ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новым гетеромультимерным белкам, способным связывать экстрадомен В фибронектина (ED-B). Кроме того, изобретение относится к составным белкам, включающим упомянутый гетеромультимерный связывающий белок, объединенный с фармацевтически и/или диагностически активным компонентом. Изобретение далее направлено на способ создания такого гетеромультимерного связывающего белка или составного белка и к фармацевтическим и/или диагностическим композициям, содержащим упомянутые гетеромультимерные связывающие белки. Дополнительно, изобретение относится к библиотекам, содержащим ДНК, кодирующие упомянутые белки.
В других вариантах осуществления изобретение направлено на полинуклеотиды, кодирующие упомянутый гетеромультимерный связывающий белок или составной белок, векторы, включающие упомянутый полинуклеотид, и клетки-хозяева, включающие упомянутый белок, составной белок, вектор и/или полинуклеотид. В одном предпочтительном варианте осуществления упомянутый гетеромультимерный связывающий белок или составной белок включен в медикамент или диагностический агент. Дополнительно, описаны способы получения упомянутого рекомбинантного белка или составного белка, а также использование упомянутых белков в способах медицинского лечения.
УРОВЕНЬ ТЕХНИКИ
Существует растущий спрос на связывающие молекулы, состоящие из аминокислот, которые не являются иммуноглобулинами. Хотя на настоящее время антитела представляют собой лучше всего изученный класс связывающих молекул, все же существует необходимость в новых связывающих молекулах, чтобы получить лиганды с высоким сродством и специфичностью, поскольку молекулы иммуноглобулинов имеют крупные недостатки. Хотя их можно легко получить и они могут быть направлены почти на любую мишень, они имеют очень сложную молекулярную структуру. Существует постоянная необходимость в замещении антител более мелкими молекулами, с которыми можно легко работать. Эти альтернативные связывающие агенты могут быть с выгодой использованы, например, в медицинских областях диагностики, профилактики и лечения заболеваний.
Белки, имеющие относительно определенные трехмерные структуры, обычно называемые белковые каркасы, могут быть использованы в качестве исходного материала для конструирования упомянутых альтернативных связывающих агентов. Эти каркасы обычно содержат один или больше областей, которые подвержены специфическому или случайному изменению последовательности, и такую рандомизацию последовательности часто выполняют для получения библиотеки белков, из которой могут быть выбраны специфические связывающие молекулы. Молекулы меньшего размера чем антитела и со сравнимым или даже более хорошим сродством к целевому антигену будут, как ожидается, лучше антител в смысле фармакокинетических свойств и иммуно-генности.
В нескольких предыдущих подходах действительно используют белковые каркасы в качестве исходного материала для связывающих белков. Например, в документе WO 99/16873 описаны разработанные модифицированные белки семейства липокалинов (так называемые антикалины), проявляющие связывающую активность для некоторых лигандов. Структуру пептидов семейства липокалинов модифицируют путем замещений аминокислот в их естественном связывающем кармане, используя способы генной инженерии. Подобно иммуноглобулинам, эти антикалины можно использовать для
идентификации или связывания молекулярных структур. Аналогично антителам, модифицируют структуры с гибкой петлей; эти модификации дают возможность распознавать лиганды, отличающиеся от естественных.
В документе WO 01/04144 описано искусственное создание связывающего домена на поверхности белка в белках со структурой бета-пласта как таковых, у которых отсутствует связывающий сайт. Посредством этого вновь созданного искусственного связывающего домена могут быть получены, например, изменения в γ-кристаллине - структурном белке хрусталика глаза - которые взаимодействуют с лигандами с высоким сродством и специфичностью. В противоположность модификации сайтов связывания, которые уже присутствуют и образованы из структур с гибкой петлей, которые упомянуты выше для антикалинов, эти домены связывания создаются заново на поверхности бета-пластов. Однако в документе WO 01/04144 описано только изменение относительно крупных белков для создания новых связывающих свойств. Из-за их размера, белки согласно WO 01/04144 могут быть модифицированы на уровне генной инженерии только способами, которые требуют некоторых усилий. Кроме того, в белках, раскрытых до сих пор, только относительно малая процентная доля всех аминокислот модифицирована для поддержания общей структуры белка. Поэтому только относительно небольшая области поверхности белка доступна для создания связывающих свойств, которые не существовали ранее. Более того, WO 01/04144 раскрывает только создание свойства связывания с γ-кристаллином.
В документе WO 04/106368 описано создание искусственных связывающих белков на основе убиквитиновых белков. Убиквитин - это небольшой, мономерный и цитозоль-ный белок, который в высокой степени сохраняется в последовательности и присутствует во всех известных эукариотических клетках от простейших до позвоночных организмов. В организме он играет очень важную роль в регулировании контролируемой деградации клеточных белков. Для этого белки, предназначенные для деградации, ковалентно связываются с убиквитиновыми или полиубиквитиновыми цепями во время их прохождения через каскад ферментов и выборочно деградируют из-за этой метки. Согласно последним результатам, убиквитин или мечение белков убиквитином, соответственно, также играет важную роль в других клеточных процессах, таких как импорт нескольких белков или их генное регулирование.
Помимо прояснения его физиологической функции, убиквитин является объектом исследований главным образом из-за его структурных и белково-химических свойств. Полипептидная цепь убиквитина состоит из 76 аминокислот, уложенных в чрезвычайно компактную α/β структуру (Виджай-Кумар (Vijay-Kumar), 1987): почти 87% этой полипептидной цепи участвует в формировании вторичных структурных элементов посредством водородных связей. Вторичными структурами являются 3,5 альфа-спиральные витки, а также антипараллельный бета-пласт, состоящий из четырех нитей. Характерное расположение этих элементов - антипараллельный бета-пласт с открытой белковой поверхностью, на задней стороне которой упакована альфа-спираль, которая лежит вертикально на ее верху - обычно считается так называемым убиквитино-подобным мотивом укладки. Еще одной структурной особенностью является выраженная гидрофобная область внутри белка между альфа-спиралью и бета-пластом.
Из-за его небольшого размера искусственное приготовление убиквитина может быть осуществлено как химическим синтезом, так и посредством биотехнологических способов. Из-за благоприятных свойств укладки убиквитин можно получать генной инженерией, используя такие микроорганизмы, как Escherichia coli, в относительно больших количествах или в цитозоли, или в периплазматическом пространстве. Из-за окисляющих условий, преобладающих в периплазме, последняя стратегия обычно резервируется для получения секреторных белков. Из-за простого и эффективного бактериального приготовления убиквитин можно использовать как партнера в объединении для других получаемых инородных белков, получение которых вызывает трудности. Посредством объединения с убиквитином можно достигнуть повышенной растворимости и, посредством этого, повышенного выхода продукта.
По сравнению с антителами или другими альтернативными каркасами, искусственные связывающие белки на основе убиквитиновых белков (также называемых Affilin®) имеют много преимуществ: небольшой размер, высокая стабильность, высокое сродство, высокая специфичность, экономичное микробное производство и регулировка сывороточного периода полураспада. Однако, все же существует необходимость в дальнейшей разработке этих белков в смысле новых терапевтических подходов с высокими степенями сродства со специфическими мишенями. Хотя в документе WO 05/05730 описано в общих чертах использование каркасов убиквитина для получения искусственных связывающих белков, там не предложено решения, как модифицировать убиквитиновый белок, чтобы получить специфическое и высокоаффинное связывание с ED-В фибронектина.
В документе WO 2008/022759 описаны рекомбинантные связывающие белки, причем домен 3 (SH3) Src гомологии FYN-киназы используется для получения новых связывающих белков. Установлено, что целевая специфичность может быть разработана путем мутации RT-петли и/или Src-петли, чтобы разработать белковую терапию и/или диагностику. Как и в липокалинах, используемых в качестве каркаса, мутагенизируе-мые аминокислотные остатки лежат в вариабельных и гибких областях петлей, имитируя принцип, лежащий в основе связывающей функции антител а/антигена. Эта общая гибкость сайта взаимодействия, которым антитела связывают эпитоп, является в основном энтальпически проходящим процессом; этот процесс, однако, приводит к неблагоприятному энтропическому вкладу потерей мобильности при связывании гибкой области, определяющей комплементарность. В противоположность этому, используя убиквитин в качестве каркаса, авторы настоящего изобретения не изменяли аминокислотные остатки в основном в гибких областях петли, а в жестких и не гибких бета-нитях области бета-пласта или рядом с бета-нитями. Преимуществом выбора аминокислотных остатков в не гибких и жестких бета-нитях или рядом с бета-нитями убиквитина в качестве областей связывания для ED-B является, помимо прочего, следующее: считается, что связывающиеся партнеры уже представляют комплементарную геометрию, подходящую для плотного связывания. Следовательно, эти взаимодействия включают комплементарность в форме, заряде и гидрофильных/гидрофобных элементах более жестких структур связывающихся партнеров. Эти взаимодействия жестких тел оптимизируют поверхность раздела и обеспечивают биологическую функцию.
Фибронектины (FN) являются важным классом высокомолекулярных внеклеточных матричных гликобелков, обильно экспрессированных в здоровых тканях и жидкостях организма. Их основная роль заключается в способствовании адгезии клеток к ряду разных внеклеточных матриксов.
Присутствие фибронектинов на поверхности нетрансформированных клеток в культуре, а также их отсутствие в случае трансформированных клеток привели к идентификации фибронектинов как важных адгезионных белков. Они взаимодействуют с многочисленными другими разными молекулами, например, коллагена, гепарансульфат-протеогликанов и фибрина и таким образом регулируют форму клеток и создание цитоскелета. Помимо этого, они отвечают за миграцию клеток и дифференциацию клеток во время эмбриогенеза. Они также играют важную роль в излечении ран, в которых они облегчают миграцию макрофагов и других иммунных клеток, и в формировании сгустков крови, позволяя адгезию тромбоцитов к поврежденным областям кровеносных сосудов.
Экстрадомен В (ED-B) фибронектина является небольшим доменом, который вводят альтернативным сплайсингом первичного транскрипта РНК в молекулу фибронектина. Эта молекула присутствует или отсутствует в молекулах фибронектина внеклеточного матрикса и представляет один из наиболее селективных маркеров, связанных с ангиогенезом и ремоделированием ткани, поскольку она обильно экспрессируется вокруг новых кровеносных сосудов, но не может быть детектирована практически во всех нормальных тканях взрослого человека (за исключением матки и яичников). Известно, что ED-B вовлечен главным образом в рак. Высокие уровни экспрессии ED-B были детектированы в областях первичного поражения, а также в местах метастаз многих твердых раковых объектов у человека, включая грудной, немелкоклеточный легочный, колоректальный, панкреатический, кожный, гепатоцеллюлярный, внутричерепную менингеому, глиобластому (Менрад и Менссен (Menrad u. Menssen), 2005). Кроме того, ED-B может быть связан с диагностическими агентами и может выгодно использоваться в качестве диагностического средства. Одним примером является его использование в молекулярной визуализации, например, атеросклеротических бляшек и детектировании рака, например, путем иммуносцинтиграфии раковых пациентов. Также возможны многие другие способы использования в диагностике.
Аминокислотная последовательность из 91 аминокислоты человеческого экстрадомена В (ED-B) фибронектина показана в SEQ ID NO: 2. Для экспрессии белка приходится добавлять стартовый метионин. ED-B обильно присутствует у млекопитающих, на пример, у грызунов, крупного рогатого скота, приматов, плотоядных, людей и т.д. Примерами животных, у которых существует 100% идентичность последовательности с ED-B человека, являются Rattus norvegicus, Bos taurus, Mus musculus, Equus caballus, Macaca mulatta, Canis lupus familiaris и Pan troglodytes.
ED-B специфически аккумулируется в неоваскулярных структурах и представляет мишень для молекулярной интервенции в рак. Некоторое количество антител или фрагментов антител к домену ED-B фибронектина известны в данной области как потенциальные терапевтические средства для рака и других показаний (смотрите, например, WO 97/45544, WO 07/054120, WO 99/58570, WO 01/62800). Фрагмент ScFvL19 антитела Fv одиночной цепи человека (также именуемый L19) является специфическим к домену ED-B фибронектина и, как подтверждено на экспериментальных моделях опухолей, так и у пациентов с раком, селективно нацелен на образование новых сосудов в опухолях. Кроме того, конъюгаты, включающие анти-ED-B антитело или анти-ED-B фрагмент антитела с цитокинами, такими как IL-12, IL-2, IL-10, IL-15, IL-24 или GM-CSF, были описаны для лекарств против клеток-мишеней для производства медикамента для ингибирования, в частности, рака, ангиогенеза роста метастаз (смотрите, например, WO 06/119897, WO 07/128563, WO 01/62298). Селективное нацеливание на образование новых сосудов в твердых опухолях с помощью анти-ED-B антител или анти-ED-B фрагментов антител, таких как L19, конъюгированньгх с подходящей функцией эффектора, такого как цитотоксический или иммуностимулирующий агент, оказалось успешным в экспериментах на животных. Для лечения рака поджелудочной железы составные белки, включающие часть интерлейкина-2 (IL-2) и часть анти-ED-B антитела, были объединены с небольшой молекулой гемцитабина (2'-деокси-2'Д'- дифторцитидин) (смотрите, например, WO 07/115837).
Вышеуказанные документы из существующего уровня техники описывают использование разных каркасов белков, включая антитела, для создания новых, связывающих ED-B белков. Таргетинг ED-B с доступными в настоящее время соединениями имеет определенные недостатки. Более малые молекулы (такие как гетеромультимерные связывающие ED-B белки настоящего изобретения на основе убиквитина) со сравнимым или даже более высоким сродством к антигену ED-B, как ожидается, будут иметь значительные преимущества перед антителами или другими связывающими белками.
Поскольку рак представляет одну из ведущих причин смертности в мире, существует растущая потребность в усовершенствованных агентах для лечения рака. Текущие химиотерапевтические агенты и лечение облучением имеют плохую селективность, и большинство химиотерапевтических агентов не аккумулируются в месте опухоли и, таким образом, не могут достигать адекватных уровней в опухоли. Существует сильная медицинская потребность в эффективном лечении рака.
Таким образом, цель настоящего изобретения заключается в том, чтобы предложить гетеромультимерные связывающие белки на основе убиквитина, способные специфически связываться с очень высоким сродством с внеклеточным доменом фибронектина (ED-B). Еще одна цель настоящего изобретения заключается в том, чтобы идентифицировать и предложить новые связывающие белки с очень высокой связывающей специфичностью к ED-B, например, для использования при лечении рака. Кроме того, должен быть предложен способ, чтобы получать упомянутые гетеромультимерные связывающие молекулы.
Вышеуказанные цели решены предметом независимых пунктов прилагаемой формулы изобретения. Предпочтительные варианты осуществления изобретения включены в зависимые пункты формулы изобретения, а также в последующее описание, примеры и фигуры чертежей.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Более конкретно, авторы изобретения предлагают белок, способный связывать ED-B фибронектина человека, включающий модифицированный гетеродимерный убиквитиновый белок, в котором два мономерных убиквитина (убиквитиновых звена) связаны между собой в расположении "голова к хвосту", причем каждый мономер упомянутого димерного белка по-разному модифицирован путем замещения по меньшей мере 6 аминокислот в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO: 1, и причем упомянутые замещения включают
(1) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно 2, или
(2) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 2, 4, 6, 62, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно 2,
и, по выбору, дальнейшие модификации, предпочтительно замещения других аминокислот, причем упомянутое модифицированное мономерное убиквитиновое звено имеет аминокислотную идентичность с SEQ ID NO: 1 по меньшей мере на одно из группы из 80%, по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 83% и по меньшей мере 90%, и причем упомянутый белок имеет специфическое связывающее сродство с упомянутым доменом ED-B фибронектина с килодальтон =10-7-10-12 М и проявляет одновалентную связывающую активность с упомянутым экстрадоменом В (ED-B) фибронектина.
В одном предпочтительном варианте осуществления, белок является рекомбинантным.
В других вариантах осуществления изобретения, 7, 8, 9 или все из аминокислот в положениях 2,4, 6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO: 1 модифицированы в каждом мономерном убиквитиновом звене. Следует понимать, что настоящее изобретение позволяет комбинацию каждой из этих вариаций в каждом мономерном звене, т.е., в первом и втором звене. Например, первое мономерное звено может включать 6 модификаций, когда второе звено включает 7 или 8 модификаций, первое звено может включать 8 модификаций и второе звено 7 модификаций и т.д. Каждая из вышеперечисленных аминокислот может быть выбрана в первом и втором звене, и затем оба звена объединяют. Предпочтительные замещения описаны ниже.
Определения важных терминов, используемых в настоящей заявке
Термин "экстрадомен В фибронектина" или сокращенно указываемый как "ED-B" включает все белки, которые показывают идентичность последовательности с SEQ ID NO: 2 по меньшей мере на 70%, по выбору на 75%, далее по выбору на 80%, 85%, 90%, 95%, 96% или 97% или больше, или на 100% и которые имеют вышеопределенную функциональность ED-B.
Термины "белок, способный связывать" или "связьшающий белок" относятся к убиквитиновому белку, включающему домен связывания с ED-B, который определен ниже. Любой такой связывающий белок на основе убиквитина может включать дополнительные белковые домены, которые не являются связьшающими доменами, такие как, например, жидкости мультимеризации, полипептидные метки, полипептидные линкеры и/или молекулы небелковых полимеров. Некоторыми примерами молекул небелковых полимеров являются гидроксиэтиловый крахмал, полиэтиленгликоль, полипропиленгликоль или полиоксиалкилен.
Антитела и их фрагменты хорошо известны специалисту в данной области. Связывающий белок изобретения не является антителом или его фрагментом, таким как фрагменты Fab или scFv. Кроме того, связывающий домен изобретения не включает иммуноглобулин, уложенный как в антителах.
В настоящем описании изобретения термины "лиганд" и "мишень" и "связывающийся партнер" используются как синонимы и могут быть взаимно заменены. Лигандом является любая молекула, способная связываться со сродством, определенным здесь для гетеромультимерного модифицированного убиквитинового белка.
Термин "убиквитиновый белок" охватывает убиквитин в соответствии с SEQ ID NO: 1 и его модификации согласно следующему определению. Убиквитин в высокой степени сохраняется в эукариотических организмах. Например, у всех млекопитающих,
исследованных до настоящего времени, убиквитин имеет идентичную аминокислотную последовательность. Особенно предпочтительными являются молекулы убиквитина от человека, грызунов, свиней и приматов. Помимо этого можно использовать убиквитин из любого другого эукаритического источника. Например, убиквитин дрожжей отличается только тремя аминокислотами от последовательности SEQ ID NO: 1. В общем, убиквитиновые белки, охватываемые упомянутым термином "убиквитиновый белок" показывают аминокислотную идентичность более чем 70%, предпочтительно более чем 75% или более чем 80%», более чем 85%», более чем 90%», более чем 95%», более чем 96% или до идентичности последовательности 97% с SEQ ID NO: 1.
Термин "модифицированный убиквитиновый белок" относится к модификациям убиквитинового белка любым одним из замещения, вставок или удалений аминокислот или их сочетания, при этом замещения являются наиболее предпочтительными модификациями, которые могут быть дополнены любой одной из модификаций, описанных выше. Число модификаций строго ограничено, так как упомянутые модифицированные мономерные убиквитиновые звенья имеют аминокислотную идентичность с SEQ ID NO: 1 по меньшей мере одну из группы из 80%, по меньшей мере 83%, по меньшей мере 85%», по меньшей мере 83% и по меньшей мере 90%. Самое большее, общее число замещений в мономерном звене поэтому ограничено 15 аминокислотами, соответствующими 80%» аминокислотной идентичности. Совокупное число модифицированных аминокислот в гетеродимерной молекуле убиквитина составляет 30 аминокислот, соответствующих 20% аминокислотных модификаций на основе гетеродимерного белка. Аминокислотная идентичность димерного модифицированного убиквитинового белка по сравнению с димерным немодифицированным убиквитиновым белком с основной мономерной последовательностью SEQ ID NO: 1 выбирается по меньшей мере как одно из группы из 80%, по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 83% и по меньшей мере 90%.
Для определения степени идентичности последовательности производного убиквитина с аминокислотной последовательностью SEQ ID NO: 1 можно использовать, например, программу SIM локального сходства (Сяокин Хуан и Уэбб Миллер (Xiaoquin Huang and Webb Miller), "Достижения прикладной математики, Том 12: 337-357,1991) или Клустал У. (Clustal, W.) (Томпсон и др. (Thompson et al.), Nucleic Acids Res., 22(22): 4673-4680,1994.).
Предпочтительно, степень идентичности последовательности модифицированного белка с SEQ ID NO: 1 определяют относительно полной последовательности SEQ ID NO: 1.
Тетеродимерным составным белком" или "гетеродимерным белком" изобретения считается белок, который включает два по-разному модифицированных мономерных убиквитиновых белка с двумя взаимодействующими связывающими доменными областями, совместно обеспечивающими одновалентное связывающее свойство (связьшающий домен) для ED-B как специфического связывающегося партнера. Гетеродимер получают путем соединения двух мономерных молекул убиквитина, причем обе эти молекулы модифицированы по-разному, как описано в настоящем документе.
Преимущество мультимеризации по-разному модифицированных мономеров убиквитина для создания гетеромультимерных связывающих белков (здесь: гетеродимерные белки) с одновалентной связывающей активностью заключается в увеличении совокупного числа аминокислотных остатков, которые могут быть модифицированы для создания нового свойства высокоаффинного связывания с ED-B. Основное преимущество заключается в том, что хотя модифицируют даже больше аминокислот, химическая целостность белка сохраняется без ухудшения общей стабильности каркаса упомянутого вновь созданного белка связывания с ED-B. Совокупное число остатков, которые могут быть модифицированы, чтобы создать новый связывающий сайт для ED-В, увеличивается, так как модифицированные остатки могут быть распределены на два мономерных убиквитиновых белка. Число модификаций поэтому может быть равно двум, соответствуя числу модифицированных молекул мономерного убиквитина. Модульная структура белка связывания с ED-B на основе убиквитина позволяет увеличить общее число модифицированных аминокислот, так как упомянутые модифицированные аминокислоты включены в две мономерных молекулы убиквитина. Данный способ обеспечивает идентификацию гетеродимерных молекул убиквитина, имеющих одну одновалентную специфичность (для одного эпитопа) для ED-B.
Таким образом, использование гетеродимеров, имеющих общий сайт связывания для связывающихся партнеров, открывает возможность вводить повышенное число модифицированных остатков, которые не влияют недолжным образом на химическую целостность белка конечной связывающей молекулы, поскольку совокупное количество этих модифицированных остатков распределено по двум мономерным звеньям, которые образуют димер. Упомянутые гетеродимерные модифицированные убиквитиновые белки, связывающиеся с ED-B, представлены в библиотеке белков.
"Одновалентный" должен пониматься как возможность того, что обе связывающих области, созданные в первом и втором мономерном звене модифицированного димерного убиквитина, вместе связывают ED-B синергистически и объединенно, т.е., обе связывающие области действуют совместно, чтобы образовать одновалентную связывающую активность. Если взять отдельно каждую связывающую область первого и второго модифицированного убиквитина в упомянутой гетеродимерной молекуле, то такая область очевидно будет связывать ED-B с намного меньшей эффективностью и сродством чем димерная молекула. Обе связывающие области образуют уникальный сайт связывания, который сформирован как смежная область аминокислот на поверхности гетеродимерного модифицированного убиквитинового белка, так что упомянутый модифицированный убиквитин способен связываться гораздо более эффективно с ED-B чем каждый мономерный белок, взятый отдельно. Особенно важно, что, согласно настоящему изобретению два мономерных белка не связаны между собой после скрининга наиболее сильных связывающих молекул убиквитина, но уже процесс скрининга выполняется в присутствии гетеродимерных убиквитинов. После получения информации о последовательности по наиболее сильно связывающим молекулам убиквитина эти молекулы могут быть получены любым другим способом, например, химическим синтезом или способами генной инженерии, например, путем связывания двух уже идентифицированных мономерных убиквитиновых звеньев вместе.
Согласно изобретению, два по-разному модифицированных мономеров убиквитина, которые связываются с одним лигандом, должны быть связаны путем соединения "го-лова-к-хвосту" друг с другом с использованием, например, генетических способов. Эти по-разному модифицированные объединенные мономеры убиквитина связываются одновалентно и эффективны только если обе "области связывающего домена" ("ОСД") действуют совместно. "Область связывающего домена" определена здесь как область на мономере убиквитина, которая имеет модифицированные аминокислоты по меньшей мере 6 аминокислот в положениях 2,4, 6, 8, 62,63, 64, 65, 66, 68 SEQ ID NO: 1, которые участвуют в связывании мишени.
Модифицированные и связанные мономеры убиквитина, которые образуют гетероди-мерный белок, связываются с тем же эпитопом через единую смежную связывающуюся область. Эта смежная область гетеромера формируется обеими определяющими связывание областями двух модулей, сформированных двумя по-разному модифицированными мономерами убиквитина.
"Соединение голова-к-хвосту" должно пониматься как соединение двух белков вместе путем соединения их в направлении N-C-N-C-, в зависимости от числа звеньев, содержащихся в димере. При этом соединении голова-к-хвосту мономеры убиквитина могут быть прямо соединены без любого линкера.
Альтернативно, соединение мономеров убиквитина может быть выполнено посредством линкеров, например, линкера, имеющего по меньшей мере аминокислотную последовательность GIG или имеющего по меньшей мере аминокислотную последовательность SGGGG или любого другого линкера, например GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG или SGGGGSGGGG. Известны из уровня техники и могут быть использованы и другие линкеры для генетического соединения двух мономеров убиквитина.
