Область техники
Группа изобретений относится к области биотехнологии и молекулярной биологии, а именно к вакцинам против инфекционных агентов, в частности, к рекомбинантным вакцинам для иммунизации против Mycobacterium tuberculosis.
Уровень техники
Туберкулез - опасное заболевание, от которого ежегодно умирает 1,5 млн человек. Это делает его основной причиной смерти в мире от инфекционных агентов, несмотря на то, что это предотвратимое и излечимое заболевание [https://www.who.int/publications/i/item/9789240052055]. В мире ежегодно регистрируется около 10 миллионов новых случаев заболеваемости туберкулезом (ТБ). Хотя туберкулез распространен во всем мире, большинство людей, заболевших туберкулезом, живут в странах с низким и средним уровнем дохода.
В настоящий момент действует Стратегия ВОЗ по борьбе с туберкулезом, предусматривающая к 2035 году снижение смертности от туберкулеза на 95% и заболеваемости туберкулезом во всем мире на 90%. Для достижения поставленных целей должна быть разработана новая вакцина, эффективная для всех возрастных групп, особенно для взрослых и подростков. Вакцины также рассматриваются как основная профилактика развития туберкулеза с множественной лекарственной устойчивостью, который становится все более распространен [https://www.who.int/teams/global-tuberculosis-programme/research-innovation/vaccines, дата обращения 17.03.2023].
В 1882 г. Роберт Кох впервые описал возбудителя туберкулеза [Die Aetiologie der Tuberculose von Dr. Robert Koch. Ausw: Berliner Klinische Wochenschrift, Band 19 (1882). Zentralbl Bakteriol Mikrobiol Hyg A Med Mikrobiol Infekt Parasitol. 1982 Mar;251(3):287-96. PMID: 6805152], что послужило основой для создания вакцин, средств диагностики и терапии. В 1890 г. Кох предпринял первую попытку вакцинации против туберкулеза и создал так называемый «туберкулин» [Koch, 1890], который не показал эффективности ни как терапия, ни как вакцина против туберкулеза, но оказался отличным диагностическим инструментом. Это была субъединичная вакцина из белковых антигенов и различных гликолипидов, которую вводили подкожно, и у инфицированных туберкулезом через 2-3 дня появлялась характерная реакция замедленной гиперчувствительности [Duthie, Reed, 2021]. Для современной диагностики туберкулеза используется усовершенствованный вариант туберкулина - кожная туберкулиновая проба на основе очищенного белкового производного.
Было предпринято много попыток создать вакцину против туберкулеза, Кальметт и Герен пришли к выводу, что субъединичные и убитые цельноклеточные вакцины не обеспечивают достаточной защиты. Вакцинный штамм БЦЖ был первоначально разработан Институтом Пастера во Франции в 1931 г. [Calmette, 1931]. Он был разработан в первую очередь для снижения младенческой смертности от туберкулеза и применялся для новорожденных. БЦЖ представляет собой живую аттенуированную вакцину, полученную путем непрерывного культивирования Mycobacterium bovis в обедненных условиях. Во время аттенуации БЦЖ потеряла кластер генов, присутствующий у М. bovis и М. tuberculosis, известный как область различия 1 (RD-1), которая кодирует основные факторы вирулентности, включая ESAT-6 и CFP-10 [Mahairas et al., 1996]
Вакцина Кальметта-Герена (БЦЖ) является единственной зарегистрированной вакциной для профилактики туберкулеза и до сих пор используется для предотвращения тяжелых форм туберкулеза у детей. В настоящее время не существует вакцины, которая была бы эффективна для профилактики туберкулеза у взрослых как до, так и после заражения туберкулезом [WHO Global Tuberculosis Reports 1997-2022]. Недавние данные показывают, что 153 страны проводят вакцинацию БЦЖ для всего населения: 87 из этих стран сообщили об охвате не менее 90% населения [WHO Global Tuberculosis Report 2020, A database of global BCG vaccination policies and practices].
Несмотря на то, что вакцина БЦЖ долгое время была единственной вакциной против туберкулеза, ее эффективность оценивается от 0% до 80% в разных регионах [Mangtani et al., 2014], причем в тропических странах защита значительно ниже, чем в более высоких широтах. Также эффективность БЦЖ ниже у жителей сельской местности [Comstock et al., 1974]. Различия в эффективности вакцины могут быть обусловлены полом привитых [Rhodes et al., 2016], риском заболевания туберкулезом в исследуемой популяции [Mangtani et al., 2017], содержанием микобактерий в окружающей среде [Palmer & Long, 1966] и другими факторами. Другим недостатком БЦЖ является гетерогенность: 22 штамма, используемые для вакцинации [Molivaet al., 2015], имеют существенные отличия [Zhang et al., 1995]. Это приводит к вариабельности иммуногенности [Oettinger et al., 1999]. Другим недостатком является отсутствие у БЦЖ ряда иммунодоминантных антигенов, таких как ESAT6, CFP10, системы ESX-1 типа VII, что обусловлено потерей участка RD1 [Brodin et al., 2006, Mostowy et al., 2003, Mahairas et al., 1996]. Также эффективность БЦЖ снижается через 20 лет после вакцинации [Nguipdop-Djomo et al., 2016, Mangtani et al., 2018], и хотя она защищает от милиарной и менингеальной форм туберкулеза у детей, БЦЖ не обеспечивает защиты от туберкулеза легких у взрослых [Rodrigues et al., 1993, Fifteen year follow up of trial of BCG vaccines, 1999], а ревакцинация БЦЖ, хотя и имеет эффективность 45,4% в профилактике инфекции M.tb [Nemes et al., 2018], не рекомендуется ВОЗ для использования. Также на эффективность БЦЖ влияет недостаточная индукция CD8+Т-клеток [Hess & Kaufmann, 1999], играющих ключевую роль в защите от туберкулеза. Другим недостатком является то, что вакцинация БЦЖ не может применяться у людей с ВИЧ, поскольку риск заболевания при вакцинации БЦЖ у ВИЧ-инфицированных повышен в несколько сотен раз [WHO Bacille Calmette-Guerin (BCG) Information sheet, 2020]. Однако до сих пор ни одна новая противотуберкулезная вакцина не прошла клинических испытаний. Таким образом, БЦЖ долгое время остается одним из наиболее экономически эффективных средств профилактики туберкулеза в странах с высокой заболеваемостью туберкулезом [Li et al., 2020].
Недостатки БЦЖ, обусловливающие ее низкую эффективность, привели к необходимости создания новых противотуберкулезных вакцин. Например, существует подход, основанный на рекомбинантных штаммах БЦЖ, экспрессирующих антигены М. tuberculosis: Ag85B, антигены из локуса RD1, слитый белок Ag85A-ESAT-6 и многие другие [Horwitz et al., 2000, Рут et al., 2003, Deng et al., 2014, Nieuwenhuizen & Kaufmann, 2018]. Также создан штамм rBCGAUre:CHly, экспрессирующий листериолизин (Hly) из Listeria monocytogenes, который позволяет повысить эффективность доставки микобактериального антигена к МНС I и приводит к усилению CD8+Т-клеточных ответов [Stetson & Medzhitov, 2006]. Существуют подходы, не связанные с БЦЖ: вирусные векторы, аттенуированные штаммы М. tuberculosis, детоксицированные, фрагментированные клетки М. tuberculosis, субъединичные вакцины и вакцины на основе нуклеиновых кислот. Многие новые вакцины уже прошли клинические испытания [Usman et al., 2017].
Терапевтические вакцины, используемые после заражения возбудителем, можно разделить на три группы: инактивированные противотуберкулезные вакцины, субъединичные вакцины и вакцины на основе нуклеиновых кислот. Терапевтические вакцины не являются «классическими» вакцинами, потому что эти вакцины должны модулировать, а не усиливать иммунный ответ.Их модулирующий эффект можно объяснить восстановлением баланса Th1/Th2 иммунного ответа. Терапевтические вакцины должны индуцировать иммунный ответ, направленный на определенные антигены возбудителя или широкий иммунный ответ для предотвращения риска избегания иммунного ответа [Doherty, 2012, Mi et al., 2021, Li & Zhu, 2006]. Большинство противотуберкулезных терапевтических вакцин применяются в сочетании с химиотерапией.
Некоторые терапевтические вакцины (М. vaccae, вакцина MIP) находятся на III фазе клинических испытаний, и обе они являются инактивированными противотуберкулезными вакцинами. Также к инактивированным вакцинам относятся ДАР-901 и РУТИ, они находятся на II фазе клинических исследований.
Четыре вакцины на основе рекомбинантных слитых белков (Mtb72f/AS01, H56/IC31, ID93/GLA-SE и АЕС/ВС02) прошли клинические испытания фазы I или фазы II, все они вызывают сильный Т-клеточный ответ [Mi et al., 2021].
Терапевтическая ДНК-вакцина против туберкулеза содержала последовательности ДНК, кодирующие антигены М. tuberculosis. В настоящее время GX-70, сконструированная из четырех антигенных плазмид М. tuberculosis, является единственной ДНК-вакциной, прошедшей клинические испытания при туберкулезе, но вакцинация GX-70 была связана с высоким риском неэффективности лечения или рецидива (NCT03159975). Химерная ДНК-вакцина Ag85a/b демонстрирует в доклинических исследованиях образование умеренных уровней антител, усиление клеточного иммунного ответа Th1 и уменьшение поражений легочной ткани [Liang et al., 2017].
Лечение туберкулеза осложняется лекарственно-устойчивыми штаммами. Существует туберкулез с множественной лекарственной устойчивостью (МЛУ-ТБ), устойчивый к двум наиболее сильнодействующим противотуберкулезным препаратам: изониазиду и рифампицину, туберкулез с широкой лекарственной устойчивостью (ШЛУ-ТБ), устойчивый к фторхинолонам и инъекционным препаратам последней линии, (амикацин, канамицин и капреомицин) и туберкулез с полной лекарственной устойчивостью (TDR-TB) [Seung et al., 2015, Maitre et al., 2017, WHO Fact Sheets. Tuberculosis, 2009, Udwadia, 2012].
Многие новые противотуберкулезные терапевтические вакцины все еще находятся на доклинической стадии, и большая их часть представляет собой ДНК-вакцины. ДНК-вакцины предпочтительнее вирусных векторов или субъединичных вакцин из-за нескольких особенностей: низкая стоимость производства, стабильность при комнатной температуре, безопасность, индукция Th1-ответа, возможность использования в качестве первичной и бустерной вакцины [Bruffaerts et al., 2014].
С момента первой ДНК-иммунизации против М. tuberculosis в 1996 г. протестировано более 60 антигенов в составе противотуберкулезных вакцин [Huygen et al., 1996, Sefidi-Heris etal., 2020], некоторые из них были предсказаны методами in silico [Moradi et al., 2017, Wang et al., 2010]. Наиболее используемыми и изученными являются конструкции на основе Ag85A, Ag85B, Hsp65, CFP10 и ESAT6 и химерных белков на их основе [Sefidi-Heris et al., 2020, Yuan et al., 2012, Yu et al., 2008]. Также для ДНК-вакцинации используют последовательности фосфат-связывающих белков [Romano et al., 2004], полипротеин слияния Mtb72F [Skeiky et al., 2004], антигены из регулона покоя DosR [Roupie et al., 2007], представители семейства PPE [Romano et al., 2008, Skeiky et al., 2000], ДНК-связывающий белок MDP1 [Suzuki et al., 2010], секретируемый липопротеин MPT83 [Kao et al., 2012] и факторы реактивации [Romano et al., 2012].
Несмотря на то, что РНК-вакцины доказали свою эффективность и безопасность [Fan & Lowrie, 2021], в настоящее время мало исследований посвящено противотуберкулезным РНК-вакцинам. Первые данные о вакцинации РНК указывали на то, что защитный эффект против М. tuberculosis был на уровне БЦЖ [Xue et al., 2004], но позже сообщалось, что она также стимулирует продукцию IFNγ и фактора некроза опухоли альфа, что указывает на Thl-ответ, характерный для ДНК-вакцин [Lorenzi et al., 2010]. Новые эксперименты с использованием РНК, кодирующей четыре белка М. tuberculosis: Rv3619 (esxV; ESAT6-подобный белок), Rv2389 (Rpffl), Rv3478 (PPE60) и Rvl886 (Ag85B), показывают снижение бактериальной нагрузки, а также гуморальный и клеточный иммунный ответ.Подходы in silico используются для разработки новых конструкций противотуберкулезных РНК-вакцин [Larsen et al., 2022; A1 Tbeishat, 2022].
