Настоящее изобретение относится к способу получения гликопротеинов в среде для культивирования клеток, которая содержит марганец, и к среде для культивирования клеток для применения согласно такому способу.
ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка находится одновременно на рассмотрении, имеет по меньшей мере одного общего автора и испрашивает приоритет в соответствии с предварительной заявкой на патент США №60/830,658, поданной 13 июля 2006 года, содержание которой включено в данную заявку посредством ссылки в полном объеме.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Белки и полипептиды становятся все более важными терапевтическими агентами. В большинстве случаев такие белки и полипептиды получают в культуре клеток из клеток, продуцирующих необычно высокое количество конкретного представляющего интерес белка или полипептида, что достигают с помощью генной инженерии и/или селекции. Контроль и оптимизация условий культивирования клеток важны для успешного коммерческого производства белков и полипептидов.
Многие белки и полипептиды, полученные в культуре клеток, представляют собой гликопротеины, содержащие ковалентно присоединенные углеводородные структуры, включающие олигосахаридные цепи. Такие олигосахаридные цепи присоединяются к белку в эндоплазматическом ретикулуме и аппарате Гольджи посредством присоединения по атому азота или кислорода (N-присоединение или О-присоединение соответственно). Олигосахаридные цепи могут составлять значительную часть массы гликопротеина. Считают, что олигосахаридные цепи играют ключевую роль в функционировании гликопротеина, в том числе облегчают правильную укладку гликопротеина, опосредуют белок-белковые взаимодействия, сообщают стабильность, фармакодинамические и/или фармакокинетические свойства, ингибируют протеолитическое расщепление, направляют гликопротеин на соответствующий секреторный путь и способствуют попаданию гликопротеина в конкретный орган или органы.
Обычно N-присоединенные олигосахаридные цепи присоединяются к образующемуся транслоцируемому белку в просвете эндоплазматического ретикулума (см. Molecular Biology of the Cell, Alberts et al., 1994, включенную в данную заявку посредством ссылки). Олигосахариды присоединяются к аминогруппе на боковой цепи остатка аспарагина, находящегося в составе консенсусной последовательности-мишени Asn-X-Ser/Thr, где X может представлять собой любую аминокислоту, кроме пролина. Исходная олигосахаридная цепь обычно укорачивается специфическими ферментами гликозидазами в эндоплазматическом ретикулуме, в результате чего образуется короткий разветвленный коровый олигосахарид из двух остатков N-ацетилглюкозамина и трех остатков маннозы.
После начальных этапов процессинга в эндоплазматическом ретикулуме гликопротеин транспортируется через малые везикулы в аппарат Гольджи, где олигосахаридная цепь подвергается дальнейшему процессингу перед секрецией на поверхность клетки. Указанная укороченная N-присоединенная олигосахаридная цепь может быть модифицирована добавлением нескольких остатков маннозы, в результате чего образуется олигосахарид с высоким содержанием маннозы. Кроме того, к коровым субъединицам маннозы может быть присоединен один или более мономер моносахарида N-ацетилглюкозамина с образованием сложных полисахаридов. К субъединицам N-ацетилглюкозамина может быть присоединена галактоза, а к субъединицам галактозы могут быть присоединены субъединицы сиаловой кислоты с образованием цепей, содержащих на конце либо сиаловую кислоту, либо галактозу, либо N-ацетилглюкозамин. Кроме того, к остатку N-ацетилглюкозамина корового олигосахарида может быть присоединен остаток фукозы. Каждая такая реакция присоединения катализируется специфичной гликозилтрансферазой.
Помимо модификации путем N-гликозилирования, гликопротеины могут также быть модифицированы путем присоединения О-присоединенных олигосахаридных цепей к конкретным остаткам серина или треонина в ходе процессинга в аппарате Гольджи. Присоединение остатков О-присоединенного олигосахарида происходит поочередно, при этом присоединение каждого остатка катализируется специфическим ферментом. В отличие от N-гликозилирования консенсусная последовательность аминокислот для О-гликозилирования является менее определенной.
На конечный рисунок и степень гликозилирования белка могут оказывать существенное влияние условия культивирования клеток. Например, при стандартном периодическом культивировании и культивировании с подпиткой обращают основное внимание на конечный уровень продуцируемого пептида, но при этом часто образующийся гликопротеин характеризуется меньшей степенью гликозилирования и/или характером гликозилирования, при котором остатки сахара олигосахаридов слабо соответствуют остаткам сахаров, присутствующим в природной форме гликопротеина. Повышение степени гликозилирования и/или изменение состава сахарных остатков для более точного соответствия уровню и составу гликозилирования, которые присутствуют в природной форме гликопротеина, могло бы потенциально способствовать получению терапевтического агента на основе гликопротеинов с большей активностью, улучшенными фармакодинамическими и/или фармокинетическими свойствами и меньшими побочными эффектами. Хотя были предприняты некоторые попытки улучшения качественных и количественных показателей гликозилирования гликопротеинов, полученных в культуре клеток, остается потребность в дополнительных усовершенствованиях. В частности, существует потребность в разработке систем для получения гликопротеинов с улучшенными характером гликозилирования в культуре клеток в среде определенным составом.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Способы и композиции согласно настоящему изобретению обеспечивают улучшенную систему для крупномасштабного получения гликопротеинов с улучшенными характеристиками гликозилирования в культуре клеток. Например, в конкретных вариантах реализации настоящее изобретение обеспечивает способы культивирования в промышленных масштабах (например, 500 л или более), в которых применяют среду, содержащую суммарную молярную концентрацию марганца приблизительно 10-600 нМ. В конкретных вариантах реализации суммарная молярная концентрация глутамина в среде составляет менее чем приблизительно 8 мМ. В некоторых вариантах реализации настоящего изобретения суммарная молярная концентрация глутамина в среде составляет менее чем приблизительно 4 мМ. Следует понимать, что термин «суммарный» при использовании в данной заявке относится к общему количеству конкретного компонента или компонентов, добавленных в процессе культивирования клеток, включая компоненты, добавленные в начале культивирования, и компоненты, добавляемые впоследствии. В конкретных вариантах реализации изобретения желательно минимизировать «подпитку» культуры с течением времени, при этом желательно максимально увеличить содержание питательных веществ в начале культивирования. Очевидно, компоненты в среде метаболизируются в процессе культивирования таким образом, что в клеточных культурах с одинаковым суммарным количеством данных компонентов будут наблюдаться различные абсолютные уровни, если указанные компоненты добавляют в различные моменты времени (например, все компоненты присутствуют на начальном этапе или же некоторые компоненты добавляют при подпитке).
Согласно настоящему изобретению применение такой питательной среды позволяет получать гликопротеины с желательным характером гликозилирования. В некоторых вариантах реализации настоящего изобретения гликопротеины могут иметь большую степень гликозилирования и/или могут иметь распределение олигосахаридных цепей, которое в большей степени соответствует распределению олигосахаридных цепей, присоединенных к гликопротеину, в природной клетке-хозяине. В некоторых вариантах реализации результатом применения системы согласно настоящему изобретению может быть продукция гликопротеина, который имеет характер гликозилирования, схожий или идентичный характеру гликозилирования, который наблюдался бы при экспрессии указанного гликопротеина эндогенной клеткой человека.
Как очевидно специалисту в данной области, состав среды согласно настоящему изобретению охватывает как среды с определенным составом, так и сложные среды. В конкретных вариантах реализации настоящего изобретения культуральная среда представляет собой среду с определенным составом, состав которой известен и контролируется.
В некоторых вариантах реализации клетки выращивают при одном или более варианте условий, описанных в предварительной заявке на патент США №60/605,097, включенной в данную заявку посредством ссылки. В некоторых вариантах реализации клетки выращивают при одном или более варианте условий, описанных в предварительной заявке на патент США №11/213,308, включенной в данную заявку посредством ссылки.
В культуры клеток согласно настоящему изобретению можно добавлять питательные вещества и/или другие компоненты среды, в том числе гормоны и/или другие факторы роста, определенные ионы (например, натрий, хлорид, кальций, магний, и фосфат), буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы (неорганические соединения, обычно присутствующие в очень низкой конечной концентрации), аминокислоты, липиды или глюкоза или другие источники энергии. В конкретных вариантах реализации настоящего изобретения может потребоваться добавление в среду химических индукторов, таких как гексаметиленбис(ацетамид) ("НМВА") и бутират натрия ("NaB"). Такие вещества можно добавлять в начале культивирования или можно добавлять на последующих этапах для пополнения истощенных запасов питательных веществ или для другой цели. В целом, желательно выбрать исходный состав среды таким образом, чтобы минимизировать добавление веществ в ходе культивирования в соответствии с настоящим изобретением.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фигуре 1 показано исследование гликозидазной активности в ретентате ультрафильтрации/диафильтрации (УФ/ДФ). Для каждого эксперимента приведены данные для различных вариантов К4 и К4', столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4′ (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 2 показано распределение вариантов К4 в rFIX, полученных в культурах, выращенных в колбах со встряхиванием. Для каждого эксперимента приведены данные для различных вариантов К4 и К4′, столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4′ (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 3 показано распределение вариантов К4 из культур, выращенных в колбах со встряхиванием с добавление различных компонентов в среду. Для каждого эксперимента приведены данные для различных вариантов К4 и К4′, столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4' (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 4 показано распределение вариантов К4 из культур, выращенных в колбах со встряхиванием в среде с добавками. Для каждого эксперимента приведены данные для различных вариантов К4 и К4′, столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4′ (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 5 показано распределение образцов К4 из культур, выращенных в колбах со встряхиванием с добавлением различных компонентов в среду. Для каждого эксперимента приведены данные для различных вариантов К4 и К4′, столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4′ (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 6 показано распределение вариантов К4 из культур, выращенных в колбах со встряхиванием с различным содержанием марганца. Для каждого эксперимента приведены данные для различных вариантов К4 и К4′, столбцы слева направо: К4 (Fuc-GlcNAc-Gal-SA), К4′ (Fuc-GlcNAc-Gal), К4′ (Fuc-GlcNAc) и К4′ (Fuc).
На Фигуре 7 представлено графическое сравнение процента общей площади пиков для G0, G1 и G2, определенных методом HPAEC-PED. Для каждого эксперимента приведены данные для сложных N-присоединенных разветвленных (biantennary) гликанов, столбцы слева направо: G0, G1 и G2.
На Фигуре 8 представлено графическое сравнение процента общей площади пиков для G0, G1 и G2, определенных методом HPAEC-PED. Для каждого эксперимента приведены данные для сложных N-присоединенных разветвленных гликанов, столбцы слева направо: G0, G1 и G2.
ОПРЕДЕЛЕНИЯ
«Аминокислота»: термин «аминокислота» при использовании в данной заявке относится к любой из двадцати природных аминокислот, которые в норме используются при образовании полипептидов, или аналогам или производным таких аминокислот, или любым неприродным аминокислотам. Аминокислоты согласно настоящему изобретению содержатся в среде для культивирования клеток. Аминокислоты, содержащиеся в среде, могут быть представлены в виде солей или в форме гидратов.
«Антитело»: термин «антитело» при использовании в данной заявке относится к молекуле иммуноглобулина или иммунологически активной части молекулы иммуноглобулина, т.е. молекуле, содержащей антигенсвязывающий сайт, специфично связывающийся с антигеном, например, Fab- или F(ab′)2 фрагменту. В некоторых вариантах реализации настоящего изобретения антитело представляет собой типичное природное антитело, известное специалистам в данной области, например гликопротеин, включающий четыре полипептидные цепи: две тяжелые цепи и две легкие цепи. В конкретных вариантах реализации антитело представляет собой одноцепочечное антитело. Например, в некоторых вариантах реализации настоящего изобретения одноцепочечное антитело включает вариант типичного природного антитела, в котором два или более компонента тяжелой и/или легкой цепей ковалентно связаны, например, посредством пептидной связи. В конкретных вариантах реализации одноцепочечное антитело представляет собой белок, имеющий структуру из двух полипептидных цепей, состоящую из тяжелой и легкой цепи, которые стабилизированы, например, внутрицепочечным линкерным пептидом, при этом такой белок обладает способностью специфично связываться с антигеном. В конкретных вариантах реализации настоящего изобретения антитело представляет собой антитело, которое содержит только тяжелые цепи, например, встречающиеся у представителей семейства Camelidae, включающего лам и верблюдов (см., например, патенты США №№6,765,087, Casterman et al., 6,015,695 Casterman et al., и 6,005,079 Casterman et al., включенные в данное описание посредством ссылки в полном объеме). Термины «моноклональные антитела» и «композиции моноклональных антител» при использовании в данной заявке обозначают популяцию молекул антител, которые содержат только один вид антигенсвязывающего сайта и, таким образом, обычно взаимодействуют только с отдельным эпитопом или конкретным антигеном. Композиции моноклональных антител, таким образом, демонстрируют одинаковую аффинность к конкретному эпитопу, с которым они связываются. Термины «поликлональные антитела» и «композиции поликлональных антител» относятся к популяциям молекул антитела, содержащим различные виды антигенсвязывающих сайтов, которые взаимодействуют с конкретным антигеном.
«Периодическое культивирование»: термин «периодическое культивирование» (butch culture) при использовании в данной заявке относится к способу культивирования клеток, при котором все компоненты, которые в конечном счете будут использованы при культивировании клеток, включая среду (см. определение «среды» ниже), а также сами клетки, обеспечивают в начале процесса культивирования. Периодическое культивирование, как правило, прекращают в некоторый момент времени и собирают клетки и/или компоненты в среде, после чего их могут подвергать очистке.
«Биореактор»: термин «биореактор» при использовании в данной заявке относится к любому сосуду, используемому для выращивания культуры клеток млекопитающего. Биореактор может быть любого размера, при условии, что его можно применять для культивирования клеток млекопитающего. Обычно такой биореактор имеет объем по меньшей мере 1 литр и может иметь объем 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000 литров или более или любой объем между данными значениями. Внутренние условия биореактора, включающие, но не ограниченные указанными параметрами: рН, растворенный кислород и температуру, обычно регулируют в ходе культивирования. Биореактор может быть изготовлен из любого материала, подходящего для содержания культуры клеток млекопитающего, ресуспендированных в среде при условиях культивирования согласно настоящему изобретению, в том числе из стекла, пластмассы или металла. Термин «промышленный биореактор» при использовании в данной заявке относится к окончательному варианту биореактора, используемого при производстве интересующего гликопротеина. Объем промышленного биореактора обычно составляет по меньшей мере 500 литров и может составлять 1000, 2500, 5000, 8000, 10000, 12000 литров или более или может равняться любому значению между указанными. Специалисту в данной области известны, и он способен выбрать подходящие биореакторы для применения при реализации настоящего изобретения на практике.
«Плотность клеток»: термин «плотность клеток» при использовании в данной заявке относится к количеству клеток, присутствующих в данном объеме в среде.
«Жизнеспособность клеток»: термин «жизнеспособность клеток» при использовании в данной заявке относится к способности клеток в культуре выживать при данных условиях культивирования или вариантах эксперимента. Данный термин при использовании в данной заявке также относится к доле клеток, жизнеспособных в конкретный момент времени относительно общего числа клеток, живых и мертвых, присутствующих в культуре в конкретный момент времени.
«Сложная среда»: термин «сложная среда» при использовании в данной заявке относится к среде, содержащей по меньшей мере один компонент, природа которого или количество либо не известно, либо не контролируется.
«Культура», «культура клеток»: данные термины при использовании в данной заявке относятся к популяции клеток, ресуспендированных в среде (см. определение «среды» ниже) при условиях, подходящих для выживания и/или роста популяции клеток. Как очевидно специалистам в данной области, в конкретных вариантах реализации данные термины при использовании в данной заявке относятся к совокупности популяции клеток и среды, в которой ресуспендирована популяция клеток. В конкретных вариантах реализации клетки в составе популяции клеток включают клетки млекопитающего.
«Среда с определенным составом»: термин «среда с определенным составом» при использовании в данной заявке относится к среде, состав которой известен и контролируется.
«Культура с подпиткой»: термин «культура с подпиткой» при использовании в данной заявке относится к способу культивирования клеток, при котором дополнительные компоненты добавляют в культуру в один или несколько моментов времени или после начала процесса культивирования. Такие дополнительные компоненты обычно включают питательные компоненты для клеток, истощенные в процессе культивирования. В качестве дополнения или альтернативы, такие дополнительные компоненты могут включать вспомогательные компоненты (см. определение «вспомогательных компонентов» ниже). В конкретных вариантах реализации дополнительные компоненты добавляют в питательную среду (см. определение «питательной среды» ниже). Культивирование с подпиткой обычно прекращают в некоторый момент времени и собирают клетки и/или компоненты в среде, после чего их можно подвергнуть очистке.
«Питательная среда»: термин «питательная среда» при использовании в данной заявке относится к раствору, содержащему питательные вещества, которые питают растущие клетки млекопитающего, который добавляют после начала культивирования клеток. Питательная среда может содержать компоненты, идентичные компонентам, содержащимся в исходной среде для культивирования клеток. Кроме того, питательная среда может содержать один или более дополнительный компонент, помимо компонентов, содержащихся в исходной среде для культивирования клеток. Кроме того или в качестве альтернативы, в питательной среде может отсутствовать один или более компонент, содержащийся в исходной среде для культивирования клеток. В некоторых вариантах реализации один или более компонент питательной среды присутствует в той же или схожей концентрации или на том же или схожем уровне, что и в исходной питательной среде. В некоторых вариантах реализации настоящего изобретения один или более компонент питательной среды присутствует в концентрации или на уровне, отличающемся от концентрации или уровня данного компонента в исходной среде для культивирования клеток. Примеры питательных сред приведены в Таблице 2, хотя настоящее изобретение не ограничивается применением данных сред. Специалисту в данной области очевидно, что можно использовать альтернативные питательные среды и/или что состав питательных сред, приведенных в качестве примеров в Таблице 2, можно варьировать. В конкретных вариантах реализации питательная среда содержит вспомогательные компоненты (см. определение «вспомогательных компонентов» ниже).
«Фрагмент»: термин «фрагмент» при использовании в данной заявке относится к полипептиду, определяемому как любая дискретная часть данного полипептида, которая является уникальной или характерной для данного полипептида. Например, данный термин при использовании в данной заявке относится к любой части данного полипептида, которая включает по меньшей мере часть с установленной последовательностью, имеющуюся в составе полноразмерного полипептида. Для определенных фрагментов указанная часть последовательности составляет по меньшей мере 4-5, 10, 15, 20, 25, 30, 35, 40, 45, 50 или более аминокислот полноразмерного полипептида. Кроме того или в качестве альтернативы, указанный термин при использовании в данной заявке относится к любой дискретной части данного полипептида, которая сохраняет по меньшей мере часть по меньшей мере одной активности полноразмерного полипептида. В конкретных вариантах реализации доля сохраняющейся активности составляет по меньшей мере 10% от активности полноразмерного полипептида. В конкретных вариантах реализации доля сохраняющейся активности составляет по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% от активности полноразмерного полипептида. В конкретных вариантах реализации доля сохраняющейся активности составляет по меньшей мере 95%, 96%, 97%, 98% или 99% от активности полноразмерного полипептида. В конкретных вариантах реализации настоящего изобретения фрагмент сохраняет 100% активности полноразмерного полипептида. В конкретных вариантах реализации фрагмент согласно настоящему изобретению содержит последовательность пептида, которая служит в качестве сайта гликозилирования. В некоторых вариантах реализации фрагмент согласно настоящему изобретению содержит часть сайта гликозилирования, таким образом, что, когда он соединяется с другим фрагментом, содержащим другую часть сайта гликозилирования, происходит восстановление функционального сайта гликозилирования.
«Ген»: термин «ген» при использовании в данной заявке относится к любой нуклеотидной последовательности, ДНК или РНК, по меньшей мере некоторая часть которой кодирует дискретный конечный продукт, который, как правило, представляет собой, но не ограничивается им, полипептид, функционирующий в некотором аспекте метаболизма или развития клетки. Ген может включать не только кодирующую последовательность, которая кодирует полипептид или другой дискретный конечный продукт, но также включает области, предшествующие кодирующей последовательности и/или следующие за ней, которые модулируют базовый уровень экспрессии (см. определение «генетического контролирующего элемента» ниже), и/или промежуточные последовательности («интроны»), расположенные между отдельными кодирующими сегментами («экзонами»).
«Генетический контролирующий элемент»: термин «генетический контролирующий элемент» при использовании в данной заявке относится к любой последовательности элемента, который модулирует экспрессию гена, с которым он связан с сохранением функциональности. Генетические контролирующие элементы могут функционировать либо увеличивая, либо снижая уровень экспрессии и могут располагаться перед, в пределах или после кодирующей последовательности. Генетические контролирующие элементы могут функционировать на любом этапе экспрессии гена, регулируя, например, инициацию, элонгацию или терминацию транскрипции, сплайсинг мРНК, редактирование мРНК, стабильность мРНК, внутриклеточную локализацию мРНК, инициацию, элонгацию или терминацию трансляции или любой другой этап экспрессии гена. Генетические контролирующие элементы могут функционировать в отдельности или в комбинации друг с другом.
