ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к молекулярной биологии и относится к рекомбинатной технологии и экспрессии белков. Более конкретно, изобретение относится к нуклеиновым кислотам для рекомбинантной экспрессии белков, содержащим последовательности, полученные из Lactococcus и подходящие в качестве промоторов. Настоящее изобретение, кроме того, относится к векторам, содержащим указанные нуклеиновые кислоты, и к клеткам-хозяевам, трансформированным ими. Изобретение также относится к применению клеток-хозяев, содержащих указанные нуклеиновые кислоты или векторы, для экспрессии гетерологичных или гомологичных белков, а также для доставки, особенно терапевтической доставки, указанных белков индивидам.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Молочнокислые бактерии становятся все более или более важными в качестве хозяев для рекомбинантной экспрессии гетерологичных полипептидов in vitro (например, патент США 5559007), а также для экспрессии in vivo или in situ и доставки антигенов и/или терапевтических полипептидов (например, WO 97/14806).
Полагают, что молочнокислые бактерии, в частности Lactococcus, являются полностью безвредными микроорганизмами (т.е. их рассматривают как в целом безопасные) и, таким образом, могут относительно просто вводиться людям и животным.
Однако для достижения устойчивого уровня гетерологичной экспрессии в молочнокислых бактериях часто требуется введение промоторов и других последовательностей, которые являются экзогенными для этих бактерий (например, смотри Wells et al., 1993A) и, следовательно, могут ставить под угрозу восприятие их как полностью безвредных.
Соответственно, существует потребность в дополнительных промоторах, которые получают из молочнокислых бактерий, более предпочтительно из Lactococcus, и которые можно успешно использовать для экспрессии белков, предпочтительно экспрессии гетерологичных белков, в молочнокислых бактериях.
Также необходимы такие промоторы, с помощью которых можно добиться высоких уровней экспрессии и таким образом получить достаточное количество экспрессированных белков для промышленного и/или терапевтического использования.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Аспекты по настоящему изобретению относятся по крайней мере к некоторым, например, одной или нескольким, указанным выше потребностям данной области.
В частности, авторы настоящего изобретения идентифицировали нуклеиновые кислоты и последовательности нуклеиновых кислот, происходящие из Lactococcus, использование которых в качестве дополнительных промоторов для рекомбинантной экспрессии, такой как предпочтительно экспрессия полипептидов, в клетках-хозяевах, предпочтительно в бактериях и более предпочтительно в Lactococcus, может иметь преимущества.
Более конкретно, авторы настоящего изобретения были намерены идентифицировать нуклеиновые кислоты и последовательности нуклеиновых кислот из Lactococcus, которые могут действовать в качестве сильных промоторов, т.е. промоторов, с помощью которых достигается высокий уровень экспрессии, для рекомбинантной экспрессии, например предпочтительно экспрессии полипептидов, в клетках-хозяевах, предпочтительно в бактериях и более предпочтительно в Lactococcus, и осуществили поставленную цель. Преимуществом стабильной экспрессии является увеличение количества продуктов экспрессии, например, полипептидов, рекомбинантно продуцируемых клетками-хозяевами, которые становятся доступными для дальнейшего использования, например, для очистки из клеток-хозяев или для терапевтической доставки в такие клетки.
Еще более неожиданно для авторов настоящего изобретения стало ясно, что нуклеиновые кислоты и их последовательности по настоящему изобретению могут действовать в качестве промоторов, которые сильней, особенно при использовании в Lactococcus, чем промоторы, ранее полученные из Lactococcus. Более конкретно, нуклеиновые кислоты и последовательности по настоящему изобретению могут так функционировать в качестве более сильных промоторов, например, более сильных конститутивных промоторов, чем промотор гена тимидилатсинтазы (thyA) из Lactococcus lactis, который, как известно авторам настоящего изобретения, является самым сильным промотором, полученным из Lactococcus, более конкретно самым сильным конструктивным промотором, полученным из Lactococcus, на сегодняшний день. Промотор thyA Lactococcus lactis является, как известно авторам настоящего изобретения, также самым сильным промотором, который известен в настоящее время, для рекомбинантной, например гетерологичной, экспрессии генов в Lactococcus и предпочтительно в Lactococcus lactis.
Неожиданно было отмечено, что комбинированный транскриптомный анализ, описанный в примерах, вместе со сложными данными протеомного анализа не обладал большой возможностью в предположении силы или активности промоторов-кандидатов. В частности, некоторые идентифицированные промоторы были слабо активными при проведении проверки. Кроме того, некоторые последовательности возможных промоторов были неактивны вне природного окружения или природной конфигурации, т.е. при поверке с гетерологичными генами.
В качестве дополнительного преимущества особенно высокая экспрессия наблюдается при использовании промоторов по настоящему изобретению, в числе прочих, для IL-10 человека, пептида YY человека (PYY), глюкагоноподобного пептида-1 человека (GLP-1), GLP-2 человека (GLP-2) и факторов «трилистника» человека (TTF) в качестве предпочтительных целей.
Следует также принимать во внимание, что нуклеиновые кислоты и их последовательности, идентифицированные авторами настоящего изобретения, были получены из Lactococcus, который утвержден как микроорганизм GRAS, т.е. «G (обычно) R (рассматриваемого) A (как) S (безопасный)». Следовательно, композиции, например, клетки-хозяева, содержащие такие промоторы, могут вводиться людям или животным с меньшим беспокойством относительно биологической безопасности, чем при введении последовательностей, полученных, например, из микроорганизмов, не являющихся GRAS, или из других источников.
Таким образом, настоящее изобретение относится к полученным из Lactococcus, которые имеют определенные преимущества, т.е. сравнительно безопасные, нуклеиновым кислотам и последовательностям, которые составляют дополнительные промоторы, более предпочтительно к дополнительным сильным промоторам и еще более предпочтительно к промоторам, которые сильнее промотора thyA, для использования для большого числа применений, включая рекомбинантную экспрессию, например, полипептидов, в клетках-хозяевах, предпочтительно в бактериях и еще более предпочтительно в Lactococcus.
Настоящее изобретение включает вышеприведенные релевантные варианты осуществления в разнообразных аспектах.
Соответственно, в одном из аспектов настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей промотор (Р), который является природным промотором вида Lactococcus, или его функциональный вариант или функциональный фрагмент, функционально связанный с одной или несколькими открытыми рамками считывания (ORF), являющимися гетерологичными для промотора (Р), отличающийся тем, что промотор (Р) сильнее промотора гена тимидилатсинтазы (thyA) Lactococcus lactis в Lactococcus.
В связанном аспекте настоящее изобретение, таким образом, также относится к рекомбинантной нуклеиновой кислоте, содержащей промотор (Р), функционально связанный с одной или несколькими открытыми рамками считывания, являющимися гетерологичными для промотора (Р), причем промотор (Р) выбран из группы, включающей или состоящей из природных промоторов генов Lactococcus для 1) бета'-субъединицы/субъединицы с М.м. 160 кДа ДНК-зависимой РНК-полимеразы (rpoC), 2) бета-субъединицы/субъединицы с М.м. 140 кДа ДНК-зависимой РНК-полимеразы (rpb2), 3) ДНК-связывающего ферритиноподобного белка (защитного средства от окислительной деградации) (dps), 4) пируваткиназы (pyk), 5) глютамил- и глютаминил-тРНК-синтетаз (glnS), 6) энолазы (eno), 7) глютаминсинтетазы (glnA), 8) регулятора транскрипции НТН-типа (glnR), 9) (Xaa-His)-дипептидазы (argE или pepV), 10) бета-субъединицы АТФ-синтазы F0F1-типа (бета-субъединицы АТФ-синтазы F1) (atpD), 11) 3-фосфоглицераткиназы (pgk), 12) глицеральдегид-3-фосфатдегидрогеназы/эритрозо-4-фосфатдегидрогеназы (gapA), 13) ацетаткиназы (ackA), 14) 3-оксоацил-[ацил-носитель-белок]-синтазы (fabB), 15) 3-оксоацил-[ацил-носитель-белок]-редуктазы (fabG), 16) альфа-субъединицы/субъединицы с М.м. 40 кДа ДНК-зависимой РНК-полимеразы (rpoА), 17) (Xaa-Pro)-аминопептидазы (pepP), 18) фруктозо/тагатозобифосфатальдолазы (tbp), 19) рибосомного белка S4 (rpsD), 20) супероксиддисмутазы (sodA), 21) рибосомного белка S12 (rpsL) и рибосомного белка S7 (rpsG), 22) рибосомного белка L18 (rplR) и рибосомного белка S5 (rpsE) и рибосомного белка L30/L7E (rpmD), 23) S-рибозилгомоцистеинлиазы (luxS), 24) рибосомного белка L19 (rplS), 25) рибосомного белка S11 (rpsK), 26) рибосомного белка L10 (rplJ), 27) рибосомного белка L7/L12 (rplL), 28) ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU (hup), 29) белка L28 50S-субъединицы рибосомы (rpmB), 30) специфичного в отношении целлобиозы компонента фосфотрансферазной системы IIB (lacE), 31) альфа-субъединицы АТФ-синтазы F0F1-типа (atpA), 32) транспортной системы сахаров АВС-типа (компонента АТФазы) (malK), 33) альфа-субъединицы компонента ацетоиндегидрогеназного комплекса Е1 (acoA), 34) белка клеточного деления (diflVA или ftsA), 35) UDP-галактопиранозомутазы (glf), 36) глютамиламинопептидазы (frvX), 37) спрогнозированного относящегося к дегидрогеназе белка (mviM), 38) рибосомного белка S2, 39) фактора 3 инициации трансляции (IF-3) (infC), 40) рибосомного белка L4 (rplD) и рибосомного белка L23 (rplW) и рибосомного белка L2 (rplB), 41) домена EMAP (ydjD), 42) фактора элонгации транскрипции (greA), 43) субъединицы АТФ-зависимой протеазы Clp (clpP), 44) рибосомного белка L15 (rplO), 45) рибосомного белка L11 (rplK), 46) рибосомного белка S8 (rpsH), 47) рибосомного белка L21 (rplU), 48) рибосомного белка S13 (rpsM), 49) рибосомного белка S19 (rpsS) и рибосомного белка L22 (rplU) и рибосомного белка L16 (rplP) и рибосомного белка L14 (rplN), 50) рибосомного белка S10 (rpsJ), 51) шаперонина GroES (Hsp10) (cpn10), 52) рибосомного белка L24 (rplX) и 53) гипотетического белка LACR_0137 (duf965), и их функциональных вариантов и функциональных фрагментов указанных природных промоторов.
Настоящее изобретение также относится к рекомбинантной нуклеиновой кислоте, причем промотор (Р) выбран из группы, состоящей из природных промоторов генов Lactococcus, предпочтительно Lactococcus lactis, для 1) бета'-субъединицы/субъединицы с М.м. 160 кДа ДНК-зависимой РНК-полимеразы (rpoC), 3) связывающего негемное железо ферритина (dpsA), 4) пируваткиназы (pyk), 5) глютаминил-тРНК-синтетаз (gltX), 6) фосфопируватгидрaтазы (eno), 9) дипептидазы PepV (pepV), 12) глицеральдегид-3-фосфатдегидрогеназы (gapB), 13) ацетаткиназы (ackA), 18) фруктозобифосфатальдолазы (fbaA), 20) супероксиддисмутазы (sodA), 21) рибосомного белка S12 (rpsL) и рибосомного белка S7 (rpsG), 22) рибосомного белка L18 (rplR) и рибосомного белка S5 (rpsE) и рибосомного белка L30/L7E (rpmD), 24) рибосомного белка L19 (rplS), 26) рибосомного белка L10 (rplJ), 28) HU-подобного ДНК-связывающего белка (hllA), 29) белка L28 50S-субъединицы рибосомы (rpmB), 30) компонента фосфотрансферазной системы IIB (ptcB), 31) альфа-субъединицы АТФ-синтазы F0F1-типа (atpA), 32) связывающего множество сахаров транспортного АТФ-связывающего белка (msmK), 33) альфа-субъединицы компонента пируватдегидрогеназы Е1 (pdhA), 34) белка клеточного деления (diflVA или ftsA), 35) UDP-галактопиранозомутазы (glf1), 36) глютамиламинопептидазы (pepA), 37) спрогнозированного относящегося к дегидрогеназе белка (llmg 0272), 38) рибосомного белка S2 (rpsB), 39) фактора 3 инициации трансляции (IF-3) (infC), 40) рибосомного белка L4 (rplD) и рибосомного белка L23 (rplW) и рибосомного белка L2 (rplB), 41) бета-цепи фенилаланил-тРНК-синтетазы (pheT), 42) фактора элонгации транскрипции GreA (greA), 43) протеолитической субъединицы АТФ-зависимой протеазы Clp (clpP), 44) рибосомного белка L15 (rplO), 45) рибосомного белка L11 (rplK), 46) рибосомного белка S8 (rpsH), 47) рибосомного белка L21 (rplU), 48) рибосомного белка S13 (rpsM), 49) рибосомного белка S19 (rpsS) и рибосомного белка L22 (rplU) и рибосомного белка L16 (rplP) и рибосомного белка L14 (rplN), 50) рибосомного белка S10 (rpsJ), 51) шаперонина GroES (Hsp10) (groES), 52) рибосомного белка L24 (rplX) и 53) предполагаемой резольвазы, осуществляющей разделение-соединение (llmg_0151), и их функциональных вариантов и функциональных фрагментов указанных природных промоторов.
В более предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, причем промотор (Р) выбран из группы, состоящей из природных промоторов генов Lactococcus, предпочтительно Lactococcus lactis, для 1) бета'-субъединицы/субъединицы с М.м. 160 кДа ДНК-зависимой РНК-полимеразы (rpoC), 3) связывающего негемное железо ферритина (dpsA), 4) пируваткиназы (pyk), 5) глютаминил-тРНК-синтетаз (gltX), 6) фосфопируватгидрaтазы (eno), 9) дипептидазы PepV (pepV), 12) глицеральдегид-3-фосфатдегидрогеназы (gapB), 13) ацетаткиназы (ackA), 18) фруктозобифосфатальдолазы (fbaA), 20) супероксиддисмутазы (sodA), 21) рибосомного белка S12 (rpsL) и рибосомного белка S7 (rpsG), 22) рибосомного белка L18 (rplR) и рибосомного белка S5 (rpsE) и рибосомного белка L30/L7E (rpmD), 24) рибосомного белка L19 (rplS), 26) рибосомного белка L10 (rplJ), 28) HU-подобного ДНК-связывающего белка (hllA), 29) белка L28 50S-субъединицы рибосомы (rpmB), 30) компонента фосфотрансферазной системы IIB (ptcB), которые определены в таблице 1.
В другом предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей промотор гена для 28) ДНК-связывающего белка бактериального ядра/HU-подобного ДНК-связывающего белка (hlla или hup); функционально связанный с одной или несколькими открытыми рамками считывания, которые являются гетерологичными для промотора. Еще более предпочтительно указанный промотор является промотором PhllA.
В другом предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей промотор гена для 3) связывающего негемное железо ферритина (dpsA или LACR_2311), промотор гена 9) дипептидазы PepV (pepV или LACR_0908) или промотор гена 20) супероксиддисмутазы (sodA или LACR_0458), соответственно; функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными для промотора. Еще более предпочтительно указанный промотор является промотором PdpsA, PpepV или PsodA.
В предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей промотор (Р), функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными для промотора (Р), причем промотор (Р) выбран из группы, включающей или состоящей из природных промоторов генов Lactococcus, перечисленных выше или в таблице 1 под номерами 1-30, предпочтительно промотора PhllA, PdpsA, PpepV или PsodA, и функциональных вариантов и функциональных фрагментов указанных природных промоторов.
В еще одном предпочтительном варианте осуществления указанные выше гены и соответствующие им природные промоторы и их функциональные варианты и функциональные фрагменты получены из Lactococcus lactis.
В связанных, приводимых в качестве примеров аспектах, также предложены векторы, содержащие рекомбинантные нуклеиновые кислоты по настоящему изобретению; интегрированные в хромосомы экспрессионные кассеты, клетки-хозяева, трансформированные рекомбинантными нуклеиновыми кислотами по настоящему изобретению или векторами, их содержащими; к применению рекомбинантных нуклеиновых кислот по настоящему изобретению для экспрессии продуктов экспрессии, предпочтительно одного или нескольких полипептидов, кодируемых указанными открытыми рамками считывания, в клетке-хозяине; к способам рекомбинантной экспрессии и выделения представляющих интерес продуктов экспрессии, предпочтительно полипептидов, используя указанные клетки-хозяева; к способам лечения, включая in situ доставку терапевтически значимых продуктов экспрессии, предпочтительно полипептидов, например, антигенов и/или невакциногенных терапевтически активных полипептидов, людям или животным с помощью таких клеток-хозяев; и связанные применения клеток-хозяев для получения лекарственных средств для облегчения указанной доставки; к фармацевтическим композициям, содержащим указанные клетки-хозяева; и тому подобное.
Эти и другие аспекты и предпочтительные варианты осуществления по настоящему изобретению описаны в следующих далее разделах и в прилагаемой формуле изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 (А-Н) представлены последовательности предпочтительных промоторов.
На фиг.2 показано электрофоретическое разделение полипептидов Lactococcus lactis MG1363.
На фиг.3 предложена стратегия клонирования.
Фиг.4. Суклонирование слитых конструкций промотор-Usp45-h[Gly2]GLP2. Р обозначает любой из следующих промоторов: P1 (Waterfield et al., 1995), PthyA (промотор тимидилатсинтазы) и PhllA (промотор ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU); usp45 означает сигнал секреции usp45 дикого типа (van Asseldonk et al., 1990) или его мутант (в том числе Usp45 N4, в котором лизин в положении 4 заменен аспарагином); Em означает селектируемый маркер - маркер устойчивости к эритромицину; ori означает точку начала репликации.
Фиг.5. Продукция и секреция h[Gly2]GLP-2 рекомбинантными штаммами L. lactis, обнаруживаемые с помощью антитела против hGLP-2. (А) секреция h[Gly2]GLP-2. В качестве положительного контроля использовали 1 мкг рекомбинантного hGLP-2. (В) Продукция и секреция h[Gly2]GLP-2 клетками. Каждая линия на блоте представляет 1 мл клеточной фракции или супернатанта культуры клеток L. lactis, полученных через три часа роста. SeeBlue® Plus2 (invitrogen) использовали в качестве маркеров молекулярной массы (Маркеры М.м.).
Фиг.6. Схематическое сравнение L. lactis MG1369 и различных экспрессирующих штаммов hIL-10, используемых в этом исследовании. Во время конструирования экспрессирующие hIL-10 кассеты интегрировали в хромосому L. lactis MG1369 путем гомологичной рекомбинации идентичных последовательностей, находящихся 5' от thyA и экспрессирующих hIL-10 кассет, а также 3' от них, соответственно. Места рекомбинации схематически представлены как «\». Это предполагает, что все последовательности ДНК вне экспрессионных кассет являются идентичными для описанных выше штаммов. Генетические элементы не масштабированы.
Фиг.7. Сравнение экспрессии hIL-10 c PthyA (промотора тимидилатсинтазы, штаммы sAGX0005 и Thy12) c экспрессией hIL-10 с (A) промотора PdpsA (ДНК-связывающего ферритиноподобного белка, SEQ ID NO: 3; sAGX0012), (B) PpepV (промотора (Xaa-His)-дипептидазы, SEQ ID NO: 9 или 158; штамм sAGX0018), (C) PsodA (промотора супероксиддисмутазы, SEQ ID NO: 20; sAGX0029) и (D) PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; штамм sAGX0037). «Промотор» означает промотор, расположенный перед геном hIL-10.
Фиг.8. Сравнение экспрессии hIL-10 в 109 клеток MG1363, sAGX0005 и sAGX0037.
Фиг.9. Схематическое сравнение L. lactis MG1363 и различных экспрессирующих hTFF штаммов, используемых в этом исследовании. Во время конструирования экспрессирующие TFF кассеты интегрировали в хромосому L. lactis MG1363 путем гомологичной рекомбинации идентичных последовательностей, находящихся 5' от thyA и экспрессирующих TFF кассет, а также 3' от них, соответственно. Места рекомбинации схематически представлены как «\». Это предполагает, что все последовательности ДНК вне экспрессионных кассет являются идентичными для описанных выше штаммов. Мутант usp45 указан *. Генетические элементы не масштабированы.
Фиг.10. Сравнение
(А) экспрессии hTFF1 c PthyA (промотора тимидилатсинтазы), связанного с сигналом секреции usp45 дикого типа и hTFF1 (штамм sAGX0041), PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28), связанного с мутантным (mut) сигналом секреции usp45 и hTFF1 (штамм sAGX0049), или PhllA, связанного с сигналом секреции usp45 дикого типа и hTFF1 (штамм sAGX0048);
(B) экспрессии hTFF3 c PthyA (промотора тимидилатсинтазы), связанного с сигналом секреции usp45 дикого типа и hTFF3 (штамм sAGX0043), промотора PdpsA (ДНК-связывающего ферритиноподобного белка, SEQ ID NO: 3), связанного с сигналом секреции usp45 дикого типа и hTFF3 (sAGX0059) и PhllA, связанного с мутантным сигналом секреции usp45 и hTFF3 (штамм sAGX0047).
«Промотор» и «сигнал секреции» означают промотор и сигнал секреции, которые расположены перед геном TFF.
Фиг.11. Анализ Вестерн-блоттинг супернатантов различных указанных штаммов. На линии, содержащие эталонные белки, указаны как «hTFF1» (эталон hTFF3) и «Маркеры М.м.» (маркеры молекулярной массы, «кДа» означает молекулярные массы). Все другие линии содержат эквивалент 0,5 мл супернатанта культуры указанных штаммов, полученного, как указано выше. Первым антителом было мышиное моноклональное антитело против hTFF1: 1/1000 (Abnova: номер в каталоге Н00007031-М02). Вторым антителом было козье антимышиное антитело, конъюгированное с АР: 1/1000 (Southern Biotech: 4050-04), и обнаружение проводили с использованием NBT/BCIP (Roche 11 697 471 0001). Маркерами М.м. является предварительно окрашенный стандарт Invitrogen SeeBlue plus2 (номер в каталоге LC5925).
