Область техники, к которой относится изобретение
Настоящее изобретение относится к способу специфического и селективного изменения нуклеотидной последовательности в специфическом сайте ДНК в целевой клетке посредством введения олигонуклеотида в эту клетку. Результатом этого является направленная замена одного или нескольких нуклеотидов, вследствие которой последовательность целевой ДНК трансформируется в последовательность олигонуклеотида там, где они различались. В более специфичном понимании изобретение относится к направленной замене нуклеотидов с использованием модифицированных олигонуклеотидов. Изобретение дополнительно относится к олигонуклеотидам и наборам. Изобретение также относится к применению способа.
Уровень техники изобретения
Генетическая модификация представляет собой процесс создания осмысленных изменений в генетическом материале живых клеток с целью модификации одного или нескольких генетически кодируемых биологических свойств этой клетки или организма, частью которого эта клетка является или в котором она может регенерироваться. Эти изменения могут иметь форму делеции частей генетического материала, добавления экзогенного генетического материала или изменений в существующей нуклеотидной последовательности генетического материала. Способы генетической модификации эукариотических организмов известны уже более 20 лет и нашли широкое применение в клетках растений, человека, животных и микроорганизмах для улучшения в таких сферах, как сельское хозяйство, здравоохранение, качество пищи и защита окружающей среды. Распространенные способы генетической модификации состоят из добавления фрагментов экзогенной ДНК к геному клетки, вследствие этого приобретающей новое свойство, благодаря чему эта клетка или организм, в составе которого она находится, улучшают и усиливают свои свойства, кодируемые существующими генами (включая такие прикладные варианты применения, в которых экспрессия существующих генов будет подавляться). Хотя многие примеры такого рода эффективны для получения желаемых свойств, тем не менее известные способы не отличаются большой точностью, поскольку они не позволяют контролировать, в какие положения генома встраиваются фрагменты экзогенной ДНК (то есть конечные уровни экспрессии непредсказуемы), поэтому желаемый эффект будет проявляться неуправляемо, относительно природных свойств, кодируемых первоначальным и хорошо сбалансированным геномом. И наоборот, способы генетической модификации, которые приводят к добавлению, делеции или изменению нуклеотидов в заранее определенных локусах генома, позволяют добиться точной модификации существующих генов.
Управляемая олигонуклеотидами направленная замена нуклеотидов (TNE, иногда ODTNE) представляет собой способ, основанный на доставке в эукариотическое клеточное ядро синтетических олигонуклеотидов (молекул, состоящих из коротких участков, организованных по типу нуклеотидных последовательностей, которые напоминают ДНК по свойствам спаривания оснований в модели Крика-Уотсона, но могут иметь химические отличия от ДНК (Alexeev and Yoon, Nature Biotechnol. 16: 1343, 1998; Rice, Nature Biotechnol. 19: 321, 2001; Kmiec, J. Clin. Invest. 112: 632, 2003). Преднамеренно конструируя ошибочный нуклеотид в гомологической последовательности олигонуклеотида, можно добиться его встраивания в последовательность геномной ДНК. Этот способ позволяет изменить один или не более чем несколько нуклеотидов в существующих локусах, но его можно применять и для создания стоп-кодонов в существующих генах, что приводит к блокированию их функции, или для создания изменений в кодонах, приводящих к тому, что в кодируемых этими генами белках меняется аминокислотная последовательность (белковая инженерия).
Направленная замена нуклеотидов (TNE) была описана в растительных, животных и дрожжевых клетках. Первые сообщения о TNE были посвящены так называемой химере, которая состояла из самокомплементарного олигонуклеотида, который сконструирован для интеркаляции в целевой хромосомный сайт. Химера содержит ошибочно спариваемый нуклеотид, который образует шаблон для введения мутации в хромосомную цель. Для того чтобы провести отбор на события TNE в большинстве исследований предпринимались попытки введения единичной замены нуклеотида в эндогенный (изначально присущий биологическому виду) ген, что приводит к появлению устойчивости к гербицидам. Первые примеры использования химер приведены в литературе для клеток человека (см. для обзора ссылку Rice et al. Nat. Biotech. 19: 321-326). Применение химер также было успешным для таких видов растений, как табак, рис и кукуруза (Beetham et al. 1999 Proc. Natl. Acad. Sci. USA 96: 8774-8778; Kochevenko et al. 2003 Plant Phys. 132: 174-184; Okuzaki et al. 2004 Plant Cell Rep. 22: 509-512). Однако было обнаружено, что активность химер воспроизводится с трудом, в связи с чем была протестирована активность однонитевых (ss) олигонуклеотидов на TNE. Более воспроизводимые результаты были получены на клетках пшеницы, дрожжей и человека (Liu et al. 2002 Nuc. Acids Res.: 30; 2742-2750; review, Parekh-Olmedo et al. 2005 Gene Therapy 12: 639-646; Dong et al. 2006 Plant Cell Rep. 25: 457-65).
Несколько групп исследователей показали, что TNE также можно выявлять, используя общие белковые экстракты клеток. Такие анализы на активность TNE называются бесклеточными анализами (Cole-Strauss et al. 1999 Nucleic Acids Res. 27: 1323-1330; Gamper et al. 2000 Nucleic Acids Res. 28, 4332-4339; Kmiec et al. 2001 Plant J. 27: 267-274; Rice et al. 2001 40: 857-868). Анализ организован следующим образом. Плазмиду, содержащую два бактериальных гена устойчивости к антибиотикам (канамицину и карбенициллину), мутируют так, что один из генов устойчивости к антибиотикам (например, к канамицину) содержит внутри рамки считывания стоп-кодон из-за замены одного нуклеотида (например, TAT на TAG). Затем эту мутировавшую плазмиду инкубируют с общим клеточным белком и однонитевым олигонуклеотидом, предназначенным для коррекции стоп-кодона в гене устойчивости к антибиотику. Белки, необходимые для TNE, присутствуют в клеточном экстракте и используют олигонуклеотид для изменения стоп-кодона в гене устойчивости к антибиотику, восстанавливая резистентный фенотип. Затем плазмидную ДНК очищают от реакционной смеси и трансформируют в E.coli. Затем бактерии высевают на питательную среду, содержащую канамицин, и число выросших бактериальных колоний отражает число событий TNE с эффектом восстановления. Эффективность электропорации вычисляют, подсчитывая число колоний, выросших на среде, содержащей карбенициллин. Эффективность TNE можно выразить количественно, рассчитав отношение восстановленных плазмид к общему количеству трансформированных плазмид.
В таком эксперименте олигонуклеотид производит замену, меняя TAG на TAC. Кроме того, для изучения возможности применения олигонуклеотидов с целью получения вставок единичного нуклеотида можно использовать бесклеточную систему. Можно создать плазмиды, с делецией одного нуклеотида в гене устойчивости к антибиотику, которая порождает сдвиг рамки считывания. В бесклеточном анализе делецию компенсируют добавлением нуклеотида, опосредованного олигонуклеотидом.
Самой большой проблемой, с которой приходится сталкиваться, применяя TNE в клетках высших организмов, например растений, является низкая эффективность, на которую указывают все опубликованные до настоящего времени данные. Для кукурузы Zhu et al. (2000 Nature Biotech. 18: 555-558) приводят частоту конверсии 1×10-4. В последующих исследованиях на табаке (Kochevenko et al. 2003 Plant Phys. 132: 174-184) и рисе (Okuzaki et al. 2004 Plant Cell Rep. 22: 509-512) приведены данные о частоте конверсии 1×10-6 и 1×10-4, соответственно. Эти частоты, по-прежнему, слишком низки для практического применения TNE.
