Настоящее изобретение относится к применению S-аденозилметионина (SAM) в комбинации с супероксиддисмутазой (SOD) для изготовления лекарственных средств для лечения болезни Альцгеймера.
Предшествующий уровень техники
Болезнь Альцгеймера (AD) - очень широко распространенная нейродегенеративная форма деменции, которая поражает значительную часть мирового населения старше 70 лет в соотношении 1:2 между мужчинами и женщинами. Заболеваемость AD постоянно возрастает, так как наиболее распространенная негенетическая форма начинается в возрасте примерно 70-75 лет, а средняя продолжительность жизни увеличивается вследствие роста уровня жизни и прогресса медицины и фармакологии. Заболевание также существует в передаваемой по наследству (генетически) форме, связанной с мутацией в некоторых локусах хромосом 14, 19 и 21 (типичной для людей, страдающих синдромом Дауна). Раннее проявление наследственной формы AD происходит в возрасте 50 лет и приводит к дегенерации мозга с последующей смертью в течение 2-3 лет. Поздняя форма заболевания также вызывает дегенерацию мозга и смерть, но в течение 10 и более лет.
В мозге обнаруживается большое количество бляшек в пространстве между нейронами и типичных нейрофибриллярных клубков в нейронах, особенно в нейронах коры головного мозга, гиппокампа и миндалевидного тела, а также в других частях мозга, отвечающих за когнитивные функции. Амилоидные бляшки, также известные как сенильные бляшки, представляют собой полимеры пептида бета-амилоида (Аβ), который является производным более крупного белка-предшественника бета-амилоида (АРР). АРР принадлежит к высококонсервативному суперсемейству трансмембранных гликопротеинов.
В последнее десятилетие учеными, занятыми изучением AD, был проведен ряд исследований с целью понимания этиологии заболевания и в особенности его молекулярных механизмов. Исследование молекулярных компонентов и их регуляции может прояснить их терапевтические и диагностические перспективы. Внимание исследователей сосредоточено на пресенилинах, чья роль в процессинге АРР и, следовательно, в образовании Ар, представляется очень значимой. Было показано, что эти белки, пресенилин 1 (PS1) и пресенилин 2 (PS2), проявляют ферментативную активность или регулируют активность других ферментов, а именно секретаз, которые расщепляют АРР в нормально распадающиеся катаболиты (альфа-секретаза) или в пептид Аβ (бета- и гамма-секретаза). В наследственной AD, мутации генов, кодирующих PS1 и PS2, приводят к избыточному образованию Аβ и, в частности, накоплению изоформы Аβ-42, которая в высокой степени амилоидогенна. Недавно эксперименты на мышах с «выключенным» геном PS1 показали сильное снижение активности гамма-секретазы и тем самым продемонстрировали, что PS1, так же, как и принадлежащий гамма-секретазе комплекс, тоже главным образом отвечает за выработку и накопление Аβ. Что касается бета-секретазы, считается, что продукт гена ВАСЕ может сам полностью выполнять бета-секретазное расщепление.
Разработка пригодных к клиническому использованию ингибиторов гамма- и бета-секретазы может стать решающим оружием против болезни Альцгеймера и в настоящее время является одним из наиболее захватывающих видов конкуренции в нейробиологии. Было ясно показано, что активность PS1 не может быть полностью подавлена, так как этот белок необходим для процессинга фактора трансдукции Notch-1, который является ключевым фактором в созревании многих стволовых клеток, например клеток, вовлеченных в эритропоэз.
