Область техники
Изобретение относится к биотехнологии и медицине, а именно направленному изменению сплайсинга в гене MARK2, кодирующем фермент MARK2, являющийся членом семейства серин/треонин-протеинкиназ, мутации в котором связаны с патогенезом онкологических заболеваний.
Уровень техники
На данный момент препараты на основе антисмысловых олигонуклеотидов, модулирующих альтернативный сплайсинг, применяются для лечения спинальной мышечной атрофии (СМА) (Eteplirsen, Golodirsen, Viltolarsen, Casimersen) и миодистрофии Дюшенна (МДД) (Nusinersen под торговой маркой Spinraza). Препарат Nusinersen был одобрен FDA США в конце 2016 года, а в 2019 году был официально зарегистрирован и в России.
Nusinersen - химически модифицированный олигонуклеотид, который способствует включению экзона 7 белка SMN2. В интроне, следующем за данным экзоном, находится сайт связывания фактора hnRNP, ингибирующего включение экзона. Олигонуклеотид частично комплементарен сайту связывания фактора и конкурирует с ним, чем способствует включению экзона.
Препараты для лечения МДД также являются химически модифицированными олигонуклеотидами и способствуют исключению экзона 51 (Eteplirsen или Exondys 51), экзона 45 (Golodirsen, Viltolarsen) или экзона 53 (Casimersen) гена DMD. Для этого они связываются с пре-мРНК в начале соответствующего экзона и препятствуют распознаванию 3’-сайта сплайсинга.
Помимо вышеперечисленных уже выпущенных на рынок препаратов для терапии СМА и МДД, за последние 10 лет запатентовано множество технологий, направленных на модуляцию сплайсинга при помощи химически модифицированных олигонуклеотидов, для лечения других заболеваний, например, болезни Альцгеймера (WO2019162692A1, US10900041B2 компании ProQR Therapeutics) и дистрофии сетчатки глаза (US2022340901A1). Перечисленные технологии влияют на включение экзонов в генах APP (экзоны 7,8 и 17) и CEP290 (экзон 36), соответственно.
Нарушения сплайсинга - как мутации факторов сплайсинга, так и аберрантный сплайсинг – при онкогенезе являются широко известными. Тем не менее, согласно недавнему обзору Nat Rev Cancer (doi: 10.1038/s41568-022-00541-7), мишенями для разработки терапевтических олигонуклеотидов являются сайты сплайсинга и участки связывания регуляторных белков, а регуляция сплайсинга дальними взаимодействиями в пре-мРНК не рассматривается. Более того, такие значимые при онкогенезе регуляторные белки как киназы, широко рассматриваются как проблема, но упоминаются лишь несколько примеров (ATM, FGFR1, PKM2) среди рассматриваемых для коррекции альтернативного сплайсинга.
Большая часть технологий, связанных с направленным изменением сплайсинга, основана на использовании модифицированных олигонуклеотидов, комплементарных участкам вокруг донорного или акцепторного сайтов сплайсинга, примыкающих к интересующему экзону. Такие олигонуклеотиды, подобно препаратам для терапии МДД, изменяют сплайсинг, блокируя распознавание сплайсосомой сайтов сплайсинга вокруг интерсующего экзона. Терапевтических модифицированных олигонуклеотидов, активирующих, а не блокирующих сплайсинг, практически не представлено, а об олигонуклеотидах, воздействующих на вторичную структуру РНК, до исследований, проведенных авторами настоящего изобретения, не сообщалось.
Раскрытие изобретения
Задачей изобретения является направленное изменение альтернативного сплайсинга пре-мРНК опухоль-ассоциированного ген MARK2, а именно, увеличение степени включения кассетного экзона 17 при помощи синтетических антисмысловых олигонуклеотидов, ингибирующих образование вторичной структуры РНК, выпетливающей этот экзон, с целью изменения свойств белкового продукта этого гена.
