Изобретение относится к микробиологии и биотехнологии и касается получения штаммов дрожжей Komagataella phaffii (Pichia pastoris), способных продуцировать фермент фитазу.
Фосфор является важным химическим элементом, необходимым для роста и развития животных. В большинстве используемых в животноводстве растительных кормов, таких как зерновые и бобовые культуры, фосфор содержится, в основном, в форме фитата (мио-инозитол гексакисфосфат) [Advanced Food Research, 1982, 28, 1-92.]. Моногастричные животные не способны усваивать фитатный фосфор из-за отсутствия в пищеварительном тракте необходимого фермента [Can. J. Anim. Sci., 2013, 93, 9-21.].
Присутствие в составе комбикорма фитата препятствует также эффективному усвоению кальция, магния, цинка, железа и других минеральных веществ и приводит к связыванию белков корма в недоступные для переваривания комплексы [Current Topics In Nutraceutical Research, 2008, 6(3), 131-144.].
Фитазы (мио-инозитол гексакисфосфат фосфогидролазы) гидролизуют фитат с отщеплением фосфатных групп. Эти ферменты с успехом используют в качестве кормовой добавки, значительно повышая усвоение фосфора [J. Sci. Food Agric, 2015, 95, 878-896.]. Они обеспечивают высвобождение не только фитат-связанного фосфора, но также белков, макро- и микроэлементов, повышая питательные свойства кормов [British Poultry Science, 2004, 45(1), 101-108.].
Природными источниками фитаз являются различные микроорганизмы: бактерии, грибы [FEMS Microbiology Reviews, 2005, 29 (1), 3-23.].
В промышленности фитазы получают путем микробиологического синтеза с использованием штаммов-продуцентов, потребность промышленности в которых постоянно растет [Afr. J. Biotechnol, 2009, 8(17), 4229-4232.]. Наиболее перспективные продуценты фитазы получены на основе рекомбинантных штаммов метилотрофных дрожжей Komagataella phaffii {Pichia pastoris) [J. Mol. Recognit., 2005, 18(2), 119-138. doi: 10.1002/jmr.687], которые обладают мощными системами экспрессии и секреции рекомбинантных белков (включая фитазы) и для которых разработаны питательные среды и отработан процесс ферментации с использованием культуры высокой плотности. Процесс культивирования метилотрофных дрожжей достаточно прост, поскольку их рост не блокируется продуктами метаболизма [FEMS Microbiol.Rev., 2000, 24:45-66, doi: 10.1111/j.1574-6976.2000.tb00532.x].
На основе дрожжей K. phaffii сконструированы продуценты следующих фитаз: Citrobacter amalonaticus [ВМС Biotechnology, 2015, 15, 88.], Escherichia coli [J. Agric. Food Chem., 2013, 61(25), 6007-6015., CN 102586294, CN 102943083], Peniophora lycii [Appl. Microbiol. Biotechnol, 2006, 72(5), 1039-47.], Citrobacter braakii [Acta Microbiologica Sinica, 46(6), 945-950.], Yersinia rohdei, Yersinia pestis, Y. Intermedia [Appl. Microbiol. Biotechnol., 2008, 80(3), 417-426.] и других.
Показано, что гены, кодирующие фитазы различного происхождения, экспрессируются в дрожжах K. phaffii с различной эффективностью, а именно при экспрессии фитаз из Yersinia rohdei, Yersinia pestis, Y. intermedia, E. coli или Aspergillus niger в дрожжах K. phaffii фитазная активность в культуральной жидкости на 4 сутки культивирования составляет соответственно 313, 19, 270, 219 и 16 ед/мл. [Appl. Microbiol. Biotechnol, 2008, 80(3), 417-426.]. При этом, фитазы различного происхождения отличаются между собой по свойствам, существенным для использования в промышленности, таким как, рабочий диапазон температуры и рН, устойчивость к действию протеолитических ферментов и другим [J. Agric. Food Chem., 2013, 61(25), 6007-6015; Amu. Rev. Anim. Biosci., 2013, 1, 283-309.].
Получение новых фитаз, обладающих промышленно-ценными свойствами и способных эффективно экспрессироваться в дрожжах K. Phaffii является актуальной задачей.
Технической проблемой, на решение которой направлено заявляемое изобретение является расширение арсенала рекомбинантных микроорганизмов, продуцирующих фитазу.
