Область изобретения
Настоящее изобретение относится к антителам к C3b и к профилактике и лечению связанного с комплементом нарушения с использованием таких антител.
Предпосылки изобретения
Система комплемента представляет собой комплексный каскад ферментов, состоящий из серии сывороточных гликопротеинов, которые в норме существуют в неактивной форме проферментов. Комплемент могут активировать два основных каскада, классический и альтернативный, которые объединяются на уровне C3, где две сходных C3-конвертазы расщепляют C3 до C3a и C3b.
Макрофаги представляют собой специализированные клетки, у которых развита присущая им способность распознавать тонкие различия в структуре экспрессируемых на клеточной поверхности идентификационных меток, так называемых молекулярных паттернов (Taylor et al., Eur J Immunol 33, 2090-2097 (2003); Taylor et al., Annu Rev Immunol 23, 901-944 (2005)). Хотя прямое распознавание этих поверхностных структур является фундаментальным аспектом врожденного иммунитета, опсонизация позволяет характерным для макрофагов рецепторам опосредовать поглощение, повышая эффективность и внося разнообразие в репертуар распознавания фагоцита (Stuart and Ezekowitz, Immunity 22, 539-550 (2005)). Процесс фагоцитоза вовлекает множество взаимодействий лиганд-рецептор, и в настоящие время очевидно, что различные опсонины, включая иммуноглобулины, коллектины и компоненты комплемента, регулируют клеточную активность, требуемую для интернализации патогена, через взаимодействие с рецепторами клеточной поверхности макрофагов (рассмотрено Aderem and Underhill, Annu Rev Immunol 17, 593-623 (1999); Underhill and Ozinsky, Annu Rev Immunol 20, 825-852 (2002)). Хотя природные иммуноглобулины, кодируемые генами зародышевой линии, могут распознавать широкое множество патогенов, большинство опсонизирующих IgG образуются путем адаптивного иммунитета, и, таким образом, эффективное выведение через Fc-рецепторы не является прямым (Carroll, Nat Immunol 5, 981-986 (2004)). Комплемент, с другой стороны, быстро распознает патогенные поверхностные молекулы и запускает поглощение частицы с помощью рецепторов комплемента (Brown, Infect Agents Dis 1, 63-70 (1991)).
Комплемент состоит из более чем 30 белков сыворотки, которые опсонизируют широкое множество патогенов для распознавания рецепторами комплемента. В зависимости от исходного пускового фактора каскада можно выделить три каскада (рассмотрены (Walport. N Engl J 344, Med 1058-1066 (2001)). Все три каскада имеют общую стадию активации центрального компонента C3, однако они отличаются способом распознавания и исходными биохимическими стадиями, ведущими к активации C3. Классический каскад активируется антителами, связанными с поверхностью патогена, которые в свою очередь связывают C1q-компонент комплемента, запуская каскад сериновых протеаз, который в конечном итоге расщепляет C3 до его активной формы, C3b. Каскад лектина активируется после распознавания углеводных мотивов белками лектинами. К настоящему времени, идентифицированы три члена этого каскада: связывающие маннозу лектины (MBL), семейство лектинов SIGN-R1 и фиколины (Pyz et al., Ann Med 38, 242-251 (2006)). Как MBL, так и фиколины ассоциированы с сериновыми протеазами, которые действуют подобно C1 в классическом каскаде, активируя компоненты C2 и C4, что ведет к центральной стадии C3. Альтернативный каскад отличается как от классического, так и от лектинового каскадов тем, что он активируется вследствие прямой реакции внутреннего сложного эфира C3 с распознаваемыми мотивами на поверхности патогена. Первоначальное связывание C3 с активирующей поверхностью приводит к быстрому усилению накопления C3b под действием протеаз альтернативного каскада, фактора B и фактора D. Важно, что C3b, накапливаемый путем либо классического, либо лектинового каскада, также может привести к усилению накопления C3b под действием факторов B и D. Во всех трех каскадах активация комплемента, главная стадия опсонизации, представляет собой превращение компонента C3 в C3b. Расщепление C3 ферментами каскадов комплемента экспонирует сложный тиоэфир для нуклеофильной атаки, позволяя ковалентное присоединение C3b к поверхности антигена через домен сложного тиоэфира. Это является начальной стадией опсонизации комплементом. Последующий протеолиз связанного C3b приводит к образованию фрагментов iC3b, C3c и C3dg, которые распознаются различными рецепторами (Ross and Medof, Adv Immunol 37, 217-267 (1985)). Это расщепление устраняет способность C3b к последующему усилению накопления C3b и активации поздних компонентов каскада комплемента, включая мембраноатакующий комплекс, способный прямо повреждать мембрану. Однако фагоцитарные рецепторы макрофагов предпочтительно распознают C3b и его фрагменты; вследствие универсальности образования сложноэфирной связи опосредуемая C3 опсонизация является центральной для распознавания патогена (Holers et al., Immunol Today 13, 231-236 (1992)), и, таким образом, рецепторы для различных продуктов деградации C3 играют важную роль в иммунном ответе хозяина.
C3 сам по себе представляет собой комплексный и гибкий белок, состоящий из 13 отдельных доменов. Центральная часть молекулы состоит из 8 так называемых макроглобулиновых (MG) доменов, которые формируют плотно упакованные α- и β-цепи C3. В эту структуру встроены CUB- (C1r/C1s, Uegf и белок морфогенеза костей-1) и TED-домены, которые содержат тиосложноэфирную связь, которая позволяет ковалентное связывание C3b с поверхностями патогенов. Остальные домены включают C3a или действуют в качестве линкеров и спейсеров центральных доменов. Сравнение структур C3b и C3c с C3 демонстрирует, что молекула претерпевает значительные конформационные перестройки при каждом протеолизе, которые экспонируют не только TED, но также дополнительные новые поверхности молекулы, которые могут взаимодействовать с клеточными рецепторами (Janssen and Gros, Mol Immunol 44, 3-10 (2007)). Для предотвращения нежелательной активации комплемента большинство клеток млекопитающих снабжены регуляторами, которые блокируют усиление комплемента на собственных клетках хозяина (Hourcade et al. Adv Immunol 45:381 (1989)). В отсутствие этих собственных регуляторов обнажение для сыворотки приводит к образованию продукта расщепления комплемента, который, в свою очередь, способствует воспалению и повреждению ткани (Oglesby et al. J Exp Med 175: 1 547 (1992) и Oglesby et al., Trans Assoc. Am. Physicians 104: 164 (1991)). Неклеточные поверхности, которые лишены собственных регуляторов комплемента, таким образом, особенно подвержены атаке комплемента и полностью зависят от защиты растворимыми регуляторами комплемента в сыворотке. Неконтролируемая активация комплемента вследствие отсутствия соответствующей регуляции комплемента ассоциирована с различными хроническими воспалительными заболеваниями и дегенеративными заболеваниями. Доминирующими в этом воспалительном каскаде являются продукты расщепления комплемента C3a и C5a, которые функционируют в качестве хемоаттрактантов и активаторов нейтрофилов и воспалительных макрофагов через рецепторы для C3a и C5a (Mollnes et al., Trends Immunol. 23:61 (2002)). Пропердин, высвобождаемый из нейтрофилов, далее усиливает воспалительный каскад через стабилизацию AP-конвертазы (Lutz and Jelezarova, Mol. Immunol. 43:2 (2006)). Было показано, что активация комплемента является важным компонентом, запускающим воспаление при опосредуемых иммунными комплексами заболеваниях, таких как мембранопролиферативный гломерулонефрит, нефротоксический нефрит и артрит (Walport, N. Engl. J. Med. 344: 1058 (2001); Thurman and Holers, J Immunol 176: 1305 (2006): Banda et al., J Immunol 171: 2109 (2003); Weisman et al., Science 249: 146 (1990); Morgan and Harris, Mol. Immunol, 40: 159 (2003)), а также связанная со старением дегенерация желтого пятна (Anderson et al. Am. J. Ophthalmol. 134: 411 (2002); Donoso et al., Surv. Ophthalmol. 51: 137 (2006); Gold et al., Natl. Genet 38: 458 (2006); Hageman et al., Proc. Natl. Acad. Sci. USA 102: 7227 (2005); Hageman et al., Ann. Med. 38: 592 (2006); Hageman et al., Prog. Retin. Eye Res. 20: 705 (2001)).
Большинство регуляторов активации комплемента действуют на уровне C3b, центрального компонента конвертаз комплемента. Эти природные регуляторы активации комплемента, как правило, имеют большие размеры (>100 кДа) и их трудно конструировать в качестве терапевтического реагента. Таким образом, существует необходимость в лекарственных средствах для профилактики и лечения ассоциированных с комплементом нарушений путем блокирования C3b.
Сущность изобретения
Настоящее изобретение относится к разработке антител, которые специфично распознают фрагменты расщепления C3 и не распознают нативный C3, таким образом, избегая действия нативного C3 в качестве "груза" для антител. Более конкретно, изобретение относится к специфичным к C3b антителам и фрагментам антител и к их применению для лечения связанных с комплементом заболеваний.
В одном аспекте изобретение относится к способу профилактики или лечения связанного с комплементом нарушения, включающему введение субъекту, нуждающемуся в эффективном количестве антагониста C3b, который является селективным ингибитором альтернативного каскада комплемента.
В одном варианте осуществления субъектом является млекопитающее. В другом варианте осуществления субъектом является человек. В следующем варианте осуществления антагонист C3b представляет собой антитело, распознающее эпитоп на активном продукте деградации C3, но не на C3.
В другом варианте осуществления антагонист C3b представляет собой антитело или фрагмент антитела, селективно связывающиеся с C3b.
В другом варианте осуществления антитело ингибирует связывание C5 с C3b. В другом варианте осуществления антитело связывается с эпитопом, включая остатки эпитопа C3b, распознаваемого антителом S77.
В другом варианте осуществления антитело связывается по существу с тем же эпитопом, что и антитело S77.
В следующем варианте осуществления антитело конкурентно ингибирует связывание антитела S77.
В другом варианте осуществления антитело связывается с эпитопом C3b, содержащим остатки, которые контактируют с антителом S77.
В дополнительном варианте осуществления антитело содержит антигенсвязывающий центр, содержащий остатки антитела S77, которые контактируют с C3b.
В предпочтительном варианте осуществления антитело содержит последовательности CDR тяжелой (SEQ ID NO:1-4) и/или легкой (SEQ ID NO:5-8) цепей антитела S77 и/или представляет собой антитело S77 или его фрагмент.
В различных вариантах осуществления антитело может быть человеческим, гуманизированным или химерным.
В других вариантах осуществления фрагмент антитела выбран из группы, состоящей из Fab, Fab', F(ab')2, scFv, (scFv)2, dAb, фрагментов определяющих комплементарность областей (CDR), линейных антител, молекул одноцепочечных антител, миниантител, антител-димеров и полиспецифических антител, образованных из фрагментов антител.
Способы по настоящему изобретению включают профилактику или лечение любого связанного с комплементом нарушения, включая воспалительные и аутоиммунные заболевания, например, такие как ревматоидный артрит (RA), острый респираторный дистресс-синдром (ARDS), отдаленное повреждение ткани после ишемии и реперфузии, активация комплемента в ходе хирургической операции с искусственным кровообращением, дерматомиозит, пемфигус, волчаночный нефрит и являющиеся его следствиями гломерулонефрит и васкулит, искусственное кровообращение, индуцируемая кардиоплегией дисфункция эндотелия коронарных артерий, мембранопролиферативный гломерулонефрит типа II, IgA-нефропатия, острая почечная недостаточность, криоглобулинемия, антифосфолипидный синдром, дегенеративные заболевания желтого пятна, такие как связанная со старением дегенерация желтого пятна (AMD), хориоидальная неоваскуляризация (CNV), увеит, диабетическая и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные с внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы, неоваскуляризация сетчатки, а также аллотрансплантация, сверхострое отторжение, гемодиализ, хронический легочный дистресс-синдром (COPD), астма и аспирационная пневмония.
В конкретном варианте осуществления связанное с комплементом нарушение представляет собой связанное с комплементом состояние глаза, такое как связанная со старением дегенерация желтого пятна (AMD) или хориоидальная неоваскуляризация (CNV).
В другом аспекте изобретение относится к антителу против C3b, селективно связывающемуся с C3b и не связывающемуся с C3 и ингибирующему связывание C5 с C3b. В одном варианте осуществления антитело связывается с эпитопом, включающим остатки эпитопа C3b, распознаваемые антителом S77.
В другом варианте осуществления антитело связывается по существу с тем же эпитопом, что и антитело S77.
В другом варианте осуществления антитело конкурентно ингибирует связывание антитела S77.
В другом варианте осуществления антитело связывается с эпитопом C3b, включающим остатки, которые контактируют с антителом S77.
В следующем варианте осуществления антитело содержит антигенсвязывающий центр, содержащий остатки антитела S77, которые контактируют с C3b.
В следующем варианте осуществления антитело содержит последовательности CDR тяжелой (SEQ ID NO:1-4) и/или легкой (SEQ ID NO:5-8) цепей антитела S77 или представляет собой антитело S77 или его фрагмент.
В различных вариантах осуществления антитело представляет собой антитело человека, гуманизированное или химерное антитело.
Фрагмент антитела может быть выбран, например, из группы, состоящей из Fab, Fab', F(ab')2, scFv, (scFv)2, dAb, фрагментов определяющих комплементарность областей (CDR), линейных антител, молекул одноцепочечных антител, миниантител, антител-димеров и полиспецифических антител, образованных из фрагментов антител.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей антагонист C3b, такой как антитело против C3b, в смеси с фармацевтически приемлемым носителем.
В конкретном варианте осуществления фармацевтическая композиция предназначена для применения при лечении связанного с комплементом нарушения.
В следующем аспекте изобретение относится к набору, содержащему контейнер, содержащий антагонисты C3b или антитело против C3b по настоящему изобретению, или фармацевтическую композицию, содержащую такой антагонист или антитело, и инструкции по введению антитела или фармацевтической композиции для лечения связанного с комплементом нарушения.
Краткое описание рисунков
Фиг.1. Результаты пэннига C3b в фаговой библиотеке антител.
Фиг. 2. Результаты конкуренции фага с различными клонами антител против C3b.
Фиг.3. Кристаллическая структура C3b в комплексе с Fab антитела YW144.2.43.S77 (в дальнейшем в настоящем документе обозначаемого как S77). Бета-цепь C3b указана зеленым цветом, альфа-цепь указана оранжевым. Тяжелая цепь (HC) и легкая цепь (EC) S77 указаны темно-зеленым и желтым соответственно. CRIg размещен на комплексе C3b:Fab исходя из структуры сокристалла C3b:CRIg и показан фиолетовым цветом.
Фиг. 4. Увеличенный масштаб связывающего взаимодействия антитела S77 с C3b. C3b показан с помощью представления поверхности, ленточная диаграмма голубого цвета соответствует наложению C3 на структуру C3b. HC и LC в S77 указаны в качестве ленточной диаграммы темно-зеленым и желтым. Поверхность C3b окрашена в соответствии с расстоянием до S77. Все атомы, расположенные ближе 4,7 Å, 4,0 Å и 3,5 Å, окрашены желтым, оранжевым и красным соответственно. Следует отметить, что LC в S77 совпадает на C3. Однако петля C3 может быть способна к движению.
Фиг. 5. Аминокислотные последовательности тяжелых (SEQ ID NO:1-4) и легких (SEQ ID NO:5-8) цепей Fab-фрагмента антитела S77. Красным указаны остатки, которые непосредственно контактируют с C3b.
Фиг. 6A и 6B. Аффинность связывания Fab исходного антитела YW144.2.43 и его варианта после созревания аффинности: Fab 144.2.43.S77 (S77 Fab).
Фиг. 7A, 7B и 7С. Сенсограммы SPR и аффинность связывания S77 с C3 и C3b.
Фиг. 8A и 8B. S77 распознает C3b, но не промолекулу C3. Очищенный C3b или C3 иммобилизовывали на микропланшетах для титрования с использованием поликлонального антитела против C3. Связывание S77 (A) или поликлонального антитела против C3 (B) с иммобилизованным C3b или C3 определяли с использованием вторичного конъюгированного с HRPO антитела. Цвет проявляли с помощью TMB (KPL), проявление останавливали 2 н. H2SO4 и поглощение считывали при 450 нм.
Фиг. 9A, 9B и 9С. Антитело IgG S77 селективно ингибирует альтернативный, но не классический каскад комплемента. Эритроциты кролика и эритроциты овцы инкубировали в сыворотке с истощением C1q и фактором B и проводили мониторинг гемолиза в присутствии возрастающей концентрации ингибитора или контрольного белка. Величину гемолиза выражали в качестве процента от максимального гемолиза в отсутствие ингибитора.
Фиг. 10. Fab S77 после созревания аффинности ингибирует альтернативный каскад комплемента.
Фиг. 11. Fab против C3b (S77) ингибирует C5-конвертазу. C5-конвертазу получали, как описано (Rawal, N и Pangburn, M. J Immunol. 2001 Feb 15; 166(4):2635-42).
Фиг. 12. Антитело IgG S77 и его Fab-фрагмент ингибируют C5-конвертазу блокированием связывания C5 с C3b, некаталитической субъединицей конвертазы. C5 в присутствии возрастающих концентраций ингибитора добавляли в планшеты, покрытые C3b. C5 связывалась с мультимерами C3b.
Фиг. 13. S77 не вызывает распад конвертазы, в противоположность фактору H. Анализ распада проводили по образованию иммобилизованной на планшете C3-конвертазы в присутствии возрастающих концентраций S77 или фактора H (положительный контроль).
Фиг. 14A и 14B. S77 ингибирует связывание профактора B с C3b и ингибирует образование C3bBb-конвертазы.
Фиг. 15A и 15B. S77 может связывать C3b в присутствии связанного fBb и не вызывает распад C3-конвертазы.
Фиг. 16 и 16B. S77 ингибирует связывание фактора H с C3b и ингибирует активность кофактора H.
Фиг. 17. S77 ингибирует связывание CR1 с C3b.
Фиг. 18A и 18B. Аминокислотные последовательности вариабельных областей легкой (SEQ ID NO:13) и тяжелой (SEQ ID NO:14) цепей антитела против HER2 rhuMAB 4D5-8.
Добавочная фиг. 1. Остатки на C3b, контактирующие с HC и LC в Fab S77 (остатки 833-839 охватывают SEQ ID NO:15; остатки 895-899 охватывают SEQ ID NO:16).
Добавочная фиг. 2. Остатки на Fab S77, контактирующие с C3b (остатки 1030-1033 охватывают SEQ ID NO:17; остатки 1098-1107 охватывают SEQ ID NO:18).
Добавочная фиг. 3. Аминокислотные последовательности C3-фактора комплемента человека (SEQ ID NO:9) и C3-фактора комплемента мыши (SEQ ID NO:10).
Подробное описание предпочтительного варианта осуществления
I. Определения
Термины "C3" и "C3-компонент комплемента" используют взаимозаменяемо, и они относятся к полипептидам C3 с нативной последовательностью.
"C3 с нативной последовательностью" представляет собой полипептид, имеющий такую же аминокислотную последовательность, что и природный C3-полипептид, независимо от способа его получения. Таким образом, C3 с нативной последовательностью может быть выделен из природного источника или он может быть получен рекомбинантными и/или синтетическими способами. Термин "C3 с нативной последовательностью" конкретно охватывает встречающиеся в природе формы вариантов (например, альтернативно сплайсированные формы) и встречающиеся в природе аллельные варианты C3, а также структурные конформационные варианты, имеющие аминокислотную последовательность, как у полипептида C3 из природного источника. Полипептиды C3 с нативной последовательностью конкретно включают C3 человека с нативной последовательностью (добавочная фигура 3, SEQ ID NO:9; также см. De Bruijn and Fey, Proc. Natl. Acad. Sci. USA 82:708-712) и полипептиды не являющихся человеком животных, включая высших приматов и других не являющихся человеком млекопитающих, такие как последовательность C3 мыши, представленная на добавочной фигуре 3, SEQ ID NO:10).
Термины "C3b" используют в настоящем документе для обозначения полипептида C3b с нативной последовательностью, образующегося из C3b после расщепления C3-конвертазой, высвобождающей фрагмент анафилатоксина C3a с N-конца α-цепи C3 и оставляющей C3b. Термин "нативная последовательность" имеет такое же значение, как определено в связи с C3, и конкретно включает C3b человека с нативной последовательностью SEQ ID NO:9.
Термин "антагонист C3b" используют в наиболее широком значении, и он включает любую молекулу, которая способна нейтрализовывать, блокировать, частично или полностью ингибировать, устранять, снижать биологическую активность C3 или препятствовать ей. Антагонисты C3b включают, но не ограничиваются ими, антитела против C3b и их антигенсвязывающие фрагменты, другие связывающие полипептиды, пептиды и непептидные низкомолекулярные соединения, которые связываются с C3b и способны нейтрализовывать, блокировать, частично или полностью ингибировать, устранять, снижать биологическую активность C3 или препятствовать биологической активности C3, такой как способность C3b участвовать в патологии связанного с комплементом нарушения. Антагонисты C3b, такие как антитела против C3b, представленные в настоящем документе, специфично распознают C3b и не распознают его предшественника, C3.
"Низкомолекулярное соединение" определяют в настоящем документе как соединение, имеющее молекулярную массу менее приблизительно 600, предпочтительно менее приблизительно 1000 дальтон.
"Активный", или "активность", или "биологическая активность" в контексте антагониста C3b, такого как антитело против C3b, по настоящему изобретению, представляет собой способность осуществлять антагонизм (частично или полностью ингибировать) биологической активности C3b. Предпочтительная биологическая активность антагониста C3b представляет собой способность достигать улучшения состояния, например при патологии, в случае связанного с C3b заболевания или состояния, например, такого как связанное с комплементом нарушение. Активность можно определять в тестах in vitro или in vivo, включая анализы связывания, с использованием соответствующей модели на животных или клинических испытаний у человека.
Термин "связанное с комплементом нарушение" используют в настоящем документе в наиболее широком значении, и он включает все заболевания и патологические состояния, патогенез которых вовлекает нарушение активации системы комплемента, например, такие как дефициты компонентов комплемента. Конкретно термин включает заболевания и патологические состояния, при которых является полезным ингибирование C3-конвертазы. Кроме того, термин включает заболевания и патологические состояния, при которых является полезным ингибирование, включая селективное ингибирование, альтернативного каскада комплемента. Связанные с комплементом нарушения включают, но не ограничиваются ими, воспалительные заболевания и аутоиммунные заболевания, например, такие как ревматоидный артрит (RA), острый респираторный дистресс-синдром (ARDS), отдаленное повреждение ткани после ишемии и реперфузии, активация комплемента в ходе хирургической операции с искусственным кровообращением, дерматомиозит, пемфигус, волчаночный нефрит и являющиеся его следствиями гломерулонефрит и васкулит, искусственное кровообращение, индуцируемая кардиоплегией дисфункция эндотелия коронарных артерий, мембранопролиферативный гломерулонефрит типа II, IgA-нефропатия, острая почечная недостаточность, криоглобулинемия, антифосфолипидный синдром, дегенеративные заболевания желтого пятна, такие как связанная со старением дегенерация желтого пятна (AMD), хориоидальная неоваскуляризация (CNV), увеит, диабетическая и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные с внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы, неоваскуляризация сетчатки, а также аллотрансплантация, сверхострое отторжение, гемодиализ, хронический легочный дистресс-синдром (COPD), астма и аспирационная пневмония.