Точная репликация ДНК является одним из ключевых критериев, опосредующих поддержание стабильности генома и обеспечивающих передачу генетической информации, содержащейся в ДНК, от поколения к поколению без мутаций. Многочисленные ошибки возникают из-за повреждения в родительской нити ДНК или могут быть вызваны агентами, реагирующими с основаниями ДНК (УФ-свет, токсины окружающей среды). Для предупреждения или исправления этих мутаций каждый организм должен поддерживать защиту. Считается, что система репарации ошибочного спаривания оснований (ММR) распознает и исправляет несогласованные или неспаренные основания, появляющиеся в процессе репликации ДНК, систематически отслеживая повреждения ДНК и предупреждая рекомбинацию между неидентичными последовательностями (Fedier and Fink, 2004 Int. J. Oncol. 2004; 24(4):1039-47), а также внося вклад в точность репликации ДНК в живых клетках.
Сущность TNE была описана во многих патентных заявках Kmiec, среди прочего в документах WO 0173002, WO 03/027265, WO 01/87914, WO 99/58702, WO 97/48714, WO 02/10364. В заявке WO 01/73002 рассматривается вопрос о том, что низкая эффективность изменения гена при использовании немодифицированных олигонуклеотидов ДНК, по-видимому, в значительной степени является результатом разрушения донорных олигонуклеотидов нуклеазами, присутствующими в реакционной смеси или в целевой клетке. Для решения этой проблемы предложено включать модифицированные нуклеотиды, придающие получаемым в конечном результате олигонуклеотидам устойчивость к воздействию нуклеаз. Типичные примеры включают нуклеотиды с фосфоротиоатными связями, 2'-O-метил-аналоги или закрытые (замкнутые) нуклеиновые кислоты (LNA). Предпочтительно, чтобы такие модификации располагались на концах олигонуклеотида, оставляя незатронутым центральный домен ДНК, окружающий целевое основание. Кроме того, в публикации оговорено, что имеют место химические взаимодействия между конвертирующим олигонуклеотидом и белками, участвующими в конверсии. Эффект таких химических взаимодействий по созданию устойчивых к нуклеазам концов при использовании других модификаций помимо LNA, фосфоротиоатных связей или включения 2'-O-метил-аналога в олигонуклеотид предсказать невозможно, поскольку белки, вовлеченные в процесс изменения, и их химическое взаимодействие с олигонуклеотидными заместителями еще неизвестны и, согласно авторам WO 0173002, не могут быть предсказаны.
Поскольку эффективность современных методов ODTNE относительно невелика (как было установлено ранее, она составляет от 10-6 до 10-4, несмотря на сообщения о высокой степени доставки олигонуклеотида, порядка 90%), в данной области знаний ощущается явная потребность в разработке более эффективных способов TNE. В соответствии с этим авторы настоящего изобретения поставили перед собой задачу улучшить существующую технологию TNE.
Описание изобретения
Авторы настоящего изобретения недавно обнаружили, что при включении в донорный олигонуклеотид для TNE таких нуклеотидов, которые способны прочнее связываться с акцепторной ДНК, чем соответствующие немодифицированные нуклеотиды A, C, T или G, можно значительно повысить частоту TNE. Не прибегая к теории, авторы настоящего изобретения полагают, что при включении в донорный олигонуклеотид модифицированных нуклеотидов этот донорный олигонуклеотид прочнее связывается с акцепторной ДНК, следовательно, повышает пропорцию TNE. Авторы настоящего изобретения обнаружили, что олигонуклеотиды, содержащие одну или несколько LNA в положениях, близких, но не прилегающих (непосредственно) к точке ошибочного спаривания, т.е. расположенные от точки ошибочного спаривания по меньшей мере на расстоянии одного нуклеотида, значительно увеличивают эффективность TNE.
С этой целью были проведены исследования, направленные на оценку влияния олигонуклеотидов, включающих одну или несколько LNA в различных положениях (в пределах олигонуклеотида), на частоту TNE в бесклеточной системе. Активность таких олигонуклеотидов в отношении TNE сравнивали с соответствующей активностью олигонуклеотидов, представляющих собой фрагменты нормальной ДНК. Было обнаружено, что олигонуклеотиды, которые содержат одну или несколько LNA в положениях, отстоящих по меньшей мере на один нуклеотид от точки ошибочного спаривания, повышают эффективность TNE как по замещениям, так и по вставкам (инсерциям) в бесклеточном анализе до такого уровня, которого до сих пор не удавалось достичь. Олигонуклеотиды, содержащие LNA, по результатам бесклеточного анализа оказались до 10 раз эффективнее по сравнению с олигонуклеотидами, полученными из нормальной ДНК. Также было обнаружено, что эффективность TNE можно улучшить при увеличении числа LNA вставок в олигонуклеотиде, если они будут отстоять друг от друга по меньшей мере на 2 нуклеотида, предпочтительнее по меньшей мере на 3, еще предпочтительнее по меньшей мере на 4. Кроме того, было обнаружено, что наблюдаемое улучшение не зависит от локуса, а это указывает на то, что олигонуклеотиды, предлагаемые изобретением, с LNA в определенных положениях по отношению к точке ошибочного спаривания, способны обеспечить повышенную частоту TNE вне зависимости от биологического вида, например, как в растительных, так и в животных клетках.
Таким образом, настоящее изобретение основано на идее, обладающей изобретательным уровнем о том, что желаемая направленная замена нуклеотидов достижима при использовании частично модифицированных при помощи LNA олигонуклеотидов (т.е. модифицированных не более чем на 75%, предпочтительнее не более чем на 50%). Расположение, тип и количество модификаций олигонуклеотида могут варьироваться в тех пределах, которые будут раскрыты здесь несколько ниже.
Таким образом, в одном из аспектов настоящее изобретение обеспечивает олигонуклеотиды, модифицированные LNA. Модифицированные LNA ss-олигонуклеотиды можно применять для введения специфических генетических изменений в растительные и животные клетки или в клетки человека. Изобретение применимо в области биомедицинских исследований, в сельском хозяйстве, а также для конструирования специфически мутированных растений и животных, включая человека. Изобретение также применимо в области медицины и генной терапии.
Последовательность олигонуклеотида, предлагаемого изобретением, гомологична целевой нити (мишени) за исключением той части, которая содержит ошибочно спаренное основание и вносит замену основания в целевую нить (мишень). В целевую последовательность вводится ошибочно спаренное основание. Можно улучшить эффективность TNE (или повысить степень успешного включения желательного нуклеотида в нужное положение дуплекса ДНК), манипулируя модификацией нуклеотидов (по сравнению с обычными основаниями A, C, T или G) или, более специфично, манипулируя расположением и количеством LNA-модификаций олигонуклеотида, который вносит ошибочно спаренное основание.
Еще один аспект изобретения состоит в способе направленного изменения родительской нити ДНК (первой нити, второй нити) посредством контактирования родительского дуплекса ДНК с олигонуклеотидом, который содержит по меньшей мере один ошибочно спаренный нуклеотид по сравнению с родительской нитью, причем донорный олигонуклеотид содержит участок, модифицированный LNA в специфических положениях для придания большей способности связывания по сравнению с родительской (акцепторной) нитью в присутствии белков, способствующих направленной замене нуклеотидов.
Таким образом, изобретательская сущность изобретения связана с улучшением способности связывания интеркалирующего олигонуклеотида (который иногда называют донорным), модифицированного LNA-нуклеотидами, по сравнению с немодифицированным интеркалирующим олигонуклеотидом, при том условии, что модификация LNA расположена в одном или нескольких положениях, не прилегающих к точке ошибочного спаривания.
