Настоящая заявка испрашивает приоритет на основании предварительной заявки на выдачу патента США № 60/740205, поданной 28 ноября 2005, полное содержание которой включено в данное описание в виде ссылки.
Государственная поддержка
В соответствии с 35 U.S.C. §202(c), подтверждается, что правительство Соединенных Штатов может иметь определенные права на описанное в данной публикации изобретение, которое было осуществлено отчасти благодаря финансированию, предоставленному Национальными Институтами Здравоохранения по гранту № GM 62134.
Область техники, к которой относится изобретение
Настоящее изобретение относится к активации каскада комплемента в организме. В частности, настоящее изобретение относится к пептидам и пептидомиметикам, способным связывать белок C3 и ингибировать активацию комплемента.
Уровень техники
Различные публикации, включая патенты, опубликованные заявки, технические статьи и научные статьи, цитируются на протяжении всего описания. Каждая из цитируемых публикаций включена в данное описание в виде ссылки в полном объеме. Полные ссылки публикаций, не полностью цитированных в описании, указаны в конце описания.
Система комплемента является первой линией иммунологической защиты от чужеродных патогенов. Ее активация посредством классического, альтернативного или лецитинового путей приводит к образованию анафилатоксических пептидов C3a и C5a и образованию мембраноатакующего комплекса C5b-9. Компонент комплемента C3 играет центральную роль в активации всех трех путей. Активация C3-конвертазами C3-пути комплемента и его последующее связывание с поверхностью мишени приводит к сборке мембраноатакующего комплекса и, в конечном счете, к повреждению или лизису клеток-мишеней. C3 является уникальным, так как он имеет сложную структуру, которая обеспечивает многообразие разных участков связывания лигандов, которые важны для путей иммунологического надзора и иммунного ответа.
Несоответствующая активация комплемента может приводить к повреждению клетки-хозяина. Комплемент вносит вклад в несколько патологических состояний, включая различные аутоиммунные заболевания, и, как было обнаружено, вносит вклад в другие клинические состояния, такие как респираторный синдром взрослых, инфаркт миокарда, отторжение после ксенотрансплантации и ожоговые поражения. Также было обнаружено, что опосредованное комплементом повреждение ткани возникает в результате ситуаций биологической несовместимости, таких как ситуации, с которыми сталкиваются пациенты, подвергающиеся диализу или искусственному кровообращению.
Опосредованные комплементом повреждения тканей прямо опосредованы мембраноатакующим комплексом и косвенно - образованием C3a и C5a. Такие пептиды индуцируют повреждение благодаря их влиянию на различные клетки, включая нейтрофилы и тучные клетки. In vivo регуляция комплемента на стадиях активации C3 и C5 обеспечивается как плазматическими, так и мембранными белками. Плазматическими белками-ингибиторами являются фактор H и C4-связывающий белок, и регуляторными мембранными белками, расположенными на поверхности клеток, являются рецепторы комплемента 1 (CR1), фактор ускорения распада (DAF) и мембранный кофакторный белок (MCP). Указанные белки ингибируют конвертазы C3 и C5 (мультисубъединичные протеазы), стимулируя диссоциацию мультисубъединичных комплексов и/или инактивируя комплексы посредством протеолиза (катализируемого фактором I). В анализе in vitro идентифицировано несколько фармакологических средств, которые регулируют или модулируют активность комплемента, но в большинстве случаев, как было показано in vivo, они обладали низкой активностью или были токсичными.
До настоящего времени не существует ингибиторов активации комплемента, одобренных для применения в клинике, хотя существуют некоторые кандидаты для клинического применения, в частности, рекомбинантная форма рецептора комплемента 1, известная как растворимый рецептор комплемента 1 (sCR1), и гуманизированное моноклональное анти-C5-антитело (5G1.1-scFv). Показано, что оба указанных вещества супрессируют активацию комплемента в животных моделях in vivo (Kalli K.R. et al., 1994; и Wang et al., 1996). Однако каждое вещество обладает тем недостатком, что является белком с высокой молекулярной массой (240 кД и 26 кД, соответственно), который трудно производить и который необходимо вводить посредством инфузии. Соответственно, в последних исследованиях внимание акцентировано на разработке меньших по размеру активных агентов, которые легче доставлять, которые являются более стабильными и требуют меньших затрат при производстве.
В патенте США №6319897 Lambris et al. описано применение комбинаторных случайных библиотек пептидов в фаговом дисплее для идентификации состоящего из 27 остатков пептида, который связывается с С3 и ингибирует активацию комплемента. Такой пептид укоротили до состоящего из 13 остатков циклического фрагмента, который полностью сохраняет активность, который назван в данной области компстатином. Компстатин ингибирует расщепление С3 до C3a и C3b С3-конвертазами. Компстатин тестировали в серии экспериментов in vitro, in vivo, ex vivo и in vivo/ex vivo и показали, что он: (1) ингибирует активацию комплемента в сыворотке человека (Sahu A. et al., 1996); (2) ингибирует индуцированную гепарином/протамином активацию комплемента у приматов без существенных побочных эффектов (Soulika A.M. et al., 2000); (3) продлевает время жизни ксенотрансплантата от свиньи к человеку, перфузируемого кровью человека (Fiane A.E. et al., 1999a; Fiane A.E. et al., 1999b; и Fiane A.E. et al., 2000); (4) ингибирует активацию комплемента в моделях искусственного кровообращения, плазмафереза и экстракорпоральных контурах при диализе (Nilsson В. et al., 1998); и (5) обладает низкой токсичностью (Furlong S.T. et al., 2000).
Компстатин представляет собой пептид, содержащий последовательность ICVVQDWGHHRCT-NH2 (SEQ ID NO:1), где Cys2 и Cys1 образуют дисульфидный мостик. Его трехмерная структуоа определена с использованием гомоядерной 2D-ЯМР-спектроскопии в комбинации с двумя отдельными экспериментально ограниченными вычислительными методиками. Первая методика включает в себя метрическую геометрию, молекулярную динамику и моделирование отжига (Morikis D. et al., 1998; WO99/13899), а вторая методика включает в себя глобальную оптимизацию (Klepeis et al., J. Computational Chern., 20: 1344-1370, 1999). В структуре компстатина выявлена молекулярная поверхность, которая содержит полярный участок и неполярный участок. Полярный участок включает в себя β-поворот типа I, а неполярный участок содержит дисульфидный мостик. Кроме того, синтезировали серию аналогов с аланиновыми заменами (сканирование аланином) и тестировали в отношении активности, при этом выявили, что четыре остатка β-поворота и дисульфидный мостик с окружающим гидрофобным кластером вносят важный вклад в ингибиторную активность компстатина (Morikis et al., 1998; WO99/13899).
Используя анализ активности комплемента, включающий в себя измерение опосредованного альтернативным путем лизиса эритроцитов, измерили IC50 компстатина, составляющую 12 мкМ. Некоторые тестированные ранее аналоги показали активность, эквивалентную или более высокую, чем активность компстатина. В опубликованной международной заявке № WO2004/026328 описаны аналоги и миметики компстатина с изменениями на N- и C-концах и в положениях 4 и 9, которые придавали повышенную активность в указанном выше анализе. Для некоторых аналогов сообщалось о повышении вплоть до 99-кратного по сравнению с компстатином (см. также Mallik et al., 2005). Разработка аналогов или миметиков компстатина с более высокой активностью может привести к существенному прогрессу в данной области.
Сущность изобретения
Настоящее изобретение относится к аналогам и миметикам ингибирующего комплемент пептида, компстатина (НООС-ICVVQDWGHHRCT-NH2; SEQ ID NO:1), которые обладают улучшенной активностью, связанной с ингибированием комплемента, по сравнению с компстатином.
В одном аспекте отличительным признаком изобретения является соединение, которое ингибирует активацию комплемента, которое включает в себя пептид, имеющий последовательность:
Xaa1-Cys-Val-Xaa2-Gln-Asp-Xaa3-Gly-Xaa4-His-Arg-Cys-Xaa5 SEQ ID NO:26); где:
Xaa1 означает Ile, Val, Leu, Ac-Ile, Ac-Val, Ac-Leu или дипептид, содержащий Gly-Ile;
Xaa2 означает Trp или аналог Trp, при этом аналог Trp имеет более гидрофобный характер по сравнению с Trp, при условии, что если Хаа3 означает Trp, то Xaa2 означает аналог Trp;
Хаа3 означает Trp или аналог Trp, содержащий химическую модификацию в своем индольном цикле, при этом химическая модификация увеличивает потенциал водородной связи индольного цикла;
Хаа4 означает His, Ala, Phe или Trp;
Хаа5 означает L-Thr, D-Thr, Ile, Val, Gly, дипептид, содержащий Thr-Asn, или дипептид, содержащий Thr-Ala, или трипептид, содержащий Thr-Ala-Asn, при этом ОН на карбоксильном конце любой из аминокислот L-Thr, D-Thr, Ile, Val, Gly или Asn необязательно заменен -NH2; и два остатка Cys связаны дисульфидной связью.
В некоторых вариантах Xaa2 принимает участие в неполярном взаимодействии с C3. В других вариантах Xaa3 принимает участие в образовании водородной связи с C3. В других вариантах Xaa2 принимает участие в неполярном взаимодействии в C3, а Xaa3 принимает участие в образовании водородной связи с C3.
В других вариантах аналог Trp Xaa2 представляет собой галогенированный триптофан, такой как 5-фтор-1-триптофан или 6-фтор-1-триптофан. В других вариантах аналог Trp в положении Xaa2 содержит низший алкокси- или низший алкильный заместитель в положении 5, например, 5-метокситриптофан или 5-метилтриптофан. В других вариантах аналог Trp в положении Xaa 2 содержит низший алкильный или низший алкеноильный заместитель в положении 1, примерные варианты включают 1-метилтриптофан или 1-формилтриптофан. В других вариантах аналог Trp Xaa3 представляет собой галогенированный триптофан, такой как 5-фтор-1-триптофан или 6-фтор-1-триптофан.
В некоторых вариантах Xaa2 содержит низший алкеноильный или низший алкильный заместитель в положении 1 триптофана, Xaa3 необязательно содержит галогенированный триптофан, и Xaa4 содержит аланин. В конкретных вариантах Xaa2 означает 1-метилтриптофан или 1-формилтриптофан, и Xaa3 необязательно содержит 5-фтор-1-триптофан. Некоторые примерные соединения согласно изобретению содержат любую из последовательностей SEQ ID NO:15-25.
В некоторых вариантах соединение содержит пептид, полученный экспрессией полинуклеотида, кодирующего пептид. В других вариантах соединение получают, по меньшей мере, частично в результате синтеза пептидов. Также можно использовать комбинацию способов синтеза.
В некоторых вариантах предлагаются аналоги компстатина, в которых соединение пегилировано, примером является соединение, содержащее последовательность SEQ ID NO:36.
В других вариантах аналог компстатина, кроме того, содержит дополнительный пептидный компонент, который продлевает сохранение соединения in vivo. Например, дополнительным пептидным компонентом может быть пептид, связывающий альбумин. Один пример конъюгата компстатинсвязывающий альбумин пептид содержит последовательность SEQ ID NO:39.
Отличительным признаком другого аспекта изобретения является соединение, которое ингибирует активацию комплемента, содержащее непептидный или частичный пептидный миметик последовательности SEQ ID NO:26, или любой из других последовательностей аналогов и конъюгатов, описанных выше. Такие непептидные или частичные пептидные миметики предназначены для связывания C3 и ингибирования активации комплемента с активностью, по меньшей мере, в 100 раз более высокой, чем пептид, содержащий последовательность SEQ ID NO:1 в эквивалентных условиях анализа.
Аналоги, конъюгаты и миметики компстатина согласно изобретению имеют практическое применение для любых целей, для которых применим компстатин, которые известны в данной области и описаны более подробно в данной публикации. Некоторые из таких применений заключаются в приготовлении соединений в виде фармацевтических композиций для введения пациенту. Такие препараты могут содержать фармацевтически приемлемые соли соединений, а также один или несколько фармацевтически приемлемых разбавителей, носителей, эксципиентов и тому подобного, которые известны специалисту в данной области.
Различные отличительные признаки и преимущества настоящего изобретения будут понятны при обращении к подробному описанию, чертежам и примерам, которые следуют далее.
Краткое описание чертежей
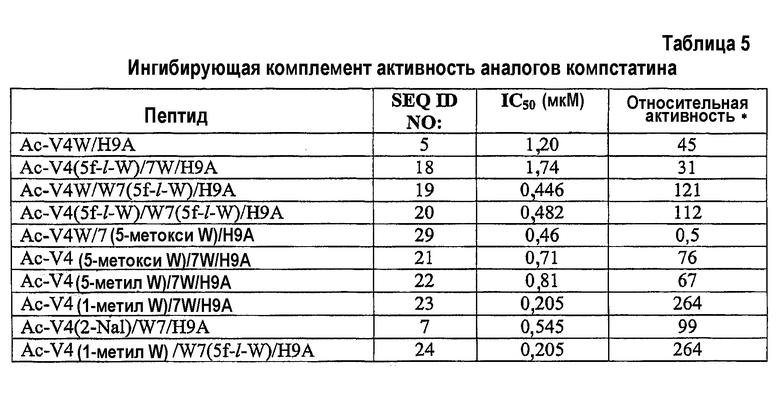
Фиг.1. Активность экспрессированного компстатина и его аналогов. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для Ac-V4W/H9A (SEQ ID NO:5) (квадраты) и экспрессированного компстатина с триптофаном (SEQ ID NO:15) (кружки), 5-фтортриптофаном (SEQ ID NO:16) (треугольники), 6-фтортриптофаном (SEQ ID NO:17 (звездочки), 5-гидрокситриптофаном (SEQ ID NO:27) (шестиугольники), 7-азатриптофаном (SEQ ID NO:28) (ромбы).
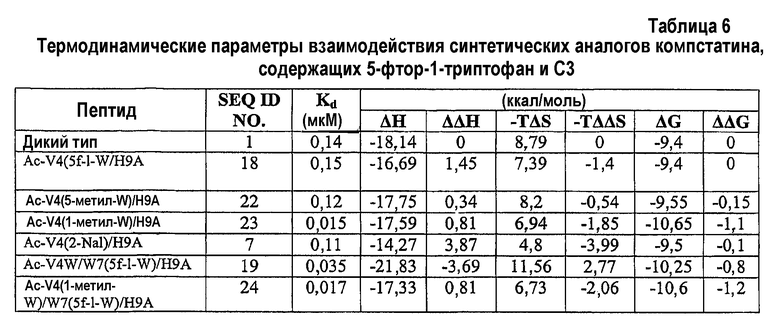
Фиг.2. Активность синтетических аналогов компстатина. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для Ac-V4W/H9A (SEQ ID NO:5) (квадраты) и аналогов компстатина с включением 5-фтор-1-триптофана в положении 4 (SEQ ID NO:18) (кружки), положении 7 (SEQ ID NO:19) (треугольники), в обоих положениях 4 и 7 (SEQ ID NO:20) (ромбы).
Фиг.3. Активность дополнительных синтетических аналогов компстатина. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для (A) Ac-V4W/H9A (SEQ ID NO:5) (треугольники) по сравнению с Ac-V4(5f-l-W)/H9A (SEQ ID NO:18) (перевернутые треугольники), Ac-V4(5-метил-W)/H9A (SEQ ID NO:22) (кружки), Ac-V4(1-метил-W)/H9A (SEQ ID NO:23) (ромбы), Ac-V4(2-Nal)/H9A (SEQ ID NO:7) (квадраты); (B) Ac-V4W/H9A (SEQ ID NO:5) (треугольники) по сравнению с Ac-V4W/W7(5f-l-W)/H9A (SEQ ID NO:19) (шестиугольники); и (C) компстатина (SEQ ID NO:1) (треугольники) по сравнению с Ac-V4(1-метил-W)/W7(5f-l-W)/H9A (SEQ ID NO:24) (треугольники, ориентированные влево).
Фиг.4. Термодинамическая характеристика взаимодействия дополнительных аналогов компстатина с C3. Данные ITC, представляющие связывание (A) Ac-V4W/H9A (SEQ ID NO:5); (B) Ac-V4(5f-l-W)/H9A (SEQ ID NO:18); (C) Ac-V4(5-метил-W)/H9A (SEQ ID NO:22); (D) Ac-V4(1-метил-W)/H9A (SEQ ID NO:23); (E) Ac-V4(2-Nal)/H9A (SEQ ID NO:7); и (F) Ac-V4W/W7(5f-l-W)/H9A (SEQ ID NO:19) с C3. Графики получали подгонкой скорректированных исходных данных к модели «один набор участков» в Origin 7.0.
Фиг.5. Графики, показывающие зависимость между гидрофобностью аналогов, обозначенных log P, и константой ингибирования (A), энтропией, обозначенной -TΔS (B), и константой связывания (C).