Поставленная задача решается путем разработки антисмыслового олигонуклеотида, способного увеличить частоту включения экзона 17 в мРНК гена MARK2 человека, когда указанная мРНК образуется путем сплайсинга из пре-мРНК в клетке, характеризующегося тем, что указанный антисмысловой олигонуклеотид имеет последовательность, 5’-CGCTCAGCGAGAC-3’, в которой каждый второй нуклеотид представляет собой заблокированную нуклеиновую кислоту (LNA) и каждая межнуклеотидная связь представляет собой фосфоротиоатную связь.
Поставленная задача также может быть решена путем разработки способа увеличения частоты включения экзона 17 в мРНК гена MARK2 в клетке человека, включающего доставку к: (i) данной клетке in vitro или ex vivo; (ii) ткани, содержащей данную клетку, in vitro или ex vivo; или (iii) человеку in vivo, антисмыслового олигонуклеотида, как описано выше, в условиях, способствующих поглощению данного антисмыслового олигонуклеотида данной клеткой, и обеспечения возможности воздействия на процесс сплайсинга.
Также поставленная задача может быть решена применением описанного выше антисмыслового олигонуклеотида для увеличения частоты включения экзона 17 в мРНК гена MARK2 человека.
В результате осуществления изобретения достигаются следующие технические результаты:
- для терапевтической практики впервые предлагается подход, направленный на изменение альтернативного сплайсинга пре-мРНК гена MARK2, основанный на увеличении степени включения кассетного экзона 17 при помощи синтетических антисмысловых олигонуклеотидов, ингибирующих образование вторичной структуры РНК, выпетливающей этот экзон, что в конечном итоге обеспечивает изменение свойств белкового продукта этого гена;
- разработана технология, обеспечивающая увеличение степени включения кассетного экзона 17 гена MARK2 в результате действия не приводящих к деградации РНК антисмысловых олигонуклеотидов на комплементарные участки, задействованные в формировании вторичной структуры молекулы пре-мРНК;
- результатом применения предлагаемой технологии является изменение паттерна сплайсинга пре-мРНК опухоль-ассоциированного гена, что обеспечивает изменение его свойств, и соответственно, коррекцию свойств клетки при патологии;
- разработан антисмысловой олигонуклеотид, способный увеличивать частоту включения экзона 17 в мРНК гена MARK2 человека;
- предлагаемое решение является основой для разработки терапевтического лечения, направленного на коррекцию свойств опухолевых клеток без использования редактирования генома, в широком классе патологий.
Краткое описание рисунков
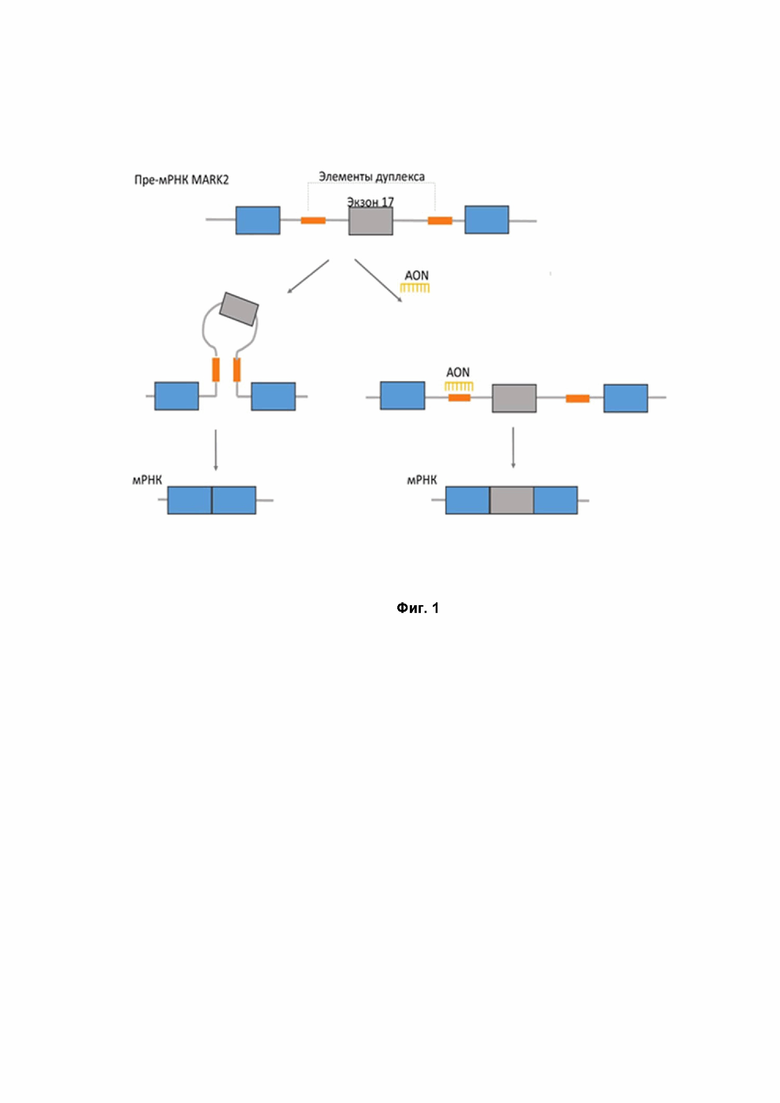
Фиг.1 - Схема изменения паттерна сплайсинга гена MARK2 при действии антисмыслового олигонуклеотида. Обозначения: большие прямоугольники синего и серого цветов – экзоны, линии серого цвета – интроны, маленькие прямоугольники оранжевого цвета – комплементарные последовательности, образующие вторичную структуру. AON – антисмысловой олигонуклеотид.
