Область техники, к которой относится изобретение
Настоящее изобретение относится к рекомбинантному связывающему белку, содержащему, по меньшей мере, одно производное домена 3 (SH3) Src-гомологии киназы FYN, где, по меньшей мере, одна аминокислота, находящаяся в Src-петле или расположенная на расстоянии до двух аминокислот рядом с Src-петлей, и/или, по меньшей мере, одна аминокислота, находящаяся в RT-петле или расположенная на расстоянии до двух аминокислот рядом с RT-петлей, заменена, делегирована или добавлена. Кроме того, данное изобретение относится к слитым белкам, содержащим связывающий белок согласно данному изобретению, соединенный с фармацевтически и/или диагностически активным компонентом. Кроме того, данное изобретение относится к нуклеотидам, кодирующим эти связывающие и/или слитые белки, а также соответствующим векторам или клеткам-хозяевам. Последнее, но не менее важное, это то, что данное изобретение относится к применению связывающих и/или слитых белков настоящего изобретения для получения лекарственного средства или диагностического средства, а также к фармацевтическим и/или диагностическим композициям, содержащим указанные связывающие и/или слитые белки.
Уровень техники
Специфические и высокоаффинные связывающие агенты являются незаменимыми инструментами в биологических и медицинских исследованиях, а также используются в медицинской диагностике, профилактике и лечении. В настоящее время моноклональные антитела являются доминирующим классом связывающих молекул, которые могут быть легко выделены с высокой аффинностью и специфичностью практически в отношении любой мишени. Однако иммуноглобулины имеют ограничения, обусловленные, главным образом, их общими биофизическими свойствами и их довольно сложной молекулярной структурой. Поэтому уже в 1990-х годах несколько исследовательских групп проводили исследование малых глобулярных белков в качестве замены антител. В основе таких соображений лежит идея переноса универсального сайта связывания со структуры антитела на альтернативные белковые несущие конструкции, так называемые каркасы. До сих пор описано более 40 каркасов, среди них два SH3-домена, SH3-домены киназ Abl и Src (см. Binz et al., Nature Biotechnology, Vol.23, No.10, 1257-1268, 2005).
SH3-домены обнаружены во многих различных белках, участвующих в трансдукции внутриклеточных сигналов и организации цитоскелета (Cohen et al., "Modular binding domains in signal transduction proteins." Cell 80(2): 237-48, 1995). Несмотря на вариабельность в их первичных структурах эти SH3-домены имеют очень сходную общую структуру и способ связывания с белками, обладая минимальной консенсусной последовательностью РххР, представляющей собой критическую детерминанту природного SH3-связывания. Важной функцией ЗН3-доменов является их участие в высокоселективных белок-белковых взаимодействиях. Erpel et al. ("Mutational analysis of the Src SH3 domain: the same residues of the ligand binding surface are important for intra - and intermolecular interactions." Embo J. 14(5): 963-75, 1995) исследовали влияние мутаций в RT- и n-Src-петлях SH3-доменов Src и показали, что мутации в обеих петлях, которые находятся рядом с гидрофобной поверхностью, могут оказывать влияние на способность этого домена участвовать в меж- и внутримолекулярных связях.
Hiipakka et al. ("SH3 domains with high affinity and engineered ligand specificities targeted to HIV-1 Net" J. Mol. Biol. 293(5): 1097-106, 1999) исследовали способность RT-петли в SH3-домене Hck действовать в качестве детерминанты разносторонней специфичности и аффинности. Авторы сконструировали фаговую библиотеку Hck-доменов, где 6 аминокислот RT-петли были рандомизированы (названные RRT-SH3). Используя эту стратегию, они идентифицировали индивидуальные RRT-SH3 домены, которые могут связываться с Nef HIV-1 максимально в 40 раз лучше, чем Hck-SH3. Авторы указывают на важность RT-петли в выборе лиганда SH3 в качестве общей стратегии создания SH3-доменов с желаемыми связывающими свойствами.
Lee et al. ("A single amino acid in the SH3 domain of Hck determines its high affinity and specificity in binding to HIV-1 Nef protein." Embo. J. 14(20): 5006-15, 1995) исследовали структурную основу различных аффинностей связывания SH3 и специфичностей Hck в отношении Nef белка HIV-1 и могли переносить связывающую способность SH3 Hck в отношении Nef на SH3-домен Fyn единственной мутацией в RT-петле SH3-домена Fyn (R96I).
Hosse et al. ("A new generation of protein display scaffolds for molecular recognition", Protein Science, 15:14-27, 2006) особо рассмотрели требования для связывающих белков, подходящих для терапевтических применений. Авторы отмечают важность некоторых свойств для терапевтически применимых связывающих белков, таких как сывороточная стабильность, проникновение в ткани, клиренс из крови, удерживание мишени и иммунный ответ. В последнем случае замечено, что терапевтические белки, выделенные не из людей, должны быть сделаны как можно более похожими на их человеческие аналоги и человеческий каркас может быть менее иммуногенным сразу же с самого начала. Эти авторы делают вывод:
"Однако, даже полностью человеческий каркас не является гарантией того, что белок не вызывает иммунную реакцию у человека, особенно если он является внутриклеточным белком. Рандомизация аминокислот во время конструирования библиотеки может потенциально вводить новые эпитопы Т-клеток. Даже единичные точковые мутации могут делать белок человека иммуногенным. Кроме того, большинство человеческих каркасов вызывают некоторый аутоиммунный ответ".
В настоящее время SH3-домены киназ AbI и Hck считают белковыми каркасами для генерирования белковых связывающих агентов с предсказуемой специфичностью, даже если до сих пор идентифицированы только связующие вещества в отношении известных лигандов, подобных белкам Nef или синтетическим пептидам (см. Binz et al. выше).
SH3-домен киназы Fyn (Fyn SH3) содержит 63 остатка (аа 83-145 последовательности, описанной Semba et al. ("yes-родственный протоонкоген, син., относится к семейству белков, относящихся к протеинтирозинкиназному семейству". Proc. Natl. Acad. Sci. USA 83(15): 5459-63, 1986) и Kawakami et al. ("Isolation and oncogenic potential of a novel human src-like gene". Mol Cell Biol. 6(12): 4195-201, 1986). Fyn является членом Src-семейства тирозинкиназ с молекулярной массой 59 кДа. В результате альтернативного сплайсинга белок Fyn существует в виде двух различных изоформ, различающихся их киназными доменами; одна форма найдена в тимоцитах, спленоцитах и некоторых гематолимфоидных клеточных линиях, в то время как вторая форма накапливается в основном в головном мозгу (Cooke and Perlmutter, "Expression of a novel form of the Fyn proto-oncogene in hematopoietic cells". New Biol. 1(1): 66-74, 1989). Биологические функции Fyn разнообразны и включают в себя передачу сигнала через рецептор Т-клеток, регуляцию функции головного мозга, а также передачу сигнала, опосредованную адгезией (Resh, M.D. "Fyn, a Src family tyrosine kinase". Int. J. Biochem. Cell Biol. 30(11): 1159-62, 1998). Fyn является внутриклеточным белком. SEQ ID NO:1 показывает последовательность Fyn SH3 (аа 83-145 киназы Fyn, как сообщено Kawakami et al. и Semba et al. в 1986, см. выше) :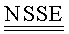
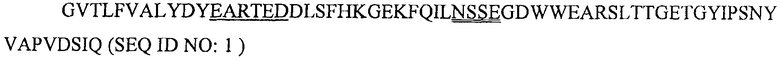
Последовательности RT-Src и n-Src петли подчеркнуты один раз и два раза, соответственно.
Аминокислотная последовательность Fyn SH3 полностью консервативна среди людей, мышей, крыс и обезьян (гиббона). Fyn SH3 цыпленка отличается в одном положении, последовательность Xenopus laevis в двух положениях аминокислот от соответствующего домена человека. Так же, как и другие SH3-домены, Fyn SH3 состоит из двух антипараллельных β-слоев и содержит две гибкие петли (называемые RT-Src и n-Src-петли) для взаимодействия с другими белками.
Таким образом, известный уровень техники считает белковые несущие конструкции, так называемые каркасы, альтернативой установленным структурам антител. Домен 3 (SH3) Src-гомологии является одним из этих, приблизительно 40 или более, каркасов. Среди многих различных SH3-доменов (приблизительно, 300 в геноме человека и нескольких тысячах, описанных до настоящего времени, в природе) Fyn SH3 является единственным, который был использован однажды ранее для выяснения специфичности связывания и аффинности SH3 в целом. Специалисту в данной области техники известно также, что внутриклеточные белки особенно склонны вызывать иммунные реакции и, следовательно, менее применимы для применений в прикладных областях in vivo, например в терапии и диагностике.
Целью настоящего изобретения является обеспечение улучшенных мишень-специфических и высокоаффинных связывающих белков, применимых в качестве исследовательских и, в частности, диагностических и медицинских агентов. Кроме того, эти связывающие белки должны быть стабильными и растворимыми в физиологических условиях, вызывать мало иммунных эффектов или вовсе не вызывать иммунных эффектов в людях, получающих их, и обеспечивать связывающую структуру, которая также доступна для больших структур-мишеней, т.е. которая не экранирована стерическими препятствиями.
Раскрытие изобретения
Неожиданно было обнаружено, что SH3-домен киназы Fyn Src-семейства обладает превосходными свойствами для конструирования рекомбинантных связывающих доменов, обладающих специфичностью и высокой аффинностью в отношении выбранных мишеней. В частности, было показано, что специфичность в отношении мишеней может быть сконструирована мутацией RT-петли и/или src-петли, приводящей к более высокой вариабельности и улучшенным связывающим свойствам в отношении многих мишеней.
