Настоящее изобретение относится к способам лечения заболеваний, опосредованных Т-клетками, путем модулирования активности Аннексина-1.
Аутоиммунные заболевания - это хронические приводящие к инвалидности патологии, вызванные неправильным функционированием иммунной системы. В большинстве случаев их причиной является неконтролируемый ответ Т-клеток на собственные антигены, презентированные в контексте молекул ГКГС антиген-презентирующих клеток (АПК). Было описано несколько факторов, вовлеченных в патогенез аутоиммунных заболеваний, в том числе факторы внешней среды, генетические и вирусные факторы, которые связаны с одной общей особенностью: гиперчувствительностью Т-клеток.
Глюкокортикоиды (ГК) часто применяют для лечения ряда хронических аутоиммунных заболеваний благодаря их способности одновременно блокировать как врожденный, так и приобретенный иммунный ответ. Исследования последних десяти лет, проведенные авторами настоящего изобретения и другими исследовательскими группами, показали, что некоторые из воспалительных эффектов, оказываемых ГК на врожденный иммунный ответ, опосредованы белком Аннексином-1 (Annexin-1, Anx-A1). Было показано, что этот белок осуществляет гомеостатическую регуляцию многих типов клеток, в том числе нейтрофилов, макрофагов и клеток эндотелия. Однако одним аспектом, которому не уделяли никакого внимания, является роль Anx-A1 в адаптивном иммунном ответе. Это вызывает удивление, с учетом того, что, как предполагается, Anx-A1 является одним из вторичных посредников фармакологического действия ГК.
Авторы данного изобретения ранее показали, что Anx-A1 играет гомеостатическую роль в Т-клетках, за счет модулировния интенсивности передачи сигналов рецептором Т-клеток (TCR) (D'Acquisto et al., Blood 109: 1095-1102, 2007).
Более того, авторы настоящего изобретения показали, что высокий уровень Anx-A1 снижает порог активации Т-клеток и способствует дифференцировке в клетки Th1, в то время как мыши, дефицитные по Anx-A1, демонстрируют нарушение активации Т-клеток и более интенсивную дифференцировку в клетки Th2 (D'Acquisto et al., Eur. J. Immunol. 37: 3131-3142, 2007).
В публикации WO 2005/027965 описано открытие механизма, за счет которого апоптотические нейтрофилы доставляют противовоспалительные сигналы к дендритным клеткам, а также идентифицированы антитела, которые препятствуют этому процессу. В частности, в публикации WO 2005/027965 Anx-1 идентифицирован в качестве сигнальной молекулы, которая предположительно экспрессируется апоптотическими нейтрофилами для подавления активации и созревания дендритных клеток. Как предполагается в публикации WO 2005/027965, антитела, называемые DAC5 (Detector of Apoptotic Cells Nr.5, Детектор Апоптотических Клеток 5), опознают и блокируют противовоспалительное действие Anx-A1, презентированного на поверхности апоптотических нейтрофилов, при фагоцитозе дендритными клетками. Таким образом, в публикации WO 2005/027965 говорится о возможности лечения различных заболеваний при помощи направленного воздействия на такие апоптотические клетки и их удаления за счет индукции воспалительного ответа, но в ней не обсуждается роль Anx-A1 в активации Т-клеток.
В публикации WO 2005/027965 заявлено, что аннексины экспрессируются на поверхности клеток, которые подвергаются апоптозу (например, см. стр.8, строки 6-7 и 29-30), и аннексины презентированы на поверхности таких клеток (например, см. стр.6, строки 10-11 и стр.8, строки 16-17). Однако в двух отдельных исследованиях (Maderna et al., J Immunol., 174: 3727-3733, 2005; Scannell et al., J Immunol., 178: 4595-4605, 2007) показано, что апоптотические клетки, включая нейтрофилы, вероятнее, секретируют аннексин-1, а не экспрессируют этот белок и презентируют его на поверхности клетки. Поскольку аннексин-1 секретируется клеткой, нельзя утверждать, что DAC5 распознает только апоптотические клетки, экспрессирующие этот белок на своей поверхности, так как указанные антитела будут также распознавать секретированный аннексин-1.
Кроме того, в публикации WO 2005/027965 указано, что совместная инкубация апоптотических нейтрофилов, экспрессирующих аннексин-1 на своей клеточной мембране, и дендритных клеток, активированных с помощью ЛПС, приводит к подавлению секреции TNF-α, и к увеличению уровня синтеза маркеров активации CD83, CD86 и HLA-DR, и что добавление DAC5 к этой культуре обращает ингибирующее действие аннексина-1, экспрессируемого апоптотическими нейтрофилами (стр.5, строка 31 до стр.6, строка 8). Данные авторов настоящего изобретения (Huggins et al., FASEB J. 2008, в печати) показывают, что дендритные клетки высвобождают Anx-A1 после стимуляции с помощью ЛПС и, таким образом, DAC5, описанные в публикации WO 2005/027965, должны связываться как с Anx-A1, находящимся на нейтрофилах, так и с аннексином-1, секретированным дендритными клетками. Кроме того, авторы настоящего изобретения обнаружили, что отсутствие аннексина-1 в дендритных клетках вызывает увеличение уровня экспрессии маркеров созревания/активации и продукцию провоспалительных цитокинов, таких как TNF-α и IL-1β и IL-12. Следовательно, антитела DAC5, описанные в публикации WO 2005/027965, должны влиять на созревание и активацию дендритных клеток и, таким образом, на дальнейшее модулирование иммунного ответа.
В поддержку этого, авторами настоящего изобретения было показано, что дендритные клетки Anx-А1-/-, при совместном культивировании с наивными Т-клетками в смешанной лимфоцитарной реакции (СЛР), демонстрируют значительное уменьшение способности индуцировать как пролиферацию Т-клеток, так и продукцию IL-2 и IFN-γ. Таким образом, агенты, блокирующие функции Anx-A1 в дендритных клетках, должны уменьшать их способность стимулировать интенсивный иммунный ответ, опосредованный Т-клетками. Антитела, описываемые в публикации WO 2005/027965, таким образом, по-видимому, не подходят для лечения заболеваний, описанных в настоящей заявке на патент.
Настоящее изобретение предусматривает применение специфично связывающихся молекул, которые связываются с Аннексином-1 (Anx-A1), при лечении заболеваний, опосредованных Т-клетками.
Согласно первому аспекту изобретения предусмотрена специфично связывающаяся молекула, которая связывается с Аннексином-1 (Anx-A1) для применения в лечении заболеваний, опосредованных Т-клетками.
Авторами настоящего изобретения ранее было показано, что Anx-A1 изменяет интенсивность сигналинга от рецептора Т-клеток (TCR) и что высокий уровень Anx-A1 снижает порог активации Т-клеток и способствует дифференцировке в клетки Th1. В настоящее время авторы идентифицировали сигнальный путь, опосредованный аннексином, и следующий за ним сигнал, в качестве мишени для блокады с целью лечения заболеваний, опосредованных Т-клетками. Такие заболевания включают заболевания, при которых наблюдается аномальная активация Т-клеток, например, многие аутоиммунные заболевания, и заболевания, при которых желательно изменить дифференцировку Т-клеток в направлении Th1 вместо Th2.
В данном изобретении применяют специфично связывающуюся молекулу, которая связывается с Аннексином-1 (Anx-А1).
Аннексины - это группа кальций- и фосфолипид-связывающих клеточных белков, также известных как липокортины. Семейство аннексинов состоит из 13 членов, включая Аннексин А1, Аннексин А2 и Аннексин А5. Аннексин А-1, также известен как Аннексин-1, и в данной заявке обозначен "Anx-A1". Аннексин-1 (Anx-A1) представляет собой белок с молекулярной массой 37 кДа, который изначально был описан как посредник действия глюкокортикоидов. За последние несколько лет появились доказательства того, что Anx-A1 выполняет гомеостатическую функцию в приобретенном иммунном ответе, в частности, в Т-клетках, за счет изменения интенсивности сигналинга от Т-клеточного рецептора (TCR). Anx-A1 действует как эндогенный ингибитор воспаления в клетках врожденной иммунной системы in vivo. На Фигуре 1А представлена ленточная диаграмма трехмерной структуры Anx-A1.
У человека существует 8 нуклеотидных последовательностей, кодирующих Anx-A1. Из них транслируются только четыре и, таким образом, существует четыре изоформы Anx-A1, обозначаемые как ANXA1-002, ANXA1-003, ANXA1-004 и ANXA1-006. Эти последовательности доступны на сайте Ensembl (www.ensembl.org) и обозначены как OTTHUMT00000052664 (ANXA1-002), OTTHUMT00000052665 (ANXA1-003), OTTHUMT00000052666 (ANXA1-004) и OTTHUMT00000052668 (ANXA1-006). Аминокислотная и нуклеотидная последовательности одной из изоформ человеческого Аннексина-1 (Anx-A1), ANXA1-003, показаны на Фигуре 2а. Аминокислотные и нуклеотидные последовательности изоформ ANXA1-002, ANXA1-004 и ANXA1-006 приведены на Фигурах 2b, 2с и 2d, соответственно. Как можно видеть на Фигуре 2, изоформы ANXA1-002, ANXA1-004 и ANXA1-006 являются короткими сплайсированными вариантами ANXA 1-003 или вариантами ANXA 1-003 с небольшим числом аминокислотных замен.
В ряде исследований показано, что N-концевой пептид Anx-A1, называемый Ас.2-26, действует как биологически активный суррогат целого белка (см., напр., Lim et al., Proc Nati Acad Sci USA 95, 14535-9, 1998).
На Фигуре 1В схематично представлены повторы в составе аннексина и локализация такой биологически активной последовательности. Пептид Ас.2-26 представляет собой апетилированный пептид с последовательностью аминокислотных остатков 2-26 из полноразмерной аминокислотной последовательности Anx-A1, показанной на Фигуре 2. Последовательность пептида Ас.2-26 показана на Фигуре 1C.
Биологическое действие Anx-A1 и его производных N-концевых биологически активные пептидов опосредованы членами семейства рецепторов формильных пептидов (FPR, formyl peptide receptor). Anx-A1 оказывает контррегуляторное действие на экстравазацию нейтрофилов и врожденный иммунитет за счет непосредственного связывания и активации одного из членов этого семейства, белка, подобного рецептору формильных пептидов-1 (Formyl Peptide Receptor-Like-1, FPRL-1). Авторами настоящего изобретения ранее было обнаружено, что стимуляция Т-клеток в присутствии hrAnx-A1 повышает уровень активации Т-клеток за счет стимуляции FPRL-1 (D'Acquisto et al., Blood 109; 1095-1102, 2007).
В настоящем изобретении применяют специфично связывающуюся молекулу, которая связывается с Аннексином-1 (Anx-A1). Anx-A1, с которым связывается указанная специфично связывающаяся молекула в основном представляет собой Anx-A1 человека, имеющий полипептидную последовательность, приведенную на Фигуре 2а, или его вариант, или его фрагмент, как, например, одну из изоформ Anx-A1 человека, включающую полипептидную последовательность, показанную на Фигуре 2b, Фигуре 2с или Фигуре 2d. Фрагмент Anx-A1 человека, с которым связывается указанная специфично связывающаяся молекула, как правило представляет собой полипептид, имеющий последовательность, показанную на Фигуре 1C. Anx-A1, с которым связывается специфично связывающаяся молекула согласно настоящему изобретению, как правило кодируется нуклеотидной последовательностью, показанной на Фигуре 2а
Используемый в настоящей заявке термин «вариант» относится к белкам, которые имеют сходные аминокислотные последовательности и/или которые сохраняют одинаковые функции. В частности, термин «вариант» охватывает белки или полипептиды, которые содержат одну и более аминокислотную вставку, делецию, замену и т.п. Аминокислотные замены как правило представляют собой консервативные замены, то есть замены одной аминокислоты на другую со сходными свойствами, таким образом, что в целом функционирование белка не подвергается значительным изменениям.
Так, аминокислоты глицин, аланин, валин, лейцин и изолейцин часто могут заменять друг друга (аминокислоты с алифатическими боковыми цепями). Среди возможных предпочтительны замены, при которых замещают друг друга глицин и аланин (поскольку они имеют относительно короткие боковые цепи) или валин, лейцин и изолейцин (они имеют большие алифатические боковые цепи, которые обладают свойством гидрофобности). Другими аминокислотами, которые часто могут заменять друг друга, являются фенилаланин, тирозин и триптофан (аминокислоты, содержащие ароматические боковые цепи); лизин, аргинин и гистидин (аминокислоты с основными боковыми цепями); аспарагин и глутамин (аминокислоты, с кислыми боковыми цепями) и цистеин и метионин (аминокислоты с серосодержащими боковыми цепями).
Для аминокислот используются следующие трехбуквенные и однобуквенные обозначения: глицин (G или Gly), аланин (А или Ala), валин (V или Val), лейцин (L или Leu), изолейцин (I или Ile), пролин (Р или Pro), фенилаланин (F или Phe), тирозин (Y или Tyr), триптофан (W или Trp), лизин (K или Lys), аргинин (R или Arg), гистидин (Н или His), аспарагиновая кислота (D или Asp), глутаминовая кислота acid (E или Glu), аспарагин (N или Asn), глутамин (Q или Gln), цистеин (С или Cys), метионин (М или Met), серин (S или Ser) и треонин (Т или Thr). В случае, если аминокислотный остаток может представлять собой аспарагиновую кислоту или асапарагин, могут использоваться обозначения Asx или В. В случае, если аминокислотный остаток может представлять собой глутаминовую кислоту или глутамин, могут использоваться обозначения Glx или Z. Аспарагиновая кислота также подразумевает аспартат, а глутаминовая кислота - глутамат, если в тексте не обозначено иначе.
Указанная выше последовательность белка также может содержать делеции или вставки аминокислот. Так, например, аминокислоты, удаление которых не оказывает значительного влияния на активность белка или по меньшей мере не уничтожает указанную активность, могут быть делетированы. Такие делеции могут быть выгодны, в случае если они уменьшают общую длину и молекулярный вес полипептида при сохранении его активности. Это может позволить уменьшить количество белка, необходимого для конкретной цели - например, могут быть уменьшены размеры доз.
Также могут осуществляться вставки аминокислот в последовательность химерного белка. Это может быть сделано для изменения свойств указанного вещества (например, облегчения его идентификации, очистки или экспрессии).
Также вышеописанная последовательность может содержать аминокислотные замены, которые можно осуществлять для изменения свойств белка (например, для облегчения его определения, очистки или экспрессии). Аминокислотные замены можно осуществлять с использованием любой подходящей техники, например, с помощью направленного мутагенеза или твердофазного синтеза.
Необходимо учитывать, что аминокислотные замены или вставки в рамках данного изобретения можно осуществлять с применением как природных, так и неприродных аминокислот. Вне зависимости от того, используется ли природная или неприродная аминокислота, предпочтительно присутствие только L-аминокислот.
Для сравнения аминокислотных последовательностей можно использовать такую программу, как CLUSTAL. Эта программа сравнивает аминокислотные последовательности и находит оптимальное выравнивание за счет внедрения пропусков в любую из последовательностей при необходимости. Возможно рассчитать идентичность или сходство (идентичность, а также сохранение типа аминокислоты) для оптимального выравнивания. Такие программы, как BLASTx, выравнивают наиболее длинный участок со сходными последовательностями и оценивают уровень идентичности. Таким образом, можно сравнить найденные области со сходными последовательностями, каждая из которых имеет свой уровень идентичности. В данном изобретении рассмотрены оба типа анализа идентичности.
