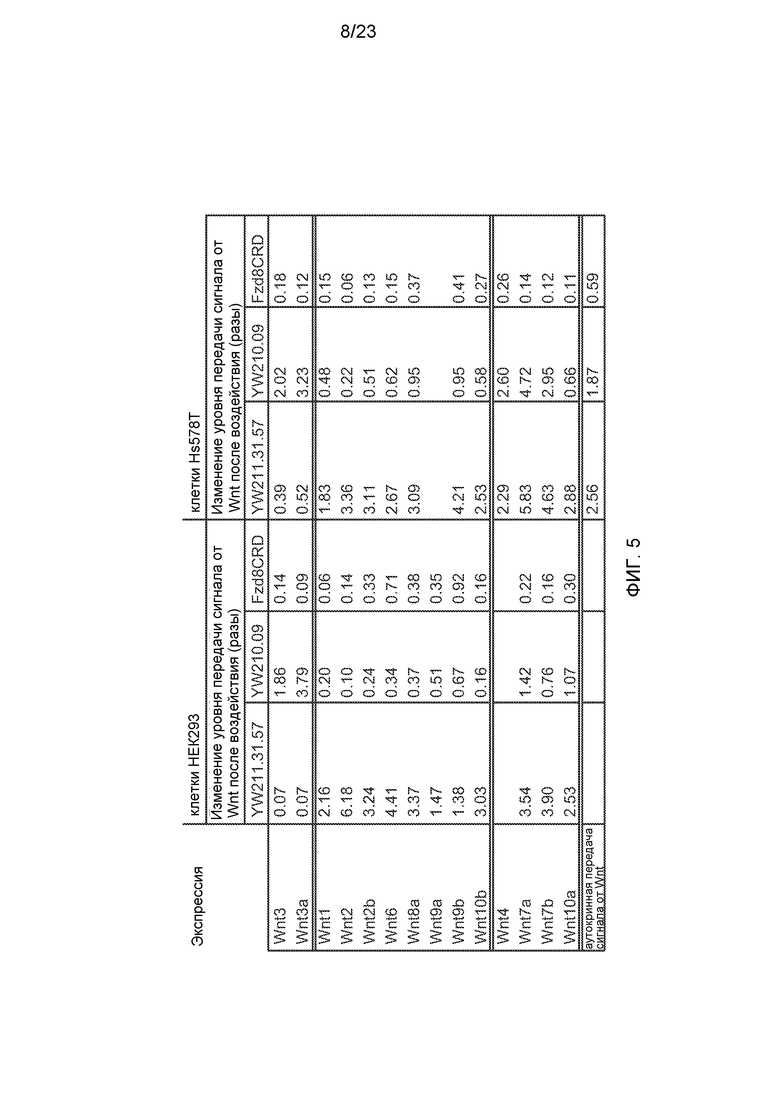
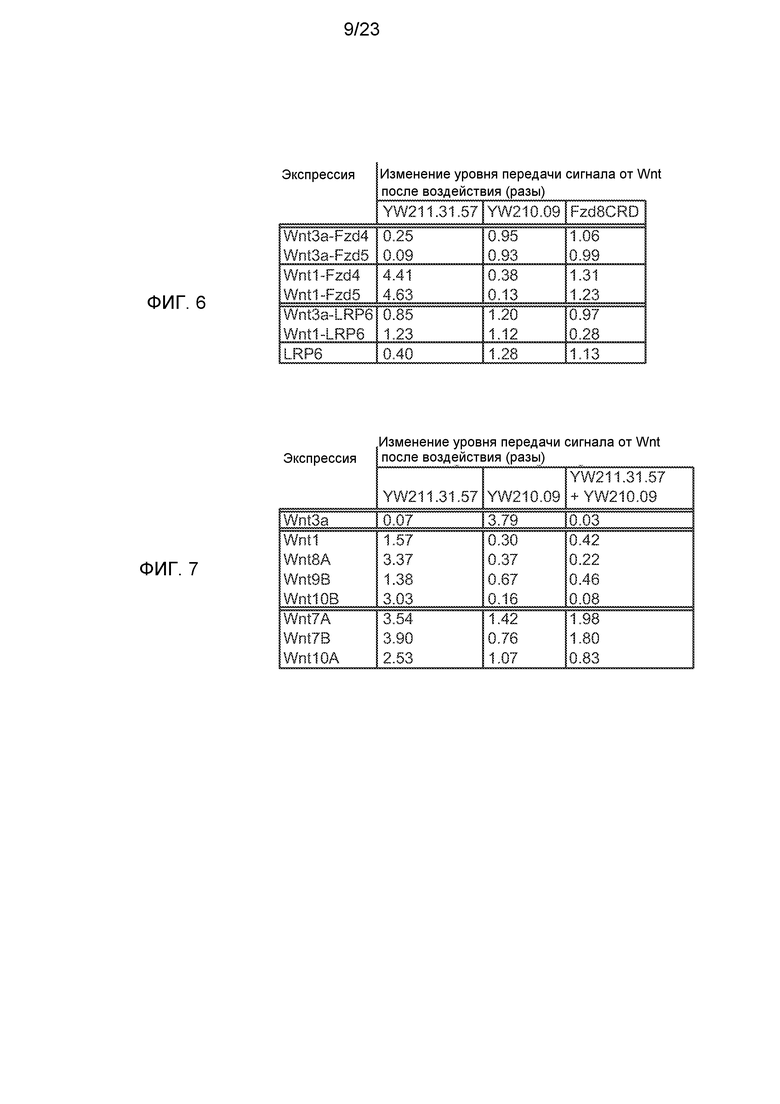
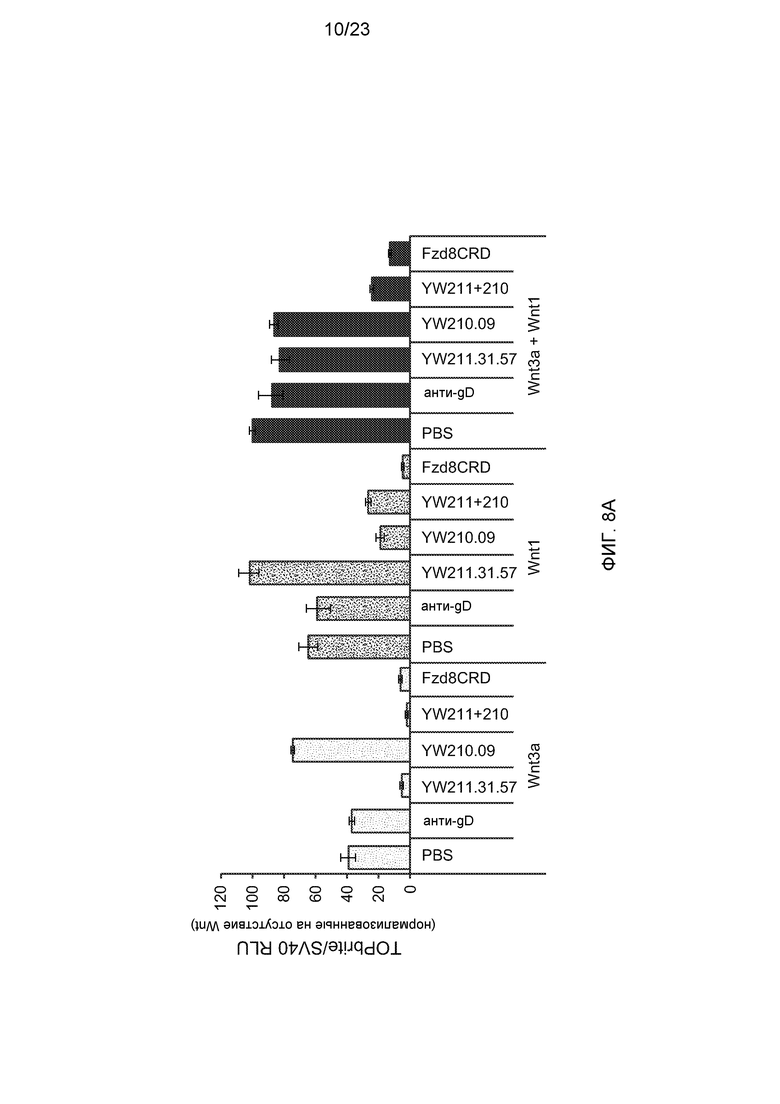
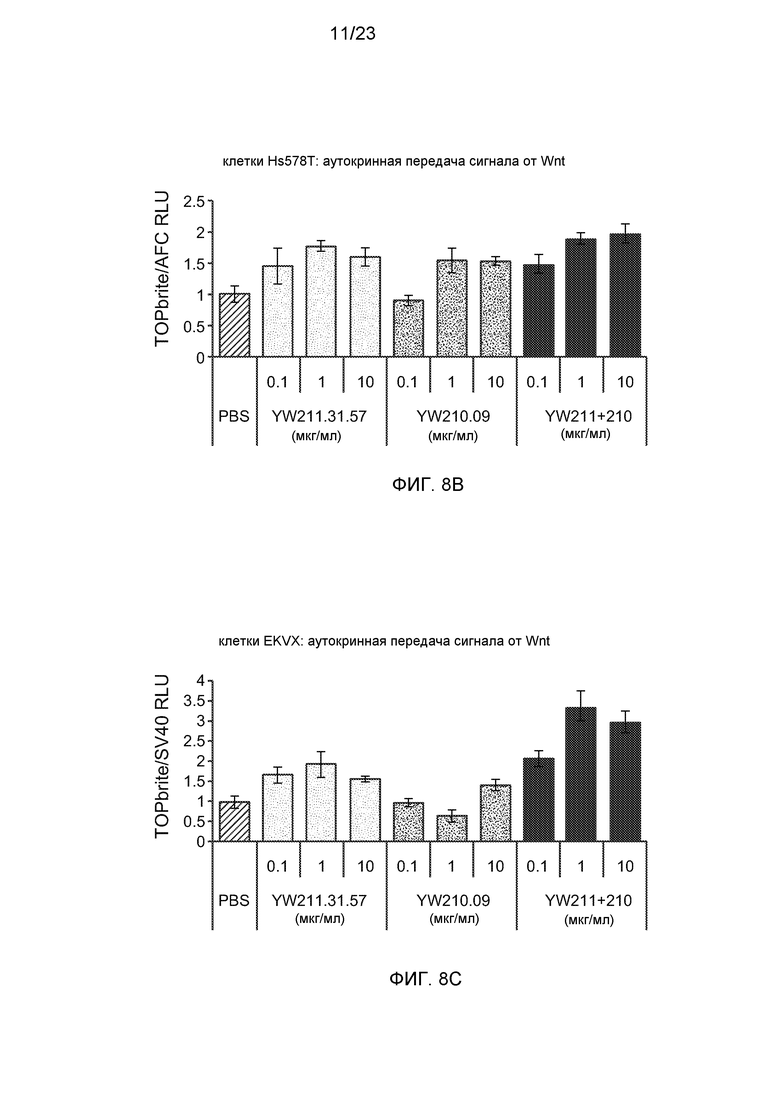
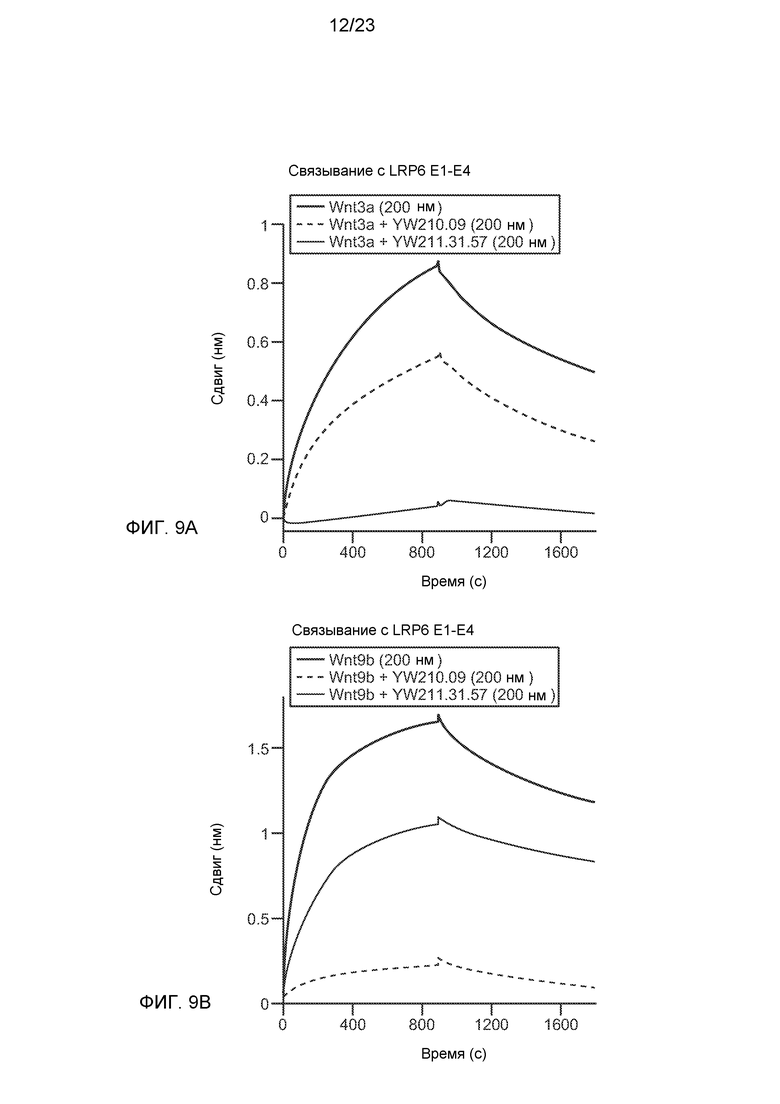
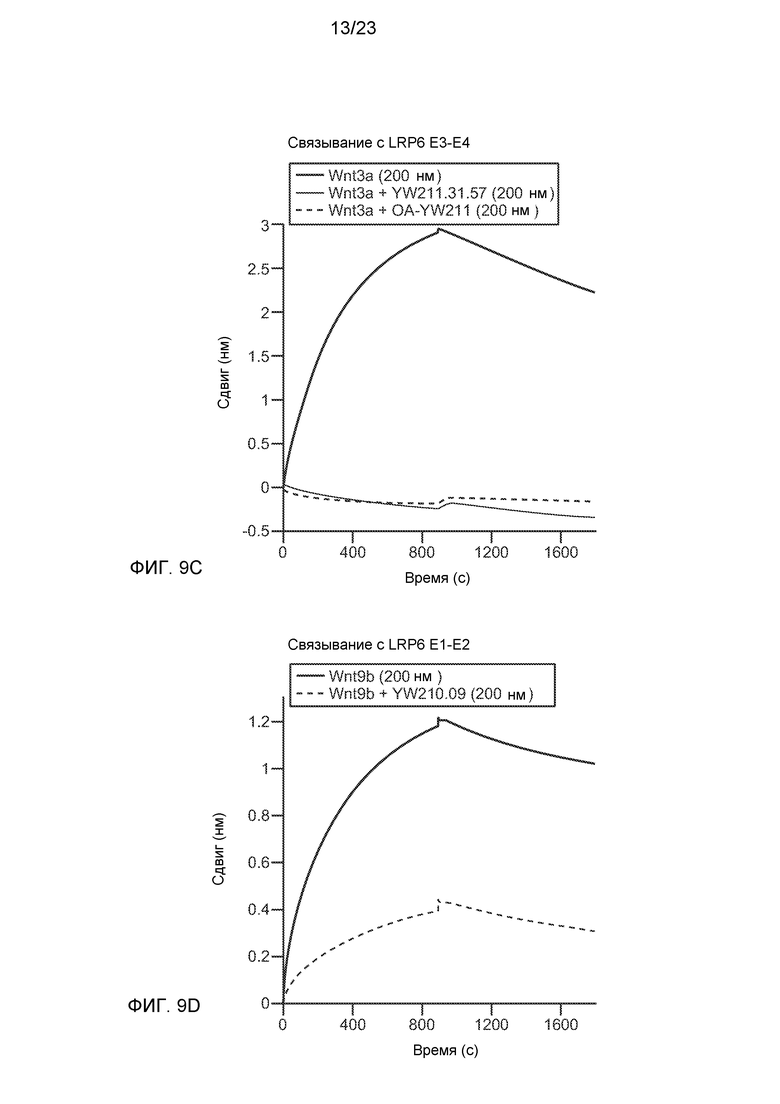
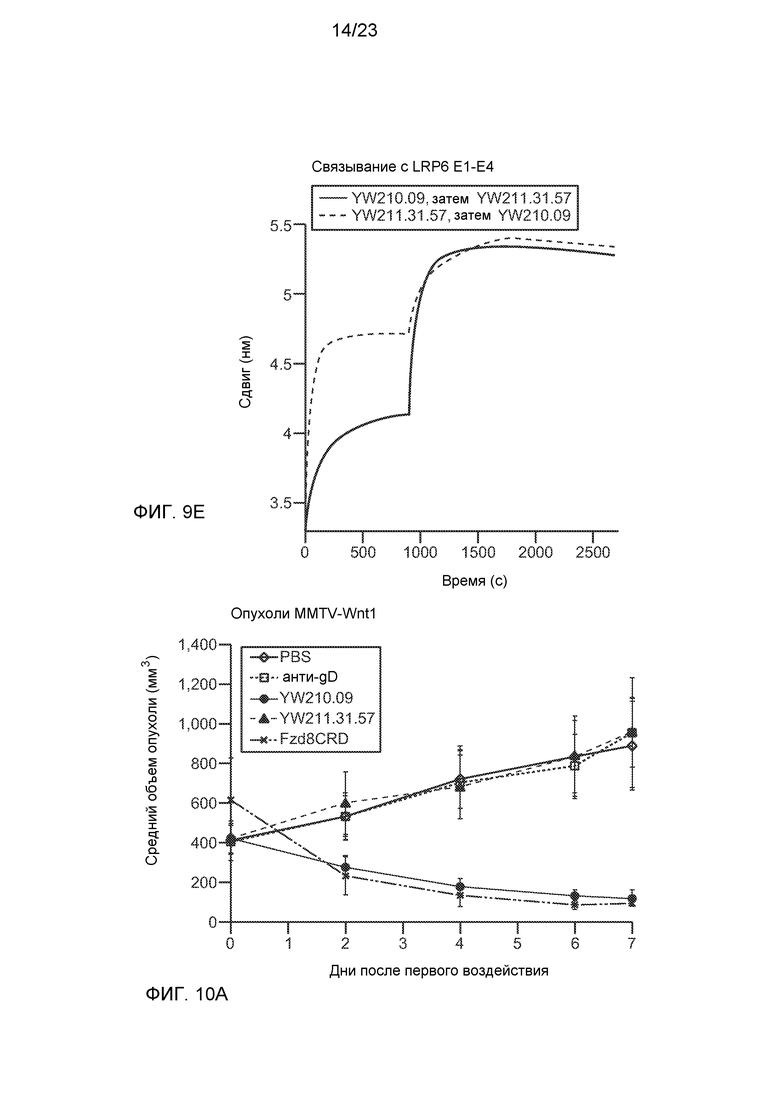
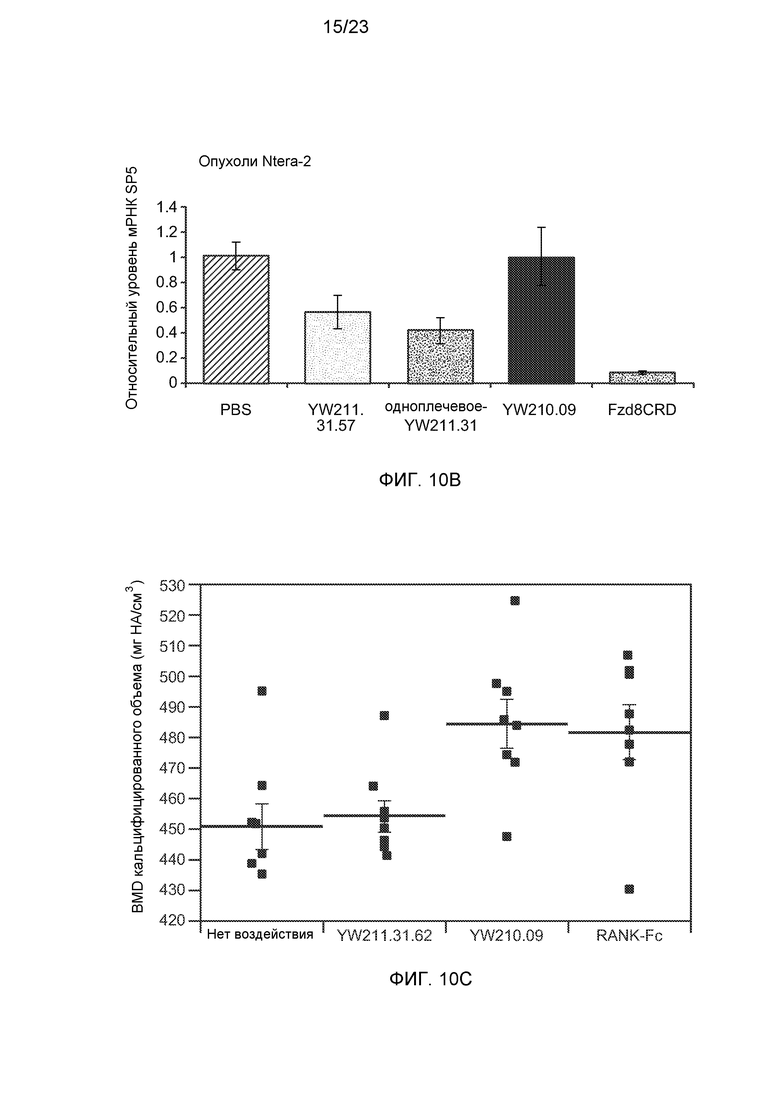
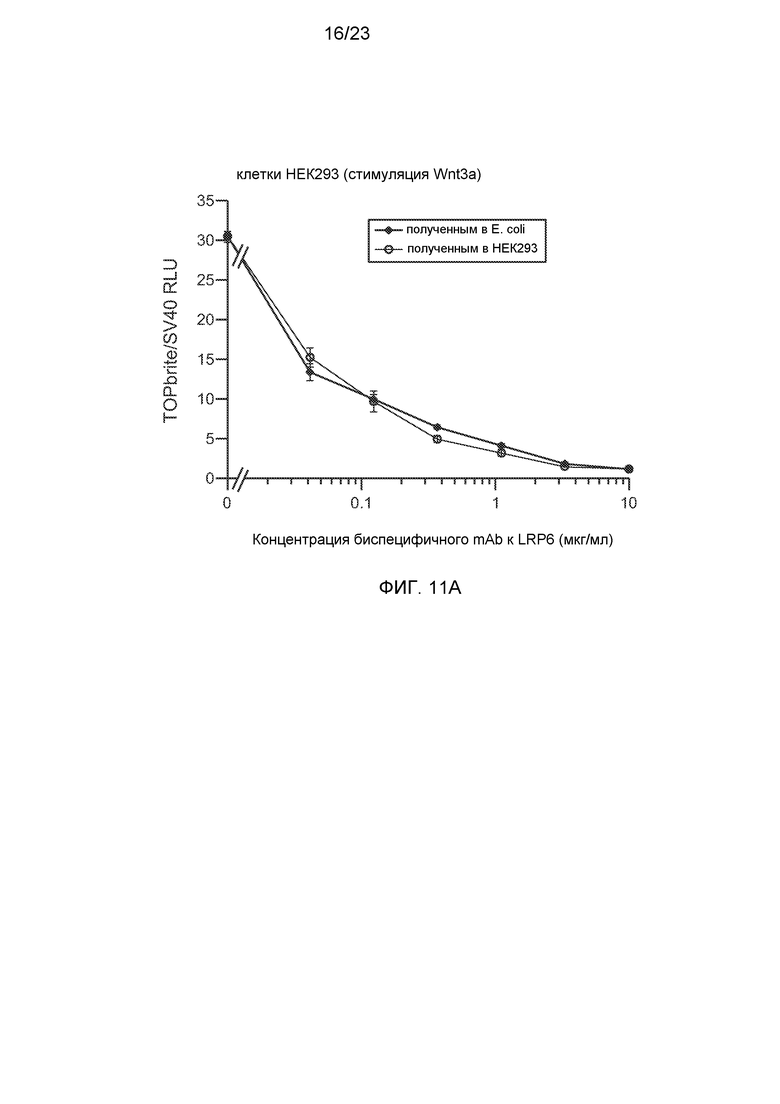
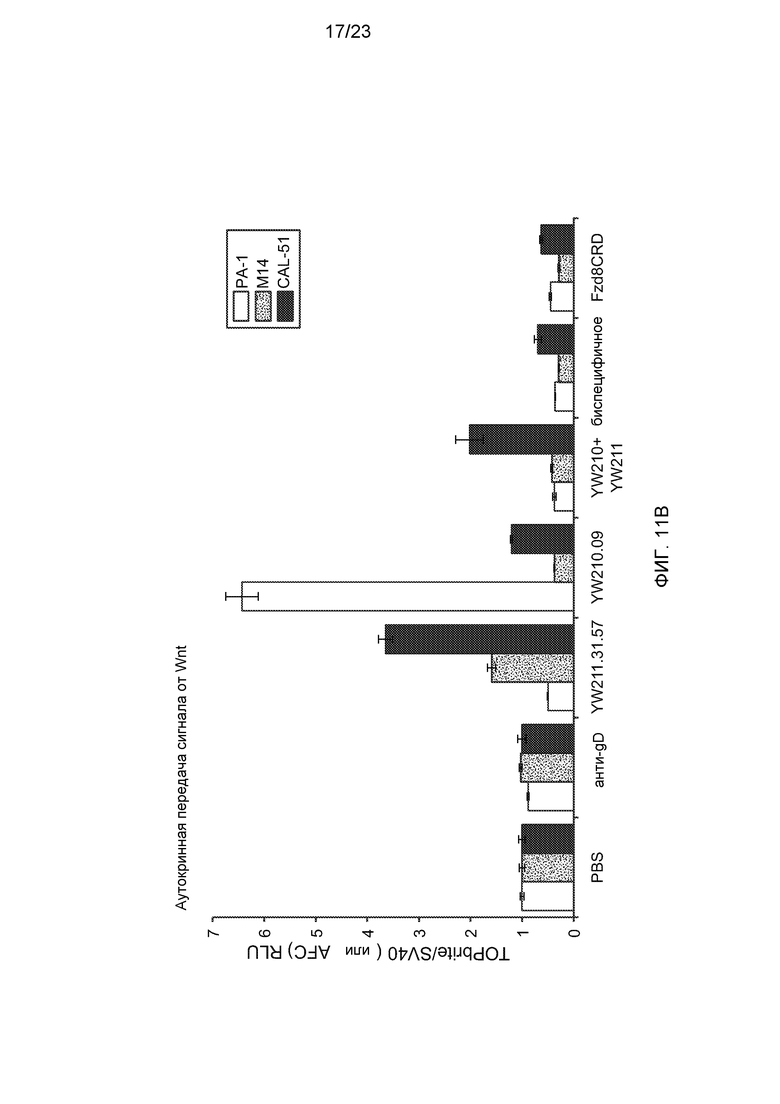
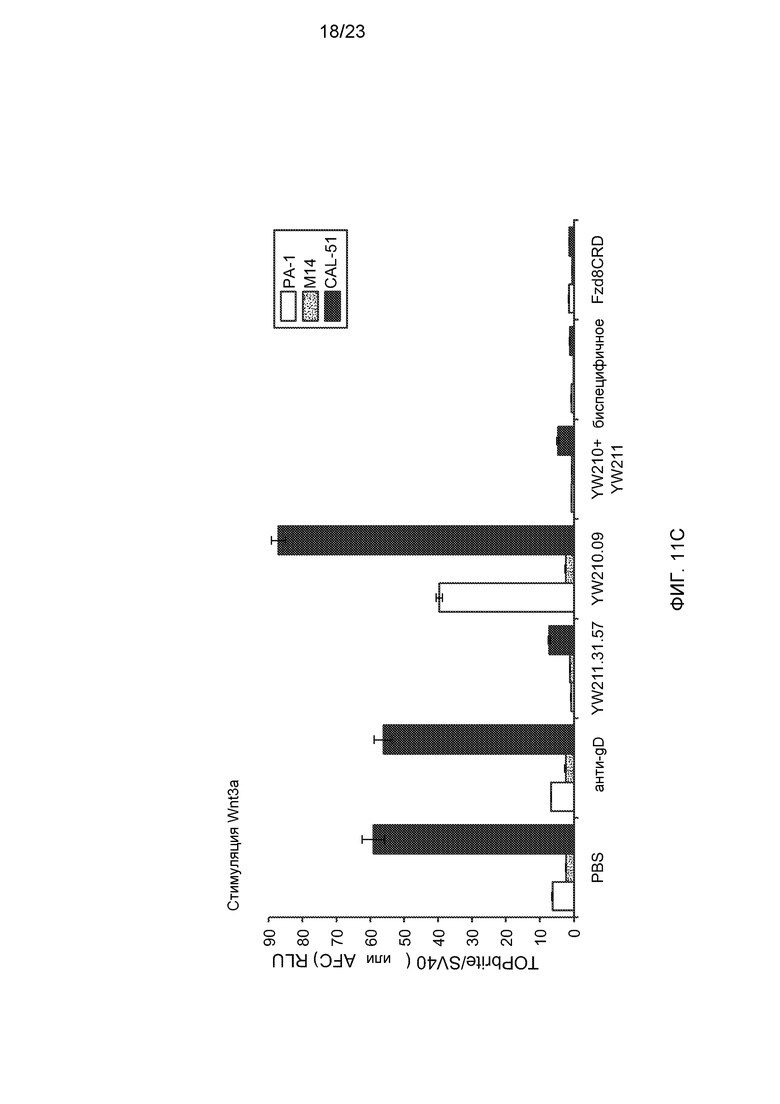
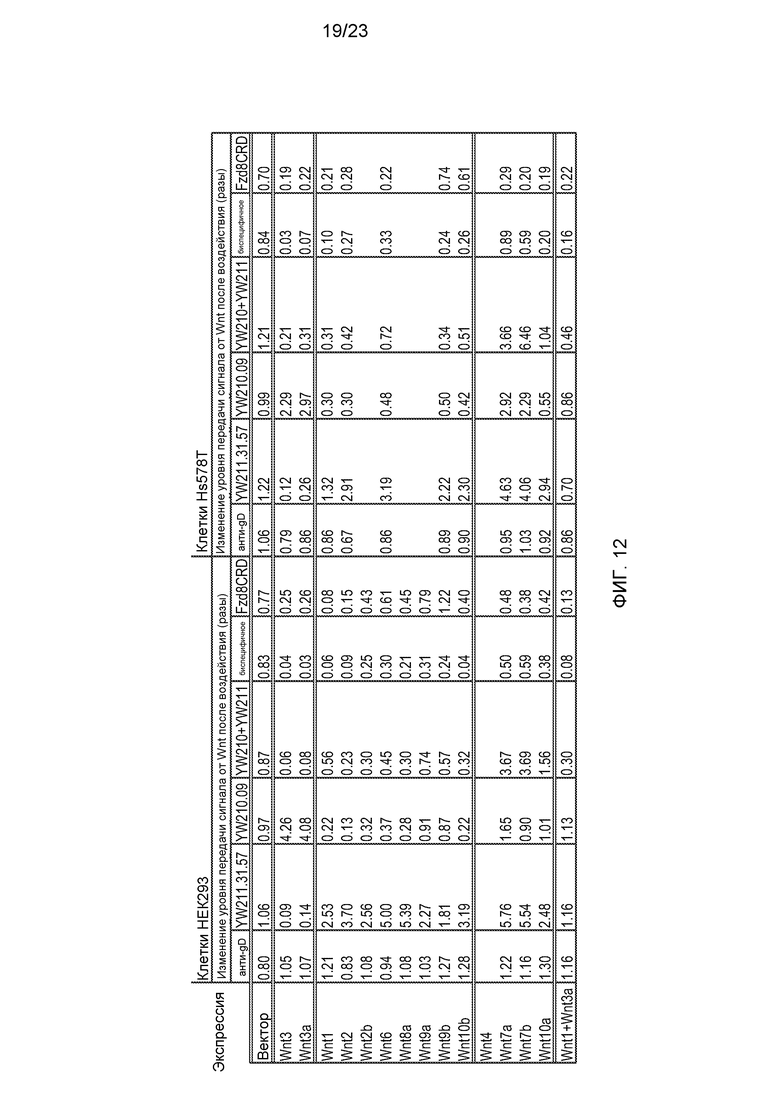
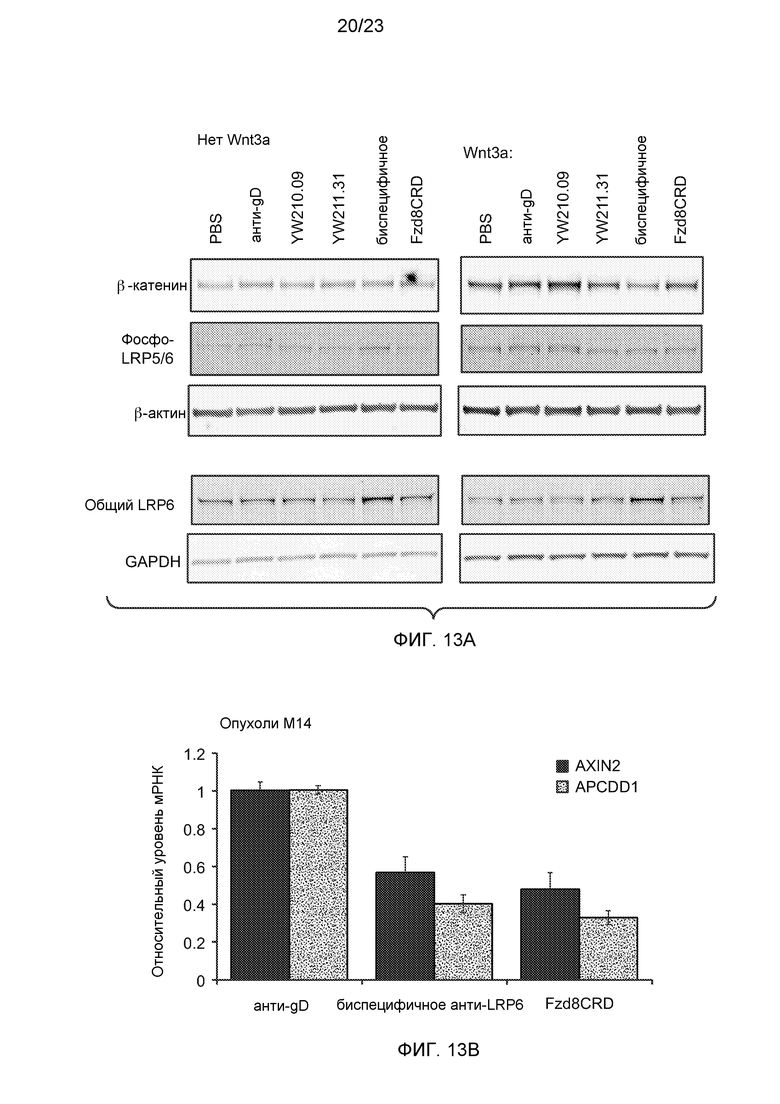
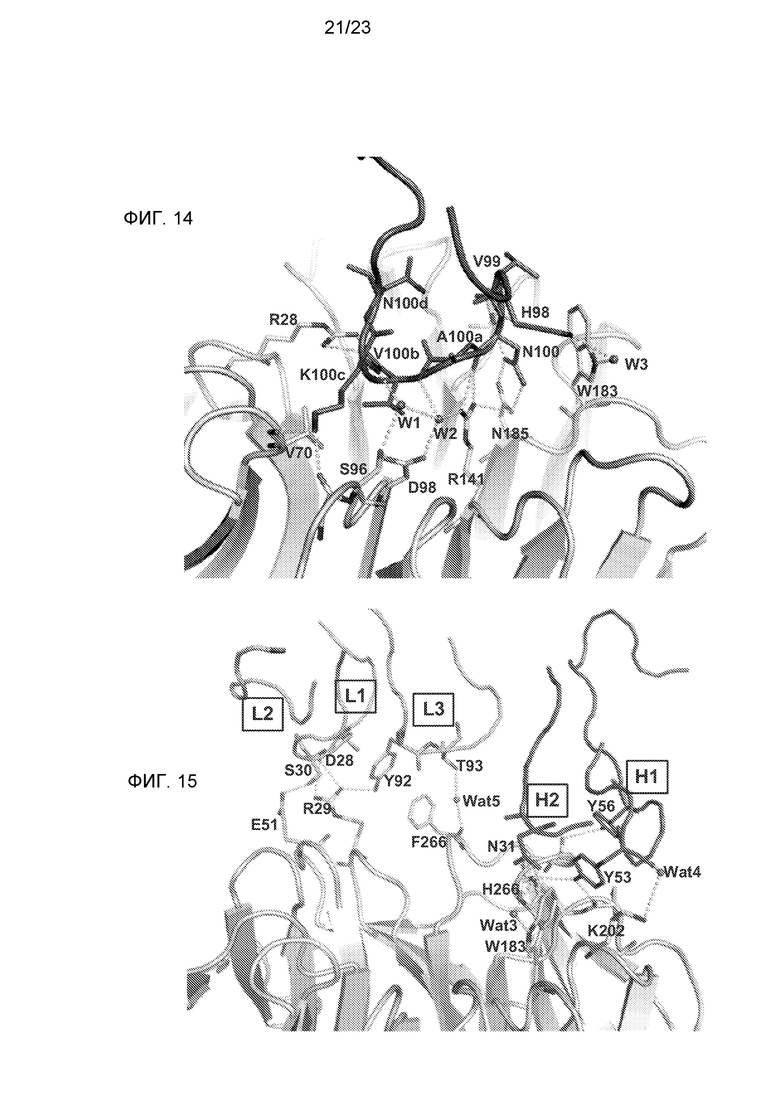
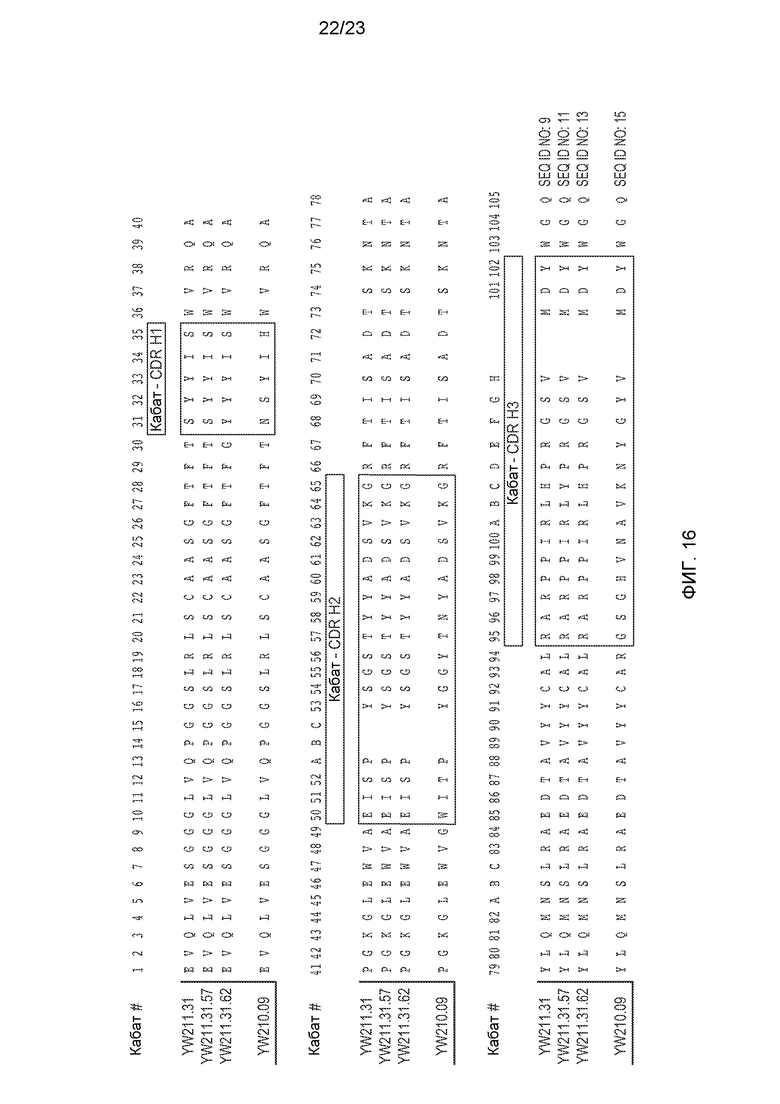
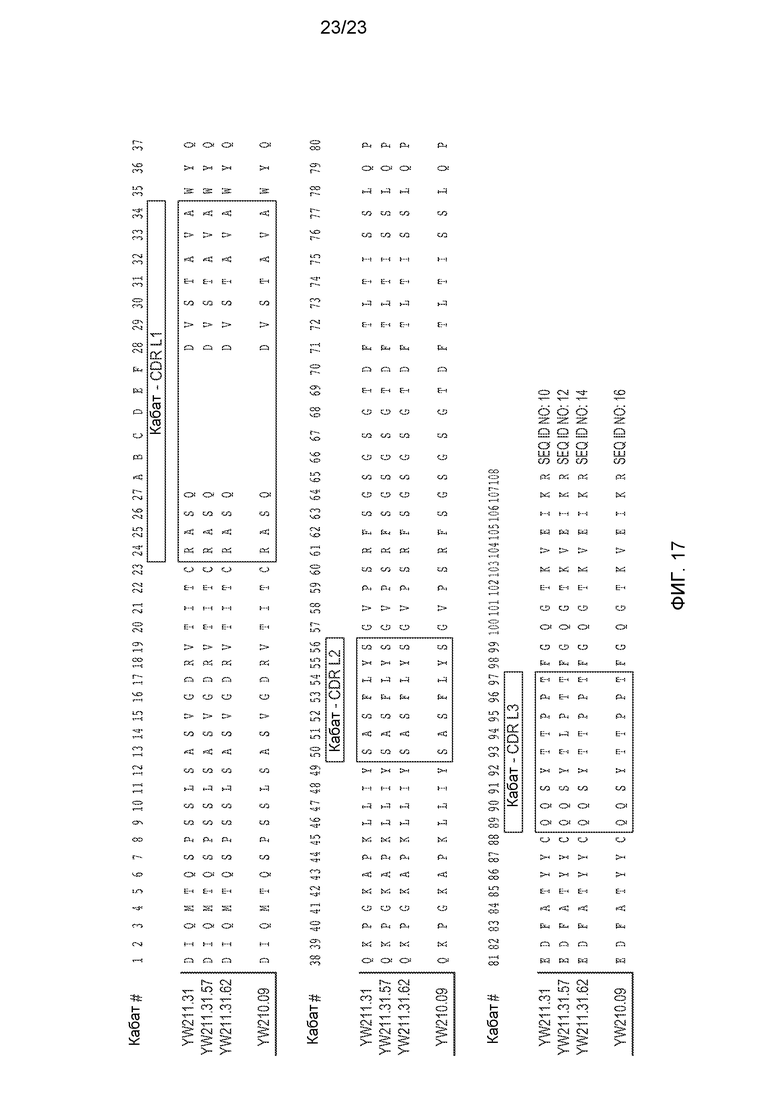












































