Изобретение относится к области биотехнологии, а именно к получению новых термостабильных фитаз методами генетической инженерии.
Фитазы представляют собой ферменты, катализирующие последовательный гидролиз фитатов до мио-инозитола и неорганического фосфата.
Известно, что более 70% фосфора, содержащегося в растительных кормах для цыплят бройлеров, находится в форме фитиновой кислоты и ее солей фитатов. Так как животные с однокамерным желудком не способны усваивать фитатный фосфор, то неусвоенные фитаты, попадая в почву и водоемы, вызывают загрязнение окружающей среды. Кроме того, фитаты уменьшают доступность белков и микроэлементов в кормах.
В связи с этим фитазы, отщепляющие фосфатные группы от фитатов, с успехом используются в животноводстве в качестве кормовой добавки, значительно повышая усвояемость фосфора.
Для практического использования препараты фитаз должны быть устойчивы к высушиванию и действию высоких температур (60-80°C) в процессе приготовления комбикормов, а также обладать высокой удельной активностью и эффективно функционировать при температуре, соответствующей физиологической температуре животных.
Известна термостабильная фитаза phyA из Aspergillus niger [Appl Environ Microbiol., 1998, v.64, p.4446-4451], которая имеет оптимальную температуру работы 55°C, высокую термостабильность (не теряет активность после воздействия на нее температуры 70°C в течение 30 минут) и способна работать в условиях низких рН в гастроэнтеральном тракте животных. Однако данная фитаза обладает низкой удельной активностью (100 ед/мг белка при 37°C).
Известна мутантная термостабильная фитаза [US 20060183213], полученная путем замены ряда аминокислот в аминокислотной последовательности фитазы Escherichia coli. Положения замен определены в ходе проведения насыщенного мутагенеза. Полученная фитаза способна работать в условиях низких рН в гастроэнтеральном тракте животных, однако имеет небольшую удельную активность (800 ед/мг белка при 37°C) и термостабильность (сохраняет 40% активности после воздействия на нее температуры 60°C в течение 30 минут).
Известна мутантная термостабильная фитаза [Appl Microbiol Biotechnol, 2008, v.79, p.69-75], полученная путем применения метода error-prone PCR к гену, кодирующему фитазу аррА Е.coli. Фитаза сохраняет 60% активности после воздействия на нее температуры 80°C в течение 10 минут и имеет хорошую удельную активность (905 ед/мг белка при 37°C), однако способна работать только в очень узком диапазоне рН (2,5-4,0).
