Область изобретения
В настоящем изобретении раскрыто применение мономиколилглицерина (MMG), его синтетического гомолога, аналога или его модифицированного варианта для получения иммуномодулятора, адъюванта, адъюванта и вакцины или системы доставки, содержащей данный адъювант.
Предшествующий уровень техники
Первые вакцины состояли из живых аттенуированных патогенов. Аттенуированные формы представляли собой или встречающиеся в природе родственные организмы, или полученные с помощью серийных пассажей в культуре. Например, в течение многих лет с туберкулезом (ТВ) человека боролись при помощи вакцинации аттенуированным штаммом Mycobacterium bovis, - вакцину М. bows BCG разработали более чем 80 лет назад. Однако, несмотря на то, что было введено более 3 миллиардов доз BCG (больше любой другой вакцины), это не всегда обеспечивает удовлетворительную устойчивость к человеческому ТВ в каждой популяции.
В настоящее время более современным подходом является использование высокоочищенных веществ, например очищенных рекомбинантных белков или пептидов. Данные вакцины являются хорошо определенными, и побочные эффекты минимизированы. К сожалению, многие высокоочищенные вещества не являются очень иммуногенными и не индуцируют достаточного иммунного ответа для обеспечения защиты. Для достижения этого антиген нуждается в некоторой помощи агентов усиления иммунного ответа, называемых адъювантами. В зависимости от патогена для защиты может требоваться преобладание или гуморального, или клеточно-опосредованного ответа. Иммунный ответ, который может быть перенесен иммунной сывороткой, называется гуморальным иммунитетом и относится к устойчивости, опосредованной антителами, которые связываются с антигенным веществом, ассоциированным с инфекционным агентом, и тем самым инициируют иммунный ответ против него. Клеточно-опосредованный иммунитет (CMI) основан на клетках иммунной системы, демонстрирующих иммунный ответ. CMI, или Т-хелперный (Th)1, иммунный ответ ассоциирован главным образом с борьбой против внутриклеточных патогенов, включая лейшманию и туберкулез, но также играет роль в борьбе против других типов инфекции, например дрожжевой инфекции Candida. Гуморальный, или Th2, иммунный ответ требуется для защиты от внеклеточных патогенов, например гельминтной инфекции.
В ряде случаев, например при гриппе, гепатите С (HCV), вирусе иммунодефицита человека (ВИЧ), хламидиозе и малярии, в зависимости от стадии инфекции, может требоваться смешанный Тh1/Тh2-ответ (Mosmann and Sad 1996). Для них требуется как Тh1, так и Th2, так как части их жизненного цикла являются внутриклеточными, но кроме того, они проходят через внеклеточные фазы, например передачу между клетками.
Развитие определенного вида иммунного ответа (гуморального или клеточно-опосредованного) можно детерминировать посредством выбора адъюванта. Например, для протективного иммунитета против внутриклеточных патогенов, таких как М. tuberculosis, требуется клеточно-опосредованный иммунный ответ, и подходящий адъювант для субъединичной вакцины, направленной против ТВ, должен усиливать ТМ-ответ (Lindblad et al. 1997).
Существует большое количество адъювантов, но большинство из них подвержено многочисленным проблемам, которые мешают их применению у людей. Только несколько адъювантов допущены для применения у человека, например адъюванты на основе алюминия (AlOH-соли) и MF-59, но они оба индуцируют Тh2-преимущественные ответы, что делает их непригодными для ТВ-вакцины и других вакцин, для которых требуется Th1-ответ (Lindblad et al. 1997).
В течение последних 20-30 лет был идентифицирован ряд новых адъювантных систем, и некоторые из них в настоящее время находятся на стадии разработки. Несмотря на это, по-прежнему признается потребность в новых адъювантных системах (Moingeon et al. 2001) и это проявляется в малочисленности альтернатив, пригодных для клинического применения.
Адъювант (от латинского «adjuvare», помогать) можно определить как любое вещество, которое при введении в вакцину служит для направления, ускорения, продления и/или усиления специфического иммунного ответа. Адъюванты были разделены на две основные категории, или системы доставки, или иммуномодуляторы/иммуностимуляторы. Система доставки может, например, представлять собой эмульсии, полистирольные частицы, ниосомы, ISCOMS (иммуностимулирующие комплексы), виросомы, микросферы, или липосомы, подобные поверхностно-активным веществам, которые представляют собой пузырьки, образованные двойными слоями липидов. Липосомы действуют как носители антигена (либо внутри пузырьков, либо присоединенных к поверхности) и могут образовывать депо в месте внесения, обеспечивая медленное, непрерывное высвобождение антигена. В течение некоторого времени после инъекции и фагоцитоза, липосомная презентация гарантирует, что конкретное количество антигена становится доступным для отдельных антиген-представляющих клеток (Gluck 1995). Иммуномодуляторы нацелены на определенные клетки или рецептор, например toll-подобные рецепторы на поверхности АРС (антигенпрезентирующие клетки). Системы доставки и иммуномодуляторы можно использовать вместе, например, как в сериях адъювантов Glaxo. Следовательно, в дополнение к доставке вакцины антигенную систему доставки также можно использовать для доставки иммуномодуляторов.
Помимо того что они представляют собой компонент вакцины, иммуномодуляторы могут быть введены без антигена(ов). При таком подходе можно активировать иммунную систему локально, например, наблюдать в виде созревания антиген-представляющих клеток, продуцирования цитокинов, которое является важным для противоопухолевой и антивирусной активности. Таким образом, введение иммуномодуляторов может, например, помогать облучению при раке и кожных заболеваниях. Примерами иммуномодуляторов, которые могут быть введены локально, являются таксаны, например таксол, лиганд toll-подобного рецептора 7/8 резиквимод, имиквимод, гардиквимод.
Диметилдиоктадециламмония бромид, хлорид, фосфат, ацетат или другие органические или неорганические соли (DDA) представляют собой липофильное четвертично-аммониевое соединение, которое образует катионные липосомы в водных растворах при температурах выше 40°С. DDA представляет собой особенно эффективную систему доставки, усиливающую захват вакцинного антигена АРС. Были описаны комбинации DDA и иммуномодулирующих агентов. Введение арквада 2НТ, который содержит DDA, людям было перспективным и не вызывало видимых побочных эффектов (Stanfield, 1973). Комбинирование DDA и TDB, или DDA и MPL показало очень отчетливый синергизм между средством доставки (DDA) и иммуномодулятором (TDB или MPL) с существенно повышенными уровнями CMI-ответа по сравнению с ответом, полученным с любым из компонентов по отдельности. Следовательно, DDA является перспективным средством доставки вакцинного антигена и иммуномодулятора, например при разработке адъювантной системы для вакцины против ТВ и других внутриклеточных патогенов.
Сообщалось, что различные соединения из микобактерий являются иммуномодулирующими. При использовании липидов, экстрагированных из М. bovis BCG, в качестве адъюванта, реакцию кожных проб на овальбумин получали на морских свинках (Hiu 1975). Липосомы, образованные при повышенных температурах из всех полярных липидов М. bovis BCG, способны вызывать гуморальный ответ на овальбумин, и вакцина, полученная из этих полярных липидов, обеспечивала защиту мышам при контрольном введении опухолевых клеток (WO 03/011336). Был изучен (Dascher et al. 2003) эффект общих липидов из М. tuberculosis H37Rv в качестве антигена в экспериментальной ТВ-вакцине для морских свинок. В этом исследовании липосомы на основе холестерина и 1,2-дистеароил-sn-глицеро-3-фосфохолина (DSPC) смешивали с экстрактом всех липидов М.tuberculosis H37Rv. После удаления растворителя липиды восстанавливали
диметилдиоктадециламмонием (DDA) в качестве адъюванта в буфере PBS. У морских свинок, иммунизированных этой вакциной, не показано значительного уменьшения бактерий, что позволяет предположить, что в данной композиции липосом, смешанных с DDA, нет сильного антигена, или что композиция микобактериальных липидов с холестерин:DSPC препятствует адъювантному действию DDA. Альтернативно, при введении смеси различных липидов, эффективные липиды могут составлять очень ограниченную часть всех липидов.
Различные очищенные компоненты из микобактерий также были изучены в отношении их адъювантной активности. Производное очищенного белка (PPD) не индуцировало реакцию гиперчувствительности замедленного типа само по себе, но при добавлении в качестве адъюванта Wax D (комплекс фрагмент микобактериальной пептидогликановой клеточной стенки-арабиногалактан-миколовая кислота) наблюдалась сильная реакция (Yamazaki S 1969). Иммуномодулятор SSM или Z-100, липид арабиноманнан, экстрагированный из М.tuberculosis, обладает противоопухолевой активностью (Suzuki F 1986). Еще одним соединением, полученным из микобактериальных клеток, является 6,6'-димиколат трегалозы (TDM) (фактор жгутообразования; миколовая кислота, содержащая гликолипид) (Saito et al, 1976). Кроме того, TDM (или синтетические аналоги) обладает иммуностимулирующими эффектами и был включен в различные адъювантные композиции (McBride et al. 1998) (Koike et al. 1998).
В статье Silva et al (1985) пять компонентов, очищенные из Mycobacterium bovis BCG, инъецировали внутривенно в виде частиц угля, покрытых липидом, и они вызывали воспалительную реакцию в легких мышей. Пять компонентов включали TDM, мономиколят трегалозы (ММТ), мономиколят глюкозы (MMGlc), мономиколят арабинозы (ММАr) и мономиколят глицерина (MMGlyc). В статье описана головная группа мономиколилглицерина, в то время как композиция миколовых кислот определена недостаточно и не представлены структурные данные. Кроме того, реакция на введение липидов описана исключительно как воспалительная активность в легких, в то время как способность усиливать специфический иммунный ответ, известная как адъювантный эффект, не описана.
Хотя иммуностимулирующая и воспалительная активность липидов, полученных из микобактерий, была признана в течение многих десятилетий с непрерывно растущей литературой о липидах, способных стимулировать иммунные ответы в животных (мышиных) моделях, до настоящего времени не был(и) идентифицирован(ы) отдельный(ые) липид(ы), способные стимулировать человеческие дендритные клетки (DC). Например, хотя было показано, что TDM является наиболее активным микобактериальным липидом с точки зрения провоспалительных ответов, не наблюдалась активация дендритных клеток при стимулировании TDM (Uehori et al, 2003). Итак, хотя TDM продемонстрировал воспалительную активность в ряде статей, у данного липида, несомненно, отсутствует способность активировать дендритные клетки, которая является ключевой для инициирования иммунного ответа. Идентификация такого липида, способного активировать человеческие дендритные клетки, предполагает, что его можно использовать в качестве части новой адъювантной системы, пригодной для применения у людей. Кроме того, отсутствие Тh1-индуцирующих адъювантов, пригодных для применения у людей, делает идентификацию отдельного липида, полученного из микобактерий, с Th1-стимулирующей способностью, важной находкой.
DC представляют собой специализированные антиген-представляющие клетки (АРС), которые играют существенную роль в управлении иммунным ответом при инфицировании патогенами, такими как М. tuberculosis. Таким образом, продуцирование IL-12 активированными DC является жизненно важной стадией контролирования инфекции М. tuberculosis, так как именно этот цитокин имеет первостепенную важность в управлении продуцированием IFN-γ Th-1-клетками, что стимулирует активацию макрофагов (Nathan et al. 1983). Кроме того, в последние годы были обнаружены свидетельства, которые указывают на то, что микобактерии также нацелены на DC в попытке модулировать иммунный ответ, и была установлена критическая роль микобактериальных липидов в данном процессе.
Вплоть до 40% сухой массы клеточной оболочки микобактерий составляют липиды (Minnikin 1982). Эти липиды долгое время были ассоциированы с особенной патогенностью этого семейства организмов и, как известно, играют существенную роль в ответе хозяина на микобактериальную инфекцию (Brennan и Goren 1979). Известными среди данных липидов являются воски фтиоцерол димикоцерозата (PDIM) (Minnikin et al. 2002), присутствие которых, как было показано, связано с патогенностью; PDIM-дефицитные мутанты М. tuberculosis демонстрируют ослабленный рост у мышей (Sirakova et al. 2003). Близкородственными PDIM являются так называемые "фенольные гликолипиды" (PGLs), подходящим примером являются 2-метил-рамнозил-фенолфтиоцерол димикоцерозаты ("микозид В"), обнаруженные у Mycobacterium bovis. В последнее время была обнаружена связь между данным моногликозилом PGL и гипервирулентностью некоторых изолятов М. tuberculosis (Reed et al. 2004).
Еще один классом особенно интересных липидов являются 6,6-димиколяты трегалозы (TDM), так называемые "факторы жгутикообразования". TDM способствуют сохранению гранулематозных поражений посредством стимулирования высвобождения провоспалительных цитокинов, таких как TNF α, IL-6 и IL-12, и Th-1-стимулирующего цитокина IFN-γ (Lima et al. 2001), и играют роль в пролонгировании выживания М tuberculosis в макрофагах посредством ингибирования слияния фагосом-лизосом (Indrigo et al. 2002). Тонкая структура миколятных компонентов TDM является важной в провоспалительной активации макрофагов при ранней инфекции (Rao et al, 2005).
