ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей непредварительной заявке, поданной по 37 CFR § 1.53(b), по 35 USC §119(e), испрашивается приоритет предварительной заявки США серийный №60/886068, поданной 22 января 2007 г., и предварительной заявки США серийный №61/013446, поданной 13 декабря 2007 г., полное содержание которых приведено здесь в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к способам очистки белков.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Крупномасштабная, экономически выгодная очистка белков является все в большей степени важной проблемой для биотехнологической промышленности. Как правило, белки продуцируют в культуре клеток, с использованием линий клеток либо млекопитающих, либо бактерий, сконструированных для продукции интересующего белка посредством введения рекомбинантной плазмиды, содержащей ген этого белка. Поскольку используемые линии клеток являются живыми организмами, их необходимо подпитывать комплексной средой для роста, содержащей сахара, аминокислоты и факторы роста, обычно дополненной препаратами из сыворотки животных. Отделение желаемого белка от смеси соединений, питательных для клеток, и от побочных продуктов самих клеток до чистоты, достаточной для использования в качестве терапевтических средств для человека, составляет сложную задачу.
Рекомбинантные терапевтические белки обычно продуцируют в нескольких линиях клеток-хозяев млекопитающих, включая клетки миеломы мыши NS0 и клетки яичника китайского хомячка (CHO) (Anderson, D.C. and Krummen, L. (2002) Curr. Opin. Biotech. 13: 117-123; Chu, L. and Robinson, D.K. (2001) Curr. Opin. Biotechnol. 12:180-187). Каждая линия клеток обладает преимуществами и недостатками с точки зрения продуктивности и характеристик белков, продуцируемых клетками. Выборы линий клеток для коммерческой продукции часто уравновешивают необходимость высокой продуктивности со способностью обеспечивать показатели качества продукта, необходимые для данного продукта. Одним важным классом терапевтических рекомбинантных белков, для которых часто необходимы способы с высоким титром, являются моноклональные антитела. Для некоторых моноклональных антител необходимы эффекторные функции, опосредованные через Fc-область, чтобы активировать их биологические функции. Примером является ритуксимаб (РИТУКСАН®, Genentech, Inc. и Biogen-Idec), химерное моноклональное антитело, которое связывается с CD-20 поверхности клеток и приводит к элиминации B-клеток (Cartron et al. (2002) Blood 99: 754-758; Idusogie et al. (2000) J. Immunol. 164: 4178-4184). Для других антител, таких как бевацизумаб (АВАСТИН™, Genentech, Inc.), гуманизированное антитело против VEGF (фактор роста эндотелия сосудов), не требуются эффекторные функции Fc для их активности.
Успехи в способах ферментации и культивирования клеток сильно увеличили титры намеченных белков в культуральной жидкости. Это увеличение эффективности на более ранних стадиях создало трудности при более поздней переработке на стадии сбора клеток. Сбор клеток или осветление собранной культуральной жидкости является важным процессом почти во всех последующих очистках продуктов на биотехнологической основе. Когда продукт является внутренним для клеток, сбор клеток используют для уменьшения жидкого объема клеток, подлежащего переработке на стадиях выделения продукта. Когда продукт является внеклеточным, сбор клеток используют для отделения продукта от клеток и клеточного дебриса, например, для выделения внеклеточного антитела из культуры клеток млекопитающих (Anthony S. Lubiniecki, Ed. (1990) Large-Scale Mammalian Cell Culture Technology, Marcel Dekker; Hansjoerg Hauser, Roland Wagner, Eds. (1997) Mammalian Cell Biotechnology in Protein Production, Walter Gruyter Publishing).
Способы для очистки белков от клеточного дебриса исходно зависят от места экспрессии белка. Некоторые белки можно заставить секретироваться непосредственно из клетки в окружающую среду для роста; другие оставляют внутри клетки. В случае последних белков первая стадия способа очистки включает в себя лизис клеток, который можно осуществлять множеством способов, включая механическое разрезание, осмотический шок или ферментативные обработки. Такое разрушение высвобождает все содержимое клетки в гомогенат, и, кроме того, образует субклеточные фрагменты, которые трудно удалить из-за их малого размера. Их, как правило, удаляют дифференциальным центрифугированием или фильтрацией. Та же самая проблема возникает, хотя и в меньшей степени, с непосредственно секретирующимися белками из-за естественной гибели клеток и высвобождения внутриклеточных белков клетки-хозяина в ходе прохождения продукции белка.
Во время очистки терапевтических антител, примеси, включая белки клетки-хозяина, варианты продукта, ДНК клетки-хозяина, малые молекулы, связанные с процессом загрязнения, эндотоксины и вирусные частицы необходимо удалить (Fahrner, R.L. et al. (2001) Biotechnol. Genet. Eng. Rev. 18:301-327). Используемые способы очистки должны являться масштабируемыми, эффективными, экономически эффективными, надежными, и соответствовать жестким требованиям чистоты конечного продукта. Современные способы очистки, как правило, включают в себя множественные хроматографические разделения, включающие в себя ортогональные режимы разделения. Типичный способ может включать в себя некоторые из следующих стадий: преципитация (US 7169908), диализ, электрофорез, ультрафильтрация, аффинная хроматография, катионообменная хроматография, анионообменная хроматография и/или хроматография гидрофобного взаимодействия. Общепринятые стадии колоночной хроматографии являются эффективными и надежными, однако, как правило, обладают низкой производительностью для продукта (переработанных кг/ч). Поскольку моноклональные антитела становятся используемыми более широко, необходима более эффективная продукция в масштабе способа.
В способах хроматографии используют химические и физические свойства белков для достижения высокой степени очистки. Эти химические и физические свойства, как правило, включают в себя размер, изоэлектрическую точку, распределение заряда, гидрофобные участки и аффинность для лигандов (Janson, J. C. and L. Ryden (eds.). (1989) Protein Purification: Principles, High Resolution Methods and Applications. VCH Publishers, Inc., New York). Различные способы разделения в хроматографии включают в себя: ионообменную хроматографию, хроматофокусировку, гель- фильтрацию (эксклюзионную), хроматографию гидрофобного взаимодействия, обращеннофазовую и аффинную хроматографию. Ионообменная хроматография (IEX), включая анионообменную и катионообменную хроматографию, разделяет аналиты (например, белки) по различиям в их суммарных поверхностных зарядах. IEX является первичным средством для отделения экспрессированных белков от клеточного дебриса и других примесей. В настоящее время IEX является одним из наиболее часто используемых способов для очистки белков, пептидов, нуклеиновых кислот и других заряженных биомолекул, предоставляя высокоразрешающие и групповые разделения с высокой емкостью загрузки. Способ способен разделять виды молекул, обладающие только незначительными различиями в свойствах их зарядов, например, два белка, различающиеся одной заряженной аминокислотой. Эти свойства делают IEX хорошо подходящей для захвата, промежуточной очистки или окончательных стадий в протоколе очистки, и способ используют от очистки и анализа в микромасштабе до очистки килограммов продукта.
Способы хроматографии являются надежными, однако емкость и производительность могут являться проблематичными для крупномасштабных применений. Общепринятые стадии колоночной хроматографии являются эффективными и надежными, однако, как правило, обладают низкой производительностью для продукта (переработанных кг/час). Поскольку рекомбинантные белки становятся более широко используемыми, необходима более эффективная продукция в масштабе процесса. Производительность стадии хроматографии, как правило, ограничена емкостью хроматографической смолы для интересующего белка. При увеличении загрузки белка на колонку отделение интересующего белка от примесей часто уменьшается.
Известно, что полиэлектролиты образуют комплексы с белками, которые принимают формы растворимых комплексов (Dellacherie, E. (1991) Am. Chem. Soc, Div. Polym. Chem. Prepr. 32(1):602), аморфных преципитатов (Mattiasson et al. (1998) Polym. Plast. Technol. Eng. 37(3):303-308; Clark et al. (1987) Biotech. Progress 3(4):241; Fisher et al. (1988) Biotechnol. Bioeng. 32:777; Shieh et al. (1991) Am. Chem. Soc, Div. Polym. Chem. Prepr. 32(1)606; Sternberg et al. (1974) Biochimica et Biophysica Acta 342:195-206; WO 2004/014942) или коацерватов (Wang et al. (1996) Biotechnol. Prog. 12:356-362; Veis, A. (1991) Am. Chem. Soc. Div. Polym. Chem. Prepr. 32(1) 596). Протеолиз папаином моноклональных антител в присутствии антигена-поликатиона (полиметакриловая кислота) образует Fab-фрагменты (Dainiak et al. (2000) Analytical Biochem. 277:58-66).
Комплексы белок-полиэлектролит образуют коацерваты, т.е. разделяются на две отдельные жидкие фазы, где коацерватная фаза содержит большинство комплексов, а другая фаза представляет собой равновесную фазу (Burgess, D.J. «Complex Coacervation: Microcapsule Formation» in Macromolecular Complexes in Chemistry and Biology, Dubin, P. L., et al. Eds. (1994) Springer-Verlag, Berlin; Dubin et al. (1994) Sep. Purif. Complexes 23:1-16). Коацервация белков полиэлектролитами представляет собой сложный процесс и не является применимой для широкого диапазона белков. Межмолекулярные связи в комплексах белок-полиэлектролит обусловлены электростатическими взаимодействиями, водородными связями и гидрофобными силами (Cooper et al. (2005) Current Opinion in Colloid & Interface Science 10:52-78; Mattison et al. (1999) Macromol. Symp. 140:53-76). В то время как является известным, что добавление полиэлектролита к раствору белка может приводить к образованию комплексов белок-полиэлектролит и более крупных кластеров, и в конечном счете, к образованию коацерватов и/или преципитации, обратный процесс может возникать при дальнейшем добавлении полиэлектролита, в результате чего происходит повторное растворение белка, препятствуя выделению или очистке белка (Carlsson et al. (2003) J. Am. Chem. Soc. 125:3140-3149). Преципитация белков с использованием полиэлектролитов может обеспечивать экономически эффективную альтернативу хроматографическому разделению. С использованием этого способа возможно использовать функциональную химию хроматографических способов для достижения сходного уровня очистки белка в растворе. В частности, производительность стадии преципитации больше не будет ограниченной емкостью конкретной хроматографической смолы.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к способам, относящимся к выделению и очистке белков, полученных из жидкостей культур клеток.
Одним аспектом изобретения является способ очистки белка посредством преципитации белка с помощью полиэлектролита, такого как полианионный полиэлектролит.
Изобретение относится также к способу очистки белка посредством преципитации примесей жидкости культуры клеток с помощью поликатионного полиэлектролита. За стадией преципитации могут следовать стадии катионообменной хроматографии, анионообменной хроматографии и другой преципитации.
Способы по изобретению включают в себя неаффинный способ для очистки антитела.
Одним аспектом изобретения является способ очистки антител, включающий в себя:
(a) регуляцию кислотности или концентрации соли в смеси, содержащей антитело, где антитело получено из собранной жидкости культуры клеток;
(b) добавление отрицательно заряженного полиэлектролита, в результате чего образуется преципитат белок-полиэлектролит;
(c) отделение преципитата белок-полиэлектролит от примесей, выбранных из агрегатов белка, фрагментов белка, белков клетки-хозяина, инсулина, гентамицина, ДНК и вымываемого белка A;
(d) выделение преципитата белок-полиэлектролит; и
(e) ресуспендирования преципитата белок-полиэлектролит в водном растворе.
Другим аспектом изобретения является способ очистки антител, включающий в себя:
(a) регуляцию кислотности или концентрации соли в смеси, содержащей антитело, где антитело получено из жидкости культуры клеток;
(b) добавление положительно заряженного, поликатионного полиэлектролита к смеси, в результате чего образуется преципитат, содержащий положительно заряженный, поликатионный полиэлектролит и примеси, выбранные из агрегатов белка, фрагментов белка, белков клетки-хозяина, инсулина, гентамицина и ДНК; и
(c) отделение преципитата от смеси, содержащей антитело.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фигуре 1 показаны стадии способа для очистки пула белка A, очищенного из собранной жидкости культуры клеток (HCCF), с последующими: левая колонка - катионообменной хроматографией (Sepharose™ fast flow, SPSFF) затем анионообменной хроматографией (QSFF); средняя колонка - QSFF; или правая колонка - преципитацией поливинилсульфоновой кислотой (PVS) при pH 7, затем QSFF.
На фигуре 2 показаны стадии способа очистки пула SPSFF, очищенного из собранной жидкости культуры клеток (HCCF), с последующими: левая колонка - QSFF; или правая колонка - преципитацией PVS при pH 7, затем QSFF.
На фигуре 3 показаны стадии способа для непосредственного захвата и очистки антитела из HCCF: левая колонка - преципитацией PVS при pH 5, с последующими QSFF, затем SPSFF; или преципитацией PVS при pH 7, с последующими QSFF, затем SPSFF.
На фигуре 4 показаны кривые растворимости rhuMab 2H7 в PVS (1800 Да) при pH 5 при 3,0 и 5,6 мСм/см (пул белка A). мСм = миллисименс, единица проводимости.
На фигуре 5 показаны кривые растворимости rhuMab 2H7 в PVS (1800 Да) при pH 7 при 0,7, 1,5, 3,0 и 4,7 мСм/см (пул белка A).
На фигуре 6 показаны кривые растворимости rhuMab 2H7 в PVS (1800 Да) при pH 7 при 0,8; и 1,5 мСм/см (пул захвата SPSFF).
На фигуре 7 показаны кривые растворимости rhuMab 2H7 в PVS (1800 Да) при pH 5 и 3,0 мСм/см; и при pH 7 и 0,7 мСм/см (HCCF).
На фигуре 8 показаны кривые растворимости, сравнивающие преципитацию PVS (1800 Да) с преципитацией PAA (1200 и 8000 Да) при pH 5, 5 мСм/см для антитела против CD20, 2H7.
На фигуре 9 показаны кривые растворимости, сравнивающие преципитацию PVS (1800 Да) с преципитацией PAA (1200 и 8000 Да) при pH 7, 1,5 мСм/см для антитела против CD20, 2H7.
На фигуре 10 показаны кривые растворимости антитела против CD20 (ритуксимаб) при pH 7, 1,5 мСм/см, сравнивающие PAA с молекулярными массами в диапазоне от 1200 до 1100000 Да.
На фигуре 11 показана таблица краткого изложения результатов последующей переработки для анти-CD20 rhuMab 2H7: (a) HCCF, пропущенная через белок A; (b) HCCF, пропущенная через SPSFF; (c) преципитация HCCF PVS.
На фигуре 12 показаны кривые растворимости Апомаб в PVS (1800 Да) при pH 5,5 и 5,1 мСм/см; pH 6,0 и 3,0 мСм/см; pH 6,0 и 5,5 мСм/см.
На фигуре 13 показаны кривые растворимости Апомаб в PVS (1800 Да) при pH 6,5 и 1,5 мСм/см; pH 6,5 и 3,2 мСм/см; и pH 7,0 и 0,7 мСм/см.
На фигуре 14 показана фотография флаконов, содержащих пулы белка A для Апомаб при pH 5,5, с содержанием: (слева) в отсутсвие PVS (0%), (в центре) 0,1% PVS (масс./об.), и (справа) 1% PVS (масс./об.).
На фигуре 15 показаны кривые растворимости анти-cMet в PVS (1800 Да) при pH 4 и 2 мСм/см; pH 5 и 1 мСм/см; pH 5 и 2,4 мСм/см; pH 6 и 0,5 мСм/см; pH 7 и 0,5 мСм/см.
На фигуре 16 показаны стадии способа для прямого захвата и очистки антитела rhuMab 2H7 из CCF: левая колонка - центрифугирование HCCF с последующей преципитацией антитела с помощью PVS, затем QSFF; правая колонка - флокуляция/преципитация примесей полиаргинином, центрифугирование HCCF, преципитация антитела с помощью PVS, затем QSFF.
На фигуре 17 показаны стадии способа для очистки антитела rhuMab 2H7 из собранной жидкости культуры клеток (HCCF) с последующими: левая колонка - Prosep vA, катионообменной хроматографией (SEPHAROSE™ fast flow, SPSFF) затем анионообменной хроматографией (QSFF); правая колонка - преципитацией полиаргинином, Prosep vA, SPSFF, затем QSFF.
На фигуре 18 показан такой же способ очистки антитела rhuMab 2H7 из собранной жидкости культуры клеток (HCCF), как на фигуре 17, с исключением стадии Prosep vA: левая колонка - SPSFF, затем QSFF; правая колонка - преципитация полиаргинином, SPSFF, затем QSFF.
На фигуре 19 показаны стадии способа для очистки антитела rhuMab 2H7 из собранной жидкости культуры клеток (HCCF) с последующими: левая колонка - преципитацией антитела с помощью PVS, затем QSFF; правая колонка - преципитацией полиаргинином, преципитацией антитела с помощью PVS, затем QSFF.
На фигуре 20 показана кривая растворимости rhuMab 2H7 CCF с полиаргинином 110 кДа.
На фигуре 21 показаны кривые растворимости rhuMab 2H7 HCCF - полиаргинин 42 кДа.
На фигуре 22 показаны кривые растворимости rhuMab 2H7 HCCF - полиаргинин 110 кДа.
На фигуре 23 показаны кривые растворимости rhuMab 2H7, анти-CD22, rhuMab C2B8 HCCF - полиаргинин 42 кДа.
На фигуре 24 показаны кривые растворимости rhuMab 2H7, анти-CD22, rhuMab C2B8 HCCF - полиаргинин 110 кДа.
На фигуре 25 показаны кривые растворимости для преципитации PVS rhuMab 2H7 CCF, флокулированной с помощью 0,075% масс./об. полиаргинина 110 кДа.
На фигуре 26 показаны кривые проскока на SPSFF для пула белка A, полученного из преципитированной полиаргинином HCCF в качестве загрузки.
На фигуре 27 показаны кривые проскока на SPSFF для HCCF, полученной при преципитации полиаргинином, в качестве загрузки.
На фигуре 28 показан гель-электрофорез для ингибирования восстановления антитела для 2H7 Mab. HCCF 4-12% BT, буфер MOPS, краситель Sypro рубиновый, загрузка 2 мкг HCCF, 5 мин 70°C, экспозиция 9 с, 2 F/T прогон 7, инкубация при RT в SS мини-флаконах, образцы по 500 мкл (микролитров), немедленно замороженные при -70°C.
На фигуре 29 показаны кривые растворимости пула белка A rhuMab C2B8 - полилизин 2,5 кДа.
На фигуре 30 показаны кривые растворимости пула белка A rhuMab C2B8 - полилизин 50 кДа.
На фигуре 31 показаны кривые растворимости rhuMab C2B8 HCCF - полилизин 50 кДа.
На фигуре 32 показаны кривые растворимости rhuMab C2B8 HCCF - полилизин 225 кДа.
На фигуре 33 показаны кривые растворимости гуманизированного анти-CD20 с образцами от 1800 Да до 1132 кДа от 0,00% до 0,20% полистиролсульфоната (PSS) при pH 7 и проводимости 12 мСм/см.
На фигуре 34 показан график Vmax емкости (измеренной в граммах антитела на площадь фильтра) в зависимости от понижения давления для каждого фильтрующего материала: A1HC, B1HC, C0HC, 45CE, 20CE, 30CE.
На фигуре 35 показан эффект изменения количества пропускаемой жидкости на выход в пересчете на объем буфера, необходимого для полного повторного растворения преципитата белок-полиэлектролит 2H7-PVS с фильтрующей среды C0HC MILLISTAK+®. LMH = литров на квадратный метр в час, измерение скорости потока.
На фигуре 36 показаны эффекты двух диапазонов интенсивности протока (потока буфера для повторного растворения в LMH) на повторное растворение белка с фильтра C0HC MILLISTAK+® (преципитат белок-полиэлектролит 2H7-PVS).
На фигуре 37 показан схематический способ поточной настройки кондиционирования HCCF/преципитации PVS.
На фигуре 38 показаны выходы выделения антитела при кондиционировании/преципитации с непрерывной подачей.
ПОДРОБНОЕ ОПИСАНИЕ ПРИМЕРНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Здесь будет сделана подробная ссылка на конкретные варианты осуществления изобретения, примеры которых проиллюстрированы в сопутствующих структурах и формулах. В то время как изобретение будет описано в соответствии с перечисленными вариантами осуществления, следует понимать, что они не предназначены для ограничения изобретения этими вариантами осуществления. И напротив, изобретение предназначено, чтобы включать все альтернативы, модификации и эквиваленты, которые можно включать в объем настоящего изобретения, как определено посредством формулы изобретения. Специалисту в данной области будут известны много методов и материалов, сходных с описанными здесь или эквивалентных им, которые можно использовать в осуществлении настоящего изобретения на практике. Настоящее изобретение никаким образом не является ограниченным описанными методами и материалами. В случае, когда одно или несколько из включенной литературы, патентов и сходных материалов отличаются от этой заявки или противоречат ей, включая в качестве неограничивающих примеров определенные термины, использование терминов, описанные способы или т.п., эта заявка имеет преимущественную силу.
ОПРЕДЕЛЕНИЯ
Если не указано иначе, следующие термины и фразы, как применяют здесь, предназначены, чтобы обладать следующими значениями.
Термин «собранная жидкость культуры клеток», также обозначаемая как HCCF, обозначает жидкость культуры прокариотических или эукариотических клеток, из которой клетки удалили средствами, включающими центрифугирование или фильтрацию. Культивирование клеток представляет собой способ, посредством которого либо прокариотические, либо эукариотические клетки выращивают в контролируемых условиях. Термин «культура клеток» относится к культивированию клеток, полученных из многоклеточных эукариот, включая клетки животных, или одноклеточных прокариот, включая бактерии и дрожжи. Культуры эукариотических клеток включают в себя клетки млекопитающих, такие как клетки яичника китайского хомячка, гибридомы и клетки насекомых. При наличии подходящего сосуда для культивирования клеток секретированные белки можно получить из субстратзависимых клеток или суспензионных линий клеток. Культуры клеток млекопитающих включают в себя клетки яичника китайского хомячка (CHO).
Термин «микробиологическая ферментация» обозначает культивирование клеток бактерий или дрожжей, которые являются генетически модифицированными для продукции химических веществ, таких как белки. Ферментацию используют для размножения клонированных бактерий и дрожжей, так же как других микроорганизмов, и продукции ценных белков. Продуктивность клеток и рост этих организмов максимизируют предоставлением конкретной среды для роста и контролем различных факторов внешней среды (таких как pH, температура, и аэрация). Жидкость после бактериальной ферментации можно получать от E. Coli.
Термин «антитело» используют здесь в самом широком смысле и конкретно распространяют на интактные моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антитела, пока они обладают желаемой биологической активностью. Антитела могут являться мышиными, человеческими, гуманизированными, химерными или полученными из других видов.
Антитело представляет собой белок, образованный иммунной системой, который является способным узнавать и связывать специфический антиген (Janeway, et al. (2001) «Immunobiology», 5th Ed., Garland Publishing, New York). Антиген-мишень, как правило, обладает многочисленными участками связывания, также называемыми эпитопами, узнаваемыми CDR на многих антителах. Каждое антитело, которое специфически связывается с отличающимся эпитопом, обладает отличающейся структурой. Таким образом, один антиген может обладать более чем одним соответствующим антителом.
Термин «антитело», как применяют здесь, также относится к полноразмерной молекуле иммуноглобулина или иммунологически активной части полноразмерной молекулы иммуноглобулина, т.е. к молекуле, содержащей антигенсвязывающий участок, который иммуноспецифически связывает антиген из интересующей мишени или ее части, где такие мишени включают в качестве неограничивающих примеров, раковую клетку или клетки, продуцирующие аутоиммунные антитела, связанные с аутоиммунным заболеванием. Иммуноглобулин, описанный здесь, может принадлежать любому типу (например, IgG, IgE, IgM, IgD и IgA), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекулы иммуноглобулина. Иммуноглобулины можно получать из любых видов. В одном аспекте, однако, иммуноглобулин происходит из человека, мыши или кролика.
«Фрагменты антител» содержат часть полноразмерного антитела, как правило, его антигенсвязывающий участок. Примеры фрагментов антител включают в себя фрагменты Fab, Fab', F(ab')2 и Fv; диатела; линейные антитела; фрагменты, продуцированные экспрессирующей библиотекой Fab, антиидиотипические (анти-Id) антитела, CDR (определяющую комплементарность область), ECD (внеклеточный домен), и эпитопсвязывающие фрагменты любого из вышеуказанного, которые иммуноспецифически связываются с антигенами раковой клетки, вирусными антигенами или микробными антигенами, одноцепочечные молекулы антител; и мультиспецифические антитела, сформированные из фрагментов антител.
