ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способу идентификации гетеромультимерного убиквитина со способностью связываться с лигандом. Кроме того, изобретение предлагает ДНК-библиотеки, ответственные за получение совокупности упомянутых гетеро-мультимерных убиквитиновых белков, а также библиотеки белков, полученных экспрессией упомянутых ДНК-библиотек, клетки и фаги, содержащие упомянутую ДНК или белки, полинуклеотиды, ответственные за получение упомянутых составных белков, и векторы, включающие упомянутые полинуклеотиды. Кроме того предложены новые связывающие белки на основе гетеромультимерных убиквитинов, способных специфически связываться с высоким сродством с выбираемыми лигандами.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Существует растущий спрос на связывающие молекулы, состоящие из аминокислот, которые не являются иммуноглобулинами. Хотя на настоящее время антитела представляют собой лучше всего изученный класс связывающих молекул, все же существует необходимость в новых связывающих молекулах, чтобы получить лиганды с высоким сродством и специфичностью, поскольку молекулы иммуноглобулинов имеют крупные недостатки. Хотя их можно легко получить и они могут быть направлены почти на любую мишень, они имеют очень сложную молекулярную структуру. Существует постоянная необходимость в замещении антител более мелкими молекулами, с которыми можно легко работать. Эти альтернативные связывающие агенты могут быть с выгодой использованы, например, в медицинских областях диагностики, профилактики и лечения заболеваний.
Белки, имеющие относительно определенные трехмерные структуры, обычно называемые белковые каркасы, могут быть использованы в качестве исходного материала для конструирования упомянутых альтернативных связывающих агентов. Эти каркасы обычно содержат один или больше областей, которые подвержены специфическому или случайному изменению последовательности, и такую рандомизацию последовательности часто выполняют для получения библиотеки белков, из которой могут быть выбраны специфические связывающие молекулы. Молекулы меньшего размера чем антитела и со сравнимым или даже более хорошим сродством к целевому антигену будут, как ожидается, лучше антител в смысле фармакокинетических свойств и иммуногенности.
В нескольких предыдущих подходах действительно используют белковые каркасы в качестве исходного материала для связывающих белков. Например, в документе WO 99/16873 описаны разработанные модифицированные белки семейства липокалинов (так называемые антикалины), проявляющие связывающую активность для некоторых лигандов. Структуру пептидов семейства липокалинов модифицируют путем замещений аминокислот в их естественном связывающем кармане, используя способы генной инженерии. Подобно иммуноглобулинам, эти антикалины можно использовать для идентификации или связывания молекулярных структур. Аналогично антителам, модифицируют структуры с гибкой петлей; эти модификации дают возможность распознавать лиганды, отличающиеся от естественных.
В документе WO 01/04144 описано искусственное создание связывающего домена на поверхности белка в белках со структурой бета-пласта как таковых, у которых отсутствует связывающий сайт.Посредством этого вновь созданного искусственного связывающего домена могут быть получены, например, изменения в γ-кристаллине - структурном белке хрусталика глаза - которые взаимодействуют с лигандами с высоким сродством и специфичностью. В противоположность модификации сайтов связывания, которые уже присутствуют и образованы из структур с гибкой петлей, которые упомянуты выше для антикалинов, эти домены связывания создаются заново на поверхности бета-пластов. Однако в документе WO 01/04144 описано только изменение относительно крупных белков для создания новых связывающих свойств. Из-за их размера, белки согласно WO 01/04144 могут быть модифицированы на уровне генной инженерии только способами, которые требуют некоторых усилий. Кроме того, в белках, раскрытых до сих пор, только относительно малая процентная доля всех аминокислот модифицирована для поддержания общей структуры белка. Поэтому только относительно небольшая области поверхности белка доступна для создания связывающих свойств, которые не существовали ранее. Более того, WO 01/04144 раскрывает только создание свойства связывания с γ-кристаллином.
В документе WO 04/106368 описано создание искусственных связывающих белков на основе убиквитиновых белков. Убиквитин - это небольшой, мономерный и цитозольный белок, который в высокой степени сохраняется в последовательности и присутствует во всех известных эукариотических клетках от простейших до позвоночных организмов. В организме он играет очень важную роль в регулировании контролируемой деградации клеточных белков. Для этого белки, предназначенные для деградации, ко-валентно связываются с убиквитиновыми или полиубиквитиновыми цепями во время, их прохождения через каскад ферментов и выборочно деградируют из-за этой метки. Согласно последним результатам, убиквитин или мечение белков убиквитином, соответственно, также играет важную роль в других клеточных процессах, таких как импорт нескольких белков или их генное регулирование.
Помимо прояснения его физиологической функции, убиквитин является объектом исследований главным образом из-за его структурных и белково-химических свойств. Полипептидная цепь убиквитина состоит из 7б аминокислот, уложенных в чрезвычайно компактную α/β структуру (Виджай-Кумар (Vijay-Kumar), 1987): почти 87% этой полипептидной цепи участвует в формировании вторичных структурных элементов посредством водородных связей. Вторичными структурами являются 3,5 альфа-спиральные витки, а также антипараллельный бета-пласт, состоящий из четырех нитей. Характерное расположение этих элементов - антипараллельный бета-пласт с открытой белковой поверхностью, на задней стороне которой упакована альфа-спираль, которая лежит вертикально на ее верху - обычно считается так называемым убиквитино-подобным мотивом укладки. Еще одной структурной особенностью является выраженная гидрофобная область внутри белка между альфа-спиралью и бета-пластом.
Из-за его небольшого размера искусственное приготовление убиквитина может быть осуществлено как химическим синтезом, так и посредством биотехнологических способов. Из-за благоприятных свойств укладки убиквитин можно получать генной инженерией, используя такие микроорганизмы, как Escherichia coli, в относительно больших количествах или в цитозоли, или в периплазматическом пространстве. Из-за окисляющих условий, преобладающих в периплазме, последняя стратегия обычно резервируется для получения секреторных белков. Из-за простого и эффективного бактериального приготовления убиквитин можно использовать как партнера в объединении для других получаемых инородных белков, получение которых вызывает трудности. Посредством объединения с убиквитином можно достигнуть повышенной растворимости и, посредством этого, повышенного выхода продукта.
По сравнению с антителами или другими альтернативными каркасами, искусственные связывающие белки на основе убиквитиновых белков (также называемых Affilin®) имеют много преимуществ: небольшой размер, высокая стабильность, высокое сродство, высокая специфичность, экономичное микробное производство и регулировка сывороточного периода полураспада. Однако, все же существует необходимость в дальнейшей разработке этих белков в смысле новых терапевтических подходов с высокими степенями сродства со специфическими мишенями. Хотя в документе WO 05/05730 описано в общих чертах использование каркасов убиквитина для получения искусственных связывающих белков, там не предложено решения, как модифицировать убиквитиновый белок, чтобы получить специфическое и высокоаффинное связывание с лигандами наподобие гаптенов и антигенов, например, белков и пептидов и их эпитопов. Способы, описанные в WO 05/05730, относятся к мономерам модифицированных убиквитиновых белков или к связанным белкам модифицированного убиквитина. Связанные формы создают путем скрининга и выбора одного, двух или больше модифицированных убиквитиновых белков с последующим их объединением генетическими или химическими способами, чтобы получить связанные формы, которые допускают, например, мультиспецифическое связывание лигандов разных типов молекулой одного связанного убиквитина. В одном примере описано сайт-направленное связывание двух идентичных белков на основе убиквитина (гомодимеров), чтобы повысить связывающее сродство по сравнению с молекулой одного модифицированного убиквитина.
Цель настоящего изобретения заключается в том, чтобы предложить способ идентификации мультимерных убиквитиновых белков с высокой способностью связываться с лигандом. Еще одна цель настоящего изобретения заключается в том, чтобы предложить способ идентификации новых связывающих белков на основе модифицированного убиквитина, способных специфически связываться с высоким сродством с выбираемыми лигандами.
Вышеуказанные цели решены предметом независимых пунктов прилагаемой формулы изобретения. Предпочтительные варианты осуществления изобретения включены в зависимые пункты формулы изобретения, а также в последующее описание, примеры и фигуры чертежей.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Более конкретно, авторы изобретения предлагают способ идентификации гетеромультимерного модифицированного убиквитина со способностью связываться с лигандом, включающий следующие этапы:
a) предоставление совокупности гетеромультимерных модифицированных убиквитиновых белков, происходящих от мономерных модифицированных убиквитиновых белков, причем упомянутая совокупность включает гетеромультимерные белки, содержащие два или больше по-разному модифицированных мономера убиквитина или по меньшей мере один мономер модифицированного убиквитина, связанные между собой по схеме голова к хвосту, причем по меньшей мере два каждых упомянутых мономера упомянутого гетеромультимерного белка по-разному модифицированы путем замещений поверхностно открытых аминокислот в по меньшей мере трех аминокислотах, расположенных в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1, причем упомянутый модифицированный мономерный белок имеет идентичность аминокислотной последовательности по меньшей мере 80%, или по меньшей мере 90%, или по меньшей мере 95% с немодифицированным убиквитиновым белком;
b) предоставление потенциального лиганда для упомянутой совокупности по-разному модифицированных белков;
c) контакт упомянутой совокупности по-разному модифицированных белков с упомянутым лигандом;
d) идентификация модифицированного гетеромультимерного белка посредством процесса скрининга, причем упомянутый модифицированный мультимерный белок связывается с упомянутым лигандом со специфическим связывающим сродством Kd в диапазоне 10-7-10-12 М и проявляется одновалентную связывающую активность в отношении упомянутого ли-ганда; и по выбору
е) изоляция упомянутого гетеромультимерного модифицированного убик-витинового белка с упомянутым связывающим сродством.
Определения важных терминов, используемых в настоящей заявке
Термин "убиквитиновый белок" охватывает убиквитин в соответствии с SEQ ID NO:1 и его модификации согласно следующему определению. Убиквитин в высокой степени сохраняется в эукариотических организмах. Например, у всех млекопитающих, исследованных до настоящего времени, убиквитин имеет идентичную аминокислотную последовательность. Особенно предпочтительными являются молекулы убиквитина от человека, грызунов, свиней и приматов. Помимо этого можно использовать убиквитин из любого другого эукаритического источника. Например, убиквитин дрожжей отличается только тремя аминокислотами от последовательности SEQ ID NO:1. В общем, убиквитиновые белки, охватываемые упомянутым термином "убиквитиновый белок" показывают аминокислотную идентичность более чем 70%, предпочтительно более чем 75% или более чем 80%, более чем 85%, более чем 90%, более чем 95%, более чем 96% или до идентичности последовательности 97% с SEQ ID NO:1.
Для определения степени идентичности последовательности производного убиквитина с аминокислотной последовательностью SEQ ID NO:1 можно использовать, например, программу SIM локального сходства (Сяокин Хуан и Уэбб Миллер (Xiaoquin Huang and Webb Miller), "Достижения прикладной математики. Том 12: 337-357, 1991) или Клустал У. (Clustal, W.) (Томпсон и др. (Thompson et al.). Nucleic Acids Res., 22(22): 4673-4680, 1994.). Предпочтительно, степень идентичности последовательности модифицированного белка с SEQ ID NO:1 определяют относительно полной последовательности SEQ ID N0:1.
В настоящем описании изобретения термины "лиганд" и "мишень" и "связывающийся партнер" используются как синонимы и могут заменять друг друга. Лигандом является любая молекула, способная связываться со сродством, которое здесь определено, с гетеромультимерным модифицированным убиквитиновым белком.
"Гетеромультимерный составной белок" или "гетеромультимерный белок" настоящего изобретения считается белком, который включает один или больше по-разному модифицированных мономерных убиквитиновых белков. "Гетеромультимером" настоящего изобретения поэтому считается слияние по меньшей мере двух по-разному модифицированных мономерных убиквитиновых белков с двумя взаимодействующими между собой доменными областями, вместе создавая свойство одновалентного связывания для специфического связывающегося партнера. Предпочтительны гетеродимеры или гетеротримеры.
Согласно изобретению, два по-разному модифицированных мономера убиквитина, которые связываются с одним лигандом, должны быть связаны путем соединения "голова-к-хвосту" друг с другом с использованием, например, генетических способов. Эти по-разному модифицированные объединенные мономеры убиквитина связываются одновалентно и эффективны только если обе "области связывающего домена" ("ОСД") действуют совместно. Эти модифицированные и связанные убиквитиновые мономеры, которые образуют гетеромерный белок, связываются с одним и тем же эпитопом посредством одной непрерывной области связывания. Эта непрерывная область гетеро-мера формируется обеими определяющими связывание областями по меньшей мере двух модулей, образованных по меньшей мере двумя по-разному модифицированными убиквитиновыми мономерами.
"Соединение голова-к-хвосту" должно пониматься как слияние двух белков путем соединения их в направлении N-C-N-C-, в зависимости от числа звеньев, содержащихся в мультимере. При этом соединении голова-к-хвосту мономеры убиквитина могут быть прямо соединены без любого линкера. Альтернативно, соединение мономеров убиквитина может быть выполнено посредством линкеров, например, линкера, имеющего по меньшей мере аминокислотную последовательность GIG или имеющего по меньшей мере аминокислотную последовательность SGGGG или любого другого линкера, например GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG или SGGGGSGGGG. Известны из уровня техники и могут быть использованы и другие линкеры для генетического соединения двух мономеров убиквитина. В общем, гетеромультимерные немодифицированные убиквитиновые белки получают путем слияния двух, трех или больше по-разному модифицированных мономерных убиквитиновых белков, чтобы получить составной белок из модифицированных мономеров убиквитина. В еще одном варианте осуществления по меньшей мере один из мономеров убиквитина не модифицирован, тогда как по меньшей мере одна из других молекул убиквитина модифицирована.
Термин ″популяция″ относится к библиотеке, которая является смесью гетерогенных полипептидов, кодированных гетерогенными нуклеиновыми кислотами. Библиотека состоит из элементов, которые имеют один полипептид, кодированный нуклеинокислотной последовательностью. Различия в последовательностях между элементами библиотеки отвечают за разнообразие, присутствующее в библиотеке. Библиотека может иметь форму простой смеси полипептидов или нуклеиновых кислот или форму организмов или клеток, например, бактерий, вирусов, животных или растительных клеток и т.п., трансформированных библиотекой нуклеиновых кислот. Предпочтительно, каждый отдельный организм или клетка содержит только один элемент библиотеки. Предпочтительно, нуклеиновые кислоты введены в экспрессирующие векторы, чтобы позволить производство полипептидов, кодированных нуклеиновыми кислотами. Поэтому в одном предпочтительном аспекте библиотека может иметь форму популяции организмов-хозяев, причем каждый организм содержит одну или больше копий экспрессирующего вектора, содержащего один элемент библиотеки в форме нуклеиновой кислоты, которая может быть экспрессирована для производства ее соответствующего полипептидного элемента. Таким образом, популяция организмов-хозяев имеет потенциал для кодирования большого спектра генетически разных вариантов полипептидов.
Такую популяцию (совокупность) гетеромультимерных убиквитиновых белков получают, например, путем генетического слияния ДНК-библиотек, каждая из которых отвечает за по-разному модифицированные мономерные белки, или в альтернативном варианте, в котором модифицирован по меньшей мере один из упомянутых мономерных убиквитиновых белков, за трансляцию ДНК в гетеромультимерные составные белки, отображая такие белки и проверяя отображенные белки на присутствие модифицированных гетеромультимерных убиквитиновых белков, содержащих мономерные убиквитиновые белки, связанные между собой по схеме голова к хвосту, причем упомянутые модифицированные гетеромультимерные убиквитиновые белки связываются с упомянутым лигандом со специфическим связывающим сродством в кД в диапазоне 10-7-10-12 М и проявляют одновалентную связывающую активность в отношении упомянутого лиганда. Для того, чтобы получить гетеродимерный убиквитиновый белок, соединяют две ДНК-библиотеки, отвечающие или за получение каждого, или по меньшей мере одного по-разному модифицированного мономерного белка, чтобы получить гетеротримерный убиквитиновый белок, соединяют три ДНК-библиотеки, отвечающие за получение каждого или по меньшей мере одного по-разному модифицированного мономерного белка и т.д. Для предоставления библиотек для скрининга могут быть использованы и другие альтернативы. Одним примером является химический синтез белков, например, по твердотельной технологии, и введение вариаций в их аминокислотный состав. Специалист может рассмотреть и счесть полезными и другие варианты. Поэтому изобретение не должно пониматься как ограниченное примерами, описанными в настоящем документе.
Изобретение, соответственно, раскрывает способ, посредством которого получают спектр полипептидов согласно функциональности, которая определяется способностью связывать лиганд, и подгруппу полипептидов, полученных в результате выбора, затем применяют для дальнейших циклов выбора согласно способности связывать целевой лиганд, чтобы аккумулировать и повышать способность связывания с лигандом.
Изобретение позволяет специалисту в данной области удалять из выбранного спектра полипептидов те, которые неспособны связываться с целевым лигандом со сродством, определенным в формуле изобретения. Изобретение позволяет специалисту в данной области расширять выбираемый спектр полипептидов теми, которые функциональны и отвечают требованиям к сродству.
Один из наиболее важных аспектов настоящего изобретения заключается в выборе модифицированных гетеромультимерных убиквитиновых белков с одновалентным связывающим сродством по отношению к мишени и в последующем определении модифицированных аминокислот, отвечающих за связывающее сродство.
Еще одно преимущество мультимеризации, предпочтительно димеризации, заключается в увеличении числа аминокислотных остатков, которые могут быть модифицированы для создания нового высокоаффинного связывающего свойства. Преимущество заключается в том, что хотя модифицируют даже больше аминокислот, белково-химическая целостность поддерживается без уменьшения общей стабильности каркаса упомянутого заново созданного связывающего белка с целью. С одной стороны, совокупное число остатков, которые могут быть модифицированы для создания нового сайта связывания для данной мишени, увеличивается, так как модифицированные остатки могут быть распределены по двум, или трем, или больше мономерным убиквитиновым белкам. Количество модификаций таким образом может быть в два или х-раз больше числа модифицируемых молекул мономерного убиквитина. В результате, модульная структура связывающего белка на основе убиквитина позволяет увеличить общее число модифицированных аминокислот, так как упомянутые модифицированные аминокислоты включены в две, или три, или больше молекул мономерного убиквитина. Настоящий способ предусматривает идентификацию молекул гетеромультимерного убиквитина, имеющих одну одновалентную специфичность (для одного отдельного эпитопа).
"Одновалентный" должен пониматься как возможность того, что обе связывающих области, созданные в первом и втором (и, по выбору, последующем) мономерном звене модифицированного димерного (по выбору гримерного или, в общем, мультимерного) убиквитина, вместе связывают ED-B синергистически и объединенно, т.е., обе связывающие области действуют совместно, чтобы образовать одновалентную связывающую активность. Если взять отдельно каждую связывающую область первого и второго модифицированного убиквитина в упомянутой гетеродимерной молекуле, то такая область очевидно будет связывать ED-B с намного меньшей эффективностью и сродством чем димерная молекула. Обе связывающие области образуют уникальный сайт связывания, который сформирован как непрерывная область аминокислот на поверхности. гетеродимерного модифицированного убиквитинового белка, так что упомянутый модифицированный убиквитин способен связываться гораздо более эффективно с ED-B чем каждый мономерный белок, взятый отдельно. Особенно важно, что, согласно настоящему изобретению два мономерных белка не связаны между собой после скрининга наиболее сильных связывающих молекул убиквитина, но уже процесс скрининга выполняется в присутствии гетеродимерных убиквитинов. После получения информации о последовательности по наиболее сильно связывающим молекулам убиквитина эти молекулы могут быть получены любым другим способом, например, химическим синтезом или способами генной инженерии, например, путем связывания двух уже идентифицированных мономерных убиквитиновых звеньев вместе. При этом понимается, что все представленные здесь примеры для димерных модифицированных убиквитиновых белков могут быть также изменены для гримерного или, в общем, мультимерного модифицированного убиквитинового белка.