«Гликопротеин»: термин «гликопротеин» при использовании в данной заявке относится к белку или полипептиду, который содержит одну или более ковалентно присоединенную цепь олигосахарида. Олигосахаридные цепи могут быть образованы из одного остатка сахара, одиночной неразветвленной цепи остатков сахара или могут быть образованы цепью остатков сахара с одним или более разветвлением. В некоторых вариантах реализации олигосахаридные цепи являются N-присоединенными. В некоторых вариантах реализации цепи являются О-присоединенными.
«Характер/степень гликозилирования»: термин «характер/степень гликозилирования» относится к наблюдаемому характеру гликозилирования данного гликопротеина или гликопротеинов. Говорят, что гликопротеин с большим числом ковалентно присоединенных остатков сахара в олигосахаридной цепи имеет повышенное гликозилирование или повышенную степень гликозилирования. И наоборот, говорят, что гликопротеин с меньшим числом ковалентно присоединенных сахарных остатков в олигосахаридной цепи имеет пониженное гликозилирование или меньшую степень гликозилирования. Термин «характер/степень гликозилирования» при использовании в данной заявке также относится к характерному распределению нескольких различных вариантов гликозилирования на конкретных гликопротеинах, экспрессируемых согласно настоящему изобретению. В этом значении увеличение степени гликозилирования означает усиление характерных вариантов гликозилирования экспрессируемых гликопротеинов.
«Клетка-хозяин»: термин «клетка-хозяин» при использовании в данной заявке относится к клетке, которой манипулируют согласно настоящему изобретению с получением гликопротеина с желаемым характером гликозилирования, как описано выше. В некоторых вариантах реализации клетка-хозяин представляет собой клетку млекопитающего.
«Гибридома»: термин «гибридома» при использовании в данной заявке относится к клетке или потомству клетки, образованной слиянием иммортализованной клетки с клеткой, продуцирующей антитела. Образующаяся в результате этого гибридома представляет собой иммортализованную клетку, продуцирующую антитела. Отдельные клетки, используемые для создания гибридомы, могут быть получены от любого млекопитающего, включая без ограничений крысу, свинью, кролика, овцу, козу и человека. В конкретных вариантах реализации гибридома представляет собой триомную линию клеток, образующуюся при слиянии клеток-потомков гетерогибридной миеломы, являющихся в свою очередь продуктом слияния клеток человека и линии клеток миеломы мыши, с плазматической клеткой. В конкретных вариантах реализации настоящего изобретения гибридома представляет собой любую иммортализованную гибридную линию клеток, продуцирующую антитела, такую как квадрома (см., например, Milstein et al., Nature, 537:3053, 1983).
«Среда», «среда для культивирования клеток», «культуральная среда»: эти термины при использовании в данной заявке относятся к раствору, содержащему питательные вещества, питающие растущие клетки млекопитающего. Обычно такие растворы обеспечивают незаменимые и заменимые аминокислоты, витамины, источники энергии, липиды и микроэлементы, требующиеся клетке для минимального роста и/или жизнеспособности. Такой раствор может также содержать вспомогательные компоненты (см. определение «вспомогательных компонентов» ниже), которые усиливают рост и/или жизнеспособность, делая их выше минимальных показателей, включающие - но не ограниченные перечисленными - гормоны и/или другие факторы роста, конкретные ионы (такие как натрий, хлорид, кальций, магний и фосфат), буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы (неорганические соединения, обычно присутствующие в очень низкой конечной концентрации), аминокислоты, липиды и/или глюкозу или другой источник энергии. В конкретных вариантах реализации среда преимущественно имеет рН и концентрацию солей, оптимальные для жизнеспособности и пролиферации клеток. Примеры культуральных сред представлены в Таблице 1, хотя настоящее изобретение не ограничено применением указанных сред. Как понятно специалисту в данной области, можно применять альтернативные культуральные среды, и/или могут быть сделаны некоторые изменения в составе культуральных сред, приведенных в качестве примеров в Таблице 1. В конкретных вариантах реализации среда представляет собой среду для подпитки, которую добавляют после начала культивирования клеток (см. определение «среды для подпитки», выше).
«Полипептид»: термин «полипептид» при использовании в данной заявке обозначает цепь последовательных аминокислот, соединенных вместе посредством пептидных связей. Термин используется для обозначения цепи из аминокислот любой длины, но, как очевидно специалисту в данной области, термин не ограничивается протяженными цепями и может относиться к минимальной цепи, включающей две аминокислоты, связанные вместе посредством пептидной связи. Как известно специалистам в данной области, полипептиды могут подвергаться процессингу и модификациям. Например, полипептид может быть гликозилирован (см. определение «гликопротеина» выше).
«Белок»: термин «белок» при использовании в данной заявке относится к одному или более полипептиду, который функционирует в виде дискретной единицы. Если отдельный полипептид представляет собой дискретную функционирующую единицу и не требует постоянной или временной физической ассоциации с другими полипептидами для формирования дискретной функционирующей единицы, термины «полипептид» и «белок» могут использоваться равнозначно. Если дискретная функциональная единица включает более одного полипептида, физически соединенных друг с другом, термин «белок» относится к совокупности полипептидов, которые физически связаны и функционируют вместе в виде дискретной единицы.
«Рекомбинантно экспрессируемый гликопротеин» и «рекомбинантный гликопротеин»: эти термины при использовании в данной заявке относятся к гликопротеину, экспрессируемому в клетке-хозяине, с которой провели манипуляции таким образом, чтобы указанная клетка экспрессировала данный гликопротеин. В конкретных вариантах реализации настоящего изобретения клетка-хозяин представляет собой клетку млекопитающего. В конкретных вариантах реализации такого рода манипуляции включают одну или более генетическую модификацию. Например, клетки-хозяева млекопитающего могут быть генетически модифицированы введением одного или более гетерологичного гена, кодирующего гликопротеин, экспрессию которого осуществляют. Гетерологичный рекомбинатно экспрессируемый гликопротеин может быть идентичным или схожим с гликопротеином, который в норме экспрессируется в клетке-хозяине млекопитающего. Гетерологичный рекомбинантно экспрессируемый гликопротеин может также быть чужеродным для указанной клетки-хозяина, то есть гетерологичным по отношению к гликопротеинам, в норме экспрессируемым указанным клеткой-хозяином млекопитающего. В конкретных вариантах реализации гетерологичный рекомбинантно экспрессируемый гликопротеин является химерным в том отношении, что он содержит последовательности аминокислот, идентичные или схожие с последовательностями гликопротеинов, в норме экспрессирующихся в клетке-хозяине млекопитающего, тогда как другие участки указанного гликопротеина являются чужеродными по отношению к клетке-хозяину. Кроме того, клетка-хозяин млекопитающего может быть генетически модифицирована путем активации или увеличения экспрессии одного или более эндогенных генов.
«Вспомогательные компоненты»: термин «вспомогательные компоненты» при использовании в данной заявке относится к компонентам, которые усиливают рост и/или жизнеспособность клеток, делая их выше минимального уровня, включающим - но не ограниченным перечисленными - гормоны и/или другие факторы роста, конкретные ионы (например, натрий, хлорид, кальций, магний и фосфат), буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы (неорганические соединения, как правило, присутствующие в очень низкой конечной концентрации), аминокислоты, липиды и/или глюкозу или другой источник энергии. В некоторых вариантах реализации вспомогательные компоненты могут быть добавлены к начальной культуре клеток. В некоторых вариантах реализации вспомогательные компоненты могут быть добавлены после начала культивирования клеток.
«Титр»: термин «титр» при использовании в данной заявке относится к общему количеству рекомбинантно экспрессируемого гликопротеина, продуцируемого культурой клеток млекопитающего в данном объеме среды. Обычно титр выражают в единицах миллиграмм гликопротеина на миллилитр среды.
ПОДРОБНОЕ ОПИСАНИЕ НЕКОТОРЫХ ВАРИАНТОВ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает улучшенные системы для получения гликопротеинов в культуре клеток. В частности, предусмотрены системы, обеспечивающие продукцию гликопротеина с желаемым характером гликозилирования. Например, гликопротеин может иметь более высокую степень гликозилирования и/или может иметь распределение олигосахаридных цепей, имеющее большую степень сходства с распределением олигосахаридных цепей, присоединенных к гликопротеину в природной клетке-хозяине. В некоторых вариантах реализации применение системы согласно настоящему изобретению может обеспечивать производство гликопротеина с характером гликозилирования, схожим или идентичным характеру гликозилирования, наблюдаемому при экспрессии гликопротеина в эндогенной клетке-хозяине. Некоторые варианты реализации настоящего изобретения рассмотрены подробно ниже. Однако, как ясно специалисту в данной области, возможны различные модификации указанных вариантов реализации в пределах объема прилагаемой формулы. Именно формула и эквиваленты определяют область настоящего изобретения, которая не ограничена и которую не следует ограничивать настоящим описанием отдельных вариантов реализации.
СОСТАВ СРЕД
В соответствии с настоящим изобретением можно применять широкий спектр сред для роста клеток млекопитающего. В конкретных вариантах реализации клетки можно выращивать в одной из ряда сред с известным определенным составом {defined media), при котором компоненты среды известны и контролируемы. В конкретных вариантах реализации клетки можно выращивать в сложной среде, в которой не все компоненты известны и/или контролируемы.
В течение последних десятилетий были разработаны и широко описаны в литературе среды для роста культур клеток млекопитающего с определенным химическим составом. Все компоненты среды с определенным составом хорошо охарактеризованы, и такие среды не содержат сложных добавок, например сыворотку или гидролизаты. Ранее состав сред разрабатывали таким образом, чтобы они обеспечивали рост клеток и поддерживали их жизнеспособность, при этом лишь незначительное внимание уделяли продукции белка или вообще ее не учитывали. В последнее время были разработаны среды, предназначенные специально для поддержания клеточных культур с высоким уровнем продукции рекомбинантного белка и/или гликопротеина.
Среда с определенным составом обычно включает около пятидесяти химических соединений в известной концентрации, растворенных в воде. Большинство из них также содержат один или более из известных белков, например инсулин, IGF-1, трансферрин или БСА, но другие среды не требуют белковых компонентов, и их называют безбелковыми средами. Химические компоненты среды разделяют на пять больших категорий: аминокислоты, витамины, неорганические соли, микроэлементы и смешанную категорию, не поддающуюся четкой классификации.
Микроэлементы состоят из разнообразных неорганических солей, присутствующих в микромолярных или более низких концентрациях. Четыре наиболее распространенных микроэлемента, входящие в состав практически всех сред с известным составом - железо, цинк, селен и медь. Железо (железистые или соли железа) и цинк, как правило, добавляют в микромолярных концентрациях, тогда как другие микроэлементы - в наномолярных концентрациях. Многочисленные менее распространенные микроэлементы обычно добавляют в наномолярных концентрациях.
Марганец часто присутствует среди микроэлементов в виде дивалентного катиона (MnCl2 или MnSO4). В ранних вариантах сред с известным составом он либо отсутствовал, либо присутствовал в высокой концентрации порядка 1 мкМ (см., например, Barnes and Sato, 1980 [Medium DMEM/F12] и Kitos et al., 1962 [Medium MD 705/1]). В состав более поздних вариантов сред марганец, как правило, входил, но в значительно более низких концентрациях, например в диапазоне 1-5 нМ (см., например, Hamilton and Ham, 1977 [Medium MCDB 301] и Cleveland and Erlanger, 1988 [среда без названия]).
Настоящее изобретение основано на обнаружении того, что гликопротеины, продуцируемые культурой клеток, выращенной в среде с определенным составом, содержащей марганец в концентрации между указанными значениями, характеризуются более высокой степенью гликозилирования, чем если бы такие клетки выращивали в стандартной среде такой, как описано выше. В конкретных вариантах реализации марганец содержится в среде в концентрации между приблизительно 10 и 600 нМ. В конкретных вариантах реализации марганец содержится в среде в концентрации между приблизительно 20 и 100 нМ. В конкретных вариантах реализации марганец содержится в среде в концентрации приблизительно 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 210, 220, 230, 240, 250, 260, 270, 280, 290, 300, 310, 320, 330, 340, 350, 360, 370, 380, 390, 400, 410, 420, 430, 440, 450, 460, 470, 480, 490, 500, 510, 520, 530, 540, 550, 560, 570, 580, 590 или 600 нМ или в любом диапазоне в пределах указанных концентраций.
Настоящее изобретение также включает открытие того, что гликопротеины, продуцируемые культурой клеток, выращенной в среде с определенным составом, содержащей сравнительно низкий уровень глутамина, характеризуются более высокой степенью гликозилирования, чем если бы клетки выращивали в стандартной среде, содержащей более высокий уровень глутамина. В конкретных вариантах реализации исходный уровень глутамина в среде составляет менее или равен приблизительно 8 мМ. В конкретных вариантах реализации исходный уровень глутамина в среде составляет менее или равен приблизительно 4 мМ.
Специалист в данной области сможет выбрать точную концентрацию марганца в пределах диапазона значений согласно настоящему изобретению на основании особенностей конкретной схемы эксперимента, в том числе типа клеток, в которых экспрессируется гликопротеин, типа экспрессируемого гликопротеина и наличия или отсутствия других компонентов в среде, в которой растут клетки. Например, различные N-присоединенные и О-присоединенные структуры или различные структуры олигосахаридов в пределах каждого из указанных широких классов могут требовать различных концентраций марганца в культуральной среде для получения олигосахаридных цепей с большей протяженностью и/или в большей степени приближенных к природным.
ГЛИКОПРОТЕИНЫ
Любой гликопротеин, экспрессируемый в клетке-хозяине, может быть получен в соответствии с замыслом настоящего изобретения. Гликопротеин может экспрессироваться с использованием гена, который является эндогенным по отношению к клетке-хозяину, или с применением гетерологичного гена, введенного в клетку-хозяин. Гликопротеин может быть природным или же может содержать последовательность, сконструированную или отобранную специалистом. Гликопротеин, который можно экспрессировать, может быть собран из полипептидных фрагментов, которые в отдельности являются природными, по меньшей мере один из которых содержит пептидную последовательность, которая служит в качестве сайта гликозилирования. Кроме того, каждый полипептидный фрагмент может содержать только часть сайта гликозилирования, который затем восстанавливается при сборке полипептидных фрагментов. Кроме того или в качестве альтернативы, полученный методом генной инженерии гликопротеин может включать один или более фрагмент, не встречающийся в природе, если при этом такой гликопротеин содержит по меньшей мере одну пептидную последовательность, которая служит в качестве сайта гликозилирования.
Гликопротеины, которые можно экспрессировать в соответствии с настоящим изобретением, часто выбирают на основании интересующей или полезной биологической или химической активности. Например, настоящее изобретение можно применять для экспрессии любого фармацевтически или коммерчески значимого фермента, рецептора, антитела, гормона, регуляторного фактора, антигена, связывающие агента и т.п. Следующий ниже список гликопротеинов, которые можно продуцировать согласно настоящему изобретению, представлен только в качестве примера, но не для ограничения. Специалисту в данной области очевидно, что любой гликопротеин можно экспрессировать в соответствии с настоящим изобретением, и специалист способен выбрать конкретный гликопротеин, который необходимо получить для конкретных задач.
Факторы свертывания крови
Как было показано, факторы свертывания крови эффективны в качестве фармацевтических и/или коммерческих агентов. С учетом важности рекомбинантных факторов свертывания крови для лечения таких заболеваний, как гемофилия, оптимизация характера гликозилирования полученных рекомбинантным способом факторов свертывания крови в соответствии с настоящим изобретением представляет особенный интерес. Например, фактор свертывания крови IX (Фактор IX или "FIX") представляет собой одноцепочечный гликопротеин, недостаток которого приводит к гемофилии В, нарушению, при котором кровь больного не способна свертываться. Таким образом, любая небольшая рана, приводящая к кровотечению, потенциально представляет угрозу для жизни.
FIX синтезируется в виде одноцепочечного зимогена, который может быть активирован с образованием двуцепочечной сериновой протеазы (Фактор IХа) при высвобождении активирующего пептида. Каталитический домен Фактора IХа локализован в тяжелой цепи (см. работу Chang et al., J.Clin. Invest, 100:4, 1997, включенную в данную заявку посредством ссылки). FIX имеет многочисленные сайты гликозилирования, включающие как N-присоединенные, так и О-присоединенные углеводы. Одно время О-присоединенная структура у серина 61 (Sia-α2,3-Gal-β1,4-GlcNAc-β1,3-Fuc-α1-O-Ser) считалась уникальной для FIX, но позднее была обнаружена в нескольких других молекулах, в том числе в белке Notch у млекопитающих и дрозофилы (Maloney et al, Journal of Biol. Chem., 275(13), 2000). FIX, синтезированные культурой клеток яичника китайского хомячка ("СНО"), демонстрируют некоторую изменчивость по олигосахаридной цепи у серина 61. Такие гликоформы и другие потенциальные гликоформы могут различаться по способности индуцировать коагуляцию при введении людям или животным и/или могут иметь различную стабильность в крови, обуславливающую менее эффективное свертывание крови.
Гемофилия А, клинически не отличимая от гемофилии В, вызвана недостатком фактора свертывания крови VIII, другого гликопротеина, синтезируемого в виде одноцепочечного зимогена, который затем преобразуется в двуцепочечную активную форму. Настоящее изобретение можно также применять для регуляции или изменения характера гликозилирования фактора свертывания крови VIII с целью изменения его коагулирующей активности. Другие гликопротеиновые факторы свертывания крови, которые могут быть получены и характер гликозилирования которых можно регулировать или изменять согласно настоящему изобретению, включают, например, тканевой фактор и фактор фон Виллебранда, но не ограничиваются ими.
Антитела
Антитела представляют собой белки, обладающие способностью специфично связываться с конкретным антигеном. С учетом большого числа антител, которые в настоящее время применяют или исследуют как фармацевтические или другие коммерческие агенты, производство антител с желательным характером гликозилирования согласно настоящему изобретению представляет особый интерес. Кроме того, антитела с отличающимся характером гликозилирования с меньшей вероятностью вызывают иммунный ответ у пациента, которому их вводят, что приводит к более эффективной схеме лечения. Кроме того или в качестве альтернативы, антитела с отличающимся характером гликозилирования в их константных областях могут демонстрировать улучшенные фармакокинетические или фармакодинамические эффекторные функции. Кроме того или в качестве альтернативы, антитела с отличающимся характером гликозилирования могут быть более стабильными при условиях культивирования клеток, в которых они синтезируются, например, могут быть более устойчивыми к протеазам или другим компонентам в культуре клеток, обеспечивая таким образом более высокий конечный титр продуцируемых антител.
Любое антитело, которое можно экспрессировать в клетке-хозяине, можно применять в соответствии с идеями настоящего описания. В некоторых вариантах реализации антитело, которое можно экспрессировать, представляет собой моноклональное антитело. В конкретных вариантах реализации моноклональное антитело представляет собой химерное антитело. Химерное антитело содержит аминокислотные фрагменты, полученные от более чем одного организма. Молекулы химерного антитела могут включать, например, антигенсвязывающий домен антитела мыши, крысы или другого вида, при этом константные области такого антитела могут быть получены от человека. Описан ряд способов получения химерных антител (см. например, Morrison et al., Proc. Natl. Acad. Sci. U.S.A. 81, 6851, 1985; Takeda et al., Nature 314, 452, 1985, Cabilly et al., патент США №4,816,567; Boss et al., патент США №4,816,397; Tanaguchi et al., публикацию европейского патента EP 171496; публикацию европейского патента 0173494, патент Великобритании GB 2177096В, включенные в данную заявку посредством ссылки).
В некоторых вариантах реализации моноклональное антитело представляет собой гуманизированное антитело. Гуманизированное антитело представляет собой химерное антитело, в котором значительное большинство аминокислотных остатков получено от антител человека, что таким образом минимизирует любые возможные иммунные реакции при его введении человеку. В гуманизированных антителах аминокислотные остатки в гипервариабельных участках заменены на остатки антитела отличного от человека вида, которые обеспечивают антигену желаемую специфичность или сродство. В конкретных вариантах реализации гуманизированное антитело имеет последовательность аминокислот, на 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 процентов или выше идентичную антителу человека. В конкретных вариантах реализации гуманизированное антитело оптимизируют путем введения консервативных замен, замен в консенсусной последовательности, замен зародышевого типа и/или обратных мутаций. Такие измененные молекулы иммуноглобулинов могут быть получены любым из нескольких способов, известных в данной области (например, Teng et al., Proc. Natl. Acad. Sci. U.S.A., 80, 7308-7312, 1983; Kozbor et al., Immunology Today, 4, 7279, 1983; Olsson et al., Meth. Enzymol., 92, 3-16.1982, включенных в данную заявку посредством ссылки). В некоторых вариантах реализации измененные молекулы иммуноглобулина получают согласно идее публикации РСТ WO92/06193 или ЕР 0239400, включенных в данную заявку посредством ссылки в полном объеме.
В конкретных вариантах реализации антитело, полученное согласно настоящему описанию, может содержать константную область или Fc-область иммуноглобулина, демонстрирующую улучшенный характер гликозилирования. Например, антитело, полученное согласно настоящему изобретению, может связываться с большей силой или с большей специфичностью с эффекторными молекулами, такими как рецепторы комплемента и/или рецептор Fc, которые могут контролировать несколько иммунных функций антитела, например активность эффекторной клетки, лизис, комплемент-опосредованную активность, выведение антитела и период полураспада антитела. Типичные рецепторы Fc, которые связываются с Fc-областью антитела (например, антитела IgG), включают, но не ограничены ими, подклассы рецепторов Fc Rl, Fc RII, и Fc RIII, и FcRn, в том числе их аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Обзор Fc-рецепторов представлен в Ravetch and Kinet, Аnnu. Rev. Immunol 9:457-92, 1991; Capel et al., Immunomethods 4:25-34,1994; и de Haas et al., J. Lab. Clin. Med. 126:330-41,1995, включенных в данную заявку посредством ссылки в полном объеме.