Термин «моноклональное антитело», как применяют здесь, относится к антителу, полученному из популяции по существу однородных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высоко специфичными, являясь направленными против отдельного антигенного участка. Кроме того, в отличие от препаратов поликлональных антител, которые содержат различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональные антитело направлено против одной детерминанты антигена. В дополнение к их специфичности, моноклональные антитела являются преимущественными в том, что их можно синтезировать без загрязнения другими антителами. Определение «моноклональное» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует рассматривать как требование получения антитела каким-либо конкретным способом. Например, моноклональные антитела для применения по настоящему изобретению можно получить способом гибридомы, первоначально описанным Kohler et al. (1975) Nature 256:495, или можно получить способами рекомбинантной ДНК (US 4816567). «Моноклональные антитела» можно выделить также из фаговых библиотек антител с применением способов, описанных, например, в Clackson et al. (1991) Nature, 352:624-628; Marks et al. (1991) J. Mol. Biol, 222:581-597.
Применимые моноклональные антитела представляют собой однородные популяции антител к конкретной антигенной детерминанте (например, антигену раковой клетки, вирусному антигену, микробному антигену, белку, пептиду, углеводу, химическому веществу, нуклеиновой кислоте или их фрагментам). Моноклональное антитело (Mab) к интересующему антигену можно получать с использованием любого способа, известного в данной области, который обеспечивает продукцию молекул антитела стабильными клеточными линиями в культуре. Они включают в себя в качестве неограничивающих примеров способ гибридомы, первоначально описанный Köhler and Milstein (1975) Nature 256:495-497), способ гибридомы B-клетки человека (Kozbor et al. (1983) Immunology Today 4:72), и способ EBV-гибридомы (Cole et al. (1985) in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96). Такие антитела могут принадлежать любому классу иммуноглобулинов, включая IgG, IgM, IgE, IgA, и IgD и любому их подклассу. Гибридому, продуцирующую Mab для применения по этому изобретению, можно культивировать in vitro или in vivo.
Терапевтические моноклональные антитела, применимые в способах по изобретению, включают в себя трастузумаб (ГЕРЦЕПТИН®, Genentech, Inc., Carter et al. (1992) Proc. Natl. Acad. Sci. USA, 89:4285-4289; US 5725856); антитела против CD20, такие как химерные анти-CD20 «C2B8» (US 5736137); ритуксимаб (РИТУКСАН®), окрелизумаб, химерный или гуманизированный вариант антитела 2H7 (US 5721108; WO 04/056312), или тозитумомаб (БЕКСАР®); анти-IL-8 (St John et al. (1993) Chest, 103:932, и WO 95/23865); антитела против VEGF, включая гуманизированные и/или аффинно зрелые антитела против VEGF, такие как гуманизированное антитело против VEGF huA4.6.1 бевацизумаб (АВАСТИН®, Genentech, Inc., Kim et al. (1992) Growth Factors 7:53-64, WO 96/30046, WO 98/45331); антитела против PSCA (WO 01/40309); антитела против CD40, включая S2C6 и его гуманизированные варианты (WO 00/75348); анти-CD11a (US 5622700; WO 98/23761; Steppe et al. (1991) Transplant Intl. 4:3-7; Hourmant et al. (1994) Transplantation 58:377-380); анти-IgE (Presta et al. (1993) J. Immunol. 151:2623-2632; WO 95/19181); анти-CD18 (US 5622700; WO 97/26912); анти-IgE, включая E25, E26 и E27 (US 5714338; US 5091313; WO 93/04173; US 5714338); антитело против рецептора Apo-2 (WO 98/51793); антитела против TNF-альфа, включая cA2 (РЕМИКЕЙД®), CDP571 и MAK-195 (US 5672347; Lorenz et al. (1996) J. Immunol. 156(4): 1646-1653; Dhainaut et al. (1995) Crit. Care Med. 23(9): 1461-1469); против тканевого фактора (TF) (EP 0 420 937 B1); против интегрина альфа-4 бета-7 человека (WO 98/06248); анти-EGFR, химеризованное или гуманизированное антитело 225 (WO 96/40210); антитела против CD3, такие как OKT3 (US 4515893); антитела против CD25 или против tac, такие как CHI-621 СИМУЛЕКТ® и ЗЕНАПАКС® (US 5693762); антитела против CD4, такие как антитело cM-7412 (Choy et al. (1996) Arthritis Rheum 39(l):52-56); антитела против CD52, такие как КАМПАТ-1H (Riechmann et al. (1988) Nature 332:323-337); антитела против рецептора Fc, такие как антитело M22, направленное против RI Fc-гамма, как в Graziano et al. (1995) J. Immunol. 155(10):4996-5002; антитела против карциноэмбрионального антигена (CEA), такие как hMN-14 (Sharkey et al. (1995) Cancer Res. 55(23Suppl): 5935s-5945s; антитела, направленные против эпителиальных клеток молочной железы, включая huBrE-3, hu-Mc 3 и CHL6 (Ceriani et al. (1995) Cancer Res. 55(23):5852s-5856s; и Richman et al. (1995) Cancer Res. 55(23 Supp): 5916s-5920s); антитела, связывающиеся с клетками карциномы толстой кишки, такие как C242 (Litton et al. (1996) Eur J. Immunol. 26(1): 1-9); антитела против CD38, например, AT 13/5 (Ellis et al. (1995) J. Immunol. 155(2):925-937); антитела против CD33, такие как Hu M195 (Jurcic et al. (1995) Cancer Res 55(23 Suppl):5908s-5910s и CMA-676 или CDP771; антитела против CD22, такие как LL2 или лимфоцид (Juweid et al. (1995) Cancer Res 55(23 Suppl):5899s-5907s); антитела против EpCAM, такие как 17-1A (ПАНОРЕКС®); антитела против GpIIb/IIIa, такие как абциксимаб или c7E3 Fab (РЕОПРО®); антитела против RSV, такие как MEDI-493 (СИНАГИС®); антитела против CMV, такие как ПРОТОВИР®; антитела против HIV, такие как PRO542; антитела против гепатита, такие как антитело против Hep B ОСТАВИР®; антитело против CA 125 оварекс; антиидиотипическое антитело против эпитопа GD3 BEC2; антитело ВИТАКСИН® против альфа-v бета-3; антитело против почечноклеточного рака человека, такое как ch-G250; ING-1; антитело против 17-1A человека (3622W94); антитело против опухоли ободочной и прямой кишки человека (A33); антитело против меланомы человека R24, направленное против ганглиозида GD3; против плоскоклеточной карциномы человека (SF-25); и антитела против лейкоцитарного антигена человека (HLA), такие как Smart ID10 и антитело против HLA DR Онколим (Lym-1).
Применимые моноклональные антитела включают в себя в качестве неограничивающих примеров, человеческие моноклональные антитела, гуманизированные моноклональные антитела, фрагменты антител или химерные моноклональные антитела человек-мышь (или другие виды). Человеческие моноклональные антитела можно получать любым из множества способов, известных в данной области (Teng et al. (1983) Proc. Natl. Acad. Sci. USA 80:7308-7312; Kozbor et al. (1983) Immunology Today 4:72-79; и Olsson et al. (1982) Methods in Enzymology 92:3-16).
Антитело может также представлять собой биспецифическое антитело. Биспецифические антитела могут обладать гибридной тяжелой цепью иммуноглобулина с первой специфичностью связывания на одном плече, и гибридной парой тяжелая цепь - легкая цепь иммуноглобулина (обеспечивающей вторую специфичность связывания) на другом плече. Эта ассиметричная структура облегчает отделение желательного биспецифического соединения от нежелательных сочетаний цепей иммуноглобулинов, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы обеспечивает легкий способ разделения (WO 94/04690; Suresh et al. (1986) Methods in Enzymology, 121:210; Rodrigues et al. (1993) J. of Immunology 151:6954-6961; Carter et al. (1992) Bio/Technology 10:163-167; Carter et al. (1995) J. of Hematotherapy 4:463-470; Merchant et al. (1998) Nature Biotechnology 16:677-681). Способы получения биспецифических антител известны в данной области (Milstein et al. (1983) Nature 305:537-539; WO 93/08829; Traunecker et al. (1991) EMBO J. 10:3655-3659). С использованием таких способов биспецифические антитела можно получить для конъюгации в качестве ADC для лечения или предупреждения заболевания, как определено здесь.
Согласно другому способу, вариабельные домены антител с желаемыми специфичностями связывания (участками объединения антитело-антиген) сливают с последовательностями константного домена иммуноглобулина. Слитый белок может обладать константным доменом тяжелой цепи иммуноглобулина, содержащей по меньшей мере часть шарнирной, CH2 и CH3 областей. Первая константная область тяжелой цепи (CH1) может содержать участок, необходимый для связывания легкой цепи, присутствующий по меньшей мере в одном из слитых белков. Нуклеиновые кислоты с последовательностями, кодирующими слитые белки с тяжелой цепью иммуноглобулина и, если желательно, с легкой цепью иммуноглобулина, вставляют в отдельные экспрессирующие векторы и котрансфицируют в подходящий организм-хозяин. Это предоставляет большую гибкость в регулировании взаимного соотношения трех полипептидных фрагментов в вариантах осуществления, когда с неравными соотношениями трех полипептидных цепей, применяемых в конструкции, достигают оптимального выхода. Однако можно вставлять кодирующие последовательности для двух или всех трех полипептидных цепей в один экспрессирующий вектор, когда экспрессия по меньшей мере двух полипептидных цепей в равных соотношениях приводит к высоким выходам, или когда соотношения не являются особенно важными.
Гибридные или бифункциональные антитела можно получать либо биологически, т.е. способами слияния клеток, либо химически, особенно с помощью перекрестно-сшивающих средств или формирующих дисульфидный мостик реагентов, и они могут содержать целые антитела или их фрагменты (EP 105360; WO 83/03679; EP 217577).
Антитело может представлять собой функционально активный фрагмент, производное или аналог антитела, которые иммуноспецифически связываются с антигенами раковой клетки, вирусными антигенами или микробными антигенами, или другими антителами, связанными с опухолевыми клетками или матриксом. В этом отношении, «функционально активный» означает, что фрагмент, производное или аналог является способным вызывать образование анти-антиидиотипических антител, которые узнают тот же самый антиген, который узнавало антитело, из которого получен фрагмент, производное или аналог. В частности, в примерном варианте осуществления антигенность идиотипа молекулы иммуноглобулина можно увеличить делецией последовательностей каркаса и CDR, которые находятся на C-конце от последовательности CDR, которая специфически узнает антиген. Чтобы определить, какие последовательности CDR связывают антиген, синтетические пептиды, содержащие последовательности CDR, можно использовать в анализах связывания с антигеном посредством любого способа анализа связывания, известного в данной области, например анализа BIA core (Kabat et al., (1991) in Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, Md; Kabat et al. (1980) J. of Immunology 125(3):961-969).
Другие применимые антитела включают в себя фрагменты антител, например, в качестве неограничивающих примеров, фрагменты F(ab')2, содержащие вариабельную область, константную область легкой цепи и домен CH1 тяжелой цепи, можно получить расщеплением пепсином молекулы антитела, и фрагменты Fab, которые можно получить восстановлением дисульфидных мостиков фрагментов F(ab')2. Другие применимые антитела представляют собой димеры тяжелой цепи и легкой цепи антител, или любой их минимальный фрагмент, такой как Fv или одноцепочечные антитела (SCA) (например, как описано в US 4946778; Bird (1988) Science 242:423-42; Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; и Ward et al. (1989) Nature 334:544-54), или любую молекулу с той же самой специфичностью, что и антитело.
Моноклональные антитела здесь конкретно включают в себя «химерные» антитела, в которых часть тяжелой и/или легкой цепи является идентичной или гомологичной соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, в то время как остаток цепи(цепей) является идентичным или гомологичным соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител, так же как фрагменты таких антител, пока они обладают желательной биологической активностью (US 4816567; и Morrison et al. (1984) Proc. Natl. Acad. Sci. USA, 81:6851-6855). Химерное антитело представляет собой молекулу, в которой различные части происходят из различных видов животных, такую как молекулы, обладающие вариабельной областью, происходящей из мышиного моноклонального иммуноглобулина, и константными областями человеческого иммуноглобулина (US 4816567; US 4816397). Химерные антитела включают в себя «приматизированные» антитела, содержащие антигенсвязывающие последовательности вариабельного домена, происходящие из не относящегося к человеку примата (например, обезьяны Старого Света, человекообразной обезьяны и т.д.), и последовательности константных областей человека.
Химерные и гуманизированные моноклональные антитела, содержащие как человеческие, так и не относящиеся к человеку части, можно получать с использованием общепринятых способов рекомбинантной ДНК (WO 87/02671; EP 184187; EP 171496; EP 173494; WO 86/01533; US 4816567; EP 12023; Berter et al. (1988) Science 240:1041-1043; Liu et al. (1987) Proc. Natl. Acad. Sci. USA 84:3439-3443; Liu et al. (1987) J. Immunol. 139:3521-3526; Sun et al. (1987) Proc. Natl. Acad. Sci. USA 84:214-218; Nishimura et al. (1987) Cancer. Res. 47:999-1005; Wood et al. (1985) Nature 314:446-449 и Shaw et al. (1988) J. Natl. Cancer Inst. 80:1553-1559; Morrison (1985) Science 229:1202-1207; Oi et al. (1986) BioTechniques 4:214; US 5225539; Jones et al. (1986) Nature 321:552-525; Verhoeyan et al. (1988) Science 239:1534 и Beidler et al. (1988) J. Immunol. 141:4053-4060; полное содержание каждого из которых приведено здесь в качестве ссылки.
Полностью человеческие антитела можно получать с использованием трансгенных мышей, которые неспособны экспрессировать эндогенные гены тяжелых и легких цепей иммуноглобулинов, но способны экспрессировать человеческие гены тяжелых и легких цепей. Трансгенных мышей иммунизируют обычным образом выбранным антигеном, например полным полипептидом по изобретению или его частью. Моноклональные антитела, направленные против антигена, можно получать с использованием общепринятого способа гибридомы. Трансгены иммуноглобулинов человека, которые несут трансгенные мыши, подвергаются реаранжировке во время дифференцировки B-клеток, и затем подвергаются переключению классов и соматической мутации. Таким образом, с использованием такого способа можно получить терапевтически применимые антитела IgG, IgA, IgM и IgE. Обзор этого способа для получения человеческих антител см. в Lonberg and Huszar (1995) Int. Rev. Immunol. 13:65-93; патентах США No. 5625126; 5633425; 5569825; 5661016; 5545806). Другие человеческие антитела можно получать из коммерческих источников, например Abgenix, Inc. (Freemont, CA) и Genpharm (San Jose, CA).
Полностью человеческие антитела, узнающие выбранный эпитоп, можно получать с использованием способа, обозначаемого «направленный отбор». В этом способе выбранное не относящееся к человеку моноклональное антитело, например мышиное антитело, используют, чтобы направлять отбор полностью человеческого антитела, узнающего тот же самый эпитоп (Jespers et al. (1994) Biotechnology 12:899-903). Человеческие антитела можно также получать с использованием различных способов, известных в данной области, включая библиотеки фагового дисплея (Hoogenboom and Winter, J. Mol. Biol. 227:381 (1991); Marks et al. (1991) J. Mol. Biol. 222:581).
Антитело может представлять собой слитый белок с антителом или его функционально активный фрагмент, например, в котором антитело сливают через ковалентную связь (например, пептидную связь), либо с N-концом, либо с C-концом аминокислотной последовательности другого белка (или его части, такой как часть белка по меньшей мере из 10, 20 или 50 аминокислот), который не является антителом. Антитело или его фрагмент может являться ковалентно связанным с другим белком на N-конце константного домена.
Антитела включают в себя аналоги и производные, которые являются каким-либо образом модифицированными, т.е. посредством ковалентного присоединения любого типа молекулы, пока такое ковалентное присоединение позволяет антителу сохранять иммуноспецифичность его связывания антигена. Например, но не в качестве ограничения, производные и аналоги антител включают в себя те, которые были дополнительно модифицированы, например, посредством гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидирования, дериватизации известными защитными/блокирующими группами, протеолитического расщепления, присоединения к клеточному комплексу антитела или другому белку и т.д. Любую из многочисленных химических модификаций можно осуществлять известными способами, включая, в качестве неограничивающих примеров, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез в присутствии туникамицина и т.д. Кроме того, аналог или производное может содержать одну или несколько неприродных аминокислот.
Примеры антител против CD20 включают в себя: «C2B8», в настоящее время ритуксимаб (РИТУКСАН®/МАБТЕРА®) (US 5736137); меченное иттрием-[90] мышиное антитело 2B8 Y2B8 или ибритутомаб тиуксетан (ЗЕВАЛИН®, Biogen Idec, Inc. US 5736137); 2B8, депонированное в ATCC под инвентарным № HB11388 22 июня 1993 г.); мышиное IgG2a «B1», называемое также тозитумомаб, необязательно, меченное 131I для получения антитела «131I-B1» или «иод 113I-тозитумомаба» (БЕКСАР®), коммерчески доступного от Corixa (US 5595721); мышиное моноклональные антитело «1F5» (например, Press et al. (1987) Blood 69(2):584-591) и его варианты, включая «обладающее частично замененным каркасом» или гуманизированное 1F5 (WO 2003/002607, Leung, S.; депозит в ATCC HB-96450); мышиное 2H7 и химерное 2H7 антитело (US 5677180); окрелизумаб, гуманизированный вариант 2H7 и другие варианты 2H7 (WO 2004/056312; US 5721108); HUMAX-CD20™, полностью человеческое, высокоаффинное антитело, нацеленное на молекулу CD20 на клеточной мембране B-клеток (Genmab, Denmark). См., например, Glennie и van de Winkel, (2003) Drug Discovery Today 8:503-510 и Cragg et al. (2003) Blood 101:1045-1052); человеческие моноклональные антитела, описанные в WO 2004/035607 и WO 2005/103081 (Teeling et al., GenМab/Medarex); антитела, обладающие комплексными N-гликозид-связанными сахарными цепями, связанными с областью Fc, описанные в US 2004/0093621 (Shitara et al.); моноклональные антитела и антигенсвязывающие фрагменты, связывающиеся с CD20 (WO 2005/000901, Tedder et al.), такие как HB20-3, HB20-4, HB20-25, и MB20-11; одноцепочечные белки, связывающиеся с CD20 (US 2005/0186216; US 2005/0202534; US 2005/0202028; US 2005/0202023); связывающиеся с CD20 молекулы, такие как серии антител AME, например антитела AME-133™ (WO 2004/103404; US 2005/0025764); и антитела против CD20 с мутациями в Fc (WO 2005/070963); связывающиеся с CD20 молекулы (WO 2005/016969; US 2005/0069545); биспецифические антитела (WO 2005/014618); гуманизированные моноклональные антитела LL2 (US 2005/0106108); химерные или гуманизированные антитела B-Ly1 против CD20 (WO 2005/044859; US 2005/0123546); = антитело A20 или его варианты, такие как химерное или гуманизированное антитело A20 (cA20, hA20, соответственно) и IMMUN-106 (US 2003/0219433); и моноклональные антитела L27, G28-2, 93-1B3, B-Cl или NU-B2, доступные из International Leukocyte Typing Workshop (Valentine et al. (1987) In: Leukocyte Typing III, McMichael, Ed., p. 440, Oxford University Press). Примерные антитела против CD20 включают в себя химерные, гуманизированные или человеческие антитела против CD20, такие как ритуксимаб, гуманизированное 2H7, химерное или гуманизированное антитело A20, такое как HUMAX-CD20™, человеческое антитело против CD20 (Genmab), и иммуноглобулины/белки, связывающиеся с CD20 (Trubion Pharm Inc.).
«Интактное» антитело представляет собой антитело, содержащее антигенсвязывающую вариабельную область, так же как константный домен легкой цепи (CL) и константные домены тяжелой цепи, CH1, CH2 и CH3. Константные домены могут представлять собой константные домены с нативной последовательностью (например, константные домены с человеческой нативной последовательностью) или вариант их аминокислотной последовательности.
Интактное антитело может обладать одной или несколькими «эффекторными функциями», которые относятся к тем видам биологической активности, которые можно приписать области Fc (нативной последовательности области Fc или варианту аминокислотной последовательности области Fc) антитела. Примеры эффекторных функций антитела включают в себя связывание C1q; комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую опосредуемую клетками цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов поверхности клеток (например B-клеточного рецептора; BCR) и т.д.
В зависимости от аминокислотной последовательности константных доменов их тяжелых цепей интактные антитела можно отнести к различным «классам». Существует пять главных классов интактных антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно далее разделить на «подклассы» (изотипы), например IgG1, IgG2, IgG3, IgG4, IgA и IgA2. Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно. Субъединичные структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны.
Термин «вариант аминокислотной последовательности» относится к полипептидам, обладающим аминокислотными последовательностями, которые до некоторой степени отличаются от нативной последовательности полипептида. Обычно варианты аминокислотной последовательности будут обладать по меньшей мере приблизительно 70% идентичностью последовательности по меньшей мере с одним связывающим рецептор доменом нативного антитела или по меньшей мере с одним связывающим лиганд доменом нативного рецептора, и предпочтительно, они будут по меньшей мере приблизительно на 80%, более предпочтительно, по меньшей мере приблизительно на 90% гомологичными по последовательности с такими связывающими рецептор или лиганд доменами. Варианты аминокислотной последовательности обладают заменами, делециями и/или вставками в конкретных положениях внутри аминокислотной последовательности нативной аминокислотной последовательности. Аминокислоты обозначены общепринятыми наименованиями, однобуквенным и трехбуквенным кодами.
«Идентичность последовательности» определяют как процент остатков в варианте аминокислотной последовательности, которые являются идентичными после выравнивания последовательностей и введения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей. Способы и компьютерные программы для выравнивания хорошо известны в данной области. Одной такой компьютерной программой является «Align 2», созданная Genentech, Inc., представленная с пользовательской документацией в United States Copyright Office, Washington, DC 20559, 10 декабря 1991 г.
ЭКСПРЕССИЯ И ПРОДУКЦИЯ БЕЛКА
Рекомбинантные белки экспрессируют посредством клонирования ДНК из векторов и способов, известных в данной области. Белки для способов очистки полиэлектролитами по изобретению можно продуцировать из подходящих клеток-хозяев, таких как клетки прокариот, дрожжей и высших эукариот. Подходящие прокариоты для этой цели включают в себя эубактерии, такие как грамотрицательные или грамположительные организмы, например Enterobacteriaceae, такие как Escherichia, например E. coli, Enterohacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, так же как Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанный в DD 266710, опубликованной 12 апреля 1989 г.), Pseudomonas, такой как P. aeruginosa, и Streptomyces. Одним из предпочтительных хозяев E. coli для клонирования является E. coli 294 (ATCC 31446), хотя пригодны другие штаммы, такие как E. coli B; E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Данные примеры являются скорее иллюстративными, чем ограничивающими.
Кроме прокариот, подходящими клонирующими или экспрессирующими хозяевами для векторов, кодирующих связывающееся с CD20 антитело, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи. Saccharomyces cerevisiae, или общераспространенные пекарские дрожжи, является общеупотребительным микроорганизмом-хозяином из низших эукариот. Однако ряд других родов, видов и штаммов являются общедоступными и применимыми здесь, такие как Schizosaccharomyces pombe; хозяева Kluyveromyces, такие как, например, K. Lactis, K. fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и мицелиальные грибы, такие как, например, Neurospora, Penicillium, Tolypocladium и хозяева Aspergillus, такие как A. nidulans и A. niger.
Подходящие клетки-хозяева для экспрессии гликозилированных антител могут происходить из многоклеточных эукариотических организмов. Примеры клеток беспозвоночных включают в себя клетки насекомых и растений. Идентифицирован ряд штаммов и вариантов бакуловирусов и соответствующих пермиссивных клеток-хозяев среди таких хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (москит), Aedes albopictus (москит), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Множество штаммов вирусов для трансфекции являются широко доступными, например вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV, и такие вирусы можно использовать в качестве вирусов по изобретению здесь, в частности для трансфекции клеток Spodoptera frugiperda.
Размножение клеток позвоночных в культуре (культуре тканей) стало общепринятым способом. Примерами применимых линий клеток хозяев-млекопитающих являются линия клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); линия клеток эмбриональной почки человека (клетки 293 или 293, субклонированные для выращивания в суспензионной культуре, Graham et al. (1977) J. Gen Virol. 36:59); клетки почки детеныша хомячка (BHK); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al. (1980) Proc. Natl. Acad. Sci. USA 77:4216) Mather (1980) Biol. Reprod. 23:243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки цервикальной карциномы человека (HELA); клетки почки собаки (MDCK); клетки печени крысы буффало (BRL 3A); клетки легкого человека (W138); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562); клетки TRI (Mather et al., Annals N.Y. Acad. Sci 383:44-68 (1982)); клетки MRC 5; клетки FS4 и линия гепатомы человека (Hep G2).