Более того, было неожиданно обнаружено, что не только нативный связывающий белок Fyn SH3, но также мутированные связывающие белки, полученные на основе Fyn SH3, также не были иммуногенными in vivo. Таким образом, рекомбинантные мутантные связывающие белки Fyn SH3 особенно применимы для создания неиммунногенных белковых терапевтических и/или диагностических средств.
В результате вышесказанного первый аспект настоящего изобретения относится к рекомбинантному связывающему белку, содержащему, по меньшей мере, одно производное домена 3 (SH3) Src-гомологии киназы Fyn, в котором
(a) по меньшей мере, одна аминокислота, находящаяся в Src-петле или расположенная на расстоянии вплоть до двух аминокислот рядом с Src-петлей, и/или
(b) по меньшей мере, одна аминокислота, находящаяся в RT-петле или расположенная на расстоянии вплоть до двух аминокислот рядом с RT-петлей, заменена, делегирована или добавлена, причем производное SH3-домена имеет аминокислотную последовательность, имеющую, по меньшей мере, 70%, предпочтительно, по меньшей мере, 80%, более предпочтительно, по меньшей мере, 90% и наиболее предпочтительно, по меньшей мере, 95%-ную идентичность относительно аминокислотной последовательности SEQ ID NO:1,
предпочтительно при условии, что этот рекомбинантный связывающий белок не содержит аминокислотной последовательности SEQ ID NO:2,
и, предпочтительно, этот рекомбинантный белок не является природным SH3-доменом, содержащим белок, существующим в природе.
Аминокислотная последовательность SEQ ID NO:2 (вариант Fyn SH3 R96I по Lee et al., см. выше) представлена ниже.
GVTLFVALYDYEAITEDDLSFHKGEKFQILNSSEGDWWEARSLTTGETGYIPSNYVAPVDSIQ (SEQ ID NO:2)
В контексте данного изобретения RT-петля киназы Fyn (иногда также обозначаемая RT-Src-петлей) состоит из аминокислот Е A R Т Е D, находящихся в положениях 12-17 в SEQ ID NO:1. Положения, подлежащие замене, делегированию или добавлению, т.е. подлежащие мутации, находящиеся внутри RT-петли или расположенные рядом с RT-петлей, представляют собой аминокислоты 10-19, предпочтительно 11-18, более предпочтительно 12-17.
В контексте данного изобретения Src-петля киназы FYN (иногда также обозначаемая как n-Src-петля) состоит из аминокислот N S S Е, находящихся в положениях 31-34 в SEQ ID NO:1. Положения, подлежащие замене, делегированию или добавлению, т.е. подлежащие мутации, находящиеся в Src-петле или рядом с Src-петлей, представляют собой аминокислоты 29-36, предпочтительно 30-35, более предпочтительно 31-34.
Рекомбинантный белок данного изобретения предпочтительно не является природным белком, содержащим SH3-домен, существующим в природных условиях или выделенным из них. Другими словами объем данного изобретения предпочтительно исключает белки, содержащие SH3-домен дикого типа. В природе имеется множество белков, содержащих SH3 домен. Такие природные SH3-белки обладают связывающей способностью в отношении их природных лигандов. Большинство, если не все из этих природных SH3-лигандов, имеют РххР мотив. Однако рекомбинантные белки данного изобретения являются генно-инженерными белками, сконструированными для проявления аффинности к неприродным мишеням, т.е. неприродным мишеням, являющимся любыми мишенями, например, находящимися в природе, предпочтительно в млекопитающих, более предпочтительно в человеке, за исключением природных (дикого типа) SH3-лигандов. Более предпочтительно, рекомбинантные белки данного изобретения по существу не имеют связывающей способности в отношении любых природных SH3-связывающих лигандов, наиболее предпочтительно не к какому-либо природному SH3-связывающему лиганду, имеющему РххР мотив.
Предпочтительно, число аминокислот, которое добавляют в одну и/или обе петли, равно 1-20, более предпочтительно 1-10 или 1-5 аминокислот и наиболее предпочтительно, когда аминокислоты не добавляют в эти петли.
В другом предпочтительном варианте осуществления части производного SH3-домена, которые лежат вне RT- и Src-петель, консервируют, насколько это возможно, чтобы не вводить иммуногенных мотивов.
Предпочтительно, чтобы рекомбинантные белки данного изобретения по существу не вызывали иммуногенной реакции в млекопитающих, предпочтительно в мышах, крысах и/или в человеке, наиболее предпочтительно в человеке. Конечно, иммуногенность полного рекомбинантного белка данного изобретения будет зависеть не только от этой части производного SH3-домена, но может зависеть от других частей полного белка.
В предпочтительном варианте осуществления этого изобретения, по меньшей мере, часть производного SH3-домена рекомбинантного белка является по существу неиммуногенной в млекопитающих, предпочтительно, в мыши, крысе и/или в человеке, наиболее предпочтительно в человеке.
Например, квалифицированный в данной области техники специалист может определить иммуногенные реакции рекомбинантного белка или части производного его SH3-домена стандартными и рутинными способами, например введением (например, внутривенной (i/v) инъекцией) представляющего интерес рекомбинантного белка или производного его SH3-домена млекопитающему, например мыши и анализом ответа иммуногенных клеток крови и/или факторов (например, интерлейкинов) по истечении соответствующего времени, необходимого для появления иммунной реакции.
В более предпочтительном варианте осуществления связывающий белок согласно настоящему изобретению представляет собой такой белок, в котором указанное производное SH3-домена имеет аминокислотную последовательность, имеющую, по меньшей мере, 70%, предпочтительно, по меньшей мере, 85%, предпочтительно, по меньшей мере, 90%, более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 98-100%-ную идентичность относительно аминокислотной последовательности домена 3 (SH3) Src-гомологии киназы FYN вне src- и RT-петель.
В предпочтительном варианте осуществления мутации вводят как в RT-, так и Src-петли.
В дополнительном более предпочтительном варианте осуществления связывающий белок настоящего изобретения содержит один или предпочтительно два измененных остатка в положениях 37 и/или 50 производного SH3-домена, предпочтительно два гидрофобных измененных остатка, более предпочтительно Trp37 и/или Tyr50, причем Trp37 и Tyr50 являются наиболее предпочтительными. Как показано на фиг.3b ниже, их рандомизация может увеличивать аффинность.
Термин "производное домена 3 Src-гомологии киназы FYN" в данном контексте означает аминокислотную последовательность, имеющую, по меньшей мере, 70%, предпочтительно, по меньшей мере, 80%, более предпочтительно, по меньшей мере, 90% и наиболее предпочтительно, по меньшей мере, 95%-ную идентичность относительно аминокислотной последовательности SEQ ID NO:1. То же самое относится и к производному SH3-домена, имеющего, по меньшей мере, 70% или, по меньшей мере, 85%, предпочтительно, по меньшей мере, 90%, более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, 98%-ную идентичность относительно домена 3 (SH3) Src-гомологии киназы FYN вне src- и RT-петель, за исключением того, что аминокислоты, образующие указанные петли, исключаются при определении идентичности последовательности.
Для определения степени идентичности последовательности производного SH3-домена Fyn относительно аминокислотной последовательности SEQ ID NO:1 может быть, например, использована программа SIM Local Similarity (Xiaoquin Huang and Webb Miller, "A Time-Efficient, Linear-Space Local Similarity Algorithm". Advances in Applied Mathematics, vol.12: 337-357, 1991), без ограничений доступная от авторов и их института (см. также www: http://www.expasy.org/tools/sim-prot.html); для анализа путем множественного выравнивания может быть использована программа Clustal W (Thompson et al., CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice". Nucleic Acids Res., 22(22): 4673-4680, 1994). Предпочтительно степень идентичности последовательности производного относительно SEQ ID NO:1 определяют относительно полной последовательности SEQ ID NO:1.
В предпочтительном варианте осуществления связывающий белок настоящего изобретения содержит, по меньшей мере, два производных SH3 домена Fyn. Более предпочтительно, он представляет собой двухвалентный связывающий белок. По меньшей мере, два производных SH3-домена могут быть одинаковыми или различными. Предпочтительно, они являются одинаковыми.
Связывающий белок настоящего изобретения может быть сконструирован таким образом, что он имеет какую-либо аффинность связывания в отношении к конкретной мишени. В предпочтительном варианте осуществления этой мишенью является мишень на основе аминокислот, такая как пептид или белок, более предпочтительно белок, содержащий мотив РххР. Конечно, только меньшая часть природных и физиологически релевантных белков-мишеней содержит мотив РххР. Примеры, приведенные ниже, показывают, что доступны связывающие белки согласно настоящему изобретению для мишеней (например, ED-B домена фибронектина) с мотивами, иными, чем РххР. Следовательно, связывающий белок настоящего изобретения ни в коей мере не ограничен мотивом РххР и может иметь специфическую аффинность связывания в отношении любой конкретной мишени, например сахаров, полипептидов и т.д.
Более предпочтительно, связывающий белок согласно этому изобретению имеет специфическую аффинность связывания в отношении мишени 10-7-10-12 М, предпочтительно 10-6-10-12 М, предпочтительно к терапевтически и/или диагностически подходящей мишени, более предпочтительно к мишени на основе аминокислот, содержащей мотив РххР.
В наиболее предпочтительном аспекте связывающий белок согласно этому изобретению имеет специфическую (in vivo и/или in vitro) аффинность связывания в диапазоне 10-7-10-12 М, предпочтительно от 10-6-10-12 М, в отношении внеклеточного домена онкофетального фибронектина (ED-B).
В предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантному связывающему белку, содержащему, по меньшей мере, одно производное домена 3 (SH3) Src-гомологии киназы FYN, где
(а) по меньшей мере, одна аминокислота, находящаяся в Src-петле или расположенная на расстоянии вплоть до двух аминокислот рядом с Src-петлей, и/или
(b) по меньшей мере, одна аминокислота, находящаяся в RT-петле или расположенная на расстоянии вплоть до двух аминокислот рядом с RT-петлей, заменена, делегирована или добавлена, причем производное SH3-домена имеет аминокислотную последовательность, имеющую, по меньшей мере, 70%, предпочтительно, по меньшей мере, 80%, более предпочтительно, по меньшей мере, 90% и наиболее предпочтительно, по меньшей мере, 95%-ную идентичность относительно аминокислотной последовательности SEQ ID NO:1,
предпочтительно при условии, что этот рекомбинантный связывающий белок не содержит аминокислотной последовательности SEQ ID NO:2,
и, предпочтительно, этот рекомбинантный белок не является природным SH3-доменом, содержащим белок, существующим в природе,
где указанный связывающий белок имеет специфическую (in vivo и/или in vitro) аффинность связывания предпочтительно 10-7-10-12 М, предпочтительно 10-6-10-12 М, в отношении внеклеточного домена онкофетального фибронектина (ED-B).
В более предпочтительном варианте осуществления указанное производное SH3-домена имеет, по меньшей мере, 85%, предпочтительно, по меньшей мере, 90%, более предпочтительно, по меньшей мере, 95%, наиболее предпочтительно, по меньшей мере, от 98 до 100% идентичности относительно домена 3 (SH3) Src-гомологии киназы FYN вне src- и RT-петель.
В другом более предпочтительном варианте осуществления вышеупомянутый ED-В-специфический связывающий белок содержит, по меньшей мере, два производных SH3-домена, предпочтительно он является двухвалентным связывающим белком.
Предпочтительно, указанный ED-B-специфический связывающий белок содержит один или несколько, предпочтительно два измененных, предпочтительно гидрофобных остатков в положениях 37 и/или 50 производного SH3-домена, в частности Trp37 и/или Tyr50, причем наиболее предпочтительными являются Trp37 и Tyr50.
Помимо специфической аффинности связывания в отношении полипептидных и белковых мишеней связывающий белок этого изобретения может также иметь специфическую аффинность связывания в отношении низкомолекулярных органических соединений или соединений не на основе аминокислот, например сахаров, олиго- или полисахаридов, жирных кислот и т.д.
Ряд слитых белков антитело-цитокин были уже исследованы в отношении применений, например, в лечении артрита или рака, часто с впечатляющими результатами. Например, антитело L19 человека, специфическое в отношении ED-B домена фибронектина (маркера ангиогенеза), использовали для доставки провоспалительных цитокинов (таких, как IL-2, IL-12 или TNF) к солидным опухолям, иногда с поразительной терапевтической пользой [в отношении обзора и соответствующих ссылок см. Neri & Bicknell, Nat. Rev. Cancer (2005) 5: 436-446, а также WO 01/62298].
В настоящее время связывающий белок настоящего изобретения используют для замены антител в слитых белках известного уровня техники, а также для конструирования новых и менее иммуногенных слитых (гибридных) для фармацевтических и диагностических применений in vivo и in vitro.
Во втором аспекте данное изобретение относится к слитому белку, содержащему связывающий белок этого изобретения, слитый с фармацевтически или диагностически активным компонентом.
Слитый белок этого изобретения может содержать неполипептидные компоненты, например непептидные линкеры, непептидные лиганды, например, для относящихся к терапии или диагностике радионуклидов.
Предпочтительно, указанный активный компонент является цитокином, предпочтительно цитокином, выбранным из группы, состоящей из IL-2, IL-12, TNF-альфа, IFN-альфа, IFN-бета, IFN-гамма, IL-10, IL-15, IL-24, GM-CSF, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, LIF1 CD80, В70, TNF-бета, LT-бета, CD-40 лиганда, Fas-лиганда, TGF-бета, IL-1-альфа и IL-1-бета.
Более предпочтительно, указанный активный компонент является токсичным соединением, предпочтительно низкомолекулярным органическим соединением или полипептидом, предпочтительно токсичным соединением, выбранным из группы, состоящей из калихеамицина, неокарциностатина, эсперамицина, динемицина, кедарцидина, мадуропептина, доксорубицина, даунорубицина, ауристатина, А-цепи рицина, модеккина, укороченного экзотоксина Pseudomonas А, дифтерийного токсина и рекомбинантного гелонина.
В другом предпочтительном варианте осуществления слитый белок согласно изобретению представляет собой белок, в котором указанный активный компонент является хемокином, предпочтительно хемокином, выбранным из группы, состоящей из IL-8, GRO-альфа, GRO-бета, GRO-гамма, ENA-78, LDGF-PBP, GCP-2, PF4, Mig, IP-10, SDF-1 альфа/бета, BUNZO/STRC33, I-TAC, BLC/BCA-1, MIP-1-альфа, MIP-1-бета, MDC, TECK, TARC, RANTES, HCC-1, HCC-4, DC-CK1, MIP-3-альфа, MIP-3-бета, MCP-1-5, эотаксина, эотаксина-2, 1-309, MPIF-1, 6Ckine, CTACK, MEC, лимфотактина и фракталкина.
В дополнительном предпочтительном варианте осуществления связывающий белок этого изобретения содержит искусственные аминокислоты.
В дополнительных предпочтительных вариантах осуществления слитого белка настоящего изобретения указанный активный компонент представляет собой флуоресцентный краситель, предпочтительно компонент выбран из группы, состоящей из Alexa Fluor или красителей Су (Berlier et al., «Quantitative Comparison of Long-wavelength Alexa Fluor Dyes to Cy Dyes: Fluorescence of the Dyes and Their Bioconjugates», J Histochem Cytochem. 51 (12): 1699-1712, 2003.); фотосенсибилизатора, предпочтительно бис(триэтаноламин)Sn(IV) хлорин е6 (SnChe6); прокоагулянтного фактора, предпочтительно тканевого фактора; фермента для активации пролекарства, предпочтительно фермента, выбранного из группы, состоящей из карбоксипептидаз, глюкуронидаз и глюкозидаз; радионуклида, либо из группы гамма-излучающих изотопов, предпочтительно, 99mTc, 123I, 111In, или из группы излучателей позитронов, предпочтительно, 18F, 64Cu, 68Ga, 86Y1 124I, или из группы бета-излучателей, предпочтительно, 131I, 90Y, 177Lu, 67Cu, или из группы альфа-излучателей, предпочтительно, 213Bi1 211At; и/или функционального Fc-домена, предпочтительно функционального Fc-домена человека.
Вышеупомянутый функциональный Fc-домен позволит направлять иммунную реакцию млекопитающего к сайту специфического связывания мишени связывающего белкового компонента слитого белка, например, в терапевтических, профилактических и/или диагностических применениях.
Дополнительный предпочтительный вариант осуществления относится к слитым белкам согласно этому изобретению, указанным выше, дополнительно содержащим компонент, модулирующий время полужизни в сыворотке, предпочтительно компонент, выбранный из группы, состоящей из полиэтиленгликоля (ПЭГ), иммуноглобулина и альбумин-связывающих пептидов.
В наиболее предпочтительном варианте осуществления гибридный белок этого изобретения, как упоминалось выше, содержит связывающий белок этого изобретения, имеющий специфическую (in vivo и/или in vitro) аффинность связывания 10-7-10-12 М, предпочтительно 10-8-10-12 М, в отношении дополнительного домена онкофетального фибронектина (ED-B). Предпочтительно, указанный ED-B-специфический связывающий белок имеет один или несколько, предпочтительно два гидрофобных остатка в положениях 37 и/или 50 производного домена SH3, в частности Trp37 и/или Tyr50, причем Trp37 и Tyr50 являются наиболее предпочтительными.
Связывающие и слитые белки согласно этому изобретению могут быть получены любым из многих общепринятых и хорошо известных способов, таких как обычные органические синтетические приемы, твердофазные синтетические способы или способа с использованием коммерчески доступных автоматических синтезаторов. С другой стороны, они могут также быть получены только общепринятыми рекомбинантными способами или в сочетании с общепринятыми синтетическими способами.
Связывающие белки настоящего изобретения также могут быть выделены путем селекции из белковой библиотеки, полученной на основе Fyn SH3, как это описано в примерах. Существует множество способов получения библиотек и они, в том числе, включают получение библиотек путем рандомизации определенных участков. Получение библиотек с помощью данной методики описано в статьях Silacci et al. ("Design, construction, and characterization of a large synthetic human antibody phage display library" Proteomics 5: 2340-50, 2005), Hoogenboom & Winter ("By-passing Immunisation Human antibodies from synthetic repertoires of germline VH gene segments rearranged in vitro" J. Mol. Biol 227: 381-88, 1992), Nissim et al. ("Antibody fragments from a 'single pot' phage display library as immunochemical reagents" The EMBO Journal 13(3); 692-98, 1994), Hoogenboom ("Designing and optimizing library selection strategies for generating high-affinity antibodies" TIBTECH 15: 62-70, 1997), Beste et al (Small antibody-like proteins with prescribed ligand specificities derived from lipocalpin fold" Proc. Natl. Sci USA 96: 1898-903, 1999) и Nygren & Skerra ("Binding proteins from alternative scaffolds" Journal of immunological methods 290: 3-28, 2004). В настоящем изобретении для получения библиотеки проводилась рандомизация участков, относящихся к Src- и RT-петле. Дополнительные аспекты настоящего изобретения относятся к (i) полинуклеотиду, кодирующему связывающий белок или слитый белок согласно этому изобретению, (ii) вектору, содержащему указанный нуклеотид, (iii) клетке-хозяину, содержащей указанный нуклеотид и/или указанный вектор.