Термин "связанное с комплементом состояние глаза" используют в настоящем документе в наиболее широком значении, и он включает все состояния и заболевания глаза, патология которых вовлекает комплемент, включая классический и альтернативный каскады, и, в частности, альтернативный каскад комплемента. Конкретно, в эту группу включены все состояния и заболевания глаза, ассоциированные с альтернативным каскадом, возникновение, развитие или прогрессирование которых можно контролировать ингибированием альтернативного каскада. Связанные с комплементом состояния глаза включают, но не ограничиваются ими, дегенеративные заболевания желтого пятна, такие как все стадии связанной со старением дегенерации желтого пятна (AMD), включая сухую и влажную (неэкссудативную и экссудативную) формы, хориоидальную неоваскуляризацию (CNV), увеит, диабетическую и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные с внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзию центральной вены сетчатки (CRVO), неоваскуляризацию роговицы, неоваскуляризацию сетчатки. Предпочтительная группа связанных с комплементом состояний глаза включает связанную со старением дегенерацию желтого пятна (AMD), включая экссудативную (влажную) и неэкссудативную (сухую или атрофическую) AMD, хориоидальную неоваскуляризацию (CNV), диабетическую ретинопатию (DR) и эндофтальмит.
Термин "воспалительное заболевание" и "воспалительное нарушение" используют взаимозаменяемо, и он означает заболевание или нарушение, при котором компонент иммунной системы млекопитающего вызывает, опосредует или иным образом вносит вклад в воспалительный ответ, приводящий к заболеваемости млекопитающего. Также к этим терминам относятся заболевания, при которых снижение воспалительного ответа оказывает улучшающий эффект на прогрессирование заболевания. В этот термин включены иммуноопосредуемые воспалительные заболевания, включая аутоиммунные заболевания.
Термин "опосредуемое T-клетками" заболевание означает заболевание, при котором T-клетки прямо или непрямо опосредуют или иным образом вносят вклад в заболеваемость у млекопитающего. Опосредуемое T-клетками заболевание может быть ассоциировано с клеточно-опосредуемыми эффектами, опосредуемыми лимфокинами эффектами и т.д. и даже эффектами, связанными с B-клетками, если B-клетки стимулируются, например, лимфокинами, секретируемыми T-клетками.
Примеры связанных с иммунной системой и воспалительных заболеваний, некоторые из которых являются опосредуемыми T-клетками, включают, но не ограничиваются ими, воспалительное заболевание кишечника (IBD), системную красную волчанку, ревматоидный артрит, ювенильный хронический артрит, спондилоартропатии, системную склеродермию (склеродермию), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (панцитопения клеток иммунной системы, пароксизмальная ночная гемоглобинурия), аутоиммунную тромбоцитопению (идиопатическая тромбоцитопеническая пурпура, иммуноопосредуемая тромбоцитопения), тиреоидит (болезнь Грэйва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), сахарный диабет, иммуноопосредуемое заболевание почек (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервной системы, такие как рассеянный склероз, идиопатическая полиневропатия, заболевания печени и желчевыводящих путей, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит и склерозирующий холингит, воспалительные и фиброзные заболевания легких (например, кистозный фиброз), глютензависимую энтеропатию, болезнь Уиппла, аутоиммунные или иммуноопосредуемые заболевания кожи, включая буллезные заболевания кожи, полиформную эритему и контактный дерматит, псориаз, аллергические заболевания легкого, такие как эозинофильная пневмония, идиопатический фиброз легких и связанный с гиперчувствительностью пневмонит, связанные с трансплантацией заболевания, включая отторжение трансплантата, реакцию "трансплантат против хозяина", болезнь Альцгеймера и атеросклероз.
"Лечение" представляет собой вмешательство, проводимое с намерением предотвращения развития или изменения патологии нарушения. Таким образом, "лечение" относится как к терапевтическому лечению, так и к профилактическим или предупредительным мерам. Субъекты, нуждающиеся в лечении, включают субъектов, уже имеющих нарушение, а также субъектов, у которых нарушение подлежит профилактике. При лечении связанного с иммунной системой заболевания лекарственное средство может прямо изменять силу ответа компонента иммунного ответа, или оно может обеспечивать повышенную восприимчивость заболевания к лечению другими лекарственными средствами, например антибиотиками, противогрибковыми средствами, противовоспалительными средствами, химиотерапевтическими средствами и т.д.
"Патология" заболевания, такого как связанное с комплементом нарушение, включает все явления, которые нарушают здоровье пациента. Она включает, но не ограничивается ими, аномальный или неконтролируемый рост клеток (нейтрофильных, эозинофильных, моноцитарных, лимфоцитарных клеток), продукцию антител, продукцию аутоантител, продукцию комплемента, препятствие нормальному функционированию соседних клеток, высвобождение цитокинов или других секретируемых продуктов на аномальных уровнях, подавление или усиление воспалительного или иммунологического ответа, инфильтрацию воспалительных клеток (нейтрофильных, эозинофильных, моноцитарных, лимфоцитарных) в клеточные пространства и т.д.
Термин "млекопитающее", как используют в настоящем документе, относится к любому животному, классифицируемому как млекопитающее, включая, но не ограничиваясь ими, человека, высших приматов, домашних и сельскохозяйственных животных, животных зоопарков, спортивных животных или комнатных животных, таких как лошади, свиньи, крупный рогатый скот, собаки, кошки, хорьки и т.д. В предпочтительном варианте осуществления изобретения млекопитающее представляет собой человека.
Введение "в сочетании с" одним или несколькими дополнительными лекарственными средствами включает одновременное (совместное) и последовательное введение в любом порядке.
"Терапевтически эффективное количество" представляет собой количество "антагониста C3b", такого как "антитело против C3b", которое требуется для достижения измеримого улучшения состояния, например при патологии, в случае определенного заболевания или состояния, например, такого как связанное с комплементом нарушение.
Термин “последовательности контроля” относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, которые пригодны для прокариот, включают, например, промотор, необязательно последовательность оператора и участок связывания рибосом. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", если она находится в функциональной связи с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторной лидерной последовательности функционально связана с ДНК полипептида, если она экспрессируется в качестве пребелка, который участвует в секреции полипептида; промотор или энхансер является функционально связанным с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосом является функционально связанным с кодирующей последовательностью, если он расположен таким образом, чтобы облегчить трансляцию. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются смежными и, в случае секреторной лидерной последовательности, являются смежными и находятся в рамке считывания. Однако энхансеры не должны быть смежными. Связывание осуществляют посредством лигирования в соответствующих участках рестрикции. Если таких участков нет, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
"Строгость" реакций гибридизации легко определяет специалист в данной области, и, как правило, она представляет собой эмпирическое вычисление, зависящее от длины зонда, температуры промывания и концентрации соли. Как правило, для надлежащего отжига более длинные зонды требуют более высоких температур, в то время как более короткие зонды требуют более низких температур. Гибридизация, как правило, зависит от способности денатурированной ДНК к повторному отжигу, когда комплементарные цепи присутствуют в окружающей среде при температуре ниже их температуры отжига. Чем более высокой является степень требуемой гомологии между зондом и гибридизуемой последовательностью, тем более высокой является относительная температура, которую можно использовать. В результате, из этого следует, что более высокие относительные температуры будут обеспечивать тенденцию к более строгим условиям реакции, в то время как более низкие температуры снижают строгость. Для дополнительных деталей и разъяснения строгости реакций гибридизации см. Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995).
"Строгие условия" или "условия высокой строгости", как определено в настоящем документе, можно определить как условия, в которых: (1) используют низкую ионную силу и высокую температуру промывания, например 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) используют в ходе гибридизации денатурирующее средство, такое как формамид, например 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% Ficoll/0,1% поливинилпирролидоном/50 мМ натрий-фосфатным буфером при pH 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°C; или (3) проводят гибридизацию в течение ночи в растворе, в котором используется 50% формамид, 5 × SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5 × раствор Денхардта, облученная ультразвуковым облучением ДНК спермы лосося (50 мкг/мл), 0,1% SDS и 10% сульфат декстрана при 42°C, с промыванием в течение 10 минут при 42°C в 0,2 × SSC (хлорид натрия/цитрат натрия) с последующим промыванием высокой строгости в течение 10 минут, состоящим из 0,1 × SSC, содержащего EDTA, при 55°C.
"Условия умеренной строгости" можно определить, как описано в Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, и они включают применение менее строгого раствора для промывания и условий гибридизации (например, температуры, ионной силы и %SDS), чем раствор для промывания и условия гибридизации, описанные выше. Примером умеренно строгих условий является инкубация в течение ночи при 37°C в растворе, содержащем: 20% формамид, 5 × SSC (150 мМ NaCl, 15 мМ трицитрат натрия), 50 мМ фосфат натрия (pH 7,6), 5 × раствор Денхардта, 10% сульфат декстрана и 20 мг/мл денатурированной расщепленной ДНК спермы лосося, с последующим промыванием фильтров в 1 × SSC при приблизительно 37-50°С. Специалисту будет понятно, каким образом корректировать температуру, ионную силу и т.д., в случае необходимости приспосабливания к таким факторам, как длина зонда и т.п.
Термин "меченный эпитопом", при применении в настоящем документе, относится к химерному полипептиду, содержащему полипептид по изобретению, слитый с "полипептидом-меткой". Полипептид-метка имеет достаточное количество остатков для обеспечения эпитопа, против которого можно получать антитело, но в то же время является достаточно коротким, чтобы не препятствовать активности полипептида, с которым он слит. Также полипептид-метка предпочтительно является совершенно уникальным, так что антитело по существу не вступает в перекрестные реакции с другими эпитопами. Пригодные полипептиды-метки, как правило, имеют по меньшей мере шесть аминокислотных остатков и, как правило, между приблизительно 8 и 50 аминокислотных остатков (предпочтительно между приблизительно 10 и 20 аминокислотных остатков).
Термин "антитело" используют в наиболее широком значении, и конкретно он включает, но не ограничивается ими, отдельные антитела, распознающие фрагмент распада C3, но не нативный C3, такие как моноклональные антитела против C3b, специфично связывающиеся с C3b, и композиции антител со специфичностью к множеству эпитопов. Термин "моноклональное антитело", как используют в настоящем документе, относится к антителу, получаемому из совокупности по существу гомогенных антител, т.е. отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах.
Как используют в настоящем документе, термин "моноклональное антитело" относится к антителу из совокупности по существу гомогенных антител, например отдельные антитела, составляющие совокупность, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут быть представлены в небольших количествах. Моноклональные антитела являются высокоспецифичными, и они направлены против единственного антигенного участка. Кроме того, в противоположность препаратам традиционных (поликлональных) антител, которые, как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной детерминанты на антигене. Определение "моноклональный" указывает на тот признак антитела, что его получают из по существу гомогенной совокупности антител, и не подразумевает того, что антитело должно быть получено каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, можно получать множеством способов, включая, например, способ гибридом, впервые описанным Kohler et al. (1975) Nature, 256: 495, или их можно получать способами рекомбинантных ДНК (см., например, патент США No. 4816567). "Моноклональные антитела" также можно выделять из фаговых библиотек антител с использованием способов, описанных, например, в Clackson et al. (1991) Nature 352: 624-628 и Marks et al. (1991) J. Mol. Biol 222: 581-597.
В настоящем документе моноклональные антитела конкретно включают "химерные" антитела (иммуноглобулины), в которых участок тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они проявляют требуемую биологическую активность (патент США No 4816567; и Morrison et al. (1984) Proc. Natl. Acad. Sci USA, 81: 6851-6855).
"Гуманизированные" формы не являющихся человеческими антител (например, грызунов) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, не являющегося человеческим. В основном, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены остатками из гипервариабельной области видов, не относящихся к человеку (донорное антитело), таких как мышь, крыса, кролик или не относящиеся к человеку приматы, которые обладают требуемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки Fv каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками, не являющимися человеческими. Более того, гуманизированные антитела могут содержать остатки, которые отсутствуют в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения параметров антитела. Как правило, гуманизированное антитело содержит по существу все по меньшей мере из одного и, как правило, из двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют гипервариабельным петлям иммуноглобулина, не являющегося человеческим, и все или по существу все FR-области представляют собой FR-области из последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно содержит по меньшей мере участок константного домена (Fc) иммуноглобулина, как правило, константного домена иммуноглобулина человека. Для более подробной информации см. Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-329; и Presta (1992) Curr. Op. Struct. Biol. 2:593-596.
"Видозависимое антитело" представляет собой антитело, которое обладает более высокой аффинностью связывания к антигену из первого вида млекопитающего, чем к гомологу этого антигена из второго вида млекопитающего. В норме, видозависимое антитело "специфично связывается" с антигеном человека (т.е. имеет величину аффинности связывания (Kd) не более чем приблизительно 1×10-7 M, предпочтительно не более чем приблизительно 1×10-8 M и наиболее предпочтительно не более чем приблизительно 1×10-9 M), но имеет аффинность связывания с гомологом антигена второго не являющегося человеком вида млекопитающего, которая по меньшей мере приблизительно в 50 раз, или по меньшей мере приблизительно в 500 раз, или по меньшей мере приблизительно раз 1000 слабее, чем его аффинность связывания с антигеном человека. Видозависимое антитело может быть любым из различных типов антител, как определено выше, однако предпочтительно оно представляет собой гуманизированное антитело или антитело человека.
Как используют в настоящем документе, "мутантное антитело" или "вариант антитела" относится к варианту по аминокислотной последовательности видозависимого антитела, где один или несколько из аминокислотных остатков видозависимого антитела являются модифицированными. Такие мутанты обладают менее чем 100% идентичностью или сходством последовательности с видозависимым антителом. В предпочтительном варианте осуществления мутантное антитело может иметь аминокислотную последовательность, обладающую по меньшей мере 75% идентичностью или сходством аминокислотной последовательности с аминокислотной последовательностью вариабельного домена либо тяжелой цепи, либо легкой цепи зависимого от вида антитела, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%. Идентичность или сходство в отношении этой последовательности определяют в настоящем документе как процент аминокислотных остатков в последовательности-кандидате, которые идентичны (т.е. являются такими же остатками) или сходны (т.е. представляет собой аминокислотный остаток из той же группы исходя из общих свойств боковых цепей, см. ниже) с остатками зависимого от вида антитела, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Ни одно из N-концевых, C-концевых или внутренних удлинений, делеций или вставок в последовательности антитела вне вариабельного домена не следует истолковывать как влияющее на идентичность или сходство последовательностей.
"Выделенное" антитело представляет собой антитело, которое идентифицировано и отделено и/или извлечено из компонента его естественных окружающих условий. Загрязняющие компоненты его естественных окружающих условий представляют собой вещества, которые препятствуют применению антитела для диагностики или лечения, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах осуществления антитело очищают (1) до более чем 95% по массе антитела, как определяют способом Лоури, и наиболее предпочтительно до более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (3) до гомогенности при SDS-PAGE в восстанавливающих или невосстанавливающих условиях с использованием кумасси синего или предпочтительно окрашивания серебром. Выделенное антитело включает антитело в рекомбинантных клетках in situ, поскольку в этом случае отсутствует по меньшей мере один компонент условий естественного окружения антитела. Однако, как правило, выделенное антитело будет получено посредством по меньшей мере одной стадии очистки.
Как используют в настоящем документе, "вариабельный домен антитела" относится к частям легкой и тяжелой цепей молекул антитела, которые включают аминокислотные последовательности определяющих комплементарность областей (CDR; т.е. CDR1, CDR2 и CDR3) и каркасные области (FR). VH относится к вариабельному домену тяжелой цепи. VL относится к вариабельному домену легкой цепи. В соответствии с используемыми способами по этому изобретению аминокислотные положения, относящиеся к CDR и FR, можно определять в соответствии с Kabat (Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, Md., 1987 и 1991)). Нумерация аминокислот антител или антигенсвязывающих фрагментов также соответствует нумерации по Kabat.
Как используют в настоящем документе, термин "определяющие комплементарность области" (CDR; т.е. CDR1, CDR2 и CDR3) относится к аминокислотным остаткам вариабельного домена антитела, наличие которых необходимо для связывания антигена. Каждый вариабельный домен, как правило, имеет три области CDR, обозначаемые как CDR1, CDR2 и CDR3. Каждая определяющая комплементарность область может содержать аминокислотные остатки из "определяющей комплементарность области" при определении по Kabat (т.е. приблизительно остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (H1), 50-65 (H2) и 95-102 (H3) в вариабельном домене тяжелой цепи; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)) и/или остатки из "гипервариабельной петли" (т.е. приблизительно остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (H1), 53-55 (H2) и 96-101 (H3) в вариабельном домене тяжелой цепи; Chothia and Lesk (1987) J. Mol. Biol. 196:901-917). В некоторых случаях определяющая комплементарность область может включать аминокислоты как из области CDR, определенной в соответствии с Kabat, так и гипервариабельную петлю. Например, CDRH1 тяжелой цепи антитела 4D5 включает аминокислоты с 26 по 35.
"Каркасные области" (в дальнейшем в настоящем документе FR) представляют собой остатки вариабельных доменов, отличные от остатков CDR. Каждый вариабельный домен, как правило, имеет четыре FR, обозначаемых как FR1, FR2, FR3 и FR4. Если CDR определены в соответствии с Kabat, остатки FR легкой цепи расположены приблизительно в области остатков 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4) и остатки FR тяжелой цепи расположены приблизительно в области остатков 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103-113 (HCFR4) в остатках тяжелой цепи. Если CDR содержат аминокислотные остатки из гипервариабельных петель, остатки FR легкой цепи расположены приблизительно в области остатков 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4) в легкой цепи и остатки FR тяжелой цепи расположены приблизительно в области остатков 1-25 (HCFR1), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4) в остатках тяжелой цепи. В некоторых случаях, когда CDR содержит аминокислоты как из CDR, определенных по Kabat, так и из CDR гипервариабельной петли, остатки FR корректируют соответствующим образом. Например, когда CDRH1 включает аминокислоты H26-H35, остатки FR1 тяжелой цепи находятся в положениях 1-25 и остатки FR2 находятся в положениях 36-49.
Как используют в настоящем документе, "набор кодонов" относится к набору различных нуклеотидных триплетных последовательностей, используемых для кодирования требуемых вариантов аминокислот. Можно синтезировать набор олигонуклеотидов, например, твердофазным синтезом, включающий последовательности, которые соответствуют всем возможным сочетаниям нуклеотидных триплетов, обеспечиваемых набором кодонов, и которые будут кодировать требуемую группу аминокислот. Стандартной формой обозначения кодонов является код IUB, который известен в данной области и описан в настоящем документе. Набор кодонов, как правило, представляют 3 заглавными буквами курсивом, например NNK, NNS, XYZ, DVK и т.п. Таким образом, "неслучайный набор кодонов", как используют в настоящем документе, относится к набору кодонов, который кодирует определенные аминокислоты, которые частично, предпочтительно полностью, удовлетворяют критериям выбора аминокислот, как описано в настоящем документе. Синтез олигонуклеотидов с определенной "вырожденностью" нуклеотидов в определенных положениях хорошо известен в данной области, например подход TRIM (Knappek et al. (1999) J. Mol. Biol. 296:57-86); Garrard & Henner (1993) Gene 128: 103). Такие наборы олигонуклеотидов, имеющие определенные наборы кодонов, можно синтезировать с использованием коммерческих устройств для синтеза нуклеиновых кислот (доступных, например, от Applied Biosystems, Foster City, CA), или их можно получать коммерчески (например, от Life Technologies, Rockville, MD). Таким образом, набор синтезированных олигонуклеотидов, имеющих конкретный набор кодонов, как правило, включает множество олигонуклеотидов с различными последовательностями, причем различия устанавливаются набором кодонов в пределах общей последовательности. Олигонуклеотиды, как используют в соответствии с изобретением, имеют последовательности, которые позволяют гибридизацию с матрицей нуклеиновой кислоты вариабельного домена и также могут, но необязательно, включать участки для ферментов рестрикции, пригодные, например, для клонирования.
Термин "фрагменты антитела" используют в настоящем документе в наиболее широком значении, и он включает, но не ограничивается ими Fab, Fab', F(ab')2, scFv, (scFv)2, dAb, фрагменты определяющих комплементарность областей (CDR), линейные антитела, молекулы одноцепочечных антител, миниантитела, антитела-димеры и полиспецифические антитела, образованные из фрагментов антител.
"Fv"-фрагмент представляет собой минимальный фрагмент антитела, который содержит полный антигенраспознающий и антигенсвязывающий центр. Этот участок состоит из димера вариабельных доменов одной тяжелой и одной легкой цепей, прочно связанных друг с другом связью, которая может быть ковалентной, например, в scFv. Именно в этой конфигурации три CDR каждого вариабельного домена взаимодействуют, образуя антигенсвязывающий центр на поверхности димера VH-VL. В совокупности, шесть CDR или их подгруппа придают антигенсвязывающую специфичность антителу. Однако даже один вариабельный домен (или половина Fv, содержащая только три CDR, специфичных к антигену) обладает способностью распознавать и связывать антиген, хотя, как правило, с более низкой аффинностью, чем целый связывающий участок.
"Fab"-фрагмент содержит вариабельный и константный домен легкой цепи и вариабельный домен и первый константный домен (CH1) тяжелой цепи. F(ab')2-фрагменты антитела содержат пару Fab-фрагментов, которые, как правило, ковалентно связаны вблизи их C-концов шарнирными остатками цистеина между ними. Также в данной области известно другое химическое связывание фрагментов антител.
"Одноцепочечные Fv" или "scFv" фрагменты антител содержат VH и VL-домены антитела, где эти домены находятся в единой полипептидной цепи. Как правило, полипептид sFv дополнительно содержит полипептидный линкер между VH и VL-доменами, который обеспечивают формирование в sFv структуры, требуемой для связывания антигена. Для обзора sFv см. Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, p. 269-315 (1994).
Термин "антитела-димеры" относится к небольшим фрагментам антител с двумя антигенсвязывающими центрами, причем эти фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH и VL). C использованием линкера, который является слишком коротким для обеспечения образования пары между двумя доменами на одной цепи, домены вынуждают образовывать пару с комплементарными доменами другой цепи и образовывать два антигенсвязывающих центра. Антитела-димеры более подробно описаны, например, в EP 404097; WO 93/11161; и Hollinger et al. (1993) Proc. Natl Acad. Sci. USA 90:6444-6448.
Выражение "линейные антитела" относится к антителам, описанным в Zapata et al. (1995 Protein Eng, 8(10): 1057-1062). В кратком изложении эти антитела содержат пару тандемных сегментов Fd (VH-CH1-VH-CH1), которые, вместе с комплементарными полипептидами легкой цепи, образуют пару антигенсвязывающих областей. Линейные антитела могут быть биспецифическими или моноспецифическими.
Как используют в настоящем документе, "библиотека" относится к множеству последовательностей антител, или фрагментов антител (например, полипептидов по изобретению), или нуклеиновых кислот, которые кодируют эти последовательности, причем последовательности отличаются сочетанием вариантов аминокислот, которые внесены в эти последовательности в соответствии со способами по изобретению.
"Фаговый дисплей" представляет собой способ, посредством которого варианты полипептидов экспонируются в качестве белков, слитых по меньшей мере с частью белка оболочки, на поверхности частиц фага, например нитевидного фага. Применимость фагового дисплея основана на том факте, что большие библиотеки рандомизированных вариантов белков можно быстро и эффективно сортировать по тем последовательностям, которые связываются с антигеном-мишенью с высокой аффинностью. Дисплей пептидных и белковых библиотек на фаге используют при скрининге миллионов полипептидов в отношении пептидов со свойствами специфичного связывания. Способы поливалентного фагового дисплея используют для дисплея небольших случайных пептидов и небольших белков посредством слияния либо с геном III, либо с геном VIII нитевидного фага. Wells and Lowman (1992) Ciirr. Opin. Struct. Biol. 3:355-362, и ссылки, цитированные в ней. В одновалентном фаговом дисплее библиотеку белков или пептидов подвергают слиянию с геном III или его частью и экспрессируют на низких уровнях в присутствии белка гена III дикого типа, так чтобы фаговые частицы экспонировали одну копию или ни одной копии слитых белков. Эффекты авидности снижаются относительно поливалентного фага, так что сортировка основана на собственной аффинности к лиганду, и используют фагмидные векторы, которые упрощают манипулирование с ДНК. Lowman and Wells (1991) Methods: A companion to Methods in Enzymology 3:205-0216.