Фиг.6. Активность дополнительного синтетического аналога компстатина. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для Ac-V4(1-метил-W)/H9A (SEQ ID NO:23) (кружочки) и Ac-V4(1-формил-W)/H9A (SEQ ID NO:25) (квадраты).
Фиг.7. Активность пегилированного аналога компстатина. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для Ac-V4(1-метил-W)/H9A (SEQ ID NO:23) (кружки) и Ac-V4(1-метил-W)/H9A-K-PEG 5000 (SEQ ID NO:36) (квадраты).
Фиг.8. Активность аналога компстатина, конъюгированного со связывающим альбумин белком. Графики зависимости ингибирования комплемента в процентах от концентрации пептида для Ac-V4(1-метил-W)/H9A (SEQ ID NO:23) (кружки) и слитого пептида (Ac- ICV(1MeW)QDWGAHRCTRLIEDICLPRWGCLWEDD-NH2) (SEQ ID NO:39) (квадраты).
Подробное описание иллюстративных вариантов
В описании и формуле изобретения используются различные термины, относящиеся к способам и другим аспектам настоящего изобретения. Таким терминам необходимо придавать их обычное в данной области значение, если не оговорено особо. Другие специально определенные термины необходимо рассматривать в таком смысле, который совместим с приведенным в данном описании определением.
Определения:
В описании и примерах могут быть использованы следующие сокращения: Ac - ацетильная группа; NH2 - амид; MALDI - лазерная десорбция-ионизация с использованием матрицы; TOF - время пролета; ITC - изотермическая титровальная калориметрия; ВЭЖХ - высокоэффективная жидкостная хроматография; NA - неактивный; dT - D-треонин; 2-Nal - 2-нафтилаланин; 1-Nal - 1-нафтилаланин; 2-Igl - 2-инданилглицин; Dht - дигидротриптофан; Bpa - 4-бензоил-L-фенилаланин; 5f-l-W - 5-фтор-l-триптофан; 6f-l-W - 6-фтор-l-триптофан; 5-OH-W - 5-гидрокситриптофан; 5-метокси-W - 5-метокситриптофан; 5-метил-W - 5-метилтриптофан; 1-метил-W - 1-метилтриптофан; для сокращенных названий аминокислот используют стандартную трех- или однобуквенную номенклатуру, например, Trp или W для триптофана.
Термин «примерно», который используют в данном описании по отношению к измеряемому значению, такому как количество, продолжительность во времени и тому подобное, охватывает вариации ±20% или ±10%, в некоторых вариантах ±5%, в некоторых вариантах ±1% и в некоторых вариантах ±0,1% от указанного значения, при этом такие вариации подходят для получения и применения описанных соединений и композиций.
Термины «фармацевтически активный» и «биологически активный» относятся к способности соединений согласно изобретению связывать C3 или его фрагменты и ингибировать активацию комплемента. Такую биологическую активность можно измерить одним или несколькими известными в данной области анализами, которые более подробно описаны в данной публикации.
В используемом в данном описании смысле «алкил» относится к необязательно замещенному насыщенному, имеющему неразветвленную цепь, разветвленную цепь или циклическому углеводороду, содержащему примерно от 1 до 10 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода), при этом предпочтительно примерно от 1 до 7 атомов углерода. Алкильные группы включают без ограничения метил, этил, н-пропил, изопропил, н-бутил, изобутил, трет-бутил, н-пентил, циклопентил, изопентил, неопентил, н-гексил, изогексил, циклогексил, циклооктил, адамантил, 3-метилпентил, 2,2-диметилбутил и 2,3-диметилбутил. Термин «низший алкил» относится к необязательно замещенному насыщенному имеющему неразветвленную цепь, разветвленную цепь или циклический углеводород, содержащий примерно от 1 до 5 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода). Низшие алкильные группы включают без ограничения метил, этил, н-пропил, изопропил, н-бутил, изобутил, трет-бутил, n-пентил, циклопентил, изопентил и неопентил.
В используемом в данном описании смысле «галоген» относится к F, Cl, Br или I.
В используемом в данном описании смысле «алканоил», который взаимозаменяемо может быть использован с термином «ацил», относится к необязательно замещенному, имеющему неразветвленную или разветвленную цепь алифатическому ацильному остатку, имеющему примерно от 1 до 10 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода), при этом предпочтительно примерно от 1 до 7 атомов углерода. Алканоильные группы включают без ограничения формил, ацетил, пропионил, бутирил, изобутирил, пентаноил, изопентаноил, 2-метилбутирил, 2,2-диметилпропионил, гексаноил, гептаноил, октаноил и тому подобные. Термин «низший алканоил» относится к необязательно замещенному, имеющему неразветвленную или разветвленную цепь алифатическому ацильному остатку, содержащему примерно от 1 до 5 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода). Низшие алканоильные группы включают без ограничения формил, ацетил, н-пропионил, изопропионил, бутирил, изобутирил, пентаноил, изопентаноил и тому подобные.
В используемом в данном описании смысле «арил» относится к необязательно замещенной, моно- или бициклической ароматической кольцевой системе, имеющей примерно от 5 до 14 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода), при этом предпочтительно примерно от 6 до 10 атомов углерода. Неограничивающие примеры включают, например, фенил и нафтил.
В используемом в данном описании смысле «аралкил» относится к алкильным радикалам, несущим арильный заместитель, и имеет примерно от 6 до 20 атомов углерода (и все комбинации и подкомбинации диапазонов и конкретных количеств атомов углерода), при этом предпочтительно примерно от 6 до 12 атомов углерода. Аралкильные группы необязательно могут быть замещены. Неограничивающие примеры включают, например, бензил, нафтилметил, дифенилметил, трифенилметил, фенилэтил и дифенилэтил.
В используемом в данном описании смысле термины «алкоксигруппа» и «алкоксил» относятся к необязательно замещенной алкил-O-группе, в которой алкил имеет ранее определенное значение. Примеры алкоксигрупп и алкоксильных групп включают наряду с другими метокси-, этокси-, н-пропокси-, изопропокси-, н-бутокси- и гептоксигруппу.
В используемом в данном описании смысле «карбоксигруппа» относится к группе -C(=O)OH.
В используемом в данном описании смысле «алкоксикарбонил» относится к группе -C(=O)O-алкил, где алкил имеет определенное ранее значение.
В используемом в данном описании смысле «ароил» относится к -C(=O)-арильной группе, в которой арил имеет определенное ранее значение. Примеры ароильных групп включают бензоил и нафтоил.
Обычно замещенные химические остатки включают один или несколько заместителей, которые замещают водород в выбранных положениях на молекуле. Примеры заместителей включают, например, галоген, алкил, циклоалкил, аралкил, арил, сульфгидрил, гидроксил (-OH), алкоксил, цианогруппу (-CN), карбоксил (-COOH), ацил (алканоил: -C(=O)R), -C(=O)O-алкил, аминокарбонил (-C(O)NH2), -N-замещенный аминокарбонил (-C(=O)NHR”), CF3, CF2CF3 и тому подобные. Что касается указанных выше заместителей, то каждый остаток R” независимо может представлять собой, например, любой из H, алкила, циклоалкила, арила или аралкила.
В используемом в данном описании смысле «L-аминокислота» относится к любой из встречающихся в природе левовращающим альфа-аминокислотам, обычно присутствующих в белках, или сложным алкиловым эфирам таких альфа-аминокислот. Термин «D-аминокислота» относится к правовращающим альфа-аминокислотам. Если не оговорено особо, то все аминокислоты, упоминаемые в данном описании, являются L-аминокислотами.
«Гидрофобное» или «неполярное» используют в данном описании как синонимы, и термины относятся к любому межмолекулярному или внутримолекулярному взаимодействию, которое не характеризуется диполем.
В используемом в данном описании смысле «пи-характер» относится к способности компстатина принимать участие в образовании пи-связи с C3. Пи-связи образуются в результате бокового перекрывания двух параллельных p-орбиталей.
В используемом в данном описании смысле «потенциал водородной связи» относится к способности компстатина принимать участие в электростатическом притяжении с C3, в которое вовлечены электроотрицательные остатки на модифицированных остатках триптофана или аналогах триптофана в компстатине и атомы водорода на C3. Неограничивающим примером такого электроотрицательного остатка является атом фтора.
«Пегилирование» относится к реакции, в которой, по меньшей мере, один остаток полиэтиленгликоля (ПЭГ), независимо от размера, химически связан с белком или пептидом с образованием конъюгата ПЭГ-пептид. «Пегилированный» означает, что, по меньшей мере, один остаток ПЭГ, независимо от размера, химически связан с пептидом или белком. Термин ПЭГ обычно сопровождается числовым индексом, который указывает приблизительную среднюю молекулярную массу полимеров ПЭГ; например, ПЭГ-8000 относится к полиэтиленгликолю, имеющему среднюю молекулярную массу около 8000.
В используемом в данном описании смысле «фармацевтически приемлемые соли» относится к производным заявленных соединений, в которых исходное соединение модифицируют, получая соли кислоты или основания. Примеры фармацевтически приемлемых солей включают без ограничения соли неорганических или органических кислот основных остатков, таких как амины; щелочные или органические соли кислотных остатков, таких как остатки карбоновых кислот; и тому подобные. Таким образом, термин «кислотно-аддитивная соль» относится к соответствующему производному в виде соли исходного соединения, которое было получено присоединением кислоты. Фармацевтически приемлемые соли включают обычные соли или соли четвертичного аммония исходного соединения, образованные, например, неорганическими или органическими кислотами. Например, такие обычные соли включают без ограничения соли, полученные с использованием неорганических кислот, таких как хлористоводородная, бромистоводородная, серная, сульфаминовая, фосфорная, азотная и тому подобные; и соли, полученные с использованием органических кислот, таких как уксусная, пропионовая, янтарная, гликолевая, стеариновая, молочная, яблочная, винная, лимонная, аскорбиновая, памовая, малеиновая, гидроксималеиновая, фенилуксусная, глутаминовая, бензойная, салициловая, сульфаниловая, 2-ацетоксибензойная, фумаровая, толуолсульфоновая, метансульфоновая, этандисульфоновая, щавелевая, изэтиновая и тому подобные. Некоторые кислотные или основные соединения согласно настоящему изобретению могут существовать в виде цвиттерионов. Все формы соединений, включая свободную кислоту, свободное основание и цвиттерионы, входят в объем настоящего изобретения.
Описание:
Согласно настоящему изобретению информация о биологических и физико-химических свойствах компстатина была использована для конструирования аналогов компстатина, имеющих в значительной степени повышенную активность по сравнению с исходным пептидом компстатина. В некоторых вариантах аналоги имеют, по меньшей мере, в 50 раз более высокую активность, чем активность компстатина. В других вариантах аналоги имеют в 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125 или 130 раз более высокую активность, чем активность компстатина. В других вариантах аналоги имеют 135-, 140-, 145-, 150-, 155-, 160-, 165-, 170-, 175-, 180-, 185-, 190-, 195-, 200-, 205-, 210-, 215-, 220-, 225-, 230-, 235-, 240-, 245-, 250-, 255-, 260-, 265-кратную или более высокую активность, чем компстатин, при сравнении с использованием анализов, описанных в примерах.
Показано, что аналоги компстатина, синтезированные согласно другим способам, обладают немного улучшенной активностью по сравнению с исходным пептидом, т.е. примерно до 99-кратной (Mallik, B. et al., 2005, supra; WO2004/026328). Аналоги, полученные согласно настоящему изобретению, обладают еще более высокой активностью, чем исходный пептид или его аналоги, полученные до настоящего времени, как продемонстрировано в анализах in vitro, которые показаны в данном описании на фигурах и в примерах.
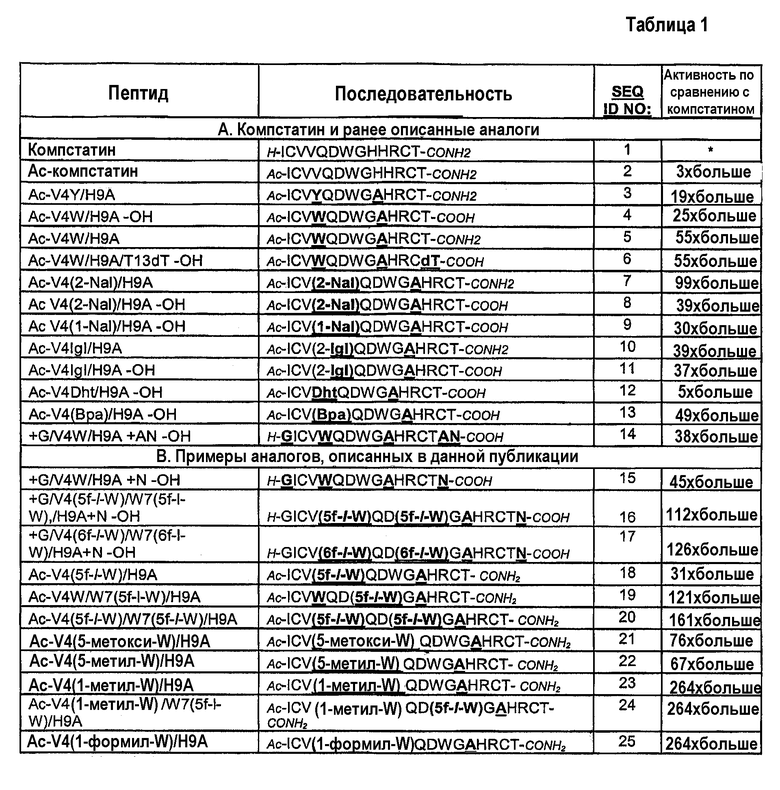
В таблице 1B показана аминокислотная последовательность и ингибирующие комплемент активности компстатина и выбранных аналогов, имеющих в значительной степени повышенную активность. Выбранные аналоги названы по конкретным модификациям требуемых положений (1-13) по сравнению с исходным пептидом, компстатином (SEQ ID NO:1) и с пептидами SEQ NO:2-14, показанными в таблице 1A, которые описаны в WO2004/026328. Пептиды SEQ ID NO:15-24 являются примерами модификаций, осуществленных согласно настоящему изобретению, приводящих к значительно более эффективным аналогам компстатина. Как описано более подробно ниже, будет понятно, что некоторые модификации, осуществленные в триптофане в положении 4, которые указаны в последовательностях SEQ ID NO:2-13, можно комбинировать с заменой аналогом триптофана в положении 7 с образованием дополнительных эффективных аналогов компстатина.
В данной таблице использованы следующие сокращения:
dT = D-треонин
2-Nal = 2-нафтилаланин
1-Nal = 1-нафтилаланин
2-Igl = 2-инданилглицин
Dht = дигидротриптофан
Bpa = 4-бензоил-L-фенилаланин
5f-l-W = 5-фтор-l-триптофан
6f-l-W = 6-фтор-l-триптофан
5-OH-W = 5-гидрокситриптофан
5-метокси-W = 5-метокситриптофан
5-метил-W = 5-метилтриптофан
1-метил-W = 1-метилтриптофан
1-формил-W = 1-формилтритофан
Модификации на N-конце. Ацетилирование N-конца обычно увеличивает ингибирующую комплемент активность компстатина и его аналогов, как можно видеть, в частности, при сравнении последовательностей SEQ ID NO:1 и SEQ ID NO:2. Соответственно, добавление ацильной группы на амино-конце пептида, включая без ограничения N-ацетилирование, является одним предпочтительным вариантом осуществления изобретения, особенно применимым, когда пептиды получают синтетически. Однако иногда преимущество имеет получение пептидов посредством экспрессии молекулы нуклеиновой кислоты, кодирующей пептид, в прокариотической или эукариотической системе экспрессии или транскрипцией и трансляцией in vitro. В случае таких вариантов можно использовать встречающийся в природе N-конец. Один пример аналога компстатина, подходящего для экспрессии in vitro или in vivo, представлен последовательностями SEQ ID NO:15-17, в которых ацетильная группа заменена немодифицированным глицином на N-конце. Последовательности SEQ ID NO:15-17, которые дополнительно содержат модификации в пептидах и на C-концах, которые обсуждаются ниже, примерно в 45 раз и примерно в 125 раз более активны, чем компстатин, в анализе ингибирования комплемента, описанного в данной публикации.