Настоящее изобретение относится к области иммунологии. Предложено выделенное биспецифическое антитело, которое связывается с двумя различными областями белка 6 рецептора липопротеинов низкой плотности (LRP6). Рассмотрен фармацевтический состав для лечения злокачественной опухоли, содержащий биспецифическое антитело по настоящему изобретению; а также применение биспецифического антитела или фармацевтического состава в качестве лекарственного средства для лечения злокачественной опухоли и иммуноконъюгат для лечения злокачественной опухоли, содержащий биспецифическое антитело и цитотоксическое средство. Описана выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по изобретению, и клетка-хозяин, содержащая указанную нуклеиновую кислоту для получения антитела. Биспецифическое антитело по настоящему изобретению способно ингибировать передачу сигнала Wnt изоформ 3, 3a и изоформ 1, 2, 2b, 6, 8a, 9a, 9b и 10b и может найти дальнейшее применение в терапии различных заболеваний, ассоциированных с сигнальным путем Wnt. 7 н. и 10 з.п. ф-лы, 4 табл., 17 ил., 13 пр.
1. Выделенное биспецифическое антитело, которое связывается с двумя различными областями белка 6 рецептора липопротеинов низкой плотности (LRP6),
где антитело содержит первый VH, содержащий аминокислотную последовательность SEQ ID NO: 13, первый VL, содержащий аминокислотную последовательность SEQ ID NO: 14, второй VH, содержащий