"Фагмида" представляет собой плазмидный вектор, имеющий бактериальный ориджин репликации, например ColE1, и копию межгенной области бактериофага. Фагмиду можно использовать в любом известном бактериофаге, включая нитевидный бактериофаг и лямбдоидный бактериофаг. Также плазмида, как правило, содержит селективный маркер для устойчивости к антибиотику. Сегменты ДНК, клонированные в эти векторы, могут размножаться в качестве плазмид. Когда клетки, обладающие этими векторами, предоставляют со всеми генами, необходимыми для продукции фаговых частиц, способ репликации плазмиды меняется на репликацию по типу "катящегося кольца" с образованием копий одной цепи плазмидной ДНК и упаковки их в фаговые частицы. Фагмида может образовывать инфекционные или неинфекционные фаговые частицы. Этот термин включает фагмиды, которые содержат ген белка оболочки фага или его фрагмент, связанный с геном гетерологичного пептида, в качестве слитых генов, так что гетерологичный полипептид экспонируется на поверхности фаговой частицы.
Термин "фаговый вектор" означает двухцепочечную репликативную форму бактериофага, содержащую гетерологичный ген и способную к репликации. Фаговый вектор имеет фаговый ориджин репликации, позволяющий репликацию фага и образование фаговых частиц. Фаг предпочтительно представляет собой нитевидный бактериофаг, такой как фаг M13, f1, fd, Pf3 или их производные, или лямбдоидный фаг, такой как, фаг лямбда, 21, phi80, phi81, 82, 424, 434 и т.д., или его производное.
Как используют в настоящем документе, "доступное для растворителя положение" относится к положению аминокислотного остатка в вариабельных участках тяжелой и легкой цепей исходного антитела или антигенсвязывающего фрагмента, которое определяют на основании структуры, совокупности структур и/или смоделированной структуры антитела или антигенсвязывающего фрагмента, в качестве потенциально доступного для доступа растворителя и/или контакта с молекулой, такой как специфический для антитела антиген. Эти положения, как правило, находятся в CDR и на наружной части белка. Доступные для растворителя положения антитела или антигенсвязывающего фрагмента, как определяют в настоящем документе, можно определять с использованием любого из множества алгоритмов, известных в данной области. Предпочтительно, доступные для растворителя положения определяют с использованием координат из 3-мерной модели антитела, предпочтительно с использованием компьютерной программы, такой как программа InsightII (Accelrys, San Diego, CA). Также доступные для растворителя положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (1971) J Mol. Biol 55, 379, и Connolly (1983) J. Appl Cryst. 16, 548). Определение доступных для растворителя положений можно проводить с использованием программного обеспечения, пригодного для моделирования белков, и полученной информации о 3-мерной структуре антитела. Программное обеспечение, которое можно использовать для этих целей, включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Как правило и предпочтительно, когда алгоритм (программа) требует введения пользователем параметра размера, "размер" зонда, который используют для вычисления, устанавливают на размер радиуса приблизительно 1,4 Å или менее. Кроме того, способы определения доступных для растворителя областей и площади с использованием программного обеспечения для персональных компьютеров описаны Pacios (1994) Comput. Chem. 18(4): 377-386.
II. Подробное описание
Система комплемента
Комплемент играет ключевую роль в защите организма и, вместе с другими компонентами иммунной системы, защищает индивида от патогенов, поступающих в организм. Однако если он активируется или контролируется не надлежащим образом, комплемент также может вызывать повреждение тканей хозяина. Ненадлежащая активация комплемента вовлечена в патогенез множества заболеваний, называемых связанными с комплементом заболеваниями или нарушениями, таких как связанные с иммунными комплексами и аутоиммунные заболевания и различные воспалительные состояния, включая опосредуемое комплементом повреждение тканей. Патология связанных с комплементом нарушений варьирует, и она может вовлекать активацию комплемента в течение длительного или короткого периода времени, активацию всего каскада, только одного из каскадов (например, классического или альтернативного каскада), только некоторые компоненты каскада и т.д. При некоторых заболеваниях биологическая активность фрагментов комплемента приводит к повреждению и заболеванию. Таким образом, ингибиторы комплемента обладают высоким терапевтическим потенциалом. Селективные ингибиторы альтернативного каскада могут быть особенно пригодными, поскольку выведение патогенов и других организмов из крови посредством классического каскада остается неизмененным.
Антитела против C3b и их применение для профилактики и лечения связанных с комплементом нарушений
Настоящее изобретение основано, по меньшей мере частично, на разработке антител, которые специфично распознают фрагменты разрушения C3 и не распознают нативный C3. В частности, изобретение относится к антителам, распознающим и специфично связывающим C3b, разработанным с использованием комбинаторных библиотек антител человека и фагового дисплея, где обогащения специфичными к C3b фагами достигали блокированием насыщающими количествами C3. С использованием этих способов авторы настоящего изобретения смогли разработать антитела, которые специфичны к активированным формам C3. Кроме того, эти антитела человека далее подвергали созреванию аффинности, таким образом, повышая их эффективность в анализах гемолиза in vitro. Fab-фрагмент получали клонированием, и было показано, что он сохраняет высокую эффективность ингибирования активации комплемента через альтернативный каскад. Совместная структура Fab (обозначаемого как S77) в комплексе с C3b была решена и были картированы остатки, вовлеченные во взаимодействие C3b-S77. Насколько известно авторам настоящего изобретения, это антитело представляет собой первое полученное из фага антитело с селективностью к C3-фрагментам, которое ингибирует альтернативный каскад комплемента.
Антитела и другие специфичные к C3b антагонисты по настоящему изобретению пригодны для профилактики и лечения связанных с комплементом нарушений. Конкретные примеры связанных с комплементом заболеваний включают, но не ограничиваются ими, ревматоидный артрит (RA), острый респираторный дистресс-синдром (ARDS), отдаленное повреждение ткани после ишемии и реперфузии, активацию комплемента в ходе хирургической операции с искусственным кровообращением, дерматомиозит, пемфигус, волчаночный нефрит и являющиеся его следствиями гломерулонефрит и васкулит, искусственное кровообращение, индуцируемую кардиоплегией дисфункцию эндотелия коронарных артерий, мембранопролиферативный гломерулонефрит типа II, IgA-нефропатию, острую почечную недостаточность, криоглобулинемию, антифосфолипидный синдром, дегенеративные заболевания желтого пятна, такие как связанная со старением дегенерация желтого пятна (AMD), хориоидальная неоваскуляризация (CNV), увеит, диабетическую и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные с внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы, неоваскуляризация сетчатки, а также аллотрансплантацию, сверхострое отторжение, гемодиализ, хронический легочный дистресс-синдром (COPD), астму и аспирационную пневмонию.
Более широкий перечень воспалительных состояний в качестве примеров связанных с комплементом заболеваний включает, например, воспалительное заболевание кишечника (IBD), системную красную волчанку, ревматоидный артрит, ювенильный хронический артрит, спондилоартропатии, системную склеродермию (склеродермию), идиопатические воспалительные миопатии (дерматомиозит, полимиозит), синдром Шегрена, системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (панцитопения клеток иммунной системы, пароксизмальная ночная гемоглобинурия), аутоиммунную тромбоцитопению (идиопатическая тромбоцитопеническая пурпура, иммуноопосредуемая тромбоцитопения), тиреоидит (болезнь Грэйва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), сахарный диабет, иммуноопосредуемое заболевание почек (гломерулонефрит, тубулоинтерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервной системы, такие как рассеянный склероз, идиопатическая полиневропатия, заболевания печени и желчевыводящих путей, такие как инфекционный гепатит (гепатит A, B, C, D, E и другие негепатотропные вирусы), аутоиммунный хронический активный гепатит, первичный биллиарный цирроз, гранулематозный гепатит и склерозирующий холингит, воспалительные и фиброзные заболевания легких (например, кистозный фиброз), глютензависимую энтеропатию, болезнь Уиппла, аутоиммунные или иммуноопосредуемые заболевания кожи, включая буллезные заболевания кожи, полиформную эритему и контактный дерматит, псориаз, аллергические заболевания легкого, такие как эозинофильная пневмония, идиопатический фиброз легких и связанный с гиперчувствительностью пневмонит, связанные с трансплантацией заболевания, включая отторжение трансплантата, реакцию "трансплантат против хозяина".
При системной красной волчанке центральным опосредующим заболевание звеном является продукция аутореактивных антител к собственным белкам/тканям и последующее возникновение иммуноопосредуемого воспаления. Антитела либо прямо, либо непрямо опосредуют повреждение тканей. Хотя не было показано, что T-лимфоциты прямо вовлечены в повреждение ткани, T-лимфоциты необходимы для образования аутореактивных антител. Возникновение заболевания, таким образом, является зависимым от T-лимфоцитов. Клинически поражаются многие органы и системы, включая почки, легкие, опорно-двигательный аппарат, кожно-слизистую систему, глаз, центральную нервную систему, сердечно-сосудистую систему, желудочно-кишечный тракт, костный мозг и кровь.
Ревматоидный артрит (RA) представляет собой хроническое системное аутоиммунное воспалительное заболевание, которое, главным образом, вовлекает синовиальную мембрану множества суставов, с повреждением в итоге суставного хряща. Патогенез является зависимым от T-лимфоцитов и ассоциирован с продукцией ревматоидных факторов, аутоантител, направленных против собственных IgG, с образованием в итоге иммунных комплексов, которые достигают высоких уровней в суставной жидкости и крови. Эти комплексы в суставе могут индуцировать выраженный инфильтрат лимфоцитов и моноцитов в синовиальную мембрану и последующие выраженные изменения синовиальной мембраны; суставная щель/жидкость инфильтрируется сходными клетками, а также множеством нейтрофилов. Пораженными тканями являются, главным образом, суставы, часто по типу симметрии. Однако также происходит внесуставное заболевание в двух основных формах. Одной из форм является развитие внесуставных очагов повреждения с продолжающимся прогрессирующим заболеванием суставов и типичными повреждениями фиброза легких, васкулита и кожных язв. Второй формой внесуставного заболевания является так называемый синдром Фелти, который возникает поздно в ходе заболевания RA, иногда после того, как заболевание суставов становится латентным, и вовлекает наличие нейтропении, тромбоцитопении и спленомегалии. Это может сопровождаться васкулитом во множестве органов с образованием инфарктов, кожных язв и гангрены. У пациентов также часто развиваются ревматоидные узелки в подкожной ткани, покрывающей пораженные суставы; узелки на поздних стадиях имеют некротические центры, окруженные смешанным инфильтратом воспалительных клеток. Другие проявления, которые могут происходить при RA, включают: перикардит, плеврит, коронарный артериит, интерстициальный пневмонит с фиброзом легких, сухой кератоконъюнктивит и ревматоидные узелки.
Ювенильный хронический артрит представляет собой хроническое идиопатическое воспалительное заболевание, которое часто начинается в возрасте 16 лет или менее. Его фенотип имеет некоторые сходства с RA; некоторых пациентов, которые являются положительными по ревматоидному фактору, классифицируют как пациентов с ювенильным ревматоидным артритом. Заболевание подразделяют на три основных категории: олигоартикулярный, полиартикулярный и системный. Артрит может быть тяжелым, и, как правило, он является деструктивным и ведет к анкилозу суставов и замедленному росту. Другие проявления могут включать хронический передний увеит и системный амилоидоз.
Спондилоартропатии представляют собой группу нарушений с некоторыми общими клиническими признаками и общей ассоциацией с экспрессией продукта гена HLA-B27. Нарушения включают: анкилозирующий спондилит, синдром Рейтера (реактивный артрит), артрит, ассоциированный с воспалительным заболеванием кишечника, спондилит, ассоциированный с псориазом, ювенильную спондилоартропатию и недифференцируемую спондилоартропатию. Отличительные признаки включают сакроилеит со спондилитом или без него; воспалительный асимметричный артрит; ассоциацию с HLA-B27 (серологически определяемый аллель локуса HLA-B класса I MHC); воспаление глаза и отсутствие аутоантител, ассоциированных с другим ревматоидным заболеванием. Клеткой, наиболее вовлеченной в индукцию заболевания в качестве ключевой, является CD8+ T-лимфоцит, клетка, которая нацелена на антиген, представленный молекулами MHC класса I. CD8+ T-клетки могут реагировать против аллеля HLA-B27 MHC класса I, как если бы он представлял собой чужеродный пептид, экспрессируемый молекулами MHC класса I. Было сделано предположение, что эпитоп HLA-B27 может имитировать бактериальный или другой микробный антигенный эпитоп и, таким образом, индуцировать CD8+ T-клеточный ответ.
Системная склеродермия (склеродермия) имеет неизвестную этиологию. Характерным признаком заболевания является уплотнение кожи; вероятно оно индуцируется воспалительным процессом. Склеродермия может быть локализованной или системной; частыми являются сосудистые повреждения, и повреждение клеток эндотелия в микроциркуляторном русле является ранним и важным событием при развитии системной склеродермии; сосудистое повреждение может быть иммуноопосредуемым. Иммунологическая основа вытекает из наличия инфильтратов мононуклеарных клеток в кожных очагах повреждения и наличия антиядерных антител у многих пациентов. Часто активируется ICAM-1 на клеточной поверхности фибробластов в очагах повреждения кожи, позволяя предположить, что взаимодействие T-клеток с этими клетками может играть роль в патогенезе заболевания. Другие вовлеченные органы включают: желудочно-кишечный тракт: атрофия и фиброз гладких мышц, приводящие к аномальной перистальтике/подвижности; почки: концентрическая субэндотелиальная интимальная пролиферация, поражающая малые дугообразные и междольковые артерии со снижением в итоге кровотока в коре почек, что приводит к протеинурии, азотемии и гипертензии; скелетные мышцы: атрофия, интерстициальный фиброз; воспаление; легкое: интерстициальный пневмонит и интерстициальный фиброз; и сердце: некроз полос сокращения, рубцевание/фиброз.
Идиопатические воспалительные миопатии, включая дерматомиозит, полимиозит и прочие, представляют собой нарушения с хроническим воспалением мышц неизвестной этиологии, приводящим к мышечной слабости. Повреждение/воспаление мышц часто является системным и прогрессирующим. С большинством форм ассоциированы аутоантитела. Эти специфичные для миозита аутоантитела направлены против компонентов, белков и РНК, вовлеченных в синтез белка, и ингибируют их функцию.
Синдром Шегрена является следствием иммуноопосредуемого воспаления и последующего функционального разрушения слезных желез и слюнных желез. Заболевание может быть ассоциировано с воспалительными заболеваниями соединительной ткани, или оно может сопровождаться ими. Заболевание ассоциировано с продукцией аутоантител против антигенов Ro и La, оба из которых представляют собой комплексы малая РНК-белок. Очаги повреждения приводят к сухому кератоконъюнктивиту, ксеростомии, с другими проявлениями или ассоциациями, включая биллиарный цирроз, периферическую или сенсорную невропатию и пальпируемую пурпуру.
Системный васкулит включает заболевания, при которых первичным очагом повреждения является воспаление и последующее разрушение кровеносных сосудов, которое приводит к ишемии/некрозу/дегенерации тканей, кровоснабжаемых пораженными сосудами, и, в некоторых случаях, конечной дисфункции органов-мишеней. Также могут встречаться васкулиты в качестве вторичного повреждения или осложнения других иммуноопосредуемых заболеваний, таких как ревматоидный артрит, системная склеродермия, и т.д., в частности при заболеваниях, также ассоциированных с образованием иммунных комплексов. Заболевания группы первичного системного васкулита включают: системный некротизирующий васкулит: узелковый полиартериит, аллергический ангиит и гранулематоз, полиангиит; гранулематоз Вегенера; лимфоматоидный гранулематоз и гигантоклеточный артериит. Прочие васкулиты включают: слизисто-кожный лимфоузелковый синдром (MLNS или болезнь Кавасаки), изолированный васкулит ЦНС, болезнь Бехчета, облитерирующий тромбангит (болезнь Бюргера) и кожный некротизирующий венулит. Полагают, что патогенный механизм большинства приведенных типов васкулита является следствием, главным образом, накопления комплексов иммуноглобулинов в стенке сосудов и последующей индукции воспалительного ответа либо через ADCC, либо через активацию комплемента, либо обоими путями.
Саркоидоз представляет собой состояние неизвестной этиологии, которое характеризуется наличием эпителиоидных гранулем практически в любой ткани организма; наиболее распространенным является вовлечение легких. Патогенез вовлекает персистенцию активированных макрофагов и лимфоидных клеток в пораженных областях с последующими хроническими осложнениями вследствие локального или системного высвобождения активных продуктов этими типами клеток.
Аутоиммунная гемолитическая анемия, включающая аутоиммунную гемолитическую анемию, панцитопению иммунных клеток и пароксизмальную ночную гемоглобинурию, является результатом продукции антител, которые реагируют с антигенами, экспрессируемыми на поверхности эритроцитов (и в некоторых случаях других клеток крови, включая также тромбоциты), и является отражением удаления этих покрытых антителами клеток через опосредуемый комплементом лизис и/или опосредуемые ADCC/Fc-рецептором механизмы.
При аутоиммунной тромбоцитопении, включая тромбоцитопеническую пурпуру и иммуноопосредуемую тромбоцитопению в других клинических условиях, происходит разрушение/удаление тромбоцитов в результате связывания либо антитела, либо комплемента, с тромбоцитами и последующего их удаления лизисом с помощью комплемента, ADCC или опосредуемыми FC-рецепторами механизмами.
Тиреоидит, включая болезнь Грэйва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит и атрофический тиреоидит, является результатом аутоиммунного ответа против антигенов щитовидной железы с продукцией антител, которые реагируют с белками, присутствующими в щитовидной железе и часто специфическими для нее. Существуют экспериментальные модели, включая спонтанные модели: крысы (крысы BUF и BB) и куры (линия кур с ожирением); индуцируемые модели: иммунизация животных либо тиреоглобулином, либо микросомальным антигеном щитовидной железы (пероксидазой щитовидной железы).
Сахарный диабет 1 типа или инсулинзависимый диабет представляет собой аутоиммунное разрушение островковых β-клеток поджелудочной железы; это разрушение опосредуется аутоантителами и аутореактивными T-клетками. Антитела к инсулину или рецептору инсулина также могут приводить к фенотипу с отсутствием ответа на инсулин.
Иммуноопосредуемые заболевания почек, включая гломерулонефрит и тубулоинтерстициальный нефрит, являются результатом опосредуемого антителами или T-лимфоцитами повреждения ткани почки либо прямо в результате продукции аутореактивных антител или T-клеток против антигенов почки, либо непрямо в результате отложения в почке антител и/или иммунных комплексов, которые являются реактивными к другим, непочечным антигенам. Таким образом, другие иммуноопосредуемые заболевания, которые приводят к образованию иммунных комплексов, также могут индуцировать иммуноопосредуемое заболевание почек в качестве непрямого последствия. Как прямой, так и непрямой иммунные механизмы приводят к воспалительному ответу, который вызывает/индуцирует развитие очага повреждения в тканях почек с нарушением в итоге функции органа и, в некоторых случаях, прогрессированием в почечную недостаточность. В патогенез очагов повреждения могут быть вовлечены как гуморальные, так и клеточные, иммунные механизмы.
Полагают, что демиелинизирующие заболевания центральной и периферической нервной системы, включая рассеянный склероз; идиопатическую демиелинизирующую полиневропатию или синдром Гийена-Барре; и хроническую воспалительную демиелинизирующую полиневропатию, имеют аутоиммунную основу и приводят к демиелинизации нервов в результате повреждения олигодендроцитов или непосредственно миелина. Существуют данные, позволяющие предположить, что при MS индукция и прогрессирование заболевания зависит от T-лимфоцитов. Рассеянный склероз представляет собой демиелинизирующее заболевание, которое является зависимым от T-лимфоцитов и имеет либо ремитирующее течение, либо хроническое прогрессирующее течение. Его этиология неизвестна; однако вклад вносят вирусные инфекции, генетическая предрасположенность, окружающая среда и аутоиммунитет. Очаги повреждения содержат инфильтраты, опосредуемые, главным образом, T-лимфоцитами, клетками микроглии и инфильтрирующими макрофагами; CD4+T-лимфоциты являются главным типом клеток в очагах повреждения. Механизм гибели олигодендроцитов и последующей демиелинизации не известен, однако вероятно, что он запускается T-лимфоцитами.
Воспалительное и фиброзное заболевание легких, включая эозинофильную пневмонию, идиопатический фиброз легких и связанный с гиперчувствительностью пневмонит, может вовлекать иммуновоспалительный ответ с нарушенной регуляцией. Ингибирование этого ответа может быть терапевтически полезным.
Аутоиммунное и иммуноопосредуемое заболевание кожи, включая буллезные заболевания кожи, полиформную эритему и контактный дерматит, опосредуются аутоантителами, и их генез является зависимым от T-лимфоцитов.
Псориаз представляет собой опосредуемое T-лимфоцитами воспалительное заболевание. Очаги повреждения содержат инфильтраты T-лимфоцитов, макрофагов и процессирующих антиген клеток и некоторые нейтрофилы. Аллергические заболевания, включая астму; аллергический ринит; атопический дерматит; пищевую гиперчувствительность и крапивницу, являются зависимыми от T-лимфоцитов. Эти заболевания, главным образом, опосредуются индуцируемым T-лимфоцитами воспалением, опосредуемым IgE воспалением или сочетанием обоих из них.
Связанные с трансплантацией заболевания, включая отторжение трансплантата и реакцию "трансплантат против хозяина" (GVHD), являются зависимыми от T-лимфоцитов; ингибирование функции T-лимфоцитов приводит к улучшению.
Антагонисты C3b, такие как антитела против C3b, по настоящему изобретению, также пригодны для профилактики и лечения связанных с комплементом состояний глаз (всех состояний и заболеваний глаз, патология которых вовлекает комплемент, включая классический и альтернативный каскады, и особенно альтернативный каскад комплемента), например, таких как дегенеративные заболевания желтого пятна, такие как все стадии связанной со старением дегенерации желтого пятна (AMD), включая сухую и влажную (неэкссудативную и экссудативную) формы, хориоидальная неоваскуляризация (CNV), увеит, диабетическую и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные с внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пята, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы и неоваскуляризация сетчатки. Предпочтительная группа связанных с комплементом состояний глаз включает связанную со старением дегенерацию желтого пятна (AMD), включая экссудативную (влажную) и неэкссудативную (сухую или атрофическую) AMD, хориоидальную неоваскуляризацию (CNV), диабетическую ретинопатию (DR) и эндофтальмит.
AMD представляет собой связанную со старением дегенерацию желтого пятна, которая является основной причиной необратимой дисфункции зрения у индивидов в возрасте свыше 60 лет. Существует два типа AMD, неэкссудативная (сухая) и экссудативная (влажная) AMD. Сухая, или неэкссудативная, форма вовлекает атрофические и гипертрофические изменения в пигментном эпителии сетчатки (RPE), лежащем под центральной зоной сетчатки (желтым пятном), а также отложения (друзы) на RPE. У пациентов с неэкссудативной AMD может происходить прогрессирование во влажную, или экссудативную, форму AMD, при которой под сетчаткой образуются аномальные кровеносные сосуды, называемые хориоидальными неоваскулярными мембранами (CNVM), из которых проникает жидкость и кровь, и это в конечном итоге приводит к вызывающему слепоту дисковидному рубцу в сетчатке и под ней. Неэкссудативная AMD, которая обычно предшествует экссудативной AMD, является более распространенной. Признаки неэкссудативной AMD варьируют; могут присутствовать твердые друзы, мягкие друзы, географическая атрофия RPE и агрегация пигмента. Компоненты комплемента накапливаются на RPE на ранних этапах AMD, и они являются основными составными элементами друз.