Модификация в пептиде. Используя вычислительные способы для оценки низкоэнергетических последовательностей, ранее было определено, что Tyr и Val являются наиболее вероятными кандидатами в положении 4 для поддержания стабильности и активности пептида (Klepeis J.L. et al., 2003). В WO2004/026328 обсуждалось, что Trp в положении 4, особенно в сочетании с Ala в положении 9, дает во много раз более высокую активность, чем активность исходного пептида (например, сравните активности последовательностей SEQ ID NO:4, 5 и 6 и активности последовательностей SEQ ID NO:2 и 3). В WO2004/026326 также описано, что все пептиды, содержащие аналоги триптофана, 2-нафтилаланин (SEQ ID NO:7, 8), 1-нафтилаланин (SEQ ID NO:9), 2-инданилглицин (SEQ ID NO:10, 11) или дигидротриптофан (SEQ ID NO:12) в положении 4 обладают повышенной ингибирующей комплемент активностью в диапазоне от 5-кратной до 99-кратной по сравнению с активностью компстатина. Кроме того, пептид, содержащий аналог фенилаланина, 4-бензоил-L-аланин в положении 4 (SEQ ID NO:13) имел в 49 раз более высокую активность, чем компстатин.
Согласно настоящему изобретению пептиды, содержащие 5-фтор-l-триптофан (SEQ ID NO:19) или 5-метокси-, 5-метил- или 1-метил-триптофан или 1-формилтриптофан (SEQ ID NO:21, 22, 23 и 25, соответственно) в положении 4 обладают в 31-264 раз более высокой активностью, чем компстатин. Включение 1-метил- или 1-формилтриптофана больше увеличивало активность и аффинность связывания по сравнению с другими аналогами. Предполагается, что опосредованная N индола водородная связь не является обязательной в положении 4 для связывания и активности компстатина. Отсутствие такой водородной связи или уменьшение полярного характера заменой водорода низшим алкилом, алканоилом или азотом индола в положении 4 усиливает связывание и активность компстатина. Не имея намерения ограничиться какой-либо конкретной теорией или механизмом действия, предполагают, что гидрофобное взаимодействие или влияние в положении 4 усиливает взаимодействие компстатина с C3. Соответственно, модификации Trp в положении 4 (например, изменение структуры боковой цепи согласно способам, известным в данной области) или замены аналогами Trp, которые сохраняют или усиливают указанное выше гидрофобное взаимодействие, предполагаются в настоящем изобретении для получения аналогов компстатина с еще более высокой активностью. Такие аналоги хорошо известны в данной области и включают без ограничения аналоги, приведенные в качестве примеров в данном описании, а также их незамещенные или альтернативно замещенные производные. Примеры подходящих аналогов можно найти в следующих публикациях и многих других: Beene, et al. (2002) Biochemistry 41: 10262-10269 (описаны наряду с прочими моногалогенированные и многократно галогенированные аналоги Trp); Babitzky and Yanofsky (1995) J. Biol. Chem. 270: 12452-12456 (описаны наряду с прочим метилированный и галогенированный Trp и другие аналоги Trp и индола); и патенты США 6214790, 6169057, 5776970, 4870097, 4576750 и 4299838. Аналоги Trp могут быть введены в пептид компстатина посредством экспрессии in vitro или in vivo или при синтезе пептидов, как известно в данной области, и более подробно описано в примерах.
В некоторых вариантах Trp в положении 4 компстатина заменяют аналогом, содержащим 1-алкильный заместитель, более конкретно - низший алкильный (например, C1-C5) заместитель, который определен выше. Такие аналоги включают без ограничения N(α)-метилтриптофан и 5-метилтриптофан. В других вариантах Trp в положении 4 компстатина заменяют аналогом, содержащим 1-алканоильный заместитель, более конкретно - низший алканоильный (например, C1-C5) заместитель, который определен выше. Кроме приведенных в качестве примеров аналогов к ним относятся без ограничения 1-ацетил-L-триптофан и L-β-гомотриптофан.
Термодинамические эксперименты показали, что включение 5-фтор-l-триптофана в положение 7 в компстатине увеличивало энтальпию взаимодействия между компстатином и C3 по сравнению с компстатином дикого типа, тогда как включение 5-фтортриптофана в положение 4 в компстатине снижало энтальпию такого взаимодействия. Не имея намерения быть связанными с каким-либо конкретным механизмом, полагают, что первые результаты показывают, что замена атомов водорода индола атомом фтора в остатке Trp в положении 7 компстатина может усиливать потенциал водородной связи индольного цикла, вводить новый потенциал водородной связи или опосредовать взаимодействие с C3 через молекулу воды в области контакта при связывании. (Katragadda M. et al., 2004). Следовательно, модификации Trp в положении 7 (например, изменение структуры боковой цепи согласно способам, известным в данной области) или замены аналогами Trp, которые сохраняют или усиливают указанный выше потенциал водородной связи, или опосредуют взаимодействие с C3 через молекулу воды в области контакта при связывании, предполагаются в настоящем изобретении для получения аналогов с еще более высокой активностью. В некоторых вариантах аналоги Trp, индольные циклы которых имеют модификации, которые приводят к повышенному потенциалу водородной связи или опосредуют взаимодействие с C3 через молекулу воды в области контакта при взаимодействии, могут быть введены в положение 7 пептида компстатина экспрессией in vitro или in vivo, или при пептидном синтезе. Обнаружено, что пептид, содержащий аналог триптофана, 5-фтортриптофан (SEQ ID NO:19) в положении 7 обладает в 121 раз повышенной активностью по сравнению с компстатином.
В другом варианте аналоги Trp вводят в оба положения 4 и 7 молекулы компстатина, а His в положении 9 компстатина необязательно заменяют Ala. Термодинамические эксперименты показали, что введение 5-фтортриптофана в положения 4 и 7 в компстатине увеличивало энтальпию взаимодействия между компстатином и C3 по сравнению с компстатином дикого типа. Соответственно, модификации Trp в положениях 4 и 7 (например, изменение боковой цепи согласно способам, хорошо известным в данной области) или замены аналогами Trp, которые сохраняют или усиливают указанное выше гидрофобное взаимодействие с C3 посредством положения 4 и сохраняют или усиливают указанный выше потенциал водородной связи с C3 посредством положения 7, или взаимодействие с C3 через молекулу воды в области контакта при связывании посредством положения 7, предполагаются в настоящем изобретении для получения аналогов компстатина с еще более высокой активностью. Такие модифицированные Trp или аналоги Trp могут быть введены в пептид компстатина в положениях 4 и 7 экспрессией in vitro или in vivo, или при синтезе пептидов. Обнаружено, что пептиды, содержащие аналог триптофана, 5-фтортриптофан (SEQ ID NO:16), и содержащие аналог триптофана, 6-фтортриптофан (SEQ ID NO:17) в положениях 4 и 7 обладают значительно повышенной активностью по сравнению с компстатином в диапазоне от 112- до 264-кратного увеличения активности. Кроме того, обнаружено, что пептиды, содержащие аналоги триптофана, 1-метилтриптофан в положении 4 и 5-фтортриптофан в положении 7 (SEQ ID NO:24) имеют 264-кратное увеличение активности по сравнению с компстатином.
Модификации на карбоксильном конце. Пептиды, полученные способами синтеза, обычно модифицируют на карбоксильном конце так, чтобы они содержали амид вместо кислоты; такую обычную модификацию можно видеть в таблице 1 в компстатине (SEQ ID NO:1) и нескольких аналогах. Действительно, в некоторых случаях определено, что пептиды, содержащие на конце амид, имеют более высокую активность, чем пептиды, содержащие на конце кислоту (сравни, например, последовательности SEQ ID NO:5 и 7 с последовательностями SEQ ID NO:4 и 8, соответственно). Соответственно, в одном предпочтительном варианте осуществления изобретения используют C-концевую амидную модификацию. Однако в некоторых условиях предпочтительно использование кислоты на C-конце. Такие условия включают без ограничения удовлетворение требований растворимости и экспрессию пептидов in vitro или in vivo с молекул нуклеиновых кислот, кодирующих пептид.
Остатком компстатина на карбоксильном конце является треонин. В некоторых вариантах осуществления настоящего изобретения C-концевой треонин заменяют одним или несколькими встречающимися в природе аминокислотами или аналогами. Например, пептид, имеющий последовательность SEQ ID NO:6, содержит D-треонин вместо L-треонина и, кроме того, имеет COOH-группу на C-конце. Указанный пептид проявляет активность, равную активности пептида SEQ ID NO:5, содержащего L-треонин и CONH2 на C-конце. Кроме того, аминокислотой Ile заменили Thr в положении 13, получив пептид с активностью в 21 раз более высокой, чем активность компстатина. Кроме того, пептиды с последовательностями SEQ ID NO:14-17, которые содержат C-концевое удлинение пептида Asn или удлинение дипептидом Ala-Asn, наряду с COOH на C-конце и неацетилированный N-конец, показали активность в 38-126 раз более высокую, чем активность компстатина. Такие пептиды также подходят для получения с использованием прокариотической или эукариотической системой, которые более подробно описаны ниже.
Аналоги компстатина согласно настоящему изобретению могут быть получены различными способами синтеза пептидов посредством конденсации одного или нескольких аминокислотных остатков согласно обычным способам синтеза пептидов. Например, пептиды синтезируют стандартным твердофазным способом, таким как способ, который может быть осуществлен с использованием синтезатора пептидов Applied Biosystems, модель 431A (Applied Biosystems, Foster City, Calif.) согласно инструкциям производителя. Другие способы синтеза пептидов или пептидомиметиков - либо твердофазными способами, либо в жидкой фазе, хорошо известны специалистам в данной области. Во время синтеза пептидов аминогруппы и карбоксильные группы разветвленной цепи при необходимости могут быть подвергнуты защите/удалению защиты с использованием общеизвестных защитных групп. Пример подходящего способа синтеза пептидов приведен в примере 3. Модификация с использованием альтернативных защитных групп пептидов и производных пептидов будет понятна специалистам в данной области.
Альтернативно некоторые пептиды согласно изобретению могут быть получены экспрессией в подходящей прокариотической или эукариотической системе. Например, конструкция ДНК может быть встроена в плазмидный вектор, приспособленный для экспрессии в бактериальной клетке (такой как E. coli) или дрожжевой клетке (такой как Saccharomyces cerevisiae), или в бакуловирусный вектор для экспрессии в клетке насекомого или вирусный вектор для экспрессии в клетке млекопитающего. Такие векторы содержат регуляторные элементы, необходимые для экспрессии ДНК в клетке-хозяине, расположенные таким образом, который обеспечивает экспрессию ДНК в клетке-хозяине. Такие регуляторные элементы, необходимые для экспрессии, включают промоторные последовательности, последовательности инициации транскрипции и необязательно энхансерные последовательности.
Пептиды SEQ ID NO:14-17 и другие, сконструированные подобным образом, подходят для получения посредством экспрессии молекулы нуклеиновой кислоты in vitro или in vivo. Конструкция ДНК, кодирующая конкатемер пептидов, причем верхний предел конкатемера зависит от используемой системы экспрессии, может быть введена в систему экспрессии in vivo. После продуцирования конкатемера осуществляют расщепление между C-концевым Asn и следующим N-концевым G воздействием на полипептид гидразина.
Пептиды, полученные генной экспрессией в рекомбинантной прокариотической или эукариотической системе, могут быть очищены способами, известными в данной области. В примерах 1 и 2 указаны способы, подходящие для применения в настоящем изобретении. В одном варианте можно использовать коммерчески доступную систему экспрессии/секреции, с помощью которой рекомбинантный пептид экспрессируется и затем секретируется из клетки-хозяина, чтобы его легко очистить от окружающей среды.
Комбинацию экспрессии генов и способов синтеза также можно использовать для получения аналогов компстатина. Например, аналог может быть получен экспрессией генов и затем подвергнут одному или нескольким способам посттрансляционного синтеза, например, чтобы модифицировать N- или C-конец или циклизовать молекулу.
Структура компстатина известна в данной области, и структуры указанных выше аналогов определяют сходным образом. После выяснения конкретной требуемой конформации короткого пептида способы конструирования пептида или пептидомиметика, соответствующего такой конформации, известны в данной области. См., например, G.R. Marshall (1993), Tetrahedron, 49: 3547-3558; Hruby and Nikiforovich (1991), in Molecular Conformation and Biological Interactions, P. Balaram and S. Ramasehan, eds., Indian Acad, of Sci., Bangalore, P.P. 429-455). Особенно важно для настоящего изобретения, что конструкция пептидных аналогов может быть дополнительно улучшена благодаря рассмотрению вклада различных боковых цепей аминокислотных остатков, который обсуждается выше (т.е. в отношении влияния функциональных групп или в отношении учитываемых стерических факторов).
Специалистам в данной области будет понятно, что пептидомиметик может служить в равной мере с пептидом в целях получения конкретной конформации остова и функциональных групп боковых цепей, необходимых для связывания с C3 и ингибирования активации комплемента. Соответственно, в объеме настоящего изобретения предполагается получение C3-связывающих, ингибирующих комплемент соединений с использованием встречающихся в природе аминокислот, производных аминокислот, аналогов или молекул, отличных от аминокислот, способных к связыванию с образованием подходящей конформации остова. Непептидный аналог или аналог, содержащий пептидный и непептидный компоненты, иногда называют в данном описании «пептидомиметиком» или «изостерическим миметиком», чтобы указать замены или отклонения в пептидах согласно изобретению, которые имеют такие же признаки конформации остова и/или другие функциональные свойства, чтобы быть в достаточной степени сходными с приведенными в качестве примера пептидами в ингибировании активации комплемента.
Применение пептидомиметиков для разработки высокоаффинных пептидных аналогов хорошо известно в данной области (см., например, Zhao B. et al., 1995; Beeley, N. 1994; и Hruby, V.J. 1993). Принимая во внимание ограничения вращения, сходные с таковыми для аминокислотных остатков в пептиде, аналоги, содержащие неаминокислотные остатки, можно проанализировать и можно проверить их конформационные мотивы с помощью диаграммы Рамачандрана (Hruby and Nikiforovich 1991) наряду с другими известными способами.
Аналоги компстатина согласно настоящему изобретению могут быть модифицированы добавлением к пептиду компонентов полиэтиленгликоля (ПЭГ). Как хорошо известно в данной области, пегилирование может увеличивать время полужизни терапевтических пептидов и белков in vivo. В одном варианте ПЭГ имеет среднюю молекулярную массу примерно от 1000 до примерно 50000. В другом варианте ПЭГ имеет среднюю молекулярную массу примерно от 1000 до примерно 20000. В другом варианте ПЭГ имеет среднюю молекулярную массу примерно от 1000 до примерно 10000. В иллюстративном варианте ПЭГ имеет среднюю молекулярную массу примерно 5000. Полиэтиленгликоль может иметь разветвленную или прямую цепь и предпочтительно прямую цепь.
Аналоги компстатина согласно настоящему изобретению могут быть ковалентно связаны с ПЭГ линкерной группой. Такие способы хорошо известны в данной области. (Обзор в Kozlowski A. et al. 2001; см. также Harris J.M. and Zalipsky S, eds. Poly(ethylene glycol), Chemistry and Biological Applications, ACS Symposium Series 680 (1997)). Неограничивающие примеры приемлемых линкерных групп включают сложноэфирную группу, амидную группу, имидную группу, карбаматную группу, карбоксильную группу, гидроксильную группу, углеводную, сукцинимидную группу (включая без ограничения сукцинимидилсукцинат (SS), сукцинимидилпропионат (SPA), сукцинимидилкарбоксиметилат (SCM), сукцинимидилсукцинамид (SSA) и N-гидроксисукцинамид (NHS)), эпоксидную группу, оксикарбонилимидазольную группу (включая без ограничения карбонилдимидазол (CDI)), нитрофенильную группу (включая без ограничения нитрофенилкарбонат (NPC) или трихлорфенилкарбонат (TPC)), трисилатную (trysylate) группу, альдегидную группу, изоцианатную группу, винилсульфоновую группу, группу тирозина, цистеиновую группу, группу гистидина или первичный амин. В некоторых вариантах линкерной группой является сукцинимидная группа. В одном варианте линкерной группой является NHS.
Аналоги компстатина согласно настоящему изобретению альтернативно могут быть непосредственно связаны с ПЭГ (т.е. без линкерной группы) посредством аминогруппы, сульфгидрильной группы, гидроксильной группы или карбоксильной группы. В одном варианте ПЭГ связывают с остатком лизина, добавленного к C-концу компстатина.
Пегилирование является одним из путей увеличения продолжительности удерживания терапевтических пептидов и белков in vivo. Клиренс пептидов in vivo также может быть снижен связыванием пептидов с некоторыми другими пептидами. Например, некоторые связывающие альбумин пептиды имеют необычно длительное время полужизни 2,3 часа при инъекции в виде внутривенного болюса кроликам (Dennis et al., 2002). Пептид такого типа, слитый с Fab D3H44 против тканевого фактора, обеспечивает возможность связывания Fab с альбумином, при этом сохраняя способность Fab связываться с тканевым фактором (Nguyen et al., 2006). Такое взаимодействие с альбумином приводило к существенно сниженному клиренсу in vivo и увеличенному времени полужизни у мышей и кроликов по сравнению с Fab D3H44 дикого типа, сопоставимым с клиренсом и временем полужизни, обнаруженным для пегилированных молекул Fab, иммуноадгезинов и слияний с альбумином. Как описано в примере 11 в данном описании, авторы изобретения синтезировали аналог компстатина, слитый со связывающим альбумин пептидом, и показали, что слитый белок является активным в ингибировании активации комплемента.