Одним из неограничивающих примеров антитела, которое можно получить согласно настоящему изобретению, является антитело к А-бета. Антитела к А-бета представляют собой особенно перспективный потенциальный способ лечения терапия болезни Альцгеймера. Болезнь Альцгеймера (БП) представляет собой прогрессирующую болезнь, приводящую к старческому сумасшествию (общее описание см. в: Selkoe, TINS 16:403, 1993; Hardy et al., WO 92/13069; Selkoe, J. Neuropathol. Exp. Neurol. 53:438, 1994; Duff et al., Nature 373:476, 1995; Games et al., Nature 373:523, 1995, включенные в данную заявку посредством ссылки). В целом, болезнь разделяют на две категории: с поздним возникновением, развивающуюся в старости (в 65 лет и более), и с ранним возникновением, которая развивается в более ранний период перед старческим, т.е. между 35 и 60 годами. Для обоих типов болезни характерны одинаковые патологические признаки, но аномалии, как правило, более серьезны и более выражены в случаях развития болезни в раннем возрасте. Болезнь характеризуется по меньшей мере двумя типами повреждений в мозге: нейрофибриллярными клубками и старческими бляшками. Нейрофибриллярные клубки представляют собой внутриклеточные отложения ассоциированного с микротрубочками белка тау, состоящего из двух филаментов, скрученных друг с другом в пары. Старческие бляшки (т.е. амилоидные бляшки) представляют собой области неорганизованного нейропиля до 150 мкм в поперечном направлении с внеклеточными амилоидными отложениями в центре, которые можно обнаружить при микроскопическом анализе срезов тканей мозга. Накопление амилоидных бляшек в пределах мозга также связано с синдромом Дауна и другими расстройствами познавательной способности.
Основным компонентом бляшек является пептид, называемый А-бета или бета-амилоидный пептид. А-бета пептид представляет собой внутренний фрагмент, имеющий массу 4 кДа и состоящий из 39-43 аминокислот, большего по размеру трансмембранного гликопротеина, известного как белок-предшественник амилоида (АРР, amyloid precursor protein). В результате протеолитического процессинга АРР с участием различных ферментов-секретаз А-бета, главным образом, представлен как в короткой форме, длиной 40 аминокислот, так и в длинной форме, длиной 42-43 аминокислот. Часть гидрофобного трансмембранного домена АРР оказывается на карбокси-конце А-бета, что может обуславливать способность А-бета агрегировать в бляшки, в особенности в случае длинной формы. Накопление амилоидных бляшек в мозге в конечном счете приводит к гибели нейронов. Физические симптомы, связанные с этим типом повреждения нервов, характеризуют болезнь Альцгеймера.
С присутствием болезни Альцгеймера коррелируют несколько мутаций в белке АРР (см., например, Goate et al., Nature 349:704, 1991 (валин717 в изолейцин); Chartier Harlan et al., Nature 353:844, 1991 (валин717 в глицин); Murrell et al., Science 254:97,1991 (валин717 в фенилаланин); Mullan et al., Nature Genet. 1:345, 1992 (двойная мутация лизин595-метионин596 в аспарагин595-лейцин596), включенные в данную заявку посредством ссылок в полном объеме). Считается, что такие мутации вызывают болезнь Альцгеймера путем повышения уровня или изменения процессинга АРР в А-бета, в частности в результате процессинга АРР с образованием большего количества А-бета длинной формы (т.е. А-бета1-42 и А-бета1-43). Предполагается, что мутации в других генах, например генах пресенилина, PS1 и PS2, косвенно влияют на процессинг АРР с образованием большего количества А-бета длинной формы (см. работу Hardy, TINS 20: 154,1997, включенную в данную заявку посредством ссылки в полном объеме).
Для определения значения амилоидных бляшек в болезни Альцгеймера успешно использовали мышиные модели (см. работы Games et al., выше; Johnson-Wood et al., Proc. Natl. Acad. Sci. USA 94:1550, 1997, включенные в данную заявку посредством ссылки в полном объеме). В частности, когда трансгенным мышам PDAPP (экспрессирующим мутантную форму АРР человека с развитием болезни Альцгеймера в молодом возрасте) вводили А-бета длинной формы, они демонстрировали как уменьшение прогрессирования болезни Альцгеймера, так и увеличение титра антител к пептиду А-бета (см. работу Schenk et al., Nature 400, 173, 1999, включенную в данную заявку посредством ссылки в полном объеме). Наблюдения, рассмотренные выше, указывают на то, что А-бета, в особенности в его длинной форме, является фактором, обуславливающим развитие болезни Альцгеймера.
А-бета пептид может существовать в растворе и может быть выявлен в ЦНС (например, в цереброспинальной жидкости, ЦСЖ) и плазме. При определенных условиях растворимый А-бета преобразуется в фибриллярную токсичную форму с бета-слоями, обнаруживаемую в невритических бляшках и церебральных кровеносных сосудах пациентов с БА. Были исследованы способы лечения, включающие иммунизацию моноклональными антителами к А-бета. На модели БА у мышей были протестированы как активная, так и пассивная иммунизация. Активная иммунизация приводила к некоторому уменьшению числа бляшек в мозге, но только при назальном введении. Также была исследована пассивная иммунизация трансгенных мышей PDAPP (работа Bard, et al.,Nat. Med. 6:916-19, 2000, включенная в данную заявку посредством ссылки в полном объеме). Было обнаружено, что антитела, узнающие аминоконцевой и центральный домены А-бета, стимулируют фагоцитоз отложений А-бета, тогда как антитела к доменам около С-конца не оказывают такого эффекта.
Механизм разрушения/выведения А-бета после пассивной или активной иммунизации продолжают исследовать. Предполагается, что в эффективное разрушение А-бета могут быть вовлечены два механизма, а именно центральная деградация и периферическая деградация. Центральный механизм деградации основан на способности антитела проходить сквозь гематоэнцефалический барьер, связываться с бляшками и индуцировать разрушение ранее существующих бляшек. Было показано, что такое разрушение обусловлено фагоцитозом, опосредованным Fc-рецептором (Bard, et al., см. выше). Механизм периферической деградации для удаления А-бета основан на нарушении динамического равновесия А-бета между мозгом, ЦСЖ и плазмой при введении антитела, приводящего к транспорту А-бета из одного отдела в другой. А-бета из центрального отдела перемещается в ЦСЖ и плазму, где происходит его деградация. Недавние исследования показали, что растворимый и несвязанный А-бета вовлечен в процесс ухудшения памяти, ассоциированный с БА, даже без уменьшения амилоидных отложений в мозге. Требуются дополнительные исследования для определения действия и/или взаимодействия указанных путей разрушения А-бета (работа Dodel, et al., The Lancet Vol.2:215, 2003, включенная в данную заявку посредством ссылки в полном объеме).
Антитела к А-бета представляют собой потенциально перспективный способ лечения БА, поскольку они могут связывать и опосредовать разрушение А-бета или других компонентов, входящих в состав амилоидных бляшек. Антитела к А-бета, полученные в соответствии с идеей настоящего описания, могут способствовать более эффективному лечению болезни Альцгеймера или других схожих болезней, например, посредством связывания и разрушения компонентов амилоидных бляшек с большей эффективностью, разрушения амилоидных бляшек с меньшими или менее серьезными побочными эффектами или предотвращения образования или отложения амилоидных бляшек. В конкретных вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, представляют собой моноклональные антитела.
В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, специфично связываются с агрегированной формой А-бета, но не связываются с растворимой формой. В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, специфично связываются с растворимой формой А-бета, при условиях, при которых они не связываются с агрегированной формой. В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, связываются как с агрегированной, так и с растворимой формами. В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, связываются с А-бета в бляшках. В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, проходят сквозь гематоэнцефалический барьер. В конкретных вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, уменьшают амилоидные скопления у объекта. В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, уменьшают невритическую дистрофию у объекта. В некоторых вариантах реализации антитела к А-бета могут поддерживать архитектуру синапса (например, синаптофизин).
Согласно некоторым вариантам реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, связываются с эпитопом в пределах остатков 13-28 А-бета (при этом первый N-терминальный остаток природного А-бета обозначен 1). В некоторых вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, связываются с эпитопом в пределах остатков 19-22 А-бета. В некоторых вариантах реализации применяют многочисленные моноклональные антитела, характеризующиеся специфическим связыванием с различными эпитопами А-бета. Например, в некоторых вариантах реализации антитело, специфичное к эпитопу в пределах остатков 19-22 А-бета, вводят совместно с антителом, специфичным к эпитопу вне остатков 19-22 А-бета. Такие антитела можно вводить последовательно или одновременно. Можно также применять антитела к компонентам амилоида, помимо А-бета (например, вводимых или вводимых совместно).
В конкретных вариантах реализации антитела к А-бета, полученные в соответствии с настоящим изобретением, связываются с эпитопом А-бета с большей силой или с большей специфичностью по сравнению с антителами к А-бета, полученными иным способом. Специфичность антитела к эпитопам можно определить известными способами, например путем создания библиотеки фагового дисплея, различные элементы которой экспонируют различные частичные последовательности А-бета. Затем можно проводить селекцию элементов библиотеки фагового дисплея, специфично связывающихся с тестируемым антителом. Изолируют семейство последовательностей. Как правило, различные элементы такого семейства содержат общую коровую последовательность и варьирующую длину фланкирующих последовательностей.
Наиболее короткая коровая последовательность, демонстрирующая специфическое связывание с антителом, как правило, соответствует эпитопу, с которым связывается антитело. В качестве альтернативы или дополнения, антитела можно протестировать на специфичность по отношению к эпитопу с помощью конкурентного анализа с антителом с ранее определенной специфичностью по отношению к эпитопу. Например, считается, что антитела, конкурирующие с антителом 15С11 за связывание с А-бета, связываются с тем же самым или аналогичным эпитопом, что и 15С11, т.е. в пределах остатков 19-22 А-бета. В конкретных вариантах реализации скрининг антител на специфичность по отношению к эпитопу можно применять для предсказания терапевтической эффективности. Например, антитело, которое, как было определено, связывается с эпитопом в пределах остатков 13-28 (например, Аβ 19-22) А-бета, вероятно, будет эффективным для предотвращения и лечения болезни Альцгеймера согласно методологии настоящего изобретения.
Антитела, которые специфично связываются с предпочтительным участком А-бета, не связывающиеся с другими участками А-бета, имеют ряд преимуществ по сравнению с моноклональными антителами, связывающимися с дополнительными участками, или с поликлональной сывороткой, полученной к целой молекуле А-бета. Между прочим, при равной массе доз дозы антител, специфично связывающихся с предпочтительными участками, содержат более высокую молярную дозу антител, эффективную для разрушения амилоидных бляшек. Кроме того, антитела, специфично связывающиеся с предпочтительными участками, могут индуцировать ответ, приводящий к разрушению амилоидных отложений, при этом не индуцируя ответ, приводящий к разрушению интактного полипептида АРР, что уменьшает потенциальные побочные эффекты.
В некоторых вариантах реализации моноклональные, химерные, одноцепочечные или гуманизированные антитела, описанные выше, могут содержать аминокислотные остатки, не встречающиеся в антителах ни у одного вида в природе. Такие чужеродные остатки можно применять, например, для придания новой или модифицированной специфичности, аффинности или эффекторной функции моноклональному, химерному, одноцепочечному или гуманизированному антителу.
Ферменты
Другой класс гликопротеинов, которые, как было показано, являются эффективными как фармацевтические и/или коммерческие агенты, включает ферменты. Ферменты могут представлять собой гликопротеины, характер гликозилирования которых влияет на ферментативную активность. Таким образом, производство ферментов с желательным характером гликозилирования в соответствии с настоящим изобретением также представляет собой особый интерес.
В качестве одного из неограничивающих примеров, недостаток глюкоцереброзидазы (GCR) вызывает патологическое состояние, известное как болезнь Гошера, обусловленное накоплением глюкоцереброзидазы в лизосомах некоторых клеток. У субъекта, страдающего болезнью Гошера, наблюдается ряд симптомов, включающих спленомегалию, гепатомегалию, нарушения скелета, тромбоцитопению и анемию. Friedman и Hayes показали, что рекомбинантная глюкоцереброзидаза GCR (rGCR), содержащая одиночную замену в первичной последовательности аминокислот, характеризовалась измененным характером гликозилирования, в частности увеличением числа остатков фукозы и N-ацетилглюкозамина, по сравнению с природной GCR (см. патент США №5,549,892).
Friedman и Hayes также показали, что такая rGCR демонстрировала улучшенные фармакокинетические свойства по сравнению с природной GCR. Например, приблизительно в два раза больше rGCR направленно транспортировалось в клетки Купффера в печени по сравнению с природной GCR. Хотя первичные последовательности аминокислот двух белков отличались по одному остатку, Friedman и Hayes выдвинули гипотезу, что измененный характер гликозилирования rGCR также мог оказать влияние на локализацию в клетках Купфера.
Специалисту в данной области известны другие примеры ферментов, демонстрирующих измененные ферментативные, фармакокинетические и/или фармакодинамические свойства, обусловленные изменением характера их гликозилирования.
Факторы роста и другие сигнальные молекулы
Другой класс гликопротеинов, которые, как было показано, эффективны как фармацевтические и/или коммерческие агенты, включает факторы роста и другие сигнальные молекулы. Таким образом, продукция рецепторов с желательным характером гликозилирования в соответствии с настоящим изобретением также представляет особый интерес. Факторы роста, как правило, представляют собой гликопротеины, которые секретируются клетками и которые связываются и активируют рецепторы на других клетках, инициируя изменения в метаболизме или в развитии клетки, содержащей рецептор.
Неограничивающие примеры факторов роста млекопитающих и других сигнальных молекул включают цитокины; эпидермальный фактор роста (EGF); тромбоцитарный фактор роста (PDGF); факторы роста фибробластов (FGF), например FGF-5; инсулиноподобный фактор роста I и -II (IGF-I и IGF-II); дез(1-3) -IGF-I (IGF-I мозга), белки, связывающие инсулиноподобный фактор роста; белки CD, например CD-3, CD-4, CD-8 и CD-19; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; морфогенетические белки кости (BMPs); интерфероны, например интерферон-альфа, -бета и -гамма; колониестимулирующие факторы (CSFs), например M-CSF, GM-CSF, и G-CSF; большинство интерлейкинов; фактор некроза опухоли бета (TNF); фолликулостимулирующий гормон; кальцитоцин; лютеинизирующий гормон; антикоагулирующие факторы, например протеин С; атриальный натрийуретический пептид; сурфактант легкого; активаторы плазминогена, например урокиназа, или уриновый, или активатор тканевого типа (t-PA) человека; гематопоэтический фактор роста и энкефалиназа. Специалисту в данной области известны другие факторы роста или сигнальные молекулы, которые могут экспрессироваться в соответствии с настоящим изобретением.
Было показано, что специфические изменения в характере гликозилирования факторов роста или других сигнальных молекул оказывают значительное влияние на их терапевтические свойства. В качестве одного неограничивающего примера распространенным способом лечения пациентов, страдающих хронической анемией, являются многократные инъекции рекомбинантного эритропоэтина человека (rHuEPO) для усиления у пациентов продукции красных кровяных клеток. Был разработан аналог rHuEPO, дарбепоэтин альфа (Aranesp®) с более продолжительным периодом существования в организме по сравнению с нормальным rHuEPO. Основное отличие дарбепоэтина альфа от rHuEPO - наличие двух дополнительных N-присоединенных олигосахаридных цепей, содержащих сиаловые кислоты.
Производство дарбепоэтина альфа осуществляли с использованием гликоинженерии in vitro (см. Elliott et al., Nature Biotechnology 21(4):414-21, 2003). Elliott с соавторами использовали мутагенез in vitro для включения дополнительных сайтов гликозилирования в полипептидный остов rHuEPO, что приводило к экспрессии аналога дарбепоэтина альфа. Дополнительные олигосахаридные цепи расположены дистально по отношению к сайту связывания с рецептором ЕРО и, очевидно, не препятствует связыванию с рецептором. Тем не менее, период полураспада дарбепоэтина альфа до трех раз выше, чем у rHuEPO, что делает его более эффективным терапевтическим агентом.
Этот пример показывает, что изменения характера гликозилирования фактора роста или другой сигнальной молекулы может оказывать значительный эффект на стабильность и/или активность терапевтического гликопротеина in vivo. Таким образом, экспрессия фактора роста или другой сигнальной молекулы, представляющей интерес, в соответствии с замыслом настоящего изобретения может приводить к экспрессии фактора роста или сигнальной молекулы с улучшенным характером гликозилирования и улучшенными терапевтическими свойствами.
Рецепторы
Другой класс гликопротеинов, которые, как было показано, эффективны как фармацевтические и/или коммерческие агенты, представляет собой рецепторы. Таким образом, продукция рецепторов с желательным характером гликозилирования в соответствии с настоящим изобретением также представляет особый интерес. Рецепторы, как правило, представляют собой трансмембранные гликопротеины, которые функционируют, распознавая внеклеточный сигнальный лиганд. Помимо домена, распознающего лиганд, у рецептора часто имеется протеинкиназный домен, который запускает сигнальный каскад, фосфорилируя внутриклеточные молекулы-мишени при связывании лиганда, что приводит к изменениям в развитии или метаболизме клетки.
В некоторых вариантах реализации рецепторный гликопротеин, продукция которого возможна в соответствии с настоящим изобретением, представляет собой рецепторную тирозинкиназу (РТК). Семейство РТК включает рецепторы, которые важны для функционирования различных типов клеток (см., например, работы Yarden and Ullrich, Ann. Rev. Biochem. 57:433-478, 1988; Ullrich and Schlessinger, Cell 61:243-254, 1990, включенные в данную заявку посредством ссылки). Неограничивающие примеры РТК включают представителей семейства рецепторов фактора роста фибробластов (FGF), представителей семейства рецепторов эпидермального фактора роста (EGF), рецепторов тромбоцитарного фактора роста (PDGF), тирозинкиназных рецепторов с иммуноглобуллиновым и EGF-гомологичным доменами-1 (TIE-1) и TIE-2 (Sato et al., Nature 376(6535)10-74, 1995) и рецептора c-Met; как предполагается, некоторые из этих рецепторов стимулируют ангиогенез, прямым или опосредованным способом (Mustonen and Alitalo, J. Cell Biol. 129:895-898, 1995). Другие неограничивающие примеры РТК включают киназу печени плода 1 FLK-1 (которую иногда называют KDR, kinase insert domain-containing receptor, рецептор с киназным доменом) (Terman et al., Oncogene 6:1677-83, 1991) или рецептор фактора роста клеток эндотелия сосудов 2 (VEGFR-2)), fms-подобную тирозинкиназу-1 (Flt-1) (DeVries et al. Science 255; 989-991, 1992; Shibuya et al., Oncogene 5:519-524, 1990), иногда называемую рецептором фактора роста клеток эндотелия сосудов 1 (VEGFR-1), нейрофилин-1, эндоглин, эндосиалин и Axl. В некоторых вариантах реализации рецепторы фактора некроза опухоли альфа и бета (TNFR-1; ЕР 417,563, с датой публикации 20 марта 1991; и TNFR-2, ЕР 417,014 с датой публикации 20 марта 1991) экспрессируют в соответствии с настоящим изобретением (обзор, см. в работе Naismith and Sprang, J Inflamm. 47(1-2):1-7, 1995-96, включенной в данную заявку посредством ссылки).
В некоторых вариантах реализации рецепторный гликопротеин, продукция которого возможна в соответствии с настоящим изобретением, представляет собой рецептор, сопряженный с G-белком (GPCR). GPCR представляют собой гликопротеины, имеющие семь трансмембранных областей. При связывании лиганда с GPCR происходит преобразование сигнала в клетке, что приводит к изменению биологического или физиологического свойства клетки. GPCR являются основной мишенью действия и разработки лекарственных препаратов. В действительности, на действии рецепторов основано более половины к настоящему времени известных лекарственных препаратов (Drews, Nature Biotechnology, 14:1516, 1996); GPCR представляют наиболее важную мишень для терапевтического вмешательства, при этом 30% клинически предписанных лекарств представляют собой либо антагонисты, либо агонисты GPCR (Milligan, G. and Rees, S., TIPS, 20: 118-124, 1999). Поскольку использование таких рецепторов в качестве терапевтических мишеней доказано и проверено временем, производство GPCR с желательным характером гликозилирования в соответствии с настоящим изобретением также представляет особый интерес. Например, внеклеточные домены GPCR с желательным характером гликозилирования, экспрессированные в соответствии с замыслом настоящего изобретения, могут функционировать как важные терапевтические агенты путем титрования или секвестрования лиганда, связывание которого с эндогенным GPCR оказывает отрицательное воздействие на организм.
Белки GPCR, наряду с G-белками и эффекторами (внутриклеточными ферментами и каналами, которые регулируются G-белками) являются компонентами модульной сигнальной системы, связывающей статус внутриклеточных вторичных посредников с внеклеточными стимулами. Гены, их кодирующие, и продукты таких генов являются потенциальными этиологическими факторами заболевания.