Полинуклеотидами могут быть ДНК, РНК, ПНК и их любые аналоги. Векторы и клетки-хозяева могут относиться к любому обычному типу, который соответствует данной цели, например получению связывающих и слитых белков этого изобретения, терапевтически применимых векторов и клеток-хозяев, например, для генной терапии. Квалифицированный специалист в данной области сможет выбрать такие полинуклеотиды, векторы и клетки-хозяева из изобилующего известного уровня техники и подтвердить их индивидуальную пригодность для желаемой цели с использованием рутинных способов и без чрезмерных затрат.
Связывающие и гибридные белки настоящего изобретения не вызывают сильной иммунной реакции и предпочтительно по существу не вызывают иммунной реакции в млекопитающих, в людях и мышах, как было показано на мышах, и по аналогии ожидается, что это будет справедливо также и для людей, потому что Fyn SH3 идентичен в обоих типах млекопитающих. Неожиданно было показано, что ни нативный домен Fyn SH3, ни мутированный домен Fyn SH3 не вызывает иммунной реакции в мышах, инъецированных любым из них. Это было неожиданно, потому что киназа Fyn является внутриклеточным белком и не участвует в неонатальной селекции В-клеток. Таким образом, произведенные из Fyn SH3 связывающие и гибридные белки со сконструированной специфичностью и аффинностью относительно клеток-мишеней особенно хорошо подходят для терапевтических, профилактических и/или диагностических применений in vivo.
Таким образом, особенно значимый аспект настоящего изобретения относится к применению связывающих или слитых белков согласно этому изобретению для получения лекарственного средства.
В дополнительном аспекте связывающий или слитый белок настоящего изобретения используют для получения диагностических средств, в частности для применения in vivo. Предпочтительно. ED-B-специфический связывающий или гибридный белок, описанный выше, используют для получения лекарственных или диагностических средств для лечения и диагностики рака.
Другой аспект настоящего изобретения относится к фармацевтической композиции, содержащей связывающий или слитый белок настоящего изобретения и необязательно фармацевтически приемлемый эксципиент.
Другой аспект настоящего изобретения относится к диагностической композиции, предпочтительно, для применений in vivo, содержащей связывающий или слитый белок настоящего изобретения и необязательно фармацевтически приемлемый эксципиент.
Предпочтительно, фармацевтическая или диагностическая композиция содержит ED-B-специфический связывающий или слитый белок настоящего изобретения и необязательно фармацевтически приемлемый эксципиент.
Фармацевтические композиции или диагностические средства для применений in vivo в настоящем изобретении обычно содержат терапевтически или диагностически эффективное количество связывающего и/или слитого белка согласно настоящему изобретению и необязательно вспомогательные вещества, такие как фармацевтически приемлемый эксципиент (эксципиенты). Указанные фармацевтические композиции готовят способами, хорошо известными в фармацевтической области. Носитель или эксципиент могут быть жидким материалом, который может служить в качестве носителя или среды для активного ингредиента. Подходящие носители или эксципиенты хорошо известны в данной области и включают в себя, например, стабилизаторы, антиоксиданты, вещества, регулирующие рН, эксципиенты для контролируемого высвобождения. Фармацевтический препарат этого изобретения может быть применен, например, для парентерального применения и может быть введен пациенту в форме растворов или т.п.
Наконец, другой аспект настоящего изобретения относится к способу лечения или диагностики, в котором эффективное количество вышеописанной фармацевтической или диагностической композиции вводят пациенту, нуждающемуся в этом, предпочтительно пациенту, страдающему или предположительно страдающему от рака и/или воспалительных заболеваний.
При проведении лечения или диагностики субъекта, страдающего от этих заболеваний, связывающий или слитый белок настоящего изобретения может быть введен в любой форме или любым способом, который делает терапевтическое или диагностическое соединение биодоступным в эффективном количестве, в том числе пероральными или парентеральным способами. Например, композиции настоящего изобретения можно вводить подкожно, внутримышечно, внутривенно и т.п. Специалист в области приготовления композиций может легко выбрать нужную форму и способ введения в зависимости от конкретных свойств выбранного продукта, заболевания или состояния, подлежащего лечению или диагностике, стадии заболевания и других релевантных обстоятельств (см., например, Remingtons Pharmaceutical Sciences, Mack Publishing Co. (1990)). Композиции настоящего изобретения можно вводить отдельно или в форме фармацевтического или диагностического препарата в комбинации с фармацевтически приемлемыми носителями или эксципиентами, соотношение и природа которых определяются растворимостью и химическими свойствами выбранного продукта, выбранным способом введения и стандартной фармацевтической и диагностической практикой. Продукты настоящего изобретения, будучи сами эффективными, могут быть приготовлены в виде смеси и введены в форме их фармацевтически приемлемых солей, например кислотно-аддитивных солей или основно-аддитивных солей, для целей стабильности, удобства кристаллизации, повышения растворимости и т.п.
Краткое описание чертежей
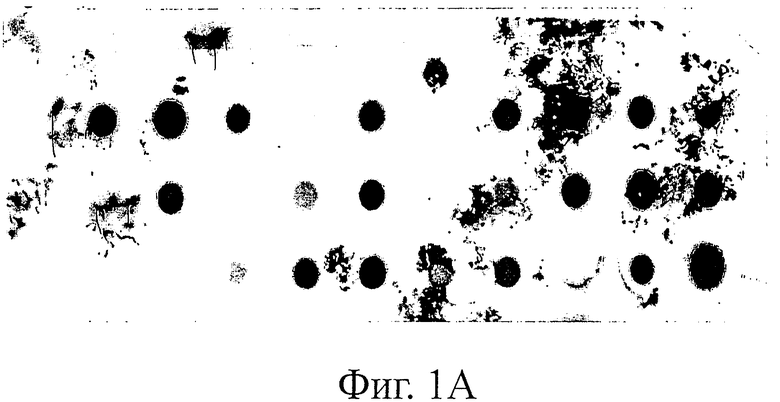
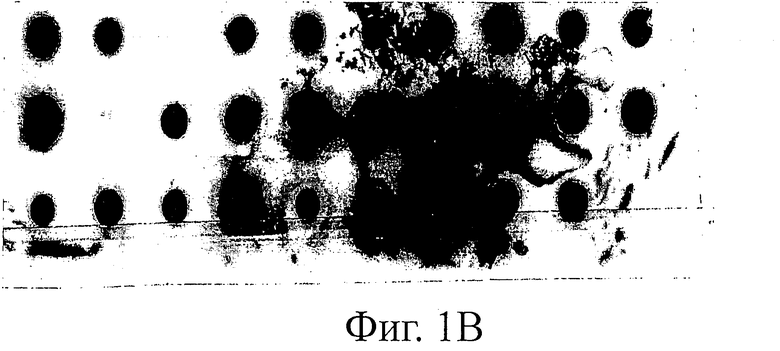
Фиг.1 иллюстрирует дот-блот анализ. Процент клонов, экспрессирующих детектируемое количество растворимых SH3-мутантов Fyn, определяли дот-блот анализом лизатов бактериальных клеток с использованием конъюгата анти-HIS-HRP антитела (Sigma) в качестве детектирующего реагента. Пероксидазную активность детектировали с использованием системы детектирования для Вестерн-блоттинга ECL plus (Amersham).
A) Fyn SH3-мутанты с рандомизированной RT-Src-петлей.
B) Fyn SH3-мутанты с удлиненной (4->6) и рандомизированной n-Src-петлей.
C) FynSH3 с RT- и n-Src-рандомизированными петлями.
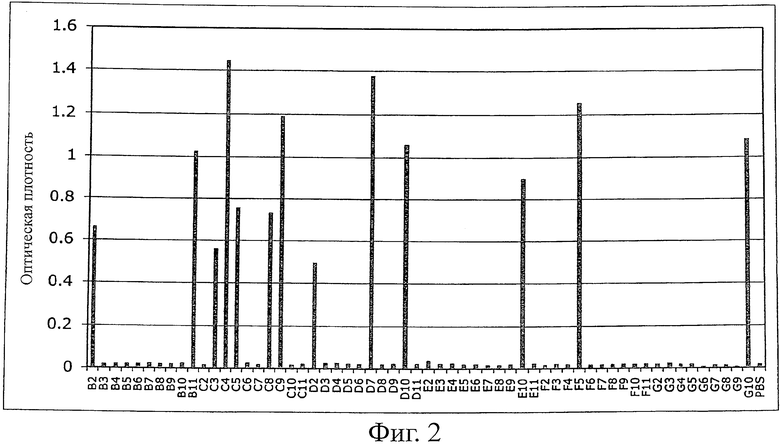
Фиг.2 иллюстрирует ELISA моноклональных фагов. После третьего цикла пэннинга против MSA моноклональные бактериальные супернатанты, содержащие фаги, представляющие Fyn SH3-мутанты, тестировали при помощи ELISA с использованием планшетов MaxiSorp (Nunc), покрытых MSA (100 мкг/мл в течение ночь, 100 мкл на лунку). Связанные фаги детектировали с использованием конъюгатов антител против М-13 с hrp (Amersham).
