Настоящее изобретение относится к способам и средствам для стимулирования фагоцитоза и, в частности, фагоцитоза апоптотических клеток.
Аполипопротеин-Е (ароЕ) наряду с аполипопротеином-В составляет основную долю белков, присутствующих в богатых триглицеридами частицах липопротеинов - ЛНП и ЛОНП.
Компонент АпоЕ является лигандом рецептора ЛНП (РЛНП) и семейства родственных рецептору ЛНП белков или ЛРБ (LRPs, LDL-receptor-related proteins) [Herz, J. and U. Beffert. 2000. Nat Rev Neurosci 1:51]. Гомозиготная делеция гена ароЕ у мышей оказывает значительный эффект на транспорт холестерина, что приводит к значительному увеличению содержания ЛНП и ЛОНП в плазме вследствие отсутствия РЛНП и ЛРБ опосредовать удаление этих липопротеинов из крови [Moghadasian, M.H. et al 2001. Faseb J 15:2623', Zhang, S.H. et al 1992. Science 258:468]. Наиболее явной фенотипической особенностью мышей, дефектных по ароЕ, является формирование липидных поражений сосудов (бляшек), напоминающих ранний атеросклероз у людей, даже при диете с нормальным содержанием жиров.
У человека встречаются три основных аллельных варианта гена ароЕ, обозначаемых ароε2, ε3 и ε4, кодирующих АроЕ2, Е3 и Е4 соответственно.
Каждый из АроЕ2 и Е4 отличается от наиболее распространенного изотипа Е3 единственной аминокислотной заменой. Присутствие хотя бы одной копии аллели ароε4 приводит к увеличению риска развития коронарной болезни сердца по сравнению с гомозиготой ароε3 дикого типа [например, см. Davignon, J., et al 1988. Arteriosclerosis 8:1; Eichner, J.E. et al. 2002. Am J Epidemiol 155:487]. Присутствие аллели арое2 связано с комплексными нарушениями метаболизма липопротеинов, например гиперлипопротеинемией III типа, и было связано как с развитием, так и с подавлением атеросклероза в зависимости от модели исследования. Конкретные молекулярные основы связи между генотипом ароЕ и развитием сердечно-сосудистых заболеваний остаются неясными.
Присутствие аллели ароε4 связано с повышенным риском развития болезни Альцгеймера [Corder, Е.Н., et al 1993 Science 261:921, Poirier, J. et al 1993. Lancet 342:697, Mahley, R.W. et al 2000. Annu Rev Genomics Hum Genet 1:507] и повышенным риском развития остеопороза [Cauley, J.A. et al 1999. J Bone Miner Res 14:1175; Salamone, L.M., et al. 2000. J Bone Miner Res 15:308.]. Действительно, связь между гаплотипом ароЕ и рядом широко распространенных заболеваний достаточно устойчива, так что ароЕ остается одним из нескольких локусов, который напрямую связан с продолжительностью жизни: аллель ароε2 широко распространена среди долгожителей, которым более 80 лет, что позволяет предположить, что данный генотип ароЕ способствует долгожительству [Schachter, F. et al 1994. Nat Genet 6:29}.
АроЕ также может регулировать локальное воспаление посредством сигнальных каскадов, опосредованных рецептором, независимых от транспорта липопротеинов. Например, АроЕ, а также связывающийся с рецептором пептид, состоящий из 14 аминокислот, который не связывает холестерин, оказались способными подавлять локальный воспалительный ответ на ишемию головного мозга в экспериментах in vivo [Lynch, J.R. et al. 2001. J Neuroimmunol 114:107 and Sheng, Н. et al 1998. J Cereb Blood Flow Metab 18:361]. Исследования на клеточных культурах позволяют предположить, что данный эффект может быть связан с подавлением функции макрофагов [Laskowitz, D.Т. et al 1997 J Neuroimmunol 76:70 and Laskowitz, D.T. et al 2001. Exp Neurol 167:74]. Исследования хронических инфекций у мышей, дефектных по ароЕ, приводят к аналогичному выводу, что АроЕ каким-то образом подавляет функцию макрофагов [Van Oosten, M. et al. 2001. J Biol Chem 276:8820 and Roselaar, S.Е. and A. Daugherty. 1998 J Lipid Res 39:1740. and de Bont, N. et al. 1999. J Lipid Res 40:680].
Настоящими изобретателями была обнаружена неожиданная роль белкового продукта гена ароЕ (аполипопротеинаЕ) в качестве регулятора уничтожения апоптотических клеток. Миметики АроЕ и другие соединения, которые стимулируют уничтожение апоптотических клеток, могут быть полезными при лечении ряда заболеваний.
В одном своем аспекте настоящее изобретение предусматривает способ выявления и/или получения соединения для лечения патологического состояния, связанного с уменьшением активности эндогенного АроЕ у индивида, включающий определение поглощения апоптотических клеток макрофагами в присутствии тестируемого соединения.
Поглощение апоптотических клеток макрофагами в присутствии тестируемого соединения можно сравнивать с поглощением в аналогичной реакционной среде и условиях, когда тестируемое соединение отсутствует. Увеличение поглощения апоптотических клеток в присутствии тестируемого соединения по сравнению со случаем, когда тестируемое соединение отсутствует, свидетельствует о том, что соединение может быть полезным при лечении патологического состояния, связанного с уменьшением активности эндогенного АроЕ.
В некоторых способах реализации макрофаги можно обработать тестируемым соединением до определения поглощения апоптотических клеток.
Патологическое состояние, связанное с уменьшением активности эндогенного АроЕ, может иметь генетическую природу (например, присутствие одной или более аллелей ароЕ4), определяться факторами окружающей среды (например, уменьшением образования АроЕ клетками под воздействием высокого уровня холестерина) или может являться болезненным состоянием, выбранным из группы, состоящей из болезни Альцгеймера, атеросклероза, инсульта и остеопороза.
В некоторых способах реализации поглощение можно определять в присутствии АроЕ. Например, способ может быть осуществлен при условиях, когда уровень АроЕ является нормальным, например с использованием макрофагов и апоптотических клеток дикого типа (в которых присутствует АроЕ) в буферном растворе с физиологическим уровнем содержания АроЕ. В таких способах реализации можно идентифицировать тестируемое соединение, которое представляет собой агонист АроЕ и которое увеличивает степень фагоцитоза относительно нормальных условий. В некоторых вариантах реализации настоящего изобретения способ может включать стадии обработки макрофагов тестируемым соединением и приведения макрофага в контакт с апоптотическим полипептидом.
В других способах реализации способ может быть осуществлен при отсутствии АроЕ, например, с применением дефектных по АроЕ макрофагов и апоптотических клеток (например, клеток, полученных от мышей, дефектных по ароЕ) при условиях, характеризующихся отсутствием АроЕ (например, с применением среды и буферном растворе, которые не содержат ароЕ). В таких способах реализации настоящего изобретения можно идентифицировать тестируемое соединение, которое воспроизводит функцию ароЕ при стимулировании фагоцитоза апоптотических клеток. Способ может включать определение поглощения дефектных по АроЕ апоптотических клеток дефектным по АроЕ макрофагом в условиях, характеризующихся отсутствием АроЕ, в присутствии тестируемого соединения.
В настоящем описании показано, что снижение эффективности уничтожения апоптотических клеток приводит к явному провоспалительному фенотипу. Способы, описанные в данной заявке, можно применять для идентификации соединений, имеющих противовоспалительные свойства. Соответствующие соединения стимулируют уничтожение апоптотических клеток, уменьшая необходимость рекрутирования большого числа фагоцитов к воспаленной ткани, уменьшая продукцию медиаторов воспаления вышеуказанными фагоцитами и тем самым оказывая системное противовоспалительное действие. В результате способы, описанные в данной заявке, можно применять для идентификации терапевтических агентов при широком спектре заболеваний с воспалительной составляющей, даже если такие заболевания явно не связаны со снижением функции АроЕ или с увеличением потребности уничтожения продуктов разрушения клеток.
В другом своем аспекте настоящее изобретение предусматривает способ идентификации и/или получения соединений для лечения или предотвращения патологических состояний, связанных с повышенной воспалительной активностью, включающий определение способности полипептида АроЕ стимулировать поглощение апоптотических клеток макрофагами в присутствии тестируемого соединения.
Стимулирование полипептидом АроЕ поглощения или фагоцитоза апоптотических клеток макрофагами в присутствии тестируемого соединения можно оценивать по сравнению с поглощением в аналогичной реакционной среде и условиях отсутствия тестируемого соединения. Увеличение поглощения апоптотических клеток в присутствии тестируемого соединения по сравнению с поглощением в его отсутствие может свидетельствовать о том, что соединение может быть полезным при лечении патологического состояния, связанного с повышенной воспалительной активностью.
Полипептид АроЕ может представлять собой полипептид АроЕ, происходящий от любого вида млекопитающих, например АроЕ мыши или АроЕ человека (например, АроЕ2, АроЕ3 или АроЕ4 человека) или может представлять собой его фрагмент или вариант, который сохраняет одну или более активностей белка дикого типа, в частности способность стимулирования фагоцитоза апоптотических клеток.
Например, полипептид АроЕ может иметь аминокислотную последовательность, которая имеет более около 30% идентичности (identity) с последовательностью АроЕ3 человека, более примерно 40%, более примерно 45%, более примерно 55%, более примерно 65%, более примерно 70%, более примерно 80%, более примерно 90% или более примерно 95%. Последовательность может иметь более около 30% сходства (similarity) с АроЕ3 человека, более примерно 40% сходства, более примерно 50% сходства, более примерно 60% сходства, более примерно 70% сходства, более примерно 80% сходства или более примерно 90% сходства.
Сходство и идентичность последовательностей обычно определяют согласно алгоритму GAP (Genetics Computer Group, Madison, WI). GAP использует алгоритм Нидлмана и Вунша (Needleman and Wunsch) для выравнивания двух полных последовательностей с максимальным числом совпадений и минимальным числом пробелов. Обычно применяют параметры по умолчанию при штрафе за создание пробела (gap creation penalty) = 12 и штрафе за продление пробела (gap extension penalty) = 4. Применение GAP может являться предпочтительным, но можно применять и другие алгоритмы, например, BLAST (который использует метод Альтшула и соавторов, Altschul et al. (1990) J. Mol. Biol. 215: 405-410), FASTA (который использует метод Пирсона и Липмана, Pearson and Lipman (1988) PNAS USA 85: 2444-2448) или алгоритм Смита-Уотермана (Smith and Waterman (1981) J. Mol Biol. 147: 195-197), или программу TBLASTN, Altschul et al. (1990) см. выше, обычно с применением заданных по умолчанию параметров. В частности, можно использовать алгоритм psi-Blast (Nucl. Acids Res. (1997) 25 3389-3402). Идентичность и сходство последовательностей можно также определить с применением программного обеспечения Genomequest™ (Gene-IT, Worcester MA USA). Сравнение последовательностей предпочтительно осуществляют относительно полноразмерной релевантной последовательности, описанной в данной заявке. Сходство допускает «консервативную изменчивость», то есть замену одного гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин, на другой, или замену одного полярного на другой, например аргинина на лизин, глутаминовой кислоты на аспарагиновую или глутамин на аспарагин.
В некоторых способах реализации настоящего изобретения полипептид АроЕ может представлять собой полипептид АроЕ млекопитающего, например АроЕ мыши, имеющий последовательность в базе данных под номером доступа ААН83351.1 или АроЕ человека, имеющий последовательность в базе данных под номером доступа Р02649 или NP_000032.1 или аллельный вариант, описанный в данной заявке.
Фрагмент полноразмерной последовательности может состоять из меньшего числа аминокислот, чем полная последовательность. Например, фрагмент может состоять из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот полноразмерной последовательности длины, но не более 290, не более 280, не более 270, не более 260, не более 250, не более 240, не более 230, не более 220, не более 210 или не более 200 аминокислот.
Патологические состояния, связанные с повышенной воспалительной активностью, могут включать рассеянный склероз, ревматоидный артрит, синдром раздраженного кишечника, болезнь Крона, инсульт, инфаркт миокарда, астму, аллергический ринит, экзему, псориаз и контактный дерматит с повышенной чувствительностью.
В некоторых способах реализации настоящего изобретения полипептид АроЕ находится на поверхности макрофага и/или апоптотических клеток. В других способах реализации настоящего изобретения полипептид АроЕ присутствует в реакционной среде.
В некоторых способах реализации настоящего изобретения способность тестируемого соединения стимулировать поглощение апоптотических клеток макрофагами можно определять в отсутствие АроЕ. Способ идентификации и/или получения соединения для лечения или предотвращения патологических состояний, связанных с повышенной воспалительной активностью, может включать определение поглощения одной или более дефектных по АроЕ апоптотических клеток макрофагами, дефектными по АроЕ, в присутствии тестируемого соединения.