Клетки-хозяева трансформируют описанными выше экспрессирующими и клонирующими векторами для продукции антитела и культивируют в общепринятой питательной среде, модифицированной соответствующим образом веществами для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих желаемые последовательности.
Клетки-хозяева, используемые для продукции антител для способов по этому изобретению, можно культивировать в разнообразных средах. Для культивирования клеток-хозяев пригодны коммерчески доступные среды для роста, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда (MEM), (Sigma), RPMI-1640 (Sigma), и среда Игла, модифицированная Дульбекко ((DMEM), Sigma). Кроме того, в качестве культуральной среды для клеток-хозяев можно использовать любую из сред, описанных в Ham et al. (1979) Meth. Enz. 58:44, Barnes et al. (1980) Anal. Biochem.102:255, US 4767704; US 4657866; US 4927762; US 4560655; US 5122469; WO 90/03430; WO 87/00195; или в патенте США Re. 30985. Любые из этих сред можно дополнить при необходимости гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид и фосфат натрия, кальция, магния), буферами (такими как MES и HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство ГЕНТАМИЦИН™), микроэлементами (определяемыми как неорганические соединения, обычно присутствующие в конечных концентрациях микромолярного диапазона) и глюкозой или эквивалентным источником энергии. Также можно добавлять любые другие необходимые добавки в соответствующих концентрациях, известных специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., представляют собой условия, которые ранее использовали для выбранных для экспрессии клеток-хозяев, и они будут очевидными обычному специалисту в данной области.
После получения собранной жидкости культуры клеток, содержащей интересующий белок, его отделение от других белков, продуцируемых клеткой, обычно предпринимают с использованием комбинации различных способов хроматографии. Этими способами разделяют смеси белков на основании их заряда, степени гидрофобности или размера. Для каждого из этих способов доступно несколько различных хроматографических смол, что позволяет осуществить более точную настройку схемы очистки для конкретного рассматриваемого белка. Сущность каждого из этих способов разделения в том, что белки можно заставить либо двигаться с различными скоростями вниз по длинной колонке, достигая физического разделения, которое увеличивается, когда они проходят дальше вниз колонки, либо избирательно связываться со средой для разделения, с последующей дифференциальной элюцией различными растворителями. В некоторых случаях желаемый белок отделяют от примесей, когда примеси специфически связываются с колонкой, а интересующий белок нет, то есть интересующий белок присутствует в «проскоке».
С использованием рекомбинантных способов можно продуцировать антитело внутри клетки, в периплазматическом пространстве, или оно может непосредственно секретироваться в среду. Если антитело продуцируется внутри клеток, в качестве первой стадии, дебрис из твердых частиц, или клеток-хозяев, или лизированных фрагментов, удаляют, например, центрифугированием или ультрафильтрацией, как для антител, которые секретируются в периплазматическое пространство E. coli (Carter et al. (1992) Bio/Technology 10:163-167). Клеточную массу можно размягчать в присутствии ацетата натрия (pH 3,5), ЭДТА и фенилметилсульфонилфторида (PMSF) в течение приблизительно 30 мин. Клеточный дебрис можно удалять центрифугированием. Когда антитело секретируется в среду, супернатанты из таких экспрессирующих систем, как правило, сначала концентрируют с использованием коммерчески доступного фильтра для концентрации белка, например установки для ультрафильтрации Amicon или Millipore Pellicon. Ингибитор протеаз, такой как PMSF, можно включать в любую из вышеуказанных стадий для ингибирования протеолиза, и антибиотики можно включать для предотвращения роста случайных контаминантов.
Общепринятые способы очистки белков, экспрессированных из клеток, включают в себя хроматографию на гидроксилапатите, ионообменную хроматографию, аффинную хроматографию, хроматографию гидрофобного взаимодействия, гель-электрофорез, диализ и их комбинации (Fahrner, RX. et al. (2001) Biotechnol. Genet. Eng. Rev. 18:301-327). Белок A является общепринятым в качестве аффинного лиганда, который можно иммобилизовать на различных подложках и который позволяет обогащение на начальной стадии собранных жидкостей культуры клеток (HCCF), содержащих экспрессированные белки. Белок A является применимым адсорбентом для аффинной хроматографии белков, таких как антитела, которые содержат область Fc. Белок A представляет собой белок клеточной стенки 41 кДа из Staphylococcus aureas, который связывается с высокой аффинностью (приблизительно 10-8 M для IgG человека) с областью Fc антител (Sulkowski, E. (1987) Protein Purification: Micro to Macro, pgs 177-195; Chadha et al. (1981) Preparative Biochemistry 11(4):467-482; Reifsnyder et al. (1996) J. Chromatography 753:73-80; US 6127526; 6333398). Белок A можно иммобилизовать на твердой фазе, такой как стекло, оксид кремния, агароза или полистирол. Твердая фаза может представлять собой колонку для очистки или дискретную фазу из дискретных частиц, например колонку из стекла с контролированным размером пор или колонку с кремниевой кислотой, или покрытую реагентом (таким как глицерин), который предназначен для предотвращения неспецифической адгезии загрязнений на твердой фазе (US 6870034). Колонка PROSEP A™, коммерчески доступная от Bioprocessing Limited, является примером стеклянной колонки с контролируемым размером пор с белком A, покрытой глицерином. Другие примеры колонок, предусматриваемых здесь, включают в себя колонку с POROS 50 A™ (полистирол) или колонку с rProtein A SEPHAROSE FAST FLOW™ (агароза). Твердую фазу для хроматографии с белком A можно уравновешивать подходящим буфером. Например, буфер для уравновешивания представляет собой 25 мМ Трис, 25 мМ NaCl, 5 мМ ЭДТА, pH 7,1.
При иммобилизации на среде для хроматографии белок A обеспечивает способ для очистки рекомбинантных антител, поскольку он может избирательно связывать антитела в сложных растворах, позволяя протекать примесям, таким как белки клетки-хозяина и малые молекулы. Пригодность белка A в качестве аффинного лиганда зависит от вида и изотипа любого домена Fc иммуноглобулина, который присутствует в антителе. Белок A можно использовать для очистки антител, основанных на тяжелых цепях γ1, γ2 или γ4 человека (Lindmark et al. (1983) J. Immunol. Meth. 62:1-13). Белок G рекомендован для всех изотипов мыши и для γ3 человека (Guss et al. (1986) EMBO J. 5:15671575). Матрица, к которой присоединен аффинный лиганд, наиболее часто представляет собой агарозу, но доступны другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол позволяют более быстрые скорости потока и более короткие периоды времени обработки, чем можно достичь с агарозой. Где антитело содержит домен CH3, смола Bakerbond ABX™ (J. T. Baker, Phillipsburg, NJ) является применимой для очистки. Другие способы очистки белка, такие как фракционирование на ионообменной колонке, преципитация этанолом, обращеннофазовая HPLC, хроматография на оксиде кремния, хроматография на гепарин-СЕФАРОЗЕ™, хроматография на анионо- или катионообменной смоле (например, на колонке с полиаспарагиновой кислотой), хроматофокусировка, SDS-PAGE и преципитация сульфатом аммония, также являются доступными, в зависимости от подлежащего выделению антитела.
ПОЛИЭЛЕКТРОЛИТЫ
Полиэлектролиты представляют собой водорастворимые полимеры, составленные из заряженных мономерных единиц. Мономерные единицы полиэлектролита по изобретению несут электролитную (заряженную функциональную) группу, которая подвергается протонной диссоциации (ионизации) в водных растворах согласно кислотности (pH). Заряженные функциональные группы полиэлектролитов включают в себя в качестве неограничивающих примеров сульфоновую кислоту, фосфоновую кислоту, карбоновую кислоту и амины, и их соответствующие ионы: сульфонат, фосфонат, карбоксилат и аммоний. Диссоциация влияет на ионную силу и электропроводность раствора.
Полиэлектролиты, применимые для способов по изобретению, могут обладать молекулярной массой, лежащей в диапазоне от приблизительно тысячи (1000) дальтон (Да) до приблизительно миллиона (1000000) дальтон. Полиэлектролиты по изобретению можно использовать в виде смеси из конкретного типа повторяющейся мономерной единицы, но с широким диапазоном длин цепей, т.е. с диапазоном молекулярной массы от приблизительно 1200 дальтон (Да) до приблизительно миллиона (1000000) дальтон. Смесь может лежать в узком диапазоне, например от приблизительно 1200 Да до приблизительно 2400 Да, или от приблизительно 4000 Да до приблизительно 8000 Да. Среднюю молекулярную массу и профиль распределения молекулярных масс можно контролировать в конкретных условиях полимеризации мономерных единиц, таких как концентрация, инициатор или катализатор полимеризации, температура или время. Среднюю молекулярную массу и профиль распределения молекулярных масс полиэлектролита можно также выбирать для конкретных способов препаративной очистки.
Отрицательно заряженные, анионные, полиэлектролиты и положительно заряженные, катионные, полиэлектролиты можно использовать в реакции преципитации. Когда pH раствора меньше, чем pI конкретного антитела, антитело является положительно заряженным. В этих условиях катионный полиэлектролит может преципитировать примеси и оставлять интересующее антитело в растворе. И наоборот, анионный полиэлектролит может преципитировать антитело, образуя преципитат белок-полиэлектролит, оставляя примеси в растворе. Другие факторы при выборе полиэлектролитов для способов по изобретению включают в себя стабильность и реакционную способность функциональной группы, молекулярную массу, плотность заряда и жесткость цепи.
Полиэлектролиты по изобретению включают в себя полиамфолиты, несущие как катионные, так и анионные заряженные функциональные группы.
Примерные полианионные полиэлектролиты по изобретению включают в себя: полиакриловую кислоту (PAA), поливинилсульфонат (PVS), полистиролсульфоновую кислоту (PSS, поли(4-винилбензолсульфонат, соль металла), полиметакрилат (PMA), полиакриламидометилпропансульфонат (PAMPS), карбоксиметилцеллюлозу (CMC), сополимер малеинового ангидрида-стирола (MAS), сополимер малеинового ангидрида-винилметилэфира (MAVE), полиаспартат, полиглутамат, декстрансульфат, пектин, альгинат и гликозаминогликаны, такие как хондроитинсульфат, гепарин/гепарансульфат и гиалуроновая кислота; и все их соли и сополимеры.
Полиакриловая кислота (PAA) и поливинилсульфоновая кислота (PVS), и их анионы, полиакрилат и поливинилсульфонат, соответственно, являются применимыми полиэлектролитами для способов по изобретению. PAA обладает сходными структурными свойствами с PVS. Обе являются полимерными и обладают линейными углеродными остовами. Два полиэлектролита различаются их функциональными группами. PVS обладает функциональной группой сульфоновой кислоты, придающей ей pKa приблизительно единица. В отличие от этого, PAA обладает функциональной группой карбоновой кислоты и pKa приблизительно пять. PVS доступна с одной молекулярной массой, определенной как в среднем 1800 Да посредством исключения по размеру, где среднее n составляет приблизительно 10-20, по детекции динамического рассеяния света. PAA является коммерчески доступной со множеством молекулярных масс, включая 1200 и 8000, где среднее n составляет приблизительно 15-20 и 100-120, соответственно. PAA и PVS закуплены у Sigma-Aldrich Co., St. Louis MO, Polysciences Inc., Warrington, PA, и Carbomer, Inc., San Diego, CA.
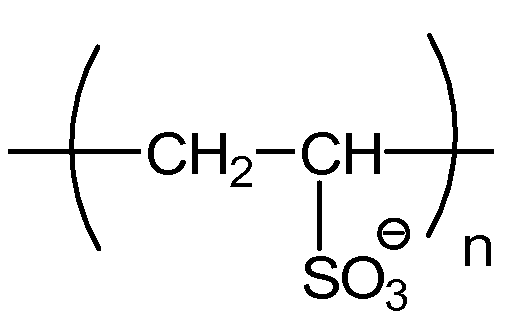
Поливинилсульфоновая кислота (PVS)
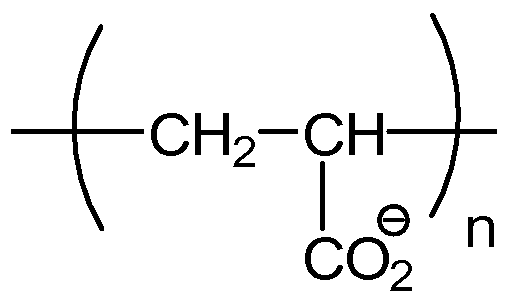
Полиакриловая кислота (PAA, полиакрилат)
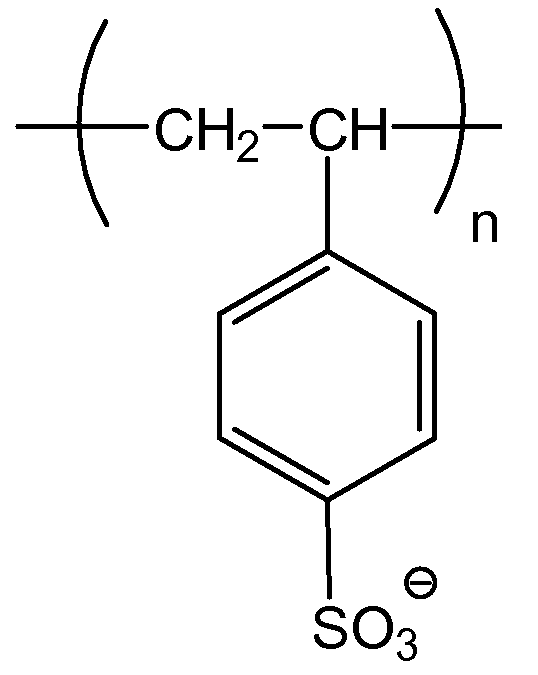
Полистиролсульфонат (PSS)
Примерные положительно заряженные, поликатионные полиэлектролиты по изобретению включают в себя: полиаргинин (PLA, включая гидрохлорид поли-L-аргинина: Sigma-Aldrich P-4463, MW 5-15 кДа, P-7762 MW 15-70 кДа, P-3892, MW >70 кДа, CAS No. 26982-20-7, и сульфат поли-L-аргинина, Sigma-Aldrich P-7637 MW 15-50 кДа, CAS No. 26700-68-5); полилизин (например, гидрохлорид поли-L-лизина, номер CAS: 26124-78-7, Sigma-Aldrich: P-2658 MW 15-30 кДа, CAS No. 26124-78-7, PLL, Jacobson, B.S. and Branton, D. (1977) Science 195, 302), полиорнитин (например, поли-L-орнитин, Sigma-Aldrich P-2533 MW 15-30 кДа, CAS No. 26982-21-8), поливинилгуанидин (поли(винилгуанидин), US 6087448), хлорид полидиаллилдиметиламмония, бромид поли(N-этил-4-винилпиридиния), хлорид полиметилакриламидопропилтриметиламмония, хлорид поливинилбензилтриметиламмония и полигистидин.
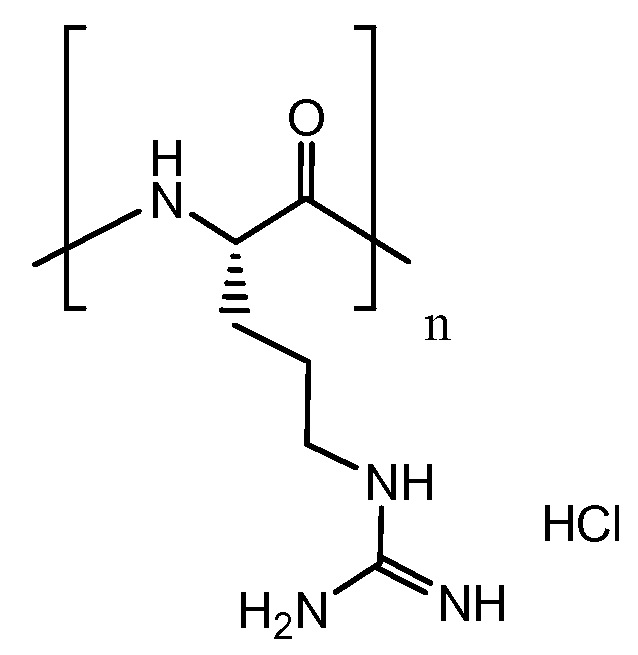
Полиаргинин (гидрохлорид поли-L-аргинина)
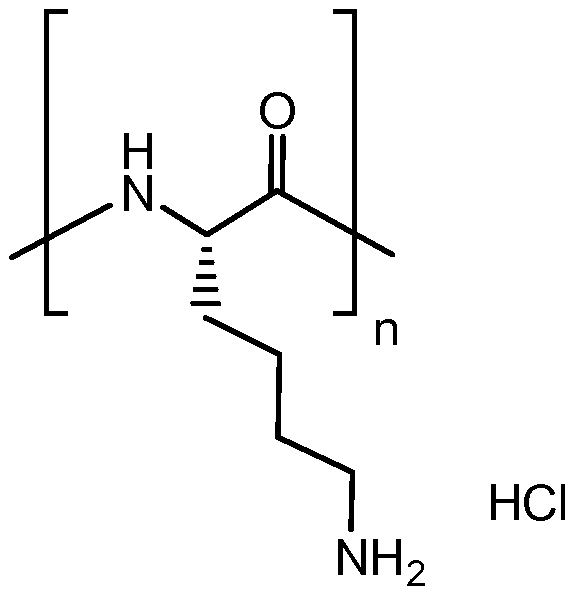
Полилизин (гидрохлорид поли-L-лизина)
ПРЕЦИПИТАЦИЯ ПОЛИЭЛЕКТРОЛИТАМИ В ОЧИСТКЕ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ
Преципитацию полиэлектролитами можно использовать для отделения антител от примесей во время очистки. Эти примеси включают в себя примеси клетки-хозяина, такие как белки CHO (CHOP) и белки E.coli (ECP), компоненты культуры клеток, такие как инсулин, гентамицин и ДНК, связанные с процессом примеси, такие как вымываемый белок A, и связанные с продуктом примеси, такие как агрегаты и фрагменты антитела. Стадия преципитации может служить заменой существующим хроматографическим разделениям. Альтернативно, преципитацию белков полиэлектролитами можно использовать в качестве стадии прямого захвата из собранной жидкости культуры клеток (HCCF) из эукариотической или прокариотической культуры, или в качестве стадии промежуточной очистки. Преципитацию полиэлектролитами можно также использовать в качестве стадии осветления при очистке белка, осветляющей жидкость культуры клеток, включая HCCF. Полиэлектролиты могут формировать хлопьевидный материал в жидкости культуры клеток, который оседает и позволяет быстрое и эффективное осветление, обогащение, преципитацию или очистку смесей, содержащих белок, такой как антитело, экспрессированный в культуре прокариотических или эукариотических клеток. Способы очистки полиэлектролитами по изобретению могут заменять операции сбора, такие как операции центрифугирования, фильтрации и хроматографии. Способы очистки полиэлектролитами по изобретению предоставляют удивительные и неожиданные преимущества в очистке белков, таких как антитела.
Условия, подлежащие оптимизации для преципитации полиэлектролитами, включают в себя pH раствора, проводимость, буферы, концентрацию белка, концентрацию полиэлектролита, так же как скорость и тип перемешивания, и скорость добавления полиэлектролита.
Преципитацию можно проводить в реакторе с перемешиванием. pH и проводимость пула антител можно доводить до намеченных условий на основании оптимальных условий, идентифицированных из кривых растворимости. Полиэлектролит добавляют и перемешивают. Скорость и тип перемешивания может влиять на эффективность преципитации и время преципитации. Преципитация следует за добавлением полиэлектролита. Преципитирующее средство отделяют от супернатанта с использованием фильтрации или центрифугирования. В случае преципитации анионным полиэлектролитом, где используют фильтрацию, преципитирующее средство отмывают буфером для промывки. Преципитирующее средство, содержащее антитело, будут затем ресуспендировать и перерабатывать далее. Возможно также проводить преципитацию в поточной линии, т.е. без сосуда с перемешиванием, где пул антитела будут доводить до намеченного pH и затем будут добавлять полиэлектролит. Полная преципитация не будет происходить, пока пул не будет разведен, что можно проводить на линии либо с аппаратом для фильтрации, либо с центрифугой. Альтернативно, после доведения пула антитела до намеченного pH, будут проводить поточное растворение с последующим добавлением полиэлектролита в поточной линии перед аппаратом для фильтрации или центрифугой.
В одном примере гормон роста человека (pI 5,2) при кристаллизации или преципитации в буфере при pH 7 будет заряжен отрицательно и, таким образом, будет взаимодействовать или образовывать комплекс с катионными полиэлектролитами. Подобным образом, моноклональные антитела, такие как ритуксимаб и трастузумаб, с pI более 9, будут способны формировать комплекс с анионными полиэлектролитами в нейтральных буферах. Оценку суммарного заряда белка можно вычислить, когда аминокислотную последовательность определят посредством общедоступных программ. Кислые белки, те белки, которые обладают более высоким содержанием аспарагиновой кислоты (pKa 4,5) и глутаминовой кислоты (pKa 4,5), как правило, обладают pI менее 6-6,4. С другой стороны, основные белки, те белки, которые обладают более высоким содержанием гистидина (pKa 6,2), лизина (pKa 10,4) и аргинина (pKa 12), как правило, обладают pI выше, чем приблизительно 7,5-8. В отличие от них обоих, нейтральные белки, те, которые, как правило, обладают сходными количествами кислых и основных аминокислотных остатков, обладают pI, которые являются нейтральными (pI составляют, как правило, приблизительно 6,5-7,4).
Хотя и не в исчерпывающем перечислении, некоторыми примерами pI для различных терапевтических белков являются следующие: рекомбинантный эритропоэтин человека (pI 4); дорназа альфа, rhДНКаза (ПУЛЬМОЗИМ®) (pI 5); этанерцепт (ЭНБРЕЛ™) (pI 5,1); инсулин (pI 5,4); гранулоцитарный колониестимулирующий фактор (pI 5,5-5,9); TNF альфа (pI 5,6); фибролаза (pI 6,7); IL-1 beta (pI 6,9); рекомбинантный тканевой активатор плазминогена (pI 6,5-8,5); ортоклон OKT3 (pI 6,7-7,2); фактор VIII (pI 7-7,6); бычий соматотропин (pI 7,4); интерлейкин 2 (pI 7,44); инсулиноподобный фактор роста-1 (pI 8,4) и апротинин (pI 10,5).
Способ преципитации анионным полиэлектролитом оценивали в качестве замены стадии катионного обмена (SPSFF) для нескольких примерных антител (анти-CD20 rhuMab 2H7, rhuMab DR5 Апомаб и анти-cMet), и также в качестве замены для стадии белка A (Prosep vA) для rhuMab 2H7 (окрелизумаб). Другие антитела и другие белки можно очищать способами преципитации полиэлектролитами, описанными здесь. Кривые растворимости использовали для идентификации эффективных или оптимальных условий преципитации (pH, проводимость и концентрация полимера) для трех антител. Для стадии преципитации полиэлектролитом продемонстрировали способность уменьшать количество белков клетки-хозяина, таких как белки яичника китайского хомячка (CHOP) и белки E. coli (ECP), вымываемый белок A, малые молекулы, такие как инсулин и гентамицин, так же как фрагменты и агрегаты антитела. Стадия преципитации полиэлектролитом не оказывает отрицательного влияния на биологическую активность 2H7, как измерено по активности CDC. Полиэлектролит PVS удаляли от преципитированного белка до уровня менее 1 мкг/мл с использованием анионообменной (QSFF) хроматографии.
Различные серии стадий способа проиллюстрированы на схемах организации производства из фигур 1-3. Стадии комбинировали и исключали, чтобы оценивать их результаты для очистки белка и эффективности способа. В одном из общепринятых способов, в настоящее время применяемых на практике в данной области для очистки экспрессированного белка из собранной жидкости культуры клеток (HCCF), используют три стадии хроматографии: (1) захват белком A (Prosep vA), (2) катионный обмен (SPSFF) и (3) анионный обмен (QSFF), показанные в виде левой колонки на фигуре 1. Предпринимали попытку очистки захватом белком A непосредственно после анионного обмена (средняя колонка, фигура 1). Стадию катионного обмена можно заменить стадией преципитации PVS при pH 7 (правая колонка, фигура 1). Собранную жидкость культуры клеток можно перерабатывать без захвата белком A посредством исходного катионного обмена со следующим непосредственно за ним анионным обменом (левая колонка, фигура 2), или посредством преципитации PVS при pH 7 с последующим анионным обменом (правая колонка, фигура 2). Преципитацию полиэлектролитом можно проводить в качестве прямого захвата из HCCF, с последующей анионообменной, затем катионообменной хроматографией (фигура 3). Преципитацию полиэлектролитом проводили при pH 5 (левая колонка, фигура 3) и при pH 7 (правая колонка, фигура 3).