Техническим результатом, достигаемым при реализации заявляемого изобретения является получение трансформанта дрожжей Komagataella phaffii - продуцента фитазы Citrobacter gillenii.
Получение заявляемого трансформанта включает введение гена phyCg, кодирующего фитазу С. gillenii, в клетки дрожжей K. phaffii с помощью экспрессионной кассеты, включающей ген phyCg, промотор, работающий в дрожжах K. phaffii, сигнальный пептид для осуществления секреции фермента в культуральную жидкость, а также терминатор, маркерный ген и, предпочтительно, сайт для гомологичной интеграции в хромосому.
Нуклеотидная последовательность гена phyCg, кодирующего фитазу С. gillenii, представлена в международной базе Genbank под номером NZ_QVEK01000009.1 (22167…23465). Так как в силу «вырожденности» генетического кода одна и та же аминокислотная последовательность может кодироваться большим числом нуклеотидных последовательностей, то для клонирования может быть использована не только сама нуклеотидная последовательность (ген phyCg), но и все ее формы, определяемые вырожденностью генетического кода.
Интеграцию осуществляют путем или гомологичной, или негомологичной рекомбинации. Трансформацию экспрессионной кассеты в клетки дрожжей K. phaffii проводят в частности, методом электропорации [http://tools.thermofisher.com/content/sfs/manuals/pich_man.pdf], методом с использованием полиэтиленгликоля или протопластов [http://www.thermofisher.com/order/catalog/product/K173001].
Конструирование экспрессионных кассет осуществляют стандартными методами генетической инженерии [Рыбчин В.Н. Основы генетической инженерии. - СПб.: СПбГТУ, 1999; Sambrook J., Maniatis Т., Fritsch Е. Molecular cloning: a laboratory manual. - N.Y.: Cold Spring Harbor Laboratory, 1989.] с использованием генетических элементов, подходящих для работы с дрожжами K. phaffii. В качестве промоторов используют АОХ1, DAS, FLD1, ICL1, PHO89, THI11, ADH1, ENO1, GUT1, GAP, TEF1, PGK1, GCW14, G1, G6 или другие [Appl. Microbiol. Biotechnol, 2014, 98, 5301-5317.]. В качестве сигнальных пептидов используют а-фактор, PHO1, SUC2, РНА-Е, KILM1, pGKL, CLY, CLY-L8, K28 pre-pro-toxin или другие [Appl. Microbiol. Biotechnol, 2014, 98, 5301-5317.]. В качестве селективных маркеров используют любые подходящие маркеры, например, гены резистентности к антибиотикам зеоцину, генетицину (G418) или бластицидину С, а также гены комплементирующие ауксотрофные мутации в геноме K. phaffii, например, HIS4, МЕТ2, ADE1, ARG4, URA3, URA5, GUT1 [Yeast, 2005, 22, 249-270.]. В качестве плечей для гомологичной интеграции используют последовательности генов АОХ1, HIS4 [http://www.thermofisher.com/order/catalog/product/V17520] или другие последовательности, гомологичные участкам хромосомы дрожжей K. phaffii.
Полученные трансформанты тестируют на наличие целевого гена в составе хромосомной ДНК и способность проявлять фитазную активность.
Изобретение проиллюстрировано следующими фигурами.