Таким образом, использование гетеромультимеров, в частности гетеродимеров, имеющих общий сайт связывания для связывающихся партнеров, открывает возможность введения повышенного числа модифицированных остатков, которые не влияют отрицательно на белково-химическую целостность конечной связывающей молекулы, поскольку общее число этих модифицированных остатков распределено по двум или больше мономерным звеньям, которые образуют ди- или мультимер. Упомянутые гетеромультимерные модифицированные убиквитиновые белки присутствуют в библиотеке белков.
После того, как, например, по меньшей мере две разных ДНК-библиотеки, ответственные за получение мономерных модифицированных убиквитиновых белков, были созданы путем разной модификации выбранных кодонов в каждом из мономерных убиквитиновых звеньев, эти библиотеки генетически объединяют для получения молекул ДНК, ответственных за получение гетеромультимерных модифицированных убиквитиновых белков. ДНК этих библиотек транслируют в белки, и полученные этим путем модифицированные гетеромультимерные белки вводят в контакт, согласно изобретению, с целевой молекулой, чтобы обеспечить связывание партнеров друг с другом, если связывающее сродство существует.Предпочтительным модифицированным убиквитином является гетеродимер.
Крайне важным аспектом настоящего изобретения является то, что процесс контакта и скрининга выполняют уже в отношении гетеромультимерного, например, гетеродимерного, модифицированного убиквитинового белка. Этот процесс позволяет выполнять скрининг на тех убиквитиновых белках, которые обеспечивают одновалентную связывающую активность с мишенью.
Особенно важно, что, согласно настоящему изобретению, мономерные модифицированные убиквитиновые белки не связывают друг с другом после выбора путем скрининга наиболее сильных связывающих молекул убиквитина, но что уже процесс скрининга выполняют в присутствии гетеромультимерных убиквитинов. Однако, следует отметить, что после получения информации о последовательности по наиболее сильным связывающим молекулам гетеромультимерного убиквитина эти молекулы могут быть получены любым другим способом, например, химическим синтезом или способами генной инженерии, например, путем связывания двух уже идентифицированных мономерных убиквитиновых звеньев вместе, чтобы сформировать гетеродимерный связывающий белок.
Согласно изобретению контакт предпочтительно выполняют посредством подходящего способа представления и выбора, такого как фаговый дисплей, рибосомный дисплей, ТАТ-фаговый дисплей, РНК дисплей или способы дисплея поверхностей клеток, дисплея поверхностей дрожжей или дисплея поверхностей бактерий, предпочтительно посредством способа фагового дисплея. Для полноты раскрытия ссылка делается также на следующие публикации: Hoess, Curr. Opin. Struct. Biol. 3 (1993), 572-579; Wells and Lowmann, Curr. Opin. Struct. Biol. 2 (1992), 597-604; Kay et al., Phage Display ofPeptides and Proteins - A Laboratory Manual (1996), Academic Press. Вышеуказанные способы известны специалистам в данной области и могут быть использованы согласно изобретению, включая их модификации.
Определение того, имеет ли модифицированный белок вычисляемое количественно связывающее сродство по отношению к предварительно определенному связывающемуся партнеру может быть выполнено, согласно изобретению, предпочтительно одним или несколькими из следующих способов: ИФА, спектроскопия методом плазменного поверхностного резонанса, флуоресцентная спектроскопия, возбужденной флуоресценцией сортированных клеток (FACS), изотермической титриметрической калориметрией и аналитическим ультрацентрифугированием. Специалист с общими знаниями может использовать и другие способы, имеющиеся в этой области.
В настоящем документе под "гетеромультимером" понимается белок, который включает по меньшей мере два разных мономерных убиквитиновых белка. "Гетеродимерами" настоящего изобретения считается слияние двух по-разному модифицированных мономерных убиквитиновых белков. Оба проявляют объединенное одновалентное связывающее свойство для специфического связывающегося партнера. Следует подчеркнуть, что модифицированный мультимерный, например, димерный, связывающий лиганд убиквитиновый белок настоящего изобретения не получают путем раздельного скрининга каждого мономерного убиквитинового белка и объединения по меньшей мере двух из них впоследствии, а путем скрининга для определения мультимерных, по выбору димерных, белков, состоящих из первого и второго или последующего мономерного звена, которые вместе проявляют одновалентную связывающую активность с упомянутым лигандом. Следует ожидать, что каждое из упомянутых субзвеньев, проявляет вполне ограниченное связывающее сродство к лиганду, тогда как только объединенный мультимерный или димерный модифицированный убиквитиновый белок будет иметь превосходные свойства связывания, описанные в настоящем документе.
В одном варианте осуществления способ относится к идентификации модифицированного гетеродимерного убиквитинового белка, в котором два мономерных убиквитиновых звена связаны между собой по схеме голова к хвосту, причем каждый мономер упомянутого димерного белка по-разному модифицирован путем замещения по меньшей мере 3, предпочтительно 6, аминокислот в положениях 2,4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1 (каждая из них поверхностно открытая)
причем упомянутые замещения включают:
(1) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 63, 64, 65 и 66;и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно в аминокислотном положении 2; или
(2) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 2, 4, 6, 62, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно в аминокислотном положении 2; и, по выбору, другие модификации, предпочтительно замещения других аминокислот, причем упомянутое звено модифицированного мономерного убиквитин имеет аминокислотную идентичность с SEQ ID NO:1 по меньшей мере одну из группы из 80%, по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 83% и по меньшей мере 90%, причем упомянутый белок имеет специфическое связывающее сродство с лигандом Kd=10-7-10-12 М и проявляет одновалентную связывающую активность в отношении упомянутого лиганда.
В других вариантах осуществления настоящего изобретения, 6, 7, 8, 9 или все аминокислоты в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1 модифицированы в каждом мономерном убиквитином звене. При этом понимается, что настоящее изобретение допускает объединение каждой из этих вариаций в каждом из мономерных звеньев, например, в первом и втором звене. Например, первое мономерное звено может включать 6 модификаций, тогда как второе звено включает 7 или 8 модификаций, первое звено может включать 8 модификаций и второе звено 7 модификаций и т.д. Каждая из вышеперечисленных аминокислот может быть выбрана в первом и втором звене, и оба звена затем объединяют. Предпочтительные замещения описаны ниже.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ Способы дисплея модифицированных гетеромультимерных убиквитиновых белков
Процедуры фагового и рибосомного дисплея, адаптированные к настоящей заявке, описаны ниже и в разделе "Примеры". Они упомянуты как примеры процедуры выбора согласно изобретению для детектирования вариаций убиквитина, которые показывают свойства связывания с потенциальным лигандом, описываемые в настоящем документе. Таким же образом могут быть применены, например, способы для представления на бактериях (бактериальный поверхностный дисплей; Daugherty et al., 1998, Protein Eng. 11(9):825-832) или дрожжевых клетках (yeast surface display; Kieke et al., 1997 Protein Eng. 10(11): 1303-10) или бесклеточные системы выбора, такие как рибосомный дисплей (Hanes and Pluckthun, 1997 Proc Natl Acad Sci USA. 94(10):4937-4942; He and Taussig, 1997_Nucleic 5 Acids Res. 25(24):5132-5134) или cis дисплей (Odegrip et al, 2004 Proc Natl Acad Sci USA. 101(9):2806-2810) или мРНК дисплей. В последнем случае временная физическая связь генотипа и фенотипа достигается путем соединения вариации белка с подходящей мРНК через рибосомы.
В описанной здесь процедуре фагового дисплея рекомбинантные вариации убиквитина представлены на нитчатом фаге, тогда как кодирующая ДНК представленной вариации представлена в то же время упакованной в однонитевую форму в фаговой оболочке. Таким образом, в рамках аффинного обогащения вариации, имеющие определенные свойства, могут быть выбраны из библиотеки, и их генетическая информация может быть амплифицирована путем инфекции подходящих бактерий или добавлена к другому циклу обогащения, соответственно. Представление мутировавшего убиквитина на поверхности фага достигается путем генетического объединения с сигнальной последовательностью с амино-окончанием, предпочтительно с Ре1 В сигнальной последовательностью, и капсид или поверхностный белок предпочтительного фага является карбокситерминальным объединением с капсидным белком рШ или его фрагментом. Кроме того, полученный составной белок может содержать другие функциональные элементы, такие как метка сродства или эпитоп антитела для детектирования и/или очистки аффинной хроматографией или последовательность распознавания протеазы для специфического расщепления составного белка в ходе аффинного обогащения. Кроме того, терминатор UAG может быть представлен, например, между геном для вариации убиквитина и кодирующей областью капсидного белка фага или его фрагмента, который не распознан во время трансляции в подходящий супрессорный штамм, частично из-за введения одной аминокислоты.
Бактериальный вектор, подходящий для процедуры выбора в контексте изоляции вариаций убиквитина со свойствами связывания с задаваемой мишенью и в который введена генная кассета для описываемого составного белка, называется фагемидом. Среди прочих, он содержит межгенную область нитчатого фага (например, М13 или fl) или ее часть, которая в случае суперинфекции бактериальной клетки, несущей фагемид посредством фагов-хелперов, таких как, например, М13К07, приводит к упаковке кова-лентно закрытой нити ДНК фагемида в капсид фага. Созданные таким образом фаговые частицы секретируются бактерией и представляют соответственную вариацию убиквитина, кодированную из-за ее объединения с капсидным белком pIII или его фрагментом, на их поверхности. Нативные капсидные белки pIII присутствуют в фаговой частице, так что ее способность реинфицировать подходящие бактериальные штаммы и, поэтому, возможность амплифицировать соответствующую ДНК сохраняется. Таким образом, обеспечивается физическая связь между фенотипом вариации убиквитина, т.е., ее потенциальное связывающее свойство, и ее генотипом.
Полученные фаговые частицы могут быть выбраны по отношению к связыванию представленной на них вариации убиквитина с любой мишенью, например, ED-B, TNFalpha, MIA-2, NGF, IgG и другими посредством способов, известных специалистам в данной области. Для этой цели представленные вариации убиквитина могут быть временно иммобилизованы на субстанции мишени, связанной, например, на микротитровальных планшетах, и могут быть специфически элюированы после отделения не связывающихся вариаций. Элюирование предпочтительно выполняют щелочными растворами, такими как, например, 100 мМ триэтиламин. Альтернативно, элюирование может быть выполнено в кислых условиях, путем протеолиза или прямого добавления инфицированных бактерий. Фаговые частицы, полученные таким образом, могут быть реамплифицированы и обогащены последовательными циклами выбора и амплификации вариаций убиквитина со свойствами связывания, например, с ED-B, TNFalpha, MIA-2, NGF, IgG или любыми другими мишенями.
Одним вариантом фагового дисплея является способ Tat-фагового дисплея (Пашке, М. и В.Хоне (Paschke, М. and W. Hohne (2005)). Gene 350(1): 79-88; см. также документ ЕР 1567643). В этом способе вариант убиквитина, который кодирован фагемидом, секре-тируется посредством системы двойной транслокации аргинина (Tat), которая экспортирует сложенные белки, которые уже достигли своей нативной конформации уже. в цитоплазме (Брюзер (Bruser) 2007 Appl Microbiol Biotechnol 76(1): 35-45). Требованием для секреции является слияние со специфическим N-терминальным сигнальным пепти-дом, который направляет этот вариант убиквитина к поре Tat. После входа в периплаз-матическое пространство N-терминальный сигнальный пептид удаляют сигнальной пептидазой. В периплазматическом пространстве вариант убиквитина затем ковалентно соединяется с капсидным белком pIII или его С-терминальным фрагментом, который получает секретированные из цитоплазмы по пути Sec, а также другие белки фага. Эта связь между убиквитином и pIII реализуется путем высокоаффинного взаимодействия зиппера лейкина Jun на N-окончании белка pIII и зиппера лейкина Fos на С-окончании варианта убиквитина. Дополнительные цистеины на окончаниях N и С каждого из зипперов лейкина позволяют осуществить ковалентную связь между обоими белками и, как следствие, они также позволяют осуществить ковалентную связь между дисплейным убиквитином и продуктом его кодирующего гена в фаговой частице.
Дальнейшая характеризация вариаций убиквитина, полученных таким образом, может быть выполнена еще в форме фагемида, т.е., объединенных с фагом, или после клонирования соответствующей генной кассеты в подходящий экспрессирующий вектор в форме растворимого белка. Подходящие способы известны специалистам в данной области или описаны в литературе. Характеризация может включать, например, определение последовательности ДНК и, таким образом, первичной последовательности изолированных вариаций. Кроме того, сродство и специфичность изолированных вариаций можно детектировать стандартными биохимическими способами, такими как ИФА или спектроскопия плазмонного поверхностного резонанса, вытеснительная хроматография, флуоресцентная анизотропия, FACS, изотермическая титриметрическая калориметрия, аналитическое ультрацентрифугирование. В свете анализа стабильности, например, спектроскопические способы в связи с химическим или физическим развертыванием известны специалистам в данной области. Другими хорошо известными способами являются КД спектроскопия, белковая флуоресцентная спектроскопия и ЯМР спектроскопия.
В еще одном варианте осуществления изобретения вариации убиквитина для процедуры рибосомного дисплея готовят посредством бесклеточной системы транскрипции/трансляции и представляют как комплекс с соответствующей мРНК, а также рибосомой. Для этой цели библиотеку ДНК, которая описана выше, используют в качестве основы, в которой гены вариаций присутствуют в форме объединений с соответствующими регуляторными последовательностями для экспрессии и биосинтеза белка. Из-за делеции терминирующего кодона в окончании 3′ генной библиотеки, а также подходящих экспериментальных условий (низкая температура, высокая концентрация Mg2+) трехчастный комплекс, состоящий из возникающего белка, мРНК и рибосомы, сохраняется во время транскрипции/трансляции in vitro.
После создания библиотеки белков, содержащей гетеродимерные модифицированные убиквитиновые белки, путем разных модификаций выбранных аминокислот в каждом из мономерных звеньев убиквитина модифицированные димерные белки вводят в контакт, согласно изобретению, с мишенью, чтобы позволить партнерам связываться друг с другом, если существует связывающее сродство. Эти библиотеки белков могут быть в форме отображения библиотеки дисплейным способом, или можно использовать любой другой способ, представляющий модифицированные белки таким образом, чтобы обеспечить контакт между модифицированными белками и белком-мишенью, причем упомянутый дисплейный способ является, по выбору, фаговым дисплеем, рибосомным дисплеем, фаговым дисплеем ТАТ, дрожжевым дисплеем, бактериальным дисплеем или дисплеем мРНК.
Потенциальные лиганды и мишени модифицированных гетеромультимерных убиквитиновых белков
Настоящее изобретение было успешно проверено на следующих представительных антигенах: ED-B, TNF-alpha, MIA-2, NGF и IgG. При этом понимается, что эти антигены были выбраны только для того, чтобы показать, что описываемые здесь способы могут быть успешно осуществлены специалистом в данной области без ненужных трудностей после получения представленной здесь информации. Изобретение не ограничено этими специфическими антигенами, и может быть выполнено на всех или по меньшей мере большинстве лигандов и молекулах-мишенях, известных в данной области. Эти мишени могут быть выбраны специалистом в пределах его общих знаний в этой области. Ниже представлены общие определения лигандов и мишеней, а также антигенов и гаптенов, а также приведены некоторые примеры других потенциальных молекул-мишеней.
Согласно изобретению, термин "антиген" относится к веществу, способному быть связанным описываемым здесь модифицированным убиквитином, функция которого сравнима с антителом. Используемыми здесь альтернативными терминами являются "лиганды", "связывающийся партнер" или "мишень". Модифицированные убиквитиновые белки настоящего изобретения дают связывающие молекулы, которые действуют также как антитело, в то же время не имея его недостатков. Термин "антиген" включает гаптены, пептиды, белки, сахара, ДНК и т.д. Из публикации Roche Lexikon Medizin (4th edition; Urban & Fischer/Elsevier GmbH) могут быть взяты следующие определения антигена и гаптена, которые также используются в данном описании.
Антиген (АГ): Обозначение любого вещества, распознанного как инородное ("не собственное") иммунной системой. Инициирует в большинстве случаев иммунную реакцию, ведущую к иммунитету (="иммуноген"), в случае аллергии (="аллерген") и атопии ("атопиген"), соответственно, эта иммунная реакция чрезмерно повышена. АГ индуцирует гуморальную (реакция антиген-антитело) и/или клеточную защитную реакцию (см. "иммунитет" ниже). Если АГ переносится иммунной системой (иммунная толерантность), он также называется "толероген". Эффективны как антиген в основном сложные и высокомолекулярные вещества (белковые тела, полисахариды, нуклеотиды и многие синтетические соединения), имеющие химически идентифицируемые функции (детерминанту), ответственные за иммунную реакцию. Классифицируется как 1) полный АГ, по большей части с высокой молекулярной массой и способный самостоятельно вызывать иммунную реакцию, 2) как гаптен низкой молекулярной массы (=полу-антиген), который действует как иммуноген только после того как он соединен с большей молекулой-носителем. Называется, например, как ксено-, алло- или изогенный, аутологический АГ; ауто-, гетеро-, трансплантационный, противоопухолевый вирусный АГ.
Гаптен: простое низкомолекулярное химическое соединение, ответственное за специфичность антигена (АГ) или способное специфически связываться с антителом из-за своей структуры (детерминанты), соответственно, но неспособное создавать аллергию: в противоположность полному АГ. Он становится полным антигеном после связывания с белковьм телом, называемым "носитель".
"Лиганд" или "мишень" или "связывающийся партнер" является молекулой, которая распознается описываемыми здесь модифицированными гетеромультимерными убик-витиновьми белками. Примеры лигандов, которые могут быть применены при практическом осуществлении настоящего изобретения, включают, но без ограничения, агонисты и антагонисты для рецепторов клеточных мембран, токсины и яды, вирусные эпи-топы, гормоны, рецепторы гормонов, полипептиды, пептиды, ферменты, субстраты ферментов, кофакторы, лекарства (например, опиаты, стероиды и т.д.), пектины, сахара, полинуклеотиды, нуклеиновые кислоты, олигосахариды, белки и моноклональные антитела.
Суммируя, в качестве связывающегося партнера для модифицированных белков, предложенных согласно изобретению, могут быть использованы все биологически и медицински активные и релевантные молекулы. Возможные связывающиеся партнеры будут описаны ниже как пример. Следует сказать, однако, что к этому списку может быть добавлено некоторое множество других возможных лигандов. Подобно отношению между антителом и антигеном, перечень потенциальных связывающихся партнеров может быть дополнен другими потенциальными лигандами.