Потенциал РНК-вакцин против М. tuberculosis остается нереализованным, и им уделяется незаслуженно мало внимания, особенно в контексте потребности в новых противотуберкулезных вакцинах.
Первая мРНК-вакцина против туберкулеза была описана в 2004 г в работе Xue с соавторами [Xue et al., 2004]. Они использовали РНК-конструкцию, кодирующую РНК-репликазу вируса Синдбис и ген М. tuberculosis МТР83, и проверили индукцию защитных иммунных ответов на мышиной модели вирулентной инфекции М. tuberculosis. В этом исследовании матричную РНК (мРНК) транскрибировали in vitro с ДНК, кодирующей антиген М. tuberculosis, очищали, и чистую мРНК четыре раза вводили мышам внутрикожно с 3-недельными интервалами. Через четыре недели после этого заражали мышей вирулентным М. tuberculosis, мыши были значительно защищены. Защита была меньше, чем у БЦЖ. Работа не получила дальнейшего развития - возможно, ввиду того, что разработка РНК-вакцин на тот момент считалась слишком сложной и дорогой [Fan & Lowrie, 2021].
В последнее время успешное создание нескольких мРНК-вакцин против SARS-Cov2 привело к заметному интересу к созданию аналогичной вакцины против туберкулеза, с поправкой на особенности развития заболевания и иммунного ответа на него.
Помимо важных для стабильности и эффективной трансляции мРНК регуляторных элементов (3'UTR, 5'UTR и поли(А) хвост), чрезвычайную роль в эффективности мРНК вакцин играет выбор антигена. В настоящее время наиболее перспективной стратегией выбора антигена для вакцин является дизайн мультиэпитопного антигена, который состоит из множества отдельных эпитопов, соединенных линкерами [Zhang, 2018]. Некоторые участки полноразмерных белков могут вызывать нежелательные иммунные последствия или просто не обладать нужной активностью, снижая таким образом эффективность вакцины. Мультиэпитопная вакцина позволяет выбрать наиболее иммуногенные и безопасные эпитопы, т.е. не вызывающие аутоиммунный, аллергический ответ и не являющиеся токсичными [Mahapatra, et al., 2021; Dey et al., 2022].
Мультиэпитопная вакцина может включать эпитопы, которые способны вызывать ответ цитотоксических Т-лимфоцитов, хелперных Т-лимфоцитов, ответ В-клеток против бактериальных или вирусных антигенов. Базовые принципы подбора антигенов заключаются в выборе наиболее распространенного штамма возбудителя и поиске кандидатных белков, из эпитопов которых формируется антиген. Как правило, в качестве таких белков рассматриваются поверхностные белки возбудителя, которые в первую очередь контактируют с компонентами иммунной системы. Характерным примером является разработка 2-х широко используемых мРНК-вакцин против SARS-CoV-2, антиген которых включает последовательности поверхностного S-белка (spike protein). Однако следует обратить внимание на то, что существуют ограничения, связанные с отсутствием аннотированного протеома для некоторых бактериальных и новых вирусных патогенов.
Интенсивное развитие биоинформатических методов и искусственного интеллекта, включающих в себя машинное обучение и нейросети, позволило создать большое количество инструментов для анализа и предсказания эффективности презентации антигенов [Parvizpour et al., 2020; Gong et al., 2022]. Можно выделить 5 базовых этапов дизайна мультиэпитопных вакцин. Во-первых, это поиск иммуногенных эпитопов среди известных белковых последовательностей инфекционного агента. Для этих целей применяют программное обеспечение Immune Epitope Database Analysis Resource (IEDB-AR), которое использует экспериментальные данные по идентификации и характеризации эпитопов и специфичных для этих эпитопов рецепторов клеток иммунной системы [Vita et al., 2015]. Во-вторых, необходимо охарактеризовать (спрогнозировать) антигенность, аллергенность и токсичность этих эпитопов, для этого часто используют базы данных Vaxijen, AllerTOP and Toxinpred [Doytchinova & Flower, 2007; Dimitrov et al., 2014; Gupta et al., 2013]. Оценку антигенности, аллергенности и токсичности проводят посредством сопоставления выбранных эпитопов с известными антигенными, аллергенными и токсичными эпитопами, после автоматического кросс-ковариантного преобразования белковых последовательностей в однородные векторы равной длины. Эти преобразования позволяют использовать подход без прямого сравнения последовательностей эпитопов, основанный на основных физико-химических свойствах белковых последовательностей, таких как гидрофобность аминокислот, размер молекулы, склонность к образованию спирали, относительное содержание аминокислот и склонность к образованию (3-цепей. Третий этап дизайна включает в себя анализ третичной структуры белка, дисульфидных связей, а также молекулярный докинг с рецепторами-мишенями лимфоцитов и молекулярно-динамическую стимуляцию. Два заключительных этапа анализа включают в себя оценку встречаемости конкретных аллелей главного комплекса гистосовместимости (МНС) 1 и 2 классов в популяции, показавших наибольшее сродство к выбранному эпитопу, и оптимизацию кодонов для достижения оптимального уровня трансляции. Однако несмотря на то, что инструменты in silico позволяют с высокой долей вероятности предсказать эффективность будущих вакцин, исследования in vivo все еще необходимы для подтверждения их эффективности и безопасности.
Помимо последовательности самих антигенных эпитопов важную роль в эффективности вакцины играет последовательность линкеров, которыми будут соединены эпитопы, а также наличие адъювантных последовательностей и/или последовательностей сигналов локализации. Линкеры AAY, GPGPG и КК широко используются для соединения Т- и В-клеточных эпитопов. Некоторые исследователи используют другие линкерные последовательности, такие как HEYGAEALERAG, GGGS или RVRR [Solanki et al., 2019; Tarrahimofrad et al., 2021].
Ранняя секретируемая антигенная мишень 6 кДа (ESAT-6) М. tuberculosis является одним из важных антигенов для создания вакцины против туберкулеза [Valizadeh et al., 2022]. Этот белок был впервые идентифицирован как Т-клеточный антиген, в настоящее время определяется как порообразующий токсин, необходимый для вирулентности М. tuberculosis. ESAT-6 секретируется через систему секреции ESX-1 (тип VII) М. tuberculosis. Он опосредует транслокацию микобактерий внутри зараженных макрофагов, разрушая мембраны фагосом. Деактивация ESAT-6 приводит к снижению вирулентности М. tuberculosis. Известно, что ESAT-6 сам по себе или в комплексе со своим шапероном CFP-10 модулирует иммунный ответ хозяина. ESAT-6/ESAT-6: CFP-10 может проникать в эндоплазматический ретикулум, где он инициирует диссоциацию бета-2-микроглобулина (р2М) и комплексов МНС I класса, снижая экспрессию комплексов МНС-1-р2М на клеточной поверхности и подавляя презентацию антигенов на МНС I класса [Refai et al., 2018]. ESAT-6 экспрессируется геном (RV3875).
Известен слитый белок М. tuberculosis, содержащий иммуногенный ESAT-6 М. tuberculosis и С-концевой участок HSP70M. tuberculosis (HSP70 (359-359- 610)) в качестве подходящего носителя и адъюванта [Tebianian et al., 2011]. Сконструированный ген встраивали в прокариотический вектор экспрессии (pQE30), экспрессировали в Escherichia coli Ml5 и очищали. Очищенный и рефолдированный белок затем использовали для иммунизации мышей, в результате которой выявляли высокие титры специфических антител, высокий уровень IFN-γ и пролиферации клеток.
Известна субъединичная вакцина на основе рекомбинантного целого ESAT6 [Brandt et al., 2000]. Полезность ESAT6 в дизайне вакцины была элегантно продемонстрирована Брандтом и его коллегами, когда после иммунизации их мультисубъединичной (цельный рекомбинантный белок) вакциной был зарегистрирован устойчивый ESAT6-специфический Т-клеточный ответ.
Baldwin и соавторы описали вакцину под названием ID93 и охарактеризовали ее эффективность на мышиной модели [Baldwin et al., 2021]. ID93 представляет собой рекомбинантную субъединичную вакцину, которая объединяет четыре антигена, принадлежащих к семействам белков М. tuberculosis, связанных с вирулентностью (Rv2608, Rv3619, Rv3620) или латентностью (Rvl813). Рекомбинантный слитый полипептид ID93 в сочетании с агонистом TLR-4 адъювантом GLA-SE, успешно завершил клиническое испытание фазы 2а [Penn-Nicholson et al., 2018]. Разработчики заявили о намерении разработать мРНК-вакцину, кодирующую полипептид ID93 [https://www.validate-network.org/development-of-an-rna-based-vaccine-against-mycobacterium-tuberculosis, дата обращения 20.03.2023].
Использование рекомбинантных белков или отдельных фрагментов пептидов в качестве вакцинных препаратов имеет ряд ограничений, связанных с высокой стоимостью наработки и очистки от примесей фрагментов бактериальных клеток. Отдельной проблемой является фолдинг белковой молекулы и наличие или отсутствие пост-трансляционных модификаций. Использование рефолдинга может привести к отличиям природного и рекомбинантного вариантов белков. Соответственно, специфический иммунитет на вакцинный препарат может быть неэффективным при встрече с патогеном.
Известна ДНК-вакцина, содержащая три Т-клеточных эпитопа ESAT-6: Esat-64-18 (QQWNFAGIEAAASAI), Esat-622-36 (VTSIHS LLDEGKQS L) и Esat-656-70 (QKWDATATELNNALQ) и участок, кодирующий FLT3L человека [Feng et al., 2013]. Для защиты эпитопов от слияния и повышения эффективности разрезания внутриклеточной протеазой использовали линкер Ala-Ala-Tyr (AAY). Описанная рекомбинантная ДНК-вакцина была покрыта наночастицами хитозана. Иммунологическую и защитную эффективность ДНК-вакцины на основе нанохитозана оценивали на мышах C57BL/6 после внутримышечной первичной вакцинации плазмидной ДНК и назальной ревакцинации пептидом Esat-6/3е (пептид, содержащий упомянутые выше три Т-клеточных эпитопа ESAT-6). Иммунизированные мыши демонстрировали усиленный Т-клеточный ответ и защиту от Mtb H37Rv. Использование ДНК вакцин имеет ряд преимуществ по сравнению с вакцинами на основе рекомбинантных белков. ДНК молекула достаточно стабильна и, поэтому экспрессия целевого антигена может сохраняться продолжительное время. Однако один из главных недостатков ДНК вакцин низкая эффективность. Для наработки целевого антигена молекуле ДНК необходимо преодолеть не только цитоплазматическую мембрану, но и ядерную.
В статье Ларсен с соавторами группа исследователей из Сиэтла в конце 2022 года представила новую платформу репликативной РНК (repRNA) в качестве гомологичной противотуберкулезной вакцины [Larsen et al., 2023]. Авторы предполагают, что репликативная способность repRNA, приводящая к более длительной экспозиции антигена, обеспечивает большее сходство с эффектом БЦЖ, чем стратегии вакцин с белковым антигеном.
Раскрыта мРНК-вакцина против М. tuberculosis и клинического изолята NTM, М. avium 2-151 smt: доклиническая вакцина-кандидат, кодирующая антиген ID91, составленный слиянием четырех эпитопов: Rv3619 (esxV; Е8АТ6-подобный белок), Rv2389 (RpfD), Rv3478 (РРЕ60) и Rvl886 (Ag85B) [Rais et al., 2023].
Самоамплифицирующиеся мРНК вакцины несомненно являются перспективным направлением в разработке мРНК вакцин, поскольку они позволяют уменьшить количество вводимого препарата в несколько раз. Тем не менее, нерешенным остается вопрос биобезопасности использования таких вакцинных препаратов, связанный как с избыточной наработкой целевой РНК, так и неспецифической активностью репликазы.
Технической проблемой, на решение которой направлено настоящее изобретение, является решение как минимум одной из вышеуказанных в уровне техники проблем.
Сущность изобретения
Одной из технических проблем, на решение которой направлено настоящее изобретение, является расширение арсенала технических средств для иммунизации против возбудителя туберкулеза Mycobacterium tuberculosis. В частности, одной из технических проблем, на решение которой направлено настоящее изобретение, являлось создание средства для иммунизации против Mycobacterium tuberculosis, способного стимулировать Т- клеточный иммунитет.