Суперсемейство белков GPCR в настоящее время включает более 250 типов паралогов, рецепторов, которые представляют собой варианты, образованные в результате дупликации генов (или других процессов), в противоположность ортологам, соответствующим одному и тому же рецептору из различных видов. Суперсемейство можно разделить на пять семейств: семейство I, рецепторы, типичными представителями которых являются родопсин и бета2-адренорецептор и в настоящее время включает свыше 200 уникальных представителей; семейство II, недавно охарактеризованное семейство рецепторов паратиреоидного гормона/кальцитонина/секретина; семейство III, семейство метаботропного глутаматного рецептора у млекопитающих; семейство IV, семейство рецептора цАМФ, играющего важную роль в хемотаксисе и в развитии D.discoideum; и семейство V, рецепторы феромона спаривания грибов, например STE2.
GPCR включают рецепторы биогенных аминов, липидных медиаторов воспаления, пептидных гормонов и сенсорных сигнальных медиаторов. GPCR активируются при связывании рецептора с его внеклеточным лигандом. «Информационные изменения GPCR, вызванные лиганд-рецепторным взаимодействием, влияют на сродство связывания G-белка с внутриклеточными доменами GPCR. Это позволяет ГТФ связываться с G-белком с большим сродством.
Активация G-белка под действием ГТФ приводит к взаимодействию α-субъединицы G-белка с аденилатциклазой или с другим комплексом, синтезирующим вторичный посредник. Это взаимодействие регулирует активность аденилатциклазы и, следовательно, продукцию молекулы вторичного посредника цАМФ. цАМФ регулирует фосфорилирует и активацию других внутриклеточных белков. В качестве альтернативы, активность GPCR может увеличивать или снижать содержание в клетке молекул других вторичных посредников, таких как цГМФ или эйкозиноидов. α-субъединица G-белка дезактивируется при гидролизе ГТФ с участием ГТФазы, и α-, β и γ-субъединицы реассоциируют. Гетеротримерный G-белок затем диссоциирует от аденилатциклазы или другого комплекса, синтезирующего молекулу внутриклеточного посредника. Активность GPCR может также регулироваться путем фосфорилирования внутри- и внеклеточных доменов или петель.
Глутаматные рецепторы формируют группу GPCR, которая играет важную роль в процессе нейротрансмиссии. Глутамат является основным нейротрансмиттером в ЦНС и считается, что он играет важную роль в пластичности нейронов, познании, памяти, изучении и некоторых неврологических нарушениях, таких как эпилепсия, инсульт и нейродегенерация (Watson, S. and S. Arkinstall, 1994. The G- Protein Linked Receptor Facts Book, Academic Press, San Diego CA, pp.130-132). Такие эффекты глутамата опосредованы двумя различными классами рецепторов, называемых ионотропными и метаботропными. Ионотропные рецепторы содержат функционирующий катионный канал, и они опосредуют быстрое возбудительное действие глутамата. Метаботропные рецепторы представляют собой модуляторы, увеличивающие возбудимость мембраны нейронов путем ингибирования кальцийзависимых проводимостей калия и как ингибирования, так и усиления действия возбудительной трансмиссии ионотропных рецепторов. Метаботропные рецепторы разделяют на пять подтипов на основании фармакологии агонистов и путей сигнальной трансдукции, и они широко распространены в тканях мозга. Было показано, что N-гликозилирование является важным для функционирования метаботропного глутаматного рецептора 1 альфа (mGlu1alpha) человека (Mody et al., Neuropharmacology 38(10):1485-92, 1999). mGlu1alpha в норме экспрессируется, по меньшей мере частично, в виде димера, состоящего из мономеров ок. 145 и 160 кДа. Путем обработки mGlu1alpha туникамицином, эффективным ингибитором N-гликозилирования, Моди с соавторами (Mody et al.) показал, что хотя экспрессия на поверхности клетки не была затронута, экспрессировался только один пептид с массой 130 кДа, предсказанной на основании его первичной последовательности аминокислот. Функционально, обработка туникамицином снижала стимулированный агонистом гидролиз фосфоинозитида приблизительно на 50%, по сравнению с необработанной популяцией клеток. Таким образом, корректировка характера гликозилирования GPCR, экспрессируемого согласно системе настоящего изобретения, может быть полезна для регуляции сигнальной функции экспрессируемого GPCR и потенциально для регуляции или изменения фармацевтических или других свойств экспрессируемого GPCR.
Семейство вазоактивного интестинального полипептида (VIP) представляет собой группу родственных полипептидов, действие которых также опосредовано GPCR. Ключевые представители этого семейства - сам VIP, секретин и фактор, высвобождающий гормон роста (GRF). VIP имеет широкий профиль физиологических действий, в том числе - расслабление гладких мышц, стимулирование или ингибирование секреции в различных тканях, модуляция активности различных иммунных клеток и различные активности возбуждения и торможения в ЦНС. Секретин стимулирует секрецию ферментов и ионов в поджелудочную железу и кишку, и он также присутствует в небольшом количестве в мозге. Гликозилирование рецептора VIP, как было показано, важно для связывания его лиганда VIP (Chochola et al., J. Biol. Chem. 268: 2312-2318, 1993). Стерическое блокирование олигосахаридной цепи путем обработки рецептора VIP агглютинином зародыша пшеницы значительно ингибировало связывание VIP дозозависимым способом и снижало стимулированный VIP цАМФ-ответ. Кроме того, мутация специфических сайтов N-гликозилирования в рецепторе VIP вызывала задержку рецептора в эндоплазматическом ретикулуме, что указывает на то, что правильное гликозилирование важно для доставки рецептора VIP на поверхность клетки (Couvineau et al., Biochemistry 35(6): 1745-52, 1996). Таким образом, корректировка характера гликозилирования GPCR, экспрессируемого согласно настоящему изобретению, может быть полезна для модулирования связывания (например, либо повышения, либо уменьшения) экспрессируемого GPCR с соответствующим ему лигандом и потенциально для регулирования или влияния на фармацевтические или другие свойства экспрессируемого GPCR.
В общих чертах, при реализации настоящего изобретения на практике выбирают интересующий гликопротеин, точная последовательность аминокислот которого известна. Способы согласно настоящему изобретению имели успешное применение как для О-присоединенных (Примеры 1 и 2), так и для N-присоединенных (Примеры 3 и 4) гликопротеинов, указывая на то, что настоящее изобретение полезно для экспрессии гликопротеинов в целом. Любой гликопротеин, который можно экспрессировать в соответствии с настоящим изобретением может иметь собственные конкретные характеристики и может влиять на плотность клеток или жизнеспособность культивируемых клеток, может быть экспрессирован на более низком уровне, чем другой гликопротеин, экспрессируемый при одинаковых условиях культивирования, и может отличаться характером гликозилирования по одному или более сайту, в зависимости от конкретных условий культивирования и осуществляемых этапов. Специалист в данной области способен соответствующим образом модифицировать стадии и композиции, применяемые для продукции конкретного гликопротеина согласно замыслу настоящего изобретения для того, чтобы оптимизировать рост клетки и продукции и/или характера гликозилирования любого конкретного экспрессируемого гликопротеина.
В конкретных вариантах реализации ингибиторы фактора некроза опухоли в виде рецепторов факторов некроза опухоли альфа и бета (TNFR-1; ЕР 417,563, с датой публикации 20 марта 1991; и TNFR-2, ЕР 417,014, с датой публикации 20 марта 1991, включенные в данную заявку посредством ссылки) экспрессируют в соответствии с системами и способами согласно настоящему изобретению (см. обзор Naismith and Sprang, J Inflamm. 47(1-2):1-7, 1995-96, включенный в данную заявку посредством ссылки в полном объеме). Согласно некоторым вариантам реализации ингибитор фактора некроза опухоли включает растворимый рецептор TNF. В конкретных вариантах реализации ингибитор фактора некроза опухоли включает растворимый TNFR-Ig. В некоторых вариантах реализации ингибиторы TNF согласно настоящему изобретению представляют собой растворимые формы TNFRI и TNFRII. В некоторых вариантах реализации ингибиторы TNF согласно настоящему изобретению представляют собой растворимые TNF-связывающие белки. В некоторых вариантах реализации ингибиторы TNF согласно настоящему изобретению представляют собой гибридные белки TNFR-Ig, например TNFR-Fc или этанерцепт. При использовании в данной заявке «этанерцепт» относится к TNFR-Fc, который представляет собой димер двух молекул внеклеточной части рецептора р75 TNF-α, при этом каждая молекула включает Fc-область IgG1 человека, состоящую из 235 аминокислот.
ВВЕДЕНИЕ ГЕНОВ ДЛЯ ЭКСПРЕССИ ГЛИКОПРОТЕИНОВ В КЛЕТКИ-ХОЗЯЕВА
Как правило, молекула нуклеиновой кислоты, введенная в клетку, кодирует гликопротеин, который можно экспрессировать согласно настоящему изобретению. Кроме того, молекула нуклеиновой кислоты может кодировать продукт гена, который индуцирует экспрессию желаемого гликопротеина в клетке. Например, введенный генетический материал может кодировать фактор транскрипции, который активирует транскрипцию эндогенного или гетерологичного гликопротеина. В качестве альтернативы или дополнения, введенная молекула нуклеиновой кислоты может усиливать трансляцию или стабильность гликопротеина, экспрессируемого клеткой.
В данной области техники известны способы, подходящие для введения нуклеиновых кислот, достаточных для обеспечения экспрессии интересующего гликопротеина, в клетки-хозяева млекопитающего. См., например, Gething et al., Nature, 293:620-625, 1981; Mantei et al, Nature, 281:40-46, 1979; Levinson et al. EP 117,060; и ЕР 117,058, включенные в данную заявку посредством ссылки. Для клеток млекопитающих распространенные способы введения генетического материала в клетки включают способ с преципитацией фосфатом кальция согласно Graham и van der Erb (Virology, 52:456-457, 1978) или способ с использованием липофектамина™ (Gibco BRL) согласно Hawley-Nelson (Focus 15:73, 1993). Общие принципы трансформации клеточных систем млекопитающих описаны в патенте США №4,399,216, Axel, выданном 16 августа 1983. Различные способы введения генетического материала в клетки млекопитающих см. в Keown et al., Methods in Enzymology, 1989, Keown et al., Methods in Enzymology, 185:527-537, 1990, и Mansour et al., Nature, 336:348-352, 1988.
В некоторых вариантах реализации нуклеиновая кислота, которую можно ввести в клетки, находится в форме «голой» молекулы нуклеиновой кислоты. Например, молекула нуклеиновой кислоты, вводимая в клетку, может состоять только из нуклеиновой кислоты, кодирующей гликопротеин, и необходимых генетических контролирующих элементов. Кроме того, нуклеиновая кислота, кодирующая гликопротеин (включая необходимые регуляторные элементы), может находиться в пределах векторной плазмиды. Неограничивающие примеры пригодных векторов для экспрессии гликопротеинов в клетках млекопитающих включают pCDNAI; pCD, см. Okayama, et al. Mol. Cell Biol. 5:1136-1142, 1985; pMCIneo Poly-A; см. Thomas, et al. Cell 51:503-512, 1987; бакуловирусный вектор, такой как рАС 373 или рАС 610; CDM8, см. Seed, В. Nature 329:840, 1987; и рМТ2РС, см. Kaufman, et al. EMBO J. 6:187-195, 1987, включенные в данную заявку посредством ссылки в полном объеме. В некоторых вариантах реализации молекула нуклеиновой кислоты, которую можно ввести в клетку, содержится в пределах вирусного вектора. Например, нуклеиновая кислота, кодирующая гликопротеин, может быть включена в геном вируса (или часть вирусного генома). Регуляторные элементы, управляющие экспрессией гликопротеина, могут быть включены в нуклеиновую кислоту, встроенную в вирусный геном (т.е. связаны с геном, встроенным в вирусный геном), или могут быть предоставлены самим вирусным геномом.
«Голая» ДНК может быть введена в клетки посредством образования преципитата, содержащего ДНК и фосфат кальция. Кроме того, «голая» ДНК может быть также введена в клетки посредством образования смеси ДНК с DEAE-декстараном и инкубирования такой смеси с клетками или посредством инкубирования совместного инкубирования клеток и ДНК в подходящем буфере и воздействия на клетки высоковольтного электрического импульса (например, путем электропорации). Другим методом введения «голой» ДНК в клетки является смешивание ДНК с суспензией липосом, содержащих катионные липиды. Затем комплекс ДНК/липосома инкубируют с клетками. «Голую» ДНК можно также непосредственно инъецировать в клетки, например, путем микроинъекции.
Кроме того, «голую» ДНК можно также вводить в клетки путем формирования комплекса ДНК с катионом, например полилизином, который связан с лигандом к рецептору на поверхности клетки (см., например Wu, G. and Wu, С.Н. J. Biol. Chem. 263:14621, 1988; Wilson et al. J. Biol. Chem. 267:963-967, 1992; и патент США №5,166,320, включенный посредством ссылки в полном объеме). Связывание комплекса ДНК-лиганд с рецептором облегчает поглощение ДНК посредством рецептор-опосредованного эндоцитоза.
Применение вирусных векторов, содержащих конкретные последовательности нуклеиновых кислот, например кДНК, кодирующую гликопротеин, является распространенным подходом для введения последовательностей нуклеиновой кислоты в клетку. Преимуществом инфицирования клеток вирусным вектором состоит в том, что большая доля клеток получает нуклеиновую кислоту, что позволяет избежать необходимости селекции клеток, получивших нуклеиновую кислоту. К тому же, молекулы, закодированные в вирусном векторе, например кДНК, содержащаяся в вирусном векторе, обычно эффективно экспрессируются в клетках, получивших нуклеиновую кислоту вирусного вектора.
Дефектные ретровирусы хорошо охарактеризованы для применения в целях передачи генов или генной терапии (см. обзор Miller, A.D. Blood 76:271, 1990). Может быть сконструирован рекомбинантный ретровирус, содержащий нуклеиновую кислоту, кодирующую интересующий гликопротеин, включенную в ретровирусный геном. Кроме того, часть ретровирусного генома может быть удалена с целью нарушения системы репликации ретровируса. Такой ретровирус с нарушенной системой репликации затем упаковывают в вирионы, которые можно применять для инфицирования клеток-мишеней с помощью вируса-помощника стандартными способами.
Геном аденовируса может быть изменен таки образом, чтобы он кодировал и экспрессировал интересующий гликопротеин, но при этом был бы неактивен с точки зрения его способности к репликации при нормальном литическом жизненном цикле вируса. См., например, Berkner et al. BioTechniques 6:616, 1988; Rosenfeld et al. Science 252:431-434, 1991; и Rosenfeld et al. Cell 68:143-155, 1992. Подходящие аденовирусные векторы, полученные из аденовирусного штамма Ad5 типа dl324 или аденовируса другого типа (например, Ad2, Ad3, Ad7 и т.п.), известные специалистам в данной области. Преимущество рекомбинантных аденовирусов состоит в том, что им не требуются делящиеся клетки для эффективной системы доставки гена и их можно применять для инфицирования разнообразных типов клеток, включая эпителий дыхательных путей (Rosenfeld et al., 1992, см. выше), клетки эндотелия (Lemarchand et al., Proc. Natl. Acad. Sci. USA 89:6482-6486, 1992), гепатоциты (Herz and Gerard, Proc. Natl. Acad. Sci. USA 90:2812-2816, 1993) и мышечные клетки (Quantin et al., Proc. Natl. Acad. Sci. USA 89:2581-2584, 1992). Кроме того, введенная аденовирусная ДНК (и содержащаяся в ней чужеродная ДНК) не интегрируется в геном клетки-хозяина, но остается в виде эписомы, таким образом избегая потенциальных проблем, которые могут произойти в результате инсерционного мутагенеза, в случаях, когда введенная ДНК встраивается в геном клетки-хозяина (например, ретровирусная ДНК). Кроме того, аденовирусный геном обладает большей вмещающей способностью для чужеродной ДНК (вплоть до 8 тысяч пар оснований) по сравнению с другими векторами для доставки гена (Berkner et al. см выше; Haj-Ahmand and Graham, J. Virol. 57:267, 1986). В большинстве используемых в настоящее время аденовирусных векторов с нарушенной системой репликации удалены целиком или частично вирусные гены Е1 и Е3, при этом сохранено до 80% генетического материала аденовируса.
Аденоассоциированный вирус (ААВ) представляет собой природный дефектный вирус, нуждающийся в другом вирусе, например аденовирусе или вирусе герпеса, в качестве вируса-помощника для осуществления его эффективной репликации и продуктивного жизненного цикла (см. обзор Muzyczka et al. Curr. Topics in Micro, and Immunol., 158:97-129, 1992). ААВ также является одним из нескольких вирусов, который может вводить свою ДНК в неделящиеся клетки и демонстрирует высокую частоту стабильной интеграции (см., например, Flotte et al., Am. J. Respir. Cell. Mol. Biol. 7:349-356, 1992; Samulski et al., J. Virol. 63:3822-3828, 1989; and McLaughlin et al., J. Virol. 62:1963-1973, 1989). Могут быть упакованы и введены в клетку векторы, содержащие всего 300 пар оснований ААВ. Размер экзогенной ДНК ограничен около 4,5 тысячами п.о. Вектор ААВ, например, описанный в работе Tratschin et al. (Mol. Cell. Biol. 5:3251-3260, 1985), можно применять для введения ДНК в клетки. С применением векторов ААВ осуществлено введение различных нуклеиновых кислот в различные типы клеток (см., например, Hermonat et al., Proc. Natl. Acad. Sci. USA 81:6466-6470, 1984; Tratschin et al., Mol. Cell. Biol. 4:2072-2081, 1985; Wondisford et al., Mol. Endocrinol. 2:32-39, 1988; Tratschin et al., J. Virol. 51:611-619, 1984; и Flotte et al., J. Biol. Chem. 268:3781-3790, 1993).
При применении данного способа введения молекул нуклеиновых кислот в популяцию клеток, который приводит к модификации большей части клеток и эффективной экспрессии гликопротеина клетками, модифицированную популяцию клеток можно в применять без дополнительного выделения или субклонирования индивидуальных клеток в пределах популяции. То есть продукция гликопротеина популяцией клеток может быть достаточной, таким образом, что не требуется дополнительной изоляции клеток, и популяцию клеток можно сразу же использовать для засева культуры клеток для продукции гликопротеина. Кроме того, может быть желательным изолировать и увеличить гомогенную популяцию клеток из нескольких клеток или единичной клетки для эффективной продукции гликопротеина.
В качестве альтернативы введению в клетки молекулы нуклеиновой кислоты, кодирующей интересующий гликопротеин, вводимая нуклеиновая кислота может кодировать другой полипептид или белок, который индуцирует или увеличивает уровень экспрессии гликопротеина, эндогенно продуцируемого клеткой. Например, клетка может быть способна экспрессировать конкретный гликопротеин, но экспрессия может не осуществляться без дополнительной обработки клетки. Аналогично, клетка может экспрессировать недостаточное количество гликопротеина для заданной цели. Таким образом, агент, который стимулирует экспрессию интересующего гликопротеина, можно применять для индукции или увеличения экспрессии данного гликопротеина клеткой. Например, введенная молекула нуклеиновой кислоты может кодировать фактор транскрипции, который активирует или увеличивает уровень транскрипции интересующего гликопротеина. Экспрессия такого фактора транскрипции в свою очередь приводит к экспрессии или более высокому уровню экспрессии интересующего гликопротеина.
В некоторых вариантах реализации нуклеиновую кислоту, обеспечивающую экспрессию гликопротеина, вводят в клетку-хозяин на постоянной основе. В некоторых вариантах реализации нуклеиновую кислоту, обеспечивающую экспрессию гликопротеина, вводят в клетку-хозяин временно. Специалист в данной области сможет легко выбрать на основании задач эксперимента, вводить ли нуклеиновую кислоту стабильно или временно.
Ген, кодирующий интересующий гликопротеин, может быть соединен с одним или более регуляторным генетическим контролирующим элементом. В некоторых вариантах реализации генетический контролирующий элемент обеспечивает конститутивную экспрессию гликопротеина. В некоторых вариантах реализации можно применять генетический контролирующий элемент, обеспечивающий индуцибельную экспрессию гена, кодирующего интересующий гликопротеин. Применение индуцибельного генетического контролирующего элемента (например, индуцибельного промотора) позволяет модулировать продукцию гликопротеина в клетке. Не ограничивающие примеры индуцибельных генетических контролирующих элементов, которые потенциально подходят для применения в эукариотических клетках, включают гормонрегулируемые элементы (например, см. Mader, S. and White, J.H., Proc. Natl. Acad. Sci. USA 90:5603-5607, 1993), элементы, регулируемые синтетическими лигандами (см., например, Spencer, D.M. et al., Science 262:1019-1024, 1993), и элементы, регулируемые ионизирующим излучением (например, см. Маnоmе, Y. et al., Biochemistry 32:10607-10613, 1993; Datta, R. et al., Proc. Natl. Acad. Sci. USA 89:10149-10153, 1992). В данной области известны дополнительные специфичные для конкретного типа клеток или другие регуляторные системы, и они могут быть использованы в соответствии с настоящим изобретением.