Регуляция экспрессии гена с помощью метилирования ДНК может быть с успехом изучена в системе культуры клеток, способной экспрессировать гены, вовлеченные в AD. В результате наших исследований была выявлена очень интересная закономерность регуляторных механизмов, связанных со старением, которая заключалась в постепенном общем возрастании гипометилирования ДНК у пожилых людей и накоплении гомоцистеина, наблюдаемом у пациентов, страдающих сенильной деменцией. Накопление гомоцистеина и гипометилирование ДНК метаболически взаимосвязаны, так как нарушение преобразования гомоцистеина в метионин обращает метаболизм в сторону синтеза S-аденозилгомоцистеина, который известен как сильный ингибитор ДНК-метилтрансферазы, тем самым вызывая гипометилирование ДНК. В соответствии с общепринятой теорией, что многие гены экспрессируются при деметилировании цитозинов специфической последовательности, этот биохимический механизм может приводить к экспрессии неэкспрессирующихся генов и сверхэкспрессии нормально экспрессирующихся генов. Это может иметь место в случае AD, потому что сверхэкспрессия PS1, т.е. гамма-секретазы, может скачком превысить активность альфа-секретазы и, следовательно, вызвать выработку пептида Аβ, который, накапливаясь, через много лет может привести к возникновению заболевания. Еще одним свидетельством возможной роли метилирования ДНК в AD является тот факт, что пациенты с AD имеют намного меньший посмертный уровень доноров метильной группы в мозге. Несомненно, снижение количества S-аденозилметионина (SAM) может приводить к изменению или увеличению экспрессии генов, вовлеченных в метаболизм АРР, в конечном итоге вызывая накопление пептида Аβ в сенильных бляшках.
Предварительные эксперименты были проведены на клетках нейробластомы линии (SK-N-SH), которая экспрессирует АРР, PS1, PS2, ВАСЕ, альфа-секретазу, другие компоненты гамма-секретазы и Notch-1. Культуры выращивали на культуральной среде, лишенной фолата, витамина В12 и витамина В6 (для изменения метаболизма гомоцистеина), в которую добавляли в различных концентрациях SAM (чтобы сбалансировать эффект недостатка витаминов). Авторы изобретения обнаружили рост экспрессии PS1 и ВАСЕ в среде, лишенной витаминов В, и заметное снижение экспрессии PS1 и ВАСЕ после введения SAM. В экспериментах, проведенных с Hpall/PCR на промоторах АРР и PS1, авторы изобретения обнаружили существенные различия в метилировании одного из CpG сайтов промотора PS1. Мы пришли к выводу, что ген PS1 может быть частично подавлен введением экзогенного SAM. Введение SAM может снижать экспрессию PS1, восстанавливая метаболический баланс в пользу альфа-секретазы. Также были проведены эксперименты с трансгенными мышами линии TgCRNDS и соответствующей контрольной группой; эти мыши характеризуются наличием мутантного человеческого гена АРР, и поэтому у них могут быстро возникать амилоидные бляшки. Эти животные получали полноценный рацион и рацион, лишенный витаминов группы В; точно так же, как и в экспериментах с клетками, наблюдалось увеличение экспрессии PS1 и ВАСЕ.
В обеих экспериментальных моделях изменение экспрессии генов приводило к возрастанию активности гамма- и бета-секретазы с последующим избыточным образованием Аβ, который накапливался, образуя сенильные бляшки, быстрее, чем у животных, которые получали контрольный рацион.
Данные, обобщенные выше, были представлены в следующих публикациях:
Fuso A., Nicolia V., Cavallaro R.A., Ricceri L, D'Anselmi F., Coluccia P., Calamandrei G. and Scarpa S. 2008. B-Vitamin Deprivation Induces Hyperhomocysteinemia and Brain S-adenosylhomocysteine, Depletes Brain S-adenosylmethionine, and Enhances PS1 and BACE Expression and Amyloid-β Deposition in Mice. Mol. Cell. Neurosci. 37: 731-746.
Fuso A., Cavallaro R.A., Zampelli A., D'Anselmi F., Piscopo P., Confaloni A. and Scarpa S. 2007. y-secretase is differentially modulated by alterations of Homocysteine cycle in neuroblastoma and glioblastoma cells. J. Alz. Dis. 11: 275-290.
