ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] Настоящая заявка испрашивает приоритет согласно предварительной заявке на патент США №62/278,493, поданной 14 января 2016 года, и предварительной заявке на патент США №62/350,472, поданной 15 июня 2016 года, каждая из которых полностью включена в настоящую заявку.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[002] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и таким образом полностью включен в настоящую заявку посредством ссылки. Упомянутая копия в формате ASCII, созданная 10 января 2017 года, названа 205350-0030-00-WO-549987_SL.txt и имеет размер 189 854 байта.
УРОВЕНЬ ТЕХНИКИ
[003] Приблизительно 10-15 миллионов человек страдают от сахарного диабета 1-го типа (T1D), который представляет собой наиболее распространенное метаболическое расстройство в младенчестве и подростковом возрасте, затрагивающее 112000 детей в возрасте до 16 лет только в Европе. T1D возникает вследствие прогрессирующего иммунно-опосредованного разрушения островковых бета-клеток поджелудочной железы, продуцирующих инсулин ("бета-клеток"), у генетически предрасположенных лиц, приводящего к хронической гипергликемии, которая вызывает микро- и макрососудистые осложнения. См., например, van Belle, T.L. et al., Physiol. Rev. 2011, 91(1): 79-118. В то время как для некоторых аутоиммунных заболеваний доступны различные варианты терапии, в настоящее время не существует одобренных вариантов терапии T1D. Пациентам с T1D требуется пожизненное введение инсулина. Более того, долгосрочная терапия требует многопрофильного подхода, включающего врачей, медсестер, диетологов и других специалистов.
[004] Единственной терапевтической стратегией при аутоиммунных заболеваниях, как в контексте краткосрочной терапии, так и при длительных схемах лечения, является блокирование аутореактивных эффекторных Т-клеток с применением генерализованной иммуносупрессии. Полагают, что активация или экспансия регуляторных Т-клеток (Treg) способна восстанавливать баланс между эффекторными Т-клетками и Treg-клетками и может обеспечить достижение той же цели без токсичности, ассоциированной с иммуносупрессией.
[005] Интерлейкин-2 ("IL-2") имеет ключевые функции в иммунной системе, в первую очередь, благодаря своему прямому воздействию на Т-клетки. В тимусе, где Т-клетки созревают, он предотвращает развитие аутоиммунных заболеваний, способствуя дифференцировке некоторых незрелых Т-клеток в регуляторные Т-клетки, которые подавляют другие Т-клетки, которые в противном случае были бы примированы для атаки на здоровые клетки в организме. При более высоких концентрациях IL-2 также способствует дифференцировке Т-клеток в эффекторные Т-клетки и в Т-клетки памяти, в случае, когда исходная Т-клетка также стимулируется антигеном, тем самым помогая организму бороться с инфекциями.
[006] Первоначально нативный IL-2 был идентифиуцирован как фактор роста лимфоцитов и сочтен в первую очередь способствующим эффекторному ответу Т-клеток in vivo, таким образом был разработан рекомбинантный IL-2 для лечения состояний, требующих стимуляции эффекторных Т-клеток, а именно, рака и инфекционных заболеваний. Однако было показано, что IL-2 отвечает за дифференциацию, выживание и функционирование эффекторных Т-клеток, поскольку у нокаутных по IL-2 мышей развивается аутоиммунное заболевание, опосредованное Т-клетками. Известно, что IL-2 является цитокином, критичным для развития, экспансии, выживания и периферической активности Treg-клеток. Дефицит продуцирования IL-2 или отсутствие ответа на IL-2 приводит к утрате функций Treg-клеток и повышению аутоиммунитета. Treg-клетки конститутивно экспрессируют тримерный высокоаффинный рецептор к IL-2 (IL-2Rαβγ) с более высокими уровнями, чем эффекторные Т-клетки CD4+ и CD8+, NK-клетки и эозинофилы. Стимулирование передачи сигналов STAT5a в Treg-клетках происходит при более низких дозах IL-2, чем в эффекторных Т-клетках. Следовательно, IL-2 в низких дозах, по-видимому, стимулирует предпочтительную активацию Treg-клеток и способствует их выживанию in vivo. См., например, Yu, A., et al., Diabetes 2015, 64: 2172-2183.
[007] В клинических исследованиях введение IL-2 вызывало иммунологические изменения, но не меняло параметров метаболизма глюкозы. См., например, Hartemann A. et al., Lancet Diabetes Endocrinol. 2013; 1:295-305; и Rosenzwajg M. et a., J Autoimmun. 2015; 58:48-58.
[008] Фармацевтические препараты IL-2 вводят путем инъекции. Хотя пероральная доставка кажется перспективной, например, в силу простоты введения, деградация в желудочно-кишечном тракте и низкий уровень абсорбции обычно делают этот путь неэффективным для доставки полипептидов. Для препаратов, основанных на полипептидах, также исследуются альтернативные способы применения, например, назальное, ректальное, легочное и глазное применение.
[009] Для доставки терапевтических молекул в ткани слизистой оболочки использовались генетически модифицированные бактерии. См., например, Steidler, L., et al., Nat. Biotechnol. 2003, 21(7): 785-789; и Robert S. and Steidler L., Microb. Cell Fact. 2014, 13 Suppl. 1: S11.
[0010] Было показано, что кишечное введение антигенов, участвующих в бета-клеточном аутоиммунитете, с помощью генетически модифицированного штамма Lactococcus lactis, угнетает T1D у NOD-мышей посредством стимуляции Foxp3+ Treg-клеток. Пероральное введение генетически измененного штамма Lactococcus lactis направляет проинсулин (PINS) человека совместно с IL-10 человека в слизистую оболочку желудочно-кишечного тракта (ЖКТ) и в сочетании с системным антителом против CD3 в низкой дозе восстанавливает долгосрочную толерантность иммунной системы почти у 60% NOD мышей с впервые выявленным диабетом. См., например, Robert, S. et a.l., Diabetes 2014, 63: 2876-2887; и Takiishi, Т. et al., J. Clin. Inv. 2012, 122(5): 1717-1725. Однако переход к клиническому применению такой антигенспецифической комбинированной терапии, включающей дополнительные иммуномодуляторы, такие как антитела против CD3, затрудняется, например, тем, что Fc-модифицированные антитела против CD3 человека не были одобрены регулирующими организациями для лечения T1D.
[0011] В данной области техники существует потребность в эффективных, направленных и контролируемых способах лечения T1D, без побочных действий и системной токсичности. Такие стратегии должны облегчать введение, повышать безопасность и в идеале улучшать эффективность и снижать терапевтически эффективные дозы. Настоящее изобретение направлено на удовлетворение этих потребностей.
ВКЛЮЧЕНИЕ ПУТЕМ ССЫЛКИ
[0012] Все публикации, патенты и патентные заявки в данном документе включены посредством ссылки в той же степени, как если бы каждая отдельная публикация, патент или патентная заявка была специально и индивидуально указана как включенная посредством ссылки. В случае противоречия между термином в настоящей заявке и термином в источнике, включенном посредством ссылки, термин в настоящей заявке имеет преимущественную силу.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0013] Соответственно, предложены композиции и способы, включающие живые молочнокислые бактерии (LAB), например, генетически модифицированные штаммы Lactococcus lactis (LL), в качестве средств доставки в слизистую оболочку IL-2 в низкой дозе, например, в комбинации с TlD-специфическими аутоантигенами, такими как проинсулин (PINS). Указанные LAB генетически модифицированы для экспрессии биоактивных полипептидов, вызывающих биологические реакции, которые, в свою очередь, блокируют дальнейшее аутоиммунное разрушение бета-клеток поджелудочной железы. Такая стратегия может способствовать регрессии сформировавшегося T1D, например, у субъектов с достаточной остаточной функцией бета-клеток и, таким образом, обеспечивать "истинное" лечение аутоиммунного диабета. Такие композиции можно вводить перорально, например, в форме фармацевтического препарата с энтеросолюбильным покрытием, который транспортирует бактерии в желудочно-кишечный тракт, например, в нижнюю часть желудочно-кишечного тракта (например, дистальные отделы толстой кишки), где они будут секретировать подходящую низкую дозу IL-2, в некоторых случаях - в комбинации с TlD-специфическим антигеном (например, PINS).
[0014] Предлагаемая композиция содержит ферментирующую молочную кислоту бактерию (LAB), содержащую экзогенную нуклеиновую кислоту, кодирующую полипептид интерлейкина-2 (IL-2), и экзогенную нуклеиновую кислоту, кодирующую полипептид специфического антигена диабета 1-го типа (T1D). В качестве альтернативы, предлагаемая композиция содержит первую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую полипептид интерлейкина-2 (IL-2), и вторую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую полипептид специфического антигена диабета 1-го типа (T1D). Эта композиция может дополнительно содержать фармацевтически приемлемый носитель. Указанная LAB может быть адаптирована для доставки в слизистую оболочку IL-2 в низкой дозе при введении субъекту-млекопитающему.
[0015] В одном аспекте указанная LAB может быть выбрана из группы, состоящей изо: вида Lactococcus, вида Lactobacillus, вида Bifidobacterium, вида Streptococcus и вида Enterococcus. Указанная LAB может представлять собой вид Lactococcus. Например, указанная LAB может представлять собой Lactococcus lactis. В качестве альтернативы, указанная LAB может быть выбрана из группы, состоящей из: Lactococcus garvieae, Lactococcus lactis, Lactococcus lactis subsp. cremoris, Lactococcus lactis subsp. hordniae, Lactococcus lactis subsp. Lactis, Lactococcus piscium, Lactococcus plantarum, Lactococcus raffinolactis, Lactobacillus acetotolerans, Lactobacillus acidophilus, Lactobacillus agilis, Lactobacillus algidus, Lactobacillus alimentarius, Lactobacillus amylolyticus, Lactobacillus amylophilus, Lactobacillus amylovorus, Lactobacillus animalis, Lactobacillus aviarius, Lactobacillus aviarius subsp. arqffinosus, Lactobacillus aviarius subsp. aviarius, Lactobacillus bavaricus, Lactobacillus bifermentans, Lactobacillus brevis, Lactobacillus buchneri, Lactobacillus bulgaricus, Lactobacillus carnis, Lactobacillus casei, Lactobacillus casei subsp. alactosus, Lactobacillus casei subsp. casei, Lactobacillus casei subsp. pseudoplantarum, Lactobacillus casei subsp. rhamnosus, Lactobacillus casei subsp. tolerans, Lactobacillus catenaformis, Lactobacillus cellobiosus, Lactobacillus collinoides, Lactobacillus confusus, Lactobacillus coryniformis, Lactobacillus coryniformis subsp. coryniformis, Lactobacillus coryniformis subsp. torquens, Lactobacillus crispatus, Lactobacillus curvatus, Lactobacillus curvatus subsp. curvatus, Lactobacillus curvatus subsp. melibiosus, Lactobacillus delbrueckii, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus delbrueckii subsp. delbrueckii, Lactobacillus delbrueckii subsp. lactis, Lactobacillus divergens, Lactobacillus farciminis, Lactobacillus fermentum, Lactobacillus fornicalis, Lactobacillus fructivorans, Lactobacillus fructosus, Lactobacillus gallinarum, Lactobacillus gasseri, Lactobacillus graminis, Lactobacillus halotolerans, Lactobacillus hamsteri, Lactobacillus helveticus, Lactobacillus heterohiochii, Lactobacillus hilgardii, Lactobacillus homohiochii, Lactobacillus iners, Lactobacillus intestinalis, Lactobacillus jensenii, Lactobacillus johnsonii, Lactobacillus kandleri, Lactobacillus kefiri, Lactobacillus kefiranofaciens, Lactobacillus kefirgranum, Lactobacillus kunkeei, Lactobacillus lactis, Lactobacillus leichmannii, Lactobacillus lindneri, Lactobacillus malefermentans, Lactobacillus mali, Lactobacillus maltaromicus, Lactobacillus manihotivorans, Lactobacillus minor, Lactobacillus minutus, Lactobacillus mucosae, Lactobacillus murinus, Lactobacillus nagelii, Lactobacillus oris, Lactobacillus panis, Lactobacillus parabuchneri, Lactobacillus paracasei, Lactobacillus paracasei subsp. paracasei, Lactobacillu sparacasei subsp. tolerans, Lactobacillus parakefiri, Lactobacillus paralimentarius, Lactobacillus paraplantarum, Lactobacillus pentosus, Lactobacillus perolens, Lactobacillus piscicola, Lactobacillus plantarum, Lactobacillus pontis, Lactobacillus reuteri, Lactobacillus rhamnosus, Lactobacillus rimae, Lactobacillus rogosae, Lactobacillus ruminis, Lactobacillus sakei, Lactobacillus sakei subsp. camosus, Lactobacillus sakei subsp. sakei, Lactobacillus salivarius, Lactobacillus salivarius subsp. salicinius, Lactobacillus salivarius subsp. salivarius, Lactobacillus sanfranciscensis, Lactobacillus sharpeae, Lactobacillus suebicus, Lactobacillus trichodes, Lactobacillus uli, Lactobacillus vaccinostercus, Lactobacillus vaginalis, Lactobacillus viridescens, Lactobacillus vitulinus, Lactobacillus xylosus, Lactobacillus yamanashiensis, Lactobacillus yamanashiensis subsp.mali, Lactobacillus yamanashiensis subsp. Yamanashiensis, Lactobacillus zeae, Bifidobacterium adolescentis, Bifidobacterium angulatum, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium catenulatum, Bifidobacterium longum, Bifidobacterium infantis, Enterococcus alcedinis, Enterococcus aquimarinus, Enterococcus asini, Enterococcus avium, Enterococcus caccae, Enterococcus camelliae, Enterococcus canintestini, Enterococcus canis, Enterococcus casseliflavus, Enterococcus cecorum, Enterococcus columbae, Enterococcus devriesei, Enterococcus diestrammenae, Enterococcus dispar, Enterococcus durans, Enterococcus eurekensis, Enterococcus faecalis, Enterococcus faecium, Enterococcus gallinarum, Enterococcus gilvus, Enterococcus haemoperoxidus, Enterococcus hermanniensis, Enterococcus hirae, Enterococcus italicus, Enterococcus lactis, Enterococcus lemanii, Enterococcus malodoratus, Enterococcus moraviensis, Enterococcus mundtii, Enterococcus olivae, Enterococcus pallens, Enterococcus phoeniculicola, Enterococcus plantarum, Enterococcus pseudoavium, Enterococcus quebecensis, Enterococcus raffinosus, Enterococcus ratti, Enterococcus rivorum, Enterococcus rotai, Enterococcus saccharolyticus, Enterococcus silesiacus, Enterococcus solitarius, Enterococcus sulfureus, Enterococcus termitis, Enterococcus thailandicus, Enterococcus ureasiticus, Enterococcus ureilyticus, Enterococcus viikkiensis, Enterococcus villorum, Enterococcus xiangfangensis, Streptococcus agalactiae, Streptococcus anginosus, Streptococcus bovis, Streptococcus canis, Streptococcus constellatus, Streptococcus dysgalactiae, Streptococcus equinus, Streptococcus iniae, Streptococcus intermedius, Streptococcus milleri, Streptococcus mitis, Streptococcus mutans, Streptococcus oralis, Streptococcus parasanguinis, Streptococcus peroris, Streptococcus pneumoniae, Streptococcus pseudopneumoniae, Streptococcus pyogenes, Streptococcus ratti, Streptococcus salivarius, Streptococcus tigurinus, Streptococcus thermophilus, Streptococcus sanguinis, Streptococcus sobrinus, Streptococcus suis, Streptococcus uberis, Streptococcus vestibularis, Streptococcus viridans и Streptococcus zooepidemicus.
[0016] В другом аспекте указанный T1D-специфический антиген может быть выбран из группы, состоящей из следующего: проинсулин (PINS), декарбоксилаза глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитическая субъединица связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP), транспортер цинка 8 (ZnT8), хромогранин А, (препро) островковый амилоидный полипептид (ppIAPP), периферии и цитруллинированный глюкозо-регулируемый белок (GRP). Например, указанный T1D-специфический антиген может представлять собой PINS.
[0017] В другом аспекте LAB может содержать указанную экзогенную нуклеиновую кислоту, кодирующую полипептид IL-2, и указанную экзогенную нуклеиновую кислоту, кодирующую полипептид T1D-специфического антигена. В качестве альтернативы, первая LAB может содержать указанную экзогенную нуклеиновую кислоту, кодирующую полипептид IL-2, а вторая LAB может содержать указанную экзогенную нуклеиновую кислоту, кодирующую полипептид T1D-специфического антигена. Указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, может быть интегрирована в хромосому указанной LAB. Указанная экзогенная нуклеиновая кислота, кодирующая полипептид T1D-специфического антигена, может быть интегрирована в хромосому указанной LAB или может присутствовать на плазмиде, содержащейся в указанной LAB. Указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая полипептид T1D-специфического антигена, может быть интегрирована в хромосому указанной LAB. Указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая полипептид T1D-специфического антигена, может быть частью полицистронной единицы экспрессии, регулируемой одним и тем же промотором.
[0018] В еще одном аспекте указанный IL-2 может представлять собой мембраносвязанную форму IL-2 или растворимую форму IL-2. Указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, может кодировать вариант полипептида IL-2. Указанный вариант полипептида IL-2 может иметь уменьшенную активность IL-2 или повышенную активность IL-2 по сравнению с соответствующим полипептидом IL-2 дикого типа. Указанный вариант полипептида IL-2 может быть выбран из группы, состоящей из: алдеслейкина, тецелейкина и биолейкина. Например, указанный вариант полипептида IL-2 содержит:
(a) первую аминокислотную замену по сравнению со зрелым IL-2 дикого типа, выбранную из группы, состоящей из L72G, L72A, L72S, L72T, L72Q, L72E, L72N, L72D, L72R и L72K; или
(b) вторую аминокислотную замену по сравнению со зрелым IL-2 дикого типа, выбранную из группы, состоящей из F42A, F42G, F42S, F42T, F42Q, F42E, F42N, F42D, F42R и F42K; или
(c) третью аминокислотную замену по сравнению со зрелым IL-2 дикого типа, выбранную из группы, состоящей из Y45A, Y45G, Y45S, Y45T, Y45Q, Y45E, Y45N, Y45D, Y45R и Y45K; или
(d) их сочетание.
[0019] Предложенная композиция может содержать Lactococcus lactis, где указанная Lactococcus lactis содержит экзогенную нуклеиновую кислоту, кодирующую полипептид IL-2, и экзогенную нуклеиновую кислоту, кодирующую PINS, и где указанная Lactococcus lactis адаптирована для доставки в слизистую оболочку IL-2 в низкой дозе при введении субъекту-млекопитающему. Указанная доставка низкой дозы IL-2 может составлять от приблизительно 0,01 млн. МЕ/сут/субъекта до приблизительно 5,4 млн. МЕ/сут/субъекта; от приблизительно 0,02 млн. МЕ/сут/субъекта до приблизительно 3,0 млн. МЕ/сут/субъекта; от приблизительно 0,1 млн. МЕ/сут/субъекта до приблизительно 3,0 млн. МЕ/сут/субъекта; или от приблизительно 0,2 млн. МЕ/сут/субъекта до 2,0 млн. МЕ/сут/субъекта.
[0020] Также предлагается применение данной композиции для лечения T1D у субъекта-млекопитающего, нуждающегося в этом. Предложенный способ лечения сахарного диабета 1-го типа (T1D) включает введение нуждающемуся в этом субъекту-млекопитающему терапевтически эффективного количества данной композиции.
[0021] В одном аспекте при данном способе лечения T1D указанному субъекту не вводят антитело против CD3. В качестве альтернативы, данный способ лечения T1D дополнительно включает введение указанному субъекту антитела против CD3. Указанное антитело против CD3 можно вводить в низкой дозе одновременно с указанной композицией указанному субъекту. Указанное антитело против CD3 можно вводить указанному субъекту внутривенно.
[0022] В другом аспекте у указанного субъекта может присутствовать остаточная функция бета-клеток. У указанного субъекта может присутствовать недавно выявленный T1D. Указанный субъект может иметь концентрацию С-пептида в крови или моче, указывающую на остаточную функцию бета-клеток. Указанный субъект может быть пациентом-человеком, имеющим концентрацию в крови натощак С-пептида менее приблизительно 1 нмоль/л, но по меньшей мере приблизительно 0,2 нмоль/л; или стимулированную концентрацию С-пептида в крови менее приблизительно 4 нмоль/л, но по меньшей мере приблизительно 0,5 нмоль/л. T1D у указанного субъекта может быть диагностирован в течение 12 месяцев до введения указанной композиции.
[0023] В следующем аспекте указанную композицию можно вводить указанному субъекту в слизистую оболочку. Указанную композицию можно вводить указанному субъекту в жидкой форме. Указанную композицию можно вводить указанному субъекту в виде пищевого продукта, пищевой добавки или суппозитория. Указанную композицию можно вводить в виде единичной лекарственной формы, содержащей от приблизительно 1×104 до приблизительно 1×1012; от приблизительно 1×106 до приблизительно 1×1012; или от приблизительно 1×109 до приблизительно 1×1012 колониеобразующих единиц (КОЕ). Указанная единичная дозированная форма может быть выбрана из группы, состоящей из: капсулы, таблетки, гранулы, суппозитория и дозированного аэрозоля. Указанная композиция может быть в форме сухого порошка или в прессованном варианте.
[0024] Кроме того, предложен генетически модифицированный микроорганизм, содержащий экзогенную нуклеиновую кислоту, кодирующую полипептид IL-2; и экзогенную нуклеиновую кислоту, кодирующую полипептид T1D-специфического антигена. Например, указанный микроорганизм может представлять собой LAB. Указанная экзогенная нуклеиновая кислота, кодирующая интерлейкин-2 и/или полипептид T1D-специфического антигена, может быть стабильно интегрирована в хромосому этого микроорганизма.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0025] Фиг. 1: нуклеотидная последовательность (SEQ ID NO: 46), кодирующая гибрид лидера секреции usp45 (SSusp45) и гена hIL-2, кодирующего интерлейкин-2 человека (hIL-2, UniProt: Р60568, аа 21-153), ниже экспрессируемого на высоком уровне гена фосфопирувата гидратазы (eno; Gene ID: 4797432; локализация: NC_009004.1 (606184..607485)), содержащая межгенную область, предшествующую экспрессированному на высоком уровне гену L30 рибосомного белка MG1363 50S L. lactis (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911, комплемент)) между еnо и SSusp45.
[0026] Фиг. 2: схематическое представление генетических локусов LL-IL-2 (ΔthyA, trePTS (ΔtrePP), otsB, ΔptcC и eno >> hIL-2) с межгенными областями и размерами продуктов амплификации ПЦР (пар нуклеотидов, п.н.).
[0027] Фиг. 3: пример плазмиды-носителя с основной цепью, состоящей из фрагмента pORI19, к которому был добавлен модуль экспрессии PhllA>>β-глюкуронидаза (uidA; Gene ID: 946149); карго-области (область встраивания), содержащей pins ниже от gapB, сцепленной межгенной областью rpmD, фланкированной кроссоверными (ХО) областями, расположенными в позициях 5'и 3' от eno>>hil-2; а также маркера отбора с помощью эритромицина: устойчивый к эритромицину ген 23S РНК метилазы (еrmС).
[0028] Фиг. 4: анализ в агарозном 1,2% геле фрагментов ПЦР, полученных из LL-IL-2.
[0029] Фиг. 5: вестерн-блоттинг, демонстрирующий присутствие hIL-2 в супернатантах культуры LL-IL-2.
[0030] Фиг. 6: нуклеотидная последовательность (SEQ ID NO: 57), кодирующая слияние лидера секреции usp45 (SSusp45), с геном pins, кодирующим проинсулин человека (PINS; UniProt: Р01308, аа 25-110), ниже экспрессируемого на высоком уровне гена глицеральдегида 3-фосфатдегидрогеназы (gapB; Gene ID: 4797877; локализация: NC_009004.1 (2492509..2493519, комплемент)), содержащая межгенную область, предшествующую экспрессированному на высоком уровне гену L30 рибосомного белка L. lactis MG1363 50S (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911, комплемент), см., например, Steidler et al., Nat. Biotechnol. 2003; 21(7):785-789) между gapB и pins.
[0031] Фиг. 7: схематическое представление соответствующих генетических локусов LL-PINS/IL-2: ΔthyA, trePTS (ΔtrePP), otsB, ptcC-, gapB>>pins и eno>>hil-2 с указанием соответствующих сайтов связывания олигонуклеотидов, сайта рестрикции EcoRI, (/усеченных/) генетических признаков, межгенных областей (IR), размеров продуктов амплификации ПЦР (п.н.).
[0032] Фиг. 8: пример плазмиды-носителя с основной цепью, которая состоит из фрагмента pORI19, к которому добавлен модуль экспрессии PhllA>>β-глюкуронидазы (uidA; Gene ID: 946149); карго-области, содержащей pins ниже gapB, сцепленной межгенной областью rpmD, и фланкированной кроссоверными (ХО) областями, расположенными в позициях 5' и 3' от gapB>>pins; а также маркера отбора с помощью эритромицина: устойчивый к эритромицину ген 23S РНК метилазы (ermC).
[0033] Фиг. 9: анализ в агарозном 1,2% геле фрагментов ПЦР, полученных из LL-PINS/IL-2.
[0034] Фиг. 10А и 10В результаты вестерн-блоттинга, показывающие (1) присутствие PINS (черная стрелка) в супернатантах культуры LL-Pins/IL-2, (Фиг. 10А) и (2) присутствие hIL-2 (незаштрихованная стрелка) в супернатантах культуры LL-PINS/IL-2(Фиг. 10В).
[0035] Фиг. 11: стабильная регрессия гипергликемии у NOD-мышей с впервые выявленным диабетом при примере антигенспецифической терапии в соответствии с настоящим изобретением. NOD-мыши с впервые выявленным диабетом получали лечение, описанное в настоящей заявке, например, в Примере 3, и концентрации глюкозы в крови наблюдали в течение 14 недель после начала лечения. Показан процент мышей, сохранявших диабетический статус при лечении доставкой в слизистую оболочку LL-PINS или доставкой в слизистую оболочку LL-PINS+LL-IL-2.
[0036] Фиг. 12: эффективность примера антигенспецифической терапии в соответствии с настоящим изобретением у мышей с недавно выявленным диабетом с начальной концентрацией глюкозы в крови более или менее 350 мг/дл. Показан процент мышей, сохранявших диабетический статус при лечении с помощью доставки в слизистую оболочку LL-PINS+LL-IL-2, описанной в настоящей заявке, например, в Примере 3.
[0037] Фиг. 13: сравнение биологической активности LL-IL-2 с рекомбинантным IL-2 человека (rhIL-2).
[0038] Фиг. 14А и 14В: концентрации живых бактерий (Фиг. 14А: КОЕ/ткань, Фиг. 14В: КОЕ/г соответственно) в разных тканях ЖКТ в разные моменты времени после введения однократной дозы (1010 КОЕ) LL-IL-2 у не страдающих ожирением мышей с диабетом. Все столбики представляют среднее значение от 3 мышей (n=3). SIP=проксимальная тонкая кишка; SID = дистальная тонкая кишка; CAE = слепая кишка; СОР = проксимальная толстая кишка; COD = дистальная толстая кишка.
[0039] Фиг. 15: структура плазмиды pAGX0053. Основная цепь плазмиды содержит фрагмент pT1NX, к которому добавлен модуль экспрессии PthyA>>SSusp45::hpins. PthyA, промотор гена тимидилатсинтазы. EmR: маркер отбора с помощью эритромицина; repD, герЕ: гены репликации.
[0040] Фиг. 16: вестерн-блоттинг, показывающий наличие полноразмерного, полученного из плазмиды, PINS в супернатантах культуры LL-PINS.
[0041] Фиг. 17: частоты ремиссии диабета у NOD-мышей с впервые выявленным диабетом, получавших различные рекомбинантные бактерии. Результаты показывают, что доставленный в слизистую оболочку LL-IL-2 (например, введение в низкой дозе IL-2), в некоторых случаях - в сочетании с примером T1D-специфического антигена (то есть PINS) в соответствии с настоящим изобретением стимулирует ремиссию диабета и вызывает стабильную регрессию гипергликемии у NOD-мышей с впервые выявленным диабетом. Мыши получали лечение в течение 6 недель, как описано в настоящей заявке, и измеряли концентрации глюкозы в крови, включая 14 недель после начала лечения. Показан процент мышей, у которых оставался диабет (логранговый критерий Мантеля-Кокса; ** p<0,01).
[0042] Фиг. 18: частоты ремиссии диабета в зависимости от начальных концентраций глюкозы в крови. Результаты показывают, что начальные концентрации глюкозы в крови способны прогнозировать успех терапии у мышей. Мышей с недавно выявленным диабетом стратифицировали исходя из начального (до лечения) уровня глюкозы в крови ниже или выше 350 мг/дл. Полученные результаты показывают, что доставка в слизистую оболочку LL-IL-2, в некоторых случаях - в сочетании с примером антигенспецифической терапии (например, PINS), в соответствии с настоящим изобретением особенно эффективна у мышей с недавно выявленным диабетом с начальной концентрацией глюкозы в крови ниже 350 мг/дл. Показан процент мышей, которые имели диабет при лечении LL-IL-2 или LL-PINS+LL-IL-2, описанным в настоящей заявке. Следует отметить, что 6 мышей, получавших только LL-IL-2 и имеющих уровень глюкозы ниже 350 мг/дл, обозначенные как "LL-IL2<350 (n=9)", были умерщвлены после 6 недель лечения, а 3 мышей наблюдали в течение всего 14-недельного периода. Точка данных для LL-IL-2>350 (n=6) (сплошной треугольник) скрыта за точкой данных для не получавших лечения Untreated>350 (n=16) (сплошной квадрат). Все получавшие лечение LL-IL-2 мыши с концентрацией глюкозы >350 мг/дл, все еще имели диабет после 6 недель лечения и после 14-недельного периода наблюдения (логранговый критерий Мантеля-Кокса; **** p<0,0001).
[0043] Фиг. 19: оценка степени инсулита в островковых бета-клетках у NOD мышей с диабетом. Полученные результаты показывают, что доставленный в слизистую оболочку LL-IL-2 (например, введение IL-2 в низкой дозе), в некоторых случаях - в сочетании с примером антигенспецифической терапии в соответствии с настоящим изобретением (например, LL-PINS+LL-IL-2) не только предотвращал ухудшение инсулита (обычно наблюдаемое при прогрессировании к длительному диабету при отсутствии лечения), но и снижал инсулит островковых бета-клеток по сравнению с мышами с недавно выявленным диабетом и не получавшими лечения мышами с длительным диабетом (например, уменьшает инсулит до степени, сравнимой с инсулитом, обнаруженным у "нормогликемических" NOD-мышей с длительным диабетом). После лечения степень тяжелого инсулита изменилась в сторону улучшения, по сравнению с не получавшими лечения мышами с длительным диабетом. Процент бета-клеток без инсулита резко возрос по сравнению с мышами с недавно выявленным диабетом и не получавшими лечения мышами с длительным диабетом. У значительного процента островков мягкий инсулит изменился в сторону улучшения до "пери-инсулита" или "отсутствия инсулита". Неожиданно это значительное снижение степени инсулита наблюдалось у всех получавших лечение мышей с недавно выявленным диабетом (с ремиссией и без ремиссии - классифицированных как "излеченные" и "неизлеченные").
[0044] Фиг. 20: Измеренные не натощак концентрации С-пептида в плазме NOD мышей с диабетом. Полученные результаты показывают, что IL-2 в низкой дозе с проинсулином или без проинсулина сохраняет функцию бета-клеток у NOD мышей с диабетом. После 6 недель лечения LL-IL-2, в некоторых случаях - в сочетании с примером антигенспецифической терапии в соответствии с настоящим изобретением (например, LL-PINS+LL-IL-2) концентрации С-пептида в плазме у мышей с недавно выявленным диабетом, показавших ремиссию (классифицированных как "излеченные") увеличивались по сравнению с мышами с недавно выявленным диабетом и не получавшими лечения мышами с длительным диабетом (Т-критерий Манна-Уитни, * p<0,05). Концентрации С-пептида в плазме получавших лечение, но "неизлеченных" мышей не были обнаружены (ND), свидетельствуя, что у таких мышей активные бета-клетки могут практически отсутствовать. Уровни С-пептида, измеренные у 3 из 4 проанализированных не получавших лечения мышей с длительным диабетом, также не были обнаружены.
[0045] Фиг. 21: Экспансия регуляторных foxp3+CTLA4+ Т-клеток в разных субпопуляциях иммунных клеток, локально (т.е. брыжеечные дренирующие лимфатические узлы, MLN), системно (т.е. селезенка и кровь) и в целевом органе (то есть лимфатические узлы поджелудочной железы) NOD мышей с диабетом, измеренная с помощью проточной цитометрией после 6 недель лечения. Результаты показывают, что доставленный в слизистую оболочку LL-IL-2, в некоторых случаях - в сочетании с примером антигенспецифической терапии в соответствии с настоящим изобретением (например, LL-PINS+LL-IL-2) повышает количество foxp3+CTLA4+ регуляторных Т-клеток в LN поджелудочной железы по сравнению с мышами с недавно выявленным диабетом. Неожиданно повышение в LN поджелудочной железы наблюдалось у мышей с ремиссией ("излеченных") и у мышей без ремиссии ("неизлеченных") мышей (Т-критерий Манна-Уитни, * p<0,05, ** p<0,01, ** * p<0,001).
[0046] Фиг. 22А и 22В: концентрации глюкозы в крови (мг/дл) у мышей с недавно выявленным диабетом, получавших только LL-IL-2 (Фиг. 22А) и получавших LL-IL-2+LL-INS (Фиг. 22В).
[0047] Фиг. 23: частота ремиссии диабета в зависимости от начальных концентраций глюкозы в крови до 350 мг/дл при начале исследования. Мышей распределяли в 5 экспериментальных групп лечения: контрольная группа не получавших лечения, LL-PINS, LL-IL2, смесь LL-PINS+LL-IL2 и смесь LL-PINS+LL-IL2 в сочетании с системным иммуномодулирующим анти-CD3, как описано в Примере 9. Показан процент мышей, сохранявших диабетический статус в различные моменты времени после лечения.
ПОДРОБНОЕ ОПИСАНИЕ
[0048] Сокращения и аббревиатуры, используемые в настоящей заявке, могут включать:
[0049] В настоящей заявке предложены композиции и способы для лечения T1D, для стимулирования Treg и/или для восстановления толерантности к T1D-специфическим антигенам (то есть аутоантигенам) у субъекта.
[0050] В настоящей заявке представлены композиции, содержащие (1) LAB, содержащую ген интерлейкина-2 (IL-2) и ген T1D-специфического антигена, или (2) первую LAB, содержащую ген интерлейкина-2 (IL-2), и вторую LAB, содержащую T1D-специфический антиген. В некоторых примерах LAB экспрессирует ген IL-2 и/или ген T1D-специфического антигена для продуцирования IL-2 и T1D-специфического антигена (например, PINS). В некоторых вариантах реализации эти композиции представляют собой фармацевтические композиции, содержащие LAB и фармацевтически приемлемый носитель. Примеры носителей описаны в настоящей заявке. В некоторых примерах фармацевтическая композиция адаптирована для доставки композиции субъекту в слизистую оболочку.
[0051] Предложены способы для лечения T1D у субъекта-млекопитающего, нуждающегося в этом. Эти способы включают введение (например, в слизистую оболочку) субъекту композиции в соответствии с настоящим изобретением. Примеры способов включают: введение субъекту терапевтически эффективного количества LAB, способных экспрессировать IL-2 и T1D-специфический антиген (например, PINS).
[0052] Неожиданно было обнаружено, что особенно хорошо реагируют на способы терапии, описанные в настоящей заявке, субъекты со значительной остаточной функцией бета-клеток. Поэтому, в некоторых вариантах реализации у субъекта-млекопитающего в описанных в настоящей заявке способах T1D был диагностирован недавно и/или указанный субъект имеет недавно возникший T1D. В некоторых примерах T1D мог быть диагностирован у субъекта в течение предыдущих 12 месяцев, предыдущих 24 месяцев или предыдущих 36 месяцев до введения композиции, содержащей LAB, описанной в настоящей заявке.
[0053] В некоторых примерах в описанных в настоящей заявке способах IL-2 и антигенные полипептиды доставляют в слизистую оболочку. Такой подход может обеспечить доставку концентраций IL-2 в низкой дозе, которые даже ниже, чем требуются для системного введения в низкой дозе. Таким образом можно избежать побочных токсических эффектов, ассоциированных с системной доставкой IL-2.
Определения
[0054] Используемые в настоящей заявке единственные формы включают ссылки на множественные формы, если контекст явно не диктует иное. Например, термин "клетка" включает в себя множество клеток, включая их смеси. Аналогичным образом, применение "соединения" для лечения или приготовления лекарственных средств, описанное в настоящей заявке, предполагает применение одного или нескольких соединений для такого лечения или приготовления, если контекст явно не диктует иное.
[0055] Используемый в настоящей заявке термин "содержащий" означает, что композиции и способы включают в себя указанные элементы, но не исключают других. "Состоящий по существу из" при применении для определения композиций и способов означает исключение других элементов любого существенного значения для данной комбинации. Таким образом, композиция, состоящая по существу из элементов, в соответствии с определением в настоящей заявке, не будет исключать следовые примеси, обусловленные способом выделения и очистки, и фармацевтически приемлемые носители, такие как физиологический раствор с фосфатным буфером, консерванты и тому подобное. "Состоящий из" подразумевает исключение любого другого компонента в виде более чем следовых количеств других ингредиентов и существенных этапов способа для введения описанных в настоящей заявке композиций. Варианты реализации, определенные каждым из этих связующих терминов, входят в объем настоящего изобретения.
[0056] Используемый в настоящей заявке термин "экспрессирующий" ген или полипептид или "продуцирующий" полипептид (например, полипептид IL-2 или полипептид T1D-специфического антигена) также включает "способный экспрессировать" и "способный продуцировать", соответственно. Например, микроорганизм, который содержит экзогенную нуклеиновую кислоту, может в достаточных условиях (например, достаточной гидратации и/или в присутствии питательных веществ) продуцировать полипептид, кодируемый этой экзогенной нуклеиновой кислотой). Однако этот микроорганизм может не всегда активно продуцировать данный кодируемый полипептид. Такую LAB (например, Lactococcus lactis) можно высушить (например, лиофилизировать), и в этом состоянии ее можно считать бездействующей (то есть не продуцирующей активно полипептид). Однако, как только LAB попадет в достаточные условия, например, будет введена субъекту и высвобождена (например, в желудочно-кишечный тракт субъекта), она может начать продуцировать полипептид. Таким образом, LAB, "экспрессирующая" ген или полипептид или "продуцирующая" полипептид по настоящему изобретению, включает LAB в ее "бездействующем" состоянии.
[0057] Термин "приблизительно" в отношении упоминаемого числового значения и его грамматические эквиваленты, используемые в настоящей заявке, могут включать в себя само это упоминаемое числовое значение и диапазон значений плюс или минус 10% от этого упоминаемого числового значения. Например, термин "приблизительно 10" включает в себя 10 и любые величины от 9 до 11 включительно. В некоторых случаях термин "приблизительно" относительно упоминаемого числового значения также может включать диапазон значений плюс или минус 10% 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от этого упоминаемого числового значения.
[0058] "Ген IL-2" обозначает ген интерлейкина-2, кодирующий "полипептид IL-2". Термин "ген IL-2" включает "варианты гена IL-2", кодирующие "варианты полипептида IL-2".
[0059] Термин "IL-2" или "полипептид IL-2" обозначает функциональный, например, полноразмерный полипептид интерлейкина-2 (например, полипептид человека IL-2), включая мембраносвязанные формы и растворимые формы, а также "варианты полипептида IL-2".
[0060] "Вариант IL-2" или "вариант полипептида IL-2" обозначает модифицированный (например, усеченный или мутантный), но функциональный полипептид IL-2, например, усеченную или мутантную версию IL-2 человека. Термин "вариант полипептида IL-2" включает полипептиды IL-2 с повышенной активностью или пониженной активностью по сравнению с соответствующим полипептидом IL-2 дикого типа. "Вариант полипептида IL-2" сохраняет по меньшей мере некоторую активность IL-2.
T1D-специфический антиген
[0061] Термины "T1D-специфический аутоантиген", "T1D-специфический антиген", "специфический для заболевания антиген", "собственный антиген", "аутоантиген" или "антиген" используются в настоящей заявке взаимозаменяемо. Эти термины используются в настоящей заявке в соответствии с принятым в данной области техники значением терминов собственный антиген или аутоантиген и обычно относятся к полипептиду/белку, происходящему из собственного организма субъекта (продуцируемого собственным организмом субъекта), где антиген распознается собственной иммунной системой субъекта и обычно порождает антитела против такого антигена. Аутоиммунные заболевания обычно ассоциированы с определенными специфическими для заболевания аутоантигенами. При T1D иммунная система субъекта может продуцировать антитела против по меньшей мере одного антигена, ассоциированного с процессом разрушения бета-клеток. Такие аутоантигены включают проинсулин (PINS), декарбоксилазу глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитической субъединицы связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP) и транспортер цинка (ZnT) 8. Клинический T1D может быть дополнительно ассоциирован с дополнительными целевыми молекулами-кандидатами, экспрессируемыми бета-клетками, такими как хромогранин А, (препро) островковый-амилоидный полипептид (ppIAPP), периферии и цитруллинированный глюкозо-регулируемый белок (GRP).
[0062] Термин "ген T1D-специфического антигена" обозначает ген, кодирующий вышеупомянутый "T1D-специфический антиген". Термин "ген T1D-специфического антигена" включает "варианты гена T1D-специфического антигена", кодирующие "варианты полипептида T1D-специфического антигена".
[0063] Термин " полипептид T1D-специфического антигена " обозначает функциональный, например, полноразмерный, полипептид, а также "варианты полипептида T1D-специфического антигена", которые могут иметь повышенную активность или пониженную активность по сравнению с соответствующим полипептидом дикого типа.
[0064] Термин "вариант T1D-специфического антигена" или "вариант полипептида T1D-специфического антигена" обозначает модифицированный (например, усеченный или мутантный), но функциональный полипептид, например, усеченную или мутантную версию PINS человека. Термин "вариант полипептида" включает полипептиды с повышенной активностью или пониженной активностью по сравнению с соответствующим полипептидом дикого типа. "Вариант полипептида" сохраняет по меньшей мере некоторую биологическую активность (функциональный полипептид). Примеры вариантов GAD65 и IA-2 включают их укороченные версии (например, GAD65370-575 и IA-2635-979 соответственно, по сравнению с полипептидами с номерами доступа NCBI NP_000809.1 (SEQ ID NO: 7) и NP_002837.1 (SEQ ID NO: 9, соответственно), сохраняющие антигенные свойства, и поэтому применимые в композициях и способах согласно настоящему изобретению, например, при стимуляции Treg и индуцировании толерантности у субъекта. Как правило, укороченные или усеченные версии T1D-специфического антигена эффективно экспрессируются и секретируются LAB (например, Lactococcus lactis).
[0065] Термин "функционально связанный" обозначает взаимное расположение, при котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать надлежащим образом. Регуляторная последовательность, "функционально связанная" с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с такими регуляторными последовательностями.
Субъект
[0066] "Субъект" представляет собой организм, который может получить пользу от введения композиции согласно настоящему изобретению, например, согласно способам согласно настоящему изобретению. Субъектом может быть млекопитающее ("субъект- млекопитающее"). Примеры субъектов-млекопитающих включают людей, сельскохозяйственных животных (таких как коровы, свиньи, лошади, овцы, козы), домашних животных (например, собаки, кошки и кролики), а также других животных, таких как мыши, крысы и приматы. В некоторых примерах субъектом-млекопитающим является пациент, представляющий собой человека.
IL-2 в низкой дозе
[0067] Термин "IL-2 в низкой дозе" обозначает дозу или концентрацию полипептида IL-2, которая может способствовать повышению компетентности и стабильности популяций регуляторных клеток Т (Treg) и/или способствует развитию наивных CD4+ Т-клеток в Treg-клетки у соответствующего субъекта, но которая ниже пороговой дозы/концентрации, которая стимулирует дифференцировку наивных Т-клеток в эффекторные Т-клетки и/или Т-клетки памяти у субъекта. Было показано, что клетки Treg имеют пороговое значение активации для IL-2 в 10-20 раз ниже, чем эффекторные Т-клетки, например, при измерении методом фосфорилирования STAT5 (pSTAT5). В звеньях сигнальных каскадов, расположенных после pSTAT5, активация многочисленных генов, важных для функционирования клеток, требует доз IL-2, которые для клеток Treg в 100 раз ниже, чем для эффекторных Т-клеток (см., например, Yu, A., et al., Diabetes 2015, 64: 2172-2183). Однако применительно к известным схемам лечения у людей минимальная доза IL-2, необходимая для стимуляции клеток Treg, установлена не была.
[0068] В некоторых вариантах реализации, например, в контексте лечения человека, "IL-2 в низкой дозе" обычно обозначает дозу полипептида IL-2 или варианта полипептида IL-2, которая может находиться в диапазоне от приблизительно 0,01 млн. МЕ/сут/субъекта до приблизительно 5,4 млн. МЕ/сут/субъекта. Низкая доза может быть в диапазоне от приблизительно 0,01 млн. МЕ/сут/субъекта до приблизительно 3,0 млн. МЕ/сут/ субъекта. Низкая доза может быть в диапазоне от приблизительно 0,02 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,03 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,04 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,05 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,06 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,07 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,08 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,09 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,1 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,2 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,3 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,4 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,5 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,6 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,7 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,8 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, от приблизительно 0,9 млн. МЕ/сут/субъекта до приблизительно 3 млн. МЕ/сут/субъекта, или от приблизительно 1,0 млн. МЕ/сут/субъекта до 3 млн. МЕ/сут/субъекта. Низкая доза может быть в диапазоне от приблизительно 0,02 млн. МЕ/сут/субъекта до приблизительно 2,5 млн. МЕ/сут/субъекта. Низкая доза также может быть в диапазоне от приблизительно 0,05 млн. МЕ/сут/субъекта до приблизительно 2,0 млн. МЕ/сут/ субъекта. Низкая доза может быть в диапазоне от приблизительно 0,1 млн. МЕ/сут/субъекта до приблизительно 1,5 млн. МЕ/сут/ субъекта. В других вариантах реализации низкая доза может быть в диапазоне от приблизительно 0,3 млн. МЕ/сут/субъекта до приблизительно 1,0 млн. МЕ/сут/ субъекта. Низкая доза может быть в диапазоне от 0,5 млн. МЕ/сут/субъекта до приблизительно 1,0 млн. МЕ/сут/субъекта.
[0069] Термин "международная единица" (ME) используется в настоящей заявке в соответствии с его принятым в данной области техники значением и представляет собой количество вещества (например, полипептида). Масса или объем, составляющие одну международную единицу, варьируют в зависимости от того, какое вещество измеряется. Всемирная организация здравоохранения (ВОЗ) определяет характеристики единиц для биоактивных полипептидов. Например, 1 ME IL-2 человека эквивалентна приблизительно 73 пг биоактивного полипептида (международный стандарт ВОЗ, NIBSC 86/500).
Анти-CD3 в низкой дозе
[0070] Термин "анти-CD3 в низкой дозе (низкодозовое)" обозначает совокупную дозу или концентрацию антитела против CD3, которая ниже стандартной дозы антитела против CD3 или одобренной регулятором дозы антитела против CD3 у людей для лечения таких заболеваний, как T1D или рак. Например, у людей терапия анти-CD3 в низкой дозе может включать дозу менее 50 мг (суммарно) антитела против CD3 у одного человека. Например, низкая доза анти-CD3 может включать от приблизительно 1 мг до приблизительно 50 мг; от приблизительно 5 мг до приблизительно 40 мг; от приблизительно 10 мг до приблизительно 30 мг; от приблизительно 15 мг до приблизительно 25 мг; от приблизительно 20 мг до приблизительно 30 мг; от приблизительно 15 мг до приблизительно 20 мг или от приблизительно 30 мг до приблизительно 35 мг совокупного лечения анти-CD антителом. Анти-CD3 в низкой дозе может включать менее приблизительно 50 мг; приблизительно 45 мг; приблизительно 40 мг; приблизительно 35 мг; приблизительно 30 мг; приблизительно 25 мг; приблизительно 20 мг; приблизительно 15 мг; приблизительно 10 мг; или приблизительно 5 мг совокупного лечения антителом против CD.
[0071] Например, в некоторых случаях совокупная доза антитела против CD3, назначаемая людям, может составлять приблизительно 34 мг или приблизительно 17 мг, вводимых в течение определенных периодов времени, например, в течение 14 дней. Это означает, что в течение 14-дневного периода вводят приблизительно 2,43 мг или 1,21 мг антитела против CD3.
[0072] Лечение анти-CD3 в низкой дозе может также содержать дозу приблизительно 80%; приблизительно 70%; приблизительно 60%; приблизительно 50%; приблизительно 40%; приблизительно 30%; приблизительно 20%; приблизительно 10%; приблизительно 5%; приблизительно 2%; приблизительно 1%; от приблизительно 80% до приблизительно 70%; от приблизительно 70% до 60%; от приблизительно 60% до 50%; от приблизительно 50% до 25%; от приблизительно 40% до 15%; или от приблизительно 30% до 5% от одобренной регулятором дозы антитела против CD3 для лечения T1D или рака.
[0073] У других млекопитающих, таких как мыши, "анти-CD3 в низкой дозе (низкодозовое)" может относиться к дозе менее приблизительно 5 мкг; 2,5 мкг; или 1 мкг. Например, для лечения мышей можно использовать приблизительно 5 мкг. В некоторых случаях для лечения мышей можно использовать приблизительно 2,5 мкг. В других случаях для лечения мышей можно использовать 1 мкг. Общая доза для мышей может составлять 12,5 мкг или 6 мкг анти-CD3.
[0074] Анти-CD3 можно вводить по меньшей мере один раз в сутки до 5 раз в сутки. Например, один раз в сутки, 2 раза в сутки, или 3 раза в сутки, пока не превышена суммарная доза. Например, если запланированная доза составляет 2,43 мг/сут, а введение происходит два раза в сутки, то можно вводить 1,215 мг на дозу.
[0075] Для реализации схемы анти-CD3 в низкой дозе, дозы можно вводить по меньшей мере один раз в сутки без перерыва в течение по меньшей мере 3 дней; 4 дней; 5 дней; 6 дней; 7 дней; 8 дней; 9 дней; 10 дней; 11 дней; 12 дней; 13 дней; 14 дней; 15 дней; 16 дней; 17 дней, 18 дней; 19 дней; 20 дней; 30 дней; или 40 дней, пока не превышена совокупная доза. Например, если назначена схема дозирования 34 мг анти-CD3 (анти-CD3 в низкой дозе) в течение 14 дней, субъекту можно вводить приблизительно 2,43 мг/сут. В некоторых случаях анти-CD в низкой дозе можно вводить по меньшей мере один раз в сутки без перерыва в течение по меньшей мере 1 месяца, 2 месяцев, 3 месяцев, 6 месяцев, 1 года или более.
[0076] Анти-CD3 в низкой дозе можно вводить внутривенно одновременно с введением композиции, описанной в настоящей заявке. В некоторых случаях анти-CD3 в низкой дозе можно вводить через 1 день; 2 дня; 3 дня; 4 дня; 5 дней; 6 дней; 7 дней; 2 недели; 3 недели; или через 1 месяц после первого введения композиции, описанной в настоящей заявке. Кроме того, в некоторых случаях анти-CD3 в низкой дозе можно вводить через 1 день; 2 дня; 3 дня; 4 дня; 5 дней; 6 дней; 7 дней; 2 недели; 3 недели; или за 1 месяц до первого введения композиции, описанной в настоящей заявке.
[0077] В некоторых случаях стандартная доза антитела против CD3 или одобренная регулятором доза антитела против CD3 у людей для лечения таких заболеваний, как T1D и рак, может быть введена пациентам до или после введения композиций, описанных в настоящей заявке.
[0078] В некоторых вариантах реализации антителом против CD3 может быть теплизумаб.
Субпопуляции пациентов
[0079] У субъекта, подвергнутого лечению с применением описанных в настоящей заявке способов, может присутствовать значимая (например, измеримая) остаточная функция бета-клеток. В таких обстоятельствах у субъекта может сохраняться ремиссия болезни, даже после прерывания или полного прекращения лечения. У пациентов с недавно диагностированным заболеванием часто на момент постановки диагноза сохраняется определенное минимальное количество островковых бета-клеток поджелудочной железы (бета-клетки), так что такие пациенты могут быть способны продуцировать определенное минимальное количество эндогенного инсулина. В такой популяции пациентов лечение композициями и способами согласно настоящему изобретению (например, IL-2 в низкой дозе и PINS-терапия) может быть особенно перспективным. Способы лечения, описанные в настоящей заявке, могут предотвратить дальнейшее разрушение бета-клеток и, таким образом, вызвать ремиссию болезни. Было обнаружено, что начальная масса бета-клеток может влиять на эффективность лечения. Например, у 57% NOD мышей с недавно выявленным диабетом, получавших композиции согласно настоящему изобретению, и с концентрацией глюкозы в крови приблизительно 350 мг/дл или менее при начале лечения удавалось добиться регрессии диабета. Регрессия болезни была достигнута только у 22% мышей с начальной концентрацией глюкозы более 350 мг/дл. Кроме того, у мышей с недавно выявленным диабетом регрессия болезни оставалась стабильной после прекращения лечения, показывая, что способы настоящего изобретения (включая доставку в слизистую оболочку биоактивных полипептидов) могут эффективно корректировать гипергликемию и восстанавливать долгосрочную толерантность к бета-клеткам. Однако, в случае если бета-клетки субъекта уничтожены, такой субъект может больше не получать пользы от описанного лечения таким же образом.
Лечение
[0080] Используемый в настоящей заявке термин "лечение", «лечить» и т.п. обозначают уменьшение интенсивности или облегчение характерных симптомов или проявлений заболевания или состояния, например T1D. Например, лечение T1D может привести к восстановлению или стимуляции антигенспецифической иммунной толерантности у субъекта. В других примерах лечение означает купирование аутоиммунного диабета или регрессию аутоиммунного диабета. Например, лечение может привести к сохранению оставшейся массы бета-клеток. В других примерах лечение T1D приводит к увеличению количества или активации Treg-клеток. В других примерах лечение может увеличивать популяцию антигенспецифических Treg-клеток (например, в тимусе) и/или стимулировать миграцию Treg-клеток в периферическую кровь. В других примерах лечение включает в себя улучшение по меньшей мере одного из клинических маркеров субъекта (пациента-человека). Например, лечение может повышать уровень С-пептида в крови и/или моче. В других примерах лечение может снизить уровень глюкозы в крови субъекта (например, пациента-человека) (например, в ответ на прием пищи или уровень глюкозы натощак); уменьшить количество инъецированного инсулина, необходимое для поддержания соответствующих уровней глюкозы в крови у субъекта, снизить уровни связанного с диабетом анти-антитела у субъекта и/или увеличить/сохранить уровни С-пептида (например, после перорального теста толерантности к глюкозе). Лечение может обозначать непрерывное/хроническое лечение или лечение, при котором у субъекта нет клинических симптомов данного заболевания или состояния в течение значительного периода времени (например, по меньшей мере 6 месяцев, по меньшей мере 1 год, по меньшей мере 2 года, по меньшей мере 3 года, по меньшей мере 4 лет или по меньшей мере 5 лет), после прекращения лечения.
[0081] Эти термины, используемые в настоящей заявке, также охватывают предотвращение или задержку начала заболевания или состояния или симптомов, ассоциированных с заболеванием или состоянием, включая снижение тяжести заболевания или состояния или ассоциированных с ним симптомов до начала явного поражения данной болезнью или состоянием. Такое предотвращение или снижение тяжести до начала явного поражения подразумевает введение описанного в настоящей заявке соединения или композиции пациенту, который на момент этого введения не имеет явного поражения этой болезнью или состоянием. "Предотвращение" также включает предотвращение рецидива или обострения заболевания или состояния или ассоциированных с ними симптомов, например, после периода улучшения.
Терапевтически эффективное количество
[0082] Используемый в настоящей заявке термин "терапевтически эффективное количество" обозначает количество непатогенного микроорганизма или композиции согласно настоящему изобретению, которые будут вызывать желаемый терапевтический эффект или реакцию при введении в соответствии с желаемой схемой лечения. Данные соединения или композиции обычно поставляются в единичной дозированной форме, например, таблетке или капсуле, которая содержит количество активного компонента, эквивалентов терапевтически эффективному количеству при введении один раз или несколько раз в сутки.
[0083] Специалисту в данной области техники понятно, что терапевтически эффективное количество рекомбинантного микроорганизма, которое требуется для достижения желаемого терапевтического эффекта (например, для эффективного лечения T1D), будет варьироваться, например, в зависимости от характера полипептида IL-2, экспрессируемого данным микроорганизмом, характера антигенного полипептида, экспрессируемого LAB, способа введения, а также возраста, массы и других характеристик реципиента.
Недавно выявленный T1D
[0084] В некоторых вариантах реализации у субъекта имеется недавно выявленный T1D. Термин "недавно выявленный T1D", "впервые выявленный T1D" или "недавно выявленная болезнь" обозначает состояние субъекта (например, пациента-человека), у которого недавно был диагностирован T1D (например, в течение приблизительно 3 предшествующих месяцев, в течение приблизительно шести предшествующих месяцев, в течение приблизительно 9 предшествующих месяцев, в течение приблизительно 12 предшествующих месяцев, в течение приблизительно 15 предшествующих месяцев, в течение приблизительно 18 предшествующих месяцев, в течение приблизительно предшествующих 24 месяцев, в течение приблизительно 30 предшествующих месяцев, в течение приблизительно 36 предшествующих месяцев, в течение приблизительно 42 предшествующих месяцев, в течение приблизительно 48 предшествующих месяцев, в течение приблизительно 54 предшествующих месяцев или в течение приблизительно 60 предшествующих месяцев).
[0085] У людей снижение функции бета-клеток, которое происходит до и после постановки диагноза T1D, может быть измерено с применением диагностических соединений-маркеров. Например, С-пептид продуцируется в равных количествах с инсулином (во время ферментативного расщепления про-инсулина) и поэтому может быть использован в качестве меры эндогенной секреции инсулина (в том числе у пациентов, получающих лечение инсулином). С-пептид ранее уже использовался в клиническом ведении пациентов с диабетом, а системы анализа для измерения уровня С-пептида известны специалистам в данной области. См., например, Jones A.G. and Hattersley A.T., Diabetic Medicine 2013, 30: 803-817; Little RR et al., Clin. Chem. 2008, 54: 1023-1026; Wiedmeyer et al., Clin. Chem. 2007, 53: 784-787.
[0086] Уровни С-пептида могут быть измерены в нмоль/л (где 1 нмоль/л составляет 1000 пмоль/л и эквивалентен приблизительно 3 нг/мл). С-пептид может быть измерен в крови или моче субъекта. Уровни С-пептида в крови могут быть определены у пациентов, находящихся в состоянии не натощак (С-пептид в произвольный момент), у субъектов в состоянии натощак (С-пептид натощак) или у субъектов, стимулированных диетическим стимулятором, таким как сбалансированная жидкая питательная смесь или глюкагон (стимулированный С-пептид). С-пептид в моче может быть измерен как общее количество С-пептида, секретируемого субъектом в течение 24 часов. Часто С-пептид, содержащийся в моче, измеряется как соотношение между С-пептидом и креатинином.
[0087] В некоторых вариантах реализации субъект (например, человек) перед введением композиции согласно настоящему изобретению (например, субъект с недавно выявленным T1D) имеет концентрацию С-пептида в крови натощак менее приблизительно 1 нмоль/л, но по меньшей мере приблизительно 0,5 нмоль/л, по меньшей мере приблизительно 0,4 нмоль/л, по меньшей мере приблизительно 0,3 нмоль/л или по меньшей мере приблизительно 0,2 нмоль/л. В других вариантах реализации субъект (например, человек) имеет стимулированную концентрацию С-пептида в крови менее приблизительно 4 нмоль/л, но по меньшей мере приблизительно 1 нмоль/л, по меньшей мере приблизительно 0,9 нмоль/л, по меньшей мере приблизительно 0,8 нмоль/л, по меньшей мере приблизительно 0,7 нмоль/л, по меньшей мере приблизительно 0,6 нмоль/л или по меньшей мере приблизительно 0,5 нмоль/л. В других вариантах реализации субъект (например, человек) с недавно выявленным T1D имеет соотношение С-пептид : креатинин в моче после приема пищи (нмоль/ммоль) менее 4, но по меньшей мере приблизительно 1, по меньшей мере приблизительно 0,9, по меньшей мере приблизительно 0,8, по меньшей мере приблизительно 0,7, по меньшей мере приблизительно 0,6, по меньшей мере приблизительно 0,5, по меньшей мере приблизительно 0,4 или по меньшей мере приблизительно 0,3.