Специалист в данной области сможет выбрать, а также, возможно, соответствующим образом модифицировать способ введения генов, обеспечивающих экспрессию интересующего гликопротеина в клетке, в соответствии с замыслом настоящего изобретения.
КЛЕТКИ
Любую клетку-хозяин, которую можно культивировать и которая может экспрессировать гликопротеин, можно применять в соответствии с настоящим изобретением. В некоторых вариантах реализации клетка хозяин представляет собой клетку млекопитающего. Не ограничивающие примеры клеток млекопитающих, которые можно применять в соответствии с настоящим изобретением, включают линию клеток миеломы мышей BALB/c (NSO/I, ЕСАСС No: 85110503); ретинобласты человека (PER.C6 (CruCell, Leiden, Нидерланды)); линию клеток печени обезьян CV1, трансформированная SV40 (COS-7, АТСС CRL 1651); линию клеток печени зародыша человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol., 36:59,1977); клетки почек детеныша хомячка (ВНК, АТСС CCL 10); клетки яичников китайского хомяка +/-DHFR (СНО, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA, 77:4216, 1980); клетки Сертоли мыши (TM4, Mather, Biol. Reprod., 23:243-251, 1980); клетки почек обезьяны (CV1 АТСС CCL 70); клетки почек африканской зеленой мартышки (VERO-76, АТСС CRL-1 587); клетки цервикальной карциномы человека (HeLa, АТСС CCL 2); клетки почек собаки (MDCK, АТСС CCL 34); клетки печени крыс линии Буффало (BRL 3А, АТСС CRL 1442); клетки легкого человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383:44-68, 1982); клетки MRC 5; клетки FS4; и линия клеток гепатомы человека (Hep G2).
Кроме того, в соответствии с настоящим изобретением можно применять любые коммерчески доступные и некоммерческие линии гибридомных клеток, экспрессирующих гликопротеины. Специалисту в данной области ясно, что линии клеток гибридомы могут нуждаться в различных питательных веществах, и/или им могут требоваться различные условия культивирования для оптимального роста и экспрессии гликопротеина, и специалист сможет модифицировать условия по необходимости.
Как упомянуто выше, во многих случаях осуществляют селекцию клеток или модифицируют их генно-инженерными методами для продукции гликопротеина на высоком уровне. Часто клетки направленно изменяют для продукции гликопротеина на высоком уровне, например путем введения гена, кодирующего интересующий гликопротеин, и/или введением генетических контролирующих элементов, которые регулируют экспрессию упомянутого гена (либо эндогенного, либо привнесенного).
Как ясно специалисту в данной области, гликопротеины, продуцируемые в различных типах клеток, могут содержать различный итоговый характер гликозилирования. Например, Przybylo с соавторами показал, что характер гликозилирования кадгеринов различается при экспрессии в незлокачественных клетках эпителия мочеточника, в клетках HCV29, трансфецированных v-raf, и промежуточноклеточном раке мочевого пузыря (см. Przybylo et al., Cancer Cell International, 2(1):6, 2002). Lifely с соавторами показал, что характер гликозилирования и биологическая активность гуманизированного антитела IgG различались при его экспрессии в СНО, линии клеток миеломы Y0 и линии клетках миеломы NS0 (см. Lifely et al., Glycobiology. 5(8):813-22, 1995). Специалист в данной области сможет выбрать необходимую линию клеток для продукции конкретного гликопротеина без проведения лишних экспериментов. Вне зависимости от выбранной линии клеток гликопротеин может быть экспрессирован в соответствии с настоящим изобретением с повышенной степенью гликозилирования.
Некоторые гликопротеины могут оказывать неблагоприятное воздействие на рост клеток, жизнеспособность клеток или некоторые другие характеристики клеток, которые в конечном счете ограничивают продукцию интересующего гликопротеина каким-либо образом. Даже среди популяции клеток одного конкретного типа, предназначенной для экспрессии специфического гликопротеина, существует изменчивость в пределах популяции клеток, такая что некоторые отдельные клетки растут лучше, продуцируют большее количество интересующего гликопротеина, продуцируют гликопротеин с повышенной степенью гликозилирования или продуцируют гликопротеин, характер гликозилирования которого более точно отражает характер гликозилирования природного гликопротеина. В конкретных вариантах реализации специалист-практик выбирает линию клеток эмпирически для интенсивного роста при конкретных условиях, выбранных для культивирования клеток. В некоторых вариантах реализации отбирают индивидуальные клетки, предназначенные для экспрессии конкретного гликопротеина, для крупномасштабной продукции на основании роста клеток, конечной плотности клеток, процента жизнеспособности клеток, титра экспрессируемого гликопротеина, протяженности и состава боковых олигосахаридных цепей или любой комбинации этих или любых других условий, важных, по мнению специалиста-практика.
КУЛЬТИВИРОВАНИЕ КЛЕТОК
Настоящее изобретение можно применять с любым способом культивирования клеток, подходящим для экспрессии гликопротеинов. Например, клетки могут быть выращены в периодических культурах или культурах с подпиткой, при этом рост культуры останавливают после того, как гликопротеин экспрессируется в достаточном количестве, и затем отбирают экспрессированный гликопротеин. Кроме того, клетки могут быть выращены в перфузионных культурах, при этом культуру не останавливают в росте и периодически или непрерывно добавляют к культуре питательные вещества и другие компоненты, а экспрессированный гликопротеин периодически или непрерывно.
Клетки могут быть выращены в любом удобном объеме, выбранном специалистом-практиком. Например, клетки могут быть выращены в сосудах для проведения реакции в небольших масштабах, объемом от нескольких миллилитров до несколько литров. Кроме того, клетки могут быть выращены в коммерческих биореакторах для крупномасштабного производства, объемом от приблизительно по меньшей мере 1 литра до 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000 литров или более, или любого объема между указанными значениями.
Температуру культивирования клеток выбирают главным образом на основании диапазона температур, при которых культура клеток остается жизнеспособной, диапазона, при котором наблюдается высокий уровень продукции гликопротеина, и/или диапазона, при котором экспрессированный гликопротеин имеет желательный характер гликозилирования. Например, клетки СНО хорошо растут и могут продуцировать гликопротеины с желательным характером гликозилирования в количествах, соответствующих коммерческим целям, приблизительно при 37°С. В целом, большинство клеток млекопитающих наиболее хорошо растут и могут продуцировать гликопротеины с желательным характером гликозилирования в количествах, соответствующих коммерческим целям, в диапазоне от 25°С до 42°С, хотя способы, излагаемые в настоящем описании, не ограничены этими температурами. Некоторые клетки млекопитающих хорошо растут и могут продуцировать гликопротеины с желательным характером гликозилирования в количествах, соответствующих коммерческим целям, в диапазоне от приблизительно 35°С до 40°С. В некоторых вариантах реализации культуру клеток выращивают при температуре 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45°С за один или более раз в течение процесса культивирования клеток. Специалист в данной области способен выбрать подходящую температуру или температуры для выращивания клеток в зависимости от конкретных потребностей клеток и конкретных практических задач, стоящих перед специалистом-практиком.
Кроме того, в процессе культивирования можно один или более раз изменять температурный режим. При смене температурного режима культивирования изменение температуры может быть сравнительно постепенным. Например, процесс изменения температуры может занимать несколько часов или дней. Альтернативно, изменение температуры может быть сравнительно внезапным. Температура может непрерывно повышаться или уменьшаться в течение процесса культивирования. Альтернативно, температура может повышаться или уменьшаться дискретно на различных этапах в процессе культивирования. Последующая температура (температуры) или диапазон (диапазоны) температур может быть ниже или выше начальной или предшествующей температуры (температур) или диапазона температур. Как понятно специалисту в данной области, различные дискретные изменения температуры включены в данный вариант реализации изобретения. Например, температура может быть изменена один раз (либо на более высокую, либо на более низкую температуру или диапазон температур), клетки содержат при такой температуре или диапазоне температур в течение некоторого промежутка времени, после чего температура может снова быть изменена на новую температуру или диапазон температур, которая/который может быть либо выше, либо ниже предшествующей температуры или диапазона температур. Температура культивирования после каждого дискретного изменения может быть постоянной или может поддерживаться в пределах некоторого диапазона температур.
Как и исходную температуру или диапазон температур, температуру или диапазон температур культивирования клеток после изменения температуры, как правило, выбирают на основании температуры (температур), при которой клеточная культура остается жизнеспособной, диапазона, при котором гликопротеин продуцируется на высоком уровне, и/или диапазона, при котором экспрессированный гликопротеин характеризуется желательным характером гликозилирования. Как правило, большинство клеток млекопитающих остаются жизнеспособными и экспрессируют гликопротеины с желательным характером гликозилирования в количествах, соответствующих коммерческим целям, в диапазоне от 25°С до 42°С, хотя способы, излагаемые в настоящем описании, не ограничиваются такими значениями температуры. В некоторых вариантах реализации клетки млекопитающих остаются жизнеспособными и экспрессируют гликопротеины с желаемым характером гликозилирования в количествах, соответствующих коммерческим целям, в диапазоне температур от 25°С до 35°С.Специалисты в данной области способны выбрать подходящую температуру (температуры) или диапазон температур (диапазоны) для выращивания клеток, в зависимости от конкретных потребностей клеток и от задач, стоящих перед специалистом-практиком. Клетки можно выращивать в течение любого времени, в зависимости от потребностей специалиста-практика и потребностей самих клеток.
В конкретных вариантах реализации периодическое культивирование или культивирование с подпиткой останавливают, когда количество экспрессированного гликопротеина достигает достаточно высокого титра и/или когда экспрессированные гликопротеины демонстрируют желательный характер гликозилирования, в соответствии с потребностями специалиста-практика. В качестве дополнения или альтернативы, реакции периодического культивирования или культивирования с подпиткой останавливают, как только клетки достигнут достаточно высокой плотности, в соответствии с потребностями специалиста-практика. Например, культивирование может быть прекращено, как только клетки достигнут 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99 процентов максимальной плотности жизнеспособных клеток. В качестве дополнения или альтернативы, периодическое культивирование или культивирование с подпиткой может быть остановлено до избыточного накопления конечных продуктов обмена веществ, например лактата и аммония.
В некоторых случаях может быть полезно добавление в культуру клеток в ходе последующей стадии производства питательных веществ или других компонентов среды, истощенных или метаболизированных клетками. В качестве неограничивающих примеров, может быть полезно добавление в культуру клеток гормонов и/или других факторов роста, неорганических ионов (таких как, например, натрий, хлорид, кальций, магний и фосфат), буферов, витаминов, нуклеозидов или нуклеотидов, микроэлементов (неорганические соединения, как правило, присутствующие в очень низкой конечной концентрации), аминокислот, липидов или глюкозы или других источников энергии. Такие вспомогательные компоненты можно добавлять к культуре клеток одновременно, или их можно добавлять в культуру клеток последовательно.
В некоторых вариантах реализации клетки выращивают согласно любому из способов культивирования клеток, описанных в предварительной заявке на патент США сер. №60/605,097, включенной в данное описание посредством ссылки. В некоторых вариантах реализации клетки выращивают при одном или более варианте условий, описанных в заявке на патент Соединенных Штатов сер. №11/213,308, включенной в данную заявку посредством ссылки.
Специалист в данной области сможет адаптировать конкретные условия культивирования клеток для оптимизации конкретных характеристик культуры клеток, включающих, но не ограниченных перечисленными, скорость роста, жизнеспособность клеток, конечная плотность клеток в культуре, конечная концентрация вредных побочных продуктов обмена веществ, например лактата и аммония, конечный титр экспрессированного гликопротеина, протяженность и состав боковых олигосахаридных цепей или любой комбинации этих или других параметров, важных по мнению специалиста-практика.
ВЫДЕЛЕНИЕ ЭКСПРЕССИРОВАННОГО ГЛИКОПРОТЕИНА
Как правило, в большинстве случаев желательно выделение и/или очистка гликопротеинов, экспрессированных согласно настоящему изобретению. В конкретных вариантах реализации экспрессированный гликопротеин секретируется в среду, и в таком случае клетки и другие твердые компоненты можно удалить, например, путем центрифугирования или фильтрации, на первом этапе процесса очистки.
Альтернативно, экспрессированный гликопротеин может быть связан с поверхностью клетки-хозяина. Например, можно удалить среду и лизировать клетки-хозяева, экспрессирующие гликопротеины, на первом этапе процесса очистки. Лизис клеток-хозяев млекопитающих можно осуществить при помощи ряда способов, известных специалистам в данной области, включая физическое разрушение с помощью стеклянных бусин и воздействие высоких значений рН.
Экспрессированный гликопротеин может быть выделен и очищен стандартными способами, включающими, но не ограниченными перечисленными, хроматографию (например, ионообменная, аффинная, эксклюзионная и гидроксиапатитная хроматография), гель-фильтрацию, центрифугирование или дифференциальную растворимость, преципитацию этанолом и/или любым другим доступным способом очистки белков (см., например, Scopes, Protein Purification Principles and Practice 2nd Edition, Springer-Verlag, New York, 1987; Higgins, S.J. and Hames, B.D. (eds.), Protein Expression: A Practical Approach, Oxford Univ Press, 1999; and Deutscher, M.P., Simon, M.I., Abelson, J.N. (eds.), Guide to Protein Purification: Methods in Enzymology (Methods in Enzymology Series, Vol.182), Academic Press, 1997, включенные в данную заявку посредством ссылки). В случае иммуноаффинной хроматографии, в частности, гликопротеин может быть выделен путем его связывания с аффинной колонкой, содержащей антитела, полученные к такому гликопротеину и фиксированные на стационарной подложке. Кроме того, для облегчения очистки при пропускании гликопротеина через аффинную колонку к гликопротеину может быть присоединена с помощью стандартных рекомбинантных способов аффинная метка, такая как последовательность оболочки вируса гриппа, полигистидин или глутатион-S-трансфераза. Для уменьшения или предотвращения деградации гликопротеина в процессе очистки на любом или на каждом этапе можно добавлять ингибиторы протеаз, например фенилметилсульфонилфторид (PMSF), леупептин, пепстатин или апротинин. Применение ингибиторов протеаз особенно полезно при необходимости лизиса клеток для выделения и очистки экспрессированного гликопротеина. В качестве дополнения или альтернативы, на любом или каждом этапе можно добавлять ингибиторы гликозидазы для уменьшения или предотвращения энзиматического отщепления ковалентно присоединенных олигосахаридных цепей.
Гликопротеины, экспрессированные согласно настоящему изобретению, могут иметь более протяженный или иным образом измененный характер гликозилирования, чем если бы такие гликопротеины продуцировали при стандартных условиях культивирования клеток. Таким образом, одно из практических преимуществ настоящего изобретения, которое можно применять на стадии очистки, состоит в том, что дополнительные и/или измененные остатки сахара на молекуле гликопротеина, полученного в соответствии с некоторыми способами согласно настоящему изобретению, могут сообщать молекуле гликопротеина различные биохимические свойства, которые может применять специалист-практик для более легкой очистки такого гликопротеина или для достижения большей степени очистки, чем если бы гликопротеин получали с использованием более традиционных способов культивирования клеток.
Специалисту в данной области очевидно, что конкретные способы очистки варьируют в зависимости от типа гликопротеина, подлежащего очистке, типа клеток, в которых экспрессируют гликопротеин, и/или состава среды, в которой выращивают клетки.
ИМУНОГЕННЫЕ КОМПОЗИЦИИ
Гликопротеины, полученные согласно настоящему изобретению, можно также применять в иммуногенных композициях, например вакцинах. В некоторых вариантах реализации улучшенный характер гликозилирования, достигаемый в результате продукции гликопротеинов в соответствии со способами согласно настоящему изобретению, может обеспечивать более эффективную иммуногенную композицию. Например, иммуногенная композиция, содержащая полученный гликопротеин, может запускать более эффективный иммунный ответ, при котором иммунная система объекта вырабатывает большее количество антител к гликопротеину и/или вырабатывает антитела, демонстрирующие большую специфичность к иммуногенному гликопротеину. В качестве дополнения или альтернативы, такой гликопротеин может запускать иммунный ответ с менее выраженными и/или менее серьезными побочными эффектами. В конкретных вариантах реализации иммуногенные композиции изобретения включают один или более гликопротеин. В качестве дополнения или альтернативы, иммуногенная композиция согласно изобретению может включать один или более физиологически приемлемый носитель.
Как правило, выбор подходящего «эффективного количества» или дозировки компонентов иммуногенной композиции (композиций) согласно настоящему изобретению основан на ряде факторов, включающих, но не ограниченных перечисленными, природу выбранного гликопротеина (гликопротеинов) в применяемой иммуногенной композиции, характер гликозилирования гликопротеина (гликопротеинов) и физического состояния субъекта, в частности, включающего общее состояние здоровья, возраст и/или вес иммунизированного субъекта. Как известно в данной области, конкретные способы и пути введения и присутствие дополнительных компонентов в иммуногенной композиции могут также повлиять на дозировку и количество композиции, включающей плазмидную ДНК. Выбор и увеличение или уменьшение эффективной дозы находится в пределах компетенции специалиста в данной области. Количество иммуногенной композиции, необходимое для запуска иммунного ответа, включающего защитный ответ, но не ограниченного им, или необходимое для оказания внешних эффектов на пациента без значительных неблагоприятных побочных эффектов, варьирует в зависимости от этих факторов. Специалисты в данной области могут легко определить подходящие дозы.
Определенные иммуногенные композиции согласно настоящему изобретению могут содержать адъювант.Адъювант представляет собой вещество, усиливающее иммунный ответ при совместном введении с иммуногеном или антигеном. Было показано, что многие цитокины или лимфокины обладают иммуномодулирущей активностью, и таким образом их можно применять в качестве адъювантов, и они включают, но не ограничены перечисленными, интерлейкины 1-α, 1-β, 2, 4, 5, 6, 7, 8, 10, 12 (см., например, патент США 5,723,127, включенный в данную заявку посредством ссылки в полном объеме), 13, 14, 15, 16, 17 и 18 (и их мутантные формы), интерфероны α, β и γ, гранулоцитарно-макрофагальный колониестимулирующий фактор (см., например, патент США №5,078,996, включенный в данную заявку посредством ссылки в полном объеме), макрофагальный колониестимулирующий фактор, гранулоцитарный колониестимулирующий фактор, GSF и факторы некроза опухоли α и β. Другие адъюванты, применимые в настоящем изобретении, включают хемокин, включающие, без ограничений, МСР-1, MIP-1α, MIP-1β и RANTES. Молекулы адгезии, такие как селектин, например L-селектин, Р-селектин и Е-селектин, также можно применять в качестве адъювантов. Другие адъюванты, которые можно применять, включают, без ограничений, муцин-подобную молекулу, например CD34, GlyCAM-1 и MadCAM-1, член семейства интегринов, например LFA-1, VLA-1, Мас-1 и р150.95, член суперсемейства иммуноглобулинов, например РЕСАМ, ICAM, например ICAM-1, ICAM-2 и ICAM-3, CD2 и LFA-3, костимулирующие молекулы, такие как CD40 и CD40L, факторы роста, в том числе фактор роста сосудов, фактор роста нервов, фактор роста фибробластов, эпидермальный фактор роста, В7.2, PDGF, BL-1 и фактор роста эндотелия сосудов, рецепторные молекулы, включающие рецептор Fas, рецептор TNF, FIt, Аро-1, р55, WSL-1, DR3, TRAMP, Аро-3, AIR, LARD, NGRF, DR4, DR5, KILLER, TRAIL-R2, TRICK2 и DR6. Еще одна молекула адъюванта включает каспазу (ICE). См. также международные публикации патентов WO98/17799 и WO99/43839, включенные в данную заявку посредством ссылки в полном объеме.
Также в качестве адъювантов можно применять холерные токсины (XT) и их мутантные формы, в том числе описанные в международной публикации заявки на патент WO 00/18434 (в которых глутаминовая кислота в позиции 29 последовательности аминокислот заменена на другую аминокислоту (отличную от аспарагиновой кислоты), предпочтительно гистидин). Схожие XT или мутантные формы описаны в международной публикации заявки на патент WO 02/098368 (в которых изолейцин в позиции 16 заменен на другую аминокислоту, либо в отдельности, либо в комбинации с заменой серина в позиции 68 на другую аминокислоту; и/или в которых валин в позиции 72 заменен на другую аминокислоту). Другие токсины XT описаны в международной публикации заявки на патент WO 02/098369 (в которых аргинин в позиции 25 заменен на другую аминокислоту; и/или содержащие вставку аминокислоты в позиции 49; и/или вставку двух аминокислот в позициях 35 и 36). Каждый из этих источников включен в данную заявку посредством ссылки в полном объеме.
В некоторых вариантах реализации иммуногенную композицию согласно настоящему изобретению вводят человеку или позвоночному, отличному от человека, с помощью различных способов, включающих, но не ограниченных перечисленными, интраназальное, пероральное, вагинальное, ректальное, парентеральное, интрадермальное, трансдермальное (см., например, международную публикацию №WO 98/20734, включенную в данную заявку посредством ссылки в полном объеме), внутримышечное, внутрибрюшинное, подкожное, внутривенное и внутриартериальное введение. Подходящий способ может быть выбран в зависимости от природы применяемой иммуногенной композиции, оценки возраста, веса, пола и общего состояния здоровья пациента, а также антигенов, присутствующих в иммуногенной композиции, и/или от других факторов, известных специалистам в данной области.