Клетки, дефектные по АроЕ, можно, например, получить от дефектной по АроЕ мыши, которая может быть получена известными способам, как описано в данной заявке.
Поглощение дефектными по АроЕ апоптотических клеток макрофагами в присутствии тестируемого соединения можно оценивать по сравнению с поглощением в аналогичной реакционное среде без АроЕ и аналогичных условиях при отсутствии тестируемого соединения.
Накопление продуктов разрушения клеток у индивида, например, в результате повреждения ткани (например, травмы головы или огнестрельной раны) или выраженном инфекционном заболевании (таком, как бактериальный сепсис, панкреатит или перикардит) может привести к значительным медицинским осложнениям.
Способы, описанные в данной заявке, могут быть полезными для идентификации и/или получения соединений, которые стимулируют уничтожение апоптотических клеток, и могут быть полезным при лечении патологического состояния, связанного с накоплением апоптотических клеток.
Способ идентификации и/или получения соединения для лечения патологических состояний, связанного с накоплением апоптотических клеток, может включать определение способности полипептида АроЕ стимулировать поглощение апоптотических клеток макрофагами в присутствии тестируемого соединения.
Стимулирование поглощения или фагоцитоза апоптотических клеток макрофагами полипептидом АроЕ в присутствии тестируемого соединения можно сравнивать с поглощением в аналогичной реакционной среде и условиях отсутствия тестируемого соединения. Более сильное стимулирование в присутствии тестируемого соединения по сравнению со случаем его отсутствия свидетельствует о том, что соединение может быть полезным при лечении патологического состояния, связанного с накоплением апоптотических клеток.
В некоторых способах реализации настоящего изобретения полипептид АроЕ может находиться на поверхности макрофага и/или апоптотической клетки. В других способах реализации настоящего изобретения полипептид АроЕ может присутствовать в реакционной среде.
Патологические состояния, связанные с накоплением апоптотических клеток, включают повреждение тканей, травматическое повреждение мозга, острую дыхательную недостаточность, бактериальный сепсис, панкреатит или перикардит.
В некоторых способах реализации настоящего изобретения способность тестируемого соединения стимулировать поглощение апоптотических клеток макрофагами может быть определена в отсутствие АроЕ.
Способ идентификации и/или получения соединения для лечения или предотвращения патологических состояний, связанных с накоплением апоптотических клеток, может включать:
приведение в контакт дефектного по АроЕ макрофага с одной или более дефектной по АроЕ апоптотической клеткой в присутствии тестируемого соединения; и
определение поглощения макрофагами апоптотических клеток.
Поглощение апоптотических клеток дефектным по АроЕ макрофагом в присутствии тестируемого соединения можно сравнить с поглощением в аналогичной реакционной среде без АроЕ и условиях отсутствия тестируемого соединения.
Макрофаги представляют собой большие одноядерные фагоцитарные клетки, которые присутствуют в крови, лимфе и других тканях и поглощают чужеродные вещества и продукты разрушения клеток, включая апоптотические клетки. Макрофаги вовлечены как в специфический, так и в неспецифический иммунные ответы.
Макрофаг для применения в способах согласно настоящему изобретению может быть культивируемым макрофагом. Культивируемые макрофаги могут быть получены при помощи одного из ряда подходящих способов, включая, например, дифференцировку in vitro свежеприготовленных моноцитов периферической крови человека (например, под действием форболовых эфиров); дифференцировку in vitro моноцитарных клеточных линий, таких как миеломоноцитарная клеточная линия ТНР-1 человека; и получение перитонеальных макрофагов путем промывки брюшины животного (такого как мышь или крыса) стерильным буфером и культивирования образующегося в результате этого брюшинного экссудата.
Апоптотическая клетка представляет собой клетку, которая претерпевает или претерпела апоптоз или программируемую клеточную гибель. Апоптотическая клетка для применения в способах согласно настоящему изобретению предпочтительно представляют собой клетку млекопитающего и может происходить из той же ткани, из того же организма или организма того же вида, что и макрофаги или может происходить из другой ткани, организма и/или вида. Для удобства в способах настоящего изобретения можно применять апоптотические тимоциты.
В данной области техники известен ряд подходящих способов для получения апоптотических клеток, включающих, например, удаление сыворотки от культивируемых первичных лимфоцитов, обычно тимоцитов; обработку культивируемых клеток индукторами апоптоза, такими как Fas-лиганд или гранзим В; и выделение клеток, претерпевающих апоптоз, in vivo с применением селективных маркеров индукции апоптоза, располагающихся на поверхности клеток (например, связывание с аннексином V). Обычно более чем 50% используемой популяции клеток находятся в состоянии апоптоза, что определяют, например, при помощи окрашивания меченным аннексином V или окрашиванием на активированную каспазу 3.
Поглощение апоптотической клетки макрофагом можно определить любым из подходящих способов. Например, к макрофагу или популяции макрофагов добавляют фиксированное количество апоптотических клеток, например в серию дублированных лунок многолуночного планшета, варьируя от 0,1 апоптотической клетки на макрофаг до 1000 апоптотических клеток на макрофаг, обычно от 1 до 100 апоптотических клеток на макрофаг. В предпочтительных способах реализации настоящего изобретения тестируемое соединение присутствует в культуральной среде в то время, когда происходит фагоцитоз.
Макрофаги можно инкубировать с апоптотическими клетками в определенный период времени, в течение которого происходит фагоцитоз. Обычно инкубацию проводят при температуре между 25°C и 40°C, как правило, при 37°C. Длительность инкубации можно выбрать такую, при которой поглощается детектируемое число апоптотических клеток, но поглощается не более чем 50% от общего числа добавленных апоптотических клеток (чтобы доступность апоптотических клеток не стала лимитирующей для дальнейшего фагоцитоза). Инкубация может занимать, например, от 10 минут до 10 часов при 37°C, как правило, от 30 минут до 2 часов.
В некоторых способах реализации настоящего изобретения можно определить число или долю апоптотических клеток, которые поглотили макрофаги путем фагоцитоза. Это удобно выполнять путем детекции апоптотических клеток, которые были добавлены к макрофагам, и различения апоптотических клеток снаружи или на внешней поверхности, макрофагов и апоптотических клеток внутри или на внутренней поверхности макрофагов.
Апоптотические клетки можно детектировать любым пригодным способом, известным в данной области техники, это удобно выполнять с помощью предварительного мечения клеток до индукции апоптоза. Ряд подходящих меток и красителей коммерчески доступен, в том числе Cell Tracker Green (Molecular Probes Inc.). Меченые апоптотические клетки можно затем идентифицировать и подсчитать путем детекции метки в подходящих условиях, например при помощи микроскопии.
Апоптотические клетки, которые находятся снаружи или внутри, можно различить путем промывки макрофагов в условиях, при которых апоптотические клетки, которые находятся снаружи от (или на внешней поверхности) макрофагов, удаляют и не учитывают, тогда как апоптотические клетки, которые находятся внутри (или на внутренней поверхности) макрофагов остаются. Можно применять любой подходящий способ промывки, при котором удаляются внешние, но не внутренние апоптотические клетки, в том числе, например, многократные промывки холодным ФСБ-Дульбекко. После промывки можно подсчитать общее число поглощенных апоптотических клеток вместе с общим числом макрофагов, участвующих в фагоцитозе.
Скорость фагоцитоза можно вычислить на основе величины фагоцитоза, происходящего в течение определенного периода времени, и для удобства ее можно представить как количество апоптотических клеток (например, тимоцитов), поглощенных макрофагом за час. Обычно эта величина колеблется от 0,1 до 100 у клеток дикого типа при отсутствии стимулятора фагоцитоза. Например, для перитонеальных макрофагов мыши, поглощающих апоптотические тимоциты, эта величина может колебаться от 2 до 4, а для макрофагов, дефектных по ароЕ, поглощающих тимоциты, дефектные по АроЕ, эта величина может колебаться от 0,5 до 1.
Эффект тестируемого соединения можно выразить в виде кратности изменения данной величины по сравнению с контрольными клетками, на которые не воздействовали данным соединением. Типичный ответ, полученный с применением рекомбинантного АроЕ человека в качестве тестируемого соединения, представлен на Фигуре 2.
Тестируемое соединение может быть добавлено в соответствующей концентрации, которую обычно определяют методом проб и ошибок, в зависимости от типа применяемого соединения, но которая обычно составляет от 10 пМ до 10 мМ, в основном от 1 нМ до 100 мкМ. Тестируемое соединение может быть добавлено в любой подходящий биосовместимый буфер, известный в данной области техники, в котором соединение достаточно хорошо растворимо, такой как ДМСО, этанол, метанол, ДМСО, ДМА, ДМФ или вода.
Обычно культивируемые макрофаги подвергают воздействию различных концентраций тестируемого соединения (включая контроли, на которых воздействуют только разбавителем), например тестируемое соединение может быть добавлено в продублированные лунки многолуночного планшета, в группах от двух до десяти лунок, как правило, от трех до пяти лунок. Клетки можно, кроме того, подвергать воздействию тестируемого соединения в течение различных временных интервалов, от нескольких минут до нескольких часов или более, обычно при 37°C.
Тестируемое соединение, пригодное для применения согласно способам настоящего изобретения, может быть малой химической молекулой, пептидом, молекулой антитела или другой молекулой, чье действие на фагоцитоз апоптотической клетки необходимо определить. Можно применять природные или синтетические химические соединения или растительные экстракты, которые содержат некоторое количество охарактеризованных или неохарактеризованных компонентов.
Подходящие тестируемые соединения могут быть соединениями, выбранными из коллекции, или сконструированными соединениями. Технология комбинаторных библиотек (Schultz, JS (1996) Biotechnol. Prog. 12:729-743) предоставляет эффективный способ тестирования потенциально большого числа различных веществ на способность стимулировать фагоцитоз апоптотических клеток.
В некоторых предпочтительных способах реализации настоящего изобретения соответствующее тестируемое соединение может быть пептидным фрагментом полипептида АроЕ млекопитающего, например АроЕ человека, или его аналогом, миметиком, производным или модифицированной формой. Например, пептидные фрагменты от 5 до 40 аминокислот, например от 6 до 10 аминокислот АроЕ, могут быть протестированы на способность стимулировать фагоцитоз апоптотических клеток.
Пептидные фрагменты АроЕ могут быть получены любым способом синтеза пептидов, известным в данной области техники, например химическим синтезом или путем экспрессии рекомбинантного гена in vitro или in vivo.
Пептиды АроЕ могут быть созданы целиком или частично путем химического синтеза. Пептид АроЕ, как здесь описано в данной заявке, может быть легко получен согласно общепринятым стандартным жидкостным или предпочтительно твердофазным способам пептидного синтеза, общее описание которых широко доступно (см, например, работу J.M.Stewart and J.D.Young, Solid Phase Peptide Synthesis, 2nd edition, Pierce Chemical Company, Rockford, Illinois (1984), работы М.Bodanzsky and A.Bodanzsky, The Practice of Peptide Synthesis, Springer Verlag, New York (1984); Applied Biosystems 430A Users Manual, ABI Inc., Foster City, California; Roberge, J.Y. et al. (1995) Science 269:202-204; Caruthers, М.H. et al. (1980) Nucl. Acids Res. Symp.Ser. 215-223; Horn, T. et al. (1980) Nucl. Acids Res. Symp. Ser. 225-232; and Merrifield J. (1963) J. Am. Chem. Soc. 85:2149-2154), или может быть получен в растворе, жидкофазным способом или при помощи любой комбинации твердофазного, жидкофазного способа или синтеза в растворе, например сначала путем получения соответствующей пептидной части и затем, если это является желательным и уместным, после удаления любой из присутствующих защитных групп, путем введения группы Х при помощи реакции с соответствующей карбоновой или сульфокислотой или их химически активным производным. Можно выполнять автоматизированный синтез, например, с применением пептидного синтезатора ABI 431 (Applied Biosystems).
Пептид АроЕ может иметь N-концевую кэпирующую (защитную) группу (например, ацетильную группу) и/или С-концевую кэпирующую группу, (например, амидную группу) для защиты концевого остатка от нежелательных химических реакций в ходе применения или для возможности осуществления последующего конъюгирования или манипуляций с пептидом. Например, модулирующие свойства пептидного фрагмента, как описано выше, можно усилить путем добавления одной из следующих групп к С-концу: хлорметилкетона, альдегида и бороновой кислоты. Эти группы являются аналогами переходного состояния [субстрата] для сериновых, цистеиновых и треониновых протеаз. N-конец пептидного фрагмента может быть блокирован карбобензилом для ингибирования аминопептидаза и увеличения стабильности (Proteolytic Enzymes 2nd Ed, Edited by R.Beynon and J.Bond Oxford University Press 2001).