[0088] В других вариантах реализации субъект с недавно выявленным T1D (например, пациент-человек) может быть определен путем измерения аутоантител к инсулину (IAA) в сыворотке или крови субъекта. В некоторых примерах субъекты дают положительный результат теста на IAA. Концентрация IAA в сыворотке также может использоваться для измерения прогрессирования заболевания или прогресса в лечении. Способы измерения аутоантител к инсулину были описаны ранее. См., например, Demeester et al., Diabetes Care 2015, 38(4): 644-651.
Слизистая оболочка
[0089] Термин "слизистая оболочка" или "мембрана слизистой оболочки" используется в настоящей заявке в соответствии с его принятым в данной области техники значением. "Слизистой оболочкой" может быть любая слизистая оболочка, находящаяся в организме, такая как слизистая оболочка полости рта, слизистая оболочка прямой кишки, слизистая оболочка желудка, слизистая оболочка кишечника, слизистая оболочка уретры, слизистая оболочка влагалища, слизистая оболочка глаз, слизистая оболочка щеки, бронхиальная или легочная слизистая оболочка и слизистая оболочка носа или обонятельной области.
[0090] Используемый в настоящей заявке термин "доставка в слизистую оболочку" используется в соответствии с его принятым в данной области техники значением, то есть подразумевает доставку к слизистой оболочке, например, посредством приведения в контакт композиции по настоящему изобретения со слизистой оболочкой. Пероральная доставка в слизистую оболочку включает трансбуккальные, сублингвальные и десневые пути доставки. Соответственно, в некоторых вариантах реализации "доставка в слизистую оболочку" включает желудочную доставку, кишечную доставку, ректальную доставку, трансбуккальную доставку, пульмональную доставку, внутриглазную доставку, назальную доставку, вагинальную доставку и пероральную доставку.
[0091] Термин "мукозная толерантность (mucosal tolerance)" относится к ингибированию специфической иммунной реакции на антиген у субъекта-млекопитающего (например, человека), после того как субъект взаимодействовал с указанным антигеном посредством слизистой оболочки. Как правило, указанная мукозная толерантность является системной толерантностью. Низкодозовая пероральная толерантность - это пероральная толерантность, вызванная низкими дозами антигенов, которая характеризуется активным иммунным подавлением, опосредованным чувствительными к циклофосфамиду регуляторными Т-клетками, которые могут передавать толерантность ранее не подвергавшимся воздействию хозяевам. Высокодозовая пероральная толерантность - это пероральная толерантность, вызванная высокими дозами антигенов, она нечувствительна к циклофосфамиду и приводит к стимуляции гипореактивности Т-клеток посредством анергии и/или уничтожению антигенспецифических Т-клеток. Разница в чувствительности к циклофосфамиду может служить для выявления различия между низкодозовой и высокодозовой толерантностью. Strobel et al., Immunology 1983, 49:451-456. Примером пероральной толерантности является низкодозовая пероральной толерантность, описанная в работе Mayer and Shao, Nature Rev. Immunol. 2004, 4:407-419.
Иммуномодулирующее соединение
[0092] В некоторых вариантах реализации настоящее изобретение предлагает способы лечения T1D, при которых субъект не получает одновременно дополнительного иммуномодулирующего соединения (то есть в дополнение к IL-2). Таким образом, субъекту вводят только T1D-специфический антиген и IL-2.
[0093] В некоторых вариантах реализации настоящее изобретение предлагает способы лечения T1D, при которых субъект одновременно получает дополнительное иммуномодулирующее соединение. Таким образом, субъекту вводят T1D-специфический антиген, IL-2 и дополнительное иммуномодулирующее соединение.
[0094] Термины "иммуномодулирующее соединение" или "иммуномодулятор" используются в настоящей заявке в соответствии с их принятым в данной области техники значениям. Иммуномодулирующим соединением может быть любое иммуномодулирующее соединение, известное специалисту в данной области. Специалист в данной области техники может предпочесть включать или не включать иммуномодулирующее соединение в описанное в настоящей заявке лечение. Решение о включении иммуномодулирующего соединения в схему лечения может быть определено по результатам описанного в настоящей заявке лечения, генетических и/или физиологических характеристик субъекта, среди прочих факторов.
[0095] В некоторых вариантах реализации иммуномодулирующее соединение представляет собой толерогенное соединение. Стимуляция толерантности может быть получена, например, путем стимулирования регуляторных Т-клеток или косвенным способом, например, путем активации незрелых дендритных клеток для получения толерантных дендритных клеток и/или ингибирования Th2-иммунного ответа, стимулирующего экспрессию факторов "ко-стимуляции" на зрелых дендритных клетках. Иммуномодулирующие и иммуносупрессивные соединения известны специалисту в данной области техники и включают, среди прочего, бактериальные метаболиты, такие как спергуалин, метаболиты грибов и стрептомицетов, такие как такролимус или циклоспорин, иммуносупрессивные цитокины, такие как IL- 4, IL-10, IFNα, TGFβ (в качестве селективного адъюванта для регуляторных Т-клеток) Flt3L, TSLP и Rank-L (в качестве селективных толерогенных индукторов DC (дендритных клеток)), антитела и/или антагонисты, такие как анти-CD40b, анти-CD25, анти-CD20, анти-IgE, анти-CD3 и белки, пептиды или слитые белки, такие как слитый белок-агонист CTL-41 g или CTLA-4. В некоторых вариантах реализации иммуномодулирующее соединение представляет собой иммуносупрессивное соединение. Иммуносупрессивное соединение может быть иммуносупрессивным цитокином или антителом. В других вариантах реализации иммуносупрессивный цитокин представляет собой усиливающий толерантность цитокин или антитело. Специалисту в данной области техники будет понятно, что термин "иммуномодулирующее соединение" также включает их функциональные гомологи. Функциональный гомолог представляет собой молекулу, имеющую, по существу, ту же или аналогичную функцию для целевого предназначения, но может отличаться структурно. В некоторых примерах иммуномодулирующее соединение представляет собой анти-CD3 или его функциональный гомолог.
LAB
[0096] В настоящем изобретении предложено применение генетически модифицированных молочнокислых бактерий (LAB). Штамм LAB может представлять собой виды Lactococcus, виды Lactobacillus, виды Bifidobacterium, виды Streptococcus или виды Enterococcus.
[0097] Используемый в настоящей заявке термин Lactococcus или Lactobacillus не ограничивается конкретным видом или подвидом, но подразумевает включение любого из видов или подвидов Lactococcus или Lactobacillus. Примеры видов Lactococcus включают Lactococcus garvieae, Lactococcus lactis, Lactococcus piscium, Lactococcus plantarum и Lactococcus raffinolactis. В некоторых примерах Lactococcus lactis представляет собой Lactococcus lactis subsp. cremoris, Lactococcus lactis subsp. hordniae или Lactococcus lactis subsp. lactis.
[0098] Примеры видов Lactobacillus включают Lactobacillus acetotolerans, Lactobacillus acidophilus, Lactobacillus agilis, Lactobacillus algidus, Lactobacillus alimentarius, Lactobacillus amylolyticus, Lactobacillus amylophilus, Lactobacillus amylovorus, Lactobacillus animalis, Lactobacillus aviarius, Lactobacillus aviarius subsp. araffinosus, Lactobacillus aviarius subsp. aviarius, Lactobacillus bavaricus, Lactobacillus bifermentans, Lactobacillus brevis, Lactobacillus buchneri, Lactobacillus bulgaricus, Lactobacillus carnis, Lactobacillus casei, Lactobacillus casei subsp. alactosus, Lactobacillus casei subsp. casei, Lactobacillus casei subsp. pseudoplantarum, Lactobacillus casei subsp. rhamnosus, Lactobacillus casei subsp. tolerans, Lactobacillus catenaformis, Lactobacillus cellobiosus, Lactobacillus collinoides, Lactobacillus confusus, Lactobacillus coryniformis, Lactobacillus coryniformis subsp. coryniformis, Lactobacillus coryniformis subsp. torquens, Lactobacillus crispatus, Lactobacillus curvatus, Lactobacillus curvatus subsp. curvatus, Lactobacillus curvatus subsp. melibiosus, Lactobacillus delbrueckii, Lactobacillus delbrueckii subsp.bulgaricus, Lactobacillus delbrueckii subsp. delbrueckii, Lactobacillus delbrueckii subsp. lactis, Lactobacillus divergens, Lactobacillus farciminis, Lactobacillus fermentum, Lactobacillus fornicalis, Lactobacillus fructivorans, Lactobacillus fructosus, Lactobacillus gallinarum, Lactobacillus gasseri, Lactobacillus graminis, Lactobacillus halotolerans, Lactobacillus hamsteri, Lactobacillus helveticus, Lactobacillus heterohiochii, Lactobacillus hilgardii, Lactobacillus homohiochii, Lactobacillus iners, Lactobacillus intestinalis, Lactobacillus jensenii, Lactobacillus johnsonii, Lactobacillus kandleri, Lactobacillus kefiri, Lactobacillus kefiranofaciens, Lactobacillus kefirgranum, Lactobacillus kunkeei, Lactobacillus lactis, Lactobacillus leichmannii, Lactobacillus lindneri, Lactobacillus malefermentans, Lactobacillus mali, Lactobacillus maltaromicus, Lactobacillus manihotivorans, Lactobacillus minor, Lactobacillus minutus, Lactobacillus mucosae, Lactobacillus murinus, Lactobacillus nagelii, Lactobacillus oris, Lactobacillus panis, Lactobacillus parabuchneri, Lactobacillus paracasei, Lactobacillus paracasei subsp. paracasei, Lactobacillus paracasei subsp. tolerans, Lactobacillus parakefiri, Lactobacillus par alimentarius, Lactobacillus paraplantarum, Lactobacillus pentosus, Lactobacillus perolens, Lactobacillus piscicola, Lactobacillus plantarum, Lactobacillus pontis, Lactobacillus reuteri, Lactobacillus rhamnosus, Lactobacillus rimae, Lactobacillus rogosae, Lactobacillus ruminis, Lactobacillus sakei, Lactobacillus sakei subsp. camosus, Lactobacillus sakei subsp. sakei, Lactobacillus salivarius, Lactobacillus salivarius subsp.salicinius, Lactobacillus salivarius subsp.salivarius, Lactobacillus sanfranciscensis, Lactobacillus sharpeae, Lactobacillus suebicus, Lactobacillus trichodes, Lactobacillus uli, Lactobacillus vaccinostercus, Lactobacillus vaginalis, Lactobacillus viridescens, Lactobacillus vitulinus, Lactobacillus xylosus, Lactobacillus yamanashiensis, Lactobacillus yamanashiensis subsp. mali, Lactobacillus yamanashiensis subsp. Yamanashiensis, Lactobacillus zeae, Bifidobacterium adolescentis, Bifidobacterium angulatum, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium catenulatum, Bifidobacterium longum и Bifidobacterium infantis. В некоторых примерах LAB представляет собой Lactococcus lactis (LL).
[0099] В других примерах бактерия выбрана из группы, состоящей из Enterococcus alcedinis, Enterococcus aquimarinus, Enterococcus asini, Enterococcus avium, Enterococcus caccae, Enterococcus camelliae, Enterococcus canintestini, Enterococcus canis, Enterococcus casseliflavus, Enterococcus cecorum, Enterococcus columbae, Enterococcus devriesei, Enterococcus diestrammenae, Enterococcus dispar, Enterococcus durans, Enterococcus eurekensis, Enterococcus faecalis, Enterococcus faecium, Enterococcus gallinarum, Enterococcus gilvus, Enterococcus haemoperoxidus, Enterococcus hermanniensis, Enterococcus hirae, Enterococcus italicus, Enterococcus lactis, Enterococcus lemanii, Enterococcus malodoratus, Enterococcus moraviensis, Enterococcus mundtii, Enterococcus olivae, Enterococcus pollens, Enterococcus phoeniculicola, Enterococcus plantarum, Enterococcus pseudoavium, Enterococcus quebecensis, Enterococcus rqffinosus, Enterococcus ratti, Enterococcus rivorum, Enterococcus rotai, Enterococcus saccharolyticus, Enterococcus silesiacus, Enterococcus solitarius, Enterococcus sulfureus, Enterococcus termitis, Enterococcus thailandicus, Enterococcus ureasiticus, Enterococcus ureilyticus, Enterococcus viikkiensis, Enterococcus villorum и Enterococcus xiangfangensis,
[00100] В других примерах бактерия выбрана из группы, состоящей из Streptococcus agalactiae, Streptococcus anginosus, Streptococcus bovis, Streptococcus canis, Streptococcus constellatus, Streptococcus dysgalactiae, Streptococcus equinus, Streptococcus iniae, Streptococcus intermedius, Streptococcus milleri, Streptococcus mitis, Streptococcus mutans, Streptococcus oralis, Streptococcus parasanguinis, Streptococcus peroris, Streptococcus pneumoniae, Streptococcus pseudopneumoniae, Streptococcus pyogenes, Streptococcus ratti, Streptococcus salivarius, Streptococcus tigurinus, Streptococcus thermophilus, Streptococcus sanguinis, Streptococcus sobrinus, Streptococcus suis, Streptococcus uberis, Streptococcus vestibularis, Streptococcus viridans и Streptococcus zooepidemicus.
[00101] В качестве примера штаммом LAB может быть Lactococcus lactis или любой из его подвидов, включая Lactococcus lactis subsp.cremoris, Lactococcus lactis subsp. hordniae, Lactococcus lactis и Lactococcus lactis subsp. lactis. В другом аспекте штамм LAB может представлять собой биологически ограниченную из соображений безопасности систему, такую как бесплазмидный штамм MG1363 Lactococcus lactis, утративший способность к нормальному росту и производству кислоты в молоке, как описано в работе Gasson, M.J. (1983) J. Bacterid. 154:1-9; или ауксотрофные по треонину и пиримидину производные штаммы L. lactis, как описано в работе Sorensen et al. (2000) Appl. Environ. Microbiol. 66:1253-1258; и Glenting et al. (2002) 68:5051-5056.
[00102] Рекомбинантная бактериальная система векторов-хозяев может быть биологически ограниченной из соображений безопасности системой. Биологическое ограничение из соображений безопасности известно специалистам в данной области техники и может быть реализовано путем введения ауксотрофной мутации, например, суицидной ауксотрофной мутации, такой как мутация ThyA, или ее эквивалентов. В качестве альтернативы, биологическое ограничение может быть реализовано на уровне плазмиды, несущей ген, кодирующий полипептид IL-2 или вариант IL-2, например, с применением нестабильной эписомальной конструкции, которая утрачивается через несколько поколений. При необходимости, для обеспечения высокого уровня ограничения можно объединить несколько уровней ограничения, таких как нестабильность плазмид и ауксотрофия.
Конструкции
[00103] Согласно настоящей заявке, LAB доставляет полипептид IL-2 и T1D-специфический антиген к целевой области, то есть, к слизистой оболочке. Например, LAB экспрессирует полипептид IL-2, после чего полипептид IL-2 экспонируется на поверхности клетки (если используется мембраносвязанная форма IL-2) или секретируется (если используется секретируемая форма IL-2). Следовательно, в частном варианте реализации, LAB, такая как L. lactis, содержит вектор экспрессии, способный экспрессировать полипептид IL-2 и T1D-специфический антиген, внутриклеточно. Например, эти полипептиды экспонируются на поверхности клетки в условиях, присутствующих в целевой слизистой оболочке, например, в желудочно-кишечном тракте. LAB может содержать векторы экспрессии, способные экспрессировать полипептид IL-2 внутриклеточно, так что полипептид IL-2 экспонируется на поверхности клетки в степени, достаточной для обеспечения IL-2 в низкой дозе, эффективного для лечения T1D у реципиента. При применении штаммов LAB, экспрессирующих более значительные количества полипептида IL-2 и T1D-специфического антигена, для лечения T1D могут потребоваться менее частые и более низкие дозы LAB. Таким образом, специалист в данной области техники может регулировать количество штамма LAB, чтобы обеспечить доставку желаемого количества полипептида IL-2 и T1D-специфического антигена.
[00104] Обычно система экспрессии будет содержать генетическую конструкцию, содержащую по меньшей мере одну нуклеотидную последовательность, кодирующую полипептид IL-2 и/или полипептид TD1-специфического антигена, обычно функционально связанный с промотором, способным направлять экспрессию этой последовательности (последовательностей) в микроорганизме-хозяине. Соответственно, экспрессируемый полипептид IL-2 и T1D-антиген могут быть кодированы последовательностью нуклеиновой кислоты, которая адаптирована к использованию предпочтительных кодонов хозяином. Эта конструкция может дополнительно содержать (все) другие подходящие элементы, включая энхансеры, последовательности инициации транскрипции, сигнальные последовательности, репортерные гены, последовательности терминации транскрипции и т.д., функциональные в выбранном хозяине, что известно специалисту в данной области.
[00105] Такая конструкция обычно выполнена в форме, подходящей для трансформации хозяина и/или в форме, которая может стабильно поддерживаться в хозяине, например, вектор, плазмида или минихромосома. Подходящие векторы, содержащие нуклеиновую кислоту для введения в штаммы LAB, например, L. lactis, могут быть выбраны или сконструированы, и содержат соответствующие регуляторные последовательности, включая промоторные последовательности, терминаторные фрагменты, энхансерные последовательности, маркерные гены и другие последовательности сообразно обстоятельствам. Векторами могут быть плазмиды, вирусные векторы, например, фаг или фагмида в зависимости от ситуации. Более подробную информацию можно найти, например, в работе Molecular Cloning: а Laboratory Manual 2nd edition, Sambrook et al., 1989, Cold Spring Harbor Laboratory Press.
[00106] Многие известные методики и протоколы для манипулирования нуклеиновой кислотой, например, для получения конструкций нуклеиновых кислот, мутагенеза, секвенирования, введения ДНК в клетки и экспрессии генов и анализа белков, подробно описаны в Short Protocols in Molecular Biology, Second Edition, Ausubel et al., eds., John Wiley & Sons, 1992. В одном варианте реализации кодирующая последовательность для полипептида IL-2 может содержаться в опероне, то есть в конструкции нуклеиновой кислоты для полицистронной экспрессии. В опероне транскрипция с промотора обеспечивает мРНК, содержащую более чем одну кодирующую последовательность, каждая из которых имеет свой собственный надлежаще расположенный сайт связывания рибосомы против хода транскрипции. Таким образом, из одной мРНК может быть транслирован более чем один полипептид. Применение оперона позволяет координировать экспрессию полипептида IL-2 и полипептида T1D-специфического антигена. Полицистронные системы экспрессии в бактериальных клетках-хозяевах описаны, например, в заявке на патент США №2014/0105863.
[00107] Для получения стабильно трансфицированных штаммов LAB, ген, кодирующий полипептид IL-2, и/или ген T1D-специфического антигена, можно интегрировать в геном LAB-хозяина. Способы создания стабильно трансфицированных штаммов LAB известны в данной области. Например, полипептид IL-2 и/или ген T1D-специфического антигена могут быть клонированы в геном хозяина посредством гомологичной рекомбинации. Как правило, незаменимый ген хозяина разрушается в результате гомологичной рекомбинации, например, делеции гена, одной или более аминокислотных замен, приводящих к неактивной форме белка, кодируемого незаменимым геном, или мутации со сдвигом рамки, приводящей к усеченной форме белка, кодируемого незаменимым геном. В одном варианте реализации незаменимым геном является ген thyA. Пример методики описан в WO 02/090551. Выбор трансформирующей плазмиды особо не ограничен, при условии, что она не дополняет разрушенный незаменимый ген, например, ген thyA. Плазмида может быть самореплицирующейся, обычно несущей один или несколько целевых генов и один или несколько маркеров устойчивости, или плазмида может быть интегративной плазмидой. В последнем случае сама интегративная плазмида может быть использована для разрушения незаменимого гена путем интеграции в локусе незаменимого гена, например, сайта thyA, в результате чего функция незаменимого гена, например, гена thyA, нарушается. Как правило, незаменимый ген, такой как ген thyA, заменяется двойной гомологичной рекомбинацией с помощью кассеты, содержащей целевой ген или гены, фланкированные целевыми последовательностями, которые нацелены на инсерцию в незаменимый ген, такой как целевой сайт thyA. Понятно, что эти целевые последовательности являются достаточно длинными и достаточно гомологичными, чтобы обеспечить интеграцию рассматриваемого гена в целевой сайт.
[00108] Генетическая конструкция, кодирующая полипептид IL-2 и/или T1D-специфический антиген, может, таким образом, присутствовать в клетке-хозяине вне хромосомы, обычно автономно реплицируясь с использованием собственной точки начала репликации или может быть интегрирована в геномную ДНК LAB, например, хромосому Lactococcus. В последнем случае может быть интегрирована одна или несколько копий нуклеиновой кислоты; интеграция может происходить в случайном участке хромосомы или, как описано выше, на ее заранее пределенном участке, например, в локусе thyA бактерии Lactococcus, например, Lactococcus lactis.
[00109] Следовательно, генетическая конструкция, кодирующая полипептид IL-2 и/или T1D-специфический антиген, может дополнительно содержать последовательности, сконфигурированные для реализации инсерции данной генетической конструкции в геном, например, хромосому, клетки LAB-хозяина.
[00110] В одном из примеров, инсерции генетической конструкции в конкретные участки в пределах генома, например, хромосому, клетки-хозяина LAB может способствовать гомологичная рекомбинация. Например, описанные в настоящей заявке генетические конструкции могут содержать одну или несколько областей гомологии с указанным сайтом интеграции в геноме, например, хромосоме, клетки-хозяина LAB. Последовательность в указанном сайте генома, например, хромосомы, может быть естественной, то есть, встречающейся в природе, или может быть экзогенной последовательностью, введенной ранее методами генной инженерии.
[00111] Например, область (области) гомологии может составлять по меньшей мере 50 п.н., 100 п.н., 200 п.н., 300 п.н., 400 п.н., 500 п.н., 600 п.н. 700 п.н., 800 п.н., 900 п.н., 1000 п.н. или более.
[00112] В одном из примеров могут быть включены две области гомологии, каждая из которых фланкирует каждую сторону соответствующих единиц экспрессии, присутствующих в описанных в настоящей заявке генетических конструкциях. Такая конфигурация может эффективно обеспечивать инсерцию соответствующих последовательностей, то есть, по меньшей мере тех, которые кодируют и осуществляют экспрессию целевого антигена, в клетки-хозяева. Способы выполнения гомологичной рекомбинации, особенно у бактериальных хозяев, и выбор рекомбинантов, как правило, известны в данной области.
[00113] Способы трансформации штаммов LAB известны специалисту в данной области; например, это трансформация протопластов и электропорация.
[00114] Высокая степень экспрессии может быть достигнута при применении гомологичной экспрессии и/или сигналов секреции на векторах экспрессии, присутствующих в LAB, например, L. lactis. Сигналы экспрессии известны специалисту в данной области техники. Вектор экспрессии может быть оптимизирован для экспрессии в зависимости от LAB, например, L. lactis, в которую он включен. Например, известны специфические векторы экспрессии, которые обеспечивают достаточные уровни экспрессии в Lactococcus, Lactobacillus lactis, casei и plantarum. Кроме того, известны системы, которые были разработаны для экспрессии гетерологичных антигенов в непатогенной, не колонизирующей, неинвазивной пищевой бактерии Lactococcus lactis (например, патент Великобритании GB 2278358 B). Пример конструкции содержит многокопийный вектор экспрессии, описанный в PCT/NL95/00135 (WO 96/32487), в котором указана нуклеотидная последовательность, кодирующая IL-2-полипептид и T1D-специфический антиген. Такая конструкция может быть подходящей для экспрессии целевого антигена в молочнокислой бактерии, в частности в Lactobacillus, при высоком уровне экспрессии, а также может быть успешно использована для направления экспрессированного продукта на поверхность бактериальной клетки. Такие конструкции (например, PCT/NL95/00135) могут характеризоваться тем, что последовательности нуклеиновой кислоты, кодирующей полипептид IL-2 и/или TlD-специфический антиген, предшествует 5'-нетранслируемая последовательность нуклеиновой кислоты, содержащая по меньшей мере минимальную последовательность, необходимую для распознавания рибосом и стабилизации РНК. За этим может следовать кодон инициации трансляции, за которым может (непосредственно) следовать фрагмент по меньшей мере пяти ко донов 5-концевой части транслируемой последовательности нуклеиновой кислоты гена молочнокислой бактерии или структурного или функционального эквивалента этого фрагмента. Этот фрагмент также может контролироваться промотором. Один из аспектов настоящего изобретения обеспечивает способ, позволяющий регулировать на высоком уровне экспрессию гетерологичных генов в хозяине и сцепление экспрессии с секрецией. В другом варианте реализации РНК-полимеразу Т7-бактериофага и ее опосредованный промотор используют для создания мощной системы экспрессии в соответствии с WO 93/17117. В одном варианте реализации плазмида экспрессии может быть получена из pTl NX.
[00115] Используемый в настоящей заявке промотор обычно конститутивно экспрессируется в бактерии. Применение конститутивного промотора позволяет избежать необходимости обеспечения индуктора или другого регулятивного сигнала для реализации экспрессии. Как правило, промотор регулирует экспрессию на уровне, при котором бактериальная клетка-хозяин остается жизнеспособной, то есть сохраняет некоторую метаболическую активность, даже если рост не поддерживается. Благодаря этому такая экспрессия может быть на низком уровне. Например, когда продукт экспрессии накапливается внутриклеточно, уровень экспрессии может приводить к накоплению продукта экспрессии в количестве менее 10% клеточного белка, в некоторых случаях - приблизительно 5% или менее приблизительно 5%, например, приблизительно 1-3%. Промотор может быть гомологичен используемой бактерии, то есть встречаться в этой бактерии в природе. Например, лактококковый промотор может быть использован в Lactococcus. Примером промотора для применения в Lactococcus lactis (или других лактококках) является "Р1", полученный из хромосомы Lactococcus lactis (Waterfield NR et al., Gene 1995, 165(1):9-15). Другим примером промотора является промотор usp45. Другие подходящие промоторы описаны в патенте США №8,759,088 и в заявке на патент США №2014/0105863.
[00116] Конструкция или конструкции нуклеиновой кислоты могут содержать секреторную сигнальную последовательность. Таким образом, в некоторых вариантах реализации нуклеиновая кислота, кодирующая IL-2 и/или T1D-специфический антиген, может обеспечивать секрецию полипептидов, например, посредством надлежащего сцепления последовательности нуклеиновой кислоты, кодирующей сигнальную последовательность, с последовательностью нуклеиновой кислоты, кодирующей полипептид). Способность бактерии, несущей нуклеиновую кислоту, секретировать антиген, может быть протестирована in vitro в условиях культивирования, поддерживающих жизнеспособность организма. Примеры секреторных сигнальных последовательностей включают последовательности, активные в штаммах LAB. Такие последовательности могут включать лидер секреции α-амилазы Bacillus amyloliquetaciens или лидер секреции фермента стафилокиназы, секретируемого некоторыми штаммами Staphylococcus, который, как известно, функционирует как в грамположительных, так и в грамотрицательных хозяевах (Rapoport, Current Opinion in Biotechnology 1990, 1: 21-27), или лидерные последовательности из множества других ферментов Bacillus или белков S-слоев (см. стр. 341-344 Harwood and Cutting, "Molecular Biological Methods for Bacillus," John Wiley & Co. 1990). В одном варианте реализации указанный сигнал секреции получают из usp45 (Van Asseldonk et al. (1993) Mol. Gen. Genet. 240:428-434). В некоторых вариантах реализации полипептид IL-2 или вариант IL-2 могут быть секретированы конститутивно.
Полипептиды IL-2
[00117] Примеры полипептидов IL-2 включают IL-2 человека дикого типа в мембраносвязанной или секретируемой форме, и любой вариант полипептида IL-2, например полипептиды, имеющие идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 90%, или по меньшей мере приблизительно на 98% с последовательностью IL-2 дикого типа или соответствующего зрелого полипептида IL-2. Пример аминокислотной последовательности IL-2 человека дикого типа представлен в SEQ ID NO: 1, тогда как пример последовательности нуклеиновой кислоты, кодирующей IL-2, представлен в SEQ ID NO: 2. Зрелый IL-2 человека дикого типа представлен в SEQ ID NO: 3.
[00118] Сигнальный пептид для IL-2 (SEQ ID NO: 4) подчеркнут и представляет собой аминокислоты 1-20 из SEQ ID NO: 1. Сигнальный пептид IL-2 может быть замещен бактериальной секреторной сигнальной последовательностью (например, SSusp45), описанной в настоящей заявке. Пример нуклеотидной последовательности согласно этому варианту реализации представлен в SEQ ID NO: 5.
[00119] Термин "вариант IL-2" включает полипептиды IL-2, характеризующиеся инсерциями, делециями, заменами и/или модификациями аминокислот в одном или нескольких сайтах нативной полипептидной цепи IL-2. В соответствии с этим определением любые такие инсерции, делеции, замены и модификации приводят к варианту полипептида IL-2, сохраняющему по меньшей мере некоторую активность связывания с IL-2RP. Примеры вариантов включают полипептиды с заменами 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот. Варианты IL-2 могут иметь консервативные модификации и замены в других положениях IL-2 (то есть такие, которые оказывают минимальное влияние на вторичную или третичную структуру варианта полипептида). Такие консервативные замены включают замены, которые описаны в Dayhoff The Atlas of Protein Sequence and Structure 5' (1978) и Argos EMBOJ., 8:779-785 (1989). Например, аминокислоты, принадлежащие к одной из следующих групп, представляют собой консервативные изменения: Группа I: ala, pro, gly, gln, asn, ser, thr; Группа II: cys, ser, tyr, thr; Группа III: val, ile, leu, met, ala, phe; Группа IV: lys, arg, his; Группа V: phe, tyr, tip, his; и Группа VI: asp, glu.
[00120] В некоторых примерах IL-2 представляет собой вариант, описанный в патенте США №4518584, в котором цистеин, обычно встречающийся в положении 125 дикого типа или нативной молекулы, был заменен нейтральной аминокислотой, такой как серии или аланин. В качестве альтернативы или дополнительно вариант IL-2 может быть таким, как описано в заявке США №06/810,656, поданной 17 декабря 1985 г., в которой метионин, обычно встречающийся в положении 104 дикого типа или нативной молекулы, был заменен на нейтральную аминокислоту, такую как аланин. В некоторых примерах у варианта IL-2 могут быть удалены одна или несколько из первых пяти N-концевых аминокислот нативного IL 2. Также были получены мутанты IL-2 с уменьшенной аффинностью связывания с CD 122 (для достижения более низкой токсичности IL-2), например, BAY 50-4798 (содержащий мутацию N88R IL 2).
[00121] Другие формы IL-2, которые могут быть использованы, включают варианты последовательности IL-2, такие как варианты, присутствующие в алдеслейкине или пролейкине (Prometheus Laboratories), тецелейкине (Roche), биолейкине (Glaxo), а также варианты, описанные в Taniguchi etal., Nature 1983, 302(5906):305-10 и Devos et al, Nucleic Acids Res. 1983, 11(13): 4307-23; Европейские патентные заявки №№. 91,539 и 88,195; Патент США №. 4,518,584; публикация патента США №2012/0244112; Патент США №. 7,569,215; Патент США №. 5,229,109; публикация патента США №2006/0269515; публикация патента ЕР №. ЕР 1730184 А2; и публикация РСТ WO 2005/086751.
[00122] В некоторых вариантах реализации вариант IL-2 имеет пониженную способность связываться с высокоаффинным рецептором IL-2, но сохраняет сродство варианта IL-2 для связывания рецептора IL-2 со средней аффинностью по сравнению с полипептидом IL-2 дикого типа. В некоторых вариантах реализации зрелый полипептид IL-2 характеризуется одной, двумя или тремя аминокислотными заменами, например, когда замещенные аминокислотные остатки выбраны из L72, F42 и Y45. В некоторых вариантах реализации вариант IL-2 характеризуется заменой L72, например, содержит первую аминокислотную замену, выбранную из группы, состоящей из L72G, L72A, L72S, L72T, L72Q, L72E, L72N, L72D, L72R и L72K. Вариант IL-2 может быть охарактеризован заменой F42, например, содержит вторую аминокислотную замену, выбранную из группы, состоящей из F42A, F42G, F42S, F42T, F42Q, F42E, F42N, F42D, F42R и F42K. В других вариантах реализации вариант IL-2 характеризуется заменой Y45, например, содержит третью аминокислотную замену, выбранную из группы, состоящей из Y45A, Y45G, Y45S, Y45T, Y45Q, Y45E, Y45N, Y45D, Y45R и Y45K. Вариант IL-2 настоящего изобретения может содержать любую комбинацию вышеуказанных первой, второй и третьей аминокислотных замен.
[00123] Варианты IL-2, описанные в настоящей заявке, могут быть приблизительно на 75%, приблизительно на 80%, приблизительно на 85%, приблизительно на 90%, приблизительно на 95%, приблизительно на 96%, приблизительно на 97%, приблизительно на 98% или приблизительно на 99% идентичными с соответствующим IL-2 дикого типа, при условии, что такой вариант полипептида IL-2 сохраняет некоторую активность IL-2 (функциональный полипептид).
[00124] Процент идентичности полипептидных последовательностей может быть рассчитан с применением коммерчески доступных алгоритмов, которые сравнивают эталонную последовательность (например, SEQ ID NO: 1 настоящего изобретения) с искомой последовательностью.
[00125] Специалисту в данной области техники будет понятно, что оптимальное количество IL-2, которое должно быть доставлено субъекту с применением способов настоящего изобретения, варьирует, например, в зависимости от LAB, экспрессирующей полипептид IL-2, и например, эффективности промотора, используемого в генетической конструкции. Обычно LAB можно вводить в количестве, эквивалентном конкретному количеству экспрессированного полипептида IL-2, или в количестве, которое обеспечивает желаемый ФК-профиль для соответствующего полипептида IL-2 у соответствующего субъекта. В качестве примера ежедневные дозы полипептида IL-2 составляют от приблизительно 10 фг до приблизительно 100 мкг активного полипептида в сутки. Другие диапазоны доз в качестве примера составляют от приблизительно 1 пг до приблизительно 100 мкг в сутки; или от приблизительно 1 нг до приблизительно 100 мкг в сутки.
[00126] Приведенные выше дозы могут быть реализованы путем введения субъекту эффективных количеств микроорганизма в сутки, причем этот микроорганизм адаптирован для экспрессии достаточного количества IL-2 для достижения желаемой дозы, такой как вышеуказанные. LAB, секретирующая полипептид IL-2, может быть доставлена в дозе от приблизительно 104 колониеобразующих единиц (КОЕ) до приблизительно 1012 КОЕ в сутки, в частности от приблизительно 106 КОЕ до приблизительно 1012 КОЕ в сутки, более конкретно, от приблизительно 109 КОЕ до приблизительно 1012 КОЕ в сутки. Количество секретируемого полипептида IL-2 в КОЕ может быть определено, например, в соответствии со способами, описанными в работе Steidler et al., Science 2000; 289(5483): 1352-1355, или с помощью ИФА. Например, LAB может секретировать по меньшей мере от 1 нг до 1 мкг активного полипептида на 109 КОЕ. На основании этого специалист может вычислить диапазон количества полипептида IL-2, секретируемого при других дозах в КОЕ.
[00127] Каждая из вышеуказанных доз/диапазонов доз может вводиться в рамках любой схемы дозирования, описанной в настоящей заявке. Ежедневную дозу можно вводить в виде 1, 2, 3, 4, 5 или 6 порций в течение суток. Кроме того, ежедневные дозы можно вводить в течение любого количества дней с любым количеством периодов отдыха между периодами введения. Например, субъекту можно вводить микроорганизм в дозе, эквивалентной приблизительно от 0,1 млн. ME до приблизительно 3 млн. МЕ/сут или через день, в течение периода, составляющего по меньшей мере приблизительно 1 неделю, по меньшей мере приблизительно 2 недели по меньшей мере приблизительно 3 недели по меньшей мере приблизительно 4 недели, по крайней мере, приблизительно 5 недель или, по крайней мере, приблизительно 6 недель. В некоторых примерах субъекту вводят LAB в дозе, эквивалентной приблизительно от 0,1 до приблизительно 5 млн. МЕ/сут, или от приблизительно 0,3 до приблизительно 3 млн. ME, например, в течение приблизительно 5 дней, приблизительно 7 дней или приблизительно 14 дней. В качестве примера дозы описаны, например, в Hartemann et al., Lancet Diabetes Endocrinol. 2013, 1(4): 295-305.
Полипептиды T1D-специфичного антигена
[00128] LAB по настоящему изобретению содержит по меньшей мере один специфический для заболевания (то есть T1D-специфический) ген аутоантигена и может экспрессировать такой ген в условиях, достаточных для экспрессии. В качестве примера T1D-специфичные антигены включают островковые антигены, ассоциированные с процессом разрушения бета-клеток. Примеры включают среди прочего: проинсулин (PINS), декарбоксилазу глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитическую субъединицу связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP) и транспортер цинка 8 (ZnT8). Другие примеры включают молекулы, экспрессируемые бета-клетками, такие как хромогранин А, (препро) островковый амилоидный полипептид (ppIAPP), периферии и цитруллинированный глюкозо-регулируемый белок (GRP).
[00129] Примеры полипептидов PINS включают PINS человека дикого типа и полипептиды, имеющие идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96% %, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью с таким PINS человека дикого типа. В качестве примера аминокислотная последовательность PINS человека дикого типа представлена SEQ ID NO: 6, тогда как пример кодирующей последовательности, кодирующей PINS, представлен в SEQ ID NO: 7 (см. CDS, содержащуюся под номером доступа NM_000207.2).
[00130] Дополнительные примеры нуклеотидных последовательностей PINS представлены кодирующими последовательностями под номерами доступа NCBI AY899304 (полная CDS, альтернативно сплайсированная, SEQ ID NO: 8); NM_000207 (вариант транскрипта 1, SEQ ID NO: 9); NM 001185097 (вариант транскрипта 2, SEQ ID NO: 10); NM_001185098 (вариант транскрипта 3, SEQ ID NO: 11); NM_001291897 (вариант транскрипта 4, SEQ ID NO: 12) и их частичные функциональные последовательности. Типичные аминокислотные последовательности PINS включают последовательности, которые кодируются любой из вышеуказанных нуклеотидных последовательностей PINS.
[00131] Может быть использована любая нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO: 6, или любая нуклеотидная последовательность, кодирующая по меньшей мере приблизительно 20, по меньшей мере приблизительно 30, по меньшей мере приблизительно 40, по меньшей мере приблизительно 50, по меньшей мере приблизительно 60, по меньшей мере приблизительно 70, по меньшей мере приблизительно 80, по меньшей мере приблизительно 90 или по меньшей мере приблизительно 100 последовательных аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью с SEQ ID NO: 6.
[00132] Дополнительные полипептиды PINS описаны, например, в базе UniProtKB-Р01308 и находящихся там ссылках. В некоторых примерах полипептид PINS представлен аминокислотными остатками 25-110 (нумерация согласно SEQ ID NO: 6).
[00133] Примеры полипептидов GAP (например, GAD65) включают GAD65 человека дикого типа и полипептиды, имеющие идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью такого GAD65 дикого типа. Пример аминокислотной последовательности GAD65 человека дикого типа представлен в SEQ ID NO: 13, тогда как пример последовательности нуклеиновой кислоты, кодирующей GAD65, представлен в SEQ ID NO: 14 (см., например, CDS под номером доступа М81882.1).
[00134] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленную в SEQ ID NO: 13, или любая нуклеотидная последовательность, кодирующая по меньшей мере приблизительно 100, по меньшей мере приблизительно 200, по меньшей мере приблизительно 300, по меньшей мере приблизительно 400 или по меньшей мере приблизительно 500 последовательных аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью в SEQ ID NO: 13.
[00135] Другие примеры последовательности глутаматдекарбоксилазы (например, GAD65) описаны, например, в UniProtKB-Q05329 и находящихся там ссылках. В одном из примеров полипептид GAD представляет собой укороченный вариант, содержащий менее приблизительно 500, менее приблизительно 400 или менее приблизительно 300 аминокислот дикого типа. В качестве примера полипептидные фрагменты (укороченные варианты GAD65) описаны, например, в работе Robert et al., Benef. Microbes 2015, 6(4): 591-601. В некоторых примерах укороченные варианты GAD эффективно экспрессируются и секретируются LAB (то есть, Lactococcus lactis). Примером укороченного варианта GAD является GAD65370-575 (нумерация аминокислот согласно последовательности с номером доступа NCBI NP_000809.1, то есть, SEQ ID NO: 13).
[00136] Другие примеры нуклеотидных последовательностей GAD представлены последовательностями со следующими номерами доступа NCBI: М81882 (GAD65; SEQ ID NO: 15); М81883 (GAD67; SEQ ID NO: 16); NM_000818 (вариант GAD2 1, SEQ ID NO: 17); и NM_001134366 (вариант GAD2 2; SEQ ID NO: 18); и открытые рамки считывания (CDS), содержащиеся там. Примеры аминокислотных последовательностей включают последовательности, кодируемые вышеуказанными нуклеотидными последовательностями под номером доступа М81882, М81883, NM 001134366 и NM_000818.
[00137] Примеры полипептидов IA-2 включают IA-2 человека дикого типа и полипептиды с идентичностью по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью с таким IA-2 дикого типа. Пример аминокислотной последовательности IA-2 человека дикого типа представлен в SEQ ID NO: 19, тогда как пример нуклеотидной последовательности, кодирующей IA-2, представлен в SEQ ID NO: 20 (см., например, открытую рамку считывания последовательности под номером доступа NM_002846.3).
[00138] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленную SEQ ID NO: 19, или любая нуклеотидная последовательность, кодирующая по меньшей мере приблизительно 100, по меньшей мере приблизительно 200, по меньшей мере приблизительно 300, по меньшей мере приблизительно 400, по меньшей мере приблизительно 500, по меньшей мере приблизительно 600 или по меньшей мере приблизительно 800 последовательных аминокислот указанной последовательности, или любая нуклеотидную последовательность, кодирующая полипептид, идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью в SEQ ID NO: 19.
[00139] Примеры нуклеотидных последовательностей IA-2 представлены под номерами доступа NCBI NM_002846 (IA-2 человека или протеинтирозинфосфатаза, рецептор типа N (PTPRN), вариант транскрипта 1, SEQ ID NO: 21); NM 001199763 (IA-2 человека или протеинтирозинфосфатаза, рецептор типа N (PTPRN), вариант транскрипта 2, SEQ ID NO: 22); NM_001199764 (IA-2 человека или протеинтирозинфосфатаза, рецептор типа N (PTPRN), вариант транскрипта 3, SEQ ID NO: 23). Примеры аминокислотных последовательностей IA-2 включают последовательности, кодированные вышеуказанными нуклеотидными последовательностями.
[00140] Другие примеры последовательности IA-2 описаны, например, в UniProtKB-Q16849 и находящихся там ссылках. В одном из примеров полипептид IA-2 может быть укороченным вариантом, содержащим менее приблизительно 700, менее приблизительно 600, менее приблизительно 500 или менее приблизительно 400 аминокислот дикого типа. В качестве примера полипептидные фрагменты (укороченные варианты IA-2) описаны, например, в работе Robert et al., Benef. Microbes 2015, 6(4): 591-601. В некоторых примерах укороченные варианты IA-2 могут эффективно экспрессироваться и секретироваться LAB (то есть Lactococcus lactis). В одном примере укороченный вариант IA-2 представляет собой IA-2б35-979 (нумерация аминокислот относительно последовательности с номером доступа NCBI NP_002837.1, т.е., SEQ ID NO: 19).
[00141] Примеры полипептидов IGRP включают IGRP человека дикого типа и полипептиды, имеющие идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью с таким IGRP дикого типа. Пример аминокислотной последовательности IGRP человека дикого типа представлен в SEQ ID NO: 24, тогда как пример кодирующей нуклеиновой кислоты, кодирующей IGRP, представлен в SEQ ID NO: 25 (см. открытую рамку считывания последовательности под номером доступа NCBI ВС113376.1).
[00142] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленную SEQ ID NO: 24, или любая нуклеотидная последовательность, кодирующая по меньшей мере 50, по меньшей мере 100, по меньшей мере 200 или по меньшей мере 300 аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью, представленной в SEQ ID NO: 24.
[00143] Дополнительные примеры нуклеотидных последовательностей представлены под следующими номерами доступа NCBI: NM_021176 (G6PC2, вариант транскрипта 1, SEQ Ш NO: 26); NM_001081686 (глюкоза-6-фосфатаза человека, каталитическая, 2, (G6PC2) вариант транскрипта 2, SEQ ID NO: 27); и NM_001270397 (G6PC, вариант транскрипта 2, SEQ ID NO: 28). Примеры аминокислотных последовательностей IGRP включают последовательности, кодируемые вышеуказанными нуклеотидными последовательностями.
[00144] Другие примеры последовательностей описаны, например, в UniProtKB -Q9NQR9 и в находящихся там ссылках, а также в работах Arden et al, Diabetes 1999; 48(3):531-542; Martin et al., J. Biol. Chem. 2001; 276(27):25197-207; и Dogra et al., Diabetologia 2006; 49(5):953-7. В некоторых примерах полипептид IGRP представляет собой укороченный вариант, содержащий менее приблизительно 300, менее приблизительно 200, менее приблизительно 100 или менее приблизительно 50 аминокислот дикого типа. В некоторых примерах укороченные варианты IGRP выбирают так, чтобы они эффективно экспрессировались и секретировались LAB (т.е., Lactococcus lactis).
[00145] Примеры полипептидов ZnT8 включают ZnT8 человека дикого типа и полипептиды, имеющие идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью такого ZnT8 дикого типа. Пример аминокислотной последовательности ZnT8 человека дикого типа представлен в SEQ ID NO: 29, тогда как пример последовательности нуклеиновой кислоты, кодирующей ZnT8, представлен в SEQ ID NO: 30 (см., например, открытую рамку считывания, содержащуюся в последовательности под номером доступа NCBI NM_173851.2).
[00146] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленную SEQ ID NO: 29, или любая нуклеотидная последовательность, кодирующая по меньшей мере 50, по меньшей мере 100, по меньшей мере 200, по меньшей мере 250 или по меньшей мере 300 аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с последовательностью, представленной в SEQ ID NO: 29.
[00147] Другие примеры нуклеотидных последовательностей ZnT8 представлены следующими номерами доступа NCBI AY212919.1 (транспортер цинка 8 человека, полная CDS, SEQ ID NO: 31); NM_173851.2 транспортер цинка 8 человека, вариант транскрипта 1, SEQ ID NO: 32); NM_001172814.1 (транспортер цинка 8 человека, вариант транскрипта 2, SEQ ID NO: 33); NM_001172811.1 (транспортер цинка 8 человека, вариант транскрипта 3, SEQ ID NO: 34); NM_001172813.1 (транспортер цинка 8 человека, вариант транскрипта 4, SEQ ID NO: 35); NM_001172815.2 (транспортер цинка 8 человека, вариант транскрипта 5, SEQ ID NO: 36) и их частичными последовательностями. Примеры аминокислотных последовательностей ZnT8 включают последовательности, которые кодируются вышеуказанными нуклеотидными последовательностями.
[00148] Другие примеры последовательностей описаны, например, в UniProtKB-Q8IWU4 и находящихся там ссылках. В некоторых примерах полипептид ZnT8 представляет собой укороченный вариант, содержащий менее приблизительно 300, менее приблизительно 200 или менее приблизительно 100 аминокислот дикого типа. В некоторых примерах укороченные варианты ZnT8 выбирают так, чтобы они эффективно экспрессировались и секретировались штаммом LAB (т.е., Lactococcus lactis).
[00149] Примеры полипептидов ppIAPP включают ppIAPP человека дикого типа и полипептиды, имеющие идентичность последовательности по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или, по меньшей мере приблизительно на 99% с таким ppIAPP дикого типа. Пример аминокислотной последовательности ppIAPP человека дикого типа представлен в SEQ ID NO: 37, тогда как пример нуклеотидной последовательности, кодирующая ppIAPP, представлен в SEQ ID NO: 38 (см., например, открытую рамку считывания с номером доступа NCBI NM_000415.2).
[00150] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленную SEQ ID NO: 37, или любая нуклеотидная последовательность, кодирующая по меньшей мере 50, по меньшей мере 100, по меньшей мере 200, по меньшей мере 250 или по меньшей мере 300 аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность последовательности по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с SEQ ID NO: 37.
[00151] Другие примеры полипептидных последовательностей ppIAPP раскрыты, например, в UniProtKB-P10997 и находящихся там ссылках. В некоторых примерах полипептид ppIAPP может быть укороченным вариантом, содержащим менее приблизительно 80, менее приблизительно 60, менее приблизительно 40 или менее приблизительно 20 аминокислот дикого типа. В некоторых примерах укороченные варианты ppIAPP выбирают так, чтобы они были эффективно экспрессировались и секретировались штаммом LAB (то есть, Lactococcus lactis).
[00152] Примеры полипептидов периферина включают периферии человека дикого типа и полипептиды, имеющие идентичность последовательности по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с таким диким типом. Пример аминокислотной последовательности периферина человека дикого типа представлена SEQ ID NO: 39, тогда как пример нуклеотидной последовательности, кодирующей периферии, представлен в SEQ ID NO: 40 (см., например, открытую рамку считывания номера доступа NCBI NM_006262.3).
[00153] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленная SEQ ID NO: 39, или любая нуклеотидная последовательность, кодирующая по меньшей мере приблизительно 100, по меньшей мере приблизительно 200, по меньшей мере приблизительно 300 или по меньшей мере приблизительно 400 последовательных аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность последовательности по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с SEQ ID NO: 39.
[00154] Другие примеры последовательности периферина раскрыты, например, в UniProtKB-P41219 и находящихся там ссылках. В некоторых примерах полипептид периферина представляет собой укороченный вариант, содержащий менее приблизительно 400, менее приблизительно 300, менее приблизительно 200 или менее приблизительно 100 аминокислот дикого типа. В некоторых примерах укороченные варианты периферии выбирают так, чтобы они эффективно экспрессировались и секретировались штаммом LAB (т.е., Lactococcus lactis).
[00155] Другие примеры нуклеотидных последовательностей представлены под номерами доступа NCBI NM_006262.3 (периферии человека, PRPH, SEQ ID NO: 41); ХМ_005269025.1 (предсказанный периферии человека, вариант транскрипта XI, SEQ ID NO: 42); XR_944623.1 (предсказанный периферии человека, вариант транскрипта Х2, SEQ ID NO: 43) и их частичные последовательности. Примеры аминокислотных последовательностей периферина включают последовательности, которые кодируются вышеуказанными нуклеотидными последовательностями.
[00156] Примеры полипептидов GRP включают GRP78/BiP человека дикого типа и полипептиды, имеющие по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95%, по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с таким GRP дикого типа. Пример аминокислотной последовательности GRP человека дикого типа представлен в SEQ ID NO: 44, тогда как пример нуклеотидной последовательности, кодирующей GRP, представлен в SEQ ID NO: 45 (см., например, открытую рамку считывания под номером доступа NCBI X87949.1).
[00157] Может быть использована любая нуклеотидная последовательность, кодирующая вышеуказанную аминокислотную последовательность, представленная SEQ ID NO: 44, или любая нуклеотидная последовательность, кодирующая по меньшей мере приблизительно 100, по меньшей мере приблизительно 200, по меньшей мере приблизительно 300, по меньшей мере приблизительно 400 или по меньшей мере приблизительно 500 последовательных аминокислот указанной последовательности, или любая нуклеотидная последовательность, кодирующая полипептид, имеющий идентичность последовательности по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% по меньшей мере приблизительно на 96%, по меньшей мере приблизительно на 97%, по меньшей мере приблизительно на 98% или по меньшей мере приблизительно на 99% с SEQ ID NO: 44.
[00158] Другие примеры последовательностей периферина раскрыты, например, в UniProtKB-P11021 и находящихся там ссылках. В некоторых примерах полипептид GRP представляет собой укороченный вариант, содержащий менее приблизительно 500, менее приблизительно 400, менее приблизительно 300 или менее приблизительно 200 аминокислот дикого типа. В некоторых примерах укороченные варианты GRP выбирают так, чтобы они эффективно экспрессировались и секретировались штаммом LAB (то есть, Lactococcus lactis).
[00159] Специалисту в данной области техники понятно, что оптимальное количество аутоантигена, которое должно быть доставлено субъекту с применением способов настоящего изобретения, варьирует, например, в зависимости от типа антигена, микроорганизма, экспрессирующего этот антиген, и генетической конструкции, например, эффективности промотора, используемого в генетической конструкции. Как правило, микроорганизм вводят в количестве, эквивалентном конкретному количеству экспрессированного антигена, или в количестве, которое обеспечивает желаемый ФК-профиль для соответствующего полипептидного антигена у соответствующего субъекта. Примеры ежедневных доз антигена составляют от приблизительно 10 фг до приблизительно 100 мкг активного полипептида в сутки. Другие диапазоны доз в качестве примера составляют от приблизительно 1 пг до приблизительно 100 мкг в сутки; или от приблизительно 1 нг до приблизительно 100 мкг в сутки.
[00160] Приведенные выше дозы антигена могут быть реализованы путем введения субъекту эффективных количеств LAB в сутки, причем LAB адаптирована для экспрессии достаточного количества биоактивного полипептида для реализации желаемой дозы, такой как вышеуказанные. LAB, секретирующая антигенный полипептид, может быть доставлена в дозе от приблизительно 104 колониеобразующих единиц (КОЕ) до приблизительно 1012 КОЕ в сутки, в частности от приблизительно 106 КОЕ до приблизительно 1012 КОЕ в сутки, или от приблизительно 109 КОЕ до приблизительно 1012 КОЕ в сутки.
[00161] Количество секретируемого антигенного полипептида в КОЕ может быть определено, например, в соответствии со способами, описанными в работе Steidler et al., Science 2000; 289(5483): 1352-1355, или с помощью ИФА. Например, LAB может секретировать по меньшей мере от 1 нг до 1 мкг активного полипептида на 109 КОЕ. На основании этого специалист может вычислить диапазон количества антигенного полипептида, секретируемого при других дозах в КОЕ.
[00162] Каждая из вышеуказанных доз/диапазонов доз можно вводить в рамках любой схемы дозирования, описанной в настоящей заявке. Ежедневную дозу можно вводить в 1, 2, 3, 4, 5 или 6 порций в течение суток. Кроме того, суточные дозы можно вводить в течение любого количества дней с любым количеством периодов отдыха между периодами введения. Например, дозу, эквивалентную приблизительно от 0,1 млн. ME до приблизительно 3 млн. МЕ/сут/субъекта можно вводить через день, в течение периода, составляющего в общей сложности 6 недель.
Составы и схемы
[00163] В способах, описанных в настоящей заявке, IL-2 и T1D могут экспрессироваться одной и той же LAB или разными LAB. Когда два полипептида экспрессируются различными микроорганизмами, их можно вводить субъекту в одном и том же (например, комбинированном) составе или вводить в виде отдельных (например, разных) составов. Отдельные составы можно вводить одновременно или в разные моменты времени. Например, продуцирующие IL-2 и T1D-специфический антиген микроорганизмы в их соответствующих составах можно вводить субъекту одновременно или последовательно, например, с периодом отдыха между введениями.
[00164] Штаммы LAB, продуцирующие IL-2 и T1D-специфический антиген, можно вводить одновременно. В некоторых примерах микроорганизм, продуцирующий IL-2, и микроорганизм, продуцирующий T1D-специфический антиген, могут быть включены в один и тот же фармацевтический состав или в несколько фармацевтических составов, принимаемых в одно и то же время. В примерах вариантах реализации два биоактивных полипептида доставляют субъекту с использованием одного штамма LAB, продуцирующего как IL-2, так и T1D-специфический антиген.
[00165] В некоторых вариантах реализации композицию, описанную в настоящей заявке, будут вводить один, два, три, четыре, пять или шесть раз в сутки, например, с использованием перорального состава. В некоторых вариантах реализации штаммы LAB вводят каждый день, через день, один раз в неделю, два раза в неделю, три раза в неделю или четыре раза в неделю. В других вариантах лечение проводят один раз в две недели. В других вариантах лечение проводят один раз в три недели. В других вариантах лечение проводят один раз в месяц.
[00166] Продолжительность цикла лечения для данного способа может составлять, например, от 7 дней до времени жизни субъекта, в зависимости от того, сколько необходимо для лечения или регрессии T1D или предотвращения рецидива. Цикл лечения может длиться от 30 дней до 2 лет. В других вариантах реализации цикл лечения у субъекта может длиться от 30 дней до 1,5 лет. В других вариантах реализации цикл лечения у субъекта может длиться от 30 дней до 1 года. В других вариантах реализации цикл лечения у субъекта может длиться от 30 дней до 11 месяцев. В других вариантах реализации цикл лечения у субъекта может длиться от 30 дней до 10 месяцев. В других вариантах реализации цикл лечения у субъекта может длиться от 30 дней до 9 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 8 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 7 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 6 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 5 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 4 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 3 месяцев. Цикл лечения у субъекта может длиться от 30 дней до 2 месяцев.
[00167] Ежедневные поддерживающие дозы можно вводить в течение периода, клинически подходящего для субъекта, например, от 1 дня до нескольких лет (например, в течение всего оставшегося времени жизни субъекта); например, от приблизительно (2, 3 или 5 дней, 1 или 2 недели или 1 месяц) и выше и/или, например, до приблизительно (5 лет, 1 год, 6 месяцев, 1 месяц, 1 неделя или 3 или 5 дней). Типичным является введение ежедневной поддерживающей дозы в течение от приблизительно 3 до приблизительно 5 дней или от приблизительно 1 недели до приблизительно 1 года. Вместе с тем, унифицированные дозы в некоторых случаях - могут вводиться от двух раз в сутки до одного раза в две недели до тех пор, пока не будет наблюдаться терапевтический эффект.
[00168] Штаммы LAB, продуцирующие полипептид IL-2 и антигенный полипептид, могут быть доставлены в виде моно- или комбинированной терапии для лечения T1D. В некоторых вариантах реализации композиции настоящего изобретения включают в себя дополнительные терапевтически активные агенты. В некоторых вариантах реализации лечение субъекта не включает другие активные компоненты, например, не включает дополнительные иммуномодулирующие вещества, такие как антитела (например, против CD3). Таким образом, в некоторых примерах фармацевтические композиции настоящего изобретения состоят существенным образом (по существу) из LAB, описанных в настоящей заявке (экспрессирующих терапевтические IL-2 и антигенные полипептиды) и фармацевтически приемлемого носителя.
Фармацевтические композиции и носители
[00169] Описанные в настоящей заявке штаммы LAB (например, L. lactis) можно вводить в чистой форме, в сочетании с другими активными ингредиентами и/или в сочетании с фармацевтически приемлемыми (то есть нетоксичными) вспомогательными веществами или носителями. Термин "фармацевтически приемлемый" используется в настоящей заявке в соответствии с его принятым в настоящей области техники значением и обозначает носители, которые совместимы с другими ингредиентами фармацевтической композиции и не являются вредными для реципиента.
[00170] Композиции, описанные в настоящей заявке, могут быть получены в любой известной или иным образом эффективной дозе или форме продукта, пригодной для применения при обеспечении системной доставки штаммов LAB (например, L. lactis) в слизистую оболочку, включая фармацевтические композиции и лекарственные формы, а также формы пищевых продуктов.
[00171] В некоторых вариантах реализации состав представляет собой пероральный состав или фармацевтическую композицию. В некоторых примерах в соответствии с этим вариантом реализации состав или фармацевтическая композиция содержит штаммы LAB в форме сухого порошка (например, в лиофилизированной форме) или в прессованной форме, в некоторых случаях - в сочетании с другими сухими носителями. Пероральные составы обычно включают в себя инертный носитель-разбавитель или съедобный носитель.
[00172] В некоторых примерах эта пероральная композиция содержит покрытие или использует стратегию капсулирования, которая облегчает доставку композиции в кишечный тракт и/или позволяет высвобождать и гидратировать микроорганизм в кишечном тракте (например, в подвздошной кишке, тонкой кишке или толстой кишке). Как только LAB высвобождается из состава и достаточно гидратируется, она начинает экспрессировать биоактивный полипептид, который затем высвобождается в окружающую среду или экспрессируется на поверхности микроорганизма. Такие стратегии покрытия и инкапсуляции (то есть стратегии с задержкой высвобождения) известны специалистам в данной области техники. См., например, патент США 5972685; WO 2000/18377; и WO 2000/22909.
[00173] Предложена фармацевтическая композиция, которая может содержать штаммы LAB в лиофилизированной или сублимированной форме, в некоторых случаях - в сочетании с другими компонентами, такими как декстраны, глутамат натрия и полиолы. Примеры лиофилизированных композиций описаны, например, в патентной публикации США. №2012/0039853. Примеры составов включают лиофилизированные бактерии (например, терапевтически эффективное количество бактерий) и фармацевтически приемлемый носитель. Лиофилизированные бактерии могут быть приготовлены в виде капсул, таблеток, гранулятов и порошков, каждое из которых можно вводить перорально. В качестве альтернативы, лиофилизированные бактерии могут быть приготовлены в виде водных суспензий в подходящих средах, или лиофилизированные бактерии могут быть суспендированы в подходящей среде, такой как напиток, непосредственно перед использованием.