Термины и определения
Если иное не оговаривается, все технические и научные термины, используемые в данной заявке, имеют то же самое значение, которое понятно для специалистов в данной области. Ссылки на методики, используемые при описании данного изобретения, относятся к хорошо известным методам, включая изменения этих методов и замену их эквивалентными методами, известными специалистам.
В документах данного изобретения термины «включает», «включающий» и т.п., а также «содержит», «содержащий» и т.п. интерпретируются как означающие «включает, помимо всего прочего» (или «содержит, помимо всего прочего»). Указанные термины не предназначены для того, чтобы их истолковывали как «состоит только из».
Термин «и/или» означает один, несколько или все перечисленные элементы.
Термин «выпетливающий» означает образующий петлю, содержащую целевой экзон, как это показано на Фиг.1.
Термин «заблокированная нуклеиновая кислота» («замкнутая (закрытая) нуклеиновая кислота», англ. locked nucleic acid, LNA) означает модифицированный нуклеотид РНК, в котором фрагмент рибозы модифицирован дополнительным этиленовым мостиком, соединяющим 2'-кислородную группу и 4' углерод.
Антисмысловые олигонуклеотиды согласно изобретению могут быть введены совместно с фармацевтически приемлемыми вспомогательными веществами, добавками, стабилизаторами, растворителями и т.п. Особенно предпочтительным в данном изобретении является использование вспомогательного вещества или реагентов для трансфекции, которые будут содействовать доставке антисмысловых олигонуклеотидов к клетке и/или в клетку, предпочтительно опухолевую клетку. Предпочтительными являются вспомогательные вещества или реагенты для трансфекции, способные образовывать комплексы, наночастицы, мицеллы, везикулы и/или липосомы, которые доставляют антисмысловые олигонуклеотиды заключенным внутри везукулы или липосомы, через клеточную мембрану. Многие из этих вспомогательных веществ известны в данной области. Подходящие вспомогательные вещества или реагенты для трансфекции включают полиэтиленимин (PEI; ExGen500 (MBI Fermentas)), LipofectAMINE 2000 (Invitrogen) или их производные, или сходные катионные полимеры, включая сополимеры полипропиленимина или полиэтиленимина (РЕС) и производные, синтетические амфифилы (SAINT-18), липофектин, но не ограничиваются ими. Специалист в данной области может выбрать и адаптировать любое из указанных выше вспомогательных веществ или другие коммерчески доступные альтернативные вспомогательные вещества и системы доставки для упаковки и доставки антисмысловых олигонуклеотидов для применения в настоящем изобретении.