Cavallaro R. A., Fuso A., D'Anselmi F. and Scarpa S. 2006. The effect of S-adenosylmethionine on CNS gene expression studied by cDNA microarrays analysis. J. Alz. Disease. 9: 415-419.
Scarpa S., Cavallaro R.A., D'Anselmi F. and Fuso A. 2006. Gene silencing through methylation: an epigenetic intervention on Alzheimer Disease. J. Alz. Disease. 9: 407-414.
Fuso A., Seminara L., Cavallaro R.A., D'Anselmi F. and Scarpa S. 2004. Homocysteine/S-adenosylmethionine Cycle Alterations Unbalance DNA Methylation Status with Consequent Up-regulation of Beta-amyloid Production. Mol. Cell. Neurosci. 28(1): 195-204.
Scarpa S., Fuso A., D'Anselrni F., Cavallaro R.A. 2003. Presenilin 1 gene silencing by S-adenosylmethionine: a treatment for Alzheimer disease? FEBS Letters 541 (1-3): 145-148.
Fuso A., Cavallaro R. A., Orru L., Buttarelli F.R. and Scarpa S. 2001. Gene silencing by S-adenosylmethionine in muscle differentiation. FEBS Letters 508 (3): 337-340.
Применение SAM для лечения AD также предлагалось в US 2002/025926 и US 2004/0048827. Последний документ описывает способность SAM влиять на бета-секретазу, пресенилин 1 и 2 и экспрессию гена белка-предшественника бета-амилоида. Исследования с меченым (тритием) SAMe, проведенные после вышеуказанной заявки на патент, продемонстрировали, как описано ниже, способность молекул достигать центральной нервной системы после перорального введения.
В US 5519058 и в CN 1099224 авторы сделали предположение о благоприятном воздействии SOD при лечении AD.
Фармацевтические композиции, содержащие SAM и SOD вместе с рядом активных ингредиентов, для применения при других, отличных от AD, болезненных состояниях, раскрыты в US 2003/129261 и в WO 2005/041996.
Описание изобретения
В настоящее время обнаружено, что активность S-аденозилметионина может быть неожиданно улучшена при введении в комбинации с супероксиддисмутазой (SOD). Доказано, что этот фермент не только способен содействовать преодолению S-аденозилметионином гематоэнцефалического барьера, но и синергетически взаимодействует с SAM в снижении экспрессии генов PS1 и ВАСЕ при их сверхэкспрессии в результате дефицита витаминов В.
Таким образом, один из аспектов изобретения относится к продуктам, содержащим S-аденозилметионин, или его производное, и супероксиддисмутазу, в форме комбинированного препарата для совместного, раздельного или последовательного введения при лечении болезни Альцгеймера.
Дозы могут варьироваться в широких пределах вследствие очень низкой токсичности SAM и SOD и зависят от ряда факторов, таких как масса тела пациента, пол и возраст. Однако, вообще говоря, они находятся в диапазоне от 200 до 2000 мг/день для SAM и от 50 до 1000 мг/день для SOD.
SAM можно вводить, преимущественно перорально, либо в чистом виде, либо в виде его стабильной соли, такой как тозилат, бутандисульфонат, дисульфат тозилат, дисульфат дитозилат и дисульфат монотозилат. Преимущественной формой введения являются клетки Saccharomyces cerevisiae, обогащенные SAM, описанные в WO 2006/131382.
SOD, полученный ферментацией из штаммов Saccharomyces cerevisiae, как описано в заявке WO 2006/131382, поданной авторами данного изобретения, можно также вводить перорально, на глиадиновой пленке в качестве носителя или с помощью других способов гастропротекции. Как альтернатива пероральному введению, SAM и SOD можно вводить парентерально, например, внутримышечно.
Примеры этих препаратов включают таблетки с пленочной оболочкой из акриловых или метакриловых полимеров, желудочно-резистентные капсулы, микрокапсулы и тому подобные лекарственные формы.