Ингибирующую активацию комплемента активность аналогов, пептидомиметиков и конъюгатов компстатина можно тестировать в различных анализах, известных в данной области. В предпочтительном варианте используют анализ, описанный в примере 4. Неисчерпывающий список других анализов приведен в патенте США 6319897, включающий без ограничения (1) связывание пептида с C3 и фрагментами C3; (2) различные гемолитические анализы; (3) измерение опосредованного C3-конвертазой расщепления C3; и (4) измерение расщепления фактора B фактором D.
Пептиды и пептидомиметики, описанные в данной публикации, практически применимы для любых целей, для которых используют сам компстатин, которые известны в данной области. Такие применения включают без ограничения: (1) ингибирование активации комплемента в сыворотке, тканях или органах пациента (человека или животного), которое может способствовать лечению некоторых заболеваний или состояний, включая без ограничения связанную с возрастом дегенерацию желтого пятна, ревматоидный артрит, повреждение спинного мозга, болезнь Паркинсона и болезнь Альцгеймера; (2) ингибирование активации комплемента, которая происходит при использовании искусственных органов или имплантатов (например посредством покрывания или иной обработки искусственного органа или имплантата пептидом согласно изобретению); (3) ингибирование активации комплемента, которая происходит во время экстракорпорального шунтирования физиологических жидкостей (крови, мочи) (например, посредством покрывания трубок, с помощью которых производят шунтирование жидкостей, пептидом согласно изобретению); и (4) скрининг библиотек малых молекул для идентификации других ингибиторов активации компстатина (например, в высокопроизводительных анализах в жидкой фазе или в твердой фазе, предназначенных для измерения способности тестируемого соединения конкурировать с аналогом компстатина за связывание с C3 или фрагментом C3).
Следующие примеры приведены для более подробного описания изобретения. Они предназначены для иллюстрации, но не для ограничения изобретения. Материалы и способы, указанные в примерах 1-5, использовали для получения результатов, описанных в примерах 6-11.
ПРИМЕР 1
Бактериальная экспрессия компстатина
Аналог компстатина, имеющий следующую последовательность NH2-GICVWQDWGAHRCTN-OH («G(-1)/V4W/H9A/N14») (SEQ ID NO:15), экспрессировали в слиянии со связывающим хитин доменом и интеином DnaB (New England Biolabs, Beverly, M.A.). Руководствуясь пептидной последовательностью и использованием кодонов в случае E. coli, использовали следующий генетический код для создания синтетического гена для данного пептида со следующей последовательностью:
5'ATTTGCGTTTGGCAGGATTGGGGTGCGCACCGTTGCACCAATTTAA3' (SEQ ID NO:29)
Чтобы клонировать синтетический ген в векторе pGEM-T, конструировали 5'-фланкирующую область, содержащую сайт SapI, и 3'-фланкирующую область, содержащую сайт PstI. Чтобы сконструировать синтетический ген, конструировали четыре перекрывающихся олигонуклеотида, показанных ниже, используя компьютерную программу DnaWorks, и синтезировали в Invitrogen Inc. (Carlsbad, CA):
5'GGTGGTGGCTCTTCCAACGGTATTTGCGTTTGGCAGGA3' (SEQ ID NO:30)
5'TTGGGGTGGCGCACCGTTGCACCAATTAACTGCAGG3' (SEQ ID NO:31)
3'CAACGTGGTTAATTGACGTCCGC5' (SEQ ID NO:32)
3'CATAAACGCAAACCGTCCTAACCCCACGCGTGG5'(SEQ ID NO:33)
Перекрывающиеся фрагменты ДНК собирали в ПЦР, как описано в Stemmer et al., 1995. Полученный, в результате, ген амплифицировали, используя следующие праймеры:
5'CGCCTGCAGTTAATTGGT3' (SEQ ID NO:34)
5'GGTGGTGCTCTTCCAACG3' (SEQ ID NO:35)
Амплифицированные в ПЦР фрагменты компстатина затем клонировали в векторе pGEM-T и полученный в результате клон расщепляли PstI и SapI. Фрагмент PstI-SapI, кодирующий аналог компстатина, затем субклонировали в экспрессирующем векторе pTWIN1, который был предварительно расщеплен PstI и SapI; последовательность клона подтверждали секвенированием ДНК.
Чтобы экспрессировать аналог компстатина, клетки ER2566 E. coli, трансформированные клоном компстатина, выращивали в среде SOB (20 г/л триптона, 5 г/л дрожжевого экстракта, 0,5 г/л NaCl, 2,5 мМ KCl, 10 мМ MgCl2) при 37°C. Когда OD600 достигала значения 0,7, экспрессию индуцировали добавлением IPTG до конечной концентрации 0,3 мМ с последующей дополнительной инкубацией при 37°C в течение 4 часов. Клетки собирали центрифугированием и лизировали обработкой ультразвуком в буфере B1 (20 мМ фосфатный буфер, pH 8,5, с 500 мМ NaCl и 1 мМ EDTA) с добавлением 0,2% твина-20. Экстракт клеток центрифугировали и растворимую фракцию наносили на колонку, связывающую хитин (New England Biolabs, Beverly, MA), предварительно уравновешенную буфером B1. Колонку промывали 100 мл буфера B1 с последующей быстрой промывкой 3 объемами колонки буфера B2 (50 мМ ацетат аммония, pH 7,0). Колонку инкубировали при комнатной температуре в течение 20 часов и пептид элюировали буфером B2, лиофилизовали и затем очищали на колонке C18 для ВЭЖХ. Очищенный пептид идентифицировали, используя MALDI-TOF-масс-спектрометрию.
ПРИМЕР 2
Экспрессии триптофановых аналогов компстатина в E. coli
Чтобы экспрессировать аналоги компстатина, содержащие производные триптофана, клоном pTWTN1-компстатин, трансформировали ауксотрофа ER2566 Trp 82. Экспрессию осуществляли в минимальной среде M9 с добавлением 1 мМ L-триптофана, который описан выше. Клетки выращивали до OD600 0,8-1,0, затем собирали центрифугированием и ресуспендировали в свежей минимальной среде, содержащей 2 мМ требуемого аналога(ов) триптофана: 5-фтортриптофана, 6-фтортриптофана, 7-азатриптофана или 5-гидрокситриптофана. Экспрессированные аналоги компстатина затем очищали, как описано в примере 1.
ПРИМЕР 3
Синтез пептидов
Синтез и очистку пептидов осуществляли, как описано в Sahu et al., 1996; Sahu et al., 2000; и Mallik et al., 2005. Коротко, пептиды синтезировали в синтезаторе пептидов Applied Biosystem (модель 431A), используя Fmoc-амидную смолу и стандартные защитные группы боковой цепи. Пептид отщепляли от смолы инкубацией в течение 3 часов при 22°C со смесью растворителей, содержащей 5% фенола, 5% тиоанизола, 5% воды, 2,5% этандитиола и 82,5% трифторуксусной кислоты (ТФУ). Реакционную смесь фильтровали через пористую стеклянную воронку, осаждали холодным эфиром, растворяли в 50% ацетонитриле, содержащем 0,1% ТФУ, и подвергали лиофилизации.
Неочищенные пептиды, полученные после отщепления, растворяли в 10% ацетонитриле, содержащем 0,1% ТФУ, и очищали, используя колонку C-18 с обращенной фазой (Waters, Milford, MA). Окисление дисульфида осуществляли способом циклизации на смоле, используя реагент трифторацетат таллия (III). Такой способ исключает стадии окисления в разбавленном растворе и последующее трудоемкое концентрирование с использованием стадий лиофилизации перед обращенно-фазовой ВЭЖХ. При использовании такого способа не происходило образования мультимеров, и получали высокий уровень (~90%) окисленного или циклизованного вещества с полным удалением защиты. Идентичность и чистоту всех пептидов подтверждали масс-спектроскопией с лазерной десорбцией и ВЭЖХ.
Для синтеза аналогов с 5-фтортриптофаном, 1-метилтриптофаном и 5-метилтриптофаном использовали Fmoc-dl-производные. Разделение энантиомерных пептидов осуществляли, как описано Meyers et al. 1978. dl-смесь каждого пептида разделяли на d- и l-изомерные пептиды на колонке C18 для обращенно-фазовой ВЭЖХ, используя 10% ацетонитрил в 0,01 М ацетате аммония, pH 4,1. Идентификацию изомеров элюированных пептидов осуществляли обработкой пептидов протеазой V8 с последующим анализом с помощью MALDI-TOF-масс-спектрометрии (MicroMass TOFspec2E).
ПРИМЕР 4
Анализы ингибирования комплемента
Ингибирующую активность компстатина и его аналогов по отношению к системе комплемента определяли, измеряя их влияние на активацию системы комплемента иммунокомплексами. Ингибирование активации комплемента оценивали посредством измерения ингибирования фиксации C3 с комплексами овальбумин - антиовальбумин в нормальной плазме человека. Лунки планшета для микротитрования покрывали 50 мкл овальбумина (10 мг/мл) в течение 2 часов при 25°C (в течение ночи при 4°C). Лунки насыщали 200 мкл раствора 10 мг/мл БСА в течение 1 часа при 25°C и затем добавляли кроличье антитело против овальбумина, чтобы образовать иммунокомплекс, которым комплемент может быть активирован. Тридцать микролитров пептидов в разных концентрациях добавляли непосредственно в каждую лунку, затем добавляли 30 мкл плазмы человека в разведении 1:80. После 30 мин инкубации связанный C3b/iC3b выявляли, используя козье конъюгированное с HRP антитело против C3 человека. Окраску проявляли добавлением субстрата пероксидазы ABTS, и оптическую плотность измеряли при 405 нм.
Данные по оптической плотности, полученные при 405 нм, переводили в значение % ингибирования на основании оптической плотности, соответствующей 100% активации комплемента. Значение % ингибирования откладывали на графике против концентрации пептида и полученный набор данных подгоняли к логистической функции доза-ответ, используя компьютерную программу Origin 7.0. Концентрация пептида, вызывающая 50% ингибирование отложения C3b/iC3b, принимали за IC50 и использовали для сравнения активностей различных пептидов. Значения IC50 получали на основании подгоняемых параметров, которые имели наименьшее значение хи-квадрат.
ПРИМЕР 5
Анализ изотермической титровальной калориметрией взаимодействия C3 с компстатином и его аналогами
Эксперименты по изометрической титровальной калориметрии осуществляли, используя калориметр Microcal VP-ITC (Microcal Inc, Northampton, MA). Для таких экспериментов использовали концентрации белка 3,5-5 мкМ и концентрации пептида 80-200 мкМ. Все титрования осуществляли в PBS (10 мМ фосфатный буфер с 150 мМ NaCl, pH 7,4). В каждом эксперименте белок-мишень, C3, вносили в ячейку, а пептид вносили в шприц. Все эксперименты осуществляли при 25°C, и в случае каждого эксперимента осуществляли инъекции пептида по 2 мкл в ячейку, содержащую белок. В каждом эксперименте исходные изотермы корректировали в отношении теплоты разбавления вычитанием изотерм, представляющих инъекции пептида в буфер. Полученные, в результате, изотермы подгоняли к различным моделям, используя компьютерную программу Origin 7.0, и модель, в которой достигается наименьшее значение хи-квадрат, считали подходящей для соответствующего набора данных. Значения аффинности связывания и энтропии наносили на графике против значений log P.
ПРИМЕР 6
Роль триптофана во взаимодействии C3-компстатин, которую оценивали, используя экспрессированные бактериями аналоги компстатина
Четыре разных аналога триптофана, которые отличаются по химической природе индольного цикла, включали в компстатин, используя систему опосредованной интеином экспрессии белков. После экспрессии пептиды очищали в одну стадию с конечным выходом 2 мг/л культуры. Аналоги триптофана: 5-фтортриптофан, 6-фтортриптофан, 7-азатриптофан и 5-гидрокситриптофан, также экспрессировали, используя ауксотроф ER2566/Trp 82, как показано с помощью профилей MALDI, и полученные, в результате, пептиды очищали до гомогенности. Нативный компстатин и аналоги циклизовали in vivo посредством дисульфидной связи, что подтверждается их неспособностью взаимодействовать с PHMB. Все пептиды дополнительно очищали на колонке C18 для ВЭЖХ с обращенной фазой.
Для активности экспрессированного аналога компстатина G(-1)/V4W/H9A/N14 (SEQ ID NO:15) получено значение IC50 1,2 мкМ, что сходно с активностью, наблюдаемой в случае аналога Ac-V4W/H9A аналог (SEQ ID NO:5). Полученные данные свидетельствуют, что глицин, расположенный на N-конце экспрессированного пептида, играет роль, сходную с ролью ацетильной группы, расположенной на N-конце аналога AC-V4W/H9A.
Обнаружено, что все экспрессированные аналоги компстатина, за исключением аналога, содержащего 7-азатриптофан, были активны при тестируемых концентрациях. Однако пептиды проявляли разные уровни активности по сравнению с аналогом Ac-V4W/H9A (фиг.1; таблица 2). Компстатин, содержащий 6-фтортриптофан и 5-фтортриптофан, а также аланин в положении 9, имел в 2,8 и 2,5 раза более высокую активность, соответственно, чем активность аналога Ac-V4W/H9A.
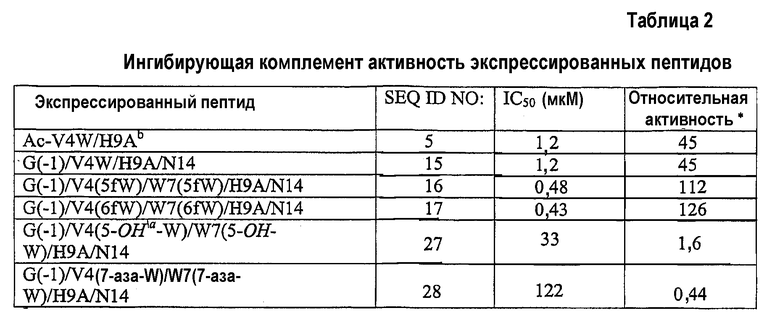
* относительно активности пептида H-I(CWQDWGHHRC)T-NH2 (компстатин, SEQ ID NO:1)
a означает гидроксигруппу
b синтетический пептид.
Не ограничиваясь каким-либо конкретным механизмом, полагают, что добавление атома фтора увеличивает активность пептида благодаря увеличению гидрофобности индольного цикла. Также исследовали включение менее гидрофобных аналогов триптофана, 5-гидрокситриптофана и 7-азатриптофана. В отличие от результатов, полученных с использованием 5-фтор- и 6-фтор-аналогов, аналоги компстатина, содержащие 5-гидрокситриптофан, проявляли в 27,5 раза меньшую активность по сравнению с аналогом Ac-V4W/H9A (SEQ ID NO:5), а пептид, содержащий 7-азатриптофан, не проявлял активности при всех тестированных концентрациях. 7-азатриптофан похож на триптофан по молекулярной структуре, за исключением того, что он имеет атом азота в положении 7 индольного кольца, а не атома углерода. Утрата активности, наблюдаемая при замене 7-азатриптофаном, свидетельствует об относительной важности данного атома углерода.
ПРИМЕР 7
Роль отдельных триптофанов во взаимодействии C3-компстатин
Твердофазный синтез пептидов использовали для создания аналогов компстатина с 5-фтортриптофаном, включенным избирательно в положении 4, положении 7 или в обоих положениях 4 и 7, с аланином в положении 9. Синтез осуществляли, используя Fmoc-5-фтор-dl-триптофан. Указанная реакция давала смесь энантиомеров пептидов, несущих 5-фтор-d-триптофан и 5-фтор-l-триптофан. Синтезировали три разных пептида: два пептида с одной заменой независимо в положении 4 или 7 и один пептид с заменами в обоих положениях 4 и 7. В то время как в случае единственных замен может образовываться смесь аналогов, содержащих 5-фтор-l-триптофан и 5-фтор-d-триптофан, в случае двойной замены возможна смесь комбинаций четырех энантиомеров. Каждую смесь пептидов затем подвергали обращенно-фазовой ВЭЖХ, чтобы разделить энантиомеры пептидов. Идентификацию энантиомеров осуществляли при расщеплении пептидов протеазой V8, и затем анализируя расщепленный продукт, используя MALDI. Протеаза V8 расщепляет с C-концевой стороны остатков Asp только тогда, когда далее следует l-аминокислота. Идентификация продуктов расщепления в масс-спектрах показала, что l-энантиомерный пептид элюируется первым, а затем следует d-форма, при этом не выявлено расщепленных фрагментов.