Несмотря на их роль в улучшении выживания микобактерий, иммуномодулирующие способности микобактериальных липидов также можно использовать для создания нового поколения Тh1 -индуцирующих адъювантов. При идентификации отдельных липидов с сильной иммуностимулирующей активностью, можно избежать проблем с токсичностью, ассоциированной с использованием убитых нагреванием целых клеток М tuberculosis, смешанных с маслом - полным адъювантом Фрейнда (CFA) - при сохранении сильной адъювантной активности. Действительно, недавно было доказано, что липосомы, образованные полярными липидами бациллы Кальмета-Герена (BCG) М.bovis, активируют дендритные клетки, полученные из костного мозга мышей (BM-DC). Было обнаружено, что большую часть этой активности можно отнести к липиду гликофосфолипидфосфатидилинозитдиманнозиду (Sprott et al. 2004).
Последние исследования в лабораториях авторов охарактеризовали новую адъювантную комбинацию микобактериального липидного экстракта, из М.bovis BCG, и диметилдиоктадециламмония бромида (DDA), который способен стимулировать комплексный и длительный иммунный ответ, с сильной как гуморальной, так и клеточно-опосредованной компонентой (Rosenkrands et al. 2005). Было обнаружено, что большую часть адъювантной активности всех липидов BCG можно отнести к неполярным липидам.
Хотя такие исследования дополнительно подтверждают способность микобактериальных липидов действовать в качестве адъювантов, оптимальным решением является идентификация одного наиболее иммуностимулирующего липида, который сам обладает сильной активностью. Он будет представлять собой даже более простой, более дешевый адъювант и, кроме того, также будет увеличивать возможность изготовления синтетических аналогов липида, обеспечивая очистительную систему, которую можно производить в больших количествах, необходимых для адъюванта, используемого в вакцинах, которые вводят во всем мире.
Краткое изложение сущности изобретения
В настоящем изобретении раскрыт иммуностимулирующий липид, мономиколилглицерин (MMG) и его синтетические гомологи, аналоги и модифицированные варианты, способные активировать DC человека. MMG получают из неполярной фракции всех липидов BCG, и он отвечает за индуцирование адъюванта и протективный эффект, связанный с данными липидами. Синтетический MMG с меньшим углеродным каркасом способен усиливать стимулирующие свойства природного MMG на человеческие DC in vitro и, кроме того, индуцировать сильный Th1-ответ in vivo, который преобразуется в длительный протективный иммунный ответ против ТВ в мышиной модели.
Подробное описание изобретения
В настоящем изобретении раскрыто применение мономиколилглицерина (MMG) или его синтетических гомологов, аналогов и модифицированных вариантов для получения иммуномодулятора, адъюванта и вакцины или системы доставки, содержащей этот адъювант, который обладает уникальной способностью стимулировать человеческие дендритные клетки.
В качестве иммуномодуляторов MMG или его синтетические гомологи, аналоги и модифицированные варианты вводят без антигена(ов). Этим способом можно активировать иммунную систему локально, например, наблюдать в виде созревания антиген-представляющих клеток, продуцирования цитокина, которое является важным для противоопухолевой и противовирусной активности.
Адъювант (от латинского adjuvare, помогать) может быть определен как любое вещество, которое при введении в вакцину служит для направления, ускорения, продления и/или усиления специфического иммунного ответа. В зависимости от природы адъюванта, он может стимулировать клеточно-опосредованный иммунный ответ, гуморальный иммунный ответ или их комбинацию. При использовании в качестве вакцинного адъюванта антигенный компонент добавляют к адъюванту. Так как усиление иммунного ответа, опосредованное адъювантами, является неспецифическим, в данной области техники известно, что один и тот же адъювант можно использовать с разными антигенами для стимулирования ответов против разных мишеней, например, с антигеном из М.tuberculosis - для стимулирования иммунитета против М.tuberculosis, или с антигеном, полученным из опухоли, - для стимулирования иммунитета против опухолей данного конкретного типа.
Предпочтительным адъювантом, раскрытым в изобретении, является адъювант, содержащий MMG или его синтетический гомолог, аналог или модифицированный вариант, который дополнительно содержит средство доставки, например эмульсии, полистирольные частицы, ниосомы, IS-COMS, виросомы, микросферы или липосомы, подобные поверхностно-активным веществам. Предпочтительными поверхностно-активными веществами являются наиболее предпочтительно катионные липиды на основе диметилдиоктадециламмония бромида или хлорида (DDA-B или DDA-C) или его сульфатной, фосфатной или ацетатной соли (DDA-X), или диметилдиоктадециниламмония бромида или хлорида (DODA-B или DODA-C) или его сульфатного, фосфатного или ацетатного соединения (DODA-X). Другие типы предпочтительных катионных липидов, используемых в данном изобретении, включают 1,2-диолеил-3-триметиламмоний-пропан (DOTAP), 1,2-димиристоил-3-триметиламмоний-пропан, 1,2-дипальмитоил-3-триметиламмоний-пропан, 1,2-дистеароил-3-триметиламмоний-пропан и диолеил-3-диметиламмоний-пропан (DODAP) и N-(1-(2,3-диолеилокси)пропил]-NXN-триметиламмоний (DOTMA), но не ограничиваются ими. Другие поверхностно-активные вещества выбирают из DXPC, DXPE, DXPG или их комбинаций, где X заменяет описание длины цепи, например Р=пальмитоил (16С), 3=стеароил (18С), А=арахидоил (20С).
Средство доставки также можно использовать для других иммуномодуляторов, таких как TLR (Toll-подобный рецептор) и he-TLR-лиганды, такие как MPL (монофосфориллипид А), полиинозиновая-полицитидиловая кислота (поли-IC), мурамилдипептид (MDP), зимозан, двухнитевая РНК (dsPHK), DC-Chol, CpG олигодезоксинуклеотиды, катионные пептиды, TDM, TDB, тамоксифен или любые аналоги любых из этих молекул. Таким образом, предпочтительный адъювант содержит MMG или его гомолог, аналог или модифицированный вариант и дополнительно содержит TLR- или не-TLR лиганды в средстве доставки.
Системы доставки, содержащие MMG или его синтетические гомологи, аналоги и модифицированные варианты, можно использовать для лечения рака, аутоиммунного расстройства, нервного расстройства, например болезни Альцгеймера, воспаления дыхательных путей, воспалительных расстройств, инфекционного заболевания, кожных расстройств, аллергии, астмы или заболевания, вызванного патогеном. MMG или его синтетические гомологи, аналоги и модифицированные варианты вводят в комбинации с одной или более вакцинами, антигенами, антителами, цитотоксическими агентами, аллергенами, антибиотиками, антисмысловыми олигонуклеотидами, TLR- и не-TLR агонистами, TLR- и не-TLR антагонистами, пептидами, белками, геннотерапевтическими векторами, ДНК-вакцинами или ко-стимулирующими молекулами.
Антигенный компонент или вещество представляет собой молекулу, которая взаимодействует с ранее сформированным антителом и/или специфическими рецепторами на Т- и В-клетках. В контексте вакцинации это молекула, которая может стимулировать развитие специфических Т- или В-клеток, приводя к образованию популяции клеток памяти иммунных клеток, которые будут стимулировать более быстрый "анамнестический" ответ, если антиген обнаруживается иммунными клетками вторично. Так как популяции клеток памяти редко являются клональными, на практике это означает, что антигеном является любая молекула или совокупность молекул, которая может стимулировать усиление иммунных ответов при повторном обнаружении иммунными клетками индивидуума, который ранее подвергался его воздействию.
Кроме того, в изобретении раскрыта вакцина для парентерального, перорального или мукозального введения или система доставки, содержащая адъювант. Вакцина может быть предназначена для парентерального, перорального, мукозального введения, сублингвального, трансдермального, местного, ингаляционного, интраназального, аэрозольного, внутриглазного, внутритрахеального, внутриректального, вагинального, с помощью генной пушки, кожного пластыря или в форме глазных капель или полоскания для рта. Предпочтительная вакцина содержит антигенный эпитоп из внутриклеточного патогена, например вирулентной микобактерий (например, слитые продукты Ag85b_TB10.4, Ag85b_ESAT-6_Rv2660, Ag85b_TB10.4_Rv2660 и Ag85a_TB10.4_Rv2660), Plasmodium falciparum (Mspl, Msp2, Msp3, Amal, GLURP, LSAI, LSA3 или CSP), Chlamydia trachomatis (e.g. CT184, CT521, CT443, CT520, CT521, CT375, CT583, CT603, CT610 или CT681), ВИЧ, вирусов гриппа или гепатита В или С. Адъювант или систему доставки также можно использовать в вакцинах для лечения рака, аллергии или аутоиммунных заболеваний.
Экстракт всех микобактериальных липидов представляет собой смесь липидов, полученных из микобактерий, например BCG, М.microti, М.tuberculosis и М.vaccae, с помощью химического или физического способа. В настоящей работе способ, используемый для экстракции, представляет собой воздействие органических растворителей (как описано ниже), но другие возможности, известные специалистам в данной области техники, являются возможными.
Фракция неполярных липидов определена как неполярные липиды. Фракцию неполярных липидов получают путем обработки микобактерий двухфазной смесью метанол/солевой раствор и петролейный эфир. Экстракт петролейного эфира составляют аполярные (неполярные) липиды. Затем получают фракцию полярных липидов добавлением хлороформа к микобактериям и остаточной водной фазе. Экстракт хлороформа содержит оставшиеся полярные липиды. Основными компонентами во фракции неполярных липидов являются фтиоцеролдимикоцерозаты, триацилглицерины, трегалозы миколипенаты и менахиноны. Основными компонентами фракции полярных липидов являются фосфолипиды, такие как фосфатидилэтанол, фосфатидилглицерин и фосфатидилинозит. Липиды с промежуточной полярностью представляют собой сульфолипиды, миколяты трегалозы, гликозилированные фенолфтиоцеролы (включая фенольные гликолипиды, PGL) и ацилированные трегалозы (Dobson et al, 1985).
MMG относится к липиду мономиколилглицерину, полученному из неполярной липидной фракции, и его производным, например альфа-MMG и кето-MMG, и его природным и синтетическим аналогам. MMG может быть выделен посредством TLC (тонкослойной хроматографии) в смеси толуол/ацетон (95:5). С помощью данного способа PGL и MMG экстрагируют вместе, но они могут быть разделены на 1-D TLC в смеси хлороформ:метанол:аммоний 0,880 (97:3:0,5). Производные MMG, альфа-MMG и кето-MMG могут быть получены при помощи нагревания в течение ночи при 100°С с 5%-ным водным ТВАН (тетрабутиламмония гидроксид) (2,5 мл) в пробирке 16×100 мм (Minnikin 1988).
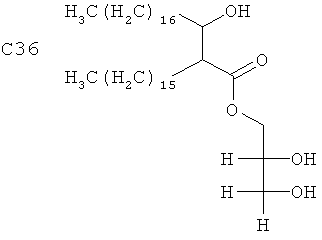
Синтетический гомолог, аналог или модифицированный вариант MMG может быть получен любым общепринятым способом химического синтеза. Аналог относится к одному из группы соединений, похожих по структуре, но различающихся по элементному составу, и гомолог относится к любому члену гомологичных рядов соединений. Такие соединения могут иметь варьирующиеся длины углеродной цепи; в частности, укороченный размер ассоциируется с пониженной токсичностью и может, следовательно, служить для ослабления любой явной токсичности аналогов. Таким образом, синтетические версии могут быть основаны на алкильных цепях, например с 8-36 атомами углерода и с 0-3 двойными связями на каждом липидном хвосте. Альтернативно, упрощенную форму можно получать путем удаления одного из липидных хвостов. Размер углеродного каркаса синтетического MMG предпочтительно составляет С8-С36, например 3-гидрокси-2-этил-гексановой кислоты 2,3-дигидроксипропиловый эфир (С8), 3-гидрокси-2-бутил-октановой кислоты 2,3-дигидроксипропиловый эфир (С12), 3-гидрокси-2-гексил-декановой кислоты 2,3-дигидроксипропиловый эфир (С16), 3-гидрокси-2-гептил-ундекановой кислоты 2,3-дигидроксипропиловый эфир (С18), 3-гидрокси-2-тетрадецил-октадекановой кислоты 2,3-дигидроксипропиловый эфир (С32) или 3-гидрокси-2-гексадецил-икозановой кислоты 2,3-дигидроксипропиловый эфир (С36) и наиболее предпочтительно С8 или С16. Модифицированный вариант может быть получен путем замещения глицеринового фрагмента другими полиольными головными группами, например полипропиленгликолем и полиэтиленглицерином. Стереохимия С2 и С3 синтетического мономиколята, а также в глицерине, может варьироваться. В дальнейшем MMG, написанный один, также означает синтетический гомолог, аналог или модифицированный вариант MMG, как описано выше.
