ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к новым растениям томата, образующим плоды, демонстрирующие модифицированное содержание Сахаров, в частности, демонстрирующие увеличенное содержание сахарозы. Настоящее изобретение также относится к семенам и частям указанных растений, например, к плодам. Настоящее изобретение дополнительно относится к способам получения и применения таких семян и растений. Настоящее изобретение также относится к новому аллелю SucMod, являющемуся модификатором содержания сахарозы, который в комбинации с аллелем TIV, отвечающим за накопление сахарозы, полученным из образца дикорастущего зеленоплодного томата, значительно изменяет долю Сахаров, содержащихся в плоде, обеспечивает увеличенное содержание сахарозы в плоде за счет гексозных Сахаров и приводит к получению плода с характерным ароматом.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Томат представляет собой хорошо известный источник витаминов, минеральных веществ и антиоксидантов, которые являются необходимыми компонентами сбалансированного здорового рациона. Также широко признается, что признаки качества, такие как цвет, аромат и плотная консистенция, будут оказывать сильное влияние на выбор потребителя при покупке данных дорогостоящих и скоропортящихся плодов сельскохозяйственных культур.
Растворимые сахара составляют примерно половину сухого вещества в спелом плоде томата, и их уровни оказывают сильное влияние на параметры, по которым может быть измерено качество плода, такие как аромат плода, сладкий вкус, предпочтение потребителя и общее содержание растворимых твердых веществ (в единицах по шкале Брикса). Согласно сложившимся представлениям культивируемые сорта томата (Solanum lycopersicum) накапливают гексозные моносахариды глюкозу и фруктозу подобно другим видам с красными, оранжевыми и желтыми плодами - Solanum cheesmaniae и Solanum pimpinellifotium, которые образуют подгруппу Eulycopersieum. В то же время они накапливают низкие уровни дисахарида сахарозы, состоящего из фруктозы и глюкозы. В отличие от этого, все зеленоплодные дикорастущие виды, которые образуют группу Eriopersicum (Solanum habrochaites (ранее Lycopersicon hirsutum), Solanum chmielewskii, Solanum pennellii и Solanum peruvianum), накапливают дисахарид сахарозу в качестве основного растворимого сахарного компонента (Davies, 1966; Manning and Maw, 1975).
Было обнаружено, что данный характерный тип накопления Сахаров обусловлен ферментом вакуолярной инвертазой томата (сокращенно VA или TIV), расщепляющим сахарозу в вакуоли на ее гексозные компоненты. В более ранних исследованиях было показано, что отдельный локус (sucr) контролирует признак накопления сахарозы/гексозы (Yelle et al., 1991; Chetelat et al., 1993; Klann et al., 1993, 1996; Hadas et al., 1995). Соответствующий ген (TIV), который кодирует фермент кислую растворимую инвертазу, катализирующий гидролиз импортируемой сахарозы до гексозы (Chetelat et al., 1993; Klann et al., 1993), был картирован на хромосоме 3 (Solyc03g083910). В процессе развития зеленоплодные виды демонстрируют прекращение экспрессии TIV во время созревания, что запускает уменьшение уровня белка и ферментативной активности, а также в конечном счете обеспечивает возможность накопления сахарозы в вакуоли. В отличие от этого, виды с красными/оранжевыми/желтыми плодами в процессе развития демонстрируют повышение экспрессии TIV, и формирующаяся в результате ферментативная активность обуславливает почти полный гидролиз сахарозы до гексозных фрагментов в виде глюкозы и фруктозы (Klann et al., 1993; Miron et al., 2002). Следовательно, введение "дикого" аллеля TIV в культивируемую линию посредством возвратного скрещивания может приводить к значительному увеличению уровней сахарозы и значительному снижению уровней глюкозы и фруктозы по сравнению с рекуррентной фоновой линией, в которой отсутствует "дикий" аллель TIV (Hadas et al., 1995, таблица 2).
Экспрессию TIV и соответствующую ей инвертазную активность можно также модулировать посттрансляционным способом с помощью белковых ингибиторов инвертазы. Действительно, исследования, проводившиеся в течение последних нескольких лет, показали, что контроль над экспрессией ингибитора инвертазы может оказывать значительное влияние на гидролиз, осуществляемый инвертазой, in planta и, следовательно, на метаболизм Сахаров. Например, сайленсинг экспрессии ингибитора инвертазы клеточной стенки (CIF) в развивающемся плоде томата приводил к повышению активности апопластной инвертазы (LIN5) и последующему увеличению акцептирующей активности и накопления Сахаров в плоде (Jin et at., 2009). Аналогичным образом, сообщалось, что очищенную вакуолярную инвертазу томата (TIV) можно было ингибировать с помощью белка Solycl299190, что указывало на то, что последний функционирует в качестве ингибитора TIV (также называемого VIF; Tauzin et at., 2014; Qin et at., 2016).
Однако относительное увеличение содержания сахарозы в культивируемых растениях томата, содержащих "дикий" аллель TIV (Klann et at., 1993, фигура 2; Hadas et at., 1995, таблица 2, данные для BC1F3) или сверхэкспрессируемый аллель VIF (Qin et al., 2016, фигура 6D), по всей видимости, происходит за счет общего содержания Сахаров, которое уменьшается или в лучшем случае поддерживается на сходном уровне, а также соотношения сахарозы и гексозы, которое быстро достигает пика при значении, составляющем около 0,50 или меньше. Таким образом, существует потребность в дополнительном повышении содержания сахарозы в плодах культивируемых растений томата при одновременном увеличении соотношения сахарозы и гексозы, а также общего содержания Сахаров, и, таким образом, обеспечении растениеводов и потребителей различающимися растениями и плодами томата.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение удовлетворяет потребность в получении новых растений томата, образующих плоды, демонстрирующие модифицированное содержание сахаров, в частности, демонстрирующие увеличенное содержание сахарозы.
В первом варианте осуществления настоящее изобретение предусматривает культивируемое растение томата, предпочтительно культивируемое растение Solanum lycopersicum, содержащее:
a) по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, характеризующегося по меньшей мере 90% идентичностью последовательности с SEQ ID NO: 1, и
b) две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата;
где указанный аллель SucMod содержит нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотид Т в положении, которое соответствует положению 498 в SEQ ID NO: 1; и
где указанное растение образует плод томата, демонстрирующий увеличенное содержание сахарозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления настоящего изобретения аллель SucMod получен из Solanum chmielewskii или Solanum pimpinellifotium.
В дополнительном варианте осуществления настоящего изобретения аллель SucMod содержит нуклеотидную последовательность под SEQ ID NO: 1.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV характеризуется по меньшей мере 98% идентичностью последовательности с SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV содержит нуклеотид А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотид А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотид Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотид С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотид Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотид С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотид А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотид G в положении, которое соответствует положению 1886 в SEQ ID NO: 6. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV получен из Solanum habrochaites.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV содержит нуклеотид С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотид G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов оуществления, где указанный аллель TIV содержит нуклеотидную последовательность под SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение содержит две копии аллеля SucMod.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV и указанный аллель SucMod могут быть получены из Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение представляет собой инбредное, дигаплоидное или гибридное растение.
В дополнительном варианте осуществления настоящее изобретение предусматривает семя, из которого образуется растение согласно любому из предыдущих вариантов осуществления.
В дополнительном варианте осуществления настоящее изобретение предусматривает способ получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающий стадии:
a) скрещивания растения согласно любому из предыдущих вариантов осуществления, содержащего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, с культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV;
b) отбора растения-потомка, образующего плоды, демонстрирующие увеличенное содержание сахарозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV;
где отбор на стадии b) проводят посредством выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотида Т в положении, которое соответствует положению 498 в SEQ ID NO: 1; и посредством выявления нуклеотида А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по предыдущему варианту осуществления, где отбор на стадии b) проводят посредством дополнительного выявления нуклеотида С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу согласно любому из предыдущих вариантов осуществления, где растение из стадии а) представляет собой Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169.
В дополнительном варианте осуществления настоящее изобретение предусматривает способ идентификации культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды, демонстрирующие увеличенное содержание сахарозы, и имеющего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата, включающий стадии:
a) выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотида Т в положении, которое соответствует положению 498 в SEQ ID NO: 1; и
b) выявления нуклеотида А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по предыдущему варианту осуществления, где стадию b) проводят посредством дополнительного выявления нуклеотида С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
Было показано, что использование аллеля SucModchm в генетическом окружении элитного культивара, а также в комбинации с "диким" аллелем TIV, таким как аллель TIVhab, как описано в данном документе, приводит к дополнительному повышению содержания сахарозы, увеличению общего содержания Сахаров и отличающемуся накоплению гексоз фруктозы и глюкозы по сравнению с накоплением сахарозы, что приводит к уникальному и выраженному ощущению аромата и вкуса плода. Таким образом, настоящее изобретение обладает потенциалом для использования в будущих программах разведения с целью улучшения аромата и вкуса плодов томата.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1. Распределение содержания сахарозы в плоде (%) в зрелых плодах томата из 15 объединенных сегрегирующих популяций F8, полученных из линии 4510 с высоким содержанием сахарозы, демонстрирующее бимодальное распределение, свидетельствующее об одиночном моногенном наследовании.
Фигура 2. Накопление сахарозы, гексозы и общего сахара (сахароза + гексоза) в развивающихся плодах (G - зелено-зрелые; BR - бланжевые; R - полностью спелые) томата линий  (обозначены квадратами) и
(обозначены квадратами) и 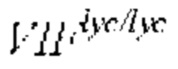 (обозначены ромбами) в генетическом окружении
(обозначены ромбами) в генетическом окружении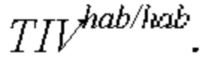
Фигура 3. (А) Общее содержание сахаров - (В) Соотношение сахарозы и гексозы в полностью спелых плодах томата линий 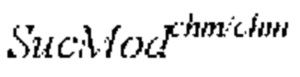 и
и  в генетическом окружении
в генетическом окружении 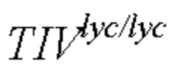 или
или  Растения выращивали в условиях пассивной защиты на произвольно взятых площадках, содержащих по шесть растений на площадку.
Растения выращивали в условиях пассивной защиты на произвольно взятых площадках, содержащих по шесть растений на площадку.
Фигура 4. Выравнивания последовательностей и матрица процентов идентичности для аллельных последовательностей SucMod/VIF из Solanum lycopersicum (Solyc12g099190), Solanum chmielewskii BD732 (SEQ ID NO: 1) и Solanum pennellii LA0716 (Solpen12g033870), а также гомологичных последовательностей из Solanum cheesmaniae LA0429 и Solanum pimpinellifolium LA1589.
Фигура 5. Выравнивания последовательностей и матрица процентов идентичности для аллельных последовательностей TIV из Solanum habrochaites (SEQ ID NO: 6), Solanum peruvianum (KY565130), Solanum pennellii (XMO15214462), Solanum chmielewskii (KY565126), Solanum lycopersicum (NM001247914), Solanum lycopersicum var cerasiforme (GU784870), Solanum cheesmaniae (KY565124) и Solanum pimpinellifolium (Z12026).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ ОПРЕДЕЛЕНИЯ
Технические термины и выражения, используемые в объеме настоящей заявки, как правило, должны иметь значение, обычно применяемое к ним в соответствующей области разведения и культивирования растений, если иное не указано в данном документе ниже.
Используемые в данном описании и прилагаемой формуле изобретения формы единственного числа включают ссылки на множественное число, если в контексте явно не указано иное. Таким образом, например, ссылка на "растение" включает одно или несколько растений, а ссылка на "клетку" включает смеси клеток, тканей и т.п.
Подразумевается, что термин "приблизительно", используемый в данном документе в отношении значения или величины массы, веса, времени, объема, концентрации или процентной доли, охватывает отклонения от указанной величины, составляющие в некоторых вариантах осуществления ± 20%, в некоторых вариантах осуществления ± 10%, в некоторых вариантах осуществления ± 5%, в некоторых вариантах осуществления ± 1%, в некоторых вариантах осуществления ± 0,5% и в некоторых вариантах осуществления ± 0,1%, поскольку такие отклонения являются подходящими для осуществления раскрытого способа.
Термин "культивируемое растение томата" в пределах объема настоящего изобретения понимают как относящийся к растению, которое больше не находится в естественном состоянии, а было выведено и одомашнено усилиями человека для сельскохозяйственного применения и/или потребления человеком, и исключает образцы дикорастущего томата, такие как Solanum chmielewskii BD732 и Solanum habrochaites LAI777. В качестве примера, в вариантах осуществления растение томата согласно настоящему изобретению способно образовывать желтые, оранжевые или красные плоды. В качестве альтернативы или дополнительно, культивируемое растение томата представляет собой гибридное растение. В качестве альтернативы или дополнительно, культивируемое растение томата представляет собой растение Solanum lycopersicum. В контексте межвидового скрещивания растения Solanum lycopersicum и образца дикорастущего томата культивируемое растение томата определяют как растение-потомок, полученное в результате указанного межвидового скрещивания, где указанное растение-потомок было подвергнуто возвратному скрещиванию с растением Solanum lycopersicum по меньшей мере три раза.
Термин "аллель" в пределах объема настоящего изобретения понимают как относящийся к альтернативным или вариантным формам различных генетических единиц, идентичным различным формам гена или идентифицируемой генетической детерминанты любого типа или ассоциированным с ними, которые являются альтернативными при наследовании, поскольку они расположены в одном и том же локусе на гомологичных хромосомах. Такие альтернативные или вариантные формы могут быть результатом однонуклеотидных полиморфизмов, вставок, инверсий, транслокаций или делеций или следствием генной регуляции, обусловленным, например, химической или структурной модификацией, регуляцией транскрипции или посттрансляционной модификацией/регуляцией. В диплоидных клетке или организме два аллеля данного гена или генетического элемента обычно занимают соответствующие локусы на паре гомологичных хромосом.
Аллель, ассоциированный с качественным признаком, может содержать альтернативные или вариантные формы различных генетических единиц, в том числе идентичные одному гену или нескольким генам или их продуктам или даже гену, нарушаемому или контролируемому генетической детерминантой, способствующей проявлению фенотипа, представленного локусом, или ассоциированные с ними.
Собственно говоря, в данном документе термин "увеличенное содержание сахарозы" понимают как означающий, что растение согласно настоящему изобретению, например, содержащее а) по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, характеризующегося по меньшей мере 90% генетическим сходством с SEQ ID NO: 1, и b) две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата, способно образовывать плоды, демонстрирующие увеличенное содержание сахарозы по сравнению с растением, в котором отсутствуют указанные аллели. В предпочтительном варианте осуществления содержание сахарозы измеряют, когда плоды томата достигают стадии полной спелости.
Термин "увеличенное содержание сахарозы" в пределах объема настоящего изобретения понимают как означающий плод томата, который характеризуется статистически значимым увеличенным содержанием сахарозы по сравнению с плодом от контрольного растения (например, как описано в разделе "Примеры"), при использовании стандартной ошибки и/или при Р<0,05 или Р<0,01 при использовании критерия Стьюдента.
Термин "контрольное растение томата" в пределах объема настоящего изобретения понимают как означающий растение томата, которое содержит такое же генетическое окружение, как и культивируемое растение томата по настоящему изобретению, при этом контрольное растение не имеет ни одного из по меньшей мере одного аллеля по настоящему изобретению, связанного с увеличенным содержанием сахарозы. В частности, контрольное растение томата представляет собой растение томата, принадлежащее к тому же сорту растений и не содержащее ни одного из по меньшей мере одного аллеля. Контрольное растение томата выращивают в течение такого же периода времени и при таких же условиях, что и культивируемое растение томата по настоящему изобретению. Сорт растения в данном документе следует понимать в соответствии с определением UPOV. Таким образом, контрольное растение томата может представлять собой почти изогенную линию, инбредную линию или гибрид при условии, что они содержат такое же генетическое окружение, как и растение томата по настоящему изобретению, за исключением того, что контрольное растение не имеет ни одного из по меньшей мере одного аллеля по настоящему изобретению, связанного с увеличенным содержанием сахарозы.
Термин "признак" относится к характеристике или фенотипу. В контексте настоящего изобретения признак содержания сахарозы представляет собой признак увеличенного содержания сахарозы. Признак может наследоваться по доминантному или рецессивному типу или по частично доминантному или неполно-доминантному типу. Признак может быть моногенным или полигенным или может быть результатом взаимодействия одного или нескольких генов с окружающей средой. Растение томата может быть гомозиготным или гетерозиготным по данному признаку.
Термины "гибрид", "гибридное растение" и "гибридное потомство" относятся к особи, полученной от генетически различных родительских организмов (например, к генетически гетерозиготной или преимущественно гетерозиготной особи).
Термин "инбредная линия" относится к генетически гомозиготной или почти гомозиготной популяции. Например, инбредная линия может быть получена посредством нескольких циклов братско-сестринского разведения или самовоспроизводства или при получении дигаплоидов.
Термин "дигаплоидная линия" относится к стабильным инбредным линиям, полученным из культуры пыльников. Некоторые пыльцевые зерна (гаплоидные), культивируемые на определенной среде и в определенных условиях, могут формировать проростки, содержащие n хромосом. Затем в проростках происходит "удвоение", и они содержат 2n хромосом. Потомство этих проростков называют "дигаплоидным", и оно по существу больше не сегрегирует (является стабильным).
Термин "культивар" или "сорт" относится к сорту, полученному методами плодоовощеводства, в отличие от встречающегося в природе сорта. В некоторых вариантах осуществления настоящего изобретения культивары или сорта обладают коммерческой ценностью.