Варианты белков и полипептидов, описанные в настоящей заявке, должны сохранять функцию исходного белка или полипептида. В качестве альтернативы или в добавление к сохранению функции исходного белка или полипептида, варианты белков или полипептидов, описанные в настоящей заявке, обычно имеют по меньшей мере 60% идентичности (как говорилось выше) с белками или полипептидами, описанными в настоящей заявке, в частности, с полипептидными последовательностями, показанными на Фигуре 1C или Фигуре 2. В большинстве случаев варианты, применяемые в настоящем изобретении, имеют по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичности с белками или полипептидами, описанными в настоящей заявке, в частности, с полипептидными последовательностями, показанными на Фигуре 1C или Фигуре 2.
Процент идентичности двух аминокислотных последовательностей или двух нуклеотидных последовательностей определяется при помощи выравнивания последовательностей с целью оптимального сравнения (например, в первую последовательность можно ввести пропуски для лучшего сравнения с другой последовательностью) и для сравнения аминокислотных остатков или нуклеотидов в позициях, соотносящихся друг с другом. «Наилучшее выравнивание» - это такое выравнивание двух последовательностей, которое приводит к максимальному проценту идентичности. Процент идентичности определяется как число одинаковых аминокислотных или нуклеотидных остатков в сравниваемых последовательностях (т.е. % идентичности = число идентичных позиций/общее число позиций × 100).
Определение процента идентичности двух последовательностей может быть усовершенствовано за счет использования математического алгоритма, известного специалистам в данной области техники. Одним из примеров математического алгоритма для сравнения двух последовательностей является алгоритм Карлина и Альтшула (Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2264-2268), модифицированный Карлином и Альтшулом в 1993 (Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5877. Программы NBLAST и XBLAST (Altschul, et al. (1990) J. Mol. Biol. 215: 403-410) включают такой алгоритм. Для получения нуклеотидных последовательностей, гомологичных нуклеиновым кислотам данного изобретения, нуклеотидный поиск BLAST может осуществляться с помощью программы NBLAST, счет = 100, длина слова = 12. Для получения аминокислотных последовательностей, гомологичных белковым молекулам данного изобретения, поиск белков BLAST может осуществляться с помощь программы XBLAST, счет = 50, длина слова = 3. Для получения выравниваний с пропусками с целью сравнения можно использовать Gapped BLAST, как описано у Альтшула и др. (Altschul et al. (1997) Nucleic Acids Res. 25: 3389-3402). В качестве альтернативы может использоваться PSI-Blast для проведения повторного поиска, который выявляет отдаленные сходства между молекулами (Id.). При использовании программ BLAST, Gapped BLAST, и PSI-Blast, могут быть использованы параметры по умолчанию в соответствующих программах (например, XBLAST и NBLAST). См. http://www.ncbi.nlm.nih.gov. Другим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Миерса и Миллера (Myers and Miller, CABIOS (1989)). Программа ALIGN (версия 2.0), которая является частью пакета программ выравнивания последовательностей CGC, включает в себя такой алгоритм. Другие алгоритмы для анализа последовательностей, известные в данной области техники, включают ADVANCE ADAM как описано у Тореллиса и Роботти (Torellis и Robotti (1994) Comput. Appl. Biosci., 10: 3-5); и FASTA, описанный у Пирсона и Липмана (Pearson и Lipman (1988) Proc. Natl. Acad. Sci. 85: 2444-8). В FASTA, параметр ktup представляет собой контрольную опцию, которая позволяет настраивать чувствительность и скорость поиска.
При использовании альтернативного подхода варианты могут представлять собой химерные белки, включающие компоненты, которые облегчают очистку, например за счет эффективного тэгирования (присоединения) нужного белка или полипептида. Иногда необходимо удаление «тэга», но в некоторых случаях возможно, что сам химерный белок сохраняет функциональность, достаточную для применения.
«Специфично связывающаяся молекула, которая связывается с Anx-A1» при использовании в настоящей заявке обозначает молекулу, которая связывается с Anx-A1 с большей афинностью, чем с другими молекулами, то есть молекула, которая специфично связывается с Anx-A1. Специфично связывающиеся молекулы, которые связываются с Anx-A1, включают антитела к Anx-A1 и аптамеры. Антитела к Anx-А, применяемые в настоящем изобретении, функционируют за счет блокирования активации Т-клеток и, таким образом, при введении, их можно применять для лечения заболеваний, опосредованных Т-клетками, причиной которых обычно является аберрантная активация Т-клеток.
Например, могут быть получены антитела к Anx-A1, направленные против Anx-A1 человека с аминокислотной последовательностью, показанной на Фигуре 2 (в большинстве случаев - с последовательностью, показанной на Фигуре 2а). В качестве альтернативы, антитела к Anx-A1 могут быть нацелены на определенный эпитоп или эпитопы человеческого Anx-A1 с аминокислотной последовательностью, показанной на Фигуре 2 (в большинстве случаев - с последовательностью, показанной на Фигуре 2а). В частности, антитела к Anx-A1 могут быть нацелены против N-концевого фрагмента Anx-A1, например, N-концевого фрагмента, содержащего по меньшей мере 188, 100, 50 или 25 аминокислотных остатков от N-конца аминокислотной последовательности, показанной на Фигуре 2а. В большинстве случаев, антитела к Anx-A1 для применения в данном изобретении представляют собой антитела против N-концевого фрагмента Anx-А, обозначаемого Ас2-26, который имеет последовательность, показанную на Фигуре 1C, или против фрагмента, состоящего по меньшей мере из 6 аминокислот из Ас2-26. Специфично связывающиеся молекулы, которые связываются с Anx-A1, включают, таким образом, антитела к Anx-A1, представляющие собой антитела против фрагмента Anx-A1 Ас2-26 с последовательностью, показанной на Фигуре 1C, или против фрагмента, включающего по меньшей мере 6 аминокислот из Ас2-26. В данном варианте реализации антитела к Anx-A1 получены против фрагмента последовательности, показанной на Фигуре 1C, которая является антигенной и способной к стимуляции продукции антител которые, при введении, можно применять для лечения заболеваний, опосредованных Т-клетками, причиной которых обычно является аберрантная активация Т-клеток.
Как говорилось выше, можно применять активный субфрагмент указанной последовательности, как было описано. Активные субфрагменты могут состоять из или включать фрагмент из по меньшей мере шести аминокислотных остатков (гексапептид) N-концевой части Anx-A1 называемой Ас2-26, с последовательностью, показанной на Фигуре 1C, включая одну или более последовательностей из:

Активные субфрагменты могут состоять из или включать фрагмент из по меньшей мере шести аминокислотных остатков (гексапептид) N-концевой части Anx-A1, называемой Ас2-26, с последовательностью, показанной на Фигуре 1C, например, фрагмент из по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23 или по меньшей мере 24 аминокислот из последовательности, представленной на Фигуре 1C.
Антитела к Anx-A1 включают моноклональные и поликлональные антитела. Обычно антитела к Anx-A1 являются моноклональными. Антитела к Anx-A1 могут быть коммерчески доступными, как, например, кроличьи поликлональные или мышиные моноклональные антитела. Обычно антитела к Anx-A1 гуманизированы, как детально описано ниже.
Моноклональные антитела можно получать с помощью гибридом. Обычно их получают за счет слияния клеток миеломы и клеток селезенки, которые производят необходимые антитела, с получением бессмертной линии клеток. Для получения моноклональных антител для применения в соответствии с настоящим изобретением можно применять широко известную методику Kohler & Milstein (Nature 256: 495-497 (1975)) или варианты, разработанные впоследствии на базе этой методики.
Поликлональные антитела могут быть получены за счет стимуляции их продукции в подходящем животном-хозяине (например, мышь, крыса, морская свинка, кролик, овца, коза или обезьяна) при помощи инъекций Anx-A1 или его варианта или фрагмента. Если это необходимо, также вместе с белком Anx-A1 назначается адъювант. Хорошо известные адъюванты включают адъювант Фрейнда (полный или неполный) и гидроксид алюминия. Антитела могут быть в последствие очищены за счет их связывания с Anx-A1.
Технологии для получения моноклональных и поликлональных антител, которые связываются с определенным полипептидом/белком, в настоящее время хорошо разработаны в данной области техники; они описаны в общеизвестных учебниках по иммунологии, например, во втором издании учебника «Иммунология» Ройта и др. (Roitt et al. Immunology second edition (1989), Churchill Livingstone, London).
В дополнение к целым антителам, настоящее изобретение включает их производные, которые способны к связыванию с Anx-A1, как описано в настоящей заявке. Таким образом, настоящее изобретение включает фрагменты антител и синтетические конструкции. Примеры фрагментов антител и синтетических конструкций даны Дугаллом и соавторами (Dougall et al. Trends BiotechnoL, 12: 372-379 (1994).
Фрагменты антител включают, например, фрагменты Fab, F(ab')2 и Fv. Фрагменты Fab описаны Ройтом с соавторами (Roitt et al [supra]). Фрагменты Fv могут быть модифицированы для получения синтетической конструкции, известной как одноцепочечная молекула Fv (single chain Fv, scFv). Она включает пептидный линкер, ковалентно соединяющий области вариабельной тяжелой цепи и вариабельной легкой цепи, который придает молекуле стабильность. Линкер может состоять из 1-20 аминокислот, как, например, из 1, 2, 3 или 4 аминокислот, 5, 10 или 15 аминокислот, или же других подходящих промежуточных значений в интервале от 1 до 20. Пептидный линкер может быть образован из любых в целом подходящих аминокислотных остатков, таких как глицин и/или серин. Одним из примеров пригодного линкера является Gly4Ser. Также можно применять мультимеры из таких линкеров, как, например, димер, тример, тетрамер или пентамер, т.е. (Gly4Ser)2, (GlySer)3, (Gly4Ser)4 или (Gly4Ser)5. Однако в других вариантах реализации пептидный линкер отсутствует, и домен VL связан с доменом VH через пептидную связь.
Специфично связывающаяся молекула может быть аналогом вариабельного одноцепочечного фрагмента (scFv). Например, scFv может быть связан с другими специфично связывающимися молекулами (например, другие фрагменты антител: scFvs, Fab и химерные антитела IgG (напр., с каркасными участками человеческого происхождения)). cFv может быть связан с другими scFv, формируя таким образом мультимер, который представляет собой мультиспецифичный связывающийся белок, например, димер, тример или тетрамер. Биспецифичные scFv иногда называют дитела, триспецифичные - тритела и тетраспецифичные - тетратела.
scFv может быть получен с помощью любой подходящей методики с применением стандартных химических или молекулярно-биологических способов. В одном из вариантов реализации изобретения аналоги моноклональных антител могут быть получены как scFv из библиотеки нативных человеческих антител фагового дисплея (McCafferty et at. Nature 348, 552-554 (1990); и как описано в международной публикации 92/01047).
Можно применять другие синтетические конструкции, включая пептиды из гипервариабельного участка (Complementarity Determining Region, CDR). Они представляют собой синтетические пептиды, состоящие из детерминант, связывающих антиген. Также можно применять пептидные миметики. Эти молекулы представляют собой конформационно ограниченные органические кольца, которые имитируют структуру петли CDR и включают боковые цепи, взаимодействующие с антигеном.
Синтетические конструкции включают химерные молекулы. Так, в группу, антител, применяемых в настоящем изобретении, входят гуманизированные антитела или их производные. Способы гуманизации антител хорошо известны в данной области техники. Антитело может быть гуманизировано за счет модификации его аминокислотной последовательности. Примером гуманизированного антитела являются антитела, имеющие каркасные области человеческого происхождения и гипервариабельные участки от грызунов (например, мышиные). Способы продукции химерных антител описаны, например, Моррисоном и др. (Morrison et al, PNAS, 81: 6851-6855 (1984) и Такеда и др. (Takeda et al. Nature, 314: 452-454 (1985). Гуманизация может осуществляться, например, так, как описано Джонсом и др. (Jones et al. Nature, 321: 522-525, 1986) Верхоэном и др; (Verhoeyen et al. Science, 239; 1534-1536); Ричманом и др. (Riechmann et al. Nature 332: 323-327, 1988). Способы уменьшения иммуногенности специфично связывающихся молекул в настоящем изобретении могут включать пришивание CDR на каркасную часть подходящего антитела или ремоделирование остатков вариабельной поверхности, например, за счет сайт-направленного мутагенеза и других часто используемых технологий молекулярной биологии. (Roguska et al. Protein Eng. 9 895-904 (1996)).
Другие применимые методы включают идентификацию возможных эпитопов для Т-клеток внутри молекулы и их последующее удаление, например, с помощью сайт-направленного мутагенеза (деиммунизация). Если необходимо, может быть проведена гуманизация участков CDR или окружающей их каркасной последовательности.
Синтетические конструкции также включают молекулы, включающие дополнительную часть, которая придает молекуле некоторые желаемые особенности в дополнение к возможности связывания с антигеном. Например, такой частью может быть метка (например, флуоресцентная или радиоактивная метка). Или же это может быть фармакологически активное вещество.
Настоящее изобретение связано с применением специфично связывающейся молекулы, которая связывается с Anx-A1, для лечения заболеваний, опосредованных Т-клетками.
Настоящее изобретение можно применять для лечения широкого круга заболеваний, опосредованных Т-клетками. В данном контексте, «заболевание, опосредованное Т-клетками» означает любое заболевание или состояние, при котором Т-клетки принимают участие в патогенезе или в развитии заболевания или патологического состояния. Заболевания, опосредованные Т-клетками, обычно вызываются аберрантной активацией Т-клеток. Соответственно, такие заболевания можно лечить, предотвращая активацию Т-клеток за счет блокирования активности Anx-A1. В большинстве случаев, заболевания, опосредованные Т-клетками, лечение которых предлагается в настоящем изобретении, - это заболевания, в которых участвуют клетки Th1.
Заболевания, опосредованные Т-клетками, включают (но не ограничены перечисленными) заболевание «трансплантат против хозяина», отторжение трансплантата, атеросклероз, ВИЧ и/или СПИД, псориаз и некоторые аутоиммунные заболевания. Аутоиммунные заболевания, которые можно лечить с помощью настоящего изобретения, включают ревматоидный артрит (РА), рассеянный склероз (PC), системную красную волчанку (СКВ), болезнь Аддисона, болезнь Грейвса (базедову болезнь), склеродермию, полимиозит, некоторые формы сахарного диабета (например, ювенильный диабет), аутоиммунный увеоретинит, язвенный колит, обыкновенную пузырчатку, воспалительную болезнь кишечника и аутоиммунный тиреоидит. Наиболее распространенные заболевания, опосредованные Т-клетками - это ревматоидный артрит, рассеянный склероз, системная красная волчанка или атеросклероз.
Типичное заболевание, опосредованное Т-клетками - это ревматоидный артрит. Считается, что при ревматоидном артрите (РА) Т-клетки распознают и взаимодействуют с антиген-презентирующими клетками в синовиальной оболочке. Будучи активированными, эти клетки продуцируют цитокины и эффекторные молекулы; эта последовательная, расширяющаяся продукция цитокинов составляет «цитокиновый каскад», который приводит к активации макрофагов и индукции воспалительного процесса, итогом которого является деградация и резорбция хрящей и костей. С течением времени может происходить эрозия костей, разрушение хряща и полная потеря целостности сустава. В конечном счете могут быть поражены многие системы органов.