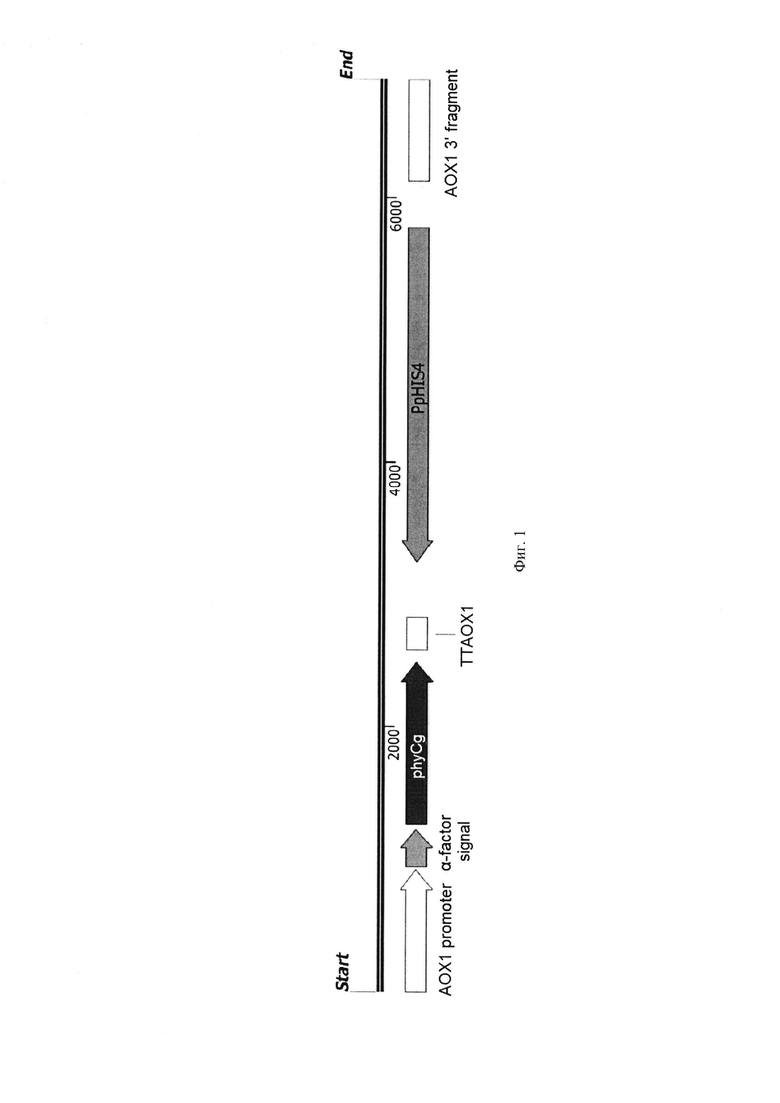
Фиг. 1. Экспрессионная кассета
Фиг. 2. Электрофореграмма фрагмента ПЦР, соответствующего гену phyCg
Фиг. 3. Активность фитазы PhyCg при различных значениях рН
Пример 1. Конструирование трансформанта дрожжей K. phaffii, продуцирующего фитазу С. gillenii
При конструировании интегративной экспрессионной кассеты используют метод "фьюжн-пцр" [Gene., 1989, 15, 77(1), 61-68.].
Последовательность гена phyCg, представленную в международной базе Genbank под номером NZ_QVEK01000009.1 (22167…23465) и кодирующую фитазу С. gillenii, синтезируют методом, описанным в [J. Microbiol. Methods, 2010, 81(2), 147-152.]. Получают ДНК последовательность, на 3'- и 5'-концах которой предусмотрены сайты рестрикции EcoRI и NotI, приведенную в перечне последовательностей под номером SEQ ID NO: 1.
Полученную последовательность ДНК встраивают в состав экспрессионной кассеты (фиг. 1), в состав которой входят следующие генетические элементы:
1. Ген phyCg, встроенный в единую рамку считывания с нуклеотидной последовательностью сигнального пептида α-фактора из Saccharomyces cerevisiae, под контролем АОХ1 промотора;
2. Терминатор транскрипции ТТАОХ1;
3. Дрожжевой селективный маркер HIS4, комплементирующий у дрожжей Komagataella phaffii мутацию в гене HIS4;
4. Область интеграции - 3' фрагмент нуклеотидной последовательности гена АОХ1.
Полученную интегративную экспрессионную кассету трансформируют в штамм K. phaffii ВКПМ Y-2837, который предварительно выращивают в жидкой питательной среде YP (мас. %: дрожжевой экстракт - 1, пептон - 2, вода - остальное) с добавлением глюкозы (2 мас. %) до концентрации 1×108 клеток на 1 мл. Клетки отделяют центрифугированием и промывают ледяной стерильной водой, а затем ледяным раствором 1 М сорбитола. Затем клетки инкубируют в 25 мМ растворе дитиотрейтола в течение 15 минут и промывают в ледяном растворе 1 М сорбитола. Далее клетки ресуспендируют в ледяном растворе 1 М сорбитола в концентрации 1-5×109 клеток на 1 мл. Аликвоту, объемом 40 мкл клеточной суспензии, переносят в охлажденный эппендорф, добавляют 400 нг ДНК экспрессионной интеграционной кассеты и инкубируют во льду 5 минут. Смесь клеток и ДНК переносят в предварительно охлажденную кювету для электропорации. Электропорацию проводят при следующих условиях: 1,5 кВ, 400 Ом, 25uF. После порации добавляют 1 мл ледяного раствора 1 М сорбитола.