В настоящей заявке предлагается полипептид для иммунизации против Mycobacterium tuberculosis, содержащий соединенные между собой посредством линкеров аминокислотные последовательности, соответствующие аминокислотам 6-20, 33-47, 65-79, 28-36, 41-51 белка ESAT-6 Mycobacterium tuberculosis, и последовательность, соответствующую аминокислотам 308-362 МНС класса I человека, при этом в качестве линкеров используют расщепляемые линкеры.
В одном из вариантов осуществления изобретения предлагается полипептид, содержащий аминокислотную последовательность Seq Id No:2.
В другом варианте осуществления изобретения предлагается нуклеиновая кислота, содержащая последовательность нуклеотидов, кодирующую полипептид по настоящему изобретению.
В еще одном варианте осуществления изобретения упомянутая нуклеиновая кислота представляет собой мРНК, в том числе для иммунизации против Mycobacterium tuberculosis, которая в частном случае реализации изобретения содержит последовательность Seq Id No: 1.
В одном из вариантов осуществления изобретения предлагается композиция для иммунизации человека против Mycobacterium tuberculosis, содержащая нуклеиновую кислоту, кодирующую полипептид по настоящему изобретению и смесь липидов. В одном из вариантов осуществления изобретения смесь липидов содержит ионизируемый катионный липид, ПЭГилированный липид, фосфолипид и холестерин. В предпочтительном варианте осуществлений изобретения смесь липидов содержит ионизируемый катионный липид ALC-0315, дистеароилфосфатидилхолин, холестерин и DMG-PEG-2000 в молярном соотношении 46,3: 9,4: 42,7: 1,6.
Группа изобретений иллюстрируется следующими графическими материалами:
На Фиг. 1А схематично представлены использованные в исследовании препараты, на Фиг. 1Б представлен дизайн эксперимента на мышах линии C57BL6/J.
На Фиг. 2 показан результат оценки Т-клеточного ответа спленоцитов иммунизированных мышей методом ELISPOT. По оси ординат количество клеток, секретирующих IFNγ в ответ на стимуляцию DC2.4., активированных белком ESAT-6. PBS - мыши, которым вводили физиологический раствор; LNP - мыши, которым вводили ЛНЧ без мРНК; ESAT-6 + LNP - мыши, которым вводили полноразмерный белок ESAT-6 (М. tuberculosis) с ЛНЧ без мРНК; мРНК- ESAT-6 - мыши, которым вводили ЛНЧ с мРНК, кодирующей полноразмерный белок ESAT-6; Мультиэпитопная мРНК- ESAT-6 мыши, которым вводили ЛНЧ с мРНК с последовательностью Seq Id No: 1, кодирующей мультиэпитопный полипептид по настоящему изобретению. Данные представлены как среднее ± ошибка среднего. По 3 животных в группе. * - р<0,05 по сравнению с группой PBS, # - р<0,05 по сравнению с группой LNP, & - р<0,05 по сравнению с группой ESAT-6+LNP.
На Фиг. 3 представлены репрезентативные данные проточной цитометрии по CD3+Т-лимфоцитам (CD8+и CD8-), продуцирующим IFNγ. PBS мыши, которым вводили физиологический раствор; ESAT-6+LNP мыши, которым вводили полноразмерный белок ESAT-6 (М. tuberculosis) с ЛНЧ без мРНК; mRNA- ESAT-6 - мыши, которым вводили ЛНЧ с мРНК, кодирующей полноразмерный белок ESAT-6; Мультиэпитопная mRNA-ESAT-6 мыши, которым вводили препарат ЛНЧ с мРНК с последовательностью Seq Id No: 1, кодирующей мультиэпитопный полипептид по настоящему изобретению. Число показывает процент клеток, которые продуцируют IFNγ, от общего пула CD3+CD8+и CD3+CD8- Т-лимфоцитов.
Детальное описание изобретения
Если не указано иначе, предполагается, что все термины, обозначения и другие научные термины, используемые в данной заявке, имеют значения, которые обычно понимают специалисты в области, к которой относится настоящее изобретение. В некоторых случаях определения терминов с общепринятыми значениями приведены в данной заявке для ясности и/или для быстрой справки и понимания, и включение таких определений в настоящее описание не должно истолковываться как наличие существенного отличия значения термина от обычно подразумеваемого в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, трансфекции и трансдукции клеток эукариот, методы молекулярной биологии, аналитические методы определения специфических белков в образце, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Реакция ПЦР и методы работы с нуклеиновыми кислотами и белками осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
В настоящем изобретении предлагается полипептид для иммунизации против Mycobacterium tuberculosis, содержащий соединенные между собой посредством линкеров аминокислотные последовательности, соответствующие аминокислотам 6-20, 33-47, 65-79, 28-36, 41-51 белка ESAT-6 Mycobacterium tuberculosis (UniProt ID: P9WNK7) и последовательность, соответствующую аминокислотам 308-362 МНС класса I человека (MHC-I). Пример последовательности аминокислот полноразмерного белка ESAT-6 Mycobacterium tuberculosis представлена в Seq Id No.: 3, последовательность аминокислот MHC-I взята из базы данных Uniprot https://www.uniprot.org/.
Под иммунизацией подразумевается формирование специфического иммунного ответа иммунной системы организма в ответ на введение иммуногена в любой доступной форме (на основе мРНК, ДНК, белка, микроорганизма и пр.). В частности, иммунизация в контексте описания изобретения выражалась в формировании специфического иммунного ответа у лабораторных животных, иммунизированных препаратом мРНК-вакцины по настоящему изобретению, а именно специфического Т-клеточного иммунного ответа.
Под эпитопом подразумевается часть макромолекулы антигена, которая распознается иммунной системой (антителами, В-лимфоцитами, Т-лимфоцитами). Аминокислотные последовательности, соответствующие аминокислотам 6-20, 33-47, 65-79, 28-36, 41- 51 белка ESAT-6 Mycobacterium tuberculosis, представляют собой наиболее вероятные эпитопы для стимуляции клеточного иммунного ответа по результатам проведенного компьютерного моделирования. Поскольку полипептид по настоящему изобретению состоит из нескольких эпитопов, по сути он представляет собой мультиэпитопный полипептид.
Последовательность, соответствующая аминокислотам 308-362 MHC-I человека представляет собой С-концевой участок молекулы MHC-I, содержащий цитоплазматический домен, отвечающий за внутриклеточный транспорт MHC-I из эндоплазматического ретикулума на поверхность клетки [Story et al., 1999]. Включение данного пептида обеспечивает эффективный транспорт синтезированного клеткой полипептида в эндосомы и его ко-локализацию с молекулами главного комплекса гистосовместимости для эффективной презентации эпитопов на клеточной мембране. Мультиэпитопный полипептид за счет транспортного сигнала ко-локализуется в эндосомах с молекулами МНС, где осуществляется его деградация протеазами по линкерам до отдельных пептидов. Эти пептиды образуют комплекс с молекулами МНС, экспрессируемыми на мембране клетки. Передача сигнала через Toll-подобные рецепторы (TLR), запускаемая продуктами трансляции мРНК по настоящему изобретению, обеспечивает высокий уровень экспрессии МНС.
Линкер по заявленному изобретению представляет собой аминокислотную последовательность, разделяющую последовательности отдельных эпитопов в полипептиде по настоящему изобретению. Выбор линкера определяется конкретными задачами, стоящими перед специалистом. В полипептиде по настоящему изобретению используют расщепляемые линкеры, которые вводятся для высвобождения свободных функциональных доменов in vivo. Этот тип линкеров может улучшать биологическую активность или обеспечивать независимое действие отдельных доменов рекомбинантных слитых белков после расщепления линкера [Chen et al., 2013; Wang et al, 2004]. После трансляции мультиэпитопный белок подвергается протеолизу по сайтам узнавания протеазами в области расщепляемых линкеров. Это приводит к тому, что отдельные эпитопы высвобождаются в цитоплазме и становятся доступными для презентации главным комплексом гистосовместимости. Примеры конкретных линкеров широко описаны в литературе, доступны и известны специалистам, выбор конкретного линкера не влияет на реализацию назначения или достижение технического результата настоящего изобретения. Эмпирические и прогнозируемые расщепляемые линкеры известны специалистам, например, из обзорной работы Chen с соавторами [Chen et al., 2013]. Линкеры AAY и КК обеспечивают эффективную деградацию синтезированного клеткой полипептида до отдельных эпитопов [Wang et al, 2004; Lei et al., 2019].
Эффективность иммунизации можно определить с использованием стандартных тестов, известных из уровня техники, например, методами ELISPOT и проточной цитометрии, которые описаны в примерах и известны в данной области техники [см., например, Ranieri et al., 2014, Flaxman & Ewer, 2018].
В одном из вариантов осуществления изобретения предлагается полипептид, содержащий аминокислотную последовательность Seq Id No:2.
Мультиэпитопный полипептид по настоящему изобретению может быть синтезирован in vitro с использованием способов, известных в данной области техники, например, с помощью прямого химического синтеза или синтеза в прокариотических или эукариотических клетках-продуцентах, трансформированных экспрессионными векторами, содержащими последовательность нуклеотидов, кодирующую мультиэпитопный полипептид по настоящему изобретению. Синтезированный мультиэпитопный полипептид может быть доставлен в организм известными из уровня техники средствами доставки лекарственных средств пептидной природы, например, описанных в обзорах Madge et al., 2021 или Torchilin, 2008.
Мультиэпитопный полипептид по настоящему изобретению может быть закодирован в последовательности нуклеиновой кислоты. В одном из вариантов осуществления настоящего изобретения предлагается нуклеиновая кислота, содержащая последовательность нуклеотидов, кодирующую мультиэпитопный полипептид по настоящему изобретению. Вакцины на основе нуклеиновых кислот известны из уровня техники и могут быть использованы для введения в организм генетического материала, экспрессирующего мультиэпитопный полипептид по настоящему изобретению. Нуклеиновые кислоты могут быть введены в клетку в составе экспрессионных векторов, известных из уровня техники. Выбор вектора определяется задачами, для которых вектор будет в дальнейшем использован и не ограничивает объем настоящего изобретения. Способы получения векторов известны из уровня техники. Векторы, содержащие нуклеиновые кислоты с последовательностью нуклеотидов, кодирующей мультиэпитопный полипептид по настоящему изобретению, могут быть получены специалистами в области генетической инженерии.
Например, мультиэпитопный полипептид по настоящему изобретению может быть закодирован в последовательности ДНК либо в последовательности РНК. При этом кодирующие его последовательности нуклеотидов могут быть созданы с использованием оптимизации кодонов, в зависимости от задач, для решения которых они создаются, и организмов, в которых предполагается экспрессия соответствующих последовательностей.
В одном из вариантов осуществления изобретения мультиэпитопный полипептид закодирован в последовательность мРНК. мРНК по настоящему изобретению может быть получена in vitro любым известным из уровня техники способом.
В частном случае реализации изобретения упомянутая мРНК содержит последовательность Seq Id No: 1, сконструированную с оптимизацией кодонов для экспрессии в эукариотических клетках.
В одном из вариантов осуществления изобретения предлагается композиция, содержащая мРНК и смесь липидов. Смеси липидов для доставки мРНК-вакцин в клетки организма человека широко известны и подробно описаны, выбор конкретных липидов зависит от целей и задач исследователей и не ограничивают объем прав настоящего изобретения. Как правило, смесь липидов, образуя липидные наночастицы (ЛНЧ) с мРНК позволяют улучшить эффективность внутриклеточной доставки мРНК, а также улучшить ее стабильность и уровень трансляции. Доставка мРНК при помощи липидных наночастиц обеспечивает попадание мРНК в цитоплазму клетки для последующей трансляции in vivo, приводящей к образованию внутри клетки мультиэпитопного полипептида по настоящему изобретению.
Внутримышечное введение мРНК, кодирующей мультиэпитопный полипептид с последовательностью Seq Id No:2, приводит к формированию у мышей устойчивого Т-клеточного иммунного ответа. Интенсивность Т-клеточного ответа иммунизированных мышей, оцениваемая по количеству спленоцитов, продуцирующих IFNγ, была выше в группе, вакцинированной упомянутой мРНК, по сравнению с группой, вакцинированной полноразмерным белком ESAT-6 или мРНК, кодирующей полноразмерный белок ESAT-6.
Сущность и промышленная применимость изобретения поясняется следующими примерами.