В другом варианте реализации типичным заболеванием, опосредованным Т-клетками, является атеросклероз. Воспаление играет ключевую роль в заболевании коронарной артерии и других проявлениях атеросклероза. Клетки иммунитета управляют ранними атеросклеротическими повреждениями; их эффекторные молекулы ускоряют развитие повреждений, а активация воспаления может вызвать острые коронарные синдромы. Приобретенный иммунитет также тесно связан с атерогенезом, поскольку было показано, что он взаимодействует с метаболическими факторами риска, вызывая, распространяя и активируя повреждения в артериальном дереве.
Две мышиные модели с гиперхолестеринемией и быстрым развитием атеросклероза, мыши АроЕ-/- и мыши-нокауты по рецептору липопротеинов низкой плотности (LDLR-/-), оказались полезными в изучении атеросклероза, поскольку в них воспроизводится клеточный состав участков повреждений у человека, в частности - по содержанию Т-лимфоцитов. Рекрутинг лимфоцитов увеличен в артериях мышей АроЕ-/-, склонных к атеросклерозу, даже до начала патологии.
Присутствие Т-лимфоцитов имеет функциональные последствия, так же, как и их полное отсутствие сокращает формирование повреждений при умеренной гиперхолестеринемии. Хелперные клетки CD4+1 типа, секретирующие IFN-γ (Th1), являются преобладающим типом Т-клеток, обнаруживаемых в бляшках, и эти Т-клетки оказывают проатерогенное действие и дестабилизацию бляшек.
В настоящее время авторами данного изобретения обнаружено, что Anx-A1 экспрессируется и в человеческих, и в мышиных атеросклеротических бляшках, и что в мышиной модели существует корреляция между экспрессией Anx-A1 и МС.
В другом варианте реализации в качестве заболевания, опосредованного Т-клетками, выступает системная красная волчанка (СКВ). Авторами данного изобретения в настоящее время обнаружено, что мРНК и белок Anx-A1 экспрессируются в Т-клетках больных СКВ на более высоком уровне, чем в Т-клетках от здоровых добровольцев.
Настоящее изобретение можно также применять в связи со способностью Anx-A1 стимулировать дифференцировку клеток Th1, например, для ограничения неконтролируемых защитных клеточных (Th1) ответов, направленных против внутриклеточных патогенов и для лечения внеклеточных инфекций (ответ Th2) за счет подавления дифференцировки Th1 и стимуляции дифференцировки Th2.
Специфично связывающуюся молекулу, которая связывается с Anx-A1, для применения обычно смешивают с фармакологически подходящим носителем, эксципиентом, вспомогательным веществом, адъювантом и/или разбавителем. Настоящее изобретение таким образом включает фармакологический препарат, содержащий специфично связывающуюся молекулу, которая связывается с Аннексином-1 (Anx-A1), для применения при лечении заболевания, опосредованного Т-клетками. Фармакологический препарат включает специфично связывающуюся молекулу, которая связывается с Аннексином-1 (Anx-A1) и подходящий по фармакологическим параметрам носитель, эксципиент, несущую основу, адъювант и/или разбавитель. Такие составы могут быть приготовлены с помощью способов, известных в области фармакологии, например, за счет смешивания активного ингредиента с носителем, эксципиентом, вспомогательным веществом, адъювантом и/или разбавителем в стерильных условиях.
Подходящие носители, вспомогательные вещества, адъюванты и/или разбавители широко известны в данной области техники и включают минеральные соли, минеральные соли с фосфатным буфером (ФСБ), карбоксиметилцеллюлозу (КМЦ), метилцеллюлозу, гидроксипропилметилцеллюлозу (ГПМЦ), декстрозу, липосомы, поливиниловый спирт, фармакологически чистый крахмал, маннитол, лактозу, стеарат магния, сахарин натрия, тальк, целлюлозу, глюкозу, сахарозу (и другие сахара), карбонат магния, желатин, масло, спирт, детергенты, эмульгаторы или воду (предпочтительно стерильную). Специфично связывающаяся молекула, которая связывается с Anx-A1, может находиться в составе жидкого препарата, который в целом будет состоять из суспензии или раствора специфично связывающейся молекулы, которая связывается с Anx-A1 в подходящем водном или неводном жидком носителе, например, в воде, этаноле, глицерине, полиэтиленгликоле (ПЭГ) или масле.
Обычно, если специфично связывающаяся молекула, которая связывается с Anx-A1 представляет собой антитело, это антитело ПЭГилировано, т.е. ковалентно присоединено к полиэтиленгликолю. Обычно это уменьшает иммуногенность и увеличивает период полужизни указанного антитела.
Специфично связывающаяся молекула, которая связывается с Anx-A1, может быть назначена отдельно или вместе с другим агентом.
Специфично связывающаяся молекула, которая связывается с Anx-A, предназначенная для применения в данном изобретении, обычно вводят объекту в терапевтически эффективном количестве. Терапевтически эффективное количество - это количество, эффективное для улучшения состояния, устранения или предотвращения одного или более симптома заболевания, опосредованного Т-клетками. Предпочтительно объектом лечения является человек. Однако настоящее изобретение в равной степени применимо и в медицине, и в ветеринарии. Например, настоящее изобретение можно применять при лечении домашних животных, таких как кошки и собаки, или «рабочих животных», таких как скаковые лошади.
Специфично связывающаяся молекула, которая связывается с Anx-A, можно вводить объекту любым подходящим способом. Специфично связывающуюся молекулу, которая связывается с Anx-A1, можно вводить системно, в частности, внутрисуставно, внутриартериально, внутриперитонеально (i.p.), внутривенно или внутримышечно. Однако специфично связывающуюся молекулу, которая связывается с Anx-A1, также можно вводить с помощью других энтеральных или парентеральных путей введения, таких как подкожный, внутрикожный, местный (включая буккальный, сублингвальный или трансдермальный), оральный (включая оральный или сублингвальный), назальный, вагинальный, анальный, пульмонарный или другие подходящие пути доставки.
Фармакологические препараты для перорального введения могут быть представлены в виде отдельных форм, таких как капсулы или таблетки; в виде порошков или гранул, в виде растворов, сиропов или суспензий (в водных или неводных жидкостях; или как съедобная пена, крем или эмульсия). Подходящие эксципиенты для таблеток или твердых желатиновых капсул включают лактозу, кукурузный крахмал или его производные, стеариновую кислоту или ее соли. Подходящие наполнители для применения в мягких желатиновых капсулах включают, например, растительные масла, воска, жиры, полутвердые или жидкие полиолы и т.п. Эксципиенты, которые могут быть использованы для приготовления растворов и сиропов, включают, например, воду, полиолы и сахара. Для приготовления суспензий можно применять масла (например, растительные масла) (для получения суспензии масла в воде или воды в масле).
Фармакологические препараты, адаптированные для трансдермального приема, могут быть представлены в виде отдельных пластырей, предназначенных для того, чтобы оставаться в тесном контакте с эпидермисом реципиента в течение продолжительного периода времени. Например, активный ингредиент может доставляться из пластыря через ионтофорез, как в целом описано в журнале «Фармакологические исследования» (Pharmaceutical Research, 3 (6): 318 (1986)).
Фармакологические препараты, адаптированные для местного приема, могут быть представлены в виде мазей, кремов, суспензий, лосьонов, порошков, растворов, паст, гелей, спреев, аэрозолей или масел. Для инфекций глаза или других наружных тканей, например, полости рта и кожи, предпочтительно локальное применение препаратов в виде мази или крема. Если активный ингредиент входит в состав мази, его можно применять с парафиновой или водорастворимой мазевой основой. В качестве альтернативы, активный ингредиент может входить в состав крема с кремовой основой в виде масла в воде или воды в масле. Фармакологические препараты, адаптированные для местной доставки в глаза, включают глазные капли, в которых активный ингредиент растворен или суспендирован в подходящем переносчике, главным образом - в водном растворителе. Фармакологические препараты, адаптированные для местной доставки в ротовую полость включают леденцы, пастилки и бальзамы для полости рта.
Фармакологические препараты, предназначенные для ректального приема, могут быть представлены в виде свечей или клизм.
Фармакологические препараты, предназначенные для приема назально с твердым носителем, включают крупнозернистый порошок, имеющий размер частиц, например, от 20 до 500 микрон, который принимается так же, как нюхательный табак, т.е. за счет быстрого вдыхания через носовые ходы порошка из контейнера, который подносят к носу. Подходящие препараты, в которых носитель является жидким, для приема в виде назального спрея или назальных капель, включают водные или масляные растворы активного ингредиента.
Фармакологические препараты, адаптированные для приема через ингаляцию, включают пудру или аэрозоли, которые могут генерироваться с помощью разнообразных типов герметичных дозированных аэрозолей, распылителей или инжекторов.
Фармакологические препараты, адаптированные для вагинальной доставки, могут быть представлены в виде маточных колец, тампонов, кремов, гелей, паст, пен или спреев.
Фармакологические препараты, адаптированные для парентеральной доставки включают водные или неводные стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостатические факторы и растворы, которые делают препарат по существу изотоничным крови данного реципиента; а также водные или неводные стерильные суспензии, которые могут включать суспензирующие агенты и загустители. Эксципиенты, которые могут быть использованы в растворах для инъекций, включают, в частности, воду, спирты, полиолы, глицерин и растительные масла. Препараты могут быть представлены в виде единичных доз или многодозовых контейнеров, например, запаянных ампул и бутылочек, и могут храниться в условиях сухой заморозки (лиофилизации); в этом случае требуется лишь добавить стерильный жидкий носитель, например, воду для инъекций, непосредственно перед применением. Растворы и суспензии инъекций для немедленного приема могут быть приготовлены из стерильных порошков, гранул и таблеток.
Фармакологические препараты могут содержать консервирующие, солюбилизирующие, смачивающие вещества, эмульгаторы, подсластители, красители, ароматизаторы, соли, буферы, вещества для оболочки или антиоксиданты. Они могут также содержать терапевтически активные вещества в дополнение к специфично связывающейся молекуле, которая связывается с Anx-A1.
Дозировку вводимого количества специфично связывающейся молекулы, которая связывается с Anx-A1, определяют в соответствии с различными параметрами, в особенности с тем, какую именно специфично связывающуюся молекулу, которая связывается с Anx-A1, применяют; а также в зависимости от возраста, веса и состояния пациента, способа введения препарата и требуемого режима.
Дозирование можно повторять так часто, как необходимо. Если развиваются побочные эффекты, количество или частота приема могут быть уменьшены, в соответствии с обычной клинической практикой.
При назначении млекопитающим, в частности, человеку, ожидается, что дневная доза активного вещества будет составлять от 1 мкг/кг веса до 10 мг/кг веса, обычно около 10 мкг - 1 мг на 1 кг веса. В любом случае, врач определяет необходимую дозу, которая лучше всего подходит для данного организма и которая зависит от возраста, веса, пола и индивидуального ответа. Дозы, представленные выше, являются характерными для среднего случая. Конечно же, возможны случаи, когда необходимы большие или меньшие дозы, и такие случаи также охвачены настоящим изобретением.
Во втором аспекте настоящего изобретения обеспечено применение специфично связывающейся молекулы, которая связывается с Anx-A1, в производстве медикаментов для лечения заболеваний, опосредованных Т-клетками.
В третьем аспекте настоящего изобретения обеспечен способ лечения заболеваний, опосредованных Т-клетками, представляющий собой введение объекту, нуждающемуся в таком лечении, терапевтического количества специфично связывающейся молекулы, которая связывается с Anx-A1. Как утверждалось выше, данный способ лечения можно применять как для человека, так и для животных, и настоящее изобретение охватывает в равной степени способы лечения, применяемые и в медицине и в ветеринарии.
Предпочтительные особенности второго и третьего аспектов изобретения - такие же, как для первого аспекта, с учетом необходимых изменений.
Настоящее изобретение в дальнейшем описано при помощи ссылок на последующие примеры и Фигуры, которые представлены в настоящей заявке лишь с целью иллюстрации и никоим образом не ограничивают настоящее изобретение. Даны ссылки на ряд Фигур, где:
Фигура 1А представляет собой ленточную диаграмму структуры Аннексина-1, демонстрирующую четыре аннексиновых повтора и N-концевой домен. Фигура 1В - это схема, показывающая аннексиновые повторы и локализацию биологически активной последовательности Аннексина-1, пептида Ас.2-26. На Фигуре 1C показана аминокислотная последовательность пептида Ас.2-26, который является ацетилированным N-концевым пептидным фрагментом Anx-A1.
На Фигуре 2а показана (i) аминокислотная последовательность и (11) нуклеотидная последовательность Аннексина-1 человека (Anx-A1), изоформы ANXA1-003. На Фигуре 2b показана аминокислотная последовательность Аннексина-1 человека (Anx-A1), изоформы ANXA1-002. На Фигуре 2с показана аминокислотная последовательность Аннексина-1 человека (Anx-A1), изоформы ANXA1-004. На Фигуре 2d показана аминокислотная последовательность Аннексина-1 человека (Anx-A1), изоформы ANXA1-006.
На Фигуре 3 показано действие рекомбинантного Аннексина-1 человека (hrAnx-А1) на активацию Т-клеток. Предварительная обработка мышиных наивных первичных клеток CD4+ hrAnx-A1c последующей активацией за счет различных концентраций анти-CD3/CD28 стимулировала пролиферацию клеток (Фигура 3А), продукцию IL-2 (Фигура 3В) и экспрессию CD25 и CD69 на клеточной поверхности (Фигуры 3С и 3D).
На Фигуре 4 показано, что эндогенный Anx-A1 влияет на пролиферацию Т-клеток. При стимуляции Т-клеток Anx-A1+/+ или Anx-A1-/- с помощью анти-CD3, анти-CD3/CD28 или ФМА/Иономицина наблюдался сниженный уровень включения 3H-тимидина (Фигуры 4А, 4В и 4С, соответственно) и продукции IL-2 (Фигура 4D) в Т-клетках, дефицитных по Anx-A1 в сравнении с контрольными нестимулированными Т-клетками.
На Фигуре 5 показана активация активаторного белка-1 (АР-1), ядерного фактора-κВ (Nuclear facror κВ, NF-κВ) и ядерного фактора активированных Т-клеток (NFAT) в присутствии или в отсутствии Anx-A1 (Фигура 5А), и сравнение активации АР-1, NF-κВ и NFAT в Т-клетках Anx-A1+/+ и Anx-/- (Фигура 5В).
На Фигуре 6А показан ФСК-анализ экспрессии FPRL-1 в Т-клетках, стимулированных анти-CD3/CD28 (5.0 мкг/мл) в течение обозначенных периодов времени. На Фигуре 6 В показана клеточная локализация Anx-A1 в Т-клетках до (контроль) или после стимуляции анти-CD3/CD28 (5.0 мкг/мл). Фигура 6С - это схематическое изображение роли системы Anx-A1/FPRL-1 в Т-клетках.