Задачей заявляемого изобретения является расширение арсенала термостабильных фитаз и рекомбинантных микроорганизмов, продуцирующих термостабильные фитазы.
Поставленная задача решена тем, что получены:
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, а аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
- Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
Штамм Pichia pastoris PhyA-CfDVG ВКПМ Y-3495 - продуцент мутантной фитазы PhyA-CfDVG, обладающей термостабильными свойствами.
Штамм Pichia pastoris PhyA-CfDV ВКПМ Y-3494 - продуцент мутантной фитазы PhyA-CfDV, обладающей термостабильными свойствами.
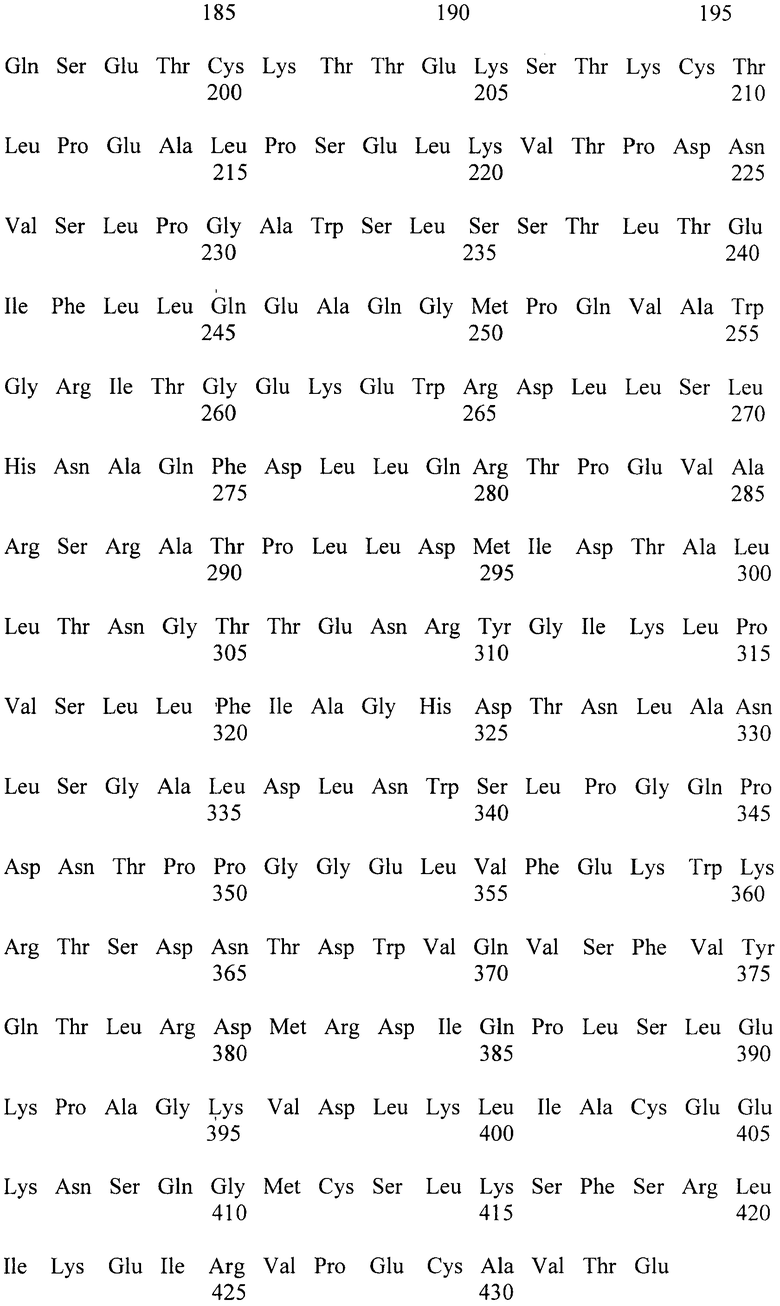
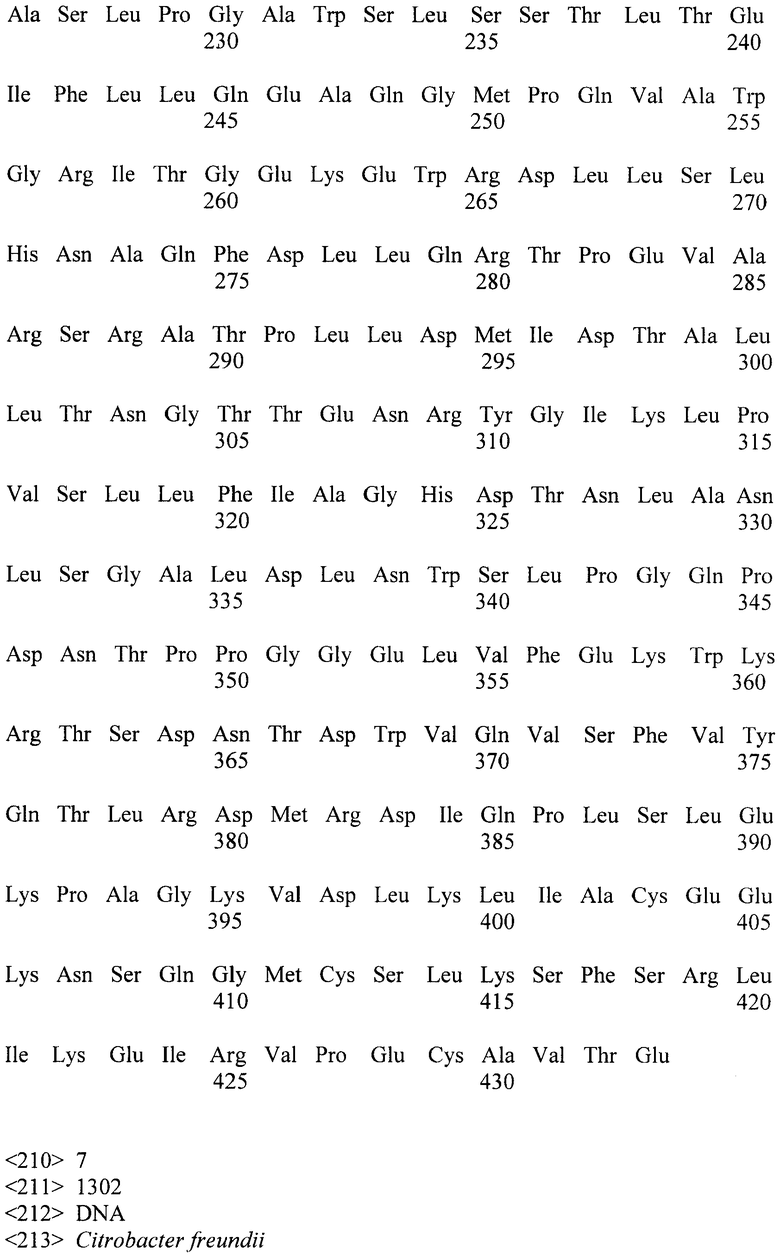
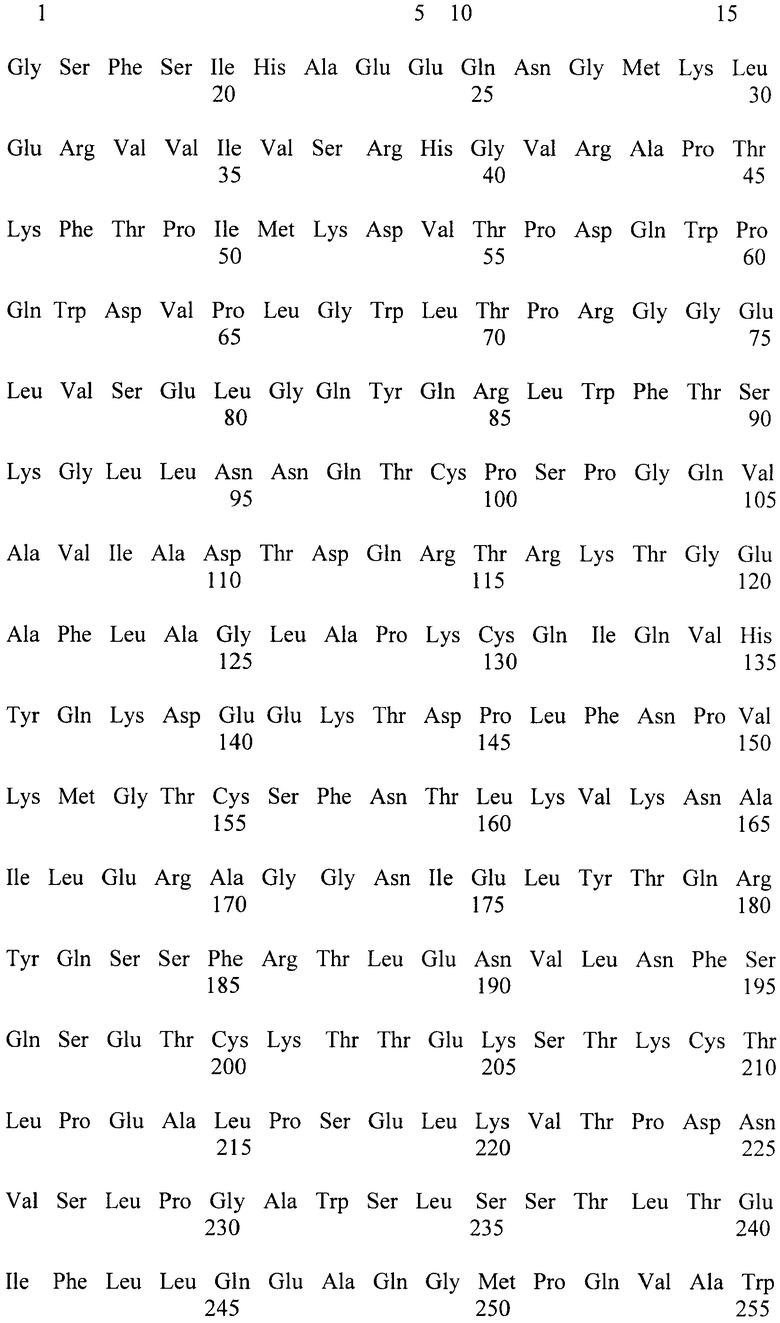
Известна [Биотехнология, 2003, №2, с.3-10] фитаза PhyA-Cf, выделенная из Citrobacter freundii и имеющая аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:1 Удельная активность фермента составляет 3000 ед/мг белка, а определение его устойчивости к действию высокой температуры показало, что после прогрева в течение 10 минут при 60°C фермент сохраняется 10% активности, а после прогрева в течение 10 минут при 70°C фермент инактивируется полностью, т.е. фермент термостабильность фермента низка.
В источниках информации не обнаружено термостабильных фитаз, полученных в результате мутагенеза фитазы из бактерий рода Citrobacter.
Известно [J. Mol. Biol., 2006, v.356, p.547-557], что термостабильность белка связана с такими структурными свойствами белка, как число водородных связей, число солевых мостиков, порядок контактов, доля остатков в белке, вовлеченных в различные элементы вторичной структуры, т.е. структурными свойствами, которые играют важную роль в сворачивании белка и приобретении им своей конечной нативной структуры.
Показано [J. Mol. Biol, 1990, v.215, p.403-410], что у термофилов наблюдается повышенное значение числа солевых мостиков (образующихся за счет электоростатического взаимодействия) и числа водородных связей на остаток. Термостабильные белки более компактны и изменение структурных свойств, приводящих к увеличению компактности белка и его стабилизации (стабилизации его нативной структуры), приводит к увеличению его термостабильности.