Термин "генетически фиксированный" относится к генетическому элементу, который был стабильно включен в состав генома растения, который обычно не содержит данный генетический элемент. Будучи генетически фиксированным, генетический элемент может передаваться другим растениям простым и прогнозируемым образом путем половых скрещиваний.
Термин "растение" или "часть растения" в дальнейшем в данном документе относится к части, органу или ткани растения, которые могут быть получены из растения томата согласно настоящему изобретению, в том числе без ограничения к листьям, стеблям, корням, цветкам или частям цветка, плодам, побегам, гаметофитам, спорофитам, пыльце, пыльникам, микроспорам, яйцеклеткам, зиготам, зародышам, участкам меристемы, каллюсной ткани, семенам, черешкам, клеточной или тканевой культуре или любым другим части или продукту растения, которые по-прежнему демонстрируют признаки содержания сахарозы согласно настоящему изобретению, в частности, при выращивании из них растения, образующего плоды.
"Растение" представляет собой любое растение на любой стадии развития.
Семя растения томата представляет собой семя, из которого вырастает растение томата согласно любому из вариантов осуществления.
"Растительная клетка" представляет собой структурную и физиологическую единицу растения, содержащую протопласт и клеточную стенку. Растительная клетка может быть представлена в виде выделенной одиночной клетки или культивируемой клетки или в виде части более высокоорганизованной единицы, такой как, например, растительная ткань, орган растения или целое растение.
"Культура растительных клеток" означает культуры растительных единиц, таких как, например, протопласты, клетки в клеточной культуре, клетки в растительных тканях, пыльца, пыльцевые трубки, семязачатки, зародышевые мешки, зиготы и зародыши на различных стадиях развития.
"Орган растения" представляет собой отдельную и визуально структурированную и дифференцированную часть растения, такую как корень, стебель, лист, цветочная почка или зародыш.
Термин "растительная ткань", используемый в данном документе, означает группу растительных клеток, организованных в структурную и функциональную единицу. Включена любая ткань растения in planta или в культуре. Данный термин включает без ограничения целые растения, органы растений, семена растений, тканевую культуру и любые группы растительных клеток, организованных в структурные и/или функциональные единицы. Использование данного термина в сочетании с любым конкретным типом растительной ткани, перечисленным выше или иным образом охватываемым данным определением, или при его отсутствии не предполагается как исключающее любой другой тип растительной ткани.
Термин "зелено-незрелая стадия" определяется как период, в котором плоды являются неспелыми и все еще увеличиваются в размерах. Данная стадия понимается как первая стадия процесса выспевания.
Термин "зелено-зрелая стадия" определяется как период, в котором плод увеличен до полного размера и является зрелым, но неспелым, который следует за "зелено-незрелой стадией" в процессе выспевания.
Термин "стадия бланжевой спелости" определяется как появление первого признака изменения цвета наружной части плода с зеленого на розовый цвет.
Термин "стадия полной спелости" определяется как период, в котором плоды являются полностью красными без признаков наличия зеленого цвета.
Термины "внутренний слой околоплодника" и "наружный слой околоплодника" в пределах объема настоящего изобретения понимают как означающие ткань плода, в которой наружный слой околоплодника представляет собой слой (толщиной примерно 2 мм), расположенный непосредственно ниже наружного слоя эпидермиса и выше слоя сосудистой ткани. Внутренний слой околоплодника располагается на глубине от примерно 3 мм до 10 мм ниже сосудистого слоя и перед внутренним слоем эпидермиса. Термин "переработанные пищевые продукты" в пределах объема настоящего изобретения понимают как означающий пищевые продукты, которые были подвергнуты изменению по сравнению с их естественным состоянием, например, томатную пасту. Способы, применяемые для переработки пищевых продуктов, включают без ограничения консервирование, замораживание, охлаждение, обезвоживание и асептическую обработку. Растения по настоящему изобретению являются особенно предпочтительными для применения в способах с использованием нагревания, которое может вызывать гидролиз сахарозы до гексозных Сахаров и, следовательно, уменьшение содержания сахарозы в плодах, образуемых данными растениями. Однако, поскольку плоды, образуемые растениями по настоящему изобретению, характеризуются увеличенным содержанием сахарозы, потеря сахарозы во время обработки будет оказывать меньшее влияние на формирующийся в результате аромат и вкус. Кроме того, увеличение общего содержания Сахаров будет сохраняться, поскольку дополнительная сахароза будет разрушаться до гексозных фрагментов. Термин "рынок свежих овощей" в пределах объема настоящего изобретения понимают как означающий реализуемые на рынке овощи, которые были подвергнуты минимальной обработке.
Используемый в данном документе термин "маркерный аллель" относится к альтернативной или вариантной форме генетической единицы, определенной в данном документе выше, при использовании в качестве маркера для определения локализации на хромосоме генетических локусов, содержащих аллели, которые способствуют изменчивости фенотипических признаков.
Используемый в данном документе термин "разведение" и его грамматические варианты относятся к любому процессу, в результате которого получают особь-потомка. Разведение может быть половым или бесполым или любой их комбинацией. Иллюстративные неограничивающие типы разведения включают варианты скрещивания, самовоспроизводство, получение двойных гаплоидных производных и их комбинации.
Используемая в данном документе фраза "устойчивая популяция для разведения" относится к совокупности потенциальных партнеров по разведению, полученных от родительских организмов и/или используемых в качестве родительских организмов в программе разведения; например, коммерческой программе разведения. Представители устойчивой популяции для разведения обычно хорошо изучены генетически и/или фенотипически. Например, некоторые фенотипические признаки, представляющие интерес, могли быть оценены, например, в разных условиях окружающей среды, в нескольких местоположениях и/или в разные моменты времени. В качестве альтернативы или в дополнение, один или несколько генетических локусов, ассоциированных с экспрессией фенотипических признаков, могли быть идентифицированы, а один или несколько представителей популяции для разведения могли быть генотипированы по одному или нескольким генетическим локусам, а также по одному или нескольким генетическим маркерам, ассоциированным с одним или несколькими генетическими локусами.
Используемая в данном документе фраза "диплоидная особь" относится к особи, у которой есть два набора хромосом, обычно по одному от каждого из двух ее родительских организмов. Однако понятно, что в некоторых вариантах осуществления диплоидная особь может получать свои "материнский" и "отцовский" наборы хромосом от одного и того же организма, как, например, при самовоспроизводстве растения для получения последующего поколения растений.
Термин "гомозиготный" в пределах объема настоящего изобретения понимают как относящийся к одинаковым аллелям в одном или нескольких соответствующих локусах на гомологичных хромосомах. В контексте настоящего изобретения растение томата, содержащее две идентичные копии определенного аллеля в определенном локусе, например, аллеля SucModchm в локусе Solyc12g099190, является гомозиготным по соответствующему локусу.
Термин "гетерозиготный" в пределах объема настоящего изобретения понимают как относящийся к неодинаковым аллелям в одном или нескольких соответствующих локусах на гомологичных хромосомах. В контексте настоящего изобретения растение томата, содержащее одну копию определенного аллеля в определенном локусе, например, аллеля SucModchm в локусе Solyc12g099190, является гетерозиготным по соответствующему локусу.
Термин "доминантный аллель" в пределах объема настоящего изобретения понимают как относящийся к аллелю, который определяет фенотип в случае его присутствия в гетерозиготном или гомозиготном состоянии.
Термин "рецессивный аллель" относится к аллелю, который определяет фенотип в случае его присутствия только в гомозиготном состоянии.
Термин "возвратное скрещивание" в пределах объема настоящего изобретения понимают как относящийся к способу, в рамках которого гибридное потомство подвергается многократному возвратному скрещиванию с одним из родительских организмов. Различные рекуррентные родительские организмы могут использоваться в последующих возвратных скрещиваниях.
Термин "локус" в пределах объема настоящего изобретения понимают как относящийся к участку на хромосоме, который содержит ген или любой другой генетический элемент или фактор, способствующий проявлению признака.
Используемый в данном документе термин "маркерный локус" относится к участку на хромосоме, содержащему нуклеотидную или полинуклеотидную последовательность, которая присутствует в геноме особи и которая ассоциирована с одним или несколькими локусами, представляющими интерес, которые могут содержать ген или любую другую генетическую детерминанту или фактор, способствующий проявлению признака. "Маркерный локус" также относится к участку на хромосоме, который содержит полинуклеотидную последовательность, комплементарную геномной последовательности, такую как последовательность нуклеиновой кислоты, используемая в качестве зондов.
Термин "генетическое сцепление" в пределах объема настоящего изобретения понимают как относящийся к ассоциации признаков при наследовании вследствие расположения генов близко друг к другу на одной и той же хромосоме, которая измеряется в процентах рекомбинации между локусами (в сантиморганидах, сМ).
Для цели настоящего изобретения термин "косегрегация" относится к тому факту, что аллель, определяющий признак, и аллель(аллели), определяющие маркер(маркеры), как правило, передаются совместно, поскольку они физически расположены близко друг к другу на одной и той же хромосоме (ввиду их физического близкого расположения между ними наблюдается сниженная рекомбинация), что приводит к неслучайной ассоциации таких аллелей в результате их близкого расположения на одной хромосоме. "Косегрегация" также относится к наличию двух или более признаков в одном растении, из которых по меньшей мере один, как известно, является генетическим, и которое нельзя объяснить простой случайностью.
Используемый в данном документе термин "генетическая архитектура в локусе количественного признака" относится к геномному участку, который характеризуется статистически значимой корреляцией с фенотипическим признаком, представляющим интерес, и представляет генетическую основу, обуславливающую фенотипический признак, представляющий интерес.
Используемые в данном документе фразы "подвергнутый половому скрещиванию" и "половое размножение" применительно к раскрытому в данном документе объекту изобретения относятся к слиянию гамет с получением потомства (например, путем оплодотворения, как, например, для получения семян путем опыления у растений). "Половое скрещивание" или "перекрестное оплодотворение" представляет собой в некоторых вариантах осуществления оплодотворение одной особи другой (например, перекрестное опыление у растений). Термин "самовоспроизводство" относится в некоторых вариантах осуществления к получению семян путем самооплодотворения или самоопыления; т.е. пыльца и семязачаток происходят из одного и того же растения.
Используемая в данном документе фраза "генетический маркер" относится к характерной особенности в геноме особи (например, нуклеотидной или полинуклеотидной последовательности, которая присутствует в геноме особи), которая ассоциирована с одним или несколькими локусами, представляющими интерес.В некоторых вариантах осуществления генетический маркер является полиморфным в популяции, представляющей интерес, или в локусе, занятом полиморфизмом, в зависимости от контекста. Генетические маркеры включают, например, однонуклеотидные полиморфизмы (SNP), инсерционно-делеционные мутации (т.е. в ставки/деле ции), простые повторяющиеся последовательности (SSR), полиморфизмы длин рестрикционных фрагментов (RFLP), случайно амплифицированные полиморфные ДНК (RAPD), маркеры рестрикционного полиморфизма амплифицированных последовательностей (CAPS), маркеры технологии микроматриц для анализа генетического разнообразия (DArT) и полиморфизмы длин амплифицированных фрагментов (AFLP), среди многих других примеров. Генетические маркеры можно, например, использовать для определения локализации на хромосоме генетических локусов, содержащих аллели, которые способствуют изменчивости фенотипических признаков. Фраза "генетический маркер" может также относиться к полинуклеотидной последовательности, комплементарной геномной последовательности, такой как последовательность нуклеиновой кислоты, используемая в качестве зондов.
"Генетический маркер" может быть физически локализован в положении на хромосоме, которое находится в пределах или вне генетического локуса, с которым он ассоциирован (т.е. является внутригенным или экстрагенным соответственно). Иными словами, хотя генетические маркеры обычно используют, если местоположение на хромосоме гена или функциональной мутации, например, в контрольном элементе вне гена, который соответствует локусу, представляющему интерес, не было идентифицировано, и существует ненулевая частота рекомбинации между генетическим маркером и локусом, представляющим интерес, в раскрытом в данном документе объекте изобретения могут также использоваться генетические маркеры, которые физически находятся в пределах генетического локуса (например, внутри геномной последовательности, которая соответствует такому гену, как, например, без ограничения, полиморфизм в интроне или экзоне гена). В некоторых вариантах осуществления раскрытого в данном документе объекта изобретения один или несколько генетических маркеров предусматривают от одного до десяти маркеров, а в некоторых вариантах осуществления один или несколько генетических маркеров предусматривают более десяти генетических маркеров.
Используемый в данном документе термин "генотип" относится к генетической конституции клетки или организма. "Генотип по набору генетических маркеров" особи включает специфические аллели в одном или нескольких локусах генетических маркеров, присутствующие в гаплотипе особи. Как известно из уровня техники, генотип может относиться к одному локусу или к нескольким локусам, независимо от того, являются ли локусы связанными или несвязанными и/или сцепленными или несцепленными. В некоторых вариантах осуществления генотип особи относится к одному или нескольким генам, которые являются связанными в том плане, что один или несколько генов участвуют в экспрессии фенотипа, представляющего интерес (например, количественного признака, определенного в данном документе). Таким образом, в некоторых вариантах осуществления генотип содержит совокупность одного или нескольких аллелей, присутствующих у особи в одном или нескольких генетических локусах, определяющих количественный признак. В некоторых вариантах осуществления генотип экспрессируется в виде гаплотипа (определенного в данном документе ниже).
Используемый в данном документе термин "зародышевая плазма" относится ко всей совокупности генотипов в популяции или другой группе особей (например, у вида). Термин "зародышевая плазма" может также относиться к растительному материалу; например, к группе растений, которые выступают в качестве вместилища для различных аллелей. Фраза "адаптированная зародышевая плазма" относится к растительным материалам с доказанным генетическим превосходством; например, для данной среды обитания или географической области, тогда как фразы "неадаптированная зародышевая плазма", "необработанная зародышевая плазма" и "экзотическая зародышевая плазма" относятся к растительным материалам с неизвестной или недоказанной генетической ценностью; например, для данной среды обитания или географической области; таким образом, фраза "неадаптированная зародышевая плазма" относится в некоторых вариантах осуществления к растительным материалам, которые не являются частью устойчивой популяции для разведения и для которых неизвестно родство с представителями устойчивой популяции для разведения.
Используемый в данном документе термин "сцепление" и его грамматические варианты относится к склонности аллелей в различных локусах на одной и той же хромосоме сегрегировать вместе чаще, чем можно было бы ожидать случайно, если бы их передача была независимой, в некоторых вариантах осуществления вследствие их физической близости.
Используемая в данном документе фраза "нуклеиновая кислота" относится к любому физическому отрезку из мономерных единиц, который может соответствовать отрезку из нуклеотидов, в том числе полимеру из нуклеотидов (например, типичному полимеру ДНК, кДНК или РНК), модифицированным олигонуклеотидам (например, олигонуклеотидам, содержащим основания, которые не являются типичными для биологической РНК или ДНК, такие как 2'-O-метилированные олигонуклеотиды) и т.п. В некоторых вариантах осуществления нуклеиновая кислота может быть однонитевой, двухнитевой, многонитевой или представлять собой их комбинацию. Если не указано иное, конкретная последовательность нуклеиновой кислоты раскрытого в данном документе объекта изобретения необязательно содержит или кодирует комплементарные последовательности в дополнение к любой явным образом указанной последовательности.
Используемый в данном документе термин "множество" относится к более чем одному объекту. Таким образом, "множество особей" относится к по меньшей мере двум особям. В некоторых вариантах осуществления термин "множество" относится к более чем половине от всего количества. Например, в некоторых вариантах осуществления "множество из популяции" относится к более чем половине представителей данной популяции.
Используемый в данном документе термин "потомство" относится к потомку(потомкам), полученным в результате конкретного скрещивания. Как правило, потомство является результатом разведения с участием двух особей, хотя некоторые виды (особенно некоторые растения и животные-гермафродиты) могут подвергаться самовоспроизводству (т.е. одно и то же растение выступает в качестве донора как мужских, так и женских гамет). Потомок(потомки) могут относиться, например, к F1, F2 или любому последующему поколению.
Используемая в данном документе фраза "количественный признак" относится к фенотипическому признаку, который может быть описан в числовом виде (т.е. количественно оценен или количественно определен). Количественный признак, как правило, демонстрирует непрерывную изменчивость между особями в популяции; то есть различия в числовом значении фенотипического признака незначительны и постепенно переходят друг в друга. Часто распределение частот количественного фенотипического признака в популяции имеет колоколообразную кривую (т.е. демонстрирует нормальное распределение между двумя крайними значениями).
"Количественный признак" обычно является результатом взаимодействия генетического локуса с окружающей средой или взаимодействия нескольких генетических локусов друг с другом и/или с окружающей средой. Примеры количественных признаков включают высоту и урожайность растения.
Для цели настоящего изобретения термин "косегрегация" относится к тому факту, что аллель, определяющий признак, и аллель(аллели), определяющие маркер (маркеры), как правило, передаются совместно, поскольку они физически расположены близко друг к другу на одной и той же хромосоме (ввиду их физического близкого расположения между ними наблюдается сниженная рекомбинация), что приводит к неслучайной ассоциации таких аллелей в результате их близкого расположения на одной хромосоме. "Косегрегация" также относится к наличию двух или более признаков в одном растении, из которых по меньшей мере один, как известно, является генетическим, и которое нельзя объяснить простой случайностью.
Используемые в данном документе термины "локус количественного признака" (QTL) и "ассоциация маркер-признак" относятся к ассоциации между генетическим маркером и хромосомным участком и/или геном, который влияет на фенотип признака, представляющего интерес. Как правило, она определяется статистически; например, с использованием одного или нескольких способов, опубликованных в литературе. QTL может представлять собой хромосомный участок и/или генетический локус с по меньшей мере двумя аллелями, которые дифференциально влияют на фенотипический признак (количественный признак либо качественный признак).