Выбор условий преципитации для антитела против CD20, rhuMab 2H7
Кривые растворимости получали для рекомбинантного гуманизированного моноклонального антитела против CD20, rhuMab 2H7, в диапазоне ионных сил. Кривые растворимости представляют собой график остаточного белка, оставшегося в супернатанте после преципитации (выраженного в процентах) в зависимости от концентрации PVS, выраженной как моль PVS/моль антитела. Кривые растворимости получены для пула белка A при pH 5 и pH 7 (фигуры 4 и 5), пула SPSFF (фигура 6) при использовании в качестве стадии прямого захвата при pH 7 и для HCCF при pH 5 и pH 7 (фигура 7). Эти кривые использовали для выбора pH, ионной силы и концентрации PVS, при которой будут проводить преципитации в препаративном масштабе.
Преципитация пула белка A (фигура 4 и фигура 5)
При увеличении концентрации PVS присутствовала незамедлительная преципитация при pH 5, как показано по уменьшению процента антитела, оставшегося в растворе - C/Co, т.е. концентрация антитела в растворе, деленная на общее антитело (фигура 4). C/Co (%) представляет собой процент (%) непреципитированного белка, деленный на исходный белок. Когда C/Co (%) составляет 0%, присутствовала полная преципитация.
Снижением ионной силы являлось возможным достигать более высокого уровня преципитации (4,7 мСм/см против 3,0 мСм/см, где мСм представляет собой миллисименс, единицу проводимости). При pH 7 требовалось разведение для достижения значительной преципитации (фигура 5). При снижении проводимости присутствовало увеличение преципитации. Полную преципитацию наблюдали при проводимости 0,7 мСм/см. При конкретной проводимости, после того как наблюдали максимальный уровень преципитации, добавление дополнительного PVS вызывало повторное растворение преципитирующего средства.
Преципитация пула SPSFF (фигура 6) и HCCF (фигура 7): Сходные тенденции наблюдали для пула SPSFF и HCCF. При снижении проводимости присутствовало увеличение преципитации. Полную преципитацию наблюдали при проводимости 0,7 мСм/см. При конкретной проводимости, после того как наблюдали максимальный уровень преципитации, добавление дополнительного PVS вызывало повторное растворение преципитирующего средства.
Оценка вклада молекулярной массы полиэлектролита
Получены также кривые растворимости, сравнивающие PVS с PAA. Кривые растворимости представляют собой график остаточного белка, оставшегося в супернатанте после преципитации (выраженного в процентах) в зависимости от концентрации полиэлектролита, выраженной как моль полиэлектролита/моль антитела. Кривые растворимости для PAA (1200 & 8000 Да) и PVS (1800 Да) в пределах диапазона эквивалентов полиэлектролита на антитело против CD20 (rhuMab-2H7, pI 9,0, 150 кДа), измерено при pH 5 (фигура 8) и pH 7 (фигура 9). Форма PAA с наиболее высокой молекулярной массой (8000 Да) приводит к большей преципитации при pH 5 и величине проводимости 5 мСм/см (фигура 8) по сравнению с полиэлектролитами - формой PAA с более низкой молекулярной массой (1200 Да) и PVS (1800 Да) (фигура 8). Сходную тенденцию наблюдали при pH 7, 1,5 мСм/см (фигура 9). PAA с более высокой молекулярной массой 8000 Да приводит к большей преципитации при pH 7 и величине проводимости 1,5 мСм/см по сравнению с полиэлектролитами - PAA с более низкой молекулярной массой 1200 Да и PVS 1800 Да. Основной единицей проводимости является сименс (S), ранее называемый мо (mho). Измерения проводимости являются зависимыми от температуры. Значения проводимости выражены как мСм/см. В обоих случаях более крупный полиэлектролит PAA обладал более узким рабочим диапазоном в отношении концентрации полиэлектролита, необходимой для достижения максимальной преципитации перед тем, как дополнительное добавление полиэлектролита вызывает повторное растворение преципитирующего средства. Увеличение молекулярной массы полиэлектролита может позволять проведение преципитации при более высоких проводимостях.
Эффект молекулярной массы полиэлектролита на преципитацию антитела против CD20 (ритуксимаба) при pH 7 и 1,5 мСм/см исследовали с PAA в пределах диапазона молекулярной массы от 1200 Да до 1100000 Да (фигура 10). Увеличение молекулярной массы приводит к увеличению преципитации в этих условиях. При pH 7 и 1,5 мСм/см не достигали полной преципитации, пока не использовали PAA с молекулярной массой более 35000 Да (35 кДа).
Эффект молекулярной массы полиэлектролита на преципитацию гуманизированного антитела против CD20 при pH 7 и 12 мСм/см исследовали также с полистиролсульфонатом (PSS) в пределах диапазона молекулярной массы от 1800 Да до 1132000 Да (фигура 33). Кривые растворимости получали для преципитации PSS гуманизированного антитела против CD20 в пределах этого диапазона молекулярной массы. Увеличение молекулярной массы приводит к увеличению преципитации в этих условиях. При pH 7 и 12 мСм/см не достигали полной преципитации, пока не использовали PSS с молекулярной массой более 220000 Да (220 кДа). Использование PSS позволяет проводить преципитацию при более высокой проводимости и минимизирует необходимость снижать проводимость/ионную силу пулов для достижения полной преципитации.
Эффект молекулярной массы полиэлектролита на очистку от примесей измеряли в условиях таблицы 1, и результаты последующей переработки показаны в таблице 2. После стадии преципитации PAA 8000 Да обладала более высокими уровнями яичника китайского хомячка (CHOP), чем PAA 1200 Да или PVS (таблица 2). С пулами QSFF, сходные уровни CHOP наблюдали для всех пулов независимо от молекулярной массы полиэлектролита, используемого для очистки. Обе молекулярные массы PAA снижали вымываемый белок A и фрагменты антитела до сходных уровней и не оказывали отрицательного влияния на уровни агрегатов.
Условия преципитации PAA
Обобщение результатов последующей переработки
Очистка антитела против CD20
Преципитацию полиэлектролитом исходно оценивали в качестве замены стадии катионообменной хроматографии. Первичной функцией стадии катионного обмена является уменьшение примесей клетки-хозяина, удаление вымываемого белка, гентамицина, ДНК и уменьшение агрегатов антитела, если они присутствуют (фигура 11). Инсулин удаляют предыдущей стадией с белком A. Стадия преципитация PVS уменьшала CHOP до сходных уровней, как стадия SPSFF. Она уменьшала также вымываемый белок A и фрагменты антитела. Присутствовало уменьшение уровня гентамицина в ходе стадии преципитации, хотя и не до такой же степени, как для стадии SPSFF. Анализ ДНК не функционирует в присутствии PVS, так что очистку PVS в ходе стадии преципитации невозможно было оценить. Добавление стадии преципитации не увеличивало агрегаты антитела или влияние на биологическую активность, как измеряли по активности CDC (фигура 11 и таблица 3).
Чтобы предоставить более сложное сырье, HCCF пропускали непосредственно через колонку SPSFF. В пуле SPSFF присутствовало в 4 раза больше CHOP, чем в пуле белка A, при использовании в качестве стадии исходного захвата (фигура 11). % фрагментов также был выше, чем для способа с захватом белком A (9,24% против 0,24%). Вероятно, что протеаза, очищаемая совместно с антителом в ходе стадии катионного обмена, приводит к расщеплению антитела. Стадия преципитации PVS уменьшала уровень фрагментов до 5,63%. CHOP уменьшался до 4,1 нг/мг в ходе способа, по сравнению с 71 нг/мг без стадии преципитации. Гентамицин также значительно уменьшали стадией преципитации (с 146 до 8 нг/мг). Инсулин и ДНК уменьшали до низких уровней посредством стадии исходного захвата SPSFF, так что очистку от них невозможно было оценить. Высокий уровень фрагментов в контрольном прогоне (SPSFF-QSFF) приводил к уменьшению активности CDC до 87% (таблица 3). В отличие от этого, прогон с преципитацией PVS обладал 100% активностью CDC.
Стадию PVS оценивали также в качестве стадии непосредственного захвата из HCCF. Стадия преципитации PVS при pH 7 удаляла инсулин (фигура 11). В отличие от этого, стадия преципитации PVS при pH 5 только частично уменьшала уровни инсулина. Преципитация PVS при pH 7 удаляла в 3 раза больше белков клетки-хозяина, чем преципитация при pH 5. Замена конечной стадии SPSFF способом ортогональной хроматографии, такой как стадия гидрофобного взаимодействия, может дополнительно уменьшать примеси клетки-хозяина. При захвате с pH 5 присутствовало 4,66% увеличение фрагментов антитела в ходе стадии катионного обмена. Этого не происходило на стадии захвата с pH 7. При этом можно предположить, что при pH 5 (наиболее вероятно, на стадии захвата SPSFF), совместно очищают кислую протеазу, что вызывает увеличение количества фрагментов антитела. Добавление стадии преципитации не влияло на биологическую активность, как измерено по активности CDC (таблица 3). Пулы Q анализировали по биологической активности/эффективности с использованием анализа комплементзависимой цитотоксичности (CDC). Этот анализ основан на измерении способности rhuMab 2H7 лизировать клетки WIL2-S в присутствии комплемента человека.
Эффект преципитации PVS на активность CDC
Очистку от полиэлектролита (удаление полиэлектролита от ресуспендированного антитела) определяли с использованием анализа FRET (резонансный перенос энергии флуоресценции) на основании ингибирования РНКазы A. PVS является сильным ингибитором РНКазы A. В анализе используют аналог РНК с флуоресцентной меткой на одном конце и тушителем на другом. При расщеплении аналога РНК РНКазой A флуоресцентная метка освобождается от тушителя, что приводит к излучению. Присутствие PVS будет ингибировать активность РНКазы A, ограничивая флуоресцентное излучение. Затем количество PVS можно определять сравнением наблюдаемой флуоресценции из тестируемого образца со стандартной кривой. Во всех случаях, PVS очищали до менее чем 1 мкг/мкл на хроматографии QSFF (таблица 4).
Количественная оценка очистки от PVS после преципитации антитела против CD20 (2H7)
Очистка rhuMab DR5 Апомаб
Способ преципитации полиэлектролитом разработали и применили для очистки антитела rhuMab DR5 Апомаб (pI 8,0, 150 кДа). Белки, содержащие домен смерти рецептора-5 (DR5), являются членами семейства рецепторов фактора некроза опухоли (TNF) (US 6872568; US 6743625). В общепринятом способе хроматографии используют смолы Prosep vA, SPSFF и QSFF. Способ преципитации полиэлектролитом модифицировали для удаления проблематичной примеси белка клетки-хозяина, глутатион-s-трансферазы (GST). Стадия Prosep vA обладает промывкой 0,5 М TMAC, pH 5, в отличие от промывки 0,4 M фосфатом калия, pH 7. На стадии SPSFF проводят элюцию в пологом градиенте, получая объем пула приблизительно 12 объемов колонки. Преципитацию PVS оценивали в качестве замены стадии SPSFF. Кривые растворимости получали в пределах диапазона pH 5,5-7,0. При увеличении pH для достижения полной преципитации требовалось разведение для уменьшения ионной силы (фигуры 12 и 13). На фигуре 12 разведение для уменьшения ионной силы с увеличением pH являлось необходимым для достижения полной преципитации. При pH 5,5 и проводимости 5,1 мСм/см наблюдали полную преципитацию. Однако при pH 6 являлось необходимым разводить пул до 3 мСм/см для достижения полной преципитации. На фигуре 13 разведение для уменьшения ионной силы при увеличении pH являлось необходимым для достижения полной преципитации. При pH 6,5 полную преципитацию наблюдали при 1,5 мСм/см. При pH 7 являлось необходимым разводить пул до 0,7 мСм/см для достижения полной преципитации.
Как только достигали полной преципитации, добавление дополнительного PVS вызывало повторное растворение преципитирующего средства. Этот эффект проиллюстрирован фотографиями на фигуре 14. На фигуре 14 проиллюстрирован эффект преципитации 0,1% PVS (масс./об.) во флаконе Апомаб в растворе и повторное растворение при 1% PVS (масс./об.). Кривые растворимости использовали для выбора условий преципитации, описанных в таблице 5. Преципитацию PVS проводили между pH 5,5 и pH 7,0. При увеличении pH присутствовало небольшое уменьшение выхода, но улучшение в очистке от CHOP и вымываемого белка A (таблица 6). Затем эти пулы пропускали через колонку QSFF и сравнивали с контрольными прогонами (таблица 6). Стадия SPSFF или стадия преципитации PVS являлась необходимой для очистки от CHOP до < 0,79 нг/мг и для очистки от вымываемого белка A до < 2 нг/мг по сравнению с 17 нг/мг CHOP и 4 нг/мг вымываемого белка A в контрольном прогоне без стадии SPSFF или преципитации. Для преципитации PVS показали также способность уменьшать уровень GST, но не до того же самого уровня, как для стадии SPSFF.
Условия преципитации Апомаб
Очистка Апомаб - Результаты последующей переработки для промежуточных пулов Апомаб (a) и пулов QSFF (b)
Очистка анти-cMet
Кривые растворимости получали для анти-cMet в пределах диапазона pH и выбранных ионных сил из-за ограниченной доступности материала. Кривые растворимости получали для пула белка A, доведенного до pH 4 - pH 7 (фигура 15). Эти кривые использовали для выбора pH, ионной силы и концентрации PVS, при которых будут проводить преципитации в препаративном масштабе. При pH 4 и проводимости 2 мСм/см анти-cMet являлось полностью преципитированным из раствора при молярном соотношении PVS:Ab более 40 (фигура 15). При pH 5 кривые растворимости получали при проводимостях 1 мСм/см и 2,4 мСм/см. При более высокой проводимости анти-cMet являлось не полностью преципитированным из раствора. При понижении проводимости раствора до 1 мСм/см анти-cMet более легко преципитировать из раствора. При этой проводимости максимальная преципитация происходила при молярных соотношениях PVS:Ab 100. При молярных соотношениях более 100 комплекс полиэлектролита обладал увеличенной растворимостью, вероятно, из-за эффектов отталкивания заряд-заряд полиэлектролита. Сравнение кривых растворимости при pH 4 и pH 5, проводимости ~2 мСм/см позволяет предполагать, что эффекты электростатического экранирования противоионами раствора влияют на pH, при котором происходит разделение объемной фазы, и что уменьшение ионной силы в растворе является необходимым при увеличении pH. При pH 6 и pH 7, и проводимостях раствора 0,5 мСм/см, анти-cMet являлось не полностью преципитированным из раствора в пределах тестируемых концентраций PVS. Максимальная преципитация 21% (C/Co = 0,79) и 12% (C/Co = 0,88) происходила при pH 6 и pH 7, соответственно. Кривая растворимости анти-cMet при pH 6 и pH 7 заметно отличалась при сравнении с полноразмерными гуманизированными антителами со сходной pI, 8,3. Например, с использованием сходных условий, pH 7,0 и раствора с проводимостью 0,7 мСм/см, Апомаб являлось полностью преципитированным (C/Co = 0) с такими же значениями R, использованными в этих экспериментах.
Способ очистки для анти-cMet (pI 8,3, 100 кДа) с преципитацией PVS сравнивали с общепринятым способом с использованием четырех стадий хроматографии, стадии захвата белком A с последующими двумя стадиями катионного обмена, проведенными в режиме связывания и элюции, и, наконец, стадией хроматографии гидрофобного взаимодействия, действующей в поточном режиме (таблица 7). Обе стадии катионного обмена и стадию гидрофобного взаимодействия заменили преципитацией PVS с последующим прогоном анионообменной хроматографии в поточном режиме. При pH 4 реакцию преципитации можно проводить при более высоких ионных силах. Однако при преципитации при pH 5 можно получать большую очистку от примесей, так как больше примесей клетки-хозяина должны являться отрицательно заряженными при этом pH и, таким образом, оставаться в растворе. Условия pH 5,0, проводимости 0,6 мСм/см и молярного отношения PVS 100 (0,1% масс./об.) выбрали для максимизации преципитации антитела, как определено из кривых растворимости. На выход выделения после преципитации PVS анти-cMet влияла растворимость осадка PVS:антитело. Осадок PVS:антитело являлся гелеобразным и трудно растворимым. После солюбилизации осадка в буфере для уравновешивания QSFF раствор являлся мутным с твердыми частицами. Раствор легко было отфильтровать с использованием вакуумного фильтра 0,4 мкм, получая прозрачный раствор, который перерабатывали в ходе анионообменной хроматографии с Q-Сефарозой.
Стадия преципитации PVS, проведенная при pH 5, 0,5 мСм/см и 0,1% PVS (масс./об.), уменьшала ECP до сходных уровней, как и стадия с CM-Сефарозой (таблица 7). Во время процесса преципитации значительного уменьшения уровня вымываемого белка A или увеличения % мономера не наблюдали.
Обобщение результатов последующей переработки анти-CMet
Выбор условий фильтрации для захвата преципитата
Объемные фильтры, обладающие адсорбирующим фильтрующим материалом, в частности, подходящие для удаления биологических загрязнений из культуры клеток, можно использовать в способах по изобретению. Объемные фильтры, как правило, используют для продукции биофармацевтических средств, которые получают из культуры клеток млекопитающих, для целей осветления различных жидкостей неочищенного продукта. Целлюлозные объемные фильтры, такие как фильтры MILLISTAK+®, коммерчески доступные от Millipore Corporation, обладают пористым неподвижным слоем из адсорбирующего материала, такого как гранулированный адсорбент, заключенным в нерастворимое в воде термопластичное связующее. Полученный композитный фильтр обеспечивает более высокое количество адсорбента с меньшими частицами адсорбента, чем общепринятые объемные фильтры. Эти композитные фильтры содержат слой плотно структурированной целлюлозной объемной среды, и их можно оптимизировать для конкретного применения, такого как удерживание коллоидных частиц и клеточного дебриса или удерживание цельных клеток и более крупного дебриса. В них комбинируют последовательные градации среды, например, посредством пакетных мембран в одном фильтрующем картридже. Такие объемные фильтры можно использовать в способах доочистки или вторичного осветления для удаления малых количеств взвешенных веществ из потоков водного продукта (белка). Фильтры могут также служить защитой или продлевать эксплуатационный ресурс более дорогих последующих процессов разделения, таких как стерильная фильтрация и аффинная хроматография, посредством удалении коллоидных загрязнений и другого клеточного дебриса. Кроме того, такие объемные фильтры являются также применимыми для защиты фильтров для очистки от вирусов посредством удаления следовых количеств агломерированных белков.
Конкретные объемные фильтры также могут задерживать, до различной степени, некоторые растворимые загрязнения, обычно обнаруживаемые в культурах клеток млекопитающих, такие как нуклеиновые кислоты, белки клетки-хозяина, липиды, поверхностно-активные вещества и т.д. Эта задерживающая способность для конкретных растворимых загрязнений основана на адсорбционных свойствах объемных фильтрующих сред. Фильтрующие среды, обычно применяемые в объемных фильтрах, включают в себя измельченные целлюлозные волокна (полученные из древесной пульпы и/или хлопка), диатомовую землю или связующие вещества из водорастворимой термоотверждающейся смолы (US 2007/0193938). Диатомитовая земля (природная форма оксида кремния, содержащая следовые количества различных силикатов) в этих композитах составляет, как правило, 40-60% по массе, адсорбируя биологический материал, такой как фрагменты клеток, органеллы и агломерированные белки, так же как различные растворимые биохимические вещества, такие как белки, липиды и нуклеиновые кислоты (ДНК и РНК).
Объемную фильтрацию оценивали в качестве стадии захвата для преципитата PVS антитела. Широкое распределение по размерам преципитата представляет задачу для захвата преципитата с использованием фильтрации. Каждую фильтрующую среду тестировали с преципитированным антителом в условиях нормальной поточной фильтрации. Проводили скрининг объемных фильтрующих сред с различными размерами пор по их способности полностью захватывать преципитат. Оптимальную фильтрующую среду выбирали на основании ее способности полностью захватывать преципитат, а также по ее способности подвергаться загрузке с высокой емкостью. Vmax использовали для оценки падения давления на фильтре, так же как для определения максимальной емкости. В таблице 8 показаны оцененные фильтрующие среды. Оцениваемые серии среды MILLISTAK+® CE (только целлюлоза) включают в себя 20CE, 35CE и 45CE. Оцениваемые серии среды MILLISTAK+® HC (целлюлоза и вспомогательный неорганический фильтр) включают в себя C0HC, B1HC и A1HC (Rathore et al. (Aug 1, 2004) BioPharm Intl.; US 2007/0193938). Картриджи и листы фильтра MILLISTAK® доступны из Millipore Corp. (Bedford, MA).
Как показано в таблице 8, среда 20CE является более открытой по сравнению с 35CE
и 45CE. 20CE обладает номинальным диапазоном размера пор 5-10 микрон, в то время как 35CE и 45CE обладают номинальным диапазоном размера пор 2-4 микрон и 0,8-2 микрон, соответственно. Эти серии фильтрующей среды выбраны для сужения диапазона размера пор, что позволяет полный захват преципитата антитело (гуманизированный вариант 2H7)-PVS. Среда 20CE приводит к захвату 45% преципитата, в то время как 35CE приводит к захвату 48% преципитата. В отличие от этого, 45CE приводит к захвату 97% преципитата. Серии среды Millistak+ HC плотнее, чем серии Millistak+ CE. Среда C0HC обладает номинальным диапазоном размера пор 0,2-2 микрон, в то время как B1HC и A1HC обладают более плотными размерами пор в диапазоне 0,05-0,7 микрон и 0,05-0,4 микрон, соответственно. Все три приводят к захвату 99% преципитата.
На фигуре 34 показан график Vmax емкости (измеренной в граммах антитела (2H7) на площадь фильтра) против падения давления для каждой фильтрующей среды (таблица 8). Устанавливали предел давления 17 psi для обозначения конца эксперимента из-за засорения фильтра. Устанавливали намеченную емкость 125 г/м2 для обозначения конца эксперимента в случае отсутствия засорения фильтра. Обе фильтрующие среды A1HC и B1HC достигли 17 psi и, таким образом, обеспечили самую низкую емкость. Емкость в конце фильтрации с использованием среды A1HC составляла 40 г/м, в то время как емкость в конце фильтрации с использованием среды B1HC составляла 68 г/м2. C0HC достигла емкости в конце 125 г/м2 с максимальным давлением 3,5 psi. Как упомянуто выше, 20CE и 30CE задерживали менее 50% преципитата и, таким образом, не выходили на намеченную емкость. 45CE достигала намеченной емкости с максимальным давлением 7 psi.
Выбор условий повторного растворения для повторного растворения преципитата внутри объемного фильтра
Оценивали ряд способов повторного растворения для выделения захваченного антитела с объемного фильтра. Целью этой стадии являлась разработка надежного способа с высоким выделением антитела. Первым оцененным способом являлось повторное растворение антитела из преципитата с использованием буфера с высокой проводимостью при высоком pH. В условиях высоких pH и проводимости, антитело является менее положительным, и больше ионов в растворе экранируют PVS и антитело, таким образом, предотвращая преципитацию. Буфер 50 мМ Трис основание/50 мМ ацетат натрия при pH в диапазоне 8-8,5 и проводимости ~6 мСм/см использовали для повторного растворения антитела. Буфер нагнетали через объемный фильтр, и полученный пул содержал антитело и PVS. Оценивали такие переменные, как скорость потока и объем буфера, необходимые для полного повторного растворения антитела. Рециркуляцию буфера через объемный фильтр также оценивали в качестве способа получения более концентрированного пула.
На фигуре 35 показан эффект изменения скорости потока на выход в переводе на объем буфера, необходимого для достижения полного повторного растворения преципитата белок-полиэлектролит 2H7-PVS из фильтрующей среды C0HC MILLISTAK+®. Для притоков в диапазоне 52-130 LMH (литров на квадратный метр в час, измерение скорости потока) и 130-261 LMH показали сходную тенденцию с обеими кривыми, приводящими к сходным конечным уровням с 600 мл буфера. Кривая при притоке 130 LMH включает ту же самую тенденцию и также приводит к сходному выходу с 600 мл буфера. Изменение скорости потока не улучшало соотношения выхода к объему буфера. Наиболее высокая концентрация антитела, полученная с использованием этого способа, составляла 1 г/л.