[00174] Для перорального введения состав может быть гастрорезистентной пероральной лекарственной формой. Например, пероральная дозированная форма (включая капсулы, таблетки, гранулы, микрогранулы, грануляты и тому подобное) может быть покрыта тонким слоем вспомогательного вещества (обычно полимеров, целлюлозных производных и/или липофильных материалов), который противостоит растворению или разрушению в желудке, но не в кишечнике, что позволяет обеспечить беспрепятственный транзит через желудок, с последующей дезинтеграцией, растворением и абсорбцией в кишечнике (например, в тонкой кишке или толстой кишке).
[00175] В некоторых примерах пероральные составы могут включать соединения, обеспечивающие контролируемое высвобождение, замедленное высвобождение или пролонгированное высвобождение микроорганизма и, таким образом, обеспечивают контролируемое высвобождение нужного белка, кодируемого в нем. Эти лекарственные формы (например, таблетки или капсулы) типично содержат обычные и хорошо известные вспомогательные вещества, такие как липофильные, полимерные, целлюлозные, нерастворимые и способные к набуханию вспомогательные вещества. Составы с контролируемым высвобождением могут также использоваться для любых других участков доставки, включая кишечную, толстокишечную, биоадгезионную или сублингвальную доставку (т.е. дентальную доставку в слизистую оболочку) и бронхиальную доставку. Когда композиции, описанные в настоящей заявке, должны вводиться ректально или вагинально, фармацевтические составы могут включать суппозитории и кремы. В этом случае клетки-хозяева суспендируют в смеси обычных вспомогательных веществ, включая липиды. Каждый из вышеупомянутых составов хорошо известен в данной области техники и описан, например, в следующих ссылках: Hansel et al. (1990, pharmaceutical dosage forms and drug delivery systems, 5th edition, William and Wilkins); Chien 1992, novel drug delivery SYSTEM, 2nd edition, M. Dekker); Prescott et al. (1989, novel DRUG delivery, J. Wiley & Sons); Cazzaniga et al., (1994, Int. J. Pharm. 108(1): 77-83).
[00176] Оральные составы и композиции, описанные в настоящей заявке, могут дополнительно включать соединения, которые способны интенсифицировать доставку в слизистую оболочку и/или поглощение слизистой оболочкой биоактивных полипептидов, экспрессированных LAB. Составы/композиции, описанные в настоящей заявке, могут также включать соединения, которые повышают жизнеспособность микроорганизма в составе и/или после высвобождения.
[00177] LAB, описанная в настоящей заявке, может быть суспендирована в фармацевтическом составе для введения человеку или животному, с заболеванием, подлежащим лечению. Такие фармацевтические составы включают среди прочего живую LAB и среду, пригодную для введения. LAB может быть лиофилизирована в присутствии обычных вспомогательных веществ, таких как лактоза, другие сахара, щелочной и/или щелочноземельный стеарат, карбонат и/или сульфат (например, стеарат магния, карбонат натрия и сульфат натрия), каолин, диоксид кремния, ароматические и вкусовые добавки. Бактерии, таким образом лиофилизированные, могут быть приготовлены в виде капсул, таблеток, гранулятов и порошков (например, порошка для полоскания рта), каждое из которых может вводиться пероральным путем. В качестве альтернативы, штаммы LAB могут быть приготовлены в виде водных суспензий в подходящих средах или лиофилизированные бактерии могут быть суспендированы в подходящей среде непосредственно перед использованием, причем такая среда включает в себя вспомогательные вещества, упомянутые в настоящей заявке, и другие вспомогательные вещества, такие как глюкоза, глицин и сахаринат натрия.
[00178] В некоторых примерах LAB локально доставляют в желудочно-кишечный тракт субъекта с применением любого подходящего способа. Например, для улучшения доставки в кишечник может использоваться система доставки в виде микросфер. Системы доставки микросфер включают микрочастицы с покрытием, обеспечивающим локализованное высвобождение в желудочно-кишечный тракт субъекта (например, составы с контролируемым высвобождением, такие как составы с энтеросолюбильным покрытием и составы для абсорбции в толстой кишке).
[00179] Для перорального введения могут быть приготовлены гастрорезистентные пероральные лекарственные формы, которые могут также включать соединения, обеспечивающие контролируемое высвобождение штаммов LAB, и, таким образом, обеспечивают контролируемое высвобождение требуемого белка, кодируемого в LAB, в разных участках пищеварения (например, IL-2). Например, пероральная дозированная форма (включая капсулы, таблетки, гранулы, грануляты, порошки) может быть покрыта тонким слоем вспомогательного вещества (например, полимеров, целлюлозных производных и/или липофильных материалов), который противостоит растворению или разрушению в желудке, но не в кишечнике, что позволяет обеспечить транзит через желудок в пользу дезинтеграции, растворения и абсорбции в кишечнике.
[00180] Пероральная лекарственная форма может быть разработана так, чтобы обеспечивать медленное высвобождение штаммов LAB и продуцируемых экзогенных белков, например, таблетки или капсулы с контролируемым высвобождением, замедленным высвобождением, пролонгированным высвобождением, замедленным действием. Эти лекарственные формы обычно содержат обычные и хорошо известные вспомогательные вещества, такие как липофильные, полимерные, целлюлозные, нерастворимые и способные к набуханию вспомогательные вещества. Такие составы описаны, например, в следующих ссылках: Hansel et al., pharmaceutical dosage FORMS AND DRUG delivery systems, 5th edition, William and Wilkins, 1990; Chien 1992, novel drug delivery system, 2nd edition, M. Dekker; Prescott et al., novel drug delivery, J.Wiley & Sons, 1989; и Cazzaniga et al., Int. J. Pharm. 108(1):77-83 (1994).
[00181] Фармацевтическая лекарственная форма (например, капсула) обычно покрыта рН-зависимыми полимерами Eudragit® для получения устойчивости к желудочному соку и для целевой доставки в терминальном отделе подвздошной кишки и толстой кишке, где полимеры растворяются при рН 6,5. Используя другие полимеры Eudragit® или различное соотношение между этими полимерами, профиль отсроченного высвобождения можно регулировать, чтобы высвободить бактерии, например, в двенадцатиперстной кишке или тощей кишке.
[00182] Фармацевтические композиции обычно содержат по меньшей мере один фармацевтически приемлемый носитель. Неограничивающие примеры подходящих вспомогательных веществ, разбавителей и носителей включают консерванты, неорганические соли, кислоты, основания, буферы, питательные вещества, витамины, наполнители и сухие разбавители, такие как крахмал, сахара, маннит и производные диоксида кремния; связывающие агенты, такие как карбоксиметилцеллюлоза и другие производные целлюлозы, альгинаты, желатин и поливинилпиролидон; увлажняющие агенты, такие как глицерин/дезинтегрирующие агенты, такие как карбонат кальция и бикарбонат натрия; агенты для замедления растворения, такие как парафин; ускорители резорбции, такие как соединения четвертичного аммония; поверхностно-активные вещества, такие как ацетиловый спирт, моностеарат глицерина; адсорбционные носители, такие как каолин и бентонит; носители, такие как пропиленгликоль и этиловый спирт, и смазывающие вещества, такие как тальк, стеарат кальция и магния, и твердые полиэтиленгликоли.
[00183] В состав композиции могут быть включены фармацевтически совместимые связывающие агенты и/или адъювантные материалы. Таблетки, пилюли, капсулы, пастилки и тому подобное могут содержать любой из следующих ингредиентов или соединений сходного характера: связующее, такое как микрокристаллическая целлюлоза, трагакантовая камедь или желатин; вспомогательное вещество, такое как крахмал или лактоза, диспергирующий агент, такой как альгиновая кислота, Примогель или кукурузный крахмал; смазывающее вещество, такое как стеарат магния; глидант, такой как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин; или вкусо-ароматическая добавка, такая как перечная мята, метилсалицилат или апельсиновый ароматизатор. Когда единичная дозированная форма представляет собой капсулу, она может содержать, помимо материала вышеуказанного типа, жидкий носитель, такой как нелетучее масло. Кроме того, единичные дозированные формы могут содержать различные другие материалы, модифицирующие физическую форму единицы дозировки, например, покрытия из сахара, шеллака или кишечнорастворимых агентов. Кроме того, сироп может содержать, помимо активных соединений, сахарозу в качестве подсластителя и некоторые консерванты, красители, красители и ароматизаторы. Понятно, что форма и характер фармацевтически приемлемого носителя определяется количеством активного ингредиента, с которым он должен быть скомбинирован, способом введения и другими хорошо известными переменными. Носитель (-и) должен быть "приемлемым" в смысле совместимости с другими ингредиентами этого состава и не вредным для его реципиента.
[00184] Альтернативные препараты для введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются диметилсульфоксид, спирты, пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло и инъекционные органические сложные эфиры, такие как этилолеат. Водные носители включают смеси спиртов и воды, буферные среды и физиологический раствор. Внутривенные носители включают жидкие и питательные пополняющие вещества, пополнители электролитов, например, основанные на декстрозе Рингера и тому подобное. Также могут присутствовать консерванты и другие добавки, такие как, например, противомикробные средства, антиоксиданты, хелатирующие агенты, инертные газы и тому подобное. Для этих способов доставки возможны различные жидкие составы, в том числе солевые, спиртовые, ДМСО-содержащие и водные растворы.
[00185] Пероральные водные составы включают вспомогательные вещества, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния и/или тому подобное фармацевтической степени чистоты. Эти композиции могут иметь форму растворов, таких как жидкости для промывания и полоскания рта, дополнительно содержащие водный носитель, такой как, например, вода, спиртовые/водные растворы, солевые растворы, парентеральные носители, такие как хлорид натрия, декстроза Рингера и тому подобное.
[00186] Водные составы для полоскания рта хорошо известны специалистам в данной области техники. Составы, относящиеся к жидкостям для промывания и полоскания рта, подробно обсуждаются, например, в патенте США 6,387,352, патенте США 6,348,187, патенте США 6,171,611, патенте США 6,165,494, патенте США 6,117,417, патенте США 5,993,785, патенте США 5,695,746, патенте США 5,470,561, патенте США 4,919,918, заявке на патент США №2004/0076590, заявке на патент США №2003/0152530 и заявке на патент США №2002/0044910.
[00187] В составах настоящего изобретения могут присутствовать другие добавки, таких как ароматизаторы, подслащивающие или окрашивающие агенты или консерванты. Примерами подходящих ароматизаторов являются мятная субстанция, например, из мяты перечной или мяты кудрявой, корица, эвкалипт, цитрус, кассии, анис и ментол. В некоторых случаях в пероральных композициях присутствуют ароматизирующие агенты в количестве от 0 до 3%; в некоторых случаях - до 2%, например, до 0,5%, в некоторых случаях - 0,2%, в случае жидких композиций.
[00188] Подсластители включают искусственные или натуральные подслащивающие агенты, такие как сахарин натрия, сахароза, глюкоза, сахарин, декстроза, левулоза, лактоза, маннит, сорбит, фруктоза, мальтоза, ксилит, тауматин, аспартам, D-триптофан, дигидрохалконы, ацесульфам, и любые их комбинации, которые могут присутствовать в количестве от 0 до 2%, в некоторых случаях - до 1% мас./мас., например от 0,05 до 0,3% мас./мас. пероральной композиции.
[00189] Красящие агенты представляют собой подходящие природные или синтетические красители, такие как диоксид титана или CI 42090, или их смеси. Красители могут присутствовать в композициях в количестве от 0 до 3%; в некоторых случаях - до 0,1%, например, до 0,05%, в некоторых случаях - приблизительно 0,005-0,0005%, в случае жидких композиций. Из обычных консервантов обычно используют бензоат натрия в концентрациях, существенно недостаточных для изменения рН композиции, в противном случае может потребоваться корректировка количества буферного агента, для достижения требуемого значения рН.
[00190] Другие необязательные ингредиенты включают увлажнители, поверхностно-активные вещества (неионные, катионные или амфотерные), загустители, камеди и связующие вещества. Увлажнитель добавляет объем к составу и сохраняет влагу в композиции для чистки зубов. Кроме того, увлажнитель помогает предотвратить деградацию микроорганизмов при хранении этого состава. Он также помогает поддерживать фазовую стабильность и обеспечивает способ создания прозрачного или полупрозрачного средства для чистки зубов.
[00191] Подходящие увлажнители включают глицерин, ксилит, глицерин и гликоли, такие как пропиленгликоль, которые могут присутствовать в количестве до 50% мас./мас. для каждого, но суммарное количество увлажнителя может составлять не более приблизительно 60-80% мас./мас. от композиции. Например, жидкие композиции могут содержать до приблизительно 30% глицерина плюс до приблизительно 5%, в некоторых случаях - приблизительно 2% мас./мас. ксилита. Поверхностно-активные вещества могут быть не анионными и могут включать полисорбат 20 или кокоамидобетаин или тому подобное в количестве до приблизительно 6%, в некоторых случаях - приблизительно 1,5-3%, мас./мас. от композиции.
[00192] Когда пероральные композиции, описанные в настоящей заявке, находятся в жидкой форме, указанные композиции обычно могут включать пленкообразующий агент в количестве до приблизительно 3% мас./мас. от пероральной композиции, например, в диапазоне от 0 до 0,1%, в некоторых случаях - приблизительно 0,001-0,01%, например приблизительно 0,005% мас./мас. от пероральной композиции. Подходящие пленкообразователи включают (в дополнение к гиалуронату натрия) те, которые продаются под торговым наименованием Гантрез (Gantrez)™.
[00193] Жидкие питательные составы для перорального или энтерального введения могут содержать одно или несколько питательных веществ, таких как жиры, углеводы, белки, витамины и минералы. Известны многие различные источники и типы углеводов, липидов, белков, минералов и витаминов, которые могут быть использованы в описанных в настоящей заявке вариантах реализации жидких питательных составов при условии, что такие питательные вещества совместимы с добавленными ингредиентами в выбранном составе, являются безопасными и эффективными для их целевого применения и не ухудшают по иным причинам характеристики продукта.
[00194] Эти питательные жидкости обычно приготовлены с достаточной вязкостью, текучестью или другими физическими или химическими характеристиками, чтобы обеспечить более эффективное и смягчающее обволакивание слизистой оболочки при выпивании или введении питательной жидкости. Эти питательные варианты реализации также могут представлять собой сбалансированный источник питания, подходящий для удовлетворения исключительных, первичных или дополнительных потребностей в питании данного лица. Неограничивающие примеры подходящих питательных жидкостей описаны, например, в патенте США 5700782; патенте США 5869118; и в патенте США 5223285.
[00195] Пищевые белки, подходящие для применения в настоящем изобретении, могут быть гидролизованы, частично гидролизованы или не гидролизованы и могут быть получены из любого известного или иного подходящего источника, такого как молоко (например, казеин, сыворотка), животные (например, мясо, рыба), зерновые (например, рис, кукуруза), растительное сырье (например, соя) или их комбинации.
[00196] Жиры или липиды, пригодные для применения в питательных жидкостях, включают, среди прочего, кокосовое масло, соевое масло, кукурузное масло, оливковое масло, сафлоровое масло, высокоолеиновое сафлоровое масло, масло МСТ (триглицериды с цепочками средней длины), подсолнечное масло, высокоолеиновое подсолнечное масло, структурированные триглицериды, пальмовые и пальмоядровые масла, пальмовый олеин, масло канолы, морские масла, хлопковые масла и их комбинации. Углеводы, пригодные для применения в питательных жидкостях, могут быть простыми или сложными, лактозосодержащими или не содержащими лактозу или их комбинациями. Неограничивающие примеры подходящих углеводов включают в себя гидролизованный кукурузный крахмал, мальтодекстрин, глюкозные полимеры, сахарозу, кукурузный сироп, сухую кукурузную патоку, углеводы, произведенные из риса, глюкозу, фруктозу, лактозу, кукурузный сироп с высоким содержанием фруктозы и неусваиваемые олигосахариды, такие как фруктоолигосахариды (FOS) и их комбинации.
[00197] Питательные жидкости, описанные в настоящей заявке, могут дополнительно содержать любой из набора витаминов, неограничивающие примеры которых включают витамин А, витамин D, витамин Е, витамин К, тиамин, рибофлавин, пиридоксин, витамин В 12, ниацин, фолиевую кислоту, пантотеновую кислоту, биотин, витамин С, холин, инозит, соли и их производные и их комбинации.
[00198] Питательные жидкости, описанные в настоящей заявке, могут дополнительно содержать любой из набора минералов, известных или иных подходящих для наших целей пациентам с риском T1D или страдающим от него, неограничивающие примеры которых включают кальций, фосфор, магниевое железо, селен, марганец, медь, йод, натрий, калий, хлорид и их комбинации.
[00199] Препараты штаммов LAB, описанных в настоящей заявке, также могут быть изготовлены в виде эликсиров или растворов для удобного перорального или ректального введения или в виде растворов, подходящих для парентерального введения, например, внутримышечным, подкожным или внутривенным способами. Кроме того, нуклеозидные производные также хорошо подходят для состава в виде лекарственных форм замедленного или пролонгированного высвобождения, включая лекарственные формы, которые высвобождают активный ингредиент только или в некоторых случаях в определенной части кишечного тракта, в некоторых случаях - в течение замедленного или пролонгированного периода времени для дальнейшего повышения эффективности. Покрытия, оболочки и защитные матрицы в таких дозированных формах могут быть изготовлены, например, из полимерных веществ или восков, хорошо известных в фармацевтической области.
[00200] Композиции, описанные в настоящей заявке, могут включать фармацевтические лекарственные формы, такие как лепешки для рассасывания, троше или пастилки. Они обычно представляют собой твердые продукты дисковидной формы, содержащие активный ингредиент в подходящей ароматизированной основе. Основой может быть твердый сахарный леденец, глицериновый желатин или комбинация сахара с достаточным количеством клейкого вещества, чтобы придать ему форму. Троше размещаются во рту, где они медленно растворяются, освобождая активный ингредиент для непосредственного контакта со слизистой оболочкой.
[00201] Варианты реализации в виде троше могут быть приготовлены, например, путем медленного добавления воды к смеси порошкообразного активного вещества, порошкообразного сахара и камеди до образования податливой массы. Для придания массе достаточной адгезии можно использовать 7% порошок гуммиарабика. Массу раскатывают, и вырезают из сплющенной массы отдельные троше, либо массу можно раскатать в цилиндр и поделить. Каждую отрезанную или отделенную часть формируют и дают высохнуть, получая лекарственную форму в виде троше.
[00202] Если активный ингредиент является термолабильным, его препарат можно изготовить в виде лепешек для рассасывания, путем прессования. Например, стадию гранулирования выполняют способом, аналогичным способу, используемому для любой прессованной таблетки. Лепешку изготавливают с использованием оборудования высокой компрессии, чтобы получить таблетку, более твердую, чем обычно, так как желательно, чтобы эта лекарственная форма медленно растворялась или распадалась во рту. Ингредиенты обычно выбирают для получения характеристик медленной растворимости.
[00203] В одном из примеров составов штаммы LAB могут быть включены в биоадгезивный носитель, содержащий предварительно желатинизированный крахмал и поперечно-сшитую поли(акриловую кислоту), для получения биоадгезивной таблетки и биоадгезивного геля, пригодного для трансбуккального применения (то есть с длительной биоадгезией и замедленной доставкой лекарства).
[00204] Порошковая смесь штаммов LAB, биоадгезивных полимеров (прежелатинизированный крахмал и поперечно-сшитая поли (акриловая кислота), совместно обработанных сушкой распылением), стеарилфумарат натрия (смазывающее вещество) и диоксид кремния (скользящее вещество), могут быть переработаны в таблетки (вес: 100 мг, диаметр: 7 мм). Способы изготовления этих таблеток хорошо известны специалисту в данной области техники и были описаны ранее для успешной разработки биоадгезивных таблеток, содержащих различные лекарственные средства (миконазол, тестостерон, фторид, ципрофлоксацин) (Bruschi M. L. and de Freitas О., Drug Development and Industrial Pharmacy, 2005 31: 293-310).
[00205] Для оптимизации состава можно варьировать содержание лекарственного средства в таблетках и соотношение между крахмалом и поли (акриловой кислотой). Исходя из предыдущих исследований, максимальное содержание лекарственного средства в совместно обработанном биоадгезивом носителе составляет приблизительно 60% (мас./мас.), а соотношение крахмала/поли (акриловой кислоты) может варьироваться от 75/25 до 95/5 (мас./мас.). Во время оптимизационного исследования биоадгезивные свойства таблеток и высвобождение лекарственного средства из таблеток являются основными параметрами, а стандартные свойства таблеток (твердость, истираемость) - вторичными критериями оценки.
[00206] Штаммы LAB могут быть включены в водную дисперсию прежелатинизированного крахмала и поперечно-сшитой поли (акриловой кислоты). Эту полимерную дисперсию готовят по стандартной методике с использованием смесителя с высоким усилием сдвига.
[00207] Аналогично таблетке, для геля может потребоваться оптимизировать содержание лекарственного средства и соотношение крахмал/поли (акриловая кислота), чтобы получить гель, имеющий оптимальную адгезию к слизистой оболочке пищевода. Для геля концентрация полимеров в дисперсии является дополнительной переменной, поскольку она определяет вязкость геля, следовательно, его мукоадгезивные свойства.
[00208] Модель для скрининга биоадгезивных свойств полимерных дисперсий относительно слизистой оболочки пищевода была подробно описана в Batchelor et al. (Int. J. Pharm., 238: 123- 32, 2002).
[00209] Другие пути и формы введения лекарства включают пищевые продукты, содержащие живые штаммы LAB. В некоторых примерах биоактивные штаммы LAB, экспрессирующие полипептид, могут быть включены в молочный продукт.
[00210] Фармацевтические композиции, описанные в настоящей заявке, могут быть изготовлены любым известным или иным эффективным способом для приготовления или производства выбранной лекарственной формы. Например, штаммы LAB могут быть приготовлены в виде состава вместе с обычными, например, фармацевтически приемлемыми носителями, такими как вспомогательные вещества и разбавители, сформированные в пероральные таблетки, капсулы, спреи, лепешки, обработанные субстраты (например, пероральные или местные тампоны, ватные диски или одноразовые, неперевариваемые субстраты, обработанные описанными в настоящей заявке композициями); пероральные жидкости (например, суспензии, растворы, эмульсии), порошки, суппозитории или любую другую подходящую лекарственную форму. В некоторых вариантах реализации настоящее изобретение предлагает способ производства фармацевтической композиции. В качестве примера способы включают в себя: приведение в контакт штаммов LAB (например, L. lactis), содержащих ген IL-2 и ген T1D-специфического антигена (или способных экспрессировать IL-2 и T1D-специфический антиген) с фармацевтически приемлемым носителем, таким образом формируя фармацевтическую композицию. В некоторых примерах такой способ может дополнительно включать: выращивание штамма LAB в среде. Способ может дополнительно включать лиофилизацию жидкости, содержащей микроорганизм, причем эта жидкость в некоторых случаях включает фармацевтически приемлемый носитель.
Единичные лекарственные формы
[00211] Настоящее изобретение дополнительно предлагает единичные лекарственные формы, содержащие определенное количество штамма LAB, в некоторых случаях - в комбинации с пищевым или фармацевтически приемлемым носителем, в которых штамм LAB содержит: ген интерлейкина-2 (IL-2); и ген специфического антигена сахарного диабета 1-го типа (T1D). В качестве примера единичные лекарственные формы содержат от приблизительно 1×103 до приблизительно 1×1014 колониеобразующих единиц (КОЕ) LAB (например, L. lactis). В качестве примера другие единичные лекарственные формы содержат от приблизительно 1×104 до приблизительно 1×1013 колониеобразующих единиц (КОЕ) LAB (например, L. lactis) или от приблизительно 1×104 до приблизительно 1×1012 колониеобразующих единиц (КОЕ) LAB (например, L. lactis). В других вариантах реализации единичная лекарственная форма содержит от приблизительно 1×105 до приблизительно 1×1012 колониеобразующих единиц (КОЕ) или от приблизительно 1×106 до приблизительно 1×1012 колониеобразующих единиц (КОЕ) LAB (например, L. lactis). В других вариантах реализации единичная лекарственная форма содержит от приблизительно 1×108 до приблизительно 1×1012 колониеобразующих единиц (КОЕ) или от приблизительно 1×109 до приблизительно 1×1012 колониеобразующих единиц (КОЕ) LAB (например, L. lactis). В других вариантах реализации единичная дозированная форма содержит от приблизительно 1×109 до приблизительно 1×1011 колониеобразующих единиц (КОЕ) или от приблизительно 1×109 до приблизительно 1×1010 колониеобразующих единиц (КОЕ) LAB (например, L. lactis). В других вариантах реализации единичная дозированная форма содержит от приблизительно 1×1010 приблизительно 1×1011 колониеобразующих единиц (КОЕ) или от приблизительно 1×108 до приблизительно 1×1010 колониеобразующих единиц (КОЕ) LAB (например, L. lactis).
[00212] В других вариантах реализации единичная лекарственная форма содержит от приблизительно 1×109 до приблизительно 1×1010 колониеобразующих единиц (КОЕ) или от приблизительно 1×109 до приблизительно 100×109 колониеобразующих единиц (КОЕ) LAB (например, L. lactis).
[00213] Такая единичная лекарственная форма может иметь любое физическое состояние или геометрическую форму. В некоторых вариантах реализации единичная лекарственная форма может быть адаптирована для перорального введения. В некоторых примерах в соответствии с этими вариантами реализации единичная лекарственная форма может быть в форме капсулы, таблетки или гранулы. Примеры капсул включают капсулы, заполненные микрогранулами. В некоторых вариантах реализации LAB (например, L. lactis), содержащаяся в лекарственной форме, находится в форме сухого порошка. Например, LAB находится в лиофилизированной порошкообразной форме, которая в некоторых случаях спрессована и покрыта оболочкой.
ПРИМЕРЫ
[00214] Следующие примеры приведены для того, чтобы продемонстрировать и дополнительно проиллюстрировать некоторые типичные варианты реализации и аспект настоящего изобретения и не должны истолковываться как ограничивающие объем настоящего описания или формулы изобретения.
Пример 1. Конструирование Lactococcus lactis, секретирующей hIL-2 (LL-IL-2)
[00215] Был сконструирован штамм Lactococcus lactis, способный секретировать IL-2 человека (LL-IL-2) по сравнению с Lactococcus lactis MG1363 (родительский штамм). См., например, Gasson MJ, J. Bacterial. 1983, 154(1):1-9. В LL-IL-2 в геном бактерий были внесены следующие модификации:
[00216] (а) Ген тимидилатсинтазы (thyA; Gene ID: 4798358; локализация: NC_009004.1 (930251..931090)) был удален, чтобы обеспечить защиту от распространения бактерии.
[00217] (b) Ген трегалозо-6-фосфатфосфорилазы (trePP; Gene ID: 4797140; локализация: NC_009004.1 (449195..451504)) был удален, чтобы обеспечить накопление экзогенной трегалозы.
[00218] (с) Ген трегалозо-6-фосфатфосфатазы (otsB; Gene ID: 1036914, метка локуса с2311) был помещен ниже не идентифицированного гена секретируемого белка 45 кДа (usp45; Gene ID: 4797218; локализация: NC_009004.1 (2462440..2463 825, комплемент)) для облегчения превращения трегалозо-6-фосфата в трегалозу.
[00219] (d) Был добавлен конститутивный промотор гена HU-подобного ДНК-связывающего белка (PhllA; Gene ID: 4797353; локализация: NC_009004.1 (490275..490550)) перед предполагаемыми генами фосфотрансферазы в опероне трегалозы (trePTS; ptsI и ptsII; LLMG_RS02300 и LLMG_RS02305; Gene ID: 4797778; локализация: NC_009004.1 (446937..447422) и Gene ID: 4797093; локализация: NC_009004.1 (447563..449128), соответственно) для усиления поглощения трегалозы.
[00220] (е) Ген, кодирующий целлобиоза-специфический компонент IIC системы PTS (Gene ID: 4796893; локализация: NC_009004.1 (430271..431608)), ptcC, был удален для увеличения удерживания трегалозы.
[00221] (f) Ген, кодирующий слияние лидера секреции usp45 (Ssusp45) с геном hIL-2, кодирующим интерлейкин-2 человека (hIL-2, UniProt: P60568, аа 21-153), был помещен ниже гена фосфопирувата гидратазы (eno; Gene ID: 4797432; локализация:
NC_009004.1 (606184..607485)), чтобы обеспечить экспрессию и секрецию hIL-2. Единица экспрессии hIl-2 была транскрипционно и трансляционно сцеплена с eno посредством межгенной области, предшествующей экспрессированному на высоком уровне гену L30 рибосомного белка L. lactis MG1363 50S (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911), комплемент)). Пример нуклеотидной последовательности, кодирующей вышеуказанное слияние Ssusp45 и hIL-2 ниже от enoA, сцепленное посредством rpmD, показан на Фиг. 1 (SEQ ID NO: 46).
[00222] На Фиг. 2 представлен схематический обзор описанных выше генетических локусов.
[00223] Эти эксперименты также включают контрольный штамм (LL-Control) с генетическими признаками, сравнимыми с LL-IL-2, за исключением того, что контрольный штамм не содержит конструкций для экспрессии IL-2. Генетические признаки штаммов LL-IL-2 и LL-Control приведены в Таблице 1 ниже.
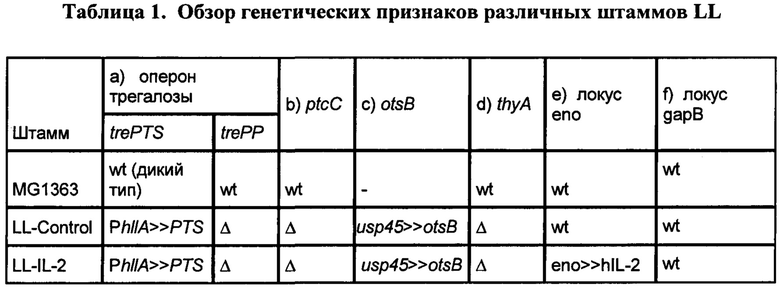
[00224] Исходя из Таблицы 1, экспрессия trePTS (на опероне трегалозы) может быть такой же, как у дикого типа (wt), или управляться промотором hllA (PhllA » PTS); trePP может быть wt или удален (Δ); ptcC может быть wt или Δ; otsB может отсутствовать (-) или локализован ниже и экпрессироваться из usp45 (usp45 >> otsB); thyA может быть wt или Δ; локус eno может быть wt (-) или содержать hIL-2. Все локусы gapB являются wt, в отличие от LL-PINS/IL-2, который несет в себе gapB >> pins, как описано в настоящей заявке ниже.
[00225] Генетические модификации выполняли с применением двойной гомологичной рекомбинации на 5'- и 3'-конце этих генетических признаков. Аналогичный способ описан для конструкции L. lactis Thy12 (см., например, Steidler L., et al., Nat. Biotechnol. 2003, 21(7):785-789), с той разницей, что плазмида-помощник pVE6007 не использовалась. Процедура включала отбор с применением эритромицина в качестве промежуточной стадии, с последующим удалением маркера отбора с помощью эритромицина. В результате LL-IL-2 практически не имеет остаточной резистентности к эритромицину.
Плазмиды-носители
[00226] Способ модификации использует плазмиды-носители, полученные из условно нерепликативного pORI19. См., например. Law J., et al., J. Bacterial. 1995; 177(24):7011-7018. Эта дефицитная по гену репликационного белка А, (repA)-плазмида, как и все ее repA- производные, не может реплицироваться в repA-L. lactis. Штамм L. lactis LL108 repA+ (см. Sanders et al., J. Bacterial. 1995, 177(18):5254-5260) был использован в качестве хозяина конструкции. Плазмиды-носители были сконструированы таким образом, что кроссоверные (ХО) области до 1 т.п.н., идентичные областям, которые фланкируют последовательность дикого типа на бактериальной хромосоме, расположены в позициях 5' и 3' переносимой плазмидами модификации. В качестве примера плазмиды используются для инсерции hIL-2 ниже от eno таким образом, что они сцеплены посредством межгенной области rpmD. Все конструкции плазмид была выполнены с применением стандартных молекулярно-биологических способов.
Хромосомные модификации
[00227] Производные плазмиды pORI19 несут маркер отбора с помощью эритромицина (ermC, ген 23S РНК-метилазы, ген ID: 1263245) и не могут реплицироваться в MG1363 или любом из его производных. После введения такой плазмиды в MG1363 к культуре применяли отбор с помощью эритромицина.
Резистентные колонии отбирали на чашках с твердым агаром, содержащих эритромицин. Из-за неспособности к репликации плазмид-носителей бактерии, резистентные к эритромицину, могут появляться только после первой гомологичной рекомбинации либо на 5'-, либо на 3'-сайте-мишени. Гомологичная рекомбинация может быть далее верифицирована посредством ПЦР.
[00228] Отбор с помощью эритромицина позволил вырезать плазмиду-носитель из бактериальной хромосомы с помощью второй гомологичной рекомбинации на 5'-, либо на 3'-сайте-мишени. Для некоторых чувствительных к эритромицину потомков вторая гомологичная рекомбинация может происходить на другом сайте-мишени по сравнению с первой гомологичной рекомбинацией. Это событие заменяет на бактериальной хромосоме дикий тип мутантом и может быть идентифицировано с помощью ПЦР. Соответствующее субкультивирование быстро уничтожит все остатки плазмид-носителей.
[00229] Наличие гена β-глюкуронидазы (uidA, Gene ID: 946149) в плазмидах-носителях, где он размножается вместе с ermC, позволяет идентифицировать чувствительные к эритромицину и резистентные к эритромицину колонии. Например, бактериальные суспензии высевали на чашки с твердым агаром, содержащие 5-бром-4-хлор-3-индолил-бета-D-глюкуроновую кислоту (X-Gluc). Экспрессирующие глюкуронидазу (GUS), (и, следовательно, резистентные к эритромицину) клоны будет выглядеть синими при превращения X-Gluc в нерастворимый синий продукт реакции дихлордибромдиниго, тогда как чувствительные к эритромицину клоны также потеряли ген uidA и, следовательно, останутся белыми. Идентификация синих и белых клонов на соответствующих этапах в описанном выше процессе значительно облегчила этот подход.
Анализ ПЦР
[00230] Колонии, демонстрирующие соответствующую гомологичную рекомбинацию на 3'-, либо в 5'-сайте-мишени, анализировали с помощью ПЦР. Фрагменты ДНК очищали с использованием набора для очистки PCR Qiagen MinElute. Полученные последовательности ДНК были идентичны предсказанным. На Фиг. 4 показан 1,2% агарозный гель, содержащий ПЦР-фрагменты, полученные с использованием олигонуклеотидов, перечисленных в Таблице 2, ДНК-полимеразы Herculase II Fusion DNA polymerase (Agilent Technologies, №600677) и соответствующих температурных циклов 50/120/30. Полученные результаты показали наличие целевых генетических признаков в LL-IL-2.

[00231] На Фиг. 4, маркеры молекулярной массы (MWM; маркер длины ДНК Invitrogen 10488-85 TrackIt 1 kb plus DNA Ladder) обозначают пары оснований: 100, 200, 300, 400, 500, 650, 850, 1000, 1650, 2000, 3000, 4000, 5000 и выше. Ожидаемые размеры фрагментов ДНК также указаны в парах оснований.
[00232] Далее секвенировали бактериальный геном LL-IL-2. Экспериментально определенные последовательности ДНК всех генетических признаков в LL-IL-2, которые отличаются от таковых родительского штамма MG1363, оказались идентичными, как и ожидалось.
Экспрессия hIL-2
[00233] Экспрессию hIL-2 с помощью LL-IL-2 измеряли методами ИФА и вестерн-блоттинга. В эксперименте ИФА (с использованием huIL-2 № DY202, R&D Systems) измерения выявили 47,1 нг/мл hIL-2 в культуральном супернатанте, тогда как контрольный штамм не продуцировал hIL-2.
[00234] На Фиг. 5 представлен результат вестерн-блоттинга, показывающий присутствие hIL-2 в супернатанте культуры LL-IL-2. Вестерн-блоттинг проводили с использованием антитела козы против IL-2 человека (1/1000 AF-202-NA, R&D Systems) в качестве первого антитела, с инкубацией с антителом кролика против антитела козы - АР (1/1000 №6160-04, Southern Biotech) в качестве идентифицирующего антитела и последующим окрашиванием NBT/BCIP (таблетки NBT/BCIP, Roche, №11697471001). Эквиваленты 1 мл бактериальных культур LL-IL-2 и контрольных штаммов загружали в белковый гель. Предварительно окрашенный стандарт SeeBlue® Plus2 Invitrogen использовали в качестве маркера молекулярной массы (MWM). Полученные данные показывают, что LL-IL-2 секретирует полноразмерный hIL-2 (то есть, такой же, как кодируемый SEQ ID NO: 46).
[00235] Бактерии культивировали в среде GM17, которая представляет собой бульон Ml 7 (Oxoid, № СМ0817) с добавкой 0,5% глюкозы, или в среде GM17T- (GM17 с добавкой 200 мкМ тимидина).
Пример 2. Lactococcus lactis, секретирующий PINS и hIL-2 (LL-PINS/IL-2)
[00236] Описаны конструирование и отбор штамма LL-PINS/IL-2, производного Lactococcus lactis (L. lactis) MG1363. LL-PINS/IL-2 включает следующие генетические признаки: (а) ген тимидилатсинтазы (thyA; Gene ID: 4798358; локализация: NC_009004.1 (930251..931090)) был удален, чтобы обеспечить защиту окружающей среды от распространения бактерии; (b) ген трегалозо-6-фосфатфосфорилазы (trePP; Gene ID: 4797140; локализация: NC_009004.1 (449195..451504)) был удален, чтобы обеспечить накопление экзогенно добавленной трегалозы; (с) ген трегалозо-6-фосфатфосфатазы (otsB; Gene ID: 1036914, метка локуса с2311) был помещен ниже неидентифицированного гена секретируемого белка 45 кДа (usp45; Gene ID: 4797218; локализация: NC_009004.1 (2462440..2463825, комплемент)) для облегчения превращения трегалозо-6-фосфата в трегалозу; (d) единица экспрессии otsB была транскрипционно и трансляционно сцеплена с gapB, посредством межгенной области, предшествующей экспрессированному на высоком уровне гену рибосомного белка L. lactis MG1363 50S (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911), комплемент)); (е) конститутивный промотор гена HU-подобного ДНК-связывающего белка (PhllA; Gene ID: 4797353; локализация: NC_009004.1 (490275..4905 50)) предшествует предполагаемым генам фосфотрансферазы в опероне трегалозы (trePTS; ptsI и ptsII; LLMG_RS02300 и LLMG_RS02305; Gene ID: 4797778; локализация: NC_009004.1 (446937..447422) и Gene ID: 4797093; локализация: NC_009004.1 (447563..449128) соответственно) для усиления поглощения трегалозы; (f) ген, кодирующий целлобиоза-специфический компонент IIC системы PTS (Gene ID: 4796893; локализация: NC_009004.1 (430271..431608)), ptcC, был разрушен (tga в положении кодона 30 из 446, tga30) для обеспечения удерживания трегалозы после накопления; (g) ген, кодирующий слияние лидера секреции usp45 (Ssusp45) с геном PINS, кодирующим проинсулин человека (PINS; UniProt: P01308, аа 25-110) помещен ниже от гена глицеральдегида 3-фосфатдегидрогеназы (gapB; Gene ID: 4797877; локализация: NC_009004.1 (2492509..2493519, комплемент)), чтобы обеспечить экспрессию и секрецию проинсулина; (h) единица экспрессии PINS была транскрипционно и трансляционно сцеплена с gapB посредством межгенной области, предшествующей экспрессированному на высоком уровне гену рибосомного белка L. lactis MG1363 50S (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911, дополнение)); и (i) ген, кодирующий слияние лидера секреции usp45 (Ssusp45) с геном hIL-2, кодирующим интерлейкин-2 человека (hIL-2, UniProt: Р60568, аа 21-153), был помещен ниже гена фосфопирувата гидратазы (eno; Gene ID: 4797432; локализация: NC_009004.1 (606184..607485)), чтобы обеспечить экспрессию и секрецию hIL-2. Единица экспрессии hIl-2 была транскрипционно и трансляционно сцеплена с eno посредством межгенной области, предшествующей экспрессированному на высоком уровне гену L30 рибосомного белка L. lactis MG1363 50S (rpmD; Gene ID: 4797873; локализация: NC_009004.1 (2316732..2316911, комплемент)).
[00237] Все генетические признаки LL-PINS/IL-2 находятся на бактериальной хромосоме. Генетический фон этого штамма обеспечивает: конститутивную секрецию PINS и hIL-2; строгую зависимость от экзогенно добавленного тимидина для роста и выживания; и способность накапливать и удерживать трегалозу для устойчивости против, например, лизиса под действием желчных кислот.
[00238] Пример нуклеотидной последовательности, кодирующей вышеуказанное слияние SSusp45 и PINS ниже гена gapB, сцепленное межгенной областью rpmD, изображена на Фиг. 6 (SEQ ID NO: 57). На Фиг. 7 представлен схематический обзор описанных выше генетических локусов. Генетические признаки вводили в бактериальный геном, как описано в Примере 1. Бактериальные штаммы выращивали и анализировали, как описано в Примере 1 выше. Олигонуклеотиды, используемые при конструировании и анализе LL-PINS/IL-2, приведены в Таблице 3 ниже.

[00239] На Фиг. 9 представлен анализ на 1,2% агарозном геле фрагментов ПЦР из LL-PINS/IL-2, указывающий на наличие целевых генетических признаков: trePTS, ptcC-, eno >> hil-2, otsB, gapB >> pins. На Фиг. 9, маркеры молекулярной массы (MWM; маркер длины ДНК Invitrogen 10488-85 Trackit 1 kb plus DNA Ladder) обозначают пары оснований: 100, 200, 300, 400, 500, 650, 850, 1000, 1650, 2000, 3000, 4000, 5000 и выше. Ожидаемые размеры фрагментов ДНК также указаны в парах оснований.
[00240] Далее секвенировали бактериальный геном LL-PINS/IL-2 Экспериментально определенные последовательности ДНК всех генетических признаков в LL-PINS/IL-2, которые отличаются от таковых родительского штамма MG1363, оказались идентичными предсказанным.
[00241] Способы гомологичной рекомбинации включали плазмиды-носители, полученные из условно нерепликативного pORI19, описанного выше. Плазмиды-носители были сконструированы таким образом, что кроссоверные (ХО) области до 1 т.п.н., идентичные областям, которые фланкируют последовательность дикого типа на бактериальной хромосоме, расположены в позициях 5' и 3' переносимой плазмидой модификации. Примером плазмиды-носителя является pAGX1145, схема которой показана на Фиг. 8. Эту плазмиду используют для инсерции PINS ниже от gapB таким образом, что они сцеплены посредством межгенной области rpmD. Аналогичную плазмиду pAGX1372 (см. Приложение: pAGX1372.gbk) используют для инсерции hil-2 ниже eno. Конструирование всех плазмид было выполнено с применением стандартных молекулярно-биологических способов.
Экспрессия PINS и hIL-2
[00242] Экспрессию PINS и hIL-2 штаммом LL-PINS/IL-2 измеряли с использованием ИФА и вестерн-блоттинга. Культуральные супернатанты из LL-PINS/IL-2 содержали 0,6 нг/мл PINS и 28,2 нг/мл hIL-2, тогда как контрольный штамм (LL-Control) не продуцировал никаких полипептидов. В качестве положительного контроля использовали бактериальный штамм MG 1363, экспрессирующий PINS из плазмидного вектора (LL-PINS). Содержимое PINS в супернатантах определяли с использованием набора кат. №10-1118-01 Mercodia, а содержание hIL-2 определяли с использованием huIL-2 № DY202, R&D Systems.
[00243] На Фиг. 10 представлен вестерн-блоттинг, показывающий присутствие PINS и hIL-2 в супернатанте культуры LL-PINS/IL-2. Эквиваленты 1 мл бактериальных культур загружали в белковый гель. Вестерн-блоттинг проводили с использованием поликлональных антител козы против инсулина В (N-20: sc-7838, Santa Cruz) и антитела козы против 1b-2 человека (1/1000 AF-202-NA, R&D Systems) в качестве первых антител к PINS и hIL-2 соответственно, с инкубацией с антителом кролика против антител козы - АР (1/1000 №6160-04, Southern Biotech) и последующим окрашиванием NBT/BCIP (таблетки NBT/BCIP, Roche, №11 697 471 001). Предварительно окрашенный стандарт SeeBlue® Plus2 Invitrogen использовали в качестве маркера молекулярной массы (MWM). Полученные данные показывают, что LL-PINS/IL-2 секретирует полноразмерный PINS и hIL-2.
[00244] Бактерии культивировали в среде GM17, представляющей собой бульон M17 (Oxoid, № СМ0817) с добавкой 0,5% глюкозы, или в среде GM17T (GM17 с добавкой 200 мкМ тимидина).
Пример 3. Фармакодинамические исследования для изучения влияния двух бактериальных штаммов на прогрессирование диабета, Lactococcus lactis (LL), секретирующей проинсулин (LL-PINS) и hIL-2 (LL-IL-2)
[00245] Бактерии культивировали согласно описанию в работе Takiishi, Т. et al., J. Clin. Inv. 2012, 122(5): 1717-1725.
[00246] Например, одиночные колонии соответствующего L. lactis инокулировали в среде GM17T (М17, Oxoid, Хемпшир, Великобритания, добавляли 0,5% глюкозы, 200 мкМ тимидина) и выращивали в течение ночи до насыщения. Разбавление этой 1/25 культуры предварительно выращивали в течение 3 часов в GM17T. Бактерии собирали центрифугированием и дополнительно инкубировали в течение 3 часов в буферной культуральной среде (1 × соли ВМ9, 0,5% казитон (Difco, BD Biosciences), 0,5% глюкозы, 25 NaHCO3, 25 mM Na2CO3, 2 mM MgSO4, 0,1 μМ CaCl2 ((среда ВМ9) (Schotte, et al. (2000) Enzyme Microb. Technol. 27(10):761-765) с добавкой 200 мкМ тимидина) (ВМ9Т). Бактерии удаляли центрифугированием и образцы супернатантов брали для анализа с помощью вестерн-блоттинга и ИФА. Для вестерн-блоттинга белки получали из неочищенных супернатантов ВМ9Т L. lactis путем осаждения диоксихолатом/ТСА (трихлоруксусная кислота)/ацетоном и растворяли в буфере для образцов SDS-PAGE. Осадок бактериальных клеток разрушали для получения внутриклеточных фракций. Культуральные супернатанты (эквивалентные 1 мл культуры) и внутриклеточные (эквивалентные 50 мкл культуры) белковые фракции разделяли с помощью SDS-12% PAGE, проводили иммуноблоттинг и проявляли антителом козы против hIL-2, а затем детектировали с помощью антитела кролика против антител козы и NBT/BCIP.
[00247] Исходные растворы всех штаммов сохраняют при -20°С в 50% глицерина в среде GM17. Бактерии культивируют в среде GM17, то есть М17 (Difco Laboratories, Детройт, Мичиган) с добавкой 0,5% глюкозы.
[00248] В экспериментальной установке использовали NOD мышей с впервые выявленным диабетом при положительной глюкозурии и двух последовательных измерениях уровня глюкозы в крови, превышающие 200 мг/дл. Мыши были распределены на три экспериментальные группы лечения: (1) не получавшие лечения контрольные мыши, (2) мыши, получавшие LL-PINS и (3) мыши, получавшие комбинацию LL-PINS и LL-IL-2 в течение 6 недель.
[00249] Этот эксперимент включал два разных штамма LL. Один штамм конститутивно экспрессирует PINS, а другой штамм конститутивно экспрессирует IL-2. Мышам вводили LL в дозе 2×109 КОЕ перорально через желудочный зонд 5 раз в неделю в течение шести (6) недель.
[00250] Мышей наблюдали в течение 42 дней (до прекращения терапии), либо 100 дней (8 недель после прекращения терапии). Помимо первоначального наблюдения на ремиссию болезни в течение 100 дней, дополнительных мышей (не получавших лечения и получавших LL-PINS + LL-IL-2) подвергали эвтаназии через 42 дня после начала лечения, и использовали их периферическую кровь и другие органы для дальнейшего анализа. Образцы сыворотки для измерения аутоантител к инсулину (IAA), воспалительных цитокинов и стимулированного глюкозой С-пептида собирали перед лечением и после прекращения терапии (день 42). Во всех экспериментальных группах (как с ремиссией болезни, так и без ремиссии) оценивали периферическую иммунную систему (фенотип и функция). Также отбирали образцы поджелудочной железы для гистологии (инсулит) и определения содержания инсулина (1C) на момент прекращения терапии (день 42).
Оценка T1D и инсулита
[00251] NOD-мышей подвергали скринингу на возникновение диабета оценкой концентрации глюкозы в моче (Clinistix, Bayer Diagnostics) и венозной крови (Accu-Chek® Aviva, Roche Diagnostics). Измерения уровня глюкозы в крови не натощак выполняли между 8 и 11 часами утра. Мышей классифицировали как имеющих диабет при наличии положительной гликозурии и двух последовательных измерений уровня глюкозы в крови, превышающих 200 мг/дл. Ремиссию диабета определяли как отсутствие гликозурии и величин гликемии <250 мг/дл в течение двух дней подряд.
[00252] Образцы поджелудочной железы фиксировали в растворе формальдегида и обрабатывали для заливки в парафин. Срезы толщиной 7 миллиметров окрашивали гематоксилином/эозином, и оценивали микроскопически степень инсулита. Повреждения островков оценивали следующим образом: 0: отсутствие инфильтрации; 1: пери-инсулит; 2: островки с инфильтрацией лимфоцитов на площади менее 50%; 3: островки с инфильтрацией лимфоцитов на площади более 50%; 4: островки полностью разрушены.
Обнаружение ауто-антител
[00253] Наличие IAA в сыворотке измеряли при начале заболевания и при прекращении терапии (день 42) с помощью анализа RIA. Было проверено, способна ли вакцинация LL-PINS + LL-IL-2 корректировать гипергликемию (ремиссию болезни) и поддерживать нормогликемию у NOD-мышей с впервые выявленным диабетом. Концентрации глюкозы в крови наблюдали в течение 14 недель после начала лечения.
Результаты - ремиссия заболевания
[00254] На Фиг. 11 показан процент мышей, которые после лечения сохраняли диабетический статус. После развития у NOD-мышей гипергликемии (2 дня подряд концентрации глюкозы в крови >200 мг/дл), они обычно прогрессировали до тяжелой гипергликемии с минимальными спонтанными ремиссиями, и большинство умирали в течение 3-6 недель (n=27).
[00255] Монотерапия инокуляцией LL-PINS (2×109 КОЕ/сутки, 5 дней в неделю в течение 6 недель, n=8) корректировала гипергликемию у 15% мышей. Примечательно, что 43% мышей с впервые выявленным диабетом (n=45), получавших комбинацию LL-PINS и LL-IL-2, быстро восстанавливали нормогликемию. Терапия LL-PINS + LL-IL-2 вызывала стабильную и постоянную ремиссию диабета, поскольку у излеченных мышей нормогликемия поддерживалась в течение дополнительного периода наблюдения 8 недель после прекращения терапии,
[00256] Клиническая эффективность терапии LL-PINS и LL-IL-2 явно зависит от концентрации глюкозы в крови на момент начала лечения. NOD-мышей с недавно выявленным диабетом стратифицировали исходя из исходных концентраций глюкозы в крови - ниже или выше 350 мг/дл. Терапия LL-PINS plus LL-IL-2 не только излечила 57% мышей с начальной гликемией ниже 350 мг/дл, но и 22% мышей с исходной гликемией выше 350 мг/дл (Фиг. 12). Эти данные впервые демонстрируют, что доставка в слизистую оболочку PINS с IL-2 рекомбинантными бактериями L. lactis эффективно корректирует гипергликемию и восстанавливает иммунную толерантность к β-клеткам у NOD мышей с манифестным недавно выявленным T1D.
[00257] Во всех кривых выживаемости Каплана-Мейера статистическую значимость между группами определяли с учетом логрангового критерия Мантеля-Кокса (*: р<0,05).
Пример 4. hIL-2, секретируемый L. lactis (LL-IL-2), обладает биологической активностью, сравнимой с рекомбинантным hIL-2
[00258] Этот эксперимент включал описанный в настоящей заявке штамм Lactococcus lactis, экспрессирующий IL-2 человека (LL-IL-2) (см., например, Пример 1), и контрольный штамм.
[00259] Биоактивность LL-IL-2 измеряли исходя из IL-2-зависимой выживаемости/пролиферации клеточной линии мышиных Т-лимфоцитов НТ2 клона А5Е. Клетки НТ2 трижды промывали средой без IL-2 и высевали с плотностью 4×103 клеток/96 лунок. Серию последовательных разбавлений рекомбинантного hIL-2 (например, № 202-IL-010, R&D systems) или супернатант из LL-IL-2 и контрольный штамм добавляли к высеянным клеткам и инкубировали в течение 24 часов при 37°С, 5% CO2 и высокой влажности. Жизнеспособность клеток измеряли с применением метода CellTiter96®AQouous One Solution (Promega № G3582). В каждую лунку добавляли 20 мкл раствора МТТ и после инкубационного периода 4 часа при 37°С, 5% CO2 и высокой влажности, планшеты считывали при длине волны 490 нм с использованием 700 нм в качестве эталона. Рекомбинантный hIL-2 (R&D Systems) и hIL-2, полученные из LL-IL-2, показывают сопоставимые дозозависимые ответы, тогда как супернатант контрольного штамма L. lactis был неактивен. Результаты показаны на Фиг. 13.
Пример 5. LL-IL-2 доставляет низкие дозы hIL-2 в желудочно-кишечный тракт не страдающих ожирением мышей с диабетом после перорального введения
[00260] Этот эксперимент включал штамм Lactococcus lactis, экспрессирующий IL-2 человека (LL-IL-2), описанный в настоящей заявке, например, в Примере 1.
Живые бактерии
[00261] Концентрации живых бактерий (КОЕ/ткань и КОЕ/г) в разных тканях ЖКТ измеряли в разные моменты времени после однократного введения 1010 КОЕ LL-IL-2 через пероральный желудочный зонд. Результаты показаны на Фиг. 14А и 14В соответственно, в которых каждый столбик представляет собой среднее значение от 3 мышей (n=3). Исходя из Фиг. 14А (КОЕ/ткань), значимые количества бактерий LL-IL-2 были обнаружены в слепой кишке (САЕ), проксимальной толстой кишке (СОР) и дистальной толстой кишке (COD) спустя 2, 4, 6 и 8 часов. Концентрация бактерий в тонком кишечнике оказалась ниже 106 КОЕ/ткань. Исходя из Фиг. 14В (КОЕ/г), концентрации бактерий LL-IL-2 были обнаружены в проксимальной тонкой кишке (SIP), дистальной тонкой кишке (SID), слепой кишке (САЕ), проксимальной толстой кишке (СОР) и дистальной толстой кишке (COD) спустя 2, 4, 6 и 8 часов, соответственно. В крови бактерий обнаружено не было.
Белок hIL-2
[00262] Концентрации белка hIL-2 (пг/ткань и пг/г) в разных тканях ЖКТ измеряли после введения однократной дозы бактерий LL-IL-2 (1010 КОЕ). Концентрации белка hIL-2 были обнаружены в слепой кишке (САЕ), проксимальной толстой кишке (СОР) и дистальной толстой кишке (COD) через 2 и 4 часа. Концентрация белка hIL-2 в тонком кишечнике оказалась ниже предела количественной оценки (LLOQ=10 пг/мл). В кровотоке тестируемых мышей белок hIL-2 обнаружен не был. Измеренные концентрации белка hIL-2 приведены в Таблице 4 и Таблице 5 ниже.
[00263] После умерщвления каждую ткань (SIP, SID, САЕ, СОР или COD) целиком взвешивали и гомогенизировали. Образец гомогената использовали для высевания на планшет (для определения КОЕ) и для ИФА (для определения hIL-2). "Суммарная ткань" в этом контексте означает, что концентрацию бактерий или белка, определенную в образце гомогената, пересчитывали в массу всей ткани.
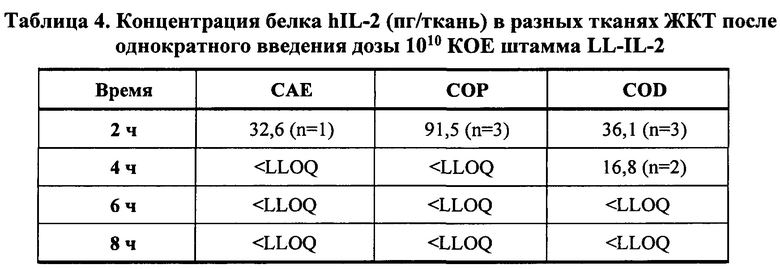
САЕ = слепая кишка; СОР = проксимальная толстая кишка; COD = дистальная толстая кишка; LLOQ = 10 пг/мл
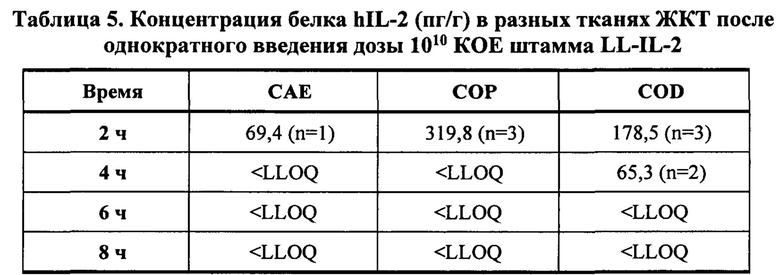
САЕ = слепая кишка; СОР = проксимальная толстая кишка; COD = дистальная толстая кишка; LLOQ = 10 пг/мл
[00264] Жизнеспособные L. lactis были обнаружены по всему ЖКТ, причем большинство бактерий находилось в проксимальной и дистальной части толстой кишки и в слепой кишке. Концентрация бактерий здесь была в 1000 раз выше, чем в дистальной и проксимальной части тонкой кишки. Это может быть объяснено большим количеством слизи и низкой перистальтикой в этих частях кишечника, приблизительно 50% вводимого L. lactis можно было выделить из дистальных частей толстой кишки через 2 часа после введения. Этот вывод является неожиданным, поскольку ранее сообщалось, что только приблизительно 10-30% перорально вводимого L. lactis выживают при прохождении двенадцатиперстной кишки (Drouault S, et al., Appl. Environ. Microbiol. 1999; 65(11): 4881-6). Было сделано предположение, что инокуляция бактерий с помощью инокуляционного буфера ВМ9 способна защитить бактерии (по крайней мере частично) от условий ЖКТ.
[00265] Согласно оценке, после дозирования приблизительно 1010 КОЕ LL-IL-2 в ткань доставляется приблизительно 90 мкг IL-2, что соответствует приблизительно 1,2 ME IL-2 (принимая 1 ME=73 пг).
Пример 6. Фармакодинамические и механистические исследования для изучения влияния клинического штамма Lactococcus lactis (LL), секретирующего как проинсулин (PINS), так и hIL-2
[00266] Этот эксперимент включает штамм Lactococcus lactis, экспрессирующий как PINS, так и IL-2 (LL-PINS/IL-2), описанный в настоящей заявке (см., например, Пример 2). Бактерии можно культивировать, как описано выше. См. Takiishi, Т. et al., J. Clin. Inv. 2012, 122(5); 1717-1725. У NOD-мышей можно проводить скрининг, лечение и анализ, как описано в Примере 3 выше. Мышам можно вводить бактерии, например, в дозе 2×109 КОЕ через желудочный зонд 5 раз в неделю в течение шести (6) недель.
Фенотипический анализ местной и периферической иммунной системы
[00267] Периферические органы (т.е. кровь, брыжеечные и поджелудочные лимфатические узлы и поджелудочная железа) можно извлечь при прекращении терапии (например, на 42-й день) и фенотипически исследовать, например, методом проточного цитометрического анализа на канонические и неканонические маркеры Treg (CD3, CD4, CD25, Foxp3, CD39, CD49b, LAG-3 и CD73). Например, совместная экспрессия CD49b и LAG-3 позволяет характеризовать высокосупрессивные Тг1-клетки, продуцирующие IL-10 (см., например, Gagliani, N. et al., Nat. Med. 2013, 19(6): 739-746), в то время как Treg, экспрессирующие как эктоферменты CD39, так и CD73, производят высокие концентрации аденозина, который считается одним из механизмов супрессии, присущих Treg. См., например, Antonioli, L., et al., Trends Mol. Med. 2013, 19(6): 355-367.