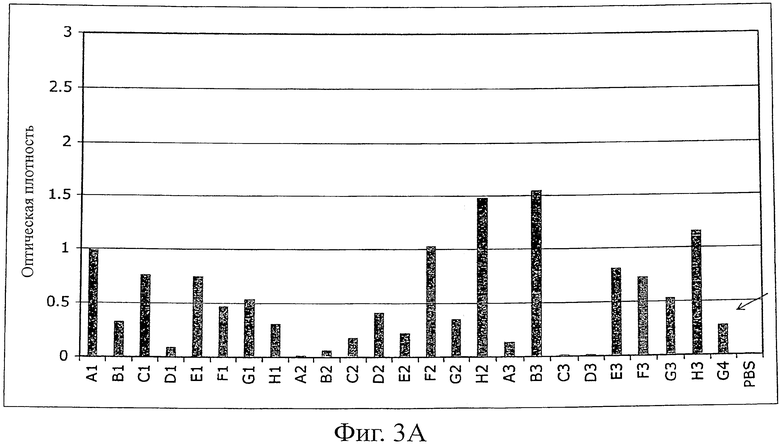
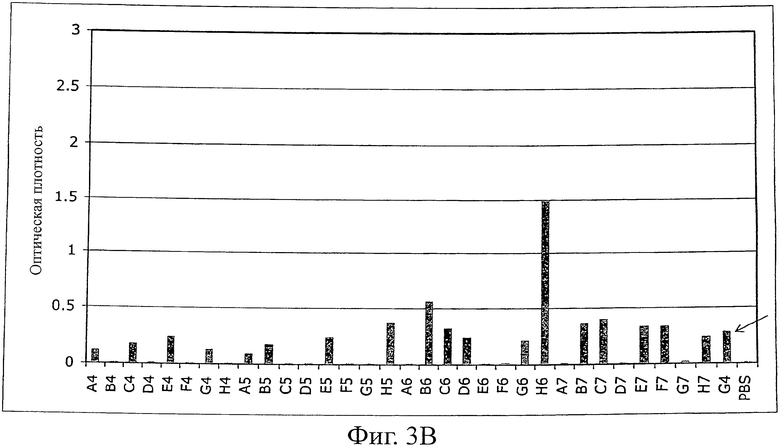
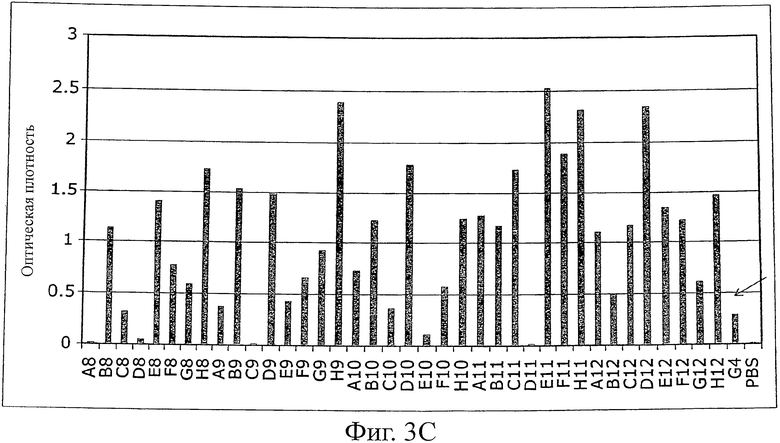
Фиг.3 иллюстрирует ELISA (против MSA) моноклональных фагов после одного цикла селекции созревания аффинности с использованием планшетов MaxiSorp (Nunc), покрытых MSA (100 мкг/мл в течение ночи, 100 мкл на лунку).
A) ELISA фагов первой суббиблиотеки G4 (рандомизированная n-Src-петля и Trp37 и Tyr50). Исходный клон G4 указан стрелкой.
B) ELISA фагов второй суббиблиотеки G4 (рандомизированная и удлиненная n-Src-петля). Исходный клон G4 указан стрелкой.
C) ELISA фагов первой и второй суббиблиотеки после одного цикла пэннинга, выполненного в условиях, благоприятствующих действию связывающих агентов с длинной koff. Исходный клон G4 указан стрелкой.
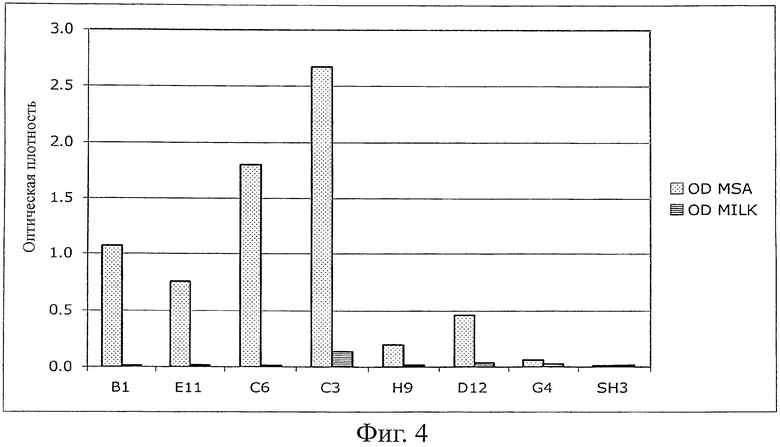
Фиг.4 показывает данные ELISA растворимых фракций (с использованием планшетов MaxiSorp (Nunc), покрытых MSA (100 мкг/мл в течение ночи, 100 мкл на лунку) нескольких MSA-связывающих клонов, после клонирования (pQE-12 вектор), экспрессии и очистки растворимого белка в соответствиями с инструкциями производителя (Qiagen, нативные условия). В качестве детектирующих агентов использовали конъюгаты антител против HIS с HRP. В качестве контроля те же связывающие белки добавляли в лунки, блокированные только 4% MPBS.
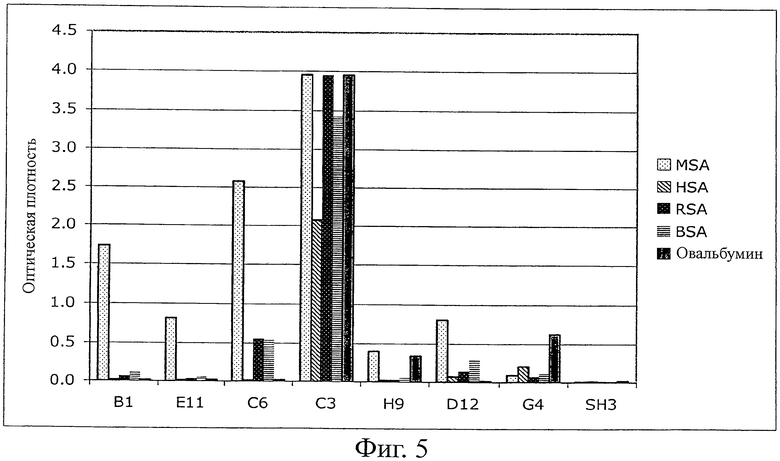
Фиг.5 - ELISA растворимого белка на специфичность. Отобранные MSA-связывающие Fyn SH3-мутанты были тестированы на связывание против сывороточного альбумина человека (HSA), сывороточного альбумина крысы (RSA), бычьего сывороточного альбумина (BSA) и овальбумина с использованием планшетов MaxiSorp (Nunc), покрытых различными альбуминами (каждым по 100 мкг/мл в течение ночи, 100 мкл на лунку).
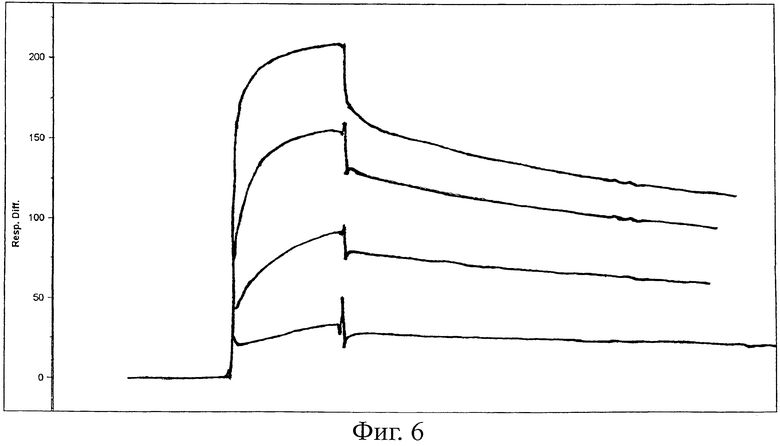
Фиг.6 - BIACore-анализ D3. Используемые концентрации: 4, 2, 1, и 0,5 мкМ (сверху вниз).
Фиг.7 - ELISA-анализ проб крови на присутствие мышиных антител.