Все пептиды, содержащие либо 5-фтор-l-триптофан или 5-фтор-d-триптофан, или оба аналога, тестировали в отношении ингибирующей комплемент активности. Синтетический пептид, замещенный 5-фтор-l-триптофаном в обоих положениях, проявлял в 2,5 раза более высокую активность, чем активность Ac-V4W/H9A (SEQ ID NO:5) (таблица 3).
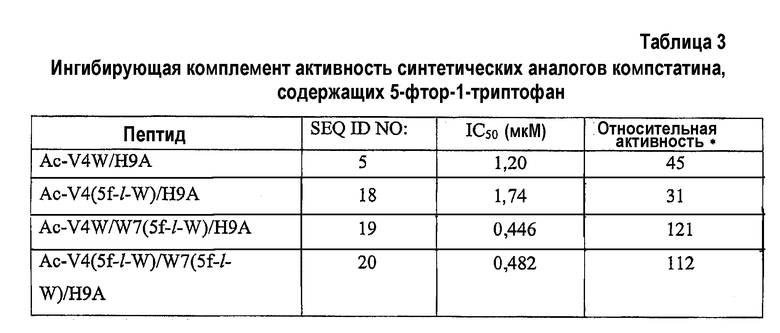
*относительно активности пептида H-I(CWQDWGHHRC)T-NH2 (компстатин, SEQ ID NO:1).
Анализы ингибирования комплемента (фиг.2; таблица 3) показали, что (a) замена 5-фтор-l-триптофаном в положении 4 отдельно делала пептид, по меньшей мере, в 1,5 раза менее активным, чем Ac-V4W/H9A (SEQ ID NO:5). Замена 5-фтор-l-триптофаном в положении 7 отдельно увеличивала активность в 2,7 раза по сравнению с Ac-V4W/H9A. Замена 5-фтор-l-триптофаном одновременно в положениях 4 и 7 также давала 2,5-кратное увеличение активности по сравнению с Ac-V4W/H9A (SEQ ID NO:5). Замена 5-фтор-d-триптофаном в любом положении 4 или 7, или в обоих положениях делала пептид неактивным.
ПРИМЕР 8
Термодинамическая основа опосредованного триптофаном узнавания компстатина компонентом C3
Изотермическую титровальную калориметрию использовали для исследования связывания пептидов с C3 и исследования термодинамической основы их активностей. Калориметрические данные, полученные для взаимодействия всех пептидов с C3, подгоняли к модели одного набора участков со стехиометрией, близкой к 1. Предполагается, что связывание указанных пептидов с C3 происходит в соотношении 1:1. Термодинамические параметры, полученные в результате таких подгонок, показаны в таблице 4. Как видно на основании значений Kd, пептид с одной заменой 5-фтор-l-триптофаном в положении 7 и двойной заменой в положениях 4 и 7 имел более тесное связывание, чем аналоги Ac-V4W/H9A (SEQ ID NO:5) и Ac-V4(5f-l-W)/H9A (SEQ ID NO:18). Полученные данные согласуются с относительными активностями, наблюдаемыми в анализе ингибирования комплемента (таблица 3), свидетельствуя, что существует корреляция между связыванием и активностью.
Все пептиды связывались с C3 с отрицательной энтальпией и положительной энтропией. Такое связывание характерно для взаимодействия компстатина с C3. Среди всех исследованных пептидов замещенный в положении 7 аналог Ac-V4W/W7(5f-l-W)/H9A (SEQ ID NO:19) имел более высокую энтальпию связывания (ΔH = -21,83, ΔΔH = -3,69), чем его эквивалент дикого типа. Замещенный в положении 4 аналог Ac-V4(5f-l-W)/H9A (SEQ ID NO:18) связывался с C3 с энтальпией - 16,69 ккал/моль, на 1,45 ккал/моль меньше, чем наблюдали в случае его эквивалента дикого типа.
Включение 5-фтортриптофана в положение 4 приводит к уменьшению энтальпии на 1,45 ккал/моль по сравнению с энтальпией при наличии триптофана в данном положении (таблица 4). Так как единственным различием между триптофаном и 5-фтортриптофаном является атом фтора у C5 индола, то такое уменьшение энтальпии можно объяснить заменой водорода фтором.
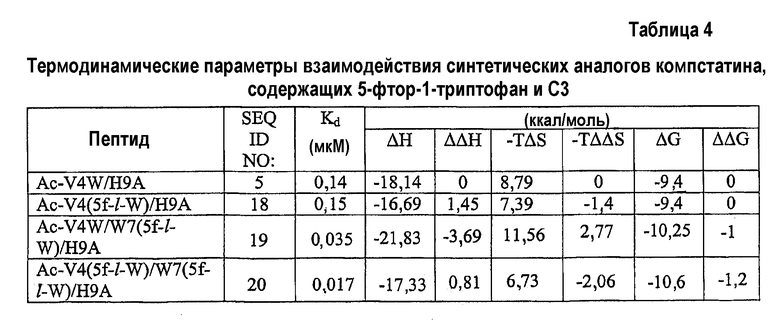
Введение 5-фтортриптофана в положение 7 увеличивало энтальпию на 3,69 ккал/моль по сравнению с диким типом (таблица 4). Не ограничиваясь каким-либо конкретным механизмом, полагают, что триптофан в положении 7 принимает участие в благоприятном с точки зрения энтальпии взаимодействии, таком как образование водородной связи. Замена одного из атомов водорода индола атомом фтора может усиливать водородные связи NH индола вследствие снижения pKa. Альтернативно атом фтора образует водородную связь в результате своей электронодонорной природы, как показано в структуре тетрадека(3-фтортирозил)глутатионтрансферазы.
Другим объяснением наблюдаемого увеличения энтальпии является то, что молекула воды образует мостик при взаимодействии между атомом фтора и акцептором водорода на C3, в таком случае должны быть образованы две водородных связи (что эквивалентно примерно 4 ккал/моль энергии). Подтверждением такой теории является снижение энтропии, наблюдаемой в случае взаимодействия замещенного в положении 7 аналога Ac-V4W/W7(5fW)/H9A (SEQ ID NO:19) по сравнению с аналогом дикого типа (таблица 4), снижение таковой может быть получено при связывании дополнительной молекулы воды в области контакта. Опосредованные водой взаимодействия между атомами фтора и другими акцепторами водородной связи наблюдали в других системах.
Связывание дважды замещенного аналога с C3 давало изменение энтальпии -19,85 ккал/моль, изменение энтропии -9,35 ккал/моль и изменение свободной энергии -10,5 ккал/моль. Полагают, что введение 5-фтортриптофана одновременно в обоих положениях отменяет влияние одиночных замен.
ПРИМЕР 9
Дополнительные аналоги компстатина
Введение аналогов триптофана в положение 4.
В примерах 5 и 6 показано, что замена валина триптофаном в положении 4 компстатина увеличивала его активность в 45 раз. Чтобы дополнительно исследовать природу взаимодействия, опосредованного остатком в положении 4, во время связывания компстатина с C3 триптофан в положении 4 заменяли аналогами и 2-нафтилаланином.
Основанные на ELISA анализы использовали для тестирования активности всех пептидных аналогов, несущих триптофан в положении 4 и аланин в положении 9. В то время как замена 1-метил триптофаном (Ac-V4(1-метил-W)/H9A) (SEQ ID NO:23) и 2-нафтилаланином (Ac-V4(2-Nal)/H9A) (SEQ ID NO:7) увеличивала активность по сравнению с компстатином в 264 и 99 раз, соответственно, замена 5-фтортриптофаном (Ac-V4(5f-l-W)/W7/H9A) (SEQ ID NO:18 и 5-метилтриптофаном (Ac-V4(5-метил-W)/H9A) (SEQ ID NO:22) приводила к более низкой активности; в 31 и 67 раз выше, чем активность, проявляемая пептидом дикого типа (таблица 5). На фиг.3 показаны кривые ингибирования, на которых изображена активность, и в таблице 5 показаны значения IC50, рассчитанные на основании кривых, и относительные активности пептидов по сравнению с активностью исходного компстатина. На фиг.5 показаны константы ингибирования (IC50) против значений log P аналогов триптофана и 2-нафтилаланина.
∗ Относительно активности H-I(CWQDWGHHRC)T-NH2 (компстатин, SEQ ID NO:1).
Связывание пептидов компстатина также исследовали, используя изотермическую титровальную калориметрию. Калориметрические данные, полученные для взаимодействия всех пептидов с C3, подгоняли к модели одного набора участков со стехиометрией, близкой к 1 (фиг.4). Результаты свидетельствуют, что связывание указанных пептидов с C3 происходит в соотношении 1:1. Термодинамические параметры, полученные в результате таких подгонок, показаны в таблице 6. Как видно на основании значений Kd, Ac-V4(1-метил-W)/H9A имел более высокую аффинность связывания (Kd=0,015 мкМ) по сравнению со всеми другими пептидами, имеющими одну замену в положении 4. Графическое изображение таких значений против значений log P аналогов показывает, что существует корреляция между аффинностью связывания и гидрофобной природой аналогов триптофана и 2-нафтилаланина. Что касается корреляции, то аффинность связывания возрастает с увеличением гидрофобности аналога, введенного в положение 4. Указанное наблюдение согласуется с корреляцией, обнаруженной между log P и константами ингибирования.
Все пептиды связывались с C3 с отрицательной энтальпией и положительной энтропией, свидетельствуя, что связывание зависит от энтальпии. Такое связывание характерно для взаимодействия компстатина с C3. Однако связывание указанных пептидов характеризуется меньшим изменением энтальпии, чем в случае дикого типа, и изменение энтропии сдвигалось в благоприятную сторону. На фиг.5B показан график log P против -TΔS, который показывает, что с увеличением гидрофобности аналогов, введенных в положение 4, энтропия становится более благоприятной, таким образом, оказывая положительное влияние на изменение свободной энергии.
Введение аналогов триптофана в положение 7. В примере 7 предполагается, что триптофан в положении 7 обеспечивает водородную связь с остатком на C3. Чтобы исследовать такую возможность дополнительно, триптофан в положении 7 заменяли аналогами триптофана, сходными с заменами в положении 4, чтобы выяснить природу взаимодействия, осуществляемого триптофаном в данном положении. Замена 5-фтортриптофаном (Ac-V4W/W7(5f-l-W)/H9A) (SEQ ID NO:19) давала в 121 раз более активный пептид (фиг.3, таблица 5). Замены триптофана 7 аналогом 5-метил-trp или 1-метил-trp делали компстатин неактивным (данные не показаны). Таким образом, не наблюдали корреляции между активностью и гидрофобностью аналогов триптофана.
Термодинамические свойства разных Trp 7-аналогов исследовали параллельно калориметрией (таблица 6). Так как не выявляли связывания в случае пептидов, содержащих 5-метил-trp или 1-метил-trp в положении 7, то параметров связывания не имеется. Только пептид Ac-V4W/W7(5f-l-W)/H9A (SEQ ID NO:19) связывался с C3. Аффинность связывания составляла 0,035 мкМ, что выше, чем аффинность связывания, наблюдаемая для всех пептидов, имеющих аналог Trp в положении 4, за исключением пептида Ac-V4(1-метил-W)/H9A (SEQ ID NO:23). В отличие от пептидов, имеющих аналог Trp в положении 4, Ac-V4W/W7(5f-l-W)/H9A (SEQ ID NO:19) связывался с C3 с высокоблагоприятным изменением энтальпии связывания (ΔH = -21,83, ΔΔH = -3,69) и неблагоприятным изменением энтропии (-TΔS = 11,56, -TΔΔS = 2,77), что свидетельствует о дополнительных благоприятных нековалентных взаимодействиях полярной природы.
Результаты показывают, что введение 5-фтортриптофана в положение 7 приводит к увеличению активности компстатина, хотя введение аналогов 5-метилтриптофана и 1-метилтриптофана делает компстатин неактивным. Утрата активности компстатина при введении 1-метилтриптофана подтверждает вывод о том, что водородная связь, опосредованная N-H Trp 7, важна для взаимодействия компстатина с C3. Кроме того, полная потеря активности компстатина при введении 5-метилтриптофана свидетельствует, что гидрофобная аминокислота мало допустима в положении 7.
Введение аналогов триптофана в оба положения 4 и 7. Так как замена триптофанов в положении 4 1-метилтриптофаном и в положении 7 5-фтортриптофаном давала аналоги компстатина, которые проявляли резкое увеличение активности, то создавали аналог компстатина, содержащий замены в положениях 4 и 7. Полученный, в результате, пептид (Ac-V4(1-метил-W)/W7(5f-l-W)/H9A) (SEQ ID NO:24) давал кривую ингибирования, сходную с кривой при одиночной замене 1-метилтриптофаном (Ac-V4(1-метил-W)/H9A) (SEQ ID NO:23) (фиг.3, таблица 5). Аффинность связывания (Kd=0,017), наблюдаемая для данного пептида в калориметре, также сходна с аффинностью связывания Ac-V4(1-метил-W)/H9A (SEQ ID NO:23). Указанные наблюдения свидетельствуют, что 5-фтортриптофан не оказывает влияния в положении 7 в присутствии 1-метилтриптофана в положении 4 в указанных экспериментальных условиях.
Введение другого аналога триптофана в положение 4. Чтобы исследовать природу взаимодействия, опосредованного остатком в положении 4, во время связывания компстатина с C3 триптофан в положении 4 заменяли аналогом триптофана 1-формилтриптофаном.
На фиг.6 показано сравнение ингибирования комплемента в процентах в зависимости от концентрации пептида для Ac-V4(1-метил-W)/H9A (SEQ ID NO:23) (кружки) и Ac-V4(1-формил-W)/H9A (SEQ ID NO:25). Как можно видеть, аналог 1-формил-W был по существу идентичен аналогу 1-метил-W по активности ингибирования комплемента.
ПРИМЕР 10
Пегилирование аналога компстатина
Пролонгированное время полужизни компстатина имеет преимущества для применения при хроническом лечении. Удлинение времени полужизни тестированных терапевтических пептидов достигали в нескольких случаях посредством пегилирования (см. Veronese et al., 2001), так как ПЭГ обладает способностью замедлять элиминирование биологических молекул из кровообращения посредством различных механизмов, включая снижение почечного клиренса, протеолиза и иммуногенности. Пегилирование заключается в ковалентном связывании полимеров ПЭГ с макромолекулами, предпочтительно с первичным амином лизина.
В настоящем примере описано получение пегилированного аналога компстатина, Ac-V4(1-метил-W)/H9A-K-ПЭГ 5000 (SEQ ID NO:36) и оценка соединения в отношении его способности ингибировать активацию комплемента.
Fmoc-NH-NHS-5000-ПЭГ приобретали из Nektar transforming therapeutics, 490 discovery Dr, Huntsville, AL 35806.
Соединение Ac-V4(1-метил-W)/H9A-K-ПЭГ 5000 (SEQ ID NO:36) синтезировали химически, используя твердофазную Fmoc-химию пептидов согласно модифицированному стандартному протоколу. Коротко, ПЭГ растворяли в 3 мл дихлорметана, вручную добавляли 2 М DIEA и перемешивали ПЭГ в течение 5 минут.
Затем ПЭГ переносили в сосуд и оставляли связываться в течение ночи. Затем снимали защиту ПЭГ 20% пиперидином в течение 20 мин.
Затем синтез продолжался согласно стандартному протоколу с включением лизина на C-конце молекулы в целях связывания ПЭГ с его боковой цепью.
Конечное отщепление пептидов осуществляли реагентом D (ТФУ:H2O:TIS:фенол, 87,5:5:2,5:5) (4 мл) при 25°C в течение 90 мин, получая требуемый продукт. Затем пептид очищали на колонке C18 для обращенно-фазовой ВЭЖХ, подвергали лиофилизации и характеризовали с помощью MALDI-TOF.
Пегилированный аналог компстатина тестировали в отношении ингибирующей комплемент активности, используя анализ in vitro, описанный в примере 4. Как показано на фиг.7, пегилированный аналог активен в ингибировании активации комплемента, однако требовалось в семь раз больше конъюгата, чтобы достичь такой же степени ингибирования, как в случае непегилированного аналога, Ac-V4(1-метил-W)/H9A (SEQ ID NO:23).