На Фигуре 7 показано, что экзогенный и эндогенный Anx-A1 оказывает влияние на дифференцировку Th1/Th2. Фигура 7А демонстрирует результаты эксперимента, в котором наивные Т-клетки лимфоузла дифференцировались in vitro в условиях Th1 (черные полоски) или в Th2 (белые полоски) в присутствии или отсутствии hrAnx-A1 и затем рестимулировались пришитыми к подложке анти-CD3 для того, чтобы измерить продукцию цитокинов у Th1 или Th2. На Фигуре 7 В показаны результаты эксперимента, в котором наивные Т-клетки лимфоузла из мышей Anx-A1+/+ или Anx-Al-/- дифференцировались in vitro в условиях Th1 (первая и вторая столбчатые диаграммы слева) или Th2 (третья и четвертая столбчатые диаграммы слева) и затем рестимулировались пришитыми к подложке анти-CD3 для измерения продукции цитокинов у Th1 или Th2.
На Фигуре 8 показан размер лапки (Фигура 8А) и клиническая оценка (Фигура 8В) мыши DBA, которую обрабатывали PBS или hrAnx-A1 в течение 12 дней в ходе фазы иммунизации в модели коллаген-индуцированного артрита (КИА). Фигура 8С демонстрирует анализ экспрессии Anx-A1 в клетках CD4+ у контрольной группы здоровых добровольцев (healthy control, НС) или у пациентов с ревматоидным артритом (РА). На Фигуре 8D показан иммуногистохимический анализ синовиальной ткани пациентов с РА.
На Фигуре 9 показано влияние полноразмерного hrAnx-A1 и N-терминального пептида Ас 2-26 на активацию Т-клеток.
Фигура 10 демонстрирует экспрессию Anx-A1 в человеческих атеросклеротических бляшках. Иммуногистохимический анализ экспрессии Anx-A1 с помощью мышиных моноклональных антител 1 В против человеческого Anx-A1 (Фигура 10А) или с помощью неиммунного IgG (Фигура 10В) в атеросклеротических бляшках сонной артерии, полученных от пациентов в ходе эндартерэктомии сонной артерии. Фотографии сделаны от одного пациента и отражают состояние шести разных пациентов со сходным нарушением.
На Фигуре 11 показана экспрессия Anx-A1 в мышиных атеросклеротических бляшках. Фигура 11 демонстрирует иммунофлуоресцентную визуализацию Anx-A1 в синусе аорты (Фигура 11А и Фигура 11В) и в плечеголовной артерии (Фигура 11С) у мышей АроЕ-/-. Срезы окрашены Dapi для локализации ядер. Проиллюстрированы результаты одного эксперимента, они являются репрезентативными для трех отдельных экспериментов. Исходное увеличение: × 200 (Фигура 11А и Фигура 11В), × 400 (Фигура 11С).
На Фигуре 12 показана экспрессия Аннексина-1 у пациентов с Системной Красной Волчанкой (СКВ). Анализ экспрессии Anx-A1 в Т-клетках от здоровых (контроль) людей и пациентов с системной красной волчанкой (СКВ), проведенный с помощью ОТ-ПЦР (верхняя панель) и Вестерн-блота (нижняя панель) Числа на фигуре обозначают объем (мкл) кДНК или количество (мкг) белка, полученных из одного и того же числа (2×106) Т-клеток, взятых у здоровых людей (контроль) или у пациентов с системной красной волчанкой (СКВ).
На Фигуре 13 показано ингибирование активации рецептора Т-клеток (TCR), измеренное в показателях продукции интерлейкина-2 (IL-2) в человеческих периферических Т-клетках от одного донора, инкубированных с нейтрализующими моноклональными антителами против человеческого рекомбинантного Аннексина-1 (анти-AnxA1 mAb1A).
На Фигуре 14 показано ингибирование активации рецептора Т-клеток (TCR), измеренное в показателях продукции интерлейкина-2 (IL-2) в человеческих периферических Т-клетках от другого донора, инкубированных с нейтрализующими моноклональными антителами против человеческого рекомбинантного Аннексина-1 (анти-AnxA1 mAb1A).
На Фигуре 15 показаны срезы спинного мозга мышей C57BL/6, иммунизированных MOG35-55 и CFA, из которых спинной мозг был удален на 12 день (оценка 0), 18 день (оценка 2) и 20 день (оценка 4). Срезы окрашены гематоксилином и эозином (haematoxylin and eosin, Н&Е, Фигура 15А) или анти-AnxA1 (Фигура 15 В). Для каждого окрашивания, на правых панелях (20Х) показано большее увеличение областей из левых панелей (4Х). Представлены результаты 3 экспериментов.
На Фигуре 16 показаны срезы спинного мозга мышей C57BL/6, иммунизированных MOG35-55 и CFA, из которых спинной мозг был удален на 20 день (оценка 4). Срезы окрашены анти-AnxA1 и анти-CD3 (А) или анти-F4/80 (В). Панели справа демонстрируют наложение двух одиночных окрашиваний. Представлены результаты 3 экспериментов.
На Фигуре 17 показаны результаты работы, в которой мыши C57BL/6 иммунизировались MOG35-55 и CFA и ежедневно наблюдались на предмет симптомов ЭАЭ (A) или потери/набора веса (В) в течение 23 дней. Результаты представлены средними значениями ± СОС (n=10/группу). **р<0,01, представлены результаты 3 экспериментов.
На Фигуре 18 показаны включение 3H-Тимидина (А) и продукция IL-2 (В) в клетках лимфатического узла, полученных из мышей AnxAl+/+ и AnxA1-/-, иммунизированных MOG35-55 CFA и убитых через 14 дней. Клетки стимулировали MOG35-55 в течение 48 часов и импульсно метили 1µCi 3H-Тимидином в течение 12 часов. Для измерения продукции IL-2 были использованы клеточные супернатанты. Результаты представлены средними значениями ±СОС (n=4/группу). *р<0,05, **р<0,01, представлены результаты 3 экспериментов.
На Фигуре 19 показано общее число клеток селезенки (А) и лимфатического узла (B), полученных из мышей AnxA1+/+ и AnxA1-/-, иммунизированных MOG35-55 CFA и убитых через 14 дней. С и D демонстрируют цитофлюориметрический анализ клеток лимфатического узла с помощью анти-CD4 FITC и анти-CD8 РЕ. Результаты представлены средними значениями ±СОС (n=10/группу). **р<0,01, представлены результаты 3 экспериментов.
На Фигуре 20 показаны уровни (A) IFN-γ, (В) IL-2, (С) TNF-α и (D) IL-17 в клеточных супернатантах клеток лимфатического узла, полученных из мышей AnxA1+/+ и AnxAl-/-, иммунизированных MOG35-55 CFA и убитых через 14 дней. Клетки стимулировали указанной концентрацией MOG35-55 в течение 4 дней и супернатанты использовали для ИФА цитокинов. Результаты представлены средними значениями ±СОС (n=4/группу). *р<0,05, **р<0,01, представлены результаты 3 экспериментов.
На Фигуре 21 показано окрашивание гематоксилином и эозином срезов спинного мозга, полученных из мышей AnxA1+/+ (А) и AnxAl-/- (В), иммунизированных MOG35-55 и CFA и убитых через 22 дня. Для каждого окрашивания на правых панелях (20Х) показано большее увеличение областей из левых панелей (4Х). Последовательные срезы были окрашены анти-CD3 (С) или анти-F4/80 (D). Фотографии являются репрезентативными для трех отдельных экспериментов со сходными результатами.
На Фигуре 22 показан ФСК-анализ CD3- (А) и F4/80 (В)-положительных мононуклеарных клеток, восстановленных с помощью градиента Перкола из гомогенатов спинного мозга, которые были получены из мышей AnxA1+/+ и AnxAl-/-, иммунизированных MOG35-55 CFA и умерщвленных через 14 дней. Показаны точечные диаграммы и гистограммы для одной мыши, и они являются репрезентативными для 2 экспериментов с количеством мышей n=4. Числа на гистограммах показывают процентную долю клеток CD3+ и F4/80+.
Примеры с 1 по 10
Материалы и Методы
Реагенты
Антитела к CD3 мыши (клон 145-2С11), CD28 мыши (клон 37,51), CD3 человека (клон ОКТ3), CD28 человека (клон CD28,2), РЕ-конъюгированные антитела к CD69 (клон H1,2F3), FITC-конъюгированные антитела к CD25 (клон РС61,5), IL-2, IL-4, IFN-γ, IL-12 мыши, антитела к IL-4 (клон 11В11) и IFN-γ мыши (клон XMG1,2) получали от eBioscience (Уэмбли, Великобритания). Человеческий рекомбинантный Anx-A1 (hrAnx-А1), очищенный от эндотоксина, готовили, как описано. В некоторых экспериментах использовали денатурированный hrAnx-A1 (инактивированный нагреванием при 95°С в течение 5 минут) в качестве положительного контроля. Если не указано иное, все другие реагенты были производства Sigma-Aldrich (Сент-Луис, Миссури).
Мыши
Самцы мышей линий BALB/C, C57/BL6 и DBA/I были получены от Charles River Laboratories (Уилмингтон, Массачусетс). Нуль-мутантную линию мышей по Аннексину 1BALB/C получали в нашей лаборатории и выращивали в среде без патогенов в нашем виварии. Возраст всех мышей, используемых в данных исследованиях, варьировал от 6 до 8 недель. Работу с животными проводили в соответствии с правилами внутренних дел Соединенного Королевства (Руководство по эксплуатации животных научно-процессуального кодекса 1986) и по директивам Европейского союза.
Получение клеток от пациентов
Мононуклеарные клетки периферической крови (МПК) получали из периферической крови путем центрифугирования в градиенте плотности фиколла (Ficoll-Paque Plus; Amersham Biosciences, Фрайбург, Германия). CD4+ клетки выделяли из периферической крови путем положительного отбора. Вкратце, периферическую кровь центрифугировали в градиенте плотности фиколла (Ficoll-Paque Plus; Amersham Biosciences). Присоединившиеся клетки отделяли от мононуклеарных клеток путем связывания с пластиком, покрытым сывороткой. Несвязавшиеся клетки инкубировали с антителами мыши к антителу CD4 человека (RFT4), промывали в буфере (фосфатно-солевой буфер [PBS], 0,5% бычьего сывороточного альбумина [BSA], 2 мМ ЭДТА рН 7,2) и инкубировали с антителами козы к иммуноглобулинам мыши, конъюгированными с магнитными шариками (Miltenyi Biotec, Оберн, Калифорния). Клетки пропускали через колонку MACS (Miltenyi Biotec) и собирали CD4+ клетки. Чистоту клеток оценивали методом проточной цитометрии. Средний процент CD3+-CD4+ клеток после 10 разведений составил 98% (в диапазоне 97%-99,3%). Остающиеся клетки ресуспендировали в лизирующем буфере (Ambion, Huntingdon, Великобритания).
Культура клеток
Первичные мышиные Т-клетки получали из лимфатических узлов путем негативной селекции. Вкратце, подмышечные, паховые и брыжеечные лимфатические узлы размельчали до суспензии отдельных клеток, затем промывали и высевали слоем на фиколле. Лимфоцитную пленку промывали 2 раза и затем инкубировали с смесью антител и магнитных шариков в соответствии с инструкциями изготовителя (набор реактивов по негативной селекции Т-клеток мыши; Dynal, Bromborough, Великобритания). В некоторых экспериментах клетки подвергали дальнейшей очистке для получения интактных CD62L4+ CD4+ Т-клеток с помощью набора реактивов по выделению CD62L+ CD4+ Т-клеток Miltenyi Biotec. Th0 условия создавали для Т-клеток в течение 4 дней в 6-луночных планшетах с иммобилизованными антителами к CD3 (5 мкг/мл) и к CD28 (5 мкг/мл) в полной среде RPMI (10% эмбриональной телячьей сыворотки [FCS], 2 мМ L-глутамина и 100 единиц/мл гентамицина), содержащей IL-2 мыши (20 ед/мл). Th1 условия создавали путем добавления IL-12 (3,4 нг/мл; eBioscience), IL-2 (20 ед/мл; eBioscience) мыши и антител к IL-4 (клон 11 ВЦ, 2 мкг/мл) мыши. Th2 условия создавали путем добавления IL-4 (3000 ед/мл; Peprotech, Роки-Хилл, Нью-Джерси), IL-2 (20 ед/мл) и антител к IFN-γ (клон XMG1,2; 2 мкг/мл). Клетки Jurkat получали от АТСС (Манассас, VA.) и культивировали в полной среде RPMI.
Анализ методом проточной цитометрии
Очищенные Т-клетки из лимфатических узлов сначала инкубировали с рекомбинантным Anx-A1 человека в течение 2 часов при 37°С в пробирках Эппендорф, а затем стимулировали антителами к CD3 и CD28, иммобилизованными на планшете, как показано на фигурах. По истечению 16 часов клетки визуализировали при помощи РЕ-конъюгированных антител к CD69 (клон H1,2F3) и FITC-конъюгированных антител к CD25 (клон РС61.5), растворенных в буфере FACS (PBS, содержащий 1% FCS и 0,02% NaN2). Интактные клетки были вычтены из обработки с помощью опции разброса вперед и в стороны и были проанализированы при помощи программного обеспечения CellQuest (Becton Dickinson, Франклин Лейке, Нью-Джерси) на проточном цитометре FACScan. Для анализа экспрессии FPRL-1, Т-клетки периферической крови человека стимулировали антителами к CD3 и CD28, иммобилизованными на планшете, в течение разных промежутков времени, и после этого визуализировали при помощи нанесения сначала антител мыши к FPRL-1 человека (клон 6С7-3-А, 5 мкг/мл), а затем FITC-конъюгированных антител.
Анализ пролиферации клеток
Очищенные Т-клетки из лимфатических узлов (105 клеток/мл) инкубировали только со средой или с различными концентрациями hrAnx-A1 в течение 2 часов при 37°С в пробирках Эппендорф. После этого аликвоты суспензии клеток по 200 мкл стимулировали антителами к CD3 и CD28, иммобилизованными на планшете в течение 24 часов в 96-луночных планшетах. По истечении 18 часов в культуры вводили импульсную метку 1 мкСi (3,7×104 Bg) [3Н]-тимидина (Amersham Pharmacia Biotech, Piscataway, Нью-Джерси) в течение 8 часов, включившуюся радиоактивную метку измеряли по автоматизированному сцинтилляционному счетчику (Packard Instruments, Пангборн, Великобритания).
Анализ изменения электроподвижности
Ядерные экстракты собирали от 3×106 до 5×106 клеток в соответствии с описанными ранее протоколами. Ядерные экстракты (от 3 мкг до 5 мкг) инкубировали с 1 мкг (для NFAT) или 2 мкг (для NF-κВ и АР-1) поли (dI:DC) в 20 мкл связывающего буфера, содержащего меченные по 32P двухцепочечные олигонуклеотидные зонды (5×105 имп/мин), и разделяли путем электрофореза в 6% полиакриламидном геле (соотношение перекрестного сшивания 29:1) в 0,5% ТВЕ в течение 2,5 часов при 150 В. NF-κВ и АР-1 связывающий буфер (10×) содержал 100 мМ Трис-HCl (рН 7,5), 500 мМ NaCl, 10 мМ ЭДТА, 50% глицерина, 10 мг/мл альбумина, 30 мМ ГТФ, 10 мМ DTT. NFAT связывающий буфер (10×) содержал 100 мМ Hepes (рН 7,9), 500 мМ KCl, 1 мМ ЭДТА, 1 мМ EGTA, 50% глицерина, 5 мг/мл альбумина, 1% Nonidet P-40, 10 мМ DTT. Использовали NF-кВ и АР-1 двухцепочечные олигонуклеотидные зонды производства Promega и NFAT производства Santa Cruz Biotechnology (Санта-Круз, Калифорния).