Модифицированные убиквитиновые белки изобретения сконструированы с новыми связывающимися сродствами к ED-B как мишени или лиганду (эти выражения
используются здесь взаимозаменяемо). Термин "замещение" также включает химическую модификацию аминокислот путем, например, замещения или добавления химических групп или остатков к исходной аминокислоте. Замещение аминокислот по меньшей мере в одной области с открытой поверхностью белка, включающей аминокислоты, расположенные по меньшей мере в одной нити бета-пласта области бета-пласта или помещение до 3 аминокислот рядом с нитью бета-пласта очень важно.
Замещение аминокислот для создания нового связывающегося домена, специфического для ED-B, может быть выполнено согласно изобретению с помощью любой желательной аминокислоты, т.е., для модификации, чтобы создать новое связывающееся свойство к ED-B, необязательно заботиться о том, чтобы аминокислоты имели какое-то конкретное химическое свойство или боковую цепь, соответственно, которые аналогичны таковым у замещаемой аминокислоты, так что для этой цели можно использовать любую желательную аминокислоту.
Этап модификации выбранных аминокислот выполняют, согласно изобретению, предпочтительно путем мутагенеза на генетическом уровне путем неспецифического мутагенеза, т.е., неспецифическим замещением выбранных аминокислот. Предпочтительно, модификацию убиквитина выполняют способами генной инженерии для изменения ДНК, принадлежащей соответствующему белку. Предпочтительно, экспрессию убиквитинового белка затем осуществляют в прокариотических или эукариотических организмах.
Замещения выполняют, в частности, в аминокислотах с открытой поверхностью четырех бета-нитей бета-пластов или аминокислотах с открытой поверхностью до 3 аминокислот рядом с нитью белка убиквитина нити бета-пласта убиквитинового белка. Каждая бета-нить обычно состоит из 5-7 аминокислот. Со ссылкой на SEQ ID N0:1, например, бета-нити обычно включают аминокислотные остатки 2-7, 12-16, 41-45 и 65-71. Области, которые могут быть дополнительно и предпочтительно модифицированы, включают положения до 3 аминокислот (т.е., 1, 2 или 3) рядом с нитью бета-пласта.
Предпочтительные области, которые могут быть дополнительно и предпочтительно модифицированы, включают, в частности, аминокислотные остатки 8-11, 62-64 и 72-75. Предпочтительные области включают бета-витки, которые связывают две бета-нити вместе. Один предпочтительный бета-виток включает аминокислотные остатки 62-64. Наиболее предпочтительной аминокислотой, которая расположена рядом с нитью бета-пласта, является аминокислота в положении 8. Кроме того, другими предпочтительными примерами замещения аминокислот являются положения 36, 44, 70 и/или 71. Например, те области, которые могут быть дополнительно и предпочтительно модифицированы, включают аминокислоты 62, 63 и 64 (3 аминокислоты), или 72, 73 (2 аминокислоты), или 8 (1 аминокислота).
В предпочтительных вариантах осуществления аминокислотные остатки изменяют путем замещений аминокислот. Однако допустимы также изъятия и вставки. Число аминокислот, которые могут быть добавлены или изъяты, ограничено 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот в мономерном субзвене убиквитина и соответственно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 аминокислот по отношению к димерному убиквитиновому белку. В одном варианте осуществления вставок аминокислот не делают. В еще одном варианте осуществления не выполняют изъятий.
При условии, что модифицированный убиквитиновый белок настоящего изобретения включает дополнительно к упомянутым замещениям, определенным в формуле изобретения и объясненным здесь, а также изъятиям и/или добавлениям одной или больше аминокислот, положения аминокислот для исходного человеческого убиквитина (SEQ ID NO: 1) должны быть выровнены с модифицированным убиквитином, чтобы выделить белки, соответствующие друг другу. В случае составных белков (смотрите ниже), нумерация (и выравнивание) каждого из субзвеньев мономерного убиквитина осуществляют одинаково, т.е., выравнивание, например, димера начинают в положении 1 аминокислоты для каждого соответственного субзвена.
В мономерном убиквитине, предпочтительно от млекопитающих, например, человека, по меньшей мере 10% аминокислот, присутствующих в бета-нитях или положениях до 3 аминокислот рядом с нитью бета-пласта, предпочтительно по меньшей мере 20%, более предпочтительно по меньшей мере 25%, могут быть модифицированы, предпочтительно замещены, согласно настоящему изобретению, чтобы создать связывающее свойство, которое ранее не существовало. Как максимум, предпочтительно приблизительно 50% аминокислот, присутствующих в бета-нитях или положениях до 3 аминокислот рядом с нитью бета-пласта, более предпочтительно, как максимум, приблизительно 40%, или приблизительно 35%, или до приблизительно 30%, или до приблизительно 25% являются модифицированными, предпочтительно замещенными. В одной бета-нити обычно модифицированы 1-4 аминокислоты. В одном варианте осуществления три из шести аминокислот предпочтительно в первой и четвертой бета-нитях, например, в области аминокислотных остатков 2-7 или 65-71, модифицированы.
Модифицированный мономерный убиквитин согласно изобретению, используемый как строительное звено для гетеродимера, насчитывает в совокупности до 20% аминокислот. Учитывая это, существует идентичность последовательности с SEQ ID NO: 1 модифицированного убиквитинового белка по меньшей мере на 80%. В других вариантах осуществления изобретения идентичность последовательности на аминокислотном уровне составляет по меньшей мере 83%, по меньшей мере 85%, по меньшей мере 87% и, кроме того, по меньшей мере 90%, по меньшей мере 92% или по меньшей мере 95% идентичности последовательности с аминокислотной последовательностью SEQ ID NO: 1. Изобретение также охватывает идентичности аминокислотной последовательности модифицированного убиквитинового белка более чем на 97%» по сравнению с аминокислотной последовательностью SEQ ID NO: 1.
В еще одном варианте осуществления изобретения убиквитин модифицирован на 3, или 4, или 5, или 6, или 7 аминокислот в положениях 2, 4, 6, 8, 62, 63, 64, 65, 66 и/или 68 SEQ ID NO: 1. В еще одном варианте осуществления убиквитин, который требуется модифицировать в этих положениях, уже предварительно модифицирован. Например, другие модификации могут включать модификации в аминокислотах 74 и 75 или в аминокислоте 45, чтобы создать повышенную стабильность или химические свойства белка. Может быть получен модифицированный мономер убиквитина, в котором в совокупности до 9, 10, 11, 12, 13, 14 и максимум 15 аминокислот убиквитина с SEQ ID NO: 1 модифицированы, предпочтительно замещены. Согласно одному примеру, может быть получен модифицированный мономерный убиквитин, имеющий 14 замещений и одно изъятие. На основании совокупного числа аминокислот убиквитина это соответствует приблизительно 20%. Это оказалось чрезвычайно удивительным и не могло ожидаться, поскольку обычно гораздо меньшего процента уже достаточно для нарушения укладки белка.
В одном варианте осуществления изобретения эти аминокислоты модифицируют для создания области, имеющей новые свойства связывания ED-B, которые образуют непрерывную область на поверхности белка. Таким образом может быть создана непрерывная область, которая имеет свойство связывания с ED-B. "Непрерывная область" согласно изобретению относится к следующему: из-за заряда, пространственной структуры и гидрофобности/гидрофильности своих боковых цепей аминокислоты взаимодействуют с окружающей их средой соответствующим образом. Этой окружающей средой может быть растворитель, обычно вода, или другие молекулы, например, пространственно близкие аминокислоты. Посредством структурной информации о белке, а также соответственного программного обеспечения можно охарактеризовать поверхность белков. Например, область поверхности раздела между атомами белка и растворителем можно визуализировать этим образом, включая информацию о том, как структурирована эта область поверхности раздела, какие части поверхности доступны для растворителя, или как на поверхности распределены заряды. Непрерывная область может быть раскрыта, например, визуализацией этого типа с использованием подходящего программного обеспечения. Такие способы известны специалистам в данной области техники. Согласно изобретению, в основном, также всю область с открытой поверхностью можно использовать в качестве непрерывной области на поверхности, которую требуется модифицировать для создания новых связывающих свойств. В одном варианте осуществления для этой цели модификация может также включать альфа-спиральную область. В гетеродимерном модифицированном убиквитиновом белке область, определяющая связывание, включает две из областей с открытой поверхностью, образующих вместе одну непрерывную область, которая включает две длины одной области, определяющей связывание.
Модификация аминокислот по меньшей мере в одной области белка с открытой поверхностью, включающей по меньшей мере одну бета-нить области бета-пласта или положения до 3 аминокислот рядом с нитью бета-пласта является очень важной. "Структура бета-пласта" определяется как в сущности пластоподобная и почти полностью растянутая. В противоположность альфа-спиралям, которые сформированы из ненарушенного сегмента полипептидной цепи, бета-пласты могут быть сформированы разными областями полипептидной цепи. Этим образом области, отстоящие друг от друга дальше в первичной структуре, могут войти в тесную близость друг с другом. Бета-нить обычно имеет длину 5-10 аминокислот (обычно 5-6 остатков в убиквитине) и имеет почти полностью растянутую конформацию. Бета-нити подходят так близко друг к другу, что образуются водородные связи между группой С-O одной нити и группой NH другой нити и наоборот. Бета-пласты могут быть сформированы из нескольких нитей и имеют пластоподобную структуру, в которой положение альфа-атомов С чередуется между положениями выше и ниже пластовой плоскости. Боковые цепи аминокислот следуют этой модели и, таким образом, альтернативно указывают вверх или вниз. В зависимости от ориентации бета-нитей, пласты классифицируют на параллельные и антипараллельные. Согласно изобретению, оба типа могут мутировать и использоваться для приготовления предложенных белков.
Для мутагенеза бета-нитей и структуры бета-пласта в убиквитине выбирают бета-нить или положения до 3 аминокислот рядом с бета-нитью (которая является нитью бета-пласта), которые близко к поверхности. Аминокислоты с открытой поверхностью могут быть идентифицированы по отношению к доступной рентгеновской кристаллографической структуре. Если кристаллическая структура не доступна, можно сделать попытки предсказать посредством компьютерного анализа области бета-пласта с открытой поверхностью и доступность индивидуальных положений аминокислот по отношению к доступной первичной структуре или смоделировать трехмерную структуру белка и получить информацию о потенциальных аминокислотах с открытой поверхностью таким образом. Дальнейшее раскрытие этого вопроса можно взять, например, в J. Mol. Biol, 1987 Apr 5; 194(3):531-44. Vijay-Kumar S, Bugg C.E., Cook W.J.
Однако также можно осуществить модификации в бета-пласте или положениях до 3 аминокислот рядом с бета-нитью, для которых отнимающий много времени предварительный выбор положений аминокислот для мутагенеза можно опустить. Области ДНК, кодирующие структуры бета-пластов или до 3 аминокислот рядом с нитью бета-пласта изолируют от их ДНК окружающей среды, подвергают неспецифическому мутагенезу и затем повторно интегрируют в ДНК кодирование для белка, из которого они были удалены ранее. За этим следует процесс выбора мутантов с желательными свойствами связывания.
В еще одном варианте осуществления изобретения бета-нити или до 3 аминокислот рядом с бета-нитью близко к поверхности выбирают как сказано выше и идентифицируют положения аминокислот для мутагенеза в этих выбранных областях. Положения аминокислот, выбранные таким образом, можно затем подвергнуть мутагенезу на уровне ДНК либо путем направленного на сайт мутагенеза, т.е., кодон, кодирующий специфическую аминокислоту замещают кодоном, кодирующим другую, ранее выбранную специфическую аминокислоту, или это замещение осуществляют в контексте неспецифического мутагенеза, в котором определяется замещаемое положение аминокислоты, но не кодон, кодирующий новую, еще не определенную аминокислоту.
"Аминокислотами с открытой поверхностью" являются аминокислоты, которые доступны для окружающего растворителя. Если доступность аминокислот в белке больше чем 8% по сравнению с доступностью аминокислоты в модельном трипептиде Gly-X-Gly, эти аминокислоты называют" поверхностно открытыми". Эти области белка или положения отдельных аминокислот, соответственно, также являются предпочтительными сайтами связывания для потенциальных связывающихся партнеров, выбор которых должен быть осуществлен согласно изобретению. Помимо этого, делается ссылка на публикации Кастера и др. (Caster et al), 1983 Science, 221, 709 - 713, и Шрейка и Рипли (Shrake & Rupley), 1973 J. Mol. Biol. 79(2):351-371, которые для полного раскрытия включены в настоящую заявку путем ссылки.
Вариации каркаса убиквитинового белка, отличающиеся замещениями аминокислот в области искусственного сайта связывания, вновь созданного из родительского белка и из друг друга, могут быть созданы путем целевого мутагенеза соответственных сегментов последовательности. В этом случае аминокислоты, имеющие некоторые свойства, такие как полярность, заряд, растворимость, гидрофобность или гидрофильность могут быть заменены или замещены, соответственно, аминокислотами с соответственными другими свойствами. Кроме замещений, термины "мутагенез" и "модифицированный" и "замененный" также включают вставки и изъятия. На белковом уровне модификации также могут быть осуществлены путем химического изменения боковых цепей аминокислот согласно способам, известным специалистам в данной области.
Способы мутагенеза убиквитина
В качестве исходной точки для мутагенеза соответственных сегментов последовательности можно использовать, например, кДНК убиквитина, которая может быть приготовлена, изменена и амплифицирована способами, известными специалистам в данной области. Коммерчески доступны реагенты и способы для сайт-специфического изменения убиквитина в относительно небольших областях первичной последовательности (приблизительно 1-3 аминокислоты) ("Quick Change", компания Stratagene; набор "Mutagene Phagemid in vitro Mutagenesis Kit", компания Biorad). Для сайт-направленного мутагенеза более крупных областей специалистам доступны специфические варианты осуществления, например, полимеразная цепная реакция (ПЦР). Для этой цели можно использовать смесь синтетических олигодеоксинуклеотидов, имеющих композиции вырожденных пар оснований в желательных положениях, например, для ввода мутации. Это также можно осуществить путем использования аналогов пар оснований, которые в естественном виде не встречаются в геномной ДНК, такие как, например, инозин.
Исходной точкой для мутагенеза одной или больше бета-нитей области бета-пласта или положений до 3 аминокислот рядом с нитью бета-пласта может быть, например, кДНК убиквитина или также геномная ДНК. Кроме того, генное кодирование для убиквитинового белка также может быть приготовлено синтетически.
Известными и доступными разными процедурами для мутагенеза являются способы сайт-специфического мутагенеза, способы неспецифического мутагенеза, мутагенез с использованием ПЦР или похожие способы.
В одном предпочтительном варианте осуществления изобретения предварительно определены положения аминокислот для мутагенеза. Выбор аминокислот для модификации осуществляют, чтобы соответствовать ограничениям пункта 1 формулы изобретения в отношении тех аминокислот, которые необходимо модифицировать. В каждом случае обычно создают библиотеку разных мутантов, которую подвергают скринингу, используя способы, известные как таковые. Обычно предварительный выбор аминокислот для модификации может быть особенно легко выполнен, поскольку для убиквитинового белка, подлежащего модификации доступно достаточно структурной информации.
Способы целевого мутагенеза, а также мутагенеза более длинных сегментов последовательности, например посредством ПЦР, химическим мутагенезом или используя бактериальные штаммы гена-мутатора также входят в уровень техники и могут быть использованы согласно изобретению.
В одном варианте осуществления изобретения мутагенез осуществляют сборкой оли-гонуклеотидов ДНК, несущих кодон NNK аминокислоты. Необходимо понимать, однако, что можно использовать и другие кодоны (триплеты). Мутации выполняют таким образом, чтобы предпочтительно сохранить структуру бета-пласта. Обычно мутагенез происходит на наружной стороне стабильной области бета-пласта, открытой на поверхности белка. Он включает и сайт-специфический и неспецифический мутагенез.
Сайт-специфический мутагенез, включающий относительно небольшую область в первичной структуре (приблизительно 3-5 аминокислот), может быть создан с использованием имеющихся в продаже наборов Stratagene® (QuickChange®) или Bio-Rad® (Mutagene® phagemid in vitro mutagenesis kit) (см. US 5,789,166; US 4,873,192).
Если сайт-специфическому мутагенезу подвергают более широкие области, должна быть подготовлена ДНК-кассета, в которой область для мутагенеза получают сборкой нуклеотидов, содержащих мутированные и неизмененные положения (Норд и др. (Nord et al), 1997 Nat. Biotechnol. 8,772-777; Макконелл и Xecc (McConell and Hoess), 1995 J. Mol. Biol. 250,460-470.). Неспецифический мутагенез может быть введен путем распространения ДНК в штаммах гена-мутатора или амплификацией ПЦР (подверженная ошибкам ПЦР) (например, Паннекоек и др. (Pannekoek et al.), 1993 Gene 128,135 140). Для этой цели используют полимеразу с повышенной частотой ошибок. Для усиления степени введенного мутагенеза или для объединения разных мутаций, соответственно, мутации в фрагментах ПЦР можно объединять посредством перестановки ДНК (Штеммер (Stemmer), 1994 Nature 370, 389-391). Обзор этих стратегий мутагенеза по отношению к ферментам представлен в обзоре Кюхнера и Арнольда (Kuchner and Arnold) (1997) TIBTECH 15, 523-530. Для осуществления этого неспецифического мутагенеза в выбранной области ДНК также должна быть сконструирована ДНК-кассета, которую используют для мутагенеза.
Неспецифическую модификацию выполняют способами, которые хорошо освоены и хорошо известны в данной области. "Неспецифически модифицированный нуклеотид или аминокислотная последовательность" является нуклеотидом или аминокислотной последовательностью, который в нескольких положениях был подвергнут вставке, изъятию или замещению нуклеотидами или аминокислотами, характер которых нельзя прогнозировать. Во многих случаях введенные неспецифические нуклеотиды (аминокислоты) или последовательности нуклеотидов (аминокислот) будут "полностью неспецифическими" (например, как следствие рандомизированного синтеза или ПЦР-опосредованного мутагенеза). Однако неспецифические последовательности также могут включать последовательности, которые имеют общую функциональную особенность (например, способность реагировать с лигандом продукта экспрессии), или неспецифические последовательности могут быть неспецифическими в том смысле, что конечный продукт экспрессии является полностью неспецифической последовательностью, например, с равномерным распределением разных аминокислот.
Для того, чтобы ввести рандомизированные фрагменты в векторы правильно, согласно изобретению предпочтительно, чтобы неспецифические нуклеотиды вводили в вектор экспрессии по принципу сайт-направленного, ПЦР-опосредованного мутагенеза. Однако специалисту известны и другие варианты, и, например, можно также вводить библиотеки синтетических неспецифических последовательностей в векторы.
Для создания мутантов или библиотек путем объединяющей ПЦР могут выполняться, например, три реакции ПЦР. Две реакции ПЦР выполняют для создания частично перекрывающихся промежуточных фрагментов. Третью реакцию ПЦР выполняют для объединения промежуточных фрагментов.
Способ конструирования библиотеки или вариантов мутантов может включать конструирование первого набора праймеров вокруг желательного рестрикционного сайта (праймер рестрикционного сайта), праймера прямой и обратной рестрикции и второго набора праймеров вокруг, например, перед и за кодоном, представляющим интерес (мутагенные праймеры), прямого и обратного мутагенного праймера. В одном варианте осуществления праймеры конструируют сразу же перед и за, соответственно, кодоном, представляющим интерес. Рестрикционные и мутагенные праймеры используют для конструирования первого промежуточного и второго промежуточного фрагментов. Две реакции ПЦР создают эти линейные промежуточные фрагменты. Каждый из этих линейных промежуточных фрагментов включает по меньшей мере один мутировавший кодон, представляющий интерес, фланкирующую нуклеотидную последовательность и сайт расщепления. Третья реакция ПЦР использует эти два промежуточных фрагмента и праймеры прямой и обратной рестрикции для производства объединенного линейного продукта. Напротив, до сих пор незакрепленные концы линейного продукта расщепляли с помощью рестрикционного фермента, чтобы создать липкие концы на линейном продукте. Липкие концы линейного продукта объединяют путем использования ДНК-лигазы, чтобы получить кольцевой продукт, например, кольцевую полинуклео-тидную последовательность.
Для конструирования промежуточных фрагментов выполняют разработку и синтез двух наборов прямых и обратных праймеров, причем первый набор содержит сайт расщепления рестрикционных ферментов вместе с его фланкирующей нуклеотидной последовательностью, и второй набор содержит по меньшей мере один вариант кодона, представляющего интерес (мутагенные праймеры). Специалисты в данной области поймут, что количество вариантов будет зависеть от количества вариантов желательных модификаций аминокислот. Автор настоящего изобретения предусматривает, что если в этом процессе будут использованы другие рестрикционные ферменты, точное местонахождение этого сайта расщепления и соответствующая последовательность прямых и обратных праймеров могут быть изменены соответственно. В уровне техники доступны другие способы, и они могут быть использованы вместо изложенного.
Кроме введения рандомизированного фрагмента продукта экспрессии в каркас в соответствии с настоящим изобретением, часто необходимо соединить неспецифическую последовательность с партнером по объединению путем объединения рандомизированной нуклеотидной последовательности с нуклеотидной последовательностью, кодирующей по меньшей мере одного партнера по объединению. Такое партнер по объединению может, например, способствовать экспрессии, и/или очистке/изоляции, и/или дальнейшей стабилизации продукта экспрессии.
Неспецифическое замещение согласно одному примеру настоящего изобретения по меньшей мере 6 аминокислот в положениях 2,4, 6, 8, 62,63, 64, 65, 66 и/или 68 мономерного убиквитина может быть выполнено особенно легко посредством ПЦР, поскольку упомянутые положения локализованы близко к амино- или карбокси-окончанию белка. Соответственно, кодоны для манипулирования находятся на 5' и 3' конце соответствующей нити кДНК. Таким образом, первый олигодеоксинуклеотид, используемый для мутагенной реакции ПЦР кроме кодонов в положениях 2,4, 6 и/или 8, которые будут мутировать, соответствует по последовательности кодирующей нити для амино-окончания убиквитина. Соответственно, второй олигодеоксинуклеотид - кроме кодонов в положениях 62,63, 64,65,66 и/или 68, которые будут мутировать, по меньшей мере частично соответствует некодирующей нити полипептидной последовательности карбокси-окончания. Посредством обоих олигодеоксинуклеотидов полиме-разная цепная реакция может быть выполнена с использованием ДНК-последовательности, кодирующей мономерный убиквитин, в качестве шаблона.
Кроме того, полученный продукт амплификации может быть добавлен к другой полимеразной цепной реакции с использованием фланкирующих олигодеоксинуклеотидов, которые вводят, например, последовательности распознавания для рестрикционных эндонуклеаз. Согласно изобретению предпочтительно вводить полученную генную кассету в векторную систему, подходящую для использования в последующей процедуре селекции для изоляции вариаций убиквитина, имеющих свойства связывания с заранее определенным гаптеном или антигеном.
Области, подлежащие модификации в убиквитине
Области для модификации могут быть в основном выбраны в зависимости от того, что они могут быть доступны для ED-B как связывающегося партнера, и от того, что общая структура белка будет предположительно проявлять толерантность к модификации.
Помимо модификаций в поверхностно открытых бета-нитях, также могут быть осуществлены модификации в других поверхностно открытых областях белка, предпочтительно в положениях до 3 аминокислот рядом с бета-нитью. Эти модифицированные области участвуют во вновь создаваемом связывании с высоким сродством с ED-B.
В еще одном варианте осуществления настоящего изобретения аминокислоты в одной или двух, предпочтительно двух, из четырех бета-нитей в белке или в положениях до 3 аминокислот рядом предпочтительно с двумя из четырех бета-нитей модифицируют, чтобы создать новое связывающее свойство. Также, по выбору, может быть осуществлена модификация в трех или четырех из четырех бета-нитей или в положениях до 3 аминокислот рядом с тремя или четырьмя из бета-нитей для создания связывания с ED-B.
В частности предпочтительно, чтобы аминокислоты в нити амино-окончания и карбок-си-окончания или в положениях до 3 аминокислот рядом с нитью амино-окончания и карбокси-окончания были модифицированы, предпочтительно замещены, чтобы создать новый сайт связывания с ED-B. В этом отношении, в частности предпочтительный, чтобы до 4 аминокислот рядом с нитью карбокси-окончания бета-пласта были модифицированы, предпочтительно замещены, и 1 аминокислота рядом с нитью амино-окончания бета-пласта была модифицирована, предпочтительно замещена. В частности предпочтительной является модификация, предпочтительно замещение, по меньшей мере в трех поверхностно открытых аминокислотах следующих положений убиквитина млекопитающего, предпочтительно убиквитина человека: 2, 4, 6, 8, 62, 63, 64, 65, 66, 68. Эти по меньшей мере четыре аминокислоты из упомянутой группы из 5 аминокислот формируют непрерывную поверхностно открытую область на поверхности убиквитина, которая, как было выяснено, в частности подходит для создания модифицированных белков, имеющих связывающее сродство, которое не существовало раньше по отношению к ED-B как связывающемуся партнеру. По меньшей мере 3 из этих аминокислотных остатков должны быть модифицированы. По выбору, 3, 4, 5, 6, 7, 8, 9 или 10 упомянутых аминокислотных остатков модифицируют, по выбору, в сочетании с дополнительными аминокислотными остатками.
После осуществления вышеуказанных модификаций авторы изобретения нашли последовательности убиквитина с модифицированными аминокислотами, описанные в примерах, которые связывают ED-B с очень высоким сродством (значения килодальтон до 10-9).
Составные белки
В еще одном предпочтительном варианте осуществления изобретение относится к составному белку, включающему связывающий белок изобретения, объединенный с фармацевтически и/или диагностически активным компонентом.
