Область техники
Настоящее изобретение относится к способу специфического и селективного изменения нуклеотидной последовательности в определенном сайте ДНК в клетке-мишени путем введения в клетку одноцепочечного ДНК-олигонуклеотида в виде мутагенного нуклеинового основания. Более конкретно, изобретение относится к способу направленного мутагенеза путем введения мутагенного нуклеинового основания в растительные протопласты с использованием полиэтиленгликоля (ПЭГ). Изобретение дополнительно относится к наборам реагентов, содержащим мутагенное нуклеиновое основание и ПЭГ. Изобретение также относится к применению ПЭГ для улучшения направленного мутагенеза.
Уровень техники
Способ специального создания изменений в генетическом материале живых клеток имеет целью модификацию одного или более из генетически кодируемых биологических свойств этой клетки или организма, часть которого образует клетка или в которую она может регенерироваться. Эти изменения могут принимать форму делеции частей генетического материала, вставку экзогенного генетического материала или изменения в существующей нуклеотидной последовательности генетического материала. Методы изменения генетического материала эукариотических организмов были известны в течение более 20 лет и обнаружили широкое применение в растительных, человеческих и животных клетках и микроорганизмах для улучшений в области сельского хозяйства, здоровья человека, качества пищи и в области защиты от неблагоприятных воздействий окружающей среды. Наиболее широко распространенные методы состоят из вставки фрагментов экзогенной ДНК в геном клетки, которые затем придадут этой клетке новое свойство или ее организму помимо и сверх свойств, кодируемых уже существующими генами (включая применения, в которых экспрессия существующих генов будет в результате подавляться). Хотя многие такие примеры эффективны в получении целевых свойств, тем не менее эти методы не очень точны из-за отсутствия контроля геномных положений, в которые вставляются фрагменты экзогенной ДНК (и, следовательно, отсутствия контроля предельного уровня экспрессии), и из-за того, что целевой эффект должен сам себя выражать относительно природных свойств, кодируемых исходным и устойчивым геномом. Напротив, методы направленного мутагенеза, которые приводят в результате к вставке, делеции или конверсии нуклеотидов в заранее определенном геномном локусе, дадут возможность точной модификации существующих генов. Кроме того, из-за точной природы направленного мутагенеза, ожидается, что новые растительные линии, полученные таким образом, будут более легко приниматься потребителями. Направленный мутагенез представляет собой метод сайт-специфического мутагенеза, который основан на доставке в эукариотическую клетку синтетических мутагенных нуклеиновых оснований (молекул, состоящих из коротких фрагментов нуклеотидо-подобных компонентов, которые напоминают ДНК по их свойствам спаривания оснований по Уотсону-Крику, но могут химически отличаться от ДНК) (Alexeev and Yoon, Nature Biotechnol. 16: 1343, 1998; Rice, Nature Biotechnol. 19: 321, 2001; Kmiec, J. Clin. Invest. 112: 632, 2003). После введения в клетку такие мутагенные нуклеиновые основания спариваются с комплементарной последовательностью в локусе-мишени. С помощью специального конструирования ошибочной пары в нуклеиновом основании ошибочная пара может представлять собой нуклеотидную конверсию в соответствующем положении в геномной последовательности-мишени. Этот метод дает возможность конверсии одного или по большей мере нескольких нуклеотидов в эндогенных локусах, но может применяться и для создания стоп-кодонов в существующих локусах, приводя в результате к нарушению их функции, или может применяться для создания изменений в кодонах, приводя в результате к получению генов, кодирующих белки с измененным аминокислотным составом (белковая инженерия).
Направленный мутагенез был описан в растительных, животных и дрожжевых клетках. В этих исследованиях использовали два различных класса синтетических мутагенных нуклеиновых оснований, химерные ДНК:РНК нуклеиновые основания или одноцепочечные нуклеиновые основания.
Химерные ДНК:РНК нуклеиновые основания (химеры) представляют собой самокомплементарные молекулы, состоящие из 25 п.о., представляющих собой только участок ДНК, и из 25 п.о. комплементарной последовательности, сделанной из 5 п.о. корового участка ДНК, фланкированного с каждой стороны участками по 10 п.о. 2'-O-метилированной РНК, которые, как считается, способствуют стабильности химеры в клетке. Коровый участок 5 п.о. включает в своем центре сконструированную ошибочную пару с нуклеотидом, который необходимо изменить в геномной последовательности-мишени. Оба эти участка связаны с помощью 4 п.о. тимидиновых шпилек. При введении в клетку химера, как считается, образует двойную D-петлю со своей последовательностью-мишенью, и ошибочная пара образуется между химерой и нуклеотидом-мишенью. Эта ошибочная пара затем исправляется с помощью эндогенных клеточных белков ДНК репарации путем конверсии геномного нуклеотида. Первые примеры направленного мутагенеза с использованием химер пришли из животных клеток (рассмотрено у Igoucheva et al. 2001 Gene Therapy 8, 391-399) и затем позднее также использовались для осуществления направленного мутагенеза в растительных клетках (Beetham et al. 1999 Proc. Natl. Acad. Sci.USA 96: 8774- 8778; Zhu et al. 1999 Proc. Natl. Acad. Sci. USA 96, 8768-8773; Zhu et al. 2000 Nature Biotech. 18, 555-558; Kochevenko et al. 2003 Plant Phys. 132: 174-184; Okuzaki et al. 2004 Plant Cell Rep. 22: 509-512). В отличие от человеческих клеток, растительная клетка, в которой происходит событие направленного мутагенеза, может регенерироваться в интактное растение и мутация перенесется в следующее поколение, что делает растительную клетку идеальным средством одновременно для исследовательского и коммерческого мутагенеза важных пищевых культур. Однако интенсивное исследование многими лабораториями показало, что частота направленного мутагенеза с использованием химер достаточно низкая и вариабельная или даже не детектируемая (Ruiter et al. 2003 Plant Mol. Biol. 53, 715-729, Van der Steege et al. (2001) Nature Biotech. 19: 305-306) и зависит от таких факторов, как транскрипционный статус мишени, положение клетки в клеточном цикле, последовательность мишени и качество химер, которые трудно синтезировать. Благодаря относительно низкой частоте направленного мутагенеза с помощью методов, известных из уровня техники, такие события могут детектироваться только, когда изменение одного нуклеотида геномной мишени приводит в результате к получению доминантного селективного фенотипа. В растительных клетках специфические точечные мутации вводились в открытую рамку считывания гена ацетолактатсинтазы (ALS, в маисе AHAS), которая катализирует исходную стадию, общую для синтеза аминокислот с разветвленными цепями, лейцина, изолейцина и валина. В табаке изменения единичного нуклеотида являются достаточными для получения кодоновых конверсий P194Q или W571L. Белок ALS, получаемый после каждой из этих кодоновых конверсий, не чувствителен к ингибированию гербицидами класса сульфонилмочевины, получая таким образом метод селекции для единичных нуклеотидных конверсий в хромосомном локусе.
