Уровень техники
1. Область техники, к которой относится изобретение
Настоящее изобретение относится к средствам, которые связывают сфингозин-1-фосфат (S1P), в частности к гуманизированным моноклональным антителам, фрагментам антител и производным антител, специфично реактивным к S1P в физиологических условиях. Такие средства можно применять в лечении и/или профилактике различных заболеваний или нарушений посредством введения фармацевтических композиций, которые содержат такие средства.
Представленное ниже описание включает информацию, которая может быть полезна для понимания настоящего изобретения. Оно не подразумевает допущения того, что любая информация, предоставленная в настоящем документе, или любая публикация, конкретно или косвенно цитируемая в настоящем документе, является предшествующим уровнем техники или даже частично относится к заявляемому настоящему изобретению.
2. Уровень техники
Биологически активные липиды каскада передачи сигнала
Липиды и их производные в настоящее время признаны важными объектами медицинских исследований, а не только простыми структурными элементами в клеточных мембранах или источником энергии для β-окисления, гликолиза или других метаболических процессов. В частности, некоторые биологически активные липиды функционируют в качестве медиаторов передачи сигнала, важных при заболеваниях животных и человека. Несмотря на то что большинство из липидов плазматической мембраны играют исключительно структурную роль, небольшая их часть вовлечена в передачу внеклеточных стимулов в клетку. "Липидная передача сигнала" относится к любому из множества клеточных каскадов передачи сигнала, в которых в качестве вторичных посредников используются липиды клеточных мембран, а также относится к прямому взаимодействию молекулы липидной передачи сигнала с ее собственным специфическим рецептором. Каскады липидной передачи сигнала активируются множеством внеклеточных стимулов, варьирующих от факторов роста до воспалительных цитокинов, и регулируют клеточную судьбу, такую как апоптоз, дифференцировка и пролиферация. Исследования передачи сигнала биологически активных липидов находятся в области интенсивного научного изучения, поскольку идентифицируется все больше и больше биологически активных липидов и охарактеризовывается их действие.
Примеры биологически активных липидов включают эйкозаноиды (включая каннабиноиды, лейкотриены, простагландины, липоксины, эпоксиэйкозатриеновые кислоты и изоэйкозаноиды), неэйкозаноидные каннабиноидные медиаторы, фосфолипиды и их производные, такие как фосфатидная кислота (РА) и фосфатидилглицерин (PG), фактор активации тромбоцитов (PAF) и кардиолипины, а также лизофосфолипиды, такие как лизофосфатидилхолин (LPC) и различные лизофосфатидные кислоты (LPA). Биологически активные липидные медиаторы передачи сигнала также включают сфинголипиды, такие как сфингомиелин, церамид, церамид-1-фосфат, сфингозин, сфингозилфосфорилхолин, сфинганин, сфинганин-1-фосфат (дигидро-S1P) и сфингозин-1-фосфат. Сфинголипиды и их производные составляют группу внеклеточных и внутриклеточных молекул передачи сигнала с плейотропыми эффектами на важные клеточные процессы. Другие примеры биологически активных липидов передачи сигнала включают фосфатидилсерин (PS), фосфатидилинозитол (PI), фосфатидилэтаноламин (PEA), диацилглицерид (DG), сульфатиды, ганглиозиды и цереброзиды.
Сфинголипиды представляют собой уникальный класс липидов, которые получили свое название вследствие их исходно загадочной природы, в честь Сфинкса. Сфинголипиды первоначально были охарактеризованы в качестве первичных структурных компонентов клеточных мембран, однако последние исследования показывают, что сфинголипиды также служат клеточными молекулами передачи сигнала и регуляторными молекулами (Hannun, et al., Adv. Lipid Res. 25:27-41, 1993; Speigel, et al., FASEB J. 10:1388-1397, 1996; Igarashi, J. Biochem 122:1080-1087, 1997; Hla, T. (2004). Semin Cell Dev Biol, 15, 513-2; Gardell, S.E., Dubin, A.E. & Chun, J. (2006). Trends Mol. Med, 12, 65-75). Сфинголипиды являются основными структурными компонентами клеточных мембран, которые также служат в качестве клеточных молекул передачи сигнала и регуляторных молекул (Hannun and Bell, Adv. Lipid Res. 25:27-41, 1993; Igarashi, J. Biochem 122:1080-1087, 1997). Сфинголипидные медиаторы передачи сигнала, церамид (CER), сфингозин (SPH) и сфингозин-1-фосфат (S1P), наиболее широко изучены, и недавно была признана их роль в сердечно-сосудистой системе, ангиогенезе и биологии опухолей (Claus, et al., Curr Drug Targets 1:185-205, 2000; Levade, et al., Circ. Res. 89:957-968, 2001; Wang, et al., J. Biol. Chem. 274:35343-50, 1999; Wascholowski and Giannis, Drug News Perspect. 14:581-90, 2001; Spiegel, S. & Milstien, S. (2003). Sphingosine-1-phosphate: an enigmatic signaling lipid. Nat Rev Mol Cell Biol, 4, 397-407).
Для обзора метаболизма сфинголипидов см. Liu, et al., Crit Rev. Clin. Lab. Sci. 36:511-573, 1999. Для обзора каскада передачи сигнала сфингомиелина см. Hannun, et al., Adv. Lipid Res. 25:27-41, 1993; Liu, et al., Crit. Rev. Clin. Lab. Sci. 36:511-573, 1999; Igarashi, J. Biochem. 122:1080-1087, 1997; Oral, et al., J. Biol. Chem. 272:4836-4842, 1997; и Spiegel et al., Biochemistry (Moscow) 63:69-83, 1998.
S1P является медиатором пролиферации клеток и защищает от апоптоза посредством активации каскадов выживания (Maceyka, et al. (2002), ВВА, vol.1585:192-201, и Spiegel, et al. (2003), Nature Reviews Molecular Cell Biology, vol.4:397-407). Было предположено, что баланс между уровнями CER/SPH и S1P обеспечивает реостатный механизм, который определяет, направить ли клетку на путь гибели или защитить от апоптоза. Ключевым регуляторным ферментом реостатного механизма является сфингозинкиназа (SPHK), ролью которой является превращение обеспечивающих гибель биологически активных липидов передачи сигнала (CER/SPH) в способствующий росту S1P. S1P имеет два исхода: S1P может деградироваться S1P-лиазой, ферментом, который расщепляет S1P до фосфоэтаноламина и гексадеканаля, или, менее часто, он гидролизуется S1P-фосфатазой до SPH.
Плейотропные виды биологической активности S1P опосредуются через семейство сопряженных с G-белком рецепторов (GPCR), исходно известных как гены дифференцировки эндотелия (EDG). Было идентифицировано пять GPCR в качестве высокоаффинных рецепторов S1P (S1PR): S1P1/EDG-1, S1P2/EDG-5, S1P3/EDG-3, S1P4/EDG-6 и S1P5/EDG-8, идентифицированный только в 1998 году (Lee, et al., 1998). Множество ответов, вызываемых S1P, сопряжено с различными гетеротримерными G-белками (Gq-, Gi, G12-13) и малыми ГТФазами семейства Rho (Gardell, et al., 2006).
У взрослых S1P высвобождается из тромбоцитов (Murata et al., 2000) и тучных клеток, создавая локальный импульс свободного S1P (достаточный для превышения Kd для S1PR) для стимуляции заживления ран и участия в воспалительном ответе. В нормальных условиях общее количество S1P в плазме является достаточно высоким (300-500 нМ); однако была предложена гипотеза о том, что большая часть S1P может быть "буферизованной" сывороточными белками, в частности липопротеинами (например, HDL>LDL>VLDL) и альбумином, так что количество биологически доступного S1P (или свободной фракции S1P) не является достаточным для существенной активации S1PR (Murata et al., 2000). Если этого не происходит, результатом будет недостаточный ангиогенез и воспаление. Также было сделано предположение о внутриклеточном действии S1P (см., например, Spiegel S, Kolesnick R (2002), Leukemia, vol.16:1596-602; Suomalainen, et al (2005), Am. J. Pathol, vol.166:773-81).
Распространенная экспрессия рецепторов S1P клеточной поверхности позволяет SIP влиять на широкий спектр клеточных ответов, включая пролиферацию, адгезию, сокращение, подвижность, морфогенез, дифференцировку и выживание. Этот спектр ответов, по-видимому, зависит от перекрывающихся или отличающихся паттернов экспрессии рецепторов S1P в клеточной и тканевой системах. Кроме того, недавно было показано перекрестное наложение каскадов передачи сигнала S1P и факторов роста, включая тромбоцитарный фактор роста (PDGF), сосудисто-эндотелиальный фактор роста (VEGF) и основной фибробластный фактор роста (bFGF) (см., например, Baudhuin, et al. (2004), FASEB J, vol.18:341-3). Регуляция различных клеточных процессов, вовлекающих S1P, оказывает конкретное влияние в числе прочих на передачу сигнала в нейронах, тонус сосудов, заживление ран, транспорт иммунных клеток, воспроизведение и сердечно-сосудистую функцию. Изменения эндогенных уровней S1P в этих системах может оказывать отрицательные эффекты, вызывая некоторые патофизиологические состояния, в том числе злокачественную опухоль, воспаление, ангиогенез, заболевание сердца; астму и аутоиммунные заболевания.
Современный новый подход к лечению различных заболеваний и нарушений, включая сердечно-сосудистые заболевания, цереброваскулярные заболевания и различные злокачественные опухоли, включает снижение уровней биологически доступного S1P, отдельно или в сочетании с другими способами лечения. Несмотря на то что были предложены стратегии лечения на основе сфинголипидов, которые нацелены на ключевые ферменты метаболического каскада сфинголипидов, такие как SPHK, подавлению непосредственно липидного медиатора S1P до настоящего времени внимания не уделяли, главным образом, вследствие сложности прямого подавления этой липидной мишени, в частности вследствие сложности сначала индукции, а затем детекции антител против SlP-мишени.
Недавно было описано получение антител, специфичных к S1P. См., например, патентную заявку США с серийным номером №20070148168; WO 2007/053447 того же заявителя. Такие антитела, которые могут, например, селективно адсорбировать S1P из сыворотки, действуют в качестве молекулярных губок для нейтрализации внеклеточного S1P. См. также патенты США 6881546 и 6858383 и патентную заявку США с серийным номером 10/029372 того же заявителя. Было показано, что SPHINGOMAB™, моноклональное антитело (mAb) мыши, разработанное Lpath, Inc. и описанное в некоторых патентах или патентных заявках, перечисленных выше, является эффективным в моделях заболеваний человека. В некоторых случаях гуманизированное антитело может быть предпочтительнее антитела мыши, в частности, для терапевтического применения у людей, поскольку может возникнуть ответ в виде антител человека против антител мыши (НАМА). Такой ответ может снижать эффективность антитела посредством нейтрализации связывающей активности и/или посредством быстрого выведения антитела из кровотока в организме. Ответ НАМА также может вызывать токсичность при последующих введениях антител мыши.
В данной работе разработано гуманизированное антитело против S1P и оно описано в настоящем документе. Предполагается, что это антитело обладает всеми преимуществами mAb мыши с точки зрения эффективности связывания S1P, нейтрализации S1P и модулирования болезненных состояний, связанных с S1P, но не обладает потенциальными недостатками mAb мыши, связанными с применением у человека. Как описано в разделе "Примеры" настоящего документа ниже, это гуманизированное антитело (обозначаемое как LT1009 или сонепцизумаб (sonepcizumab)) в действительности демонстрирует активность, превышающую активность исходного антитела (мыши) в моделях заболевания на животных.
3. Определения
Перед подробным описанием настоящего изобретения будут определены некоторые термины, используемые в контексте настоящего изобретения. В дополнение к этим терминам, при необходимости, другие термины определены в других частях настоящего описания. Если в настоящем документе не указано иначе, термины, относящиеся к данной области, используемые в данном описании, будут иметь их признанные в данной области значения. В случае противоречий следует руководствоваться настоящим описанием, включая определения.
"Иммунная молекула" включает любое антитело (Ab) или иммуноглобулин (Ig) и относится к любой форме пептида, полипептида, образованного, смоделированного на основе гена иммуноглобулина или кодируемого геном иммуноглобулина, или к фрагменту такого пептида или полипептида, который способен связывать антиген или эпитоп (см., например, Immunobiology, 5th Edition, Janeway, Travers, Walport, Shlomchiked. (editors), Garland Publishing (2001)). Для настоящего изобретения антиген представляет собой молекулу биологически активного липида.
"Антитело против S1P" или "иммунная молекула, реактивная к S1P" относится к любому антителу или образованной из антитела молекуле, которая связывает S1P. Как будет понятно из этих определений, антитела или иммунные молекулы могут быть поликлональными или моноклональными, и их можно получать различными способами и/или их можно получать из животного, включая человека.
"Биологически активный липид" относится к липидной молекуле передачи сигнала. Как правило, биологически активный липид, когда он оказывает свои эффекты передачи сигнала, находится не в биологической мембране, что означает, что, хотя такой тип липида может существовать в некоторый момент времени в биологической мембране (например, клеточной мембране, мембране клеточной органеллы и т.д.), когда он связан с биологической мембраной, он представляет собой не "биологически активный липид", а вместо этого молекулу "структурного липида". Биологически активные липиды отличаются от структурных липидов (например, мембраносвязанных фосфолипидов) тем, что они опосредуют внеклеточную и/или внутриклеточную передачу сигнала и, таким образом, вовлечены в контроль функции многих типов клеток посредством модулирования дифференцировки, миграции, пролиферации, секреции, выживания и других процессов. In vivo биологически активные липиды могут встречаться во внеклеточных жидкостях, где они могут находиться в комплексе с другими молекулами, например сывороточными белками, такими как альбумин и липопротеины, или в "свободной" форме, т.е. не в комплексе с другими типами молекул. В качестве внеклеточных медиаторов некоторые биологически активные липиды изменяют клеточную передачу сигнала посредством активации мембраносвязанных ионных каналов или сопряженных с G-белком рецепторов, которые, в свою очередь, активируют комплексные системы передачи сигнала, что приводит к изменению клеточной функции или выживанию. В качестве внутриклеточных медиаторов биологически активные липиды могут осуществлять свое действие посредством прямого взаимодействия с внутриклеточными компонентами, такими как ферменты и ионные каналы. Иллюстративные примеры биологически активных липидов включают LPA и S1P.
Термин "лекарственное средство" означает средство, подавляющее ангиогенез и/или неоваскуляризацию, например созревание CNV и BV, отек, сосудистую проницаемость и фиброз, фиброгенез и рубцевание, связанные с основной патологией заболеваний и состояний глаза или являющиеся ее частью.
Термин "комбинированная терапия" относится к терапевтической схеме, которая включает предоставление по меньшей мере двух различных способов терапии для достижения указанного терапевтического эффекта. Например, комбинированная терапия может включать введение двух или более химически отличающихся активных ингредиентов, например антитела против LPA и антитела против S1P. Альтернативно комбинированная терапия может включать введение иммунной молекулы, реактивной к биологически активному липиду, и введение одного или нескольких других химиотерапевтических средств. Альтернативно комбинированная терапия может включать введение антитела против липида вместе с проведением другого лечения, такого как лучевая терапия и/или хирургическая операция. Кроме того, комбинированная терапия может включать введение антитела против липида вместе с одним или несколькими другими биологическими средствами (например, средство против VEGF, TGFβ, PDGF или bFGF), химиотерапевтическими средствами и другими способами лечения, такими как лучевая терапия и/или хирургическая операция. В контексте комбинированной терапии с использованием двух или более химически отличающихся активных ингредиентов, понятно, что активные ингредиенты можно вводить в качестве части одной и той же композиции или в качестве различных композиций. При введении в качестве отдельных композиций композиции, содержащие различные активные ингредиенты, можно вводить в одно и то же время или в разное время, одним и тем же способом или разными способами, с использованием одной и той же схемы дозирования или различных схем дозирования, в зависимости от конкретной потребности и как определит лечащий врач. Аналогично, когда один или несколько типов антител против липидов, например антитело против LPA, отдельно или вместе с одним или несколькими химиотерапевтическими средствами, комбинируют, например, с лучевой терапией и/или хирургической операцией, лекарственное средство(а) можно вводить до или после хирургической операции или лучевой терапии.
"Средство против S1P" относится к любому средству, которое является специфично реактивным к S1P, и включает антитела или образованные из антитела молекулы или не образованные из антитела молекулы, которые связывают S1P и его варианты.
"Гаптен" относится к молекуле, адаптированной для конъюгации с гаптеном, делая, таким образом, гаптен иммуногенным. Иллюстративным неограничивающим классом молекул гаптенов являются белки, примеры которых включают альбумин, гемоцианин лимфы улитки, гемаглютинин, столбнячный и дифтерийный токсоид. Другие классы и примеры молекул гаптенов, пригодных для применения в соответствии с изобретением, известны в данной области. Эти гаптены, а также природные и синтетические гаптены, которые будут открыты или изобретены впоследствии, могут быть адаптированы для применения в соответствии с изобретением.
Термин "химиотерапевтическое средство" означает средства против злокачественной опухоли и другие антигиперпролиферативные средства. Проще говоря, "химиотерапевтическое средство" относится к химическому веществу, предназначенному для уничтожения клеток и тканей. Такие средства включают, но не ограничиваются ими: (1) средства, повреждающие ДНК, и средства, которые ингибируют синтез ДНК: например, антрациклины (например, доксорубицин, донорубицин, эпирубицин), алкилирующие средства (например, бендамустин, бусульфан, карбоплатин, кармустин, цисплатин, хлорамбуцил, циклофосфамид, дакарбазин, гексаметилмеламин, ифосфамид, ломустин, мехлорэтамин, мелфалан, митотан, митомицин, пипоброман, прокарбазин, стрептозоцин, тиотепа и триэтиленмеламин), производные платины (например, цисплатин, карбоплатин, цис-диаминдихлорплатина), ингибиторы теломеразы и топоизомеразы (например, камптосар), (2) средства, деполимеризующие тубулин: например, таксоиды (например, паклитаксел, доцетаксел, BAY 59-8862), (3) антиметаболиты, такие как капецитабин, хлордезоксиаденозин, цитарабин (и его активированная форма, ara-СМР), цитозин арабинозид, дакарбазин, флоксуридин, флударабин, 5-фторурацил, 5-DFUR, гемцитабин, гидроксимочевина, 6-меркаптопурин, метотрексат, пентостатин, триметрексат и 6-тиогуанин, (4) антиангиогенные средства (например, авастин, талидомид, сунитиниб, леналидомид), средства, разрушающие сосуды (например, флавоноиды/флавоны, DMXAA, производные комбретастатина, такие как CA4DP, ZD6126, AVE8062A, и т.д.), (5) биологические вещества, такие как антитела или фрагменты антител (например, Герцептин, Авастин, Панорекс, Ритуксан, Зевалин, Милотарг, Кэмпас, Бексар, Эрбитукс, Луцентис), и (6) эндокринную терапию: например, ингибиторы ароматазы (например, 4-гидроандростендион, экземестан, аминоглютетимид, анастрозол, летозол), антиэстрогены (например, Тамоксифен, Торемифен, Ралоксифен, Фазлодекс), стероиды, такие как дексаметазон, (7) иммуномодуляторы: например, цитокины, такие как IFN-бета и IL2, ингибиторы интегринов, других белков адгезии и матриксных металлопротеиназ, (8) ингибиторы дезацетилазы гистонов, (9) ингибиторы передачи сигнала, такие как ингибиторы тирозинкиназ, такие как иматиниб (Gleevec), (10) ингибиторы белков теплового шока, (11) ретиноиды, такие как полностью транс-ретиноевая кислота, (12) ингибиторы рецепторов факторов роста или самих факторов роста, (13) антимитотические соединения, такие как навелбин, Паклитаксел, таксотер, винбластин, винкристин, виндезин и винорелбин, (14) противовоспалительные средства, такие как ингибиторы СОХ, и (15) регуляторы клеточного цикла, например регуляторы контрольной точки и ингибиторы теломеразы.
Термин "сфинголипид", как используют в настоящем документе, относится к классу соединений, известных в данной области как сфинголипиды, включая, но не ограничиваясь ими, следующие соединения (см. http//www.lipidmaps.org в качестве сайта, содержащего ссылки, указанные посредством заключенных в скобки буквенно-цифровых строк, ниже, причем эти ссылки содержат химические формулы, структурную информацию и т.д. для соответствующих соединений):
Сфингоидные основания [SP01]
Сфинг-4-енины (сфингозины) [SP0101]
Сфинганины [SP0102]
4-Гидроксисфинганины (фитосфингозины) [SP0103]
Гомологи и варианты сфингоидных оснований [SP0104]
1-фосфаты сфингоидных оснований [SP0105]
Лизосфингомиелины и лизогликосфинголипиды [SP0106]
N-метилированные сфингоидные основания [SP0107]
Аналоги сфингоидных оснований [SP0108]
Церамиды [SP02]
N-ацилсфингозины (церамиды) [SP0201]
N-ацилсфинганины (дигидроцерамиды) [SP0202]
N-ацил-4-гидроксисфинганины (фитоцерамиды) [SP0203]
Ацилцерамиды [SP0204]
1-фосфаты церамидов [SP0205]
Фосфосфинголипиды [SP03]
Фосфохолины церамидов (сфингомиелины) [SP0301]
Фосфоэтаноламины церамидов [SP0302]
Фосфоинозитолы церамидов [SP0303]
Фосфоносфинголипиды [SP04]
Нейтральные гликосфинголипиды [SP05]
Простые серии Glc (GlcCer, LacCer и т.д.) [SP0501]
GalNAcb1-3Gala1-4Galb1-4Glc- (серия Globo) [SP0502]
GalNAcb1-4Galb1-4Glc- (серия Ganglio) [SP0503]
Galb1-3GlcNAcb1-3Galb1-4Glc- (серия Lacto) [SP0504]
Galb1-4GlcNAcb1-3Galb1-4Glc- (серия Neolacto) [SP0505]
GalNAcb1-3Gala1-3Galb1-4Glc- (серия Isoglobo) [SP0506]
GlcNAcb1-2Mana1-3Manb1-4Glc- (серия Mollu) [SP0507]
GalNAcb1-4GlcNAcb1-3Manb1-4Glc- (серия Arthro) [SP0508]
Gal- (серия Gala) [SP0509]
Другие [SP0510]
Кислые гликосфинголипиды [SP06]
Ганглиозиды [SP0601]
Сульфогликосфинголипиды (сульфатиды) [SP0602]
Глюкуроносфинголипиды [SP0603]
Фосфогликосфинголипиды [SP0604]
Другие [SP0600]
Основные гликосфинголипиды [SP07]
Амфотерные гликосфинголипиды [SP08]
Арсеносфинголипиды [SP09]
Настоящее изобретение относится к средствам против сфинголипидов S1P, которые пригодны для лечения или профилактики гиперпролиферативных нарушений, таких как злокачественная опухоль, и сердечно-сосудистых или цереброваскулярных заболеваний и нарушений и различных глазных нарушений, как более подробно описано ниже. В частности, изобретение направлено на S1P и его варианты, включая без ограничения сфингозин-1-фосфат [сфинген-1-фосфат; D-эритросфингозин-1-фосфат; сфинг-4-енин-1-фосфат; (Е,2S,3R)-2-амино-3-гидроксиоктадек-4-енокси]фосфоновая кислота (AS 26993-30-6), DHS1P определяют как дигидросфингозин-1-фосфат [сфинганин-1-фосфат; [(2S,3R)-2-амино-3-гидроксиоктадекокси]фосфоновая кислота; D-эритродигидро-D-сфингозин-1-фосфат (CAS 19794-97-9)]; SPC представляет собой сфингозилфосфорилхолин, лизосфингомиелин, сфингозилфосфохолин, сфингозин-фосфорилхолин, этанаманий; 2-((((2-амино-3-гидрокси-4-октадекенил)окси)гидроксифосфинил)окси)-N,N,N-триметилхлорид, (R-(R*,S*-(Е))), хлорид 2-[[(E,2R,3S)-2-амино-3-гидроксиоктадек-4-енокси]гидроксифосфорил]оксиэтил-1-триметилазания (CAS 10216-23-6).
Термин "эпитоп" или "антигенная детерминанта" при использовании в настоящем документе, если нет иных указаний, относится к участку S1P, к которому средство против S1P является реактивным.
Термин "гиперпролиферативное нарушение" относится к заболеваниям и нарушениям, связанным с неконтролируемой пролиферацией клеток, включая, но не ограничиваясь ими, неконтролируемый рост клеток органов и тканей, приводящий к злокачественным опухолям или новообразованиям и доброкачественным опухолям. Гиперпролиферативные нарушения, связанные с эндотелиальными клетками, могут приводить к заболеваниям, связанным с ангиогенезом, таким как ангиомы, эндометриоз, ожирение, старческая дегенерация желтого пятна и различные ретинопатии, а также к пролиферации эндотелиальных клеток и гладкомышечных клеток, которая вызывает рестеноз после стентирования при лечении атеросклероза. Гиперпролиферативные нарушения, вовлекающие фибробласты (например, фиброгенез), включают, но не ограничиваются ими, нарушения с избыточным рубцеванием (например, фиброзом), такие как старческая дегенерация желтого пятна, ремоделирование сердца и недостаточность, связанная с инфарктом миокарда, избыточным заживлением ран, которое обычно происходит после хирургической операции или повреждения, келоидами и фиброидными опухолями и стентированием.
Композиции по изобретению применяют в способах терапии на основе сфинголипидов. "Терапия" относится к профилактике и/или лечению заболеваний, нарушений или физической травмы.
"Сердечно-сосудистая терапия" включает сердечную терапию, а также профилактику и/или лечение других заболеваний, связанных с сердечно-сосудистой системой, таких как заболевание сердца. Термин "заболевание сердца" включает любой тип заболевания, нарушения, травмы или хирургического лечения, которое включает ткань сердца или миокарда. Особый интерес представляют заболевания, которые связаны с гипоксией и/или ишемией ткани миокарда и/или с сердечной недостаточностью. Одним типом заболевания сердца, которое может быть следствием ишемии, является реперфузионное повреждение, например, которое может произойти при использовании в терапии антикоагулянтов, тромболитических средств или средств против стенокардии, или когда сосуды сердца хирургически открывают посредством ангиопластики или посредством пересадки коронарной артерии. Другим типом заболевания сердца, на которое направлено изобретение, является болезнь коронарных артерий (CAD), которая может возникать вследствие артериосклероза, в частности атеросклероза, основной причины ишемии. Симптомами CAD являются стабильная или нестабильная стенокардия, и она может приводить к инфарктам миокарда (MI) и внезапной сердечной смерти. Состояния, представляющие особый интерес, включают, но не ограничиваются ими, ишемию миокарда; острый инфаркт миокарда (AMI); болезнь коронарных артерий (CAD); острый коронарный синдром (ACS); повреждение клеток и тканей сердца, которое может происходить в процессе или вследствие перкутанной реваскуляризации (ангиопластики коронарных артерий) со стентированием или без него; аорто-коронарное шунтирование (CABG) или другие хирургические или медицинские процедуры или способы терапии, которые могут вызывать ишемическое или ишемическое/реперфузионное повреждение у людей; и сердечно-сосудистую травму. Термин "сердечная недостаточность" охватывает острый инфаркт миокарда, миокардит, кардиомиопатию, застойную сердечную недостаточность, септический шок, сердечную травму и идиопатическую сердечную недостаточность. Диапазон ишемических состояний, которые приводят к сердечной недостаточности, называют острым коронарным синдромом (ACS).
Термин "кардиотерапевтическое средство" относится к средству, которое является терапевтическим в отношении заболеваний и нарушений, вызываемых заболеваниями и нарушениями сердца и миокарда или связанных с ними.
"Цереброваскулярная терапия" относится к терапии, направленной на профилактику и/или лечение заболеваний и нарушений, связанных с ишемией и/или гипоксией головного мозга. Особый интерес представляет ишемия и/или гипоксия головного мозга вследствие общей ишемии при заболевании сердца, включающем, но не ограничивающимся этим, сердечную недостаточность.
Термин "метаболит сфинголипида" относится к соединению, из которого получен сфинголипид, а также к соединению, которое образуется при деградации конкретного сфинголипида. Иными словами, "метаболит сфинголипида" представляет собой соединение, которое вовлечено в метаболические каскады сфинголипидов. Метаболиты включают метаболические предшественники и продукты метаболизма. Термин "метаболические предшественники" относится к соединениям, из которых образуются сфинголипиды. Метаболические предшественники, представляющие особый интерес, включают, но не ограничиваются ими, SPC, сфингомиелин, дигидросфингозин, дигидроцерамид и 3-кетосфинганин. Термин "продукты метаболизма" относится к соединениям, которые образуются при деградации сфинголипидов, таким как фосфорилхолин (например, фосфохолин, фосфат холина), жирные кислоты, включая свободные жирные кислоты и гексадеканаль (например, пальмитальдегид).
Как используют в настоящем документе, термин "терапевтический" охватывает полный спектр способов лечения заболевания или нарушения. "Терапевтическое" средство по изобретению может действовать профилактическим или превентивным образом, включая случаи, которые охватывают процедуры, предназначенные для индивидов, которые могут быть идентифицированы, как подверженные риску (фармакогенетика); или смягчающим или излечивающим образом; или оно может действовать, замедляя скорость или степень прогрессирования по меньшей мере одного симптома заболевания или нарушения, подлежащего лечению; или может действовать, минимизируя требуемое время, возникновение или степень дискомфорта или боли, или физические ограничения, связанные с восстановлением после заболевания, нарушения или физической травмы; или их можно использовать в качестве адъювантов для других лекарственных средств и способов лечения.
"Лечение" относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. Индивиды, нуждающиеся в лечении, включают индивидов, которые уже имеют нарушение, а также индивидов, у которых нарушение подлежит профилактике.
Термин "комбинированная терапия" относится к терапевтической схеме, которая включает предоставление по меньшей мере двух различных способов лечения для достижения намеченного терапевтического эффекта. Например, комбинированная терапия может вовлекать введение двух или более химически отличающихся активных ингредиентов, например быстродействующего химиотерапевтического средства и антитела против липида. Альтернативно комбинированная терапия может вовлекать введение антитела против липида и/или одного или нескольких химиотерапевтических средств, отдельно или вместе с предоставлением другого способа лечения, такого как лучевая терапия и/или хирургическая операция. Кроме того, комбинированная терапия может вовлекать введение антитела против липида вместе с одним или несколькими другими биологическими средствами (например, средством против VEGF, TGFβ, PDGF или bFGF), химиотерапевтическими средствами и другим способом лечения, таким как лучевая терапия и/или хирургическая операция. В контексте введения двух или более химически отличающихся активных ингредиентов, понятно, что активные ингредиенты можно вводить в качестве части одной и той же композиции или в качестве различных композиций. При введении в качестве отдельных композиций композиции, содержащие отличающиеся активные ингредиенты, можно вводить одновременно или в разное время, посредством одного и того же или разных способов введения, с использованием одинаковых или различных схем дозирования, в зависимости от потребностей в конкретных условиях и как определяет лечащий врач. Аналогично, когда один или несколько типов антител против липида, например антитело против LPA, отдельно или совместно с одним или несколькими химиотерапевтическими средствами, комбинируют, например, с лучевой терапией и/или хирургической операцией, лекарственное средство(а) можно доставлять до или после хирургической операции или лучевой терапии.
"Монотерапия" относится к схеме лечения на основе доставки одного терапевтически эффективного соединения, вводимого в качестве одной дозы или нескольких доз с течением времени.
"Новообразование" или "злокачественная опухоль" относится к аномальному и неконтролируемому клеточному росту. "Новообразование", или опухоль или злокачественная опухоль, представляет собой аномальную, нерегулируемую и дезорганизованную пролиферацию при росте клеток и, как правило, относится к злокачественной опухоли. Новообразование может быть доброкачественным или злокачественным. Новообразование является злокачественным, или раковым, если оно обладает свойствами деструктивного роста, инвазивности и метастазирования. Инвазивность относится к локальному распространению новообразования посредством инфильтрации или разрушения окружающей ткани, как правило, прорывающегося через базальные мембраны, которые определяют границы тканей, таким образом, часто проникающего в кровеносную систему организма. Метастазы, как правило, относятся к распространению опухолевых клеток через лимфатические или кровеносные сосуды. Метастазы также относятся к миграции опухолевых клеток посредством прямого распространения через серозные полости или субарахноидальное или другие пространства. Посредством процесса метастазирования миграция опухолевых клеток в другие области организма приводит к развитию новообразований в областях, отдаленных от области исходного возникновения.
"Млекопитающее" для целей лечения относится к любому животному, классифицируемому как млекопитающее, включая человека, домашних и сельскохозяйственных животных, и животных зоопарков, спортивных животных или комнатных животных, таких как собаки, лошади, кошки, коровы и т.д. Предпочтительно млекопитающим является человек.
"Нативные антитела" и "нативные иммуноглобулины", как правило, представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 дальтон, которые состоят из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, хотя количество дисульфидных связей варьирует среди тяжелых цепей различных изотипов иммуноглобулинов. Каждая тяжелая и легкая цепи также обладает дисульфидными мостиками, расположенными внутри цепи с равными интервалами. Каждая тяжелая цепь на одном конце обладает вариабельным доменом (VH), за которым следует ряд константных доменов. Каждая легкая цепь обладает вариабельным доменом (VL) на одном конце и константным доменом на другом конце; константный домен легкой цепи расположен параллельно первому константному домену тяжелой цепи, а вариабельный домен легкой цепи расположен параллельно вариабельному домену тяжелой цепи. Полагают, что область контакта между вариабельными доменами легкой цепи и тяжелой цепи формируют конкретные аминокислотные остатки.
Термин "вариабельная" область включает каркасную область и CDR (иначе известные как гипервариабельные области) и относится к тому факту, что последовательности определенных участков вариабельных доменов значительно отличаются среди антител и они задействованы в связывании и специфичности каждого конкретного антитела в отношении его конкретного антигена. Однако вариабельность не является равномерно распределенной на протяжении вариабельных доменов антител. Она сконцентрирована в трех сегментах, называемых гипервариабельными областями в вариабельных доменах как легких цепей, так и тяжелых цепей. Наиболее высококонсервативные участки вариабельных доменов называются каркасными областями (FR). Вариабельные домены нативных тяжелых и легких цепей содержат четыре FR (FR1, FR2, FR3 и FR4 соответственно), главным образом, приминающих конфигурацию β-слоев, соединенных тремя гипервариабельными областями, которые формируют петли, соединяющие структуры β-слоев и в некоторых случаях формирующие их часть. Гипервариабельные области в каждой цепи расположены вместе в непосредственной близости от FR и совместно с гипервариабельными областями другой цепи участвуют в формировании антигенсвязывающего центра антител (см. Kabat et al., Sequences of Protein of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991), стр.647-669). Константные домены не вовлечены непосредственно в связывание антитела с антигеном, но они проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (ADCC).
Как используют в настоящем документе, термин "гипервариабельная область" относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из "определяющей комплементарность области"' или "CDR" (например, остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (Н2) и 95-102 (Н3) в вариабельном домене тяжелой цепи; Kabat et al. (1991), выше), и/или остатки из "гипервариабельной петли" (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) в вариабельном домене тяжелой цепи; Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). Остатки "каркасной области" или "FR" представляют собой такие остатки вариабельного домена, которые отличаются от остатков гипервариабельной области, как определено в настоящем документе.
Расщепление антител папаином приводит к образованию двух идентичных антигенсвязывающих фрагментов, называемых "Fab"-фрагментами, каждого с отдельным антигенсвязывающим центром, и остающегося "Fc"-фрагмента, название которого отражает способность легко кристаллизоваться. Обработка пепсином приводит к образованию F(ab′)2-фрагмента, который обладает двумя антигенсвязывающими центрами и по-прежнему способен перекрестно связывать антиген.
"Fv" представляет собой минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий центр. Этот участок состоит из димера, состоящего из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи с прочной нековалентной связью. Именно в данной конфигурации три гипервариабельные области каждого вариабельного домена взаимодействуют и определяют антигенсвязывающий центр на поверхности димера VH-VL. В совокупности шесть гипервариабельных областей обеспечивают антигенсвязывающую специфичность антитела. Однако даже отдельный вариабельный домен (или половина Fv, содержащая только три гипервариабельные области, специфичные в отношении антигена) обладает способностью распознавать и связывать антиген, хотя и с меньшей аффинностью, чем полный связывающий участок.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab′-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конце тяжелой цепи домена СН1, в том числе одного или нескольких остатка(ов) цистеина из шарнирной области антитела. В настоящем документе Fab′-SH представляет собой обозначение для Fab′, в котором остаток(ки) цистеина константных доменов содержат свободную тиольную группу. F(ab′)2-фрагменты антител исходно были получены в качестве пары Fab′-фрагментов, которые имеют между ними шарнирные остатки цистеина. Также известны другие химические соединения фрагментов антител.
"Легкие цепи" антител любого вида позвоночных могут быть отнесены к одному из двух типов, называемых каппа (к) и лямбда (λ), отчетливо различающихся на основе аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотной последовательности константного домена тяжелых цепей, иммуноглобулины могут быть отнесены к различным классам. В настоящее время существует пять основных классов полноразмерных антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть далее разделены на "подклассы" (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA и IgA2. Константные домены тяжелых цепей, которые соответствуют различным классам антител, называются альфа, дельта, эпсилон, гамма и мю соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
Как используют в настоящем документе, термин "антитело" используют в наиболее широком смысле, и конкретно он включает моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, полиспецифичные антитела (например, биспецифичные антитела), фрагменты антител и связывающие агенты, в которых используются CDR (или их варианты, которые сохраняют антигенсвязывающую активность) исходного антитела. Антитела определяют в настоящем документе, как сохраняющие по меньшей мере один требуемый вид активности исходного антитела. Требуемые виды активности могут включать способность специфично связывать антиген, способность ингибировать пролиферацию in vitro, способность ингибировать ангиогенез in vivo и способность изменять профиль цитокинов in vitro. "Фрагменты антител" содержат участок полноразмерного антитела, предпочтительно содержащий его антигенсвязывающую или вариабельную область. Примеры фрагментов антител включают фрагменты Fab, Fab′, F(ab′)2 и Fv; димерные антитела; линейные антитела; молекулы одноцепочечных антител и полиспецифичные антитела, образованные фрагментом(ами) антитела.
Как используют в настоящем документе, термин "моноклональное антитело" относится к антителу, полученному из совокупности по существу гомогенных антител, например, отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных природных мутаций, которые могут быть представлены в небольших количествах. Моноклональные антитела являются высокоспецифичными и направлены против одного антигенного участка. Более того, в отличие от препаратов традиционных (поликлональных) антител, которые, как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение "моноклональный" указывает на тот признак антитела, что его получают из по существу гомогенной совокупности антител, и не подразумевает того, что антитело должно быть получено каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, могут быть получены способом гибридом, впервые описанным Kohler et al., Nature, 256:495 (1975), или они могут быть получены способами рекомбинантных ДНК (см., например, патент США №4816567). "Моноклональные антитела" также могут быть получены из фаговых библиотек антител с использованием способов, описанных, например, в Clackson et al., Nature, 352:624-628 (1991) и Marks et al., J. Mol. Biol, 222:581-597 (1991).
В настоящем документе моноклональные антитела конкретно включают "химерные" антитела (иммуноглобулины), в которых участок тяжелой и/или легкой цепи является идентичным или гомологичным соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (патент США №4816567; и Morrison et al., Proc. Natl. Acad. Sci USA, 81:6851-6855 (1984)).
"Гуманизированные" формы не являющихся человеческими антител (например, грызунов) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. В основном гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области видов, не относящихся к человеку (донорное антитело), таких как мышь, крыса, кролик или не относящиеся к человеку приматы, которые обладают требуемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками, не являющимися человеческими. Более того, гуманизированные антитела могут содержать остатки, которые отсутствуют в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения параметров антитела. Как правило, гуманизированное антитело содержит по существу все по меньшей мере из одного и, как правило, из двух вариабельных доменов, в которых все или по существу все гипервариабельные области соответствуют гипервариабельным областям иммуноглобулина, не являющегося человеческим, и все или по существу все FR представляют собой FR из последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно содержит по меньшей мере участок константного домена (Fc) иммуноглобулина, как правило, константного домена иммуноглобулина человека. Для более подробной информации см. Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992) и Hansen, WO 2006105062.
"Одноцепочечные Fv" или "scFv"-фрагменты антител содержат VH и VL-домены антитела, и эти домены находятся в единой полипептидной цепи. Как правило, полипептид Fv также необязательно содержит полипептидный линкер между VH- и VL-доменами, который обеспечивает возможность формирования в Fv структуры, требуемой для связывания антигена. Для обзора sFv см. Pluckthun in The Pharmacology of Monoclonal Antibodies, vol.113, Rosenburg and Moore eds. Springer-Verlag, New York, pp.269-315 (1994).
Термин "димерные антитела" относится к небольшим фрагментам антител с двумя антигенсвязывающими центрами, где фрагменты содержат 'вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH-VL). С использованием линкера, который является слишком коротким для возможности спаривания двух доменов одной цепи, инициируют спаривание доменов с комплементарными доменами другой цепи и формирование двух антигенсвязывающих центров. Димерные антитела более подробно описаны, например, в европейском патенте 404097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci USA, 90:6444-6448 (1993).
Выражение "линейные антитела", как используют в настоящем документе, относится к антителам, описанным в Zapata, et al. Protein Eng. 8(10):1057-1062 (1995). В кратком изложении, эти антитела содержат пару тандемных Fd-сегментов (VH-CH1-VH-CH1), которые образуют пару антигенсвязывающих участков. Линейные антитела могут быть биспецифичными или моноспецифичными.
В настоящем документе "вариант" антитела против сфинголипида относится к молекуле, аминокислотная последовательность которой отличается от аминокислотной последовательности "исходного" антитела против сфинголипида вследствие вставки, делеции и/или замены одного или нескольких аминокислотного остатка(ов) в последовательности исходного антитела и которая сохраняет по меньшей мере один требуемый вид активности исходного связывающего антитела. Требуемые виды активности могут включать способность специфично связываться с антигеном, способность ингибировать пролиферацию in vitro, способность ингибировать ангиогенез in vivo и способность изменять профиль цитокинов in vitro. В одном варианте осуществления вариант содержит одну или несколько аминокислотную замену (замен) в одной или нескольких гипервариабельной области(ях) исходного антитела. Например, вариант может содержать по меньшей мере одну, например от приблизительно одной до приблизительно десяти и предпочтительно от приблизительно двух до приблизительно пяти, замен в одной или нескольких гипервариабельных областях исходного антитела. Как правило, вариант будет обладать аминокислотной последовательностью, обладающей по меньшей мере приблизительно 50% идентичностью аминокислотной последовательности с последовательностями вариабельного домена тяжелой или легкой цепи исходного антитела, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью последовательности. Идентичность или гомологию в отношении этой последовательности определяют в настоящем документе как процентное количество аминокислотных остатков в последовательности-кандидате, которые идентичны остаткам исходного антитела, после выравнивания последовательностей и внесения разрывов, если необходимо, для достижения максимальной процентной идентичности последовательностей. Ни одно из N-концевых, С-концевых или внутренних удлинений, делеций или вставок в последовательности антитела не следует истолковывать, как влияющие на идентичность или гомологию последовательности. Вариант сохраняет способность связывать сфинголипид и предпочтительно обладает требуемыми видами активности, которые превосходят виды активности исходного антитела. Например, вариант может обладать более сильной аффинностью связывания, усиленной способностью снижать ангиогенез и/или останавливать прогрессирование опухоли. Для анализа таких требуемых свойств (например, сниженной иммуногенности, более длительного времени полужизни, усиленной стабильности, усиленной эффективности) следует сравнивать, например, форму Fab варианта с формой Fab исходного антитела или полноразмерную форму варианта с полноразмерной формой исходного антитела, поскольку было выявлено, что формат антитела против сфинголипида влияет на его активность в анализах биологической активности, описанных в настоящем документе. Особый интерес может представлять собой вариант антитела, который проявляет по меньшей мере приблизительно 5%, предпочтительно по меньшей мере приблизительно 10%, 25%, 59% или более по меньшей мере одного требуемого вида активности. Предпочтительный вариант представляет собой вариант, который обладает улучшенными биофизическими свойствами при измерении in vitro или улучшенными биологическими видами активности при измерении in vitro или in vivo по сравнению с исходным антителом.
"Исходное" антитело в настоящем документе представляет собой антитело, которое кодируется аминокислотной последовательностью, используемой для получения варианта. Предпочтительно исходное антитело обладает каркасной областью человека и, при наличии, обладает константной областью(ями) антитела человека. Например, исходное антитело может представлять собой гуманизированное антитело или антитело человека.
"Выделенное" антитело представляет собой антитело, которое идентифицировано и выделено и/или извлечено из компонента его естественных окружающих условий. Загрязняющие компоненты его естественных окружающих условий представляют собой вещества, которые препятствуют применению антитела для диагностики или лечения, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело очищают (1) до более чем 95% по массе антитела, как определяют способом Лоури, и наиболее предпочтительно до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности при SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием кумасси голубого или предпочтительно окрашивания серебром. Выделенное антитело включает антитело в рекомбинантных клетках in situ, поскольку в этом случае отсутствует по меньшей мере один компонент условий естественного окружения антитела. Однако, как правило, выделенное антитело будет получено посредством по меньшей мере одной стадии очистки.
Как используют в настоящем документе, "метка" относится к поддающемуся детекции соединению или композиции, которые конъюгированы прямо или непрямо с антителом. Метка может быть детектируемой самостоятельно (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментной метки, она может катализировать химическое изменение соединения или композиции субстрата, которое является детектируемым.
Под "твердой фазой" подразумевают неводную матрицу, на которую антитело по настоящему изобретению может прикрепляться или на которой антитело или другой связывающий реагент против S1P могут быть иным образом иммобилизованы. Примеры твердой фазы, охватываемые в настоящем документе, включают твердую фазу, образованную частично или полностью из стекла (например, стекла с контролируемым размером пор), полисахаридов (например, агарозы), полиакриламидов, полистирола, поливинилового спирта и силиконов. В определенных вариантах осуществления, в зависимости от контекста, твердая фаза может включать лунку планшета для анализа, в то время как в других вариантах осуществления она представляет собой колонку для очистки (например, колонку для аффинной хроматографии). Также этот термин включает дисперсную твердую фазу из дискретных частиц, такую как твердая фаза, описанная в патенте США №4275149.
"Липосома" представляет собой небольшую везикулу, состоящую из различных типов липидов, фосфолипидов и/или поверхностно-активного вещества, которые пригодны для доставки лекарственного средства (такого как антитела против сфинголипида, описанные в настоящем документе, и, необязательно, химиотерапевтическое средство) у млекопитающего. Компоненты липосомы обычно образуют двухслойную структуру, сходную с размещением липидов биологических мембран. "Выделенная" молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая идентифицирована и отделена по меньшей мере от одной загрязняющей молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в природном источнике нуклеиновой кислоты антитела. Выделенная молекула нуклеиновой кислоты находится в другой форме или условиях, чем форма и условия, в которых она встречается в природе. Выделенные молекулы нуклеиновой кислоты, таким образом, отличаются от молекулы нуклеиновой кислоты, существующей в клетках в природе. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно экспрессируют антитело, где, например, молекула нуклеиновой кислоты находится в области хромосомы, отличающейся от области хромосомы клеток в природе.
Выражение "последовательности контроля" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной с ними кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, которые пригодны для прокариот, например, включают промотор, необязательно последовательность оператора и участок связывания рибосом. Известно, что в эукариотических клетках используются промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота "функционально связана", когда она помещена в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторной лидерной последовательности функционально связана с ДНК полипептида, если она экспрессируется в качестве пребелка, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосом функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Как правило, "функционально связанный" означает, что молекулы нуклеиновой кислоты, подлежащие связыванию, являются соседними, и в случае секреторной лидерной последовательности соседними и находящимися в рамке считывания. Однако энхансеры не обязательно должны быть соседними. Связывание проводят лигированием в удобных участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой.
Как используют в настоящем документе, выражения "клетка", "клеточная линия" и "клеточная культура" используют взаимозаменяемо, и все такие определения включают потомство. Таким образом, слова "трансформанты" и "трансформированные клетки" включают первичную клетку и культуры, образованные из нее, независимо от количества пересевов. Также понятно, что все потомство может не быть точно идентичным с точки зрения содержащейся ДНК вследствие намеренных или случайных мутаций. Включается мутантное потомство, которое обладает той же функцией или биологической активностью, в отношении которой проводят скрининг в исходно трансформированной клетке. Когда подразумеваются отличающиеся определения, это будет очевидно из контекста.
"Патентоспособная" композиция, процесс, устройство или изделие в соответствии с изобретением означает, что объект удовлетворяет всем предусмотренным законодательством требованиям патентоспособности во время проведения анализа. Например, в отношении новизны, неясности или сходных с ними, если последующее исследование выявит, что один или несколько пунктов формулы изобретения охватывает один или несколько вариантов осуществления, которые будут опровергать новизну, неочевидность, и т.д., пункт(ы) формулы изобретения, ограниченные определением "патентоспособных" вариантов осуществления, конкретно исключают непатентоспособный вариант(ы) осуществления. Также прилагаемую формулу изобретения следует интерпретировать как обеспечивающую наиболее широкий разумный объем, а также как сохраняющую ее действительность. Более того, формулу изобретения следует интерпретировать таким образом, чтобы (1) сохранялась ее действительность и (2) обеспечивалась наиболее широкая разумная интерпретация в обстоятельствах, если одно или несколько предусматриваемых законом требований в отношении патентоспособности изменены или если меняются стандарты, касающиеся оценки того, является ли конкретное предусматриваемое законом требование в отношении патентоспособности удовлетворительным, с того времени, когда данная заявка подана или опубликована в качестве патента до того времени, когда рассматривается применимость одного или нескольких из прилагаемых пунктов формулы изобретения.
Термин "фармацевтически приемлемая соль" относится к солям, которые сохраняют биологическую эффективность и свойства веществ и соединений по этому изобретению и которые не являются биологически или иным образом нежелательными. Во многих случаях вещества и соединения по этому изобретению способны образовывать кислые и/или основные соли благодаря присутствию заряженных групп, например заряженных аминогрупп и/или карбоксильных групп или групп, сходных с ними. Фармацевтически приемлемые кислотно-аддитивные соли можно получать из неорганических и органических кислот, в то время как фармацевтически приемлемые основно-аддитивные соли можно получать из неорганических и органических оснований. Для обзора фармацевтически приемлемых солей см. Berge, et al. (1977) J. Pharm. Sci., vol.66, 1-19.
"Множество" означает более одного.
Термины "отделенный", "очищенный", "выделенный" и т.п. означают, что один или несколько компонентов образца, содержащегося в емкости для содержания образца, физически удаляют, или физически удалены, или разбавлены в присутствии одного или нескольких других компонентов образца, присутствующих в емкости. Компоненты образца, которые можно удалять или разбавлять в ходе стадии разделения или очистки, включают продукты химических реакций, не вступившие в реакцию химические вещества, белки, углеводы, липиды и не связавшиеся молекулы.
Термин "тип" используют в настоящем документе в различных контекстах, например в контексте конкретного типа химиотерапевтического средства. В каждом контексте термин относится к совокупности химически неопределенных молекул разновидности, к которой относится конкретный контекст.
"Специфично ассоциировать" и "специфичная ассоциация" и т.п. относятся к специфичному неслучайному взаимодействию между двумя молекулами, которое зависит от наличия структурных, гидрофобных/гидрофильных и/или электростатических признаков, которые обеспечивают соответствующие химические или молекулярные взаимодействия между молекулами.
"Индивид" или "пациент" относится к животному, нуждающемуся в лечении, которое может быть обеспечено молекулами по изобретению. Животные, которых можно лечить в соответствии с изобретением, включают позвоночных, причем особенно предпочтительными их примерами являются млекопитающие, такие как быки, собаки, лошади, кошки, овцы, свиньи и приматы (включая человека и не относящихся к человеку приматов).
"Терапевтически эффективное количество" (или "эффективное количество") относится к количеству активного ингредиента, например вещества по изобретению, достаточному для обеспечения лечения при введении субъекту или пациенту. Таким образом, терапевтически эффективное количество композиции по изобретению может легко определить специалист в данной области. В контексте глазной терапии "терапевтически эффективное количество" представляет собой количество, которое приводит к объективно определяемому изменению одного или нескольких параметров, связанных с лечением заболевания или состояния глаз, включая повышение или снижение экспрессии одного или нескольких генов, коррелирующей с заболеванием или состоянием глаз, индукцию апоптоза или других каскадов клеточной гибели, клиническое улучшение симптомов, снижение нарушенной неоваскуляризации или воспаления и т.д. Безусловно, терапевтически эффективное количество будет варьировать, в зависимости от конкретного индивида и состояния, подлежащего лечению, массы и возраста индивида, тяжести болезненного состояния, конкретного выбранного соединения, соблюдаемой схемы дозирования, времени введения, способа введения и сходных с ними, все из которых может легко определить специалист в данной области. Будет понятно, что в контексте комбинированной терапии терапевтически эффективное количество конкретного активного ингредиента может отличаться от терапевтически эффективного количества активного ингредиента при введении в качестве монотерапии (т.е. терапевтической схемы, в которой используется только одно химическое соединение в качестве активного ингредиента).
Термин "лечение" или "проведение лечения" заболевания или нарушения включает предотвращение заболевания или нарушения или защиту от него (т.е. обеспечение того, что клинические симптомы не развиваются); ингибирование заболевания или нарушения (т.е. остановку или подавление развития клинических симптомов); и/или смягчение заболевания или нарушения (т.е. обеспечение регрессии клинических симптомов). Как будет понятно, не всегда можно отличить "профилактику" и "подавление" заболевания или нарушения, поскольку конечное индуктивное событие или события могут быть неизвестными или латентными. Таким образом, будут подразумевать, что термин "профилактика" составляет тип "лечения", которое охватывает как "предотвращение", так и "подавление". Таким образом, термин "лечение" включает "профилактику".
Термин "терапевтическая схема" означает любое лечение заболевания или нарушения с использованием химиотерапевтических лекарственных средств, лучевой терапии, хирургической операции, генной терапии, ДНК-вакцин и терапии, терапии на основе антисмысловых молекул, включая терапию siRNA, антиангиогенной терапии, иммунотерапии, трансплантации костного мозга, аптамеров и других биологических веществ, таких как антитела и варианты антитела, рецепторы-ловушки и другие лекарственные средства на основе белков.