В еще одном аспекте изобретение относится к составному белку, включающему гетеродимерный связывающий белок изобретения, объединенный с фармацевтически и/или диагностически активным компонентом. Составной белок изобретения может включать не полипептидные компоненты, например, не пептидные линкеры, не пептидные лиганды, например, для терапевтически или диагностически релевантных радионуклеотидов. Он также может включать небольшие органические или не аминокислотные соединения, например, сахар, олиго- или полисахарид, жирную кислоту и т.д. В одном предпочтительном варианте осуществления изобретения молекула на основе гетеромерного убиквитина, связывающая ED-B, ковалентно или не ковалентно конъюгирована с белком или пептидом, имеющим терапевтически или диагностически релевантные свойства.
Ниже приведены некоторые примеры того, как получить составные белки на основе убиквитина со способностью связывать ED-B.
a) Конъюгация белка через лизиновые остатки, присутствующие в убиквитине.
b) Конъюгация связывающего белка на основе гетеродимерного убиквитина через цистеиновые остатки - могут быть расположены в С-окончании или в любом другом положении (например, аминокислотный остаток 24 или 57); конъюгация с малеимидными выбираемыми компонентами.
c) Пептидные или протеиногенные конъюгации - генетические объединения (предпочтительно С- или N-окончание).
d) Объединения на основе "меток" - белок или пептид, расположенный в С- или N-окончании белка-мишени ED-B. "Метки" объединения, например, полигистидин (в частности релевантный для радиомечения).
Эти и другие способы ковалентного и не ковалентного прикрепления представляющего интерес белка к подложке хорошо известны в данной области и поэтому более подробно здесь не описаны.
По выбору, упомянутый активный компонент является цитокином, предпочтительно цитокином, выбираемым из группы, состоящей из факторов некроза опухолей (например, TNF-альфа, TNF-бета), интерлейкинов (например, IL-2, IL-12, IL-10, IL-15, IL-24, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, IL-8, IL-1-альфа, IL-1-бета), интерферонов (например, IFN-альфа, IFN-бета, IFN-гамма), GM-CSF, GRO (GRO-альфа, GRO-бета, GRO-гамма,), MIP (MIP-1-альфа, MIP-1-бета, MIP-3-альфа, MIP-3-бета), TGF-бета LIF1 CD80, CD-40 лиганд, В70, LT-бета, Fas-лиганд, ENA-78, LDGF-PBP, GCP-2, PF4, Mig, IP-10, SDF-1 альфа/бета, BUNZO/STRC33,l-TAC, BLC/BCA-1, MDC, ТЕСК, TARC, RANTES, НСС-1, НСС-4, DC-CK1, МСР-1-5, Эотаксин, Эотаксин-2,1-309, MPIF-1, 6Ckine, СТАСК, МЕС, лимфотактин, фракталкин и др.
Одним из наиболее предпочтительных цитокинов для использования в настоящем изобретении является TNF-альфа. Воспалительный цитокин TNF имеет многочисленные виды активности в организме млекопитающего, включая противоопухолевый эффект, который в настоящее время клинически иррелевантен из-за неприемлемой токсичности эффективных доз для человека. В настоящее время TNF терапевтически используют в сочетании с цитостатическими субстанциями, такими как мелфалан.
Далее по выбору, упомянутый активный компонент, который может быть конъюгирован с гетеромультимерным связывающим белком на основе убиквитина, является токсическим соединением, предпочтительно, небольшим органическим соединением или полипептидом, по выбору токсическим соединением, например, выбираемым из группы, состоящей из сапорина, усеченного синегнойного экзотоксина А, рекомбинантного гелонина, цепи рицина-А, каличеамицина, неокарциностатина, эсперамицина, динеми-цина, кедарцидина, мадуропептина, доксорубицина, даунорубицина, ауристатина, холерного токсина, модеццина, дифтерийного токсина.
В еще одном варианте осуществления изобретения гетеромультимерный связывающий белок на основе убиквитина согласно изобретению может содержать искусственные аминокислоты.
В других вариантах осуществления составного белка настоящего изобретения упомянутый активный компонент является флуоресцентным красителем, предпочтительно компонентом, выбираемым из группы радионуклида, или из группы гамма-излучающих изотопов, предпочтительно 99Tc, 123i, 111In, или из группы излучателей позитронов, предпочтительно 18F, 64Cu, 68Ga, 86Y, 124I или из группы бета-излучателя, предпочтительно 131I 90Y, 177Lu, 67Cu, или из группы альфа-излучателя, предпочтительно 213Bi, 211At; красителями Alexa Fluor или циановыми (Берлиер и др. (Berlier et al.), J Histochem Cytochem. 51 (12): 1699-1712, 2003); фотосенсибилизатором; прокоа-гулянтным фактором, предпочтительно тканевым фактором (например, tTF-усеченным тканевым фактором); ферментом для активации пролекарства, предпочтительно ферментом, выбираемым из группы, состоящей из карбоксипептидаз, глюкуронидаз и глюкозидаз; и/или функциональным Fc доменом, предпочтительно функциональным Fc доменом человека.
Еще один вариант осуществления относится к составным белкам согласно изобретению, кроме того включающим компонент, модулирующий период полураспада сыворотки, предпочтительно компонентом, выбираемым из группы, состоящей из полиэти-ленгликоля, альбумин-связывающих пептидов и иммуноглобулина.
Специфичности связывания (константы диссоциации)
Специфичности связывания составных белков согласно изобретению такие же, как определены выше для не составного белка, выраженные в килодальтонах. В соответствии с изобретением, термин "килодальтон" определяет специфическое связывающее сродство, которое в соответствии с изобретение находится в диапазоне от 10-7 до 10-12 М. Значение 10-5 М и ниже может считаться выражаемым количественно связывающим сродством. В зависимости от применения, значение 10-7 М-10-11 М предпочтительно, например, для хроматографических применений или 10-7-10-12 М например, для диагностических или терапевтических применений. Другие предпочтительные значения связывающего сродства лежат в диапазоне от 10-7 до 10-10 М, предпочтительно до 10-11 М.
Способы определения значений связывающего сродства известны как таковые и могут быть выбраны, например, из следующих: ИФА, технология на основе поверхностного плазмонного резонанса (ППР) (предлагаемая, например, Biacore®), флуоресцентная спектроскопия, изотермическая титриметрическая калориметрия (ИТК), аналитическое ультрацентрифугирование, возбужденная флуоресценция сортированных клеток.
После осуществления вышеуказанных модификаций авторы изобретения нашли, что последовательности убиквитина с модифицированными аминокислотами, описанные в примерах связывают их мишени с очень высоким сродством (значения в килодальтонах до 10-10 М).
Димеризация убиквитина
В настоящем изобретении "димером" считается белок, который включает два мономерных убиквитиновых белка. Если димер включает два по-разному модифицированных мономера, он называется "гетеромерный димер" или "гетеродимер". Таким образом, "гетеродимер" изобретения считается объединением двух по-разному модифицированных мономерных убиквитиновых белков, имеющих объединенное одновалентное связывающее свойство для специфического связывающегося партнера ED-B.
Необходимо подчеркнуть, что модифицированный гетеродимерный связывающий ED-B убиквитиновый белок изобретения нельзя получить путем раздельного скрининга каждого мономерного убиквитинового белка и объединения двух из них впоследствии, но можно получить путем скрининга на гетеродимерные белки, состоящие из первого и второго мономерного звена, которые вместе проявляют одновалентную связьшающую активность с упомянутым лигандом ED-B. Следует ожидать, что каждое из упомянутых субзвеньев проявляет вполне ограниченное связывающее сродство к ED-B, тогда как только объединенный димерный модифицированный убиквитиновый белок будет иметь превосходные связывающие свойства, описанные в настоящем документе (смотрите, например, Фиг.4).
Согласно изобретению два по-разному модифицированных мономера убиквитина, генетически связанные путем объединения "голова к хвосту", связываются с одним и тем же эпитопом ED-B и эффективны только, если обе области связывающегося домена действуют вместе. ОСД мономеров образуют одну непрерывную область связывания.
Таким образом, убиквитиновый белок, модифицированный в соответствии с изобретением, димеризован для эффективного связывания ED-B фибронектина. Мономеры могут быть соединены непосредственно или через линкеры, как сказано выше. Можно использовать многие известные линкеры.
Каждый мономерный убиквитин имеет модификации по меньшей мере в шести аминокислотах 2, 4, 6, 8, 62, 63, 64, 65, 66, 68. Мономерные белки генетически объединены друг с другом. Связывание с мишенью опосредовано упомянутыми ОСД совместно, т.е., ОСД взаимодействуют между собой и формируют одну общую область связывания, способную связываться с упомянутым доменом ED-B фибронектина одновалентным образом.
Гетеродимеры модифицированного убиквитина связываются с ED-B Гетеродимер убиквитина согласно изобретению, связывающийся с ED-B с кД=10-7-10-2 М и проявляющий активность одновалентного связывания в отношении упомянутого экстрадомен В (ED-B) фибронектина, выбирают из двух следующих альтернатив:
(1) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 63, 64, 65 и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно 2, и
(2) в первом мономерном звене замещения по меньшей мере в аминокислотных положениях 2,4, 6, 62, 63, 64, 65, и 66; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; по выбору дополнительно 2.
В одном варианте осуществления составной белок является генетически объединенным гетеродимером упомянутого мономера убиквитина, имеющего замещения в аминокислотах в положениях 6, 8, 63-66 первого мономера убиквитина и замещения в аминокислотных остатках в положениях 6, 8, 62-66, и по выбору в положении 2 второго мономера убиквитина, предпочтительно:
- в первом мономере убиквитина замещения
лизина (K) на триптофан (W) или фенилаланин (F) в положении 6,
лейкина (L) на триптофан или фенилаланин (W, F) в положении 8, 30,
лизина (K) на аргинин (R) или гистидин (Н) в положении 63,
глютаминовую кислоту (Е) на лизин (K), аргинин (R) или гистидин (Н) в положении 64,
серии (S) на фенилаланин (F) или триптофан (W) в положении 65 и треонин (Т) на пролин (P) в положении 66;
- во втором мономере убиквитина предпочтительны замещения
лизина (K) на треонин (Т), аспарагин (N), серии (S) или глутамин (Q) в положении 6, лейкина (L) на глутамин (Q) или треонин (Т) или аспарагин (N) или серии (S) в положении 8,
глутамин (Q) на триптофан (W) или фенилаланин (F) в положении 62,
лизина (K) на серии (S), треонин (Т), аспарагин (N) или глутамин (Q) в положении 63,
глютаминовую кислоту (Е) на аспарагин (N), серии (S), треонин (Т), или глутамин (Q) в положении 64,
серии (S) на фенилаланин (F) или триптофан (W) в положении 65, и
треонин (Т) на глютаминовую кислоту (Е) или аспарагиновую кислоту (D) в положении 66, и
по выбору, глутамин (Q) на аргинин (R), гистидин (Н) или лизин (К) в положении 2.
Эти альтернативные замещения в каждом мономере могут быть объединены друг с другом без каких-либо ограничений при условии, что получаемые гетеродимеры модифицированного убиквитина имеют специфическую активность связывания с упомянутым экстрадоменом В (ED-B) фибронектина кД=10-7-10-12 М и проявляют активность одновалентного связывания в отношении упомянутого экстрадомена В (ED-B) фибронектина, и при условии, что структурная стабильность убиквитинового белка не разрушена или не затронута.
Наиболее предпочтительными являются следующие замещения:
(1) в первом мономерном звене по меньшей мере K6W, L8W, K63R, E64K, S65F и Т66Р;
и во втором мономерном звене по меньшей мере K6T, L8Q, Q62W, K63S, E64N, S65W, и Т66Е; по выбору дополнительно Q2R, или
(2) в первом мономерном звене по меньшей мере Q2T, F4W, K6H, Q62N, K63F, E64K, S65L и T66S;
и во втором мономерном звене по меньшей мере K6X, L8x, Q62X, K63X, Е64Х, S65X, и Т66Х; по выбору дополнительно Q2X, где X может быть любой аминокислотой (смотрите Фиг.2).
В частности, предпочтительными являются следующие замещения в первом мономере убиквитина для создания белков связывания с ED-B
2: Q→T, 4: F→W, 6: K→Н, 62: Q→N, 63: K→F, 64: Е→K, 65: S→L, 66: T→S
Для соединения двух мономеров "голова к хвосту" можно использовать или не использовать линкер. Предпочтительными являются линкеры SEQ ID NO: 32 или последовательность GIG, или SGGGGIG, или SGGGGSGGGGIG.
В одном предпочтительном варианте осуществления, гетеродимер убиквитина с двумя областями, определяющими связывание (ОСД) и совместно действующими для связывания ED-B, включает аминокислотную последовательность SEQ ID NO: 33 или 34. Еще один предпочтительный белок представлен следующей последовательностью, в которой ХХХХ может быть любой аминокислотой (SEQ ID NO: 47). Здесь в качестве линкера использован SGGGGSGGGGIG. При этом понимается, что также возможен другой вид линкеров или, альтернативно, отсутствие линкера.
:MTIWVHTLTGKTITLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNINFKLSLHLVLRLRGGSGGGGSGGGGIG
MQIFVXTXTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNINXXXXXLHLVLRLRGG
Консенсусные последовательности примеров белков с этими последовательностями показаны на Фиг.2.
Предпочтительный составной белок изобретения, включающий TNF-альфа в качестве фармацевтически активного компонента, имеет последовательность SEQ ID NO: 35 или 36.
В еще одном аспекте настоящее изобретение также охватывает полинуклеотиды, которые отвечают за создание белка или составного белка, как сказано выше. Дополнительно, изобретение охватывает векторы, включающие упомянутый полинуклеотид.
В дополнительном аспекте настоящего изобретения охвачены клетки-хозяева, которые содержат белок или составной белок, описанный здесь, и/или полинуклеотид для создания упомянутого рекомбинантного белка или составного белка изобретения или вектор, содержащий упомянутый полинуклеотид.
Использование белков изобретения, например, связывающих белков на основе гетеродимерного убиквитина, специфически для объединения ED-B с эффектором, таким как TNF-альфа
Белки связывания модифицированного убиквитина с ED-B согласно изобретению должны использоваться, например, для приготовления диагностических средств для использования in vitro или in vivo, а также терапевтических средств. Белки согласно изобретению могут использоваться, например, как молекулы прямого эффектора (модулятора, антагониста, агониста) или антиген-распознающих доменов. Примеры опухолей с обильным проявлением ED-B антигена показаны в таблице на Фиг.1.
В зависимости от выбранного партнера для объединения, фармацевтическая композиция изобретения адаптирована для направленности на лечение рака, например, груди и колоректальных раков, или на любые другие опухолевые заболевания с обильным проявлением ED-B (смотрите примеры на Фиг.1).
Композиции адаптированы для содержания терапевтически эффективной дозы. Величина назначаемой дозы зависит от организма пациента, типа заболевания, возраста и массы тела пациента, а также других факторов, известных как таковые.
Композиции содержат фармацевтически или диагностически приемлемый носитель и, по выбору, могут содержать другие вспомогательные агенты и наполнители, известные как таковые. Они включают, например, но без ограничения, стабилизирующие агенты, поверхностно-активные вещества, соли, буферы, подкрашивающие агенты и т.д.
Фармацевтическая композиция может быть в форме жидкого препарата, крема, лосьона для топикального применения, аэрозоли, порошков, гранул, таблеток, суппозиториев или капсул, эмульсии или липосомного препарата. Композиции предпочтительно стерильные, непирогенные и изотонные и содержат фармацевтически традиционные и приемлемые добавки, известные как таковые. Дополнительно, делается ссылка на правила Фармакопеи США или публикацию "Фармацевтические науки Ремингтона", Мае Publishing Company (1990).
В области человеческой и ветеринарной терапии и профилактики фармацевтически эффективные медикаменты, содержащие по меньшей мере один гетеромерный убиквитиновый белок, модифицированный в соответствии с изобретением, для связывания ED-B может быть приготовлен способами, известными как таковые. В зависимости от галенового препарата, эти композиции могут быть введены парэнтерально путем инъекции или инфузии, системно, ректально, интраперитонеально, внутримышечно, подкожно, чрезкожно или другими традиционно применяемыми способами. Тип фармацевтического препарата зависит от типа заболевания, тяжести заболевания, пациента и других факторов, известных специалистам в области медицины.
В одном варианте осуществления фармацевтическая композиция содержит белок или составной белок изобретения или их сочетание и кроме того содержит один или больше химиотерапевтических агентов, предпочтительно выбираемых из следующей таблицы:
В одном предпочтительном варианте осуществления, химиотерапевтический агент выбирают из мелфалана, доксорубицина, циклофосфамида, дактиномицина, фтордезок-сиурацила, цисплатина, паклитаксела и гемцитабина; или из группы ингибиторов ки-назы.
"Фармацевтическая композиция" согласно изобретению может присутствовать в форме композиции, в которой разные активные ингредиенты и разбавители и/или носители смешаны друг с другом, или может принимать форму объединенного препарата, в котором активные ингредиенты присутствуют в частично или полностью раздельной форме. Примером такой композиции или объединенного препарата является набор.
"Композиция" согласно настоящему изобретению включает по меньшей мере два фармакологически активных соединения. Эти соединения могут быть введены одновременно или отдельно с временным интервалом от одной минуты до нескольких суток. Соединения могут быть введены одинаковым способом или по-разному; например, возможно пероральное введение одного активного соединения и парэнтеральное введения другого соединения. Также, активные соединения могут быть в одном медикаменте, например, в одном растворе для инфузии или как набор, содержащий оба соединения отдельно. Также возможно, чтобы оба соединения присутствовали в двух или больше упаковках.
В частности, предпочтительной комбинацией является составной белок согласно изобретению и мелфалан и/или (липосомный) доксорубицин. Кроме неопластических агентов из класса АТС L01, TNF-составной белок изобретения может быть объединен с другими антинеопластическими веществами, включая цитокины и их производные, радиофармацевтическими препаратами, клеточными терапевтическими препаратами и наночастицами.
Из-за его активности проникновения в опухоль, TNF-составной белок изобретения (а также другие рекомбинантные белки/составные белки настоящего изобретения) может быть объединен с антинеопластическими агентами, которые указаны в рубрике L01 Системы классификации анатомико-терапевтических химических веществ (АТС), предоставляемой Всемирной организацией здравоохранения.
Авторы изобретения неожиданно выяснили, что составной белок из гетеродимера убиквитина, объединенного с TNF-альфа, причем составной белок предпочтительно имеет последовательность SEQ ID NO: 35 или 36, может быть с выгодой применен в терапии. TNF-альфа является высоко токсичным веществом и, таким образом, может назначаться только в малых дозах, которые обычно лежат ниже минимального терапевтического порога (и, таким образом, терапевтически неактивны). Из-за токсичности TNF-альфа, чтобы достигнуть терапевтически эффективной концентрации, в настоящее время при использовании TNF-альфа выбирают подход перфузии в изолированную конечность. Перфузия в конечность является медицинским приемом, который можно использовать для доставки противораковых лекарств непосредственно в руку или ногу. Поток крови в конечность и из нее временно прекращают с помощью кровоостанавливающего жгута и противораковые лекарства вводят непосредственно в кровь конечности. Это позволяет пациенту получать большую дозу TNF-альфа в той области, где имеет место рак.
Однако, путем применения TNF-альфа составных белков настоящего изобретения, можно вводить TNF-альфа в не токсичной, но все же терапевтически эффективной концентрации. Поскольку TNF-альфа соединен с (связывающим) составным белком настоящего изобретения, он может быть непосредственно активным в месте заболевания (например, месте опухоли) и, таким образом, количество "свободного" TNF-альфа может быть резко сокращено.
Системные побочные эффекты TNF-альфа могут быть заметно снижены путем введения TNF-альфа как составного белка согласно настоящему изобретению. Путем использования TNF-альфа составного белка изобретения, общая доза TNF-альфа для достижения терапевтического эффекта таким образом может быть значительно уменьшена и может с выгодой использоваться для лечения системной опухоли (без необходимости и ограничений перфузии в конечность), в частности в сочетании с химиотерапевтическими агентами (смотрите выше).
В еще одном варианте осуществления фармацевтическая композиция находится в форме набора из частей, предусматривающего отдельные позиции для рекомбинантного убиквитинового белка/составного белка изобретения и для одного или больше химиотерапевтических агентов.
Способ получения гетеродимерных, связывающих ED-B белков изобретения
Белки, связывающие ED-B согласно изобретению, могут быть приготовлены любым из многих традиционных и хорошо известных способов, таких как простые стратегии органического синтеза, способы твердофазного синтеза или в имеющихся в продаже автоматизированных синтезаторах. С другой стороны, они также могут быть приготовлены традиционными рекомбинантными способами как таковыми или в сочетании с традиционными способами синтеза.
В еще одном аспекте настоящего изобретения предложен способ создания рекомби-нантного модифицированного убиквитинового белка. Этот способ включает по меньшей мере следующие этапы:
а) предоставление популяции по-разному модифицированных димерных убиквитиновых белков, происходящих из мономерных убиквитиновых белков, причем упомянутая популяция включает димерные убиквитиновые белки, включающие два модифицированных мономера убиквитина, связанных вместе в расположении "голова к хвосту", причем каждый мономер упомянутого димерного белка по-разному модифицирован путем замещения по меньшей мере 6 аминокислот в положениях 2,4,6, 8, 62, 63, 64, 65, 66 и 68 SEQ ID NO: 1
причем упомянутые замещения включают:
В одном аспекте изобретения упомянутую популяцию по-разному модифицированных белков получают путем генетического объединения двух библиотек ДНК, каждая из которых отвечает за получение по-разному модифицированных мономерных убиквитиновых белков.
В еще одном аспекте упомянутый способ адаптирован так, чтобы объединить упомянутый модифицированный гетеродимерный убиквитиновый белок с фармацевтически активным компонентом, по выбору цитокином, предпочтительно TNF-альфа, или диагностическим компонентом, или в котором упомянутый рекомбинантный модифицированный гетеродимерный убиквитиновый белок сформирован посредством упомянутого фармацевтически активного компонента, которым, по выбору, является TNF-альфа, или посредством упомянутого диагностического компонента.
Согласно изобретению, модифицированный белок также может быть приготовлен химическим синтезом. В этом варианте осуществления этапы с) - d) из пункта 1 формулы изобретения тогда выполняют в один этап.
В еще одном аспекте настоящее изобретение направлено на библиотеку, содержащую ДНК, отвечающую за получение модифицированных мономерньгх убиквитиновых белков, как сказано выше, которые формируют основу для получения гетеродимерных убиквитиновых белков изобретения.
В еще одном аспекте изобретения представлена библиотека объединений, содержащая ДНК, полученные путем объединения двух библиотек, как сказано выше, причем каждая библиотека отвечает за получение звеньев по-разному модифицированных мономерных убиквитиновых белков, чтобы получить гетеродимерные составные белки убиквитина, и причем их мономерные звенья связаны между собой в расположении "голова к хвосту", и упомянутая библиотека отвечает за гетеродимерные составные белки убиквитина, проявляющие одновалентную связывающую активность в отношении упомянутого экстрадомена В (ED-B) фибронектина. Упомянутое связывание между собой выполняют или используя любой из линкеров, известный специалисту в данной области, или линкер, описанный в настоящем документе. В одном варианте осуществления изобретения TNF-альфа используют как линкер, одновременно действующий как фармацевтически активное соединение.
В Примере 1 описано получение сложной библиотеки. Однако необходимо проявлять осторожность в отношении качества такой библиотеки. Качество библиотеки в каркасной технологии стоит первым в зависимости от ее сложности (количества отдельных вариантов), а также функциональности (структурная и белково-химическая целостность получаемых кандидатов). Обе характеристики, однако, могут оказывать отрицательное влияние друг на друга: повышение сложности библиотеки путем увеличения числа модифицированных положений на каркасе может привести к ухудшению белково-химических характеристик вариантов. Это может привести к ухудшенной растворимости, аггрегации и/или низкому выходу продукта. Причиной этого является повышенное отклонение от исходных каркасов, имеющих энергетически благоприятную упаковку белка.
Поэтому при конструировании такой библиотеки каркасов требуется подходящий баланс между крайними положениями введения максимально возможного числа вариаций в оригинальную последовательность, чтобы оптимизировать ее для мишени и, с другой стороны, сохранить оригинальную первичную последовательность в максимальной степени, чтобы избежать отрицательных белково-химических воздействий.
Следует сказать, что настоящее раскрытие также охватывает каждую возможную комбинацию описанных здесь признаков в плане аспектов или вариантов осуществления изобретения.
Выбор модифицированных убиквитиновых белков со связывающим сродством в отношении мишени ED-B и определение модифицированных аминокислот, отвечающих за связывающее сродство После, например, создания по меньшей мере двух разных ДНК-библиотек, отвечающих за получение гетеродимерных модифицированных убиквитиновых белков, путем разных модификаций выбранных аминокислот в каждом из мономерных звеньев убиквитина эти библиотеки генетически объединяют, например, по линкерной технологии, чтобы получить молекулы ДНК, отвечающие за получение гетеродимерных модифицированных убиквитиновых белков. ДНК этих библиотек экспрессируют в белки, и полученные таким образом модифицированные димерные белки вводят в контакт согласно изобретению с ED-B, чтобы, по выбору, позволить связывание партнеров друг с другом, если связывающее сродство существует.
Очень важный аспект изобретения заключается в том, что процесс контакта и скрининга выполняют уже по отношению к гетеродимерному убиквитиновому белку. Этот процесс позволяет выполнять скрининг на тех убиквитиновых белках, которые обладают активностью одновалентного связывания с ED-B.
Согласно изобретению контакт предпочтительно выполняют посредством подходящего способа представления и выбора, такого как фаговый дисплей, рибосомный дисплей, мРНК дисплей или способы дисплея поверхностей клеток, дисплея поверхностей дрожжей или дисплея поверхностей бактерий, предпочтительно посредством способа фагового дисплея. Для полноты раскрытия ссылка делается также на следующие публикации: Hoess, Curr. Opin. Struct. Biol. 3 (1993), 572-579; Wells and Lowmann, Curr. Opin. Struct. Biol. 2 (1992), 597-604; Kay et al., Phage Display of Peptides and Proteins - A Laboratory Manual (1996), Academic Press. Вышеуказанные способы известны специалистам в данной области и могут быть использованы согласно изобретению, включая их модификации.