Антигенный компонент или вещество может представлять собой полипептид или часть полипептида, которое вызывает иммунный ответ у животного или человека и/или в биологическом образце, определенный с помощью любого из биологических анализов, описанных в данном описании изобретения. Иммуногенная часть полипептида может представлять собой Т-клеточный эпитоп или В-клеточный эпитоп. С целью идентификации релевантных Т-клеточных эпитопов, которые распознаются во время иммунного ответа, возможно использование способа "грубой силы": Так как Т-клеточные эпитопы являются линейными, делеционные мутанты полипептида будут, если они сконструированы систематично, показывать, какие участки полипептида являются основными в иммунологическом распознавании, например, подвергая эти делеционные мутанты, например, анализу гамма-INF (интерферона), описанному в данном описании изобретения. В еще одном способе используются перекрывающиеся олигопептиды (предпочтительно синтетические, имеющие длину, например, 20 аминокислотных остатков), полученные из полипептида. Данные пептиды могут быть испытаны в биологических анализах (например, анализ гамма-IFN, как описано в данном описании изобретения), и некоторые из них будут давать положительный ответ (и таким образом являться иммуногенными) как свидетельство присутствия Т-клеточного эпитопа в пептиде. Линейные В-клеточные эпитопы могут быть определены с помощью анализа распознавания В-клеток в отношении перекрывающихся пептидов, охватывающих интересующий полипептид, как, например, описано у Harboe et al, 1998.
Хотя показано, что минимальная длина Т-клеточного эпитопа составляет по меньшей мере 6 аминокислот, нормально, чтобы такие эпитопы были составлены из более длинных участков аминокислот.Таким образом, предпочитают, чтобы полипептидный фрагмент по изобретению имел длину по меньшей мере 7 аминокислотных остатков, например по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 12, по меньшей мере 14, по меньшей мере 16, по меньшей мере 18, по меньшей мере 20, по меньшей мере 22, по меньшей мере 24 и по меньшей мере 30 аминокислотных остатков. Таким образом, в важных воплощениях способа по изобретению предпочтительно, чтобы полипептидный фрагмент имел длину не больше 50 аминокислотных остатков, например не более 40, 35, 30, 25, и 20 аминокислотных остатков. Ожидается, что пептиды, имеющие длину от 10 до 20 аминокислотных остатков, окажутся наиболее эффективными в качестве диагностических средств, и, следовательно, особенно предпочтительными длинами полипептидного фрагмента, используемого в способе по изобретению, являются 18, а также 15, 14, 13, 12 и даже 11 аминокислот.
В частности, антигенное вещество может быть произведено из культуры метаболизирующих Mycobacterium tuberculosis, Mycobacterium bovis и других природных микобактерий, таких как, например, Mycobacterium avium. В частности, интересующие вещества из фильтрата таких микобактерий представляют собой белки семейства генов ESAT-6 (такие как ESAT6 и ТВ 10.4), а также другие ранние антигены, такие как Ag85A, Ад85 В, ORF2c, Rv 1036 и Rv0285, которые являются основными мишенями клеточно-опосредованного иммунитета на ранней фазе туберкулеза у ТВ-пациентов и в разных животных моделях. Кроме того, другие антигены, такие как Rv2653, Rv2655, Rv2656, Rv2657, Rv2658, Rv2659, Rv2660, которые являются основными мишенями при более поздних стадиях ТВ-инфекции, являются релевантными. Их иммуногенность сама по себе является низкой, но в комбинации с адъювантными комбинациями по настоящему изобретению они оказываются сильными кандидатами для провоцирования сильного и стойкого иммунитета против туберкулеза, как демонстрируется в следующей подробной части данного описания изобретения.
Белки генного семейства ESAT-6, а также многие другие антигены, применимые в комбинации с адъювантными комбинациями по настоящему изобретению, сегодня могут быть получены искусственно, например синтетически или с помощью генетических рекомбинантных методов.
Слитые белки признаны особенно хорошо подходящими в качестве антигенных веществ в вакцинах, например, слитые продукты Ag85b_TB10.4, Ag85b_ESAT-6_Rv2660, Ag85b_TB10.4_Rv2660 и Ag85a_TB10.4_Rv2660 признаны особенно эффективными против ТВ.
Вакцина определена как суспензия мертвых, аттенуированных или другим способом модифицированных микроорганизмов (бактерий, вирусов или риккетсии) или их частей для введения с целью создания иммунитета к заболеванию. Вакцина может быть введена или профилактически для предупреждения заболевания, или в качестве терапевтической вакцины для борьбы с уже имеющимися заболеваниями, такими как рак или скрытые инфекционные заболевания, а также в связи с аллергией и аутоиммунными заболеваниями. Вакцина может быть эмульгирована в подходящем адъюванте для усиления иммунного ответа.
Вакцины вводят способом, соответствующим дозированной композиции, и в таком количестве, которое будет терапевтически эффективным и иммуногенным. Количество, подлежащее введению, зависит от субъекта, подлежащего лечению, включая, например, способность иммунной системы индивидуума закреплять иммунный ответ, и степени желательной защиты. Пригодные диапазоны доз составляют порядка нескольких сотен микрограммов активного ингредиента на вакцинацию с предпочтительным диапазоном от примерно 0,1 мкг до 1000 мкг, например, в диапазоне от примерно 1 мкг до 300 мкг, и особенно в диапазоне от примерно 1 мкг до 50 мкг. Пригодные схемы первичного введения и бустер-инъекций антигена также варьируются, но они типизируются по первичному введению с последующими повторными иммунизациями или другими введениями.
Способ использования может широко варьироваться. Любой из общепринятых способов введения вакцины является пригодным. Как предполагается, они включают пероральное или мукозальное применение на твердой физиологически приемлемой основе или в физиологически приемлемой дисперсии, парентерально, посредством инъекции или подобное. Доза вакцины зависит от пути введения и варьируется в соответствии с возрастом человека, подлежащего вакцинации и, в меньшей степени, габаритов человека, подлежащего вакцинации.
Вакцины обычно вводят парентерально, с помощью инъекции, например, или подкожно, или внутримышечно. Дополнительные композиции, которые пригодны для других способов введения, включают суппозитории и, в некоторых случаях, пероральные или мукозальные композиции. Для суппозиториев традиционные связывающие вещества и носители могут включать, например, полиалкаленгликоли или триглицериды; такие суппозитории могут быть образованы из смесей, содержащих активный ингредиент в диапазоне от 0,5% до 10%, предпочтительно 1-2%. Пероральные композиции включают такие обычно используемые эксципиенты как, например, фармацевтические марки маннитола, лактозы, крахмала, стеарата магния, сахарината натрия, целлюлозы, карбоната магния и тому подобного. Такие композиции принимают форму растворов, суспензий, таблеток, пилюль, капсул, препаратов с замедленным высвобождением или порошков и предпочтительно содержат 10-95% активного ингредиента, предпочтительно 25-70%.
Предпочтительные вакцины могут представлять собой, например:
Белковую вакцину: вакцинная композиция, содержащая полипептид (или по меньшей мере одну иммуногенную его часть), смесь пептидов или слитый полипептид.
Живые рекомбинантные вакцины: экспрессия необходимого антигена в вакцине в непатогенном микроорганизме или вирусе. Хорошо известными примерами таких микроорганизмов являются Mycobacterium bovis BCG, Salmonella и Pseudomonas, и примерами вирусов является вирус коровьей оспы и аденовирус.
Для всех этих вакцинных конструкций добавление подходящего адъюванта приводило к улучшенной эффективности вакцин (Brandt et al, 2000), (van Rooij et al, 2002), (Bennekov et al, 2006).
Липосомы (или липидные пузырьки) представляют собой водные ячейки, окруженные липидным бислоем. Липидные компоненты обычно представляют собой фосфолипиды или другие амфифилы, такие как поверхностно-активные вещества, часто с добавлением холестерина и других заряженных липидов. Липосомы способны задерживать водо- и липидорастворимые соединения, таким образом позволяя липосоме действовать в качестве носителя. Липосомы использовали в качестве систем доставки в фармакологии и медицине, таких как иммуноадъюванты, в лечении инфекционных заболеваний и воспалений, терапии рака и генной терапии {Gregoriadis et al, 1995}. Факторами, которые могут оказывать влияние на адъювантный эффект липосом, являются размер липосом, липидная композиция и поверхностный заряд. Кроме того, также может быть важным локализация антигена (например, он адсорбирован или ковалентно связан с поверхностью липосомы, либо он инкапсулирован в липосомные водные ячейки). Дендритные клетки можно использовать в качестве средств доставки антигена. Показано, что загрузка антигена к антиген-представляющим клеткам, таким как дендритные клетки, является эффективным способом генерации активных Т-клеток, играющих роль в противоопухолевом иммунитете.
Соединения четвертичного аммония, например
диметилдиоктадециламмония бромид, -хлорид или другие его органические или неорганические соли (DDA-B, DDA-C или DDA-X), диметилдиоктадециниламмония хлорид, -бромид или другие его органические или неорганические соли (DODA-C, DODA-B или DODA-X), или 1,2-диолеил-3-триметиламмоний-пропан (DOTAP), 1,2-димироил-3-триметиламмоний-пропан, 1,2-дипальмитоил-3-триметиламмония-пропан, 1,2-дистеароил-3-триметиламмоний-пропан и диолеил-3-диметиламмоний-пропан (DO-DAP) и N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмоний (DOTMA), обладают способностью образовывать липидные агрегаты, такие как липидные бислои, липосомы всех типов, как однослойные, так и многослойные, мицеллы и тому подобное, при диспергировании в водный среде. Липидные мембраны таких структур представляют превосходную матрицу для включения других амфифильных соединений, таких как гликолипиды, например MMG или 6,6'-дибегенат альфа,альфа'-трегалозы (TDB), которые, как показано, стабилизируют дисперсии пузырьков (Davidsen et al, PCT/DK2005/000467).
Комбинация MMG и системы доставки может действовать синергически в отношении усиления иммунного ответа, например, при введении одного DDA. Таким образом, DDA способствует низкому уровню продуцирования IFN-γ, однако совместно с MMG резко усиливает продуцирование IFN-γ.
Липосомы по данному изобретению могут быть изготовлены рядом способов, хорошо известных в данной области техники (Davidsen et al, PCT/DK2005/000467). Включение MMG в липосомы/системы доставки может быть осуществлено рядом способов, хорошо известных в данной области техники, включая простое смешивание липосом и MMG. В частности, включение MMG в липосомы может быть осуществлено, как описано в Davidsen et al, PCT/DK2005/000467.
В дополнение к обеспечению иммунитета к заболеваниям адъювантные комбинации по настоящему изобретению также можно использовать для продуцирования антител против соединений, которые сами по себе являются слабыми иммуногенными веществами, и такие антитела можно использовать для обнаружения и количественной оценки интересующих соединений, например, в медицине и аналитической химии.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
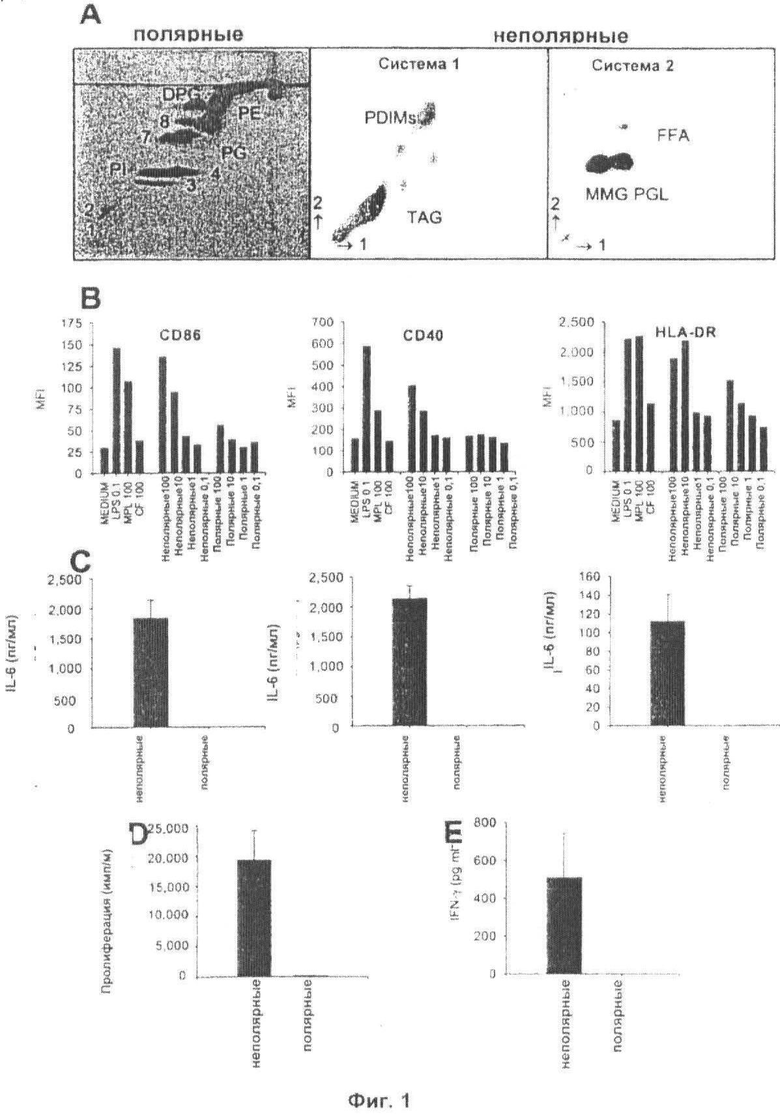
Фиг.1. Выделение и количественная оценка иммуностимулирующей активности неполярных и полярных липидов из М.bovis BCG. Полярные и неполярные липиды, экстрагированные из М.bovis BCG Copenhagen, анализировали с помощью 2-D TLC. Во фракции полярных липидов 1-4 представляют собой маннозиды фосфатидилинозита, PI представляет собой фосфатидилинозит, РЕ представляет собой фосфатидилэтаноламин, и DPG представляет собой дифосфатидилглицерин (кардиолипид). PG представляет собой L-альфа-фосфатидил-DL-глицерин, в то время как 5 и 6 представляют собой неизвестные фосфолипиды. В неполярной фракции TAG представляет собой триацилглицерин, PDIM представляет собой фтиоцерола димикоцерозат А, В и С, MMG представляет собой мономиколилглицерин, PGL представляет собой фенольные гликолипиды, и FFA представляют собой свободные жирные кислоты (Панель А). iDС инкубировали в течение 24 ч в присутствии одной лишь среды, LPS (0,1 мкг/мл), MPL (100 мкг/мл), фактора жгутикообразования (CF) (100 мкг/мл), неполярных липидов (0,1-100 мкг/мл) или полярных липидов (0,1-100 мкг/мл). Показаны средние геометрические значения интенсивности флуюресценции (MFI) уровней поверхностных маркеров на DC после обработки. Данные получали из одного типичного эксперимента из трех, используя трех разных доноров (панель В). Супернатанты культуры, полученные после обработки 100 мкг/мл неполярных или полярных липидов, анализировали с помощью ELISA на присутствие цитокинов IL-6, TNF-α и IL-12. Представлены данные, полученные от одного типичного эксперимента из трех, выполненных в трех параллелях с использованием трех разных доноров (± s.e.m (стандартная ошибка среднего)) (Панель С). Пролиферация (Панель D) и высвобождение IFN-γ (Панель Е) Т-клетками от PPD-негативных доноров после инкубирования с DC, обработанными в течение 24 часов 100 мкг/мл неполярных или полярных липидов, в анализе MLR. Представлены данные, полученные от одного типичного эксперимента из трех, выполненных в трех повторах с использованием трех разных доноров (± s.e.m).