Сущность изобретения
Это изобретение относится к патентоспособным гуманизированным средствам против сфинголипидов, включая антитела и варианты антител против сфинголипидов с требуемыми свойствами с точки зрения терапевтической и/или диагностической перспективы, включая высокую аффинность связывания со сфинголипидами, способность связывать и нейтрализовывать сфингозин-1-фосфат (S1P), в частности, в физиологическом контексте (например, в живой ткани, крови и т.д.) и в физиологических условиях, а также к изоформам, вариантам, изомерам и родственным соединениям. В частности, изобретение относится к антителам, в частности моноклональным антителам, более конкретно к гуманизированным моноклональным антителам и их вариантам, направленным против S1P. Такие антитела и варианты предпочтительно включены в фармацевтические композиции, пригодные для введения индивидам, о которых известно или предполагают, что они нуждаются в лечении такими соединениями. В дополнение к композициям изобретение также относится к наборам, включающим такие композиции, способам получения таких антител против S1P и вариантов и способам лечения с использованием таких средств.
В одном варианте осуществления предусмотрены гуманизированные антитела или их антигенсвязывающие фрагменты, которые являются специфично реактивными к S1P и содержат по меньшей мере одну тяжелую цепь и по меньшей мере одну легкую цепь. Каждая тяжелая цепь содержит вариабельный домен тяжелой цепи, содержащий: последовательность аминокислотных остатков, имеющую аминокислотную последовательность SEQ ID NO:27; или аминокислотную последовательность, имеющую по меньшей мере 50% идентичность последовательности, предпочтительно по меньшей мере 80% идентичность последовательности с ней, и каждая легкая цепь содержит вариабельный домен легкой цепи, содержащий последовательность аминокислотных остатков, имеющую аминокислотную последовательность SEQ ID NO:29; или аминокислотную последовательность, имеющую по меньшей мере 50% идентичность последовательности, предпочтительно по меньшей мере 80% идентичность последовательности с ней.
Каждая тяжелая цепь этих гуманизированных антител или их антигенсвязывающих фрагментов может содержать первую последовательность аминокислотных остатков последовательности DHTIH (SEQ ID NO:13), вторую последовательность аминокислотных остатков AISPRHDITKYNEMFRG (SEQ ID NO:18) и/или третью последовательность аминокислотных остатков GGFYGSTIWFDF (SEQ ID NO:15), а каждая легкая цепь содержит первую последовательность аминокислотных остатков последовательности ITTTDIDDDMN (SEQ ID NO:10), вторую последовательность аминокислотных остатков последовательности EGNILRP (SEQ ID NO:11) и/или третью последовательность аминокислотных остатков последовательности LQSDNLPFT (SEQ ID NO:12).
Гуманизированные антитела по изобретению могут содержать полноразмерную аминокислотную последовательность тяжелой цепи SEQ ID NO:31 и полноразмерную аминокислотную последовательность легкой цепи SEQ ID NO:32 или последовательности, имеющие по меньшей мере 80% идентичность им, включая аминокислотную последовательность тяжелой цепи SEQ ID NO:35, в которой отсутствует С-концевой лизин.
В соответствии с изобретением гуманизированное антитело может иметь те же последовательности вариабельных доменов тяжелой и легкой цепей, что и последовательности вариабельных доменов тяжелой и легкой цепей, кодируемые вектором рАТН1009 в АТСС под регистрационным №РТА-8421 или РТА-8422.
Кроме того, предусмотрены фармацевтические композиции, содержащие гуманизированные антитела или антигенсвязывающие фрагменты по изобретению, нуклеиновые кислоты, кодирующие гуманизированные антитела или антигенсвязывающие фрагменты по изобретению, клетки-хозяева, трансфицированные указанными нуклеиновыми кислотами, и способы лечения заболеваний или нарушений, связанных с аномальным уровнем S1P, с применением композиций по изобретению.
Эти и другие аспекты и варианты осуществления изобретения более подробно рассмотрены в разделах, которые следуют далее. Указанные выше и другие аспекты изобретения станут более очевидными из следующего подробного описания, прилагаемых рисунков и формулы изобретения. Несмотря на то что способы и материалы, сходные или эквивалентные способам и материалам, описанным в настоящем документе, можно использовать на практике или при тестировании настоящего изобретения, пригодные способы и материалы описаны ниже. Кроме того, материалы, способы и примеры, представленные ниже, являются только иллюстративными и не предназначены для ограничения.
Краткое описание чертежей
Данная заявка содержит по меньшей мере одну фигуру, выполненную в цвете. Копии данной заявки с цветным рисунком(ами) будут предоставлены при запросе и оплате необходимого взноса. Краткое описание каждой из фигур представлено ниже.
Фиг.1. Фиг.1 имеет две панели, А и В. На панели А графически проиллюстрированы результаты конкурентного ELISA для S1P, SPH, LPA, SPC и других структурно сходных биологических липидов, конкурирующих за конъюгированное с биотином моноклональное антитело против S1P. Эти результаты указывают на то, что антитело является специфичным и чувствительным к S1P и не распознает структурно сходные биологически активные липиды. Как описано в примере 1, ниже, связанное антитело было детектировано вторым антителом, специфичным к IgG мыши или человека, конъюгированным с HRP. Хромогенные реакции определяли и записывали в качестве оптической плотности (OD). Концентрация липидов, используемых для конкуренции, указана на оси X. Нельзя было выявить взаимодействия вторичного антитела с матрицей, покрытой S1P отдельно (данные не представлены). На панели В представлены структуры биологически активных липидов, сходных с S1P, которые приведены на панели А.
Фиг.2. На данной фигуре представлены связывающие свойства нескольких химерных и рекомбинантных вариантов гуманизированного антитела против S1P. Связывание с S1P химерного антитела (рАТН10 + рАТН50) сравнивали в анализе связывания ELISA с двумя вариантами гуманизированного моноклонального антитела против S1P (рАТН201 + рАТН308) и (рАТН201 + рАТН309). рАТН308 представляет собой гуманизированную легкую цепь с пятью возвратными мутациями мыши, и рАТН309 представляет собой гуманизированную легкую цепь с тремя возвратными мутациями в каркасной области. Гуманизированная тяжелая цепь (рАТН201) содержит только одну возвратную мутацию мыши в каркасной области.
На фиг.3 представлен график, на котором показано, что SPHINGOMAB является высокоспецифичным к S1P. На графике, данные для которого были получены с использованием конкурентного ELISA, продемонстрирована специфичность SPHINGOMAB к S1P по сравнению с другими биологически активными липидами. SPHINGOMAB не демонстрировало перекрестной реактивности к сфингозину (SPH), прямому метаболическому предшественнику S1P, или лизофосфатидной кислоте (LPA), важной молекуле внеклеточной передачи сигнала, которая структурно и функционально сходна с S1P. SPHINGOMAB не распознавал другие структурно сходные липиды и метаболиты, включая церамид-1-фосфат (G1P), дигидросфингозин (DH-SPH), фосфатидилсерин (PS), фосфатидилэтаноламин (РЕ) или сфингомиелин (SM). SPHINGOMAB не реагировал перекрестно с дигидросфингозин-1-фосфатом (DH-S1P) и, в меньшей степени, сфингозилфорилхолином (SPC). Аффинность (Kd) SPHINGOMAB к S1P составляет менее 100 пМ, что значительно превышает большинство терапевтических антител, в частности других молекулярных губок.
Фиг.4. Фиг.4 имеет две части, А и В. Эксперименты, в которых получены данные, представленные на этой фигуре, подробно описаны в примере 4, ниже. В кратком изложении, эти данные показывают, что SPHINGOMAB снижает CNV и рубцевание при глазных повреждениях. Мышам вводили SPHINGOMAB или совпадающее по изотипу неспецифическое моноклональное антитело. Повреждения CNV индуцировали лазерной перфорацией мембраны Бруха. Представлены графики и репрезентативные изображения очагов повреждения из каждой группы лечения, окрашенные конъюгированным с родамином агглютинином I R.communis в отношении васкуляризации (А) или трихроматичной окраской Массона в отношении образования коллагеновых рубцов (В). На фиг.4А показано, что в модели образования повреждения CNV на мышах SPHINGOMAB значительно ослабляет хориоидальную неоваскуляризацию через 14 и 28 суток после индуцированной лазером перфорации мембран Бруха. На фиг.4В показано, что SPHINGOMAB значительно снижает фиброз, связанный с образованием повреждения CNV через 28 суток после индуцированной лазером перфорации мембраны Бруха.
Фиг.5. Фиг.5 имеет две панели, А и В. На панели А показано, что S1P обеспечивает неоваскуляризацию посредством индукции образования трубки HUVEC и миграции, которые снижаются посредством SPHINGOMAB. На панели 5А показаны четыре микрофотографии HUVEC, посеянные на матригель и инкубированные в течение 6 ч для оценки образования трубки. На панели 5В представлены данные для HUVEC, которые обрабатывали 1 мкМ S1P ± SPHINGOMAB (1 мкг/мл) в течение 6 ч в камере для инвазии из матригеля. Количество клеток, которые мигрировали в мембрану из матригеля, подсчитывали в пяти независимых полях.
Фиг.6. Фиг.6 содержит несколько фотографий (А) и графиков (В и С) для экспериментов, описанных в примере 6, ниже, которые проводили с использованием SPHINGOMAB. SPHINGOMAB нейтрализует индуцируемую посредством SIP, VEGF и bFGF неоваскуляризацию. На фиг.6А представлены фотографии нескольких репрезентативных окрашенных FITC кровеносных сосудов из срезов вставок матригеля±указанные факторы роста. На фиг.6В показано, что S1P стимулирует инфильтрацию эндотелиальных клеток (ЕС). На фиг.6С представлено количественное определение относительной флуоресценции из вставок матригеля, стимулированных VEGF или bFGF, в качестве индикатора неоваскуляризации. Эффекты S1P, VEGF и bFGF ингибировались, когда мышам системно вводили 1 или 25 мг/кг SPHINGOMAB.
Фиг.7. На фиг.7 представлено 5 графиков, обозначенных А-Е, и две цветные фотографии. Эти данные были получены с использованием моноклонального антитела против S1P SPHINGOMAB. См. пример 7, ниже, для подробного описания эксперимента. В кратком изложении, эти данные показывают, что SPHINGOMAB нейтрализует стимулированное S1P рубцевание. В этих экспериментах фибробласты не подпитывали сывороткой, а затем обрабатывали 0, 0,1, 0,5 или 1 мкм S1P +/- 1 мкг/мл SPHINGOMAB в течение 12-24 ч. Данные демонстрируют стимулированную S1P пролиферацию фибробластов Swiss 3Т3, измеренную по включению 3Н-тимидина (А), миграцию фибробластов сердца мыши в скарификационном анализе. (В), экспрессию гена коллагена (относительная флуоресценция) в выделенных фибробластах сердца из трансгенных мышей, экспрессирующих коллаген-GFP (С), и дифференцировку клеток WI-38 в миофибробласты, определяемую по сниженной клеточной пролиферации и повышенной экспрессии α-SMA (D). SPHINGOMAB нейтрализовало каждый из этих эффектов S1P. SPHINGOMAB снижало околососудистый фиброз in vivo в модели длительного инфаркта миокарда на мышах (Е).
Фиг.8. Фиг.8 имеет три панели, 8А, 8В и 8С. Эти данные показывают, что S1P обеспечивает трансформацию глазных эпителиальных клеток и фибробластов в сократимые образующие рубцовую ткань миофибробласты. Как описано в примере 8, ниже, исследовали эффекты S1P на трансформацию в миофибробласты нескольких глазных клеточных линий человека. Было выявлено, что S1P стимулирует продукцию α-гладкомышечного актина (α-SMA; маркера миофибробластов) в клетках пигментного эпителия сетчатки человека (фиг.8А) и фибробластах конъюнктивы человека (фиг.8В). Эти данные впервые демонстрируют, что S1P относится к факторам, которые обеспечивают трансформацию эпителиальных клеток глаза и фибробластов в сократимые продуцирующие рубцовую ткань миофибробласты. Также исследовали эффекты S1P на экспрессию ингибитора активатора плазминогена (PAI-1) в фибробластах конъюнктивы человека. Повышенная экспрессия PAI-1 коррелирует со снижением протеолитической деградации соединительной ткани и активируется в связи с некоторыми фиброзными заболеваниями, которые вовлекают усиленное рубцевание. Как показано на фиг.8С, S1P стимулирует экспрессию PAI-1 зависимым от дозы образом.
Фиг.9. На фиг.9 представлены две столбиковые диаграммы, А и В, на которых показаны экспериментальные данные, полученные с использованием моноклонального антитела против S1P, называемого SPHINGOMAB. SPHINGOMAB снижал инфильтрацию иммунных клеток в рану in vivo. Мышей подвергали MI, вводили физиологический раствор или 25 мг/кг SPHINGOMAB через 48 после хирургической операции, а затем умерщвляли на 4 сутки. SPHINGOMAB снижал инфильтрацию макрофагов (А) и тучных клеток (В) в рану. Данные представлены в качестве кратности снижения относительно значений при введении физиологического раствора.
Фиг.10. Фиг.10 имеет две панели, 10А и 10В. На каждой панели показана карта клонирующего вектора для экспрессии VL- и VH-доменов моноклонального антитела мыши против S1P. На фиг.10А представлена карта вектора pKN100 для клонирования VL-домена. На фиг.10B представлена карта вектора pG1D200 для клонирования VH-домена.
Фиг.11. На фиг.11 представлены данные, показывающие связывающие свойства нескольких антител мыши против S1P, химерных и рекомбинантных гуманизированных антител против S1P. Связывание S1P с антителом мыши (muMAbS1P; кривая, образованная частными значениями в виде прямоугольников) и химерным антителом (chMAbS1P; кривая, образованная частными значениями в виде вертикального прямоугольника) сравнивали в анализе связывания ELISA с первым вариантом гуманизированного антитела (рАТН200 + рАТН300; кривая, образованная частными значениями в виде перевернутых треугольников).
Фиг.12. Фиг.12 имеет две панели, А и В, на которых показаны данные клеточных анализов in vitro, проведенных с использованием нескольких вариантов гуманизированного моноклинального антитела. На панели А показано, что гуманизированное mAb способно предотвращать защиту с помощью S1P клеток SKOV3 от индуцированного посредством Taxol апоптоза. Как описано в примере 16, ниже, клетки SKOV3 обрабатывали в течение 48 ч 500 нМ Taxol (Tax) в присутствии или в отсутствие 500 нМ S1P с huMAbHCLC3 (309), huMAbHCLC5 (308), muMAbSlP (muMAb) или неспецифическим IgG1 (NS) в концентрации 1 мкг/мл. Значения представляют собой средние значения ±SEM (n=3), и для каждого частного значения проводили три исследования. "NT" означает "без обработки" и "Veh" означает "только носитель". На панели В представлена секреция IL-8 в клетках рака яичника (OVCAR3), обработанных S1P и одним из нескольких различных моноклональных антител против S1P или контрольным моноклональным антителом. В экспериментах, подробно описанных в примере 16, ниже, 100000 OVCAR3 клеток/лунка подвергали голоданию в течение ночи и к культуральной среде добавляли 1 мкМ S1P отдельно или предварительно инкубированный с 1 мкг/мл неспецифического антитела (NS), рАТН201 + рАТН309 (LC3), рАТН201 + рАТН308 (LC5), рАТН207 + рАТН309 (cysLC3), pATH207 + pATH308 (cysLC5) и 0,1 мкг/мл (М0,1), 1 мкг/мл (Ml) или 10 мкг/мл (М10) антитела мыши против S1P. После инкубации в течение 22 часов клеточные супернатанты собирали и секрецию IL-8 измеряли посредством ELISA с использованием набора R&D system Quantikine human CXCL8/IL-8. На фигуре "NT" относится к необработанным клеткам.
Фиг.13. На фиг.13 представлена эффективность in vivo нескольких вариантов моноклонального антитела человека по сравнению с моноклональным антителом мыши против S1P и контролями в модели CNV на животных. Как описано в примере 17, ниже, в этих экспериментах мышам вводили 0,5 мкг два раза (сутки 0 и сутки 6) моноклонального антитела мыши против S1P (Mu), несколько гуманизированных вариантов моноклонального антитела против S1P (т.е. варианты LC3, LC5, HCcysLC3 и HCcysLC5), или неспецифического моноклонального антитела (NS) посредством введения в стекловидное тело, а затем проводили перфорацию их мембраны Бруха. Мышей умерщвляли через 14 суток после лазерной хирургической операции. Комплексы склера-RPE-сосудистая оболочка вырезали и окрашивали конъюгированным с родамином антителом против агглютинина I R.communis. Объемы повреждений CNV представлены в качестве средних значений ±SEM.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Соединения
Настоящее изобретение относится к определенным средствам против S1P, в частности к средствам, которые представляют собой иммунные молекулы, включая антитела, которые специфично реагируют с биологически активным липидом S1P; иными словами, биологически активным липидом, с которым реагирует средство против S1P, является S1P.
Молекулы антител или иммуноглобулины представляют собой крупные молекулы гликопротеинов с молекулярной массой приблизительно 150 кДа, как правило, состоящие из двух различных типов полипептидной цепи. Одна полипептидная цепь, называемая "тяжелой" цепью (Н), имеет массу приблизительно 50 кДа. Другой полипептид, называемый "легкой" цепью (L), обладает массой приблизительно 25 кДа. Каждая молекула иммуноглобулина, как правило, состоит из двух тяжелых цепей и двух легких цепей. Две тяжелые цепи связаны друг с другом дисульфидными связями, количество которых варьирует между тяжелыми цепями различных изотипов иммуноглобулинов. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью. В любой природной молекуле антитела две тяжелые цепи и две легкие цепи являются идентичными, обладая двумя идентичными антигенсвязывающими областями, и, таким образом, их называют двухвалентными, т.е. обладающими способностью одновременно связываться с двумя идентичными молекулами.
"Легкие" цепи молекул антитела любых видов позвоночных могут быть отнесены к двум четко отличающимся типам, каппа (k) и лямбда (l), на основе аминокислотных последовательностей их константных доменов. Соотношение двух типов легких цепей варьирует от вида к виду. В качестве примера, среднее соотношение k и l составляет 20:1 у мышей, в то время как у человека оно составляет 2:1, и у крупного рогатого скота оно составляет 1:20.
"Тяжелые" цепи молекул антитела из любых видов позвоночных могут быть отнесены к одному из пяти четко отличающихся типов, называемых изотипами, на основе аминокислотных последовательностей их константных доменов. Некоторые изотипы имеют несколько подтипов. Пять основных классов иммуноглобулинов представляют собой иммуноглобулин М (IgM), иммуноглобулин D (IgD), иммуноглобулин G (IgG), иммуноглобулин A (IgA) и иммуноглобулин Е (IgE). IgG представляет собой наиболее широко распространенный изотип и он имеет несколько подклассов (IgG1, 2, 3 и 4 у человека). Fc-фрагмент и шарнирные области отличаются в антителах различных изотипов, таким образом определяя их функциональные свойства. Однако общая организация доменов сходна во всех изотипах.
Термин "вариабельная область" относится к N-концевому участку молекулы антитела или ее фрагмента. Как правило, каждая из четырех цепей имеет вариабельную (V) область в ее N-концевой части, которая обеспечивает антигесвязывающую область, и константную (С) область, которая определяет изотип. Легкие цепи связаны с тяжелыми цепями посредством множества нековалентных взаимодействий и дисульфидных связей, и V-области тяжелой и легкой цепей образуют пары в каждом плече молекулы антитела с образованием двух идентичных антигенсвязывающих областей. Полагают, что некоторые аминокислотные остатки образуют поверхность контакта между вариабельными доменами легкой и тяжелой цепи [см. Kabat, et al. (1991), Sequences of Protein of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, Md. и Clothia et al. (1985), J. Mol. Biol, vol.186:651].
Известно, что вариабельность неравномерно распределена на протяжении вариабельных доменов антител, а сконцентрирована в трех сегментах, называемых "определяющими комплементарность областями" (CDR) или "гипервариабельными областями" в вариабельных доменах как легкой цепи, так и тяжелой цепи. Более высококонсервативные участки вариабельных доменов называют "каркасной областью" (FR). Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR-области, соединенные тремя CDR. CDR в каждой цепи расположены рядом вблизи FR-областей, и с CDR из других цепей формируют антигенсвязывающую область антител [см. Kabat, et al. (1991), Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, Md.]. В совокупности, 6 CDR обеспечивают связывающие свойства молекулы антитела в отношении антигена. Однако даже один вариабельный домен (или половина Fv, содержащей три CDR, специфичных к антигену) обладает способностью распознавать и связывать антиген [см. Pluckthun (1994), в The Pharmacology of Monoclonal Antibodies, vol.113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315].
Термин "константный домен" относится к С-концевому участку тяжелой и легкой цепи антитела. Как правило, константные домены не прямо вовлечены в связывающие свойства молекулы антитела в отношении антигена, а проявляют различные эффекторные функции, такие как преципитация антитела при антителозависимой клеточной токсичности. Здесь "эффекторные функции" относятся к различным физиологическим эффектам антител (например, опсонизации, лизису клеток, дегрануляции тучных клеток, базофилов и эозинофилов и к другим процессам), опосредуемым привлечением иммунных клеток посредством молекулярного взаимодействия между Fc-доменом и белками иммунной системы. Изотип тяжелой цепи определяет функциональные свойства антитела. Его характерные функциональные свойства обеспечиваются С-концевыми участками тяжелых цепей, где они не связаны с легкими цепями.
Как используют в настоящем документе, "фрагмент антитела" относится к участку интактного антитела, который включает антигенсвязывающую область или вариабельные участки интактного антитела, где участок может не содержать константных доменов тяжелой цепи (например, СН2, СН3 и СН4) Fc-области интактного антитела. Альтернативно участки константных доменов тяжелой цепи (например, СН2, СН3 и СН4) могут быть включены во "фрагмент антитела". Примерами фрагментов антител являются фрагменты, которые сохраняют связывание антигена и включают фрагменты Fab, Fab′, F(ab′)2, Fd и Fv; димерные антитела; тримерные антитела; молекулы одноцепочечных антител (sc-Fv); мини-антитела, наноантитела и полиспецифичные антитела, образованные фрагментами антител. В качестве примера, Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи.
Термин "вариант" относится к аминокислотной последовательности, которая отличается от нативной аминокислотной последовательности антитела по меньшей мере одним аминокислотным остатком или модификацией. Нативная или исходная аминокислотная последовательность дикого типа относится к аминокислотной последовательности антитела, встречающегося в природе. "Вариант" молекулы антитела включает, но не ограничивается ими, изменения в вариабельном участке или константном участке легкой цепи и/или тяжелой цепи, включая гипервариабельную область или CDR-область, Fc-область, Fab-область, CH1-домен, СН2-домен, СН3-домен и шарнирную область.
Термин "специфичный" относится к селективному связыванию антитела с его эпитопом-мишенью. Специфичность связывания молекул антитела можно тестировать посредством сравнения связывания антитела с требуемым антигеном и связывания антитела с не его собственным антигеном или аналогичным антигеном или смесью антигенов при данном наборе условий. Предпочтительно антитело по изобретению будет лишено существенного связывания с не его собственными антигенами или даже аналогами антигена-мишени. В настоящем документе термин "антиген" относится к молекуле, которая распознается и связывается молекулой антитела или иммунной молекулой, которая связывается с антигеном. Определенный участок антигена, который связывается антителом, называют "эпитопом". "Гаптен" относится к низкомолекулярному соединению, которое может, в большинстве условий, вызывать иммунный ответ (т.е. действовать в качестве антигена), только когда он связан с молекулой носителя, например белком, полиэтиленгликолем" (PEG), коллоидным золотом, силиконовыми гранулами и сходными с ними. Носитель может представлять собой носитель, который также самостоятельно не вызывает иммунный ответ.
Термин "антитело" используют в наиболее широком смысле, и он охватывает моноклональное, поликлональное, полиспецифичное (например, биспецифичное, где каждое плечо антитела реагирует с отличающимся эпитопом или с одним и тем же или отличающимся антигеном) антитело, мини-антитело, гетероконъюгатное антитело, димерное антитело, тримерное антитело, химерные и синтетические антитела, а также фрагменты антител, которые специфично связывают антиген с требуемым свойством связывания и/или биологической активностью.
Термин "моноклональное антитело" (mAb) относится к антителу или совокупности сходных антител, полученных из совокупности по существу гомогенных антител, и его не следует истолковывать, как требующий продукции антитела каким-либо конкретным способом. Например, моноклональные антитела можно получать способом гибридом, впервые описанным Kohler and Milstein (1975), Nature, vol.256:495-497 или способами рекомбинантных ДНК.
Термин "химерное" антитело (или иммуноглобулин) относится к молекуле, содержащей тяжелую и/или легкую цепь, которая идентична или гомологична соответствующим последовательностям в антителах конкретных видов или относящихся к конкретному классу или подклассу антител, в то время как оставшаяся цепь(и) идентична или гомологична соответствующим последовательностям в антителах другого вида или относящихся к другому классу или подклассу антител, а также к фрагментам таких антител, при условии, что они проявляют требуемую биологическую активность [Cabilly et al. (1984), ниже; Morrison et al., Proc. Natl. Acad. Sci. U.S.A. 81:6851].
Термин "гуманизированное антитело" относится к формам антител, которые содержат последовательности из не являющихся человеческими антител (например, антител мыши), а также антител человека. Гуманизированное антитело может включать консервативные аминокислотные замены или неприродные остатки из одного или различных видов, которые существенно не изменяют его связывание и/или биологическую активность. Такие антитела представляют собой химерные антитела, которые содержат минимальную последовательность из не являющихся человеческими иммуноглобулинов. По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки определяющей комплементарность области (CDR) реципиента заменены остатками CDR не относящегося к человеку вида (донорное антитело), такого как мышь, крыса, верблюд, крупный рогатый скот, коза или кролик, обладающие требуемыми свойствами. Более того, гуманизированные антитела могут содержать остатки, которые не встречаются ни в реципиентном антителе, ни в последовательностях импортных CDR или каркасной области. Эти модификации проводят для дальнейшего усовершенствования и максимизации характеристик антитела. Таким образом, как правило, гуманизированное антитело будет содержать все по меньшей мере из одного и, в одном аспекте, двух вариабельных доменов, в которых все или по существу все из гипервариабельных петель соответствуют гипервариабельным петлям иммуноглобулина человека и все или по существу все из FR-областей представляют собой FR-области из последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно будет содержать по меньшей мере часть константного участка (Fc) иммуноглобулина или константный участок (Fc) иммуноглобулина человека. См., например, Cabilly et al., патент США №4816567; Cabilly et al., патент Европы №0125023 B1; Boss et al., патент США №4816397; Boss et al., европейский патент № 0120694 B1; Neuberger, M.S. et al., WO 86/01533; Neuberger, M.S. et al., европейский патент №0194276 B1; Winter, патент США №5225539; Winter, европейский патент №0239400 B1; Padlan, E. A. et al., европейскую патентную заявку №0519596 A1; Queen et al. (1989) Proc. Natl. Acad. Sci. USA, vol.86:10029-10033).
Термин "биспецифичное антитело" может относиться к антителу, или моноклональному антителу, имеющему свойства связывания в отношении по меньшей мере двух различных эпитопов. В одном варианте осуществления эпитопы представляют собой эпитопы одного антигена. В другом варианте осуществления, эпитопы представляют собой эпитопы двух различных антигенов. Способы получения биспецифичных антител известны в данной области. Например, биспецифичные антитела можно получать рекомбинантными способами с использованием коэкспрессии двух пар тяжелая цепь/легкая цепь иммуноглобулинов. Альтернативно биспецифичные антитела можно получать с использованием образования химической связи. Биспецифичные антитела включают биспецифичные фрагменты антител.
Термин "гетероконъюгатное антитело" может относиться к двум ковалентно связанным антителам. Такие антитела можно получать с использованием известных способов в химии синтетических белков, включая использование поперечно-сшивающих агентов. Как используют в настоящем документе, термин "конъюгат" относится к молекулам, образованным ковалентным присоединением одного или нескольких фрагмента(ов) антитела или связывающих групп к одной или нескольким молекуле(ам) полимера.
Термин "биологически активный" относится к антителу или фрагменту антитела, который способен связывать требуемый эпитоп и осуществлять некоторый биологический эффект. Биологические эффекты включают, но не ограничиваются ими, модулирование сигнала роста, модулирование антиапоптотического сигнала, модулирование апоптотического сигнала, модулирование каскада эффекторной функции и модулирование других взаимодействий лиганда.
Термин "рекомбинантная ДНК" относится к нуклеиновым кислотам и продуктам генов, экспрессируемых с них, которые сконструированы, созданы или модифицированы человеком. "Рекомбинантные" полипептиды или белки представляют собой полипептиды или белки, полученные способами рекомбинантных ДНК, например, из клеток, трансформированных конструкцией экзогенной ДНК, кодирующей требуемый полипептид или белок. "Синтетические" полипептиды или белки представляют собой полипептиды и белки, полученные химическим синтезом.
Термин "экспрессирующая кассета" относится к нуклеотидной молекуле, способной влиять на экспрессию структурного гена (т.е. последовательности, кодирующей белок, такой как антитело по изобретению) в хозяине, совместимом с такими последовательностями. Экспрессирующие кассеты включают по меньшей мере промотор, функционально связанный с последовательностью, кодирующей полипептид, и необязательно с другими последовательностями, например сигналами терминации транскрипции. Также можно использовать дополнительные регуляторные элементы, необходимые или полезные для достижения экспрессии, например энхансеры. Таким образом, экспрессирующие кассеты включают плазмиды, экспрессирующие векторы, рекомбинантные вирусы, любую форму рекомбинантного вектора в виде "голой ДНК" и т.п.
Источники антитела не ограничены источником, проиллюстрированным в настоящем документе (например, антитело мыши и гуманизированное антитело мыши). Антитела можно индуцировать во многих видах, включая виды млекопитающих (например, мышь, крысу, верблюд, быка, козу, лошадь, морскую свинку, хомяка, овцу и кролика) и птиц (утку, курицу). Источником индуцированных антител могут быть виды, отличающиеся от вида животного, у которого они индуцированы. Например, в XenoMouse™ (Abgenix, Inc., Fremont CA) продуцируются полностью человеческие моноклональные антитела. Для определенных целей можно использовать нативные антитела человека, такие как аутоантитела к S1P, полученные от индивидов, у которых может быть показан титр такого аутоантитела к S1P. Альтернативно для создания антител, содержащих последовательность человека, можно использовать библиотеку последовательностей антител человека.
2. Применение
Изобретение относится к композициям и способам для лечения или профилактики определенных заболеваний и состояний с использованием одного или нескольких лекарственных средств, которые изменяют активность или концентрацию одного или нескольких нежелательных биологически активных липидов, или их предшественников или метаболитов. Терапевтические способы и композиции по изобретению действуют посредством изменения эффективной концентрации, т.е. абсолютной, относительной, эффективной и/или доступной концентрации и/или видов активности определенных нежелательных биологически активных липидов. Можно сказать, что снижение эффективной концентрации биологически активного липида "нейтрализует" липид-мишень или его нежелательные эффекты, включая последующие эффекты. Здесь "нежелательный" относится к биологически активному липиду, который является нежелательным, поскольку он вовлечен в процессы заболевания, например, в качестве молекулы передачи сигнала, или к нежелательному количеству биологически активного липида, которое приводит к заболеванию, когда присутствует в избытке.
Без связи с какой-либо конкретной теорией, полагают, что ненадлежащая концентрация S1P и/или его метаболитов или последующих эффекторов может вызывать развитие различных заболеваний и нарушений или участвовать в нем. По существу композиции и способы можно использовать для лечения этих заболеваний и нарушений, в частности, посредством снижения эффективной концентрации in vivo конкретного липида-мишини, например S1P или его вариантов. В частности, полагают, что композиции и способы по изобретению пригодны для лечения заболеваний, характеризующихся, по меньшей мере частично, аномальной неоваскуляризацией, ангиогенезом, фиброгенезом, фиброзом, рубцеванием, воспалением и иммунным ответом.
Примеры нескольких классов заболеваний, которые можно лечить в соответствии с изобретением, описаны ниже. Будет понятно, что множество заболеваний и состояний характеризуется, по меньшей мере частично, множеством патологических процессов (например, как патологическая неоваскуляризация, так и рубцевание) и что классификации, представленные в настоящем документе, представлены для удобства описания и не ограничивают изобретение.
S1P и гиперпролиферативные нарушения
Один аспект изобретения относится к способам лечения гиперпролиферативного нарушения. Эти способы включают введение млекопитающему (например, быкам, собакам, лошадям, овцам или свиньям, в частности человеку), о котором известно или предполагают, что он страдает связанным с S1P гиперпролиферативным нарушением, терапевтически эффективного количества композиции, содержащей средство, которое препятствует активности S1P, предпочтительно в приемлемом фармацевтическом или ветеринарном носителе, в зависимости от того, что требуется для предполагаемого применения. Связанные с S1P гиперпролиферативные нарушения включают новообразования, нарушения, связанные с пролиферацией эндотелиальных клеток, и нарушения, связанные с фиброгенезом. Наиболее часто новообразование представляет собой злокачественную опухоль. Типичными нарушениями, связанными с пролиферацией эндотелиальных клеток, являются зависимые от ангиогенеза нарушения, например злокачественные опухоли, вызываемые солидными опухолями, гематологическими опухолями, и старческая дегенерация желтого пятна. Нарушения, связанные с фиброгенезом, включают нарушения, которые вовлекают аномальное ремоделирование сердца, такие как сердечная недостаточность.
Существует множество известных гиперпролиферативных нарушений, при которых клетки различных тканей и органов проявляют аномальные паттерны роста, пролиферации, миграции, передачи сигнала, старения и гибели. Несмотря на то что для некоторых из этих заболеваний был разработан ряд способов лечения, многие из них тем не менее остаются в большой степени не поддающимися лечению существующими технологиями, в то время как в других случаях, несмотря на то, что доступны способы лечения, они часто являются не оптимальными и редко приводят к излечению.
Злокачественная опухоль представляет собой, возможно, наиболее широко признанный класс гиперпролиферативных нарушений. Злокачественные опухоли представляют собой класс истощающих заболеваний, и в совокупности показатель смертности от них является вторым после только сердечно-сосудистых заболеваний. Множество злокачественных опухолей не полностью понятны на молекулярном уровне. В результате злокачественная опухоль является основным объектом программ исследований и разработки как для правительства США, так и для и фармацевтических компаний. Результатом были беспрецедентные усилия R&D и продукция множества важных лекарственных средств для помощи в борьбе против злокачественной опухоли.
К сожалению, огромного количества исследований злокачественной опухоли было не достаточно для преодоления значительного вреда, вызываемого злокачественной опухолью. Ежегодно только в США все еще диагностируется свыше одного миллиона новых случаев злокачественной опухоли и более пяти тысяч смертей. Это является впечатляющей демонстрацией того, что даже, несмотря на огромные усилия, приложенные для открытия новых лекарственных средств от злокачественной опухоли, эффективные лекарственные средства для борьбы с заболеванием остаются труднодостижимыми.
Злокачественную опухоль в настоящее время сначала лечат одним или сочетанием трех типов терапии: хирургической операцией, лучевой терапией и химиотерапией. Хирургическая операция включает удаление массы пораженной ткани. Несмотря на то что хирургическая операция иногда является эффективной в отношении удаления опухолей, расположенных в определенных областях, например в молочной железе, толстом кишечнике и коже, ее нельзя использовать для лечения опухолей, расположенных в других областях, таких как позвоночник, а также для лечения диссеминированных неопластических состояний, таких как лейкоз. Лучевая терапия вовлекает воздействие на живую ткань ионизирующего облучения, вызывающего гибель или повреждение подвергаемых воздействию клеток. Побочные эффекты лучевой терапии могут быть острыми и временными, в то время как другие могут быть необратимыми. Химиотерапия включает нарушение репликации клеток или метаболизма клеток.
Дополнительный отрицательный фактор состоит в том, что современные лекарственные средства обычно обладают существенными недостатками в форме токсичности и тяжелых побочных эффектов у пациента. Таким образом, многие группы недавно начали поиск новых подходов в борьбе со злокачественной опухолью. Эти новые так называемые "инновационные способы лечения" включают генную терапию и терапевтические белки, такие как моноклональные антитела.
Первым моноклональным антителом, использованным в клинике для лечения злокачественной опухоли, был Rituxan (ритуксимаб), который был выпущен в 1997 году и который продемонстрировал применимость биоспецифичных моноклональных антител в качестве лекарственных средств. Таким образом, не удивительно, что с тех пор для применения в клинике было одобрено шестнадцать других моноклональных антител, включая шесть антител, которые назначают при злокачественной опухоли. Успех этих продуктов, а также сниженная стоимость и время для разработки моноклональных антител по сравнению с низкомолекулярными соединениями сделали лекарственные средства на основе моноклональных антител второй из наиболее крупных категорий лекарственных средств-кандидатов после низкомолекулярных соединений. Кроме того, было показано, что высокая специфичность антител по сравнению с лекарственными средствами на основе низкомолекулярных соединений обладает большими преимуществами с точки зрения как эффективности, так и токсичности. Только для злокачественной опухоли в настоящее время существует более 270 промышленных проектов R&D для антител, и в разработку новых лекарственных средств против злокачественной опухоли на основе антител вовлечено более чем 50 компаний. Впоследствии моноклональные антитела стали основными участниками лечения злокачественной опухоли и согласно оценкам они занимают возрастающую долю на рынке лекарственных средств против злокачественной опухоли.
Идентификация внеклеточных медиаторов, которые обеспечивают рост и выживание опухоли, является критической стадией в разработке терапевтических вмешательств, которые снизят заболеваемость злокачественной опухолью и смертность от нее. Как описано ниже, сфингозин-1-фосфат (S1P), ключевой компонент каскада передачи сигнала сфинголипидов, считают плейотропным туморогенным фактором роста. S1P обеспечивает рост опухоли посредством стимуляции пролиферации клеток, выживания клеток и метастазирования. Также S1P обеспечивает ангиогенез опухоли посредством поддержания миграции и выживания зндотелиальных клеток, поскольку они образуют новые сосуды в опухолях. В совокупности S1P инициирует пролиферативную, проангиогенную и антиапоптотическую последовательность событий, приводящую к прогрессии злокачественной опухоли. Таким образом, лекарственные средства, которые модулируют и, в частности, снижают уровни S1P in vivo, будут эффективными для лечения злокачественной опухоли.
Исследования показали, что сфингозинкиназа (SPHK) представляет собой недавно подтвержденный онкоген, который продуцирует молекулу внеклеточной передачи сигнала сфинголипидов, сфингозин-1-фосфат (S1P), который обеспечивает рост опухоли. Рост опухоли обеспечивается как прямо, так и непрямо посредством действия S1P в качестве фактора роста, связанного с пролиферацией и метастазированием опухолевых клеток, а также проангиогенных эффектов S1P. Заявитель получил биоспецифичное моноклональное антитело против S1P (mAb против S1P), которое можно использовать в качестве терапевтической молекулярной губки для селективной абсорбции S1P, таким образом, снижая внеклеточные концентрации этого фактора роста опухоли с ожидаемым снижением объема опухоли и метастатического потенциала, а также одновременно блокируя образование новых кровеносных сосудов, которые в ином случае будут питать растущую опухоль. Ожидаемый успех концепции молекулярной абсорбции будет представлять собой инновационный подход для лечения злокачественной опухоли. Как показано в представленных ниже абзацах, заявитель разработал mAb против важного фактора роста опухоли, сфингозин-1-фосфата (S1P). Заявитель полагает, что это антитело может быть эффективным для снижения пролиферации, метастатического потенциала и ангиогенеза, связанных со многими типами злокачественных опухолей, и, таким образом, для уменьшения злокачественной опухоли в целом, а также ангиогенеза злокачественной опухоли, которые сопровождают заболевание.
Нейтральная форма сфингомиелиназы (nSMase) представляет собой ключевой ранний компонент каскада передачи сигнала сфинголипидов (Chatterjee, Adv. Lipid Res. 26:25-46, 1993; Liu, Obein, and Hannun, Semin. Cell Dev. Biol. 8:311-322, 1997). nSMase представляет собой только один по меньшей мере из пяти классов SMase, которые были идентифицированы, включая щелочные, кислые, кислые цинк-зависимые, нейтральные магний-зависимые и нейтральные магний-независимые (Liu, Obein, и Hannun, Semin. Cell Dev. Biol. 8:311-322, 1997). Класс nSMase обычно связан с поверхностными мембранами (Das, Cook, and Spence, Biochim Biophys Acta 777:339-342, 1984; Dobrowsky, Cell Signal 12:81-90, 2000), и он может активироваться множеством стимулов, вызывающих апоптоз, такими как провоспалительный цитокин, фактор некроза опухоли альфа (TNFα) (Segui, et al., J. Clin. Invest. 108:143-151, 2001), Т-клеточный рецептор (Tonnetti, et al., J. Exp. Med 189:1581-1589, 1999), ионизирующее излучение (Haimovitz-Friedman, et al., J. Exp.Med 180:525-535, 1994) и средства против новообразований на основе антрациклина (Andrieu-Abadie, et al., FASEB J. 13:1501-1510, 1999). Фактор некроза опухоли альфа (TNFα) представляет собой хорошо известный активатор nSMase (Adam, et al., J. Biol. Chem 271:14617-14622, 1996; Dressier, Mathias, and Kolesnick, Science 255:1715-1718, 1992; Kim, et al., J. Biol. Chem. 266:1:484-489, 1991; Kronke, Chem Phys Lipids 102:157-66, 1999; Yanaga and Watson, FEBS Letters 314:297-300, 1992), продукции CER (Kronke, Chem Phys Lipids 102:157-66, 1999) и апоптоза (Rath и Aggarwal, J. Clin. Immunol. 19:350-364, 1999; Robaye, et al., Am. J. Pathol 138:447-453, 1991; Takeda et al., Int. Immunol. 5:691-694, 1993) во многих типах клеток, включая злокачественные клеточные линии (Andrieu-Abadie, et al., FASEB J. 13:1501-1510, 1999; Hannun and Obein, Trends in Biol. Sci. 20:72-76, 1995; Kolesnick, trends Biochem Sci 24:224-5, 1999; Obeid, et al., Science 259:1769-1771, 1993), и было показано, что активация nSMase является критической для индуцируемого TNFα апоптоза (Luberto, et al., J. Biol. Chem. 277:41128-41139, 2002; Segui, et al., J. Clin. Invest. 108:143-151, 2001). Вследствие этого nSMase также была предложена в качестве мишени для разработки лекарственных средств (Wascholowski and Giannis, Drug News Perspect. 14:581-90, 2001).
Молекула передачи сигнала сфинголипидов, S1P, образуется из SPH под действием сфингозинкиназы (SPHK). Идентифицировано две изоформы киназы, SPHK1 и SPHK2 (Liu, J. Biol. Chem. 275:19513-20, 2000; Nava, et al., Exp Cell Res 281:115-127, 2002). В то время как CER и SPH обычно ассоциированы с апоптозом, S1P, напротив, является медиатором клеточной пролиферации и активации каскадов выживания (An, Ann N. Y. Acad Sci 905:25-33, 2000; Maceyka, et al., BBA 1585:193-201, 2002; Zhang, et al., J. Cell Biol. 114:155-167, 1991). Недавно он был признан в качестве внеклеточного медиатора, который может активировать ряд сопряженных с G-белком рецепторов (GPCR), относящихся к семейству рецепторов S1P/LPA, ранее известных как рецепторы Edg (An, Ann N Y Acad Sci 905:25-33, 2000; An, Goetzl and Lee, J. cell biochem 30/31:147-157, 1998; Lee, et al., Science 279:1552-1555, 1998; Okamoto, et al., Biochem. Biophys. Res. Commun. 260:203-208, 1999); однако также было сделано предположение о внутриклеточных действиях SIP (Van Brooklyn, et al., J. Cell Biol. 142:229-240, 1998). Более того, было предположено, что баланс между уровнями CER/SPH и S1P обеспечивают реостатный механизм, который определяет, направить ли клетку на каскад гибели или защитить от апоптоза (Kwon, et al., J. Biol. Chem. 276:10627-10633, 2001; Maceyka, et al., BBA 1585:193-201, 2002; Pyne, Biochem. J. 349:385-402, 2000). Ключевым регуляторным ферментом реостатного механизма является SPHK, функцией которой является превращение запускающих гибель сфинголипидов (CER/SPH) в запускающий рост S1P.
Знаковое исследование, в котором впервые SPHK была предложена в качестве онкогена, было опубликовано группой из Adelaide, показавшей, что фибробласты NIH-3T3, стабильно трансфицированные киназой, проявляли усиленную клеточную пролиферацию, сопровождающуюся повышенной продукцией SIP (Vadas and Gamble, Circ. Res. 79:1216-1217, 1996; Xia et al., Curr Biol. 10:1527-1530, 2000). Кроме того, клетки, сверхэкспрессирующие SPHK, избегали контактного ингибирования, свойства, обычно проявляемого трансформированными клетками. Это наблюдение согласуется с недавним отчетом, в котором показано, что S1P усиливает метастатический потенциал некоторых злокачественных клеточных линий человека (Igarashi, Ann. N. Y. Acad. Sci. 845:19-31, 1998; Takuwa, Biochim Biophys Acta. 1582:112-120, 2002). Более того, трансфектанты образовывали опухоли при инъекции их мышам NOD/SCID. Эти результаты недавно были подтверждены в исследовании, показавшем, что низкомолекулярный ингибитор SPHK, вводимый внутрибрюшинно, может снижать объем опухоли у мышей SCID, которым проводят подкожные инъекции клеток аденокарциномы молочной железы JC (French, et al., Cancer Res 63:5962-5969, 2003). Существенно, что концепция о том, что SPHK может быть новым онкогеном, была подкреплена открытием того, что SPHK сверхэкспрессируется во многих солидных опухолях, таких как опухоли молочной железы, толстого кишечника, легкого, яичника, желудка, матки, почки и прямой кишки (French et al. (2003), выше). Кроме того, было показано, что несколько полученных из опухоли человека клеточных линий могут направляться на апоптоз при лечении низкомолекулярными ингибиторами SPHK и что их эффективность может быть объяснена их способностью снижать уровни S1P. В совокупности эти данные демонстрируют важную концепцию о том, что S1P представляет собой фактор роста, вероятно, продуцируемый самими опухолевыми клетками и что снижение концентрации S1P может вызывать апоптоз, наблюдаемый при устранении фактора роста.
S1P и ангиогенез опухоли
Ангиогенез представляет собой процесс, посредством которого из существующих сосудов образуются новые кровеносные сосуды. Ангиогенез играет критическую роль в некоторых физиологических процессах, и он вовлечен в патогенез множества нарушений, включая рост опухоли, инвазию и метастазирование. Процесс ангиогенеза, ассоциированный с солидными опухолями и опухолями кровеносной системы (ангиогенез опухоли), считают ключевым компонентом образования опухоли и прогрессии заболевания, поскольку новые кровеносные сосуды обеспечивают преимущество для роста опухолевых клеток по сравнению с незлокачественными клетками. Таким образом, клинический контроль ангиогенеза является критическим компонентом для лечения злокачественной опухоли и других зависимых от ангиогенеза заболеваний. Антиангиогенные лекарственные средства являются особенно привлекательными, поскольку эндотелиальные клетки (ЕС) сосудов не легко поддаются мутации, как злокачественные клетки; следовательно, ЕС с меньшей вероятностью приобретут устойчивость к длительной терапии, что делает их хорошими потенциальными мишенями для лекарственных средств.
В злокачественный ангиогенез вовлечено несколько факторов роста. Было выявлено, что биолипид сфингозин-1-фосфат (S1P) является медиатором множества клеточных процессов, важных для злокачественной опухоли. S1P оказывает большинство из его эффектов в качестве специфического лиганда для семейства сопряженных с G-белком рецепторов, обозначаемых SIP1-5. Эти рецепторы регулируют ангиогенез и созревание сосудов, движение клеток и миграцию лимфоцитов. В противоположность S1P, предшественники S1P, сфингозин и церамид, ассоциированы с остановкой роста и апоптозом. Наконец, существует комплексное взаимное влияние между S1P и другими ангиогенными факторами роста, такими как VEGF, EGF, PDGF, bFGF и IL-8. S1P посредством связывания с рецептором S1P1 трансактивирует рецепторную тирозинкиназу фактора роста, такую как находится на VEGFR, EGFR и PDGFR. Важность S1P в зависимых от ангиогенеза опухолях делает S1P превосходной мишенью для лечения злокачественной опухоли. Исходя из этих наблюдений, подход на основе антител для нейтрализации внеклеточного S1P может привести к выраженному снижению прогрессии злокачественной опухоли у человека в результате ингибирования образования кровеносных сосудов с сопутствующим снижением количества питательных веществ и кислорода, необходимых для поддержания роста опухоли. Более того, недавние исследования позволяют предположить, что многие ингибиторы ангиогенеза также могут действовать в качестве антиинвазивных и антиметастатических соединений, которые также могут способствовать подавлению распространения злокачественной опухоли в области, отдаленные от исходной опухоли.
Источником возрастающей совокупности последних данных, вовлекающих S1P в качестве одного из наиболее эффективных проангиогенных средств, являются исследования, непосредственно сравнивающие S1P с такими средствами, как VEGF и bFGF. S1P стимулирует синтез ДНК и хемотаксическую подвижность эндотелиальных клеток вен человека (HUVEC) и в то же время индуцирует дифференцировку многоклеточных структур, все из которых говорят о роли S1P в раннем образовании кровеносных сосудов (Argraves, et al., 2004; Lee et al., 1999; Liu, et al., 2000). Также S1P обеспечивает миграцию образованных в костном мозге предшественников ЕС в области неоваскуляризации (Annabi, et al., 2003). Клетки, которые сверхэкспрессируют S1P1, являются устойчивыми к антиангиогенным средствам талидомиду и Неовастату (Annabi et al., 2003). Кроме того, было показано, что существует существенное взаимное влияние между S1P и другими проангиогенными факторами роста, такими как VEGF, EGF, PDGF, bFGF и IL-8. Например, S1P трансактивирует рецепторы EGF (Shida, et al., 2004) и VEGF2 (Spiegel & Milstien, 2003), и VEGF активирует экспрессию рецептора SlP1 (Igarashi, et al., 2003). Также S1P, действующий через S1P1 и "линию VEGF", требуется для ангиогенеза и неоваскуляризации задних конечностей (Сhае, et al., 2004).
Антиангиогенный подход к злокачественной опухоли значительно продвинулся вследствие недавнего одобрения FDA антиангиогенного лекарственного средства, бевацизумаба (Avastin®, Genentech) для лечения рака толстого кишечника в качестве дополнения к цитотоксической химиотерапии.
Недавно было разработано МАb мыши против SIP, LT1002 с высокой аффинностью связывания и специфичностью к S1P. Было показано, что LT1002 значительно замедляет прогрессию опухоли и связанный с ней ангиогенез в нескольких моделях злокачественной опухоли человека на животных. Кроме того, LT1002 ослабляло образование очага хориоидальной неоваскуляризации (CNV) в общепринятой модели ангиогенеза при старческой дегенерации желтого пятна (AMD). CNV происходит при заболеваниях, при которых существуют аномалии мембраны Бруха и пигментного эпителия сетчатки. Наиболее распространенным заболеванием этого типа является AMD, самая распространенная причина потери зрения у пожилых пациентов. Эти результаты позволяют предположить, что LT1002 имеет несколько механизмов действия, включая: (1) прямые эффекты на рост опухолевых клеток, (2) непрямой антиангиогенный эффект на эндотелиальные клетки сосудов и (3) непрямой антиангиогенный эффект в виде препятствования высвобождению и действию других проангиогенных факторов роста.
Источником наиболее прямого доказательства in vivo того, что S1P приводит к ангиогенезу опухоли, является недавняя публикация авторов настоящего изобретения, которая сфокусирована на моноклональном антителе (mAb) мыши, предназначенном для нейтрализации внеклеточного S1P посредством молекулярной абсорбции (Visentin, et al., 2006). В различных анализах in vitro с использованием HUVEC mAb против SIP нейтрализовывало образование трубки, миграцию эндотелиальных клеток сосудов и защиту от клеточной гибели, все из которых индуцируются посредством S1P. S1P усиливал рост новых капилляров во вставках матригеля, имплантированных мышам, и этот эффект нейтрализовывало системное введение mAb против S1P. mAb по существу нейтрализовывало индуцируемый bFGF и VEGF ангиогенез в анализе вставки матригеля у мышей, и антитело подавляло стимулированное S1P высвобождение проангиогенных цитокинов (VEGF, IL-8, IL-6) из опухолевых клеток in vitro и in vivo. Важно, что у мышей, которым провели ксенотрансплантацию ортотопно помещенных злокачественных клеток человека, было показано существенное замедление прогрессии опухоли при введении mAb против S1P. Это было показано в моделях на мышах для рака молочной железы, яичника и легкого человека и в одной модели с аллотрансплантатом меланомы мыши (Visentin, et al., 2006).
Применение моноклональных антител (mAb) в качестве терапевтического лечения множества заболеваний и нарушений быстро возрастает, поскольку было показано, что они являются безопасными и эффективными лекарственными средствами. Одобренные терапевтические mAb включают Avastin®, Erbitux® и Rituxan®. Дополнительные mAb находятся в различных фазах клинической разработки для различных заболеваний, и большинство из них нацелены на различные формы злокачественной опухоли. Как правило, моноклональные антитела получают от не относящихся к человеку млекопитающих. Однако терапевтическая применимость моноклональных антител мыши ограничена, главным образом, вследствие того факта, что у людей возникает их собственный антительный ответ на антитела мыши. Этот ответ, так называемый ответ НАМА (антитела человека против антител мыши), приводит к последующей нейтрализации и быстрому устранению mAb мыши. Это ограничение было преодолено посредством разработки процесса, называемого "гуманизацией" антител мыши. Гуманизация в значительной степени снижает развитие иммунного ответа против введенного терапевтического MAb и, таким образом, предотвращает снижение времени полужизни и терапевтической эффективности вследствие НАМА. Главным образом, процесс гуманизации состоит в пересадке определяющих комплементарность областей (CDR) мыши в каркасную область (FR) иммуноглобулина человека. Эту стратегию называют "пересадкой CDR". Часто для восстановления аффинности, утраченной в исходной конструкции после пересадки, требуются "возвратные мутации" выбранных остатков в FR человека на аминокислотные остатки мыши.
Получение mAb представляет собой комплексный процесс, который обусловлен вариабельностью самого белка. Вариабельность mAb может быть локализована в белковом остове и/или в углеводной группе. Гетерогенность может быть связана с образованием альтернативных дисульфидных пар, дезамидацией и образованием изоаспартильных остатков, окислением метионина и цистеина, циклизацией N-концевых остатков глутамина до пироглутамата и неполным ферментативным отщеплением С-концевых остатков лизина карбоксипептидазами млекопитающих. Обычно для улучшения свойств молекул антител, таких как повышенная стабильность, устойчивость к протеазам, характер агрегации и усиление уровня экспрессии в гетерологичных системах, используют способы инженерии.
В настоящем документе описана гуманизация MAb мыши против S1P. В целом стратегия состоит в пересадке шести CDR из LT1002 в каркасную область человека. Дополнительные модификации осуществляли для дополнительного усовершенствования и оптимизации характеристик антитела. Гуманизированное MAb обладало теми же характеристиками, что и LT1002, и, таким образом, оно является пригодным для тестирования в клинических испытаниях.
S1P и фиброз
Фибробласты, в частности миофибробласты, представляют собой ключевые клеточные элементы при рубцевании в ответ на повреждение клеток и воспаление [Tomasek, et al. (2002), Nat Rev Mol Cell Biol, vol.3:349-63, и Virag and Murry (2003), Am. J. Pathol, vol.163:2433-40]. Экспрессия гена коллагена миофибробластами является признаком ремоделирования и необходима для рубцевания [Sun and Weber (2000), Cardiovasc Res, vol.46:250-6, и Sun and Weber (1996), J. Mol. Cell Cardiol, vol.28:851-8]. SIP обеспечивает заживление ран посредством активации миграции и пролиферации фибробластов с одновременным усилением продукции коллагена [Sun, et al. (1994), J. Biol. Chem, vol.269:16512-7]. SIP, продуцируемый локально поврежденными клетками, может быть ответственным за ненадлежащее заживление ран, связанное с ремоделированием и рубцеванием. Таким образом, полагают, что ингибиторы S1P пригодны при заболеваниях или состояниях, характеризующихся, по меньшей мере частично, аномальным фиброгенезом или фиброзом. В настоящем документе "фиброгенез" определяют как избыточную активность или количество фибробластов, а "фиброз" определяют как избыточную активность или количество фибробластов, которые приводят к избыточной или ненадлежащей продукции коллагена и рубцеванию, разрушению физиологической структуры ткани и/или к ненадлежащему уплотнению матрикса, что приводит к таким патологиям, как отслоение сетчатки, или к другим процессам, приводящим к нарушению функции органа.
S1P и экспрессия коллагена фибробластами: S1P обеспечивает дифференцировку покоящихся фибробластов в активные миофибробласты, которые проявляют усиленную экспрессию коллагена в ходе рубцевания [Urata, et al. (2005), Kobe J. Med Sci, vol.51:17-27]. Одновременно с пролиферацией и миграцией фибробластов в зону рубцевания миофибробласты образуют временную гранулярную сеть, состоящую главным образом из остеопонтина и фибронектина [Sun and Weber (2000), Cardiovasc Res, vol.46:250-6]. По мере продолжения ремоделирования временный матрикс абсорбируется и устанавливается коллагеновая сеть [Sun and Weber (2000), Cardiovasc Res, vol.46:250-6]. Авторы настоящего изобретения продемонстрировали, что S1P запускает продукцию коллагена миофибробластами. Было показано, что TGFβ, хорошо известный медиатор фиброза, активирует некоторые профибротические белки, превращает фибробласты в миофибробласты и стимулирует экспрессию воспалительных белков, вероятно, посредством действия SIP [Squires, et al. (2005), J. Mol. Cell Cardiol, vol.39:699-707 и Butt, Laurent and Bishop (1995), Eur J. Cell Biol, vol.68:330-5]. Активация TIMP1, молекулы передачи сигнала, вовлеченной в стимулируемую TGFβ дифференцировку фибробластов в миофибробласты, блокируется посредством siRNA против SPHK1 [Yamanaka, et al., J. Biol. Chem. 2004 Dec 24; 279(52):53994-4001], позволяя предположить, что гуманизированный вариант антитела против S1P может подавлять профибротические эффекты TGFβ, а также подавлять фиброгенные эффекты самого S1P.