Определение того, имеет ли модифицированный белок вычисляемое количественно связывающее сродство по отношению к предварительно определенному связывающемуся партнеру может быть выполнено, согласно изобретению, предпочтительно одним или несколькими из следующих способов: ИФА, спектроскопия методом плазмонного поверхностного резонанса, флуоресцентная спектроскопия, возбужденной флуоресценцией сортированных клеток (FACS), изотермической титриметрической калориметрией и аналитическим ультрацентрифугированием.
Способ выбора фаговым дисплеем
Один тип процедуры фагового дисплея, адаптированный к настоящей заявке, описан далее как пример выбора согласно изобретению по отношению к вариациям убиквитина, которые проявляют связывающие свойства. Таким же образом могут быть применены, например, способы для представления на бактериях (бактериальный поверхностный дисплей; Daugherty et al., 1998, Protein Eng. 11(9):825-832) или дрожжевых клетках (yeast surface display; Kieke et al, 1997 Protein Eng. 10(11): 1303-10) или бесклеточные системы выбора, такие как рибосомный дисплей (Hanes and Pluckthun, 1997 Proc Natl Acad Sci USA. 94(10):4937-4942; He and Taussig, 1997_Nucleic 5 Acids Res. 25(24):5132-5134) или cis дисплей (Odegrip et al, 2004 Proc Natl Acad Sci USA. 101 (9):2806-2810) или мРНК дисплей. В последнем случае временная физическая связь генотипа и фенотипа достигается путем соединения вариации белка с подходящей мРНК через рибосому.
В описанной здесь процедуре фагового дисплея рекомбинантные вариации убиквитина представлены на нитчатом фаге, тогда как кодирующая ДНК представленной вариации представлена в то же время упакованной в однонитевую форму в фаговой оболочке. Таким образом, в рамках аффинного обогащения вариации, имеющие определенные свойства, могут быть выбраны из библиотеки, и их генетическая информация может быть амплифицирована путем инфекции подходящих бактерий или добавлена к другому циклу обогащения, соответственно. Представление мутировавшего убиквитина на поверхности фага достигается путем генетического объединения с сигнальной последовательностью с амино-окончанием, предпочтительно с PelB сигнальной последовательностью, и капсид или поверхностный белок предпочтительного фага является кар-бокситерминальным объединением с капсидным белком рШ или его фрагментом.
Кроме того, полученный составной белок может содержать другие функциональные элементы, такие как метка сродства или эпитоп антитела для детектирования и/или очистки аффинной хроматографией или последовательность распознавания протеазы для специфического расщепления составного белка в ходе аффинного обогащения. Кроме того, терминатор UAG может быть представлен, например, между геном для вариации убиквитина и кодирующей областью капсидного белка фага или его фрагмента, который не распознан во время трансляции в подходящий супрессорный штамм, частично из-за введения одной аминокислоты.
Бактериальный вектор, подходящий для процедуры выбора в контексте изоляции вариаций убиквитина со свойствами связывания с ED-B и в который введена генная кассета для описываемого составного, называется фагемидом. Среди прочих, он содержит межгенную область нитчатого фага (например, М13 или f1) или ее часть, которая в случае суперинфекции бактериальной клетки, несущей фагемид посредством фагов-
хелперов, таких как, например, M13K07, приводит к упаковке закрытой нити ДНК фагемида в капсид фага. Созданные таким образом фагемиды секретируются бактериями и представляют соответственную вариацию убиквитина, кодированную из-за ее объединения с капсидным белком pIII или его фрагментом, на их поверхности. Нативные капсидные белки pIII присутствуют в фагемиде, так что его способность реинфицировать подходящие бактериальные штаммы и, поэтому, возможность амплифицировать соответствующую ДНК сохраняется. Таким образом, обеспечивается физическая связь между фенотипом вариации убиквитина, т.е., ее потенциальное связывающее свойство, и ее генотипом.
Полученные фагемиды могут быть выбраны по отношению к связыванию представленной на них вариации убиквитина с ED-B способами, известными специалистам в данной области. Для этой цели представленные вариации убиквитина могут быть временно иммобилизованы на субстанции мишени, связанной, например, на микротитровальных планшетах, и могут быть специфически элюированы после отделения не связывающихся вариаций. Элюирование предпочтительно выполняют щелочными растворами, такими как, например, 100 мМ триэтиламин. Альтернативно, элюирование может быть выполнено в кислых условиях, путем протеолиза или прямого добавления инфицированных бактерий. Фагемиды, полученные таким образом, могут быть реамплифицированы и обогащены последовательными циклами выбора и амплификации вариаций убиквитина со свойствами связывания с ED-B.
Дальнейшая характеризация вариаций убиквитина, полученных таким образом, может быть выполнена в форме фагемида, т.е., объединенных с фагом, или после клонирования соответствующей генной кассеты в подходящий экспрессирующий вектор в форме растворимого белка. Подходящие способы известны специалистам в данной области или описаны в литературе. Характеризация может включать, например, определение последовательности ДНК и, таким образом, первичной последовательности изолированных вариаций. Кроме того, сродство и специфичность изолированных вариаций можно детектировать стандартными биохимическими способами, такими как ИФА или спектроскопия плазмонного поверхностного резонанса, флуоресцентная спектроскопия, FACS, изотермическая титриметрическая калориметрия, аналитическое ультрацентрифугирование и др.. В свете анализа стабильности, например, спектроскопические способы в связи с химическим или физическим развертыванием известны специалистам в данной области.
Способ выбора рибосомным дисплеем
В еще одном варианте осуществления изобретения вариации убиквитина для процедуры рибосомного дисплея готовят посредством бесклеточной системы транскрипции/трансляции и представляют как комплекс с соответствующей мРНК, а также рибосомой. Для этой цели библиотеку ДНК, которая описана выше, используют в качестве основы, в которой гены вариаций присутствуют в форме объединений с соответствующими регуляторными последовательностями для экспрессии и биосинтеза белка. Из-за изъятия терминирующего кодона в окончании 3' генной библиотеки, а также подходящих экспериментальных условий (низкая температура, высокая концентрация Mg) трехчастный комплекс, состоящий из возникающего белка, мРНК и рибосомы, сохраняется во время транскрипции/трансляции in vitro.
После создания библиотеки белков, содержащей гетеродимерные модифицированные убиквитиновые белки, путем разных модификаций выбранных аминокислот в каждом из мономерных звеньев убиквитина модифицированные димерные белки вводят в контакт, согласно изобретению, с ED-B, чтобы позволить партнерам связываться друг с другом, если существует связывающее сродство. Эти библиотеки белков могут быть в форме отображения библиотеки дисплейным способом, или можно использовать любой другой способ, представляющий модифицированные белки таким образом, чтобы обеспечить контакт между модифицированными белками и белком-мишенью ED-B, причем упомянутый дисплейный способ является, по выбору, фаговым дисплеем, рибосомным дисплеем, дисплеем фага ТАТ, дрожжевым дисплеем, бактериальным дисплеем или дисплеем мРНК.
Выбор модифицированных вариаций убиквитина по отношению к их активности связывания с ED-B 20 при специфическом связывающем сродстве в кД в диапазоне 10-7-10-12М может быть выполнен способами, известными специалистам в данной области. Для этой цели вариации убиквитина, представленные, например, на рибосомных комплексах, могут быть временно иммобилизованы на субстанции мишени, связанной, например, на микротитровальных планшетах или могут быть связаны с магнитными частицами после связывания в растворе, соответственно. После отделения несвязываю-щихся вариаций генетическая информация вариаций со связывающей активностью может быть специфически элюирована в форме мРНК путем деструкции рибосомного комплекса. Элюирование предпочтительно осуществляют с 50 мМ ЭДТА. Полученная таким образом мРНК может быть изолирована и обратно транскрибирована в ДНК с использованием подходящих способов (реакция обратной транскриптазы), и ДНК, полученная таким образом, может быть реамплифицирована.
Посредством последовательных циклов in vitro транскрипции/трансляции, выбора и амплификации вариации убиквитина со свойствами связывания с определенным гаптеном или антигеном могут быть обогащены.
Характеризация белков, связывающихся с EDB
Дальнейшая характеризация вариаций убиквитина, полученных таким образом, может быть выполнена в форме растворимого белка, как подробно сказано выше, после клонирования соответствующей генной кассеты в подходящий экспрессирующий вектор. Подходящие способы известны специалистам в данной области или описаны в литературе.
Предпочтительно, за этапом детектирования белков, имеющих связывающее сродство в отношении определенного связывающегося партнера, следует этап изоляции и/или обогащения детектированного белка.
После экспрессии убиквитинового белка, модифицированного согласно изобретению, он может быть далее очищен и обогащен способами, известными как таковые. Выбираемые способы зависят от нескольких факторов, известных как таковые специалистам в данной области, например, используемого экспрессирующего вектора, организма-хозяина, намеченной области использования, размера белка и других факторов. Для упрощения очистки белок, модифицированный согласно изобретению, может быть объединен с другими пептидными последовательностями, имеющими повышенное сродство с материалами разделения. Предпочтительно, выбирают такие объединения, которые не оказывают вредного воздействия на функциональность убиквитинового белка или могут быть отделены после очистки из-за введения специфических сайтов расщепления протеазы. Такие способы также известны как таковые специалистам в данной области.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг.1 показана таблица ч указанием встречаемости ED-B в разных опухолях.
на Фиг.2 показаны консенсусные положении и замещения аминокислот в еще 16 последовательностях, которые, как было выяснено, имеют удивительно сильное связывающее сродство с ED-B. Консенсусными положениями аминокислот в первой мономерной области, определяющей связывание, являются 2,4,6, 62,63, 64, 65, 66, а консенсусными замещениями аминокислот являются Q2T, F4W, K6H, Q62N, K63F, E64K, S65L и T66S. Как можно понять из Фиг.2, 4 семейства последовательностей могут быть обогащены (консенсусные последовательности, буквы соответствуют частоте встречаемости аминокислот). Положениями 85 и 87 являются положения в гетеродимерном белке; в отношении второго мономера соответствующими положениями являются 6 и 8; 141-145 соответствуют положениям 62-64). TWH NFKLS, отображенная в темно-синем цвете, происходит из 1071-С 12. Остатки, помеченные красным цветом, относятся к одному из упомянутых четырех семейств последовательностей. Остатки, помеченные красным, были преимущественно обогащены (178/457 последовательностей) и включают, согласно ИФА, наиболее сильные связывающие молекулы.
На Фиг.3 показано, что тетрамеризация приводит к повышению сродства.
В таблице показаны значения в кД модифицированных мономеров убиквитина по сравнению с тетрамерами, состоящими из модифицированных мономеров убиквитина. Как примеры, показаны варианты 5Е1 и 1Н4 убиквитина. Связывание ED-B сравнивается со связыванием с c-FN (клеточный фибронектин). Эти Фигуры демонстрируют значительно более высокое сродство в связывании тетрамерного варианта (например, 56 нМ для 5Е1 или 1,4 нМ для 1Н4) с мишенью ED-B по сравнению с мономером (4,51 мкМ для 5Е1 или 9,98 мкМ для 1Н4).
На Фиг.4 показано, что рекомбинация переднего (первого) модифицированного мономера убиквитина (имеющего BDR1) с другим, модифицированным задним (вторым) мономером убиквитина (имеющим BDR2) для создания гетеродимера приводит к значительному увеличению сродства, а также специфичности. Молекулы модифицированного убиквитина проанализированы посредством Biacore, флуоресцентной анизотропии, связывания на клетках и срезах тканей. Показаны зависящие от концентрации результаты ИФА (конц. - ИФА) связывания нескольких вариантов с ED-B человека.
На Фиг.4А показано связывающее сродство кД=9,45 мкМ для мономера 41В10.
На Фиг.4В показано, что связывающее сродство кД=131 нМ для 41 В10, объединенного с другим, вторым мономером, приводит к 46Н9.
На Фиг.5 показаны специфические варианты, объединенные с цитокином (например, TNF-альфа). Эти составные белки тримеризуют модифицированный мономер убиквитина и являются биологически активными молекулами.
На Фиг.5 А приведен схематический чертеж составного белка - эффектора, связывающего ED-B на основе модифицированного убиквитина; зеленый (структура наверху) - эффектор, например, цитокин, предпочтительно TNF-альфа; коричневый: светло-коричневый: структура модифицированных мономеров убиквитина (Affilin®).
На Фиг.5 В показано, что конъюгат эффектора модифицированного убиквитина 5Е1-TNF-конъюгат имеет про-апоптическую активность (измеренную в анализе апоптоза L929).
На Фиг.5 С показано высокоаффинное связывание 1Н4-тар-альфа-объединения с ED-В (кД=15,1 нМ) (закрашенные круги, соединенные подогнанной линией). Связывание с BSA нанесено на график как контрольное (закрашенные круги не соединены линией).
На Фиг.6 показано сродство и активность молекулы гетородимера, связывающейся с ED-B, на основе модифицированного убиквитина, объединенной с цитокином, например, TNF-альфа.
- Индуцирующая апоптоз активность объединения цитокина, связывающегося с ED-B, на основе модифицированного убиквитина: ЕС50 0,78±0,24 пМ
- Индуцирующая апоптоз активность свободного цитокина: ЕС50 3,14±3,59 пМ
На Фиг.6А показано сродство гетеродимера 24Н12 (кД 50,7 нМ), связывающего ED-B, на основе модифицированного убиквитина.
На Фиг.6В показано сродство гетеродимера 24Н12, связывающего ED-B, на основе модифицированного убиквитина, генетически объединенного с цитокином TNF-альфа, чтобы привести к мультимеризации гетеродимера 24Н12 (кД=5,6 нМ).
На Фиг.6С показан анализ примеров кандидатов из выбранной библиотеки гетероди-мерного модифицированного убиквитина, например, клоны 9Е12, 22D1, 24Н12, 41В10 гетеродимера. Значения ИФА в килодальтонах увеличиваются для мишени ED-B по сравнению с цитозольным фибронектином, используемым в качестве контрольного, подтверждая специфическое связывание с мишенью.
На Фиг.6D показаны результаты анализа молекулы 9Е12 модифицированного гетеро-димерного убиквитина посредством анализов взаимодействия без меток с использованием Biacore®. Были проанализированы разные концентрации вариантов гетеродимерного убиквитина (смотрите легенду к Фигуре: 0-15 мкМ 9Е12) для связывания с ED-В, иммобилизованном на чипе (Biacore), чтобы проанализировать взаимодействие между гетеродимерным вариантом 9Е12 и ED-B. Значение в килодальтонах нельзя определить из анализа кривых ассоциации и диссоциации.
На Фиг.6Е показаны результаты анализа молекулы 41 В10 модифицированного гетеродимерного убиквитина посредством анализов взаимодействий без меток с использованием Biacore®. Были проанализированы разные концентрации вариантов гетероди-мерного убиквитина (смотрите легенду к Фигуре: 0-15 мкМ 41 В10) для связывания с ED-B, иммобилизованном на чипе (Biacore), чтобы проанализировать взаимодействие между гетеродимерным вариантом 41В10 и ED-B. Анализ кривых ассоциации и диссоциации дал значение кД 623 нМ (623×10-9 М, 6,2×10-7 М).
На Фиг.7 показан вклад разных вариантов на основе модифицированного убиквитина в связывающее сродство и специфичность. Эти разные варианты разделяют модули общей последовательности, которые отмечены буквами нижнего регистра. Варианты были проанализированы в отношении их связывания с ED-B. На Фиг.3 показаны другие комбинации мономеров, дающие гетеродимеры модифицированного убиквитина. Гетеродимерные варианты 46-А5, 50-G11 и 46-Н4 имеют одинаковый первый (передний) модифицированный мономер с BDR1 (помечен буквой "а" на этой Фигуре), но второй (задний) мономер убиквитина модифицирован BDR2 в разных положениях. Варианты 52-D10 и 52-В3 имеют другой первый (передний) модифицированный мономер по сравнению с 46-Н9 с BDR1, но одинаковый второй (задний) мономер убиквитина с BDR2 (помечен буквой "е").
Гетеродимеры модифицированного убиквитина имеют следующие последовательности:
46-Н4: SEQ IDNO: 25, 45-Н9: SEQ IDNO: 26, 46-А5: SEQ IDNO: 27, 50-G11: SEQ ID NO: 28, 52-В3: SEQ ID NO: 29, 52-D10: SEQ ID NO: 30
Вышеописанные последовательности были модифицированы в ходе экспериментов путем добавления His-Tag с последовательностью LEHHHHHH (SEQ ID NO: 31).
Как можно видеть из Фиг.7,46-Н4 имеет превосходное связывающее сродство с ED-B (кД=189 нМ); 46-А5 и 52-D10 не имеют связывающей активности, тогда как другие модифицированные убиквитиновые белки имеют небольшую связывающую активность с ED-B по сравнению с 46-Н4. Таким образом можно сделать вывод, что оба мс номера в гетеродимерном варианте требуются для высокоаффинного связывания с мишенью; оба мономера проявляют одновалентное связывание с мишенью.
Гетеродимер модифицированного убиквитина с высокой активностью связывания с ED-B, названный 46Н9, идентифицируется следующими заменами аминокислот в об(их областях связывающегося домена в двух мономерах по сравнению с мономерами убиквитина дикого типа:
в первом модуле (BDR1) (a) Q2G, F4V, K6R, Q62P, K63H, Е64А, S65T, T66L во втором модуле (BDR2) (е) К6Н, L8M, Q62K, K63P, E64I, S65A, Т66Е
50G11
в первом модуле (46Н9)(а) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L во втором модуле (с) K6M L8R, Q62M, K63N, Е64А, S65R, T66L
46Н4
в первом модуле (46Н9)(а) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L во втором модуле (d) K6G, L8W, Q62T, K63Q, E64Q, S65T, T66R
52В3
в первом модуле (g) Q2R, F4P, K6Y, Q62P, K63P, E64F, S65A, T66R во втором модуле (46Н9) K6H, L8M, Q62K, K63P, E64I, S65A, Т66Е
52D10 (не связывается с ED-B)
в первом модуле Q2V, F4C, K6R, Q62T, K63A, Е64Р, S65G, T66D
во втором модуле (46Н9) (е) K6H, L8M, Q62K, K63P, E64I, S65A, Т66Е
46А5 (не связывается с ED-B)
в первом модуле (46Н9)(а) Q2G, F4V, K6R, Q62P, K63H, Е64Р, S65T, T66L во втором модуле (b) K6L, L8M, Q62L, k63 A, E64F, S65A,
На Фиг.8 показано выравнивание последовательности. Линия 1: два мономера убиквитинового белка дикого типа (1-я линия) связаны с 12-аминокислотньгм линкером SGGGGSGGGGIG, начинающимся в положении 77 и заканчивающимся в положении 88; второй мономер с BDR2 начинается в положении 89 с метионином. Этот димерный убиквитиновый белок дикого типа выровнен с гетеродимерным вариантом 46-Н9 модифицированного убиквитина (2-я линия) с разными модификациями в первом и во втором мономерах, давая две ОСД. Обе ОСД действуют совместно в связывании мишени из-за одновалентного связывания с мишенью.
На Фиг.9 показано выравнивание последовательности гетеродимерного варианта 1041-D11 (1-я линия) модифицированного убиквитина с "Ub2_TsX9" (убиквитин, модифицированный в положении 45 в обоих мономерах в триптофан, показывая линкер GIG между двумя мономерами (положения 77 - 79; второй мономер начинается с метионина в положении 80), и замена с глицина на аланин в последних с-терминальных аминокислотах 2-го мономера. Третья линия показывает "Уби-димер дт", в качестве димера убиквитин дикого типа; не показывая линкерного выравнивания (таким образом, второй мономер начинается в положении 77 с метионином). 4-я линия показывает "Уби-мономер дт", который является человеческим убиквитином дикого типа.
На Фиг.10 показан зависящий от концентрации ИФА связывания варианта 1041-D11 гетеродимерного убиквитина с ED-B человека. Вариант 1041-D11 проявляет очень высокоаффинное связывание с ED-B (кД=6,9 нМ=6,9×10-9 М). Закрытые точки показывают сродство связывания варианта 1041-D11 гетеродимерного убиквитина с ED-B, содержащим фрагмент фибронектина (названный 67В89-tO), по сравнению с отсутст вием связывания этого варианта с отрицательным контролем (названным 6789-tO) (незакрашенные круги).
На Фиг.11 показаны результаты конкурентного, зависящего от концентрации ИФА связывания варианта 1041-D11 гетеродимерного убиквитина с иммобилизованным ED-В, содержащим фрагмент фибронектина (67В89), в присутствии увеличивающихся количеств свободной мишени. Вариант 1041-D11 гетеродимерного убиквитина проявляет очень высокоаффинное связывание с ED-B (IC50=140 нМ).
На Фиг.12 показан результат анализа молекулы 1041-D11 модифицированного гетеродимерного убиквитина в анализах взаимодействий без меток с использованием Biacore®. Были проанализированы разные концентрации этого варианта гетеродимерного убиквитина (смотрите легенду на Фигуре: 0-200 нМ 1041-D11) для связывания с ED-B, содержащим фрагмент фибронектина (названный 67 В89), иммобилизованным на чипе SA (Biacore). Анализ кривых ассоциации и диссоциации дал значение кД 1 нМ (1×10-9 М) и показатель koff 7,7×10-4 с-1, который показывает длительное время полураспада комплекса 1041-D11 и ED-B.
На Фиг.13 показано связывание варианта 1041-D11 гетеродимерного убиквитина с ED-B в зависящем от концентрации ИФА при одновременном анализе сывороточной стабильности связывающей активности. Показаны разные условия, такие как предварительная инкубация в течение 1 часа при 37°С этого варианта в сыворотке мыши или крысы и в PBST как контроле. Значения в кД находятся в интервале от 10 до 20 нМ. Таким образом, можно сделать вывод, что на связывание гетеродимера 1041-D11 с ED-В сыворотка крови влияет незначительно.
На Фиг.14 показан анализ формирования комплекса варианта 1041-D11 гетеродимерного убиквитина с фрагментами фибронектина за счет СО-ЖХВР.
На Фиг.14А показано формирование комплекса 1041-D11 с ED-B. Три хроматограммы ЖХВР наложены друг на друга: синий пик с временем удержания 21,651 мин происходит из чистого 1041 -Dl 1; черный пик с временем удержания 26,289 мин представляет фрагмент фибронектина 67 В89; смесь 1041-D11 и 67В89 дает красный пик с временем удержания 21,407 мин после СО-ЖХВР. Сдвиг пика 1041-D11 на меньшее время удержания, а также исчезновение пика 67В89 указывает на формирование комплекса 1041-D11 и растворимого ED-B.
На Фиг.14 В показано наложение трех хроматограмм СО-ЖХВР 1041-D11 (синий, 21,944 мин), фрагмент фибронектина 6789 без ED-B (черный, 26,289 мин) и смесь 1041-D11 и 6789 (красная линия с пиками в 21,929 мин и 26,289 мин). Почти не наблюдается сдвиг пика 1041-D11. Этот факт вместе с отсутствием исчезновения пика 6789 указывает на незначительное связывание свободного фрагмента фибронектина 6789 ED-B.
На Фиг.15 показано связывание варианта 1041-D11 гетеродимерного убиквитина с клетками клеточной культуры.
На Фиг.15А показано связывание варианта 1041-D11 гетеродимерного убиквитина на фибробластных клетках (Wi38) эмбрионального легкого человека, которые были зафиксированы. Первая колонка на Фиг.15 показывает контроль, использующий анти-ED-B антитела, вторая колонка показывает инкубацию этого варианта при концентрации белка 58,7 нМ, третья колонка показывает десятикратно более высокую концентрацию белка 1041-D11 (587 нМ), четвертая колонка является отрицательным контролем с физраствором с фосфатным буфером. В первом ряду фибробластные клетки Wi38 человека показаны в фазовом контрасте; второй ряд показывает иммунофлуорес-ценцию, и третий ряд показывает мечение ядер с помощью DAPI. Можно сделать вывод, что вариант 1041-D11 связывается с Wi38 с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.Был выполнен контрольный анализ с использованием клеток NHDF, которые экспрессируют низкий уровень ED-B (данные не показаны). Эти варианты не связываются с этими клетками.
На Фиг.15В показано связывание на фибробластных клетках (Wi38) жизнеспособного эмбрионального легкого человека. Отрицательные контрольные клетки TnnaNHDF являются первичными нормальными фибробластными клетками, которые экспрессируют низкие уровни EDB-фибронектина. Первая и третья линии показывают этот вариант при разной концентрации белка и отрицательный контроль. Вторая и четвертая линии показывают инкубацию контроля с использованием антител к EDB. Первые 2 линии показывают вариант и положительный контроль на клеточной линии Wi38. Третья и четвертая линии показывают инкубацию NHDF - клеток. Можно видеть, что вариант 1041-D11 связывается с Wi38 с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.
На Фиг.15С показано связывание на фиксированных мьппиных клетках Balb 3Т3. Были проверены три концентрации белка (1,10, 50 нМ) этого варианта. Первый ряд показывает этот вариант (SPVF-28-1041-41 1-TsX9) на клетках, второй ряд показывает положительный контроль (антитела к Fv28-EDB-15), третий ряд показывает инкубацию с отрицательным контролем (UB2_TsS9; немодифицированный убиквитин, соответствующий SEQ ID NO:1). Можно видеть, что вариант 1041-D11 связывается с мьшшными клетками Balb ЗТЗ с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.