Термины «лечение», «терапия» охватывают лечение патологических состояний у пациента и включают как излечение, так и блокирование (приостановку) течения заболевания или облегчение тяжести заболевания.
Под «пациентом» в рамках настоящего изобретения понимается человек.
Под «эффективным количеством» («терапевтически эффективным количеством») активного вещества в фармацевтической композиции понимается количественное содержание активного вещества в композиции, достаточное для достижения терапевтического (и/или профилактического) эффекта у пациента. Данные об эффективном количестве активных ингредиентов в составе композиции могут быть получены на основании результатов экспериментов.
Подробное описание изобретения
Технология антисмысловых олигонуклеотидов является эффективным средством для модуляции экспрессии одного или нескольких специфических генных продуктов, в том числе продуктов альтернативного сплайсинга, и, несомненно, может использоваться в терапевтических, диагностических и исследовательских целях. Принцип антисмысловой технологии заключается в том, что антисмысловой олигонуклеотид, который гибридизуется с целевой нуклеиновой кислотой, изменяет доступность расположенных на ней цис-регуляторных элементов и, тем самым, модулирует процессы, отвечающие за экспрессию генов, такие как транскрипция, сплайсинг или трансляция.
Основное отличие технологии, предлагаемой авторами, от используемых в терапевтической практике ранее, состоит в том, что предлагаемые модифицированные олигонуклеотиды комплементарны не сайтам сплайсинга или участкам внутри самого исключаемого в процессе аберрантного сплайсинга экзона, а участкам в интронах вокруг интересующего экзона, которые задействованы в формировании вторичной структуры молекулы пре-мРНК. При этом авторами показано, что блокирование формирующих структуру взаимодействий значимо влияет на уровень включения экзонов других генов (Kalinina M., Skvortsov D., Kalmykova S., Ivanov T., Dontsova O., Pervouchine D. Multiple competing RNA structures dynamically control alternative splicing in the human ATE1 gene // Nucleic Acids Res. 2021 Jan 11;49(1):479-490. doi: 10.1093/nar/gkaa1208).
Таким образом, в настоящем изобретении предлагается изменение сплайсинга гена MARK2 с использованием подхода, никогда ранее не применявшемся к изменению сплайсинга в терапевтических целях. Принципиальная схема изменения направления сплайсинга MARK2 при действии антисмыслового олигонуклеотида показана на Фиг.1.
Наиболее близкое по механизму действия семейство технологий - метод изменения уровня включения экзона 51 гена DMD для терапии МДД, предложенный ACADEMISCH ZIEKENHUIS LEIDEN (см., например, US10100304B2). Цитируемое изобретение включает в себя метод определения участка пре-мРНК (экзона и окружающей области), который формирует структуру, т.е. гибридизуется с другой частью РНК (закрытая структура), и участка, который не гибридизуется (открытая структура). Также изобретение включает в себя модифицированные олигонуклеотиды, которые по крайней мере частично комплементарны закрытой структуре и хотя бы частично комплементарны открытой структуре. Комплементарный олигонуклеотид по изобретению способен образовывать двухцепочечную структуру с последовательностью экзона и, таким образом, влиять на уровень включения экзона.
Стоит отметить, что в отличие от подавляющего большинства примеров на текущем уровне развития науки и техники, направленных на подавление включения экзона (например, тем или иным вариантом блокирования сплайс-сайтов), предложенная технология позволяет стимулировать включение экзона.
Технология согласно настоящему изобретению представляет собой систему направленного изменения сплайсинга в гене MARK2. Ген MARK2 кодирует фермент, являющийся членом семейства серин/треониновых протеинкиназ, локализованный на клеточных мембранах, который является важным регулятором клеточной полярности в эпителиальных и нейрональных клетках, а также контролирует стабильность микротрубочек посредством фосфорилирования и инактивации нескольких белков, ассоциированных с микротрубочками.