Полагают, что минимизация ненадлежащего рубцевания применима для лечения фиброзных заболеваний и состояний, включая, но не ограничиваясь ими, глазные и сердечно-сосудистые заболевания, заживление ран и склеродермию.
Антитела против S1P для лечения склеродермии
Композиции и способы по изобретению будут пригодны для лечения нарушений и заболеваний, характеризующихся, по меньшей мере частично, аномальной неоваскуляризацией, ангиогенезом, фиброгенезом, фиброзом, рубцеванием, воспалением и иммунным ответом. Одним таким заболеванием является склеродермия, которую также называют системной склеродермией.
Склеродермия представляет собой аутоиммунное заболевание, которое вызывает рубцевание или утолщение кожи и иногда включает другие области организма, такие как легкие, сердце и/или почки. Склеродермия характеризуется образованием рубцовой ткани (фиброзом) в коже и органах организма, которое приводит к утолщению и уплотнению вовлеченных областей, с последующим снижением функции. Согласно Scleroderma Foundation в настоящее время склеродермию имеют приблизительно 300000 американцев. Одна треть или менее из пораженных лиц имеет распространенное заболевание, в то время как остальные две трети имеют кожные симптомы. Когда заболевание поражает легкие и вызывает рубцевание, дыхание может стать ограниченным вследствие того, что легкие более не могут расширяться должным образом. Для измерения способности дышать врачи используют устройство, которое оценивает форсированную жизненную емкость (FVC). У людей с FVC, составляющей менее 50 процентов от ожидаемых показателей, уровень смертности в течение 10 лет от связанного со склеродермией заболевания легкого составляет приблизительно 42 процента. Одна из причин того, что смертность является настолько высокой, состоит в том, что в настоящее время не доступно эффективное лечение.
Как описано в разделе "Примеры" настоящей заявки, существующие данные указывают на то, что S1P представляет собой профибротический фактор роста, который может приводить к активации, пролиферации фибробластов и, наконец, к повышенной активности фибробластов, связанной с ненадлежащим рубцеванием и ремоделированием. Более того, показаны потенциальные роли для S1P в активности кожных и других типов фибробластов. Например, было показано, что биологически активные липиды стимулируют миграцию фибробластов кожи мыши (Hama, et al., J. Biol. Chem. 2004 Apr 23; 279 (17):17634-9), а фибробласты кожи человека экспрессируют несколько подтипов рецепторов S1P (Zhang, et al., Blood. 1999 May 1; 93(9):2984-90). В дополнение ко многим прямым эффектам S1P на активность фибробластов, S1P также может обладать множеством потенциальных непрямых эффектов на активность фибробластов. Например, S1P может способствовать действию других хорошо известных профибротических факторов, таких как TGF-β и тромбоцитарный фактор роста (PDGF). TGF-β представляет собой один из наиболее широко исследованных и признанных участников фиброза (Desmouliere, et al., J. Cell. Biol. 122:103-111, 1993). TGF-β активирует экспрессию и активность SphKl, что приводит к повышенной экспрессии тканевых ингибиторов металлопротеиназы 1 (TIMP-1), белка, который ингибирует деградацию ЕСМ (Yamanaka, et al., J. Biol. Chem. 279:53994-54001, 2004). Повышенная экспрессия TIMP-1 связана с фиброзом кишечника и диастолической дисфункцией у пациентов с сердечной недостаточностью (Heymans, et al., Am. J. Pathol. 166:15-25, 2005). Напротив, SIP стимулирует экспрессию и высвобождение TGF-β (Norata, et al., Circulation 111:2805-2811, 2005). Также существуют явные доказательства взаимного влияния между S1P и PDGF. S1P прямо стимулирует экспрессию PDGF (Usui, et al., J. Biol. Chem. 279:12300-12311, 2004). Кроме того, рецептор S1P1 и рецептор PDGF связываются друг с другом, и их ассоциация необходима для активации посредством PDGF последующей передачи сигнала, которая приводит к пролиферации и миграции различных типов клеток (Long, et al., Prostaglandins Other Lipid Mediat 80:74-80, 2006; Baudhuin et al., Faseb J. 18:341-343, 2004). По существу эффекты TGF-β и PDGF на фиброз частично могут быть следствием взаимного влияния с каскадом передачи сигнала S1P. По существу композиции и способы по изобретению можно использовать для лечения склеродермии, в частности, посредством снижения эффективной концентрации in vivo конкретного липида-мишени, например S1P.
Полагают, что системная склеродермия усиливается стимулирующими аутоантителами против рецепторов PDGF (Baroni, et al., N Engl J. Med. 2006 v.354 (25):2667-76), и рецепторы PDGF активируются в фибробластах при склеродермии в ответ на TGF-β (Yamakage, et al., J. Exp. Med. 1992 May 1; 175(5):1227-34). Вследствие существенного взаимного влияния между системами передачи сигнала S1P, PDGF и TGF-β, блокирование биологической активности S1P средством против S1P (например, mAb против S1P) может непрямо подавлять просклеротические эффекты PDGF и TGF-β. Более того, лечение таким средством против S1P может оказать благоприятный эффект у пациентов со склеродермией посредством подавления прямых эффектов S1P на кожные и другие формы фибробластов, которые приводят к прогрессии заболевания.
S1P и глазные заболевания и состояния
Патологический или аномальный ангиогенез/неоваскуляризация, аномальное ремоделирование, фиброз, и рубцевание, и воспаление происходят в связи с заболеваниями сетчатки и глазными заболеваниями, такими как старческая дегенерация желтого пятна (AMD), диабетическая ретинопатия (DR) и ретролентальная фиброплазия (ROP) и другие связанные с развитием нарушения [Eichler, et al. (2006), Curr Pharm Des, vol.12:2645-60], a также в результате инфекций и механического повреждения глаза [Ciulla, et al. (2001), Curr Opin Ophthalmol, vol.12:442-9 и Dart et al. (2003), Eye, vol.17:886-92]. Полагают, что антитела против S1P будут пригодны для лечения глазных заболеваний, компонентом которых является патологический или аномальный ангиогенез/неоваскуляризация, аномальное ремоделирование, фиброз и рубцевание или воспаление.
Ангиогенез/неоваскуляризация глаза:
Патологический глазной ангиогенез является основной причиной слепоты при множестве клинических состояний. Хориоидальная неоваскуляризация (CNV) происходит при ряде глазных заболеваний, наиболее распространенными из которых является экссудативная или "влажная" форма AMD. В результате все в большей степени возрастной популяции AMD в настоящее время является эпидемической и представляет собой основную причину слепоты в западном мире у пациентов старше 60 лет. Несмотря на эпидемическую потерю зрения, вызываемую AMD, только немногие способы лечения, главным образом, направленные против VEGF, могут замедлить прогрессию AMD, и еще меньшее их количество может привести к восстановлению после потери зрения [Bylsma and Guymer (2005), Clin Exp Optom, vol.88:322-34, Gryziewicz (2005), Adv Drug Deliv Rev, vol.57:2092-8, и Liu and Regillo (2004), Curr Opin Ophthalmol, vol.15:221-6]. Таким образом, разработка новых способов лечения патологической неоваскуляризации является крайне важной.
AMD используют в настоящем документе только для иллюстративных целей при описании глазных состояний, связанных с аномальным ангиогенезом/неоваскуляризацией, аномальным ремоделированием, фиброзом, и рубцеванием, и воспалением, и эти состояния встречаются при других глазных заболеваниях и нарушениях, как описано и заявлено в настоящем документе. AMD включает связанные с возрастом патологические изменения [Tezel, Bora, and Kaplan (2004), Trends Mol Med, vol.10:417-20 и Zarbin (2004), Arch Ophthalmol, 122:598-614]. Существует множество теорий, однако точная этиология и патогенез AMD все еще остаются непонятными. Старение связано с накапливающимся окислительным повреждением, утолщением мембраны Бруха и образованием друз. Окислительный стресс приводит к повреждению клеток пигментного эпителиалия сетчатки (RPE) и, в некоторых случаях, хориокапилляров [Zarbin (2004), Arch Ophthalmol, vol.122:598-614, и Gorin, et al. (1999), Mol Vis, vol.5:29]. Повреждение RPE, вероятно, приводит к хроническому воспалительному ответу в мембране Бруха и сосудистой оболочке [Johnson et al. (2000), Exp.Eye Res, vol.70:441-9]. Это повреждение и воспаление обеспечивает и усиливает повреждение сетчатки посредством стимуляции CNV и атрофии [Zarbin (2004), Arch Ophthalmol, vol.122:598-614, и Witmer, et al. (2003), Prog Retin Eye Res, vol.22:1-29]. CNV приводит к дефектным и проницаемым кровеносным сосудам (BV), которые, вероятно, распознаются как рана [Kent and Sheridan (2003), Mol. Vis, vol.9:747-55]. В сосудистой оболочке возникает заживление раны и вторгается в субретинальное пространство через мембрану Бруха и RPE. Ответы в виде заживления ран характеризуются типичным ранним воспалительным ответом, выраженным ангиогенным ответом и образованием ткани с последующим конечным созреванием всех вовлеченных элементов. Ремоделирование раны может необратимо нарушить фоторецепторы и RPE, таким образом, обосновывая необходимость в лечении CNV не только антиангиогенными лекарственными средствами [La Cour, Kiilgaard, and Nissen (2002), Drugs Aging, vol.19:101-33,12].
Изменения в нормальной ретинальной и субретинальной структуре в результате связанного с CNV фиброза, отека и воспаления, отдельно или в совокупности, приводят к связанной с AMD потере зрения [Tezel and Kaplan (2004), Trends Mol Med, vol.10:417-20, и Ambati, et al. (2003), Surv Ophthalmol, vol.48:257-93]. Множество клеточных и цитокиновых взаимодействий, которые связаны с экссудативной AMD, значительно осложняют поиск эффективных способов лечения. Несмотря на то что CNV и отек частично поддаются лечению лекарственными средствами против VEGF, потенциальные способы лечения для смягчения рубцевания и воспаления еще не были адекватно рассмотрены [Bylsma and Guymer (2005), Clin Exp Optom, vol.88:322-34, и Pauleikhoff (2005), Retina, vol.25:1065-84]. Пока неоваскулярный комплекс остается интактным, как, по-видимому, происходит в случае пациентов, которых лечат средствами против VEGF, сохраняется потенциал в отношении субретинального фиброза и последующей потери зрения.
Лекарственные средства против VEGF-A представляют собой современное существенное достижение в лечении экссудативной AMD. Однако испытания фазы III VISION с PEGAPTANIB, высокоаффинным аптамером, который селективно ингибирует изоформу 165 VEGF-A, продемонстрировали, что в среднем пациент продолжает терять зрение, и только у небольшой доли улучшалось зрение [Gragoudas, et al. (2004), N Engl J. Med, vol.351:2805-16]. Ингибирование всех изоформ VEGF-A (общее ингибирование VEGF) фрагментом антитела RANIBIZUMAB привело к значительно более впечатляющим результатам [Brown, et al., N Eng Med. (2006), vol.355:1432-44, Rosenfeld, et al. N Eng J. Med. (2006), vol.355:1419-31]. Испытание в течение 2 лет MARINA и испытание в течение 1 года ANCHOR продемонстрировали, что приблизительно у 40% пациентов происходит некоторое улучшение зрения. Несмотря на то что эти результаты представляют собой значительное достижение в способности лечить экссудативную AMD, они также показывают, что 60% пациентов не имеют улучшения зрения. Более того, эти пациенты должны были удовлетворять строго определенным критериям включения и исключения. Результаты в более крупной популяции пациентов могут быть менее высокими.
Все еще существует явная необходимость в разработке дополнительных лекарственных средств, которые нацелены на другие стадии при развитии CNV и процессы, которые в конечном итоге приводят к разрушению фоторецепторов. Во-первых, рост хориоидальных BV вовлекает организованное взаимодействие множества медиаторов, а не только VEGF, дающих возможность модулировать или ингибировать весь процесс [Gragoudas, et al. (2004), N Engl J. Med, vol.351:2805-16]. Во-вторых, экссудативная AMD состоит из сосудистых и внесосудистых компонентов. Сосудистый компонент вовлекает эндотелиальные клетки (ЕС) сосудов, предшественники ЕС и перициты. Внеклеточный компонент, который, как оказалось, с точки зрения объема является наиболее крупным компонентом, состоит из клеток воспалительного, глиального и ретинального пигментного эпителия (RPE) и фибробластов. Повреждение тканей может быть следствием любого компонента. Эти другие аспекты патологического процесса не охватываются современными способами лечения против VEGF. Нацеливание на дополнительные элементы ангиогенного каскада, ассоциированного с AMD, может обеспечить более эффективный и синергичный подход к лечению [Spaide, RF (2006), Am. J. Ophthalmol, vol.141:149-156].
Воспаление при заболевании глаз:
Существуют накапливающиеся данные о том, что воспаление, конкретно, макрофаги и система комплемента [Klein, et al. (2005), Science, vol.308:385-9; и Hageman, et al. (2005), Proc. Natl. Acad Sci USA, vol.102:7227-32], играют важную роль в патогенезе экссудативной AMD. Гистопатология хирургически вырезанных хориоидальных неоваскулярных мембран демонстрирует, что макрофаги представлены практически повсеместно [Grossniklaus, et al. (1994), Ophthalmology, vol.101:1099-111, и Grossniklaus, et al. (2002), Mol. Vis, vol.8:119-26]. Существуют накапливающиеся доказательства того, что макрофаги могут играть активную роль в опосредовании образования и распространения CNV [Grossniklaus, et al. (2003), Mol. Vis, vol.8:119-26; Espinosa-Heidmann, et al. (2003), Invest Ophthalmol Vis Sci, vol.44:3586-92; Oh, et al. (1999), Invest Ophthalmol Vis Sci, vol.40:1891-8; Cousins, et al. (2004), Arch Ophthalmol, vol.122:1013-8; Forrester (2003), Nat Med, vol. 9:1350-1, и Tsutsumi, et al. (2003), J. Leukoc Biol, vol.74:25-32] посредством множества эффектов, включающих секрецию ферментов, которые могут разрушать клетки и осуществлять деградацию мембраны Бруха, а также высвобождать проангиогенные цитокины [Otani, et al. (1999), Ophthalmol Vis Sci, vol.40:1912-20, и Amin, Puklin, and Frank (1994), Invest Ophthalmol Vis Sci, vol.35:3178-88]. В области повреждения макрофаги проявляют микроморфологические признаки активации, такие как дегрануляция [Oh, et al. (1999), Invest Ophthalmol Vis Sci, vol.40:1891-8, и Trautmann et al. (2000), J. Pathol, vol.190:100-6]. Таким образом, полагают, что молекула, которая ограничивает инфильтрацию макрофагов в хориоидальный неоваскулярный комплекс, может способствовать ограничению образования CNV.
Хориоидальная неоваскуляризация и созревание кровеносных сосудов при заболевании глаз:
Ангиогенез представляет собой необходимый компонент нормального заживления раны, поскольку он обеспечивает доставку кислорода и питательных веществ воспалительным клеткам и способствует удалению дебриса [Lingen (2001), Arch Pathol Lab Med, vol.125:67-71]. Прогрессирующий ангиогенез состоит из двух различных процессов: стадия I: миграция сосудистых ЕС в ответ на близлежащие стимулы к концам капилляров, где они пролиферируют и образуют имеющие просвет структуры; и стадия II: укорочение сосудистой сети и оптимизация сосудов [Guo, et al. (2003), Am. J. Pathol, vol.162:.1083-93].
Стадия I: неоваскуляризация. Ангиогенез наиболее часто способствует заживлению ран. Однако новые сосуды, в отсутствие контроля, обычно являются дефектными и обеспечивают проницаемость, геморрагию и воспаление. Снижение дисфункциональных и проницаемых BV посредством нацеливания на проангиогенные GF показало некоторую способность замедлять прогрессию AMD [Pauleikhoff (2005), Retina, vol.25:1065-84,14, и van Wijngaarden, Coster, and Williams (2005), JAMA, vol.293:1509-13].
Стадия II: Созревание кровеносных сосудов и снижение чувствительности к лекарственному средству. Как оказалось, общее ингибирование VEGF оказывает благоприятный эффект, главным образом посредством действия, направленного против проницаемости, что приводит к устранению внутри- и субретинального отека, поскольку фактическое повреждение CNV по существу не подвергается инволюции. Отсутствие существенной инволюции CNV частично может быть результатом созревания вновь образованных сосудов вследствие покрытия перицитами. Перициты играют критическую роль в развитии и поддержании сосудистой ткани. Наличие перицитов, по-видимому, придает устойчивость к средствам против VEGF и препятствует их способности ингибировать ангиогенез [Bergers and Song (2005), Neuro-oncol, vol.7:452-64; Yamagishi and Imaizumi (2005), Int J. Tissue React, vol.27:125-35; Armulik, Abramsson and Betsholtz (2005), Circ Res, vol.97:512-23; Ishibashi et al. (1995), Arch Ophthalmol, vol.113:227-31]. Средство, которое обладает ингибиторным эффектом на привлечение перицитов, вероятно, будет нарушать сборку сосудистых каналов и созревание хориоидальных неоваскулярных каналов, таким образом, сохраняя их чувствительность к антиангиогенным средствам.
Ремоделирование сосудистой сети вовлекает коррекцию плотности кровеносных сосудов (BV) для удовлетворения потребности в питательных веществах [Gariano and Gardner (2005), Nature, 438:960-6]. Периоды незрелости BV соответствуют периоду, в ходе которого новые сосуды функционируют, но еще не приобрели оболочку из перицитов [Benjamin, Hemo, and Keshet (1998), Development, 125:1591-8, и Gerhardt and Betsholtz (2003), Cell Tissue Res, 2003, 314:15-23]. Это замедление является необходимым для обеспечения окна пластичности для точной регуляции развивающихся сосудов согласно потребностям сетчатки или сосудистой оболочки в питательных веществах.
Биологически активный липид сфингозин-1-фосфат (S1P), VEGF, PDGF, ангиопоэтины (Ang) и другие факторы роста (GF) усиливают рост кровеносных сосудов и привлекают гладкомышечные клетки, (SMC) и перициты к новым сосудам, которые обеспечивают ремоделирование появляющихся сосудов [Allende and Proia (2002), Biochim Biophys Acta, vol.582:222-7; Gariano and Gardner (2005), Nature, vol.438:960-6; Grosskreutz, et al. (1999), Microvasc Res, vol.58:128-36; Nishishita, and Lin (2004), J. Cell Biochem, vol.91:584-93, и Erber, et al. (2004), FASEB J, vol.18:338-40,32]. Перициты, наиболее вероятно, образующиеся посредством дифференцировки in situ мезенхимных предшественников во время прорастания ЕС или вследствие миграции и дедифференцировки артериальных гладкомышечных клеток, тесно объединяются и оборачивают ЕС, что приводит, в целом, к созреванию и выживанию сосудов [Benjamin, Hemo, and Keshet (1998), Development, vol.125:1591-8]. Недавние исследования продемонстрировали, что SIP и рецептор S1P1 вовлечены в транспорт на клеточной поверхности и активацию молекулы межклеточной адгезии N-кадгерина [Paik, et al. (2004), Genes Dev, vol.18:2392-403]. N-кадгерин необходим для взаимодействий между ЕС, перицитами и клетками стенки, которые обеспечивают развитие стабильного сосудистого ложа [Gerhardt and Betsholtz (2003), Cell Tissue Res, vol.314:15-23]. Глобальная делеция гена SIP1 приводит к аномальному оборачиванию клеток стенки появляющихся BV, требуемому для стабилизации BV в ходе эмбрионального развития [Allende and Proia (2002), Biochim Biophys Acta, vol.1582:222-7].
Локальная инъекция siRNA к S1P1 подавляет стабилизацию сосудов в моделях ксенотрансплантатов опухолей [Сhае, et al. (2004), J. Chin Invest, vol.114:1082-9]. Исследования на трансгенных мышах показали, что VEGF и PDGF-B обеспечивают созревание и стабилизацию новых BV [Guo, et al. (2003), Am J. Pathol, 162:1083-93, и Gariano and Gardner (2005), Nature, vol.438:960-6,50]. VEGF активирует Ang-1 (мРНК и белок) [Asahara, et al. (1998), Circ Res, vol.83:233-40]. Ang-1 играет основную роль в привлечении перицитов и поддержании перицитами периэндотелиальной опоры [Asahara, et al. (1998), Circ Res, vol.83:233-40]. Внутриглазная инъекция VEGF ускоряла оборачивание перицитами сплетения ЕС [Benjamin, Hemo, and Keshet (1998), Development, vol.125:1591-8]. Эмбрионы мышей с дефицитом PDGF-B лишены микрососудистых перицитов, что приводит к отеку, микроаневризмам и летальным кровоизлияниям [Lindahl, et al. (1997), Science, vol.277:242-5]. Пренатальные исследования на мышах показали, что для полной стимуляции посредством VEGF и PDGF созревания сосудистого ложа требуются дополнительные сигналы. Основываясь на трансактивации S1P, указанной выше, этим фактором может быть S1P [Erber et al. (2004), FASEB J, vol.18:338-40]. Стабилизация и созревание сосудов ассоциированы с потерей пластичности и отсутствием регрессии при устранении VEGF и других GF и устойчивостью к антиангиогенной терапии [Erber, et al. (2004), FASEB J, vol.18:338-40, и Hughes and Chan-Ling (2004), Invest Ophthalmol Vis Sci, vol.45:2795-806]. Устойчивость BV к ингибиторам ангиогенеза обеспечивается перицитами, которые исходно стабилизируют созревшие сосуды, и перицитами, которые привлекаются к незрелым сосудам при терапии [Erber, et al. (2004), FASEB J, vol.18:338-40]. После оборачивания незрелых ЕС перициты экспрессируют компенсаторные факторы выживания (Ang-1 и PDGF-B), которые защищают ЕС от проапоптотических средств.
Отек и проницаемость сосудов при заболевании глаз:
Мембраны CNV состоят из фенестрированных сосудистых ЕС, которые имеют тенденцию к выходу их внутрисосудистого содержимого в окружающее пространство, что приводит к субретинальной геморрагии, экссудатам и накоплению жидкости [Gerhardt and Betsholtz (2003), Cell Tissue Res, vol.14:15-23]. В течение многих лет непосредственно CNV ткани и позднее интранеоваскуляризация сетчатки предполагались как ответственные за снижение остроты зрения, ассоциированное с AMD. Однако в настоящее время полагают, что основную роль в потере зрения, ассоциированной с AMD и другими заболеваниями глаз, включая слепоту, связанную с диабетом, играет отек желтого пятна, вызываемый повышением проницаемости сосудов (VP) и последующим разрушением гематоретинального барьера (BRB). [Hughes and Chan-Ling (2004), Invest Ophthalmol Vis Sci, vol.45:2795-806; Felinski and Antonetti (2005), Curr Eye Res, vol.30:949-57; Joussen, et al. (2003), FASEB J, vol.17:76-8, и Strom, et al. (2005), Invest Ophthalmol Vis Sci, vol.46:3855-8]. В частности, диабетическая ретинопатия (DR) и диабетический отек желтого пятна (DME) являются распространенными микрососудистыми осложнениями у пациентов с диабетом и являются наиболее распространенными причинами связанной с диабетом слепоты. DME является следствием повышенной микрососудистой проницаемости. Joussen, et al. (2003), FASEB J, vol.17:76-8. Совместно они представляют собой наиболее распространенную причину вновь возникающей слепоты в популяции трудоспособного возраста. Полагают, что соединения, такие как антитела, которые нацелены на S1P, будут терапевтически полезны при таких состояниях.
Примеры нескольких классов глазных заболеваний, которые можно лечить в соответствии с изобретением, описаны ниже. Будет понятно, что множество заболеваний и состояний характеризуются, по меньшей мере частично, множеством патологических процессов (например, как патологической неоваскуляризацией, так и рубцеванием) и что классификации, представленные в настоящем документе, представлены для удобства описания и не ограничивают изобретение.
а. Ишемические ретинопатии, связанные с патологической неоваскуляризацией, и заболевания, характеризующиеся образованием эпиретинальной и субретинальной мембраны.
Ишемические ретинопатии (IR) представляют собой группу разнообразных нарушений, характеризующуюся нарушенным кровотоком в сетчатке. Примеры IR включают диабетическую ретинопатию (DR), ретролентальную фиброплазию (ROP), серповидно-клеточную ретинопатию и окклюзивное заболевание вен сетчатки. Все из этих нарушений могут быть связаны с запускаемой VEGF пролиферацией с патологической неоваскуляризацией сетчатки, которая в конечном итоге может приводить к внутриглазному кровоизлиянию, образованию эпиретинальной мембраны и тракционному отслоению сетчатки. Идиопатические эпиретинальные мембраны (ERM), также называемые складками желтого пятна или циллофановой ретинопатией, могут вызвать снижение зрения, вторичное по отношению к деформации структуры сетчатки. Эти мембраны иногда рецидивируют, несмотря на хирургическое удаление, и иногда они связаны с ишемией сетчатки. VEGF и его рецепторы локализованы на ERM. Присутствие VEGF в мембранах связано с пролиферативной диабетической ретинопатией, пролиферативной витреоретинопатией, и складки желтого пятна далее подтверждают, что этот цитокин играет важную роль в ангиогенезе при ишемическом заболевании сетчатки и в росте мембраны при пролиферативных витреоретинальных нарушениях. Кроме того, на клетках в ERM также идентифицированы рецепторы VEGF VEGFR1 и VEGFR2. Эти данные показывают, что VEGF может быть аутокринным и/или паракринным стимулятором, который может приводить к прогрессии сосудистой и бессосудистой ERM. PDGF и его рецепторы [Robbins, et al. (1994), Invest OphthalmolVis Sci; vol.35:3649-3663] описаны при пролиферативных заболеваниях сетчатки глаз [Cassidy, et al. (1998), Br J. Ophthamol; vol.82:181-85, и Freyberger, et al. (2000), Exp. Clin. Endocrinol. Diabetes, vol.108:106-109]. Эти открытия позволяют предположить, что лиганды и рецепторы PDGF широко распространены в пролиферативных мембранах сетчатки различного происхождения, и подтверждают, что в патогенез ERM может быть вовлечена аутокринная и паракринная стимуляция PDGF. Трансформирующий фактор роста-β (TGF-β) вовлечен в образование ERM [Pournaras, et al. (1998), Klin Monatsbl Augenheilkd, vol.212:356-358], как показано по окрашиванию по TGF и иммунореактивности. Кроме того, рецептор II для TGF-β экспрессируется в миофибробластах ERM диабетиков и мембранах PVR. Эти результаты позволяют предположить, что TGF-β, продуцируемый во множестве типов клеток в сетчатке и ERM, является привлекательной мишенью для лечения PVR, диабетических и вторичных ERM. Было описано, что уровень интерлейкина-6 (IL-6) повышен в стекловидном теле человека при пролиферативной диабетической ретинопатии (PDR) [La Heij, et al. (2002), Am J. Ophthal, 134:367-375], и в одном исследовании 100% исследованных диабетических ERM экспрессировали белок IL-6 [Yamamoto, et al. (2001) Am J. Ophthal, vol.132:369-377].
Было показано, что экзогенное введение основного фибробластного фактора роста (bFGF) индуцирует пролиферацию эндотелия и экспрессию VEGF [Stavri, et al. (1995), Circulation, vol.92:11-14]. Согласно этим наблюдениям концентрация bFGF возрастает в образцах стекловидного тела от пациентов с PDR [Sivalingam, et al. (1990), Arch Ophthalmol, vol.108:869-872, и Boulton, et al. (1997), Br J. Ophthalmol, vol.81:228-233]. bFGF также вовлечен в образование ERM [Hueber, et al. (1996), Int. Ophthalmol, vol.20:345-350], и bFGF продемонстрирован в 8 из 10 исследованных мембран PDR. Более того, эти исследователи выявили положительное окрашивание по соответствующему рецептору, FGFR1. Иммунореактивность к bFGF также была показана в несосудистых идиопатических ERM. Эти результаты указывают на вовлечение bFGF в образование как сосудистых, так и бессосудистых ERM. Harada, et al. (2006), Prog in Retinal и Eye Res, vol.25:149-164. Также повышенный уровень bFGF был выявлен в сыворотке пациентов с ROP (Becerril, et al. (2005), Ophthalmology, vol.112, 2238].
С учетом известных плейотропных эффектов S1P и его взаимодействий с VEGF, bFGF, PDGF, TGF-β и IL-6 полагают, что средство, которое связывает, осуществляет антагонизм, ингибирует эффекты или продукцию S1P, будет эффективным в отношении подавления патологической неоваскуляризации сетчатки при ишемической ретинопатии и заболеваниях задних сегментов, характеризующихся образованием сосудистой или бессосудистой ERM. Другие глазные состояния, характеризующиеся, по меньшей мере частично, аномальной неоваскуляризацией или ангиогенезом, включают старческую дегенерацию желтого пятна, отторжение трансплантата роговицы, неоваскулярную глаукому, изнашивание контактных линз, инфекции роговицы, включая инфекцию вирусом простого герпеса, опоясывающего герпеса и протозойную инфекцию, птеригий, инфекционный увеит, хроническое отслоение сетчатки, повреждение лазером, серповидно-клеточную ретинопатию, окклюзивное заболевание вен, хориоидальную неоваскуляризацию, ретинальную ангиоматозную пролиферацию и идиопатическую полипоидальную хориоидальную васкулопатию.
b. Пролиферативная витреоретинопатия (PVR)
PVR наблюдают после спонтанного регматогенного отслоения сетчатки и после травматического отслоения сетчатки. Она является основной причиной неуспешной хирургической операции по поводу отслоения сетчатки. Она характеризуется ростом и уплотнением клеточных мембран на обеих сторонах сетчатки, на задней поверхности стекловидного тела и на основании стекловидного тела. Это избыточное развитие рубцовой ткани в глазу может привести к развитию тракционного отслоения сетчатки, и, таким образом, способы лечения, направленные на предотвращение или ингибирование пролиферативной витреоретинопатии (PVR), являются логическим принципом лечения отслоения сетчатки. Гистопатологически PVR характеризуется избыточной продукцией коллагена, уплотнением и пролиферацией клеток [Michels, Retinal Detachment 2nd Edition. Wilkinsin CP, Rice ТА Eds, Complicated types of retinal detachment, pp.641-771, Mosby St Louis 1997]. Типы клеток, идентифицированные в мембранах PVR, включают, главным образом, клетки пигментного эпителия сетчатки, фибробласты, макрофаги и эндотелиальные клетки сосудов [Jerdan, et al. (1989), Ophthalmology, vol.96:801-10, и Vidinova, et al. (2005), Klin Monatsbl Augenheilkd; vol.222:568-571]. Патофизиология этой реакции в виде избыточного рубцевания, по-видимому, опосредуется рядом цитокинов, включая тромбоцитарный фактор роста (PDGF), трансформирующий фактор роста (TGF) бета, основный фибробластный фактор роста (bFGF), интерлейкин-6 (IL)-6 и интерлейкин-8 (IL)-8 [Nagineni, et al. (2005), J. Cell Physiol, vol.203:35-43; La Heij, et al. (2002), Am J. Ophthalmol, 134:367-75; Planck, et al. (1992), Curr Eye Res; vol. 11:1031-9; Canataroglu et al. (2005) Ocul Immunol. Inflamm; vol.13:375-81, и Andrews, et al. (1999), Ophthalmol Vis Sci; vol.40:2683-9]. Ингибирование этих цитокинов может помочь предотвратить развитие PVR при своевременном введении или ограничить ее тяжесть [Akiyama, et al. (2006), J. Cell Physiol, vol.207:407-12, и Zheng, et al. (2003), Jpn J. Ophthalmolm, vol.47:158-65].
Сфингозин-1-фосфат (SIP) представляет собой биологически активный лизолипид с плейотропными эффектами. Он является проангиогенным, провоспалительным (стимулирует привлечение макрофагов и тучных клеток) и профибротическим (стимулирует рубцевание). Главным образом S1P стимулирует пролиферацию и миграцию клеток и является антиапоптотическим. S1P обеспечивает эти биологически разнообразные функции посредством его взаимодействий со множеством цитокинов и факторов роста. Было показано, что ингибирование S1P моноклональным антителом (SPHINGOMAB) блокирует функции сосудисто-эндотелиального фактора роста (VEGF), bFGF, IL-6 и IL-8 [Visentin, В et al. (2006), Cancer Cell, vol.9:1-14]. Связывание SIP с рецептором SIP1 может также повысить продукцию PDGF; таким образом, можно ожидать, что средство, которое связывает S1P, также снизит продукцию PDGF [Milstien and Spiegel (2006), Cancer Cell, vol.9:148-150]. Как показано в разделе "Примеры" ниже, в настоящее время продемонстрировано, что in vitro SIP трансформирует клетки RPE человека в миофибробласт-подобный фенотип, сходный с типом, наблюдаемым при PVR. С учетом патофизиологии, которая в конечном итоге приводит к избыточному рубцеванию, наблюдаемому при PVR, и известных эффектов S1P на эти же ключевые медиаторы полагают, что средство, которое связывает, осуществляет антагонизм или ингибирует эффекты или продукцию S1P, будет эффективным в отношении устранения или минимизации развития PVR и его вызывающих тяжелое повреждение эффектов на глаз.
с. Увеит
Увеит представляет собой воспалительное нарушение увеального тракта глаза. Он может поражать лицевую (переднюю) часть или обратную (заднюю) часть глаза, или обе из них. По этиологии он может быть идиопатическим или инфекционным, и он может угрожать потерей зрения. Идиопатический увеит ассоциировали с повышенной экспрессией CD4+ в передней камере [Calder, et al. (1999), Invest Ophthalmol Vis Sci, vol.40:2019-24]. Также данные позволяют сделать предположение о патологической роли Т-лимфоцитов и их хемоаттрактанта IP-10 в патогенезе увеита [Abu El-Asrar (2004), Am J. Ophthalmol, vol.138:401-11]. Другие хемокины при остром переднем увейте включают воспалительные белки макрофагов, хемоаттрактантный белок-1 и IL-8. Эти цитокины, вероятно, играют критическую роль в привлечении лейкоцитов при остром переднем увейте. Verma, et al. (1997), Curr Eye Res; vol.16:1202-8. С учетом сильных и плейотропных эффектов каскада передачи сигнала S1P полагают, что SPHINGOMAB и другие иммунные молекулы, которые снижают эффективную концентрацию биологически активного липида, могут служить в качестве эффективного способа снижения или модулирования внутриглазного воспаления, связанного с увеитом.
d. Рефракционная хирургия
Ответ в виде заживления ран роговицы имеет особое значение для рефракционных хирургических процедур, поскольку он является основным определяющим фактором безопасности и эффективности. Эти процедуры проводят для лечения миопии, гиперопии и астигматизма. Лазерный кератомилез in situ (LASIK) и фоторефракционная кератэктомия (PRK) являются наиболее распространенными рефракционными процедурами, однако в попытках преодолеть осложнения были разработаны другие процедуры. Эти осложнения включают в числе прочих гиперкоррекцию, недостаточную коррекцию, регрессию и стромальное помутнение. Ряд распространенных осложнений связан с ответом в виде заживления, и они имеют корни в биологическом ответе на хирургическую операцию. Одной из наиболее важных задач биологии роговицы является обеспечение репарации тканей через регенерацию, а не через фиброз. Полагают, что выбор между регенерацией и фиброзом основан на контроле активации фибробластов [Stramer, et al. (2003), Invest Ophthalmol Vis Sci; vol.44:4237-4246, и Fini (1999) Prog Retin Eye Res, vol.18:529-551]. Клетки, называемые миофибробластами, могут появляться в субэпителиальной строме через 1-2 недели после хирургической операции или повреждения. Миофибробласты предположительно образуются из кератоцитов под влиянием TGF-β [Jester, et al. (2003), Exp Eye Res, vol.77:581-592]. Помутнение роговицы и стромальное рубцевание характеризуются сниженной прозрачностью роговицы и могут быть ассоциированы с образованием фибробластов и миофибробластов. Исследования in situ и in vitro позволили предположить, что TGF-β и PDGF важны для стимуляции дифференцировки миофибробластов [Folger, et al. (2001), Invest Ophthalmol Vis Sci; 42:2534-2541]. Помутнение отмечалось в определенных обстоятельствах в центральной границе после LASIK. Оно включает диффузный ламеллярный кератит, лоскут, в форме пончика, и удержание эпителиального дебриса на границе. Вероятно, что каждый из этих случаев связан с повышенным доступом TGF-β из эпителиальных клеток в активированные кератоциты [Netto, et al. (2005), Cornea, vol.24:509-522]. Регрессия, наиболее вероятно, является следствием усиленных эпителиально-стромальных взаимодействий при заживлении ран, таких как повышенная продукция фибробластами роговицы и/или миофибробластами факторов роста, модулирующих эпителий [Netto, et al. (2005), выше]. Было показано, что ингибирование связывания TGF-β с рецепторами посредством антитела против TGF-β для местного применения снижает помутнение, индуцируемое PRK [Jester, et al. (1997), Cornea, vol.16:177-187]. С учетом известных эффектов антитела против биологически активного липида на фибротические процессы и TGF-β авторы настоящего изобретения полагают, что оно может способствовать лечению некоторых из осложнений рефракционной хирургической операции, таких как помутнение, стромальное рубцевание и регрессия.
е. Модулирование фильтрационной хирургической операции по поводу глаукомы
Традиционно полагают, что глаукома является заболеванием, при котором повышенное внутриглазное давление вызывает повреждение оптического нерва и в конечном итоге нарушает поле зрение или остроту зрения. Существуют другие формы глаукомы, где повреждение оптического нерва может происходить в условиях нормального давления или так называемой "нормотензивной глаукомы". У многих пациентов способы лечения способны контролировать их заболевание, однако для других требуется фильтрационная хирургическая операция по поводу глаукомы, при которой в глазу создают фистулу для обеспечения оттока жидкости. Ее можно проводить посредством трабекулоэктомии, имплантации медицинского устройства или других способов хирургического вмешательства. Неуспех фильтрационной хирургической операции по поводу глаукомы связан с процессом заживления ран, характеризующимся пролиферацией фибробластов и конечным рубцеванием. Антиметаболиты, такие как 5-фторурацил и митомицин С, могут снизить последующее рубцевание; однако даже при применении этих лекарственных средств длительное наблюдение показывает, что неуспех хирургической операции все еще остается серьезной клинической проблемой [Mutsch and Grehn (2000), Graefes Arch Clin. Exp. Ophthalmol; vol.238:884-91, и Fontana, et al. (2006), Ophthalmology, vol.113:930-936]. Исследования фибробластов теноновой капсулы демонстрируют, что они обладают способностью синтезировать bFGF и PDGF и TGF-β и что эти факторы роста вовлечены в процесс репарации тканей после фильтрационной хирургической операции по поводу глаукомы, который приводит к неуспеху процедуры. Trpathi, et al. (1996), Exp Eye Res, vol.63:339-46. Дополнительные исследования также вовлекают эти факторы роста в постфильтрационный раневой ответ [Denk, et al. (2003), Curr Eye Res; vol.27:35-44], и было сделано заключение, что различные изоформы PDGF являются главными стимуляторами пролиферации фибробластов теноновой капсулы после хирургической операции по поводу глаукомы, в то время как TGF-β необходим для трансформации фибробластов теноновой капсулы в миофибробласты. Авторы настоящего изобретения продемонстрировали, что S1P присутствует в фибробластах теноновой капсулы/конъюнктивы и что S1P строго экспрессируется при ответе в виде заживления ран. S1P также стимулирует профибротическую функцию множества фибробластных типов клеток и трансформацию в миофибробластный фенотип и продукцию коллагена. С учетом определенных плейотропных эффектов S1P и его известных взаимодействий с bFGF, PDGF и TGF-бета полагают, что средство, которое связывает, осуществляет антагонизм, ингибирует эффекты или продукцию S1P, будет эффективным для модулирования заживления ран и/или фибротического ответа, который приводит к неуспеху хирургической операции при глаукоме и будет представлять собой эффективный терапевтический способ повышения количества успешных исходов хирургической операции. Предусматривается, что средство можно вводить, например, посредством инъекции в стекловидное тело или субъконъюнктивальной инъекции или местно.
f. Трансплантация роговицы
Трансплантация роговицы (сквозная кератопластика (РК)) представляет собой наиболее успешную операцию трансплантации тканей у человека. Хотя в США ежегодно проводят 47000 трансплантаций роговицы, отторжение аллотрансплантата роговицы все еще является ведущей причиной неуспеха трансплантации роговицы [Ing, et al. (1998), Ophthalmology, vol.105:1855-1865]. В настоящее время способность предотвращать отторжение трансплантата является недостаточной, хотя иммуносупрессия и иммуномодулирование могут быть перспективным подходом. Недавно было выявлено, что при отторжении аллотрансплантатов роговицы CD4(+) Т-клетки функционируют также прямо, как и эффекторные клетки, а не как хелперные клетки. Hegde, et al. (2005), Transplantation, vol.79:23-31. Исследования на мышах показали повышенные количества нейтрофилов, макрофагов и тучных клеток в строме роговицы, подвергающейся отторжению. Макрофаги были основным инфильтрирующим типом клеток, за которым следовали Т-клетки, тучные клетки и нейтрофилы. При трансплантации роговицы с высоким риском происходила ранняя экспрессия хемокинов IL-8 (воспалительный белок макрофагов-2) и хемотаксического белка моноцитов-1 (МСР-1) [Yamagami, et al. (2005), Mol. Vis, vol.11, 632-40].
FTY720 (FTY) представляет собой новое иммуносупрессивное лекарственное средство, которое действует, изменяя движение лимфоцитов, что приводит к лимфопении периферической крови и повышенному количеству лимфоцитов в лимфатических узлах. FTY опосредует его иммуномодулирующие эффекты посредством связывания с некоторыми из рецепторов S1P, экспрессированных на лимфоцитах [Bonier, et al. (2005), Transplantation, vol.79:492-5]. Лекарственное средство вводят перорально, и единичная пероральная доза снижает количество лимфоцитов на 30-70%. FTY снижал количество Т-клеток подгруппы CD4(+) в большей степени, чем подгруппы клеток CD8(+). Bohler, et al. (2004), Nephrol Dial Transplant, vol.19:702-13. У мышей, которым вводили FTY, было показано существенное продление выживания ортотопного трансплантата роговицы при пероральном введении. Zhang, et al. (2003), Transplantation, vol.76:1511-3. Пероральное введение FTY также существенно замедляло и снижало его тяжесть в модели ксенотрансплантации роговицы от крысы к мыши [Sedlakova, et al. (2005), Transplantation, vol.79, 297-303]. С учетом известного патогенеза отторжения аллотрансплантата в сочетании с данными, позволяющими предположить, что модулирование эффектов передачи сигнала S1P может повысить выживание трансплантата роговицы, полагают, что иммунные молекулы, которые снижают эффективную концентрацию биологически активных липидов, например SPHINGOMAB, также будут пригодны для лечения иммунологических состояний, таких как отторжение аллотрансплантата, например, посредством подавления иммунного ответа, и, таким образом, вероятно, повысят выживание трансплантата роговицы после PK. Лекарственное средство также может обладать дополнительным преимуществом, состоящим в том, что в дополнение к системному введению возможно местное введение, например, посредством местной окологлазной или внутриглазной доставки.
Другие глазные заболевания с воспалительным или иммунным компонентом включают хронический витрит, инфекции, включая инфекцию вирусом простого герпеса, опоясывающего герпеса, и протозойные инфекции и гистоплазмоз глаз.
g. Заболевания переднего сегмента, характеризующиеся рубцеванием
Полагают, что лечение антителом, нацеленным на биологически активный липид, также является полезным при некоторых состояниях, характеризующихся рубцеванием в передней области глаза. Они включают следующие состояния:
i. Травма
Роговица, как наиболее передняя структура глаза, подвергается воздействию различных вредных факторов, варьирующих от переносимых воздухом частиц до тупой травмы, которая может привести к механической травме. Роговица и передняя поверхность глаза также могут подвергаться воздействию других форм травмы вследствие хирургических и химических, таких как кислоты и щелочи, повреждений. Результаты этих типов повреждений могут быть изнуряющими и часто приводят к образованию рубцового симблефарона роговицы и конъюнктивы. Кроме того, результатом может быть неоваскуляризация роговицы. Нейтрофилы накапливаются, а высвобождение ими лейкотриенов и присутствие интерлейкина-1 и интерлейкина-6 служат для привлечения последующей волны воспалительных клеток [Sotozono, et al. (1997), Curr Eye Res, vol.19:670-676], которые инфильтрируют роговицу и высвобождают протеолитические ферменты, которые приводят к дальнейшему повреждению и разрушению ткани роговицы и плавлению роговицы. Кроме того, фибробласты роговицы и конъюнктивы становятся активированными и вторгаются и вызывают отложение коллагена и фиброз. Нежелательные эффекты избыточного воспаления и рубцевания обеспечиваются посредством TGF-β. Saika, et al. (2006), Am J Pathol vol.168:1848-60. Этот процесс приводит к потере прозрачности роговицы и нарушению зрения. Снижение воспаления, включая снижение инфильтратов нейтрофилов и снижение фиброза, приводили к более быстрому и более полному заживлению в модели ожога роговиц щелочью на мышах [Ueno, et al. (2005), Ophthalmol. Vis Sci, vol.46:4097-106].
ii. Глазной рубцовый пемфигоид (ОСР)
ОСР представляет собой хроническое рубцовое (образующее рубцы) аутоиммунное заболевание, которое главным образом поражает конъюнктиву. Заболевание является непременно прогрессирующим, и его прогноз является очень плохим. На его последней стадии рубцевание в роговице и связанная с ним кератопатия приводят к билатеральной слепоте. Гистологически на конъюнктиве видны подслизистое рубцевание и хроническое воспаление, в котором область тучных клеток является неожиданно большой [Yao, et al. (2003), Ocul Immunol I nflanun, vol.11:211-222]. Аутоантигены приводят к образованию аутоантител. Связывание. аутоантитела с аутоантигеном запускает комплексную серию событий с инфильтрацией Т-лимфоцитов, где количество CD4-клеток (хелперы) значительно превосходит количество CD8-клеток (супрессоров). Также результатом является инфильтрация макрофагов и тучных клеток, также как и высвобождение провоспалительных и профибротических цитокинов. Индуцируемая цитокинами пролиферация и активация фибробластов конъюнктивы приводят к субэпителиальному фиброзу (см. примеры в настоящем документе ниже). В исследованиях была показана роль TGF-β и IL-1 в фиброзе конъюнктивы у пациентов с ОСР [Razzaque, et al. (2004), Invest Ophthalmol. Vis Sci, vol.45:1174-81].
iii. Синдром Стивенса-Джонсона (SJS) и токсический эпидермальный некролиз (TEN)
SJS и TEN представляют собой опасные для жизни побочные реакции на лекарственные средства. Глазные осложнения этих двух связанных состояний могут быть тяжелыми и включают изменения конъюнктивы глазного яблока и конъюнктивы век, век и роговицы. Лекарственные средства и инфекции являются наиболее распространенными ускоряющими факторами. Хронические глазные признаки включают рубцевание, образование симблефарона и рубцевание конъюнктивы в результате исходного воспалительного процесса. Это приводит к образованию энтропиона, трихиазу и нестабильности слезной пленки. Повреждение поверхности глаза приводит к рубцеванию, неоваскуляризации и, в тяжелых случаях, к кератинизации. Как в случае ОСР, происходит субэпителиальный фиброз конъюнктивы. Полагают, что в развитии SJS/TEN участвует сильный аутоиммунный ответ лимфоцитов на лекарственное средство или инфекцию. Harilaos, et al. (2005), Erythema Multiforme, Stevens Johnson Syndrome, and Toxic Epidermal Necrolysis, Cornea 2nd edition. Krachmer, Mannis, Holland eds. Elesevier Mosby Philadelphia. Инфильтрирующая клеточная популяция при SJS включает макрофаги, СD4-положительные Т-клетки и CD8-положительные Т-клетки. Клеточная популяция сходна с популяцией, наблюдаемой при химическом повреждении. Kawasaki, et al. (2000), J. Ophthalmol, vol.84:1191-3.
iv. Птеригий
Клинически птеригий выглядит как мясистая сосудистая масса, которая появляется в щели между веками. Тело птеригия представляет собой мясистую сосудисто-волокнистую массу. Активный птеригий характеризуется существенным переполнением и прогрессивным ростом сосудов. Они прочно прикреплены к глазному яблоку. В запущенных случаях птеригий вторгается в роговицу и может вызывать снижение зрения, вторичное по отношению к потере прозрачности роговицы в зрительной оси или к неправильному астигматизму. Симптоматически, пациенты могут испытывать ощущение чужеродного тела, слезоотделение и неясное зрение. На гистопатологии заметна гиалинизация субэпителиальной соединительной ткани собственного вещества, повышенное количество фибробластов и повышенное количество тучных клеток. Butrus, et al. (1995), Am J. Ophthalmol, vol.119:236-237. Лечение птеригия остается проблематическим. Часто проводят хирургическое удаление, однако частота рецидива является высокой [Krag, et al. (1992), Acta Ophthalmol, vol.70:530]. Для способствования снижению частоты рецидивов птеригия используют различные фармакологические адъюванты, такие как митомицин-С и даунорубицин. Хотя они могут быть полезными, данные за длительный период ограничены, и они могут быть связаны с истончением склеры и плавлением роговицы. Dougherty, et al. [(1996), Cornea, vol.15:537-540, и Lee, et al. (2001), Cornea, vol.20:238-42] впервые продемонстрировали, что VEGF может играть важную роль в развитии птеригия, и идентифицировали VEGF и оксид азота в эпителии птеригия. Эти исследователи сделали предположение о том, что эти, а также другие цитокины ответственны за сосудисто-волокнистое прорастание, характеризующее птеригий. Было продемонстрировало присутствие основного FGF и TGF-бета 1 как при первичном, так и при рецидивирующем птеригий [Kira, et al. (1998), Graefes Arch Clin Exp. Ophthalmol, vol.236:702-8], и опубликованные морфометрические и иммуногистохимические данные далее подтверждают представление о том, что ангиогенез может участвовать в образовании птеригия [Marcovich, et al. (2002), Curr Eye Res, vol.25:17-22]. Другие исследования вовлекали IL-6 и IL-8, а также VEGF в качестве медиаторов, которые могут иметь отношение к развитию птеригия [Di Girolamo, et al. (2006), Invest Ophthalmol. Vis Sci, vol.47:2430-7]. Эффективное средство против образования и роста птеригия может уменьшить необходимость в хирургическом вмешательстве или снизить частоту рецидивов. Другие глазные заболевания и состояния с компонентом в виде фиброгенеза, фиброза или рубцевания включают AMD, диабетическую ретинопатию, ретролентальную фиброплазию, серповидно-клеточную ретинопатию, ишемическую ретинопатию, окклюзивое заболевание вен сетчатки и переутомление контактными линзами.
В заключение избыточное рубцевание является основополагающим компонентом патофизиологии многих глазных и неглазных заболеваний и состояний. Биологически активные липиды, такие как S1P, участвуют в этом процессе, и лечение антителами для снижения концентраций этих веществ, вероятно, приведет к терапевтической пользе у пациентов, получающих лечение. В одном варианте осуществления полагают, что ингибиторы биологически активных липидов, в частности моноклональные антитела, направленные против S1P и/или его вариантов, пригодны для модулирования ответов в виде заживления хирургических и травматических ран.
Фиброз, фиброгенез и рубцевание:
Образование субретинального фиброза приводит к необратимому повреждению фоторецепторов и необратимому снижению зрения. При условии, что неоваскулярный комплекс остается интактным, как происходит в случае пациентов, которым вводят средства против VEGF, остается потенциал в отношении субретинального фиброза и последующего снижения зрения. В дополнение исследования PRONTO для RANIBIZUMAB (Lucentis®), было открыто, что пациенты, которые потеряли зрение, потеряли его в результате либо субретинального фиброза, либо разрыва RPE. Средство, которое может снижать степень инфильтрации фибробластов и депонирования коллагена, может быть полезным.
Фибробласты, в частности миофибробласты, представляют собой ключевые клеточные элементы при рубцевании в ответ на клеточное повреждение и воспаление [Tomasek, et al. (2002), Nat Rev Mol Cell Biol, vol.3:349-63, и Virag and Murry (2003), Am J. Pathol, vol.163:2433-40]. Экспрессия генов коллагена миофибробластами является признаком ремоделирования и необходима для рубцевания [Sun and Weber (2000), Cardiovasc Res, vol.46:250-6, и Sun and Weber (1996), J. Mol. Cell Cardiol, vol.28:851-8]. S1P обеспечивает заживление ран посредством активации миграции и пролиферации фибробластов при повышении продукции коллагена [Sun, et al. (1994), J. Biol Chem, vol.269:16512-7]. S1P, продуцируемый локально поврежденными клетками, может быть ответственен за ненадлежащее заживление ран, связанное с ремоделированием и рубцеванием. Таким образом, полагают, что ингибиторы S1P пригодны при заболеваниях или состояниях, характеризующихся, по меньшей мере частично, нарушенным фиброгенезом или фиброзом.
Образование субретинального фиброза приводит к необратимому повреждению фоторецепторов и необратимому снижению зрения. При условии, что неоваскулярный комплекс остается интактным, как происходит в случае пациентов, которых лечат средствами против VEGF, остается потенциал в отношении субретинального фиброза и снижения зрения в будущем.
Минимизация ненадлежащего рубцевания посредством нейтрализации S1P может быть полезной и может предотвратить необратимое снижение остроты зрения посредством ограничения степени субретинального фиброза и последующего повреждения фоторецепторов. Возрастающие доказательства позволяют предположить, что S1P может участвовать как в ранней, так и в поздней стадиях ненадлежащего ремоделирования сетчатки, связанного с экссудативной AMD. S1P имеет выраженный не VEGF-зависимый проангиогенный эффект. S1P также стимулирует миграцию, пролиферацию и выживание множества типов клеток, включая фибробласты, ЕС, перициты и воспалительные клетки - те же клетки, которые участвуют во множестве неадекватных процессов при экссудативной AMD и других глазных нарушениях. S1P связан с продукцией и активацией VEGF, bFGF, PDGF и других факторов роста (GF), вовлеченных в патогенез экссудативной AMD. В заключение S1P может модулировать созревание незрелых сосудов, и этот процесс приводит к потере чувствительности к антиангиогенным средствам. Ингибирование действия S1P может быть эффективным медикаментозным лечением экссудативной AMD, которое может обеспечить значительные преимущества над направленными исключительно против VEGF подходами или может действовать синергично с ними для нацеливания на комплексные процессы и множество стадий, которые в конечном итоге приводят к связанной с AMD потере зрения.
Предпочтительные в настоящее время способы лечения AMD включают Lucentis® и неутвержденное применение Avastin® (Genentech, Inc.), оба из которых нацелены на один фактор роста (VEGF-A) и, по-видимому, оказывают большую часть их благоприятного эффекта через направленное против проницаемости действие, приводящее к устранению субретинального и интраретинального отека, поскольку фактическое хориоидальное неоваскулярное (CNV) повреждение не подвергается инволюции. Однако связанное с экссудативной AMD снижение зрения происходит вследствие не индуцируемого CNV субретинального или интраретинального отека. Патологическое разрушение и ремоделирование ретинальной и субретинальной структуры, вызываемое совместно CNV, субретинальным фиброзом, отеком и воспалением, приводит к снижению остроты зрения, связанному с AMD. Доступные лекарственные средства, включая Lucentis™, не направлены на эти различные причины. Таким образом, лекарственное средство, посредством которого можно воздействовать на множество механизмов, которые вызывают снижение зрения, может иметь большую ценность либо в качестве монотерапии, либо в сочетании с другим средством, таким как средство против VEGF (например, Lucentis® или Avastin®).
Таким образом, без связи с какой-либо конкретной теорией, полагают, что уровень нежелательных сфинголипидов, таких как S1P и/или один или несколько их метаболитов, вызывает развитие или приводит к развитию различных глазных заболеваний и нарушений, где в патогенез заболевания вовлечены ненадлежащее воспаление, фиброз и/или ангиогенез. Заболевания и состояния глаза, при которых антитела против S1P, вероятно, будут клинически полезными, включают диабетическую ретинопатию, ретролентальную фиброплазию, диабетический отек желтого пятна, PVR, заболевания переднего сегмента и старческий отек желтого пятна, как влажный, так и сухой, и состояния после операций, таких как трабекулоэктомия или имплантация клапана при глаукоме.
Антитела против S1P для лечения склеродермии
Композиции и способы по изобретению будут пригодны для лечения нарушений и заболеваний, характеризующихся, по меньшей мере частично, ненадлежащей неоваскуляризацией, ангиогенезом, фиброгенезом, фиброзом, рубцеванием, воспалением и иммунным ответом. Одним таким заболеванием является склеродермия, которую также называют системной склеродермией.
Склеродермия представляет собой аутоиммунное заболевание, которое вызывает рубцевание или утолщение кожи, и иногда включает другие области организма, такие как легкие, сердце и/или почки. Склеродермия характеризуется образованием рубцовой ткани (фиброзом) в коже и органах организма, которое может приводить к утолщению и уплотнению вовлеченных областей с последующим снижением функции. Согласно Scleroderma Foundation в настоящее время приблизительно 300000 американцев имеют склеродермию. Одна треть или менее из пораженных лиц имеет распространенное заболевание, в то время как остальные две трети имеют кожные симптомы. Когда заболевание поражает легкие и вызывает рубцевание, дыхание может стать ограниченным вследствие того, что легкие более не могут расширяться должным образом. Для измерения способности дышать врачи используют устройство, которое оценивает форсированную жизненную емкость (FVC). У людей с FVC, составляющей менее 50 процентов от ожидаемых показателей, уровень смертности в течение 10 лет от связанного со склеродермией заболевания легкого составляет приблизительно 42 процента. Одна из причин того, что смертность является настолько высокой, состоит в том, что в настоящее время не доступно эффективное лечение.
Без связи с какой-либо конкретной теорией, полагают, что ненадлежащие концентрации липидов, таких как S1P и/или его метаболиты, вызывают развитие склеродермии или участвуют в ее развитии. По существу композиции и способы по изобретению можно использовать для лечения склеродермии, в частности, посредством снижения эффективной концентрации in vivo конкретного липида-мишени, например S1P.
Как описано в настоящей заявке, существующие данные указывают на то, что S1P является профибротическим фактором роста, который может приводить к активации, пролиферации фибробластов и к конечной повышенной активности фибробластов, связанной с ненадлежащим рубцеванием и ремоделированием. Полагают, что биологическая активность S1P посредством средства против S1P (например, mAb против S1P) может непрямо подавлять просклеротические эффекты PDGF и TGF-β. Более того, лечение средством против S1P может оказать благоприятное действие у пациентов со склеродермией посредством подавления прямых эффектов S1P на кожные и другие формы фибробластов, которые приводят к прогрессии заболевания.
Сердечно-сосудистые и цереброваскулярные нарушения
Без связи с какой-либо конкретной теорией, уровень нежелательных сфинголипидов, таких как CER, SPH или S1P, и/или одного или нескольких их метаболитов, может быть непосредственно ответственным за дисфункцию сердца, в процессе или после ишемии сердца, например, в ходе реперфузионного повреждения и конечного ремоделирования сердца и сердечной недостаточности.
Поскольку сфинголипиды, такие как S1P, вовлечены в фиброгенез и заживление ран ткани печени (Davaille, et al., J. Biol. Chem. 275:34268-34633, 2000; Ikeda, et al., Am J. Physiol. Gastrointest. Liver Physiol. 279:G304-G310, 2000), заживление поврежденных сосудов (Lee, et al., Am. J. Physiol. Cell Physiol. 278:C612-C618, 2000) и другие болезненные состояния или события, связанные с такими заболеваниями, такими как злокачественная опухоль, ангиогенез и воспаление (Pyne, et al., Biochem. J. 349:385-402, 2000), композиции и способы по изобретению можно применять для лечения не только этих заболеваний, но также заболеваний сердца.
Это позволяет предположить, что сфинголипиды из сердечных источников или других нецеребральных источников могут приводить к инсульту. Следовательно, препятствование продукции и/или действию сфинголипидов может быть благоприятным при подавлении инсульта, в частности инсульта, вызванного заболеванием периферических сосудов, атеросклерозом и сердечными нарушениями. Полученные недавно данные позволяют предположить, что экзогенно вводимый S1P пересекает гематоэнцефалический барьер и обеспечивает сужение сосудов в головном мозге (Tosaka, et al., Stroke 32:2913-2919, 2001).