На Фиг.15D показано связывание на фиксированных мьппиных клетках ST-2. Были проверены три разных концентрации белка (1,10, 50 нМ) этого варианта. Первый ряд показывает вариант (SPVF-28-1041-411-TsX9) на клетках, второй ряд показывает положительный контроль (антитела к Fv28-EDB), третий ряд показывает инкубацию с отрицательным контролем (UB2_TsS9; немодифицированный убиквитин, соответствующий SEQ ID NO: 1). Можно видеть, что вариант 1041-D11 гетеродимерного убиквитина связывается с мышиными клетками Balb ST-2 с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.
На Фиг.16 А показана специфичность варианта 1041-D11 гетеродимерного убиквитина с мишенью в срезах тканей млекопитающих. Были оценены опухолевые ткани F9 из семи проб. Иммуногистохимия с разными концентрациями от 10 нМ до 100 нМ варианта 1041-D11 гетеродимерного убиквитина привела к специфическому васкулярному окрашиванию ED-B на опухолях F9 от мышей. ED-B является высоко специфическим маркером для опухолевой сосудистой сети. Белок-мишень ED-B расположен на аблюминальной стороне сосудов. Вариант 1041-D11 специфически декорирует сосудистую сеть в срезах тканей из опухолей F9. Полученные результаты сравнимы с фрагментом L19 антитела. Дополнительно были проверены 48 тканей; никакого неспецифического окрашивания в любой из 48 тканей в релевантной панели FDA не наблюдалось.
На Фиг.16 В показана аккумуляция 1041-D11 в опухолевой ткани в сравнении с убиквитином дикого типа (на этой Фигуре, Ub2 (NCP2). Опухолевые ткани F9 проанализировали на присутствие 1041-D11 и убиквитин дикого типа в разные моменты времени между 30 мин и 16 ч. Наибольшая аккумуляция 1041-D11 в опухолевой ткани наблюдалась через 30 мин и 16 ч после введения, тогда как аккумуляция убиквитина дикого типа в опухолевых тканях F9 была низкой. Этот вариант обогащен в опухолях, экспрессирующих ED-B, по сравнению с убиквитином дикого типа. Это является доказательством направленности 1041-D11 на опухолевые ткани. Кроме того, отношение опухоль-кровь 1041-D11 в модели рака четко демонстрирует in vivo активность варианта 1041-D11 у животных (данные не показаны).
На Фиг.17 показана высокая селективность и специфичность составного белка 1041-D11-TNF-альфа для ED-B.
Фиг.17А и 17В: Индуцирующая апоптоз активность TNF-альфа из составного белка 1041-D11-TNF-альфа была проверена в клеточном анализе (клетки L929). Эти Фигуры четко показывают, что составной белок 1041-D11-TNF-альфа (Фиг.17В) является таким же активным, как свободный TNF-альфа (Фиг.17В) в клеточной культуре.
Фиг.17С демонстрирует высокую селективность составного белка гетеродимерного убиквитина 1041-D11-TNF-альфа с мишенью ED-B. Домен 67В89 фибронектина ED-B человека связывается с явным значением кД 1,8 нМ с вариантом 1041-D11 (закрашенные круги), показывая высокое сродство для этой мишени. Человеческий фибронектин без домена ED-B (h6789) не связывается вариантом 1041-D11-TNF-альфа (незакрашенные круги).
На Фиг.17D+E показан анализ связывания составного белка 1041-D11-TNF-альфа на основе модифицированного убиквитина с ED-B с помощью анализов Biacore. Результаты демонстрируют высокое сродство составного белка 1041-D11-TNF-альфа с значением кД=1,13 нМ.
На Фиг.17F показана высокая специфичность связывания, наблюдавшаяся с вариантом 1041-D11, в клеточной культуре, которая сохраняется, когда 1041-D11 объединен с TNF-альфа. Этот составной белок специфически связывается с экспрессирующими клетками EDB. Таким образом, составной белок 1041-D11-TNF-альфа связывается с очень высоким сродством и специфичностью с мишенью ED-B ("мишень(+)"). В сыворотке без ED-B ("мишень(-)"), перекрестной реакции не наблюдается.
На Фиг.18 показан относительный рост опухоли in vivo во время лечения мышей в течение 7 суток вариантом 1041-D11, объединенным с TNF-альфа в сочетании с мелфа-ланом. Данные четко показывают, что вариант 1041-D11-TNF-альфа в сочетании с цитостатическим агентом мелфаланом уменьшает относительный рост опухоли более эффективно чем mTNF-альфа в сочетании с мелфаланом или чем только мелфалан. Кинетика роста опухоли в течение 7 суток после лечения показывает эффективное сокращение опухолей вариантом 1041-D11-mTNFa. Это является четким доказательством эффективности лечения опухолей составным белком 1041-D11-TNF-альфа в сочетании с мелфаланом. ED-B идентичен в нескольких видах млекопитающих, включая мышей и человека, и, таким образом, эти результаты прогнозируют действие варианта 1041-D11-TNF-альфа для людей.
ПРИМЕРЫ
Следующие Примеры представлены для дальнейшей иллюстрации изобретения. Изобретение, в частности, продемонстрировано по отношению к модификации убиквитина в качестве примера. Изобретение, однако, этим не ограничено, и последующие Примеры просто показывают осуществимость изобретения на практике на основе вышеприведенного описания. Для полного раскрытия изобретения ссылка также делается на публикации, упомянутые в данной заявке и в приложении, которые все включена в полном объеме в данную заявку путем ссылки.
Пример 1. Идентификация гетеродимерных белков, связывающихся с ED-B, на основе модифицированных убиквитиновых белков
Конструирование и клонирование библиотеки
Если не указано иное, использовались хорошо известные рекомбинантные генетические способы, которые описаны, например, в публикации Сэмбрука и др. (Sambrook et al). Неспецифическая библиотека гетеродимеров человеческого убиквитина с высокой сложностью была подготовлена путем согласованного мутагенеза в совокупности 15 выбранных положений аминокислот.Модифицированные аминокислоты, которые были замещены триплетами NNK, включали по меньшей мере 3 аминокислоты, выбираемые из положений 2, 4, 6, 8, 62, 63, 64, 65, 66, 68 в проксимальном (первом) мономере убиквитина, и по меньшей мере 3 аминокислоты, выбираемые из положений 2,4, 6, 8, 62, 63, 64, 65, 66, 68 в дистальном (втором) мономере убиквитина. Оба мономера убиквитина были генетически связаны (голова к хвосту) линкером глицин/серин по меньшей мере с последовательностью GIG или линкером глицин/серин по меньшей мере с последовательностью SGGGG, например GIG, SGGGG, SGGGGIG, SGGGGSGGGGIG (SEQ ID NO: 32) или SGGGGSGGGG, но возможен любой другой линкер.
Выбор фагового дисплея TAT
Библиотека гетеродимерного убиквитина была обогащена относительно использования мишени, например, фагового дисплея ТАТ в качестве системы выбора. Могут быть использованы другие способы выбора, известные в данной области. Мишень может быть иммобилизована неспецифически на связывающихся поверхностях белка или через биотинилированные остатки, которые были ковалентно соединены с белком. Предпочтительна иммобилизация посредством биотина на гранулах стрептавидина или полосках нейтравидина. Фаги, связывающиеся с мишенью, выбирают или в растворе, или на иммобилизованной мишени; например, биотинилированную и иммобилизованную мишень с фагом инкубировали путем промывки фагов, связанных с матриксом, и путем элюирования фагов, связанных с матриксом. В каждом цикле после инкубации мишени гранулы отделяли от раствора магнитным способом и промывали несколько раз. В первом цикле выбора биотинилированная мишень была иммобилизована на полосках нейтравидина, тогда как в циклах 2-4 выполняли выборы в растворе, после чего следовала иммобилизация комплексов мишень-фаг на покрытых стрептавидином гранулах Dynabeads® (компания Invitrogen). После промывки в первых двух циклах выбора фаги связывающегося с мишенью модифицированного убиквитина высвобождали путем элюирования кислым раствором. В циклах выбора 3 и 4 элюирование фагов осуществляли путем конкурентного элюирования с избыточной мишенью. Элюированные фаги были реамплифицированы. Для направления специфичности связующих во время выбора можно включить белок, сходный с мишенью.
Альтернативно выбору фагового дисплея ТАТ: выборрибосомного дисплея
Библиотека убиквитина была обогащена относительно использования мишени, например, рибосомным дисплеем в качестве системы выбора (Занд и др. (Zahnd et al.), 2007), Охаси и др. (Ohashi et al.), 2007). Могут быть использованы другие способы выбора, известные в данной области. Мишень была биотинилирована согласно стандартным способам и иммобилизована на покрытых стрептавидином гранулах Dynabeads® (компания Invitrogen). Троичные комплексы, включающие рибосомы, мРНК и возни кающий убиквитиновый полипептид, были ассемблированы с использованием набора PURExpressTM In Vitro для синтеза белка (NEB). Были выполнены два первичных цикла выбора, в которых инкубировали троичные комплексы, после чего выли выполнены два подобных цикла выбора. В каждом цикле после инкубации мишени гранулы отделяли от раствора магнитным способом и промывали буфером рибосомного дисплея с повышающейся тщательностью. После промывки в первых двух циклах выбора гранулы снова отделяли от раствора магнитным способом, и мРНК молекул модифицированного убиквитина, связывающихся с мишенью, освободили от рибосом путем добавления 50 мМ ЭДТА. В циклах 3 и 4 выбора элюирование мРНК осуществляли конкурентным элюированием с избыточной мишенью (Липовсек и Плактун (Lipovsek and Pluckthun), 2004). После каждого цикла выполняли очистку РНК и синтез кДНК с использованием набора "RNeasy MinElute Cleanup Kit" (компания Qiagen, Германия), набора "Turbo DNA-free Kit" (компания Applied Biosystems, США) и обратной транскриптазы транскриптора (компания Roche, Германия).
Клонирование обогащенных пулов
После четвертого цикла выбора синтезированная кДНК была амплифицирована посредством ПЦР через праймеры F1
(GGAGACCACAACGGTTTCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAG ATATACATATG) (SEQ ID NO: 9) и
WUBI(co)RD_xho (AAAAAAAAACTCGAGACCGCCACGCAGACGCAGAACCAG) (SEQ ID NO: 10), вырезана с помощью рестрикционных нуклеаз Ndel и Xhol (компания Promega, США) и лигирована в экспрессирующий вектор рЕТ-20b(+) (компания Merck, Германия) через совместимые липкие концы.
Анализ совпадений клонированных колоний
После трансформации в клетки NovaBlue(DE3) (компания Merck, Германия) резистентные к ампициллину клонированные колонии выращивали в течение 6 ч при 37°С в 200 мкл среды SOB AG (среда SOB, содержащая 100 мкг/мл ампициллина и 20 г/л глюкозы), экспрессия модифицированного убиквитина, связывающегося с ED-B, была достигнута путем культивирования в течение 16 ч при 37°С в 96-луночных глубоких планшетах (компания Genetix, Великобритания) с использованием 500 мкл автоиндукционной среды ZYM-5052 (Штудиер (Studier), 2005). Клетки собрали путем центрифугирования в течение 15 мин при 4°С и 3600 g и затем лизировали путем инкубации в течение 30 мин при 37°С с 300 мкл буфера лизиса на лунку, содержащую 0,2× BugBuster® (компания Merck, Германия), 0,3 мг/мл лизоцима (компания VWR, Германия) 0,2 мМ фенилметилсульфонилфторида (компания Roth, Германия), 3 мМ MgCl2 и 0,2 ед./мл бензоназы (компания VWR, Германия) в 50 мМ NaH2PO4, 300 мМ NaCl, рН 8. После центрифугирования в течение 30 мин при 4°С и 3600 g полученные супернатанты подвергли скринингу путем ИФА, используя планшеты Nunc MediSorp (компания Thermo Fisher Scientific, США), покрытые 4 мкг/мл ED-B и убиквитин-специфическим фрагментом Fab, конъюгированным с пероксидазой хрена. В качестве детектирующего реагента использовали TMB-Plus (компания Biotrend, Германия), и желтый цвет получили, используя 50 мкл/лунка 0,2 М раствора H2SO4, измерили в спектрофотометре для чтения планшетов на 450 нм против 620 нм.
Обычно выполняли несколько, например, четыре цикла дисплея выбора против ED-B. В двух последних циклах выбора связывающие молекулы элюировали с избытком свободного ED-B. Были идентифицированы, помимо прочих, следующие связывающие ED-B варианты.
Последовательность 46Н9
MGIVVRTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSD YNIPHPTLLHLVLRLRGGSGGGGSGGGGIGMQIFVHTMTGKTITLEVEPSDTIENVKA KIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNIKPIAELHLVLRLRGG (SEQ ID NO: 6)
Последовательность 9Е12
MRIPVYTLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSD YNIPPFARLHLVLRLRGGSGGGGSGGGGIGMQIFVMTRTGKTITLEVEPSDTIENVKA KIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNIMNARLLHLVLRLRGG (SEQ ID NO: 7)
Последовательность 22D1
MLILVRTLTDKTITLEVEPSDTIGNVKLAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSD YNISVGAMLHLVLRLRGGSGGGGSGGGGIGMQIFVLTWTGKTITLEVEPSDTIENVKA KIQDKEGIPPDQQRLIWAGKQLEDGRTLSD YNIRRLPPLHLVLRLRGG (SEQ ID NO: 8)
Выравнивание последовательности мономера убиквитина дикого типа (Уби-мономер дт) с димером убиквитина дикого типа (Уби-димер дт) и убиквитиновым белком (Ub2-TsX на Фиг.9 с заменой в положении 45 каждого мономера и с двумя замещениями в С-окончании) с гетеродимерным вариантом 1041-D11 модифицированного убиквитина показано на Фиг.9. В Ub2-TsX замещения в С-окончании (GG на АА) мономера повышают стабильность в сыворотке, поскольку деубиквитиназы расщепляются за GG убиквитина, но не за АА. Вторичная структура убиквитина дикого типа по сравнению с убиквитином с этими замещениями в С-окончании почти идентична.
Модифицированные убиквитины с превосходной активностью связывания с ED-B, указанные как 1041-D11 (показаны на Фиг.9; SEQ ID NO: 36) или 1045-D10, идентифицируются по следующим заменам аминокислот по сравнению с диким типом: в первом модуле: K6W, L8W, K63R, E64K, S65F, Т66Р; во втором модуле: K6T, L8Q, Q62W, K63S, E64N, S65W, Т66Е; по выбору Q2R (в варианте 1041-D11, но не в варианте 1045-D10).Подходящими предпочтительными линкерами для этого составного белка являются таковые с SEQ ID NO: 32 или последовательностью GIG. Однако есть много известных линкеров, которые можно использовать вместо их.
В качестве еще одного предпочтительного примера белок представлен следующей последовательностью, где ХХХХ может быть любой аминокислотой (SEQ ID NO: 47). В качестве линкера здесь использовали SGGGGSGGGGIG (выделен курсивом). При этом понимается, что осуществимыми альтернативами является другой вид линкеров или отсутствие линкера.
MTIWVHTLTGKTITLTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNINFKLSLHLVLRLRGGSGGGGSGGGGIG
MQIFVXTXTGKTITLEVEPSDTIENVKAKIQDKEGIPPDQQRLIWAGKQLEDGRTLSDYNINXXXXXLHLVLRLRGG
Консенсусные последовательности примеров белков с этими последовательностями показаны на Фиг.2.
Пример 2. Получение составных белков из вариантов модифицированного убиквитина, связывающих ED-B, и TNF-альфа человека (hTNFa)
Эти варианты экспрессированы как составные белки из модифицированного убиквитина, например, гетеродимерного варианта 1041-D11, и мышиного или человеческого TNFa в E.coli. Анализ составного белка включает: экспрессию и чистоту белка, отсутствие потенциала аггрегации, активность TNFa в клеточной культуре, сродство для белка-мишени ED-B, селективность, специфическое связывание в клеточной культуре. Предпосылкой для эксперимента на животных по индукции сокращения опухоли у мышей с опухолью F9 является объединение с мышиным TNFa.
Этап 1: Получение вектора для клонирования составных белков (pETSUMO-TNFa)
pETSUMOadapt является модифицированным вектором pETSUMO (компания Invitrogen), который модифицирован путем вставки дополнительного сайта множественного клонирования (MCS). Начиная с TNF-альфа, клонированного в pETSUMOadapt, были введены рестрикционные сайты для вставки вариантов модифицированного убиквитина, связывающих ED-B. Полученный конструкт имеет структуру His6-SUMO-TNFa со следующей ДНК-последовательностью (SEQ ID NO: 11):
ATGGGCAGCAGCCATCATCATCATCATCACGGCAGCGGCCTGGTGCCGCGCGGC
AGCGCTAGCATGTCGGACTCAGAAGTCAATCAAGAAGCTAAGCCAGAGGTGAAG
CCAGAAGTCAAGCCTGAGACTCACATCAATTTAAAGGTGTCCGATGGATCTTCAG
AGATCTTCTTCAAGATCAAAAAGACCACTCCTTTAAGAAGGCTGATGGAAGCGTi
CGCTAAAAGACAGGGTAAGGAAATGGACTCCTTAAGATTCTTGTACGACGGTAT
TAGAATTCAAGCTGATCAGACCCCTGAAGATTTGGACATGGAGGATAACGATAT
TATTGAGGCTCACAGAGAACAGATTGGTGGTGTGCGTAGCAGCAGCCGTACCCC
GAGCGATAAACCGGTGGCGC ATGTGGTGGCGAATCCGC AGGCGGAAGGCC
AGCTGCAGTGGCTGAACCGTCGTGCGAATGCGCTGCTGGCCAACGGCGTGGAAC
TGCGTGATAATCAGCTGGTTGTGCCGAGCGAAGGCCTGTATCTGATTTATAGCCA
GGTGCTGTTTAAAGGCCAGGGCTGCCCGAGCACCCATGTGCTGCTGACCCATACC
ATTAGCCGTATTGCGGTGAGCTATCAGACCAAAGTGAACCTGCTGTCTGCGATTA
AAAGCCCGTGCCAGCGTGAAACCCCGGAAGGCGCGGAAGCGAAACCGTGGTAT
GAACCGATTTATCTGGGCGGCGTGTTTCAGCTGGAAAAAGGCGATCGTCTGAGC
GCGGAAATTAACCGTCCGGATTATCTGGATTTTGCGGAAAGCGGCCAGGTGTATT
TTGGCATTATTGCGCTGTAATAA
Эта последовательность TNF-альфа была амплифицирована посредством ПЦР путем введения сайта BamHI- и сайта XhoI. Использованные праймеры:
SUMO-EDB-TNFa-rw (SEQ ID NO: 12): TTT TTT GGA ТСС GTG CGTAGCAGC AGC
SUMO-EDB-TNFa-rev (SEQ ID NO: 13): CTT GTC TCT CGA GGC GGC CGC TTA TTAC
Праймер rw (прямой) (SEQ ID NO: 12) распознает первые 15 пар оснований TNFa (подчеркнутая область) и имеет BamHI-последовательность (выделена жирным шрифтом). Праймер rev (обратный) (SEQ ID NO: 13) содержит последнюю пару оснований TNFa, терминирующие кодоны (подчеркнуты) и рестрикционный сайт Xhol (выделен жирным шрифтом).
Смесь для реакции ПЦР ПОР икл):
84,5 мкл Н2О; 10 мкл 10× Pwo буфер+Mg; 2 мкл 10 мМ дезоксинуклеозидтрифосфата (=200 мкМ); по 0,5 мкл 100 мкМ праймеров прямого/обратного (=каждый 0,5 мкМ); 2 мкл ДНК (=0,25 мкг); 0,5 мкл полимеразы Pwo (=2,5 Ед.; компания Roche)
ПЦР-программа:
3 мин 94°С, 30 с 94°С, 30 с 60°С, 2 мин 72°С (этапы 2-4: 30 циклов), 5 мин 72°С, после 4°С, после очистка продукта ПЦР с помощью набора Qiagen-MinElute (элюирование в 10 мкл ЕВ). Продукт ПЦР ввели в MCS вектора pETSUMOadapt посредством BamHI-XhoI-рестрикции и лигирования.
Смесь для рестрикции (100 мкл):
Вектор: 83 мкл Н2О; 10 мкл 10× буфера NE 3; 1 мкл ЮОх BSA; 3 мкл BamHI (=30 Ед.; NEB), 1,5 мкл Xhol (=30 Ед.; NEB); 1,65 мкл вектора; инкубация в течение 3 ч при 37°С.
Продукт ПЦР: 76,5 мкл Н2О; 10 мкл 10× буфера NE 3; 1 мкл 100× BSA; 3 мкл BamHI (=30 Ед.; NEB), 1,5 мкл Xhol (=30 Ед.; NEB); 8 мкл вставки; инкубация в течение 3 ч при 37°С.
Отделение рестрикции в 1% агарозном геле (100 В цикл 60 мин); отрезание фрагмента вектора (5659 пар оснований) и вставка (491 пара оснований); очистка с помощью набора Qiagen для экстракции геля (элюирование в 30 мкл ЕВ).
Лигирование (20 мкл"):
15,2 мкл Н2О; 2 мкл 10× Т4-ДНК лигазного буфера; 2,26 мкл вектора (200 нг); 0,54 мкл вставки (40 нг) инкубация 5 мин при 65°С; охлаждение до 16°С; добавление 1 мкл Т4-ДНК лигазы (=3 Ед.; NEB); инкубация 6 ч при 16°С.
NaAc/изопропанол-осаждение: смесь для лигирования (20 мкл)+2,2 мкл 3М NaAc (рН 5,0)+22,2 мкл изопропанола; 30 мин при -20°С; 15 мин при 4°С 13000 об/мин;
повторное суспендирование гранулы в 500 мкл 70% EtOH; вращение; повторное суспен-дирование гранулы в 10 мкл Н2О.
Трансформация:
Смешивание электро-компетентных клеток Novablue(DE3) (40 мкл-аликвота) с 10 мкл продукта лигирования; перенос в кювету ОД см для электрополяции; вибрация в элек-трополяторе (1,8 кВ, 50 мкФ, 100 Ом); раствор для инкубации с 1 мл SOC-среды 45 мин при 37°С 220 об/мин; 100 мкл на LB-планшете с канамицином; инкубация в течение ночи при 37°С.
Этап 2: Клонирование EDB-составных белков на основе модифицированного убиквитина
Для получения слияний связывающих EDB вариантов на основе модифицированного убиквитина и TNFa, представляющую интерес последовательность на основе модифицированного убиквитина, связывающую EDB, амплифицировали из pET20b-вектора посредством ПЦР; ввели рестрикционные сайты Bsal и BamHI. Этот способ подходит для мономерных и димерных вариантов, связывающих EDB, на основе модифицированного убиквитина. Праймер для мономерного ДТ-Убиквитина (Wubi):
Прямой линкер SUMO-EDB-WUBI (SEQ ID NO: 14):: GTT CCA AGG TCT CAT GGT ATG CAG АТС TTC GTG
Обратный линкер SUMO-EDB (SEQ ID NO: 15):: GTG GTG GGA ТСС ACC GCC ACC ACC AGA ACC GCC ACG CAG ACG
Прямой праймер (SEQ ID NO: 14) распознает первые 15 пар оснований модифицированного убиквитина (подчеркнутая область) и имеет Bsal-последовательность (выделена жирным шрифтом). Обратный праймер (SEQ ID NO: 15) распознает последние 15 пар оснований модифицированного убиквитина и вставляет аминокислотный линкер (последовательность SGGGG) и BamHI-рестрикционньш сайт (выделен жирным шрифтом). Для каждого варианта на основе модифицированного убиквитина исполь зовали специфический прямой праймер. Праймеры для мономерных связывающих EDB вариантов на основе модифицированного убиквитина 1Н4, 5Е1 и 4В10:
1Н4 (MWIKV...): праймер (8иМО-ЕБВ-1Н4-прямой) (SEQ ID NO: 16): GTT CCA AGG TCT CAT GGT ATG TGG АТС AAG GTG
4B10 (MLILV): праймер (SUMO-EDB-4B10-npflMoft) (SEQ ID NO: 17): GTT CCA AGG TCT CAT GGT ATG TTG АТС CTG GTG
5E1 (MVINV...): праймер (8иМО-ЕОВ-5Е1-прямой) (SEQ ID NO: 18): GTT CCA AGG TCT CAT GGT ATG GTT АТС AAT GTG
Обратный праймер использовали для всех мономерных вариантов на основе модифицированного убиквитина. Обратный праймер для димерных вариантов на основе модифицированного убиквитина:
Димер-t0a-обратный (SEQ ID NO: 19): GTG GTG GGA ТСС ACC GCC ACC ACC AGA ACC ACC ACG TAA ACG
Прямой праймер для клонирования димерных ДТ-убиквитинов (WubiHubi) и для димерных связывающих EDB вариантов на основе модифицированного убиквитина:
WT (MQIFV…) праймер (SUMO-EDB-WUBI-прямой) (SEQ ID NO: 20): GTT CCA AGG TCT CAT GGT ATG CAG АТС TTC GTG
(Примечание: прямой праймер для димерного ДТ-убиквитина идентичен прямому праймеру для мономерного ДТ-убиквитина.)