Подробное описание изобретения
В одном из аспектов изобретение относится к олигонуклеотиду для направленного изменения последовательности двухнитевой ДНК, где двухнитевая последовательность ДНК содержит первую последовательность ДНК и вторую последовательность ДНК, которая комплементарна первой последовательности ДНК, олигонуклеотид содержит домен, способный гибридизироваться с первой последовательностью ДНК, где домен содержит по меньшей мере одну ошибку спаривания по отношению к первой последовательности ДНК, и где олигонуклеотид содержит по меньшей мере один участок, который содержит по меньшей мере один модифицированный нуклеотид, обладающий большей способностью связывания по сравнению с природными A, C, T или G, и где предпочтительно по меньшей мере один модифицированный нуклеотид более сильно связывается с нуклеотидом в противоположном положении в первой последовательности ДНК, по сравнению с природным нуклеотидом, комплементарным нуклеотиду в противоположном положении первой последовательности ДНК, где по меньшей мере один модифицированный нуклеотид представляет собой LNA, расположенную на расстоянии по меньшей мере одного нуклеотида от по меньшей мере одной точки ошибочного спаривания, и предпочтительно, где олигонуклеотид содержит по меньшей мере приблизительно 75% модифицированных нуклеотидов.
В одном из аспектов изобретение относится к олигонуклеотиду, модифицированному LNA, для направленного изменения последовательности двухнитевой ДНК. Двухнитевая (дуплексная) ДНК содержит первую последовательность ДНК и вторую последовательность ДНК. Вторая последовательность ДНК комплементарна первой последовательности ДНК и спаривается с ней, образуя дуплекс. Олигонуклеотид содержит домен, в котором имеется по меньшей мере одно ошибочное спаривание оснований по отношению к двухнитевой последовательности ДНК, подлежащей изменению. Предпочтительно, чтобы домен был частью олигонуклеотида, который комплементарен первой нити, включая по меньшей мере одно неправильно спаренное основание.
Предпочтительно, чтобы точка ошибочного спаривания в домене имела отношение к первой последовательности ДНК. Олигонуклеотид содержит участок, который модифицирован по меньшей мере одной LNA для приобретения большей аффинности связывания, чем соответствующий участок второй последовательности ДНК. Предпочтительно, чтобы по меньшей мере один модифицированный нуклеотид представлял собой LNA и был расположен на расстоянии по меньшей мере одного нуклеотида от по меньшей мере одной точки ошибочного спаривания, более предпочтительно, чтобы олигонуклеотид содержал не более 75% нуклеотидов, модифицированных LNA.
Домен, который содержит ошибочное спаривание, и участок, содержащий модифицированный нуклеотид (нуклеотиды), могут перекрываться. Таким образом, в некоторых вариантах осуществления изобретения домен, содержащий ошибочное спаривание, расположен в другом положении в пределах олигонуклеотида, чем участок, который предполагается модифицировать. В некоторых вариантах осуществления изобретения домен включает этот участок. В некоторых вариантах осуществления изобретения участок может включать домен. В некоторых вариантах осуществления изобретения домен и участок, подлежащий модификации, расположены в одном и том же положении в пределах олигонуклеотида и имеют одинаковую длину, то есть они совпадают и по длине, и по положению. В некоторых вариантах осуществления изобретения в домене может содержаться более одного участка.
Для настоящего изобретения это означает, что часть олигонуклеотида, содержащая ошибочное спаривание, которое должно быть включено в дуплекс ДНК, может располагаться в другом или смещенном положении по отношению к модифицированной части олигонуклеотида. В частности, в некоторых вариантах осуществления изобретения система репарации клетки или по меньшей мере белки, вовлеченные в эту систему, или по меньшей мере белки, вовлеченные в TNE, определяют, какая нить ДНК содержит ошибку спаривания, а какая нить должна быть использована как шаблон для коррекции ошибки.
В некоторых вариантах осуществления изобретения олигонуклеотид содержит участок, который содержит по меньшей мере один, предпочтительнее по меньшей мере 2, еще предпочтительнее по меньшей мере 3 модифицированных нуклеотида. В некоторых вариантах осуществления изобретения участок олигонуклеотида может содержать более чем 4, 5, 6, 7, 8, 9 или 10 LNA-модифицированных нуклеотидов.
В некоторых вариантах осуществления изобретения по меньшей мере одна LNA расположена на расстоянии не более 10 нуклеотидов, предпочтительнее не более 8 нуклеотидов, еще предпочтительнее не более 6 нуклеотидов, но еще предпочтительнее не более 4, 3 или 2 нуклеотидов от точки ошибочного спаривания. В более предпочтительном варианте осуществления изобретения по меньшей мере одна LNA расположена на расстоянии 1 нуклеотида от точки ошибочного спаривания, то есть между точкой ошибочного спаривания и LNA находится один нуклеотид. В некоторых вариантах осуществления изобретения, относящихся к таким олигонуклеотидам, которые содержат более одной LNA, каждая LNA расположена на расстоянии по меньшей мере одного нуклеотида от точки ошибочного спаривания. В предпочтительном варианте осуществления изобретения LNA расположены не рядом, а отделены друг от друга по меньшей мере одним нуклеотидом, предпочтительнее двумя или тремя нуклеотидами. В некоторых вариантах осуществления изобретения, когда олигонуклеотид модифицирован двумя или более (четным числом) LNA, эти модификации расположены (приблизительно) на одинаковом расстоянии от точки ошибочного спаривания. Иначе говоря, предпочтительно, чтобы модификации LNA располагались симметрично по отношению к точке ошибочного спаривания. Например, в предпочтительном варианте осуществления изобретения две модификации LNA расположены симметрично по отношению к точке ошибочного спаривания и отделены от нее одним нуклеотидом с каждой стороны (то есть отделены друг от друга тремя нуклеотидами).
В некоторых вариантах осуществления изобретения не более 50% модифицированных нуклеотидов олигонуклеотида представляют собой производные LNA, т.е. обычные A, C или G в олигонуклеотиде замещены соответствующими аналогами LNA. Предпочтительно, чтобы доля таких нуклеотидов составляла не более 40%, предпочтительнее не более 30%, еще предпочтительнее не более 20% и наиболее предпочтительно не более 10%.
В некоторых вариантах осуществления изобретения может быть введено более одной ошибки спаривания либо одновременно, либо последовательно. Олигонуклеотид может вместить более одной ошибки спаривания, расположенных или по соседству друг с другом, или в разных положениях (в пределах олигонуклеотида). С этой целью олигонуклеотид можно адаптировать для вмещения второго набора LNA в соответствии с изложенными здесь принципами, но при условии, что они не будут мешать друг другу в отношении улучшенной способности связывания вследствие специфической конформации LNA в олигонуклеотиде, т.е. предпочтительно, чтобы они были расположены поблизости от точки ошибочного спаривания (на расстоянии 1 нуклеотида от нее). В некоторых вариантах осуществления изобретения олигонуклеотид может включать два, три, четыре или более ошибочно спаренных нуклеотида, которые могут располагаться как по соседству, так и на некотором расстоянии друг от друга (не прилегая). Для вмещения этого олигонуклеотид может содержать дополнительные домены и сегменты, в частности он может содержать несколько сегментов. В некоторых вариантах осуществления изобретения олигонуклеотид может включать потенциальную вставку для инсерции в акцепторную нить. Такая вставка может иметь разную длину (более чем от пяти и до ста нуклеотидов). Подобным образом в некоторых вариантах осуществления изобретения могут быть введены делеции примерно такой же длины (от 1 до 100 нуклеотидов).