[00268] Для окрашивания внутриклеточных цитокинов иммунные клетки можно повторно стимулировать, например, 1 мкг/мл фосфор-миристиновой кислоты (РМА, Sigma-Aldrich) и 0,5 мкг/мл иономицина (Sigma-Aldrich) в течение 4 часов в присутствии 1 мкл/мл GolgiPlug™ (BD). После окрашивания поверхности клеток можно проводить внутриклеточное окрашивание, например, с использованием набора Cytofix/Cytoperm™ (BD) (то есть CD3, CD4, CD8, IL-2, IL-4, IL-17 и IFN-y).
[00269] Для обнаружения pSTAT-5 клеточные суспензии можно быстро фиксировать после умерщвления или после культивирования in vitro, например, в 10 объемах раствора PBS 1,5% формальдегида в течение 10 минут при комнатной температуре. Клетки можно промыть, например, в растворе PBS, содержащем 0,2% BSA, и пермеабилизировать, например, 100% метанолом в течение 10 минут на льду. Клетки можно дополнительно промыть, например, PBS 0,2% BSA, и затем инкубировать с фосфоспецифическим антителом в сочетании с интересующими антителами (например, анти-CD3, CD4, CD8, CD25, CD69, CD44, CD122), например, в течение 30 минут в темноте при комнатной температуре. В некоторых случаях можно добавлять антитело против Ki67 вместе с антителом против Foxp3. Отрицательный порог для pSTAT5 можно определить на нестимулированных клетках или на клетках, окрашенных всеми флуоресцентными антителами минус pSTAT5.
[00270] Многопараметрический анализ может быть выполнен, например, с использованием приборов FACS Gallios (Beckman Coulter), FACS Canto II (Becton Dickinson (BD)) или FACS Fortessa (BD), а результаты проанализированы с помощью программного обеспечения FlowJo® (Tree Star). Мертвые клетки (селективное окрашивания живых и мертвых клеток красителем желтый 405) и дублеты могут быть исключены из всех анализов.
Анализ in vitro поликлональной супрессии и детектирование IFN-γ
[00271] Супрессирующую функцию периферических Treg, выделенных из селезенки и лимфатических узлов (в идеале - выделенных из hCD2.Foxp3 NOD-мышей), можно оценить, например, в описанном анализе поликлональной супрессии in vitro. См., например, Takiishi, Т., et al., J Clin. Invest. 2012. 122(5):1717-1725. IFN-γ измеряют в бесклеточных супернатантах.
Результаты
[00272] Ожидается, что лечение LL-PINS/IL-2 будет стимулировать и рекрутировать Treg и показывать биологическую активность, сравнимую с лечением LL-IL-2+LL-PINS, описанным в настоящей заявке, например, в Примерах 3 и 8.
Пример 7. Конструирование Lactococcus lactis, секретирующей проинсулин (LL-PINS)
[00273] Последовательность ДНК, кодирующую проинсулин (hpins) человека, получали из GenBank (номер доступа NM_00207.2). Последовательность ДНК hpins удлиняли на 3'-конце стоп-кодоном ТАА и сайтом рестрикции SpeI. ДНК-фрагмент синтезировали с помощью ПЦР-сборки 40-мерных олигонуклеотидов (от OAGX0362 до oAGX0377) и ДНК-полимеразы Accu Prime для амплификации. Амплифицированный фрагмент сливали с сигналом секреции usp45 (SSusp45), ниже промотора thyA лактококка (PthyA), который был удлинен на 5'-конце сайтом рестрикции EcoRI. Амплифицированный продукт, имеющий 5' EcoRI-конец и 3' SpeI-конец, вставляли между сайтами рестрикции EcoRI и SpeI плазмиды pTINX (номер доступа GenBank НМ 585371.1) и лигировали. Лигирование вводили в L. lactis MG1363 путем электропорации, и колонии подвергали скринингу с помощью ПЦР-анализа. Полученную плазмиду обозначали pAGX0053 (Фиг. 15).
[00274] Из MG1363 [pAGX0053] был изготовлен фрагмент ПЦР, содержащий модуль экспрессии PthyA >> SSusp45::hpins, с использованием OAGX0169 и OAGX0170. После очистки этого фрагмента было подтверждено, что последовательность ДНК штамма MG1363 [pAGX0053] идентична предсказанной последовательности. Конструирование плазмиды проводили с применением стандартных молекулярно-биологических способов.
[00275] Экспрессию PINS тестировали на супернатанте культуры (SN) из [MG1363] pAGX0053 методом ИФА и вестерн-блоттингом, MG1363 [pAGX0053] секретирует 2,47 нг/мл PINS, по данным Pro-Insulin Elisa (Mercodia №10-1118-01). Образцы неочищенного SN готовили для вестерн-блоттинга. Эквиваленты 1 мл бактериальной культуры [MG1363] pAGX0053 и контрольных штаммов MG1363 [pTINX] и SAGX0037 загружали на белковый гель. Образцы инкубировали с поликлональным антителом козы против инсулина В (Santa Cruz N-20: sc-7838, 1/500). Детектирование проводили путем инкубации с антителом кролика против антител козы АР (Southern Biotech №6160-04, 1/1000) и последующим окрашиванием NBT/BCIP (таблетки Roche NBT/BCIP №11 697 471 001, согласно инструкциям изготовителя). Предварительно окрашенный стандарт Invitrogen SeeBlue® Plus-2 использовали в качестве маркера молекулярной массы (MWM). Данные представлены на Фиг. 16, показывающий секрецию полноразмерного PINS LL-PINS.
[00276] Штаммы Lactococcus lactis, содержащие экзогенные нуклеиновые кислоты, кодирующие T1D-специфические антигены, отличные от PINS, такие как GAD65, IA-2, IGRP, ZnT8, ppIAPP, периферии, хромогранин А и GRP, могут быть получены в соответствии с вышеописанной процедурой с применением соответствующих нуклеиновых кислот вместо hpins.
Пример 8. Фармакокинетическое профилирование LL-IL-2, перорально вводимого отдельно по сравнению с LL-PINS+LL-IL-2
Бактериальная культура
[00277] Бактерии культивировали, как описано ранее у Takiishi, Т. et al., J. Clin. Inv. 2012, 122(5): 1717-1725. Например, LL-pT1NX и LL-PINS культивировали в среде GM17TE (бульон М17 с добавлением 0,5% глюкозы, 200 мкМ тимидина и 5 мкг/мл эритромицина). LL-IL-2 культивировали в среде GM17T (бульон М17 с добавлением 0,5% глюкозы и 200 мкМ тимидина). Исходные суспензии штаммов LL хранили при -80°С в глицерине. Остатки суспензии разбавляли 1/1000 в среде роста (GM17TE или GM17T соответственно) и инкубировали в течение 16 часов при 30°С, достигая плотности насыщения 2×109 КОЕ/мл. Бактерии собирали центрифугированием в течение 10 минут при 4°С и 2900 об/мин и концентрировали 10-кратно в среде ВМ9Т (соли 5 × М9, 10% казитон, 10% глюкоза, 0,5 М NaHCO3, 0,5 М Na2COs, 1 M MgCl2, 100М CaCl2 и 100 мМ тимидина) для внутрижелудочных инокуляций. Лечение состояло из 100 мкл этой суспензии для LL-pT1NX, LL-PINS и LL-IL-2. Комбинацию LL-PINS+LL-IL-2 получали смешиванием равных частей суспензий LL-PINS и LL-IL-2. Затем лечение состояло из 200 мкл этой суспензии.
График введения и дозировка
[00278] NOD-мыши с впервые выявленным диабетом (2 измерения подряд уровня глюкозы в крови более 200 мг/дл и положительная глюкозурия) получали 2×109 колониеобразующих единиц (КОЕ) живых генетически модифицированных бактерий Lactococcus lactis 5 раз в неделю (в будние дни) в течение 6 недель. Мыши в группе лечения LL-PINS+LL-IL-2 получали 2×109 КОЕ LL-PINS и 2×109 КОЕ LL-IL-2. Мыши в контрольной группе 1 не получали никакого лечения, а мыши в контрольной группе 2 получали бактерии, несущие пустой вектор (LL-pT1NX). После 6-недельного периода лечения некоторые мыши были зарезервированы для дальнейшего анализа через 14 недель после начала лечения.
[00279] Нормогликемические NOD-мыши (возраст 22 недели) получали одну дозу 2×109 КОЕ LL-IL-2. Через два, четыре, шесть или восемь часов после введения дозы мышей подвергали эвтаназии и собирали цельную кровь и сыворотку. Проксимальную тонкую кишку (PSI), дистальную тонкую кишку (DSI), слепую кишку (САЕ), Проксимальную толстую кишку (РСО) и дистальную толстую кишку (DCO) забирали в буфере для гомогенизации (10 × соли М9, 0,5 М NaHCO3, 0,5 М Na2CO3, 10% бычий сывороточный альбумин, дистиллированная вода) при концентрации 100 мг/мл и механически диссоциировали. Гомогенаты тканей кишечника и цельной крови высевали в последовательных разведениях на планшетах GM17 для количественной оценки выделения бактерий. Концентрацию IL-2 в сыворотке и гомогенатах измеряли с помощью ИФА.
Забор органов
[00280] Через шесть недель после начала лечения мышей подвергали эвтаназии с помощью CO2. Кровь забирали путем сердечной пункции гепаринизированной иглой. Кровь разделяли на аликвоты (200 мкл) для переработки в отдельные клетки для проточной цитометрии и для разделения плазмы (центрифугировали в течение 10 мин при 2000 g при комнатной температуре). Поджелудочные железы забирали для гистологического анализа и хранили в 5% параформальдегиде (PFA). Следующие лимфоидные органы извлекали для анализа проточной цитометрией; селезенку (SPL), брыжеечные лимфатические узлы (MLN) и лимфоузлы поджелудочной железы (PLN).
Гистология поджелудочной железы и градация инсулита
[00281] Из поджелудочных желез, фиксированных с помощью PFA и залитых в парафин, готовили срезы толщиной 6 мкм, забранные на расстоянии 100 мкм, затем окрашивали гематоксилином и эозином. Парафин удаляли с использованием ксилола с последующей дегидратацией растворами этанола 100%-90%-70%-50%. Срезы регидратировали водопроводной водой и дистиллированной водой. Срезы окрашивали в течение 3 минут в гематоксилин и промывали водопроводной водой. Срезы кратковременно промывали в кислом этаноле 3 раза, а затем тщательно промывали водопроводной водой. Образцы помещали в насыщенный водный Li2CO3 и промывали в водопроводной воде. Затем образцы помещали в раствор эозина Y (0,5% водный раствор) и промывали в водопроводной воде. Срезы дегидратировали 50% этанолом, 70% этанолом, 90% этанолом, дважды 100% этанолом и дважды ксилолом в течение 10 секунд на каждой стадии. Препараты накрывали покровным стеклом, и наблюдали островки под световым микроскопом при 20× или 40× увеличении и сортировали по виду. Инфильтрацию островков оценивали следующим образом: 0 - отсутствие инфильтрации; 1 - пери-инсулит; 2 - островки с инфильтрацией на менее 50% площади; 3 - островки с инфильтрацией на более 50% площади; 4 - полностью разрушенные островки/тяжелый инсулит.
ИФА для определения С-пептида
[00282] Для определения уровней С-пептида в плазме использовали серийно производимый набор ИФА для определения С-пептида крысы/мыши (EZRMCP2-21K, EMD Millipore, Сент-Чарльз, Миссури). В кратком изложении, 96-луночный планшет три раза промывали 1 × HRP промывочным буфером (50 мМ Трис-солевой буфер, содержащий Tween-20, разбавленный 1:10 дистиллированной водой). Матричный раствор (сывороточная матрица, содержащая 0,008% азида натрия) добавляли в лунки с холостым раствором, стандартом и образцами контроля качества. Затем во все лунки добавляли буфер для анализа (0,05 М фосфосалина, 0,025 М этилен-диамин-тетрауксусной кислоты (ЭДТА), 0,08% азида натрия, 1% альбумина бычьей сыворотки (БСА), рН 7,4). Стандарты и образцы контроля качества, содержащие известные уровни С-пептида крысы, добавляли в соответствующие лунки и добавляли неразбавленную плазму мыши. Добавляли смесь раствора антитела (смесь 1:1 предварительно титрованного иммобилизованного и биотинилированного детектирующего антитела к С-пептиду) и инкубировали планшет при комнатной температуре в течение 2 часов на шейкере для микротитрационного планшета, установленном на умеренную скорость. Лунки промывали 3 раза 1 × HRP-промывочным буфером, и в каждую лунку добавляли раствор фермента (предварительно титрованный комплекс стрептавидина с пероксидазой хрена) для конъюгации пероксидазы хрена с иммобилизованными биотинилированными антителами. Планшет инкубировали в течение 30 минут при комнатной температуре на шейкере для микротитрационного планшета, установленном на умеренную скорость. Лунки тщательно промывали 1 × HRP промывочным буфером. В каждую лунку добавляли раствор субстрата, содержащий 3,3,5,5'-тетраметилбензидин, и планшет встряхивали на шейкере 15 минут. Активность фермента останавливали стоп-раствором (0,3 М HCl) и измеряли оптическую плотность при 450 нм в течение 5 минут на спектрофотометре VICTOR (Perkin Elmer).
Выделение лимфоцитов из лимфоидных органов
[00283] Органы извлекали и помещали в ледяную промывочную среду (среда RPMI 1640 (Life Technologies/Invitrogen), с добавлением 4,5% антибиотиков (сульфат G418) и 2% фетальной телячьей сыворотки (FCS). Органы протирали через 70 мкм сито с промывочной средой. Клетки осаждали центрифугированием. Для селезенки добавляли NH4C1 на 3 минуты при 37°С с последующей промывкой PBS. Клетки ресуспендировали в 150 мкл FACS-буфера (1 × PBS, 0,1% BSA, 2 мМ ЭДТА). Для окрашивания использовали следующие меченные флуоресцином антитела: CD3-PerCP-Cy5.5 (145-2С11), CD4-APC-H7 (GK1.5, BD), CD8α-eFluor450 (53-6.7), CD25-PE-Cy7 (PC61.5), CTLA4-PE (UC10-489), Foxp3-APC (FJK-165). Все антитела были получены от eBioscience, если не указано иное. Использовали антимышиный CD16/CD32 (93, eBioscience) для блокирования рецепторов Fc-γ II и III для уменьшения неспецифического связывания меченных флуоресцином антител. Для окрашивания мертвых клеток использовали желтый краситель Zombie Yellow Fixable Viability Dye (BioLegend) согласно спецификациям производителя.
Окрашивание клеток Treg
[00284] Клетки перед окрашиванием промывали 1 × PBS. Приблизительно 1×109 клеток осаждали в 96-луночный планшет и ресуспендировали в 50 мкл красителя Zombie Yellow Fixable Viability Dye, разбавленного 1/500 в 1 × PBS. Клетки инкубировали при комнатной температуре в темноте в течение 20 минут. Затем клетки промывали 200 мкл FACS-буфера и инкубировали с антителами против внеклеточных эпитопов, разбавленными в буфере FACS, в темноте в течение 30 минут при 4°С. Клетки промывали 200 мкл FACS-буфера. Клетки фиксировали и пермеабилизировали в растворе для фиксации/пермеабилизация (Foxp3/String Buffer Set, eBioscience) в течение 30 минут при комнатной температуре. Антитела против внутриклеточных эпитопов разбавляли в 1 × пермеабилизационном буфере и инкубировали с клетками в темноте в течение 30 минут при 4°С. Клетки промывали и ресуспендировали в FACS-буфере и фильтровали для регистрации данных. Следующие антитела использовали в следующих разведениях: CD3 (PerCP-Су5.5; 1/100); CD25 (РЕ-Су7; 1/625); CD4 (АРС-Н7; 1/160); CD8a (eFluor450; 1/300); CTLA4 (РЕ; 1/200); Foxp3 (APC 1/200).
Проточная цитометрия
[00285] Данные проточной цитометрии получали на установке BD Canto II с программным обеспечением FACSDiva и анализировали с помощью программного обеспечения FlowJo® (TreeStar). Для калибровки использовали микросферы UltraComp eBeads™ (eBioscience).
Статистический анализ
[00286] Статистический анализ проводили с применением программного обеспечения GraphPad Prism 6.
Результаты
[00287] Результаты в целом указывают на то, что (a) IL2 в низкой дозе (вводимый в слизистую оболочку посредством LL-IL-2) с проинсулином или без проинсулина в качестве аутоантигена (например, вводимого в виде LL-PINS) способен безопасно вызывать регрессию недавно выявленного диабета у мышей, (b) стимуляция и активация Treg (CD4+CD25+ Foxp3+CTLA4+) являются возможными механизмами действия терапии LL-IL-2 и LL-IL-2+LL-PINS, и (с) исходные концентрации глюкозы в крови являются прогностическим фактором для терапевтического успеха у мышей.
Доставка в слизистую оболочку LL-IL-2 вызывает длительную ремиссию диабета у NOD-мышей
[00288] Диагноз диабета ставили при результате глюкозы в крови у мышей более 200 мг/дл в течение двух дней подряд, в сочетании с положительной глюкозурией. Успех лечения ("ремиссия") определялся как наличие двух подряд результатов измерения уровня глюкозы в крови ниже 200 мг/дл и полного отсутствия положительной глюкозурии.
[00289] Не получавшие лечения NOD-мыши с диабетом сохраняли гипергликемию и были подвергнуты эвтаназии, когда они теряли более 20% от своей первоначальной массы тела. Лечение LL-pT1NX не восстанавливало нормогликемию у мышей с диабетом. Доставка в слизистую оболочку LL-PINS вызвала регрессию диабета у 12% мышей с недавно выявленным диабетом. Неожиданно, после 6 недель применения, введение одного LL-IL-2 вызывало регрессию диабета (т.е. приводило к восстановленной нормогликемии) у приблизительно 27% мышей с недавно выявленным диабетом. Комбинированная терапия, состоящая из доставки в слизистую оболочку LL-IL-2 в сочетании с LL-PINS (LL-IL-2+LL-PINS), восстановила нормогликемию после диабета у приблизительно 30% мышей. LL-IL-2 и LL-IL-2+LL-PINS вызывали долговременную ремиссию диабета в течение по меньшей мере 8 дополнительных недель после завершения лечения. По сравнению с терапией одним LL-IL-2, комбинированная терапия LL-IL-2+LL-PINS способна ускорить регрессию диабета. Результаты показаны на Фиг. 17.
Концентрация глюкозы в крови в начале терапии влияет на терапевтический успех
[00290] Терапия LL-IL-2 и LL-IL-2+LL-PINS в течение 6 недель приводила к регрессии диабета у приблизительно 45% и приблизительно 50% соответственно, у мышей с начальными уровнями глюкозы в крови ниже 350 мг/дл. Для сравнения, только приблизительно 8% мышей с начальными уровнями глюкозы в крови выше 350 мг/дл достигли ремиссии. Эти результаты показывают, что остаточная масса бета-клеток при начале лечения может позволить прогнозировать терапевтический успех. Эти результаты показаны на Фиг. 18. На Фиг. 22А и 22В показаны концентрации глюкозы в крови (мг/дл) у мышей с недавно выявленным диабетом, получавших только LL-IL-2 (Фиг. 22А) и LL-IL-2+LL-PINS (Фиг. 22В), в период лечения и в период наблюдения.
Доставка в слизистую оболочку LL-IL-2 сохраняет массу функциональных бета-клеток
[00291] Показатель С-пептида в плазме способен отражать содержание инсулина поджелудочной железы (см., например, Suarez-Pinzon WL et al., Combination therapy with glucagon-like peptide-1 and gastrin restores normoglycemia in diabetic NOD mice. Diabetes 2008; 57:3281-8.). Уровни С-пептида у мышей, получавших и излеченных LL-IL-2 (113,23±28,16 пМ, n=3) и LL-IL-2+LL-PINS (167,63±64,38 пмоль, n=9), составляли 19%, а 28% от значений С-пептида, измеренных у нормогликемических мышей NOD (602,93±293,43 пМ, n=6) и приблизительно в два раза выше, чем значения, обнаруженные у не получавших лечения NOD-мышей с длительным диабетом (n=4; 58,23±100,86 мМ). Уровни С-пептида могут быть мерой количества эндогенного инсулина, произведенного оставшимися островками. Когда у NOD мышей возникает диабет, уровни С-пептида падают, а у не получавших лечения мышей с длительным диабетом уровни С-пептида должны быть очень низкими или не обнаружимыми. Это фактически и наблюдалось у 3 из 4 проанализированных мышей. Только у одной мыши были обнаружимые уровни С-пептида. Это наблюдение свидетельствует, что функциональные бета-клетки все еще присутствуют при диагнозе диабета и сохраняются терапией. Мыши, получавшие LL-IL-2+LL-PINS и проявляющие диабетическую ремиссию, имели статистически достоверно более высокие уровни С-пептида по сравнению с мышами с недавно выявленным диабетом (р<0,05). У неизлеченных мышей были необнаружимые значения С-пептида, сходные с теми, что наблюдались у большинства мышей с длительным диабетом. Эти результаты показывают, что у животных с недавно выявленным диабетом осталась какая-то функция бета-клеток. Излеченные мыши имели уровни С-пептида, сравнимые с уровнем, обнаруженным у мышей с недавно выявленным диабетом. Неизлеченные животные утратили функцию бета-клеток, что проявляется в не обнаружимых уровнях С-пептида. Эти результаты показаны на Фиг. 19.
Доставка в слизистую оболочку LL-IL-2 замедляет прогрессирование инсулита
[00292] Гистологический анализ поджелудочной железы после начала диабета выявил островки поджелудочной железы, сильно инфильтрированные лейкоцитами, и лишь немногие островков с сохранившимися бета-клетками. LL-IL-2 и LL-IL-2+LL-PINS останавливали инсулит (т.е. предотвращали его ухудшение). Ухудшение инсулита обычно наблюдается при прогрессировании от недавно возникшего к давнему диабету. Неожиданно, оба препарата улучшили степень инсулита по сравнению с инсулитом, обнаруженным у нормогликемических NOD мышей с длительным диабетом. Хотя на процент островков с тяжелым инсулитом существенного влияния не было, оба препарата кардинально увеличили площадь, свободную от инсулита. Еще более неожиданно это улучшение наблюдалось и у животных, не достигших нормогликемии ("не излеченных" животных). Результаты показаны на Фиг. 20.
[00293] Окрашивание гематоксилином и эозином не позволило определить, какие именно иммунные клетки проникают в поджелудочную железу, то есть эффекторные Т-клетки (Teff) или клетки Treg.
Доставка в слизистую оболочку LL-IL-2 стимулирует экспансию CD4+Foxp3+CTLA4+Treg-клеток
[00294] Влияние LL-IL-2 на разные субпопуляции иммунных клеток, как локально (то есть в MLN), системно (то есть в селезенке и крови), так и в целевом органе (то есть в PLN) измеряли с применением проточного цитометрического анализа. Поскольку в низкой дозе IL-2, вводимый систематически, способен стимулировать экспрессию белков, ассоциированных с клетками Treg, включая Foxp3, CD25 и CTLA4, определяли относительные количества CD4+, CD8+ и CD4+ Foxp3+ клеток или их статус активации (CD44, CD62L).
[00295] Были оценены популяции CD4+ и CD8+ Т-клеток среди живых CD3+ Т-клеток. Небольшое снижение CD4+ Т-клеток в MLN было обнаружено при терапии LL-IL-2 и LL-IL-2+LL-PINS у неизлеченных мышей по сравнению с излеченными животными (р=0,044 и р=0,068 соответственно). В исследованных периферических органах (то есть крови и селезенке) и органе-мишени (то есть PLN) статистически значимых изменений в количестве этих лейкоцитов не наблюдалось. Различия в количествах CD4+ Т-клеток были ограничены участками, локально подверженными воздействию LL-IL-2 и LL-IL-2+LL-PINS. И наоборот, популяция CD8+ Т-клеток была увеличена в MLN не излеченных терапией LL-IL-2+LL-PINS мышей, по сравнению с вылеченными мышами (р=0,012). Эта тенденция также присутствовала при терапии LL-IL-2. Кроме того, получавшие LL-IL-2 и LL-IL-2+LL-PINS неизлеченные мыши имели значительно более высокие уровни CD8+ Т-клеток в селезенке (р=0,025 и р=0,011 соответственно) по сравнению с NOD-мышами с впервые выявленным диабетом. Это системное изменение могло быть следствием прогрессирования заболевания, а не терапии, содержащей L. lactis.
[00296] Присутствие клеток Treg оценивали также с использованием проточной цитометрии. Через 6 недель после начала терапии PLN мышей, излеченных LL-IL-2 и LL-IL-2+LL-PINS, показывали тенденцию к увеличению количества Foxp3+CTLA-4+Treg-клеток по сравнению с мышами с впервые выявленным диабетом (в обеих группах приблизительно 17% по сравнению с приблизительно 10% у мышей с впервые выявленным диабетом). Эта тенденция наблюдалась также у получавших LL-IL-2 и LL-IL-2+LL-PINS неизлеченных мышей (приблизительно 14% и приблизительно 17% соответственно).
[00297] В селезенке у неизлеченных мышей наблюдалось уменьшение этого компартмента клеток Treg по сравнению с излеченными мышами и по сравнению с получавшими лечения мышами с впервые выявленным диабетом. В крови оба вида терапии стимулировали тенденцию к увеличению циркулирующих Foxp3+CTLA-4+Treg-клеток по сравнению с мышами с впервые выявленным диабетом. Опять же, это увеличение не зависело от результата терапии и у неизлеченных мышей, получавших LL-IL-2+LL-PINS, это увеличение было значительным (р=0,009). В MLN у не излеченных LL-IL-2+LL-PINS мышей также наблюдались значительно более высокие количества Foxp3+CTLA4+Treg-клеток по сравнению с мышами с впервые выявленным диабетом (р=0,002). Вышеуказанные результаты проиллюстрированы на Фиг. 21.
[00298] Компартмент Treg клеток дополнительно определяли измеряя экспрессию CD25. Значительное количество Foxp3+CTLA4+Treg-клеток не экспрессировали CD25 (CD25-), но терапия LL-IL-2 и LL-IL-2+LL-PINS по-прежнему вызывала изменения в этой популяции. CD25-Foxp3+CTLA-4+Treg-клетки были относительно многочисленны в крови и повышены у не излеченных LL-IL-2+LL-PINS мышей (р=0,004) и у излеченных LL-IL-2 мышей. В селезенке тенденция к уменьшению этой популяции наблюдалась у получавших LL-IL-2 и LL-IL-2+LL-PINS не излеченных мышей по сравнению с вылеченными мышами (р=0,06 и р=0,06 соответственно). В брыжеечных лимфатических узлах наблюдалось мало влияния LL-IL-2 или LL-IL-2+LL-PINS на CD25-Foxp3+CTLA-4+Treg-клетки. В целевых лимфатических узлах (то есть, PLN) эта популяция Treg-клеток была увеличена у излеченных LL-IL-2 и LL-IL-2+LL-PINS мышей по сравнению с мышами с впервые выявленным диабетом.
[00299] Наблюдались следующие тенденции для CD25+ клеток: в MLN клетки CD25+Foxp3+CTLA-4+Treg были повышены у не излеченных мышей, получавших LL-IL-2 и LL-IL-2+LL-PINS, по сравнению с мышами с впервые выявленным диабетом (12% и 14% против 7% соответственно). Это было статистически значимо в последней группе (р=0,001). В PLN наблюдалась заметная тенденция к увеличению этой популяции в группах, получавших LL-IL-2 и LL-IL-2+LL-PINS.
[00300] Картина распределения Foxp3+CTLA-4+Treg-клеток, наблюдаемая в крови и селезенке, была сходной при дальнейшей классификации, основанной на экспрессии CD25. В локальных и целевых лимфатических узлах (т.е. MLN и PLN) повышение Foxp3+CTLA-4+Treg-клеток наблюдалось после обоих видов терапии, LL-IL-2 и LL-PINS+IL-2, что было в значительной степени обусловлено компартментом CD25+ Treg-клеток.
[00301] Размер селезенки у NOD мышей с впервые выявленным диабетом, получавших LL-PINS+IL-2, оценивали в общей сложности в течение 6 недель (n=3). Никакой спленомегалии не наблюдалось. Лечение LL-IL-2, как и LL-IL-2+LL-PINS было безопасным и хорошо переносилось.
Заключение
[00302] Терапия LL-IL-2 и LL-IL-2+LL-PINS является безопасной, хорошо переносимой и стимулирует ремиссию диабета у NOD мышей с впервые выявленным диабетом. Оба вида терапии обладают полезными метаболическими и иммунными эффектами. Неожиданно, некоторый эффект присутствует и у не излеченных мышей, что указывает на потенциальную выгоду для субъектов, которые не достигли нормогликемии в ходе экспериментов, или для которых лечение происходит слишком поздно, поскольку на момент начала лечения оставшаяся масса бета-клеток была ниже определенного порогового значения (у NOD-мышей процесс потери бета-клеток идет быстрее по сравнению с людьми). Таким образом, иммунный эффект возможно наблюдался у всех получавших лечение животных, но этот эффект транслировался в определенный терапевтический эффект только у животных, имеющих достаточную массу бета-клеток, оставшихся на момент начала лечения.
[00303] Препараты LL-IL-2 и LL-IL-2+LL-PINS останавливали прогрессирование инсулита, возможно, посредством механизма, включающего экспансию Treg-клеток. Различия в проценте Foxp3+CTLA4+ Treg-клеток между получавшими и не получавшими лечение мышами с впервые выявленным диабетом были обнаружены в PLN после терапии LL-IL-2 и LL-IL-2+LL-PINS. Увеличение числа клеток Treg, стимулированное этими препаратами, может быть результатом улучшения выживаемости Treg-клеток (см., например. Tang Q, Bluestone JA. Nat. Immunol. 2008; 9(3):239-244). В качестве альтернативы, увеличенное количество клеток Treg может быть результатом преобразования эффекторных Т-клеток (Teff) в стимулированные Treg-клетки (Zheng Y, Rudensky AY. Nat. Immunol. 2007; 8(5): 457-462), или увеличением рекрутинга Treg-клеток в PLN (Grinberg-Bleyer Y. et al., J. Exp. Med. 2010; 207(9): 1871-1878).
Пример 9. Эффект введения антитела против CD3 на частоту ремиссии диабета
[00304] Было проведено исследование по подбору дозы антитела против CD3 у мышей. Мышам с начальным уровнем глюкозы в крови более 200 мг/дл вводили 2,5 мкг/сут; 5 мкг/сут; 10 мкг/сут; 25 мкг/сут; или 50 мкг/сут антитела против CD3 (теплизумаб) путем в/в инъекций. Дозировки 25 мкг/сут или 50 мкг/сут проявляли некоторую токсичность. Лечение проводили в течение 5-дневного периода, т.е. совокупная доза антитела против CD3 составляла 12,5 мкг; 25 мкг; 50 мкг; 125 мкг; или 250 мкг. В группе лечения, получавшей 10 мкг/сут, максимальный процент ремиссии составлял приблизительно 50%. Однако, чтобы уменьшить токсичность в течение более длительного периода лечения, была выбрана доза 2,5 мкг/сут (12,5 мкг суммарно), которая считается субтерапевтической дозой (низкодозовое лечение). Введение 2,5 мкг/сут антитела против CD3 было менее эффективным, чем при более высокой дозе 10 мкг/сут, и приводила к стимуляции ремиссии у приблизительно 20%.
[00305] Как и в Примере 8, LL-PINS, LL-IL2, LL-PINS+LL-IL2 вводили путем внутрижелудочной инокуляции (2×109 КОЕ/д), 5 раз в неделю в течение 6 недель мышам с впервые выявленным диабетом. Кроме того, в течение 5 дней подряд внутримышечно вводили (2,5 мкг/сут) моноклональное антитело хомяка против CD3 мыши (mAb) (145-2C11, BioXCell, Нью-Гемпшир, США) мышам, получавшим инокуляцию LL-PINS+LL-IL2. У всех исследуемых мышей с впервые выявленным диабетом начальная концентрация глюкозы в крови была ниже 350 мг/дл. Массу и гликемию измеряли 3 раза в неделю. Ремиссия диабета определялась как отсутствие гликозурии и величина гликемии <200 мг/дл в течение двух дней подряд.
[00306] Как показано на Фиг. 23, лечение LL-PINS (n=5), LL-IL2 (n=13) и смесью LL-PINS+LL-IL2 (n=31) корректировало гипергликемию у 20%, 30,8% и 45,2% мышей, соответственно. Среди мышей с впервые выявленным диабетом (n=3), получавших комбинацию LL-PINS+LL-IL2 и анти-CD3, нормогликемия восстановилась у 66,7% мышей. Эти данные свидетельствуют о том, что дополнительное иммуномодулирующее вещество, например, антитело против CD3, улучшает исход терапии LL-IL-2+LL-PINS у субъектов с более низкой начальной концентрацией глюкозы в крови.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ИНТРЕКСОН АКТОБИОТИКС Н.В.
<120> КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ДИАБЕТА 1 ТИПА
<130> 205350-0030-00-WO-549987
<140>
<141>
<150> 62/350,472
<151> 2016-06-15
<150> 62/278,493
<151> 2016-01-14
<160> 61
<170> PatentIn версия 3.5
<210> 1
<211> 153
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
20 25 30
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
35 40 45
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe
50 55 60
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
65 70 75 80
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
85 90 95
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
100 105 110
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
115 120 125
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
130 135 140
Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150
<210> 2
<211> 462
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(459)
<400> 2
atg tac agg atg caa ctc ctg tct tgc att gca cta agt ctt gca ctt 48
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
gtc aca aac agt gct cca act tca tca tca act aaa aaa act caa ttg 96
Val Thr Asn Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
20 25 30
caa ctt gaa cac ttg ctt ttg gat ctt caa atg atc ttg aac ggt atc 144
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
35 40 45
aac aac tac aaa aac cca aaa ctt act cgt atg ttg act ttt aaa ttt 192
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe
50 55 60
tac atg cca aaa aaa gct act gaa ctt aaa cac ttg caa tgt ctt gaa 240
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
65 70 75 80
gaa gaa ttg aaa cca ctt gaa gaa gtt ttg aac ctt gct caa tca aaa 288
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
85 90 95
aac ttt cac ttg cgt cca cgt gat ctt atc tca aac atc aac gtt atc 336
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
100 105 110
gtt ttg gaa ctt aaa ggt tca gaa act act ttt atg tgt gaa tac gct 384
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
115 120 125
gat gaa act gct act atc gtt gaa ttt ttg aac cgt tgg atc act ttt 432
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
130 135 140
tgt caa tca atc atc tca act ttg act taa 462
Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150
<210> 3
<211> 133
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 4
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser
20
<210> 5
<211> 483
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
полинуклеотид"
<400> 5
atgaaaaaaa agattatctc agctatttta atgtctacag tgatactttc tgctgcagcc 60
ccgttgtcag gtgtttacgc cgctccaact tcatcatcaa ctaaaaaaac tcaattgcaa 120
cttgaacact tgcttttgga tcttcaaatg atcttgaacg gtatcaacaa ctacaaaaac 180
ccaaaactta ctcgtatgtt gacttttaaa ttttacatgc caaaaaaagc tactgaactt 240
aaacacttgc aatgtcttga agaagaattg aaaccacttg aagaagtttt gaaccttgct 300
caatcaaaaa actttcactt gcgtccacgt gatcttatct caaacatcaa cgttatcgtt 360
ttggaactta aaggttcaga aactactttt atgtgtgaat acgctgatga aactgctact 420
atcgttgaat ttttgaaccg ttggatcact ttttgtcaat caatcatctc aactttgact 480
taa 483
<210> 6
<211> 110
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu
1 5 10 15
Trp Gly Pro Asp Pro Ala Ala Ala Phe Val Asn Gln His Leu Cys Gly
20 25 30
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe
35 40 45
Phe Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly
50 55 60
Gln Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu
65 70 75 80
Ala Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys
85 90 95
Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105 110
<210> 7
<211> 333
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(330)
<400> 7
atg gcc ctg tgg atg cgc ctc ctg ccc ctg ctg gcg ctg ctg gcc ctc 48
Met Ala Leu Trp Met Arg Leu Leu Pro Leu Leu Ala Leu Leu Ala Leu
1 5 10 15
tgg gga cct gac cca gcc gca gcc ttt gtg aac caa cac ctg tgc ggc 96
Trp Gly Pro Asp Pro Ala Ala Ala Phe Val Asn Gln His Leu Cys Gly
20 25 30
tca cac ctg gtg gaa gct ctc tac cta gtg tgc ggg gaa cga ggc ttc 144
Ser His Leu Val Glu Ala Leu Tyr Leu Val Cys Gly Glu Arg Gly Phe
35 40 45
ttc tac aca ccc aag acc cgc cgg gag gca gag gac ctg cag gtg ggg 192
Phe Tyr Thr Pro Lys Thr Arg Arg Glu Ala Glu Asp Leu Gln Val Gly
50 55 60
cag gtg gag ctg ggc ggg ggc cct ggt gca ggc agc ctg cag ccc ttg 240
Gln Val Glu Leu Gly Gly Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu
65 70 75 80
gcc ctg gag ggg tcc ctg cag aag cgt ggc att gtg gaa caa tgc tgt 288
Ala Leu Glu Gly Ser Leu Gln Lys Arg Gly Ile Val Glu Gln Cys Cys
85 90 95
acc agc atc tgc tcc ctc tac cag ctg gag aac tac tgc aac tag 333
Thr Ser Ile Cys Ser Leu Tyr Gln Leu Glu Asn Tyr Cys Asn
100 105 110
<210> 8
<211> 396
<212> ДНК
<213> Homo sapiens
<400> 8
gccatcaagc aggtctgttc caagggcctt tgcgtcagat cactgtcctt ctgccatggc 60
cctgtggatg cgcctcctgc ccctgctggc gctgctggcc ctctggggac ctgacccagc 120
cgcagccttt gtgaaccaac acctgtgcgg ctcacacctg gtggaagctc tctacctagt 180
gtgcggggaa cgaggcttct tctacacacc caagacccgc cgggaggcag aggacctgca 240
ggtggggcag gtggagctgg gcgggggccc tggtgcaggc agcctgcagc ccttggccct 300
ggaggggtcc ctgcagaagc gtggcattgt ggaacaatgc tgtaccagca tctgctccct 360
ctaccagctg gagaactact gcaactagac gcagcc 396
<210> 9
<211> 469
<212> ДНК
<213> Homo sapiens
<400> 9
agccctccag gacaggctgc atcagaagag gccatcaagc agatcactgt ccttctgcca 60
tggccctgtg gatgcgcctc ctgcccctgc tggcgctgct ggccctctgg ggacctgacc 120
cagccgcagc ctttgtgaac caacacctgt gcggctcaca cctggtggaa gctctctacc 180
tagtgtgcgg ggaacgaggc ttcttctaca cacccaagac ccgccgggag gcagaggacc 240
tgcaggtggg gcaggtggag ctgggcgggg gccctggtgc aggcagcctg cagcccttgg 300
ccctggaggg gtccctgcag aagcgtggca ttgtggaaca atgctgtacc agcatctgct 360
ccctctacca gctggagaac tactgcaact agacgcagcc cgcaggcagc cccacacccg 420
ccgcctcctg caccgagaga gatggaataa agcccttgaa ccagcaaaa 469
<210> 10
<211> 495
<212> ДНК
<213> Homo sapiens
<400> 10
agccctccag gacaggctgc atcagaagag gccatcaagc aggtctgttc caagggcctt 60
tgcgtcagat cactgtcctt ctgccatggc cctgtggatg cgcctcctgc ccctgctggc 120
gctgctggcc ctctggggac ctgacccagc cgcagccttt gtgaaccaac acctgtgcgg 180
ctcacacctg gtggaagctc tctacctagt gtgcggggaa cgaggcttct tctacacacc 240
caagacccgc cgggaggcag aggacctgca ggtggggcag gtggagctgg gcgggggccc 300
tggtgcaggc agcctgcagc ccttggccct ggaggggtcc ctgcagaagc gtggcattgt 360
ggaacaatgc tgtaccagca tctgctccct ctaccagctg gagaactact gcaactagac 420
gcagcccgca ggcagcccca cacccgccgc ctcctgcacc gagagagatg gaataaagcc 480
cttgaaccag caaaa 495
<210> 11
<211> 648
<212> ДНК
<213> Homo sapiens
<400> 11
agccctccag gacaggctgc atcagaagag gccatcaagc aggtctgttc caagggcctt 60
tgcgtcaggt gggctcagga ttccagggtg gctggacccc aggccccagc tctgcagcag 120
ggaggacgtg gctgggctcg tgaagcatgt gggggtgagc ccaggggccc caaggcaggg 180
cacctggcct tcagcctgcc tcagccctgc ctgtctccca gatcactgtc cttctgccat 240
ggccctgtgg atgcgcctcc tgcccctgct ggcgctgctg gccctctggg gacctgaccc 300
agccgcagcc tttgtgaacc aacacctgtg cggctcacac ctggtggaag ctctctacct 360
agtgtgcggg gaacgaggct tcttctacac acccaagacc cgccgggagg cagaggacct 420
gcaggtgggg caggtggagc tgggcggggg ccctggtgca ggcagcctgc agcccttggc 480
cctggagggg tccctgcaga agcgtggcat tgtggaacaa tgctgtacca gcatctgctc 540
cctctaccag ctggagaact actgcaacta gacgcagccc gcaggcagcc ccacacccgc 600
cgcctcctgc accgagagag atggaataaa gcccttgaac cagcaaaa 648
<210> 12
<211> 529
<212> ДНК
<213> Homo sapiens
<400> 12
agccctccag gacaggctgc atcagaagag gccatcaagc aggtctgttc caagggcctt 60
tgcgtcaggt gggctcagga ttccagggtg gctggacccc agatcactgt ccttctgcca 120
tggccctgtg gatgcgcctc ctgcccctgc tggcgctgct ggccctctgg ggacctgacc 180
cagccgcagc ctttgtgaac caacacctgt gcggctcaca cctggtggaa gctctctacc 240
tagtgtgcgg ggaacgaggc ttcttctaca cacccaagac ccgccgggag gcagaggacc 300