аминокислотную последовательность SEQ ID NO: 15, и второй VL, содержащий аминокислотную последовательность SEQ ID NO: 14.
2. Выделенное биспецифическое антитело, которое связывается с двумя различными областями белка 6 рецептора липопротеинов низкой плотности (LRP6),
где антитело содержит
первый домен VH, содержащий
(a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 20;
(b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 18; и
(c) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 19,
первый домен VL, содержащий
(g) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 25;
(h) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26; и
(i) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27,
второй домен VH, содержащий
(d) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 22;
(e) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 23; и
(f) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 24,
и второй домен VL, содержащий
(g) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 25;
(h) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 26; и
(i) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 27.
3. Биспецифическое антитело по п.2, где антитело ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt3 и Wnt3a, и ингибирует передачу сигнала, индуцированную изоформой Wnt, выбранной из группы, состоящей из Wnt 1, 2, 2b, 6, 8а, 9а, 9b и 10b.
4. Биспецифическое антитело по п.2, где антитело ингибирует аутокринную передачу сигнала от Wnt.
5. Антитело по п.4, где один из двух эпитопов содержит аминокислотные остатки R28, Е51, D52, V70, S71, Е73, L95, S96, D98, Е115, R141 и N185 белка 6 рецептора липопротеинов низкой плотности (LRP6) c последовательностью SEQ ID NO: 29.
6. Антитело по п.5, где один из двух эпитопов дополнительно содержит R29, W188, К202, Р225, Н226, S243 и F266 белка 6 рецептора липопротеинов низкой плотности (LRP6) с последовательностью SEQ ID NO: 29.
7. Антитело по любому из пп.1-6, где антитело представляет собой моноклональное антитело.
8. Антитело по любому из пп.1-6, где антитело представляет собой химерное антитело, антитело человека или гуманизированное антитело.
