ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области биохимической инженерии. Более конкретно, настоящее изобретение относится к генетически модифицированным клеткам-хозяевам и способам получения в них полипептидов.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Терапевтические полипептиды и белки могут экспрессироваться в различных клетках-хозяевах, включая бактериальные клетки, клетки E.coli, клетки грибов или дрожжевые клетки, клетки микроорганизмов, клетки насекомых и клетки млекопитающих. Грибковые хозяева, такие как метилотрофные дрожжи Pichia pastoris, имеют определенные преимущества в отношении терапевтических белков, например, они не секретируют большого количества эндогенных белков, они имеют сильный индуцируемый промотор, их можно выращивать на определенной химической среде, и они могут продуцировать рекомбинантные белки с высоким титром (Cregg et al., Mol. Biotech. 16:23-52 (2000)). И дрожжи и нитевидные грибы успешно используются для получения рекомбинантных белков, как внутриклеточно, так и в секретируемом виде (Cereghino, J. L. and J. M. Cregg 2000 FEMS Microbiology Reviews 24(1): 45-66; Harkki, A., et al. 1989 Bio-Technology 7(6): 596; Berka, R. M., et al. 1992 Abstr. Papers Amer. Chem.Soc. 203: 121-BIOT; Svetina, M., et al. 2000 J. Biotechnol. 76(23): 245-251). S. cerevisiae представляют собой замечательные клетки-хозяева для экспрессии рекомбинантного сывороточного альбумина человека (HSA). Однако экспрессия других терапевтических полипептидов, включая полипептиды, генетически слитые с HSA, сталкивается с техническим препятствием в виде низкого титра рекомбинантных белков. Поэтому существует потребность в клетках-хозяевах, в частности штаммах S. cerevisiae , которые способны продуцировать гетерологичные пептиды, полипептиды и/или белки с высоким титром рекомбинантного белка.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте настоящего изобретения предлагаются генетически модифицированные клетки-хозяева, содержащие по меньшей мере один выделенный полинуклеотид, кодирующий протеазу Killer Expression (Kex2p) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Kex2p, и по меньшей мере один выделенный полинуклеотид, кодирующий протеин-дисульфидизомеразу (Pdi1) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Pdi. Также в настоящем изобретении предложены генетически модифицированные клетки-хозяева, содержащие по меньшей мере один выделенный полинуклеотид, кодирующий протеазу Killer Expression (Kex2p) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Kex2p, по меньшей мере один выделенный полинуклеотид, кодирующий протеин-дисульфидизомеразу (Pdi1) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Pdi, и по меньшей мере один выделенный полинуклеотид, кодирующий оксидоредуктин эндоплазматического ретикулума (Ero1) или его фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Ero1.
В другом аспекте настоящее изобретение относится к генетически модифицированным клеткам-хозяевам, которые экспрессируют или сверхэкспрессируют по меньшей мере один генный продукт по меньшей мере одного выделенного полинуклеотида, кодирующего белок или его фрагмент и/или его вариант, который имеет по меньшей мере одну функциональную активность указанного белка, выбранного из: Kex2p, Pdi1 или Ero1, когда указанные генетически модифицированные клетки-хозяева выращиваются в культуре. Другой аспект настоящего изобретения предлагает генетически модифицированные клетки-хозяева, которые сверхэкспрессируют по меньшей мере два белка или их фрагмента и/или варианта, которые имеют по меньшей мере одну функциональную активность указанных по меньшей мере двух белков, выбранных из: Kex2p, Pdi1 или Ero1, когда указанные генетически модифицированные клетки-хозяева выращиваются в культуре, по сравнению с клеткой-хозяином дикого типа, причем указанная клетка-хозяин дикого типа относится к тому же виду и выращивается в таких же условиях культивирования, но не сверхэкспрессирует по меньшей мере два генных продукта, выбранных из Kex2p, Pdi1 и Ero1. Клетки-хозяева могут быть прокариотическими или эукариотическими. Примеры клеток-хозяев могут включать в себя, но не ограничиваются ими: HeLa, СНО, COS, HEK293, THPI, дрожжи и клетки насекомых. В конкретных вариантах осуществления изобретения клеткой млекопитающего является клетка хомячка, человека или мыши. В конкретном варианте осуществления изобретения клеткой является клеточная линия CHO, клеточная линия НЕК 293 или клеточная линия ВНК.
Также в настоящем документе предложены способы получения рекомбинантного полипептида, включающие культивирование клетки-хозяина по настоящему изобретению. В другом аспекте настоящее изобретение относится к рекомбинантным полипептидам, полученным способами по настоящему изобретению. Также в настоящем документе предложены фармацевтические композиции, содержащие рекомбинантные полипептиды, изготовленные способами по настоящему изобретению. В другом аспекте настоящего изобретения предложены способы лечения пациента, нуждающегося в этом, включающие введение терапевтически эффективного количества фармацевтической композиции по настоящему изобретению.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1. Создание Предварительного главного клеточного банка штаммов-продуцентов альбиглютида. Стадии от ПЦР-продукта экспрессионной кассеты KEX2-KanMX до штамма-продуцента BXP10KEX2PDIERO1 представляют собой последовательное встраивание экспрессионных кассет для создания штамма-хозяина Процесса IV, BXP10_KEX2_PDI_ERO1. После трансформации плазмидой pCID3610 был отобран финальный штамм-продуцент и использован для изготовления предварительного главного клеточного банка (РСВ).
