В настоящем изобретении описана роль микроэлементов в клеточном росте и гликозилировании белков при производстве рекомбинантных гликопротеинов путем ферментации.
Предпосылки создания изобретения
Рекомбинантные гликопротеины, как правило, получают с помощью ферментативного процесса производства с использованием эукариотических систем экспрессии. Пост-трансляционное гликозилирование белков имеет важное значение для физико-химических и биологических свойств и функций, таких как растворимость, стабильность, клиренс, иммуногенность и иммунные эффекторные функции белков. Статус гликозилирования гликопротеина строго регулируется и даже небольшие различия в гликозилировании могут оказывать значительные воздействия. Однако из-за большого количества ферментов, участвующих, прежде всего в N-гликозилировании, этот путь представляет собой наиболее сложную пост-трансляционную модификацию, осуществляемую в эукариотических клетках. В гликопротеинах сахара присоединены либо к атому азота амида в боковой цепи аспарагина (N-связь), либо к атому кислорода в боковой цепи серина или треонина (О-связь). Гликозилирование начинается с образования N-связей в эндоплазматическом ретикулуме, так называемого «корового гликозилирования». После этого полипептиды транспортируются в аппарат Гольджи, в котором происходит добавление О-связанных сахаров и модификация N-связанных сахаров многочисленными различными путями, например, посредством удаления остатков маннозы, добавления N-ацетилглюкозамина, галактозы и/или фукозы, и/или остатков сиаловой кислоты.
Металлы и их ионы играют очень важную роль в биологии (Yannone и др., Current Opinion in Biotechnology 23.1, 2013, cc. 89-95). Описано, что из 39 имеющих решающее значение биологических элементов 26 представляют собой металлы. Кроме того, существует значительная вариация в активных концентрациях указанных металлов, требуемая для осуществления их биологической функции. Имеющие решающее значение металлы можно классифицировать как макронутриенты типа кальция, калия или магния, и микронутриенты типа металлических микроэлементов, таких как железо, цинк, марганец или медь, и ультрамикроэлементы типа ванадия и вольфрама. Хотя эти имеющие решающее значение металлы играют ключевую роль для биологического гомеостаза, они могут обладать токсичностью в концентрациях, превышающих требуемые для их биологической функции. Таким образом, строгая регуляция системной концентрации металлов является жизненно важной для биологического гомеостаза.
Переходные металлы медь, железо, цинк и марганец классифицируют в соответствии с биологически активной концентрацией в качестве микроэлементов, и они играют ключевую роль в клеточного гомеостазе. Как установлено, они являются активными сайтами многих ферментов, что описано у Fraga и Fraga (Mol. Aspects Med. 26.4-5, 2005, cc. 235-244).
К настоящему времени в течение многих лет изучалась роль микроэлементов в клеточном росте с помощью экспериментов по кормлению животных и на клеточных культурах. Необходимые для успешного культивирования клеток микроэлементы были определены Ham с соавторами (Proc. Natl. Acad. Sci. USA 53, 1965, cc. 288-293). Большой интерес представляет собой также изучение роли микроэлементов в гликозилировании белков, однако детальный вклад различных микроэлементов в ферментативные процессы и взаимодействия катионов металлов в пути гликозилирования остаются относительно мало изученными.
В целом, активность принимающих участие в гликозилировании ферментов зависит от доступности двухвалентных микроэлементов (таких как Zn2+, Mn2+, Са2+, Mg2+), значении рН среды и доступности нуклеозидфосфатов (УТФ, ГТФ, ЦТФ) и соответствующих сахарных производных (УДФ-Gal, УДФ-GlcNAc, ГДФ-Fuc, ЦМФ-сиаловая кислота).
Kaminska с соавторами (Glyconj. J. 15.8, 1998, cc. 783-788) изучали влияние двухвалентных катионов на активность гликопротеин-6-α-L-фукозилтрансферазы и обнаружили, что, в то время как двухвалентные катионы, такие как Mg2+ и Са2+, активировали этот фермент, Cu2+, Zn2+ и Ni2+ сильно ингибировали активность указанного фермента. Witsell с соавторами (J. Biol. Chem. 265.26, 1990, cc. 15731-15737) описали роль конкретных двухвалентных катионов в активации галактозилтрансферазы в нативных пузырьках Гольджи в молочной железе. Crowell с соавторами (Biotechnol. Bioeng. 96.3, 2007, cc. 538-549) описали, что микроэлементы, прежде всего марганец, взаимодействуют с другими связанными с культивированием факторами, такими как доступность аминокислот, на поздних фазах процесса в подвергнутых стрессу культурах, регулируя гликозилирование белков. Gawlitzek с соавторами (Biotechnol. Bioeng. 103.6, 2009, cc. 1164-1175) установили, что условия культивирования клеток, такие как концентрации марганца и железа, удельная продуктивность энхансера бутирата (промежуточные продукты, участвующие в превращении пирувата в бутират), тиреоидные гормоны и значение рН культуры, могут контролировать оккупацию сайта N-гликозилирования рекомбинантных гликопротеинов, которые экспрессируются в СНО-клетках.
Как изложено в US 2007/0161084, для ряда ферментов, участвующих в гликозилировании, различные катионы играют роль кофакторов. В указанной заявке на патент описано, что сиалилирование гликопротеинов, т.е. добавление концевого остатка сиаловой кислоты к углеводной цепи гликопротеина, и, в частности, сиалилирование эритропоэтина, может улучшать путем выращивания клеток-хозяев млекопитающих, которые экспрессируют гликопротеин, в среде, содержащей нетоксичное количество марганца. Марганец может присутствовать в среде для начального роста или его можно добавлять после фазы быстрого клеточного роста, или его можно добавлять после одного или двух циклов сбора.
Роль марганца и его активной концентрации в клеточном росте является спорной. Существуют данные о том, что марганец является важным для нормального клеточного роста и развития, но одновременно он является токсичным в высоких концентрациях (Au и др., Neurotoxicology 29.4, 2008, cc. 569-576). Например, установлено, что марганец связывается с локализованными на клеточной поверхности интегринами и активирует МАР-киназы, ERK1 и ERK2, обе эти субстанции являются важными «переключателями» клеточного роста (Roth и др., Neurotoxicology 23.2, 2002, cc. 147-57). С другой стороны, Roth с соавторами установили также, что марганец принимает участие в гибели и апоптозе клеток путем включения многих факторов апоптоза, которые, в конце концов, приводят к клеточной гибели путем разрушения функции митохондрий и индукции утраты АТФ. Токсическое действие марганца является особенно сильным, когда из среды для культивирования клеток удаляют железо, поскольку железо и марганец конкурируют за один и тот же клеточный мембранный белок-транспортер (шаттл) DMT1 (Roth и др.).
В US 2008/0081356 описан метод крупномасштабного производства гликопротеинов с измененными схемами гликозилирования в промышленной культуре клеток путем использования среды, содержащей марганец в кумулятивной молярной концентрации от 10 до 600 нМ, и глутамин в кумулятивной молярной концентрации менее чем примерно 8 мМ.
В WO 2012/149197 описаны методы модулирования профиля гликозилирования рекомбинантно экспрессируемых белков и, прежде всего антител, в линиях СНО- и NSO-клеток путем внесения марганца и/или галактозы в среды для производства.
Важность Mn2+ в качестве кофактора β-1,4-галактозил-трансферазной активности описана ранее у Ramakrishnan с соавторами (J. Mol. Biol. 357.5, 2006, cc. 1619-1633).
Известно, что цинк имеет решающее значение для пролиферации и дифференцировки клеток, прежде всего для регуляции синтеза ДНК и митоза. Цинк является структурным компонентом огромного количества белков, включая факторы транскрипции и ферменты, участвующие в клеточных путях передачи сигналов, таких как метаболизм вторичных мессенджеров, активность протеинкиназ и протеинфосфатаз (Beyersmann и др., Biometals 14.3-4, 2001, cc. 331-341). Таким образом, биодоступный цинк ассоциирован с клеточной пролиферацией.
Железо играет ключевую роль в регуляции клеточного роста. Ранее было установлено, что добавление железа или его транспортирующего белка трансферрина в имеющую определенный химический состав культуральную среду является жизненно важным для образования биомассы. Например, установлено, что железо стимулирует клеточный рост линий раковых клеток глиомы С6 и лейкоза L1210 в бессывороточной среде (Basset и др., Carcinogenesis 6.3, 1985, cc. 355-359). Фундаментальная роль железа в клеточном росте дополнительно продемонстрирована с помощью хелаторов, которые могут пересекать плазматическую мембрану, ограничивающих его биодоступность путем связывания металла внутри клетки. Такие агенты, как десферриоксамин и десферритиоцин, ингибируют рост различных опухолевых клеток в культуре (Reddel и др., Exp. Cell Res. 161.2, 1985, cc. 277-284 и Basset и др., выше) и значительно снижают Т-клеточную пролиферацию (Poison и др., Immunology 71.2, 1990, cc. 176-181 и Pattanapanyasat и др., Br. J. Haematol. 82.1, 1992, cc. 13-19). Так, возможный ингибирующий рост клеток механизм представляет собой депривацию железа, ассоциированную с пониженной активностью рибонуклеотид-редуктазы и пониженным количеством дезоксирибонуклеотидов. Это в результате приводит к прекращению митоза на S-фазе. Добавление железа в среду приводит к реверсии ингибирования роста (Lederman и др., Blood 64.3, 1984, cc. 748-753).
В WO 98/08934 описано применение среды, дополненной цинком и железом (среди других компонентов), в суспензионной культуре для поддержания роста при высокой плотности клеток млекопитающих.
Медь играет очень важную роль в клеточном гемостазе, хотя метаболические «судьбы» меди и железа тесно связаны (Arredondo и др., Molecular Aspects of Medicine 26.4, 2005, cc. 313-327). Системный дефицит меди приводит к дефициту клеточного железа, что обусловливает фенотипически сходные действия типа пониженного клеточного роста. Кроме того, снижение биодоступности с помощью хелатирования меди связано с пониженной способностью к стимулируемому факторами роста фосфорилированию рецепторов и, в конце концов, к ингибированию путей клеточной пролиферации в опытах на культуре клеток (Turski и др., J. Biol. Chem. 284.4, 2009, cc. 717-721). С другой стороны, установлено, что медь стимулирует образование кровеносных сосудов, и снижение количества меди с помощью диеты или хелаторов меди снижает способность опухоли поддерживать ангиогенный ответ (Harris и др., Nutr. Rev. 62.2, 2004, cc. 60-64) и прогрессирование рака (Gupte и др., Cancer Treat. Rev. 35.1, 2009, cc. 32-46).
В US 2009/0068705 описаны методы крупномасштабного производства белков и/или полипептидов в культуре клеток, в которой культуральная среда содержит медь и/или глутамин и в которой полипептид имеет более обширную или более желательную схему гликозилирования.
Хотя исследования в данной области сфокусированы на активности металлических кофакторов в стимулировании гликоферментов, различные авторы, такие как Kaminska с соавторами, выше, продемонстрировали также, что эти же или другие металлические кофакторы могут ингибировать активность другого гликофермента (например, Ca2+ активирует маннозилолигосахарид-1,2-α-маннозидазу и одновременно ингибирует α-1,3-маннозилгликопротеин-2-β-N-ацетилглюкозаминилтрансферазу). Информацию, касающуюся металлических кофакторов, которые in vitro требуются для активации или для структурной организации белка, и металлических ингибиторах ряда ферментов, участвующих в гликозилировании, можно почерпнуть в базе данных BRENDA (http://www.brenda-enzymes.org). Указанная двойная активность различных металлических кофакторов подчеркивает необходимость в строгой регуляции микроэлементов, применяемых в производстве гликопротеинов.
Установлено, что гликоинженерия, т.е. изменение ассоциированных с белком углеводов посредством рекомбинантной коэкспрессии эндогенных или гетерологичных гликоферментов, влияет на фармакокинетические свойства белков, такие как молекулярная стабильность, растворимость, биологическая активность in vivo и иммуногенность. В частности, для антител установлено, что повышенный уровень галактозилирования влияет на эффекторные функции антитела, такие как CDC (Raju, Curr. Opin. Immunol. 20.4, 2008, cc. 471-478) или антитело-обусловленная клеточнозависимая цитотоксичность (ADCC), например, галактозилирование моноклональных человеческих анти-D антител IgG-типа влияет на опосредуемую Fc-рецептором функциональную активность (Kumpel и др., Hum. Antib. Hyb. 5.3-4, 1994, cc. 143-151); или продемонстрировано, что повышение уровня галактозилирования усиливает блокирование комплементопосредованного воспаления с помощью содержащих IgG иммунных комплексов (Karsten и др., Nat. Med. 18.9, 2012, cc. 1401-1406). N-гликозилирование Fc-областей антител является важным для связывания с Fc-рецепторами, которые вовлечены в эффекторные функции антител. Фукоза представляет собой коровый остаток в N-гликанах многих антител, однако установлено, что нефукозилированные формы обладают повышенной ADCC по сравнению с их фукозилированными копиями. Таким образом, также требуются методы повышения производства нефукозилированных белков и/или влияния на фукозилирование антител.
Таким образом, в данной области сохраняется необходимость в методах повышения эффективности стратегий улучшения культивирования клеток, включая повышение клеточного роста и образования биомассы, и/или для управления гликозилированием и, как следствие, созреванием гликопротеинов.
Краткое изложение сущности изобретения
При создании изобретения определяли роль биоактивных микроэлементов в эффективности культивирования клеток и, в частности, их влияние на клеточный рост и образование гликанов в гликопротеинах и, как следствие, на созревание гликопротеинов. Таким образом, настоящее изобретение относится к способам выбора между условиями, которые влияют на клеточный рост или образование биомассы, и условиями, которые влияют на созревание N-гликанов в экспрессируемом гликопротеине, который продуцируется клетками. Так, в способах, предлагаемых в настоящем изобретении, продуцирующие гликопротеин клетки культивируют в среде, которую приспосабливают для достижения требуемого конечного результата.
В настоящем изобретении предложен способ получения рекомбинантного гликопротеина в эукариотической клетке в условиях ферментационной культуры, заключающийся в том, что регулируют концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде в процессе культивирования для того, чтобы воздействовать на образование биомассы и/или зрелость N-гликанов в экспрессируемом гликопротеине.
Настоящий способ заключается в том, что регулируют концентрации железа, меди, цинка и марганца так, чтобы влиять на образование биомассы и/или созревание N-гликанов в экспрессируемом гликопротеине. Под регуляцией концентраций железа, меди, цинка и марганца подразумевается, что условия в клетке должны благоприятствовать образованию биомассы или должны влиять на созревание экспресируемого гликопротеина. В контексте настоящего описания понятие «благоприятствовать» означает, что активность клетки должна проявляться в требуемом направлении по сравнению с клеткой, которую выращивали в таких же условиях, но в которой концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, не регулировали указанным образом. Так, когда предпочтительным является образование биомассы, то в клетке процессы должны в большей степени направляться в сторону размножения, чем в сторону производства гликопротеина. В результате должно усиливаться или повышаться образование биомассы. Аналогично этому, когда посредством регуляции концентрации железа, меди, цинка и марганца или только цинка и марганца воздействуют на созревание N-гликанов экспрессируемого гликопротеина, то в клетке процессы должны в большей степени направляться в сторону производства указанных гликопротеинов, чем в сторону, например, образования биомассы или производства других типов гликопротеинов. В результате должно усиливаться или повышаться производство гликопротеина требуемого типа.
Способы, предлагаемые в изобретении, воздействуют на созревание N-гликанов экспрессируемого гликопротеина, например, приводя к производству гликопротеина с требуемым уровнем созревания. Так, способы позволяют воздействовать на производство зрелых N-гликозилированных гликопротеинов и/или незрелых N-гликозилированных гликопротеинов, зрелых нефукозилированных гликопротеинов и/или незрелых нефукозилированных гликопротеинов. Способы, предлагаемые в настоящем изобретении, включают повышение зрелости экспрессируемых N-гликозилированных гликопротеинов, повышение производство зрелых N-гликозилированных гликопротеинов, повышение производства зрелых нефукозилированных гликопротеинов и/или повышение производства незрелых нефукозилированных гликопротеинов. Одновременно с повышением производства зрелых N-гликозилированных гликопротеинов может понижаться производство зрелых и/или незрелых нефукозилированных видов гликопротеинов и незрелых N-гликозилированных гликопротеинов. Одновременно с повышением производства незрелых нефукозилированных гликопротеинов может понижаться производство зрелых нефукозилированных гликопротеинов и/или зрелых или незрелых N-гликозилированных гликопротеинов.
В некоторых вариантах осуществления изобретения концентрации железа, меди, цинка и марганца в культуральной среде можно регулировать для того, чтобы благоприятствовать образованию биомассы, что предусматривает усиление роста клеток и тем самым повышение уровня биомассы. В других вариантах осуществления изобретения можно регулировать концентрации железа, меди, цинка и марганца в культуральной среде для повышения зрелости экспрессируемого гликопротеина. В одном из вариантов осуществления изобретения можно регулировать концентрации железа, меди, цинка и марганца или только цинка и марганца в культуральной среде для того, чтобы благоприятствовать, т.е. повышать производство зрелых N-гликозилированных гликопротеинов. В другом варианте осуществления изобретения можно регулировать концентрации железа, меди, цинка и марганца в культуральной среде для того, чтобы благоприятствовать, т.е. повышать производство незрелых нефукозилированных гликопротеинов. В следующем варианте осуществления изобретения можно регулировать концентрации железа, меди, цинка и марганца в культуральной среде сначала для повышенного роста и, как следствие, для увеличения биомассы, а затем вновь для воздействия на созревание гликопротеина, т.е. для повышения зрелости экспрессируемых гликопротеинов, увеличения производства зрелых N-гликозилированных гликопротеинов или увеличения производства зрелых или незрелых нефукозилированных гликопротеинов.
Таким образом, изобретение позволяет:
- благоприятствовать образованию биомассы путем повышения концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде;
- благоприятствовать повышению зрелости экспрессируемых N-гликозилированных гликопротеинов путем повышения концентрации каждого металла из группы, включающей цинк и марганец, и необязательно понижения концентрации железа и меди в культуральной среде; и
- благоприятствовать производству незрелых нефукозилированных гликопротеинов либо (I) путем понижения концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде, либо (II) путем повышения концентраций каждого металла из группы, включающей медь и железо, и понижения концентраций каждого металла из группы, включающей цинк и марганец, в культуральной среде.
Один из объектов изобретения включает описанные выше способы, в которых для того, чтобы благоприятствовать образованию биомассы, концентрации железа, меди, цинка и марганца в культуральной среде регулируют так, чтобы они составляли:
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ.
Одним из объектов изобретения являются описанные выше способы, в которых для того, чтобы благоприятствовать повышению зрелости экспрессируемых N-гликозилированных гликопротеинов, концентрации железа, меди, цинка и марганца [указанные ниже в подпункте (I)] или концентрации цинка и марганца [указанные ниже в подпункте (II)] в культуральной среде регулируют так, чтобы они составляли:
(I)
(а) железо: от 0 до 25 мкМ;
(б) медь: от 0 до 0,1 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ; или
(II)
(а) цинк: от 20 до более чем 50 мкМ и
(б) марганец: от 0,01 до более чем 3 мкМ.
Одним из объектов изобретения являются описанные выше способы, в которых для того, чтобы благоприятствовать производству незрелых нефукозилированных гликопротеинов концентрации железа, меди, цинка и марганца в культуральной среде регулируют так, чтобы они составляли:
(I)
(а) железо: от 0 до 35 мкМ;
(б) медь: от 0 до 1 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ; либо
(II)
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ.
Одним из объектов изобретения являются способы, описанные выше, в которых гликопротеин является экзогенным или эндогенным для эукариотической клетки, в которых необязательно гликопротеин представляет собой структурный гликопротеин, гормон, антитело или фермент.
Одним из объектов изобретения являются способы, описанные выше, в которых гликопротеин представляет собой антитело, в которых необязательно антитело представляет собой терапевтическое или диагностическое антитело, необязательно химерное, гуманизированное или человеческое антитело.
Одним из объектов изобретения являются способы, описанные выше, в которых эукариотическая клетка представляет собой клетку млекопитающего, клетку дрожжей или клетку насекомых.
Одним из объектов изобретения являются способы, описанные выше, в которых концентрации железа, меди, цинка и марганца регулируют в процессе фаз роста и/или образования продуктов в культуре.
Одним из объектов изобретения являются способы, описанные выше, в которых повышение концентрации любого металла из группы, включающей железо, медь, цинк и марганец, или их всех в культуральной среде регулируют путем добавления в среду, в которой культивируют клетки, любого металла из группы, включающей железо, медь, цинк и марганец, или их всех и/или путем расщепления клеток с помещением их в свежую среду, дополненную любым металлом из группы, включающей железо, медь, цинк и марганец, или ими всеми.
Одним из объектов изобретения являются способы, описанные выше, в которых для снижения биодоступной концентрации любого металла из группы, включающей железо, медь, цинк и марганец, или их всех в культуральной среде применяют получение комплексов железа, меди, цинка и марганца с хелатором и/или достигают путем посева клеток в свежую среду, содержащую пониженную концентрацию любого металла из группы, включающей железо, медь, цинк и марганец, или их всех по сравнению со средой, применяемой на непосредственно предшествующей фазе культивирования.
Одним из объектов изобретения являются способы, описанные выше, в которых концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде регулируют в процессе культивирования сначала для того, чтобы благоприятствовать образованию биомассы, а затем для того, чтобы благоприятствовать либо производству зрелых N-гликозилированных гликопротеинов, либо производству незрелых нефукозилированных гликопротеинов.
Также в настоящем изобретении предложена среда, пригодная для производства рекомбинантного гликопротеина в эукариотической клетке в условиях ферментационной культуры, где среда содержит каждый металл из группы, включающей железо, медь, цинк и марганец, в концентрации, благоприятствующей образованию биомассы и/или воздействующей на зрелость N-гликанов экспрессируемого гликопротеина.
Согласно одному из объектов изобретения среда содержит железо, медь, цинк и марганец в следующих концентрациях:
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ.
Применение указанной среды для производства рекомбинантного гликопротеина в эукариотической клетке в условиях ферментационной культуры благоприятствует образованию биомассы.
Согласно одному из объектов изобретения среда содержит железо, медь, цинк и марганец или цинк и марганец в следующих концентрациях:
(I)
(а) железо: от 0 до 25 мкМ;
(б) медь: от 0 до 0,1 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ, или
(II)
(а) цинк: от 20 до более чем 50 мкМ и
(б) марганец: от 0,01 до более чем 3 мкМ.
Применение среды, содержащей железо, медь, цинк и марганец или цинк и марганец в концентрациях, указанных в подпункте (I) или (II) соответственно, для производства рекомбинантного гликопротеина в эукариотической клетке в условиях ферментационных культур благоприятствует повышению зрелости экспрессируемых N-гликозилированных гликопротеинов.
Согласно одному из объектов изобретения среда содержит железо, медь, цинк и марганец в концентрациях, составляющих либо:
(I)
(а) железо: от 0 до 35 мкМ;
(б) медь: от 0 до 1 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ, либо:
(II)
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ.
Применение среды, содержащей железо, медь, цинк и марганец или цинк и марганец в концентрациях, указанных выше в подпункте (I) или (II), для производства рекомбинантного гликопротеина в эукариотической клетке в условиях ферментационных культур благоприятствует производству незрелых нефукозилированных гликопротеинов.
Описание чертежей
На чертежах показано:
на фиг. 1 - Стадии процесса ферментативного получения. Замороженные флаконы, например, из главного банка посевного материала (PSB), подвергали оттаиванию в среде для подготовки посевного материала (затравки). В ней клетки культивировали в течение примерно 3 недель для восстановления после связанного с оттаиванием стресса (нормализация времени удвоения клеток). Затем клетки переносили в среды для подготовки к инокуляции n-1 и n-2 для размножения культуры. Через примерно 1 неделю клетки переносили в среду для крупномасштабного производства посредством периодического ферментативного процесса с подпиткой в течение 2 недель;
на фиг. 2 - Периодические процессы с подпиткой, отличающиеся двумя фенотипически различными фазами. Фаза производства может быть разделена на два сегмента (фазы), на которых доминирует либо клеточный рост, либо производство целевого (требуемого) белка;
на фиг. 3 - Различные подходы к планированию экспериментов (DoE). В зависимости от требуемого результата эксперимента можно применять различные подходы к DoE для оптимизации процесса. Например, при оптимизации трех факторов, как правило, их варьируют между минимальным (-1), средним (0) и максимальным (+1) уровнями для оптимизации DoE (иллюстрация получена из http://www.gmpua.com и модифицирована);
на фиг. 4 - Воздействие микроэлементов на независимое от клона МАт галактозилирование. Маточные растворы солей алюминия, молибдена, бария, хрома, брома, йода, меди, марганца, рубидия, серебра, цинка, олова и циркония добавляли в периодическую культуру с подпиткой клона 1 (n=4) и клона 3 (n=3), используя в начале эксперимента запатентованную платформу А для культуры клеток (среда и процесс). Воздействие микроэлементов на галактозилирование МАт анализировали в дни 7, 12 и 13 путем моделирования уровней G1 (А, Б) и G2 (В, Г) на основе существующих экспериментальных данных. Каждая точка на графиках сравнения фактических и предсказанных (actual vs predicted) значений (А, В) соответствовала одному эксперименту по культивированию. Статистическую значимость воздействия каждого микроэлемента на уровни G1 и G2 центрировали относительно среднего значения и масштабировали с помощью половинного диапазона для G1 (Б) и G2 (Г);
на фиг. 5 - Измерение в зависимости от времени регуляции галактозилирования МАт с помощью марганца. Значимость воздействия марганца на образование гликанов G1 (А) и G2 (Б) клона 3 (планшеты 5, 6 и 7) анализировали с помощью инструмента, представляющего собой прогностический профайлер программы JMP. Положительное воздействие марганца на G1 и G2 различалось на ранней (день 7) и поздней фазах процесса (день 13) вплоть до 100% (крутизна наклона кривой, полученной с помощью прогностического профайлера). Эксперимент осуществляли в трех повторностях (n=3);
на фиг. 6 - Различные воздействия совместного добавления цинка, железа, меди и марганца на клеточный рост, галактозилированные и нефукозилированные виды гликопротеина. Эксперименты с периодической подпиткой, проведенные с клоном 2, осуществляли с добавлением цинка, железа, меди и марганца в различных концентрациях в DoE-эксперименте. Анализировали воздействия на максимальную плотность жизнеспособных клеток (А-В), конечный интеграл от количества клеток по времени (Г-Е), G1-виды (Ж-И) и сумму маннозилированных видов w/o Fuc (К-М). С помощью прогностических моделей (А, Г, Ж, К) и оценки параметров сортировки (Б, Д, З, Л) продемонстрированы качественные и количественные воздействия микроэлементов для каждой конкретной модели на значимые факторы в отношении клеточного роста и гликозилирования соответственно. Оптимальные концентрации (обозначенные серым цветом числа: стандартизованная концентрация, пунктирная горизонтальная линия: максимальное значение) цинка, железа, меди и марганца в периодических культурах с подпиткой для оптимального клеточного роста (В, Е), оптимального галактозилирования гликопротеина (И) высоко нефукозилированных (афукозилированных) видов гликопротеинов (М), определяли с использованием инструмента максимизации профайлера программы JMP. Примечание: при применении таких микроэлементов как цинк и медь, обнаружены обратные воздействия в отношении гликозилирования;
на фиг. 7 - Воздействие цинка и меди на нефукозилированные виды гликопротеинов. С использованием в качестве инструмента профайлера JMP анализировали воздействия уровней цинка на корреляцию с уровнями меди в отношении маннозилированных видов гликанов. Исследовали средние уровни цинка и меди (А), сбалансированные уровни цинка и меди (Б), высокие уровни цинка и средние уровни меди (В), низкие уровни цинка и низкие уровни меди (Г), низкие уровни цинка и средние уровни меди (Д), высокие уровни цинка и низкие уровни меди (Е), средние уровни цинка и высокие уровни меди (Ж), высокие уровни цинка и высокие уровни меди (З), средние уровни цинка и низкие уровни меди (И) и уровень цинка -0,2 (т.е. 25,416 мкМ) при высоких концентрациях меди (К);
на фиг. 8 - Оптимальные концентрации цинка, железа, меди и марганца для клеточного роста, формирования галактозилированных и нефукозилированных видов гликопротеинов. С использованием одной валидной прогностической модели продемонстрировали воздействие различных вариантов уровней цинка, железа, меди и марганца на VCD (плотность жизнеспособных клеток), CTI (интеграл от количества клеток по времени), уровень галактозилирования МАТ и гликанов с высоким содержанием маннозы (Man5);
на фиг. 9 - Модуляция клеточного роста с помощью целевых концентраций микроэлементов. Предсказанные концентрации микроэлементов, благоприятствующие клеточному роста и гликозилированию МАт, тестировали в трех экспериментах в биореакторах с периодической подпиткой, на стадиях, обозначенных как «Афукозилирование» (треугольники), «Рост» (квадрат) и «Галактозилирование» (окружности), применяя клон 2 и запатентованную среду, и платформу Б для процесса (А). Анализировали воздействие на плотность жизнеспособных клеток (Б), интеграл от количества клеток по времени (Б) и жизнеспособность клеток (Г). Афукозилирование обозначает то же самое, что нефукозилирование;
на фиг. 10 - Различный клеточный метаболизм в зависимости от добавления меди. Процесс «галактозилирования» на фазе «Гликозилирование» (дни d6-d14) снижал конечные уровни аммония (А) и повышал уровни лактата (Б). Оценка уровня ЛДГ, служащего маркером лизированных клеток, продемонстрировала сниженный на 50% уровень по сравнению с контролем и стадией процесса «Рост»;
на фиг. 11 - Модуляция галатозилированных и нефукозилированных видов с помощью целевых концентраций микроэлементов. Предсказанные концентрации микроэлементов, благоприятствующие гликозилированию МАт, тестировали в трех экспериментах в биореакторах с периодической подпиткой на стадиях «Афукозилирование» (треугольники), «Рост» (квадрат) и «Галактозилирование» (окружности), применяя клон 2 и запатентованную среду и платформу Б для процесса (А). Анализировали воздействие модуляций целевых микроэлементов на относительное содержание Man5- (A), G0- (Б) и G1 (В)-видов, а также на кумулятивное содержание нефукозилированных видов (Г);
на фиг. 12 - Роль металлических микроэлементов в клеточном росте и галактозилировании/нефукозилировании клона 1 - эксперимент с периодической подпиткой в биореакторе. Предсказанные концентрации микроэлементов, благоприятствующие клеточному росту и модуляции зрелости гликозилирования МАт, тестировали в трех экспериментах в реакторах с периодической подпиткой, комбинируя хороший и плохой рост клеток во время стадии подготовки к инокулированию (фазы n-2/n-1) и галактозилирование, и нефукозилирование МАт (образование незрелых гликанов) при периодической подпитке (фаза n). Условия эксперимента должны были специфически модулировать клеточный рост, галактозилирование и незрелое гликозилирование белка в процессе подготовки к инокулированию и на фазе производства (А). Различные концентрации микроэлементов, необходимые для осуществления эксперимента указаны в (Б). В биореакторе, условия, в котором обозначены как «GG» (ромбы), поддерживался клеточный рост и галактозилирование белка, в биореакторе, условия в котором обозначены как «GA» (квадраты), поддерживался клеточный рост и нефукозилирование МАт, а в биореакторе, условия в котором обозначены как «АА» (треугольники), подавлялся клеточный рост, и запускалось нефукозилирование белка. Анализировали воздействие на плотность жизнеспособных клеток (В), интеграл от количества клеток по времени (Г), созревание МАт (сумма G1- и G2-видов) в дни 6 и 14 (Д) и незрелость МАт (сумма Man6, Man5 и G0-GlcNAc) в дни 6 и 14 (Е);
на фиг. 13 - Измеренные концентрации железа, цинка, марганца и меди в эксперименте в биореакторе 1. Представлены фактические концентрации меди (A), железа (Б), марганца (В) и цинка (Г), измеренные с помощью ICP-MC-анализа (масс-спектрометрия с индуктивно связанной плазмой) в супернатанте клеточной культуры. Постепенное повышение теоретической концентрации цинка, железа, марганца и меди являлось результатом болюсного введения в дни 3, 5 и 9. В день 6 культуру расщепляли;
на фиг. 14 - Концентрации железа, цинка, марганца и меди для калькуляции среды в эксперименте в биореакторе 2. Представлены фактические концентрации цинка (А), марганца (Б), железа (В) и меди (Г) для репрезентативных вариантов периодического культивирования с подпиткой, включая фазы n-2, n-1, n, для клона 1, клона 2, клона 3, клона 4 (n>3), измеренные с помощью ICP-MC-анализа в супернатанте клеточной культуры;
на фиг. 15 - Воздействие цинка и марганца на галактозилированные виды гликопротеина. С использованием в качестве инструмента профайлера JMP анализировали воздействие уровней цинка на корреляцию с уровнями меди в отношении галактозилированных видов G1-гликанов для клона 2. Изучали низкие уровни цинка и высокие уровни марганца (А), низкие уровни цинка и низкие уровни марганца (Б), высокие уровни цинка и низкие уровни марганца (B), высокие уровни цинка и высокие уровни марганца (Г);
на фиг. 16 - Воздействие цинка и марганца на галактозилированные виды гликопротеина в зависимости от времени. С использованием в качестве инструмента профайлера JMP анализировали воздействие уровней цинка на корреляцию с уровнями меди в отношении галактозилированных видов G1- и С2-гликанов для клона 1 (n=4) и клона 3 (n=3). Изучали высокие уровни цинка и высокие уровни марганца (А), высокие уровни цинка и низкие уровни марганца (Б), низкие уровни цинка и низкие уровни марганца (В), низкие уровни цинка и высокие уровни марганца (Г).
Определения
«Регуляция» в контексте настоящего описания означает повышение или понижение концентрации элемента в культуральной среде. Повышение или понижение концентрации элемента оценивают относительно концентрации элемента в среде на фазе культивирования, непосредственно предшествующей регулированию. Например, если регуляция приводит к повышению концентрации микроэлементов и требуется в начале фазы производства, то это означает повышение концентрации указанных микроэлементов относительно концентрации указанных микроэлементов, включенных в среду на фазе, непосредственно предшествующей фазе роста.
«Регуляция концентрации» в контексте настоящего описания означает изменение измеренной или поддающейся измерению или рассчитанной или поддающейся расчету фактической концентрации элемента в среде, окружающей клетку, в данный момент времени.
«Воздействие» в контексте настоящего описания означает действие, которое приводит к изменению процессов в клетке, представляющих собой либо образование биомассы, либо созревание N-гликанов. Полученный в результате эффект может представлять собой, например, повышение образования биомассы или повышение производства гликопротеина с требуемой схемой гликозилирования.
«И/или» в контексте настоящего описания используют для конкретного обозначения каждой из двух специфических характеристик или специфических компонентов в сочетании друг с другом или не в сочетании друг с другом. Например, «А и/или Б» является конкретным обозначением каждого из понятий (I) А, (II) Б и (III) А и Б, как, если бы каждое из этих понятий было указано индивидуально.
«Антитело» в контексте настоящего описания обозначает молекулу иммуноглобулина или иммунологически активный участок молекулы иммуноглобулина, т.е. молекулу, которая содержит антигенсвязывающий центр, такую как Fab- или F(аb’)2-фрагмент, вне зависимости от того, встречается ли она в естественных условиях или получают ли ее частично или полностью путем синтеза. Понятие «антитело» используют в его наиболее широком смысле, и оно включает моноклональные антитела (включая полноразмерные антитела, которые имеют Fc-область иммуноглобулина, или интактные моноклональные антитела), композиции антител с полиэпитопной специфичностью, поликлональные антитела, многовалентные антитела (как правило, сконструированные так, что они имеют три или большее количество антигенсвязывающих центров), мультиспецифические антитела (например, биспецифические антитела), полученные по меньшей мере из двух интактных антител, димерные антитела (диабоди) и одноцепочечные молекулы, такие как scFv-молекулы, а также фрагменты антител (например, Fab, F(ab')2 и Fv). Под определение антитела подпадают конъюгаты антител, такие как конъюгаты антитела и лекарственного средства (ADC), или антитела, конъюгированные, например, с элементами-метками.
Понятие «биомасса» в контексте настоящего описания относится к количеству или массе культивируемых клеток в культуральной среде. Биомассу можно измерять непосредственно или опосредовано путем определения плотности жизнеспособных клеток, плотности всех клеток, интеграла от количества клеток по времени (на основе плотности жизнеспособных и всех клеток), интеграла от объема клеток по времени (на основе плотности жизнеспособных и всех клеток), объема упакованных клеток, массы в сухом состояния или массы во влажном состоянии.
«Биореактор» в контексте настоящего описания означает любой сосуд, применяемый для выращивания культуры клеток млекопитающих. Как правило, объем биореактора должен составлять по меньшей мере 1 л и он может составлять 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000 л или более, или представлять собой любой объем, находящийся между указанными величинами. Внутренние условия биореактора, включая (но, не ограничиваясь только ими) рН, уровень растворенного кислорода и температуру, как правило, контролируют в течение периода культивирования. Биореактор может быть изготовлен из любого материала, включая стекло, пластик или металл, пригодного для поддержания культур клеток млекопитающих, которые суспендированы в средах, в условиях культивирования, предлагаемых в настоящем изобретении.
Понятия «клетка» и «клеточная линия» в контексте настоящего описания используют взаимозаменяемо, и все указанные понятия включают потомство.
«Плотность клеток» в контексте настоящего описания означает количество клеток, присутствующих в данном объеме среды.
«Жизнеспособность клеток» в контексте настоящего описания означает способность клеток в культуре выживать при данных принятых условиях культивирования или экспериментальных вариациях. Понятие в контексте настоящего описания означает также ту часть клеток, которая остается живой в конкретный момент времени относительно общего количества клеток, живых или мертвых, в культуре в данный момент времени.
«Хелатор» в контексте настоящего описания означает соединение, обладающее способностью подавлять химическую активность посредством образования хелата, т.е. путем связывания иона металла.
«Концентрация» в контексте настоящего описания касательно микроэлементов, таких как железо, медь цинк и марганец, означает количество каждого из микроэлементов, содержащееся в культуральной среде. Концентрация может представлять измеренную или поддающуюся измерению или рассчитанную или поддающуюся расчету фактическую концентрацию элемента в среде, окружающей клетку в данный момент времени. Методы измерения концентрации указанных микроэлементов в среде известны в данной области. Примеры таких методов включают ICP-MC (фирма Agilent, Беблинген, Германия). Таким образом, понятие «концентрация» относится также к количеству каждого из микроэлементов, содержащихся в окружающей клетку культуральной среде в процессе культивирования, и, таким образом, означает фактическую концентрацию соответствующего микроэлемента в данный момент времени. Указанную концентрацию можно определять аналитически, и она обусловлена, например, внесением элемента в культуру (внесением отвешенного количества, переносом клеток и среды из предкультуры, внесением примесей, выщелачиванием и т.д.), высвобождением из клеток (например, в результате гибели клеток или в результате активной секреции), поглощением клетками и другими факторами.
«Медь» в контексте настоящего описания означает катион Cu2+.
«Культура» или «клеточная культура» в контексте настоящего описания означает клеточную популяцию, суспендированную в среде в условиях, пригодных для выживания и/или роста клеточной популяции. Указанные понятия относятся также к комбинации среды и суспендированной в ней клеточной популяции.
Понятия «условия культивирования» и «условия ферментации» в контексте настоящего описания используют взаимозаменяемо, и они обозначают такие условия, которые должны удовлетворять требованиям успешного культивирования клеток. Как правило, эти условия включают наличие соответствующей среды, а также контроль, например, температуры, которая должна составлять примерно 37°С, но могут включать также сдвиг температуры в процессе культивирования (например, с 37°С до 34°С) и значения рН, которое, как правило, находится между 6,8 и 7,2, а также наличие кислорода и диоксида углерода. Указанные условия включают также метод, с помощью которого культивируют клетки, например, культивирование на шейкере или роботизированное культивирование.
«Понижение, уменьшение» в контексте настоящего описания касательно концентраций микроэлементов, таких как железо, медь, цинк или марганец, означает понижение концентрации указанных микроэлементов в культуральной среде относительно их концентрации в среде, в которой клетки культивировали непосредственно на предыдущей фазе или частичной фазе.
«Благоприятствовать» в контексте настоящего описания означает, что активность клетки в указанном направлении можно повышать по сравнению с клеточным ростом в таких же условиях, но в которых концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, не регулировали с помощью методов, предлагаемых в настоящем изобретении. Например, когда требуется благоприятствовать образованию биомассы, регуляция концентрации микроэлементов в среде должна приводить к тому, что клетки в большей степени направлены на размножение, чем на производство гликопротеина. В этом случае результатом является повышенное образование биомассы. В контексте настоящего описания синонимами понятия «благоприятствовать» являются понятия «повышать» и «усиливать».
Понятие «периодическое культивирование с подпиткой» в контексте настоящего описания относится к методу культивирования клеток, в котором дополнительные компоненты вносятся в культуру однократно или в несколько моментов времени после начала процесса культивирования. Процесс периодического культивирования с подпиткой, как правило, прекращают в некоторый момент времени и клетки и/или компоненты в среде собирают и необязательно очищают.
«Галактозилированный» в контексте настоящего описания касательно гликопротеинов относится к гликопротеину, который содержит один или несколько остатков галактозы в G1- и G2-гликоструктурах.
Понятие «ген» в контексте настоящего описания относится к любой из таких субстанций как нуклеотидная последовательность, ДНК или РНК, по меньшей мере часть которых кодирует полипептид. Необязательно ген содержит не только кодирующую последовательность полипептида, но содержит также области, расположенные перед и/или после кодирующей последовательности, которые модулируют основной уровень экспрессии, и/или интроны между кодирующими сегментами или экзоны.
Понятие «гликоформа» в контексте настоящего описания относится к любой из нескольких различных форм гликопротеина, имеющим различные присоединенные сахариды.
Понятие «гликопротеин» в контексте настоящего описания относится к белку или полипептиду, который содержит одну или несколько ковалентно связанных олигосахаридных цепей. Олигосахаридные цепи могут состоять из одного сахарного остатка, одной неразветвленной цепи или сахарных остатков или могут состоять из цепи сахарных остатков, которые имеют одно или несколько разветвлений. Олигосахаридные цепи могут представлять собой N-связанные или О-связанные цепи.