Термин "реципиентное растение томата" используется в данном документе для обозначения растения томата, которое должно получить ДНК, полученную из донорного растения томата, содержащего аллель, отвечающий за увеличенное содержание сахарозы. Указанное "реципиентное растение томата" может уже содержать или может не содержать один или несколько аллелей, отвечающих за содержание сахарозы, и в этом случае термин обозначает растение, которое должно получить дополнительный аллель в другом локусе.
Термин "естественное генетическое окружение" используется в данном документе для обозначения исходного генетического окружения аллеля. Такое генетическое окружение может, например, представлять собой геном дикорастущего образца томата. Например, аллели по настоящему изобретению были обнаружены в определенных местоположениях на хромосомах 3 и 12 в Solanum habrochaites и Solanum chmielewskii соответственно. В качестве примера, в Solanum chmielewskii представлено естественное генетическое окружение аллеля SucModchm на хромосоме 12 Solanum chmielewskii. В то же время способ, который предусматривает перенос ДНК, содержащей данный аллель, из хромосомы 12 Solanum chmielewskii в такое же положение на хромосоме 12 другого вида томата, предпочтительно культивируемого растения томата, еще более предпочтительно растения Solanum lycopersicum, приведет к тому, что данный аллель не будет находиться в своем естественном генетическом окружении.
Термин "донорное растение томата" в пределах объема настоящего изобретения понимают как означающий растение томата, которое предоставляет по меньшей мере один аллель, связанный с увеличенным содержанием сахарозы.
Используемая в данном документе фраза "качественный признак" относится к фенотипическому признаку, который контролируется одним или несколькими генами, демонстрирующими основные фенотипические эффекты. Ввиду этого наследование качественных признаков, как правило, является простым. Примеры у растений включают без ограничения цвет цветка и несколько известных видов устойчивости к заболеваниям, такие как, например, устойчивость к грибковой пятнистости или устойчивость к вирусу мозаики томатов.
Термин "отбор на основе маркеров" в пределах объема настоящего изобретения понимают как относящийся, например, к применению генетических маркеров для выявления одной или нескольких нуклеиновых кислот у растения, где нуклеиновая кислота ассоциирована с требуемым признаком, для идентификации растений, несущих гены желательных (или нежелательных) признаков, так чтобы эти растения можно было применять (или избегать их применения) в программе селекционного разведения.
Термин "микросателлитные или SSR-маркеры (простые повторяющиеся последовательности)" в пределах объема настоящего изобретения понимают как относящийся к типу генетического маркера, который состоит из многочисленных повторов коротких последовательностей оснований ДНК, обнаруживаемых в локусах по всему геному растения и с определенной вероятностью являющихся высокополиморфными.
Однонуклеотидный полиморфизм (SNP), представляющий собой вариацию в одном сайте ДНК, является наиболее частым типом вариации в геноме. Однонуклеотидный полиморфизм (SNP) представляет собой вариацию последовательности ДНК, имеющую место в том случае, когда один нуклеотид - А, Т, С или G - в геноме (или другой общей последовательности) отличается между представителями биологического вида или между парными хромосомами у особи. Например, два секвенированных фрагмента ДНК от разных особей - AAGCCTA и AAGCTTA - содержат различие в одном нуклеотиде. В данном случае имеются два аллеля: С и Т. Основные принципы матрицы для выявления SNP являются такими же, как для ДНК-микроматрицы. Это взаимопроникновение гибридизации ДНК, флуоресцентной микроскопии и захвата ДНК. Тремя компонентами матриц для выявления SNP являются матрица, которая содержит последовательности нуклеиновых кислот (т.е. амплифицированные последовательности или мишени), один или несколько меченых аллель-специфических олигонуклеотидных зондов и система выявления, которая регистрирует и интерпретирует сигнал гибридизации.
Присутствие или отсутствие требуемого аллеля может быть определено с помощью ПЦР в режиме реального времени с использованием красителей для двухнитевой ДНК или способа с использованием флуоресцентного репортерного зонда.
Термин "ПЦР (полимеразная цепная реакция)" в пределах объема настоящего изобретения понимают как относящийся к способу получения относительно больших количеств специфических участков ДНК или сегмента(сегментов) генома, за счет чего обеспечивается возможность проведения различных анализов с использованием этих участков.
Термин "праймер для ПЦР" в пределах объема настоящего изобретения понимают как относящийся к относительно коротким фрагментам однонитевой ДНК, используемым в ПЦР-амплификации специфических участков ДНК.
Термин "фенотип" в пределах объема настоящего изобретения понимают как относящийся к различимой(различимым) характеристике(характеристикам) генетически контролируемого признака.
Используемая в данном документе фраза "фенотипический признак" относится к внешнему виду или другой выявляемой характеристике особи, являющейся результатом взаимодействия ее генома, протеома и/или метаболома с окружающей средой.
Термин "полиморфизм" в пределах объема настоящего изобретения понимают как относящийся к наличию в популяции двух или более различных форм гена, генетического маркера, или наследственного признака, или продукта гена, которые могут быть получены, например, посредством альтернативного сплайсинга, метилирования ДНК и т.д.
Термин "селекционное разведение" в пределах объема настоящего изобретения понимают как относящийся к программе разведения, в которой в качестве родительских организмов используют растения, которые обладают желательными признаками или проявляют их.
Термин "тестерное растение" в пределах объема настоящего изобретения понимают как относящийся к растению рода Solanum, используемому для определения генетических характеристик признака растения, подлежащего тестированию. Как правило, растение, подлежащее тестированию, скрещивают с "тестерным" растением, и подсчитывают соотношение сегрегации признака у потомства, полученного в результате скрещивания.
Термин "зонд", используемый в данном документе, относится к группе атомов или молекул, которая способна распознавать конкретную молекулу-мишень или клеточную структуру-мишень и связываться с ней и, следовательно, позволяет выявлять молекулу-мишень или структуру-мишень. В частности, "зонд" относится к меченой последовательности ДНК или РНК, которую можно использовать для выявления присутствия и количественной оценки комплементарной последовательности путем молекулярной гибридизации.
Используемый в данном документе термин "гибридизироваться" относится к традиционным условиям гибридизации, предпочтительно к условиям гибридизации, в которых 5х SSPE, 1% SDS, 1х раствор Денхардта используют в качестве рабочего раствора, и/или температуры гибридизации составляют от 35°С до 70°С, предпочтительно 65°С. После гибридизации отмывку предпочтительно проводят сперва с использованием 2х SSC, 1% SDS, а затем с использованием 0,2х SSC при температурах от 35°С до 75°С, в частности от 45°С до 65°С, но особенно при 59°С (в отношении определения SSPE, SSC и раствора Денхардта см. Sambrook et al., в цитируемом месте). Условия гибридизации высокой жесткости, как, например, описанные в Sambrook et al., выше, являются особенно предпочтительными. Особенно предпочтительные жесткие условия гибридизации присутствуют, например, если гибридизация и отмывка происходят при 65°С, как указано выше. Нежесткие условия гибридизации, например, с гибридизацией и отмывкой, проводимыми при 45°С, являются менее предпочтительными, а при 35°С еще менее предпочтительными.
Согласно настоящему изобретению термин "указанное положение, соответствующее положению X", где X представляет собой любое число, которое можно найти в соответствующем контексте настоящей заявки, не только включает соответствующее положение в SEQ ID NO, упоминающееся впоследствии, но также включает любую последовательность, кодирующую аллель SucMod или TIV, где после выравнивания с эталонной SEQ ID NO соответствующее положение может иметь другой номер, но соответствует указанному в эталонной SEQ ID NO. Выравнивание последовательностей аллелей SucMod или TIV можно осуществлять путем целесообразного применения различных инструментов для выравнивания и, например, путем применения инструментов, описанных ниже.
Термины "генетическое сходство" или "идентичность последовательностей" используются в данном документе взаимозаменяемо. Термины "идентичный" или "процент идентичности" применительно к двум или более последовательностям нуклеиновой кислоты или белка относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или имеют определенную процентную долю аминокислотных остатков или нуклеотидов, которые являются одинаковыми, при сравнении и выравнивании для достижения максимального соответствия согласно измерению с помощью одного из следующих алгоритмов сравнения последовательностей или при визуальном осмотре. Если две последовательности, подлежащие сравнению друг с другом, отличаются по длине, идентичность последовательностей предпочтительно относится к процентной доле нуклеотидных остатков в более короткой последовательности, которые идентичны нуклеотидным остаткам в более длинной последовательности. Как используется в данном документе, процент идентичности/гомологии между двумя последовательностями зависит от количества идентичных положений, общих для последовательностей (т.е. % идентичности = количество идентичных положений/общее количество положений × 100), с учетом количества гэпов и длины каждого гэпа, которые необходимо ввести для оптимального выравнивания этих двух последовательностей. Сравнение последовательностей и определение процента идентичности двух последовательностей можно проводить с помощью математического алгоритма, описанного в данном документе ниже. Например, идентичность последовательностей можно определить традиционно с использованием компьютерных программ, таких как программа BestFit (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Мадисон, Висконсин 53711). В BestFit используется алгоритм поиска локальной гомологии согласно Smith and Waterman, Advances in Applied Mathematics 2 (1981), 482-489 с целью выявления сегмента, характеризующегося наивысшей идентичностью последовательностей между двумя последовательностями. В ходе применения BestFit или другой программы для выравнивания последовательностей с целью определения того, является ли конкретная последовательность, например, на 95% идентичной эталонной последовательности согласно настоящему изобретению, параметры предпочтительно корректируют таким образом, чтобы процент идентичности рассчитывался по всей длине эталонной последовательности, и чтобы допускались гэпы для выявления гомологии, составляющие до 5% от общего числа нуклеотидов в эталонной последовательности. В ходе применения BestFit для так называемых необязательных параметров предпочтительно оставляют их предварительно установленные ("по умолчанию") значения. Отклонения, появляющиеся при сравнении данной последовательности и вышеописанных последовательностей по настоящему изобретению, могут быть вызваны, например, добавлением, делецией, заменой, вставкой или рекомбинацией. Такое сравнение последовательностей предпочтительно также можно проводить с помощью программы "fasta20u66" (версия 2.0u66, сентябрь 1998 г., авторы William R. Pearson и Виргинский университет; см. также W.R. Pearson (1990), Methods in Enzymology 183, 63-98, прилагаемые примеры и http://workbench.sdsc.edu/). Для этой цели можно использовать установленные параметры "по умолчанию".
Другим показателем того, что две последовательности нуклеиновой кислоты являются по существу идентичными, является то, что две молекулы гибридизируются друг с другом в жестких условиях. Фраза "специфично гибридизируется с" относится к связыванию, образованию дуплекса или гибридизации молекулы только с конкретной нуклеотидной последовательностью в жестких условиях, когда такая последовательность присутствует в сложной смеси (например, общих клеточных) ДНК или РНК. "Связываются(связывается) в значительной степени" относится к комплементарной гибридизации между нуклеиновой кислотой-зондом и нуклеиновой кислотой-мишенью и охватывает незначительные несовпадения, которые могут быть компенсированы путем снижения жесткости среды для гибридизации, чтобы достичь необходимого выявления последовательности нуклеиновой кислоты-мишени.
"Жесткие условия гибридизации" и "жесткие условия гибридизации и отмывки" применительно к экспериментам по гибридизации нуклеиновых кислот, таким как саузерн- и нозерн-блот-гибридизация, зависят от последовательности и отличаются при разных параметрах окружающей среды. Более длинные последовательности специфично гибридизируются при более высоких температурах. Обширное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, part I, chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier, New York. Как правило, условия гибридизации и отмывки высокой жесткости выбирают так, чтобы температура была приблизительно на 5°С ниже температуры точки плавления для конкретной последовательности при определенных ионной силе и рН. Как правило, при "жестких условиях" зонд будет гибридизироваться со своей подпоследовательностью-мишенью, но не с другими последовательностями.
"Температура точки плавления" представляет собой температуру (при определенных ионной силе и рН), при которой 50% последовательности-мишени гибридизируется с абсолютно совпадающим зондом. Для очень жестких условий выбирают температуру, равную температуре плавления (Tm) для конкретного зонда. Примером жестких условий гибридизации для гибридизации комплементарных нуклеиновых кислот, которые имеют более 100 комплементарных остатков на фильтре при саузерн- или нозерн-блоттинге, является 50% формамида с 1 мг гепарина при 42°С, при этом гибридизацию проводят в течение ночи. Примером условий отмывки высокой жесткости является использование 0,1 5 М NaCl при 72°С в течение приблизительно 15 минут. Примером жестких условий отмывки является отмывка с использованием 0,2 × SSC при 65°С в течение 15 минут (описание SSC-буфера см. в Sambrook ниже). Часто с целью устранения фонового сигнала зонда отмывке в условиях высокой жесткости предшествует отмывка в условиях низкой жесткости. Примером отмывки в условиях средней жесткости для дуплекса, имеющего, например, более 100 нуклеотидов, является использование 1 × SSC при 45°С в течение 15 минут. Примером отмывки в условиях низкой жесткости для дуплекса, имеющего, например, более 100 нуклеотидов, является 4-6 × SSC при 40°С в течение 15 минут. Для коротких зондов (например, длиной от приблизительно 10 до 50 нуклеотидов) жесткие условия, как правило, предусматривают концентрации солей, составляющие менее чем приблизительно 1,0 М ионов Na, как правило, концентрацию, составляющую от приблизительно 0,01 до 1,0 М ионов Na (или других солей) при рН 7,0-8,3, а также температуру, как правило, составляющую по меньшей мере приблизительно 30°С. Жестких условий также можно достичь путем добавления дестабилизирующих средств, таких как формамид. В целом соотношение сигнал-шум, которое в 2 раза (или больше) превышает наблюдаемое для неродственного зонда в конкретном гибридизационном анализе, указывает на выявление специфичной гибридизации. Нуклеиновые кислоты, которые не гибридизируются друг с другом в жестких условиях, по-прежнему являются по существу идентичными, если белки, которые они кодируют, являются по существу идентичными. Например, это происходит в том случае, когда копия нуклеиновой кислоты создана с использованием максимальной вырожденности кодонов, допускаемой генетическим кодом.
РАСТЕНИЯ, СЕМЕНА, ПЛОДЫ
В первом варианте осуществления настоящее изобретение предусматривает культивируемое растение томата, предпочтительно культивируемое растение Solanum lycopersicum, содержащее:
a) по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, характеризующегося по меньшей мере 90% генетическим сходством с SEQ ID NO: 1, и
b) две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата;
где указанное растение образует плод томата, демонстрирующий увеличенное содержание сахарозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления аллель SucMod содержит нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотид Т в положении, которое соответствует положению 498 в SEQ ID NO: 1.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где содержание сахарозы в плодах томата, достигающих стадии полной спелости, увеличено на 50%, более предпочтительно на 75%, еще более предпочтительно на 100%, в частности на 200%, по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В альтернативном или дополнительном варианте осуществления растение по настоящему изобретению представляет собой растение согласно любому из предыдущих вариантов осуществления, где указанное растение образует плоды томата, демонстрирующие содержание сахарозы, составляющее по меньшей мере приблизительно 10 мг⋅г-1 сырого веса, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие содержание сахарозы, составляющее по меньшей мере приблизительно 12 мг⋅г-1 сырого веса, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие содержание сахарозы, составляющее по меньшей мере приблизительно 15 мг⋅г-1 сырого веса, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие содержание сахарозы, составляющее по меньшей мере приблизительно 20 мг⋅г-1 сырого веса, при достижении стадии полной спелости.
В альтернативном или дополнительном варианте осуществления настоящее изобретение предусматривает культивируемое растение томата, предпочтительно культивируемое растение Solanum lycopersicum, содержащее:
a) по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, характеризующегося по меньшей мере 90% генетическим сходством с SEQ ID NO: 1, и
b) две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата;
где указанный аллель SucMod содержит нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотид Т в положении, которое соответствует положению 498 в SEQ ID NO: 1, и
где указанное растение образует плод томата, демонстрирующий увеличенное соотношение сахарозы и гексозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где соотношение сахарозы и гексозы в плодах томата, достигающих стадии полной спелости, увеличено на 50%, более предпочтительно на 75%, еще более предпочтительно на 100%, в частности на 200%, по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В альтернативном или дополнительном варианте осуществления растение по настоящему изобретению представляет собой растение согласно любому из предыдущих вариантов осуществления, где указанное растение образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 0,5, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 1, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 1,5, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 2, при достижении стадии полной спелости. В дополнительном варианте осуществления растение по настоящему изобретению образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 3, при достижении стадии полной спелости.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod получен из Solanum chmielewskii или Solanum pimpinellifolium. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod получен из Solanum chmielewskii. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod получен из образца BD732 Solanum chmielewskii или образца LA1589 Solanum pimpinellifolium. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod получен из образца BD732 Solanum chmielewskii.
В альтернативном или дополнительном варианте осуществления любой аллель SucMod или VIF, полученный из дикорастущего вида томата, может успешно применяться в контексте настоящего изобретения, при условии, что аллель SucMod или VIF из указанного дикорастущего вида томата экспрессируется на более высоком уровне, чем аллель TIV у того же указанного дикорастущего вида томата.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod содержит нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и где указанный нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1, может быть выявлен с помощью ПЦР путем амплификации фрагмента ДНК с использованием пары олигонуклеотидных праймеров: прямого праймера под SEQ ID NO: 2 и обратного праймера под SEQ ID NO: 3, а также зонда для благоприятного аллеля под SEQ ID NO: 4.