Фигура 2. Анализ методом Саузерн-блота PDI1 и KEX2 в штаммах-хозяевах. Эндогенные гены KEX2 и PDI1 расположены на хромосомах XIV и III, соответственно, в единственной копии (дикий тип). Планируемый участок встраивания на хромосоме XII изображен ниже дикого типа. Зонды для обнаружения каждого гена показаны в виде сплошного прямоугольника.
Фигура 3. Анализ методом Вестерн-блоттинга Pdi1 и Kex2p из штаммов-хозяев. Образцы нанесены в следующем порядке, дорожки 1-5: 5 клонов штамма BXP10-KEX2-PDI1; PDI: BXP10, сверхэкспрессирующий Pdi1; KEX2: BXP10, сверхэкспрессирующий Kex2p; и BXP10: штамм-хозяин в качестве контроля. Наносили равное количество белка.
Фигура 4. Электрофорез в денатурирующем полиакриламидном геле (электрофорез в ДСН-ПААГ) 12 образцов супернатанта после роста в планшете при встряхивании. Дорожки в геле: L: предварительно окрашенный белковый маркер SeeBlue2 (Invitrogen); RS. Эталон белка pCID3610; 1-12: 12 субклонов, экспрессирующих pCID3610.
Фигура 5. Анализ титра (А) и качества (В) белка pCID3610, получаемого ферментацией в DasGip. Получаемый титр в супернатанте и уровень 6-AA (в %) для белка pCID3610 сравнивали с BXP10-KEX2-PDI1 в качестве контроля, который представляет собой BXP10, сверхэкспрессирующий Kex2p и Pdi1.
Фигура 6. Кривые роста, полученные для клеток из ампулы Исследовательского клеточного банка.
Фигура 7. Кривые роста, полученные для клеток из Предварительного главного клеточного банка.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
«Клетка-хозяин (клетки-хозяева)» в контексте настоящего изобретения относится к клетке, в которую была введена (например, путем трансформации, инфекции или трансфекции) или в которую можно ввести (например, путем трансформации, инфекции или трансфекции) выделенную полинуклеотидную последовательность. Клетки-хозяева по настоящему изобретению могут включать, но не ограничиваются ими, бактериальные клетки, клетки грибов, дрожжевые клетки, клетки микроорганизмов, клетки насекомых и клетки млекопитающих. Клетки-хозяева по настоящему изобретению, имеющие свое происхождение от дрожжей и/или нитевидных грибов, могут включать, но не ограничиваются ими, следующие семейства, роды и виды: Pichia pastoris, Pichia finlandica, Pichia trehalophila, Pichia koclamae, Pichia membranaefaciens, Pichia methanolica, Pichia minuta (Ogataea minuta, Pichia lindneri), Pichia opuntiae, Pichia thermotolerans, Pichi salictaria, Pichia guercum, Pichia pijperi, Pichia stiptis, Pichia sp., Saccharomyces castelii, Saccharomyces cerevisiae, Saccharomyces kluyveri, Saccharomyces sp., Schizosaccharomyces pombe, Schizosaccharomyces japonicus, Schizosaccharomyces octosporus, Schizosaccharomyces cryophilus, Schizosaccharomyces sp., Hansenula polymorpha, Kluyveromyces sp., Kluyveromyces lactis, Candida albicans, Candida sp., Aspergillus fumigatus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Trichoderma reesei, Chrysosporium lucknowense, Fusarium sp., Fusarium gramineum, Fusarium venenatum, Physcomitrella patens, Yarrowia lipolytica, Arxula adeninivorans, Schwanniomyces occidentalis и Neurospora crassa.
«Трансформированный» в данной области представляет собой направленное изменение генома или эписомы организма путем введения внешней ДНК или РНК или любое другое стабильное введение внешней ДНК или РНК.
«Трансфицированный» в данной области представляет собой введение внешней ДНК или РНК в микроорганизм, включая, но не ограничиваясь ими, рекомбинантную ДНК или РНК.