Вестерн-блот анализ
Т-клетки из лимфатических узлов инкубировали, как указано на фигурах. После инкубации при температуре 37°С в течение различных промежутков времени, клетки лизировали в ледяном лизирующем буфере (1% NP-40, 20 мМ Трис рН 7,5, 150 мМ NaCl, 1 мМ MgCl2, 1 мМ EGTA, 0,5 мМ PMSF, 1 мкМ апротинин, 1 мкМ леупептин, 1 мкМ пепстатин, 50 мМ NaF, 10 мМ Na4P2O7 и 1 мМ NaVO4, 1 мМ β-глицерофосфат). Клеточный лизат центрифугировали при 13/226 g (13000 оборотов в мин) в течение 5 минут при 4°С, супернатант собирали и разделяли методом электрофореза в SDS-10% полиакриламидном геле. После переноса, мембраны инкубировали в течение ночи с антителами, разведенными в трис-буферном солевом растворе, содержащем Твин-20 (TTBS: 0,13 М NaCl, 2,68 мМ KCl; 0,019 М Трис-HCl; 0,001% об./об. Твин-20, рН 7,4) и 5% обезжиренное сухое молоко при температуре 4°С. Для экспериментов с антителами к pERK1/2 и к Akt, в буфер TTBS добавляли 50 мМ NaF, а вместо молока использовали бычий сывороточный альбумин (5%). Для каждого опыта использовали экстракт, полученный из равного количества клеток. Иммунноблоттинг и визуализацию белков по хемилюминесценции (ECL; Amersham Pharmacia Biotech) проводили в соответствии с инструкциями производителя. Для получения цитозольной и мембранной фракций клетки сначала собирали и промывали в ледяном PBS, а затем кратко центрифугировали в течение 2 минут при 300 g. Полученный осадок клеток лизировали в лизирующем буфере (20 мМ Трис-HCl, рН 7,5; ингибиторы протеаз, перечисленные в лизирующем буфере) и пропускали через иглу номера 25 по крайней мере 5 раз, чтобы достигнуть эффективного лизиса. Затем суспензию центрифугировали в течение 2 минут при 300 g, супернатант собирали, и снова центрифугировали в течение 45 минут при 800 g (4°С). На данном этапе супернатант (питозольную фракцию) собирали, а осадок (мембранную фракцию) ресуспендировали в лизирующем буфере, содержащем 1% (об./об.) Тритона Х-100. Все фракции держали на льду в течение эксперимента.
Твердофазный иммунноферментный анализ цитокинов
Для анализа выработки Th1/Th2 цитокинов, Th0/Th1/Th2 клетки (106/mL), полученные после 4-дневной дифференцировки в условиях задержки и 1 дня отдыха в полной среде RPMI, стимулировали антителами к CD3 (5 мкг/мл), иммобилизованными на планшете, в течение 8 ч в 24-луночных планшетах. Культуральную жидкость собирали и анализировали на содержание IFN-γ, IL-2, IL-4 и IL-10 с помощью набора реагентов для ИФА Th1/Th2 (eBioscience). Набор реагентов для ИФА IL-13 также приобретали у eBioscience.
Пример 1 - Влияние человеческого рекомбинантного аннексина-1 (hrAnx-A1) на активацию Т-клеток
Интактные Т-клетки из лимфатических узлов мышей стимулировали в течение 24 часов 5,0 (ν), 2,5 (σ) или 1,25 (τ) мкг/мл антител к CD3/CD28 в отсутствие или присутствии различных концентраций hrAnx-A1 24 часа, а затем вводили импульсную метку 3H-тимидина для оценки пролиферации. Результаты представлены на Фигуре 3А.
Фигура 3В показывает продукцию IL-2 первичными интактными Т-клетками из лимфатических узлов мыши, стимулированных антителами к CD3/CD28 (1,25 мкг/мл) в отсутствие или присутствии различных концентраций hrAnx-A1 в течение 24 часов.
Интактные Т-клетки из лимфатических узлов мышей стимулировали антителами к CD3/CD28 в концентрации 1,25 мкг/мл (левая колонка), 2,5 мкг/мл (средняя колонка) и 5,0 мкг/мл (правая колонка) в отсутствие (верхние панели) или присутствии (нижние панели) hrAnx-A1 (600 нМ) в течение 12 часов, а затем анализировали на экспрессию CD25 и CD69 методом FACS. Результаты приведены на Фигуре 3С.
На Фигуре 3D интактные Т-клетки из лимфатических узлов мышей стимулировали указанными концентрациями антител к CD3/CD28 в присутствии 150 (X), 300 (Ω) и 600 (µ) нМ hrAnx-A1 в течение 12 часов, а затем анализировали на экспрессию CD25 (левый график) и CD69 (правый график) методом FACS.
Во всех экспериментах значения приведены как среднее ± стандартная погрешность для n=3-4 мышей. *Р<0,05; **Р<0,01.
Результаты показывают, что предварительная обработка интактных первичных CD4+ клеток мышей hrAnx-A1 с последующей активацией различными концентрациями антител к CD3/CD28 усиливает пролиферацию клеток (Фигура 3А), выработку IL-2 (Фигура 3В) и экспрессию CD25 и CD69 на клеточной поверхности (Фигура 3С и D).
Пример 2 - Эндогенный Anx-A1 модулирует пролиферацию Т-клеток
Фигура 4 показывает, что: (А) антитела к CD3 (5,0 мкг/мл) (В) антитела к CD3/CD28 (5,0 мкг/мл) или (С) РМА (20 нг/мл) и Иономицин (2 нг/мл) индуцируют пролиферацию Т-клеток дикого типа и клеток, дефектных по Anx-A1, выраженную в процентах от включения 3H-тимидина по сравнению с контрольными нестимулированными Т-клетками. В некоторых экспериментах клетки активировали также рекомбинантным IL-2 мыши (20 нг/мл). Значения приведены как среднее ± стандартная погрешность для n=4-5††Р<0,01 против IL-2 стимулированных Anx-Al+/+ клеток; **Р<0,01 против стимулированных антителами к CD3 или антителами к CD3/CD28 или РМА/Иономицином Anx-Al+/+ клеток; §§Р<0,01 против стимулированных антителами к CD3 или антителами к CD3/CD28 или РМА/Иономицином Anx-A1-/- клеток.
Фигура 4D показывает выработку IL-2 интактными Т-клетками из лимфатических узлов, стимулированных антителами к CD3, антителами к CD3/CD28 (5,0 мкг/мл) или РМА (20 нг/мл) и Иономицином (2 нг/мл) в течение 24 часов. Значения приведены как среднее ± стандартная погрешность для n=4-5 мышей. **Р<0,01.
Результаты показывают, что стимуляция Anx-A1+/+ или Anx-Al-/- Т-клеток антителами к CD3, антителами к CD3/CD28 или РМА/Иономицином приводит к снижению скорости включения 3H-тимидина (Фигура 4А, 4В и 4С, соответственно) и выработки IL-2 (Фигура 4D) Т-клетками, дефектными по Anx-A1, по сравнению с контрольными нестимулированными Т-клетками.
Пример 3 - Активация АР-1, NF-κВ и NFAT в присутствии или отсутствии Anx-A1
Исследования проводили с целью выяснения, как экзогенные и эндогенные Anx-A1 модулируют активацию Т-клеток. Анализировали три основных активатора транскрипции Т-клеток, а именно Activator Protein-1 (АР-1), Nuclear Factor-кВ (NF--κВ) и Nuclear Factor of Activated T cells (NFAT), в клетках, стимулированных hrAnx-A1.
Фигура 5А показывает анализ изменения электрофоретической подвижности активации АР-1, NF-κВ и NFAT в Т-клетках, стимулированных антителами к CD3/CD28 (1,25 мкг/мл) в присутствии или отсутствии указанных концентраций hrAnx-A1. Фигура 5В показывает сравнение активации АР-1, NF--κВ и NFAT в Anx-Al+/+ или Anx-A1-/- Т-клетках, стимулированных антителами к CD3/CD28 (5,0 мкг/ мл).
Результаты показали повышенную активацию всех трех факторов транскрипции (Фигура 5А). Наоборот, Anx-A1-/- Т-клетки показали пониженную активацию этих транскрипционных факторов по сравнению с контрольными клетками (Фигура 5В).
Пример 4 - Экстернализация FPRL-1 и Anx-A1 в Т-клетках
Мы исследовали, экспрессируют ли Т-клетки рецептор для Anx-A1, Formyl Peptide Receptor Like-1 (FPRL-1). FACS-окрашивание нестимулированных Т-клеток периферической крови человека (ПКТ) специфическими моноклональными антителами к FPRL-1 не выявило экспрессии рецептора. Однако стимуляция антителами к CD3/CD28 индуцировала экстернализацию FPRL-1 в течение 1 часа с последующей стабильной экспрессией на поверхности клетки (Фигура 6А). Интересно, что аналогичный паттерн наблюдался для Anx-A1. Таким образом, анализ распределения Anx-A1 в ПКТ человека показал, что белок равномерно распределяется между цитозолем и мембраной. Однако когда клетки стимулировали антителами к CD3/CD28, наблюдалось накопление Anx-A1 на мембране.
Затем белок экспортируется к наружной стороне мембраны и высвобождается во внеклеточную среду. В соответствии с этой моделью, когда мы иммунопреципитировали Anx-A1 из культуральной среды ПКТ человека, стимулированных антителами к CD3/CD28, мы наблюдали увеличение высвобождения Anx-A1 сравнению с контрольными нестимулированными клетками (Фигура 6В). В совокупности эти наблюдения показывают, что путь передачи сигнала через TCR увеличивает высвобождение Anx-A1, одновременно со стимуляцией экспрессии его рецепторов.
В физиологических условиях, Anx-A1/FPRL-1 входят в состав TCR для модулирования силы передачи сигнала через TCR. Однако, в патологических условиях, таких как в РА или системной красной волчанки (неопубликованные данные), когда белок экспрессируется на более высоком уровне, это может привести к повышению активации Т-клеток в связи с более низким пороговым значением передачи сигнала через TCR (Фигура 6С).
Пример 5 - Экзогенные и эндогенные Anx-A1 модулируют дифференцировку Th1/Th2
Недавние исследования позволяют предположить, что сила передачи сигнала через TCR влияет на коммитацию линии Т клеток на Th1- или Th2-эффекторные клетки. Учитывая увеличение или уменьшение передачи сигнала через TCR в Т-клетках, обработанных hrAnx-A1 (Фигуры 3 и 5А) или Anx-A1-/- клетках (Фигуры 4 и 5В), мы стремились определить, могут ли различные уровни Anx-A1 влиять на дифференцировку Т клеток в Th1 или Th2 клетки.
Интактные Т-клетки из лимфатических узлов дифференцировали in vitro в Th1 (черные столбики) или Th2 (белые столбики) в условиях присутствия или отсутствия hrAnx-A1 (600 нМ), а затем повторно стимулировали антителами к CD3 (5 мкг/мл), иммобилизованными на планшете, в течение 8 часов для измерения выработки Th1 или Th2 цитокинов. Результаты представлены на Фигуре 7А. Значения приведены как среднее±стандартная погрешность для n=4-5 мышей. **Р<0,01.
Как показано на Фигуре 7А, при дифференцровке интактных Т-клеток (CD44lo, CD62Lhi) в Th1 (антитела к CD3/CD28, IL2, IL12 и антитела к IL-4) или Th2 (антитела к CD3/CD28, IL-2, IL-4 и антитела к IFNγ) условиях в присутствии hrAnx-A1 увеличивалась продукция IL-2 и IFNγ с сопутствующим снижением высвобождения IL-4 и IL10 после повторной стимуляции антителами к CD3.
Интактные Т-клетки из лимфатических узлов Anx-A1+/+ или Anx-A1-/- мышей подвергали дифференцировки in vitro в Th1 (первый и второй графики слева) и Th2 (третий и четвертый графики слева) условиях, а затем повторно стимулировали антителами к CD3 (5 мкг/мл), иммобилизованными на планшете в течение 8 часов для измерения продукции Th1 или Th2 цитокинов. Результаты приведены на Фигуре 7В. Значения приведены как среднее ± стандартная погрешность для n=4-5 мышей. **Р<0,01.
Как показано на Фигуре 7В, аналогичные результаты были получены и в отношении эндогенных белков: анализ продукции цитокинов Th1/Th2 в дифференцированных Th1/Th2 клетках Anx-A1+/+ или Anx-A1-/- мышей показал более высокие уровни IL2 и IFNγ у мышей дикого типа по сравнению с нокаутной линией, для продукции IL-4 и IL13 был показан противоположный профиль.
Пример 6 - Anx-A1 и ревматоидный артрит
Чтобы доказать, что hrAnx-A1 усиливает активацию Т-клеток in vivo, мы использовали модель хронического аутоиммунного заболевания, коллаген-индуцированного артрита (КИА), на линии мышей DBA. Мышам ежедневно в течение 12 дней после иммунизации коллагеном вводили hrAnx-A1 (время, за которое интактные клетки дифференцируются в клетки Th эффекторные клетки), а затем анализировали развитие заболевания, вызванное введением антигена. Фигура 8 показывает объем конечности (Фигура. 8а) и клиническую оценку (Фигура 8В) мышей, которым вводили PBS (100 мкл) или hrAnx-A1 (1 мкг подкожно два раза в день). Синхронизация заболевания была получена за счет повторной иммунизации коллагеном на 21 день; клинические признаки были очевидны, начиная с 22 дня (1-й день от начала заболевания). Значения приведены как среднее ± стандартная погрешность для n=6-8 мышей. Группы сравнивали с использованием критерия Манна-Уитни. *Р<0,01.
Как видно на Фигурах 8А и 8В, обработка мышей hrAnx-A1 усугубляет признаки и симптомы артрита по сравнению с мышами, которым вводили PBS, что подтверждает, что высокое содержание Anx-A1 влияет на активацию и дифференцировку Т клеток и, что эти эффекты влияют на развитие болезни в модели РА на мышах.
Для исследования клинической значимости этих исследований экспрессия Anx-A1 была проанализирована у CD4+ периферических Т клеток и синовиальных CD3+ клеток, полученных от пациентов с РА. Фигура 8С показывает результаты. Средние значения показаны горизонтальными линиями, также приведены р значения теста Манна-Уитни.* Р<0,01. Как показано на Фигуре 8С, CD4+ клетки, полученные от пациентов с РА, экспрессируют мРНК и белок Anx-A1 на высоком уровне (данные не приводятся) по сравнению с контрольными клетками, полученными от здоровых добровольцев (НС).
Также применяли метод флуоресцентной иммуногистохимии с использованием вторичной сыворотки, конъюгированной с зеленой и красной флуоресцентной меткой, как показано на каждой панели Фигуры 8D. Иммуногистохимический анализ экспрессии Anx-A1 в синовиальной ткани пациентов с РА выявил высокую степень ко-локализации с CD3+ клетками. Поэтому, учитывая, что CD4 клетки у пациентов с РА экспрессируют более высокие уровни Anx-A1, можно сделать вывод, что нарушение регуляции экспрессии этого белка может способствовать развитию этого заболевания.