В дополнительном варианте осуществления аллель SucMod характеризуется по меньшей мере 92% генетическим сходством с SEQ ID NO: 1. В дополнительном варианте осуществления аллель SucMod характеризуется по меньшей мере 95% генетическим сходством с SEQ ID NO: 1. В дополнительном варианте осуществления аллель SucMod характеризуется по меньшей мере 97% генетическим сходством с SEQ ID NO: 1. В дополнительном варианте осуществления аллель SucMod характеризуется по меньшей мере 98% генетическим сходством с SEQ ID NO: 1. В дополнительном варианте осуществления аллель SucMod характеризуется по меньшей мере 99% генетическим сходством с SEQ ID NO: 1.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель SucMod содержит нуклеотидную последовательность под SEQ ID NO: 1. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение содержит SEQ ID NO: 1.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV получен из образца томата, относящегося к группе Eriopersicum.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV характеризуется по меньшей мере 98% генетическим сходством с SEQ ID NO: 6. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV характеризуется по меньшей мере 99% генетическим сходством с SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV содержит нуклеотид А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотид А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотид Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотид С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотид Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотид С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотид А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотид G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV получен из Solanum habrochaites. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV получен из образца LA1777 Solanum habrochaites.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV содержит нуклеотид С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотид G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В альтернативном или дополнительном варианте осуществления любой аллель TIV, полученный из дикорастущего вида томата, может успешно применяться в контексте настоящего изобретения, при условии, что аллель TIV из указанного дикорастущего вида томата экспрессируется на менее высоком уровне, чем аллель SucMod или VIF у того же указанного дикорастущего вида томата.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV имеет нуклеотидную последовательность под SEQ ID NO: 6. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение содержит SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение предусматривает культивируемое растение томата, предпочтительно культивируемое растение Solanum lycopersicum, содержащее:
a) по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, характеризующегося по меньшей мере 90% генетическим сходством с SEQ ID NO: 1, и
b) две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата;
где указанный аллель SucMod получен из дикорастущего вида томата, где указанный аллель SucMod экспрессируется на более высоком уровне, чем аллель TIV у того же указанного дикорастущего вида томата, и
где указанный аллель TIV, полученный из образца дикорастущего зеленоплодного томата, экспрессируется на менее высоком уровне, чем аллель SucMod у того же указанного дикорастущего вида томата, и
где указанное растение образует плод томата, демонстрирующий увеличенное содержание сахарозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления настоящее изобретение предусматривает культивируемое растение Solanum lycopersicum, содержащее по меньшей мере одну копию аллеля SucMod из Solanum chmielewskii и две копии аллеля TIV из Solanum habrochaites, где указанное растение образует плод томата, демонстрирующий увеличенное содержание сахарозы по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение содержит две копии аллеля SucMod. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение является гомозиготным по аллелю SucMod. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение является гомозиготным по аллелю SucModchm. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение является гомозиготным по аллелю SucModchm под SEQ ID NO: 1.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанный аллель TIV и указанный аллель SucMod могут быть получены, являются полученными или происходят из Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение получено посредством скрещивания Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка с растением томата, в котором отсутствуют указанные аллели SucMod и TIV. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомок или предок представляет собой источник аллелей SucMod и TIV по настоящему изобретению. В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанные аллели SucMod и TIV по настоящему изобретению интрогрессированы из Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка.
В дополнительном варианте осуществления настоящее изобретение предусматривает растение согласно любому из предыдущих вариантов осуществления, где указанное растение представляет собой гаплоидное, дигаплоидное, инбредное или гибридное культивируемое растение томата.
В другом варианте осуществления растение согласно настоящему изобретению обладает мужской стерильностью. В другом варианте осуществления растение согласно настоящему изобретению обладает цитоплазматической мужской стерильностью.
В другом варианте осуществления растение согласно настоящему изобретению образует зрелые плоды томата, где цвет плодов является желтым, красным или оранжевым.
В дополнительном варианте осуществления предусмотрены часть, орган или ткань растения, которые могут быть получены из культивируемого растения томата, предпочтительно культивируемого Solanum lycopersicum согласно любому из предыдущих вариантов осуществления, в том числе без ограничения листья, стебли, корни, цветки или части цветка, плоды, побеги, гаметофиты, спорофиты, пыльца, пыльники, микроспоры, яйцеклетки, зиготы, зародыши, участки меристемы, каллюсная ткань, семена, черешки, клеточные или тканевые культуры или любые другие часть или продукт растения, которые по-прежнему демонстрируют признак увеличенного содержания сахарозы согласно настоящему изобретению, в частности, при выращивании из них растения, образующего плоды.
В дополнительном варианте осуществления настоящее изобретение предусматривает плод томата, образованный растением томата согласно любому из предыдущих вариантов осуществления.
В дополнительном варианте осуществления настоящее изобретение предусматривает семя томата, из которого образуется растение томата согласно любому из предыдущих вариантов осуществления.
АЛЛЕЛИ, МАРКЕРЫ
Настоящее изобретение дополнительно направлено на аллели SucMod и TIV, управляющие экспрессией признака содержания сахарозы у растения томата или контролирующие ее. В дополнительном варианте осуществления аллели по настоящему изобретению локализованы на хромосомах 3 и 12 соответственно. В дополнительном варианте осуществления настоящего изобретения аллели SucMod и TIV по настоящему изобретению могут быть получены, являются полученными или происходят из донорного растения, которое имеет генетическое окружение Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка и содержит указанные аллели SucMod и TIV по настоящему изобретению.
В дополнительном варианте осуществления настоящее изобретение относится к выделенной нуклеотидной последовательности, содержащей SEQ ID NO: 1.
В дополнительном варианте осуществления аллели по настоящему изобретению генетически или физически сцеплены с 3 маркерными локусами, которые косегрегируют с признаком содержания сахарозы и представляют собой маркерный локус ST3226 для аллелей SucMod и маркерные локусы ST3472 и ST3478 для аллеля TIVhab, или любым смежным маркером, который характеризуется статистически значимой корреляцией с признаком содержания сахарозы и, таким образом, косегрегирует с ним.
В другом варианте осуществления указанные аллели SucMod и TIV по настоящему изобретению или их функциональные части генетически сцеплены с 3 маркерными локусами соответственно, где:
i. маркерный локус ST3226 может быть идентифицирован с помощью ПЦР путем амплификации фрагмента ДНК с использованием пары олигонуклеотидных праймеров: прямого праймера под SEQ ID NO: 2 и обратного праймера под SEQ ID NO: 3, а также зонда для благоприятного аллеля под SEQ ID NO: 4,
ii. маркерный локус ST3472 может быть идентифицирован с помощью ПЦР путем амплификации фрагмента ДНК с использованием пары о лиго нуклеотид ных праймеров: прямого праймера под SEQ ID NO: 7 и обратного праймера под SEQ ID NO: 8, а также зонда для благоприятного аллеля под SEQ ID NO: 9,
iii. маркерный локус ST3478 может быть идентифицирован с помощью ПЦР путем амплификации фрагмента ДНК с использованием пары о лиго нуклеотид ных праймеров: прямого праймера под SEQ ID NO: 11 и обратного праймера под SEQ ID NO: 12, а также зонда для благоприятного аллеля под SEQ ID NO: 13.
В настоящем изобретении раскрыт набор для выявления локуса признака содержания сахарозы в культивируемом растении томата, в частности в культивируемом растении Solanum lycopersicum, где указанный набор содержит по меньшей мере одну пару олигонуклеотидных праймеров и зонд для ПЦР, выбранные из:
a. пары праймеров, представленной прямым праймером под SEQ ID NO: 2 и обратным праймером под SEQ ID NO: 3, а также зондов под SEQ ID NO: 4 и 5; или
b. пары праймеров, представленной прямым праймером под SEQ ID NO: 7 и обратным праймером под SEQ ID NO: 8, а также зондов под SEQ ID NO: 9 и 10; или
c. пары праймеров, представленной прямым праймером под SEQ ID NO: 11 и обратным праймером под SEQ ID NO: 12, а также зондов под SEQ ID NO: 13 и 14; или
другого праймера или пары праймеров, представленных для смежного маркера, который характеризуется статистически значимой корреляцией с признаком содержания сахарозы и, таким образом, косегрегирует с ним.
В дополнение к SNP-маркеру ST3226 в примере 7 и на фигуре 4 раскрыт другой SNP-маркер, ассоциированный с аллелями SucMod/VIF по настоящему изобретению, по меньшей мере с аллелем VIFpimp по настоящему изобретению.
Кроме того, в дополнение к двум SNP-маркерам ST3272 и ST3274, ассоциированным с аллелем TIVhab по настоящему изобретению, в примере 8 и на фигуре 5 раскрыты 8 SNP-маркеров, ассоциированных с дополнительными аллелями TIV по настоящему изобретению.
Специалист в данной области способен разработать соответствующие праймеры и зонды по своему усмотрению и без каких-либо трудностей на основании информации о последовательностях, раскрытой в данном документе.
В настоящем изобретении также раскрыто применение некоторых или всех из этих SNP-маркеров согласно настоящему изобретению для диагностического отбора и/или генотипирования локуса признака содержания сахарозы у культивируемого растения томата, в частности культивируемого растения Solanum lycopersicum.
В настоящем изобретении дополнительно раскрыто применение некоторых или всех из этих SNP-маркеров для идентификации у растения томата, в частности культивируемого растения томата, более конкретно, растения Solanum lycopersicum согласно настоящему изобретению, наличия локуса признака содержания сахарозы и/или для мониторинга интрогрессии локуса признака содержания сахарозы у культивируемого растения томата, в частности растения Solanum lycopersicum согласно настоящему изобретению, описанного в данном документе.
В настоящем изобретении дополнительно раскрыт полинуклеотид (продукт амплификации), который может быть получен в ходе ПЦР-реакции, предусматривающей по меньшей мере один олигонуклеотидный праймер или пару олигонуклеотидных праймеров для ПЦР, выбранных из группы, состоящей из SEQ ID NO: 2 и SEQ ID NO: 3; SEQ ID NO: 7 и SEQ ID NO: 8; SEQ ID NO: 11 и SEQ ID NO: 12; и осуществление реакции с зондами для благоприятных аллелей, выбранными из группы, включающей SEQ ID NO: 4, SEQ ID NO: 9 или SEQ ID NO: 13, или с помощью другого праймера, представленного для смежного маркера, который характеризуется статистически значимой корреляцией с признаком содержания сахарозы или с одним из раскрытых маркеров и, таким образом, косегрегирует с ним, при этом данный продукт амплификации соответствует продукту амплификации, который может быть получен из Solanum lycopersicum линии TIPC18-61141, депонированного в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка, содержащего аллели SucMod и TIV по настоящему изобретению, в ходе ПЦР-реакции с идентичными праймерами или парами праймеров при условии, что соответствующий маркерный локус по-прежнему присутствует у указанного растения томата и/или может считаться его аллелем.
Также в данном документе рассматривается полинуклеотид, который характеризуется по меньшей мере 90%, в частности по меньшей мере 95%, в частности по меньшей мере 96%, в частности по меньшей мере 97%, в частности по меньшей мере 98%, в частности по меньшей мере 99%, идентичностью последовательности с последовательностью указанного продукта амплификации, и/или полинуклеотид, имеющий нуклеотидную последовательность, которая гибридизируется с нуклеотидными последовательностями указанного продукта амплификации, который может быть получен в ходе указанной выше ПЦР-реакции.
Продукт амплификации согласно настоящему изобретению, описанный в данном документе выше, затем можно применять для получения или разработки новых праймеров и/или зондов, которые можно использовать для идентификации локуса признака содержания сахарозы.
Таким образом, настоящее изобретение в одном варианте осуществления дополнительно относится к производным маркерам, в частности к производным праймерам или зондам, разработанным на основе продукта амплификации согласно настоящему изобретению, описанного в данном документе выше, с помощью способов, известных из уровня техники, при этом данные производные маркеры генетически сцеплены с локусом признака содержания сахарозы.
СПОСОБЫ РАЗВЕДЕНИЯ
В другом варианте осуществления настоящее изобретение относится к способу получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, части или семени растения, где указанный способ включает следующие стадии:
а) скрещивание 1-го растения согласно любому из предыдущих вариантов осуществления со 2-м растением томата, в котором отсутствуют аллели SucMod и TIV по настоящему изобретению,
b) получение растения томата, являющегося потомком, и
c) необязательно отбор растения из указанного потомства, характеризующегося тем, что указанное растение образует плоды, демонстрирующие увеличенное содержание сахарозы.
В дополнительном варианте осуществления настоящее изобретение предусматривает способ получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающий стадии:
a) скрещивания 1-го растения согласно любому из предыдущих вариантов осуществления, содержащего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, со 2-м культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV;
b) отбора растения-потомка, образующего плоды, демонстрирующие увеличенное содержание сахарозы;
где отбор на стадии b) проводят посредством выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или посредством выявления нуклеотида Т в положении, которое соответствует положению 498 в SEQ ID NO: 1; и посредством выявления нуклеотида А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по любому из предыдущих вариантов осуществления, где отбор на стадии b) проводят посредством дополнительного выявления нуклеотида С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по любому из предыдущих вариантов осуществления, где 1-е растение из стадии а) представляет собой Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169.
В другом варианте осуществления настоящее изобретение относится к способу по любому из предыдущих вариантов осуществления, где указанное растение-потомок демонстрирует содержание сахарозы, составляющее по меньшей мере приблизительно 10, предпочтительно 12, более предпочтительно 15, еще более предпочтительно 15 мг⋅г-1 сырого веса, при достижении стадии полной спелости.
В дополнительном варианте осуществления настоящее изобретение предусматривает способ получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное соотношение сахарозы и гексозы, включающий стадии:
a) скрещивания 1-го растения согласно любому из предыдущих вариантов осуществления, содержащего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, со 2-м культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV;
b) отбора растения-потомка, образующего плоды, демонстрирующие увеличенное содержание сахарозы;
где отбор на стадии b) проводят посредством выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или посредством выявления нуклеотида Т в положении, которое соответствует положению 498 в SEQ ID NO: 1, и посредством выявления нуклеотида А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по любому из предыдущих вариантов осуществления, где отбор на стадии b) проводят посредством дополнительного выявления нуклеотида С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В другом варианте осуществления настоящее изобретение относится к способу по предыдущему варианту осуществления, где указанное растение из стадии с) образует плоды томата, демонстрирующие соотношение сахарозы и гексозы, составляющее по меньшей мере приблизительно 0,5, предпочтительно по меньшей мере приблизительно 1, предпочтительно по меньшей мере приблизительно 1,2, более предпочтительно по меньшей мере приблизительно 1,5, еще более предпочтительно по меньшей мере 2,0, при достижении стадии полной спелости.
В другом варианте осуществления настоящее изобретение относится к способу получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающему стадии:
a) скрещивания 1-го растения согласно любому из предыдущих вариантов осуществления со 2-м растением томата, в котором отсутствуют аллели SucMod и TIV по настоящему изобретению,
b) получения культивируемого растения томата, являющегося потомком, и
c) необязательно отбора растения из указанного потомства, характеризующегося тем, что указанное растение образует плоды, демонстрирующие содержание сахарозы, увеличенное на 50%, более предпочтительно на 75%, еще более предпочтительно на 100%, в частности на 200%, по сравнению с таким же культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV.
В дополнительном варианте осуществления рассматривается способ по любому из предыдущих вариантов осуществления, где 1-е растение томата из стадии а) представляет собой Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка.
В другом варианте осуществления рассматривается способ получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающий следующие стадии:
a) получение семян растения томата согласно любому из предыдущих вариантов осуществления,
b) проращивание указанных семян и выращивание из них зрелого фертильного растения,
c) индуцирование самоопыления у указанного растения из а), выращивание плодов и сбор из них фертильных семян, и
d) выращивание растений из семян, собранных на с), и отбор растения с увеличенным содержанием сахарозы.
В дополнительном варианте осуществления настоящее изобретение относится к способу получения культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающему стадии:
a. отбора растения томата, содержащего 3 маркерных локуса, которые косегрегируют с признаком содержания сахарозы и представляют собой маркерный локус ST3226 для аллелей SucMod и маркерные локусы ST3472 и ST3478 для аллеля TIVhab, или любой смежный маркер, который характеризуется статистически значимой корреляцией с признаком содержания сахарозы и, таким образом, косегрегирует с ним,
b. скрещивания указанного растения из стадии а) с культивируемым растением томата, предпочтительно культивируемым растением Solanum lycopersicum, которое не содержит указанных аллелей SucMod и TIV, и
c. отбора растения томата, являющегося потомком, полученного в результате указанного скрещивания, которое характеризуется признаком содержания сахарозы и демонстрирует ассоциацию с указанными 3 маркерными локусами из стадии а), а также образует плоды томата, демонстрирующие увеличенное содержание сахарозы.
В дополнительном варианте осуществления настоящего изобретения предусмотрен способ увеличения содержания сахарозы в плодах, образуемых растением томата, включающий стадии:
а) отбора растения томата, содержащего 3 маркерных локуса, которые косегрегируют с признаком содержания сахарозы и представляют собой маркерный локус ST3226 для аллелей SucMod и маркерные локусы ST3472 и ST3478 для аллеля TIVhab, или любой смежный маркер, который характеризуется статистически значимой корреляцией с признаком содержания сахарозы и, таким образом, косегрегирует с ним,
b) скрещивания указанного растения томата из стадии а), которое характеризуется признаком содержания сахарозы, с реципиентным культивируемым растением томата, которое не характеризуется признаком содержания сахарозы, и
c) отбора потомства, полученного в результате указанного скрещивания, которое демонстрирует увеличенное содержание сахарозы по сравнению с реципиентным растением из стадии b) и демонстрирует ассоциацию увеличенного содержания сахарозы с 3 маркерными локусами ST3226, ST3472 и ST3478 из стадии а).