SAM и SOD могут входить в состав одной и той же дозированной лекарственной формы или же вводиться раздельно в составе отдельных препаратов; в этом случае может поставляться набор, содержащий две лекарственные формы в отдельных дозированных формах вместе с инструкцией по их последовательному, совместному или раздельному применению.
Описание фигур
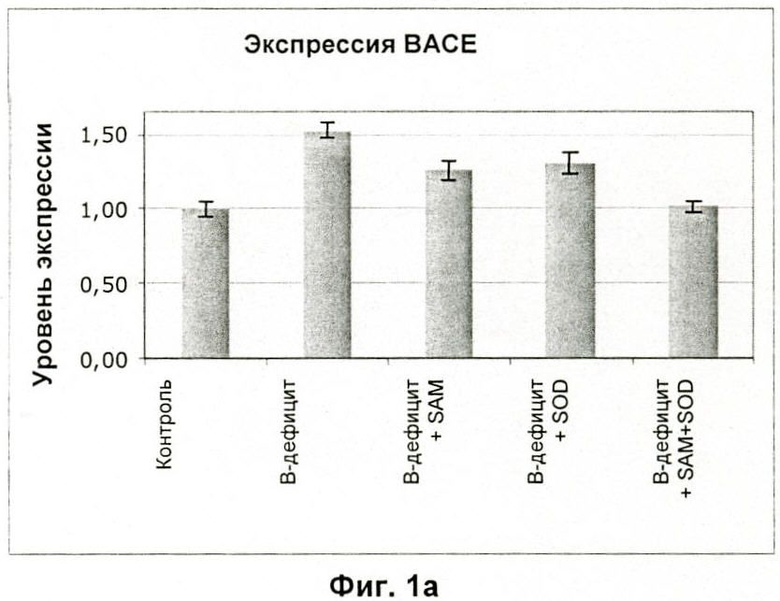
На Фиг.1а и 1b показано влияние введенных мышам SAM и SOD на сверхэкспрессию PS1 и ВАСЕ, индуцированную рационом, лишенным витаминов В.
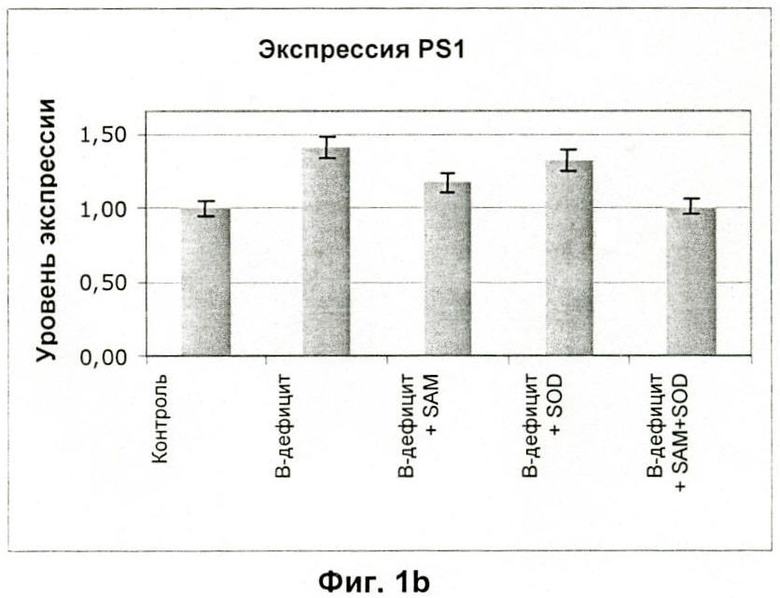
На Фиг.2 показано влияние SOD и глутатиона (GSH) на сверхэкспрессию PS1 и ВАСЕ в клетках нейробластомы линии SK-N-BE, отдельно и в комбинации с SAM.