9Е12 (MRIPV…): праймер (9ЕШ0а-прямой) (SEQ ID NO: 21): GTT CCA AGG TCT CAT GGT ATG CGTATC CCTGTG
24H12 (MVIKV…): праймер (24Н12-Юа-прямой) (SEQ ID NO: 22):GTT CCA AGG TCT CAT GGT ATG GTT АТС AAG GTG
15G7 (MEIGV…): праймер (15G7-t0a-np*Moft) (SEQ ID NO: 23): GTT CCA AGG TCT CAT GGT ATG GAG АТС GGT GTG
22D1 (MLILV…): праймер (2201-Юа-прямой) (SEQ ID NO: 24): GTT CCA AGG TCT CAT GGT ATG CTT АТС TTG GTG
ПЦР-смесь (100 мкл):
84,5 мкл Н2О; 10 мкл 1 Ох Pwo-буфер+Mg; 2 мкл 10 мМ дезоксинуклеозидтрифосфата (=200 мкМ); по 0,5 мкл 100 мкМ праймера прямого/обратного (=je 0,5 мкМ); 2 мкл ДНК (зависит от варианта); 0.5 мкл Pwo-полимеразы (=2,5 Ед.; компания Roche)
ПЦР-программа;
1. Змин94°С
2. 30 с94°С
3. 30 с60°С
4. 2 мин 72°С (этапы 2-4: 30 циклов)
5. 5 мин 72°С, после 4°С
Очистка ПЦР-продуктов в агарозном геле, отрезание требуемой полосы и очистка с помощью набора Qiagen для экстракции геля. Клонирование ПЦР-продукта посредством BsaI-BamHI-рестрикции (в pETSUMO-TNFa)
Рестрикция (100 мкл): 75 мкл Н2О; 10 мкл 10х буфер NEB 3; 1 мкл ЮОх бычьего сывороточного альбумина (БСА); 3 мкл Bsal (=30 Ед.; NEB); 8 мкл ДНК (вектор или ПЦР-продукт) инкубация 2 ч 50°С, 10 мин 65°С, добавление 3 мкл BamHI (=30 Ед.; NEB), 2 ч 37°С отделение рестрикции в 1% агарозном геле; отрезание фрагмента вектора и вставка; очистка с помощью набора Qiagen для экстракции геля (элюирование в 30 мкл ЕВ).
Дотирование (20 мкл):
12,5 мкл Н2О; 2 мкл 10х Т4-ДНК лигазный буфер; 5 мкл вектор (66 нг); 0,5 мкл вставка (вариабельная), инкубация 5 мин 65°С; охлаждение до 16°С; добавление 1 мкл Т4-ДНК лигазы (=3 Ед.; NEB); инкубация 16 ч 16°С
Осаждение NaAc/изопропанолом (смотрите Этап 1)
Трансформация в электрокомпетентньгх клетках Novablue(DE3), как сказано выше. Результатом является следующий конструкт слияния: EDB- модифицированный убиквитин и TNFa в pETSUMOadapt с der His6-SUMO- модифицированный убиквитин-SGGGG-TNFa (359 аминокислот с мономерным модифицированным убиквитином, 447 аминокислот с димерным модифицированным убиквитином)
Пример 3: Экспрессия и очистка составных белков на основе убиквитина-Т№-альфа
Анализ ДНК-последовательности показал правильность последовательностей составных белков SUMO-TNFa. Для экспрессии вариантов клоны культивировали в колбе шейкера путем разбавления прекультуры 1:100 LB/канамицином и перемешивания культуры с частотой вращения 200 об/мин и 37°С до оптической плотности при 600 нм (OD600) 0,5. Экспрессию индуцировали путем добавления IPTG (конечная концентрация 1 мМ). Культивирование продолжали в течение 4 часов при 30°С и 200 об/мин. Клетки бактерий собрали центрифугированием при 4°С, 6000× g в течение 20 мин. Клеточную гранулу суспендировали в 30 мл буфера NPI-20, включающего бензоназу и лизоцим. Клетки деструктировали ультразвуком (3×20 секунд) на льду. Супернатант, содержащий растворимые белки, был получен после центрифугирования суспензии: при 4°С и 40000× g в течение 30 мин. Оба белка были очищены аффинной хроматографией при комнатной температуре. Одна колонка Ni-агароза (5 мл, компания GE Healthcare) была уравновешена 50 мл NPI-20. Супернатант, содержащий эти растворимые белки, был добавлен в колонку, за чем последовал этап промывки NPI-20. Связанный белок элюировали с линейным градиентом до 50% NPI-500 в 100 мл. Фракции анализировали денатурирующим электрофорезом в полиакриламидном геле в отношении их чистоты. Подходящие фракции объединяли и подавали в гель-фильтрационную колонку (Superdex 75, 1,6×60 см, компания GE Healthcare), уравновешенную SUMO-гидролазным буфером для расщепления (50 мМ Трис, 300 мМ NaCl, рН 8,0) с расходом 1 мл/мин.
Реакцию расщепления выполняли согласно инструкции изготовителя (компания Invitrogen). После расщепления белок наносили на Ni-агарозную колонку (5 мл, компания GE Healthcare). His-помеченные SUMO-гидролаза и SUMO связывались с колонкой, а правильный составной белок проходил колонку (без His-метки). Чистоту белков подтвердили анализом жидкостной хроматографией высокого разрешения и гель-электрофорезом. Правильность молекулярной массы тримера (посредством TNFa) подтвердили, используя аналитический SEC-анализ (10/30 Superdex G75, компания GE Healthcare).
Пример 4: Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина с ED-B человека
Пример 4А. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем ИФА, зависящего от концентрации.
Связывание вариантов на основе убиквитина с ED-B человека анализировали путем ИФА, зависящего от концентрации. Увеличивающиеся количества очищенного белка наносили на планшеты NUNC-medisorp, покрытые ED-B человека, БСА и клеточным фибронектином (cFN). Антигенное покрытие 50 мкл (10 мкг/мл) на лунку выполнили при 4°С в течение ночи. После промывки планшетов фосфатно-солевым буфером ФСБ), 0,1% Tween-20 рН 7,4 (ФСБТ) лунки блокировали, используя блокирующий раствор (ФСБ рН 7,4; 3% БСА; 0,5% Tween-20) при 37°С в течение 2 ч. Лунки снова три раза промыли ФСБТ. Разные концентрации белка, связывающего ED-B на основе модифицированного убиквитина затем инкубировали в лунках при комнатной температуре в течение 1 ч (объем 50 мкл) (на Фиг.10, как стартовую концентрацию использовали 500 нМ белка 1041-D11). После промывки лунок ФСБТ нанесли конъюгат перокси-дазы хрена фрагмента анти-Ubi fab (AbyD) в подходящем разбавлении (например, 1:2000 или 1:6500) в ФСБТ. Планшет промыли три раза 300 мкл буфера ФСБТ на лунку. В каждую лунку добавили 50 мкл раствора субстрата ТМВ (компания KEM-EN-Тес) и инкубировали в течение 15 мин. Реакцию прекратили путем добавления 50 мкл 0,2 М H2SO4 на лунку. Планшеты ИФА считывали, используя устройство для чтения ИФА TECAN Sunrise. Фотометрические измерения спектральной поглощательной спо собности выполнили на 450 нм, используя 620 нм в качестве контрольной длины волны. На Фиг.1 четко показано специфическое связывание 1Н4 с ED-B с истинным кД-значением 11 нМ. Вариант 5Е1 показал истинное кД-значение 7,7 мкМ и 4В10 280 нМ соответственно. Фиг.10 показывает очень высокоаффинное связывание варианта 104] D11 с ED-B (кД=6,9 нМ). Таким образом, только немного модификаций (до 8 замеще ний в каждом мономере) в убиквитине дикого типа приводят к очень высокоаффинному связыванию с ED-B.
Пример 4 В. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем конкурентного ИФА, зависящего от концентрации.
Был выполнен конкурентный, зависящий от концентрации ИФА, для анализа связывания варианта 1041-30 D11 убиквитина с иммобилизованным ED-B, содержащим фрагмент фибронектина (67В89), в присутствии увеличивающихся количеств свободной мишени. Условия ИФА были как описано, например, для Примера 5А, за тем исключе нием, что белок 1041-D11 был предварительно инкубирован с ED-B (67В89) (0 мкМ -10 мкМ) или также с отрицательным контролем 6789 (0 мкМ-10 мкМ) в течение 1 ч и затем эту смесь с мишенью 67В89 поместили на планшет Medisorp; после этого этот вариант булл детектирован соответствующим антителом (анти-Убиквитин-Fab-пероксидаза хрена; разбавление 1:6500). На Фиг.11 показано, что вариант 1041-D11 имеет очень высокоаффинное связывание с ED-B (IC50=140 нМ). Результат, показанный на Фиг.10 подтвержден; только немного модификаций (до 8 замещений в каждой мономере) в убиквитине дикого типа приводят к очень высокоаффинному связывании с ED-B.
Пример 4С. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем конкурентного ИФА, зависящего от концентрации, с одновременным анализом стабильности связывающей активности в сыворотке.
ИФА выполнили, используя способы, хорошо известные в данной области, которые описаны выше (Пример 5А и 5В). ED-B (здесь названный 67В89) нанесли на микро-титровальные планшеты, этот вариант связали с ED-B и детектировали антителом, специфическим к убиквитину (анти-Ubi-Fab-пероксидаза хрена). Вариант в этом анализе обрабатывали разными путями: вариант инкубировали в сыворотке мышей в течение 1 ч при 37°С (смотрите на Фиг.13, синие крути); вариант инкубировали в сыворотке крыс в течение в течение 1 ч при 37°С (на Фиг.13 красные круги), или вариант инкубировали в ФСБ в течение 1 ч при 37°С (на Фиг.13 черные круги). Фиг.13 показывает, что все кД варианта 1041-D11 составляли от 10,3 нМ (в ФСБ) до 20,74 нМ (в сыворотке мышей).
Пример 4D. Анализ связывания вариантов, связывающих ED-B, на основе модифицированного убиквитина путем анализов Biacore.
Были проанализированы разные концентрации варианта (например, 0-200 нМ варианта, предпочтительно 1041-D11) для связывания с ED-B, содержащим фрагмент фибронектина (названный 67В89), иммобилизованным на чипе СМ5 (Biacore), используя способы, известные специалистам в данной области. Полученные данные обрабатывали посредством программного обеспечения BIAevaluation и 1:1 - Langmuir-подгонкой. Молекулярная масса варианта 1041-D11 составляла 1,0 нМ, как показано на Фиг.12. Кинетические константы связывания составили kon=7,6×105; koff=7,7×10-4 с-1. KD составного белка 1041-D11 - TNF-альфа составила 1,13 нМ, как показано на Фиг.17D. Кинетические константы связывания составили kon=4,5×105 M-1V-1; koff=5,0×10-4 с-1.
Пример 4Е. Анализ формирования комплекса вариантов, связывающих ED-B, на основе модифицированного убиквитина путем СО-ЖХВР.
Для анализа формирования комплекса использовали колонки Tricorn Superdex 755/150 GL (компания GE-Healthcare) (объем=3 мл), количество вводимого белка составило 50 мкл. Другие условия: буфер: 1× ФСБ, рН 7,3, расход: 0,3 мл/мин, цикл: 45 мин (инъекция пробы: через 15 мин). Условие: 0,72 нмоли белка 1041-D11+0,72 нмоли ED-B (здесь назван 67В89 или также отрицательного контроля 6789) инкубировали в течение 1 ч при комнатной температуре; затем наносили на колонку для анализа формирования комплекса. На Фиг.14 только вариант показан черным, только мишень ED-B показана синим, связывание варианта, формирующее комплекс с ED-B, показано розовым. Фиг.14А показывает ED-B с этим вариантом; Фиг.14 В показывает вариант без ED-B. Эта Фигура показывает, что вариант 1041-D11 формирует комплекс с ED-B (67В89), но не формирует комплекса с 6789.
Пример 5: Биологический анализ TNF-альфа
Физиологическую активность TNF-альфа слияний, связывающих ED-B, на основе TNF-альфа-модифицированного убиквитина определяли, используя анализ L929 апоп-тоза (Флик и др. (Flick et al), 1984 J. Immunol. Methods. 68:167-175). В этом анализа TNF-альфа эффективно стимулирует смерть клеток в клетках, сенсибилизированных актиномицином Д при значениях ЕС50 в пикомолярном диапазоне.
Клетки повторно суспендировали в среде, содержащей ФСБ и антибиотики. Клеточную суспензию в объеме 100 мкл с плотностью 3,5 х 105 клеток/мл посеяли в лунки 96-луночного стандартного планшета для культивирования клеток, после чего инкубировали в течение ночи в инкубаторе с увлажненным CO2. После этого культуральную среду удалили и добавили 50 мкл среды, содержащей ФСБ, актиномицин Д и антибиотики, в каждую лунку, после чего инкубировали еще 30 мин. После этого добавили 50 мкл проверяемых объектов, слияний, связывающих ED-B, на основе TNF-альфа-модифицированного убиквитина или, как контроль, рекомбинантного TNF-альфа человека в подходящем диапазоне концентраций от 10-7 до 10-18 М. После еще 48 ч инкубации определили метаболическую активность как меру выживания клеток, используя реагент WST-1 (компания Roche).
Для каждого проверяемого объекта были выполнены по меньшей мере три независимых эксперимента, каждый из них три раза. Каждая проверка составных белков, связывающих ED-B, на основе TNF-альфа-модифицированного убиквитина проходила параллельно с проверкой диапазона дозы рекомбинантного TNF-альфа человека, чтобы получить информацию о вариабельности между анализами.
Количественная оценка основана на значении ЕС50, т.е., значении концентрации проверяемого объекта, способствующему выживанию половины клеток.
Из слияния, связывающего ED-B, на основе TNF-альфа-модифицированного убиквитина были проанализированы один мономер убиквитина (Wubi) и три конструкта ди-мера убиквитина. В зависимости от варианта на основе модифицированного убиквитина, связывающего ED-B и соединенного с частью TNF-альфа, связанная с TNF-альфа активность возрастала (SPWF-28_24-H12_STNF-альфа) или убывала (SPWF-28_22-D1_TNF-альфа, Wubi-Hubi-TNF-альфа) приблизительно на один порядок. Смотрите Фиг.17 по анализу варианта 1041-D11 TNF-альфа.
Пример 6. Анализ связывания вариантов убиквитина в пробах клеточной культуры
Проверили связывание варианта 1041-D11 с клетками клеточной культуры. Были проанализированы клетки разных клеточных культур, включая нормальные фибробластные клетки эмбрионального легкого человека, имеющие высокие уровни экспрессии ED-B (клетки Wi38), клеточную линию эмбрионального фибробласта мышей (Balb 3Т3); линию стромальных клеток, полученную из моноцитов/макрофагов (RAW264.7) костного мозга мышей (ST-2), клетки NHDF и мышиные фибробластные клетки (LM).
Вариант 1041-D11 (разные концентрации) или антитело, специфическое к ED-B (500 нМ FV28 CH4/F1 1× ФСБ были инкубированы (1 ч, 37°С) с клетками Wi38 (60000 клеток/мл; из АТСС), после чего выполнили фиксацию метанолом (5 мин, -20°С), блокировку (5% хрена/ФСБ, 1 ч); инкубацию с a-Strep-Tag-IgG кролика (полученного от компании GenScript А00875,1:500) в течение 1 ч и инкубацию с a-rabbit-IgG*А1еха488-АК кролика (полученного ото компании Invitrogen Al 1008,1:1000) в течение 1 ч. Ядра были окрашены DAPI. Первая колонка на Фиг.15А показывает контроль, использующий антитела kEDB, вторая колонка показывает инкубацию варианта при концентрации белка 58,7 нМ, третья колонка показывает вдесятеро большую концентрацию белка 1041-D11 (587 нМ), четвертая колонка показывает отрицательный контроль с ФСБ. В первом ряду фибробластные клетки Wi38 человека показаны в фазовом контрасте, второй ряд показывает иммунофлуоресценцию и третий ряд показывает окрашивание DAPI. Из этих изображений можно сделать вывод, что вариант 1041-D11 связывается с фиксированными клетками Wi38 с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.Отрицательные контрольные клетки типа NHDF являются первичными нормальными фибробластньгми клетками, которые экспрессируют низкие уровни EDB-фибронектина (данные не показаны). Эти варианты не связываются с этими клетками.
На Фиг.15 В показан анализ варианта 1041-D11 на жизнеспособных клетках Wi38. Отрицательные контрольные клетки типа NHDF являются первичными нормальными фибробластными клетками, которые экспрессируют низкие уровни EDB-фибронектина. Клетки поместили в слайды камеры (NUNC, 60000 клеток/мл). Для анализа потенциала связывания клетки фиксировали с помощью 100% МеОН в течение 5 мин при -20°С.Для блокировки неспецифического связывания клетки инкубировали с 5% лошадиной сывороткой в течение 1 ч при 37°С. Клетки проверили с вариантом 1041-D11, антителом FV28 CH4/F1, специфическим к ED-B, в качестве положительного контроля или UB2 в качестве отрицательного контроля в разных концентрациях в течение 1 ч при комнатной температуре. Проверку провели путем инкубации с a-Strep-Tag-IgG кролика (полученного от компании GenScript А00875,1:500) в течение 1 ч и инкубации с a-rabbit-IgG*Alexa488-AK кролика (полученного от компании Invitrogen Al 1008, 1:1000) в течение 1 ч. Ядра окрасили DAPI. Первая и третья линии на Фиг.15 В показывает этот вариант при разной концентрации белка и отрицательном контроле. Вторая и четвертая линии показывают инкубацию контроля с использованием антител к EDB. Первые две линии показывают вариант и положительный контроль на клеточной линии Wi3 8. Третья и четвертая линии показывают инкубацию клеток NHDF. Из изображений можно видеть, что вариант 1041-D11 связывается с жизнеспособными клетками Wi38 с высокой специфичностью к ED-B, содержащему внеклеточный матрикс.Был выполнен контроль с использованием клеток NHDF, которые не содержат низкий уровень EDB (данные не показаны). Варианты не связываются с этими клетками.
Подобные эксперименты были выполнены с использованием разных типов клеток, например, Balb3T3 (АТСС, кат.№30-2002), Raw (Lonza, кат. №ВЕ12-115F/U1), ST-2 (Lonza, кат.№BE 12-115F/U1). На Фиг.15С и D показано, что связывание ED-B является высоко специфическим к мышиным клеткам Balb3T3 и ST-2. Не наблюдалось связывания с моноцитами/макрофагами (необработанными) (данные не показаны).
Как сказано выше, Фиг.16А показывает специфичность 1041-D11 в срезах тканей. Были оценены ткани опухолей F9 из семи проб. Иммуногистохимия с 500 нМ 1041-D11 дала специфическое окрашивание сосудов ED-B на опухолях F9 от мышей. ED-B является высоко специфическим маркером для сосудистой сети опухоли. Белок-мишень EDB расположен на аблюминальной стороне сосудов. 1041-D11 специфически декорирует сосудистую сеть в срезах тканей из опухолей F9. Полученные результаты сравнимы с тканевой специфичностью фрагмента L19 антитела. Кроме того были проверены 48 тканей; не наблюдалось неспецифического окрашивания в любой из 48 тканей в релевантной панели FDA. Фиг.16В показывает аккумуляцию 1041-D11 в клетках опухоли по сравнению с убиквитином дикого типа. Таким образом, составные белки на основе модифицированного убиквитина, специфически связывающиеся с ED-B, подходят для целевой раковой терапии на основе ED-B.
Пример 7: Исследование in vivo эффективности варианта 1041D11-TNF-альфа
Для того, чтобы установить терапевтическую эффективность варианта 1041-D11-TNF-альфа, это соединение проверили на тератоме F9 (смотрите, Бореи и др. (Borsi et al.), 2003 Blood 102, 4384-4392) в мьппиных моделях. Экспрессия ED-B у мышей сравнима с человеком в ситуации in vivo и подходит для оценки терапевтического воздействия 1041-D11-mTNF-альфа на рак, предпочтительно в сочетании с цитотоксическим соединением, таким как мелфалан. Тератома F9 является агрессивной опухолью с высокой плотностью сосудов. Бореи и др. описали, что нацеливание мышиного TNF-альфа через антитела к EDB повышают эффективность мелфалана, что демонстрируется замедлением роста опухоли. График экспериментов для исследования эффективности был взят у Бореи, 2003.
На стадии 1 определяли фармакологически активную и переносимую дозу с конечными точками, относящимися к отношению опухоли к массе тела, потере массы тела и выживанию. Авторы изобретения выяснили, что 1041D11-TNF-альфа переносим при самой высокой дозе (6,75 пмоль/г), но не оказывает подавляющего действия на рост опухоли (>10% массы тела спустя 3,4 и 8 суток → затем животные были умерщвлены), тогда kak104D11-TNF-альфа в самой низкой дозе (0,25 пмоль/г), как кажется, замедляет рост опухоли. Использованные далее группы дозировок снижали с 2,25 пмоль/г 1041D11-TNF-альфа.
На стадии 2 исследования определяли зависящую от дозы эффективность с мелфаланом при конечной точке замедления роста опухоли (потеря массы тела животным >10%, опухоль >10% массы тела, изъязвление опухоли). В данном исследовании были проверены 1041D11/mTNFa и мышиный TNFa в сочетании с мелфаланом. Использовали 168 животных, 14 групп дозировок (по 8 мышей в группе, брали, когда опухоли F9 достигали 300-400 мм3); за внутривенным введением проверяемой пробы следовала интраперитонеальная инъекция мелфалана через 24 часа. График дозировки показан в таблице 1.
На Фиг.18 показан относительный рост опухоли во время лечения (7 суток). Фиг.18а четко показывает, что наше соединение 1041-D11-TNF-альфа в сочетании с мелфала-ном сокращает относительный рост опухоли более эффективно чем mTNF-альфа в сочетании с мелфаланом или чем только мелфалан. Кинетика роста опухоли через 7 суток после лечения показывает значительное сокращение опухолей при воздействии 1041-D11-mTNFa. Это является явным доказательством эффективности в сочетании с мелфаланом.
1. Birchler, M., F.Viti, L.Zardi, B.Spiess, and D.Neri. 1999. Selective targeting and photocoagulation of ocular angiogenesis mediated by a phage-derived human antibody fragment. Nat Biotechnol 17:984-8.
2. Brenmoehl, J., M.Lang, M.Hausmann, S.N.Leeb, W.Falk, J.Scholmerich, M.Goke, and G.Rogler. 2007. Evidence for a differential expression of fibronectin splice forms ED-A и ED-B in Crohn's disease (CD) mucosa. Int J Colorectal Dis 22:611-23.
3. Dubin, D., J.H.Peters, L.F.Brown, B.Logan, K.C.Kent, B.Berse, S.Berven, В.Cercek, B.G.Sharifi, R.E.Pratt, et al. 1995. Balloon catheterization induced arterial expression of embryonic fibronectins. Arterioscler Thromb Vase Biol 15:1958-67.
4. Goodsell, D.S. 2001. FUNDAMENTALS OF CANCER MEDICINE: The Molecular Perspective: Antibodies. The Oncologist 6:547-548.
5. Kaczmarek, J., P.Castellani, G.Nicolo, B.Spina, G.Allemanni, and L.Zardi.
1994. Distribution of oncofetal fibronectin isoforms in normal, hyperplastic and neoplastic human breast tissues. Int J Cancer 59:11 -6.
6. Menrad, A., and H.D.Menssen. 2005. ED-B fibronectin as a target for antibody-based cancer treatments. Expert Opin Ther Targets 9:491-500.
7. Pujuguet, P., A.Hammann, M.Moutet, J.L.Samuel, F.Martin, and M.Martin.
1996. Expression of fibronectin ED-A+and ED-B+isoforms by human and experimental colorectal cancer. Contribution of cancer cells and tumor-associated myofibroblasts. Am J Pathol 148:579-92.
8. Trachsel, E., M.Kaspar, F.Bootz, M.Detmar, and D.Neri. 2007. A human mAb specific to oncofetal fibronectin selectively targets chronic skin inflammation in vivo. J Invest Dermatol 127:881 -6.
9. Van Vliet, A., H.J.Baelde, L.J.Vleming, E. de Heer, and J.A.Bruijn. 2001.
Distribution of fibronectin isoforms in human renal disease. J Pathol 193:256-62.
10. Lipovsek, D., and Pluckthun, A. (2004). In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 290,51-67.
11. Ohashi, H., Shimizu, Y„ Ying, B.W., and Ueda, T. (2007). Efficient protein selection based on ribosome display system with purified components. Biochem Biophys. Res. Commun. 352,270-276.