Понятие «клетка-хозяин» в контексте настоящего описания означает любой тип клеточной системы, которую можно конструировать для образования гликопротеинов.
«Повышение, возрастание» в контексте настоящего описания касательно концентраций микроэлементов, таких как железо, медь, цинк или марганец, означает повышение концентрации указанных микроэлементов в культуральной среде относительно их концентрации в среде, в которой клетки культивировали непосредственно на предыдущей фазе или частичной фазе.
«Железо» в контексте настоящего описания означает либо катион Fe(III) (Fe3+), либо катион Fe(II) (Fe2+).
«Марганец» в контексте настоящего описания означает катион Mn2+.
«Зрелость (созревание)» и «зрелость (созревание) гликопротеина» в контексте настоящего описания относится к схеме гликозилирования рекомбинантно продуцируемых гликопротеинов. Все N-гликозилированные белки обычно имеют пентасахаридное ядро (ядро представляет собой GlcNAc2 Man3) и концевое гликозилирование, которое имеет место в комплексе Гольджи, что приводит к огромной структурной диверсификации из-за различных комбинаций олигосахаридов, добавляемых к указанному ядру. Таким образом, понятие «повышение зрелости гликопротеина» относится к добавлению к ядру и/или последующую модификацию олигосахаридных звеньев. Таким образом, понятие «воздействие на зрелость N-гликанов» включает повышение зрелости гликопротеина относительно коровой структуры или относительно непосредственно предшествующей углеводной структуры. Таким образом, зрелость гликопротеина возрастает, когда коровую структуру модифицируют путем добавления олигосахаридных звеньев или путем удаления, например, остатков маннозы из незрелой гибридной структуры. Таким образом, повышение зрелости включает, например, производство гликопротеинов с G0-струтурой, например, из Man3-5-структуры или из коровой структуры. Полностью зрелые виды гликопротинов могут представлять собой галактозилированные виды, содержащие один или два галактозных остатка, или сиалилированные виды, которые содержат один или несколько остатков сиаловой кислоты, такие как GlcNAc3Man3GlcNAc2Gal1, GlcNAc3Man3GlcNAc2Gal2, GlcNAc3Man3GlcNAc2Gal1Sia1, GlcNAc3Man3GlcNAc2Gal2Sia1 и GlcNAc3Man3GlcNAc2Gal2Sia2. Незрелые гликопротеины могут представлять собой виды с высоким содержание маннозы, содержащие от 4 до 9 остатков маннозы, или незрелые гибридные структуры, такие как GlcNAc3Man5GlcNAc, GlcNAc3Man4GlcNAc и GlcNAc3Man3GlcNAc. В некоторых случаях требуется получать гликопротеины, в которых сахарные остатки, которые обычно входят в их состав, не включены соответствующим образом. Под объем изобретения подпадают частично или полностью нефукозилированные гликопротеины, в которых отсутствуют коровые остатки фукозы, например, G0-F, G1-F и G2-F. Зрелые и незрелые виды гликопротеинов могут быть нефукозилированными, т.е. в них могут отсутствовать коровые остатки фукозы.
Понятия «среда», «среда для культивирования клеток» и «культуральная среда» в контексте настоящего описания применяют взаимозаменяемо, и они относятся к содержащему питательные вещества раствору, который поддерживает ост клеток млекопитающих. Как правило, указанные растворы содержат незаменимые и заменимые аминокислоты, витамины, источники энергии, липиды и микроэлементы, которые требуются для минимального роста и/или выживания клеток. Указанный раствор может содержать также дополнительные компоненты, которые усиливают рост и/или выживание, превышающие минимальный уровень, которые включают (но не ограничиваясь только ими) гормоны и/или другие факторы роста, конкретные ионы, такие как натрий, хлорид, кальций, магний и фосфат, буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы, аминокислоты, липиды и/или глюкозу или другой источник энергии. Среду целесообразно приготавливать так, чтобы значение рН и концентрация солей были оптимальными для выживания и пролиферации клеток. Среда может представлять собой среду с пониженным содержанием сыворотки или бессывороточную среду, т.е. среду с содержанием сыворотки примерно 1-5% или среду, практически свободную от какой-либо сыворотки млекопитающих (например, фетальной бычьей сыворотки) соответственно. Под практически свободной средой подразумевают среду, которая содержит 0-5% сыворотки, предпочтительно при 0-1% сыворотки и наиболее предпочтительно примерно 0-0,1% сыворотки. Указанную бессывороточную среду можно применять, когда идентичность и концентрация каждого из компонентов среды является известной. Среда может представлять собой безбелковую среду, т.е. среду, которая не должна содержать белок, но может содержать неидентифицированные пептиды, например, из растительных гидролизатов. Среды могут включать человеческий сывороточный альбумин и человеческий трансферрин и инсулин и липиды потенциально животного происхождения, или свободную от ксенобиотиков среду, содержащую человеческий сывороточный альбумин, человеческий трансферрин, человеческий инсулин и липиды установленного химического состава. В альтернативном варианте среда может представлять собой среду определенного химического состава, т.е. среду, в которой все субстанции установлены и присутствуют в определенных концентрациях. Указанные среды могут содержать только рекомбинантные белки и/или гормоны, или безбелковую среду определенного химического состава, т.е. содержащую только низкомолекулярные компоненты и при необходимости синтетические пептиды/гормоны. Среды определенного химического состава могут также быть практически свободны от какого-либо белка.
«Нефукозилированный гликопротеин» представляет собой зрелый или незрелый гликопротеин, лишенный одного или нескольких коровых N-фукозных остатков. Указанные структуры могут быть нефукозилированными сами по себе, в этом случае гликопротеин в естественных условиях не содержит фукозных остатков, они могут быть нефукозилированными в результате отсутствия фукозного остатка, который, как предполагается, в естественных условиях должен присутствовать в гликопротеине. В контексте настоящего описания понятия «нефукозилированный» и «афукозилированный» применяют взаимозаменяемо.
Понятие «перфузионное культивирование» в контексте настоящего описания относится к методу культивированная клеток, заключающемуся в том, что выращивают клетки в среде на основе инокулята, когда клетки достигают требуемой плотности заменяют истощенную среду свежей средой. Перфузия может представлять собой либо непрерывную, либо прерывистую перфузию и может включать введение по меньшей одной болюсной подпитки в клеточную культуру. После перфузионного культивирования можно осуществлять периодическое культивирование с подпиткой.
Понятие «полипептид» в контексте настоящего описания относится к последовательной цепи аминокислот, сцепленных друг с другом через пептидные связи. Никакое ограничение по длине не накладывается на указанную аминокислотную цепь, которая может содержать от 2 до множества аминокислот. Полипептиды можно процессировать и/или модифицировать, например, путем гликозилирования.
Понятие «белок» в контексте настоящего описания относится к одному или нескольким полипептидам, которые функционируют в качестве дискретной единицы. Когда белок содержит только один функциональный полипептид, то понятия полипептид и белок применяют взаимозаменяемо.
Понятия «рекомбинантный гликопротеин» или «рекомбинантно экспрессируемый гликопротеин» в контексте настоящего описания применяют взаимозаменяемо для обозначения гликопротеина, экспрессируемого из клетки-хозяина, которую подвергали манипуляции для достижения указанной экспрессии. Манипуляция включала одну или несколько генетических модификаций, таких как интродукция одного или нескольких гетерологичных генов, кодирующих подлежащий экспрессии гликопротеин. Гетерологичный ген может кодировать гликопротеин, который либо экспрессируется в норме в указанной клетке, либо является чужеродным для клетки-хозяина. Альтернативно этому, манипуляция может обусловливать повышающую или понижающую регуляцию одного или несколько эндогенных генов.
Под «расщеплением» в контексте настоящего описания подразумевают также процесс, известный как пересев или субкультивирование клеток. Оно включает перенос небольшого количества клеток в свежую среду, посредством чего расщепленные клетки засевают новую культуру. В суспензионных культурах небольшое количество культуры, содержащее несколько клеток, разводят в большем объеме свежей среды.
Понятие «титр» в контексте настоящего описания относится к общему количеству рекомбинантно экспрессируемого гликопротеина, продуцируемого культурой клеток млекопитающего, в данном объеме среды. Титр, как правило, выражают в таких единицах как миллиграммы гликопротеина на миллилитр среды.
«Цинк» в контексте настоящего описания означает катион Zn2+.
В контексте настоящего описания применяли следующие сокращения:
Asn аспарагин
ADCC антитело-обусловленная клеточнозависимая
цитотоксичность
CTI интеграл от количества клеток по времени
CDC комплементзависимая цитотоксичность
ЦМФ цитидинмонофосфат
ЦТФ цитидинтрифосфат
ГДФ гуанозиндифосфат
ГТФ гуанозинтрифосфат
ICP-MC масс-спектрометрия с индуктивно связанной плазмой
ЛДГ лактатдегидрогеназа
УДФ уридиндифосфат
УТФ уридинтрифосфат
МАт моноклональное антитело
PSB главный банк посевного материала
PSE псевдо-стандартная ошибка
RSME плановая стандартная ошибка
Fuc L-фукоза
Gal D-галактоза
GlcNAc N-ацетилглюкозамин
NANA N-ацетилнейраминовая кислота
Man D-манноза
Man5 GlcNAc2 Man5
Man6 GlcNAc2 Man6
высокое содержание
маннозы GlcNAc2 Man5-8
ядро
(коровая структура) GlcNAc2 Man3
G0 GlcNAc Fuc GlcNAc Man3 GlcNAc2
G0-F GlcNAc GlcNAc Man3 GlcNAc2
G1 GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal
G1-F GlcNAc GlcNAc Man3 GlcNAc2 Gal
G2 GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal2
G2 1SA GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal2 NANA1
комплекс GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal0-2
комплекс-F GlcNAc GlcNAc Man3 GlcNAc2 Gal0-2
Подробное описание изобретения
Настоящее изобретение относится к способам и средам для производства рекомбинантного гликопротеина в условиях ферментационной культуры в эукариотической клетке, где способ заключается в том, что регулируют концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде в процессе культивирования для воздействия на образование биомассы и/или зрелости N-гликанов экспрессируемого гликопротеина.
Согласно одному из объектов изобретения способ, предлагаемый в изобретении, заключается в том, что регулируют концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде так, чтобы благоприятствовать образованию биомассы. В результате этого должны повышаться количество или масса культивируемых клеток в культуральной среде. Биомассу можно измерять, определяя плотность жизнеспособных клеток, общую плотность клеток, интеграл от количества клеток по времени (на основе плотности жизнеспособных и всех клеток), интеграл от объема клеток по времени (на основе плотности жизнеспособных и всех клеток), объем упакованных клеток, массу в сухом состоянии или массу во влажном состоянии, с помощью методов, известных в данной области.
Согласно одному из объектов изобретения способ, предлагаемый в настоящем изобретении, заключается в том, что регулируют концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде так, чтобы воздействовать на зрелость N-гликанов в экспрессируемом гликопротеине. Понятие «зрелость N-гликанов» относится к схеме гликозилирования, согласно которой гликопротеин может содержать либо все, либо практически все, либо меньшее количество по сравнению с общим количеством определяемых генетическими факторами гликановых остатков, т.е. гликановых остатков, добавленных с помощью эндогенных генетически кодируемых гликоферментов.
В предпочтительном варранте осуществления изобретения регуляция концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, или только цинка и марганца в культуральной среде должна повышать зрелость экспрессируемых N-гликопротеинов. Примерами углеводных схем в таких экспрессируемых гликопротеинах являются: GlcNAc Fuc GlcNAc Man3 GlcNAc2 (G0); GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal (G1) и GlcNAc Fuc GlcNAc Man3 GlcNAc2 Gal2 (G2). К ним относятся также зрелые нефукозилированные гликопротеины, такие как GlcNAc GlcNAc Man3 GlcNAc2 (G0-F) и GlcNAc GlcNAc Man3 GlcNAc2 Gal0-2 (G1-F, G2-F). В наиболее предпочтительном варианте осуществления изобретения благоприятствующее воздействие направлено на образование гликопротеинов с G0-, G1- и/или G2-структурой углеводов.
В альтернативном предпочтительном варианте осуществления изобретения регуляция концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде должна влиять на производство гликопротеинов, представляющих собой незрелые нефукозилированные гликопротеины. Примерами схемы гликозилирования в указанных незрелых нефукозилированных гликопротеинах являются гликопротеины с высоким содержанием маннозы, т.е. те, которые содержат Man5, Man6 и Man7, такие как GlcNAc2 Man5-8, например, GlcNAc2 Man5 и GlcNAc2 Man6.
Настоящее изобретение относится к производству рекомбинантного гликопротеина. Оно не ограничено природой гликопротеина, если его можно получать с помощью ферментационной культуры и экспрессировать в эукариотической клетке. Гликопротеины, которые можно получать с помощью указанного способа, включают секретируемые и связанные с мембраной гликопротеины и/или гликопротеины, которые могут являться экзогенными или эндогенными для эукариотической клетки, включая, например, структурные гликопротеины, гормоны, антитела, ферменты и т.п.
В предпочтительном варианте осуществления изобретения гликопротеин представляет собой антитело, как правило, терапевтическое или диагностическое антитело, и в дополнительном варианте осуществления изобретения антитело представляет собой химерное, гуманизированное или человеческое антитело.
Когда гликопротеин представляет собой антитело, то антитело может представлять собой терапевтически эффективное антитело, и оно может связываться с любым белком, включая представителя семейства ангиопоэтинов, такого как Ang1, Ang2, Ang3 и Ang4, и антитела могут представлять собой антитела, обладающие двойной специфичностью в отношение представителя семейства ангиопоэтинов и, например, VEGF, такие как Ang2/VEGF; антитело может связываться с представителем семейства рецепторов HER, таким как HER1 (EGFR), HER2, HER3 и HER4; CD-белками, такими как CD3, CD4, CD8, CD18, CD19, CD20, CD21, CD22, CD25, CD33, CD34, CD38, CD40, CD44 и CD52; молекулами клеточной адгезии, такими как LFA-1, VLA04, ICAM-1, VCAM и интегрин, включая либо α-, либо β-субъединицу (например, антитела к CD11а, к D18 или к CD11b); факторами роста, такими как сосудистый эндотелиальный фактор роста (VEGF); рецепторами цитокинов, такими как рецептор тимусного стромального лимфопоэтина (TSLP-R); IgE; антигенами группы крови; flk2/flt3-рецептором; рецептором ожирения (ОВ) и белком С. Другие примеры белков включают гормон роста (GH), включая человеческий гормон роста (hGH) и бычий гормон роста (bGH); релизинг-фактор гормона роста; паратиреоидный гормон, гормон, стимулирующий щитовидную железу; липопротеины; α-1-антитрипсин; цепь А инсулина; цепь В инсулина; проинсулин, фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания, такие как фактор VIIIC; тканевый фактор (TF); фактор фон Виллебранда; предсердный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или активатор плазминогена тканевого типа (t-PA), бомбазин, тромбин, факторы некроза опухоли -α и -β; энкефалиназу; RANTES (хемокин, экспрессируемый и секретируемый нормальными Т-клетками при активации); человеческий макрофагальный воспалительный белок (MIP-1-α); сывороточный альбумин, такой как человеческий сывороточный альбумин (HSA);
муллерианингибирующую субстанцию; цепь А релаксина, цепь В релаксина; прорелаксин; мышиный гонадотропин-ассоциированный пептид; ДНКазу; ингибин; активин; рецепторы для гормонов или факторов роста; белок А или D; фибробласт-активирующий белок (FAP); карциноэмбриональный антиген (СЕА); ревматоидные факторы; нейротропный фактор, такой как нейротропный фактор, образующийся в костном мозге (BDNF); нейротрофин-3, -4, -5 или -6 (NT-3, NT-4, NT-5 или NT-6) или фактор роста нервов, такой как NGF-β; плацентарный фактор роста (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF) и рецептор эпидермального фактора роста (EGFR); трансформирующий фактор роста (TGF) такой как TGF-α и TGF-β, включая TGF-β1, TGF-β2, TGF-β3, TGF-β4 или TGF-β5; инсулиноподобный фактор роста -I и -II (IGF-I и IGF-II); дез(1-3)-IGF-1 (IGF-I головного мозга); белки, связывающие инсулиноподобный фактор роста (IGFBP); эритропоэтин (ЕРО); тромбопоэтин (ТРО); остеоиндуктивные факторы; иммунотоксины; белок, участвующий в остеогенезе (BMP); интерферон (интерферон-α, -β или -γ); колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, IL-1-IL-10 и IL-17; супероксиддисмутазу; Т-клеточные рецепторы; BlyS (Br3)-рецептор; иммуноадгезин Вr3-Fc; Аро-2-рецептор; Fc-рецептор; поверхностные мембранные белки; фактор, ускоряющий распад (DAF); вирусный антиген, такой, например, как часть оболочки вируса СПИДа; транспортные белки; хоминг-рецепторы; аддерессины; регуляторные белки; иммуноадгезины; и биологически активные фрагменты или варианты любого из указанных выше белков. Альтернативно этому, антитело может представлять собой антитело, направленное против эпителиальных клеток молочной железы или связывающееся с клетками карциномы ободочной кишки, антитела к ЕрСАМ, антитела к GpIIb/IIIa, антитела к RSV, антитела к CMV, антитела к ВИЧ, антитела к вирусу гепатита, антитела к СА 125, антитела к αvβ3, антитела против человеческой почечноклеточной карциномы, антитела к человеческому 17-1А, антитела против человеческой колоректальной опухоли, антитело R24 против человеческой меланомы, мишенью которого является ганглиозид GD3, антитела против человеческой плоскоклеточной саркомы, антител к человеческому лейкоцитарному антигену (HLA), антитела к HLA DR.
Согласно способу, предлагаемому в настоящем изобретении, рекомбинантный гликопротеин получают в эукариотический клетке. Согласно настоящему изобретению можно применять любую эукариотическую клетку, которая пригодна для выращивания в культуре клеток и для экспрессии гликопротеинов. Эукариотическая клетка предпочтительно представляет собой линию эукариотической клетки, которая обладает способностью расти и выживать при помещении в суспензионную культуру в среду, содержащую соответствующие питательные вещества и факторы роста, и которая, как правило, обладает способностью экспрессировать и секретировать в больших количествах конкретный представляющий интерес гликопротеин в культуральной среде.
В предпочтительном варианте осуществления эукариотическая клетка представляет собой клетку млекопитающего, дрожжевую клетку или клетку насекомого.
Когда эукариотическая клетка представляет собой клетку млекопитающего, то она может представлять собой, например, клеточную линию мышиной миеломы NSO, линию клеток почки обезьяны CVI, трансформированную SV40 (COS-7, АТСС® CRL 1651); линию клеток почки человеческого эмбриона 293S (Graham и др., J. Gen. Virol. 36, 1977, с. 59); клетки почки детеныша хомяка (ВНK, АТСС® CCL 10); клетки Сертоли мыши (ТМ4, Mather, Biol. Reprod. 23, 1980, с. 243); клетки почки обезьяны (CVI-76, АТСС® CCL 70); клетки почки африканской зеленой мартышки (VERO-76, АТСС® CRL 1587); клетки карциномы шейки матки человека (HELA, АТСС® CCL 2); клетки почки собаки (MDCK, АТСС® CCL 34); клетки печени бычьей крысы (BRL 3А, АТСС® CRL 1442); клетки легкого человека (W138, АТСС® CCL 75); клетки печени человека (Hep G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, АТСС® CCL 5I); клетки гепатомы крысы (НТС, MI.54, Baumann и др., J. Cell Biol., 85, 1980, с. 1) и TR-1-клетки (Mather и др., Annals N.Y. Acad. Sci. 383, 1982, с. 44), линию клеток PER.C6 (Percivia LLC) и клеточные линии гибридом.
Для воплощения на практике изобретения предпочтительными клеточными линиями являются клетки яичника китайского хомячка (СНО, Urlaub и Chasin Р. N. A. S. 77, 1980, с. 4216) или PER.С6. Известные производные СНО, которые можно применять согласно настоящему изобретению, включают, например клетки CHO/-DHFR (Uriah и Chasin, выше), CHOK1SV (фирма Lonza), СНО-K1 DUC B11 (Simonsen и Levinson Р. N. A. S. 80, 1983, сс. 2495-2499) и DP12 СНО (ЕР 307247), а также клетки СНО DG44 (Derouazi и др., Biochem. Biophys. Res. Commun. 340, 2006, cc. 1069-1077).
Когда эукариотическая клетка представляет собой дрожжевую клетку, то она может представлять собой, например, клетку Saccharomyces cerevisiae или Pichia pastoris.
Когда эукариотическая клетка представляет собой клетку насекомого, то она может представлять собой, например, Sf-9-клетку.
Эукариотические клетки, применяемые в настоящем изобретении, могут представлять собой созданную с помощью гликоинженерии клетку, которую конструировали таким образом, чтобы модифицировать профиль гликозилирования клетки. Указанное конструирование включает, например, либо «выключение», либо «включение» генов, участвующих в синтезе N-гликанов.
Согласно настоящему изобретению наиболее предпочтительно клетка представляет собой СНО-клетку, необязательно созданную с помощью гликоинженерии СНО-клетку.
Эукариотическую клетку, применяемую согласно настоящему изобретению, выбирают или подвергают манипуляции с целью получения рекомбинантного гликопротеина. Манипуляция включает одну или несколько генетических модификаций, направленных на интродукцию одного или нескольких гетерологичных генов, которые кодируют подлежащий экспрессии гликопротеин. Гетерологичный ген может кодировать гликопротеин, который либо в норме экспрессируется в клетке, либо является чужеродным для клетки-хозяина. Посредством манипуляции можно добавлять или в альтернативном варианте осуществлять повышающую или понижающую регуляцию одного или несколько эндогенных генов. Часто клетки подвергают манипуляции с целью получения рекомбинантного гликопротеина путем, например, интродукции гена, кодирующего гликопротеин, и/или путем интродукции контролирующих элементов, которые регулируют экспрессию гена, кодирующего представляющий интерес гликопротеин. Гены, кодирующие рекомбинантные гликопротеины и/или контролирующие элементы, можно интродуцировать в клетку-хозяина с помощью векторов, таких как плазмидный, фаговый или вирусный вектор. Некоторые векторы обладают способностью к автономной репликации в клетке-хозяине, в которую они интродуцированы, в то время как другие векторы могут интегрироваться в геном клетки-хозяина и в результате реплицироваться совместно с геномом хозяина. Различные векторы являются публично доступными, и точная природа векторов не является принципиально важной для настоящего изобретения. Как правило, к компонентам векторов относятся одна или несколько сигнальных последовательностей, сайт инициации репликации, один или несколько маркерных генов, промотор и последовательность терминатора транскрипции. Указанные компоненты описаны в WO 97/25428.
Для получения биомассы и экспрессии гликопротеина из эукариотических клеток применяют способ, предлагаемый в изобретении, заключающийся в том, культивируют клетки в условиях ферментации. Согласно настоящему изобретению можно применять любой метод ферментационного культивирования клеток или любую систему, которые обеспечивают рост клеток с образованием биомассы и экспрессию гликопротеинов. Например, клетки можно выращивать в периодической культурах, периодических культурах с подпиткой или периодических культурах с расщеплением, в которых культивирование прекращают после достижения достаточного уровня экспрессии гликопротеина, после чего гликопротеин собирают и при необходимости очищают. Если применяют периодическое культивирование с подпиткой, то подпитку культуры можно осуществлять один или несколько раз в процессе культивирования. Когда используют несколько подпиток, то для этой цели можно использовать одинаковые или различные подпитывающие растворы. Альтернативно этому, клетки можно выращивать в перфузионных (проточных) культурах, в которых не осуществляют прекращение культивирования и новые питательные вещества и компоненты добавляют в культуру периодически или постоянно и экспрессируемый гликопротеин удаляют либо периодически, либо постоянно. В предпочтительном варианте осуществления изобретения применяемый в настоящем изобретении метод культивирования клеток представляет собой периодическое культивирование с подпиткой или периодическое культивирование с расщеплением или комбинацию обоих методов.
Реакторы, температуры и другие условия ферментационного культивирования клеток для образования биомассы и производства гликопротеинов, такие как концентрация кислорода и значения рН, известны в данной области. Можно выбирать любые условия, приемлемые для культивирования отобранной эукариотической клетки, используя информацию, известную в данной области. Условия культивирования, такие как температура, рН и т.п., как правило, идентичны применявшимся ранее для экспрессии в отобранной клетке-хозяине, и должны быть очевидны специалисту в данной области. При необходимости температуру и/или рН, и/или CO2 можно изменять в процессе культивирования для того, чтобы повышать выход и/или повышать относительное количество гликопротеина требуемого качества.
Среда, в которой культивируют клетки и в которой регулируют концентрации микроэлементов, таких как железо, медь, цинк и марганец, с помощью способа, предлагаемого в настоящем изобретении, может представлять собой любую из широкого разнообразия сред, известных в данной области. При необходимости среда может представлять собой среду определенного химического состава, в которой компоненты среды являются известными и контролируются, или среда может представлять комплексную среду, в которой не все компоненты известны и/или контролируются.
Среды определенного химического состава, включая среды для культивирования клеток млекопитающих, являются широко изученными, и данные о них опубликованы в современных руководствах. Все компоненты сред определенного состава хорошо охарактеризованы, и такие среды не содержат комплексных добавок, таких как сыворотка и гидролизаты. Как правило, такие среды включают в определенных количествах очищенные факторы роста, белки, липопротеины и другие субстанции, источником которых в противном случае могла бы являться добавка сыворотки или экстракта. Указанные среды создавали исключительно с целью поддержания высокопродуктивных клеточных культур. Некоторые среды определенного состава можно относить к средам с низким содержанием белков, или они могут представлять собой безбелковые среды, если в них не включены типичные компоненты сред с низким содержанием белков инсулин и трансферрин. Альтернативно этому, в способах, предлагаемых в настоящем изобретении можно применять бессывороточные среды. Такие среды, как правило, не содержат сыворотку или белковые фракции, но могут содержать неустановленные компоненты.
Примеры поступающих в продажу культуральных сред включают среду Хэма F10 (фирма Sigma), минимальную поддерживающую среду (MEM, фирма Sigma), RPMI-1640 (фирма Sigma) и модифицированную по методу Дульбекко среду Игла (DMEM, фирма Sigma), и среды определенного химического состава и подпитывающие добавки, которые поступают в продажу от фирмы Life Technologies. Любые указанные среды можно дополнять при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста); солями (такими как хлорид натрия, кальций, магний и фосфат), буферами (такими как HEPES); нуклеозидами (такими как аденозин и тимидин), антибиотиками (такими как GENTAMYCIN™) и глюкозой или другим эквивалентным источником энергии.
Необходимые для конкретной клеточной линии питательные вещества и факторы роста для среды, включая их концентрации, определяют эмпирически и без обширных экспериментов согласно методу, описанному, например, в Mammalian Cell Culture, Mather (изд-во Plenum Press: NY 1984); Barnes и Sato, Cell 22, 1980, c. 649 или Mammalian Cell Biotechnology: A Practical Approach M. Butler (изд-во IRL Press, 1991). Приемлемая среда содержит основной компонент среды, такой как композиция на основе DMEM/среды Хэма 12 с модифицированными концентрациями некоторых компонентов, таких как аминокислоты, соли, сахара, и витамины, и необязательно содержит глицин, гипоксантин, тимидин, рекомбинантный человеческий инсулин, гидролизованный пептон, такой как PRIMATONE HS™ или PRIMATONE RL™ (фирма Sheffield, Великобритания) или его эквивалент, защищающий клетки агент, такой как PLURONIC F68™ или эквивалентный полиол Плюроник и GENTAMYCIN™.
Ниже в таблице 1 представлены сведения о различных количествах таких микроэлементов, как железо, медь, цинк и марганец, в различных поступающих в продажу средах для культивирования клеток, любую из которых можно применять для клеточного культивирования.
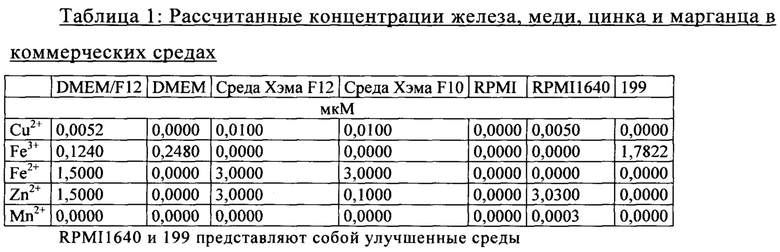
Рассчитанные концентрации железа, меди, цинка и марганца, которые должны присутствовать в указанных выше коммерческих средах, можно изменять, если дополнять эти среды комплексными ингредиентами, такими как сыворотки или пептоны.
Когда способ, предлагаемый в изобретении, предусматривает регуляцию концентраций железа, меди, цинка и марганца в культуральной среде путем добавления указанных микроэлементов в среду, или среда содержит отрегулированные концентрации указанных микроэлементов, микроэлементы можно добавлять, или они присутствуют в среде в форме соли металла. Для производства рекомбинантного гликопротеина для включения в культуральную среду можно применять любую соль железа, меди, цинка и марганца. Как правило, предпочтительно, чтобы соль металла находилась в форме соответствующего сульфата, галогенида, оксида, нитрата, цитрата, ацетата или фосфата металла, в гидратированной или безводной форме, или, чтобы ион металла был связан с хелатором, таким как трансферрин или лактоферрин.
Как правило, соли железа (II) и (III), которые можно применять согласно настоящему изобретению, включают цитрат Fe(III), FeSO4, FeCl2, FeCl3, Fe(NO3)3 и FePO4, а также железо, связанное с трансферрином или лактоферрином.
Как правило, соли меди (II), которые можно применять согласно настоящему изобретению, включают CuSO4, СuСl2, и ацетат Cu.
Как правило, соли цинка (II), которые можно применять согласно настоящему изобретению, включают ZnSO4 и ZnCl2.
Как правило, соли марганца, которые можно применять согласно настоящему изобретению, включают MnSO4, MnCl2, MnF2 и MnI2.
В настоящем изобретении предложено культивирование клеток в условиях ферментационной культуры. Оно, как правило, представляет собой многостадийную процедуру культивирования, в которой клетки культивируют с использованием несколько стадий или фаз. Согласно указанной предпочтительной процедуре процесс ферментации культуры, например, из замороженных флаконов с клетками, как правило, включает три различные фазы, например, представленные на фиг. 1, а именно:
I) фазу подготовки затравки для восстановления клеток после связанного с оттаиванием стресса и для нормализация времени удвоения клеток, которая может продолжаться от 14 до, например, более 60 дней, в зависимости от скорости восстановления клеток и масштаба производства. На фиг.1 она изображена как продолжающаяся 21 день;
II) фазу(ы) роста или подготовки для инокуляции, обозначенную(ые) как n-x-фазы (n обозначает фазу производства), где x, как правило, обозначает 1-5, предпочтительно 1 или 2. На фиг. 1 проиллюстрированы фазы n-1 и n-2. Эти фазы можно обозначать также как фаза(ы) роста, на которой(ых) клетками инокулируют среду, пригодную для ускорения роста и образования биомассы. Так, n-x-фазы, как правило, предназначены для размножения культуры для последующего использования в более крупных форматах культивирования и вымывания отобранного соединения. Когда n-x-фазы состоят из фазы n-1 и n-2, то каждая из стадий n-1 и n-2 продолжается, например, от 2 до 7 дней, как правило, каждая продолжается 3 или 4 дня; и
III) фазу производства или n-фазу, предназначенную для производства рекомбинантного гликопротеина в соответствующем количестве и/или соответствующего качества. Продолжительность этой фазы может зависеть, например, от природы рекомбинантной клетки, а также от количестве и/или качества экспрессируемого гликопротеина. Как правило, это фаза может продолжаться от примерно 11 до примерно 20 дней. На фиг. 1 эта фаза изображена как продолжающаяся 14 дней.
Клетки можно поддерживать на фазе подготовки затравки или на фазе роста в течение требуемого периода времени, например, путем добавления при необходимости свежей среды или питательной добавки к существующей среде.
Как правило, для фазы производства или n-фазы характерны две различные фазы, представленные на фиг. 2. Первая из них представляет собой фазу роста, предназначенную для образования в достаточном количестве жизнеспособной биомассы для производства специфического для клеток белка, а вторая из них представляет собой фазу, предназначенную для образования рекомбинантного гликопротеина, но без существенного роста клеток.
Любая или все из указанных фазы подготовки затравки, фазы роста и фазы производства может быть непрерывной, или клетки, полученные на одной из фаз можно применять для инокуляции следующей фазы.
Для выделения экспрессируемого гликопротеина в течение или в конце периода культивирования, предпочтительно фазы производства, можно применять методы, известные в данной области. Гликопротеин предпочтительно выделяют из культуральной среды в виде секретированного полипептида, хотя его можно выделять из лизатов клеток-хозяев, когда он продуцируется непосредственно без секреторного сигнала. Если гликопротеин связан с мембраной, то его можно высвобождать из мембраны, используя раствор приемлемого детергента (например, Тритон-Х 100), или его внеклеточную область можно высвобождать путем ферментативного расщепления. Экспрессированный гликопротеин можно выделять и/или при необходимости очищать с помощью методик, известных в данной области.
В способе, предлагаемом в настоящем изобретении, концентрации железа, меди, цинка и марганца регулируют на любой или на всех фазах роста или производства в процессе ферментации. В предпочтительном варианте осуществления изобретения концентрации железа, меди, цинка или марганца регулируют в начале или в течение фазы роста и/или в начале или в течение фазы производства. Наиболее предпочтительно регулировать концентрации железа, меди, цинка и марганца в начале фазы роста и в начале фазы производства или регулировать концентрации железа, меди, цинка и марганца в любом или во всех следующих случаях: в начале фазы роста, в течение фазы роста, в начале фазы производства и в течение фазы производства.
Концентрации железа, меди, цинка и марганца регулируют путем повышения или понижения концентрации указанных микроэлементов в культуральной среде. Согласно настоящему изобретению, когда концентрацию указанных элементов повышают или понижают, то указанное повышение или понижение концентрации определяют по повышению или понижению относительно концентрации указанных элементов в среде на непосредственно предшествующей фазе культивирования. Таким образом, если имеет место повышение концентрации, например, каждого металла из группы, включающей железо, медь, цинк и марганец, в среде в начале фазы производства, это означает, что имеет место повышение концентрации указанных микроэлементов относительно концентрации указанных микроэлементов в среде на непосредственно предшествующей фазе роста. Аналогично этому, если имеет место повышение концентрации, например, каждого металла из группы, включающей железо, медь, цинк и марганец, в среде в течение фазы производства, это означает, что имеет место повышение концентрации указанных микроэлементов относительно концентрации указанных микроэлементов в среде на непосредственно предшествующей стадии фазы производства. Аналогично этому, если имеет место понижение концентрации, например, каждого металла из группы, включающей железо, медь, цинк и марганец, или всех их, в начале или в течение любой из фаз роста или производства, это означает, что имеет место понижение концентрации указанных микроэлементов относительно концентрации указанных микроэлементов в среде на непосредственно предшествующей фазе культивирования. Аналогично этому, если имеет место понижение концентрации, например, железа и меди, и повышение концентрации, например, цинка и марганца, в среде в начале или в течение фазы производства, то означает, что имеет место понижение концентрации железа и меди и повышение концентрации цинка и марганца по сравнению с концентрациями указанных микроэлементов в среде на непосредственно предшествующей фазе культивирования.
В целом, предпочтительно, чтобы, когда регулируют концентрации в среде каждого металла из группы, включающей железо, медь, цинк и марганец, или всех их, то концентрации всех указанных микроэлементов регулируют одновременно. Однако способ, предлагаемый в настоящем изобретении, предусматривает также регуляцию концентраций железа, меди, цинка и марганца индивидуально, попарно или регуляцию трех из указанных микроэлементов одновременно и последующую регуляцию четвертого из них или наоборот. А частности, если концентрацию двух из микроэлементов требуется повышать, а концентрацию двух микроэлементов требуется понижать, то регуляция повышения и понижения может иметь место в одно и то же время или в различные моменты времени. В этом случае предпочтительно, что регуляция, приводящая к повышению концентрации двух микроэлементов и к понижению концентрации двух элементов имела место в один и тот же момент времени или в той же самой среде.
Согласно способу, предлагаемому в настоящем изобретении, регуляцию концентраций железа, меди, цинка и марганца можно обеспечивать с помощью любой методики, пригодной для использования в условиях ферментации. Метод, с помощью которого регулируют концентрации указанных микроэлементов, не имеет решающего значения для настоящего изобретения, и соответствующие методы известны в данной области. Таким образом, регуляцию концентрации микроэлемента можно осуществлять либо путем добавки с среду (в случае необходимости повышения концентрации любого или всех микроэлементов), в которой культивируют клетки, либо путем переноса всех или части клеток (т.е. расщепления) в свежую среду, которая содержит микроэлементы в требуемых концентрациях. При необходимости можно использовать комбинацию двух указанных методов.
Таким образом, регуляция концентрации железа, меди, цинка и марганца можно осуществлять непрерывно, в течение всего или части периода культивирования, или прерывисто, например, в виде реакции на предполагаемую, рассчитанную или измеренную концентрацию микроэлемента(ов) в культуральной среде. В изобретении определены диапазоны, в которых регулируют концентрацию микроэлементов. Если фактическое измерение или результаты расчета концентрации каждого или всех микроэлементов в течение определенного периода культивирования, например, в течение фазы роста или производства, свидетельствует о том, что концентрация каждого или всех микроэлементов находится в пределах диапазонов, указанных в настоящем описании, то, несмотря на это может иметь место регуляция указанной концентрации, для того, чтобы концентрация микроэлементов оставалась в указанном диапазоне.
При необходимости для измерения фактических концентраций микроэлементов в культуральной среде перед осуществлением регуляции можно применять известные методики. Они включают онлайн- и оффлайн-анализы концентрации соответствующего микроэлемента.
Так, если применяют условия периодической ферментации, то для достижения повышения концентраций микроэлементов можно, например, осуществлять посев в свежую среду, которая содержит или дополнена соответствующими микроэлементами в концентрациях, превышающих те, которые присутствуют в существующей культуральной среде, или осуществлять перенос (расщепление) клеток в среду, которая содержит или дополнена соответствующими микроэлементами в концентрациях, превышающих те, которые присутствуют в существующей культуральной среде. Если применяют условия периодической ферментации с подпиткой, то для достижения повышения концентраций микроэлементов можно, например, осуществлять посев в свежую среду, которая содержит или дополнена соответствующими микроэлементами в повышенных концентрациях, проводя одну или несколько болюсных или непрерывных подпиток культуральной среды соответствующими микроэлементами; определять уровень подпитки на основе количества клеток или осуществлять расчет на основе известных метаболических моделей, суррогатных маркеров метаболизма и т.д., или осуществлять перенос (расщепление) культуры в среду, которая содержит или дополнена соответствующими микроэлементами в повышенных концентрациях. Если пополнение культуры осуществляют с помощью болюсной или непрерывной подпитки, то она может включать другие питательные вещества/компоненты, необходимые для культивирования, в дополнение к любому или всем микроэлементам, таким как железо, медь, цинк и марганец. Если применяют условия перфузионной ферментации, то для достижения повышения концентраций микроэлементов можно, например, применять постоянное или прерывистое добавление микроэлементов в реактор либо одновременно с другими питательными веществами/компонентами, которые добавляют в перфузионную культуру, либо отдельно от них.
Если требуется понижение концентраций любого металла из группы, включающей железо, цинк, медь и марганец, или всех их, то для достижения этого можно осуществлять посев клеток в свежую среду, в которой концентрации микроэлементов понижены по сравнению с концентрацией указанных микроэлементов в среде, которая используется на непосредственно предшествующей фазе культивирования. В альтернативном или дополнительном варианте понижения концентраций любого металла из группы, включающей железо, цинк, медь и марганец, или их всех, можно достигать, например, в существующей или свежей среде путем комплексообразования иона металла, например, путем добавления хелатора иона металла в культуру и вымывания содержащих ион металла комплексов. Можно применять любой хелатор иона металла, который обладает способностью образовывать комплекс с соответствующим ионом металла, приводя к понижению биодоступной концентрации указанного иона металла в культуральной среде. Приемлемые для железа и меди указанные хелаторы включают, например, малые молекулы или связывающие металлы белки, такие, например, как ЭДТК, ЭГТК, сидерофоры, такие как дефероксамин (десфериоксамин), десферитиоцин, тетрациклин, хинолон, фосфонаты, полифенолы, белки, такие как трнсферрин или церулопламин, полисахариды, органические кислоты, такие как малеат, супризон, батокупроина сульфонат, батофенантролина сульфонат и D-пеницилламин. Кроме того, эффект снижения концентрации микроэлементов можно имитировать путем ингибирования/активации ферментов гликозилирования с помощью альтернативных микроэлементов, с помощью целевой конкуренции с клеточными транспортерами или с помощью целевой модуляции активности клеточного транспортера.
Конкретные величины пониженных или повышенных концентраций металла базируются либо на фактических измерениях металла в культуральной среде, либо на теоретических концентрациях металла в культуральном растворе, который окружает клетки. Практикующему специалисту в данной области должно быть очевидно, что может присутствовать некоторая концентрация микроэлементов, интродуцированная, например, с примесями или в результате выщелачивания, и что требуется учитывать это при расчете пониженной или повышенной концентрации микроэлементов, предлагаемых в изобретении.
В способах, предлагаемых в настоящем изобретении, предпочтительно, чтобы понижение концентрации любого или всех микроэлементов, если это возможно, достигалось путем посева клеток в свежую среду, в которой концентрация микроэлементов понижена по сравнению с концентрацией указанных микроэлементов в среде, которая используется на непосредственно предшествующей фазе культивирования.
В некоторых вариантах осуществления настоящего изобретения концентрации железа, меди, цинка и марганца можно регулировать в культуральной среде так, чтобы благоприятствовать росту клеток и тем самым увеличению биомассы. В других вариантах осуществления изобретения концентрации железа, меди, цинка и марганца или концентрации только цинка и марганца можно регулировать в культуральной среде так, чтобы повышать зрелость экспрессируемых N-гликопротеинов и производство гликопротеинов со зрелой схемой гликозилирования. В другом варианте осуществления изобретения концентрации железа, меди, цинка и марганца можно регулировать в культуральной среде так, чтобы благоприятствовать повышению производства незрелых нефукозилированных гликопротеинов. В следующем варианте осуществления изобретения концентрации железа, меди, цинка и марганца в культуральной среде можно регулировать сначала для повышения роста, а затем вновь для воздействия на зрелость гликопротеина, включая повышение зрелости экспрессируемых N-гликозилированных гликопротеинов или повышение производства незрелых нефукозилированных гликопротеинов.
В способе, предлагаемом в настоящем изобретении, используя микроэлементы в различных концентрациях в начале или в течение фаз роста и/или производства, можно оказывать прямое воздействие на образование биомассы и зрелость N-гликозилирования экспрессируемого гликопротеина, что проиллюстрировано ниже в таблице 2. Регуляция концентраций железа, меди, цинка и марганца в течении указанных фаз культивирования приводит к переключению между образованием биомассы и зрелости N-гликозилирования экспрессируемых гликопротеинов. «Афукозилирование» в таблице 2 является синонимом нефукозилирования.
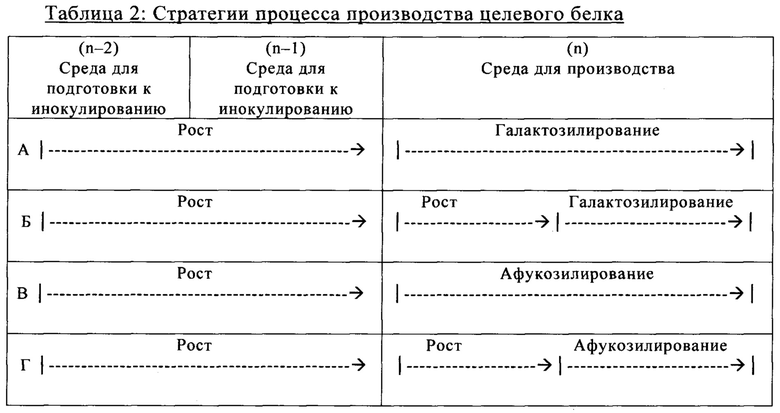