Активность гена MARK2 существенна при прогрессировании ряда опухолей, а повышение уровня его экспрессии коррелирует с опухолевой трансформацией в некоторых типах неоплазий. MARK2 также может участвовать в активации клеточного цикла и репарации ДНК (Hubaux R. el al. Microtubule affinity-regulating kinase 2 is associated with DNA damage response and cisplatin resistance in non-small cell lung cancer // Int J Cancer. 2015 Nov 1;137(9):2072-82. doi: 10.1002/ijc.29577. Epub 2015 May 6). С другой стороны, MARK2 ингибирует рост клеток HeLa и обращает эпителиально-мезенхимальный переход (Ge Xu et al. MARK2 inhibits the growth of HeLa cells through AMPK and reverses epithelial-mesenchymal transition // Oncol Rep. 2017 Jul;38(1):237-244. doi: 10.3892/or.2017.5686. Epub 2017 May 30). Высокие уровни экспрессии этой киназы коррелируют с устойчивостью к цисплатину и паклитакселу (Hubaux R. et al. 2015 doi: 10.1002/ijc.29577; Stauffer S. el al. Cyclin-dependent kinase 1-mediated AMPK phosphorylation regulates chromosome alignment and mitotic progression // J Cell Sci . 2019 Oct 28;132(20):jcs236000. doi: 10.1242/jcs.236000). Ген MARK2 содержит 19 экзонов, и некоторые из них подвергаются альтернативному сплайсингу. В частности, экзон 17 подвергается альтернативному сплайсингу и является кассетным.
Нижеследующие примеры приведены в целях раскрытия характеристик настоящего изобретения, и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения.
Номера нуклеотидных последовательностей (SEQ ID NO) в тексте описания соответствуют таковым в Перечне последовательностей согласно Стандарту ST.26, являющемуся частью настоящего описания изобретения. В случае наличия разночтений в структуре последовательностей между текстом описания и соответствующей последовательностью в Перечне последовательностей согласно Стандарту ST.26, приоритетными являются данные, представленные в тексте описания. При этом сконструированные в результате проведенной работы антисмысловые олигонуклеотиды (AON) не представлены в Перечне последовательностей, поскольку Перечень последовательностей согласно Стандарту ST.26 не должен включать никаких последовательностей с менее чем десятью немодифицированными («специально определёнными», согласно терминологии Стандарта ST.26) нуклеотидами, как последовательность со своим собственным присвоенным идентификационным номером последовательности.
Пример 1.
В рамках проведенного эксперимента было показано, что фланкирующие интроны экзона 17 гена MARK2 содержат консервативные комплементарные области, взаимодействие которых между собой выпетливает экзон 17 и, тем самым, определяет протекание альтернативного сплайсинга.
Для этого был сконструирован миниген, охватывающий фрагмент гена MARK2, заключенный между экзонами 16 и 18. Затем в миниген были введены две мутации (заменяющие последовательность фрагмента структуры в пре-мРНК на обратную), изменяющие нуклеотидную последовательность таким образом, что каждая из них по отдельности разрушает, а две мутации, введенные одновременно, восстанавливают вторичную структуру РНК. Эффекты этих мутаций на альтернативный сплайсинг минигена MARK2, а именно на включение/исключение экзона 17, оценивали с помощью ОТ-ПЦР.