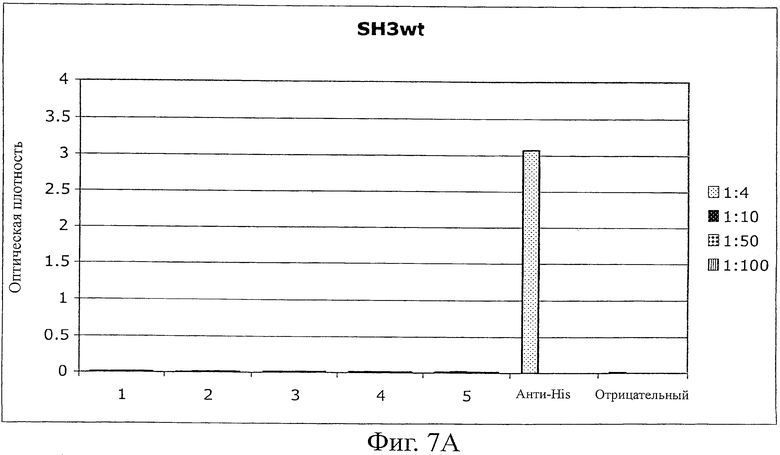
A) Планшеты MaxiSorp (Nunc) покрывали Fyn SH3 (20 мкг/мл в течение ночи, 100 мкл на лунку). Пробы крови (75-200 мкл) каждой из 5 мышей применяли в серии разведений (от 1:4 до 1:100). Детектирование антител выполняли с использованием конъюгата антитела к мышиному IgG с HRP (Sigma). В качестве контроля эффективности покрытия использовали конъюгаты анти-HIS антитела с HRP (Sigma).
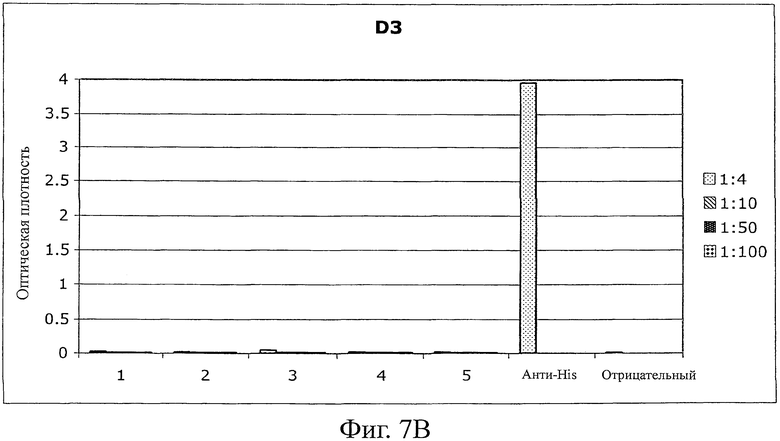
B) Планшеты MaxiSorp (Nunc) покрывали Fyn SH3 D3 (20 мкг/мл в течение ночи, 100 мкл на лунку). Пробы крови (75-200 мкл) каждой из 5 мышей использовали в серии разведений (от 1:4 до 1:100). Детектирование антител проводили с использованием конъюгата антитела к мышиному IgG с HRP (Sigma). В качестве контроля эффективности покрытия использовали конъюгаты анти-HIS-антитела с HRP (Sigma).
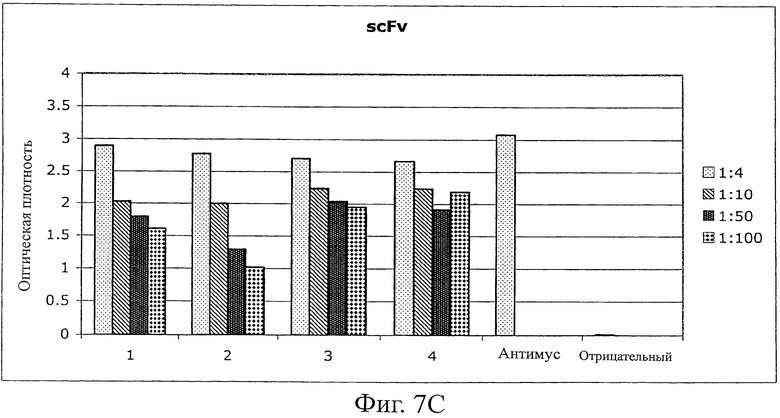
C) Планшеты MaxiSorp (Nunc) покрывали scFv (60 мкг/мл в течение ночи, 100 мкл на лунку). Пробы крови (75-200 мкл) каждой из 4 мышей использовали в серии разведений (от 1:4 до 1:100). Детектирование антител проводили с использованием конъюгата антитела к мышиному IgG с HRP (Sigma). В качестве контроля эффективности покрытия использовали конъюгаты анти-myc антитела с HRP (Roche).
Фиг.8 показывает иммунофлуоресценцию D3 (фиг.8A), соответствующий негативный контроль (8B), анти-CD31-окрашивание (фиг.8C) и соответствующий негативный контроль (8D) на гистологических срезах мышиной тератокарциномы.
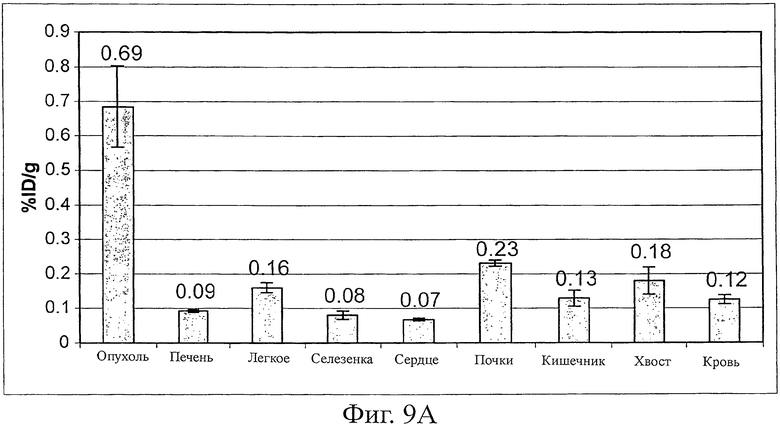
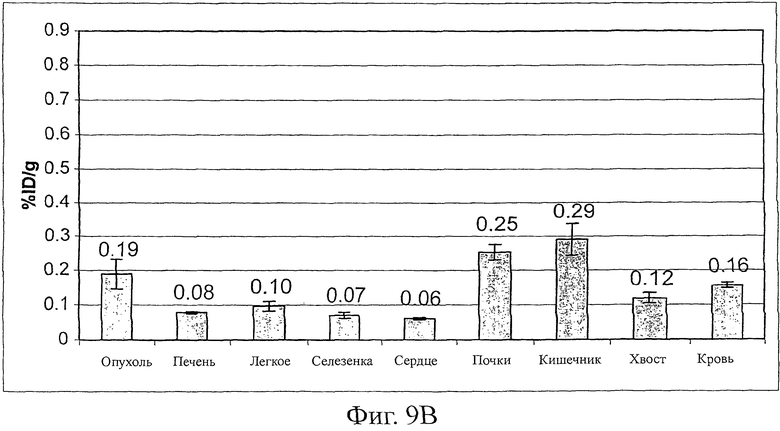
Фиг.9 показывает удерживание опухолью Fyn SH3-D3 (фиг.9A), в то время как для Fyn SH3wt не наблюдали аккумуляции в опухоли (фиг.9B). Результаты нацеливания выражены в виде % инъецированной дозы 125I-меченого белка, удерживаемого на грамм ткани (% ID/г).
Осуществление изобретения
Далее предмет настоящего изобретения будет описан более подробно со ссылкой на конкретные варианты осуществления, которые не следует рассматривать как ограничивающие объем настоящего изобретения.
Примеры
Пример 1: Экспрессия Fyn SH3-мутантов
Для оценки экспрессии Fyn SH3-мутантов проводят дот-блот-анализ трех различных Fyn SH3-суббиблиотек (фиг.1): в первой библиотеке была рандомизирована только RT-петля, во второй была рандомизирована Src-петля и удлинена до 6 остатков и в третьей были одновременно рандомизированы RT- и Src-петля, причем последняя петля была удлинена с 4 до 6 остатков. Процент экспрессированных Fyn SH3-мутантов находится в диапазоне 59-90%.
Таблица 1
1. Библиотека
2. RT- и n-Src
3. Экспрессированные мутанты
4. Число тестируемых клонов
Пример 2: Отборы с использованием фагового дисплея против мышиного сывороточного альбумина
Создавали библиотеку из 107 различных Fyn SH3 (рандомизирована только RT-петля) и клонировали в фагмидный вектор pHEN1 (Hoogenboom et al. "Multi-subunit proteins on the surface of filamentous phage: methodologies for displaying antibody (Fab) heavy and light chains". Nucleic Acids Res, 19(15): 4133-7, 1991). Библиотеку представляли на фагах и выполняли 3 цикла пэннинга против мышиного сывороточного альбумина (MSA). После третьего цикла выполняли скрининг на связывающие белки при помощи ELISA для моноклональных фагов; детектировали 13 положительных клонов (фиг.2). Секвенирование 13 клонов показывает, что две различные последовательности, обозначенные как G4 и С4, были обогащены.
Однако после субклонирования и экспрессии G4 в векторе pQE-12 (Qiagen, экспрессия и очистка в соответствии с руководством изготовителя в нативных условиях) связывание белков с MSA не могло быть обнаружено ELISA (фиг.4) из-за низкой аффинности (анализ ELISA для фагов является более чувствительным, чем ELISA растворимого белка). Поэтому для двух различающихся по созреванию аффинности библиотек (размер: 107 клонов на каждую библиотеку) использовали последовательность G4. В первой библиотеке были рандомизированы 4 остатка n-Src-петли и остатки Trp37 (SEQ ID NO:1) и Tyr50 (SEQ ID NO:1), во второй была удлинена n-Src-петля с 4 до 6 рандомизированных остатков. После одного цикла пэннинга несколько клонов обеих суббиблиотек давали более сильные сигналы в Фаг-ELISA в сравнении с исходным клоном G4 (фиг.3). После субклонирования и экспрессии нескольких клонов связывание растворимого белка подтверждали при помощи ELISA (фиг.4). Кажущиеся константы диссоциации находились в диапазоне 100 нМ (определенные при помощи BIAcore). Некоторые из этих клонов перекрестно реагировали с другими сывороточными альбуминами (анализировали: сывороточный альбумин человека (HSA), сывороточный альбумин крысы (RSA), бычий сывороточный альбумин (BSA) и овальбумин), в то время как другие клоны были высокоспецифическими в отношении MSA, что указывает на то, что можно выделить высокоспецифические связывающие белки (фиг.5).