Известно также [J. Mol. Biol, 1998, v.278, p.279-289], что вклад аминокислоты в формирование третичной структуры фермента определяется не только ее природой, но и ее положением в аминокислотной последовательности и, следовательно, осуществляя точечные замены аминокислот, находящихся в значимых положениях аминокислотной последовательности, можно добиться изменения структурных свойств белка (и его термостабильности).
Проведенные нами исследования показали, что для фитазы PhyA-Cf из бактерий Citrobacter freundii такими значимыми позициями являются аспарагиновая кислота в положении 144, аминокислота валин в положении 226 и аминокислота глицин в положении 343.
Механизм, приводящий к изменению термостабильности, различен для каждого случая, а именно;
- замена отрицательно заряженной аспарагиновой кислоты в положении 144 на аминокислоту, содержащую в нециклической боковой R-группе положительно заряженную аминогруппу (аспарагин, глутамин, аргинин, лизин), приводит к образованию дополнительных электростатических взаимодействий;
- замена аминокислоты валин в положении 226 на аминокислоту с меньшим объемом R-группы (аланин, глицин) уменьшает стерические препятствия и облегчает взаимодействие между соседними аминокислотами;
- замена неполярной аминокислоты глицин в положении 343 на высокополярную аминокислоту (аспаргиновую, глутаминовую, лизин, аргинин, аспарагин) приводит к образованию дополнительного мостика.
Для получения мутантной фитазы к гену phyA, имеющему нуклеотидную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:2 [AY390262, GenBank], и кодирующему фитазу PhyA-Cf, выделенную из Citrobacter freundii, применяют метод сайт-направленного мутагенеза [Archives of Biochemistry and Biophysics, 2000, v.382, №1, p.105-112] и получают ДНК, кодирующую фитазу с выбранными аминокислотными заменами.
Полученную ДНК клонируют в вектор, подходящий для ее трансформации в клетки Pichia pastoris. Так как в силу «вырожденности» генетического кода одна и та же аминокислотная последовательность может кодироваться большим числом нуклеотидных последовательностей, то для клонирования может быть использована не только сама вышеуказанная нуклеотидная последовательность, но и все ее формы, определяемые вырожденностью генетического кода.
Полученной рекомбинантной плазмидой осуществляют трансформацию штамма-реципиента. Трансформанты тестируют на способность проявлять фитазную активность по методу Фиске-Субарроу [J. Biol. Chem., v.66, p.376-400] и отбирают клон, обладающий максимальной фитазной активностью, получая штамм, продуцирующий мутантную фитазу.
Штаммы Pichia pastoris PhyA-CfDVG и Pichia pastoris PhyA-CfDV, продуцирующие термостабильные фитазы с наиболее интересными характеристиками, депонированы во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ).
Штамм Pichia pastoris PhyA-CfDVG - продуцент термостабильной фитазы PhyA-CfDVG депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Y-3495.
Штамм хранят в среде с глицерином при температуре -70°C или в лиофильно-высушенном виде при температуре +4°C.
Штамм характеризуется следующими признаками:
Морфологические признаки
При росте на среде «Malt agar» [The yeast of toponomic study. Ed.N.J.W.Kreger-van Rij, Amsterdam, Elsevier Sci. Publ.B.V., 1984, p.421] в течение 48 часов при температуре 30°C клетки образуют гладкие, круглые колонии с матовой поверхностью светло-кремового цвета, после 3-х дней культивирования при температуре 30°C культура образует клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
При росте на среде «Malt extract» [The yeast of toponomic study. Ed.N.J.W.Kreger-van Rij, Amsterdam, Elsevier Sci. Publ.B.V., 1984, p.421] в течение 48 часов при температуре 30°С клетки образуют интенсивную ровную суспензию. Культура имеет характерный запах дрожжей.
Культуральные признаки
Клетки хорошо растут на полной органической среде YEPD следующего состава (мас.%): пептон - 2, дрожжевой экстракт - 1, глюкоза - 2 (или глицерин - 1), вода - остальное.
Кроме того клетки хорошо растут на минимальной среде М9 (Т.Маниатис, Э.Фрич, Дж.Сэмбрук, Молекулярное клонирование, Москва, "Мир", 1984, с.390), а также на других синтетических средах для дрожжей, содержащих в качестве источника углерода глюкозу или глицерин.
Клетки штамма отличаются слабым ростом на средах, содержащих в качестве источника углерода метанол.
Физиолого-биохимические признаки
Клетки растут при температуре от 4°C до 37°C. Оптимальной температурой выращивания является 30°C.
Оптимум рН для роста составляет 4,5-6,5.
В качестве источника углерода клетки используют глюкозу, глицерин, метанол. В качестве источника азота - минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Штамм не нуждается в гистидине для своего роста.
Штамм Pichia pastoris PhyA-CfDV - продуцент термостабильной фитазы PhyA-CfDV депонирован во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Y-3494.
Штамм хранят в среде с глицерином при температуре -70°C или в лиофильно-высушенном виде при температуре +4°C.
Штамм характеризуется следующими признаками:
Морфологические признаки
При росте на среде «Malt agar» в течение 48 часов при температуре 30°C клетки образуют гладкие, круглые колонии с матовой поверхностью светло-кремового цвета, после 3-х дней культивирования при температуре 30°C культура образует клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками.
При росте на среде «Malt extract» в течение 48 часов при температуре 30°C клетки образуют интенсивную ровную суспензию. Культура имеет характерный запах дрожжей.
Культуральные признаки
Клетки хорошо растут на полной органической среде YEPD, на минимальной среде М9, а также на других синтетических средах для дрожжей, содержащих в качестве источника углерода глюкозу или глицерин.
Клетки штамма отличаются слабым ростом на средах, содержащих в качестве источника углерода метанол.
Физиолого-биохимические признаки
Клетки растут при температуре от 4°C до 37°C. Оптимальной температурой выращивания является 30°C.
Оптимум рН для роста составляет 4,5-6,5.
В качестве источника углерода клетки используют глюкозу, глицерин, метанол. В качестве источника азота - минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному и анаэробному росту.
Штамм не нуждается в гистидине для своего роста.
Изобретение проиллюстрировано следующими фигурами.
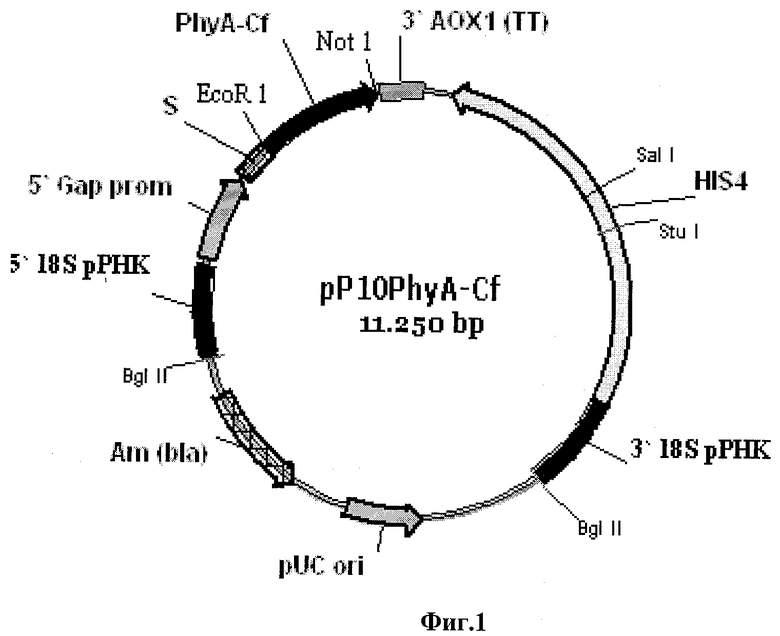
Фиг.1 - Схема рекомбинантной плазмида pP10PhyA-Cf.