Конструирование минигена производили рестриктазно-лигазным клонированием по одноименным «липким» концам на основе вектора pRK5. Фрагмент генома человека от 16 до 18 экзона гена MARK2 (3452 пар нуклеотидов) амплифицировали по методу гнездового ПЦР с помощью высокоточной ДНК полимеразы Q5 High-Fidelity DNA Polymerase (New England Biolab) и праймеров (прямой: 5’-CATCCGCCCACAACATCAGCAG-3’ (SEQ ID NO: 1), обратный: 5’-TGGCAGAAGGAAGAGAGGAGCAG-3’ (SEQ ID NO: 2)). Участок ДНК внутри ПЦР-продукта первой реакции реамплифицировали с помощью праймеров с введенными сайтами узнавания эндонуклеаз рестрикции (EcoRI: 5’-TAAGCAGAATTCCCAGACCGAACTAACTTCCC-3’ (SEQ ID NO: 3), SalI: 5’-TGCTTAGTCGACCTGAGCGTCTCCACTCGGTC-3’ (SEQ ID NO: 4)). В качестве матрицы использовали геномную ДНК клеточной линии аденокарциномы легкого A549. Линеаризацию плазмиды pRK5 и обработку фрагмента гена MARK2 проводили с помощью эндонуклеаз рестрикции EcoRI и SalI (Thermo Fisher Scientific). pRK5 вектор и фрагмент гена MARK2 лигировали с помощью T4 ДНК-лигазы (Evrogen), после чего лигазной смесью трансформировали штамм E.coli DH5α. Путем сайт-специфического мутагенеза на основе ПЦР получен мутантный вариант минигена с заменой последовательности, которая нарушает комплементарное взаимодействие (прямой праймер: 5’-CAGAGCGACTCGCCGACTTCCGAAAATGGGATGACCC-3’ (SEQ ID NO: 5), обратный праймер: 5’-TAGAGTCCCAGGCTTTCAGACC-3’ (SEQ ID NO: 6)). Последовательность полученной плазмиды и фрагментов минигена после проведения мутагенеза проверяли путем секвенирования.
В результате проведенного эксперимента было показано, что введение одиночных мутаций приводит к увеличению числа транскриптов с включенным экзоном 17, а в компенсаторном двойном мутанте, в котором спаривание оснований восстановлено, степень включения экзона 17 уменьшается до уровней, близких к исходному.
Пример 2.
Поскольку мутагенез эндогенных транскриптов неприменим в терапевтических задачах, для воздействия на вторичную структуру РНК в эндогенном транскрипте MARK2 применялись синтетические антисмысловые олигонуклеотиды, комплементарные участкам вторичной структуры и не приводящие к деградации РНК. Антисмысловые олигонуклеотиды представляли собой миксмеры LNA/DNA, которые были разработаны с использованием данных о коррекции альтернативного сплайсинга гена SMN2 (Touznik A. et al. LNA/DNA mixmer-based antisense oligonucleotides correct alternative splicing of the SMN2 gene and restore SMN protein expression in type 1 SMA fibroblasts // Sci Rep. 2017 Jun 16;7(1):3672. doi: 10.1038/s41598-017-03850-2).
Сконструированные в результате проведенной работы антисмысловые олигонуклеотиды (AON) против MARK2 представляют собой последовательности, состоящие из модифицированных нуклеотидов (миксмеры), а именно, чередующихся через один фосфотиоатного и LNA-фосфотиоатного нуклеотидов (т.е. основанные на заблокированной нуклеиновой кислоте (LNA) с заменой ДНК на каждом втором нуклеотиде и имеющие фосфонотиоированные скелеты для защиты от клеточных нуклеаз). Синтез миксмеров LNA/DNA осуществлялся ЗАО «Синтол» (Москва, Россия). Последовательности полученных AON представлены в Таблице 1.
Таблица 1. Сконструированные антисмысловые олигонуклеотиды.
каждый второй нуклеотид представляет собой заблокированную нуклеиновую кислоту (LNA),
при этом каждая межнуклеотидная связь представляет собой фосфоротиоатную связь
каждый второй нуклеотид представляет собой заблокированую нуклеиновую кислоту (LNA),
при этом каждая межнуклеотидная связь представляет собой фосфоротиоатную связь
Обозначения: G,A,T,C – немодифицированный нуклеотид;
+G, +A, +T, +C - заблокированная нуклеиновая кислота (LNA);
* - фосфоротиоатные (тиофосфатные) межнуклеотидные связи.