В дальнейшем аспекте изобретения конструкция олигонуклеотида может быть достигнута посредством:
- определения последовательности акцепторной нити или по меньшей мере участка последовательности по соседству с подлежащим замене нуклеотидом. В типичном случае это может быть фрагмент, состоящий по меньшей мере из 10, предпочтительнее из 15, 20, 25 или 30 нуклеотидов, прилегающих к точке ошибочного спаривания, предпочтительно с обеих сторон (например, GGGGGGXGGGGGG, где X является ошибочно спаренным основанием);
- конструирования донорного олигонуклеотида, комплементарного одному или обоим участкам, прилегающим к точке ошибочного спаривания и содержащим желаемый нуклеотид для замены (например, CCCCCCYCCCCCC);
- получения (например, посредством синтеза) донорного олигонуклеотида с модификациями LNA в желаемых положениях. Модификации могут широко варьироваться в зависимости от обстоятельств. Примерами могут быть CCCmCCmCYCCmCCCmC, CCCmCCCYCCCmCCC, CCCCCCYCCCmCmCmCm, CmCmCmCmCmCYCCCCCC, CCCCCmCYCCCmCCCCC и так далее, где Cm означает модифицированный LNA нуклеотидный остаток. Для другой акцепторной последовательности, например, ATGCGTACXGTCCATGAT, можно сконструировать соответствующие донорные олигонуклеотиды, например, TACGCALGYCLGGTACTA (L=LNA) с таким же варьированием модификаций, как описано выше.
- контактирования ДНК, подлежащей модификации, с донорным олигонуклеотидом в присутствии белков, способных провести направленную замену нуклеотида, например и в частности, белков, вовлеченных в механизм репарации ошибок ДНК в клетке.
Не связываясь с теорией, авторы полагают, что улучшенная аффиность (сродство) связывания повышает вероятность того, что олигонуклеотид находит свою цель и остается связанным с ней, усиливая эффективность TNE. Лучшую аффинность связывания могут придавать многие химические модификации сахарного скелета или основания. Однако настоящее изобретение сфокусировано на олигонуклеотидах, модифицированных LNA, и основано на том обстоятельстве, что активность таких модификаций в отношении TNE зависит от их положения в пределах нуклеотида.
Закрытая (замкнутая) нуклеиновая кислота (LNA) является аналогом ДНК с очень интересными свойствами для применения в антисмысловой генотерапии. LNA представляют собой аналоги бициклических и трициклических нуклеозидов и нуклеотидов, а также олигонуклеотиды, которые содержат такие аналоги. Основные структурные и функциональные характеристики LNA и родственных аналогов раскрыты в различных публикациях и патентах, включая WO 99/14226, WO 00/56748, WO 00/66604, WO 98/39352, патент США № 6043060 и патент США № 6268490 (все перечисленные документы включены сюда в качестве ссылки во всей своей полноте).
В частности, LNA сочетает способность различать правильные и неправильные цели (высокую специфичность) с очень высокой биологической стабильностью (низкий круговорот) и с беспрецедентной аффинностью (очень большая сила связывания с целью). Фактически, увеличение аффинности, зарегистрированное в связи с применением LNA, оставляет далеко позади все известные до сих пор показатели аффинности, переводя их в диапазон низких и умеренных.
LNA представляет собой аналог РНК, в котором рибоза структурно связана метиленовым мостиком между атомами 2'-кислорода и 4'-углерода. Этот мостик ограничивает пластичность кольца рибофуранозы и запирает структуру в жесткое бициклическое образование. Этот так называемый N-тип (или 3'-эндо) конформации приводит к увеличению Tm LNA-содержащих дуплексов и, вследствие этого, к более высокой аффинности связывания и к более высокой специфичности. Спектральные исследования методом ЯМР фактически продемонстрировали закрытую конформацию N-типа для сахара LNA, но, кроме того, позволяли обнаружить, что мономеры LNA способны скручивать соседние немодифицированные нуклеотиды, перевод их в конформацию N-типа. Важно, что благоприятные характеристики LNA происходят не за счет утраты других важных свойств, что часто наблюдается у других аналогов нуклеиновых кислот.
LNA может свободно перемешиваться с любыми другими химическими составами, входящими в генеральную совокупность аналогов ДНК. Основания LNA могут встраиваться в олигонуклеотиды как короткие последовательности чистой LNA или как более длинные химеры типа LNA/ДНК. Вставки LNA могут находиться в разных положениях: внутри олигонуклеотида, на 3'- или на 5'-конце. Однако вследствие жесткой бициклической конформации остатки LNA иногда нарушают спиралевидный виток нитей нуклеиновой кислоты. Поэтому в целом менее предпочтительно конструировать олигонуклеотид с двумя и более прилегающими друг к другу остатками LNA. Предпочтительно, чтобы остатки LNA были разделены по меньшей мере одним (модифицированным) нуклеотидом, который не нарушает спиралевидный виток, например обычным нуклеотидом (A, C, T или G).
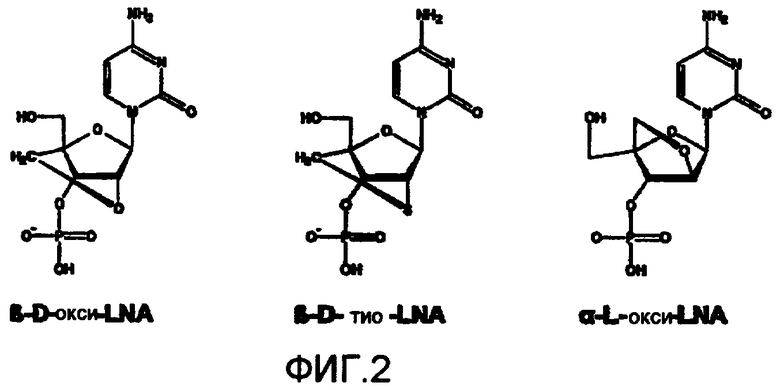
Первоначально разработанный и предпочтительный мономер LNA (β-D-окси-LNA) был модифицирован в новые мономеры LNA. Новый мономер α-L-окси-LNA демонстрирует исключительно высокую стабильность против активности 3'-эндонуклеазы, кроме того, он обладает большей мощностью и большей гибкостью по сравнению с β-D-окси-LNA в конструировании сильнодействующих антисмысловых олигонуклеотидов. Кроме того, можно использовать ксило-LNA и L-рибо-LNA, как это раскрыто в документах WO 9914226, WO 00/56748, WO 00/66604. В настоящем изобретении любая LNA вышеуказанных типов эффективна в достижении целей, на которые направлено это изобретение, т.е. в повышении эффективности TNE с предпочтительным использованием аналогов β-D-LNA.
В уровне техники по TNE модификации LNA вошли в перечень возможных олигонуклеотидных модификаций как альтернатива применению химерных молекул для TNE. Однако в данной области техники до сих пор нет указаний, по которым можно было бы предположить, что модифицированные LNA однонитевые олигонуклеотиды ДНК значительно повышают эффективность TNE, доводя ее до такой степени, которая была обнаружена в настоящем изобретении при позиционировании LNA на расстоянии по меньшей мере одного нуклеотида от точки ошибочного спаривания и/или в том случае, когда олигонуклеотид содержит не более приблизительно 75% LNA (с округлением до ближайшего целого числа нуклеотидов).
Доставка олигонуклеотида может быть обеспечена посредством электропорации или других традиционных методик, позволяющих осуществлять доставку или в ядро, или в цитоплазму. Тестирование in vitro способа, предлагаемого настоящим изобретением, можно провести, используя бесклеточную систему, как это описано среди прочего в документах WO 01/87914, WO 03/027265, WO 99/58702, WO 01/92512.