Селекцию трансформантов ведут в течение 5 суток при температуре 30°С на агаризованной среде следующего состава (мас. %): Na2HPO4 - 0,6; KH2PO4 - 0,3; NaCl - 0,05; NH4Cl - 0,1; MgSO4 7H2O - 0,065; агар - 2; глюкоза - 2; CaCl2 - 0,07; биотин, мг - 0,0002; кальций пантотенат - 0,04; фолиевая кислота - 0,0002; ниацин - 0,04; р-аминобензойная к-та - 0,02; пиридоксин гидрохлорид - 0,04; рибофлавин - 0,02; тиамин гидрохлорид - 0,04; борная кислота - 0,05; CuSO4 - 0,004; KJ - 0,01; FeCl3 - 0,02; натрий молибдат - 0,02; ZnSO4 - 0,04, вода - остальное.
Наличие целевого гена в составе хромосомной ДНК полученных трансформантов подтверждают методом ПЦР с использованием праймеров PhyCg-F (5'-aagaattcgatgaacagagcggaatgcag-3'), PhyCg-R (5'-aagcggccgcttatttctcagcacattcggaca-3').
Режим реакции ПЦР: 95°С - 3 мин - 1 цикл; 30 циклов: 95°С - 30 с, 56°С - 30 с, 72°С - 90 с; 72°С - 5 мин. - 1 цикл.
Для контроля величины амплифицированного фрагмента ДНК при электрофорезе используют молекулярный маркер GeneRuler 1 kb DNA Ladder (Fermentas) (линия 2, фиг. 2, размер фрагментов снизу вверх 10000, 8000, 6000, 5000, 4000, 3500, 3000, 2500, 2000, 1500, 1000, 750, 500, 250 п.н.).
Наличие ПЦР-фрагмента размером 1233 п. н. (линия 1 фиг. 2) подтверждает присутствие гена phyCg, кодирующего фитазу из Citrobacter gillenii, в составе хромосомы полученных трансформантов.
Пример 2. Оценка уровня фитазной активности трансформантов дрожжей Komagataella phaffii
Отбор наиболее продуктивного трансформанта проводят при культивировании в жидкой питательной среде по следующей схеме.
Посевную культуру каждого из трансформантов в отдельности выращивают в пробирках (50 мл) с 5 мл жидкой питательной среды YP с добавлением глюкозы (2 мас. %) при 30°С в течение 24 ч на качалке (250 об/мин). Засев ферментационной среды осуществляют в соотношении 1/10 для каждого трансформанта в отдельности.
Ферментацию проводят при 30°С на качалке (250 об/мин) в питательной среде YP с добавлением 1% глюкозы в течение 24 ч. Экспрессию гена фитазы индуцируют путем добавления метанола до конечной концентрации 1% каждые 24 ч в течение 4 суток. Продуктивность каждого из трансформантов Komagataella phaffii оценивают по активности фитазы, накопившейся в культуральной жидкости. Для этого клетки трансформанта осаждают центрифугированием, а супернатант используют для определения фитазной активности модифицированным методом Фиске-Субарроу [J. Microbiol. Methods, 1999, 39(1), 17-22.].

Представленные в таблице значения фитазной активности свидетельствуют о достаточно высоком уровне продуктивности по фитазе Citrobacter gillenii полученных трансформантов Komagataella phaffii, а также об эффективной экспрессии фитазы PhyCg С. gillenii в дрожжах K. phaffii.
Отобранный по результатам ферментации наиболее продуктивный трансформант №3, который синтезирует фитазу в количестве 279 ед/мл, депонируют в Биоресурсном центре Всероссийская Коллекция Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» - ГосНИИгенетика (117545 Москва, 1-ый Дорожный проезд, д. 1) как Komagataella phaffii PCG3 ВКПМ Y-4839.
Штамм характеризуется следующими признаками
Культурально-морфологические признаки
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YP следующего состава (мас. %: пептон - 2, дрожжевой экстракт -1, агар - 2, вода - остальное) с добавлением глюкозы (2 мас. %), клетки имеют овальную форму, 3-4 мкм в диаметре. Клетки почкуются, при этом почкование истинное, многостороннее. Истинного мицелия не образуют. Споруляция происходит при инкубации культуры на агаризованной среде следующего состава (мас. %): хлорид калия - 1.0, ацетат натрия - 0.5, глюкоза - 1.0, агар - 2.0, вода - остальное. Аски имеют тетраэдрическую форму, включают 4 аскоспоры.