Пример 7 - Влияние полноразмерного белка hrAnx-A1 и N-концевого пептида Ас 2-26 на активацию Т-клеток.
Исследовали влияние N-концевого пептида hrAnx-A1 (пептид Ас 2-26) и полноразмерного белка hrAnx-A1 на активацию Т-клеток. Выработка IL-2 интактными Т-клетками из лимфатических узлов мыши была стимулирована антителами к CD3/CD28 в концентрации 0,6, 1,25 или 2,5 мкг/мл при присутствии или отсутствии полноразмерного белка hrAnx-A1 (300 нМ) или N-концевого пептида Ас.2-26 (100 мкМ), производного от Anx-A1, в течение 24 часов.
Было установлено, что N-концевой пептид Ас 2-26 сохраняет большую часть биологической активности полноразмерного белка, то есть усиливает выработку IL-2 (Фигура 9) и пролиферацию Т-клеток (данные не приведены).
Пример 8 - Anx-A1 и атеросклероз
Для исследования, экспрессируется ли Anx-A1 в атеросклеротических бляшках человека, срезы атеросклеротических бляшек сонной артерии, удаленные у пациентов во время каротидной эндартерэктомии, обрабатывали моноклональными антителами мыши к Anx-A1 человека (mAbIB). Производство этих антител описано в Pepinsky et al. FEBS Letters 261: 247-252, 1990. Вкратце, BALB/с мышей иммунизировали путем введения аннексина-1 (ссылка на липокортин-1 в Pepinsky et al.) в полном адъюванте Фрейнда в брюшную полость. Животным повторно вводили аннексин-1 на 14 и 28 дни в неполном адъюванте Фрейнда. По истечении 6 недель, брали образцы крови и исследовали их на антитела, которые блокируют активность аннексина-1. Клетки селезенки мышей, противосыворотка которых проявила активность против аннексина, сливали с SP3 Х Ag8 клетками для получения гибридомы. Культуральную жидкость оТ-клеток гибридомы исследовали на наличие антител, которые могут связывать аннексин-1 с радиоактивной меткой; гибридомы, производящие антитела, которые связывают более 50% от введенной метки, пассировали путем предельного разведения. Наиболее перспективные линии были выращены как асциты в мышах, подвергнутых воздействию пристана; моноклональные антитела аффинно очищали на сефарозе с белком А, используя системы связывания Пирса и элюирующего буфера.
Как показано на Фигуре 10, в атеросклеротической бляшке наблюдается компактное и четкое окрашивание на Anx-A1, подтверждающее, что воспалительный инфильтрат в этих тканях экспрессирует высокий уровень Anx-A1.
Аналогичное исследование проводили на АроЕ-/- линии мышей. Локализацию Anx-A1 в аортальном синусе и брахиоцефальной артерии (БЦА) у 10-месячных мышей линии АроЕ-/- проводили методом конфокальной микроскопии для определения экспрессии и пространственного распределения Anx-A1. В ткани артерии, не затронутой атеросклерозом, не наблюдали иммунореактивного Anx-A1 (данные не приведены). Напротив, в атеросклеротических бляшках из аортального синуса и БЦА наблюдали яркое окрашивание на Anx-A1 (Фигура 11).
Четкую иммунореактивность Anx-A1 обнаружили в непосредственной близости от фиброзного утолщения аортального синуса (Фигура 11А) и БЦА (Фигура 11В), а также в непосредственной близости от некротического ядра бляшки в аортальном синусе (Фигура 11В). Эти результаты показывают, что Anx-A1 экспрессируется в человеческих и мышиных атеросклеротических бляшках и позволяют предположить, что его экспрессия может потенциально влиять на стабильность бляшек.
Пример 9 - Anx-A1 и системная красная волчанка (СКВ)
Клинические исследования биологических функций Аннексина-1 связали наличие аутоиммунных антител против этого белка с развитием аутоиммунных заболеваний, включая системную красную волчанку (СКВ), ревматоидный артрит и воспалительные заболевания кишечника. В свете этих выводов мы предположили, что выработка аутоиммунных антител может быть связана с неконтролируемой экспрессией Аннексина-1 у пациентов. Для проверки этой гипотезы, мы проанализировали уровень экспрессии аннексина-1 в Т-клетках, полученных от здоровых добровольцев и больных СКВ. мРНК Аннексина-1 и белок экспрессировались на значительно более высоком уровне в Т-клетках, полученных от пациентов с СКВ (Фигура 12). Таким образом, эти результаты подтверждают гипотезу, что повышение уровня экспрессии Аннексина-1 в Т-клетках пациентов с СКВ, и, следовательно, в Т-клетках у пациентов с другими аутоиммунными заболеваниями, может быть ответственно за повышение уровня Th1 цитокинов, описанного для этих заболеваний, и, таким образом, представляет фактор риска для развития аутоиммунных заболеваний.
Пример 10 - Ингибирование активации Т-клеток антителами к Anx-A1
Т-клетки очищенной периферической крови человека инкубировали со смесью антител к CD3 и к CD28 (5 мкг/мл) для активации рецептора Т-клеток (TCR): активация произошла, как показано на Фигуре 13, судя по значительной выработке интерлейкина-2 (IL-2), основного цитокина активации и дифференцировки Т клеток.
Затем клетки инкубировали с различными концентрациями (1,0, 0,1, 0,01 и 0,001 мкг/мл) нейтрализующих моноклональных антител мыши к рекомбинантному аннексину-1 человека (mAb1A). Производство этих антител описано в Pepinsky et al. FEBS Letters 261: 247-252, 1990 и в Примере 8.
Обработка mAb1A вызывала зависимое от концентрации ингибирование выработки IL-2 (Фигура 13) и пролиферации клеток (данные не приведены). В качестве контроля использовали IgG в концентрациях 1,0, 0,1, 0,01 и 0,001 мкг/мл; при всех концентрациях не было отмечено какого-либо эффекта. Результаты показали, что блокирование аннексина-1, по-видимому, является более эффективным при более низкой концентрации специфических моноклональных антител mAb1A.
Во всех случаях данные приведены как среднее ± стандартная погрешность для трех измерений. *Р<0,01.
Затем Т-клетки очищенной периферической крови человека от различных доноров инкубировали со смесью антител к CD3 и к CD28 для активации рецептора Т-клеток (TCR) при различных концентрациях (1 мкг/мл). Как и в предыдущем случае, судя по выработке интерлейкина-2 (IL-2), активация произошла, как показано на Фигуре 14, но на более низком уровне из-за более низкой концентрации примененных антител к CD3 и к CD28.
Как и в предыдущем случае, клетки инкубировали с различными концентрациями (1,0, 0,1, 0,01 и 0,001 мкг/мл) нейтрализующих моноклональных антител мыши к рекомбинантному аннексину 1 человека (mAb1A). Обработка mAb1A вызывала зависимое от концентрации ингибирование продукции IL-2 (Фигура 14). В качестве контроля использовали IgG в концентрации 1,0 мкг/мл; не было отмечено какого-либо эффекта. Как и в предыдущем случае, результаты показали, что блокирование аннексина-1, по-видимому, происходит более эффективно при более низких концентрациях специфических моноклональных антител mAb1A, за исключением концентрации 1,0 мкг/мл mAb1A.
Во всех случаях данные приведены как среднее ± стандартная погрешность для трех измерений. *Р<0,01.
Эти эксперименты показали, что эндогенный аннексин 1 способствует активации Т-клеток в присутствии специфичного стимула. Кроме того, блокирование пути через аннексин 1 ослабляло активацию Т-клеток до 50%. Тот факт, что ингибирование достигло максимума около 50%, позволяет предположить, что "нормальный» иммунитет и иммунитет «домашнего хозяйства» не будет затронут при лечении Т-клеточных заболеваний с использованием молекулы, которая специфично связывается с Anx-A1, что и требовалось показать.
Пример 11 - Anx-A1 и рассеянный склероз (PC)
Материалы и методы
Реагенты
Пептид гликопротеина миелина олигодендроцитов (MOG)33-55 (MEVGWYRSPFSRVVHLYRNGK) был синтезирован и очищен в Cambridge Research Biochemicals (Биллингем, Великобритания). Полный адъювант Фрейнда, содержащий Mycobacterium tuberculosis Н37а получали от Difco, а токсин Bordetella pertussis - от Sigma-Aldrich Co (Пула, Великобритания). Если не указано иное, все другие реагенты были производства Sigma-Aldrich Со.
Мыши
Самцов нулевой мутантной по AnxA1 линии мышей (возраст 9-11 недель), получали, как описано ранее (Hannon et al., Faseb J, 17: 253-255, 2003; Roviezzo et al., J. Physiol Pharmacol 53: 541-553, 2002), возвратно скрещивали с мышами линии C57BL/6 в течение >10 поколений и разводили в учреждении по уходу за животными В & К (Халл, Великобритания). Соответствующие по возрасту и полу мыши контрольной линии C57BL/6 были использованы в качестве контроля во всех экспериментах. Животных содержали в стандартных условиях при температуре 22±1°С и поддерживали световой режим 12 ч/12 ч в соответствии с правилами внутренних дел Соединенного Королевства (Закон о животных 1986) и по директивам Европейского союза.
Индукция ЭАЭ
В день 0 мышей иммунизировали путем введения подкожно 300 мкл эмульсии, состоящей из 300 мкг MOG35-55 в PBS в сочетании с равным объемом полного адъюванта Фрейнда, содержащего 300 мкг инактивированных нагреванием М. tuberculosis H37Ra. Эмульсию вводили в оба бока, а затем внутрь брюшной полости вводили токсин В. pertussis (500 нг/100 мкл) в 100 мкл физиологического раствора на 0 и 2 день. Мышей осматривали ежедневно на признаки ЭАЭ и потерю веса. Тяжесть заболевания оценивали по 6-балльной шкале: 0 = нет болезни; 1 = частично вялый хвост; 2 = полностью вялый хвост; 3 = гипотония задних конечностей; 4 = частичный паралич конечностей задние; 5 = полный паралич задних конечностей; 6 = умирающие или мертвые животные.
Анализ пролиферации клеток.
Клетки из лимфатических узлов (106 клеток/200 мкл), полученные от мышей, иммунизированных MOG35-55 в течение 14 дней, в течение 48 часов стимулировали MOG35-55 (50-100 мкг/200 мкл) в 96-луночных планшетах. В течение последних 12 часов в культуры клеток вводили импульсную метку 1 мкСi [3H]-тимидина (Amersham Pharmacia Biotech, Бакингемшир, Великобритания), включившуюся радиоактивную метку измеряли по автоматизированному сцинтилляционному счетчику (Packard Instrument Company, Inc, штат Иллинойс, США).
Твердофазный иммунноферментный анализ цитокинов
Клетки из лимфатических узлов (10 клеток/мл), полученные от мышей, иммунизированных MOG35-55 в течение 14 дней, стимулировали MOG35-55 (100 мкг/мл) в течение 4 дней. Культуральную среду собирали и анализировали на содержание IFN-γ, IL-2, IL-17A и TNF-α с помощью набора реактивов для ИФА (eBioscience, Дорсет, Великобритания) в соответствии с инструкциями производителя.
Выделение клеток воспаления из спинного мозга
Мыши были умерщвлены с использованием СО2. Спинной мозг извлекали путем применения гидростатического давления, впрыскивая PBS в позвоночник с помощью шприца с иглой 21 размера. Ткань нарезали на маленькие куски и пропускали через фильтр для клеток (70 нм; BD Falcon), используя поршень стерильного 1 мл шприца. Суспензию отдельных клеток центрифугировали в течение 10 мин при 390×g, ресуспендировали в 20 мл PBS, содержащего 30% перколла (Sigma) и добавляли к 10 мл PBS, содержащего 70% перколла. После центрифугирования при 390×g в течение 20 мин, мононуклеарные клетки выделяли из интерфазы, промывали и ресуспендировали в буфере FACS (PBS, содержащий 1% FCS и 0,02% NaN2) для дальнейшего анализа.
Проточная цитометрия
Образцы клеток из очищенных перколлом тканей спинного мозга или очищенных фиколлом тканей лимфатических узлов ресуспендировали в FACS буфере, содержащем блокирующие антитела к CD16/CD32 FcγIIR (клон 93; eBioscience), в течение 30 мин при температуре 4°. После этого клеточные суспензии гибиридизовали с FITC-конъюгированными антителами к CD3 (1:100; клон 145 2С11) или с антителами к F4/80 (1:100; клон ВМТ), а клетки лимфатических узлов гибридизовали с антителами к CD4, конъюгированными с FITC (1:500; клон L3T4), и антителами к CD8 (1:1000; клон Ly-2) в течение 30 мин при 4°С, с последующим анализом на проточном цитофлуориметре FACSCalibur использованием программного обеспечения CellQuest (Becton Dickinson). По крайней мере, анализировали 104 клеток на образец, определение положительных и отрицательных популяций проводили на основе развившегося окрашивания, не относящегося к изотипам IgG.
Гистология
Ткани спинного мозга разрезали и фиксировали в 4% нейтральном буферном формалине в течение 48 часов, а затем инкубировали с декальцифицирующим раствором, содержащим ЭДТА (0,1 мМ в PBS) в течение 14 дней до заключения в парафин. Гистологическое исследование проводили на парафиновых срезах, образцы которых брали в различные моменты времени в зависимости от тяжести заболевания. Из срезов спинного мозга (5 мкм) ксилолом удаляли парафин и окрашивали гематоксилином и эозином (Н & Е) для оценки воспаления. Окрашивание на AnxA1 проводили на замороженных срезах с использованием антител к AnxA1 (разведение 1:500; Zymed, Invitrogen) и антител к иммуноглобулинам кролика, конъюгированных с пероксидазой хрена (HRP) (разведение 1:500; Dako). Двойное окрашивание на AnxA1 и CD3 или F4/80 проводили, как описано ранее, с использованием FITC-конъюгированных антител к CD3 (1:100; клон 145 2С11) или антител к F4/80 (1:100; клон ВМТ). Срезы также подкрашивали гематоксилином. Во всех случаях минимальное проанализированное число срезов на животное ≥3. Цифровые изображения в режиме фазового контраста получали с использованием программного пакета анализа изображений Image Pro.
Статистический анализ
Для всех статистических тестов использовали программное обеспечение Prism (GraphPad). Статистическую оценку частот клеток, пролиферации и продукции цитокинов проводили с использованием t-критерия Стюдента. Дисперсионный анализ применяли для анализа клинической классификации ЭАЭ. Значение р<0,05 считали статистически значимым. Р-значения ниже 0,05 считали достоверными. Данные представлены как среднее ± стандартная погрешность для n образцов в каждой группе.