tgcaggtggg gcaggtggag ctgggcgggg gccctggtgc aggcagcctg cagcccttgg 360
ccctggaggg gtccctgcag aagcgtggca ttgtggaaca atgctgtacc agcatctgct 420
ccctctacca gctggagaac tactgcaact agacgcagcc cgcaggcagc cccacacccg 480
ccgcctcctg caccgagaga gatggaataa agcccttgaa ccagcaaaa 529
<210> 13
<211> 585
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Met Ala Ser Pro Gly Ser Gly Phe Trp Ser Phe Gly Ser Glu Asp Gly
1 5 10 15
Ser Gly Asp Ser Glu Asn Pro Gly Thr Ala Arg Ala Trp Cys Gln Val
20 25 30
Ala Gln Lys Phe Thr Gly Gly Ile Gly Asn Lys Leu Cys Ala Leu Leu
35 40 45
Tyr Gly Asp Ala Glu Lys Pro Ala Glu Ser Gly Gly Ser Gln Pro Pro
50 55 60
Arg Ala Ala Ala Arg Lys Ala Ala Cys Ala Cys Asp Gln Lys Pro Cys
65 70 75 80
Ser Cys Ser Lys Val Asp Val Asn Tyr Ala Phe Leu His Ala Thr Asp
85 90 95
Leu Leu Pro Ala Cys Asp Gly Glu Arg Pro Thr Leu Ala Phe Leu Gln
100 105 110
Asp Val Met Asn Ile Leu Leu Gln Tyr Val Val Lys Ser Phe Asp Arg
115 120 125
Ser Thr Lys Val Ile Asp Phe His Tyr Pro Asn Glu Leu Leu Gln Glu
130 135 140
Tyr Asn Trp Glu Leu Ala Asp Gln Pro Gln Asn Leu Glu Glu Ile Leu
145 150 155 160
Met His Cys Gln Thr Thr Leu Lys Tyr Ala Ile Lys Thr Gly His Pro
165 170 175
Arg Tyr Phe Asn Gln Leu Ser Thr Gly Leu Asp Met Val Gly Leu Ala
180 185 190
Ala Asp Trp Leu Thr Ser Thr Ala Asn Thr Asn Met Phe Thr Tyr Glu
195 200 205
Ile Ala Pro Val Phe Val Leu Leu Glu Tyr Val Thr Leu Lys Lys Met
210 215 220
Arg Glu Ile Ile Gly Trp Pro Gly Gly Ser Gly Asp Gly Ile Phe Ser
225 230 235 240
Pro Gly Gly Ala Ile Ser Asn Met Tyr Ala Met Met Ile Ala Arg Phe
245 250 255
Lys Met Phe Pro Glu Val Lys Glu Lys Gly Met Ala Ala Leu Pro Arg
260 265 270
Leu Ile Ala Phe Thr Ser Glu His Ser His Phe Ser Leu Lys Lys Gly
275 280 285
Ala Ala Ala Leu Gly Ile Gly Thr Asp Ser Val Ile Leu Ile Lys Cys
290 295 300
Asp Glu Arg Gly Lys Met Ile Pro Ser Asp Leu Glu Arg Arg Ile Leu
305 310 315 320
Glu Ala Lys Gln Lys Gly Phe Val Pro Phe Leu Val Ser Ala Thr Ala
325 330 335
Gly Thr Thr Val Tyr Gly Ala Phe Asp Pro Leu Leu Ala Val Ala Asp
340 345 350
Ile Cys Lys Lys Tyr Lys Ile Trp Met His Val Asp Ala Ala Trp Gly
355 360 365
Gly Gly Leu Leu Met Ser Arg Lys His Lys Trp Lys Leu Ser Gly Val
370 375 380
Glu Arg Ala Asn Ser Val Thr Trp Asn Pro His Lys Met Met Gly Val
385 390 395 400
Pro Leu Gln Cys Ser Ala Leu Leu Val Arg Glu Glu Gly Leu Met Gln
405 410 415
Asn Cys Asn Gln Met His Ala Ser Tyr Leu Phe Gln Gln Asp Lys His
420 425 430
Tyr Asp Leu Ser Tyr Asp Thr Gly Asp Lys Ala Leu Gln Cys Gly Arg
435 440 445
His Val Asp Val Phe Lys Leu Trp Leu Met Trp Arg Ala Lys Gly Thr
450 455 460
Thr Gly Phe Glu Ala His Val Asp Lys Cys Leu Glu Leu Ala Glu Tyr
465 470 475 480
Leu Tyr Asn Ile Ile Lys Asn Arg Glu Gly Tyr Glu Met Val Phe Asp
485 490 495
Gly Lys Pro Gln His Thr Asn Val Cys Phe Trp Tyr Ile Pro Pro Ser
500 505 510
Leu Arg Thr Leu Glu Asp Asn Glu Glu Arg Met Ser Arg Leu Ser Lys
515 520 525
Val Ala Pro Val Ile Lys Ala Arg Met Met Glu Tyr Gly Thr Thr Met
530 535 540
Val Ser Tyr Gln Pro Leu Gly Asp Lys Val Asn Phe Phe Arg Met Val
545 550 555 560
Ile Ser Asn Pro Ala Ala Thr His Gln Asp Ile Asp Phe Leu Ile Glu
565 570 575
Glu Ile Glu Arg Leu Gly Gln Asp Leu
580 585
<210> 14
<211> 1758
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(1755)
<400> 14
atg gca tct ccg ggc tct ggc ttt tgg tct ttc ggg tcg gaa gat ggc 48
Met Ala Ser Pro Gly Ser Gly Phe Trp Ser Phe Gly Ser Glu Asp Gly
1 5 10 15
tct ggg gat tcc gag aat ccc ggc aca gcg cga gcc tgg tgc caa gtg 96
Ser Gly Asp Ser Glu Asn Pro Gly Thr Ala Arg Ala Trp Cys Gln Val
20 25 30
gct cag aag ttc acg ggc ggc atc gga aac aaa ctg tgc gcc ctg ctc 144
Ala Gln Lys Phe Thr Gly Gly Ile Gly Asn Lys Leu Cys Ala Leu Leu
35 40 45
tac gga gac gcc gag aag ccg gcg gag agc ggc ggg agc caa ccc ccg 192
Tyr Gly Asp Ala Glu Lys Pro Ala Glu Ser Gly Gly Ser Gln Pro Pro
50 55 60
cgg gcc gcc gcc cgg aag gcc gcc tgc gcc tgc gac cag aag ccc tgc 240
Arg Ala Ala Ala Arg Lys Ala Ala Cys Ala Cys Asp Gln Lys Pro Cys
65 70 75 80
agc tgc tcc aaa gtg gat gtc aac tac gcg ttt ctc cat gca aca gac 288
Ser Cys Ser Lys Val Asp Val Asn Tyr Ala Phe Leu His Ala Thr Asp
85 90 95
ctg ctg ccg gcg tgt gat gga gaa agg ccc act ttg gcg ttt ctg caa 336
Leu Leu Pro Ala Cys Asp Gly Glu Arg Pro Thr Leu Ala Phe Leu Gln
100 105 110
gat gtt atg aac att tta ctt cag tat gtg gtg aaa agt ttc gat aga 384
Asp Val Met Asn Ile Leu Leu Gln Tyr Val Val Lys Ser Phe Asp Arg
115 120 125
tca acc aaa gtg att gat ttc cat tat cct aat gag ctt ctc caa gaa 432
Ser Thr Lys Val Ile Asp Phe His Tyr Pro Asn Glu Leu Leu Gln Glu
130 135 140
tat aat tgg gaa ttg gca gac caa cca caa aat ttg gag gaa att ttg 480
Tyr Asn Trp Glu Leu Ala Asp Gln Pro Gln Asn Leu Glu Glu Ile Leu
145 150 155 160
atg cat tgc caa aca act cta aaa tat gca att aaa aca ggg cat cct 528
Met His Cys Gln Thr Thr Leu Lys Tyr Ala Ile Lys Thr Gly His Pro
165 170 175
aga tac ttc aat caa ctt tct act ggt ttg gat atg gtt gga tta gca 576
Arg Tyr Phe Asn Gln Leu Ser Thr Gly Leu Asp Met Val Gly Leu Ala
180 185 190
gca gac tgg ctg aca tca aca gca aat act aac atg ttc acc tat gaa 624
Ala Asp Trp Leu Thr Ser Thr Ala Asn Thr Asn Met Phe Thr Tyr Glu
195 200 205
att gct cca gta ttt gtg ctt ttg gaa tat gtc aca cta aag aaa atg 672
Ile Ala Pro Val Phe Val Leu Leu Glu Tyr Val Thr Leu Lys Lys Met
210 215 220
aga gaa atc att ggc tgg cca ggg ggc tct ggc gat ggg ata ttt tct 720
Arg Glu Ile Ile Gly Trp Pro Gly Gly Ser Gly Asp Gly Ile Phe Ser
225 230 235 240
ccc ggt ggc gcc ata tct aac atg tat gcc atg atg atc gca cgc ttt 768
Pro Gly Gly Ala Ile Ser Asn Met Tyr Ala Met Met Ile Ala Arg Phe
245 250 255
aag atg ttc cca gaa gtc aag gag aaa gga atg gct gct ctt ccc agg 816
Lys Met Phe Pro Glu Val Lys Glu Lys Gly Met Ala Ala Leu Pro Arg
260 265 270
ctc att gcc ttc acg tct gaa cat agt cat ttt tct ctc aag aag gga 864
Leu Ile Ala Phe Thr Ser Glu His Ser His Phe Ser Leu Lys Lys Gly
275 280 285
gct gca gcc tta ggg att gga aca gac agc gtg att ctg att aaa tgt 912
Ala Ala Ala Leu Gly Ile Gly Thr Asp Ser Val Ile Leu Ile Lys Cys
290 295 300
gat gag aga ggg aaa atg att cca tct gat ctt gaa aga agg att ctt 960
Asp Glu Arg Gly Lys Met Ile Pro Ser Asp Leu Glu Arg Arg Ile Leu
305 310 315 320
gaa gcc aaa cag aaa ggg ttt gtt cct ttc ctc gtg agt gcc aca gct 1008
Glu Ala Lys Gln Lys Gly Phe Val Pro Phe Leu Val Ser Ala Thr Ala
325 330 335
gga acc acc gtg tac gga gca ttt gac ccc ctc tta gct gtc gct gac 1056
Gly Thr Thr Val Tyr Gly Ala Phe Asp Pro Leu Leu Ala Val Ala Asp
340 345 350
att tgc aaa aag tat aag atc tgg atg cat gtg gat gca gct tgg ggt 1104
Ile Cys Lys Lys Tyr Lys Ile Trp Met His Val Asp Ala Ala Trp Gly
355 360 365
ggg gga tta ctg atg tcc cga aaa cac aag tgg aaa ctg agt ggc gtg 1152
Gly Gly Leu Leu Met Ser Arg Lys His Lys Trp Lys Leu Ser Gly Val
370 375 380
gag agg gcc aac tct gtg acg tgg aat cca cac aag atg atg gga gtc 1200
Glu Arg Ala Asn Ser Val Thr Trp Asn Pro His Lys Met Met Gly Val
385 390 395 400
cct ttg cag tgc tct gct ctc ctg gtt aga gaa gag gga ttg atg cag 1248
Pro Leu Gln Cys Ser Ala Leu Leu Val Arg Glu Glu Gly Leu Met Gln
405 410 415
aat tgc aac caa atg cat gcc tcc tac ctc ttt cag caa gat aaa cat 1296
Asn Cys Asn Gln Met His Ala Ser Tyr Leu Phe Gln Gln Asp Lys His
420 425 430
tat gac ctg tcc tat gac act gga gac aag gcc tta cag tgc gga cgc 1344
Tyr Asp Leu Ser Tyr Asp Thr Gly Asp Lys Ala Leu Gln Cys Gly Arg
435 440 445
cac gtt gat gtt ttt aaa cta tgg ctg atg tgg agg gca aag ggg act 1392
His Val Asp Val Phe Lys Leu Trp Leu Met Trp Arg Ala Lys Gly Thr
450 455 460
acc ggg ttt gaa gcg cat gtt gat aaa tgt ttg gag ttg gca gag tat 1440
Thr Gly Phe Glu Ala His Val Asp Lys Cys Leu Glu Leu Ala Glu Tyr
465 470 475 480
tta tac aac atc ata aaa aac cga gaa gga tat gag atg gtg ttt gat 1488
Leu Tyr Asn Ile Ile Lys Asn Arg Glu Gly Tyr Glu Met Val Phe Asp
485 490 495
ggg aag cct cag cac aca aat gtc tgc ttc tgg tac att cct cca agc 1536
Gly Lys Pro Gln His Thr Asn Val Cys Phe Trp Tyr Ile Pro Pro Ser
500 505 510
ttg cgt act ctg gaa gac aat gaa gag aga atg agt cgc ctc tcg aag 1584
Leu Arg Thr Leu Glu Asp Asn Glu Glu Arg Met Ser Arg Leu Ser Lys
515 520 525
gtg gct cca gtg att aaa gcc aga atg atg gag tat gga acc aca atg 1632
Val Ala Pro Val Ile Lys Ala Arg Met Met Glu Tyr Gly Thr Thr Met
530 535 540
gtc agc tac caa ccc ttg gga gac aag gtc aat ttc ttc cgc atg gtc 1680
Val Ser Tyr Gln Pro Leu Gly Asp Lys Val Asn Phe Phe Arg Met Val
545 550 555 560
atc tca aac cca gcg gca act cac caa gac att gac ttc ctg att gaa 1728
Ile Ser Asn Pro Ala Ala Thr His Gln Asp Ile Asp Phe Leu Ile Glu
565 570 575
gaa ata gaa cgc ctt gga caa gat tta taa 1758
Glu Ile Glu Arg Leu Gly Gln Asp Leu
580 585
<210> 15
<211> 2400
<212> ДНК
<213> Homo sapiens
<400> 15
agctcgcccg cagctcgcac tcgcaggcga cctgctccag tctccaaagc cgatggcatc 60
tccgggctct ggcttttggt ctttcgggtc ggaagatggc tctggggatt ccgagaatcc 120
cggcacagcg cgagcctggt gccaagtggc tcagaagttc acgggcggca tcggaaacaa 180
actgtgcgcc ctgctctacg gagacgccga gaagccggcg gagagcggcg ggagccaacc 240
cccgcgggcc gccgcccgga aggccgcctg cgcctgcgac cagaagccct gcagctgctc 300
caaagtggat gtcaactacg cgtttctcca tgcaacagac ctgctgccgg cgtgtgatgg 360
agaaaggccc actttggcgt ttctgcaaga tgttatgaac attttacttc agtatgtggt 420
gaaaagtttc gatagatcaa ccaaagtgat tgatttccat tatcctaatg agcttctcca 480
agaatataat tgggaattgg cagaccaacc acaaaatttg gaggaaattt tgatgcattg 540
ccaaacaact ctaaaatatg caattaaaac agggcatcct agatacttca atcaactttc 600
tactggtttg gatatggttg gattagcagc agactggctg acatcaacag caaatactaa 660
catgttcacc tatgaaattg ctccagtatt tgtgcttttg gaatatgtca cactaaagaa 720
aatgagagaa atcattggct ggccaggggg ctctggcgat gggatatttt ctcccggtgg 780
cgccatatct aacatgtatg ccatgatgat cgcacgcttt aagatgttcc cagaagtcaa 840
ggagaaagga atggctgctc ttcccaggct cattgccttc acgtctgaac atagtcattt 900
ttctctcaag aagggagctg cagccttagg gattggaaca gacagcgtga ttctgattaa 960
atgtgatgag agagggaaaa tgattccatc tgatcttgaa agaaggattc ttgaagccaa 1020
acagaaaggg tttgttcctt tcctcgtgag tgccacagct ggaaccaccg tgtacggagc 1080
atttgacccc ctcttagctg tcgctgacat ttgcaaaaag tataagatct ggatgcatgt 1140
ggatgcagct tggggtgggg gattactgat gtcccgaaaa cacaagtgga aactgagtgg 1200
cgtggagagg gccaactctg tgacgtggaa tccacacaag atgatgggag tccctttgca 1260
gtgctctgct ctcctggtta gagaagaggg attgatgcag aattgcaacc aaatgcatgc 1320
ctcctacctc tttcagcaag ataaacatta tgacctgtcc tatgacactg gagacaaggc 1380
cttacagtgc ggacgccacg ttgatgtttt taaactatgg ctgatgtgga gggcaaaggg 1440
gactaccggg tttgaagcgc atgttgataa atgtttggag ttggcagagt atttatacaa 1500
catcataaaa aaccgagaag gatatgagat ggtgtttgat gggaagcctc agcacacaaa 1560
tgtctgcttc tggtacattc ctccaagctt gcgtactctg gaagacaatg aagagagaat 1620
gagtcgcctc tcgaaggtgg ctccagtgat taaagccaga atgatggagt atggaaccac 1680
aatggtcagc taccaaccct tgggagacaa ggtcaatttc ttccgcatgg tcatctcaaa 1740
cccagcggca actcaccaag acattgactt cctgattgaa gaaatagaac gccttggaca 1800
agatttataa taaccttgct caccaagctg ttccacttct ctaggtagac aattaagttg 1860
tcacaaactg tgtgaatgta tttgtagttt gttccaaagt aaatctattt ctatattgtg 1920
gtgtcaaagt agagtttaaa aattaaacaa aaaagacatt gctcctttta aaagtccttt 1980
cttaagttta gaatacctct ctaagaattc gtgacaaaag gctatgttct aatcaataag 2040
gaaaagctta aaattgttat aaatacttcc cttactttta atatagtgtg caaagcaaac 2100
tttattttca cttcagacta gtaggactga atagtgccaa attgcccctg aatcataaaa 2160
ggttctttgg ggtgcagtaa aaaggacaaa gtaaatataa aatatatgtt gacaataaaa 2220
actcttgcct ttttcatagt attagaaaaa aatttctaat ttacctatag caacatttca 2280
aatgtattta aatacatata attttacaaa aggaaaatat atatattaaa aaagatatcc 2340
tattttgtaa catatagatt tttattttat ataggttata caaactgcgg gggcggaatt 2400
<210> 16
<211> 3610
<212> ДНК
<213> Homo sapiens
<400> 16
gaattcttcg taggaattat cttttccctc ctctcacccg acagcctgcc tatttccaaa 60
ggaaaaaaaa aaagcgtgtt gagtacgttc tggattactc ataagacctt ttttttttcc 120
ttccgggcgc aaaaccgtga gctggattta taatcgccct ataaagctcc agaggcggtc 180
aggcacctgc agaggagccc cgccgctccg ccgactagct gcccccgcga gcaacggcct 240
cgtgatttcc ccgccgatcc ggtccccgcc tccccactct gcccccgcct accccggagc 300
cgtgcagccg cctctccgaa tctctctctt ctcctggcgc tcgcgtgcga gagggaacta 360
gcgagaacga ggaagcagct ggaggtgacg ccgggcagat tacgcctgtc agggccgagc 420
cgagcggatc gctgggcgct gtgcagagga aaggcgggag tgcccggctc gctgtcgcag 480
agccgagcct gtttctgcgc cggaccagtc gaggactctg gacagtagag gccccgggac 540
gaccgagctg atggcgtctt cgaccccatc ttcgtccgca acctcctcga acgcgggagc 600
ggaccccaat accactaacc tgcgccccac aacgtacgat acctggtgcg gcgtggccca 660
tggatgcacc agaaaactgg ggctcaagat ctgcggcttc ttgcaaagga ccaacagcct 720
ggaagagaag agtcgccttg tgagtgcctt cagggagagg caatcctcca agaacctgct 780
ttcctgtgaa aacagcgacc gggatgcccg cttccggcgc acagagactg acttctctaa 840
tctgtttgct agagatctgc ttccggctaa gaacggtgag gagcaaaccg tgcaattcct 900
cctggaagtg gtggacatac tcctcaacta tgtccgcaag acatttgatc gctccaccaa 960
ggtgctggac tttcatcacc cacaccagtt gctggaaggc atggagggct tcaacttgga 1020
gctctctgac caccccgagt ccctggagca gatcctggtt gactgcagag acaccttgaa 1080
gtatggggtt cgcacaggtc atcctcgatt tttcaaccag ctctccactg gattggatat 1140
tattggccta gctggagaat ggctgacatc aacggccaat accaacatgt ttacatatga 1200
aattgcacca gtgtttgtcc tcatggaaca aataacactt aagaagatga gagagatagt 1260
tggatggtca agtaaagatg gtgatgggat attttctcct gggggcgcca tatccaacat 1320
gtacagcatc atggctgctc gctacaagta cttcccggaa gttaagacaa agggcatggc 1380
ggctgtgcct aaactggtcc tcttcacctc agaacagagt cactattcca taaagaaagc 1440
tggggctgca cttggctttg gaactgacaa tgtgattttg ataaagtgca atgaaagggg 1500
gaaaataatt ccagctgatt ttgaggcaaa aattcttgaa gccaaacaga agggatatgt 1560
tcccttttat gtcaatgcaa ctgctggcac gactgtttat ggagcttttg atccgataca 1620
agagattgca gatatatgtg agaaatataa cctttggttg catgtcgatg ctgcctgggg 1680
aggtgggctg ctcatgtcca ggaagcaccg ccataaactc aacggcatag aaagggccaa 1740
ctcagtcacc tggaaccctc acaagatgat gggcgtgctg ttgcagtgct ctgccattct 1800
cgtcaaggaa aagggtatac tccaaggatg caaccagatg tgtgcaggat atctcttcca 1860
gccagacaag cagtatgatg tctcctacga caccggggac aaggcaattc agtgtggccg 1920
ccacgtggat atcttcaagt tctggctgat gtggaaagca aagggcacag tgggatttga 1980
aaaccagatc aacaaatgcc tggaactggc tgaatacctc tatgccaaga ttaaaaacag 2040
agaagaattt gagatggttt tcaatggcga gcctgagcac acaaacgtct gtttttggta 2100
tattccacaa agcctcaggg gtgtgccaga cagccctcaa cgacgggaaa agctacacaa 2160
ggtggctcca aaaatcaaag ccctgatgat ggagtcaggt acgaccatgg ttggctacca 2220
gccccaaggg gacaaggcca acttcttccg gatggtcatc tccaacccag ccgctaccca 2280
gtctgacatt gacttcctca ttgaggagat agaaagactg ggccaggatc tgtaatcatc 2340
cttcgcagaa catgagttta tgggaatgcc ttttccctct ggcactccag aacaaacctc 2400
tatatgttgc tgaaacacac aggccatttc attgagggaa aacataatat cttgaagaat 2460
attgttaaaa ccttacttaa agcttgtttg ttctagttag caggaaatag tgttcttttt 2520
aaaaagttgc acattaggaa cagagtatat atgtacagtt atacatacct ctctctatat 2580
atacatgtat agtgagtgtg gcttagtaat agatcacggc atgtttcccg ctccaagaga 2640
attcacttta ccttcagcag ttaccgagga gctaaacatg ctgccaacca gcttgtccaa 2700
caactccagg aaaactgttt ttcaaaacgc catgtcctag gggccaaggg aaatgctgtt 2760
ggtgagaatc gacctcactg tcagcgtttc tccacctgaa gtgatgatgg atgagaaaaa 2820
acaccaccaa atgacaagtc acaccctccc cattagtatc ctgttagggg aaaatagtag 2880
cagagtcatt gttacaggtg tactatggct gtattttaga gattaatttg tgtagattgt 2940
gtaaattcct gttgtctgac cttggtggtg ggaggggaga ctatgtgtca tgatttcaat 3000
gattgtttaa ttgtaggtca atgaaatatt tgcttattta tattcagaga tgtaccatgt 3060
taaagaggcg tcttgtattt tcttcccatt tgtaatgtat cttatttata tatgaagtaa 3120
gttctgaaaa ctgtttatgg tattttcgtg catttgtgag ccaaagagaa aagattaaaa 3180
ttagtgagat ttgtatttat attagagtgc ccttaaaata atgatttaag cattttactg 3240
tctgtaagag aattctaaga ttgtacatga cataagttat agtaatcatg gcaaatcctg 3300
ttacttaaat agcatctgct cttctcttac gctctctgtc tggctgtacg tctggtgttc 3360
tcaatgcttt tctagcaact gttggataat aactagatct cctgtaattt tgtagtagtt 3420
gatgaccaat ctctgtgact cgcttagctg aaacctaagg caacatttcc gaagaccttc 3480
tgaagatctc agataaagtg accaggctca caactgtttt tgaagaaggg aaattcacac 3540
tgtgcgtttt gagtatgcaa gaagaatata aataaataaa atatctcatg gagattgaca 3600
aaaaaaaaaa 3610
<210> 17
<211> 2824
<212> ДНК
<213> Homo sapiens
<400> 17
gcggccgccc gcacttcccg cctctggctc gcccgaggac gcgctggcac gcctcccacc 60
ccctcactct gactccagct ggcgtgcatg gtctgcctcg catcctcacg actcagctcc 120
ctccctctct cgtgtttttt tcctccgccg ccccctcatt catccccact gggctccctt 180
tccctcaaat gctctggggc tctccgcgct ttcctgagtc cgggctccga ggacccttag 240
gtagtcccgg tctcttttaa agctccccgg cttccaaagg gttgccacgt ccctaaaccc 300
tgtctccagc tcgcatacac acacgcacag acacgcacgt tttctgttcc tgcgtgacac 360
ccgccctcgc cgctcggccc cgccggtccc cgcgcggtgc cctcctcccg ccacacgggc 420
acgcacgcgc gcgcagggcc aagcccgagg cagctcgccc gcagctcgca ctcgcaggcg 480
acctgctcca gtctccaaag ccgatggcat ctccgggctc tggcttttgg tctttcgggt 540
cggaagatgg ctctggggat tccgagaatc ccggcacagc gcgagcctgg tgccaagtgg 600
ctcagaagtt cacgggcggc atcggaaaca aactgtgcgc cctgctctac ggagacgccg 660
agaagccggc ggagagcggc gggagccaac ccccgcgggc cgccgcccgg aaggccgcct 720
gcgcctgcga ccagaagccc tgcagctgct ccaaagtgga tgtcaactac gcgtttctcc 780
atgcaacaga cctgctgccg gcgtgtgatg gagaaaggcc cactttggcg tttctgcaag 840
atgttatgaa cattttactt cagtatgtgg tgaaaagttt cgatagatca accaaagtga 900
ttgatttcca ttatcctaat gagcttctcc aagaatataa ttgggaattg gcagaccaac 960
cacaaaattt ggaggaaatt ttgatgcatt gccaaacaac tctaaaatat gcaattaaaa 1020
cagggcatcc tagatacttc aatcaacttt ctactggttt ggatatggtt ggattagcag 1080
cagactggct gacatcaaca gcaaatacta acatgttcac ctatgaaatt gctccagtat 1140
ttgtgctttt ggaatatgtc acactaaaga aaatgagaga aatcattggc tggccagggg 1200
gctctggcga tgggatattt tctcccggtg gcgccatatc taacatgtat gccatgatga 1260
tcgcacgctt taagatgttc ccagaagtca aggagaaagg aatggctgct cttcccaggc 1320
tcattgcctt cacgtctgaa catagtcatt tttctctcaa gaagggagct gcagccttag 1380
ggattggaac agacagcgtg attctgatta aatgtgatga gagagggaaa atgattccat 1440
ctgatcttga aagaaggatt cttgaagcca aacagaaagg gtttgttcct ttcctcgtga 1500
gtgccacagc tggaaccacc gtgtacggag catttgaccc cctcttagct gtcgctgaca 1560
tttgcaaaaa gtataagatc tggatgcatg tggatgcagc ttggggtggg ggattactga 1620
tgtcccgaaa acacaagtgg aaactgagtg gcgtggagag ggccaactct gtgacgtgga 1680
atccacacaa gatgatggga gtccctttgc agtgctctgc tctcctggtt agagaagagg 1740
gattgatgca gaattgcaac caaatgcatg cctcctacct ctttcagcaa gataaacatt 1800
atgacctgtc ctatgacact ggagacaagg ccttacagtg cggacgccac gttgatgttt 1860
ttaaactatg gctgatgtgg agggcaaagg ggactaccgg gtttgaagcg catgttgata 1920
aatgtttgga gttggcagag tatttataca acatcataaa aaaccgagaa ggatatgaga 1980
tggtgtttga tgggaagcct cagcacacaa atgtctgctt ctggtacatt cctccaagct 2040
tgcgtactct ggaagacaat gaagagagaa tgagtcgcct ctcgaaggtg gctccagtga 2100
ttaaagccag aatgatggag tatggaacca caatggtcag ctaccaaccc ttgggagaca 2160
aggtcaattt cttccgcatg gtcatctcaa acccagcggc aactcaccaa gacattgact 2220
tcctgattga agaaatagaa cgccttggac aagatttata ataaccttgc tcaccaagct 2280
gttccacttc tctagagaac atgccctcag ctaagccccc tactgagaaa cttcctttga 2340
gaattgtgcg acttcacaaa atgcaaggtg aacaccactt tgtctctgag aacagacgtt 2400
accaattatg gagtgtcacc agctgccaaa atcgtaggtg ttggctctgc tggtcactgg 2460
agtagttgct actcttcaga atatggacaa agaaggcaca ggtgtaaata tagtagcagg 2520
atgaggaacc tcaaactggg tatcattttg cacgtgctct tctgttctca aatgctaaat 2580
gcaaacactg tgtatttatt agttaggtgt gccaaactac cgttcccaaa ttggtgtttc 2640
tgaatgacat caacattccc ccaacattac tccattacta aagacagaaa aaaataaaaa 2700
cataaaatat acaaacatgt ggcaacctgt tcttcctacc aaatataaac ttgtgtatga 2760
tccaagtatt ttatctgtgt tgtctctcta aacccaaata aatgtgtaaa tgtggacaca 2820
tctc 2824
<210> 18
<211> 2419
<212> ДНК
<213> Homo sapiens
<400> 18
gcggccgccc gcacttcccg cctctggctc gcccgaggac gcgctggcac gcctcccacc 60
ccctcactct gactccagct ggcgtgcatg gtctgcctcg catcctcacg actcagctcc 120
ctccctctct cgtgtttttt tcctccgccg ccccctcatt catccccact gggctccctt 180
tccctcaaat gctctggggc tctccgcgct ttcctgagtc cgggctccga ggacccttag 240
gtagtcccgg tctcttttaa agctccccgg cttccaaagg gttgccacgt ccctaaaccc 300
tgtctccagc tcgcatacac acacgcacag acacgcacgt tttctgttcc tgcgtgacac 360
ccgccctcgc cgctcggccc cgccggtccc cgcgcggtgc cctcctcccg ccacacgggc 420
acgcacgcgc gcgcagggcc aagcccgagg cagctcgccc gcagctcgca ctcgcaggcg 480
acctgctcca gtctccaaag ccgatggcat ctccgggctc tggcttttgg tctttcgggt 540
cggaagatgg ctctggggat tccgagaatc ccggcacagc gcgagcctgg tgccaagtgg 600
ctcagaagtt cacgggcggc atcggaaaca aactgtgcgc cctgctctac ggagacgccg 660
agaagccggc ggagagcggc gggagccaac ccccgcgggc cgccgcccgg aaggccgcct 720
gcgcctgcga ccagaagccc tgcagctgct ccaaagtgga tgtcaactac gcgtttctcc 780
atgcaacaga cctgctgccg gcgtgtgatg gagaaaggcc cactttggcg tttctgcaag 840
atgttatgaa cattttactt cagtatgtgg tgaaaagttt cgatagatca accaaagtga 900
ttgatttcca ttatcctaat gagcttctcc aagaatataa ttgggaattg gcagaccaac 960
cacaaaattt ggaggaaatt ttgatgcatt gccaaacaac tctaaaatat gcaattaaaa 1020
cagggcatcc tagatacttc aatcaacttt ctactggttt ggatatggtt ggattagcag 1080
cagactggct gacatcaaca gcaaatacta acatgttcac ctatgaaatt gctccagtat 1140
ttgtgctttt ggaatatgtc acactaaaga aaatgagaga aatcattggc tggccagggg 1200
gctctggcga tgggatattt tctcccggtg gcgccatatc taacatgtat gccatgatga 1260
tcgcacgctt taagatgttc ccagaagtca aggagaaagg aatggctgct cttcccaggc 1320
tcattgcctt cacgtctgaa catagtcatt tttctctcaa gaagggagct gcagccttag 1380
ggattggaac agacagcgtg attctgatta aatgtgatga gagagggaaa atgattccat 1440
ctgatcttga aagaaggatt cttgaagcca aacagaaagg gtttgttcct ttcctcgtga 1500
gtgccacagc tggaaccacc gtgtacggag catttgaccc cctcttagct gtcgctgaca 1560
tttgcaaaaa gtataagatc tggatgcatg tggatgcagc ttggggtggg ggattactga 1620
tgtcccgaaa acacaagtgg aaactgagtg gcgtggagag ggccaactct gtgacgtgga 1680
atccacacaa gatgatggga gtccctttgc agtgctctgc tctcctggtt agagaagagg 1740
gattgatgca gaattgcaac caaatgcatg cctcctacct ctttcagcaa gataaacatt 1800
atgacctgtc ctatgacact ggagacaagg ccttacagtg cggacgccac gttgatgttt 1860
ttaaactatg gctgatgtgg agggcaaagg ggactaccgg gtttgaagcg catgttgata 1920
aatgtttgga gttggcagag tatttataca acatcataaa aaaccgagaa ggatatgaga 1980
tggtgtttga tgggaagcct cagcacacaa atgtctgctt ctggtacatt cctccaagct 2040
tgcgtactct ggaagacaat gaagagagaa tgagtcgcct ctcgaaggtg gctccagtga 2100
ttaaagccag aatgatggag tatggaacca caatggtcag ctaccaaccc ttgggagaca 2160
aggtcaattt cttccgcatg gtcatctcaa acccagcggc aactcaccaa gacattgact 2220
tcctgattga agaaatagaa cgccttggac aagatttata ataaccttgc tcaccaagct 2280
gttccacttc tctaggtaga caattaagtt gtcacaaact gtgtgaatgt atttgtagtt 2340
tgttccaaag taaatctatt tctatattgt ggtgtcaaag tagagtttaa aaattaaaca 2400
aaaaagacat tgctccttt 2419
<210> 19
<211> 979
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Met Arg Arg Pro Arg Arg Pro Gly Gly Leu Gly Gly Ser Gly Gly Leu
1 5 10 15
Arg Leu Leu Leu Cys Leu Leu Leu Leu Ser Ser Arg Pro Gly Gly Cys
20 25 30
Ser Ala Val Ser Ala His Gly Cys Leu Phe Asp Arg Arg Leu Cys Ser
35 40 45
His Leu Glu Val Cys Ile Gln Asp Gly Leu Phe Gly Gln Cys Gln Val
50 55 60
Gly Val Gly Gln Ala Arg Pro Leu Leu Gln Val Thr Ser Pro Val Leu
65 70 75 80
Gln Arg Leu Gln Gly Val Leu Arg Gln Leu Met Ser Gln Gly Leu Ser
85 90 95
Trp His Asp Asp Leu Thr Gln Tyr Val Ile Ser Gln Glu Met Glu Arg
100 105 110
Ile Pro Arg Leu Arg Pro Pro Glu Pro Arg Pro Arg Asp Arg Ser Gly
115 120 125
Leu Ala Pro Lys Arg Pro Gly Pro Ala Gly Glu Leu Leu Leu Gln Asp
130 135 140
Ile Pro Thr Gly Ser Ala Pro Ala Ala Gln His Arg Leu Pro Gln Pro
145 150 155 160
Pro Val Gly Lys Gly Gly Ala Gly Ala Ser Ser Ser Leu Ser Pro Leu
165 170 175
Gln Ala Glu Leu Leu Pro Pro Leu Leu Glu His Leu Leu Leu Pro Pro
180 185 190
Gln Pro Pro His Pro Ser Leu Ser Tyr Glu Pro Ala Leu Leu Gln Pro
195 200 205
Tyr Leu Phe His Gln Phe Gly Ser Arg Asp Gly Ser Arg Val Ser Glu
210 215 220
Gly Ser Pro Gly Met Val Ser Val Gly Pro Leu Pro Lys Ala Glu Ala
225 230 235 240
Pro Ala Leu Phe Ser Arg Thr Ala Ser Lys Gly Ile Phe Gly Asp His
245 250 255
Pro Gly His Ser Tyr Gly Asp Leu Pro Gly Pro Ser Pro Ala Gln Leu
260 265 270
Phe Gln Asp Ser Gly Leu Leu Tyr Leu Ala Gln Glu Leu Pro Ala Pro
275 280 285
Ser Arg Ala Arg Val Pro Arg Leu Pro Glu Gln Gly Ser Ser Ser Arg
290 295 300
Ala Glu Asp Ser Pro Glu Gly Tyr Glu Lys Glu Gly Leu Gly Asp Arg
305 310 315 320
Gly Glu Lys Pro Ala Ser Pro Ala Val Gln Pro Asp Ala Ala Leu Gln
325 330 335
Arg Leu Ala Ala Val Leu Ala Gly Tyr Gly Val Glu Leu Arg Gln Leu
340 345 350
Thr Pro Glu Gln Leu Ser Thr Leu Leu Thr Leu Leu Gln Leu Leu Pro
355 360 365
Lys Gly Ala Gly Arg Asn Pro Gly Gly Val Val Asn Val Gly Ala Asp
370 375 380
Ile Lys Lys Thr Met Glu Gly Pro Val Glu Gly Arg Asp Thr Ala Glu
385 390 395 400
Leu Pro Ala Arg Thr Ser Pro Met Pro Gly His Pro Thr Ala Ser Pro
405 410 415
Thr Ser Ser Glu Val Gln Gln Val Pro Ser Pro Val Ser Ser Glu Pro
420 425 430
Pro Lys Ala Ala Arg Pro Pro Val Thr Pro Val Leu Leu Glu Lys Lys
435 440 445
Ser Pro Leu Gly Gln Ser Gln Pro Thr Val Ala Gly Gln Pro Ser Ala
450 455 460
Arg Pro Ala Ala Glu Glu Tyr Gly Tyr Ile Val Thr Asp Gln Lys Pro
465 470 475 480
Leu Ser Leu Ala Ala Gly Val Lys Leu Leu Glu Ile Leu Ala Glu His
485 490 495
Val His Met Ser Ser Gly Ser Phe Ile Asn Ile Ser Val Val Gly Pro
500 505 510
Ala Leu Thr Phe Arg Ile Arg His Asn Glu Gln Asn Leu Ser Leu Ala
515 520 525
Asp Val Thr Gln Gln Ala Gly Leu Val Lys Ser Glu Leu Glu Ala Gln
530 535 540
Thr Gly Leu Gln Ile Leu Gln Thr Gly Val Gly Gln Arg Glu Glu Ala
545 550 555 560
Ala Ala Val Leu Pro Gln Thr Ala His Ser Thr Ser Pro Met Arg Ser
565 570 575
Val Leu Leu Thr Leu Val Ala Leu Ala Gly Val Ala Gly Leu Leu Val
580 585 590
Ala Leu Ala Val Ala Leu Cys Val Arg Gln His Ala Arg Gln Gln Asp
595 600 605
Lys Glu Arg Leu Ala Ala Leu Gly Pro Glu Gly Ala His Gly Asp Thr
610 615 620
Thr Phe Glu Tyr Gln Asp Leu Cys Arg Gln His Met Ala Thr Lys Ser
625 630 635 640
Leu Phe Asn Arg Ala Glu Gly Pro Pro Glu Pro Ser Arg Val Ser Ser
645 650 655
Val Ser Ser Gln Phe Ser Asp Ala Ala Gln Ala Ser Pro Ser Ser His
660 665 670
Ser Ser Thr Pro Ser Trp Cys Glu Glu Pro Ala Gln Ala Asn Met Asp
675 680 685
Ile Ser Thr Gly His Met Ile Leu Ala Tyr Met Glu Asp His Leu Arg
690 695 700
Asn Arg Asp Arg Leu Ala Lys Glu Trp Gln Ala Leu Cys Ala Tyr Gln
705 710 715 720
Ala Glu Pro Asn Thr Cys Ala Thr Ala Gln Gly Glu Gly Asn Ile Lys
725 730 735
Lys Asn Arg His Pro Asp Phe Leu Pro Tyr Asp His Ala Arg Ile Lys
740 745 750
Leu Lys Val Glu Ser Ser Pro Ser Arg Ser Asp Tyr Ile Asn Ala Ser
755 760 765
Pro Ile Ile Glu His Asp Pro Arg Met Pro Ala Tyr Ile Ala Thr Gln
770 775 780
Gly Pro Leu Ser His Thr Ile Ala Asp Phe Trp Gln Met Val Trp Glu
785 790 795 800
Ser Gly Cys Thr Val Ile Val Met Leu Thr Pro Leu Val Glu Asp Gly
805 810 815
Val Lys Gln Cys Asp Arg Tyr Trp Pro Asp Glu Gly Ala Ser Leu Tyr
820 825 830
His Val Tyr Glu Val Asn Leu Val Ser Glu His Ile Trp Cys Glu Asp
835 840 845
Phe Leu Val Arg Ser Phe Tyr Leu Lys Asn Val Gln Thr Gln Glu Thr
850 855 860
Arg Thr Leu Thr Gln Phe His Phe Leu Ser Trp Pro Ala Glu Gly Thr
865 870 875 880
Pro Ala Ser Thr Arg Pro Leu Leu Asp Phe Arg Arg Lys Val Asn Lys
885 890 895
Cys Tyr Arg Gly Arg Ser Cys Pro Ile Ile Val His Cys Ser Asp Gly
900 905 910
Ala Gly Arg Thr Gly Thr Tyr Ile Leu Ile Asp Met Val Leu Asn Arg
915 920 925
Met Ala Lys Gly Val Lys Glu Ile Asp Ile Ala Ala Thr Leu Glu His
930 935 940
Val Arg Asp Gln Arg Pro Gly Leu Val Arg Ser Lys Asp Gln Phe Glu
945 950 955 960
Phe Ala Leu Thr Ala Val Ala Glu Glu Val Asn Ala Ile Leu Lys Ala
965 970 975
Leu Pro Gln
<210> 20
<211> 2940
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(2937)
<400> 20
atg cgg cgc ccg cgg cgg cct ggg ggt ctc ggg gga tcc ggg ggt ctc 48
Met Arg Arg Pro Arg Arg Pro Gly Gly Leu Gly Gly Ser Gly Gly Leu
1 5 10 15
cgg ctg ctc ctc tgc ctc ctg ctg ctg agc agc cgc ccg ggg ggc tgc 96
Arg Leu Leu Leu Cys Leu Leu Leu Leu Ser Ser Arg Pro Gly Gly Cys
20 25 30
agc gcc gtt agt gcc cac ggc tgt cta ttt gac cgc agg ctc tgc tct 144
Ser Ala Val Ser Ala His Gly Cys Leu Phe Asp Arg Arg Leu Cys Ser
35 40 45
cac ctg gaa gtc tgt att cag gat ggc ttg ttt ggg cag tgc cag gtg 192
His Leu Glu Val Cys Ile Gln Asp Gly Leu Phe Gly Gln Cys Gln Val
50 55 60
gga gtg ggg cag gcc cgg ccc ctt ttg caa gtc acc tcc cca gtt ctc 240
Gly Val Gly Gln Ala Arg Pro Leu Leu Gln Val Thr Ser Pro Val Leu
65 70 75 80
caa cgc tta caa ggt gtg ctc cga caa ctc atg tcc caa gga ttg tcc 288
Gln Arg Leu Gln Gly Val Leu Arg Gln Leu Met Ser Gln Gly Leu Ser
85 90 95
tgg cac gat gac ctc acc cag tat gtg atc tct cag gag atg gag cgc 336
Trp His Asp Asp Leu Thr Gln Tyr Val Ile Ser Gln Glu Met Glu Arg
100 105 110
atc ccc agg ctt cgc ccc cca gag ccc cgt cca agg gac agg tct ggc 384
Ile Pro Arg Leu Arg Pro Pro Glu Pro Arg Pro Arg Asp Arg Ser Gly
115 120 125
ttg gca ccc aag aga cct ggt cct gct gga gag ctg ctt tta cag gac 432
Leu Ala Pro Lys Arg Pro Gly Pro Ala Gly Glu Leu Leu Leu Gln Asp
130 135 140
atc ccc act ggc tcc gcc cct gct gcc cag cat cgg ctt cca caa cca 480
Ile Pro Thr Gly Ser Ala Pro Ala Ala Gln His Arg Leu Pro Gln Pro
145 150 155 160
cca gtg ggc aaa ggt gga gct ggg gcc agc tcc tct ctg tcc cct ctg 528
Pro Val Gly Lys Gly Gly Ala Gly Ala Ser Ser Ser Leu Ser Pro Leu
165 170 175
cag gct gag ctg ctc ccg cct ctc ttg gag cac ctg ctg ctg ccc cca 576
Gln Ala Glu Leu Leu Pro Pro Leu Leu Glu His Leu Leu Leu Pro Pro
180 185 190
cag cct ccc cac cct tca ctg agt tac gaa cct gcc ttg ctg cag ccc 624
Gln Pro Pro His Pro Ser Leu Ser Tyr Glu Pro Ala Leu Leu Gln Pro
195 200 205
tac ctg ttc cac cag ttt ggc tcc cgt gat ggc tcc agg gtc tca gag 672
Tyr Leu Phe His Gln Phe Gly Ser Arg Asp Gly Ser Arg Val Ser Glu
210 215 220
ggc tcc cca ggg atg gtc agt gtc ggc ccc ctg ccc aag gct gaa gcc 720
Gly Ser Pro Gly Met Val Ser Val Gly Pro Leu Pro Lys Ala Glu Ala
225 230 235 240
cct gcc ctc ttc agc aga act gcc tcc aag ggc ata ttt ggg gac cac 768
Pro Ala Leu Phe Ser Arg Thr Ala Ser Lys Gly Ile Phe Gly Asp His
245 250 255
cct ggc cac tcc tac ggg gac ctt cca ggg cct tca cct gcc cag ctt 816
Pro Gly His Ser Tyr Gly Asp Leu Pro Gly Pro Ser Pro Ala Gln Leu
260 265 270
ttt caa gac tct ggg ctg ctc tat ctg gcc cag gag ttg cca gca ccc 864
Phe Gln Asp Ser Gly Leu Leu Tyr Leu Ala Gln Glu Leu Pro Ala Pro
275 280 285
agc agg gcc agg gtg cca agg ctg cca gag caa ggg agc agc agc cgg 912
Ser Arg Ala Arg Val Pro Arg Leu Pro Glu Gln Gly Ser Ser Ser Arg
290 295 300
gca gag gac tcc cca gag ggc tat gag aag gaa gga cta ggg gat cgt 960
Ala Glu Asp Ser Pro Glu Gly Tyr Glu Lys Glu Gly Leu Gly Asp Arg
305 310 315 320
gga gag aag cct gct tcc cca gct gtg cag cca gat gcg gct ctg cag 1008
Gly Glu Lys Pro Ala Ser Pro Ala Val Gln Pro Asp Ala Ala Leu Gln
325 330 335
agg ctg gcc gct gtg ctg gcg ggc tat ggg gta gag ctg cgt cag ctg 1056
Arg Leu Ala Ala Val Leu Ala Gly Tyr Gly Val Glu Leu Arg Gln Leu
340 345 350
acc cct gag cag ctc tcc aca ctc ctg acc ctg ctg cag cta ctg ccc 1104
Thr Pro Glu Gln Leu Ser Thr Leu Leu Thr Leu Leu Gln Leu Leu Pro
355 360 365
aag ggt gca gga aga aat ccg gga ggg gtt gta aat gtt gga gct gat 1152
Lys Gly Ala Gly Arg Asn Pro Gly Gly Val Val Asn Val Gly Ala Asp
370 375 380
atc aag aaa aca atg gag ggg ccg gtg gag ggc aga gac aca gca gag 1200
Ile Lys Lys Thr Met Glu Gly Pro Val Glu Gly Arg Asp Thr Ala Glu
385 390 395 400
ctt cca gcc cgc aca tcc ccc atg cct gga cac ccc act gcc agc cct 1248
Leu Pro Ala Arg Thr Ser Pro Met Pro Gly His Pro Thr Ala Ser Pro
405 410 415
acc tcc agt gaa gtc cag cag gtg cca agc cct gtc tcc tct gag cct 1296
Thr Ser Ser Glu Val Gln Gln Val Pro Ser Pro Val Ser Ser Glu Pro
420 425 430
ccc aaa gct gcc aga ccc cct gtg aca cct gtc ctg cta gag aag aaa 1344
Pro Lys Ala Ala Arg Pro Pro Val Thr Pro Val Leu Leu Glu Lys Lys
435 440 445
agc cca ctg ggc cag agc cag ccc acg gtg gca gga cag ccc tca gcc 1392
Ser Pro Leu Gly Gln Ser Gln Pro Thr Val Ala Gly Gln Pro Ser Ala
450 455 460
cgc cca gca gca gag gaa tat ggc tac atc gtc act gat cag aag ccc 1440
Arg Pro Ala Ala Glu Glu Tyr Gly Tyr Ile Val Thr Asp Gln Lys Pro
465 470 475 480
ctg agc ctg gct gca gga gtg aag ctg ctg gag atc ctg gct gag cat 1488
Leu Ser Leu Ala Ala Gly Val Lys Leu Leu Glu Ile Leu Ala Glu His
485 490 495
gtg cac atg tcc tca ggc agc ttc atc aac atc agt gtg gtg gga cca 1536
Val His Met Ser Ser Gly Ser Phe Ile Asn Ile Ser Val Val Gly Pro
500 505 510
gcc ctc acc ttc cgc atc cgg cac aat gag cag aac ctg tct ttg gct 1584
Ala Leu Thr Phe Arg Ile Arg His Asn Glu Gln Asn Leu Ser Leu Ala
515 520 525
gat gtg acc caa caa gca ggg ctg gtg aag tct gaa ctg gaa gca cag 1632
Asp Val Thr Gln Gln Ala Gly Leu Val Lys Ser Glu Leu Glu Ala Gln
530 535 540
aca ggg ctc caa atc ttg cag aca gga gtg gga cag agg gag gag gca 1680
Thr Gly Leu Gln Ile Leu Gln Thr Gly Val Gly Gln Arg Glu Glu Ala
545 550 555 560
gct gca gtc ctt ccc caa act gcg cac agc acc tca ccc atg cgc tca 1728
Ala Ala Val Leu Pro Gln Thr Ala His Ser Thr Ser Pro Met Arg Ser
565 570 575
gtg ctg ctc act ctg gtg gcc ctg gca ggt gtg gct ggg ctg ctg gtg 1776
Val Leu Leu Thr Leu Val Ala Leu Ala Gly Val Ala Gly Leu Leu Val
580 585 590
gct ctg gct gtg gct ctg tgt gtg cgg cag cat gcg cgg cag caa gac 1824
Ala Leu Ala Val Ala Leu Cys Val Arg Gln His Ala Arg Gln Gln Asp
595 600 605
aag gag cgc ctg gca gcc ctg ggg cct gag ggg gcc cat ggt gac act 1872
Lys Glu Arg Leu Ala Ala Leu Gly Pro Glu Gly Ala His Gly Asp Thr
610 615 620
acc ttt gag tac cag gac ctg tgc cgc cag cac atg gcc acg aag tcc 1920
Thr Phe Glu Tyr Gln Asp Leu Cys Arg Gln His Met Ala Thr Lys Ser
625 630 635 640
ttg ttc aac cgg gca gag ggt cca ccg gag cct tca cgg gtg agc agt 1968
Leu Phe Asn Arg Ala Glu Gly Pro Pro Glu Pro Ser Arg Val Ser Ser
645 650 655
gtg tcc tcc cag ttc agc gac gca gcc cag gcc agc ccc agc tcc cac 2016
Val Ser Ser Gln Phe Ser Asp Ala Ala Gln Ala Ser Pro Ser Ser His
660 665 670
agc agc acc ccg tcc tgg tgc gag gag ccg gcc caa gcc aac atg gac 2064
Ser Ser Thr Pro Ser Trp Cys Glu Glu Pro Ala Gln Ala Asn Met Asp
675 680 685
atc tcc acg gga cac atg att ctg gca tac atg gag gat cac ctg cgg 2112
Ile Ser Thr Gly His Met Ile Leu Ala Tyr Met Glu Asp His Leu Arg
690 695 700
aac cgg gac cgc ctt gcc aag gag tgg cag gcc ctc tgt gcc tac caa 2160
Asn Arg Asp Arg Leu Ala Lys Glu Trp Gln Ala Leu Cys Ala Tyr Gln
705 710 715 720
gca gag cca aac acc tgt gcc acc gcg cag ggg gag ggc aac atc aaa 2208
Ala Glu Pro Asn Thr Cys Ala Thr Ala Gln Gly Glu Gly Asn Ile Lys
725 730 735
aag aac cgg cat cct gac ttc ctg ccc tat gac cat gcc cgc ata aaa 2256
Lys Asn Arg His Pro Asp Phe Leu Pro Tyr Asp His Ala Arg Ile Lys
740 745 750
ctg aag gtg gag agc agc cct tct cgg agc gat tac atc aac gcc agc 2304
Leu Lys Val Glu Ser Ser Pro Ser Arg Ser Asp Tyr Ile Asn Ala Ser
755 760 765
ccc att att gag cat gac cct cgg atg cca gcc tac ata gcc acg cag 2352
Pro Ile Ile Glu His Asp Pro Arg Met Pro Ala Tyr Ile Ala Thr Gln
770 775 780
ggc ccg ctg tcc cat acc atc gca gac ttc tgg cag atg gtg tgg gag 2400
Gly Pro Leu Ser His Thr Ile Ala Asp Phe Trp Gln Met Val Trp Glu
785 790 795 800
agc ggc tgc acc gtc atc gtc atg ctg acc ccg ctg gtg gag gat ggt 2448
Ser Gly Cys Thr Val Ile Val Met Leu Thr Pro Leu Val Glu Asp Gly
805 810 815
gtc aag cag tgt gac cgc tac tgg cca gat gag ggt gcc tcc ctc tac 2496
Val Lys Gln Cys Asp Arg Tyr Trp Pro Asp Glu Gly Ala Ser Leu Tyr
820 825 830
cac gta tat gag gtg aac ctg gtg tcg gag cac atc tgg tgc gag gac 2544
His Val Tyr Glu Val Asn Leu Val Ser Glu His Ile Trp Cys Glu Asp
835 840 845
ttt ctg gtg cgg agc ttc tac ctg aag aac gtg cag acc cag gag acg 2592
Phe Leu Val Arg Ser Phe Tyr Leu Lys Asn Val Gln Thr Gln Glu Thr
850 855 860
cgc acg ctc acg cag ttc cac ttc ctc agc tgg ccg gca gag ggc aca 2640
Arg Thr Leu Thr Gln Phe His Phe Leu Ser Trp Pro Ala Glu Gly Thr
865 870 875 880
ccg gcc tcc acg cgg ccc ctg ctg gac ttc cgc agg aag gtg aac aag 2688
Pro Ala Ser Thr Arg Pro Leu Leu Asp Phe Arg Arg Lys Val Asn Lys
885 890 895
tgc tac cgg ggc cgc tcc tgc ccc atc atc gtg cac tgc agt gat ggt 2736
Cys Tyr Arg Gly Arg Ser Cys Pro Ile Ile Val His Cys Ser Asp Gly
900 905 910
gcg ggg agg acc ggc acc tac atc ctc atc gac atg gtc ctg aac cgc 2784
Ala Gly Arg Thr Gly Thr Tyr Ile Leu Ile Asp Met Val Leu Asn Arg
915 920 925
atg gca aaa gga gtg aag gag att gac atc gct gcc acc ctg gag cat 2832
Met Ala Lys Gly Val Lys Glu Ile Asp Ile Ala Ala Thr Leu Glu His
930 935 940
gtc cgt gac cag cgg cct ggc ctt gtc cgc tct aag gac cag ttt gaa 2880
Val Arg Asp Gln Arg Pro Gly Leu Val Arg Ser Lys Asp Gln Phe Glu
945 950 955 960
ttt gcc ctg aca gcc gtg gcg gag gaa gtg aat gcc atc ctc aag gcc 2928
Phe Ala Leu Thr Ala Val Ala Glu Glu Val Asn Ala Ile Leu Lys Ala
965 970 975
ctg ccc cag tga 2940
Leu Pro Gln
<210> 21
<211> 3801
<212> ДНК
<213> Homo sapiens
<400> 21
aggtaggtcc cccgctccga cagggctctt acgccatatg tgacgcaaat tagcttacga 60
gccgggggtg gaggtgtcgt cagcacccgc ctgcggttgg ccgcgttgca gcgagagggg 120
catgtagacc ccgcctcgta gggaggtggg gagcggcaag ccccgcctca gcccctctgg 180
caggctcccg ccagcgtcgc tgcggctccg gcccgggagc gagcgcccgg agctcggaaa 240
gatgcggcgc ccgcggcggc ctgggggtct cgggggatcc gggggtctcc ggctgctcct 300
ctgcctcctg ctgctgagca gccgcccggg gggctgcagc gccgttagtg cccacggctg 360
tctatttgac cgcaggctct gctctcacct ggaagtctgt attcaggatg gcttgtttgg 420
gcagtgccag gtgggagtgg ggcaggcccg gccccttttg caagtcacct ccccagttct 480
ccaacgctta caaggtgtgc tccgacaact catgtcccaa ggattgtcct ggcacgatga 540
cctcacccag tatgtgatct ctcaggagat ggagcgcatc cccaggcttc gccccccaga 600
gccccgtcca agggacaggt ctggcttggc acccaagaga cctggtcctg ctggagagct 660