В некоторых вариантах реализации настоящего изобретения иммуногенные композиции вводят многократно. Порядок введения иммуногенной композиции и временные интервалы между отдельными введениями могут быть выбраны специалистом в данной области на основании соответствующих факторов, известных специалистам в данной области, включающих, но не ограниченных перечисленными, физические характеристики и конкретные ответные реакции хозяина на применение указанного способа.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
В конкретных вариантах реализации настоящего изобретения полученные гликопротеины обладают фармакологической активностью и применимы для приготовления фармацевтических препаратов. Композиции согласно настоящему изобретению, как описано выше, можно вводить субъекту, или сначала они могут быть сформулированы для доставки любым доступным способом, включающим, но не ограниченным перечисленными, парентеральный (например, внутривенный), интрадермальный, подкожный, пероральный, носовой, бронхиальный, глазной, трансдермальный (локальный), трансмукозальный, ректальный и вагинальный способы. Фармацевтические композиции согласно настоящему изобретению обычно включают очищенный гликопротеин, экспрессированный в линии клеток млекопитающего, агент для доставки (т.е. катионный полимер, пептидный молекулярный транспортер, сурфактант и т.п., как описано выше) в комбинации с фармацевтически приемлемым носителем. При использовании в данной заявке выражение «фармацевтически приемлемый носитель» включает растворитель, диспергатор, компонент оболочки, антибактериальный и противогрибковый агент, изотонический агент и агент, замедляющий абсорбцию, и т.п, совместимый с фармацевтическим введением. Вспомогательные активные соединения могут также входить в состав композиции. Например, гликопротеин, полученный согласно настоящему изобретению, может быть дополнительно конъюгирован с другим лекарственным средством для системной фармакотерапии, например с токсином, низкомолекулярным цитотоксическим средством, веществом, модифицирующим биологический ответ, и радионуклидом (см., например, Kunz et al., Calicheamicin derivative-carrier conjugates, US20040082764 A1).
В качестве альтернативы или дополнения, белок или полипептид, полученный согласно настоящему изобретению, можно вводить в комбинации (либо одновременно, либо последовательно) с одним или более дополнительным фармацевтически активным агентом. Перечень примеров таких фармацевтически активных агентов можно найти в Настольном справочнике врача Physicians' Desk Reference, 55 издание, изданный Medical Economics Co., Inc., Montvale, NJ, 2001, включенном в данную заявку посредством ссылки. Для большинства из таких агентов известны фармацевтически эффективные дозировки и режимы, и многие из них указаны в самом Настольном справочнике врача.
Фармацевтическая композиция преимущественно составлена таким образом, что она является совместимой с ее предполагаемым способом введения. Растворы или суспензии, подходящие для парентерального, интрадермального или подкожного применения, могут включать следующие компоненты: стерильный растворитель, такой как вода для инъекции, солевой раствор, нелетучее масло, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, например бензиловый спирт или метилпарабены; антиоксиданты, например аскорбиновую кислоту или бисульфит натрия; хелатирующие агенты, например этилендиаминтетрауксусная кислота; буферы, такие как ацетатный, цитратный или фосфатный, и агенты для поддержания тоничности, например хлорид натрия или декстрозу. рН может быть скорректирован добавлением кислоты или основания, например соляной кислоты или гидроксида натрия. Препараты для парентерального введения могут быть заключены в ампулы, одноразовые шприцы или во флаконы с многократными дозами, сделанные из стекла или пластмассы.
Фармацевтические композиции, подходящие для введения путем инъекции, обычно включают стерильные водные растворы (если компоненты водорастворимые) или коллоидные растворы и стерильные порошки для экстемпорального приготовления стерильных растворов или коллоидных растворов для введения. Для внутривенного введения подходящие носители включают физиологический солевой раствор, бактериостатическую воду, Cremophor EL™ [BASF, Parsippany, NJ) или фосфатно-солевой буфер (ФСБ). Во всех случаях композиция должна быть достаточно жидкой для возможности инъекции. Некоторые фармацевтические композиции согласно настоящему изобретению стабильны при условиях производства и хранения и должны быть устойчивы к заражению микроорганизмами, такими как бактерии и грибы. В целом, подходящий носитель может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерол, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их соответствующие смеси. Необходимая степень текучести может поддерживаться, например, с помощью оболочки, например лецитина, использованием частиц необходимого размера в случае коллоидного раствора, использованием сурфактантов. Устойчивость к действию микроорганизмов может обеспечиваться различными антибактериальными и противогрибковыми агентами, например парабенами, хлорбутанолом, фенолом, аскорбиновой кислотой, тимерозалом и т.п. Во многих случаях будет полезно включение в состав композиции изотонических агентов, например сахаров, полиспиртов, например маннитола, сорбитола или хлорида натрия. Замедленная абсорбция композиций, вводимых путем инъекции, может обеспечиваться включением в композицию агента, замедляющего абсорбцию, например моностеарата алюминия и желатина.
Стерильные растворы для инъекции могут быть приготовлены путем включения очищенного гликопротеина в требующемся количестве в подходящем растворителе в отдельности или в комбинации с компонентами, перечисленными выше, по необходимости, с последующей стерилизацией путем фильтрации. Как правило, коллоидные растворы получают путем включения очищенного гликопротеина, экспрессированного в линии клеток млекопитающего, в стерильный наполнитель, который содержит основную дисперсионную среду и другие необходимые компоненты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекции способы приготовления включают, например, вакуумную сушку и лиофилизацию, в результате которых получают порошок активного компонента, а также любой дополнительный необходимый компонент, раствор такого компонента, предварительно стерилизованный путем фильтрации.
Композиции для перорального введения обычно включают инертный растворитель или пищевой носитель. Для терапевтического перорального введения очищенный гликопротеин можно включать в состав с композиции с наполнителем и применять в форме таблеток, пастилок или капсул, например желатиновых капсул. Композиции для перорального введения можно также приготовить с использованием жидкого носителя для использования в виде полоскания для рта. В состав композиции могут быть включены фармацевтически совместимые связывающие агенты и/или адъюванты. Таблетки, пилюли, капсулы, пастилки и т.п. могут содержать любой из следующих компонентов или соединения аналогичной природы: связующее вещество, например микрокристаллическую целлюлозу, камедь трагаканта или желатин; наполнитель (эксципиент), например крахмал или лактозу, дезинтегрирующий агент, например альгиновую кислоту, Primogel или зерновой крахмал; смазывающее вещество, например стеарат магния или Sterotes; глидант, например коллоидный диоксид кремния; подсластитель, например сахарозу или сахарин; или ароматизатор, например перечную мяту, метилсалицилат или апельсиновый ароматизатор. Препараты для перорального введения могут также включать агенты для увеличения стабильности в желудочно-кишечном тракте и/или увеличения абсорбции.
Для введения путем ингаляции композиции согласно настоящему изобретению, включающие очищенный гликопротеин, экспрессированный в линии клеток млекопитающего, и агент для доставки, преимущественно доставляют в форме распыляемый аэрозоль из контейнера под давлением или диспенсера, который содержит подходящий пропеллент, например газ, такой как углекислый газ, или распылителя. Настоящее изобретение, в частности, предусматривает доставку композиций с использованием назального аэрозоля, ингалятора или другого средства для прямой доставки в верхние и/или нижние дыхательные пути. Было показано, что введение интраназальных вакцин, направленных против вирусов гриппа, индуцирует ответ CD8 Т-клеток, что указывает на то, что, по меньшей мере, некоторые клетки в дыхательных путях могут поглотить ДНК при доставке таким способом, и агенты для доставки согласно изобретению повышают поглощение композиции клетками. Согласно некоторым вариантам реализации композиции, включающие очищенный гликопротеин, экспрессированный линией клеток млекопитающего, и агент для доставки, входят в состав препарата в виде больших пористых частиц для введения с помощью аэрозоля.
Системное введение также можно осуществлять трансмукозальным или трансдермальным способом. Для трансмукозального или трансдермального введения в препаратах применяют смачивающий реагент, способный проходить через соответствующий барьер. Такие смачивающие реагенты в целом известны в данной области и включают, например, для трансмукозального введения, детергенты, соли желчных кислот и производные фусидовой кислоты. Трансмукозальное введение можно осуществлять с помощью назальных спреев или суппозиториев. Для трансдермального введения очищенные гликопротеины и агенты для доставки входят в состав препаратов в виде мазей, бальзамов, гелей или кремов, как в целом известно в данной области.
Композиции могут также входить в состав препаратов в виде суппозиториев (например, со стандартными основами для суппозиториев, таких как масло какао и другие глицериды) или удерживающих клизм для ректальной доставки.
В конкретных вариантах реализации фармацевтические композиции согласно изобретению содержат дополнительный наполнитель, например местный анестетик, пептид, липид, в том числе катионный липид, липосому, липидную частицу, поликатион, например полилизин, разветвленный, трехмерный поликатион, например дендример, углевод, катионное амфифильное вещество, детергент, сурфактант бензиламмоний или другое соединение, облегчающее доставку полинуклеотидов в клетки. Такие вспомогательные агенты включают местные анестетики бупивакаин или тетракаин (см., например, патенты США №№5,593,972; 5,817,637; 5,380,876; 5,981,505 и 6,383,512 и международную публикацию патента №WO98/17799, включенные в данную заявку посредством ссылки в полном объеме).
В конкретных вариантах реализации композиции получают с применением носителей, защищающих гликопротеин от быстрого удаления из организма, например препаратов с контролируемым высвобождением, в том числе имплантатов и систем для доставки на основе микрокапсул. Можно применять поддающиеся биологическому разложению и биологически совместимые полимеры, например этиленвинилацетат, полиангидриды, полигликолиевую кислоту, коллаген, полиортоэфиры и полимолочную кислоту. Способы получения таких препаратов известны специалистам в данной области. Вещества можно также приобрести у компаний Alza Corporation и Nova Pharmaceuticals, Inc. В качестве фармацевтически приемлемых носителей можно также применять суспензии с использованием липосом (включающие липосомы, направленно воздействующие на инфицированные клетки за счет моноклональных антител к вирусным антигенам). Такие суспензии можно также приготовить согласно способам, известным специалистам в данной области, например, описанным в патенте США №4,522,811, включенном в данную заявку посредством ссылки в полном объеме.
Для удобства введения и равномерного дозирования композиции для перорального или парентерального введения входят в состав препаратов в дозированной форме. Дозированная форма при использовании в данной заявке относится к физически дискретным элементам, которые можно применять в качестве единичных доз для субъекта, подвергаемого лечению, при этом каждый такой элемент содержит установленное количество активного гликопротеина, которое согласно расчетам оказывает желаемый терапевтический эффект совместно с необходимым фармацевтическим носителем.
Гликопротеин, экспрессированный согласно настоящему изобретению, можно вводить на различных сроках или через различные промежутки времени, по необходимости, например раз в неделю в течение примерно от 1 до 10 недель, от 2 до 8 недель, примерно от 3 до 7 недель, примерно 4, 5 или 6 недель и т.п. Как очевидно специалисту в данной области, различные факторы могут влиять на дозы и временные интервалы, необходимые для эффективного лечения субъекта, включающие, но не ограниченные перечисленными, тяжесть заболевания или нарушения, предшествующего лечения, общее состояния здоровья и/или возраст субъекта и наличие других заболеваний. Как правило, лечение субъекта гликопротеином, как описано в данной заявке, может включать однократную процедуру/воздействие или, во многих случаях, может включать серию процедур/воздействий. Очевидно, что величина соответствующей дозы может зависеть от эффективности гликопротеина, и ее можно также адаптировать для конкретного реципиента, например, посредством введения возрастающих доз до достижения предварительно выбранного желаемого ответа. Кроме того, очевидно, что конкретная величина дозы для любого конкретного животного субъекта может зависеть от ряда факторов, включающих активность конкретного применяемого гликопротеина, возраст, вес тела, общее состояние здоровья, пол и режим питания субъекта, время введения, способ введения, скорость выведения, любой комбинации лекарственных препаратов и/или степени экспрессии или активности, которую требуется модулировать.
Настоящее изобретение включает применение композиций согласно изобретению для лечения животных, отличных от человека. Соответственно, дозы и способы введения могут быть выбраны согласно известными принципам ветеринарной фармакологии и медицины. Их описание можно найти, например, в Adams, R. (ed.), Veterinary Pharmacology and Therapeutics, 8th edition, Iowa State University Press; ISBN: 0813817439; 2001.
Фармацевтические композиции согласно изобретению могут быть заключены в контейнер, упаковку или диспенсер вместе с инструкциями для введения.
Предшествующее описание следует рассматривать только как иллюстрирующее, но не ограничивающее изобретение. Специалисту очевидны альтернативные способы и вещества для осуществления изобретения, а также дополнительные варианты применения, которые охвачены прилагаемой формулой.
ПРИМЕРЫ
ПРИМЕР 1: СОСТАВ СРЕДЫ
Настоящее изобретение включает обнаружение того факта, что гликопротеины, продуцируемые культурой клеток, выращенной в культуральной среде, содержащей марганец в одной или более концентрации согласно настоящему изобретению, характеризуются увеличенной степенью гликозилирования, чем если бы клетки выращивали в стандартной культуральной среде. Марганец можно добавлять к любой культуральной среде, способной обеспечивать рост клеток. Примеры культуральных сред, к которым может быть добавлен марганец, в пределах любой из концентраций согласно изобретению, приведены в Таблице 1, хотя настоящее изобретение не ограничено применением таких культуральных сред. Как очевидно специалистам в данной области, для выращивания клеток можно применять другие среды и/или в составах сред, приведенных в качестве примеров в Таблице 1, могут быть сделаны некоторые изменения.
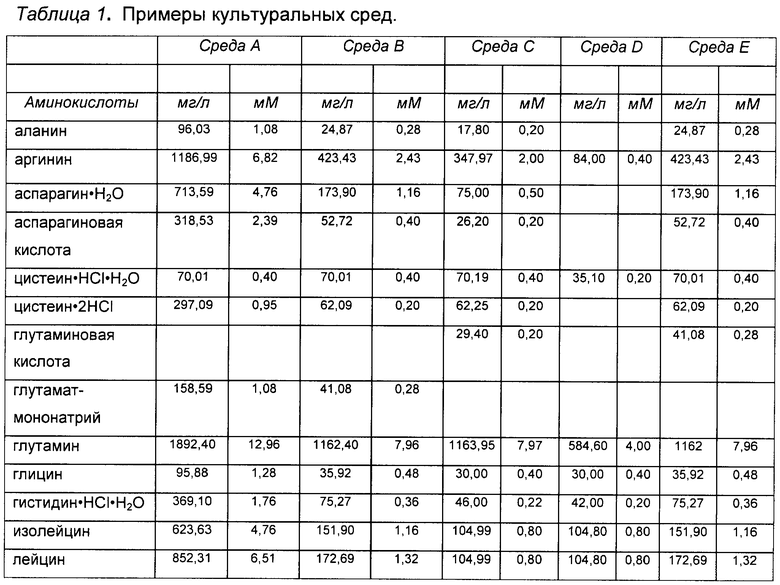
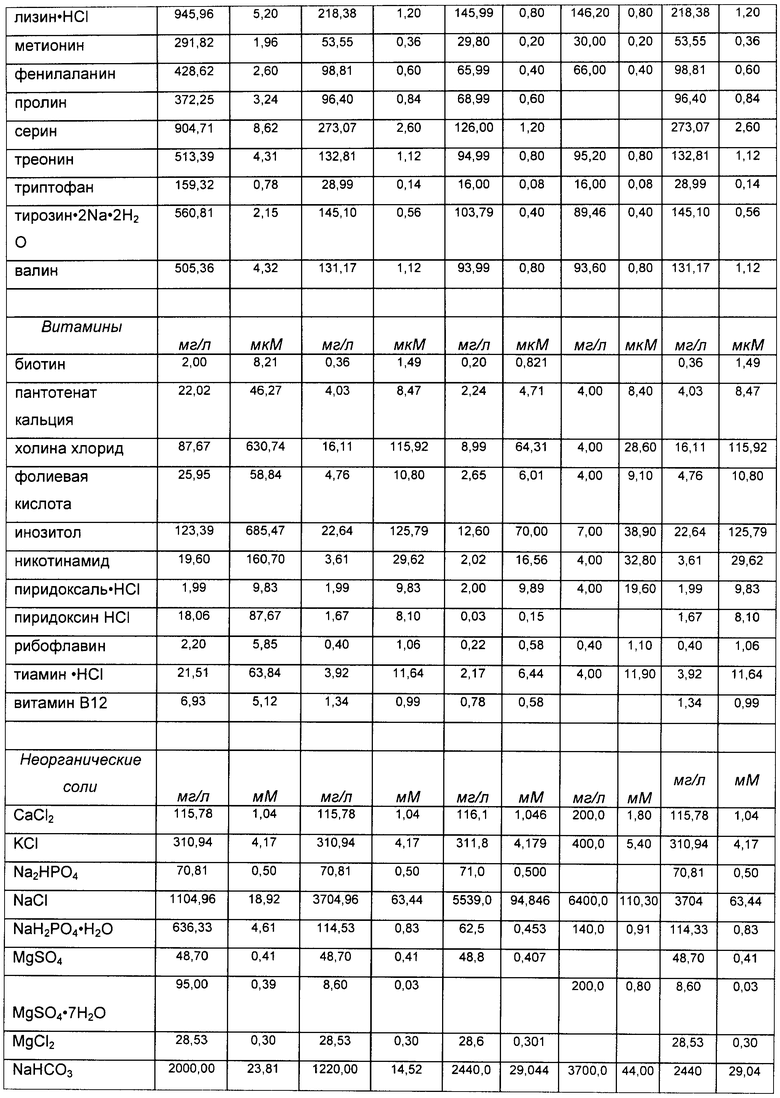
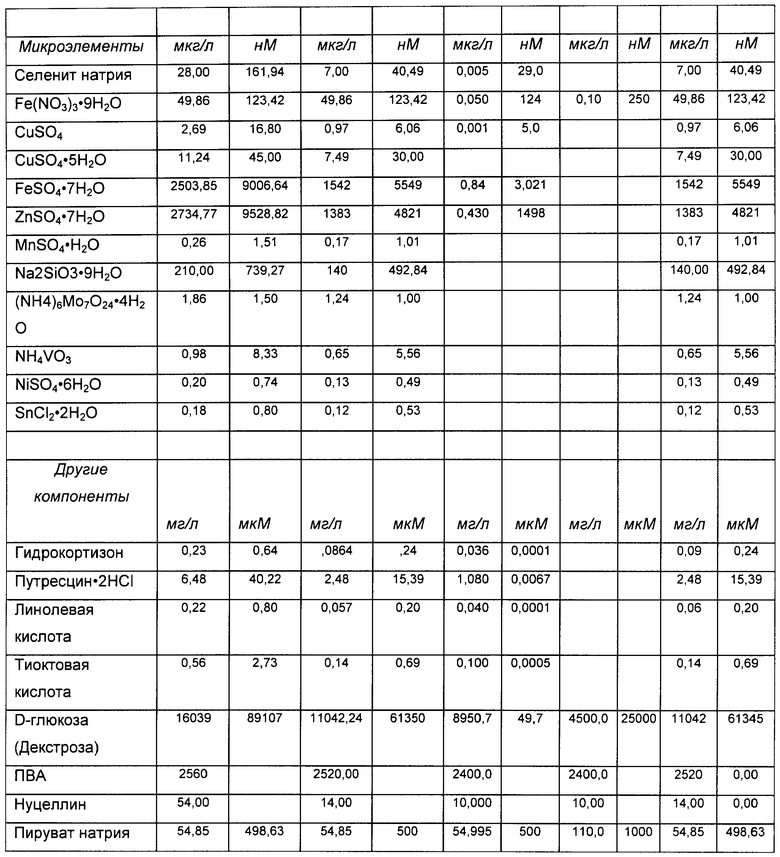
В конкретных вариантах реализации к клеткам добавляют питательную среду в один или более моментов времени после начала культивирования. Примеры питательных сред представлены в Таблице 2, хотя настоящее изобретение не ограничено применением таких питательных сред. Как очевидно специалистам в данной области, можно применять другие питательные среды для выращивания клеток и/или в состав приведенных в качестве примеров питательных сред, представленных в Таблице 2, могут быть внесены некоторые изменения. Например, концентрация одного или более компонента такой питательной среды может быть увеличена или снижена для достижения желаемой концентрации таких компонентов. В конкретных вариантах реализации концентрацию каждого компонента питательной среды увеличивают или уменьшают на один и тот же коэффициент. Например, концентрация каждого компонента питательной среды может быть увеличена или уменьшена в 1х, 2х, 3х, 4х, 5х, 6х, 7х, 8х, 9х, 10х, 11х, 12х, 13х, 14х, 15х, 16х, 17х, 18х, 19х, 20х, 25х, 30х, 35х, 40х, 45х, 50х или более.