Пример 1. Выбор эпитопов
Аминокислотные последовательности антигенов М. tuberculosis были получены из базы данных Uniprot (https://www.uniprot.org/, дата обращения 01.03.2023). Мы анализировали белок ESAT-6 М. tuberculosis. Этот белок является одним из немногих секретируемых белков М. tuberculosis и играет ключевую роль в патогенезе туберкулеза. Первый механизм его действия связан с тем, что ESAT-6 необходим для проникновения бактерий в цитозоль из фаголизосомальных компартментов, что приводит к возможности выхода бактерий из клетки, дальнейшему распространению бактерий в организме [van der Wei N et al., 2007]. Второй механизм связан со способностью ESAT-6 индуцировать образование пор на мембранах что приводит к смерти мононуклеарных фагоцитов [Smith et al., 2008; Peng & Sun, 2016].
Для предсказания эпитопов белка ESAT-6, которые будут презентоваться комплексами MHC-I, был использован пакет MHCFlurry 2.0 версии 2.0.4 [O'Donnell et al., 2020]. Количественные характеристики частоты аллелей был получены из статьи [Weiskopf et al., 2013].
Для предсказания эпитопов белка ESAT-6, которые будут презентоваться комплексами МНС-II, был использован пакет NetMHC-IIpan 4.1 [Reynisson et al., 2020]. Мы использовали следующие опции для расчета эпитопов: -length 13,14,15,16,17 для указания длины возможных эпитопов и -ВА для того, чтобы включить режим предсказания аффинности для пары эпитоп-МНС.Частота аллелей МНС была получена из статьи [Greenbaum et al., 2011].
С помощью такого алгоритма было отобрано 2 эпитопа для MHC-I и 3 для МНС-II (Таблица 1).

Пример 2. Оценка и ранжирование эпитопов
На следующем этапе была проведена оценка и ранжирование эпитопов. Полученные предсказания были отфильтрованы в среде Python 3.9 (Python Software Foundation. Python Language Reference, version 3.9. Доступен по адресу http://www.python.org. дата обращения 01.03.2023). Каждая пара эпитоп-МНС должна взаимодействовать с аффинностью, соответствующей IC50 не более 500 нМ, каждый эпитоп должен связываться с по меньшей мере 5 МНС, кодируемыми различными аллелями. Для эпитопов, которые удовлетворяют указанным выше условиям, были рассчитаны медиана, среднее и максимальная аффинность эпитопа к тем МНС, с которыми он взаимодействует.
Эпитопы, презентируемые на молекулах МНС I и II классов, были проранжированы по отдельности. Для каждого класса МНС были отобраны те эпитопы, которые не пересекаются по своим последовательностям для максимизации покрытия различных участков антигена.
В результате компьютерного моделирования была получена конструкция, представляющая собой слитый полипептид из 5 эпитопов белка ESAT-6 Mycobacterium tuberculosis и сигнального пептида MHC-I человека с аминокислотной последовательностью:
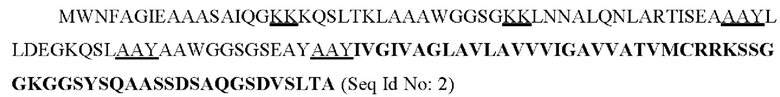
Подчеркнуты последовательности линкеров (КК или AAY), соединяющих различные эпитопы белка ESAT-6 Mycobacterium tuberculosis и участок MHC-I человека. Жирным шрифтом выделен сигнальный пептид: последовательность цитоплазматического и транс мембранного домена МНС класса I, который обеспечивает ко-локализацию целевого белка с комплексами гистосовместимости в различных эндоцитарных компартментах (аппарат Гольджи, эндоплазматический ретикулум, эндосомы) и на поверхности клеточной мембраны, что значительно усиливает презентацию антигена [Kreiter et al., 2008].
Пример 3. Синтез олигонуклеотидов, клонирование в плазмиду pSmart
Для синтеза ДНК, кодирующей мультиэпитопный полипептид по настоящему изобретению, была проведена оптимизация кодонов для экспрессии в эукариотических клетках, в результате была получена последовательность нуклеотидов Seq Id No:4.
Для получения конструкций для последующей транскрипции РНК in vitro в коммерческий вектор pSmart (Lucigen, США) встраивали кассету, содержащую последовательность 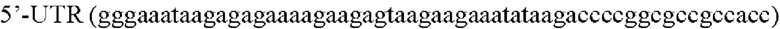 с последовательностью полноразмерного белка ESAT-6 М. tuberculosis или мультиэпитопного полипептида по настоящему изобретению и с 3'-UTR (gctggagcctcggtggcctagcttcttgccccttgggcctccccccagcccctcctccccttcctgcacccgtacccccgtgtctttgaataa agtctgagtgggcggca). Затем полученную кассету обрабатывали рестриктазами EcoRI и BglII, очищали на агарозном геле и осуществляли лигирование с аналогично подготовленным коммерческим вектором pSmart, (Lucigen, США). Химически компетентные клетки NEB-stable (New England Biolabs, Соединенное Королевство) трансформировали лигазной смесью и высевали на LB агар, содержащий ампициллин в концентрации 100 мкг/мл. Колонии на наличие вставки проверяли методом ПЦР. Верификацию плазмид осуществляли секвенированием по Сэнгеру, анализ хроматограмм секвенирования проводили с помощью программы Ugene v38.1. Е. Coli NEB-stable с вектором культивировали при 30°С и 180 оборотах в минуту. Выделение и очистку плазмидной ДНК проводили с использованием набора QIAGEN Plasmid Maxi Kit (Qiagen, США). Для получения линеаризованной плазмиды препарат обрабатывали рестриктазой Spel по уникальному сайту рестрикции, расположенному после PolyA хвоста.
с последовательностью полноразмерного белка ESAT-6 М. tuberculosis или мультиэпитопного полипептида по настоящему изобретению и с 3'-UTR (gctggagcctcggtggcctagcttcttgccccttgggcctccccccagcccctcctccccttcctgcacccgtacccccgtgtctttgaataa agtctgagtgggcggca). Затем полученную кассету обрабатывали рестриктазами EcoRI и BglII, очищали на агарозном геле и осуществляли лигирование с аналогично подготовленным коммерческим вектором pSmart, (Lucigen, США). Химически компетентные клетки NEB-stable (New England Biolabs, Соединенное Королевство) трансформировали лигазной смесью и высевали на LB агар, содержащий ампициллин в концентрации 100 мкг/мл. Колонии на наличие вставки проверяли методом ПЦР. Верификацию плазмид осуществляли секвенированием по Сэнгеру, анализ хроматограмм секвенирования проводили с помощью программы Ugene v38.1. Е. Coli NEB-stable с вектором культивировали при 30°С и 180 оборотах в минуту. Выделение и очистку плазмидной ДНК проводили с использованием набора QIAGEN Plasmid Maxi Kit (Qiagen, США). Для получения линеаризованной плазмиды препарат обрабатывали рестриктазой Spel по уникальному сайту рестрикции, расположенному после PolyA хвоста.
Для получения конструкции, для наработки полноразмерного белка ESAT-6 использовали вектор рЕТ30 (Agilent, США). Последовательность ESAT-6 с дополнительной последовательностью для 6 гистидинов на N-конце была собрана из олигонуклеотидов с помощью ПЦР. Затем полученную кассету обрабатывали рестриктазами EcoRI и NdeI, очищали на агарозном геле и осуществляли лигирование с аналогично подготовленным коммерческим вектором рЕТ30 (Agilent, США). Для трансформации использовали клетки BL21DE3 (Agilent, США). Верификацию плазмид осуществляли секвенированием по Сэнгеру.
Сборку целевой последовательности осуществляли из олигонуклеотидов путем сшивки фрагментов методом ПЦР с перекрывающимися праймерами. Затем полученный фрагмент инкубировали с эндонуклеазами рестрикции ЕсоШ и BglU, очищали на агарозном геле и осуществляли лигирование с аналогично подготовленным коммерческим вектором pSmart (Lucigen, США). Вектор содержит участок полиаденилирования длиной 110 остатков адениозинмонофосфата. Для трансформации использовали штамм Е. coli NEB-stable (New England Biolabs, Соединенное Королевство). Клоны отбирали методом ПЦР с колоний и подтверждали последовательность вставки секвенированием. Культивирование Е. coli для наработки верифицированной плазмиды проводили на шейкере-инкубаторе при 30°С и 180 оборотах в минуту. Затем с помощью набора реагентов QIAGEN Plasmid Maxi Kit (Qiagen, США) проводили выделение плазмидной ДНК из бактериальных клеток. Полученный препарат плазмиды линеаризовали по уникальному сайту рестрикции Spel с последующей визуализацией в агарозном геле.
Пример 4. In vitro транскрипция мРНК
In vitro транскрипцию проводили как описано ранее [Kirshina et al., 2022]. Вкратце, использовали 5 мкг линеаризованной плазмиды pSmart-ESAT-6, буфер (20 мМ DTT, 2 мМ спермидина, 80 мМ HEPES-KOH рН 7.4, 24 мМ MgC12), 500 единиц Т7 РНК-полимеразы (Биолабмикс, Новосибирск), 200 единиц ингибитора рибонуклеаз RiboCare (Евроген, Москва) и 1 мкл смеси ферментов из набора RiboMAX Large Scale RNA Production System (Promega, США) в качестве источника неорганической пирофосфатазы. Реакционная смесь также содержала 12 мМ аналога кэпа ARCA (Биолабмикс, Новосибирск) и по 3 мМ каждого из рибонуклеозидтрифосфатов (Биосан, Новосибирск). Реакцию проводили 2 часа при температуре 37°С, после чего в реакцию добавляли еще по 3 мМ каждого из рибонуклеозидтрифосфатов и инкубировали в течение еще 2 часов. ДНК гидролизовали при помощи нуклеазы RQ1 (Promega, США), РНК осаждали добавлением LiCl до концентрации 0,32М и EDTA рН 8,0 до концентрации 20 мМ с последующей инкубацией на льду в течение часа. Далее раствор центрифугировали в течение 15 минут (25000g, 4°С). Осадок РНК промывали 70% этанолом, растворяли в ультрачистой воде и еще раз осаждали спиртом по стандартной методике. Концентрацию РНК определяли спектрофотометрически по поглощению света при длине волны 260 нм. Длину и гомогенность синтезированных молекул РНК оценивали с помощью капиллярного электрофореза на приборе TapeStation (Agilent, USA).
Пример 5. Создание липидных наночастиц (ЛНЧ), содержащих мРНК
Капсулирование мРНК в липидные наночастицы проводили путем смешивания водного раствора мРНК в 10 мМ цитратном буфере (рН 3,0) со спиртовым раствором смеси липидов. Смешивание проводили в микрофлюидном картридже на приборе The NanoAssemblr™ Benchtop (Precision Nanosystems, США). Липидная смесь содержала следующие компоненты: ионизируемый епьтонный липид ALC-0315 (BroadPharm, США), дистеароилфосфатидилхолин (ДСФХ или DSPC) (Avanti Polar Lipids, США), холестерин (Sigma-Aldrich, США), DMG-PEG-2000 (BroadPharm, США) в молярном соотношении 46,3: 9,4: 42,7: 1,6. Массовая доля мРНК в ЛНЧ составляла 0,04% масс.Для формирования частиц водную и спиртовую фазы смешивали в соотношении 3: 1 по объему с общей скоростью смешивания 10 мл/мин. После смешивания фаз полученную водно-спиртовую суспензию частиц диализировали в 300 объемах фосфатно-солевого буфера (рН 7,4, 18 ч, +15°С). После диализа суспензию частиц концентрировали, используя фильтры Amicon Ultra-4 с отсечением MW10 000.
Затем частицы фильтровали в стерильных условиях через полиэфирсульфоновую мембрану с диаметром пор 0,22 мкм (Merck, США) и при необходимости хранили при 4°С. После фильтрации качество полученных частиц анализировали по двум параметрам: размер частиц (Zetasizer Nano ZSP, Malvern Panalitycal, США) и количество инкапсулированной мРНК. Количество инкапсулированной в липидные наночастицы мРНК определяли по разнице значений уровня флуоресцентного сигнала при окрашивании реагентом RiboGreen (Thermo Fischer Scientific, США) суспензии частиц до их разрушения и после. Для разрушения частиц использовали детергент Triton Х-100 (Sigma-Aldrich, США).