ПРИМЕР 11
Конъюгат аналога компстатина со связывающим альбумин белком
Dennis et al.(2002) идентифицировали серию пептидов, имеющих коровую последовательность DICLPRWGCLW (SEQ ID NO:37), которые специфично связывают сывороточный альбумин среди множества видов с высокой аффинностью. Такие пептиды связываются с альбумином в стехиометрическом соотношении 1:1 в сайте, отличном от известных сайтов связывания малых молекул. Пептид SA21 (AcRLIEDICLPRWGCLWEDDNHE; SEQ ID NO:38) имеет необычно длительное время полужизни, составляющее 2,3 часа, при инъекции в виде внутривенного болюса кроликам. Как указано в подробном описании, родственная последовательность, слитая с Fab D3H44 против тканевого фактора, делала возможным связывание Fab с альбумином с аффинностью, сходной с аффинностью SA21, сохраняя при этом способность Fab связывать тканевой фактор 1 (Nguyen et al. 2006). Указанное взаимодействие с альбумином приводило к пониженному клиренсу in vivo в 25 и 58 раз у мышей и кроликов, соответственно, по сравнению с Fab D3H44 дикого типа. Время полужизни удлинялось в 37 раз до 32,4 часа у кроликов и в 26 раз до 10,4 часа у мышей, достигая 25-43% времени полужизни альбумина у указанных животных. Такое время полужизни превышало время полужизни Fab 2 и сравним со временем полужизни, наблюдаемым в случае пегилированных молекул Fab, иммуноадгезинов и слияний с альбумином.
В настоящем примере описан синтез аналога компстатина, слитого со связывающим альбумин пептидом, и его активность в анализах ингибирования комплемента in vitro.
Соединение 4(1MeW)-ABP синтезировали химически, используя твердофазную Fmoc-химию пептидов согласно стандартным протоколам. N- и C-концы пептида защищали ацетильной и амидной группами. Пептид дополнительно очищали на колонке C18 для обращенно-фазовой ВЭЖХ, подвергали лиофилизации и характеризовали, используя MALDI-масс-спектрометрию.
Для циклизации пептид-смолу (загрузка 0,10 ммоль/г на основании аминокислотного анализа) подвергали набуханию в дихлорметане (ДХМ) (2 мл) в течение 5 мин, фильтровали и обрабатывали смесью 94:1:5 ДХМ/ТФУ/TIS (5 мл) при 25°C 3 раза × 2 мин, чтобы избирательно удалить защиту S-Mmt-защитными группами, удаляя растворитель под давлением N2. Полученные промежуточные продукты бис(тиол), бис(Acm)-пептид-смола промывали CH2Cl2, ДМФА и NMP (каждым 5 раз × 2 мин, 2 мл), далее подвергали набуханию в NMP (2 мл) в течение 5 мин и затем обрабатывали Et3N (2 эквивалента) в NMP при 25°C в течение 4 часов. Затем связанные пептид-смолу промывали ДМФА и CH2Cl2 ( каждым 5 раз × 2 мин, 2 мл). После образования связанной со смолой первой петли пептид-смолу снова промывали ДМФА (5 раз × 2 мин, 2 мл) и подвергали набуханию в ДМФА (2 мл) в течение 5 мин, фильтровали и обрабатывали Tl(tfa)3 (1,5 эквивалента) в смеси ДМФА-анизол (4 мл), чтобы циклизовать вторые дисульфидные петли. После осторожного встряхивания при 25°C в течение 4 часов таллиевые реагенты удаляли, используя ДМФА (8 раз × 2 мин, 2 мл) и пептид-смолу дополнительно промывали CH2Cl2 (5 раз × 2 мин, 2 мл). Конечное отщепление бициклического пептида осуществляли реагентом D (ТФУ:H2O:TIS:фенол, 87,5:5:2,5:5) (4 мл) при 25°C в течение 90 мин, получая требуемый продукт.
Полученный в результате конъюгированный пептид (SEQ ID NO:39) показан ниже.

Альбуминсвязывающий пептид-компстатин тестировали в отношении активности ингибирования комплемента, используя анализ in vitro, описанный в примере 4. Как показано на фиг.8, конъюгат был активным в ингибировании активации комплемента, однако требовалось в семь раз больше конъюгата, чтобы достичь такой же степени ингибирования, как в случае неконъюгированного аналога, Ac-V4(1-метил-W)/H9 A (SEQ ID NO:23).
CСЫЛКИ
Babitzke P, and Yanofsky C. (1995) Structural features of L-tryptophan required for activation of TRAP, the trp RNA-binding attenuation protein of Bacillus subtilis. J. Biol. Chem. 270:12452-6.
Beeley N. (1994) Peptidomimetics and small-molecule drug design: towards improved bioavailability and in vivo stability. Trends Biotechnol. 12:213-6.
Beene D.L., Brandt G.S., Zhong W, Zacharias N.M., Lester H.A., and Dougherty D.A. (2002) Cation-pi interactions in ligand recognition by serotonergic (5-HT3A) and nicotinic acetylcholine receptors: the anomalous binding properties of nicotine. Biochemistry. 41:10262-9.
Dennis M.S., Zhang M, Meng Y.G., Kadkhodayan M, Kirchhofer D, Combs D, Damico L.A. (2002) Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J Biol Chem. 277:35035-43.
Fiane A.E., Mollnes Т.Е., Videm V, Hovig T, Hogasen K, Mellbye O.J., Spruce L, Moore W.T., Sahu A, and Lambris J.D. (1999a) Prolongation of ex vivo-perfused pig xenograft survival by the complement inhibitor Compstatin. Transplant. Proc. 31:934-5.