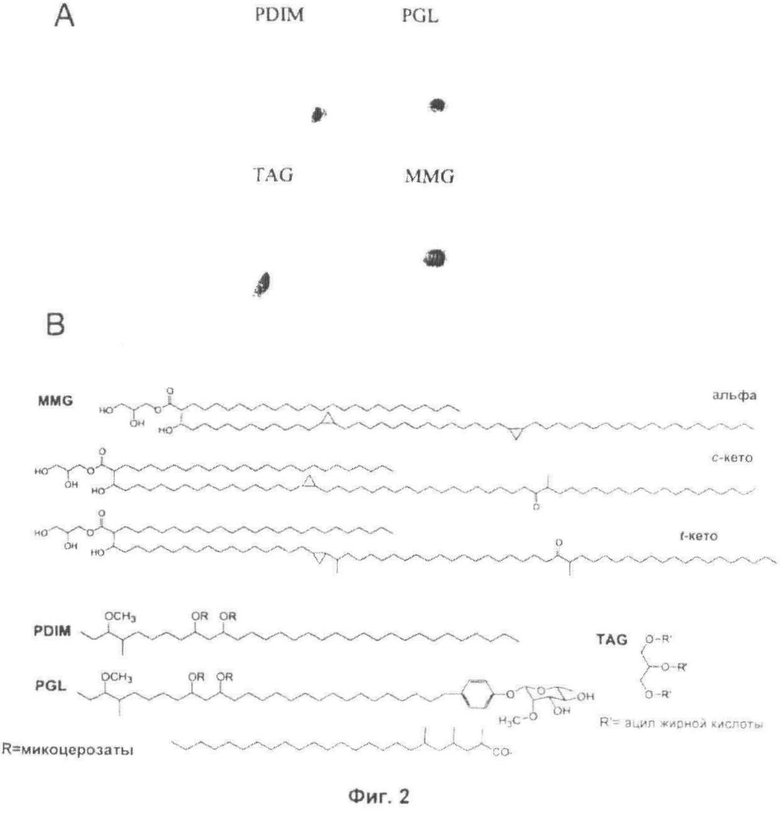
Фиг.2. Структуры MMG, TAG, PDIM и PGL. Анализ 2-D TLC отдельных липидов PDIM, TAG, PGL и MMG (Панель А) и типичные структуры MMG, PGL, PDIM и TAG (Панель В).
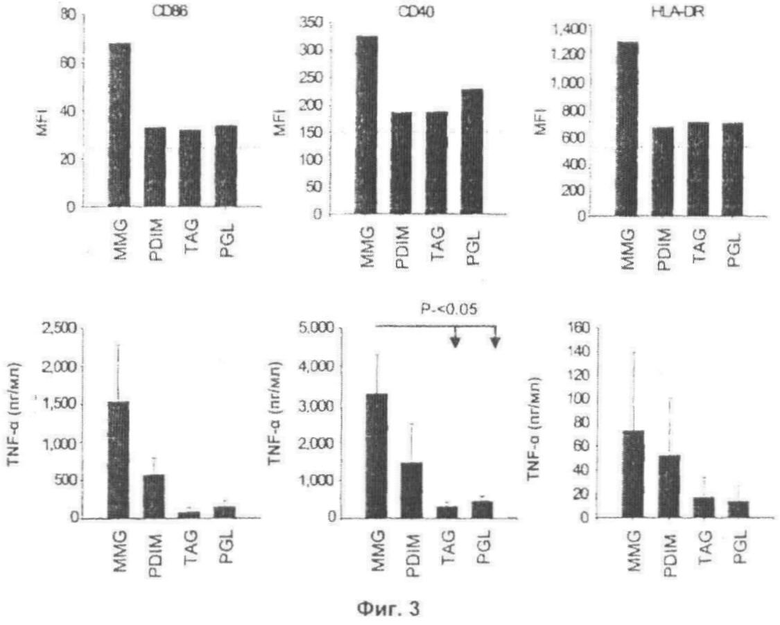
Фиг.3. Активирование человеческих дендритных клеток посредством MMG. iDС инкубировали в течение 24 ч в присутствии одной лишь среды (пунктирная линия) или MMG, PDIM, PGL или TAG (10 мкг/мл). Обнаружено, что липидные препараты не содержат эндотоксинового загрязнения (<0,00 нг LPS/мкг липида). Показаны MFI уровней поверхностных маркеров на DC для одного типичного донора из шести после обработки (Панель А). Супернатанты культур, полученные после обработки MMG, PDIM А, PGL или TAG (10 мкг/мл), анализировали с помощью ELISA на присутствие цитокинов IL-6, TNF-α и IL-12 (Панель В). Представлено среднее значение (± s.e.m.) данных, полученных из трех или четырех экспериментов с использованием разных доноров, выполненное в трех повторах. Данные анализировали, используя тест Тьюки.
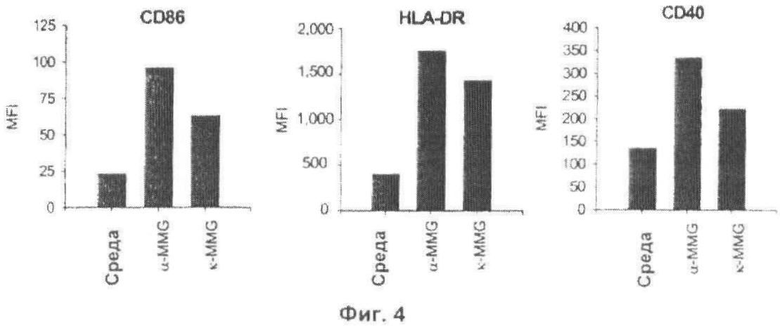
Фиг.4. Альфа - и кетомиколяты MMG представляют собой иммуностимуляторы. iDC инкубировали в течение 24 ч в присутствии только одной среды и альфа- или кетомиколята MMG (10 мкг/мл). Показаны MF1 поверхностных маркеров на DC после обработки.
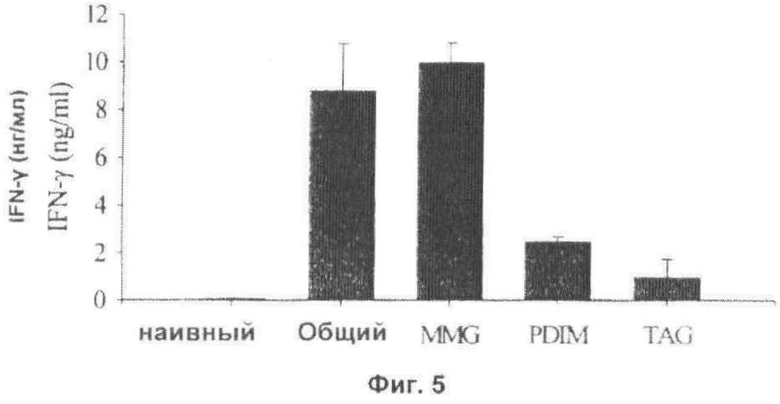
Фиг.5. Высвобождение IFN-γ, индуцированное MMG, выделенным из М.bovis BCG Copenhagen. Мышей C57BL/6 иммунизировали при помощи Ag85B-ESAT-6 в комбинации с адъювантами на липидной основе, выделенными из BCG Copenhagen, включенными в DDA липосомы. Измеряли высвобождение IFN-γ в РВМС, выделенных из дренирующих лимфатических узлов через сутки после вакцинации.
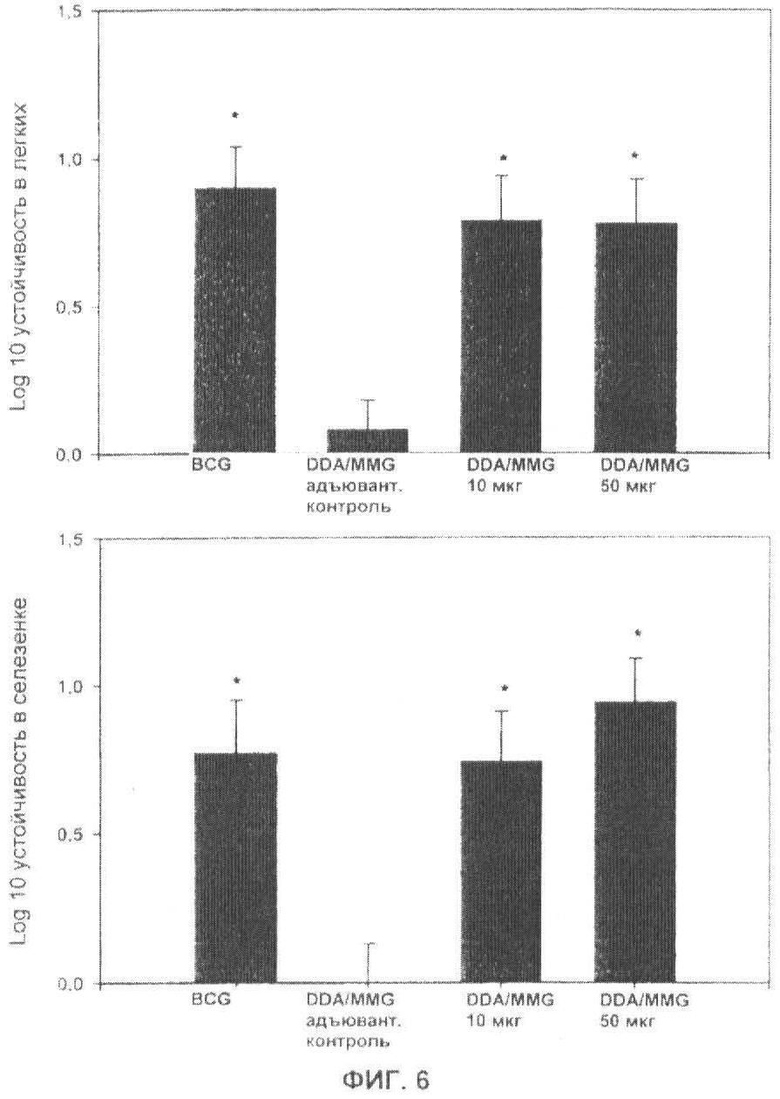
Фиг.6. Защита против вирулентной ТВ-инфекции посредством адъювантов MMG. Мышей C57BL/6 иммунизировали три раза Ag85B-ESAT-6 в комбинации с адъювантами на основе DDA и 10 или 50 мкг MMG. Через шесть недель после последней вакцинации мыши получали аэрозольную провокацию М.tuberculosis. Число бактерий измеряли в легких и селезенке через шесть недель. Мышей, получающих стандартную вакцинацию BCG, включали в качестве положительного контроля, и мышей, иммунизированных посредством DDA/MMG (10 мкг) без антигена, в качестве отрицательного контроля. Протективную эффективность экспериментальных вакцин выражают в виде Log10 уменьшения бактериальной нагрузки в легких, по сравнению с неиммунизированными мышами. Результаты представляют собой средние значения для шести мышей в каждой группе ±SEM. Значения, значительно отличающиеся от неиммунизированного контроля, помечены *Р<0,05.