Пример 6. Наработка и очистка белка ES АТ-6
Для наработки белкам, coli BL21DE3 с вектором pET30-ESAT-6 культивировали при 37°С и помешивании с частотой 180 об./мин до оптической плотности бактериальной культуры OD(600)=0,65, затем добавляли ИПТГ до конечной концентрации 1 мМ и культивировали клетки 6 часов при 30°С.Клетки ресуспендировали в буфере, содержащем 50 мМ Трис-HCl рН 8,0 и 300 мМ хлорида натрия. Далее проводили лизис клеток ультразвуком с длительностью импульса 15 с и промежутком между импульсами 15 с. Амплитуда ультразвуковых колебаний составляла 50% от максимальной. После лизиса образцы центрифугировали в течение 15 минут при 20000 g, 4°С. рН полученного после центрифугирования супернатанта доводили до 7,5 путем добавления раствора 1 М Tris-OH. Далее испытуемый образец подвергали фильтрации через полиэфирсульфоновую мембрану с диаметром пор 0,22 мкм.
Первый этап очистки белка ESAT-6 осуществляли с использованием металлохелатного аффинного сорбента со средним размером частиц 30 мкм и иммобилизованными на них через хелатирующий лиганд нитрилотриуксусную кислоту ионами Ni2+. Затем фракции очищенного белка диализовали против буфера, содержащего 25 мМ Tris-HCl. Далее, диализованный образец наносили на сильный катионообменный сорбент со средним размером частиц 45 мкм и иммобилизованными на них функциональными сульфогруппами (-SO3-). Молекулярную массу полученного белка проверяли как с помощью классического метода электрофореза в полиакриламидном геле, так и с помощью масс-спектрометра maXis 4G ETD (Bruker, США).
Таким образом, был получен полноразмерный белок ESAT-6, который был использован в дальнейшем для стимуляции созревания дендритных клеток DC2.4 и оценке уровня активации Т-клеток вакцинированных животных.
Пример 7. Дизайн экспериментального исследования
В эксперименте были использованы самцы мышей линии C57BL6/J, имеющие статус SPF (specific pathogen free, свободные от патогенной флоры), в возрасте 8-9 недель и массой 19-21 грамм, полученные из Центра генетических ресурсов лабораторных животных Института цитологии и генетики СО РАН. Мышам вводили внутримышечно препарат ЛНЧ с мРНК, кодирующей полноразмерный белок ES АТ-6, в количестве 50 мкг мРНК на дозу, либо препарат ЛНЧ с мРНК, содержащей последовательность Seq Id No: 1, кодирующей мультиэпитопный полипептид по настоящему изобретению, в количестве 50 мкг мРНК на дозу, либо 25 мкг полноразмерного белка ESAT-6 (М. tuberculosis) с липидными наночастицами без мРНК. В качестве контроля использовали животных, которым вводили липидные наночастицы без мРНК, или фосфатный буфер в том же объеме, что и экспериментальные инъекции (Фиг. 1А). Количество ЛНЧ без мРНК на одну дозу при введении с белком ESAT-6 или без него соответствовало количеству ЛНЧ в одной дозе препарата ЛНЧ с мРНК. Через 4 недели после первой инъекции животным повторно вводили те же препараты в той же дозе. Забой мышей и выделение селезенки осуществляли через 4 недели после второй инъекции (Фиг. 1Б).
Пример 8. Оценка уровня Т-клеточного ответа после вакцинации
Уровень сформированного Т-клеточного ответа у вакцинированных мышей определяли по количеству спленоцитов, секретирующих IFNγ в ответ на белок ESAT-6 М. tuberculosis, с помощью метода Elispot.
Селезенки вакцинированных и контрольных мышей забирали на 28 сутки после повторной вакцинации. Из селезенок выделяли спленоциты протиранием через фильтр с размером пор 70 мкм. Спленоциты вносили в 96-луночные планшеты Elispot с мембраной из ПВДФ (BD, США) в количестве 300 тыс.клеток на лунку. Предварительно в лунки планшета вносили питательную среду RPMI-1640 (ПанЭко, Россия) с 10% FCS из расчета общего объема жидкости в лунке 200 мкл.
В планшете формировали группы стимуляции спленоцитов: спленоциты от каждой мыши без стимуляции (отрицательный контроль), спленоциты от каждой мыши, стимулированные белком ESAT-6 (50 мкг/мл) и дендритными клетками линии DC2.4 (Merck, США) (30 тыс. на 300 тыс.спленоцитов) (DC2.4 + ESAT-6). Каждая экспериментальная лунка ставилась в двух технических повторах. Далее спленоциты культивировали 18 часов в СО2-инкубаторе при 5% СО2 и 37°С).
По истечении срока культивирования спленоциты, секретирующие IFNγ, выявляли с помощью наборов Mouse IFNg ELISPOT Set (BD, США) и AEC Substrate Set (BD, США), согласно инструкции производителя. Подсчет точек, соответствующих спленоцитам, секретирующим IFNγ, производили на приборе S6 Ultra (CTL, США).
Анализ Т-клеточного ответа методом ELISPOT показал влияние вакцинации на IFNγ-секретирующие клетки [F(10,4)=6.02, р=0.012]. Количество IFNγ-секретирующих клеток в ответ на стимуляцию DC2.4+ESAT-6 было выше в спленоцитах мышей, вакцинированных мультиэпитопным вариантом мРНК вакцины (mRNA mEpitope) по сравнению с контрольными группами (PBS, LNP) или спленоцитами мышей, иммунизированных белком ESAT-6 с наночастицами без РНК (ESAT-6+LNP) (Фиг. 2). Количество IFNγ-секретирующих клеток среди спленоцитов мышей, иммунизированных мРНК вакциной, кодирующей полноразмерный белок ESAT-6, было также выше, чем в контроле (64+-2.7 vs 2.5+-1.0), однако эти изменения были статистически не достоверны. Следует отметить, что спленоциты мышей, иммунизированных белком ESAT-6 с ЛНЧ без мРНК, демонстрировали лишь незначительное увеличение IFNγ-секретирующих клеток по сравнению со спленоцитами после иммунизации мРНК вакцинами (ESAT-6: 5.7+-1.8, mRNA ESAT-6: 64.0+-2.7, мультиэпитопная mRNA ESAT-6: 126.0+- 46.2).
Пример 9. Цитометрическое определение клеток, продуцирующих IFNγ
Для цитометрического определения клеток, продуцирующих IFNγ, спленоциты вакцинированных мышей вносили в 200 мкл среды RPMI-1640 в культуральный 96-луночный планшет в количестве 300 тыс.на лунку и стимулировали белком ESAT-6 или оставляли нестимулированными. Через 2 часа после начала стимуляции в лунки добавляли Брефелдин A (Abeam, США) в концентрации 10 мкг/мл. Далее клетки культивировали ночь в СО2-инкубаторе. По окончании инкубации клетки окрашивали мечеными антителами к CD3 и CD8 (BD, США), фиксировали и пермеабилизировали коммерческим набором (BD, США) согласно инструкции производителя, после чего внутриклеточно окрашивали антителами к IFNγ. Полученные таким образом клетки анализировали на проточном цитометре LSR Fortessa(BD, США). Полученные данные обрабатывали в программе Flow Jo (США).
Цитометрическое исследование показало, что процент CD3+CD8+и CD3+CD8- Т-лимфоцитов, продуцирующих IFNγ, выше в спленоцитах мышей, иммунизированных мРНК-вакцинами (Фиг. 3). Таким образом, данные по цитометрии подтверждают, что повышенная секреция IFNγ в спленоцитах мышей, вакцинированных мРНК, кодирующей мультиэпитопный пептид по настоящему изобретению, обеспечивается в том числе за счет Т-лимфоцитов, что говорит о формировании Т-клеточного ответа на антигены микобактерии туберкулеза у вакцинированных мышей.
Пример 10. Статистический анализ
Статистическую обработку данных проводили, используя однофакторный дисперсионный анализ ANOVA и Turkey's тест в качестве апостериорного (post hoc) анализа. Различия между экспериментальными группами считали статистически значимыми при р<0.05. Анализ данных производили с помощью пакета программ Statistica 6.0 (США).
Все публикации, патенты и заявки на патенты включены в настоящий документ посредством ссылки. Хотя в вышеприведенном описании это изобретение было описано в отношении некоторых предпочтительных вариантов его осуществления, и многие детали были изложены в целях иллюстрации, для специалистов в данной области техники будет очевидно, что изобретение допускает дополнительные варианты осуществления и что некоторые детали, описанные в данном документе, могут значительно изменяться без отклонения от сущности изобретения.
Использование терминов в единственном числе в контексте описания изобретения должно толковаться как охватывающее как единственное, так и множественное число, если иное не указано в данном документе или явно не противоречит контексту. Термины «состоящий из», «имеющий», «включающий» и «содержащий» следует толковать как неограничивающие термины, т.е. означающие «включая, но не ограничиваясь», если не указано иное. Перечисление диапазонов значений в данном документе просто предназначено для использования в качестве сокращенного способа индивидуальной ссылки на каждое отдельное значение, попадающее в этот диапазон, если здесь не указано иное, и каждое отдельное значение включено в спецификацию, как если бы оно было отдельно изложено в данном документе. Все способы, описанные в данном документе, могут выполняться в любом подходящем порядке, если иное не указано в данном документе или иным образом явно не противоречит контексту. Использование любых и всех примеров или иллюстративного языка (например, «такой как»), представленных в данном документе, предназначено просто для лучшего описания изобретения и не налагает ограничения на объем изобретения, если иное не заявлено. Никакие формулировки в описании не следует истолковывать как указывающие на какой-либо не заявленный элемент как существенный для практического применения изобретения.
Здесь описаны варианты осуществления этого изобретения, включая лучший из известных изобретателям способа осуществления изобретения. Разновидности этих вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения предшествующего описания. Авторы ожидают, что квалифицированные специалисты будут использовать такие варианты в зависимости от обстоятельств, и авторы предполагают, что изобретение будет реализовано на практике иначе, чем конкретно описано в данном документе. Соответственно, это изобретение включает в себя все модификации и эквиваленты признаков, изложенных в прилагаемой формуле изобретения, как это разрешено действующим законодательством. Более того, любая комбинация вышеописанных признаков во всех их возможных вариациях охватывается изобретением, если иное не указано в данном документе или иным образом явно не противоречит контексту.
Заявитель просит рассмотреть представленные материалы заявки «Мультиэпитопный полипептид для иммунизации против Mycobacterium tuberculosis» на предмет выдачи патента на изобретение.
Список литературы
NCT03159975 Safety and hnmunogenicity Study of GX-70 in ТВ Patient, 2017, Yonsei University, https://clinicaltrials.gov/ct2/show/NCT03159975. дата обращения 17.03.2023.
A1 Tbeishat H. Novel In Silico mRNA vaccine design exploiting proteins of M. tuberculosis that modulates host immune responses by inducing epigenetic modifications. Sci Rep.2022 Mar 17;12(1):4645. doi: 10.1038/s41598-022-08506-4. PMID: 35301360; PMCID: PMC8929471.
A database of global BCG vaccination policies and practices, http://www.bcgatlas.org/. дата обращения 17.03.2023.
Baldwin SL, Reese VA, Larsen SE, et al. Prophylactic efficacy against Mycobacterium tuberculosis using ID93 and lipid-based adjuvant formulations in the mouse model. PLoS One. 2021 Mar 11;16(3):e0247990. doi: 10.1371/journal.pone.0247990. PMID: 33705411; PMCID: PMC7951850.
Brandt L, Elhay M, Rosenkrands I, et al. ESAT-6 subunit vaccination against Mycobacterium tuberculosis. Infect Immun. 2000 Feb;68(2):791-5. doi: 10.1128/IAI.68.2.791-795.2000. PMID: 10639447; PMCID: PMC97206.
Brodin P, Majlessi L, Marsollier L, et al. Dissection of ESAT-6 system 1 of Mycobacterium tuberculosis and impact on immunogenicity and virulence. Infect Immun. 2006 Jan;74(l):88-98. doi: 10.1128/IAI.74.1.88-98.2006. PMID: 16368961; PMCID: PMC1346617.
Bruffaerts N, Huygen K, Romano M. DNA vaccines against tuberculosis. Expert Opin Biol Ther. 2014 Dec;14(12):1801-13. doi: 10.1517/14712598.2014.951630. Epub 2014 Aug 22. PMID: 25145964.