gcttttacag gacatcccca ctggctccgc ccctgctgcc cagcatcggc ttccacaacc 720
accagtgggc aaaggtggag ctggggccag ctcctctctg tcccctctgc aggctgagct 780
gctcccgcct ctcttggagc acctgctgct gcccccacag cctccccacc cttcactgag 840
ttacgaacct gccttgctgc agccctacct gttccaccag tttggctccc gtgatggctc 900
cagggtctca gagggctccc cagggatggt cagtgtcggc cccctgccca aggctgaagc 960
ccctgccctc ttcagcagaa ctgcctccaa gggcatattt ggggaccacc ctggccactc 1020
ctacggggac cttccagggc cttcacctgc ccagcttttt caagactctg ggctgctcta 1080
tctggcccag gagttgccag cacccagcag ggccagggtg ccaaggctgc cagagcaagg 1140
gagcagcagc cgggcagagg actccccaga gggctatgag aaggaaggac taggggatcg 1200
tggagagaag cctgcttccc cagctgtgca gccagatgcg gctctgcaga ggctggccgc 1260
tgtgctggcg ggctatgggg tagagctgcg tcagctgacc cctgagcagc tctccacact 1320
cctgaccctg ctgcagctac tgcccaaggg tgcaggaaga aatccgggag gggttgtaaa 1380
tgttggagct gatatcaaga aaacaatgga ggggccggtg gagggcagag acacagcaga 1440
gcttccagcc cgcacatccc ccatgcctgg acaccccact gccagcccta cctccagtga 1500
agtccagcag gtgccaagcc ctgtctcctc tgagcctccc aaagctgcca gaccccctgt 1560
gacacctgtc ctgctagaga agaaaagccc actgggccag agccagccca cggtggcagg 1620
acagccctca gcccgcccag cagcagagga atatggctac atcgtcactg atcagaagcc 1680
cctgagcctg gctgcaggag tgaagctgct ggagatcctg gctgagcatg tgcacatgtc 1740
ctcaggcagc ttcatcaaca tcagtgtggt gggaccagcc ctcaccttcc gcatccggca 1800
caatgagcag aacctgtctt tggctgatgt gacccaacaa gcagggctgg tgaagtctga 1860
actggaagca cagacagggc tccaaatctt gcagacagga gtgggacaga gggaggaggc 1920
agctgcagtc cttccccaaa ctgcgcacag cacctcaccc atgcgctcag tgctgctcac 1980
tctggtggcc ctggcaggtg tggctgggct gctggtggct ctggctgtgg ctctgtgtgt 2040
gcggcagcat gcgcggcagc aagacaagga gcgcctggca gccctggggc ctgagggggc 2100
ccatggtgac actacctttg agtaccagga cctgtgccgc cagcacatgg ccacgaagtc 2160
cttgttcaac cgggcagagg gtccaccgga gccttcacgg gtgagcagtg tgtcctccca 2220
gttcagcgac gcagcccagg ccagccccag ctcccacagc agcaccccgt cctggtgcga 2280
ggagccggcc caagccaaca tggacatctc cacgggacac atgattctgg catacatgga 2340
ggatcacctg cggaaccggg accgccttgc caaggagtgg caggccctct gtgcctacca 2400
agcagagcca aacacctgtg ccaccgcgca gggggagggc aacatcaaaa agaaccggca 2460
tcctgacttc ctgccctatg accatgcccg cataaaactg aaggtggaga gcagcccttc 2520
tcggagcgat tacatcaacg ccagccccat tattgagcat gaccctcgga tgccagccta 2580
catagccacg cagggcccgc tgtcccatac catcgcagac ttctggcaga tggtgtggga 2640
gagcggctgc accgtcatcg tcatgctgac cccgctggtg gaggatggtg tcaagcagtg 2700
tgaccgctac tggccagatg agggtgcctc cctctaccac gtatatgagg tgaacctggt 2760
gtcggagcac atctggtgcg aggactttct ggtgcggagc ttctacctga agaacgtgca 2820
gacccaggag acgcgcacgc tcacgcagtt ccacttcctc agctggccgg cagagggcac 2880
accggcctcc acgcggcccc tgctggactt ccgcaggaag gtgaacaagt gctaccgggg 2940
ccgctcctgc cccatcatcg tgcactgcag tgatggtgcg gggaggaccg gcacctacat 3000
cctcatcgac atggtcctga accgcatggc aaaaggagtg aaggagattg acatcgctgc 3060
caccctggag catgtccgtg accagcggcc tggccttgtc cgctctaagg accagtttga 3120
atttgccctg acagccgtgg cggaggaagt gaatgccatc ctcaaggccc tgccccagtg 3180
agaccctggg gccccttggc gggcagccca gcctctgtcc ctctttgcct gtgtgagcat 3240
ctctgtgtac ccactcctca ctgccccacc agccacctct tgggcatgct cagcccttcc 3300
tagaagagtc aggaagggaa agccagaagg ggcacgcctg cccagcctcg catgccagag 3360
cctggggcat cccagagccc agggcatccc atgggggtgc tgcagccagg aggagaggaa 3420
aggacatggg tagcaattct acccagagcc ttctcctgcc tacattccct ggcctggctc 3480
tcctgtagct ctcctggggt tctgggagtt ccctgaacat ctgtgtgtgt ccccctatgc 3540
tccagtatgg aagaatgggg tggagggtcg ccacacccgg ctccccctgc ttctcagccc 3600
cgggcctgcc tctgactcac acttgggcgc tctgccctcc ctggcctcac gcccagcctc 3660
ctcccaccac cctcccacca tgcgctgctc aacctctctc cttctggcgc aagagaacat 3720
ttctagaaaa aactactttt gtaccagtgt gaataaagtt agtgtgttgt ctgtgcagct 3780
gcaaaaaaaa aaaaaaaaaa a 3801
<210> 22
<211> 3712
<212> ДНК
<213> Homo sapiens
<400> 22
aggtaggtcc cccgctccga cagggctctt acgccatatg tgacgcaaat tagcttacga 60
gccgggggtg gaggtgtcgt cagcacccgc ctgcggttgg ccgcgttgca gcgagagggg 120
catgtagacc ccgcctcgta gggaggtggg gagcggcaag ccccgcctca gcccctctgg 180
caggctcccg ccagcgtcgc tgcggctccg gcccgggagc gagcgcccgg agctcggaaa 240
gatgcggcgc ccgcggcggc ctgggggtct cgggggatcc gggggtctcc ggctgctcct 300
ctgcctcctg ctgctgagca gccgcccggg gggctgcagc gccgttagtg cccacggctg 360
tctatttgac cgcaggctct gctctcacct ggaagtctgt attcaggatg gcttgtttgg 420
gcagtgccag gtgggagtgg ggcaggcccg gccccttttg caagtcacct ccccagttct 480
ccaacgctta caaggtgtgc tccgacaact catgtcccaa ggattgtcct ggcacgatga 540
cctcacccag tatgtgatct ctcaggagat ggagcgcatc cccaggcttc gccccccaga 600
gccccgtcca agggacaggt ctggcttggc acccaagaga cctggtcctg ctggagagct 660
gcttttacag gacatcccca ctggctccgc ccctgctgcc cagcatcggc ttccacaacc 720
accagtgggc aaaggtggag ctggggccag ctcctctctg tcccctctgc aggctgagct 780
gctcccgcct ctcttggagc acctgctgct gcccccacag cctccccacc cttcactgag 840
ttacgaacct gccttgctgc agccctacct gttccaccag tttggctccc gtgatggctc 900
cagggtctca gagggctccc cagggatggt cagtgtcggc cccctgccca aggctgaagc 960
ccctgccctc ttcagcagaa ctgcctccaa gggcatattt ggggaccacc ctggccactc 1020
ctacggggac cttccagggc cttcacctgc ccagcttttt caagactctg ggctgctcta 1080
tctggcccag gagttgccag cacccagcag ggccagggtg ccaaggctgc cagagcaagg 1140
gagcagcagc cgggcagagg actccccaga gggctatgag aaggaaggac taggggatcg 1200
tggagagaag cctgcttccc cagctgtgca gccagatgcg gctctgcaga ggctggccgc 1260
tgtgctggcg ggctatgggg tagagctgcg tcagctgacc cctgagcagc tctccacact 1320
cctgaccctg ctgcagctac tgcccaaggg tgcaggaaga aatccgggag gggttgtaaa 1380
tgttggagct gatatcaaga aaacaatgga ggggccggtg gagggcagag acacagcaga 1440
gcttccagcc cgcacatccc ccatgcctgg acaccccact gccagcccta cctccagtga 1500
agtccagcag gtgccaagcc ctgtctcctc tgagcctccc aaagctgcca gaccccctgt 1560
gacacctgtc ctgctagaga agaaaagccc actgggccag agccagccca cggtggcagg 1620
acagccctca gcccgcccag cagcagagga atatggctac atcgtcactg atcagaatgt 1680
ggtgggacca gccctcacct tccgcatccg gcacaatgag cagaacctgt ctttggctga 1740
tgtgacccaa caagcagggc tggtgaagtc tgaactggaa gcacagacag ggctccaaat 1800
cttgcagaca ggagtgggac agagggagga ggcagctgca gtccttcccc aaactgcgca 1860
cagcacctca cccatgcgct cagtgctgct cactctggtg gccctggcag gtgtggctgg 1920
gctgctggtg gctctggctg tggctctgtg tgtgcggcag catgcgcggc agcaagacaa 1980
ggagcgcctg gcagccctgg ggcctgaggg ggcccatggt gacactacct ttgagtacca 2040
ggacctgtgc cgccagcaca tggccacgaa gtccttgttc aaccgggcag agggtccacc 2100
ggagccttca cgggtgagca gtgtgtcctc ccagttcagc gacgcagccc aggccagccc 2160
cagctcccac agcagcaccc cgtcctggtg cgaggagccg gcccaagcca acatggacat 2220
ctccacggga cacatgattc tggcatacat ggaggatcac ctgcggaacc gggaccgcct 2280
tgccaaggag tggcaggccc tctgtgccta ccaagcagag ccaaacacct gtgccaccgc 2340
gcagggggag ggcaacatca aaaagaaccg gcatcctgac ttcctgccct atgaccatgc 2400
ccgcataaaa ctgaaggtgg agagcagccc ttctcggagc gattacatca acgccagccc 2460
cattattgag catgaccctc ggatgccagc ctacatagcc acgcagggcc cgctgtccca 2520
taccatcgca gacttctggc agatggtgtg ggagagcggc tgcaccgtca tcgtcatgct 2580
gaccccgctg gtggaggatg gtgtcaagca gtgtgaccgc tactggccag atgagggtgc 2640
ctccctctac cacgtatatg aggtgaacct ggtgtcggag cacatctggt gcgaggactt 2700
tctggtgcgg agcttctacc tgaagaacgt gcagacccag gagacgcgca cgctcacgca 2760
gttccacttc ctcagctggc cggcagaggg cacaccggcc tccacgcggc ccctgctgga 2820
cttccgcagg aaggtgaaca agtgctaccg gggccgctcc tgccccatca tcgtgcactg 2880
cagtgatggt gcggggagga ccggcaccta catcctcatc gacatggtcc tgaaccgcat 2940
ggcaaaagga gtgaaggaga ttgacatcgc tgccaccctg gagcatgtcc gtgaccagcg 3000
gcctggcctt gtccgctcta aggaccagtt tgaatttgcc ctgacagccg tggcggagga 3060
agtgaatgcc atcctcaagg ccctgcccca gtgagaccct ggggcccctt ggcgggcagc 3120
ccagcctctg tccctctttg cctgtgtgag catctctgtg tacccactcc tcactgcccc 3180
accagccacc tcttgggcat gctcagccct tcctagaaga gtcaggaagg gaaagccaga 3240
aggggcacgc ctgcccagcc tcgcatgcca gagcctgggg catcccagag cccagggcat 3300
cccatggggg tgctgcagcc aggaggagag gaaaggacat gggtagcaat tctacccaga 3360
gccttctcct gcctacattc cctggcctgg ctctcctgta gctctcctgg ggttctggga 3420
gttccctgaa catctgtgtg tgtcccccta tgctccagta tggaagaatg gggtggaggg 3480
tcgccacacc cggctccccc tgcttctcag ccccgggcct gcctctgact cacacttggg 3540
cgctctgccc tccctggcct cacgcccagc ctcctcccac caccctccca ccatgcgctg 3600
ctcaacctct ctccttctgg cgcaagagaa catttctaga aaaaactact tttgtaccag 3660
tgtgaataaa gttagtgtgt tgtctgtgca gctgcaaaaa aaaaaaaaaa aa 3712
<210> 23
<211> 3570
<212> ДНК
<213> Homo sapiens
<400> 23
aagagaattg ggctctgggg aaaattgcag gttcgggaga aggacacggg gtgcatgtgg 60
gcagagggac ttagagaccg gctgcgagac cacaggagaa gccacttgct ccgaggcgcc 120
tgggaggctg tctatttgac cgcaggctct gctctcacct ggaagtctgt attcaggatg 180
gcttgtttgg gcagtgccag gtgggagtgg ggcaggcccg gccccttttg caagtcacct 240
ccccagttct ccaacgctta caaggtgtgc tccgacaact catgtcccaa ggattgtcct 300
ggcacgatga cctcacccag tatgtgatct ctcaggagat ggagcgcatc cccaggcttc 360
gccccccaga gccccgtcca agggacaggt ctggcttggc acccaagaga cctggtcctg 420
ctggagagct gcttttacag gacatcccca ctggctccgc ccctgctgcc cagcatcggc 480
ttccacaacc accagtgggc aaaggtggag ctggggccag ctcctctctg tcccctctgc 540
aggctgagct gctcccgcct ctcttggagc acctgctgct gcccccacag cctccccacc 600
cttcactgag ttacgaacct gccttgctgc agccctacct gttccaccag tttggctccc 660
gtgatggctc cagggtctca gagggctccc cagggatggt cagtgtcggc cccctgccca 720
aggctgaagc ccctgccctc ttcagcagaa ctgcctccaa gggcatattt ggggaccacc 780
ctggccactc ctacggggac cttccagggc cttcacctgc ccagcttttt caagactctg 840
ggctgctcta tctggcccag gagttgccag cacccagcag ggccagggtg ccaaggctgc 900
cagagcaagg gagcagcagc cgggcagagg actccccaga gggctatgag aaggaaggac 960
taggggatcg tggagagaag cctgcttccc cagctgtgca gccagatgcg gctctgcaga 1020
ggctggccgc tgtgctggcg ggctatgggg tagagctgcg tcagctgacc cctgagcagc 1080
tctccacact cctgaccctg ctgcagctac tgcccaaggg tgcaggaaga aatccgggag 1140
gggttgtaaa tgttggagct gatatcaaga aaacaatgga ggggccggtg gagggcagag 1200
acacagcaga gcttccagcc cgcacatccc ccatgcctgg acaccccact gccagcccta 1260
cctccagtga agtccagcag gtgccaagcc ctgtctcctc tgagcctccc aaagctgcca 1320
gaccccctgt gacacctgtc ctgctagaga agaaaagccc actgggccag agccagccca 1380
cggtggcagg acagccctca gcccgcccag cagcagagga atatggctac atcgtcactg 1440
atcagaagcc cctgagcctg gctgcaggag tgaagctgct ggagatcctg gctgagcatg 1500
tgcacatgtc ctcaggcagc ttcatcaaca tcagtgtggt gggaccagcc ctcaccttcc 1560
gcatccggca caatgagcag aacctgtctt tggctgatgt gacccaacaa gcagggctgg 1620
tgaagtctga actggaagca cagacagggc tccaaatctt gcagacagga gtgggacaga 1680
gggaggaggc agctgcagtc cttccccaaa ctgcgcacag cacctcaccc atgcgctcag 1740
tgctgctcac tctggtggcc ctggcaggtg tggctgggct gctggtggct ctggctgtgg 1800
ctctgtgtgt gcggcagcat gcgcggcagc aagacaagga gcgcctggca gccctggggc 1860
ctgagggggc ccatggtgac actacctttg agtaccagga cctgtgccgc cagcacatgg 1920
ccacgaagtc cttgttcaac cgggcagagg gtccaccgga gccttcacgg gtgagcagtg 1980
tgtcctccca gttcagcgac gcagcccagg ccagccccag ctcccacagc agcaccccgt 2040
cctggtgcga ggagccggcc caagccaaca tggacatctc cacgggacac atgattctgg 2100
catacatgga ggatcacctg cggaaccggg accgccttgc caaggagtgg caggccctct 2160
gtgcctacca agcagagcca aacacctgtg ccaccgcgca gggggagggc aacatcaaaa 2220
agaaccggca tcctgacttc ctgccctatg accatgcccg cataaaactg aaggtggaga 2280
gcagcccttc tcggagcgat tacatcaacg ccagccccat tattgagcat gaccctcgga 2340
tgccagccta catagccacg cagggcccgc tgtcccatac catcgcagac ttctggcaga 2400
tggtgtggga gagcggctgc accgtcatcg tcatgctgac cccgctggtg gaggatggtg 2460
tcaagcagtg tgaccgctac tggccagatg agggtgcctc cctctaccac gtatatgagg 2520
tgaacctggt gtcggagcac atctggtgcg aggactttct ggtgcggagc ttctacctga 2580
agaacgtgca gacccaggag acgcgcacgc tcacgcagtt ccacttcctc agctggccgg 2640
cagagggcac accggcctcc acgcggcccc tgctggactt ccgcaggaag gtgaacaagt 2700
gctaccgggg ccgctcctgc cccatcatcg tgcactgcag tgatggtgcg gggaggaccg 2760
gcacctacat cctcatcgac atggtcctga accgcatggc aaaaggagtg aaggagattg 2820
acatcgctgc caccctggag catgtccgtg accagcggcc tggccttgtc cgctctaagg 2880
accagtttga atttgccctg acagccgtgg cggaggaagt gaatgccatc ctcaaggccc 2940
tgccccagtg agaccctggg gccccttggc gggcagccca gcctctgtcc ctctttgcct 3000
gtgtgagcat ctctgtgtac ccactcctca ctgccccacc agccacctct tgggcatgct 3060
cagcccttcc tagaagagtc aggaagggaa agccagaagg ggcacgcctg cccagcctcg 3120
catgccagag cctggggcat cccagagccc agggcatccc atgggggtgc tgcagccagg 3180
aggagaggaa aggacatggg tagcaattct acccagagcc ttctcctgcc tacattccct 3240
ggcctggctc tcctgtagct ctcctggggt tctgggagtt ccctgaacat ctgtgtgtgt 3300
ccccctatgc tccagtatgg aagaatgggg tggagggtcg ccacacccgg ctccccctgc 3360
ttctcagccc cgggcctgcc tctgactcac acttgggcgc tctgccctcc ctggcctcac 3420
gcccagcctc ctcccaccac cctcccacca tgcgctgctc aacctctctc cttctggcgc 3480
aagagaacat ttctagaaaa aactactttt gtaccagtgt gaataaagtt agtgtgttgt 3540
ctgtgcagct gcaaaaaaaa aaaaaaaaaa 3570
<210> 24
<211> 355
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Met Asp Phe Leu His Arg Asn Gly Val Leu Ile Ile Gln His Leu Gln
1 5 10 15
Lys Asp Tyr Arg Ala Tyr Tyr Thr Phe Leu Asn Phe Met Ser Asn Val
20 25 30
Gly Asp Pro Arg Asn Ile Phe Phe Ile Tyr Phe Pro Leu Cys Phe Gln
35 40 45
Phe Asn Gln Thr Val Gly Thr Lys Met Ile Trp Val Ala Val Ile Gly
50 55 60
Asp Trp Leu Asn Leu Ile Phe Lys Trp Ile Leu Phe Gly His Arg Pro
65 70 75 80
Tyr Trp Trp Val Gln Glu Thr Gln Ile Tyr Pro Asn His Ser Ser Pro
85 90 95
Cys Leu Glu Gln Phe Pro Thr Thr Cys Glu Thr Gly Pro Gly Ser Pro
100 105 110
Ser Gly His Ala Met Gly Ala Ser Cys Val Trp Tyr Val Met Val Thr
115 120 125
Ala Ala Leu Ser His Thr Val Cys Gly Met Asp Lys Phe Ser Ile Thr
130 135 140
Leu His Arg Leu Thr Trp Ser Phe Leu Trp Ser Val Phe Trp Leu Ile
145 150 155 160
Gln Ile Ser Val Cys Ile Ser Arg Val Phe Ile Ala Thr His Phe Pro
165 170 175
His Gln Val Ile Leu Gly Val Ile Gly Gly Met Leu Val Ala Glu Ala
180 185 190
Phe Glu His Thr Pro Gly Ile Gln Thr Ala Ser Leu Gly Thr Tyr Leu
195 200 205
Lys Thr Asn Leu Phe Leu Phe Leu Phe Ala Val Gly Phe Tyr Leu Leu
210 215 220
Leu Arg Val Leu Asn Ile Asp Leu Leu Trp Ser Val Pro Ile Ala Lys
225 230 235 240
Lys Trp Cys Ala Asn Pro Asp Trp Ile His Ile Asp Thr Thr Pro Phe
245 250 255
Ala Gly Leu Val Arg Asn Leu Gly Val Leu Phe Gly Leu Gly Phe Ala
260 265 270
Ile Asn Ser Glu Met Phe Leu Leu Ser Cys Arg Gly Gly Asn Asn Tyr
275 280 285
Thr Leu Ser Phe Arg Leu Leu Cys Ala Leu Thr Ser Leu Thr Ile Leu
290 295 300
Gln Leu Tyr His Phe Leu Gln Ile Pro Thr His Glu Glu His Leu Phe
305 310 315 320
Tyr Val Leu Ser Phe Cys Lys Ser Ala Ser Ile Pro Leu Thr Val Val
325 330 335
Ala Phe Ile Pro Tyr Ser Val His Met Leu Met Lys Gln Ser Gly Lys
340 345 350
Lys Ser Gln
355
<210> 25
<211> 1068
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(1065)
<400> 25
atg gat ttc ctt cac agg aat gga gtg ctc ata att cag cat ttg cag 48
Met Asp Phe Leu His Arg Asn Gly Val Leu Ile Ile Gln His Leu Gln
1 5 10 15
aag gac tac cga gct tac tac act ttt cta aat ttt atg tcc aat gtt 96
Lys Asp Tyr Arg Ala Tyr Tyr Thr Phe Leu Asn Phe Met Ser Asn Val
20 25 30
gga gac ccc agg aat atc ttt ttc att tat ttt cca ctt tgt ttt caa 144
Gly Asp Pro Arg Asn Ile Phe Phe Ile Tyr Phe Pro Leu Cys Phe Gln
35 40 45
ttt aat cag aca gtt gga acc aag atg ata tgg gta gca gtc att ggg 192
Phe Asn Gln Thr Val Gly Thr Lys Met Ile Trp Val Ala Val Ile Gly
50 55 60
gat tgg tta aat ctt ata ttt aaa tgg ata tta ttt ggt cat cga cct 240
Asp Trp Leu Asn Leu Ile Phe Lys Trp Ile Leu Phe Gly His Arg Pro
65 70 75 80
tac tgg tgg gtc caa gaa act cag att tac cca aat cac tca agt cca 288
Tyr Trp Trp Val Gln Glu Thr Gln Ile Tyr Pro Asn His Ser Ser Pro
85 90 95
tgc ctt gaa cag ttc cct act aca tgt gaa aca ggt cca gga agt cca 336
Cys Leu Glu Gln Phe Pro Thr Thr Cys Glu Thr Gly Pro Gly Ser Pro
100 105 110
tct ggc cat gca atg ggc gca tcc tgt gtc tgg tat gtc atg gta acc 384
Ser Gly His Ala Met Gly Ala Ser Cys Val Trp Tyr Val Met Val Thr
115 120 125
gct gcc ctg agc cac act gtc tgt ggg atg gat aag ttc tct atc act 432
Ala Ala Leu Ser His Thr Val Cys Gly Met Asp Lys Phe Ser Ile Thr
130 135 140
ctg cac aga ctg acc tgg tca ttt ctt tgg agt gtt ttt tgg ttg att 480
Leu His Arg Leu Thr Trp Ser Phe Leu Trp Ser Val Phe Trp Leu Ile
145 150 155 160
caa atc agt gtc tgc atc tcc aga gta ttc ata gca aca cat ttt cct 528
Gln Ile Ser Val Cys Ile Ser Arg Val Phe Ile Ala Thr His Phe Pro
165 170 175
cat caa gtt att ctt gga gta att ggt ggc atg ctg gtg gca gag gcc 576
His Gln Val Ile Leu Gly Val Ile Gly Gly Met Leu Val Ala Glu Ala
180 185 190
ttt gaa cac act cca ggc atc caa acg gcc agt ctg ggc aca tac ctg 624
Phe Glu His Thr Pro Gly Ile Gln Thr Ala Ser Leu Gly Thr Tyr Leu
195 200 205
aag acc aac ctc ttt ctc ttc ctg ttt gca gtt ggc ttt tac ctg ctt 672
Lys Thr Asn Leu Phe Leu Phe Leu Phe Ala Val Gly Phe Tyr Leu Leu
210 215 220
ctt agg gtg ctc aac att gac ctg ctg tgg tcc gtg ccc ata gcc aaa 720
Leu Arg Val Leu Asn Ile Asp Leu Leu Trp Ser Val Pro Ile Ala Lys
225 230 235 240
aag tgg tgt gct aac ccc gac tgg atc cac att gac acc acg cct ttt 768
Lys Trp Cys Ala Asn Pro Asp Trp Ile His Ile Asp Thr Thr Pro Phe
245 250 255
gct gga ctc gtg aga aac ctt ggg gtc ctc ttt ggc ttg ggc ttt gca 816
Ala Gly Leu Val Arg Asn Leu Gly Val Leu Phe Gly Leu Gly Phe Ala
260 265 270
atc aac tca gag atg ttc ctc ctg agc tgc cga ggg gga aat aac tac 864
Ile Asn Ser Glu Met Phe Leu Leu Ser Cys Arg Gly Gly Asn Asn Tyr
275 280 285
aca ctg agc ttc cgg ttg ctc tgt gcc ttg acc tca ttg aca ata ctg 912
Thr Leu Ser Phe Arg Leu Leu Cys Ala Leu Thr Ser Leu Thr Ile Leu
290 295 300
cag ctc tac cat ttc ctc cag atc ccg act cac gaa gag cat tta ttt 960
Gln Leu Tyr His Phe Leu Gln Ile Pro Thr His Glu Glu His Leu Phe
305 310 315 320
tat gtg ctg tct ttt tgt aaa agt gca tcc att ccc cta act gtg gtt 1008
Tyr Val Leu Ser Phe Cys Lys Ser Ala Ser Ile Pro Leu Thr Val Val
325 330 335
gct ttc att ccc tac tct gtt cat atg tta atg aaa caa agc gga aag 1056
Ala Phe Ile Pro Tyr Ser Val His Met Leu Met Lys Gln Ser Gly Lys
340 345 350
aag agt cag taa 1068
Lys Ser Gln
355
<210> 26
<211> 3096
<212> ДНК
<213> Homo sapiens
<400> 26
tcgacatgcc acaaaggcac agtataaaaa cggtgggaat cagagcactt cagctccaat 60
tgctctatgt ttagaattgc ctctttttca agatggattt ccttcacagg aatggagtgc 120
tcataattca gcatttgcag aaggactacc gagcttacta cacttttcta aattttatgt 180
ccaatgttgg agaccccagg aatatctttt tcatttattt tccactttgt tttcaattta 240
atcagacagt tggaaccaag atgatatggg tagcagtcat tggggattgg ttaaatctta 300
tatttaaatg gatattattt ggtcatcgac cttactggtg ggtccaagaa actcagattt 360
acccaaatca ctcaagtcca tgccttgaac agttccctac tacatgtgaa acaggtccag 420
gaagtccatc tggccatgca atgggcgcat cctgtgtctg gtatgtcatg gtaaccgctg 480
ccctgagcca cactgtctgt gggatggata agttctctat cactctgcac agactgacct 540
ggtcatttct ttggagtgtt ttttggttga ttcaaatcag tgtctgcatc tccagagtat 600
tcatagcaac acattttcct catcaagtta ttcttggagt aattggtggc atgctggtgg 660
cagaggcctt tgaacacact ccaggcatcc aaacggccag tctgggcaca tacctgaaga 720
ccaacctctt tctcttcctg tttgcagttg gcttttacct gcttcttagg gtgctcaaca 780
ttgacctgct gtggtccgtg cccatagcca aaaagtggtg tgctaacccc gactggatcc 840
acattgacac cacgcctttt gctggactcg tgagaaacct tggggtcctc tttggcttgg 900
gctttgcaat caactcagag atgttcctcc tgagctgccg agggggaaat aactacacac 960
tgagcttccg gttgctctgt gccttgacct cattgacaat actgcagctc taccatttcc 1020
tccagatccc gactcacgaa gagcatttat tttatgtgct gtctttttgt aaaagtgcat 1080
ccattcccct aactgtggtt gctttcattc cctactctgt tcatatgtta atgaaacaaa 1140
gcggaaagaa gagtcagtag agtggtgcct agagttagtg ctctgtgtca cagatcaccc 1200
ttctccatcc accagtagag ccacagagta ggcacagacc agaggcttct aatccgactt 1260
cacagaatag cggcacaggc cccattcccc atagagatgt ttagtttggc cttcgcactg 1320
gtcttttttt ttaatccttc agttaccaat atttagatac aagaatattt gacataaaaa 1380
tcggaagttc tgtatttctt gaaaaatctg atagtatgac aacacagagc ctgcatcccc 1440
agctggagac aactgaccag agctgcatac taacagtccc cagtaggagg caaggactcc 1500
attttctcac agtcttcagc atcccagcag gagccccact atgattcctt tatcttctta 1560
aggccaggct gcatctgatt cctgttgaca ttttagtggg gaccacagcc atatccagtt 1620
tcagttttca gatgaggaaa tggaagccta tttaggtaaa agaacttgcc tggagtcacg 1680
ccaccttcag agcaggaatt agaacccaag gcttctgaca catatcccca ttacactgtg 1740
tgtttgagtg tgcacacatg cacatgcttt ttgtttgtat gtttcctttt tagaaccagg 1800
gacttgctct gttgcccagg ctggagtgca gtggtggata gcggctcact gcagcctcaa 1860
actcctggct caaatgatcc tccctcctca gcctcccatt agctaagact acaggtgtgc 1920
accaccatgc ctggctaatt tttgaatatt ttttagagcc agcttcttac tatatttgcc 1980
caggctggtc ttgaactcct ggcctcaagg agttcccttg aggatgttcc acccatgtca 2040
gcctcccaca gtgctgggat tgcaggcttg agccattgta ttaatagttg gcctacactg 2100
tcttttgttt cttcatttac aagaggtaaa aatcagtaag aatgaatgct ttcatttaga 2160
ttctatgatg actgccatat aaatcagcta ccttttcaga aatgacattg aaatactgca 2220
tcctctttga cttatacccc acacatacat gcagggtcta ggtgggacca acagtggctc 2280
cagatgtata tacacacggg tccagggaca acagaacagg ccaggctaca gtatggactt 2340
gaacctctgc ttcacttctg gtctctgttt tacagtagtg ctcaccaata tttcaataaa 2400
ttctactgaa cttactctaa tagaacatta ttcaaccccc aaagacttat ctcttcacta 2460
tgacatctcc atactttatt tttaggagac agactttcaa aaccagagaa atcaggtgcc 2520
ttcctcaagg tcatgctcca acccaggcca actattaaat gcttgcatct gttagctgga 2580
ttagtctcta tgtatactga actgtgatga aaatctatag ctttgtttta gaaaattatt 2640
gttgatggac tattaatatt atattaacaa tttctcagta agtgtgtttt ttcctcattg 2700
aatctaggaa tgctgggctt taagttgata actgtgtcat ttcaatcaag tacaaggatt 2760
ttgaggcaga ttttgctaga tatcttagta atcccccaca atgttttatg taactcttct 2820
cagaatatca atacattaat tattttagat gacatattaa ataatctatg aatattaatg 2880
aaaacaatac agttgaagtg agtgttgttt aacatgatag tagctgagga taacaaacct 2940
caaaaaatca aagtattaat tactcctttc caagtatatg tatagagcat gtgtcattgc 3000
ttttataaaa cgcacttaat agctttcttt ctaaaaggca actgaacttt ctaaaaggta 3060
aataaactga acttgatatt aaaaaaaaaa aaaaaa 3096
<210> 27
<211> 2980
<212> ДНК
<213> Homo sapiens
<400> 27
tcgacatgcc acaaaggcac agtataaaaa cggtgggaat cagagcactt cagctccaat 60
tgctctatgt ttagaattgc ctctttttca agatggattt ccttcacagg aatggagtgc 120
tcataattca gcatttgcag aaggactacc gagcttacta cacttttcta aattttatgt 180
ccaatgttgg agaccccagg aatatctttt tcatttattt tccactttgt tttcaattta 240
atcagacagt tggaaccaag atgatatggg tagcagtcat tggggattgg ttaaatctta 300
tatttaaatg gatattattt ggtcatcgac cttactggtg ggtccaagaa actcagattt 360
acccaaatca ctcaagtcca tgccttgaac agttccctac tacatgtgaa acaggtccag 420
gaagtccatc tggccatgca atgggcgcat cctgtgtctg gtatgtcatg gtaaccgctg 480
ccctgagcca cactgtctgt gggatggata agttctctat cactctgcac aggcatgctg 540
gtggcagagg cctttgaaca cactccaggc atccaaacgg ccagtctggg cacatacctg 600
aagaccaacc tctttctctt cctgtttgca gttggctttt acctgcttct tagggtgctc 660
aacattgacc tgctgtggtc cgtgcccata gccaaaaagt ggtgtgctaa ccccgactgg 720
atccacattg acaccacgcc ttttgctgga ctcgtgagaa accttggggt cctctttggc 780
ttgggctttg caatcaactc agagatgttc ctcctgagct gccgaggggg aaataactac 840
acactgagct tccggttgct ctgtgccttg acctcattga caatactgca gctctaccat 900
ttcctccaga tcccgactca cgaagagcat ttattttatg tgctgtcttt ttgtaaaagt 960
gcatccattc ccctaactgt ggttgctttc attccctact ctgttcatat gttaatgaaa 1020
caaagcggaa agaagagtca gtagagtggt gcctagagtt agtgctctgt gtcacagatc 1080
acccttctcc atccaccagt agagccacag agtaggcaca gaccagaggc ttctaatccg 1140
acttcacaga atagcggcac aggccccatt ccccatagag atgtttagtt tggccttcgc 1200
actggtcttt ttttttaatc cttcagttac caatatttag atacaagaat atttgacata 1260
aaaatcggaa gttctgtatt tcttgaaaaa tctgatagta tgacaacaca gagcctgcat 1320
ccccagctgg agacaactga ccagagctgc atactaacag tccccagtag gaggcaagga 1380
ctccattttc tcacagtctt cagcatccca gcaggagccc cactatgatt cctttatctt 1440
cttaaggcca ggctgcatct gattcctgtt gacattttag tggggaccac agccatatcc 1500
agtttcagtt ttcagatgag gaaatggaag cctatttagg taaaagaact tgcctggagt 1560
cacgccacct tcagagcagg aattagaacc caaggcttct gacacatatc cccattacac 1620
tgtgtgtttg agtgtgcaca catgcacatg ctttttgttt gtatgtttcc tttttagaac 1680
cagggacttg ctctgttgcc caggctggag tgcagtggtg gatagcggct cactgcagcc 1740
tcaaactcct ggctcaaatg atcctccctc ctcagcctcc cattagctaa gactacaggt 1800
gtgcaccacc atgcctggct aatttttgaa tattttttag agccagcttc ttactatatt 1860
tgcccaggct ggtcttgaac tcctggcctc aaggagttcc cttgaggatg ttccacccat 1920
gtcagcctcc cacagtgctg ggattgcagg cttgagccat tgtattaata gttggcctac 1980
actgtctttt gtttcttcat ttacaagagg taaaaatcag taagaatgaa tgctttcatt 2040
tagattctat gatgactgcc atataaatca gctacctttt cagaaatgac attgaaatac 2100
tgcatcctct ttgacttata ccccacacat acatgcaggg tctaggtggg accaacagtg 2160
gctccagatg tatatacaca cgggtccagg gacaacagaa caggccaggc tacagtatgg 2220
acttgaacct ctgcttcact tctggtctct gttttacagt agtgctcacc aatatttcaa 2280
taaattctac tgaacttact ctaatagaac attattcaac ccccaaagac ttatctcttc 2340
actatgacat ctccatactt tatttttagg agacagactt tcaaaaccag agaaatcagg 2400
tgccttcctc aaggtcatgc tccaacccag gccaactatt aaatgcttgc atctgttagc 2460
tggattagtc tctatgtata ctgaactgtg atgaaaatct atagctttgt tttagaaaat 2520
tattgttgat ggactattaa tattatatta acaatttctc agtaagtgtg ttttttcctc 2580
attgaatcta ggaatgctgg gctttaagtt gataactgtg tcatttcaat caagtacaag 2640
gattttgagg cagattttgc tagatatctt agtaatcccc cacaatgttt tatgtaactc 2700
ttctcagaat atcaatacat taattatttt agatgacata ttaaataatc tatgaatatt 2760
aatgaaaaca atacagttga agtgagtgtt gtttaacatg atagtagctg aggataacaa 2820
acctcaaaaa atcaaagtat taattactcc tttccaagta tatgtataga gcatgtgtca 2880
ttgcttttat aaaacgcact taatagcttt ctttctaaaa ggcaactgaa ctttctaaaa 2940
ggtaaataaa ctgaacttga tattaaaaaa aaaaaaaaaa 2980
<210> 28
<211> 4092
<212> ДНК
<213> Homo sapiens
<400> 28
atagcagagc aatcaccacc aagcctggaa taactgcaag ggctctgctg acatcttcct 60
gaggtgccaa ggaaatgagg atggaggaag gaatgaatgt tctccatgac tttgggatcc 120
agtcaacaca ttacctccag gtgaattacc aagactccca ggactggttc atcttggtgt 180
ccgtgatcgc agacctcagg aatgccttct acgtcctctt ccccatctgg ttccatcttc 240
aggaagctgt gggcattaaa ctcctttggg tagctgtgat tggagactgg ctcaacctcg 300
tctttaagtg gattctcttt ggacagcgtc catactggtg ggttttggat actgactact 360
acagcaacac ttccgtgccc ctgataaagc agttccctgt aacctgtgag actggaccag 420
ggaaagataa agccgaccta cagatttcgg tgcttgaatg tcattttgtg gttgggattc 480
tgggctgtgc agctgaatgt ctgtctgtca cgaatctacc ttgctgctca ttttcctcat 540
caagttgttg ctggagtcct gtcaggcatt gctgttgcag aaactttcag ccacatccac 600
agcatctata atgccagcct caagaaatat tttctcatta ccttcttcct gttcagcttc 660
gccatcggat tttatctgct gctcaaggga ctgggtgtag acctcctgtg gactctggag 720
aaagcccaga ggtggtgcga gcagccagaa tgggtccaca ttgacaccac accctttgcc 780
agcctcctca agaacctggg cacgctcttt ggcctggggc tggctctcaa ctccagcatg 840
tacagggaga gctgcaaggg gaaactcagc aagtggctcc cattccgcct cagctctatt 900
gtagcctccc tcgtcctcct gcacgtcttt gactccttga aacccccatc ccaagtcgag 960
ctggtcttct acgtcttgtc cttctgcaag agtgcggtag tgcccctggc atccgtcagt 1020
gtcatcccct actgcctcgc ccaggtcctg ggccagccgc acaagaagtc gttgtaagag 1080
atgtggagtc ttcggtgttt aaagtcaaca accatgccag ggattgagga ggactactat 1140
ttgaagcaat gggcactggt atttggagca agtgacatgc catccattct gccgtcgtgg 1200
aattaaatca cggatggcag attggagggt cgcctggctt attcccatgt gtgactccag 1260
cctgccctca gcacagactc tttcagatgg aggtgccata tcacgtacac catatgcaag 1320
tttcccgcca ggaggtcctc ctctctctac ttgaatactc tcacaagtag ggagctcact 1380
cccactggaa cagcccattt tatctttgaa tggtcttctg ccagcccatt ttgaggccag 1440
aggtgctgtc agctcaggtg gtcctctttt acaatcctaa tcatattggg taatgttttt 1500
gaaaagctaa tgaagctatt gagaaagacc tgttgctaga agttgggttg ttctggattt 1560
tcccctgaag acttacttat tcttccgtca catatacaaa agcaagactt ccaggtaggg 1620
ccagctcaca agcccaggct ggagatccta actgagaatt ttctacctgt gttcattctt 1680
accgagaaaa ggagaaagga gctctgaatc tgataggaaa agaaggctgc ctaaggagga 1740
gtttttagta tgtggcgtat catgcaagtg ctatgccaag ccatgtctaa atggctttaa 1800
ttatatagta atgcactctc agtaatgggg gaccagctta agtataatta atagatggtt 1860
agtggggtaa ttctgcttct agtatttttt ttactgtgca tacatgttca tcgtatttcc 1920
ttggatttct gaatggctgc agtgacccag atattgcact aggtcaaaac attcaggtat 1980
agctgacatc tcctctatca cattacatca tcctccttat aagcccagct ctgctttttc 2040
cagattcttc cactggctcc acatccaccc cactggatct tcagaaggct agagggcgac 2100
tctggtggtg cttttgtatg tttcaattag gctctgaaat cttgggcaaa atgacaaggg 2160
gagggccagg attcctctct caggtcactc cagtgttact tttaattcct agagggtaaa 2220
tatgactcct ttctctatcc caagccaacc aagagcacat tcttaaagga aaagtcaaca 2280
tcttctctct tttttttttt ttttgagaca gggtctcact atgttgccca ggctgctctt 2340
gaattcctgg gctcaagcag tcctcccacc ctaccacagc gtcccgcgta gctgggacta 2400
caggtgcaag ccactatgtc cagctagcca actcctcctt gcctgctttt cttttttttt 2460
ctttttttga gacggcgcac ctatcaccca ggctggagtg gagtggcacg atcttggctc 2520
actgcaacct cttcctcctg gttcaagcga ttctcatgtc tcagcctcct cagtagctag 2580
gactaccggc gtgcaccacc atgccaggct aatttttata tttttagaat tttagaagag 2640
atgggatttc atcatgttgg ccaggctggt ctcgaactcc tgacctcaag tgatccacct 2700
gccttggcct cccaaggtgc taggattaca ggcatgagcc accgcaccgg gccctccttg 2760
cctgtttttc aatctcatct gatatgcaga gtatttctgc cccacccacc taccccccaa 2820
aaaaagctga agcctattta tttgaaagtc cttgtttttg ctactaatta tatagtatac 2880
catacattat cattcaaaac aaccatcctg ctcataacat ctttgaaaag aaaaatatat 2940
atgtgcagta ttttattaaa gcaacatttt atttaagaat aaagtcttgt taattactat 3000
attttagatg caatgtgatc tgaagtttct aattctggcc caactaaatt tctagctctg 3060
tttccctaaa caaataattt ggtttctctg tgcctgcatt ttccctttgg agaagaaaag 3120
tgctctctct tgagttgacc gagagtccca ttagggatag ggagacttaa atgcatccac 3180
aggggcacag gcagagttga gcacataaac ggaggcccaa aatcagcata gaaccagaaa 3240
gattcagagt tggccaagaa tgaacattgg ctaccagacc acaagtcagc atgagttgct 3300
ctatggcatc aaattgcaac ttgagagtag atgggcaggg tcactatcaa attaagcaat 3360
cagggcacac aagttgcagt aacacaacaa gactaggcca gctctggaat ccagtaactc 3420
agtgtcagca aggttttggg ttatagttca agaaagtcta aacagagcca gtcacagcac 3480
caaggaatgc tcaagggagc tattgcaggt ttctctgcta agagatttat ttcatcctgg 3540
gtgcagggtt cgacctccaa aggcctcaaa tcatcaccgt atcaatggat ttcctgaggg 3600
taagctccgc tatttcacac ctgaactccg gagtctgtat attcagggaa gattgcattc 3660
tcctactgga tttgggctct cagagggcgt tgtgggaacc aggcccctca cagaatcaaa 3720
tggtcccaac cagggagaaa gaaaatagtc tttttttttt ttttaataga gatgggggtc 3780
tcactatgct gcccaggctg gtcttgaact cctgggttca agtgatcctc ctgcctcagc 3840
ctcccaaagt gctgggatta cagtgtgagc cactgcgctt ggccagaaat ggttttgatc 3900
tgtctgaact gaaccctact gcttaggcat agccccatcc ttgataatct atttgctccc 3960
aaggaccaag tccaagatcc ttacaagaaa ggtctgccag aaagtaaata ctgcccccac 4020
tccctgaagt ttatgaggtt gataagaaaa cataacagat aaagtttatt gagtgctaac 4080
tttaaaaaaa aa 4092
<210> 29
<211> 369
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Met Glu Phe Leu Glu Arg Thr Tyr Leu Val Asn Asp Lys Ala Ala Lys
1 5 10 15
Met Tyr Ala Phe Thr Leu Glu Ser Val Glu Leu Gln Gln Lys Pro Val
20 25 30
Asn Lys Asp Gln Cys Pro Arg Glu Arg Pro Glu Glu Leu Glu Ser Gly
35 40 45
Gly Met Tyr His Cys His Ser Gly Ser Lys Pro Thr Glu Lys Gly Ala
50 55 60
Asn Glu Tyr Ala Tyr Ala Lys Trp Lys Leu Cys Ser Ala Ser Ala Ile
65 70 75 80
Cys Phe Ile Phe Met Ile Ala Glu Val Val Gly Gly His Ile Ala Gly
85 90 95
Ser Leu Ala Val Val Thr Asp Ala Ala His Leu Leu Ile Asp Leu Thr
100 105 110
Ser Phe Leu Leu Ser Leu Phe Ser Leu Trp Leu Ser Ser Lys Pro Pro
115 120 125
Ser Lys Arg Leu Thr Phe Gly Trp His Arg Ala Glu Ile Leu Gly Ala
130 135 140
Leu Leu Ser Ile Leu Cys Ile Trp Val Val Thr Gly Val Leu Val Tyr
145 150 155 160
Leu Ala Cys Glu Arg Leu Leu Tyr Pro Asp Tyr Gln Ile Gln Ala Thr
165 170 175
Val Met Ile Ile Val Ser Ser Cys Ala Val Ala Ala Asn Ile Val Leu
180 185 190
Thr Val Val Leu His Gln Arg Cys Leu Gly His Asn His Lys Glu Val
195 200 205
Gln Ala Asn Ala Ser Val Arg Ala Ala Phe Val His Ala Leu Gly Asp
210 215 220
Leu Phe Gln Ser Ile Ser Val Leu Ile Ser Ala Leu Ile Ile Tyr Phe
225 230 235 240
Lys Pro Glu Tyr Lys Ile Ala Asp Pro Ile Cys Thr Phe Ile Phe Ser
245 250 255
Ile Leu Val Leu Ala Ser Thr Ile Thr Ile Leu Lys Asp Phe Ser Ile
260 265 270
Leu Leu Met Glu Gly Val Pro Lys Ser Leu Asn Tyr Ser Gly Val Lys
275 280 285
Glu Leu Ile Leu Ala Val Asp Gly Val Leu Ser Val His Ser Leu His
290 295 300
Ile Trp Ser Leu Thr Met Asn Gln Val Ile Leu Ser Ala His Val Ala
305 310 315 320
Thr Ala Ala Ser Arg Asp Ser Gln Val Val Arg Arg Glu Ile Ala Lys
325 330 335
Ala Leu Ser Lys Ser Phe Thr Met His Ser Leu Thr Ile Gln Met Glu
340 345 350
Ser Pro Val Asp Gln Asp Pro Asp Cys Leu Phe Cys Glu Asp Pro Cys
355 360 365
Asp
<210> 30
<211> 1110
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(1107)
<400> 30
atg gag ttt ctt gaa aga acg tat ctt gtg aat gat aaa gct gcc aag 48
Met Glu Phe Leu Glu Arg Thr Tyr Leu Val Asn Asp Lys Ala Ala Lys
1 5 10 15
atg tat gct ttc aca cta gaa agt gtg gaa ctc caa cag aaa ccg gtg 96
Met Tyr Ala Phe Thr Leu Glu Ser Val Glu Leu Gln Gln Lys Pro Val
20 25 30
aat aaa gat cag tgt ccc aga gag aga cca gag gag ctg gag tca gga 144
Asn Lys Asp Gln Cys Pro Arg Glu Arg Pro Glu Glu Leu Glu Ser Gly
35 40 45
ggc atg tac cac tgc cac agt ggc tcc aag ccc aca gaa aag ggg gcg 192
Gly Met Tyr His Cys His Ser Gly Ser Lys Pro Thr Glu Lys Gly Ala
50 55 60
aat gag tac gcc tat gcc aag tgg aaa ctc tgt tct gct tca gca ata 240
Asn Glu Tyr Ala Tyr Ala Lys Trp Lys Leu Cys Ser Ala Ser Ala Ile
65 70 75 80
tgc ttc att ttc atg att gca gag gtc gtg ggt ggg cac att gct ggg 288
Cys Phe Ile Phe Met Ile Ala Glu Val Val Gly Gly His Ile Ala Gly
85 90 95
agt ctt gct gtt gtc aca gat gct gcc cac ctc tta att gac ctg acc 336
Ser Leu Ala Val Val Thr Asp Ala Ala His Leu Leu Ile Asp Leu Thr
100 105 110
agt ttc ctg ctc agt ctc ttc tcc ctg tgg ttg tca tcg aag cct ccc 384
Ser Phe Leu Leu Ser Leu Phe Ser Leu Trp Leu Ser Ser Lys Pro Pro
115 120 125
tct aag cgg ctg aca ttt gga tgg cac cga gca gag atc ctt ggt gcc 432
Ser Lys Arg Leu Thr Phe Gly Trp His Arg Ala Glu Ile Leu Gly Ala
130 135 140
ctg ctc tcc atc ctg tgc atc tgg gtg gtg act ggc gtg cta gtg tac 480
Leu Leu Ser Ile Leu Cys Ile Trp Val Val Thr Gly Val Leu Val Tyr
145 150 155 160
ctg gca tgt gag cgc ctg ctg tat cct gat tac cag atc cag gcg act 528
Leu Ala Cys Glu Arg Leu Leu Tyr Pro Asp Tyr Gln Ile Gln Ala Thr
165 170 175
gtg atg atc atc gtt tcc agc tgc gca gtg gcg gcc aac att gta cta 576
Val Met Ile Ile Val Ser Ser Cys Ala Val Ala Ala Asn Ile Val Leu
180 185 190
act gtg gtt ttg cac cag aga tgc ctt ggc cac aat cac aag gaa gta 624
Thr Val Val Leu His Gln Arg Cys Leu Gly His Asn His Lys Glu Val
195 200 205
caa gcc aat gcc agc gtc aga gct gct ttt gtg cat gcc ctt gga gat 672
Gln Ala Asn Ala Ser Val Arg Ala Ala Phe Val His Ala Leu Gly Asp
210 215 220
cta ttt cag agt atc agt gtg cta att agt gca ctt att atc tac ttt 720
Leu Phe Gln Ser Ile Ser Val Leu Ile Ser Ala Leu Ile Ile Tyr Phe
225 230 235 240
aag cca gag tat aaa ata gcc gac cca atc tgc aca ttc atc ttt tcc 768
Lys Pro Glu Tyr Lys Ile Ala Asp Pro Ile Cys Thr Phe Ile Phe Ser
245 250 255
atc ctg gtc ttg gcc agc acc atc act atc tta aag gac ttc tcc atc 816
Ile Leu Val Leu Ala Ser Thr Ile Thr Ile Leu Lys Asp Phe Ser Ile
260 265 270
tta ctc atg gaa ggt gtg cca aag agc ctg aat tac agt ggt gtg aaa 864
Leu Leu Met Glu Gly Val Pro Lys Ser Leu Asn Tyr Ser Gly Val Lys
275 280 285
gag ctt att tta gca gtc gac ggg gtg ctg tct gtg cac agc ctg cac 912
Glu Leu Ile Leu Ala Val Asp Gly Val Leu Ser Val His Ser Leu His
290 295 300
atc tgg tct cta aca atg aat caa gta att ctc tca gct cat gtt gct 960
Ile Trp Ser Leu Thr Met Asn Gln Val Ile Leu Ser Ala His Val Ala
305 310 315 320
aca gca gcc agc cgg gac agc caa gtg gtt cgg aga gaa att gct aaa 1008
Thr Ala Ala Ser Arg Asp Ser Gln Val Val Arg Arg Glu Ile Ala Lys
325 330 335
gcc ctt agc aaa agc ttt acg atg cac tca ctc acc att cag atg gaa 1056
Ala Leu Ser Lys Ser Phe Thr Met His Ser Leu Thr Ile Gln Met Glu
340 345 350
tct cca gtt gac cag gac ccc gac tgc ctt ttc tgt gaa gac ccc tgt 1104
Ser Pro Val Asp Gln Asp Pro Asp Cys Leu Phe Cys Glu Asp Pro Cys
355 360 365
gac taa 1110
Asp
<210> 31
<211> 1955
<212> ДНК
<213> Homo sapiens
<400> 31
ctcggcctcg cgttataaaa agcggtgggg cagggccggc gagacaatct gggaggcggg 60
taccgggcct cacggatccg cgccgcgccc cccacctgtg gctgcgcgcg gggtgggctg 120
cgctcccctg ggcggcgccg ggcgcccggg gctggtggcg agatgggccg ctactctggc 180
aagacgtgcc ggctgctctt catgctggtg ctcaccgtcg ccttcttcgt ggcggagctg 240
gtctccggct acctgggcaa ctccatcgcg ctgctctccg actccttcaa catgttctcc 300
gacctgatct cgctgtgcgt gggcctgagc gccggctaca tcgcccggcg ccccacccgg 360
ggcttcagcg ccacctacgg ctacgcccgc gccgaggtgg tgggcgcgct gagcaacgcg 420
gtcttcctca ccgcgctctg cttcaccatc ttcgtggagg ccgtgctgcg cctggcccgg 480
cccgagcgca tcgatgaccc cgagctggtg ctcatcgtcg gcgtcctggg gctgttggtc 540
aacgtggtgg ggctgctcat cttccaggac tgcgccgcct ggttcgcgtg ctgcctccgg 600
ggacgcagtc gccgcctgca gcagcggcag cagctggcgg agggctgtgt ccccggcgct 660
ttcggggggc ctcagggcgc ggaggacccg cggcgcgcgg cggacccgac agccccaggc 720
tcggactcgg ccgtaaccct ccgggggacc tcggtggaaa ggaagcggga gaagggggcg 780
accgtgttcg caaacgtagc aggtgattcc ttcaacaccc agaatgagcc agaagacatg 840
atgaaaaaag agaaaaagtc tgaagctctg aatatcagag gtgtactttt gcatgtgatg 900
ggagatgccc tggggtccgt ggttgtggtc atcacggcca tcatattcta tgtgcttccc 960
ctgaagagtg aggacccgtg taactggcag tgttacattg accccagcct gactgtcctc 1020
atggtcatca tcattttgtc atctgccttc ccgcttatca aggagaccgc tgccattctg 1080
ctacagatgg tcccaaaagg agtcaacatg gaagagctga tgagtaaact ctctgctgtg 1140
cctggaatta gcagtgtaca tgaagtgcac atctgggaac ttgtaagtgg aaagattatt 1200
gccaccctgc acatcaagta tcctaaggac aggggatatc aagatgccag cacaaaaatt 1260
cgagaaatct tccaccatgc gggaatccac aatgtgacca tccagtttga aaatgtggac 1320
ttgaaggaac ccctggagca gaaggactta ctgttgctct gcaactcacc ctgcatctcc 1380