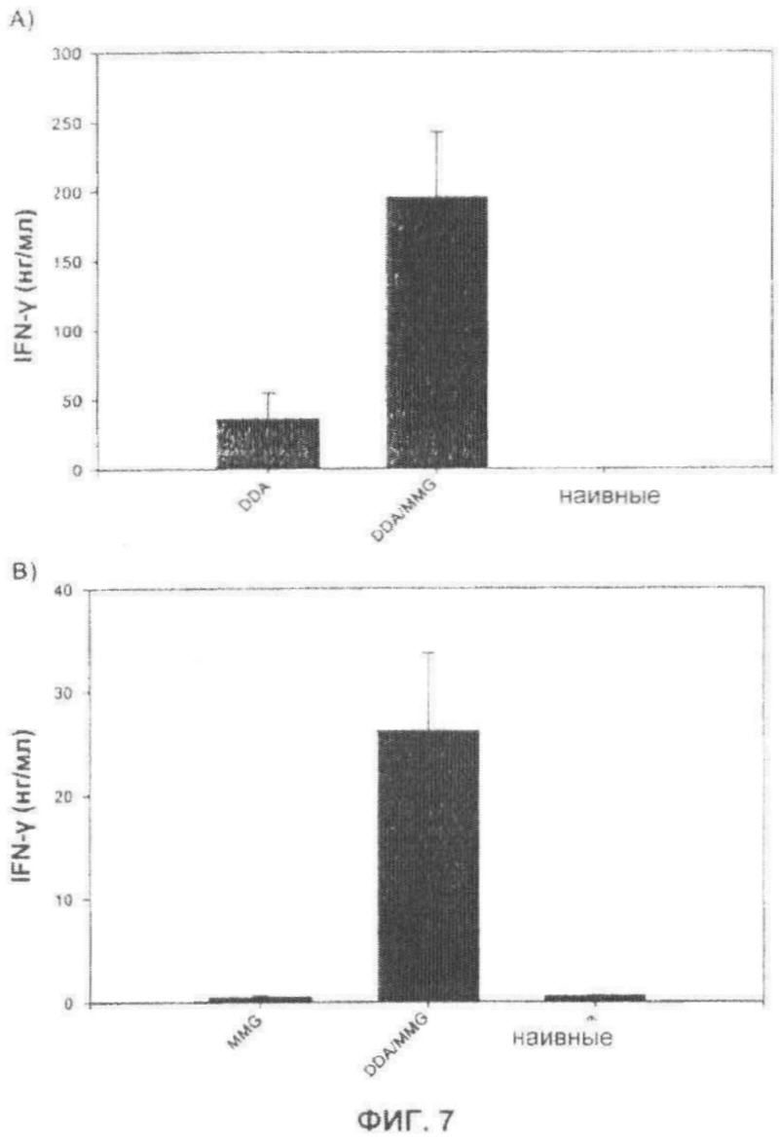
Фиг.7. Высвобождение IFN-γ улучшают посредством комбинирования MMG и DDA. В двух экспериментах мышей C57BL/6 иммунизировали при помощи Ag85B-ESAT-6 в DDA или DDA/MMG (панель А), или MMG, или DDA/MMG (панель В). Через 3 недели после последней вакцинации измеряли высвобождение IFN-γ в РВМС, выделенных из крови (панель А) или в селезенках (панель В).
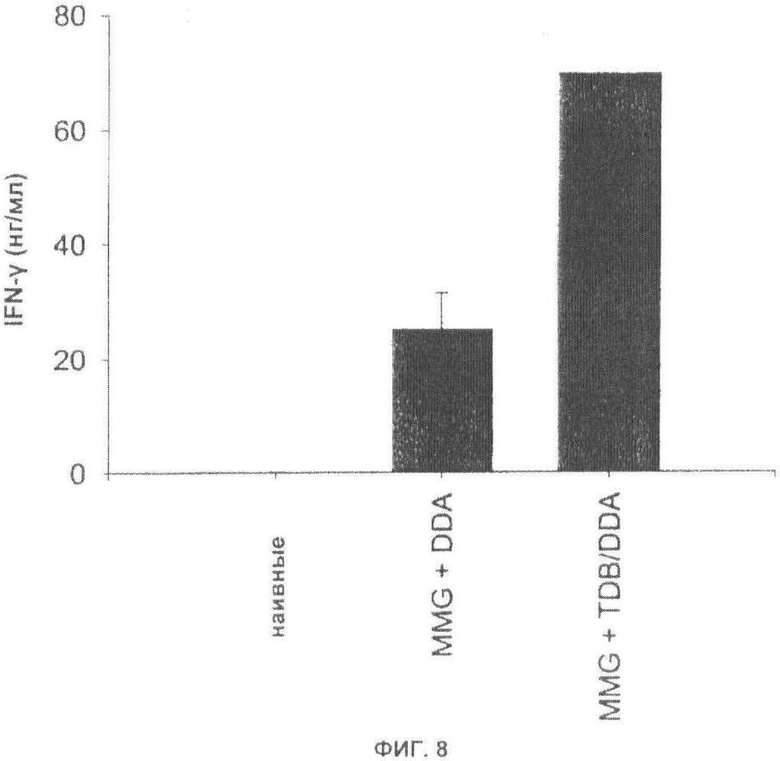
Фиг.8. Высвобождение IFN-γ улучшают посредством добавления TDB к комбинации MMG/DDA. Мышей C57BL/6 иммунизировали при помощи Ag85B-ESAT-6, введенного в MMG, включенный в DDA липосомы, или липосомы DDA, содержащие TDB. Через 5 месяцев после вакцинации измеряли высвобождение IFN-γ в РВМС, выделенных из крови.
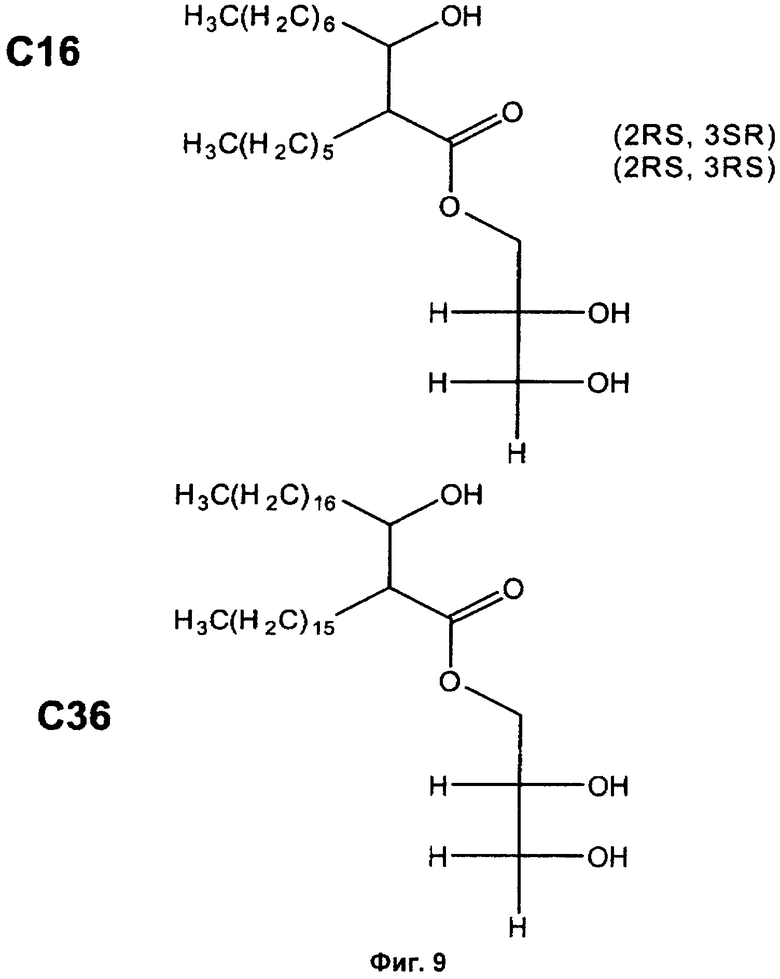
Фиг.9. Примеры структур синтетических аналогов MMG.
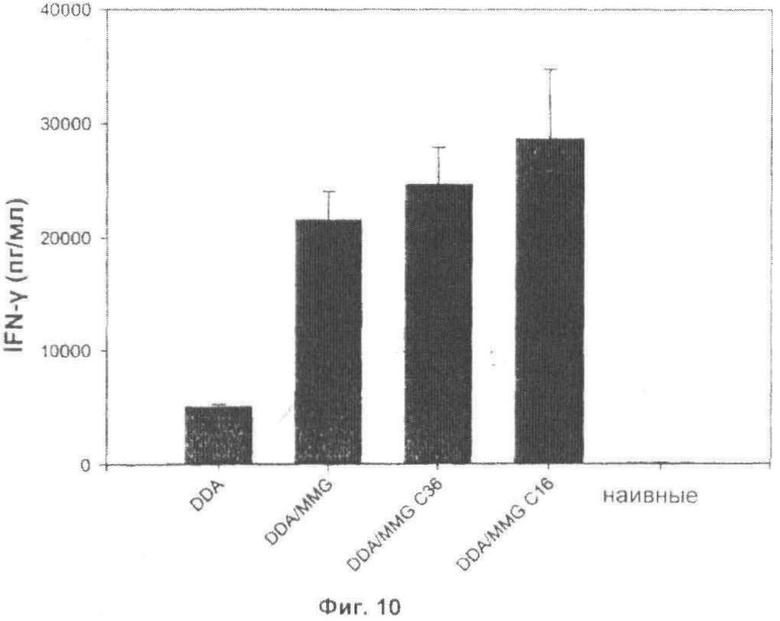
Фиг.10. Иммунные ответы сравнимы с природными и синтетическими аналогами MMG. Мышей C57BL/6 иммунизировали при помощи Ag85B-ESAT-6 в DDA, DDA/ MMG (10 мкг), DDA/MMG С36 (10 мкг) или DDA/MMG С16 (10 мкг). Измеряли высвобождение IFN-γ в РВМС, выделенных из крови, через одну неделю после последней иммунизации.
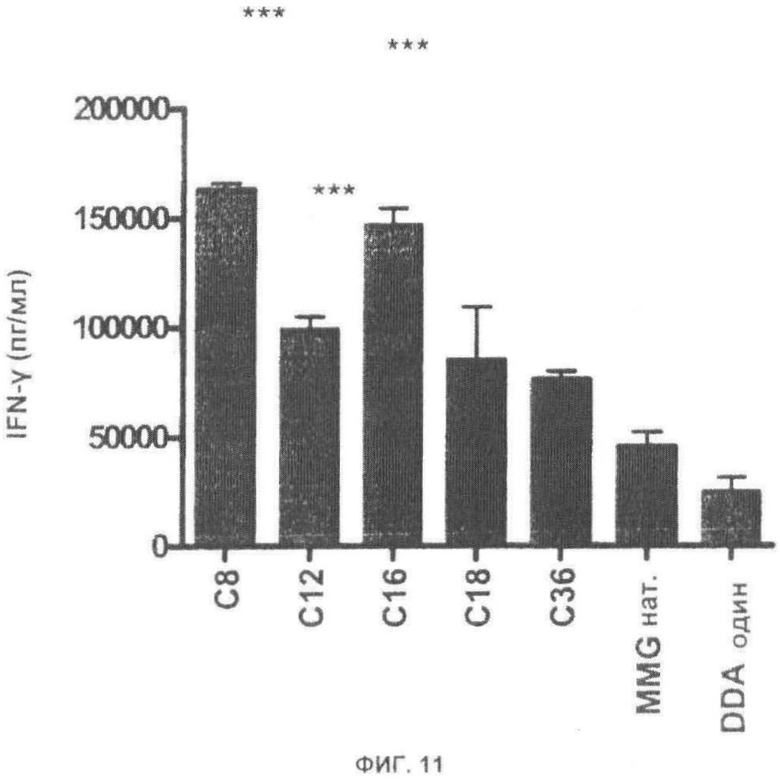
Фиг.11. Более высокие иммунные ответы при более короткой длине цепи. Мышей C57BL/6 иммунизировали при помощи Ag85B-ESAT-6 в DDA или DDA с различными аналогами MMG с длиной цепи, меняющейся от 8 до 36 (1 мкг/доза). Измеряли высвобождение IFN-γ в РВМС, выделенных из крови, через три недели после последней иммунизации.