Calmette A. Preventive Vaccination Against Tuberculosis with BCG. Proc R Soc Med. 1931 Sep;24(11):1481-90. PMID: 19988326; PMCID: PMC2182232.
Chen X, Zaro JL, Shen WC. Fusion protein linkers: property, design and functionality. Adv Drug Deliv Rev. 2013 Oct;65(10): 1357-69. doi: 10.1016/j.addr.2012.09.039. Epub 2012 Sep 29. PMID: 23026637; PMCID: PMC3726540.
Comstock GW, Livesay VT, Woolpert SF. Evaluation of BCG vaccination among Puerto Rican children. Am J Public Health. 1974 Mar;64(3):283-91. doi: 10.2105/ajph.64.3.283. PMID: 4811772; PMCID: PMC1775430.
Deng YH, He HY, Zhang BS. Evaluation of protective efficacy conferred by a recombinant Mycobacterium bovis BCG expressing a fusion protein of Ag85A-ESAT-6. J Microbiol Immunol Infect. 2014 Feb;47(1):48-56. doi: 10.1016/j.jmii.2012.11.005. Epub 2013 Jan 26. PMID: 23357605.
Dey J, Mahapatra SR, Lata S, et al. Exploring Klebsiella pneumoniae capsule polysaccharide proteins to design multiepitope subunit vaccine to fight against pneumonia. Expert Rev Vaccines. 2022;21(4):569-587.
Dimitrov I, Bangov I, Flower DR, Doytchinova I. AllerTOP v.2~a server for in silico prediction of allergens. J Mol Model. 2014;20(6):2278.
Doherty TM. Immunotherapy for ТВ. Immunotherapy. 2012 Jun;4(6):629-47. doi: 10.2217/imt. 12.52. PMID: 22788130.
Doytchinova IA, Flower DR. VaxiJen: a server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinformatics. 2007;8:4.
Duthie MS, Reed SG. Skin tests forthe detection of Mycobacterial infections: achievements, current perspectives, and implications for other diseases. Appl Microbiol Biotechnol. 2021 Jan;105(2):503-508. doi: 10.1007/s00253-020-11062-4. Epub 2021 Jan 4. PMID: 33394146; PMCID: PMC7780083.
Fan XY, Lowrie DB. Where are the RNA vaccines for ТВ? Emerg Microbes Infect. 2021 Dec;10(l):1217-1218. doi: 10.1080/22221751.2021.1935328. PMID: 34036874; PMCID: PMC8216257.
Feng G, Jiang Q, Xia M, et al. Enhanced immune response and protective effects of nano-chitosan-based DNA vaccine encoding T cell epitopes of Esat-6 and FL against Mycobacterium tuberculosis infection. PLoS One. 2013 Apr 23;8(4):e61135. doi: 10.1371/journal.pone.0061135. PMID: 23637790; PMCID: PMC3634041.
Fifteen year follow up of trial of BCG vaccines in south India for tuberculosis prevention. Tuberculosis Research Centre (ICMR), Chennai. Indian J Med Res. 1999 Aug; 110:56-69. PMID: 10573656.
Flaxman A, Ewer KJ. Methods for Measuring T-Cell Memory to Vaccination: From Mouse to Man. Vaccines (Basel). 2018 Jul 21;6(3):43. doi: 10.3390/vaccines6030043. PMID: 30037078; PMCID: PMC6161152.
Gong W, Pan C, Cheng P, et al. Peptide-Based Vaccines for Tuberculosis. Front Immunol. 2022;13:830497.
Greenbaum J, Sidney J, Chung J, et al. Functional classification of class II human leukocyte antigen (HLA) molecules reveals seven different supertypes and a surprising degree of repertoire sharing across supertypes. hnmunogenetics. 2011;63(6):325-35.
Gupta S, Kapoor P, Chaudhary K, et al. hi silico approach for predicting toxicity of peptides and proteins. PLoS One. 2013;8(9):e73957.
Hess J, Kaufmann SH. Live antigen carriers as tools for improved anti-tuberculosis vaccines. FEMS Immunol Med Microbiol. 1999 Feb;23(2): 165-73. doi: 10.1111/j.l 574-695X.1999.tb01236.x. PMID: 10076914.
Horwitz MA, Harth G, Dillon BJ, et al. Recombinant bacillus calmette-guerin (BCG) vaccines expressing the Mycobacterium tuberculosis 30-kDa major secretory protein induce greater protective immunity against tuberculosis than conventional BCG vaccines in a highly susceptible animal model. Proc Natl Acad Sci USA. 2000 Dec 5;97(25): 13853-8. doi: 10.1073/pnas.250480397. PMID: 11095745; PMCID: PMC17665.
Huygen K, Content J, Denis O, et al. hnmunogenicity and protective efficacy of a tuberculosis DNA vaccine. Nat Med. 1996 Aug;2(8):893-8. doi: 10.1038/nm0896-893. PMID: 8705859.
Kao FF, Mahmuda S, Pinto R, et al. The secreted lipoprotein, MPT83, of Mycobacterium tuberculosis is recognized during human tuberculosis and stimulates protective immunity in mice. PLoS One. 2012;7(5):e34991. doi: 10.1371/journal.pone.0034991. Epub 2012 May 2. PMID: 22567094; PMCID: PMC3342273.
Kirshina AS, Kazakova AA, Kolosova ES, et al. Effects of various mRNA-LNP vaccine doses on neuroinflammation in BALB/c mice. Bulletin of RSMU. 2022; (6): 119 25. DOI: 10.24075/brsmu.2022.068
Koch R. Weitere Mittheilungen uber ein Heilmittel gegen Tuberculose. Dtsch. Med. Wochenschr. 1890. 16 1029-1032. 10.1055/s-0029-1207546
Kreiter S, Selmi A, Diken M, et al. Increased antigen presentation efficiency by coupling antigens to MHC class I trafficking signals. J Immunol. 2008;180(1):309-318.
Larsen SE, Erasmus JH, Reese VA, et al. RNA-based vaccine demonstrates prophylactic efficacy against Mycobacterium tuberculosis challenge in a mouse model. bioRxiv 2022.02.23.481669; doi: https://doi.org/10.1101/2022.02.23.481669
Larsen SE, Erasmus JH, Reese VA, et al. An RNA-Based Vaccine Platform for Use against Mycobacterium tuberculosis. Vaccines (Basel). 2023 Jan 5; 11(1): 130. doi: 10.3390/vaccinesl 1010130. PMID: 36679975; PMCID: PMC9862644.
Lei Y, Zhao F, Shao J, et al. Application of built-in adjuvants for epitope-based vaccines. PeerJ. 2019 Jan 14;6:e6185. doi: 10.7717/peerj.6185. PMID: 30656066; PMCID: PMC6336016.
Li JM, Zhu DY. Therapeutic DNA vaccines against tuberculosis: a promising but arduous task. Chin Med J (Engl). 2006 Jul 5;119(13): 1103-7. PMID: 16834929.
Li J, Zhao A, Tang J, et al. Tuberculosis vaccine development: from classic to clinical candidates. Eur J Clin Microbiol Infect Dis. 2020 Aug;39(8): 1405-1425. doi: 10.1007/sl0096-020-03843-6. Epub 2020 Feb 15. PMID: 32060754; PMCID: PMC7223099.
Liang Y, Zhang X, Bai X, et al. Immunogenicity and therapeutic effects of a Mycobacterium tuberculosis rv2190c DNA vaccine in mice. BMC Immunol. 2017 Feb 27; 18(1): 11. doi: 10.1186/sl2865-017-0196-x. PMID: 28241799; PMCID: PMC5327546.
Lorenzi JC, Trombone AP, Rocha CD, et al. Intranasal vaccination with messenger RNA as a new approach in gene therapy: use against tuberculosis. BMC Biotechnol. 2010 Oct 20; 10:77. doi: 10.1186/1472-6750-10-77. PMID: 20961459; PMCID: PMC2972232.
Madge HYR, Stephenson RJ and Toth I. Nanocarrier-based vaccine delivery systems for synthetic peptide vaccines. Handbook of nanotechnology applications: environment, energy, agriculture and medicine. Edited by Woei Jye Lau, Kajornsak Faungnawakij, Kuakoon Piyachomkwan, and Uracha Rungsardthong Ruktanonchai. Amsterdam, Netherlands: Elsevier. 2021; 509-535.https://doi.org/10.1016/b978-0-12-821506-7.00020-x
Mahairas GG, Sabo PJ, Hickey M J, et al. Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M. bovis. J Bacteriol. 1996 Mar;178(5): 1274-82. doi: 10.1128/jb. 178.5.1274-1282.1996. PMID: 8631702; PMCID: PMC177799.
Mahapatra SR., Dey J, Kaur T, et al. Immunoinformatics and molecular docking studies reveal a novel Multi-Epitope peptide vaccine against pneumonia infection. Vaccine. 2021;39(42):6221-6237.
Maitre T, Aubry A, Jarlier V, et al. Multidrug and extensively drug-resistant tuberculosis. Med Mai Infect. 2017 Feb;47(l):3-10. doi: 10.1016/j.medmal.2016.07.006. PMID: 27637852.
Mangtani P, Abubakar I, Ariti C, et al. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin Infect Dis. 2014 Feb;58(4):470-80. doi: 10.1093/cid/cit790. Epub 2013 Dec 13. PMID: 24336911.
Mangtani P, Nguipdop-Djomo P, Keogh RH, et al. Observational study to estimate the changes in the effectiveness of bacillus Calmette-Guerin (BCG) vaccination with time since vaccination for preventing tuberculosis in the UK. Health Technol Assess. 2017 Jul;21(39): 1-54. doi: 10.3310/hta21390. PMID: 28738015; PMCID: PMC5534974.
Mangtani P, Nguipdop-Djomo P, Keogh RH, et al. The duration of protection of school-aged BCG vaccination in England: a population-based case-control study, hit J Epidemiol. 2018 Feb 1;47(1):193-201. doi: 10.1093/ije/dyxl41. PMID: 29025083.
Mi J, Liang Y, Liang J, et al. The Research Progress in Lnmunotherapy of Tuberculosis. Front Cell Infect Microbiol. 2021 Nov 15;11:763591. doi: 10.3389/fcimb.2021.763591. PMID: 34869066; PMCID: PMC8634162.
Moliva Л, Turner J, Torrelles JB. Prospects in Mycobacterium bovis Bacille Calmette et Guerin (BCG) vaccine diversity and delivery: why does BCG fail to protect against tuberculosis? Vaccine. 2015 Sep 22;33(39):5035-41. doi: 10.1016/j.vaccine.2015.08.033. Epub 2015 Aug 28. PMID: 26319069; PMCID: PMC4577463.
Moradi J, Tabrizi M, Izad M, et al. Designing a novel multi-epitope DNA-based vaccine against tuberculosis: in silico approach. Jundishapur J Microbiol. 2017; 10:e43950. 10.5812/jjm.43950
Mostowy S, Tsolaki AG, Small PM, et al. The in vitro evolution of BCG vaccines. Vaccine. 2003 Oct 1;21 (27-30):4270-4. doi: 10.1016/s0264-410x(03)00484-5. PMID: 14505909.
Nemes E, Geldenhuys H, Rozot V, et al. Prevention of M. tuberculosis Infection with H4:IC31 Vaccine or BCG Revaccination. N Engl J Med. 2018 Jul 12;379(2):138-149. doi: 10.1056/NEJMoal714021. PMID: 29996082; PMCID: PMC5937161.
Nguipdop-Djomo P, Heldal E, Rodrigues LC, et al. Duration of BCG protection against tuberculosis and change in effectiveness with time since vaccination in Norway: a retrospective
population-based cohort study. Lancet Infect Dis. 2016 Feb;16(2):219-26. doi: 10.1016/S1473-3099(15)00400-4. Epub 2015 Nov 19. PMID: 26603173.
Nieuwenhuizen NE, Kaufmann SHE. Next-Generation Vaccines Based on Bacille Calmette-Guerin. Front Immunol. 2018 Feb 5;9:121. doi: 10.3389/fimmu.2018.00121. PMID: 29459859; PMCID: PMC5807593.
O'Donnell TJ, Rubinsteyn A, Laserson U. MHCflurry 2.0: Improved Pan-Allele Prediction of MHC Class I-Presented Peptides by Incorporating Antigen Processing. Cell Syst. 2020;ll(l):42-48 e7.
Oettinger T, fergensen M, Ladefoged A, et al. Development of the Mycobacterium bovis BCG vaccine: review of the historical and biochemical evidence for a genealogical tree. Tuber Lung Dis. 1999;79(4):243-50. doi: 10.1054/tuld. 1999.0206. PMID: 10692993.