Из-за трудностей работы с химерами велись поиски более надежной альтернативы олигонуклеотидного дизайна. Некоторые лаборатории исследовали возможность использования одноцепочечных (ss) нуклеиновых оснований для осуществления направленного мутагенеза. Было обнаружено, что они дают более воспроизводимые результаты, будучи при этом более просто синтезируемыми, а также они могут включать модифицированные нуклеотиды для улучшения представленности мутагенного основания в клетке (Liu et al. 2002 Nucl. Acids Res. 30: 2742-2750; обзор, Parekh-Olmedo et al. 2005 Gene Therapy 12: 639-646; Dong et al. 2006 Plant Cell Rep. 25: 457-65; De Piedoue et al. 2007 Oligonucleotides 27: 258-263).
Направленный мутагенез был описан в различных патентных заявках Kmiec, среди прочего в WO 01/73002, WO 03/027265, WO 01/87914, WO 99/58702, WO 97/48714, WO 02/10364. В WO 01/73002 предположили, что низкая эффективность генных изменений, получаемых с использованием немодифицированных нуклеиновых оснований в значительной степени, как считается, определяется результатом их деградации нуклеазами, присутствующими в реакционной смеси клетки-мишени. Для устранения этой проблемы предложили включать модифицированные нуклеотиды, которые делают полученные в результате нуклеиновые основания устойчивыми против нуклеаз. Типичные примеры таких модифицированных нуклеотидов включают фосфоротиоатные связи или 2'-О-метил-аналоги. Эти модификации предпочтительно локализованы на концах нуклеинового основания, покидающего центральный немодифицированный домен, окружающий основание-мишень. В поддержку этого предположения, в патентной заявке WO 02/26967 продемонстрировали, что определенные модифицированные нуклеотиды, увеличивающие внутриклеточную продолжительность существования нуклеинового основания, усиливают эффективность направленного мутагенеза в in vitro тест-системах, а также в хромосомной мишени млекопитающих. Не только устойчивость к нуклеазам, но также аффинность связывания мутагенного одноцепочечного нуклеинового основания с его комплементарной ДНК-мишенью обладает значительным эффектом усиления частоты направленного мутагенеза. Одноцепочечное нуклеиновое основание, содержащее модифицированные нуклеотиды, которые усиливают его аффинность связывания, может более эффективно находить свою комплементарную мишень в сложном геноме и/или оставаться связанным со своей мишенью в течение более продолжительного периода времени и иметь меньшую вероятность удаления белками, регулирующими ДНК-транскрипцию и репликацию. Для тестирования множества модифицированных нуклеотидов для улучшения эффективности процесса мутагенеза использовали in vitro анализ направленного мутагенеза. Закрытые нуклеиновые кислоты (LNA) и C5-пропин-пиримидины имеют модификации сахарного компонента и основания соответственно, которое стабилизирует образование дуплекса и повышает температуру плавления дуплекса. Когда эти модифицированные нуклеотиды включают в одноцепочечное нуклеиновое основание, они улучшают эффективность направленного мутагенеза до 13 раз выше, чем эффективность, получаемая с использованием немодифицированного нуклеинового основания в той же последовательности. См. в связи с этим WO 2007073166 и WO 2007073170 от имени заявителей настоящего изобретения.
Авторы настоящего изобретения попытались улучшить частоту направленного мутагенеза в растительных клетках путем оптимизации метода, используемого для введения мутагенных нуклеиновых оснований в растительные клетки. Наиболее широко используемый метод трансформации растительных клеток, Agrobacterium-опосредованная трансформация переносит секцию ее опухоле-индуцирующей (Ti) плазмиды, так называемой T-ДНК, в растительные клетки, где она эффективно интегрируется в геном растения в случайное положение. T-ДНК фланкируется с каждого конца "пограничными" последовательностями длиной до 22 п.о., с происхождением из Ti-плазмиды, которые не имеют гомологии с последовательностью-мишенью. Поскольку для направленного мутагенеза используется короткая длина мутагенных одноцепочечных нуклеиновых оснований, пограничные последовательности будут мешать процессу. Таким образом, направленный мутагенез может быть достигнут только в растительных клетках посредством прямого переноса ДНК с использованием химических или физических методов. В публикациях сообщалось о некоторых таких методах переноса ДНК, которые включают электропорацию, обработку полиэтиленгликолем (ПЭГ) протопластов, биолистичекую бомбардировку каллюсного материала растения и микроинъекцию ДНК в индивидуальные протопласты или ткань. В уровне техники нет никаких указаний относительно предпочтительного метода переноса одноцепочечных нуклеиновых оснований для направленного мутагенеза, конкретно, для переноса ДНК в растения или растительные протопласты.
С целью достижения настолько высокой эффективности направленного мутагенеза в растениях, насколько это возможно, авторы настоящего изобретения в ходе своих исследований идентифицировали четыре фактора, которые следует оптимизировать. Во-первых, мутагенное нуклеиновое основание предпочтительно вводится с высокой эффективностью трансформации, т.е. вводится в настолько большое количество растительных клеток, насколько это возможно. Во-вторых, обработка предпочтительно не летальна для большинства клеток, гарантируя, что настолько, насколько возможно много трансформированных клеток также будет выживать при процедуре трансформации (эффективность выживания). В-третьих, метод трансформации предпочтительно не является вредным для последующих делений трансформированных растительных клеток при образовании микрокаллюсов (регенерация/эффективность культивирования) и, наконец, предпочтительной является возможность идентификации индивидуальных регенерированных растений, произошедших после событий направленного мутагенеза без применения селекции (эффективность идентификации). Большинство методов трансформации ДНК в индивидуальные растительные клетки использует протопласты, выделенные непосредственно из листьев (мезофильные протопласты) или из клеточных суспензий (рассмотрено в Sheen, J. (2001) Plant Phys. 127: 1466-1475). Протопласты могут использоваться для исследований транзиторной экспрессии, в случае которой экспрессия гена или локализация белка могут оцениваться вскоре после трансформации, или для получения стабильно трансформированных растений при росте протопластов на среде для усиления образования каллюса и для органогенеза.