При использовании в настоящем изобретении способность донорного олигонуклеотида повлиять на TNE зависит от типа, расположения и числа или относительного содержания модифицированных нуклеотидов, встроенных в донорный олигонуклеотид. Эту способность можно выразить количественно, например, нормализуя аффинность связывания (или энергию связывания (свободную энергию Гиббса)) между обычными нуклеотидами к 1, то есть для связок как AT, так и GC, аффинность связывания нормализована к 1. Для олигонуклеотидов, предлагаемых настоящим изобретением, относительная аффинность связывания (RBA)
каждого модифицированного нуклеотида составляет >1. Это иллюстрируется следующей формулой:
Здесь RBA означает общую относительную аффинность связывания, RBA (модифицированная) представляет собой суммарную аффинность связывания модифицированного олигонуклеотида длиной n нуклеотидов, а RBA (немодифицированная) соответствует суммарной относительной аффиности связывания немодифицированного олигонуклеотида длиной m нуклеотидов. Например, олигонуклеотид длиной 100 bp (пар оснований) содержит 10 модификаций, причем каждая из них имеет относительную аффинность связывания 1,1. В этом случае общий показатель RBA составит: RBA=[(10×1,1)+(90×1,0)]-(100×1,0)=1.
Обратите внимание на то, что определение RBA в принципе не зависит от длины сравниваемых нуклеотидных нитей. Однако при сравнении RBA для различных нитей предпочтительно, чтобы эти нити имели приблизительно одинаковую длину, или для сравнения были взяты участки, сопоставимые по длине. Обратите внимание на то, что показатель RBA не принимает во внимание, сгруппированы ли модификации в пределах нити вместе. Таким образом, более высокая степень модификации некой нити A по сравнению с нитью B означает, что RBA(A)>RBA(B). Для участков, расположенных выше и ниже, можно определить и использовать соответствующие (локальные) значения RBA. Для поправки на эффект положения модифицированного нуклеотида в величину RBA можно ввести взвешивающий коэффициент. Например, эффект модифицированного нуклеотида в донорном олигонуклеотиде, прилегающего к точке ошибочного спаривания, может быть больше по сравнению с эффектом модифицированного нуклеотида, отделенного от точки ошибочного спаривания пятью нуклеотидами. В контексте настоящего изобретения RBA (донора)>RBA (акцептора).
В некоторых вариантах осуществления изобретения величина RBA донора может превышать величину RBA акцептора по меньшей мере на 0,1. В некоторых вариантах осуществления изобретения величина RBA донора может превышать величину RBA акцептора по меньшей мере на 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,5, 2,0 или 2,5. Значения RBA можно получить при обычном анализе модифицированной аффинности связывания нуклеотида, например, при молекулярном моделировании, термодинамическом измерении и т.д. В альтернативном варианте их можно определить, измеряя различия Tm между модифицированной и немодифицированной нитями. По-другому RBA можно выражать в виде разности Tm между немодифицированной и модифицированной нитями, либо измеряя эти показатели, либо вычисляя их, пользуясь традиционными формулами для расчета Tm набора нуклеотидов, либо комбинируя измерения и вычисления.
В соответствии с изобретением донорные олигонуклеотиды могут содержать дополнительные модификации, направленные на улучшение характеристик гибридизации, например, чтобы донор проявлял повышенную аффинность к целевой нити ДНК, а интеркаляция донора проходила как можно проще. Донорный олигонуклеотид также может быть дополнительно модифицирован для приобретения большей устойчивости к нуклеазам, для стабилизации триплексной или квадруплексной структуры. Дополнительная модификация модифицированных LNA донорных олигонуклеотидов, предлагаемых изобретением, может включать фосфоротиоатную модификацию, замещения 2-OMe, применение добавочных LNA на 3'- и/или 5'-конце олигонуклеотида, PNA (пептидных нуклеиновых кислот), рибонуклеотидов и других оснований, которые модифицируют, предпочтительно усиливая, стабильность гибрида между олигонуклеотидом и акцепторной нитью.
Особенно полезными среди таких модификаций считаются PNA, которые представляют собой аналоги олигонуклеотидов с замещением дезоксирибозного каркаса олигонуклеотида пептидным каркасом. Один из таких пептидных каркасов сконструирован из повторных единиц N-(2-аминоэтил) глицина, соединенных амидными связями. Каждая субъединица пептидного каркаса прикреплена к нуклеиновому основанию (также обозначаемому как "основание"), которое может представлять собой природное основание, основание, не встречающееся в природе, или модифицированное основание. Олигомеры PNA специфически связывают свою последовательность с комплементарной ДНК или РНК, причем с более высокой аффинностью, чем обычные ДНК или РНК. В соответствии с этим результирующие дуплексы PNA/ДНК или PNA/РНК имеют более высокую температуру плавления (Tm). В дополнение к этому, Tm дуплексов PNA/ДНК или PNA/РНК намного менее чувствительна к концентрации соли, чем у дуплексов ДНК/ДНК или ДНК/РНК. Полиамидный каркас PNA также более устойчив к ферментативному разрушению. Синтез PNA описан, например, в документах WO 92/20702 и WO 92/20703, содержание которых включено сюда в качестве ссылки во всей полноте. Другие PNA проиллюстрированы, например, в документе WO 93/12129 и в патенте США № 5539082, выданном 23 июля 1996 г., и содержание этих документов включено сюда в качестве ссылки во всей полноте. В дополнение к этому, многие научные публикации описывают синтез PNA, а также их свойства и применение. См., например, ссылки Patel, Nature, 1993, 365, 490; Nielsen et al., Science, 1991, 254, 1497; Egholm, J. Am. Chem. Soc, 1992, 114, 1895; Knudson et al., Nucleic Acids Research, 1996, 24, 494; Nielsen et al., J. Am. Chem. Soc, 1996, 118, 2287; Egholm et al., Science, 1991, 254, 1497; Egholm et al., J. Am. Chem. Soc, 1992, 114, 1895 и Egholm et al., J. Am. Chem. Soc, 1992, 114, 9677.
Дополнительные полезные модификации олигонуклеотидов LNA, имеющие отношение к настоящему изобретению, также известны под названиями Super A и Super T и могут быть получены от компании Epoch Biosciences Germany. Эти модифицированные нуклеотиды содержат дополнительный заместитель, который залипает в большую бороздку спирали ДНК, где он, как полагают, улучшает стекинг оснований в дуплексе ДНК.
В дальнейших вариантах осуществления изобретения можно достичь полезных результатов, если в дополнение к олигонуклеотидам, модифицированным LNA (в соответствии с настоящим изобретением), в эти олигонуклеотиды вводятся дополнительные модификации, которые еще больше усиливают аффиность олигонуклеотида к акцепторной нити. Так, например, было обнаружено, что олигонуклеотид, модифицированный LNA в соответствии с настоящим изобретением, который дополнительно содержит пиримидин, модифицированный C5-пропином, и/или пурины, модифицированные C7 пропинилом, значительно повышает эффективность TNE.
Донорные нуклеотиды, предлагаемые настоящим изобретением, также могут быть химерными, т.е. содержать сегменты ДНК, РНК, LNA, PNA или их комбинации.
Так, в некоторых вариантах осуществления изобретения предлагаемый олигонуклеотид дополнительно содержит другие, необязательно не метилированные модифицированные нуклеотиды.
В некоторых вариантах осуществления изобретения олигонуклеотид устойчив к нуклеазам. Это выгодно потому, что олигонуклеотид защищен от разрушения нуклеазами, в связи с чем повышается вероятность того, что донорный олигонуклеотид сможет найти свою цель (акцепторную молекулу).