Fiane A.E., Mollnes Т.Е., Videm V, Hovig T, Hogasen K, Mellbye O.J., Spruce L, Moore W.T., Sahu A, and Lambris J.D. (1999b) Compstatin, a peptide inhibitor of C3, prolongs survival of ex vivo perfused pig xenografts. Xenotransplantation. 6:52-65.
Fiane A.E., Videm V, Lambris J.D., Geiran O.R., Svennevig J.L., and Mollnes Т.Е. (2000) Modulation of fluid-phase complement activation inhibits hyperacute rejection in a porcine-to-human xenograft model. Transplant, Proc. 32:899-900.
Furlong S.T., Dutta A.S., Coath M.M., Gormley J.J., Hubbs S.J., Lloyd D, Mauger R.C., Strimpler A.M., Sylvester M.A., Scott C.W., and Edwards P.D. (2000) C3 activation is inhibited by analogs of compstatin but not by serine protease inhibitors or peptidyl alpha-ketoheterocycles. Immunopharmacology. 48:199-212.
Hruby V.J. (1993) Conformational and topographical considerations in the design of biologically active peptides. Biopolyniers. 33:1073-82.
Kalli K.R., Hsu P, Fearon and D.T. (1994) Therapeutic uses of recombinant complement protein inhibitors. Springer Semin, Immunopathol. 15:417-31.
Katragadda M, Morikis D, and Lambris J.D. (2004) Thermodynamic studies on the interaction of the third complement component and its inhibitor, compstatin. J. Biol. Chem, 279:54987-95.
Klepeis J.L., Floudas C.A., Morikis D, Tsokos C.G., Argyropoulos E, Spruce L, and Lambris J.D. (2003) Integrated computational and experimental approach for lead optimization and design of compstatin variants with improved activity. J. Am. Chem, Soc. 125:8422-3.
Kozlowski A, Charles S.A., and Harris J.M. (2001) Development of pegylated interferons for the treatment of chronic hepatitis C. BioDrugs. 15:419-29.
Mallik B, Katragadda M, Spruce L.A., Carafides C, Tsokos C.G., Morikis D, and Lambris J.D. (2005) Design and NMR Characterization of Active Analogs of Compstatin Containing Non-Natural Ammo Acids. J. Med. Chem. 48:274-286.
Morikis D, Assa-Munt N, Sahu A, and Lambris J.D. (1998) Solution structure of Compstatin, a potent complement inhibitor. Protein Sci. 7:619-27.
Nguyen A, Reyes A.E. 2nd, Zhang M, McDonald P, Wong W.L., Damico L.A., Dennis M.S. (2006). The pharmacokinetics of an albumin-binding Fab (AB.Fab) can be modulated as a function of affinity for albumin. Protein Eng Des Sel. 19:291-7.
Nilsson B, Larsson R, Hong J, Elgue G, Ekdahl K.N., Sahu A, and Lambris J.D. (1998) Compstatin inhibits complement and cellular activation in whole blood in two models of extracorporeal circulation. Blood. 92:1661-7.
Sahu A, Kay B.K., and Lambris J.D. (1996) Inhibition of human complement by a С3-binding peptide isolated from a phage-displayed random peptide library. J. Immunol. 157:884-91.
Soulika A.M., Khan M.M., Hattori T, Bowen F.W., Richardson B.A., Hack C.E., Sahu A, Edmunds L.H. Jr, and Lambris J.D. (2000) Inhibition of heparin/protamine complex-induced complement activation by Compstatin in baboons. Clin. Immunol. 96:212-21.
Spruce L., E. Argyropoulos, D. Mastellos, G. Sfyroera, and J.D. Lambris (2002) Chemical synthesis of small complement proteins and protein modules. International Immunopharmacology. 2: 1320-1321.
Veronese F.M. (2001) Peptide and protein PEGylation: a review of problems and solutions. Biomaterials 22:405-417.
Wang Y, Hu Q, Madri J.A., Rollins S.A., Chodera A, and Matis L.A. (1996) Amelioration of lupus-like autoimmune disease in NZB/WP1 mice after treatment with a blocking monoclonal antibody specific for complement component C5, Proc. Natl. Acad. Sci. USA. 93:8563-8.
Zhao B, Helms L.R., DesJarlais .L., Abdel-Meguid S.S., and Wetzel R. (1995) A paradigm for drug discovery using a conformation from the crystal structure of a presentation scaffold, Nat. Struct.Biol. 2:1131-7.
The present invention is not limited to the embodiments described and exemplified above, but is capable of variation and modification within the scope of the appended claims.
Настоящее изобретение не ограничено вариантами, описанными и приведенными в примерах выше, и возможно изменение и модификация в объеме прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭФФЕКТИВНЫЕ АНАЛОГИ КОМПСТАТИНА | 2012 |
|
RU2656102C2 |
| КЛЕТОЧНО-РЕАКТИВНЫЕ АНАЛОГИ КОМПСТАТИНА, АНАЛОГИ КОМПСТАТИНА ДЛИТЕЛЬНОГО ДЕЙСТВИЯ ИЛИ АНАЛОГИ КОМПСТАТИНА НАЦЕЛЕННОГО ДЕЙСТВИЯ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2653439C9 |
| ПРОЛОНГИРОВАННАЯ ДОСТАВКА АНАЛОГОВ КОМПСТАТИНА ИЗ ГЕЛЕЙ | 2008 |
|
RU2505311C2 |
| СХЕМЫ ДОЗИРОВАНИЯ И СООТВЕТСТВУЮЩИЕ КОМПОЗИЦИИ И СПОСОБЫ | 2018 |
|
RU2779495C2 |
| КЛЕТОЧНО-РЕАКТИВНЫЕ АНАЛОГИ КОМПСТАТИНА, АНАЛОГИ КОМПСТАТИНА ДЛИТЕЛЬНОГО ДЕЙСТВИЯ ИЛИ АНАЛОГИ КОМПСТАТИНА НАЦЕЛЕННОГО ДЕЙСТВИЯ И СВЯЗАННЫЕ С НИМИ КОМПОЗИЦИИ И СПОСОБЫ | 2013 |
|
RU2705215C2 |
| СХЕМЫ ВВЕДЕНИЯ | 2016 |
|
RU2770099C2 |
| МОДУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА | 2015 |
|
RU2670988C2 |