В настоящем изобретении примерами связывающихся партнеров для гетеродимерных убиквитинов являются экстрадомен В фибронектина (ED-B), цитокин (фактор некроза опухоли альфа) (TNF-α), MIA-2, иммуноглобулин или его часть, например, целое антитело, (например, иммуноглобулинО), и фактор роста (например, NGF, например, фактор роста нервов человека). Ниже приведены краткие описания этих лигандов. Однако следует подчеркнуть, что все эти лиганды хорошо известны в данной области в течение многих лет и известны специалистам в соответствующих областях техники. Поэтому приведенные ниже описания только вкратце излагают некоторые важные параметры этих белков, аминокислотные последовательности которых также известны.
Экстрадомен В (ED-B) фибронектина является небольшим доменом, который вводят альтернативным сплайсингом первичного транскрипта РНК в молекулу фибронектина. Известно, что ED-B вовлечен главным образом в рак и псориаз. Что удивительно, высокие уровни экспрессии ED-B были детектированы в областях первичного поражения, а также в местах метастаз многих твердых раковых объектов у человека, включая грудной, немелкоклеточный легочный, колоректальный, панкреатический, кожный, гепато-целлюлярный, внутричерепную менингеому, глиобластому (Менрад и Менссен (Menrad u. Menssen), 2005). Кроме того, ED-B может быть связан с диагностическими агентами и может выгодно использоваться в качестве диагностического средства. Одним примером является его использование в молекулярной визуализации, например, атеросклеротических бляшек и детектировании рака, например, путем иммуносцинтиграфии раковых пациентов. Также возможны многие другие способы использования в диагностике.
Аминокислотная последовательность из 91 аминокислоты человеческого экстрадомена. В (ED-B) фибронектина показана в SEQ ID NO:2. Для экспрессии белка приходится добавлять стартовый метионин. ED-B обильно присутствует у млекопитающих, например, у грызунов, крупного рогатого скота, приматов, плотоядных, людей и т.д. Примерами животных, у которых существует 100% идентичность последовательности с ED-B человека, являются Rattus norvegicus, Bos taurus, Mus musculus, Equus caballus, Macaca mulatto, Canis lupus familiaris и Pan troglodytes.
Белок MIA ("активность, ингибирующая меланому", также называется CD-RAP "полу-. чаемый из хряща, чувствительный к ретиноевой кислоте белок") экспрессируется в хондроцитах и был первоначально изолирован из-за его антипролиферативных свойств in vitro. Изначально он был детектирован в супернатанте клеточной культуры клеток меланомы и оттуда изолирован. После очистки и частичного секвенсирования этого белка, фрагмент кДНК MIA человека был изолирован с помощью вырожденных праймеров и RT-PCR (полимеразной цепной реакции обратной транскриптазы). Сейчас известны последовательности для MIA человека, мыши, быка, крысы и полосатой перцины. Родственный белок, MIA-2, описан в документах ЕР 1410803 В1 и US-2010/0212037. Эти документы включены в настоящий документ путем ссылки.
Фактор некроза опухоли альфа (TNF-alpha), плейотропный цитокин, продуцируется в основном макрофагами, но его продуцируют и другие типы клеток. TNF-alpha демонстрирует полезную, а также патологическую активность. Он имеет как эффекты, стимулирующие рост, так и свойства, ингибирующие рост, помимо этого являясь саморегулируемым. Полезные функции TNF-alpha включают поддержание гомеостаза путем регулирования циркадного ритма организма, создание иммунной реакции на бактериальные, вирусные, грибковые и паразитарные инфекции, замену или реконструкцию поврежденной ткани путем стимуляции роста фибробласта и, как подразумевает само название, ликвидацию некоторых опухолей. TNF-alpha вовлечен как посредник в большое число заболеваний.
Фактор роста нервов (NGF) является секретируемым белком, который был открыт свыше 50 назад как молекула, которая способствует выживанию и дифференциации чувствительных и симпатических нейронов. NGF является членом семейства нейро-трофических факторов, известных как нейротрофины. NGF связывается с высоким сродством с киназой рецептора тропомиозина, известной как TrkA. NGF также способен связывать рецептор, известный как р75, член суперсемейства рецепторов фактора некроза опухолей, который также взаимодействует с другими нейротрофинами. Бета-цепь NGF отвечает за активность NGF, стимулирующую рост нервов. Эта бета-цепь гомодимеризуется и входит в более крупный белковый комплекс. Структура и функция NGF рассмотрена, например, в следующих публикациях: Софроньев М.В. и др. (Sofroniew, M.V. et al.) (2001) Annu. Rev. Newosci. 24:1217-1281; Вайсман К и Девос (Weismann, С.and de Vos), A.M. (2001) Cell. Mol. Life Sci. 58:748-759; Фанесток М. (Fahnestock, М.) (1991) Curr. Top.Microbiol. Immunol. 165:1-26.
Антитела IgG являются крупными молекулами приблизительно 150 kDa, состоящими из 4 пептидных цепей. Они содержат 2 идентичных тяжелых цепи приблизительно по 50 kDa и 2 идентичные легкие цепи приблизительно по 25 kDa, имея, таким образом, тетрамерную четвертичную структуру. Две тяжелых цепи связаны друг с другом и с легкой цепью дисульфидными связями. Результирующий тетрамер имеет две идентичные половины, которые совместно образуют Y-образную форму. Каждый конец этой вилки содержит идентичный сайт связывания антигена. Области Fc в IgG имеют высоко консервативный сайт N-гликозилирования. N-гликаны, прикрепленные к этому сай-ту, являются главным образом корфукозилированными диантеннальньши структурами сложного типа. Помимо этого, небольшие количества этих N-гликанов также несут биссекторные GlcNAc и α-2,6 связанные остатки сиаловой кислоты.
Способы выбора, обогащения и характеризации дисплейных белков
Выбор гетеромультимерных модифицированных убиквитинов в отношении их активности связывания с определенным лигандом при специфическом связывающем сродстве Kd в диапазоне 10-7-10-12 М может быть выполнен способами, известными специалистам в данной области. Для этой цели представленные вариации убиквитина могут быть временно иммобилизованы на субстанции мишени, связанной, например, на микротитровальных планшетах, или могут быть связаны с магнитными частицами после связывания в растворе, соответственно. После отделения несвязывающихся вариаций генетическая информация по вариациям со связывающей активностью может быть специфически элюирована в форме мРНК путем деструкции рибосомного комплекса. Элюирование предпочтительно выполняют с помощью ЭДТА. Полученную таким образом мРНК можно изолировать и обратно транскрибировать в ДНК, используя подходящие способы (реакция обратной транскриптазы), и полученную таким образом ДНК можно реамплифицировать.
Посредством последовательных циклов транскрипции/трансляции, выбора и амплификации вариации можно обогащать вариации убиквитина со свойствами связывания с заданным гаптеном или антигеном.
Дальнейшая характеризация вариаций убиквитина, полученных таким образом, может быть выполнена в форме растворимого белка, как подробно сказано выше, после клонирования соответствующей генной кассеты в подходящий экспрессирующий вектор. Подходящие способы известны специалистам в данной области или описаны в литературе.
Предпочтительно, за этапом детектирования белков, имеющих связывающее сродство в отношении определенного связывающегося партнера, следует этап изоляции и/или обогащения детектированного белка.
После экспрессии убиквитинового белка, модифицированного согласно изобретению, он может быть далее очищен и обогащен способами, известными как таковые. Выбираемые способы зависят от нескольких факторов, известных как таковые специалистам в данной области, например, используемого экспрессирующего вектора, организма-хозяина, намеченной области использование размера белка и других факторов. Для упрощения очистки белок, модифицированный согласно изобретению, может быть объединен с другими пептидными последовательностями, имеющими повышенное сродство с материалами разделения. Предпочтительно, выбирают такие объединения, которые не оказывают вредного воздействия на функциональность убиквитинового белка или могут быть отделены после очистки из-за введения специфических сайтов расщепления протеазы. Такие способы также известны как таковые специалистам в данной области.
Немодифицированные и модифицированные убиквитиновые белки как исходная точка для мутагенеза
Термины "белок, способный связывать" или "связывающий белок" относятся к убиквитиновому белку, имеющему одну связывающую область, которая описана ниже. Связывающая область может относиться по меньшей мере к двум областям, определяющим связывание ("ООС"), Каждый мономер имеет по меньшей мере одну область, определяющую связывание; по меньшей мере два мономера образуют мультимер, имеющий по меньшей мере две области, определяющие связывание, которые образуют одну область связывания для одного антигена. Любой такой связывающий белок на основе убиквитина может включать дополнительные белковые домены, которые не являются связывающими доменами, такие как, например, жидкости мультимеризации, полипептидные метки, полипептидные линкеры и/или молекулы небелковых полимеров. Некоторыми примерами молекул небелковых полимеров являются гидроксиэтиловый крахмал, полиэтиленгликоль, полипропиленгликоль или полиоксиалкилен.
Дальнейшая мультимеризация гетеромультимерных модифицированных убиквитиновых белков может быть также выполнена путем посттрансляционного слияния гетеромультимерного модифицированного убиквитинового белка с молекулами эффектора, имеющими домен мультимеризации (например, TNF-α). В еще одном варианте осуществления дальнейшую мультимеризацию выполняют путем использования полиэти-ленгликольного (PEG) линкера. В еще одном варианте осуществления упомянутый домен мультимеризации также действует как фармацевтически активный компонент; одним примером является TNF-alpha, действующий и как домен мультимеризации, и как фармацевтический компонент.
Модифицированные гетеромультимерные убиквитиновые белки
Термин "модифицированный убиквитиновый белок" относится к модификациям убиквитинового белка любым одним из замещения, вставок или удалений аминокислот или их сочетания, при этом замещения являются наиболее предпочтительными модификациями, которые могут быть дополнены любой одной из модификаций, описанных выше. Число модификаций строго ограничено, так как упомянутые модифицированные мономерные убиквитиновые звенья имеют аминокислотную идентичность с SEQ ID NO:1 по меньшей мере одну из группы из 80%, по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 83% и по меньшей мере 90%. Самое большее, общее число замещений в мономерном звене поэтому ограничено 15 аминокислотами, соответствующими 80% аминокислотной идентичности. Другими альтернативами являются 13, 12,11, 10, 9, 8, 7, 6 или 5 модифицированных аминокислот. Совокупное число модифицированных аминокислот в гетеродимерной молекуле убиквитина составляет 30 аминокислот, соответствующих 20% аминокислотных модификаций на основе гетеродимерного белка. Другими альтернативами являются 28, 26, 24, 22, 20, 18, 16, 14, 13, 12, 11, 10, 9, 8, 7, 6 или 5 модифицированных аминокислот в молекуле димерного убиквитина. Аминокислотная идентичность димерного модифицированного убиквитинового белка по сравнению с димерным убиквитином, состоящим из двух немодифицированных мономерных убиквитиновых белков с основной мономерной последовательностью SEQ ID NO:1 выбирается по меньшей мере как одно из группы из 80%, по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 83% и по меньшей мере 90%.
Модифицированные убиквитиновые белки, полученные способом настоящего изобретения, сконструированы с новыми связывающимися сродствами к молекуле-мишени или лиганду или связывающейся молекуле (эти выражения используются здесь взаимозаменяемо).
Термин ″замещение″ также включает химическую модификацию аминокислот путем, например, замещения или добавления химических групп или остатков к исходной аминокислоте. Замещение аминокислот по меньшей мере в одной области с открытой поверхностью белка, включающей аминокислоты, расположенные по меньшей мере в одной нити бета-пласта области бета-пласта или помещение до 3 аминокислот рядом с нитью бета-пласта очень важно.
Модификацию выполняют способами, которые хорошо освоены и хорошо известны в данной области. ″Неспецифически модифицированный нуклеотид или аминокислотная последовательность″ является нуклеотидом или аминокислотной последовательностью, который в нескольких положениях был подвергнут вставке, делеции или замещению нуклеотидами или аминокислотами, характер которых нельзя прогнозировать. Во многих случаях введенные неспецифические нуклеотиды (аминокислоты) или последовательности нуклеотидов (аминокислот) будут ″полностью неспецифическими″ (например, как следствие рандомизированного синтеза или ПЦР-опосредованного мутагенеза). Однако неспецифические последовательности также могут включать последовательности, которые имеют общую функциональную особенность (например, способность реагировать с лигандом продукта экспрессии), или неспецифические последовательности могут быть неспецифическими в том смысле, что конечный продукт экспрессии является полностью неспецифической последовательностью, например, с равномерным распределением разных аминокислот.
Для того, чтобы ввести рандомизированные фрагменты в векторы правильно, согласно изобретению предпочтительно, чтобы неспецифические нуклеотиды вводили в вектор экспрессии по принципу сайт-направленного, ПЦР-опосредованного мутагенеза. Однако специалисту известны и другие варианты, и, например, можно также вводить библиотеки синтетических неспецифических последовательностей в векторы.
Для создания мутантов или библиотек путем объединяющей ПЦР могут выполняться, например, три реакции ПЦР. Две реакции ПЦР выполняют для создания частично перекрывающихся промежуточных фрагментов. Третью реакцию ПЦР выполняют для объединения промежуточных фрагментов.
Способ конструирования библиотеки или вариантов мутантов может включать конструирование первого набора праймеров вокруг желательного рестрикционного сайта (праймер рестрикционного сайта), праймера прямой и обратной рестрикции и второго набора праймеров вокруг, например, перед и за кодоном, представляющим интерес (мутагенные праймеры), прямого и обратного мутагенного праймера. В одном варианте осуществления праймеры конструируют сразу же перед и за, соответственно, кодоном, представляющим интерес. Рестрикционные и мутагенные праймеры используют для конструирования первого промежуточного и второго промежуточного фрагментов. Две реакции ПЦР создают эти линейные промежуточные фрагменты. Каждый из этих линейных промежуточных фрагментов включает по меньшей мере один мутировавший кодон, представляющий интерес, фланкирующую нуклеотидную последовательность и сайт расщепления. Третья реакция ПЦР использует эти два промежуточных фрагмента и праймеры прямой и обратной рестрикции для производства объединенного линейного продукта. Напротив, до сих пор незакрепленные концы линейного продукта расщепляли с помощью рестрикционного фермента, чтобы создать липкие концы на линейном продукте. Липкие концы линейного продукта объединяют путем использования ДНК-лигазы, чтобы получить кольцевой продукт, например, кольцевую полинуклеотидную последовательность.
Для конструирования промежуточных фрагментов выполняют разработку и синтез двух наборов прямых и обратных праймеров, причем первый набор содержит сайт расщепления рестрикционных ферментов вместе с его фланкирующей нуклеотидной последовательностью, и второй набор содержит по меньшей мере один вариант кодона,. представляющего интерес (мутагенные праймеры). Специалисты в данной области поймут, что количество вариантов будет зависеть от количества вариантов желательных модификаций аминокислот. Автор настоящего изобретения предусматривает, что если в этом процессе будут использованы другие рестрикционные ферменты, точное местонахождение этого сайта расщепления и соответствующая последовательность прямых и обратных праймеров могут быть изменены соответственно. В уровне техники доступны другие способы, и они могут быть использованы вместо изложенного.
Кроме введения рандомизированного фрагмента продукта экспрессии в каркас в соответствии с настоящим изобретением, часто необходимо соединить неспецифическую последовательность с партнером по объединению путем объединения рандомизированной нуклеотидной последовательности с нуклеотидной последовательностью, кодирующей по меньшей мере одного партнера по объединению. Такое партнер по объединению может, например, способствовать экспрессии, и/или очистке/изоляции, и/или дальнейшей стабилизации продукта экспрессии.
Для целей очистки партнер в объединении может включать метку очистки, такую как метка His6, метка туе, целевую последовательность для биотинилирования BSP, BirA, метку flu, lacZ и GST. Кроме того, партнер в объединении может включать сигнал сортировки или последовательность против мишени.
Замещение аминокислот для создания нового связывающегося домена, специфического для молекулы-мишени, может быть выполнено согласно изобретению с помощью любой желательной аминокислоты, т.е., для модификации, чтобы создать новое связывающееся свойство к молекуле-мишени, необязательно заботиться о том, чтобы аминокислоты имели какое-то конкретное химическое свойство или боковую цепь, соответственно, которые аналогичны таковым у замещаемой аминокислоты, так что для этой цели можно использовать любую желательную аминокислоту.
Этап модификации выбранных аминокислот выполняют, согласно изобретению, предпочтительно путем мутагенеза на генетическом уровне путем неспецифического мутагенеза, т.е., неспецифическим замещением выбранных аминокислот. Предпочтительно, модификацию убиквитина выполняют способами генной инженерии для изменения ДНК, принадлежащей соответствующему белку. Предпочтительно, экспрессию убик-витинового белка затем осуществляют в прокариотических или эукариотических организмах.
Замещения выполняют, в частности, в аминокислотах с открытой поверхностью четырех бета-нитей бета-пластов или аминокислотах с открытой поверхностью до 3 аминокислот рядом с нитью белка убиквитина нити бета-пласта убиквитинового белка. Каждая бета-нить обычно состоит из 5-7 аминокислот. Со ссылкой на SEQ ID NO:1, например, бета-нити обычно включают аминокислотные остатки 2-7, 12-16, 41-45 и 65-71. Области, которые могут быть дополнительно и предпочтительно модифицированы, включают положения до 3 аминокислот (т.е., 1, 2 или 3) рядом с нитью бета-пласта. Предпочтительные области, которые могут быть дополнительно и предпочтительно модифицированы, включают, в частности, аминокислотные остатки 8-11, 62-64 и 72-75. Предпочтительные области включают бета-витки, которые связывают две бета-нити вместе. Один предпочтительный бета-виток включает аминокислотные остатки 62-64. Наиболее предпочтительной аминокислотой, которая расположена рядом с нитью бета-пласта, является аминокислота в положении 8. Кроме того, другими предпочтительными примерами замещения аминокислот являются положения 36, 44, 70 и/или 71. Например, те области, которые могут быть дополнительно и предпочтительно модифицированы, включают аминокислоты 62, 63 и 64 (3 аминокислоты), или 72, 73 (2 аминокислоты), или 8 (1 аминокислота).
Число аминокислот, которые могут быть добавлены или изъяты, ограничено 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или больше аминокислот в мономерном субзвене убиквитина и 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 18, 20, 22, 24, 26 или 28 аминокислот по отношению к димерному убиквитиновому белку, обычно в х раз больше числа модификаций в мономерном белке. Обычно число вставок в мономерной молекуле включает 1-10 аминокислот и/или 1-7 делеций аминокислот. Число замещений составляет по меньшей мере 6 и самое большее 14 замещений аминокислот в мономерной молекуле. Димерная молекула включает в общем по меньшей мере 12 и самое большее 28 замещений, и/или в общем по меньшей мере одну и самое большее 20 вставок, и/или по меньшей мере одно и самое большее 14 делеций. Можно использовать все числа в пределах этих диапазонов, и они охватываются изобретением, и все сочетания количеств делеций, вставок и замещений возможны при том условии, что общая структурная целостность молекулы сохраняются. В одном варианте осуществления настоящего изобретения сохраняется структура бета-пласта.