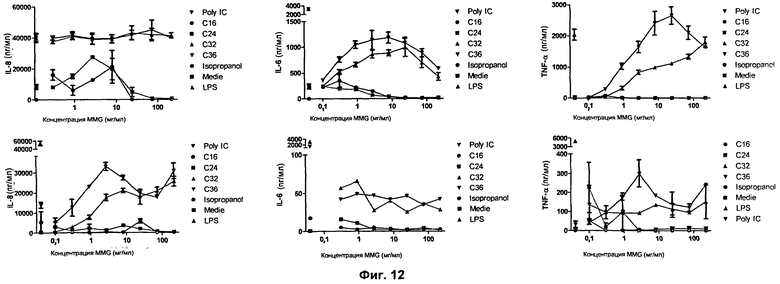
Фиг.12. Оценка иммуностимулирующей активности MMG с разной длиной цепи. Представлены результаты in vitro теста (ELISA) на человеческих макрофагах, стимулированных синтетическими MMG с разной длиной цепи. Показано, что все синтетические MMG с разной длиной цепи стимулируют иммунный ответ, причем иммунный ответ значительно сильнее при добавлении поверхностно-активного вещества, например DDA. [На Фиг.12 Isopropanol означает изопропанол].
Примеры
Вещества и методы:
ЭКСТРАКЦИЯ НЕПОЛЯРНЫХ И ПОЛЯРНЫХ ЛИПИДОВ ИЗ М.BOVIS BCG
Mycobacterium bovis BCG (Copenhagen) культивировали в модифицированной среде Сотона. Микобактерии собирали через 2-3 недели, суспендировали в PBS и убивали посредством инкубирования в течение 1,5 часов при 60°С. Неполярные и полярные липиды экстрагировали в соответствии со стандартными протоколами (Dobson et al, 1985), (Rosenkrands et al, 2005).
Для экстракции неполярных липидов смесь метанол:0,3% NaCl (440 мл) и 440 мл петролейного эфира добавляли к 20 г микобактерий (масса во влажном состоянии), и смесь перемешивали в течение 2 ч. После центрифугирования, верхний слой удаляли, и нижний слой реэкстрагировали 440 мл петролейного эфира. Супернатантные фазы от обеих экстракций объединяли и упаривали с получением неполярных липидов.
Для экстракции полярных липидов метанольный солевой раствор, содержащий биомассу, нагревали в кипящей водяной бане в течение 10 мин при 100°С, затем охлаждали в течение 10 мин при 37°С. Добавляли хлороформ:метанол:0-3% NaCl (9:10:3) объемом 520 мл, и смесь перемешивали в течение ночи. Итоговую смесь пропускали через воронку из спеченного стекла; осадок на фильтре собирали и реэкстрагировали дважды при помощи 170 мл смеси хлороформ:метанол:0-3% NaCl (5:10:4). Все три водно-метанольно-хлороформные фазы объединяли, и добавляли 580 мл смеси хлороформ:0,3% водный NaCl (1:1) при перемешивании в течение 10 минут. Фазы оставляли разделяться, после чего верхний водный слой удаляли и отбрасывали. Нижний органический слой упаривали досуха с получением полярных липидов.
ОЧИСТКА ОТДЕЛЬНЫХ НЕПОЛЯРНЫХ ЛИПИДОВ
PDIM и TAG выделяли, используя TLC, выполненную в смеси петролейный эфир/ацетон (98:2); извлекали только основной компонент PDIM, на основе фтиоцерола A. PGL и MMG выделяли вместе при помощи TLC, выполненной в смеси толуол/ацетон (95:5). PGL и MMG разделяли на 1-D TLC в смеси хлороформ:метанол:аммоний 0,880 (97:3:0,5). PDIM, TAG, PGL и MMG подвергали воздействию 500 МГц 1Н и 13С ядерного магнитного резонанса (ЯМР) (Bruker drx 500) и MALDI-TOF (матричная лазерная десорбция/ионизация) масс-спектроскопии (MS) (Bruker Biflex IV). Образцы (10 мкл) регидратированных липидных экстрактов (1 мг/мл) анализировали с помощью SDS-PAGE (электрофорез на полиакриламидном геле с додецилсульфатом натрия) (Laemmli et al, 1970} и окрашивания серебром (Blum et al, 1987) на содержание остаточного белка. Было обнаружено, что липидные препараты не содержат эндотоксинового загрязнения (<0,00 нг LPS/мкг липида).
ГИДРОЛИЗ МОНОМИКОЛИЛГЛИЦЕРИНА
Мономиколилглицерин нагревали в течение ночи при 100°С с 5% водным ТВАН (2,5 мл) в 16×100 мм пробирке (Minnikin et al, 1988). После охлаждения смесь разбавляли водой (2 мл) и добавляли 10%-ный раствор иодометана (3 мл) в дихлорметане, и пробирки помещали в ротатор на 1 ч. Верхний водный слой отбрасывали, нижний слой промывали эквивалентными объемами 1 М HCl и воды и упаривали досуха с получением остатка, содержащего, как показано при помощи TLC (петролейный эфир:ацетон 95:5), альфа- и кетомиколят-метиловые эфиры. В результате препаративной TLC (как показано выше) получали альфа-метилмиколят и кетометилмиколят.
Получение триметилсилильных производных использовали для разделения альфа-MMG и кето-MMG. Смесь MMG и 600 мл реагента TRI-SIL (Pierce) нагревали при 75°С в течение 20 мин. Охлажденный раствор затем сушили в потоке азота и загружали на пластинку для препаративной TLC и разгоняли в смеси петролейный эфир:толуол (50:50), зоны альфа-MMG и кето-MMG визуализировали, используя 0,01%-ный родамин, и рассматривали при длинноволновой флуоресценции. Соответствующие зоны соскребали с TLC-пластинки, и альфа-MMG, и кето-MMG экстрагировали с силикагеля три раза, используя диэтиловый эфир (3×5 мл). Объединенные экстракты сушили, и триметилсилильные группы удаляли путем добавления смеси гептан:метанол (1:1) и нескольких кристаллов лара-толуол-сульфоновой кислоты и смешивали в течение 1 ч. Гептановый слой восстанавливали и упаривали досуха с получением очищенных альфа-MMG и кето-MMG.
СИНТЕЗ MMG, ДЕМОНСТРИРУЮЩИЙ ПОЛУЧЕНИЕ С36
3-Гидрокси-2-тетрадецил-октадекановую кислоту (синтетическая С32 кориномиколовая кислота, синтезированная согласно Datta et al, 1991) (100 мг, 0,20 ммоль, 1 экв.) и 4-пирролидинопиридин (100 мг, 3 экв.) помещали в 50 мл круглодонную колбу, и добавляли 50 мкл раствора 2,2-диметил-4-гидроксиметил-1,3-диоксолана (sn-изопропилиденглицерина) в дихлорметане (500 мкл), а также молекулярные сита 4Å. Смесь доводили до полного высушивания в глубоком вакууме при комнатной температуре, и добавляли N,N'-дициклогексилкарбодиимидазол (DCC) (15 мл, 0,1 М DCC в DCM, 5 экв.), и реакционную смесь оставляли перемешиваться при комнатной температуре в течение ночи. Молекулярные сита удаляли фильтрацией, реакционную смесь восстанавливали досуха в вакууме, и остаток очищали, используя колоночную флэш-хроматографию (Fluka 60741 Silica Gel 60), элюируя от гексана до смеси гексан:этилацетат (8:2) с 5%-ными приростами, с получением чистого изопропилиден-защищенного соединения (3-гидрокси-2-тетрадецил-октадекановой кислоты 2,2-диметил-[1,3]-диоксолан-4-илметиловый эфир), с выходом 56% (68 мг). 1Н ЯМР (CDCl3, 300 МГц) δН 0.90 (t, 6Н, СН3), 1.20 (s, 54Н, СН2), 1.40 (s, 3Н, СН3), 1.45 (s, 3Н, СН3), 2.50 (m, 1Н, СН), 4.05-4.40 (m, 5Н, СН2, СН); 13С ЯМР (CDCl3, 75 МГц) δс 15.0 (СН3), 22.1, 28.8, 28.9, 29.0, 31.4 (СН2), 29.1 (СН3), 52.1 (СН(СН2) I3CH3), 63.2 (СН2ОСО), 69.3 (СН2O), 73.4 (СН(СН2),4СН3), 174.3 (С=O); m/z (El) 633.55 [M+Na''] (100%); HRMS, рассчитано для C38H74O5Na [M+Na''] 633,5536; обнаружено 633,5527.
3-Гидрокси-2-тетрадецил-октадекановой кислоты 2,2-диметил-[1,3]-диоксолан-4-илметиловый эфир (68 мг, 1 экв.) растворяли в 6 мл раствора трифторуксусная кислота:тетрагидрофуран:вода (8:17:3 по объему) и перемешивали при комнатной температуре в течение ночи. Раствор нейтрализовали насыщенным водным бикарбонатом натрия, и смесь экстрагировали дважды хлороформом. Органический экстракт промывали водой и рассолом, сушили и восстанавливали в вакууме с получением неочищенного продукта в виде белого твердого вещества, которое очищали при помощи колоночной флэш-хроматографии с 10 г силикагелевым картриджем Varian Bond Elut 12256026, элюируя от гексана до смеси гексан: этилацетат (7:3) с 5%-ным приростом, с получением указанного в заголовке соединения в виде белого твердого вещества с выходом 49% (32 мг). Точка плавления 72-74°С. 1Н ЯМР (CDCl3, 300 МГц) δн 0.90 (t, 6Н, СН3), 1.25 (s, 54Н, СН2), 2.50 (m, 1Н, СН), 3.45-3.85 (m, 3Н, СН, СН2), 4.25 (m, 2Н, СН2); 13С ЯМР (CDCl3, 75 МГц) δс 15.0 (СН3), 26.3, 30.9, 31.3, 33.5 (СН2), 47.5 (СН(СН2)13СН3), 68.4 (СН2), 69.5 (СН(СН2)14СН3), 72.5 (СН2O), 76.4 (СН), 175.4 (C-l); m/z (El) 593.50 [M+Na"] (100%); HRMS, рассчитано для C35H70O5Na [M+Na~], 593,5121; обнаружено 593,5143.
АНАЛИЗЫ ДЕНДРИТНЫХ КЛЕТОК
DC из человеческих РВМС, получали согласно модифицированному способу Romani et al., 1994. Периферическую кровь получали из светлого слоя кровяного сгустка. В кратком изложении, моноциты выделяли центрифугированием с фиколл-гипак (Lymphoprep 1077 средней плотности, Nycomed, Oslo, Norway) с последующим отделением CD 14-положительных клеток, используя анти-СD-меченные магнитные шарики (MACS; Miltenyi Biotech, Bergesh Gladbach, Germany). Моноциты культивировали в полной RPMI 1640, с добавлением 10% FCS (плодной сыворотки теленка), 50 мкМ 2-меркаптоэтанола, 100 Ед./мл пенициллина, 100 мкг/мл стрептомицина, 2 мМ L-глютамина (все Gibco) (СМ), и в присутствии 100 нг/мл человеческих рекомбинантных GM-CSF (гранулоцитарно-макрофагальные колониестимулирующие факторе) (Prepotech, Rocky Hill, NJ, USA) и 50 нг/мл человеческих рекомбинантных IL-4 (Becton Dickinson (BD)) в течение 7 суток при 37°С, 5% СO2.
На 7 сутки iDC (1×105 клетки/мл) культивировали в течение еще 24 часов с липополисахаридом (LPS) {Escherichia coli O127:В8) (Sigma-Aldrich, Brondby, Denmark) или липидами из М.bovis BCG. Липидные экстракты получали путем повторного растворения сухого липидного вещества М.bovis в смеси хлороформ:метанол (2:1) и последующего выпаривания растворителя и обработки пробы ультразвуком в СМ. Липиды добавляли к незрелым DC в концентрации от 0,1 до 100 мкг/мл.
ПРОТОЧНЫЙ ЦИТОМЕТРИЧЕСКИЙ АНАЛИЗ
DC окрашивали для определения поверхностных маркеров путем инкубирования сначала с релевантным mAb (BD Pharmingen) (30 мин, 4°С), затем с разбавленным 1/20 FITC-конъюгированным антителом козы против lg мыши (Jackson ImmunoResearch Laboratories, West Grove, PA) (30 мин, 4°C). Неспецифическое связывание Ab блокировали 10%-ным раствором плодной сыворотки теленка (15 мин, 4°С), затем добавляли подходящие первичные человеческие mAb. Окрашенные клетки сразу же исследовали с помощью проточной цитометрии, используя проточный цитометр FACScan (BD) и анализировали при помощи программы CellQuest.
ИЗМЕРЕНИЯ ЦИТОКИНОВ
Супернатанты DC культуры собирали и хранили при -20°С. Секретированные IL-12p70, IL-6 и TNF-α измеряли с помощью ELISA (BD), согласно инструкциям производителя.