Было сделано предположение, что ранним событием в ходе ишемии сердца (т.е. отсутствия доставки крови к сердцу) является избыточная продукция сердечной мышцей встречающегося в природе соединения сфингозина и что другие метаболиты, в частности S1P, также продуцируются либо самой сердечной тканью, либо компонентами крови, как следствие продукции сфинголипидов в сердце и последующего их превращения в крови. Настоящее изобретение относится к способам нейтрализации S1P специфичными гуманизированными моноклональными антителами. Таким образом, настоящее изобретение относится к гуманизированным антителам против сфинголипида и связанным с ними композициям и способам для снижения уровней в крови и тканях ключевого сфинголипида, S1P. Такие антитела пригодны, например, для связывания нежелательных сфинголипидов в цельной крови и, таким образом, снижения их эффективной концентрации.
Терапевтические способы и композиции по изобретению называют "основанными на сфинголипидах", чтобы указать, что эти способы терапии могут изменять относительную, абсолютную или доступную концентрацию(и) определенных нежелательных, токсических или кардиотоксических сфинголипидов. "Токсический сфинголипид" относится к любому сфинголипиду, который может вызывать или усиливать некроз и/или апоптоз клеток или иным образом нарушать функцию органа или ткани (например, посредством избыточного фиброза), включая в некоторых случаях конкретные типы клеток, которые находятся в конкретных тканях или органах. "Кардиотоксические сфинголипиды" представляют собой токсические сфинголипиды, которые прямо или непрямо обеспечивают сердечную недостаточность через ненадлежащее рубцевание (фиброгенез) и вызывают негативное инотропное состояние или вызывают или усиливают некроз и/или апоптоз клеток, находящихся в сердце, или ассоциированных с ним, включая, но не ограничиваясь ими, кардиомиоциты, нейроны сердца и т.п., и/или могут вызывать снижение сердечной функции вследствие негативных инотропных аритмических эффектов сфинголипидов и/или их метаболитов в виде сужения/спазма коронарных сосудов. "Нежелательные сфинголипиды" включают токсические и кардиотоксические сфинголипиды, а также метаболиты, в частности метаболические предшественники, токсических и кардиотоксических сфинголипидов. Представляющие особый интерес нежелательные кардиотоксические и/или токсические сфинголипиды включают, но не ограничиваются ими, церамид (CER), церамид-1-фосфат (С1Р), сфингозин-1-фосфат (S-1-P), дигидро-S1P (DHS1P), сфингозилфосфорилхолин (SPC), сфингозин (SPH; D(+)-эритро-2-амино-4-трансоктадецен-1,3-диол или сфинганин) и различные метаболиты.
Известно, что одним из наиболее ранних ответов сердечных миоцитов на гипоксию и реоксигенацию является активация нейтральной сфингомиелиназы и накопление церамида. Hernandez, et al. (2000), Circ. Res. 86:198-204, 2000. Согласно утверждениям SPH опосредует ранние события передачи сигнала при апоптотической клеточной гибели во множестве типах клеток (Ohta, et al., FEBS Letters 355:267-270, 1994; Ohta, et al., Cancer Res. 55:691-697, 1995; Cuvlilier, et al., Nature 381:800-803, 1996). Постулируется, что кардиотоксические эффекты гипоксии частично могут быть следствием продукции сфинголипидов и/или ненадлежащей продукции других метаболитов (например, протонов, кальция и некоторых свободных радикалов) или молекул передачи сигнала (например, МАР-киназ, каспаз), которые неблагоприятно влияют на сердечную функцию.
S1P запасается в тромбоцитах и является нормальным компонентом плазмы и сыворотки человека (Yatomi, et al., J. Biochem. 121:969-973, 1997). S1P представляет собой коронарное сосудосуживающее средство, и он оказывает биологические эффекты на сердце собак. Sugiyama, et al. (2000), Cardiovascular Res. 46:119-125. Была постулирована роль SIP в атеросклерозе (Siess, et al., IUBMB Life 49:161-171, 2000). Это было подтверждено другими данными, включая данные о том, что защитный эффект HDL является следствием блокирования продукции SIP (Xia, et al., PNAS 95:14196-14201, 1988; Xia, et al., J. Biol Chem. 274:33143-33147, 1999).
Описано, что уровень сфингомиелина, метаболического предшественника церамида, повышен у экспериментальных животных, подвергнутых гипоксии (Sergeev, et al., Kosm. Biol. Aviakosm. Med. (Russian) 15:71-74, 1981). В других исследованиях было описано, что внутренние мембраны мышечных клеток содержат высокие количества SPH и сфингомиелина (Sumnicht, et al., Arch. Biochem. Biophys. 215:628-637, 1982; Sabbadini, et al., Biochem. Biophys. Res. Comm. 193752-758, 1993). Введение экспериментальным животным грибковых токсинов фумонизинов В приводит к повышенным уровням SPH и DHSPH в сыворотке (S1P не измеряли) с одновременными негативными инотропными эффектами на сердце (Smithe, et al., Toxicological Sciences 56:240-249, 2000).
Другие заболевания или состояния
Вследствие вовлечения передачи сигнала биологически активных липидов во множество процессов, включая неоваскуляризацию, ангиогенез, аномальный фиброгенез, фиброз и рубцевание, и воспалительные и иммунные ответы, полагают, что ингибиторы этих биологически активных липидов будут полезны при различных заболеваниях и состояниях, связанных с одним или несколькими из этих процессов. Такие заболевания и состояния могут быть системными (например, системная склеродермия) или локализованными в одной или нескольких конкретных системах, частях или органах организма (например, в коже, легком, сердечно-сосудистой системе или глазу).
Одним способом контроля количества нежелательных сфинголипидов у пациента является предоставление композиции, которая содержит одно или несколько гуманизированных антител против сфинголипидов, для связывания одного или нескольких сфинголипидов, таким образом, действующих как терапевтические "губки", которые снижают уровень нежелательных сфинголипидов. Когда соединение называют "свободным", соединение никоим образом не ограничено от достижения области или областей, где оно оказывает его нежелательные эффекты. Как правило, свободное соединение присутствует в крови и ткани, которая либо является областью(ями) действия свободного соединения либо содержит ее (их) или из которой соединение может свободно мигрировать в его область(и) действия. Свободное соединение также может быть доступным для действия на него фермента, который превращает соединение в нежелательное соединение.
Без связи с какой-либо конкретной теорией, полагают, что уровень нежелательных сфинголипидов, таких как SPH или S1P, и/или одного или нескольких их метаболитов вызывает развитие или участвует в развитии заболеваний и нарушений сердца и миокарда.
Поскольку сфинголипиды вовлечены в фиброгенез и заживление ран ткани печени (Davaille, et al., J. Biol. Chem. 275:34268-34633, 2000; Ikeda, et al., Am J. Physiol. Gastrointest. Liver Physiol 279:G304-G310, 2000), заживление поврежденных сосудов (Lee, et al., Am. J. Physiol. Cell Physiol. 278:C612-C618, 2000) и другие болезненные состояния или события, связанные с такими заболеваниями или нарушениями, например злокачественной опухолью, ангиогенезом, различными глазными заболеваниями, связанными с избыточным фиброзом и воспалением (Pyne, et al., Biochem. J. 349:385-402, 2000), композиции и способы по изобретению можно применять для лечения этих заболеваний и нарушений, а также заболеваний и нарушений сердца и миокарда.
Одна из форм терапии на основе сфинголипидов включает манипулирование метаболическими каскадами сфинголипидов в целях снижения истинной, относительной и/или доступной концентрации in vivo нежелательных токсических сфинголипидов. Изобретение относится к композициям и способам для лечения или профилактики заболеваний, нарушений или физической травмы, в которых гуманизированные антитела против сфинголипида вводят пациенту, чтобы связать нежелательные токсические сфинголипиды или их метаболиты.
Такие гуманизированные антитела против сфинголипида могут быть изготовлены в виде фармацевтической композиции, которая пригодна для множества целей, включая лечение заболеваний, нарушений или физической травмы. Фармацевтические композиции, содержащие одно или несколько гуманизированных антител против сфинголипида по изобретению, могут быть включены в наборы и медицинские устройства для такого лечения. Медицинские устройства можно использовать для введения фармацевтических композиций по изобретению пациенту, нуждающемуся в этом, и в соответствии с одним вариантом осуществления изобретения предусмотрены наборы, которые включают такие устройства. Такие устройства и наборы могут быть предназначены для общепринятых способов введения, включая самостоятельное введение, фармацевтических композиций по изобретению. Такие устройства и наборы также могут быть предназначены для неотложного применения, например, в машинах скорой помощи или комнатах оказания экстренной помощи, или в процессе хирургической операции, или в местах, где возможно повреждение, но где полная медицинская помощь может не последовать незамедлительно (например, туристический поход и отдых в палатках или в боевой обстановке).
Способы введения
Лечение заболеваний и состояний, рассмотренных в настоящем документе, можно проводить посредством введения средств и композиций по изобретению различными способами с использованием различных составов и устройств. Пригодные фармацевтически приемлемые разбавители, носители и эксципиенты хорошо известны в данной области. Специалист в данной области поймет, что количества, подлежащие введению в ходе одного конкретного протокола лечения, можно легко определить. Можно ожидать, что пригодные количества попадут в диапазон от 10 мкг/доза до 10 г/доза, предпочтительно от 10 мг/доза до 1 г/доза.
Лекарственные вещества можно вводить способами, известными в данной области, включая, но не ограничиваясь ими, системное, подкожное, внутрикожное введение, введение на слизистую оболочку, включая введение посредством ингаляции, и местное введение. Слизистая оболочка относится к эпителиальной ткани, которая выстилает внутренние полости организма. Например, слизистую оболочку содержит пищевой канал, включая полость рта, пищевод, желудок, кишечник и анальное отверстие; дыхательные пути, включая носовые ходы, трахею, бронхи и легкие; и половые органы. Для целей данного описания слизистая оболочка также включает наружную поверхность глаза, т.е. роговицу и конъюнктиву. Местное введение (в противоположность системному введению) может быть преимущественным, поскольку этот подход может ограничить потенциальные системные побочные эффекты, но все еще обеспечивать терапевтический эффект.
Фармацевтические композиции, используемые в настоящем изобретении, включают, но не ограничиваются ими, растворы, эмульсии и содержащие липосомы составы. Эти композиции можно получать из различных компонентов, которые включают, но не ограничиваются ими, предварительно подготовленные жидкости, самоэмульгирующиеся твердые вещества и самоэмульгирующиеся полутвердые вещества.
Фармацевтические составы, используемые в настоящем изобретении, можно получать общепринятыми способами, хорошо известными в фармацевтической промышленности. Такие способы включают стадию объединения активных ингредиентов с фармацевтическим носителем(ями) или эксципиентом(ами). Предпочтительные носители включают носители, которые являются фармацевтически приемлемыми, в частности, когда композиция предназначена для терапевтического применения у человека. Для терапевтических применений не у человека (например, при лечении животных-компаньонов, домашнего скота, рыбы или птиц) можно использовать приемлемые в ветеринарии носители. Как правило, составы получают объединением в равномерную и однородную массу активных ингредиентов с жидкими носителями или высокодисперсными твердыми носителями, или и с теми и с другими, а затем, при необходимости, приданием продукту формы.
Композиции по настоящему изобретению могут быть изготовлены в виде множества возможных дозированных форм, таких как, но не ограничиваясь ими, таблетки, капсулы, жидкие сиропы, мягкие гели, суппозитории и клизмы. Композиции по настоящему изобретению также могут быть изготовлены в качестве суспензий в водных, неводных или смешанных носителях. Водные суспензии могут дополнительно содержать вещества, которые повышают вязкость суспензии, включая, например, карбоксиметилцеллюлозу натрия, сорбит и/или декстран. Также суспензия может содержать стабилизаторы.
В одном варианте осуществления фармацевтические композиции можно изготавливать и использовать в виде пен. Фармацевтические пены включают, но не ограничиваются ими, такие составы, как эмульсии, микроэмульсии, кремы, желе и липосомы.
Несмотря на по существу сходные свойства, эти составы варьируют с точки зрения компонентов и консистенции конечного продукта. Технология изготовления таких композиций и составов главным образом известна специалистам в области фармацевтики и изготовления, ее можно применять для изготовления композиций по настоящему изобретению.
В одном варианте осуществления иммунную молекулу можно доставлять в глаз, например, посредством капель или мази для местного применения, окологлазной инъекции, внутрикамерной доставки в переднюю камеру или стекловидное тело, посредством имплантированных депо или системно посредством инъекции или перорального введения. Используемое количество антитела может легко определить специалист в данной области.
Традиционные подходы к доставке лекарственных средств в глаз включают местное применение, перераспределение в глаз после системного введения или прямые внутриглазные/окологлазные инъекции [Sultana, et al. (2006), Current Drug Delivery, vol.3:207-217; Ghate and Edelhauser (2006), Expert Opinion, vol.3:275-287; и Kaur and Kanwar (2002), Drug Develop Industrial Pharmacy, vol.28:473-493]. Лекарственные средства против S1P или другие лекарственные средства против биологически активных липидов на основе антител, вероятно, можно будет использовать в любом из этих подходов, хотя все они обладают определенными известными преимуществами и недостатками. Капли для местного применения являются удобными, но они смываются, главным образом, вследствие носослезного дренажа, часто доставляя менее 5% нанесенного лекарственного средства в передний отдел глаза и еще меньшую долю этой дозы в задний сегмент глазного яблока. Помимо капель, другим способом местного введения является введение с помощью аэрозолей. Третьим способом является применение глазных мазей или эмульсий, которые можно использовать для продления времени контакта состава с поверхностью глаза, хотя неясность зрения и покрытие глазных яблок могут причинять затруднения. Такие местные подходы, тем не менее, являются предпочтительными, поскольку системное введение лекарственных средств для лечения глазных нарушений подвергает весь организм воздействию потенциальной токсичности лекарственного средства.
Лечение заднего сегмента глаза имеет медицинское значение, поскольку старческая дегенерация желтого пятна, диабетическая ретинопатия, задний увеит и глаукома являются основными причинами снижения зрения в США и других развитых странах. Myles, et al. (2005), Adv Drug Deliv Rev; 57:2063-79. Наиболее эффективным способом доставки лекарственного средства в задний сегмент является инъекция в стекловидное тело через плоскую часть. Однако прямые инъекции для обеспечения доставки требуют помощи квалифицированного медицинского специалиста и могут вызывать ограничивающее лечение беспокойство у многих пациентов. Окологлазные инъекции, подход, который включает субконъюнктивальные, ретробульбарные, перибульбарные инъекции и инъекции в заднее субтеноново пространство, являются в некоторой степени менее инвазивными, чем инъекции в стекловидное тело. Многократные и продолжительные инъекции в стекловидное тело могут вызывать осложнения, такие как кровоизлияние в стекловидное тело, отслоение сетчатки или эндофтальмит.
Лечение антителом против биологически активного липида также можно проводить с использованием одной из новых систем для доставки в глаз [Sultana, et al. (2006), Current Drug Delivery, vol.3:207-217; и Ghate and Edelhauser (2006), Expert Opinion, vol.3:275-287], включая системы для замедленного и контролируемого высвобождения, такие как (а) глазные вкладыши (растворимые, размываемые, неразмываемые или на основе гидрогеля), накладки на роговицу, например повязка на основе коллагена и контактные линзы, которые обеспечивают контролируемую доставку лекарственного средства в глаз, (b) системы, образующие гель in situ, которые обеспечивают простоту введения в качестве капель, которые превращаются в форму геля в глазу, обеспечивая, таким образом, некоторый замедленный эффект лекарственного средства в глазу, (с) везикулярные системы, такие как липосомы, ниосомы/дискомы и т.д., которые обеспечивают преимущества направленной доставки, биосовместимости и отсутствия неясности зрения, (d) мукоадгезивные системы, которые обеспечивают лучшее удержание в глазу, (е) пролекарства, (f) средства, усиливающие проникновение, (g) системы лиофилизированных носителей, (h) макрочастицы, (i) субмикронные эмульсии, (j) ионтофорез, (k) дендримеры, (l) микросферы, включая биоадгезивные микросферы, (m) наносферы и другие наночастицы, (n) колласомы и (о) системы для доставки лекарственных средств, которые сочетают одну или несколько из указанных выше систем для обеспечения аддитивного или даже синергического благоприятного эффекта. Большинство из этих подходов нацелены на передний сегмент глаза и могут быть полезными при лечении заболеваний переднего сегмента. Однако, тем не менее, один или несколько из этих подходов может быть пригоден для влияния на концентрации биологически активного липида в задней области глаза, поскольку относительно низкая молекулярная масса липидов, вероятно, позволит значительное продвижение липида в пределах глаза. Кроме того, антитело, введенное в переднюю область глаза, может быть способным мигрировать через глаз, особенно если оно изготовлено в виде варианта антитела с более низкой молекулярной массой, такого как Fab-фрагмент. Также можно использовать системы для замедленной доставки лекарственного средства в задний сегмент, такие как одобренные или разрабатываемые системы (см. ссылки, выше).
Как упоминалось ранее, лечение заболевания задней сетчатки, сосудистой оболочки и желтого пятна имеет очень большое медицинское значение. В этом отношении важным достижением является транссклеральный ионтофорез [Eljarrat-Binstock and Domb (2006), Control Release, 110:479-89], и он может обеспечивать эффективный способ доставки антител в задний сегмент глаза.
Также в состав антитела можно добавлять различные эксципиенты для улучшения качества терапии, что делает терапию более удобной, или для того, чтобы четко обеспечить применение антитела в составе только для цели, для которой он предназначен и одобрен. Примеры эксципиентов включают химические реагенты, контролирующие рН, противомикробные средства, консерванты для предотвращения потери эффективности антитела, красители для идентификации состава только для глазного применения, солюбилизирующие средства для повышения концентрации антитела в составе, средства, усиливающие проникновение, и средства для коррекции изотоничности и/или вязкости. Например, для продления времени полужизни антитела можно добавлять ингибиторы протеазы. В одном варианте осуществления антитело доставляют в глаз посредством инъекции в стекловидное тело в растворе, содержащем фосфатно-солевой буфер при пригодных для глаза значениях рН.
Средство против S1P (например, гуманизированное антитело) также можно химически модифицировать для получения пролекарства, которое вводят в одном из составов или устройств, описанных выше. Затем активная форма антитела высвобождается под действием эндогенного фермента. Возможными глазными ферментами, рассматриваемыми для этого применения, являются различные цитохромы р450, альдегидредуктазы, кетонредуктазы, эстеразы или N-ацетил-β-глюкозоамидазы. Другие химические модификации антитела могут повышать его молекулярную массу и в результате повышать время нахождения антитела в глазу. Примером такой химической модификации является пегилирование [Harris and Chess (2003), Nat Rev Drug Discov; 2:214-21], процесс, который может быть общим или специфичным для функциональной группы, такой как дисульфид [Shaunak, et al. (2006), Nat Chem Biol; 2:312-3] или тиол [Doherty, et al. (2005), Bioconjug Chem; 16:1291-8].
В представленных в настоящем документе, ниже, примерах описана продукция гуманизированных антител и вариантов антитела против сфинголипида с требуемыми свойствами с точки зрения перспективы лечения, включая высокую аффинность связывания со сфинголипидами. В частности, изобретение направлено на S1P и его варианты, которые могут включать сам S1P, определенный как сфингозин-1-фосфат [сфинген-1-фосфат; D-эритросфингозин-1-фосфат; сфинг-4-енин-1-фосфат; (E,2S,3R)-2-амино-3-гидроксиоктадек-4-енокси]фосфоновую кислоту (AS 26993-30-6)], DHS1P определенный как дигидросфингозин-1-фосфат [сфинганин-1-фосфат; [(2S,3R)-2-амино-3-гидроксиоктадекокси] фосфоновую кислоту; D-эритродигидро-D-сфингозин-1-фосфат (CAS 19794-97-9)]; SPC представляет собой сфингозилфосфорилхолин, лизосфингомиелин, сфингозилфосфохолин, сфингозинфосфорилхолин, этанаминий; 2-((((2-амино-3-гидрокси-4-октадекенил)окси)гидроксифосфинил)окси)-N,N,N-триметил-, хлорид, (R-(R*,S*-(E))), хлорид 2-[[(E,2R,3S)-2-амино-3-гидроксиоктадек-4-енокси]гидроксифосфорил]оксиэтил-1-триметилазания (CAS 10216-23-6).
Получение и охарактеризация антител
Аффинность антитела можно определять, как описано в разделе "Примеры" настоящего документа ниже. Предпочтительные гуманизированные антитела или варианты антител представляют собой антитела, которые связывают сфинголипид со значением Кd не более чем приблизительно 1×10-7 M, предпочтительно не более чем приблизительно 1×10-8 М и наиболее предпочтительно не более чем приблизительно 5×10-9 М.
Помимо антител с сильной аффинностью связывания со сфинголипидами, также желательно отбирать гуманизированные антитела или варианты антител, которые имеют другие благоприятные свойства с точки зрения терапевтической перспективы. Например, антитело может представлять собой антитело, которое снижает ангиогенез и изменяет прогрессию опухоли. Предпочтительно антитело имеет значение эффективной концентрации 50 (ЕС50) не более чем приблизительно 10 мкг/мл, предпочтительно не более чем 1 мкг/мл и наиболее предпочтительно не более чем приблизительно 0,1 мкг/мл, при измерении в прямом анализе связывания ELISA. Предпочтительно антитело имеет значение эффективной концентрации не более чем приблизительно 10 мкг/мл, предпочтительно не более чем приблизительно 1 мкг/мл и наиболее предпочтительно не более чем приблизительно 0,1 мкг/мл, при измерении в клеточных анализах в присутствии 1 мкМ S1P, например, в этих концентрациях антитело способно ингибировать индуцируемое сфинголипидом высвобождение IL-8 in vitro no меньшей мере на 10%. Предпочтительно антитело имеет значение эффективной концентрации не более чем приблизительно 10 мкг/мл, предпочтительно не более чем приблизительно 1 мкг/мл и наиболее предпочтительно не более чем приблизительно 0,1 мкг/мл, при измерении в модели CNV на животных после лазерного ожога, например, в этих концентрациях антитело способно ингибировать индуцируемую сфинголипидом неоваскуляризацию in vivo по меньшей мере на 50%.
Способы анализа для определения активности антител против сфинголипидов по изобретению включают анализы ELISA, как показано в разделе "Примеры" настоящего документа ниже.
Предпочтительно гуманизированное антитело или вариант антитела не вызывает иммуногенный ответ при введении терапевтически эффективного количества антитела пациенту-человеку. Если иммуногенный ответ индуцируется, предпочтительно ответ будет таким, что антитело, тем не менее, обеспечит терапевтическую пользу у пациента, которого им лечат.
В соответствии с одним вариантом осуществления изобретения гуманизированные антитела против сфинголипидов связывают "эпитоп", как определено в настоящем документе. Для скрининга антител, которые связывают эпитоп на сфинголипиде, связываемый представляющим интерес антителом (например, антител, которые блокируют связывание антитела со сфинголипидом), можно проводить общепринятый анализ перекрестного блокирования, как описано в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow и David Lane (1988). Альтернативно для определения того, связывает ли антитело представляющий интерес эпитоп, можно проводить эпитопное картирование, например, как описано в Champe, et al. [J. Biol. Chem. 270:1388-1394 (1995)].
Антитела по изобретению имеют вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, соответствующую формуле: FR1-CDRH1-FR2-CDRH2-FR3-CDRH3-FR4, где "FR1-4" соответствуют четырем каркасным областям и "CDRH1-3" соответствуют трем гипервариабельным областям вариабельного домена тяжелой цепи антитела против сфинголипида. FR1-4 могут быть образованы "консенсусной последовательностью" (например, наиболее часто встречающимися аминокислотами класса, подкласса или подгруппы тяжелых или легких цепей иммуноглобулинов человека), как в разделе "Примеры", ниже, или они могут быть образованы каркасной областью отдельного антитела человека или сочетанием различных последовательностей каркасных областей. Множество последовательностей каркасной области антител человека представлено, например, в Kabat, et al., выше. В одном варианте осуществления FR вариабельной области тяжелой цепи представлены консенсусной последовательностью подгруппы иммуноглобулинов человека, представленной в Kabat, et al., выше. Предпочтительно подгруппа иммуноглобулина человека представляет собой подгруппу III тяжелой цепи человека (например, как в SEQ ID NO:16).
Последовательность FR вариабельной области тяжелой цепи человека предпочтительно имеет в ней одну или несколько замен, например, где остаток FR человека заменен соответствующим остатком не человека (под "соответствующим остатком не человека" подразумевают остаток не человека с той же нумерацией положения по Kabat, что и представляющий интерес остаток человека, после выравнивания последовательностей человека и не человека), однако замена остатком не человека не является необходимой. Например, замену остатка FR, отличную от замены соответствующим остатком не человека, можно выбирать посредством фагового дисплея. Иллюстративные остатки FR вариабельной области тяжелой цепи, которые можно заменять, включают любой один или несколько из остатков FR с номерами: 37Н, 49Н, 67Н, 69Н, 71Н, 73Н, 75Н, 76Н, 78Н и 94Н (здесь использована нумерация остатков по Kabat). Предпочтительно заменяют по меньшей мере два, или по меньшей мере три, или по меньшей мере четыре из этих остатков. Особенно предпочтительное сочетание замен FR представляет собой: 49Н, 69Н, 71Н, 73Н, 76Н, 78Н и 94Н. В отношении гипервариабельных областей тяжелых цепей они предпочтительно имеют аминокислотные последовательности, приведенные в таблице 2, ниже.
Антитела предпочтительных вариантов осуществления, описанных в настоящем документе, имеют вариабельный домен легкой цепи, содержащий аминокислотную последовательность, соответствующую формуле: FR1-CDRL1-FR2-CDRL2-FR3-CDRL3-FR4, где "FR1-4" соответствуют четырем каркасным областям и "CDRL1-3" соответствуют трем гипервариабельным областям вариабельного домена тяжелой цепи антитела против сфинголипида. FR1-4 могут быть образованы "консенсусной последовательностью" (например, наиболее часто встречающиеся аминокислоты класса, подкласса или подгруппы тяжелых или легких цепей иммуноглобулинов человека), как в разделе "Примеры", ниже, или они могут быть образованы каркасной областью отдельного антитела человека или сочетанием различных последовательностей каркасных областей. В одном предпочтительном варианте осуществления FR вариабельной области легкой цепи представлен консенсусной последовательностью подгруппы иммуноглобулинов человека, представленной в Kabat, et al., выше. Предпочтительно подгруппа иммуноглобулинов человека представляет собой подгруппу I легких цепей каппа человека (например, как в SEQ ID NO:17).
Последовательность FR вариабельной области легкой цепи человека имеет замены в ней, например, где остаток FR человека заменен соответствующим остатком мыши, но замена остатком не человека не является обязательной. Например, замену остатка, отличную от замены соответствующим остатком не человека, можно отбирать посредством фагового дисплея. Иллюстративные остатки FR вариабельной области легкой цепи, которые можно заменять, включают любой один или несколько из номеров остатков FR, включая, но не ограничиваясь ими, F4, Y36, Y49, G64, S67.
В отношении CDR они предпочтительно имеют аминокислотные последовательности, приведенные в таблице 2, ниже.
Способы получения гуманизированных представляющих интерес антител против сфинголипидов, описанных в настоящем документе, более подробно рассмотрены ниже.
А. Получение антител
Способы гуманизации антител против сфинголипида не человека и получения вариантов антител против сфинголипида описаны в разделе "Примеры" ниже. Для гуманизации антитела против сфинголипида получают исходный материал в виде антитела не человека. Когда намереваются получить вариант, получают исходное антитело. Иллюстративные способы получения такого исходного материала в виде антитела не человека и исходные антитела будут описаны в следующих разделах.
(i) Получение антигена
Сфинголипидный антиген, подлежащий применению для продукции антител, может представлять собой, например, интактный сфинголипид или часть сфинголипида (например, фрагмент сфинголипида, содержащий "эпитоп"). Другие формы антигенов, пригодные для получения антител, будут очевидны специалистам в данной области. Сфинголипидный антиген, используемый для получения антитела, описан в разделе "Примеры" ниже. В одном варианте осуществления антиген представляет собой производное сфинголипида, и он может быть связан с белком-переносчиком.
(ii) Поликлональные антитела
Поликлональные антитела предпочтительно индуцируют у животных посредством многократных подкожных (sc) или внутрибрюшинных (ip) инъекций соответствующего антигена и адъюванта. Может быть пригодной конъюгация соответствующего антигена с белком, который является иммуногенным у вида, подлежащего иммунизации, например гемоцианином лимфы улитки, сывороточным альбумином, бычьим тироглобулином или ингибитором трипсина бобов сои, с использованием бифункционального или образующего производное вещества, например сложного эфира малеимидобензоилсульфосукцинимида (конъюгация через остатки цистеина), N-гидроксисукцинимида (через остатки лизина), глутаральдегида, янтарного ангидрида, SOC12, или R1N=C=NR, где R и R1 представляют собой отличающиеся алкильные группы.
Животных иммунизируют против антигена, иммуногенных конъюгатов или производных посредством комбинирования, например, 100 мкг или 5 мкг белка или конъюгата (для кроликов или мышей соответственно) с тремя объемами полного адъюванта Фрейнда, и инъекции раствора внутрикожно в различные области. Через один месяц животным проводят вторичную иммунизацию исходным количеством пептида или конъюгата, умноженным на 0,1-0,2, в полном адъюванте Фрейнда посредством подкожной инъекции в несколько областей. Через от семи до 14 суток после этого животным пускают кровь и сыворотку оценивают в отношении титра антител. Животным проводят повторные иммунизации до достижения титрами плато. Предпочтительно животным проводят повторную иммунизацию конъюгатом того же антигена с другим белком и/или посредством другого поперечно-сшивающего реагента. Также конъюгаты можно продуцировать в рекомбинантной культуре клеток в качестве слитых белков. Также для усиления иммунного ответа можно пригодным образом использовать вызывающие агрегацию средства, такие как квасцы.
(iii) Моноклональные антитела
Моноклональные антитела можно получать с использованием способа гибридом, впервые описанным Kohler, et al., Nature, 256:495 (1975), или другими пригодными способами, включая способы рекомбинантных ДНК (см., например, патент США №4816567). В способе гибридом мышь или другое пригодное животное-хозяина, такого как хомяк или макак-резус, иммунизируют, как описано в настоящем документе выше, для индукции лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфично связываться с белком, используемым для иммунизации. Альтернативно лимфоциты можно иммунизировать in vitro. Затем лимфоциты подвергают слиянию с клетками миеломы с использованием пригодного средства для слияния, такого как полиэтиленгликоль, с получением гибридомных клеток (Goding, Monoclonal Antibodies: Principles and Practice, pp.59-103 (Academic Press, 1986)).
Клетки гибридомы, полученные таким образом, высевают и выращивают в пригодной культуральной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост и выживание неслитых родительских клеток миеломы. Например, если в родительских клетках миеломы отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT или HPRT), культуральная среда для гибридомы, как правило, будет включать гипоксантин, аминоптерин и тимидин (среда HAT), которые препятствуют росту клеток с дефицитом HGPRT.
Предпочтительные клетки миеломы представляют собой клетки, которые поддаются эффективному слиянию, поддерживают стабильную продукцию антитела на высоком уровне отобранными антителопродуцирующими клетками и являются чувствительными к среде, такой как среда HAT. Среди них, предпочтительные клеточные линии миеломы представляют собой линии миеломы мыши, такие как линии, образованные из опухолей мыши МОР-21 и М.С.-11, доступных от Salk Institute Cell Distribution Center, San Diego, Calif. USA, и клетки SP-2 или X63-Ag8-653, доступные от American Type Culture Collection, Rockville, Md. USA. Также для продукции моноклональных антител человека описаны клеточные линии миеломы человека и гетеромиеломы мыши-человека (Kozbor, J. Immunol., 133:3001 (1984); Brodeur, et al., Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой выращивают клетки гибридомы, оценивают в отношении продукции моноклональных антител, направленных против антигена. Предпочтительно специфичность связывания моноклональных антител, продуцируемых клетками гибридомы, определяют посредством иммунопреципитации или анализа связывания in vitro, такого как радиоиммунный анализ (RIA) или твердофазный иммуноферментный анализ (ELISA).
Аффинность связывания моноклонального антитела можно определять, например, анализом Скэтчарда, Munson, et al., Anal. Biochem., 107:220 (1980).
После идентификации гибридомных клеток, которые продуцируют антитела с требуемой специфичностью, аффинностью и/или активностью, клоны можно субклонировать способами лимитирующего разведения и выращивать стандартными способами (Goding, Monoclonal: Principles and Practice, pp.59-103 (Academic Press, 1986)). Пригодные для этой цели культуральные среды включают, например, среду D-MEM или RPMI-1640. Кроме того, клетки гибридомы можно выращивать in vivo в качестве асцитных опухолей у животного.
Моноклональные антитела, секретируемые субклонами, пригодным образом выделяют из культуральной среды, асцитной жидкости или сыворотки с помощью общепринятых способов очистки иммуноглобулинов, например, таких как хроматография на белке А-Sepharose, хроматография с гидроксилапатитом, гель-электрофорез, диализ или аффинная хроматография.
ДНК, кодирующую моноклональные антитела, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител). Клетки гибридомы служат в качестве предпочтительного источника такой ДНК. После выделения ДНК можно встраивать в экспрессирующие векторы, которые затем трансфицируют в клетки-хозяева, такие как клетки Е.coli, клетки обезьяны COS, клетки яичника китайского хомяка (СНО) или клетки миеломы, которые в ином случае не продуцируют белок иммуноглобулина, обеспечивая синтез моноклональных антител в рекомбинантных клетках-хозяевах. Рекомбинантная продукция антител будет более подробно описана ниже.
(iv) Гуманизация и варианты аминокислотной последовательности
В примере 12, ниже, описаны способы гуманизации антитела против сфинголипида. Общие способы гуманизации описаны, например, в US 5861155, US 19960652558, US 6479284, US 20000660169, US 6407213, US 19930146206, US 6639055, US 20000705686, US 6500931, US 19950435516, US 5530101, US 5585089, US 19950477728, US 5693761, US 19950474040, US 5693762, US 19950487200, US 6180370, US 19950484537, US 2003229208, US 20030389155, US 5714350, US 19950372262, US 6350861, US 19970862871, US 5777085, US 19950458516, US 5834597, US 19960656586, US 5882644, US 19960621751, US 5932448, US 19910801798, US 6013256, US 19970934841, US 6129914, US 19950397411, US 6210671, US 6329511, US 19990450520, US 2003166871, US 20020078757, US 5225539, US 19910782717, US 6548640, US 19950452462, US 5624821 и US 19950479752. В определенных вариантах осуществления может быть желательным получение вариантов аминокислотной последовательности этих гуманизированных антител, в частности, где они повышают аффинность связывания или улучшают другие биологические свойства гуманизированного антитела. В примере 12 описаны технологии получения вариантов аминокислотной последовательности антитела против сфинголипида с усиленной аффинностью относительно исходного антитела.
Варианты аминокислотной последовательности антитела против сфинголипида получают внесением соответствующих нуклеотидных замен в ДНК антитела против сфинголипида или посредством пептидного синтеза. Такие варианты включают, например, делеции, и/или вставки, и/или замены остатков в аминокислотных последовательностях антител против сфинголипида, представленных в примерах, приведенных в настоящем документе. Для получения конечной конструкции проводят любое сочетание из делеции, вставки и замены, при условии, что конечная конструкция обладает требуемыми характеристиками. Аминокислотные замены также могут изменять посттрансляционные процессы гуманизированного антитела против сфинголипида или варианта антитела против сфинголипида, например они могут изменять количество или положение участков гликозилирования.
Пригодный способ идентификации определенных остатков или участков антитела против сфинголипида, которые являются предпочтительными участками для мутагенеза, называют "сканирующим аланином мутагенезом", как описано Cunningham and Wells, Science, 244:1081-1085 (1989). В данном случае выявляют остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяют на нейтральную или отрицательно заряженную аминокислоту (наиболее предпочтительно аланин или полиаланин) для нарушения взаимодействия аминокислот с антигеном. Те положения аминокислот, которые демонстрируют функциональную чувствительность к замещению, затем подтверждают посредством внесения дополнительных или других вариантов в участки замещения, или вместо них. Таким образом, поскольку участок для внесения изменения в аминокислотную последовательность является предопределенным, нет необходимости в непосредственной предопределенности характера мутации. Например, для анализа действия мутации в данной области в кодоне-мишени или участке-мишени проводят сканирующий ala или случайный мутагенез и экспрессированные варианты антитела против сфинголипида подвергают скринингу в отношении наличия требуемой активности. Вставки в аминокислотную последовательность включают N- и/или С-концевые слитые последовательности, обладающие длиной в диапазоне от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или нескольких аминокислотных остатков внутрь последовательности. Примеры концевых вставок включают антитело против сфинголипида с N-концевым метионильным остатком или антитело, слитое с эпитомной меткой. Другие варианты вставок в молекулу антитела против сфинголипида включают слияние N- или С-конца антитела против сфинголипида с ферментом или полипептидом, который повышает время полужизни антитела в сыворотке.
Другим типом варианта является вариант с аминокислотной заменой. Эти варианты имеют по меньшей мере один остаток аминокислоты в молекуле антитела против сфинголипида, который удален, и другой остаток, встроенный вместо него. Участки, представляющие наибольший интерес для проведения мутагенеза с замещением, включают гипервариабельные области, но также предусматриваются изменения FR. Консервативные замены являются предпочтительными заменами. Если результатом таких замен является изменение биологической активности, тогда можно вносить более значительные изменения, обозначенные "иллюстративными" заменами, приведенными ниже, или как описано далее в отношении классов аминокислот, и продукты подвергать скринингу.
Существенные модификации биологических свойств антитела проводят, выбирая замены, которые значительно отличаются по их эффекту на поддержание (а) структуры полипептидного остова в области замены, например, такой как конформация в виде слоя или спирали, (b) заряда или гидрофобности молекулы в участке-мишени или (с) объема боковой цепи. Встречающиеся в природе остатки можно разделить на основе общих свойств их боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gin, his, lys, arg;
(5) остатки, влияющие на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены вовлекают замену члена одного из этих классов членом другого класса.
Любой остаток цистеина, не вовлеченный в поддержание надлежащей конформации гуманизированного антитела против сфинголипида или варианта антитела против сфинголипида, также можно заменять для повышения устойчивости молекулы к окислению и предотвращения ошибочного поперечного сшивания. Напротив, к антителу можно добавлять цистеиновую связь(и) для повышения его стабильности (в частности, когда антитело представляет собой фрагмент антитела, такой как Fv-фрагмент).
Один тип варианта с заменой включает замену одного или нескольких остатков гипервариабельной области исходного антитела (например, гуманизированного антитела или антитела человека). Как правило, полученный вариант(ы), отобранный для дальнейшей разработки, обладает улучшенными свойствами относительно исходного антитела, из которого он получен. Удобным способом получения таких вариантов с заменами является созревание аффинности с использованием фагового дисплея. В кратком изложении, в несколько участков гипервариабельной области (например, в 6-7 участков) вносят мутации для получения всех возможных замен аминокислот в каждом участке. Варианты антитела, полученные таким образом, экспонируют в одновалентной форме в частицах нитевидных фагов в качестве антител, слитых с продуктом гена III M13, упакованных в каждой частице. Затем проводят скрининг вариантов, экспонированных посредством фагов, в отношении их биологической активности (например, аффинности связывания), как описано в настоящем документе. В целях выявления участков гипервариабельной области, являющихся кандидатами для модификации, можно проводить сканирующий аланином мутагенез для выявления остатков гипервариабельной области, в значительной степени участвующих в связывании антигена. Альтернативно, или дополнительно, может быть целесообразным проведение анализа кристаллической структуры комплекса антиген-антитело для определения участков контакта между антителом и его сфинголипидом. Такие образующие контакт остатки и соседние остатки являются кандидатами для замены в соответствии с представленными в настоящем документе способами. После получения таких вариантов панель вариантов подвергают скринингу, как описано в настоящей заявке, и для дальнейшей разработки могут отбирать антитела с наилучшими свойствами в одном или нескольких подходящих анализов.
В другом типе аминокислотного варианта антитела изменен исходный характер гликозилирования антитела. Под изменением подразумевают удаление одной или нескольких углеводных групп, находящихся на антителе, и/или добавление одного или нескольких участков гликозилирования, которые не присутствуют в антителе.
Гликозилирование антител, как правило, является либо N-связанным или О-связанным. N-связанное относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где X представляет собой любую аминокислоту, за исключением пролина, представляют собой последовательности, распознаваемые для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. О-связанное гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, обычно к серину или треонину, хотя также могут быть использованы 5-гидроксипролин или 5-гидроксилизин.
Добавление участков гликозилирования в антитело удобно проводить посредством изменения аминокислотной последовательности таким образом, чтобы она содержала одну или несколько из описанных выше трипептидных последовательностей (в случае участков N-связанного гликозилирования). Также изменение можно осуществлять добавлением одного или нескольких остатков серина или треонина в последовательность исходного антитела или заменой одним или несколькими остатками серина или треонина в последовательности исходного антитела (в случае участков O-связанного гликозилирования).
Молекулы нуклеиновых кислот, кодирующие варианты аминокислотных последовательностей антитела против сфинголипида, получают множеством способов, известных в данной области. Эти способы включают, но не ограничиваются ими, выделение из природного источника (в случае природных вариантов аминокислотных последовательностей) или получение посредством олигонуклеотид-направленного (или сайт-направленного) мутагенеза, мутагенеза посредством ПЦР и кассетного мутагенеза ранее полученного измененного или не измененного варианта антитела против сфинголипида.
(v) Антитела человека
В качестве альтернативы гуманизации можно получать антитела человека. Например, в настоящее время возможным является получение трансгенных животных (например, мышей), которые способны, при иммунизации, продуцировать полный набор антител человека в отсутствие эндогенной продукции иммуноглобулинов. Например, было описано, что гомозиготная делеция гена участка соединения тяжелых цепей антител (JH) у химерных мышей и мутантных мышей зародышевой линии приводит к полному ингибированию эндогенной продукции антител. Перенос системы генов иммуноглобулинов зародышевой линии человека в таких мутантных мышей зародышевой линии приводит к продукции антител человека при антигенной стимуляции. См., например, Jakobovits et al., Proc. Natl. Acad. Sci USA, 90:2551 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggermann et al., Year in Immunol., 7:33 (1993); и патенты США №5591669, 5589369 и 5545807. Также антитела человека можно получать из библиотек фагового дисплея (Hoogenboom, et al., J. Mol. Biol, 227:381 (1991); Marks, et al., J. Mol. Biol, 222:581-597 (1991); и патенты США №5565332 и 5573905). Как рассмотрено выше, антитела человека также можно получать с помощью активированных В-клеток in vitro (см., например, патенты США №5567610 и 5229275) или другими пригодными способами.
(vi) Фрагменты антител
В определенных вариантах осуществления гуманизированное антитело против сфинголипида или вариант антитела против сфинголипида представляет собой фрагмент антитела. Для получения фрагментов антител были разработаны различные способы. Традиционно, эти фрагменты получали посредством протеолитического расщепления интактных антител (см., например, Morimoto et al., Journal of Biochemical and Biophysical Methods 24:107-117 (1992); и Brennan et al., Science, 229:81 (1985)). Однако эти фрагменты в настоящее время можно получать непосредственно с помощью рекомбинантных клеток-хозяев. Например, фрагменты Fab′-SH можно выделять непосредственно из Е.coli и химически связывать с получением фрагментов F(ab′)2 (Carter et al., Bio/Technology. 10:163-167 (1992)). В другом варианте осуществления F(ab′)2 получают с использованием лейциновой молнии GCN4 для обеспечения сборки молекулы F(ab′)2. В соответствии с другим подходом фрагменты Fv, Fab или F(ab′)2 можно выделять непосредственно из рекомбинантной культуры клеток-хозяев. Другие способы получения фрагментов антител будут очевидны квалифицированному специалисту.
(vii) Полиспецифичные антитела
В некоторых вариантах осуществления может быть желательным получение полиспецифичных (например, биспецифичных) гуманизированных антител или вариантов антител против сфинголипида, имеющих специфичность связывания по меньшей мере по отношению к двум различным эпитопам. Иллюстративные биспецифичные антитела могут связываться с двумя различными эпитетами белка сфинголипида. Альтернативно плечо против сфинголипида можно объединять с плечом, которое связывается с другой молекулой. Биспецифичные антитела можно получать в качестве полноразмерных антител или фрагментов антител (например, биспецифичные антитела F(ab′)2).
В соответствии с другим подходом для получения биспецифичных антител для максимального увеличения процентного количества гетеродимеров, выделяемых из рекомбинантных клеточных культур, можно конструировать область контакта между парой молекул антител. Предпочтительная область контакта содержит по меньшей мере часть CH3-домена константного домена антитела. В этом способе одну или несколько небольших боковых цепей аминокислот из области контакта первой молекулы антитела заменяют боковыми цепями большего размера (например, тирозина или триптофана). В области контакта второй молекулы антитела создают компенсаторные "полости" идентичного или сходного с крупной боковой цепью(ями) размера посредством замены крупных боковых цепей аминокислот цепями меньших размеров (например, аланина или треонина). Это обеспечивает механизм для повышения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры. См., например, патент США №5731168.
Биспецифичные антитела включают поперечно-связанные антитела или "гетероконъюгаты" антител. Например, одно из антител в гетероконъюгате может быть соединено с авидином, а другое - с биотином. Гетероконъюгаты антител можно получать с использованием любого пригодного способа поперечного связывания. Пригодные вещества для поперечного связывания хорошо известны в данной области и описаны, например, в патенте США №4676980, совместно с рядом способов поперечного связывания.
Способы получения биспецифичных антител из фрагментов антител также описаны в литературе. Например, биспецифичные антитела можно получать с использованием образования химических связей. В Brennan et al., Science, 229:81 (1985) описан способ, где полноразмерные антитела протеолитически расщепляют с получением F(ab′)2-фрагментов. Эти фрагменты восстанавливают в присутствии вещества, образующего комплексы с дитиолами, арсенита натрия, для стабилизации соседних дитиолов и предотвращения образования межмолекулярного дисульфида. Затем полученные Fab′-фрагменты превращают в производные с тионитробензоатом (TNB). Одно из производных Fab′-TNB затем повторно превращают в Fab′-тиол восстановлением меркаптоэтиламином и смешивают с эквимолярным количеством другого производного Fab′-TNB с получением биспецифичного антитела. Полученные биспецифичные антитела можно использовать в качестве средств для селективной иммобилизации ферментов. В другом варианте осуществления фрагменты Fab′-SH, прямо выделенные из Е.coli, можно химически связывать in vitro с получением биспецифичных антител. Shalaby et al., J. Exp. Med., 175:217-225 (1992).
Также были описаны различные способы получения и выделения фрагментов биспецифичных антител непосредственно из рекомбинантной клеточной культуры. Например, были получены биспецифичные антитела с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5):1547-1553 (1992). Пептиды лейциновой молнии белков Fos и Jun соединяли с Fab′-участками двух различных антител посредством слияния генов. Гомодимеры антител восстанавливали в шарнирной области с получением мономеров, а затем повторно окисляли с получением гетеродимеров антител. Этот способ также можно использовать для получения гомодимеров антител. Технология "димерных антител", описанная Hollinger, et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993), обеспечила альтернативный механизм получения биспецифичных фрагментов антител. Фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким для возможности спаривания двух доменов одной цепи. Таким образом, домены VH и VL одного фрагмента вынуждены образовывать пару с комплементарными доменами VL и VH другого фрагмента, таким образом, формируя два антигенсвязывающих центра. Также описана другая стратегия для получения фрагментов биспецифичных антител с использованием димеров одноцепочечных Fv (sFv). См. Gruber et al., J. Immunol., 152:5368 (1994). Альтернативно биспецифичное антитело может представлять собой "линейное антитело", продуцируемое, как описано в Zapata et al. Protein Eng. 8 (10):1057-1062 (1995).
Также предусмотрены антитела более чем с двумя валентностями. Например, можно получать триспецифичные антитела. Tutt et al., J. Immunol. 147:60 (1991).
Антитело (или полимер или полипептид) по изобретению, содержащее один или несколько участков связывания на плечо, или его фрагмент называют в настоящем документе "поливалентным" антителом. Например, "бивалентное" антитело по изобретению содержит два участка связывания на Fab или его фрагмент, в то время как "тривалентный" полипептид по изобретению содержит три участка связывания на Fab или его фрагмент. В поливалентном полимере по изобретению два или более участка связывания на Fab могут связываться с одинаковыми или отличающимися антигенами. Например, два или более участка связывания в поливалентном полипептиде по изобретению могут быть направлены против одного антигена, например против одинаковых частей или эпитопов указанного антигена или против двух или более одинаковых или отличающихся частей или эпитопов указанного антигена; и/или они могут быть направлены против различных антигенов; или их сочетания. Таким образом, бивалентный полипептид по изобретению, например, может содержать два идентичных участка связывания, может содержать первый участок, направленный против первой части или эпитопа антигена, и второй участок связывания, направленный против той же части или эпитопа указанного антигена или против другой части или эпитопа указанного антигена; или он может содержать первый участок связывания, направленный против первой части или эпитопа антигена, и второй участок связывания, направленный против другого антигена. Однако, как будет очевидно из описания, представленного в настоящем документе выше, изобретение не ограничиваются ими, в том смысле, что поливалентный полипептид по изобретению может содержать любое количество участков связывания, направленных против одного или различных антигенов.
Антитело (или полимер, или полипептид) по изобретению, которое содержит по меньшей мере два участка на Fab, или его фрагмент, в котором по меньшей мере один участок связывания направлен против первого антигена и второй участок связывания направлен против второго антигена, отличающегося от первого антигена, также будет называться "полиспецифичным". Таким образом, "биспецифичный" полимер содержит по меньшей мере один участок, направленный против первого антигена, и по меньшей мере один второй участок, направленный против второго антигена, в то время как "триспецифичный" представляет собой полимер, который содержит по меньшей мере один участок связывания, направленный против первого антигена, по меньшей мере один дополнительный участок связывания, направленный против второго антигена, и по меньшей мере один дополнительный участок связывания, направленный против третьего антигена, и т.д. Таким образом, в его наиболее простой форме, биспецифичный полипептид по изобретению представляет собой двухвалентный полипептид (на Fab) по изобретению. Однако, как будет очевидно из описания, представленного в настоящем документе выше, изобретение не ограничивается этим, в том смысле, что полиспецифичный полипептид по изобретению может содержать любое количество участков связывания, направленных против двух или более различных антигенов.
(viii) Другие модификации
Предусмотрены другие модификации гуманизированного антитела против сфинголипида или варианта антитела против сфинголипида. Например, изобретение также относится к иммуноконъюгатам, содержащим антитело, описанное в настоящем документе, конъюгированное с цитотоксическим веществом, таким как токсин (например, ферментативно активный токсин бактерий, грибов, растений или животных или его фрагменты), или радиоактивный изотоп (например, радиоконъюгат). Конъюгаты получают с использованием различных бифункциональных средств для связывания белков, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как 2,6-диизоцианат толуола) и вторичные активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол).
Антитела против сфинголипидов, описанные в настоящем документе, также можно изготавливать в качестве иммунолипосом. Липосомы, содержащие антитело, получают способами, известными в данной области, такими как описаны в Epstein et al., Proc. Natl. Acad. Sci. USA 82:3688 (1985); Hwang, et al., Proc. Natl Acad. Sci. USA 77:4030 (1980); и патентах США №№4485045 и 4544545. Липосомы с повышенным временем циркуляции описаны в патенте США №5013556. Например, липосомы можно получать способом обращенно-фазового выпаривания с липидной композицией, содержащей фосфатидилхолин, холестерин и производное фосфатидилэтаноламина с PEG (PEG-РЕ). Липосомы экструдируют через фильтры с определенным размером пор с получением липосом с требуемым диаметром. Fab′-фрагменты антитела по настоящему изобретению можно конъюгировать с липосомами, как описано в Martin et al., J. Biol. Chem. 257:286-288 (1982), посредством реакции дисульфидного обмена. Другой активный ингредиент необязательно содержится в липосоме.
Ферменты или другие полипептиды можно ковалентно связывать с антителами против сфинголипидов способами, хорошо известными в данной области, такими как применение гетеробифункциональных поперечно-сшивающих реагентов, рассмотренных выше. Альтернативно можно конструировать слитые белки, содержащие по меньшей мере антигенсвязывающую область антитела по изобретению, связанные по меньшей мере с функционально активной частью фермента по изобретению, с использованием способов рекомбинантных ДНК, хорошо известных в данной области (см., например, Neuberger, et al., Nature 312:604-608 (1984)).
В некоторых вариантах осуществления изобретения может быть желательным использование фрагмента антитела, а не интактного антитела, например, для повышения проникновения в ткани-мишени и клетки-мишени. В этом случае может быть желательной модификация фрагмента антитела для повышения его времени полужизни в сыворотке. Этого можно достигать, например, посредством включения во фрагмент антитела связывающего рецептор спасения эпитопа (например, посредством мутации соответствующего участка во фрагменте антитела или посредством включения эпитопа в пептидный маркер, который затем подвергают слиянию с фрагментом антитела на любом конце или в середине, например, с помощью синтеза ДНК или пептидов). См., например, патент США №6096871.
Также в объем этого изобретения включены ковалентные модификации гуманизированного антитела против сфинголипида или варианта антитела против сфинголипида. Их можно проводить химическим синтезом или ферментативным или химическим расщеплением антитела, если это применимо. Другие типы ковалентных модификаций антитела вносят в молекулу реакцией аминокислотных остатков-мишеней антитела с органическим образующим производное веществом, которое способно реагировать с выбранными боковыми цепями или N- или С-концевыми остатками. Иллюстративные ковалентные модификации полипептидов описаны в патенте США №5534615, конкретно включенном в настоящий документ в качестве ссылки. Предпочтительный тип ковалентной модификации антитела включает связывание антитела с одним из множества небелковых полимеров, например с полиэтиленгликолем, полипропиленгликолем или полиоксиалкиленами, так, как указано в патентах США №№4640835; 4496689; 4301144; 4670417; 4791192 или 4179337.
В. Векторы. Клетки-хозяева и рекомбинантные способы