В дополнительном варианте осуществления настоящего изобретения предусмотрен способ получения растений томата, образующих плоды, демонстрирующие увеличенное содержание сахарозы, посредством введения в растение томата нуклеотидной последовательности под SEQ ID NO: 1. В дополнительном варианте осуществления предусмотрен способ по предыдущему варианту осуществления, где нуклеотидную последовательность под SEQ ID NO: 6 дополнительно вводят в указанное растение томата.
Аллели, отвечающие за содержание сахарозы, также могут быть введены посредством мутагенеза, например, посредством химического мутагенеза, например, посредством мутагенеза, индуцированного EMS. В качестве альтернативы, аллели, отвечающие за содержание сахарозы, также могут быть идентифицированы и/или введены посредством применения методик TILLING.
Аллели, отвечающие за содержание сахарозы, также могут быть введены посредством целенаправленного мутагенеза, например, с помощью гомологичной рекомбинации, нуклеаз с "цинковыми пальцами", индукции мутаций с использованием олигонуклеотидов, эффекторных нуклеаз, подобных активаторам транскрипции (TALEN), систем коротких палиндромных повторов, регулярно расположенных группами (CRISPR), или любой альтернативной методики редактирования генома.
В качестве альтернативы, аллели, отвечающие за содержание сахарозы, также могут быть введены с помощью трансгенных или цисгенных способов посредством нуклеотидной конструкции, которая может содержаться в векторе.
СПОСОБЫ ОТБОРА
В дополнительном варианте осуществления настоящее изобретение предусматривает способ идентификации культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды, демонстрирующие увеличенное содержание сахарозы, и имеющего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата, включающий стадии:
a) выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и/или нуклеотида Т в положении, которое соответствует положению 498 в SEQ ID NO: 1; и
b) выявления нуклеотида А в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида Т в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида С в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида А в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
В дополнительном варианте осуществления настоящее изобретение относится к способу по предыдущему варианту осуществления, где стадию b) проводят посредством дополнительного выявления нуклеотида С в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
В настоящем изобретении дополнительно раскрыты способы идентификации культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, характеризующегося признаком содержания сахарозы, включающие стадии:
a) получения популяции, сегрегирующей по содержанию сахарозы,
b) скрининга сегрегирующей популяции на наличие представителя, характеризующегося признаком содержания сахарозы, где указанный признак может быть идентифицирован по наличию 3 маркерных локусов ST3226, ST3472 и ST3478,
с) отбора одного представителя сегрегирующей популяции, где указанный представитель характеризуется признаком содержания сахарозы.
В настоящем изобретении дополнительно раскрыты способы идентификации культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, образующего плоды, демонстрирующие увеличенное содержание сахарозы, и имеющего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата, включающие стадии:
a) получения популяции, сегрегирующей по содержанию сахарозы,
b) скрининга сегрегирующей популяции на наличие представителя, характеризующегося признаком содержания сахарозы, где указанный признак может быть идентифицирован по наличию 3 маркерных локусов, при этом данные маркерные локусы расположены на хромосомах 3 и 12 соответственно и косегрегируют с признаком содержания сахарозы, и может быть идентифицирован с помощью олигонуклеотидного праймера для ПЦР или пары олигонуклеотидных праймеров для ПЦР, выбранных из группы пары праймеров, представленных прямым праймером под SEQ ID NO: 2 и обратным праймером под SEQ ID NO: 3, идентифицирующими маркерный локус ST3226; пары праймеров, представленных прямым праймером под SEQ ID NO: 7 и обратным праймером под SEQ ID NO: 8, идентифицирующими маркерный локус ST3472; и пары праймеров, представленных прямым праймером под SEQ ID NO: 11 и обратным праймером под SEQ ID NO: 12, идентифицирующими маркерный локус ST3478,
c) отбора одного представителя сегрегирующей популяции, где указанный представитель характеризуется признаком содержания сахарозы.
В настоящем изобретении дополнительно раскрыт способ выявления у растения томата генотипа, связанного с фенотипом увеличенного содержания сахарозы, включающий стадии:
a) выявления посредством генотипирования у растения томата набора молекулярных маркеров, включающего маркерные локусы ST3226, ST3472 и ST3478, связанные с увеличенным содержанием сахарозы;
b) отбора указанного выявленного растения томата, содержащего набор молекулярных маркеров, включающий маркерные локусы ST3226, ST3472 и ST3478, связанные с увеличенным содержанием сахарозы; и
c) скрещивания указанного отобранного растения томата для получения растения томата, являющегося потомком, содержащего набор молекулярных маркеров, включающий маркерные локусы ST3226, ST3472 и ST3478, связанные с увеличенным содержанием сахарозы.
Способ по любому из предыдущих вариантов осуществления, где указанный набор молекулярных маркеров может быть выявлен посредством применения SEQ ID NO: 1-14.
Способ по любому из предыдущих вариантов осуществления, где выявляемые маркерные локусы характеризуются генотипом, имеющим один аллель в аллельном состоянии G в маркерном локусе ST3226, один аллель в аллельном состоянии С в маркерном локусе ST3472 и один аллель в аллельном состоянии G в маркерном локусе ST3478.
ПУТИ ПРИМЕНЕНИЯ
В другом варианте осуществления настоящее изобретение относится к применению культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, части или семени растения согласно любому из предыдущих вариантов осуществления, для получения и сбора плодов томата.
В другом варианте осуществления настоящее изобретение относится к применению культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum согласно любому из предыдущих вариантов осуществления, для получения плодов томата, демонстрирующих увеличенное содержание сахарозы, для реализации в свежем виде или для переработки пищевых продуктов. Особый интерес представляет то, что увеличенное содержание сахарозы сохраняется даже после переработки пищевых продуктов, такой как консервирование или замораживание. В дополнительном варианте осуществления настоящее изобретение предусматривает переработанные пищевые продукты, изготовленные из плода томата, образованного культивируемым растением томата, предпочтительно культивируемым растением Solanum lycopersicum, согласно любому из предыдущих вариантов осуществления. В другом варианте осуществления настоящее изобретение относится к применению культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, части или семени растения согласно любому из предыдущих вариантов осуществления, где культивируемое растение томата, предпочтительно культивируемое растение Solanum lycopersicum, часть или семя растения, представляет собой Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка. В дополнительном варианте осуществления настоящее изобретение относится к применению культивируемого растения томата, предпочтительно культивируемого растения Solanum lycopersicum, части или семени растения согласно любому из предыдущих вариантов осуществления, для посева в поле, оранжерее или теплице с пленочным покрытием.
Настоящее изобретение также относится к применению материала для размножения, характеризующегося содержанием сахарозы, который может быть получен из растения томата согласно любому из предыдущих вариантов осуществления, для выращивания растения томата с целью получения растений томата, образующих плоды томата, демонстрирующие увеличенное содержание сахарозы, где указанное увеличенное содержание сахарозы может быть оценено в ходе стандартного анализа, в частности анализа, описанного в примере 9 ниже.
Настоящее изобретение также предусматривает способ получения плодов томата, при этом способ включает посадку семени согласно любому из предыдущих вариантов осуществления, культивирование растения томата, полученного из него, и сбор плода томата, образованного указанным растением томата.
В дополнительном варианте осуществления настоящее изобретение относится к применению аллелей, генетически или физически сцепленных с 3 маркерными локусами, которые косегрегируют с признаком содержания сахарозы и представляют собой маркерные локусы ST3226, ST3472 и ST3478 соответственно, для придания признака увеличенного содержания сахарозы растению томата, в котором отсутствуют указанные аллели.
Настоящее изобретение дополнительно относится к применению растения томата согласно любому из предыдущих вариантов осуществления для интрогрессии признака содержания сахарозы в растение томата, в котором отсутствует указанный признак содержания сахарозы.
Настоящее изобретение дополнительно относится к применению SEQ ID NO: 1-14 для отбора растений с помощью маркеров. Настоящее изобретение дополнительно относится к применению SEQ ID NO: 1-14 для интрогрессии в растения.
На основе описания настоящего изобретения специалисту в данной области, имеющему в распоряжении Solanum lycopersicum линии TIPC18-61141, депонированный в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169, или его потомка или предка, содержащих аллели SucMod и TIV по настоящему изобретению, описанные в данном документе, будет нетрудно перенести указанные аллели по настоящему изобретению в другие растения томата различных типов с применением методик разведения, хорошо известных из уровня техники, с помощью маркерных локусов, раскрытых в данном документе.
ПОДРОБНОСТИ В ОТНОШЕНИИ ДЕПОНИРОВАНИЯ СЕМЯН
Заявитель произвел депонирование 2500 семян Solanum lycopersicum линии TIPC18-61141 в NCIMB 20 августа 2018 года под номером доступа в NCIMB 43169.
Для вынесения экспертного заключения и удовлетворения запросов эксперта заявителем было принято решение о выдаче депонированного материала только эксперту в соответствии с Правилом 32(1) ЕРС или соответствующим законодательством и правилами, принятыми в других странах, или договорами (условиями соглашения с привлеченным экспертом) вплоть до публикации извещения о выдаче патента или в течение 20 лет от даты подачи, если патентная заявка была отклонена, отозвана или считается отозванной.
ПРИМЕРЫ
Пример 1. Идентификация нового аллеля, связанного с увеличенным содержанием сахарозы
У Solanum lycopersicum линии 2927, несущего интрогрессированный TIV (TIVhab) Solanum habrochaites на хромосоме 3 (полученный из Solanum habrochaites LA1777), наблюдалось, что накопление сахарозы происходило до уровней в диапазоне приблизительно 10-20% от общего содержания Сахаров (что называется умеренным уровнем накопления сахарозы), а не до более высоких уровней, составляющих 60-80% (высокий уровень накопления сахарозы), обнаруживаемых у самих соответствующих дикорастущих видов (таблица 1, строки, выделенные жирным шрифтом).
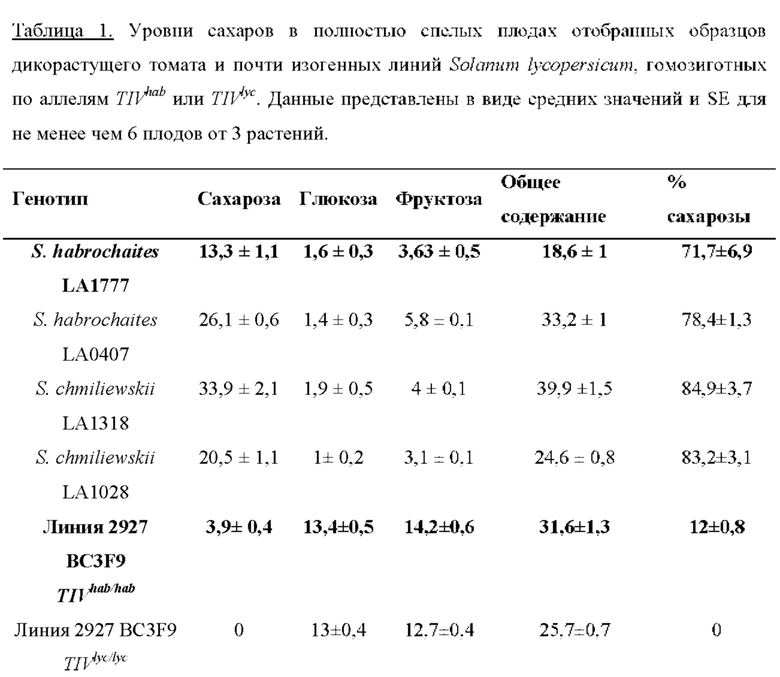
С целью изучения данного явления разрабатывали сегрегирующие популяции на основе скрещивания интрогрессионной линии томата TIVhab/hab (линии 2927) с интрогрессионной линией, несущей аллель TIV из дикорастущего вида Solanum chmielewskii BD732, которая также характеризуется накоплением высоких уровней сахарозы (линией 2928). С целью исключения возможных эффектов двух аллелей TIV - TIVhab и TIVchm - разрабатывали популяции, полученные в результате возвратного скрещивания, в которых проводили отбор посредством генотипирования по наличию гомозиготного генотипического окружения TIVhab/hab и высокому уровню накопления сахарозы.
Результаты для 260 растений, полученных из сегрегирующих гетерозиготных популяций F9 (идентифицированных как 4510 и 4511), указывали на то, что отдельный ген определяет умеренный либо высокий уровень накопления сахарозы в присутствии генотипа TIVhab/hab, и высокий уровень накопления сахарозы, по-видимому, является доминантным признаком (фигура 1).
Пример 2. Идентификация гена, лежащего в основе признака высокого уровня накопления сахарозы
С целью идентификации гена, отвечающего за признак высокого уровня накопления сахарозы, который был назван модификатором содержания сахарозы (SucMod), осуществляли картирование QTL с последующим тонким картированием на уровне отдельных генов.
В качестве первой стадии сегрегирующие популяции F9, состоящие из 260 растений, описанных в примере 1, подвергали скринингу в отношении накопления сахарозы, и отбирали 20 растений из популяций F10, полученных от каждого из 20 растений F9, характеризующихся умеренным уровнем накопления сахарозы (предположительно гомозиготных по рецессивному аллелю Solanum lycopersicum, отвечающему за умеренный уровень накопления сахарозы), и 20 растений из популяций F10, полученных от каждого из 20 растений F9, характеризующихся высоким уровнем накопления сахарозы (как гетерозиготных, так и гомозиготных по доминантному аллелю Solanum chmielewskii, отвечающему за высокий уровень накопления сахарозы). На основании результатов теста на гомозиготность и в соответствии с сегрегацией по уровням накопления сахарозы среди 20 растений из каждого семейства 15 гомозиготных семейств F10 с умеренным уровнем накопления сахарозы и 15 гомозиготных семейств F10 с высоким уровнем накопления сахарозы анализировали с помощью массового сегрегационного анализа, разделяя их на 3 группы, каждая из которых состояла из 5 семейств F10, и генотипируя с помощью системы Illumina. Признак определенно был локализован в дистальном участке хромосомы 12 от SL2.40ch12:64339465 до дистального конца (на основании генетической карты Heinz версии 6) - участке размером около 1 млн. о.
Интересно, что данный участок несет три тандемных гена, кодирующих ингибиторы инвертазы, на участке размером 10 т.о. от SL2.40ch12:64769000 до SL2.40ch12:64779000. Однако полный участок размером ~ 1 млн. п.о. содержит также дополнительные гены-кандидаты, в том числе транспортеры Сахаров, факторы транскрипции и гены с неизвестной функцией. Следовательно, осуществляли стратегию тонкого картирования, и 10000 растений F10, полученных от гетерозиготных растений F9, генотипировали с помощью платформы Illumina с маркерами в 4 положениях вдоль участка, представляющего интерес, и в общей сложности отбирали 327 гомозиготных рекомбинантов, которые выращивали для получения спелых плодов для анализа Сахаров. Результаты анализа Сахаров в зрелых плодах данных рекомбинантов позволили ограничить интрогрессируемый участок, представляющий интерес, до участка размером 440 т.о. между SL2.40ch12:64479000 и SL2.40ch12:64919000. Сорок шесть информативных рекомбинантов, представляющих растения, характеризующиеся как высоким, так и умеренным уровнем накопления сахарозы, использовали для поэтапного тонкого картирования, основанного на ПЦР-клонировании и секвенировании восьми дополнительных участков. Среди рекомбинантов два являлись информативными и позволили ограничить участок с обеих сторон до отдельного гена Solyc12g099190 и исключить два дополнительных нижерасположенных гена ингибиторов инвертазы в качестве кандидатов для QTL SucMod. Рекомбинант SM335 характеризовался уровнем сахарозы, составлявшим 55%, и его событие рекомбинации имело место в участке выше промоторного участка локуса Solyc12g099190, кодирующего ингибитор вакуолярной инвертазы (VIF). Рекомбинант SM79 характеризовался уровнем сахарозы, составлявшим 22%, и его событие рекомбинации имело место в участке ниже 3'-участка гена.
С целью определения того, могут ли отличия в транскрипции генов объяснять дифференциальный эффект аллелей SucMod/VIF, проводили анализ методом секвенирования РНК с использованием развивающихся плодов томата от почти изогенных линий томата TIVhab/hab, отличающихся по интрогрессии VIF и, таким образом, несущих аллель SucModchm либо VIFlyc. Результаты анализа дифференциальной экспрессии генов в участке четко демонстрируют весьма значительную повышенную экспрессию SucModchm по сравнению с VIFluc в ходе всего развития плода (таблица 2). На экспрессию аллеля TIVhad не влияла принадлежность аллелей SucMod/VIF.