Вновь синтезированный пептид может быть в значительной степени очищен препаративной высокоэффективной жидкостной хроматографией (Creighton, Т. (1983) Proteins, Structures and Molecular Principles, W H Freeman and Co., New York, N.Y.) или при помощи других аналогичных способов, доступных в данной области техники. Состав синтетических пептидов можно подтвердить анализом аминокислотного состава или секвенированием (например, при помощи деградации по методу Эдмана).
Получение рекомбинантного пептида АроЕ in vivo или in vitro можно осуществить путем экспрессии нуклеиновой кислоты, содержащей кодирующую нуклеотидную последовательность, с применением стандартных рекомбинантных способов.
Другие подходящие соединения могут быть основаны на моделировании 3-мерной структуры АроЕ и применении рационального дизайна лекарств, который обеспечивает потенциально активные соединения с конкретными характеристиками формы, размера и заряда молекулы. Способы и средства рационального дизайна лекарств хорошо известны в данной области техники.
Эффект соединения, идентифицированного описанным выше способом, можно оценить при вторичном скрининге. Например, действие соединения на один или более симптомов патологического состояния, описанного выше, можно определить in vivo на модельном животном.
Контрольные эксперименты могут быть выполнены в соответствии со способами, описанными в данной заявке. Постановка соответствующих контролей находится в пределах компетенции и способности специалиста в данной области техники.
Метод, как описано в данной заявке, может включать идентификацию тестируемого соединения как агента, стимулирующего, или повышающего, или увеличивающего фагоцитоз апоптотической клетки.
Идентифицированное соединение может быть изолировано и/или очищено. В некоторых способах реализации настоящего изобретения соединение может быть получено, синтезировано и/или произведено с применением стандартных способов синтеза.
Соединения, идентифицированные как агенты, стимулирующие фагоцитоз апоптотических клеток, с применением способа, описанного в данной заявке, факультативно могут быть модифицированы или подвергнуты технике рационального дизайна лекарств для оптимизации активности или обеспечения других полезных характеристик, таких как увеличение периода полувыведения или уменьшение побочных действий при введении индивиду.
Кроме того, можно проводить дальнейшую оптимизацию или модификацию для получения одного или более конечных соединений либо для in vivo или при клинических испытаниях.
Соединение, полученное способами согласно настоящему изобретению, описанными выше, может входить в состав композиции, например лекарственного препарата, фармацевтической композиции или лекарства, с фармацевтически приемлемым наполнителем, носителем, буфером, стабилизатором или другими веществами, хорошо известными специалисту в данной области техники.
Фармацевтически приемлемый наполнитель или другое вещество должен быть нетоксичным и не должен препятствовать действию активного компонента. Точная природа носителя или другого материала зависит от способа введения, который может быть пероральным, или путем инъекции, например, кожной, подкожной или внутривенной.
Способ создания фармацевтической композиции для применения при лечении патологического состояния, описанного в данной заявке, может включать;
идентификацию и/или получение соединения, стимулирующего фагоцитоз апоптотических клеток, с применением способа, описанного в данной заявке, и
смешивание идентифицированного таким образом соединения с фармацевтически приемлемым носителем.
Как описано выше, соединение может быть модифицировано для оптимизации его фармацевтических свойств.
Способ получения фармацевтической композиции для лечения патологического состояния, описанного в данной заявке, может включать;
i) идентификацию и/или получение соединения, стимулирующего фагоцитоз апоптотических клеток, например, с применением способа, описанного в данной заявке,
ii) синтез идентифицированного соединения и
iii) включение соединения в фармацевтическую композицию.
Фармацевтическая композиция может содержать в дополнение к соединению, идентифицированному в качестве стимулятора фагоцитоза апоптотических клеток, фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие вещества, хорошо известные специалисту в данной области техники.
Фармацевтические композиции для перорального введения могут быть в форме таблеток, капсул, порошка или в жидкой форме. Таблетка может включать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические композиции обычно включают жидкий носитель, такой как вода, углеводороды, животные или растительные масла, минеральное масло или синтетическое масло. В состав композиции могут входить физиологический солевой раствор, раствор декстрозы или другого сахарида или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной, кожной или подкожной инъекции или инъекции в область повреждения композиция может быть в форме приемлемого для парентерального введения водного раствора, который является апирогенным и имеет подходящие pH, изотоничность и стабильность. Специалисты в данной области техники способны подготовить соответствующие растворы с применением, например, изотонических разбавителей, таких как физиологический раствор для инъекций, раствор Рингера для инъекций или лактат Рингера для инъекций. В состав композиции при необходимости могут входить консерванты, стабилизаторы, буферы, антиоксиданты и/или другие вспомогательные вещества.
Изобретение охватывает соединение, идентифицированное и/или полученное с применением способа, описанного выше, в качестве агента, который может быть полезен при лечении патологического состояния, описанного выше, фармацевтическую или ветеринарную композицию, лекарственный препарат, лекарство или другой состав, включающий такое соединение, способ, включающий введение такой композиции индивиду, например человеку или животному, не являющемуся человеком, для лечения (которое может включать профилактическое лечение патологического состояния, описанного выше, применение такого соединения для производства композиции для введения, например для лечения патологического состояния, описанного выше, и способ получения фармацевтической или ветеринарной композиции, включающий смешивание такого соединения с наполнителем, разбавителем или носителем, например фармацевтически приемлемым наполниетелем, разбавителем или носителем, и, возможно, с другими компонентами.
Введение соединения предпочтительно осуществляют в «профилактически эффективном количестве» или «терапевтически эффективном количестве» (в зависимости от обстоятельств, хотя профилактику можно считать терапией), которое при этом является достаточным для оказания полезного действия на индивида. Фактическое вводимое количество, а также скорость и периодичность введения зависят от природы и тяжести заболевания, которое подвергают лечению. Назначение лечения, например определение дозы и т.п., находится в рамках обязанностей врачей общей практики и врачей других специальностей.
Другие аспекты изобретения относятся к применению полипептидов АроЕ и их миметиков для стимулирования уничтожения апоптотических клеток, например, при лечении патологического состояния, связанного с накоплением апоптотических клеток.
Способ лечения патологического состояния, связанного с накоплением апоптотических клеток у индивида, может включать: увеличение экспрессии и/или активности АроЕ у вышеуказанного индивида.
Патологические состояния, связанные с накоплением апоптотических клеток, включают повреждение ткани, травматическое повреждение мозга, острую дыхательную недостаточность, бактериальный сепсис, панкреатит или перикардит.
Активность АроЕ можно повысить, например, при введении полипептида АроЕ или его миметика указанному индивиду.
Полипептиды АроЕ описаны более подробно выше.
Миметики АроЕ представляют собой соединения, сохраняющие активность АроЕ при стимулировании фагоцитоза апоптотических клеток. Миметики АроЕ могут быть получены, например, при определении конкретных частей соединения, которые необходимы и/или важны для обладания требуемым свойством. Это можно сделать, например, путем систематичного изменения аминокислотных остатков в пептиде, например путем поочередного изменения каждого остатка. Эти части или остатки, составляющие активную область АроЕ, известны как «фармакофор».
После обнаружения фармакофора создают модель его структуры в соответствии с его физическими свойствами, например стереохимией, связями, размером и/или зарядом, с применением данных из различных источников, например, полученных способами спектроскопии, рентгенодифракционным методом и ЯМР. В процессе такого моделирования можно применять компьютерный анализ, подобное отображение (моделирующее заряд и/или объем фармакофора, но не связи между атомами) и другие способы.
Затем выбирают молекулу-матрицу, к которой можно присоединить химические группы, имитирующие фармакофор. Молекулу-матрицу и химические группы, присоединяемые к ней, удобно выбирать такими, чтобы модифицированное соединение можно было легко синтезировать, чтобы оно, насколько это возможно, было фармакологически приемлемым и не разрушалось in vivo и при этом сохраняло способность стимулировать фагоцитоз апоптотических клеток. Миметики АроЕ, полученные с помощью такого подхода, можно затем скринировать с применением способов, описанных в данной заявке, чтобы обнаружить, стимулируют ли они или насколько стимулируют фагоцитоз апоптотических клеток.
Другие аспекты изобретения обеспечивают полипептид АроЕ или его миметик для применения при лечении патологического состояния, связанного с накоплением апоптотических клеток, и применение полипептида АроЕ или его миметика для получения лекарственного препарата для лечения патологического состояния, связанного с накоплением апоптотических клеток.
В других аспектах настоящее изобретение относится к стимулированию уничтожения апоптотических клеток, например, при лечении патологического состояния, связанного с уменьшением эндогенного АроЕ или повышенной воспалительной активностью.
Способ лечения патологического состояния, связанного с уменьшением содержания эндогенного АроЕ или повышенной воспалительной активностью у индивида, может включать введение соединения, стимулирующего фагоцитоз апоптотических клеток.
Соединение может быть фрагментом пептида АроЕ человека или его аналогом или производным. Соответствующие соединения описаны более подробно выше.
Патологическое состояние, связанное с уменьшением эндогенного ароЕ, может быть выбрано из группы, состоящей из болезни Альцгеймера, атеросклероза, инсульта и остеопороза
Патологическое состояние, связанное с повышенной воспалительной активностью, может быть выбрано из группы, состоящей из рассеянного склероза, ревматоидного артрита, синдрома раздраженного кишечника, болезни Крона, инсульта, инфаркта миокарда, астмы, аллергического ринита, экземы, псориаза и контактного дерматита с повышенной чувствительностью.
В других аспектах настоящее изобретение предусматривает соединение, стимулирующее фагоцитоз апоптотических клеток, для лечения патологического состояния, связанного со сниженной активностью эндогенного АроЕ или повышенной воспалительной активностью, и применение соединения, стимулирующего фагоцитоз апоптотических клеток, для производства лекарственного препарата для лечения патологического состояния, связанного со сниженной активностью эндогенного АроЕ или повышенной воспалительной активностью.
Различные дополнительные аспекты и способы реализации очевидны для специалиста в данной области техники на основании настоящего описания. Содержание всех документов, упомянутых в данном описании, включено посредством ссылки в полном объеме.
Изобретение охватывает все комбинации и частичные комбинации характерных черт, описанных выше.
Определенные аспекты и способы реализации настоящего изобретения проиллюстрированы посредством примеров и со ссылками на фигуры, описанные ниже.
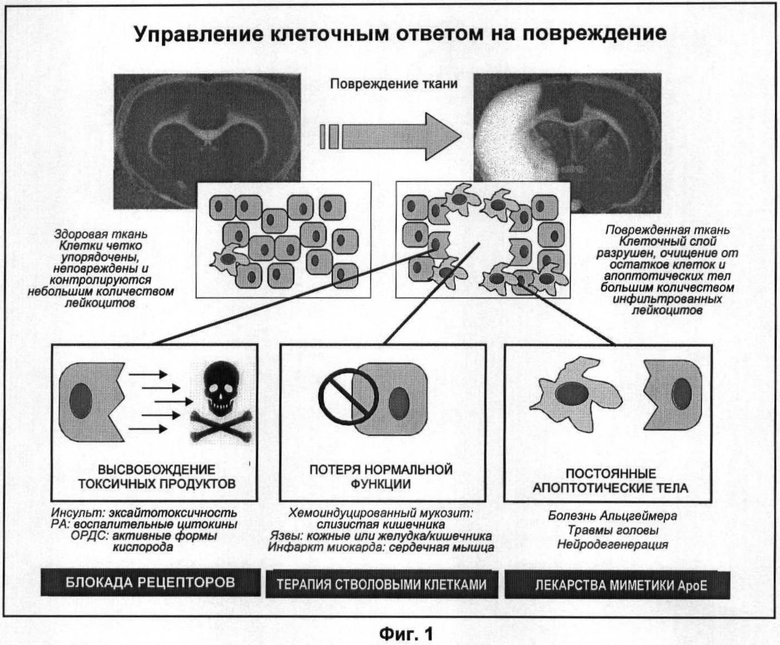
На Фигуре 1 показаны три пути, составляющие ответ ткани на повреждение. После повреждения ткани (такого как церебральный инфаркт во время инсульта, показанный сверху на изображениях мозга в норме (слева) и церебрального инфаркта (справа), окрашенных для обнаружения поврежденной области (белым)), организм должен справиться с выделяющимися цитокинами и другими продуктами поврежденных клеток с потерей функции погибшей ткани и должен уничтожить продукты разрушения и апоптотические клетки. Эта третья составляющая ответа может быть слабоэффективной у некоторых индивидов (например, у имеющих определенный полиморфизм по гену ароЕ).