О трансформации растительных протопластов с использованием протопластов сообщалось ранее (Fromm et al. (1985) Proc. Natl. Acad. Sci. USA 82: 5824-5828; Nishiguchi et al. (1986) Plant Cell Rep. 5: 57-60; Ou-Lee et al. (1986) Proc. Natl. Acad. Sci. USA 83: 6815-6819; Hauptmann et al. (1987) Plant Cell Rep. 6: 265-270; Jones et al. (1989) Plant Mol. Biol. 13: 503-511). Как правило, интенсивность поля (В/см), дающая наибольшую эффективность трансформации, приводит в результате к выживанию протопластов <50% (Jones et al. (1989) Plant Mol.Biol. 13: 503-511). При исследованиях электропорации табака нами было обнаружено, что только приблизительно 10% от всех протопластов табака в образце трансформируется плазмидой, экспрессирующей GFP, и эта относительно низкая эффективность трансформации также наблюдалась после электропорации протопластов Arabidopsis (Miao et al. (2007) Nature Protocols 10: 2348-2353). Вообще, оптимальные условия электропорации должны определяться эмпирически для каждого вида растений, и они также могут варьироваться в зависимости от типа прибора для электропорации, метода и буферов, используемых для выделения протопластов. В то время как электропорация успешно применяется для многих видов растений, она остается трудным методом с несколькими серьезными ограничениями (как обсуждается в: http://genetics.mgh.harvard.edu/sheenweb/faq.html), конкретно, в том, что касается воспроизводимости. Следовательно, использование электропорации менее желательно для улучшения эффективности в целом для направленного мутагенеза TNE (направленная замена нуклеотидов).
Прямой перенос генов с использованием биолистической доставки очень успешно использовался при генерировании трансгенных растительных культур и регулярно используется для стабильной интеграции трансгенов. Клеточные суспензии переносят на твердую среду для индукции каллюса и этот материал бомбардируют золотыми частицами, получаемыми с помощью газового источника высокого давления. Сообщалось, что частота трансформации низкая, ~0,01% от общего числа трансформируемых клеток. Из-за низкой эффективности трансформации, выживание трансформированных клеток трудно оценить. Напротив, эффективность регенерации после бомбардировки, вероятно, является высокой благодаря сильному делению обработанного материала. Однако так как TNE встречается в одной клетке одного каллюса, такое событие легко потерять, если оно не селектируется или, альтернативно, регенерированные растения будут химерными для события направленного мутагенеза. Таким образом, бомбардировка не практична для осуществления направленного мутагенеза в не селектируемых локусах. Напротив, возможно извлечь события мутагенеза с использованием протопластов, поскольку каждый микрокаллюс выделяют из одного протопласта.
Индуцированные однонуклеотидные конверсии с помощью химер в ALS продемонстрировали, что направленный мутагенез может детектироваться в клетках табака, кукурузы и риса. Бомбардировку использовали для табака (Beetham et al. 1999 Proc.Natl.Acad.Sci. USA 96: 8774-8778; Kochevenko et al. 2003 Plant Phys. 132: 174-184), кукурузы (Zhu et al. 1999 Proc. Natl. Acad. Sci. USA 96, 8768- 8773) и риса (Okuzaki et al. 2004 Plant Cell Rep. 22: 509-512). Beetham et al. (1999) сообщили, что частота устойчивости к гербициду после прямого переноса химер повышается в 20 раз по сравнению с фоновой частотой мутаций (по оценкам 10-7-10-8). Kochevenko et al. также использовали электропорацию для осуществления экспериментов направленного мутагенеза в мезофильных протопластах табака. Авторы настоящего изобретения смогли получить гербицидоустойчивый каллюс табака с частотой 0,0001% по сравнению с частотой, полученной Beetham et al. Это предполагает, что при работе с одинаковыми видами растений и одинаковой нуклеотидной мишенью, как в этом случае, метод прямой доставки ДНК не имеет большого воздействия на эффективность направленного мутагенеза, который остается на нежелательно низком уровне. Однако Ruiter et al. (2003 Plant Mol. Biol. 53, 715-729) проводили оба эксперимента бомбардировки и электропорации в обоих типах растений, в табаке, и рапсе и не смогли детектировать никакого эффекта химер.
Сообщали об эффективности бомбардировки каллюсов обоих растений, кукурузы и риса, с эффективностью 0,01% трансформированных клеток (Zhu et al. (1999) Proc. Natl. Acad. Sci USA 96, 8768-8773; Okuzaki et al. (2004) Plant Cell Rep. 22: 509-512). Однако это было осуществимо только в селектируемых локусах.
ПЭГ-опосредованная трансформация протопластов известна уже с 1985. В первом методе трансформации протопластов применяли ПЭГ (Krens et al. (1982) Nature 296: 72-74; Potyrykus et al. (1985) Plant Mol. Biol. Rep. 3: 117-128; Negrutiu et al. (1987) Plant Mol.Biol. 8: 363-373). Этот метод применим к протопластам из множества различных растений (Rasmussen et al. (1993) Plant Sci. 89: 199-207). Считается, что ПЭГ стимулирует трансформацию путем осаждения ДНК в присутствии двухвалентных катионов на поверхности протопластов, где она затем интернализируется (Maas & Werr (1989) Plant Cell Rep. 8: 148-151). ПЭГ-трансформация представляет собой наиболее часто используемый метод трансформации протопластов Arabidopsis (http://genetics.mgh.harvard.edu/sheenweb/faq.html) (Mathur et al. Methods in molecular biology, vol. 82, 267-276) и хорошо подходит четырем требованиям для эффективной TNE. Когда протопласты табака обрабатывают с помощью ПЭГ, меченный биотином одноцепочечный олигонуклеотид может детектироваться во всех исследуемых клетках. Выживание, по оценкам с помощью прижизненного окрашивания с использованием флюоресцеин диацетата, составляет >90% после обработки ПЭГ. Не все протопласты сохраняют способность деления из микрокаллюсов. В типичном выделении не обработанных протопластов табака приблизительно 25% приходится на микрокаллюсы. Обработка ПЭГ оказывает незначительное воздействие на эффективность регенерации, которая падает приблизительно на 10%, но это не очень значительно по сравнению с другими методами трансформации. Ни в одном из вышеописанных методов, известных их уровня техники, не предполагалось использование ПЭГ-трансформации для сайт-направленного мутагенеза, конкретно TNE.
