ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к области эпигенетической регуляции экспрессии генов. В частности, данное изобретение относится к композициям и способам, включающим гистон-метилтрансферазную активность, которые нацелены на контроль экспрессии генов in vivo и in vitro.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Эпигенетика относится к передаче информации от клетки или многоклеточного организма к ее или его потомкам при отсутствии кодирования этой информации в нуклеотидной последовательности генов. Эпигенетический контроль обычно осуществляется через химическую модификацию структуры ДНК или хроматина. Экспрессию гена можно замедлять, например, метилированием и ацетилированием гистонов, которые ассоциируются с геномной ДНК. Метилирование и ацетилирование гистонов, как правило, осуществляются в "хвосте" гистонов, домене, который находится на поверхности и имеет результирующий положительный заряд вследствие распространенности таких аминокислотных остатков, как аргинин (R) и лизин (K). Химическая модификация "хвоста" гистона замедляется ферментом, имеющим гистон-метилтрансферазную активность (HMTase) и гистон-ацетилтрансферазную активность (HAT).
Эпигенетическая модификация может происходить в различные периоды нормального развития организма, а также в ходе превращения нормальных клеток в раковые клетки. Такие модификации часто приводят к сайленсингу или к активации некоторых генов. Документальными доказательствами убедительно подтверждено, что при раковом заболевании большинство опухолевых клеток имеют аномальные эпигенетические импринты ДНК (Feinberg АР & Vogelstein В, (1983) Nature 1(5895): 89-92).
Стволовые клетки представляют собой клетки, способные как к интенсивному самообновлению, так и к дифференцировке в клетки-предшественники. Стволовые клетки сами по себе являются также возможной первопричиной (возможным первоисточником) возникновения раковых заболеваний. Стволовые клетки могут иметь большую продолжительность жизни, в течение которой они приобретают генетические мутации и эпигенетические модификации, которые могут повышать склонность к злокачественному новообразованию. Предполагают, что так как стволовые клетки занимают нишу, которая так тонко балансирует между участием в конкурентной пролиферации и дифференцировке, то небольшие, но принципиальные эпигенетические изменения могут резко сдвинуть равновесие в сторону фенотипа раковых стволовых клеток. Понимание того, почему и как регулируются эпигенетические модификации, является решающим для понимания, обнаружения и лечения рака и, в частности, лечения (обработки) раковых стволовых клеток. На самом деле полагают, что одной из особенностей рецидивирующих и агрессивных раковых заболеваний, которые трудно поддаются лечению, является то, что опухоли могут содержать раковые стволовые клетки, которые не реагируют на обычные лекарственные средства.
Перенос ядра соматической клетки (SCNT) используют для получения животных в животноводстве (для клонирования или для терапии стволовыми клетками), биопроизводства белков и для моделирования заболевания (Wilmut I, Beaujean N, de Sousa PA, Dinnyes A, King TJ, Paterson LA, Wells DN, Young LE. (2002) Nature. Oct 10; 419(6907): 583-6). Одной из проблем, связанных с эффективностью и результативностью попыток SCNT, является то, что геном соматических клеток содержит интенсивные и устойчивые эпигенетические метки, которые могут препятствовать успешному перепрограммированию. Кроме того, реципиентная яйцеклетка может содержать факторы, вызывающие эпигенетический эффект, которые могут также вносить вклад в неудачу процесса. Следовательно, необходимо создать композиции и способы для повышения эффективности SCNT и, значит, тех применений в биопроцессинге, которые упрощаются с помощью этой операции.
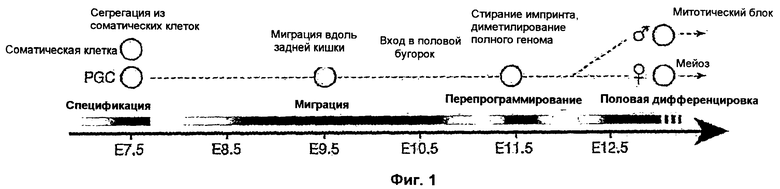
Спецификация примордиальных (первичных половых) зародышевых клеток (ПЗК, PGC) у мыши предоставляет привлекательную экспериментальную модель для анализа влияния эпигенетических модификаций in vivo. Популяцию клеток-"основательниц" (специальных клеток, founder-cells), примерно, из 45 PGC (ПЗК) сначала обнаруживают в Е7.5 в эмбрионах мыши (Ginsburg, М., Snow, М.Н. & McLaren, А. (1990) Development 110, 521-8). Затем эти PGC мигрируют и входят в половые бугорки из Е10.5 впереди, где продолжается дальнейшее экстенсивное эпигенетическое программирование зародышевых клеток. К Е13.5 зародышевые клетки входят в профазу мейоза.
Значительные эпигенетические модификации происходят сразу же после спецификации PGC, включая метилирование и ацетилирование "хвостов" гистонов с помощью HMTase (НМТаз) и HAT, соответственно (Surani et al., 2004 (CSH Symposium); Seki et al., 2004; Lachner, M., O'Sullivan, R.J. & Jenuwein, T. (2003) J Cell Sci 116, 2117-24, и Vaquero, A., Loyola, A. & Reinberg, D. (2003) Sci Aging Knowledge Environ 2003, RE4). Среди кандидатных генов, предположительно, участвующих в регуляции этих эпигенетических изменений в PGC, имеются HMTase (НМТазы), которые относятся к семейству белков с консервативным SET/PR доменом.
Следовательно, существует потребность в создании реагентов и способов улучшения контроля эпигенетических регуляторных механизмов в клетке. В частности, необходимы композиции (составы) и способы, позволяющие лучше контролировать экспрессию генов таким образом, чтобы влиять на решение судьбы клеток в стволовых клетках и в раковых клетках.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Первый аспект изобретения включает выделенный полипептидный комплекс, содержащий, по меньшей мере, первый домен, имеющий сайт - специфическую ДНК-связывающую активность, и, по меньшей мере, второй домен, имеющий аргинин-метилтрансферазную активность, причем второй домен способен метилировать аргининовый остаток, локализованный в хвостовой области гистона Н2А.
В специфическом варианте изобретения второй домен имеет аргинин-метилтрансферазную активность, которая обеспечивает симметричное NG,N'G-диметилирование аргининового остатка, надлежащим образом локализованного в положении 3 в хвостовой области гистона Н2А (H2AR3). Необязательно, второй домен способен, кроме того, метилировать аргининовый остаток, локализованный в области "хвоста" гистона Н4, аргининовый остаток, надлежащим образом локализованный в положении 3 в хвостовой области гистона Н4 (H4R3). В одном варианте изобретения аргинин-метилтрансферазная активность содержится в Prmt5 аргинин-метилтрансферазном домене, или в его производном, или в его гомологе.
Согласно изобретению первый домен можно специфически направлять на связывание с одной или более консенсусных последовательностей в геномной ДНК млекопитающего, которые участвуют в контроле экспрессии генов. Обычно, но не исключительно, такие сайты могут находиться в некодирующих промоторных областях, нетранслируемых областях или интронах. В специфическом варианте изобретения ДНК-связывающий домен по изобретению способен связываться с сайтом связывания типа PRDI/Blimp1, имеющим консенсусную последовательность из четырех мотивов GGGAAAG, два находятся в 5' промоторной области целевого гена, а два расположены 3' от сайта начала транскрипции. Соответственно, ДНК-связывающий домен содержит белок Blimp1, ДНК-связывающий участок полипептида PRDI/Blimp1 или его гомолог или производное.
Второй аспект изобретения включает нуклеотидную экспрессирующую векторную конструкцию, которая применима для индукции экспрессии полипептидного комплекса в клетке млекопитающего, причем вектор содержит:
одну или более кодирующих последовательностей, функционально связанных с промоторной последовательностью,
где одна или более кодирующих последовательностей кодируют, по меньшей мере, первый полипептидный домен, имеющий сайт-специфическую ДНК-связывающую активность и, по меньшей мере, второй полипептидный домен, имеющий аргинин-метилтрансферазную активность, при этом первый домен специфически направлен на связывание с одной или более консенсусных последовательностей в геномной ДНК млекопитающего, которые участвуют в контроле экспрессии генов, а второй домен имеет аргинин-метилтрансферазную активность, которая обеспечивает симметричное NG,N'G-диметилирование аргининового остатка, локализованного в полипептидном субстрате.
Согласно особому варианту изобретения полипептидный субстрат представляет собой гистон. Необязательно, второй полипептидный домен способен метилировать аргининовый остаток, локализованный в положении 3 в "хвостовой" области гистона Н2А (H2AR3), а также аргининовый остаток, находящийся в "хвостовой" области гистона Н4.
Подходящие экспрессирующие векторы включают плазмиды, космиды, вирусные векторы и искусственные хромосомы, такие как YAC. Необязательно, промоторную последовательность можно выбирать либо из конститутивного промотора, либо из индуцибельного промотора. Подходящие индуцибельные промоторы включают хорошо описанные Tet- или Тамоксифен-регулируемые системы. Альтернативные системы могут включать промоторы, чувствительные к тепловому шоку.
В одном варианте изобретения экспрессирующий вектор содержит кассету экспрессии, в которой первая кодирующая последовательность кодирует первый полипептидный домен, а вторая кодирующая последовательность кодирует второй полипептидный домен. Необязательно, первая кодирующая последовательность кодирует полипептид PRDI/Blimp1, а вторая кодирующая последовательность кодирует полипептид Prmt5. В особых вариантах изобретения может быть предпочтительно, чтобы первая и вторая кодирующие последовательности были разделены одной или более промежуточными последовательностями. Одна или более промежуточных последовательностей может содержать, по меньшей мере, один участок внутренней посадки рибосом (IRES) с тем, чтобы содействовать бицистронной экспрессии первой и второй кодирующих последовательностей в клетке. Экспрессирующие векторы по изобретению могут также содержать одну или более нуклеотидных последовательностей, которые кодируют полипептид, выбранных из: селективного маркера; маркера устойчивости к антибиотикам; и гена-репортера.
Третий аспект изобретения включает способ контроля экспрессии генов в клетке млекопитающего, заключающийся в индукции образования в клетке полипептидного комплекса, содержащего, по меньшей мере, первый домен, имеющий сайт-специфическую ДНК-связывающую активность, и, по меньшей мере, второй домен, имеющий аргинин-метилтрансферазную активность, причем второй домен способен метилировать аргининовый остаток, локализованный в "хвостовой" области гистона Н2А. В особом варианте изобретения образование полипептидного комплекса индуцируется в клетке индукцией экспрессии полипептида PRDI/Blimp1, или его гомолога или производного, в клетке.
В особом варианте изобретения экспрессия полипептида PRDI/Blimp1 индуцируется в клетке с помощью трансфекции клетки экспрессирующим вектором, который кодирует полипептид Blimp1, или его производное или гомолог. Необязательно, экспрессия полипептида PRDI/Blimp1 индуцируется в клетке с помощью трансфекции клетки экспрессирующим вектором, описанным ранее. Обычно клетка млекопитающего представляет собой человеческую клетку ткани или представлена в виде линии клеток. В особом варианте изобретения клетка млекопитающего представляет собой опухолевую или раковую клетку или линию клеток. Способ по данному аспекту изобретения можно осуществлять in vitro или in vivo.