Рециркуляцию буфера через объемный фильтр оценивали в качестве способа, который потенциально может приводить к более концентрированному пулу. Резервуар, содержащий буфер, служил как входным контейнером для буфера, так и выходным контейнером для пула. Как показано на фигуре 36, два диапазона протока (потока буфера для повторного растворения в LMH) тестировали для определения оптимального времени переработки, во время которой авторы настоящего изобретения выделяют большинство исследуемого ими белка с фильтра в форме преципитата белок-полиэлектролит 2H7-PVS. С притоком в диапазоне 130-261 LMH достигали такого же выхода 84%, как с притоком в диапазоне 261-522 LMH через 60 минут. В пересчете на концентрацию пула, способ с рециркуляцией приводил к более концентрированному пулу при сравнении с однократным пропусканием буфера через объемный фильтр. Наиболее высокая концентрация антитела, полученная с использованием способа рециркуляции, составляла 2 г/л.
Операция реакции преципитации и захвата объемной фильтрацией в непрерывном режиме
Преципитация PVS основана на ионном взаимодействии между сильно анионной молекулой PVS и только слабо катионными молекулами антитела в растворе. Разбавление исходных подаваемых белков для снижения проводимости раствора до приемлемого диапазона для преципитации PVS может приводить к большим увеличениям объема подаваемого сырья. Чтобы избежать возможных ограничений по объему в производственном масштабе, необходимо разработать схему для поточного кондиционирования подаваемого сырья. Поточное разведение можно использовать для уменьшения проводимости потока подаваемого сырья без необходимости резервуара для перемешивания. Посредством объединения потока PVS с потоком кондиционированного подаваемого сырья можно осуществлять поточную преципитацию PVS-антитело и исключать необходимость большого резервуара для преципитация в масштабе. На фигуре 37 изображена схема поточной настройки кондиционирования HCCF/преципитации PVS с целями: (i) долгого времени удержания и низкой скорости жидкости, чтобы способствовать агрегации частиц PVS-Mab; (ii) большой площади поперечного сечения поточного реактора для поддержания высокой производительности способа; и (iii) реализации в лабораторном масштабе.
В то время как преципитация PVS-антитело может легко происходить при поточном перемешивании PVS и кондиционированных потоков подаваемого сырья, процесс агрегации частиц преципитата PVS-антитело является очень чувствительным к турбулентности потока, времени запаздывания и средней линейной скорости жидкости. Период времени запаздывания с низким перемешиванием является необходимым для агрегации частиц образца, необходимой для формирования частиц преципитата, достаточно больших для захвата фильтрующей средой. Высокая скорость потока может предупреждать формирование более крупного агрегата преципитата PVS-Ab, создавая поток очень маленьких частиц преципитата, которые могут являться сложными для захвата фильтрующей средой.
Трубопровод с поточными, непрерывными кондиционированием подаваемого сырья/преципитацией PVS представлен в лабораторном масштабе в сочетании со способом захвата объемной фильтрацией, перерабатывающей непрерывно преципитируемый поток подаваемого сырья. Наблюдали выходы выделения антитела в диапазоне 71%-60% (графики выхода антитела на фигуре 38) для способа, схематически показанного на фигуре 37 с очисткой от CHOP, в 9 раз больше по сравнению со стадией поточной преципитации PVS.
Выбор условий преципитации катионным полиэлектролитом
Кривые растворимости получали с использованием двух молекулярных масс полиаргинина (поли-L-аргинина) в CCF и HCCF. Кривые растворимости представляют собой график остаточного белка, оставшегося в супернатанте после преципитации (выраженного в процентах) в зависимости от концентрации полиэлектролита, выраженной как масса полиэлектролита (г)/объем раствора (мл).
Кривые растворимости получены для rhuMab 2H7 в CCF (Фигура 20) и в HCCF (обычно при pH 7) в диапазоне ионных сил (фигуры 21 и 22). Эти кривые использовали для выбора оптимальных условий преципитации CHOP, при которых будут проводить преципитации в препаративном масштабе, а именно ионной силы, концентрации и молекулярной массы полиаргинина. Кривые растворимости получены также для HCCF
с анти-CD22 и rhuMab C2B8 (фигуры 23 и 24), чтобы убедиться в надежности и стабильности способа преципитации. Для оценки осуществимости очистки антитела посредством двух стадий преципитации, преципитации катионным полиэлектролитом с последующей преципитацией анионным полиэлектролитом кривые растворимости получили для преципитации PVS rhuMab HCCF, полученной в результате флокуляции CCF с помощью 0,075% масс./об. 110 кДа полиаргинина (фигура 25). При увеличении концентрации полиаргинина присутствовала преципитация CHOP, как показано по уменьшению остаточного CHOP (нг/мг) в супернатанте. Оптимальная преципитация CHOP происходила при концентрации полиаргинина 0,2% масс./об. с уменьшением уровня CHOP в 22 раза, с выходом 95%. Дополнительное увеличение концентрации полиаргинина не приводило к значительному уменьшению уровня CHOP, но присутствовало уменьшение выхода.
Преципитация CHOP, по-видимому, является функцией ионной силы и молекулярной массы полиэлектролита, как видно на кривых растворимости для rhuMab 2H7 HCCF (фигуры 21 и 22). При уменьшении проводимости присутствовало увеличение преципитации. Как правило, увеличение молекулярной массы полиэлектролита, по-видимому, оказывает влияние на преципитацию CHOP. При сходных концентрациях полиаргинина полиэлектролит с более высокой молекулярной массой (110 кДа) преципитировал больше CHOP по сравнению с полиэлектролитом с более низкой молекулярной массой (42 кДа). При конкретной проводимости (за исключением 3 мСм/см), добавление полиаргинина более 0,1% масс./об. не приводило к дополнительному значительному уменьшению уровня CHOP. При концентрации полиаргинина 0,1% масс./об. 10-кратное и 13-кратное уменьшение уровня CHOP получили для молекулярных масс 42 кДа и 110 кДа, соответственно, и выходы составляли >95% для обеих молекулярных масс.
Сходные тенденции наблюдали для HCCF с анти-CD22 и rhuMab C2B8, как видно на кривых растворимости для HCCF с rhuMab 2H7, анти-CD22, rhuMab C2B8 (фигура 23 и 24). Преципитация анти-CD22 HCCF 0,1% масс./об. 110 кДа полиаргинином приводила к 25-кратному уменьшению уровня CHOP и 95% выходу. Оптимальная преципитация CHOP для rhuMab C2B8 присутствовала для концентрации 0,15% масс./об. 110 кДа полиаргинина, что приводило к 20-кратному уменьшению уровня CHOP и 95% выходу.
При увеличении концентрации PVS присутствовала полная преципитация антитела в подаваемом сырье, флокулированном с помощью полиаргинина, при обоих тестированных pH и проводимостях, показанная по уменьшению выхода (%) (фигура 25). Это указывает на минимальное влияние присутствия полиаргинина на преципитацию антитела PVS в тестированных условиях. При pH 7 и проводимости 1,5 мСм/см, после того как наблюдали максимальный уровень преципитации, добавление дополнительного PVS вызывало повторное растворение преципитирующего средства.
Последующая переработка rhuMab 2H7
В общепринятом способе в настоящее время используют три стадии хроматографии, стадию захвата белком A с последующей стадией катионного и анионного обмена. Флокуляция CCF с помощью 0,075% масс./об. 110 кДа полиаргинина и переработка HCCF с помощью преципитации антитела PVS с последующей стадией QSFF показана как схема организации производства a), таблица 9). Первичной функцией этой стадии флокуляции является осветление CCF удалением твердых частиц (клеточного дебриса), таким образом, уменьшая мутность, и преципитация примесей клетки-хозяина, таких как CHOP и ДНК. Полиаргинин являлся способным флокулировать твердые частицы и компоненты среды для культивирования клеток, и уменьшать уровень CHOP до 23480 нг/мг (уменьшение в 4 раза) и полностью удалять ДНК. Если концентрацию полиаргинина 110 кДа увеличивали до 0,2% масс./об., получали уменьшение уровня CHOP вплоть до 22 раз, как показано кривыми растворимости (фигура 20). Преципитация PVS флокулированной HCCF дополнительно уменьшала CHOP до 212 нг/мг, что являлось значительно меньшим, чем CHOP в контрольных пулах HCCF-PVS. Контрольные пулы HCCF-PVS-Q обладали почти в два раза большим количеством CHOP, чем пулы поли(Arg) HCCF-PVS-Q. Анализ ДНК не функционирует в присутствии PVS, так что очистку PVS в ходе стадии преципитации невозможно было оценить. Выходы в ходе всех стадий превышали 95%, за исключением стадий преципитации PVS (таблица 9). Эти низкие выходы для стадий PVS можно отнести к способу преципитации и манипуляциям в препаративном масштабе, а не к механизму и условиям преципитации (фигура 25). Процент мономера посредством SEC, инсулин и гентамицин не оценивали для этих пулов.
Чтобы исследовать вклад последующих переработок, HCCF преципитировали с помощью 0,1% масс./об. полиаргинина 110 кДа и проводили через различные схемы очистки. Выход для антитела составлял 97%, и CHOP уменьшали в 18 раз, и ДНК полностью удаляли преципитацией полиаргинином. Инсулин и гентамицин не очищали этой стадией преципитации. Перед последующей переработкой преципитированной HCCF получали кривые проскока для оценки влияния полиаргинина на емкость загрузки смол SPSFF. Для обеих загрузок, преципитированной полиаргинином HCCF и пулом после преципитации белком A, 5% проскок на SPSFF присутствовал при 60 мг антитела на мл смолы, что сходно с контролем (без преципитации полиаргинином). Это показывает, что полиаргинин не оказывал отрицательного влияния на емкость связывания SPSFF (фигура 26 и 27).
Общепринятый способ для антитела (схема организации производства b), Таблица 9): преципитированную полиаргинином HCCF пропускали через белок A с последующей ионообменной хроматографией. Пул полиаргинин-белок A обладал уровнями CHOP, сходными с контрольным белком A, даже несмотря на то, что уровни CHOP в загружаемом подаваемом сырье являлись очень разными. Гентамицин и инсулин уменьшали до низких уровней посредством стадии с белком A. CHOP уменьшали в 490 раз до 3 нг/мг в ходе SPSFF по сравнению с 307 нг/мг в отсутствие стадии преципитации. ДНК, вымываемый белок A, инсулин и гентамицин также уменьшали посредством способа с полиаргинином-белком A-SP до приемлемых уровней, сравнимых с контрольным способом (Белок A-SPSFF-QSFF) (таблица 9). Поскольку уровни примесей являлись приемлемыми в пулах SPSFF после преципитации полиаргинином, не является необходимым перерабатывать их на QSFF.
Способ неаффинной очистки антитела (схема организации производства c), Таблица 9): преципитированную полиаргинином HCCF пропускали через SPSFF с последующей QSFF. Пул после преципитации SPSFF обладал 63 нг/мг CHOP (уменьшение в 76 раз) по сравнению с 6731 нг/мг в отсутствие преципитации. SPSFF полностью удаляла инсулин и частично уменьшала уровень гентамицина. Очистка от CHOP являлась лучшей в способе с полиаргинином-SP, в то время как очистка от ДНК, инсулина и гентамицина являлись сравнимыми с контрольным способом (SPSFF-QSFF) (фигура 28). После преципитации полиаргинином уровни CHOP в пуле SPSFF являются сходными с пулом QSFF, что позволяет предполагать, что стадия QSFF является излишней в этом способе.
Способ очистки с двумя стадиями преципитации/хроматографии
Преципитированную полиаргинином HCCF затем подвергали преципитации антитела PVS и затем перерабатывали на QSFF (схема организации производства d, таблица 9). Пул PVS обладал 49 нг/мг CHOP (уменьшение в 99 раз) по сравнению с 1517 нг/мг в отсутствие преципитации полиаргинином. Стадия преципитации PVS полностью удаляла инсулин и частично уменьшала уровень гентамицина. Очистка от CHOP являлась лучшей в способе с полиаргинином-PVS, в то время как очистка от ДНК и инсулина являлись сравнимыми с контрольным способом (SPSFF-QSFF). Очистка от гентамицина являлась в 2 раза меньшей в контрольном способе, в присутствии стадии QSFF. После преципитации полиаргинином уровни CHOP в пуле SPSFF являлись сходными с пулом QSFF, что позволяет предполагать, что стадия QSFF является излишней в этом способе.
Обобщение результатов последующей переработки для rhuMab 2H7
Ингибирование восстановления антитела
HCCF, содержащую rhuMab 2H7, обрабатывали полиаргинином и содержали в минифлаконах из нержавеющей стали в течение 48 часов, и образцы отбирали через равномерные промежутки времени и анализировали. Антитело являлось интактным и не восстановленным в течение всего периода времени в преципитированной полиаргинином HCCF в отличие от контроля, где антитело начинает восстанавливаться через 10 часов (фигура 28). Полиаргинин ингибирует восстанавливающие антитело агенты посредством их преципитации вместе с другими отрицательно заряженными примесями.
Оценка других катионных полиэлектролитов
Оптимизация условий преципитации для дополнительного увеличения удаления примесей может включать в себя использование других полиэлектролитов, так же как оптимизацию других условий, таких как скорость и тип перемешивания, и скорость добавления полиэлектролита. Другую основную поли(аминокислоту), полилизин, выбрали для исходного сравнения с полиаргинином. Полиаргинин обладает катионной функциональной группой гуанидиния, придающей pKa приблизительно 12,5. Полилизин обладает катионом амина в качестве функциональной группы, придающей pKa приблизительно 10,5. Как полиаргинин, так и полилизин являются коммерчески доступными со множеством молекулярных масс.
Кривая растворимости пула белка A rhuMab C2B8
Преципитация CHOP положительно заряженным поликатионным полиэлектролитом коррелирует с ионной силой и молекулярной массой полиэлектролита (фигуры 29 и 30). Полилизин с более низкой молекулярной массой (2,5 кДа) не преципитировал CHOP в диапазоне проводимости 3-12 мСм/см. Однако полилизин 50 кДа приводил к 4-кратному и 2-кратному уменьшениям уровня CHOP при 3 мСм/см и 6мСм/см, соответственно. Не наблюдали значительной преципитации CHOP при 12 мСм/см. Как правило, при уменьшении проводимости, присутствовало увеличение преципитации CHOP, и увеличение молекулярной массы полиэлектролита, по-видимому, оказывало влияние на преципитацию CHOP. При более низких проводимостях, после того как получали максимальную преципитацию CHOP, дополнительное добавление полилизина приводило к повторному растворению преципитата. Выходы превышали 95% для всех тестированных условий.
Кривая растворимости пула rhuMab C2B8 HCCF
Сходные тенденции наблюдали с rhuMab C2B8 HCCF (фигуры 31 и 32). При проводимостях 3 мСм/см и 6 мСм/см полилизин как 50 кДа, так и 225 кДа приводил к уменьшению CHOP в 9 раз. Однако не получили значительного уменьшения количества CHOP при 12 мСм/см даже с 225 кДа полилизином. Это показывает, что преципитация CHOP при высоких проводимостях требует высоко заряженного полиэлектролита, подобного полиаргинину, чтобы преодолеть ионную интерференцию. Выходы превышали 90% для всех тестированных условий.
Кривые растворимости получены для трех антител (rhuMab 2H7, анти-CD22 и rhuMab C2B8) с использованием полиаргинина (фигуры 20-25, 29-32). Преципитация CHOP являлась функцией pKa, молекулярной массы и концентрации полиэлектролита, так же как ионной силы раствора. Свойства антитела, такие как плотность заряда и изоэлектрическая точка, также могут играть роль в отделении примесей от антител. Примеси, такие как CHOP и ДНК, удаляли посредством управления концентрацией полиэлектролита и ионной силой. Можно изменять свойства полиэлектролита, такие как функциональная группа, молекулярная масса, плотность заряда и гидрофобность. Полиаргинин можно использовать в качестве флокулирующего средства в CCF или в качестве преципитирующего примеси средства в HCCF. Преципитация является зависимой от проводимости. Наблюдали 10-25-кратное удаление CHOP, и ДНК не поддавалась детекции (таблица 9). Полиаргинин не оказывал отрицательного влияния на емкость связывания SPSFF и являлся совместимым с преципитацией антитела с помощью анионных полиэлектролитов и катионообменных смол. Добавление полиаргинина к HCCF обладало также удивительными и неожиданными преимуществами, включая ингибирование восстановления антитела, возможно, посредством преципитации восстанавливающего агента вместе с другими примесями.
ПРИМЕРЫ
Чтобы проиллюстрировать изобретение, включены следующие примеры.
Однако следует понимать, что эти примеры не ограничивают изобретение и предназначены только для того, чтобы предложить способы практического осуществления изобретения. Специалисту в данной области будет понятно, что описанные примерные методы, протоколы, способы, реагенты и аппараты можно легко адаптировать для практического осуществления альтернативных способов по этому изобретению, подразумевая, что они входят в объем этого изобретения.
Пример 1 Хроматография с белком A
В колонке с белком A использовали смолу Prosep vA, и ее использовали для очистки rhuMab 2H7, присутствующего в HCCF. Диаметр колонки составлял 2,5 см, и высота наполнителя составляла 14 см. Колонкой управляли при скорости потока 40 CV/h (объемов колонки в час). Колонку Prosep vA использовали в нескольких циклах (2 цикла). После уравновешивания HCCF наносили на колонку, и rhuMab 2H7 задерживалось на колонке. Затем колонку промывали буфером для уравновешивания, затем буфером для промывки и затем снова буфером для уравновешивания. После окончания этих промывок на колонку наносили буфер для элюции. Пулирование начинали на основании поглощения при 280 нм (0,5 OD) и останавливали после 2 объемов колонки. Затем на колонку наносили буфер для регенерации. Состав буфера и продолжительности фаз указаны в таблице 10.
Состав буфера и продолжительности фаз для Prosep vA
Собранную жидкость культуры клеток (HCCF), полученную от рекомбинантных клеток-хозяев, можно наносить на уравновешенную твердую фазу с использованием буфера для нанесения, который может являться таким же, как буфер для уравновешивания. При протекании загрязненного препарата через твердую фазу белок связывается с иммобилизованным белком A, а другие загрязнения (такие как белки яичника китайского хомячка, CHOP, когда белок продуцируют в клетке CHO) могут неспецифически связываться с твердой фазой. Следующая стадия, проводимая последовательно, вызывает удаление загрязнений, связанных с твердой фазой, антителом и/или белком A, посредством промывки твердой фазы на стадии промежуточной промывки. После загрузки твердую фазу можно уравновешивать буфером для уравновешивания до начала стадии промежуточной промывки. Буфер для промежуточной промывки может содержать соль и дополнительное соединение, где дополнительное соединение представляет собой (a) детергент (например, полисорбат 20 или полисорбат 80); (b) растворитель (например, гексиленгликоль); и (c) полимер (например, PEG). Применяемую соль можно выбирать на основании интересующего белка, однако, предпочтительно, она представляет собой ацетат (например, ацетат натрия). Количества соли и дополнительного соединения в композиции являются такими, что объединенное количество элюирует загрязнение(загрязнения), по существу, без удаления интересующего белка. Концентрации соли в таких буферах для промывки могут лежать в диапазоне от приблизительно 0,1 до приблизительно 2 M, или от приблизительно 0,2 M до приблизительно 0,6 M. Применимые концентрации детергента составляют от приблизительно 0,01 до приблизительно 5%, или приблизительно 0,1-1%, или приблизительно 0,5%, например, когда детергент представляет собой полисорбат. Примерные концентрации растворителя составляют приблизительно от 1% до 40%, или приблизительно от 5 до приблизительно 25%. Когда дополнительное соединение представляет собой полимер (например, PEG 400 или PEG 8000), его концентрация может, например, составлять от приблизительно 1% до приблизительно 20%, предпочтительно, от приблизительно 5% до приблизительно 15%.
В другом варианте осуществления стадия промежуточной промывки включает в себя использование высоко концентрированного буферного раствора, например, буфера в концентрации более приблизительно 0,8 M, например, вплоть до приблизительно 2 M, и предпочтительно, в диапазоне от приблизительно 0,8 M до приблизительно 1,5 M, наиболее предпочтительно, приблизительно 1 M. В этом варианте осуществления буфер предпочтительно представляет собой Трис-буфер, такой как Трис-ацетат. pH буфера для промежуточной промывки может составлять от приблизительно 4 до приблизительно 8, или от приблизительно 4,5 до приблизительно 5,5, или приблизительно 5,0. В другом варианте осуществления pH составляет приблизительно 7,0. После стадии промежуточной промывки из предыдущего раздела интересующий белок выделяют с колонки. Этого обычно достигают с использованием подходящего буфера для элюции. Белок можно, например, элюировать с колонки с использованием буфера для элюции, обладающего низким pH, например, в диапазоне от приблизительно 2 до приблизительно 5, и предпочтительно, в диапазоне от приблизительно 2,5 до приблизительно 3,5. Примеры буферов для элюции для этих целей включают в себя цитратные или ацетатные буферы. В ходе фазы элюции колонки с белком A любой неспецифически связанный CHOP может элюироваться совместно с интересующим белком, подвергая риску чистоту пула продукта. Для удаления CHOP перед фазой элюции, примерную стадию промежуточной промывки с использованием хлорида тетраметиламмония (TMAC) можно проводить для удаления CHOP (US 6870034; US 6127526; US 6333398).
Пример 2 Хроматография SPSFF
В катионообменной колонке применяли смолу SP Sepharose Fast Flow (SPSFF) в режиме связывания и элюции с градиентной элюцией. Колонкой SPSFF управляли при скорости потока 150 см/ч. Во всех случаях высота слоя составляла 30 см. При переработке пулов промежуточной хроматографии (пулы Prosep vA и QSFF) использовали колонку 0,66 см i.d., и колонку загружали до 40 г/л. При переработке HCCF использовали колонку диаметром 0,6-2,2 см, и колонку загружали до 10-40 г/л. Во всех случаях загрузку кондиционировали до pH 5,0 и проводимости приблизительно 5,5 мСм/см. После загрузки колонку затем промывали буфером для уравновешивания, затем буфером для промывки и затем снова буфером для уравновешивания. После завершения этих промывок rhuMab 2H7 элюировали в градиенте от 50 мМ ацетата до 350 мМ ацетата на протяжении 15 объемов колонки. Пулирование начинали и останавливали на основании поглощения при 280 нм (0,5 OD). Колонку регенерировали и дезинфицировали 0,5 н. гидроксидом натрия и хранили в 0,1 н. гидроксиде натрия. Состав буфера и продолжительности фаз указаны в таблице 11.
Состав буфера и продолжительности фаз для SPSFF
B: 350 мМ ацетат NA pH 5,5
Пример 3 Анионообменная хроматография
Использовали анионообменную колонку со смолой Q Sepharose Fast Flow (QSFF) и управляли в проточном режиме. Колонкой QSFF управляли при скорости потока 150 см/час. Диаметр колонки составлял 0,66 см, и высота слоя составляла 20 см. Колонку уравновешивали и загружали при pH 8,0. Антитело RhuMab 2H7 пропускали через колонку, которую затем промывали буфером для уравновешивания. Пулирование начинали и останавливали на основании поглощения при 280 нм (0,5 OD). Колонку регенерировали и дезинфицировали 0,5 н. гидроксидом натрия и хранили в 0,1 н. гидроксиде натрия. Состав буфера и продолжительности фаз указаны в таблице 12.
Состав буфера и продолжительности фаз для QSFF
Пример 4a Очистка анионным полиэлектролитом
Пулы антител доводили до pH и проводимостей, указанных в таблице 13. HCCF доводили до pH 6 с помощью 1 М уксусной кислоты и до проводимости 2,0 мСм/см с помощью WFI (вода для орошения). Проводили поточное добавление PVS к кондиционированной HCCF для достижения конечной концентрации 0,05% масс./об. в течение периода 20 минут. При перемешивании PVS добавляли к кондиционированному пулу до конечных концентраций, указанных в таблице 13. Преципитированный PVS пул центрифугировали при 4000 об./мин (4657 g) при 10°C в течение 30 минут с использованием центрифуги Sorval R3-CB. Супернатант удаляли и осадок промывали 25 мМ MOPS, pH 7,1. Осадок ресуспендировали в 50 мМ Трис, 50 мМ ацетате, и pH 8,0 (приблизительно 6,5 мСм/см). Ресуспендированный осадок затем подвергали хроматографии QSFF.