12. Studier, F.W. (2005). Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41,207-234.
13. Zahnd, C, Amstutz, P., and Pluckthun, A. (2007). Ribosome display: selecting and evolving proteins in vitro that specifically bind to a target. Nat. Methods 4, 269-279.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИДЕНТИФИКАЦИИ ГЕТЕРОМУЛЬТИМЕРНЫХ МОДИФИЦИРОВАННЫХ УБИКВИТИНОВЫХ БЕЛКОВ СО СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С ЛИГАНДАМИ | 2010 |
|
RU2553333C2 |
| СПЕЦИФИЧЕСКИЕ И ВЫСОКОАФИННЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ МОДИФИЦИРОВАННЫЕ SH3-ДОМЕНЫ КИНАЗЫ Fyn | 2007 |
|
RU2478707C2 |
| НОВЫЕ IL-17-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2550272C2 |
| TRAIL R2-СПЕЦИФИЧЕСКИЕ МУЛЬТИМЕРНЫЕ СКАФФОЛДЫ | 2011 |
|
RU2628699C2 |
| КОМПОЗИЦИИ IL-12, НАЦЕЛЕННЫЕ НА EDB | 2019 |
|
RU2758143C1 |
| РЕЦЕПТОР ED-ДОМЕНА ФИБРОНЕКТИНА | 2001 |
|
RU2280254C2 |
| МУТЕИНЫ ЛИПОКАЛИНА 2 ЧЕЛОВЕКА (LCN2, HNGAL) С АФФИННОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ МИШЕНИ | 2010 |
|
RU2564125C2 |
| АНТАГОНИСТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2005 |
|
RU2401842C2 |
| АНТИТЕЛА | 2018 |
|
RU2798399C2 |
| АНТИТЕЛА ПРОТИВ ПОЛИУБИКВИТИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2603093C2 |
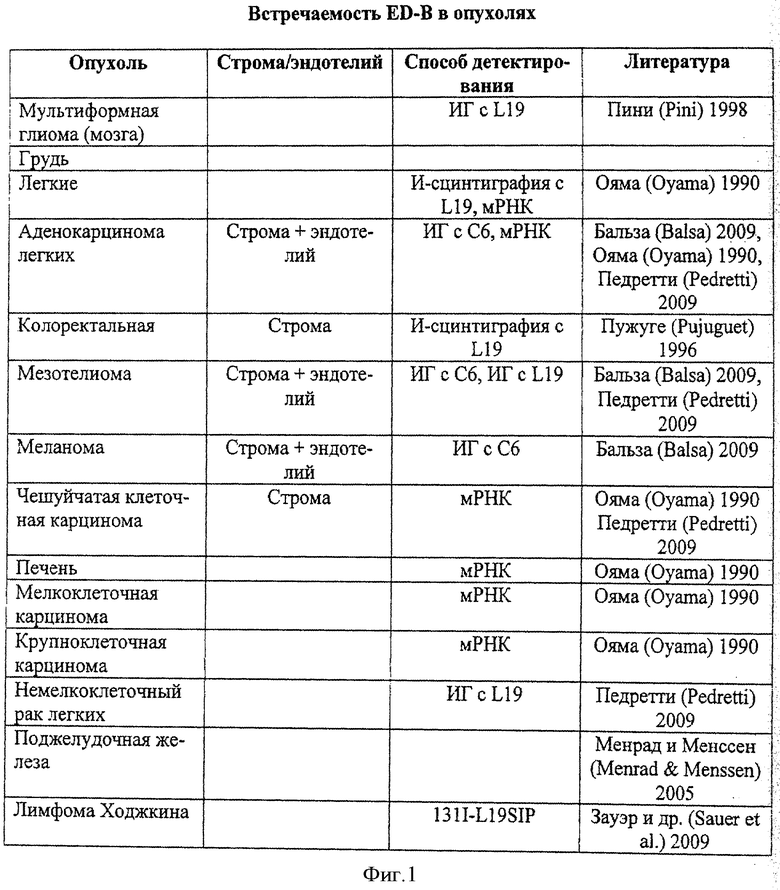
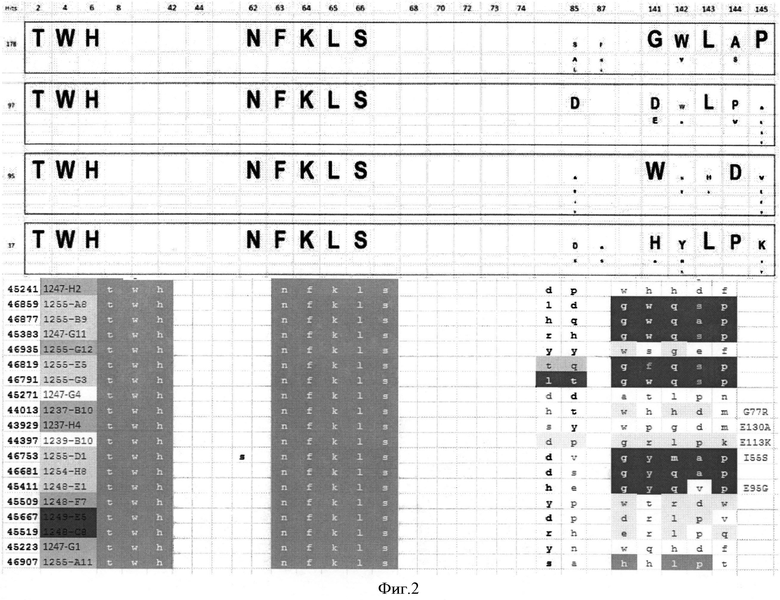
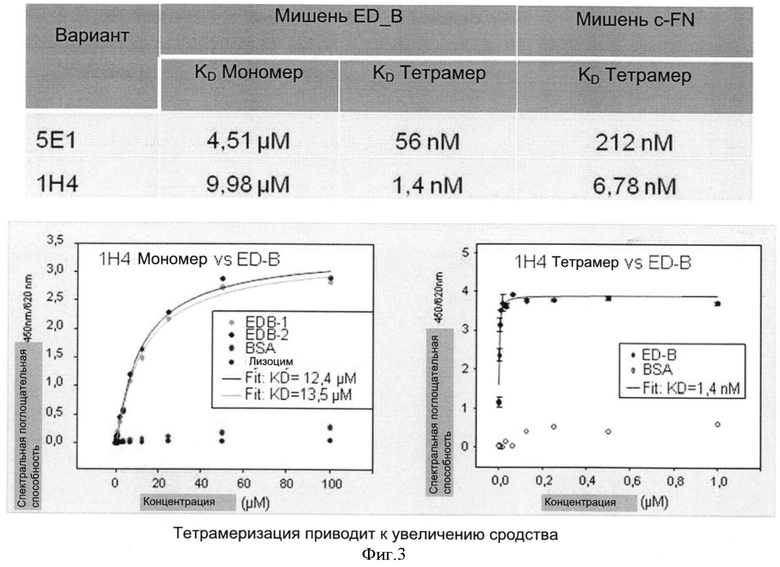

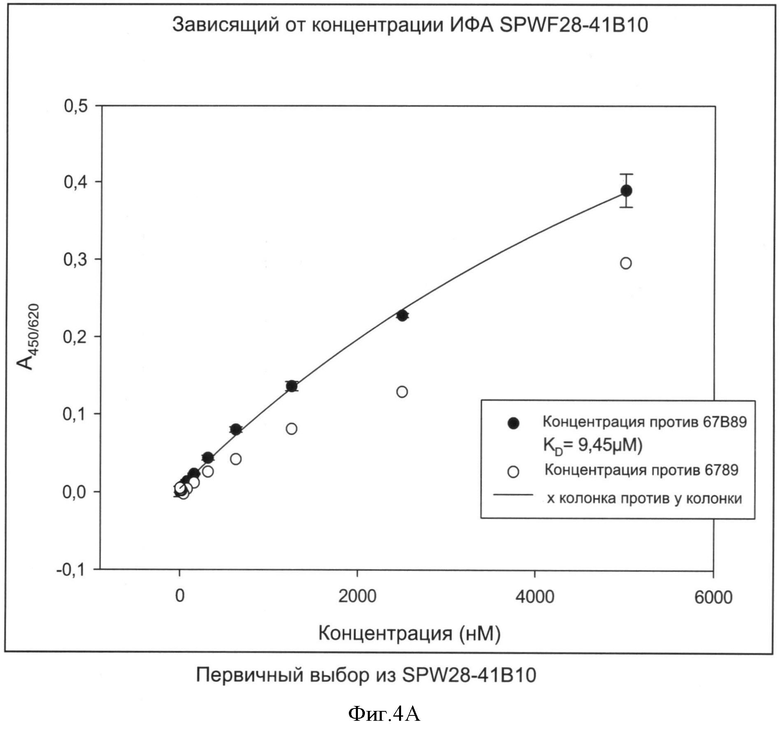
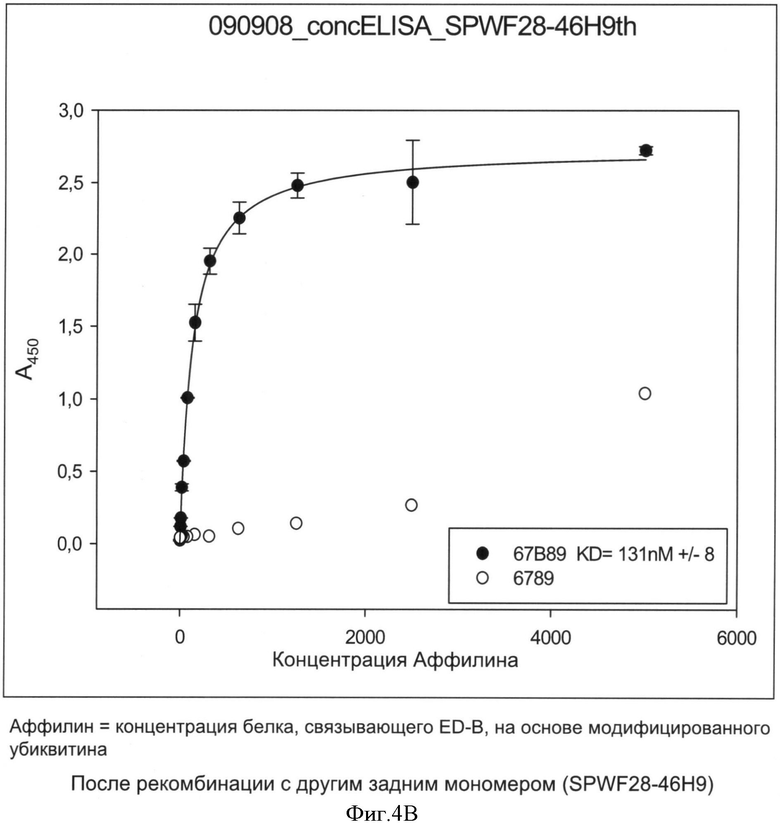

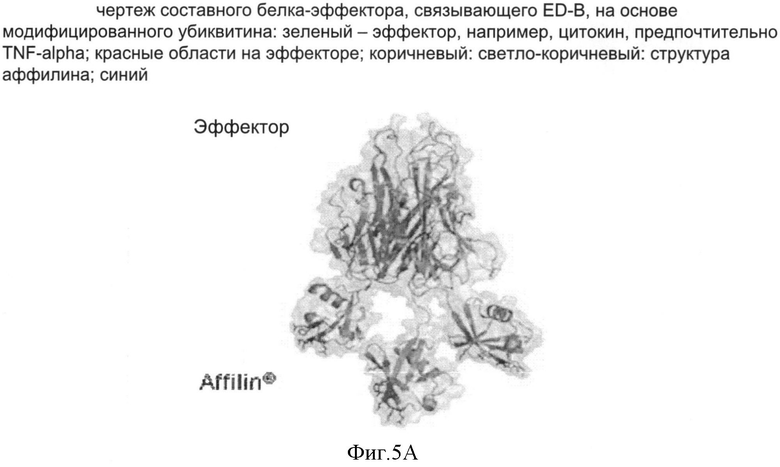

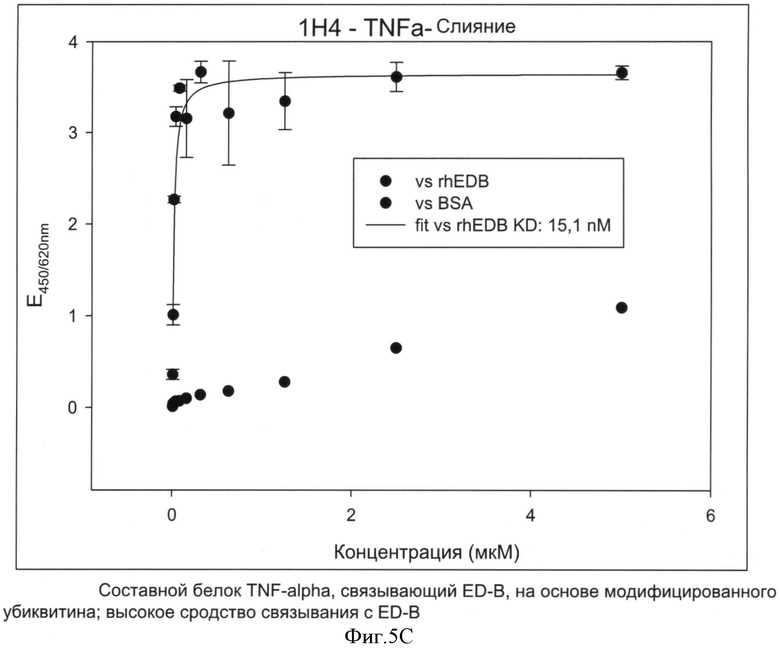
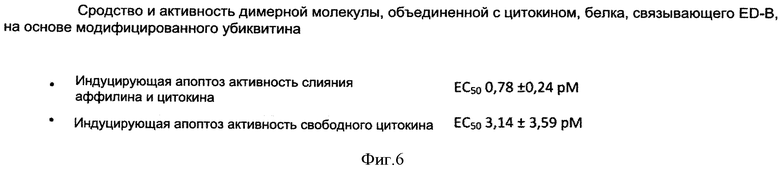
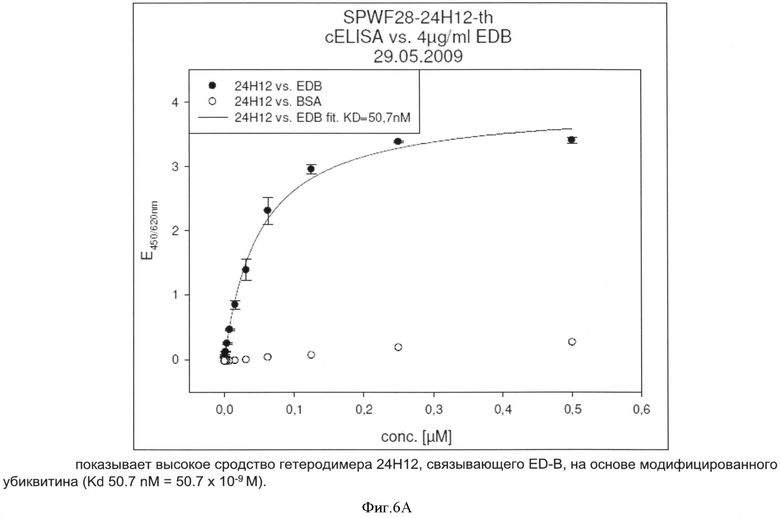
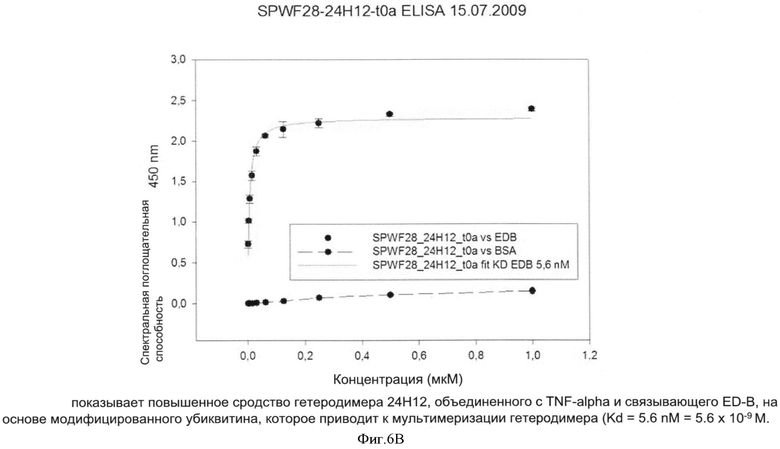
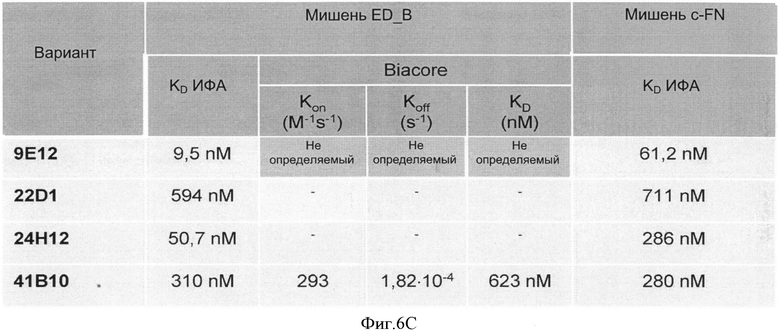
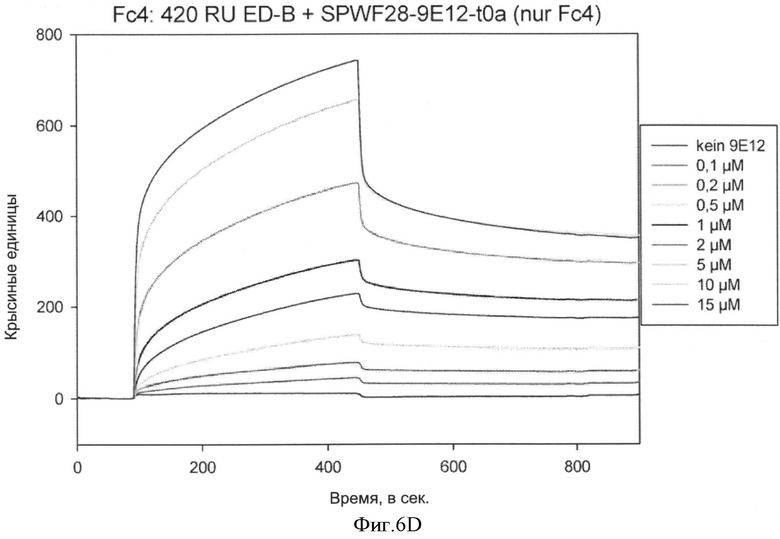
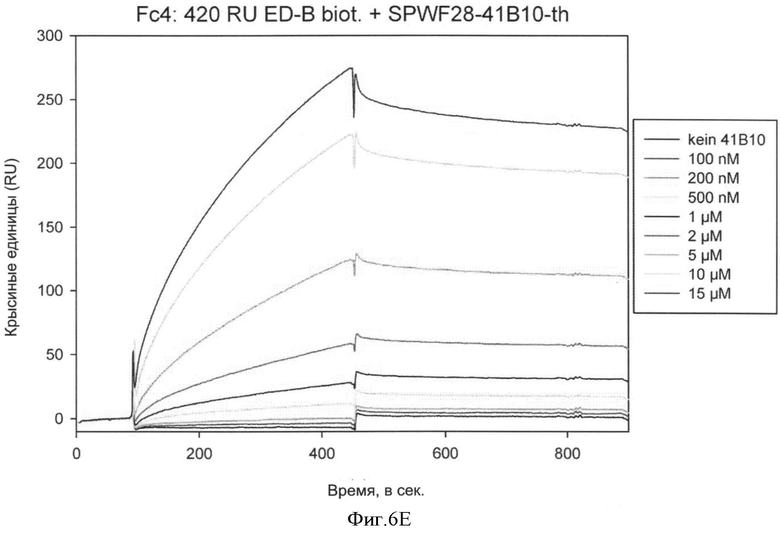
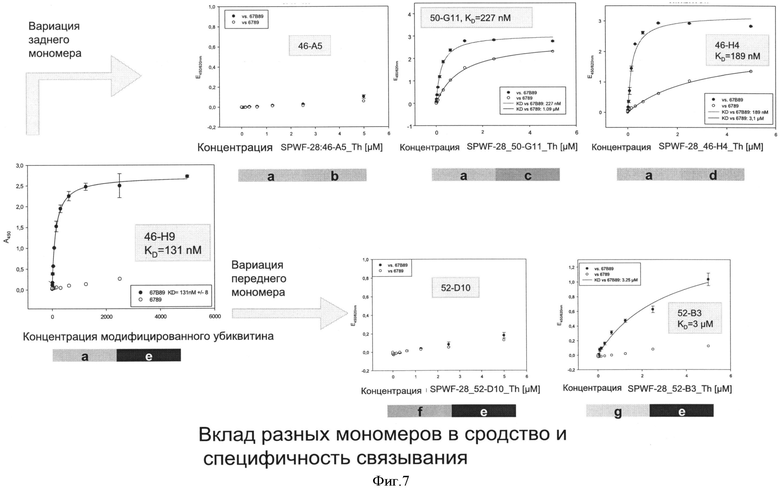


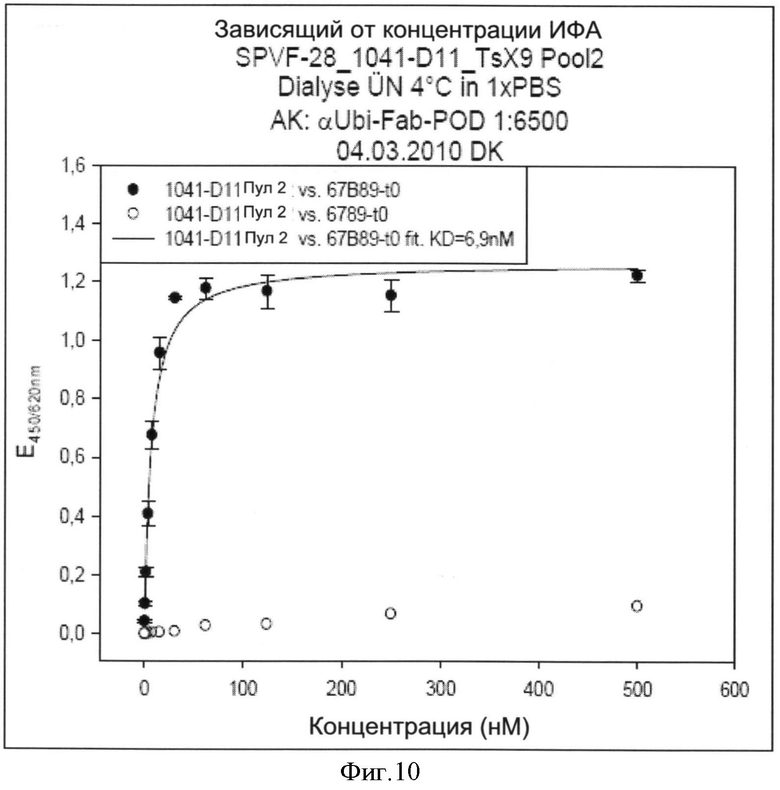
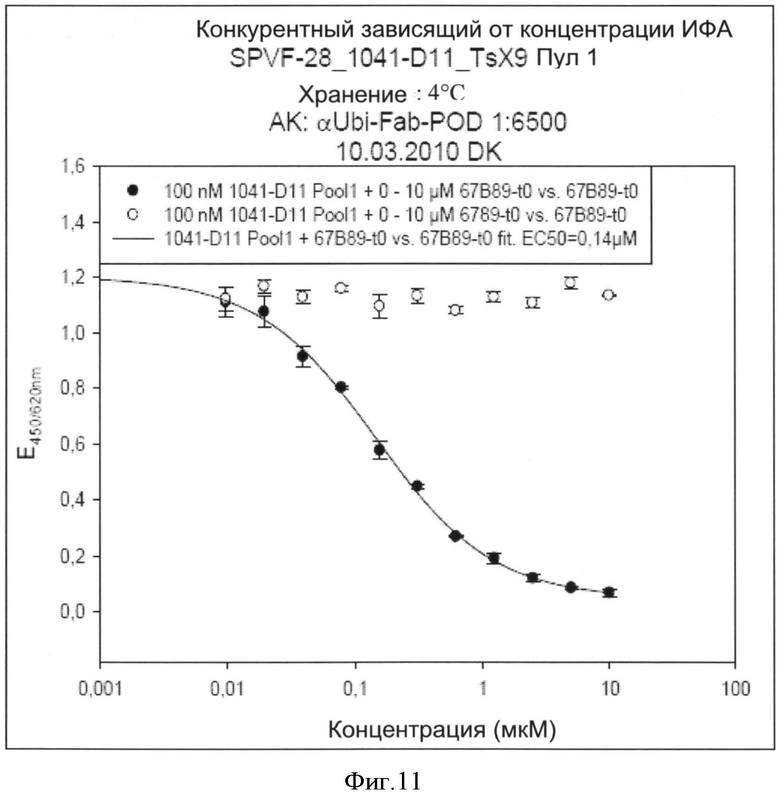

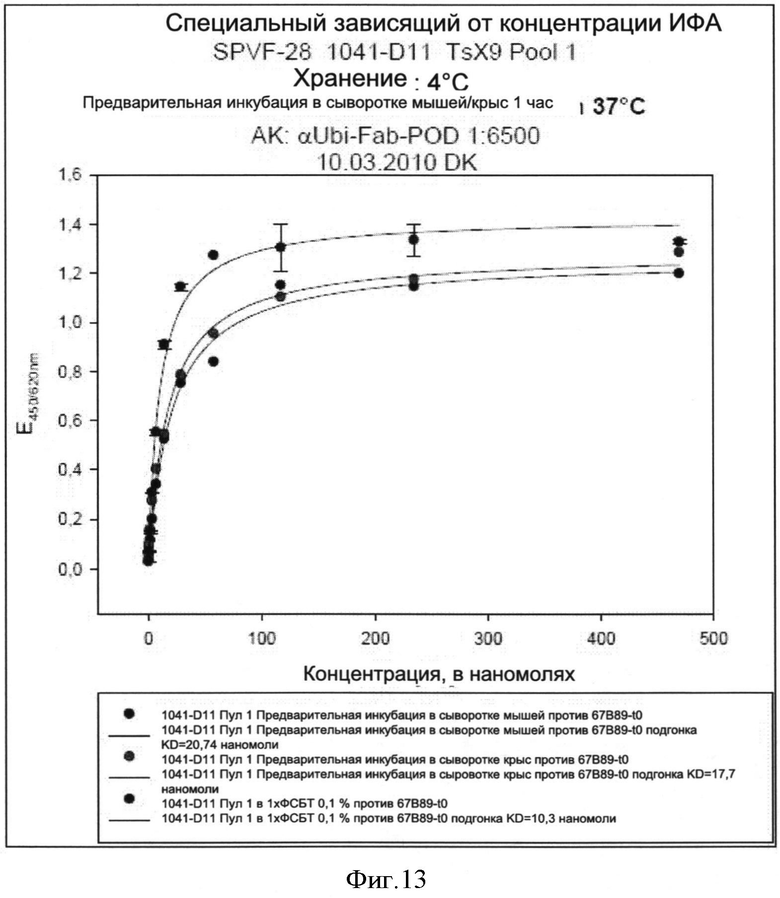

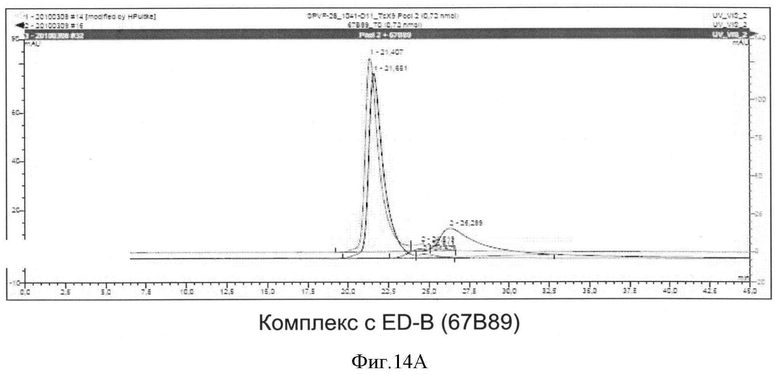
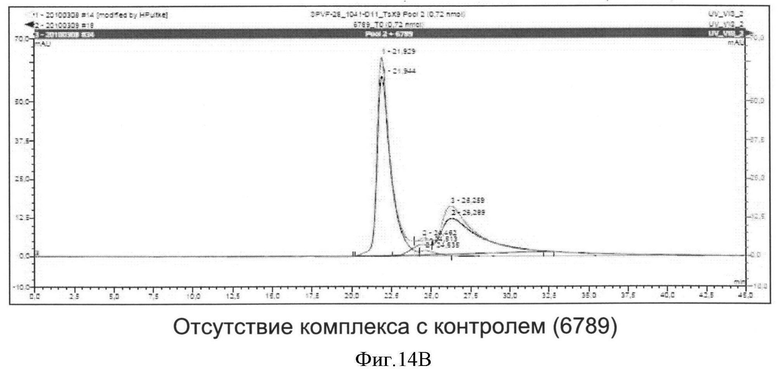

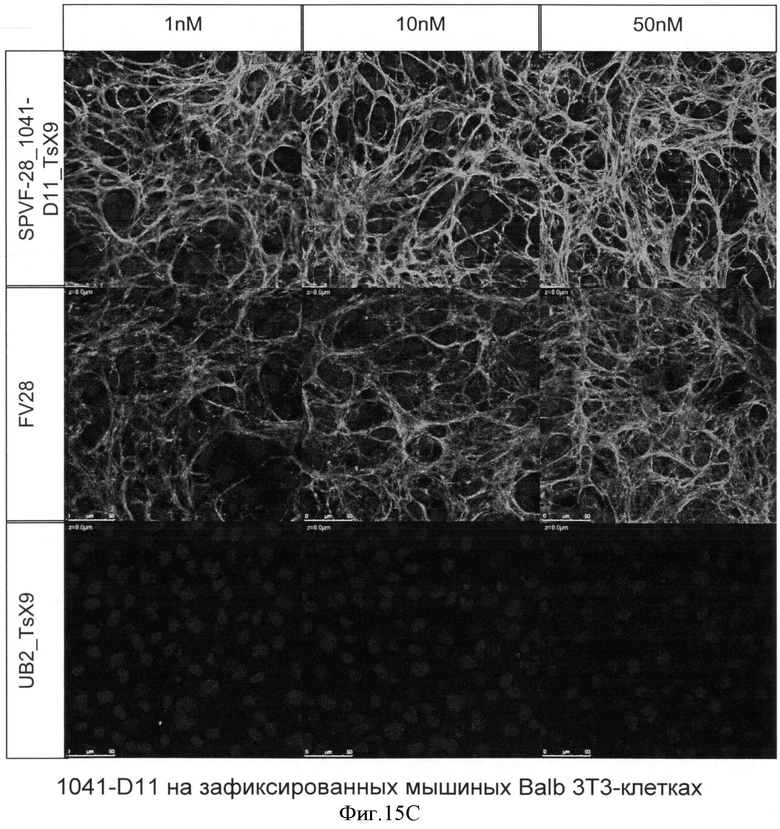
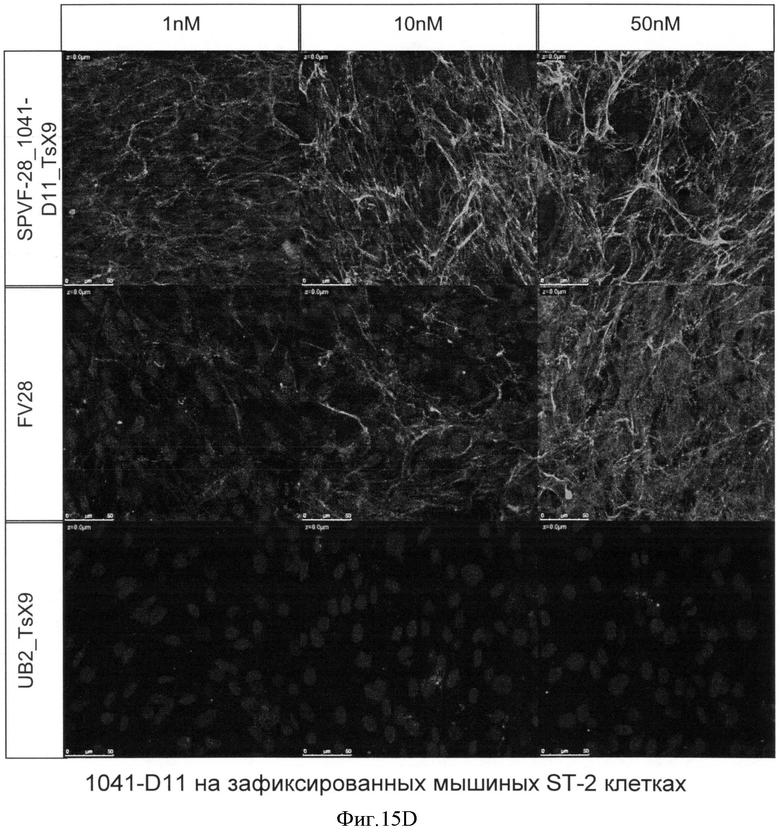

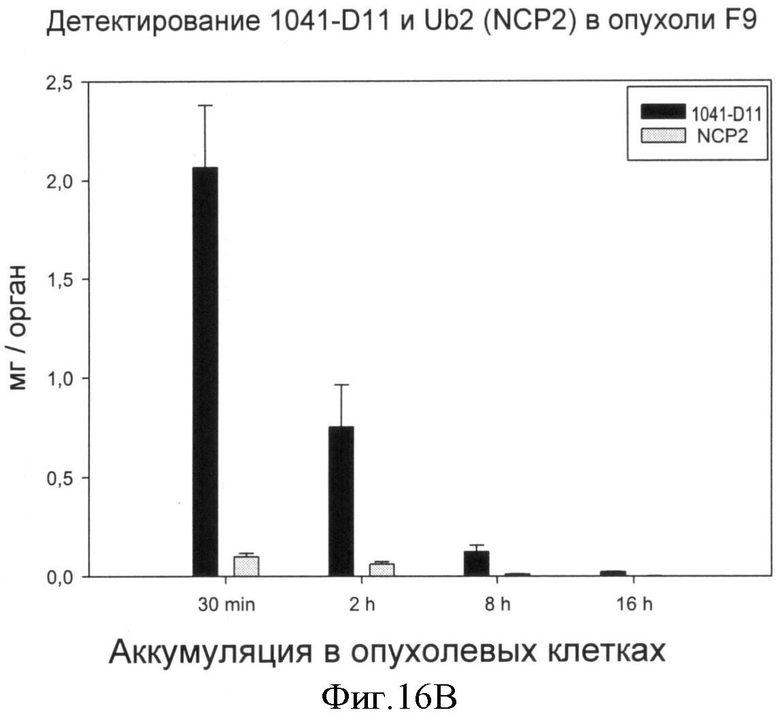
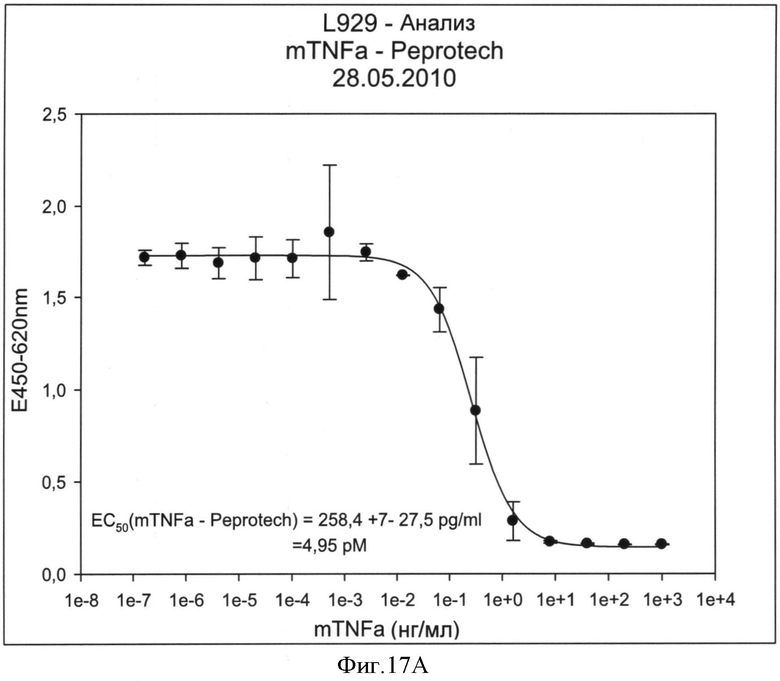

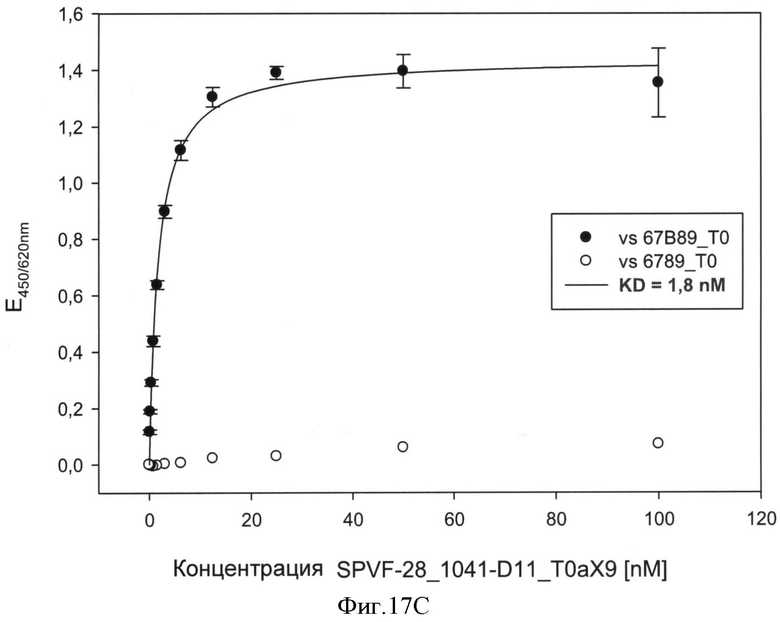
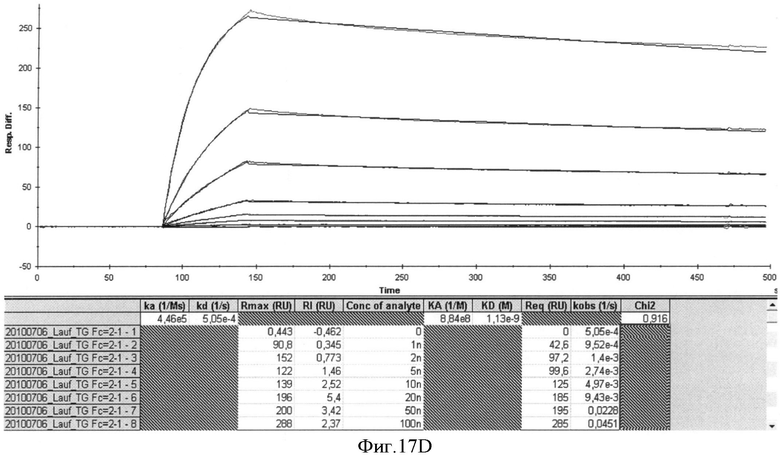
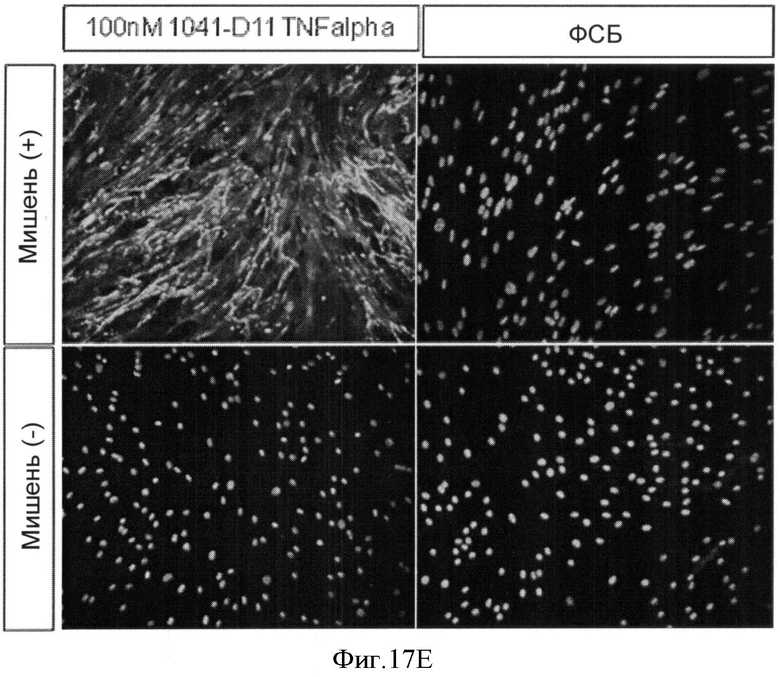
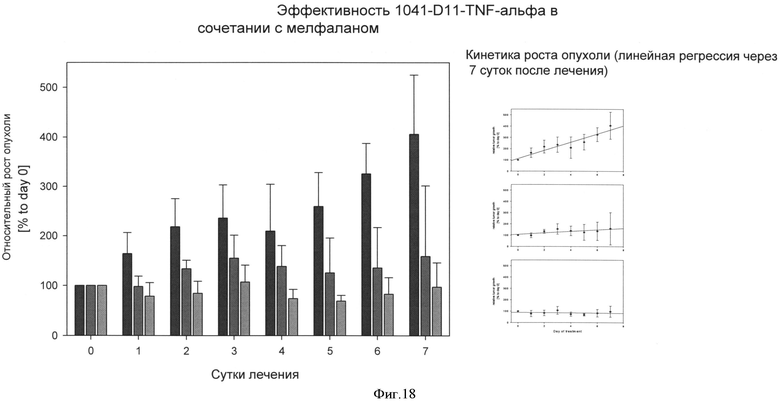
Изобретение относится к области биотехнологии, конкретно к новым гетеромультимерным белкам, полученным из модифицированного убиквитина, и может быть использовано в медицине для лечения или диагностики заболеваний, ассоциированных с гиперпродукцией экстрадомена В фибронектина (ED-B). Белок включает два мономерных убиквитиновых звена, по-разному модифицированных путем замещений по меньшей мере 6 аминокислот в положениях 4, 6, 8, 62, 63, 64, 65 и 66 SEQ ID NO: 1. При этом в 1-м мономерном звене замещения включают: F4W, K6(Н,W или F), Q62N, Е64(K,R или Н), S65(L,F или W), Т66(S или Р), а во 2-м мономерном звене: K6(Т,N,S или Q), L8(Q,Т,N или S), Q62(W или F), K63(S,Т,N или Q), Е64(N, S, Т или Q), S65(F или W), Т66(Е или D). Изобретение позволяет получить модифицированный гетеродимерный белок убиквитина, способный связывать ED-B с высоким сродством. 20 н. и 8 з.п. ф-лы, 18 ил., 3 табл., 7 пр.
1. Белок, способный связывать экстрадомен В (ED-B) фибронектина, состоящий из модифицированного гетеродимерного убиквитинового белка, в котором два мономерных убиквитиновых звена связаны между собой в расположении "голова к хвосту" непосредственно или посредством линкера, отличающийся тем, что каждый мономер упомянутого димерного белка по-разному модифицирован путем замещений по меньшей мере 6 аминокислот в положениях 4, 6, 8, 62, 63, 64, 65 и 66 SEQ ID NO: 1, причем упомянутые замещения включают:
в первом мономерном звене замещения в аминокислотных положениях 4, 6, 62, 64, 65, и 66 по SEQ ID NO: 1,
отличающийся тем, что замещения осуществляют следующим образом:
в первом мономерном звене замещения в аминокислотных позициях 4, 6, 62, 64, 65 и 66, причем в положении 4 F замещается на W, в положении 6 K замещается на Н, W или F, в положении 62 Q замещается на N, в положении 64 Е замещается на K, R или Н, в положении 65 S замещается на L, F или W, а в положении 66 Т замещается на S или Р;
и
во втором мономерном звене замещения в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; где K в положении 6 замещается на Т, N, S или Q, L в положении 8 замещается на Q, Т, N или S, Q в положении 62 замещается на W или F, K в положении 63 замещается на S, Т, N или Q, Е в положении 64 замещается на N, S, Т или Q, S в положении 65 замещается на F или W, Т в положении 66 замещается на Е или D; где мономерное звено необязательно включает дополнительные аминокислотные замены, не влияющие на активность белка,
и причем упомянутый белок имеет специфическое связывающее сродство с упомянутым доменом ED-B фибронектина КД=10-7-10-12 М и проявляет одновалентную связывающую активность по отношению к упомянутому экстрадомену В (ED-B) фибронектина.
2. Белок по п. 1, отличающийся тем, что оба мономера убиквитина связаны посредством линкера, имеющего по меньшей мере последовательность GIG или по меньшей мере SGGGG, предпочтительно или SGGGGIG, или SGGGGSGGGGIG (SEQ ID NO: 32).
3. Конъюгат для лечения заболеваний, ассоциированных с гиперпродукцией ED-В, включающий белок по любому из пп. 1 и 2, объединенный с фармацевтически активным компонентом, отличающийся тем, что упомянутый фармацевтически активный компонент является цитокином, хемокином, цитотоксическим соединением или ферментом.
4. Конъюгат по п. 3, который включает гетеродимер убиквитина с SEQ ID NO: 33, или 34, или 47 или имеет аминокислотную идентичность по меньшей мере 90% или 95% с последовательностью SEQ ID NO: 33, или 34, или 47.
5. Конъюгат по п. 3 или 4, имеющий белковую природу.
6. Конъюгат по п. 3, отличающийся тем, что упомянутым фармацевтически активным компонентом является TNF-альфа или его производное, предпочтительно отличающийся тем, что упомянутый составной белок имеет последовательность SEQ ID NO: 35 или 36, или отличающийся тем, что SEQ ID NO: 47 объединена с TNF-альфа или его производным или имеет аминокислотную идентичность по меньшей мере 90% или 95% с последовательностью SEQ ID NO: 35 или 36, или с SEQ ID NO: 47, объединенной с TNF-альфа или его производным.