Авторы настоящего изобретения попытались улучшить метод прямого переноса ДНК для получения эффективного направленного мутагенеза в растительных клетках. Авторы настоящего изобретения обнаружили, что из числа методов трансформации, описанных где-либо в настоящем документе, ПЭГ-трансформация протопластов значительно улучшает общую эффективность направленного мутагенеза по сравнению с электропорацией и биолистикой. Это оказалось неожиданным, поскольку методы направленного мутагенеза в растениях к настоящему моменту, как представляется, предпочитали использование электропорации, несмотря на ассоциированные низкие эффективности. Кроме того, большинство улучшений в методе были направлены на улучшение мутагенных нуклеиновых оснований, а не на систему доставки мутагенного нуклеинового основания в мишень геномной ДНК.
Для сравнения авторы настоящего изобретения использовали мутагенное одноцепочечное нуклеиновое основание для получения конверсии P194Q в локусе ALS, ведущей к устойчивости к гербицидам. Идентичные мутагенные одноцепочечные нуклеиновые основания вводили в мезофильные протопласты табака с использованием или ПЭГ-опосредованной трансформации, или электропорации и селектировали клетки, устойчивые к гербицидам, с использованием условий селекции. Таким образом, авторы настоящего изобретения обнаружили, что ПЭГ-опосредованная трансформация растительных клеток является наиболее эффективным методом для осуществления направленного мутагенеза в растительных клетках по сравнению с известными методами трансформации.
В одном аспекте изобретение относится к способу направленного изменения дуплексной акцепторной ДНК-последовательности в протопласте растительной клетки, включающему объединение дуплексной акцепторной ДНК-последовательности с мутагенным одноцепочечным нуклеиновым основанием, где дуплексная акцепторная ДНК-последовательность содержит первую ДНК-последовательность и вторую ДНК-последовательность, которая комплементарна первой ДНК-последовательности, и где донорное мутагенное одноцепочечное нуклеиновое основание включает по меньшей мере одну ошибочную пару по отношению к дуплексной акцепторной ДНК-последовательности, которую необходимо изменить, предпочтительно, по отношению к первой ДНК-последовательности, где способ дополнительно включает стадию введения мутагенного одноцепочечного нуклеинового основания в протопласты клеток с использованием полиэтиленгликоль (ПЭГ)-опосредованной трансформации.
Мутагенное одноцепочечное нуклеиновое основание вводят в контакт с протопластами растения, трансформируемыми с использованием метода на основе ПЭГ-трансформации. Метод ПЭГ-опосредованной трансформации сам по себе является широко известным, и по необходимости специалист может произвести небольшие изменения в конкретных протоколах, не выходя за рамки сущности настоящего изобретения.
Мутагенное одноцепочечное нуклеиновое основание, используемое в настоящем изобретении, имеет длину, которая соответствует другому (химерному) мутагенному одноцепочечному нуклеиновому основанию, используемому в направленном мутагенезе, т.e. обычно между 10-60 нуклеотидов, предпочтительно 20-55 нуклеотидов, более предпочтительно 25-50 нуклеотидов. В определенных воплощениях изобретения, мутагенное одноцепочечное нуклеиновое основание, используемое в настоящем изобретении, может быть модифицировано, например, с помощью LNA и/или пропиниловых модификаций, как описано в заявках WO 2007073166 и WO 2007073170. Таким образом, в некоторых вариантах осуществления, мутагенное одноцепочечное нуклеиновое основание содержит по меньшей мере одну LNA, локализованную в положении, взятом из ошибочной пары-мишени и предпочтительно две LNA, локализованные на месте по меньшей мере одного нуклеотида, удаленного с каждой стороны от ошибочной пары. Кроме того, эти LNA представляют собой по меньшей мере 3, 4 или 5 нуклеотидов, удаленных с 5'- и/или 3'-конца мутагенного одноцепочечного нуклеинового основания. В некоторых вариантах осуществления, мутагенное одноцепочечное нуклеиновое основание может включать одну или более пропиновых замен, по существу, как описано в WO 2007073166 и WO 2007073170. В некоторых вариантах осуществления, донорное мутагенное одноцепочечное нуклеиновое основание может быть конъюгировано с белком, таким как сигнал ядерной локализации. В этом варианте осуществления, олигонуклеотид, используемый в настоящем изобретении, соединяется посредством подходящего (линкерного) метода с сигналом ядерной локализации, таким как известный (NLS) пептид большого Т-антигена SV40, GATA транскрипционный фактор 11, хеликаза XBP1 ДНК-репарации, белок светоопосредованного развития DET1, транскрипционный фактор ERF, PR-родственный транскрипционный аткиватор PTI6 и ядерный спиральный белок, по существу, как описано в заявках на стадии рассмотрения PCT/NL2007/000279. Конъюгат олигонуклеотида и сигнала ядерной локализации может использоваться в способе трансформации на основе ПЭГ, описанном в настоящем описании.
Изменение, полученное с помощью способа по настоящему изобретению, представляет собой делецию, замену или вставку по меньшей мере одного нуклеотида. Предпочтительно, изменение представляет собой замену. Больше нуклеотидов может быть заменено на один нуклеотид, но ожидается, что эффективность уменьшится, следовательно, существует предпочтение для изменения одного нуклеотида. ДНК-мишень (или дуплексная акцепторная ДНК) может быть из любого источника, но предпочтительно, ДНК-мишень берут из растения. Предпочтительно, ДНК-мишень берут из геномной ДНК, линейной ДНК, искусственных хромосом, ядерной хромосомной ДНК, хромосомной ДНК органелл, эписомной ДНК. Способ согласно изобретению может использоваться для изменения клетки, коррекции мутации путем восстановления дикого типа, индуцирования мутации, инактивации фермента путем нарушения кодирующей области, модификации биоактивности фермента путем нарушения кодирующей области, модификации белка путем нарушения кодирующей области.