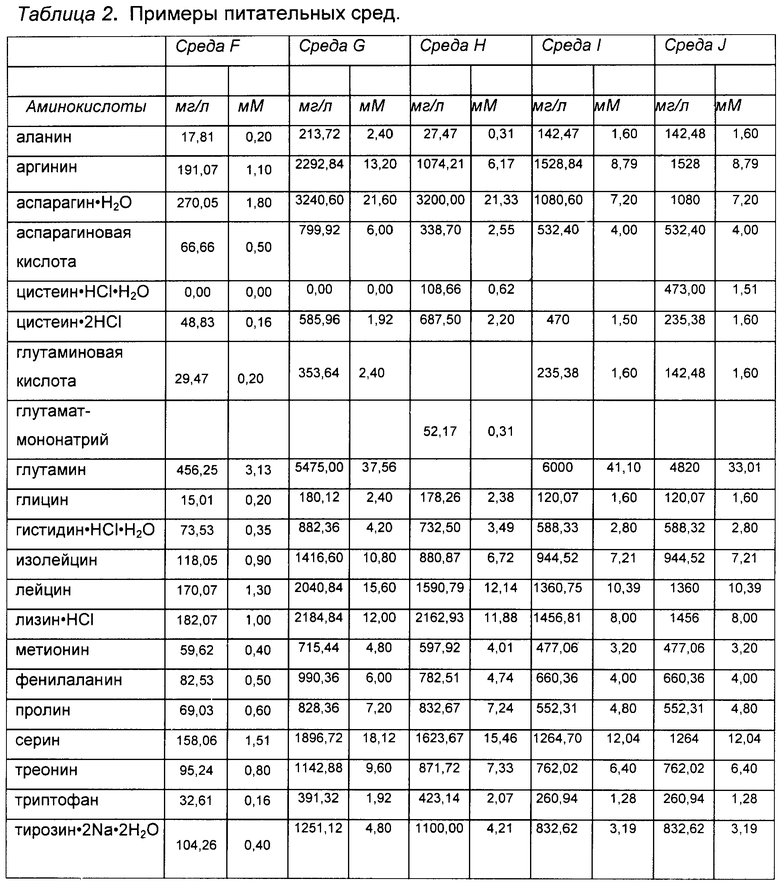
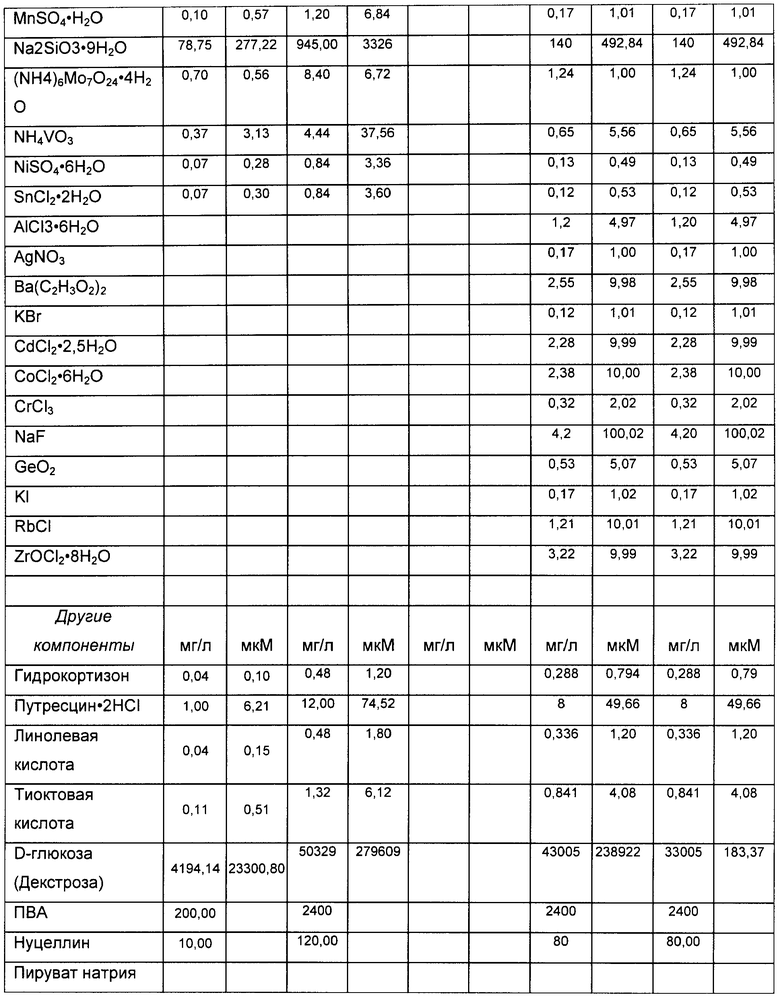
ПРИМЕР 2: МЕЛКОМАСШТАБНОЕ ИССЛЕДОВАНИЕ КАРТИРОВАНИЯ ПЕПТИДА rFIX
Введение: В ходе продукции сырьевого вещества лекарственной формы (СВЛФ) рекомбинантного Фактора свертывания крови IX человека ("rFIX") наблюдали различия в величинах отношений площадей пика («relative peak area ratio, RPAR») пептида K4 на пептидной карте. Пробы готовили путем ферментативного гидролиза с помощью лизил-эндопептидазы из Achromobacter lyticus "AchroK", Wako номер в каталоге #129-02541), с последующим разделением с помощью обращенно-фазовой ВЭЖХ. Величины RPAR К4 имеют значения меньше нижнего предела 82% от контрольного вещества-эталона в некоторых партиях и достигают минимума 78% от контрольного образца. Баланс веществ был обнаружен в пике К4'. Различие между двумя пиками обусловлено исключительно степенью гликозилирования Ser-61. В варианте К4 к серину присоединен тетрасахарид Sia-α23-Gal-β1, 4-GlcNAc-β1, 3-Fuc-α1-O, тогда как в варианте К4' с пропорциональным увеличением - только фукоза.
Полномасштабный эксперимент с культурой клеток проводили в тот период, когда формировались различия в карте (гликозилирования) пептида. Клетки выращивали в культуральной среде, в которую добавляли FeSO4, CuSO4 и холина хлорид в 2Х, 7Х и 2Х соответственно. СВЛФ, полученное в результате мелкомасштабной очистки экспериментальных партий, демонстрировало повышенные значения RPAR К4, тогда как контрольное вещество, очищенное таким же способом - нет. Большинство СВЛФ соответствовало требованиям карты пептида (≥82% от контрольного образца), хотя они не достигали уровня, наблюдаемого для вещества-эталона. Это указывает на то, что различия, наблюдаемые на картах RPAR, являются результатом процесса культивирования клеток и могут быть обусловлены недостатком питательных веществ.
Анализ проб в процессе производства: Клетки удаляли из среды для культивирования клеток с помощью микрофильтрации («МФ») и ультрафильтрации/диафильтрации («УФ/ДФ»), в результате чего получали достаточно чистое вещество из отдельных биореакторов, доступных для анализа.
Для исследования того, происходит ли деградация варианта К4 обратно в К4′ (содержащий только фукозу) после секреции из клетки, проводили модифицированный анализ проб из ретентата УФ/ДФ. Образец ретентата разделяли на три равных аликвоты. Очистку одной из них проводили сразу же путем мелкомасштабной очистки на колонке; этот образец служил отрицательным контролем. Две другие аликвоты инкубировали в течение ночи при 37°С перед очисткой аналогичным способом. К одной из них добавляли сиалидазу для удаления концевой сиаловой кислоты из варианта К4 на случай, если сиаловая кислота блокирует активность некоторых других гликозидаз. После мелкомасштабной очистки во всех трех образцах анализировали К4, как описано выше. На Фигуре 1 показано, что в образцах не наблюдали деградации, кроме деградации, катализируемой сиалидазой. Это убедительно свидетельствует о том, что проблема, связанная с картой пептида, является анаболической, что означает, что «отсутствующие» остатки сахара в положении Ser-61 ранее не были присоединены к синтезированной цепи. Сохраняется вероятность, хотя это крайне маловероятно, что гликозидазная активность, ответственная за удаление таких остатков сахара, была инактивирована или удалена на этапах МФ и УФ/ДФ. Результаты также показывают возможность применения мелкомасштабной системы очистки для анализа образцов на стадии до использования Q Сефарозы.
Мелкомасштабное моделирование: Мелкомасштабные культуры для производства rFIX, выращенные во встряхиваемых колбах на качалке, использовали как модель для оценки влияния различных сред и добавок на варианты К4. В каждом случае очищали непосредственно кондиционированную среду из встряхиваемых колб (без УФ/ДФ) через колонку для мелкомасштабной очистки с отбором пиков на основе объема и оценивали распределение вариантов К4 в образце.
Возможность применения мелкомасштабной модели была продемонстрирована с использованием тех же добавок, которые использовали и в полномасштабном эксперименте. Также исследовали добавление марганца, поскольку при анализе литературных источников было обнаружено, что Mn++ необходим для аналогичного фермента с гликозилирующей активностью у дрозофилы (см. Moloney et al., J. Biol. Chem. 275(13): 9604-9611, 2000; Bruckner et al., Nature 406: 411-415, 2000). Проводили сравнение более четырех пассажей во встряхиваемых колбах, результаты показаны на Фигуре 2. Показаны инъекции в двух повторах для каждого образца. "Р#" указывает номер пассажа для каждого варианта условий. Для Р1-2 концентрация Mn составляла 1 нМ; для Р3-4 - 10 нМ. Различие между распределениями вариантов в К4 в контрольной среде и среде с добавками сравнимо с различиями, обнаруженными при производстве в биореакторах. Различия проявлялись после одного пассажа, и многократные пассажи не показали каких-либо изменений.
Поскольку добавки включали три компонента (FeSO4, CuSO4 и холина хлорид), следующий эксперимент был направлен на выявление того, какой из этих компонентов ответственен за увеличение распределения вариантов К4. Три компонента добавляли к трем культурам во встряхиваемых колбах, продуцируемых rFIX. Компоненты добавляли попарно для выявления какого-либо синергического эффекта. Другие культуры включали положительный (все три компонента) и отрицательный (без добавок) контроли.
На Фигуре 3 показано, что добавкой, способствующей увеличению распределения вариантов К4, был FeSO4. Показаны инъекции в двух повторах для каждого образца. Добавки имели экспериментальные концентрации. Оказалось, что добавление CuSO4 и холина хлорида, при отсутствии FeSO4, могло способствовать даже уменьшению распределения. Следует отметить, что по неизвестным причинам десиалированная форма К4' начинала появляться в большом количестве как в анализируемых образцах, так, эталонном веществе. Такая тенденция видна на Фигуре 3 и остается в последующих экспериментах.
Спектроскопический анализ с индуктивно-связанной плазмой («ИСП») содержания железа в порошке среды показал, что в порошке присутствует соответствующее количество железа. Это позволило предположить, что эффект, вызываемый добавлением FeSO4, в действительности обусловлен микропримесями указанного исходного вещества. Партия FeSO4, использованная в исходной среде, была проанализирована ИСП-анализом, и были обнаружены несколько микропримесей, присутствующих в концентрациях, превышающих предел детекции. После исключения из рассмотрения известных ингибиторов и компонентов среды для культивирования клеток, продуцирующих rFIX, список был сужен до следующих девяти потенциально полезных элементов: Sb, Bi, В, Со, Ge, Mn, Mo, Ni и V.
Затем проводили эксперименты по мелкомасштабному моделированию для изучения некоторых других возможных добавок, которые могли дополнить исходные добавки для среды. Дополнительно были протестированы CuSO4, ZnSO4 и MnSO4 (до 10 нМ). ZnSO4 был включен в анализ, поскольку цинк может конкурентно ингибировать поглощение других бивалентных катионов. По неизвестным причинам контрольные и экспериментальные условия давали очень схожие распределения вариантов К4, которые в большей степени напоминали наблюдаемые ранее для контрольных условий (см. Фигуру 4, показаны инъекции каждого образца в двух повторах). Однако добавление MnSO4 в экспериментальные условия отчетливо увеличивало распределение вариантов К4. Как было показано, добавление ZnSO4 может уменьшить распределение вариантов К4, но значимость таких различий не достоверна.
Как было отмечено выше, на Фигуре 3 наблюдали увеличение уровня пика десиалированного варианта К4'. Это явление сохранялось и имело тенденцию к возрастанию в ходе описанных мелкомасштабных исследований. Такая тенденция не изменяет интерпретацию представленных результатов.
Вывод: Подробное изучение элюатов, полученных при очистке в процессе производства и мелкомасштабной очистке на колонках, убедительно показало, что различия в порошке среды обусловили сдвиг RPAR К4, что в свою очередь привело к различиям в пептидной карте. Поскольку добавление конкретных компонентов в среду для культивирования клеток делало такой сдвиг обратимым, в частности, вероятно, что различие в порошке среды заключается в том, уменьшалось содержание одного или более компонента. Поскольку было показано, что наиболее эффективной добавкой является FeSO4 и поскольку согласно ИСП-анализу порошок среды содержал соответствующее количество Fe, было выдвинуто предположение, что микропримеси FeSO4 обуславливают надлежащее гликозилирование Ser-61. Согласно ИСП-анализу FeSO4 примесь в среде представляла собой не определенный компонент порошка среды, но скорее была побочным питательным веществом, ранее также присутствующим в среде.
ПРИМЕР 3: МЕЛКОМАСШТАБНЫЕ ИССЛЕДОВАНИЯ ВЛИЯНИЯ ДОБАВОК В СРЕДЕ НА ПЕПТИДНУЮ КАРТУ rFIX
Введение: Пример 2 показал, что различия между партиями, связанные со степенью гликозилирования по Ser-61, понаблюдали в различных партиях rFIX, что проявлялось в виде сдвига в распределении популяции вариантов пептида К4. Все партии rFIX имели распределение цепей различной длины, в котором преобладал полноразмерный тетрасахарид (Sia-α2,3-Gal-β1,4-GlcNAc-β1,3-Fuc-α1), но некоторые партии характеризовались необычно высокой долей форм, содержащих только фукозу.
Пример 2 также показал, что в биореакторах наблюдали сдвиг в распределении К4 и что он был тесно связан с изменением партии порошка среды. Результаты Примера 2 убедительно подтверждают предположение о том, что изменение распределения гликоформ обусловлено анаболической функцией, но не катаболической. Кроме того, эти эксперименты показали, что добавление FeSO4 могло частично обратить сдвиг, но ИСП-анализ показал отсутствие каких-либо значимых различий в концентрации FeSO4 между различными партиями порошка среды. Таким образом, было выдвинуто предположение о том, что другой неидентифицированный следовой компонент FeSO4, присутствующий на различных уровнях в разных партиях порошка среды, ответственен за такой сдвиг.
Аддитивный эффект: Как обсуждалось в Примере 2, ИСП-анализ показал измеримые количества девяти следовых элементов в партии FeSO4, использованной для экспериментальных условий культивирования. При сравнении этих следовых элементов с опубликованными прописями сред исключили необходимость добавления Sb или Bi. На основании составов сред и ИСП-анализа FeSO4 была приготовлена смесь пяти соединений для добавления к культурам, продуцируемым rFIX (указана конечная концентрация в среде): 1 нМ (NH4)6Mo7O24, 10 нМ CoCl2, 5,5 нМ NH4VO3, 1,5 нМ NiSO4 и 20 нМ H3BO3. MnSO4 добавляли отдельно, поскольку обзор более ранней литературы выявил, что марганец может быть важен для гликозилтрансферазной деятельности (см. Breton and Imberty, Curr. Opinion in Structural Biol. 9: 563-571, 1999; Bruckner et al., Nature 406: 411-415, 2000). В том же эксперименте было исследовано добавление FeSO4 (2Х или 4Х), для определения того, мог ли он дополнительно увеличивать относительное содержание тетрасахарида. В каждом случае добавки использовали дополнительно к добавляемым компонентам, описанным в Примере 2.
Результаты экспериментов с использованием таких добавок представлены на Фигуре 5. Определяли распределение вариантов К4, и представленные значения представляют собой отношения площадей каждого из четырех пиков К4 к их суммарному значению. На Фигуре также приведены данные для вещества-эталона, контроля (без добавок) и культуры с добавлением компонентов (как в Примере 2). Показаны инъекции в двух повторах для каждого образца. В каждом ряду столбцов самый левый столбец соответствует доле молекул с тетрасахаридом, присоединенным по Ser-61, а самый правый столбец соответствует доле молекул, содержащих только фукозу. Наблюдаемые различия между культурами, соответствующими положительному и отрицательному контролям (с добавлением компонентов и без добавления соответственно), были меньше, чем наблюдали ранее. Вне зависимости от этого Фигура 5 отчетливо демонстрирует, что смесь следовых элементов (микроэлементов) не оказывала влияния на распределение вариантов К4, тогда как добавление как FeSO4, так и MnSO4 улучшало распределение вариантов К4. При включении в добавки 15 нМ MnSO4 наблюдали приблизительно такой же эффект на распределение вариантов К4, как и при добавлении 12 мкМ FeSO4.
После обнаружения отчетливого ответа на добавление марганца были проведены эксперименты по нахождению оптимальной концентрации марганца в среде для культивирования клеток, продуцирующих rFIX. Как показано на Фигуре 6, результаты этого эксперимента согласуются с результатами, представленными на Фигуре 5, поскольку все культуры, в которые добавляли марганец, характеризовались меньшим содержанием вариантов К4, содержащих только фукозу, по сравнению с культурами с другими добавками или контрольной культурой. Фактически, культуры с концентрацией марганца 40 нМ или более характеризовались примерно таким же содержанием вариантов К4, содержащих только фукозу, как и эталонное вещество. Однако при концентрации марганца 100 или 500 нМ наблюдали большую долу трисахаридов, чем при 40 нМ. Таким образом, было определено, что 40 нМ неожиданно является эффективной концентрацией марганца для более протяженного гликозилирования FIX по Ser-61.
Применимость мелкомасштабной модели: Одна из необычных особенностей таких мелкомасштабных экспериментов и экспериментов, представленных в Примере 2, заключалась в варьировании уровня трисахаридов или десиалированных вариантов от эксперимента к эксперименту. Поскольку такие варианты варьировали аналогичным образом при анализе эталонного вещества, можно считать, что это явление является артефактом, вызванным ферментативным гидролизом в одном сосуде/реакторе. Поскольку все образцы в данном эксперименте подвергались гидролизу в одно и то же время при использовании одного и того же сырья, можно предположить, что такая вариабельность не влияет на результаты анализа, представленные в данном Примере или в Примере 2.
Вывод: Эти эксперименты показали, что добавление 40 нМ MnSO4 в среду для культивирования клеток, продуцирующих rFIX, увеличивает распределение вариантов К4.
ПРИМЕР 4: АНАЛИЗ N-ПРИСОЕДИНЕННЫХ ОЛИГОСАХАРИДОВ В ОБРАЗЦАХ СРЕДЫ ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК, ПРОДУЦИРУЮЩИХ АНТИТЕЛА К А-БЕТА
Введение: Был исследован характер распределения N-присоединенных олигосахаридов в клетках СНО, экспрессирующих гуманизированное моноклональное антитело IgG1 к пептиду А-бета («клетки, продуцирующие антитела к А-бета»), при четырех вариантах условий культивирования. Характеристики образца и соответствующие данные представлены в Таблице 3.
Процедура: Культуру, продуцирующую антитела к А-бета, №1 выращивали с периодическим добавлением питательной среды. В культуру, продуцирующую антитела к А-бета, №2 добавляли микроэлементы Е в начале культивирования. Состав Микроэлементов Е указан в Таблице 4. Культуру, продуцирующую антитела к А-бета, №3 выращивали в условиях, идентичных культуре №2, за исключением того, что начальный уровень глутамина был 4 мМ. Культуру, продуцирующую антитела к А-бета, №4 выращивали в условиях, идентичных культуре №3, за исключением того, что в культуру не добавляли Микроэлементов Е и добавляли глутамат в концентрации 2 г/л.
Распределение гликоформ каждого образца определяли путем расщепления с помощью пептид-N-глкизидазы F (PNGase F) с последующей анионообменной хроматографией при высоком рН с анализом путем импульсной электромеханической детекции (HPAEC-PED). Вкратце, осуществляли ионный обмен образцов с буфером в 50 мМ муравьинокислом аммонии, забуференном при рН 7,3 с использованием концентратора белка Amicon Ultra 30000 MWCO. После восстановления каждый образец подвергали ферментативному гидролизу с 5 мкл PNGase F (без глицерина) и инкубировали в течение ночи при 37°С. Затем образцы высушивали с помощью скоростного вакуумного центрифугирования и восстанавливали в очищенной воде. Затем переносили образцы во флаконы с автодозатором для анализа путем HPAEC-PED. Система HPAEC-PED оснащена устройством Dionex CarboPac РА100, аналитической колонкой (2×250 мм) и детектором ED-40. Использовали линейный градиент ацетата, включающий два элюента: элюент А, содержащий 100 мМ NaOH, и элюент В, содержащий 100 мМ NaOH /500 мМ ацетат натрия.
Анализ данных: Три типа комплексов N-присоединенных двуветвистых гликанов, ассоциированных с антителом к А-бета, содержат ноль ("G0"), один ("G1") или два (G2") остатка галактозы на внешних ветвях N-присоединенных двуветвистых гликанов. Во всех образцах показано присутствие трех типичных пиков гликоформ G0, G1 и G2. На Фигуре 7 представлено графическое сравнение процента общей площади пика для пиков G0, G1 и G2, определенных при анализе каждого образца методом HPAEC-PED. В профилях всех рассмотренных образцов наблюдали присутствие дополнительных небольших пиков. Наблюдаемые пики соответствуют низкому содержанию моно- и ди-сиалированных гликоформ.