В конкретном варианте изобретения контроль экспрессии генов представляет собой контроль экспрессии одного или более генов, выбранных из группы, состоящей из: с-Мус; Dhx38; Pcdh7; Q8C9T7; Xylt1; DnaH1; Baip2; Nek7; Dusp2; ENSMUSG00000027041; Sirt4; и Blimp1. Индукция полипептидного комплекса в клетке может привести к снижению экспрессии одного или более из этих генов.
Четвертый аспект изобретения включает способ стимуляции самообновления и ингибирования дифференцировки стволовой клетки, заключающийся в ингибировании образования комплекса Blimp1/Prmt5 в стволовой клетке. Необязательно, стволовая клетка представляет собой стволовую клетку млекопитающего, предпочтительно, человеческую стволовую клетку. В особых вариантах изобретения стволовую клетку выбирают из группы, состоящей из взрослой стволовой клетки; стволовой клетки-предшественника; и плюрипотентной стволовой клетки. Понятно, что изобретение никоим образом не относится к репродуктивному клонированию человека или к манипуляции с человеческими эмбрионами и их использованию для репродуктивного клонирования человека.
В специфическом варианте этого аспекта изобретения ингибирование образования комплекса Blimp1/Prmt5 в стволовой клетке осуществляется экспозицией клетки с соединением, ингибирующим (ингибитором) Blimp1, с соединением, ингибирующим Prmt5, и/или с соединением, ингибирующим комплекс Blimp1/Prmt5. Предпочтительно, ингибитор можно выбирать из низкомолекулярного ингибитора; молекулы siPHK (малой интерферирующей РНК), которая связывается с мРНК Blimp1 или Prmt5; антисмыслового олигонуклеотида, который связывается с мРНК Blimp1 или Prmt5; и доминантно-негативного варианта полипептида Blimp1 или Prmt5.
Следующий аспект изобретения включает способ контроля локализации Prmt5 в клетке, обычно локализации эндогенного Prmt5, заключающийся в индукции экспрессии в клетке полипептида Blimp1, тем самым индукции образования комплекса Blimp1/Prmt5 в клетке. Blimp1, индуцируемый в клетке, может быть либо экзогенным Blimp1, либо эндогенным Blimp1. Предпочтительно, клетка может быть стволовой клеткой млекопитающего.
Еще один аспект изобретения относится к применению полипептидных комплексов по изобретению для лечения рака и к клеткам - предпочтительно, клеткам млекопитающего/человека, - содержащим описанные выше экспрессирующие векторные конструкции.
ОПИСАНИЕ ФИГУР
На Фиг.1 представлены (а) краткое изложение основных событий в процессе спецификации и развития мышиной зародышевой клетки от Е7.5 до Е12.5, и (b) анализ экспрессии кандидатных генов домена SET/PR с помощью ПЦР кДНК единичной клетки от 2 репрезентативных клеток-основательниц (специальных клеток, founder cells) PGC (серые) и 2 соматических клеток (белые). Черные области показывают детекцию экспрессии в PGC и соматических клетках.
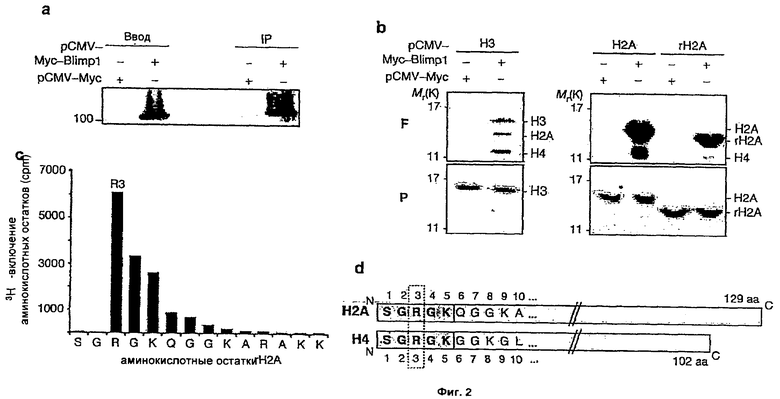
На Фиг.2 показано, что осажденный иммунопреципитацией комплекс мышиного Blimp1 проявляет аргинин-метилтрансферазную активность, (а) Меченный Myc мышиный Blimp1 или соответствующий контроль экспрессируют как указано в клетках 293Т. Мус иммунопреципитаты анализируют Вестерн-блоттингом, используя антитела против белка Мус; (b) те же самые иммунопреципитаты используют в HMTase анализах в отношении очищенного гистона Ну; Н2А и рекомбинантного Н2А (rН2А). Для каждого показана флуорограмма (F) и мембрана, окрашенная раствором Понсо (Р); (с) микросеквенирование меченного радиоизотопной меткой rН2А, по оси x показаны аминокислоты 1-14 rН2А, по оси у показано [3Н]- включение отдельных аминокислотных остатков в виде числа импульсов в минуту (cmp); и (d) выравнивание, показывающее сохранение последовательности крайнего N-конца Н4 и Н2А.1;
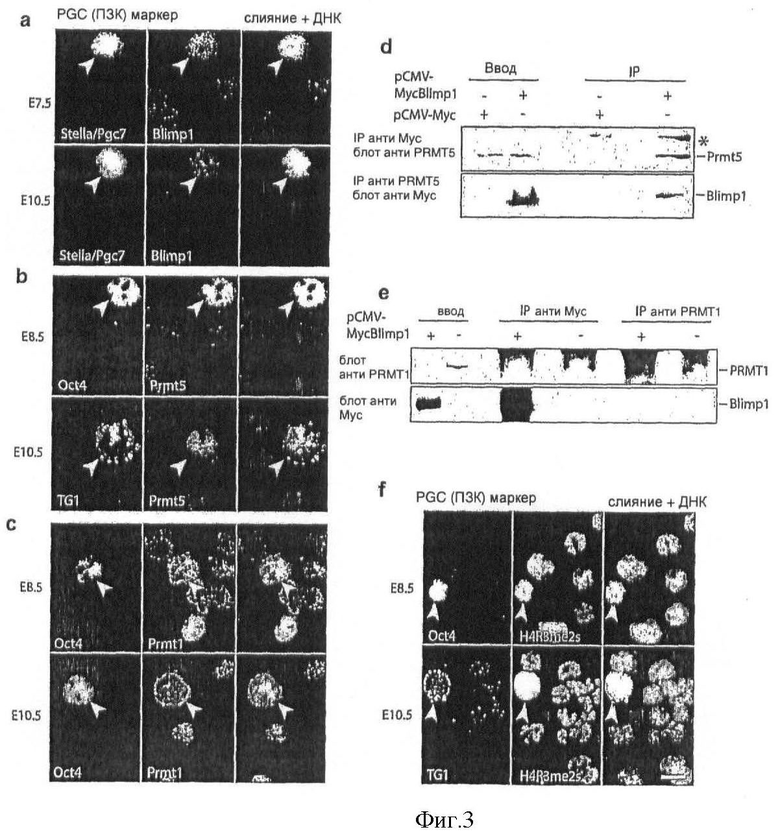
На Фиг.3 показано, что перекрывающаяся экспрессия Blimp1 и Prmt5 в зародышевых клетках приводит к специфическому паттерну H2A/H4R3me, (a, b, c) Паттерн экспрессии Blimp1, Prmt5 и Prmt1 в PGC на различных стадиях обнаруживают иммуноокрашиванием антителами, специфическими к Blimp1 (a), Prmt5 (b) и Prmt1, (с) зародышевые клетки детектируют как показано, используя антитела против stella/PGC7, Oct4 или TG1/SSЕА1, объединенные (слитые) изображения показаны с ДНК, окрашенной с помощью DAPI; (d, е) Меченный Myc мышиный Blimp1 или соответствующие контроли экспрессируют как указано в клетках 293Т, Мус, Prmt5 или Prmt1 иммунопреципитаты анализируют Вестерн-блоттингом, используя антитела против Prmt5, Мус или Prmt1, звездочкой показан неспецифический сигнал; и (f) Метилирование Н2А/Н4 R3 в зародышевых клетках оценивают иммуноокрашиванием в клетках PGC на различных указанных стадиях развития H4R3me2s антителами, зародышевые клетки совместно окрашивают антителами против стадиеспецифических маркеров, а именно, Oct4 или TG1/SSEA1, объединенные (слитые) изображения показаны с ДНК, окрашенной с помощью DAPI, масштаб: 10 мкм (масштаб идентичен во всех рисунках).
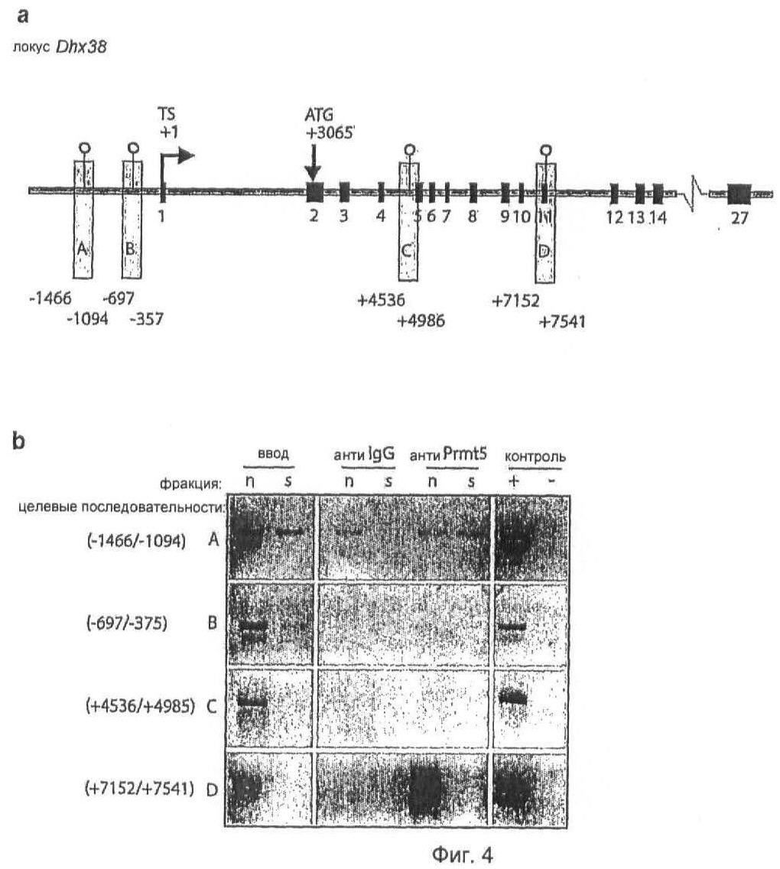
На Фиг.4 показана in vivo идентификация Blimp1/Prmt5 связывающих элементов с геномным локусом Dhx38, (а) положения предполагаемых сайтов связывания Blimp1 близ Dhx38 начала транскрипции (TS) и стартового кодона (ATG), также показаны амплифицируемые последовательности для ChIP анализа (А, В, С, D), (b) Взаимодействие эндогенного Prmt5 с геномной ДНК локуса Dhx38 с помощью ChIP анализа, фракцию супернатанта (s) или ядерную фракцию (n) клеточных экстрактов выделенных клеток половых бугорков Е10.5 эмбрионов осаждают иммунопреципитацией антителами либо к Prmt5, либо к IgG, в качестве контроля используют геномную ДНК "хвоста" (+) и воду (-).