В одном аспекте изобретение относится к применению ПЭГ-опосредованной трансформации для улучшения эффективности направленного мутагенеза в растительных протопластах. Не имея ограничений по части теории, считается, что использование ПЭГ-опосредованной трансформации осаждает ДНК на клеточной мембране протопласта. Осажденная ДНК инкапсулируется клеточной мембраной и вводится в протопласт в защищенной форме. Протопласт в ходе его обычного клеточного цикла непосредственно после этого образования с удалением клеточной стенки начнет свой обычный процесс регенерации клеточной стенки. Клеточное деление обычно начинается позже (от нескольких часов до нескольких дней). Направленное нуклеотидное изменение, как правило, имеет место во время клеточного деления с использованием клеточного механизма репарации. В период времени между введением донорной ДНК в протопласт и стартом клеточного деления, донорная ДНК подвергается воздействию со стороны механизма защиты клетки, такого как экзонуклеазы, и, вероятно, дегенерирует, и, следовательно, становится неэффективной для TNE. При использовании метода ПЭГ-опосредованной трансформации, донорная ДНК инкапсулируется посредством эндоцитоза и таким путем, по меньшей мере, временно защищается от дегенеративного действия эндонуклеаз. Когда ДНК высвобождается из своей инкапсулированной формы, она имеет повышенные шансы присутствия в сам момент или приблизительно в момент клеточного деления, во время которого ДНК (т.e. мутагенное одноцепочечное нуклеиновое основание) становится доступно для поиска комплемента в ДНК акцепторной клетки и для замены нуклеотида, как в обычных механизмах направленного мутагенеза.
Примеры
Сравнение частот направленного мутагенеза с использованием или ПЭГ-опосредованной трансформации, или электропорации
Выделение протопластов
In vitro культуры ростков Nicotiana tabacum cv Petit Havana линии SR1 поддерживали на среде MS20 с 0,8% агаром Difco в высоких стеклянных колбах в течение 16/8-часового светового дня при освещенности 2000 люкс при 25°C и 60-70% относительной влажности. Среда MS20 представляет собой основную среду Murashige и Skoog's medium (Murashige, T. and Skoog, F., Physiologia Plantarum, 15: 473-497, 1962), содержащую 2% (мас./об.) сахарозу, без добавления гормонов, и 0,8% агар Difco. Собирали полностью распустившиеся листья 3-6-недельных культур ростков. Листья нарезали полосками толщиной 1 мм, которые затем переносили в большие чашки Петри (100 мм×100 мм), содержащие 45 мл базальной среды MDE для преплазмолизисной обработки в течение 30 минут. Базальная среда MDE содержит 0,25 г KCl, 1 г MgSO4·7H2O, 0,136 г KH2PO4, 2,5 г поливинилпирролидона (MW 10000), 6 мг нафталинуксусной кислоты и 2 мг 6-бензиламинопурина в общем объеме 900 мл. Осмоляльность раствора регулировали до 600 мОсм·кг-1 сорбитом, pH до 5,7. Затем добавляли 5 мл базового фермента SR1. Базовый фермент состоит из 750 мг целлюлазы Onozuka R10, 500 мг дризелазы (driselase) и 250 мг мацерозима R10 на 100 мл, отфильтрованные через ватманскую бумагу и стерилизованные посредством фильтрации. Гидролизу давали пройти в течение ночи в темноте при 25°C. Гидролизованные листья фильтровали через нейлоновый фильтр 50 мкм в стерильной пробирке. Равный объем холодной промывочной среды KCl для промывания фильтра и объединенной с протопластами суспензии. Промывочная среда KCl состоит из 2 г CaCl2·2H2O на литр и достаточного количества KCl для доведения осмоляльности до 540 мОсм·кг-1. Суспензию переносили в 10-мл пробирки, и протопласты осаждали в течение 10 минут при 85xg при 4°C. Супернатант удаляли и осажденные протопласты тщательно ресуспендировали в 5 мл холодной промывочной среды MLm, которая представляет собой макронутриенты среды MS (Murashige, T. and Skoog, F., Physiologia Plantarum, 15: 473-497, 1962) в половине обычной концентрации, 2,2 г CaCl2·2H2O на литр и количество маннита для доведения осмоляльности до 540 мОсм·кг-1. Содержимое 2 пробирок объединяли и центрифугировали в течение 10 минут при 85xg при 4°C. Супернатант удаляли и осажденные протопласты тщательно ресуспендировали в 5 мл холодной промывочной среды MLs, которая представляет собой среду MLm, в которой маннит заменен на сахарозу. Содержимое 2 пробирок объединяли и осторожно добавляли 1 мл промывочной среды KCl поверх раствора сахарозы, чтобы не разрушить нижнюю фазу. Протопласты центрифугировали в течение 10 минут при 85xg при 4°C. Интерфазу между растворами сахарозы и KCl, содержащую живые протопласты, тщательно собирали. Добавляли равный объем промывочной среды KCl и тщательно перемешивали. Плотность протопластов измеряли с помощью гемоцитометра.
ПЭГ-трансформация
Суспензию протопластов центрифугировали при 85xg в течение 10 минут при 5°C. Супернатант удаляли и осажденные протопласты ресуспендировали в промывочной среде KCl до конечной концентрации 106·мл-1. В 10-мл пробирке мягко, но тщательно перемешивали 250 мкл суспензии протопластов, 1,6 нмоль мутагенного одноцепочечного нуклеинового основания и 250 мкл ПЭГ-раствора. Через 20 минут инкубации при комнатной температуре капельно добавляли 5 мл холодного раствора 0,275 M Ca(NO3)2. Суспензию протопластов центрифугировали при 85xg в течение 10 минут при 4°C. Супернатант удаляли и осажденные протопласты тщательно ресуспендировали в 1,25 мл культуральной среды T0, содержащей 50 мкг·мл-1 цефотаксима и 50 мкг·мл-1 ваномицина, культуральной среды для мутагенного одноцепочечного нуклеинового основания, содержащей (на 1 литр, pH 5,7) 950 мг KNO3, 825 мг NH4NO3, 220 мг CaCl2·2H2O, 185 мг MgSO4·7H2O, 85 мг KH2PO4, 27,85 мг FeSO4·7H2O, 37,25 мг Na2EDTA·2H2O, микронутриенты согласно среде Heller's medium (Heller, R., Ann Sci Nat Bot Biol Veg 14: 1-223, 1953), витамины согласно среде Morel и Wetmore's medium (Morel, G. and R.H. Wetmore, Amer. J. Bot. 38: 138-40, 1951), 2% (мас./об.) сахарозы, 3 мг нафталинуксусной кислоты, 1 мг 6-бензиламинопурина и количество маннита для доведения осмоляльности до 540 мОсм·кг-1. Суспензию переносили в 35-мм чашку Петри. Добавляли равный объем агарозной среды T0 и мягко перемешивали agarose. Образцы инкубировали при 25°C в темноте и далее культивировали, как описано ниже.