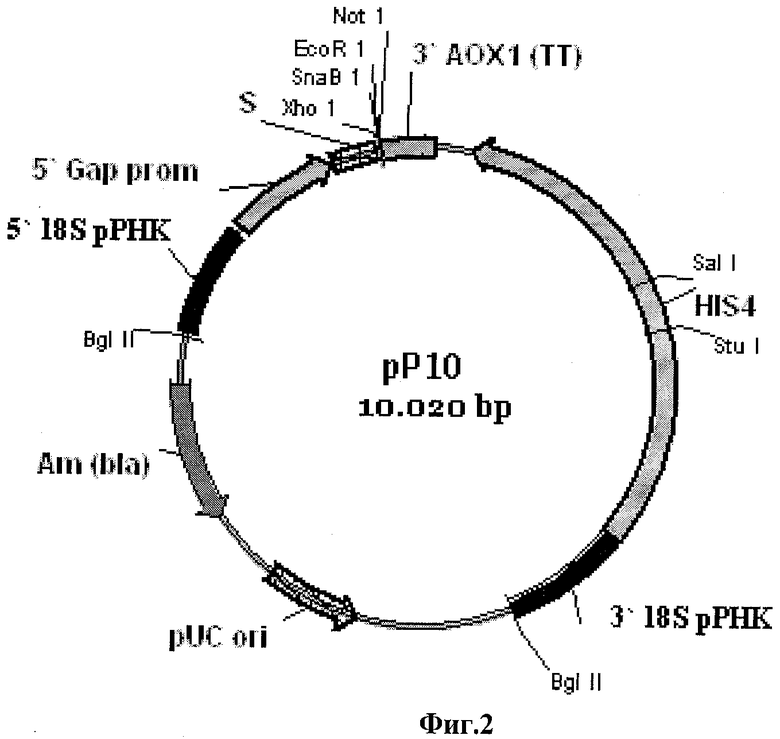
Фиг.2 - Схема вектора рР10.
Следующие примеры иллюстрируют изобретение, но не исчерпывают его.
Пример 1. Конструирование рекомбинантной плазмиды pP10phyA-CfD
1. Проведение мутагенеза
Мутантный ген phyA-CfD получают применяя метод сайт-направленного мутагенеза к гену гену phyA, кодирующему фитазу PhyA-Cf.
Для этого ПЦР методом синтезируют два фрагмента ДНК, включающих нуклеотидную замену в положениях 430 и 432 нуклеотидного сиквенса SEQ ID NO:2 таким образом, что кодон gat (кодирующий аспарагиновую кислоту) заменяют на кодон аас (кодирующий аспарагин).
Для ПЦР-синтеза используют следующие праймеры (нуклеотидные замены в приведенных последовательностях подчеркнуты):
1. Для синтеза первого фрагмента
прямой - pic3638D (5'-aggaattcGAAGAGCAGAACGGTATGA-3')
обратный - D/Nobr (5'-ATTAAAAAGAGG G T T AGTTT-3')
2. Для синтеза второго фрагмента
прямой - D/Npr (5'-aagaaaaaact a a c cctct-3')
обратный - pic3638R (5'-agcggccgcTTACTTATTCCGTAACTG-3').
В качестве матрицы для проведения полимеразной цепной реакции используют плазмиду pP10PhyA-Cf [Заявка РФ №2009126296 от 10.07.2009, приор. 21.08.2008], имеющую размер 11250 п.о. и состоящую из следующих элементов (фиг.1):
1) фрагмент 5' 18S рРНК: 1-269 п.о.;
2) промотор GAP: 270-1026 п.о.;
3) α'-амилазный сигнальный пептид: 1029-1295 п.о.;
4) ген фитазы PhyA-Cf: 1304-2534 п.о.;
5) терминатор транскрипции 3' АОХ1: 2552-2885 п.о.;
6) HIS4 ORF: 3279-5813 п.о.;
7) фрагмент 3' 18S рРНК: 6085-6774 п.о.;
8) pUC ori: 7974-8648 п.о.;
9) ген резистентности к ампициллину: 9448-10308 п.о.
Для проведения полимеразной цепной реакции используют 100 мкл реакционной смеси, содержащей 0,5 нг ДНК-матрицы, по 1 мкМ соответствующего прямого и обратного праймеров, 2,5 ед. Pfu-полимеразы, 10 мкл 10х Pfu-буфера, 0,8 мМ dNTP. Реакцию осуществляют по следующей схеме: 94°C - денатурирование (1 мин), 57°C - отжиг (1 мин), 72°C - полимеризация (50 сек). Всего проводят 30 циклов амплификации. Амплифицированные фрагменты ДНК очищают в агарозном геле и затем при помощи «DNA extraction KIT» (Fermentas).
Наработанные фрагменты имеют следующие размеры:
- первый фрагмент 378 пар оснований;
- второй фрагмент 908 пар оснований.
Мутантный ген phyA-CfD синтезируют методом полимеразной цепной реакции из двух вышеописанных фрагментов. Для проведения ПЦР используют 100 мкл реакционной смеси, содержащей 0,2 нг каждого ДНК-фрагмента, по 2 мкМ соответствующего прямого pic3638D и обратного pic3638R праймеров, 2,5 ед. Pfu-полимеразы, 10 мкл 10х Pfu-буфера, 0,8 мМ dNTP. Реакцию осуществляют по следующей схеме: 94°C - денатурирование (1 мин), 57°C - отжиг (1 мин), 72°C - полимеризация (1 мин). Всего проведено 30 циклов амплификации. Из агарозного геля выделен амплифицированный фрагмент ДНК, размером 1256 пар оснований, который очищен при помощи «DNA extraction KIT».
2. Конструирование плазмиды
Полученный фрагмент ДНК phyA-CfD гидролизуют эндонуклеазами рестрикции EcoR1 и Not1 (Fermentas), очищают в агарозном геле и лигируют с вектором рР10 (Заявка РФ №2008134196, приор, от 21.08.2008), который в качестве сайта интеграции содержит последовательность ДНК, кодирующую область 18S рРНК, в качестве селективного маркера для отбора трансформантов в клетках E.coli содержит селективный маркер bla, в качестве сайта начала репликации содержит pUC ori, в состав экспрессионной кассеты входит промотор GAP, терминатор транскрипции АОХ1, селективный маркер HIS4, а в качестве сигнального пептида вектор содержит α-амилазный сигнальный пептид (фиг.2).