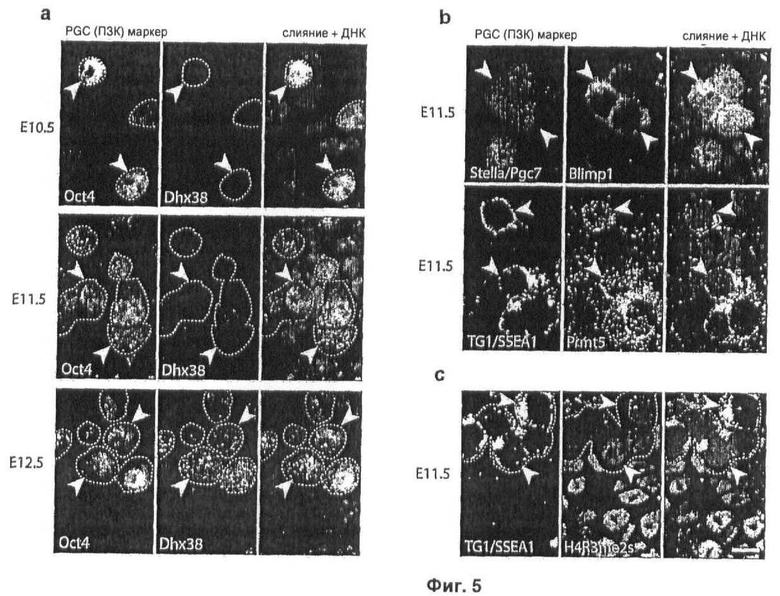
На Фиг.5 показано, что экспрессия Dhx38 активируется в зародышевых клетках при транслокации Blimp1 и Prmt5 из ядра в цитоплазму, что приводит к снижению уровней модификации H2A/H4R3me2s, иммуноокрашивание (a) Dhx38, (b) Blimp1 и Prmt5, и (с) H2A/H4R3me2s осуществляют на криосрезах половых бугорков на указанных стадиях развития, зародышевые клетки детектируют, используя специфические антитела: stella/Pgc7, Oct4 или TG1/SSEA1, объединенные изображения показаны с ДНК, окрашенной с помощью DAPI, масштаб: 10 мкм (масштаб идентичен во всех рисунках).
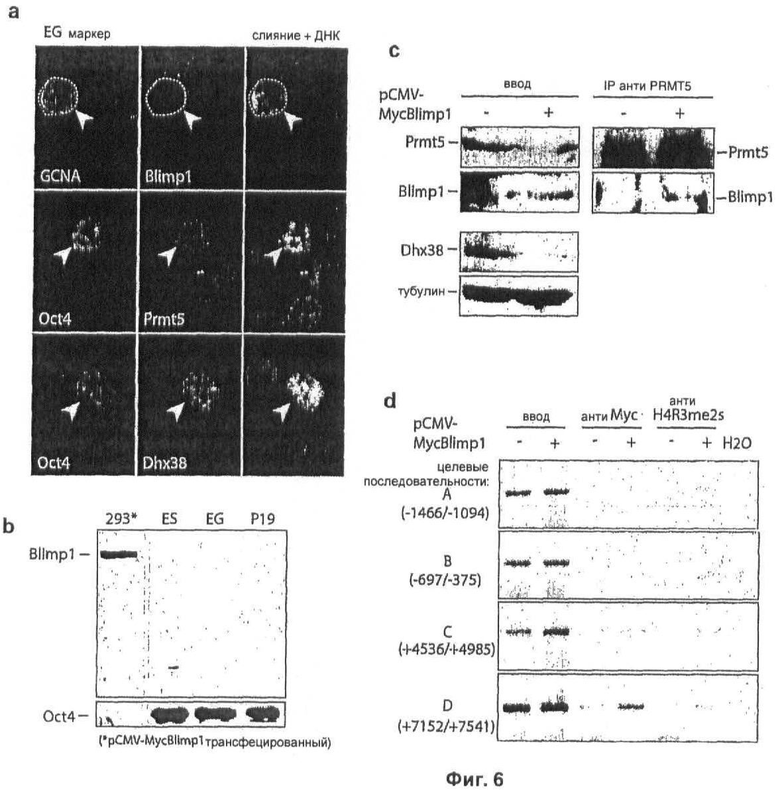
На Фиг.6 показан анализ Blimp1, Prmt5 и Dhx38 в плюрипотентных EG клетках и клетках эмбриональной карциномы (ЕС) (а) Иммуноокрашивание на Blimp1, Prmt5 и Dhx38 осуществляют на EG клетках, объединенные изображения показаны с ДНК, окрашенной DAPI, обратите внимание на обратное соотношение между экспрессией Dhx38 и Blimp1; (b) Вестерн-блоттинг ES, EG или ЕС (Р19) экстрактов для Blimp1 и Oct4; (с) Меченный Myc мышиный Blimp1 экспрессируют как указано в Р19 плюрипотентных ЕС клетках, Prmt5 иммунопреципитаты анализируют Вестерн-блоттингом, используя антитела к Prmt5, Blimp1 или Dhx38, причем уровни тубулина, показывающие равную нагрузку на дорожке ввода, обнаруживают, используя антитело против тубулина, следует отметить репрессию Dhx38, когда Blimp1 вводят в ЕС (Р19) клетки; (d) Повышенные уровни H4R3me2s на Dhx38 локусе в Myc-Blimp1 трансфецированных ЕС (Р19) клетках анализируют с помощью ChIP, клеточные экстракты Р19 клеток осаждают иммунопреципитацией антителами либо к Myc, либо к H4R3me2s, А, В, С, D относятся к областям в Dhx38 локусе, содержащем сайты связывания Blimp1, как объясняется на Фиг.4.
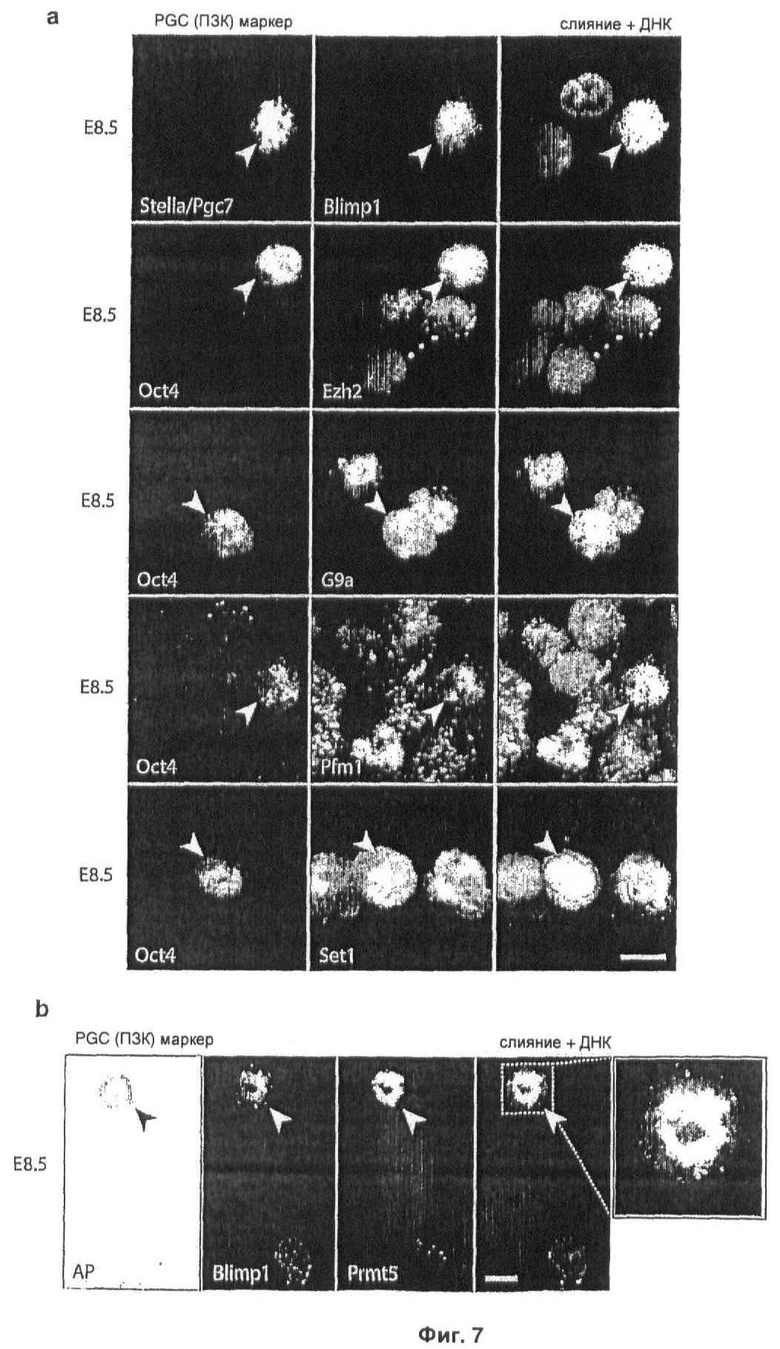
На Фиг.7 показаны (а) Иммунофлуоресцентный анализ кандидатных генов домена SET/PR при использовании скрининга экспрессии методом ПНР в Е7.5, показанном на Фиг.1; иммуноокрашивание выделенных клеток из Е8.5 эмбрионов специфическими антителами против специфических гистоновых метилтрансфераз, как указано, зародышевые клетки детектируют, используя антитела, специфические к зародышевым клеткам, Oct4 или Stella/PGC7; (b) совместное иммуноокрашивание Blimp1 и Prmt5 с применением соответствующих антител в Е8.5 PGC, зародышевые клетки метят с применением экспрессии тканенеспецифической щелочной фосфатазы (АР), объединенные изображения показаны с ДНК, окрашенной с помощью DAPI, масштаб: 10 мкм (масштаб идентичен во всех рисунках).