Пример 4b Очистка катионным полиэлектролитом
Преципитация жидкости культуры клеток (CCF): преципитацию примесей проводили с помощью полиаргинина, молекулярная масса 110 кДа. Полиаргинин добавляли к CCF в биореакторе в течение периода 5 минут для достижения конечной концентрации 0,075% масс./об. при перемешивании при 100 об./мин. Затем флокулированную CCF центрифугировали при 10000 g и фильтровали в стерильных условиях через фильтр 0,2 мкм.
Преципитация собранной жидкости культуры клеток (HCCF): Полиаргинин (поли-L-аргинин) добавляли к HCCF в биореакторе в течение периода 5 минут для достижения конечной концентрации 0,1% масс./об. при перемешивании при 100 об./мин. Затем преципитированную HCCF центрифугировали при 5000 g и фильтровали в стерильных условиях через фильтр 0,2 мкм. Фильтрованное подаваемое сырье перерабатывали посредством различных способов очистки.
Условия преципитации PVS для 2H7
Пример 5 Определение концентрации белка:
Концентрацию антитела в пулах определяли по поглощению при 280 нм (с вычитанием поглощения при 320 нм для поправки на рассеяние света), с использованием спектрофотометра 8453 с проточной кюветой с длиной пути 10 мм (Agilent). Применяли коэффициент поглощения 1,75 мл/(мг см). Концентрацию антитела вычисляли с использованием следующего уравнения:
Пример 6 Эксклюзионная хроматография
Эксклюзионную хроматографию использовали для мониторирования гетерогенности по размеру rhuMab 2H7 в нативных условиях. В анализе применяли колонку TSK-GEL G3000SWXL (7,8 мм × 300 мм, Tosohaas) для разделения агрегатов, мономера и фрагментов. Колонкой управляли при скорости потока 0,3 мл/мин с использованием рабочего буфера 0,20 M калий фосфат, 0,25 M хлорид калия, pH 6,2. Колонкой управляли при температуре окружающей среды. Образцы разводили в рабочем буфере, и для каждого образца инъецировали 20 мкг. Поглощение при 280 нм использовали для мониторирования уровней агрегатов, мономера и фрагментов.
Пример 7 ELISA CHOP - Y:42
Твердофазный иммуноферментный анализ (ELISA) с использованием антител козы против CHOP использовали для определения концентраций CHOP во всех пулах. Для ELISA аффинно очищенные антитела козы против CHOP иммобилизовали в лунках микропланшета для титрования. Разведения пулированных образцов инкубировали в лунках с последующей инкубацией с конъюгированными с пероксидазой антителами козы против CHOP. Ферментативную активность пероксидазы хрена оценивали количественно с помощью o-фенилендиамина, образующего колориметрический сигнал. Проводили серийные разведения образцов в разбавителе для анализа, так чтобы считывание поглощения попадало в диапазон анализа (5-320 нг/мл).
Пример 8 ELISA белка A - Y:80
Уровень белка A определяли посредством сэндвич-ELISA для белка A. Антитела курицы против белка A стафилококка иммобилизовали в лунках микропланшета для титрования. Образцы разводили до 0,2 мг/мл антитела и наносили в лунки. Белок A, если он присутствовал в образце, связывался с покрытым антителом. Связанный белок A детектировали с помощью антител против белка, конъюгированных с пероксидазой хрена. Ферментативную активность пероксидазы хрена оценивали количественно с помощью o-фенилендиамина, образующего колориметрический сигнал.
Пример 9 ELISA инсулина - Y:64
ELISA с использованием поликлональных антител морской свинки против инсулина использовали для определения концентрации инсулина во всех пулах. Для ELISA аффинно очищенные антитела против инсулина иммобилизовали в лунках микропланшета для титрования. Разведения пулированных образцов инкубировали в лунках с последующей инкубацией с конъюгированными с пероксидазой хрена антителами морской свинки против инсулина. Ферментативную активность пероксидазы хрена оценивали количественно с помощью тетраметилбензидина. Проводили серийные разведения образцов в разбавителе для анализа, так чтобы считывание поглощения попадало в диапазон анализа (0,094-3,000 нг/мл).
Пример 10 ELISA для гентамицина - Y:81
Конкурентный ELISA использовали для определения концентраций гентамицина во всех пулах. Поликлональные антитела козы против гентамицина-BSA иммобилизовали в лунках микропланшета для титрования. Гентамицин конкурировал с биотинилированным гентамицином за связывание с антителами. Количество связанного меченного биотином гентамицина детектировали с помощью добавления пероксидазы хрена-стрептавидина и субстрата o-фенилендиамина. Проводили серийные разведения образцов в разбавителе для анализа, так чтобы считывание поглощения попадало в диапазон анализа (0,37-90 нг/мл).