Фиг.12. Схематический обзор структуры pT1NX и различных экспрессирующих hPYY G9 (3-36) плазмид, используемых в этом исследовании. Экспрессионные плазмиды pAGX0211, pAGX0212 и pAGX0213 получены встраиванием соответствующих экспрессионных кассет в виде EcoRI-SpeI-фрагментов в плазмиду pT1NX, открытую расщеплением ферментами EcoRI и SpeI. Как таковые, структура и положение всех последовательностей ДНК вне экспрессионных кассет, таких как точка начала репликации (ori) и маркер устойчивости к эритромицину (EmR), идентичны для всех плазмид. Генетические элементы не масштабированы.
Фиг.13. Сравнение экспрессии hPYY G9 (3-36) c P1 (Waterfield et al., 1995) (плазмида pAGX0211), PthyA (промотора тимидилатсинтазы, плазмида pAGX0212) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; плазмида pAGX0213), связанных с сигналом секреции ups45 и hPYY G9 (3-36). Все плазмиды присутствовали в L. lactis MG1369. «Промотор» означает промотор, который находится в направлении 3'-5' от гена hPYY G9 (3-36) или присутствует в эквивалентном сайте в pT1NX.
Фиг.14. Схематический обзор структуры pT1NX и различных экспрессирующих hGLP-1 G8 (7-36) плазмид, используемых в этом исследовании. Экспрессионные плазмиды pAGX0233 и pAGX0234 получены встраиванием соответствующих экспрессионных кассет в виде EcoRI-SpeI-фрагментов в плазмиду pT1NX, открытую расщеплением ферментами EcoRI и SpeI. Как таковые, структура и положение всех последовательностей ДНК вне экспрессионных кассет, таких как точка начала репликации (ori) и маркер устойчивости к эритромицину (EmR), идентичны для всех плазмид. Генетические элементы не масштабированы.
Фиг.15. Сравнение экспрессии hGLP-1 G8 (7-36) c P1 (Waterfield et al., 1995) (плазмида pAGX0211), PthyA (промотора тимидилатсинтазы, плазмида pAGX0212) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; плазмида pAGX0213), связанных с сигналом секреции ups45 и hGLP-1 G8 (7-36). Все плазмиды присутствовали в L. lactis MG1369. «Промотор» означает промотор, который находится в направлении 3'-5' от гена hGLP-1 G8 (7-36) или присутствует в эквивалентном сайте в pT1NX.
Фиг.16. Схематическое сравнение L. lactis MG1363 и различных экспрессирующих hIL-10 штаммов, используемых в этом исследовании. Во время конструирования экспрессирующие hIL-10 кассеты подвергают интеграции в хромосому L. lactis MG1363 с помощью гомологичной рекомбинации между идентичными последовательностями, находящимися 5' от thyA и экспрессирующих hIL-10 кассет, а также 3' от них, соответственно. Места рекомбинации схематически представлены «\». Это предполагает, что все последовательности ДНК вне экспрессионных кассет являются идентичными для описанных выше штаммов. В настоящем описании «промотором» является любой один из промоторов, представленных в штамме «sAGX00xx» (таблица 11). Генетические элементы не масштабированы.
Фиг.17. Сравнение экспрессии hIL-10 c PthyA (промотора тимидилатсинтазы, штамм sAGX0005) c экспрессией hIL-10 с любого другого промотора ряда штаммов (смотри фиг.16 и таблицу 11), в которых эти промоторы Lactococcus помещены 5' от слитого гена usp45-hIL-10.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Используемые в настоящем описании формы единственного числа подразумевают как единичное, так и множественное число, если из контекста явно не следует другое. Например, «клетка» относится к одной клетке или к более, чем одной клетке.
Используемые в настоящем описании термины «содержит» и «содержащий» являются синонимами терминов «включающий» и «включает», являются «содержащими» или «открытыми» формами, не исключают дополнительные, не указанные члены, элементы или стадии способов.
Перечисление численных диапазонов с помощью конечных точек включает все числа и доли, находящиеся в пределах этого диапазона, включая указанные конечные значения.
Под термином «приблизительно» понимают, если этот термин используют в описании в отношении измеряемой величины, такой как параметр, количество, период времени и тому подобные отклонения, составляющие +/-20% или меньше, предпочтительно +/-10% или меньше, более предпочтительно +/-5% или меньше, еще более предпочтительно +/-1% или меньше или даже еще более предпочтительно +/-0,1% или меньше, от определенной величины до таких пределов, при которых такие отклонения подходят для осуществления раскрытого изобретении.
Все приведенные в настоящем описании документы включены в качестве ссылки в полном объеме. В частности, описания всех документов, на которые в описании дана ссылка, включены в качестве ссылки.
Кроме случаев оговоренных особо, значения всех терминов, используемых при раскрытии по настоящему изобретению, в том числе технических и научных термины, такие же, как значения, обычно понимаемые средним специалистом в данной области, к которой относится это изобретение. Посредством дополнительного определения далее приведены определения для лучшего понимания сущности по настоящему изобретению.
Используемый в настоящем описании термин «нуклеиновая кислота» означает полимер любой длины, по существу состоящий из нуклеотидов, например, дезоксирибонуклеотидов и/или рибонуклеотидов. Нуклеиновые кислоты могут включать пуриновые и/или пиримидиновые основания и/или другие природные (например, ксантин, инозин, гипоксантин), химически или биохимически модифицированные (например, метилированные), неприродные или производные нуклеотидные основания. Остов нуклеиновых кислот может включать сахара и фосфатные группы, которые обычно встречаются в РНК или ДНК, и/или один или несколько модифицированных или замещенных сахаров и/или одну или несколько модифицированных или замещенных фосфатных групп. Термин «нуклеиновая кислота», кроме того, предпочтительно включает ДНК, РНК или гибридные молекулы ДНК/РНК, конкретно, включая яРНК человека, пре-мРНК, мРНК, кДНК, геномную ДНК, продукты амплификации, олигонуклеотиды и синтетическую (например, химически синтезированную) ДНК, РНК или гибриды ДНК/РНК. «Нуклеиновая кислота» может быть двухцепочечной, частично двухцепочечной или одноцепочечной. Если нуклеиновая кислота является одноцепочечной, то она может быть смысловой цепью или антисмысловой цепью. Кроме того, нуклеиновая кислота может быть кольцевой или линейной.
В предпочтительном варианте осуществления нуклеиновая кислота, содержащая промотор по настоящему изобретению, является ДНК или РНК, более предпочтительно ДНК.
Термин «рекомбинантная нуклеиновая кислота» обычно относится к нуклеиновой кислоте, которая состоит из сегментов, соединенных вместе, используя технологию рекомбинантных ДНК. Если рекомбинантная нуклеиновая кислота подвергается репликации в организме-хозяине, то нуклеиновые кислоты потомства также входят в определение термина «рекомбинантная нуклеиновая кислота».
Стандартные ссылки, в которых описаны общие принципы технологии рекомбинантных ДНК, включают Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989; Current Protocols in Molecular Biology, ed. Ausubel et al., Greene Publishing and Wiley-Interscience, New York, 1992 (с периодическими обновлениями) (“Ausubel et al. 1992”); Innis et al., PCR Protocols: A Guide to Methods and Applications, Academic Press: San Diego, 1990. Основные принципы микробиологии изложены, например, в Davis, B.D. et al. В Microbiology, 3rd edition, Harper & Row, издательство Philadelphia, PA (1980).
Под промотором обычно понимают участок на молекуле нуклеиновой кислоты, предпочтительно на молекуле ДНК, с которым связывается РНК-полимераза, инициируя транскрипцию. Промотор предпочтительно, но не обязательно, располагается выше, т.е. в направлении 5' от последовательности, транскрипцию которой он контролирует. В настоящем изобретении конкретные промоторы обозначают термином «Р», за которым следует название гена, из которых они получены. Например, «PthyA» обозначает промотор гена thyA, а «PhllA» обозначает промотор гена hllA.
Термин «природный промотор» относится к промотору, нуклеотидная последовательность которого идентична нуклеотидной последовательности природного промотора, например, в клетке или организме в природе. Термин «природный промотор», таким образом, относится к последовательности промотора и не должен истолковаться как требование того, чтобы промотор был получен или продуцирован каким-либо конкретным способом. Наример, но не ограничиваясь, термин таким образом включает промоторы природных хозяев, выделенных из них, клонированных и репродуцированных с использованием технологии рекомбинантных ДНК, продуцированных способом амплификации или полученных синтетическим способом и тому подобное, если последовательность таких промоторов остается одинаковой последовательностям их природных копий (смотри, например, таблицу 1).
Специалисту в данной области будет понятно, что природная последовательность промотора данного гена может отличаться у различных видов Lactococcus и/или различных подвидов внутри одного вида Lactococcus и/или различных штаммов внутри одного вида или подвида Lactococcus из-за природной генетической дивергенции указанных видов, подвидов и/или штаммов. Таким образом, такие дивергентные, но обнаруживаемые в природе последовательности промоторов могли бы считаться природными.
Специалист в данной области обычно может спрогнозировать и идентифицировать природные бактериальные промоторы, такие как промоторы Lactococcus. Тем не менее, в качестве дополнительного руководства природный промотор можно, как правило, идентифицировать путем анализа геномной последовательности или ее части из бактерии, предпочтительно из вида Lactococcus; идентификации в ней открытой рамки считывания, т.е. последовательности кодирующих нуклеотидных триплетов, начинающейся с кодона инициации трансляции (предпочтительно ATG) и заканчивающейся кодоном терминации трансляции (например, TAA, TAG или TGA), и не содержащей какого-либо внутреннего, находящегося в рамке кодона терминации трансляции, и анализа последовательности, находящейся в направлении 5' от указанного кодона инициации трансляции, для того, чтобы локализовать самый верхний кодон инициации трансляции, в направлении 5' от которого наблюдается в рамке кодон терминациии трансляции. Предпочтительно транскрипцию идентифицированной таким образом открытой рамки считывания можно подтвердить экспериментально, например с помощью Норзерн-блоттинга или ОТ-ПЦР, а сайт инициации транскрипции (например, прилегающий к самому верхнему и/или, возможно, одному или нескольким из более нижних кодонов инициации трансляции) можно определить, используя, например, способ быстрой амплификации 5'-концов кДНК (5'-RACE).
Как правило, последовательности, прилегающие и ближайшие в направлении 3'-5' к самому верхнему кодону инициации трансляции (и/или, если экспериментальные данные указывают на это, одному или нескольким из более нижних кодонов инициации трансляции) могут включать природный промотор, ответственный за транскрибирование указанной ORF. В качестве предпочтительного примера, если первый нуклеотид кодона инициации трансляции обозначают «+1» (например, нуклеотид А кодона ATG - +1), а нуклеотид, находящийся непосредственно в направлении 5' от него, обозначают «-1», то термин «природный промотор» может относиться к последовательности приблизительно от «-500» до приблизительно «+50», например приблизительно от «-500» до приблизительно «+20», приблизительно от «-500» до приблизительно «+10», приблизительно от «-500» до приблизительно «+5», приблизительно от -«500» до приблизительно «+2» или приблизительно от «-500» до приблизительно «-1», предпочтительно приблизительно от «-400» до приблизительно «+50», например, в предпочтительных примерах приблизительно от «-400» до приблизительно «+20», например приблизительно от «-400» до приблизительно «+10», приблизительно от «-400» до приблизительно «+5», приблизительно от «-400» до приблизительно «+2» или приблизительно от «-400» до приблизительно «-1», более предпочтительно приблизительно от «-300» до приблизительно «+50», например, в предпочтительных примерах приблизительно от «-300» до приблизительно «+20», например приблизительно от «-300» до приблизительно «+10», приблизительно от «-300» до приблизительно «+5», приблизительно от «-300» до приблизительно «+2» или приблизительно от «-300» до приблизительно «-1», такой как, например, в предпочтительных примерах приблизительно от «-200» до приблизительно «+50» или дальнейших предпочтительных примерах приблизительно от «-200» до приблизительно «+20», например приблизительно от «-200» до приблизительно «+10», приблизительно от «-200» до приблизительно «+5», приблизительно от «-200» до приблизительно «+2» или приблизительно от «-200» до приблизительно «-1», или такой как, например, в других предпочтительных примерах приблизительно от «-100» до приблизительно «+50» и в дальнейших предпочтительных примерах приблизительно от «-100» до приблизительно «+20», например приблизительно от «-100» до приблизительно «+10», приблизительно от «-100» до приблизительно «+5», приблизительно от «-100» до приблизительно «+2» или приблизительно от «-100» до приблизительно «-1», если указанная последовательность демонстрирует промоторную активность.
Также предусматривается использование функциональных вариантов природных промоторов Lactococcus в рекомбинантных нуклеиновых кислотах по настоящему изобретению. Термин «вариант» относится к последовательности, которая по существу идентична (т.е. в основном, но не полностью идентична) соответствующей природной последовательности, например последовательности соответствующего природного промотора Lactococcus. «По существу идентична» обозначает идентичность по крайней мере на 60%, предпочтительно по крайней мере на 70%, более предпочтительно по крайней мере на 80%, например по крайней мере на 85%, более предпочтительно по крайней мере на 90%, например по крайней мере на 91%, 92%, еще более предпочтительно по крайней мере на 93%, например, 94%, еще более предпочтительно по крайней мере на 95%, например по крайней мере на 96%, более предпочтительно по крайней мере на 97%, например по крайней мере на 98%, и наиболее предпочтительно по крайней мере на 99%. Выравнивание последовательностей и определение идентичности последовательностей можно осуществить, например, используя инструмент Basic Local Alignment Search Tool (BLAST), первоначально описанный Altschul et al. (1990), такой как алгоритм «Blast 2 sequences», описанный Tatusova и Madden (1999).
Изобретение также относится к использованию функциональных фрагментов природных промоторов Lactococcus в рекомбинантных нуклеиновых кислотах по настоящему изобретению. Используемый в настоящем описании термин «фрагмент» относится к последовательности, которая имеет 5' и/или 3' делецию одного или нескольких нуклеотидов по сравнению с природной последовательностью, например, природным промотором Lactococcus или его вариантом, но в которой остальная последовательность нуклеиновой кислоты фрагмента идентична соответствующим положениям в последовательности природного промотора Lactococcus или его варианта. Остальная последовательность фрагмента может представлять предпочтительно по крайней мере 30%, например по крайней мере 40%, более предпочтительно по крайней мере 50%, например по крайней мере 60%, даже более предпочтительно по крайней мере 70%, например по крайней мере 80% или по крайней мере 85%, и еще более предпочтительно по крайней мере 90%, например по крайней мере 95% или более от последовательности нуклеиновой кислоты соответствующего природного промотора Lactococcus или его варианта, например, идентифицированного способами по настоящему изобретению, например, приведенными конкретными последовательностями SEQ ID NO: в таблице 1.
Термин «функциональный» относительно вариантов и фрагментов промоторов, указанных выше, относится к тому факту, что у конкретных вариантов и фрагментов, по крайней мере частично сохраняется промоторная активность, т.е. способность связывать РНК-полимеразу и инициировать транскрипцию, соответствующего природного промотора. Предпочтительно такие функциональные варианты или функциональные фрагменты будут сохранять по крайней 50% активности соответствующего природного промотора, например по крайней мере 60%, более предпочтительно по крайней мере 70%, например по крайней мере 80%, еще более предпочтительно по крайней мере 85%, например по крайней мере 86%, по крайней мере 87%, по крайней мере 88% или 89%, еще более предпочтительно по крайней мере 90%, например по крайней мере 91%, по крайней мере 92%, по крайней мере 93%, по крайней мере 94% и наиболее предпочтительно по крайней мере 95%, например по крайней мере 96%, по крайней мере 97% и наиболее предпочтительно по крайней мере 98% или по крайней мере 99% активности соответствующего природного промотора. Также предпочтительно такие функциональные варианты или функциональные фрагменты могут иметь еще более высокую активность, чем активность соответствующего природного промотора.
Специалисту также будет понятно, что в вариантах осуществления рекомбинантные нуклеиновые кислоты по настоящему изобретению могут, кроме того, включать более одного промотора (Р), и/или функционального варианта, и/или функционального фрагмента по настоящему изобретению. Например, указанные промоторы, функциональные варианты или функциональные фрагменты, которые могут быть одинаковыми или различными, могут быть функционально связаны и контролировать экспрессию различных транскрипционных единиц в пределах указанных нуклеиновых кислот. Альтернативно или дополнительно, экспрессия одной транскрипционной единицы может контролироваться более чем одним промотором(ами), функциональным вариантом(ами) и/или функциональным фрагментом(ами), функционально связанной с нею, которые могут быть одинаковыми или различными. Например, функциональная связь более чем одного из указанных выше элементов, имеющих промоторную активность, с одной транскрипционной единицей может дополнительно увеличить уровень транскрипции указанной единицы. Например, но им не ограничиваясь, такие промоторы, функциональные варианты и/или функциональные фрагменты могут располагаться последовательно, например последовательно в направлении 3'-5' от соответствующей транскрипционной единицы.
В еще одном альтернативном варианте осуществления настоящее изобретение также относится к рекомбинантным нуклеиновым кислотам, содержащим химерные промоторы, включающие две или более частей, полученных из различных промоторов, функциональных вариантов и/или функциональных фрагментов по настоящему изобретению и вместе составляющих новый промотор.
Специалисту хорошо известны способы определения активности промоторов. Например, последовательность нуклеиновой кислоты, активность которой в качестве промотора пытаются определить, может быть встроена в рекомбинантную репортерную конструкцию при условии, чтобы она была функционально связана с последовательностью репортера, предпочтительно с последовательностью, кодирующей репортер, такой как, например, зеленый флуоресцентный белок (GFP) или хлорамфениколацетилтрансфераза (САТ) и тому подобное, и оценивают экспрессию и/или накопление мРНК репортера (например, с помощью Норзерн-блоттинга, количественной ОТ-ПЦР и тому подобное) и/или белка (например, с помощью Вестерн-блоттинга, ELISA, измерения флуоресценции или ферментативной активности и тому подобное), если указанная репортерная конструкция вводится в представляющие интерес клетки-хозяева или организмы.
В приведенном в качестве примера предпочтительном варианте осуществления можно измерить экспрессию белка, гетерологичного для организма, в котором измеряют экспрессию, например, гетерологичного для бактерий, предпочтительно гетерологичного для Lactococcus, еще более предпочтительно гетерологичного для Lactococcus lactis. Например, в предпочтительном варианте осуществления можно оценить экспрессию в бактериях, предпочтительно в Lactococcus, еще более предпочтительно в Lactococcus lactis, гена, кодирующего полипептид эукариотического происхождения, еще более предпочтительно любого гена, кодирующего терапевтически значимые полипептиды, предназначенные для экспрессии, используя нуклеиновые кислоты по настоящему изобретению (указанные в этом описании), предпочтительно GLP-2, GLP-1, PYY и TFF и еще более предпочтительно любой из иммуномодулирующих полипептидов, цитокинов, факторов роста или интерлейкинов, например, очень предпочтительно hIL-10. Таким образом, определенные значения для исследованных последовательностей нуклеиновых кислот указывают активность или силу возможных промоторов, включенных в такие последовательности. Специалисту также понятно, что для обеспечения сравнительного характера таких анализов промоторной активности условия, отличные от условий исследуемых последовательностей нуклеиновых кислот, должны оставаться приблизительно одинаковыми или в идеальном случае одинаковыми для различных анализов. Такие условия могут включать, например, количество репортерной конструкции, вводимое в клетки, способ трансформации, используемый для такого введения, число и сайт интеграции таких репортерных конструкций в геноме исследуемых реципиентных клеток и/или состояние (например, фазу роста, например предпочтительно экспоненциальную фазу роста) реципиентных клеток-хозяев во время измерения и тому подобное. Приводимый в качестве примера способ измерения силы промотора в реципиентных клетках Lactococcus lactis, используя hIL-10 в качестве экспрессируемого гена, показан в примере 2. Дополнительный примерный способ измерения силы промотора в реципиентных клетках Lactococcus lactis и Lactobacillus casei, используя GLP2 в качестве экспрессируемого гена, показан в примерах 3 и 4. Другие дополнительные примерные способы измерения силы промотора в реципиентных клетках Lactococcus lactis, используя GLP1, hPYY, hIL-10 и TFF в качестве экспрессируемых генов, показаны в примерах 5-9. Эксперименты, в которых используются Lactobacillus casei в качестве реципиентных клеток, приводят к схожим результатам (непредставленным).
Соответственно, промотор (1), который указывают как «более сильный» по сравнению с другим промотором (2), должен демонстрировать значительно большую активность, оцениваемую подходящими анализами, например анализами, описанными в предыдущем абзаце, например анализом, приведенным в качестве примера в примерах 2-9. Значительно большая активность относится к статистически значимому обнаружению большей активности промотора (1), например предпочтительно с р<0,5 или р<0,05.
Активность промотора (1) может быть больше активности промотора (2) на любую величину, и предпочтительно активность промотора (1) может быть больше активности промотора (2) по крайней мере на 1% от величины активности промотора (2), например по крайней мере на 2%, по крайней мере на 3% или по крайней мере на 4%, более предпочтительно по крайней мере на 5%, например по крайней мере на 6%, по крайней мере на 7%, по крайней мере на 8% или по крайней мере на 9%, даже более предпочтительно по крайней мере на 10%, например по крайней мере на 15%, еще более предпочтительно по крайней мере на 20%, например по крайней мере на 30% или по крайней мере на 40%, даже более предпочтительно по крайней мере на 50%, например по крайней мере на 60%, по крайней мере на 70%, по крайней мере на 80% или по крайней мере на 90%, и в дальнейших очень предпочтительных примерах по крайней мере на 100%, по крайней мере на 150%, по крайней мере на 200%, по крайней мере на 250%, по крайней мере на 300%, по крайней мере на 400% или по крайней мере на 500% от величины активности промотора (2), или в других предпочтительных примерах активность промотора (1) может быть больше по крайней мере в 10 раз активности промотора (2), например по крайней мере приблизительно в 50 раз, по крайней мере приблизительно в 100 раз, по крайней мере приблизительно в 500 раз или по крайней мере приблизительно в 1000 раз.
Соответственно, для получения функциональных вариантов или фрагментов промоторов, особенно промоторов, раскрытых в настоящем изобретении, специалист должен бы знать, как получить такие варианты (например, с помощью направленного или неспецифического мутагенеза) или фрагменты (например, с помощью 5' и/или 3' усечений, например, с помощью расщепления ферментами рестрикции или ПЦР) и исследовать такие варианты или фрагменты на их промоторную активность, как указано выше. Тем не менее, в качестве дополнительного руководства, но им не ограничиваясь, было отмечено, что бактериальные промоторы часто включают консенсусные последовательности, прилегающие к положениям -10 и -35. Соответственно, функциональные фрагменты бактериальных, например Lactococcus, промоторов могут предпочтительно включать по крайней мере последовательности, соответствующие положениям природных промоторов приблизительно от -10 до приблизительно -35, более предпочтительно приблизительно от -8 до приблизительно -40, более предпочтительно приблизительно от -5 до приблизительно -40 или приблизительно от -5 до приблизительно -45 и еще более предпочтительно приблизительно от -2 или -1 до приблизительно -50. Кроме того, функциональные варианты бактериальных, например Lactococcus, промоторов могут предпочтительно включать консенсусные последовательности, прилегающие к положениям -10 и -35, представленные в природных аналогичных промоторах или известные в данной области.
«Функциональная связь» является связью, с помощью которой регуляторные последовательности ДНК и последовательность ДНК, которую пытаются экспрессировать, соединяются так, что обеспечивают экспрессию. Например, говорят, что последовательности ДНК, такие как, например, предпочтительно промотор, и гетерологичная открытая рамка считывания функционально связаны, если характер связи между этими последовательностями (1) не приводит к введению мутации со сдвигом рамки, (2) не мешает промотору управлять транскрипцией открытой рамки считывания или (3) не мешает транскрипции открытой рамки считывания с помощью последовательности промоторного района.
В приводимом в качестве примера предпочтительном варианте осуществления указанный промотор может быть расположен выше, т.е. в направлении 5' от, открытой рамки (рамок) считывания, с которой он функционально связан.
Конкретная природа регуляторных участков, требуемых для экспрессии, может изменяться в зависимости от организма, но указанные участки должны включать промоторный участок, который, в прокариотах, содержит как промотор (который управляет инициацией транскрипции РНК), так и последовательности ДНК, которые после транскрибирования в РНК будут подавать сигнал к инициации синтеза белка. Такие участки обычно включают такие 5'-некодирующие последовательности, которые вовлечены в инициацию транскрипции и трансляции, такие как рамка Прибноу (подтвержденная ТАТА-рамка), последовательность Шайна-Дальгарно и тому подобное.
Преимущественно соответствующий кодон инициации трансляции, с которым заданный промотор по настоящему изобретению обычно связан в природе, может быть не включен в рекомбинантные нуклеиновые кислоты по настоящему изобретению, например, для предотвращения инициации трансляции с таких кодонов. Например, 3'-конец промотора может быть усечен в положении -1 и выше этого положения; альтернативно, кодон инициации трансляции может быть мутирован (например, ATG в другой кодон) и тому подобное. В еще одном альтернативном случае может присутствовать указанный природный кодон инициации трансляции и возможно также наличие нескольких (например, предпочтительно ≤ 20, более предпочтительно ≤ 10, еще более предпочтительно ≤ 5, например максимально 1, 2, 3 или 4) дополнительных кодонов открытой рамки считывания, связанных с заданным промотором в природе, и гетерологичная открытая рамка считывания, представляющая интерес, может быть слита с ним в рамке считывания, продуцируя таким образом слитый продукт.
Термин «открытая рамка считывания» или «ORF» относится к последовательности кодирующих нуклеотидных триплетов, начинающейся с кодона инициации трансляции (предпочтительно ATG), заканчивающейся кодоном терминации трансляции (например, TAA, TAG или TGA) и не содержащей какого-либо внутреннего находящегося в рамке кодона терминации трансляции и потенциально способной кодировать полипептид. Следовательно, этот термин может быть синонимом термина «кодирующая последовательность, используемого в данной области. В рекомбинантной нуклеиновой кислоте по настоящему изобретению кодоны инициации трансляции одной или нескольких ORF могут быть, как правило, связаны с регуляторными последовательностями, контролирующими инициацию трансляции, например, с последовательностью Шайна-Дальгарно. Известно, что в бактериях, в том числе Lactococcus, мультицистронные единицы, содержащие две или более последовательных ORF, контролируемых общим расположенным выше в последовательности промотором, можно получить путем соединения находящихся ниже в последовательности кодонов инициации трансляции с указанными последовательностями, контролирующими такую инициацию трансляции.
Термин «гетерологичный», при указании взаимоотношения между данной ORF и промотором, означает, что указанный промотор обычно не связан с указанной ORF в природе, т.е. промотор обычно не контролирует ее транскрипцию. Другими словами, связь создается методами рекомбинантных ДНК в рекомбинантных нуклеиновых кислотах по настоящему изобретению.
Термин «Lactococcus» обычно относится к роду Lactococcus и включает любой таксон (например, вид, подвид, штамм), отнесенный к такому роду в данной области. В качестве примера, Lactococcus включает виды Lactococcus garvieae, Lactococcus lactis, Lactococcus piscium, Lactococcus plantarum и Lactococcus raffinolactis и любые их подвиды (subsp.) и штаммы.
В предпочтительных вариантах осуществления настоящего изобретения Lactococcus является Lactococcus lactis. Lactococcus lactis включает, но ими не ограничивается, Lactococcus lactis subsp. cremoris, Lactococcus lactis subsp. hordniae, Lactococcus lactis subsp. lactis, Lactococcus lactis subsp. bv. diacetylactis.
В дополнительных предпочтительных вариантах осуществления настоящего изобретения Lactococcus lactis является Lactococcus lactis subsp. cremoris или Lactococcus lactis subsp. lactis, более предпочтительно Lactococcus lactis subsp. lactis, и включает любой его штамм, такой как, например, Lactococcus lactis subsp. cremoris SK11 или Lactococcus lactis subsp. lactis MG 1363.
В предпочтительных вариантах осуществления промотор (Р) получают из Lactococcus, как определено выше, более предпочтительно из предпочтительных таксонов Lactococcus, определенных выше, особенно в двух предшествующих абзацах.
Если говорят, что промотор (Р) сильнее промотора thyA Lactococcus lactis в Lactococcus, то это означает, что он сильнее по крайней мере в одном и возможно в большем числе таксонов или во всех таксонах (например, видах, подвидах или штаммах) Lactococcus. Предпочтительно промотор (Р) может быть таким образом сильнее по крайней мере в Lactococcus lactis. Также предпочтительно промотор (Р) может быть таким образом сильнее по крайней мере в Lactococcus lactis subsp. cremoris или Lactococcus lactis subsp. lactis, более предпочтительно по крайней мере в Lactococcus lactis subsp. lactis.
Термин «тимидилатсинтаза» относится к ферменту ЕС 2.1.1.45, а «ген тимидилатсинтазы (thyA) Lactococcus lactis» обозначает ген, кодирующий указанный фермент в Lactococcus lactis. Последовательность гена thyA из нескольких таксонов Lactococcus lactis, таких как, например, Lactococcus lactis subsp. lactis MG 1363 (Ross et al., 1990; Steidler et al., 2003; входящий № в Genbank: AF462070), Lactococcus lactis subsp. lactis IL1403 (Bolotin et al., 2001; идентификатор гена в Genbank: 1115198) и Lactococcus lactis subsp. cremoris SK 11 (входящий № в Genbank: NC_008527, локус LACR_1631, идентификатор гена в Genome: 4434110) была идентифицирована. Специалисту в данной области не составит труда идентифицировать и выделить гомологи гена thyA из дополнительных таксонов Lactococcus lactis.
Промотор гена thyA таким образом относится к природному промотору гена thyA, определенному в настоящем описании. В качестве примера, промотор thyA может относиться к последовательности нуклеиновой кислоты приблизительно от «-500» до приблизительно «+50» соответствующего гена thyA (при этом «+1» обозначает первый нуклеотид кодона инициации трансляции данного гена thyA), и в качестве дальнейших предпочтительных примеров приблизительно от «-500» до приблизительно «+20», приблизительно от «-500» до приблизительно «+10», приблизительно от «-500» до приблизительно «+5», приблизительно от «-500» до приблизительно «+2» или приблизительно от «-500» до приблизительно «-1», приблизительно от «-400» до приблизительно «+50», приблизительно от «-400» до приблизительно «+20», приблизительно от «-400» до приблизительно «+10», приблизительно от «-400» до приблизительно +5, приблизительно от «-400» до приблизительно «+2» или приблизительно от «-400» до приблизительно «-1», приблизительно от «-300» до приблизительно «+50», приблизительно от «-300» до приблизительно «+20», приблизительно от «-300» до приблизительно «+10», приблизительно от «-300» до приблизительно «+5», приблизительно от «-300» до приблизительно «+2» или приблизительно от «-300» до приблизительно «-1», приблизительно от «-200» до приблизительно «+50», приблизительно от «-200» до приблизительно «+20», приблизительно от «-200» до приблизительно «+10», приблизительно от «-200» до приблизительно «+5», приблизительно от «-200» до приблизительно «+2» или приблизительно от «-200» до приблизительно «-1», или приблизительно от «-100» до приблизительно «+50», приблизительно от «-100» до приблизительно «+20», приблизительно от «-100» до приблизительно «+10», приблизительно от «-100» до приблизительно «+5», приблизительно от «-100» до приблизительно «+2» или приблизительно от «-100» до приблизительно «-1» до тех пор, пока указанная последовательность демонстрирует силу, приблизительно одинаковую и предпочтительно одинаковую с силой промотора thyA в природе.
Предпочтительно промотор thyA, использующийся в качестве эталонного в настоящем изобретении, является промотором гена thyA Lactococcus lactis subsp. lactis MG 1363, и промотор по настоящему изобретению сильнее указанного промотора, как определено выше.
В приводимом в качестве примера анализе, описанном в примере 2 и основанном на анализе Steidler et al. 2003 (там же), промотор thyA Lactococcus lactis subsp. lactis MG 1363 контролирует экспрессию IL-10 человека при интеграции в одной копии в локус thyA в штамме MG 1363, составляющей 6,5 нг/1×109 бактерий. Соответственно, с помощью промотора по настоящему изобретению, который сильнее промотора thyA, может, в указанном анализе, достигаться экспрессия hIL-10, превышающая 6,5 нг/1×109 бактерий, предпочтительно ≥ 7 нг/1×109 бактерий, например ≥ 8 нг/1×109 или ≥ 9 нг/1×109 бактерий, даже более предпочтительно ≥ 10 нг/1×109 бактерий, например ≥ 11 нг/1×109 бактерий, ≥ 12 нг/1×109 бактерий, ≥ 13 нг/1×109 бактерий или ≥ 14 нг/1×109 бактерий, все еще более предпочтительно ≥ 15 нг/1×109 бактерий, еще более предпочтительно ≥ 20 нг/1×109 бактерий, например ≥ 25 нг/1×109 бактерий, ≥ 30 нг/1×109 бактерий, ≥ 35 нг/1×109 бактерий или ≥ 40 нг/1×109 бактерий, и наиболее предпочтительно ≥ 50 нг/1×109 бактерий или больше, например ≥ 100 нг/1×109 бактерий, и в дальнейших очень предпочтительных примерах ≥ 200 нг/1×109 бактерий, например ≥ 500 нг/1×109 бактерий, или ≥ 1000 нг/1×109 бактерий, или ≥ 2000 нг/1×109 бактерий, или ≥ 5000 нг/1×109 бактерий.
В дополнительных приводимых в качестве примеров анализах, например, описанных в примерах 3 и 4, штамм MG 1363 или L. casei включает вектор pT1NX, в котором промотор thyA L. lactis subsp. lactis MG 1363 контролирует экспрессию гена GLP-2 человека. Соответственно, промотор по настоящему изобретению, например промотор PhllA, сильнее промотора thyA, если при использовании эквивалентов 1 мл культур, полученных в конце логарифмической фазы, загруженных в SDS-PAGE для электрофореза и проанализированных с помощью Вестерн-блоттинга, количество GLP-2, экспрессированного посредством промотора по настоящему изобретению, превышает количество GLP-2, экспрессированного посредством промотора thyA. Дополнительные приводимые в качестве примеров анализы описываются в примерах 5-9.
В связанном варианте осуществления настоящее изобретение, таким образом, также относится к рекомбинантной нуклеиновой кислоте, включающей промотор (Р), функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными относительно промотора (Р), например гена GLP-2, гена hIL-10, гена GLP-1, гена hPYY или гена TTF, причем промотор (Р) выбран из группы, включающей или состоящей из природных промоторов генов Lactococcus, перечисленных под номерами 1) - 53) в разделе сводных данных, предпочтительно природных промоторов перечисленных генов Lactococcus lactis, причем промотор (Р) выбран из группы, состоящей из природных промоторов генов 1) бета'-субъединицы/субъединицы с М.м. 160 кДа ДНК-зависимой РНК-полимеразы (rpoC), 3) связывающего негемное железо ферритина (dpsA), 4) пируваткиназы (pyk), 5) глютаминил-тРНК-синтетаз (gltX), 6) фосфопируватгидрaтазы (eno), 9) дипептидазы PepV (pepV), 12) глицеральдегид-3-фосфатдегидрогеназы (gapB), 13) ацетаткиназы (ackA), 18) фруктозобифосфатальдолазы (fbaA), 20) супероксиддисмутазы (sodA), 21) рибосомного белка S12 (rpsL) и рибосомного белка S7 (rpsG), 22) рибосомного белка L18 (rplR) и рибосомного белка S5 (rpsE) и рибосомного белка L30/L7E (rpmD), 24) рибосомного белка L19 (rplS), 26) рибосомного белка L10 (rplJ), 28) HU-подобного ДНК-связывающего белка (hllA), 29) белка L28 50S-субъединицы рибосомы (rpmB), 30) компонента фосфотрансферазной системы IIB (ptcB), которые определены в таблице 1 под номерами 1)-53) в таблице 1, даже более предпочтительно промоторов, включенных в список в таблице 1 под номерами 1), 3)-6), 9), 12), 13), 18), 20)-22), 24), 26) и 28)-30), даже более предпочтительно промотора PhllA, включенного в список под номером 28), промотора PdpsA, включенного в список под номером 3), промотора PpepV, включенного в список под номером 9), или промотора PsodA, включенного в список под номером 20), и функциональных вариантов и функциональных фрагментов указанных природных промоторов. Также предпочтительно когда промотор (Р) выбран из группы, включающей или состоящей из нуклеиновых кислот, приведенных в SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-54, 160, 163-165, 167, 169, 171-180, и функциональных вариантов и функциональных фрагментов указанных природных промоторов.
Указанные названия понятны специалисту в данной области и определяют конкретные гены различных таксонов (например, видов, подвидов и штаммов) Lactococcus, таких как предпочтительные таксоны Lactococcus, описанные выше. Несмотря на это, для дополнительного руководства в таблице 1 ниже указаны уникальные номера - идентификаторы генов, которые обозначают указанные гены, выделенные из Lactococcus lactis subsp. cremoris SK11 (входящий номер в Genbank для полноразмерной геномной последовательности SK11: NC_008525) и subsp. lactis MG1363. Номера - идентификаторы генов уникальным образом обозначают указанные гены в базе данных «Entrez Gene» NCBI (www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene), как описано Maglott et al. 2005. На их основе специалист в данной области может идентифицировать и выделить гомологи указанных генов из дополнительных таксонов Lactococcus, отличных от SK11 и/или MG1363, например предпочтительных видов, подвидов и штаммов, о которых в настоящем описании говорится. Как используются в настоящем описании, «гомологи» относятся к последовательностям, в частности последовательностям генов, из двух или более различных таксонов, которые схожи (например, предпочтительно могут быть в значительной степени идентичны, как определено в настоящем описании) в результате происхождения из общего предка. Гомологи отмеченных выше генов из SK11 и/или MG1363 предпочтительно выполняют одну и ту же функцию в других таксонах Lactococcus.
Специалист в данной области может идентифицировать и выделить природные промоторы, контролирующие экспрессию генов, перечисленных в таблице 1, из SK11 и/или MG1363 или их гомологи из дополнительных таксонов Lactococcus (в том числе промоторы, которые контролируют экспрессию генов, обнаруживаемых в мультицистронных транскрипционных единицах и, следовательно, могут не содержать промотор непосредственно выше в последовательности их кодона инициации трансляции; например, в случае SK11 и/или MG1363, с целью иллюстрации для rpsL-rpsG с промотором, находящимся выше ORF rpsL, происходит контроль транскрипции обеих ORF; или для rplR-rpsE-rpmD с промотором, находящимся в направлении 5' от rplR, происходит контроль транскрипции всех ORF; или для rplD-rplW-rplB с промотором, находящимся выше rplD, происходит контроль всех ORF; или для rpsS-rplU-rplP-rplN с промотором, находящимся выше rpsS, происходит контроль всех ORF; или для rpsM с промотором, находящимся выше гена infA, кодирующего фактор 1 инициации трансляции (IF-1), управляющим транскрипцией rpsM), следуя рекомендациям этого описания и используя общие знания в данной области.
В предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, включающей промотор (Р), функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными относительно промотора (Р), причем промотор (Р) выбран из группы, включающей или состоящей из природных промоторов генов Lactococcus, перечисленных в списке под номерами 1)-30) в разделе сводных данных (т.е. первых 30 генов в таблице 1), даже более предпочтительно промоторов, включенных в список в таблице 1 под номерами 1), 3)-6), 9), 12), 13), 18), 20)-22), 24), 26) и 28)-30), даже более предпочтительно промотора PhllA, включенного в список под номером 28), промотора ДНК-связывающего ферритиноподобного белка (средства защиты от окислительной деградации) (dps), включенного в список под номером 3), промотора (Xaa-His)-дипептидазы (argE или pepV), включенного в список под номером 9), или промотора супероксиддисмутазы (sodA), включенного в список под номером 20), и функциональных вариантов и функциональных фрагментов указанных природных промоторов.
В предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, включающей промотор (Р), функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными для промотора (Р), причем промотор (Р) выбран из группы, включающей или состоящей из нуклеиновых кислот, приведенных в SEQ ID NO: 1-54 и 157-180, и их гомологов, даже более предпочтительно SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-54, 160, 163-165, 167, 169, 171-180 и функциональных вариантов и функциональных фрагментов указанных природных промоторов, более предпочтительно SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-30, 160, 163-165, 167, 169, 171, и функциональных вариантов и функциональных фрагментов указанных природных промоторов, предпочтительно SEQ ID NO: 28, 3, 9, 158 и/или 20, и их функциональных вариантов и функциональных фрагментов.
Указанные последовательности SEQ ID NO: 1-54 и 157-180 являются неограничивающими примерами природных промоторов, связанных с экспрессией генов, перечисленных под номерами 1)-53) выше (о которых говорится подробнее в таблице 1), в Lactococcus lactis subsp. lactis MG 1363 и/или Lactococcus lactis subsp. cremoris SK11. Соответственно, эти промоторы и их гомологи (особенно из таксонов Lactococcus, отличных от MG1363 и/или SK11), и их функциональные варианты и функциональные производные могут использоваться в рекомбинантных нуклеиновых кислотах по настоящему изобретению для осуществления экспрессии эффективных открытых рамок считывания в клетках-хозяевах, предпочтительно в бактериальных клетках-хозяевах, более предпочтительно в Lactococcus, еще более предпочтительно в Lactococcus lactis, еще более предпочтительно в Lactococcus lactis subsp. lactis, например, в предпочтительном примере в штамме MG 1363.
В другом предпочтительном варианте осуществления промотор (Р) выбран из группы, включающей или состоящей из нуклеиновых кислот, приведенных в SEQ ID NO: 1-54 и 157-180, более предпочтительно SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-54, 160, 163-165, 167, 169, 171-180, более предпочтительно SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-30, 160, 163-165, 167, 169, 171, наиболее предпочтительно SEQ ID NO: 28, 3, 9, 158 и/или 20, и их гомологов, их функциональных вариантов и/или функциональных фрагментов.
В еще одном предпочтительном варианте осуществления промотор (Р) выбран из группы, включающей или состоящей из нуклеиновых кислот, приведенных в SEQ ID NO: 1-30 и 157-171, более предпочтительно SEQ ID NO: 28, 3, 9, 158 и/или 20, и их функциональных вариантов и функциональных фрагментов.
В другом предпочтительном варианте осуществления рекомбинантная нуклеиновая кислота по настоящему изобретению, кроме того, включает последовательность терминатора транскрипции, находящуюся в направлении 3' от указанных одной или нескольких открытых рамок считывания. Например, если присутствует одна открытая рамка считывания, то последовательность терминатора транскрипции может преимущественно находиться ниже, т.е. в направлении 3' от нее. Если рекомбинантная нуклеиновая кислота содержит две или более ORF, например, последовательно упорядоченных и образующих вместе мультицистронную транскрипционную единицу, то терминатор транскрипции может преимущественно находиться в направлении 3' от самой нижней ORF.
Термин «терминатор транскрипции» относится к элементу последовательности, находящейся в конце транскрипционной единицы, который подает сигнал к терминации транскрипции. Предпочтительно терминатор транскрипции, использующийся в настоящем изобретении, будет подавать сигнал к терминации транскрипции в клетках-хозяевах, предназначенных для использования с рекомбинантными нуклеиновыми кислотами по настоящему изобретению, таких как, например, бактериальные клетки-хозяева, более предпочтительно Lactococcus, еще более предпочтительно Lactococcus lactis. Методы идентификации и выделения подходящих терминаторов, например, терминаторов из Lactococci, известны в данной области (например, смотри van der Vossen et al., 1995), примеры терминаторов приведены, например, в van der Vossen et al. (1992) и тому подобное
В другом предпочтительном варианте осуществления рекомбинантная нуклеиновая кислота по настоящему изобретению, кроме того, включает оператор, сконструированный для контроля транскрипции с промотора (Р). Используемый в настоящем описании термин «оператор» относится к нуклеотидной последовательности, предпочтительно последовательности ДНК, которая контролирует инициацию и/или поддержание транскрипции последовательности с промотора.
Как правило, оператор может размещаться между промотором и находящейся в направлении 5'-3' от него последовательности, транскрипцию которой промоторы контролируют. Обычно оператор способен связывать полипептид-репрессор, тем самым уменьшая транскрипцию с указанного промотора. Подходящий репрессор может чередовать состояние, в котором он связывает оператор, с состоянием, в котором он не связывает оператор, и такое чередование может преимущественно контролироваться внешними условиями, например, внешним веществом или конкретным метаболитом. Соответственно, в клетках-хозяевах, включающих совместимый репрессор, введение оператора в нуклеиновую кислоту по настоящему изобретению может обечпечить контроль активности промотора и его экспрессию. Приводимые в качестве примеров системы операторов-репрессоров включают, например, систему lac и смотри также, например, Nauta et al. (1996) или систему биосинтеза гистидина (смотри, например, Delorme et al., 1999). Последовательности операторов можно, как правило, получить из бактериальных хромосом.
В другом предпочтительном варианте осуществления рекомбинантные нуклеиновые кислоты по настоящему изобретению, кроме того, включают последовательности, сконструированные для осуществления вставки указанных нуклеиновых кислот в геном, например, хромосому, клетки-хозяина.
В особенно предпочтительном примере вставке нуклеиновых кислот по настоящему изобретению в конкретные сайты внутри генома, например, хромосомы, клетки-хозяина может способствовать гомологичная рекомбинация. Например, рекомбинантные нуклеиновые кислоты по настоящему изобретению могут включать один или несколько участков гомологии с указанным сайтом интеграции внутри генома, например, хромосомы, клетки-хозяина. Последовательность в указанном сайте генома, например, хромосомы, может быть природной, т.е. встречающейся в природе, или может быть экзогенной последовательностью, введенной с помощью предшествующей генной инженерии.
Например, размер указанного участка(ов) гомологи может составлять по крайней мере 50 п.о., предпочтительно по крайней мере 100 п.о., например, по крайней мере 200 п.о., более предпочтительно по крайней мере 300 п.о., например, по крайней мере 400 п.о., более предпочтительно по крайней мере 500 п.о., например, по крайней мере 600 п.о. или по крайней мере 700 п.о., еще более предпочтительно по крайней мере 800 п.о., например, по крайней мере 900 п.о. или по крайней мере 1000 п.о. или более.
В предпочтительном примере могут быть включены два участка гомологии, при этом они фланкируют каждый из концов экспрессионной единицы (единиц), присутствующей в нуклеиновых кислотах по настоящему изобретению. С помощью такой конфигурации могут преимущественно встраивать релевантные последовательности, т.е. последовательности, позволяющие осуществлять экспрессию представляющих интерес открытых рамок считывания с промоторов по настоящему изобретению, в клетках-хозяевах. Способы осуществления гомологичной рекомбинации, особенно в бактериальных клетках, и отбора рекомбинантов общеизвестны в данной области. Приводимым в качестве примера и предпочтительным способом является, например, способ Steidler et al. (2003) введения экзогенных последовательностей в локус thyA Lactococcus.
Следовательно, в предпочтительном варианте осуществления настоящее изобретение также относится к рекомбинантной нуклеиновой кислоте после интеграции в геном, например, хромосому, предпочтительно бактериальную хромосому, более предпочтительно хромосому Lactococcus. Такой интеграции можно добиться в клетке-хозяине, трансформированной указанной рекомбинантной нуклеиновой кислотой или вектором, включающим такую НК, как описано еще где-нибудь в этом описании.
В предпочтительном варианте осуществления одна или несколько открытых рамок считывания, связанных с промотором по настоящему изобретению, в нуклеиновых кислотах по настоящему изобретению кодируют полипептид.
Понятно, что сущностью настоящего изобретения, главным образом, являются новые промоторы и их применение, а природа указанных продуктов экспрессии, предпочтительно полипептидов, никаким образом не ограничена.
Тем не менее, были описаны клетки-хозяева, включающие контролируемые промоторами Lactococcus открытые рамки считывания, в качестве средства для продукции in vitro (смотри, например, патент США 5559007; Steidler et al., 1995; Wells et al., 1993B) и доставки in vivo (смотри, например, WO 97/14806; WO 01/02570; Steidler et al., 2003; Steidler et al., 2000) релевантных полипептидов вирусного, прокариотического или эукариотического происхождения, в том числе полипептидов, подходящих для профилактики или терапии заболеваний у людей или животных.
Соответственно, в одном из вариантов осуществления указанные одна или несколько открытых рамок считывания рекомбинантных нуклеиновых кислот по настоящему изобретению могут кодировать продукт экспрессии, предпочтительно полипептид, способный вызывать терапевтический ответ у индивида, предпочтительно у человека или животного.
В особенно эффективном, приводимом в качестве неограничивающего примера, варианте осуществления указанные одна или несколько открытых рамок считывания рекомбинантных нуклеиновых кислот по настоящему изобретению могут кодировать антиген и/или невакциногенный терапевтически активный полипептид.
Используемый в настоящем описании термин «антиген» обычно относится к веществу, которое индуцирует иммунный ответ, включая гуморальный иммунный ответ и/или клеточный иммунный ответ, и которое способно связываться с продуктом, например, антителом или Т-клеткой, иммунного ответа. Следовательно, в предпочтительном примере для антигена требуется функционирующая иммунная система индивида, которому он вводится, для индукции физиологической реакции у такого индивида. «Антиген» по настоящему изобретению также включает «аутоантигены», которые не вызывают иммунный ответ у здорового индивидуума, но могли бы вызвать такой ответ у индивидуума, страдающего аутоиммунным заболеванием, т.е. организм которого не способен распознавать собственные составные части (отказ на субмолекулярном уровне) в качестве «собственных», что приводит к иммунному ответу против его собственных клеток и тканей. Любое заболевания, являющееся следствием такого аберрантного иммунного ответа, называют аутоиммунным заболеванием. Соответственно, «антиген» по настоящему изобретению также включает (физиологически активный) белок, который не мог бы вызывать иммунный ответ у здорового индивидуума, но мог бы вызывать такой ответ у индивидуума с генетическим дефектом в таком белке. Кроме того, «антиген» по настоящему изобретению также включает аллерген, который не может вызывать иммунный ответ у здорового индивидуума, но мог бы вызывать такой ответ у индивидуума, страдающего аллергическим заболеванием.
Антиген по настоящему изобретению может происходить из любого полипептида, против которого иммунный ответ у человека или животного мог бы быть терапевтически эффективен, например, из патогена, например, из вирусного, прокариотического (например, бактериального) или эукариотического патогена, из не являющегося физиологическим белка (например, белка, происходящего из раковой ткани), из аллергена (например, для вызова иммунологической толерантности) и тому подобное. Антиген мог бы также быть метаболитом белка. В качестве примера, антиген мог бы быть полисахаридом, липидом или другим веществом. Сильные промоторы, описанные в настоящем описании, могли бы запускать экспрессию ферментов, необходимых для синтеза или сборки указанного полисахарида, липида или другого вещества.
Термин «невакциногеннный терапевтически активный полипептид» относится обычно к полипептиду, который у человека или животного, которому его вводят, не вызывает иммунный ответ против себя самого и с помощью которого можно добиваться терапевтического эффекта. Следовательно, можно было бы предполагать, что терапевтический эффект такого полипептида непосредственно связан с его собственным природным биологическим функционированием, посредством чего с его помощью можно добиться конкретных эффектов в организме индивида, а не с вызовом терапевтического эффекта посредством действия в качестве иммуногенного и/или иммунопротективного антигена у индивида. Следовательно, невакциногенный терапевтически активный полипептид должен быть биологически активным в своей экспрессированной форме или, по крайней мере, должен превращаться в биологически активную форму после высвобождения из экспрессирующей клетки-хозяина. Предпочтительно такая биологически активная форма указанного полипептида может демонстрировать вторичную и предпочтительно также третичную конформацию, которая является одинаковой или близко аналогичной его природной конфигурации.
Предпочтительно невакциногенный терапевтически активный полипептид является также нетоксичным и непатогенным.
В предпочтительном варианте осуществления невакциногенный терапевтически активный полипептид может быть получен у человека или животного и может предпочтительно соответствовать тому же таксону, что и человек или животное, которому его вводят.
Неограничивающие примеры подходящих невакциногенных терапевтически активных полипептидов включают невакциногенные терапевтически активные полипептиды, которые способны функционировать местно или системно, например, являются полипептидом, способным влиять на эндокринные функции, воздействуя на местный метаболизм или метаболизм всего организма, и/или биологически активный полипептид(ы) является полипептидом(ами), который способен регулировать активности клеток, принадлежащих иммуногемопоэтической системе, и/или один или несколько биологически активных полипептидов являются полипептидами, которые способны оказывать влияние на жизнеспособность, рост и дифференциацию множества нормальных или раковых клеток в организме или оказывать влияние на иммунорегуляцию или индукцию острых воспалительных реакций на повреждение и инфицирование, и/или один или несколько биологически активных полипептидов являются полипептидами, которые способны усиливать или индуцировать устойчивость к инфицированию клеток и тканей, опосредуемую хемокинами, действующими на свои рецепторы клеток-мишеней, или пролиферацию эпителиальных клеток, или стимулирование заживление ран, и/или один или несколько биологически активных полипептидов модулируют экспрессию или продукцию веществ клетками в организме.
Конкретные примеры таких полипептидов включают, но ими не ограничиваются, инсулин, гормон роста, пролактин, кальцитонин, лютеинизирующий гормон, паратиреоидный гормон, соматостатин, тиреостимулирующий гормон, вазоактивный интестинальный полипептид, цитокины, такие как IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-10, IL-11, IL-12, IL-13, любой из IL-14 - IL-32, GM-CSF, M-CSF, SCF, IFN, EPO, G-CSF, LIF, OSM, CNTF, GH, PRL, TNF-семейство цитокинов, например, лиганды TNFα, TNFβ, CD40, CD27 или FAS, IL-1-семейство цитокинов, семейство факторов роста фибробластов, тромбоцитарные факторы роста, трансформирующие факторы роста и факторы роста нервов, семейство цитокинов - факторов роста эпидермиса, связанные с инсулином цитокины и тому подобное. В альтернативном случае терапевтически активный полипептид может быть рецептором или антагонистом терапевтически активных полипептидов, определенных выше. В альтернативном случае терапевтически активный полипептид может быть нейтрализующим антителом или ему подобным. Другие конкретные примеры таких подходящих полипептидов перечислены, например, в заявке WO 96/11277, стр. 14, строки 1-30, включенной в настоящее описание посредством ссылки; в заявке WO 97/14806, со стр. 12, строки 1, до включительно стр. 13, строки 27, включенной в настоящее описание посредством ссылки; или патенте США 5559007, с колонки 8, строки 31, до включительно колонки 9, строки 9, включенном в настоящее описание посредством ссылки. Предпочтительно подходящим полипептидом является пептид IL-10, например, IL-10 человека (hIL-10), например, приведенный в качестве примера в примере 2, 5 или 9.
В предпочтительном варианте осуществления подходящим полипептидов является пептид «трилистника», такой как TFF1, TFF2 и/или TFF3. Три члена этого семейства пептидов «трилистника» были определены у людей и сначала обозначены: pS2 (индуцибельный ген эстрогена рака молочной железы, Lefebvre, 1993), SP (плазмолитический пептид) и ITF (интестинальный фактор «трилистника»). В используемой в настоящее время номенклатуре pS2 переименован в TFF1, SP в TFF2 и ITF в TFF3 (смотри, например, Wong et al., 1999). Эта новая номенклатура используется по всему тексту настоящего изобретения. У людей, мышей и крыс TFF1 и TFF2, главным образом, выявляются в желудке, в то время как TFF3, главным образом, выявляется в двенадцатиперстной кишке и ободочной кишке. Полагают, что TFF1 действует через рецептор на поверхности клеток (Tan et al., 1997). Wong et al. (1999) недавно сделали обзор пептидов «трилистника». Содержание этой статьи включено посредством ссылки в настоящее описание. Пептиды «трилистника» секретируются эпителиальными продуцирующими слизь клетками и стабильны в кислотном окружении. Эти пептиды вносят вклад в защиту слизистой оболочки (образование геля над эпителием) и, вероятно, вовлечены в восстановление поврежденной слизистой оболочки путем стимуляции миграции эпителиальных клеток (Playford et al., 1996). Продукция пептидов «трилистника» увеличена местно в районах, в которых происходит повреждение, таких как язвы слизистой мембраны, выстилающей желудок, и колит (Wright et al., 1990). Babyatsky et al. (1996) установили, что введение рекомбинантных пептидов «трилистника» уменьшает повреждение в этих местах. В большинстве исследований установлено, что, в противоположность большинству других белков, которые важны для защиты слизистой оболочки (таких как фактор роста эпидермиса), пептиды «трилистника» не вызывают пролиферацию или вызывают небольшую пролиферацию (Playford et al., 1996).
В другом предпочтительном варианте осуществления подходящим пептидом является пептид PYY, например PYY человека (hPYY) и даже более предпочтительно Gly9 вариант (3-36) hPYY, приведенный в качестве примера в примере 7.
В другом предпочтительном варианте осуществления подходящим пептидом является глюкагоноподобный пептид-1 (GLP-1 или GLP1), например GLP-1 человека (hGLP-1) и даже более предпочтительно Gly8 вариант (7-36) hGLP-1, приведенный в качестве примера в примере 8. В предпочтительном варианте осуществления подходящим пептидом является глюкагоноподобный пептид-2 (GLP-2 или GLP2). Ген проглюкагона человека (входящий № в GenBank - NM_002054) кодирует препробелок, который расщепляется на четыре различных зрелых пептида, включающих глюкагоноподобный пептид-2 (GLP-2). Внешне вводимый GLP-2 вызывает у индивидуумов, например людей, ряд эффектов, включающих интестинальный рост, усиление функции кишечника, уменьшение разрушения кости и нейропротекцию. GLP-2 может действовать эндокринно для осуществления связи интестинального роста и метаболизма с поглощением питательных веществ. GLP-2 и родственные аналоги могут представлять лечения для синдрома укороченной тонкой кишки, болезни Крона, остеопороза и использоваться в качестве адъювантной терапии во время химиотерапии рака. Предпочтительно ген GLP-2 адаптирован к использованию предпочтительных кодонов в Lactococcus, например, L. lactis. Предпочтительно ген кодирует аминокислотную последовательность HADGSFSDEMNTILDNLAARDFINWLIQTKITD.
В другом предпочтительном варианте осуществления ген кодирует аналог зрелого GLP-2 человека с заменой аланина на глицин в положении 2, которая, как было установлено, уменьшает чувствительность к деградации дипептидилпептидазой-IV (Booth et al., 2004), например, ген кодирует аминокислотную последовательность HGDGSFSDEMNTILDNLAARDFINWLIQTKITD.
В более предпочтительном варианте осуществления ген GLP-2 адаптирован к использованию предпочтительных кодонов в Lactococcus и включает замену аланина на глицин в положении 2, например, нуклеотидную последовательность h[Gly2]GLP-2:
CACGGTGATGGTTCATTTTCAGATGAAATGAACACTATCCTTGATAACCTTGCTGCTCGTGATTTTATCAACTGGCTTATCCAAACTAAAATCACTGATTAA.
Соответственно в одном из вариантов осуществления рекомбинантная нуклеиновая кислота по настоящему изобретению кодирует антиген и/или невакциногенный терапевтически активный полипептид, причем указанный антиген способен вызывать иммунный ответ, предпочтительно протективный иммунный ответ, у являющегося человеком или животным индивида, и/или указанный невакциногенный терапевтически активный полипептид способен вызывать терапевтический эффект у являющегося человеком или животным индивида.
В заявке WO 97/14806, кроме того, описывается, в частности, коэкспрессия антигенов со стимулирующими иммунный ответ молекулами, такими как, например, интерлейкины, например, IL-2 или IL-6, бактериями. Соответственно, также предусматривается такая коэкспрессия, используя промоторы по настоящему изобретению.
В другом предпочтительном варианте осуществления открытая рамка считывания в соответствии с настоящим изобретением, кроме того, включает последовательность, кодирующую сигнал секреции в рамке с полипептидом, кодируемым ORF. Это преимущественно делает возможным секрецию экспрессированного полипептида из клетки-хозяина и, тем самым, может облегчить, например, выделение или доставку.
Как правило, последовательность сигнала секреции представляет сегмент из приблизительно 16 - приблизительно 35 аминокислот, который обычно содержит гидрофобные аминокислоты, которые встраиваются в липидную двухслойную мембрану и тем самым делают возможной секрецию сопутствующей последовательности белка или пептида из клетки-хозяина, и который обычно отщепляется от этого белка или пептида. Предпочтительно последовательность сигнала секреции может быть таким образом активна в клетке-хозяине, намеченной для использования с нуклеиновой кислотой, включающей указанную сигнальную последовательность, например, в бактериальной клетке-хозяине, предпочтительно молочнокислой бактерии, более предпочтительно Lactococcus, более предпочтительно Lactococcus lactis.
Последовательности сигналов секреции, активные в подходящих клетках-хозяевах, известны в данной области; примерные сигнальные последовательности Lactococcus включают сигнальные последовательности usp45 (смотри патент США 5559007) и другие, смотри, например, Perez-Martinez et al. (1992); Sibakov et al. (1991). Предпочтительно сигнальная последовательность расположена между последовательностью промотора и ORF, т.е. сигнальная последовательность расположена 3' от последовательности промотора и предшествует ORF представляющего интерес полипептида. В предпочтительном варианте осуществления сигнальная последовательность кодирует аминокислотную последовательность MKKKIISAIL MSTVILSAAA PLSGVYA (usp45).
Авторы настоящего изобретения неожиданно обнаружили, что комбинация промотора PhllA с мутированной сигнальной последовательностью usp45 приводит к дополнительно контролируемой продукции и секреции представляющего интерес полипептида. В частности, мутант включает аспарагин (N) в положении 4 вместо лизина (К). В предпочтительном варианте осуществления сигнальная последовательность кодирует аминокислотную последовательность MKKNIISAIL MSTVILSAAA PLSGVYADTN.
В другом предпочтительном варианте осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, определенной в настоящем описании, причем указанная рекомбинантная нуклеиновая кислота включает:
(a) PdpsA, usp45 и hIL-10 (sAGX0012); PdpsA, usp45N4 и hIL-10;
PpepV, usp45 и hIL-10 (sAGX0018); PpepV, usp45N4 и hIL-10;
PsodA, usp45 и hIL-10 (sAGX0029); PsodA, usp45N4 и hIL-10;
PhllA, usp45 и hIL-10 (sAGX0037); PhllA, usp45N4 и hIL-10;
(b) PdpsA, usp45N4 и hTFF1; PdpsA, usp45 и hTFF1;
PpepV, usp45N4 и hTFF1; PpepV, usp45 и hTFF1;
PsodA, usp45N4 и hTFF1; PsodA, usp45 и hTFF1;
PhllA, usp45N4 и hTFF1 (sAGX0048); PhllA, usp45 и hTFF1 (sAGX0049);
(c) PdpsA, usp45N4 и hTFF3; PdpsA, usp45 и hTFF3 (sAGX0048);
PpepV, usp45N4 и hTFF3; PpepV, usp45 и hTFF3;
PsodA, usp45N4 и hTFF3; PsodA, usp45 и hTFF3;
PhllA, usp45N4 и hTFF3 (sAGX0057); PhllA, usp45 и hTFF3;
(d) PdpsA, usp45N4 и hPYY; PdpsA, usp45 и hPYY (sAGX0048);
PpepV, usp45N4 и hPYY; PpepV, usp45 и hPYY;
PsodA, usp45N4 и hPYY; PsodA, usp45 и hPYY;
PhllA, usp45N4 и hPYY (sAGX0057); PhllA, usp45 и hPYY; PhllA, usp45 и hPYY G9 (3-36) (sAGX0213);
(e) PdpsA, usp45N4 и GLP-1; PdpsA, usp45 и GLP-1;
PpepV, usp45N4 и GLP-1; PpepV, usp45 и GLP-1;
PsodA, usp45N4 и GLP-1; PsodA, usp45 и GLP-1;
PhllA, usp45N4 и GLP-1; PhllA, usp45 и GLP-1;
(f) PdpsA, usp45N4 и GLP-2; PdpsA, usp45 и GLP-2;
PpepV, usp45N4 и GLP-2; PpepV, usp45 и GLP-2;
PsodA, usp45N4 и GLP-2; PsodA, usp45 и GLP-2;
PhllA, usp45N4 и GLP-2; или PhllA, usp45 и GLP-2.
В другом аспекте настоящее изобретение относится к вектору, включающему рекомбинантную нуклеиновую кислоту по настоящему изобретению.
Как в настоящем описании используется, «вектор» относится к молекуле нуклеиновой кислоты, обычно ДНК, в которую фрагменты нуклеиновых кислот могут быть встроены и клонированы, т.е. репродуцированы. Следовательно, вектор будет, как правило, содержать один или несколько уникальных рестрикционных сайтов и может быть способен к автономной репликации в определенном хозяине или организме-носителе из условия, чтобы клонированная последовательность была репродуцируемой. Векторы могут включать, без ограничения, плазмиды, фагемиды, бактериофаги, происходящие из бактериофагов векторы, РАС, ВАС, линейные нуклеиновые кислоты, например, линейная ДНК, и тому подобное, в соответствующих случаях (смотри, например, Sambrook et al., 1989; Ausubel 1992).
Факторы, важные при выборе конкретного вектора, например плазмиды, включают, среди прочих, легкость, с которой реципиентные клетки, содержащие вектор, можно распознать и отделить от тех реципиентных клеток, которые не содержат вектор; число копий вектора, которое желательно в конкретном хозяине; и то, желательна ли возможность «челночного перемещения» вектора между клетками-хозяевами различных видов. Предпочтительные прокариотические векторы включают плазмиды, такие как плазмиды, способные реплицироваться в E.coli (такие как, например, pBR322, ColE1, pSC101, pUC19 и тому подобное). Такие плазмиды описываются, например, Sambrook et al., 1989; Ausubel 1992. Особенно предпочтительными векторами могут быть векторы, способные реплицироваться в E.coli (и других грамотрицательных бактериях), а также в другой представляющей интерес клетке-хозяине, например, в грамположительной бактерии, молочнокислой бактерии, предпочтительно Lactococcus, более предпочтительно Lactococcus lactis (смотри, например, Kok et al. (1984)). Другими предпочтительными векторами могут быть векторы, способные реплицироваться и/или осуществлять челночное перемещение между одной или несколькими грамположительными бактериями, но не в грамотрицательных бактериях. В предпочтительном варианте осуществления вектором является pT1NX, описанный Steidler et al. (1998), публикация которых включена в описание в качестве ссылки.
В другом аспекте настоящее изобретение относится к клетке-хозяину, трансформированной рекомбинантной нуклеиновой кислотой и/или вектором по настоящему изобретению. Например, такая трансформация может быть пригодна для репродукции и сохранения указанной нуклеиновой кислоты или векторов.
Альтернативно или дополнительно и преимущественно трансформированная клетка-хозяин будет способна транскрибировать открытую рамку(и) считывания нуклеиновой кислоты по настоящему изобретению, используя промоторы по настоящему изобретению, и предпочтительно экспрессировать продукты экспрессии, предпочтительно один или несколько полипептидов, кодируемых указанными открытыми рамками считывания.
Следовательно, в другом аспекте настоящее изобретение относится к применению рекомбинантной нуклеиновой кислоты или вектора по настоящему изобретению для достижения экспрессии продуктов экспрессии, предпочтительно одного или нескольких полипептидов, кодируемых указанными открытыми рамками считывания, в клетке-хозяине. Предпочтительно продукты экспрессии, например, полипептиды, могут быть гетерологичными, т.е. экзогенными, для указанной клетки-хозяина.
Предпочтительно клеткой-хозяином будет прокариотическая клетка, например бактериальная клетка, более предпочтительно непатогенная и/или не осуществляющая инвазию бактерия, еще более предпочтительно грамположительная бактерия, более предпочтительно молочнокислая бактерия (например, подвиды Lactococcus, подвиды Streptococcus, подвиды Lactobacillus, подвиды Leuconostos, подвиды Pediococcus, подвиды Brevibacterium, подвиды Propionibacterium или подвиды Bifidobacterium), очень предпочтительно бактерия Lactococcus и наиболее предпочтительно бактерия Lactococcus lactis, например, определенная выше, если промоторы по настоящему изобретению соответствующим образом активны в указанных клетках-хозяевах.
Рекомбинантная нуклеиновая кислота или вектор по настоящему изобретению могут присутствовать в клетке-хозяине вне хромосомы, предпочтительно автономно реплицируясь, используя собственную точка начала репликации, или может быть интегрирована в бактериальную геномную ДНК, например бактериальную хромосому, например хромосому Lactococcus. В последнем случае может быть интегрирована одна или множество копий указанной нуклеиновой кислоты, предпочтительно одна копия; интеграция может происходить в случайном сайте хромосомы или, как описано выше, в заранее определенном ее сайте, предпочтительно в заранее определенном сайте, таком как, в предпочтительном неограничивающем примере, в локусе thyA Lactococcus, например, Lactococcus lactis, например, как описано Steidler et al. (2003), публикация которых специально включена в настоящее описание посредством ссылки.
Соответственно, в другом аспекте настоящее изобретение относится к клетке-хозяину, включающему рекомбинантную нуклеиновую кислоту, определенную выше, причем указанный промотор (Р) присутствует в хромосоме указанной клетки-хозяина, и указанный промотор (Р) функционально связан с одной или несколькими открытыми рамками считывания, гетерологичными относительно указанного промотора (Р), более предпочтительно указанный промотор (Р), кроме того, включает сигнальную последовательность, предпочтительно usp45 или usp45N4, более предпочтительно указанный промотор (Р), кроме того, включает оператор, сконфигурированный для контроля транскрипции с указанного промотора (Р), и более предпочтительно промотор (Р) выбран из группы, состоящей из нуклеиновых кислот, приведенных в SEQ ID NO: 1, 3-6, 9, 12, 13, 18, 20-22, 24, 26, 28-54, 160, 163-165, 167, 169, 171-180, и их гомологов и их функциональных вариантов и функциональных фрагментов.
В дополнительном аспекте настоящее изобретение относится к клетке-хозяину, определенной в настоящем описании, причем указанные одна или несколько открытых рамок считывания кодируют полипептид, способный вызывать терапевтическую реакцию или иммунный ответ у индивида, предпочтительно у человека или животного, предпочтительно указанные одна или несколько открытых рамок считывания кодируют антиген и/или невакциногенный терапевтически активный полипептид, более предпочтительно указанный антиген способен индуцировать иммунный ответ, предпочтительно иммунологическую толерантность, у человека или животного, и/или указанный невакциногенный терапевтически активный полипептид способен вызывать терапевтический эффект у человека или животного. В другом предпочтительном аспекте настоящее изобретение относится к клетке-хозяину, определенной в настоящем описании, причем указанным невакциногенным терапевтически активным полипептидом является hIL-10, GLP-2, GLP-1, TFF или hPYY. В другом дальнейшем предпочтительном аспекте настоящим изобретением предоставляется клетка-хозяин, определенная в настоящем описании, причем указанный антиген способен вызывать иммунный ответ и используется в качестве вакцины для человека или животного.
В другом предпочтительном варианте осуществления настоящее изобретение относится клетке-хозяину, определенной в настоящем описании, причем указанная клетка включает:
(a) PdpsA, usp45 и hIL-10 (sAGX0012); PdpsA, usp45N4 и hIL-10;
PpepV, usp45 и hIL-10 (sAGX0018); PpepV, usp45N4 и hIL-10;
PsodA, usp45 и hIL-10 (sAGX0029); PsodA, usp45N4 и hIL-10;
PhllA, usp45 и hIL-10 (sAGX0037); PhllA, usp45N4 и hIL-10;
(b) PdpsA, usp45N4 и hTFF1; PdpsA, usp45 и hTFF1;
PpepV, usp45N4 и hTFF1; PpepV, usp45 и hTFF1;
PsodA, usp45N4 и hTFF1; PsodA, usp45 и hTFF1;
PhllA, usp45N4 и hTFF1 (sAGX0048); PhllA, usp45 и hTFF1 (sAGX0049);
(c) PdpsA, usp45N4 и hTFF3; PdpsA, usp45 и hTFF3 (sAGX0048);
PpepV, usp45N4 и hTFF3; PpepV, usp45 и hTFF3;
PsodA, usp45N4 и hTFF3; PsodA, usp45 и hTFF3;
PhllA, usp45N4 и hTFF3 (sAGX0057); PhllA, usp45 и hTFF3;
(d) PdpsA, usp45N4 и hPYY; PdpsA, usp45 и hPYY (sAGX0048);
PpepV, usp45N4 и hPYY; PpepV, usp45 и hPYY;
PsodA, usp45N4 и hPYY; PsodA, usp45 и hPYY;
PhllA, usp45N4 и hPYY (sAGX0057); PhllA, usp45 и hPYY; PhllA, usp45 и hPYY G9 (3-36) (sAGX0213);
(e) PdpsA, usp45N4 и GLP-1; PdpsA, usp45 и GLP-1;
PpepV, usp45N4 и GLP-1; PpepV, usp45 и GLP-1;
PsodA, usp45N4 и GLP-1; PsodA, usp45 и GLP-1;
PhllA, usp45N4 и GLP-1; PhllA, usp45 и GLP-1;
(f) PdpsA, usp45N4 и GLP-2; PdpsA, usp45 и GLP-2;
PpepV, usp45N4 и GLP-2; PpepV, usp45 и GLP-2;
PsodA, usp45N4 и GLP-2; PsodA, usp45 и GLP-2;
PhllA, usp45N4 и GLP-2; или PhllA, usp45 и GLP-2.
В другом аспекте настоящее изобретение относится к способу рекомбинантной экспрессии представляющего интерес продукта экспрессии, предпочтительно полипептида, включающему
а) культивирование клетки-хозяина, включающей рекомбинантную нуклеиновую кислоту или вектор по настоящему изобретению, в которой указанные одна или несколько открытых рамок считывания кодируют представляющий интерес продукт экспрессии, предпочтительно полипептид, и
b) выделение представляющего интерес продукта экспрессии, предпочтительно полипептида, продуцируемого клеткой-хозяином при указанном культивировании.
Специалист в данной области обычно знает и может дополнительно оптимизировать условия культивирования, например, температуру, присутствие и концентрацию необходимых питательных веществ, насыщение кислородом, перемешивание, инокуляцию и тому подобное, которые позволяют экспрессировать полипептиды в клетках-хозяевах, предпочтительно клетках-хозяевах по настоящему изобретению.
Специалист в данной области также знает и может дополнительно оптимизировать способы очистки для выделения продуктов экспрессии, таких как предпочтительно полипептиды, экспрессируемые указанными клетками-хозяевами. Например, подходящие методы выделения белков могут включать лизис организма-хозяина (например, когда полипептид аккумулируется внутриклеточно) путем, например, механического или ферментативного разрушения клеточной стенки и удаление клеточного дебриса или собирание супернатанта (например, когда полипептиды секретируются), удаление небелковых компонентов, таких как ДНК и липополисахариды, преципитацию сульфатом аммония, гель-хроматографию, например, для обогащения желаемым белком, аффинную хроматографию и тому подобное.
В другом аспекте настоящее изобретение относится к применению клетки-хозяина, трансформированной рекомбинантной нуклеиновой кислотой и/или вектором по настоящему изобретению для получения лекарственного средства, для доставки продукта 9(ов) экспрессии, предпочтительно полипептида(ов), кодируемого одной или несколькими открытыми рамками считывания, введенными в указанную рекомбинантную нуклеиновую кислоту, человеку или животному.
В связанном аспекте настоящее изобретение относится к способу доставки полипептида, кодируемого одной или несколькими открытыми рамками считывания, включенными в рекомбинантную нуклеиновую кислоту по настоящему изобретению, человеку или животному, включающему введение указанному человеку или животному терапевтически эффективного количества клеток-хозяев, трансформированных указанной рекомбинантной нуклеиновой кислотой и/или вектором по настоящему изобретению. Животным может быть, предпочтительно млекопитающее, такое как, например, домашние животные, сельскохозяйственные животные, животные зоопарков, спортивные животные, комнатные животные и экспериментальные животные, такие как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, крупный рогатый скот, коровы; приматы, такие как обезьяны, мартышки, орангутанги и шимпанзе; псовые, такие как собаки и волки; кошачьи, такие как кошки, львы и тигры; лошадиные, такие как лошади, ослы и зебры; продовольственные животные, такие как коровы, свиньи и овцы; копытные, такие как олени и жирафы; грызуны, такие как мыши, крысы, хомяки и морские свинки, и т.д.
Используемые в настоящем описании термины «лечить» или «лечение» относятся как к терапевтическому лечению, так и к профилактическим или предупредительным мерам, при которых у индивида предотвращается или замедляется (уменьшается) нежелательное физиологическое изменение или нарушение. «Человек или животное, которым показано лечение» включает человека или животного, у которых лечение данного состояния могло бы быть эффективным.
Термин «терапевтически эффективное количество» относится к количеству терапевтического вещества или композиции, эффективному для лечения заболевания или нарушения у индивида, например, человека или животного, т.е. для получения местного или системного эффекта и действия. Например, терапевтически эффективное количество бактерий может включать по крайней мере 1 бактерию или по крайней мере 10 бактерий, или по крайней мере 102 бактерий, или по крайней мере 103 бактерий, или по крайней мере 104 бактерий, или по крайней мере 105 бактерий, или по крайней мере 106 бактерий, или по крайней мере 107 бактерий, или по крайней мере 108 бактерий, или по крайней мере 109, или по крайней мере 1010, или по крайней мере 1011, или по крайней мере 1012, или по крайней мере 1013, или по крайней мере 1014, или по крайней мере 1015 или более клеток-хозяев, например, бактерий, например, в однократной или повторной дозе.
Клетки-хозяева по настоящему изобретению могут вводиться отдельно или в комбинации с одним или несколькими активными соединениями. Последние могут вводиться до, после или одновременно с введением указанных клеток-хозяев.
Существует ряд описаний предшествующего уровня техники, касающихся доставки антигенов и/или терапевтически активных полипептидов, и должно быть понятно, что такие описания могут быть далее преимущественно модифицированы сильными промоторами по настоящему изобретению. В качестве примера, а не ограничения, бактериальная доставка интерлейкинов, в частности, IL-10, для лечения колитов (например, смотри WO 00/23471), доставка антигенов в качестве вакцин (например, WO 97/14806), доставка GLP-2 и родственных аналогов может использоваться для лечения синдрома укороченной тонкой кишки, болезни Крона, остеопороза и в качестве адъювантной терапии во время химиотерапии рака, и тому подобное. Кроме того, бактериальная доставка пептидов «трилистника» может использоваться для лечения заболеваний пищеварительного тракта (например, смотри WO 01/02570). В частности, применение белков или пептидов «трилистника» для лечения нарушений и повреждения пищеварительного тракта, включающего ротовое отверстие, пищевод, желудок и толстую и тонкую кишку, а также для защиты и лечения тканей, выстилающих снаружи пищеварительный тракт, описывается в WO 97/38712 и WO 92/14837. Эти белки могут использоваться или для лечения поражений в этих областях, или для подавления образования поражений. Эти поражения, включающие, но ими не ограничивающие, неязвенную диспепсию, гастрит, пептическую язву или язву двенадцатиперстной кишки, рак желудка, лимфому MALT, болезнь Менетрие, желудочно-пищеводный рефлюкс, болезнь Крона, неспецифический язвенный колит и острый колит химического, бактериального или неясного происхождения, могут быть вызваны лучевой терапией или химиотерапий для лечения рака, любым другим лекарственным средством, включающим этиловый спирт, который повреждает пищеварительный тракт, случайным подверганием облучению и воздействию едкого вещества, инфекцией, нарушением пищеварения. Пептиды «трилистника» особенно эффективны для лечения острого колита. Возможны дополнительные терапевтические применения, используя промоторы и клетки-хозяева по настоящему изобретению.
Другие неограничивающие примеры типов заболеваний, поддающихся лечению у людей или животных путем доставки терапевтических полипептидов в соответствии с настоящим изобретением, включают, но ими не ограничиваются, например, воспалительные заболевания кишечника, например болезнь Крона и неспецифический язвенный колит (лечение которых эффективно с помощью IL-Ira или IL-10 или пептидами «трилистника»); аутоиммунные заболевания, включающие, но ими не ограничивающиеся, псориаз, ревматоидный артрит, красную волчанку (лечение которых эффективно, например, с помощью IL-Ira или IL-10 или релевантным аутоантигеном); аллергические заболевания, включающие, но без ограничения, астму, пищевые аллергии (лечение которых эффективно релевантным аллергеном); глютеновую болезнь (лечение которой эффективно глютеновыми аллергенами); неврологические нарушения, включающие, но без ограничения, болезнь Альцгеймера, болезнь Паркинсона и болезнь Шарко (лечение которых эффективно, например, нейротропным фактором головного мозга и цилиарным нейротропным фактором); рак (лечение которого эффективно, например, с помощью IL-1, колониестимулирующими факторами или интерфероном-W); остеопороз (лечение которого эффективно, например, трансформирующим фактором роста f3); диабет (лечение которого эффективно, например, инсулином); сердечно-сосудистое заболевание (лечение которого эффективно, например, тканевым активатором плазминогена); атеросклероз (лечение которого эффективно, например, цитокинами и антагонистами цитокинов); гемофилию (лечение которой эффективно, например, факторами свертывающей системы крови); дегенеративное заболевание печени (лечение которого эффективно, например, фактором роста гепатоцитов или интерфероном-α); легочные заболевание, такие как муковисцидоз (лечение которого эффективно, например, альфа-антитрипсином); ожирение; инфицирования патогенными микроорганизмами, например, вирусные или бактериальные инфекции (лечение которых эффективно любым рядом вышеупомянутых композиций или антигенов); и тому подобное.
Специалисту в данной области должно быть понятно, что специально перечисленные заболевания приведены только в качестве примеров, и никоим образом не подразумевается, что их перечисление ограничивает применение средств, предоставляемых настоящим изобретением, например, промоторов, нуклеиновых кислот, векторов и клеток-хозяев по настоящему изобретению, этими конкретными заболеваниями. Взамен являющийся специалистом в данной области читатель понимает, что средства по настоящему изобретению могут использоваться для экспрессии, по существу, любых представляющих интерес продуктов экспрессии, предпочтительно полипептидов, которые могут иметь терапевтическую значимость не только для перечисленных заболеваний, но также для различных дополнительных заболеваний или состояний людей и животных. Следовательно, после того как подходящий продукт экспрессии, предпочтительно полипептид, например, антиген и/или невакциногенный терапевтически активный полипептид, выбран или определен для данного недомогания, специалист в данной области мог бы успешно выполнить его экспрессию выделение и/или доставку, используя средства по настоящему изобретению.
Настоящее изобретение также относится к лечению заболеваний у других животных, включая собак, лошадей, кошек и птиц. Заболевания собак включают, но ими не ограничиваются, чумку собак (парамиксовирус), гепатит собачьих (аденовирус Cav-1), питомниковый кашель или ларинготрахеит (аденовирус Cav-1), инфекционный энтерит собачьих (коронавирус) и геморрагический энтерит (парвовирус).
Заболевания кошек включают, но ими не ограничиваются, вирусный ринотрахеит (вирус герпеса), калицивирусное заболевание кошачьих (калицивирус), инфекционный перитонит кошачьих (парвовирус) и лейкоз у кошек (вирус лейкоза кошачьих). Также предусматриваются другие вирусные заболевания лошадей и птиц в качестве поддающихся лечению, используя способы и композиции по настоящему изобретению. Для этой цели особенно предпочтительным будет применение микроорганизмов, экспрессирующих рекомбинантные интерфероны.
В другом аспекте настоящее изобретение также относится к фармацевтической композиции, содержащей клетку-хозяина, трансформированную нуклеиновой кислотой и/или вектором по настоящему изобретению.
Предпочтительно такая композиция содержит фармацевтически эффективное количество клеток-хозяев по настоящему изобретению и фармацевтически приемлемый носитель, т.е. одно или несколько фармацевтически приемлемых веществ-носителей и/или добавок, например, буферов, носителей, наполнителей, стабилизаторов и тому подобное.
Используемый в настоящем описании термин «фармацевтически приемлемый» не противоречит термину, известному в данной области, и означает совместимый с другими ингредиентами фармацевтической композиции и не вредный для реципиента.
Рекомбинантные клетки-хозяева по настоящему изобретению могут быть суспендированы в фармацевтической композиции для введения человеку или животному, страдающему заболеванием, на которое направлено лечение. Такие фармацевтические композиции включают, но ими не ограничиваются, живые клетки-хозяева и среду, подходящую для введения. Рекомбинантные клетки-хозяева могут быть лиофилизированы в присутствии обычных наполнителей, таких как лактоза, другие сахара, стеарат, карбонат и/или сульфат щелочного и/или щелочно-земельного металла (например, магния стеарат, натрия карбонат и натрия сульфат), каолин, диоксид кремния, коррригенты и ароматизирующие вещества.
Лиофизированные таким образом клетки-хозяева могут быть получены в форме капсул, таблеток, гранул и порошков, и вводиться перорально.
Альтернативно, некоторые рекомбинантные бактерии могут быть получены в виде водных суспензий в подходящих средах, или лиофизированные бактерии могут быть суспендированы в подходящей среде непосредственно перед применением, при этом такая среда включает наполнители, указанные в настоящем описании, и другие наполнители, такие как глюкоза, глицин и натрия сахаринат.
Для перорального введения могут быть получены устойчивые к желудочному соку пероральные лекарственные формы, которые могут также включать соединения, обеспечивающие контролируемое высвобождение клеток-хозяев, и, таким образом, обеспечивают контролируемое высвобождение желаемого белка, кодируемого в них. Например, пероральная лекарственная форма (в том числе таблетки, пилюли, гранулы, порошки) может быть покрыта тонким слоем наполнителя (обычно полимеров, целлюлозных производных и/или липофильных материалов), который препятствует растворению или разрушению в желудке, но растворяется в кишечнике, тем самым делая возможным прохождение через желудок, получая определенные преимущества дезинтеграции, растворения и абсорбции в кишечнике.
Пероральная лекарственная форма может быть составлена так, чтобы обеспечивать медленное высвобождение клеток-хозяев и их рекомбинантного белка, например, в виде таблеток или капсул с контролируемым высвобождением, непрерывным высвобождением, пролонгированным высвобождением, непрерывного действия. Такие лекарственные формы обычно содержат общепринятые и широко известные наполнители, такие как липофильные, полимерные, целлюлозные, нерастворимые, способные к набуханию наполнители. Композиции с контролируемым высвобождением также можно использовать для любых других мест доставки, включая доставку в кишечник, ободочную кишку, биоадгезию или сублингвальную доставку (т.е. доставку к зубам и слизистой оболочке) и бронхиальную доставку. Если композиции по настоящему изобретению вводят ректально или вагинально, то фармацевтические композиции могут включать суппозитории и кремы. В этом случае клетки-хозяева суспендируют в смеси обычных наполнителей, также включающих липиды. Все вышеупомянутые композиции широко известны в данной области и описаны, например, в следующих ссылочных документах: Hansel et al. (1990), Chien (1992), Prescott et al. (1989) и Cazzaniga et al. (1994).
Для ректального введения предпочтительно может использоваться композиция для клизм. Термин «клизма» включает жидкие препараты, предназначенные для ректального введения. Клизма может, как правило, поставляться в контейнерах для однократного введения и содержит одно или несколько активных веществ, растворенных или диспергированных в воде, глицерине или макроголах или других подходящих растворителях.
Таким образом, в соответствии с настоящим изобретением в предпочтительном варианте осуществления рекомбинантные клетки-хозяева, кодирующие желаемый ген, могут вводиться животному или человеку через слизистые оболочки, например, пероральный, назальный, ректальный, вагинальный или бронхиальный путь введения с помощью любой из современных композиций, используемых для конкретного пути. Дозы клеток-хозяев для введения будут изменяться в зависимости от ряда факторов, включая тип бактерий и гена, колируемого ими, типа и тяжести заболевания, подвергаемого лечению, и пути введения, который должен использоваться.
Таким образом, для каждого и любого варианта осуществления настоящего изобретения невозможно определить точную дозу, но она будет очевидна специалисту в данной области на основании настоящего изобретения. Дозу можно было бы так или иначе определить в каждом случае путем измерения концентраций на уровне сыворотки рекомбинантного белка после введения заранее определенного числа клеток, используя широко известные способы, такие как способы, известные как ELISA или Biacore (смотри примеры). Анализ кинетического профиля и полупериода существования доставляемого рекомбинантного белка обеспечивает достаточную информацию для того, чтобы определить эффективный диапазон доз для трансформированных клеток-хозяев.
В одном из вариантов осуществления, если клетки-хозяева экспрессируют антиген, то настоящее изобретение может, таким образом, также относиться к вакцине.
Термин «вакцина» обозначает фармацевтически приемлемую композицию, которая, после введения в эффективном количестве животным или человеку, способна индуцировать выработку антител к иммуногену, введенному в вакцины, и/или вызывать защитный иммунитет у индивида.
Вакцина по настоящему изобретению может включать клетки-хозяева, трансформированные нуклеиновыми кислотами или векторами по настоящему изобретению, и, кроме того, необязательно, наполнитель. Такие вакцины могут также включать адъювант, т.е. соединение или композицию, которые усиливают иммунный ответ на антиген. Адъюванты включают, но ими не ограничиваются, полный адъювант Фрейнда, неполный адъювант Фрейнда, сапонин, минеральные гели, такие как гидроокись алюминия, поверхностно-активные вещества, такие как лизолецитин, плюроники полиолы, полианионы, пептиды, масляные или углеводородные эмульсии, и потенциально пригодные, фармацевтически приемлемые для человека адъюванты, такие БЦЖ (бацилла Кальметта-Герена) и Corynebacterium parvum.
В других предпочтительных вариантах осуществления настоящее изобретение относится к применению клетки-хозяина, определенной в настоящем описании для получения лекарственного средства, для доставки полипептида, кодируемого указанными одной или несколькими открытыми рамками считывания рекомбинантной нуклеиновой кислоты, человеку или животному, предпочтительно указанным полипептидом является антиген и/или невакциногенный терапевтически активный полипептид, предпочтительно hIL-10, GLP-2, GLP-1, TFF или hPYY, как определено выше. Настоящее изобретение, кроме того, относится к фармацевтической композиции, содержащей клетку-хозяина, определенную в настоящем описании. Кроме того, настоящее изобретение относится к клетке-хозяину, определенной в настоящем описании, для применения в качестве лекарственного средства.
Настоящее изобретение ниже проиллюстрировано с помощью примеров, которые не следует рассматривать как ограничивающие.
ПРИМЕРЫ
Материалы, используемые в примерах
GM17 - питательная среда М17 (Oxoid CM0817), приготовленная в соответствии с инструкциями производителя, но без добавления каких-либо сахаров.
- 20% глюкоза (Merck 1.08337), стерилизованная фильтрованием (Stericup-GV 0,22 мкм PVDF Millipore CGVU05RE).
- питательная среда GM17: М17, дополненная 0,5% глюкозы.
GM17T представляет собой GM17, дополненную 200 мкМ тимидином (Sigma # T9250).
GM17E представляет собой GM17, дополненную 5 мкг/мл эритромицина (Sigma # E6376) путем разведения из маточного раствора 25 мг/мл эритромицина в 96% этаноле.
BM9 - 10% казитон (Difco 225930), подвергнутый автоклавированию в течение 15 мин при 121°С.
- 0,5 М NaHCO3 (Merck 1.06329), стерилизованный фильтрованием (Stericup-GV 0,22 мкм PVDF Millipore CGVU05RE).
- 0,5 М Na2CO3 (Merck 1.06392), стерилизованный фильтрованием (Stericup-GV 0,22 мкм PVDF Millipore CGVU05RE).
- 1 М MgSO4 (Merck 1.05886), подвергнутый автоклавированию в течение 15 мин при 121°С.
- 100 мМ CaCl2 (Merck 1.02382), подвергнутый автоклавированию в течение 15 мин при 121°С.
- 10х соли М9 (Difco 248510), подвергнутые автоклавированию в течение 15 мин при 121ºС. Развести в 10х в стерильной воде для получения солей М9.
- 10 мл ВМ9: добавить различные компоненты в порядке, приведенном ниже:
* 1 мл 10х солей М9
* 500 мкл 10% казитона
* 250 мкл 20% глюкозы
* 7,75 мл воды
* 500 мкл 0,5 М NaHCO3
* 500 мкл 0,5 М Na2CO3
Перемешать надлежащим образом и добавить следующие компоненты:
* 20 мкл 1 М MgSO4
* 10 мкл 100 мМ CaCl2
BM9T представляет собой BM9, дополненную 200 мкМ тимидином (Sigma # T9250).
BM9E представляет собой BM9, дополненную 5 мкг/мл эритромицина (Sigma # E6376) путем разведения из маточного раствора 25 мг/мл эритромицина в 96% этаноле.
ELISA
IL-10 - набор II для ELISA IL-10 человека BD OptEIA; номер в каталоге 550613; BD Biosciences; http://www.bdbiosciences.com/
TFF-1 - сэндвич-ELISA
- антитело для покрытия: мышиное моноклональное антитело против TFF-1 (М02), клон 3Н5 (Abrova: номер в каталоге Н00007031-М02)
- антитело для детекции: кроличье поликлональное антитело против hTFF-1 (Alpha Diagnostics: номер в каталоге TFF12-A; www.4adi.com)
- конъюгат: антикроличье антитело, конъюгированное с пероксидазой хрена (HRP) (Southern Biotech: номер в каталоге 4050-05; http://www.southernbiotech.com/)
- субстрат: ТМВ
TFF-3 - сэндвич-ELISA
- антитело для покрытия: мышиное моноклональное антитело 4408 против TFF-3 (R&D Systems: номер в каталоге MAB4408; www.rndsystems.com)
- антитело для детекции: биотинилированное собственными силами мышиное моноклональное антитело 15С6 против TFF-3 человека (Calbiochem: номер в каталоге 585350; www.calbiochem.com)
- конъюгат: стрептавидин-HRP (номер в каталоге 554066; BD Biosciences; http://www.bdbiosciences.com/)
- субстрат: ТМВ
PYY - имеющийся в продаже набор для ELISA пептидов YY (PYY) в целом; номер в каталоге DSL-10-33600; Diagnostic System Laboratories, Inc., www.dslabs.com.
GLP-1 - имеющийся в продаже набор для ELISA глюкагоноподобного пептида-1 (активного); номер в каталоге EGLP-35K; Linco Research; www.millipore.com.
Пример 1. Выделение сильных промоторов из Lactococcus lactis.
Используя способ Antelmann et al. (2000), в Lactococcus lactis subsp. lactis MG 1363 были идентифицированы в высокой степени экспрессирующиеся белки.
Вкратце, в высокой степени экспрессирующиеся белки идентифицировали по изобильным белковым зонам после одномерного электрофореза в геле (фиг.2). Указанные белковые полосы расщепляли трипсином, и смеси пептидов анализировали в режиме LC-MS/MS в масс-спектрометрической ионной ловушке (Esquire HCT, Bruker). Спектры (только спектры 250 MS/MS для каждого выполнения сохраняли для анализа данных) анализировали с помощью модульного подхода к разработке, эксплуатации и тестированию программных средств (MASCOT) с использованием банка данных из белков Lactococcus lactis subsp. cremoris sk11 (Genbank NC_008527). Пептиды с показателем MASCOT, превышающим пороговый показатель идентичности, идентифицируют с вероятностью, составляющей 95%. Чем больше имеется пептидов, происходящих из одного и того же белка, тем более надежной является идентификация.
С помощью этого анализа был получен перечень в высокой степени распространенных белков и соответствующих генов (таблица 1 и фиг.1). На этой основе авторы по настоящему изобретению идентифицировали и выделили последовательности, находящиеся в направлении 3'-5' от указанных генов (SEQ ID NO: 1-54 и 157-180) с их собственными последовательностями Шайна-Дальгарно (SD). Для большинства промоторов это было осуществлено с помощью ПЦР на хромосомной ДНК Lactococcus lactis MG 1363. Например, праймеры, использованные для амплификации последовательностей промоторов, соответствующих генам, перечисленным под номерами 1)-30), продемонстрированы в таблице 2. Также подходящие праймеры можно выбрать на основе имеющейся геномной информации о промоторах, соответствующих генам, перечисленным под номерами 31)-53) выше. Промоторы для eno, tbp и lacE были также синтезированы, используя олигонуклеотиды (олигонуклеотиды в таблице 3).
Пример 2. Активность промотора
Была разработана стратегия, позволяющая субклонировать различные промоторы впереди гена способного к секреции hIL-10 (фиг.3). Субклонирование выполняли таким образом, чтобы экспрессирующие hIL-10 кассеты были фланкированы последовательностями-мишенями. Такая структура делает возможной двойную гомологичную рекомбинацию вокруг гена thyA и, таким образом, интеграцию в хромосомы, по существу, как описано в Steidler et al. 2003, выше и в WO 02/090551.
Схематически стратегия очерчена со ссылкой на фиг.3.
А) Используя соответствующие праймеры, промоторы выделяют с помощью ПЦР с хромосомной ДНК L. lactis MG1363; В) промоторы соединяют с релевантными фланкирующими районами (частичным 3' районом мишени, частичной ORF hIL-10) с помощью ПЦР с отжигом; С) и D) продукты присоединения субклонируют в условно нерепликативную плазмиду с помощью использования соответствующим образом расположенных сайтов для эндонуклеаз рестрикции (NcoI + AflIII или альтернативно NcoI + BstEII). Это приводит к получению как полного 3' района мишени, так и полной ORF hIL-10. Е) Промоторные конструкции вводят в L. lactis MG1363 с помощью последующей 5' и 3' рекомбинации. Интеграцию в хромосому выполняют благодаря двухстадийной процедуре. После введения нерепликативной плазмиды в исходный штамм L. lactis MG1363 первую гомологичную рекомбинацию между 5'-последовательностями-мишенями, или 3'-последовательностями-мишенями можно отобрать на содержащих эритромицин селективных средах. Вторую гомологичную рекомбинации между альтернативными мишенями отбирают по отсутствию thyA. F) Конечной структурой является хромосомная структура, в которой thyA заменен hIL-10.
Интегрированные в хромосомы трансгены оценивают в отношении уровней экспрессии способного к секреции hIL-10 (т.е. hIL-10, обеспеченного в рамке с последовательностью сигнала секреции Lactococcus, например, usp45 или схожей последовательности) относительно других штаммов Lactococcus, по существу как у Steidler et al. 2003, в заявке WO 02/090551, озаглавленной «Self-containing Lactococcus strain», и вкратце описано в настоящем описании: посев различных штаммов Lactococcus lactis осуществляют в виде штриховых линий до отдельной колонии, и предварительную культуру готовят инокулированием 1 колонии в GM17 (M17, Oxoid, Hampshire, UK, дополненную 0,5% глюкозы) и инкубируют в течение 16 ч при 30οС. Эти предварительные культуры инокулируют 1/25 в 5 мл GM и инкубируют в течение 4 ч при 30οС. Клетки собирают центрифугированием и ресуспендируют в 5 мл свежей ВМ9 (состав в таблице 4). Эти суспензии клеток инкубируют в течение 3 ч при 30οС. Бактериальные клетки удаляют центрифугированием, а супернатанты переносят в чистые пробирки. Содержание hIL-10 в супернатантах определяют с помощью ELISA, и белки hIL-10 визуализируют анализом эквивалента 1 мл культуры с помощью Вестерн-блоттинга.
Даже после повторных попыток и использования различных стратегий клонирования было невозможно субклонировать различные последовательности промоторов. Успешно субклонированные последовательности промоторов суммированы в таблице 12.
Пример 3. Сила промотора
После идентификации нескольких промоторов авторы по настоящему изобретению проверили отдельные промоторные конструкции в дальнейшей серии экспериментов. Кроме того, для того чтобы установить, является активность промотора независимой от репортерного гена, авторы по настоящему изобретению разработали репортерные конструкции, включающие ген GLP-2.
Ген проглюкагона человека (входящий № в GenBank: NM_002054) кодирует препробелок, который расщепляется на четыре различных зрелых пептида, включающих глюкагоноподобный пептид-2 (GLP-2). Авторы по настоящему изобретению сконструировали ген для h[Gly2]GLP-2, кодируемый при использовании предпочтительных кодонов L. lactis. Этот ген кодирует аналог зрелого GLP-2 человека с заменой аланина на глицин в положении 2, который, как было установлено, уменьшает чувствительность к деградации дипептидилпептидазой-IV (Booth et al., 2004). Синтетический ген собирали, используя методологию уровня техники, по существу как описано Stemmer et al. (1995), публикация которых включена в настоящее описание посредством ссылки.
Получаемый в результате фрагмент сливали с сигналом секреции usp45 (van Asseldonk, et al., 1990). Конструкцию слияния помещали 3' от ряда промоторов Lactococcus: P1 (Waterfield, et al., 1995), PthyA (промотора тимидилатсинтазы) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28).
С помощью этой стратегии были получены фрагменты, которые можно было субклонировать в виде EcoRI-XbaI-фрагментов для P1 (pT1GLP2) или EcoRI-SpeI-фрагментов (для всех других конструкций) в плазмиду pT1NX, открытую расщеплением EcoRI и SpeI (Steidler et al., 1998; фиг.4).
Обзор различных плазмид приведен в таблице 5. Последовательности всех плазмид были подтверждены, после чего их трансформировали в L. lactis MG1363, используя способ Gasson (1983).
Экспрессию и секрецию GLP-2 документировали с использованием экстрактов белков различных штаммов, получаемых параллельно. Вкратце, культуры с предельно высокой плотностью клеток разводили в 25 раз, выращивали в подходящим образом забуференной среде для роста и собирали в конце логарифмической фазы. Эквиваленты 1 мл культуры загружали в SDS-ПААГ для электрофореза и анализировали с помощью Вестерн-блоттинга. GLP-2 детектировали с помощью иммуноблоттинга с использованием поликлонального кроличьего антитела против GLP-2 (Abcam, Cambridge, UK). Вторым антителом был козий антикроличий IgG (Southern Biotechnology Associates, Birmingham, AL), связанный со щелочной фосфатазой. Ферментативную активность выявляли с использованием субстрата NBT/BCIP (Roche Diagnostics, Basel, Швейцария).
На основании данных авторов по настоящему изобретению можно заключить, что промотор PhllA является очень сильным.
Пример 4. Влияние сигнальной последовательности
Для того чтобы контролировать уровни экспрессии, в том числе продукцию и секрецию, авторы настоящего изобретения сконструировали ряд мутантов usp45, которые были сначала проверены в соединении с промотором PhllA 28).
Неожиданно, мутант usp45, посредством чего лизин в положении 4 был заменен аспарагином, (usp45 N4), приводил к значительному увеличению продукции и секреции h[Gly2)GLP-2 (PhllAN4GLP2) относительно управляемой промотором thyA и промотором P1 экспрессии (фиг.5А), Более того, эти трансформанты были очень стабильными. Конструкция, интегрированная в геном L. lactis, приводит также к в значительной степени увеличенной продукции и экспрессии. Экспрессия репортерной конструкция в Lactobacillus casei дает по существу такие же результаты, как и в L. lactis.
Затем авторы настоящего изобретения ввели эту мутацию в usp45 3' от промотора Р1 и промотора thyA, создавая плазмиды pT1N4GLP2 и pThyAN4GLP2, соответственно. Неожиданно, эта мутация имела незначительный эффект на продукцию h[Gly2)GLP-2, регулируемую промотором Р1, но полностью отменяла продукцию h[Gly2)GLP-2 под транскрипционным контролем промотора thyA (фиг.5В).
На основе этих данных очевидно, что промотор PhllA в соединении с сигнальной последовательностью uspN4 очень хорошо подходит для контролирования экспрессии генов.
Пример 5. Сила промотора, проверяемая по экспрессии интерлейкина-10 человека (hIL-10)
Авторы по настоящему изобретению проверили несколько промоторных конструкций в ряде экспериментов. Более того, для того чтобы установить, является ли активность промотора независимой от репортерного гена, авторы по настоящему изобретению сконструировали репортерные конструкции, в которых были сформированы экспрессионные кассеты, которые содержали исследуемые промоторы впереди сигнала секреции usp45 (van Asseldonk et al., 1990), слитого с синтетическим геном hIL-10 (смотри также пример 2). В этом случае конструкцию слияния помещали 3' от ряда промоторов Lactococcus: PthyA (промотора тимидилатсинтазы, штаммы sAGX0005 и Thy12), промотора PdpsA (ДНК-связывающего ферритиноподобного белка, SEQ ID NO: 3; штамм sAGX0012), PpepV (промотора (Xaa-His)-дипептидазы, SEQ ID NO: 158; штамм sAGX0018), PsodA (промотора супероксиддисмутазы, SEQ ID NO: 20; штамм sAGX0020) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; штамм sAGX0037).
Все экспрессионные кассеты были интегрированы в локус thyA хромосомы L. lactis MG1363 с помощью двойной гомологичной рекомбинации, тем самым ген thyA был удален (стратегией, схожей со стратегией, используемой для конструирования Thy12 (Steidler et al., 2003), но без использования хелперной плазмиды pVE6007, как показано на фиг.3). Это предполагает, что все последовательности ДНК вне экспрессирующих hIL-10 кассет являются идентичными. Обзор структуры и положения экспрессирующих hIL-10 кассет в различных штаммах приведен на фиг.6. Были подтверждены последовательности экспрессирующих hIL-10 кассет (промотора, сигнала секреции usp45, hIL-10) описанных в настоящем описании штаммов.
Посев различных проверяемых штаммов осуществляли в виде штриховых линий до отдельной колонии на чашки с твердым агаром GM17T. Отдельные колонии инокулировали в GM17T и выращивали в течение ночи до предельно высокой плотности. Соответствующие разведения этих культур предварительно выращивали в GM17T в течение 4 ч. Бактерии собирали центрифугированием и далее инкубировали в течение 3 ч в забуференной среде для культивирования (ВМ9Т). Бактериальные клетки удаляли центрифугированием, а из этого осветленного супернатанта получали образцы для анализа. Образцы анализировали с помощью ELISA, специфичной для hIL-4. Уровни экспрессии приведены на фиг.7. Все данные представлены в виде среднеарифметического трех отдельных измерений. Относительная сила промотора приведена в таблице 6.
Для исключения влияния различных между штаммами характеристик роста авторы настоящего изобретения определяли колониеобразующие единицы (CFU) в конце эксперимента по экспрессии, как описано выше, и рассчитывали продукцию hIL-10 в 109 CFU для различных штаммов. Этот эксперимент показал, что не наблюдаются в значительной степени различные скорости роста и что, по оценке экспрессии hIL-10, PhllA приблизительно в 4х сильнее PthyA (фиг.8 и таблица 7).
На основании данных авторов настоящего изобретения можно заключить, что PdpsA, PpepV, PsodA и PhllA являются очень сильными и очень компетентными для управления экспрессией гетерологичных генов.
Пример 6. Сила промотора, проверяемая по экспрессии фактора «трилистника» человека (TFF)
После идентификации потенциально сильных промоторов авторы по настоящему изобретению проверили несколько промоторных конструкций в ряде экспериментов. Более того, для того чтобы установить, является ли активность промотора независимой от репортерного гена, авторы по настоящему изобретению сконструировали репортерные конструкции, в которых были сформированы экспрессионные кассеты, которые содержали исследуемые промоторы впереди сигнала секреции usp45 дикого типа (van Asseldonk et al., 1990) или его мутанта (mut; в том числе Usp45 N4, в котором лизин в положении 4 был заменен аспарагином), слитого с синтетическим геном hTFF1 или hTFF3. Конструкции слияния помещали 3' от ряда промоторов Lactococcus: PthyA (промотора тимидилатсинтазы, штаммы sAGX0041 и sAGX0043), промотора PdpsA (ДНК-связывающего ферритиноподобного белка, SEQ ID NO: 3; штамм sAGX0059) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; штаммы sAGX0048, sAGX0049 и sAGX0057). Все экспрессионные кассеты были интегрированы в локус thyA хромосомы L. lactis MG1363 с помощью двойной гомологичной рекомбинации, тем самым ген thyA был удален (стратегией, схожей со стратегией, используемой для конструирования Thy12 (Steidler et al., 2003), но без использования хелперной плазмиды pVE6007, как показано на фиг.3). Это предполагает, что все последовательности ДНК вне экспрессирующих TFF кассет являются идентичными. Обзор структуры и положения экспрессирующих TFF кассет в различных штаммах приведен на фиг.9 и в таблице 8. Были подтверждены последовательности экспрессирующих TFF кассет (промотора, сигнала секреции, TFF) описанных в настоящем описании штаммов.
Посев различных проверяемых штаммов осуществляли в виде штриховых линий до отдельной колонии на чашки с твердым агаром GM17T. Отдельные колонии инокулировали в GM17T и выращивали в течение ночи до предельно высокой плотности. Соответствующие разведения этих культур предварительно выращивали в GM17T в течение 4 ч. Бактерии собирали центрифугированием и далее инкубировали в течение 3 ч в забуференной среде для культивирования (ВМ9Т). Бактериальные клетки удаляли центрифугированием, а из этого осветленного супернатанта получали образцы для анализа. Образцы анализировали с помощью ELISA, специфичной для hTFF. Уровни экспрессии приведены на фиг.10. Образцы штаммов-экспрессоров hTFF1 также анализировали с помощью Вестерн-блоттинга, специфичного для hTFF1 (фиг.11). Все данные представлены в виде среднеарифметического трех отдельных измерений. Относительная сила промотора приведена в таблице 8.
Согласуясь с примером 4, данные фиг.10 показывают, что мутант Usp45 N4 не несет ответственность за увеличенную экспрессию hTFF3 штаммом sAGX0057, но обеспечивает дополнительный уровень контроля экспрессии.
PdpsA, PpepV и PsodA, помещенные впереди экспрессирующих hTFF1 и hTFF3 конструкций, демонстрируют также увеличенную экспрессию относительно PthyA.
На основании данных авторов по настоящему изобретению можно заключить, что PdpsA, PpepV, PsodA и PhllA являются очень сильными и очень компетентными для управления экспрессией гетерологичных генов.
Пример 7. Сила промотора, проверяемая по экспрессии аминокислот 3-36 варианта Gly9 пептида YY человека (hPYY G9 (3-36))
После идентификации потенциально сильных промоторов авторы по настоящему изобретению проверили несколько промоторных конструкций в дальнейшем ряде экспериментов. Более того, для того чтобы установить, является ли активность промотора независимой от репортерного гена, авторы по настоящему изобретению сконструировали репортерные конструкции, в которых были сформированы экспрессионные кассеты, которые содержали исследуемые промоторы впереди сигнала секреции usp45 (van Asseldonk et al., 1990), слитого с синтетическим геном hPYY G9 (3-36). Конструкцию слияния помещали 3' от ряда промоторов Lactococcus: P1 (Waterfield et al., 1995) (плазмида pAGX0211); PthyA (промотора тимидилатсинтазы, плазмида pAGX0212) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; плазмида pAGX0213). Все экспрессионные кассеты были встроены в виде EcoRI-SpeI-фрагментов в плазмиду pT1NX (Schotte et al., 2003). Это предполагает, что все последовательности ДНК вне экспрессионных кассет, в том числе точка начала репликации и маркер устойчивости к эритромицину, являются идентичными для вышеуказанных плазмид. Обзор структуры различных экспрессионных плазмид приведен на фиг.12 и в таблице 9. Были подтверждены последовательности экспрессирующих hPYY кассет (промотора, сигнала секреции usp45, hPYY) описанных в настоящем описании штаммов.
Пустой экспрессионный вектор pT1NX, а также все экспрессирующие hPYY G9 (3-36) плазмиды были трансформированы в L. lactis MG1363. Посев полученных в результате штаммов осуществляли в виде штриховых линий до отдельной колонии на чашки с твердым агаром GM17E. Отдельные колонии инокулировали в GM17E и выращивали в течение ночи до предельно высокой плотности. Соответствующие разведения этих культур предварительно выращивали в GM17E в течение 4 ч. Бактерии собирали центрифугированием и далее инкубировали в течение 3 ч в забуференной среде для культивирования (ВМ9E). Бактериальные клетки удаляли центрифугированием, а из этого осветленного супернатанта получали образцы для анализа. Образцы анализировали с помощью ELISA, специфичной для hPYY. Уровни экспрессии приведены на фиг.13. Относительная сила промотора приведена в таблице 9.
PdpsA, PpepV и PsodA, помещенные впереди сигнала секреции usp45 (van Asseldonk et al., 1990), слитого с синтетическим геном hPYY G9 (3-36), демонстрируют также увеличенную экспрессию относительно экспрессии, управляемой PthyA.
На основании данных авторов по настоящему изобретению можно заключить, что PdpsA, PpepV, PsodA и PhllA являются очень сильными и очень компетентными для управления экспрессией гетерологичных генов.
Пример 8. Сила промотора, проверяемая по экспрессии аминокислот 7-36 варианта Gly8 глюкагоноподобного пептида-1 человека (hGLP-1 G8 (7-36))
После идентификации потенциально сильных промоторов авторы по настоящему изобретению проверили несколько промоторных конструкций в дальнейшем ряде экспериментов. Более того, для того чтобы установить, является ли активность промотора независимой от репортерного гена, авторы по настоящему изобретению сконструировали репортерные конструкции, в которых были сформированы экспрессионные кассеты, которые содержали исследуемые промоторы впереди сигнала секреции usp45 (van Asseldonk et al., 1990), слитого с синтетическим геном hGLP-1 G8 (7-36). Вариант GLP-1 Gly8 демонстрирует сниженную чувствительность к протеолитическому расщеплению дипептидилпептидазой VI (Deacon et al., 1998). Конструкцию слияния помещали 3' от двух промоторов Lactococcus: PthyA (промотора тимидилатсинтазы, плазмида pAGX0233) и PhllA (промотора ДНК-связывающего белка бактериального ядра/ДНК-связывающего белка HU; SEQ ID NO: 28; плазмида pAGX0234). Все экспрессионные кассеты были встроены в виде EcoRI-SpeI-фрагментов в плазмиду pT1NX (Schotte et al., 2003). Это предполагает, что все последовательности ДНК вне экспрессионных кассет, в том числе точка начала репликации и маркер устойчивости к эритромицину, являются идентичными для вышеуказанных плазмид. Обзор структуры различных экспрессионных плазмид приведен на фиг.14 и в таблице 10. Были подтверждены последовательности экспрессирующих GLP-1 Gly8 кассет (промотора, сигнала секреции usp45, GLP-1 Gly8) описанных в настоящем описании штаммов. Пустой экспрессионный вектор pT1NX, а также все экспрессирующие hGLP-1 G8 (7-36) плазмиды были трансформированы в L. lactis MG1363. Посев полученных в результате штаммов осуществляли в виде штриховых линий до отдельной колонии на чашки с твердым агаром GM17E. Отдельные колонии инокулировали в GM17E и выращивали в течение ночи до предельно высокой плотности. Соответствующие разведения этих культур предварительно выращивали в GM17E в течение 4 ч. Бактерии собирали центрифугированием и далее инкубировали в течение 3 ч в забуференной среде для культивирования (ВМ9E). Бактериальные клетки удаляли центрифугированием, а из этого осветленного супернатанта получали образцы для анализа. Образцы анализировали с помощью ELISA, специфичной для hGLP-1. Уровни экспрессии приведены на фиг.15. Относительная сила промотора приведена в таблице 10.
Пример 9. Сила промотора, проверяемая по экспрессии интерлейкина-10 человека (hIL-10)
Авторы по настоящему изобретению проверили несколько промоторных конструкций в последующем ряде экспериментов. Более того, для того чтобы установить, является ли активность промотора независимой от репортерного гена, авторы по настоящему изобретению сконструировали репортерные конструкции, в которых были сформированы экспрессионные кассеты, которые содержали исследуемые промоторы впереди сигнала секреции usp45 (van Asseldonk et al., 1990), слитого с синтетическим геном hIL-10. Конструкцию слияния помещали 3' от ряда промоторов Lactococcus (фиг.16 и таблица 11).
Все экспрессионные кассеты были интегрированы в локус thyA хромосомы L. lactis MG1363 с помощью двойной гомологичной рекомбинации, тем самым ген thyA был удален (стратегией, схожей со стратегией, используемой для конструирования Thy12 (Steidler et al., 2003), но без использования хелперной плазмиды pVE6007). Это предполагает, что все последовательности ДНК вне экспрессирующих hIL-10 кассет являются идентичными. Обобщенный обзор структуры и положения экспрессирующих hIL-10 кассет, присутствующих в различных штаммах (обычно обозначаемых «sAGX00xx»), приведен на фиг.16. Были подтверждены последовательности экспрессирующих hIL-10 кассет (промотора, сигнала секреции usp45, hIL-10) описанных в настоящем описании штаммов.
Посев различных проверяемых штаммов осуществляли в виде штриховых линий до отдельной колонии на чашки с твердым агаром GM17T. Отдельные колонии инокулировали в GM17T и выращивали в течение ночи до предельно высокой плотности. Соответствующие разведения этих культур предварительно выращивали в GM17T в течение 4 ч. Бактерии собирали центрифугированием и далее инкубировали в течение 3 ч в забуференной среде для культивирования (ВМ9Т). Бактериальные клетки удаляли центрифугированием, а из этого осветленного супернатанта получали образцы для анализа. Образцы анализировали с помощью ELISA, специфичной для hIL-10. Уровни экспрессии приведены на фиг.17. Все данные представлены в виде среднеарифметического трех отдельных измерений. Относительная сила промотора приведена в таблице 11.
В отличие от рассчитанного протеомного и транскриптомного анализа сила и компетентность конкретного промотора, управляющего экспрессией гетерологичных генов, осталась непредвиденной.
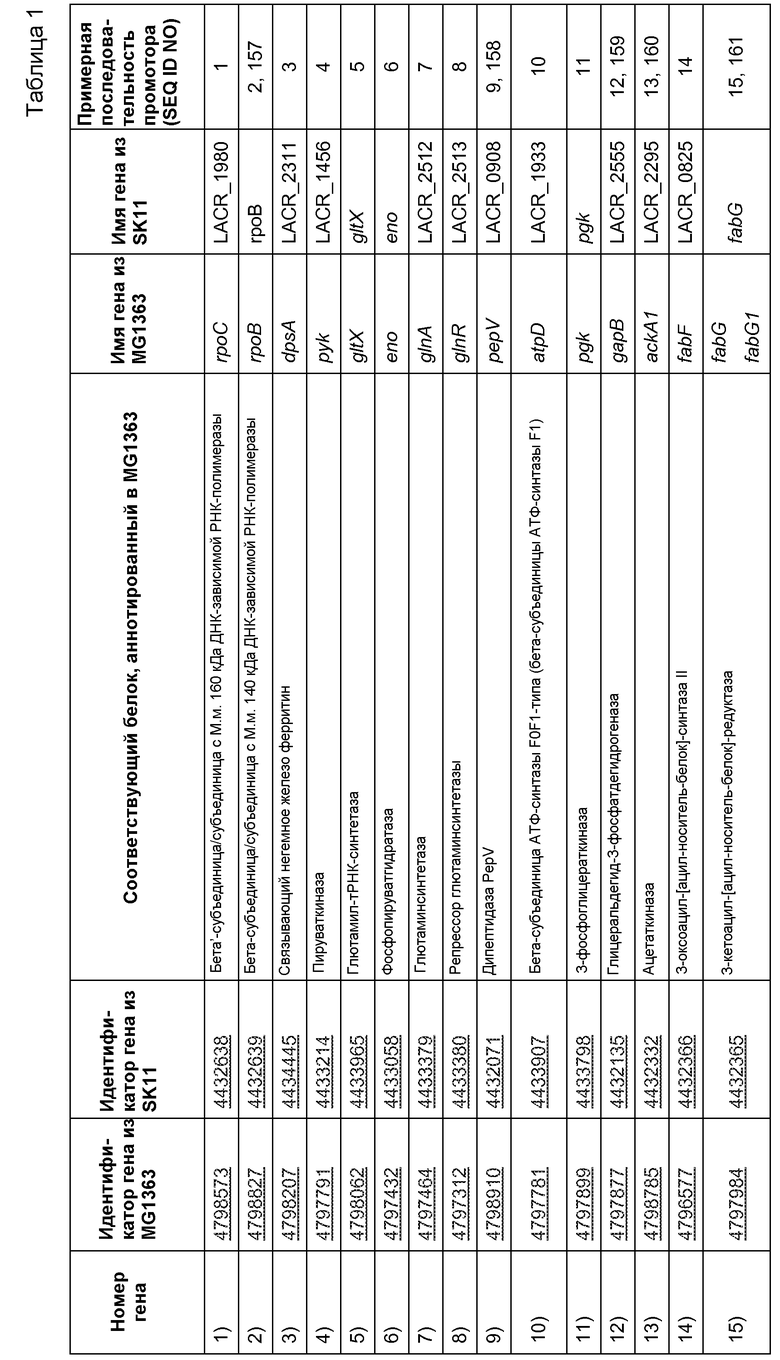

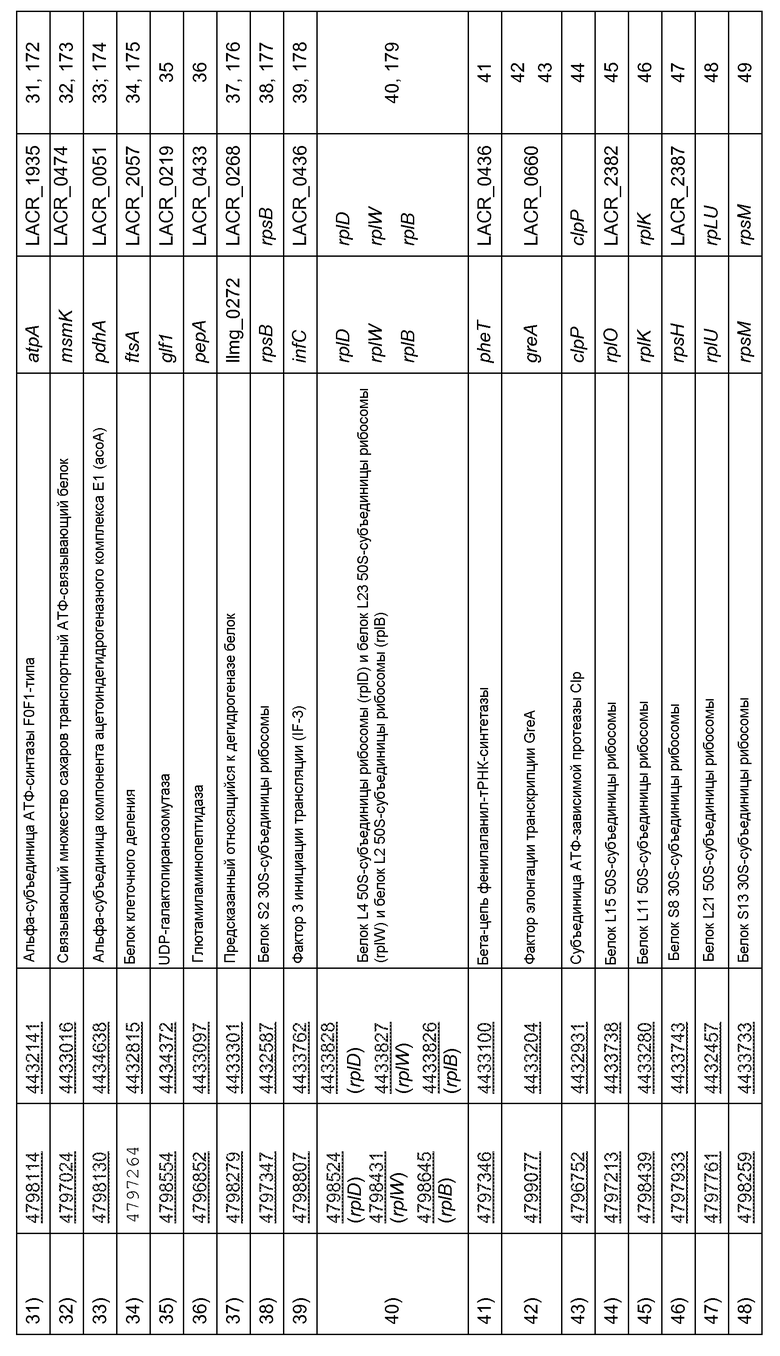




10х соли М9 содержат на литр: 60 г Na2CO3, 30 г KH2PO4, 10 г NH4Cl, 5 г NaCl.

sAGX0005
Эта работа

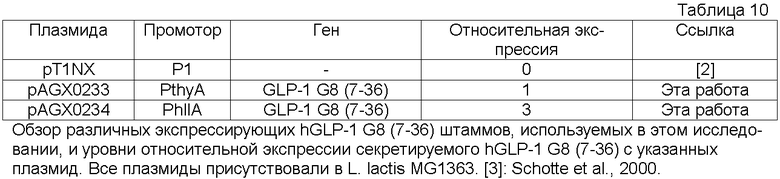
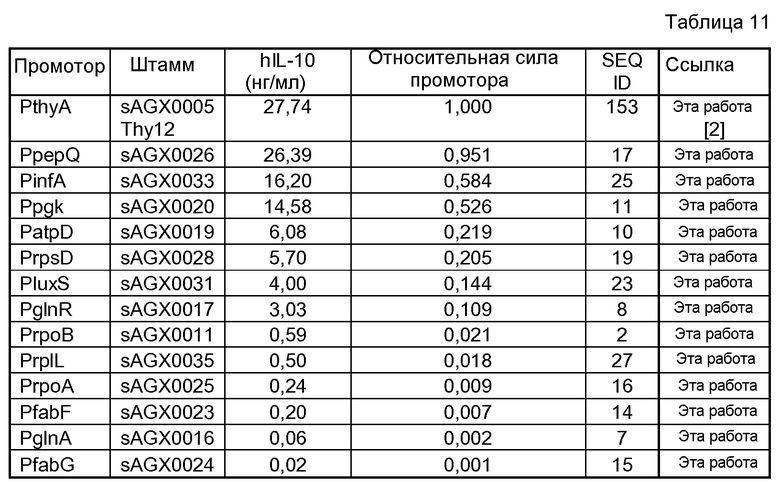

Клинический штамм, состав и их получение
Идентификация клинического штамма
L.lactis SAGX0085 представляет собой модифицированный штамм L.lactis, полученный из подвида L.lactis cremoris MG1363. В MG1363 кассета экспрессии тимидилатсинтазы (thyA) [PthyA>>thyA] была заменена на кассету экспрессии hTFF1 [PhllA>>SSusp45>>htff1], в которой промотор гена hllA MG1363 (PhllA) расположен перед слитой конструкцией лидера секреции лактококка (SSusp45) и гена htff1. Этот штамм назван sAGX0085 или AG013 L.lactis.
Получение
Банк базовых клеток cGMP был получен из штамма sAGX0085.
Существующий ферментативный процесс представляет собой двустадийный процесс и включает стадию предкультивирования и стадию рН-контролируемой ферментации партий в 1000 литровом биореакторе.
Ферментов инокулировали 580 мл PCGM, используя дополнительный стерилизуемый паром вход. После ферментизации накопленную биомассу концентрировали и промывали чистой водой. Стадию концентрирования и промывания биомассы проводили путем центрифугирования в стерилизованном CIP-SIP (121°С/20 мин), охлаждали (4°С) в стековом дисковом устройстве. Степень концентрирования составляла 20 раз. Для последующей стадии лиофилизации стабилизацию бактерий обеспечивали путем добавления криозащитного раствора в биомассу, полученную на стадии концентрирования и промывки. После стадии замораживания лиофилизированный остаток измельчали в порошок, а затем фильтровали. После фильтрации порошок становился гомогенным и образцы, взятые из полученной окончательной партии DS, помещали в мешки Alu (экструзионная ламинированная пленка OPA-PE-ALU-PE), каждый содержал 0,95-1,05 кг DS, термически герметизировали и хранили при -20°С.
Получение партий GLP и GMP и их применение (основное доклиническое и клиническое применение).
Партии, полученные для доклинического и клинического испытания AG013, по шкале располагались следующим образом:
- 200 л (2 партии GMP AG013 DS) Synco BioPartners (Голландия);
- 800 л (1 партия не-GMP AG013 DS) Lactosan (Австрия);
- 1000 л (3 партии не-GMP и 3 партии GMP AG013 DS) Lactosan (Австрия).
Клинический состав
- AG013-DP состоит из AG013-DS, смешанного с подходящим эксципиентом, и находится в виде порошка для восстановления. Клинический состав AG013 предназначен для перорального и местного применения в форме MR. Эта суспензия MR получена путем восстановления AG013-DP в выбранном растворе для восстановления (SR).
- AG013 или подходящее плацебо (AG013-плацебо) предназначено для перорального введения в состав MR. Для этой цели AG013-DP и AG013-плацебо должны быть восстановлены в SR упакованных индивидуальных стеклянных бутылочках с получением суспензии MR до введения.
- Как AG013-DP, так и соответствующее плацебо находятся в 30 мл-овых прозрачных стеклянных пробирках. SR вводят в 15 мл-овые темные стеклянные пробирки. Конечная концентрация AG013 составляет 2,0×1011 CFU L.lactis sAGX0085 на 15 мл MR.
Фаза Ib клинических испытаний у человека
AG013 исследовали на людях в фазе Ib клинических испытаний (AG013-ODOM-101). Фаза Ib - это многоцелевое, одинарное слепое, контролируемое плацебо исследование с последовательной эскалации дозы для оценки безопасности, переносимости и профиля РК AG013 у индивидов с ОМ во время индукционной химиотерапии (СТ) для лечения рака головы и шеи. Также оценивалась способность AG013 усиливать эффективность курса и уменьшать тяжесть ОМ.
Было зарегистрировано 25 индивидов (8 получали плацебо и 17 получали AG013) в 6 штатах США для исследования в фазе Ib.
Дозы увеличивали в группах, в которых был отмечен эффект, путем увеличения частоты приема от одного раза в день (сутки) до 3 раз в день (три раза в сутки) и до 6 раз в день, что дало ежедневную дозу 2×1011 КОЕ/день, 6×1011 КОЕ/день и 1,2×1012 КОЕ/день соответственно.
Исследование достигло своей основной цели, показав, что AG013 в целом является безопасным и хорошо переносится.
Несколько эффективных конечных точек были оценены как исследовательские конечные точки. У индивидов, получивших AG013, было отмечено уменьшение процента дней, в течение которых наблюдался язвенный мукозит, а у индивидов, которые получали AG013 при любом графике дозирования, было отмечено отсутствие язвенного мукозита вообще или наличие только в течение 1 дня, по сравнению с пациентами, получавшими плацебо. Кроме того, индивиды, которые получали AG013, реже обращались в центр и в скорую помощь за внеплановой помощью по сравнению с пациентами, получавшими плацебо.
ССЫЛКИ


| название | год | авторы | номер документа |
|---|---|---|---|
| МУКОАДГЕЗИВНЫЙ МИКРООРГАНИЗМ | 2017 |
|
RU2762940C2 |
| СИСТЕМА ПОЛИЦИСТРОННОЙ ЭКСПРЕССИИ ДЛЯ БАКТЕРИЙ | 2012 |
|
RU2644346C2 |
| СИСТЕМА ПОЛИЦИСТРОННОЙ ЭКСПРЕССИИ ДЛЯ БАКТЕРИЙ | 2012 |
|
RU2766157C2 |
| МОДИФИЦИРОВАННЫЕ ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2773435C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ БАКТЕРИИ, СТАБИЛЬНО ЭКСПРЕССИРУЮЩИЕ ИЛ-10 И ИНСУЛИН | 2017 |
|
RU2768027C2 |
| МОДИФИЦИРОВАННЫЕ ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2653757C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ДИАБЕТА 1 ТИПА | 2017 |
|
RU2760997C2 |
| РЕКОМБИНАНТНЫЕ ПРОБИОТИЧЕСКИЕ БАКТЕРИИ | 2015 |
|
RU2723324C2 |
| ХЛОРИД-ИНДУЦИРУЕМАЯ ПРОКАРИОТИЧЕСКАЯ СИСТЕМА ЭКСПРЕССИИ | 2019 |
|
RU2812766C2 |
| КЛЕТКИ-ХОЗЯЕВА И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2014 |
|
RU2663587C2 |



































































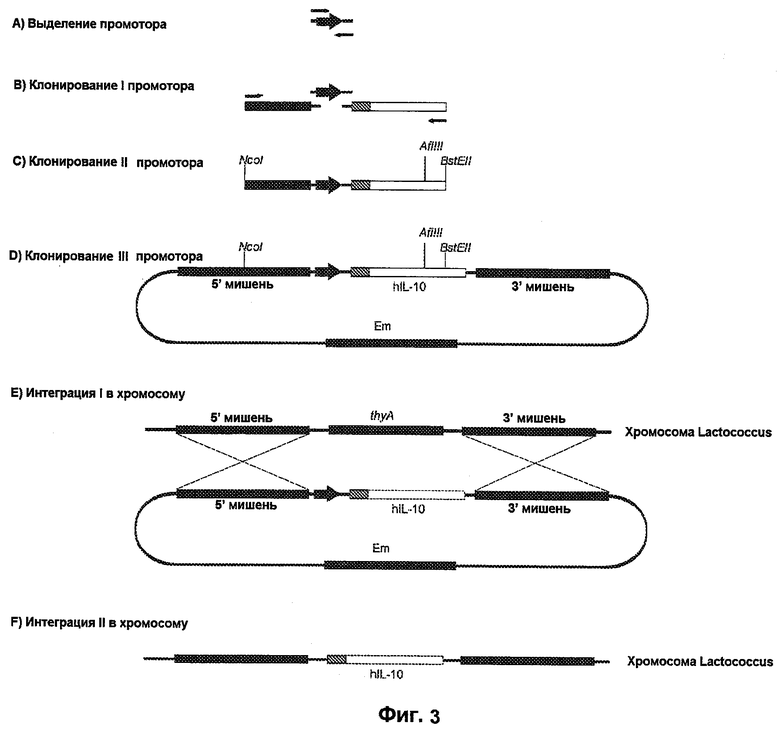


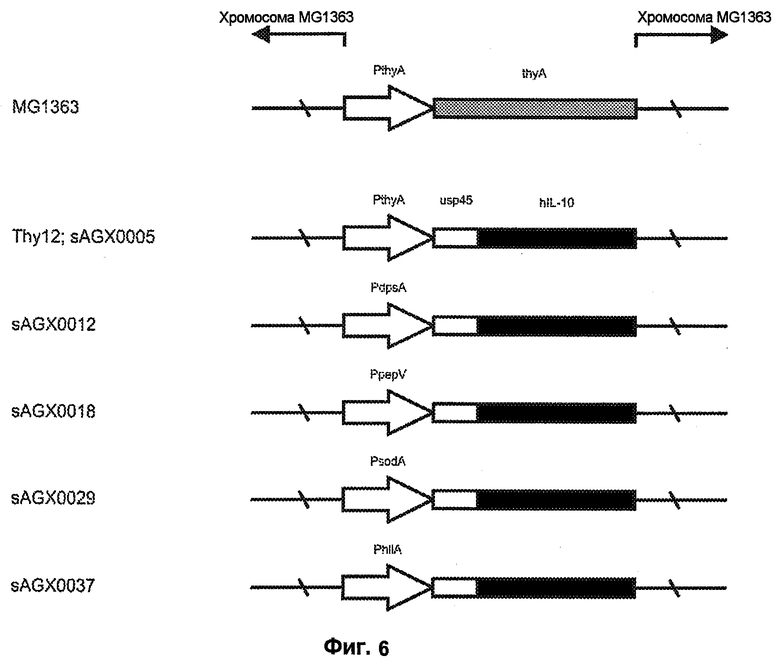
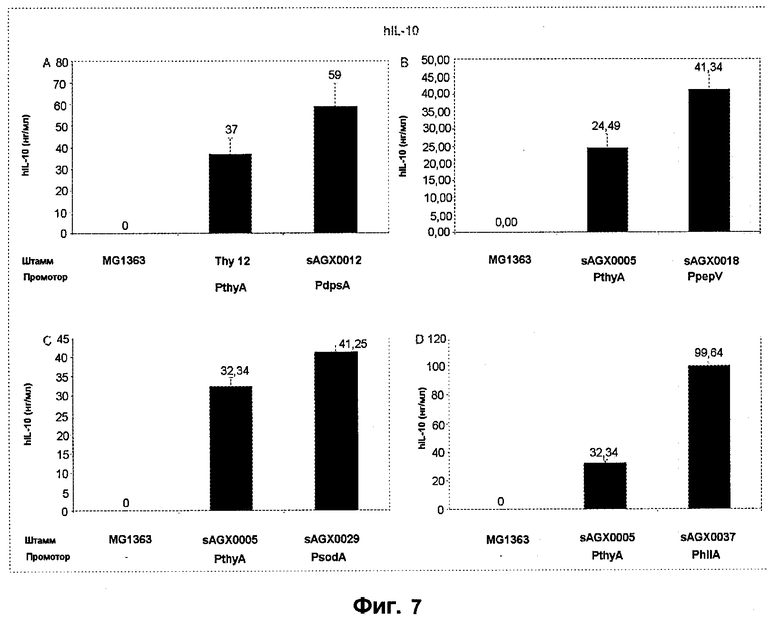
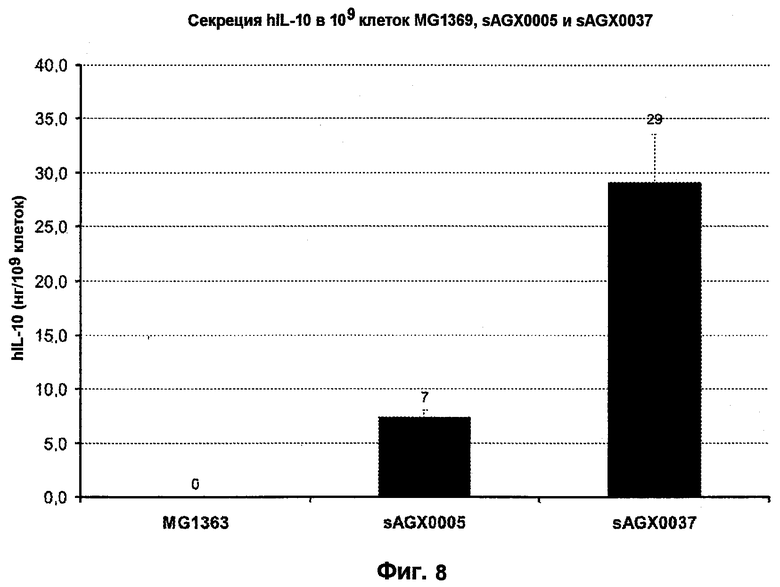


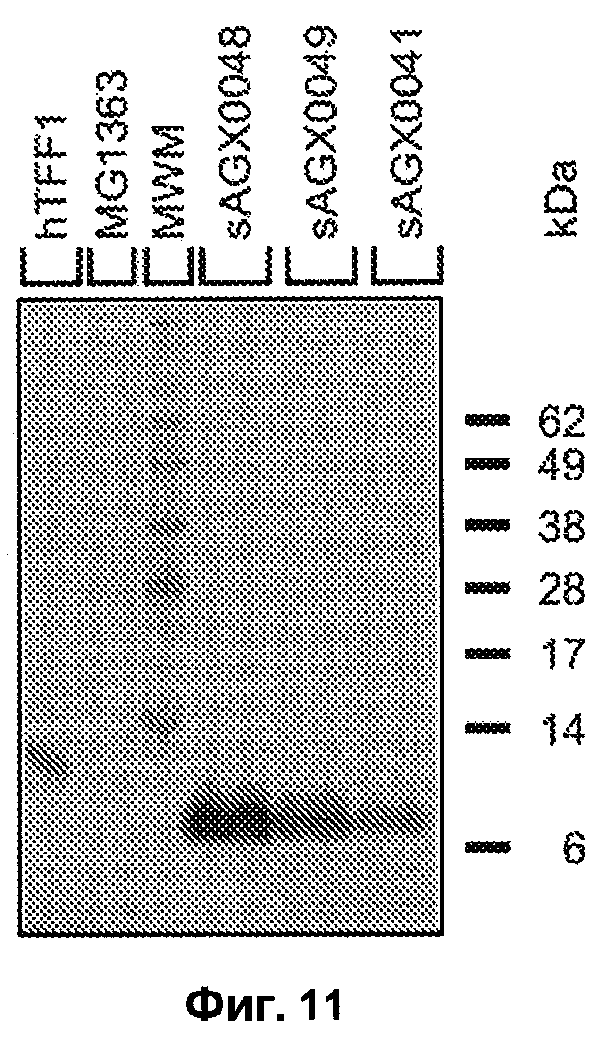
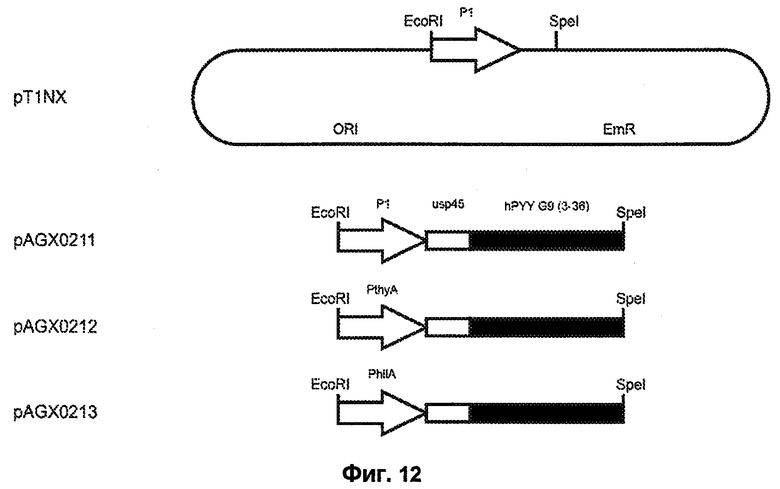


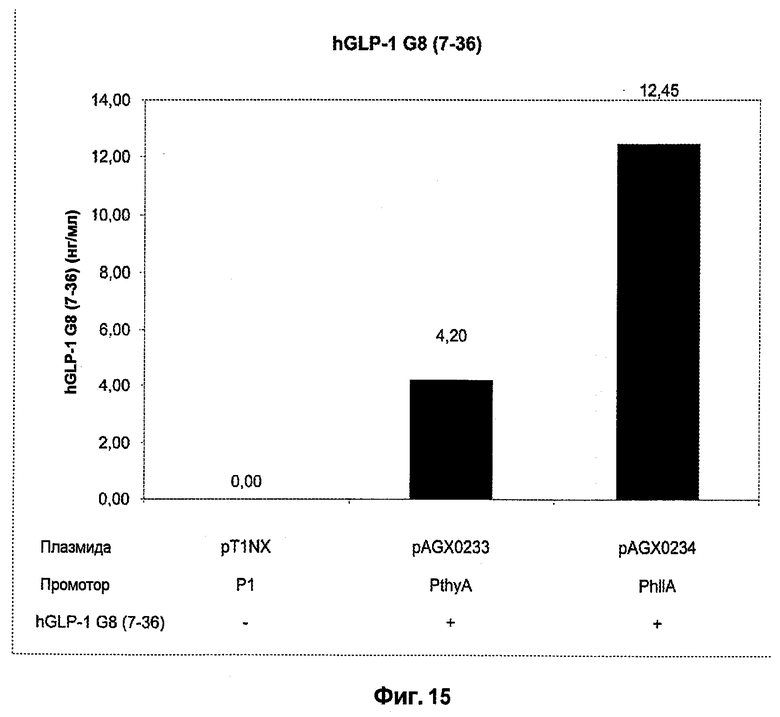

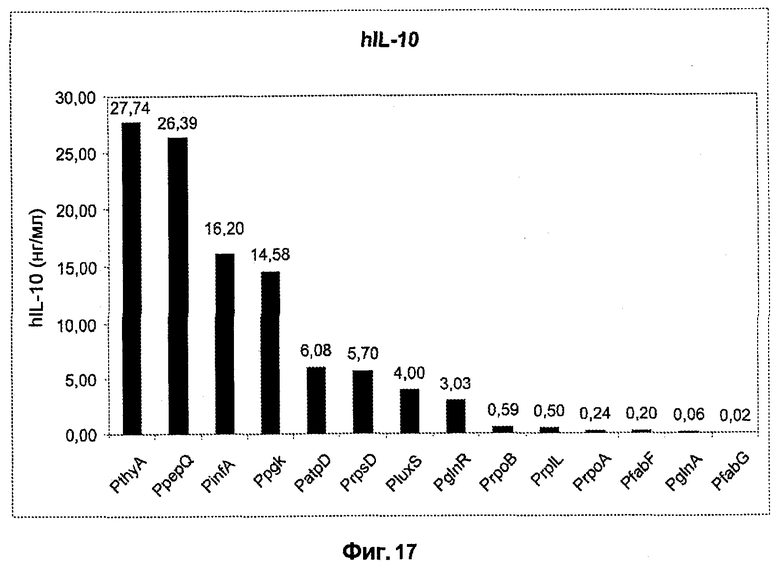
Изобретение относится к области биотехнологии, а именно к рекомбинантной нуклеиновой кислоте, экспрессирующей один или несколько представляющих интерес полипептидов, вектору экспрессии и бактерии, которые содержат данную рекомбинантную нуклеиновую кислоту, и их применению, а также к фармацевтической композиции для доставки одного или нескольких представляющих интерес полипептидов человеку или животному. Рекомбинантная нуклеиновая кислота содержит природный промотор гена HU-подобного ДНК-связывающего белка (PhIIA) вида Lactococcus с последовательностью SEQ ID NO:28, или его гомологичный или функциональный вариант, который по меньшей мере на 95% идентичен промотору с последовательностью SEQ ID NO:28, функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными для промотора PhIIA, где промотор PhIIA расположен выше одной или нескольких открытых рамок считывания. Вектор экспрессии содержит вышеуказанную рекомбинантную нуклеиновую кислоту, предпочтительно, указанный вектор получают из pT1NX. Бактерия содержит вышеуказанную рекомбинантную нуклеиновую кислоту или вышеуказанный вектор. Предложенное изобретение позволяет повысить уровень экспрессии представляющих интерес генов полипептидов и таким образом получить достаточное количество экспрессированных белков. 11 н. и 8 з.п.ф-лы, 26 ил., 12 табл., 9 пр.
1. Рекомбинантная нуклеиновая кислота, экспрессирующая один или несколько представляющих интерес полипептидов, содержащая природный промотор гена HU-подобного ДНК-связывающего белка (PhIIA) вида Lactococcus с последовательностью SEQ ID NO:28, или его гомологичный или функциональный вариант, который по меньшей мере на 95% идентичен промотору с последовательностью SEQ ID NO:28, функционально связанный с одной или несколькими открытыми рамками считывания, гетерологичными для промотора PhIIA, где промотор PhIIA расположен выше одной или нескольких открытых рамок считывания.
2. Рекомбинантная нуклеиновая кислота по п.1, дополнительно содержащая:
- последовательность терминатора транскрипции, расположенную в направлении 3' от указанной одной или нескольких открытых рамок считывания;
- оператор, сконструированный для контроля транскрипции с указанного PhIIA промотора;
- последовательности, сконструированные для вставки указанной рекомбинантной нуклеиновой кислоты в хромосому клетки-хозяина предпочтительно с помощью гомологичной рекомбинации; и/или
- сигнальную последовательность, предпочтительно usp45 или usp45N4.
3. Рекомбинантная нуклеиновая кислота по п.1, в которой указанные одна или несколько открытых рамок считывания кодируют:
- полипептид, способный вызывать терапевтическую реакцию или иммунный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид.
4. Рекомбинантная нуклеиновая кислота по п.3, где:
- указанный антиген способен индуцировать иммунный ответ, предпочтительно иммунологическую толерантность, у человека или животного,
- указанный невакциногенный терапевтически активный полипептид способен вызывать терапевтический эффект у человека или животного;
- указанный антиген способен индуцировать иммунный ответ и используется в качестве вакцины для человека или животного;
- указанный невакциногенный терапевтически активный полипептид представляет собой hIL-10, GLP-2, GLP-1, TFF или hPYY; и/или
- указанная рекомбинантная нуклеиновая кислота содержит:
(a) PhIIA, usp45 и hIL-10 или PhIIA, usp45N4 и hIL-10; или
(b) PhIIA, usp45N4 и hTFF1 или PhIIA, usp45 и hTFF1; или
(c) PhIIA, usp45N4 и hTFF3 или PhIIA, usp45 и hTFF3; или
(d) PhIIA, usp45N4 и hPYY или PhIIA, usp45 и hPYY; PhIIA, usp45 и hPYY G9 (3-36) (sAGX0213);
(e) PhIIA, usp45N4 и GLP-1 или PhIIA, usp45 и GLP-1; или
(f) PhIIA, usp45N4 и GLP-2 или PhIIA, usp45 и GLP-2.
5. Вектор экспрессии, содержащий рекомбинантную нуклеиновую кислоту по любому из пп.1-4, предпочтительно указанный вектор получают из pT1NX.
6. Бактерия, экспрессирующая один или несколько представляющих интерес полипептидов, содержащая рекомбинантную нуклеиновую кислоту по любому из пп.1-4, или вектор по п.5.
7. Бактерия, экспрессирующая один или несколько представляющих интерес полипептидов, содержащая рекомбинантную нуклеиновую кислоту по п.1, в которой указанный промотор PhIIA присутствует в хромосоме указанной клетке-хозяине, и указанный промотор PhIIA функционально связан с одной или несколькими открытыми рамками считывания, гетерологичными для указанного промотора PhIIA, где промотор PhIIA расположен выше одной или нескольких открытых рамок считывания.
8. Бактерия по п.7, в которой указанный промотор PhIIA дополнительно содержит:
- сигнальную последовательность, предпочтительно usp45 или usp45N4; и/или
- оператор, сконструированный для контроля транскрипции с указанного промотора PhIIA.
9. Бактерия по п.6, в которой указанный один или несколько представляющих интерес полипептидов представляют собой:
- полипептид, способный вызывать терапевтическую реакцию или иммунный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид.
10. Бактерия по п.9, в которой:
- указанный антиген способен индуцировать иммунный ответ, предпочтительно иммунологическую толерантность, у человека или животного,
- указанный невакциногенный терапевтически активный полипептид способен вызывать терапевтический эффект у человека или животного;
- указанный невакциногенный терапевтически активный полипептид представляет собой hIL-10, GLP-2, GLP-1, TFF или hPYY,
- указанный антиген способен индуцировать иммунный ответ и используется в качестве вакцины для человека или животного;
- и/или указанная бактерия содержит:
(a) PhIIA, usp45 и hIL-10 или PhIIA, usp45N4 и hIL-10; или
(b) PhIIA, usp45N4 и hTFF1 или PhIIA, usp45 и hTFF1; или
(c) PhIIA, usp45N4 и hTFF3 (sAGX0057) или PhIIA, usp45 и hTFF3; или
(d) PhIIA, usp45N4 и hPYY или PhIIA, usp45 и hPYY; PhIIA, usp45 и hPYY G9 (3-36); или
(e) PhIIA, usp45N4 и GLP-1 или PhIIA, usp45 и GLP-1; или
(f) PhIIA, usp45N4 и GLP-2 или PhIIA, usp45 и GLP-2.
11. Бактерия по п.6, являющаяся непатогенной и неинвазивной бактерией, предпочтительно грамположительной бактерией, более предпочтительно молочнокислой бактерией, еще более предпочтительно бактерией Lactococcus или бактерий Lactobacillus и наиболее предпочтительно бактерией Lactococcus lactis или Lactobacillus casei.
12. Применение рекомбинантной нуклеиновой кислоты по любому из пп.1-4 для экспрессии одного или нескольких представляющих интерес полипептидов в бактерии.
13. Применение вектора по п.5 для экспрессии одного или нескольких представляющих интерес полипептидов в бактерии.
14. Способ рекомбинантной экспрессии представляющего интерес полипептида, включающий:
a) культивирование бактерии по любому из пп.6-11, в которой указанные одна или несколько открытых рамок считывания кодируют один или несколько представляющих интерес полипептидов, и
b) выделение представляющего интерес полипептида, продуцированного бактерией при указанном культивировании.
15. Применение бактерии по п.11 для доставки указанного одного или нескольких представляющих интерес полипептидов человеку или животному, где указанный один или несколько представляющих интерес полипептидов представляют собой:
- полипептид, способный вызывать терапевтический или иммуногенный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид.
16. Применение бактерии по п.11 для получения лекарственного средства, где один или несколько представляющих интерес полипептидов представляют собой:
- полипептид, способный вызывать терапевтический или иммуногенный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид.
17. Применение по п.15, где указанным полипептидом является антиген и/или невакциногенный терапевтически активный полипептид, предпочтительно hIL-10, GLP-2, GLP-1, TFF или hPYY.
18. Фармацевтическая композиция для доставки одного или нескольких представляющих интерес полипептидов человеку или животному, содержащая бактерию по п.11, и фармацевтически приемлемый носитель, где указанный один или несколько представляющих интерес полипептидов представляют собой:
- полипептид, способный вызывать терапевтический или иммуногенный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид.
19. Бактерия по п.11, где указанный один или несколько представляющих интерес полипептидов представляют собой:
- полипептид, способный вызывать терапевтический или иммуногенный ответ у индивида, предпочтительно у человека или животного;
- антиген и/или
- невакциногенный терапевтически активный полипептид,
использующаяся в качестве медикамента.
| СОЛЬ НАТРИЯ β -ФЕНИЛИЗОСЕРИНА-(2R, 3S) И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 1992 |
|
RU2090551C1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Приспособление к токарному полуавтомату для подачи обрабатываемых колец в зажимной патрон | 1955 |
|
SU102570A1 |
| CHATEL J M et al | |||
| Induction of mucosal immune response after intranasal or oral inoculation of mice with Lactococcus lactis producing bovine beta-lactoglobulin, CLINICAL AND DIAGNOSTIC LABORATORY IMMUNOLOGY, 2001, v | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Музыкальный инструмент со звучащими металлическими пластинками и ручной клавиатурой | 1922 |
|
SU545A1 |
| TERMONT S | |||
| et al | |||
| Intracellular accumulation of | |||