Результаты
Экспрессия AnxA1 коррелирует с тяжестью экспериментального аутоиммунного энцефаломиелита (ЭАЭ)
Корреляция между уровнями AnxA1 в содержимом спинного мозга и степенью проникновения мононуклеарных клеток в ЦНС были оценены на мышиной модели PC, индуцированного иммунизацией MOG35-55, ЭАЭ. MOG35-55-индуцированный ЭАЭ является моделью для аутоиммунной демиелинизации ЦНС и широко используется для исследования патогенных механизмов, ответственных за развитие PC. С этой целью, собирали спинной и головной мозг мышей дикого типа, иммунизированных пептидом MOG35-55 на разных стадиях заболевания, т.е. на день 12 (оценка 0), день 18 (оценка 2) и день 20 (оценка 4), и проводили иммуногистохимическое выявление AnxA1, параллельно с окрашиванием гематоксилином и эозином.
Как показано на Фигуре 15, ткани спинного мозга, собранные во время фазы индукции, без признаков заболевания мышей, показали слабое окрашивание на AnxA1 (оценка 0, Фигуры 15А и В, соответственно). Однако, с появлением клинических признаков и воспалительного инфильтрата в ЦНС, дискретные участки иммуноокрашивания на AnxA1 наблюдались повсеместно в оболочке головного мозга (оценка 2, Фигура 15А и В, соответственно). По мере развития заболевания наблюдали увеличение числа AnxA1-положительных участков клеточного инфильтрата (оценка 4, Фигуры 15А и В, соответственно), что позволяет предположить, что проникновение воспалительных клеток, экспрессирующих AnxA1 на высоком уровне, коррелирует с тяжестью заболевания.
Чтобы определить клеточные источники иммунореактивности AnxA1 в спинном мозгу, проводили двойное иммунофлуоресцентное окрашивание срезов антителами к AnxA1 и или антителами к CD3 (маркер Т-клеток), или антителами к F4/80 (маркер для макрофагов). Обнаружили большое количество проникших Т-клеток и макрофагов на срезах спинного мозга мышей на пике развития ЭАЭ (Фигура 16А и В, средние панели, соответственно). Однако окрашивание на AnxA1 на тех же срезах показало частичную ко-локализацию как с Т-клетками, так и с макрофагами без особенного сродства к одному или другому типу клеток (Фигура 16А и В, правые панели, соответственно).
У мышей AnxA1-/- симптомы ЭАЭ развиваются с нарушениями
Поскольку экспрессия AnxA1 на пике развития ЭАЭ была повышена, исследовали роль этого белка в развитие ЭАЭ. AnxA1+/+ и AnxA1-/- мышей подкожно иммунизировали пептидом MOG35-55 в CFA в день 0, а затем вводили внутривенно токсин В. pertussis в день 0 и на 2-й день. У обеих линий мышей AnxA1+/+и AnxA1-/- симптомы ЕАЕ начали развиваться на 12-й день после иммунизации, достигая пика заболевания на 20-й день. При этом у мышей AnxA1-/- заболевание протекало легче по сравнению с AnxA1+/+ (Фигура 17А). Интересно, что различия становились очевидными и значительными только на поздней стадии заболевания, т.е. с 18 по 23 день и далее.
Исследование ЭАЭ на животных моделях показало, что острая фаза заболевания совпадает с потерей веса, возможно, из-за анорексии и недостаточного потребления жидкости. Измерение веса иммунизированных мышей коррелировало с тяжестью клинических оценок и показало уменьшение потери веса - с 18 дня и далее - у AnxAl-/- мышей по сравнению с AnxA1+/+ контролем (Фигура 17В). Дальнейшее сравнение развития ЭАЭ у AnxA1+/+ и AnxA1-/- мышей показало снижение смертности и максимальной оценки заболевания, без различия по росту заболеваемости или времени начала заболевания (Таблица 1).
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО К АННЕКСИНУ А1 | 2011 |
|
RU2596403C2 |
| КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА | 2008 |
|
RU2492234C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИММУННЫХ ЗАБОЛЕВАНИЙ | 2000 |
|
RU2203682C2 |
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| НОВАЯ СУБПОПУЛЯЦИЯ CD8CD45RC КЛЕТОК TREG И ЕЕ ПРИМЕНЕНИЯ | 2016 |
|
RU2766691C2 |
| СПОСОБ ДИАГНОСТИКИ АЛЛЕРГИЧЕСКИХ РЕАКЦИЙ | 2009 |
|
RU2536291C2 |
| РНК ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2790447C2 |
| АНТИТЕЛО ПРОТИВ БЕЛКА ЗАПРОГРАММИРОВАННОЙ СМЕРТИ КЛЕТКИ 1 (PD-1) И ЕГО ПРИМЕНЕНИЕ | 2015 |