Предварительно вектор рР10 расщепляют ферментами рестрикции по сайтам EcoR1-Not1. Лигазной смесью трансформируют компетентные клетки E.coli C600 [Т.Маниатис, Э.Фрич, Дж.Сэмбрук, Молекулярное клонирование, Москва, "Мир", 1984, с.84], приготовленные накануне обработкой хлористым кальцием. После стандартной процедуры трансформации (0°C - 40 мин, 42°C - 2 мин, 0°C - 5 мин) клетки разводят в 10 раз средой LB [Т.Маниатис, Э.Фрич, Дж.Сэмбрук, Молекулярное клонирование, Москва, "Мир", 1984, с.84], подращивают в течение одного часа и высевают на агар LB, содержащий ампициллин в концентрации 50 мкг/мл. Посевы инкубируют при 37°C.
На следующие сутки выросшие устойчивые к ампициллину колонии тестируют с помощью вышеприведенных праймеров и отбирают позитивные клоны, из которых выделяют плазмидную ДНК по стандартной методике (Т.Маниатис, Э.Фрич, Дж.Сэмбрук, Молекулярное клонирование, Москва, "Мир", 1984, с.89). Выделенные плазмидные ДНК гидролизуют эндонуклеазами рестрикции EcoR1 и Not1, размер полученных фрагментов определяют с помощью гель-электрофореза. Затем отбирают клон, плазмидная ДНК которого содержит последовательность, соответствующую размеру последовательности кодирующей renphyA-CfD.
Из отобранного клона выделяют рекомбинантную плазмиду pP10phyA-CfD. Ген phyA-CfD секвенируют. Нуклеотидная последовательность гена соответствует последовательности, приведенной в перечне последовательностей под номером SEQ ID NO:3, и кодирует аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:4, отличающуюся от исходной заменой в позиции 144.
Пример 2. Создание рабочего штамма Pichia yastoris PhyA-CfD для получения и определения свойств фитазы PhyA-CfD
1. Получение рабочего штамма-продуцента
Для получения штамма Pichia pastoris PhyA-CfD, клетки штамма Pichia pastoris GS115 (his4) ВКПМ Y-2837 трансформируют плазмидой pP10phyA-CfD, которую предварительно гидролизуют рестриктазой BglII и электроэлюируют в 1% агарозном геле.
1 мл ночной культуры клеток Pichia pastoris Y-2837 выращивают в 100 мл среды YEPD при 30°C до достижения культурой оптической плотности, соответствующей 2 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30°C в течение 30 мин. К 50 мкл полученной суспензии клеток добавляют 1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 мин при 100°C), и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 мин при 30°C и 20 минут при 42°C, помещают на 15 секунд в ледяную баню и отделяют клетки центрифугированием в течение 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду М9 с добавлением 0,3 об.% микроэлементов. Клоны трансформантов выращивают 2 суток.
Полученные трансформанты высевают в жидкую среду YEPD и выращивают при 30°C до стационарной фазы роста в течение 2 суток. Далее клетки осаждают центрифугированием при 10000 об/мин в течение 5 минут, отбирают 100 мкл супернатанта и определяют фитазную активность по накоплению в реакционной среде свободного фосфат-иона, детектируемого методом Фиске-Субарроу. Одна единица активности фермента соответствует высвобождению 1 мкмоль фосфата за 1 минуту. Активность фермента измеряют при помощи спектрофотометра при длине волны 700 нм.
Трансформант, показавший наибольшую фитазную активность культуральной жидкости, выбран для наработки фермента фитазы с целью определения ее термостабильности.
2. Синтез фитазы PhyA-CfD штаммом-продуцентом Pichia pastoris PhyA-CfD
Посевную культуру штамма-продуцента Pichia pastoris PhyA-CfD выращивают в 5 мл жидкой среды YEPD при 30°C в течение ночи. Полученную ночную культуру переносят в колбу Эрленмейера, объемом 750 мл, содержащую 100 мл среды YEPD, в соотношении 1:100. Культуру инкубируют при 30°C и интенсивном встряхивании (240 об/мин) до достижения стационарной фазы в течение 2 суток. Фитазную активность штамма измеряют, как описано выше. Активность составляет 50 ед/мл культуральной жидкости.
Для определения удельной фитазной активности [Can J Microbiol., 2000, v.46, №1, p.59-71] фитазу PhyA-CfD очищают, используя метод высокоскоростной жидкостной хроматографии (ВСЖХ). Образцы готовят с помощью диализа против буфера (50mM Tris-HCl, рН 7.0, 1М NaCl) в течение ночи и наносят на колонку Superdex 75-HR, уравновешенную при помощи того же буфера. Колонку предварительно калибруют с использованием белковых маркеров #SM0431 (Fermentas). Количество белка на выходе колонки измеряют при помощи УФ-детектора при длине волны 280 нм. Фитаза PhyA-CfD обнаружена во фракциях, соответствующих молекулярной массе 45 кДа. От солей буфера образец фитазы очищают диализом против воды в течение ночи.
Затем отбирают аликвоту, содержащую 30 мкг очищенного фермента, и измеряют его фитазную активность по методу Фиске-Субарроу.
Активность отобранного образца составила 80 единиц, т.е. удельная активность фермента PhyA-CfD равна 2400 ед/мг белка.
Для определения устойчивости к действию высокой температуры образец фитазы PhyA-CfD инкубируют при температуре 80°C в течение 10 мин, а затем при 5°C в течение 30 мин с целью ренатурации белка. Остаточная удельная активность составляет 216 ед/мг белка, т.е. мутантная термостабильная фитаза PhyA-CfD после прогрева при температуре 80°C в течение 10 минут сохраняет 9% активности.
Пример 3. Конструирование рекомбинантной плазмиды pP10phyA-CfV
1. Проведение мутагенеза
Мутантный ген phyA-CfV получают, применяя метод сайт-направленного мутагенеза к гену phyA, кодирующему фитазу PhyA-Cf.
Для этого ПЦР методом синтезируют два фрагмента ДНК, включающих нуклеотидную замену в положениях 677 и 678 нуклеотидного сиквенса SEQ ID NO:2 таким образом, что кодон gta (кодирующий валин) заменяют на кодон gct (кодирующий аланин).
Для ПЦР-синтеза используют следующие праймеры (нуклеотидные замены в приведенных последовательностях подчеркнуты):
1. Для синтеза первого фрагмента
прямой - pic3638D (5'-aggaattcGAAGAGCAGAACGGTATGA-3')
обратный - V/Aobr (5'-ACCAGGTAATGA AG CATTGT-3')
2. Для синтеза второго фрагмента
прямой - V/Apr (5'- actcctgacaatgcttcatta-3')
обратный - pic3638R (5'-agcggccgcTTACTTATTCCGTAACTG-3').
В качестве матрицы для проведения полимеразной цепной реакции используют плазмиду pP10PhyA-Cf.
Полимеразную цепную реакцию осуществляют, как описано в примере 1.
Наработанные фрагменты имеют следующие размеры:
- первый фрагмент 620 пар оснований;
- второй фрагмент 665 пар оснований.