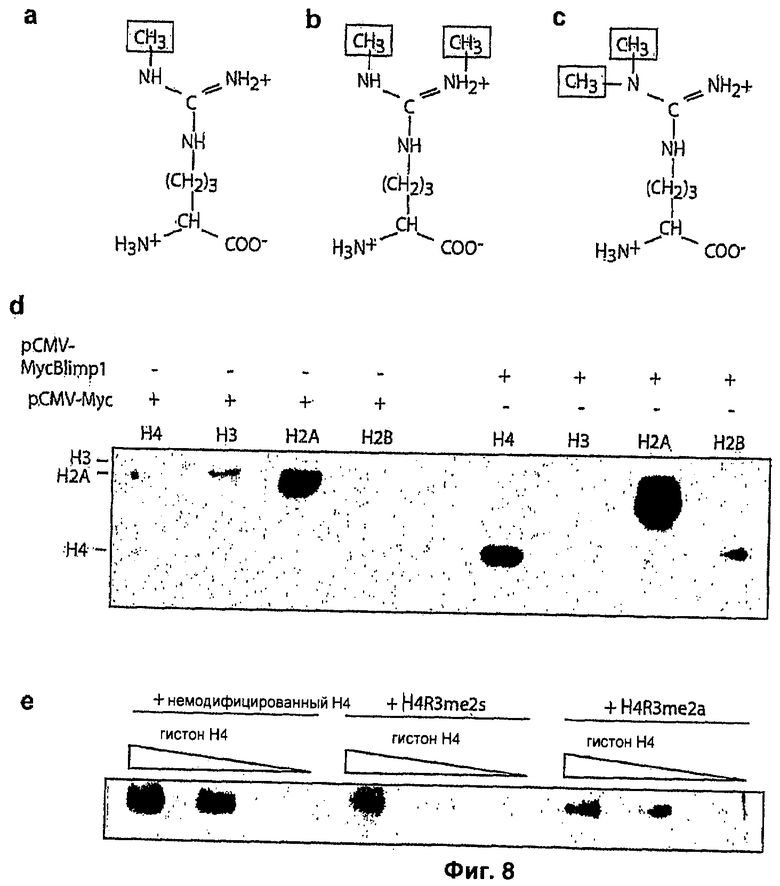
На Фиг.8 дается характеристика Н4 R3me2s антител, аргинин может быть модифицирован единственной метальной группой (а), или двумя метильными группами, которые расположены симметрично (b) или асимметрично (с); антитело против Н4 R3me2s (Abeam™) сначала получают, используя Н4 синтетический пептид с R3 симметричным ди(е?)метилированием, для проверки его специфичности проводят Вестерн-блоттинг; (d) против гистонов из тимуса теленка (Н4, Н3, Н2А, Н2В) инкубируют в присутствии и в отсутствие осажденного иммунопреципитацией Myc-Blimp1, и (е) согласно конкурентному анализу в отношении Н4 пептидов, включая немодифицированные, R3me2s и R3me2a, антитело четко узнает симметрично диметилированный пептид.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Перед тем, как давать подробное описание изобретения, приводится ряд определений, которые помогут понять изобретение. Все ссылочные материалы, цитируемые в данном описании, вводятся ссылками во всей полноте. Если не указано иначе, все технические и научные термины, используемые в данном описании, имеют то значение, которое оно обычно имеет для рядовых специалистов в той области техники, к которой относится данное изобретение.
Термин "перепрограммирование" по данному описанию относится к стадии изменения или удаления эпигенетических модификаций из ядра клетки. Перепрограммирование способствует редукции при коммитировании клеток и, следовательно, состоянию дифференцировки клетки в целом и, в частности, ядра. По сути перепрограммирование состоит из возврата ядра соматической дифференцированной или коммитированной клетки к экспрессии гена, эпигенетическому и функциональному состоянию, характерному для эмбриональной, зародышевой или стволовой клетки. Перепрограммирование ядер соматических клеток представляет собой предпочтительную первую стадию в таких методах, как SCNT, но оно также представляет интерес для других методов, в которых важен контроль дифференцировки клеток, т.е. способность к дифференцировке, дифференцировочные потенции.
Термин "рак" применяется в данном описании для обозначения ткани, локализованной в новообразовании (неоплазме, опухоли) или обладающей свойствами, ассоциированными с новообразованием (неоплазмой, опухолью). Новообразования обычно обладают характеристиками, которые отличают их от нормальной ткани и нормальных клеток. Такие характеристики включают, но без ограничения: степень анаплазии, изменения морфологии клеток, неправильность формы, пониженную адгезивность, способность к метастазированию, повышенные уровни ангиогенеза, повышенная инвазивность клеток, пониженные уровни клеточного апоптоза и, как правило, повышенную злокачественность клеток. Термины, соответствующие термину "рак", а часто являющиеся его синонимами, включают саркому, карциному, опухоль, эпителиому, лейкоз (лейкемию), лимфому, полип, перерождение, неоплазму (новообразование) и т.п.
Термин "эпигенетическая модификация" относится к химическому мечению (маркировке) генома. Эпигенетические метки могут включать метилирование ДНК (импринты), а также метилирование и ацетилирование белков, ассоциированных с ДНК, таких как гистоны. Экспрессию генов, специфических в отношении родных родителей (либо из материнской, либо из отцовской хромосомы), часто наблюдается у млекопитающих и является результатом эпигенетических модификаций. В родительских зародышевых линиях эпигенетическая модификация может привести к стабильному сайленсингу или к стабильной активации генов.
"Биопроцессинг" относится к методам, в которых живые клетки или их компоненты используют для получения нужного конечного продукта. В контексте настоящего изобретения эпигенетические модификации в клетках можно использовать для повышения этой способности клеток с целью применения в биопроцессинге. Обычно методы биопроцессинга включают SCNT.
Термины "производные или гомологи" доменов ДНК-связывания и/или аргинин-метилтрансфераз по данному описанию относятся к мРНК и полипептидам, которые обладают идентичностью последовательностей, практически аналогичной молекулам по изобретению, соответственно. Полагают, что производные и гомологи включают ортологи последовательностей из других видов и мутанты, у которых, тем не менее, наблюдается высокий уровень функциональной эквивалентности. Под практически аналогичной (сходной) идентичностью последовательностей понимают уровень сходства последовательностей, примерно, от 50%, 60%, 70%, 80%, 90%, 95%, примерно, до 99% идентичности. Процент идентичности последовательностей можно определить, используя обычные методы (Henikoff and Henikoff, Pros. Natl. Acad. Sci. USA 1992; 89: 10915, и Altschul et al. Nucleic Acids Res. 1997; 25: 3389-3402). Или же гомологи полипептидов по изобретению могут представлять собой такие последовательности, которые могут демонстрировать способность гибридизоваться с последовательностями по данному описанию в условиях высокой, средней или низкой жесткости.
Термин "экспрессирующий (экспрессионный) вектор" ("вектор экспрессии") применяют для обозначения молекулы ДНК, являющиеся либо линейной, либо кольцевой, в которую может быть интегрирован фрагмент последовательности другой ДНК. Такой (такие) фрагмент(ы) ДНК могут включать дополнительные сегменты, которые обеспечивают транскрипцию гена, кодированного фрагментом последовательности ДНК. Дополнительные сегменты могут включать, но без ограничения: промоторы, терминаторы транскрипции, энхансеры, участки внутренней посадки рибосом, нетранслируемые области, сигналы полиаденилирования, селективные маркеры, ориджины репликации и тому подобное. Экспрессирующие векторы часто получают из плазмид, космид, вирусных векторов и искусственных хромосом дрожжей; векторы часто представляют собой рекомбинантные молекулы, содержащие последовательности ДНК из нескольких источников.
Выражение "функционально связанный", в применении к последовательностям ДНК, например, в экспрессирующем векторе, указывает, что последовательности расположены таким образом, что для достижения определенных целей они действуют совместно, а именно, промоторная последовательность содействует инициации транскрипции, которая происходит с помощью связанной кодирующей последовательности до сигнала терминации транскрипции.
"Полинуклеотид" представляет собой одно- или двухнитевую (тяжевую, цепочечную) ковалентно связанную последовательность нуклеотидов, в которых 3' и 5' концы каждого нуклеотида соединены фосфодиэфирными связями. Полинуклеотид может состоять из дезоксирибонуклеотидных оснований или рибонуклеотидных оснований. Полинуклеотиды включают ДНК и РНК и могут быть получены синтетически in vitro или выделены из природных источников. Размеры полинуклеотидов обычно выражаются в виде числа пар (нуклеотидных) оснований (пар нуклеотидов, п.н., bp) для двухнитевых полинуклеотидов, или, в случае однонитевых полинуклеотидов, в виде числа нуклеотидов (nt). Одна тысяча п.н. или nt равна килобазе (kb). Полинуклеотиды длиной менее примерно 40 нуклеотидов обычно называют "олигонуклеотидами".
Термин "промотор" по данному описанию обозначает область в гене, с которой факторы транскрипции и/или РНК полимераза могут (может) связываться таким образом, чтобы контролировать экспрессию ассоциированной кодирующей последовательности. Обычно, но не всегда, промоторы локализованы в 5' некодирующих областях генов, 5' от кодона инициации трансляции. Промоторная область гена может содержать одну или более консенсусных последовательностей, которые действуют как распознаваемые сайты связывания для специфических в отношении последовательности ДНК-связывающих доменов ДНК-связывающих белков. Тем не менее, такие сайты связывания могут также быть локализованы в областях вне промотора, например, в энхансерных областях, расположенных в интронах или 3' от кодирующей последовательности.
Термин "выделенный", в применении к полипептиду или комплексу (объединению) полипептидов, обозначает полпептид, который выделен из организма, который является его природным источником. Предпочтительно, чтобы выделенный полипептид практически не содержал других полипептидов, нативных для протеома организма - источника. Наиболее предпочтительно, чтобы чистота выделенного полипептида составляла, по меньшей мере, 95%, более предпочтительно, более 99%. В данном контексте предполагается, что термин "выделенный" включает один и тот же полипептид в альтернативных физических формах, либо в нативной форме, в форме денатурированного белка, в димерной/мультимерной форме, в форме гликозилированного, кристаллизованного, либо дериватизированного белка. Ссылка на "комплекс" ("объединение") в данном описании включает примеры, в которых первый и второй полипептидные домены находятся в единой полипептидной цепи, а также в которых первый и второй домены входят в состав отдельных полипептидных цепей, которые нековалентной связью связаны друг с другом, а также в которых образуются посттрансляционные ковалентные связи, связывающие отдельные домены в один объединенный функциональный элемент (единицу).
В одном варианте настоящего изобретения предусматривается новый комплекс между Blimp1 и Prmt5, способный регулировать экспрессию генов в клетках млекопитающих по механизму эпигенетического контроля.