С помощью способа, предлагаемого в настоящем изобретении, продуцирующие гликопротеин клетки культивируют в среде, специально приспособленной для получения требуемого конечного результата. Таким образом, как проиллюстрировано ниже в таблице 3, среда для роста прежде всего усиливает образование биомассы, среда для галактозилирования прежде всего повышает зрелость экспрессируемых N-гликопротеинов и производство зрелых гликопротеинов, а среда, обеспечивающая отсутствие фукозилирования, прежде всего усиливает образование незрелых нефукозилированных гликопротеинов. Могут иметь место некоторые взаимосвязанные воздействия каждой среды, что может в определенной степени влиять и на другие результаты, как это продемонстрировано ниже, однако при этом оказывается благоприятствующее действие на один требуемый результат.

В предпочтительном варианте осуществления настоящего изобретения предложено повышение концентрации каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде для того, что благоприятствовать клеточному росту/образованию биомассы. В этом варианте осуществления изобретения концентрацию каждого металла из группы, включающей железо, медь, цинк и марганец, повышают в начале и/или в течение фазы роста и необязательно также в начале и/или в течение фазы производства. Указанный вариант осуществления изобретения является наиболее пригодным для клеток с низкой способностью к росту, клеточных линий, в которых образование биомассы и продуктивность взаимосвязаны, и в том случае, когда требуются низкие уровни зрелости галактозилирования гликопротеина.
В альтернативном предпочтительном варианте осуществления настоящего изобретения предложено повышение концентрации каждого металла из группы, включающей цинк и марганец, и необязательно снижение концентрации железа и меди в культуральной среде для повышения зрелости экспрессируемых N-гликопротеинов, приемлемого производства гликопротеинов со зрелой схемой гликозилирования и, как следствие, снижения производства незрелых гликопротеинов, включая незрелые нефукозилированные гликопротеины. Согласно этому варианту осуществления изобретения концентрацию каждого металла из группы, включающей железо, медь, цинк и марганец, или только цинка и марганца регулирую в начале фазы роста и/или в начале, и/или в течение фазы производства. Наибольшее благоприятное действие в отношении повышенной зрелости гликопротеинов и производства гликопротеинов со зрелой схемой гликозилирования путем регуляции концентраций микроэлементов в течение фаз роста и производства имеет место, когда желательным является низкий роста клеток, и прежде всего, когда экспрессируемый гликопротеин представляет собой комплекс с несколькими нежелательными побочными продуктами. Благоприятное действие в отношении повышенной зрелости гликопротеинов и производства гликопротеинов со зрелой схемой гликозилирования путем регуляции концентраций микроэлементов в среде в начале фазы производства, является наиболее ценным, когда в процессе фазы роста достигается хороший рост. Благоприятное действие в отношении повышенной зрелости гликопротеинов и производства гликопротеинов со зрелой схемой гликозилирования путем регуляции концентраций микроэлементов в среде в процессе фазы производства является наиболее ценным, когда концентрации микроэлементов регулировали на фазе роста и в начале фазы производства для гарантии эффективного образования биомассы.
Альтернативным предпочтительным вариантом осуществления настоящего изобретения является повышение концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде для того, чтобы оказывать благоприятное воздействие на производство незрелых нефукозилированных гликопротеинов. В этом варианте осуществления изобретения концентрацию каждого металла из группы, включающей железо, медь, цинк и марганец, регулируют в начале фазы роста и/или в начале, и/или в течение фазы производства. Наибольшее благоприятное действие в отношении производства незрелых нефукозилированных гликопротеинов путем регуляции концентраций микроэлементов в начале фаз роста и производства имеет место, когда рост клеток является наиболее высоким. Благоприятное действие в отношении производства незрелых нефукозилированных гликопротеинов путем регуляции концентраций микроэлементов в начале фазы производства является наиболее ценным в том случае, когда в течение фазы роста достигался хороший рост. Благоприятное действие в отношении производства незрелых нефукозилированных гликопротеинов путем регуляции концентраций микроэлементов в среде в течение фазы производства является наиболее ценным, когда концентрации микроэлементов регулировали на фазе роста и в начале фазы производства для гарантии высокого образования биомассы.
Альтернативным предпочтительным вариантом осуществления настоящего изобретения является повышение концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде в начале фазы роста и необязательно в начале фазы производства для того, чтобы оказывать благоприятное действие на клеточный рост/образование биомассы, и дополнительно повышение концентрации каждого металла из группы, включающей цинк и марганец, и повышение концентрации железа и меди в культуральной среде в начале или в течение фазы производства для повышения зрелости гликопротеинов, например, для благоприятного воздействия на производство гликопротеинов со зрелой схемой N-гликозилирования по сравнения с производством незрелых гликопротеинов, включая незрелые нефукозилированные гликопротеины. В этом варианте осуществления изобретения, когда концентрации железа, меди, цинка и марганца повышают в начале фазы производства для того, чтобы оказывать благоприятное воздействие на образование массы, то повышение зрелости гликопротеинов, например, благоприятное воздействие на производство гликопротеинов со зрелой схемой N-гликозилирования, получают путем дополнительной регуляции концентраций указанных микроэлементов в процессе фазы производства.
Альтернативным предпочтительным вариантом осуществления настоящего изобретения является повышение концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде в начале фазы роста и необязательно в начале фазы производства для того, чтобы оказывать благоприятное действие на клеточный рост/образование биомассы, и дополнительно понижение концентрации каждого металла из группы, включающей цинк и марганец, и повышение концентраций каждого металла из группы, включающей железо, медь, цинк и марганец, в культуральной среде в начале или в течение фазы производства для благоприятного воздействия на производство незрелых нефукозилированных гликопротеинов. В этом варианте осуществления изобретения, когда концентрации железа, меди, цинка и марганца повышают в начале фазы производства для того, чтобы оказывать благоприятное воздействие на образование массы, благоприятное воздействие на производство незрелых нефукозилированных гликопротеинов получают путем дополнительной регуляции концентраций указанных микроэлементов в процессе фазы производства.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на образование биомассы, предпочтительно регулировать концентрацию железа в форме Fe2+ или Fe3+ в культуральной среде так, чтобы она составляла от 15 до более чем 80 мкМ, предпочтительно от 20 до более чем 60 мкМ и наиболее предпочтительно от 25 до более чем 50 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для повышения зрелости экспрессируемых N-гликопротеинов, например, чтобы оказывать благоприятное воздействие на производство зрелых видов N-гликозилированного гликопротеина, предпочтительно регулировать концентрацию железа в форме Fe2+ или Fe3+ в культуральной среде так, чтобы она составляла от 0 до 25 мкМ, предпочтительно от 0 до 20 мкМ и наиболее предпочтительно от 0 до 16 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на производство незрелых нефукозилированных видов гликопротеина, предпочтительно регулировать концентрацию железа в форме Fe2+ или Fe3+ в культуральной среде так, чтобы она составляла от 0 до 35 мкМ, предпочтительно от 0 до 30 мкМ и наиболее предпочтительно от 0 до 25 мкМ, или составляла от 15 до более чем 80 мкМ, предпочтительно от 20 до более чем 60 мкМ и наиболее предпочтительно от 25 до более чем 50 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на образование биомассы, предпочтительно регулировать концентрацию меди в форме Cu2+ в культуральной среде так, чтобы она составляла от 0,3 до более чем 2,5 мкМ, предпочтительно от 0,5 до более чем 1,5 мкМ и наиболее предпочтительно от 0,3 до более чем 1 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для повышения зрелости экспрессируемых N-гликопротеинов, например, чтобы оказывать благоприятное воздействие на производство зрелых видов N-гликозилированного гликопротеина, предпочтительно регулировать концентрацию меди в форме Cu2+ в культуральной среде так, чтобы она составляла от 0 до 0,1 мкМ, предпочтительно от 0 до 0,08 мкМ и наиболее предпочтительно от 0 до 0,06 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на производство незрелых нефукозилированных видов гликопротеина, предпочтительно регулировать концентрацию меди в форме Cu2+ в культуральной среде так, чтобы она составляла от 0 до 1 мкМ, предпочтительно от 0 до 0,5 мкМ и наиболее предпочтительно от 0 до 0,3 мкМ, или составляла от 0,3 до более чем 2,5 мкМ, предпочтительно от 0,5 до более чем 1,5 мкМ и наиболее предпочтительно от 0,3 до более чем 1 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на образование биомассы, предпочтительно регулировать концентрацию цинка в форме Zn2+ в культуральной среде так, чтобы она составляла от 20 до более чем 50 мкМ, предпочтительно от 25 до более чем 45 мкМ и наиболее предпочтительно от 28 до более чем 43 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для повышения зрелости экспрессируемых N-гликопротеинов, например, чтобы оказывать благоприятное воздействие на производство зрелых видов N-гликозилированного гликопротеина, предпочтительно регулировать концентрацию цинка в форме Zn2+ в культуральной среде так, чтобы она составляла от 20 до более чем 50 мкМ, предпочтительно от 25 до более чем 45 мкМ и наиболее предпочтительно от 28 до более чем 43 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на производство незрелых нефукозилированных видов гликопротеина, предпочтительно регулировать концентрацию цинка в форме Zn2+ в культуральной среде так, чтобы она составляла от 0 до 20 мкМ, предпочтительно от 0 до 15 мкМ и наиболее предпочтительно от 0 до 13 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на образование биомассы, предпочтительно регулировать концентрацию марганца в форме Mn2+ в культуральной среде так, чтобы она составляла от 0,01 до более чем 3 мкМ, предпочтительно от 0,05 до более чем 2 мкМ и наиболее предпочтительно от 0,1 до более чем 1 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для повышения зрелости экспрессируемых N-гликопротеинов, например, чтобы оказывать благоприятное воздействие на производство зрелых видов N-гликозилированного гликопротеина, предпочтительно регулировать концентрацию марганца в форме Mn2+ в культуральной среде так, чтобы она составляла от 0,01 до более чем 3 мкМ, предпочтительно от 0,05 до более чем 2 мкМ и наиболее предпочтительно от 0,1 до более чем 1 мкМ.
В предпочтительном варианте осуществления настоящего изобретения для того, чтобы оказывать благоприятное воздействие на производство незрелых нефукозилированных видов гликопротеина, предпочтительно регулировать концентрацию марганца в форме Mn2+ в культуральной среде так, чтобы она составляла от 0 до 0,01 мкМ, предпочтительно от 0 до 0,05 мкМ и наиболее предпочтительно от 0 до 0,07 мкМ.
Предпочтительными концентрациями комбинаций железа, меди, цинка и марганца в культуральной среде для получения требуемых согласно настоящему изобретению результатов являются:
A. для того, чтобы благоприятствовать образованию биомассы:
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ.
Б. для того, чтобы повышать зрелость экспрессируемых гликопротеинов, например для того, чтобы благоприятствовать производству зрелых N-гликозилированных гликопротеинов:
(I)
(а) железо: от 0 до 25 мкМ;
(б) медь: от 0 до 0,1 мкМ;
(в) цинк: от 20 до более чем 50 мкМ и
(г) марганец: от 0,01 до более чем 3 мкМ или
(II)
(а) цинк: от 20 до более чем 50 мкМ и
(б) марганец: от 0,01 до более чем 3 мкМ.
B. для того, чтобы благоприятствовать производству незрелых нефукозилированных гликопротеинов, либо:
(I)
(а) железо: от 0 до 35 мкМ;
(б) медь: от 0 до 1 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ, либо:
(II)
(а) железо: от 15 до более чем 80 мкМ;
(б) медь: от 0,3 до более чем 2,5 мкМ;
(в) цинк: от 0 до 20 мкМ и
(г) марганец: от 0 до 0,01 мкМ.
Наиболее предпочтительные комбинации концентраций железа, меди, цинка и марганца в культуральной среде для того, чтобы благоприятствовать образованию биомассы, для того, чтобы повышать зрелость экспрессируемых N-гликозилированных гликопротеинов, например для производства гликопротеинов со зрелой схемой N-гликозилирования, и/или для того, чтобы благоприятствовать производству незрелых нефукозилированных гликопротеинов, представлены ниже в таблице 4.