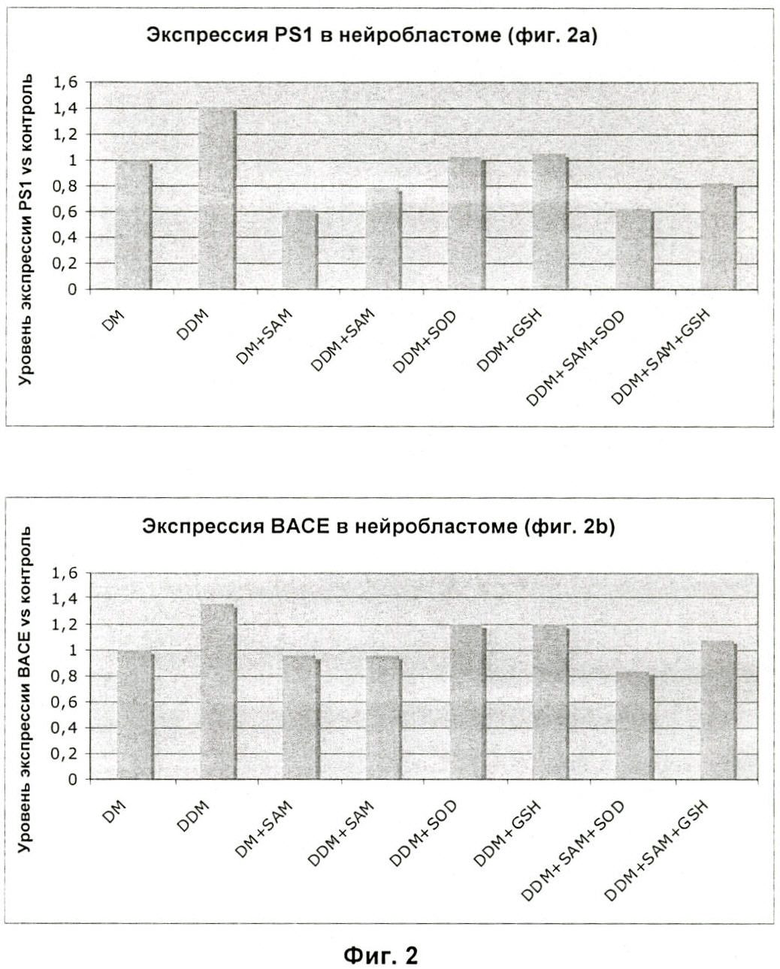
На Фиг.3 показаны уровни меченого тритием SAM в мозге мышей после перорального введения 400 мкг/день SAM.
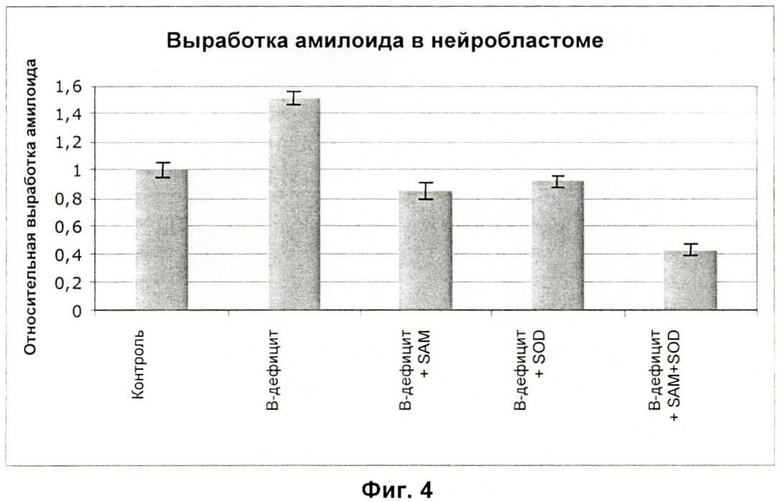
На Фиг.4 показана степень продукции бета-амилоида клетками нейробластомы человека после одной недели применения соединений.