В других вариантах осуществления аминокислотные остатки изменяют путем замещений аминокислот. Однако также допустимы делеций и вставки. Число аминокислот, которые могут быть добавлены или изъяты, ограничено 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот в мономерном субзвене убиквитина и соответственно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 аминокислот по отношению к димерному убиквитиновому белку. В одном варианте осуществления вставок аминокислот не делают. В еще одном варианте осуществления не выполняют делеций.
При условии, что модифицированный убиквитиновый белок настоящего изобретения включает дополнительно к упомянутым замещениям, определенным в формуле изобретения и объясненным здесь, а также изъятиям и/или добавлениям одной или больше аминокислот, положения аминокислот для человеческого убиквитина немутантного типа (SEQ ID NO:1) должны быть выровнены с модифицированным убиквитином, чтобы выделить белки, соответствующие друг другу. В случае составных белков (смотрите ниже), нумерация (и выравнивание) каждого из субзвеньев мономерного убиквитина осуществляют одинаково, т.е., выравнивание, например, димера начинают в положении 1 аминокислоты для каждого соответственного субзвена.
В мономерном убиквитиновом белке, предпочтительно от млекопитающих, например, человека, по меньшей мере 10% аминокислот, присутствующих в бета-нитях или положениях до 3 аминокислот рядом с нитью бета-пласта, предпочтительно по меньшей мере 20%, более предпочтительно по меньшей мере 25%, могут быть модифицированы, предпочтительно замещены. Как максимум, предпочтительно приблизительно 50% аминокислот, присутствующих в бета-нитях или положениях до 3 аминокислот рядом с нитью бета-пласта, более предпочтительно, как максимум, приблизительно 40%, или приблизительно 35%, или до приблизительно 30%, или до приблизительно 25% являются модифицированными, предпочтительно замещенными. В одной бета-нити обычно модифицированы 1-4 аминокислоты. В одном варианте осуществления две из шести аминокислот предпочтительно в первой и четвертой бета-нитях, например, в области аминокислотных остатков 2-7 или 65-71, модифицированы.
Модифицированный мономерный убиквитин согласно изобретению, используемый как строительное звено для гетеромультимера, насчитывает в совокупности до 20% аминокислот, которые должны быть модифицированы. Учитывая это, существует идентичность последовательности с SEQ ID NO:1 модифицированного убиквитинового белка по меньшей мере на 80%. В других вариантах осуществления изобретения идентичность последовательности на аминокислотном уровне составляет по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 87% и, кроме того, по меньшей мере 90%, по меньшей мере 92% или по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO:1. Изобретение также охватывает идентичности аминокислотной последовательности модифицированного убиквитинового белка более чем на 97% по сравнению с аминокислотной последовательностью SEQ ID NO:1.
В еще одном варианте осуществления настоящего изобретения уже предварительно модифицированный убиквитин (в котором модифицированы 3, или 4, или 5, или 6, или 7 аминокислот в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и/или 68 SEQ ID NO:1) используется как исходная точка для дальнейших модификаций, чтобы создать свойство связывания с мишенью, и может быть получен убиквитин, в котором в совокупности до 9, 10, 11, 12, 13, 14 и максимум 15 аминокислот убиквитина SEQ ID NO:1 модифицированы, предпочтительно замещены. Например, дальнейшие модификации могут включать модификации в аминокислотах 74 и 75 или в аминокислоте 45, чтобы создать повышенную стабильность или улучшенные белково-химические свойства. Согласно одному примеру, модифицированный мономерный убиквитин как строительное звено для гетеромультимерного белка может быть получен таким образом с 14 замещениями и делециями от совокупного числа аминокислот убиквитина, что соответствует приблизительно 20%. Это было полностью неожиданно, поскольку обычно гораздо меньшего процента уже достаточно, чтобы нарушить укладку белка.
В одном варианте осуществления изобретения эти аминокислоты модифицируют для создания области, имеющей новые свойства связывания, которые образуют непрерывную область на поверхности белка. Таким образом может быть создана непрерывная область, которая имеет свойство связывания с лигандом-мишенью. ″Непрерывная область″ согласно изобретению относится к следующему: из-за заряда, пространственной структуры и гидрофобности/гидрофильности своих боковых цепей аминокислоты взаимодействуют с окружающей их средой соответствующим образом. Этой окружающей средой может быть растворитель, обычно вода, или другие молекулы, например, пространственно близкие аминокислоты. Посредством структурной информации о белке, а также соответственного программного обеспечения можно охарактеризовать поверхность белков. Например, область поверхности раздела между атомами белка и растворителем можно визуализировать этим образом, включая информацию о том, как структурирована эта область поверхности раздела, какие части поверхности доступны для растворителя, или как на поверхности распределены заряды. Непрерывная область может быть раскрыта, например, визуализацией этого типа с использованием подходящего программного обеспечения. Такие способы известны специалистам в данной области техники. Согласно изобретению, в основном, также всю область с открытой поверхностью можно использовать в качестве непрерывной области на поверхности, которую требуется модифицировать для создания новых связывающих свойств. В одном варианте осуществления для этой цели модификация может также включать альфа-спиральную область. В гетеродимерном модифицированном убиквитиновом белке область, определяющая связывание, включает две из областей с открытой поверхностью, образующих вместе одну непрерывную область, которая включает две длины одной области, определяющей связывание.
Модификация аминокислот по меньшей мере в одной области белка с открытой поверхностью, включающей по меньшей мере одну бета-нить области бета-пласта или положения до 3 аминокислот рядом с нитью бета-пласта является очень важной. "Структура бета-пласта" определяется как в сущности пластоподобная и почти полностью растянутая. В противоположность альфа-спиралям, которые сформированы из ненарушенного сегмента полипептидной цепи, бета-пласты могут быть сформированы разными областями полипептидной цепи. Этим образом области, отстоящие друг от друга дальше в первичной структуре, могут войти в тесную близость друг с другом. Бета-нить обычно имеет длину 5-10 аминокислот (обычно 5-6 остатков в убиквитине) и имеет почти полностью растянутую конформацию. Бета-нити подходят так близко друг к другу, что образуются водородные связи между группой С-0 одной нити и группой NH другой нити и наоборот. Бета-пласты могут быть сформированы из нескольких нитей и имеют пластоподобную структуру, в которой положение альфа-атомов С чередуется между положениями выше и ниже пластовой плоскости. Боковые цепи аминокислот следуют этой модели и, таким образом, альтернативно указывают вверх или вниз. В зависимости от ориентации бета-нитей, пласты классифицируют на параллельные и антипараллельные. Согласно изобретению, оба типа могут мутировать и использоваться для приготовления предложенных белков.
Для мутагенеза бета-нитей и структуры бета-пласта в убиквитине выбирают бета-нить. или положения до 3 аминокислот рядом с бета-нитью (которая является нитью бета-пласта), которые близко к поверхности. Аминокислоты с открытой поверхностью могут быть идентифицированы по отношению к доступной рентгеновской кристаллографической структуре. Если кристаллическая структура не доступна, можно сделать попытки предсказать посредством компьютерного анализа области бета-пласта с открытой поверхностью и доступность индивидуальных положений аминокислот по отношению к доступной первичной структуре или смоделировать трехмерную структуру белка и получить информацию о потенциальных аминокислотах с открытой поверхностью таким образом. Дальнейшее раскрытие этого вопроса можно взять, например, в J. Мо1. BioL, 1987 Apr 5; 194(3):531-44. Vijay-Kumar S, Bugg C.E., Cook W.J.
Однако также можно осуществить модификации в бета-пласте или положениях до 3 аминокислот рядом с бета-нитью, для которых отнимающий много времени предварительный выбор положений аминокислот для мутагенеза можно опустить. Области ДНК, кодирующие структуры бета-пластов или до 3 аминокислот рядом с нитью бета-пласта изолируют от их ДНК окружающей среды, подвергают неспецифическому мута-генезу и затем повторно интегрируют в ДНК кодирование для белка, из которого они были удалены ранее. За этим следует процесс выбора мутантов с желательными свойствами связывания.
В еще одном варианте осуществления изобретения бета-нити или до 3 аминокислот рядом с бета-нитью близко к поверхности выбирают как сказано выше и идентифицируют положения аминокислот для мутагенеза в этих выбранных областях. Положения аминокислот, выбранные таким образом, можно затем подвергнуть мутагенезу на уровне ДНК либо путем направленного на сайт мутагенеза, т.е., кодон, кодирующий специфическую аминокислоту замещают кодоном, кодирующим другую, ранее выбранную специфическую аминокислоту, или это замещение осуществляют в контексте неспецифического мутагенеза, в котором определяется замещаемое положение аминокислоты, но не кодон, кодирующий новую, еще не определенную аминокислоту.
"Аминокислотами с открытой поверхностью" являются аминокислоты, которые доступны для окружающего растворителя. Если доступность аминокислот в белке больше чем 8% по сравнению с доступностью аминокислоты в модельном трипептиде Gly-X-Gly, эти аминокислоты называют " поверхностно открытыми". Эти области белка или положения отдельных аминокислот, соответственно, также являются предпочтительными сайтами связывания для потенциальных связывающихся партнеров, выбор которых должен быть осуществлен согласно изобретению. Помимо этого, делается ссылка на публикации Кастера и др. (Caster et al), 1983 Science, 221, 709-713, и Шрейка и Рипли (Shrake & Rupley), 1973 J. Mol. Biol. 79(2):351-371, которые для полного раскрытия включены в настоящую заявку путем ссылки.
Вариации убиквитина, отличающиеся замещениями аминокислот в области искусственного сайта связывания, вновь созданного из родительского белка и из друг друга, могут быть созданы путем целевого мутагенеза соответственных сегментов последовательности. В этом случае аминокислоты, имеющие некоторые свойства, такие как полярность, заряд, растворимость, гидрофобность или гидрофильность могут быть заменены или замещены, соответственно, аминокислотами с соответственными другими свойствами. Кроме замещений, термины ″мутагенез″ и ″модифицированный″ и ″замененный″ также включают вставки и делеции. На белковом уровне модификации также могут быть осуществлены путем химического изменения боковых цепей аминокислот согласно способам, известным специалистам в данной области.
Способы мутагенеза убиквитина
В качестве исходной точки для мутагенеза соответственных сегментов последовательности можно использовать, например, кДНК убиквитина, которая может быть приготовлена, изменена и амплифицирована способами, известными специалистам в данной области. Коммерчески доступны реагенты и способы для сайт-специфического изменения убиквитина в относительно небольших областях первичной последовательности (приблизительно 1-3 аминокислоты) (″Quick Change″, компания Stratagene; набор ″Mutagene Phagemid in vitro Mutagenesis Kit″, компания Biorad). Для сайт-направленного мутагенеза более крупных областей специалистам доступны специфические варианты осуществления, например, полимеразная цепная реакция (ПЦР). Для этой цели можно использовать смесь синтетических олигодеоксинуклеотидов, имеющих композиции вырожденных пар оснований в желательных положениях, например, для ввода мутации. Это также можно осуществить путем использования аналогов пар оснований, которые в естественном виде не встречаются в геномной ДНК, такие как, например, инозин.
Исходной точкой для мутагенеза одной или больше бета-нитей области бета-пласта или положений до 3 аминокислот рядом с нитью бета-пласта может быть, например, кДНК убиквитина или также геномная ДНК. Кроме того, генное кодирование для убиквити-нового белка также может быть приготовлено синтетически.
В одном варианте осуществления изобретения мутагенез осуществляют сборкой олигонуклеотидов ДНК, несущих кодон NNK аминокислоты. Необходимо понимать, однако, что можно использовать и другие кодоны (триплеты). Мутации выполняют таким образом, чтобы предпочтительно сохранить структуру бета-пласта. Обычно мутагенез происходит на наружной стороне стабильной области бета-пласта, открытой на поверхности белка. Он включает и сайт-специфический и неспецифический мутагенез. Сайт-специфический мутагенез, включающий относительно небольшую область в первичной структуре (приблизительно 3-5 аминокислот), может быть создан с использованием имеющихся в продаже наборов Stratagene® (QuickChange®) или Bio-Rad® (фагемид) Mutagene® в наборе для мутагенеза in vitro) (см. US 5,789,166; US 4,873,192). Если сайт-специфическому мутагенезу подвергают более широкие области, должна быть подготовлена ДНК-кассета, в которой область для мутагенеза получают сборкой нуклеотидов, содержащих мутированные и неизмененные положения (Норд и др. (Nord et al), 1997 Nat. Biotechnol. 8, 772-777; Макконелл и Хесс (McConell and Hoess), 1995 J. Mol. Biol. 250, 460-470.). Неспецифический мутагенез может быть введен путем распространения ДНК в штаммах гена-мутатора или амплификацией ПЦР (подверженная ошибкам ПЦР) (например, Паннекоек и др. (Pannekoek et al.), 1993 Gene 128, 135 140). Для этой цели используют полимеразу с повышенной частотой ошибок. Для усиления степени введенного мутагенеза или для объединения разных мутаций, соответственно, мутации в фрагментах ПЦР можно объединять посредством перестановки ДНК (Штеммер (Stemmer), 1994 Nature 370, 389-391). Обзор этих стратегий мутагенеза по отношению к ферментам представлен в обзоре Кюхнера и Арнольда (Kuchner and Arnold) (1997) TIBTECH 15, 523-530. Для осуществления этого неспецифического мутагенеза в выбранной области ДНК также должна быть сконструирована ДНК-кассета, которую используют для мутагенеза.
Разные процедуры, известные как таковые и доступные для мутагенеза, являются способами для сайт-специфического мутагенеза, способами для неспецифического мутагенеза, мутагенеза с использованием ПЦР или им подобными способами.
В одном предпочтительном варианте осуществления настоящего изобретения предварительно определяют положения аминокислот для мутагенеза. Выбор аминокислот для модификации осуществляют в соответствии с ограничениями пункта 1 формулы изобретения в отношении тех аминокислот, которые должны быть модифицированы. В каждом случае обычно создают библиотеку разных мутантов, которую подвергают скринингу, используя способы, известные как таковые. Обычно предварительный выбор аминокислот для модификации можно особенно легко выполнить, когда достаточная структурная информация имеет по убиквитиновому белку, подлежащему модификации.
Способы для целевого мутагенеза, а также мутагенеза более длинных сегментов последовательностей, например, посредством ПЦР, химическим мутагенезом или с использованием штаммов бактерий гена-мутатора также известны из уровня техники и могут быть использованы согласно изобретению.
В одном варианте осуществления изобретения мутагенез осуществляют сборкой олигонуклеотидов ДНК, несущих кодон NNK. аминокислоты. Необходимо понимать, однако, что можно использовать и другие кодоны (триплеты). Мутации выполняют таким образом, чтобы предпочтительно сохранить структуру бета-пласта. Обычно мутагенез происходит на наружной стороне стабильной области бета-пласта, открытой на поверхности белка. Он включает и сайт-специфический и неспецифический мутагенез. Сайт-специфический мутагенез, включающий относительно небольшую область в первичной структуре (приблизительно 3-5 аминокислот), может быть создан с использованием имеющихся в продаже наборов Stratagene® (QuickChange®) или Bio-Rad® (фагемид Mutagene® в наборе для мутагенеза in vitro) (см. US 5,789,166; US 4,873,192).
Если сайт-специфическому мутагенезу подвергают более широкие области, должна быть подготовлена ДНК-кассета, в которой область для мутагенеза получают сборкой нуклеотидов, содержащих мутированные и неизмененные положения (Норд и др. (Nord et al), 1997 Nat. Biotechnol. 8, 772-777; Макконелл и Хесс (McConell and Hoess), 1995 J. Mol. Biol. 250, 460-470.). Неспецифический мутагенез может быть введен путем распространения ДНК в штаммах гена-мутатора или амплификацией ПЦР (подверженная ошибкам ПЦР) (например, Паннекоек и др. (Pannekoek et al.), 1993 Gene 128, 135 140).
Для этой цели используют полимеразу с повышенной частотой ошибок. Для усиления степени введенного мутагенеза или для объединения разных мутаций, соответственно, мутации в фрагментах ПЦР можно объединять посредством перестановки ДНК (Штеммер (Stemmer), 1994 Nature 370, 389-391). Обзор этих стратегий мутагенеза по отношению к ферментам представлен в обзоре Кюхнера и Арнольда (Kuchner and Arnold) (1997) TIBTECH 15, 523-530. Для осуществления этого неспецифического мутагенеза в выбранной области ДНК также должна быть сконструирована ДНК-кассета, которую используют для мутагенеза.
Неспецифическое замещение согласно одному примеру настоящего изобретения по меньшей мере 6 аминокислот в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и/или 68 мономерного убиквитина может быть выполнено особенно легко посредством ПЦР, поскольку упомянутые положения локализованы близко к амино- или карбокси-окончанию белка. Соответственно, кодоны для манипулирования находятся на 5' и 3' конце соответствующей нити кДНК. Таким образом, первый олигодеоксинуклеотид, используемый для мутагенной реакции ПЦР кроме кодонов в положениях 2, 4, 6 и/или 8, которые будут мутировать, соответствует по последовательности кодирующей нити для амино-окончания убиквитина. Соответственно, второй олигодеоксинуклеотид - кроме кодонов в положениях 62, 63, 64, 65, 66 и/или 68, которые будут мутировать, по меньшей мере частично соответствует некодирующей нити полипептидной последовательности карбокси-окончания. Посредством обоих олигодеоксинуклеотидов полимеразная цепная реакция может быть выполнена с использованием ДНК-последовательности, кодирующей мономерный убиквитин, в качестве шаблона.
Кроме того, полученный продукт амплификации может быть добавлен к другой полимеразной цепной реакции с использованием фланкирующих олигодеоксинуклеотидов, которые вводят, например, последовательности распознавания для рестрикционных эндонуклеаз. Согласно изобретению предпочтительно вводить полученную генную кассету в векторную систему, подходящую для использования в последующей процедуре селекции для изоляции вариаций убиквитина, имеющих свойства связывания с заранее определенным гаптеном или антигеном.
Замещение аминокислот для создания нового связывающегося домена, специфического для выбранного лиганда, может быть выполнено согласно изобретению с помощью любой желательной аминокислоты, т.е., для модификации, чтобы создать новое свойство связывания с этим выбранным лигандом, необязательно заботиться о том, чтобы аминокислоты имели какое-то конкретное химическое свойство или боковую цепь, соответственно, которые аналогичны таковым у замещаемой аминокислоты, так что для этой цели можно использовать любую желательную аминокислоту.
Этап модификации выбранных аминокислот выполняют, согласно изобретению, предпочтительно путем мутагенеза на генетическом уровне путем неспецифического мутагенеза, т.е., неспецифическим замещением выбранных аминокислот. Предпочтительно, модификацию убиквитина выполняют способами генной инженерии для изменения ДНК, принадлежащей соответствующему белку. Предпочтительно, экспрессию убиквитинового белка затем осуществляют в прокариотических или эукариотических организмах.
Согласно изобретению, модифицированный убиквитиновый белок может быть кроме того предпочтительно приготовлен путем химического синтеза. В предпочтительных вариантах осуществления аминокислотные остатки изменяют путем замещений аминокислот.Однако также допустимы делеции и вставки. По выбору, число аминокислот для вставки или делеции составляет 1-10, 1-5, 2, 3 или 4 аминокислоты. В одном варианте осуществления вставку аминокислот не осуществляют. В еще одном варианте осуществления не осуществляют делеций.