Пример 3.
Анализ эффективности действия сконструированных AON проводился на клеточной линии аденокарциномы легкого человека A549. Клетки линии культивировались в среде DMEM/F-12, содержащей 10% эмбриональной бычьей сыворотки, 50 ЕД/мл пенициллина и 0,05 мг/мл стрептомицина (все продукты Thermo Fisher Scientific) при 37°C и 5% CO2. Трипсин-ЭДТА (Thermo FisherScientific) использовали для субкультивирования клеток в соответствии с рекомендациями ATCC. Аутентификацию клеточной линии подтверждали с помощью анализа коротких тандемных повторов. Линию клеток тестировали на отсутствие микоплазмы с использованием набора MycoReport (Евроген). Тестирование AON проводили с использованием Lipofectamine RNAiMAX (Invitrogen) на 50–70% конфлюэнтных клетках в 12-луночном планшете при концентрации АОN 10 - 100 нМ. Процедуру трансфекции проводили в соответствии с протоколом производителя. Через 48 часов после трансфекции клетки разрушали с помощью буфера для лизиса EasyRNA (Евроген), выделяли РНК и оценивали соотношение изоформ с помощью ОТ-ПЦР.
В результате проведенного эксперимента было установлено, что обработка антисмысловым олигонуклеотидом MARK2_1 в концентрации 100 нМ приводит значимому увеличению частоты включения экзона 17 в эндогенном транскрипте гена MARK2 в 2.5 раза (см. Таблицу 2). Также увеличение использования данного экзона в клетках, обработанных в концентрации 100 нМ, детектируется на электрофорезе продуктов ОТ-ПЦР и при анализе ОТ-кПЦР отдельных изоформ. Таким образом, в гене MARK2 человека наблюдается увеличение частоты включения экзона 17 при блокировании вторичной структуры РНК, выпетливающией экзон 17, антисмысловым олигонуклеотидом MARK2_1.
В то же время, обработка антисмысловым олигонуклеотидом MARK2_2 не приводила к увеличению частоты включения экзона 17 (см. Таблицу 2).
Таблица 2. Эффективность AON на степень включения экзона 17.
Проведенный анализ различий в действии антисмысловых олигонуклеотидов MARK2_1 и MARK2_2 позволяет предположить, что причина различий объясняется котранскрипционным протеканием сплайсинга в этом гене. Так, биоинформатический анализ данных отклика транскриптома на замедление полимеразы RNAPII (мутант R749H) показал, что степень включения экзона 17 значимо увеличивается при замедлении скорости элонгации транскрипции (Fong N. et al. Pre-mRNA splicing is facilitated by an optimal RNA polymerase II elongation rate // Genes Dev. 2014 Dec 1;28(23):2663-76. doi: 10.1101/gad.252106.114). Таким образом, при котранскрипционном сплайсинге действие антисмыслового олигонуклеотида на последовательность в направлении 3’-конца ослабляется конкуренцией со вторичной структурой РНК.
В результате проведенной работы разработана технология направленного изменения альтернативного сплайсинга гена MARK2 через ингибирование образования вторичной структуры РНК, выпетливающей экзон 17, при помощи синтетических антисмысловых олигонуклеотидов. Разработанная технология обеспечивает увеличение степени включения кассетного экзона 17 гена MARK2 в результате действия не приводящего к деградации РНК антисмыслового олигонуклеотида MARK2_2 на комплементарные участки, задействованные в формировании вторичной структуры молекулы пре-мРНК. Предлагаемое решение является основой для разработки терапевтического лечения, направленного на коррекцию свойств опухолевых клеток путем направленного изменения свойств белкового продукта гена MARK2 без использования редактирования генома.