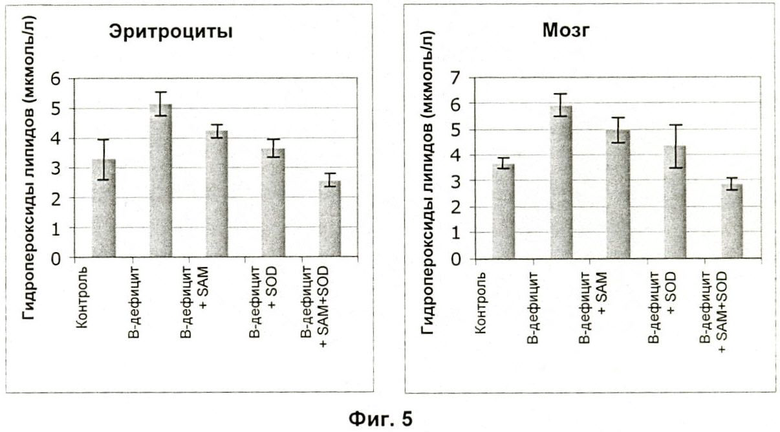
На Фиг.5 показано влияние SAM и SOD на уровень окисления в эритроцитах и мозге мышей, которые получали лишенный витаминов В рацион.
Далее изобретение представлено подробно посредством фармакологических испытаний, описанных в следующих примерах.
Пример 1 - Экспрессия генов
В частности, воздействие SAM проверяли при фармакологических концентрациях (400 мкг/день) на TgCRND8 мышах и соответствующем контроле дикого типа.
Пути экспрессии генов
РНК экстрагировали из культур клеток и гомогенизированных тканей мозга и синтезировали кДНК. Всего использовали 0,5 мг кДНК для каждой реакции в реальном масштабе времени на Opticon2 DNA Engine (MJ Research) с SYBR-Green реагентами. Эффективность амплификации для каждой пары праймеров предварительно определяли с помощью амплификации калибровочной линейной кривой. Для определения количества специфической кДНК, присутствующей в стандартной реакции, экспериментальные образцы сравнивали с калибровочной кривой специфического гена. Стандарты получали из продуктов ПЦР (полимеразной цепной реакции) высокой очистки, амплифицированных с помощью позитивного контроля. Общие уровни кДНК нормализовали по р-актин контролю (ген "домашнего хозяйства").
Результаты показывают, что SAM также обращает сверхэкспрессию PS1 и ВАСЕ, вызванную дефицитным рационом in vivo (Фиг.1) и даже снижает ее до уровня ниже, чем в контроле с полноценным рационом.
Так как метаболизм гомоцистеина вовлечен как в реакции метилирования, так и в окислительно-восстановительные реакции, авторы проверяли влияние различных антиоксидантов на экспрессию этих двух генов. Первые данные, полученные на клетках нейробластомы линии SK-N-BE, показали, что как SOD, так и глутатион (GSH) ингибируют сверхэкспрессию PS1 и ВАСЕ, вызванную дефицитом витаминов, хотя и в меньшей степени, чем SAM. Однако интересно отметить, что при совместном введении SOD и SAM, они проявляют синергетический эффект (Фиг.2), который вызывает дальнейшее снижение экспрессии двух генов до более низких уровней, чем наблюдали при введении одного только SAM (ниже на 15-20%).
Пример 2 - Захват SAM
Для подтверждения того, что SAM преодолевает гематоэнцефалический барьер, проводили эксперименты на клетках и мышах.
Для культур клеток добавляли 100 мкмоль/л SAM к среде культуры клеток F14 (полноценной и лишенной витаминов В, согласно плану эксперимента) и культивирование останавливали после 96 часов.