Настоящее изобретение, конкретно, относится к лечению AMD высокого риска, включая AMD 3 категории и 4 категории. AMD 3 категории характеризуется отсутствием развернутой AMD в обоих глазах, причем по меньшей мере один глаз имеет остроту зрения 20/32 или лучше, и по меньшей мере одной большой друзой (например, 125 мкм), обширной (как измеряют по площади друзы) промежуточной друзой, или географической атрофией (GA), которая не вовлекает центр желтого пятна, или любым их сочетанием. AMD 3 категории (которую все еще считают "сухой" AMD) имеет высокий риск перехода в хориоидальную неоваскуляризацию (CNV).
AMD высокого риска 4 категории (классифицируемая как "влажная" AMD) характеризуется остротой зрения 20/32 или лучше и отсутствием развернутой AMD (GA, вовлекающий центр желтого пятна или признаки хориоидальной неоваскуляризации) в служащем показателем глазу. Второй глаз характеризуется развернутой AMD, или остротой зрения менее 20/32, свойственной макулопатии при AMD. Как правило, AMD высокого риска без лечения быстро прогрессирует в хориоидальную неоваскуляризацию (CNV), со скоростью, приблизительно в 10-30 раз превышающей скорость прогрессирования для AMD 1 или 2 категории (невысокого риска).
Антагонисты C3b также являются применимыми для профилактики прогрессирования AMD (в частности, AMD 3 категории или 4 категории) в CNV и/или для профилактики развития/прогрессирования AMD или CNV в непораженном или менее пораженном втором глазу. В этом контексте термин "профилактика" используют в наиболее широком значении, и он включает полное или частичное блокирование и замедление прогрессирования заболевания, а также замедление начала более тяжелой формы заболевания. Пациентам с высоким риском развития или прогрессирования в AMD высокого риска (категория 4) или CMV особенно полезен этот аспект изобретения.
Известно, что полиморфизм фактора комплемента H (CFH) ассоциирован с риском развития у индивида AMD и/или CNV. Мутации в CFH могут активировать комплемент, что, в свою очередь, может привести к AMD/CNV. Недавно было описано, что полиморфизм фактора комплемента H (CFH) объясняет 50% имеющегося риска AMD (Klein et al., Science 308:385-9 (2005)). Было выявлено, что распространенный гаплотип CFH (HF1/CFH) предрасполагает индивидов к связанной с возрастом дегенерации желтого пятна (Hageman et al., Proc. Natl. Acad. Sci. USA, 102(2):7227-7232 (2005)). AMD была выделена в качестве аутосомно-доминантного признака, и локус заболевания был картирован на хромосоме 1q25-q31 между маркерами D1S466 и D1S413, с максимальным показателем lod приблизительно 3,20 (Klein et al., Arch Opthalmol 116(8): 1082-9 (1998); Majewski et al., Am. J. Hum. Genet. 73(3):540-50 (2003); Seddon et al., Am. J. Hum. Genet. 73(4):780-90 (2003); Weeks et al., Am. J. Ophthalmol. 132(5):682-92 (2001); Iyengar et al., Am. J. Hum. Genet. 74(1):20-39 (2004)); на хромосоме 2q3/2q32 между маркерами D12S1391 и D2S1384, с максимальным показателем lod 2,32/2,03 (Seddon et al., выше); 3pl3, между маркерами D12S1300 и D12S1763, с максимальным показателем lod 2,19 (Majewski et al., выше; Schick et al., Am. J. Hum. Genet. 72(6): 1412-24 (2003)); 6q14, между маркерами D6S1056 и DS249, с максимальным показателем lod 3,59/3,17 (Kniazeva et al., Am. J. Ophthlmol. 130(2): 197-202 (2000)); 9q33, в области маркера D9S934, с максимальным показателем lod 2,06 (Mejwski et al., выше); 10q26, в области маркера D10S1230, с максимальным показателем lod 3,06 (Majewski et al., выше; Iyengar et al., выше; Kenealy et al., Mol. Vis. 10: 57-61 (2004)); 17q25, в области маркера D17S928, с максимальным показателем lod 3,16 (Weeks et al., выше); и 22ql2, в области маркера D22S1045, с максимальным показателем lod 2,0 (Seddon et al., выше). Таким образом, генетический скрининг является важной частью выявления пациентов, которые являются особенно подходящими кандидатами для профилактического лечения, включая профилактику прогрессирования заболевания в более тяжелую форму, например от AMD до CNV.
Получение и селекция антител против C3b
Изобретение, представленное в настоящем документе, включает продукцию и применение антител, которые распознают C3b, но не распознают его неактивный предшественник C3. Иллюстративные способы получения антител более подробно описаны в следующих разделах.
Антитела против C3b антитела отбирают с использованием полипептида C3b вида млекопитающего. Предпочтительно полипептид представляет собой C3b человека. Однако также в качестве антигена-мишени можно использовать полипептиды C3b других видов, такие как C3b мыши. Антигены C3b различных видов млекопитающих можно выделять из природных источников. В других вариантах осуществления антиген получают рекомбинантными способами или изготавливают с использованием других способов синтеза, известных в данной области.
Выбранное антитело в норме может иметь достаточно высокую аффинность связывания с антигеном C3b. Например, антитело может связывать C3b человека со значением Kd не более чем приблизительно 5 нМ, предпочтительно не более чем приблизительно 2 нМ и более предпочтительно не более чем приблизительно 500 пМ. Аффинность антитела можно определять, например, анализом на основе поверхностного плазмонного резонанса (такого, как анализ BIAcore, описанный в разделе "Примеры"); твердофазным иммуноферментным анализом (ELISA) и конкурентными анализами (например, RIA).
Также, антитело можно подвергать другим анализам биологической активности, например, для оценки его эффективности в качестве лекарственного средства. Такие анализы известны в данной области и зависят от антигена-мишени и предполагаемого применения антитела. Примеры включают анализ ингибирования HUVEC (как описано в разделе "Примеры", ниже) и анализы in vitro и in vivo, описанные ниже, для идентификации антител, которые селективно блокируют альтернативный каскад и демонстрируют активность в отношении профилактики и/или лечения по меньшей мере одного связанного с комплементом нарушения.
Для скрининга антител, которые связываются с конкретным эпитопом на представляющем интерес антигене, можно проводить общепринятый анализ перекрестного блокирования, такой как описан в Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory, Ed Harlow and David Lane (1988). Альтернативно можно проводить картирование эпитопа, например, как описано в Champe et al. (1995) J. Biol. Chein. 270: 1388-1394, для определения наличия связывания антитела с представляющим интерес эпитопом.
В предпочтительном варианте осуществления антитела против C3b по настоящему изобретению отбирают с использованием уникального подхода фагового дисплея. Подход вовлекает создание фаговых библиотек синтетических антител человека на основе матрицы каркасной области, внесение достаточного разнообразия в вариабельные домены, дисплей полипептидов, имеющих разнообразные вариабельные домены, и селекцию антител-кандидатов с высокой аффинностью к антигену-мишени C3b. Обогащения специфическими к C3b фагами, кодирующими антитела, селективно блокирующими C3b, но не C3, можно достигать, например, блокированием насыщающими количествами C3, как описано в разделе "Примеры", ниже.
Подробное описание способов фагового дисплея может быть найдено, например, в WO 03/102157, опубликованной 11 декабря 2003 года. В одном аспекте библиотеки антител можно создавать внесением мутаций в доступные для растворителя и/или высоко разнообразные положения по меньшей мере в одной CDR вариабельного домена антитела. С использованием способов, представленных в настоящем документе, можно вносить мутации в некоторые или все из CDR. В некоторых вариантах осуществления может быть предпочтительным создание библиотек разнообразных антител внесением мутаций в положения в CDRH1, CDRH2 и CDRH3 с образованием единой библиотеки, или внесением мутаций в положения в CDRL3 и CDRH3 с образованием единой библиотеки, или внесением мутаций в положения в CDRL3 и CDRH1, CDRH2 и CDRH3 с образованием единой библиотеки.
Можно создавать библиотеку вариабельных доменов, например, имеющих мутации в доступных для растворителя и/или имеющих высокое разнообразие положениях CDRH1, CDRH2 и CDRH3. Можно создавать другую библиотеку, имеющую мутации в CDRL1, CDRL2 и CDRL3. Эти библиотеки также можно использовать вместе для создания связывающих молекул с требуемой аффинностью. Например, после одного или нескольких раундов селекции библиотек тяжелых цепей в отношении связывания с антигеном-мишенью можно заменять библиотеку легких цепей в совокупности связывающих молекул с тяжелой цепью для последующих раундов селекции для повышения аффинности связывающих молекул.
Предпочтительно, библиотеку создают заменой исходных аминокислот вариантами аминокислот в области CDRH3 вариабельного участка последовательности тяжелой цепи. Полученная библиотека может содержать множество последовательностей антител, где разнообразие последовательности, главным образом, находится в области CDRH3 тяжелой последовательности.
В одном аспекте создают библиотеку применительно к последовательности гуманизированного антитела 4D5 или последовательности каркасных аминокислот в последовательности гуманизированного антитела 4D5. Предпочтительно, библиотеку создают заменой по меньшей мере остатков 95-100a тяжелой цепи аминокислотами, кодируемыми набором кодонов DVK, где набор кодонов DVK используют для кодирования набора вариантов аминокислот для каждого из этих положений. Пример набора олигонуклеотидов, который пригоден для создания этих замен, включает последовательность (DVK)7. В некоторых вариантах осуществления библиотеку создают заменой остатков 95-100a аминокислотами, кодируемыми как набором кодонов DVK, так и набором кодонов NNK. Пример набора олигонуклеотидов, который пригоден для внесения этих замен, включает последовательность (DVK) 6 (NNK). В другом варианте осуществления библиотеку создают заменой по меньшей мере остатков 95-100a аминокислотами, кодируемыми как набором кодонов DVK, так и набором кодонов NNK. Пример набора олигонуклеотидов, который пригоден для внесения этих замен, включает последовательность (DVK) 5 (NNK). Другой пример набора олигонуклеотидов, который пригоден для внесения этих замен, включает последовательность (NNK) 6. Другие примеры пригодных олигонуклеотидных последовательностей может определить специалист в данной области в соответствии с критериями, описанными в настоящем документе.
В другом варианте осуществления используют различные конструкции CDRH3 для выделения высокоаффинных связывающих молекул и для выделения связывающих молекул для различных эпитопов. Диапазон длин CDRH3, создаваемых в этой библиотеке, составляет от 11 до 13 аминокислот, хотя также можно создавать CDRH3 с длинами, отличными от этих. Разнообразие H3 можно расширять с использованием наборов кодонов NNK, DVK и NVK, а также более ограниченного разнообразия на N- и/или C-конце.
Разнообразие также можно вносить в CDRH1 и CDRH2. Конструкции для разнообразия CDR-H1 и H2 согласуются со стратегией нацеливания для имитации природного набора антител, как описано, с модификацией, которая фокусируется на разнообразии, более сходном с природным разнообразием, чем предыдущие конструкции. Для разнообразия в CDRH3 можно конструировать множество библиотек отдельно с различными длинами H3, а затем комбинировать для селекции связывающих молекул для нацеливания на антигены. Множество библиотек можно объединять и сортировать с использованием селекции на твердой подложке и способов сортировки в растворе, как описано ранее и в настоящем документе ниже. Можно использовать множество стратегий сортировки. Например, один из ее вариантов вовлекает сортировку на мишени, связанной с твердой подложкой, а затем сортировку по метке, которая может присутствовать на слитом полипептиде (например, метка анти-gD), и затем другую сортировку на мишени, связанной с твердой подложкой. Альтернативно библиотеки сначала можно сортировать на мишени, связанной с твердой поверхностью, затем элюированные связывающие молекулы сортировать с использованием связывания в фазе раствора со снижающимися концентрациями антигена-мишени. Применение сочетаний различных способов сортировки обеспечивает минимизацию селекции не только высокоэкспрессируемых последовательностей, но и обеспечивает селекцию ряда различных высокоаффинных клонов.
Связывающие молекулы, высокоаффинные к антигену-мишени C3b, можно выделять из библиотек. Ограничение разнообразия в области H1/H2 снижает вырожденность приблизительно от 104 до 105 раз, и обеспечение большего разнообразия H3 обеспечивает более высокоаффинные связывающие молекулы. Применение библиотек с различными типами разнообразия в CDRH3 (например, с использованием DVK или NVT) обеспечивает выделение связывающих молекул, которые могут связываться с различными эпитопами антигена-мишени.
В другом варианте осуществления создают библиотеку или библиотеки с разнообразием в областях CDRH1, CDRH2 и CDRH3. В этом варианте осуществления разнообразие в CDRH3 создают с использованием различных длин областей H3 и с использованием, главным образом, наборов кодонов XYZ и NNK или NNS. Библиотеки можно создавать с использованием отдельных олигонуклеотидов и объединять или можно объединять олигонуклеотиды с образованием группы библиотек. Библиотеки этого варианта осуществления можно сортировать против мишени, связанной с твердой поверхностью. Клоны, выделенные при множестве сортировок, можно подвергать скринингу в отношении специфичности и аффинности с использованием анализов ELISA. Для специфичности клоны можно подвергать скринингу против требуемых антигенов-мишеней, а также против других не являющихся мишенями антигенов. Эти связывающие молекулы к антигену-мишени C3b затем можно подвергать скринингу в отношении аффинности в конкурентном анализе ELISA связывания в растворе или конкурентном спот-анализе. Высокоаффинные связывающие молекулы можно выделять из библиотеки с использованием наборов кодонов XYZ, полученных, как описано выше. Эти связывающие молекулы можно легко получать в качестве антител или антигенсвязывающих фрагментов с высоким выходом в клеточной культуре.
В некоторых вариантах осуществления может быть желательным создание библиотек с более высоким разнообразием длин области CDRH3. Например, может быть желательным создание библиотек с областями CDRH3 в диапазоне приблизительно от 7 до 19 аминокислот.
Высокоаффинные связывающие молекулы, выделенные из библиотек этих вариантов осуществления, легко получать в бактериальной и эукариотической клеточной культуре с высоким выходом. Можно конструировать векторы для легкого удаления последовательностей, таких как метки gD, последовательность белкового компонента оболочки вируса, и/или для добавления в константную область последовательностей для обеспечения продукции полноразмерных антител или антигенсвязывающих фрагментов с высоким выходом.
Библиотеку с мутациями в CDRH3 можно комбинировать с библиотекой, содержащей версии вариантов других CDR, например CDRL1, CDRL2, CDRL3, CDRH1 и/или CDRH2. Таким образом, например, в одном варианте осуществления библиотеку CDRH3 комбинируют с библиотекой CDRL3, созданной применительно к последовательности гуманизированного антитела 4D5 с вариантами аминокислот в положениях 28, 29, 30, 31 и/или 32 с использованием заданных наборов кодонов. В другом варианте осуществления библиотеку с мутациями CDRH3 можно комбинировать с библиотекой, содержащей вариабельные домены тяжелой цепи с вариантами CDRH1 и/или CDRH2. В одном варианте осуществления создают библиотеку CDRH1 с помощью последовательности гуманизированного антитела 4D5 с вариантами аминокислот в положениях 28, 30, 31, 32 и 33. Можно создавать библиотеку CDRH2 с последовательностью гуманизированного антитела 4D5 с вариантами аминокислот в положениях 50, 52, 53, 54, 56 и 58 с использованием заданных наборов кодонов.
Антитело против C3b, полученное из фаговых библиотек, можно далее модифицировать для получения мутантов антитела с улучшенными физическими, химическими и/или биологическими свойствами относительно исходного антитела. Когда необходимым анализом является анализ биологической активности, мутант антитела предпочтительно имеет биологическую активность в выбираемом анализе, которая по меньшей мере приблизительно в 10 раз лучше, предпочтительно по меньшей мере приблизительно в 20 раз лучше, более предпочтительно по меньшей мере приблизительно в 50 раз лучше и иногда по меньшей мере приблизительно в 100 раз или 200 раз лучше, чем биологическая активность исходного антитела в этом анализе. Например, мутант антитела против C3b предпочтительно имеет аффинность связывания с C3b, которая по меньшей мере приблизительно в 10 раз выше, предпочтительно по меньшей мере приблизительно в 20 раз выше, более предпочтительно по меньшей мере приблизительно в 50 раз выше, и иногда по меньшей мере приблизительно в 100 раз или 200 раз выше, чем аффинность связывания исходных антител против C3b, таких как антитело S77.
Для получения мутанта антитела вносят одно или несколько изменений (например, замен) аминокислот в один или несколько из гипервариабельных участков исходного антитела. Альтернативно или дополнительно, можно вносить одно или несколько изменений (например, замен) в остатки каркасной области исходного антитела, где это приводит к улучшению аффинности связывания мутанта антитела с антигеном из второго вида млекопитающих. Примеры остатков каркасной области для модификации включают остатки, которые нековалентно связывают антиген прямо (Amit et al. (1986) Science 233: 747-753); взаимодействуют с конформацией CDR /влияют на нее (Chothia et al. (1987) J. Mol. Biol 196: 901-917); и/или участвуют в границе контакта VL-VH (EP 239400 B1). В определенных вариантах осуществления модификация одного или нескольких таких остатков каркасной области приводит к усилению аффинности связывания антитела с антигеном второго вида млекопитающих. Например, в этом варианте осуществления изобретения можно изменять от приблизительно одного до приблизительно пяти остатков каркасной области. Иногда, для получения мутанта антитела, пригодного для применения в доклинических испытаниях, может быть достаточным, чтобы даже ни один из остатков гипервариабельной области не изменяли. Однако в норме мутант антитела содержит дополнительное изменение(я) гипервариабельной области.
Остатки гипервариабельной области, которые изменены, можно менять случайным образом, особенно когда первоначальная аффинность связывания исходного антитела является такой, что такие полученные случайным образом мутанты антитела можно легко подвергать скринингу.
Один пригодный способ для получения таких мутантов антител называют "сканирующим аланином мутагенезом" (Cunningham and Wells (1989) Science 244: 1081-1085). Здесь, один или несколько остаток(ов) гипервариабельной области заменяют аланином или полиаланиновым остатком(ами) для обеспечения взаимодействия аминокислот с антигеном из второго вида млекопитающих. Тот остаток(и) гипервариабельной области, который демонстрирует функциональную чувствительность к заменам, затем можно выявлять внесением другой или других мутаций в эти участки или вместо этих участков для замены. Таким образом, хотя участок внесения вариации аминокислотной последовательности является заданным, тип мутации сам по себе не должен быть заданным. Полученные этим способом мутанты по ala подвергают скринингу в отношении их биологической активности, как описано в настоящем документе.
Обычно начинают с консервативных замен, таких как замены, представленные ниже под заголовком "предпочтительные замены". Если такие замены приводят к изменению биологической активности (например, аффинности связывания), тогда вносят более существенные изменения, обозначаемые "иллюстративными заменами" в представленной ниже таблице, или как дополнительно описано ниже в отношении классов аминокислот, и продукты подвергают скринингу. Предпочтительные замены приведены в таблице ниже.
Еще более существенные модификации биологических свойств антител проводят, выбирая замены, которые значительно отличаются по их эффекту на поддержание (a) структуры полипептидного остова в области замены, например, такой как конформация в виде слоя или спирали, (b) заряда или гидрофобности молекулы в участке-мишени или (c) объема боковой цепи. Встречающиеся в природе остатки можно разделить на группы на основе общих свойств их боковых цепей:
гидрофобные: норлейцин, met, ala, val, leu, ile;
нейтральные гидрофильные: cys, ser, thr, asn, gln;
кислые: asp, glu;
основные: his, lys, arg;
остатки, которые влияют на ориентацию цепи: gly, pro; и
ароматические: trp, tyr, phe.
Неконсервативные замены вовлекают замену члена одного из этих классов членом из другого класса.
В другом варианте осуществления участки, выбранные для модификации, подвергают созреванию аффинности с использованием фагового дисплея (см. выше).
Молекулы нуклеиновых кислот, кодирующие мутанты по аминокислотной последовательности, получают различными способами, известными в данной области. Эти способы включают, но не ограничиваются ими, олигонуклеотид-опосредуемый (или сайт-направленный) мутагенез, мутагенез посредством ПЦР и кассетный мутагенез ранее полученного мутанта или немутантного варианта исходного антитела. Предпочтительным способом получения мутантов является сайт-направленный мутагенез (см., например, Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488).
В определенных вариантах осуществления мутант антитела имеет только один замененный остаток гипервариабельной области. В других вариантах осуществления заменены два или более из остатков гипервариабельной области исходного антитела, например от приблизительно двух до приблизительно десяти замен в гипервариабельной области.
Обычно мутант антитела с улучшенными биологическими свойствами имеет аминокислотную последовательность, имеющую по меньшей мере 75% идентичность или сходство аминокислотной последовательности с аминокислотной последовательностью вариабельного домена либо тяжелой, либо легкой цепи исходного антитела, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%. Идентичность или сходство в отношении этой последовательности определяют в настоящем документе, как процент аминокислотных остатков в последовательности-кандидате, которые идентичны (т.е. являются такими же) или сходны (т.е. представляют собой аминокислотный остаток из той же группы на основе общих свойств боковых цепей, см. выше) с остатками исходного антитела, после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Ни одну из N-концевых, C-концевых или внутренних удлинений, делеций или вставок в последовательность антитела вне вариабельного домена не следует считать влияющей на идентичность или сходство последовательностей.
После продукции мутанта антитела определяют биологическую активность этой молекулы относительно исходного антитела. Как указано выше, это может вовлекать определение аффинности связывания и/или других видов биологической активности антитела. В предпочтительном варианте осуществления изобретения получают панель мутантов антитела и проводят их скрининг в отношении аффинности связывания с антигеном, таким как C3b или его фрагмент. Один или несколько из мутантов антител, выбранных при этом первичном скрининге, необязательно подвергают одному или нескольким дополнительным анализам биологической активности для подтверждения того, что мутант(ы) антитела с повышенной аффинностью связывания действительно пригодны, например, для доклинических испытаний.
Мутант(ы) антитела, отобранные таким образом, можно подвергать дополнительным модификациям, часто в зависимости от предполагаемого применения антитела. Такие модификации могут вовлекать дополнительное изменение аминокислотной последовательности, слияние с гетерологичным полипептидом(ами) и/или ковалентные модификации, такие как модификации, представленные ниже. В отношении изменений аминокислотной последовательности иллюстративные модификации приведены выше. Например, любой остаток цистеина, не вовлеченный в поддержание надлежащей конформации мутанта антитела, также можно заменять, главным образом, серином, для повышения окислительной стабильности молекулы и для предотвращения неправильного поперечного сшивания. Напротив, цистеиновый мостик(и) можно добавлять в антитело для повышения его стабильности (в частности, где антитело представляет собой фрагмент антитела, такой как Fv-фрагмент). Другой тип с мутантными аминокислотами имеет измененный паттерн гликозилирования. Этого можно достигать удалением одной или нескольких углеводных групп, находящихся в антителе, и/или добавлением одного или нескольких участков гликозилирования, которые отсутствуют в антителе. Гликозилирование антител и других полипептидов, как правило, является либо N-связанным, либо O-связанным. N-связанное гликозилирование относится к присоединению углеводной группы к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин, где X представляет собой любую аминокислоту, за исключением пролина, представляют собой последовательности, распознаваемые для ферментативного присоединения углеводной группы к боковой цепи аспарагина. Таким образом, наличие любой из этих трипептидных последовательностей в полипептиде создает потенциальный участок гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, обычно, к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин. Добавление участков гликозилирования можно проводить, изменяя аминокислотную последовательность, чтобы она содержала одну или несколько из описанных выше трипептидных последовательностей (для участков N-связанного гликозилирования). Также изменение можно проводить посредством добавления к последовательности или замены посредством одного или нескольких остатков серина или треонина в последовательности исходного антитела (для участков O-связанного гликозилирования).
Дополнительные детали получения, селекции, обогащения и созревания аффинности антител против C3b путем фагового дисплея представлены в разделе "Примеры", ниже.