В некоторых вариантах осуществления изобретения может быть модифицирован тот нуклеотид в олигонуклеотиде, который находится в положении ошибочного спаривания. Ответ на вопрос о том, может или не может быть модифицировано ошибочное спаривание, во многом зависит от точного механизма направленной замены нуклеотида или от механизма репарации ДНК в клетке с использованием различий в аффинности между донорной и акцепторной нитями. То же самое относится к точной локализации других модифицированных положений вблизи или по соседству с точкой ошибочного спаривания. Однако, основываясь на раскрытых здесь сведениях, такой олигонуклеотид можно легко сконструировать и протестировать, принимая во внимание процедуры тестирования для подходящих олигонуклеотидов, как это описано здесь в другом месте. В некоторых вариантах осуществления изобретения нуклеотид в положении ошибочного спаривания не модифицирован. В некоторых вариантах осуществления изобретения модификация находится на расстоянии одного нуклеотида от точки ошибочного спаривания, предпочтительно 2, 3, 4, 5, 6 или 7 нуклеотидов от точки ошибочного спаривания. В некоторых вариантах осуществления изобретения модификация находится в положении (репликации) ниже точки ошибочного спаривания. В некоторых вариантах осуществления изобретения модификация находится в положении (репликации) выше точки ошибочного спаривания. В некоторых вариантах осуществления изобретения модификация расположена на расстоянии от 10 пар оснований до 10 т.п.о. от точки ошибочного спаривания, предпочтительно от 50 до 5000 пар оснований, более предпочтительно от 100 до 500 пар оснований.
Олигонуклеотиды, используемые в качестве доноров, могут варьироваться по длине, обычно от 10 до 500 нуклеотидов, предпочтительно от 11 до 100 нуклеотидов, предпочтительнее от 15 до 90, еще предпочтительнее от 20 до 70 и наиболее предпочтительно от 30 до 60 нуклеотидов.
В одном из аспектов изобретение относится к способу направленного изменения последовательности дуплекса акцепторной ДНК, включающему комбинирование последовательности дуплекса акцепторной ДНК с донорным олигонуклеотидом, причем последовательность дуплекса акцепторной ДНК содержит первую последовательность ДНК и вторую последовательность ДНК, которая комплементарна первой последовательности ДНК, где донорный олигонуклеотид содержит домен, который содержит по меньшей мере одно ошибочное спаривание по отношению к последовательности дуплекса акцепторной ДНК, подлежащей изменению, предпочтительно по отношению к первой последовательности ДНК, где участок донорного олигонуклеотида модифицирован LNA для экспрессии более высокой степени аффинности к первой последовательности ДНК по сравнению с немодифицированным нуклеотидом в этом же положении (в пределах олигонуклеотида), а также в присутствии белков, способствующих направленной замене нуклеотида, причем LNA находится на расстоянии по меньшей мере одного нуклеотида по отношению к точке ошибочного спаривания.
Изобретение в своей самой широкой форме, в общем, применимо ко всем разновидностям организмов, включая человека, животных, растения, рыб, рептилий, насекомых, грибы, бактерии и т.д. Изобретение применимо для модификации любых типов ДНК, например ДНК, извлеченной из геномной ДНК, линейной ДНК, искусственных хромосом, ядерной хромосомной ДНК, хромосомной ДНК органелл, BAC, YAC. Изобретение может быть реализовано как in vivo, так и ex vivo.
Изобретение в своей самой широкой форме применимо во многих целях для изменения клетки, исправления мутаций с возвратом к дикому типу, индуцирования мутаций, инактивирующих фермент за счет разрушения кодирующего региона, модифицирования биологической активности фермента за счет изменения кодирующей области, модифицирования белка за счет разрушения кодирующей области.
Изобретение также относится к применению олигонуклеотидов, главным образом, в соответствии с приведенными здесь описаниями, для изменения клеток, для исправления мутаций с возвратом к дикому типу, для индуцирования мутаций, инактивирующих фермент за счет разрушения кодирующего региона, для модифицирования биологической активности фермента за счет изменения кодирующей области, для модифицирования белка за счет разрушения кодирующей области, для репарации ошибок спаривания, для направленного изменения генетического материала (растений), включая генные мутации, целевую репарацию генов и генной нокаут.
Изобретение дополнительно относится к наборам, включающим один или более нуклеотидов, как это определено здесь в другом месте, по выбору, в комбинации с белками, способными индуцировать MRM и, в особенности, способными осуществлять TNE.
Изобретение дополнительно относится к модифицированному генетическому материалу, полученному способом, который предлагается настоящим изобретением, к клеткам и организмам, которые включают модифицированный генетический материал, к растениям и частям растений, которые получены таким способом.
В частности, изобретение относится к применению способа TNE с использованием олигонуклеотидов, модифицированных LNA, которые предложены этим изобретением, для создания у растений устойчивости к гербицидам. В частности, изобретение относится к растениям, которые приобрели устойчивость к гербицидам, особенно к гербицидам на основе глифосата и/или сульфонилмочевины, таким как хлорсульфорон.
Описание рисунков
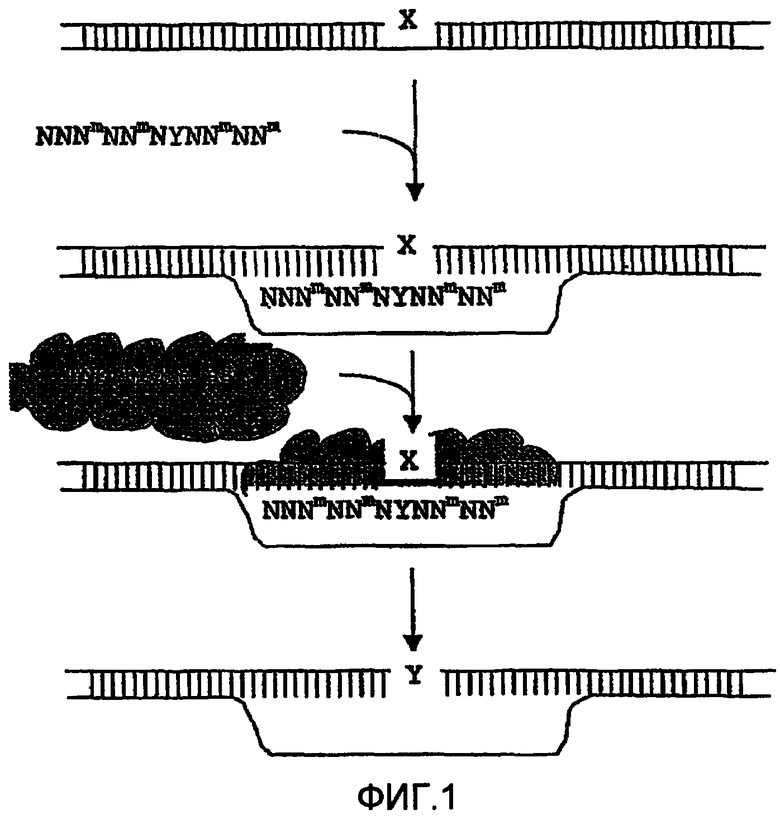
Фиг.1: Схематическое изображение направленной замены нуклеотида. Акцепторную двойную нить ДНК, содержащую нуклеотид, который подлежит замене (X), вводят в контакт с донорным олигонуклеотидом, модифицированным LNA (схематически представленным как NNNmNNNmYNNmNNm) и содержащим нуклеотид, который будет вставлен в целевую последовательность (Y). Триплексную структуру подвергают контактированию (или вводят в контакт) с окружающей средой, способствующей TNE, или по меньшей мере с белками, которые способны осуществлять TNE, например, с бесклеточной смесью ферментов или с бесклеточным экстрактом (см., среди прочего, WO 99/58702, WO 01/73002).