После выполнения вышеописанных модификаций авторы изобретения нашли аминокислотные последовательности модифицированного убиквитина, описанные в примерах, которые связывают свои мишени с очень высоким сродством (значения Kd до 10-10 М).
Области, подлежащие модификации в убиквитине
Области для модификации могут быть в основном выбраны в зависимости от того, что они могут быть доступны для выбранного связывающегося партнера, и от того, что общая структура белка будет предположительно проявлять толерантность к модификации.
Помимо модификаций в поверхностно открытых бета-нитях, также могут быть осуществлены модификации в других поверхностно открытых областях белка, предпочтительно в положениях до 3 аминокислот рядом с бета-нитью. Эти модифицированные области участвуют во вновь создаваемом связывании с высоким сродством с мишенью.
Согласно еще одному предпочтительному варианту осуществления настоящего изобретения по меньшей мере 3, или 4, или 6, по выбору по меньшей мере 8, 10, 12 и максимум 15 поверхностно открытых аминокислот убиквитина, предпочтительно убиквитина млекопитающего или человека, могут быть модифицированы в мономерном убиквитине, в котором в качестве модификации предпочтительно замещение. Это включает модификацию 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 поверхностно открытых аминокислот убиквитина. Эти по меньшей мере 3 и максимум 15 поверхностно открытых модифицированных аминокислот затем образуют область со связывающим сродством к предварительно определенному связывающемуся партнеру. Эта область определена здесь как" область связывающегося домена" ("ОСД"). В этом отношении особенно предпочтительно, чтобы по меньшей мере 2, по выбору по меньшей мере 4, далее по выбору по меньшей мере 6, 8, 10, 12 и максимум 15 поверхностно открытых аминокислот находились в области бета-пласта, т.е., в нити бета-пласта, или были распределена по нескольким бета-нитям или положениям до 3 аминокислот рядом с нитью бета-пласта. Кроме того предпочтительно, чтобы по меньшей мере 3 из всех модифицированных, предпочтительно замещенных, аминокислот были расположены непосредственно рядом друг с другом в первичной последовательности.
В еще одном варианте осуществления настоящего изобретения аминокислоты в одной или двух, предпочтительно двух, из четырех бета-нитей в белке или в положениях до 3 аминокислот рядом предпочтительно с двумя из четырех бета-нитей модифицируют, чтобы создать новое связывающее свойство. Также, по выбору, может быть осуществлена модификация в трех или четырех из четырех бета-нитей или в положениях до 3 аминокислот рядом с тремя или четырьмя из бета-нитей для создания связывания с выбранной мишенью или лигандом.
В частности предпочтительно, чтобы аминокислоты в нити амино-окончания и карбокси-окончания или в положениях до 3 аминокислот рядом с нитью амино-окончания и карбокси-окончания были модифицированы, предпочтительно замещены, чтобы создать новый сайт связывания с лигандом или мишенью. В этом отношении, в частности предпочтительно, чтобы до 3 аминокислот рядом с нитью карбокси-окончания бета-пласта были модифицированы, предпочтительно замещены, и 1 аминокислота рядом с нитью амино-окончания бета-пласта была модифицирована, предпочтительно замещена.
Согласно изобретению убиквитин модифицирован в его аминокислотах, предпочтительно путем замещения, в по меньшей мере трех аминокислотах в следующих положениях убиквитина млекопитающего, предпочтительно убиквитина человека: 2, 4, 6, 8, 62, 63, 64, 65, 66, 68. Эти по меньшей мере три аминокислоты из упомянутой группы аминокислот формируют непрерывную поверхностно открытую область на поверхности убиквитина, которые, как было установлено, являются особенно подходящими для создания модифицированных белков, имеющих связывающее сродство, которое раньше не существовало по отношению к специфическому связывающемуся партнеру, например, ED-B, TNFalpha, NGF, IgG, MIA-2 или любой другой мишени. По меньшей мере три из этих аминокислотных остатков должны быть модифицированы. По выбору, 3, 4, 5, 6, 7, 8, 9 или 10 упомянутых аминокислотных остатков модифицированы, предпочтительно замещены, по выбору, в сочетании с дополнительными аминокислотными остатками.
Для определения степени идентичности последовательности производного убиквитина с аминокислотной последовательностью SEQ ID NO:1 можно использовать, например, программу SIM локального сходства (Сяокин Хуан и Уэбб Миллер (Xiaoquin Huang and Webb Miller), "Достижения прикладной математики. Том 12: 337-357, 1991) или Клустал У. (Clustal, W.) (Томпсон и др. (Thompson et al.). Nucleic Acids Res., 22(22): 4673-4680, 1994), которая доступна бесплатно от авторов и их института для анализа множественного выравнивания. Предпочтительно, степень идентичности последовательности производного с SEQ ID NO:1 определяют относительно полной последовательности SEQ ID NO:1.
Способы определения значений связывающего сродства известны как таковые и могут быть выбраны, например, из следующих: ИФА, технология на основе поверхностного плазменного резонанса (ППР) (предлагаемая, например, Biacore®), вытеснительная хроматография, флуоресцентная спектроскопия и изотермическая титриметрическая калориметрия (ИТК).
В еще одном аспекте изобретение относится к составному белку, включающему гете-ромультимерный связывающий белок настоящего изобретения, объединенный с фармацевтически и/или диагностически активным компонентом; ссылка делается, например, на документ US 7,838,629, все содержание которого включено путем ссылки.
Составной белок настоящего изобретения может включать не полипептидные компоненты, например, не пептидные линкеры, не пептидные лиганды, например, для терапевтически или диагностически релевантных радионуклеотидов. Он также может включать небольшие органические или не аминокислотные соединения, например, сахар, олиго- или полисахарид, жирную кислоту и т.д. В одном предпочтительном варианте осуществления изобретения связывающаяся молекула на основе гетеромерного убиквитина связана пептидным, имеющим терапевтически или диагностически релевантные свойства линкером на основе аминокислоты, или лигандом, или белком, имеющим терапевтически или диагностически релевантные свойства.
Специфичности связывания (константы диссоциации)
Специфичности связывания составных белков согласно изобретению такие же, как определены выше для не составного белка, выраженные в килодальтонах. В соответствии с изобретением, термин "килодальтон" определяет специфическое связывающее сродство, которое в соответствии с изобретение находится в диапазоне от 10-7 до 10-12 М. Значение 10-5 М и ниже может считаться выражаемым количественно связывающим сродством. В зависимости от применения, значение 10-7 М - 10-11 М предпочтительно, например, для хроматографических применений или 10-9-10-12 М например, для диагностических или терапевтических применений. Другие предпочтительные значения связывающего сродства лежат в диапазоне от 10-7 до 10-10 М, предпочтительно до 10-11 М.
Мультимеризация убиквитина
Согласно изобретению два по-разному модифицированных мономера убиквитина, генетически связанные путем объединения "голова к хвосту", связываются с одним и тем же эпитопом молекулы-мишени, например, ED-B, TNFalpha, IgG, Mia-2, NGF или любой другой молекулой-мишенью, и эффективны только, если обе области связывающегося домена действуют вместе. Другими словами, они связываются с одним и тем же эпитопом через непрерывную область связывания, которая сформирована совместным действием обеих связывающих областей двух мономеров.
Мономеры могут быть соединены непосредственно или через линкеры. Подходящими предпочтительными линкерами являются линкеры SEQ ID NO:32 или имеющие по меньшей мере последовательность GIG, или имеющие по меньшей мере последовательность SGGGG, или любой другой линкер, например GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG или SGGGGSGGGG. Однако, существует много известных линкеров, которые можно использовать вместо этих.
Библиотеки
В еще одном аспекте настоящее изобретение направлено на библиотеку, содержащую ДНК, ответственную за получение модифиц ированных мономерных убиквитиновых белков, которые определены выше, которая формирует основу для предоставления гетеромультимерных, предпочтительно гетеродимерных, убиквитиновых белков настоящего изобретения.
В еще одном аспекте изобретения представлена библиотека объединений, содержащая ДНК, полученные путем объединения двух библиотек, как сказано выше, причем каждая библиотека отвечает за получение звеньев по-разному модифицированных мономерных убиквитиновых белков, чтобы получить гетеродимерные составные белки убиквитина, и причем их мономерные звенья связаны между собой в расположении "голова к хвосту". Упомянутая библиотека отвечает за получение гетеродимерных составных белков убиквитина, проявляющих одновалентную связывающую активность в отношении заданной мишени. Упомянутое связывание между собой выполняют или используя любой из линкеров, известный специалисту в данной области, или линкер, описанный в настоящем документе. "По-разному модифицированных" также включает альтернативу одной немодифицированной молекулы, присутствующей в гетеродимерном белке.
В Примере 1 описано получение сложной библиотеки. Однако необходимо проявлять осторожность в отношении качества такой библиотеки. Качество библиотеки в каркасной технологии стоит первым в зависимости от ее сложности (количества отдельных вариантов), а также функциональности (структурная и белково-химическая целостность получаемых кандидатов). Обе характеристики, однако, могут оказывать отрицательное влияние друг на друга: повышение сложности библиотеки путем увеличения числа модифицированных положений на каркасе может привести к ухудшению белково-химических характеристик вариантов. Это может привести к ухудшенной растворимости, агрегации и/или низкому выходу продукта. Причиной этого является повышенное отклонение от исходных каркасов, имеющих энергетически благоприятную упаковку белка.
Поэтому при конструировании такой библиотеки каркасов требуется подходящий баланс между крайними положениями введения максимально возможного числа вариаций в оригинальную последовательность, чтобы оптимизировать ее для мишени и, с другой стороны, сохранить оригинальную первичную последовательность в максимальной степени, чтобы избежать отрицательных белково-химических воздействий.
Специфические модификации в гетеродимерных убиквитиновых белках
Гетеродимер убиквитина согласно изобретению, связывающийся с лигандом с Kd=10-7-10-12 М и проявляющий одновалентную связывающую активность в отношении упомянутого лиганда, выбирают из двух следующих альтернатив:
(1) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору, дополнительно 2, и
(2) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 2, 4, 6, 62, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору, дополнительно 2.
В одном варианте осуществления составной белок является генетически объединенным гетеродимером упомянутого мономера убиквитина, имеющего замещения в аминокислотах в положениях 6, 8,63-66 первого мономера убиквитина и замещения в аминокислотных остатках в положениях 6, 8, 62-66, и по выбору в положении 2 второго мономера убиквитина, предпочтительно
- в первом мономере убиквитина замещения
лизина (K) на триптофан (W) или фенилаланин (F) в положении 6,
лейкина (L) на триптофан или фенилаланин (W, F) в положении 8,
лизина (K) на аргинин (R) или гистидин (Н) в положении 63,
глютаминовую кислоту (Е) на лизин (K), аргинин (R) или гистидин (Н) в положении 64,
серии (S) на фенилаланин (F) или триптофан (W) в положении 65 и
треонин (Т) на пролин (Р) в положении 66;
- во втором мономере убиквитина предпочтительны замещения лизина (K) на треонин (Т), аспарагин (N), серин (S) или глутамин (Q) в положении 6,
лейкина (L) на глутамин (Q) или треонин (Т) или аспарагин (N) или серин (S) в положении 8,
глутамин (Q) на триптофан (W) или фенилаланин (F) в положении 62,
лизина (K) на серин (S), треонин (Т), аспарагин (N) или глутамин (Q) в положении 63,
глютаминовую кислоту (Е) на аспарагин (N), серин (S), треонин (Т), или глутамин (Q) в положении 64,
серин (S) на фенилаланин (F) или триптофан (W) в положении 65, и
треонин (Т) на глютаминовую кислоту (Е) или аспарагиновую кислоту (D) в положении 66, и
по выбору, глутамин (Q) на аргинин (R), гистидин (Н) или лизин (K) в положении 2.
Эти альтернативные замещения в каждом мономере могут быть объединены друг с другом без каких-либо ограничений при условии, что получаемые гетеродимеры модифицированного убиквитина имеют специфическую активность связывания с упомянутым лигандом кД=10-7-10-12 М и проявляют активность одновалентного связывания в отношении упомянутого лиганда, и при условии, что структурная стабильность убиквитинового белка не разрушена или не затронута.
Наиболее предпочтительными являются следующие замещения:
(1) в первом мономерном звене по меньшей мере K6W, L8W, K63R, Е64К, S65F и Т66Р;
и во втором мономерном звене по меньшей мере K6T, L8Q, Q62W, K63S, E64N, S65W и Т66Е; по выбору дополнительно Q2R, или
(2) в первом мономерном звене по меньшей мере Q2T, F4W, K6H, Q62N, K63F, Е64К, S65LHT66S;
и во втором мономерном звене модификации по меньшей мере в положениях 6, 8, 62, 63, 64, 65 и 66, кроме того, по выбору,
во втором мономерном звене по меньшей мере K6X, L8X, Q62X, K63X, Е64Х, S65X и Т66Х; по выбору дополнительно Q2X, где Х может быть любой аминокислотой.
В частности, предпочтительными являются следующие замещения в первом мономере убиквитина для создания белков связывания с ED-B:
2: Q□T, 4: F□W, 6: K□H, 62: Q□N, 63: K□F, 64: E□K, 65: S□L, 66: T□S.
Для соединения двух мономеров "голова к хвосту" можно использовать или не использовать линкер. Предпочтительными являются линкеры SEQ ID NO:32 или последовательность GIG, или SGGGGIG, или SGGGGSGGGGIG.
В одном предпочтительном варианте осуществления, гетеродимер убиквитина с двумя областями, определяющими связывание и совместно действующими для связывания лиганда ED-B, включает аминокислотную последовательность SEQ ID NO:33 или 34. Еще один предпочтительный составной белок настоящего изобретения, включающий TNF-alpha в качестве фармацевтически активного компонента, имеет последовательность SEQ ID NO:35 или 36. В еще одном варианте осуществления гетеродимер убиквитина с двумя областями, определяющими связывание и действующими совместно для связывания лиганда ED-B, включает аминокислотную последовательность с Фиг. 11, соответствующую SEQ ID NO:XX.
Еще один предпочтительный белок представлен следующей последовательностью, в которой ХХХХ может быть любой аминокислотой (SEQ ID NO:47).
MTIWVHTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSD
YNINFKLSLHLVLRLRGGSGGGGSGGGGIG
MQIFVXTXTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSD
UNIXXXXXLHLVLRLRGG
Примеры белков с этими последовательностями показаны на Фиг. 11. В качестве линкера здесь использован SGGGGSGGGGIG. При этом понимается, что осуществимыми альтернативами также является другой тип линкеров или отсутствие линкера.
Полинуклеотиды, клетки-хозяева, векторы настоящего изобретения
В еще одном аспекте настоящее изобретение также охватывает полинуклеотиды, которые отвечают за создание белка или составного белка, как сказано выше. Дополнительно, изобретение охватывает векторы, включающие упомянутый полинуклеотид.
В дополнительном аспекте настоящего изобретения охвачены клетки-хозяева, которые включают описанный здесь белок или составной белок и/или полинуклеотидное кодирование для упомянутого рекомбинантного белка или составного белка настоящего изобретения или вектор, содержащий упомянутый полинуклеотид.
Использование молекул модифицированного гетеромультимерного убиквитина
Модифицированные убиквитиновые белки настоящего изобретения, способные связывать лиганд с высоким сродством, должны использоваться, например, для приготовления диагностических средств для использования in vitro или in vivo, а также терапевтических средств. Белки согласно изобретению можно использовать, например, в качестве молекул прямого эффектора (модулятора, антагониста, агониста) или антиген-распознающих доменов.
В области человеческой и ветеринарной терапии и профилактики фармацевтически эффективные медикаменты, содержащие по меньшей мере один гетеромерный убиквитиновый белок, модифицированный в соответствии с изобретением, для связывания ED-B может быть приготовлен способами, известивши как таковые. В зависимости от галенового препарата, эти композиции могут быть введены парэнтерально путем инъекции или инфузии, системно, ректально, интраперитонеально, внутримышечно, подкожно, чрезкожно или другими традиционно применяемыми способами. Тип фармацевтического препарата зависит от типа заболевания, тяжести заболевания, пациента и других факторов, известных специалистам в области медицины.
В зависимости от выбранного партнера для объединения, фармацевтическая композиция изобретения адаптирована для направленности на лечение рака, например, груди и колоректальных раков, или на любые другие опухолевые заболевания с обильным проявлением мишени.
Композиции адаптированы для содержания терапевтически эффективной дозы. Величина назначаемой дозы зависит от организма пациента, типа заболевания, возраста и массы тела пациента, а также других факторов, известных как таковые.
Композиции содержат фармацевтически или диагностически приемлемый носитель и, по выбору, могут содержать другие вспомогательные агенты и наполнители, известные как таковые. Они включают, например, но без ограничения, стабилизирующие агенты, поверхностно-активные вещества, соли, буферы, подкрашивающие агенты и т.д.
Фармацевтическая композиция может быть в форме жидкого препарата, крема, лосьона для топикального применения, аэрозоли, порошков, гранул, таблеток, суппозиториев или капсул, эмульсии или липосомного препарата. Композиции предпочтительно стерильные, непирогенные и изотонные и содержат фармацевтически традиционные и приемлемые добавки, известные как таковые. Дополнительно, делается ссылка на правила Фармакопеи США или публикацию "Фармацевтические науки Ремингтона", Мае Publishing Company (1990).
"Фармацевтическая композиция" согласно изобретению может присутствовать в форме композиции, в которой разные активные ингредиенты и разбавители и/или носители смешаны друг с другом, или может принимать форму объединенного препарата, в котором активные ингредиенты присутствуют в частично или полностью раздельной форме. Примером такой композиции или объединенного препарата является набор.
"Композиция" согласно настоящему изобретению включает по меньшей мере два фармакологически активных соединения. Эти соединения могут быть введены одновременно или отдельно с временным интервалом от одной минуты до нескольких суток. Соединения могут быть введены одинаковым способом или по-разному; например, возможно пероральное введение одного активного соединения и парэнтеральное введения другого соединения. Также, активные соединения могут быть в одном медикаменте, например, в одном растворе для инфузии или как набор, содержащий оба соединения отдельно. Также возможно, чтобы оба соединения присутствовали в двух или больше упаковках.
В еще одном варианте осуществления фармацевтическая композиция находится в форме набора из частей, предусматривающего отдельные позиции для рекомбинантного убиквитинового белка/составного белка изобретения и для одного или больше химиотерапевтических агентов.
Модифицированные убиквитиновые белки согласно изобретению могут быть приготовлены многими традиционными и хорошо известными способами, такими как стратегии простого органического синтеза, способы твердофазного синтеза или в имеющихся в продаже автоматизированных синтезаторах. С другой стороны, они также могут быть приготовлены традиционными рекомбинантными способами как таковыми или в сочетании с традиционными способами синтеза.
По выбору, модификации могут быть выполнены путем генетической инженерии на уровне ДНК и экспрессии модифицированного белка в прокариотических или эукариотических организмах или in vitro.
В еще одном варианте осуществления упомянутый этап модификации включает этап химического синтеза.
В одном аспекте настоящего изобретения упомянутую совокупность по-разному модифицированных белков получают путем генетического объединения двух ДНК-библиотек, каждая из которых отвечает за по-разному модифицированные мономерные убиквитиновые белки.