На агаризованной среде YP с добавлением глюкозы (2 мас. %) колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией. При росте в жидкой среде YP с добавлением глюкозы (2 мас. %), при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок не образует.
Физиолого-биохимические признаки
Способен к росту как в аэробных, так и в анаэробных условиях.
В качестве единственного источника углерода способен использовать метанол, этанол, глюкозу, глицерин, лактат, сукцинат, не способен ассимилировать мальтозу, сахарозу, ацетат, крахмал, лактозу.
При культивировании в присутствии метанола способен синтезировать фитазу.
Пример 3. Рабочий диапазон рН фитазы PhyCg, синтезированной заявляемым трансформантом.
По окончании процесса ферментации клетки заявляемого трансформанта осаждают центрифугированием, а супернатант используют для определения фитазной активности при различных значениях рН модифицированным методом Фиске-Субарроу [J. Microbiol, Methods, 1999, 39(1), 17-22.]. Определение рабочего диапазона рН фермента осуществляют с использованием следующих буферных растворов: глицин-HCl (рН 2,0-3,5), Na-ацетатный (4,0-5,5) и трис-HCl (рН 6,0-8,0). Реакционную смесь, содержащую 100 мкл супернатанта и 900 мкл 2%-ного раствора фитата натрия в соответствующем буфере инкубируют при 37°С в течение 25 мин. В образец добавляют 1000 мкл красителя (свежеприготовленная смесь, состоящая из четырех объемов 1,5%-ного молибдата аммония в 5,5%-ном растворе серной кислоты и 1 объема 2,5%-ного водного раствора сульфата железа (II)). Смесь выдерживают 5 мин и спектрофотометрически измеряют оптическую плотность при 700 нм. Единицу активности фитазы определяют, как количество фермента, катализирующего высвобождение 1 мкмоль неорганического фосфата из фитата натрия в минуту при 37°С.
На фиг. 3 представлена относительная активность фитазы PhyCg заявляемого трансформанта при различных значениях рН (за 100% принимается активность при оптимальном значении рН 4,5). Из представленных результатов видно, что фитаза, продуцируемая заявляемым трансформантом сохраняет более 50% активности в интервале рН от 3,0 до 6,0, Такой диапазон рН выгодно отличают полученную с помощью заявляемого трансформанта фитазу от большинства известных. В частности, коммерческая фитаза из Citrobacter braakii сохраняет около 60% активности при рН 3,0, около 50% активности при рН 5,5, и около 30% при рН 6,0 [Biotechnology Letters, 2003,25,1231-1234.].
Высокая активность фитазы PhyCg при рН 6 делает ее привлекательной для промышленного применения, в частности, для использования в качестве кормовой добавки при получении аквакультуры.
--->
Перечень последовательностей
<110>Федеральное государственное бюджетное учреждение "Государственный
научно-исследовательский институт генетики и селекции промышленных
микроорганизмов Национального исследовательского центра "Курчатовский
институт" (НИЦ «Курчатовский институт» - ГосНИИгенетика).