Согласно способам, предлагаемым в настоящем изобретении, повышение концентрационных уровней железа, меди, цинка и марганца в среде для того, чтобы благоприятствовать, стимулировать или повышать образование биомассы в процессе культивирования в условиях ферментации с получением рекомбинантного гликопротеина, может приводить к повышению содержания биомассы по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 15%, 20%, 25%, 30%, 40%, 45%, 50%, 55% или 60% по сравнению с сопоставимой культурой, в которой концентрационные уровни железа, меди, цинка и марганца не регулировали. Методы измерения и количественной оценки повышении клеточного роста известны в данной области. Как правило, для этой цели можно анализировать максимальную плотность жизнеспособных клеток и конечный интеграл от количества клеток по времени.
Как должно быть очевидно специалисту в данной области, при производстве рекомбинантного гликопротеина повышение биомассы в течение одной стадии клеточного культивирования должно приводить к повышению количества гликопротеина, продуцируемого на более поздней стадии клеточного культивирования.
Согласно способам, предлагаемым в настоящем изобретении, целенаправленная регуляция концентрационных уровней железа, меди, цинка и марганца или только цинка и марганца в среде в процессе культивирования в условиях ферментации для повышения зрелости экспрессируемых N-гликопротеинов, например, для того, чтобы благоприятствовать, стимулировать или повышать производство гликопротеинов со зрелой схемой N-гликозилирования, может приводить к повышению производства зрелых N-гликозилированных гликопротеинов по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 15%, 20%, 25%, 30%, 40%, 45%, 50%, 55% или 60% по сравнению с сопоставимой культурой, в которой концентрационные уровни железа и меди и/или цинка и марганца не регулировали в процессе культивирования.
Согласно способам, предлагаемым в настоящем изобретении, целенаправленная регуляция концентрационных уровней железа, меди, цинка и марганца или только цинка и марганца в среде в процессе культивирования в условиях ферментации для того, чтобы благоприятствовать, стимулировать или повышать производство гликопротеинов с G0-, G1- или С2-схемой, может приводить к повышению производства гликопротеинов с G0-, G1- или G2-схемой по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 15%, 20%, 25%, 30%, 40%, 45%, 50%, 55% или 60% по сравнению с сопоставимой культурой, в которой концентрационные уровни железа и меди и/или цинка и марганца не регулировали в процессе культивирования.
Согласно способам, предлагаемым в настоящем изобретении, целенаправленная регуляция концентрационных уровней железа, меди, цинка и марганца в среде в процессе культивирования в условиях ферментации для того, чтобы благоприятствовать, стимулировать или повышать производство незрелых нефукозилированных гликопротеинов, может приводить к повышению производства незрелых нефукозилированных гликопротеинов по меньшей мере на 5%, предпочтительно по меньшей мере на 10%, 15%, 20%, 25%, 30%, 40%, 45%, 50%, 55% или 60% по сравнению с сопоставимой культурой, в которой концентрационные уровни железа, меди, цинка и марганца не регулировали в процессе культивирования.
Приведенные ниже примеры служат для иллюстрации изобретения.
Примеры
Материалы и химические субстанции
Клеточные линии
Для описанного исследования при создании изобретения применяли 9 созданных в лаборатории заявителей полученных рекомбинантным путем линий клеток СНО-K1: клон 1 и клон 2, которые оба экспрессируют одно и то же моноклональное антитело, клон 3, который экспрессирует другое моноклональное антитело, и клон А, клон Б, клон В, клон Г, клон Д и клон Е, которые экспрессируют созданное с помощью гликоинженерии антитело. По сравнению с клоном 1 для клона 2 характерны высокие уровни видов гликанов с высоким содержанием маннозы (Man5). Все клоны культивировали с использованием запатентованной созданной в лаборатории заявителей безбелковой среды химически определенного состава и платформ процесса, обозначенных ниже как платформа А и платформа Б.
Контроль при осуществлении процесса - клеточный рост и анализы метаболитов
Рост и жизнеспособность клеток анализировали по вытеснения трипанового синего (Strober, Curr. Protoc. Immunol., приложение 3, 2001) и с использованием автоматического устройства CedexHiRes (фирма Roche Innovatis, Билефельд, Германия). Для количественной оценки метаболитов, таких как лактат и аммоний, жидкую культуру клеток центрифугировали для отделения клеток и анализировали с помощью системы Cobas Integra 400 plus (фирма Roche, Маннгейм, Германия). Титр продукта количественно оценивали с помощью системы Cobas Integra 400 plus (фирма Roche, Маннгейм, Германия) или метода ЖХВР PorosA, описанного у Zeck и др., PLoS. One 7.7, 2012, е40328.
Контроль при осуществлении процесса - микроэлементы
Концентрации микроэлементов анализировали, используя не содержащий клеток супернатант культуры и разработанный в лаборатории заявителей метод для ICP-MC-системы (фирма Agilent, Беблинген, Германия).
DoE-эксперименты
Статистическое планирование экспериментов с помощью DoE (планирование экспериментов) представляет собой эффективный и хорошо известный метод, применяемый, прежде всего, для оптимизации биологических и химических взаимодействий, в которых на конечный результат оказывают влияние независимо друг от друга много факторов. Например, DoE успешно применяли для оптимизации сред для культивирования клеток (Zhang и др., Cytotechnology 65.3, 2013, cc. 363-378), процессов ферментации (Fu и др., Biotechnology Progress 28.4, 2012, cc. 1095-1105), процессов очистки белков (Pezzini и др., J. Chromatogr. A 1218.45, 2011, cc. 8197-8208) и условий культивирования клеток (Chen и др., Tissue Eng. Part С. Methods 17.12, 2011, cc. 1211-1221).
В зависимости от требуемого результата можно применять различные DoE-подходы к процессу, описанному в настоящем изобретении (фиг. 3). Для систем с огромным количеством факторов влияния часто применяют фракционные факторные планы (фракционные факториальные дизайны), что позволяет идентифицировать значимые принципиальные модуляторы в процессе. С другой стороны, планы экспериментов типа полного факторного плана, факторного плана Бокса-Бенкена или центрального композиционного плана, не только позволяют идентифицировать имеющие решающее значение модуляторы, но также и количественно оценивать их дискретные оптимальные концентрации или задаваемые параметры. Для этого применяют так называемые инструменты прогностического профайлера, которые выдают результаты эксперимента, когда концентрации или задаваемые параметры имеющих решающее значение модуляторов варьируют в интерактивном режиме in silico в диапазоне от минимального (-1) до максимального (1) состояния. Для расчета оптимальных концентраций следует определять предсказанный оптимальный уровень (значение от -1 до +1) путем увеличения количества задаваемых применяемых концентраций.
В этом исследовании все DoE-подходы планировали и анализировали, используя в качестве инструмента статистическую программу JMP (фирма SAS Institute GmbH,  , Германия). Эксперименты планировали как варианты периодического культивирования с подпиткой для моделирования условий, сходных с процессом производства. Все спланированные с помощью DoE эксперименты по периодическому культивированию с подпиткой осуществляли, используя шейкер или роботизированные системы культивирования и запатентованные среды для культивирования и платформы подпитки А и Б. Применяемые для культивирования среды представляли собой бессывороточные и безбелковые среды определенного химического состава
, Германия). Эксперименты планировали как варианты периодического культивирования с подпиткой для моделирования условий, сходных с процессом производства. Все спланированные с помощью DoE эксперименты по периодическому культивированию с подпиткой осуществляли, используя шейкер или роботизированные системы культивирования и запатентованные среды для культивирования и платформы подпитки А и Б. Применяемые для культивирования среды представляли собой бессывороточные и безбелковые среды определенного химического состава
Анализ гликозилированных видов
Для анализа гликозилирования МАт собранную жидкость, в которой культивировали клетки, центрифугировали для отделения клеток и фильтровали, используя фильтрационную мембрану с размером пор 0,2 мкМ.
Гликозилированные виды МАт анализировали согласно методу, описанному у Reusch и др. (Anal. Biochem. 432.2, 2013, cc. 82-89), применяя разработанный заявителями протокол капиллярного электрофореза.
Пример 1
Обратная регуляция галактозилирования марганцем и медью
Для тестирования в целом влияния указанных микроэлементов на характеристики клеточной культуры при создании изобретения применяли две созданные в лаборатории заявителей рекомбинантные линии клеток, а именно клон 1 и клон 3, экспрессирующие два различных моноклональных антитела соответственно. Используя фракционный факторный DoE-план (таблица 5), можно анализировать статистическую значимость различных микроэлементов (используя для этой цели пополнение базальной среды специфическими маточными растворами). Подход на основе фракционного факторного DoE применяли для каждого клона и всех повторностей (четырех для клона 1, трех для клона 3), используя полностью контролируемую роботизированную систему культивирования.