Пример 3. Эффект аллелей SucMod/VIF в отношении содержания сахарозы и сахара
С целью определения стадии развития, в ходе которой может наблюдаться отличие в накоплении сахарозы, обусловленное аллелями SucMod/VIF, уровни Сахаров измеряли в ходе развития почти изогенной линии F14 (на основе первоначального скрещивания линий 2928 и 2927), сегрегирующей по аллелям, отвечающим за умеренный (TIVhab/hab VIFlyc/lyc) и высокий (TIVhab/hab SucModchm/chm) уровень накопления сахарозы. В ходе развития накопление сахарозы у линий как с умеренным, так и с высоким уровнем накопления сахарозы начинается до стадии бланжевой спелости, но скорость накопления сахарозы является значительно более высокой для генотипа SucModchm/chm, чем для генотипа VIFlyc/lyc (фигура 2). Уже на стадии бланжевой спелости можно увидеть небольшие, но значительные отличия в уровнях сахарозы, обусловленные генотипом VIF, и эти отличия увеличиваются по мере выспевания (фигура 2В). Увеличение уровней сахарозы также сочетается с увеличением общего содержания сахаров (фигура 2А). Что касается абсолютных значений, наблюдалось чистое увеличение уровней сахарозы, составлявшее 18 мг⋅г-1 сырого веса, при этом общее содержание сахаров увеличивалось на 13 мг⋅г-1 сырого веса. Таким образом, приблизительно 60% чистого увеличения содержания сахарозы выражается в чистом увеличении общего содержания сахаров, что обусловлено тем фактом, что уровни гексоз у растений, характеризующихся умеренным накоплением, являются более высокими, и на них приходится более высокая доля в общем содержании сахаров (фигура 2С). Тем не менее, фенотип высокого содержания сахарозы сопровождается чистым увеличением общего содержания сахаров.
С целью подтверждения данного наблюдения разрабатывали популяции, полученные в результате возвратного скрещивания, несущие две копии аллеля TIVhab и сегрегирующие по аллелям SucModchm и VIFluc. Измеряли содержание растворимых сахаров и сухих веществ в градусах Брикса в полностью спелых плодах, и результаты представлены в таблице 3. Можно видеть, что общее содержание Сахаров увеличивается параллельно с увеличением уровней сахарозы. Кроме того, растения, которые характеризовались наиболее высоким уровнем накопления общего сахара, также характеризовались наиболее высоким уровнем накопления сахарозы. Наконец, линии, несущие по меньшей мере одну копию аллеля SucModchm, демонстрировали намного более высокое соотношение сахарозы и гексозы.
Результаты также демонстрируют, что аллель SucModchm характеризуется полным доминированием, и что гетерозиготное состояние SucModchm/VIFlyc и гомозиготное состояние SucModchm/chm были неразличимы с точки зрения уровней сахарозы и гексозы.

Пример 4. Подтверждение эффекта аллелей SucMod/VIF в отношении содержания сахарозы и сахаров в другом генетическом окружении и другом местоположении
С целью подтверждения эффекта аллелей SucMod/VIF измеряли уровни сахаров в линиях томата F6, полученных от родительских линий томата Ikram (Solanum lycopersicum), сегрегирующих по аллелям TIVlyc/lyc, TIVhab/hab, VIFlyc/lyc и SucModchm/chm. Результаты представлены в таблице 4 и на фигуре 3 и демонстрируют, что накопление сахарозы и соотношение сахарозы и гексозы являются значительно более высокими в случае комбинации генотипа SucModchm/chm с генотипом TIVhab/hab.


Пример 5. Информация о последовательности для аллелей VIF и TIV, включающая иллюстративные ассоциированные SNP-маркеры
В таблице 5 изложена нуклеотидная последовательность аллеля SucModchm (SEQ ID NO: 1) и аллеля TIVhab (SEQ ID NO: 6) по настоящему изобретению. Кроме того, в таблице 5 раскрыты иллюстративные молекулярные SNP-маркеры, специфически ассоциированные с аллелем SucModchm (маркер ST3226) и аллелем TIVhab (маркеры ST3472 и ST3478). Для каждого маркера в таблице показаны местоположение на хромосоме, праймеры (прямой и обратный) для амплификации маркерного фрагмента ДНК и зонды для благоприятных/рекуррентных аллелей, обеспечивающие идентификацию целевого генотипа. Благоприятный SNP-аллель подчеркнут и выделен жирным шрифтом как в последовательности аллеля, так и в зонде для благоприятного аллеля, а также указано его положение относительно последовательности аллеля.

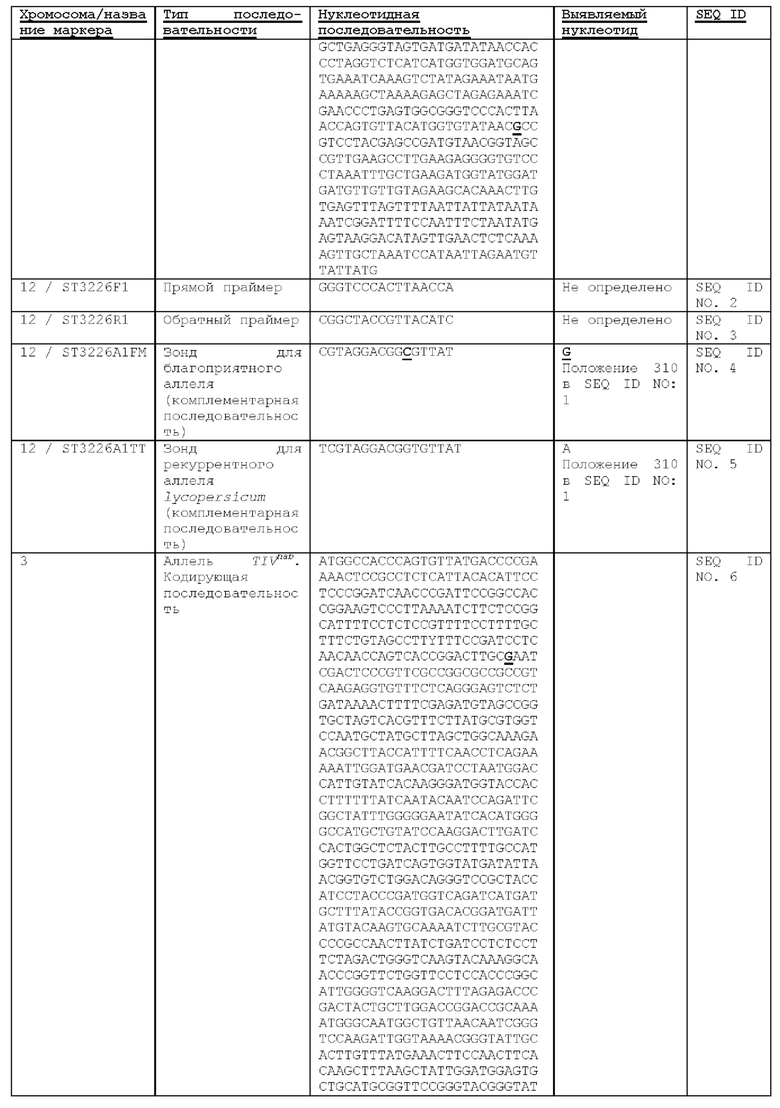

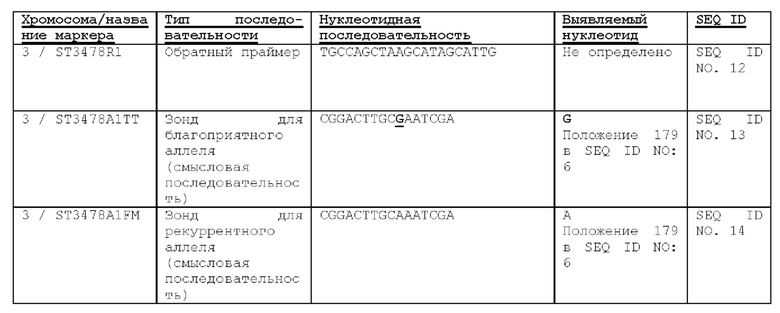
Пример 6. Идентификация дополнительных аллелей SucMod/VIF у других дикорастущих видов томата
С целью идентификации потенциальных новых генетических источников ортологичных аллелей SucMod/VIF измеряли уровни растворимых сахаров в зрелых плодах дикорастущих и культивируемых видов томата. Результаты раскрыты в таблице 6 и подчеркивают то, что образцы Solanum видов habrochaites, pennellii, peruvianum и chmielewskii образуют подгруппу, в которой содержание сахарозы превышает 10 мг⋅г-1 сырого веса, и соотношение сахарозы и гексозы составляет приблизительно 2 или больше. В свою очередь, образцы Solanum видов lycopersicum, cheesmaniae и pimpinellifolium образуют другую подгруппу, в которой содержание сахарозы составляет приблизительно 3 мг⋅г-1 сырого веса или меньше, и соотношение сахарозы и гексозы составляет менее 0,2. Природа последней подгруппы дополнительно подтверждается данными об экспрессии соответствующих этим образцам аллелей TIV и VIF из образцов видов habrochaites, pennellii, peruvianum и chmielewskii. Все образцы в подгруппе демонстрируют высокий уровень экспрессии аллеля VIF и низкий уровень экспрессии аллеля TIV. Напротив, по меньшей мере образцы видов lycopersicum и cheesmaniae демонстрируют противоположный профиль экспрессии: аллель TIV экспрессируется на высоком уровне, и аллель VIF экспрессируется на незначительном уровне. С точки зрения уровня экспрессии образцы вида pimpinellifolium, по-видимому, образуют отдельную третью подгруппу, в которой аллель TIV экспрессируется на высоком уровне, как и аллель VIF.
Эти данные указывают на то, что по меньшей мере образцы видов habrochaites, pennellii, peruvianum и chmielewskii, а также образцы вида pimpinellifolium могут применяться в качестве генетических источников дополнительных аллелей SucMod/VIF, при условии, что их применяют в комбинации с аллелями TIV из образца дикорастущего зеленоплодного томата, для увеличения содержания сахарозы, общего содержания Сахаров и соотношения сахарозы и гексозы.
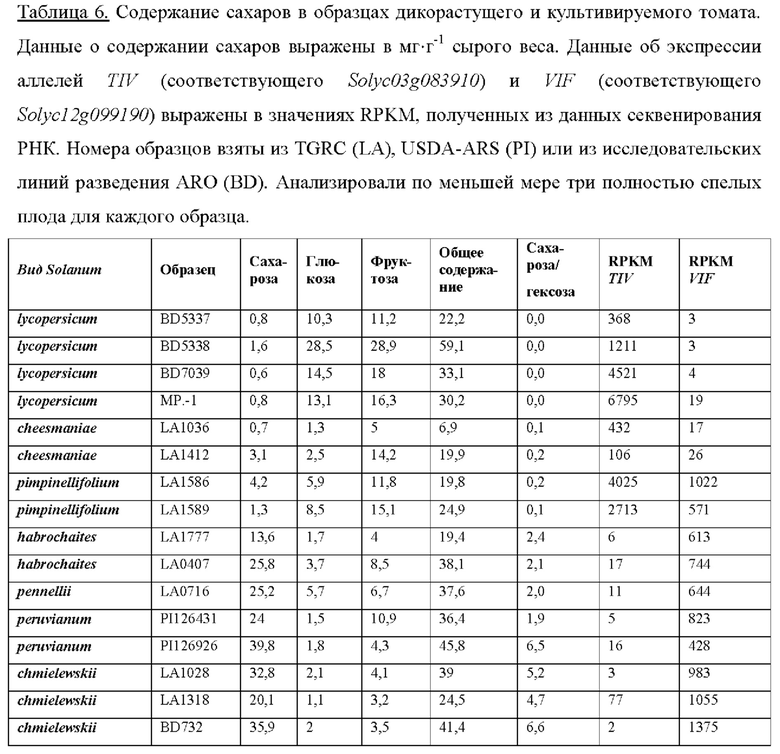
Для дополнительного изучения того, могут ли дополнительные аллели SucMod/VIF быть получены из источников в других дикорастущих видах томата, проводили выравнивания последовательностей с помощью Clustal Omega с использованием последовательностей аллелей SucMod/VIF из Solanum lycopersicum, Solanum chmielewskii BD732 и Solanum pennellii LA0716, а также гомологичных последовательностей из Solanum cheesmaniae LA0429 и Solanum pimpinellifolium LA1589.
На фигуре 4 показано, что аллель VIF из Solanum pimpinellifolium LA1589 (VIFpimp) демонстрирует такой же полиморфизм по SNP-маркеру ST3226, который был идентифицирован для аллеля SucModchm, а именно нуклеотид G в положении, которое соответствует положению 310 в SEQ ID NO: 1 (см. также таблицу 5). Кроме того, аллель SucModchm и аллель VIFpimp имеют еще один общий SNP, а именно нуклеотид Т в положении, которое соответствует положению 498 в SEQ ID NO: 1.
Наконец, аллель VIFpimp и аллель SucModchm обладают 98,86% генетической идентичностью по их полным последовательностям.
Следовательно, предполагается, что по меньшей мере аллель VIFpimp из образца LA1589 Solanum pimpinellifolium обеспечивает эффекты в отношении накопления сахарозы, общего содержания Сахаров и соотношения сахарозы и гексозы, подобные наблюдаемым для аллеля SucModchm.
Пример 7. Идентификация дополнительных аллелей TIV у других дикорастущих видов томата
Результаты, раскрытые в таблице 6 и обсуждаемые в примере 7, также позволяют предположить, что образцы зеленоплодного Solanum видов habrochaites, pennellii, peruvianum и chmielewskii также могут применяться в качестве генетических источников дополнительных аллелей TIV, которые в комбинации с аллелями SucMod/VIF по настоящему изобретению способствуют увеличению содержания сахарозы.
Для дополнительного изучения того, действительно ли дополнительные аллели TIV могут быть получены из источников в других дикорастущих видах томата, проводили выравнивания последовательностей с помощью Clustal Omega с использованием последовательностей аллелей TIV из Solanum habrochaites (SEQ ID NO: 6), Solanum peruvianum, Solanum pennellii, Solanum chmielewskii, Solanum lycopersicum, Solanum lycopersicum var cerasiforme, Solanum cheesmaniae и Solanum pimpinellifolium, которые показаны на фигуре 5.
На фигуре 5 показано, что аллели TIV из, Solanum habrochaites (SEQ ID NO: 6), Solanum peruvianum, Solanum pennellii и Solanum chmielewskii демонстрируют идентичные полиморфизмы в 8 положениях, которые выделены жирным шрифтом и серым затушевыванием, что тем самым обеспечивает 8 SNP-маркеров для распознавания аллелей TIV из подгруппы habrochaites/peruvianum/pennellii/chmielewskii. Два разных SNP-маркера демонстрируют полиморфизм, специфичный для единственного аллеля TIVhab из обряда LA1777 Solanum habrochaites, и они также выделены жирным шрифтом и серым затушевыванием. Последние из SNP-маркеров - ST3472 и ST3478, соответствующая дополнительная информация о последовательностях которых может быть найдена в таблице 5 - эффективно применялись для распознавания аллеля TIVhad. Кроме того, аллели TIV из зеленоплодных Solanum habrochaites (SEQ ID NO: 6), Solanum peruvianum, Solanum pennellii и Solanum chmielewskii обладают по меньшей мере 98% генетической идентичностью по их полным последовательностям.
Пример 8. Иллюстративный протокол анализа сахаров в плодах томата
Образцы тканей околоплодника плодов томата (приблизительно 1 г сырого веса FW) экстрагировали 3 раза в 5 мл 80% (об.:об.) этанола в течение 45 мин при 70°С, и три экстракта объединяли. Раствор сахаров затем выпаривали досуха при 75°С и повторно растворяли в 2 мл дистиллированной воды. Анализ растворимых Сахаров осуществляли с помощью высокоэффективной жидкостной хроматографии (HPLC, Shimadzu, Япония) с использованием колонки Alltech 700СН для анализа углеводов (Alltech Associates, номер по каталогу 70057) и рефрактометрического детектора (RID-10A, Shimadzu, Япония), как было описано ранее у Miron и Schaffer, 1991.
В качестве альтернативы, анализ сахарозы и гексозных Сахаров можно выполнять с помощью доступных методик с использованием UV-излучения, таких как анализ ENZYTEC для D-глюкозы/D-фруктозы/сахарозы, доступный от R-Biopharm AG. Концентрации сахаров количественно определяли по изменениям показателей поглощения UV-излучения растворами сахаров с течением времени в присутствии коммерчески доступных наборов ферментов, содержащих β-фруктозидазу, гексокиназу, фосфоглюкоизомеразу и глюкозо-6-фосфатдегидрогеназу.
СПИСОК ЛИТЕРАТУРЫ
• Davies J.N., 1966, Occurrence of sucrose in the fruit of some species of Lycopersicon, Nature 209, p. 640-641.
• Manning K. and Maw G.A., 1975, Distribution of acid invertase in the tomato plant, Phytochemistry 14(9), p. 1965-1969.
• Yelle S. et al., 1991, Sink metabolism in tomato fruit: IV. Genetic and biochemical analysis of sucrose accumulation, Plant Physiol. 95(4), p. 1026-1035.
• Stommel J.R., 1992, Enzymic Components of Sucrose Accumulation in the Wild Tomato Species Lycopersicon peruvianum, Plant Physiol. 99(1), p. 324-328.
• Klann E. et al., 1993, Expression of acid invertase gene controls sugar composition in tomato (lycopersicon) fruit, Plant Physiol. 103(3), p. 863-870.
• Chetelat R.T. et al., 1993, Inheritance and genetic mapping of fruit sucrose accumulation in Lycopersicon chmielewskii, Plant J. 4, p. 643-650.
• Hadas R. et al., 1995, PCR-generated molecular markers for the invertase gene and sucrose accumulation in tomato, Theor. Appl. Genet. 90(7-8), p. 1142-1148.
• Miron D. et al., 2002, Sucrose uptake, invertase localization and gene expression in developing fruit of Lycopersicon esculentum and the sucrose-accumulating Lycopersicon hirsutum, Physiol. Plant. 115(1), p. 35-47.
• Jin Y. et al, 2009, Posttranslational elevation of cell wall invertase activity by silencing its inhibitor in tomato delays leaf senescence and increases seed weight and fruit hexose level, Plant Cell 21, p. 2072-2089.