Пример 3: Отбор с использованием фагового дисплея против дополнительного домена В фибронектина (ED-B)
ED-B выбрали в качестве белка-мишени для демонстрации возможности отбора произведенных из Fyn SH3-связывающих агентов против фармацевтически релевантного белка. ED-B является доменом гомологии Типа III, состоящим из 91 аминокислоты, который инсертируется в молекулу фибронектина по механизму альтернативного сплайсинга на уровне первичного транскрипта всегда, когда происходит ремоделирование ткани (Zardi et al., "Transformed human cells produce a new fibronectin isoform by preferential alternative splicing of a previously unobserved exon". Embo J. 6(8): 2337-42, 1987). Он является хорошим маркером качества ангиогенеза, который сверхэкспрессируется в большинстве солидных опухолей (например, почечно-клеточной карциноме, колоректальной, печеночно-клеточной карциноме, высокоэффективной астроцитоме, опухолях головы и шеи, раке мочевого пузыря и т.д.), но является практически недетектируемым в нормальной ткани взрослого человека (за исключением эндометрия в пролиферативной фазе и некоторых сосудов в яичниках). (В отношении более подробного освещения действия ED-B в качестве мишени см. Menrad and Menssen, "ED-B fibronectin as a target for antibody-based cancer treatments". Expert Opin. Ther. Targets 9(3): 491-500, 2005.)
Готовили библиотеку из более чем 1 биллиона Fyn SH3-мутантов и представляли ее на фагах (одновременная рандомизация RT-Src- и n-Src-петель). После трех циклов пэннинга против ED-B 3 связывающих клона идентифицировали для фагов ELISA. Секвенирование выявило две различные последовательности (клоны, названные B11 и D3). Константу диссоциации D3 определяли анализом, производимом в реальном времени, данных резонанса поверхностного плазменного с использованием прибора BIAcore3000, и она оказалась равной величине 8.5×10-8 М (фиг.6).
D3 (SEQ ID NO:3)
GVTLFVALYDYHAQSGADLSFHKGEKFQILKFGRGKGDWWEARSLTTGETGYIPSNYVAPVDSIQ
Пример 4: Иммуногенность
Иммуногенность белков является одним из главных недостатков терапии, связанной с использованием белков, особенно для лечения, включающего в себя повторяющееся введение лекарственного средства. Вследствие сохранения Fyn SH3-последовательности в мышах и людях иммуногенный потенциал Fyn SH3-белка дикого типа (Fyn SH3wt) и Fyn SH3-мутанта (Fyn SH3D3, связывающий агент в отношении ED-B) исследовали in vivo повторяемым инъецированием 5 мышам двух белков. Мышам инъецировали 4 раза (каждый третий день) 20 мкг белка. Через день после 4-й инъекции мышей умерщвляли и отбирали пробы крови для исследования присутствия или отсутствия мышиных анти-Fyn SH3wt- и анти-Fyn SH3D3-антител. В качестве положительного контроля 4 мышам инъецировали (одинаковые промежутки времени между инъекциями и одинаковые дозы, равные 60 мкг) человеческое антитело в формате одноцепочечного Fv (scFv). Однако одна мышь scFv-группы умерла через 20 минут после третьей инъекции, а 3 другие были близки к смерти, поэтому пробы крови брали уже после третьей инъекции. Фиг.7A и B показывают, что не было детектируемых антител против Fyn SH3wt и Fyn SH3D3, в то время как наблюдали сильные сигналы для контрольной группы (фиг.7C).
Пример 5: Иммуногистофлуоресценция
Для выяснения вопроса, узнает ли Fyn SH3-D3 (D3, связывающий агент против ED-В) свою мишень в нативной конформации в ткани, выполняли иммунофлуоресценцию срезов F9-тератокарциномы. Фиг.8 показывает, что D3 связывался со стромой опухоли вокруг кровеносных сосудов (фиг.8.A). Детектирование проводили с использованием конъюгата антитела анти-His - Alexa488. В негативном контроле не добавляли D3-белок (фиг.8.B). Для визуализации кровеносных сосудов те же срезы одновременно окрашивали крысиным антителом против CD31 мыши, а в качестве вторичного антитела использовали конъюгат ослиного антитела против крысиного Ig - Alexa594 (фиг.8C). Негативный контроль проводили с использованием вторичного антитела без первичного антитела (фиг.8D).
Пример 6: Количественное биораспределение in vivo
Эффективность нацеливания in vivo Fyn SH3-D3 (связывающего агента против ED-В) и Fyn SH3 дикого типа (агента, не связывающего агента ED-B, оценивали в экспериментах по биораспределению в мышах, несущих трансплантированную s.c. тератокарциному F9. Поскольку ED-B идентичен в мышах и людях, результаты исследования нацеливания на опухоли должны быть прогнозирующими в отношении активности D3 в людях. 125I-меченый D3 и SH3wt инъецировали внутривенно и через 24 часа, животных умерщвляли, органы извлекали, взвешивали и определяли радиоактивность. Фиг.9A показывает, что D3 селективно накапливается в опухоли (соотношения опухоль:орган находятся в диапазоне от 3:1 до 10:1), в то время как никакого обогащения не наблюдали для белка Fyn SH3 дикого типа (фиг.9B).


| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ IL-17-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2550272C2 |
| НОВЫЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ С ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2013 |
|
RU2627185C1 |
| МОДИФИЦИРОВАННЫЕ УБИКВИТИНОВЫЕ БЕЛКИ СО СПЕЦИФИЧЕСКОЙ СВЯЗЫВАЮЩЕЙ АКТИВНОСТЬЮ ДЛЯ ЭКСТРАДОМЕНА "В" ФИБРОНЕКТИНА | 2010 |
|
RU2556820C2 |
| КОНЪЮГАТЫ АНТИТЕЛО-ФИНОМЕР | 2015 |
|
RU2732226C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ ГЕТЕРОМУЛЬТИМЕРНЫХ МОДИФИЦИРОВАННЫХ УБИКВИТИНОВЫХ БЕЛКОВ СО СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С ЛИГАНДАМИ | 2010 |
|
RU2553333C2 |
| TRAIL R2-СПЕЦИФИЧЕСКИЕ МУЛЬТИМЕРНЫЕ СКАФФОЛДЫ | 2011 |
|
RU2628699C2 |
| СПОСОБЫ И СОСТАВЫ ДЛЯ ПРОИЗВОДСТВА ЛИМФОЦИТОВ И ИХ РЕГУЛИРУЕМОГО УВЕЛИЧЕНИЯ | 2017 |
|
RU2755059C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2642310C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2727452C1 |
| АНГИОПОЭТИН-2-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ | 2005 |
|
RU2404992C2 |




Настоящее изобретение относится к области биотехнологии и может быть использовано в медицине, в частности в онкологии. Предложен способ создания библиотеки рекомбинантных белков, представленных мутантными по src-петле и RT-петле формами домена SH3 киназы Fyn, с помощью которого получена библиотека фагового дисплея, содержащая более 1 млрд мутантных производных природной последовательности SH3-домена данного фермента. В результате скрининга полученной библиотеки на наличие связывающих интересующий лиганд рекомбинантных белков получен белок, характеризующийся высокой афинностью связывания с доменом В фибронектина (ED-B). На основе нового белка сконструированы слитые белки, а также химические производные, которые, наряду со связывающим белком по изобретению, содержат фармацевтически и/или диагностически активные компоненты. Предлагаются различные варианты применения нового связывающего белка и его производных в качестве диагностических и лекарственных средств, в частности предназначенных для диагностики и лечения раковых заболеваний. 17 н. и 11 з.п.ф-лы, 1 табл., 19 ил., 6 пр.
1. Способ получения библиотеки рекомбинантных связывающих белков, состоящей из производных домена 3 (SH3) Src-гомологии киназы Fyn с мутациями в src и RT петлях, в которых
(a) по меньшей мере, одна аминокислота, находящаяся в src-петле, и
(b) по меньшей мере, одна аминокислота, находящаяся в RT-петле, заменена, делетирована или добавлена,
причем производное SH3 домена имеет, по меньшей мере, 80% идентичности относительно аминокислотной последовательности SEQ ID NO:1 и при условии, что этот рекомбинантный связывающий белок не является белком, содержащим природный SH3 домен;
включающий стадии:
(i) генерирование полинуклеотидов, кодирующих указанные рекомбинантные белки;
(ii) составление библиотек, экспрессирующих указанные рекомбинантные белки.
2. Способ по п.1, в котором генерирование полинуклеотидов на стадии (i) осуществляют одновременной рандомизацией, по меньшей мере, одной аминокислоты, находящейся в src-петле, и случайной заменой, делетированием или добавлением, по меньшей мере, одной аминокислоты, находящейся в RT-петле.
3. Способ по п.1, в котором библиотека на стадии (ii) представляет собой библиотеку фагового дисплея.
4. Способ по п.1, дополнительно включающий стадию (iii) подвергания указанных библиотек селекционному скринингу.