Затем для каждого состояния обобщали теоретические эффективные концентрации алюминия, молибдена, бария, хрома, брома, йода, меди, марганца, рубидия, серебра, цинка, олова и циркония (сTE effective) в начале DoE-эксперимента с периодической подпиткой (таблица 6). Теоретические cTE effective для указанных микроэлементов в начале эксперимента рассчитывали с помощью уравнения 1:
(уравнение 1)
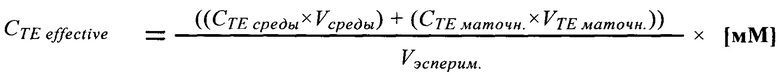

Возможное влияние алюминия, молибдена, бария, хрома, брома, йода, меди, марганца, рубидия, серебра, цинка, олова и циркония на галактозилирование анализировали на основе относительных уровней G1- и G2-гликанов для обоих клонов (клон 1 и клон 3, каждый из которых экспрессирует различные МАт) и, применяя образцы, полученные в дни 7, 12 и 13, для идентификации общих, независящих от клона и времени воздействий.
Используя в качестве инструмента JMP DoE-анализ была определена хорошая линейная модель, описывающая влияние микроэлементов на галактозилирование МАт. Так, обнаружено сильное положительное воздействие марганца на уровни G1 и G2 (фиг. 4А-Г). Кроме того, для цинка обнаружена тенденция к положительному воздействию на относительное содержание G1 и G2. С другой стороны, для меди обнаружено статистически значимое отрицательное влияние на образование G1-гликанов (фиг. 4А-Б), свидетельствующее о том, что обнаруженное in vitro ингибирующее действие (подробно описанное у  J и др., Glycoconj J. 15.8, 1998, cc. 783-788) удалось подтвердить в экспериментах in vivo на культуре клеток.
J и др., Glycoconj J. 15.8, 1998, cc. 783-788) удалось подтвердить в экспериментах in vivo на культуре клеток.
Важно отметить, что добавление цинка в комбинации с марганцем приводило к синергетическому действию в отношении образования галактозилированных гликанов, и это действие, вероятно, зависело от продолжительности процесса. Для количественной оценки действия при создании изобретения моделировали образование G1 и G2 только с воздействием цинка и марганца в высоких и низких концентрациях (фиг. 16). Было спрогнозировано получение самого большого количества G1- и G2-видов при использовании марганца в высокой концентрации в комбинации с высокими уровнями цинка.
Используя балльные оценки в скрининговых исследованиях (фиг. 4Б, Г) данные об оптимальных для галактозилирования концентрациях можно обобщить следующим образом:
марганец: >0,0963 мкМ, поскольку на линейной модели продемонстрированы наилучшие результаты при наиболее высокой концентрации → оптимальная концентрация еще не достигнута),
медь: <0,3142 мкМ, поскольку на линейной модели продемонстрированы наилучшие результаты при наиболее низкой концентрации → оптимальная концентрация еще не достигнута).
При создании изобретения анализировали также зависимость воздействия указанных выше важных элементов на галактозилирование от времени при использовании клона 3 в качестве модельной клеточной линии. Путем сравнения данных, полученных в день 7 с данными, полученными в день 13 с использованием инструмента прогностического профайлера количественно оценивали роль марганца на ранней и поздней фазе производства. Воздействие марганца как на G1, так и на G2 снижалось на поздних фазах процесса (день 13). Уменьшение крутизны наклона полученной с помощью прогностического профайлера кривой свидетельствует о том, что биодоступность марганца не является столь же высокой, как на ранних фазах процессе (день 7) (фиг. 5).
Пример 2
Обратные воздействия меди и железа на клеточный рост и гликозилирование
Для выявления индивидуальных и объединенных воздействий микроэлементов, ответственных за модуляцию клеточного роста и гликозилирования, при создании изобретения анализировали воздействия цинка, меди, железа и марганца, которые известны, как обладающие высокой биологической активностью микроэлементы, на клеточный рост и гликозилирование МАт с помощью DoE-эксперимента полного факторного плана, используя рекомбинантный клон 2 (таблица 7). Клон 2 применяли в качестве репортерной клеточной линии, поскольку он имел эндогенный повышенный уровень формирования структур гликанов с высоким содержанием маннозы.


В этом эксперименте теоретические эффективные концентрации цинка, железа, меди и марганца (сTE effective) в начале DoE-эксперимента с периодической подпиткой обобщали для каждого состояния (таблица 8). Теоретические сTE effective для цинка, железа, меди и марганца в начале эксперимента рассчитывали согласно описанному выше методу.

Наиболее выраженное воздействие на клеточный рост, которое анализировали по максимальной плотности жизнеспособных клеток (фиг. 6А-В) и конечного интеграла от количества клеток по времени (фиг. 6Г-Е), обнаружено при добавлении железа. В целом, для высоких концентраций цинка, марганца и меди, применяемых в эксперименте, обнаружена тенденция обусловливать более высокий клеточный рост и образование биомассы (фиг. 6В, Е). С другой стороны, целевая комбинация цинка, меди, железа и марганца, модулировала количество галактозилированных или нефукозилированных видов. Марганец в высоких концентрациях повышал галактозилирование, что продемонстрировано в настоящем описании для G1 (фиг. 6Ж-И), в то время как совместное действие цинка и меди и отсутствие марганца приводило к  количествам незрелых гликозилированных видов МАт (фиг. 6К-М).
количествам незрелых гликозилированных видов МАт (фиг. 6К-М).
Для меди и цинка обнаружено выраженное воздействие на образование незрелых гликанов. Поэтому при создании изобретения дополнительно детально анализировали указанное воздействие на нефукозилированные виды гликанов (гликопротеина с высоким содержанием маннозы), используя в качестве инструмента прогностический профайлер JMP. В целом, для цинка при его применении в средних уровнях (уровень 0 в эксперименте) (уровень 0 означает 28,295 мкМ) обнаружена отрицательная, а для меди (уровень 0 означает 0,330 мкМ) положительная корреляция с количеством гликанов с высоким содержанием маннозы (фиг. 7А). Это подтверждается тем фактом, что отрицательный уровень цинка -0,2 (означает 25,416 мкМ) и положительный уровень меди 0,6 (означает 0,349 мкМ) уже не оказывает влияния на сумму маннозилированных видов гликанов (фиг. 7Б). Затем моделировали различные «сценарии» важности таких модуляторов как цинк, медь и марганец, в формировании видов гликанов с высоким содержанием маннозы:
- средние и высокие уровни (от -0,2 до 1, которые соответствуют концентрациям от 25,416 до 43,301 мкМ) цинка индуцируют положительное воздействие меди (фиг. 7В, Е, Ж, З, И),
- низкие уровни (от -1 до -0,2, которые соответствуют концентрациям от 13,900 до 25,416 мкМ) цинка индуцируют отрицательное воздействие меди (фиг. 7Г, Д),
- средний уровень (-0,2, который соответствует концентрации 25,416 мкМ) цинка аннулирует воздействие меди (фиг. 7Б, К),
- отрицательная роль марганца не зависела от уровней цинка и меди.
При создании изобретения в качестве инструмента применяли прогностический профайлер JMP для определения оптимальных концентраций цинка, меди, железа и марганца для совместной оптимизации клеточного роста и формирования гликанов МАт. При создании изобретения применяли одну валидную модель, учитывающую все параметры микроэлементов (квадратичный эффект железа).
Согласно результатам, полученным с помощью прогностического профайлера, цинк, медь и марганец при применении в наиболее высоких концентрациях и железо при применении на уровнях >0,42 (т.е. >36,704 мкМ) могут улучшать клеточный рост (фиг. 8А, В). Применение цинка, железа и марганца в высоких концентрациях и меди в низкой концентрации может повышать количество галактозилированных видов МАт (фиг. 8Г). С другой стороны, применение в низких концентрациях марганца, уровень -1 (т.е. 0,071 мкМ), меди (т.е. 0,312 мкМ) и цинка (т.е. 13,900 мкМ), а железа на уровнях -0,3 (т.е. 11,288 мкМ) параллельно повышает количество нефукозилированных видов МАт (фиг. 8Е).
Минимальные показатели VCD и CTI имели место, когда выбирали высокий уровень цинка и меди и низкие уровни железа и марганца (фиг. 8Б). Низкогалактозилированные виды гликанов образовывались при выборе низких концентраций всех четырех элементов (фиг. 8Г). Минимальный уровень маннозилированных гликанов получали при высоких уровнях цинка и марганца и низких уровнях железа и меди (фиг. 8Ж).
И в этом случае также при создании изобретения тестировали вариант, в котором добавляли цинк в комбинации только с марганцем для повышенного образования галактозилированных гликанов с помощью моделирования данных с использованием JMP. Как продемонстрировано ранее, цинк и марганец оказывают синергетическое действие на образование галактозилированных гликанов. Для количественной оценки этого действия моделировали образование G1 при применении в качестве эффекторов цинка и марганца в высоких и низких концентрациях (фиг. 15). Добавление высоких уровней цинка в сочетании с высокими уровнями марганца могло увеличивать количество G1-видов на 17% (фиг. 15А, Г).
Касательно модуляции степени зрелости видов гликанов сделаны прогнозы в отношении оптимальных уровней микроэлементов для следующей комбинации G1 и видов с высоким содержанием маннозы (фиг. 8Г-Ж):
- для зрелых гликанов: оптимальные уровни, определенные для высокого содержания G1 и низкого содержания Man5,
- для незрелых гликанов: оптимальные уровни, определенные для высокого содержания Man5 и низкого содержания G1.
Пример 3
Целевое производство галактозилированных и нефукозилированных видов гликанов путем модуляции меди, железа, цинка и марганца: эксперимент в биореакторе 1
Клон 2 СНО-K1 выращивали в течение 14 дней в 2-литровых контролируемых системах для ферментации типа Quad (фирма Sartorius, Геттинген, Германия), применяя запатентованную среду и процесс болюсной подпитки с использованием платформы Б. Концентрированную питательную подпитку вносили в объеме, составляющем 10% от объема в начала культивирования, в дни 3, 6 и 9. Требуемую модуляцию микроэлементов осуществляли путем добавления специфической среды, которая благоприятствует гликозилированию МАт, в день 6.
Соотношения и требуемые концентрации цинка, меди, железа и марганца, идентифицированные в предыдущих экспериментах, обобщены в таблице 9. Указанные концентрации и соотношения цинка, меди, железа и марганца тестировали в проведенном с целью верификации эксперименте с биореакторе с периодической подпиткой, используя клон 2, для которого характерны эндогенные высокие уровни Man5-видов гликанов. При создании изобретения модулировали соотношения микроэлементов, используя стратегию культивирования с расщеплением и пополнение требуемыми добавками для поддержания клеточного роста на первой фазе периодического процесса с подпиткой (дни d0-d5) и для созревания гликозилирования МАт на второй фазе периодического процесса с подпиткой (дни d6-d14), применяя две различные среды для культивирования, предназначенные для фаз образования биомассы («Биомасса») и гликозилирования («Гликозилирование»), которые описаны в таблице 9А и таблице 9Б соответственно.
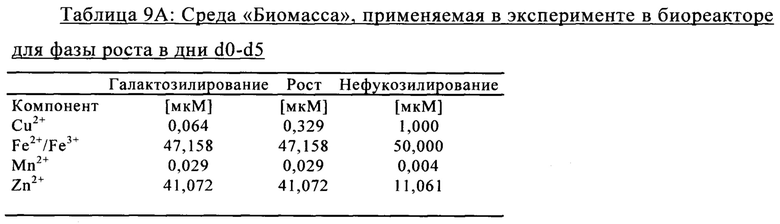
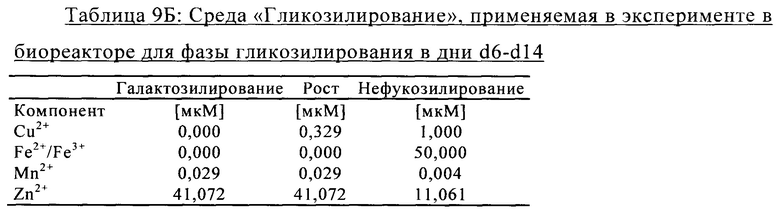
По сравнению с условиями, созданными для того, чтобы благоприятствовать формированию незрелых гликанов, что обозначено как «Нефукозлирование», в которых в течение всего процесса применяли постоянные концентрации цинка, меди, железа и марганца, при создании изобретении на стадиях, обозначенных в дальнейшем как «Рост» и «Галактозилирование» (фиг. 9А), тестировали микроэлементы в концентрациях, которые, как прогнозировалось, должны влиять на клеточный рост и созревание гликозилирования МАт, изменяя концентрацию меди между двумя уровнями. Такое решение было принято в связи с тем, что медь, вероятно, оказывает более сильное действие на гликозилирование, чем на максимальную плотность жизнеспособных клеток (фиг. 8).
В приведенной ниже таблице (таблица 10) проиллюстрированы теоретические концентрации, применяемые в эксперименте, проведенном в биореакторе 1.

Используя указанные условия, при создании изобретения доказано, что вплоть до дня 6 способность клеток расти можно модулировать путем баланса микроэлементов, при применении которого продемонстрированы наиболее высокая плотность жизнеспособных клеток и CTI на стадии, обозначенной как «Рост», после чего осуществляли процессы, обозначенные как «Нефукозилирование» и «Галактозилирование» (фиг. 9Б, В). Последующее переключение на условия, благоприятствующие гликозилированию, и приводящие к снижению клеточного роста, начиная со дня 6, четко продемонстрировано для процесса, обозначенного как «Галактозилирование» по плотности жизнеспособных клеток и интегралу от количества клеток по времени. Важно отметить, что модуляция соотношений микроэлементов оказывала сильное воздействие на жизнеспособность клеток на поздних фазах процесса (фиг. 9Г). При снижении концентрации меди обнаружена тенденция к повышению конечной плотности жизнеспособных клеток почти на 30% (для процесса, обозначенного как «Галактозилирование») по сравнению с процессом, обозначенным как «Рост». Воздействие снижения концентрации меди на повышение жизнеспособности клеток можно продемонстрировать также с помощью анализа высвобождения ЛДГ (фиг. 10В).
С другой стороны, медь, как было установлено, оказывает положительное воздействие на реметаболизацию лактата на поздних фазах процесса (Luo и др., Biotechnol. Bioeng. 109.1, 2012, cc. 146-156). В описанном выше эксперименте по верификации было продемонстрировано участие меди в реметаболизации лактата, поскольку на стадии, обозначенной как «Галактозилирование», обнаружена повышенная, но не имеющая решающего значения (→ высокая жизнеспособность клеток), концентрация лактата на поздних фазах процесса по сравнению со стадиями, обозначенными как «Нефукозилирование» и «Рост», на которых вплоть до дня 14 почти весь лактат поглощался (фиг. 10Б). Исходя из того, что как известно, высокие уровни лактата могут благоприятствовать детоксикации аммония (Li и др., Biotechnol. Bioeng. 109.5, 2012, cc. 1173-1186), при создании изобретения удалось подтвердить для процесса «Галактозилирования» обратную корреляцию между накоплением лактата и повторной ассимиляцией аммония (фиг. 10А).
Для изучения воздействия модуляции микроэлементов на кинетику гликозилирования МАт при создании изобретения анализировали образцы, которые отбирали в процессе осуществления культивирования в биореакторе, соответствующие нефукозилированным и галактозилированным видам. Содержание видов с высоким содержанием маннозы (Man5) (фиг. 11А), а также нефукозилированных видов в целом (фиг. 11Г) можно снижать (вплоть до 30% в случае Man5) с помощью спрогнозированной модуляции микроэлементов в зависимости от концентрации меди (концентрация меди: «Нефукозилирование» > «Рост» > «Галактозилирование»). Модуляция микроэлементов приводила к сдвигу от видов с высоким содержанием маннозы к G0-видам (вплоть до 30%), а затем к G1-видами (вплоть до 30% в день 13) по мере переключения условий процесса с условий, применяемых на стадии роста, к условиям, применяемым на стадии гликозилирования (фиг. 11А-В). Снижение G1-видов на стадии, обозначенной как «Галактозилирование», в день 14 могут обусловливаться ограничением биодоступности Mn2+ и Zn2+ на поздних фазах процесса.
Пример 4
Целевая модуляция клеточного роста и гликозилирования МАт с помощью железа, меди, цинка и марганца: эксперимент в биореакторе 2
Клон 2 СНО-K1 выращивали во встряхиваемых колбах для размножения культуры и 14 дней в 2-литровых контролируемых системах для ферментации типа Quad (фирма Sartorius, Геттинген, Германия) для осуществления периодического культивирования с подпитой с целью оценки влияния железа, меди, цинка и марганца на клеточный рост и гликозилирование белка. Применяли запатентованную среду и платформу Б процесса. Концентрированную питательную подпитку вносили непрерывно в объеме, составляющем 2,73% (об./об.) от объема в начале культивирования, в дни 3, 6 и 9. Требуемую модуляцию микроэлементов осуществляли путем использования конкретных концентраций металлов в среде для культивирования и непрерывной подпитки (таблицы 11-13) и путем расщепления культуры в процессе подготовки для инокуляции (фазы n-2 и n-1) и переносили в среду для производства (фаза n). Соответствующие концентрации железа, цинка, меди и марганца рассчитывали на основе специфических для клеток и системы культивирования скоростей (фиг. 13 и 14) и, учитывая сбалансированный объем в процессе расщепления и культивирования (например, осуществляя добавление корректирующих агентов, таких как маточный раствор глюкозы, противовспенивающее средство, основание). Фактические концентрации железа, цинка, меди и марганца можно измерять с помощью ICP-MC (фирма Agilent, Беблинген, Германия).
Условия тестирования создавали так, чтобы либо поддерживать клеточный рост в процессе размножения культуры, начальное образование биомассы во время периодического процесса с подпиткой и галактозилирование белка на фазе производства во время периодического процесса с подпиткой (дни 6-14), либо поддерживать клеточный рост в процессе размножения культуры, начальное образование биомассы во время периодического процесса с подпиткой и нефукозилирование белка на фазе производства во время периодического процесса с подпиткой (дни 6-14), либо препятствовать клеточному росту в процессе размножения культуры, начальному образованию биомассы во время периодического процесса с подпиткой и поддерживать нефукозилирование белка на фазе производства во время периодического процесса с подпиткой (дни 6-14) (фиг. 12А).

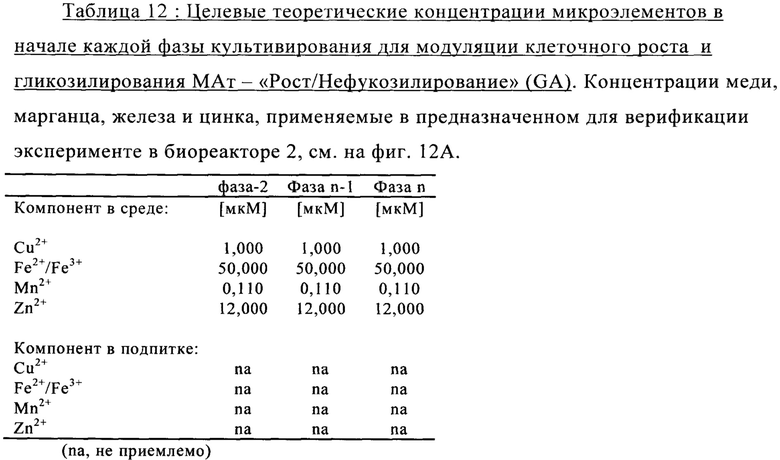
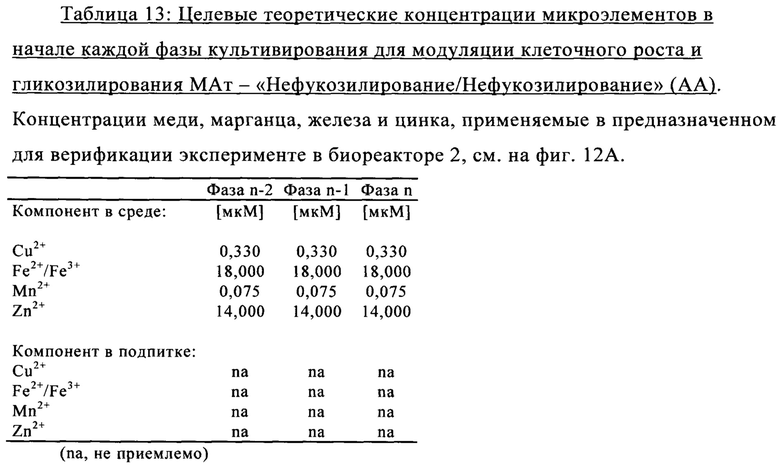
Как и предполагалось, тестируемый вариант «АА» заметно препятствовал клеточному росту в процессе подготовки для инокуляции и на фазе производства, что продемонстрировано по плотности жизнеспособных клеток и интеграла от количества клеток по времени на фазе n (фиг. 12В-Г). С другой стороны, никаких различий в образовании биомассы не обнаружено в тестируемых вариантах «GG» и «GA» в процессе размножения клеточной культуры и на фазе клеточного роста при использовании периодического процесса с подпиткой. Воздействие на созревание МАт анализировали путем измерения количества гликанов. Для зрелых и незрелых гликозилированнных видов МАт объединяли G1 и G2 или Man6, Man5 и G0-GlcNAc. Как и предполагалось, тестируемый вариант «GG» благоприятствовал образованию зрелых видов гликанов (увеличение в 1,4-7 раз по сравнению с вариантами «АА» и «GA») (фиг. 12Д), а тестируемые варианты «GA» и «АА» поддерживали образование незрелых гликанов (2-3-кратное увеличение по сравнению с вариантом «GG») (фиг. 12Е).
В целом, описанная процедура позволят модулировать гликопротеины и прежде всего созревание гликозилирования МАт, с помощью целевого баланса микроэлементов с использованием биотехнологического процесса производства. Путем переключения условий с тех, которые применяют на стадии, обозначенной как «Биомасса», на условия, которые применяют на стадии, обозначенной как «Гликозилирование», с использованием стратегий модуляции концентраций и соотношения цинка, меди, железа и марганца, способ удовлетворяет требованиям экономичности процесса, обеспечивающего высокие титры.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ГЛИКОПРОТЕИНОВ | 2007 |
|
RU2463345C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ГИАЛУРОНИДАЗЫ | 2021 |
|
RU2822202C1 |
| Способ контроля гликозилирования рекомбинантного гликопротеина | 2015 |
|
RU2670889C9 |
| СПОСОБ ПОЛУЧЕНИЯ ЦЕЛЕВОГО АНТИТЕЛА С МОДУЛИРОВАННЫМ ГАЛАКТОЗИЛИРОВАНИЕМ (ВАРИАНТЫ) И СПОСОБ МОДУЛИРОВАНИЯ ГАЛАКТОЗИЛИРОВАНИЯ ЦЕЛЕВОГО АНТИТЕЛА (ВАРИАНТЫ) ПУТЕМ ОПТИМИЗАЦИИ КУЛЬТУРАЛЬНОЙ СРЕДЫ | 2015 |
|
RU2642285C1 |
| СПОСОБ ПОЛУЧЕНИЯ СЫРЬЯ ИЗ ARTHROSPIRA PLATENSIS, ОБОГАЩЕННОЙ МИКРОЭЛЕМЕНТАМИ В ОРГАНИЧЕСКОЙ ФОРМЕ | 2024 |
|
RU2837132C1 |
| КОНСТРУКЦИИ СЛИЯНИЯ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ АНТИТЕЛ С ПОВЫШЕННЫМИ АФФИННОСТЬЮ СВЯЗЫВАНИЯ Fc-РЕЦЕПТОРА И ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2010 |
|
RU2623167C2 |
| СПОСОБЫ, СВЯЗАННЫЕ С ГЛИКОЗИЛИРОВАНИЕМ КЛЕТОЧНОЙ ПОВЕРХНОСТИ | 2008 |
|
RU2475751C2 |
| Способ культивирования метанокисляющих микроорганизмов | 2023 |
|
RU2811437C1 |
| ШТАММ ГРИБА TRAMETES PUBESCENS C-23 - ПРОДУЦЕНТ ЭРГОСТЕРИНА И ПРЕПАРАТ, ПОЛОЖИТЕЛЬНО ВЛИЯЮЩИЙ НА ТКАНЕВЫЙ ОБМЕН, СТИМУЛИРУЮЩИЙ ИММУНОГЕНЕЗ И СПОСОБСТВУЮЩИЙ ВОССТАНОВЛЕНИЮ НАРУШЕННОЙ ОКСИДАЗ-СМЕШАННОЙ ФУНКЦИИ ПЕЧЕНИ | 2005 |
|
RU2323966C2 |
| ПОЛУЧЕНИЕ РЕКОМБИНАНТНОГО ФАКТОРА VIII В СРЕДЕ, НЕ СОДЕРЖАЩЕЙ БЕЛКА | 1998 |
|
RU2222547C2 |