Обычно соматические клетки развиваются по пути дифференцировки, развиваясь от состояния менее специализированной клетки к состоянию более специализированной или коммитированной клетки. Менее специализированные клетки могут демонстрировать способность вести себя как стволовые клетки-предшественники, дающие начало некоторым различным типам клеток. Количество этих различных типов клеток, для которых данная стволовая клетка может служить в качестве предшественника, обычно называют "потентностью" этой стволовой клетки. Следовательно, плюрипотентные стволовые клетки могут вести себя как предшественники очень многих различных типов дифференцированных клеток. Если клетка может дифференцироваться во все клетки организма, она является тотипотентной стволовой клеткой, если клетка может дифференцироваться в большинство типов клеток, она является плюрипотентной стволовой клеткой. Эмбриональные стволовые клетки обычно называют плюрипотентными, так как они могут генерировать большинство типов клеток млекопитающих, за исключением экстраэмбриональных тканей (т.е. трофэктодермы). Настоящее изобретение включает способ контроля выбора судьбы клетки на уровне контроля экспрессии генов. Белковый комплекс по изобретению, будучи экспрессирован или иным способом введен в клетки млекопитающих, сдвинуть судьбу клетки в другую сторону от выбора плюрипотентности. Напротив, ингибирование активности белкового комплекса по изобретению, или даже одного компонента, Blimp1, в стволовых клетках может сдвинуть судьбу клетки в сторону выбора плюрипотентности и самообновления.
Другой родственной областью полезности настоящего изобретения является терапия рака. Большинство, если не все, раковые заболевания подвергаются эпигенетическим изменениям, включая в значительной степени деактивацию и сайленсинг генов-супрессоров опухолей и активацию онкогенов. Реактивация генов-супрессоров опухолей может уменьшить интенсивность фенотипа рака, поскольку может подавить (негативно регулировать) онкогены. Следовательно, способ контроля экспрессии генов и выбора судьбы клеток in vivo представляет очень обещающее направление в терапии рака.
Созревание белка (Blimp1), индуцированное В-лимфоцитами
Blimp1 представляет собой белок длиной 100 кДа, который содержит пять ДНК-связывающих мотивов цинковых пальцев (GENBANK регистрационный по: NM_007548). Человеческий гомолог Blimp1 называется либо PRDI-BF1, либо PRDM1 (GENBANK регистрационный по: NM_000198). кДНК Blimp1 сначала выделяли вычитательным (субстрактивным) скринингом линии клеток В-клеточной лимфомы (BCL1) после обработки цитокинами IL-2 и IL-5. эктопической экспрессии Blimp1 достаточно для того, чтобы вызвать конечную дифференцировку BCL1 клеток. Считается, что Blimp1 является 'мастер-регулятором' (общим регулятором) конечного развития В-клеток (Yu J. et al. (2000) Mol. Cell. Biol. 20(7): 2592-2603).
У человека PRDI-BF1, человеческий ортолог мышиного Blimp1, способен образовывать комплекс с Н3 лизин-метилтрансферазой G9a. Показано, что этот комплекс способен вызвать сайленсинг гена человеческого интерферона-β (IFN-β) по опосредованному хроматином механизму в промоторной области гена (Gyory I. et al. (2004) Nat. Imm. 5: 299-308).
Показано, что Blimp1 образует комплекс с факторами, которые принимают участие в эпигенетических модификациях. Полагают, что комплекс, содержащий Blimp1 и гистонную деацетилазу (HDAC), изменяет структуру нуклеосомы и ингибирует транскрипцию генов за счет деацетилирования лизиновых остатков на "хвостах" гистона. Так как ацетилирование лизиновых остатков эффективно нейтрализует их положительный заряд, деацетилирование возвращает заряд и приводит к модификации структуры нуклеосомы вследствие стерических и других эффектов.
Известные мишени Blimp1 включают c-Myc, IFN-β, CD23, CD22, ГКГ (МНС) класса II, BSAP (Pax 5), фактор ранних В-клеток и CIITA. Все эти гены претерпевают транскрипционную регрессию под действием Blimp1. Транскрипция гена c-Myc подавляется Blimp1 в процессе дифференцировки В-клеток и является важной мишенью Blimp1 в клетках лимфомы BCL1. Для того чтобы Blimp1 подавлял промотор c-Myc, нужны различные области молекулы Blimp1, включая N-концевой кислый домен и область между аа 90 и 464 (Yu J. et al., см. выше).
Онкобелок c-Myc важен для контроля регуляторов роста, апоптоза и/или дифференцировки, а его дисрегуляция также является причиной широкого ряда новообразований. Дисрегуляция экспрессии c-Myc в В-клетках часто вызывает опухоли. Хромосомные транслокации гена с-Мус в локусы гена Ig присутствует в большинстве лимфом Беркитта и мышиных плазмацитом (Lin K.I. et al. (2000) Mol. Cell Biol. (20)23: 8684-8695).
Мышиные клетки ES можно сохранять как плюрипотентную, самообновляющуюся популяцию с помощью LIF/STAT3-зависимой передачи сигнала. Было показано, что STAT3 регулирует экспрессию транскрипционного фактора (фактора транскрипции) Myc. Полагают также, что для взрослых стволовых клеток требуется активация Myc. Методом RT-ПЦР (RT-PCR) анализа показано, что транскрипция Myc повышается в ES клетках. Обычно для ES клеток требуется культуральная среда, которая включает LIF, иначе ES клетки имеют тенденцию скорее к дифференцировке, нежели к самообновлению. Обнаружено, что уровни Myc в клетках ES быстро разрушаются после удаления LIF из культуры, это указывает, что он может быть необходим для дифференцировки ES клеток. Эксперименты наводят на мысль, что один Myc может поддерживать состояние клеток ES (т.е. плюрипотентный фенотип) на уровне, сопоставимом с LIF, а когда Myc инактивируется, пул стволовых клеток уменьшается (Cartwright P. et al. (2005) Development 132: 885-896).
Белок Аргинин-Метилтрансфераза 5 (Prmt5)
Известно два типа гистонных метилтрансфераз (HMTase), которые включают любой домен SET (Suvar3-9, Энхансер Zeste, Trithorax), обнаруженный сначала в белках, обладающих лизин-специфической метилазной активностью, или домен с аргинин-специфической метилазной каталитической активностью, обнаруженный в PRMT. PRMT делятся на типы I и II: PRMT типа I катализируют монометилирование и асимметрическое ди(е?)метилирование аргининовых остатков, в то время как PRMT типа II катализируют образование монометилированных и симметрично диметилированых аргининовых остатков. Из шести известных PRMT только PRMT5 (GENBANK регистрационные No: NM_006109 (человеческий); NM_013768 (мышиный)) ведут себя как PRMT типа II, которые могут нацеливаться на гистоны. Конкретно, показано, что PRMT5 нацелен на конкретные остатки аргинина в Н3 и Н4 N-концевых хвостах.
PRMT5 может ассоциироваться с комплексами ремоделирования хроматина на основе BRG1 и hBRM hSWI/SNF. В таких комплексах PRMT5 способен стимулировать рост клеток и независимый от заякоривания рост, метилируя остаток аргинина 8 гистона Н3 (H3R8), и тем самым снижая экспрессию генов, таких как ST7 и NM23, которые, как известно, принимают участие в супрессии опухолей (Richard S. et al. (2005) Biochem. J. 388: 379-386). PRMT5 также принимает участие в транскрипционной регрессии CYCLIN Е и САD.
Комплекс Blimp1/Prmt5
Заявитель настоящего изобретения приступили к идентификации возможной активности домена Blimp1 SET/PR, так как никакой специфической гистон-метилтрансферазной активности не было отнесено к самому Blimp1. Заявитель показал, что Blimp1 могут образовывать новый комплекс с PRMT5, комплекс, который in vivo сохраняется в линии дифференцировки мышиных зародышевых клеток до входа PGC в половые бугорки, это наводит на мысль, что Blimp1 постоянно принимает участие в последовательности поколений ранних зародышевых клеток млекопитающих. Дальнейший анализ, более подробно описанный ниже, показывает, что новый комплекс Blimp1/Prmt5 сообщает уникальные характерные эпигенетические признаки благодаря метилированию гистонов Н2А и Н4. Полагают, что это первый пример, в котором метилирование хвоста гистона Н2А показано как эпигенетический регуляторный механизм. Далее, эксперименты, более подробно описанные ниже, показывают, что в то время как Prmt5 присутствует в плюрипотентных EG и ES клетках, Blimp1 в них отсутствует.
Экспрессия Blimp1 в линии плюрипотентных клеток ЕС Р19 приводит к репрессии гена, который, как известно, экспрессируются в плюрипотентных клетках на высоком уровне. Эксперименты наводят на мысль, что комплекс Blimp1/Prmt5 является важным регулятором экспрессии генов и может оказывать заметное влияние на выбор судьбы клеток, в частности, в отношении плюрипотентности и дифференцировки. По этой причине один вариант настоящего изобретения включает механизм контроля генов, основанный на использовании различной биологической активности, на примере комплекса Blimp1/Prmt5 in vitro или in vivo.
Не связывая себя какой-либо теорией, полагают, что в этом комплексе Blimp1 обеспечивает функцию таргетирования генов, так как он имеет пять ДНК-связывающих доменов в виде цинковых пальцев. Prmt5 обеспечивает аргинин-метилтрансферазную активность, которая служит в качестве HMTase при локализации на ДНК, что приводит к репрессии экспрессии генов. Известно, что человеческий ортолог Blimp1 связывается с сайтом PRDI в промоторной области человеческого IFN-β (Keller A.D. & Maniatis T. (1991) Genes Dev. 5: 868-879). Как указывалось выше, известно также, что Blimp1 способен репрессировать экспрессию c-Myc. Согласно данному изобретению идентифицирован новый поднабор генов, которые, как показано, являются специфическими мишенями комплекса Blimp1/Prmt5 и экспрессия которых может регулироваться новым комплексом. Этот поднабор включает гены, имеющие различные функции, но, как полагают, играющие важную роль в регуляции потентности, клеточного цикла, дифференцировки, адгезии клеток, эпигенетического перепрограммирования и, возможно, также супрессии опухоли.
Согласно изобретению репрессорный комплекс Blimp1/Prmt5 коррелирует с высокими уровнями H2A/H4R3me2s в зародышевых клетках. Однако, в ходе дифференцировки В клеток в плазматические клетки Blimp1 управляет G9a-зависимым H3K9me2. Таким образом, Blimp1 способен управлять выбором судьбы и свойствами различных клеток за счет ассоциации с различными связывающими партнерами, и ремоделирование (структуры) хроматина может являться самым главным для этих процессов. Помимо очевидной важности Blimp1 на ранних стадиях спецификации зародышевых клеток у мышей, настоящее изобретение демонстрирует дополнительное участие Blimp1 в образовании уникального характерного эпигенетического признака хроматина, наблюдаемого в зародышевых клетках после спецификации PGC (ПЗК). Однако, Blimp1/Prmt5 существует благодаря ядрам PGC (ПЗК) и входит в цитоплазму после Е10.5, когда экстенсивное программирование полного (всего) генома детектируют в зародышевых клетках. Таким образом, понимание связи между комплексом Blimp1/Prmt5, симметричным метилированием аргинина 3 на Н2А/Н4 и последующим эпигенетическим перепрограммированием полного генома в зародышевых клетках позволяет проникнуть в суть механизма (понять механизм), лежащего в основе этого важного процесса, и понять, как ядерное перепрограммирование контролируется внутри клетки.