Рекомбинантная продукция антител против C3b
Антитела против C3b по изобретению можно получать рекомбинантными способами, с использованием легко получаемых технологий и материалов.
Для рекомбинантной продукции антитела против C3b нуклеиновую кислоту, кодирующую его, выделяют и встраивают в реплицируемый вектор для последующего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую моноклональное антитело, легко выделяют и секвенируют с использованием общепринятых способов (например, с использованием олигонуклеотидных зондов, которые способны специфично связываться с генами, кодирующими тяжелую и легкую цепи антитела). Доступно множество векторов. Компоненты векторов, как правило, включают, но не ограничиваются ими, один или несколько из следующих компонентов: сигнальная последовательность, ориджин репликации, один или несколько генов селективных маркеров, энхансерный элемент, промотор и последовательность терминации транскрипции.
(i) Компонент - сигнальная последовательность
Антитело по изобретению можно получать рекомбинантными способами не только прямо, но также в качестве полипептида, слитого с гетерологичным полипептидом, который предпочтительно представляет собой сигнальную последовательность или другой полипептид, обладающий участком для специфичного расщепления на N-конце зрелого белка или полипептида. Выбранная гетерологичная сигнальная последовательность предпочтительно представляет собой последовательность, которая распознается и преобразуется (т.е. расщепляется сигнальной пептидазой) в клетке-хозяине. В случае прокариотических клеток-хозяев, которые не распознают и не преобразуют нативную сигнальную последовательность антитела, сигнальную последовательность можно заменять прокариотической сигнальной последовательностью, выбранной, например, из группы из лидерных последовательностей щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II. В случае секреции в дрожжах нативную сигнальную последовательность можно заменять, например, лидерной последовательностью инвертазы дрожжей, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора Saccharomyces и Kluyveromyces) или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы C. albicans или сигнальной последовательностью, описанной в WO 90/13646. Для экспрессии в клетках млекопитающих доступны сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например сигнальная последовательность gD вируса простого герпеса. ДНК такого участка-предшественника лигируют в рамке считывания с ДНК, кодирующей антитело.
(ii) Компонент - ориджин репликации
Как экспрессирующие, так и клонирующие векторы содержат последовательность нуклеиновой кислоты, которая обеспечивает репликацию вектора в одной или нескольких выбранных клетках-хозяевах. Как правило, в векторах для клонирования эта последовательность представляет собой последовательность, которая обеспечивает репликацию вируса независимо от хромосомной ДНК хозяина, и включает ориджины репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для множества бактерий, дрожжей и вирусов. Ориджин репликации из плазмиды pBR322 является пригодным для большинства грамотрицательных бактерий, плазмидный ориджин 2µ пригоден для дрожжей, и различные вирусные ориджины пригодны для клонирования векторов в клетках млекопитающих. Как правило, для экспрессирующих векторов млекопитающих компонент, представляющий собой ориджин репликации, не требуется (ориджин SV40, главным образом, можно использовать только потому, что он содержит ранний промотор).
(iii) Компонент - селективный ген
Экспрессирующие и клонирующие векторы могут содержать ген для селекции, также называемый селективным маркером. Типичные гены для селекции кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефекты ауксотрофов или (c) обеспечивают основные питательные вещества, которые не доступны из комплексных сред, например ген, кодирующий D-аланинрацемазу для Bacilli.
В одном примере схемы селекции используют лекарственное средство для остановки роста клетки-хозяина. Клетки, которые успешно трансформированы гетерологичным геном, продуцируют белок, придающий устойчивость к лекарственному средству, и, таким образом, выживают после режима селекции. Примерами такого доминантного вида селекции является применение лекарственных средств метотрексата, неомицина, гистидинола, пуромицина, микофеноловой кислоты и гигромицина.
Другим примером пригодных селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентификацию клеток, способных захватывать нуклеиновую кислоту, кодирующую антитело, такие как гены DHFR, тимидинкинизы, металлотеонеина-I и -II, предпочтительно гены металлотеонеина приматов, аденозиндезаминазы, орнитиндекарбоксилазы и т.д.
Например, клетки, трансформированные геном для селекции DHFR, сначала идентифицируют культивированием всех трансформантов в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. При использовании DHFR дикого типа пригодные клетки-хозяева представляют собой клеточную линию яичника китайского хомяка (CHO), дефицитную по активности DHFR (например, ATCC CRL-9096).
Альтернативно, клетки-хозяева (в частности, хозяева дикого типа, которые содержат эндогенный DHFR), трансформированные или совместно трансформированные с последовательностями ДНК, кодирующими антитело по изобретению, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), можно отбирать выращиванием клеток на среде, содержащей вещество для селекции по селективному маркеру, такое как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США No. 4965199.
Пригодным геном для селекции для применения в дрожжах является ген trp1, находящийся в дрожжевой плазмиде YRp7 (Stinchcomb et al, Nature (1979) 282:39). Ген trp1 представляет собой селективный маркер для мутантного штамма дрожжей, у которого отсутствует способность расти в отсутствии триптофана, например ATCC No. 44076 или PEP4-1. Jones (1977), Genetics, 85: 12. Затем наличие повреждения trp1 в геноме дрожжевой клетки-хозяина обеспечивает эффективные условия для определения трансформации по росту в отсутствие триптофана. Аналогично, дефицитные по Leu2 штаммы дрожжей (ATCC 20622 или 38626) комплементируют известными плазмидами, обладающими геном Leu2.
Кроме того, векторы, полученные из кольцевой плазмиды pKD1 1,6 мкм, можно использовать для трансформации дрожжей Kluyveromyces. Альтернативно описаны экспрессирующие системы для крупномасштабной продукции рекомбинантного химозина телят для K. lactis. Van den Berg (1990) Bio/Technology, 8: 135. Также описаны стабильные мультикопийные экспрессирующие векторы для секреции зрелого рекомбинантного сывороточного альбумина человека посредством промышленных штаммов Kluyveromyces. Fleer et al. (1991) Bio/Technology, 9: 968-975.
(iv) Компонент - промотор
Экспрессирующие и клонирующие векторы, как правило, содержат промотор, который распознается организмом-хозяином и функционально связан с нуклеиновой кислотой, кодирующей антитело. Промоторы, пригодные для применения в прокариотических хозяевах, включают промотор phoA, промоторные системы β-лактамазы и лактозы, промотор щелочной фосфатазы, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако пригодными являются другие бактериальные промоторы. Промоторы для применения в бактериальных системах также содержат последовательность Шайна-Дальгарно (SD), функционально связанную с ДНК, кодирующей антитело по изобретению.
Известны промоторные последовательности для эукариот. Практически все эукариотические гены обладают AT-богатой областью, расположенной приблизительно от 25 до 30 оснований выше участка инициации транскрипции. Другая последовательность, находящаяся от 70 до 80 оснований выше точки начала транскрипции множества генов, представляет собой участок CNCAAT, где N может представлять собой любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может представлять собой сигнал для добавления поли-A хвоста к 3'-концу кодирующей последовательности. Все из этих последовательностей можно встраивать в эукариотические экспрессирующие векторы.
Примеры пригодных промоторных последовательностей для применения в дрожжевых хозяевах включают промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, таких как енолаза, глицеральдегид-3-фосфат дегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкоза-6-фосфатизомераза, 3-фосфоглицеромутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкоизомераза и глюкокиназа.
Другие дрожжевые промоторы, которые представляют собой индуцибельные промоторы, обладающие дополнительным преимуществом контролируемой условиями выращивания транскрипции, представляют собой промоторные участки алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, ферментов деградации, связанных с метаболизмом азота, металлотеонеина, глицеральдегид-3-фосфат дегидрогеназы и ферментов, ответственных за употребление мальтозы и галактозы. Кроме того, пригодные векторы и промоторы для применения для экспрессии в дрожжах описаны в EP 73657. Также предпочтительно с дрожжевыми промоторами используют дрожжевые энхансеры.
Транскрипция антитела с векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами, получаемыми из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус (такой, как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита-B и наиболее предпочтительно вирус обезьяны 40 (SV40), гетерологичными промоторами млекопитающих, например промотором актина или промотором иммуноглобулина, промоторами теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Ранние и поздние промоторы вируса SV40 обычно получают в качестве рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Предранний промотор цитомегаловируса человека обычно получают в качестве рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в хозяевах, относящихся к млекопитающим, с использованием вируса папилломы крупного рогатого скота в качестве вектора описана в патенте США No. 4419446. Модификация этой системы описана в патенте США No. 4601978. См. также Reyes et al. (1982) Nature 297:598-601 в отношении экспрессии кДНК β-интерферона человека в клетках мышей под контролем промотора тимидинкиназы из вируса простого герпеса. Альтернативно в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
(v) Компонент - энхансерный элемент
Транскрипцию ДНК, кодирующей антитело по изобретению, у высших эукариот часто усиливают посредством встраивания в вектор энхансерной последовательности. Известно много энхансерных последовательностей из генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако, как правило, используют энхансер из вируса эукариотической клетки. Их примеры включают энхансер SV40 на поздней стороне ориджина репликации (п.о. 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне ориджина репликации и энхансеры аденовируса. См. также Yaniv, Nature 297: 17-18 (1982) в отношении энхансеров для активации эукариотических промоторов. Энхансер может быть присоединен в векторе к кодирующей антитело последовательности в положении 5' или 3', однако предпочтительно он расположен в участке, расположенном в 5'-направлении от промотора.
(vi) Компонент для терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (в клетках дрожжей, грибов, насекомых, растений, животных, человека или содержащих ядро клетках из других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно находятся на 5'- и, иногда, на 3'-нетранслируемых участках эукариотических или вирусных ДНК или кДНК. Эти участки содержат нуклеотидные фрагменты, транскрибируемые в качестве полиадинилированных фрагментов в нетранслируемом участке мРНК, кодирующей антитело. Одним пригодным компонентом для терминации транскрипции является участок полиаденилирования бычьего гормона роста. См. WO 94/11026 и описанный в ней экспрессирующий вектор.
(vii) Селекция и трансформация клеток-хозяев.
Пригодные клетки-хозяева для клонирования или экспрессии ДНК в векторах, представленных в настоящем документе, представляют собой клетки прокариот, дрожжей или высших эукариот, описанных выше. Пригодные для этой цели прокариоты включают эубактерии, такие как грамотрицательные или грамположительные организмы, например Enterobacteriaceae, такие как Escherichia, например E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанные в DD 266710, опубликованной 12 апреля 1989 года), Pseudomonas, такие как P. aeruginosa и Streptomyces. Одним предпочтительным хозяином E. coli для клонирования является E. coli 294 (ATCC 31446), хотя также пригодными являются другие штаммы, такие как E. coli B, E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Эти примеры являются иллюстративными, но не ограничивающими.
В дополнение к прокариотам пригодными хозяевами для клонирования или экспрессии кодирующих антитело векторов являются эукариотические микроорганизмы, такие как нитевидные грибы или дрожжи. Saccharomyces cerevisiae, или обычные пекарские дрожжи, наиболее распространены среди низших эукариотических микроорганизмов-хозяев. Однако широко доступным и пригодным здесь является ряд других родов, видов и штаммов, таких как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K. lactis, K.fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитевидные грибы, такие как, например, хозяева Neurospora, Penicillium, Tolypocladium и Aspergillus, такие как A. nidulans и A. niger.
Пригодные клетки-хозяева для экспрессии гликозилированного антитела получают из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки растений и насекомых. Выявлен ряд штаммов и вариантов бакуловирусов и соответствующих пермиссивных клеток-хозяев среди хозяев, таких как Spodoptera frugiperda (гусеница), Aedes aegypti (москит), Aedes albopictus (москит), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Множество штаммов вирусов для трансфекции являются широко доступными, например вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы можно использовать здесь в качестве вирусов в соответствии с настоящим изобретением, в частности, для трансфекции клеток Spodoptera frugiperda. Также в качестве хозяев можно использовать растительные клеточные культуры хлопка, кукурузы, картофеля, сои, петунии, томата и табака, ряски и другие клетки растений.
Однако наибольший интерес представляют клетки позвоночных, и размножение клеток позвоночных в культуре (культуре тканей) стало общепринятым способом. Примерами пригодных клеточных линий млекопитающих являются линия почки обезьяны CV1, трансформированная SV40 (COS-7, ATCC CRL 1651); линия почки эмбриона человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol. 36:59 (1977)); клетки почки детенышей хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы буффало (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al, Annals N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC 5; клетки FS4; и линия гепатомы человека (Hep G2).
Для продукции антитела клетки-хозяева трансформируют описанными выше векторами для экспрессии и клонирования и для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих требуемые последовательности, их культивируют в общепринятой среде с соответствующими питательными веществами.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продукции антитела по изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев пригодными являются коммерчески доступные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда ((MEM), (Sigma)), RPMI-1640 (Sigma), среда Игла, модифицированная способом Дульбекко, ((DMEM), Sigma). Кроме того, для клеток-хозяев в качестве культуральной среды можно использовать любую из сред, описанных в Ham et al. (1979) Meth. Enz. 58:44, Barnes et al. (1980) Anal Biochem. 102:255, патентах США No. 4767704; 4657866; 4927762; 4560655 или 5122469; WO 9003430; WO 87/00195; или патенте США Re. 30985. Любые из этих сред можно дополнить при необходимости гормонами и/или другими факторами роста (такими, как инсулин, трансферрин или эпидермальный фактор роста), солями (такими, как хлорид натрия, кальция, магния и фосфат), буферами (такими, как HEPES), нуклеотидами (такими, как аденозин и тимидин), антибиотиками (такими, как лекарственное средство GENTAMYCIN™), микроэлементами (определяемыми как неорганические соединения, обычно представленные в конечных концентрациях микромолярного диапазона) и глюкозой или эквивалентными источниками энергии. Также можно добавлять любые другие необходимые дополнительные вещества в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, которые ранее использовали для выбранных для экспрессии клеток-хозяев, и они будут понятны специалисту в данной области.
(ix) Очистка антитела
При использовании рекомбинантных способов антитело может продуцироваться внутриклеточно, в периплазматическом пространстве, или непосредственно секретироваться в среду. Если такие молекулы продуцируются внутриклеточно, на первой стадии, дебрис в виде частиц, либо клеток-хозяев, либо лизированных фрагментов удаляют, например, посредством центрифугирования или ультрафильтрации. В Carter et al. (1992) Bio/Technology 10:163-167 описан способ выделения антител, которые секретируются в периплазматическое пространство E. coli. В кратком изложении клеточную массу размораживают в присутствии ацетата натрия (pH 3,5), EDTA и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 мин. Клеточный дебрис можно удалять центрифугированием. Когда антитело секретируется в среду, супернатанты из таких экспрессирующих систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например элемента для ультрафильтрации Amicon или Millipore Pellicon. На любой из предшествующих стадий можно включать ингибитор протеаз, такой как PMSF, для ингибирования протеолиза и для предотвращения роста ненужных контаминирующих организмов можно добавлять антибиотики.
Очистку антитела можно проводить с использованием, например, хроматографии с гидроксилапатитом, гель-электрофореза, диализа и аффинной хроматографии, причем аффинная хроматография является предпочтительным способом очистки. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, который присутствует в антителе. Белок A можно использовать для очистки антител, которые основаны на тяжелых цепях γ1, γ2 или γ4 человека (Lindmark et al. (1983), J. Immunol. Meth. 62: 1-13). Белок G рекомендован для всех изотипов мыши и для γ3 человека (Guss et al. (1986), EMBO J. 5: 15671575). Матрицей, к которой присоединяют аффинный лиганд, наиболее часто является агароза, однако доступны другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, позволяют более быстрые скорости потока и более короткое время обработки, чем скорости потока и время обработки, которые можно достигать посредством агарозы. Когда антитело содержит CH3-домен, для очистки пригодна смола Bakerbond ABXTM (J. T. Baker, Phillipsburg, NJ). Также доступны другие способы очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на heparin-SEPHAROSETM, хроматография на анионо- или катионообменной смоле (такая, как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония, в зависимости от подлежащего выделению антитела.
После предварительной стадии(ий) очистки смесь, содержащую представляющее интерес антитело и примеси, можно подвергать хроматографии гидрофобного взаимодействия с низким значением pH с использованием буфера для элюирования при pH между приблизительно 2,5-4,5, предпочтительно проводимой при низких концентрациях соли (например, от приблизительно 0-0,25 M соли).
Скрининговые анализы и модели на животных для идентификации антител против C3b и других антагонистов C3b
Антитела против C3b и другие антагонисты C3b можно оценивать во множестве анализов in vitro и in vivo в отношении их способности селективно ингибировать альтернативный каскад комплемента и предупреждать и лечить ассоциированные с комплементом нарушения.
Анализы in vitro, такие как анализы связывания и конкурентного связывания, анализы гемолиза, описаны в разделе "Примеры".
Терапевтическую активность in vivo антагонистов C3b, таких как антитела против C3b, описанные в настоящем документе, можно тестировать в соответствующих моделях на животных. Таким образом, например, можно создавать способами инженерии модели на рекомбинантных (трансгенных) животных путем введения кодирующей части представляющих интерес генов в геном представляющих интерес животных, с использованием стандартных способов получения трансгенных животных. Животные, которые могут служить мишенью для трансгенных манипуляций, включают, но не ограничиваются ими, мышей, крыс, кроликов, морских свинок, овец, коз, свиней и не являющихся человеком приматов, например бабуинов, шимпанзе и других обезьян. Известные в данной области способы введения трансгена таким животным включают микроинъекцию в пронуклеус (Hoppe and Wanger, патент США No. 4873191); опосредуемый ретровирусом перенос гена в зародышевые линии (например, Van der Putten et al., Proc. Natl. Acad. Sci. 82 USA 6148-615 [1985]); генное нацеливание в эмбриональных стволовых клетках (Thompson et al., Cell. 56, 313-321 [1989]); электропорацию эмбрионов (Lo, Mol Cell Biol. 3, 1803-1814 [1983]); опосредуемый семенной жидкостью перенос (Lavitrano et al., Cell 57, 717-73 [1989]). Для обзора см., например, патент США No. 4736866.
Для целей настоящего изобретения трансгенные животные включают животных, которые несут трансген только в части их клеток ("мозаичные животные"). Трансген можно встраивать либо в качестве отдельного трансгена, либо, в конкатамерах, например, повторами типа "голова к голове" или "голова к хвосту". Селективное введение трансгена в конкретный тип клеток также возможно, например, в соответствии со способом Lasko et al., Proc. Natl. Acad. Sci. USA 89, 623-636 (1992). Мониторинг экспрессии трансгена у трансгенных животных можно проводить стандартными способами. Например, для подтверждения встраивания трансгена можно использовать саузерн-блот анализ или амплификацию путем ПЦР. Затем уровень экспрессии мРНК можно анализировать с использованием способов, таких как гибридизация in situ, нозерн-блот анализ, ПЦР или иммуноцитохимия.
Кроме того, животных можно исследовать в отношении признаков патологии иммунного заболевания, например путем гистологического исследования, для определения инфильтрации иммунных клеток в определенные ткани.
Модели на рекомбинантных (трансгенных) животных можно создавать путем введения кодирующей части представляющих интерес генов в геном представляющих интерес животных, с использованием стандартных способов создания трансгенных животных. Животные, которые могут служить мишенью для трансгенных манипуляций, включают, но не ограничиваются ими, мышей, крыс, кроликов, морских свинок, овец, коз, свиней и не являющихся человеком приматов, например бабуинов, шимпанзе и других обезьян. Известные в данной области способы введения трансгена таким животным включают микроинъекцию в пронуклеус (Hoppe and Wanger, патент США No. 4873191); опосредуемый ретровирусом перенос гена в зародышевые линии (например, Van der Putten et al., Proc. Natl. Acad. Sci. 82 USA 6148-615 [1985]); генное нацеливание в эмбриональных стволовых клетках (Thompson et al., Cell. 56, 313-321 [1989]); электропорацию эмбрионов (Lo, Mol Cell Biol. 3, 1803-1814 [1983]); опосредуемый семенной жидкостью перенос (Lavitrano et al., Cell 57, 717-73 [1989]). Для обзора см., например, патент США No. 4736866.
Эффективность для профилактики и/или лечения артрита можно оценивать, например, в модели индуцируемого коллагеном артрита (Terato et al. Brit. J. Rheum. 35:828-838 (1966)). Потенциальные профилактические/терапевтические средства против артрита также можно подвергать скринингу в модели опосредуемого антителом артрита, индуцируемого внутривенной инъекцией коктейля из четырех моноклональных антител, как описано Terato et al., J. Immunol. 148:2103-8 (1992), и Terato et al., Autoimmunity 22: 137-47 (1995). Кандидаты в средства для профилактики и/или лечения артрита также можно исследовать в моделях на трансгенных животных, например, таких как трансгенные по TNF-α мыши (Taconic). Эти животные экспрессируют фактор некроза опухоли человека (TNF-α), цитокин, который вовлечен в патогенез ревматоидного артрита человека. Экспрессия TNF-α у этих мышей приводит к тяжелому хроническому артриту передних и задних лап и обеспечивает простую модель воспалительного артрита на мышах.
В последние годы также были разработаны модели псориаза на животных. Таким образом, Asebia (ab), "шершавая кожа" (fsn) и "хронический пролиферативный дерматит" (cpd) являются спонтанными мутациями у мышей с подобными псориазу изменениями кожи. Также для исследования in vivo псориаза и идентификации лекарственных средств для лечения псориаза можно использовать трансгенных мышей со сверхэкспрессией в коже цитокинов, таких как интерферон-γ, интерлейкин-1α, фактор роста кератиноцитов, трансформирующий фактор роста-α, интерферон-6, сосудисто-эндотелиальный фактор роста или белок 6 морфогенеза кости. Также подобные псориазу очаги повреждения были описаны у мышей с гипоморфным β2-интегрином, подвергнутых возвратному скрещиванию с линией PL/J, и у трансгенных по β1-интегрину мышей, мышей scid/scid, восстановленных с помощью CD4+/CD45RBhi T-лимфоцитов, а также у трансгенных крыс HLA-B27/hβ2m. Также известны модели с ксенотрансплантацией с использованием кожи человека, пересаженной иммунодефицитным мышам. Таким образом, антитела и другие антагонисты C3b по изобретению можно тестировать в модели на мышах scid/scid, описанной Schon, M. P. et al., Nat. Med. (1997) 3: 183, при которых у мышей выявляют гистопатологические повреждения кожи, сходные с псориазом. Другой пригодной моделью является химера кожа человека/мышь scid, полученная, как описано Nickoloff, B. J. et al., Am. J. Path. (1995) 146:580. Для дополнительных деталей см., например, Schon, M.P., J Invest Dermatology 112:405-410 (1999).
Описана модель астмы, в которой индуцируемую антигеном гиперреактивность дыхательных путей, эозинофилию и воспаление легких индуцируют сенсибилизацией животного овальбумином, а затем нагрузкой животного тем же белком, доставляемым с помощью аэрозоля. Несколько моделей на животных (морская свинка, крыса, не являющийся человеком примат) демонстрируют симптомы, сходные с атопической астмой у человека при нагрузке антигенами в виде аэрозоля. Модели на мышах имеют многие признаки астмы человека. Пригодные способы тестирования агонистов CRIg и CRIg в отношении активности и эффективности при лечении астмы описаны Wolyniec, W.W. et al., Am. J. Respir. Cell Mol. Biol. (1998) 18: 777, и ссылках, цитированных в ней.
Контактная гиперчувствительность представляет собой простой анализ in vivo клеточноопосредуемой иммунной функции. В этом способе эпидермальные клетки подвергают воздействию экзогенных гаптенов, которые приводят к реакции гиперчувствительности замедленного типа, которую выявляют и количественно определяют. Контактная чувствительность вовлекает исходную фазу сенсибилизации с последующей фазой оценки. Фазу оценки проводят, когда эпидермальные клетки встречают антиген, с которым они ранее имели контакт. Происходит набухание и воспаление, что делает ее превосходной моделью аллергического контактного дерматита человека. Пригодный способ подробно описан в Current Protocols in Immunology, Eds. J. E. Cologan, A. M. Kruisbeek, D. 11. Margulies, E. M. Shevach and W. Strober, John Wiley & Sons, Inc., 1994, раздел 4.2. См. также Grabbe, S. and Schwarz, T, Immun. Today 19 (1): 37-44 (1998).