Мышам вводили перорально 400 мкг/день SAM с помощью зонда-иглы для кормления и животных умерщвляли через 2 месяца; животные получали полноценный рацион или рацион, лишенный витаминов В, согласно плану эксперимента. Уровни SAM анализировали с помощью HPLC (высокоэффективная жидкостная хроматография) на системе Varian HPLC. Клетки и гомогенизированные ткани мозга лизировали в дистиллированной воде, и макромолекулы осаждали 1,5 моль/л РСА (хлорная кислота). Калибровочные кривые SAM рассчитывали до и после отбора экспериментальных образцов. Для анализа меченого тритием SAM, клетки обрабатывали, как описано выше, тогда как мышей подвергали воздействию в течение 4 дней, чтобы свести к минимуму облучение радиоактивным препаратом. Клетки и гомогенизированные ткани мозга лизировали в дистиллированной воде. Часть лизированных тканей мозга разрушали ультразвуком и центрифугировали, чтобы отделить мембраны и клеточные органеллы; другую часть после разрушения ультразвуком для отделения макромолекул обрабатывали хлорной кислотой (РСА). Усвоение радиоактивного SAM измеряли с помощью сцинтилляционного счетчика бета-частиц.
Было обнаружено, что внутриклеточные уровни SAM возросли с 1 мкмоль/л (контроль) до 2,5-3 мкмоль/л. Чтобы определить, обусловлен ли этот рост захватом экзогенного SAM или же он вызван увеличением эндогенных резервов в результате повышенных внеклеточных концентраций молекул, был проведен эксперимент с радиоактивным SAM. 100 мкмоль/л меченого тритием SAM (SAM[3H]) добавляли к культурам клеток SK-N-BE и определяли захват с помощью лизиса клеток и измерения радиоактивности. Уровень радиоактивности клеточных лизатов соответствовал 1,5 мкмоль/л SAM; это значение сравнимо с обнаруженным ростом в пределах от 1 до 2,5 раз для немеченого SAM и ясно показывает, что повышение внутриклеточного уровня обусловлено захватом экзогенного SAM.
Похожий рост уровней SAM обнаружили в лизатах тканей мозга мышей, которым перорально вводили 400 мкг/день SAM; аналогично проводили дополнительный эксперимент с SAM[3H] в концентрации 400 и 800 мкг/день. Возрастание радиоактивности в полном лизате тканей мозга соответствует 0,5 нг SAM на мозг у мышей, которым вводили 400 мкг SAM, и 1 нг на мозг у мышей, которым вводили 800 мкг SAM (Фиг.3).
Часть полного лизата тканей мозга также разрушали ультразвуком и центрифугировали для получения очищенного лизата (цитоплазмы), и еще одну часть этого очищенного лизата осаждали хлорной кислотой (РСА) для устранения белков (растворимый лизат). Интересно отметить, что растворимый лизат мышей, которым вводили 800 мкг SAM, показывает уровень радиоактивности, соответствующий 0,5 нг SAM (тогда как полный и очищенный лизат демонстрирует более высокие уровни), это свидетельствует о том, что избыточные метильные группы, связанные с экзогенным SAM, были конъюгированы с другими молекулами клетки.
Пример 3 - Продукция амилоида в линиях клеток нейробластомы
Способы: среда и культуры клеток
Линию клеток нейробластомы человека SK-N-BE выдерживали в среде F14 с 10% FCS (фетальная телячья сыворотка) и переносили в полную дифференциальную среду (контрольная среда с 1% FCS и 10 мкмоль/л ретиноевой кислоты) или в дифференциальную среду, лишенную фолата, витамина В12 и витамина В6 (В-дефицитная среда). В культуры вносили питательную среду каждые двое суток и останавливали рост через 96 часов.