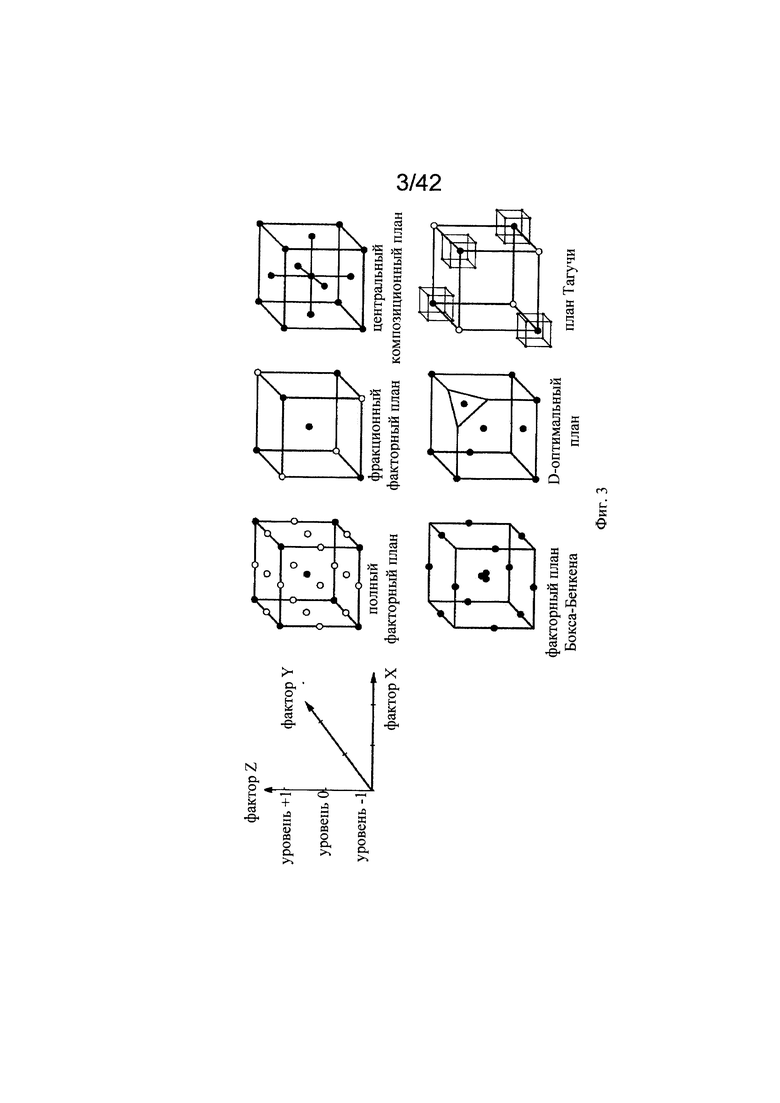
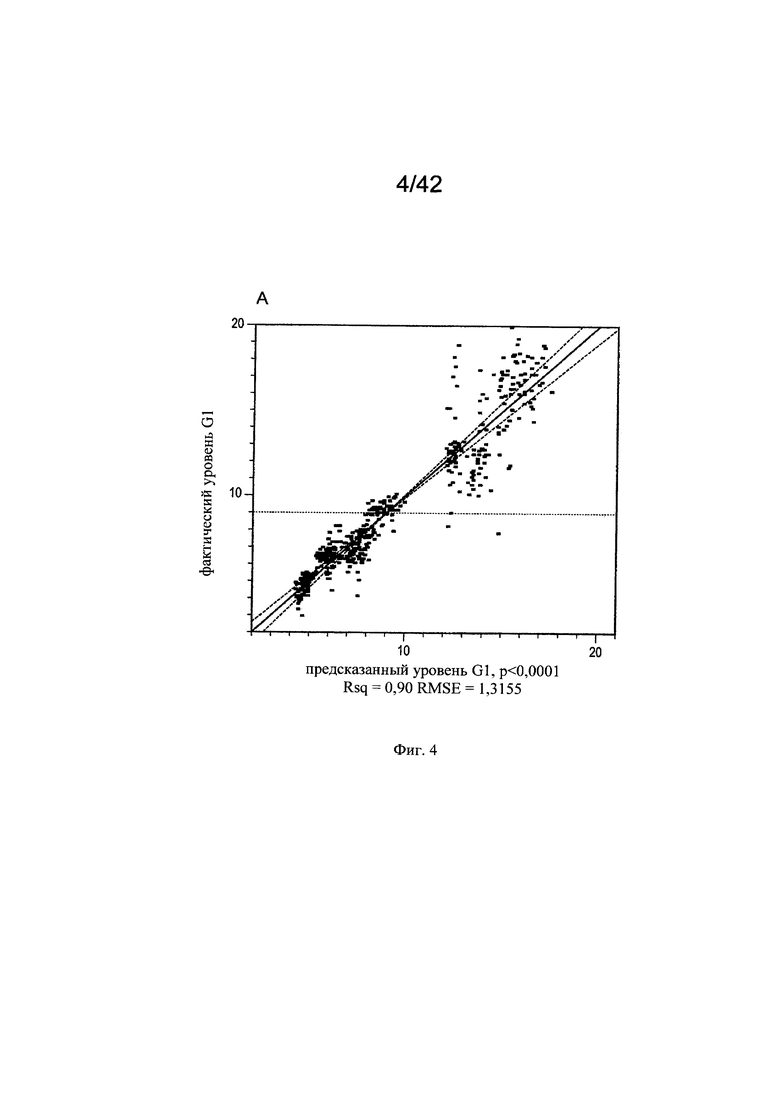

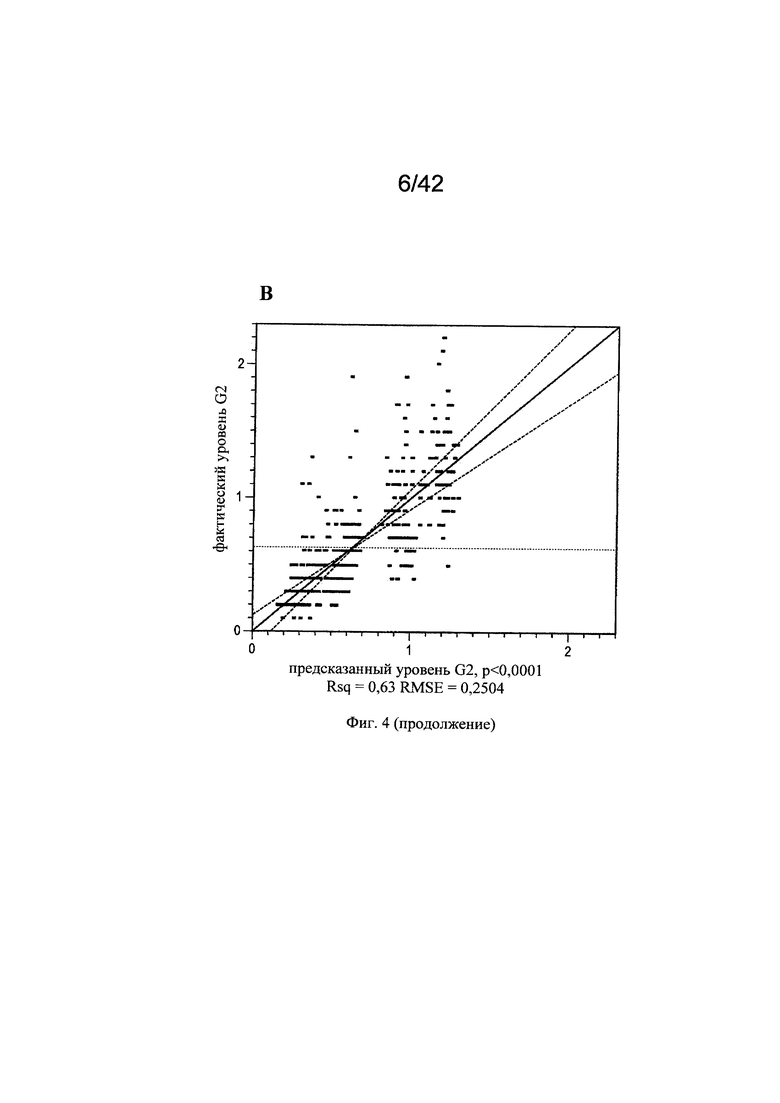
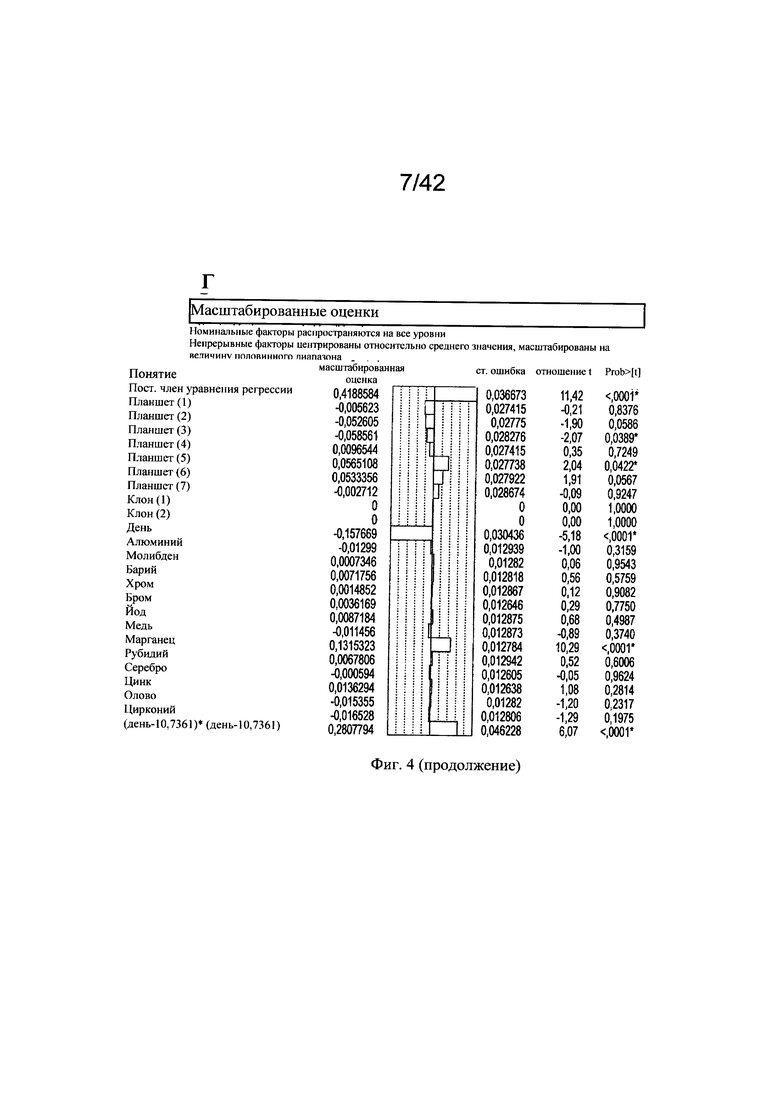
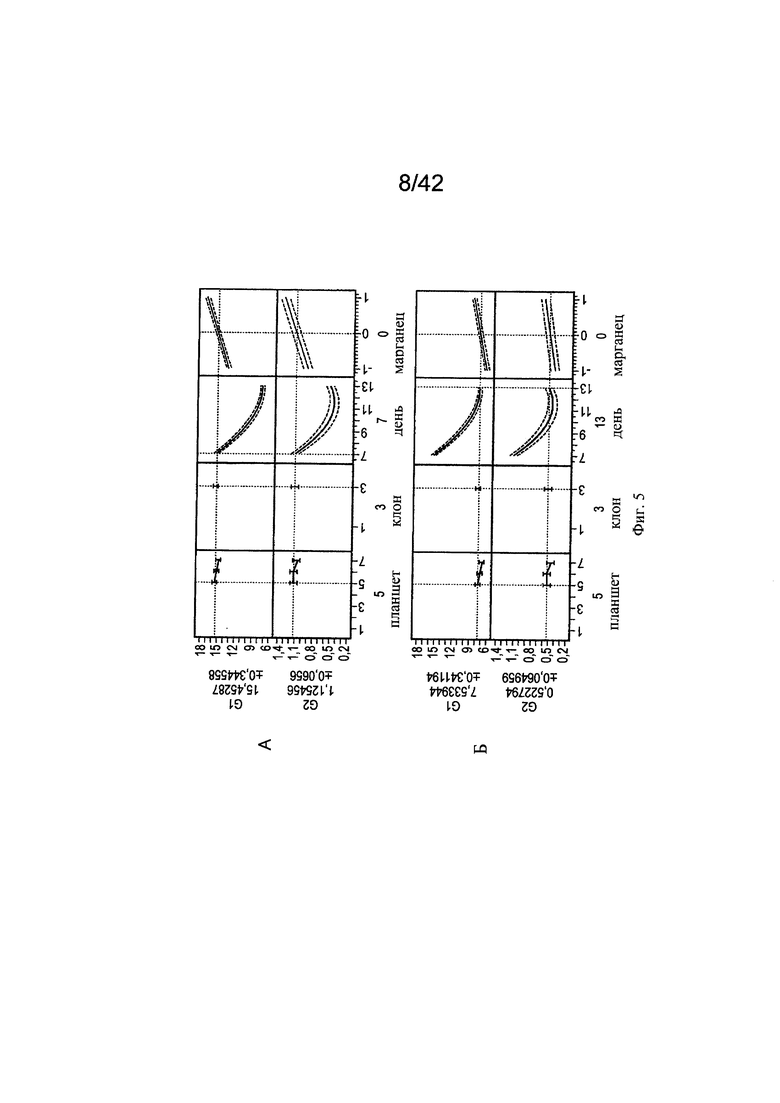

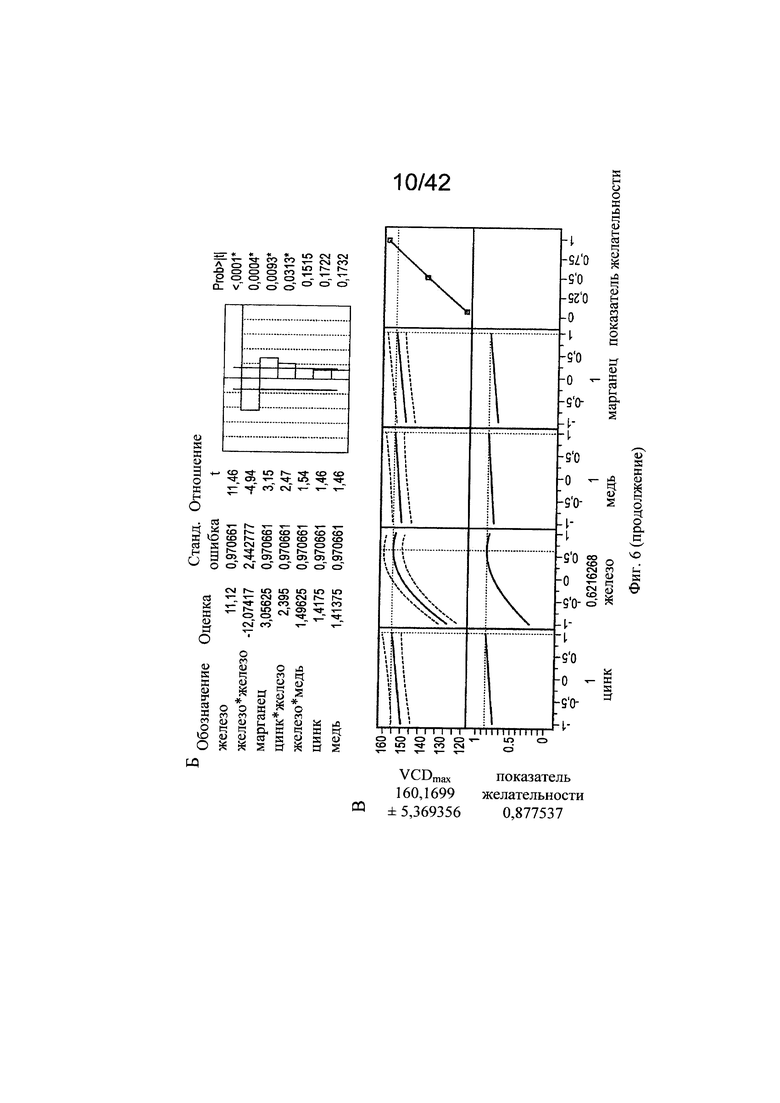

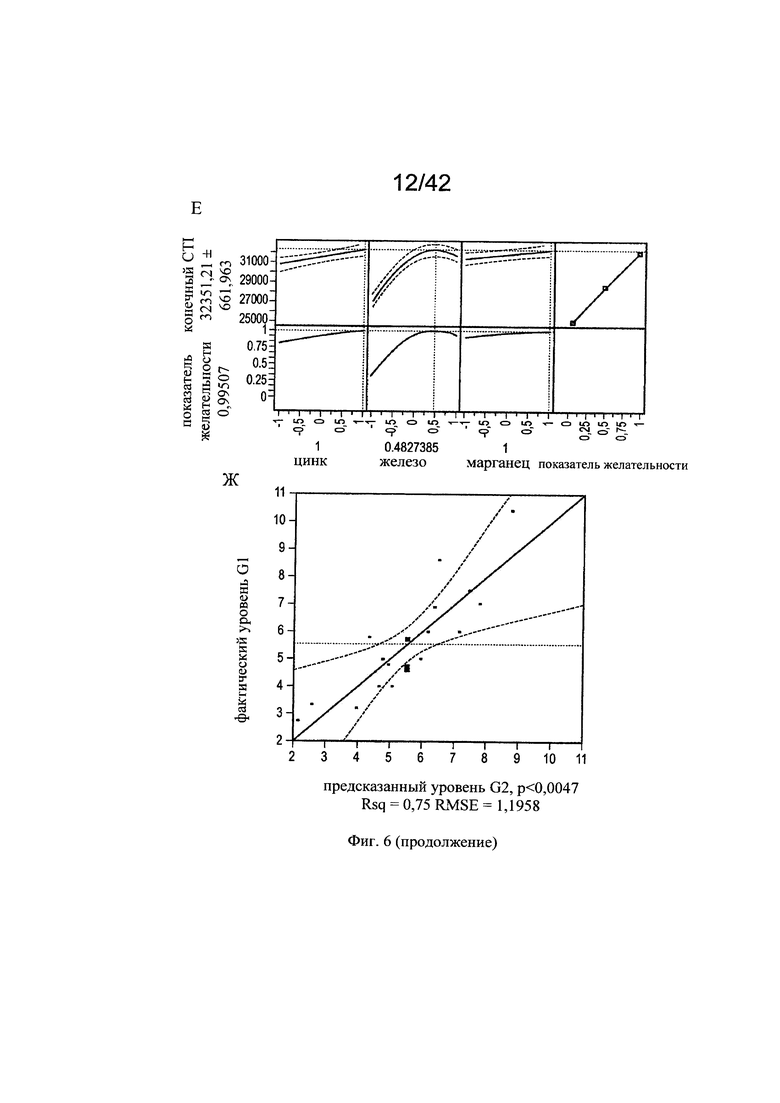
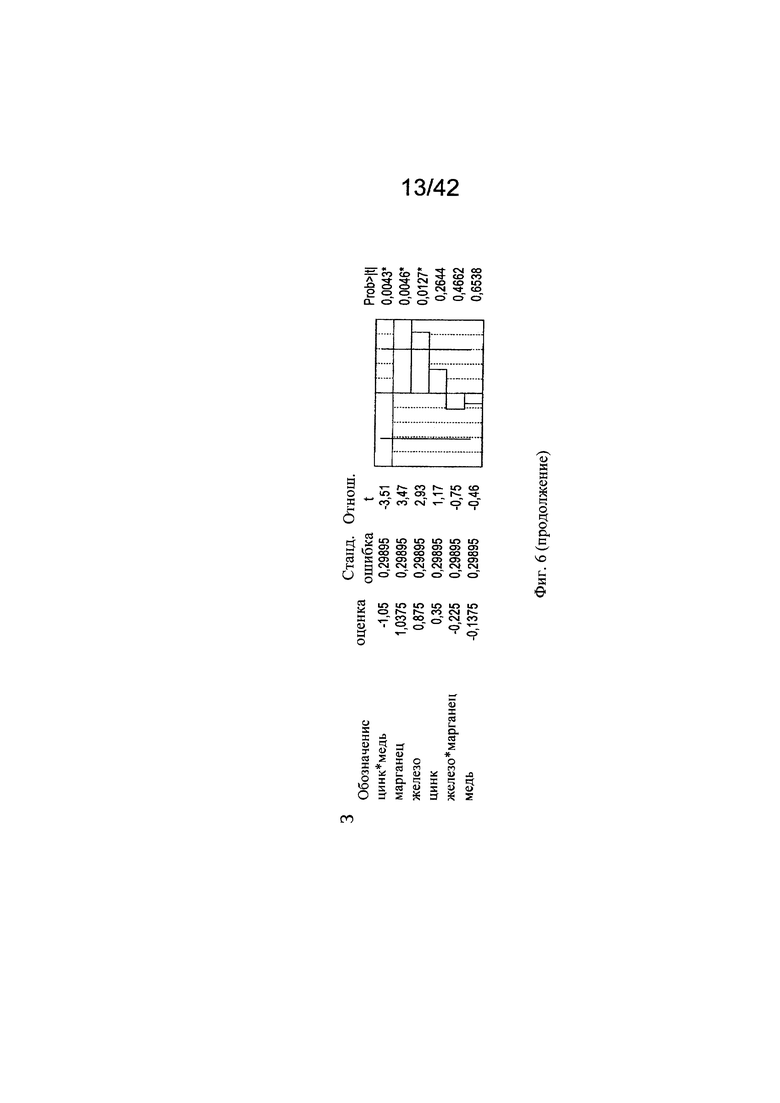
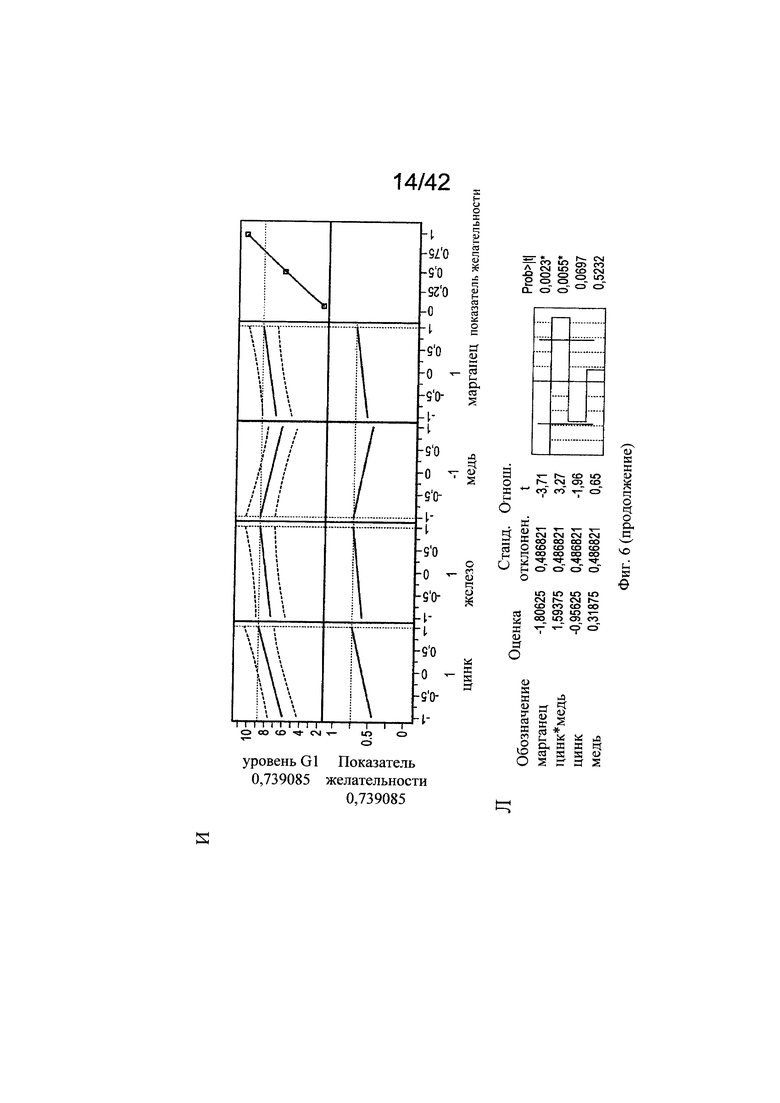
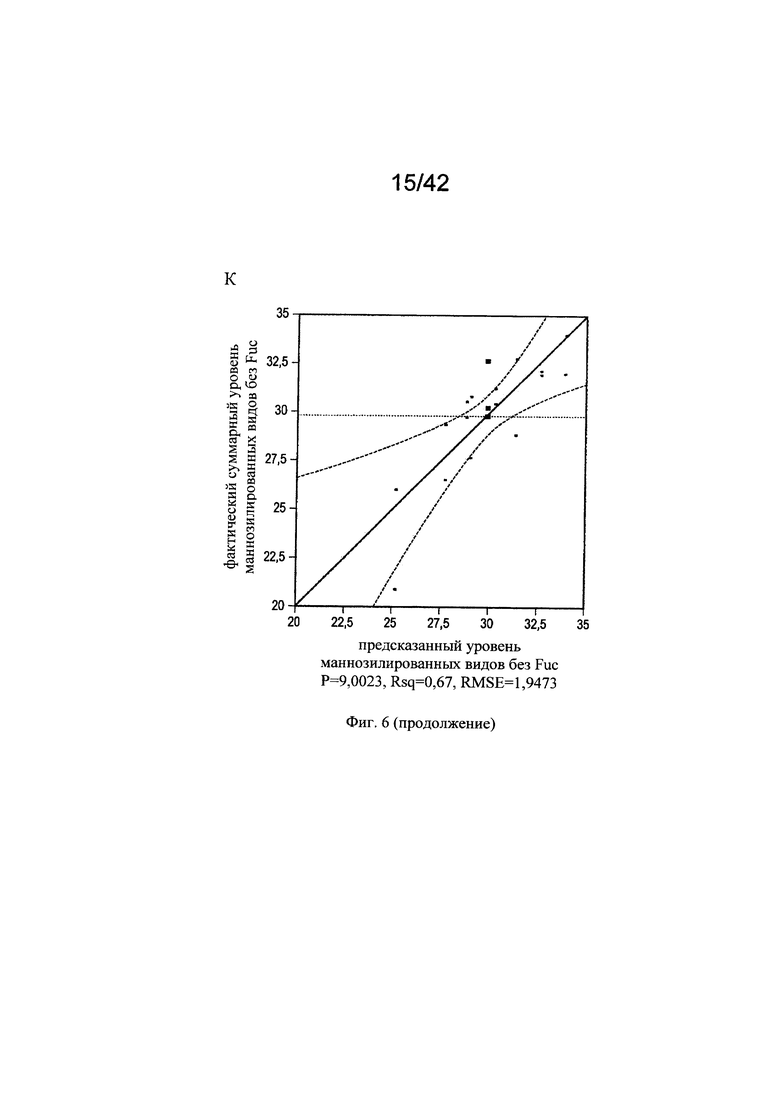