АНАЛИЗ СМЕШАННОЙ РЕАКЦИИ ЛЕЙКОЦИТОВ (MLR)
iDС для анализа смешанной реакции лейкоцитов (MLR) получали из моноцитов, как указано выше. Полученные клетки культивировали в течение 24 часов в той же самой среде (iDС) или в среде, содержащей липиды (10 или 100 мкг/мл). Титры DC от 0,125×105 до 2×105 инкубировали при 37°С/5% СO2 с аллогенными Т-клетками (105 клеток/лунка) от PPD-отрицательного донора в 96-луночных титрационных микропланшетах с плоским дном. Т-клетки выделяли, используя набор для выделения клеток Раn-Т (Miltenyi) в соответствии с инструкциями производителя. Т-клеточные сокультуры аллогенных DC инкубировали в течение 6 суток. Надосадочную жидкость собирали и хранили при -20°С до измерения секретированных IFN-γ с помощью ELISA (BD) в соответствии с инструкциями производителя. Затем оба анализа импульсно обрабатывали средой, содержащей 1 мкКи/лунка [3Н] тимидина, в течение последних 18 ч культивирования. Клетки собирали, и Т-клеточную пролиферацию измеряли с помощью жидкостно-сцинтиляционного подсчета (Microbeta Systems). Все анализы выполняли в трех повторах, используя по меньшей мере три разных донора.
АНТИГЕНЫ
Слитый белок Аg85 В и ESAT-6 (в дальнейшем обозначается Аg85В-ESAT-6) получали в виде рекомбинантных белков, как описано ранее (Olsen et al, 2001).
ЖИВОТНЫЕ
Самок мышей BALB/c или C57BL/6 в возрасте от 8 до 12 недель получали от Bomholtgaard (Ry, Denmark) или Harlan Scandinavia (Denmark). Инфицированных мышей содержали в клетках в защитном кожухе с ламинарным потоком BL-3.
ИММУНИЗАЦИИ
Мышей иммунизировали подкожно (s.c.) в основание хвоста вплоть до трех раз с двухнедельным перерывом между каждой из иммунизаций. Вакцины (0,2 мл/мышь) состояли из 2 мкг слитого белка Ag85B-ESAT-6, эмульгированного в 250 мкг DDA, и 10 мкг регидратированного липидного экстракта, если не указано иное. В некоторых случаях в DDA липосомы включали 11 мол. % TDB (Davidsen et al, PCT/DK2005/000467). В качестве положительного контроля в экспериментах с участием инфекции М. tuberculosis одна группа мышей получала одну дозу BCG Danish 1331, инъецируемую подкожно в основание хвоста. Общие или отдельные липидные экстракты получали путем регидратирования сухого липидного вещества М.bovis водой Milli Q в количестве 1 или 5 мг/мл и затем обработки пробы ультразвуком. Стандартные липидные вакцины получали смешиванием антигена с солевым раствором, с последующим добавлением регидратированного липидного экстракта и DDA и перемешиванием на вортексе. Вакцины оставляли в течение ночи для обеспечения адсорбции антигена.
КУЛЬТУРЫ ЛИМФОЦИТОВ
Образцы крови или паховые лимфатические узлы брали от мышей через 7-150 суток после последней иммунизации и готовили, как описано ранее (Rosenkrands et al, 2005). Культирование клеток выполняли в трех повторах в круглодонных микротитровальных лунках, содержащих 2×105 клеток в объеме 200 мкл RPMI с добавлением 2-меркаптоэтанола, глютамина, пенициллин-стрептомицина, HEPES (N-2-гидроксиэтил-пиперазин-N-2-этансульфоновой кислоты) и 10%-ной плодовой сыворотки теленка. Антиген для повторной стимуляции использовали в концентрации 5 мкг/мл. Клетки, содержащие только среду или 5 мкг/мл СоnА, включали во все эксперименты в качестве отрицательного и положительного контролей, соответственно. Супернатанты культур собирали от параллельных культур через 72 часа инкубирования в присутствии антигена, и количество IFN-γ определяли при помощи ферментного иммуносорбентного анализа (Brandt et al, 2000).
ЭКСПЕРИМЕНТАЛЬНЫЕ ИНФЕКЦИИ
Для оценки эффективности вакцины мышам вводили через 2,5 месяца после первой иммунизации аэрозольным путем с помощью системы ингаляционного воздействия Glas-Col, калиброванной для отложения приблизительно 25 CFU вирулентной М. tuberculosis Erdman в легкие. Бактериальную нагрузку в селезенке и легких определяли через шесть недель путем нанесения последовательных разведений на агар Middlebrook 7HI 1, с добавлением 2 мкл гидразида 2-тиофен-карбоновой кислоты на миллилитр для селективного подавления роста BCG. Колонии подсчитывали через 2-3 недели инкубации при 37°С.
СТАТИСТИЧЕСКИЙ АНАЛИЗ
Различия в количестве колоний у инфицированных мышей и контрольных мышей оценивали с помощью дисперсионного анализа. Если показаны значительные эффекты, различия между средними значениями оценивали с помощью теста Даннетта. Для анализов человеческих DC различия в высвобождении цитокинов в ответ на разные липиды оценивали с помощью дисперсионного анализа, и если показаны значительные эффекты, различие между средними значениями, оценивали с помощью теста Тьюки.
ПРИМЕР 1
ВЫДЕЛЕНИЕ И ИММУНОСТИМУЛИРУЮЩАЯ АКТИВНОСТЬ НЕПОЛЯРНЫХ ЛИПИДОВ ИЗ М.BOVIS BCG
Общие липиды М. bovis BCG разделяли на полярные и неполярные фракции. В полярной фракции липиды, которые можно было идентифицировать, представляли собой маннозиды фосфатидилинозита (1-4), фосфатидилинозит (PI), фосфатидилэтаноламин (РЕ), дифосфатидилглицерин (DPG) и L-альфа-фосфатидил-DL-глицерин (PG). Также был идентифицирован ряд неизвестных фосфолипидов (7 и 8) (Фиг.1А). В неполярной фракции большая часть идентифицированных липидов представляла собой фтиоцерола димикоцерозаты (PDIMs), триацилглицерины (TAGs), фенольный гликолипид (PGL) и мономиколилглицерин (MMG) (Фиг.1). FFA представляет собой свободные жирные кислоты.
Сравнительную иммуностимулирующую активность неполярных и полярных липидов изучали, используя незрелые DC (iDС) из моноцитов периферической крови человека (Фиг.1). Обработка неполярными липидами приводила к дозозависимому увеличению уровней активационных маркеров CD86, CD40 и HLA-DR по сравнению с необработанными контролями (Фиг.2В). Доза неполярных липидов 100 мкг/мл приводила к активации DC, сравнимой с активацией, наблюдаемой для сильной иммуностимулирующей молекулы LPS (0,1 мкг/мл), и более высокой по сравнению с наблюдаемой для микобактериального фактора жгутикообразования (TDM) и MPL. Активация этих молекул сопровождалась секрецией провоспалительных медиаторов фактора некроза опухоли-α (TNF-α), интерлейкина (IL)-6 и IL-12 (Фиг.2С). Уровни таких провоспалительных цитокинов в супернатантах iDС, обработанных полярными липидами, были ниже пределов определения с использованием данного анализа. Наконец, авторы изобретения использовали смешанную реакцию лейкоцитов (MLR), используя аллогенные Т-клетки от PPD-негативного донора (Фиг.2D-E) в качестве другого показателя активации DC. Кроме того, поддерживая состояние высокой активации DC, обработанных неполярными липидами, авторы изобретения обнаружили высокие уровни пролиферации и высвобождения IFN-γ, в то время как MLR не индуцировался полярной фракцией.
ПРИМЕР 2
ВЫДЕЛЕНИЕ ОТДЕЛЬНОГО ЛИПИДА ИЗ НЕПОЛЯРНОГО ЛИПИДНОГО ЭКСТРАКТА М. BOVIS BCG И ХАРАКТЕРИСТИКА MMG
Липиды из иммуностимулирующей неполярной фракции выделяли, используя препаративную TLC, с получением чистых образцов фтиоцерола димикоцерозата A (PDIM A), TAG, PGL и MMG; минорные количества фтиоцерола В и фтиодиолона А также были определены, но не были извлечены посредством используемых препаративных способов (Фиг.2А). Структура и идентичность липидов была подтверждена с помощью 1Н ядерно-магнитного резонанса (ЯМР) и масс-спектроскопии (MS) (Фиг.2 В и данные не показаны). Компонент MMG демонстрировал 1Н и 13С ЯМР-спектры, характерные для 1-моноацилглицерина (Gunstone et al, 1991). Массспектроскопия MALDI-TOF вместе с данными ЯМР общей фракции MMG (Таблица 1) выявили присутствие альфа-миколята и кето-миколята в цис- и транс-форме. Приблизительное соотношение главных компонентов составляло 1,00:0,29:0,24 соответственно.
ПРИМЕР 3
АКТИВИРОВАНИЕ ЧЕЛОВЕЧЕСКИХ ДЕНДРИТНЫХ КЛЕТОК С ПОМОЩЬЮ MMG
Очищенные MMG, PDIM A, PGL и TAG оценивали в отношении их способности активировать человеческие iDС. В этих анализах постоянно обнаруживалось, что MMG является наиболее сильным индуктором активации DC, приводящей к четко выраженной положительной регуляции CD86, CD40 и HLA-DR (Фиг.3А). MMG активировал DC даже в большей мере, чем PDIM А, липид, который долгое время ассоциировался с патогенностью микобактерий (Сох et al, 1999), который выступал как второй наиболее активный липид, в то время как PGL и TAG вызывали меньшую активацию. У шести отдельных доноров порядок активации наблюдали как MMG>PDIM A>PGL>TAG, и средняя кратность увеличения уровней CD86 над уровнями, ассоциированными с необработанным iDС, составляла 1,91±0,29, 1,82±0,43, 1,52±0,26 и 1,32±0,14, соответственно. Индукция цитокина следовала той же общей тенденции (Фиг.3В), причем MMG выделялся как наиболее сильный иммуностимулирующий липид. IL-6 высвобождался DC, подверженными воздействию MMG, с значительно более высокими уровнями (Р<0,05), чем индуцированный PGL или TAG. Не было отмечено значительных различий между индукциями цитокинов другими липидами. MMG можно, таким образом, классифицировать как наиболее сильный иммуностимулирующий липид в неполярной липидной фракции М. bovis BCG.
ПРИМЕР 4
ИММУНОСТИМУЛИРУЮЩАЯ АКТИВНОСТЬ АЛЬФА- И KETO-MMG
В данном примере авторы изобретения хотели дополнительно изучить стимулирующие свойства MMG и идентифицировать активный компонент, ответственный за его сильную иммуностимулирующую способность. Альфа-MMG и кето-MMG разделяли после получения триметилсилиловых эфиров MMG, препаративной TLC и последующего гидролиза защитных групп триметилсилильного эфира с получением альфа-MMG и кето-MMG. Структуры альфа- и кето-миколятов представлены на Фиг.2В. При оценке их способности активировать человеческие iDС, альфа- и кето-MMG стимулировали в порядке 2-3-кратного увеличения в уровнях активационных маркеров (Фиг.4). Таким образом, два субкомпонента MMG также проявляют выраженную способность стимулировать человеческие DC.
ПРИМЕР 5
ИНДУЦИРОВАНИЕ Th1 ИММУННОГО ОТВЕТА ПОСРЕДСТВОМ MMG, ВЫДЕЛЕННОГО ИЗ М.BOVIS BCG
Для изучения адъювантной активности MMG, выделенные липиды испытывали на их способность индуцировать продуцирование IFN-γ у мышей. Мышам C57BL/6 вводили 10 мкг общих или отдельных липидов. In vivo, DDA служит в качестве средства для доставки липидов. Таким образом, 2 мкг слитого белка Ag85B-ESAT-6 и 10 мкг регидратированного липидного экстракта, эмульгированного в 250 мкг DDA, вводили подкожным путем. Доза 10 мкг MMG, введенных в DDA липосомы, приводила к уровням IFN-γ 10 нг/мл при повторной стимуляции РВМС, выделенных из дренирующих лимфатических узлов; уровень, который сравним с зарегистрированным для общих липидов в DDA при эквивалентной дозе (Фиг.5). PDIM-A также индуцировали продуцирование IFN-γ, хотя и на более низком уровне, в то время как DDA липосомы с включенными TAG или PGL стимулировали очень небольшое высвобождение IFN-γ (Фиг.5, данные не представлены). Следует заметить, что данные отдельные липиды проявляют активность в качестве адъювантов, так как не наблюдался повторный ответ при повторной стимуляции с помощью или экстракта общих липидов, или отдельных липидов (данные не представлены). Таким образом, MMG также был идентифицирован как наиболее активный неполярный липид in vivo, и ему одному можно приписать большую часть адъювантной активности общих липидов из BCG.