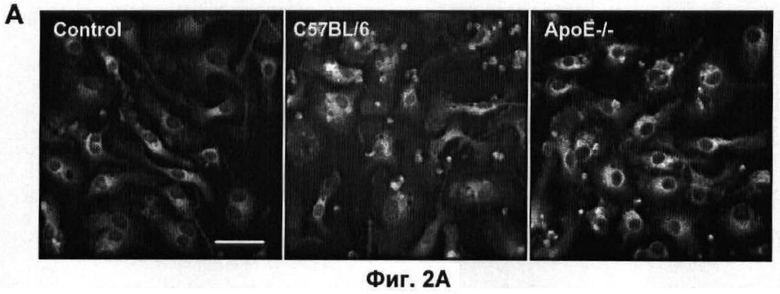
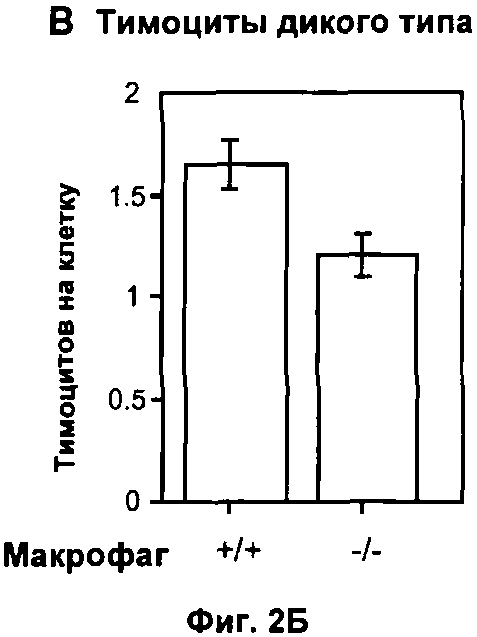
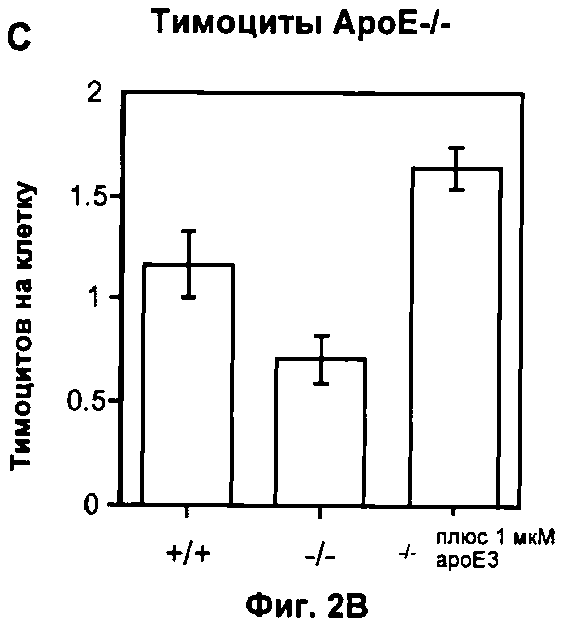
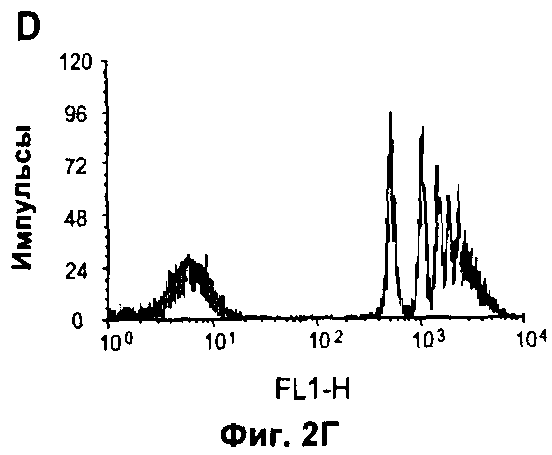
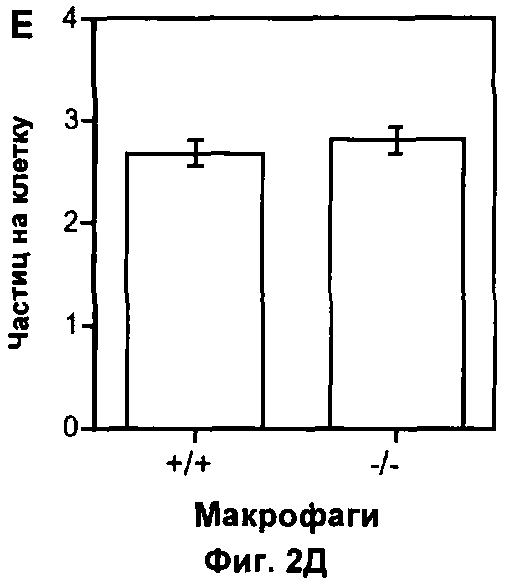
На Фигуре 2 показано влияние дефекта АроЕ на фагоцитоз in vitro: (A) Флуоресцентные микрофотографии макрофагов после поглощения меченых апоптотических тимоцитов в течение 30 минут при 37°C. Контрольное изображение получено без добавления тимоцитов. Изображения получали при высоком коэффициенте усиления, так что видны немеченые макрофаги из-за автофлуоресценции в дополнение к меченым тимоцитам (показаны стрелками). Масштаб 25 мкм. (Б) Количественная оценка тимоцитов дикого типа, поглощенных одним макрофагом, на изображениях, таких как представлены на (А). Общее число тимоцитов в десяти полях зрения делили на общее число макрофагов. Значения представлены в виде среднего ±СО по трем лункам в каждом из трех экспериментов, (В) Количественная оценка дефектных по АроЕ тимоцитов, поглощенных одним макрофагом. Также показан эффект преинкубации с 1 мкМ растворимого рекомбинантного АроЕ3 человека. (Г) Типичная гистограмма проточной цитометрии макрофагов после поглощения флуоресцентно меченных латексных частиц в течение 30 минут при 37°C. Клетки, не поглотившие частицы, имеют низкую интенсивность флуоресценции (FL1-H), тогда клетки, поглотившие 1, 2, 3 или больше частиц, постепенно смещаются в область более высокой интенсивности флуоресценции. Типичная гистограмма для макрофагов дикого типа показана черным, и для макрофагов, дефектных по ароЕ - красным цветом. (Д) Количественный анализ латексных частиц, поглощенных макрофагом из гистограмм проточной цитометрии, таких, как показаны на (Г). Значения представлены в виде среднего ±СО по трем лункам в каждом из трех экспериментов.
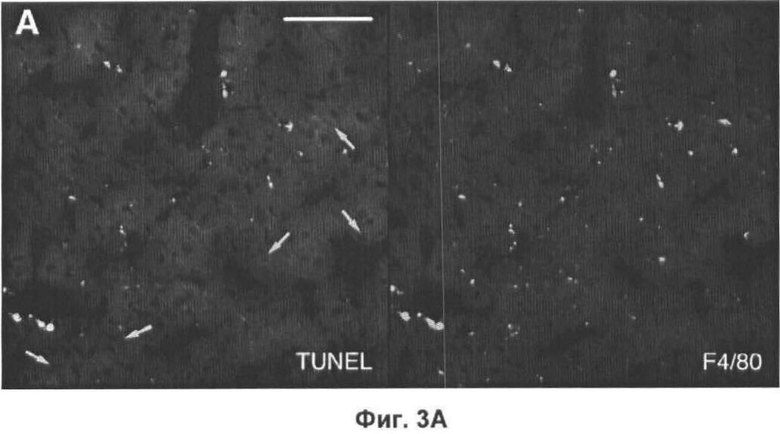
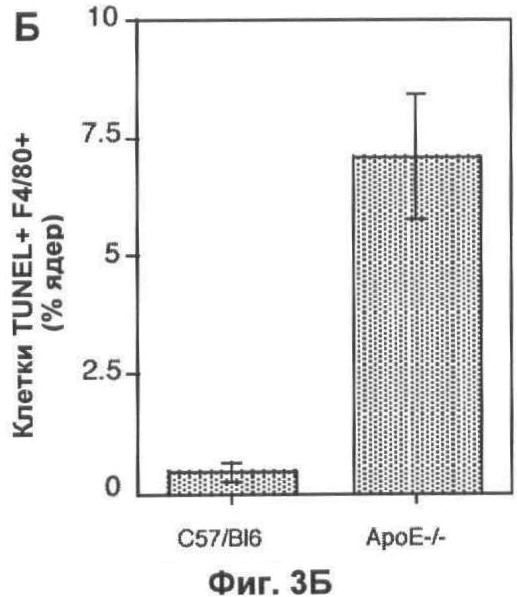
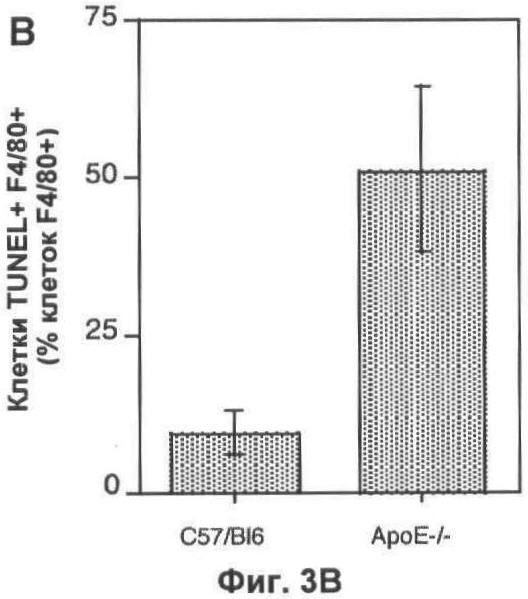
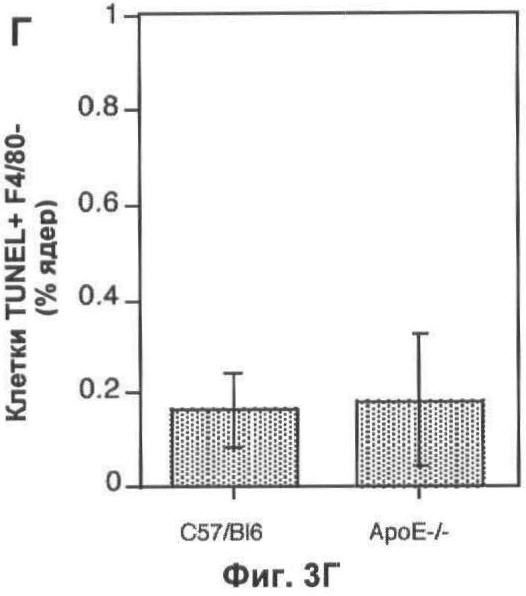
На Фигуре 3 представлено TUNEL-окрашивание печени мыши. (А) Две флуоресцентные микрофотографии с одного и того же поля зрения типичного среза печени мыши, дефектной по АроЕ. Макрофаги (главным образом, клетки Купфера) окрашены красным (правая часть) при помощи моноклонального антитела F4/80. Остатки клеток и погибающие клетки окрашены зеленым с применением метода TUNEL (левая часть). Все TUNEL+ клетки на данном изображении также являются F4/80+, но около 50% F4/80+ клеток являются TUNEL- (белые стрелки). Масштаб 25 мкм. (Б) Оценка абсолютного количества TUNEL+ F4/80+ клеток в 10 срезах печени каждой из 6 мышей каждого генотипа, выраженная в виде процента от общего числа проанализированных ядер. (В) Количество TUNEL+ F4/80+ клеток, выраженное в виде процента от количества F4/80+ клеток, на тех же срезах, которые анализировали в (Б). (Г) Количество TUNEL+ F4/80- клеток (то есть остатки клеток и погибающие клетки, не являющиеся макрофагами), выраженное в виде процента от общего числа проанализированных ядер. В (Б). (Г) значения представлены в виде среднего ±СО для шести животных.