Реакция "трансплантат против хозяина" происходит, когда иммунокомпетентные клетки трансплантируют пациентам с иммуносупрессией или иммунной толерантностью. Донорные клетки распознают и отвечают на антигены хозяина. Ответ может варьировать от угрожающего жизни тяжелого воспаления до мягких случаев диареи и снижения массы тела. Модели реакции "трансплантат против хозяина" обеспечивают способ оценки реактивности T-клеток против антигенов MHC и минорных антигенов трансплантата. Пригодный способ подробно описан в Current Protocols in Immunology, выше, раздел 4.3.
Модели на животных для отторжения аллотрансплантата кожи представляют собой средство для тестирования способности T-клеток опосредовать разрушение тканей in vivo, что указывает на их роль в противовирусном и противоопухолевом иммунитете и является его мерой. В наиболее распространенных и признанных моделях используют трансплантаты кожи хвоста мыши. Неоднократные эксперименты показали, что отторжение аллотрансплантата кожи опосредуется T-клетками, хелперными T-клетками и киллерными-эффекторными T-клетками, но не антителами. Auchincloss, H. Jr. and Sachs, D. H., Fundamental Immunology, 2nd ed., W. E. Paul ed., Raven Press, NY, 1989, 889-992. Пригодный способ подробно описан в Current Protocols in Immunology, выше, раздел 4.4. Другие модели отторжения трансплантата, которые можно использовать для тестирования агонистов CRIg и CRIg, представляют собой модели аллогенного трансплантата сердца, описанные Tanabe, M. et al., Transplantation (1994) 58:23 и Tinubu, S. A. et al., J. Immunol. (1994) 4330-4338.
Модели на животных для гиперчувствительности замедленного типа также обеспечивают анализ клеточноопосредуемой иммунной функции. Реакции гиперчувствительности замедленного типа представляют собой опосредуемый T-клетками иммунный ответ in vivo, характеризующийся воспалением, которое не достигает пика до тех пор, пока не пройдет период времени после нагрузки антигеном. Эти реакции также происходят при тканеспецифических аутоиммунных заболеваниях, таких как рассеянный склероз (MS) и экспериментальный аутоиммунный энцефаломиелит (EAE, модель для MS). Пригодный способ подробно описан в Current Protocols in Immunology, выше, раздел 4.5.
EAE представляет собой опосредуемое T-клетками аутоиммунное заболевание, характеризующееся воспалением, связанным с T-клетками и мононуклеарными клетками, и последующей демиелинизацией аксонов в центральной нервной системе. EAE, как правило, считают подходящей моделью на животных для MS у человека. Bolton, C, Multiple Sclerosis (1995) 1: 143. Были разработаны как острые, так и ремитирующие модели. CRIg и его агонисты и антагонисты можно тестировать в отношении стимулирующей или ингибирующей T-клетки активности против иммуноопосредуемого демиелинизирующего заболевания с использованием протокола, описанного в Current Protocols in Immunology, выше, разделы 15.1 и 15.2. Также см. модели для связанного с миелином заболевания, при котором олигодендроциты или шванновские клетки пересаживают в центральную нервную систему, как описано в Duncan, I. D. et al., Molec. Med. Today (1997) 554-561.
Модели ишемии-реперфузии миокарда можно выполнять у мышей или крыс. Животным проводят трахеостомию и их вентилируют вентилятором для небольших животных. Во внутреннюю сонную артерию и яремную вену помещают полиэтиленовые катетеры для измерения среднего артериального кровяного давления. Ишемическую реперфузию миокарда начинают лигированием левой передней нисходящей артерии (LAD) швом 6-O. Ишемии достигают обратимым стягиванием лигатуры вокруг LAD до полной окклюзии сосуда. Через 30 мин лигатуру удаляют и сердце подвергают перфузии в течение 4 часов. Агонисты CRIg и CRIg можно тестировать в отношении их эффективности измерением размера инфаркта сердца, активности сердечных креатинкиназ, активности миелопероксидазы и иммуногистохимией с использованием антител против C3.
Модель диабетической ретинопатии вовлекает введение мышам или крысам стрептозотоцина. Агонисты CRIg и CRIg можно тестировать в отношении их эффекта на расширение венул, микрососудистые нарушения в сетчатке и неоваскуляризацию сетчатки и полости стекловидного тела.
Модель мембранопролиферативного гломерулонефрита можно получать следующим образом: самок мышей иммунизируют внутрибрюшинно 0,5 мг контрольного IgG кролика в CFA (-7 сутки). Через семь суток (0 сутки) 1 мг антитела кролика против базальной мембраны мыши (GBM) инъецируют внутривенно через хвостовую вену. Повышение антитела против IgG кролика в сыворотке измеряют с помощью ELISA. Образцы мочи мышей через 24 ч собирают в метаболических клетках и оценивают почечную функцию мыши измерением белка в моче, а также азота мочевины в крови.
Модель на животных для связанной со старением дегенерации желтого пятна (AMD) включает мышей с нуль-мутацией в генах Ccl-2 или Ccr-2. У этих мышей развиваются основные признаки AMD, включая накопление липофусцина в пигментном эпителии сетчатки (RPE) и друзы под ним, атрофию фоторецепторов и хориоидальную неоваскуляризацию (CNV). Эти признаки развиваются в возрасте более 6 месяцев. Агонисты CRIg и CRIg можно тестировать в отношении образования друз, атрофии фоторецепторов и хориоидальной неоваскуляризации.
CNV можно тестировать в различных моделях индуцируемой лазером хориоидальной неоваскуляризации. Таким образом, например, CNV можно индуцировать у крыс и яванских макаков интенсивной фотокоагуляцией лазером, которая приводит к хориоидальной неоваскуляризации. Прогрессирование и лечение этого состояния можно оценивать, например, посредством ангиографии с флуоресцеином, гистопатологической и иммуногистохимической оценки и путем анализа фармакокинетики, гемолиза, скринингового анализа антитела и анализа активации комплемента в сыворотке, полученной от животных до и после лечения, с различными промежутками времени. Мониторинг эффективности профилактического введения можно проводить сходными способами, включая мониторинг сосудистой проницаемости ангиографией с флуоресцеином, ингибирование накопления комплемента в области ожога лазером, обследованием глаза, фотографированием глаза, сбором тканей стекловидного тела и сетчатки и т.п. Дополнительные детали представлены в разделе "Примеры", ниже.
Способы лечения
Для профилактики, лечения или снижения тяжести связанного с комплементом нарушения пригодная дозировка соединения по изобретению зависит от типа подлежащего лечению нарушения, как определено выше, тяжести и течения нарушения, введения средства в профилактических или терапевтических целях, предшествующего лечения, клинического анамнеза пациента и ответа на соединение и решения лечащего врача. Соединение пригодным образом вводят пациенту за один раз или в ходе серии введений. Предпочтительно, перед тестированием у человека желательно определить кривую доза-эффект и фармацевтическую композицию по изобретению сначала in vitro, а затем в пригодных моделях на животных.
Например, в зависимости от типа и тяжести заболевания, исходной дозой-кандидатом для введения пациенту является приблизительно от 1 мкг/кг до 15 мг/кг (например, 0,1-20 мг/кг) антитела против C3b или другого антагониста C3b, например, посредством либо одного введения, либо разделенных введений, либо непрерывной инфузии. Типичная суточная дозировка может варьировать приблизительно от 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. Для повторяющихся введений через несколько суток или более, в зависимости от состояния, лечение продолжают до возникновения желательных симптомов подавления заболевания. Однако могут быть пригодными другие режимы дозирования. Мониторинг этой терапии легко проводить общепринятыми способами и анализами.
Эффективность лечения связанных с комплементом состояний глаза, таких как AMD или CNV, можно определять по различным конечным результатам, обычно используемым для оценки внутриглазных заболеваний. Например, можно оценивать снижение остроты зрения. Снижение остроты зрения можно оценивать, например, но не ограничиваясь ими, измерением среднего изменения максимально корригированной остроты зрения (BCVA) по сравнению с исходным уровнем до требуемого момента времени (например, когда BCVA основана на карте для оценки остроты зрения Early Treatment Diabetic Retinopathy Study (ETDRS) и оценке на тестируемом расстоянии 4 метра), определением доли субъектов, которые потеряли менее 15 букв в остроте зрения в требуемый момент времени по сравнению с исходным уровнем, определением доли субъектов, у которых прибавилось 15 букв или более в требуемый момент времени по сравнению с исходным уровнем, определением доли субъектов с эквивалентом остроты зрения Snellen, составляющим 20/2000 или хуже в требуемый момент времени, определением NE1 Visual Functioning Questionnaire, определением размера CNV и величины проницаемости при CNV в требуемый момент времени, например, с помощью ангиографии с флуоресцеином и т.д. Можно проводить оценку глаз, например, которая включает, но не ограничиваются ими, например, проведение обследования глаза, измерение внутриглазного давления, оценку остроты зрения, измерение давления на щелевой лампе, оценку внутриглазного воспаления и т.д.
Фармацевтические композиции
Антитела против C3b и другие антагонисты C3b по настоящему изобретению можно вводить для лечения связанных с комплементом нарушений в форме фармацевтических композиций.
Терапевтические составы антитела против C3b или другого антагониста по изобретению получают для хранения смешиванием активной молекулы, имеющей требуемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharnaceutical Sciences 16th edition, Osol, A. Ed. [1980]), в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие, как хлорид октадецилдиметилбензиламмония; хлорид гексаметэтония; хлорид бензалкония, хлорид бензэтония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как TWEEN®, PLURONICS® или полиэтиленгликоль (PEG).
Для доставки полипептида, антитела или фрагмента антитела в клетки можно использовать липофекцию или липосомы. Когда используют фрагменты антитела, предпочтительным является наименьший фрагмент, который специфично связывается со связывающим доменом белка-мишени. Например, на основе последовательностей вариабельных участков антитела можно конструировать пептидные молекулы, которые сохраняют способность связывать белковую последовательность-мишень. Такие пептиды можно синтезировать химически и/или продуцировать технологией рекомбинантных ДНК (см., например, Marasco et al., Proc. Natl. Acad. Sci. 90 USA 7889-7893 [1993]).
Активные молекулы также могут быть заключены в микрокапсулы, полученные, например способами коацервации или межфазной полимеризации, например микрокапсулы на основе гидроксиметилцеллюлозы или желатина и микрокапсулы на основе полиметилметакрилата соответственно, в коллоидные системы для доставки лекарственного средства (например, липосомы, микросферы на основе альбумина, микроэмульсии, наночастицы и нанокапсулы) или в микроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980).
Составы, подлежащие применению для введения in vivo, должны быть стерильными. Этого легко достигают фильтрацией через стерильные фильтрационные мембраны.
Можно получать препараты с замедленным высвобождением. Пригодные примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащие антитело, которые представлены в виде имеющих форму изделий, например пленок или микрокапсул. Примеры матриц с замедленным высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США No. 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочная кислота-гликолевая кислота, такие как LUPRON DEPOT® (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата леупролида), и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этилен-винилацетат и молочная кислота-гликолевая кислота, обеспечивают высвобождение молекул в течение 100 суток, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Когда инкапсулированные антитела остаются в организме длительно, они могут денатурировать или агрегировать в результате воздействия увлажнения при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Для стабилизации можно разработать целесообразные стратегии, в зависимости от вовлеченного механизма. Например, если выявлено, что механизм агрегации представляет собой внутримолекулярное образование связей S-S через тио-дисульфидный обмен, стабилизации можно достигать модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, применением соответствующих добавок и разработкой определенных композиций полимерных матриксов.
Соединения по изобретению для профилактики или лечения заболевания или состояния глаз, как правило, вводят путем глазной, внутриглазной инъекции и/или инъекции в стекловидное тело. Также можно использовать другие способы введения, которые включают, но не ограничивается ими, местное, парентеральное, подкожное, внутрибрюшинное, внутрилегочное, интраназальное и проводимое внутрь очага поражения введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение.
Составы для глазного, внутриглазного введения или введения в стекловидное тело можно получать способами и с использованием ингредиентов, известных в данной области. Главным требованием для эффективного лечения является надлежащее проникновение через глаз. В отличие от заболеваний передней части глаза, где лекарственные средства можно доставлять местно, заболевания сетчатки требуют более область-специфического подхода. Глазные капли и мази редко проникают в заднюю часть глаза, а гематоофтальмический барьер препятствует проникновению системно вводимых лекарственных средств в ткань глаза. Таким образом, как правило предпочтительным для выбора способом доставки лекарственного средства для лечения заболевания сетчатки, такого как AMD и CNV, является прямая инъекция внутрь стекловидного тела. Инъекции внутрь стекловидного тела обычно повторяют с интервалами, которые зависят от состояния пациента и от свойств и времени полужизни доставляемого лекарственного средства. Для внутриглазного проникновения (например, внутрь стекловидного тела), как правило, предпочтительными являются молекулы меньшего размера.
Следующие примеры предоставлены только для иллюстративных целей и не предназначены для ограничения объема настоящего изобретения никоим образом. Коммерчески доступные реагенты, указанные в примерах, использовали в соответствии с инструкциями изготовителя, если нет иных указаний. Источником клеток, указанных в представленных примерах и на протяжении описания под регистрационными номерами ATCC, является American Type Culture Collection, 10801 University Boulevard, Manassas, VA 201 10-2209.
Аминокислотные остатки в аминокислотных последовательностях антитела пронумерованы в соответствии с Kabat (Kabat et al., Sequences of proteins of immunological interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). Используют однобуквенные сокращения аминокислот. Вырожденность ДНК представлена с использованием кода IUB (N = A/C/G/T, D = A/G/T, V = A/C/G, B=C/G/T, H= A/C/T, K = G/T, M = A/C, R = A/G, S = G/C, W= A/T, Y = C/T).
Пример 1
Антитела, образованные из фага, кодирующего гипервариабельные участки исходного антитела
Последовательности нуклеиновых кислот VL- и VH-доменов антитела против HER2 rhuMAB 4D5-8 HERCEPTIN® (Genentech, Inc.) (фиг. 18A и 18B) использовали в качестве исходной последовательности для мутагенеза HVR и фаговой селекции в отношении связывания с C3b человека. Антитело 4D5 представляет собой гуманизированное антитело, специфичное к ассоциированному со злокачественной опухолью антигену, известному как Her-2 (erbB2). Антитело включает вариабельные домены, имеющие консенсусные каркасные области, где несколько положений были обратно заменены на последовательность мыши в процессе повышения аффинности гуманизированного антитела. Последовательность и кристаллическая структура гуманизированного антитела 4D5 описана в патенте США No. 6054297, Carter et al., PNAS 89: 4285 (1992), кристаллическая структура представлена в Carter et al., J. Mol. Biol. 229:969 (1993) и он-лайн на www/ncbi/nih/gov/structure/mmdb(MMDB#s-990-992), полное описание которых включено в настоящий документ в качестве ссылки.
VL- и VH-домены HERCEPTIN® содержат консенсусный VL-домен каппа-I человека и вариант консенсусного VH-домена человека III подгруппы. Вариант VH-домена имеет 3 замены относительно консенсусной последовательности человека: R71A, N73T и L78A.
Фагмида, использованная для этой работы, представляла собой одновалентный вектор для дисплея Fab-g3 (pV0350-2B), имеющий 2 открытые рамки считывания под контролем промотора phoA, по существу, как описано в Lee et al., J. Mol. Biol. (2004), 340(5): 1073-93. Первая открытая рамка считывания состоит из двух сигнальных последовательностей stII, слитых с VL- и CH1-доменами акцепторной легкой цепи, а вторая состоит из сигнальной последовательности stII, слитой с VH- и CH1-доменами акцепторной тяжелой цепи, за которыми следует укороченный минорный белок оболочки фага P3. См. Lee et al., выше.
Антитела, полученные мутагенезом HVR тяжелой цепи
Клон Fab YW 144.2.43 получили мутагенезом HVR-H1, H2 и H3 тяжелой цепи huMAb 4D5-8 (антитело против HER2 HERCEPTIN®, Genentech, Inc.) и селекцией против слитого белка C3b человека. В HVR-H1 постоянными оставляли положения по Kabat 26 (G), 27 (F), 28 (T), 29 (I), 34 (I) и 35 (H) и аминокислоты в положениях 30-33 варьировали. В HVR-H2 постоянными оставляли положения по Kabat 51 (I), 52a (P), 55 (G), 57 (T), 59 (Y), 60 (A), 61 (D), 62 (S), 63 (V), 64 (K) и 65 (G) и положения 49, 50, 52, 53, 54, 56 и 58 варьировали. В HVR-H3 постоянными оставляли положения по Kabat 93 (A) и 102 (Y) и положения 94-100, 100a-h и 101 варьировали. Легкая цепь YW 144.2.43 представляла собой модифицированную последовательность huMAb 4D5-8 (модифицированную в положениях 30, 66 и 91), HVR которой не изменяли в ходе фаговой селекции. В каждый гипервариабельный участок вносили разнообразие последовательностей мутагенезом выбранных аминокислотных положений с использованием стандартных способов мутагенеза.
Создание фаговых библиотек
Пулы рандомизированных олигонуклеотидов, сконструированных для каждой гипервариабельной области, фосфорилировали по отдельности в шести реакционных смесях объемом 20 мкл, содержащих 660 нг олигонуклеотида, 50 мМ Tris pH 7,5, 10 мМ MgCl2, 1 мМ ATP, 20 мМ DTT и 5 Е полинуклеотидкиназы в течение 1 ч при 37°С. Затем шесть пулов фосфорилированных олигонуклеотидов комбинировали с 20 мкг матрицы Kunkel в 50 мМ Tris pH 7,5, 10 мМ MgCl2 в конечном объеме 500 мкл, что приводило к соотношению олигонуклеотида и матрицы, составляющему 3. Смесь подвергали отжигу при 90°С в течение 4 мин, при 50°С в течение 5 мин, а затем охлаждали на льду. Избыток не подвергнутого отжигу олигонуклеотида удаляли набором для очистки продуктов ПЦР QIAQUICKTM (набор Qiagen 28106) с использованием модифицированного протокола для предотвращения чрезмерной денатурации подвергнутой отжигу ДНК. К 500 мкл подвергнутой отжигу смеси добавляли 150 мкл PB и смесь разделяли между двумя 2 колонками на диоксиде кремния. После промывания каждой колонки 750 мкл PE и дополнительного вращения для высушивания колонок каждую колонку элюировали 110 мкл 10 мМ Tris, 1 мМ EDTA, pH 8. Затем подвергнутую отжигу и очищенную матрицу (220 мкл) достраивали добавлением 1 мкл 100 мМ ATP, 10 мкл 25 мМ dNTP (по 25 мМ каждого из dATP, dCTP, dGTP и dTTP), 15 мкл 100 мМ DTT, 25 мкл 10x буфера TM (0,5 M Tris pH 7,5, 0,1 M MgCl2), 2400 Е T4-лигазы и 30 Е T7-полимеразы в течение 3 ч при комнатной температуре.
Достроенный продукт анализировали на гелях Tris-ацетат-EDTA/агароза (Sidhu et al., Methods in Enzymology 328:333-363 (2000)). Обычно были видимы три полосы: нижняя полоса представляет собой точно достроенный и лигированный продукт, средняя полоса представляет собой достроенный, но не лигированный продукт, и верхняя полоса представляет собой продукт, полученный вытеснением цепи. Верхняя полоса образуется внутренней побочной активностью T7-полимеразы, и ее трудно избежать (Lechner el al., J. Biol Chem. 258: 11174-11184 (1983)); однако эта полоса трансформируется в 30 раз менее эффективно, чем нижняя полоса, и обычно приводит к получению библиотеки. Средняя полоса является следствием отсутствия 5'-фосфата для конечной реакции лигирования; эта полоса эффективно трансформируется и дает, главным образом, последовательность дикого типа.
Затем достроенный продукт очищали и вводили путем электропорации в клетки SS320 и размножали в присутствии фага-помощника M13/KO7, как описано Sidhu et al., Methods in Enzymology 328: 333-363 (2000). Размеры библиотек варьировали в диапазоне 1-2 × 109 независимых клонов. Случайные клоны из исходных библиотек секвенировали для оценки качества библиотеки.
Фаговая селекция
В качестве антигенов для селекции использовали белки C3b человека. C3b человека наносили на микропланшеты для титрования MaxiSorp (Nunc) в количестве 10 мкг/мл в PBS и инкубировали в течение ночи при 4°С. Для первого раунда селекции использовали 12 лунок. Лунки блокировали в течение 1 ч при КТ с использованием блокирующего фаг буфера (1% BSA, 0,05% Tween 20, PBS). Фаговые библиотеки представляли собой осадок с PEG из замороженных в глицерине исходных смесей, ресуспендированный в блокирующем фаг буфере и инкубированный в течение 1 ч при КТ. Затем фаговые библиотеки добавляли в блокированные планшеты с антигеном и инкубировали в течение ночи при КТ. После связывания в течение ночи несвязанный/неспецифический фаг удаляли из планшетов с антигеном промыванием буфером для промывания (PBS, 05% Tween 20). Связанный фаг оценивали инкубацией лунок с 50 мМ HCl, 0,5 M KCl в течение 30 мин. Фаг амплифицировали с использованием клеток XL-I Blue и фага-помощника M13/KO7 и выращивали в течение 36 ч при 30°С в 2YT, 50 мкг/мл карбанециллина, 50 мкг/мл канамицина, 10 мкг/мл тетрациклина. Затем амплифицированный фаг выделяли с использованием модифицированного протокола осаждения с PEG (Monaci, P., Cortese, R., Screening phage libraries with sera, In: Phage display - A practical approach, Clackson and Lowman, eds., 2004, p. 193-215). Титры фага, элюированного с покрытой мишенью лунки, сравнивали с титрами фага, выделенными из не покрытой мишенью лунки, для оценки обогащения. Четыре раунда фаговой селекции завершали с количеством лунок-мишеней, снижающимся до 4 (раунд 2) и 2 (раунды 3 и 4). Для раундов 2 и 4 в качестве блокирующего реагента для планшетов с антигеном и фагом использовали блокирующий буфер с казеином (Pierce). В раундах селекции 2-4 использовали период связывания фаг-антиген, составляющий 3-4 часа, и возрастающую строгость промывания. В случае пэннига C3b человека также добавляли C3 человека (>1 мкМ) в раундах селекции 2-4 в ходе инкубации фаг-антиген в качестве обратной селекции против фаговых антител, которые также могли связывать C3 человека. Отобрали клон фага для человека YW144.2.43. Результаты пэннинга C3b представлены на фигуре 1. Характеристики связывания C3b определяли, как описано в примере 3.
Пример 2
Антитела, созданные вариацией HVR H1, H2, H3 и L3
Клон YW144.2.43 получали мутагенезом HVR-H1, H2, H3 и L3 вариабельного домена тяжелой цепи huMAb 4D5-8 (антитело против HER2 HERCEPTIN®, Genentech, Inc.) и модифицированного вариабельного домена легкой цепи huMAb 4D5-8. В HVR-H1 положения по Kabat 26 (G), 28 (T), 29 (F), 30 (S), 31 (S) и 35 (S) оставляли постоянными и варьировали аминокислоты в положениях 27, 32-34. В HVR-H2 положения по Kabat 49 (S), 51 (I), 55 (G), 57 (T), 59 (Y), 60 (A), 61 (D), 62 (S), 63 (V), 64 (K) и 65 (G) оставляли постоянными и варьировали положения 50, 52, 52a, 53, 54, 56 и 58. В HVR-H3 положения по Kabat 93 (A), 94 (R), 100f-g (делеция) оставляли постоянными и варьировали положения 95-100, 100a-e, 100h и 102. В HVR-L3 положения по Kabat 89 (Q), 90 (Q), 95 (P) и 97 (T) оставляли постоянными и варьировали положения 91-94 и 96. Последовательность HVR-L1 оставляли постоянной как RASQSISSYLA (SEQ ID NO:11) и последовательность HVR-L2 оставляли постоянной как GASSRAS (SEQ ID NO:12). Разнообразие последовательностей вносили в каждый гипервариабельный участок мутагенезом выбранных аминокислотных положений с использованием стандартных способов мутагенеза. Клоны антитела против C3v подвергали селекции и секвенировали.