Электропорация
Протопласты центрифугировали при 85xg в течение 10 минут при 5°C. Супернатант удалили и осадок ресуспендировали в ледяном буфере электропорации, состоящем из 10 мМ HEPES, 80 мМ NaCl, 0,04 мМ CaCl2, 0,4 M маннита, pH 5,7 регулировали до 540 мОсм·кг-1 с помощью маннита до конечной концентрации 106 мл-1. Протопласты держали на льду на протяжении всей процедуры. В широкую 0,4-см кювету для электропорации добавляли 4,5 нмоль мутагенного одноцепочечного нуклеинового основания и 700 мкл суспензии протопластов. Однократный пульс экспоненциального распада доставляется к клеточной суспензии с использованием системы электропорации Biorad GenePulser XceII, оборудованной PC- и CE-модулем согласно следующим параметрам:
интенсивность поля 500 В·см-1,
емкость 950 мкФ.
При этих условиях сопротивление образца составляет приблизительно 30 Ом, и полученная в результате постоянная времени составляет приблизительно 30 мс. Эти параметры выбирали в качестве параметров, дающих наибольший уровень транзиторной экспрессии GFP в протопластах табака через 24 часа после электропорации. После подачи импульса, протопластам давали возможность восстановиться в кювете при комнатной температуре в течение 30 минут. Протопласты затем восстанавливали в 1 мл культуральной среды T0 и переносили в 10-мл пробирку. Кювету промывали с помощью дополнительных 5 мл культуральной среды T0, которую объединяли с суспензией протопластов. После основательного, но мягкого перемешивания добавляли 50 мкг·мл-1 цефотаксима и 50 мкг·мл-1 ванкомицина и 1,25 мл суспензии протопластов переносили в 35-мм чашку Петри. Добавляли равный объем агарозной среды T0 и смесь мягко гомогенизировали. Образцы инкубировали при 25°C в темноте и далее культивировали, как описано ниже.
Культивирование протопластов
Через 10 дней культивирования, агарозную пластинку разрезали на 6 равных частей и переносили в чашку Петри, содержащую 22,5 мл среды MAP1AO, содержащей 20 нМ хлорсульфурона. Эта среда состоит из (на 1 литр, pH 5,7) 950 мг KNO3, 825 мг NH4NO3, 220 мг CaCl2·2H2O, 185 мг MgSO4·7H2O, 85 мг KH2PO4, 27,85 мг FeSO4·7H2O, 37,25 мг Na2EDTA·2H2O, микронутриенты согласно среде Murashige и Skoog's medium (Murashige, T. and Skoog, F., Physiologia Plantarum, 15: 473-497, 1962) в одной десятой от исходной концентрации, витамины согласно среде Morel и Wetmore's medium (Morel, G. and R.H. Wetmore, Amer. J. Bot. 38: 138-40, 1951), 6 мг пирувата, 12 мг каждого из соединений, фумаровой кислоты и лимонной кислоты, 3% (ма./об.) сахарозы, 6% (мас./об.) маннита, 0,03 мг нифталинуксусной кислоты и 0,1 мг 6-бензиламинопурина. Образцы инкубировали при 25°C при слабом освещении в течение 6-8 недель. Растущие каллюсы затем переносили в среду MAPI и давали им возможность развиться в течение еще 2-3 недель. Среда MAP1 имеет тот же состав, что и среда MAP1AO, однако содержит 3% (мас./об.) манит, вместо 6%, и 46,2 мг·л-1 гистидина (pH 5,7). Она затвердевает с помощью 0,8% (мас./об.) агара Difco. Каллюсы затем переносили в среду RP с использованием стерильных пинцетов. Среда RP состоит из (на 1 литр, pH 5,7) 273 мг KNO3, 416 мг Ca(NO3)2·4H2O, 392 мг Mg(NO3)2·6H2O, 57 мг MgSO4·7H2O, 233 мг (NH4)2SO4, 271 мг KH2PO4, 27,85 мг FeSO47H2O, 37,25 мг Na2EDTA·2H2O, микронутриенты согласно среде Murashige и Skoog's medium в одной пятой от опубликованной концентрации, витамины согласно среде Morel и Wetmore's medium (Morel, G. and R.H. Wetmore, Amer. J. Bot. 38: 138-40, 1951), 0,05% (мас./об.) сахарозы, 1,8% (мас./об.) маннита, 0,25 мг зеатина и 41 нМ хлорсульфурона, и затвердевает с помощью 0,8% (мас./об.) агара Difco. Зрелые ростки переносили в субстрат для выращивания растений через 2-3 недели.
Мутагенные одноцепочечные нуклеиновые основания
Все мутагенные одноцепочечные нуклеиновые основания синтезировали с помощью Eurogentec (Seraing, Belgium), очищали с использованием HPLC с обращенной фазой и ресуспендировали в стерильной воде milliQ. Перед использованием мутагенное одноцепочечное нуклеиновое основание нагревали до 95°C в течение 5 минут. Мутагенное одноцепочечное нуклеиновое основание 06Q262 сконструировали для введения одной ошибочной пары (подчеркнутый нуклеотид) в ген ALS табака (регистрационный номер X07644) в положении кодона P194, которое приводит в результате к конверсии CCA на CAA (P194Q). Мутагенное одноцепочечное нуклеиновое основание 06Q261 является точной парой для последовательности гена ALS табака и служит в качестве отрицательного контроля. Мутагенное одноцепочечное нуклеиновое основание 06Q263 состоит из случайной комбинации 40 нуклеотидов и служит в качестве отрицательного контроля.
06Q261 5'TCAGTACCTATCATCCTACGTGGCACTTGACCTGTTATAG [SEQ ID 1]
06Q262 5'TCAGTACCTATCATCCTACGTTGCACTTGACCTGTTATAG [SEQ ID 2]
06Q263 5'ATCGATCGATCGATCGATCGATCGATCGATCGATCGATCG [SEQ ID 3]
Выживание протопластов при обработке
Выживание протопластов после обеих процедур, ПЭГ-трансформации и электропорации, оценивали с помощью активности эстеразы с использованием флуоресцентного красителя для прижизненной окраски флюоресцеиндиацетата (FDA) через 24 часа после трансформации. Добавляли два мкл 5 мг·мл-1 базового раствора FDA в ацетоне к 1 мл трансформированных протопластов. Пропорцию флуоресцентных протопластов во всей популяции считали на гемоцитометре. Наблюдения проводили с помощью прямого эпифлуоресцентного микроскопа Nikon Eclipse E600, оборудованного набором фильтров GFP LP (EX480/40, DM505, BA510). Возбуждение обеспечивали с помощью ртутной 100 Вт лампы сверхвысокого давления. Обнаруживали изображения с использованием камеры DS-2MBWc CCD, присоединенной к контроллеру DS-U 1, присоединенному к PC-модулю, выполняющего программу NIS Element image по сбору данных/анализу изображений.