| ПЕПТИДЫ, ИЛИ ИХ ФАРМАЦЕВТИЧЕСКИ ПРИЕМЛЕМЫЕ КИСЛОТЫ, ИЛИ СОЛИ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ОБЛАДАЮЩАЯ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТЬЮ ПО ОТНОШЕНИЮ К БОМБЕЗИНУ, СПОСОБ ЛЕЧЕНИЯ РАКА У МЛЕКОПИТАЮЩИХ | 1994 |
|
RU2114118C1 |
| МОДУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА | 2015 |
|
RU2778514C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА D И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799044C2 |

















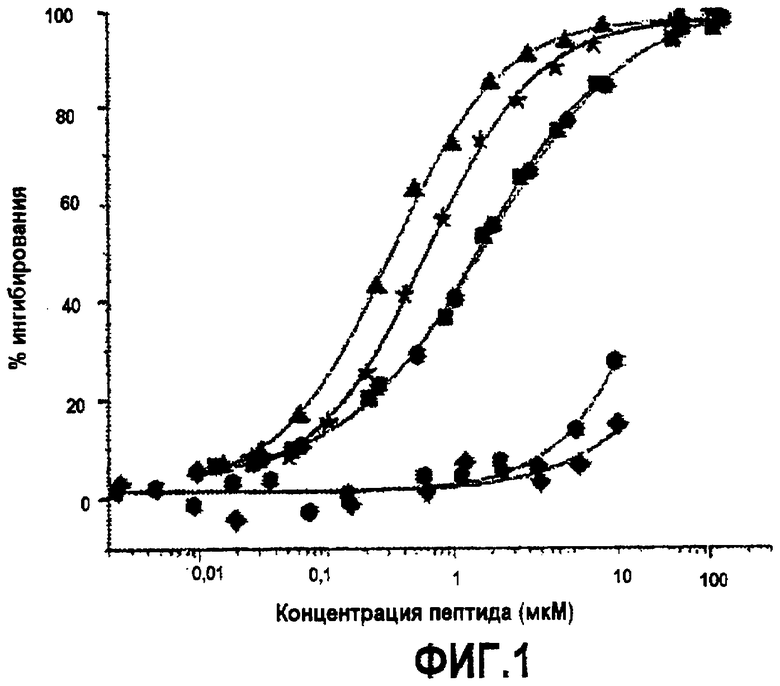
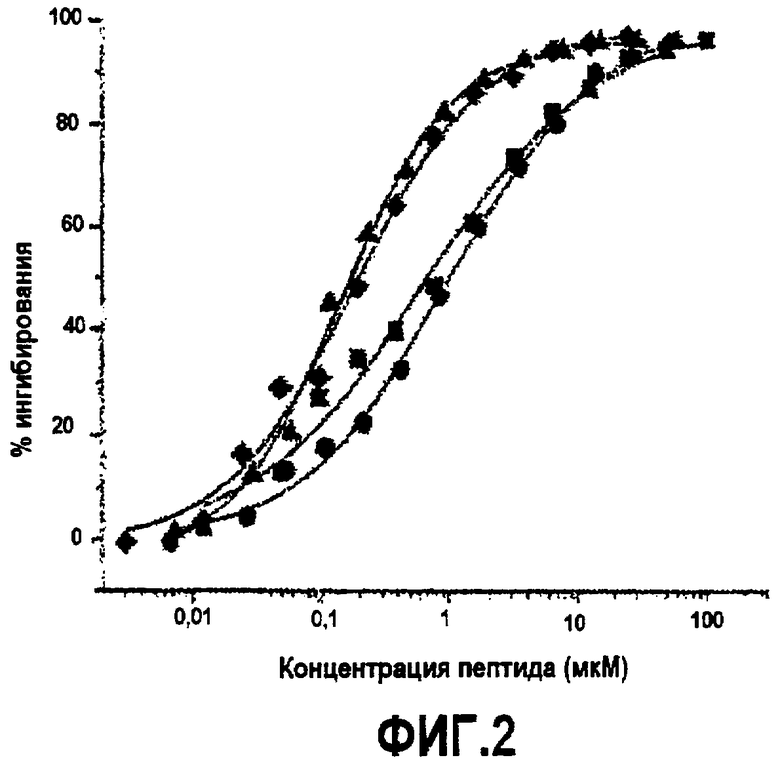
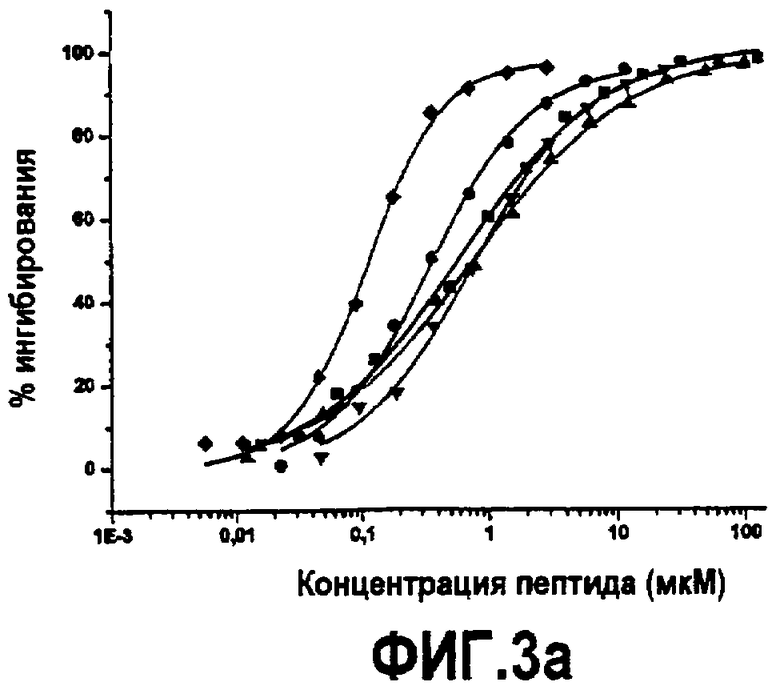
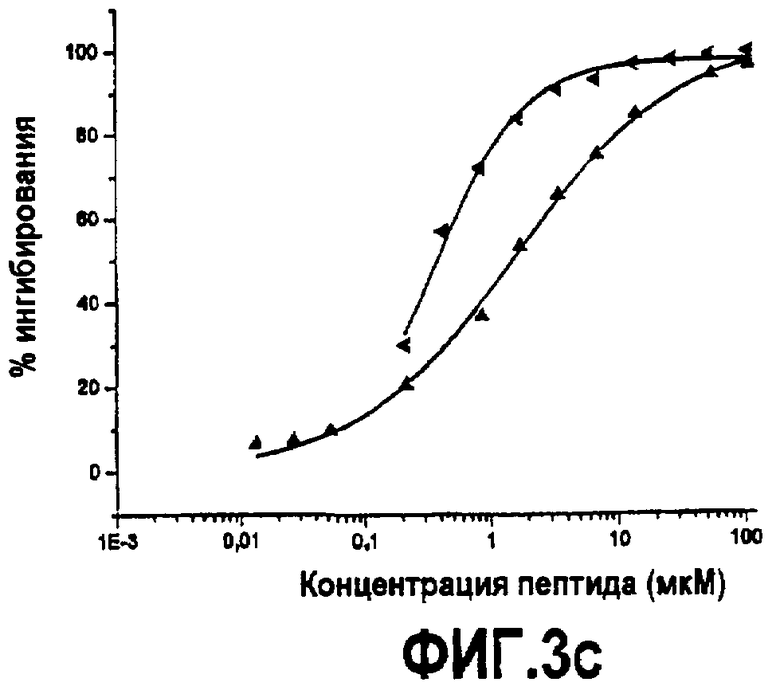
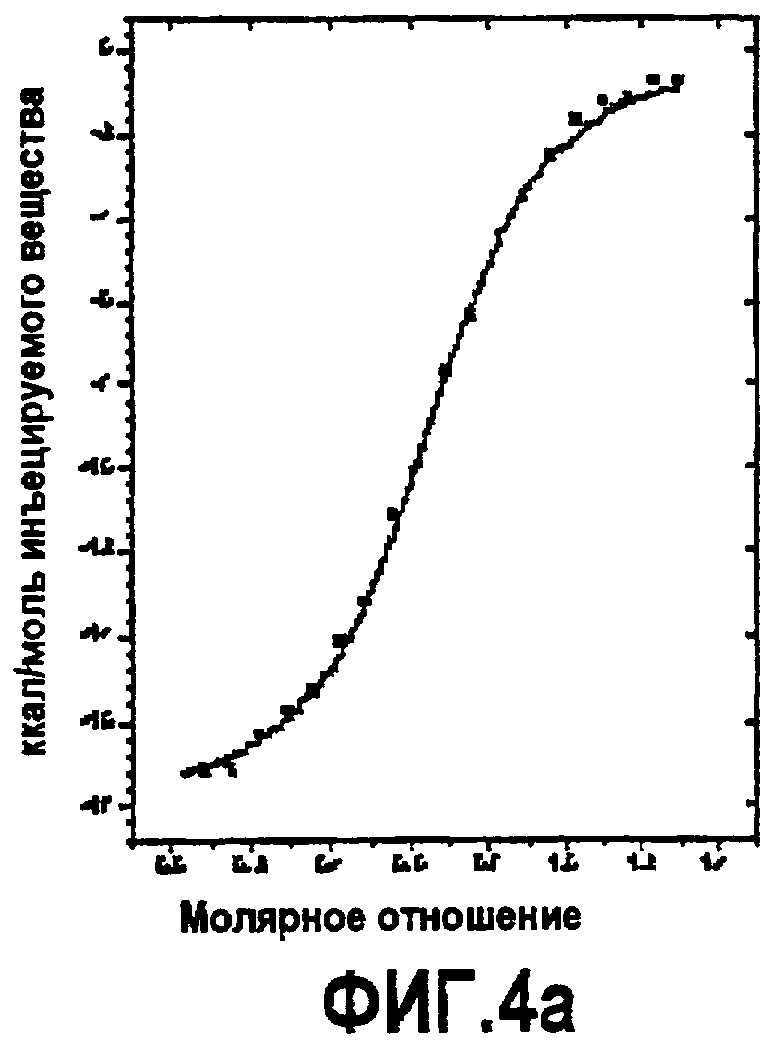
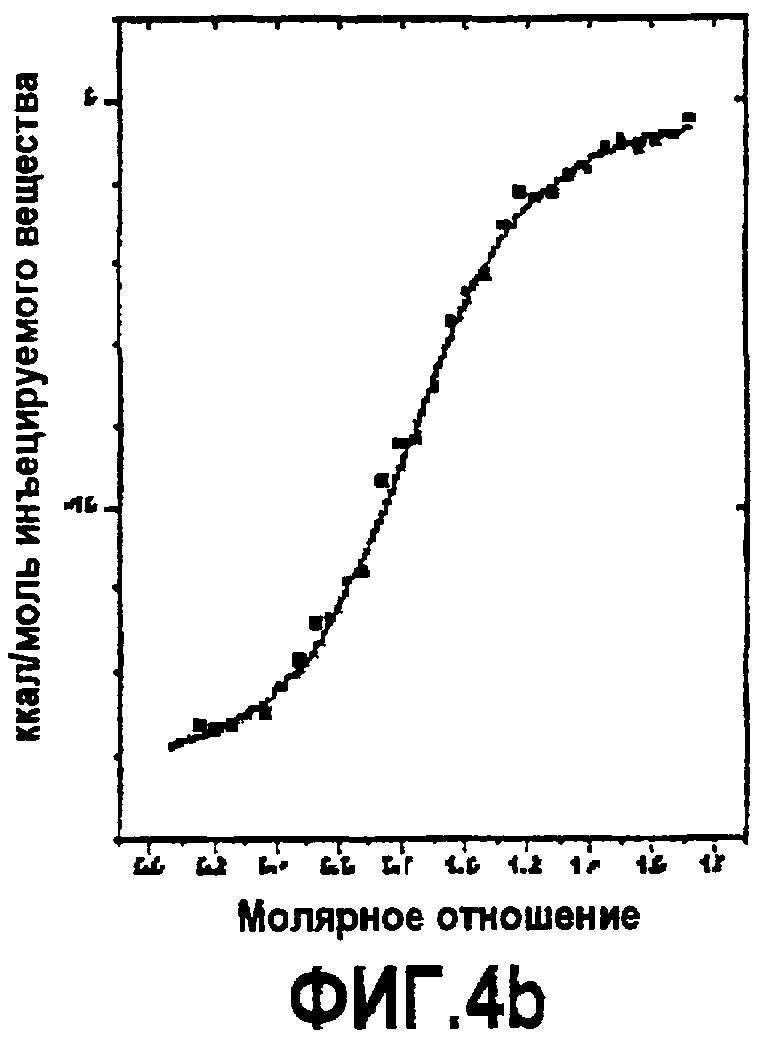
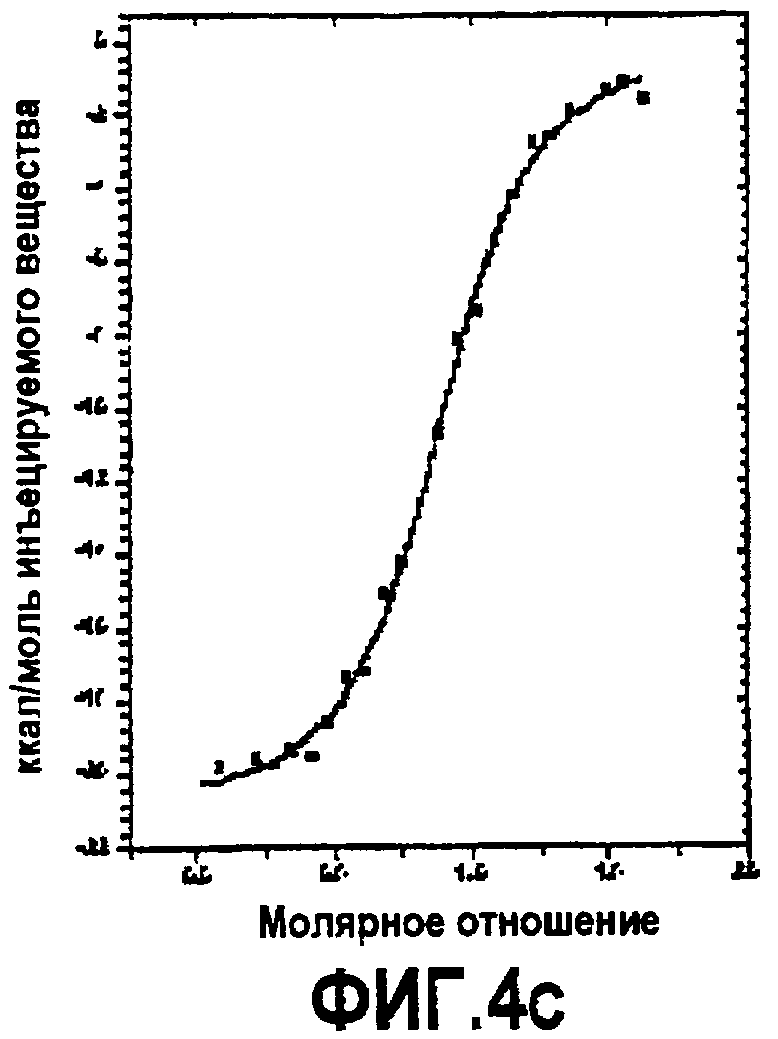
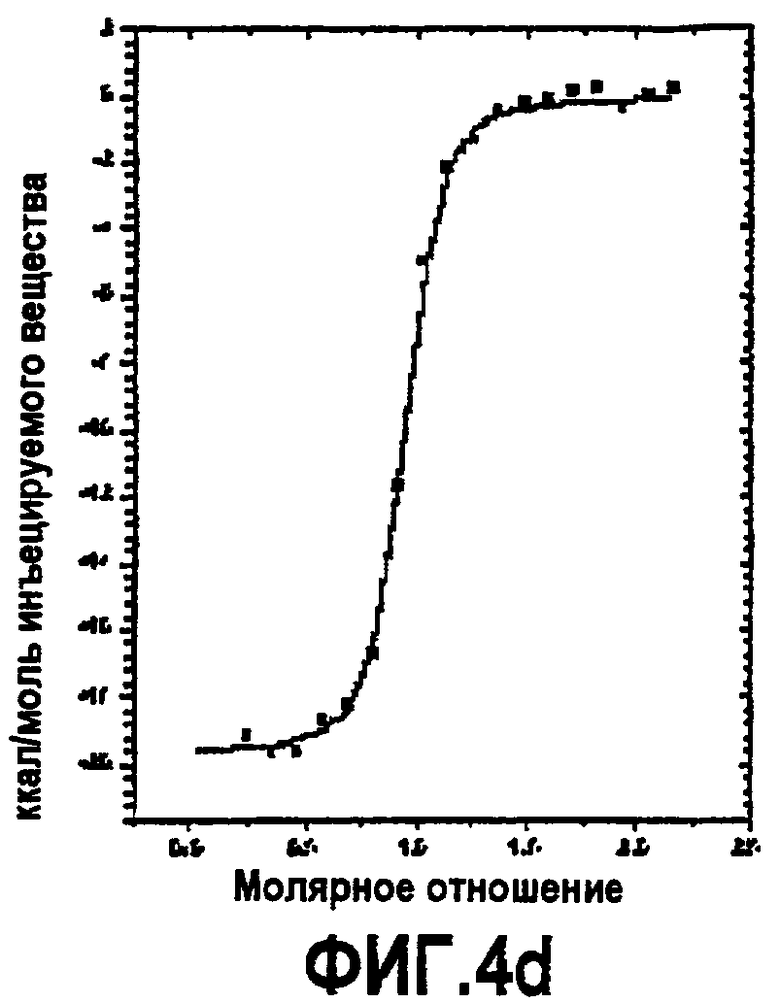
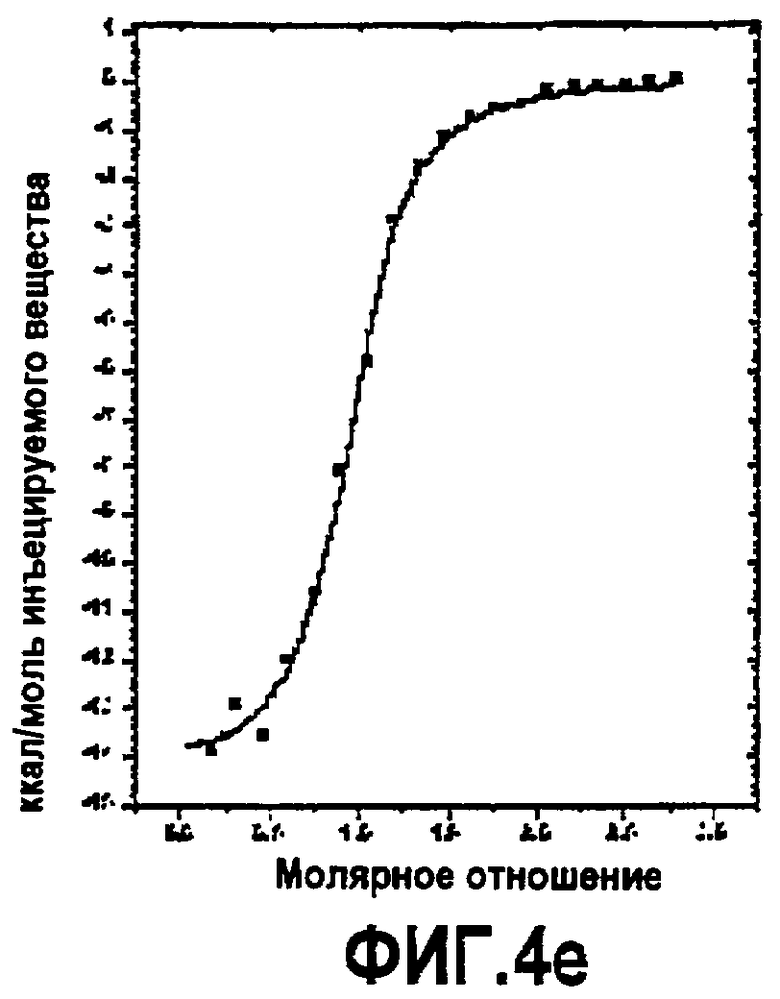
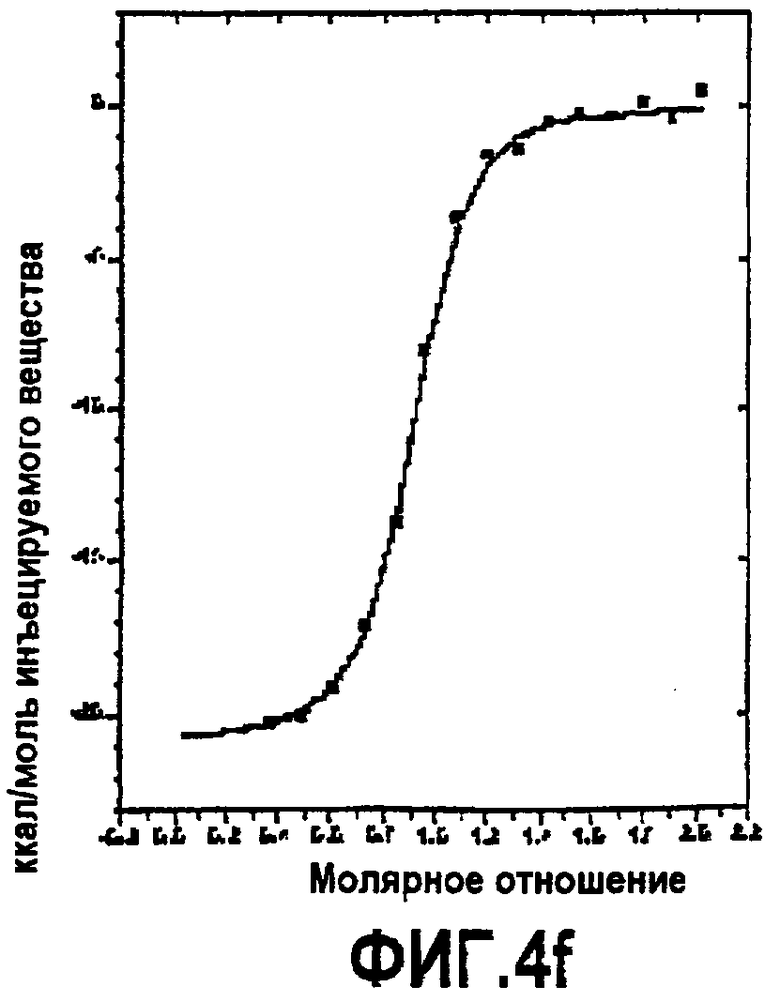
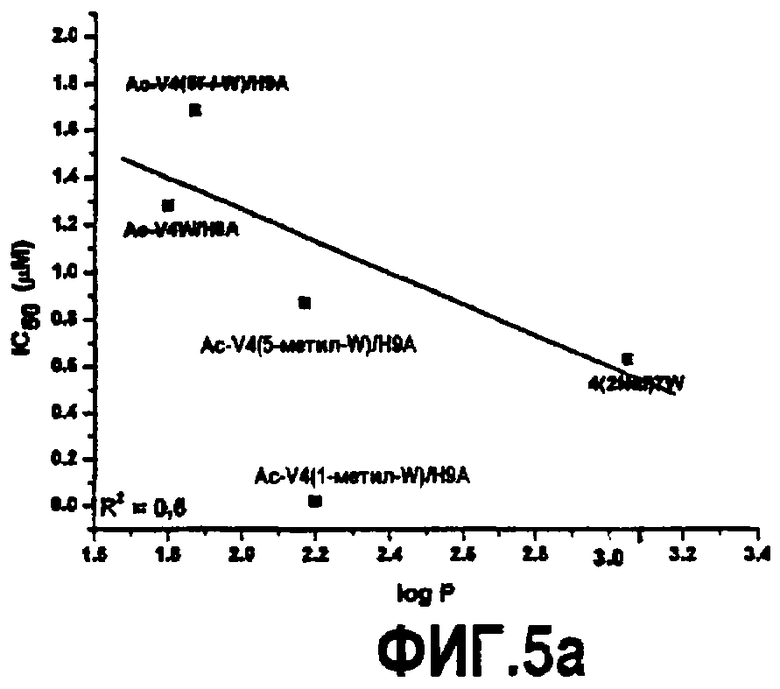
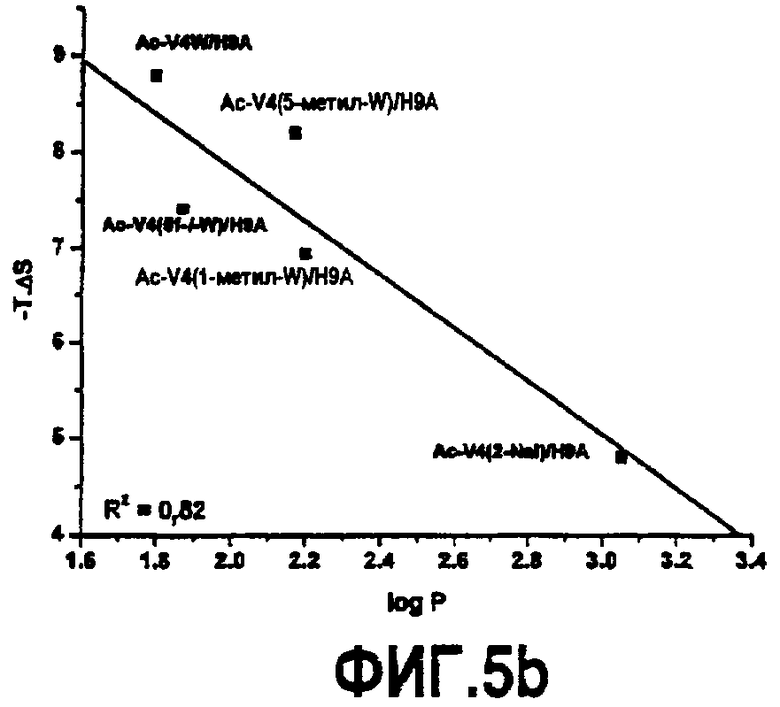
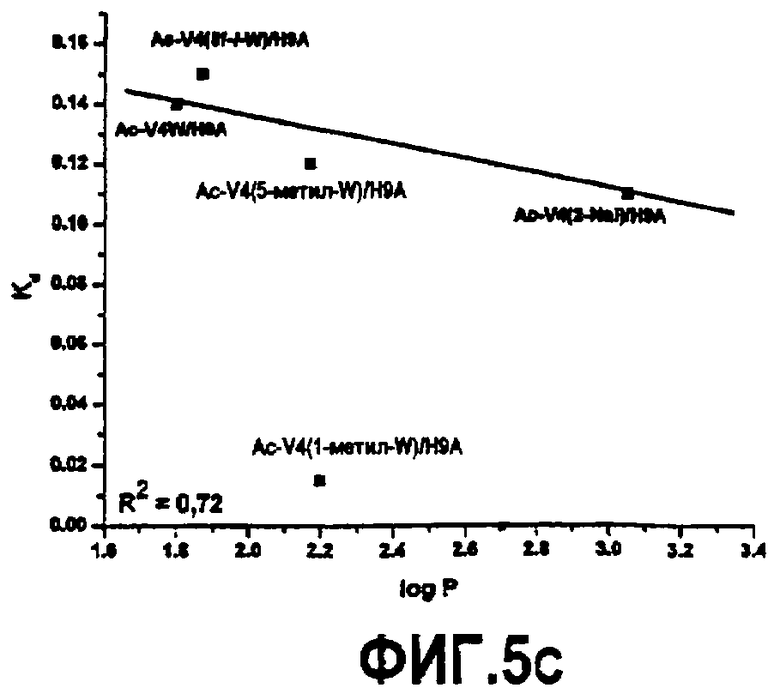
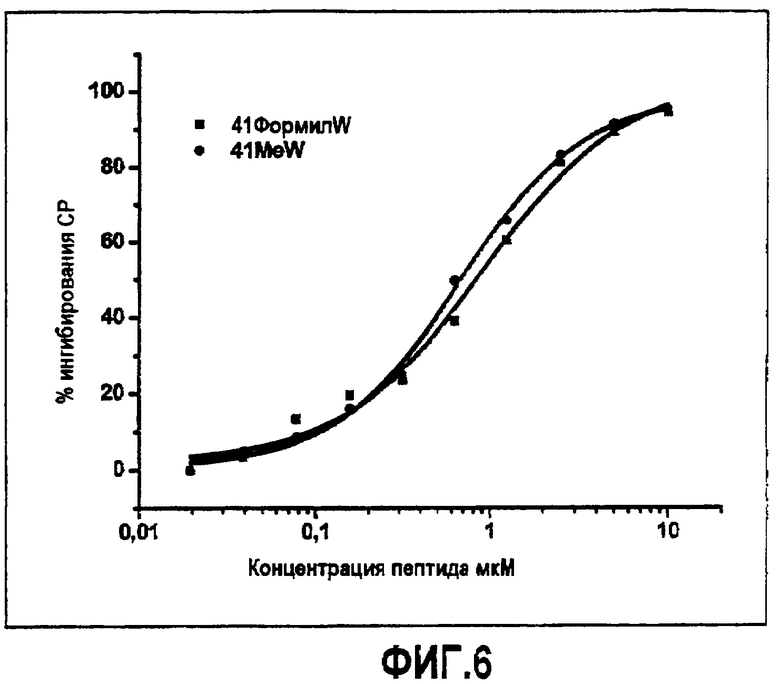
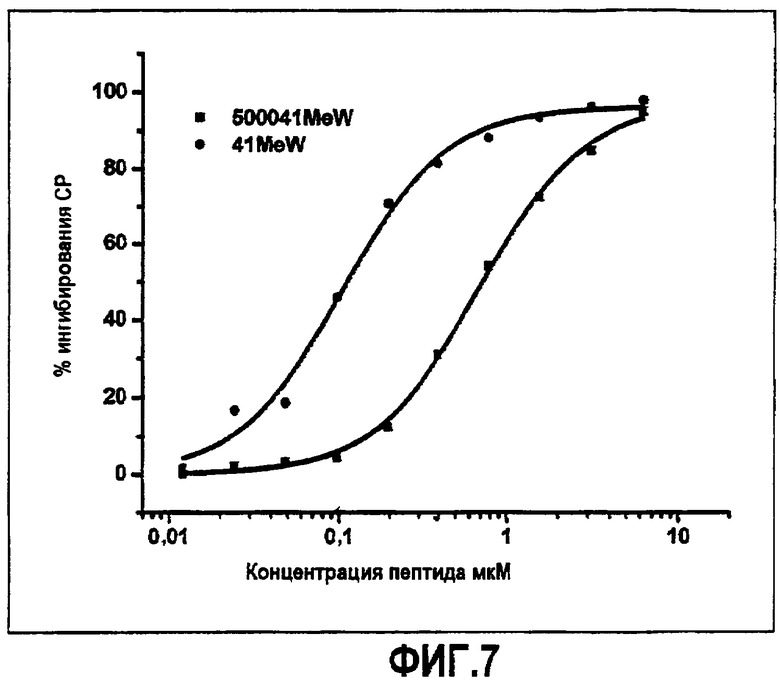
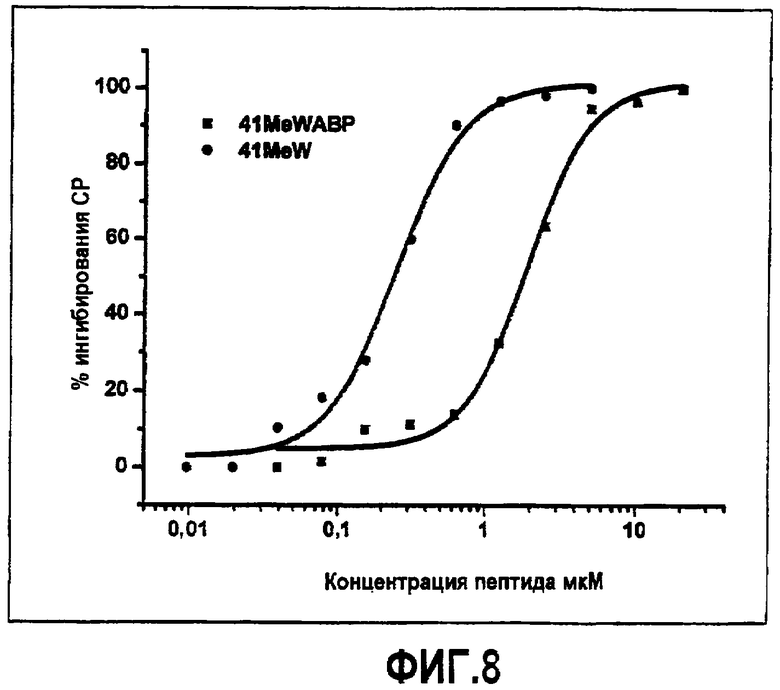
Изобретение относится к соединениям, содержащим пептиды и пептидомиметики, способные связывать белок С3 и ингибировать активацию комплемента. Указанные соединения проявляют значительно повышенную активность в ингибировании активации комплемента по сравнению с имеющимися в настоящее время соединениями. 15 з.п. ф-лы, 8 ил., 6 табл., 11 пр.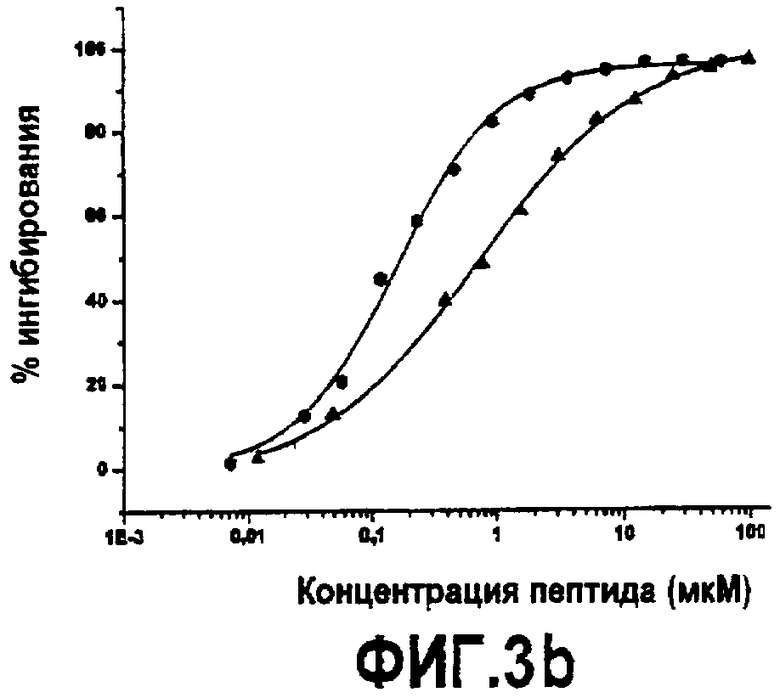
1. Соединение, которое ингибирует активацию комплемента, которое представляет собой пептид следующей последовательности:
Xaa1-Cys-Val-Xaa2-Gln-Asp-Xaa3-Gly-Xaa4-His-Arg-Cys-Xaa5 (SEQ ID NO:26);
где:
Xaa1 представляет собой Ilе, Val, Leu, Ac-Ile, Ac-Val, Ac-Leu или дипептид Gly-Ile;
Xaa2 представляет собой Trp или аналог Trp, выбранный из 5-фтор-l-триптофана, 6-фтор-l-триптофана или аналога с низшим алкокси, низшим алканоильным или низшим алкильным заместителем в положении 1 или 5 триптофана, при условии, что если Хаа3 представляет собой Trp, то Xaa2 представляет собой аналог Trp;
Хаа3 представляет собой Trp или аналог Trp, который является галогенированным триптофаном;
Хаа4 представляет собой His, Ala, Phe или Trp;
Хаа5 представляет собой L-Thr, D-Thr, Ile, Val, Gly, дипептид Thr-Asn или Thr-Ala, или трипептид Thr-Ala-Asn, где -OH на карбоксильном конце любой аминокислоты L-Thr, D-Thr, Ile, Val, Gly или Asn необязательно заменен -NH2; и
два остатка Cys связаны дисульфидной связью;
где соединение, необязательно, может быть пегилировано или может быть слито с альбумин-связывающим белком, включающим SEQ ID NO:37.
2. Соединение по п.1, в котором аналог Trp в Хаа2 представляет собой 5-фтор-l-триптофан или 6-фтор-l-триптофан.
3. Соединение по п.1, в котором аналог Trp в Хаа2 имеет низший алкокси- или низший алкильный заместитель в положении 1 или 5 триптофана.
4. Соединение по п.3, в котором аналог Trp в Хаа2 представляет собой 5-метокситриптофан или 5-метилтриптофан или 1-метилтриптофан.
5. Соединение по п.1, в котором аналог Trp в Хаа2 представляет собой 1-формилтриптофан.
6. Соединение по п.4, в котором аналог Trp в Хаа2 представляет собой 1-метилтриптофан.
7. Соединение по п.1, в котором галогенированный триптофан Хаа3 представляет собой 5-фтор-l-триптофан или 6-фтор-l-триптофан.
8. Соединение по п.1, в котором Хаа4 означает Ala.
9. Соединение по п.1, в котором Хаа2 имеет низший алканоильный или низший алкильный заместитель в положении 1 триптофана, Хаа3 необязательно представляет собой галогенированный триптофан, и Хаа4 представляет собой Ala.
10. Соединение по п.9, в котором Хаа2 означает 1-метилтриптофан или 1-формилтриптофан, и Хаа3 необязательно представляет собой 5-фтор-l-триптофан.
11. Соединение по п.1, имеющее любую из последовательностей SEQ ID NO:16-25.
12. Соединение по п.1, которое представляет собой пептид, полученный экспрессией полинуклеотида, кодирующего этот пептид.
13. Соединение по п.1, которое получено, по меньшей мере частично, пептидным синтезом.
14. Соединение по п.11, имеющее любую из последовательностей SEQ ID NO:23, 24 или 25.
15. Соединение по п.1, имеющее последовательность SEQ ID NO:36.
16. Соединение по п.1, имеющее последовательность SEQ ID NO:39.
| WO 2004026328 A1, 01.04.2004 | |||
| ПРОТЕИН, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ К ОБРАЗОВАНИЮ С3-КОНВЕРТАЗЫ, ДНК, ВЕКТОР, КОНЪЮГАТ И ИХ ИСПОЛЬЗОВАНИЕ | 1997 |
|
RU2238322C2 |
| KATRAGADDA MADAN ET AL: "Hydrophobic effect and hydrogen bonds account for the improved activity of a complement inhibitor, compstatin." JOURNAL OF MEDICINAL CHEMISTRY, 2006, vol.49, no.15, pages 4616-4622 | |||
| KATRAGADDA ET AL: "Structure-activity-based design of potent compstatin analogs" | |||