Мутантный ген phyA-CfV синтезируют из двух вышеописанных фрагментов методом полимеразной цепной реакции, как описано в примере 1. Получают амплифицированный фрагмент ДНК, размером 1256 пар оснований.
2. Конструирование плазмиды
Конструирование плазмиды pf10phyA-CfV осуществляют, как описано в примере 1, используя амплифицированный ген phyA-CfV.
После получения рекомбинантной плазмиды ген phyA-CfV секвенируют. Нуклеотидная последовательность гена соответствует последовательности, приведенной в перечне последовательностей под номером SEQ ID NO:5, и кодирует аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:6, отличающуюся от исходной заменой в позиции 226.
Полученную плазмиду pP10phyA-CfV гидролизуют рестриктазой BglII, электроэлюируют в 1% агарозном геле и используют для трансформации клеток дрожжей Pichia pastoris GS115 (his4) ВКПМ Y2837, как описано в примере 4.
Пример 4. Создание рабочего штамма Pichia pastoris PhyA-CfV для получения и определения свойств фитазы PhyA-CfV
1. Получение рабочего штамма-продуцента
Для получения штамма Pichia pastoris PhyA-CfV, клетки штамма Pichia pastoris GS115 (his4) ВКПМ Y-2837 трансформируют плазмидой pP10phyA-CfV, как описано в примере 2.
Трансформант, показавший наибольшую фитазную активность культуральной жидкости, выбран для наработки фермента фитазы с целью определения ее термостабильности.
2. Синтез фитазы PhyA-CfV штаммом-продуцентом Pichia pastoris PhyA-CfV
Подготовку посевной культуры и культивирование штамма-продуцента Pichia pastoris PhyA-CfV осуществляют, как описано в примере 2.
Фитазная активность штамма, измеренная в соответствии с примером 2, составляет 60 ед/мл культуральной жидкости.
Очистку фитазы, определение удельной фитазной активности и определение устойчивости фитазы к кратковременному действию высокой температуры осуществляют по примеру 2.
Активность отобранного образца, содержащего 36 мкг фермента, составляет 76 единиц, т.е. удельная активность фермента PhyA-CfV равна 2736 ед/мг белка.
Остаточная удельная активность составляет 278 ед/мг белка, т.е. мутантная термостабильная фитаза PhyA-CfV после прогрева при температуре 80°C в течение 10 минут сохраняет 10% активности.
Пример 5. Конструирование рекомбинантной плазмиды pP10phyA-CfG
1. Проведение мутагенеза
Мутантный ген phyA-CfG получают применяя метод сайт-направленного мутагенеза к гену phyA, кодирующему фитазу PhyA-Cf.
Для этого ПЦР методом синтезируют два фрагмента ДНК, включающих нуклеотидную замену в положении 1028 нуклеотидного сиквенса SEQ ID NO:2 таким образом, что кодон ggt (кодирующий глицин) заменяют на кодон gat (кодирующий аспарагиновую кислоту).
Для ПЦР-синтеза используют следующие праймеры (нуклеотидные замены в приведенных последовательностях подчеркнуты):
1. Для синтеза первого фрагмента
прямой - pic3638D (5'-aggaattcGAAGAGCAGAACGGTATGA-3')
обратный - G/Dobr (5'-ATTATCGGGTTGA T CGGGTA-3')
2. Для синтеза второго фрагмента
прямой - G/Dpr (5'-actggtcgctacccg a tcaa-3')
обратный - pic3638R (5'-agcggccgcTTACTTATTCCGTAACTG-3').
В качестве матрицы для проведения полимеразной цепной реакции используют плазмиду pP10PhyA-Cf.
Полимеразную цепную реакцию осуществляют, как описано в примере 1.
Наработанные фрагменты имеют следующие размеры:
- первый фрагмент 1021 пар оснований;
- второй фрагмент 265 пар оснований.
Мутантный ген phyA-CfG синтезируют из двух вышеописанных фрагментов методом полимеразной цепной реакции, как описано в примере 1. Получают амплифицированный фрагмент ДНК, размером 1256 пар оснований.
2. Конструирование плазмиды
Конструирование плазмиды pP10phyA-CfG осуществляют, как описано в примере 1, используя амплифицированный ген phyA-CfG.
После получения рекомбинантной плазмиды ген phyA-CfG секвенируют.
Нуклеотидная последовательность гена соответствует последовательности, приведенной в перечне последовательностей под номером SEQ ID NO:7, и кодирует аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:8, отличающуюся от исходной заменой в позиции 343.
Полученную плазмиду pf10phyA-CfG гидролизуют рестриктазой BglII, электроэлюируют в 1% агарозном геле и используют для трансформации клеток дрожжей Pichia pastoris GS115 (his4) ВКПМ Y2837, как описано в примере 6.
Пример 6. Создание рабочего штамма Pichia pastoris PhyA-CfG для получения и определения свойств фитазы PhyA-CfG
1. Получение рабочего штамма-продуцента
Для получения штамма Pichia pastoris PhyA-CfG, клетки штамма Pichia pastoris GS115 (his4) ВКПМ Y-2837 трансформируют плазмидой pP10phyA-CfD, как описано в примере 2.
Трансформант, показавший наибольшую фитазную активность культуральной жидкости, был выбран для наработки фермента фитазы с целью определения ее термостабильности.
2. Синтез фитазы PhyA-CfG штаммом-продуцентом Pichia pastoris PhyA-CfG
Подготовку посевной культуры и культивирование штамма-продуцента Pichia pastoris PhyA-CfG осуществляют, как описано в примере 2.
Фитазная активность штамма измеренная в соответствии с примером 2, составляет 57 ед/мл культуральной жидкости.
Очистку фитазы, определение удельной фитазной активности и определение устойчивости фитазы к кратковременному действию высокой температуры осуществляют по примеру 2.
Активность отобранного образца, содержащего 32 мкг фермента, составляет 62 единицы, т.е. удельная активность фермента PhyA-CfG составляет 1984 ед/мг белка.
Остаточная удельная активность составляет 238 ед/мг белка, т.е. мутантная термостабильная фитаза PhyA-CfD после прогрева при температуре 80°C в течение 10 минут сохраняет 12% активности.
Пример 7. Конструирование рекомбинантной плазмиды pP10phyA-CfDVG
1. Проведение мутагенеза
Мутантный ген phyA-CfDVG получают, применяя метод сайт-направленного мутагенеза к гену phyA, кодирующему фитазу PhyA-Cf.
Для этого ПЦР методом синтезируют четыре фрагмента ДНК, включающих нуклеотидные замены в следующих положениях нуклеотидного сиквенса SEQ ID NO:2:
- 430 и 432 таким образом, что кодон gat (кодирующий аспарагиновую кислоту) заменяют на кодон аас (кодирующий аспарагин);
- 677 и 678 таким образом, что кодон gta (кодирующий валин) заменяют на кодон gct (кодирующий аланин);
- 1028 таким образом, что кодон ggt (кодирующий глицин) заменяют на кодон gat (кодирующий аспарагиновую кислоту).