Результаты
Суммирование результатов трансформации с использованием обеих процедур, ПЭГ-трансформации и электропорации, представлено в таблице 1. При использовании ПЭГ-трансформации степень выживания протопластов была значительно выше по сравнению с использованием электропорации. Природа электропорации сама по себе является более вредной для выживания протопластов, чем природа ПЭГ-трансформация, которая приводит в результате к гораздо более высокой степени восстановления/выживания, а также к более высокой эффективности направленного мутагенеза. Эффективность направленного мутагенеза оценивали после инкубирования протопластов в присутствии хлорсульфурона.
Таблица 1: Сравнение ПЭГ-трансформации и электропорации в отношении степени выживания протопластов, выраженное в процентном содержании флуоресцентных протопластов после окрашивания FDA в восстановленной популяции протопластов. Результаты представляют собой среднее значение 3 независимых повторов экспериментов ± Станд.Откл.
ПЦР-амплификация ALS и секвенирование
ДНК выделяли из устойчивых к хлорсульфурону микроколоний табака с использованием набора реагентов DNeasy kit (Qiagen) и использовали в качестве матрицы в реакции ПЦР. Конверсии кодонов-мишеней в гене ALS табака детектировали с использованием праймеров 5'GGTCAAGTGCCACGTAGGAT [SEQ ID 4] и 5'GGGTGCTTCACTTTCTGCTC [SEQ ID 5], которые амплифицируют фрагмент 776 п.о. этого гена, включая кодон 194. Нуклеотидную конверсию в устойчивом к гербициду каллюсе табака подтверждали с помощью клонирования ПЦР-продуктов в вектор pCR2.1::TOPO (Invitrogen) и секвенирования индивидуальных плазмид. Табак содержит 2 аллеля ALS (SurA и SurB). Нуклеотидная конверсия кодона P194 в каждом из этих локусов является достаточной для придания устойчивости к хлорсульфурону. Поскольку табак представляет собой аллотетраплоидный вид, существует восемь возможных мишеней в табаке, в которых может встретиться TNE. В соответствии с этим, необходимо секвенировать >10 плазмидных клонов, содержащих ПЦР-продукт для детектирования одного с конверсией CCA на CAA. Это предполагает, что в каждом устойчивом каллюсе только 1 из 8 аллелей ALS подвергся нуклеотидной конверсии, опосредованной направленным мутагенезом. Для всех каллюсов, полученных в настоящем исследовании, мы наблюдали ожидаемую нуклеотидную конверсию CCA на CAA.
Эффективность направленного мутагенеза увеличивается, если вместо электропорации используется ПЭГ-опосредованная трансформация.
Были проведены эксперименты по сравнению эффективности двух способов трансформации протопластов для направленного мутагенеза, электропорации и полиэтиленгликоль-опосредованной трансформации. В качестве мишени был выбран ген ацетолактатсинтазы табака (ALS) (регистрационный номер X07644), поскольку конверсия пролина 194 в глутамин (конверсия P194Q) дает в результате фермент, не чувствительный к гербициду хлорсульфурону. Впоследствии протопласты, в которые была успешно внедрена мутация, приводящая к такой аминокислотной замене, будут нормально развиваться в культуральной среде, дополненной гербицидом хлорсульфуроном, в то время как немутированные протопласты погибнут, что позволит провести сравнение различных методов доставки.
Протокол
Протопласты выделяли, как описано в настоящей заявке, и трансформировали используя 20 мкг олигонуклеотида 06Q262 (5'TCAGTACCTATCATCCTACGTTGCACTTGACCTGTTATAG), разработанного для получения C->A трансверсии в положении кодона P194 в кодирующей последовательности гена ALS табака, в результате которой пролин 194 превращается в глутамин. ПЭГ-опосредованную трансформацию и электропорацию выполняли используя параметры, приведенные в таблице 2. Трансформированные протопласты восстанавливали в культуральной среде и выращивали до получения конечной плотности 105/мл. Протопластам давали возможность развиваться в течение 10 дней в темноте при 25°C, а затем переносили в культуральную среду, дополненную 20 нМ хлорсульфурона. Через 4-6 недель культивирования устойчивые каллюсы собирали и генотипировали.
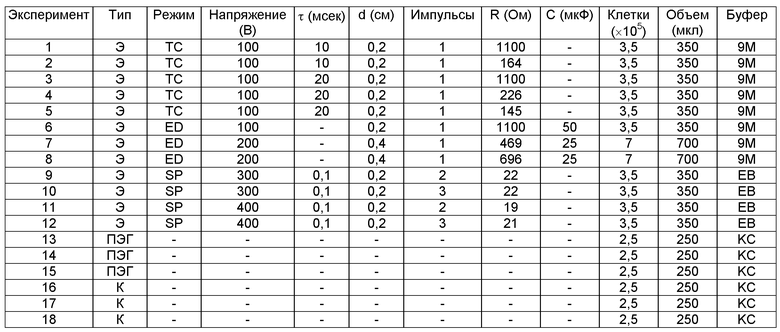
Таблица 2: Условия проведения экспериментов посредством электропорации и ПЭГ-трансформации протопластов табака с помощью мутагенных олигонуклеотидов. Протопласты были трансформированы 20 мкг олигонуклеотида 06Q262.
Тип: Э=электропорация; ПЭГ=ПЭГ-трансфекция, K=контроль.
Режим: TC=постоянное время; ED=экспоненциальный спад, SP=прямоугольные импульсы.
Буфер: 9M=9 г маннита в 100 мл, фильтр стерилизовать, хранить при температуре 4°C; KC=7 мМ CaCl2·2H2O, 260 мМ KCl, pH 5,7, фильтр стерилизовать, хранить при температуре 4°C; EB=10 мМ HEPES, pH 6,7, 50 мМ KCl, 5 мМ CaCl2, фильтр стерилизовать.
Для каждого условия, электропорацию или ПЭГ-трансфекцию выполняли в трех экземплярах, и образцы объединяли до внесения в культуру.