КРАТКОЕ ОПИСАНИЕ ФИГУР ЧЕРТЕЖЕЙ
На Фиг. 1 показано, показано, что рекомбинация переднего (первого) модифицированного мономера убиквитина с областью, определяющей связывание, упоминаемой как ООС1, с по-другому модифицированным задним (вторым) мономером убиквитина с областью, определяющей связывание, упоминаемой как ООС2, для создания гетеродимера приводит к значительному увеличению сродства, а также специфичности связывания. Молекулы модифицированного убиквитина проанализированы посредством Biacore, флуоресцентной анизотропии, связывания на клетках и срезах тканей. Показаны зависящие от концентрации результаты ИФА (конц. - ИФА) связывания нескольких вариантов гетеродимерного убиквитина с ED-B человека.
На Фиг. 1А показано связывающее сродство Kd=9.4 мкМ =9,45×10-6 М для мономера 41В10 (здесь: SPWF28-41B10-th). Закрашенные круги показывают связывание первого мономера 41В10 с фрагментом 67В89, который представляет экстрадомен В(ED-В) фибронектина. Контрольный фрагмент 6789 не содержит ED-B и показан незакрашенными кругами.
На Фиг. 1В показано связывающее сродство гетеродимерного убиквитина. Гетеродимер содержит первый мономер 41В10, объединенный с другим, вторым мономером, что дает вариант 46Н9 (здесь: SPWF28-46H9-th). Связывающее сродство 46Н9 намного повышено по сравнению с мономером, показанным на Фиг. 1А из-за одновалентного связывания обоих мономеров с мишенью ED-B (Kd=131 нМ =1,3×10-7 М; здесь 67В89, закрашенные круги). Контрольный фрагмент 6789 не содержит ED-B и показан незакрашенными кругами.
На Фиг. 2 показано сродство и активность объединенной с цитокином молекулы гетеродимера, связывающего ED-B, на основе модифицированного убиквитина.
На Фиг. 2А показано высокое сродство связывающего ED-B гетеродимера 24Н12 (Kd 50,7 нМ=5×10-8 М) на основе модифицированного убиквитина. Закрашенные круги показывают связывание 24Н12 с EDB; в качестве отрицательного контроля использовали связывание 24Н12 с БСА (бычий сывороточный альбумин) (незакрашенные круги).
На Фиг. 2В показано повышенное сродство связывающего ED-B гетеродимера 24Н12 на основе модифицированного убиквитина, объединенного с цитокином TNFalpha для мультимеризации гетеродимера 24Н12 (Kd=5,6 нМ=5,6×10-9 М).
На Фиг. 2С показан анализ примеров кандидатов, выбранных из библиотеки гетеродимерного модифицированного убиквитина, например, варианты 9Е12, 22D1, 24Н12, 41В10 гетеродимера. Значения ИФА в килодальтонах увеличиваются для мишени ED-B по сравнению с цитозольным фибронектином (c-FN), используемым в качестве контрольного, подтверждая специфическое связывание с мишенью.
На Фиг. 2D показаны результаты анализа молекулы 9Е12 модифицированного гетеродимерного убиквитина посредством анализов взаимодействия без меток с использованием Biacore®. Были проанализированы разные концентрации вариантов гетеродимерного убиквитина (смотрите легенду к Фигуре: 0-15 мкМ 9Е12) для связывания с ED-B, иммобилизованном на чипе (Biacore), чтобы проанализировать взаимодействие между гетеродимерным вариантом 9Е12 и ED-B. Значение в килодальтонах нельзя определить из анализа кривых ассоциации и диссоциации.
На Фиг. 2Е показаны результаты анализа молекулы 41 В10 модифицированного гетеродимерного убиквитина посредством анализов взаимодействий без меток с использованием Biacore®. Были проанализированы разные концентрации вариантов гетеродимерного убиквитина (смотрите легенду к Фигуре: 0-15 мкМ 41В10) для связывания с ED-В, иммобилизованном на чипе (Biacore), чтобы проанализировать взаимодействие между гетеродимерным вариантом 41 В10 и ED-B. Анализ кривых ассоциации и диссоциации дал значение кД 623 нМ (6,2×10-7 М).
На Фиг. 3 показан вклад разных вариантов на основе модифицированного убиквитина в связывающее сродство и специфичность. Эти разные варианты разделяют модули общей последовательности, которые отмечены буквами нижнего регистра. Варианты были проанализированы в отношении их связывания с ED-B. На Фиг. 3 показаны другие комбинации мономеров, дающие гетеродимеры модифицированного убиквитина. Гетеродимерные варианты 46-А5, 50-G11 и 46-Н4 имеют одинаковый первый (передний) модифицированный мономер с ОСД1 (помечен буквой "а" на этой Фигуре), но второй (задний) мономер убиквитина модифицирован ОСД2 в разных положениях. Варианты 52-D10 и 52-В3 имеют другой первый (передний) модифицированный мономер по сравнению с 46-Н9 с ООС1, но одинаковый второй (задний) мономер убиквитина с ОСД2 (помечен буквой "е").
Гетеродимеры модифицированного убиквитина имеют следующие последовательности:
46-Н4: SEQ ID NO:25, 45-Н9: SEQ ID NO:26, 46-А5: SEQ ID NO:27, 50-G11: SEQ ID NO:28, 52-В3: SEQ ID NO:29, 52-D10: SEQ ID NO:30
Вышеуказанные последовательности были модифицированы в ходе экспериментов путем добавления метки His-Tag с последовательностью LEHHHHHH (SEQ ID NO:31).
Как можно видеть из Фиг. 3, 46-Н4 имеет превосходное связывающее сродство с ED-B (кД=189 нМ); 46-А5 и 52-D10 не имеют связывающей активности, тогда как другие модифицированные убиквитиновые белки имеют небольшую связывающую активность с ED-B по сравнению с 46-Н4. Таким образом можно сделать вывод, что оба мономера в гетеродимерном варианте требуются для высокоаффинного связывания с мишенью: оба мономера проявляют одновалентное связывание с мишенью.
Гетеродимер модифицированного убиквитина с высокой активностью связывания с ED-B, названный 46Н9, идентифицируется следующими заменами аминокислот в обеих областях связывающегося домена в двух мономерах по сравнению с мономерами убиквитина дикого типа:
в первом модуле (ОСД1) (a) Q2G, F4V, K6R, Q62P, K63H, Е64А, S65T, T66L
во втором модуле (ОСД2) (е) K6H, L8M, Q62K, K63P, E64I, S65A, Т66Е
50G11
в первом модуле (46Н9)(а) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L
во втором модуле (с) K6M L8R, Q62M, K63N, Е64А, S65R, T66L
46Н4
в первом модуле (46Н9) (a) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L
во втором модуле (d) K6G. L8W, Q62T, K63Q, E64Q, S65T, T66R
52В3
в первом модуле (g) Q2R, F4P, K6Y, Q62P, K63P, E64F, S65A, T66R
во втором модуле (46Н9) K6H, L8M, Q62K, K63P, E64I, S65A, Т66Е
52D10 (не связывается с ED-B)
в первом модуле Q2V, F4C, K6R, Q62T, K63A, Е64Р, S65G, T66D
во втором модуле (46Н9) (е) K6H, L8M, Q62K, K63P, E64I, S65A, Т66Е
46А5 (не связывается с ED-B)
в первом модуле (46Н9)(а) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L
во втором модуле (b) K6L, L8M, Q62L, K63A, E64F, S65A,
На Фиг. 4 показано выравнивание последовательности. Линия 1: два мономера убиквитинового белка дикого типа (1-я линия) связаны с 12-аминокислотным линкером SGGGGSGGGGIG, начинающимся в положении 77 и заканчивающимся в положении 88; второй мономер с ОСД2 начинается в положении 89 с метионином. Этот димерный убиквитиновый белок дикого типа выровнен с гетеродимерным вариантом 46-Н9 модифицированного убиквитина (2-я линия) с разными модификациями в первом и во втором мономерах, давая две ОСД. Обе ОСД действуют совместно в связывании мишени из-за одновалентного связывания с мишенью.
На Фиг. 5 показано выравнивание последовательности гетеродимерного варианта 1041-D11 (1-я линия) модифицированного убиквитина с "Ub2_TsX9" (убиквитин, модифицированный в положении 45 в обоих мономерах в триптофан, показывая линкер GIG между двумя мономерами (положения 77-79; второй мономер начинается с метионина в положении 80), и замена с глицина на аланин в последних с-терминальных аминокислотах 2-го мономера. Третья линия показывает "Уби-димер дт", в качестве димера убиквитин дикого типа; не показывая линкерного выравнивания (таким образом, второй мономер начинается в положении 77 с метионином). 4-я линия показывает "Уби-мономер дт", который является человеческим убиквитином дикого типа.
На Фиг. 6 показан зависящий от концентрации ИФА связывания варианта 1041-D11 гетеродимерного убиквитина с ED-B человека. Вариант 1041-D11 проявляет очень высокоаффинное связывание с ED-B (кД=6,9 нМ=6,9×10-9 М). Закрытые точки показывают сродство связывания варианта 1041-D11 гетеродимерного убиквитина с ED-B, содержащим фрагмент фибронектина (названный 67B89-tO), по сравнению с отсутствием связывания этого варианта с отрицательным контролем (названным 6789-tO) (незакрашенные круги).
На Фиг. 7 показаны результаты конкурентного, зависящего от концентрации ИФА связывания варианта 1041-D11 гетеродимерного убиквитина с иммобилизованным ED-В, содержащим фрагмент фибронектина (67 В89), в присутствии увеличивающихся количеств свободной мишени. Вариант 1041-D11 гетеродимерного убиквитина проявляет очень высокоаффинное связывание с ED-B с IC50=140 нМ растворимого 67 В89, указывая, что связывание 1041-D11 не является артефактом ухудшения структурных качеств ED-B из-за иммобилизации на гиброфобной поверхности, используемой в настройках конкурентного ИФА.
На Фиг. 8 показан результат анализа молекулы 1041-D11 модифицированного гетеродимерного убиквитина в анализах взаимодействий без меток с использованием Biacore®. Были проанализированы разные концентрации этого варианта гетеродимерного убиквитина (смотрите легенду на Фигуре: 0-200 нМ 1041-D11) для связывания с ED-B, содержащим фрагмент фибронектина (названный 67 В89), иммобилизованным на чипе SA (Biacore). Анализ кривых ассоциации и диссоциации дал значение кД 1 нМ (1×10-9 М) и показатель koff 7,7×10-4 с-1, который показывает длительное время полураспада комплекса 1041-D11 и ED-B.
На Фиг. 9 показано связывание варианта 1041-D11 гетеродимерного убиквитина с ED-В в зависящем от концентрации ИФА при одновременном анализе сывороточной стабильности связывающей активности. Показаны разные условия, такие как предварительная инкубация в течение 1 часа при 37°С этого варианта в сыворотке мыши или крысы и в PBST как контроле. Значения в кД находятся в интервале от 10 до 20 нМ. Таким образом, можно сделать вывод, что на связывание гетеродимера 1041-D11 с ED-В сыворотка крови влияет незначительно.
На Фиг. 10 показан анализ формирования комплекса варианта 1041-D11 гетеродимерного убиквитина с фрагментами фибронектина за счет СО-ЖХВР.
На Фиг. 10А показано формирование комплекса 1041-D11 с ED-B. Три хроматограммы ЖХВР наложены друг на друга: синий пик с временем удержания 21,651 мин происходит из чистого 1041-D11; черный пик с временем удержания 26,289 мин представляет фрагмент фибронектина 67 В 8 9; смесь 1041-D11 и 67 В 8 9 дает красный пик с временем удержания 21,407 мин после СО-ЖХВР. Сдвиг пика 1041-D11 на меньшее время удержания, а также исчезновение пика 67 В89 указывает на формирование комплекса 1041-D11 и растворимого ED-B.
На Фиг. 10 В показано наложение трех хроматограмм СО-ЖХВР 1041-D11 (синий, 21,944 мин), фрагмент фибронектина 6789 без ED-B (черный, 26,289 мин) и смесь 1041-D11 и 6789 (красная линия с пиками в 21,929 мин и 26,289 мин). Почти не наблюдается сдвиг пика 1041-D11. Этот факт вместе с отсутствием исчезновения пика 6789 указывает на незначительное связывание свободного фрагмента фибронектина 6789 ED-B.
На Фиг. 11 показаны консенсусные положения и замещения аминокислот в вариантах связывания ED-B. Показаны 16 представительных последовательностей гетеродимеров, которые, как было выяснено, имеют удивительно сильное связывающее сродство с ED-В. Консенсусными положениями аминокислот в первой мономерной области, определяющей связывание, являются 2, 4, 6, 62, 63, 64, 65, 66, а консенсусными замещениями аминокислот являются Q2T, F4W, K6H, Q62N, K63F, E64K, S65L и T66S.
На Фиг. 12 показано выравнивание последовательности шести гетеродимерных белков на основе убиквитина, связывающих MIA2. Второй мономер убиквитина начинается с метионина в положении 89 (1111-В4,1111-С9) или в положении 80 (1111-Е10, 1111-F6, 1П1-Н12,1111-Н2).
На Фиг. 13 показано выравнивание области, определяющей связывание ОСД1 и ОСД2, а также линкеры связывающих MIA2 гетеродимерных белков на основе убиквитина с Фиг. 12. Также показаны дополнительные замены аминокислот в последовательности убиквитина.
На Фиг. 14 показан зависящий от концентрации ИФА связывания варианта 1111-Е 10 с Фиг. 12 с биотинилированным MIA-2 (biot. MIA2), Kd=2,6 мкмоль (закрашенные круги); контрольный человеческий сывороточный альбумин (HSA) (незакрашенные круги).
Фиг. 15А. Модификации были выполнены в аминокислотных остатках в серии молекул первого и второго мономерных убиквитиновых звеньев, и выравнивания последовательностей были выполнены для оценки наиболее сильных сайтов связывания. Часть А показывает информацию по последовательности для первого, и Часть В для второго мономерного звена модифицированного убиквитина.
Фиг. 15В. Модификации в положениях 2, 4, 6, 62-66, 68 первого мономера убиквитина и в положениях 6, 8, 62-66 во втором мономере. Линкер между двумя убиквитиновыми мономерами: SGGGGSGGGGIG.:
Фиг. 15С.Показан зависящий от концентрации ИФА связывания варианта SPWF-15_6-А12 гетеродимерного убиквитина с человеческим TNFalpha. Связывающий белок SPWF-15_6-A12 показывает связывание с очень высоким сродством с TNFalpha (Kd=12 нМ=1,2×10-8 M). На этой фигуре показано связывание с высоким сродством к человеческому TNFalpha (закрашенные круги); контрольный БСА (незакрашенные круги).
Фиг. 15D. Последовательность связывающего белка SPWF-15_16-D4_Th гетеродимерного убиквитина со специфичностью к TNFalpha. Модификации в положениях 2, 4, 6, 62-66 первого мономера убиквитина и в положениях 6, 8, 62-66 во втором мономере. Линкер между двумя мономерами убиквитина: SGGGGSGGGGIG.
Фиг. 15Е. Показан зависящий от концентрации ИФА связывания варианта SPWF-15_16-D4_Th гетеродимерного убиквитина с человеческим TN Falpha. Связывающий белок SPWF-15_6-A12 показывает очень высокое сродство с TNFalpha (Kd=1,7 нМ=1,7×10-9 M). На этой фигуре показано связывание с человеческим TNFalpha (закрашенные круги); контроль: бычий сывороточный альбумин (БСА) (незакрашенные круги).
На Фиг. 16 показано связывание NGF гетеродимерами на основе модифицированного убиквитина.
Фиг. 16А. Последовательность связывающего белка SPWF9-1B7-th со специфичностью для NGF. Модификации в положениях 2, 4, 6, 62-66 и в положении 51 первого мономера убиквитина и в положениях 6, 8, 62-66 во втором мономере. Линкер между двумя мономерами убиквитина: SGGGGSGGGGIG.
Фиг. 16В. Зависящий от концентрации ИФА определяет связывание с высоким сродством Kd 0,9 мкМ=9×10-7 М) с NGF. На фигуре показано связывание с рекомбинант-ным человеческим NGF (rhNGF; закрашенные круги); контроль: БСА (незакрашенные круги).
Фиг. 16С. Последовательность связывающего белка SPWF9-6A2-th гетеродимерного убиквитина со специфичностью для NGF. Модификации в положениях 2, 4, 6, 62-66 первого мономера убиквитина и в положениях 6, 8, 62, 64-66 во втором мономере. Линкер между двумя мономерами убиквитина: SGGGGSGGGGIG.
Фиг. 16D. Зависящий от концентрации ИФА определяет связывание с высоким сродством Kd 180 нМ=1,8×10-7 М) с NGF. На фигуре показано связывание с рекомбинант-ные человеческим NGF (rhNGF; закрашенные круги); контроль: БСА (незакрашенные круги).
Фиг. 17 Гетеродимерные белки, связывающие IgG.
Фиг. 17А. Последовательность связывающего белка SPVF4-16B2-ts гетеродимерного убиквитина со специфичностью для IgG. Модификации в положениях 6, 62, 63, 65, 66 первого мономера убиквитина и в положениях 6, 62-66 во втором мономере. Линкер между двумя мономерами убиквитина: SGGGGSGGGGIG. Фиг. 17 В. Зависящий от концентрации ИФА определяет связывание со сродством Kd 3,8 мкМ с IgG. На фигуре показано связывание с IgG (закрашенные черные круги); контроль: БСА-1, БСА-2 и Энбрель (красные, зеленые и синие круги без подогнанной линии). Энбрель несет часть Fe человеческого IgG1. Слабое связывание с Энбрель указывает на связывание SPVF4-16B2-ts с частью Fab IgG.
Фиг.17С. Последовательность связывающего белка SPVF4-9C6-ts гетеродимерного убиквитина со специфичностью для IgG. Модификации в положениях 6, 8, 62-66 первого мономера убиквитина и в положениях 6, 8,62-66 во втором мономере. Линкер между двумя мономерами убиквитина: SGGGGSGGGGIG. Фиг. 17D. Зависящий от концентрации ИФА определяет связывание со сродством Kd 4,1 мкМ с IgG. На фигуре показано связывание с IgG (закрашенные черные круги); контроль: БСА (незакрашенные круги) и Этанерцептом (фирменное наименование Энбрель) (красные круги, без подогнанной линии). Энбрель несет часть Fc человеческого IgG1. Слабое связывание с Энбрель указывает на связывание SPVF4-9C6-ts с частью Fab IgG.
ПРИМЕРЫ
Следующие Примеры представлены для дальнейшей иллюстрации изобретения. Изобретение, в частности, продемонстрировано по отношению к модификации убиквитина в качестве примера. Изобретение, однако, этим не ограничено, и последующие Примеры просто показывают осуществимость изобретения на практике на основе вышеприведенного описания. Для полного раскрытия изобретения ссылка также делается на публикации, упомянутые в данной заявке и в приложении, которые все включена в полном объеме в данную заявку путем ссылки.
Пример 1. Идентификация гетеродимерных белков, связывающихся с ED-B, на основе модифицированных убиквитиновых белков
Конструирование и планирование библиотеки
Если не указано иное, использовались хорошо известные рекомбинантные генетические способы, которые описаны, например, в публикации Сэмбрука и др. (Sambrook et al).