Обсуждение: В ходе таких экспериментов исследовали относительное распределение пиков G0:G1:G2 в образцах культуры клеток, продуцирующих антитела к А-бета, с добавлением питательной среды при различных экспериментальных условиях. Культуры, в которые добавляли Микроэлементы Е, демонстрировали снижение уровня G0 с соответствующим увеличением уровня G1 и G2, по сравнению с условиями культивирования при отсутствии Микроэлементов Е (Фигура 7). Культивирование при низком уровне глутамина производило аналогичный эффект, и такие эффекты были аддитивными. В культурах с низким уровнем глутамина (4 мМ) с добавлением Микроэлементов Е демонстрировали значительный сдвиг в распределении N-присоединенных гликоформ, практически с равным соотношением G0 и G1, при этом G2 представляли приблизительно 10% общей площади пика (Фигура 7). Культуры, в которые добавляли Микроэлементы Е, содержали марганец в концентрации 156 нМ. Однако, следует отметить, что такие культуры также имели более высокий уровень других металлов. Таким образом, возможно, что, помимо марганца, улучшению наблюдаемого характера гликозилирования способствовали и другие условия культивирования.
Вывод: Различия в распределениях гликозилирования, наблюдаемые в образцах антител к А-бета, наиболее вероятно обусловлены соответствующими изменениями в условиях культивирования. Наши данные убедительно демонстрируют, что низкое содержание глутамина (4 мМ) и/или добавление Микроэлементов Е, содержащих 100 мМ MnSO4, приводило к значительному изменению в распределении процентного содержания различных N-присоединенных гликоформ антител к А-бета. Эти эффекты, как было показано, являются независимыми и аддитивными.
ПРИМЕР 5: АНАЛИЗ N-ПРИСОЕДИНЕННЫХ ОЛИГОСАХАРИДОВ В ОБРАЗЦАХ АНТИТЕЛ К А-БЕТА ПРИ ДОБАВЛЕНИИ МАРГАНЦА
Введение: Пример 4 показал, что увеличение распределения гликозилирования в образцах антител к А-бета может быть обусловлено добавлением Микроэлементов Е в среды для культивирования и поддержанием низкого уровня глутамина. В данном примере исследовано, может ли добавление одного только марганца в среды для культивирования вызвать аналогичное увеличение распределения гликозилирования.
Процедура: Культуры, продуцирующие антитела к А-бета, выращивали в культуральной среде либо с добавлением, либо без добавления 40 мМ марганца. К культурам добавляли в качестве подпитки 40% общего объема питательной среды. Образцы собирали и анализировали согласно способу, описанному в Примере 4.
Анализ данных: Проанализированные образцы сравнивали по наличию пиков и проценту общей площади пика для каждого пика. На Фигуре 8 показано графическое сравнение процента общей площади пика для пиков G0, G1 и G2 для каждого образца, определенных методом HPAEC-PED.
Обсуждение: Три типа комплексов N-присоединенных разветвленных гликанов, ассоциированных с антителом к А-бета, представляют собой структуры G0, G1 и G2, которые соответственно содержат ноль, один или два остатка галактозы на внешних ветвях N-присоединенных разветвленных гликанов. Образцы, полученные от клеток, выращенных в среде либо с отсутствием, либо с добавлением 40 мМ марганца, демонстрировали наличие всех трех пиков, характерных для гликоформ G0, G1 и G2. Общая площадь пика G0 уменьшалась от 68% в контрольном образце до 53% в образце, полученном из среды, содержащей дополнительный марганец (см. Фигуру 8). Также в образце, полученном от среды, содержащей марганец, наблюдали увеличение процентов G1 и G2 от общей площади пиков. Процент G1 общей площади пика составил 26% в контрольном образце и 39% в образце среды с добавлением марганца. Процент G2 общей площади пика составил 6% в контрольном образце и 9% в образце среды с добавлением марганца (Фигура 8).
Вывод: Полученные данные указывают на то, что добавление только марганца в среду приводило к формированию повышенной степени гликозилирования, о чем свидетельствует сдвиг в процентном соотношении распределения G0:G1:G2 в этих образцах.
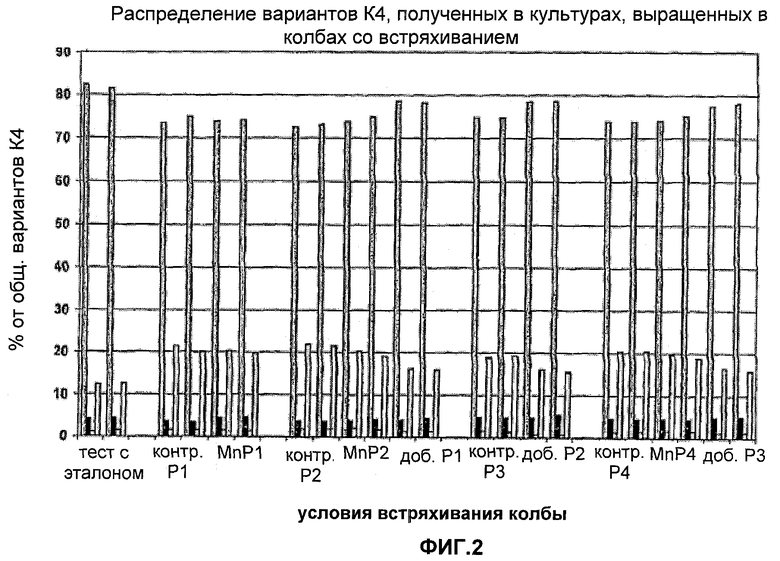
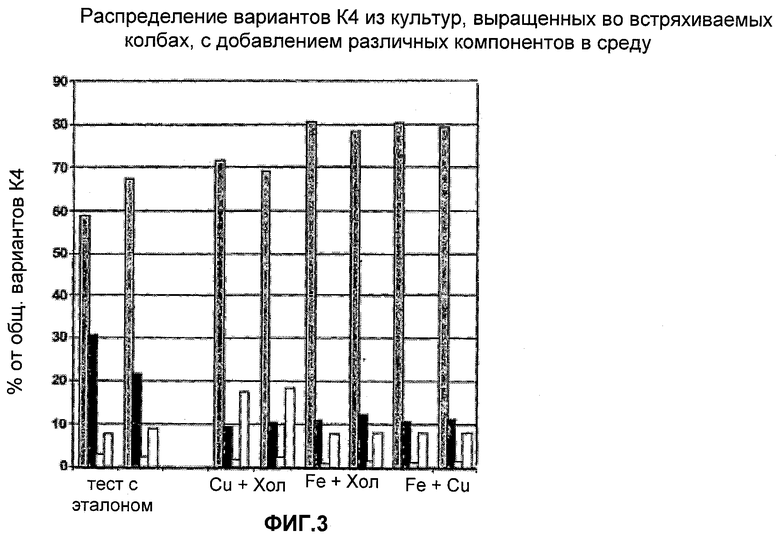
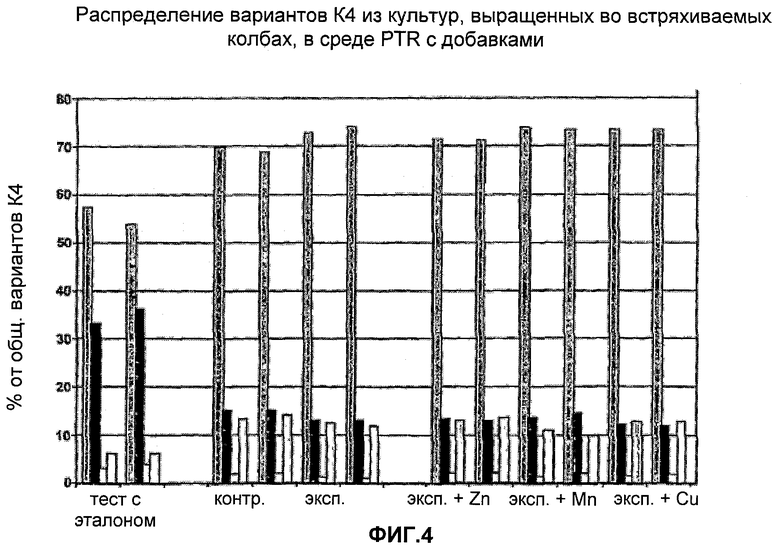
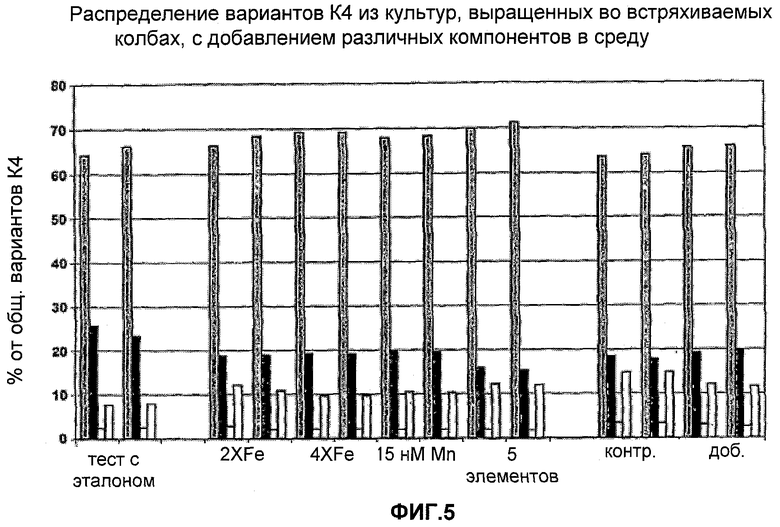
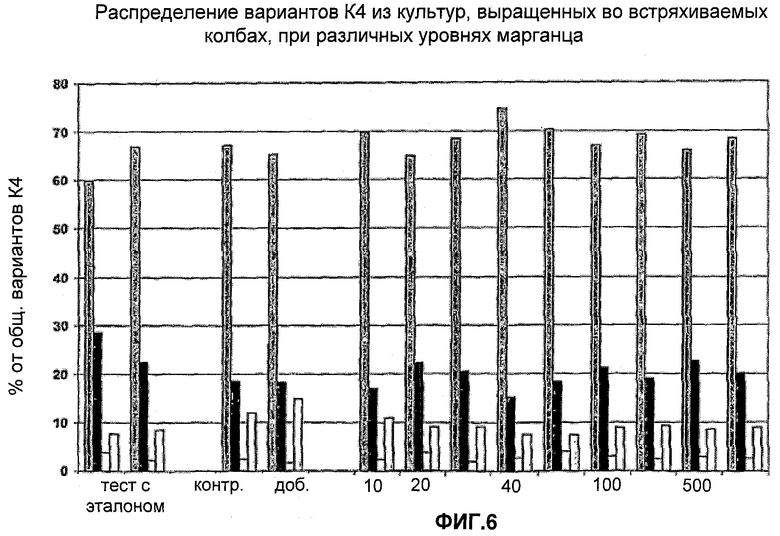
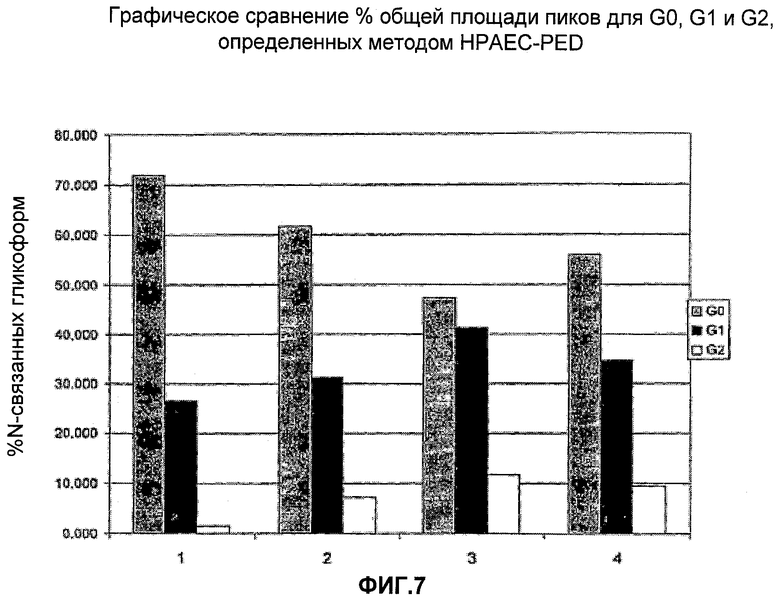
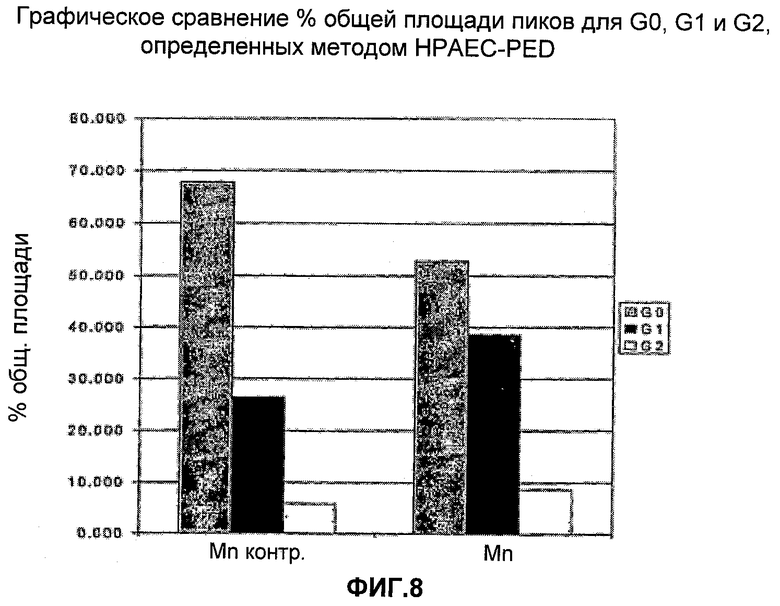
Изобретение относится к области биотехнологии, конкретно к получению гликопротеинов, и может быть использовано для крупномасштабного получения фактора свертывания крови IX в культуре клеток. Способ включает культивирование клеток млекопитающего, содержащих ген, кодирующий фактор свертывания крови IX, в среде для культивирования клеток, которая содержит приблизительно от 10 нМ до 600 нМ марганца при условиях и в течение времени, достаточного для осуществления экспрессии фактора свертывания крови IX. Изобретение позволяет получить экспрессированный фактор свертывания крови IX с повышенной степенью гликозилирования по сравнению со степенью гликозилирования, наблюдаемой при использовании таких же условий и такой же среды, но в отсутствие марганца. 2 н. и 23 з.п. ф-лы, 8 ил., 4 табл., 5 пр.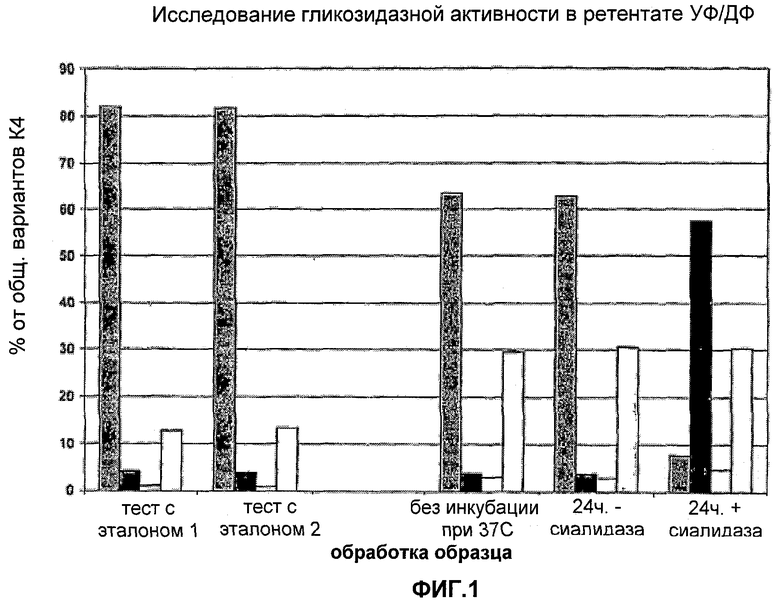
1. Способ получения фактора свертывания крови IX в культуре клеток, включающий:
культивирование клеток млекопитающего, содержащих ген, кодирующий фактор свертывания крови IX, в среде для культивирования клеток, которая содержит приблизительно от 10 нМ до 600 нМ марганца при условиях и в течение времени, достаточного для осуществления экспрессии фактора свертывания крови IX, при этом экспрессированный фактор свертывания крови IX имеет повышенную степень гликозилирования, по сравнению со степенью гликозилирования, наблюдаемой при использовании таких же условий и такой же среды, но в отсутствие марганца.
2. Способ по п.1, включающий стадии:
культивирования клеток млекопитающего, содержащих ген, кодирующий фактор свертывания крови IX, в среде для культивирования клеток, которая содержит приблизительно от 10 нМ до 600 нМ марганца поддерживание культуры при первом диапазоне температур в течение первого промежутка времени, достаточного для воспроизводства клеток до достижения плотности жизнеспособных клеток в диапазоне приблизительно 20-80% от максимально возможной плотности жизнеспособных клеток, который можно достигнуть при поддерживании культуры при первом температурном диапазоне;
перевод культуры во второй диапазон температур, такой что по меньшей мере одно значение температуры второго диапазона температур ниже самого низкого значения температуры первого диапазона температур; поддерживание культуры в течение второго промежутка времени при условиях и в течение времени, достаточного для осуществления экспрессии фактора свертывания крови IX, при этом экспрессированный фактор свертывания крови IX имеет повышенную степень гликозилирования по сравнению со степенью гликозилирования, наблюдаемой при использовании таких же условий и такой же среды, но в отсутствие марганца.
3. Способ по п.2, отличающийся тем, что первый диапазон температур включает диапазон температур приблизительно 30-42°С.
4. Способ по п.3, отличающийся тем, что первый диапазон температур включает температуру приблизительно 37°С.
5. Способ по любому из пп.2-4, отличающийся тем, что второй диапазон температур включает диапазон температур приблизительно 25-41°С.
6. Способ по п.5, отличающийся тем, что второй диапазон температур включает температуру приблизительно 31°С.
7. Способ по п.2, дополнительно включающий вторую смену температурного режима после указанной первой смены температурного режима, которая представляет собой перевод культуры на третье значение температуры или в третий диапазон температур, при этом по меньшей мере одно значение температуры указанного третьего диапазона температур ниже самого низкого значения температуры указанного второго диапазона температур.
8. Способ по п.7, отличающийся тем, что указанный третий диапазон температур включает диапазон температур приблизительно 25-40°С.
9. Способ по п.1, отличающийся тем, что среда для культивирования клеток содержит приблизительно 20-200 нМ марганца.
10. Способ по п.9, отличающийся тем, что среда для культивирования клеток содержит приблизительно 40 нМ марганца.
11. Способ по п.1, отличающийся тем, что среда для культивирования клеток включает глутамин, начальная концентрация которого меньше или равна приблизительно 8 мМ.
12. Способ по п.11, отличающийся тем, что начальная концентрация глутамина в среде для культивирования клеток меньше или равна приблизительно 4 мМ.
13. Способ по п.1, отличающийся тем, что объем культуры клеток составляет по меньшей мере около 500 л.
14. Способ по п.1, отличающийся тем, что в культуру клеток дополнительно после начала культивирования клеток добавляют питательную среду.
15. Способ по п.1, отличающийся тем, что в культуру клеток дополнительно добавляют вспомогательные компоненты.
16. Способ по п.15, отличающийся тем, что указанные вспомогательные компоненты выбраны из группы, включающей гормоны и/или другие факторы роста, неорганические ионы, буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы, аминокислоты, липиды, глюкозу или другой источник энергии и их комбинации.
17. Способ по п.16, отличающийся тем, что в культуру клеток добавляют глюкозу в количестве приблизительно 2 г/л.
18. Способ по п.1, отличающийся тем, что среда для культивирования клеток характеризуется определенным составом.
19. Способ по любому из пп.1-17, отличающийся тем, что фактор свертывания крови IX представляет собой рекомбинантный фактор свертывания крови IX человека.
20. Среда для культивирования клеток для применения в способе по п.1, которая содержит приблизительно 10-600 нМ марганца.
21. Среда для культивирования клеток по п.20, которая содержит приблизительно 20-200 нМ марганца.
22. Среда для культивирования клеток по п.21, которая содержит приблизительно 40 нМ марганца.
23. Среда для культивирования клеток по любому из пп.20-22, отличающаяся тем, что указанная среда для культивирования клеток содержит глутамин, начальная концентрация которого менее или равна приблизительно 8 мМ.
24. Среда для культивирования клеток по п.23, отличающаяся тем, что начальная концентрация глутамина менее или равна приблизительно 4 мМ.
25. Среда для культивирования клеток по п.20, отличающаяся тем, что указанная среда характеризуется определенным составом.
| Станок для гибки монтажных петель и анкеров | 1982 |
|
SU1096017A1 |
| WO 9639488 А1, 12.12.1996 | |||
| RESTELLIV | |||
| et al., The Effect of Cell Culture Parameters on Protein Glycosylation, Cell Engineering, 2002, v.3, p.61-92 | |||
| ANDERSEN D.C | |||
| et al., Multiple cell culture factors can affect the glycosylation of Asn-184 in CHO-produced tissue-type plasminogen activator, Biotechnol | |||
| Bioeng., 2000, v.70, | |||