Фиг.2: Химические структуры разных LNA.
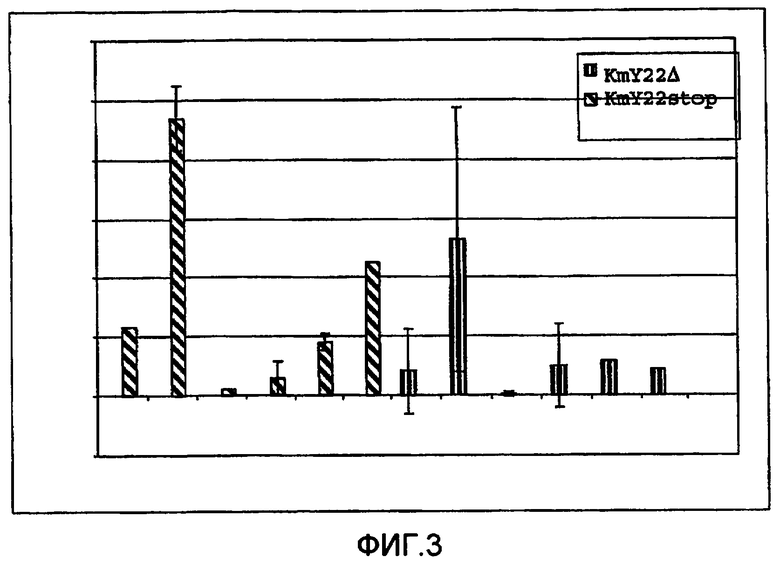
Фиг.3: Эффективность репарации TNE при использовании олигонуклеотидов, содержащих модифицированные LNA нуклеотиды по результатам измерений в бесклеточном анализе. В этом эксперименте сначала была определена эффективность репарации с применением олигонуклеотида нормальной ДНК, величину которой приняли за 1. Кратность увеличения указывает на возрастание репарации, наблюдаемое при применении олигонуклеотидов, содержащих β-D-LNA, по сравнению с эффективностью репарации при использовании олигонуклеотида нормальной ДНК.
Пример
Материалы и методы
Олигонуклеотиды, содержащие нуклеотиды LNA, были закуплены в компании Eurogentec. Последовательности использованных олигонуклеотидов представлены ниже. Плазмида, использованная в экспериментах, представляла собой производную pCR2.1 (Invitrogen), в которой содержались гены, придающие устойчивость к канамицину и карбенициллину. В ORF (открытую рамку считывания) канамицина и карбенициллина были введены внутрирамочные стоп-кодоны и делеции, как это было описано ранее (Sawano et al. 2000 Nucleic Acids Res. 28: e78). Плазмида KmY22stop несет мутацию TAT→TAG в кодоне Y22 внутри ORF канамицина. В плазмиде KmY22 третий нуклеотид кодона Y22 (TAT) был делетирован, давая сдвиг рамки.
Дефектные гены канамицина и олигонуклеотиды, использованные в бесклеточной системе:
Показаны релевантные последовательности открытых рамок считывания канамицина и закодированные аминокислоты. Были введены мутации одного нуклеотида, продуцирующие стоп-кодон (TAG, *), как это описано ранее (Sawano et al. 2000 Nucleic Acids Res. 28: e78). Последовательности использованных в эксперименте олигонуклеотидов LNA показаны прописными буквами. Олигонуклеотиды L1-L6 были использованы для конвертирования мутаций KmY22stop и KmY22Δ в альтернативный кодон, кодирующий тирозин (TAC). Ошибочно спаренный нуклеотид в каждом олигонуклеотиде выделен подчеркиванием. Области связывания в олигонуклеотиде для ORF канамицина выделены подчеркиванием. Олигонуклеотиды L1-L6 комплементарны кодирующей последовательности канамицина.
Бесклеточные анализы проводили следующим образом. Были собраны цветочные почки растения Arabidopsis thaliana (экотип Col-0), которые сохраняли в азотной атмосфере. Затем добавляли 200 мкл буфера для выделения белков (20 мМ HEPES pH 7,5, 5 мМ KCl, 1,5 мМ MgCl2, 10 мМ DTT, 10% (объем/объем) глицерина, 1% (вес/объем) PVP). Остатки растительного материала осаждали центрифугированием на скорости 14 тысяч оборотов в минуту в течение 30 минут, а надосадочную жидкость хранили при температуре -80°C. Концентрацию белков измеряли при помощи набора NanoOrange (Molecular Probes, Inc.). Типичное выделение приводило к концентрации белка приблизительно 3-4 мкг/мкл. Бесклеточные реакции содержали следующие компоненты. 1 мкг плазмидной ДНК (KmY22stop или KmY22Δ), 100 нг олигонуклеотида, 30 мкг общего растительного белка, 4 мкл ДНК денатурированной спермы лосося, (3 мкг/мкл), 2 мкл смеси ингибитора протеазы (50× концентрации: таблетки коктейля полного ингибитора протеаз без ЭДТА, Roche Diagnostics), 50 мкл 2× бесклеточного реакционного буфера (400 мМ трис pH 7,5, 200 мМ MgCl2, 2 мМ DTT, 0,4 мМ спермидина, 50 мМ АТФ, по 2 мМ CTP, GTP, UTP, по 0,1 мМ каждого dNTP и 10 мМ NAD) при доведении общего объема водой до 100 мкл. Смесь инкубировали при 37°C в течение 1 часа. Затем плазмидную ДНК выделяли следующим образом. К каждой реакции добавляли 100 мкл H2O для увеличения объема с последующим добавлением 200 мкл щелочного забуференного фенола (pH 8-10). Материал коротко перемешивали вихревым движением, а затем центрифугировали на скорости 13 тысяч оборотов в минуту в течение 3 минут. Верхнюю водную фазу переносили в новую пробирку, куда затем добавляли 200 мкл хлороформа. Материал коротко перемешивали вихревым движением, вращали в центрифуге со скоростью 13 тысяч оборотов в минуту в течение 3 минут, после чего водную фазу переносили в новую пробирку. ДНК осаждали, добавляя 0,7 объема 2-пропанола, а полученный осадок ресуспендировали в TE. Для удаления любых совместно очищенных олигонуклеотидов ДНК пропускали через очистительную колонку Qiagen PCR, а плазмидную ДНК элюировали в конечном объеме 30 мкл. 2 мкл плазмидной ДНК электропорировали в 18 мкл электрокомпетентных клеток DH10B (Invitrogen). После электропорации клеткам давали восстановиться в среде SOC в течение l часа при 37°C. По истечении указанного времени добавляли канамицин до концентрации 100 мкг/мл, а клетки инкубировали еще 3 часа. Твердые среды содержали 100 мкг/мл канамицина или карбенициллина. В экспериментах с KmY22stop и KmY22 число событий TNE определяли на среде с канамицином, а эффективность электропорации вычисляли, подсчитывая количество колоний, полученных в разведениях электропорации 10-4 и 10-5 после высева на среду с карбенициллином. Эффективность TNE вычисляли делением числа событий TNE на общее число трансформированных клеток.
Результаты
Олигонуклеотиды были сконструированы для осуществления замены одного нуклеотида (KmY22stop) или инсерции (KmY22Δ) в стоп-кодоне (TAG), введенном в открытую рамку считывания канамицина, чтобы этот кодон вновь начал кодировать правильную аминокислоту.