State Research Institute for Genetics and Selection of Industrial
Microorganisms of National Research Center «Kurchatov Institute»
(NRC «Kurchatov Institute» – GOSNIIGENETIKA)
<120>Трансформант дрожжей Komagataellaphaffii – продуцент фитазы
Citrobactergillenii
<130> 20-03
<160>1
<210>1
<211>1251
<212> DNA
<213>Citrobactergillenii
<220>
<223>ген phyCg, кодирующий фитазу PhyCg избактерий Citrobactergillenii
<400>1
aagaattcgatgaacagagcggaatgcagcttgagcgtgttgtcatcgtcagtcgtcatg 60
gcgtcagggcaccgacaaagttcacgccgcttatgcagcaagtcactcccgaccgctggc 120
cgcaatgggacgttcctctggggtggttgactcctcgcggcggggcactcattactgaat 180
taggacggtatcaacgtttacgcctggcggacaaaggtctgctggataataaaacgtgtc 240
caacggcagggcaggtcgcggtcattgccgatagcgatcaacgtacccgtaaaacgggtg 300
aagcattcctggcaggactggccccgaaatgtaaagtacaggttcattatcaacaagata 360
agtcaaaatctgatcccctttttaatcccatcaaggcggggcagtgttcgctgaacacat 420
cgcaggtgaaagaggccatcctgacccgggctggcggaagtcttgatgagtacacgcgcc 480
actaccaacccgcatttcaagccctggaacgggtgttaaatttctcccagtcagaaaagt 540
gtcaagcagctgggcagtctgcacagtgtacgctaaccgacgtcttacctgctgaactca 600
aggtctctccagaaaatatatcgttgtcaggctcatggggactggcttcaaccctgacgg 660
aaatcttcctgctgcaacaagcacaagggatgtcgcaggtggcctgggggcgtattcatg 720
gcgataaagaatggcgtacattattaagtctgcacaatgcgcagtttgaccttctgcaga 780
aaaccccggaggttgcccgtagcagggccacaccgttacttgatttgatacgtacagcgc 840
tcgtaacacagggggcaacagaaaataaatacgcaattcagttgcccgtctctttgttgt 900
ttattgcggggcatgacaccaatcttgccaatatcagcggggcattaggccttaacgtgt 960
ctctgcccggtcagccagataatacgccgccgggtggagagtttgttttcgaaaggtgga 1020
aacgggttagcgatcattctgattgggtgcaggtttcttttatgtatcagacattgcagg 1080
aaatgcgtgatatgcaacctttgtcgttgcaatcgcctcccggaaaaattgtgctgccct 1140
tagcggcctgcgatgagaaaaatacgcagggaatgtgctcattaaaaagtttttctacac 1200
tcattgattccgttcgcgtgtccgaatgtgctgagaaataagcggссgctt 1251
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Трансформант дрожжей Komagataella phaffii, продуцирующий фитазу Cronobacter turicensis | 2021 |
|
RU2756330C1 |
| Штамм дрожжей Komagataella phaffii с инактивированным геном HIS4 - реципиент для конструирования безмаркерных штаммов-продуцентов гетерологичных белков | 2022 |
|
RU2787584C1 |
| Штамм дрожжей Komagataella phaffii с инактивированным геном LEU2 - реципиент для конструирования штаммов-продуцентов гетерологичных белков | 2022 |
|
RU2788528C1 |
| Интегративная кассета для экспрессии генов и ее использование при конструировании трансформанта дрожжей Komagataella mondaviorum | 2023 |
|
RU2836330C1 |
| Трансформант дрожжей Pichia pastoris, продуцирующий фитазу | 2019 |
|
RU2708446C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент фитазы | 2018 |
|
RU2701498C1 |
| Трансформант дрожжей Pichia pastoris, продуцирующий фитазу Escherichia coli | 2019 |
|
RU2737621C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris с увеличенной продукцией фитазы Escherichia coli | 2019 |
|
RU2737623C1 |
| Рекомбинантный штамм дрожжей Pichia pastoris - продуцент фитазы Escherichia coli | 2019 |
|
RU2751595C2 |
| Трансформант дрожжей Ogataea haglerorum - продуцент фитазы Escherichia coli | 2021 |
|
RU2771079C1 |


Изобретение относится к биотехнологии. Предложен трансформант дрожжей Komagataella phaffii, продуцирующий фитазу Citrobacter gillenii. Указанный трансформант содержит нуклеотидную последовательность гена, приведенную в перечне последовательностей под номером SEQ ID NO:1, кодирующего фитазу C. gillenii, или ее формы, определяемые вырожденностью генетического кода. Изобретение обеспечивает расширение арсенала рекомбинантных микроорганизмов, продуцирующих фитазу. 3 ил., 1 табл., 3 пр.
Трансформант дрожжей Komagataella phaffii - продуцент фитазы Citrobacter gillenii, содержащий нуклеотидную последовательность гена, приведенную в перечне последовательностей под номером SEQ ID NO:1, кодирующего фитазу C. gillenii, или ее формы, определяемые вырожденностью генетического кода.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФИТАЗЫ (ВАРИАНТЫ), ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФИТАЗЫ (ВАРИАНТЫ) | 2009 |
|
RU2409670C1 |
| база данных Genbank, NZ_QVEK01000009.1, 16.04.2020 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Найдено онлайн: https://www.ncbi.nlm.nih.gov/nuccore/1468439911?sat=49&satkey=35749075 Дата обращения 15.06.2021 | |||
| US 20100261259 A1, 14.10.2010 | |||
| KARBALAEI M | |||
| ET AL | |||