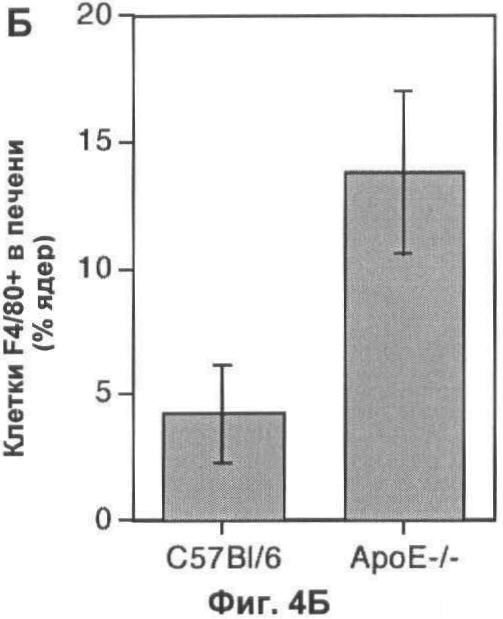
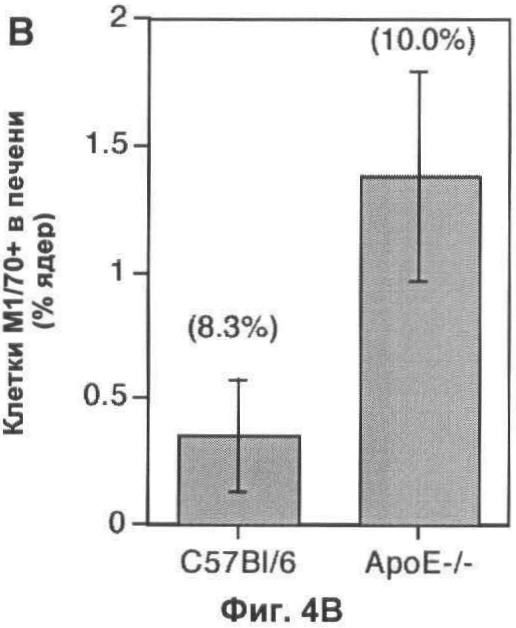
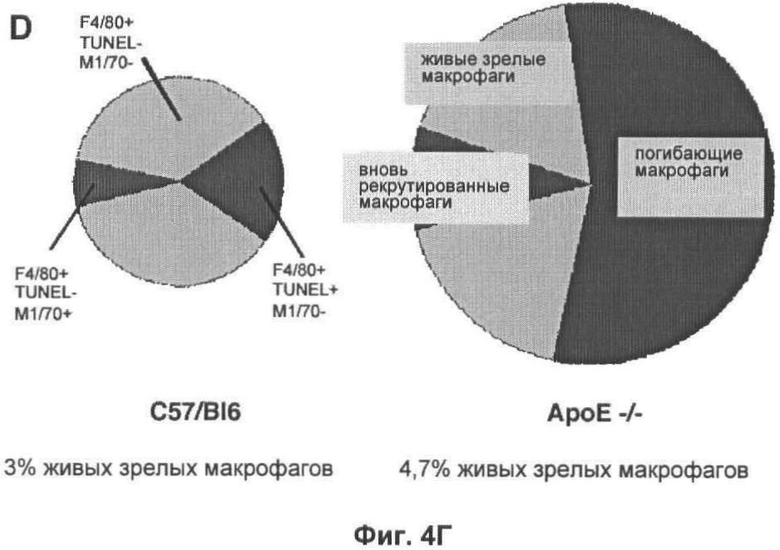
На Фигуре 4 показана динамика популяции макрофагов в печени мыши. (А) Флуоресцентные микрофотографии при слабом увеличении типичных срезов печени мышей дикого типа и мышей, дефектных по АроЕ, окрашенных с помощью моноклональных антител F4/80. При данном увеличении (масштаб 25 мкм), макрофаги видны в виде белых пятен, которые, как было подтверждено при высоком увеличении, представляют собой специфическое окрашивание клеток (вставка; масштаб 10 мкм). (Б) Оценка количества F4/80+ клеток на 10 срезах печени каждой из 6 групп мышей каждого генотипа, выраженная в виде процента от общего числа проанализированных ядер. (В) Оценка количества М1/70+ клеток (с высоким уровнем экспрессии интегрина CD11b, представляющих собой недавно рекрутированные макрофаги) на тех же срезах, что и (Б). Абсолютное количество М1/70+ клеток представлено в виде процента от общего числа проанализированных ядер, и числа над каждым отрезком представляют число М1/70+ клеток в виде доли от числа F4/80+ клеток в том же срезе. В (Б) и (В) значения представлены в виде среднего ±СО для шести животных. (Г) Общая характеристика динамики популяции макрофагов у мыши дикого типа (левая схема) и дефектной по АроЕ (правая схема). Площадь под графиком пропорциональна общему числу F4/80+ клеток и разделена на сегменты, показывающие доли популяции макрофагов, которые являются вновь рекрутированными (М1/70+ TUNEL-; зеленый), зрелыми (М1/70- TUNEL-; серый) или погибающими (М1/70- TUNEL+; красный). Также показано количество живых зрелых макрофагов (М1/70- F4/80+ TUNEL- клетки), выраженное в виде процента от общего числа ядер.
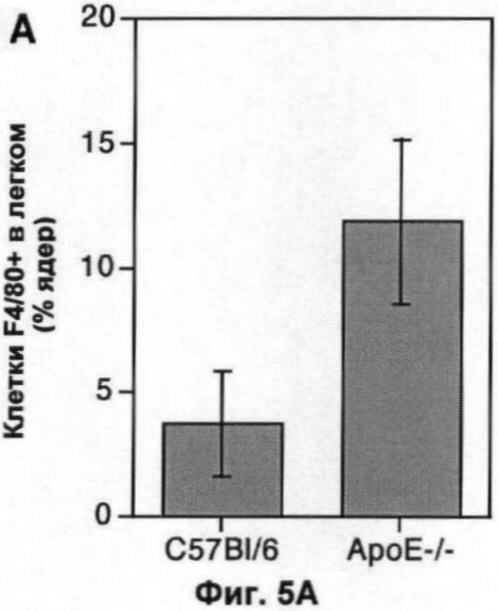
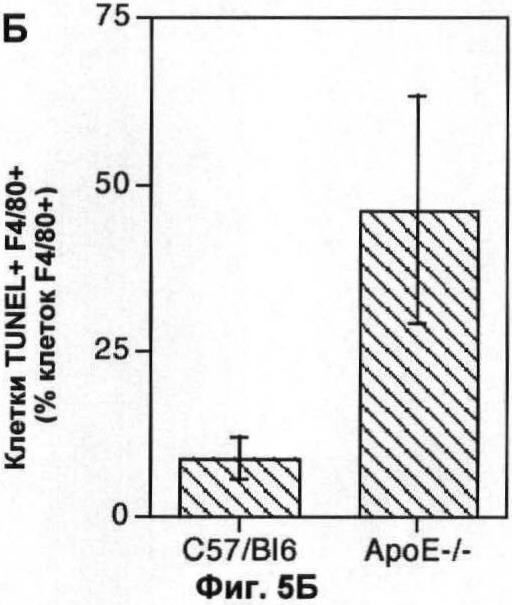
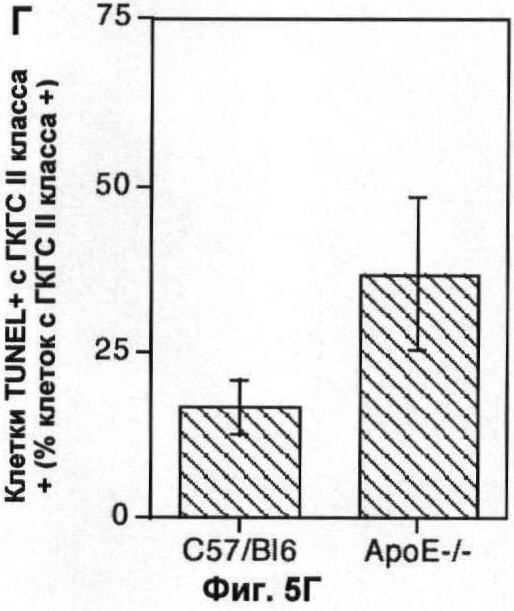
На Фигуре 5 показаны популяции макрофагов в легком и мозге мыши. (А) Количество F4/80+ клеток (по большей части, альвеолярные макрофаги) в 10 срезах легких 6 мышей каждого генотипа, выраженное в виде процента от общего числа проанализированных ядер. (Б) Количество TUNEL+ F4/80+ клеток в той же секции, что и в (А), выраженное в виде процента от числа F4/80+ клеток. (В) Количество ГКГС II класса + клеток (преимущественно микроглия) в 10 срезах мозга 6 мышей каждого генотипа, выраженное в виде процента от общего числа проанализированных ядер. (Г) Количество TUNEL+II класса + клеток в тех же срезах, что и (В), выраженное в виде процента от числа ГКГС II класса + клеток. В каждом случае значения представляют собой среднее ±СО для 6 животных.
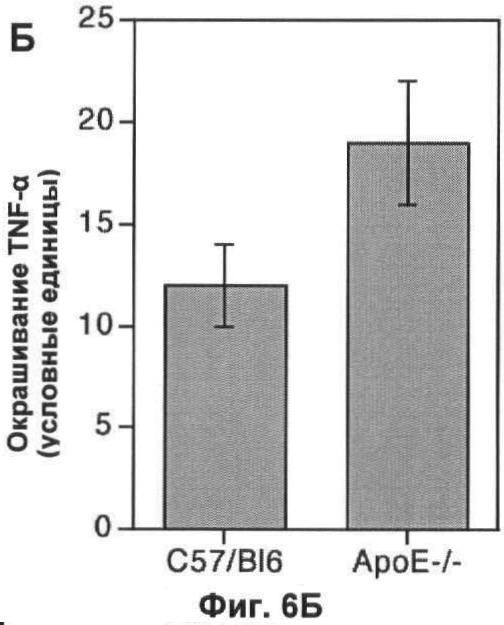
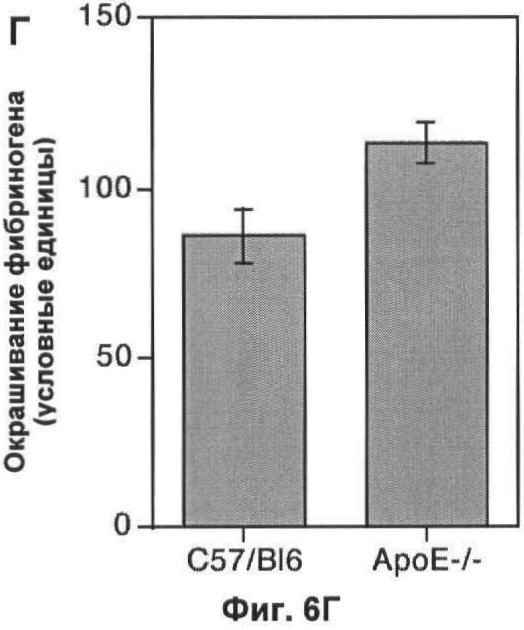
На Фигуре 6 показаны маркеры воспаления в печени мыши. (А) Флуоресцентные микрофотографии типичного среза печени мыши дикого типа, окрашенного на TNF-α. Большинство областей среза содержат слабо детектируемый TNF-α или не детектируемый TNF-α, а клетки в области небольшого кровяного сосуда окрашены интенсивно. Масштаб 25 мкм. (Б) Окрашивание по TNF-α в 10 срезах печени каждой из 6 мышей каждого генотипа, измеренное методами количественной иммунофлуоресценции, как описано ранее. (В) Флуоресцентные микрофотографии типичного среза печени мыши дикого типа, окрашенного на фибриноген. Высокая степень окрашивания видна в центральных венах каждой функциональной единицы печени, а также в синусоидных капиллярах Масштаб 25 мкм. (Г) Окрашивание на фибриноген 10 срезов печени каждой из 6 мышей каждого генотипа, измеренное методами количественной иммунофлуоресценции, как описано ранее. На каждом графике значения представляют собой среднее±СО для 6 мышей.