Созревание аффинности YW144.2.43
Для повышения аффинности антитела против C3b YW144.2.43 было создано три библиотеки фагового дисплея на фоне YW 144.2.43, каждая из которых нацелена на множество HVR для мутагенеза с мягкой рандомизацией, как описано в Lee et al., J. MoL Biol. (2004), 340(5): 1073-93. Для избежания повторной селекции YW144.2.43 из потенциально высокого фона матрицы, перед созданием каждой библиотеки в HVR, подлежащие мутации, вносили стоп-кодоны. Для усиления эффективности процесса селекции на основе аффинности использовали способ сортировки в растворе. Посредством манипулирования концентрацией биотинилированной мишени, снижения времени захвата фага для более низкого фона и добавления небиотинилированной мишени для устранения клонов с более высокими скоростями диссоциации можно умело отбирать клоны с высокой аффинностью. Lee et al., J. Mol. Biol. (2004), 340(5): 1073-93. После первого раунда селекции наблюдали обогащение (захват зависимого от мишени фага), что позволило предположить, что в каждой библиотеке присутствовало большое количество клонов с довольно высокой аффинностью к C3b человека. Строгость селекции возрастала при последующих раундах. После 5 раундов селекции клоны из каждой библиотеки анализировали. Наблюдали новые последовательности, нацеленные на каждую из шести HVR. Выбранные клоны подвергали скринингу фаговым ELISA, а затем экспрессировали в качестве белка IgG и их аффинность охарактеризовывали с использованием анализа связывания BiacoreTM.
Фаговые библиотеки клонов после созревания аффинности сортировали с использованием способа сортировки в твердом состоянии/в растворе. C3b человека биотинилировали смешиванием 500 мкл 3,6 мг/мл C3b человека в PBS и 10 мкл 1 M фосфата калия, pH 8, с 20 мкл 4 мМ Sulfo-NHS-LC-биотина (Pierce). Для 1 раунда селекции биотинилированные C3b наносили на микропланшеты для титрования MaxiSorp (Nunc) в количестве 10 мкг/мл в PBS и инкубировали в течение ночи при 4°С. Для первого раунда селекции использовали 16 лунок с мишенью. Лунки блокировали в течение 1 ч при КТ с использованием SuperBlock (Pierce). Фаговые библиотеки после созревания разбавляли в буфере SuperBlock и инкубировали в течение 1 ч при КТ. Затем в блокированные планшеты с антигеном, инкубированные в течение 2 ч при КТ, добавляли фаговые библиотеки. После связывания несвязанный/неспецифический фаг удаляли из планшетов с антигеном промыванием буфером для промывания (PBS, 05% Tween20). Связанный фаг элюировали инкубацией лунок с 50 мМ HCl, 0,5 M KCl в течение 30 мин. Фаг амплифицировали с использованием клеток XL-1 Blue и фага-помощника M13/KO7 и выращивали в течение 36 ч при 30°С в 2YT, 50 мкг/мл карбанециллина, 50 мкг/мл канамицина, 1 мкг/мл тетрациклина. Затем амплифицированный фаг выделяли с использованием модифицированного протокола осаждения с PEG (Monaci, P., Cortese, R., выше). Титры фага, элюированного с покрытой мишенью лунки, сравнивали с титрами фага, выделенного из не покрытой мишенью лунки, для оценки обогащения. Для раундов 2-5 селекции выполняли протокол сортировки в растворе. На лунки микропланшета для титрования наносили 10 мкг/мл нейтравидина в PBS в течение ночи при 4°С, а затем блокировали в течение 1 ч с использованием SuperBlock (Pierce). Выделенные фаговые библиотеки суспендировали в SuperBlock и смешивали с 50 нМ b-Robo4-His в течение 1 ч. Фаг, связанный с b-C3b, иммобилизовывали на покрытых нейтравидином лунках в течение 30 мин и несвязанный фаг смывали буфером для промывания. Фаг элюировали с использованием 50 мМ HCl, 500 мМ KCl в течение 30 мин, нейтрализовывали и размножали в клетках XL1 blue (Stratagene) в присутствии фага-помощника KO7 (New England Biolabs). Последующие раунды сортировки проводили аналогично со следующими исключениями: в раунде 2 конечная концентрация b-C3b составляла 50 нМ, в раунде 3 конечная концентрация b-C3b составляла 25 нМ, в раунде 4 конечная концентрация b-C3b составляла 5 нМ и в раунде 5 конечная концентрация b-C3b составляла 0,5 нМ, с добавлением 50 нМ небиотинилированного C3b к смеси за 1 ч до иммобилизации на нейтравидине.
Несколько клонов после созревания аффинности были отобраны для связывания с C3b человека и отсеквенированы. Аминокислотные последовательности Fab-фрагментов тяжелой и легкой цепей антитела после созревания аффинности YW144.2.43.S77 (сокращенно S77) представлены на фигуре 5.
Пример 3
Охарактеризация отобранного клона антитела против C3b
Фаговый ELISA - Фаговые конкурентные анализы связывания проводили для приближенного определения аффинности связывания (определяемой как фаговая IC50) экспонированных на фаге Fab для C3b. Анализы проводили следующим образом. Очищенные супернатанты фагов от каждого клона получали с использованием модифицированного протокола осаждения с PEG, как описано выше. Очищенные супернатанты фагов подвергали серийному разведению в блокирующем фаг буфере, затем инкубировали на планшетах, покрытых C3b (1 мкг/мл) в течение 15 минут. Планшеты промывали буфером для промывания и инкубировали в течение 30 минут с конъюгатом пероксидаза хрена/антитело против M13 (разбавленным 1:5000 в буфере PBS) (Amersham Pharmacia Biotech). Планшеты промывали, проявляли субстратом метилбензидина (TMB) (Kirkegaard и Perry Laboratories) и гасили 0,1 н. H2SO4. Поглощение измеряли спектрофотометрически при 450 нм для определения концентрации фага, дающей приблизительно 50% сигнала при насыщении. Фиксированную субнасыщающую концентрацию фага разбавляли в блокирующем фаг буфере, содержащем двукратные серийные разведения белка C3b от 350 нМ C3b до 5нМ C3b. Смеси инкубировали в течение одного часа при осторожном встряхивании при комнатной температуре, переносили в планшеты, покрытые C3b (1 мкг/мл) и планшеты инкубировали в течение 20 минут. Планшеты промывали и обрабатывали, как описано выше. Аффинности связывания оценивали в качестве значений IC50 (определенных как концентрация антигена, которая блокировала 50% связывания фага с иммобилизованным антигеном). Результаты конкуренции фага за C3b представлены на фигуре 2.
Продукция IgG и определение аффинности - Для экспрессии белка IgG для охарактеризации аффинности стоп-кодон вносили между тяжелой цепью и g3 в векторе для фагового дисплея. Клоны трансформировали в клетки E. coli 34B8 и выращивали в среде AP5 при 30°С (Presta et al. Cancer Res. 57: 4593-4599 (1997)). Клетки собирали центрифугированием, суспендировали в 10 мМ Tris, 1 мМ EDTA, pH 8 и разрушали с использованием микрофлюидизатора. Fab очищали аффинной хроматографией с белком G. Аффинность связывания образованного из фага антитела против C3b YW144.2.43 и его варианта после созревания аффинности YW144.2.43S77 (Fab-фрагменты) к C3b и C3 человека определяли измерением поверхностного плазмонного резонанса с использованием системы BIACORE® 3000 (Biacore, Inc., Piscataway, NJ). Тестированные Fab-фрагменты антитела представляли собой YW144.2.43 и YW144.2.43S7. В кратком изложении проточные ячейки 1 и 2 на карбоксиметилированных декстрановых биосенсорных чипах (CM5, Biacore Inc.) активировали 0,2 M гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и 0,05 M N-гидроксисукцинимидом (NHS) со скоростью потока 5 мкл/мин в течение 7 мин. В качестве отрицательного контроля проточные ячейки оставляли непокрытыми. Эти активированные чипы покрывали Fab против C3b разведением до 5 мкг/мл 10 мМ ацетатом натрия, pH 4,8, перед инъекцией со скоростью потока 5 мкл/минута до достижения приблизительно 50 единиц ответа (RU) связанного антитела. Далее инъецировали 1 M этаноламин для блокирования не вступивших в реакцию групп. Для измерения кинетики двукратные серийные разведения растворимого антигена C3b или C3 человека (от приблизительно 100 нМ до приблизительно 3 нМ для C3b, от ~1 мкМ до 50 нМ для C3) инъецировали в PBS с 0,05% Tween 20 при 25°C со скоростью потока 35 мкл/мин. После каждой инъекции чип регенерировали с использованием 20 мМ HCl. Связывающий ответ корректировали вычитанием RU из пустой проточной ячейки. Константы ассоциации (kon) и константы диссоциации (kоff) вычитали с использованием простой модели связывания Ленгмюра "один к одному" (BIAevaluation Software версии 3.2). Равновесную константу диссоциации (KD) вычисляли в качестве соотношения kдиссоциации/kассоциации. Аффинность связывания представлена на фигурах 6 и 7.
В другом эксперименте очищенный C3b или C3 иммобилизовывали на микропланшетах для титрования с использованием поликлонального C3 антитела. Связывание S77 (A) или поликлонального антитела против C3 (B) с иммобилизованным C3b или C3 определяли с использованием вторичного конъюгированного с HRPO антитела. Цвет проявляли TMB (KPL), проявление останавливали 2 н. H2SO4 и считывали поглощение при 450 нм.
ELISA C3b для тестирования специфичности антитела против C3b S77. Двадцать пять мкл антитела для иммобилизации (YW144.2.45.S77 после созревания xC3/C3b (Genentech) 2 мкг/мл), разбавленного в PBS, добавляли в лунки микропланшета для титрования и инкубировали в течение ночи при 4°С. Планшет промывали 3x буфером для промывания (PBS/0,05% Tween 20 (20x исходный раствор; Media Prep; каталожный номер A3355)). В лунки добавляли пятьдесят мкл блокирующего буфера и планшет инкубировали в течение 1-3 часов при осторожном встряхивании (комнатная температура) и промывали 3x буфером для промывания. Стандартный исходный раствор (C3b, Complement Technology Inc.; каталожный номер A114, хранящийся при 100x в -20°С) изготавливали в буфере Magic Buffer (I X PBS pH 7,4, 0,5% BSA, 0,05% Tween 20, 0,2% BgG, 15PPM Proclin (Media Prep; Cat. A3381 ) + 0,35M NaCl. Также для получения образцов для анализа использовали Magic Buffer + 0,35M NaCl. Двадцать пять мкл стандартов/образцов добавляли в предназначенные для них лунки. Образцы инкубировали в течение ~2 часов (+/- 0,5 ч) при КТ при осторожном встряхивании и промывали 3x буфером для промывания. Планшет переворачивали на 180 градусов и стадию промывания повторяли. Антитело для детекции (конъюгированное с пероксидазой F(ab')2 козы против C3 человека (Protos Immunoresearch; каталожный номер 765)) разбавляли 1:7K разбавителем для анализа и инкубировали в планшете в течение 1-2 часов при КТ при осторожном встряхивании. Планшет промывали 3x буфером для промывания и переворачивали на 180 градусов и повторяли стадию промывания. Изготавливали раствор 50/50 TMB. Планшет для ELISA промывали 3x буфером для промывания. Планшет переворачивали на 180 градусов и стадию промывания повторяли. В лунки добавляли 25 мкл TMB. Проявляли цвет при комнатной температуре. Время проявления: 10 минут для обоих планшетов. Проявление цвета останавливали добавлением в лунки 25 мкл 1,0 M фосфорной кислоты. Проводили считывание OD планшета (450/630 нм). Результаты представлены на фигуре 8, панель A.
Анализ общего C3, используемого в качестве положительного контроля для детекции C3, проводили аналогично анализу ELISA C3b, описанному выше, используемому для тестирования специфичности S77 за исключением того, что фракцию IgG против C3 человека (Cappel 55033) использовали в качестве антитела для иммобилизации. Результаты представлены на фигуре 8, панель B.
Как представлено на фигуре 8, S77 распознает C3b, но не промолекулу C3.
Пример 4
Антитела против C3b специфично ингибируют альтернативный каскад комплемента
Анализ гемолиза - Для определения активности альтернативного каскада эритроциты кролика (Er, Colorado Serum) промывали 3x в GVB и ресуспендировали до 2×109/мл. Ингибиторы (50 мкл) и 20 мкл суспензии Er смешивали 1:1 с GVB/0,1 M EGTA/0,1 M MgCl2. Активацию комплемента начинали добавлением сыворотки человека с истощенным C1q (Quidel; 30 мкл, разбавленная 1:3 в GVB). После инкубации в течение 30 минут при комнатной температуре добавляли 200 мкл GVB/10 мМ EDTA для остановки реакции и образцы центрифугировали в течение 5 мин при 500 g. Гемолиз определяли в 200 мкл супернатанта измерением поглощения при 412 нм. Данные выражали в качестве % от гемолиза, индуцированного в отсутствие ингибитора. Для определения эффекта CRIg на классический каскад комплемента следовали сходному способу за исключением того, что Er заменяли покрытыми IgM эритроцитами овцы (E-IgM, CompTech) и анализ проводили в сыворотке человека с дефицитом фактора B в GVB++.
Как представлено на фигурах 9 и 10, антитело S77 с созреванием аффинности против C3b ингибирует альтернативный каскад комплемента и не ингибирует классический каскад.
Пример 5
Антитела против C3b ингибируют связывание C5 с C5-конвертазой
Конкурентный анализ C5 - C3b наносили на микропланшет для титрования с 3 мкг/мл C3b в PBS в течение ночи при 4°C. Планшет блокировали 1% BSA в PBS и инкубировали с возрастающими концентрациями антитела, смешанного с 0,4 мкМ C5 в 20 мМ Tris/20 мМ Ca/20 мМ Mg/150 мМ NaCl/0,05% Tween/1 % BSA. Детекцию связывания C5 проводили инкубацией с антителом против C5 человека (клон 7D12, Genentech) в течение 30 мин при КТ, а затем с антителом осла против антител мыши с HRP (Jackson) 1:5000. Как представлено на фигурах 11 и 12, антитело после созревания аффинности S77 против C3b ингибирует C5-конвертазу.
Пример 6
Антитела против C3b не проявляют активности распада
Активность ускорения распада - На микропланшеты для титрования наносили в течение ночи 3 мкг/мл C3b в PBS. Планшеты промывали 2 раза в PBST (PBS/0,1 %-Tween), блокировали в течение 2 ч при 37°C PBST, содержащим 4% BSA. Планшеты инкубировали в течение 2 ч при комнатной температуре в вероналовом буфере, содержащем 400 нг/мл фактора B, 25 нг/мл фактора D и 2 мМ NiCl2, 25 мМ NaCl, 0,05% Tween 20 и 4% BSA, а затем инкубировали в течение 15 мин с фактором H или S77 в PBST. Детекцию фактора Bb проводили последовательными инкубациями в течение 1 ч с поликлональным антителом козы против фактора B человека (Kent), разбавленным 1:5000, в PBST и антителом осла против антител козы, конъюгированным с HRPO (Caltag), разбавленным 1:5000, в PBST. Цвет проявляли TMB (KPL), проявление останавливали в 2 н. H2SO4 и считывали поглощение при 450 нм. Активность кофактора для опосредуемого фактором I расщепления C3b измеряли инкубацией 0,8 мкМ C3b и 80 нМ фактора I с 80 нМ фактором H или варьирующими концентрациями S77 в 30 мл GVB. Смесь инкубировали в течение 60 мин при 37°С и образцы анализировали гель-электрофорезом, как описано для анализа C3-конвертазы.
Как представлено на фигуре 13, S77 не проявляет активности ускорения распада.
Пример 7
С77 ингибирует связывание профактора В с С3b и образование С3bBb-конвертазы
Протокол
На планшет MaxiSorp наносили в течение 4 ч при КТ 3 мкг/мл С3b (PUR13420) в PBS (20 мкл/лунка). Промывание проводили 6х посредством 100 мкл PBS 0,1% Tween (PBST) (устройство для промывания BioTek EL405). Планшеты блокировали в течение 2 ч при КТ 4% BSA/0,05% Tween/PBS, а затем блокированный раствор удаляли встряхиванием в раковину. Добавляли 20 мкл буфера для конвертазы АР в течение 2 ч при КТ, а затем промывали 6х PBST. Добавляли 20 мкл Аb в течение 45 мин при КТ и лунки промывали 6х PBST. Для детекции фактора В/Вb, лунки инкубировали с антителом козы против fB (Kent Labs) 1:7000 в течение 30 мин при КТ. Для детекции S77 и CR1 в лунки добавляли PBST, их затем промывали 6х PBST и инкубировали в течение 30 мин с антителом осла против IgG козы-HRPO (Jackson) 1:7000 в PBST++. После инкубации в течение 30 мин при КТ с антителом против 6х-His (SEQ ID NO:19) (R&D) 1:100 в PBST++ проводили промывание 6х PBST. Проявление проводили 20 мкл субстрата ТМВ и реакцию останавливали 10 мкл 2 н. серной кислоты. Считывание планшетов проводили при 450 нм.
"Буфер для конвертазы АР»: 4% BSA, 0,1% Tween 20, 2 мМ NiCl2, 25 мМ NaCl, 25 нг/мл фактора D, 400 нг/мл фактора В (CompTech).
В соответствии с указанным выше протоколом С3b наносили на микропланшеты для титрования. Добавляли С77, контрольный Fab или CR1-фрагмент (LHRA-C), а через 1 ч добавляли фактор В. Детекцию связывания фактора B с C3b проводили с помощью конъюгированного с HRPO вторичного антитела и поглощение считывали при 450 нм. Результаты представлены на фигуре 14, панель A.
Аналогично, следуя указанному выше протоколу, C3b наносили на микропланшеты для титрования, а затем добавляли S77, контрольный Fab или CR1-фрагмент (LHRA-C). Образования C3-конвертазы достигали добавлением фактора D и фактора B. Образование конвертазы определяли с использованием первичного антитела, которое распознает фактор Bb, и вторичного конъюгированного с HRPO антитела. Цвет проявляли посредством TMB (KPL), проявление останавливали в 2 н. H2SO4 и поглощение считывали при 450 нм. Результаты представлены на фигуре 14, панель B.
Как представлено на фигуре 14, антитело S77 ингибирует связывание профактора B с C3B и ингибирует образование C3bBb-конвертазы.
Пример 8
S77 связывает C3b в присутствии связанного fBb и не разрушает C3-конвертазу
С использованием протокола, описанного в примере 7, C3b наносили на микропланшеты для титрования. C3-конвертазу получали добавлением фактора D и фактора B. В планшет добавляли S77, контрольный Fab или CR1-фрагмент (LHRA-C) и определяли связывание этих молекул со вторичными антителами, конъюгированными с HRPO. Результаты представлены на фигуре 15, панель A.
Аналогично, следуя протоколу, описанному в примере 7, на микропланшеты для титрования наносили 3 мкг/мл C3b. Планшеты инкубировали с фактором B и фактором D, а затем инкубировали с CR1 (LHRA-C), S77 или контрольным Fab. Детекцию фактора Bb проводили с антителом козы против фактора B человека и антителом осла против антител козы, конъюгированным с HRPO. Цвет проявляли с помощью TMB (KPL), проявление останавливали в 2 н. H2SO4 и поглощение считывали при 450 нм. Результаты представлены на фигуре 15, панель B.
Результаты, указанные на фигуре 15, демонстрируют, что S77 может связывать C3b в присутствии связанного fBb и не вызывает распада C3-конвертазы.
Пример 9
S77 ингибирует связывание фактора H с C3b и ингибирует активность кофактора для фактора H
Протокол 1 (Фиг. 15, панель A) - На планшеты MaxiSorp наносили в течение 3 ч при КТ 3 мкг/мл C3b (PUR13420) в PBS (20 мкл/лунка). Планшеты промывали 6х 100 мкл PBS 0,1% Tween (PBST) (устройство для промывания BioTek EL405) и блокировали в течение 2 ч при КТ 4% BSA/0,05% Tween/PBS. Планшеты инкубировали в течение 30 мин при КТ при встряхивании с блокирующими Ab, 20 мкл, с последующей инкубацией в течение 1 ч при КТ с 0,33 мкМ fH (CompTech) и добавлением 10 мкл 1 мкМ fH. Планшеты промывали 6x PBST в устройстве для промывания планшетов (BioTek EL405), инкубировали в течение 30 мин с антителом осла против IgG мыши (H+L)-HRPO (Jackson) 1:7000 и промывали 6x PBST в устройстве для промывания планшетов (BioTek EL405). Проявление проводили с помощью 20 мкл субстрата TMB. Реакцию останавливали 10 мкл 2 н. серной кислоты и считывание планшетов проводили при 450 нм.
Протокол 2 (Фиг. 15, панель B) - Все разведения проводить в GVB++(1 мМ MgCl, 0,15 мМ CaCl). Добавить в пробирку eppendorf 10 мкл 1,6 мкМ C3b (конечная концентрация 0,4 мкМ C3b). Добавить 10 мкл Fab против C3b, контрольный Fab или CR1. Инкубировать 20 мин при КТ. Добавить 10 мкл 0,08 мкМ fI (конечная концентрация 20 мкМ fI). Инкубировать в течение 60 мин при 37°С. Добавить 40 мкл буфера Laemmeli + 2-bME, кипятить 3 мин. Разделять на 8% геле Invitrogen, 25 мкл/лунка, 125 мВ в течение 1,5 часов. Промыть гель 3x в течение 5 мин H2O. Окрашивать Simply Blue (Invitrogen) 60 мин при КТ при качании. Промыть 3x 5 мин ddH2O. Промыть в течение ночи ddH2O в большой форме для выпечки на устройстве для качания, закрыть пластмассой. Реагенты: C3b PUR13240, fI от Complement Technologies, fH от Complement Technologies, GVB++ от BioWhittaker.
Как описано выше, на планшеты наносили C3b. Добавляли фактор H в присутствии возрастающих концентраций контрольного Fab или S77. Связывание фактора H с C3b определяли с использованием антитела против фактора H и вторичного конъюгированного с HRPO антитела против антител мыши. Цвет проявляли с помощью TMB (KPL), проявление останавливали в 2 н. H2SO4 и поглощение считывали при 450 нм. Результаты представлены на фигуре 16, панель A.
Активность кофактора для опосредуемого фактором I расщепления C3b измеряли инкубацией 0,8 мкМ C3b и 80 нМ фактора I с 80 нМ фактором H или различными концентрациями S77 в 30 мл GVB. Смесь инкубировали в течение 60 мин при 37°С и образцы анализировали гель-электрофорезом, как описано для анализа C3-конвертазы. Результаты представлены на фигуре 16, панель B.
Как представлено на фигуре 16, антитело S77 ингибирует связывание фактора H с C3b, а также ингибирует активность кофактора для фактора H.