aagggctgtg ctaagcagct gtgttgtccc cccggggcac tgcctctggc tcacgtcaat 1440
ggctgtgctg agcacttcct ctgtcacgtc aatggctgtg ctgagcacaa tggtgggccc 1500
tctctagaca catacggaag tgatggcctc agtagaagag acgcaagaga agtggctatt 1560
gaagtgtctt tggatagctg tctgagtgac cacggacaaa gtcttaacaa aactcaggag 1620
gaccaatgtt atgtcaacag aacgcatttt taatctggta ctcacataat cagaccatat 1680
agacgaggca ctttggaacc acaagcttgg ctcacaaaaa gagctttcct gggttgtagg 1740
cccagactag acttgcagca tgcatgctct gtgttcacta ggggttggct gtttgggatt 1800
ttagttaaac gtgtctgtga atttttatgt aactaactcc tttccattcc cctgggtgtc 1860
tcatgctgct ctttgactgt ttcagcttga acatgcattt tctaaagcaa actgcactag 1920
tgtatatatc agggtttgaa gctcatgggc tctct 1955
<210> 32
<211> 5373
<212> ДНК
<213> Homo sapiens
<400> 32
agcagttttt gtaggtgaaa acaatgaagc caggtaatat tgcaaggagg ctgtaatttt 60
agcagaccta ccaacaacac tgatgtagga agctcattat tttaatttct ggagcctttt 120
aattttttct ttagaaagtg tataaataat tgcagtgctg ctttgcttcc aaaactgggc 180
agtgagttca acaacaacga caacaacagc cgcagctcat cctggccgtc atggagtttc 240
ttgaaagaac gtatcttgtg aatgataaag ctgccaagat gtatgctttc acactagaaa 300
gtgtggaact ccaacagaaa ccggtgaata aagatcagtg tcccagagag agaccagagg 360
agctggagtc aggaggcatg taccactgcc acagtggctc caagcccaca gaaaaggggg 420
cgaatgagta cgcctatgcc aagtggaaac tctgttctgc ttcagcaata tgcttcattt 480
tcatgattgc agaggtcgtg ggtgggcaca ttgctgggag tcttgctgtt gtcacagatg 540
ctgcccacct cttaattgac ctgaccagtt tcctgctcag tctcttctcc ctgtggttgt 600
catcgaagcc tccctctaag cggctgacat ttggatggca ccgagcagag atccttggtg 660
ccctgctctc catcctgtgc atctgggtgg tgactggcgt gctagtgtac ctggcatgtg 720
agcgcctgct gtatcctgat taccagatcc aggcgactgt gatgatcatc gtttccagct 780
gcgcagtggc ggccaacatt gtactaactg tggttttgca ccagagatgc cttggccaca 840
atcacaagga agtacaagcc aatgccagcg tcagagctgc ttttgtgcat gcccttggag 900
atctatttca gagtatcagt gtgctaatta gtgcacttat tatctacttt aagccagagt 960
ataaaatagc cgacccaatc tgcacattca tcttttccat cctggtcttg gccagcacca 1020
tcactatctt aaaggacttc tccatcttac tcatggaagg tgtgccaaag agcctgaatt 1080
acagtggtgt gaaagagctt attttagcag tcgacggggt gctgtctgtg cacagcctgc 1140
acatctggtc tctaacaatg aatcaagtaa ttctctcagc tcatgttgct acagcagcca 1200
gccgggacag ccaagtggtt cggagagaaa ttgctaaagc ccttagcaaa agctttacga 1260
tgcactcact caccattcag atggaatctc cagttgacca ggaccccgac tgccttttct 1320
gtgaagaccc ctgtgactag ctcagtcaca ccgtcagttt cccaaatttg acaggccacc 1380
ttcaaacatg ctgctatgca gtttctgcat catagaaaat aaggaaccaa aggaagaaat 1440
tcatgtcatg gtgcaatgca cattttatct atttatttag ttccattcac catgaaggaa 1500
gaggcactga gatccatcaa tcaattggat tatatactga tcagtagctg tgttcaattg 1560
caggaatgtg tatatagatt attcctgagt ggagccgaag taacagctgt ttgtaactat 1620
cggcaatacc aaattcatct cccttccaat aatgcatctt gagaacacat aggtaaattt 1680
gaactcagga aagtcttact agaaatcagt ggaagggaca aatagtcaca aaattttacc 1740
aaaacattag aaacaaaaaa taaggagagc caagtcagga ataaaagtga ctctgtatgc 1800
taacgccaca ttagaacttg gttctctcac caagctgtaa tgtgattttt ttttctactc 1860
tgaattggaa atatgtatga atatacagag aagtgcttac aactaatttt tatttacttg 1920
tcacattttg gcaataaatc cctcttattt ctaaattcta acttgtttat ttcaaaactt 1980
tatataatca ctgttcaaaa ggaaatattt tcacctacca gagtgcttaa acactggcac 2040
cagccaaaga atgtggttgt agagacccag aagtcttcaa gaacagccga caaaaacatt 2100
cgagttgacc ccaccaagtt gttgccacag ataatttaga tatttacctg caagaaggaa 2160
taaagcagat gcaaccaatt cattcagtcc acgagcatga tgtgagcact gctttgtgct 2220
agacattggg cttagcattg aaactataaa gaggaatcag acgcagcaag tgcttctgtg 2280
ttctggtagc aactcaacac tatctgtgga gagtaaactg aagatgtgca ggccaacatt 2340
ctggaaatcc tatgtcaatg ggtttggttt ggaacctgga cttctgcatt tttaaaagtt 2400
acccagagat gcttctaaag atgagccata gtctagaaga ttgtcaacca caggagttca 2460
ttgagtggga cagctagaca catacattgg cagctacaat agtatcatga attgcaatga 2520
tgtagtgggg tataaaagga aagcgatgga tattgccgga tgggcatggc cagtgatgtt 2580
tcacgtcatt gaggtgacag ctctgctgga ctttgaatta catatggagg ctctccagga 2640
agacgaagaa gagaaggaca ttctaggcaa aaagaagact aggcacaagg cacacttatg 2700
tttgtctgtt agcttttagt tgaaaaagca aaatacatga tgcaaagaaa cctctccacg 2760
ctgtgatttt taaaactaca tactttttgc aactttatgg ttatgagtat tgtagagaac 2820
aggagatagg tcttagatga tttttatgtt gttgtcagac tctagcaagg tactagaaac 2880
ctagcaggca ttaataattg ttgaggcaat gactctgagg ctatatctgg gccttgtcat 2940
tatttatcat ttatatttgt atttttttct gaaatttgag ggccaagaaa acattgactt 3000
tgactgagga ggtcacatct gtgccatctc tgcaaatcaa tcagcaccac tgaaataact 3060
acttagcatt ctgctgagct ttccctgctc agtagagaca aatatactca tcccccacct 3120
cagtgagctt gtttaggcaa ccaggattag agctgctcag gttcccaacg tctcctgcca 3180
catcgggttc tcaaaatgga aagaatggtt tatgccaaat cacttttcct gtctgaagga 3240
ccactgaatg gttttgtttt tccatatttt gcataggacg ccctaaagac taggtgactt 3300
ggcaaacaca caagtgttag tataattctt tgcttctgct tctttttgaa aatcatgttt 3360
agatttgatt ttaagtcaga aattcactga atgtcaggta atcattatgg agggagattt 3420
gtgtgtcaac caaagtaatt gtcccatggc cccagggtat ttctgttgtt tccctgaaat 3480
tctgcttttt tagtcagcta gattgaaaac tctgaacagt agatgtttat atggcaaaat 3540
gcaagacaat ctacaaggga gattttaagg attttgagat gaaaaaacag atgctactca 3600
ggggctttat gaaccatcca tcaattctga agttctgact ctcccattac cctttccctg 3660
gtgtggtcag aactccaggt cactggaagt tagtggaatc atgtagttga attctttact 3720
tcaagacatt gtattctctc cagctatcaa aacattaatg atcttttatg tctttttttt 3780
gttattgtta tactttaagt tctggggtac atgtgcggaa catgtaggtt tgttacatag 3840
gtatacatgt gccatggtgg tttgctgcac tcatcaacct gtcatctaca ttcttttatg 3900
tctgtctttc aaagcaacac tctgttcttc tgagtagtga aatcaggtca actttaccac 3960
cagcctccat ttttaatatg cttcaccatc atccagcacc tacttaagat ttatctaggg 4020
ctctgtggtg atgttaggac ccataaaaga aatttatgcc ttccatatgt ttggttacag 4080
atgggaaatg ggaatgttga aggacatgaa agaaaggatg tttacacatt aagcatcagt 4140
tctgaagcta gattgtctga gtttgaatct tagctcttcc ctttattagc tctgtgacct 4200
cgagctagtt acttaaatgc tctgatcctc tatttcctga tcagtgaaac ctccctattc 4260
aaatgtgtga gagtttaata aattaggaca cttaaaaatg ttggagcagt gcatagcatg 4320
tagtgttcag tacatgttaa atgttgtttt ttattatgta caaacatgag tgggcacaga 4380
attttaaatc atctcaactt ttgagaaatt ttgagttatc aacaccgttc ccacaagaca 4440
gtggcaaaat tattggtgag aattaaacag ctgtttctca gaggaagcaa tggaggcttg 4500
ctgggataaa ggcatttact gagaggctgt tacctagtga gagtgatgaa ttaattaaaa 4560
tagtcgaatc cctttctgac tgtctctgaa agcttccgct tttatctttg aagagcagaa 4620
ttgtcactcc aaggacattt attaataaaa agaacaactg tccagtgcaa tgaaggcaaa 4680
gtcataggtc tcccaagtct taccccattc ctgtgaaata tcaagttctt ggcttttctc 4740
tgtcatgtag cctcaacttt ctctgaccgg gtgcatttct ttctctggtt tctaaattgc 4800
cagtggcaaa tttggatcac ttacttaata tctgttaaat tttgtgaccc aacaaagtct 4860
tttagcactg tggtgtcaaa aagaaaaaca cctcccaggc atatacattt tatagattcc 4920
tggagaatgt tgctctccag ctccatcccc acccaatgaa atatgatcca gagagtcttg 4980
caaagagaca agcctcattt tccacaatta gctctaaagt gcctccagga aatgattttc 5040
tcagctcatc tctctgtatt ccctgttttg gatcacaggg caatctgttt aaatgactaa 5100
ttacagaaat cattaaaggc accaagcaaa tgtcatctct gaatacacac atcccaagct 5160
ttacaaatcc tgcctggctt gacagtgatg aggccactta acagtccagc gcaggcggat 5220
gttaaaaaaa ataaaaaggt gaccatctgc ggtttagttt tttaactttc tgatttcaca 5280
cttaacgtct gtcattctgt tactgggcac ctgtttaaat tctattttaa aatgttaatg 5340
tgtgttgttt aaaataaaat caagaaagag aga 5373
<210> 33
<211> 5403
<212> ДНК
<213> Homo sapiens
<400> 33
agcagttttt gtaggtgaaa acaatgaagc caggtaatat tgcaaggagg ctgtaatttt 60
agcagaccta ccaacaacac tgatgtagga agctcattat tttaatttct ggagcctttt 120
aattttttct ttagaaagtg tataaataat tgcagtgctg ctttgcttcc aaaactgggc 180
agtgagttca acaacaacga caacaacagc cgcagctcat cctggccgtc atggagtttc 240
ttgaaagaac gtatcttgtg aatgataaag ctgccaagat gtatgctttc acactagaaa 300
gaaggagctg caaatgaaca cttcatagca atgtggaact ccaacagaaa ccggtgaata 360
aagatcagtg tcccagagag agaccagagg agctggagtc aggaggcatg taccactgcc 420
acagtggctc caagcccaca gaaaaggggg cgaatgagta cgcctatgcc aagtggaaac 480
tctgttctgc ttcagcaata tgcttcattt tcatgattgc agaggtcgtg ggtgggcaca 540
ttgctgggag tcttgctgtt gtcacagatg ctgcccacct cttaattgac ctgaccagtt 600
tcctgctcag tctcttctcc ctgtggttgt catcgaagcc tccctctaag cggctgacat 660
ttggatggca ccgagcagag atccttggtg ccctgctctc catcctgtgc atctgggtgg 720
tgactggcgt gctagtgtac ctggcatgtg agcgcctgct gtatcctgat taccagatcc 780
aggcgactgt gatgatcatc gtttccagct gcgcagtggc ggccaacatt gtactaactg 840
tggttttgca ccagagatgc cttggccaca atcacaagga agtacaagcc aatgccagcg 900
tcagagctgc ttttgtgcat gcccttggag atctatttca gagtatcagt gtgctaatta 960
gtgcacttat tatctacttt aagccagagt ataaaatagc cgacccaatc tgcacattca 1020
tcttttccat cctggtcttg gccagcacca tcactatctt aaaggacttc tccatcttac 1080
tcatggaagg tgtgccaaag agcctgaatt acagtggtgt gaaagagctt attttagcag 1140
tcgacggggt gctgtctgtg cacagcctgc acatctggtc tctaacaatg aatcaagtaa 1200
ttctctcagc tcatgttgct acagcagcca gccgggacag ccaagtggtt cggagagaaa 1260
ttgctaaagc ccttagcaaa agctttacga tgcactcact caccattcag atggaatctc 1320
cagttgacca ggaccccgac tgccttttct gtgaagaccc ctgtgactag ctcagtcaca 1380
ccgtcagttt cccaaatttg acaggccacc ttcaaacatg ctgctatgca gtttctgcat 1440
catagaaaat aaggaaccaa aggaagaaat tcatgtcatg gtgcaatgca cattttatct 1500
atttatttag ttccattcac catgaaggaa gaggcactga gatccatcaa tcaattggat 1560
tatatactga tcagtagctg tgttcaattg caggaatgtg tatatagatt attcctgagt 1620
ggagccgaag taacagctgt ttgtaactat cggcaatacc aaattcatct cccttccaat 1680
aatgcatctt gagaacacat aggtaaattt gaactcagga aagtcttact agaaatcagt 1740
ggaagggaca aatagtcaca aaattttacc aaaacattag aaacaaaaaa taaggagagc 1800
caagtcagga ataaaagtga ctctgtatgc taacgccaca ttagaacttg gttctctcac 1860
caagctgtaa tgtgattttt ttttctactc tgaattggaa atatgtatga atatacagag 1920
aagtgcttac aactaatttt tatttacttg tcacattttg gcaataaatc cctcttattt 1980
ctaaattcta acttgtttat ttcaaaactt tatataatca ctgttcaaaa ggaaatattt 2040
tcacctacca gagtgcttaa acactggcac cagccaaaga atgtggttgt agagacccag 2100
aagtcttcaa gaacagccga caaaaacatt cgagttgacc ccaccaagtt gttgccacag 2160
ataatttaga tatttacctg caagaaggaa taaagcagat gcaaccaatt cattcagtcc 2220
acgagcatga tgtgagcact gctttgtgct agacattggg cttagcattg aaactataaa 2280
gaggaatcag acgcagcaag tgcttctgtg ttctggtagc aactcaacac tatctgtgga 2340
gagtaaactg aagatgtgca ggccaacatt ctggaaatcc tatgtcaatg ggtttggttt 2400
ggaacctgga cttctgcatt tttaaaagtt acccagagat gcttctaaag atgagccata 2460
gtctagaaga ttgtcaacca caggagttca ttgagtggga cagctagaca catacattgg 2520
cagctacaat agtatcatga attgcaatga tgtagtgggg tataaaagga aagcgatgga 2580
tattgccgga tgggcatggc cagtgatgtt tcacgtcatt gaggtgacag ctctgctgga 2640
ctttgaatta catatggagg ctctccagga agacgaagaa gagaaggaca ttctaggcaa 2700
aaagaagact aggcacaagg cacacttatg tttgtctgtt agcttttagt tgaaaaagca 2760
aaatacatga tgcaaagaaa cctctccacg ctgtgatttt taaaactaca tactttttgc 2820
aactttatgg ttatgagtat tgtagagaac aggagatagg tcttagatga tttttatgtt 2880
gttgtcagac tctagcaagg tactagaaac ctagcaggca ttaataattg ttgaggcaat 2940
gactctgagg ctatatctgg gccttgtcat tatttatcat ttatatttgt atttttttct 3000
gaaatttgag ggccaagaaa acattgactt tgactgagga ggtcacatct gtgccatctc 3060
tgcaaatcaa tcagcaccac tgaaataact acttagcatt ctgctgagct ttccctgctc 3120
agtagagaca aatatactca tcccccacct cagtgagctt gtttaggcaa ccaggattag 3180
agctgctcag gttcccaacg tctcctgcca catcgggttc tcaaaatgga aagaatggtt 3240
tatgccaaat cacttttcct gtctgaagga ccactgaatg gttttgtttt tccatatttt 3300
gcataggacg ccctaaagac taggtgactt ggcaaacaca caagtgttag tataattctt 3360
tgcttctgct tctttttgaa aatcatgttt agatttgatt ttaagtcaga aattcactga 3420
atgtcaggta atcattatgg agggagattt gtgtgtcaac caaagtaatt gtcccatggc 3480
cccagggtat ttctgttgtt tccctgaaat tctgcttttt tagtcagcta gattgaaaac 3540
tctgaacagt agatgtttat atggcaaaat gcaagacaat ctacaaggga gattttaagg 3600
attttgagat gaaaaaacag atgctactca ggggctttat gaaccatcca tcaattctga 3660
agttctgact ctcccattac cctttccctg gtgtggtcag aactccaggt cactggaagt 3720
tagtggaatc atgtagttga attctttact tcaagacatt gtattctctc cagctatcaa 3780
aacattaatg atcttttatg tctttttttt gttattgtta tactttaagt tctggggtac 3840
atgtgcggaa catgtaggtt tgttacatag gtatacatgt gccatggtgg tttgctgcac 3900
tcatcaacct gtcatctaca ttcttttatg tctgtctttc aaagcaacac tctgttcttc 3960
tgagtagtga aatcaggtca actttaccac cagcctccat ttttaatatg cttcaccatc 4020
atccagcacc tacttaagat ttatctaggg ctctgtggtg atgttaggac ccataaaaga 4080
aatttatgcc ttccatatgt ttggttacag atgggaaatg ggaatgttga aggacatgaa 4140
agaaaggatg tttacacatt aagcatcagt tctgaagcta gattgtctga gtttgaatct 4200
tagctcttcc ctttattagc tctgtgacct cgagctagtt acttaaatgc tctgatcctc 4260
tatttcctga tcagtgaaac ctccctattc aaatgtgtga gagtttaata aattaggaca 4320
cttaaaaatg ttggagcagt gcatagcatg tagtgttcag tacatgttaa atgttgtttt 4380
ttattatgta caaacatgag tgggcacaga attttaaatc atctcaactt ttgagaaatt 4440
ttgagttatc aacaccgttc ccacaagaca gtggcaaaat tattggtgag aattaaacag 4500
ctgtttctca gaggaagcaa tggaggcttg ctgggataaa ggcatttact gagaggctgt 4560
tacctagtga gagtgatgaa ttaattaaaa tagtcgaatc cctttctgac tgtctctgaa 4620
agcttccgct tttatctttg aagagcagaa ttgtcactcc aaggacattt attaataaaa 4680
agaacaactg tccagtgcaa tgaaggcaaa gtcataggtc tcccaagtct taccccattc 4740
ctgtgaaata tcaagttctt ggcttttctc tgtcatgtag cctcaacttt ctctgaccgg 4800
gtgcatttct ttctctggtt tctaaattgc cagtggcaaa tttggatcac ttacttaata 4860
tctgttaaat tttgtgaccc aacaaagtct tttagcactg tggtgtcaaa aagaaaaaca 4920
cctcccaggc atatacattt tatagattcc tggagaatgt tgctctccag ctccatcccc 4980
acccaatgaa atatgatcca gagagtcttg caaagagaca agcctcattt tccacaatta 5040
gctctaaagt gcctccagga aatgattttc tcagctcatc tctctgtatt ccctgttttg 5100
gatcacaggg caatctgttt aaatgactaa ttacagaaat cattaaaggc accaagcaaa 5160
tgtcatctct gaatacacac atcccaagct ttacaaatcc tgcctggctt gacagtgatg 5220
aggccactta acagtccagc gcaggcggat gttaaaaaaa ataaaaaggt gaccatctgc 5280
ggtttagttt tttaactttc tgatttcaca cttaacgtct gtcattctgt tactgggcac 5340
ctgtttaaat tctattttaa aatgttaatg tgtgttgttt aaaataaaat caagaaagag 5400
aga 5403
<210> 34
<211> 5316
<212> ДНК
<213> Homo sapiens
<400> 34
gggctgccag catgctgtca cctctcaata ggagatcagt taatgcatac tgaaggaagg 60
cttgttggag aagaatcctc tcctgaaccc tgtggagact ctagaatcct gcggatgtct 120
ctctccctaa gtaagagatg ttacttcctg gagggaatgc agtgttggga atctgaagac 180
ccagctttga gctgaatttg ctttgtgata cctgaaggag ctgcaaatga acacttcata 240
gcaatgtgga actccaacag aaaccggtga ataaagatca gtgtcccaga gagagaccag 300
aggagctgga gtcaggaggc atgtaccact gccacagtgg ctccaagccc acagaaaagg 360
gggcgaatga gtacgcctat gccaagtgga aactctgttc tgcttcagca atatgcttca 420
ttttcatgat tgcagaggtc gtgggtgggc acattgctgg gagtcttgct gttgtcacag 480
atgctgccca cctcttaatt gacctgacca gtttcctgct cagtctcttc tccctgtggt 540
tgtcatcgaa gcctccctct aagcggctga catttggatg gcaccgagca gagatccttg 600
gtgccctgct ctccatcctg tgcatctggg tggtgactgg cgtgctagtg tacctggcat 660
gtgagcgcct gctgtatcct gattaccaga tccaggcgac tgtgatgatc atcgtttcca 720
gctgcgcagt ggcggccaac attgtactaa ctgtggtttt gcaccagaga tgccttggcc 780
acaatcacaa ggaagtacaa gccaatgcca gcgtcagagc tgcttttgtg catgcccttg 840
gagatctatt tcagagtatc agtgtgctaa ttagtgcact tattatctac tttaagccag 900
agtataaaat agccgaccca atctgcacat tcatcttttc catcctggtc ttggccagca 960
ccatcactat cttaaaggac ttctccatct tactcatgga aggtgtgcca aagagcctga 1020
attacagtgg tgtgaaagag cttattttag cagtcgacgg ggtgctgtct gtgcacagcc 1080
tgcacatctg gtctctaaca atgaatcaag taattctctc agctcatgtt gctacagcag 1140
ccagccggga cagccaagtg gttcggagag aaattgctaa agcccttagc aaaagcttta 1200
cgatgcactc actcaccatt cagatggaat ctccagttga ccaggacccc gactgccttt 1260
tctgtgaaga cccctgtgac tagctcagtc acaccgtcag tttcccaaat ttgacaggcc 1320
accttcaaac atgctgctat gcagtttctg catcatagaa aataaggaac caaaggaaga 1380
aattcatgtc atggtgcaat gcacatttta tctatttatt tagttccatt caccatgaag 1440
gaagaggcac tgagatccat caatcaattg gattatatac tgatcagtag ctgtgttcaa 1500
ttgcaggaat gtgtatatag attattcctg agtggagccg aagtaacagc tgtttgtaac 1560
tatcggcaat accaaattca tctcccttcc aataatgcat cttgagaaca cataggtaaa 1620
tttgaactca ggaaagtctt actagaaatc agtggaaggg acaaatagtc acaaaatttt 1680
accaaaacat tagaaacaaa aaataaggag agccaagtca ggaataaaag tgactctgta 1740
tgctaacgcc acattagaac ttggttctct caccaagctg taatgtgatt tttttttcta 1800
ctctgaattg gaaatatgta tgaatataca gagaagtgct tacaactaat ttttatttac 1860
ttgtcacatt ttggcaataa atccctctta tttctaaatt ctaacttgtt tatttcaaaa 1920
ctttatataa tcactgttca aaaggaaata ttttcaccta ccagagtgct taaacactgg 1980
caccagccaa agaatgtggt tgtagagacc cagaagtctt caagaacagc cgacaaaaac 2040
attcgagttg accccaccaa gttgttgcca cagataattt agatatttac ctgcaagaag 2100
gaataaagca gatgcaacca attcattcag tccacgagca tgatgtgagc actgctttgt 2160
gctagacatt gggcttagca ttgaaactat aaagaggaat cagacgcagc aagtgcttct 2220
gtgttctggt agcaactcaa cactatctgt ggagagtaaa ctgaagatgt gcaggccaac 2280
attctggaaa tcctatgtca atgggtttgg tttggaacct ggacttctgc atttttaaaa 2340
gttacccaga gatgcttcta aagatgagcc atagtctaga agattgtcaa ccacaggagt 2400
tcattgagtg ggacagctag acacatacat tggcagctac aatagtatca tgaattgcaa 2460
tgatgtagtg gggtataaaa ggaaagcgat ggatattgcc ggatgggcat ggccagtgat 2520
gtttcacgtc attgaggtga cagctctgct ggactttgaa ttacatatgg aggctctcca 2580
ggaagacgaa gaagagaagg acattctagg caaaaagaag actaggcaca aggcacactt 2640
atgtttgtct gttagctttt agttgaaaaa gcaaaataca tgatgcaaag aaacctctcc 2700
acgctgtgat ttttaaaact acatactttt tgcaacttta tggttatgag tattgtagag 2760
aacaggagat aggtcttaga tgatttttat gttgttgtca gactctagca aggtactaga 2820
aacctagcag gcattaataa ttgttgaggc aatgactctg aggctatatc tgggccttgt 2880
cattatttat catttatatt tgtatttttt tctgaaattt gagggccaag aaaacattga 2940
ctttgactga ggaggtcaca tctgtgccat ctctgcaaat caatcagcac cactgaaata 3000
actacttagc attctgctga gctttccctg ctcagtagag acaaatatac tcatccccca 3060
cctcagtgag cttgtttagg caaccaggat tagagctgct caggttccca acgtctcctg 3120
ccacatcggg ttctcaaaat ggaaagaatg gtttatgcca aatcactttt cctgtctgaa 3180
ggaccactga atggttttgt ttttccatat tttgcatagg acgccctaaa gactaggtga 3240
cttggcaaac acacaagtgt tagtataatt ctttgcttct gcttcttttt gaaaatcatg 3300
tttagatttg attttaagtc agaaattcac tgaatgtcag gtaatcatta tggagggaga 3360
tttgtgtgtc aaccaaagta attgtcccat ggccccaggg tatttctgtt gtttccctga 3420
aattctgctt ttttagtcag ctagattgaa aactctgaac agtagatgtt tatatggcaa 3480
aatgcaagac aatctacaag ggagatttta aggattttga gatgaaaaaa cagatgctac 3540
tcaggggctt tatgaaccat ccatcaattc tgaagttctg actctcccat taccctttcc 3600
ctggtgtggt cagaactcca ggtcactgga agttagtgga atcatgtagt tgaattcttt 3660
acttcaagac attgtattct ctccagctat caaaacatta atgatctttt atgtcttttt 3720
tttgttattg ttatacttta agttctgggg tacatgtgcg gaacatgtag gtttgttaca 3780
taggtataca tgtgccatgg tggtttgctg cactcatcaa cctgtcatct acattctttt 3840
atgtctgtct ttcaaagcaa cactctgttc ttctgagtag tgaaatcagg tcaactttac 3900
caccagcctc catttttaat atgcttcacc atcatccagc acctacttaa gatttatcta 3960
gggctctgtg gtgatgttag gacccataaa agaaatttat gccttccata tgtttggtta 4020
cagatgggaa atgggaatgt tgaaggacat gaaagaaagg atgtttacac attaagcatc 4080
agttctgaag ctagattgtc tgagtttgaa tcttagctct tccctttatt agctctgtga 4140
cctcgagcta gttacttaaa tgctctgatc ctctatttcc tgatcagtga aacctcccta 4200
ttcaaatgtg tgagagttta ataaattagg acacttaaaa atgttggagc agtgcatagc 4260
atgtagtgtt cagtacatgt taaatgttgt tttttattat gtacaaacat gagtgggcac 4320
agaattttaa atcatctcaa cttttgagaa attttgagtt atcaacaccg ttcccacaag 4380
acagtggcaa aattattggt gagaattaaa cagctgtttc tcagaggaag caatggaggc 4440
ttgctgggat aaaggcattt actgagaggc tgttacctag tgagagtgat gaattaatta 4500
aaatagtcga atccctttct gactgtctct gaaagcttcc gcttttatct ttgaagagca 4560
gaattgtcac tccaaggaca tttattaata aaaagaacaa ctgtccagtg caatgaaggc 4620
aaagtcatag gtctcccaag tcttacccca ttcctgtgaa atatcaagtt cttggctttt 4680
ctctgtcatg tagcctcaac tttctctgac cgggtgcatt tctttctctg gtttctaaat 4740
tgccagtggc aaatttggat cacttactta atatctgtta aattttgtga cccaacaaag 4800
tcttttagca ctgtggtgtc aaaaagaaaa acacctccca ggcatataca ttttatagat 4860
tcctggagaa tgttgctctc cagctccatc cccacccaat gaaatatgat ccagagagtc 4920
ttgcaaagag acaagcctca ttttccacaa ttagctctaa agtgcctcca ggaaatgatt 4980
ttctcagctc atctctctgt attccctgtt ttggatcaca gggcaatctg tttaaatgac 5040
taattacaga aatcattaaa ggcaccaagc aaatgtcatc tctgaataca cacatcccaa 5100
gctttacaaa tcctgcctgg cttgacagtg atgaggccac ttaacagtcc agcgcaggcg 5160
gatgttaaaa aaaataaaaa ggtgaccatc tgcggtttag ttttttaact ttctgatttc 5220
acacttaacg tctgtcattc tgttactggg cacctgttta aattctattt taaaatgtta 5280
atgtgtgttg tttaaaataa aatcaagaaa gagaga 5316
<210> 35
<211> 5561
<212> ДНК
<213> Homo sapiens
<400> 35
ttttaagaag catcaagaaa gcctgcctgt ctaactttgg aaatatcacc ctcatgctgt 60
cttcccagga tgtctctctc cctaagtaag agatgttact tcctggaggg aatgcagtgt 120
tgggaatctg aagacccagc tttgagctga atttgctttg tgatacctga gacagcattt 180
ccccatgttg gccaggctgg tcttgaactc ctgacctcaa gtgatccgcc cgcctcggcc 240
tcccaaagtg ctgggtttac aagcgtgagc caccgcgccc cgggggagtc agaaagtgta 300
taaataattg cagtgctgct ttgcttccaa aactgggcag tgagttcaac aacaacgaca 360
acaacagccg cagctcatcc tggccgtcat ggagtttctt gaaagaacgt atcttgtgaa 420
tgataaagct gccaagatgt atgctttcac actagaaaga aggagctgca aatgaacact 480
tcatagcaat gtggaactcc aacagaaacc ggtgaataaa gatcagtgtc ccagagagag 540
accagaggag ctggagtcag gaggcatgta ccactgccac agtggctcca agcccacaga 600
aaagggggcg aatgagtacg cctatgccaa gtggaaactc tgttctgctt cagcaatatg 660
cttcattttc atgattgcag aggtcgtggg tgggcacatt gctgggagtc ttgctgttgt 720
cacagatgct gcccacctct taattgacct gaccagtttc ctgctcagtc tcttctccct 780
gtggttgtca tcgaagcctc cctctaagcg gctgacattt ggatggcacc gagcagagat 840
ccttggtgcc ctgctctcca tcctgtgcat ctgggtggtg actggcgtgc tagtgtacct 900
ggcatgtgag cgcctgctgt atcctgatta ccagatccag gcgactgtga tgatcatcgt 960
ttccagctgc gcagtggcgg ccaacattgt actaactgtg gttttgcacc agagatgcct 1020
tggccacaat cacaaggaag tacaagccaa tgccagcgtc agagctgctt ttgtgcatgc 1080
ccttggagat ctatttcaga gtatcagtgt gctaattagt gcacttatta tctactttaa 1140
gccagagtat aaaatagccg acccaatctg cacattcatc ttttccatcc tggtcttggc 1200
cagcaccatc actatcttaa aggacttctc catcttactc atggaaggtg tgccaaagag 1260
cctgaattac agtggtgtga aagagcttat tttagcagtc gacggggtgc tgtctgtgca 1320
cagcctgcac atctggtctc taacaatgaa tcaagtaatt ctctcagctc atgttgctac 1380
agcagccagc cgggacagcc aagtggttcg gagagaaatt gctaaagccc ttagcaaaag 1440
ctttacgatg cactcactca ccattcagat ggaatctcca gttgaccagg accccgactg 1500
ccttttctgt gaagacccct gtgactagct cagtcacacc gtcagtttcc caaatttgac 1560
aggccacctt caaacatgct gctatgcagt ttctgcatca tagaaaataa ggaaccaaag 1620
gaagaaattc atgtcatggt gcaatgcaca ttttatctat ttatttagtt ccattcacca 1680
tgaaggaaga ggcactgaga tccatcaatc aattggatta tatactgatc agtagctgtg 1740
ttcaattgca ggaatgtgta tatagattat tcctgagtgg agccgaagta acagctgttt 1800
gtaactatcg gcaataccaa attcatctcc cttccaataa tgcatcttga gaacacatag 1860
gtaaatttga actcaggaaa gtcttactag aaatcagtgg aagggacaaa tagtcacaaa 1920
attttaccaa aacattagaa acaaaaaata aggagagcca agtcaggaat aaaagtgact 1980
ctgtatgcta acgccacatt agaacttggt tctctcacca agctgtaatg tgattttttt 2040
ttctactctg aattggaaat atgtatgaat atacagagaa gtgcttacaa ctaattttta 2100
tttacttgtc acattttggc aataaatccc tcttatttct aaattctaac ttgtttattt 2160
caaaacttta tataatcact gttcaaaagg aaatattttc acctaccaga gtgcttaaac 2220
actggcacca gccaaagaat gtggttgtag agacccagaa gtcttcaaga acagccgaca 2280
aaaacattcg agttgacccc accaagttgt tgccacagat aatttagata tttacctgca 2340
agaaggaata aagcagatgc aaccaattca ttcagtccac gagcatgatg tgagcactgc 2400
tttgtgctag acattgggct tagcattgaa actataaaga ggaatcagac gcagcaagtg 2460
cttctgtgtt ctggtagcaa ctcaacacta tctgtggaga gtaaactgaa gatgtgcagg 2520
ccaacattct ggaaatccta tgtcaatggg tttggtttgg aacctggact tctgcatttt 2580
taaaagttac ccagagatgc ttctaaagat gagccatagt ctagaagatt gtcaaccaca 2640
ggagttcatt gagtgggaca gctagacaca tacattggca gctacaatag tatcatgaat 2700
tgcaatgatg tagtggggta taaaaggaaa gcgatggata ttgccggatg ggcatggcca 2760
gtgatgtttc acgtcattga ggtgacagct ctgctggact ttgaattaca tatggaggct 2820
ctccaggaag acgaagaaga gaaggacatt ctaggcaaaa agaagactag gcacaaggca 2880
cacttatgtt tgtctgttag cttttagttg aaaaagcaaa atacatgatg caaagaaacc 2940
tctccacgct gtgattttta aaactacata ctttttgcaa ctttatggtt atgagtattg 3000
tagagaacag gagataggtc ttagatgatt tttatgttgt tgtcagactc tagcaaggta 3060
ctagaaacct agcaggcatt aataattgtt gaggcaatga ctctgaggct atatctgggc 3120
cttgtcatta tttatcattt atatttgtat ttttttctga aatttgaggg ccaagaaaac 3180
attgactttg actgaggagg tcacatctgt gccatctctg caaatcaatc agcaccactg 3240
aaataactac ttagcattct gctgagcttt ccctgctcag tagagacaaa tatactcatc 3300
ccccacctca gtgagcttgt ttaggcaacc aggattagag ctgctcaggt tcccaacgtc 3360
tcctgccaca tcgggttctc aaaatggaaa gaatggttta tgccaaatca cttttcctgt 3420
ctgaaggacc actgaatggt tttgtttttc catattttgc ataggacgcc ctaaagacta 3480
ggtgacttgg caaacacaca agtgttagta taattctttg cttctgcttc tttttgaaaa 3540
tcatgtttag atttgatttt aagtcagaaa ttcactgaat gtcaggtaat cattatggag 3600
ggagatttgt gtgtcaacca aagtaattgt cccatggccc cagggtattt ctgttgtttc 3660
cctgaaattc tgctttttta gtcagctaga ttgaaaactc tgaacagtag atgtttatat 3720
ggcaaaatgc aagacaatct acaagggaga ttttaaggat tttgagatga aaaaacagat 3780
gctactcagg ggctttatga accatccatc aattctgaag ttctgactct cccattaccc 3840
tttccctggt gtggtcagaa ctccaggtca ctggaagtta gtggaatcat gtagttgaat 3900
tctttacttc aagacattgt attctctcca gctatcaaaa cattaatgat cttttatgtc 3960
ttttttttgt tattgttata ctttaagttc tggggtacat gtgcggaaca tgtaggtttg 4020
ttacataggt atacatgtgc catggtggtt tgctgcactc atcaacctgt catctacatt 4080
cttttatgtc tgtctttcaa agcaacactc tgttcttctg agtagtgaaa tcaggtcaac 4140
tttaccacca gcctccattt ttaatatgct tcaccatcat ccagcaccta cttaagattt 4200
atctagggct ctgtggtgat gttaggaccc ataaaagaaa tttatgcctt ccatatgttt 4260
ggttacagat gggaaatggg aatgttgaag gacatgaaag aaaggatgtt tacacattaa 4320
gcatcagttc tgaagctaga ttgtctgagt ttgaatctta gctcttccct ttattagctc 4380
tgtgacctcg agctagttac ttaaatgctc tgatcctcta tttcctgatc agtgaaacct 4440
ccctattcaa atgtgtgaga gtttaataaa ttaggacact taaaaatgtt ggagcagtgc 4500
atagcatgta gtgttcagta catgttaaat gttgtttttt attatgtaca aacatgagtg 4560
ggcacagaat tttaaatcat ctcaactttt gagaaatttt gagttatcaa caccgttccc 4620
acaagacagt ggcaaaatta ttggtgagaa ttaaacagct gtttctcaga ggaagcaatg 4680
gaggcttgct gggataaagg catttactga gaggctgtta cctagtgaga gtgatgaatt 4740
aattaaaata gtcgaatccc tttctgactg tctctgaaag cttccgcttt tatctttgaa 4800
gagcagaatt gtcactccaa ggacatttat taataaaaag aacaactgtc cagtgcaatg 4860
aaggcaaagt cataggtctc ccaagtctta ccccattcct gtgaaatatc aagttcttgg 4920
cttttctctg tcatgtagcc tcaactttct ctgaccgggt gcatttcttt ctctggtttc 4980
taaattgcca gtggcaaatt tggatcactt acttaatatc tgttaaattt tgtgacccaa 5040
caaagtcttt tagcactgtg gtgtcaaaaa gaaaaacacc tcccaggcat atacatttta 5100
tagattcctg gagaatgttg ctctccagct ccatccccac ccaatgaaat atgatccaga 5160
gagtcttgca aagagacaag cctcattttc cacaattagc tctaaagtgc ctccaggaaa 5220
tgattttctc agctcatctc tctgtattcc ctgttttgga tcacagggca atctgtttaa 5280
atgactaatt acagaaatca ttaaaggcac caagcaaatg tcatctctga atacacacat 5340
cccaagcttt acaaatcctg cctggcttga cagtgatgag gccacttaac agtccagcgc 5400
aggcggatgt taaaaaaaat aaaaaggtga ccatctgcgg tttagttttt taactttctg 5460
atttcacact taacgtctgt cattctgtta ctgggcacct gtttaaattc tattttaaaa 5520
tgttaatgtg tgttgtttaa aataaaatca agaaagagag a 5561
<210> 36
<211> 5430
<212> ДНК
<213> Homo sapiens
<400> 36
ttttaagaag catcaagaaa gcctgcctgt ctaactttgg aaatatcacc ctcatgctgt 60
cttcccagga tgtctctctc cctaagtaag agatgttact tcctggaggg aatgcagtgt 120
tgggaatctg aagacccagc tttgagctga atttgctttg tgatacctgg agagaagacg 180
tgttttcttg acaacagcac agtacctagt gagttcaaca acaacgacaa caacagccgc 240
agctcatcct ggccgtcatg gagtttcttg aaagaacgta tcttgtgaat gataaagctg 300
ccaagatgta tgctttcaca ctagaaagaa ggagctgcaa atgaacactt catagcaatg 360
tggaactcca acagaaaccg gtgaataaag atcagtgtcc cagagagaga ccagaggagc 420
tggagtcagg aggcatgtac cactgccaca gtggctccaa gcccacagaa aagggggcga 480
atgagtacgc ctatgccaag tggaaactct gttctgcttc agcaatatgc ttcattttca 540
tgattgcaga ggtcgtgggt gggcacattg ctgggagtct tgctgttgtc acagatgctg 600
cccacctctt aattgacctg accagtttcc tgctcagtct cttctccctg tggttgtcat 660
cgaagcctcc ctctaagcgg ctgacatttg gatggcaccg agcagagatc cttggtgccc 720
tgctctccat cctgtgcatc tgggtggtga ctggcgtgct agtgtacctg gcatgtgagc 780
gcctgctgta tcctgattac cagatccagg cgactgtgat gatcatcgtt tccagctgcg 840
cagtggcggc caacattgta ctaactgtgg ttttgcacca gagatgcctt ggccacaatc 900
acaaggaagt acaagccaat gccagcgtca gagctgcttt tgtgcatgcc cttggagatc 960
tatttcagag tatcagtgtg ctaattagtg cacttattat ctactttaag ccagagtata 1020
aaatagccga cccaatctgc acattcatct tttccatcct ggtcttggcc agcaccatca 1080
ctatcttaaa ggacttctcc atcttactca tggaaggtgt gccaaagagc ctgaattaca 1140
gtggtgtgaa agagcttatt ttagcagtcg acggggtgct gtctgtgcac agcctgcaca 1200
tctggtctct aacaatgaat caagtaattc tctcagctca tgttgctaca gcagccagcc 1260
gggacagcca agtggttcgg agagaaattg ctaaagccct tagcaaaagc tttacgatgc 1320
actcactcac cattcagatg gaatctccag ttgaccagga ccccgactgc cttttctgtg 1380
aagacccctg tgactagctc agtcacaccg tcagtttccc aaatttgaca ggccaccttc 1440
aaacatgctg ctatgcagtt tctgcatcat agaaaataag gaaccaaagg aagaaattca 1500
tgtcatggtg caatgcacat tttatctatt tatttagttc cattcaccat gaaggaagag 1560
gcactgagat ccatcaatca attggattat atactgatca gtagctgtgt tcaattgcag 1620
gaatgtgtat atagattatt cctgagtgga gccgaagtaa cagctgtttg taactatcgg 1680
caataccaaa ttcatctccc ttccaataat gcatcttgag aacacatagg taaatttgaa 1740
ctcaggaaag tcttactaga aatcagtgga agggacaaat agtcacaaaa ttttaccaaa 1800
acattagaaa caaaaaataa ggagagccaa gtcaggaata aaagtgactc tgtatgctaa 1860
cgccacatta gaacttggtt ctctcaccaa gctgtaatgt gatttttttt tctactctga 1920
attggaaata tgtatgaata tacagagaag tgcttacaac taatttttat ttacttgtca 1980
cattttggca ataaatccct cttatttcta aattctaact tgtttatttc aaaactttat 2040
ataatcactg ttcaaaagga aatattttca cctaccagag tgcttaaaca ctggcaccag 2100
ccaaagaatg tggttgtaga gacccagaag tcttcaagaa cagccgacaa aaacattcga 2160
gttgacccca ccaagttgtt gccacagata atttagatat ttacctgcaa gaaggaataa 2220
agcagatgca accaattcat tcagtccacg agcatgatgt gagcactgct ttgtgctaga 2280
cattgggctt agcattgaaa ctataaagag gaatcagacg cagcaagtgc ttctgtgttc 2340
tggtagcaac tcaacactat ctgtggagag taaactgaag atgtgcaggc caacattctg 2400
gaaatcctat gtcaatgggt ttggtttgga acctggactt ctgcattttt aaaagttacc 2460
cagagatgct tctaaagatg agccatagtc tagaagattg tcaaccacag gagttcattg 2520
agtgggacag ctagacacat acattggcag ctacaatagt atcatgaatt gcaatgatgt 2580
agtggggtat aaaaggaaag cgatggatat tgccggatgg gcatggccag tgatgtttca 2640
cgtcattgag gtgacagctc tgctggactt tgaattacat atggaggctc tccaggaaga 2700
cgaagaagag aaggacattc taggcaaaaa gaagactagg cacaaggcac acttatgttt 2760
gtctgttagc ttttagttga aaaagcaaaa tacatgatgc aaagaaacct ctccacgctg 2820
tgatttttaa aactacatac tttttgcaac tttatggtta tgagtattgt agagaacagg 2880
agataggtct tagatgattt ttatgttgtt gtcagactct agcaaggtac tagaaaccta 2940
gcaggcatta ataattgttg aggcaatgac tctgaggcta tatctgggcc ttgtcattat 3000
ttatcattta tatttgtatt tttttctgaa atttgagggc caagaaaaca ttgactttga 3060
ctgaggaggt cacatctgtg ccatctctgc aaatcaatca gcaccactga aataactact 3120
tagcattctg ctgagctttc cctgctcagt agagacaaat atactcatcc cccacctcag 3180
tgagcttgtt taggcaacca ggattagagc tgctcaggtt cccaacgtct cctgccacat 3240
cgggttctca aaatggaaag aatggtttat gccaaatcac ttttcctgtc tgaaggacca 3300
ctgaatggtt ttgtttttcc atattttgca taggacgccc taaagactag gtgacttggc 3360
aaacacacaa gtgttagtat aattctttgc ttctgcttct ttttgaaaat catgtttaga 3420
tttgatttta agtcagaaat tcactgaatg tcaggtaatc attatggagg gagatttgtg 3480
tgtcaaccaa agtaattgtc ccatggcccc agggtatttc tgttgtttcc ctgaaattct 3540
gcttttttag tcagctagat tgaaaactct gaacagtaga tgtttatatg gcaaaatgca 3600
agacaatcta caagggagat tttaaggatt ttgagatgaa aaaacagatg ctactcaggg 3660
gctttatgaa ccatccatca attctgaagt tctgactctc ccattaccct ttccctggtg 3720
tggtcagaac tccaggtcac tggaagttag tggaatcatg tagttgaatt ctttacttca 3780
agacattgta ttctctccag ctatcaaaac attaatgatc ttttatgtct tttttttgtt 3840
attgttatac tttaagttct ggggtacatg tgcggaacat gtaggtttgt tacataggta 3900
tacatgtgcc atggtggttt gctgcactca tcaacctgtc atctacattc ttttatgtct 3960
gtctttcaaa gcaacactct gttcttctga gtagtgaaat caggtcaact ttaccaccag 4020
cctccatttt taatatgctt caccatcatc cagcacctac ttaagattta tctagggctc 4080
tgtggtgatg ttaggaccca taaaagaaat ttatgccttc catatgtttg gttacagatg 4140
ggaaatggga atgttgaagg acatgaaaga aaggatgttt acacattaag catcagttct 4200
gaagctagat tgtctgagtt tgaatcttag ctcttccctt tattagctct gtgacctcga 4260
gctagttact taaatgctct gatcctctat ttcctgatca gtgaaacctc cctattcaaa 4320
tgtgtgagag tttaataaat taggacactt aaaaatgttg gagcagtgca tagcatgtag 4380
tgttcagtac atgttaaatg ttgtttttta ttatgtacaa acatgagtgg gcacagaatt 4440
ttaaatcatc tcaacttttg agaaattttg agttatcaac accgttccca caagacagtg 4500
gcaaaattat tggtgagaat taaacagctg tttctcagag gaagcaatgg aggcttgctg 4560
ggataaaggc atttactgag aggctgttac ctagtgagag tgatgaatta attaaaatag 4620
tcgaatccct ttctgactgt ctctgaaagc ttccgctttt atctttgaag agcagaattg 4680
tcactccaag gacatttatt aataaaaaga acaactgtcc agtgcaatga aggcaaagtc 4740
ataggtctcc caagtcttac cccattcctg tgaaatatca agttcttggc ttttctctgt 4800
catgtagcct caactttctc tgaccgggtg catttctttc tctggtttct aaattgccag 4860
tggcaaattt ggatcactta cttaatatct gttaaatttt gtgacccaac aaagtctttt 4920
agcactgtgg tgtcaaaaag aaaaacacct cccaggcata tacattttat agattcctgg 4980
agaatgttgc tctccagctc catccccacc caatgaaata tgatccagag agtcttgcaa 5040
agagacaagc ctcattttcc acaattagct ctaaagtgcc tccaggaaat gattttctca 5100
gctcatctct ctgtattccc tgttttggat cacagggcaa tctgtttaaa tgactaatta 5160
cagaaatcat taaaggcacc aagcaaatgt catctctgaa tacacacatc ccaagcttta 5220
caaatcctgc ctggcttgac agtgatgagg ccacttaaca gtccagcgca ggcggatgtt 5280
aaaaaaaata aaaaggtgac catctgcggt ttagtttttt aactttctga tttcacactt 5340
aacgtctgtc attctgttac tgggcacctg tttaaattct attttaaaat gttaatgtgt 5400
gttgtttaaa ataaaatcaa gaaagagaga 5430
<210> 37
<211> 89
<212> БЕЛОК
<213> Homo sapiens
<400> 37
Met Gly Ile Leu Lys Leu Gln Val Phe Leu Ile Val Leu Ser Val Ala
1 5 10 15
Leu Asn His Leu Lys Ala Thr Pro Ile Glu Ser His Gln Val Glu Lys
20 25 30
Arg Lys Cys Asn Thr Ala Thr Cys Ala Thr Gln Arg Leu Ala Asn Phe
35 40 45
Leu Val His Ser Ser Asn Asn Phe Gly Ala Ile Leu Ser Ser Thr Asn
50 55 60
Val Gly Ser Asn Thr Tyr Gly Lys Arg Asn Ala Val Glu Val Leu Lys
65 70 75 80
Arg Glu Pro Leu Asn Tyr Leu Pro Leu
85
<210> 38
<211> 267
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(267)
<400> 38
atg ggc atc ctg aag ctg caa gta ttt ctc att gtg ctc tct gtt gca 48
Met Gly Ile Leu Lys Leu Gln Val Phe Leu Ile Val Leu Ser Val Ala
1 5 10 15
ttg aac cat ctg aaa gct aca ccc att gaa agt cat cag gtg gaa aag 96
Leu Asn His Leu Lys Ala Thr Pro Ile Glu Ser His Gln Val Glu Lys
20 25 30
cgg aaa tgc aac act gcc aca tgt gca acg cag cgc ctg gca aat ttt 144
Arg Lys Cys Asn Thr Ala Thr Cys Ala Thr Gln Arg Leu Ala Asn Phe
35 40 45
tta gtt cat tcc agc aac aac ttt ggt gcc att ctc tca tct acc aac 192
Leu Val His Ser Ser Asn Asn Phe Gly Ala Ile Leu Ser Ser Thr Asn
50 55 60
gtg gga tcc aat aca tat ggc aag agg aat gca gta gag gtt tta aag 240
Val Gly Ser Asn Thr Tyr Gly Lys Arg Asn Ala Val Glu Val Leu Lys
65 70 75 80
aga gag cca ctg aat tac ttg ccc ctt 267
Arg Glu Pro Leu Asn Tyr Leu Pro Leu
85
<210> 39
<211> 470
<212> БЕЛОК
<213> Homo sapiens
<400> 39
Met Ser His His Pro Ser Gly Leu Arg Ala Gly Phe Ser Ser Thr Ser
1 5 10 15
Tyr Arg Arg Thr Phe Gly Pro Pro Pro Ser Leu Ser Pro Gly Ala Phe
20 25 30
Ser Tyr Ser Ser Ser Ser Arg Phe Ser Ser Ser Arg Leu Leu Gly Ser
35 40 45
Ala Ser Pro Ser Ser Ser Val Arg Leu Gly Ser Phe Arg Ser Pro Arg
50 55 60
Ala Gly Ala Gly Ala Leu Leu Arg Leu Pro Ser Glu Arg Leu Asp Phe
65 70 75 80
Ser Met Ala Glu Ala Leu Asn Gln Glu Phe Leu Ala Thr Arg Ser Asn
85 90 95
Glu Lys Gln Glu Leu Gln Glu Leu Asn Asp Arg Phe Ala Asn Phe Ile
100 105 110
Glu Lys Val Arg Phe Leu Glu Gln Gln Asn Ala Ala Leu Arg Gly Glu
115 120 125
Leu Ser Gln Ala Arg Gly Gln Glu Pro Ala Arg Ala Asp Gln Leu Cys
130 135 140
Gln Gln Glu Leu Arg Glu Leu Arg Arg Glu Leu Glu Leu Leu Gly Arg
145 150 155 160
Glu Arg Asp Arg Val Gln Val Glu Arg Asp Gly Leu Ala Glu Asp Leu
165 170 175
Ala Ala Leu Lys Gln Arg Leu Glu Glu Glu Thr Arg Lys Arg Glu Asp
180 185 190
Ala Glu His Asn Leu Val Leu Phe Arg Lys Asp Val Asp Asp Ala Thr
195 200 205
Leu Ser Arg Leu Glu Leu Glu Arg Lys Ile Glu Ser Leu Met Asp Glu
210 215 220
Ile Glu Phe Leu Lys Lys Leu His Glu Glu Glu Leu Arg Asp Leu Gln
225 230 235 240
Val Ser Val Glu Ser Gln Gln Val Gln Gln Val Glu Val Glu Ala Thr
245 250 255
Val Lys Pro Glu Leu Thr Ala Ala Leu Arg Asp Ile Arg Ala Gln Tyr
260 265 270
Glu Ser Ile Ala Ala Lys Asn Leu Gln Glu Ala Glu Glu Trp Tyr Lys
275 280 285
Ser Lys Tyr Ala Asp Leu Ser Asp Ala Ala Asn Arg Asn His Glu Ala
290 295 300
Leu Arg Gln Ala Lys Gln Glu Met Asn Glu Ser Arg Arg Gln Ile Gln
305 310 315 320
Ser Leu Thr Cys Glu Val Asp Gly Leu Arg Gly Thr Asn Glu Ala Leu
325 330 335
Leu Arg Gln Leu Arg Glu Leu Glu Glu Gln Phe Ala Leu Glu Ala Gly
340 345 350
Gly Tyr Gln Ala Gly Ala Ala Arg Leu Glu Glu Glu Leu Arg Gln Leu
355 360 365
Lys Glu Glu Met Ala Arg His Leu Arg Glu Tyr Gln Glu Leu Leu Asn
370 375 380
Val Lys Met Ala Leu Asp Ile Glu Ile Ala Thr Tyr Arg Lys Leu Leu
385 390 395 400
Glu Gly Glu Glu Ser Arg Ile Ser Val Pro Val His Ser Phe Ala Ser
405 410 415
Leu Asn Ile Lys Thr Thr Val Pro Glu Val Glu Pro Pro Gln Asp Ser
420 425 430
His Ser Arg Lys Thr Val Leu Ile Lys Thr Ile Glu Thr Arg Asn Gly
435 440 445
Glu Val Val Thr Glu Ser Gln Lys Glu Gln Arg Ser Glu Leu Asp Lys
450 455 460
Ser Ser Ala His Ser Tyr