Животные и рационы
В примерно трехнедельном возрасте мышей систематично распределяли либо в группу с контрольным рационом, либо в группу с дефицитным рационом; животные получали корм и воду ad libitum. Контрольный (AIN-93M; рацион А: фолат 1,98 мг; витамин В12 0,025 мг; витамин В6 7 мг) и опытный рационы (AIN-93M В; рацион В, лишенный фолата, витамина В12 и витамина В6) приобретали в Mucedola (Италия). Оба рациона содержали 1% сульфатиазола для того, чтобы ингибировать образование фолата бактериями кишечника и гарантировать, что единственным источником фолата является рацион. Кроме того, другие три группы животных получали SAM (800 мкг/день) и SOD (10 ED/день) или их комбинацию (SAM 400 мкг/день и SOD 5 Е/день). Эксперимент проводили в течение одной недели, затем мышей анестезировали и умерщвляли для получения мозга и крови. Кровь получали пункцией сердца, собирали в пробирку, содержащую 2 г/дл ЭДТА, и немедленно центрифугировали для отделения плазмы и эритроцитов, затем хранили при -80°С. Мозг перфузировали PBS и удаляли.
Анализ амилоида
Гомогенизированные ткани мозга лизировали раствором следующего состава: 50 ммоль/л трис-HCl рН 7,4, 150 ммоль/л NaCl, 0,2% Nonidet P-40, 1% CHAPS, 2 ммоль/л ЭДТА, PMSF (200 мкмоль/л), леупептин (1 мкмоль/л), пепстатин А (1 мкмоль/л) и ингибитор кальпаина I (5 мкмоль/л). Экстракт белка анализировали с помощью ELISA (твердофазный иммуноферментный метод) и иммунодиагностического набора Аβ 1-42 (BioSource International, Бельгия); этот набор гарантирует хорошую линейную чувствительность до 10 пг/мл. Все измерения проводили трижды.
Статистический анализ
Все представленные здесь оценки значимости (р<0.05) различий рассчитывали с помощью однофакторного дисперсионного анализа и апостериорного критерия Бонферрони.
Результаты
Исходя из данных, представленных на Фиг.4, синергизм между SAM 400 и SOD выглядит несомненным и подтверждается нижеследующим статистическим анализом, в котором все обозначения ссылаются на Фиг.4.
В-дефицит vs Контроль: р<0,001
В-дефицит + SAM и В-дефицит + SOD vs В-дефицит: р<0,001
В-дефицит + SAM + SOD vs В-дефицит: р<0,001
В-дефицит + SAM + SOD vs В-дефицит + SAM и В-дефицит + SOD:
р<0,001.
Группа изобретений относится к медицине и предназначена для лечения болезни Альцгеймера. Используют S-аденозилметионин (SAM) в комбинации с супероксиддисмутазой (SOD). SAM и SOD могут применяться совместно, раздельно или последовательно. Группа изобретений позволяет уменьшить продукцию амилоида в клетках нейробластомы и снизить уровень окисления в тканях мозга и эритроцитов. 2 н. и 3 з.п. ф-лы, 3 пр., 5 ил.
1. Применение S-аденозилметионина (SAM) в комбинации с супероксиддисмутазой (SOD) для изготовления лекарственных средств для лечения болезни Альцгеймера.
2. Применение по п.1, где лекарственное средство ингибирует сверхэкспрессию PS1 (пресенилина 1) и ВАСЕ (бета-секретазы).
3. Применение по любому из пп.1 или 2, где лекарственные средства вводят перорально.
4. Комбинация для совместного, раздельного или последовательного введения при лечении болезни Альцгеймера, содержащая в качестве единственных активных ингредиентов (1) S-аденозилметионин или его производное и (2) супероксиддисмутазу.
5. Комбинация по п.4, где SAM находится в форме тозилата, бутандисульфоната, дисульфата тозилата, дисульфата дитозилата или дисульфата монотозилата или содержится в клетках Saccharomyces cerevisiae, обогащенных SAM.
| СПОСОБ ИЗГОТОВЛЕНИЯ ТАБЛЕТОК, СОДЕРЖАЩИХ S-АДЕНОЗИЛМЕТИОНИН | 2002 |
|
RU2313348C2 |
| WO 2005041996, 12.05.2005 | |||
| US 2002025926, 28.02.2002 | |||
| SCARPA S | |||
| et al | |||
| Gene silencing through methylation: an epigenetic intervention on Alzheimer disease J Alzheimers Dis | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| SCARPA S | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