Также изобретение относится к выделенной нуклеиновой кислоте, кодирующей гуманизированное антитело против сфинголипида или вариант антитела против сфинголипида, векторам и клеткам-хозяевам, содержащим нуклеиновую кислоту, и рекомбинантным способам продукции антитела.
Для рекомбинантной продукции антитела нуклеиновую кислоту, кодирующую его, можно выделять и встраивать в реплицируемый вектор для последующего клонирования (амплификации ДНК) или для экспрессии. В другом варианте осуществления антитело можно получать посредством гомологичной рекомбинации, например, как описано в патенте США №5204244. ДНК, кодирующую моноклональное антитело, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелую и легкую цепи антитела). Доступно множество векторов. Компоненты векторов, как правило, включают, но не ограничиваются ими, один или несколько из следующих компонентов: сигнальная последовательность, ориджин репликации, один или несколько генов селективных маркеров, энхансерный элемент, промотор и последовательность терминации транскрипции, как описано, например, в патенте США №5534615.
Пригодные клетки-хозяева для клонирования или экспрессии ДНК в векторах, представленных в настоящем документе, представляют собой клетки прокариот, дрожжей или высших эукариот, описанных выше. Пригодные для этой цели прокариоты включают эубактерии, такие как грамотрицательные или грамположительные организмы, например Enterobacteriaceae, такие как Escherichia, например Е.coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, а также Bacilli, такие как В.subtilis и В.licheniformis (например, В.licheniformis 41P), Pseudomonas, такие как P. aeruginosa и Streptomyces. Одним предпочтительным хозяином Е. coli для клонирования является Е.coli 294 (АТСС 31446), хотя также пригодными являются другие штаммы, такие как Е.coli В, Е.coli X1776 (АТСС 31537) и Е.coli W3110 (АТСС 27325). Эти примеры являются иллюстративными, но не ограничивающими.
В дополнение к прокариотам пригодными хозяевами для клонирования или экспрессии кодирующих антитело против сфинголипида векторов являются эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи. Saccharomyces cerevisiae, или обычные пекарские дрожжи, наиболее распространены среди низших эукариотических микроорганизмов-хозяев. Однако широко доступным и пригодным здесь является ряд других родов, видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K.lactis, K.fragilis (ATCC 12424), K.bulgaricus (ATCC 16045), К.wickeramii (ATCC 24178), K.waltii (ATCC 56500), К. drosophilarum (ATCC 36906), K.thermotolerans и K.marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитевидные грибы, например, такие как хозяева Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A.nidulans и A.niger.
Пригодные клетки-хозяева для экспрессии гликозилированного антитела против сфинголипида получают из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки растений и насекомых. Выявлен ряд штаммов и вариантов бакуловирусов и соответствующих пермиссивных клеток-хозяев среди хозяев, таких как Spodoptera frugiperda (гусеница), Aedes aegypti (москит), Aedes albopictus (москит), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Множество штаммов вирусов для трансфекции являются широко доступными, например вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы можно использовать здесь в качестве вирусов в соответствии с настоящим изобретением, в частности, для трансфекции клеток Spodoptera frugiperda. Также в качестве хозяев можно использовать растительные клеточные культуры хлопка, кукурузы, картофеля, сои, петунии, томата и табака.
Однако наибольший интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (культуре тканей) стало общепринятым способом. Примерами пригодных хозяев, представляющих собой клеточные линии млекопитающих, являются линия почки обезьяны CV1, трансформированная SV40 (COS-7, АТСС CRL 1651); эмбриональная линия почки человека (клетки 293 или 293, субклонированные для выращивания в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977); клетки почки детеныша хомяка (ВНК, АТСС CCL 10); клетки яичника китайского хомяка / -DHFR (СНО, Urlaub, et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (ТМ4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 АТСС CCL 70); клетки почки африканской зеленой мартышки (VERO-76, АТСС CRL-1587); клетки цервикальной карциномы человека (HELA, АТСС CCL 2); клетки почки собаки (MDCK, АТСС CCL 34); клетки печени крысы буффало (BRL 3А, АТСС CRL 1442); клетки легкого человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линия гепатомы человека (Hep G2).
Для продукции антитела против сфинголипида клетки-хозяева трансформируют описанными выше векторами для экспрессии и клонирования и культивируют в общепринятой среде, модифицированной соответствующим образом для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности.
Клетки-хозяева, используемые для продукции антитела против сфинголипида по изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев пригодными являются коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда ((MEM), (Sigma)), RPMI-1640 (Sigma) и среда Игла, модифицированная способом Дульбекко, ((DMEM), Sigma). Кроме того, для клеток-хозяев в качестве культуральной среды можно использовать любую из сред, описанных в Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal Biochem. 102:255 (1980), патентах США №№4767704; 4657866; 4927762; 4560655 или 5122469; WO9003430; WO 87/00195; или патенте США Re. 30985. Любые их этих сред можно дополнять, при необходимости, гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство GENTAMYCIN™), микроэлементами (определяемыми как неорганические соединения, обычно представленные в конечных концентрациях микромолярного диапазона) и глюкозой или эквивалентными источниками энергии. Также можно добавлять любые другие необходимые дополнительные вещества в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, рН и т.п., представляют собой условия, которые ранее использовали для выбранных для экспрессии клеток-хозяев, и они будут понятны специалисту в данной области.
При использовании рекомбинантных способов антитело может продуцироваться внутриклеточно, в периплазматическом пространстве, или оно может непосредственно секретироваться в среду. При внутриклеточной продукции антитела в качестве первого этапа удаляют частицы дебриса, либо клетки-хозяева, либо лизированные фрагменты, например, посредством центрифугирования или ультрафильтрации. В Carter, et al., Bio/Technology 10:163-167 (1992) описан способ выделения антител, которые секретируются в периплазматическое пространство Е. coli. В кратком изложении, клеточную массу размораживают в присутствии ацетата натрия (рН 3,5), ЭДТА и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 мин. Клеточный дебрис можно удалять центрифугированием. Когда антитело секретируется в среду, супернатанты таких экспрессирующих систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например установка для ультрафильтрации Amicon или Millipore Pellicon. На любой из упомянутых выше стадий можно включать ингибитор протеаз, такой как PMSF, для ингибирования протеолиза, и антибиотики можно включать для предотвращения роста ненужных загрязняющих организмов.
Композицию антитела, полученную из клеток, можно очищать с использованием, например, хроматографии с гидроксилапатитом, катионной или анионной ионообменной хроматографии и аффинной хроматографии, причем аффинная хроматография является предпочтительным способом очистки. Пригодность белка А в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который находится в антителе. Белок А можно использовать для очистки антител, в основе которых лежат тяжелые цепи человека (Lindmark et al., J. Immunol. Meth. 62:1-13 (1983)). Белок G рекомендован для всех изотипов мыши и для γ3 человека (Guss et al., EMBO J. 5:1567-1575 (1986)). Матрица, к которой присоединен аффинный лиганд, наиболее часто представляет собой агарозу, однако доступными являются и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и более короткое время обработки, чем те, которых достигают с помощью агарозы. В случае, когда антитело содержит домен СН3, для очистки пригодна смола Bakerbond ABX™ (J.Т.Baker, Phillipsburg, NJ). Также в зависимости от антитела, подлежащего выделению, пригодны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на SEPHAROSE™ с гепарином, хроматография на анионной или катионной обменной смоле (такая как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
После предварительной стадии(й) очистки смесь, содержащую представляющее интерес антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия с низким значением рН с хроматографией с использованием буфера для элюирования с рН между приблизительно 2,5-4,5, предпочтительно проводимой при низких концентрациях солей (например, от приблизительно 0-0,25 М соль).
С. Фармацевтические составы
Терапевтические составы антитела или иммунной молекулы по изобретению получают для хранения смешиванием антитела, обладающего требуемой степенью чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (см., например, Remington′s Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензэтония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как TWEEN™, PLURONICS™ или полиэтиленгликоль (PEG).
При необходимости, для конкретного показания к лечению состав, представленный в настоящем документе, также может содержать более одного активного соединения, предпочтительно соединения с дополняющими видами активности, которые не оказывают неблагоприятного эффекта друг на друга. Такие молекулы пригодным образом присутствуют в сочетании в количествах, которые эффективны для предполагаемой цели.
Активные ингредиенты также можно заключать в микрокапсулу, полученную, например способами коацервации или межфазной полимеризации, например, микрокапсулы на основе гидроксиметилцеллюлозы или желатина и микрокапсулы на основе полиметилметацилата, соответственно, в коллоидных системах для доставки лекарственного средства (например, липосомы, микросферы на основе альбумина, микроэмульсии, наночастицы и нанокапсулы) или в микроэмульсии. Такие способы описаны в Remington′s Pharmaceutical Sciences, 16th edition, Osol, A., Ed., (1980).
Составы, подлежащие применению для введения in vivo, должны быть стерильными. Этого легко достигают фильтрацией через стерильные фильтрационные мембраны.
Можно получать препараты с замедленным высвобождением. Пригодные примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, причем матрицы находятся в форме изделий определенной формы, например пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочная кислота-гликолевая кислота, такие как Lupron Depot™ (инъецируемые микросферы, состоящие из сополимера молочная кислота-гликолевая кислота и ацетата леупролида), и поли-D-(-)-3-гидроксимасляная кислота. В то время как полимеры, такие как этиленвинилацетат и молочная кислота - гликолевая кислота, обеспечивают высвобождение молекулы в течение свыше 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. При инкапсулировании антитела остаются в организме в течение длительного времени, они могут подвергаться денатурации или образовывать агрегаты под воздействием влажности при 37°С, что приводит к потере биологической активности и возможным изменениями иммуногенности. В зависимости от вовлеченного механизма можно разрабатывать рациональные стратегии для стабилизации. Например, если открыто, что механизмом агрегации является межмолекулярное образование S-S связи через тиодисульфидный обмен, стабилизации можно достигать модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, использованием соответствующих добавок и разработки специфичных полимерных композиций матриц.
D. Нетерапевтическое применение антител
Антитела по изобретению можно применять в качестве средств для аффинной очистки. В этом процессе антитела иммобилизуют на твердой фазе, такой как смола Sephadex или фильтровальная бумага, с использованием способов, хорошо известных в данной области. Иммобилизованное антитело подвергают контактированию с образцом, содержащим сфинголипид, подлежащий очистке, и впоследствии подложку промывают пригодным растворителем, который будет удалять по существу любой материал из образца, за исключением сфинголипида, который связан с иммобилизованным антителом. Наконец, подложку промывают другим пригодным растворителем, таким как глициновый буфер, например, с рН между 3 и 5,0, который будет высвобождать сфинголипид от антитела.
Антитела против сфинголипида также могут быть пригодны в диагностических анализах сфинголипидов, например, для детекции его экспрессии в конкретных клетках, тканях (таких как образец биоптатов) или жидкостях организма. Такие диагностические способы могут быть пригодны для диагностики сердечно-сосудистого или цереброваскулярного заболевания или нарушения.
Для диагностических применений антитело, как правило, будет мечено поддающейся детекции группой. Доступно множество меток, которые главным образом группируют в следующие категории:
(а) Радиоизотопы, такие как 35S, 14C, 125I, 3Н и 131I. Антитело можно метить радиоизотопом с использованием способов, описанных, например, в Current Protocols in Immunology, Volumes 1 and 2, Coligen et al., Ed. Wiley-Interscience, New York, N.Y., Pubs. (1991), и радиоактивность можно измерять с использованием подсчета на сцинтилляционном счетчике.
(b) Доступными являются флуоресцентные метки, такие как хелатные соединения редкоземельных металлов (хелатные соединения европия) или флуоресцеин и его производные, родамин и его производные, дансил, лиссамин, фикоэритрин и техасский красный. Флуоресцентные метки можно конъюгировать с антителом с использованием способов, описанных, например, в Current Protocols in Immunology, выше. Флуоресценцию можно количественно определять с использованием флуориметра.
(c) Доступны различные фермент-субстратные метки. Например, в патенте США №4275149 представлен обзор некоторых из них. Фермент главным образом катализирует химическое изменение хромогенного субстрата, которое можно измерять с использованием различных способов. Например, фермент может катализировать изменение цвета в субстрате, которое можно измерять спектрофотометрически. Альтернативно фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Способы количественного определения изменения флуоресценции описаны выше. Хемилюминесцентный субстрат становится электронно-возбужденным посредством химической реакции, и затем он может испускать свет, который можно измерять (с использованием, например, хемилюминометра), или передавать энергию флуоресцентному акцептору. Примеры ферментных меток включают люциферазы (например, люциферазу плодовой мушки и бактериальную люциферазу; патент США №4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, мочевиназу, пероксидазу, такую как пероксидаза хрена (HRPO), щелочную фосфатазу, бета-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы (например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу), гетероциклооксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.п. Способы конъюгации ферментов с антителами описаны в О′Sullivan, et al., Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (ed J. Langone & H. Van Vunakis), Academic press, New York, 73:147-166 (1981).
Примеры фермент-субстратных сочетаний включают, например:
(i) пероксидазу хрена (HRPO) с водородпероксидазой в качестве субстрата, где водородпероксидаза окисляет предшественник красителя (например, ортофенилендиамин (OPD) или 3,3′,5,5′-тетраметилбензидингидрохлорид (ТМВ));
(ii) щелочную фосфатазу (АР) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и
(iii) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, п-нитрофенил-β-D-галактозидазой) или флуорогенным субстратом 4-метилумбеллиферил-β-D-галактозидазы.
Множество других фермент-субстратных сочетаний доступно специалистам в данной области. Для их общего обзора см. патенты США №№4275149 и 4318980.
Иногда метка непрямо конъюгирована с антителом. Квалифицированному специалисту известны различные способы для достижения этого. Например, антитело можно конъюгировать с биотином, и любую из трех широких категорий меток, упомянутых выше, можно конъюгировать с авидином, или наоборот. Биотин селективно связывается с авидином, и, таким образом, метка может быть конъюгирована с антителом таким непрямым образом. Альтернативно, для достижения непрямой конъюгации метки с антителом антитело конъюгируют с небольшим гаптеном (например, дигоксином), и один из отличающихся типов меток, упомянутых выше, конъюгируют с антителом против гаптена (например, антителом против дигоксина). Таким образом, можно достигать непрямой конъюгации метки с антителом.
В другом варианте осуществления изобретения антитело против сфинголипида не обязательно должно быть меченым, и его присутствие можно выявлять с использованием меченого антитела, которое связывается с антителом против сфинголипида.
Антитела по настоящему изобретению можно использовать в любом известном способе анализа, таком как конкурентные анализы связывания, прямые и непрямые сэндвич-анализы и анализы иммунопреципитации. См., например, Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc. 1987).
Конкурентные анализы связывания основаны на способности меченого стандарта конкурировать с анализируемым компонентом тестируемого образца за связывание с ограниченным количеством антитела. Количество сфинголипида в тестируемом образце обратно пропорционально количеству стандарта, который становится связанным с антителами. Для упрощения определения количества стандарта, который становится связанным, антитела, как правило, являются нерастворимыми до или после конкуренции, так что стандарт и анализируемый компонент, которые связываются с антителами, можно удобно отделить от стандарта или анализируемого компонента, которые остаются несвязанными.
Сэндвич-анализы вовлекают применение двух антител, каждое из которых способно связываться с отличающимся иммунногенным участком, или эпитопом, белка, подлежащего детекции. В сэндвич-анализе анализируемый компонент тестируемого образца связывают первым антителом, которое иммобилизовано на твердой подложке, и впоследствии второе антитело связывается с анализируемым компонентом, таким образом, образуя нерастворимый комплекс из трех частей. См., например, патент США №4376110. Второе антитело само по себе может быть мечено поддающейся детекции группой (прямые сэндвич-анализы), или его можно определять с использованием антитела против иммуноглобулина, которое мечено поддающейся детекции группой (непрямой сэндвич-анализ). Например, одним типом сэндвич-анализа является анализ ELISA, в случае которого поддающаяся детекции группа представляет собой фермент. Для иммуногистохимии образец крови или ткани может быть свежим или замороженным или он может быть погруженным в парафин и фиксированным консервантом, например, таким как формалин.
Антитела также можно использовать для диагностических анализов in vivo. Как правило, антитело метят радионуклидом (таким как 111In, 99Tc, 14С, 131I, 125I, 3Н, 32Р или 35S), так что локализацию связанной молекулы-мишени можно определять с использованием иммуносцинтиллографии.
Е. Диагностические наборы
Для удобства антитело по настоящему изобретению может быть предоставлено в наборе, например, в виде упакованного сочетания реагентов в заданных количествах с инструкциями по проведению диагностического анализа. Когда антитело мечено ферментом, набор будет включать субстраты и кофакторы, требуемые для фермента (например, предшественник субстрата, который образует поддающийся детекции хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или лизирующий буфер) и т.п. Относительные количества различных реагентов могут широко варьировать для обеспечения концентраций в растворе реагентов, которые по существу оптимизируют чувствительность анализа. В частности, реагенты могут быть предоставлены в качестве сухих порошков, обычно лиофилизированных, включающих эксципиенты, которые при растворении будут приводить к раствору реагента, имеющему соответствующую концентрацию.
F. Терапевтическое применение антитела
Для терапевтического применения антитела против сфинголипидов по изобретению вводят млекопитающему, предпочтительно человеку, в фармацевтически приемлемой дозированной форме, такой как формы, описанные выше, включая формы, которые можно вводить внутривенно человеку в качестве болюса или в виде непрерывной инфузии в течение периода времени, посредством внутримышечного, внутрибрюшинного, интрацереброспинального, подкожного, внутрисуставного, интрасиновиального, интратекального, перорального, местного или ингаляционного способов.
Для профилактики или лечения заболевания соответствующая дозировка антитела будет зависеть от типа заболевания, подлежащего лечению, как определено выше, тяжести и течения заболевания, от профилактических или терапевтических целей введения молекулы, предшествующего лечения, клинического анамнеза пациента и ответа на антитело и от решения лечащего врача. Антитело пригодным образом вводят пациенту за один раз или на протяжении серии введений.
В зависимости от типа и тяжести заболевания исходная дозировка для введения пациенту составляет от приблизительно 1 мкг/кг до приблизительно 50 мг/кг (например, 0,1-20 мг/кг) антитела, например, либо посредством одного или нескольких отдельных введений, либо посредством непрерывной инфузии. Типичная суточная или недельная дозировка может варьировать от приблизительно 1 мкг/кг до приблизительно 20 мг/кг или более, в зависимости от упомянутых выше факторов. В случае многократных введений в течение нескольких суток или более, в зависимости от состояния, введение повторяют до возникновения требуемого подавления симптомов заболевания. Однако могут быть пригодны другие схемы дозирования. Мониторинг хода этой терапии легко проводить посредством общепринятых способов и анализов, включая, например, радиографическую визуализацию.
В соответствии с другим вариантом осуществления изобретения эффективность антитела в отношении профилактики или лечения заболевания можно повышать посредством серийного введения антитела или введения в сочетании с другим средством, которое эффективно для этих целей, например, такого как химиотерапевтические лекарственные средства против злокачественной опухоли. Такие другие средства могут присутствовать во вводимой композиции, или их можно вводить по отдельности. Антитело пригодным образом вводят серийно или в сочетании с другим средством.
G. Изделия
В другом варианте осуществления этого изобретения предусмотрено изделие, содержащее материалы, пригодные для лечения требуемого состояния. Изделие содержит контейнер и этикетку. Пригодные контейнеры включают, например, бутыли, флаконы, шприцы и пробирки для анализа. Контейнеры могут быть изготовлены из множества материалов, таких как стекло или пластмасса. Контейнер содержит композицию, которая эффективна для лечения состояния, и может иметь стерильное отверстие для доступа (например, контейнер может представлять собой мешок для внутривенного раствора или флакон, имеющий пробку, проницаемую для иглы для подкожных инъекций). Активное средство в композиции представляет собой антитело против сфинголипида. На этикетке на контейнере, или прилагаемой к нему, указано, что композицию применяют для лечения выбранного состояния. Изделие далее может содержать второй контейнер, содержащий фармацевтически приемлемый контейнер, такой как фосфатно-солевой буфер, раствор Рингера или раствор декстрозы для разбавления лиофилизированного состава антитела. Кроме того, он может включать другие материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями для применения.
Изобретение будет более понятным с помощью следующих примеров, которые предназначены только для иллюстрации наилучшего известного в настоящее время способа осуществления на практике изобретения. Объем изобретения не следует считать ограниченным ими.
ПРИМЕРЫ
Пример 1
Моноклональное антитело мыши к S1P (Sphingomab™; LT1002)
Один тип терапевтического антитела специфично связывает нежелательные сфинголипиды для достижения благоприятных эффектов, например, таких как (1) снижение эффективной концентрации нежелательных токсических сфинголипидов (и/или концентрации их метаболических предшественников), которые могут обеспечить нежелательный эффект, такой как кардиотоксический, туморогенный или ангиогенный эффект; (2) ингибирование связывания нежелательных токсических, туморогенных или ангиогенных сфинголипидов с клеточным рецептором, и/или, таким образом, снижения концентрации сфинголипида, который доступен для связывания с таким рецептором. Примеры таких терапевтических эффектов включают, но не ограничиваются ими, применение антител против S1P для снижения эффективной концентрации доступного S1P в сыворотке in vivo, таким образом, блокируя или по меньшей мере ограничивая туморогенные и ангиогенные эффекты S1P и его роль в сердечной недостаточности после инфаркта миокарда, злокачественной опухоли или фиброгенных заболеваниях.
Синтезировали тиолированный S1P, чтобы он содержал реакционно-способную группу, способную поперечно сшивать основные структурные элементы S1P с молекулой носителя, такой как KLH. Перед иммунизацией аналог тио-S1P конъюгировали поперечным сшиванием через IOA или SMCC с белковыми носителями (например, KLH) с использованием стандартных протоколов. SMCC представляет собой гетеробифункциональное поперечно-сшивающее вещество, которое реагирует с первичными аминами и сульфгидрильными группами, и он является предпочтительным поперечно-сшивающим веществом.
Мышей Swiss Webster или BALB-C иммунизировали четыре раза на протяжении двух месяцев посредством 50 мкг иммуногена (конъюгат тиолированного S1P и KLH через SMCC) на инъекцию. Образцы сыворотки собирали через две недели после второй, третьей и четвертой иммунизаций и подвергали скринингу посредством прямого ELISA в отношении присутствия антител против S1P. После этого селезенки от животных, которые обладали высокими титрами антитела, использовали для получения гибридом стандартными способами слияния. Полученные гибридомы выращивали до смыкания монослоя, после чего клеточный супернатант собирали для анализа ELISA. Среди 55 мышей, которых иммунизировали, 8 отвечали хорошо, демонстрируя существенные сывороточные титры антител, реагирующих с S1P. Слияние впоследствии проводили с использованием селезенок этих мышей и клеток миеломы в соответствии с общепринятыми способами. Затем полученные 1500 гибридом подвергали скринингу посредством прямого ELISA, с получением 287 положительных гибридом. Из этих 287 гибридом, подвергнутых скринингу посредством прямого ELISA, 159 показали значительные титры. Затем каждую из 159 гибридом распределяли в 24-луночные планшеты. Затем кондиционированные клетками среды от выросших гибридом подвергали повторному скринингу для идентификации стабильных гибридом, способных секретировать представляющие интерес антитела. Для 60 стабильных гибридом с наиболее высоким титром проводили конкурентные ELISA.
Среди 55 мышей и практически 1500 гибридом, подвергнутых скринингу, была выявлена одна гибридома, которая проявляла характеристики, которые оправдывали клонирование способом ограничивающих разведений, требуемым для получения в итоге истинного моноклонального антитела. В результате этого процесса были получены 47 клонов, большинство из которых были признаны положительными в отношении продукции антител против S1P. Среди этих 47 клонов 6 выращивали в 24-луночных планшетах и после этого подвергали скринингу посредством конкурентного ELISA. Из 4 клонов, которые оставались положительными, один клон был выбран для начала крупномасштабной продукции моноклонального антитела против S1P. Мышам SCID инъецировали эти клетки и полученные асцитные жидкости подвергали очистке посредством белка А (50% выход) и анализировали в отношении уровней эндотоксина (<3 МЕ/мг). Для одного раунда получения асцитных жидкостей проводили инъекции 50 мышам с получением всего 125 мл асцитной жидкости. Антитела были изотипированы как IgG1 каппа, и их определили как >95% чистые посредством ВЭЖХ. Антитело получали в 20 мМ фосфате натрия с 150 мМ хлоридом натрия (рН 7,2) и хранили при -70°С. Это антитело было обозначено как LT1002 или Sphingomab™.
Положительный клон гибридомы (обозначенный как клон 306D326.26) был депонирован в АТСС (номер ячейки для хранения SD-5362), и он представляет собой первое mAb мыши, направленное против S1P. Также клон содержит вариабельные участки тяжелой и легкой цепей антитела, которые можно использовать для получения "гуманизированного" варианта антитела, а также информацию о последовательности, требуемую для конструирования химерного антитела.
Скрининг сыворотки и клеточного супернатанта в отношении S1P-специфичных антител проводили посредством прямого ELISA с использованием в качестве антигена тиолированного аналога S1P. Стандартный ELISA проводили, как описано ниже, за исключением того, что в ходе первичной инкубации 50 мкл образца (сыворотки или клеточного супернатанта) разбавляли равным объемом PBS/0,1% Tween-20 (PBST). ELISA проводили в 96-луночных планшетах для ELISA высокого связывания (Costar), покрытых 0,1 мкг химически синтезированного тиолированного S1P, конъюгированного с BSA в буфере для связывания (33,6 мМ Na2CO3, 100 мМ NaHCO3; рН 9,5). Тиолированный-S1P-BSA инкубировали при 37°С в течение 1 ч при 4°С в течение ночи в лунках планшета для ELISA. Затем планшеты промывали четыре раза PBS (137 мМ NaCl, 2,68 мМ KCl, 10,14 мМ Na2HPO4, 1,76 мМ KH2PO4; рН 7,4) и блокировали PBST в течение 1 ч при комнатной температуре. Для первоначальной стадии инкубации 75 мкл образца (содержащего S1P, подлежащий измерению), инкубировали с 25 мкл mAb против S1P в концентрации 0,1 мкг/мл, разбавленного в PBST, и добавляли в лунку планшета для ELISA. Каждый образец анализировали в трех лунках. После инкубации в течение 1 ч при комнатной температуре планшеты для ELISA промывали четыре раза посредством PBS и инкубировали со 100 мкл на лунку вторичного антитела козы против антител мыши с HRP (Jackson Immunoresearch) в концентрации 0,1 мкг/мл в течение 1 ч при комнатной температуре. Затем планшеты промывали четыре раза PBS и обрабатывали тетраметилбензидином (Sigma) в течение 1-10 минут. Реакцию детекции останавливали добавлением равного объема 1М H2SO4. Оптическую плотность образцов определяли посредством измерения при 450 нм с использованием устройства для считывания планшетов ELISA EL-X-800 (Bio-Tech).
В отношении перекрестной реактивности проводили конкурентный ELISA, как описано выше, за исключением следующих изменений. Первичная инкубация состояла из конкурента (S1P, SPH, LPA и т.д.) и конъюгированного с биотином mAb против S1P. Биотинилирование очищенного моноклонального антитела проводили с использованием набора EZ-Link Sulfo-NHS-Biotinylation (Pierce). Включение биотина определяли в соответствии с протоколом набора, и оно варьировало от 7 до 11 молекул биотина на антитело. Конкурент получали следующим образом: исходные растворы липида облучали ультразвуковым излучением и сушили в атмосфере аргона, а затем разбавляли в DPBS/BSA [1 мг/мл, не содержащий жирную кислоту BSA (Calbiochem) в DPBS (Invitrogen 14040-133)]. Очищенное mAb против S1P разбавляли при необходимости в PBS/0,5% Triton X-100. Растворы конкурента и антитела смешивали для получения 3 частей конкурента на 1 часть антитела. Для получения сигнала использовали конъюгированное с HRP вторичное антитело против стрептавидина (Jackson Immunoresearch).
Другой аспект данных конкурентного ELISA (представленного на фиг.1, панель А) состоит в том, что он демонстрирует, что mAb против S1P неспособно отличить тиолированный аналог S1P от природного S1P, который добавляли в конкурентном эксперименте. Также он демонстрирует, что антитело не распознает какие-либо продукты окисления, поскольку аналог конструировали без каких-либо двойных связей. Также mAb против S1P тестировали против природного продукта, содержащего двойную связь, которому позволяли оседать при комнатной температуре в течение 48 часов. Обращенно-фазовую ВЭЖХ природного S1P проводили в соответствии со способами, описанными ранее (Deutschman, et al. (July 2003), Am Heart J., vol.146(1):62-8), и результаты показали отсутствие различий по времени удержания. Кроме того, сравнение характеристик связывания моноклонального антитела с различными липидами, представленными на фиг.1, панель А, указывает на то, что эпитоп, распознаваемый антителом, не включает углеводородную цепь в области двойной связи природного S1P. С другой стороны, эпитоп, распознаваемый моноклональным антителом, представляет собой область, содержащую аминоспирт на основном остове сфингозина и свободный фосфат. Если свободный фосфат был связан с холином (как в случае SPC), то связывание в некоторой степени снижалось. Если аминогруппа была этерифицирована с жирной кислотой (как в случае С1Р), связывания антителом не наблюдалось. Если аминоспиртовой остов сфингозина заменяли глициновым остовом (как в случае LPA), тогда S1P-специфичное моноклональное антитело не проявляло связывания. Эти данные по картированию эпитопа указывают на то, что существует только один эпитоп на S1P, распознаваемый моноклональным антителом, и что этот эпитоп определяется уникальной полярной концевой группой S1P.
В сходном эксперименте с использованием показателей ELISA оценивали пригодные контрольные материалы, чтобы убедиться, что это моноклональное антитело против S1P не распознавало белок-переносчик или поперечно-сшивающее вещество. Например, нормальное вещество для поперечного сшивания SMCC заменяли на IOA при конъюгации тиолированного S1P с BSA в качестве материала для нанесения в ELISA. Когда использовали IOA, характеристики связывания антитела были практически идентичными случаю, когда использовали BSA-SMCC-тиолированный S1P. Аналогично BSA заменяли на KLH в качестве белка, который находился в комплексе с тиолированным S1P в качестве материала для нанесения. В этом эксперименте также отсутствовало значимое отличие в характеристиках связывания антитела.
Кинетика связывания: кинетика связывания S1P с его рецептором или другими молекулами традиционно является проблематичной вследствие структуры липидов. Многие проблемы связаны с нерастворимостью липидов. Для измерения посредством BIAcore эти проблемы были решены прямой иммобилизацией S1P на чип BIAcore. Затем для определения характеристик связывания антитела с S1P антитело пропускали над поверхностью чипа и измеряли изменение оптической плотности. Чтобы преодолеть двухвалентную природу связывания антител, S1P наносили на чип с низкой плотностью. Кроме того, на чип с различной плотностью наносили S1P (7, 20 и 1000 RU) и данные по связыванию антитела в целом приводили в соответствие с моделью взаимодействия 1:1. Результаты, представленные на фиг.2, демонстрируют изменения оптической плотности вследствие связывания моноклонального антитела с S1P при трех различных плотностях S1P. В целом аффинность моноклонального антитела к S1P определили как очень высокую, в диапазоне приблизительно от 88 пикомоль (пМ) Д9 99 нМ, в зависимости от того, одновалентную или двухвалентную модель связывания использовали для анализа данных связывания.
Пример 2
Анализы ELISA
1. Количественные ELISA
На микропланшеты для титрования ELISA (Costar, каталожный №3361) наносили антитело кролика против IgG мыши, специфичное к F(ab′)2-фрагменту антитело (Jackson, 315-005-047), разбавленное в 1 М карбонатном буфере (рН 9,5) при 37°С в течение 1 ч. Планшеты промывали PBS и блокировали PBS/BSA/Tween-20 в течение 1 ч при 37°С. Для первичной инкубации к лункам добавляли разведения неспецифичного IgG мыши или IgG человека, целые молекулы (используемые для калибровочной кривой), и образцы, подлежащие измерению. Планшеты промывали и инкубировали со 100 мкл на лунку конъюгированного с HRP антитела козы против антител мыши (H+L), разбавленного 1:40000 (Jackson, каталожный №115-035-146) в течение 1 ч при 37°С. После промывания проводили детекцию ферментативной реакции посредством тетраметилбензидина (Sigma, каталожный №Т0440) и останавливали добавлением 1 М H2SO4. Оптическую плотность (OD) измеряли при 450 нм с использованием Thermo Multiskan EX. Полученные данные переносили в программное обеспечение GraphPad для анализа.
2. Прямые ELISA
На микропланшеты для титрования ELISA (Costar, каталожный №3361) наносили LPA-BSA, разбавленный в 1 М карбонатном буфере (рН 9,5) при 37°С в течение 1 ч. Планшеты промывали PBS (137 мМ NaCl, 2,68 мМ KCl, 10,1 мМ Na2HPO4, 1,76 мМ KH2PO4; рН 7,4) и блокировали посредством PBS/BSA/Tween-20 в течение 1 ч при комнатной температуре или в течение ночи при 4°С. Образцы, подлежащие тестированию, разбавляли до 0,4 мкг/мл, 0,2 мкг/мл, 0,1. мкг/мл, 0,05 мкг/мл, 0,0125 мкг/мл и 0 мкг/мл и в каждую лунку добавляли по 100 мкл. Планшеты промывали и инкубировали со 100 мкл на лунку конъюгированного с HRP антитела козы против антител мыши (разведение 1:20000) (Jackson, каталожный №115-035-003) в течение 1 ч при комнатной температуре. После промывания проводили детекцию ферментативной реакции посредством тетраметилбензидина (Sigma, каталожный №Т0440) и реакцию останавливали добавлением 1 М H2SO4. Оптическую плотность (OD) измеряли при 450 нм с использованием Thermo Multiskan EX. Полученные данные переносили в программное обеспечение GraphPad для анализа.
3. Конкурентные анализы
Специфичность mAb тестировали в анализах ELISA. На микропланшеты для титрования ELISA (Costar, каталожный №3361) наносили 18:0 LPA-BSA, разбавленного в 1 М карбонатном буфере (рН 9,5) при 37°С в течение 1 ч. Планшеты промывали посредством PBS (137 мМ NaCl, 2,68 мМ KCl, 10,1 мМ Na2HPO4/ 1,76 мМ KH2PO4; рН 7,4) и блокировали посредством PBS/BSA/Tween-20 при 37°С в течение 1 ч или в течение ночи при комнатной температуре. Для первичной инкубации в лунки планшетов для ELISA добавляли 0,4 мкг/мл mAb против LPA и заданные количества (14:0, 16:0, 18:0, 18:1, 18:2 и 20:4) LPA, DSPA, 18:1 LPC (лизофосфатидилхолина), S1P, церамида и церамид-1-фосфата и инкубировали при 37°С в течение 1 ч. Планшеты промывали и инкубировали со 100 мкл на лунку конъюгированного с HRP антитела козы против антител мыши (разведение 1:20000) (Jackson, каталожный №115-035-003) или конъюгированного с HRP антитела козы против антитела человека (H+L), разведенного 1:50000 (Jackson, каталожный №109-035-003) при 37°С в течение 1 ч. После промывания проводили детекцию ферментативной реакции посредством тетраметилбензидина и реакцию останавливали добавлением 1 М H2SO4. Оптическую плотность (OD) измеряли при 450 нм с использованием Thermo Multiskan EX. Полученные данные переносили в программное обеспечение GraphPad для анализа.
Пример 3
mAb мыши SPHINGOMAB является высокоспецифичным к S1P
Конкурентный ELISA демонстрирует специфичность SPHINGOMAB у S1P по сравнению с другими биологически активными липидами. SPHINGOMAB демонстрирует отсутствие перекрестной реактивности в отношении сфингозина (SPH), прямого метаболического предшественника S1P, или лизофосфатидной кислоты (LPA), важной молекулы внеклеточной передачи сигнала, которая структурно и функционально сходна с S1P. SPHINGOMAB не распознавало другие структурно сходные липиды и метаболиты, включая церамид-1-фосфат (ClP), дигидросфингозин (DH-SPH), фосфатидилсерин (PS), фосфатидилэтаноламин (РЕ) или сфингомиелин (SM). SPHINGOMAB не реагировало перекрестно с дигидросфингозин-1-фосфатом (DH-SlP) и, в меньшей степени, сфингозилфорилхолином (SPC) (фиг.3).
Пример 4
SPHINGOMAB значительно уменьшает CNV и рубцевание в модели CNV на мышах
Самок мышей C57BL6/J подвергали индуцируемой лазером перфорации мембраны Бруха и вводили либо 0,5 мкг Sphingomab, либо совпадающее по изотипу неспецифичное (NS) антитело, разбавленное в 2 мкл физиологического раствора. Мышей умерщвляли через 14 и 28 суток после лазерной перфорации.
Для индукции повреждений CNV зрачок расширяли офтальмологическим тропикамидом (0,5%) и фенилэфрином (2,5%). На глаз помещали покровное стекло. Для перфорации мембраны Бруха в трех квадрантах правого глаза, расположенных приблизительно на расстоянии 50 мкм от оптического диска в относительных положениях в области 9, 12 и 3 часов, использовали Oculight GL 532 nm (Iridex Corporation, Mountain View, CA), соединенный с щелевой лампой, установленный для доставки 100-мсек импульсов при 150 мВт с размером пятна 50 мкм. Левый глаз служил в качестве неповрежденного контроля во всех случаях. Любые повреждения, не связанные с пузырьками пара или повреждениями, которые сливались, исключали из анализа.
Для измерения очага повреждения CNV получали хориоидальные плоские срезы комплекса склера-хориоид-RPE и окрашивали на сосуды (агглютинин I R.communis; красный) и перициты (CD 140b; зеленый). Цифровые изображения получали с использованием эпифлуоресцентного Zeiss Axioplan 2 с цифровой камерой высокого разрешения RGB Spot и лазерным сканирующим конфокальным микроскопом (BioRad MRC 1024, BioRad Corporation, Temecula, CA). Для волюметрического анализа использовали захват z-серии и для получения объема повреждения сумму площадей повреждения на протяжении z-серии умножали на толщину z (4 мкм).
Для оценки отложения коллагена комплекс склера-хориоид-RPE окрашивали трихроматическим красителем Массона. Комплекс склера-хориоид-RPE погружали в парафин, а затем получали серийные срезы толщиной 6 микрон. Оценивали приблизительно 30 срезов на очаг повреждения. Количественное определение объема отложения коллагена вычисляли таким же образом, как описано для объема повреждений CNV.
Полученные цифровые изображения оценивали морфометрически с использованием программного обеспечения ImageJ (Research Services Branch, National Institutes of Health, Bethesda, MD). На фиг.4А показано, что SPHINGOMAB значительно ослабляет хориоидальную неоваскуляризацию через 14 и 28 суток после индуцированной лазером перфорации мембраны Бруха. На фиг.4В показано, что SPHINGOMAB значительно снижает фиброз, связанный с образованием очага повреждения CNV, через 28 суток после индуцированной лазером перфорации мембраны Бруха.
Пример 5
SPHINGOMAB ингибирует неоваскуляризацию посредством множества механизмов, включая ингибирование миграции эндотелиальных клеток и образования трубки
S1P обеспечивает миграцию эндотелиальных клеток пупочной вены человека (HUVEC) и в матригеле и в других анализах образование de novo BV in vitro; SPHINGOMAB может нейтрализовывать эти эффекты S1P. Эксперименты проводили, как описано Visentin et al. (Cancer Cell 2006 Mar; 9(3):225-38). Данные на фиг.5А позволяют предположить, что HUVEC, посеянные на GF-восстановленный матригель, образовывали множество подобных капиллярам структур в присутствии S1P и не образовывали подобные капиллярам структуры в отсутствие S1P или при совместной инкубации со SPHINGOMAB и S1P. Данные на фиг.5В демонстрируют способность 0,1-1 мкМ S1P к сильной стимуляции миграции HUVEC, в 2-2,5 раза превышающую необработанные HUVEC, или HUVEC, совместно инкубированные со SPHINGOMAB в анализе хемоинвазии в матригеле. В совокупности эти исследования демонстрируют, что SPHINGOMAB может эффективно подавлять проангиогенные эффекты S1P на ЕС.
Пример 6
SPHINGOMAB ингибирует неоваскуляризацию посредством множества механизмов, включая подавление эффектов S1P, VEGF и bFGF in vivo
Исходя из исследований in vivo, показывающих, что S1P повышал эндотелиальный рост капилляров в подкожно имплантированные вставки матригеля, авторы настоящего изобретения предположили, что SPHINGOMAB может снижать образование de novo BV in vivo. Для исследования этого авторы настоящего изобретения использовали анализ неоваскуляризации во вставке матригеля in vivo. В одной серии экспериментов матригель дополняли либо 1 мкМ S1P, 0,5 мкМ/мл bFGF, либо 1 мкг/мл VEGF, a затем инъецировали внутрибрюшинно мышам (n=4). Через 10 суток мышей гепаринизировали и инъецировали им флуоресцентный лектин, Isolectin B4-FITC, который связывается с молекулами адгезии, экспрессируемыми сосудистыми ЕС, которые образуют растущие BV. Затем вставки извлекали, замораживали в ОСТ, разрезали и оценивали в отношении FITC-окрашенных BV. Данные на фиг.6А позволяют предположить, что S1P является более сильным стимулятором неоваскуляризации in vivo, чем bFGF или VEGF [Lee, et al., (1999), Biochem Biophys Res Commun, vol.264:743-50], на что указывает большое количество окрашенных FITC BV во вставках, содержащих S1P, по сравнению со вставками, содержащими bFGF или VEGF.
Затем срезы вставок окрашивали гемотоксилином и эозином для оценки инфильтрации ЕС (фиг.6В). Инфильтрация ЕС является критической стадией при неоваскуляризации. Вставки, содержащие S1P, имели повышенную в 3 раза инфильтрацию ЕС по сравнению со вставками только матригеля. Инфильтрация клеток предположительно происходит посредством ЕС, хотя авторы настоящего изобретения признают, что также могут окрашиваться другие типы клеток, такие как иммунные клетки. Мыши, которым системно вводили SPHINGOMAB каждые 48 ч (начиная с 1 суток перед имплантацией вставки), демонстрировали сниженный уровень инфильтрации ЕС, даже когда S1P добавляли к вставкам матригеля. Эти результаты демонстрируют способность SPHINGOMAB ингибировать инфильтрацию ЕС in vivo.
Эндогенный S1P из крови и окружающей ткани могут поставлять в рану проангиогенные стимулы. Исследовали способность SPHINGOMAB снижать эндогенный S1P в ране. Оптимально стимулированные вставки (матригель, дополненный 0,5 мкг/мл bFGF или 10 мг/мл VEGF) имплантировали мышам. Мышам проводили внутрибрюшинные инъекции SPHINGOMAB в количестве 25 мг/кг или физиологического раствора каждые 48 ч, начиная с 1 суток перед имплантацией матригеля. Каждая группа по введению (матригель, матригель плюс GF или матригель плюс GF и вводимый SPHINGOMAB) состояла минимум из 6 мышей. Через 10 суток мышам вводили гепарин, инъецировали Isolectin B4-FITC, вставки вырезали, погружали в среду для замораживания ОСТ и получали срезы. Плотность микрососудов количественно оценивали по окрашенным лектином-FITC сосудам, как представлено на фиг.6С. Окрашивание BV было спорадическим в контрольных (необработанных) вставках, в то время как вставки, содержащие bFGF или VEGF, демонстрировали признаки значительной васкуляризации. Вставки из мышей, которым вводили SPHINGOMAB, демонстрировали значительное снижение образования BV по сравнению со вставками с bFGF или VEGF из мышей, которым вводили физиологический раствор. Количественное определение окрашенных сосудов показало снижение неоваскуляризации в от 5 до 8,5 раз в содержащих VEGF или bFGF оставках, соответственно, из животных, которым вводили SPHINGOMAB, по сравнению с животными, которым вводили физиологический раствор (фиг.6С). Эта оценка далее демонстрирует способность эндогенного сывороточного и тканевого S1P усиливать микроваскуляризацию, а также способность SPHINGOMAB нейтрализовывать проангиогенные эффекты эндогенного S1P.
Пример 7
SPHINGOMAB ингибирует рубцевание in vivo
S1P вносит значительный вклад в заживление раны посредством активации миграции, пролиферации фибробластов и продукции ими коллагена; SPHINGOMAB нейтрализует эти эффекты. В нескольких исследованиях с использованием нескольких типов фибробластов была подтверждена способность S1P обеспечивать заживление ран: 1) S1P повышал пролиферацию фибробластов Swiss-3Т3 при измерении по включению 3Н-тимидина с использованием стандартных способов (фиг.7А); 2) S1P обеспечивал миграцию фибробластов сердца в стандартном анализе заживления царапин (фиг.7В); 3) S1P запускал экспрессию коллагена фибробластами сердца, выделенными из трансгенных мышей, обладающих репортером GFP коллагена 1а, как показано посредством иммунофлуоресцентной микроскопии (фиг.7С); и 4) S1P индуцировал дифференцировку фибробластов легких WI-38 в миофибробласты, клетки, которые являются активными при ремоделировании рубца, как показано по повышенной экспрессиии маркерного белка миофибробластов, актина гладких мышц, с использованием иммуноблот-анализа (фиг.7Б). В каждом из этих анализов SPHINGOMAB нейтрализовывал S1P. Ожидается, что глазные фибробласты будут сходным образом отвечать на S1P и SPHINGOMAB. Было замечено сходство между сердечно-сосудистым заболеванием и неоваскулярными очагами повреждения при AMD, включая ремоделирование рубца и последующее ненадлежащее образование фиброзной ткани (Vine, et al. (2005), Ophthalmology, vol.112:2076-80 и Seddon and Chen (2004), Int Ophthalmol Clin., vol.44:17-39); таким образом, полагают, что SPHINGOMAB может иметь эффекты на неоваскуляризацию и рубцевание в глазу, сходные с эффектами, демонстрируемыми в сердечно-сосудистой системе.
В исследованиях оценивали эффективность SPHINGOMAB в отношении снижения рубцевания в сердце после длительного инфаркта миокарда (MI) посредством лигирования левой нисходящей коронарной артерии у мышей. Системное введение 25 мг/кг SPHINGOMAB или физиологического раствора начинали через 48 ч после хирургической операции. Введение антитела через 48 ч было выбрано для обеспечения нормального образования репаративного рубца в ходе ранней фазы ремоделирования и обеспечения полезного S1P-стимулируемого ангиогенеза сразу после MI. Через две недели после инфаркта мышей умерщвляли и фиброз оценивали посредством окрашивания сердечной ткани трихроматическим красителем Массона. У животных, которым вводили SPHINGOMAB, происходила практически полная отмена периваскулярного фиброза (фиг.7, фотографии). В качестве контроля каких-либо неспецифических ответов в отношении заживления ран животных для симуляции подвергали трахеотомии без лигирования коронарной артерии (фиг.7Е).
Пример 8
S1P обеспечивает трансформацию эпителиальных клеток глаза и фибробластов в сокращающиеся образующие рубцовую ткань миофибробласты
Патологический фиброз ткани (рубцевание) представляет собой основной фактор, способствующий ряду глазных нарушений, включая старческую дегенерацию желтого пятна, диабетическую ретинопатию, ретролентальную фиброплазию, пролиферативную витреоретинопатию и последствия хирургической операции по поводу глаукомы.
При многих из этих нарушений циркулирующие факторы роста и хемокины обеспечивают трансформацию нормальных клеток глаза в фибросократимые продуцирующие рубцовую ткань клетки, которые были названы "миофибробластами". В норме, миофибробласты ответственны за репарацию ткани в качестве части ответа на повреждение в виде заживления ран. Однако измененное количество и функция миофибробластов вовлечены в заболевания, характеризующиеся образованием патологической рубцовой ткани в печени, коже, легком, почках, сердце и глазах. В глазу трансформация клеток пигментного эпителия (RPE) в миофибробластный фенотип связана с образованием фиброзно-сократимых мембран, которые вызывают отслоение сетчатки и последующее нарушение зрения. Кроме того, трансформация миофибробластов в глазные фибробласты может приводить к аномальной продукции рубцовой ткани после повреждения глаза, что приводит к последующей потере зрения. Несмотря на то что было идентифицировано множество из циркулирующих белковых факторов в глазу, которые обеспечивают образование миофибробластов, ничего не известно в отношении о роли в этом процессе лизолипидов, таких как S1P. Таким образом, авторы настоящего изобретения исследовали эффекты S1P на миофибробластную трансформацию нескольких глазных клеточных линий человека. Как представлено на фиг.8, S1P стимулирует продукцию α-гладкомышечного актина (α-SMA; маркера миофибробластов) в клетках пигментного эпителия сетчатки человека (фиг.8А) и в фибробластах конъюнктивы человека (фиг.8В). Эти данные демонстрируют впервые, что S1P относится к среде циркулирующих химических факторов, которые запускают трансформацию глазных эпителиальных клеток и фибробластов в сократимые продуцирующие рубцовую ткань миофибробласты, которые могут приводить к отслоению сетчатки, глазному фиброзу и последующему нарушению зрения.
В этих экспериментах способность S1P обеспечивать экспрессию α-SMA в клетках пигментного эпителия сетчатки и фибробластах конъюнктивы отличалась зависимым от концентрации образом. Как показано, значительное повышение экспрессии α-SMA наблюдалось в эпителиальных клетках при концентрации 0,001 мкМ, которая затем снижалась до базальных уровней при концентрации 10 мкМ. В противоположность этому, значительное повышение экспрессии α-SMA в фибробластах конъюнктивы наблюдали только в концентрации 10 мкМ. Полагают, что это различие является следствием повышенной экспрессии рецептора S1P в эпителиальных клетках по сравнению с фибробластами. Вследствие повышенных уровней экспрессии рецептора S1P клетки пигментного эпителия сетчатки, вероятно, являются более чувствительными к S1P при низких концентрациях. В противоположность этому, при высоких уровнях S1P рецепторы становятся сенсибилизированными или возможно даже интернализуются, что приводит к сниженной стимуляции посредством S1P.
Коллаген является одним из основных структурных белков, который поддерживает все ткани в организме, и он представляет собой один из главных компонентов рубцовой ткани. В непатологических условиях общее содержание коллагена в ткани поддерживается посредством баланса между продукцией коллагена фибробластами и деградацией определенными ферментами. Ряд нарушений, которые включают повышенные уровни рубцовой ткани, являются следствием, частично, физиологических и молекулярных процессов, которые ингибируют деградацию коллагена, которая необходима для рубцевания. Было предположено, что способность S1P обеспечивать образование рубцовой ткани может быть следствием его способности ингибировать деградацию коллагена, таким образом, приводя к суммарному увеличению рубцовой ткани в органах. Таким образом, оценивали эффекты S1P на экспрессию ингибитора активатора плазминогена (PAI-1) в фибробластах конъюнктивы человека. Повышенная экспрессия PAI-1 коррелирует со снижением протеолитической деградации соединительной ткани и активируется в связи с несколькими фиброзными заболеваниями, которые включают повышенное рубцевание. Как представлено на фиг.8С, S1P стимулирует экспрессию PAI-1 дозозависимым образом. Эти данные подтверждают, что S1P также может обеспечивать образование рубцовой ткани посредством стимуляции экспрессии белков, которые ингибируют ее деградацию, что позволяет предположить, что для обеспечения и поддержания патологического рубцевания, связанного с глазными заболеваниями, S1P функционирует посредством множества механических каскадов.
Пример 9
SPHINGOMAB ингибирует инфильтрацию воспалительных и иммунных клеток
Воспаление является первым ответом в процессе ремоделирования. Оно запускается как ишемией, так и клеточным повреждением и приводит к активации экспрессии цитокинов, которые стимулируют миграцию макрофагов и нейтрофилов в поврежденную область для фагоцитоза погибших клеток и для дальнейшей активации воспалительного ответа [Jordan, et al. (1999), Cardiovasc Res., vol.43:860-78]. Тучные клетки также являются важными клеточными медиаторами воспалительного ответа. S1P, высвобождаемый из тучных клеток, отвечает за множество неблагоприятных ответов, наблюдаемых в экспериментальных моделях воспаления на животных [Jolly, et al. (2004), J. Exp. Med., vol.199:959-70 и Jolly et al. (2005), Blood., vol.105:4736-42].
На основе сходства иммунного и воспалительного ответов при CNV и CVD оценивали эффективность SPHINGOMAB в отношении подавления инфильтрации иммунных клеток в рану в модели инфаркта на мышах, в качестве показателя потенциальных эффектов SPHINGOMAB в отношении подавления этих повреждений в ходе AMD [Vine, et al. (2005), Ophthalmology., vol.112:2076-80; и Seddon and Chen (2004), Int Ophthalmol Clin., vol.44:17-39]. Через четверо суток после MI инфильтрацию макрофагов и тучных клеток оценивали с использованием антител МАС-1 и MCG35, соответственно, в пределах области риска. SPHINGOMAB значительно снижал плотность воспалительных макрофагов (фиг.9А) и тучных клеток (фиг.9В), позволяя предположить, что SPHINGOMAB может нейтрализовывать иммунные и воспалительные повреждения в ходе AMD.
Пример 10
Клонирование и охарактеризация вариабельных доменов моноклонального антитела против S1P мыши (LT1002; Sphingomab)
В этом примере описано клонирование mAb мыши против S1P. В целом стратегия состояла из клонирования вариабельных доменов как легкой цепи (VL), так и тяжелой'цепи (VH) мыши. Консенсусная последовательность 306D VH показывает, что фрагмент константного участка соответствует изотипу гамма 2b. Вариабельные домены мыши клонировали совместно с константным доменом легкой цепи (CL) и с константным доменом тяжелой цепи (CH1, CH2 и СН3), что привело к химерной конструкции антитела.
1. Клонирование mAb мыши
Клон из гибридомной клеточной линии против S1P 306D326.1 (ATCC#SD-5362) выращивали в DMEM (модифицированной способом Дульбекко среде Игла с GlutaMAX™ I, 4500 мг/л D-глюкозы, пируватом натрия; Gibco/Invitrogen, Carlsbad, CA, 111-035-003), 10% FBS (Sterile Fetal Clone I, Perbio Science), и lx глутамине/пенициллине/стрептомицине (Gibco/Invitrogen). Тотальную РНК выделяли из 107 гибридомных клеток с использованием способа на основе набора RNeasy Mini (Qiagen, Hilden Germany). РНК использовали для получения первой цепи кДНК в соответствии с протоколом изготовителей (набор для синтеза 1 цепи, Amersham Biosciences).
кДНК вариабельного участка (VH) тяжелой цепи иммуноглобулина амплифицировали посредством ПЦР с использованием праймера MHV7 (MHV7: 5′-ATGGRATGGAGCKGGRTCTTTMTCTT-3′ [SEQ ID NO:1]) в сочетании со смесью праймеров для константного участка IgG2b MHCG1/2a/2b/3 (MHCG1: 5′-CAGTGGATAGACAGATGGGGG-3′ [SEQ ID NO:2]; MHCG2a: 5′-CAGTGGATAGACCGATGGGGC-3 [SEQ ID NO:3]; MHCG2b: 5′-CAGTGGATAGACTGATGGGGG-3′ [SEQ ID NO:4]; MHCG3: 5′-CAAGGGATAGACAGATGGGGC-3′ [SEQ ID NO:5]). Продукт реакции лигировали в вектор pCR2.1®-TOPO® (Invitrogen) с использованием набора ТОРО-ТА cloning® и секвенировали. Затем вариабельный домен тяжелой цепи амплифицировали посредством ПЦР с этого вектора и встраивали в качестве фрагмента Hind III и Ара I, и лигировали в экспрессирующий вектор pG1D200 (см. патент США №7060808) или pG4D200 (ниже), содержащий промотор HCMVi, лидерную последовательность, и константный участок гамма-1 с получением плазмиды pG1D200306DVH (фиг.10). Консенсусная последовательность 306D VH (представленная ниже) показала, что фрагмент константной области соответствовал изотипу гамма 2b.
Аналогично вариабельный участок (VК) цепи каппа иммуноглобулина амплифицировали с использованием праймера MKV 20 (5′-GTCTCTGATTCTAGGGCA-3′ [SEQ ID NO:6]) в сочетании с праймером для константной области МКС (5′-ACTGGATGGTGGGAAGATGG-3′ [SEQ ID NO:7]). Продукт этой реакции лигировали в вектор pCR2.1®-ТОРО® с использованием набора ТОРО-ТА cloning® и секвенировали. Затем вариабельный домен легкой цепи амплифицировали посредством ПЦР, а затем встраивали в качестве фрагмента Bam HI и Hind III в экспрессирующий вектор pKN100 (см. патент США №7060808), содержащий промотор HCMVi, лидерную последовательность и константный домен каппа человека, с получением плазмиды PKN100306DVK.
Плазмиды тяжелой и легкой цепей pGlD200306DVH и pKN100306DVK трансформировали в бактерии DH4a и запасали в глицерине. Крупномасштабную плазмидную ДНК получали, как описано изготовителем (Qiagen, набор MAXIPREP™ без эндотоксина). Образцы ДНК, очищенные с использованием набора Qiagen QIAprep Spin Miniprep Kit или набора EndoFree Plasmid Mega/Maxi Kit, секвенировали с использованием автоматического секвенатора ABI 3730×1, который также транслирует флуоресцентные сигналы в их соответствующую последовательность нуклеиновых оснований. Праймеры были сконструированы на 5′- и 3′-концах, чтобы полученная последовательность перекрывалась. Длина праймеров составляла 18-24 оснований, и предпочтительно они обладали 50% содержанием GC и не обладали предсказанными димерами или вторичной структурой. Аминокислотные последовательности для доменов VH и VL мыши из Sphingomab™ представляют собой SEQ ID NO:8 и 9 соответственно (таблица 2). Остатки CDR (см. Kabat, EA (1982), Pharmacol Rev, vol.34:23-38) подчеркнуты в таблице 2 и показаны отдельно в таблице 3, ниже.