Palmer CE, Long MW. Effects of infection with atypical mycobacteria on BCG vaccination and tuberculosis. Am Rev Respir Dis. 1966 Oct;94(4):553-68. doi: 10.1164/arrd. 1966.94.4.553. PMID: 5924215.
Parvizpour S, Pourseif MM, Razmara J, et al. Epitope-based vaccine design: a comprehensive overview of bioinformatics approaches. Drug Discov Today. 2020;25(6): 1034-1042.
Peng X, Sun J. Mechanism of ESAT-6 membrane interaction and its roles in pathogenesis of Mycobacterium tuberculosis. Toxicon. 2016 Jun 15;116:29-34. doi: 10.1016/j.toxicon.2015.10.003. Epub 2015 Oct 9. PMID: 26456678; PMCID: PMC4973572.
Penn-Nicholson A, Tameris M, Smit E, et al. Safety and immunogenicity of the novel tuberculosis vaccine ID93+GLA-SE in BCG-vaccinated healthy adults in South Africa: a randomised, double-blind, placebo-controlled phase 1 trial. Lancet Respir Med. 2018 Apr;6(4):287-298. doi: 10.1016/S2213-2600(18)30077-8. PMID: 29595510.
Pym AS, Brodin P, Majlessi L, et al. Recombinant BCG exporting ESAT-6 confers enhanced protection against tuberculosis. Nat Med. 2003 May;9(5):533-9. doi: 10.1038/nm859. Epub 2003 Apr 14. PMID: 12692540.
Rais M, Abdelaal H, Reese VA, et al. Immunogenicity and protection against Mycobacterium avium with a heterologous RNA prime and protein boost vaccine regimen. Tuberculosis (Edinb). 2023 Jan;138:102302. doi: 10.1016/j.tube.2022.102302. Epub 2022 Dec 27. PMID: 36586154.
Ranieri E, Popescu I, Gigante M. CTL ELISPOT assay. Methods Mol Biol. 2014;1186:75-86. doi: 10.1007/978-1-4939-1158-5 6. PMID: 25149304.
Refai A, Gritli S, Barbouche MR, Essafi M. Mycobacterium tuberculosis Virulent Factor ESAT-6 Drives Macrophage Differentiation Toward the Pro-inflammatory Ml Phenotype and
Subsequently Switches It to the Anti-inflammatory M2 Phenotype. Front Cell Infect Microbiol. 2018 Sep 18;8:327. doi: 10.3389/fcimb.2018.00327. PMID: 30283745; PMCID: PMC6157333.
Reynisson B, Barra C, Kaabinejadian S, et al. Improved Prediction of МНС II Antigen Presentation through Integration and Motif Deconvolution of Mass Spectrometry MHC Eluted Ligand Data. J Proteome Res. 2020;19(6):2304-2315.
Rhodes SJ, Knight GM, Fielding K, et al. Individual-level factors associated with variation in mycobacterial-specific immune response: Gender and previous BCG vaccination status. Tuberculosis (Edinb). 2016 Jan;96:37-43. doi: 10.1016/j.tube.2015.10.002. Epub 2015 Nov 11. PMID: 26786653.
Rodrigues LC, Diwan VK, Wheeler JG. Protective effect of BCG against tuberculous meningitis and miliary tuberculosis: a meta-analysis, hit J Epidemiol. 1993 Dec;22(6): 1154-8. doi: 10.1093/ije/22.6.1154. PMID: 8144299.
Romano M, Denis O, D'Souza S, et al. Induction of in vivo functional Db-restricted cytolytic T cell activity against a putative phosphate transport receptor of Mycobacterium tuberculosis. J Immunol. 2004 Jun 1;172(11):6913-21. doi: 10.4049/jimmunol.l72.11.6913. PMID: 15153510.
Romano M, Rindi L, Korf H, et al. Immunogenicity and protective efficacy of tuberculosis subunit vaccines expressing PPE44 (Rv2770c). Vaccine. 2008 Nov 11;26(48):6053-63. doi: 10.1016/j.vaccine.2008.09.025. Epub 2008 Sep 24. PMID: 18822333.
Romano M, Aryan E, Korf H, et al. Potential of Mycobacterium tuberculosis resuscitation-promoting factors as antigens in novel tuberculosis sub-unit vaccines. Microbes Infect. 2012 Jan;14(l):86-95. doi: 10.1016/j.micinf.2011.08.011. Epub 2011 Aug 31. PMID: 21920450.
Roupie V, Romano M, Zhang L, et al. Immunogenicity of eight dormancy regulon-encoded proteins of Mycobacterium tuberculosis in DNA-vaccinated and tuberculosis-infected mice. Infect Immun. 2007 Feb;75(2):941-9. doi: 10.1128/IAI.01137-06. Epub 2006 Dec 4. PMID: 17145953; PMCID: PMC1828490.
Sefidi-Heris Y, Jahangiri A, Mokhtarzadeh A, et al. Recent progress in the design of DNA vaccines against tuberculosis. Drug Discov Today. 2020 Sep 11:S 1359-6446(20)30345-7. doi: 10.1016/j.drudis.2020.09.005. Epub ahead of print. PMID: 32927065.
Seung KJ, Keshavjee S, Rich ML. Multidrug-Resistant Tuberculosis and Extensively Drug-Resistant Tuberculosis. Cold Spring Harb Perspect Med. 2015 Apr 27;5(9):a017863. doi: 10.1101/cshperspect.a017863. PMID: 25918181; PMCID: PMC4561400.
Skeiky YA, Ovendale PJ, Jen S, et al. T cell expression cloning of a Mycobacterium tuberculosis gene encoding a protective antigen associated with the early control of infection. J Immunol. 2000 Dec 15;165(12):7140-9. doi: 10.4049/jimmunol. 165.12.7140. PMID: 11120845.
Skeiky YA, Alderson MR, Ovendale PJ, et al. Differential immune responses and protective efficacy induced by components of a tuberculosis polyprotein vaccine, Mtb72F, delivered as naked DNA or recombinant protein. J Immunol. 2004 Jun 15;172(12):7618-28. doi: 10.4049/jimmunol. 172.12.7618. PMID: 15187142.
Smith J, Manoranjan J, Pan M, et al. Evidence for pore formation in host cell membranes by ESX-1-secreted ESAT-6 and its role in Mycobacterium marinum escape from the vacuole. Infect Immun. 2008 Dec;76(12):5478-87. doi: 10.1128/IAI.00614-08. Epub 2008 Oct 13. PMID: 18852239; PMCID: PMC2583575.
Solanki V, Tiwari M, Tiwari V. Prioritization of potential vaccine targets using comparative proteomics and designing of the chimeric multi-epitope vaccine against Pseudomonas aeruginosa. Sci Rep.2019;9(1):5240.
Stetson DB, Medzhitov R. Recognition of cytosolic DNA activates an IRF3-dependent innate immune response. Immunity. 2006 Jan;24(l):93-103. doi: 10.1016/j.immuni.2005.12.003. PMID: 16413926.
Story CM, Furman MH, Ploegh HL. The cytosolic tail of class I MHC heavy chain is required for its dislocation by the human cytomegalovirus US2 and US 11 gene products. Proc Natl Acad Sci U S A. 1999 Jul 20;96(15):8516-21. doi: 10.1073/pnas.96.15.8516. PMID: 10411907; PMCID: PMC 17548.
Suzuki D, Nagata T, Eweda G, et al. Characterization of murine T-cell epitopes on mycobacterial DNA-binding protein 1 (MDP1) using DNA vaccination. Vaccine. 2010 Feb 23;28(8):2020-5. doi: 10.1016/j.vaccine.2009.10.062. PMID: 20188258.
Tarrahimofrad H, Rahimnahal S, Zamani J, et al. Designing a multi-epitope vaccine to provoke the robust immune response against influenza A H7N9. Sci Rep.2021;11(1):24485.
Tebianian M, Hoseini AZ, Ebrahimi SM, et al. Cloning, expression, and immunogenicity of novel fusion protein of Mycobacterium tuberculosis based on ESAT-6 and truncated C-terminal fragment of HSP70. Biologicals. 2011 May;39(3): 143-8. doi: 10.1016/j.biologicals.2011.02.002. Epub 2011 Mar 8. PMID: 21388826.
Torchilin V. Intracellular delivery of protein and peptide therapeutics. Drug Discov Today Technol. 2008 Autumn;5(2-3):e95-el03. doi: 10.1016/j.ddtec.2009.01.002. PMID: 24981097.
Udwadia ZF. MDR, XDR, TDR tuberculosis: ominous progression. Thorax. 2012 Apr;67(4):286-8. doi: 10.1136/thoraxjnl-2012-201663. PMID: 22427352.
Usman MM, Ismail S, Teoh TC. Vaccine research and development: tuberculosis as a global health threat. Cent Eur J Immunol. 2017;42(2): 196-204. doi: 10.5114/ceji.2017.69362. Epub 2017 Aug 8. PMID: 28867962; PMCID: PMC5573893.
Valizadeh A, Lnani Fooladi AA, Sedighian H, et al. Evaluating the Performance of PPE44, HSPX, ESAT-6 and CFP-10 Factors in Tuberculosis Subunit Vaccines. Curr Microbiol. 2022 Jul 19;79(9):260. doi: 10.1007/s00284-022-02949-8. PMID: 35852636; PMCID: PMC9295111.
van der Wei N, Hava D, Houben D, et al. M. tuberculosis and M. leprae translocate from the phagolysosome to the cytosol in myeloid cells. Cell. 2007 Jun 29; 129(7): 1287-98. doi: 10.1016/j.cell.2007.05.059. PMID: 17604718.
Vita R, Overton JA, Greenbaum JA, Pet al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 2015;43(Database issue):D405-12.
Wang J, Zhang H, Wang H. Analysis of predicted CD8+T cell epitopes from proteins encoded by the specific RD regions of Mycobacterium tuberculosis for vaccine development and specific diagnosis. Mol Biol Rep.2010 Apr;37(4): 1793-9. doi: 10.1007/sl 1033-009-9613-4. Epub 2009 Jul 17. PMID: 19609720.
Wang QM, Sun SH, Hu ZL, et al. Epitope DNA vaccines against tuberculosis: spacers and ubiquitin modulates cellular immune responses elicited by epitope DNA vaccine. Scand J Immunol. 2004 Sep;60(3):219-25. doi: 10.1111/j.0300-9475.2004.01442.x. PMID: 15320877.
Weiskopf D, Angelo MA, de Azeredo EL, et al. Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+T cells. Proc Natl Acad Sci USA. 2013;110(22):E2046-53.
WHO Bacille Calmette-Guerin (BCG) Information sheet, 2020. Geneva: World Health Organization, https://www.who.int/publications/m/item/bcg-information-sheet, дата обращения 17.03.2023.
WHO Fact Sheets. Tuberculosis, 2009. Geneva: World Health Organization, https://www.who.int/europe/news-room/fact-sheets/item/tuberculosis, дата обращения 17.03.2023.
WHO Global Tuberculosis Reports 1997-2022. Geneva: World Health Organization, https://www.who.int/teams/global-tuberculosis-programme/tb-reports, дата обращения 17.03.2023.
WHO Global Tuberculosis Report 2020. (2020). WHO Global Tuberculosis Report 2020. Geneva: World Health Organization, https://www.who.int/publications/i/item/978924001313 L дата обращения 17.03.2023.Xue T, Stavropoulos E, Yang M, et al. RNA encoding the MPT83 antigen induces protective immune responses against Mycobacterium tuberculosis infection. Infect Immun. 2004 Nov;72(11):6324-9. doi: 10.1128/IAI.72.11.6324-6329.2004. PMID: 15501761; PMCID: PMC523008.
Yu DH, Hu XD, Cai H. Efficient tuberculosis treatment in mice using chemotherapy and immunotherapy with the combined DNA vaccine encoding Ag85B, MPT-64 and MPT-83. Gene Ther. 2008 May;15(9):652-9. doi: 10.1038/gt.2008.13. Epub 2008 Feb 21. PMID: 18288210.
Yuan W, Dong N, Zhang L, et al. hnmunogenicity and protective efficacy of a tuberculosis DNA vaccine expressing a fusion protein of Ag85B-Esat6-HspX in mice. Vaccine. 2012 Mar 23;30(14):2490-7. doi: 10.1016/j.vaccine.2011.06.029. Epub 2011 Jun23. PMID: 21704108.