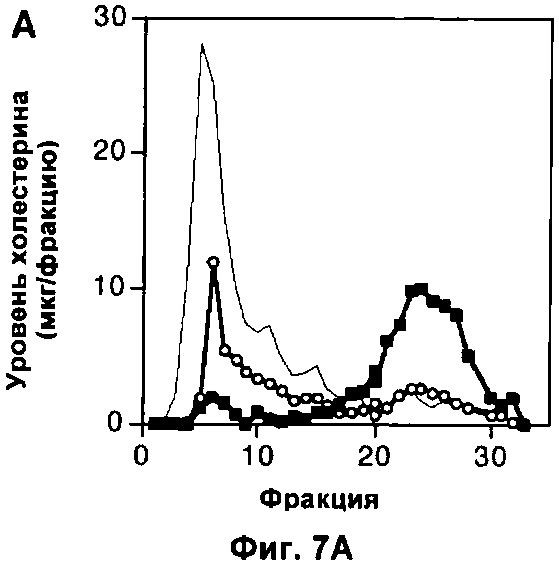
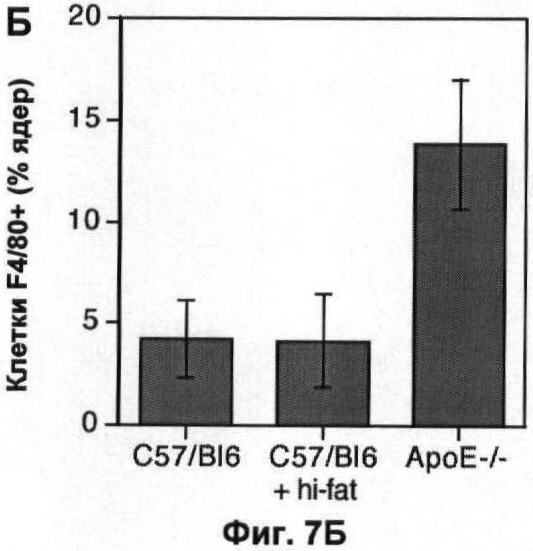
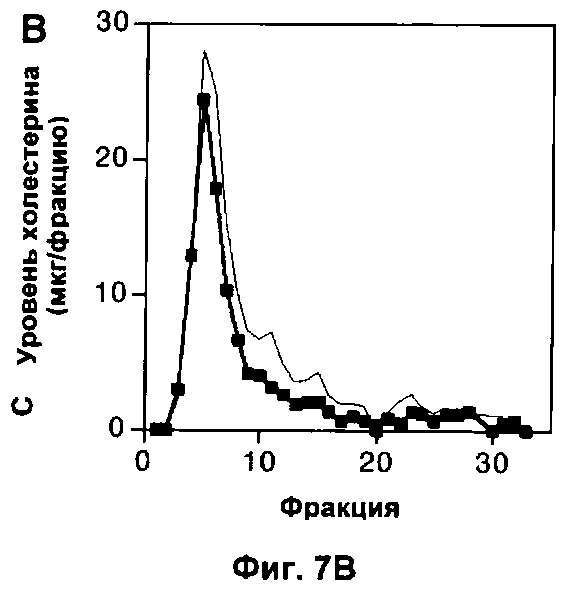
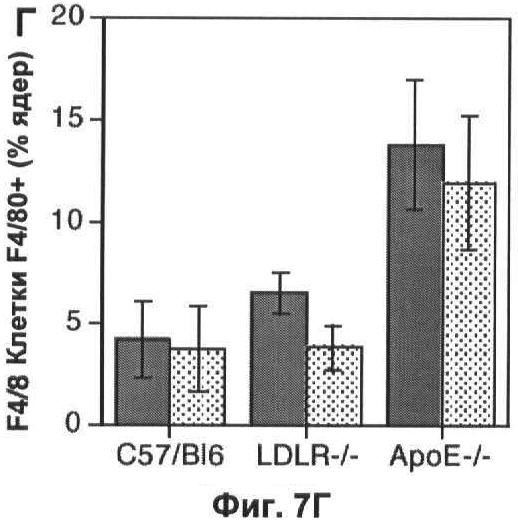
На Фигуре 7 показано влияние измененного метаболизма липопротеинов на популяцию макрофагов. (А) Профиль липопротеинов объединенных сывороток 6 мышей дикого типа, умерщвленных в возрасте 22 недель, которым давали корм либо с высоким содержанием жиров (неокрашенные кружки), либо нормальную пищу (окрашенные квадратики) в течение 10 недель. Профиль липопротеинов объединенных сывороток 6 мышей, дефектных по АроЕ, представлен для сравнения (простая линия). В данном протоколе ЛОНП элюировали до фракции 10, ЛНП между фракциями 10 и 20 и ЛВП после фракции 20. (Б) Количество F4/80+ клеток, выраженное в виде процента от общего числа проанализированных ядер, в 10 срезах печени каждой из 6 мышей в каждой группе. (В) Профиль липопротеинов объединенных сывороток 6 мышей, дефектных по рецептору, которым давали нормальный корм в ходе эксперимента. Профиль липопротеинов объединенных сывороток 6 мышей, дефектных по ароЕ, показан (простая линия) для сравнения. (Г) Количество F4/80+ клеток, выраженное в виде процента от общего числа проанализированных ядер, в 10 срезах печени (темные столбики) или легкого (светлые столбики) каждой из 6 мышей каждого генотипа. На каждом графике значения представляют собой среднее ±СО для 6 мышей.
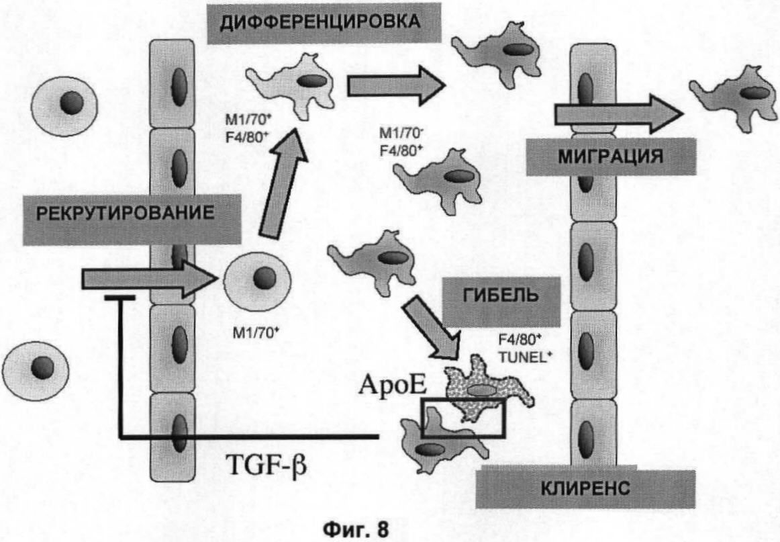
На Фигуре 8 показана модель динамики популяции макрофагов в печени мыши. Процессы увеличения равновесной популяции зрелых макрофагов (F4/80+) включают рекрутирование моноцитарных предшественников через эндотелий сосудов, дифференцировку через М1/70+ F4/80+ вновь рекрутированных макрофагов и пролиферацию. Процессы уменьшения равновесной популяции зрелых макрофагов включают миграцию через эндотелий в лимфу или кровь и гибель либо путем апоптоза, либо некроза (TUNEL+ F4/80+ клетки; розовая клетка, обозначенная пунктиром) с последующим их уничтожением с помощью фагоцитоза. Мы показали, что нарушение АроЕ уменьшает уничтожение остатков апоптотических клеток.
Эксперименты
Методы и материалы
Анализы фагоцитоза in vitro
Перитонеальные макрофаги мышей использовали во всех способах анализа фагоцитоза. Макрофаги асептически выделяли из мышей либо C57BI/6 (дикий тип), либо ароЕ-/- путем промывания брюшины мышей ледяным стерильным раствором Хэнкса (Sigma). Вымытые перитонеальные клетки хранили в предварительно охлажденных стерильных стеклянных пробирках, а затем промывали дважды средой RPMI (осаждали при 300g в течение 10 мин). Осадок клеток в конце ресуспендировали в RPMI + 10% ФТС (фетальная сыворотка телят) и рассевали из расчета 5×105 клеток/мл на пластиковых чашках для культур тканей (Nunc). Макрофагам позволяли адгезировать в течение 2 часов при 37°C, затем промывали холодным ФСБ Дульбекко перед инкубацией в RPMI + 10% ФТС до тех пор, пока не использовали в анализах фагоцитоза через 4-10 дней после выделения. Все клетки для анализа фагоцитоза тщательно промывали и инкубировали в бессывороточных условиях для исключения возможных эффектов, обусловленных присутствием АроЕ быка в ФТС, применяемой для культивирования клеток.
Анализ фагоцитоза с помощью латексных частиц проводили согласно Ichinose с изменениями [Ichinose, M. et al 1994 Cell Immunol 156:508]. Кратко, 2-мкм флуоресцентные микросферы (модифицированные карбоксилатом, Molecular Probes) в дистиллированной воде в концентрации 4,5×109 частиц/мл покрывали 1%-ным БСА, а затем разрушали ультразвуком в течение 5 минут. Монослои макрофагов промывали забуференным HEPES раствором солей pH 7,5 и предварительно инкубировали в течение 30 минут в 250 мкл этого же раствора. Покрытые БСА латексные частицы добавляли до концентрации 2,5×106 частиц на лунку, а затем инкубировали макрофаги в течение 1 часа при 37°С, в течение этого времени происходил фагоцитоз. Нефагоцитированные частицы затем удаляли путем пятикратной тщательной промывки ледяным ФСБ Дульбекко. Макрофаги затем освобождали добавлением 0,25% трипсина (Тип IIS из поджелудочной железы свиньи, Sigma) в течение 2-х часов при 37°С и фиксировали добавлением глутаральдегида (конечная концентрация 0,5%). Количество фагоцитированных частиц измеряли проточной цитометрией с использованием FACStar (Becton Dickinson), анализируя по 10000 клеток в пробирке, и высчитывали среднее количество фагоцитированных каждым макрофагом частиц (показатель фагоцитоза), используя программное обеспечение FCSPress.
Анализ фагоцитоза апоптотических тимоцитов проводили аналогично, по методике, описанной ранее, с изменениями [Scott, R.S. et al 2001. Nature 411:207]. Тимоциты получали из мышей C57BI/6 или АроЕ-/- путем изолирования тимуса в RPMI + 10% ФТС, забуференного 20 мМ Трис-HCl, pH 7,2 и пропускания суспензии через клеточный фильтр 40 меш (Sigma). Получившуюся суспензию клеток осаждали (500 g × 5 мин) и ресуспендировали в RPMI + 10% ФТС до 107 клеток/мл. За день перед использованием клеток в анализефагоцитоза (от 3 до 9 дней после помещения их в культуру) тимоциты промывали три раза стерильным ФСБ Дульбекко и доводили до концентрации 5×106 клеток/мл, а затем метили Cell Tracker Green (Molecular Probes; конечная концентрация 2 мкМ) в течение 30 минут при 37°C. Меченые клетки затем промывали ФСБ и инкубировали в свежей бессывороточной среде RPMI в течение 30 минут при 37°C, снова промывали ФСБ, а затем инкубировали бессывороточной среде RPMI в течение ночи. Удаление сыворотки приводили к более чем 70%-ному уровню апоптотических тимоцитов согласно измерению путем окрашивания аннексином V и проточной цитометрии. Анализы фагоцитоза проводили в соответствии с таковыми для латексных частиц за исключением того, что добавляли 2×106 апоптотических тимоцитов на лунку. Подсчитывали поглощенные тимоциты, фиксируя макрофаги на стеклянной подложке 1%-ной уксусной кислотой в 70%-ном этаноле в течение 90 минут и анализируя на флуоресцентном микроскопе (Provis AX; Olympus), соединенным с системой анализа изображения. Количество поглощенных тимоцитов и общее количество макрофагов подсчитывали в каждом из 18 зон обзора на лунку и высчитывали средний показатель фагоцитоза.
Приготовление препаратов тканей
Взрослых самцов мыши (мыши либо C57/BI6, либо АроЕ-/-, либо LDLR-/- [Ishibashi, S. et al 1994. J Clin Invest 93:1885]; по шесть животных в каждой группе) умерщвляли с помощью асфиксии CO2 и откачивали кровь для получения сыворотки с помощью сердечной пункции. Ткани (левую долю печени, левое легкое и левое полушарие мозга) быстро препарировали в ледяном физиологическом растворе, затем заливали в среде для заливки ОСТ и замораживали на -80°C. Для животных, стимулированных высоким содержанием липидов, заменяли обычную пищу пищей с высоким содержанием липидов (1,25% холестерина, 7,5% насыщенных жиров) в течение 10 недель перед умерщвлением. Все другие животные находились на обычной диете и имели неограниченный доступ к воде.
Срезы замороженных клеток (4 мкм) получали затем из каждой ткани, собирали на подложках, покрытых поли-L-лизином, и фиксировали в ледяном ацетоне в течение 90 с, высушивали на воздухе и замораживали на -20°C до анализа. Поперечные срезы делали с точки, находящейся примерно посередине ткани по переднезадней оси, через расстояние в 4 мм. Для каждого антигена (или группы антигенов в экспериментах по иммунофлуоресценции с 2-я или 3-я цветами) использовали по 16 срезов, сделанных через каждые 4 мм (каждые 250 мкм), где 10 срезов обрабатывали первичным антителом, а 6 срезов (случайно выбранных из 16), служивших в качестве контролей, не подвергали взаимодействию с первичным антителом, как было рекомендовано Mosedale с соавторами [Mosedale, D.E. et al. 1996. J Histochem Cytochem 44:1043].