9. Антитело по п.8, где антитело представляет собой химерное антитело.
10. Антитело по п.8, где антитело представляет собой антитело человека.
11. Антитело по п.8, где антитело представляет собой гуманизированное антитело.
12. Фармацевтический состав, содержащий биспецифическое антитело в концентрации от 0,5 мг/кг до 15 мг/кг по любому из пп.1-11 и фармацевтически приемлемый носитель, для лечения злокачественной опухоли.
13. Применение биспецифического антитела по любому из пп.1-11 или фармацевтического состава по п.12 в качестве лекарственного средства для лечения злокачественной опухоли.
14. Применение фармацевтического состава по п.13, где злокачественная опухоль представляет собой немелкоклеточный рак легких, рак молочной железы, рак поджелудочной железы, рак яичников, рак почки или рак предстательной железы.
15. Иммуноконъюгат, содержащий биспецифическое антитело по любому из пп.1-11 и цитотоксическое средство, для лечения злокачественной опухоли.
16. Выделенная нуклеиновая кислота, кодирующая биспецифическое антитело по любому из пп.1-11.
17. Клетка-хозяин, содержащая нуклеиновую кислоту по п.16 для получения антитела по любому из пп.1-11.
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| BINNERTS M.E | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| BOURHIS E | |||
| et al | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |

