Область техники настоящего изобретения
Изобретение относится к области биотехнологии и медицине, в частности к получению и использованию наноантител для блокирования инфекции, вызванной патогенными микроорганизмами, в частности Mycoplasma hominis.
Предшествующий уровень техники настоящего изобретения
Микоплазмы (класс Mollicutes) - мельчайшие патогенные бактерии, характерной особенностью которых является отсутствие клеточной стенки. Представители данного семейства являются причиной различных заболеваний респираторного и урогенитального тракта, а также острых и хронических заболеваний суставов у человека и животных [Прозоровский С.В., Раковская И.В., 1995]. Одной из особенностей микоплазм является их способность персистировать in vivo в течение длительного периода без каких-либо клинических проявлений. Такая способность микоплазм обусловлена рядом особенностей, которые позволяют им эффективно «ускользать» от иммунной системы хозяина. Для микоплазм свойственна молекулярная мимикрия и фенотипическая пластичность, которые приводят к тому, что микоплазмы не эффективно распознаются и элиминируются иммунной системой хозяина. Это позволяет микоплазмам сохраняться в организме хозяина в течение длительного времени, что часто может являться причиной хронических заболеваний [Barile М., 1993, Razin S., 1991, 1998]. Другой характерной особенностью микоплазм является их способность к длительному персистированию in vitro - в культурах эукариотических клеток. Являясь облигатными мембранными паразитами, они сохраняются в клеточных культурах в течение десятков лет.
В настоящее время показано, что из 14 видов микоплазм, естественным хозяином которых является человек, пять видов (M.hominis, M.pneumoniae, M.genitalium, M.fermentans и U.urealyticum) ассоциированы с различными заболеваниями человека. Борьба с этими инфекциями представляет большие трудности в связи с развивающейся устойчивостью к антибиотикам и особенностями ответных реакций организма [Прозоровский, 1995].
Клинические проявления микоплазменной инфекции неспецифичны и весьма разнообразны. Во многих случаях микоплазмы вызывают латентную инфекцию. Под влиянием различных провоцирующих факторов латентная инфекция может перейти в хроническую рецидивирующую или острую форму [Прозоровский, 1995]. В настоящее время считается, что патогены, вызывающие хронические заболевания, ассоциированные с хроническим воспалением, являются одним из ключевых факторов, влияющих на опухолевую прогрессию [Coussens L., 2002, DeMarzo А., 2004, 2007, Gupta S., 2004, Karin M., 2006, Nelson W., 2003, Platz E., 2004, Sfanos K., 2008, Sutcliffe S., 2008, Wagenlehner F., 2007].
Mycoplasma hominis вызывает около половины негонококковых урогенитальных инфекций. Кроме этого, у 70% больных гонококковыми инфекциями обнаруживается M.hominis. M.hominis ассоциирована с патологиями урогенитального тракта у мужчин и верхней части урогенитального тракта у женщин. Например, M.hominis считается основным возбудителем негонококковых уретритов, урепростатитов, вагинитов, эндометритов, воспалений тазовой области, цервицитов, бесплодия, недостаточного веса новорожденных (Cassell et al. (1991) Clin. Perinatol. 18:241-262; Cassell et al. (1984) Adv. Exp. Med. Biol. 224:93-115; and Cassell et al. (1983) Sex. Transm. Dis. 10:294-302).
M.hominis относится к труднокультивируемым микроорганизмам, что делает ее диагностику бактериологическими методами длительной (от 2 до 6 дней) и дорогостоящей.
Ввиду обозначенных проблем желательно разработать способ лечения микоплазменной инфекции, не связанный с применением антибиотиков, а также способ специфической детекции микоплазм, отличный от классического бактериологического метода.
В качестве ближайших аналогов технического решения, составляющих основу настоящего изобретения, можно привести:
1) человеческие моноклональные антитела против бактериальных токсинов, описанные в патенте ("Human Monoclonal Antibodies Against Bacterial Toxins - US Patent 4689299"). Патент посвящен получению и продукции гибридных клеточных линий, секретирующих моноклональные антитела против бактериальных токсинов, и методу использования полученных моноклональных антител для защиты от бактериальных (дифтерийного и столбнячного) токсинов;
2) моноклональные антитела против Mycoplasma pneumoniae (Monoclonal antibodies against Mycoplasma pneumoniae, hybridomas producing these, methods for the preparation thereof, and the use thereof - US Patent 5641638). В патент описаны моноклональные антитела способные специфически связывать антиген Р2 микоплазмы M.pneumoniae, а также клетки гибридомы, продуцирующие эти моноклональные антитела.
Данные технические решения как наиболее близкие к заявляемому по составу действующего вещества фармацевтической композиции и способу его использования выбраны авторами настоящего изобретения за прототип.
Недостатками прототипа являются:
1) отсутствие высокоэффективного способа генерирования и селекции антител;
2) большой размер, что влечет за собой пониженную проницаемость;
3) структурные особенности не позволяют выявлять «скрытые» эпитопы антигенов;
4) ограниченность генно-инженерных манипуляций, адаптации для конкретных задач, возможность создания многовалентных и многофункциональных производных.
Таким образом, в уровне техники существует острая потребность в разработке эффективных специфичных антител против микоплазмы хоминис (M.hominis).
Задачей настоящего изобретения явилось создание новых антител, способных эффективно связывать антигены микоплазмы M.hominis и блокировать размножение этой бактерии.
Поставленая задача решается за счет того, что создают наноантитело, специфически связывающее липид-ассоциированный антиген M.hominis и имеющее аминокислотную последовательность SEQ ID NO:2. Создают также наноантитело, специфически связывающее липид-ассоциированный антиген M.hominis и имеющее аминокислотную последовательность SEQ ID NO:4. При этом наноантитело, имеющее аминокислотную последовательность SEQ ID NO:2, подавляет инфекцию, вызванную микоплазмой M.hominis, и также наноантитело, имеющее аминокислотную последовательность SEQ ID NO:4, подавляет инфекцию, вызванную микоплазмой M.hominis. Способ лечения инфекции млекопитающего, вызванной микоплазмой M.hominis, заключается во введении указанному млекопитающему терапевтически эффективного количества наноантитела, имеющего аминокислотную последовательность SEQ ID NO:2. Способ лечения инфекции млекопитающего, вызванной микоплазмой M.hominis, заключается во введении указанному млекопитающему терапевтически эффективного количества наноантитела, имеющего аминокислотную последовательность SEQ ID NO:4.
В основе настоящего изобретения находятся не классические бивалентные антитела, которые рассматриваются в качестве прототипа, а малые наноантитела, которые имеют ряд преимуществ по сравнению с классическими моноклональными антителами для практического применения в области терапии заболеваний. Поскольку наноантитела (молекулярная масса около 12-15 кДа) на порядок меньше по размеру традиционных антител, они приобретают ряд новых положительных качеств, имеющих практическую значимость. Существует эффективные способы получения (и селекции) таких антител в отношении различных (в том числе низкоиммуногенных) антигенов микоплазм, а благодаря своей низкой иммуногенности наноантитела могут применятся для лечения инфекций, вызванных патогенами данной группы.
Полным эквивалентом термина "наноантитела" для целей настоящего изобретения является вошедшее в широкое употребление обозначение «нанотело», введенное фирмой ABLYNX (NANOBODYTM), а также «однодоменное мини-антитело» и «однодоменное наноантитело».
Рекомбинантные наноантитела получают на основе особых неканонических одноцепочечных антител, существующих в норме наряду с классическими антителами у животных семейства верблюдовых (и у некоторых видов хрящевых рыб). Эти особые антитела состоят из димера только одной укороченной (без первого константного района СН1) тяжелой цепи иммуноглобулина и полнофункциональны в отсутствие легкой цепи иммуноглобулина. Для собственно специфического узнавания и связывания антигена при этом необходим и достаточен лишь один вариабельный домен (VHH, «наноантитело», «nanobody» или однодоменное наноантитело) этого антитела. Организация вариабельных доменов (VHH) неканонических антител в значительной степени подобна той, что у вариабельных доменов (VH) классических антител (у человека VH-домены иммуноглобулинов подкласса IgG3 имеют особо выраженную гомологию с VH и VHH верблюдовых). В обоих случаях V-домены состоят из четырех консервативных каркасных участков (FR, «framework regions»), окружающих три гипервариабельных участка (определяющие комплементарность, CDR, от «complementarity determining regions»). В обоих случаях домены формируют типичную для V-домена иммуноглобулина пространственную структуру из двух бета-слоев (-листов), один - из четырех аминокислотных цепочек, и второй - из пяти [Padlan Е.А. X-Ray crystallography of antibodies. Adv. Protein Chem. 1996; 49: 57-133. Muyldermans S., Cambillau C., Wyns L. Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains./ «Узнавание антигенов фрагментами однодоменных антител: избыточная роскошь спаренных доменов». TIBS 2001; 26: 230-235]. В этой структуре все три гипервариабельных участка кластеризуются с одной стороны V-домена (где они участвуют в узнавании антигена) и располагаются в петлях, соединяющих бета-структуры. Однако имеются и важные отличия, связанные с функционированием VHH в формате одного домена. Так, гипервариабельные участки CDR1 и CDR3 заметно увеличены в случае VHH. Часто в гипервариабельных участках VHH обнаруживаются цистеиновые остатки, причем присутствующие сразу в двух участках (чаще всего в CDR1 и CDR3, реже - в CDR2 и CDR3). При исследовании кристаллических структур VHH было показано, что эти цистеиновые остатки формируют дисульфидные связи, что приводит к дополнительной стабилизации структуры петель данного антигена. Наиболее явным и воспроизводимым отличительным признаком VHH являются четыре замены гидрофобных аминокислотных остатков на гидрофильные во втором каркасном участке (Val37Phe, Gly44Glu, Leu45Arg, Trp47Gly, согласно нумерации Кабат). Этот каркасный участок в случае VH домена является высококонсервативным, обогащен гидрофобными аминокислотными остатками и особо важен для образования связи с вариабельным доменом VL легкой цепи. VHH-домен в этом плане сильно отличается: указанные замены гидрофобных аминокислот на гидрофильные делают невозможной ассоциацию VHH и VL. Эти замены также объясняют высокую растворимость VHH, наноантитела, когда его получают в виде рекомбинантного белка [Тиллиб С.В. «Верблюжьи наноантитела» - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология 2011; 45(1): 77-85].
По сравнению с традиционными и чисто рекомбинантными антителами верблюжьи наноантитела обладают рядом преимуществ, что позволяет предполагать большой потенциал их будущего использования в различных исследованиях и при создании новых биотехнологических устройств, а также в клинических целях для диагностики и лечения заболеваний.
Характерными особенностями наноантител, определяющими большой потенциал их использования для самых разнообразных практических приложений в иммунобиотехнологии, являются следующие [см. обзор: Тиллиб С.В. «Верблюжьи наноантитела» - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология 2011; 45(1): 77-85]:
1) наличие высокоэффективного способа генерирования и селекции наноантител;
2) малый размер, ~2×4 нм, 13-15 кДа (улучшенная проницаемость клеток);
3) структурные особенности (способность образовывать необычные для классических антител паратопы, позволяющие связываться с углублениями и активными центрами белков); могут использоваться для выявления «скрытых» эпитопов или эпитопов, которые не могут быть узнаны существенно более крупными обычными антителами;
4) высокий экспрессионный выход, экономичность наработки в больших количествах. Обычно наноантитела нарабатывают в периплазме бактерий E.coli (в количестве 1-10 мг из 1 литра культуры). Продемонстрирована возможность их эффективной наработки в дрожжах, растениях и клетках-млекопитающих;
5) простота всевозможных генно-инженерных манипуляций, адаптации для конкретных задач, возможность создания многовалентных и многофункциональных производных;
6) низкая иммуногенность; возможность экономично «гуманизировать» антитела без заметной потери их специфической активности.
Возможность получения рекомбинантных наноантител с заданной специфичностью определяется существованием у представителей семейства Camelidae функциональных и обладающих достаточно широким спектром узнавания неканонических антител. Неканонические антитела состоят из димера только одной укороченной тяжелой цепи иммуноглобулина (без легких цепей), специфичность узнавания которых определяется лишь одним вариабельным доменом [Hamers-Casterman C, Atarhouch Т, Muyldermans S, et al. Naturally occurring antibodies devoid of light chains./ «Существующие в природе антитела без легких цепей». Nature 1993; 363:446-448]. Техническая реализация отбора наноантител, являющихся генно-инженерными производными антиген-распознающих доменов одноцепочечных антител верблюда, основана на высокоэффективной процедуре селекции антиген-узнающих полипептидов, экспонированных на поверхности частицы нитчатого фага («фаговый дисплей»).
Метод фагового дисплея является весьма эффективной и широко используемой технологией для функционального отбора из больших рекомбинантных библиотек последовательностей ДНК, кодирующих пептиды и белки, обладающие заданными свойствами и экспрессирующиеся в составе поверхностного белка нитчатых фагов [Brissette R & Goldstein NI. The use of phage display peptide libraries for basic and translational research. /«Использование пептидных фагово-дисплейных библиотек для фундаметальных исследований и исследований трансляции». Methods Mol Biol. 2007; 383:203-13; Sidhu SS & Koide S. Phage display for engineering and analyzing protein interaction interfaces./ «Фаговый дисплей для конструирования и анализа взаимодействий белковых доменов». Curr Opin Struct Biol. 2007; 17:481-7]. Одно из особо важных приложений этой технологии - генерирование специфических рекомбинантных антител для самых различных антигенов [Hoogenboom HR. Selecting and screening recombinant antibody libraries. / «Селекция и анализ библиотек рекомбинантных антител». Nat Biotechnol. 2005; 23:1105-16]. Обычно вместо больших целых молекул классических антител для экспонирования на поверхности фага используют гибридные рекомбинантные одноцепочечные белки, представляющие собой случайные комбинации клонированных последовательностей вариабельных районов тяжелой и легкой цепей иммуноглобулинов, соединенные короткой серин/глицин-богатой линкерной последовательностью. Такая химерная молекула, в случае правильного сочетания доменов, способна сохранять специфичность исходного иммуноглобулина, несмотря на введенные по сравнению с нативной молекулой антител изменения. Одной из проблем традиционных рекомбинантных технологий является необходимость работы с очень большими библиотеками рекомбинантных антител, в которых должны быть представлены всевозможные комбинации двух случайных вариабельных районов (тяжелой и легкой цепей иммуноглобулинов), соединенных линкерной последовательностью. Помимо проблемы представленности здесь также очевидна и проблема формирования правильной относительной конформации этих двух доменов, а также проблема растворимости индивидуальных вариабельных доменов, которые часто имеют тенденцию к агрегации. Упомянутых проблем можно избежать при использовании наноантител, так как практически каждый клонированный вариабельный домен одноцепочечных антител будет в этом случае обладать определенной антиген-узнающей специфичностью, соответствующей одному из антител иммунизированного животного, и можно эффективно проводить селекцию из относительно небольших библиотек таких доменов.
Наноантитела с заданной специфичностью или их производные могут использоваться, как и классические антитела, в различных приложениях, включающих в себя, но не ограниченных, детекцию антигенов (как в исследовательских, так и в диагностических целях), блокирование активности белка-антигена, специфическую доставку за счет связывания с антигеном желаемых молекул, конъюгированных с антителом. Также наноантитела могут быть исходными модулями (блоками) более сложных многомодульных препаратов. Возможно объединение в одном мультивалентном производном двух, трех и более моновалентных первичных наноантител. Эти объединяемые в одну конструкцию наноантитела могут связываться как с одним и тем же эпитопом антигена-мишени, так и с его разными эпитопами или даже с различными антигенами-мишенями. Возможно также комбинированное объединение в одну конструкцию наноантител и других молекул или лекарств с получением многофункциональных препаратов [Conrath KE, Lauwereys М, Wyns L, Muyldermans S. Camel single-domain antibodies as modular building units in bispecific and bivalent antibody constructs. / «Верблюжьи однодоменные антитела в качестве модулярныйх строительных единиц в биспецифичных и бивалентных конструкциях антител». J Biol Chem. 2001 Mar 9; 276 (10): 7346-50; Zhang J, Tanha J, Hirama T, Khieu NH, To R, Tong-Sevinc H, Stone E, Brisson JR, MacKenzie CR. Pentamerization of single-domain antibodies from phage libraries: a novel strategy for the rapid generation of high-avidity antibody reagents. / «Пентамеризация однодоменных антител из фаговых библиотек: новая стратегия быстрого получения реагентов антител с высокой авидностью». J Mol Biol. 2004 Jan 2; 335 (1): 49-56; Cortez-Retamozo V, Backmann N, Senter PD, Wernery U, De Baetselier P, Muyldermans S, Revets H. Efficient cancer therapy with a nanobody-based conjugate. / «Эффективная раковая терапия конъюгатами на основе нанотел». Cancer Res. 2004 Apr 15; 64 (8): 2853-7; Baral TN, Magez S, Stijlemans B, Conrath K, Vanhollebeke B, Pays E, Muyldermans S, De Baetselier P. Experimental therapy of African trypanosomiasis with a nanobody-conjugated human trypanolytic factor. / «Экспериментальная терапия африканской трипаносомии с помощью человеческого трипанолитического фактора, конъюгиованного с нанотелом». Nat. Med. 2006 May; 12 (5): 580-4; Coppieters K, Dreier T, Silence K, Haard HD, Lauwereys M, Casteels P, Beirnaert E, Jonckheere H, Wiele CV, Staelens L, Hostens J, Revets H, Remaut E, Elewaut D, Rottiers P. Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis. / «Форматированные анти-TNFalpha VHH-белков, выделенные из верблюдовых, демонстрируют высокое сродство к воспаленным суставам в мышиной модели коллаген-индуцированного артрита». Arthritis Rheum. 2006 Jun; 54 (6): 1856-66]; мультимеризация с помощью введения дополнительных аминокислотных последовательностей взаимодействующих белковых доменов, таких как лейциновые зипперы [Harbury Р.В., Zhang Т., Kim P.S., et al. A switch between two-, three- and four-stranded coiled coils in GCN4 leucine zipper mutants. / «Перключение между двух-, трех- и четырех-цепочечными спиральными структурами в мутантах GCN-«лейциновой молнии»». Science, 1993, 262:1401-1407; Shirashi T., Suzuyama k., Okamoto H. et al. Increased cytotoxicity of soluble Fas ligand by fusing isoleucine zipper motif. / «Усиленная цитотоксичность растворимого Fas-лиганда вследствие его соединения с мотивом «изолейциновой молнии»». Biochem. Biophys. Res. Communic. 2004, 322: 197-202; Chenchik A., Gudkov A., Komarov A., Natarajan V. Reagents and methods for producing bioactive secreted peptides. / «Реагенты и методы для получения биоактивных секретируемых пептидов». 2010. US Patent Application 20100305002] или последовательностей небольших белков, образующих стабильные комплексы [Deyev SM, Waibel R, Lebedenko EN, Schubiger AP, Plückthun A. Design of multivalent complexes using the barnase*barstar module. / «Дизайн мультивалентных комплексов используя модуль барназа-барстар». Nat Biotechnol. 2003, 21(12):1486-92].
Также было показано [Vincke C., Loris R., Saerens D., et al. // J. Biol. Chem. 2009. V.284. №5. P.3273-3284], что можно «гуманизировать» такие верблюжьи наноантитела без заметной потери их специфической активности, проведя небольшое число точечных замен аминокислот. Это открывает потенциальную возможность широкого использования наноантител в качестве средств пассивной иммунизации для предотвращения развития различных опасных инфекционных заболеваний [Wesolowski J., Alzogaray V., Reyelt J. et al. Single domain antibodies: promising experimental and therapeutic tools in infection and immunity. / «Однодоменные антитела: многообещающие экспериментльные и терапевтические инструменты в области инфекции и иммунитета». Med. Microbiol. Immunol. 2009; 198, 157-174].
Раскрытие настоящего изобретения
Способ получения наноантител, связывающих антигены микоплазмы M.hominis, проводят на основании селекции методом фагового дисплея, генно-инженерных модификаций кодирующих эти антитела последовательностей и использования в качестве продуцента действующего вещества (антитела) бактерии E.coli. Наноантитела (молекулярная масса около 12-15 кДа) на порядок меньше по размеру традиционных антител и являются полнофункциональными антиген-узнающими единицами, обладающими рядом новых положительных качеств практической значимости. Наноантитело представляет собой однодоменный белок, который хорошо растворим и обладает повышенной стабильностью (в широком диапазоне температур и pH). Это позволяет избегать проблем с растворимостью и правильным сворачиванием молекул антител при продукции прокариотическими клетками и ведет к существенному снижению затрат на их производство по сравнению с традиционными способами получения терапевтических моноклональных антител в эукариотических системах экспрессии. Существенно облегчается также процедура сохранения и транспортировки антител по сравнению с заметно менее стабильными традиционными антителами. В силу меньшего размера наноантитела характеризуются лучшей способностью проникать в ткани. Наконец, наноантитела позволяют легко проводить генно-инженерные манипуляции с целью последующей продукции биспецифических наноантител или химер, в состав которых помимо наноантитела входит другой белок с желаемыми свойствами.
Несмотря на то что, главным образом, в иллюстративных целях авторами настоящего изобретения делается акцент на продукции антител с использованием бактерии E.coli, среднему специалисту в данной области техники будет очевидно, что под объем настоящего изобретения подпадают и другие варианты систем реализации настоящего изобретения, прямо не заявленные в настоящем документе. Например, в качестве эукариотического продуцента могут быть использованы дрожжевые культуры или любые другие системы, очевидные в данной области техники.
Способ получения наноантител описан в примерах 1 и 2. Выбор путей реализации с целью получения наноантител с заявляемыми свойствами из прокариотической системы экспрессии в соответствии с одним из предпочтительных вариантов осуществления настоящего изобретения обусловлен следующими факторами:
1) высокий экспрессионный выход, экономичность наработки в больших количествах, обеспечивающиеся экспрессией наноантител в периплазму бактерий E.coli (в количестве 1-10 мг из 1 литра культуры);
2) простота всевозможных генно-инженерных манипуляций, адаптации для конкретных задач, возможность создания многовалентных и многофункциональных производных;
3) высокая экономическая рентабельность производства. Авторы настоящего изобретения исходят из того, что, как известно среднему специалисту в данной области техники, первичные, исходно получаемые последовательности наноантител могут быть затем адаптированы или «форматированы» различным образом для последующего практического использования.
Так, наноантитела могут быть исходными модулями (блоками) более сложных многомодульных препаратов. Возможно объединение в одном мультивалентном производном двух, трех и более моновалентных первичных наноантител. Эти объединяемые в одну конструкцию наноантитела могут связываться как с одним и тем же эпитопом антигена-мишени, так и с его разными эпитопами или даже с различными антигенами-мишенями. Возможно также комбинированное объединение в одну конструкцию наноантител и других молекул или лекарств с получением многофункциональных препаратов; мультимеризация с помощью введения дополнительных аминокислотных последовательностей, взаимодействующих белковых доменов, таких как лейциновые зипперы, или последовательностей небольших белков, образующих стабильные комплексы. Для модулирования свойств препарата наноантитела, например увеличения времени жизни или совершенствования способа очистки, в состав конечного соединения могут быть введены дополнительные аминокислотные последовательности. Среднему специалисту в данной области техники будет очевидно, что такие модификации и прочие варианты антител, лежащих в основе настоящего изобретения, подпадают под объем настоящего изобретения, поскольку являются структурными и функциональными вариантами наноантител. Таким образом, авторы настоящего изобретения понимают под термином "наноантитела" как первичные, исходно получаемые, "минимальные" аминокислотные последовательности наноантител, так и их модификации, полученные в результате упомянутых адаптаций или «форматирования» и их варианты. Термин «вариант антитела» для целей настоящего изобретения означает полипептид, который содержит изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности белка, например как минимум 10% от активности исходного наноантитела. Ряд изменений в варианте белка зависит от положения или от типа аминокислотного остатка в трехмерной структуре белка. Количество изменений может составлять, например, от 1 до 30, более предпочтительно, от 1 до 15, и наиболее предпочтительно, от 1 до 5 изменений в последовательности исходного наноантитела. Эти изменения могут иметь место в областях полипептида, которые не являются критичными для его функции. Это становится возможным благодаря тому, что некоторые аминокислоты обладают высокой гомологией друг с другом, и поэтому третичная структура или активность белка не нарушаются при таком изменении. Поэтому в качестве варианта белка может выступать белок, который характеризуется гомологией не менее 70%, предпочтительно, не менее 80%, более предпочтительно, не менее 90%, и наиболее предпочтительно, не менее 95% по отношению к аминокислотной последовательности исходного наноантитела при условии сохранения активности полипептида. Гомология между аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например с помощью выравнивания последовательностей в компьютерной программе BLAST 2.0, которая вычисляет три параметра: счет, идентичность и сходство.
Замена, делеция, вставка, добавление или замена одного или нескольких аминокислотных остатков будут представлять собой консервативную мутацию или консервативные мутации при условии, что активность белка при этом сохраняется. Примером консервативной мутацей(ями) является консервативная замена(ы). "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток заменяется аминокислотным остатком, имеющим сходную боковую цепь. В данной области техники определены семейства аминокислот, имеющих сходные боковые цепи. Эти семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Поскольку гипервариабельные районы наноантител определяют их специфическое взаимодействие с антигеном, то именно гомологичные замены аминокислот в этих участках могут приводить к получению несколько различающихся по последовательности наноантител, которые обладают идентичными или близкими свойствами. Таким образом, среднему специалисту в данной области техники будет очевидно, что под объем настоящего изобретения подпадают не только указанные в приложении последовательности наноантител, но и те, которые могут быть получены путем замен аминокислот в гипервариабельных участках (указанных в перечне последовательностей как CDR) на другие, но очень близкие по свойствам, аминокислоты (консервативных замен).
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК, кодирующего исходное наноантитело, например посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид исходного наноантитела, могут быть получены путем экспрессирования фрагментов ДНК, имеющих мутацию, описанную выше, и установления активности экспрессируемого продукта.
Замена, делеция, вставка или добавление нуклеотидов, описанных выше, также включают мутации, которые имеют место в природе и, например, обусловлены изменчивостью.
Полипептиды наноантител согласно настоящему изобретению могут кодироваться большим множеством молекул нуклеиновых кислот, что является результатом хорошо известного в данной области техники явления вырожденности генетического кода. Суть феномена состоит в том, что любая аминокислота (за исключением триптофана и метионина), входящая в состав природных пептидов, может кодироваться более чем одним триплетным нуклеотидным кодоном. Любая из этих вырожденных кодирующих молекул нуклеиновых кислот может входить в состав кассет, экспрессирующих антитела, заявленные в соответствии с настоящим изобретением и подпадающие под объем настоящего изобретения.
Получение функционально-активных наноантител, распознающих антигены M.hominis, показано в примерах 3, 4, 5.
Эффективность лечения микоплазменной инфекции выбранным фармацевтически активным количеством антител доказана в примере 6.
Краткое описание чертежей
На фиг.1 представлены нуклеотидные и аминокислотные последовательности двух отобранных наноантител, aMh1 и aMh2, полученные параллельно одним и тем же способом и обладающие искомыми свойствами: способностью специфически связывать Mycoplasma hominis. Нуклеотидные последовательности кДНК, кодирующие два отобранных наноантитела, были определены (для aMh1 - SEQ ID NO: 1; для aMh2 - SEQ ID NO: 3), и из них были выведены соответствующие аминокислотные последовательности отобранных наноантител (для aMh1 - SEQ ID NO: 2; для aMh2 - SEQ ID NO: 4). В указанных последовательностях подчеркнуты соответственно (слева направо, от N- к С-концу) гипервариабельные участки CDR1, CDR2 и CDR3 антигенузнающих последовательностей отобранных наноантител.
На фиг.2 представлена элелекторофореграмма полиакриламидного геля с антигенным материалом белков микоплазмы хоминис (M.hominis) штамм Н-34, использованным для иммунизации верблюда.