Неспецифическая библиотека гетеродимеров человеческого убиквитина с высокой сложностью была подготовлена путем согласованного мутагенеза в совокупности 15 выбранных положений аминокислот.Модифицированные аминокислоты, которые были замещены триплетами NNK, включали по меньшей мере 3 аминокислоты, выбираемые из положений 2, 4, 6, 8, 62, 63, 64, 65, 66, 68 в проксимальном (первом) мономере убиквитина, и по меньшей мере 3 аминокислоты, выбираемые из положений 2, 4, 6, 8, 62, 63, 64, 65, 66, 68 в дистальном (втором) мономере убиквитина. Оба мономера убиквитина были генетически связаны (голова к хвосту) линкером глицин/серин по меньшей мере с последовательностью GIG или линкером глицин/серин по меньшей мере с последовательностью SGGGG, например GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG (SEQ ID NO:32) или SGGGGSGGGG, но возможен любой другой линкер.
Выбор фагового дисплея ТАТ
Библиотека гетеродимерного убиквитина была обогащена относительно использования мишени, например, фагового дисплея ТАТ в качестве системы выбора. Могут быть использованы другие способы выбора, известные в данной области. Мишень может быть иммобилизована неспецифически на связывающихся поверхностях белка или через биотинилированные остатки, которые были ковалентно соединены с белком. Предпочтительна иммобилизация посредством биотина на гранулах стрептавидина или полосках нейтравидина. Фаги, связывающиеся с мишенью, выбирают или в растворе, или на иммобилизованной мишени; например, биотинилированную и иммобилизованную мишень с фагом инкубировали путем промывки фагов, связанных с матриксом, и путем элюирования фагов, связанных с матриксом. В каждом цикле после инкубации мишени гранулы отделяли от раствора магнитным способом и промывали несколько раз. В первом цикле выбора биотинилированная мишень была иммобилизована на полосках нейтравидина, тогда как в циклах 2-4 выполняли выборы в растворе, после чего следовала иммобилизация комплексов мишень-фаг на покрытых стрептавидином гранулах Dynabeads® (компания Invitrogen). После промывки в первых двух циклах выбора фаги связывающегося с мишенью модифицированного убиквитина высвобождали путем элюирования кислым раствором. В циклах выбора 3 и 4 элюирование фагов осуществляли путем конкурентного элюирования с избыточной мишенью. Элюированные фаги были реамплифицированы. Для направления специфичности связующих во время выбора можно включить белок, сходный с мишенью.
Альтернативно выбору фагового дисплея ТАТ: выбор рибосомного дисплея
Библиотека убиквитина была обогащена относительно использования мишени, например, рибосомным дисплеем в качестве системы выбора (Занд и др. (Zahnd et al.), 2007), Охаси и др. (Ohashi et al.), 2007). Могут быть использованы другие способы выбора, известные в данной области. Мишень была биотинилирована согласно стандартным способам и иммобилизована на покрытых стрептавидином гранулах Dynabeads®. (компания Invitrogen). Троичные комплексы, включающие рибосомы, мРНК и возникающий убиквитиновый полипептид, были ассемблированы с использованием набора PURExpress™ In Vitro для синтеза белка (NEB). Были выполнены два первичных цикла выбора, в которых инкубировали троичные комплексы, после чего выли выполнены два подобных цикла выбора. В каждом цикле после инкубации мишени гранулы отделяли от раствора магнитным способом и промывали буфером рибосомного дисплея с повышающейся тщательностью. После промывки в первых двух циклах выбора гранулы снова отделяли от раствора магнитным способом, и мРНК молекул модифицированного убиквитина, связывающихся с мишенью, освободили от рибосом путем добавления. 50 мМ ЭДТА. В циклах 3 и 4 выбора элюирование мРНК осуществляли конкурентным элюированием с избыточной мишенью (Липовсек и Плактун (Lipovsek and Pluckthuh), 2004). После каждого цикла выполняли очистку РНК и синтез кДНК с использованием набора "RNeasy MinElute Cleanup Kit" (компания Qiagen, Германия), набора "Turbo DNA-free Kit" (компания Applied Biosystems, США) и обратной транскриптазы транс-криптора (компания Roche, Германия).
Клонирование обогащенных пулов
После четвертого цикла выбора синтезированная кДНК была амплифицирована посредством ПЦР согласно известному способу, вырезана с помощью подходящих рест-рикционных нуклеаз и лигирована в экспрессирующий вектор рЕТ-20b(+) (компания Merck, Германия) через совместимые липкие концы.
Анализ совпадений клонированных колоний
После трансформации в клетки NovaBlue(DE3) (компания Merck, Германия) резистентные к ампициллину клонированные колонии выращивали в течение 6 ч при 37°С в 200 мкл среды SOBAG (среда SOB, содержащая 100 мкг/мл ампициллина и 20 г/л глюкозы), экспрессия модифицированного убиквитина, связывающегося с ED-B, была достигнута путем культивирования в течение 16 ч при 37°С в 96-луночных глубоких планшетах (компания Genetix, Великобритания) с использованием 500 мкл автоиндукционной среды ZYM-5052 (Штудиер (Studier), 2005). Клетки собрали и затем лизирова-ли. После центрифугирования полученные супернатанты подвергли скринингу путем ИФА, используя планшеты Nunc MediSorp (компания Thermo Fisher Scientific, США), покрытые 4 мкг/мл ED-B и убиквитин-специфическим фрагментом Fab, конъюгиро-ванным с пероксидазой хрена. В качестве детектирующего реагента использовали TMB-Plus (компания Biotrend, Германия), и желтый цвет получили, используя 50 мкл/лунка 0,2 М раствора H2S04, измерили в устройстве для чтения планшетов на 450 нм против 620 нм.
Обычно выполняли несколько циклов дисплея выбора против мишени. В двух последних циклах выбора связывающие молекулы элюировали с избытком свободной мишени.
Например, были идентифицированы гетеродимерные модифицированные убиквитино-вые связывающие белки против мишени ED-B, такие как 46Н9 (SEQ ID NO:6), 9Е12 (SEQ ID NO:7), 22D1 (SEQ ID NO:8), 1041-D11 на Фиг. 5 (SEQ ID NO:33), 1045-D10 (SEQ ID NO:34). Например, были идентифицированы гетеродимерные модифицированные убиквитиновые связывающие белки против другой мишени, например, против белка 1111-Е10 Фиг. 12 (SEQ ID NO:53), связывающего мишень MIA-2, против белков SPWF-15_6-A12 Фиг. 15 В (SEQ ID NO:57) и SPWF-15J6-D4 Фиг. 15D(SEQ ID NO:90), связывающих мишень TNFalpha, против белков SPWF9-1B7-th Фиг. 16А (SEQ ID NO:91) и SPWF9-6A2-th Фиг. 16С (SEQ ID NO:92), связывающих мишень NGF и против белков SPVF4-16B2-ts Фиг. 17А (SEQ ID NO:93) и SPVF4-9C6-ts Фиг. 17С (SEQ ID NO:94), связывающих мишень IgG.
Выравнивание последовательности мономера убиквитина дикого типа (Уби-мономер дт) с димером убиквитина дикого типа (Уби-димер дт) и убиквитиновым белком (Ub2-TsX на Фиг. 5 с заменой в положении 45 каждого мономера и с двумя замещениями в С-окончании) с гетеродимерным вариантом 1041-D11 модифицированного убиквитина показано на Фиг. 5. В Ub2-TsX замещения в С-окончании (GG на АА) мономера повышают стабильность в сыворотке, поскольку деубиквитиназы расщепляются за GG убиквитина, но не за АА. Вторичная структура убиквитина дикого типа по сравнению с убиквитином с этими замещениями в С-окончании почти идентична.
Модифицированные убиквитины с превосходной активностью связывания с ED-B, указанные как 1041-D11 (показаны на Фиг. X; SEQ ID NO:36) или 1045-D10, идентифицируются по следующим заменам аминокислот по сравнению с диким типом: в первом модуле: K6W, L8W, K63R, Е64К, S65F, Т66Р; во втором модуле: КбТ, L8Q, Q62W, K.63S, E64N, S65W, Т66Е; по выбору Q2R (в варианте 1041-D11, но не в варианте 1045-D10). Подходящими предпочтительными линкерами для этого составного белка являются линкеры, имеющие по меньшей мере последовательность GIG или имеющие по меньшей мере последовательность SGGGG, или любой другой линкер, например, GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG или SGGGGSGGGG. Однако, есть много известных линкеров, которые можно здесь использовать. Другие вещества, связывающие EDB, с их консенсусной последовательностью в первой мономерной области, определяющей связывание, показаны на Фиг. 11.
Модифицированные убиквитины с превосходной активностью связывания MIA-2 показаны на Фиг. 12-14.
Модифицированные убиквитины с превосходной активностью связывания NGF показаны на Фиг. 16.
Модифицированные убиквитины с превосходной активностью связывания TNFalpha показаны на Фиг. 15.
Модифицированные убиквитины с превосходной активностью связывания IgG показаны на Фиг. 17.
Пример 2: Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина с человеческой мишенью
Пример 2А. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем ИФА, зависящего от концентрации.
Связывание вариантов на основе убиквитина с мишенью человека анализировали путем ИФА, зависящего от концентрации. Увеличивающиеся количества очищенного белка наносили на планшеты NUNC-medisorp, покрытые мишенью человека, БСА или сывороточным альбумином человека (САЧ) и возможными другими контролями, такими как клеточный фибронектин (cFN), если в качестве мишени использовали ED-B. Антигенное покрытие с 50 мкл (10 мкг/мл) раствора белка на лунку выполнили при 4°С в течение ночи. После промывки планшетов фосфатно-солевым буфером (ФСБ), 0,1% Tween-20 pH 7,4 (ФСБТ) лунки блокировали, используя блокирующий раствор (ФСБ рН 7,4; 3% БСА; 0,5% Tween-20) при комнатной температуре в течение 2 ч. Лунки снова три раза промыли ФСБТ и затем три раза ФСБ. Покрытые лунки инкубировали с разными концентрациями белка, связывающего мишень, при комнатной температуре в течение 1 ч. После промывки лунок ФСБТ нанесли конъюгаты фрагмента анти-Ubi fab (AbyD) POD в подходящем разбавлении в ФСБТ. Планшет промыли три раза буфером ФСБТ. В каждую лунку добавили 50 мкл раствора субстрата ТМВ (компания KEM-EN-Тес) и инкубировали в течение 15 мин. Реакцию прекратили путем добавления 0,2 М H2SO4. Планшеты ИФА считывали, используя устройство для чтения ИФА TECAN Sunrise. Фотометрические измерения спектральной поглощательной способности выполнили на 450 нм, используя 620 нм в качестве контрольной длины волны. На Фиг. 6 показано связывание с высоким сродством варианта 1041-D11 с ED-B (Kd=6,9 нМ). Это подтверждено в отношении других молекул-мишеней MIA-2, TNFalpha, NGF и IgG результатами, показанными на Фиг. 14, 15, 16 и 17, соответственно. Таким образом, только несколько модификаций (до 8 замещений в каждом мономере) в убиквитине дикого типа дают сродство с данными мишенями в низком микромолярном диапазоне.
Пример 2В. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем конкурентного ИФА, зависящего от концентрации.
Здесь описан анализ связывания для мишени ED-B, но без дальнейших экспериментов, его можно использовать для любой другой мишени. Был выполнен конкурентный, зависящий от концентрации ИФА, для анализа связывания варианта 1041-30 D11 убиквитина с иммобилизованным ED-B, содержащим фрагмент фибронектина (67 В 8 9), в присутствии увеличивающихся количеств свободной мишени. Условия ИФА были как описано, например, для Примера 2А, за тем исключением, что белок 1041-D11 был предварительно инкубирован с ED-B (67 В 89) (0 мкМ - 10 мкМ) или также с отрицательным контролем 6789 (0 мкМ - 10 мкМ) в течение 1 ч и затем эту смесь с мишенью 67 В89 поместили на планшет Medisorp; после этого этот вариант был детектирован соответствующим антителом (анти-Убиквитин-Fab-пероксидаза хрена; разбавление 1:6500).
На Фиг. 7 показано, что вариант 1041-D11 имеет очень высокоаффинное связывание с ED-B (IC50=140 нМ). Результат, показанный на Фиг. 6 подтвержден; только немного модификаций (до 8 замещений в каждом мономере) в убиквитине дикого типа приводят к очень высокоаффинному связыванию с ED-B.
Пример 2С. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем ИФА, зависящего от концентрации, с одновременным анализом стабильности связывающей активности в сыворотке.
ИФА выполнили, используя способы, хорошо известные в данной области, которые описаны выше (Пример 2А и 2В). ED-B (здесь названный 67 В89) нанесли на микротит-ровальные планшеты, этот вариант связали с ED-B и детектировали антителом, специфическим к убиквитину (анти-Ubi-Fab-пероксидаза хрена). Вариант в этом анализе обрабатывали разными путями: вариант инкубировали в сыворотке мышей в течение 1 ч при 37°С (смотрите на Фиг. 9, синие круги); вариант инкубировали в сыворотке крыс в течение в течение 1 ч при 37°С (на Фиг. 13Х красные круги), или вариант инкубировали в ФСБ в течение 1 ч при 37°С (на Фиг. 9 черные круги). Фиг. 13 показывает, что все кД варианта 1041-D11 составляли от 10,3 нМ (в ФСБ) до 20,74 нМ (в сыворотке мышей).
Пример 2D. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем анализов Biacore.
Были проанализированы разные концентрации варианта (например, 0 - 200 нМ варианта, предпочтительно 1041-D11) для связывания с ED-B, содержащим фрагмент фибро-нектина (названный 67 В89), иммобилизованным на чипе СМ5 (Biacore), используя способы, известные специалистам в данной области. Полученные данные обрабатывали посредством программного обеспечения BIAevaluation и 1:1-Langmuir-подгонкой. Молекулярная масса варианта 1041-D11 составляла 1,0 нМ, как показано на Фиг. 8. Кинетические константы связывания составили kon=7,6*105 М-1с-1; koff=7,7*10-4 c-1. kd составного белка 1041-D11 - TNF-альфа составила 1,13 нМ. Кинетические константы связывания составили kon=4,5*105 М-1с-1; koff=5,0*10-4 c-1.
Пример 2Е. Анализ формирования комплекса вариантов, связывающих ED-B, на основе модифицированного убиквитина путем СО-ЖХВР.
Для анализа формирования комплекса использовали колонки Tricorn Superdex 75 5/150 GL (компания GE-Healthcare) (объем =3 мл), количество вводимого белка составило 50 мкл. Другие условия: буфер: 1×ФСБ, pH 7,3, расход: 0,3 мл/мин, цикл: 45 мин (инъекция пробы: через 15 мин). Условие: 0,72 нмоли белка 1041-D11+0,72 нмоли ED-B (здесь назван 67В89 или также отрицательного контроля 6789) инкубировали в течение 1 ч при комнатной температуре; затем наносили на колонку для анализа формирования комплекса. На Фиг. 10 только вариант показан черным, только мишень ED-B показана синим, связывание варианта, формирующее комплекс с ED-B, показано розовым. Фиг. 10А показывает фибронектин, содержащий ED-B, с этим вариантом; Фиг. 10 В показывает вариант с фибронектином без ED-B(6789). Эта Фигура показывает, что вариант 1041-D11 формирует комплекс с ED-B(67В89), но не формирует комплекса с фибронектином (6789), подтверждая специфичность.
Пример 3: Гетеродимерные связывающие белки на основе убиквитина с улучшенным связыванием с TNF-alpha
Гетеродимерные связывающие белки на основе убиквитина, специфические для TNF-alpha, были выбраны согласно способу настоящего изобретения, т.е., была создана фаговая библиотека, которая включала совокупность модифицированных гетеродимерных убиквитиновых связывающих белков, которые были подвергнуты скринингу на их потенциал связывания с TNF-alpha. Были выполнены следующие модификации:
в первом мономере: в одной или нескольких аминокислотах в положениях 2, 4, 6, 62-66, по выбору дополнительно в одном или нескольких из положений 68, 70, 72-74, по выбору в дополнительных положениях;
во втором мономере: модификации in одной или нескольких аминокислотах в положениях 6, 8, 62-66.
В качестве линкера в большинстве случаев использовали SGGGGSGGGGIG, за исключением 1144-D11 (SEQ ID NO:79) и 1144-Е9 (SEQ ID NO:80). Линкер не использовали для 1144-D11 и 1144-Е9 между первым и вторым мономером убиквитина. Положения 75 и 76 или АА, или GG. Линкер показан в части А на Фиг. 15. Связывающее сродство показано на Фиг. 15В-Е.
Пример 4: Создание гетеродимерных связывающих белков на основе убиквитина с улучшенным связыванием с MIA2
MIA2 является диагностическим и терапевтическим маркером, помимо прочего в контексте цирроза, фиброза и рака печени. Подробную информацию по этому маркеру можно найти в документе US 2004076965.
Белком-мишенью для модифицированных убиквитиновых связывающих белков настоящего изобретения является стабильная основная область MIA-2 из 101 аминокислоты, упоминаемая здесь как SPR30-3. SPR30-3 является структурированной частью MIA-2. Она гомологична MIA (CD-RAP), OTOR, TANGO, исключая сигнальный пептид. Ее молекулярная масса составляет 11569,198 Да.
Основная аминокислотная область MIA-2 следующая (SEQ ID NO:95):
MLESTKLLADLKKCGDLECEALINRVSAMRDYRGPDCRYLNFTKGEEISVYVKLAGE REDLWAGSKGKEFGYFPRDAVQIEEVFISEEIQMSTKESDFLCL
Гетеродимерные связывающие белки на основе убиквитина, специфические для MIA2, были выбраны согласно способу настоящего изобретения, т.е., была создана фаговая библиотека, которая включала совокупность модифицированных гетеродимерных убиквитиновых связывающих белков, которые были подвергнуты скринингу на их потенциал связывания с MIA2. Результаты следующие.
На Фиг. 13 показано выравнивание гетеродимерных белков, связывающих MIA2, на основе убиквитина.
Вариант 1111-Е10 показывает сродство в микромолярном диапазоне на биотинилиро-ванной мишени и формирование комплекса в вытеснительной хроматографии. Наиболее сильное связывающее вещество обозначено 1111-Е10 с замещениями аминокислот в положениях 6, 8, 62, 63, 64, 65, 66 в первом мономерном звене убиквитина (ОСД1) и разными замещениями в положениях 6, 8, 62, 63, 64, 65, 66 во втором мономерном звене убиквитина (ОСД2).
Первое мономерное звено убиквитина (ОСД1) показывает те же замещения, что и в 1111-H2 и 1111-Н12. Варианты 111 1-H2 и 1111-Н12 могут поэтому считаться сочетанием ОСД1 и ОСД2, отличающиеся только одной замещенной аминокислотой.