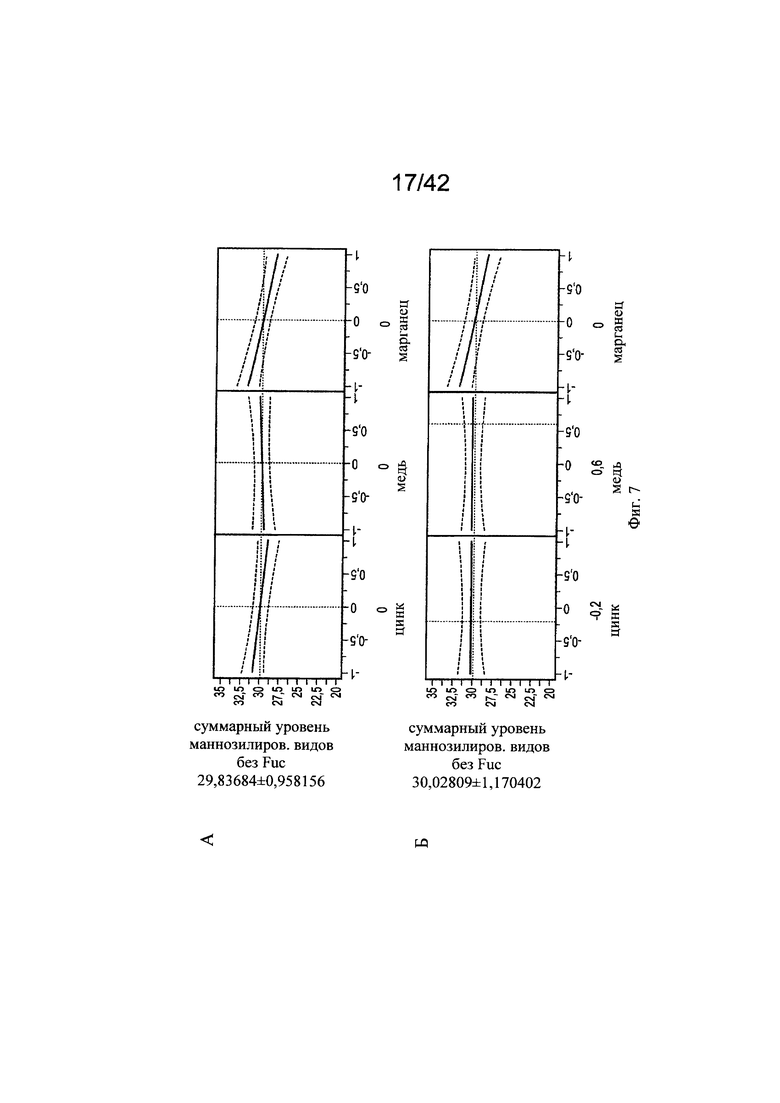


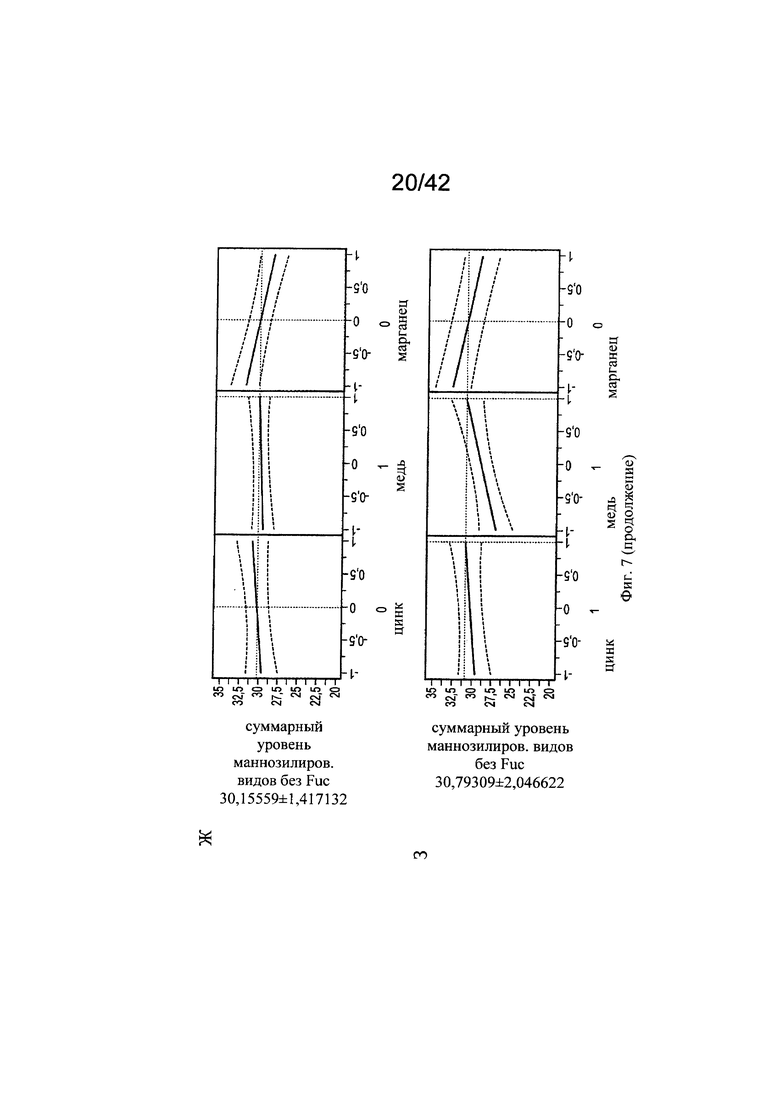
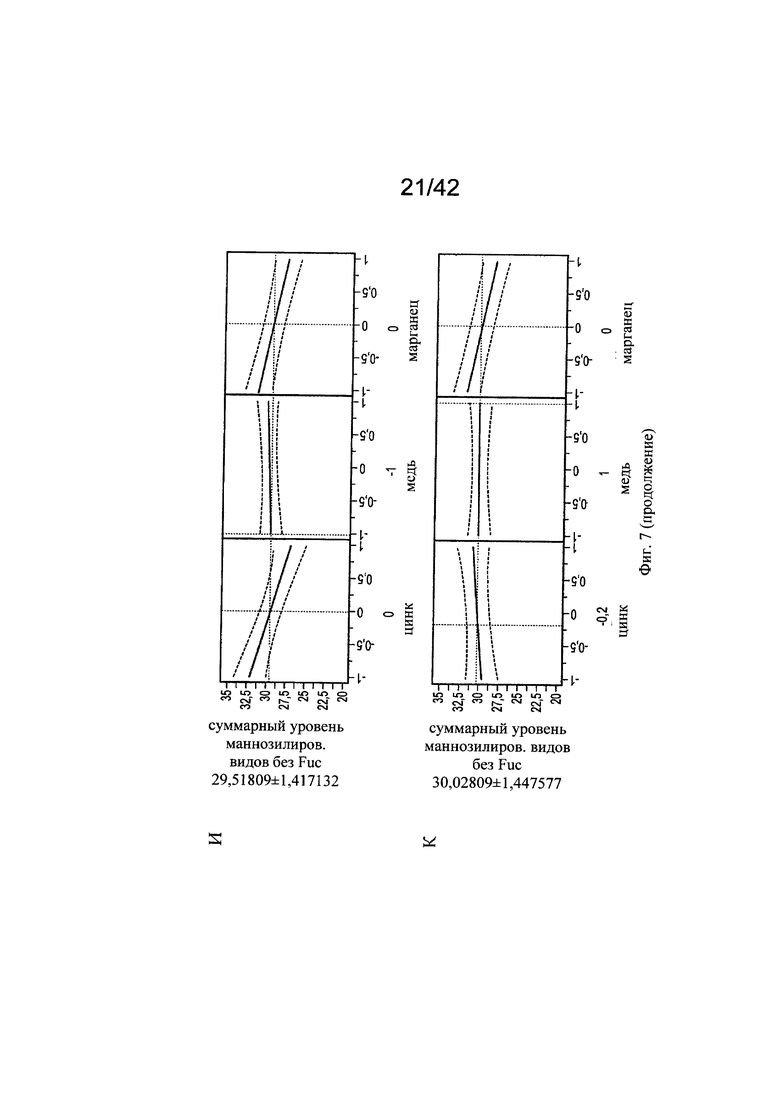


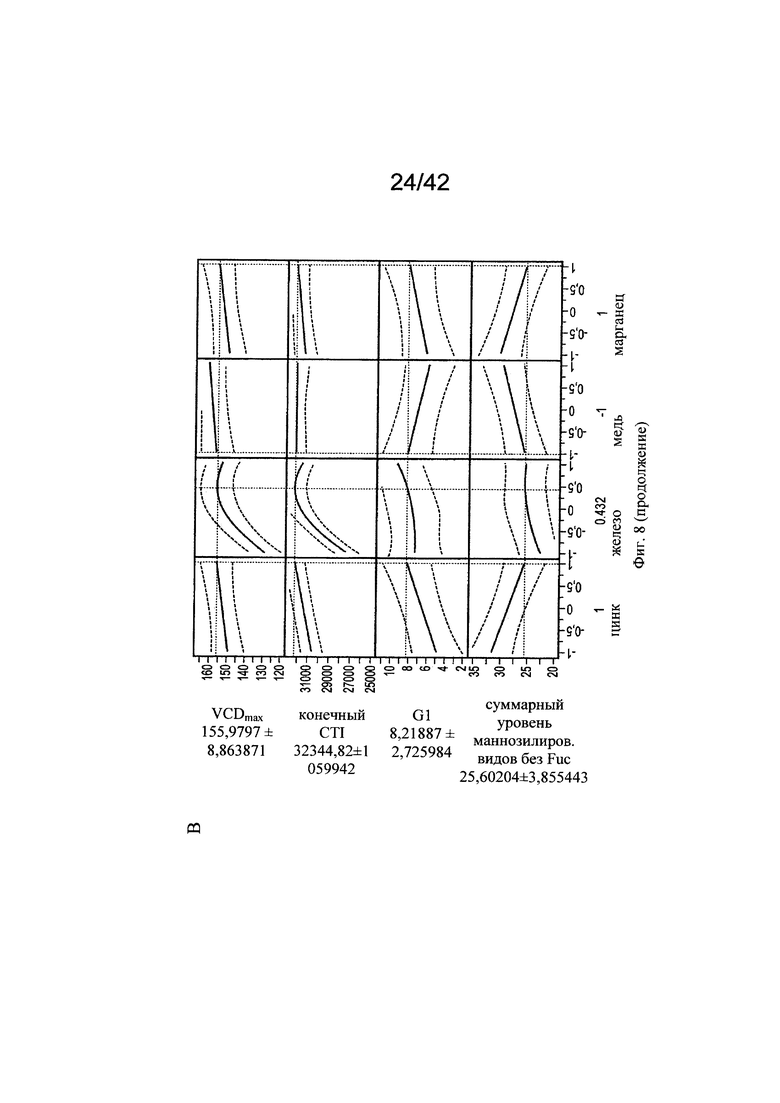
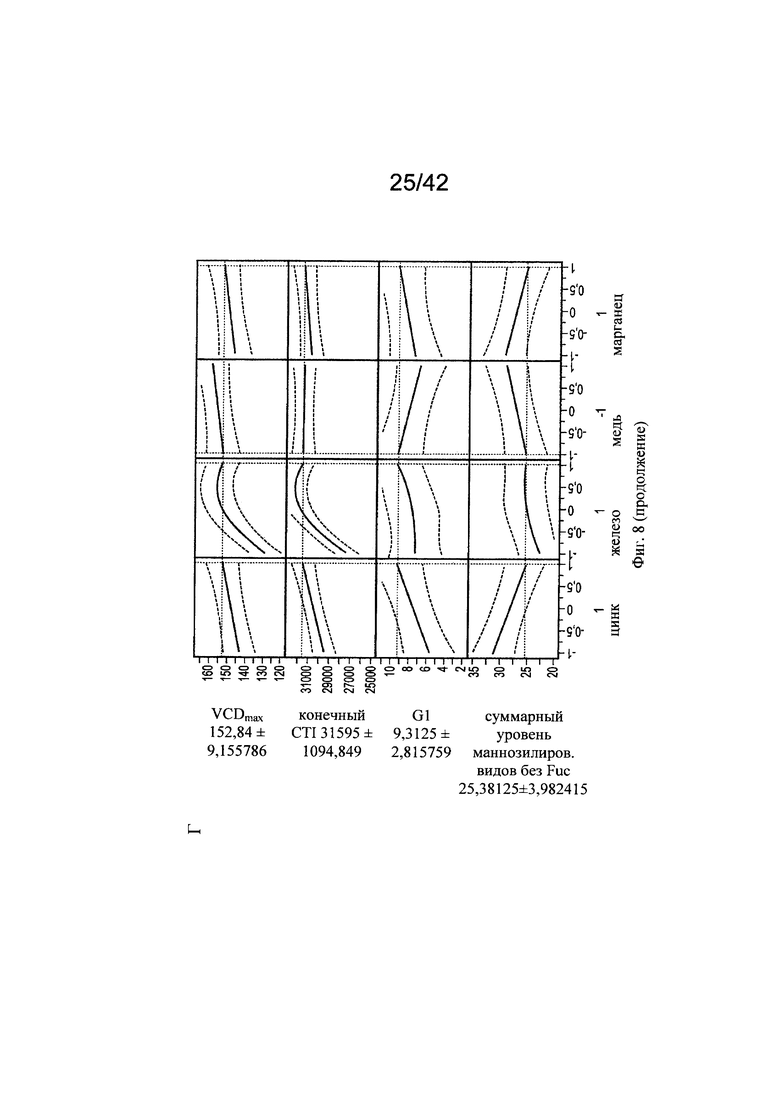
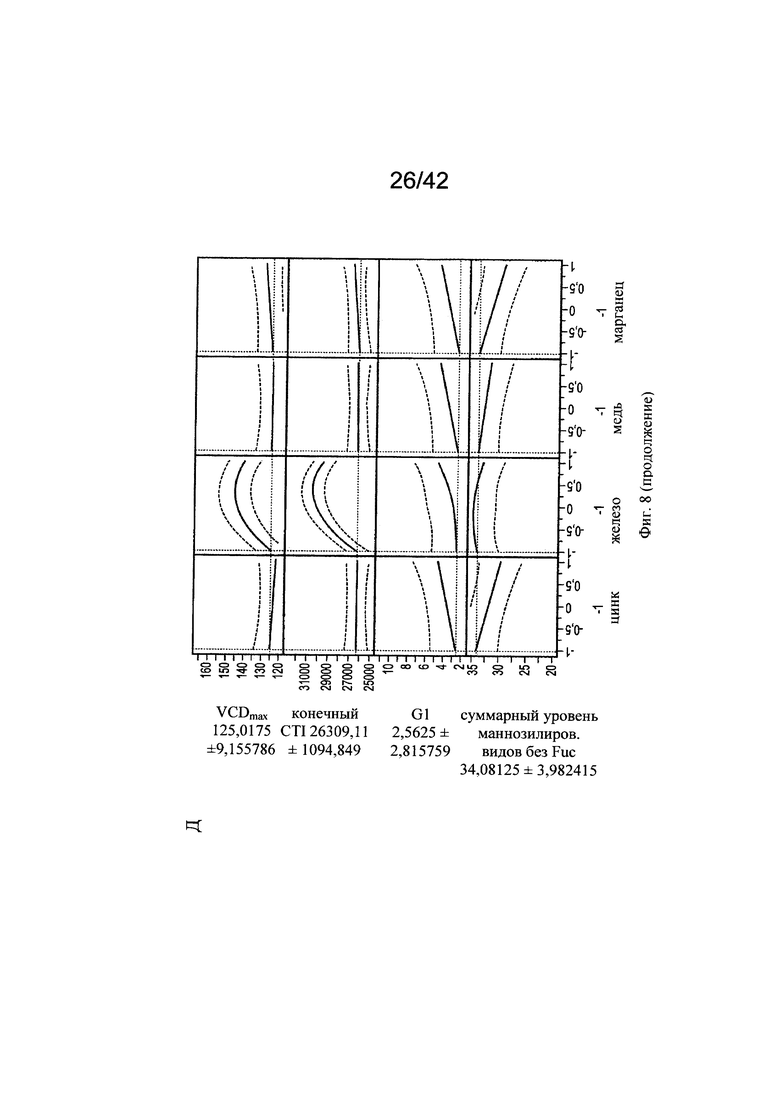
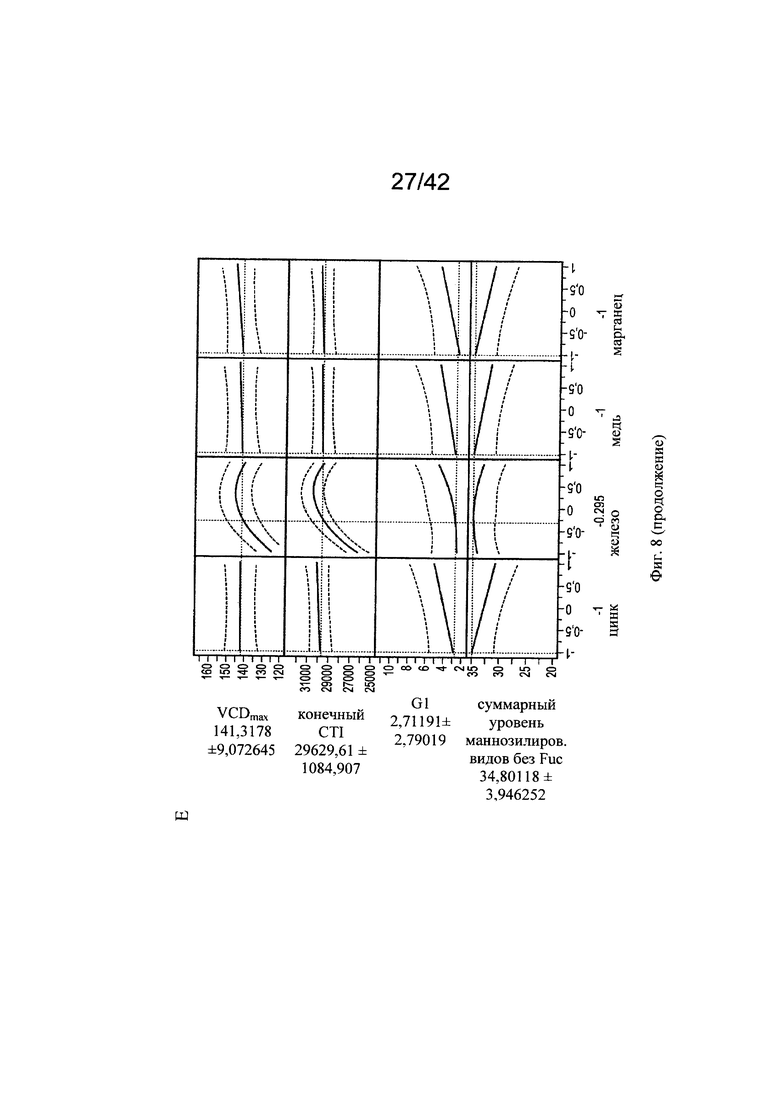