В каждом эксперименте параллельно использовали немодифицированные олигонуклеотиды ДНК и олигонуклеотиды, модифицированные LNA. В каждом эксперименте эффективность TNE, полученную при использовании олигонуклеотида нормальной ДНК, произвольно принимали за 1, а эффективность TNE при использовании олигонуклеотидов LNA впоследствии выражали как кратность увеличения по сравнению с олигонуклеотидом нормальной ДНК. Авторы обнаружили, что увеличение эффективности репарации зависит от положения LNA в олигонуклеотиде. Олигонуклеотид L1 содержал нуклеотиды LNA, фланкирующие ошибочно спаренный нуклеотид, и такая конструкция не продемонстрировала никаких преимуществ перед олигонуклеотидом нормальной ДНК. Однако олигонуклеотид L2 с нуклеотидами LNA, отделенными одним нуклеотидом от точки ошибочного спаривания, продемонстрировал среднее увеличение (эффективности TNE)
в 5 раз. Введение добавочных нуклеотидов LNA (L3 и L4) уменьшало эффективность репарации до уровня ниже олигонуклеотида нормальной ДНК, доводя ее до фонового состояния, то есть указанные олигонуклеотиды оказались биологически неактивными. Это подтверждено данными, полученными при использовании L5 и L6. В L5 помимо LNA, фланкирующих ошибочно спаренные нуклеотиды (как в L2), также содержались добавочные нуклеотиды LNA. В этом случае увеличение репарации, наблюдавшееся при использовании L2, отсутствовало, предположительно, из-за ингибиторного эффекта добавочных нуклеотидов LNA. L6 также демонстрирует улучшение репарации при использовании KmY22stop. Такая же тенденция наблюдалась, когда эксперименты проводились с использованием KmY22Δ. Известно, что репарация посредством вставок одного нуклеотида менее эффективна по сравнению с репарацией на основе замещений, и именно этот эффект наблюдался в наших экспериментах. L2 демонстрирует увеличение частоты репарации, а это свидетельствует о том, что для инсерций положение нуклеотидов LNA относительно точки ошибочного спаривания является важным признаком в отношении частоты репарации.
Эксперименты настоящего изобретения с применением анализа TNE in vitro и бесклеточной системы демонстрируют, что олигонуклеотиды, содержащие нуклеотиды LNA, обеспечивают более высокий уровень TNE по сравнению с эффективностью TNE, наблюдаемой при использовании олигонуклеотидов нормальной ДНК. Это увеличение может быть пятикратным. Эффект в значительной степени зависит от положения нуклеотидов LNA относительно ошибочно спаренного нуклеотида. В дополнение следует отметить, что процесс репарации очень чувствителен к числу нуклеотидов LNA в олигонуклеотиде, и если оно достигает 50%, олигонуклеотид экспрессирует в анализах сниженную биологическую активность. Еще одна форма нуклеотидов LNA, 2'-амино-LNA аналоги (Rosenbohm et al. (2003) Org Biomol Chem. 1, 655-663), может быть дополнительно функционализирована при добавлении к нуклеотиду других химических групп. В дополнение к повышенной аффинности связывания такие нуклеотиды LNA обладают дополнительными химическими группами, которые могут взаимодействовать с ДНК многими способами, что дополнительно увеличивает аффинность связывания (Sorensen et al. (2003) Chem ComMun (Camb) 17, 2130-2131). Такие 2'-амино-LNA могут увеличивать репарацию более 5-кратного уровня, который уже достигнут и продемонстрирован.





Изобретение относится к области молекулярной биологии и может быть использовано для направленной модификации последовательности дуплексной ДНК (днДНК). Предложен донорный олигонуклеотид, последовательность которого по всей ее длине комплементарна подлежащей изменению последовательности акцепторной ДНК, за исключением одного нуклеотида, формирующего с изменяемой последовательностью днДНК ошибочное спаривание, определяющее характер предполагаемой замены. При этом олигонуклеотид по изобретению содержит два модифицированных нуклеотида, которые представляют собой LNA и располагаются на расстоянии одного нуклеотида по обе стороны от точки ошибочного спаривания. Описан способ эффективного изменения последовательности днДНК с использованием нового олигонуклеотида, раскрыты условия и средства, необходимые для его осуществления. 4 н. и 10 з.п. ф-лы, 3 ил., 1 пр.
1. Способ направленного изменения последовательности дуплексной акцепторной ДНК, включающий а) контактирование последовательности дуплексной акцепторной ДНК с донорным олигонуклеотидом, который содержит одно ошибочное спаривание по отношению к последовательности дуплексной акцепторной ДНК, подлежащей изменению, и два модифицированных нуклеотида, представляющих собой LNA, расположенные на расстоянии одного нуклеотида по обе стороны от точки ошибочного спаривания, и b) инкубацию ДНК и олигонуклеотида в присутствии белков и в условиях, необходимых для протекания реакции замены нуклеотидов.
2. Способ по п.1, в котором ошибочное спаривание расположено в центре олигонуклеотида.
3. Способ по п.1, в котором изменение осуществляют внутри клетки, предпочтительно выбранной из группы, которая состоит из растительной клетки, клетки гриба, клетки грызуна, клетки примата, клетки человека или дрожжевой клетки.
4. Способ по п.1, в котором белки получены из клеточного экстракта.
5. Способ по п.4, в котором клеточный экстракт выбран из группы, состоящей из экстракта растительных клеток, экстракта клеток грибов, экстракта клеток грызунов, экстракта клеток приматов, экстракта клеток человека и экстракта дрожжевых клеток.
6. Способ по п.1, в котором изменение представляет собой делецию, замещение или инсерцию по меньшей мере одного нуклеотида.
7. Способ по любому из предшествующих пунктов, в котором клетка является эукариотической клеткой, растительной клеткой, клеткой млекопитающего, отличного от человека, или клеткой человека.
8. Способ по любому из пп.1-6, в котором ДНК-мишень происходит от грибов, бактерий, растений, млекопитающих или человека.
9. Способ по любому из пп.1-6, в котором дуплексная ДНК происходит из геномной ДНК, искусственных хромосом млекопитающих, бактериальных искусственных хромосом, дрожжевых искусственных хромосом, растительных искусственных хромосом, ядерной хромосомной ДНК, хромосомной ДНК органелл или эписомной ДНК.
10. Способ по любому из пп.1-6, предназначенный для исправления мутации с возвратом к дикому типу, индуцирования мутации, инактивирующей фермент за счет разрушения кодирующей области, модифицирования биологической активности фермента за счет изменения кодирующей области или модифицирования белка за счет разрушения кодирующей области.
11. Олигонуклеотид для направленного изменения последовательности дуплексной ДНК, который содержит одно ошибочное спаривание по отношению к подлежащей изменению последовательности ДНК и два модифицированных нуклеотида, представляющие собой LNA, расположенные на расстоянии одного нуклеотида по обе стороны от точки ошибочного спаривания.
12. Олигонуклеотид по п.11, в котором ошибочное спаривание расположено в центре олигонуклеотида.
13. Применение олигонуклеотида, определенного по п.11, для улучшенного направленного изменения последовательности дуплексной ДНК.
14. Применение олигонуклеотида, определенного по п.11, в способе создания у растений устойчивости к гербицидам.
| PAREKH-OLMEDO H | |||
| ET AL., Chem | |||
| Biol., v.9, no.10, 1073-1084, 2002 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| PETERSEN M | |||
| ET AL., J | |||
| Mol | |||
| Recognit., 13(1), 44-53, 2000. | |||