|
RU2661678C1 |
| ПРЕДОТВРАЩЕНИЕ АЛЛЕРГИЧЕСКОЙ СЕНСИБИЛИЗАЦИИ | 2008 |
|
RU2429881C2 |
| МЕЧЕНЫЕ ХИМЕРНЫЕ ЭФФЕКТОРНЫЕ МОЛЕКУЛЫ И ИХ РЕЦЕПТОРЫ | 2014 |
|
RU2729463C2 |









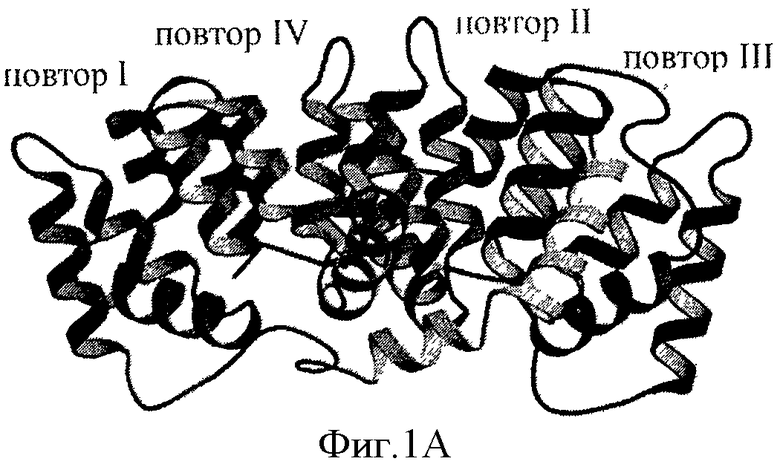
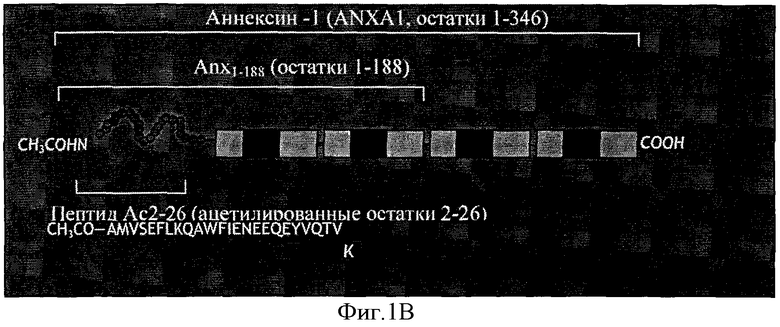
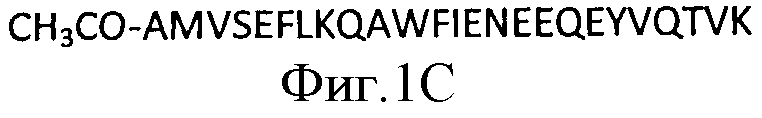




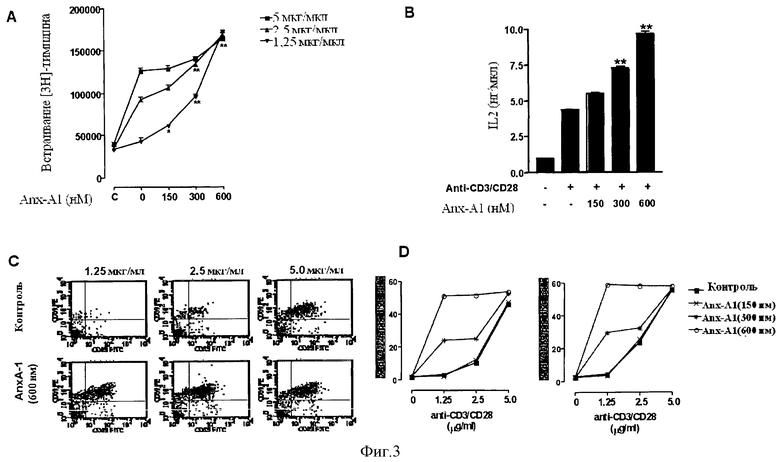
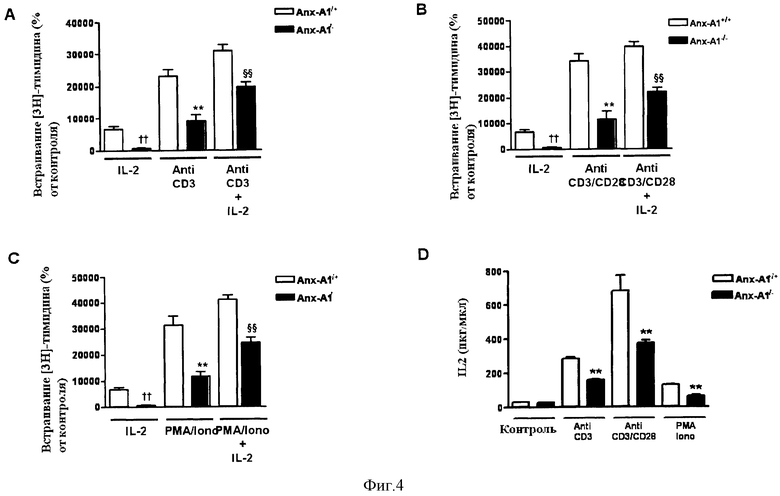
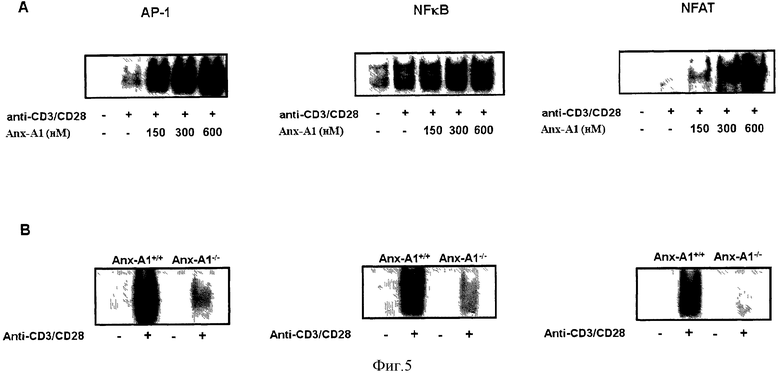
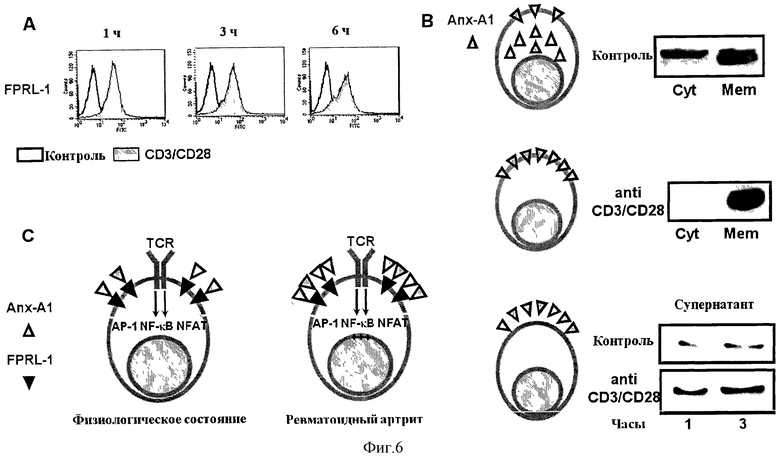
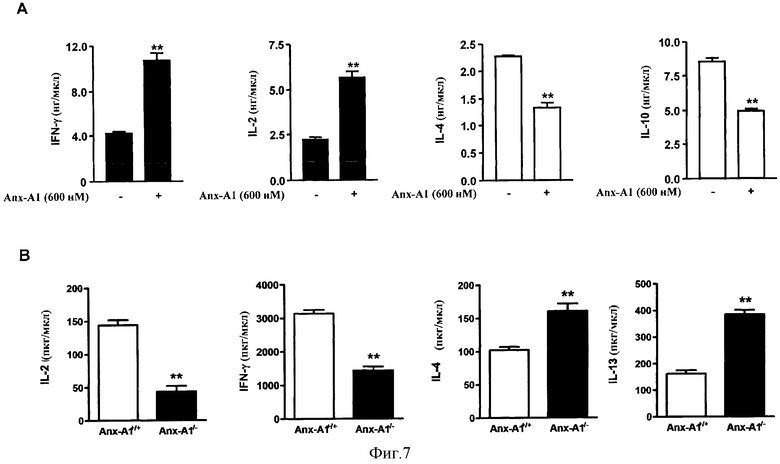
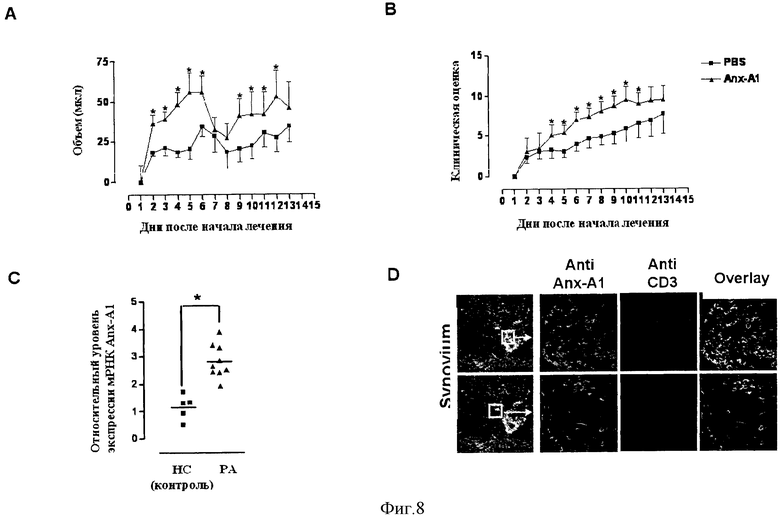
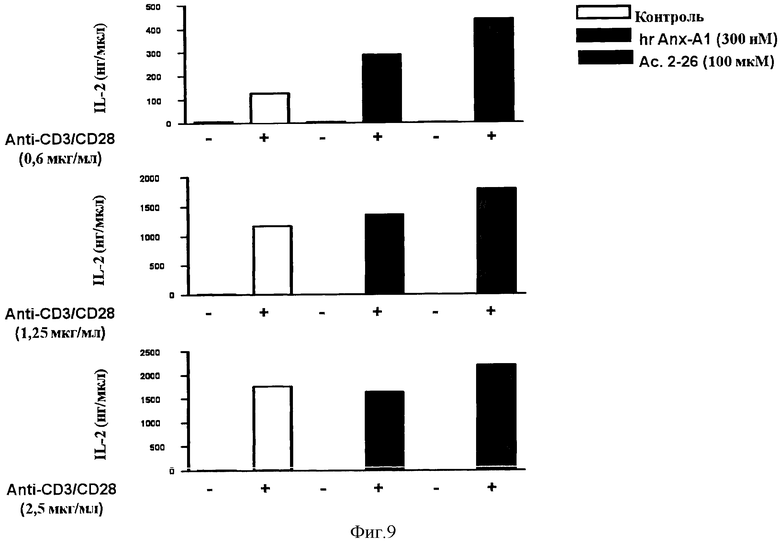

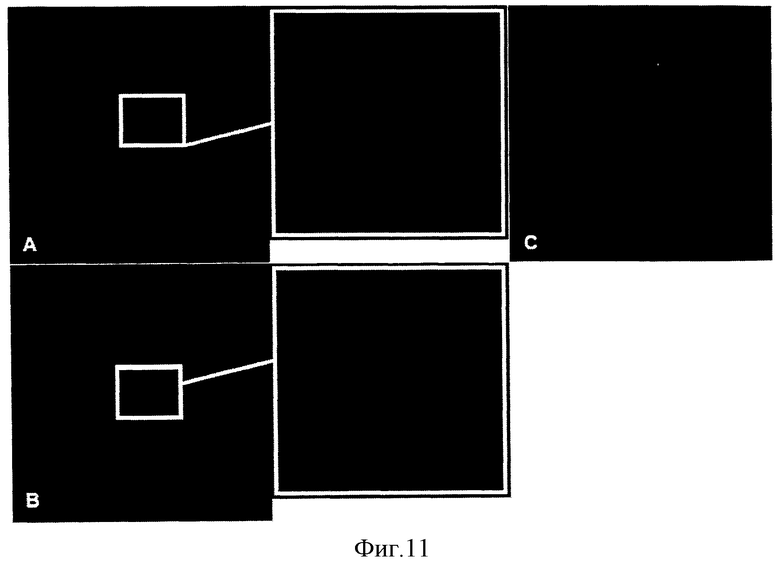
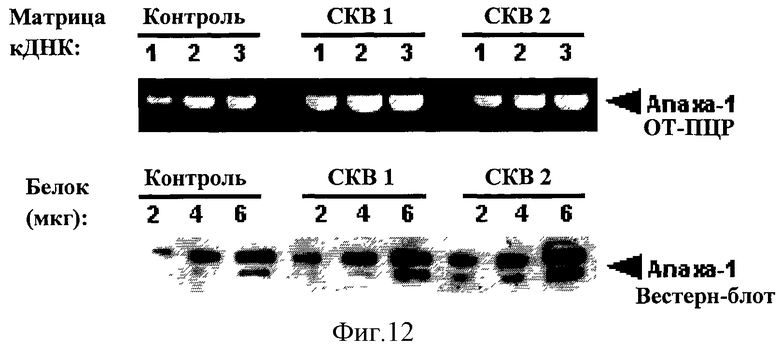
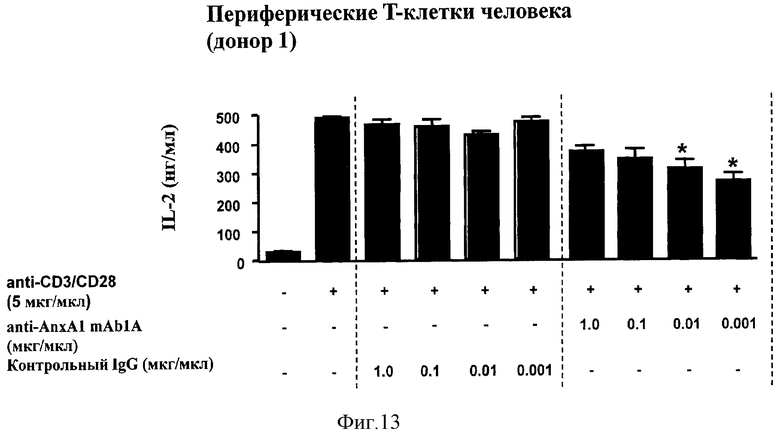
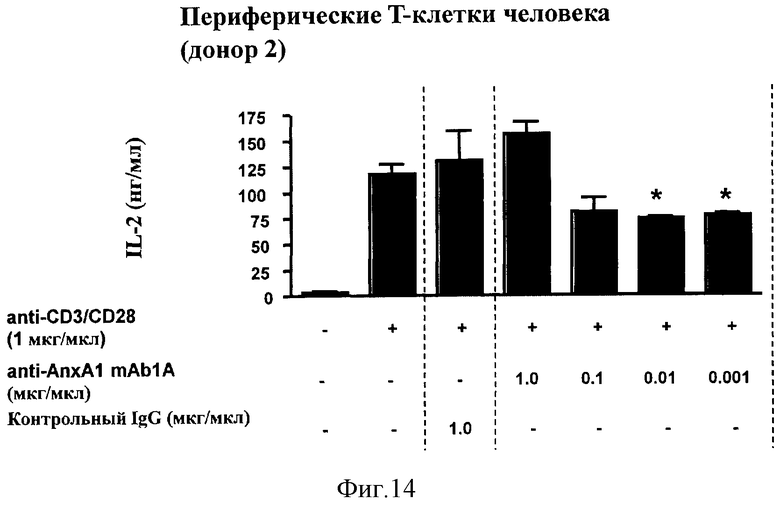
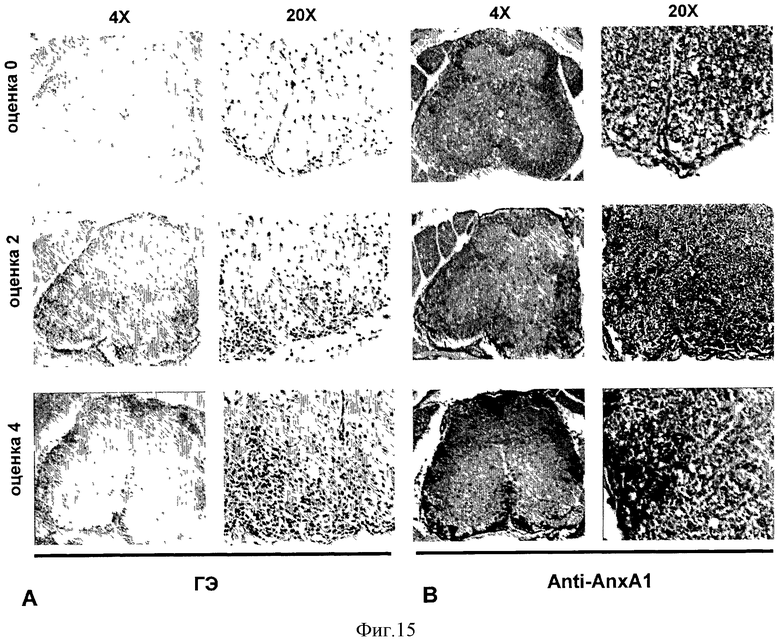

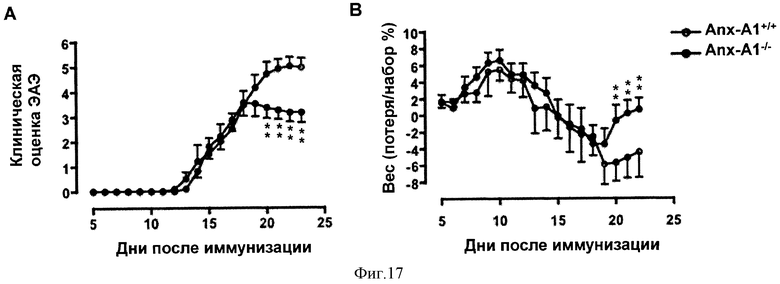
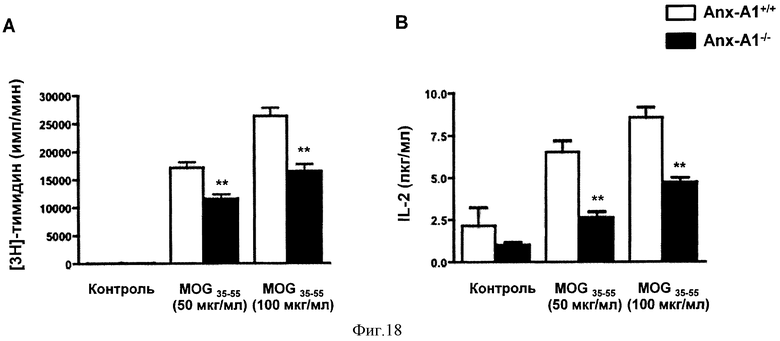
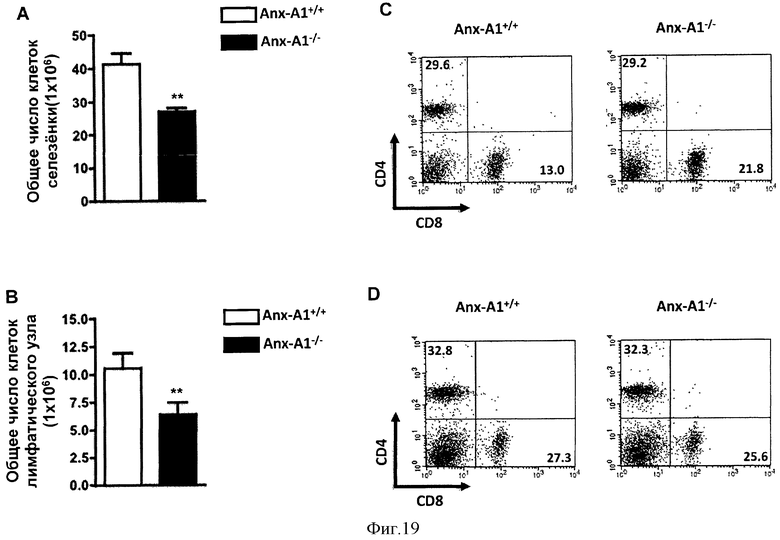
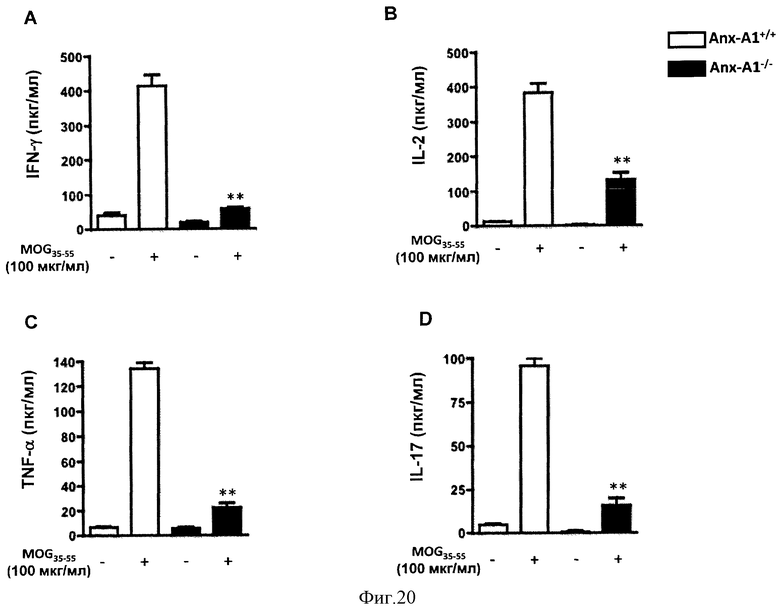

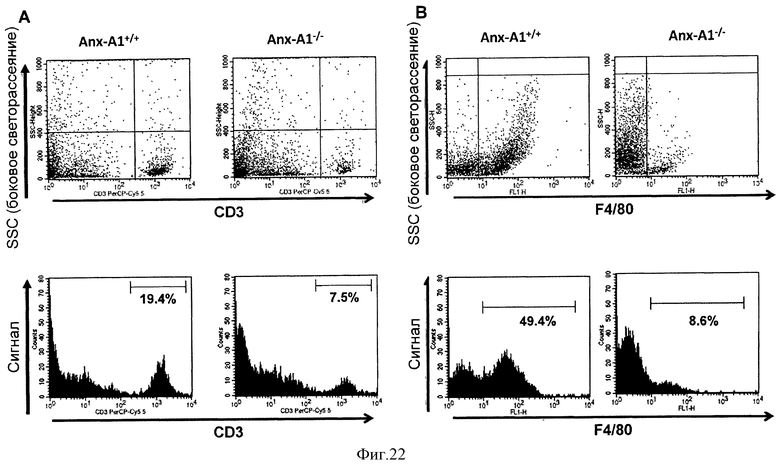
Группа изобретений относится к медицине и касается применения антитела к Аннексину-1 (Anx-А1) человека, которое имеет последовательность SEQ ID NO: 23, для лечения заболевания, вызванного аномальной активацией Т-клеток. Группа изобретений также касается способа лечения заболевания, вызванного аномальной активацией Т-клеток, включающего введение субъекту, нуждающемуся в таком лечении, терапевтического количества указанного антитела; применения указанного антитела в производстве лекарственного средства для лечения заболевания, вызванного аномальной активацией Т-клеток. Группа изобретений обеспечивает лечение заболевания, вызванного аномальной активацией Т-клеток. 3 н. и 6 з.п. ф-лы, 11 пр., 22 ил.
1. Применение антитела к Аннексину-1 (Anx-А1) человека, которое имеет последовательность SEQ ID NO: 23, для лечения заболевания, вызванного аномальной активацией Т-клеток.
2. Применение по п. 1, отличающееся тем, что указанное антитело к Anx-A1 представляет собой моноклональное антитело или его фрагмент.
3. Применение по п. 2, отличающееся тем, что моноклональное антитело является гуманизированным.
4. Применение по п. 1, отличающееся тем, что указанное заболевание выбрано из группы, включающей реакцию «трансплантат против хозяина», отторжение трансплантата, атеросклероз, ВИЧ, СПИД, псориаз и аутоиммунные заболевания.
5. Применение по п. 4, отличающееся тем, что указанное аутоиммунное заболевание выбрано из группы, включающей ревматоидный артрит (РА), рассеянный склероз (PC), системную красную волчанку (СКВ), болезнь Аддисона, болезнь Грейвса, склеродермию, полимиозит, сахарный диабет, аутоиммунный увеоретинит, язвенный колит, обыкновенную пузырчатку, воспалительную болезнь кишечника и аутоиммунный тиреоидит.
6. Применение по п. 5, отличающееся тем, что указанное аутоиммунное заболевание выбрано из группы, включающей ревматоидный артрит (РА), рассеянный склероз (РС) и системную красную волчанку (СКВ).
7. Применение по п. 4, отличающееся тем, что указанное заболевание выбрано из группы, включающей атеросклероз, ВИЧ, СПИД и псориаз.
8. Применение антитела к Аннексину-1 (Anx-A1) человека, которое имеет последовательность SEQ ID NO: 23, в производстве лекарственного средства для лечения заболевания, вызванного аномальной активацией Т-клеток.
9. Способ лечения заболевания, вызванного аномальной активацией Т-клеток, включающий введение субъекту, нуждающемуся в таком лечении, терапевтического количества антитела к Аннексину-1 (Anx-A1) человека, которое имеет последовательность SEQ ID NO: 23.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| D’ACQUISTO et al., Impaired T cell activation and increased Th2 lineage commitment in Annexin-1-deficient T cells | |||
| Eur J Immunol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| O’KENNEDY R., et al., Antibody engineering: an overview | |||
| Essays Biochem | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| LIM L.H.K | |||
| et al., Promoting detachment of neutrophils adherent | |||