465 470
<210> 40
<211> 1413
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(1410)
<400> 40
atg agc cac cac ccg tcg ggc ctc cgg gcc ggc ttc agc tcc acc tca 48
Met Ser His His Pro Ser Gly Leu Arg Ala Gly Phe Ser Ser Thr Ser
1 5 10 15
tac cgc cgt acc ttc ggt cca ccg ccc tca cta tcc ccc ggg gcc ttc 96
Tyr Arg Arg Thr Phe Gly Pro Pro Pro Ser Leu Ser Pro Gly Ala Phe
20 25 30
tcc tac tcg tcc agc tcc cgc ttc tcc agc agc cgc ctg ctg ggc tcc 144
Ser Tyr Ser Ser Ser Ser Arg Phe Ser Ser Ser Arg Leu Leu Gly Ser
35 40 45
gcg tcc ccg agc tcc tcg gtg cgc ctg ggc agc ttc cgt agc ccc cga 192
Ala Ser Pro Ser Ser Ser Val Arg Leu Gly Ser Phe Arg Ser Pro Arg
50 55 60
gcg gga gcg ggc gcc ctc ctg cgc ctg ccc tcg gag cgc ctc gac ttc 240
Ala Gly Ala Gly Ala Leu Leu Arg Leu Pro Ser Glu Arg Leu Asp Phe
65 70 75 80
tcc atg gcc gag gcc ctc aac cag gag ttc ctg gcc acg cgc agc aac 288
Ser Met Ala Glu Ala Leu Asn Gln Glu Phe Leu Ala Thr Arg Ser Asn
85 90 95
gag aag cag gag ctg cag gag ctc aac gac cgc ttc gcc aac ttc atc 336
Glu Lys Gln Glu Leu Gln Glu Leu Asn Asp Arg Phe Ala Asn Phe Ile
100 105 110
gag aag gta cgc ttt ctg gag cag cag aac gcg gcc ctg cgc ggg gag 384
Glu Lys Val Arg Phe Leu Glu Gln Gln Asn Ala Ala Leu Arg Gly Glu
115 120 125
ctg agc caa gcc cgg ggc cag gag ccg gcg cgc gcc gac cag ctg tgc 432
Leu Ser Gln Ala Arg Gly Gln Glu Pro Ala Arg Ala Asp Gln Leu Cys
130 135 140
cag cag gag ctg cgc gag ctg cgg cga gag ctg gag ctg ttg ggc cgc 480
Gln Gln Glu Leu Arg Glu Leu Arg Arg Glu Leu Glu Leu Leu Gly Arg
145 150 155 160
gag cgt gac cgg gtg cag gtg gag cgc gac ggg ctg gcg gag gac ctg 528
Glu Arg Asp Arg Val Gln Val Glu Arg Asp Gly Leu Ala Glu Asp Leu
165 170 175
gcg gcg ctc aag cag agg ttg gag gag gag acg cgc aag cgg gag gac 576
Ala Ala Leu Lys Gln Arg Leu Glu Glu Glu Thr Arg Lys Arg Glu Asp
180 185 190
gcg gag cac aac ctc gtg ctc ttc cgc aag gac gtg gac gat gcc act 624
Ala Glu His Asn Leu Val Leu Phe Arg Lys Asp Val Asp Asp Ala Thr
195 200 205
ctg tcc cgc ctg gaa cta gag cgc aag att gag tct ctg atg gat gag 672
Leu Ser Arg Leu Glu Leu Glu Arg Lys Ile Glu Ser Leu Met Asp Glu
210 215 220
att gag ttc ctc aag aag ctg cac gag gag gag ctg cga gac ctg cag 720
Ile Glu Phe Leu Lys Lys Leu His Glu Glu Glu Leu Arg Asp Leu Gln
225 230 235 240
gtg agt gtg gag agc cag cag gtg cag cag gtg gag gtg gaa gcc acg 768
Val Ser Val Glu Ser Gln Gln Val Gln Gln Val Glu Val Glu Ala Thr
245 250 255
gtg aag ccc gag ctg acg gca gcg ctg agg gac atc cgc gcg cag tac 816
Val Lys Pro Glu Leu Thr Ala Ala Leu Arg Asp Ile Arg Ala Gln Tyr
260 265 270
gag agc atc gcc gcg aag aac ctg cag gag gcg gag gag tgg tac aag 864
Glu Ser Ile Ala Ala Lys Asn Leu Gln Glu Ala Glu Glu Trp Tyr Lys
275 280 285
tcc aag tac gcg gac ctg tcc gac gct gcc aac cgg aac cac gag gcc 912
Ser Lys Tyr Ala Asp Leu Ser Asp Ala Ala Asn Arg Asn His Glu Ala
290 295 300
ctg cgc cag gcc aag cag gag atg aac gag tcc cga cgc cag atc cag 960
Leu Arg Gln Ala Lys Gln Glu Met Asn Glu Ser Arg Arg Gln Ile Gln
305 310 315 320
agt cta acg tgc gag gtg gac ggg ctg cgc ggc acg aac gag gcg ctg 1008
Ser Leu Thr Cys Glu Val Asp Gly Leu Arg Gly Thr Asn Glu Ala Leu
325 330 335
ctc agg cag ttg aga gag ctg gag gag cag ttc gcc ctg gag gcg ggg 1056
Leu Arg Gln Leu Arg Glu Leu Glu Glu Gln Phe Ala Leu Glu Ala Gly
340 345 350
ggc tac cag gcg ggc gct gcg cgg ctc gag gag gag ctg cga cag cta 1104
Gly Tyr Gln Ala Gly Ala Ala Arg Leu Glu Glu Glu Leu Arg Gln Leu
355 360 365
aaa gag gag atg gcg cgg cac ctg agg gag tac cag gag ctc ctc aac 1152
Lys Glu Glu Met Ala Arg His Leu Arg Glu Tyr Gln Glu Leu Leu Asn
370 375 380
gtc aag atg gcc ctg gac atc gag atc gcc acc tac cgc aag ctg ctg 1200
Val Lys Met Ala Leu Asp Ile Glu Ile Ala Thr Tyr Arg Lys Leu Leu
385 390 395 400
gag ggc gag gag agc cgg atc tcc gtg ccc gtc cat tct ttt gcc tcc 1248
Glu Gly Glu Glu Ser Arg Ile Ser Val Pro Val His Ser Phe Ala Ser
405 410 415
tta aat ata aag acg act gtg cct gag gtg gag cct ccc cag gac agc 1296
Leu Asn Ile Lys Thr Thr Val Pro Glu Val Glu Pro Pro Gln Asp Ser
420 425 430
cac agc cgg aag acg gtt ctg atc aag acc att gag acc cgg aat ggg 1344
His Ser Arg Lys Thr Val Leu Ile Lys Thr Ile Glu Thr Arg Asn Gly
435 440 445
gag gtg gtg aca gag tcc cag aag gag cag cgc agt gag ctg gac aag 1392
Glu Val Val Thr Glu Ser Gln Lys Glu Gln Arg Ser Glu Leu Asp Lys
450 455 460
tct tct gcc cac agt tac taa 1413
Ser Ser Ala His Ser Tyr
465 470
<210> 41
<211> 1833
<212> ДНК
<213> Homo sapiens
<400> 41
cctcgcagcg gtctgcggct ccttcccagc ccccggccta gctctgcgaa cggtgactgc 60
ccatccttgg ccgcaatgag ccaccacccg tcgggcctcc gggccggctt cagctccacc 120
tcataccgcc gtaccttcgg tccaccgccc tcactatccc ccggggcctt ctcctactcg 180
tccagctccc gcttctccag cagccgcctg ctgggctccg cgtccccgag ctcctcggtg 240
cgcctgggca gcttccgtag cccccgagcg ggagcgggcg ccctcctgcg cctgccctcg 300
gagcgcctcg acttctccat ggccgaggcc ctcaaccagg agttcctggc cacgcgcagc 360
aacgagaagc aggagctgca ggagctcaac gaccgcttcg ccaacttcat cgagaaggta 420
cgctttctgg agcagcagaa cgcggccctg cgcggggagc tgagccaagc ccggggccag 480
gagccggcgc gcgccgacca gctgtgccag caggagctgc gcgagctgcg gcgagagctg 540
gagctgttgg gccgcgagcg tgaccgggtg caggtggagc gcgacgggct ggcggaggac 600
ctggcggcgc tcaagcagag gttggaggag gagacgcgca agcgggagga cgcggagcac 660
aacctcgtgc tcttccgcaa ggacgtggac gatgccactc tgtcccgcct ggaactagag 720
cgcaagattg agtctctgat ggatgagatt gagttcctca agaagctgca cgaggaggag 780
ctgcgagacc tgcaggtgag tgtggagagc cagcaggtgc agcaggtgga ggtggaagcc 840
acggtgaagc ccgagctgac ggcagcgctg agggacatcc gcgcgcagta cgagagcatc 900
gccgcgaaga acctgcagga ggcggaggag tggtacaagt ccaagtacgc ggacctgtcc 960
gacgctgcca accggaacca cgaggccctg cgccaggcca agcaggagat gaacgagtcc 1020
cgacgccaga tccagagtct aacgtgcgag gtggacgggc tgcgcggcac gaacgaggcg 1080
ctgctcaggc agttgagaga gctggaggag cagttcgccc tggaggcggg gggctaccag 1140
gcgggcgctg cgcggctcga ggaggagctg cgacagctaa aagaggagat ggcgcggcac 1200
ctgagggagt accaggagct cctcaacgtc aagatggccc tggacatcga gatcgccacc 1260
taccgcaagc tgctggaggg cgaggagagc cggatctccg tgcccgtcca ttcttttgcc 1320
tccttaaata taaagacgac tgtgcctgag gtggagcctc cccaggacag ccacagccgg 1380
aagacggttc tgatcaagac cattgagacc cggaatgggg aggtggtgac agagtcccag 1440
aaggagcagc gcagtgagct ggacaagtct tctgcccaca gttactgaac cccttggtcc 1500
ggagccttga ctctgcccta ggcctgctca aagcccaaac cctaagacca ctcctgaatt 1560
gtctcctctc cctctgcatg tgtctaaaag gtggtaccag gcatcccttt cctggcttat 1620
ggccaagccc tacccggcca gcagtcgctg ggcctctccc tgccctgaca cttgatgtga 1680
cctatgtgct tcccttttca tgtcccgata agaagccaat gatcccccct caggacaaat 1740
ctactccagc cacgatgaga agtgggtgag ccagggtctg agtttcacat ttgaaccaaa 1800
taaaatgctg tcaagagaaa actctccagt gca 1833
<210> 42
<211> 1949
<212> ДНК
<213> Homo sapiens
<400> 42
tgttccgcag gggtggggca tccccctccc catacaaccc ccctccagcg ggccatcagg 60
ccagtgggag gagctgcccg tgccccccct gagaccgcag ggctataaag ccgcctcgca 120
gcggtctgcg gctccttccc agcccccggc ctagctctgc gaacggtgac tgcccatcct 180
tggccgcaat gagccaccac ccgtcgggcc tccgggccgg cttcagctcc acctcatacc 240
gccgtacctt cggtccaccg ccctcactat cccccggggc cttctcctac tcgtccagct 300
cccgcttctc cagcagccgc ctgctgggct ccgcgtcccc gagctcctcg gtgcgcctgg 360
gcagcttccg tagcccccga gcgggagcgg gcgccctcct gcgcctgccc tcggagcgcc 420
tcgacttctc catggccgag gccctcaacc aggagttcct ggccacgcgc agcaacgaga 480
agcaggagct gcaggagctc aacgaccgct tcgccaactt catcgagaag gtacgctttc 540
tggagcagca gaacgcggcc ctgcgcgggg agctgagcca agcccggggc caggagccgg 600
cgcgcgccga ccagctgtgc cagcaggagc tgcgcgagct gcggcgagag ctggagctgt 660
tgggccgcga gcgtgaccgg gtgcaggtgg agcgcgacgg gctggcggag gacctggcgg 720
cgctcaagca gaggttggag gaggagacgc gcaagcggga ggacgcggag cacaacctcg 780
tgctcttccg caaggacgtg gacgatgcca ctctgtcccg cctggaacta gagcgcaaga 840
ttgagtctct gatggatgag attgagttcc tcaagaagct gcacgaggag gagctgcgag 900
acctgcaggt gagtgtggag agccagcagg tgcagcaggt ggaggtggaa gccacggtga 960
agcccgagct gacggcagcg ctgagggaca tccgcgcgca gtacgagagc atcgccgcga 1020
agaacctgca ggaggcggag gagtggtaca agtccaagta cgcggacctg tccgacgctg 1080
ccaaccggaa ccacgaggcc ctgcgccagg ccaagcagga gatgaacgag tcccgacgcc 1140
agatccagag tctaacgtgc gaggtggacg ggctgcgcgg cacgaacgag gcgctgctca 1200
ggcagttgag agagctggag gagcagttcg ccctggaggc ggggggctac caggcgggcg 1260
ctgcgcggct cgaggaggag ctgcgacagc taaaagagga gatggcgcgg cacctgaggg 1320
agtaccagga gctcctcaac gtcaagatgg ccctggacat cgagatcgcc acctaccgca 1380
agctgctgga gggcgaggag agccggatct ccgtgcccgt ccattctttt gcctccttaa 1440
atataaagac gactgtgcct gaggtggagc ctccccagga cagccacagc cggaagacgg 1500
ttctgatcaa gaccattgag acccggaatg gggagcaggt ggtgacagag tcccagaagg 1560
agcagcgcag tgagctggac aagtcttctg cccacagtta ctgaacccct tggtccggag 1620
ccttgactct gccctaggcc tgctcaaagc ccaaacccta agaccactcc tgaattgtct 1680
cctctccctc tgcatgtgtc taaaaggtgg taccaggcat ccctttcctg gcttatggcc 1740
aagccctacc cggccagcag tcgctgggcc tctccctgcc ctgacacttg atgtgaccta 1800
tgtgcttccc ttttcatgtc ccgataagaa gccaatgatc ccccctcagg acaaatctac 1860
tccagccacg atgagaagtg ggtgagccag ggtctgagtt tcacatttga accaaataaa 1920
atgctgtcaa gagaaaactc tccagtgca 1949
<210> 43
<211> 1628
<212> ДНК
<213> Homo sapiens
<400> 43
tgttccgcag gggtggggca tccccctccc catacaaccc ccctccagcg ggccatcagg 60
ccagtgggag gagctgcccg tgccccccct gagaccgcag ggctataaag ccgcctcgca 120
gcggtctgcg gctccttccc agcccccggc ctagctctgc gaacggtgac tgcccatcct 180
tggccgcaat gagccaccac ccgtcgggcc tccgggccgg cttcagctcc acctcatacc 240
gccgtacctt cggtccaccg ccctcactat cccccggggc cttctcctac tcgtccagct 300
cccgcttctc cagcagccgc ctgctgggct ccgcgtcccc gagctcctcg gtgcgcctgg 360
gcagcttccg tagcccccga gcgggagcgg gcgccctcct gcgcctgccc tcggagcgcc 420
tcgacttctc catggccgag gccctcaacc aggagttcct ggccacgcgc agcaacgaga 480
agcaggagct gcaggagctc aacgaccgct tcgccaactt catcgagaag gtacgctttc 540
tggagcagca gaacgcggcc ctgcgcgggg agctgagcca agcccggggc caggagccgg 600
cgcgcgccga ccagctgtgc cagcaggagc tgcgcgagct gcggcgagag ctggagctgt 660
tgggccgcga gcgtgaccgg gtgcaggtgg agcgcgacgg gctggcggag gacctggcgg 720
cgctcaagca gaggtcaggg ggcagggctg ggccgctgcc gtcgaggcga ggtcgaagcg 780
gccgtcgagg cggctgctct tgcctcccct cgcttcccct ctccatcagc agcccaaggg 840
tgtggctccc cttaccaacc caggttggag gaggagacgc gcaagcggga ggacgcggag 900
cacaacctcg tgctcttccg caaggacgtg gacgatgcca ctctgtcccg cctggaacta 960
gagcgcaaga ttgagtctct gatggatgag attgagttcc tcaagaagct gcacgaggag 1020
gagctgcgag acctgcaggt gagtgtggag agccagcagg tgcagcaggt ggaggtggaa 1080
gccacggtga agcccgagct gacggcagcg ctgagggaca tccgcgcgca gtacgagagc 1140
atcgccgcga agaacctgca ggaggcggag gagtggtaca agtccaagta cgcggacctg 1200
tccgacgctg ccaaccggaa ccacgaggcc ctgcgccagg ccaagcagga gatgaacgag 1260
tcccgacgcc agatccagag tctaacgtgc gaggtggacg ggctgcgcgg cacgaacgag 1320
gcgctgctca ggcagttgag agagctggag gagcagttcg ccctggaggc ggggggctac 1380
caggcgggcg ctgcgcggct cgaggaggag ctgcgacagc taaaagagga gatggcgcgg 1440
cacctgaggg agtaccagga gctcctcaac gtcaagatgg ccctggacat cgagatcgcc 1500
acctaccgca agctgctgga gggcgaggag agccggatct ccgtgcccgt ccattctttt 1560
gcctccttaa atataaagac gactgtgcct gaggtggagc ctccccagga cagccacagc 1620
cggaagac 1628
<210> 44
<211> 653
<212> БЕЛОК
<213> Homo sapiens
<400> 44
Met Lys Leu Ser Leu Val Ala Ala Met Leu Leu Leu Leu Ser Ala Ala
1 5 10 15
Arg Ala Glu Glu Glu Asp Lys Lys Glu Asp Val Gly Thr Val Val Gly
20 25 30
Ile Asp Leu Gly Thr Thr Tyr Ser Cys Val Gly Val Phe Lys Asn Gly
35 40 45
Arg Val Glu Ile Ile Ala Asn Asp Gln Gly Asn Arg Ile Thr Pro Ser
50 55 60
Tyr Val Ala Phe Thr Pro Glu Gly Glu Arg Leu Ile Gly Asp Ala Ala
65 70 75 80
Lys Asn Gln Leu Thr Ser Asn Pro Glu Asn Thr Val Phe Asp Ala Lys
85 90 95
Arg Leu Ile Gly Arg Thr Trp Asn Asp Pro Ser Val Gln Gln Asp Ile
100 105 110
Lys Phe Leu Pro Phe Lys Val Val Glu Lys Lys Thr Lys Pro Tyr Ile
115 120 125
Gln Val Asp Ile Gly Gly Gly Gln Thr Lys Thr Phe Ala Pro Glu Glu
130 135 140
Ile Ser Ala Met Val Leu Thr Lys Met Lys Glu Thr Ala Glu Ala Tyr
145 150 155 160
Leu Gly Lys Lys Val Thr His Ala Val Val Thr Val Pro Ala Tyr Phe
165 170 175
Asn Asp Ala Gln Arg Gln Ala Thr Lys Asp Ala Gly Thr Ile Ala Gly
180 185 190
Leu Asn Val Met Arg Ile Ile Asn Glu Pro Thr Ala Ala Ala Ile Ala
195 200 205
Tyr Gly Leu Asp Lys Arg Glu Gly Glu Lys Asn Ile Leu Val Phe Asp
210 215 220
Leu Gly Gly Gly Thr Phe Asp Val Ser Leu Leu Thr Ile Asp Asn Gly
225 230 235 240
Val Phe Glu Val Val Ala Thr Asn Gly Asp Thr His Leu Gly Gly Glu
245 250 255
Asp Phe Asp Gln Arg Val Met Glu His Phe Ile Lys Leu Tyr Lys Lys
260 265 270
Lys Thr Gly Lys Asp Val Arg Lys Asp Asn Arg Ala Val Gln Lys Leu
275 280 285
Arg Arg Glu Val Glu Lys Ala Lys Ala Leu Ser Ser Gln His Gln Ala
290 295 300
Arg Ile Glu Ile Glu Ser Phe Tyr Glu Gly Glu Asp Phe Ser Glu Thr
305 310 315 320
Leu Thr Arg Ala Lys Phe Glu Glu Leu Asn Met Asp Leu Phe Arg Ser
325 330 335
Thr Met Lys Pro Val Gln Lys Val Leu Glu Asp Ser Asp Leu Lys Lys
340 345 350
Ser Asp Ile Asp Glu Ile Val Leu Val Gly Gly Ser Thr Arg Ile Pro
355 360 365
Lys Ile Gln Gln Leu Val Lys Glu Phe Phe Asn Gly Lys Glu Pro Ser
370 375 380
Arg Gly Ile Asn Pro Asp Glu Ala Val Ala Tyr Gly Ala Ala Val Gln
385 390 395 400
Ala Gly Val Leu Ser Gly Asp Gln Asp Thr Gly Asp Leu Val Leu Leu
405 410 415
His Val Cys Pro Leu Thr Leu Gly Ile Glu Thr Val Gly Gly Val Met
420 425 430
Thr Lys Leu Ile Pro Ser Asn Thr Val Val Pro Thr Lys Asn Ser Gln
435 440 445
Ile Phe Ser Thr Ala Ser Asp Asn Gln Pro Thr Val Thr Ile Lys Val
450 455 460
Tyr Glu Gly Glu Arg Pro Leu Thr Lys Asp Asn His Leu Leu Gly Thr
465 470 475 480
Phe Asp Leu Thr Gly Ile Pro Pro Ala Pro Arg Gly Val Pro Gln Ile
485 490 495
Glu Val Thr Phe Glu Ile Asp Val Asn Gly Ile Leu Arg Val Thr Ala
500 505 510
Glu Asp Lys Gly Thr Gly Asn Lys Asn Lys Ile Thr Ile Thr Asn Asp
515 520 525
Gln Asn Arg Leu Thr Pro Glu Glu Ile Glu Arg Met Val Asn Asp Ala
530 535 540
Glu Lys Phe Ala Glu Glu Asp Lys Lys Leu Lys Glu Arg Ile Asp Thr
545 550 555 560
Arg Asn Glu Leu Glu Ser Tyr Ala Tyr Ser Leu Lys Asn Gln Ile Gly
565 570 575
Asp Lys Glu Lys Leu Gly Gly Lys Leu Ser Ser Glu Asp Lys Glu Thr
580 585 590
Met Glu Lys Ala Val Glu Glu Lys Ile Glu Trp Leu Glu Ser His Gln
595 600 605
Asp Ala Asp Ile Glu Asp Phe Lys Ala Lys Lys Lys Glu Leu Glu Glu
610 615 620
Ile Val Gln Pro Ile Ile Ser Lys Leu Tyr Gly Ser Ala Gly Pro Pro
625 630 635 640
Pro Thr Gly Glu Glu Asp Thr Ala Glu Lys Asp Glu Leu
645 650
<210> 45
<211> 1962
<212> ДНК
<213> Homo sapiens
<220>
<212> Кодирующая последовательность
<222> (1)..(1959)
<400> 45
atg aag ctc tcc ctg gtg gcc gcg atg ctg ctg ctg ctc agc gcg gcg 48
Met Lys Leu Ser Leu Val Ala Ala Met Leu Leu Leu Leu Ser Ala Ala
1 5 10 15
cgg gcc gag gag gag gac aag aag gag gac gtg ggc acg gtg gtc ggc 96
Arg Ala Glu Glu Glu Asp Lys Lys Glu Asp Val Gly Thr Val Val Gly
20 25 30
atc gac ttg ggg acc acc tac tcc tgc gtc ggc gtg ttc aag aac ggc 144
Ile Asp Leu Gly Thr Thr Tyr Ser Cys Val Gly Val Phe Lys Asn Gly
35 40 45
cgc gtg gag atc atc gcc aac gat cag ggc aac cgc atc acg ccg tcc 192
Arg Val Glu Ile Ile Ala Asn Asp Gln Gly Asn Arg Ile Thr Pro Ser
50 55 60
tat gtc gcc ttc act cct gaa ggg gaa cgt ctg att ggc gat gcc gcc 240
Tyr Val Ala Phe Thr Pro Glu Gly Glu Arg Leu Ile Gly Asp Ala Ala
65 70 75 80
aag aac cag ctc acc tcc aac ccc gag aac acg gtc ttt gac gcc aag 288
Lys Asn Gln Leu Thr Ser Asn Pro Glu Asn Thr Val Phe Asp Ala Lys
85 90 95
cgg ctc atc ggc cgc acg tgg aat gac ccg tct gtg cag cag gac atc 336
Arg Leu Ile Gly Arg Thr Trp Asn Asp Pro Ser Val Gln Gln Asp Ile
100 105 110
aag ttc ttg ccg ttc aag gtg gtt gaa aag aaa act aaa cca tac att 384
Lys Phe Leu Pro Phe Lys Val Val Glu Lys Lys Thr Lys Pro Tyr Ile
115 120 125
caa gtt gat att gga ggt ggg caa aca aag aca ttt gct cct gaa gaa 432
Gln Val Asp Ile Gly Gly Gly Gln Thr Lys Thr Phe Ala Pro Glu Glu
130 135 140
att tct gcc atg gtt ctc act aaa atg aaa gaa acc gct gag gct tat 480
Ile Ser Ala Met Val Leu Thr Lys Met Lys Glu Thr Ala Glu Ala Tyr
145 150 155 160
ttg gga aag aag gtt acc cat gca gtt gtt act gta cca gcc tat ttt 528
Leu Gly Lys Lys Val Thr His Ala Val Val Thr Val Pro Ala Tyr Phe
165 170 175
aat gat gcc caa cgc caa gca acc aaa gac gct gga act att gct ggc 576
Asn Asp Ala Gln Arg Gln Ala Thr Lys Asp Ala Gly Thr Ile Ala Gly
180 185 190
cta aat gtt atg agg atc atc aac gag cct acg gca gct gct att gct 624
Leu Asn Val Met Arg Ile Ile Asn Glu Pro Thr Ala Ala Ala Ile Ala
195 200 205
tat ggc ctg gat aag agg gag ggg gag aag aac atc ctg gtg ttt gac 672
Tyr Gly Leu Asp Lys Arg Glu Gly Glu Lys Asn Ile Leu Val Phe Asp
210 215 220
ctg ggt ggc gga acc ttc gat gtg tct ctt ctc acc att gac aat ggt 720
Leu Gly Gly Gly Thr Phe Asp Val Ser Leu Leu Thr Ile Asp Asn Gly
225 230 235 240
gtc ttc gaa gtt gtg gcc act aat gga gat act cat ctg ggt gga gaa 768
Val Phe Glu Val Val Ala Thr Asn Gly Asp Thr His Leu Gly Gly Glu
245 250 255
gac ttt gac cag cgt gtc atg gaa cac ttc atc aaa ctg tac aaa aag 816
Asp Phe Asp Gln Arg Val Met Glu His Phe Ile Lys Leu Tyr Lys Lys
260 265 270
aag acg ggc aaa gat gtc agg aag gac aat aga gct gtg cag aaa ctc 864
Lys Thr Gly Lys Asp Val Arg Lys Asp Asn Arg Ala Val Gln Lys Leu
275 280 285
cgg cgc gag gta gaa aag gcc aag gcc ctg tct tct cag cat caa gca 912
Arg Arg Glu Val Glu Lys Ala Lys Ala Leu Ser Ser Gln His Gln Ala
290 295 300
aga att gaa att gag tcc ttc tat gaa gga gaa gac ttt tct gag acc 960
Arg Ile Glu Ile Glu Ser Phe Tyr Glu Gly Glu Asp Phe Ser Glu Thr
305 310 315 320
ctg act cgg gcc aaa ttt gaa gag ctc aac atg gat ctg ttc cgg tct 1008
Leu Thr Arg Ala Lys Phe Glu Glu Leu Asn Met Asp Leu Phe Arg Ser
325 330 335
act atg aag ccc gtc cag aaa gtg ttg gaa gat tct gat ttg aag aag 1056
Thr Met Lys Pro Val Gln Lys Val Leu Glu Asp Ser Asp Leu Lys Lys
340 345 350
tct gat att gat gaa att gtt ctt gtt ggt ggc tcg act cga att cca 1104
Ser Asp Ile Asp Glu Ile Val Leu Val Gly Gly Ser Thr Arg Ile Pro
355 360 365
aag att cag caa ctg gtt aaa gag ttc ttc aat ggc aag gaa cca tcc 1152
Lys Ile Gln Gln Leu Val Lys Glu Phe Phe Asn Gly Lys Glu Pro Ser
370 375 380
cgt ggc ata aac cca gat gaa gct gta gcg tat ggt gct gct gtc cag 1200
Arg Gly Ile Asn Pro Asp Glu Ala Val Ala Tyr Gly Ala Ala Val Gln
385 390 395 400
gct ggt gtg ctc tct ggt gat caa gat aca ggt gac ctg gta ctg ctt 1248
Ala Gly Val Leu Ser Gly Asp Gln Asp Thr Gly Asp Leu Val Leu Leu
405 410 415
cat gta tgt ccc ctt aca ctt ggt att gaa act gta gga ggt gtc atg 1296
His Val Cys Pro Leu Thr Leu Gly Ile Glu Thr Val Gly Gly Val Met
420 425 430
acc aaa ctg att cca agt aat aca gtg gtg cct acc aag aac tct cag 1344
Thr Lys Leu Ile Pro Ser Asn Thr Val Val Pro Thr Lys Asn Ser Gln
435 440 445
atc ttt tct aca gct tct gat aat caa cca act gtt aca atc aag gtc 1392
Ile Phe Ser Thr Ala Ser Asp Asn Gln Pro Thr Val Thr Ile Lys Val
450 455 460
tat gaa ggt gaa aga ccc ctg aca aaa gac aat cat ctt ctg ggt aca 1440
Tyr Glu Gly Glu Arg Pro Leu Thr Lys Asp Asn His Leu Leu Gly Thr
465 470 475 480
ttt gat ctg act gga att cct cct gct cct cgt ggg gtc cca cag att 1488
Phe Asp Leu Thr Gly Ile Pro Pro Ala Pro Arg Gly Val Pro Gln Ile
485 490 495
gaa gtc acc ttt gag ata gat gtg aat ggt att ctt cga gtg aca gct 1536
Glu Val Thr Phe Glu Ile Asp Val Asn Gly Ile Leu Arg Val Thr Ala
500 505 510
gaa gac aag ggt aca ggg aac aaa aat aag atc aca atc acc aat gac 1584
Glu Asp Lys Gly Thr Gly Asn Lys Asn Lys Ile Thr Ile Thr Asn Asp
515 520 525
cag aat cgc ctg aca cct gaa gaa atc gaa agg atg gtt aat gat gct 1632
Gln Asn Arg Leu Thr Pro Glu Glu Ile Glu Arg Met Val Asn Asp Ala
530 535 540
gag aag ttt gct gag gaa gac aaa aag ctc aag gag cgc att gat act 1680
Glu Lys Phe Ala Glu Glu Asp Lys Lys Leu Lys Glu Arg Ile Asp Thr
545 550 555 560
aga aat gag ttg gaa agc tat gcc tat tct cta aag aat cag att gga 1728
Arg Asn Glu Leu Glu Ser Tyr Ala Tyr Ser Leu Lys Asn Gln Ile Gly
565 570 575
gat aaa gaa aag ctg gga ggt aaa ctt tcc tct gaa gat aag gag acc 1776
Asp Lys Glu Lys Leu Gly Gly Lys Leu Ser Ser Glu Asp Lys Glu Thr
580 585 590
atg gaa aaa gct gta gaa gaa aag att gaa tgg ctg gaa agc cac caa 1824
Met Glu Lys Ala Val Glu Glu Lys Ile Glu Trp Leu Glu Ser His Gln
595 600 605
gat gct gac att gaa gac ttc aaa gct aag aag aag gaa ctg gaa gaa 1872
Asp Ala Asp Ile Glu Asp Phe Lys Ala Lys Lys Lys Glu Leu Glu Glu
610 615 620
att gtt caa cca att atc agc aaa ctc tat gga agt gca ggc cct ccc 1920
Ile Val Gln Pro Ile Ile Ser Lys Leu Tyr Gly Ser Ala Gly Pro Pro
625 630 635 640
cca act ggt gaa gag gat aca gca gaa aaa gat gag ttg tag 1962
Pro Thr Gly Glu Glu Asp Thr Ala Glu Lys Asp Glu Leu
645 650
<210> 46
<211> 700
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
полинуклеотид"
<400> 46
aactgaagat tcaacaatct cagacatcgc tgttgcaact aacgctggtc aaatcaaaac 60
tggttcactt tcacgtacag accgtatggc taaatacaac caattgcttc gtattgaaga 120
ccaattggct gaagttgctc aatacaaagg tcttaaagca ttctacaacc ttaaaaaata 180
aggaggaaaa aatgaaaaaa aagattatct cagctatttt aatgtctaca gtgatacttt 240
ctgctgcagc cccgttgtca ggtgtttacg ccgctccaac ttcatcatca actaaaaaaa 300
ctcaattgca acttgaacac ttgcttttgg atcttcaaat gatcttgaac ggtatcaaca 360
actacaaaaa cccaaaactt actcgtatgt tgacttttaa attttacatg ccaaaaaaag 420
ctactgaact taaacacttg caatgtcttg aagaagaatt gaaaccactt gaagaagttt 480
tgaaccttgc tcaatcaaaa aactttcact tgcgtccacg tgatcttatc tcaaacatca 540
acgttatcgt tttggaactt aaaggttcag aaactacttt tatgtgtgaa tacgctgatg 600
aaactgctac tatcgttgaa tttttgaacc gttggatcac tttttgtcaa tcaatcatct 660
caactttgac ttaaggttta gatggtttta attagcaata 700
<210> 47
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 47
aatccaatga cggcacttct tc 22
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 48
cttgtcgtta aagcctattc 20
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 49
cgtaaccatg taaaagcact tctg 24
<210> 50
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 50
gtaattctaa tgctggtggg 20
<210> 51
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 51
attacgccat ctaaatcaaa c 21
<210> 52
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 52
catcgctgaa gctatcatcg 20
<210> 53
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 53
gatggctgaa gctccaactc 20
<210> 54
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 54
gcatggaaga ggacaaagag 20
<210> 55
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 55
aacctgtggg agggcgaaag 20
<210> 56
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 56
tgggtcgtga atacttcc 18
<210> 57
<211> 500
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
полинуклеотид"
<400> 57
tcagctaacg gagctcaact tgttaaaact gtatcttggt acgataacga aatgtcatac 60
acttcaaacc ttgttcgtac acttgcatac ttcgctaaaa tcgctaaata aggaggaaaa 120
aatgaagaag aaaatcatta gtgccatctt aatgtctaca gtgattcttt cagctgcagc 180
tcctttatca ggcgtttatg catttgtgaa ccaacacctg tgcggctcac acctggtgga 240
agctctctac ctagtgtgcg gggaacgagg cttcttctac acacccaaga cccgccggga 300
ggcagaggac ctgcaggtgg ggcaggtgga gctgggcggg ggccctggtg caggcagcct 360
gcagcccttg gccctggagg ggtccctgca gaagcgtggc attgtggaac aatgctgtac 420
cagcatctgc tccctctacc agctggagaa ctactgcaac taattttccg attttaacgg 480
tataaaaacc agtcttcggg 500
<210> 58
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 58
aaccgctttc agaagaaggg 20
<210> 59
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 59
caccgaatta acacgcatta tgactt 26
<210> 60
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 60
tttcgctggg aaagcacac 19
<210> 61
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтезированный
олигонуклеотид"
<400> 61
gcgtgtccaa gcaatagatg 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУКОАДГЕЗИВНЫЙ МИКРООРГАНИЗМ | 2017 |
|
RU2762940C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ БАКТЕРИИ, СТАБИЛЬНО ЭКСПРЕССИРУЮЩИЕ ИЛ-10 И ИНСУЛИН | 2017 |
|
RU2768027C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| СРЕДСТВА НА ОСНОВЕ КОНСТАНТНОЙ ОБЛАСТИ ИММУНОГЛОБУЛИНА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР | 2008 |
|
RU2729829C2 |
| ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ ЛИЗИН-ПРОТИВОМИКРОБНЫЙ ПЕПТИД (AMP), ЛИЗИНЫ, ВЫДЕЛЕННЫЕ ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ, А ТАКЖЕ ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2803121C2 |
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |
| СПОСОБ ПОЛУЧЕНИЯ ХИМИЧЕСКИХ ПРОДУКТОВ ТОНКОГО СИНТЕЗА С ПОМОЩЬЮ CORYNEBACTERIUM, СЕКРЕТИРУЮЩЕЙ МОДИФИЦИРОВАННЫЕ α-1,6-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2763317C2 |

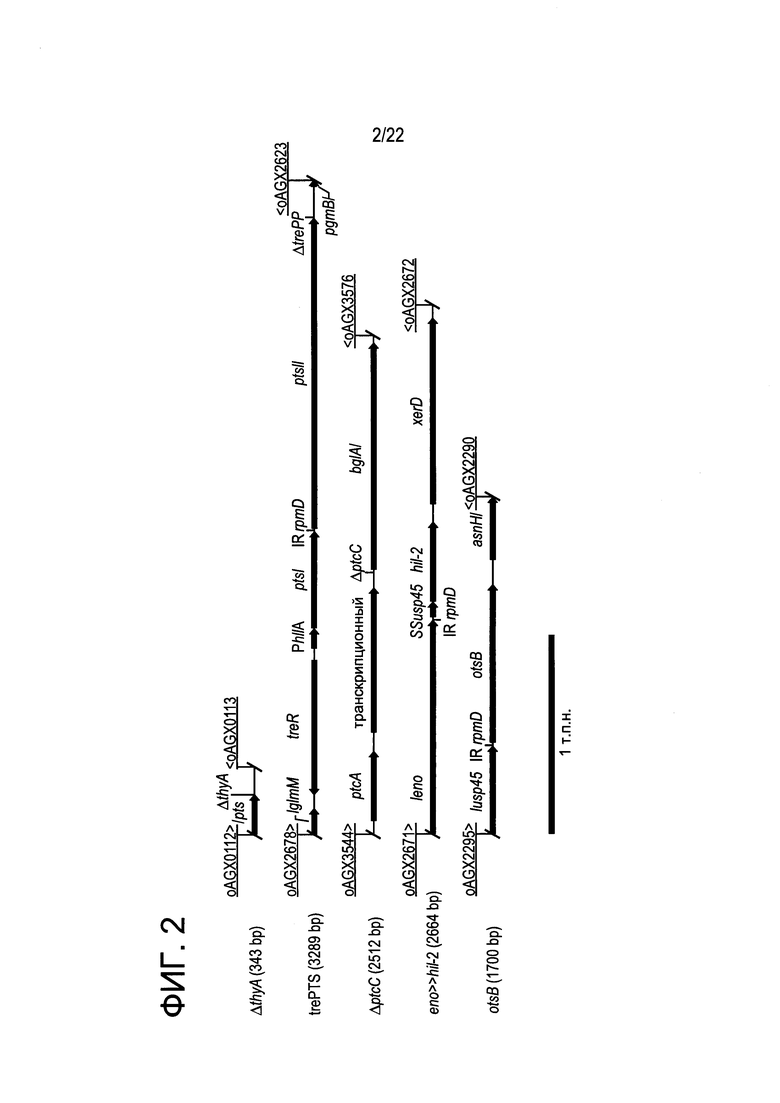
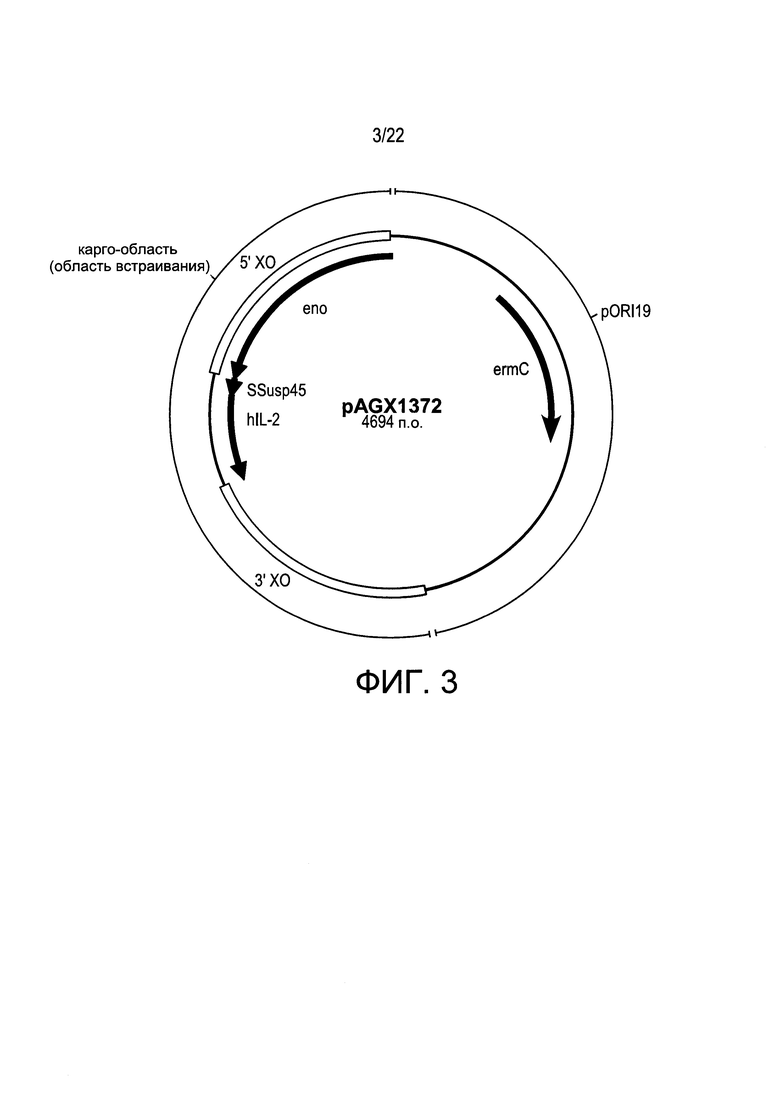
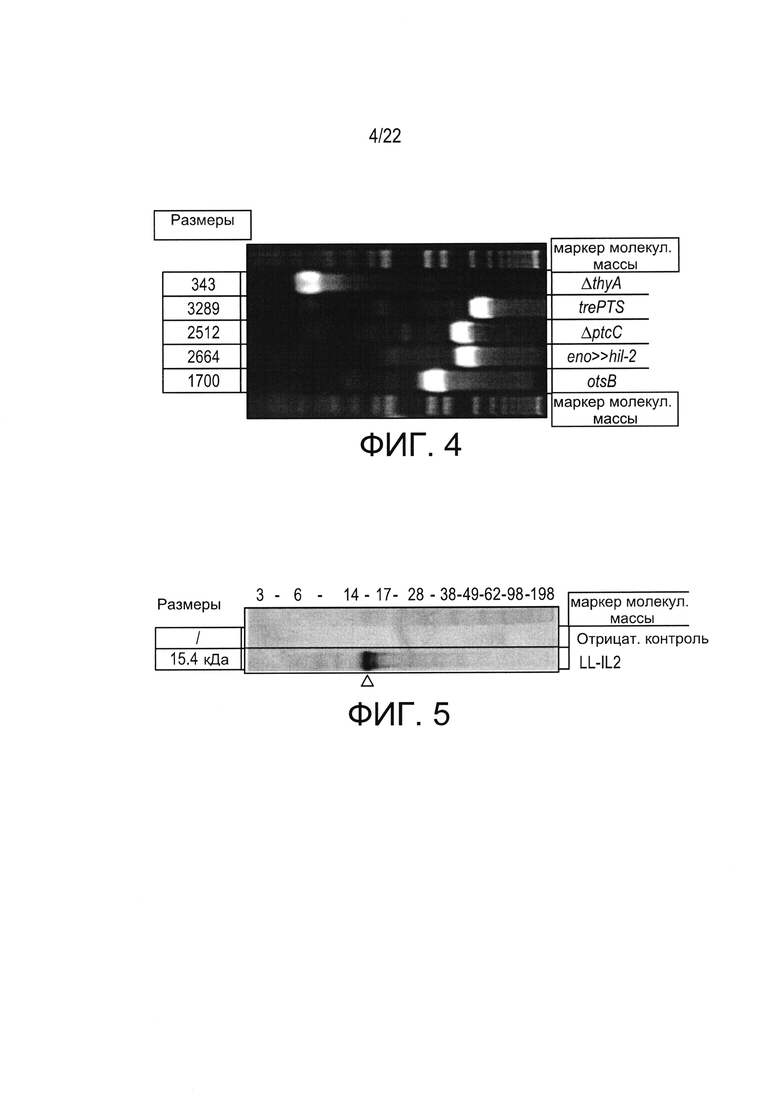


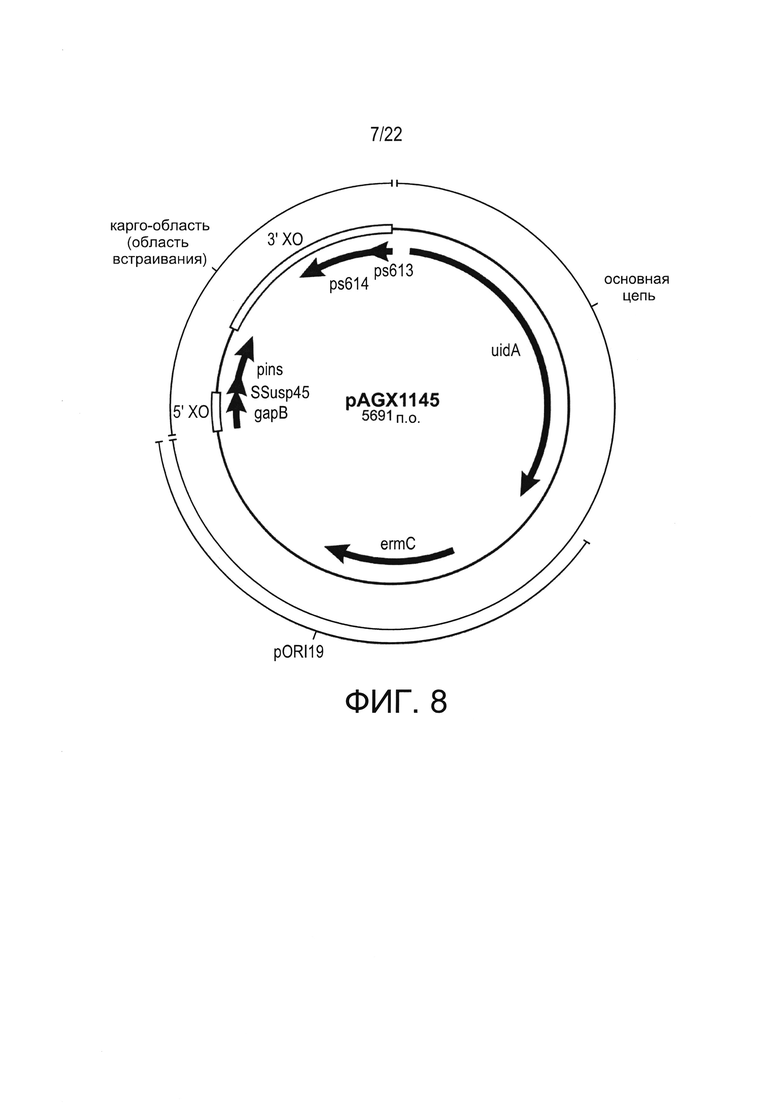

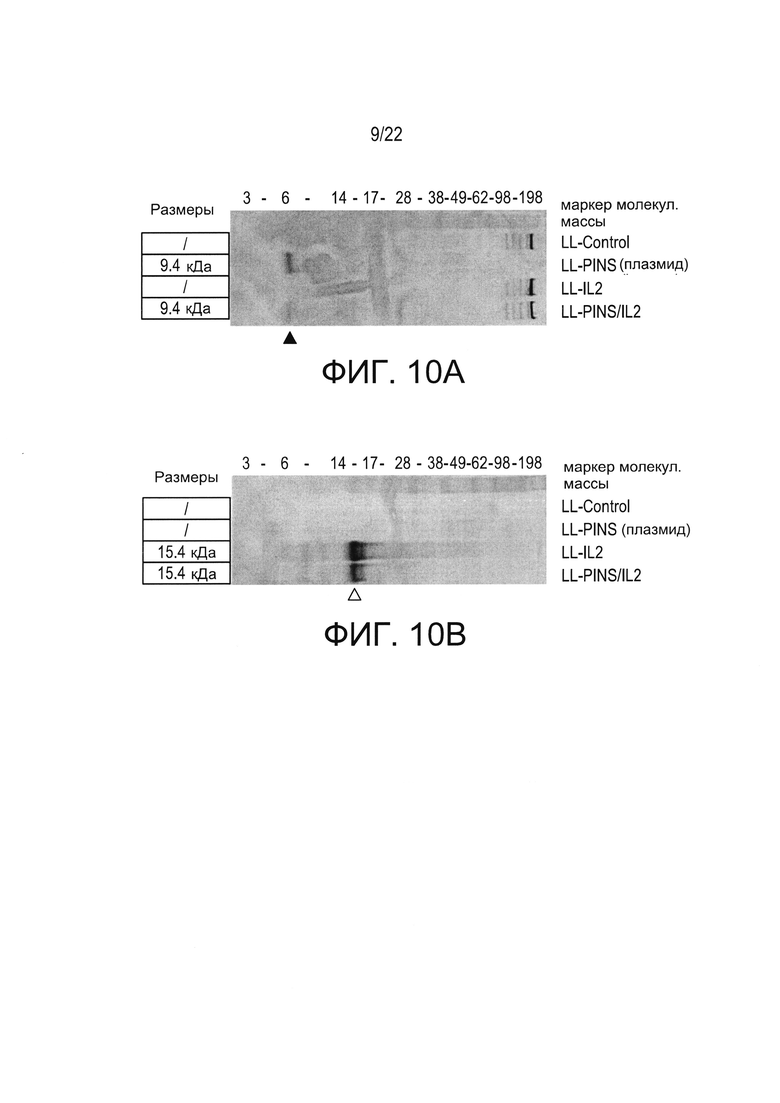
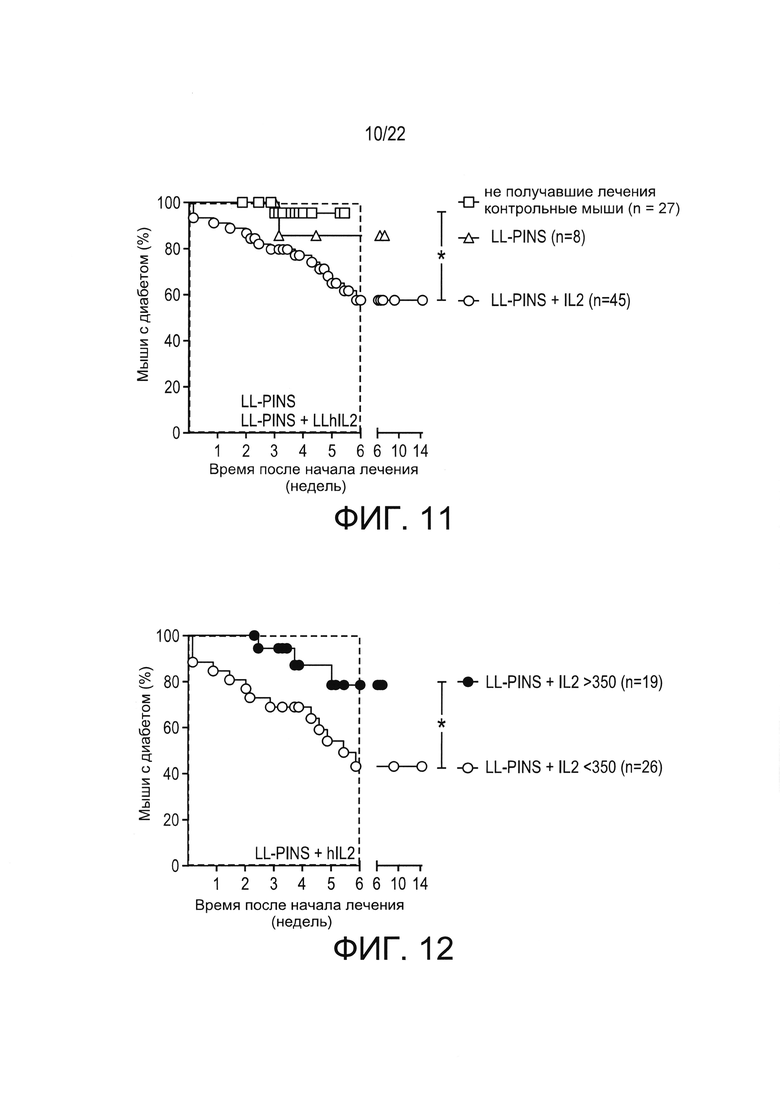
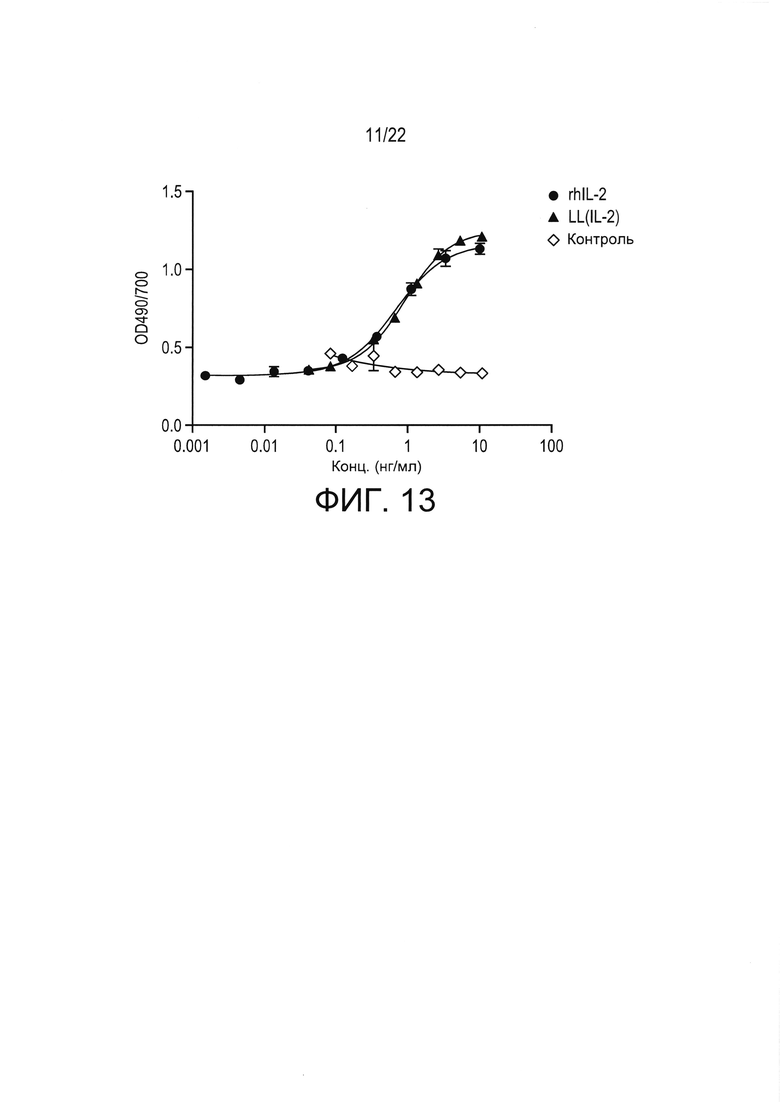
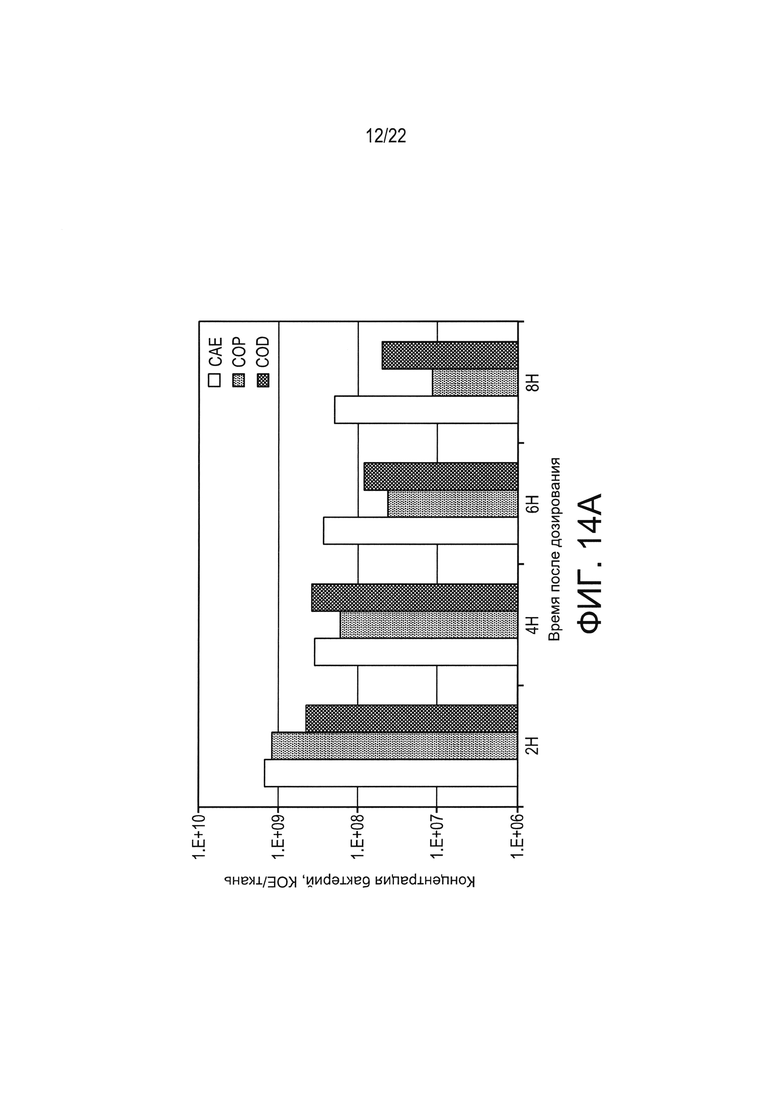
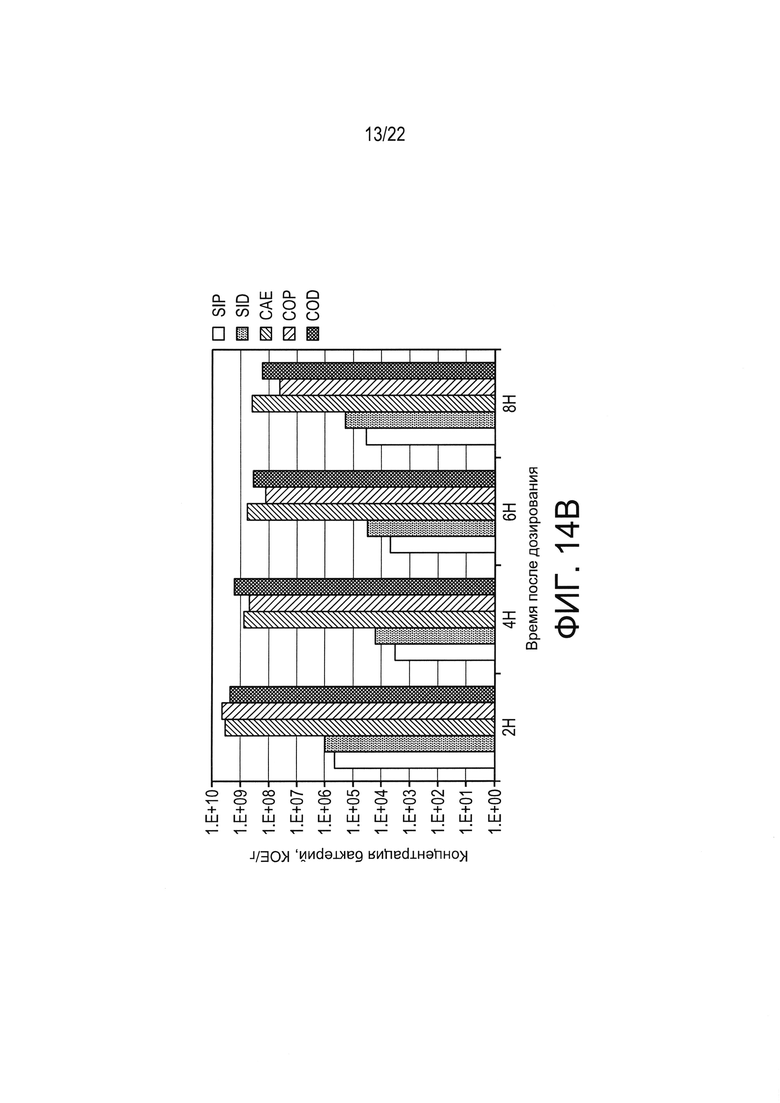
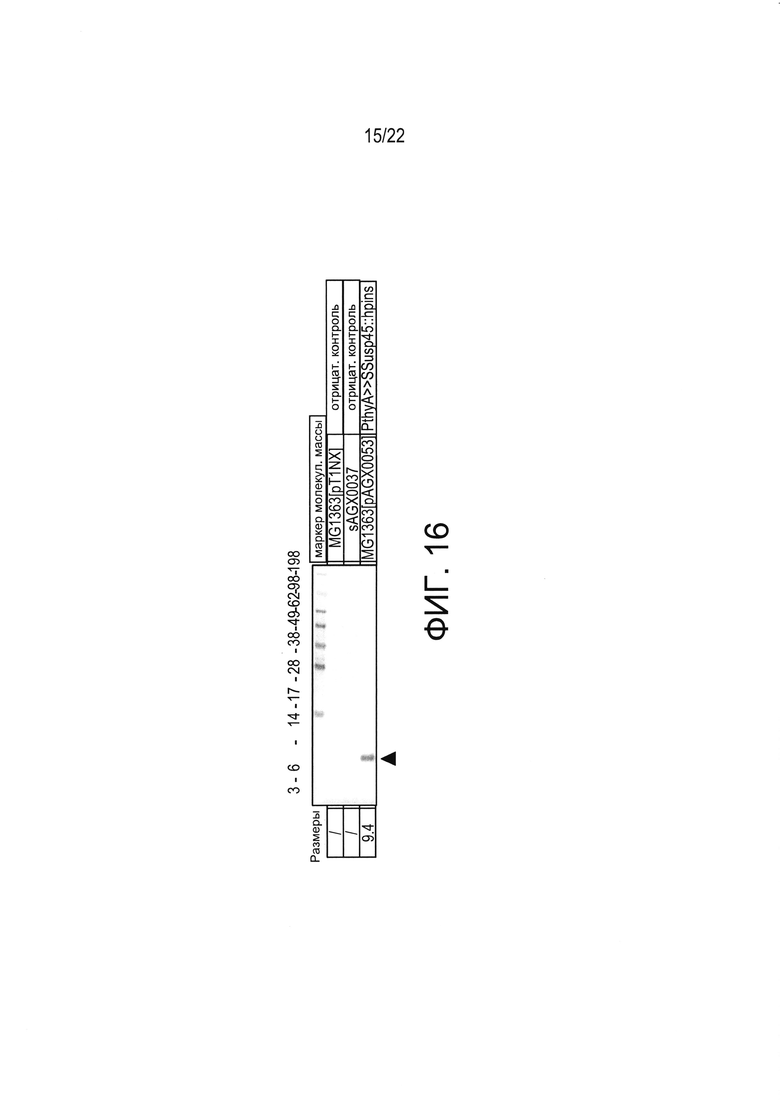
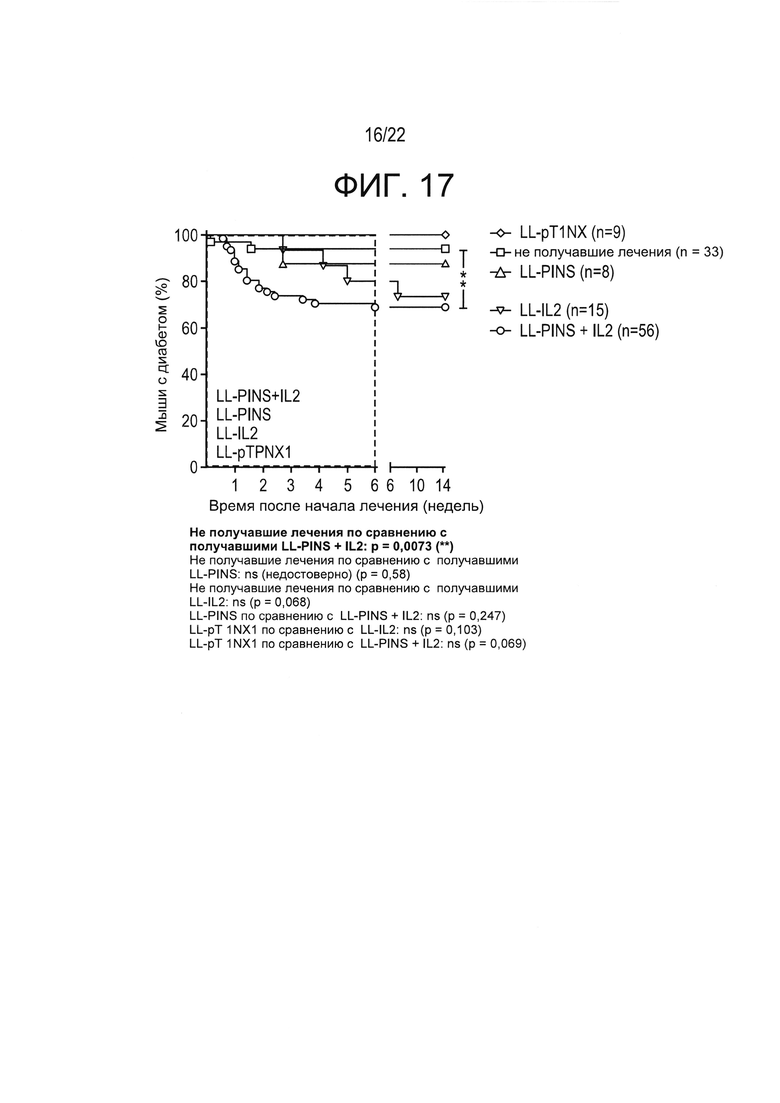
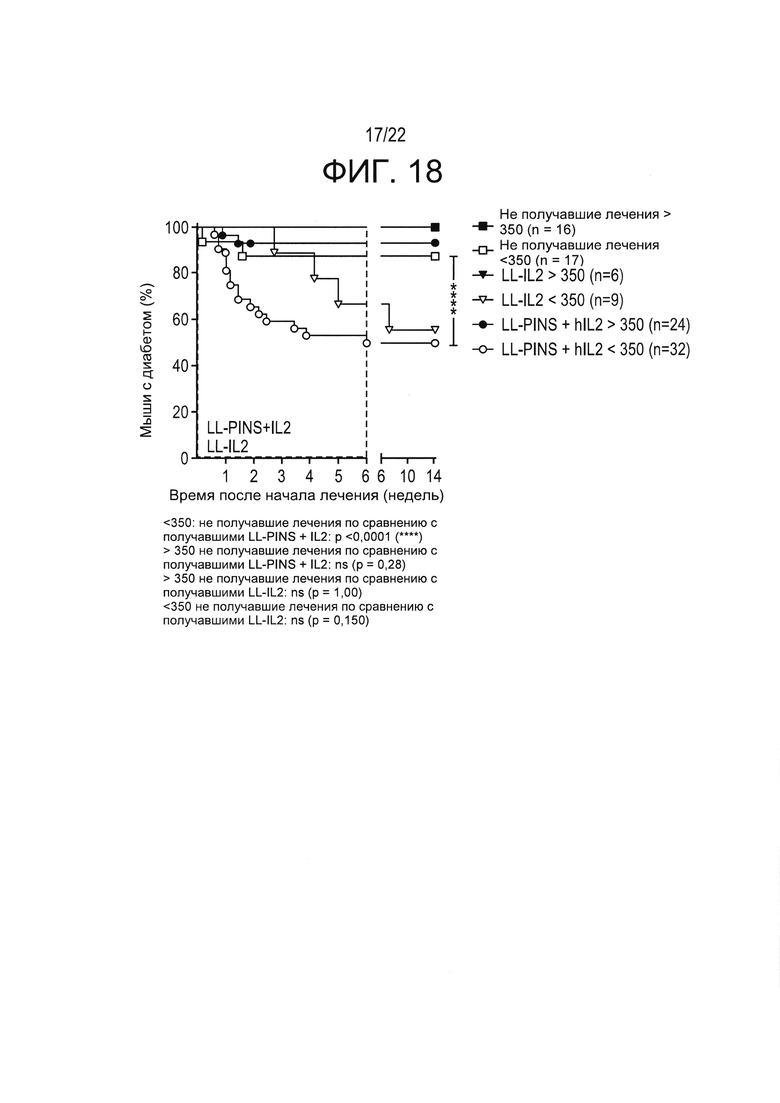
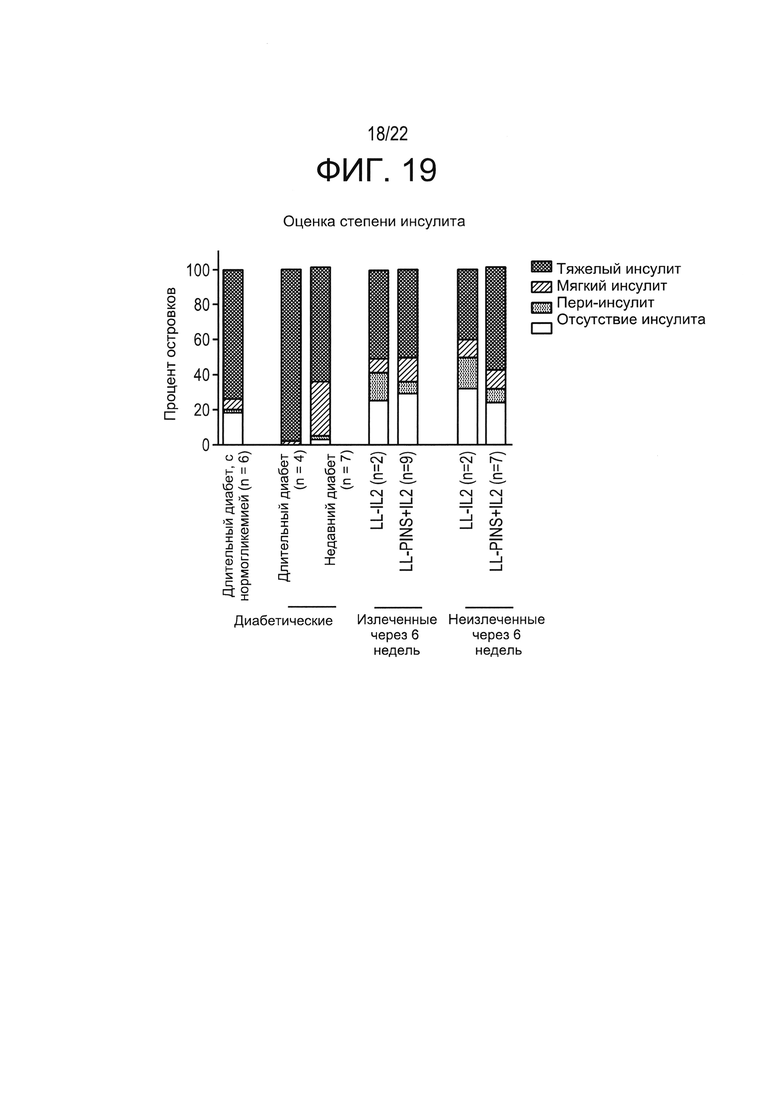
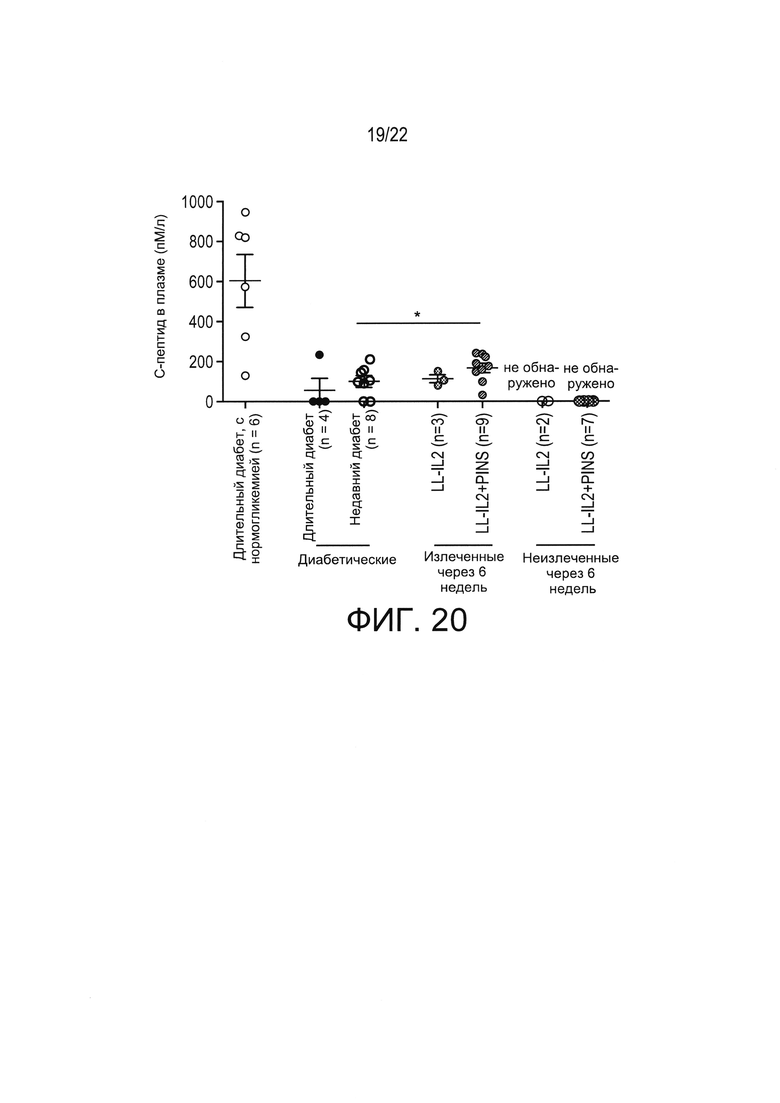
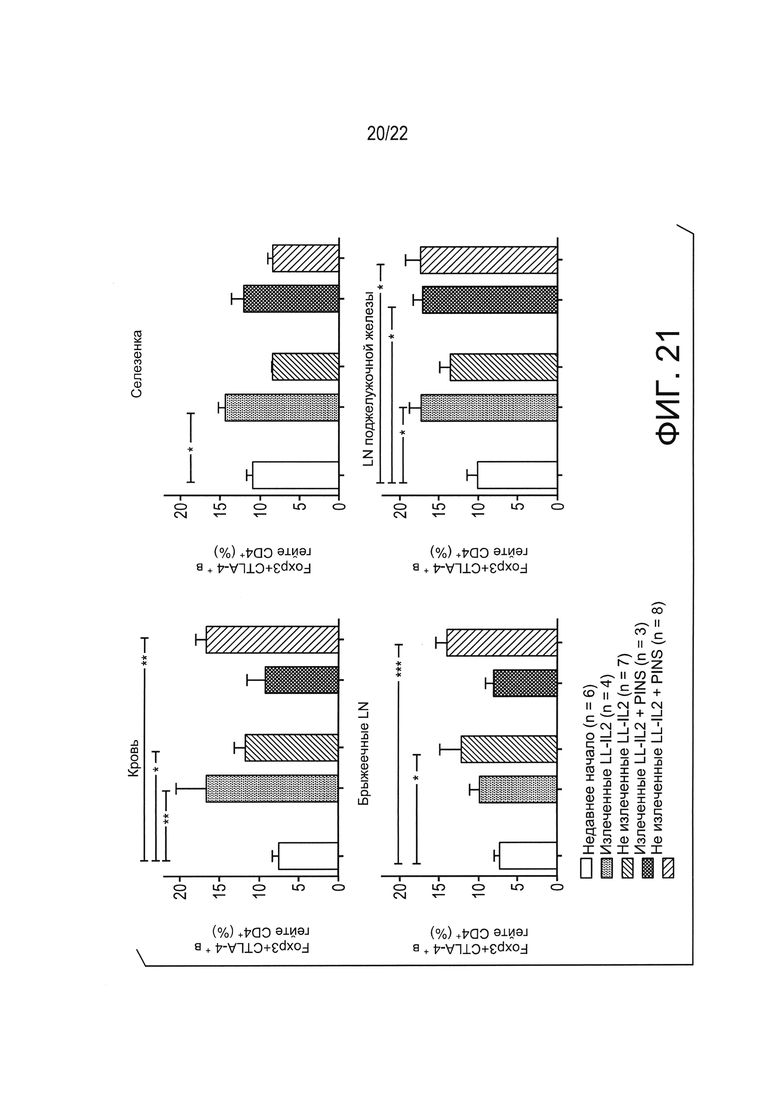
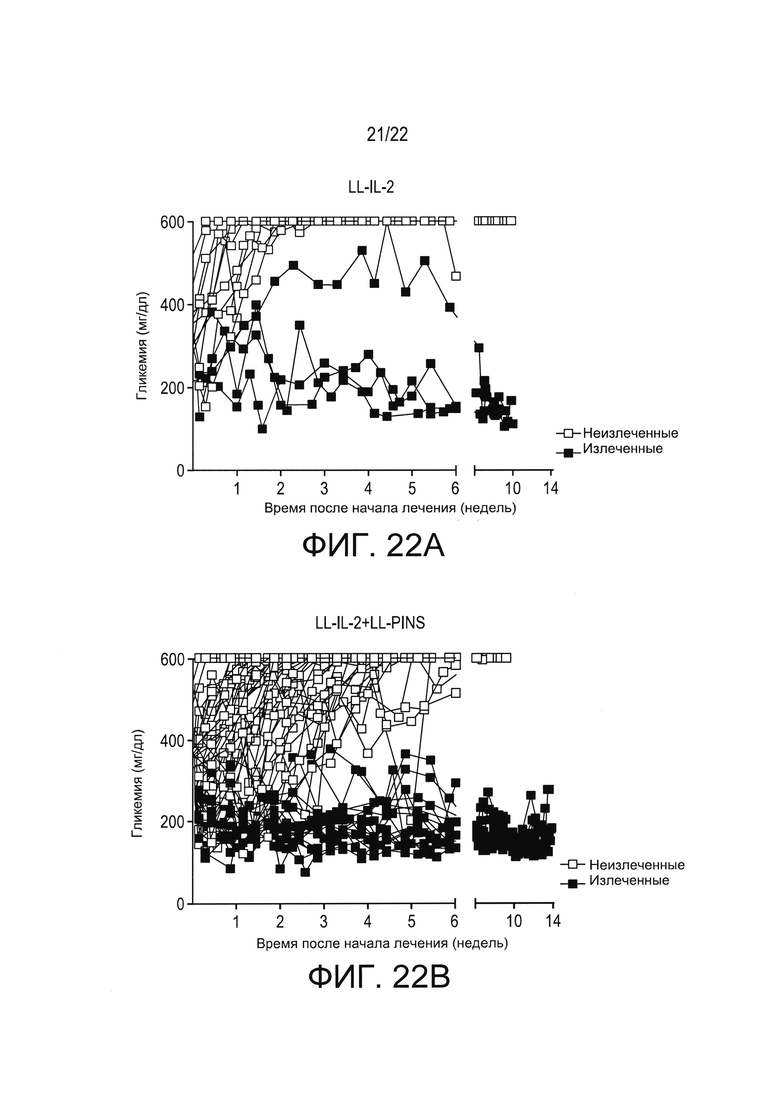
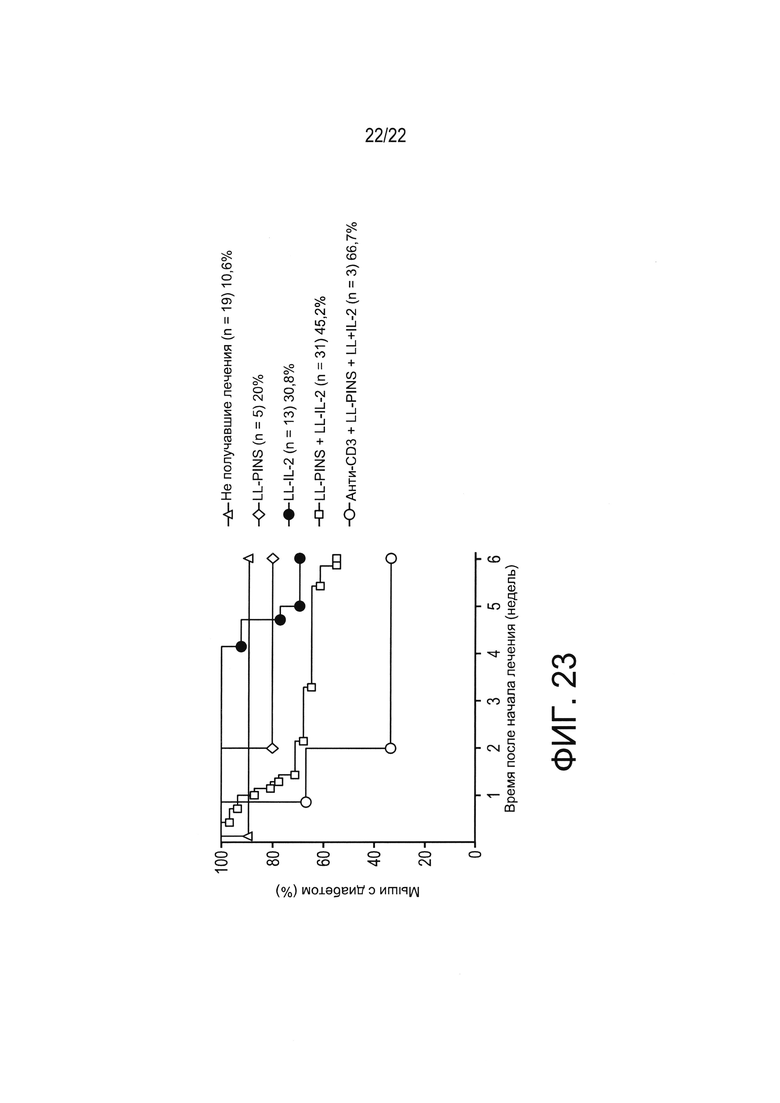
Группа изобретений относится к лечению сахарного диабета 1-го типа (T1D). Предложена молочнокислая бактерия (LAB) для лечения T1D посредством доставки в слизистую оболочку полипептида интерлейкина-2 (IL-2) и полипептида T1D-специфического антигена, где указанная LAB содержит экзогенную нуклеиновую кислоту, кодирующую указанный полипептид IL-2, и экзогенную нуклеиновую кислоту, кодирующую указанный полипептид T1D-специфического антигена. При этом указанная LAB способна продуцировать указанный полипептид IL-2 и указанный полипептид T1D-специфического антигена. При этом указанный полипептид T1D-специфического антигена представляет собой: проинсулин (PINS), декарбоксилазу глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитическую субъединицу связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP), транспортер цинка 8 (ZnT8), хромогранин A, (препро) островковый амилоидный полипептид (ppIAPP), периферин или цитруллинированный глюкозорегулируемый белок (GRP). Предложена также композиция для лечения T1D посредством доставки в слизистую оболочку полипептида IL-2 и полипептида T1D-специфического антигена, содержащая в эффективном количестве (а) первую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую указанный полипептид IL-2 и способную продуцировать указанный полипептид IL-2, и (б) вторую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую указанный полипептид T1D-специфического антигена и способную продуцировать указанный полипептид T1D-специфического антигена. Указанные LAB или композиция используются в составе фармацевтической композиции, а также применяются в способе лечения T1D у нуждающегося в этом субъекта-млекопитающего. Группа изобретений обеспечивает стимуляцию и активацию Treg клеток. 5 н. и 35 з. п. ф-лы, 23 ил., 5 табл., 9 пр.
1. Молочнокислая бактерия (LAB) для лечения сахарного диабета 1-го типа (T1D) посредством доставки в слизистую оболочку полипептида интерлейкина-2 (IL-2) и полипептида T1D-специфического антигена, где указанная LAB содержит экзогенную нуклеиновую кислоту, кодирующую указанный полипептид IL-2, и экзогенную нуклеиновую кислоту, кодирующую указанный полипептид T1D-специфического антигена, при этом указанная LAB способна продуцировать указанный полипептид IL-2 и указанный полипептид T1D-специфического антигена, причём указанный полипептид T1D-специфического антигена представляет собой: проинсулин (PINS), декарбоксилазу глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитическую субъединицу связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP), транспортер цинка 8 (ZnT8), хромогранин A, (препро) островковый амилоидный полипептид (ppIAPP), периферин или цитруллинированный глюкозорегулируемый белок (GRP).
2. LAB по п. 1, отличающаяся тем, что указанная LAB принадлежит к виду Lactococcus, виду Lactobacillus, виду Bifidobacterium, виду Streptococcus или виду Enterococcus.
3. LAB по п. 2, отличающаяся тем, что указанная LAB представляет собой вид Lactococcus.
4. LAB по п. 3, отличающаяся тем, что указанная LAB представляет собой Lactococcus lactis.
5. LAB по любому из пп. 1-4, отличающаяся тем, что указанный полипептид T1D-специфического антигена представляет собой PINS.
6. LAB по любому из пп. 1-5, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид специфического антигена сахарного диабета 1-го типа (T1D), содержит в направлении от 5’ к 3’ промотор gapB (PgapB), gapB и лидер секреции usp45, транскрипционно- и трансляционно-связанные с pins.
7. LAB по любому из пп. 1-6, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид IL-2, включает в себя в направлении от 5’ к 3’ промотор eno (Peno) и лидер секреции usp45, транскрипционно- и трансляционно-связанные с последовательностью, кодирующей полипептид IL-2.
8. LAB по любому из пп. 1-7, отличающаяся тем, что указанная LAB дополнительно содержит по меньшей мере одну из следующих генетических модификаций:
а) инактивацию thyA;
b) инактивацию trePP;
c) инактивацию ptcC;
d) встраивание экзогенного otsB; и
e) экспрессию ptsI и ptsII под контролем конститутивного промотора.
9. LAB по любому из пп. 1-8, отличающаяся тем, что указанная LAB представляет собой Lactoccus lactis и дополнительно содержит следующие генетические модификации:
а) удаление thyA;
b) удаление trePP;
c) нарушение ptcC;
d) встраивание и экспрессию экзогенного otsB; и
e) экспрессию ptsI и ptsII под контролем конститутивного промотора.
10. LAB по любому из пп. 1-9, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая полипептид T1D-специфического антигена, интегрированы в хромосому указанной LAB.
11. LAB по любому из пп. 1-10, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид T1D-специфического антигена, каждая являются частью полицистронной единицы экспрессии.
12. LAB по любому из пп. 1-11, отличающаяся тем, что указанный полипептид IL-2 секретируется.
13. LAB по любому из пп. 1-12, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид IL-2, кодирует вариант полипептида IL-2.
14. LAB по любому из пп. 1-13, отличающаяся тем, что указанная LAB обеспечивает доставку указанного IL-2 и указанного полипептида специфического антигена сахарного диабета 1-го типа (T1D) в слизистую оболочку при введении указанной композиции в организм субъекта-млекопитающего.
15. Композиция для лечения сахарного диабета 1-го типа (T1D) посредством доставки в слизистую оболочку полипептида интерлейкина-2 (IL-2) и полипептида T1D-специфического антигена, содержащая в эффективном количестве:
(а) первую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую указанный полипептид IL-2 и способную продуцировать указанный полипептид IL-2, и
(б) вторую LAB, содержащую экзогенную нуклеиновую кислоту, кодирующую указанный полипептид T1D-специфического антигена и способную продуцировать указанный полипептид T1D-специфического антигена, причём указанный полипептид T1D-специфического антигена представляет собой: проинсулин (PINS), декарбоксилазу глутаминовой кислоты (GAD65), инсулинома-ассоциированный белок 2 (IA-2), каталитическую субъединицу связанного белка островково-специфической глюкозо-6-фосфатазы (IGRP), транспортер цинка 8 (ZnT8), хромогранин A, (препро) островковый амилоидный полипептид (ppIAPP), периферин или цитруллинированный глюкозорегулируемый белок (GRP).
16. Композиция по п. 15, отличающаяся тем, что указанная LAB принадлежит к виду Lactococcus, виду Lactobacillus, виду Bifidobacterium, виду Streptococcus или виду Enterococcus.
17. Композиция по п. 16, отличающаяся тем, что указанная LAB представляет собой вид Lactococcus.
18. Композиция по п. 17, отличающаяся тем, что указанная LAB представляет собой Lactococcus lactis.
19. Композиция по любому из пп. 15-18, отличающаяся тем, что указанный полипептид T1D-специфического антигена представляет собой PINS.
20. Композиция по любому из пп. 15-19, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид специфического антигена сахарного диабета 1-го типа (T1D), содержит в направлении от 5’ к 3’ промотор gapB (PgapB), gapB и лидер секреции usp45, транскрипционно- и трансляционно-связанные с pins.
21. Композиция по любому из пп. 15-20, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, включает в себя в направлении от 5’ к 3’ промотор eno (Peno) и лидер секреции usp45, транскрипционно- и трансляционно-связанные с последовательностью, кодирующей полипептид IL-2.
22. Композиция по любому из пп. 15-21, отличающаяся тем, что указанная LAB дополнительно содержит по меньшей мере одну из следующих генетических модификаций:
а) инактивацию thyA;
b) инактивацию trePP;
c) инактивацию ptcC;
d) встраивание экзогенного otsB; и
e) экспрессию ptsI и ptsII под контролем конститутивного промотора.
23. Композиция по любому из пп. 15-22, отличающаяся тем, что указанная LAB представляет собой Lactoccus lactis и дополнительно содержит следующие генетические модификации:
а) удаление thyA;
b) удаление trePP;
c) нарушение ptcC;
d) встраивание и экспрессию экзогенного otsB; и
e) экспрессию ptsI и ptsII под контролем конститутивного промотора.
24. Композиция по любому из пп. 15-23, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая полипептид T1D-специфического антигена, интегрированы в хромосому указанной LAB.
25. Композиция по любому из пп. 15-24, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая полипептид IL-2, и указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид T1D-специфического антигена, каждая являются частью полицистронной единицы экспрессии.
26. Композиция по любому из пп. 15-25, отличающаяся тем, что указанный полипептид IL-2 секретируется.
27. Композиция по любому из пп. 15-26, отличающаяся тем, что указанная экзогенная нуклеиновая кислота, кодирующая указанный полипептид IL-2, кодирует вариант полипептида IL-2.
28. Композиция по любому из пп. 15-27, отличающаяся тем, что указанная LAB обеспечивает доставку указанного полипептида IL-2 и указанного полипептида специфического антигена сахарного диабета 1-го типа (T1D) в слизистую оболочку, при введении указанной композиции в организм субъекта-млекопитающего.
29. Фармацевтическая композиция для лечения сахарного диабета 1-го типа (T1D), содержащая (i) эффективное количество LAB по любому из пп. 1-14 и фармацевтически приемлемый носитель или (ii) эффективное количество композиции по любому из пп. 15-28 и фармацевтически приемлемый носитель.
30. Применение LAB по любому из пп. 1-14, композиции по любому из пп.15-28 или фармацевтической композиции по п. 29 для лечения сахарного диабета 1-го типа (T1D) у субъекта-млекопитающего, нуждающегося в этом.
31. Способ лечения сахарного диабета 1-го типа (T1D) у нуждающегося в этом субъекта-млекопитающего, включающий:
пероральное введение указанному субъекту-млекопитающему терапевтически эффективного количества LAB по любому из пп. 1-14, композиции по любому из пп.15-28 или фармацевтической композиции по п. 29.
32. Способ по п. 31, отличающийся тем, что указанному субъекту не вводят антитело против CD3.
33. Способ по п. 31, дополнительно включающий введение указанному субъекту антитела против CD3.
34. Способ по п. 33, отличающийся тем, что указанное антитело против CD3 вводят указанному субъекту одновременно с указанной LAB, указанной композицией или указанной фармацевтической композицией.
35. Способ по любому из пп. 31-34, отличающийся тем, что указанный субъект имеет концентрацию C-пептида в крови или моче, указывающую на остаточную функцию бета-клеток.
36. Способ по любому из пп. 31-35, отличающийся тем, что указанным субъектом является пациент, представляющий собой человека, имеющий концентрацию С-пептида в крови натощак от приблизительно 0,2 нмоль/л до приблизительно 1 нмоль/л или имеющий стимулированную концентрацию C-пептида в крови от приблизительно 0,5 нмоль/л до приблизительно 4 нмоль/л.
37. Способ по любому из пп. 31-36, отличающийся тем, что у указанного субъекта был диагностирован T1D в течение предшествующих 12 месяцев до введения указанной LAB, указанной композиции или указанной фармацевтической композиции.
38. Способ по любому из пп. 31-37, отличающийся тем, что указанную LAB, указанную композицию или указанную фармацевтическую композицию вводят в виде единичной лекарственной формы, содержащей от приблизительно 1 × 104 до приблизительно 1 × 1012; от приблизительно 1 × 106 до приблизительно 1 × 1012 или от приблизительно 1 × 109 до приблизительно 1 × 1012 колониеобразующих единиц (КОЕ).
39. Способ по п. 38, отличающийся тем, что указанная единичная лекарственная форма представляет собой капсулу, таблетку, гранулу, суппозиторий или дозированный аэрозоль.
40. Способ по любому из пп. 31-39, отличающийся тем, что указанная LAB, указанная композиция или указанная фармацевтическая композиция находится в форме сухого порошка или его спрессованном варианте.
| TAKIISHI Т | |||
| ET AL | |||
| Reversal of autoimmune diabetes by restoration of antigen-specific tolerance using genetically modified Lactococcus lactis in mice | |||
| J Clin Invest | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| STEIDLER L | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