• Sievers F. et al, 2011, Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega, Mol. Syst. Biol. 7:539. https://www.ebi.ac.uk/Tools/msa/clustalo/
• Tauzin A.S. et al., 2014, Functional characterization of a vacuolar invertase from Solanum lycopersicon: post-translational regulation by N-glycosylation and a proteinaceous inhibitor, Biochimie 101, p. 39-49.
• Qin G. et al., 2016, A tomato vacuolar invertase inhibitor mediates sucrose metabolism and influences fruit ripening, Plant Physiol. 172, p. 1596-1611.
• Miron D. and Schaffer A.A., 1991, Sucrose Phosphate Synthase, Sucrose Synthase, and Invertase Activities in Developing Fruit of Lycopersicon esculentum Mill, and the Sucrose Accumulating Lycopersicon hirsutum Humb. and Bonpl., Plant Physiol. 95(2), p. 623-627.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СИНГЕНТА ПАРТИСИПЙШНС АГ
<120> РАСТЕНИЯ ТОМАТА, ОБРАЗУЮЩИЕ ПЛОДЫ С МОДИФИЦИРОВАННЫМ
СОДЕРЖАНИЕМ
САХАРОВ В ПЛОДАХ
<130> 81697-EP
<160> 25
<170> PatentIn версия 3.5
<210> 1
<211> 527
<212> ДНК
<213> Solanum chmielewskii
<400> 1
atgagaaatt tattccccat attgatgtta atcactaatt tggcactcaa caacgataac 60
aacaacaaca acaacatcat acacgcaacg tgtagggaga ctccatacta ctccctatgt 120
ctctcagtcc tagaatccga tccacgtagc tacaaggctg agggtagtga tgatataacc 180
accctaggtc tcatcatggt ggatgcagtg aaatcaaagt ctatagaaat aatgaaaaag 240
ctaaaagagc tagagaaatc gaaccctgag tggcgggtcc cacttaacca gtgttacatg 300
gtgtataacg ccgtcctacg agccgatgta acggtagccg ttgaagcctt gaagaggggt 360
gtccctaaat ttgctgaaga tggtatggat gatgttgttg tagaagcaca aacttgtgag 420
tttagtttta attattataa taaatcggat tttccaattt ctaatatgag taaggacata 480
gttgaactct caaaagttgc taaatccata attagaatgt tattatg 527
<210> 2
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 2
gggtcccact taacca 16
<210> 3
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 3
cggctaccgt tacatc 16
<210> 4
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для благоприятного аллеля
<400> 4
cgtaggacgg cgttat 16
<210> 5
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для рекуррентного аллеля
<400> 5
tcgtaggacg gtgttat 17
<210> 6
<211> 1911
<212> ДНК
<213> Solanum habrochaites
<400> 6
atggccaccc agtgttatga ccccgaaaac tccgcctctc attacacatt cctcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcttct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttytttccg atcctcaaca accagtcacc ggacttgcga 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctctgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcagaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcggct 420
atttggggga atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcagtgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac ggatgattat 600
gtacaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagtaca aaggcaaccc ggttctggtt cctccacccg gcattggggt caaggacttt 720
agagacccga ctactgcttg gaccggaccg caaaatgggc aatggctgtt aacaatcggg 780
tccaagattg gtaaaacggg tattgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tgtctactaa aaaaacaaat gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgacaaagaa caaatggaca cccgacaacc cggaattkga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
caacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctatt
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagtctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg ttggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccgtttg gtgtcatagt gattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttctacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctcag aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacattcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat agcatcgcga atttacccaa caaaggcagc aaatggagca
1800
gcacgactct tcgttttcaa caatgctaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg actcagctaa tattcgatcc ctccctttgc aagacttgta a 1911
<210> 7
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 7
agtctcaacc caattccaca atcc 24
<210> 8
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 8
ggttccgggt acgggtatg 19
<210> 9
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для благоприятного аллеля
<400> 9
aattccgggt tgtcg 15
<210> 10
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для рекуррентного аллеля
<400> 10
aattccgggt tatcgg 16
<210> 11
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер
<400> 11
tgctttctgt agccttcttt cc 22
<210> 12
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер
<400> 12
tgccagctaa gcatagcatt g 21
<210> 13
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для благоприятного аллеля
<400> 13
cggacttgcg aatcga 16
<210> 14
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд для рекуррентного аллеля
<400> 14
cggacttgca aatcga 16
<210> 15
<211> 527
<212> ДНК
<213> Solanum lycopersicum
<400> 15
atgagaaatt tattccccat attgatgtta ctcactaatt tgtcactcaa catcgataac 60
aacaacaaca acaacatcat acgcgcaacg tgtagggaga ctccatacta ctccctatgt 120
ctctcagtcc tagaatccga tccacgtagc tacaaggctg agggtagtga tgatataacc 180
accctaggcc tcatcatggt ggatgcggtg aaatcaaagt ctatagaaat aatgaaaaag 240
ctaaaagagc tagagaaatc gaaccctgag tggcgggtcc cacttaacca gtgttacatg 300
gtgtataaca ccgtcctacg agccgatgta acggtagccg ttgaagcctt gaagaggggt 360
gtccctaaat ttgctgaaga tggtatggat gatgttgttg tagaagcaca aacttgtgag 420
tttagtttta attattataa taaatcggat tttccaattt ctaatatgag taaggacata 480
gttgaactct caaaagtcgc taaatccata attagaatgt tattatg 527
<210> 16
<211> 527
<212> ДНК
<213> Solanum pennellii LA0716
<400> 16
atgagaaatt tattccccat attgatgtta atcactaatt tatcactcaa caacgataac 60
aacaacaaca acaacatcat acgcgcaacg tgtagggaga ctccatacta ctccctatgt 120
ctctcaatcc tagaatccga tccacgtagc tacgaggctg agggtagtga tgatataact 180
accctaggcc tcatcatggt ggatgcagtg aaatcaaagt ctatagaaat aatgaaaaag 240
ctaaaagagc tagaaaaatc gaaccctgag tggcgggtcc cacttaacca gtgttacatg 300
gtgtataaca ccgtcctacg agccgatgta acggtagccg ttgaagcctt gaagaggggt 360
gtccctaaat ttgctgaaga tggtatggat gatgttgttg tagaagcaca aacttgtgag 420
tttagtttta attattataa taaatcggat tttccaattt ctaatatgag taaggacata 480
attgaactct caaaagtcgc taaatccata attagaatgt tattatg 527
<210> 17
<211> 527
<212> ДНК
<213> Solanum cheesmaniae LA0429
<400> 17
atgagaaatt tattccccat attgatgtta ctcactaatt tgtcactcaa catcgataac 60
aacaacaaca acaacatcat acgcgcaacg tgtagggaga ctccatacta ctccctatgt 120
ctctcagtcc tagaatccga tccacgtagc tacaaggctg agggtagtga tgatataacc 180
accctaggcc tcatcatggt ggatgcggtg aaatcaaagt ctatagaaat aatgaaaaag 240
ctaaaagagc tagagaaatc gaaccctgag tggcgggtcc cacttaacca gtgttacatg 300
gtgtataaca ccgtcctacg agccgatgta acggtagccg ttgaagcctt gaagaggggt 360
gtccctaaat ttgctgaaga tggtatggat gatgttgttg tagaagcaca aacttgtgag 420
tttagtttta attattataa taaatcggat tttccaattt ctaatatgag taaggacata 480
gttgaactct caaaagtcgc taaatccata attagaatgt tattatg 527
<210> 18
<211> 527
<212> ДНК
<213> Solanum pimpinellifolium LA1589
<400> 18
atgagaaatt tattccccat attgatgtta ctcactaatt tgtcactcaa catcgataac 60
aacaacaaca acaacatcat acgcgcaacg tgtagggaga ctccatacta ctccctatgt 120
ctctcagtcc tagaatccga tccacgtagc tacaaggctg agggtagtga tgatataacc 180
accctaggcc tcatcatggt ggatgcggtg aaatcaaagt ctatagaaat aatgaaaaag 240
ctaaaagagc tagagaaatc gaaccctgag tggcgggtcc cacttaacca gtgttacatg 300
gtgtataacg ccgtcctacg agccgatgta acggtagccg ttgaagcctt gaagaggggt 360
gtccctaaat ttgctgaaga tggtatggat gatgttgttg tagaagcaca aacttgtgag 420
tttagtttta attattataa taaatcggat tttccaattt ctaatatgag taaggacata 480
gttgaactct caaaagttgc taaatccata attagaatgt tattatg 527
<210> 19
<211> 1911
<212> ДНК
<213> Solanum peruvianum KY565130
<400> 19
atggccactc agtgttatga ccccgaaaac tccgcctctc attacacatt cctcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagtaca aaggcaaccc ggttctggtt cctccacccg gcattggtgt caaggacttt 720
agggacccga ctactgcttg gaccggaccg caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaagcggg tattgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaat gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat ggcaataagc aagatcatta tgctattggt
1020
acgtatgact tggcaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa taagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctatt
1320
gttaaggaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagtctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gaagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgaattaac gccagtttac
1560
ttctacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctcag aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacactcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt agatggagca
1800
gcacgactct tcgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcaattg agtcagctaa tattcgatcc ttccctttgc aagacttgta a 1911
<210> 20
<211> 1911
<212> ДНК
<213> Solanum pennellii XM015214462
<400> 20
atggccaccc agtgttatga ccccgaaaac tccgcctctc actacacatt cctcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg ggctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaactgctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gactgggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagtaca aaggcaaccc ggttctggtt cctccacccg gcattgctgt caaggacttt 720
agagacccga ctactgcttg gaccggaccg caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tattgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaat gggttggaca catcatataa cgggccgggt 960
gtaaagcatg ttttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgacaaagaa caaatggacg cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
caacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctatt
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagtctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttctacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctcag aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacactcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt aaatggagca
1800
gcacgactct tcgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg agtcagctaa tattcgatcc ttccctttgc aagacttgta a 1911
<210> 21
<211> 1908
<212> ДНК
<213> Solanum chmielewskii KY565126
<400> 21
atggccactc agtgttatga ccccgaaaac tccgcctctc attacacatt actcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttgtgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg tcatggttcc tgatcaatgg tatgatatta atggtgtctg gactgggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagtaca aaggcaaccc ggttctggtt cctccacccg gcattggggt caaggacttt 720
agagacccga ctactgcttg gaccggaccg caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tgttgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgcgtggac 900
ttttacccgg tatctactaa aaaaacaaat gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggg
1020
acgtatgact tggcaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcaccaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaag tattccaagg acagtgcttt acgacaagaa gacagggaca
1260
catctacttc agtggccagt ggaagaaatt gaaagcttaa gagtgggtga tcctattgtt
1320
aagcaagtcg atcttcaacc aggctcaatt gagctactcc gtgttgactc agctgcagag
1380
ttggatatag aagtctcatt tgaagtggac aaagtcgcgc ttcagggaat aattgaagca
1440
gatcatgtag gtttcagttg ctctactagt ggaggtgctg ctagcagagg cattttggga
1500
ccatttggtg tcatagtaat tgctgatcaa acgctatctg agctaacgcc agtttacttc
1560
tacatttcta aaggagctga tggtcgtgca gagactcact tctgtgctga tcaaactaga
1620
tcctcagagg ctccgggagt tggtaaacaa gtttatggta gttcagtacc tgtgttggac
1680
ggtgaaaaac attcaatgag attattggtg gatcactcaa ttgtggagag ctttgctcaa
1740