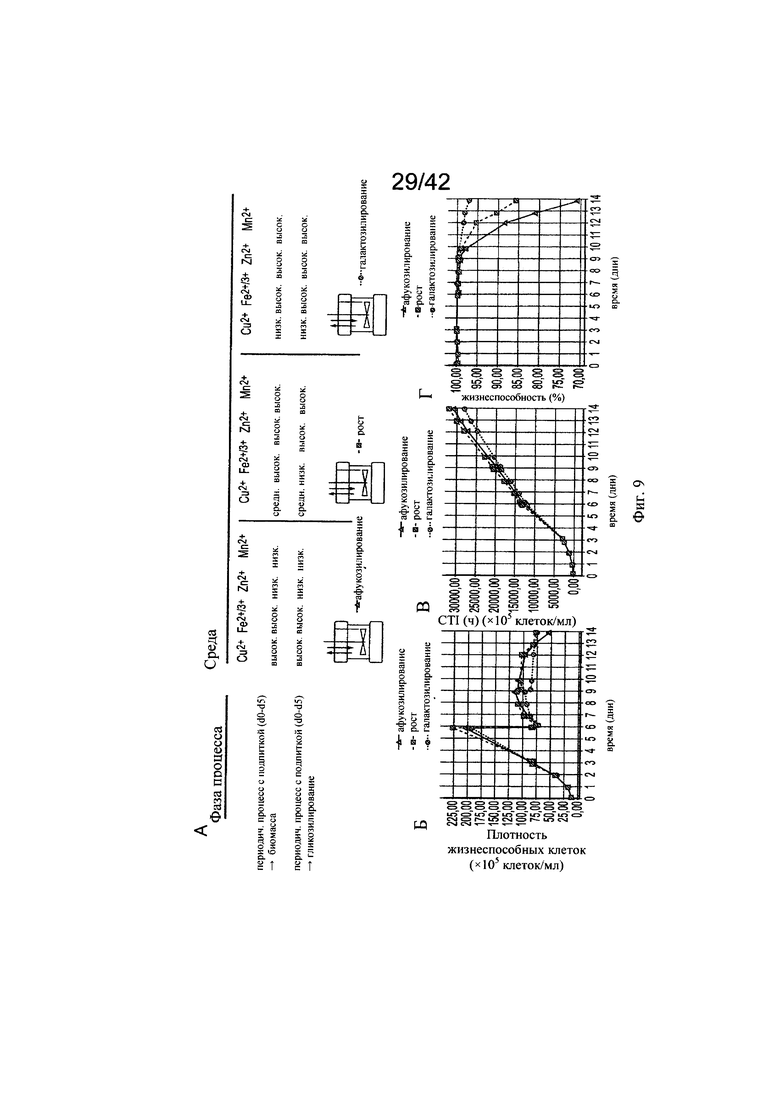
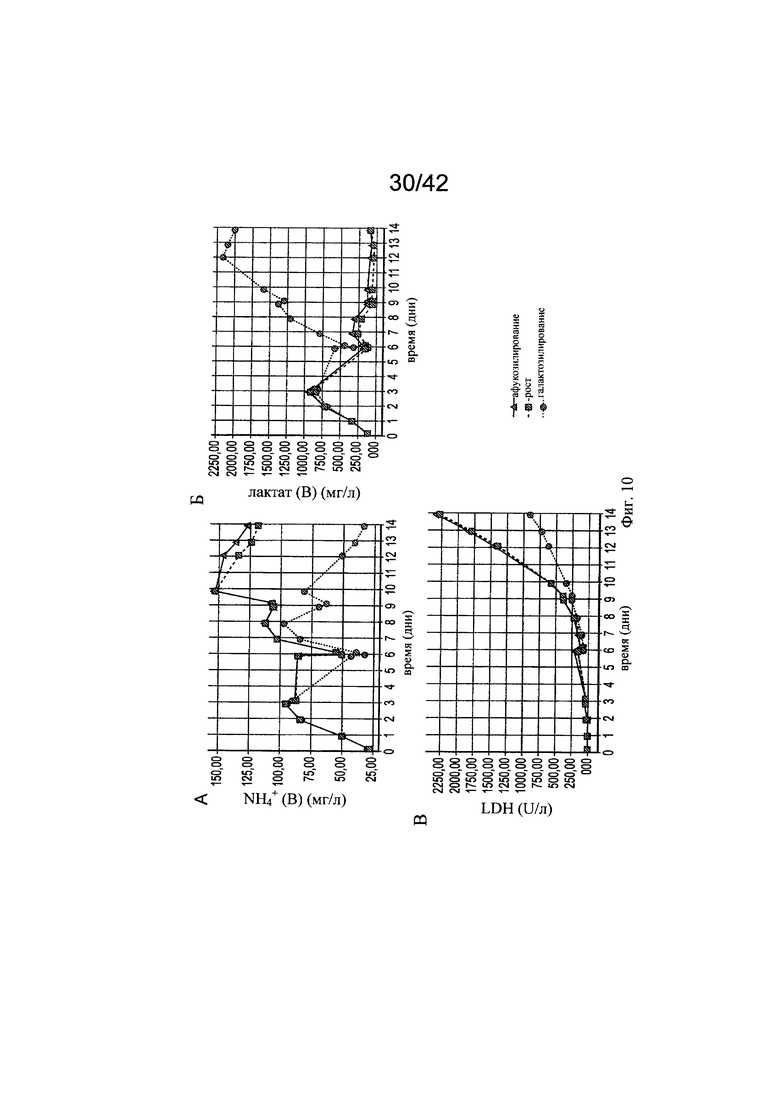
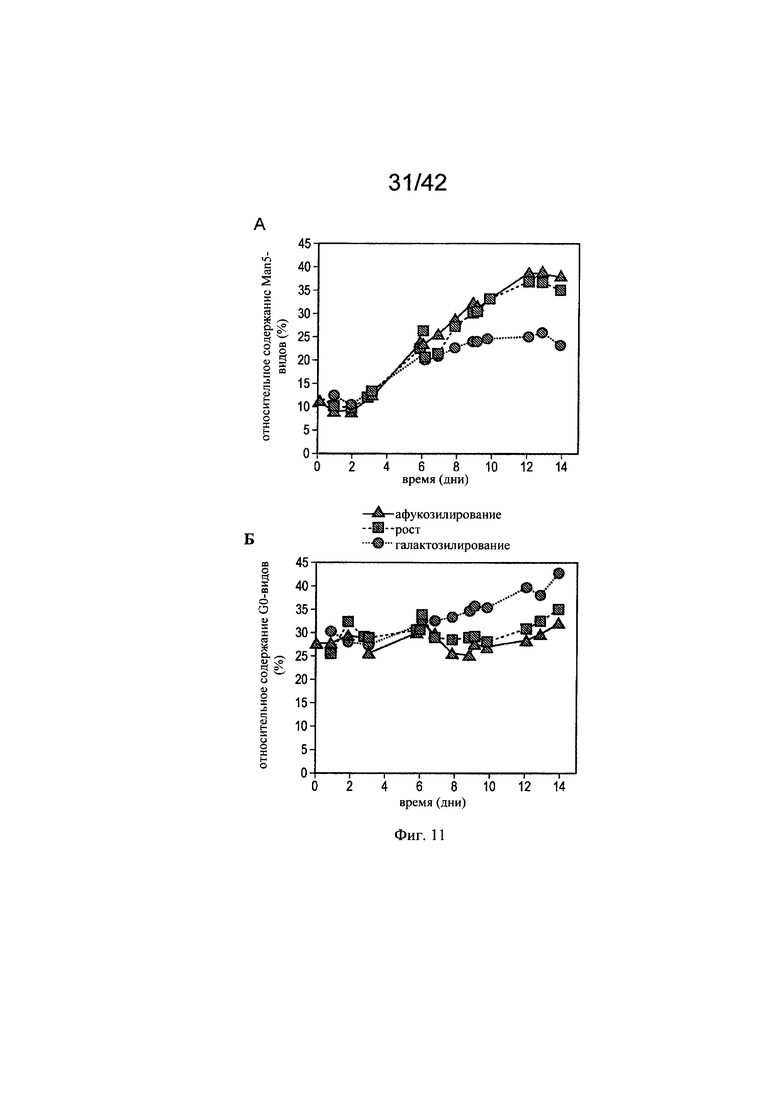
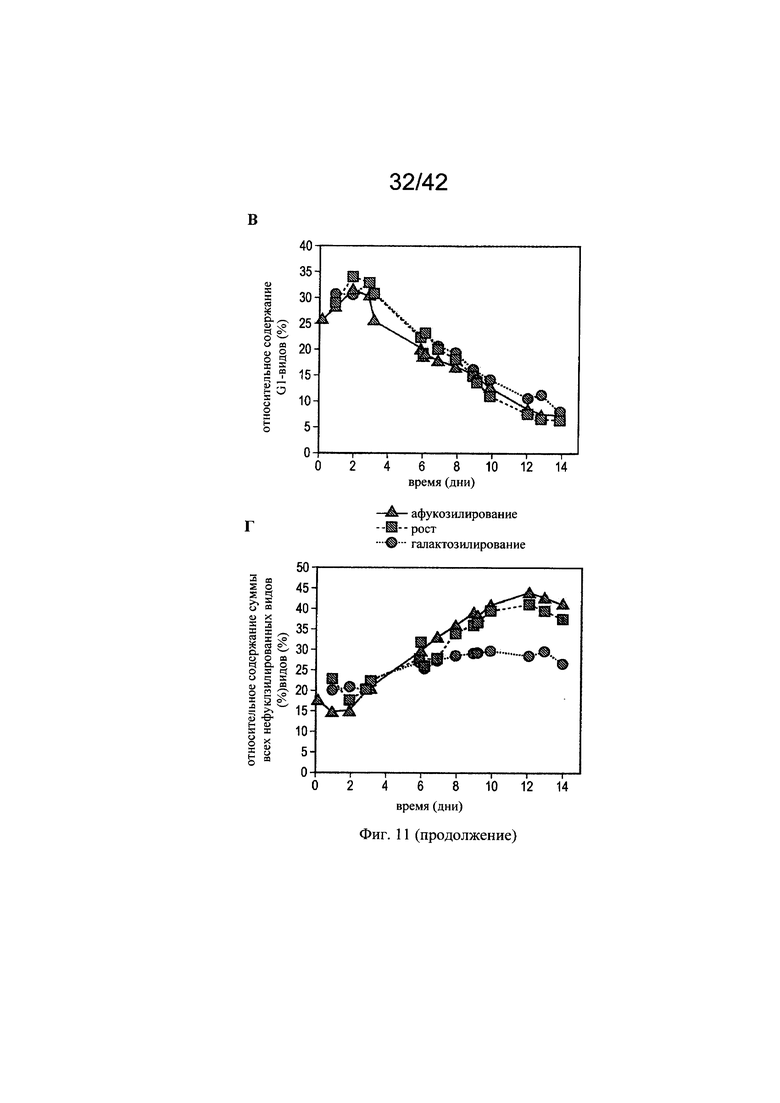

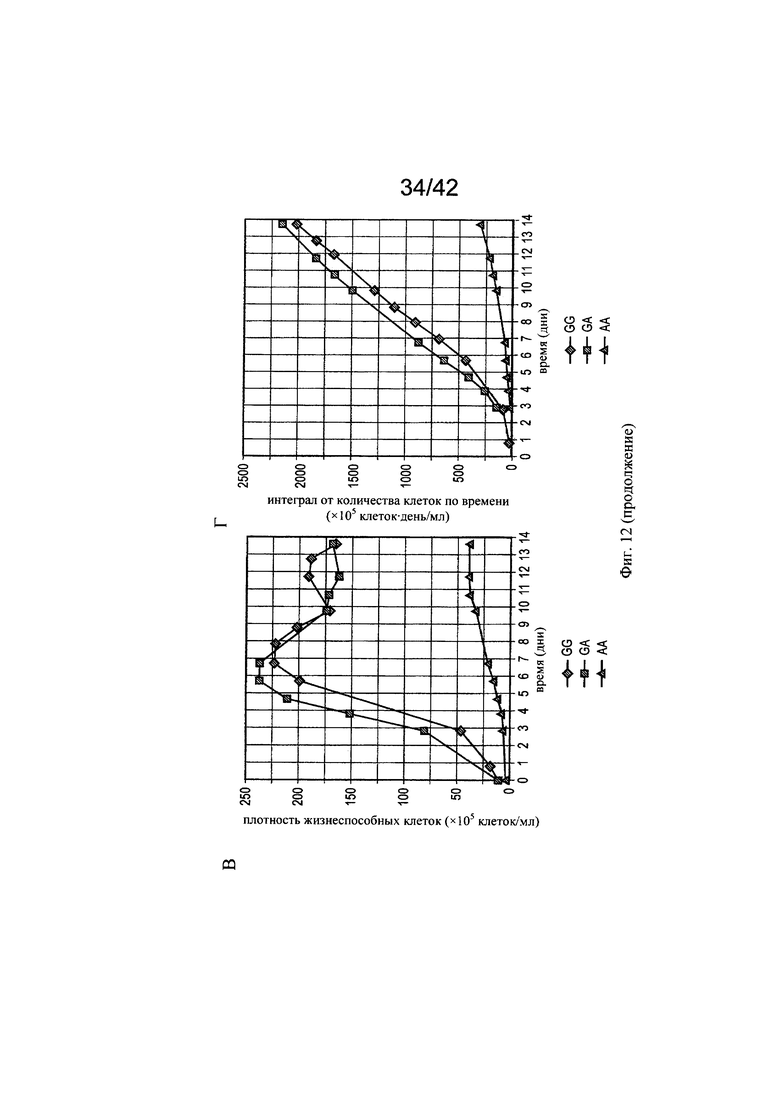

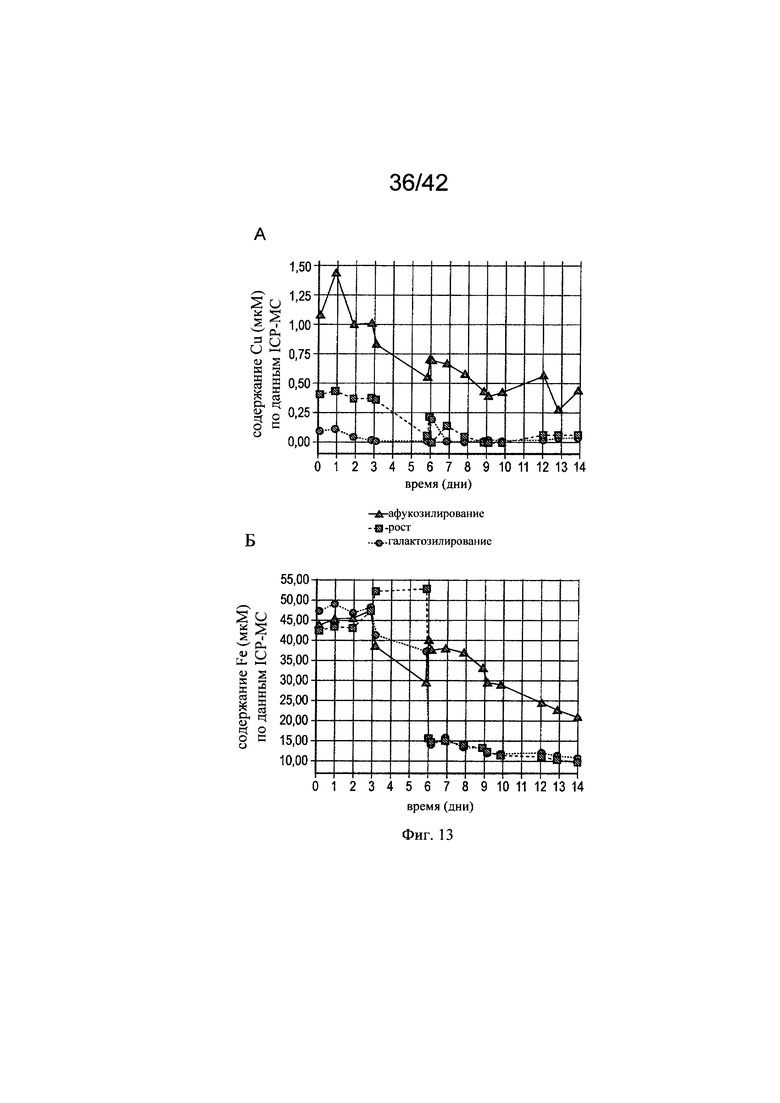
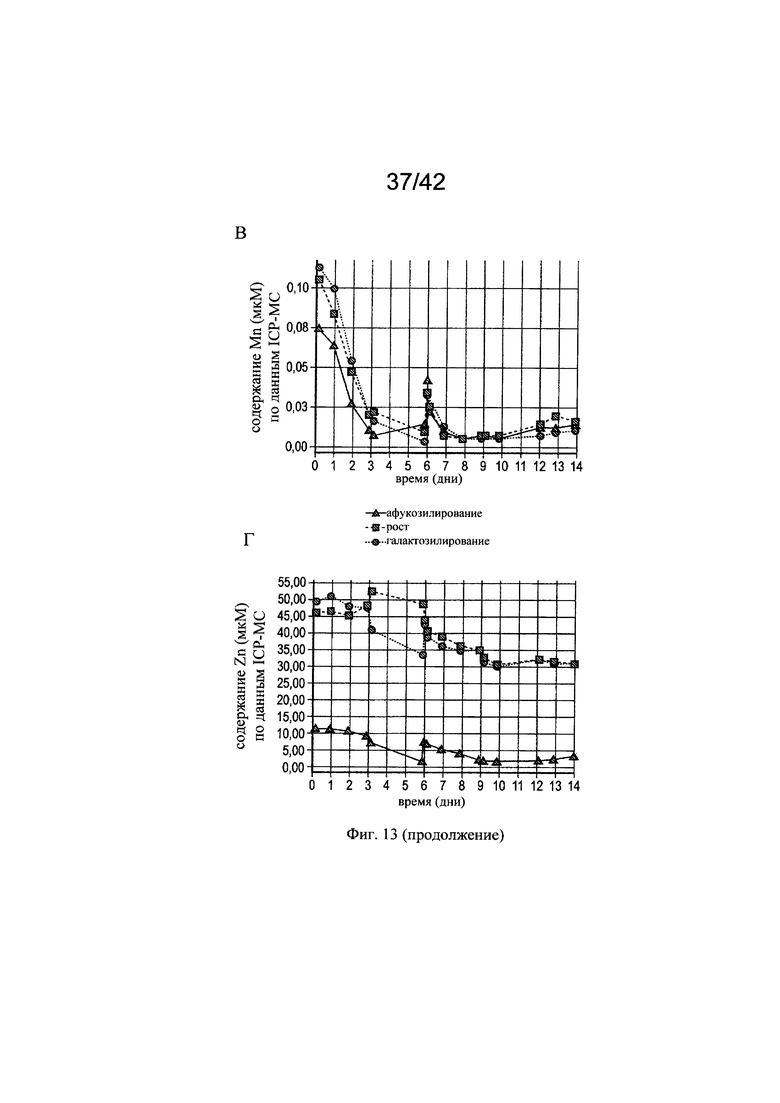

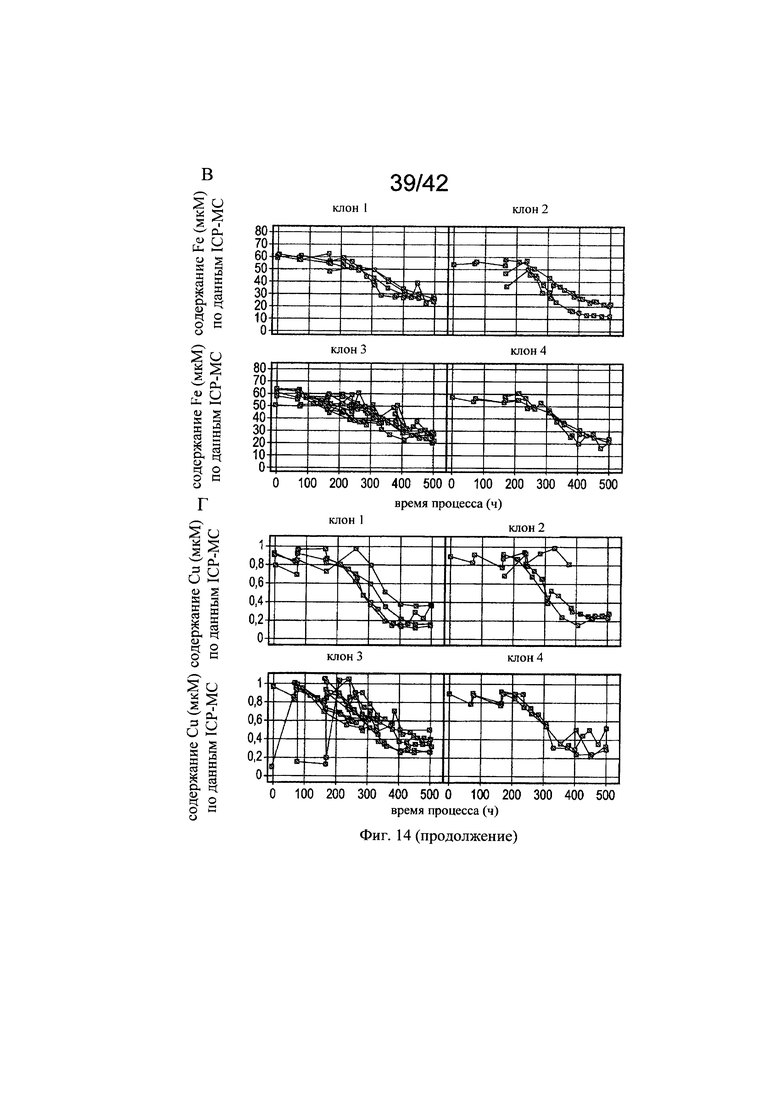



Группа изобретений относится к биотехнологии, в частности к способам получения рекомбинантного гликопротеина в условиях ферментационной культуры в СНО-клетке путем регулирования концентраций железа, меди, цинка и марганца в среде. В одном варианте увеличивают концентрацию железа, меди, цинка и марганца в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации для увеличения образования биомассы. Во втором варианте увеличивают концентрацию цинка и марганца и необязательно уменьшают концентрацию железа и меди в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта для увеличения продукции G0, G1 или G2 гликоформы рекомбинантного гликопротеина. В третьем варианте (A) уменьшают концентрацию железа, меди, цинка и марганца или (B) увеличивают концентрацию железа и меди и уменьшают концентрацию цинка и марганца в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации для увеличения продукции гликоформы рекомбинантного гликопротеина с высоким содержанием маннозы. Изобретения позволяют воздействовать на производство зрелых N-гликозилированных гликопротеинов и/или незрелых N-гликозилированных гликопротеинов, зрелых нефукозилированных гликопротеинов и/или незрелых нефукозилированных гликопротеинов и/или благоприятствовать образованию биомассы. 3 н. и 11 з.п. ф-лы, 16 ил., 13 табл., 4 пр.
1. Способ получения рекомбинантного гликопротеина в условиях ферментационной культуры в СНО-клетке, включающий увеличение концентрации каждого из металлов, таких как железо, медь, цинк и марганец, в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации для увеличения образования биомассы, где увеличение концентрации каждого из металлов, таких как железо, медь, цинк и марганец, в культуральной среде относится к соответствующей концентрации железа, меди, цинка и марганца в культуральной среде, применяемой непосредственно на предшествующей фазе культивирования, и где после увеличения концентрация каждого из металлов, таких как железо, цинк и марганец, в культуральной среде составляет:
(а) железо: по меньшей мере 15 мкМ и приблизительно до 80 мкМ;
(б) медь: по меньшей мере 0,3 мкМ и приблизительно до 2,5 мкМ;
(в) цинк: по меньшей мере 20 мкМ и приблизительно до 50 мкМ; и
(г) марганец: по меньшей мере 0,01 мкМ и приблизительно до 3 мкм;
где объем биомассы, полученной в культуральной среде, увеличен по меньшей мере на 5% по сравнению с процессом ферментации, при котором концентрации каждого из металлов, таких как железо, медь, цинк и марганец, в культуральной среде не увеличены в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации.
2. Способ по п. 1, в котором экспрессируемый гликопротеин представляет собой зрелый N-гликозилированный гликопротеин, зрелый нефукозилированный гликопротеин или незрелый нефукозилированный гликопротеин.
3. Способ по п. 1, в котором углеводный фрагмент экспрессируемого гликопротеина имеет G0-, G1- или G2-структуру.
4. Способ по п. 1, в котором для увеличения концентрации биомассы концентрации регулируют так, чтобы они составляли:
(а) железо: от 15 мкМ до 80 мкМ;
(б) медь: от 0,3 мкМ до 2,5 мкМ;
(в) цинк: от 20 мкМ до 50 мкМ; и
(г) марганец: от 0,01 мкМ до 3 мкМ.
5. Способ получения рекомбинантного гликопротеина в условиях ферментационной культуры в СНО-клетке, где способ включает увеличение концентрации каждого из металлов, таких как цинк и марганец, и необязательное уменьшение концентрации железа и меди в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации для увеличения продукции G0, G1 или G2 гликоформы рекомбинантного гликопротеина, где увеличение концентраций каждого из металлов, таких как цинк и марганец, и необязательное уменьшение концентрации железа и меди в культуральной среде относится к соответствующей концентрации железа, меди, цинка и марганца в культуральной среде, применяемой непосредственно на предшествующей фазе культивирования, и где после увеличения концентрации цинка и марганца их концентрация в культуральной среде составляет:
(а) цинк: по меньшей мере 20 мкМ и приблизительно до 50 мкМ; и
(б) марганец: по меньшей мере 0,01 мкМ и приблизительно до 3 мкМ; и
где после необязательного уменьшения концентрации железа и меди их концентрация в культуральной среде составляет:
(в) железо: менее чем 25 мкМ; и
(г) медь: менее чем 0,1 мкМ;
где G0, G1 или G2 гликоформа экспрессируемого гликопротеина увеличена по меньшей мере на 5% по сравнению с процессом ферментации, при котором концентрация каждого из металлов, таких как цинк и марганец, не увеличена и необязательно концентрация каждого из металлов, таких как железо и медь, не уменьшена в культуральной среде в начале или в процессе как фазы роста, так и фазы образования продукта процесса ферментации.
6. Способ по п. 5, в котором для повышения зрелости экспрессируемого гликопротеина концентрации регулируют так, чтобы они составляли:
(а) железо: от 0 мкМ до 25 мкМ;
(б) медь: от 0 мкМ до 0,1 мкМ;
(в) цинк: от 20 мкМ до 50 мкМ; и
(г) марганец: от 0,01 мкМ до 3 мкМ.
7. Способ по п. 5, в котором для повышения зрелости экспрессируемого гликопротеина концентрации регулируют так, чтобы они составляли:
(а) цинк: от 20 мкМ до 50 мкМ; и
(б) марганец: от 0,01 мкМ до 3 мкМ.
8. Способ получения рекомбинантного гликопротеина в условиях ферментационной культуры в СНО-клетке, где способ включает:
(A) уменьшение концентрации каждого из металлов, таких как железо, медь, цинк и марганец; или
(B) увеличение концентраций железа и меди и уменьшение концентраций цинка и марганца;
в свободной от сыворотки культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации для увеличения продукции гликоформы рекомбинантного гликопротеина с высоким содержанием маннозы, где уменьшение концентрации в культуральной среде согласно подпункту (А) и увеличение и уменьшение концентраций в культуральной среде согласно подпункту (В) относится к соответствующей концентрации железа, меди, цинка и марганца в культуральной среде, применяемой непосредственно на предшествующей фазе культивирования, и где:
для (А) после уменьшения концентрации железа, цинка и марганца концентрация в культуральной среде составляет:
(а) железо: менее чем 35 мкМ;
(б) медь: менее чем 1 мкМ;
(а) цинк: менее чем 20 мкМ; и
(б) марганец: менее чем 0,01 мкМ; или
для (В) после увеличения концентрации железа и меди и уменьшения концентрации цинка и марганца концентрация в культуральной среде составляет:
(а) железо: по меньшей мере 15 мкМ и приблизительно до 80 мкМ;
(б) медь: по меньшей мере 0,3 мкМ и приблизительно до 2,5 мкМ;
(в) цинк: менее чем 20 мкМ; и
(г) марганец: менее чем 0,01 мкМ;
где гликоформа экспрессируемого гликопротеина с высоким содержанием маннозы увеличена по меньшей мере на 5% по сравнению с процессом ферментации, при котором концентрация каждого из металлов, таких как железо, медь, цинк или марганец, не уменьшена в соответствии с подпунктом (А) в культуральной среде или концентрации железа и меди не увеличены и концентрации цинка и марганца не уменьшены в соответствии с подпунктом (В) в культуральной среде в начале и/или в процессе как фазы роста, так и фазы образования продукта процесса ферментации.
9. Способ по п. 8, в котором для повышения получения незрелых нефукозилированных гликопротеинов концентрации регулируют так, чтобы они составляли: либо
(I)
(а) железо: от 0 мкМ до 35 мкМ;
(б) медь: от 0 мкМ до 1 мкМ;
(в) цинк: от 0 мкМ до 20 мкМ и
(г) марганец: от 0 мкМ до 0,01 мкМ, или
(II)
(а) железо: от 15 мкМ до 80 мкМ;
(б) медь: от 0,3 мкМ до 2,5 мкМ;
(в) цинк: от 0 мкМ до 20 мкМ и
(г) марганец: от 0 мкМ до 0,01 мкМ.
10. Способ по одному из пп. 1-9, в котором гликопротеин является экзогенным или эндогенным для эукариотической клетки, в котором необязательно гликопротеин представляет собой структурный гликопротеин, гормон, антитело или фермент.
11. Способ по п. 10, в котором гликопротеин представляет собой антитело, в котором необязательно антитело представляет собой терапевтическое или диагностическое антитело, необязательно химерное, гуманизированное или человеческое антитело.
12. Способ по одному из пп. 1-11, в котором повышение концентрации любого металла, а именно железа, меди, цинка и марганца или их всех, в культуральной среде обеспечивают путем дополнения среды, в которой культивируют клетки, и/или путем расщепления клеток посредством переноса в свежую среду, дополненную любым металлом, а именно железом, медью, цинком и марганцем, или ими всеми.
13. Способ по одному из пп. 1-11, в котором снижение биодоступной концентрации любого металла, а именно железа, меди, цинка и марганца или их всех, обеспечивают путем образования комплекса железа, меди, цинка и марганца с хелатором и/или путем посева клеток на свежую среду, содержащую любой металл, а именно железо, медь, цинк и марганец, или их все, в пониженной концентрации по сравнению со средой непосредственно на предшествующей фазе культивирования.
14. Способ по одному из пп. 1-13, в котором концентрации каждого металла, а именно железа, меди, цинка и марганца, в культуральной среде регулируют в процессе культивирования сначала так, чтобы оказывать благоприятное воздействие на образование биомассы, а затем так, чтобы повышать зрелость экспрессируемых N-гликопротеинов или повышать получение незрелых нефукозилированных гликопротеинов.
| Станок для гибки монтажных петель и анкеров | 1982 |
|
SU1096017A1 |
| HAN Y | |||
| et al | |||
| "Cultivation of Recombinant Chinese hamster ovary cells grown as suspended aggregates in stirred vessels" | |||
| Journal of Bioscience and Bioengineering, 2006, 102(5), 430-435 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| ПОЛУЧЕНИЕ ГЛИКОПРОТЕИНОВ | 2007 |
|
RU2463345C2 |
| GAWLITZEK M | |||
| Estacio M, Furch T, Kiss R | |||
| Identification of cell culture conditions | |||