Иммунофлуоресцентное окрашивание
Все реакции иммунофлуоресцентного окрашивания выполняли в соответствии с описанным Mosedale et al. при условиях, оптимизированных для количественной иммунофлуоресценции [Mosedale et al., см. выше]. Кратко, срезы регидратировали в буфере ФСБ, содержащем 3% БСА (бычий сывороточный альбумин) без жирных кислот, в течение 30 минут, затем подвергали взаимодействию с первичным антителом в ФСБ/3% БСА в течение 18 часов. После трехкратного промывания в ФСБ в течение 3 минут каждое препараты подвергали взаимодействию с соответствующим вторичным флуоресцентно меченным антителом в концентрации 25 мкг/мл в ФСБ/3% БСА в течение 6 часов, за исключением тех случаев, когда использовали меченые первичные антитела. После 3-х дополнительных промываний в ФСБ препараты промывали водой, высушивали на воздухе и готовили для изучения на Citifluor AF-1, затем замораживали на -20°C до исследования с помощью системы анализа изображений. Следующие имеющиеся в продаже первичные антитела были использованы: к антигену F4/80 макрофагов (МСАР497; Serotec; 25 мкг/мл); к CD11b (M1/70 (MCA74G); Serotec; 25 мкг/мл); к ГКГ класса II (I-Ab) (MCA1500F; Serotec; 20 мкг/мл); к фибрин(оген)у (4440-8004; Biogenesis; 20 мкг/мл); к мышиному TNF-α (AB-410-NA; R&D Systems; 50 мкг/мл); к PCNA (М0879; Dako; 12 мкг/мл). Все вторичные антитела представляли собой антитела осла с минимальной перекрестной активностью (Jackson Immunoresearch). Все растворы антител также содержали Хехст 33342 (конечная концентрация 1 мкг/мл) для контрастирующей окраски ядер, видимой в диапазоне синего света.
Определение апоптотических и некротических клеток
Погибающие клетки детектировали с использованием реакции TUNEL в соответствии с описанным ранее [Gavrieli, Y. et al. 1992. J Cell Biol 119:493], применяя набор «In Situ Cell Death Detection kit» (Roche) согласно инструкциям производителя. Срезы, предварительно обработанные ДНКазой I быка (Roche; конечная концентрация 10 мкг/мл в 50 мМ Трис pH 7,5, 1 мМ MgCl2), были использованы как положительные контроли; отсутствие флуоресцентно меченного дУТФ использовали в качестве отрицательного контроля. В печени мыши примерно 70% клеток TUNEL+ также окрашивались по активированной каспазе-3 (используя антитела СМ-1) и демонстрировали признаки конденсации в ядре при флуоресценции Хехст 33342, в результате чего можно предположить, что большинство клеток, определяемых как TUNEL+ с помощью указанного набора, подвергались апоптозу, в противоположность некрозу, согласно определениям Stadelmann и Lassmann [Stadelmann, С., and H. Lassmann. 2000. Cell Tissue Res 301:19], как заявлено производителями.
Анализ профиля липопротеинов
Объединенные образцы сыворотки подвергали гель-хроматографии с применением колонки Sepharose 6B в точности, как было описано ранее [Grainger, D.J. et al. 1995. Nat Med 1:1067]. Холестерин детектировали в получающихся фракциях, используя способ анализа с холестерин-оксидазой, и относили к разным классам липопротеинов на основании профилей элюирования аполипопротеинов, анализированных с помощью гель-электрофореза и Вестерн-блота, в соответствии с описанным ранее [Yokode, М. et al. (1990) Science 250:1273].
Результаты
Эффект мутаций по ароЕ на поглощение макрофагом апоптотических клеток in vitro
Перитонеальные макрофаги получали из мышей, дефектных по АроЕ (Е-/-), и мышей дикого типа (WT) в качестве контроля. После 96 часов культивирования клеток Е-/- и WT осуществляли их взаимодействие с флуоресцентно меченными апоптотическими тимоцитами WT, которых они фагоцитировали в течение 1 часа при 37°C. Количество тимоцитов, поглощенных макрофагом, измеряли с помощью иммунофлуоресцентной микроскопии (Фиг.2А) как показатель способности макрофагов уничтожать апоптотические тельца. В течение этого времени каждый макрофаг WT поглощал 1,65±0,12 апоптотических тимоцитов WT, но фагоцитоза не наблюдали при применении живых тимоцитов (данные не показаны), подтверждая тем самым, что этот анализ является специфичным для фагоцитоза апоптотических телец. В отличие от этого макрофаги Е-/- фагоцитировали 25% или менее апоптотических тимоцитов в течение того же времени (p<0.05, непарный Т-критерий Стьюдента; Фиг.2Б).
Проточная цитометрия с использованием ПТС-меченных антител к мышиному ароЕ показало, что тимоциты экспрессируют ароЕ, хотя на более низком уровне, чем макрофаги. Поэтому мы повторили эксперимент с использованием тимоцитов Е-/-. Интересно, что макрофаги WT не фагоцитируют тимоциты Е-/- так же эффективно, как и тимоциты WT (p<0.05, t-тест Стьюдента; Фиг.2В), и даже более значительное нарушение в фагоцитозе апоптотических телец наблюдали в случае, когда и фагоцитирующие макрофаги, и апоптотические тимоциты являлись дефектными по ароЕ, при этом фагоцитировалось примерно на 60% или менее апоптотических клеток по сравнению с системой дикого типа (p<0,05, ANOVA; Фиг.2В). Таким образом, эти результаты показывают, что хотя фагоцитоз апоптотических телец может иметь место при полном отсутствии ароЕ, процесс заметно ослаблен.
Поскольку фагоцитоз апоптотических телец уменьшен в сходной степени нарушениями по ароЕ у взаимодействующего макрофага или тимоцита, мы проанализировали, может ли экзогенное добавление ароЕ восстанавливать нормальную функцию. Добавление человеческого ароЕ3 в концентрации 1 мкМ (схожей с концентрацией в плазме крови) изменяло фагоцитоз тимоцитов, вызванный нарушениями по ароЕ (p<0.05 без добавления ароЕ; р=0.98 для клеток дикого типа, непарный t-тест Стьюдента; Фиг.2 В). Мы сделали вывод, что ароЕ является важным модулятором фагоцитоза апоптотических телец, но, скорее всего, не функционирует в паре рецептор: лиганд, участвуя в прямом взаимодействии клеток во время фагоцитоза, но имеет более комплексную регуляторную роль, возможно, включающую сигналинг на клеточной поверхности.
Далее мы проанализировали, является ли ароЕ модулятором широкого спектра макрофагального фагоцитоза или имеет специфичность для фагоцитоза апоптотических клеток. Фагоцитоз флуоресцентно меченных латексных частиц, проанализированный с помощью проточной цитометрии (Фиг.2Г), макрофагами Е-/- не отличался от такового макрофагами WT (р=0.86, непарный t-тест Стьюдента; Фиг.2Д), показывая тем самым, что влияние ароЕ на фагоцитоз является специфичным для захвата апоптотических телец.
Эффект нарушений по ароЕ на уничтожение апоптотических телец
Проанализировали количество апоптотических телец, присутствующих in vivo в мышах, дефектных по АроЕ, и мышах дикого типа, являющихся одним пометом в качестве контроля. Срезы замороженных клеток получали через интервалы 250 мкм через 4 мм левой доли печени, клетки убивали и остатки клеток детектировали с помощью TUNEL-метода, в котором метятся разрывы ДНК. Соокрашивание для общего маркера макрофагов F4/80 показало, что большинство (>98%) TUNEL-положительных клеток в печени как дикого типа, так и дефектных по ароЕ мышей было F4/80+ (большинство TUNEL-положительных клеток были макрофагами). Следует отметить, тем не менее, что мечение TUNEL окрашивает не только апоптотические клетки, но может окрашивать некротические клетки или даже здоровые клетки в зависимости от способа приготовления ткани. Однако на наших срезах многие TUNEL+ клетки имели характеристики апоптотических клеток при флуоресценции Хехст (то есть очевидные конденсацию хроматина и вакуоляризацию ядер). Более того, 60-70% TUNEL+ клеток также окрашивались по активной каспазе-3. Таким образом, эти наблюдения показывают, что окрашивание TUNEL является полезным как показатель клеточной гибели макрофагов в печени, что согласуется с открытием Stadelmann и Lassmann [см. выше].
Количество погибших макрофагов (F4/80+ TUNEL+) в печени значительно увеличивается у мышей, дефектных по ароЕ, по сравнению с контрольными мышами дикого типа того же помета (Фиг.3А-В). Как абсолютное количество апоптотических макрофагов (p<0.01, непарный t-тест Стьюдента; Фиг.3Б), так и доля популяции макрофагов, которые были TUNEL+ (p<0.05, непарный t-тест Стьюдента; Фиг.3 В), увеличивалось. У животных дикого типа примерно 10% клеток F4/80+ были также TUNEL+, тогда как у животных, дефектных по ароЕ, более 50% были TUNEL+. Мы также проанализировали влияние нарушений по ароЕ на определенном количестве апоптотических гепатоцитов (клетки F4/80-TUNEL+). Однако уровень гибели клеток в немакрофагальных клеточных компартментах был значительно ниже по сравнению с макрофагальным компартментом (<2% всех клеток TUNEL+ были F/480-), поэтому данный эксперимент был мало чувствителен для определения влияния нарушений по ароЕ на эту популяцию (Фиг.3Г).
Наблюдаемое увеличение апоптотических макрофагов в печени у мышей, дефектных по АроЕ, может быть следствием либо повышенного уровня гибели макрофагов, либо пониженного уровня уничтожения апоптотических телец. Для того чтобы разделить эти два варианта, авторы проанализировали популяцию макрофагов более полно, чтобы получить картину динамики популяции макрофагов в печени мыши. Сначала мы оценили размер популяции макрофагов в печени путем выражения общего количества клеток F4/80+ как процент общего количества ядер в печени (Фиг.4А, Б). Несмотря на значительное увеличение количества и доли макрофагов TUNEL+ у мышей, дефектных по ароЕ, общая популяция макрофагов в печени была значительно больше у животных, дефектных по ароЕ (р<0.05; непарный t-тест Стьюдента; Фиг.3Б). Существует два простых объяснения этого очевидного парадокса: либо уничтожение апоптотических телец значительно менее эффективно у мышей, дефектных по ароЕ, что согласуется с наблюдениями авторов in vitro, либо рекрутирование макрофагов увеличивается в отсутствие ароЕ до степени, большей, чем увеличение гибели макрофагов.
Для того чтобы исследовать второй вариант, авторы использовали наблюдение, касающееся того, что интегрин CD11b (выявляемый моноклональным антителом М1/70), экспрессирующийся на высоком уровне на моноцитах, негативно регулируется последующим рекрутированием в популяцию тканевых макрофагов. Как результат, количество клеток М1/70+ F4/80+ в печени может быть использовано как косвенный показатель рекрутирования макрофагов. Нарушение по ароЕ увеличивает абсолютное количество макрофагов М1/70+ F4/80+ в печени более чем в 2 раза (p<0,05, непарный t-тест Стьюдента; Фиг.4В), но это лишь небольшое увеличение доли макрофагов, заново рекрутированных (10,0% у мышей, дефектных по ароЕ, по сравнению с 8,3% у мышей C57/BI6). Таким образом, увеличение уровня рекрутирования макрофагов сильно меньше более значительного увеличения погибших или погибающих макрофагов. Поскольку пролиферация популяции макрофагов является очень редким событием (не были выявлены клетки PCNA+F4/80+ в анализируемых срезах печени), авторы сделали вывод, что повышенное рекрутирование макрофагов, скорее всего, не является причиной увеличенной популяции макрофагов у мышей, дефектных по АроЕ.
Краткий анализ динамики популяции макрофагов в печени мышей дикого типа и дефектных по АроЕ показан на Фиг.4Г. Нарушения по ароЕ оказывали значительное негативное влияние на гомеостаз популяции макрофагов, что приводило к небольшому, но статистически значимому увеличению рекрутирования макрофагов, более значительному увеличению количества живых макрофагов, но наиболее значительный эффект представлял собой сильное увеличение количества апоптотических телец. Учитывая результат этого анализа, авторы сделали вывод, что уничтожение апоптотических телец снижено в печени у мышей, дефектных по АроЕ, как in vivo, так и in vitro.
Динамика популяции макрофагов в других тканях
Срезы замороженных клеток были сделаны из мозга (левое полушарие) и из левого легкого той же мыши, у которой брали образцы печени. Как и в печени, нарушения по ароЕ увеличивали размер альвеолярной популяции макрофагов (Фиг.5А), при этом главный фактор, влияющий на это увеличение, представляет собой накопление погибших или погибающих клеток (Фиг.5Б). Параметры эффектов, наблюдаемых в легком, были сходными с таковыми, наблюдаемыми в печени.
Было невозможно использовать те же маркерные антитела для характеристики популяции макрофагов в ткани мозга (микроглия), поскольку они лишь слабо окрашивались при помощи моноклонального антитела F4/80. Тем не менее, антитело к молекулам главного комплекса гистосовместимости (ГКГС) II класса (которое только окрашивает субпопуляцию клеток F4/80+ в печени и в легком) окрашивало микроглию. Нарушение по АроЕ увеличивало количество клеток ГКГС II класса + в мозге (Фиг.5В), хотя в меньшей степени, чем наблюдалось для популяции F4/80+ в легком и в печени. Однако в соответствии с наблюдениями авторов для других тканей как абсолютное количество, так и доля клеток ГКГС II класса +, которые были TUNEL+, заметно увеличивалось (Фиг.5Г). Сходные результаты были получены с использованием антител к CD14 для детекции популяции микроглиальных клеток, хотя по CD14 можно селективно детектировать заново рекрутированные макрофаги или активированные микроглиальные клетки.