Сконструированный в рамках изобретения антисмысловой олигонуклеотид может быть использован для корректировки возможной дерегуляции сплайсинга в гене MARK2 и, соответственно, для производства лекарственных средств, направленных на такую корректировку. Биоинформатический анализ показал, что значительная дерегуляция сплайсинга наблюдается при онкогенезе, и, например, включение экзона 17 гена MARK2 значительно понижено в опухолях головы и шеи, а также в раке груди. Свидетельством функциональной значимости кодируемого этим экзоном фрагмента белка, является его вероятное участие в локализации и находящиеся в нем множественные сайты фосфорилирования.
Несмотря на то, что изобретение описано со ссылкой на раскрываемые варианты воплощения, для специалистов в данной области должно быть очевидно, что конкретные подробно описанные эксперименты приведены лишь в целях иллюстрирования настоящего изобретения, и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения. Должно быть понятно, что возможно осуществление различных модификаций без отступления от сути настоящего изобретения.
--->
Перечень последовательностей
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Система
направленного изменения сплайсинга в гене MARK2.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-09-14">
<ApplicationIdentification>
<IPOfficeCode></IPOfficeCode>
<ApplicationNumberText>-</ApplicationNumberText>
<FilingDate></FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>498297</ApplicantFileReference>
<ApplicantName languageCode="ru">АВТОНОМНАЯ НЕКОММЕРЧЕСКАЯ
ОБРАЗОВАТЕЛЬНАЯ ОРГАНИЗАЦИЯ ВЫСШЕГО ОБРАЗОВАНИЯ «СКОЛКОВСКИЙ ИНСТИТУТ
НАУКИ И ТЕХНОЛОГИЙ»</ApplicantName>
<ApplicantNameLatin>SKOLKOVO INSTITUTE OF SCIENCE AND TECHNOLOGY
(SKOLTECH))</ApplicantNameLatin>
<InventionTitle languageCode="ru">Система направленного изменения
сплайсинга в гене MARK2</InventionTitle>
<SequenceTotalQuantity>6</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>catccgcccacaacatcagcag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>23</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..23</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tggcagaaggaagagaggagcag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>32</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..32</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>taagcagaattcccagaccgaactaacttccc</INSDSeq_sequenc
e>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>32</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..32</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q13">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tgcttagtcgacctgagcgtctccactcggtc</INSDSeq_sequenc
e>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>37</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..37</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q10">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cagagcgactcgccgacttccgaaaatgggatgaccc</INSDSeq_se
quence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q12">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tagagtcccaggctttcagacc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к биотехнологии и медицине. Предложен антисмысловой олигонуклеотид, способный увеличивать частоту включения экзона 17 в мРНК гена MARK2 человека в процессе образования указанной мРНК путем сплайсинга из пре-мРНК в клетке. Антисмысловой олигонуклеотид имеет последовательность 5'-CGCTCAGCGAGAC-3'Ф, в которой каждый второй нуклеотид представляет собой заблокированную нуклеиновую кислоту (LNA) и каждая межнуклеотидная связь представляет собой фосфоротиоатную связь. Изобретение обеспечивает направленное изменение сплайсинга в гене MARK2 и может быть использовано для корректировки возможной дерегуляции сплайсинга в гене MARK2 и, соответственно, для производства лекарственных средств, направленных на такую корректировку. 1 ил., 2 табл., 3 пр.
Антисмысловой олигонуклеотид, способный увеличивать частоту включения экзона 17 в мРНК гена MARK2 человека в процессе образования указанной мРНК путем сплайсинга из пре-мРНК в клетке, характеризующийся тем, что указанный антисмысловой олигонуклеотид имеет последовательность 5'-CGCTCAGCGAGAC-3', в которой каждый второй нуклеотид представляет собой заблокированную нуклеиновую кислоту (LNA) и каждая межнуклеотидная связь представляет собой фосфоротиоатную связь.
| US 10100304 B2, 16.10.2018 | |||
| KALININA M., et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Nucleic Acids Res | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| WO 2020210537 A1, 15.10.2020 | |||
| Прибор для измерения температуры | 1928 |
|
SU13375A1 |