* в объединенных 3 реакциях электропорации (1,05×106 протопластов)
Результаты
В образцах, подвергнутых электропорации, нельзя было выявить каллюсы, устойчивые к хлорсульфурону (см. таблицу 3). Во всех образцах протопластов, подвергнутых ПЭГ-трансформации, протопласты развились в микроколонии. После переноса в селективную среду развивались каллюсы, устойчивые к хлорсульфурону (см. таблицу 3), которые были собраны для генотипирования. Наличие мутации было подтверждено во всех устойчивых микроколониях каллюсов с помощью Sange последовательности.
Заключение
Количественное сравнение эффективности направленного мутагенеза, выполненного либо ПЭГ-опосредованной трансформацией, либо электропорацией, в качестве способа доставки олигонуклеотида, показало, что ПЭГ-опосредованная трансформация значительно превосходит электропорацию (см. таблицу 3). Начиная с сопоставимых популяций протопластов, полученный увеличенный показатель доставки олигонуклеотида позволяет успешно идентифицировать мутантов, устойчивых к хлорсульфурону, в то время как в образцах с электропорацией такие мутанты не могли быть выявлены. Учитывая все факты, эти результаты демонстрируют, что ПЭГ-трансформация в протопластах приводит к более высокой эффективности направленного мутагенеза по сравнению с электропорацией.
Таблица 3: Количественное сравнение эффективности направленного мутагенеза, выполненного либо ПЭГ-опосредованной трансформацией, либо электропорацией. Эффективность направленного мутагенеза увеличивается, если вместо электропорации используется ПЭГ-опосредованная трансформация.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |
| БЫСТРЫЙ НАПРАВЛЕННЫЙ АНАЛИЗ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР ДЛЯ ОПРЕДЕЛЕНИЯ ДОНОРНОЙ ВСТАВКИ | 2014 |
|
RU2668819C2 |
| ТОЛЕРАНТНОЕ К СТРЕССУ ТРАНСГЕННОЕ РАСТЕНИЕ ПШЕНИЦЫ | 2005 |
|
RU2376377C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ ZEA MAYS L., УСТОЙЧИВЫХ К ПОВРЕЖДЕНИЯМ, ВЫЗЫВАЕМЫМ НАСЕКОМЫМИ | 1988 |
|
RU2126047C1 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2010 |
|
RU2556376C2 |
| ФРАГМЕНТ ДНК (ВАРИАНТЫ), СЛИТЫЙ ФРАГМЕНТ ДНК ДЛЯ ЭКСПРЕССИИ В РАСТЕНИЯХ И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ И ЕГО ПОТОМКОВ | 1989 |
|
RU2130491C1 |
| СПОСОБ ПОЛУЧЕНИЯ УСТОЙЧИВЫХ К СУЛЬФОНИЛМОЧЕВИННЫМ ГЕРБИЦИДАМ ДВУДОЛЬНЫХ РАСТЕНИЙ | 1987 |
|
RU2099422C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ МОДИФИКАЦИИ ЗАДАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ-МИШЕНИ | 2012 |
|
RU2663354C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2017 |
|
RU2664865C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2010 |
|
RU2612156C2 |


Изобретение относится к области биохимии, в частности к способам направленного изменения дуплексной акцепторной ДНК-последовательности и улучшения эффективности направленного мутагенеза в растительных протопластах. Заявленные способы включают введение в протопласты растительных клеток одноцепочечного мутагенного олигонуклеотида, имеющего в своем составе по меньшей мере один ошибочный нуклеотид по отношению к дуплексной акцепторной ДНК-последовательности, которую необходимо изменить, с использованием ПЭГ-опосредованной трансформации. Изобретение позволяет повысить частоту направленного мутагенеза. 2 н. и 8 з.п. ф-лы, 3 табл.
1. Способ направленного изменения дуплексной акцепторной ДНК-последовательности в протопласте растительной клетки, включающий объединение дуплексной акцепторной ДНК-последовательности с донорным одноцепочечным мутагенным олигонуклеотидом, где дуплексная акцепторная ДНК-последовательность содержит первую ДНК-последовательность и вторую ДНК-последовательность, которая комплементарна первой ДНК-последовательности, и где донорный мутагенный олигонуклеотид имеет в своем составе по меньшей мере один ошибочный нуклеотид по отношению к дуплексной акцепторной ДНК-последовательности, которую необходимо изменить, предпочтительно, по отношению к первой ДНК-последовательности, где способ включает стадию введения мутагенного олигонуклеотида в протопласты клеток с использованием полиэтиленгликоль (ПЭГ)-опосредованной трансформации.
2. Способ по п.1, где одноцепочечный мутагенный олигонуклеотид имеет длину от 10 до 60 нуклеотидов.
3. Способ по п.1, где одноцепочечный мутагенный олигонуклеотид включает замены закрытых нуклеиновых кислот (LNA).
4. Способ по п.1, где одноцепочечный мутагенный олигонуклеотид включает пропиновые замены.
5. Способ по п.1 для изменения клетки, коррекции мутации путем восстановления дикого типа, индуцирования мутации, инактивации фермента путем нарушения кодирующей области, модификации биоактивности фермента путем изменения кодирующей области, модификации белка путем нарушения кодирующей области.
6. Способ улучшения эффективности направленного мутагенеза в растительных протопластах, включающий стадию ПЭГ-опосредованной трансформации, в котором используют одноцепочечный мутагенный олигонуклеотид, где донорный мутагенный олигонуклеотид имеет в своем составе по меньшей мере один ошибочный нуклеотид по отношению к дуплексной акцепторной ДНК-последовательности, которую необходимо изменить.
7. Способ по п.6, в котором одноцепочечный мутагенный олигонуклеотид имеет длину от 10 до 60 нуклеотидов.
8. Способ по п.7, где одноцепочечный мутагенный олигонуклеотид включает LNA-замены.
9. Способ по п.6, где одноцепочечный мутагенный олигонуклеотид включает пропиновые замены.
10. Способ по п.6 для изменения клетки, коррекции мутации путем восстановления дикого типа, индуцирования мутации, инактивации фермента путем нарушения кодирующей области, модификации биоактивности фермента путем изменения кодирующей области или модификации белка путем нарушения кодирующей области.
| WO 2007073154 A, 28.06.2007 | |||
| HOHE ANNETTE et al., An improved and highly standardised transformation procedure allows efficient production of single and multiple targeted gene-knockouts in a moss, Physcomitrella patens, Current Genetics, 2004, Volume 44, Issue 6, pp | |||
| Ручной ткацкий станок | 1922 |
|
SU339A1 |
| WO2001087914 A, 22.11.2001 | |||
| RU 2003137841 A, 10.02.2005 | |||