Были оценены следующие другие сильные связывающие молекулы: 1111-C9, 1111-B4 и 1111-F6. Эти связывающие молекулы или были нерастворимы, или не показали связывания с SPR30-3 MIA2 в ИФА и в хроматографии со ступенчатым вытеснением. Варианты 1111-Е10 и 1111- С9, соответственно, и 1111-В4 были обогащены (дополнительное замещение Т9А в 1111-В4 произошло несколько раз). Вариант 111 1-F6 не был обогащен, но казался подходящим кандидатом из-за его высокого сигнала в ИФА; однако этот вариант кажется был нерастворимым.
На ФИГ.14 показан зависящий от концентрации ИФА с вариантом 1111-Е10, связывающимся с биотинилированным MIA-2 (биот.MIA2), Kd=2,6 мкМ (закрашенные -круги); контроль САЧ (незакрашенные круги). Этот вариант 1111-E10 оказался лучшей молекулой, связывающейся с MIA2. Последовательность следующая:
MQIFVETFTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSDY
NIGWHPELHLVLRLRGGGIGMQIFVRTETGKTITLEVEPSDTIENVKAKIQDKEGIPPD
QQRLIWAGKQLEDGRTLSDYNILMGYVLHLVLRLRAA
(SEQ ID NO:53)
Использованные линкеры показаны в прилагаемом листинге последовательности как:
ПУБЛИКАЦИИ
1. Birchler, M., F.Viti, L.Zardi, B.Spiess, and D.Neri. 1999. Selective targeting and photocoagulation of ocular angiogenesis mediated by a phage-derived human antibody fragment. Nat Biotechnol 17:984-8.
2. Brenmoehl, J., M.Lang, M.Hausmann, S.N.Leeb, W.Falk, J. Scholmerich, M. Goke, and G. Rogler. 2007. Evidence for a differential expression offibronectin splice forms ED-A and ED-B in Crohn's disease (CD) mucosa. Int J Colorectal Dis 22:611-23.
3. Dubin, D., J. H.Peters, L.F.Brown, B.Logan, K.C.Kent, B.Berse, S.Berven, B.Cercek, B.G.Sharifi, R.E.Pratt, and et al. 1995. Balloon catheterization induced arterial expression of embryonic fibronectins. Arterioscler Thromb Vase Biol 15:1958-67.
4. Goodsell, D.S. 2001. FUNDAMENTALS OF CANCER MEDICINE: The Molecular Perspective: Antibodies. The Oncologist 6:547-548.
5. Kaczmarek, J., P.Castellani, G.Nicolo, B.Spina, G.Allemanni, and L.Zardi. 1994. Distribution ofoncofetal fibronectin isoforms in normal, hyperplastic and neoplastic human breast tissues. Int J Cancer 59:11-6.
6. Menrad, A., and H.D.Menssen. 2005. ED-B fibronectin as a target for antibody-based cancer treatments. Expert Opin Ther Targets 9:491-500.
7. Pujuguet, P., A.Hammann, M.Moutet, J.L.Samuel, F.Martin, and M. Martin. 1996. Expression offibronectin ED-A+ и ED-B+ isoforms by human and experimental colorectal cancer. Contribution of cancer cells and tumor-associated myofibroblasts. Am J Pathol 148:579-92.
8. Trachsel, E., M.Kaspar, F.Bootz, M.Detmar, and D.Neri. 2007. A human mAb specific to oncofetal fibronectin selectively targets chronic skin inflammation in vivo. J Invest Dermatol 127:881-6.
9. Van Vliet, A., H.J.Baelde, L.J.VIeming, E.de Heer, and J.A.Bruijn. 2001. Distribution offibronectin isoforms in human renal disease. J Pathol 193:256-62.
10. Lipovsek, D., and Pluckthun, A. (2004). In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 290, 51-67.
11. Ohashi, H., Shimizu, Y., Ying, B.W., and Ueda, T. (2007). Efficient protein selection based on ribosome display system with purified components. Biochem Biophys. Res. Commun. 352,270-276.
12. Studier, F.W. (2005). Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41, 207-234.
13. Zahnd, С., Amstutz, P., and Pliickthun, A. (2007). Ribosome display: selecting and evolving proteins in vitro that specifically bind to a target. Nat. Cnoco6s 4, 269-279.
14. Paschke, M. and W.Hohne (2005). Gene 350(1): 79-88
15. Briiser 2007 Appl Microbiol Biotechnol 76(1): 35-45

































































| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ УБИКВИТИНОВЫЕ БЕЛКИ СО СПЕЦИФИЧЕСКОЙ СВЯЗЫВАЮЩЕЙ АКТИВНОСТЬЮ ДЛЯ ЭКСТРАДОМЕНА "В" ФИБРОНЕКТИНА | 2010 |
|
RU2556820C2 |
| АНТИТЕЛА ПРОТИВ ПОЛИУБИКВИТИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2603093C2 |
| TRAIL R2-СПЕЦИФИЧЕСКИЕ МУЛЬТИМЕРНЫЕ СКАФФОЛДЫ | 2011 |
|
RU2628699C2 |
| БИСПЕЦИФИЧЕСКИЕ АСИММЕТРИЧНЫЕ ГЕТЕРОДИМЕРЫ, СОДЕРЖАЩИЕ АНТИ-CD3 КОНСТРУКЦИИ | 2013 |
|
RU2650868C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (LCN2, HNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2564125C2 |
| CD40L-СПЕЦИФИЧНЫЕ КАРКАСНЫЕ СТРУКТУРЫ, ПРОИСХОДЯЩИЕ ИЗ Tn3, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2704992C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К АМИЛОИДУ БЕТА | 2008 |
|
RU2567151C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ АМИЛОИДА-БЕТА ПРИ ГЛАЗНЫХ ЗАБОЛЕВАНИЯХ | 2008 |
|
RU2571859C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ЛЕГОЧНОЙ ГИПЕРТЕНЗИИ | 2017 |
|
RU2748278C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ АМИЛОИДА БЕТА ПРИ ГЛАЗНЫХ ЗАБОЛЕВАНИЯХ | 2008 |
|
RU2542967C2 |














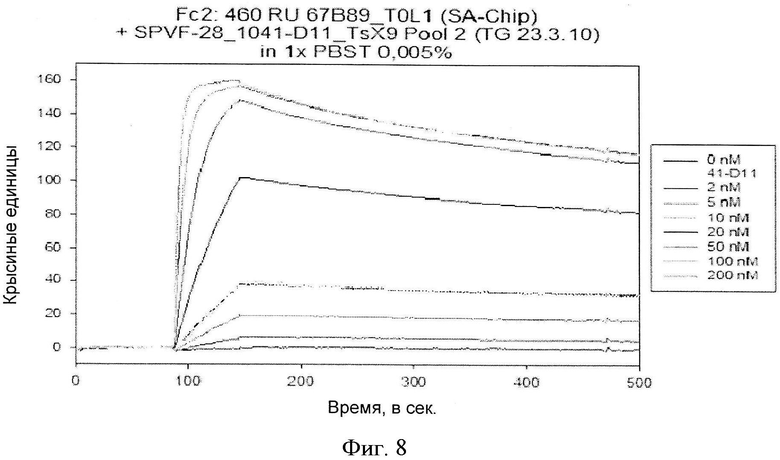

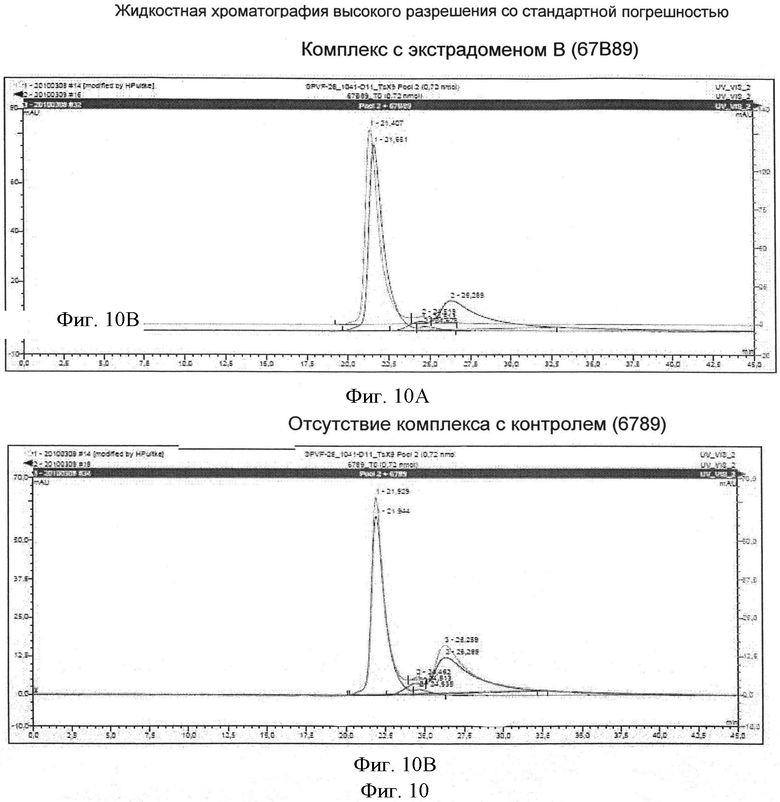
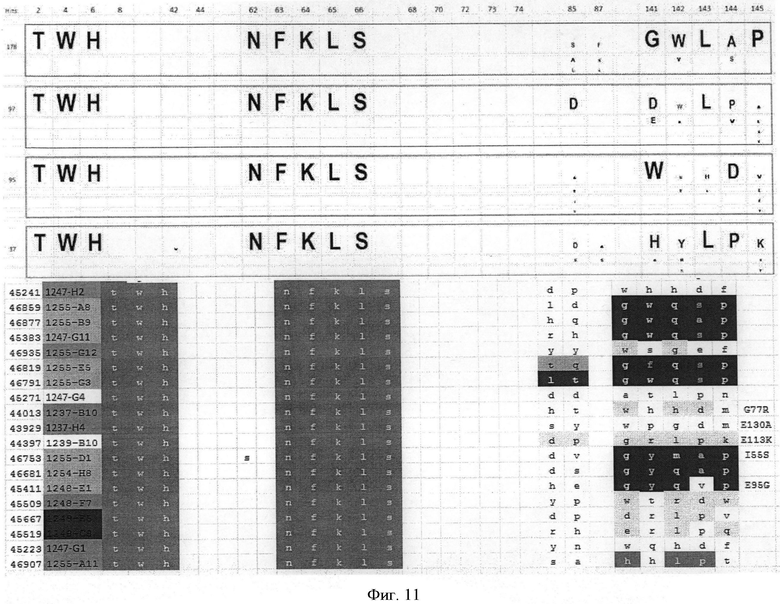
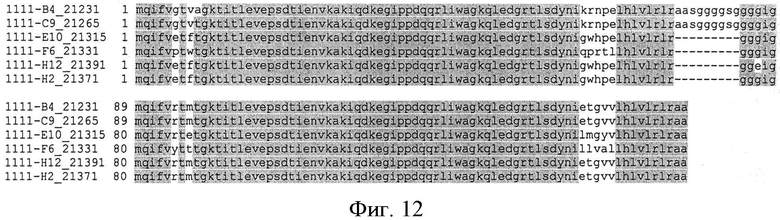
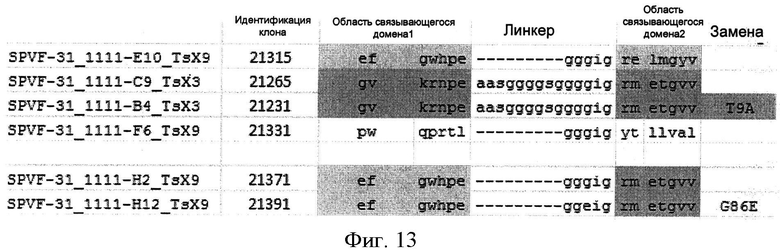





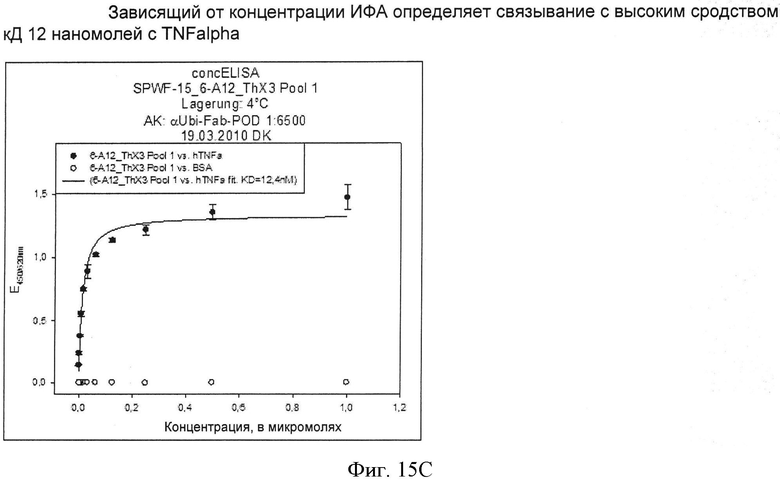






Изобретение относится к области биотехнологии и может быть использовано для идентификации гетеромультимерных убиквитинов со способностью связываться с лигандом-антигеном. Способ включает контактирование совокупности гетеродимерных модифицированных убиквитинов, включающих два мономера убиквитина, связанных между собой по схеме голова к хвосту, с потенциальным лигандом дисплейным способом. При этом каждый из упомянутых мономеров модифицирован по-разному и содержит 5-8 замещений в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1. Далее идентифицируют гетеродимерный модифицированный белок, связавшийся с лигандом с сродством связывания Kd в диапазоне 10-7-10-12 М и одновалентной связывающей активностью. Предложены ДНК-библиотеки, отвечающие за получение популяции упомянутых гетеромультимерных убиквитинов, а также библиотеки белков, полученные путем экспрессии упомянутых ДНК-библиотек. Изобретение позволяет получить новые связывающие белки на основе гетеромультимерного убиквитина, способные специфически связываться с высоким сродством с выбранными лигандами. 3 н. и 5 з.п. ф-лы, 17 ил., 4 пр.
1. Способ идентификации гетеродимерного модифицированного убиквитина со способностью связываться с лигандом-антигеном, включающий следующие этапы:
a) предоставление совокупности гетеродимерных модифицированных убиквитинов, происходящих от мономерных модифицированных убиквитиновых белков, включающей два мономера убиквитина, связанных между собой по схеме голова к хвосту, в которой каждый из упомянутых мономеров содержит 5-8 замещений в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1, причем упомянутый модифицированный мономерный белок имеет идентичность аминокислотной последовательности по меньшей мере 80%, или по меньшей мере 90%, или по меньшей мере 95% с немодифицированным убиквитиновым белком;
b) предоставление потенциального лиганда для упомянутой совокупности по-разному модифицированных белков;
c) контакт упомянутой совокупности по-разному модифицированных белков с упомянутым лигандом дисплейным способом;
d) идентификация гетеродимерного модифицированного белка процессом скрининга, причем упомянутый модифицированный гетеродимерный белок связывается с упомянутым лигандом со специфическим сродством связывания
Kd в диапазоне 10-7-10-12 М и проявляет одновалентную связывающую активность в отношении упомянутого лиганда; и, по выбору,
e) изоляция упомянутого гетеродимерного модифицированного убиквитина с упомянутым сродством связывания,
причем упомянутые мономеры по-разному модифицированы путем замещений поверхностно открытых аминокислот в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1.
2. Способ по п. 1, отличающийся тем, что упомянутый модифицированный мономерный белок включает от 1 до 10 вставок аминокислот в мономерной молекуле и/или от 1 до 7 делеций аминокислот в мономерной молекуле.
3. Способ по любому из предшествующих пунктов, отличающийся тем, что упомянутый модифицированный мономерный убиквитиновый белок получают путем генетической инженерии ДНК, отвечающей за получение убиквитина, и экспрессии упомянутого белка в прокариотических или эукариотических организмах или in vitro.
4. Способ по п. 3, отличающийся тем, что упомянутый димерный белок получают способом фагового дисплея, рибосомного дисплея, фагового дисплея ТАТ, дрожжевого дисплея, бактериального дисплея, дисплея поверхности клетки или способом дисплея мРНК.
5. Способ по п. 1, отличающийся тем, что еще от 1 до 7 дополнительных аминокислот замещают по меньшей мере в одном из мономерных убиквитиновых белков, которые выбирают из одной или нескольких аминокислот в положениях 8, 36, 44, 62, 63,64, 70, 71, 72 и 73 SEQ ID NO:1.
6. Способ по п. 1, отличающийся тем, что упомянутую совокупность гетеродимерных составных белков убиквитина получают путем генетического слияния двух ДНК-библиотек, каждая из которых отвечает за получение по-разному модифицированных мономерных белков, трансляции этой ДНК в гетеродимерные составные белки, дисплей упомянутых белков и скрининг дисплейных белков на присутствие модифицированных гетеродимерных убиквитиновых белков, включающих мономерные убиквитиновые белки, соединенные между собой по схеме голова к хвосту, и отличающийся тем, что упомянутые модифицированные гетеродимерные убиквитиновые белки связываются с упомянутым лигандом со специфическим сродством связывания Kd в диапазоне 10-7-10-12 М и проявляют одновалентную связывающую активность в отношении упомянутого лиганда, или отличающийся тем, что упомянутую совокупность гетеромультимерных составных белков убиквитина получают химическим синтезом белков.
7. ДНК-библиотека для использования в способе по п. 1, отвечающая за получение совокупности гетеродимерных убиквитиновых составных белков, происходящих из мономерных убиквитинов, причем каждый димерный белок включает два модифицированных мономера убиквитина, соединенных между собой по схеме голова к хвосту, отличающаяся тем, что упомянутые мономеры по-разному модифицированы путем замещений поверхностно открытых аминокислот в 5-8 аминокислотах, расположенных в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO:1, причем упомянутый модифицированный мономерный белок имеет идентичность аминокислотной последовательности по меньшей мере 80%, или по меньшей мере 90%, или по меньшей мере 95% с немодифицированным белком.
8. Библиотека белков для использования в способе по п. 1, полученная путем экспрессии ДНК-библиотеки по п. 7.
| WO 2008059011 A1, 22.05.2008 | |||
| US 2006099686 A1, 11.05.2006 | |||
| RAASI S | |||
| et al., Binding of polyubiquitin chains to ubiquitin-associated (UBA) domains of HHR23A, J.Mol.Biol., 2004, v.341, p.1367-1379 | |||
| ДИМЕР МОЛЕКУЛЯРНОГО ВАРИАНТА АПО-ЛИПОПРОТЕИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 1992 |
|
RU2134696C1 |
| ELLISON M.J | |||
| et al., Epitope-tagged Ubiquitin, The journal of biological chemistry, 1991, v.266, n | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| АВТОМАТИЧЕСКОЕ УСТРОЙСТВО К ПАРОВОЗНЫМ ВОДОПОДОГРЕВАТЕЛЯМ ДЛЯ УСТРАНЕНИЯ ПИТАНИЯ КОТЛА ЧЕРЕЗ ВОДОПОДОГРЕВАТЕЛЬ ПРИ ЕЗДЕ С ЗАКРЫТЫМ РЕГУЛЯТОРОМ | 1930 |
|
SU21150A1 |