Пример 11 Анализ ПЦР ДНК CHO - Y:77
ДНК оценивали количественно амплификацией ПЦР ДНК CHO. Сначала ДНК из образцов и контролей выделяли с использованием мининабора для вирусной РНК Qiagen. Затем экстракты и стандартную кривую, вместе с готовой смесью для ПЦР, содержащей праймеры и зонд, загружали в формате 96-луночного планшета в коммерческую систему детекции последовательности (Applied Biosystems), где ДНК CHO оценивали количественно с использованием ПЦР с детекцией в реальном времени. Для TaqMan® ПЦР применяли праймеры и зонд, сконструированные специфическими для последовательности-мишени ДНК CHO. Продукт амплификации измеряли с использованием флуорогенного зонда, меченного репортерным красителем на 5'-конце, подавляемого тушащим красителем на 3'-конце. Taq-полимераза начинает амплификацию ДНК-мишени и при достижении зонда ее 5'-нуклеазная активность замещает зонд, высвобождая репортерный краситель. Когда репортерный краситель больше не находится поблизости от тушащего красителя, репортер флуоресцирует и измеряют увеличение интенсивности излучения. Число циклов, при котором ДНК является амплифицированной выше порога (Ct), вычисляют для стандартной кривой, по сравнению с которой количественно оценивают неизвестные концентрации в образцах.
Пример 12 Активность rhuMab 2H7
Активность rhuMab 2H7 определяли с использованием анализа комплементзависимой цитотоксичности (CDC). Этот анализ основан на измерении способности rhuMab 2H7 лизировать клетки WIL2-S в присутствии комплемента человека.
Пример 13 Анализ ингибирования РНКазы A для PVS
PVS является сильным ингибитором РНКазы A. В анализе используют аналог РНК с флуоресцентной меткой на одном конце и тушителем на другом. При расщеплении аналога РНК РНКазой A флуоресцентная метка освобождается от тушителя, что приводит к излучению. Присутствие PVS будет ингибировать активность РНКазы A, ограничивая флуоресцентное излучение. Затем количество PVS можно определять сравнением наблюдаемой флуоресценции из тестируемого образца со стандартной кривой.
Предшествующее описание рассматривают только как иллюстративное для принципов изобретения. Кроме того, поскольку многочисленные модификации и изменения будут ясно очевидными специалистам в данной области, им не желают ограничивать изобретение до точных конструкций и процессов, показанных, как описано выше. Соответственно, все подходящие модификации и эквиваленты можно рассматривать как попадающие в объем изобретения, как определено следующей ниже формулой изобретения.
Слова «содержит», «содержащий», «включать», «включая» и «включает» при использовании в этом описании и в следующей ниже формуле изобретения предназначены для обозначения присутствия указанных признаков, чисел, компонентов или стадий, однако они не препятствует присутствию или добавлению одного или нескольких других признаков, чисел, компонентов, стадий или их групп.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ОЧИСТКИ ПОЛИПЕПТИДОВ | 2011 |
|
RU2594163C2 |
| ОЧИСТКА АНТИТЕЛ С ПОМОЩЬЮ КАТИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2008 |
|
RU2498991C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2793783C1 |
| УЛУЧШЕННАЯ ОЧИСТКА БЕЛКА ПОСРЕДСТВОМ МОДИФИЦИРОВАННОГО ЭЛЮИРОВАНИЯ БЕЛКА А | 2010 |
|
RU2571929C2 |
| СПОСОБ ОЧИСТКИ БЕЛКА СЛИЯНИЯ | 2015 |
|
RU2698654C2 |
| СПОСОБЫ И КОМПОЗИЦИИ, ВКЛЮЧАЮЩИЕ ОЧИЩЕННЫЕ РЕКОМБИНАНТНЫЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2671481C2 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ЭТАПОВ ОЧИСТКИ БЕЛКА, НАХОДЯЩИХСЯ НИЖЕ ПО ПОТОКУ, С ИСПОЛЬЗОВАНИЕМ МЕМБРАННОЙ ИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2012 |
|
RU2648999C2 |
| СПОСОБЫ ИНАКТИВАЦИИ ВИРУСА С ИСПОЛЬЗОВАНИЕМ ЭКОЛОГИЧНЫХ ДЕТЕРГЕНТОВ | 2014 |
|
RU2707951C1 |
| СПОСОБЫ ОЧИСТКИ ГЕТЕРОДИМЕРНЫХ ПОЛИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2019 |
|
RU2820588C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ УМЕНЬШЕНИЯ АГРЕГАЦИИ МАКРОМОЛЕКУЛ ПРИ ФИЗИОЛОГИЧЕСКИХ УСЛОВИЯХ | 2009 |
|
RU2563823C2 |
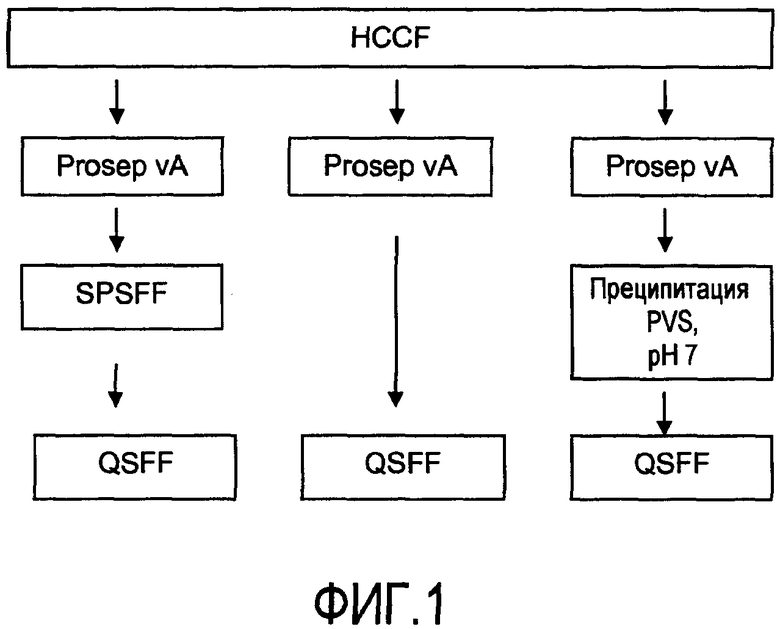
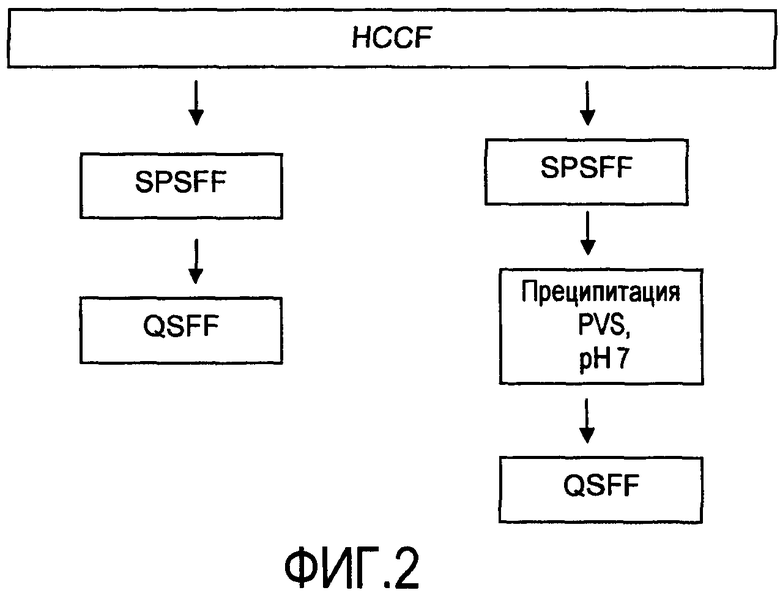
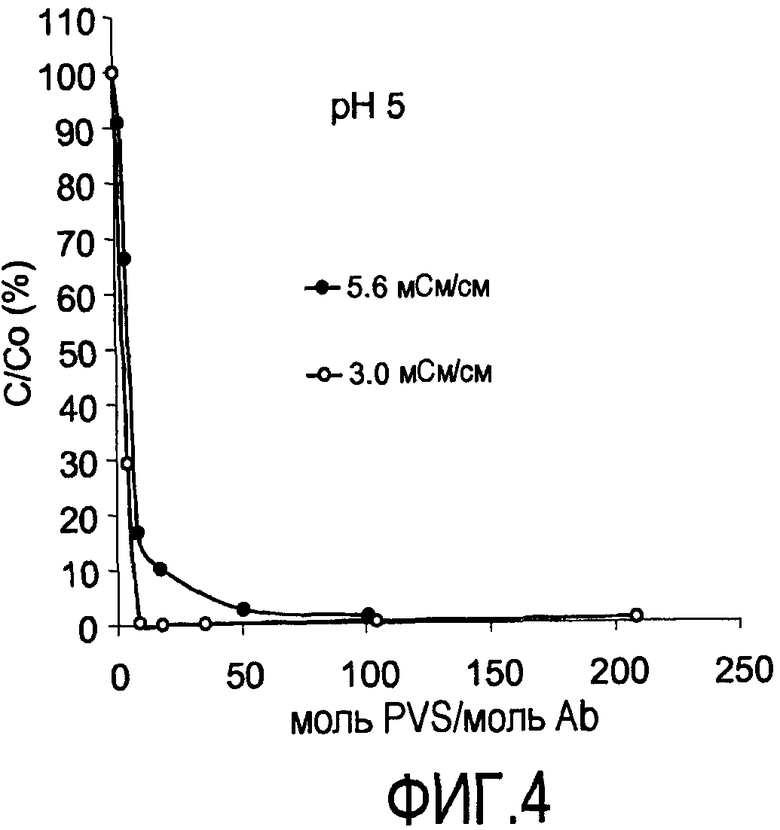
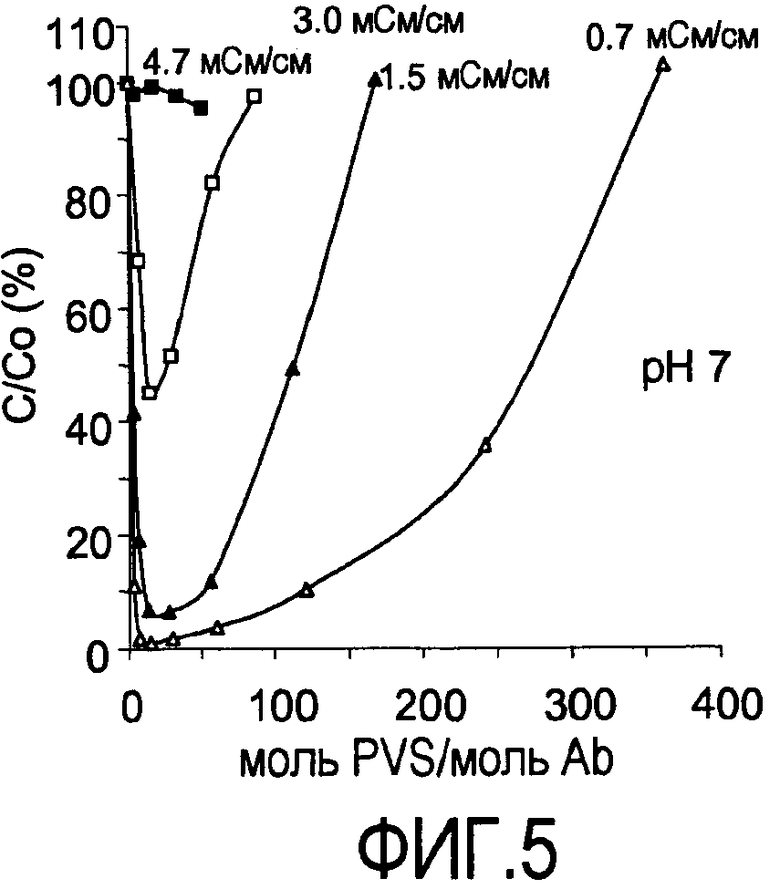
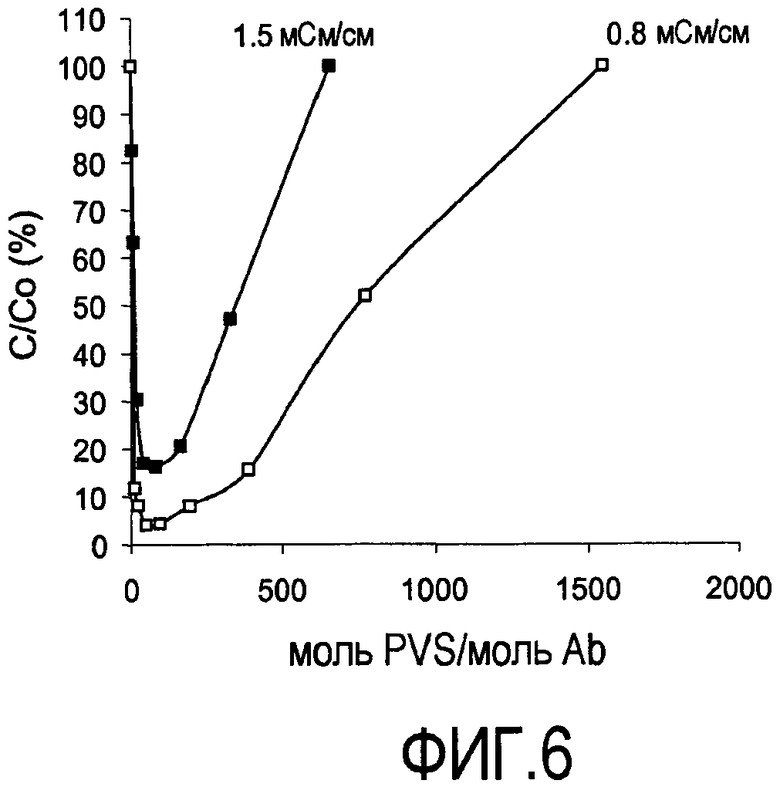
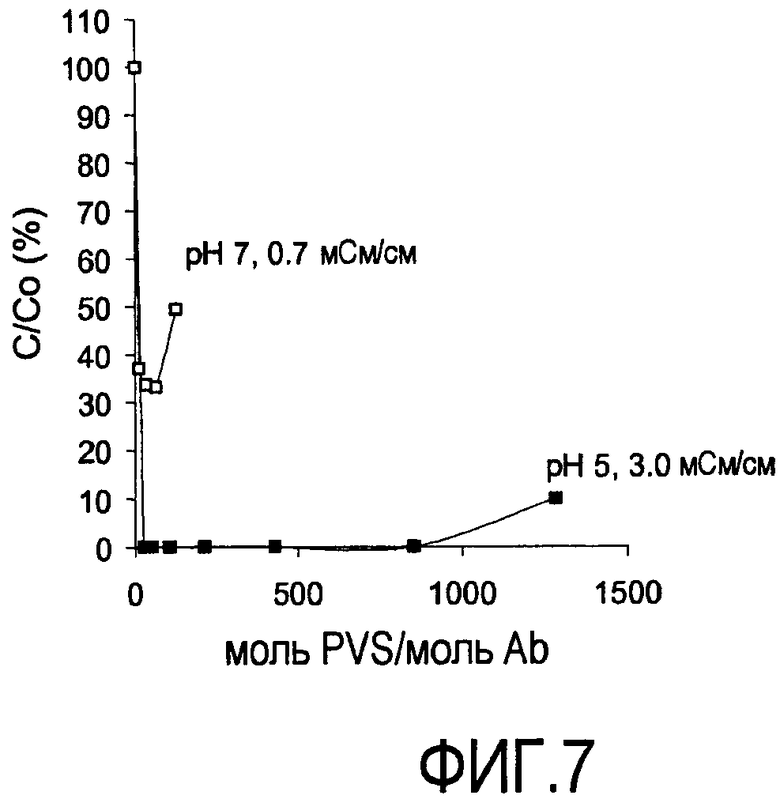
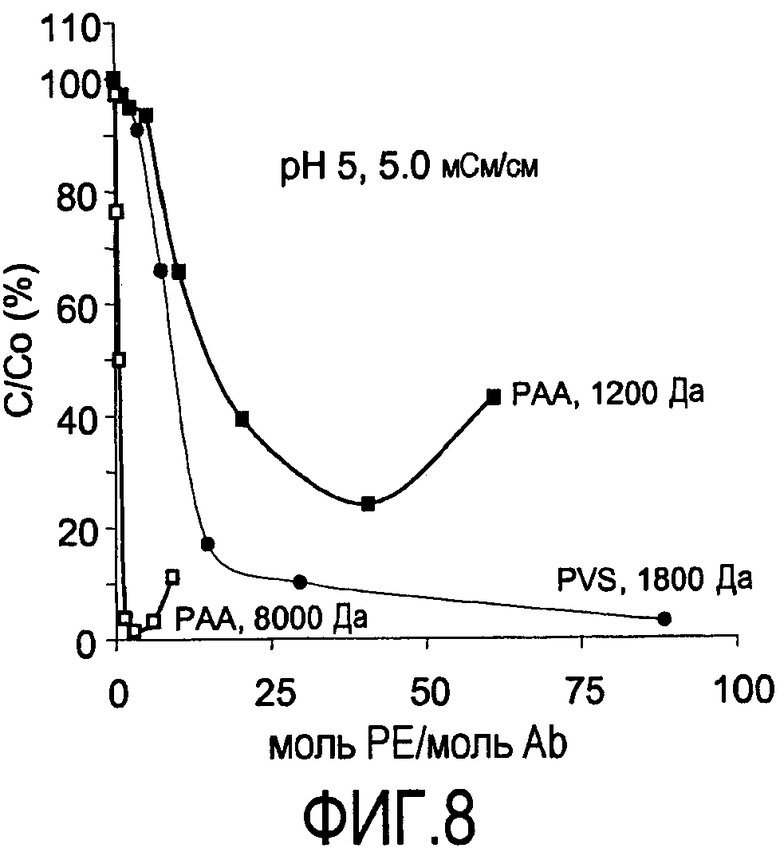
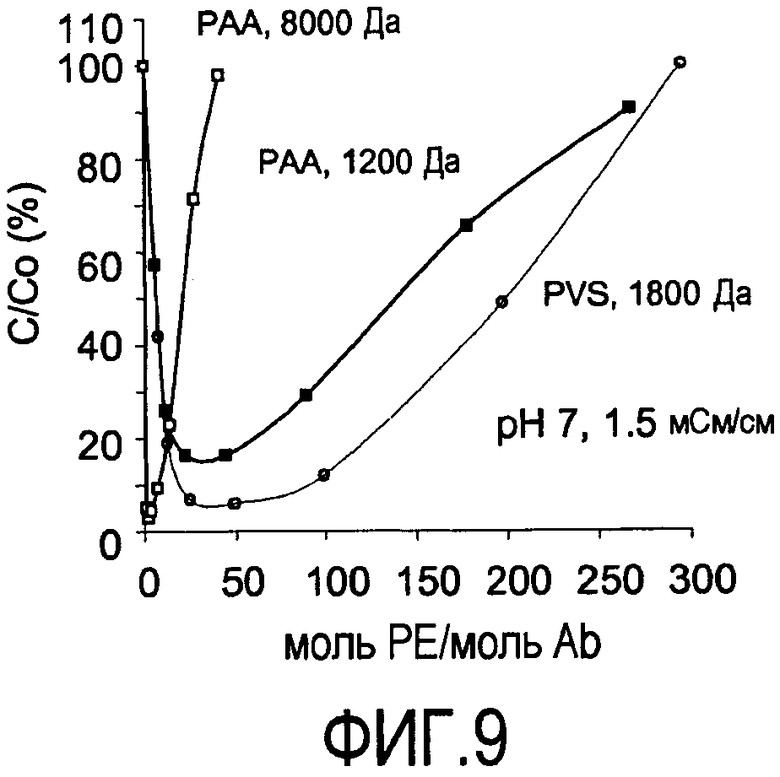
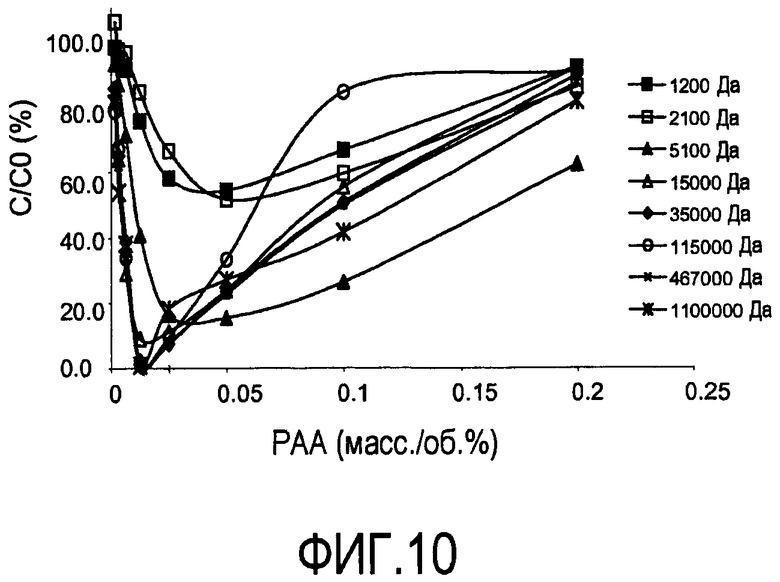
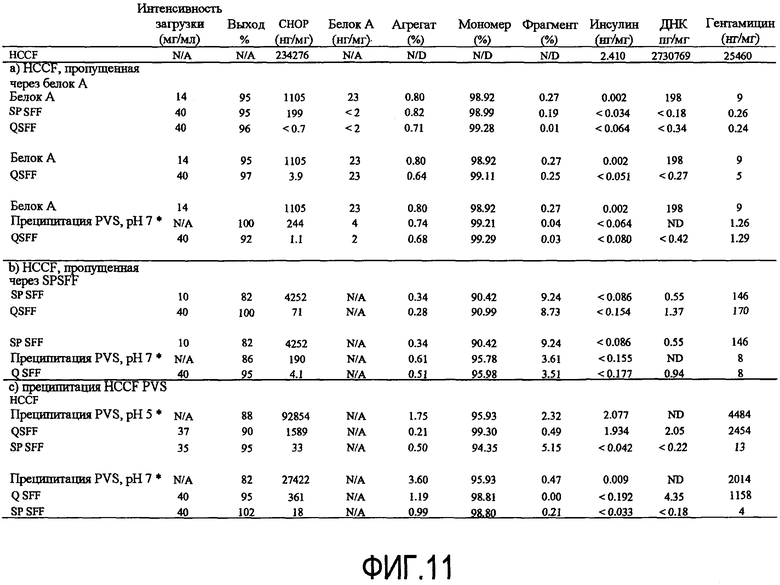
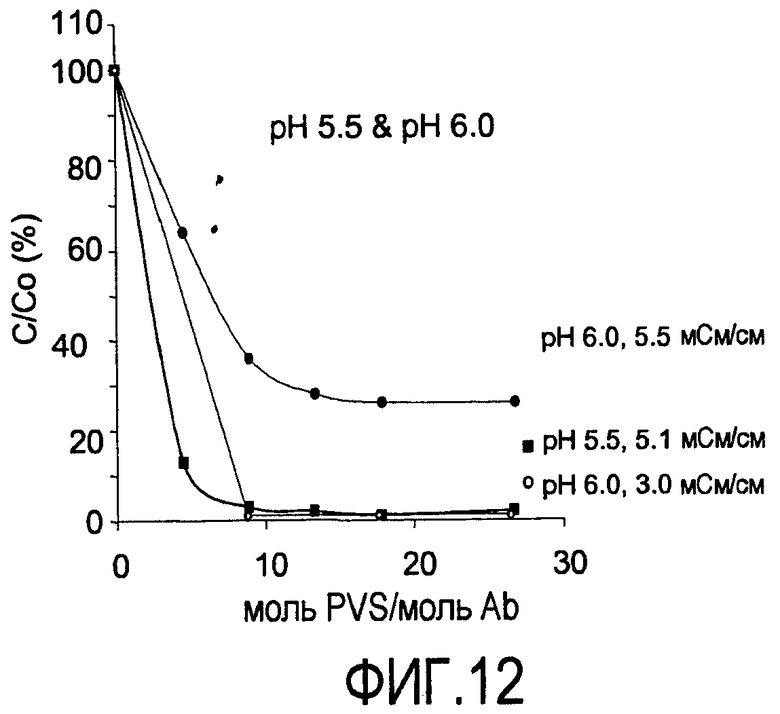
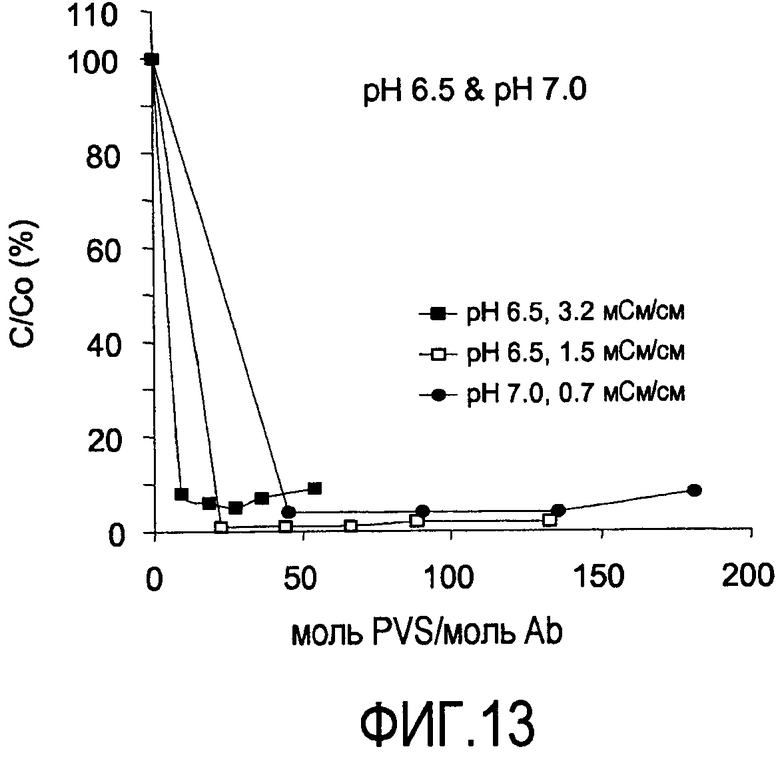

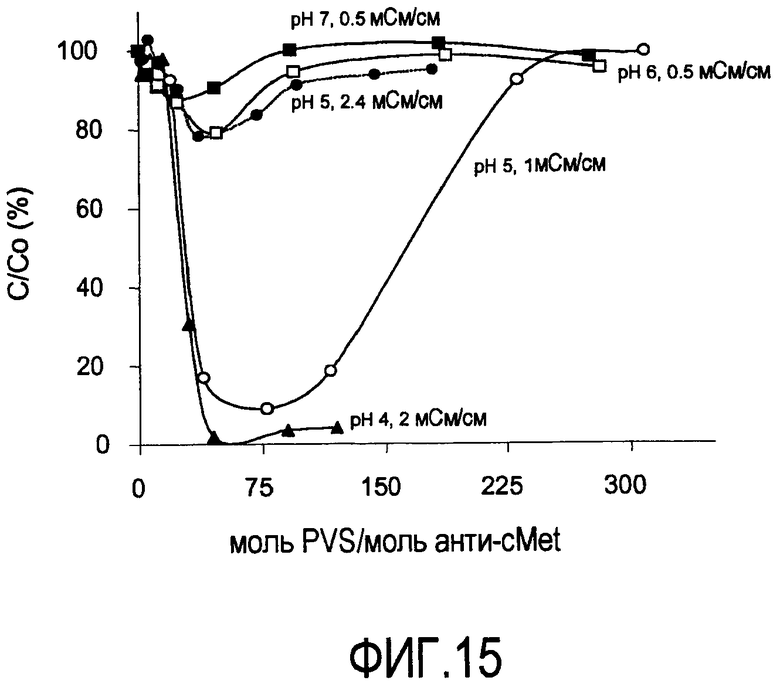