7. Конъюгат для диагностики заболеваний, ассоциированных с гиперпродукцией ED-B, включающий белок по любому из пп. 1 и 2, объединенный с диагностически активным компонентом, отличающийся тем, что упомянутый диагностически активный компонент является флуоресцентным соединением, фотосенсибилизатором или радионуклидом.
8. Конъюгат по п. 7, который включает гетеродимер убиквитина с SEQ ID NO: 33, или 34, или 47 или имеет аминокислотную идентичность по меньшей мере 90% или 95% с последовательностью SEQ ID NO: 33, или 34, или 47.
9. Фармацевтическая композиция для лечения заболеваний, ассоциированных с гиперпродукцией ED-B, содержащая белок, способный связывать домен ED-B фибронектина, согласно любому из пп. 1 и 2 в фармацевтически эффективном количестве, дополнительно содержащая еще один или более химиотерапевтических агентов и один или больше фармацевтически приемлемых носителей или наполнителей.
10. Фармацевтическая композиция для лечения заболеваний, ассоциированных с гиперпродукцией ED-B, содержащая конъюгат по любому из пп. 3-6 в фармацевтически эффективном количестве и дополнительно содержащая еще один или больше химиотерапевтических агентов и один или больше фармацевтически приемлемых носителей или наполнителей.
11. Фармацевтическая композиция по п. 9 или 10, отличающаяся тем, что упомянутые химиотерапевтические агенты выбирают из мелфалана, доксорубицина, циклофосфамида, дактиномицина, фтордезоксиурацила, цисплатина, паклитаксела и гемцитабина, или из группы ингибиторов киназы, или из радиофармацевтических препаратов.
12. Фармацевтическая композиция по п. 11, которая находится в форме объединенного препарата или в форме набора отдельных компонентов.
13. Полинуклеотид, кодирующий рекомбинантный белок согласно любому из пп. 1 и 2.
14. Полинуклеотид, кодирующий конъюгат по п 5.
15. Вектор экспрессии, включающий полинуклеотид по п. 13.
16. Вектор экспрессии, включающий полинуклеотид по п. 14.
17. Клетка-хозяин для экспрессии белка согласно любому из пп. 1 и 2, включающая полинуклеотид по п. 13.
18. Клетка-хозяин для экспрессии конъюгата по п. 5, включающая полинуклеотид по п. 14.
19. Клетка-хозяин для экспрессии белка согласно любому из пп. 1 и 2, содержащая вектор экспрессии по п. 15.
20. Клетка-хозяин для экспрессии конъюгата согласно п. 5, содержащая вектор экспрессии по п. 16.
21. Композиция для диагностики ED-B-асоциированных заболеваний, включающая эффективное количество белка согласно любому из пп. 1 и 2 с диагностически приемлемым носителем.
22. Композиция для диагностики ED-B-ассоциированных заболеваний, включающая эффективное количество конъюгата согласно любому из пп. 7 и 8 с диагностически приемлемым носителем.
23. Способ создания белка согласно любому из пп. 1 и 2, включающий следующие этапы:
а) обеспечение совокупности по-разному модифицированных димерных убиквитиновых белков, происходящих из мономерных убиквитиновых белков, причем упомянутая совокупность состоит из димерных убиквитиновых белков, включающих два модифицированных
мономера убиквитина, связанных между собой в расположении "голова к хвосту", причем каждый мономер упомянутого димерного белка по-разному модифицирован путем замещения по меньшей мере 6 аминокислот в положениях 4, 6, 8, 62, 63, 64, 65 и 66 из SEQ ID NO: 1,
причем упомянутые замещения включают:
в первом мономерном звене замещения в аминокислотных положениях 4, 6, 62, 64, 65 и 66, причем в положении 4 F замещается на W, в положении 6 K замещается на Н, W или F, в положении 62 Q замещается на N, в положении 64 Е замещается на K, R или Н, в положении 65 S замещается на L, F или W, а в положении 66 Т замещается на S или Р; и
во втором мономерном звене замещения по меньшей мере в аминокислотных положениях 6, 8, 62, 63, 64, 65 и 66; где K в положении 6 замещается на Т, N, S или Q, L в положении 8 замещается на Q, Т, N или S, Q в положении 62 замещается на W или F, K в положении 63 замещается на S, Т, N или Q, Е в положении 64 замещается на N, S, Т или Q, S в положении 65 замещается на F или W, Т в положении 66 замещается на Е или D,
где мономерное звено необязательно включает дополнительные аминокислотные замены, не влияющие на активность белка;
b) предоставление экстрадомена В (ED-B) фибронектина в качестве потенциального лиганда;
c) контакт упомянутой совокупности по-разному модифицированных белков с упомянутым экстрадоменом В (ED-B) фибронектина;
d) идентификация модифицированного димерного убиквитинового белка способом скрининга, причем упомянутый модифицированный димерный убиквитиновый белок связывается с упомянутым экстрадоменом В (ED-B) фибронектина со специфическим связывающим сродством при значении КД в диапазоне 10-7-10-12 М и проявляет активность одновалентного связывания по отношению к упомянутому экстрадомену В (ED-B) фибронектина.
24. Способ по п. 21, отличающийся тем, что упомянутую совокупность по-разному модифицированных белков получают путем генетического объединения двух библиотек ДНК, каждая из которых отвечает за получение по-разному модифицированных мономерных убиквитиновых белков.
25. Способ создания конъюгата согласно любому из пп. 3-6, отличающийся тем, что белок согласно любому из пп. 1 и 2 конъюгирован с фармацевтически активным компонентом, отличающийся тем, что упомянутый фармацевтически активный компонент является цитокином, хемокином, цитотоксическим соединением или ферментом.
26. Способ создания конъюгата согласно любому из пп. 7 и 8, отличающийся тем, что белок согласно любому из пп. 1 и 2 конъюгирован с диагностически активным компонентом, отличающийся тем, что упомянутый диагностически активный компонент является флуоресцентным соединением, фотосенсибилизатором или радионуклидом.
27. Применение рекомбинантного белка по любому из пп. 1 и 2 или конъюгата согласно любому из пп. 3-6 для лечения заболеваний, ассоциированных с гиперпродукцией ED-B.
28. Применение рекомбинантного конъюгата согласно любому из пп. 7 и 8 для диагностики заболеваний, ассоциированных с гиперпродукцией ED-B.
| WO2008059011 A1, 22.05.2008 | |||
| RAASI S | |||
| et al., Binding of polyubiquitin chains to ubiquitin-associated (UBA) domains of HHR23A, J.Mol.Biol., 2004, v.341, p.1367-1379 | |||
| WO2004106368 A1, 09.12.2004 | |||
| ДИМЕР МОЛЕКУЛЯРНОГО ВАРИАНТА АПО-ЛИПОПРОТЕИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 1992 |
|
RU2134696C1 |
| WO2006040129 A2, 20.04.2006 | |||
| WO2005087797 A1, 22.09.2005 | |||
| МОЛДОГАЗИЕВА Н.Т | |||
| и др, Структурно-функциональные взаимосвязи в | |||