Zhang Y, Wallace RJ Jr, Mazurek GH. Genetic differences between BCG substrains. Tuber Lung Dis. 1995 Feb;76(1):43-50. doi: 10.1016/0962-8479(95)90579-0. PMID: 7718847.
Zhang L. Multi-epitope vaccines: a promising strategy against tumors and viral infections. Cell Mol Immunol. 2018; 15(2): 182-184.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="2023110496_перечень.xml" softwareName="WIPO Sequence"
softwareVersion="2.2.0" productionDate="2023-04-24">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023110496</ApplicationNumberText>
<FilingDate>2023-04-24</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">Автономная некоммерческая
образовательная организация высшего образования
"Научно-Технологический Университет
"Сириус"</ApplicantName>
<ApplicantNameLatin>Sirius University of Science and
Technology</ApplicantNameLatin>
<InventionTitle languageCode="ru">Мультиэпитопный полипептид для
иммунизации против Mycobacterium tuberculosis</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>402</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..402</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgtggaacttcgccggcatagaggccgccgccagcgccatacagggca
agaagaagcagagcctgaccaagctggccgccgcctggggcgggagcggcaagaagctgaacaacgccct
gcagaacctggcccgcaccataagcgaggccgccgcctacctgctcgacgagggcaagcagagcctggcc
gcctacgccgcctggggcgggagcggcagcgaggcctacgccgcctacatagtgggcatagtggccggcc
tggccgtgctgatagtggtcgtgataggcgccgtggtcgccaccgtgatgtgccgcaggaagagctccgg
cgggaagggcgggagctacagccaggccgccagctccgacagcgcccagggcagcgacgtgagcctgacc
gcc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>134</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..134</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MWNFAGIEAAASAIQGKKKQSLTKLAAAWGGSGKKLNNALQNLARTISE
AAAYLLDEGKQSLAAYAAWGGSGSEAYAAYIVGIVAGLAVLAVVVIGAVVATVMCRRKSSGGKGGSYSQA
ASSDSAQGSDVSLTA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>95</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..95</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MTEQQWNFAGIEAAASAIQGNVTSIHSLLDEGKQSLTKLAAAWGGSGSE
AYQGVQQKWDATATELNNALQNLARTISEAGQAMASTEGNVTGMFA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>402</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..402</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgtggaacttcgccggcatagaggccgccgccagcgccatacagggca
agaagaagcagagcctgaccaagctggccgccgcctggggcgggagcggcaagaagctgaacaacgccct
gcagaacctggcccgcaccataagcgaggccgccgcctacctgctcgacgagggcaagcagagcctggcc
gcctacgccgcctggggcgggagcggcagcgaggcctacgccgcctacatagtgggcatagtggccggcc
tggccgtgctgatagtggtcgtgataggcgccgtggtcgccaccgtgatgtgccgcaggaagagctccgg
cgggaagggcgggagctacagccaggccgccagctccgacagcgcccagggcagcgacgtgagcctgacc
gcc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА | 2007 |
|
RU2443773C2 |
| ПОЛИАНТИГЕННАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ И ВСПОМОГАТЕЛЬНОГО ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА | 2019 |
|
RU2724896C1 |
| РЕКОМБИНАНТНАЯ BCG, СВЕРХЭКСПРЕССИРУЮЩАЯ PhoP-PhoR | 2017 |
|
RU2760580C2 |
| ШТАММ Mycobacterium tuberculosis BN ДЛЯ МОДЕЛИРОВАНИЯ ЛАТЕНТНОЙ ТУБЕРКУЛЕЗНОЙ ИНФЕКЦИИ | 2021 |
|
RU2760751C1 |
| ВАКЦИНА ПРОТИВ ТУБЕРКУЛЕЗА | 2017 |
|
RU2647831C1 |
| Способ детекции штаммов Mycobacterium bovis BCG в формате реального времени | 2018 |
|
RU2689801C1 |
| СПОСОБ ОЦЕНКИ АКТИВНОСТИ ТУБЕРКУЛЕЗНОЙ ИНФЕКЦИИ У ДЕТЕЙ И ПОДРОСТКОВ | 2015 |
|
RU2586279C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422525C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
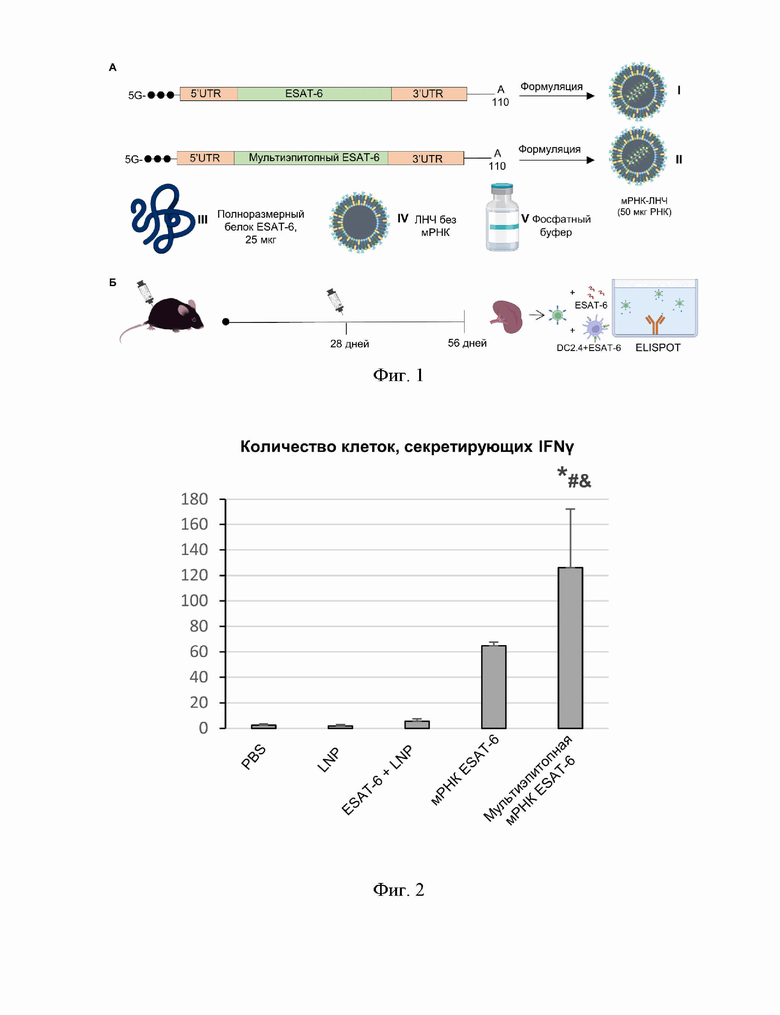
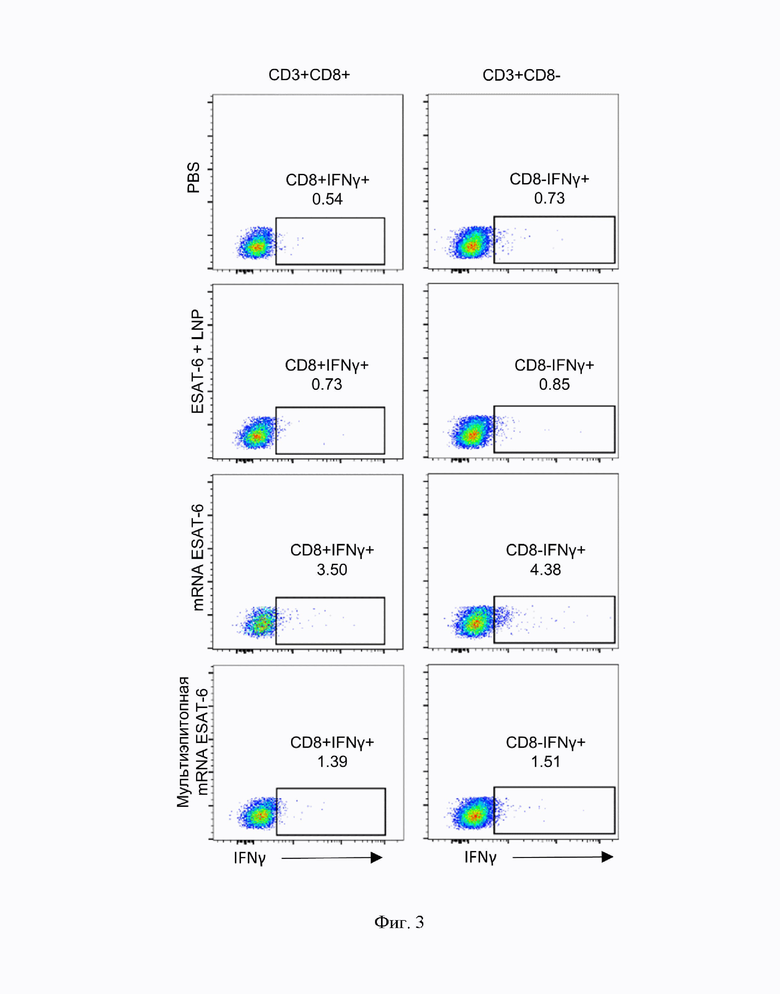
Изобретение относится к области биотехнологии и молекулярной биологии, а именно к вакцинам против инфекционных агентов, в частности к рекомбинантным вакцинам для иммунизации против Mycobacterium tuberculosis. Раскрыт полипептид для иммунизации против Mycobacterium tuberculosis, содержащий соединенные между собой посредством расщепляемых линкеров аминокислотные последовательности, соответствующие аминокислотам 6-20, 33-47, 65-79, 28-36, 41-51 белка ESAT-6 Mycobacterium tuberculosis, и последовательность, соответствующую аминокислотам 308-362 MHC I класса человека. Использование изобретения приводит к формированию устойчивого специфического Т-клеточного иммунного ответа против Mycobacterium tuberculosis. Изобретение может быть использовано при производстве вакцины от туберкулеза. 3 н. и 6 з.п. ф-лы, 3 ил., 1 табл., 10 пр.
1. Полипептид для иммунизации против Mycobacterium tuberculosis, содержащий соединенные между собой посредством линкеров аминокислотные последовательности, соответствующие аминокислотам 6-20, 33-47, 65-79, 28-36, 41-51 белка ESAT-6 Mycobacterium tuberculosis, и последовательность, соответствующую аминокислотам 308-362 MHC класса I человека, при этом упомянутая последовательность MHC класса I человека расположена на C-конце упомянутого полипептида, а в качестве линкеров используют расщепляемые линкеры.
2. Полипептид по п. 1, содержащий аминокислотную последовательность Seq Id No: 2.
3. Нуклеиновая кислота, кодирующая полипептид по любому из пп. 1 или 2.
4. Нуклеиновая кислота по п. 3, представляющая собой мРНК.
5. Нуклеиновая кислота по любому из пп. 3 или 4, представляющая собой мРНК для иммунизации против Mycobacterium tuberculosis.
6. Нуклеиновая кислота по любому из пп. 4 или 5, содержащая последовательность Seq Id No: 1.
7. Композиция для иммунизации человека против Mycobacterium tuberculosis, содержащая нуклеиновую кислоту по любому из пп. 3-6 и смесь липидов.
8. Композиция по п. 7, в которой смесь липидов содержит ионизируемый катионный липид, ПЭГилированный липид, фосфолипид и холестерин.
9. Композиция по любому из пп. 7 или 8, в которой смесь липидов содержит ионизируемый катионный липид ALC-0315, дистеароилфосфатидилхолин, холестерин и DMG-PEG-2000 в молярном соотношении 46,3:9,4:42,7:1,6.
| АРАКЕЛОВ С.А., ВАКЦИНЫ ПРОТИВ ТУБЕРКУЛЕЗА - НОВЫЙ ВЕК, Вопросы практической педиатрии, 2015, Том 10, Номер 3, стр | |||
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| OLSEN A | |||
| W | |||
| et al., Efficient protection against Mycobacterium tuberculosis by vaccination with a single subdominant epitope from the ESAT-6 antigen, Eur J Immunol., 2000, Volume 30(6), pp | |||
| Телефонная трансляция с катодным реле | 1920 |
|
SU1724A1 |
| ВАКЦИНА РЕКОМБИНАНТНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ И АДЪЮВАНТ ДЛЯ НЕЕ | 2017 |
|
RU2665817C1 |