Изобретение также дает понимание роли Blimp1/Prmt5 в плюрипотентных эмбриональных (EG) клеток гонад, образование которых из PGC несомненно ассоциируется с утратой Blimp1. Настоящее изобретение предоставляет прямое доказательство того, что Blimp1 ключевым для репрессии образующихся и мигрирующих PGC от приобретения явно плюрипотентного фенотипа, подобного стволовым клеткам. Поэтому агонисты и антагонисты активности Blimp1/Prmt5 могут играть важную роль в сдерживании выбора судьбы клеток либо в сторону от плюрипотентного фенотипа, либо в сторону к плюрипотентному фенотипу. Кроме того, экспрессия Blimp1 в клетках предоставляет механизм блокирования аргинин-метилтрансферазной активности Prmt5 в ядре и вдали от потенциальных цитоплазматических субстратов. Такой эффект может быть целесообразным в ходе дифференцировки стволовых клеток и в ходе всего клеточного цикла в заданной клетке или линии клеток.
Конкретные малые нуклеотидные молекулы, которые применяются в изобретении в качестве ингибиторов Blimp1 и/или Prmt5, представляют собой короткие фрагменты двухнитевой РНК, известные как малые (короткие) интерферирующие РНК (siRNA). Технология интерферирующих РНК (iPHK, RNAi) позволяет осуществлять селективную активацию генной функции vivo. В настоящем изобретении iPHK (RNAi) можно использовать для того, чтобы задавить экспрессию Blimp1 и/или Prmt5 в клетках. В этом процессе двухнитевые мРНК распознаются и расщепляются разрезающей РНКазой, в результате получают фрагменты iPHK (RNAi) длиной 21-23 нуклеотида. Эти RNAi включают в индуцируемый РНК комплекс сайленсинга (RISC) и раскручивают с помощью этого комплекса. Затем единичный антисмысловой тяж направляет RISC на мРНК, содержащую комплементарную последовательность, что приводит к эндонуклеолитическому расщеплению мРНК (Elbashir et al. (2001) Nature 411; 494-498). Следовательно, эта технология дает метод для нацеливания на и расщепления мРНК Blimp1 и/или Prmt5 в соматических клетках, предназначенных для применения в биопроцессинге или в тех случаях, когда нужно стимулирование самообновления плюрипотентного фенотипа. Методы получения siPHK для Prmt5 описаны в уровне техники (Richard S., см. выше). Примеры подходящих последовательностей siPHK для таргетирования человеческого PRMT5 включают:
5' CTCATTTGCTGACAATGAA 3' [SEQ ID NO:1]
5' GGACCTGAGAGATGATATA 3' [SEQ ID NO:2]
5' GTTTCAAGAGGGAGTTCAT 3' [SEQ ID NO:3]
Комплекс Blimp1/Prmt5 можно использовать для идентификации других белков и полипептидов, которые взаимодействуют с ним в клеточной среде. Обычные методы определения белок-белковых взаимодействий, такие как двухгибридный скрининг в дрожжах, можно использовать для идентификации возможных агонистов и антагонистов интерактивности комплекса Blimp1/Prmt5. Также в сферу изобретения входит идентификация малых молекул, которые ингибируют ассоциацию Blimp1 с Prmt5, например, экранируя или нарушая взаимодействие с теми доменами Blimp1, о которых известно, что они непосредственно взаимодействуют с другими белками (Yu J. et al. см. выше). Белок- белковые взаимодействия или взаимодействия белок-малая молекула, в которых участвуют Blimp1, Prmt5 и/или комплекс Blimp1/Prmt5, можно изучать, используя такие методы, как BIAcore®, который изучает молекулярные взаимодействия с помощью поверхностно-плазменного резонанса (BIAcore, Inc., Piscataway, NJ; см. также www.biacore.com).
Скрининг молекул и белков на связывание с комплексом Blimp1/Prmt5 можно осуществлять методами автоматического высокопроизводительного скрининга. Поэтому изобретение включает методы идентификации молекул, взаимодействующих с комплексом Blimp1/Prmt5, путем обнаружения позитивного взаимодействия связывания между комплексом Blimp1/Prmt5 и молекулой-мишенью. Можно применять дополнительные стадии скрининга, чтобы определить, действительно ли идентифицированное позитивное взаимодействие связывания имеет фармакологическое значение - т.е. действительно ли молекула-мишень (целевая молекула) способна снижать (уменьшать) биологическую активность или функцию Blimp1, Prmt5 и/или комплекса Blimp1/Prmt5. Если идентифицирована молекула с позитивным снижающим эффектом, молекулу классифицируют как 'hit' (попадание или хит), а затем ее можно оценить как потенциальное кандидатное лекарство. В это время, или ранее, можно принять во внимание дополнительные факторы, такие как например, всасывание (адсорбция), распределение, метаболизм и выделение (ADME), профили биодоступности и токсичности молекулы. Если молекула потенциального лекарства удовлетворяет фармакологическим требованиям, то считают, что она является фармацевтически совместимой. Можно приготовить соответствующие композиции для проверки активности in-vitro и in-vivo в соответствии со стандартными методами, известными в уровне техники.
ПРИМЕРЫ
Реагенты и методы
Выделение из эмбрионов. Примордиальные зародышевые клетки выделяют из эмбрионов аутбредных MF1 мышей на различных стадиях развития. День введения вагинального тампона обозначают Е0.5. кДНК-библиотеки из единичных клеток взяты из ранее опубликованной работы (Saitou M., Barton S.С. & Surani M.А. (2002) Nature 418, 293-300).
Иммуноокрашивание. Препарированные эмбрионы, содержащие зародышевые клетки, обрабатывают трипсином для получения суспензии одиночных клеток. Затем клетки оставляют осаждаться на покрытых поли-L-лизином срезах, фиксируют с помощью 2% PFA (параформальдегида), трижды отмывают PBS и обрабатывают далее. Целые эмбриональные половые бугорки в Е11.5 срезают, отмывают в PBS, фиксируют в течение двух часов при 4°C в 4% PFA, отмывают в PBS и оставляют осаждаться в 20% растворе сахарозы при 4°C. Их заключают в О.С.Т (BDH) и делают криосрезы. Одиночные клетки или срезы пермеализуют в IF буфере (PBS; 0,1% тритона; 10 мг/мл BSA). Инкубацию первичных антител проводят при 4°C, а затем трижды отмывают в IF буфере и инкубируют со вторичными антителами (Alexa 564, Alexa 488; Молекулярные зонды) в течение двух часов при комнатной температуре и отмывают в PBS. Затем срезы заключают в среду Vectashields с DAPI (Vector laboratories). Иммунофлуоресценцию наблюдают с помощью конфокального микроскопа BioRad radiance 2000. Используют следующие антитела и разведения: PGC7 (от T.Nakano; 1: 2500), Oct4 (BD Transduction Laboratories; 1: 200), TG1 (специфическое в отношении мышиных зародышевых клеток моноклональное антитело против SSEA1; 1:1), Blimp1 (от K.Calame; 1:10), Prmt5 (фирма Upstate; 1:250), Prmt1 (Upstate, 1:200), H4R3me2s (Abeam; 1:1000), Dhx38/Prp16 (Proteintech Group; 1:200), Ezh2 (Upstate; 1:50), G9a (Abeam™; 1:100), Pfm1 (Abeam™; 1:50), Set1 (от W. Herr; 1:200).
Иммунопреципитационные анализы и приготовление ядерных экстрактов. Кодирующую область мышиного Blimp1 амплифицируют с помощью RT-ПЦР и продукт клонируют в pcDNA3-MycHisA, получают конструкцию pCMV-MycBlimp1. Клетки 293Т или Р19, трансфецированные либо оригинальным вектором (называемым pCMV-Myc), либо pCMV-MycBlimp1, отмывают PBS и лизируют в IP буфере, содержащем 150 мМ NaCl, 1% NP40, 0,1% тритона, 50 мМ Трис (Tris) pH 8.0 и полный протеазный ингибирующий коктейль (Roche).
В типичной реакции иммунопреципитации используют 2×107 клеток. Экстракт цельных клеток инкубируют с 2 мкг Myc антител (New England Biolabs). Или же, используют антитела либо к Prmt5, либо к Prmt1 (Upstate) в течение ночи при 4°C. Затем прибавляют 30 мкл гранул белок A/G сефароза и выдерживают 2 часа при 4°C. Гранулы отмывают пять раз в IP буфере. Связанные белки элюируют при кипячении в буфере Лэмли для образцов. Ядерные экстракты клеток ES, EG или Р19 готовят в соответствии с инструкцией производителя (набор ядерных экстрактов; Active Motif), и 25 мкг ядерной фракции используют на нагрузку.
In vitro анализы с метилтрансферазой. Гранулы от иммунопреципитационных анализов 293Т клеток трансфецируют либо pCMV-Myc или pCMV-MycBlimp1, обрабатывают, дополнительно отмывая дважды в HMTase буфере (25 мМ NaCl, 25 мМ Tris pH 8.8), и используют HMTase анализе, описанном в 18, с небольшими модификациями. Вкратце, 1 мкг Н3 или Н2А (Roche) или рекомбинантного Н2А (любезный подарок от A. Brehm), в качестве субстрата, и 2 мкКи S-аденозил-L-[метил-3H]метионина ([3H]SAM; Amersham Biosciences), в качестве донора метильной группы, инкубируют в смеси 20 мкл буфера для HMTase анализа в течение трех часов при 37°C. Белки разделяют в 18% геле для SDS-PAGE, переносят на PDVF мембрану и визуализируют окрашиванием Понсо и флуорографией. In vitro метилированный rН2А микросеквенируют (определяют первичную последовательность короткого фрагмента) ступенчатой деградацией по Эдману (Protein and Nucleic acid Chemistry Facility, Cambridge University, UK), затем определяют 3H включение отдельных аминокислот подсчетом сцинтилляций. Для изучения Н2А/Н4 специфического метилирования Н4, Н3, Н2А и Н2В (Roche) инкубируют в присутствии иммуноосажденного комплекса Blimp1 и SAM как описано выше и проводят Вестерн-блоттинг, используя H4R3me2s антитела (Abeam™, Cambridge, UK), которые проверяют на их специфичность конкурентным анализом (см. приложение, Фиг.8).