ggaggaagaa cagtcataac atcgcgaatt tacccaacaa aggcagtaaa tggagcagca
1800
cgactctttg ttttcaacaa tgccacaggg gctagcgtta ctgcctccgt caagatttgg
1860
ccacttgagt cagctaatat tcgatccttc cctttgcaag acttgtaa 1908
<210> 22
<211> 1911
<212> ДНК
<213> Solanum lycopersicum NM001247914
<400> 22
atggccactc agtgttatga ccccgaaaac tccgcctctc gttacacatt actcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagttca aaggcaaccc ggttctggtt cctccacccg gcattggtgt caaggacttt 720
agagacccga ctactgcttg gaccggacca caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tgttgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaac gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgggaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctact
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagcctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttttacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctctg aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacattcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt aaatggagca
1800
gcacgactct ttgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg agtcagctaa tattcaatcc ttccctttgc aagacttgta a 1911
<210> 23
<211> 1911
<212> ДНК
<213> Solanum lycopersicum var. cerasiforme GU784870
<400> 23
atggccactc agtgttatga ccccgaaaac tccgcctctc gttacacatt actcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atccccaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagttca aaggcaaccc ggttctggtt cctccacccg gcattggtgt caaggacttt 720
agagacccga ctactgcttg gaccggacca caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tgttgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaac gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgggaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctact
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagcctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttttacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctctg aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacattcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt aaatggagca
1800
gcacgactct ttgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg agtcagctaa tattcaatcc ttccctttgc aagacttgta a 1911
<210> 24
<211> 1911
<212> ДНК
<213> Solanum cheesmaniae KY565124
<400> 24
atggccactc agtgttatga ccccgaaaac tccgcctctc gttacacatt actcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagttca aaggcaaccc ggttctggtt cctccacccg gcattggtgt caaggacttt 720
agagacccga ctactgcttg gaccggacca caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tgttgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaac gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgggaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctact
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagcctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttttacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctctg aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacattcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt aaatggagca
1800
gcacgactct ttgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg agtcagctaa tattcaatcc ttccctttgc aagacttgta a 1911
<210> 25
<211> 1911
<212> ДНК
<213> Solanum pimpinellifolium Z12026
<400> 25
atggccactc agtgttatga ccccgaaaac tccgcctctc gttacacatt actcccggat 60
caacccgatt ccggccaccg gaagtccctt aaaatcatct ccggcatttt cctctccgtt 120
ttccttttgc tttctgtagc cttctttccg atcctcaaca accagtcacc ggacttgcaa 180
atcgactccc gttcgccggc gccgccgtca agaggtgttt ctcagggagt ctccgataaa 240
acttttcgag atgtagccgg tgctagtcac gtttcttatg cgtggtccaa tgctatgctt 300
agctggcaaa gaacggctta ccattttcaa cctcaaaaaa attggatgaa cgatcctaat 360
ggaccattgt atcacaaggg atggtaccac cttttttatc aatacaatcc agattcagct 420
atttggggaa atatcacatg gggccatgct gtatccaagg acttgatcca ctggctctac 480
ttgccttttg ccatggttcc tgatcaatgg tatgatatta acggtgtctg gacagggtcc 540
gctaccatcc tacccgatgg tcagatcatg atgctttata ccggtgacac tgatgattat 600
gtgcaagtgc aaaatcttgc gtaccccgcc aacttatctg atcctctcct tctagactgg 660
gtcaagttca aaggcaaccc ggttctggtt cctccacccg gcattggtgt caaggacttt 720
agagacccga ctactgcttg gaccggacca caaaatgggc aatggctgtt aacaatcggg 780
tctaagattg gtaaaacggg tgttgcactt gtttatgaaa cttccaactt cacaagcttt 840
aagctattgg atggagtgct gcatgcggtt ccgggtacgg gtatgtggga gtgtgtggac 900
ttttacccgg tatctactaa aaaaacaaac gggttggaca catcatataa cgggccgggt 960
gtaaagcatg tgttaaaagc aagtttagat gacaataagc aagatcatta tgctattggt
1020
acgtatgact tgggaaagaa caaatggaca cccgataacc cggaattgga ttgtggaatt
1080
gggttgagac tagactatgg gaaatattat gcatcaaaga ctttttatga cccgaagaaa
1140
gaacgaagag tactgtgggg atggattggg gaaactgaca gtgaatctgc tgacctgcag
1200
aagggatggg catctgtaca gagtattcca aggacagtgc tttacgacaa gaagacaggg
1260
acacatctac ttcagtggcc agtggaagaa attgaaagct taagagtggg tgatcctact
1320
gttaagcaag tcgatcttca accaggctca attgagctac tccgtgttga ctcagctgca
1380
gagttggata tagaagcctc atttgaagtg gacaaagtcg cgcttcaggg aataattgaa
1440
gcagatcatg taggtttcag ttgctctact agtggaggtg ctgctagcag aggcattttg
1500
ggaccatttg gtgtcatagt aattgctgat caaacgctat ctgagctaac gccagtttac
1560
ttttacattt ctaaaggagc tgatggtcgt gcagagactc acttctgtgc tgatcaaact
1620
agatcctctg aggctccggg agttggtaaa caagtttatg gtagttcagt acctgtgttg
1680
gacggtgaaa aacattcaat gagattattg gtggatcact caattgtgga gagctttgct
1740
caaggaggaa gaacagtcat aacatcgcga atttacccaa caaaggcagt aaatggagca
1800
gcacgactct ttgttttcaa caatgccaca ggggctagcg ttactgcctc cgtcaagatt
1860
tggtcacttg agtcagctaa tattcaatcc ttccctttgc aagacttgta a 1911
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ ТОМАТОВ С УЛУЧШЕННЫМИ ПРИЗНАКАМИ | 2019 |
|
RU2821992C2 |
| РАСТЕНИЯ ТОМАТА С УЛУЧШЕННОЙ УСТОЙЧИВОСТЬЮ К БОЛЕЗНЯМ | 2015 |
|
RU2721952C2 |
| РАСТЕНИЯ SOLANUM LYCOPERSICUM С ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ | 2015 |
|
RU2696827C2 |
| РАСТЕНИЯ ТОМАТА, ОБЛАДАЮЩИЕ ПОВЫШЕННЫМИ УРОВНЯМИ УСТОЙЧИВОСТИ К BOTRYTIS | 2007 |
|
RU2469094C2 |
| РАСТЕНИЕ АРБУЗА, ХАРАКТЕРИЗУЮЩЕЕСЯ ПРОДУКТИВНЫМ ЦВЕТЕНИЕМ | 2017 |
|
RU2792674C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ СЕМЯН, ИМЕЮЩИХ В ЗАРОДЫШЕ ПОЛНЫЙ ДИПЛОИДНЫЙ ГЕНОМ, КОМПЛЕМЕНТАРНЫЙ МАТЕРИНСКОМУ | 2006 |
|
RU2438297C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2008 |
|
RU2522477C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2014 |
|
RU2617955C2 |
| КЛОНИРОВАНИЕ И ИСПОЛЬЗОВАНИЕ ФУНКЦИОНАЛЬНОГО R-ГЕНА ИЗ SOLANUM CHACOENSE | 2010 |
|
RU2586509C2 |
| ФАРНЕЗЕН-СИНТАЗА | 2009 |
|
RU2546877C2 |
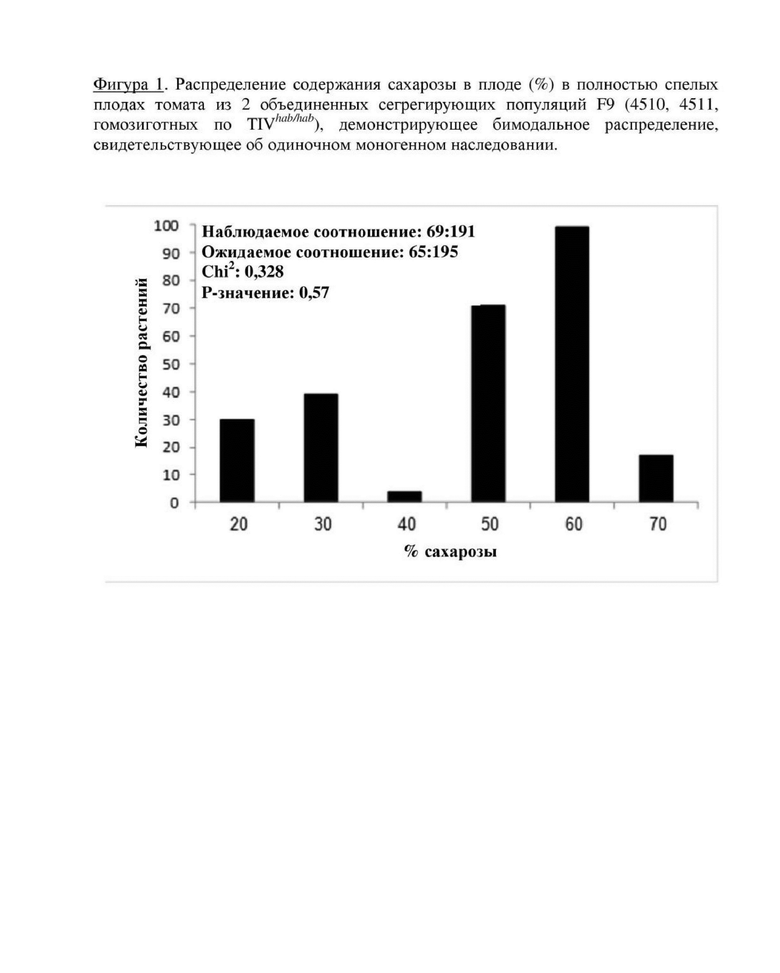
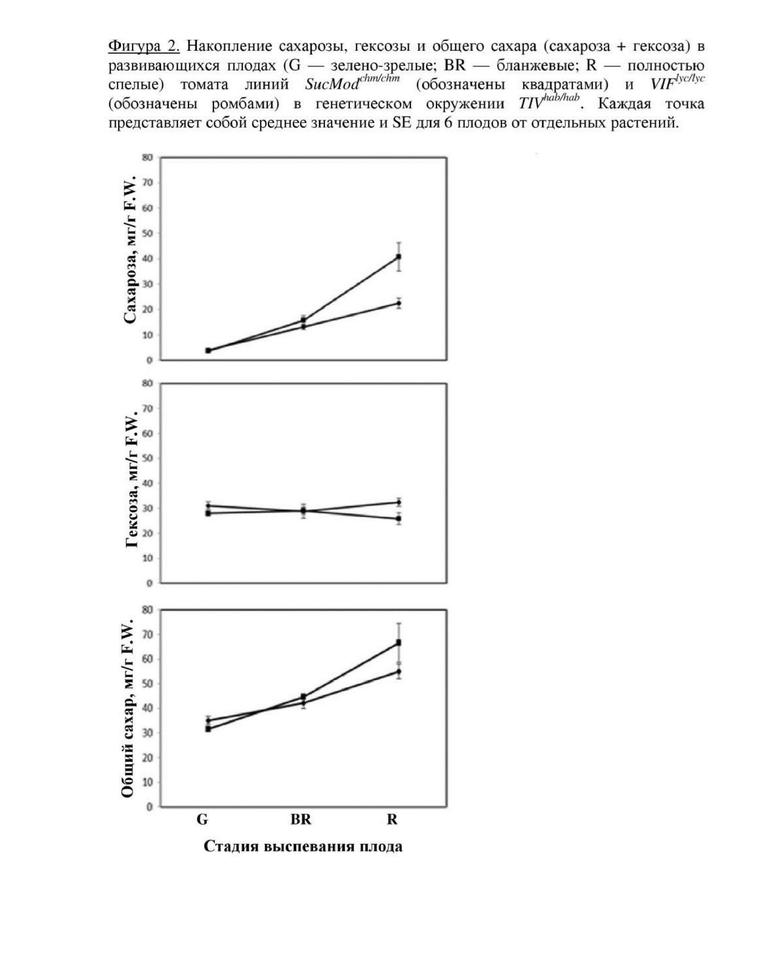

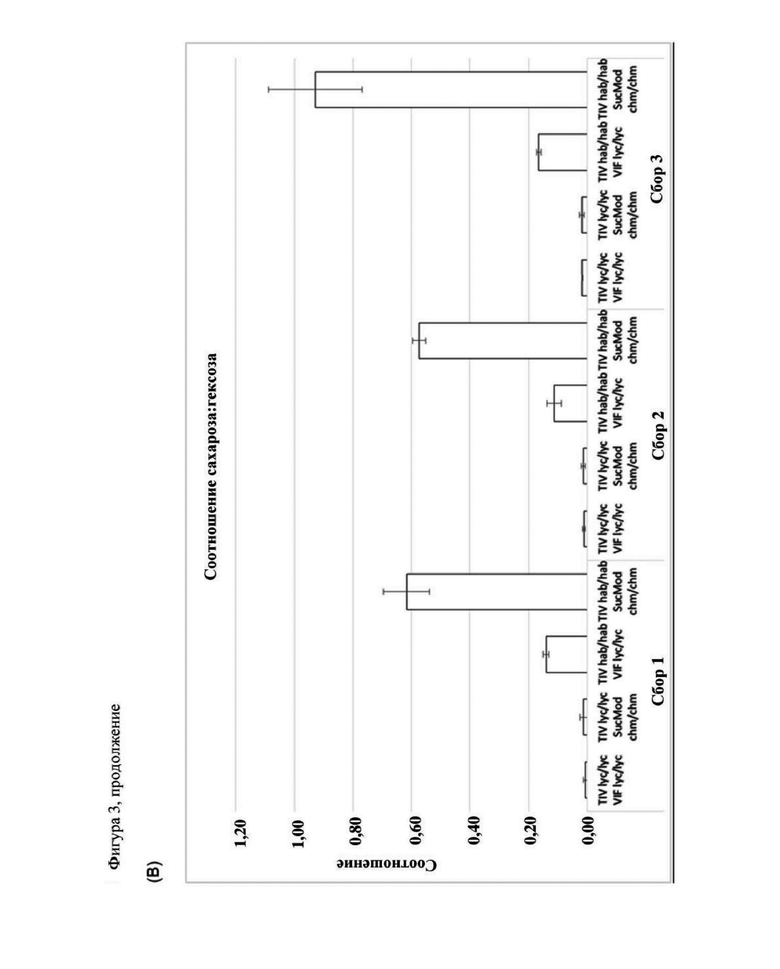

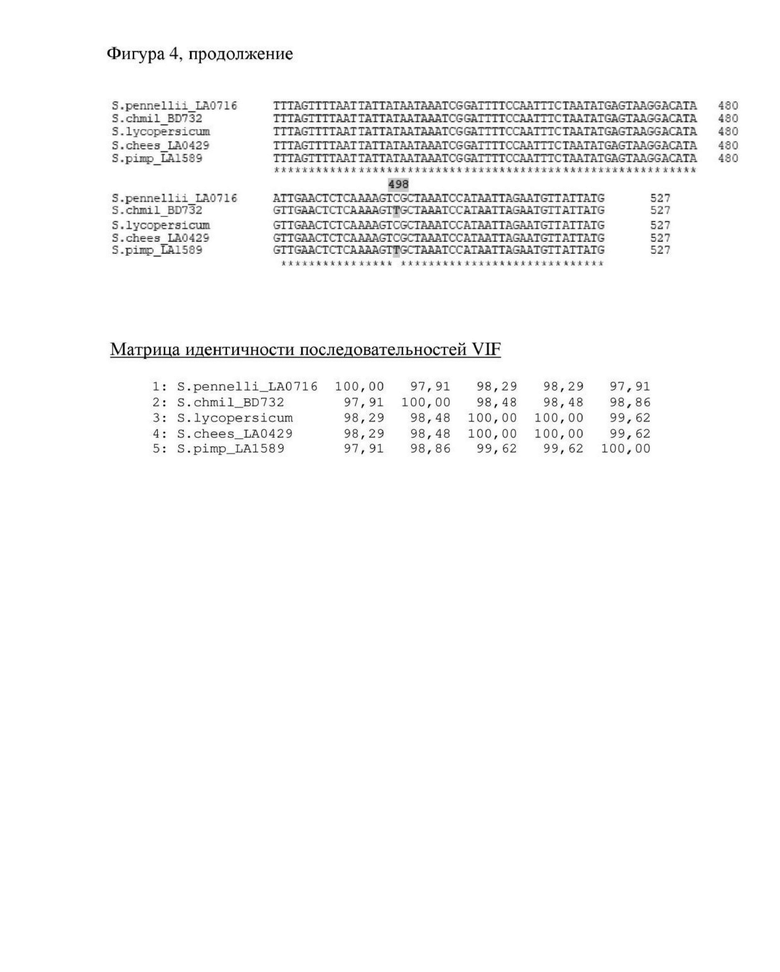



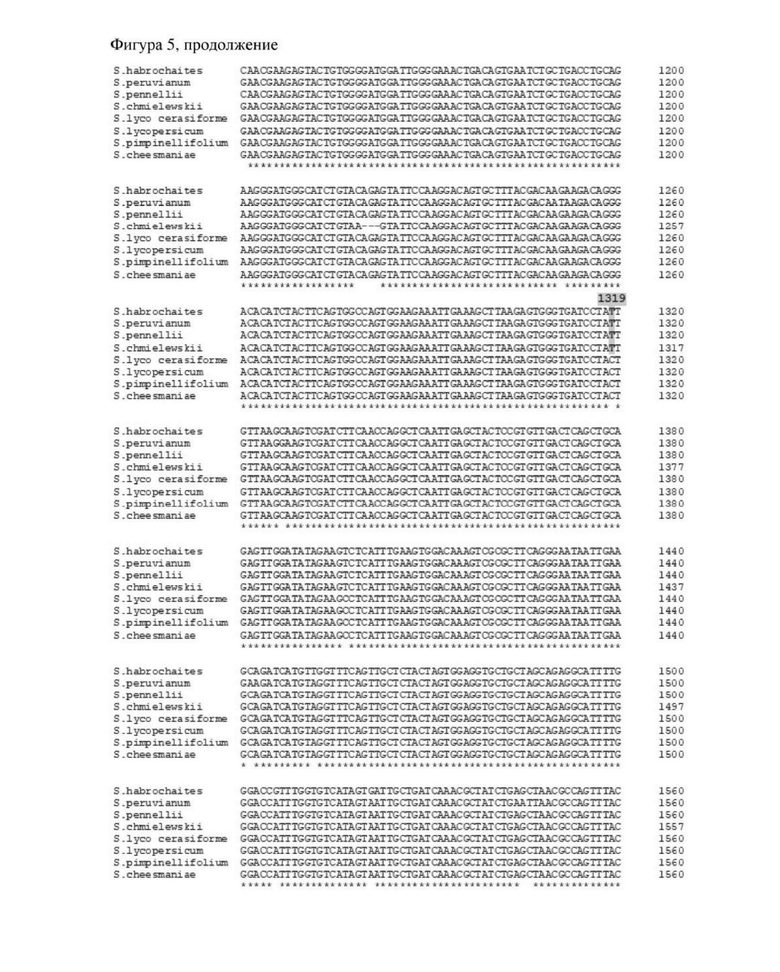


Изобретение относится к области биохимии, в частности к способу получения культивируемого растения томата. Также раскрыт способ идентификации растения томата. Изобретение позволяет эффективно культивировать растения томата Solanum lycopersicum. 3 н. и 3 з.п. ф-лы, 6 табл., 5 ил., 8 пр.
1. Способ получения культивируемого растения томата, культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающий стадии:
a) скрещивания первого растения, содержащего:
(i) по меньшей мере одну копию аллеля модификатора сахарозы SucMod, по меньшей мере на 90% идентичного SEQ ID NO: 1 и содержащего нуклеотид G в положении, которое соответствует положению 310 SEQ ID NO: 1, и нуклеотид Т в положении, которое соответствует положению 498 SEQ ID NO: 1, и
(ii) две копии аллеля TIV, отвечающего за накопление сахарозы, последовательность которых по меньшей мере на 98% идентична SEQ ID NO: 6; с культивируемым растением томата, в котором отсутствуют указанные аллели SucMod и TIV;
б) отбора растения-потомка, образующего плоды, демонстрирующие увеличенное содержание сахарозы;
где отбор на стадии б) проводят посредством выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и нуклеотида T в положении, которое соответствует положению 498 в SEQ ID NO: 1; и посредством выявления нуклеотида A в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида A в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида T в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида C в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида T в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида C в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида A в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
2. Способ по п. 1, где отбор на стадии b) проводят посредством дополнительного выявления нуклеотида C в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
3. Способ по п. 1 или 2, в котором первое растение этапа а) представляет собой линию Solanum lycopersicum TIPC18-61141, депонированную в NCIMB 20 августа 2018 под номером NCIMB № 43169.
4. Способ идентификации культивируемого растения томата, культивируемого растения Solanum lycopersicum, образующего плоды, демонстрирующие увеличенное содержание сахарозы, и имеющего по меньшей мере одну копию аллеля SucMod, являющегося модификатором содержания сахарозы, и две копии аллеля TIV, отвечающего за накопление сахарозы, полученного из образца дикорастущего зеленоплодного томата, включающий стадии:
a) выявления нуклеотида G в положении, которое соответствует положению 310 в SEQ ID NO: 1, и нуклеотида T в положении, которое соответствует положению 498 в SEQ ID NO: 1; и
b) выявления нуклеотида A в положении, которое соответствует положению 41 в SEQ ID NO: 6; и/или нуклеотида A в положении, которое соответствует положению 668 в SEQ ID NO: 6; и/или нуклеотида T в положении, которое соответствует положению 930 в SEQ ID NO: 6; и/или нуклеотида C в положении, которое соответствует положению 1034 в SEQ ID NO: 6; и/или нуклеотида T в положении, которое соответствует положению 1319 в SEQ ID NO: 6; и/или нуклеотида C в положении, которое соответствует положению 1563 в SEQ ID NO: 6; и/или нуклеотида A в положении, которое соответствует положению 1629 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 1886 в SEQ ID NO: 6.
5. Способ по п. 4, где стадию b) проводят посредством дополнительного выявления нуклеотида C в положении, которое соответствует положению 1056 в SEQ ID NO: 6; и/или нуклеотида G в положении, которое соответствует положению 179 в SEQ ID NO: 6.
6. Способ продуцирования культивируемого растения томата, культивируемого растения Solanum lycopersicum, образующего плоды томата, демонстрирующие увеличенное содержание сахарозы, включающий стадии:
а) обеспечения семенами растения томата, содержащего:
(i) по меньшей мере одну копию аллеля модификатора сахарозы SucMod, по меньшей мере на 90% идентичного SEQ ID NO: 1 и содержащего нуклеотид G в положении, которое соответствует положению 310 SEQ ID NO: 1, и нуклеотид Т в положении, которое соответствует положению 498 SEQ ID NO: 1, и
(ii) две копии аллеля TIV, отвечающего за накопление сахарозы, последовательность которых по меньшей мере на 98% идентична SEQ ID NO: 6;
б) проращивание указанных семян и выращивание из них взрослых фертильных растений;
в) индукцию самоопыления указанных растений при а), выращивание фруктов и сбор у них фертильных семян; и
г) производство растений из собранных на стадии в) семян и отбор растений с повышенным содержанием сахарозы.
| US 5658773 A, 19.08.1997 | |||
| WO 2012095841 A1, 19.07.2021 | |||
| РАСТЕНИЯ ТОМАТА, ОБЛАДАЮЩИЕ ПОВЫШЕННЫМИ УРОВНЯМИ УСТОЙЧИВОСТИ К BOTRYTIS | 2007 |
|
RU2469094C2 |