Пример 10
S77 ингибирует связывание CR1 с C3b
Протокол - Наносить на планшет MaxiSorp в течение ночи при 4°С 3 мкг/мл C3b (PUR13420) в PBS (100 мкл/лунка). Промыть 3x 100 мкл PBS 0,1% Tween (PBST) (устройство для промывания BioTek EL405). Блокировать в течение 2 ч при КТ 4% BSA/0,1 % Tween/PBS. Инкубировать в течение 30 мин при КТ при встряхивании с блокирующими Ab, 20 мкл. Инкубировать 1 ч при КТ с 50 нМ CR1 LHR-AC. Промыть 3х PBST в устройстве для промывания планшетов (BioTek EL405). Инкубировать в течение 45 мин при КТ с 1:10 mIgG1 против hCD35-FITC (Pharmingen) в PBST. Промыть 3x PBST в устройстве для промывания планшетов (BioTek EL405). Инкубировать 30 мин с антителом осла против IgG мыши-HRPO (Jackson) 1:7000. Промыть 6x PBST в устройстве для промывания планшетов (BioTek EL405). Проявить 20 мкл субстрата TMB. Остановить реакцию 10 мкл 2 н. серной кислотой. Считывать планшет при 450 нм.
Как показано на фигуре 17, антитело S77 ингибирует связывание CR1 с C3b.
Пример 11
Кристаллизация и уточнение данных
Эксперименты с висящей каплей проводили с использованием способа паровой диффузии с 2-мкл каплями, состоящими из раствора белка и резервуарного раствора в соотношении 1:1. Раствор белка содержал комплекс C3b:S7714 в концентрации 10 мг/мл в 25 мМ Tris, 50 мМ NaCl при рН7, и резервуарный раствор содержал 10% PEG 4000, 0,2 М MgCl2 в 0,1 М Hepes при рН 7,2. Кристаллы появлялись через две недели. Кристаллы инкубировали в резервуарном растворе, дополненном 20% глицерином, перед быстрым замораживанием. Данные собирали от отдельного замороженного кристалла с помощью beam line 5.0.1 от the Advanced Light Source (Berkeley) и обрабатывали с использованием программ DENZO и SCALEPACK. Кристаллы относились к пространственной группе С2 с параметрами ячеек а=216,4 Å, b=180,4 Å, с=154,6 Å и β=115,73 Å с 2 комплексами, каждый из которых состоял из одной молекулы C3b, связанной с одной молекулой Fab, в асимметричном элементе. Структуру решали путем молекулярного замещения с использованием программы Phaser и координат С3b, константных доменов и вариабельных доменов Fab-фрагмента. Модель вручную корректировали с использованием программы О и уточнение проводили с помощью программы REFMAC с использованием узких 2-кратных ограничений некристаллографической симметрии. R и Rfree уточненной модели составляют 22,5% и 29,0% соответственно.
Кристаллическая структура C3b в комплексе с антителом S77 представлена на фигуре 3. На фигуре 4 представлено приближение связывающего взаимодействия антитела S77 с C3b. С использованием данных кристаллизации остатки в последовательности Fab тяжелой цепи С77, находящиеся в тесном контакте с С3b, показаны красным цветом.
Кроме того, на добавочной фигуре 1 представлены остатки на C3b, которые контактируют с С77. На добавочной фигуре 2 представлены остатки Fab C77, которые контактируют с C3b.
Нацеливание на C3b, компонент, центральный для активации комплемента, обеспечивает мощный подход для ингибирования каскада комплемента на уровне как C3-, так и C5-конвертаз. В исследованиях, описанных в представленных выше примерах, использовали фаговую технологию для получения антител, которые селективно распознают C3b, но не его промолекулу C3. Кристаллическая структура C3b в комплексе с Fab-фрагментом специфического антитела (S77) указывает на то, что антитело распознает эпитоп на домене MG7, экспонируемом после расщепления C3 до C3b. S77 блокирует связывание фактора B и C5 с C3b, что приводит к эффективному ингибированию C3- и C5-конвертаз альтернативного, но не классического, каскада комплемента. Кроме того, S77 ингибирует связывание fH и активность кофактора, а также связывание CR1 с C3b, указывая на то, что участок связывания S77 с доменом MG7 C3b является "горячей" точкой для регуляции активации комплемента. Взятые вместе результаты этого исследования иллюстрируют молекулярную основу активации и ингибирования комплемента на уровне C3- и C5-конвертаз альтернативного каскада и демонстрируют применимость фагового дисплея и других технологий дисплея для получения селективных антител с перспективным терапевтическим потенциалом.
Предполагается, что указанное выше письменное описание является достаточным для того, чтобы специалист в данной области смог осуществить на практике это изобретение. Различные модификации по изобретению в дополнение к модификациям, представленным и описанным в настоящем документе, станут очевидными специалистам в данной области из представленного выше описания и находятся в объеме прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОФИЛАКТИКА И ЛЕЧЕНИЕ ПАТОЛОГИЧЕСКИХ СОСТОЯНИЙ ГЛАЗ, ВЫЗВАННЫХ КОМПЛЕМЕНТОМ | 2008 |
|
RU2522976C2 |
| ПРИМЕНЕНИЕ ИНГИБИТОРОВ ПУТИ КОМПЛЕМЕНТА ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2006 |
|
RU2417099C2 |
| ВАРИАНТЫ CRIG С СОЗРЕВШЕЙ АФФИННОСТЬЮ | 2009 |
|
RU2553517C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ MASP-1, И/ИЛИ MASP-2, И/ИЛИ MASP-3 ДЛЯ ЛЕЧЕНИЯ ПАРОКСИЗМАЛЬНОЙ НОЧНОЙ ГЕМОГЛОБИНУРИИ | 2013 |
|
RU2655299C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ MASP-1, И/ИЛИ MASP-2, И/ИЛИ MASP-3 ДЛЯ ЛЕЧЕНИЯ РАЗЛИЧНЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2013 |
|
RU2709351C2 |
| ПРИМЕНЕНИЕ ИНГИБИТОРОВ ПУТИ КОМПЛЕМЕНТА ДЛЯ ЛЕЧЕНИЯ ГЛАЗНЫХ БОЛЕЗНЕЙ | 2010 |
|
RU2583927C2 |
| КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ MASP-2-ЗАВИСИМОЙ АКТИВАЦИИ КОМПЛЕМЕНТА | 2012 |
|
RU2636038C2 |
| ОПТИМИЗИРОВАННЫЕ ВАРИАНТЫ АНТИ-VEGF АНТИТЕЛ | 2016 |
|
RU2763916C2 |
| Способы лечения состояний, связанных с masp-2 зависимой активацией комплемента | 2012 |
|
RU2662563C2 |
| НОВЫЕ АНТИТЕЛА | 2008 |
|
RU2490277C2 |






















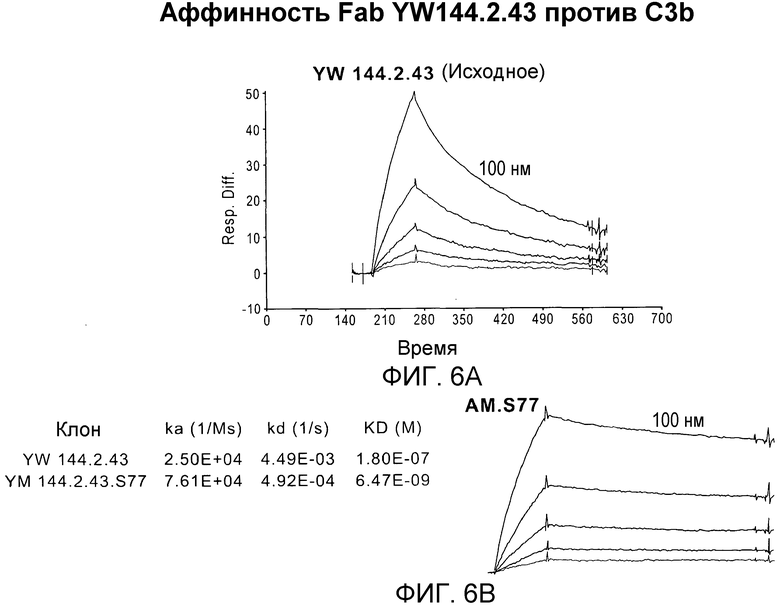


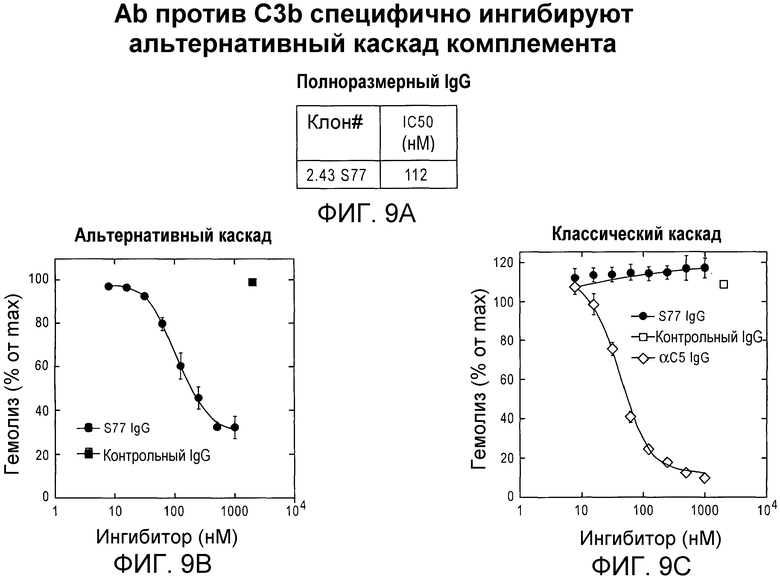

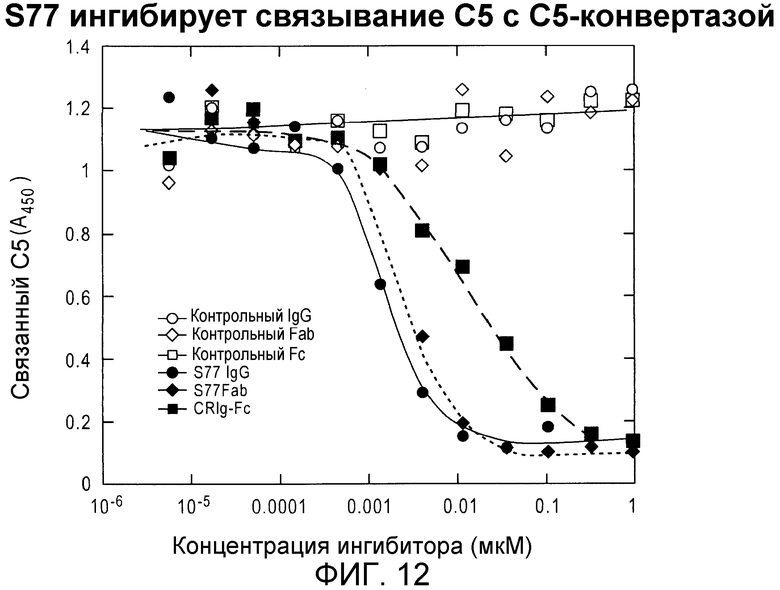
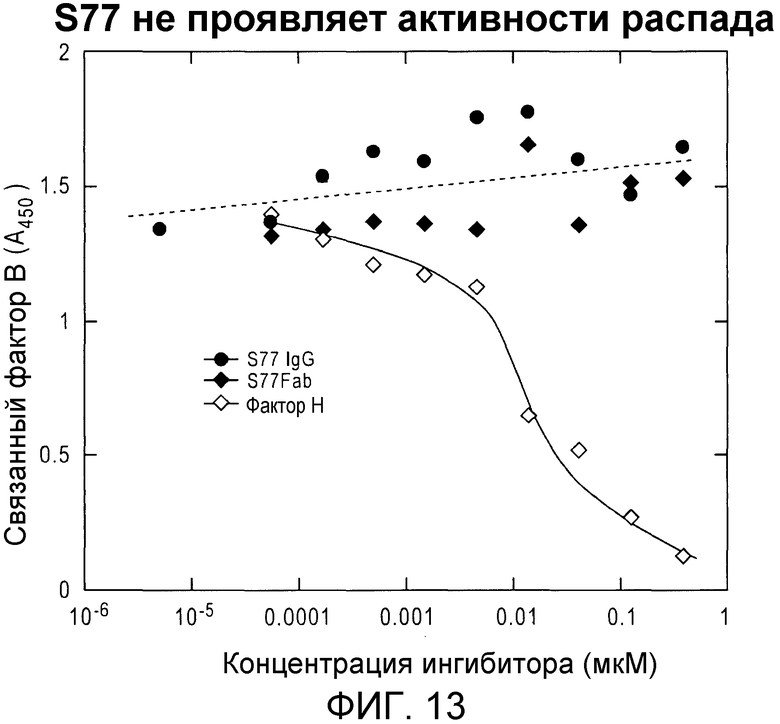




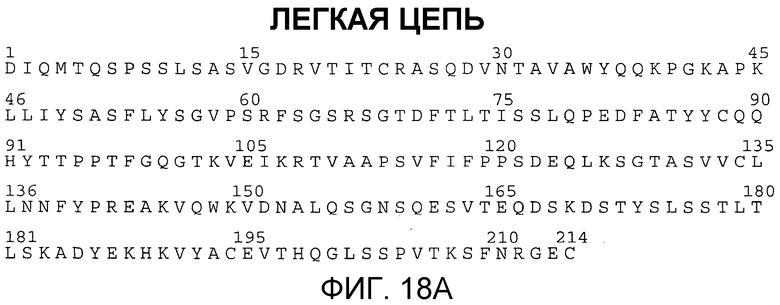



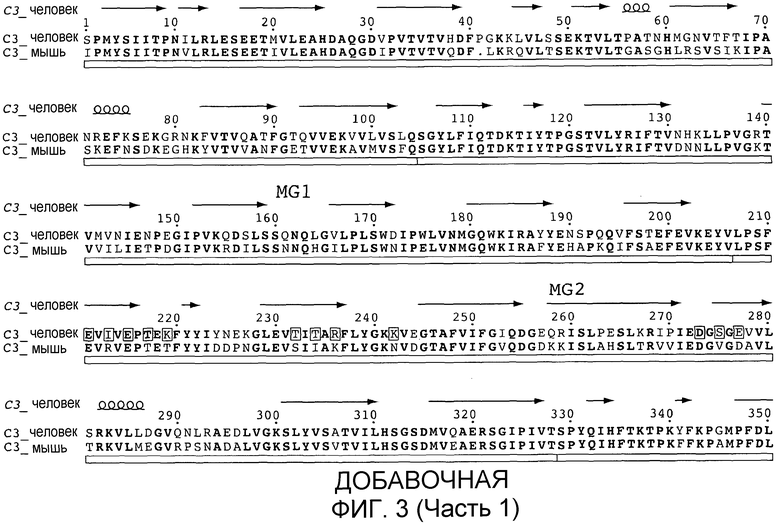



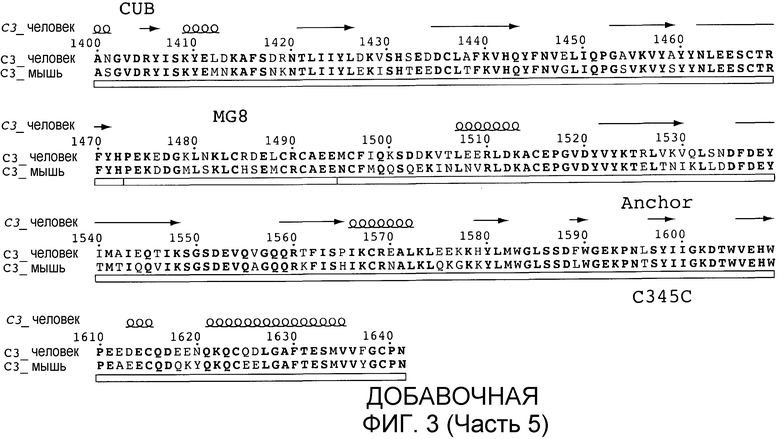
Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты антител против С3b, каждое из которых характеризуется наличием шести CDR и не связывает С3. Описана фармацевтическая композиция на основе антитела и набор, используемые для профилактики или лечения комплемент-зависимого заболевания. Использование изобретения обеспечивает новые антитела против С3b, Fab-фрагмент которых ингибирует альтернативный каскад комплемента с IС50 около 100 нМ, что может найти применение в медицине для профилактики и лечения связанного с комплементом нарушения, например воспалительного или дегенеративного заболевания. 4 н. и 25 з.п. ф-лы, 21 ил., 1 табл., 11 пр.
1. Антитело против С3b или его функциональный фрагмент, которое представляет собой селективный ингибитор альтернативного каскада комплемента и которое ингибирует связывание С5 с С3b, где указанное антитело содержит последовательности тяжелой цепи CDRH1, CDRH2 и CDRH3, SEQ ID NO:2, 3 и 4, соответственно, и последовательности легкой цепи CDRL1, CDRL2 и CDRL3, SEQ ID NO:6, 7 и 8, соответственно, для применения в профилактике или лечении связанного с комплементом нарушения у субъекта.
2. Антитело по п.1, где указанным субъектом является млекопитающее.
3. Антитело по п.2, где указанным субъектом является человек.
4. Антитело по п.3, где указанное антитело распознает эпитоп на активном продукте деградации С3, но не на С3.
5. Антитело по п.4, где указное антитело против С3b селективно связывается с С3b.
6. Антитело по п.5, где указанное антитело связывается с эпитопом, включающим остатки эпитопа С3b, распознаваемые антителом с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
7. Антитело по п.5, где указанное антитело связывается по существу с тем же эпитопом, что и антитело с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи SEQ ID NO:5-8.
8. Антитело по п.5, где указанное антитело конкурентно ингибирует связывание антитела с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи SEQ ID NO:5-8.
9. Антитело по п.5, где указанное антитело связывается с эпитопом С3b, содержащим остатки, которые контактируют с антителом с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
10. Антитело по п.5, где указанное антитело содержит антигенсвязывающий центр, содержащий остатки антитела с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8, которые контактируют с С3b.
11. Антитело по п.5, которое представляет собой антитело человека, гуманизированное или химерное антитело.
12. Антитело по п.5, где указанный фрагмент антитела выбран из группы, состоящей из Fab, Fab′, F(ab′)2, scFv, (scFv)2, dAb, фрагментов, определяющих комплементарность областей (CDR), молекул одноцепочечных антител, мини-антител и диател.
13. Антитело по п.12, где указанный фрагмент антитела представляет собой Fab-, Fab′-, F(ab′)2-, scFv или (scFv)2-фрагмент.
14. Антитело по п.3, где указанное связанное с комплементом нарушение представляет собой воспалительное заболевание или аутоиммунное заболевание.
15. Антитело по п.3, где указанное связанное с комплементом нарушение выбрано из группы, состоящей из ревматоидного артрита (RA), острого респираторного дистресс-синдрома (ARDS), отдаленного повреждения ткани после ишемии и реперфузии, активации комплемента в ходе хирургической операции с искусственным кровообращением, дерматомиозита, пемфигуса, волчаночного нефрита и являющихся его следствиями гломерулонефрита и васкулита, искусственного кровообращения, индуцируемой кардиоплегией дисфункции эндотелия коронарных артерий, мембранопролиферативного гломерулонефрита типа II, IgA-нефропатии, острой почечной недостаточности, криоглобулинемии, антифосфолипидного синдрома, дегенеративных заболеваний желтого пятна, таких как связанная со старением дегенерация желтого пятна (AMD), хориоидальная неоваскуляризация (CNV), увеит, диабетическая и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы, неоваскуляризация сетчатки, а также аллотрансплантации, сверхострого отторжения, гемодиализа, хронического легочного дистресс-синдрома (COPD), астмы и аспирационной пневмонии.
16. Антитело по п.3, где указанное связанное с комплементом нарушение представляет собой связанное с комплементом состояние глаза.
17. Антитело по п.16, где связанное с комплементом состояние глаза представляет собой связанную со старением дегенерацию желтого пятна (AMD) или хориоидальную неоваскуляризацию (CNV).
18. Антитело против С3b, селективно связывающееся с С3b и не связывающееся с С3 и ингибирующее связывание С5 с С3b, где указанное антитело содержит последовательности тяжелой цепи CDRH1, CDRH2 и CDRH3, SEQ ID NO:2, 3, и 4, соответственно, и последовательности легкой цепи CDRL1, CDRL2 и CDRL3, SEQ ID NO:6, 7, и 8, соответственно, для применения в профилактике или лечении связанного с комплементом нарушения у индивида.
19. Антитело по п.18, где указанное антитело связывается с эпитопом, включающим остатки эпитопа С3b, распознаваемые антителом с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
20. Антитело по п.18, где указанное антитело связывает по существу тот же эпитоп, что и антитело с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
21. Антитело по п.18, где указанное антитело конкурентно ингибирует связывание антитела с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
22. Антитело по п.18, где указанное антитело связывается с эпитопом С3b, содержащим остатки, которые контактируют с антителом с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8.
23. Антитело по п.18, где указанное антитело содержит антигенсвязывающий центр, содержащий остатки антитела с последовательностью вариабельного домена тяжелой цепи, SEQ ID NO:1-4, и последовательностью вариабельного домена легкой цепи, SEQ ID NO:5-8, которые контактируют с С3b.
24. Антитело по п.18, которое представляет собой антитело человека, гуманизированное или химерное антитело.
25. Антитело по п.18, где указанный фрагмент антитела выбран из группы, состоящей из Fab, Fab′, F(ab′)2, scFv, (scFv)2, dAb, фрагментов определяющих комплементарность областей (CDR), молекул одноцепочечных антител, мини-антител и диател.
26. Антитело по п.18, где указанный фрагмент антитела представляет собой Fab-, Fab′-, F(ab′)2-, scFv или (sсFv)2-фрагмент.
27. Фармацевтическая композиция, использующаяся для профилактики или лечения комплемент-зависимого заболевания, содержащая эффективное количество антитела по пп.18-26 в смеси с фармацевтически приемлемым носителем.
28. Фармацевтическая композиция по п.27, где указанное связанное с комплементом нарушение выбрано из группы, состоящей из ревматоидного артрита (RA), острого респираторного дистресс-синдрома (ARDS), отдаленного повреждения ткани после ишемии и реперфузии, активации комплемента в ходе хирургической операции с искусственным кровообращением, дерматомиозита, пемфигуса, волчаночного нефрита и являющихся его следствиями гломерулонефрита и васкулита, искусственного кровообращения, индуцируемой кардиоплегией дисфункции эндотелия коронарных артерий, мембранопролиферативного гломерулонефрита типа II, IgA-нефропатии, острой почечной недостаточности, криоглобулинемии, антифосфолипидного синдрома, дегенеративных заболеваний желтого пятна, таких как связанная со старением дегенерация желтого пятна (AMD), хориоидальная неоваскуляризация (CNV), увеит, диабетическая и другие связанные с ишемией ретинопатии, эндофтальмит и другие связанные внутриглазной неоваскуляризацией заболевания, такие как диабетический отек желтого пятна, патологическая миопия, болезнь фон Гиппеля-Линдау, гистоплазмоз глаза, окклюзия центральной вены сетчатки (CRVO), неоваскуляризация роговицы, неоваскуляризация сетчатки, а также аллотрансплантации, сверхострого отторжения, гемодиализа, хронического легочного дистресс-синдрома (COPD), астмы и аспирационной пневмонии.
29. Набор для профилактики или лечения комплемент-зависимого заболевания, содержащий контейнер, антитело или фрагмент антитела по п.18 или фармацевтическую композицию по п.27, и инструкцию по применению указанного антитела или фрагмента антитела для лечения комплемент-зависимого заболевания.
| WO 2006012621 А2, 02.02.2006 | |||
| WO 2006063150 А2, 15.06.2006 | |||
| US 20050107319 A1, 19.05.2005 | |||
| PORTEU F, et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Immunol | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| МАШИНА ДЛЯ ПЕРЕРАБОТКИ ТОРФЯНОЙ МАССЫ | 1923 |
|
SU629A1 |
| NIEBOER С., et al | |||
| "Immunofluorescence patterns in sun-exposed and not-sun-exposed skin of healthy | |||