Иммунопреципитация хроматина (ChIP)-клонирование и ChIP. Суспензии одиночных клеток сшивают в 1% формальдегиде 10 мин при комнатной температуре. Затем клетки разрушают и ядра лизируют в 50 мМ Tris pH 8.0, 10 мМ EDTA, 1% SDS с последующим центрифугированием. Экстракт подвергают иммунопреципитации как описано выше, добавляя очищенный IgG (Santa Cruz) в качестве негативного контроля. Затем гранулы дважды элюируют в 50 мМ NaHCO3, 1% SDS и супернатанты обрабатывают протеиназой K в течение 5 ч при 65°C с последующей очисткой смесью фенол/хлороформ и осаждением (преципитацией) спиртом. Затем PRMT5 (Upstate) или H4R3m32 (Abeam™) ChIP образцы анализируют стандартной реакцией ПЦР, используя следующие праймеры (1for (прямой) ccaggaggggtttcatcaactg [SEQ ID NO:4] и 1rev (обратный) tgttaccgtctcacttggtgtttg [SEQ ED NO:5]; 2for acctcacaactgctgggattac [SEQ ID NO:6] и 2rev ttcgttttctgcgtccgtg [SEQ ID NO:7]; 3for tttgtcgcagtgtcttatcgtaac [SEQ ID NO:8] и 3rev taggaaggtgttggggaggg [SEQ ID NO:9]; 4for atgaggtttgagaagtgtggc [SEQ ID NO:10] и 4rev atcagcggtggtggtgacagc [SEQ ID NO:11]). В случае анализа "ChIP cloning" проводят два последовательных цикла иммунопреципитации, используя антитела против Myc. Далее проводят преципитацию (осаждение) ДНК, которую "затупляют" по концам Т4 ДНК-полимеразой, а затем лигируют к отожженным JW102 и JW103 25. Затем продукты лигирования ПЦР-амплифицируют с JW102 (один цикл 55°C 2 мин; 72°C 5 мин; 94°C 2 мин и 20 циклов 94°C 30 с; 55°C 30 с; 72°C 1 мин и, наконец, один цикл 72°C 5 мин). ПЦР-продукты клонируют в pGEM-T (Promega), колонии проверяют на инсерции с помощью ПЦР и продукты секвенируют стандартными методами. Затем проводят программный поиск, используя он-лайн источники сведений по биоинформатике, такие как Ensembl (http://www.ensembl.org).
Пример 1: Определение Blimp1 активности
Значительные эпигенетические модификации происходят непосредственно после спецификации мышиных PGC (ПЗК), включая метилирование и ацетилирование гистонных концов метилтрансферазами (HMTase) и ацетилтрансферазами (HAT), соответственно. Среди кандидатных генов, которые могут участвовать в регуляции эпигенетических изменений в PGC, имеются HMTase, которые принадлежат к семейству белков с консервативным SET/PR доменом. Проводят анализ экспрессии двадцати пяти генов, содержащих домен SET/PR, в Е7.5 PGC и в соседних соматических клетках и обнаруживают, что Blimp1, G9a, Set1, Ezh2 и Pfm1 экспрессируются в эмбриональной области, содержащей PGC (Фиг.1b и Фиг.7). Однако, только экспрессия Blimp1 ограничивается PGC в Е7.5, и его экспрессия сохраняется затем в зародышевых клетках.
Для изучения активности домена Blimp1 SET/PR сначала устанавливают, что меченный Myc мышиный Blimp1, после транзиторной экспрессии в клетках 293Т, можно подвергнуть эффективной иммунопреципитации, используя антитела против Myc (Фиг.2а). Затем иммунопреципитат можно подвергнуть стандартному радиоактивному анализу гистон-метилтрансферазной активности по гистону Н3 (Rea, S. et al. (2000) Nature 406, 593-9). Наблюдают сравнительно слабый сигнал, соответствующий Н3, но неожиданно также обнаруживают заметные полосы, соответствующие гистонам Н2А и Н4 (Фиг.2b), причем, по определению Вестерн-блоттингом, последняя присутствует в виде небольшой примеси (низкие уровни) в препарате Н3 (данные не приводятся). Был сделан вывод, что слабый сигнал на Н3 может быть вызван Blimp1-ассоциированным G9a, как сообщалось ранее для В клеток (Gyory et al., см. выше). Однако, зародышевые клетки не показывают заметного диметилирования лизина 9 в гистоне Н3 (Н3K9me2), которое является основной модификацией, приписываемой G9a, и на спецификацию PGC не оказывает значительного влияния утрата G9a функции. Поэтому было решено сосредоточиться на наблюдаемой активности получаемого иммунопреципитацией Blimp1 на Н2А и Н4 гистонах.
Ввиду новизны открытия метилирования "хвоста" Н2А было решено сначала проверить метилтрансферазную активность иммунопреципитата на препаратах тимуса теленка и рекомбинантного Н2А, и было найдено, что он обладает сильной метилирующей активностью к этому гистону (Фиг.2b). Для идентификации целевого (целевых) аминокислотного(ых) остатка(ов) на Н2А "хвосте"меченный радиоактивной меткой рекомбинантный белковый продукт анализа метилтрансферазной активности подвергают ступенчатой деградации по Эдману. Подсчетом сцинтилляций во фракции освобожденных аминокислот обнаруживают радиоактивное мечение rH2A R3 (Фиг.2c). Те же самые результаты получают на препаратах тимуса теленка и рекомбинантного Н4 (данные не показаны). Это логичный результат, если принять во внимание сохранение аминокислотной последовательности первых N-концевых остатков гистонов Н2А и Н4 (Фиг.2d). Известно, что метилирование Н4 R3 играет важную роль в регуляции транскрипции. Однако, эти результаты предсказывают существование дополнительного нового метилирования R3 на гистоне Н2А.
Пример 2: Идентификация комплекса Blimp1/Prmt5
Так как SET/PR домены ассоциируются с гистон-метилтрансферазной активностью только на лизиновых остатках, было резонно предположить, что аргинин-метилирующая активность, обнаруженная выше, могла быть вызвана не Blimp1, a должна быть связана с другой HMTase, присутствующей в иммунопреципитате. Ранее сообщалось, что два белка аргинин-метилтрансферазы, Prmt1 и Prmt5, опосредуют метилирование гистона Н4 R3. Prmt1 представляет собой класса I аргинин-метилтрансферазу, приводящую в результате к NG-монометиларгинину (Rme1) и асимметричному NG,N'G-диметиларгинину (Rme2a) на различных типах субстратов. Как указывалось выше, Prmt5 относится к класса II аргинин-метилтрансферазе, которая отвечает за монометилирование аргинина (Rme1) и симметричное NG,N'G-диметилирование (Rme2s) (Фиг.8) 20. Чтобы проверить, действительно ли эти белки могут ассоциироваться с Blimp1 in vivo, их экспрессию анализируют в образующихся PGC и соседних соматических клетках, используя библиотеки кДНК одиночных клеток, описанные выше. Найдено, что Prmt1 исключается из PGC в Е7.5, тогда как Prmt5 присутствует как в PGC, так и в соматических клетках (данные не показаны). Однако, на уровне белка Prmt5 показывает ядерное окрашивание и становится высокообогащенным PGC по сравнению с соматическими клетками от Е8.5 и далее (Фиг.3а, b; Фиг.S1b и см. ниже). С другой стороны, на этих стадиях Prmt1 обнаруживается, главным образом, в цитоплазме зародышевых клеток (Фиг.3c).
Затем решено было проверить, какой именно фермент, Prmt5 или Prmt1, взаимодействует Blimp1. Действительно, найдено, что меченный Myc Blimp1 может подвергаться эффективной совместной иммунопреципитации с эндогенным Prmt5 в 293Т клетках (Фиг.3d). Верно и обратное, Prmt5 может также тянуть за собой меченный Myc Blimp1 (Фиг.3d). В отличие от этого нельзя обнаружить никакого взаимодействия между меченным Myc Blimp1 и эндогенным Prmt1 (Fig.3e). Эти эксперименты подтверждают, что Blimp1 и Prmt5 могут образовывать комплекс в 293Т клетках. Принимая во внимание перекрывающуюся экспрессию этих белков в PGC (Фиг.3а, b; Фиг.S1b), логично представить, что Blimp1 и Prmt5 являются частями одного и того же белкового комплекса в PGC и что этот комплекс может встречаться где-либо в другом месте в процессе развития или в нормальных, а также опухолевых взрослых тканях.
Пример 3: Активность комплекса Blimp1/Prmt5 in vivo
Проверяют специфичность антитела к H4R3me2s и показывают, что оно эффективно узнает оба гистона Н2А и Н4 из тимуса теленка (Фиг.8b, левый рисунок, Н2А и Н4 присутствуют также в качестве примесей в препаратах Н3 и Н2В). В конкурентном анализе это антитело эффективно титруется Н4 (1-9С) пептидом, содержащим R3me2s (Фиг.S2c). Кроме того, это антитело также распознает продукт Blimp1/Prmt5 HMTase анализа как на Н4, так и на Н2А (Фиг.52b, правый рисунок), тем самым давая дополнительное доказательство того, что комплексу присуща именно симметричная диметилирующая активность Prmt5, а не асимметричная диметилирующая активность Prmt1.
При использовании антитела, специфического к модификации H2/H4R3me2s, PGC, выделенные из ранних эмбрионов, анализируют иммуногистохимическими методами. В Е8.5 H2A/H4R3me2s явно наблюдается в PGC и соматических клетках (Фиг.3f), но в Е10.5 значительно более высокая аккумуляция Н2А/Н4 R3me2s наблюдается преимущественно в зародышевых клетках (Фиг.3f). Однако, когда используют другое антитело, которое распознает моно- и/или асимметричное диметилирование H4R3 (H4R3me1 и H4R3me2a), наблюдается, что эта модификация в PGC находится в Е8.5, но не в Е10.5 (данные не приведены). Совместные результаты с этими двумя антителами показывают рост H2A/H4R3me2s в PGC от Е8.5 к Е10.5. Эти результаты демонстрируют наличие специфического характерного свойства хроматина в ходе развития зародышевых клеток, которое, как полагают, определяется совместным присутствием Prmt5 и Blimp1. Важно отметить, что существует множество дополнительных модификаций "хвостов" гистонов, которые вносят вклад в специфическую для зародышевых клеток хроматиновую структуру. Недавно было показано, что утрата функции Blimp1 ведет к неправильному (аберрантному) развитию PGC-подобных клеток-основателей, которые прекращают пролиферировать (Ohinata, Y. et al. (2005) Nature 5, 5).
Пример 4: Идентификация in vivo мишеней комплекса Blimp1/Prmt5
Как было показано ранее, Blimp1 может направлять генную регуляцию в процессе дифференцировки клеток через рекрутинг (привлечение, пополнение) факторов взаимодействия к специфическим сайтам. Для идентификации предполагаемых мишеней Blimp1 применяют метод клонирования с хроматин-идентификацией, в котором меченный Myc вариант Blimp1 сначала сверхэкспрессируют в 293Т клетках. Затем проводят иммунопреципитацию ядерных экстрактов из этих клеток 293Т антителами к Myc, а иммунопреципитированную ДНК экстрагируют, очищают, сшивают по тупым концам с помощью линкеров и ПЦР-амплифицируют и клонируют. Несколько клонов отбирают и анализируют секвенированием с последующим BLAST анализом для картирования клонированных инсертов. Из 32 клонов 11 соответствует областям, локализованным вблизи регуляторных последовательностей или в подходящих регуляторных последовательностях известных генов (см. Таблицу 1).