Для ПЦР-синтеза используют праймеры, содержащие вышеописанные нуклеотидные замены (в приведенных последовательностях подчеркнуты):
1. Для синтеза первого фрагмента
прямой - pic3638D (5'-aggaattcGAAGAGCAGAACGGTATGA-3')
обратный - D/Nobr (5'-ATTAAAAAGAGG G T T AGTTT-3')
2. Для синтеза второго фрагмента
прямой - D/Npr (5'-aagaaaaaact a a c cctct-3')
обратный - V/Aobr (5'-ACCAGGTAATGA AG CATTGT-3')
3. Для синтеза третьего фрагмента
прямой - V/Apr (5'-actcctgacaatg ct tcatta-3')
обратный - G/Dobr (5'-ATTATCGGGTTGA T CGGGTA-3')
4. Для синтеза четвертого фрагмента
прямой - G/Dpr (5'-actggtcgctacccg at caa-3')
обратный - pic3638R (5'-agcggccgcTTACTTATTCCGTAACTG-3').
На 5-конце праймеров pic3638D и pic3638R находится сайт рестрикции для эндонуклеазы, образующей липкие концы: EcoR1 для прямого праймера и Not1 - для обратного (в приведенных последовательностях выделены жирным шрифтом).
В качестве матрицы для проведения полимеразной цепной реакции используют плазмиду pP10PhyA-Cf.
Полимеразную цепную реакцию осуществляют, как описано в примере 1.
Наработанные фрагменты имеют следующие размеры:
- первый фрагмент 378 пар оснований;
- второй фрагмент 272 пары оснований;
- третий фрагмент 378 пар оснований;
- четвертый 265 пар оснований.
Мутантный ген phyA-CfDVG синтезируют из четырех вышеописанных фрагментов методом полимеразной цепной реакции, как описано в примере 1. Получают амплифицированный фрагмент ДНК, размером 1256 пар оснований.
2. Конструирование плазмиды
Конструирование плазмиды pP10phyA-CfDVG осуществляют, как описано в примере 1, используя амплифицированный ген phyA-CfDVG.
После получения рекомбинантной плазмиды ген phyA-CfDVG секвенируют. Нуклеотидная последовательность гена соответствует последовательности, приведенной в перечне последовательностей под номером SEQ ID NO:9, и кодирует аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:10, отличающуюся от исходной заменами в позиции 144 и 226 и 343.
Полученную плазмиду pP10phyA-CfDVG гидролизуют рестриктазой BglII, электроэлюируют в 1% агарозном геле и используют для трансформации клеток дрожжей Pichia pastoris GS115 (his4) ВКПМ Y2837, как описано в примере 8.
Пример 8. Получение штамма Pichia pastoris PhyA-CfDVG ВКПМ Y-3495 - продуцента фитазы Phy-CfDVG и синтез фитазы PhyA-CfDVG
1. Получение штамма-продуцента
Для получения штамма Pichia pastoris PhyA-CfDVG - продуцента фитазы PhyA-CfDVG, клетки штамма Pichia pastoris GS115 (his4) ВКПМ Y-2837 трансформируют плазмидой pP10phyA-CfDVG, как описано в примере 2.
Трансформант с наибольшей фитазной активностью культуральной жидкости заложен на хранение и депонирован в ВКПМ под номером ВКПМ Y-3495.
В отличие от исходного штамма заявляемый штамм не нуждается в гистидине для своего роста.
2. Синтез фитазы PhyA-CfDVG штаммом-продуцентом Pichia pastoris PhyA-CfDVG ВКПМ Y-3495
Подготовку посевной культуры и культивирование штамма-продуцента Pichia pastoris PhyA-CfDVG осуществляют, как описано в примере 2.
Фитазная активность штамма измеренная в соответствии с примером 2, составляет 100 ед/мл культуральной жидкости.
Очистку фитазы, определение удельной фитазной активности и определение устойчивости фитазы к кратковременному действию высокой температуры осуществляют по примеру 2.
Активность отобранного образца, содержащего 25 мкг очищенного фермента, составляет 37,5 единиц, т.е. удельная активность фермента PhyA-CfDVG составляет 1500 ед/мг белка.
Остаточная удельная активность составляет 1050 ед/мг белка, т.е. мутантная термостабильная фитаза PhyA-CfDVG после прогрева при температуре 80°C в течение 10 минут сохраняет 70% активности.
Пример 9. Конструирование рекомбинантной плазмиды pP10phyA-CfDV
1. Проведение мутагенеза
Мутантный ген phyA-CfDV получают, применяя метод сайт-направленного мутагенеза к гену phyA-CfDVG.
Для этого ПЦР методом синтезируют два фрагмента ДНК, включающих нуклеотидную замену в положении 1028 нуклеотидного сиквенса SEQ ID NO:9 таким образом, что кодон gat (кодирующий аспарагиновую кислоту) заменяют на кодон ggt (кодирующий глицин).
Для ПЦР-синтеза используют следующие праймеры (нуклеотидные замены в приведенных последовательностях подчеркнуты):
1. Для синтеза первого фрагмента
прямой - pic3638D (5'-aggaattcGAAGAGCAGAACGGTATGA-3')
обратный - D/Gobr (5'-ATTATCGGGTTGA C CCGGGTA-3')
2. Для синтеза второго фрагмента
прямой - D/Gpr (5'-actggtcgctacccg g tcaa-3')
обратный - pic3638R (5'-agcggccgcTTACTTATTCCGTAACTG-3').
В качестве матрицы для проведения полимеразной цепной реакции используют плазмиду pP10phyA-CfDVG.
Полимеразную цепную реакцию осуществляют, как описано в примере 1.
Наработанные фрагменты имеют следующие размеры:
- первый фрагмент 1021 пар оснований;
- второй фрагмент 265 пар оснований.
Мутантный ген phyA-CfDV синтезируют из двух вышеописанных фрагментов методом полимеразной цепной реакции, как описано в примере 1. Получают амплифицированный фрагмент ДНК размером 1256 пар оснований.
2. Конструирование плазмиды
Конструирование плазмиды pP10phyA-CfDV осуществляют, как описано в примере 1, используя амплифицированный ген phyA-CfDV.
После получения рекомбинантной плазмиды ген phyA-CfDV секвенируют. Нуклеотидная последовательность гена соответствует последовательности, приведенной в перечне последовательностей под номером SEQ ID NO:11, и кодирует аминокислотную последовательность, приведенную в перечне последовательностей под номером SEQ ID NO:12, отличающуюся от исходной заменами в позиции 144 и 226.
Полученную плазмиду pP10phyA-CfDV гидролизуют рестриктазой BglII, электроэлюируют в 1% агарозном геле и используют для трансформации клеток дрожжей Pichia pastoris GS115 (his4) ВКПМ Y2837, как описано в примере 10.
Пример 10. Получение штамма Pichia pastoris PhyA-CfDV ВКПМ Y-3494 - продуцента фитазы PhyA-CfDV и синтез фитазы PhyA-CfDV
1. Получение штамма-продуцента
Для получения штамма Pichia pastoris PhyA-CfDV - продуцента фитазы PhyA-CfDV, клетки штамма Pichia pastoris GS115 (his4) ВКПМ Y-2837 трансформируют плазмидой pf10phyA-CfDV, как описано в примере 2.