Исходя из этого данные наблюдения показали, что ослабление поглощения апоптотических телец в отсутствие АроЕ, наблюдаемое in vitro, приводило к общему накоплению неуничтоженных апоптотических телец при равновесии у мышей, дефектных по АроЕ, и что имело место небольшое, но статистически значимое увеличение рекрутирования макрофагов в ряде тканей, вероятно, в ответ на нарушение в уничтожении апоптотических телец.
Эффект нарушения по АроЕ на маркеры воспаления в печени
Использовали количественную иммунофлуоресценцию для измерения относительного уровня двух маркеров воспаления в печени от мышей, дефектных по АроЕ, и их сибсов дикого типа. Цитокин TNF-α является положительно регулируемым при процессе острого воспаления, но в норме присутствует на очень низком уровне в здоровой печени, преимущественно в периваскулярных областях (Фиг.6А). Окрашивание по TNF-α было интенсивнее в 1,5 раза у мышей, дефектных по ароЕ, по сравнению с сибсами дикого типа (p<0,05; U-тест Манна-Уитни; Фиг.6Б). Хотя уровень этого цитокина был низок в обеих группах, нарушение по АроЕ приводило к статистически и, возможно, биологически значимому изменению уровня TNF-α.
Окрашивание по продукту гепатоцитов фибриногену (Фиг.6В) было также интенсивнее при дефектах АроЕ (p<0,05; непарный t-тест Стьюдента; Фиг.6Г). Хотя первоначально открытый в процессе свертывания крови фибриноген известен как положительно регулируемый участник острой фазы (то есть ген, уровень продукта которого увеличен во время островоспалительной ответной реакции). Уровень этих двух неродственных маркеров общего воспаления (TNF-α и фибриногена) увеличен у мышей, дефектных по АроЕ, по сравнению с их контролями.
Эффект концентрации холестерина в плазме на динамику популяции макрофагов
In vivo дефект АроЕ приводит к значительному нарушению регуляции метаболизма липопротеинов. Для того чтобы определить, могут ли изменения в метаболизме липопротеинов косвенно воздействовать на динамику популяции макрофагов, авторы использовали две стратегии, независящие от делеции ароЕ, для изменения метаболизма липопротеинов. Во-первых, авторы проанализировали влияние корма с высоким содержанием холестерина при кормлении мышей дикого типа в течение 10 недели, приводящего к увеличению концентрации холестерина в плазме как ЛНП, так и ЛОНП (липопротеинов очень низкой плотности) и к уменьшению концентрации холестерина ЛВП (Фиг.7А), так что профиль липопротеинов мышей C57/BI6, которых кормили жирной пищей, очень сильно походил на аналогичный профиль у мышей, дефектных по АроЕ, который был установлен предварительно. В конце этого периода не наблюдали разницы в популяции макрофагов в печени (р=0,94; непарный t-тест Стьюдента при сравнении с мышами дикого типа при нормальной диете; Фиг.7Б). Аналогично делеция гена рецептора ЛНП оказывала незначительное влияние на популяцию макрофагов в печени (р=0,07; непарный t-тест Стьюдента; Фиг.7Г) и не оказывала влияния на популяции в мозге и легком (p=0,88, t-тест Стьюдента; Фиг.7Г), несмотря на то что вызывало нарушение метаболизма липопротеинов, сравнимое с таковым при делеции ароЕ (Фиг.7В), которое было установлено предварительно. Авторы сделали вывод, что общие изменения в метаболизме липопротеинов, скорее всего, не вызывают изменений динамики популяции макрофагов, какие наблюдаются у мышей, дефектных по АроЕ, и, более того, взаимодействие ароЕ с рецептором ЛНП, скорее всего, не опосредуют ослабление уничтожения апоптотических телец, которое наблюдали авторы.
Показано, что полное отсутствие белка АроЕ у макрофагов специфично ослабляет фагоцитоз апоптотических клеток in vitro без нарушения общей функции фагоцитоза. Это нарушение приводит к заметному увеличению накопления апоптотических клеток и частей в ряде тканей у мышей, дефектных по АроЕ, in vivo, а также к более крупной популяции живых макрофагов в этих тканях. Это, в свою очередь, связано с общим увеличением провоспалительных маркеров, включая TNF-α и фибриноген. Хотя генная делеция ароЕ, как ранее сообщалось, способствует рекрутированию макрофагов у стенок кровеносных сосудов [например, Lessner, S. et а1 2002. Am J Pathol 160:2145 and Reckless, J., J.C. et al 1997. Circulation 95:1542], можно предположить, что это скорее непрямой ответ на местное отложение липидов, чем непосредственно результат нарушения по АроЕ. Показано, что изменения метаболизма липидов не являются причиной этих эффектов и эти данные являются прямым подтверждением общего влияния АроЕ на тканевое рекрутирование макрофагов (Фиг.8), которое не зависит от метаболизма липопротеинов и обусловлено ослабленным захватом остатков апоптотических клеток.
Эти результаты представляют прямое подтверждение главного физиологического механизма, лежащего в основе связи между генотипом ароЕ и спектром заболеваний, имеющих макрофагальную воспалительную оставляющую. АроЕ требуется для эффективного уничтожения апоптотических телец, и различные генотипы по ароЕ связаны с небольшими различиями в скорости уничтожения апоптотических телец. В динамике по времени in vivo даже это небольшое уменьшение уничтожения апоптотических телец приводит к увеличенному потоку через моноцитарно-макрофагальный путь и способствует общему провоспалительному изменению в фенотипе. Одно из последствий такого изменения представляет собой тенденцию к фиброзу, что приводит к потере функциональной архитектуры ткани, что, в свою очередь, является признаком болезни Альцгеймера и атеросклероза.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДУЛЯТОР АКТИВНОСТИ | 2018 |
|
RU2773957C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2012 |
|
RU2577978C2 |
| Способ лечения и профилактики гипергликемии и инсулинорезистентности и связанных с ними заболеваний и функциональных нарушений органов и систем и средство для его осуществления | 2023 |
|
RU2823334C1 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2020 |
|
RU2819731C2 |
| ГУМАНИЗИРОВАННЫЕ M-CSF МЫШИ | 2012 |
|
RU2730643C2 |
| ЛЕЧЕНИЕ АУТОИММУННОГО ЗАБОЛЕВАНИЯ ПУТЕМ МОДУЛИРОВАНИЯ АКТИВНОСТИ АННЕКСИНА-1 (ЛИПОКОРТИНА-1) | 2009 |
|
RU2554801C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ НЕВРОЛОГИЧЕСКИХ НАРУШЕНИЙ | 2005 |
|
RU2387453C2 |
| СПОСОБ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ГЕПАТОЦИТОВ, СПОСОБ ИНГИБИРОВАНИЯ КЛЕТОЧНОЙ АДГЕЗИИ ГЕПАТОЦИТОВ И СПОСОБ ИНГИБИРОВАНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ ANGPTL4 В ГЕПАТОЦИТАХ ИЛИ ПРЕДШЕСТВЕННИКАХ ГЕПАТОЦИТОВ | 2005 |
|
RU2380411C2 |
| ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, С НАРУШЕНИЕМ В ЛОКУСЕ C9ORF72 | 2016 |
|
RU2725737C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕЧЕСКОМУ РОДУ ЖИВОТНЫЕ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2775425C2 |




Изобретение относится к области медицины, а именно медицинской иммунологии. Сущность способа идентификации соединения для лечения состояния, связанного с накоплением апоптотических клеток состоит в том, что проводят определение способности белкового продукта гена ароЕ к стимулированию фагоцитоза апоптотических клеток макрофагом в присутствии тестируемого соединения. Увеличение фагоцитоза апоптотических клеток в присутствии тестируемого соединения по отношению к случаю его отсутствия указывает на то, что соединение может быть полезным при лечении состояния, связанного с пониженной активностью эндогенного АроЕ. Способ получения соединения для лечения состояния, связанного с накоплением апоптотических клеток, включает определение способности полипептида АроЕ стимулировать поглощение апоптотических клеток макрофагом в присутствии тестируемого соединения, стимулирующего фагоцитоз апоптотических клеток. Увеличение поглощения апоптотических клеток в присутствии тестируемого соединения по отношению к случаю его отсутствия указывает, что соединение является полезным в лечении состояния и может быть выделено. Способ лечения состояния включает повышение экспрессии и/или активности АроЕ у указанного индивида путем введения полипептида АроЕ или его миметика указанному индивиду. Использование заявленного изобретения позволяет оптимизировать способы и средства стимулирования фагоцитоза. 4 н. и 12 з.п.ф-лы., 8 ил., 1 пр.
1. Способ идентификации и соединения для лечения состояния, связанного с накоплением апоптотических клеток, включающий:
определение способности полипептида АроЕ стимулировать поглощение апоптотических клеток макрофагом в присутствии тестируемого соединения,
при этом увеличение поглощения апоптотических клеток в присутствии тестируемого соединения по отношению к случаю его отсутствия указывает, что указанное соединение может быть полезным в лечении состояния, связанного с накоплением апоптотических клеток.
2. Способ по п.1, отличающийся тем, что указанное состояние выбрано из группы, включающей травму ткани, травматическое повреждение головного мозга, острый респираторный дистресс-синдром, бактериальный сепсис, панкреатит и перикардит.
3. Способ по любому из пп.1 или 2, отличающийся тем, что указанное тестируемое соединение представляет собой фрагмент пептида АроЕ человека или его аналог.
4. Способ по любому из пп.1 или 2, включающий идентификацию тестируемого соединения в качестве агента, стимулирующего фагоцитоз апоптотических клеток.
5. Способ получения соединения для лечения состояния, связанного с накоплением апоптотических клеток, включающий:
определение способности полипептида АроЕ стимулировать поглощение апоптотических клеток макрофагом в присутствии тестируемого соединения, стимулирующего фагоцитоз апоптотических клеток,
при этом увеличение поглощения апоптотических клеток в присутствии тестируемого соединения по отношению к случаю его отсутствия указывает, что указанное соединение может быть полезным в лечении состояния, связанного с накоплением апоптотических клеток, и выделение указанного тестируемого соединения.
6. Способ по п.5, отличающийся тем, что указанное состояние выбрано из группы, включающей травму ткани, травматическое повреждение головного мозга, острый респираторный дистресс-синдром, бактериальный сепсис, панкреатит и перикардит.
7. Способ по любому из пп.5 или 6, отличающийся тем, что указанное тестируемое соединение представляет собой фрагмент пептида АроЕ человека или его аналог.
8. Способ по любому из пп.5 или 6, включающий синтез и/или производство тестируемого соединения.
9. Способ по любому из пп.5 или 6, включающий модификацию соединения для оптимизации его фармацевтических свойств.
10. Способ по любому из пп.5 или 6, включающий обеспечение соединения в виде композиции с фармацевтически приемлемым эксципиентом.
11. Способ лечения состояния, связанного с накоплением апоптотических клеток у индивида, который включает:
повышение экспрессии и/или активности АроЕ у указанного индивида посредством введения полипептида АроЕ или его миметика указанному индивиду.
12. Способ по п.11, отличающийся тем, что полипептид АроЕ представляет собой пептидный фрагмент АроЕ человека.
13. Способ по любому из пп.11 или 12, отличающийся тем, что указанное состояние, связанное с накоплением апоптотических клеток, выбрано из группы, включающей травму ткани, травматическое повреждение головного мозга, острый респираторный дистресс-синдром, бактериальный сепсис, панкреатит или перикардит.
14. Применение полипептида АроЕ или его миметика для производства лекарственного средства для применения в лечении состояния, связанного с накоплением апоптотических клеток.
15. Применение по п.14, отличающееся тем, что полипептид АроЕ представляет собой пептидный фрагмент АроЕ человека.
16. Применение по любому из пп.14 или 15, отличающееся тем, что указанное состояние выбрано из группы, включающей: травму ткани, травматическое повреждение головного мозга, острый респираторный дистресс-синдром, бактериальный сепсис, панкреатит и перикардит.
| APRAHAMIAN T | |||
| et al Impaired clearance of apoptotic cells promotes synergy between atherogenesis and autoimmune disease, 2004, v.199, №8, p.1121-31, PubMed, PMID: 15096538 | |||
| BELLOSTA S | |||
| et al Macrophage-specific expression of human apolipoprotein E reduces atherosclerosis in hypercholesterolemic apolipoprotein E-null mice, 1995 v.96, №5, p.2170-9, PubMed, PMID: 7593602. |