Пример 5: Исследование контроля экспрессии гена Dhx38 с применением комплекса Blimp1/Prmt5
Среди возможных идентифицированных мишеней Blimp1 для изучения выбирают Dhx38. Dhx38 представляет собой консервативный ген, кодирующий содержащую DEAH-бокс РНК-геликазу (также известную как Prp16), которая необходима в C.elegans для посттрансляционной регуляции пол-детерминирующих генов при переключении сперматозоидов на ооциты (Graham, P. L. & Kimble, J. (1993). Genetics 133, 919-31). При детальном изучении Dhx38 локуса обнаружено, что он содержит четыре мотива GGGAAAG, соответствующих консенсусному сайту связывания Blimp1: два в 5' области и два в 3' (downstream) области старт-сайта транскрипции (Фиг.4а). Хотя имеющееся в настоящее время антитело против Blimp1 работает в методах иммуноокрашивания, его нельзя применять для эффективной иммунопреципитации Blimp1 (данные не показаны). Поэтому используют Prmt5, чтобы выяснить, является ли Dhx38 мишенью для комплекса Blimp1/Prmt5 в зародышевых клетках. ChIP анализ с применением Prmt5 антител проводят на PGC, содержащихся в суспензиях клеток половых бугорков из Е10.5 эмбрионов, фазы, когда Blimp1 и Prmt5 коэкспрессируются в ядрах зародышевых клеток (см. выше). Действительно, найдено, что Prmt5 могут специфически оттягивать нуклеотиды, перекрывающие последовательность от +7152 до +7541, охватывающую экзон 11 гена Dhx38, который содержит Blimp1 консенсус-связывающий сайт (Фиг.4b). Другие три предполагаемых сайта связывания не ассоциируются с Prmt5 в нашем анализе ChIP. Эти результаты показывают, что Prmt5 рекрутируются в мишени Blimp1, такие как Dhx38, тем самым наводя на мысль, что комплекс Blimp1/Prmt5 регулирует экспрессию таких целевых генов в зародышевых клетках.
Экспрессию/репрессию Dhx38 в PGC рассматривают с учетом того, что Dhx38 является мишенью комплекса Blimp1/Prmt5 в зародышевых клетках. Обнаружено, что Dhx38 не детектируется в Е10.5 и Е11.5 (Фиг.5а). Однако, к Е12.5 Dhx38 активируется как в женских, так и в мужских PGC (Фиг.5а), что поразительным образом существует параллельно с делокализацией Prmt5 и Вlimp1 из ядра в цитоплазму в зародышевых клетках B Е11.5 (Фиг.5b). В Е12.5 как Prmt5, так и Blimp1 отсутствуют в ядрах PGC (данные не показаны). Активация Dhx38 непосредственно предшествует блокаде мейоза и митоза женских и мужских зародышевых клеток, соответственно. Следовательно, существует обратная связь между экспрессией Blimp1/Prmt5 и Dhx38. Эти результаты показывают, что Blimp1 и Prmt5 могут функционировать при репрессии транскрипции генов-мишеней, таких как Dhx38, в клетках зародышевой линии. Эта репрессия, по-видимому, снимается, когда зародышевые клетки входят в половой бугорок и как Blimp1, так и Prmt5 претерпевают транслокацию из ядра в цитоплазму. Следует отметить, что метилирование аргинина в гистоне ассоциируется в основном с активацией транскрипции, хотя недавно ассоциированную с Prmt5 активность H3R8me связывали с пониженной экспрессией гена (Pal S. et al., см. выше). Далее, так как ранее сообщалось, что и Blimp1, и Prmt5 являются репрессорами транскрипции, возможно, что симметричное диметилирование аргининового остатка, ассоциированное с комплексом Blimp1/Prmt5 в зародышевых клетках, представленное в данном описании, будет способствовать репрессии генов.
Пример 6: Комплекс Blimp1-Prmt5 в плюрипотентных стволовых клетках
Чтобы лучше понять роль комплекса Blimp1/Prmt5, было решено изучить плюрипотентные эмбриональные зародышевые (EG) клетки. EG можно получать из выделенных PGC in vitro и, следовательно, их можно рассматривать как ближайшую линию клеток, эквивалентную PGC. Найдено, что EG клетки являются позитивными по отношению к Prmt5, но в отличие от PGC, в них отсутствует Blimp1 (Фиг.6а, b). В соответствии с этими результатами было найдено, что EG клетки также позитивны в отношении Dhx38, это наводит на мысль, что экспрессия этого гена может быть вызвана отсутствием Blimp1 (Фиг.6а, b). Для восстановления комплекса репрессии Blimp1/Prmt5 меченный Myc Blimp1 сверхэкспрессируют в EG клетках. Однако, эти попытки приводят к сильной цитотоксичности уже через 12 ч трансфекции клеток, тогда как жизнеспособность трансфецированных при использовании pCMV-Myc контрольных EG клеток не нарушается (данные не показаны). Аналогичные результаты получены также с плюрипотентными эмбриональными стволовыми (ES) клетками (данные не приводятся).
Выбирают плюрипотентные клетки эмбриональной карциномы (ЕС) мыши линии Р19, которые также имеют признаки плюрипотентности, аналогичные клеткам EG и ES cells. Действительно, в клетках Р19, подобно клеткам ES/EG, наблюдается экспрессия Prmt5 и Dhx38, но не Blimp1 (Фиг.6b, с). Найдено, однако, что эти клетки толерантны к экспрессии Blimp1, и поэтому используют преципитацию, которая действительно подтвердила, что сверхэкспрессированный Myc-Blimp1 взаимодействует с Prmt5 в мышиных ЕС клетках (Фиг.6c). Следует отметить, что этот неустойчивый (временный) комплекс Blimp1/Prmt5 затем вызывает даун-регуляцию Dhx38 в клетках Р19 (Фиг.6c). Анализ ChIP подтверждает, что эта даун-регуляция Dhx38 сопровождается повышенными уровнями H2A/H4R3me2s в Dhx38 локусе (Фиг.6d). Эти результаты подтверждают мнение, что комплекс Blimp1/Prmt5 отвечает за репрессию генов-мишеней, таких как Dhx38, которые также вероятны в PGC.
Хотя конкретные варианты изобретения подробно раскрываются в данном описании, это делается с помощью примеров и только с целью иллюстрации. Вышеприведенные варианты изобретения не предполагают ограничивать объем прилагаемой формулы изобретения, представленной ниже. Заявители полагают, что различные замены, изменения и модификации можно вносить в изобретение, не отступая от сущности и объема изобретения в соответствии с формулой изобретения.



Изобретение относится к области биотехнологии и молекулярной генетики. Выделенный полипептидный комплекс участвует в эпигенетическом контроле экспрессии генов у млекопитающего. При этом комплекс содержит белок Blimp 1 или его гомолог PRD1, имеющий сайт-специфическую ДНК-связывающую активность, и белок PRMT5. Комплекс может регулировать экспрессию генов в клетках, в частности в стволовых клетках млекопитающих, за счет контроля метилирования R3 в С-концевых областях гистонов Н2А и Н4. 3 н. и 8 з.п. ф-лы, 8 ил., 1 табл., 5 пр.
1. Выделенный полипептидный комплекс, который участвует в эпигенетическом контроле экспрессии генов у млекопитающего, содержащий белок Blimpl или его гомолог PRD1, имеющий сайт-специфическую ДНК-связывающую активность, и белок PRMT5, имеющий аргинин-метилтрансферазную активность, причем белок PRMT5 способен метилировать аргининовый остаток, локализованный в положении 3 в N-концевой области гистона Н2А.
2. Способ контроля экспрессии генов в клетке млекопитающего, заключающийся в индукции образования в клетке полипептидного комплекса, который участвует в эпигенетическом контроле экспрессии генов у млекопитающего, содержащего белок Blimpl или его гомолог PRD1, имеющий сайт-специфическую ДНК-связывающую активность, и белок PRMT5, имеющий аргинин-метилтрансферазную активность, причем белок PRMT5 способен метилировать аргининовый остаток, локализованный в положении 3 в N-концевой области гистона Н2А, причем контроль экспрессии генов приводит к контролю экспрессии одного или более генов, содержащихся в группе, состоящей из: с-Мус; Dhx38; Pcdh7; Q8C9T7; Xylt1; DnaH1; Baip2; Nek7; Dusp2; ENSMUSG00000027041; Sirt4; и Blimp1.
3. Способ по п.2, отличающийся тем, что образование полипептидного комплекса индуцируют в клетке индукцией экспрессии нуклеиновой кислоты, кодирующей полипептид PRDI/Blimp1, в клетке.
4. Способ по п.3, отличающийся тем, что экспрессию нуклеиновой кислоты, кодирующей полипептид PRDI/Blimp1, индуцируют в клетке трансфекцией клетки экспрессирующим вектором, который кодирует полипептид Blimp1.
5. Способ по п.2, отличающийся тем, что образование полипептидного комплекса индуцируют в клетке трансфекцией клетки нуклеотидным экспрессирующим вектором, содержащим:
одну или более кодирующую последовательность, функционально связанную с промоторной последовательностью, причем одна или более кодирующих последовательностей кодируют, по меньшей мере, первый полипептид, включающий белок Blimp1 или его гомолог PRD1, имеющий сайт-специфическую ДНК-связывающую активность, и, по меньшей мере, второй полипептид, включающий белок PRJV1T5, имеющий аргинин-метилтрансферазную активность.
6. Способ по любому из пп.2-5, отличающийся тем, что клетка млекопитающего представляет собой человеческую клетку.
7. Способ по любому из пп.2-6, отличающийся тем, что клетка млекопитающего представляет собой опухолевую или раковую клетку.
8. Способ по любому из пп.2-7, отличающийся тем, что способ осуществляют in vitro.
9. Способ по любому из пп.2-7, отличающийся тем, что способ осуществляют in vivo.
10. Способ по любому из пп.2-9, отличающийся тем, что индукция полипептидного комплекса в клетке приводит к снижению экспрессии одного или более генов, представленных в п.2.
11. Способ идентификации молекулы, способной оказывать антагонистическое действие на биологическую активность полипептидного комплекса по п.1, включающий определение того, демонстрирует ли соединение-кандидат связывающее взаимодействие с PRDI/Blimp1, PRMT5 и/или комплексом PRDI/Blimp-PRMT5, и идентификацию такого соединения в качестве молекулы, взаимодействующей с комплексом PRDI/Blimp1-PRMT5, отличающийся тем, что соединение, которое связывается с PRDI/Blimp1, PRMT5 и/или комплексом PRDI/Blimp-PRMT5 и которое ингибирует биологическую активность или функцию комплекса PRDI/Blimp1-PRMT5 по п.1, идентифицируют в качестве антагонистической молекулы.
| WO 2005060722 (PRESIDENT AND FELLOWS OF HARDV; GLIMCHER LH; MOWEN K) Garcia JF et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Fabbrizio E et al | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| EMBO Rep | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Немцова M.B | |||
| Нарушения | |||