Аминокислотные последовательности нескольких вариабельных доменов химерного антитела (VH и VL) сравнивают в таблице 4. Эти варианты клонировали в экспрессионные векторы после лидерных последовательностей зародышевой. линии. Лидерные последовательности зародышевой линии подчеркнуты в таблице 4 на последовательностях рАТН200 (первые 19 аминокислот) и рАТН300 (первые 22 аминокислоты). CDR показаны полужирным шрифтом. Аминокислоты, которые следуют после С-конца каждой из последовательностей тяжелой и легкой цепей в таблице 4, показаны курсивом. Они представляют собой первые несколько аминокислот константного домена, и они не являются частью вариабельного домена.
Следует отметить, что, несмотря на то, что серийные номера рАТН200 и рАТН300 обычно относятся к вектору, содержащему конкретную последовательность варианта вариабельного домена, для удобства эту номенклатуру можно использовать в настоящем документе для обозначения и отличия непосредственно вариантов вариабельных доменов.
Последовательности VH- и VL-доменов мыши использовали для конструирования молекулярной модели для определения того, какие остатки каркасной области следует включать в гуманизированное антитело.

2. Экспрессия и связывающие свойства химерного антитела
Плазмиды тяжелой и легкой цепей как pG1D200306DVH, так pKN100306DVK трансформировали в бактерии DH4a и запасали в глицерине. Крупномасштабную плазмидную ДНК получали, как описано изготовителем (Qiagen, набор MAXIPREP™ без эндотоксина каталожный №12362).
Для экспрессии антитела в не являющейся человеческой системе млекопитающих плазмиды трансфицировали в клеточную линию фибробластов почки африканской зеленой мартышки COS 7 посредством электропорации (0,7 мл с 107 клеток/мл) с использованием 10 мкг каждой плазмиды. Трансфицированные клетки выращивали в 8 мл среды для роста в течение 4 суток. Химерное антитело 306DH1 × 306DVK-2 экспрессировалось в количестве в 1,5 мкг/мл в среде, кондиционированной временно котрансфицированными клетками COS. Связывание этого антитела с S1P определяли с использованием ELISA для S1P.
Уровень экспрессии химерного антитела определяли в количественном ELISA следующим образом. На микропланшеты для титрования (Nunc MaxiSorp immunoplate, Invitrogen) наносили 100-мкл аликвоты 0,4 мкг/мл антитела козы против IgG человека (Sigma, St. Louis, МО), разбавленного в PBS, и инкубировали в течение ночи при 4°С. Затем планшеты промывали три раза посредством 200 мкл/лунка буфера для промывания (1×PBS, 0,1% TWEEN). Аликвоты объемом 200 мкл каждого разбавленного образца сыворотки или супернатанта для слияния переносили в покрытые токсином планшеты и инкубировали при 37°С в течение 1 ч. После 6 промываний буфером для промывания в каждую лунку добавляли конъюгат антитела козы против легкой цепи каппа человека с пероксидазой (Jackson Immunol. Research) в разведении 1:5000. Реакцию проводили в течение 1 ч при комнатной температуре, планшеты промывали 6 раз буфером для промывания и 150 мкл субстрата K-BLUE (Sigma) добавляли в каждую лунку, инкубированную в темноте при комнатной температуре в течение 10 мин. Реакцию останавливали добавлением 50 мкл раствора RED STOP (SkyBio Ltd.) и поглощение определяли при 655 нм с использованием Microplater Reader 3550 (Bio-Rad Laboratories Ltd.). Результаты анализов связывания антитела представлены на фиг.11.
3. Экспрессия в 293F
Плазмиды тяжелой и легкой цепей трансформировали в Е.coli Тор 10 (химически компетентные клетки Е. coli One Shot Top 10 (Invitrogen, C4040-10)) и запасали в глицерине. Крупномасштабную плазмидную ДНК получали, как описано изготовителем (Qiagen, набор MAXIPREP™ без эндотоксина каталожный №12362).
Для экспрессии антитела в системе человека плазмиды трансфицировали в клеточную линию почки эмбриона человека 293F (Invitrogen) с использованием 293fectin (Invitrogen) и с использованием сред 293F-FreeStyle Media (Invitrogen) для культивирования. Плазмиды как легкой, так и тяжелой цепей трансфицировали в количестве 0,5 г/мл. Трансфекции проводили при клеточной плотности 106 клеток/мл. Супернатанты собирали центрифугированием при 1100 об/мин в течение 5 минут при 25°С через 3 суток после трансфекции. Уровни экспрессии количественно определяли посредством количественного ELISA (см. предыдущие примеры), и они варьировали от ~0,25 до 0,5 г/мл для химерного антитела.
4. Очистка антитела
Моноклональные антитела очищали из культуральных супернатантов, пропуская культуральные супернатанты над колонками с белком A/G (Pierce, каталожный №53133) со скоростью 0,5 мл/мин. Подвижные фазы состояли из IX буфера для связывания IgG Pierce (каталожный №21001) и 0,1 М глицина, рН 2,7 (Pierce, Elution Buffer, каталожный №21004). Коллекции антител в 0,1 М глицине разбавляли до 10% (об./об.) посредством 1 М фосфатного буфера, рН 8,0, для нейтрализации рН. Коллекции IgGi объединяли и подвергали тщательному диализу против IX PBS (Pierce Slide-A-Lyzer Cassette, 3500 MWCO, каталожный №66382). Элюаты концентрировали с использованием Centricon YM-3 (10000 MWCO Amicon каталожный №4203) посредством центрифугирования в течение 1 ч при 2500 об/мин. Концентрацию антитела определяли посредством количественного ELISA, как.описано выше, с использованием в качестве стандарта коммерческого исходного раствора миеломного IgG1. Изотипы тяжелых цепей mAb определяли посредством ELISA с использованием набора для изотипирования моноклональных антител (Sigma, ISO-2).
5. Сравнительное связывание вариантов антитела с S1P
В таблице 5, ниже, представлен сравнительный анализ мутантных форм с химерным антителом. Для получения этих результатов проводили детекцию связывания антитела вторичным антителом, специфичным к IgG мыши или человека, конъюгированным с HRP. Хромогенную реакцию измеряли и представляли в качестве оптической плотности (OD). Концентрация панели антител составляла 0,1 мкг/мл. Не было выявлено взаимодействия второго антитела с покрытой S1P матрицей отдельно.
6. Определение кинетики связывания посредством поверхностного плазмонного резонанса (SPR)
Все данные связывания получали на оптическом биосенсоре Biacore 2000 (Biacore АВ, Uppsala Sweden). S1P связывали с сенсорным чипом на основе малеимида СМ5. Сначала чип СМ5 активировали смесью равных количеств NHS/EDC в течение семи минут с последующей стадией блокирования этилдиамином в течение 7 минут. Далее над поверхностями пропускали сульфо-MBS (Pierce Со.) в 0,5-мМ концентрации в буфере для анализа HBS (10 мМ HEPES, 150 мМ NaCl, 0,005% р20, рН 7,4). S1P разбавляли в буфере для анализа HBS до концентрации 0,1 мМ и инъецировали в течение различных периодов времени с получением 2 поверхностей с различной плотностью S1P (305 и 470 RU). Далее, данные связывания для mAb получали с использованием серии 3-кратных разведений, начиная с 16,7 нМ, 50,0 нМ, 50,0 нМ, 16,7 нМ и 16,7 нМ для антител мыши, 201308, 201309 и 207308 соответственно.
Каждую концентрацию тестировали в двух экземплярах. Поверхности регенерировали посредством 50 мМ NaOH. Все данные получали при 25°С. Данные об ответе обрабатывали с использованием поверхности для сравнения, а также пустых инъекций. Наборы данных (ответы от двух поверхностей и каждого варианта, тестированного дважды) приводили в соответствие с моделями взаимодействия с получением параметров связывания. Данные для различных концентраций mAb в целом приводили в соответствие с использованием модели взаимодействия 1:1 (мышь) или 1:2 (варианты) для определения кажущихся констант скорости связывания. Число в скобках указывает на ошибку в последней цифре.
Пример 11
Химерное mAb к S1P
Как используют в настоящем документе, термин "химерное" антитело (или "иммуноглобулин") относится к молекуле, содержащей тяжелую и/или легкую цепь, которая идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретных видов или принадлежащих конкретному классу или подклассу антител, в то время как оставшаяся цепь(и) идентична или гомологична соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также к фрагментам таких антител, при условии, что они проявляют требуемую биологическую активность (Cabilly, et al., выше; Morrison et al., Proc. Natl. Acad. Sci. U.S.A. 81:6851 (1984)).
Химерное антитело к SIP получали с использованием вариабельных участков (Fv), содержащих активные связывающие S1P участки антитела мыши из конкретной гибридомы (номер ячейки для хранения депонированного образца в АТСС SD-5362) с Fc-областями иммуноглобулина IgG1 человека. Fc-области содержали домены CL, ChL и Ch3 антитела человека. Не ограничиваясь конкретным способом, химерные антитела также можно было получить из Fc-областей IgG1, IgG2, IgG3, IgG4, IgA или IgM. Как будет понятно специалистам в данной области, "гуманизированные" антитела можно получать посредством пересадки определяющих комплементарность участков (CDR, например CDR1-4) mAb мыши против S1P в каркасные области антитела человека (например, Fr1, Fr4 и т.д.), такие как каркасные области IgG1. На фиг.11 представлено связывание химерных и полностью мышиных mAb при прямом определении посредством ELISA с использованием тиолированного S1P в качестве материала для нанесения.
В прямых экспериментах ELISA, представленных на фиг.11, химерное антитело к S1P имело сходные характеристики связывания с моноклональным антителом полностью мыши. ELISA проводили в 96-луночных планшетах для ELISA с высоким связыванием (Costar), на которые нанесен 0,1 мкг химически синтезированного тиолированного S1P, конъюгированного с BSA в буфере для связывания (33,6 мМ Na2CO3, 100 мМ NaHCO3; рН 9,5). Тиолированный S1P-BSA инкубировали при 37°С в течение 1 ч или при 4°С в течение ночи в планшете для ELISA. Затем планшеты промывали четыре раза посредством PBS (137 мМ NaCl, 2,68 мМ KCl, 10,14 мМ Na2HPO4, 1,76 мМ KH2PO4; рН 7,4) и блокировали посредством PBST в течение 1 ч при комнатной температуре. Для первоначальной стадии инкубирования 75 мкл образца (содержащего подлежащий измерению S1P) инкубировали с 25 мкл 0,1 мкг/мл моноклонального антитела против S1P, разбавленного в PBST, и добавляли в лунку планшета для ELISA.
Анализ каждого образца проводили в трех лунках. После инкубации в течение 1 ч при комнатной температуре планшеты для ELISA промывали четыре раза посредством PBS и инкубировали со 100 мкл на лунку 0,1 мкг/мл вторичного антитела козы против антител мыши с HRP (Jackson Immunoresearch) в течение 1 ч при комнатной температуре. Затем планшеты промывали четыре раза посредством PBS и подвергали воздействию тетраметилбензидина (Sigma) в течение 1-10 минут. Реакцию детекции останавливали посредством добавления равного объема 1М H2SO4. Оптическую плотность образцов определяли посредством измерения при 450 нм с использованием устройства для считывания планшетов ELISA EL-X-800 (Bio-Tech).
Также предпочтительный способ измерения титра антитела либо в сыворотке иммунизированного животного, либо в кондиционированных клетками средах (например, супернатанте) антителопродуцирующей клетки, такой как гибридома, включает покрытие планшета для ELISA лигандом-мишенью (например, тиолированным аналогом SIP, LPA и т.д.), который ковалентно связан с белком-носителем, таким как BSA.
Не ограничиваясь конкретным способом или примером, химерные антитела можно получать против других липидных мишеней, таких как LPA, PAF, церамиды, сульфатиды, цереброзиды, кардиолипины, фосфотидилсерины, фосфотидилинозитолы, фосфатидные кислоты, фосфотидилхолины, фосфатидилэтаноламины, эйкозиноиды и другие лейкотриены и т.д. Кроме того, если желательно, многие из этих липидов также могут быть гликозилированными и/или ацетилированными.
Пример 12
Получение и охарактеризация гуманизированного моноклонального антитела против S1P LT1009 (Sonepcizumab)
Было показано, что моноклональное антитело мыши против S1P 306D (LT1002; Sphingomab™), которое специфично связывает S1P, эффективно подавляет ангиогенез и рост опухоли в различных моделях на животных. Как рассмотрено ниже, LT1002 гуманизировали с использованием поиска идентичности и гомологии последовательностей для каркасных областей человека, в которые подлежит пересаживать CDR мыши, и создаваемой компьютером модели, указывающей на некоторые возвратные мутации каркасной области. Два варианта, НuМАbНСLС3 (LT1004) (с 3 возвратными мутациями в легкой цепи) и HuMAbHCLC5 (LT1006) (с 5 возвратными мутациями в легкой цепи) обладали аффинностью связывания в наномолярном диапазоне. Дополнительное конструирование проводили в попытках улучшить биофизические и биологические свойства гуманизированных вариантов. Гуманизированные варианты HuMAbHCCysAlaLC3 (LT1007) и HuMAbHCCysAlaLC5 (LT1009), в которых свободный остаток цистеина в HCDR2 заменен аланином, проявляли аффинность связывания в пикомолярном диапазоне. Все гуманизированные варианты ингибировали ангиогенез в модели хориоидальной неоваскуляризации (CNV) при связанной со старением дегенерации желтого пятна (AMD), при этом HuMAbHCCysAlaLC5 (LT1009) проявлял лучшую стабильность и эффективность in vivo no сравнению с исходным антителом мыши. Вариант huMAbHCCysAlaLC5 (LT1009) был назван Sonepcizumab™.
а. Схема гуманизации для антитела против S1P
Вариабельные домены mAb мыши LT1002 (Sphingomab™) гуманизировали посредством пересадки CDR (патент США №5225539, Winter). Остатки CDR идентифицировали на основе гипервариабельности последовательности, как описано Kabat et al. 1991.
В этом исследовании пригодные акцепторные структуры выбирали, исходя из поиска гомологии антител человека в базах данных IMGT и Kabat с использованием программы для структурного выравнивания (SR v7.6). Исходной стадией был запрос этих баз данных вариабельных последовательностей тяжелой цепи (VH) и вариабельных последовательностей легкой цепи (VL) человека с последовательностями VH и VL белка LT1002 соответственно для идентификации каркасных областей человека (FR) с высокой идентичностью последовательностей в FR, в остатках Vernier (Foote, J. & Winter, G. Antibody framework residues affecting the conformation of the hypervariable loops. J. Mol. Biol. 224, 487-499 (1992)), канонических остатках (Morea, et al., Antibody modeling: implications for engineering and design, Methods 20, 267-279 (2000)) и остатках границы контакта VH-VL (Chothia, C, Novotny J., Bruccoleri, R., & Karplus, M. Domain association in immunoglobulin molecules. The packing of variable domains. J. Mol. Biol. 186, 651-663 (1985)) и с CDR идентичного канонического класса и/или длины. Идентичность каждого члена этой библиотеки отдельным выровненным остаткам антитела мыши вычисляли с использованием программы. Идентифицировали последовательности человека с последовательностью FR, наиболее идентичной FR мыши, получая исходный короткий список "акцепторных" последовательностей человека. Также вычисляли последовательности с наибольшей идентичностью антителу мыши в остатках Vernier, канонических остатках и остатках границы контакта VH-VL (VCI). Различия в этих положениях у человека и мыши классифицировали на консервативные и неконсервативные замены, чтобы выбранная наилучшая каркасная область имела наименьшее количество неконсервативных отличий VCI от LT1002. Петли CDR L3 и H1 LT1002 можно классифицировать как канонические структуры. Эти структуры L3 и HI использовали для отбора FR антитела человека с идентичными каноническими структурами. Для неклассифицированных CDR была предпринята попытка отобрать каркасные области человека с длиной CDR, идентичной антителу мыши. Обоснованием этого является то, что структуры петель CDR зависят не только от самой последовательности петли CDR, но также от основных остатков каркасной области (канонических остатков). Таким образом каркасная область человека с совпадающими каноническими структурами CDR и/или длиной CDR, вероятно, будет удерживать пересаженные CDR мыши в наиболее подходящей ориентации для сохранения аффинности связывания антигена. Этого достигали для всех CDR, за исключением Н3 CDR, посредством выбора последовательностей каркасной области человека. Кроме того, каркасные области с необычными остатками цистеина или пролина исключали, где это возможно. Эти вычисления проводили по отдельности для последовательностей тяжелой и легкой цепей. Наконец, в наиболее совпадающих последовательностях сравнивали различия отдельных последовательностей на протяжении каркасной области. Среди антител человека, которые наиболее соответствовали указанным выше сравнительным вычислениям, антитела AY050707 и AJ002773 были отобраны как наиболее пригодные антитела для каркасной области человека легкой цепи и тяжелой цепи соответственно. Каркасная область AY050707 была описана van den Brink, et al. (Blood, 15 April 2002, Vol.99, №8, pp.2828-2834), и ее последовательность доступна через Genbank (регистрационный №AY050707, неполная cds мРНК вариабельной области легкой цепи иммуноглобулина клона WR3VL Homo Sapiens; внесенная 13 ноября 2001 года, последний пересмотр 8 апреля 2002 года).
Аналогично каркасная область антитела (AJ002773) была описана Snow et al. [Eur. J. Immunol. 28 (10), 3354-3361 (1998)], и ее последовательность доступна через Genbank (регистрационный №AJ002773, мРНК Homo Sapiens для вариабельной области 5 тяжелой цепи иммуноглобулина G4 пациента 2,2; внесенная 6 ноября 1998 года, последний пересмотр 16 октября 2006 года).
Последовательности как AY050707 (легкая цепь), так и AJ002773 (тяжелая цепь) также находятся в IMGT/LIGM, многоцелевой базе данных нуклеотидных последовательностей иммуноглобулинов (IG) и Т-клеточных рецепторов (TR) человека и других видов позвоночных. Эта база данных была создана в 1989 году, Marie-Paule Lefrane, LIGM, Montpellier, France, и она интерактивно доступна с июля 1995 года.
Второй стадией было создание молекулярной модели вариабельных участков LT1002 и идентификация остатков FR, которые могут влиять на связывание антигена, но не были включены в группу остатков Vernier, канонических остатков и остатков границы контакта. Для лучшего понимания того, каким образом различные остатки FR влияют на конформацию петель CDR и наоборот, исследовали множество структурных признаков пересаженных донорных и акцепторных вариабельных доменов. Неконсервативные остатки FR в LT1002, которые, вероятно, влияют на CDR, были идентифицированы, исходя из определений остатков Vernier и канонических остатков (см. выше), и таким образом в нескольких остатках FR человека были восстановлены исходные аминокислоты мыши (подвергнуты возвратной мутации).
b. Мутагенез
Мутации в последовательностях вариабельного домена создавали с использованием набора QuikChange Site-Directed Mutagenesis Kit (Stratagene, каталожный #200524). Отдельные реакции проводили с 50 нг двухцепочечной ДНК-матрицы, 2,5 Ед ДНК-полимеразы PfuUltre HF и ее соответствующим буфером (Stratagene, каталожный №200524), 10 мМ смеси dNTP и 125 нг каждого из мутагенных олигонуклеотидов, ресуспендированных в 5 мМ Tris-HCl (pH 8,0), и 0,1 мМ EDTA. Исходную денатурацию проводили при 95°С в течение 30 секунд, с последующими 16 циклами амплификации: 95°С в течение 30 секунд, 55°С в течение 60 секунд и 68°С в течение 8 мин. После температурных циклов конечную реакционную смесь расщепляли веществом для расщепления DpnI при 37°С в течение 1 часа для удаления метилированной исходной ДНК. Полученную мутантную форму трансформировали в компетентные E.coli XL1-Blue и высевали на LB-агар, содержащий 50 мкг/мл ампициллина. Затем колонии проверяли секвенированием. Затем каждую из мутантных форм культивировали во встряхиваемых колбах объемом 1 литр и очищали с использованием набора EndoFree Plasmid Purification Kit от Qiagen, каталожный №12362.
с. Получение вариантов гуманизированного антитела
Химерное антитело мыши-человека (chMAb S1P) конструировали посредством клонирования вариабельных доменов LT1002 в вектор, который содержал константные участки каппа-цепи и тяжелой цепи человека для обеспечения экспрессии полноразмерного антитела в клетках млекопитающих. Гуманизированную тяжелую цепь получили пересадкой CDR 1, 2 и 3 по Kabat из VH LT1002 в акцепторную каркасную область AJ002773. Ближайшим геном зародышевой линии к AJ002773 был VH5-51, лидерная последовательность которого была включена, в качестве лидерной последовательности, в гуманизированный вариант тяжелой цепи. Белковая последовательность рАТН200, первого гуманизированного варианта LT1002 VH, с лидерной последовательностью VH5-51, представлена в таблице 4. В случае VH-домена LT1002 остатки в положении 2, 27, 37, 48, 67 и 69 представляли собой остатки Vernier или остатки на границе контакта VH- и VL-доменов, и, вероятно, они влияют на ориентацию CDR. Положение 37, по-видимому, является критическим для границы контакта между VH- и VL-доменами. Остатки в этих положениях в каркасной области человека подвергали возвратной мутации посредством остатка мыши, находящегося в соответствующем положении. Мутации V37M, M48I и Y27F тестировали по-отдельности. Один вариант (рАТН205) содержал 3 мутации совместно с V67A и I69L, а другой вариант (рАТН206) содержал все 5 мутаций и V2A.
Гуманизированную легкую цепь получали пересадкой CDR 1, 2 и 3 по Kabat из LT1002 VL в акцепторную каркасную область AY050707. Ближайшим геном зародышевой линии к AY050707 был L11, лидерная последовательность которого была включена в гуманизированную конструкцию легкой цепи. Последовательность белка рАТН300 (легкая цепь LT1002) показана в таблице 4. Лидерные последовательности зародышевой линии указаны в таблице 4 подчеркиванием. В случае VL, четыре неконсервативных положения Vernier 4, 36, 49, 64 были отобраны для возвратной мутации на остатки мыши, поскольку они вовлечены в поддержание структуры петель CDR. Исследование молекулярной модели LT1002 подтвердило, что Tyr 67 расположен вблизи поверхности CDR и ориентирован в направлении связывающей антиген плоскости и может взаимодействовать с S1P. Таким образом, в более поздние гуманизированные варианты также добавляли возвратную мутацию S67Y. Две мутации были внесены по-отдельности с получением двух вариантов, содержащих либо Y49S, либо Y36F. Было получено несколько вариантов со следующими сочетаниями мутаций: (Y49S, F4V), (Y49S, Y36F), (Y49S, Y36F, F4V), (Y49S, G64S), (Y49S, Y36F, F4V, G64S), (Y49S, Y36F, F4V, G64S, S67Y), (Y49S, G64S, S67Y).
d. Селекция основных гуманизированных кандидатов
Вариабельные участки основных пересаженных вариантов (рАТН200 и рАТН300) и все варианты, содержащие возвратные мутации, клонировали в экспрессирующие векторы, содержащие константные участки VH или VL человека. Все гуманизированные варианты получали в клетках млекопитающих в тех же условиях, что и химерное (chMAb) антитело, и тестировали в отношении связывания с S1P посредством ELISA. Выход составлял приблизительно 10-20 мг/л для гуманизированных вариантов и 0,3-0,5 мг/л для chMAb против SIP. SDS-PAGE в восстанавливающих условиях показал две полосы на уровне 25 кДа и 50 кДа с высокой чистотой (>98%), что согласуется с ожидаемыми массами легкой и тяжелой цепей. При невосстанавливающих условиях наблюдали одну полосу с ожидаемой массой ~150 кДа. В качестве стандарта в анализах связывания гуманизированного антитела использовали chMAb, поскольку оно содержало те же вариабельные участки, что и исходное антитело мыши, и имело те же константные участки, что и гуманизированные антитела, и, таким образом, его можно было выявить с использованием того же протокола ELISA.
Исходное гуманизированное антитело, в котором шесть CDR мыши были пересажены в немутантные каркасные области человека, не демонстрировало какого-либо поддающегося детекции связывания с S1P (фиг.11). Легкая цепь каппа, содержащая 4 возвратные мутации (Y49S, Y36F, F4V и G64S), совместно с химерной тяжелой цепью, проявляла субоптимальное связывание с S1P при измерении посредством ELISA. Включение дополнительной мутации в положение Y67 значительно повышало связывание. Как вариант рАТН308, который содержал возвратные мутации Y49S, Y36F, F4V, G64S и S67Y, так и вариант рАТН308, который содержал возвратные мутации Y49S, G64S и S67Y, совместно с химерной тяжелой цепью, приводили к антителам, которые связывали S1P аналогично химерному антителу, при определении посредством ELISA. 2 мутации, Y36F и F4V, не считали необходимыми возвратными мутациями с точки зрения связывания S1P. Для восстановления активности требовалось конструирование от 3 до 5 возвратных мутаций в каркасной области VL.
Включение возвратной мутации Vernier V37M в каркасную область человека тяжелой цепи, совместно с химерной легкой цепью, было достаточным для восстановления связывающего поведения, сходного с химерным антителом (фиг.11).
В общих словах, гуманизация VH-домена LT1002 требовала только одной аминокислоты из последовательности каркасной области мыши, в то время как в VL-домене каркасной области мыши для достижения связывания, эквивалентного исходному LT1002 мыши, необходимо было сохранить три или пять остатков мыши.
е. Оптимизация гуманизированного основного кандидата
Антитело мыши против S1P содержит свободный остаток цистеина в CDR2 (Cys50) тяжелой цепи, который может потенциально вызывать некоторую нестабильность молекулы антитела. С использованием сайт-направленного мутагенеза создавали варианты рАТН201 с заменой остатка цистеина аланином (huMAbHCCysAlaLC3) (рАТН207), глицином (huMAbHCCysAlaLC3), серином (huMAbHCCysSerLC3) и фенилаланином (huMAbHCCysPheLC3). Мутантную тяжелую цепь с цистеином также тестировали с гуманизированной легкой цепью (рАТН308), содержащей 5 возвратных мутаций (huMAbHCCysAlaLC5=LT1009). Варианты экспрессировали в клетках млекопитающих, а затем охарактеризовывали в панели анализов in vitro. Важно, что уровень экспрессии гуманизированных вариантов был значительно выше, чем для chMAb против S1P.
f. Углубленная охарактеризация основного гуманизированного кандидата
i. Специфичность. Гуманизированные варианты тестировали в отношении специфичности в конкурентном анализе ELISA (фиг.1) против S1P и нескольких других биологических липидов. Этот анализ имел дополнительное преимущество, позволяя эпитопное картирование. Гуманизированное антитело LT1009 демонстрировало отсутствие перекрестной реактивности к сфингозину (SPH), прямому метаболическому предшественнику S1P, или LPA (лизофосфатидной кислоте), важной внеклеточной молекуле передачи сигнала, которая структурно и функционально сходна с S1P. Более того, rhuMAb против S1P не распознавало другие структурно сходные липиды и метаболиты, включая церамид (CER), церамид-1-фосфат (С1Р). Однако, как и ожидалось, LT1009 перекрестно реагировал с сфингозилфосфохолином (SPC), липидом, в котором свободная фосфатная группа S1P связана с остатком холина. Важно, что все гуманизированные варианты проявляли профиль специфичности, сравнимый с антителом мыши.
ii. Аффинность связывания. Измерение посредством Biacore связывания IgG с чипом, покрытым S1P, показало, что варианты LT1004 или LT1006 проявляли аффинность связывания в низком наномолярном диапазоне, сходную с chMAb против S1P, как показано на фиг.11. Гуманизированные варианты LT1007 и LT1009, в которых остаток цистеина был заменен аланином, проявляли аффинность связывания в пикомолярном диапазоне, сходную с исходным LT1002 мыши (Sphingomab™).
iii. Стабильность. Гуманизированные варианты тестировали в отношении стабильности после нагрузки при высокой температуре. Для каждого гуманизированного варианта определяли приблизительные средние точки термических конформационных переходов с устранением сворачивания (TM), подвергая супернатанты воздействию температур в диапазоне от 60 до 74°С. Эти температуры были выбраны, исходя из профиля денатурации, наблюдаемого для молекулы антитела мыши после термической нагрузки в широком диапазоне температур между 50 и 80°С. Связывающие свойства каждого варианта определяли до и после температурной нагрузки. Антитело мыши проявляло TM 65°С. Вариант huMAbHCCysAlaLC5 (LT1009) проявлял TM, превосходящую все другие варианты. В таблице 6 представлены основные гуманизированные кандидаты и их характеристики.
Указано количество мутаций в тяжелой и легкой цепях. В столбце "описание" приведена идентичность тяжелой и легкой цепей.
iv. Последовательности
Как в случае природных антител, LT1009 включает три определяющих комплементарность области (каждую "CDR") в каждом из двух полипептидов легкой цепи и в каждом из двух полипептидов тяжелой цепи, которые содержат каждую молекулу антитела. Аминокислотные последовательности для каждой из этих шести CDR представлены непосредственно ниже ("VL" обозначает вариабельный участок легкой цепи иммуноглобулина, в то время как "VH" обозначает вариабельный участок тяжелой цепи иммуноглобулина):
CDR1 VL: ITTTDIDDDMN [SEQ ID NO:10]
CDR2 VL: EGNILRP [SEQ ID NO:11]
CDR3 VL: LQSDNLPFT [SEQ ID NO:12]
CDR1 VH: DHTIH [SEQ ID NO:13]
CDR3 VH: GGFYGSTIWFDF [SEQ ID NO:15]
CDR2 VH: AISPRHDITKYNEMFRG [SEQ ID NO:18]
Пример 13
Продукция и очистка гуманизированного mАb против S1P
В этом примере описана продукция рекомбинантного гуманизированного моноклонального антитела. (LT1009; Sonepcizumab™), которое связывается с высокой аффинностью с биологически активным липидом сфингозин-1-фосфатом (SIP). LT1009 представляет собой полноразмерное антитело изотипа IgG1/каппа, состоящее из двух идентичных легких цепей и двух идентичных тяжелых цепей с общей молекулярной массой приблизительно 150 кДа. Тяжелая цепь содержит N-связанный участок гликозилирования. Природа олигосахаридной структуры еще не определена, однако ожидается, что она представляет собой комплексную биантеннальную структуру с сердцевиной в виде фукозы. Природа гликоформы, которая будет преобладающей, не известна на этой стадии. Ожидается некоторая С-концевая гетерогенность вследствие присутствия остатков лизина в константном домене тяжелой цепи. Две тяжелые цепи ковалентно связаны друг с другом через две межцепочечные дисульфидные связи, которые согласуются со структурой IgG1 человека.
LT1009 исходно было получено из моноклонального антитела мыши (LT1002; Sphingomab™), которое было получено с использованием гибридом, полученным из мышей, иммунизированных S1P. Гуманизация антитела мыши вовлекала встраивание шести CDR мыши вместо CDR каркасной области антитела человека, отобранного вследствие его структурного сходства с исходным антителом мыши. Для создания гуманизированного антитела в каркасную область была внесена серия замен. Эти замены называют возвратными мутациями, и они представляют собой замены остатков человека остатками мыши, которые играют существенную роль во взаимодействии антитела с антигеном. Конечный гуманизированный вариант содержит одну возвратную мутацию мыши в каркасной области вариабельного домена тяжелой цепи человека и пять возвратных мутаций мыши в каркасной области вариабельного домена легкой цепи человека. Кроме того, один остаток, присутствующий в CDR2 тяжелой цепи, заменяли остатком аланина. Было показано, что эта замена повышает стабильность и эффективность молекулы антитела.
Гуманизированные вариабельные домены (как легкой, так и тяжелой цепи) клонировали в систему для экспрессии генов GS Lonza с получением плазмиды рАТН1009. Эта экспрессирующая система состоит из экспрессирующего вектора, имеющего константные домены генов антитела и селективный маркер глутаминсинтетазу (GS). GS представляет собой фермент, ответственный за биосинтез глутамина из глутамата и аммиака. Вектор, имеющий как гены антитела, так и селективный маркер, трансфицируют в запатентованную хозяйскую клеточную линию яичника китайского хомяка (CHOK1SV), адаптированную для роста в бессывороточной среде, и он обеспечивает достаточное количество глутамина для выживания клетки без экзогенного глутамина. Кроме того, в среду добавляют специфичный ингибитор GS, метионинсульфоксимин (MSX), для ингибирования активности эндогенной GS, чтобы могли выживать только клеточные линии с активностью GS, обеспечиваемой вектором. Полученная клеточная линия СНО, трансфицированная рАТН1009, называется LH1.
Следует отметить, что природные лидерные последовательности зародышевой линии, описанные в указанных выше примерах, заменены лидерными последовательностями в остове экспрессирующего вектора GS, используемого для получения плазмиды рАТН1009. Последние лидерные последовательности можно видеть как 19 аминокислот, начинающихся с "mewswv", на N-конце тяжелой цепи LT1009 (SEQ ID NO:19 и 24), а лидерная последовательность LC представляет собой 20 аминокислот, начинающихся с "msvpt" (как показано на N-конце SEQ ID NO:20 и 26).
Трансфицированные клетки СНО LH1 отбирали по их способности расти в не содержащей глутамина среде в присутствии MSX, и отбирали изоляты (клоны) для высокого уровня секреции активного LT1009. LH1 275 представляет собой название, данное основному клону клеточной линии СНО LH1, содержащему вектор рАТН1009, отобранный для создания банка основных клеток (МСВ) для продукции всех партий продукта в виде антитела LT1009. Затем получали материал для токсикологических исследований и клинической разработки.
Депонированные образцы АТСС: Е.coli StB 12, содержащий плазмиду рАТН1009, был депонирован в American Type Culture Collection (номер депонированного образца РТА-8421). Клеточная линия СНО LH1 275, которая содержит вектор рАТН1009, также была депонирована в American Type Culture Collection (номер депонированного образца РТА-8422). АТСС расположена по адресу 10801 University Blvd., Manassas, VA 20110-2209.
Последовательности:
Нуклеотидные и аминокислотные последовательности для полипептидов тяжелой и легкой цепей LT1009 приведены непосредственно ниже, лидерные последовательности (из экспрессирующего вектора Lonza GS) подчеркнуты; CDR. указаны полужирным шрифтом.
Аминокислотная последовательность вариабельного домена LC LT1009 [SEQ ID NO:19]:

Аминокислотная последовательность вариабельного домена НС LT1009 [SEQ ID NO:20]:

Соответствующие нуклеотидные последовательности, кодирующие вариабельные домены тяжелой и легкой цепей, приведены непосредственно ниже. Лидерные последовательности (из экспрессирующего вектора Lonza GS) подчеркнуты; последовательности, предшествующие лидерной последовательности, представляют собой сайт разрезания HindIII (aagctt) и консенсусную последовательность Козака (gccgccacc), которая играет основную роль в инициации процесса трансляции; CDR указаны полужирным шрифтом:
Нуклеотидная последовательность вариабельного домена НС LT1009 [SEQ ID NO:21]:

Нуклеотидная последовательность вариабельного домена LC LT1009 [SEQ ID NO:22]:

Полноразмерная нуклеотидная последовательность (кДНК) НС LT1009 [SEQ ID NO:23], где CDR указаны полужирным шрифтом и лидерная область подчеркнута; шарнирная область показана курсивом. Последовательности, предшествующие лидерной последовательности, представляют собой сайт разрезания HindIII (aagctt) и консенсусную последовательность Козака (gccgccacc):

Аминокислотная последовательность НС LT1009 с лидерной последовательностью (подчеркнута) и без шарнирной области. CDR показаны полужирным шрифтом. [SEQ ID NO:24]:

Полноразмерная нуклеотидная последовательность LC LT1009 [SEQ ID NO:25], где лидерная последовательность подчеркнута, а CDR указаны полужирным шрифтом; последовательности, предшествующие лидерной последовательности, представляют собой сайт разрезания HindIII (aagctt) и консенсусную последовательность Козака (gccgccacc):

Аминокислотная последовательность LC LT1009, где лидерная последовательность подчеркнута, a CDR указаны полужирным шрифтом [SEQ ID NO:26]:

Последовательности тяжелой и легкой цепей LT1009 без лидерных последовательностей (и без предшествующих участков разрезания нуклеазой и последовательностей Козака) являются следующими. CDR указаны полужирным шрифтом.
Аминокислотная последовательность вариабельного домена НС LT1009 [SEQ ID NO:27]:

Соответствующая нуклеотидная последовательность НС LT1009, кодирующая вариабельный домен [SEQ ID NO:28]:

Аминокислотная последовательность вариабельного домена LC LT1009 [SEQ ID NO:29]:

Соответствующая нуклеотидная последовательность LC LT1009, кодирующая вариабельный домен [SEQ ID NO:30]:

Аминокислотные последовательности полноразмерных тяжелой и легкой цепей LT1009 без лидерных последовательностей являются следующими (CDR указаны полужирным шрифтом):
Полноразмерная аминокислотная последовательность тяжелой цепи LT1009 без лидерной последовательности (и без предшествующего сайта расщепления нуклеазой и последовательности Козака), включающая шарнирную область (подчеркнута). (SEQ ID NO:31):

Полноразмерная аминокислотная последовательность легкой цепи LT1009 без лидерной последовательности. [SEQ ID NO:32]:

Соответствующие нуклеотидные последовательности (без лидерных последовательностей или предшествующего сайта расщепления нуклеазой и участка Козака) представлены ниже. Понятно, что вследствие вырожденности генетического кода также практически любую данную аминокислотную последовательность могут кодировать альтернативные нуклеотидные последовательности.
Полноразмерная нуклеотидная последовательность (кДНК) тяжелой цепи LT1009 [SEQ ID NO:33]:

Полноразмерная нуклеотидная последовательность легкой цепи LT1009 [SEQ ID NO:34]:

С-концевой лизин на тяжелой цепи LT1009 не всегда может присутствовать на зрелом белке тяжелой цепи.
Несмотря на то что в нуклеотидных и аминокислотных последовательностях для тяжелой цепи LT1009 показан лизин в качестве последнего (самого С-концевого) аминокислотного остатка белка, при экспрессии, например в клетках СНО клона LH1 275, LT1009 не содержит С-концевой лизин. Это показано посредством пептидного картирования, и без связи с теорией полагают, что это является следствием посттрансляционной модификации белка в системах млекопитающих. Также без связи с теорией полагают, что в других экспрессирующих системах, в частности в системах не млекопитающих, в зрелой тяжелой цепи LT1009 присутствует С-концевой лизин.
Аминокислотная последовательность тяжелой цепи LT1009, экспрессируемая в клетках СНО (т.е. без лидерных последовательностей и без С-концевого лизина), представлена ниже (CDR указаны полужирным шрифтом, шарнирная область указана курсивом) [SEQ ID NO:35]

Пример нуклеотидной последовательности, которая может кодировать эту аминокислотную последовательность, представлен ниже в качестве SEQ ID NO:36. Будет понятно, что вследствие вырожденности генетического кода множество нуклеотидных последовательностей могут кодировать ту же аминокислотную последовательность, и по этой причине признается, что эти и другие нуклеотидные последовательности, представленные в настоящем документе в качестве кодирующих аминокислотные последовательности, представлены для целей иллюстрации. CDR указаны полужирным шрифтом, шарнирная область указана курсивом:


Пример 14
Изготовление гуманизированного mAb
Как правило, процесс получения включает три стадии: серию посевов, серию инокуляций и получение культуры. Во всех стадиях используют бессывороточные реагенты и среду для роста клеточной культуры с низким содержанием белка. Для начала серии посевов используют клетки из рабочего банка клеток; клетки субкультивируют каждые от трех до четырех суток и после назначенного периода в культуре серийных посевов начинают серию инокуляций. Для размножения клеток в целях включения в стадию продукции предпочтительно используют неселективную среду (не содержащую МТХ среду). Клетки размножают серийным субкультивированием в емкостях возрастающего объема. Через определенное количество суток в серии инокуляций начинают стадию продукции. Культивирование для продукции проводят в биореакторе объемом 200 л, 400 л, 2000 л или 20000 л. Пример биореактора описан ниже.
Пример биореактора: Увеличение масштаба от 2-л биореакторов будут продолжать сначала до 15-л перемешиваемой емкости Applikon, а затем до биореактора объемом 50 л, биореактора объемом 200 л и в конце биореактора объемом 2000 л (все созданы до одного масштаба). Характеристики этих емкостей являются следующими:
Изготовитель: АВЕС, Inc.
Изготовлен для ASME, Section VIII Pressure Vessel Code. Контактные поверхности представляют собой 316L SS.
Нижнее приводное устройство, конструкция АВЕС.
Импеллер с малым усилием сдвига 316LSS, шлифованный до 15-20 микродюймов и пассивированный. Диаметр ~1/2 диаметра емкости.
Устройства управления: Allen Bradley Control Logic PLC, с пультом оператора Versa view.
Возбуждение: датчик Allen Bradley, контроль А-В PLC выхода для VFI для контроля RPM.
Температура: Двойной контроль платинового детектора RTD 100 Ом, контрольные тепловой, холодовой и паровой клапаны А-В PLC w/рециркуляционный насос. Цикл автоматической стерилизации с пустым биореактором.
рН: Датчик Ingold, заполненный гелем, поддающимся сжатию, контроль СО2 для разбрызгивания посредством А-В PLC.
Растворенный кислород: полярографический электродный датчик Ingold, контроль разбрызгивания О2 посредством А-В PLC.
Поток воздуха и газа: Датчики представляют собой Four Brooks Thermal Mass Flowmeters для распыления воздуха, О2, N2 и СО2, также предоставляется тепловой массовый Brooks для наружного слоя воздуха, контроль посредством А-В PLC потока газа для автоматического контроля рН, автоматического контроля DO или ручного контроля всего потока газа через А-В PLC.
Давление в емкости: Датчик представляет собой гигиенический трансдуктор диафрагмального типа Rosemount, контроль представляет собой контроль А-В PLC трансдуктора с возвратным контрольным клапаном давления.
Программируемый логический контроллер (PLC): Allen Bradley Control Logix System для последовательного циклического управления процессом, как указано. Программное обеспечение: программирование PLC использует программное обеспечение Rockwell Software (Allen Bradley) RS Logix 5000.
Интерфейс "пользователь-машина" (HMI): Локальный пульт оператора на компьютере Allen Bradley HMI Verso View Industrial со встроенным входом FPD/сенсорного экрана, сообщающегося с PLC через Ethernet. Программное обеспечение представляет собой программное обеспечение RS View 32.
Пример процесса продукции.
Биореактор из нержавеющей стали с перемешиванием с контролем температуры растворенного кислорода и рН.
Для оптимального выхода определяют плотность посева.
Как правило, используют бессывороточную среду.
Как правило, процесс с подпиткой.
Как правило, с переключением температуры.
Ожидается длительность культивирования в биореакторе от 8 до 14 суток.
Во время сбора жизнеспособность подлежит определению.
Сбор будут осветлять посредством фильтрации.
Сбор будут хранить при 2-8°С после осветления и перед очисткой.
Пример 15
Крупномасштабная очистка гуманизированного mAb
Процесс очистки лекарственного вещества, как правило, состоит из четырех стадий: хроматография с белком А, анионообменная хроматография (Q-сефароза), катионообменная хроматография (СМ-сефароза) и ультрафильтрация/диафильтрация (UF/DF). Аффинная колонка, как правило, является первой стадией после сбора и осветления. В этой колонке, как правило, используется смола с иммобилизованным белком А. Эта аффинная стадия очищает антитело от белков и ДНК клеток-хозяев. Для инактивации потенциальных вирусов элюат, как правило, подвергают процессу инактивации вируса с последующей стадией анионобменной хроматографии для уменьшения количества белков, ДНК клеток-хозяев, белка А и потенциальных вирусов. Далее, как правило, используют стадию катионообменной хроматографии для последующего снижения количеств белков клеток-хозяев и агрегатов антитела. В конце пул подвергают диафильтрации и далее концентрируют.
Репрезентативный процесс очистки:
Сбор можно концентрировать и подвергать замене буфера перед колонкой с белком А. Следующей стадией в процессе является аффинная хроматография на колонке с белком А. Связанное антитело элюируют буфером с низким значением рН. Элюат белка А выдерживают в течение периода времени для инактивации вирусов.
Следующей стадией в процессе может быть ионообменная хроматография на колонке Q(+) в условиях, при которых через нее протекает продукт антитела, и примеси, такие как ДНК и белки клетки-хозяина, связываются со смолой колонки.
Следующей стадией в процессе может быть вторая ионообменная хроматография на колонке S(-) в условиях, при которых примеси проходят через колонку. Вместо стадии на колонке S(-) можно использовать стадию на колонке гидрофобного взаимодействия. Следующей стадией в процессе, вероятно, является стадия удаления вируса нанофильтрацией с использованием фильтра DV20 или Planova. Продукт протекает через фильтр. Конечными стадиями в процессе является диафильтрация в буфер конечного состава лекарственного вещества и ультрафильтрация для достижения намеченной концентрации белка.
Пример 16
Биологическая активность гуманизированных вариантов антитела мыши против S1P
Клеточные анализы in vitro
Гуманизированные антитела тестировали в отношении их способности изменять выживаемость опухолевых клеток в присутствии химиотерапевтических средств, как показано на фиг.12. Опухолевые клетки SKOV3 подвергали воздействию Taxol, химиотерапевтического средства, которое индуцирует гибель опухолевых клеток посредством активации апоптотического уничтожающего агента, каспазы-3. S1P был способен снижать индуцируемую посредством Taxol активацию каспазы-3 и/или клеточную гибель по сравнению с контрольными необработанными клетками. Анализы апоптоза проводили, как рекомендовано изготовителем (Promega, каталожный №G7792). В кратком изложении, клетки А549 (2500 клеток на лунку) высевали в 96-луночные планшеты и перед обработкой им позволяли расти до 80% смыкания монослоя. Затем клетки обрабатывали с 0,1-1 мкМ Paclitaxel (Sigma, каталожный №Т 7409) или без него, 0,1-1 мкМ S1P и 1 мкг/мл mAb против S1P, в средах Маккоя в течение 48 ч. Через 48 часов к клеткам добавляли буфер для анализа каспазы. Активность каспазы-3 в супернатанте измеряли с помощью набора Apo-One Homogeneous Caspase-3/7 Assay kit (Promega, кат. №G7792) в соответствии с протоколом изготовителя. Активность каспазы-3/7 выражают в качестве количества раз, в которое возрос флуоресцентный сигнал относительно обработанных носителем клеток.
Активация каспазы-3 возрастала при добавлении mAb против S1P в присутствии S1P, что позволяет предположить, что защитный антиапоптотический эффект S1P устранялся посредством селективного поглощения S1P антителом. Оба гуманизированных варианта антитела, huMAbHCLC3 (LT1004) и huMAbHCLC5 (LT1006), проявляли активность, превосходящую LT1002. Параллельно все варианты тестировали в отношении их эффектов на индуцируемое S1P высвобождение цитокинов из злокачественных клеток. Известно, что S1P вызывает значительное высвобождение IL-8 в кондиционированные клетками среды от злокачественных клеток. Добавление контрольного mAb мыши против S1P снижало высвобождение IL-8 из клеток рака яичника зависимым от концентрации образом. Два гуманизированных варианта huMAbHCCysAlaLC3 (LT1007) и huMAbHCCysAlaLC5 (LT1009) проявляли более высокое снижение высвобождения IL-8 по сравнению с HuMAbHCLC3 (LT1004) и huMAbHCLC5 (LT1006).
Пример 17
Эффективность in vivo mAb мыши (Sphingomab) против гуманизированного mAb (Sonepcizumab) в моделях неоваскуляризации на животных
Хориоидальная неоваскуляризация (CNV) относится к росту новых кровеносных сосудов, источником которых является сосудистая оболочка при разрушении мембраны Бруха, в субретинальный пигментный эпителий (sub-RPE) или субретинальное пространство глаза. CNV является основной причиной потери зрения при дегенерации желтого пятна и других глазных состояниях. Для оценки mAb против S1P в этом примере используют модель CNV на мышах.
Гуманизированные варианты антитела и антитело мыши сравнивали по их способности ингибировать неоваскуляризацию в моделях на животных CNV при AMD, как показано на фиг.13. Мышам дважды (0 сутки и 6 сутки) вводили 0,5 мкг антитела мыши (Мu; LT1002), гуманизированных вариантов [LC3 (LT1004), LC5 (LT1006), HCCysAlaLC3 (LT1007) и HCCysAlaLC5 (LT1009)] или неспецифического mAb (NS) посредством введения в стекловидное тело, а затем подвергали лазерной перфорации мембраны Бруха. Мышей умерщвляли через 14 суток после лазерной хирургической операции. Контрольным мышам вводили водный буфер (PBS) или совпадающее по изотипу неспецифическое антитело. Три из гуманизированных вариантов ингибировали ангиогенез, по существу эквивалентно антителу мыши, как оценивают по измерению площади CNV. Объемы повреждений CNV представлены в качестве средних значений ±SEM. Гуманизированный вариант, содержащий 5 возвратных мутаций в легкой цепи и мутацию цистеина в CDR2 тяжелой цепи (huMAbHCCysAlaLC5; LT1009), значительно подавлял неоваскуляризацию. Это отличие было статистически высокозначимым.
Для индукции CNV мышам проводили анестезию смесью кетамина (14 мг/кг) и ксилазина (30 мг/кг) в стерильном физиологическом растворе, вводимом внутрибрюшинно в дозе 5 мкл на 20 г массы тела. Затем проводили расширение их зрачков посредством одной капли каждого из офтальмологического тропикамида (0,5%) и фенилэфрина (2,5%). Затем для перфорации мембраны Бруха в трех квадрантах правого глаза, расположенных на расстоянии приблизительно 50 мкм от оптического диска в относительных положениях 9, 12 и 3 часов, будут использовать аргоновый зеленый офтальмологический лазер (Oculight GL 532 нм, Iridex Corporation, Mountain View, CA), соединенный с щелевой лампой, установленной для доставки 100-мсек импульсов при 150 мВ с размером пятна 50 мкм. Левый глаз служил в качестве неповрежденного контроля во всех случаях.
Морфометрические и волюметрические показатели очагов повреждения CNV измеряли следующим образом. Через две недели после лазерной индукции CNV животных умерщвляли передозировкой смеси кетамина-ксилазина, а затем проводили перфузию всего тела посредством пункции сердца 6 мл 4% параформальдегида в PBS, рН 7,5, (фиксаж), как описано ранее (Sengupta et al., 2003). Затем глаза будут извлекать, пунктировать иглой 27 г на 1 мм позади края и погружать в фиксаж на 1 ч при комнатной температуре, а затем промывать 2Х погружением в PBS в течение 30 мин. Затем глаза будут разрезать для извлечения заднего сегмента, состоящего из пигментного эпителия сетчатки, хориоидальных капилляров и склеры. Затем повышали проницаемость этой ткани и ее подвергали реакции с конъюгированным с родамином агглютинином I R.communis (Vector Laboratories, Burlingame, CA) для детекции очага повреждения CNV, как описано ранее (Sengupta et al., 2003; Sengupta et al., 2005). Затем задние чашки разрезали на 4-7 радиальных срезов и помещали в плоском виде на покровные стекла микроскопа с каплей среды против обесцвечивания Vectashield (Vector Laboratories, Burlingame, VT) для получения цифрового изображения с помощью эпифлуоресцентного Zeiss Axioplan 2 с цифровой камерой высокого разрешения RGB Spot) и лазерной сканирующей конфокальной микроскопии (BioRad MRC 1024, BioRad Corporation, Temecula, CA).
Полученные цифровые изображения оценивали морфометрически с использованием программного обеспечения ImageJ (Research Services Branch, National Institutes of Health, Bethesda, MD). Изображения разделяли на отдельные каналы RGB для анализа красного и зеленого каналов следующим образом: 1) калибровку для конкретного объектива и микроскопа применяли, устанавливая соотношение пикселей к длине; 2) применяли порог с использованием алгоритма Otsu; 3) изображения будут сделаны бинарными; 4) представляющую интерес область (ROI) обрисовывали, включая полную область повреждения; 5) проводили гранулометрический анализ для количественного определения площадей пикселей над пороговым уровнем в ROI. Для волюметрического анализа процесс был сходным с процессом, описанным выше, за исключением того, что использовали захват z-серии. Затем сумму площади очага повреждения на протяжении z-серии умножали на толщину z (как правило, 4 мкм) с получением объема повреждения.
Лекарственные продукты, тестированные в этой модели, представляли собой LT1002 (mAb мыши к SIP; Sphingomab™); LT1004 (гуманизированное mAb), LT1006 (гуманизированное mAb), LT1007 (гуманизированное mAb) и LT1009 (гуманизированное mAb; Sonepcizumab™). Также были включены носитель в виде физиологического раствора и контроли в виде неспецифического антитела (NSA). Как представлено на фиг.13, как mAb LT1002 мыши (Sphingomab™), так и гуманизированное mAb LT1009 (Sonepcizumab™) специфично снижали область повреждения в этой модели CNV на мышах. Все тестированные mAb показали приблизительно 80-98% снижение размера очага повреждения, которое было значимым (р<0,001 против физиологического раствора) во всех случаях. Кроме того, LT1007 и LT1009 также показали значимое ингибирование (р<0,05) по сравнению с неспецифическим контрольным антителом. Процентное ингибирование размера очага повреждения составляло приблизительно 80% для LT1002 (мышь), 82% LT1004 (гуманизированное), 81% для LT1006 и 99% для LT1009. Таким образом, LT1009 представлял собой гуманизированный вариант mAb, наиболее активный в этой модели неоваскуляризации in vivo.
Пример 18
Определение эффекта дозы Sonepcizumab
Мышам (n=10) проводили однократную билатеральную инъекцию в стекловидное тело возрастающих доз sonepcizumab (0,085, 0,85, 1,7 или 5,0 мкг/глаз) или высокой дозы неспецифического (NS) антитела (3,0 мкг/глаз) за одни сутки до индуцируемой лазером перфорации мембраны Бруха. Через четырнадцать суток после лазерной перфорации мышей подвергали анестезии и осуществляли перфузию меченным флуоресцеином декстраном и получали плоские препараты сосудистой оболочки для анализа размера повреждения CNV.
В этом исследовании изучали эффект величины дозы sonepcizumab и интервала дозы на ингибирование CNV с использованием другого валидированного способа количественного определения площади CNV, в котором животным проводили перфузию меченным флуоресцеином декстраном непосредственно перед умерщвлением. Sonepcizumab индуцировал дозозависимое снижение площади CNV, что приводило к максимальному ингибированию приблизительно 50%, в дозе 5 мкг/глаз. Это снижение было значимым (р<0/0001 по сравнению с неспецифическим контрольным антителом с использованием непарного t-теста). В исследовании частоты дозирования сходную эффективность наблюдали в группах, в которых вводили Sonepcizumab в один момент времени (0 сутки) или в несколько моментов времени (0 и 7 сутки) в ходе исследования в течение 14 суток.
Максимальное ингибирование, составляющее приблизительно 50%, наблюдаемое при введении Sonepcizumab (5 мкг/глаз), сравнимо подходящим образом с ранее опубликованными данными для той же модели и полученными тем же исследователем, демонстрирующими снижение площади CNV посредством VEGF-Trap (4,92 мкг/глаз). Saishin, et al. J. Cell Physiol, 2003. 195(2): p.241-8. "Ловушки" (Regeneron Pharmaceuticals, Inc.) представляют собой слитые молекулы двух различных компонентов рецептора и Fc-области молекулы антитела, называемой Fc-областью, и VEGF-Trap изготавливаются для лечения заболеваний глаз и злокачественной опухоли компанией Regeneron. Сравнение этих двух независимых исследований показало, что снижение очага повреждения CNV посредством Sonepcizumab было на 20 процентных пунктов выше, чем снижение, наблюдаемое посредством VEGF-Trap. Таким образом, эти данные не только подтверждают предварительные данные авторов настоящего изобретения в отношении способности терапии против S1P снижать образование очага в модели CNV на мышах, однако они также демонстрируют повышенную эффективность гуманизированного антитела, sonepcizumab, в отношении ингибирования образования очага повреждения CNV и обеспечивают понимание эффекта, направленного против проницаемости.
Пример 19
Эффективность Sonepcizumab в отношении снижения развития неоваскуляризации сетчатки в модели ретролентальной фиброплазии на мышах
Мышей C57BL/6 (n=7) помещали в 75% кислород на 7 сутки жизни и на 12 сутки жизни возвращали в комнатный воздух, и проводили внутриглазную инъекцию 5 мкг sonepcizumab в один глаз и носителя в противоположный глаз. На 17 сутки мышам проводили внутриглазную инъекцию антитела против РЕСАМ, меченного FITC, и через 8 часов мышей умерщвляли, и глаза извлекали и фиксировали в забуференном PBS формалине при комнатной температуре в течение 5 часов. Сетчатки вырезали и промывали фосфатно-солевым буфером, содержащим 0,25% Triton X-100, и получали их целый препарат. Предметные стекла рассматривали с помощью флуоресцентного микроскопа Nikon и область ретинального NV на сетчатку измеряли посредством анализа изображения.
В соответствии со снижением CNV, наблюдаемым в модели лазерной перфорации на мышах, авторы настоящего изобретения также наблюдали значительное снижение CNV в модели на мышах ретролентальной фиброплазии (ROP). Введение в стекловидное тело Sonepcizumab (5,0 мкг/глаз) привело практически к 4-кратному снижению неоваскуляризации сетчатки по сравнению с контролем в виде физиологического раствора. Эти данные подтверждают эффективность sonepcizumab в отношении ингибирования патологического ангиогенеза в глазу как в ретинальном, так и в хориоидальном сосудистом ложах, индуцированного либо ишемией, либо перфорацией мембраны Бруха.
Пример 20
Эффект Sonepcizumab на индуцируемый VEGF ангиогенез в анализе вставки матригеля
Неоваскуляризацию in vivo осуществляли с использованием анализа вставки матригеля GFR, как описано в Staton, et al., Int J. Exp Pathol, 2004. 85(5): p.233-48. Мышам nu/nu в возрасте 4-6 недель инъецировали в левый бок 500 мкл ледяного матригеля GFR. Матригель GFR инъецировали либо отдельно (контроль), либо после добавления 10 мкг/мл VEGF, дополненного 100 мкг/мл гепарина. Группы для введения контроля и sonepcizumab состояли из 3 животных. Животным вводили физиологический раствор или sonepcizumab (10 мг/кг) за 1 сутки до имплантации матригеля GFR и дозы вводили внутрибрюшинно каждые 72 ч в ходе эксперимента. Через 12 суток животных умерщвляли; вставки вырезали и сразу фиксировали в фиксаже, не содержащем цинка и формалина, в течение ночи, погружали в парафин и получали срезы (5 мкм). Затем погруженные в парафин срезы окрашивали в отношении CD31 (Pharmingen). Изображения (9 изображений на срез, 3 среза на вставку) получали посредством цифровой камеры при 20х увеличении, а затем количественно определяли CD31-положительное окрашивание с помощью программы PhotoShop 6,0 и выражали его в качестве показателя ангиогенеза (пиксел2) посредством ImageJ.
Антиангиогенные эффекты sonepcizumab были очевидными в этом анализе вставки матригеля. Как и ожидалось, во вставках матригеля, дополненных 10 мкг/мл VEGF, индуцировалась обширная неоваскуляризация (приблизительно 5,75х относительно необработанного контроля, лишенного VEGF или sonepcizumab). Важно, что системное внутрибрюшинное введение sonepcizumab перед инъекцией матригеля предотвращало приблизительно 80% этого стимулированного VEGF повышения насыщенности клетками и плотности микрососудов. Это снижение является значимым (р<0,05 по сравнению с VEGF отдельно) и подтверждает мощную антиангиогенную активность sonepcizumab при системном введении животным и строго подтверждает, что sonepcizumab способен значительно ингибировать индуцированный VEGF ангиогенез. Это открытие согласуется с данными онкологической программы Lpath, на основании которых это антитело против S1P снижало сывороточные уровни нескольких ангиогенных факторов, включая VEGF, в модели ортотопного рака молочной железы мыши.
Основным компонентом роста кровеносных сосудов, связанным с AMD, является привлечение перицитов, которые оборачивают и поддерживают рост эндотелиальной трубки. Jo, et al., Am J. Pathol, 2006, 168(6): p.2036-53. Исследования на трансгенных мышах показали, что VEGF и PDGF-B являются основными факторами, которые стимулируют инфильтрацию и дифференцировку перицитов, что приводит к созреванию и стабилизации кровеносных сосудов. Guo, et al., Am J. Pathol, 2003, 162(4): p.1083-93; Benjamin, L.E., I.Hemo, and E.Keshet, Development, 1998, 125(9):.p.1591-8. Важно, что SIP обеспечивает трансактивацию VEGF и PDGF. Таким образом, способность sonepcizumab непрямо нейтрализовывать эти факторы роста позволяет предположить, что sonepcizumab может предотвращать аномальный рост кровеносных сосудов при AMD.
Пример 21
Sonepcizumab значимо снижает проницаемость сосудов после лазерной перфорации мембраны Бруха
Эффективность sonepcizumab, вводимого для ингибирования сосудистой проницаемости (в дополнение к ингибированию неоваскуляризации, как показано выше), оценивали в модели лазерной перфорации мембраны Бруха на мышах.
Мышей C57BL/6 (n=10) подвергали лазерной перфорации мембраны Бруха в 3 областях каждого глаза и проводили внутриглазную инъекцию 5 мкг sonepcizumab в один глаз и носителя в противоположный глаз. Через одну неделю после лазерной перфорации мышам проводили внутрибрюшинную инъекцию 12 мкл/г массы тела 1% флуоресцеина натрия и умерщвляли через 5 минут. Глаза извлекали и фиксировали в забуференном PBS формалине при комнатной температуре в течение 5 часов. Затем сетчатки разрезали, промывали и инкубировали с первичным антителом против РЕСАМ-1. Затем сетчатки промывали, инкубировали с вторичным антителом (антитело козы против IgG крысы, конъюгированное с родамином), а затем делали плоский препарат.
Количественное определение области повреждения CNV измеряют посредством окрашивания на РЕСАМ-1. Количественное определение сосудистой проницаемости определяют посредством окрашивания флуоресцеином натрия. Общая площадь проникновения из CNV = CNV + проникновение (зеленый) - площадь CNV (красный). Значения соответствуют среднему значению ±SEM для n=10 мышей/группа. Площадь хориоидальной неоваскуляризации (окрашенная посредством РЕСАМ-1) составляла приблизительно 0,015 мм2 для животных, которым вводили LT1009, и приблизительно 0,03 мм2 для животных, которым вводили физиологический раствор. Это представляет собой 50% снижение неоваскуляризации (р-0,018). Площадь проникновения при хориодальной неоваскуляризации (окрашенной флуоресцеином) составляла приблизительно 0,125 мм2 для животных, которым вводили LT1009, и приблизительно 0,2 мм2 для контрольных животных, которым вводили физиологический раствор. Это представляет собой приблизительно 38% снижение (р-0,017) проницаемости кровеносных сосудов.
Репрезентативные иммуногистохимические изображения снижения хориоидальной неоваскуляризации и сосудистой проницаемости у мышей, которым вводили 5,0 мкг/глаз Sonepcizumab или контроля в виде PBS, согласуются с этими результатами. Таким образом, в дополнение к снижению CNV, sonepcizumab значительно снижал отек сетчатки вследствие сосудистой проницаемости после лазерной перфорации мембраны Бруха, который играет основную роль в потере остроты зрения, ассоциированной с: (i) проницаемостью хориоидальных новых сосудов при AMD и (ii) разрушением гематоретинального барьера при диабете. Gerhardt, H. and С.Betsholtz, Cell Tissue Res, 2003, 314(1): p.15-23. Sonepcizumab снижает образование патологических кровеносных сосудов в глазу, а также сосудистую проницаемость, которая приводит к отеку сетчатки. Эти открытия согласуются с данными, полученными в эксперименте по количественному определению площади CNV, в котором мышам проводили перфузию меченного флуоресцеином декстрана. На количественное определение CNV этим способом, безусловно, влияет сосудистая проницаемость. Высокоположительные результаты говорят в пользу эффекта против проницаемости в хориоидальном сосудистом ложе. С учетом этих данных, авторы настоящего изобретения полагают, что sonepcizumab имеет потенциал в отношении монотерапии. Также существует возможность синергического эффекта с сопутствующими общими блокирующими VEGF-A средствами.
Пример 22
Снижение инфильтрации макрофагов в сетчатку после введения антитела к S1P
Старческая дегенерация желтого пятна (AMD) представляет собой заболевание, связанное со старением, которое постепенно нарушает остроту центрального зрения. Существуют два основных типа дегенерации желтого пятна: сухая или атрофическая, которая составляет 85-90% случаев AMD, и влажная форма AMD, характеризующаяся ростом аномальных кровеносных сосудов. Сухую дегенерацию желтого пятна диагностируют, когда из скоплений или дебриса повреждающейся ткани начинают накапливаться желтоватые пятна, известные как друзы, главным образом в области желтого пятна. Может происходить постепенное снижение центрального зрения. Эффективное лечение наиболее преобладающей атрофической (сухой) формы AMD отсутствует. Атрофическая AMD запускается аномалиями в пигментном эпителии сетчатки (RPE), который расположен позади фоторецепторных клеток и в норме обеспечивает основную метаболическую поддержку для этих светочувствительных клеток. Вторично относительно дисфункции RPE, палочки и колбочки желтого пятна дегенерируют, что приводит к необратимой потере зрения. Патологическими процессами, вовлеченными в патогенез атрофической AMD, являются окислительный стресс, ишемия, образование друз, накопление липофусцина, местное воспаление и реактивный глиоз. Среди этих процессов воспаление происходит в качестве основного участника повреждения тканей. Было показано, что инфильтрация макрофагов в желтое пятно пациентов с сухой AMD является важным компонентом повреждающего воспалительного ответа. Таким образом, средство, которое может подавлять инфильтрацию макрофагов, может быть ценным лекарственным средством, поскольку ингибирование инфильтрации макрофагов, вероятно, снизит повреждение ткани желтого пятна. Такое средство также может снижать скорость, с которой сухая AMD превращается во влажную AMD.
В настоящее время в модели ишемической и воспалительной ретинопатии показано 55% ингибирование инфильтрации макрофагов после введения антитела против S1P. Эти данные были получены с использованием общепринятой модели индуцируемой кислородом ретинопатии на мышах (также известной, как модель ретролентальной фиброплазии (ROP)). Конкретно, мышей C57BL/6 помещали в 75% кислород на 7 сутки жизни и на 12 сутки жизни возвращали в комнатный воздух и проводили им внутриглазную инъекцию 5 мкг гуманизированного антитела против SIP (LT1009, Sonepcizumab™) в один глаз и носителя во второй глаз. На 17 сутки жизни мышам проводили внутриглазную инъекцию меченного FITC антитела к F4/80 (общий маркер макрофагов) и через 8 часов мышей умерщвляли. Глазные яблоки извлекали и фиксировали в забуференном PBS формалине при комнатной температуре в течение 5 часов. Сетчатки вырезали и промывали фосфатно-солевым буфером, содержащим 0,25% Triton X-100, и делали их целый препарат. Предметные стекла рассматривали с помощью флуоресцентного микроскопа Nikon, и область ретинального NV на сетчатку измеряли посредством анализа изображения, и количественно определяли макрофаги. Результаты представлены в таблице 7, ниже.
На основании этих данных и известной роли макрофагов в патогенезе сухой AMD полагают, что антитела против S1P представляют собой эффективное лекарственное средство для лечения сухой AMD.
Пример 23
Ответ колоректального трансплантата опухоли SC COLO205 у мышей Nude NCr на лечение 25-75 мг/кг LT1009 отдельно и в сочетании с Avastin или Paclitaxel
Целью этого исследования было определение эффективности LT1009, отдельно и в сочетании с другими средствами против злокачественной опухоли, в отношении замедления прогрессии опухолей колоректальной карциномы человека (COLO205), пересаженных подкожно (sc) и установившихся у самок мышей Ncr (nu/nu).
Мышам Nude имплантировали подкожно вблизи правого бока один фрагмент опухоли COLO205 на мышь из пассажа in vivo. Все введения начинали в день, когда у 60 мышей в каждом эксперименте устанавливалась опухоль, варьирующая по размеру приблизительно от 100 до 200 мм3. Затем мышам (n=10 на группу) вводили либо 25 мг/кг LT1009, либо 50 мг/кг LT1009, либо 40 мг/кг Avastin, либо 50 мг/кг LT1009 плюс 40 мг/кг Avastin, 15 мг/кг Paclitaxel, либо носитель (физиологический раствор). В ходе эксперимента 25 или 50 мг/кг LT1009 и физиологический раствор вводили внутрибрюшинно один раз в q3d в объеме 0,1 мл/20 г массы тела. Avastin вводили внутривенно в дозировке 40 мг/кг/доза по схеме q7d, инъецируя в объеме 0,1 мл/20 г массы тела. Paclitaxel (положительный контроль) вводили внутривенно в дозировке 15 мг/кг/доза по схеме q1d×5, инъецируя в объеме 0,1 мл/10 г массы тела. На 21 сутки в ходе исследования дозу 25 мг/кг LT1009 повышали до 75 мг/кг LT1009.
Животных осматривали каждые сутки в отношении смертности. Размеры опухоли и массу тела определяли два раза в неделю, начиная с первых суток введения и включая сутки завершения исследования. Когда средний размер опухоли в контрольной группе, в которой вводили носитель, в каждом исследовании достигал приблизительно 4000 мг, исследование завершали. Опухоли каждой мыши извлекали, записывали полную массу, опухоли обрабатывали для определения плотностей микрососудов (MVD) посредством окрашивания CD-31. Массы опухолей (мг) вычисляли с использованием уравнения для эллипсоидной сферы (l×w2)/2=мм3, где l и w относятся к большему и меньшему размерам, полученным при каждом измерении, и подразумевают единицу плотности (1 мм3=1 мг).
50 мг/кг LT1009 по существу ингибировали прогрессию опухоли (р<0,018), как измеряют по конечной массе опухоли, на 32% по сравнению с опухолями из животных, которым вводили физиологический раствор. 25/75 мг/кг LT1009 также были эффективными в отношении снижения конечной массы опухоли на 20%; однако это снижение не было статистически значимым. 50 мг/кг LT1009 были настолько же эффективными в отношении снижения конечной массы опухоли, как и Avastin, (снижение на 32% против 35% соответственно). Сочетание LT1009 и Avastin было более эффективным, чем каждое средство отдельно, демонстрируя 48% снижение массы опухоли по сравнению с животными, которым вводили физиологический раствор. Таким образом, эффекты LT1009 и Avastin, по-видимому, являются аддитивными. Положительный контроль, Paclitaxel, полностью устранял предварительно развившиеся опухоли.
Пример 24
Ответ колоректального трансплантата опухоли SC HT29 у мышей Nude NCr на лечение 50 мг/кг LT1009 отдельно и в сочетании с Avastin и 5-FU
Целью этого исследования является оценка противоопухолевой эффективности LT1009, отдельно и в сочетании с другими средствами против злокачественной опухоли, против ксенотрансплантатов опухоли толстого кишечника НТ29, имплантированных подкожно самкам бестимусных мышей NCr-nu/nu.
Мышам Nude имплантировали подкожно вблизи правого бока один фрагмент опухоли НТ29 на мышь из пассажа in vivo. Все введения начинали в день, когда у 60 мышей в каждом эксперименте устанавливалась опухоль, варьирующая по размеру приблизительно от 100 до 200 мм3. В каждой группе было десять мышей. В ходе эксперимента внутрибрюшинно вводили 50 мг/кг LT1009 и физиологический раствор q2d в объеме 0,1 мл/20 г массы тела. 75 мг/кг 5-FU и 20 мг/кг Avastin вводили внутрибрюшинно и внутривенно в дозировке 75 мг/кг/доза и 20 мг/кг/доза соответственно q4d, инъецируемой в объеме 0,1 мл/10 г массы тела. Первая доза LT1009 состояла из 100 мг/кг, вводимых внутривенно.
Животных осматривали каждые сутки в отношении смертности. Размеры опухоли и массу тела определяли два раза в неделю, начиная с первых суток введения и включая сутки завершения исследования. Когда средний размер опухоли в контрольной группе, в которой вводили носитель, в каждом исследовании достигал приблизительно 4000 мг, исследование завершали. Опухоли каждой мыши извлекали, записывали полную массу, опухоли обрабатывали для определения плотностей микрососудов (MVD) посредством окрашивания CD-31. Массы опухолей (мг) вычисляли с использованием уравнения для эллипсоидной сферы (l×w2)/2=мм3, где l и w относятся к большему и меньшему размерам, полученным при каждом измерении, и подразумевают единицу плотности (1 мм3=1 мг).
50 мг/кг LT1009 снижали прогрессию опухоли, при определении по массе опухоли, на 13%, в то время как Avastin снижал массу опухолей на 30% по сравнению с опухолями из животных, которым вводили физиологический раствор. Сочетание LT1009 и Avastin было более эффективным, чем каждое средство отдельно, демонстрируя статистически значимое снижение массы опухоли, составляющее 41%, по сравнению с животными, которым вводили физиологический раствор. Введение 5-FU снижало массу опухоли на 28%. 5-FU показал минимальный аддитивный эффект с LT1009, демонстрируя ингибирование конечной массы опухоли на 29%.
Пример 25
Ответ ксенотрансплантата опухоли предстательной железы SC DU145 у мышей Nude NCr на введение 50 мг/кг LT1009, отдельно или в сочетании с Avastin или Paclitaxel
Целью этого исследования было определение эффективности LT1009, отдельно и в сочетании с другими средствами против злокачественной опухоли, в отношении замедления прогрессии опухолей карциномы предстательной железы человека (DU145), пересаженных подкожно (sc) и установленных у самок мышей Ncr (nu/nu).
Мышам Nude имплантировали подкожно вблизи правого бока один фрагмент опухоли DU145 на мышь из пассажа in vivo. Все введения начинали в день, когда у 60 мышей в каждом эксперименте устанавливалась опухоль, варьирующая по размеру приблизительно от 100 до 200 мм3. Затем в ходе эксперимента мышам (n=10/группа) вводили либо 50 мг/кг LT1009, либо 20 мг/кг Avastin, либо 7,5 мг/кг Paclitaxel, либо 50 мг/кг LT1009 плюс 20 мг/кг Avastin, 50 мг/кг LT1009 плюс 7,5 мг/кг Paclitaxel или носитель (физиологический раствор). 50 мг/кг LT1009 и физиологический раствор вводили внутрибрюшинно q2d в объеме 0,1 мл/20 г массы тела. Paclitaxel и Avastin вводили внутривенно и внутрибрюшинно в дозировке 7,5 мг/кг/доза и 20 мг/кг/доза, q1d ×5 и q4d соответственно, инъецируя в объеме 0,1 мл/10 г массы тела. Первая доза LT1009 состояла из 100 мг/кг, вводимых внутривенно.
В ходе исследования мониторинг роста опухоли проводили посредством измерения подкожных опухолей по трем осям и вычисления объема. В конце исследования определяли конечную массу и объемы опухоли, а затем мышей умерщвляли и опухоли извлекали. Затем определяли плотность микрососудов (MVD) опухолей посредством окрашивания по CD-31.
50 мг/кг LT1009 значимо (р<0,00) снижали прогрессию опухоли, при измерении по конечной массе опухоли, на 28%. Avastin и Paclitaxel также значимо (р<0,00) снижали конечную массу опухоли на 80% по сравнению с опухолями животных, которым вводили физиологический раствор. LT1009 не повышал значимо противоопухолевую активность Avastin или Paclitaxel при измерении по конечным объемам опухолей.
Пример 26
Ответ ксенотрансплантата опухоли миеломы человека RPMI 8226 у мышей СВ17 SCID на лечение посредством 25 мг/кг или 50 мг/кг LT1009, отдельно и в сочетании с Bortezomib
Целью этого исследования является оценка противоопухолевой эффективности LT1009, отдельно и в сочетании со средством против злокачественной опухоли Bortezomib, в отношении подкожных ксенотрансплантатов опухоли миеломы человека RPMI у самок мышей СВ17 SCID.
Мышам Nude (CB17 SCID, в возрасте 4-5 недель, массой 18-22 г, самки мышей, полученные от Harlan) подкожно инъецировали клетки RPMI 8226, собранные из культуры ткани (~1×107 клеток/мышь). Когда опухоли выросли приблизительно до 100 мм по размеру, животных попарно подбирали по размеру опухоли в группу введения и контроля (10 мышей на группу). Исходное дозирование начали на 1 сутки после подбора пар. Животным во всех группах проводили дозирование по массе (0,01 мл на грамм; 10 мл/кг). LT1009 в носителе вводили посредством внутрибрюшинной (IP) инъекции один раз каждые трое суток до завершения исследования (Q3D до конца). Bortezomib вводили посредством внутривенной инъекции через хвостовую вену один раз каждые трое суток в течение шести введений (Q3D×6). В качестве отрицательного контроля вводили носитель для LT1009 (0,9% физиологический раствор) внутрибрюшинно на Q3D до конца схемы.
Отдельные средние объемы опухоли ±SEM и в группах записывают два раза в неделю до завершения исследования, начиная на 1 сутки. В конце исследования записывают конечный средний объем опухоли ±SEM для каждой группы; животных, у которых происходили частичные или полные регрессии опухолей, или животных, испытавших техническую или связанную с лекарственным средством смерть, исключали из этих вычислений.
Все композиции и способы, описанные и заявленные в настоящем документе, можно получать и выполнять без излишнего экспериментирования с учетом настоящего описания. Несмотря на то что композиции и способы по этому изобретению описаны с точки зрения предпочтительных вариантов осуществления, специалистам в данной области будет очевидно, что в композиции и способы можно вносить изменения. Все такие сходные замены и модификации, очевидные специалистам в данной области, считают относящимися к сущности и объему изобретения, как определено прилагаемой формулой изобретения.
Все патенты, патентные заявки и публикации, упомянутые в описании, являются характерными для уровня специалиста в области, к которой относится изобретение. Все патенты, патентные заявки и публикации, включая патенты, патентные заявки и публикации, по которым испрашивается приоритет или другое преимущество, включены в настоящий документ в качестве ссылок в такой же степени, как если бы каждая отдельная публикация была конкретно и индивидуально указана как включенная в качестве ссылки.
Изобретение, иллюстративно описанное в настоящем документе, пригодным образом можно применять на практике в отсутствие любого элемента(ов), конкретно не описанного в настоящем документе. Таким образом, например, в каждом случае в настоящем документе любой из терминов "содержащий", "состоящий по существу из" и "состоящий из" можно заменить любым из двух других терминов. Термины и выражения, которые использованы, используют в качестве терминов описания, а не для ограничения, и при применении таких терминов и выражений не предполагается исключение любых эквивалентов представленных и описанных признаков или их частей, однако признается, что в объеме заявленного изобретения возможны различные модификации. Таким образом, следует понимать, что, несмотря на то, что настоящее изобретение конкретно описано посредством предпочтительных вариантов осуществления и необязательных признаков, специалисты в данной области могут прибегать к модификации и отклонению от концепций, описанных в настоящем документе, и что такие модификации и отклонения считают находящимися в объеме этого изобретения, как определено прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ АНТИТЕЛА | 2008 |
|
RU2490277C2 |
| ХИМЕРНЫЕ И ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ИНТЕГРИНА α5β1, КОТОРЫЕ МОДУЛИРУЮТ АНГИОГЕНЕЗ | 2003 |
|
RU2346701C2 |
| СОСТАВЫ АНТИТЕЛ | 2014 |
|
RU2694055C2 |
| СПОСОБЫ ИНГИБИРОВАНИЯ ГЛАЗНОГО АНГИОГЕНЕЗА | 2009 |
|
RU2530583C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ АЛЬФА5БЕТА1 | 2007 |
|
RU2465282C2 |
| АНТИАНГИОГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАНЕЕ ПОДВЕРГАВШЕГОСЯ ЛЕЧЕНИЮ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2010 |
|
RU2576027C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТФР-БЕТА-АНТИТЕЛА | 2005 |
|
RU2386638C2 |
| ВАРИАНТЫ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2536937C2 |
| АНТИТЕЛО ПРОТИВ panNrp (ВАРИАНТЫ) | 2007 |
|
RU2454427C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ПРИМЕНЕНИЕМ АНТАГОНИСТОВ АКСИАЛЬНОГО СВЯЗЫВАНИЯ PD-1 И VEGF АНТАГОНИСТОВ | 2013 |
|
RU2689760C2 |






























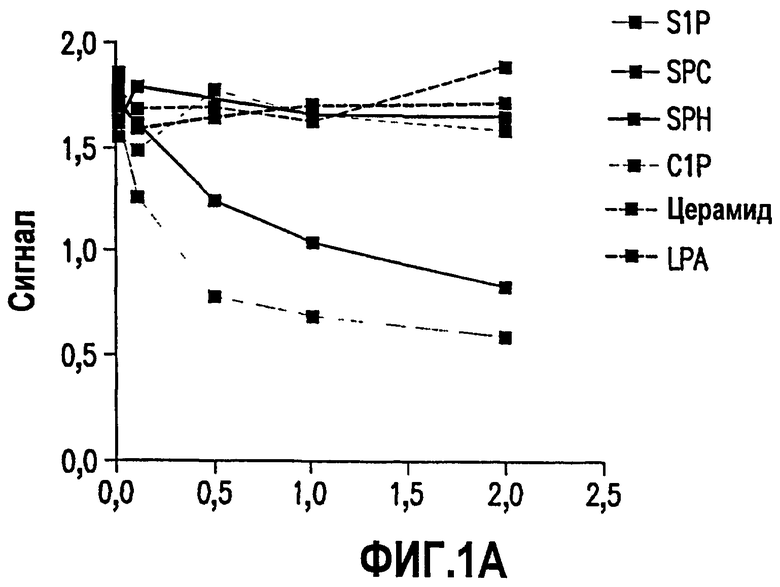

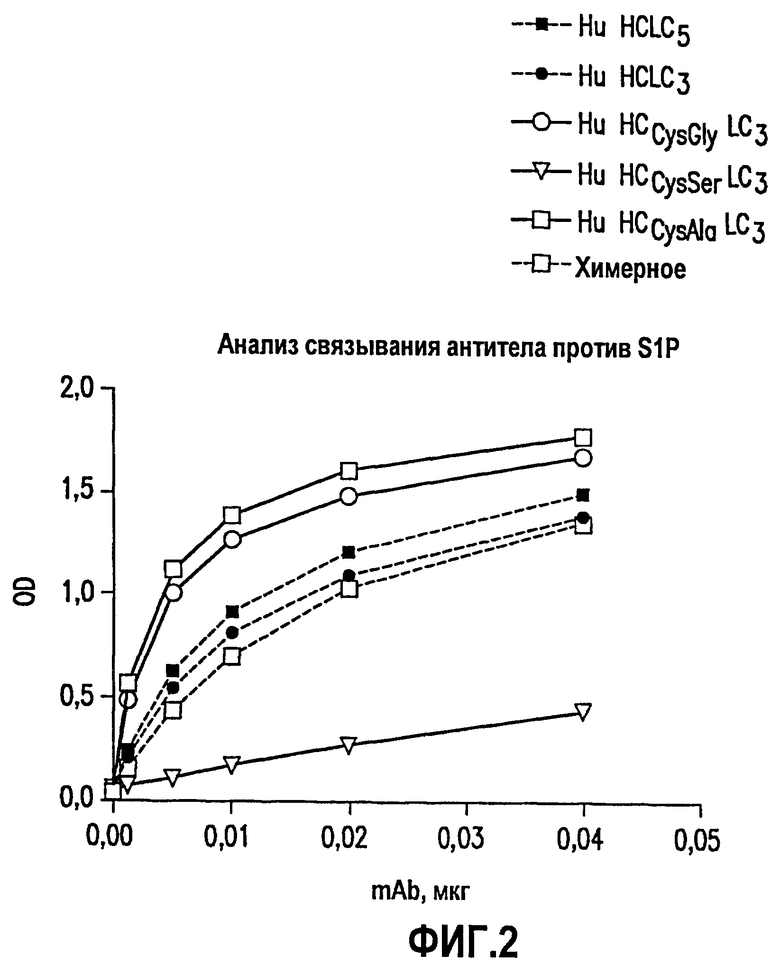
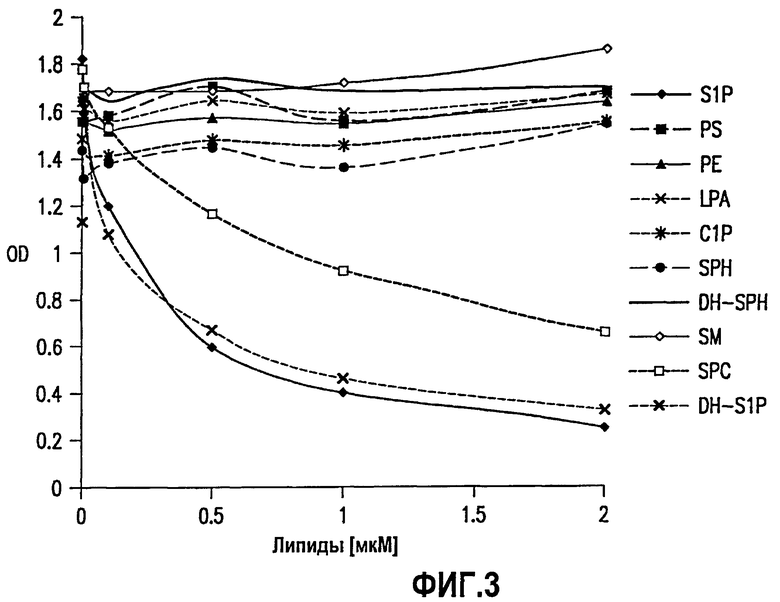
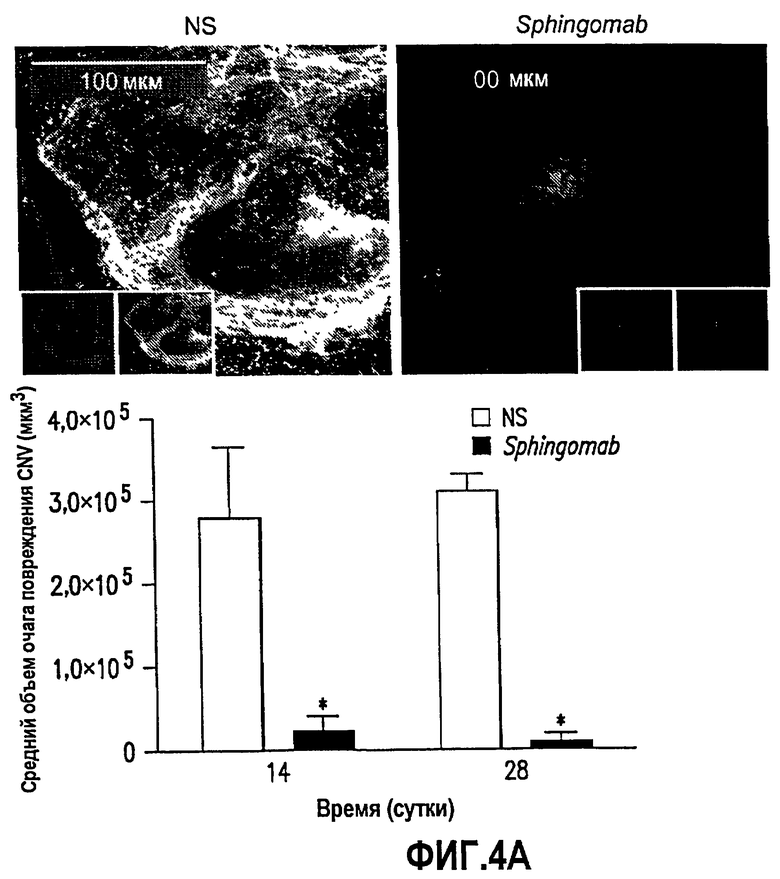










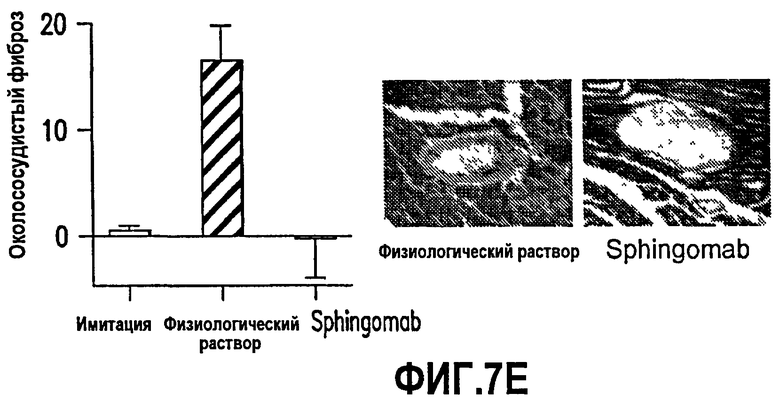
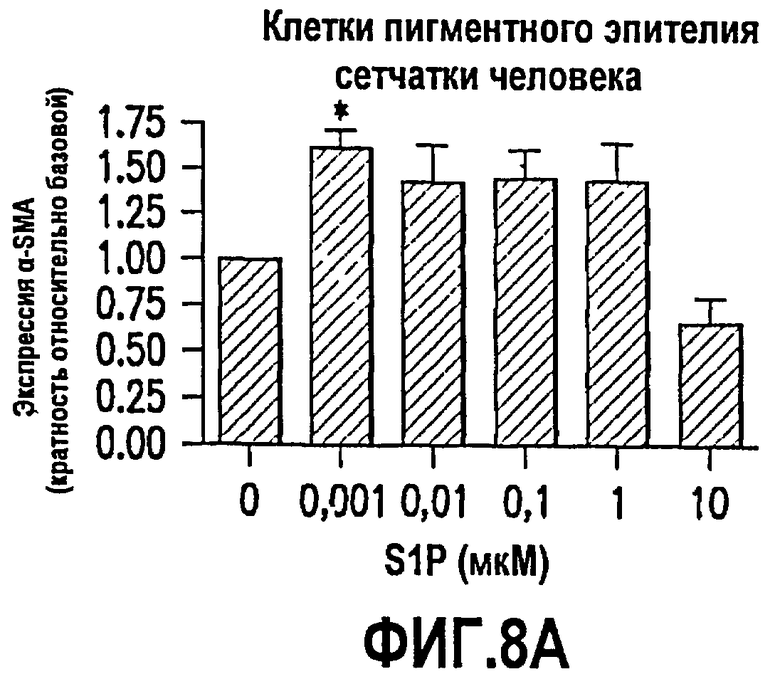
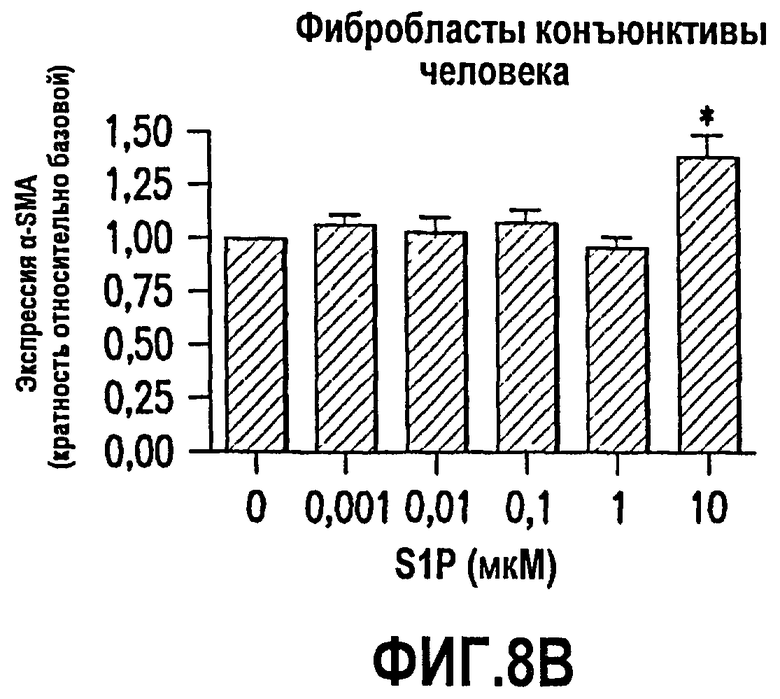
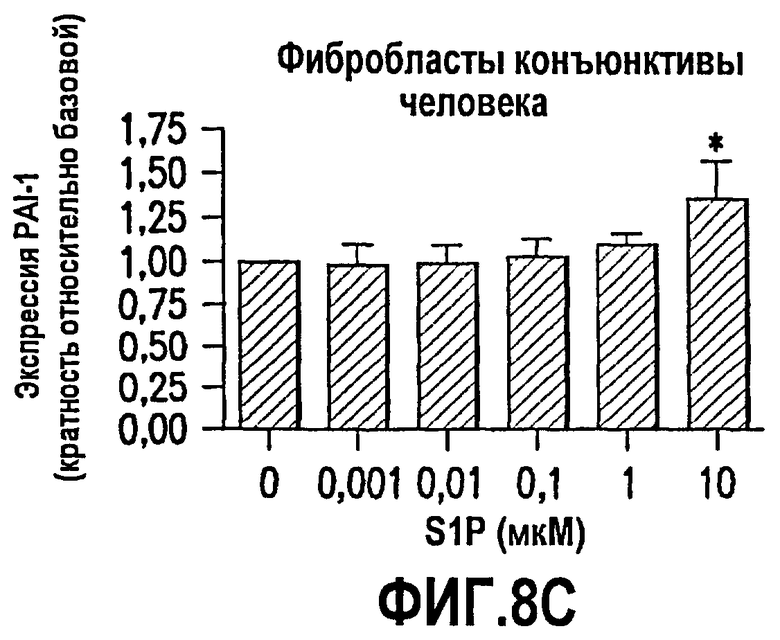


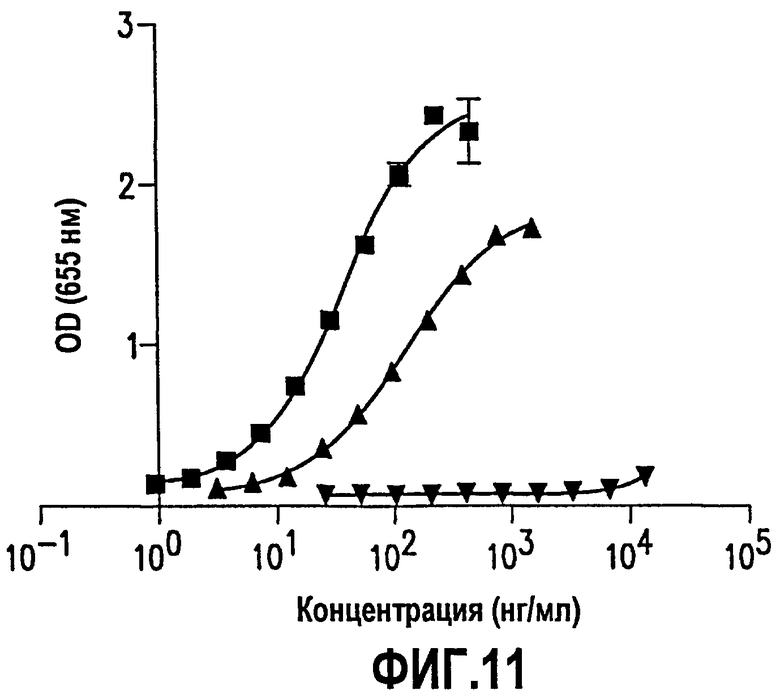

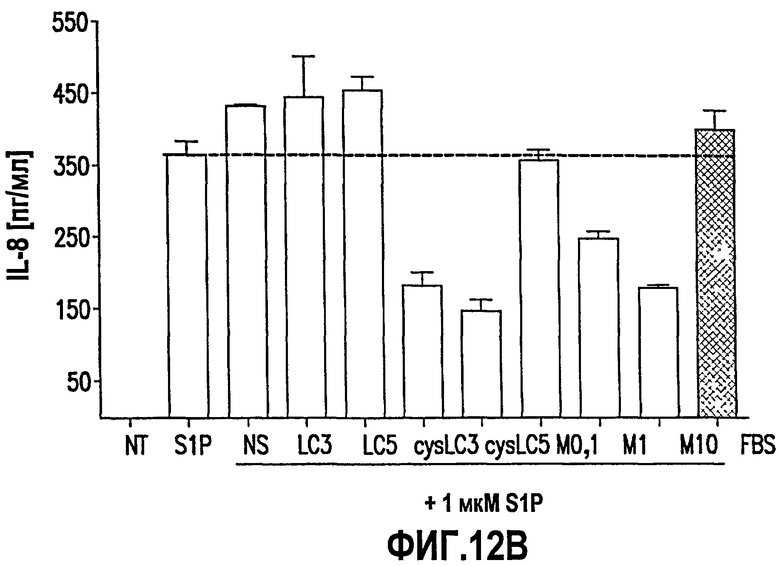
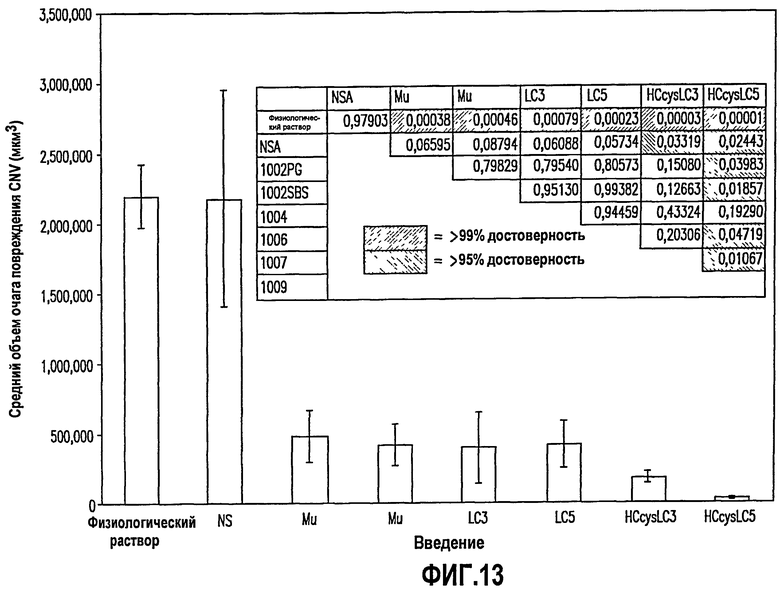
Настоящее изобретение относится к области иммунологии. Рассмотрены антитело, которое специфически связывает сфингозин-1-фосфат (S1P), и его антигенсвязывающий фрагмент. Предложены фармацевтическая композиция, вектор для экспрессии антитела, клетка-хозяин, а также способ лечения заболевания или нарушения, связанного с S1P. Использование изобретения может найти дальнейшее применение в лечении заболеваний, связанных с S1P. 10 н. и 7 з.п. ф-лы, 26 пр., 11 табл., 13 ил.
1. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент, которое специфично связывает сфингозин-1-фосфат (S1P), и которое содержит по меньшей мере один вариабельный домен тяжелой цепи и по меньшей мере один вариабельный домен легкой цепи, в котором:
A. каждый вариабельный домен тяжелой цепи содержит:
(i) первую последовательность аминокислотных остатков последовательности DHTIH [SEQ ID NO:13];
(ii) вторую последовательность аминокислотных остатков AISPRHDITKYNEMFRG [SEQ ID NO:18]; и
(iii) третью последовательность аминокислотных остатков последовательности GGFYGSTIWFDF [SEQ ID NO:15]; и
B. каждый вариабельный домен легкой цепи содержит:
(i) первую последовательность аминокислотных остатков последовательности ITTTDIDDDMN [SEQ ID NO:10];
(ii) вторую последовательность аминокислотных остатков последовательности EGNILRP [SEQ ID NO:11]; и
(iii) третью последовательность аминокислотных остатков последовательности LQSDNLPFT [SEQ ID NO:12].
2. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.1, в котором:
A. каждый вариабельный домен тяжелой цепи содержит последовательность аминокислотных остатков, имеющую аминокислотную последовательность

B. каждый вариабельный домен легкой цепи содержит последовательность аминокислотных остатков, имеющую аминокислотную последовательность

3. Выделенное гуманизированное антитело по п.1, в котором:
A. каждая тяжелая цепь содержит последовательность аминокислотных остатков, имеющую аминокислотную последовательность

B. каждая легкая цепь содержит последовательность аминокислотных остатков, имеющую аминокислотную последовательность


4. Выделенное гуманизированное антитело по п.3, в котором каждая тяжелая цепь содержит последовательность аминокислотных остатков, имеющую аминокислотную последовательность

5. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.1, в котором:
A. каждая тяжелая цепь содержит такую же аминокислотную последовательность вариабельного домена тяжелой цепи, что и вариабельный домен тяжелой цепи, кодируемый геном тяжелой цепи в векторе рАТН1009 из АТСС с регистрационным №РТА-8421; и
B. каждая легкая цепь содержит такую же аминокислотную последовательность вариабельного домена легкой цепи, что и вариабельный домен легкой цепи, кодируемый геном легкой цепи в векторе рАТН1009 из АТСС с регистрационным №РТА-8421.
6. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.1, в котором по меньшей мере один аминокислотный остаток антитела является гликозилированным.
7. Фармацевтическая композиция для лечения или профилактики состояний, заболеваний или нарушений, связанных со сфингозин-1-фосфатом (SIP), содержащая терапевтически эффективное количество выделенного гуманизированного антитела или его антигенсвязывающего фрагмента по п.1 и фармацевтически приемлемый носитель, где выделенное гуманизированное антитело или его антигенсвязывающий фрагмент связывают внеклеточный S1P.
8. Вектор для экспрессии антитела по п.1 или антигенсвязывающего фрагмента такого антитела, где вектор содержит:
A. первую экспрессирующую кассету, которая содержит промотор и ген, который кодирует полипептид(ы) тяжелой цепи гуманизированного антитела или его антигенсвязывающего фрагмента; и
B. вторую экспрессирующую кассету, которая содержит промотор и ген, который кодирует полипептид(ы) легкой цепи гуманизированного антитела или его антигенсвязывающего фрагмента.
9. Вектор по п.8, где вектор представляет собой плазмиду, содержащую первую и вторую экспрессирующие кассеты.
10. Рекомбинантная клетка-хозяин, трансфицированная композицией рекомбинантной нуклеиновой кислоты по п.8, необязательно клетка-хозяин, депонированная под регистрационным номером АТСС РТА-8421.
11. Способ лечения заболевания или нарушения у индивида, включающий введение индивиду, нуждающемуся в таком лечении, выделенного гуманизированного антитела или его антигенсвязывающего фрагмента по п.1, где указанное заболевание или нарушение коррелирует с аномальным уровнем S1P.
12. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент по п.1, в котором:
А. каждая тяжелая цепь содержит такую же тяжелую цепь, что и тяжелая цепь антитела, экспрессируемого клеточной линией СНО LH1 275, депонированной под регистрационным номером АТСС РТА-8422; и
В. каждая легкая цепь содержит такую же легкую цепь, что и легкая цепь антитела, экспрессируемого клеточной линией СНО LH1 275, депонированной под регистрационным номером АТСС РТА-8422.
13. Выделенное гуманизированное антитело или его антигенсвязывающий фрагмент, которое специфично связывает сфингозин-1-фосфат (S1P) и которое содержит гуманизированный вариабельный домен легкой цепи, включающий гипервариабельные определяющие комплементарность области (CDR), характеризуемые как CDRL1 (SEQ ID NO:10), CDRL2 (SEQ ID NO:11) и CDRL3 (SEQ ID NO:12), и гуманизированный вариабельный домен тяжелой цепи, включающий гипервариабельные CDR, характеризуемые как CDRH1 (SEQ ID NO:13), CDRH2 (SEQ ID NO:18) и CDRH3 (SEQ ID NO:15).
14. Вектор для экспрессии антитела по п.1 или антигенсвязывающего фрагмента такого антитела, где вектор содержит первый ген, который кодирует вариабельный домен легкой цепи, который включает первый, второй и третий пептиды CDR легкой цепи SEQ ID NO:10, SEQ ID NO:11 и SEQ ID NO:12 соответственно, и второй ген, который кодирует ген тяжелой цепи, который кодирует вариабельный домен тяжелой цепи, который включает первый, второй и третий пептиды CDR тяжелой цепи SEQ ID NO:13, SEQ ID NO:18 и SEQ ID NO:15 соответственно.
15. Плазмида, содержащая вектор по п.14, где указанная плазмида содержит ген тяжелой цепи, включающий последовательность нуклеиновой кислоты SEQ ID NO:33, и ген легкой цепи, включающий последовательность нуклеиновой кислоты легкой цепи SEQ ID NO:34, которая при экспрессии в линии эукариотических клеток продуцирует гуманизированное антитело или антигенсвязывающий фрагмент такого антитела, которые специфично связывают сфингозин-1-фосфат (S1P).
16. Рекомбинантная клетка-хозяин, трансфицированная плазмидой по п.15 и способная экспрессировать указанное антитело, где указанная клетка-хозяин необязательно представляет собой клетку CHOK1SV, необязательно клеточную линию, имеющую обозначение LH1 275 и депонированную под регистрационным номером АТСС РТА-8422.
17. Плазмида, содержащая вектор по п.14, где указанная плазмида содержит ген тяжелой цепи, который кодирует последовательность аминокислотных остатков, содержащую аминокислотную последовательность SEQ ID NO:31 или SEQ ID NO:35, и ген легкой цепи, который кодирует последовательность аминокислотных остатков, содержащую аминокислотную последовательность SEQ ID NO:32, которая при экспрессии в линии эукариотических клеток продуцирует гуманизированное антитело или антигенсвязывающий фрагмент такого антитела, которые специфично связывают сфингозин-1-фосфат (S1P).
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Visentin В | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| МОИСЕЕНКО B.M | |||
| Моноклональные антитела в лечении злокачественных опухолей, Практическая онкология (2003), т.4, 3:148-156. | |||