Трансформант, показавший наибольшую фитазную активность культуральной жидкости, заложен на хранение и депонирован в ВКПМ под номером Y-3494.
2. Синтез фитазы PhyA-CfDV штаммом-продуцентом Pichia pastoris PhyA-CfDV ВКПМ Y-3494
Подготовку посевной культуры и культивирование штамма-продуцента Pichia pastoris PhyA-CfDV осуществляют, как описано в примере 2.
Фитазная активность штамма, измеренная в соответствии с примером 2, составляет 150 ед/мл культуральной жидкости.
Очистку фитазы, определение удельной фитазной активности и определение устойчивости фитазы к кратковременному действию высокой температуры осуществляют по примеру 2.
Активность отобранного образца, содержащего 30 мкг фермента, составляет 51 единицу, т.е. удельная активность фермента PhyA-CfDV составляет 1700 ед/мг белка.
Остаточная удельная активность составляет 731 ед/мг белка, т.е. мутантная термостабильная фитаза PhyA-CfDV после прогрева при температуре 80°С в течение 10 минут сохраняет 43% активности.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФИТАЗЫ (ВАРИАНТЫ), ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФИТАЗЫ (ВАРИАНТЫ) | 2009 |
|
RU2409670C1 |
| Мутантная рекомбинантная термостабильная фитаза | 2018 |
|
RU2706086C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ ФИТАЗЫ | 2012 |
|
RU2504579C2 |
| ИНТЕГРАТИВНЫЙ ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГЕНОВ В ДРОЖЖАХ | 2008 |
|
RU2388823C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент фитазы | 2018 |
|
RU2701498C1 |
| Трансформант дрожжей Pichia pastoris, продуцирующий фитазу | 2019 |
|
RU2708446C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент фитазы Escherichia coli | 2019 |
|
RU2751595C2 |
| Рекомбинантный штамм дрожжей Pichia pastoris с увеличенной продукцией фитазы Escherichia coli | 2019 |
|
RU2737623C1 |
| Способ оценки продуктивности рекомбинантных трансформантов дрожжей Pichia pastoris, секретирующих фитазы, относящиеся к кислым гистидиновым фосфатазам | 2017 |
|
RU2665836C1 |
| Трансформант дрожжей Komagataella phaffi - продуцент фитазы Citrobacter gillenii | 2020 |
|
RU2771582C1 |














Изобретение относится к области биотехнологии. Предложены варианты мутантных рекомбинантных фитаз PhyA-Cf из бактерий Citrobacter freundii: 1) вариант, содержащий аминокислотную замену D144N или D144R, или D144Q, или D144K; 2) вариант, содержащий аминокислотную замену V226A или V226G; 3) вариант, содержащий аминокислотную замену G343D или G343E, или G343K, или G343R, или G343N; 4) вариант, содержащий аминокислотные замены в положении D 144 на N, R, Q или K, и в положении V226 на А или G; 5) вариант, содержащий аминокислотные замены в положении D 144 на N, R, Q или K, и в положении G343 на D, Е, K, R или N; 6) вариант, содержащий аминокислотные замены в положении V226 на А или G, и в положении G343 на D, Е, K, R или N; 7) вариант, содержащий аминокислотные замены в положении D 144 на N, R, Q или K, и в положении V226 на А или G, и в положении G343 на D, Е, K, R или N. Представлены фрагменты ДНК, кодирующие указанные рекомбинанатные фитазы. Описаны штаммы-продуценты рекомбинанатных фитаз Pichia pastoris PhyA-CfDVG и Pichia pastoris PhyA-CfDV, депонированные во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под номерами ВКПМ Y-3495 и ВКПМ Y-3494 соответственно. Изобретение позволяет получить фитазы, обладающие термостабильными свойствами. 11 н. и 2 з.п. ф-лы, 2 ил., 10 пр.
1. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин.
2. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин.
3. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
4. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, а аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин.
5. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
6. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
7. Мутантная рекомбинантная фитаза, характеризующаяся аминокислотной последовательностью, соответствующей аминокислотной последовательности фитазы PhyA-Cf из бактерий Citrobacter freundii, в которой аспарагиновая кислота в положении 144 заменена на аминокислоту аспарагин, или аминокислоту аргинин, или аминокислоту глутамин, или аминокислоту лизин, аминокислота валин в положении 226 заменена на аминокислоту аланин или аминокислоту глицин, а аминокислота глицин в положении 343 заменена на аспаргиновую кислоту или глутаминовую кислоту, или аминокислоту лизин, или аминокислоту аргинин, или аминокислоту аспарагин.
8. Мутантная рекомбинантная фитаза по п.4, отличающаяся тем, что аспарагиновая кислота в положении 144 заменена аминокислотой аспарагин и аминокислота валин в положении 226 заменена аминокислотой аланин.
9. Мутантная рекомбинантная фитаза по п.7, отличающаяся тем, что аспарагиновая кислота в положении 144 заменена аминокислотой аспарагин и аминокислота валин в положении 226 заменена аминокислотой аланин, а аминокислота глицин в положении 343 заменена аспарагиновой кислотой.
10. Фрагмент ДНК, кодирующий мутантную фитазу по п.9, характеризующийся тем, что имеет нуклеотидную последовательность, указанную в списке последовательностей под номером SEQ ID NO:9, в которой нуклеотиды в положении 430-432 образуют кодон, кодирующий аспарагин, в положении 676-678 образуют кодон, кодирующий аланин, а в положении 1027-1029 образуют кодон, кодирующий аспарагиновую кислоту, или последовательность, отличающуюся от последовательности SEQ ID NO:9, вследствие вырожденности генетического кода.
11. Фрагмент ДНК, кодирующий мутантную фитазу по п.8, характеризующийся тем, что имеет нуклеотидную последовательность, указанную в списке последовательностей под номером SEQ ID NO:11, в которой нуклеотиды в положении 430-432 образуют кодон, кодирующий аспарагин, а в положении 676-678 образуют кодон, кодирующий аланин, или последовательность, отличающуюся от последовательности SEQ ID NO:11, вследствие вырожденности генетического кода.
12. Штамм Pichia pastoris PhyA-CfDVG ВКПМ Y-3495 - продуцент мутантной фитазы по п.9, обладающей термостабильными свойствами.
13. Штамм Pichia pastoris PhyA-CfDV ВКПМ Y-3494 - продуцент мутантной фитазы по п.8, обладающей термостабильными свойствами.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Паровая машина с периодическим парообразователем | 1922 |
|
SU676A1 |
| YOUNG-OK KIM, et al | |||
| "Molecular cloning of the phytase gene from Citrobacter braakii and its expression in Saccharomyces cerevisiae", Biotechnology Letters (2006) 28: pp.33-38 | |||
| ЗИНИН Н.В., САМСОНОВ В.В | |||
| и др | |||
| Фитазная активность некоторых групп бактерий | |||
| - Биотехнология, 2003, №2, с.3-10. | |||