5. Библиотека рекомбинантных белков, состоящая из производных домена 3 (SH3) Src-гомологии киназы Fyn с мутациями в src и RT петлях, в которых
(a) по меньшей мере, одна аминокислота, находящаяся в src-петле, и
(b) по меньшей мере, одна аминокислота, находящаяся в RT-петле, заменена, делетирована или добавлена,
причем производное SH3 домена имеет, по меньшей мере, 80% идентичности относительно аминокислотной последовательности SEQ ID NO:1 и при условии, что этот рекомбинантный связывающий белок не является белком, содержащим природный SH3 домен.
6. Библиотека по п.5, содержащая более 1 миллиарда связывающих белков.
7. Библиотека рекомбинантных белков, состоящая из производных домена 3 (SH3) Src-гомологии киназы Fyn с мутациями в src и RT петлях, в которых
(a) по меньшей мере, одна аминокислота, находящаяся в src-петле, и
(b) по меньшей мере, одна аминокислота, находящаяся в RT-петле, заменена, делетирована или добавлена,
причем производное SH3 домена имеет, по меньшей мере, 80% идентичности относительно аминокислотной последовательности SEQ ID NO:1 и при условии, что этот рекомбинантный связывающий белок не является белком, содержащим природный SH3 домен, где
как минимум один рекомбинантный связывающий белок содержит аминокислотную последовательность SEQ ID NO:3.
8. Рекомбинантный белок, связывающий дополнительный домен В фибронектина (ED-B), имеющий аминокислотную последовательность SEQ ID NO:3.
9. Слитый белок, содержащий рекомбинантный белок, охарактеризованный в п.8, слитый с фармацевтически активным компонентом.
10. Слитый белок по п.9, где указанный компонент выбран из группы, состоящей из
(i) цитокинов, предпочтительно цитокинов, выбранных из группы, состоящей из IL-2, IL-12, TNF-альфа, IFN-альфа, IFN-бета, IFN-гамма, IL-10, IL-15, IL-24, GM-CSF, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, LIF, CD80, B70, TNF-бета, LT-бета, лиганда CD-40, Fas-лиганда, TGF-бета, IL-1-альфа и IL-1-бета;
(ii) токсичных соединений, выбранных из полипептидов, предпочтительно токсичных соединений, выбранных из группы, состоящей из калихеамицина, неокарциностатина, эсперамицина, динемицина, кедарцидина, мадуропептина, доксорубицина, даунорубицина, ауристатина, А-цепи рицина, модеккина, укороченного экзотоксина А Pseudomonas, дифтерийного токсина и рекомбинантного гелонина;
(iii) хемокинов, предпочтительно хемокинов, выбранных из группы, состоящей из IL-8, GRO-альфа, GRO-бета, GRO-гамма, ENA-78, LDGF-РВР, GCP-2, PF4, Mig, IP-10, SDF-1 альфа/бета, BUNZO/STRC33, I-TAC, BLC/BCA-1, MIP-1-альфа, MIP-1-бета, MDC, TECK, TARC, RANTES, HCC-1, HCC-4, DC-CK1, MIP-3-альфа, MIP-3-бета, MCP-1-5, эотаксина, эотаксина-2, I-309, MPIF-1, 6Ckine, CTACK, MEC, лимфотактина и фракталкина.
(iv) прокоагулянтных факторов, предпочтительно тканевых факторов;
(v) ферментов для активации пролекарств, предпочтительно ферментов, выбранных из группы, состоящей из карбоксипептидаз, глюкуронидаз и глюкозидаз;
(vi) функциональных Fc-доменов, предпочтительно функциональных Fc-доменов человека.
11. Слитый белок по п.9, дополнительно содержащий компонент, модулирующий время полужизни в сыворотке, предпочтительно компонент, выбранный из группы, состоящей из полиэтиленгликоля (ПЭГ), иммуноглобулина и альбуминсвязывающих пептидов.
12. Конструкт, содержащий рекомбинантный белок, охарактеризованный в п.8, слитый с фармацевтически активным компонентом, где фармацевтически активный компонент выбран из
(i) токсичных соединений, предпочтительно низкомолекулярных органических соединений;
(ii) флуоресцентных красителей, предпочтительно компонента, выбранного из Alexa Fluor или красителей Су;
(iii) фотосенсибилизаторов, предпочтительно
бис(триэтаноламин)Sn(IV)хлорин е6(SnСhе6);
(iv) радионуклидов, либо из группы гамма-излучающих изотопов, предпочтительно 99mТс, 123I, 111In, либо из группы излучателей позитронов, предпочтительно 18F, 64Cu, 68Ga, 86Y, 124I, либо из группы бета-излучателей, предпочтительно 131I, 90Y, 177Lu, 67Cu, либо из группы альфа-излучателей, предпочтительно 213Bi, 211At.
13. Слитый белок, содержащий рекомбинантный белок, охарактеризованный в п.8, слитый с диагностически активным компонентом.
14. Слитый белок по п.13, где указанный компонент выбран из группы, состоящей из
(i) цитокинов, предпочтительно, цитокинов, выбранных из группы, состоящей из IL-2, IL-12, TNF-альфа, IFN-альфа, IFN-бета, IFN-гамма, IL-10, IL-15, IL-24, GM-CSF, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, LIF, CD80, B70, TNF-бета, LT-бета, лиганда CD-40, Fas-лиганда, TGF-бета, IL-1-альфа и IL-1-бета;
(ii) токсичных соединений, выбранных из полипептидов, предпочтительно токсичных соединений, выбранных из группы, состоящей из калихеамицина, неокарциностатина, эсперамицина, динемицина, кедарцидина, мадуропептина, доксорубицина, даунорубицина, ауристатина, А-цепи рицина, модеккина, укороченного экзотоксина А Pseudomonas, дифтерийного токсина и рекомбинантного гелонина;
(iii) хемокинов, предпочтительно хемокинов, выбранных из группы, состоящей из IL-8, GRO-альфа, GRO-бета, GRO-гамма, ENA-78, LDGF-РВР, GCP-2, PF4, Mig, IP-10, SDF-1 альфа/бета, BUNZO/STRC33, I-TAC, BLC/BCA-1, MIP-1-альфа, MIP-1-бета, MDC, TECK, TARC, RANTES, HCC-1, HCC-4, DC-CK1, MIP-3-альфа, MIP-3-бета, MCP-1-5, эотаксина, эотаксина-2, I-309, MPIF-1, 6Ckine, CTACK, MEC, лимфотактина и фракталкина.
(iv) прокоагулянтных факторов, предпочтительно тканевых факторов;
(v) ферментов для активации пролекарств, предпочтительно ферментов, выбранных из группы, состоящей из карбоксипептидаз, глюкуронидаз и глюкозидаз;
(vi) функциональных Fc-доменов, предпочтительно функциональных Fc-доменов человека.
15. Слитый белок по п.13, дополнительно содержащий компонент, модулирующий время полужизни в сыворотке, предпочтительно компонент, выбранный из группы, состоящей из полиэтиленгликоля (ПЭГ), иммуноглобулина и альбуминсвязывающих пептидов.
16. Конструкт, содержащий рекомбинантный белок, охарактеризованный в п.8, слитый с диагностически активным компонентом, где диагностически активный компонент выбран из
(i) токсичных соединений, предпочтительно низкомолекулярных органических соединений;
(ii) флуоресцентных красителей, предпочтительно компонента, выбранного из Alexa Fluor или красителей Су;
(iii) фотосенсибилизаторов, предпочтительно бис(триэтаноламин)Sn(IV)хлорин е6(SnСhе6);
(iv) радионуклидов либо из группы гамма-излучающих изотопов, предпочтительно 99mТе, 123I, 111In, либо из группы излучателей позитронов, предпочтительно 18F, 64Cu, 68Ga, 86Y, 124I, либо из группы бета-излучателей, предпочтительно 131I, 90Y, 177Lu, 67Cu, либо из группы альфа-излучателей, предпочтительно 213Bi, 211At.
17. Полинуклеотид, кодирующий рекомбинантный белок, охарактеризованный в п.8.
18. Полинуклеотид, кодирующий слитый белок по любому из пп.9-11 или 13-15.
19. Экспрессионный вектор, содержащий полинуклеотид по п.17.
20. Экспрессионный вектор, содержащий полинуклеотид по п.18.
21. Клетка-хозяин для экспрессии, содержащая полинуклеотид по п.17 или вектор по п.19.
22. Клетка-хозяин для экспрессии, содержащая полинуклеотид по п.18 или вектор по п.20.
23. Применение рекомбинантного белка, охарактеризованного в п.8, для получения лекарственного средства или диагностического средства.
24. Применение по п.23, где лекарственное средство или диагностическое средство предназначено для лечения или диагностики рака.
25. Фармацевтическая композиция для лечения рака, содержащая терапевтически эффективное количество рекомбинантного белка, охарактеризованного в п.8, или слитого белка по любому из пп.9-11, или конструкта по п.12.
26. Фармацевтическая композиция по п.25, дополнительно содержащая фармацевтически приемлемый эксципиент.
27. Диагностическая композиция для диагностики рака, содержащая диагностически эффективное количество рекомбинантного белка, охарактеризованного в п.8, или слитого белка по любому из пп.13-15, или конструкта по п.16.
28. Диагностическая композиция по п.27, дополнительно содержащая фармацевтически приемлемый эксципиент.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| PANNI S | |||
| et al | |||
| J | |||
| Biol | |||
| Chem | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| ERPEL Т | |||
| et al | |||
| EMBO J | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| BORCHERT Т | |||
| et al | |||
| EMBO J | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| РЕЦЕПТОР ED-ДОМЕНА ФИБРОНЕКТИНА | 2001 |
|
RU2280254C2 |