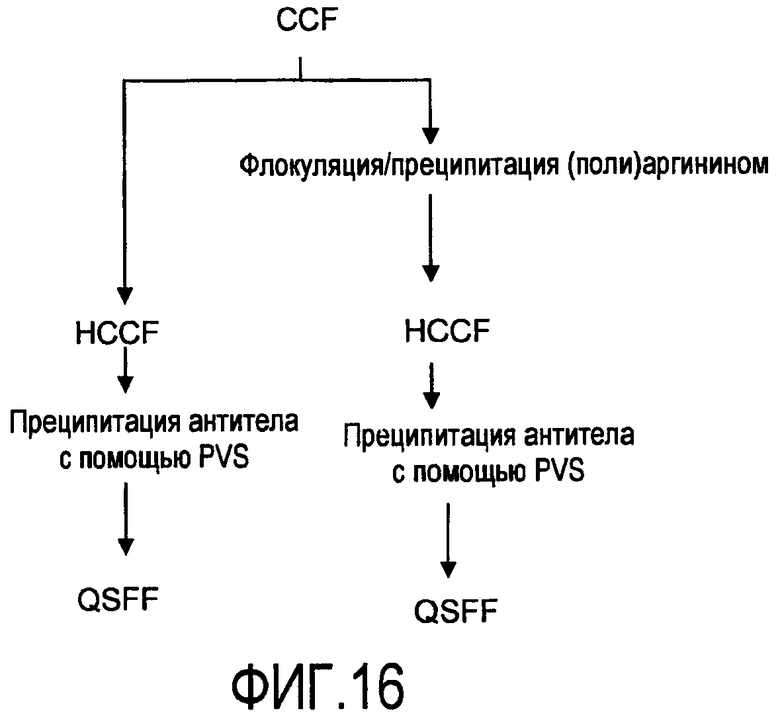
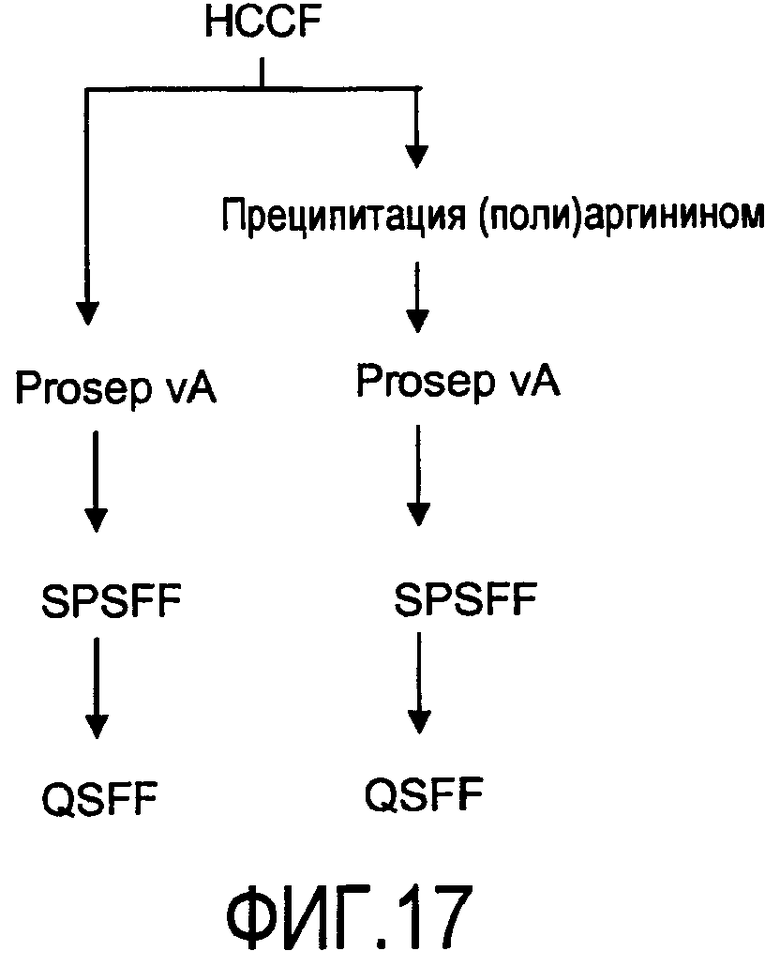
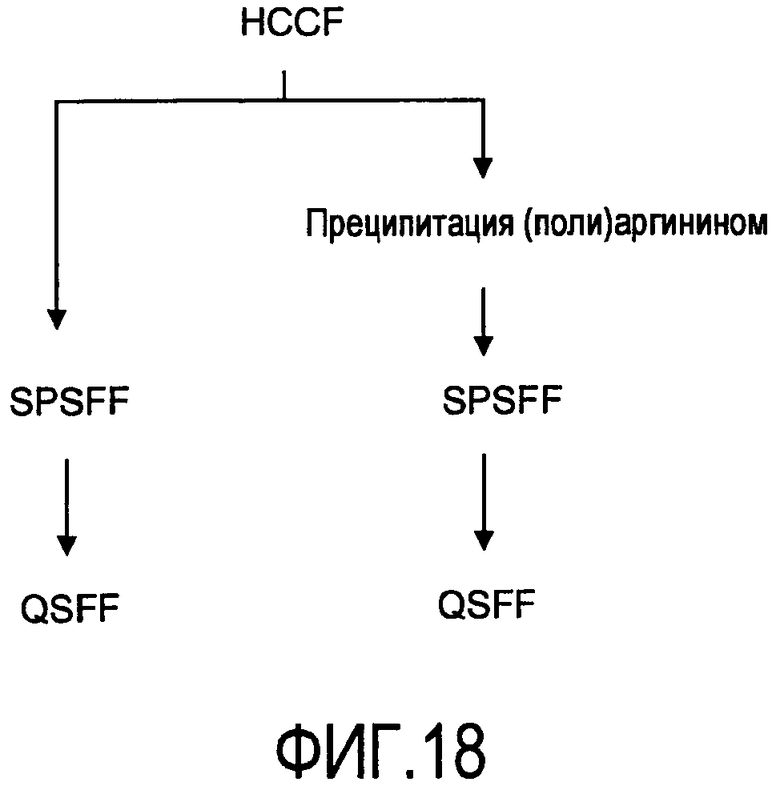
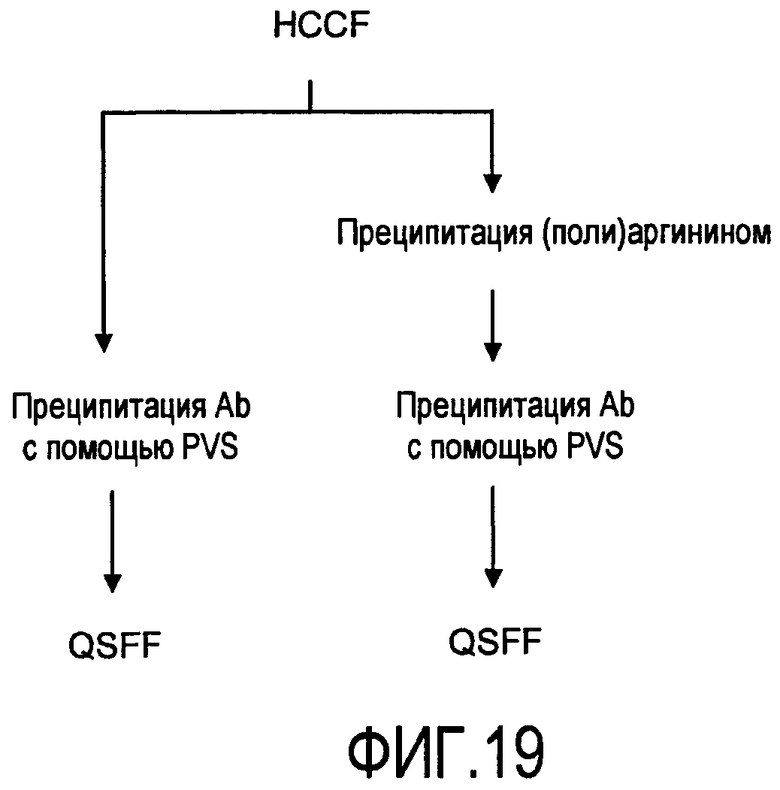
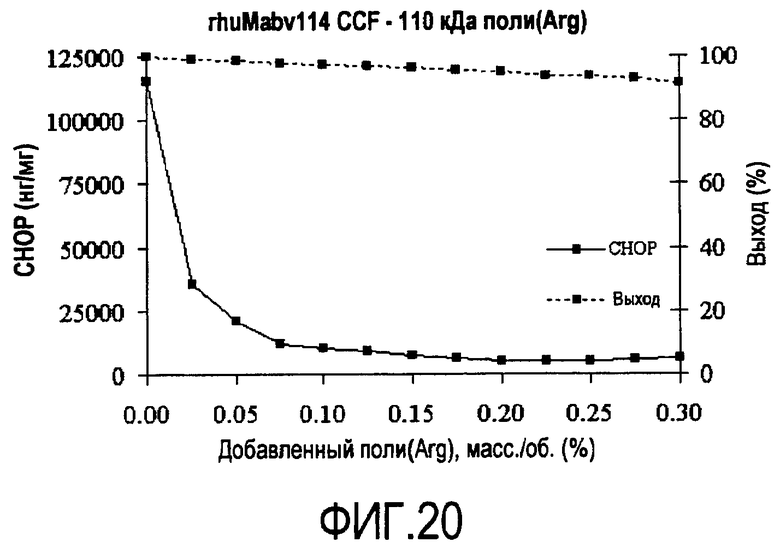
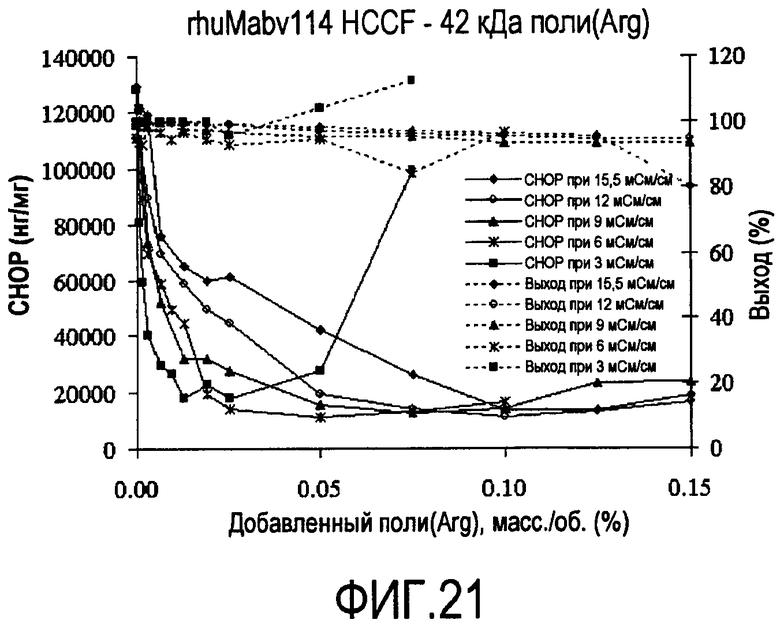
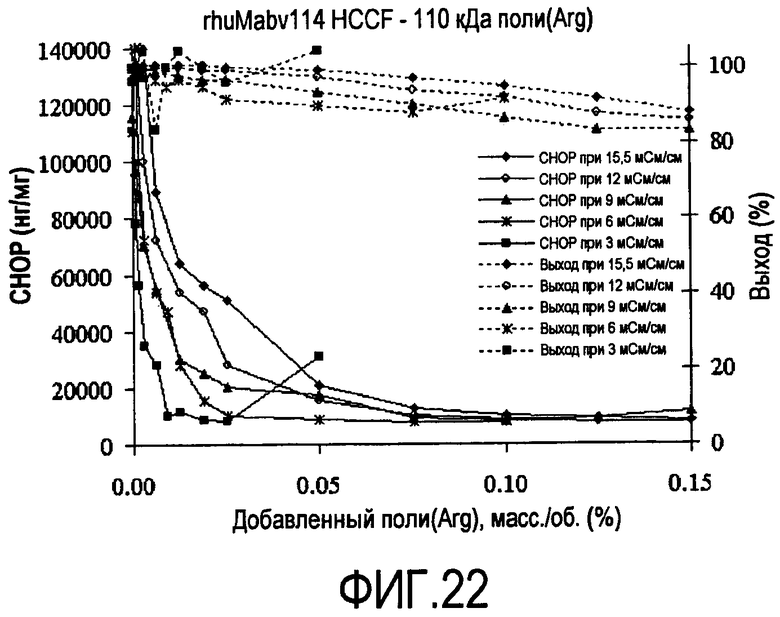
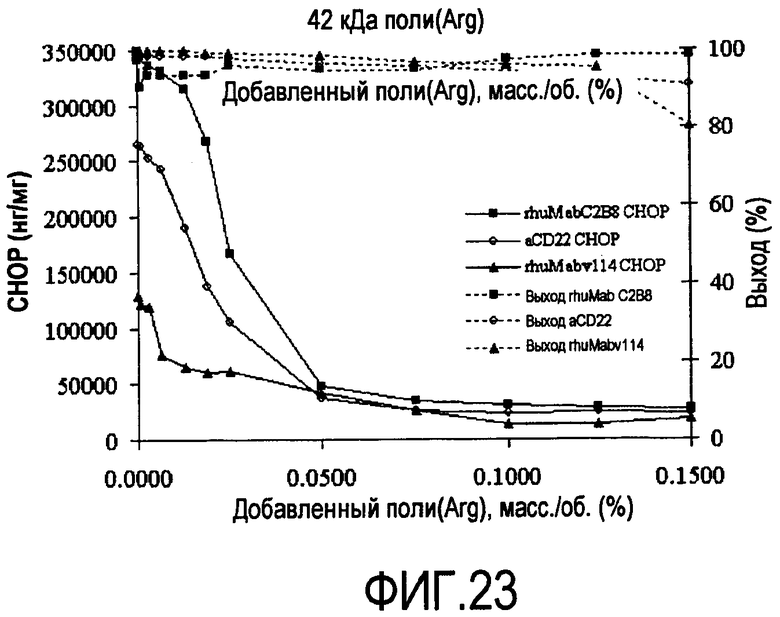
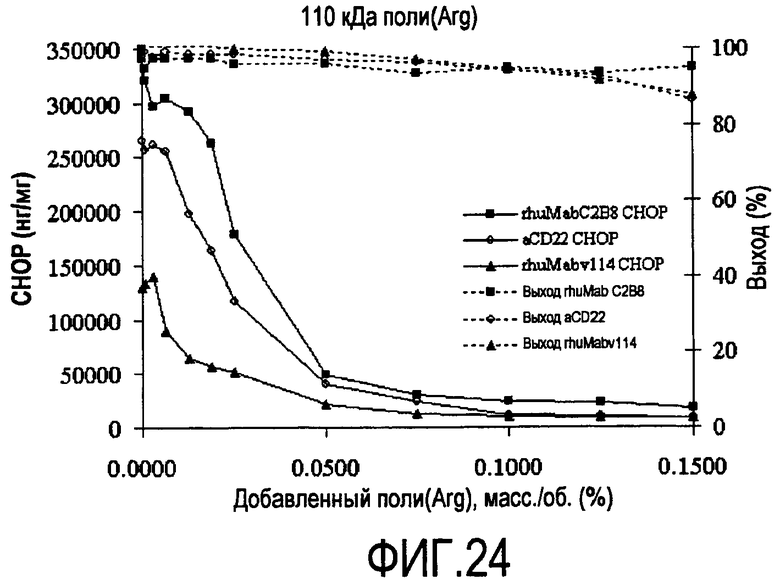
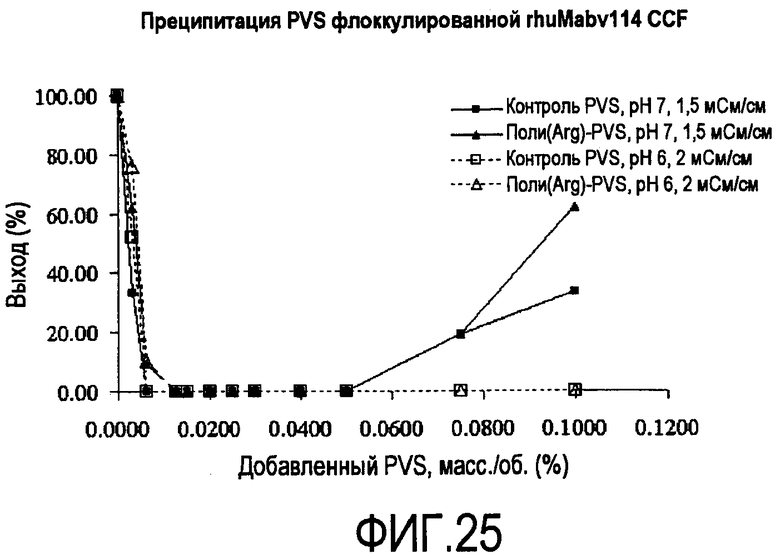
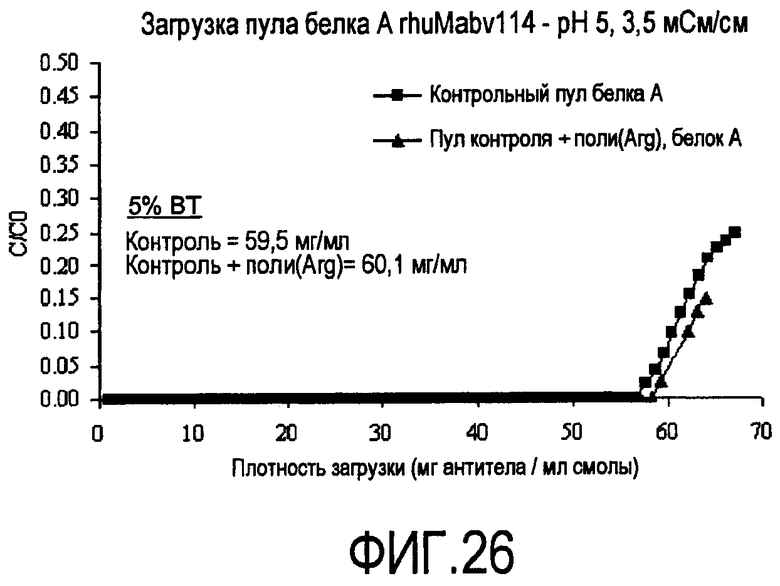
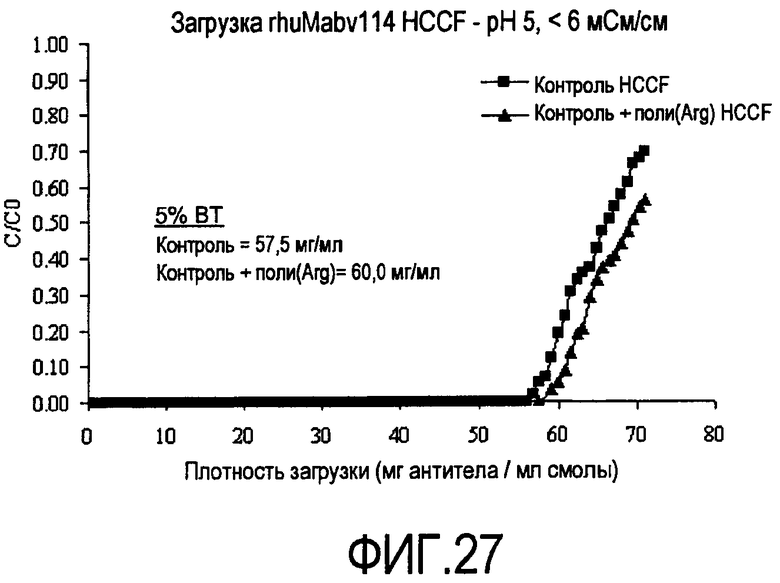
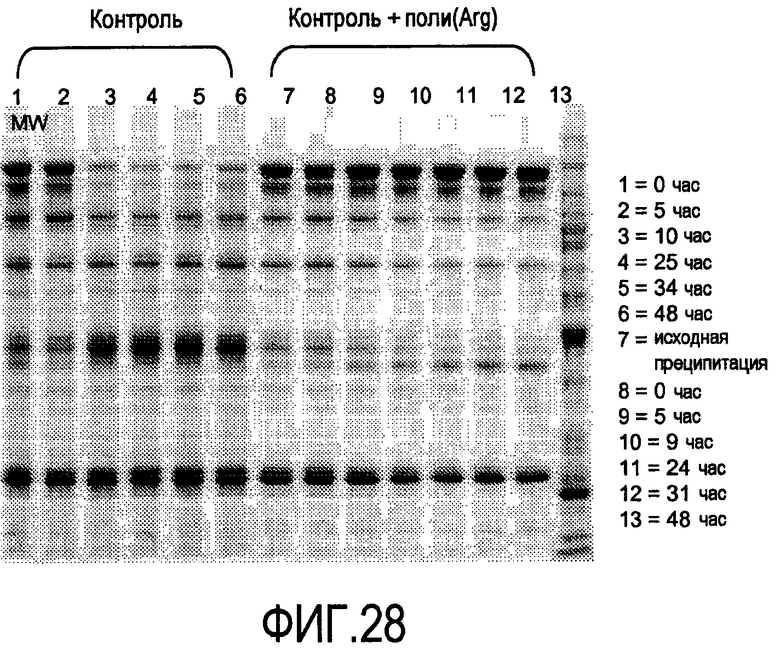
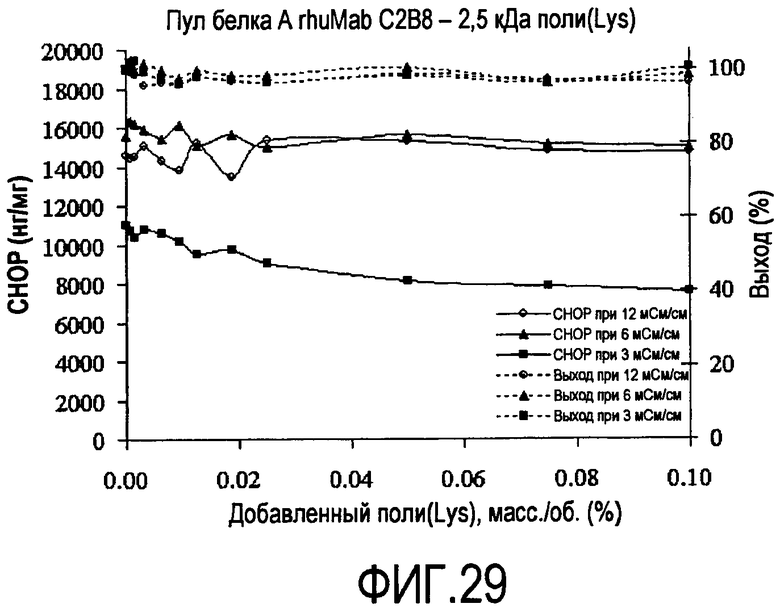
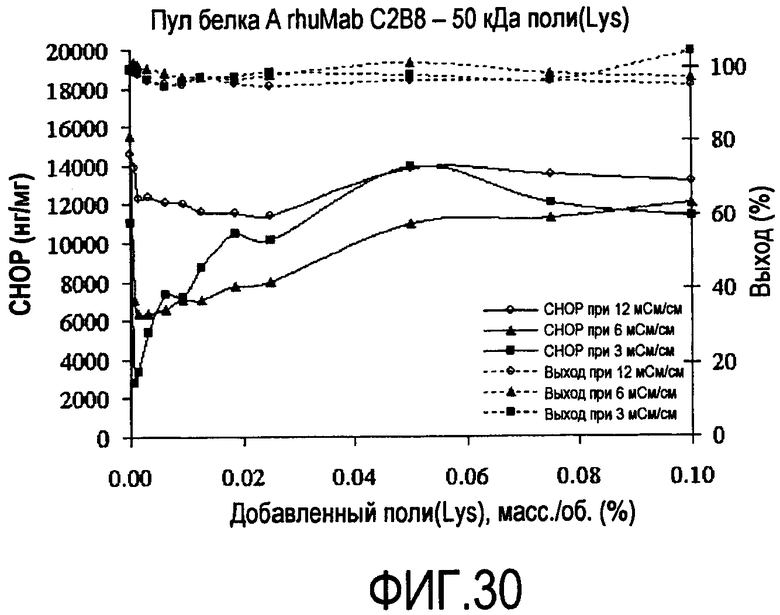
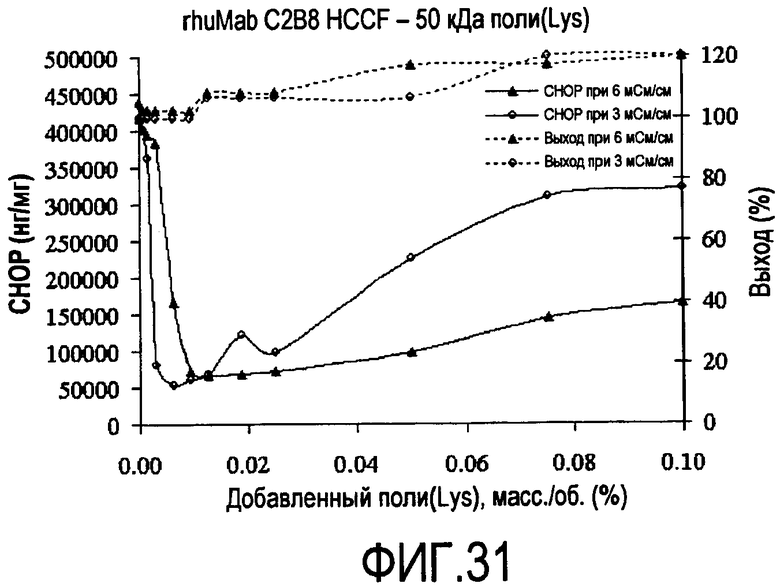
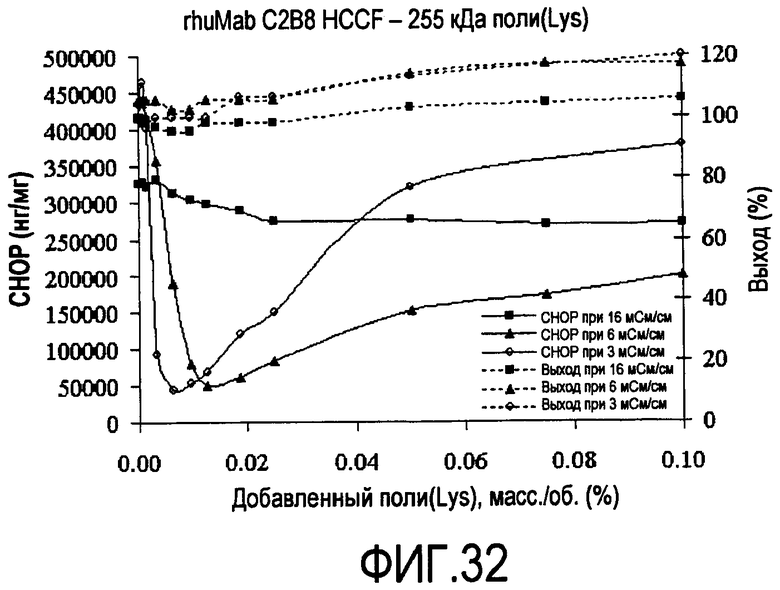
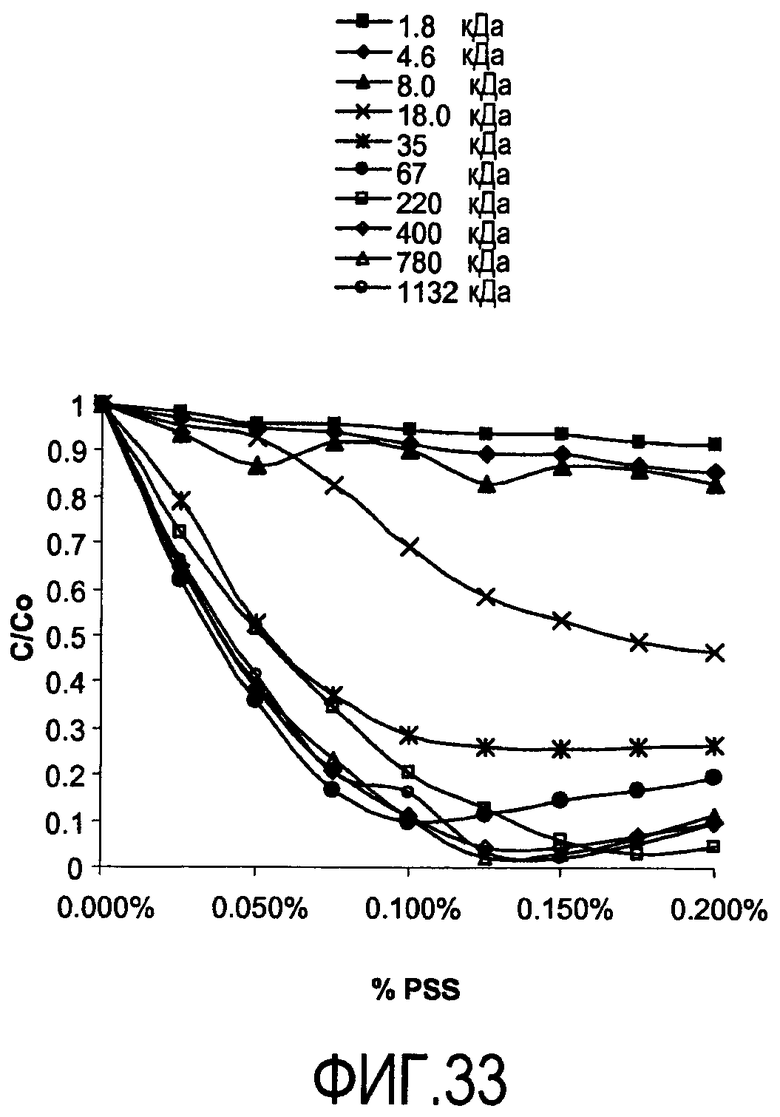
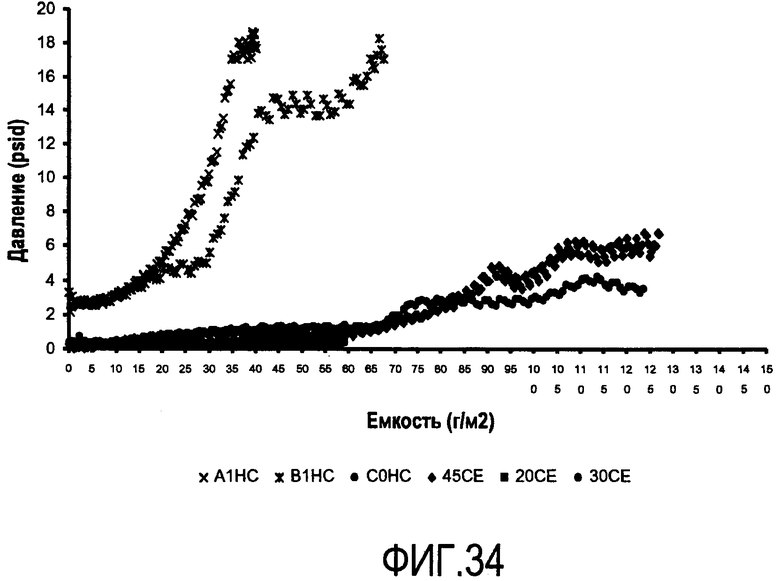
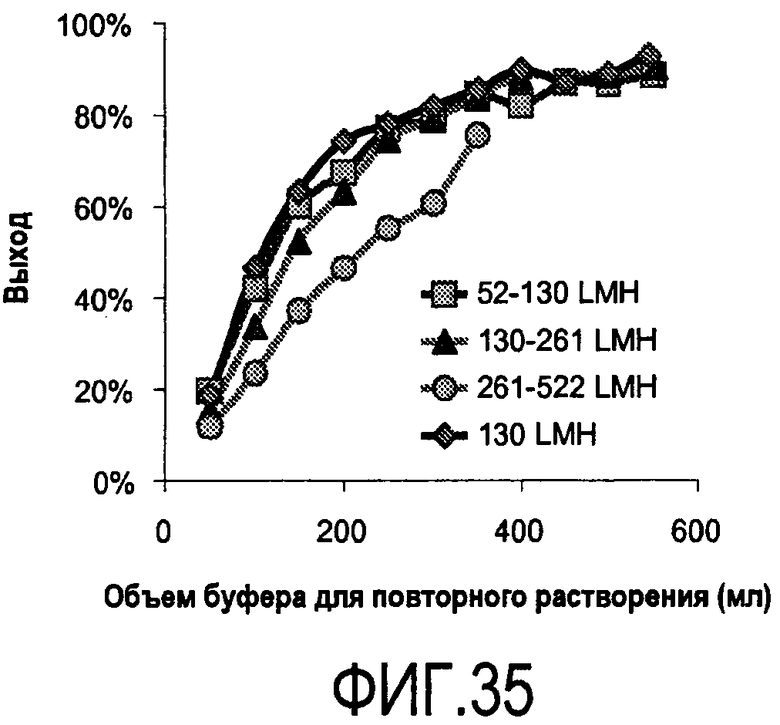
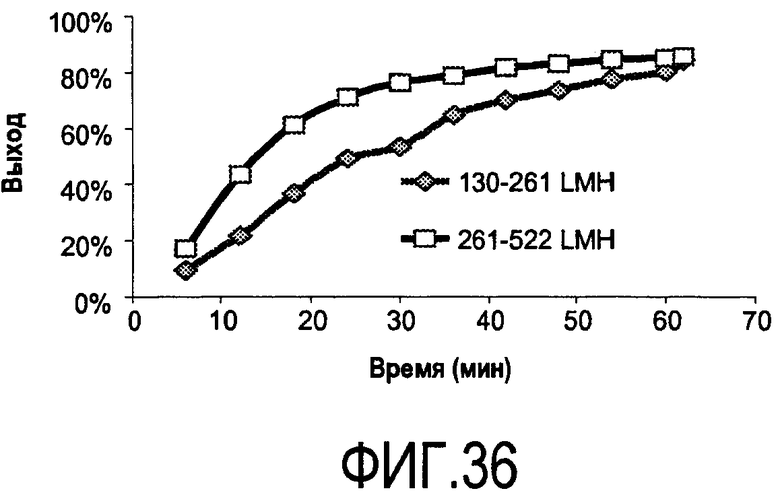
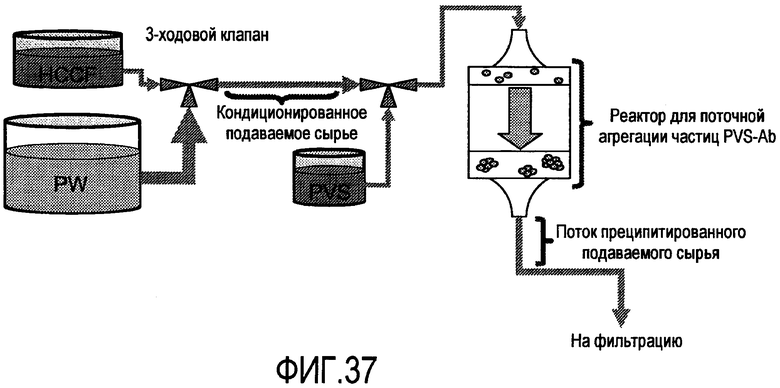
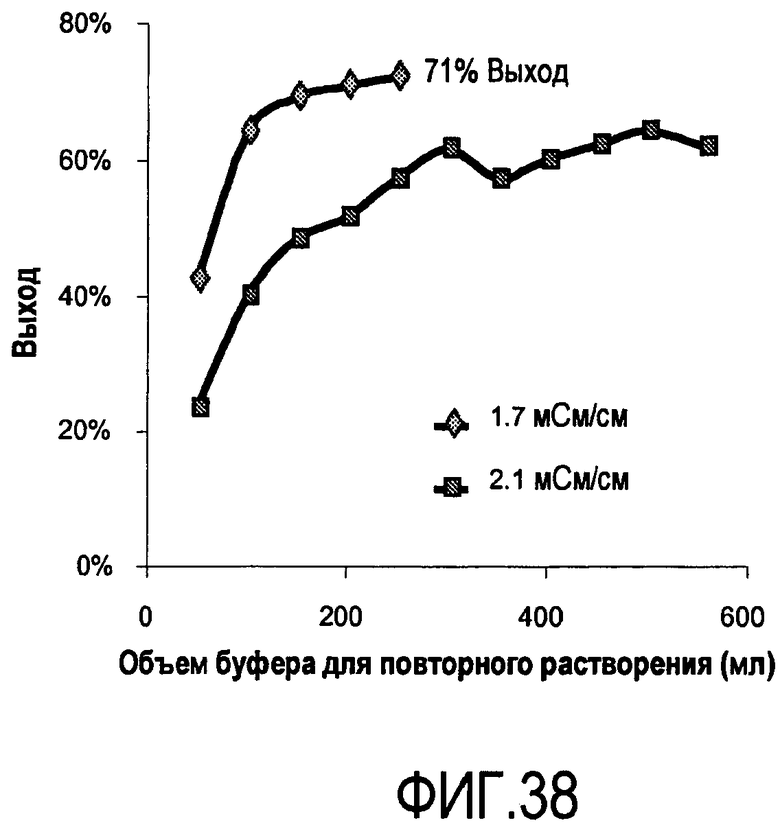
Настоящее изобретение относится к области биохимии. Предложен способ очистки антител посредством добавления отрицательно заряженного полиэлектролита, такого как поливинилсульфоновая кислота, поливинилсульфонат, полистиролсульфоновая кислота или полиакриловая кислота, к смеси, содержащей антитело, и выделения получаемого преципитата, содержащего антитело. Использование изобретения может найти дальнейшее применение в технологии выделения антител из биологических жидкостей и их очистки. 12 з.п. ф-лы, 13 табл., 38 ил., 13 пр.
1. Способ очистки антител, включающий:
(a) регуляцию кислотности до pH 4-7 или концентрации соли до 0,1-2 М в смеси, содержащей антитело, где антитело получено из собранной жидкости культуры клеток, и собранная жидкость культуры клеток получена из культуры клеток млекопитающих;
(b) добавление отрицательно заряженного полиэлектролита, выбранного из поливинилсульфоновой кислоты, поливинилсульфоната, полистиролсульфоновой кислоты и полиакриловой кислоты, и где отрицательно заряженный полиэлектролит обладает молекулярной массой от 1000 до 1000000 Да, посредством чего образуется преципитат белок-полиэлектролит, содержащий отрицательно заряженный полиэлектролит и антитело, до тех пор, пока преципитат не начнет повторно растворяться;
(c) отделение преципитата белок-полиэлектролит от примесей, выбранных из агрегатов белка, фрагментов белка, белков клетки-хозяина, инсулина, гентамицина, ДНК и вымываемого белка А;
(d) выделение преципитата белок-полиэлектролит; и
(e) ресуспендирование преципитата белок-полиэлектролит в водном растворе, где способ необязательно дополнительно включает стадии очистки с помощью хроматографии на основе белка А, катионообменной и/или анионообменной хроматографии до и/или после стадий (а)-(е).
2. Способ по п.1, где антитело представляет собой моноклональное антитело, необязательно выбранное из антитела против CD20, антитела против DR5 и антитела против СМЕТ.
3. Способ по п.1 или 2, где антитело получено из собранной жидкости культуры клеток посредством иммобилизации белка на адсорбенте с белком А.
4. Способ по п.1 или 2, где антитело получено из собранной жидкости культуры клеток посредством катионообменной хроматографии.
5. Способ по п.1 или 2, где антитело захватывают непосредственно из собранной жидкости культуры клеток.
6. Способ по п.1, где в смесь добавляют более одного отрицательно заряженного полиэлектролита.
7. Способ по п.1, где преципитат белок-полиэлектролит выделяют центрифугированием.
8. Способ по п.1, где преципитат белок-полиэлектролит выделяют фильтрацией.
9. Способ по п.1, где преципитат белок-полиэлектролит выделяют объемной фильтрацией.
10. Способ по п.1, где белок повторно растворяют с помощью однократного пропускания буфера для повторного растворения через фильтр.
11. Способ по п.1, где белок ресуспендируют посредством рециркуляции буфера для повторного растворения через фильтр.
12. Способ по п.1, где полиэлектролит отделяют от ресуспендированного преципитата белок-полиэлектролит посредством анионообменной очистки.
13. Способ по п.1, где культура клеток млекопитающих представляет собой культуру клеток яичника китайского хомячка.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| US 3737377 A, 05.06.1973 | |||
| DUBIN P.L., GAO J., MATTISON K., "Protein purification by Selective Phase Separation with Polyelectrolytes", Separation and Purification Methods (1994), 23(1): 1-16 | |||
| COOPER CL et al., "Polyelectrolyte-protein complexes", Current Opinion in Colloid & Interface Science (2005), 10(1-2):52-78. | |||