ПРИМЕР 6
ПРОТЕКТИВНАЯ ЭФФЕКТИВНОСТЬ АДЪЮВАНТОВ НА ОСНОВЕ MMG
Чтобы оценить способность адъювантов на основе MMG обеспечивать защиту против ТВ инфекции, мышей C57BL/6 иммунизировали Ag85B-ESAT-6, доставленным в MMG (две разные дозы) и DDA. Группы мышей, получающие вакцинацию BCG и один адъювант, были включены в качестве положительного и отрицательного контролей, соответственно. Через шесть недель после последней вакцинации, мышам вводили живой М.tuberculosis аэрозольным путем. Способность вакцины уменьшать бактериальную нагрузку определяли в легких и селезенке через шесть недель. Эти данные показали значительные уровни защиты с MMG/DDA в качестве адъюванта и протективные уровни, сравнимые с BCG (Фиг.6). Как ожидалось, данный эффект был специфическим, так как у мышей, вакцинированных адъювантом без антигена, не подавлялся бактериальный рост.
ПРИМЕР 7
УЛУЧШЕННЫЙ ЭФФЕКТ ПРИ КОМБИНИРОВАНИИ MMG И PDA
Чтобы оценить эффект комбинирования иммуномодуляторов (MMG) и системы доставки (DDA), мышей C57BL/6 вакцинировали одним DDA или комбинацией DDA/MMG (эксперимент 1, Фиг.7А), или в эксперименте 2 (Фиг.7В) одним MMG или комбинацией DDA/MMG. Из этих экспериментов видно, что иммунный ответ значительно усиливается при комбинировании DDA и MMG.
ПРИМЕР 8
УСИЛЕНИЕ ИММУННОГО ОТВЕТА ПУТЕМ ВВЕДЕНИЯ ТDВ/ТРЕТЬЕГО КОМПОНЕНТА В MMG И PDA ЛИПОСОМЫ
С целью изучения влияния на адъювантную активность MMG при комбинировании с другими иммуностимулирующими компонентами, оценивали иммунный ответ у мышей C57BL/6 после подкожной иммунизации при помощи Ag85B-ESAT-6 и 10 мкг MMG, включенного в DDA липосомы или липосомы DDA с включенными иммуномодуляторами TDB {Davidsen et al, PCT/DK2005/000467} через 5 месяцев после первой вакцинации. Хотя комбинация MMG, включенного в липосомы DDA, приводила к уровням IFN-γ ~25 нг/мл при повторной стимуляции РВМС, выделенных из крови, высвобождение IFN-γ значительно возрастало, если использовали DDA липосомы с включенным TDB (Фиг.8). Таким образом, наблюдался синергический эффект между MMG, DDA и TDB, указывающий на то, что добавление третьего компонента к комбинации MMG и DDA может дополнительно усиливать адъювантную активность.
ПРИМЕР 9
АДЪЮВАНТНАЯ АКТИВНОСТЬ АНАЛОГОВ MMG СРАВНИМА С АДЪЮВАНТНОЙ АКТИВНОСТЬЮ ПРИРОДНОГО MMG
Чтобы оценить иммунологический эффект синтетических аналогов MMG, мышей C57BL/6 иммунизировали с помощью Ag85B-ESAT-6 в DDA с природным MMG, синтетическим аналогом MMG с 16 атомами углерода (как показано на Фиг.9) и синтетическим аналогом MMG с 36 атомами углерода (как показано на Фиг.9) (все 10 мкг/DDA/MMG). Иммунный ответ измеряли в крови через одну неделю после последней вакцинации, и были показаны сравнимые уровни ответов с тремя адъювантами на основе MMG, в то время как DDA сам по себе опять показал более слабый эффект.
ПРИМЕР 10
БОЛЕЕ ВЫСОКИЕ ИММУННЫЕ ОТВЕТЫ ПРИ БОЛЕЕ КОРОТКОЙ ДЛИНЕ ЦЕПИ
Чтобы оценить иммунологический эффект синтетических аналогов MMG с более короткой длиной цепи, мышей C57BL/6 иммунизировали с помощью Ag85B-ESAT-6 в одном DDA, DDA с природным MMG, DDA с разными синтетическими аналогами с диапазоне от С8 до С36 (все 1 мкг/доза). Иммунный ответ измеряли в крови через три недели после последней вакцинации, и было продемонстрировано, что синтетические аналоги MMG активизируют даже при уровнях доз 1 мкг. Кроме того, эти результаты также показывают, что синтетические аналоги MMG с более короткими (от 16 С или менее) цепями являются более эффективными в сравнении с природной MMG. *** Р<0,001 (Фиг.11).
Источники информации




| название | год | авторы | номер документа |
|---|---|---|---|
| РАСШИРЕНИЕ СПЕКТРА Т КЛЕТОК ДЛЯ ВКЛЮЧЕНИЯ СУБДОМИНАНТНЫХ ЭПИТОПОВ ПУТЕМ ВАКЦИНАЦИИ С АНТИГЕНАМИ, ДОСТАВЛЕННЫМИ КАК БЕЛКОВЫЕ ФРАГМЕНТЫ ИЛИ ПЕПТИДНЫЕ КОКТЕЙЛИ | 2007 |
|
RU2490024C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
| НОВЫЕ АДЪЮВАНТНЫЕ КОМПОЗИЦИИ | 2009 |
|
RU2510280C2 |
| СТИМУЛЯЦИЯ ИММУННОГО ОТВЕТА ЭНАНТИОМЕРАМИ КАТИОННЫХ ЛИПИДОВ | 2009 |
|
RU2530555C2 |
| Мультиэпитопный полипептид для иммунизации против Mycobacterium tuberculosis | 2023 |
|
RU2824195C1 |
| КОМПОЗИЦИИ ЛИПИДНЫХ ВЕЗИКУЛ И СПОСОБЫ ПРИМЕНЕНИЯ | 2011 |
|
RU2595872C2 |
| СОСТАВ, СОДЕРЖАЩИЙ АГОНИСТ TLR, И СПОСОБЫ ПРИМЕНЕНИЯ | 2017 |
|
RU2761870C2 |
| ИММУНОЛОГИЧЕСКИЙ АДЪЮВАНТ НА ОСНОВЕ НАНОЧАСТИЦ ДЛЯ ВАКЦИН ПРОТИВ ВЫСОКОПАТОГЕННЫХ ШТАММОВ ВИРУСА ГРИППА | 2013 |
|
RU2529959C1 |
| КОМПЛЕКС, СОДЕРЖАЩИЙ ОЛИГОНУКЛЕОТИД, ОБЛАДАЮЩИЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2723943C2 |
| ВАКЦИННЫЕ КОМПОЗИЦИИ С КАТИОННЫМИ ЛИПИДАМИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2013 |
|
RU2649365C2 |
Группа изобретений относится к медицине и может быть использована для изготовления адъюванта или иммуностимулятора. Для этого применяют синтетический мономиколилглицерин (MMG) на основе алкильных цепей с 8-36 атомами углерода. Наиболее предпочтительно с алкильной цепью из 8-16 атомов углерода. Также предложены адъювант для усиления иммунного ответа, иммуномодулятор для стимуляции дендритных клеток, синтетический MMG, вакцина и система доставки. Группа изобретений обеспечивает стимулирование и активирование человеческих дендритных клеток (DC) в низких дозах, что является эффективным в лечении рака, аутоиммунных расстройств, нервных расстройств, например болезни Альцгеймера, воспаления дыхательных путей, инфекционных заболеваний, кожных расстройств, аллергии или астмы. 6 н. и 13 з.п. ф-лы, 12 ил., 1 табл., 10 пр.
1. Применение синтетического мономиколилглицерина (MMG) на основе алкильных цепей с 8-36 атомами углерода, наиболее предпочтительно с алкильной цепью из 8-16 атомов углерода для изготовления адъюванта или иммуномодулятора.
2. Применение по п.1 синтетического MMG на основе алкильных цепей с 8-36 атомами углерода, имеющего формулу
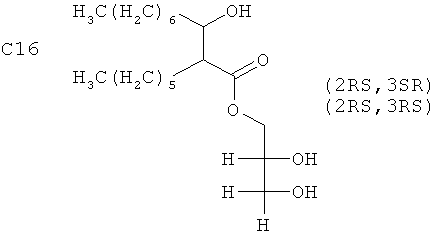
или
 .
.
3. Адъювант для усиления иммунного ответа, включающий синтетический мономиколилглицерин (MMG) на основе алкильных цепей с 8-36 атомами углерода, наиболее предпочтительно с алкильной цепью из 8-16 атомов углерода.
4. Адъювант по п.3, дополнительно содержащий поверхностно-активное вещество.
5. Адъювант по п.4, где поверхностно-активное вещество представляет собой DDA-B (диметилдиоктадециламмония бромид), DDA-C (диметилдиоктадециламмония хлорид), DDA-X (диметилдиоктадециламмония сульфатная, фосфатная или ацетатная соль), DODA-B (диметилдиоктадециниламмония бромид), DODA-C (диметилдиоктадециниламмония хлорид), DODA-X (диметилдиоктадециниламмония сульфатная, фосфатная или ацетатная соль), DO-ТАР (1,2-диолеил-3-триметиламмоний-пропан), DODAP (диолеил-3-диметиламмоний-пропан), DOTMA (N-[l-(2,3-диолеилокси)пропил]-N,N,N-триметиламмоний), DXPC, DXPE, DXPG или их комбинации.
6. Адъювант по любому из пп.3-5, содержащий дополнительный иммуномодулятор, где иммуномодулятор представляет собой не-TLR(Toll-подобный рецептор)-лиганды, например TDB (дибегенат альфа,альфа'-трегалозы) и MDP (мурамилдипептид), или TLR-лиганды, например Poly 1:С.
7. Иммуномодулятор для стимуляции дендритных клеток, включающий синтетический мономиколилглицерин (MMG) на основе алкильных цепей с 8-36 атомами углерода, наиболее предпочтительно с алкильной цепью из 8-16 атомов углерода.
8. Иммуномодулятор по п.7, дополнительно содержащий поверхностно-активное вещество.
9. Иммуномодулятор по п.8, где поверхностно-активное вещество представляет собой DDA-B (диметилдиоктадециламмония бромид), DDA-С (диметилдиоктадециламмония хлорид), DDA-X (диметилдиоктадециламмония сульфатная, фосфатная или ацетатная соль), DODA-B (диметилдиоктадециниламмония бромид), DODA-C (диметилдиоктадециниламмония хлорид), DODA-X (диметилдиоктадециниламмония сульфатная, фосфатная или ацетатная соль), DO-ТАР (1,2-диолеил-3-триметиламмоний-пропан), DODAP (диолеил-3-диметиламмоний-пропан), DOTMA (N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмоний), DXPC, DXPE, DXPG или их комбинации.
10. Иммуномодулятор по любому из пп.7-9, содержащий дополнительный иммуномодулятор, где иммуномодулятор представляет собой He-TLR(Toll-подобный рецептор)-лиганды, например TDB (дибегенат альфа,альфа'-трегалозы) и MDP (мурамилдипептид), или TLR-лиганды, например Poly I:C.
11. Синтетический MMG на основе алкильных цепей с 8-36 атомами углерода, имеющий формулу
или
.
12. Вакцина, содержащая адъювант или иммуномодулятор, содержащий синтетический мономиколилглицерин на основе алкильных цепей с 8-36 атомами углерода, имеющий формулу
или
,
и антигенный компонент.
13. Вакцина по п.12, где антигенный компонент содержит антигенный эпитоп внутриклеточного патогена.
14. Вакцина по п.13, где антигенный компонент содержит антигенный эпитоп вирулентной микобактерии, например Mycobacterium tuberculosis, M.bovis или M.africanum.
15. Вакцина по п.14, где антигенный компонент выбран из слитых продуктов Ag85b_TB10.4, Ag85b_ESAT-6_Rv2660, Ag85b_TB10.4_Rv2660 и Ag85a_TB10.4_Rv2660.
16. Вакцина по п.13, где антигенный компонент содержит антигенный эпитоп из Plasmodium falciparum, Chlamydia trachomatis, ВИЧ (вирус иммунодефицита человека), гриппа или гепатитов В или С.
17. Вакцина по п.13 для лечения рака, аутоиммунного расстройства, нервных расстройств, например болезни Альцгеймера, воспаления дыхательных путей, воспалительных расстройств, инфекционного заболевания, кожных расстройств, аллергии или астмы.
18. Система доставки, содержащая адъювант или иммуномодулятор, содержащий синтетический мономиколилглицерин на основе алкильных цепей с 8-36 атомами углерода, имеющий формулу
или
,
для лечения рака, аутоиммунного расстройства, нервных расстройств, например болезни Альцгеймера, воспаления дыхательных путей, воспалительных расстройств, инфекционного заболевания, кожных расстройств, аллергии или астмы.
19. Система доставки по п.18, где MMG вводят в комбинации с одной или более вакцинами, антигенами, антителами, цитотоксическими агентами, аллергенами, антибиотиками, антисмысловыми олигонуклеотидами, TLR- и не-TLR-агонистами, TLR- и He-TLR-антагонистами, пептидами, белками, генотерапевтическими векторами, ДНК-вакцинами, иммуномодуляторами или костимуляторными молекулами.
| JP 59089632 А, 23.05.1984 | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| SILVA C.L | |||
| Inflammation induced by mycolic acid-containing glycolipids of Mycobacterium Bovis (BCG) // Brazilian J | |||
| med | |||
| Biol | |||
| Res., 1998, v.18, pp.327-335. | |||