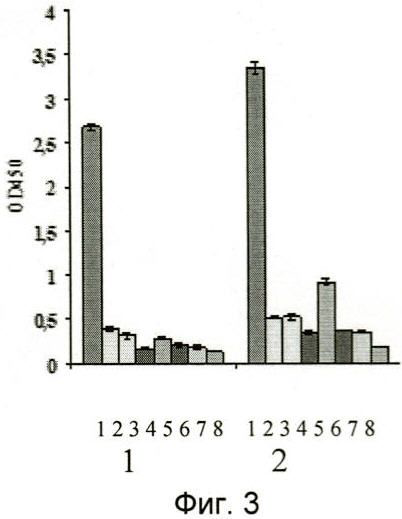
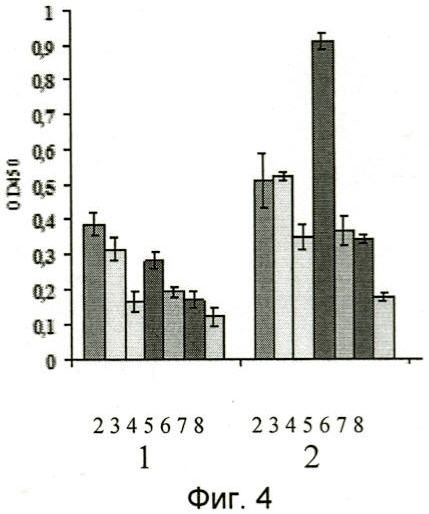
На фиг.3 и 4 представлены результаты иммуноферментного анализа узнавания отобранными наноантителами aMh1, aMh2 иммобилизованных в лунках иммунологической плашки липид-ассоциированных мембранных белков микоплазм. В качестве контрольных использовали ячейки с иммобилизованным белком овальбумином кур. Видно, что антитела специфически связываются с антигенами микоплазмы хоминис M.hominis, и, с меньшей аффинностью (фиг.4), микоплазмы орале M.orale и микоплазмы аргинини M.arginini и не взаимодействуют с липид-ассоциированными мембранными белками других видов микоплазм.
Цифрами от 1 до 8 обозначены столбцы, соответствующие различным видам микоплазм и отрицательному контролю эксперимента:
1 - Mycoplasma hominis (микоплазма хоминис),
2 - Mycoplasma orale (микоплазма орале),
3 - Mycoplasma arginini (микоплазма аргинини),
4 - Mycoplasma arthritidis (микоплазма артритидис),
5 - Mycoplasma pneumoniae (микоплазма пневмониа),
6 - Mycoplasma fermentans (микоплазма ферментанс),
7 - Ureaplasma urealiticum (уреаплазма уреалитикум),
8 - отрицательный контроль, цифрами 1 и 2 обозначены соответственно наноантитела aMh1, aMh2.
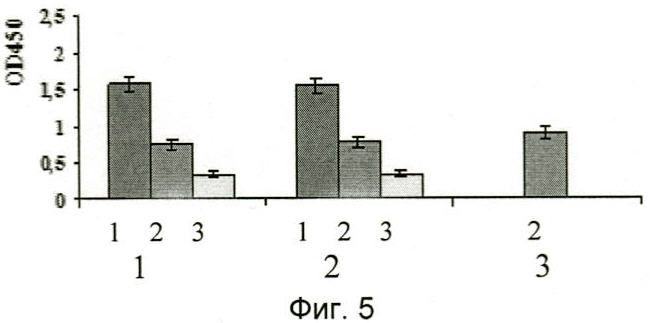
На фиг.5 проиллюстрированы результаты иммуноферментного анализа узнавания отобранными наноантителами aMh1, aMh2 M.hominis в фиксированной параформальдегидом зараженной культуре клеток. Видно, что антитела специфически связываются с M.hominis в зараженной культуре и не взаимодействуют с контрольными незараженными клетками. Цифрами 1 и 2 обозначены соответственно наноантитела aMh1, aMh2, цифре 3 соответствует отрицательный контроль, цифрами от 1 до 3 обозначены столбцы, соответстветствующие зараженным клеткам, незараженным клеткам и неинкубированным с антителами клеткам.
Примеры осуществления настоящего изобретения
Пример 1
Получение библиотеки вариабельных доменов одноцепочечных антител
Иммунизация
Двугорбого верблюда Camelus bactrianus последовательно иммунизировали 5 раз путем подкожного введения антигенного материала, смешанного с равным объемом полного (при первой инъекции) или неполного (при остальных инъекциях) адъюванта Фрейнда. В качестве антигена использовали препарат липид-ассоциированных белков микоплазмы хоминис (M.hominis), штамм Н-34 (фиг.2), выделенных стандартым методом фракционирования белков в ТХ-114 (Bordier C. Phase separation of integral membrane proteins in Triton X-114 solution., J Biol Chem. 1981 Feb 25; 256(4): 1604-7). Вторую иммунизацию проводили через 3 недели после первой, затем с интервалом в две недели проводили еще три иммунизации. Взятие крови (150 мл) проводили через 5 дней после последней инъекции. Для предотвращения свертывания взятой крови добавляли 50 мл стандартного солевого раствора (PBS), содержащего гепарин (100 ед./мл) и ЭДТА (3 мМ).
Кровь разводили в 2 раза стандартным солевым раствором (PBS), содержащим 1 мМ ЭДТА. 35 мл разбавленного раствора крови наслаивали на ступеньку специальной среды (Histopaque-1077, Sigma) с плотностью 1,077 г/мл объемом 15 мл и проводили центрифугирование в течение 20 мин при 800×g. Мононуклеарные клетки (лимфоциты и моноциты) отбирали из интерфазной зоны плазма/Histopaque, после чего промывали раствором PBS, содержащим 1 мМ ЭДТА.
Суммарную РНК из В-лимфоцитов выделяли с помощью реагента TRIzol (Invitrogen). Затем, на колонке с олиго(dT)-целлюлозой из тотальной РНК очищали поли(А)-содержащую РНК. Концентрацию РНК определяли с помощью биофотометра (Eppendorf) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле с формальдегидом.
Реакцию обратной транскрипции проводили по стандартному протоколу [Sambrook et al., 1989] с использованием обратной транскриптазы H-M-MuLV и праймера олиго(dT)15 в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции и полученные продукты амплификации клонировали по сайтам Ncol(Pstl) и Notl в фагмидный вектор, как описано ранее [Hamers-Casterman et al., 1993; Nguyen et al., 2001; Saerens et al., 2004; Rothbauer et al., 2006]. Процедедуру селекции проводили также аналогично тем, что в указанных работах. Она базировалась на методе фагового дисплея, в которой в качестве фага-помощника использовали бактериофаг M13KO7 (New England Biolabs, США).
Пример 2
Селекция наноантител, специфически узнающих M.hominis
Селекцию наноантител проводили методом фагового дисплея с использованием препарата липид-ассоциированных белков M.hominis, штамм Н-34, иммобилизованного на дне лунок 96-луночного ИФА-планшета. Использовали полистироловые иммунологические плашки с высокой сорбцией MICROLON 600 (Greiner Bio-One). Для блокировки использовали 1% БСА (Sigma-Aldrich, США) и/или 1% обезжиренное молоко (Bio-Rad, США) в PBS. Процедуру селекции и последующей амплификации отбираемых фаговых частиц (содержащих ген наноантитела внутри, а экспрессирующееся наноантитело - в составе поверхностного фагового белка pIII) повторяли, как правило, последовательно три раза. Все манипуляции проводили, как описано в публикациях Тиллиб С.В., Иванова Т.И., Васильев Л.А. 2010. Фингерпринтный анализ селекции «наноантител» методом фагового дисплея с использованием двух вариантов фагов-помощников. Acta Naturae 2010; 2 (3): 100-108; Hamers-Casterman C., Atarhouch T., Muyldermans S. et al. Nature 1993; 363: 446-448.; Nguyen V.K., Desmyter A., Muyldermans S. Adv. Immunol. 2001; 79: 261-296; Saerens D., Kinne J., Bosmans E., Wernery U., Muyldermans S., Conrath K. J Biol Chem. 2004; 279: 51965-51972; Rothbauer U., Zolghadr K., Tillib S., et al. Nature Methods 2006; 3: 887-889].
Последовательности клонов отобранных наноантител группировали согласно схожести их фингерпринтов, получаемых при электрофоретическом разделении продуктов гидролиза амплифицированных последовательностей наноантител параллельно тремя частощепящими рестрикционными эндонуклеазами (Hinfl, Mspl, Rsal). Последовательности кДНК наноантител (SEQ ID NO: 1 и 3) были определены (фиг.1). В указанных последовательностях подчеркнуты соответственно (слева направо) гипервариабельные участки CDR1, CDR2 и CDR3 антиген-узнающих последовательностей отобранных наноантител.
Продукция нанонтител
Последовательности кДНК отобранного наноантитела, переклонировали в экспрессионный плазмидный вектор - модифицированный вектор pHEN6 [Conrath KE, Lauwereys M, Gallenj M, Matagne A, Frère JM, Kinne J, Wyns L, Muyldermans S. Beta-lactamase inhibitors derived from single-domain antibody fragments elicited in the Camelidae. / «Бэта-лактомазные ингибиторы, происходящие из фрагментов однодоменных антител, индуцированных у Верблюдовых». Antimicrob Agents Chemother. 2001; 45:2807-12], позволяющий присоединение к С-концу наноантитела (His)6-эпитопа (сразу вслед за НА-эпитопом, кодируемым в векторе pHEN6). Благодаря наличию на N-конце экспрессируемой последовательности сигнального пептида (peIB) нарабатываемый рекомбинантный белок (наноантитело) накапливается в периплазме бактерий, что позволяет эффективно его выделять методом осмотического шока, не разрушая собственно бактериальные клетки. Продукцию наноантител проводили в Е.coli (штамм BL21). Экспрессию индуцировали добавлением 1 мМ индолил-бета-D-галактопиранозида и клетки инкубировали при интенсивном перемешивании в течение 7 часов при 37°C или в течение ночи при 29°C. Наноантитело выделяли из периплазматического экстракта с использованием аффинной хроматографии на Ni-NTA-агарозе с использованием системы для очистки QIAExpressionist (QIAGEN, США).
Пример 3
Детекция специфических антигенов с использованием наноантител, специфически узнающих M.hominis.
Демонстрация связывания наноантител с липид-ассоциированными белками M.hominis
Способность наноантител связывать липид-ассоциированные белки M.hominis проверяли с использованием метода иммуноферментного анализа с иммобилизованными липид-ассоциированными белками M.hominis, по стандартному протоколу. В качестве контроля использовали лунки с иммобилизованным белком овальбумином кур или бычьим сывороточным альбумином (неспецифические белки). Детекцию связавшихся aMh1 и aMh2 проводили с помощью анти-НА антител мыши, вторичных антител к иммуноглобулинам мыши, конъюгированных с пероксидазой хрена, и хромогенного субстрата ТМБ (3,3',5,5'-тетраметилбензидин, Sigma).
На фиг.3 представлены результаты анализа, из которых следует, что наноантитела специфически связываются с иммобилизованными липид-ассоциированными белками M.hominis, но не связываются с овальбумином, использованным в качестве блокирующего агента.
Пример 4
Демонстрация специфичности связывания наноантител с липид-ассоциированными белками M.hominis, M.pneumoniae, M.orale и M.arginini
Способность наноантител связывать липид-ассоциированные белки М.hominis проверяли с использованием метода иммуноферментного анализа с иммобилизованными липид-ассоциированными белками M.hominis, M.arginini, M.arthritidis, M.orale, M.fermentans, M.pneumoniae, U.urealiticum по стандартному протоколу. В качестве контроля использовали лунки с иммобилизованным белком овальбумином кур или бычьим сывороточным альбумином (неспецифические белки). Детекцию связавшихся aMh2, aMh1 проводили с помощью анти-НА антител мыши, вторичных антител к иммуноглобулинам мыши, конъюгированных с пероксидазой хрена, и хромогенного субстрата ТМБ (3,3',5,5'-тетраметилбензидин, Sigma).
На фиг.3 и 4 представлены результаты анализа, из которых следует, что наноантитела специфически связываются с иммобилизованными липид-ассоциированными белками микоплазмы хоминис (M.hominis) и, с меньшей афинностью, микоплазмы пневмонии (M.pneumoniae), микоплазмы орале (M.orale) и микоплазмы аргинини (M.arginini), но не связываются с липид-ассоциированными белками других видов микоплазм.
Пример 5
Демонстрация связывания наноантител эукариотическими клетками, инфицированными микоплазмой хоминис (M.hominis), in vitro
Эукариотическая культура клеток MCF7 была заражена M.hominis путем добавления культуры M.hominis в культуральную среду и инкубацией клеток MCF7 на этой среде. Затем клетки высевали на культуральный планшет и фиксировали 0,8% параформальдегидом по стандартному протоколу. Способность наноантител aMh3, aMh1 связываться с M.hominis, персистирующей на поверхности эукариотических клеток, проверяли с использованием метода иммуноферментного анализа по стандартному протоколу. В качестве контроля использовали лунки с фиксированными незараженными клетками. Детекцию связавшихся aMh1, aMh2 проводили с помощью анти-НА антител мыши, вторичных антител к иммуноглобулинам мыши, конъюгированных с пероксидазой хрена, и хромогенного субстрата ТМБ (3,3',5,5'-тетраметилбензидин, Sigma).
На фиг.5 представлены результаты анализа, из которых следует, что наноантитела специфически связываются с M.hominis, персистирующей на поверхности эукариотических клеток, но не связываются с незараженными клетками.
Таким образом, заявленные в соответствии с настоящим изобретением фармацевтические композиции доказали свою применимость для детекции M.hominis in vitro.
Пример 6
Для демонстрации эффективности терапевтического действия наноантител для лечения микоплазменной инфекции мыши линии BALB/C (18-20 g) с синхронизированным овуляционным циклом были заражены внутривагинально M.hominis линии MY17386, в дозе 1*10^7 на мышь, как описано ранее (Taylor-Robinson D, Furr РМ. Further observations on the murine model of Mycoplasma hominis infection. / «Дальнейшие наблюдения на мышиной модели инфекции Mycoplasma hominis». J Med Microbiol. 2010 Aug; 59 (Pt 8):970-5. Epub 2010 May 6). Спустя неделю после заражения мыши получали ежедневные внутривагинальные спринцевания объемом 100 мкл ФБР с наноантителами aMh1, 2 в количестве 70 мкг/мышь/день (минимально активное количество антител, способных блокировать микоплазменную инфекцию) в течение 5 дней, доза наноантител была выбрана в соответствии с литературными данными, согласно которым в терапевтических целях антитела используют в дозах от 30 до 100 мкг на мышь (Xiao X, Zhu Z et al. Human anti-plague monoclonal antibodies protect mice from Yersinia pestis in a bubonic plague model.PLoS One. 2010 Oct 13; 5(10):e13047; Chen Z, Moayeri M, Purcell Monoclonal Antibody Therapies against Anthrax. RToxins (Basel). 2011 Aug; 3(8): 1004-19. Epub 2011 Aug 15). Контрольным группам мышей спринцевание проводили 100 мкл ФБР. Спустя 2 дня после последнего спринцевания органы половой системы мышей были гомогенизированы с использованием гомогенизатора с принципом бисерной мельницы, гомогенаты использованы для определения титра микоплазм методом культурального высева.
В табл.1 и 2 представлены результаты культуральных высевов M.hominis из гомогенатов половых органов мышей. Из представленных результатов следует, что наноантитела aMh1 и aMh2 оказывают достоверное ингибирующее действие на микоплазменную внутривагинальную инфекцию, что приводит к снижению титра микоплазм в гомогенатах органов половой системы. Таким образом, продемонстрирована эффективность терапии микоплазменной инфекции с использованием наноантител aMh1, 2. В таблице 1 представлена демонстация эффективности применения наноантитела aMh1 для лечения микоплазменной инфекции, вызванной M.hominis.
В таблице 2 представлена демонстация эффективности применения наноантитела aMh2 для лечения микоплазменной инфекции, вызванной M.hominis.
Таким образом, в указанных примерах раскрыто: создание новых антител, способных эффективно связывать антигены микоплазмы M.hominis; способ их получения; способ лечения урогенитального микоплазмоза, вызванного у млекопитающего микоплазмой M.hominis, из чего очевидно, что поставленная задача достигнута.

Изобретение относится к области биотехнологии и медицины. Предложены антитела, специфически связывающие липид-ассоциированный антиген M.hominis, с охарактеризованными аминокислотными и нуклеотидными последовательностями, а также способ лечения инфекции, вызванной микоплазмой M.hominis, включающий введение указанному млекопитающему терапевтически эффективного количества указанных наноантител. Настоящее изобретение может найти дальнейшее применение в терапии микоплазменной инфекции. 4 н. и 2 з.п. ф-лы, 2 табл., 6 пр., 5 ил.
1. Наноантитело, специфически связывающее липид-ассоциированный антиген M.hominis и имеющее аминокислотную последовательность SEQ ID NO:2.
2. Наноантитело, специфически связывающее липид-ассоциированный антиген M.hominis и имеющее аминокислотную последовательность SEQ ID NO:4.
3. Наноантитело по п.1, отличающееся тем, что оно подавляет инфекцию, вызванную микоплазмой M.hominis.
4. Наноантитело по п.2, отличающееся тем, что оно подавляет инфекцию, вызванную микоплазмой M.hominis.
5. Способ лечения инфекции млекопитающего, вызванной микоплазмой M.hominis, включающий в себя введение указанному млекопитающему терапевтически эффективного количества наноантитела по п.1.
6. Способ лечения инфекции млекопитающего, вызванной микоплазмой M.hominis, включающий в себя введение указанному млекопитающему терапевтически эффективного количества наноантитела по п.2.
| ТИЛЛИБ С.В | |||
| Верблюжьи Наноантитела" - Эффективный инструмент для исследований, диагностики и терапии"// Молекулярная Биология | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| MORRISON-PLUMMER J et al | |||
| Biological effects of anti-lipid and anti-protein monoclonal antibodies on Mycoplasma pneumoniae// Infect Immun | |||
| Пневматический водоподъемный аппарат-двигатель | 1917 |
|
SU1986A1 |
| RAZIN S et al. | |||