«Идентичность» в данной области представляет собой связь между двумя или несколькими полипептидными последовательностями или двумя или несколькими полинуклеотидными последовательностями (в зависимости от обстоятельств), определенную путем сравнения последовательностей. В данной области «идентичность» также означает степень родства по последовательности между полипептидными или полинуклеотидными последовательностями (в зависимости от обстоятельств), определенную путем сравнения цепей этих последовательностей. «Идентичность» может быть легко вычислена известными способами, включающими, но не ограниченными ими, описанные в (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; а также Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073 (1988)). Способы определения идентичности разработаны, чтобы достигнуть наибольшего совпадения между исследуемыми последовательностями. Более того, способы определения идентичности входят в общедоступные компьютерные программы. Компьютерные программные способы определения идентичности между двумя последовательностями включают, но не ограничиваются ими, программный пакет GCG (Devereux, J., et al, Nucleic Acids Research 12(1): 387 (1984)), BLASTP, BLASTN и FASTA (Altschul, S.F. et al., J. Molec. Biol. 215: 403-410 (1990)). Программа BLAST X общедоступна в NCBI и других источниках (BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215: 403-410 (1990)). Также для определения идентичности может использоваться хорошо известный алгоритм Смита-Уотермана.
Параметры для сравнения полипептидных последовательностей включают следующие:
Алгоритм: Needleman and Wunsch, J. Mol Biol. 48: 443-453 (1970); Матрица сравнения: BLOSSUM62 от Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89: 10915-10919 (1992)
Штраф за разрыв: 12
Штраф за длину разрыва: 4
Программой с этими параметрами является общедоступная программа «gap» от Genetics Computer Group, Madison WI. Вышеуказанные параметры является параметрами «по умолчанию» для сравнения пептидов (вместе с отсутствием штрафа за окончание разрывов).
Параметры для сравнения полинуклеотидных последовательностей включают следующие: алгоритм: Needleman and Wunsch, J. Mol Biol. 48: 443-453 (1970)
Матрица сравнения: совпадения = +10, несовпадения = 0
Штраф за разрыв: 50
Штраф за длину разрыва 3
Доступны в программе «gap» от Genetics Computer Group, Madison WI. Они является параметрами «по умолчанию» для сравнения нуклеиновых кислот.
Смысл «идентичности» для полинуклеотидов и полипептидов, в зависимости от обстоятельств, приведен в пп.(1) и (2) ниже.
(1) Полинуклеотидные варианты осуществления дополнительно включают выделенный полинуклеотид, содержащий полинуклеотидную последовательность, имеющую по меньшей мере 50, 60, 70, 80, 85, 90, 95, 97 или 100% идентичности с эталонной последовательностью, например, SEQ ID NO: 3, причем указанная полинуклеотидная последовательность может быть идентична эталонной последовательности SEQ ID NO: 3 или может включать в себя до некоторого целого числа нуклеотидных изменений по сравнению с эталонной последовательностью, где указанные изменения выбраны из группы, состоящей по меньшей мере из одной нуклеотидной делеции, замены, включая транзицию и трансверсию, или вставки, и где указанные изменения могут происходить в 5'- или 3'-концевых позициях эталонной нуклеотидной последовательности или в любом месте между этими концевыми позициями, расположенные либо индивидуально среди нуклеотидов в эталонной последовательности, либо в одной или нескольких непрерывных группах в эталонной последовательности, и где указанное число нуклеотидных изменений определяется умножением общего числа нуклеотидов в SEQ ID NO:3 на целое число, определяющее процент идентичности, деленное на 100, а затем вычитанием этого результата из указанного общего числа нуклеотидов в SEQ ID NO: 3, или:
nn ≤ xn - (xn • y),
где nn представляет собой число нуклеотидных изменений, xn является общим числом нуклеотидов в SEQ ID NO:3, y является 0,95 для 95%, 0,97 для 97% или 1,00 для 100%, а • является символом для оператора умножения, и где любой нецелочисленный результат xn и y округляется до ближайшего целого числа до вычитания его из xn. Изменения полинуклеотидной последовательности, кодирующей полипептид, может создать нонсенс, миссенс или мутации со сдвигом рамки в этой кодирующей последовательности и тем самым изменять полипептид, кодируемый полинуклеотидом после таких изменений.
(2) Полипептидные варианты осуществления дополнительно включают выделенный полипептид, содержащий полипептид, имеющий по меньшей мере 50, 60, 70, 80, 85, 90, 95, 97 или 100% идентичности с эталонной последовательностью полипептида, например, SEQ ID NO: 1, причем указанная полипептидная последовательность может быть идентична эталонной последовательности или может включать в себя до некоторого целого числа аминокислотных изменений по сравнению с эталонной последовательностью, где указанные изменения выбраны из группы, состоящей по меньшей мере из одной аминокислотной делеции, замены, включая консервативную и неконсервативную замену, или вставки, и где указанные изменения могут происходить в амино- или карбокси-концевых позициях эталонной полипептидной последовательности или в любом месте между этими концевыми позициями, расположенные либо индивидуально среди аминокислот в эталонной последовательности, либо в одной или нескольких непрерывных группах в эталонной последовательности, и где указанное число аминокислотных изменений определяется умножением общего числа аминокислот на целое число, определяющее процент идентичности, деленное на 100, а затем вычитанием этого результата из указанного общего числа аминокислот, или:
nа ≤ xa - (xa • y),
где nа представляет собой число аминокислотных изменений, xа является общим числом аминокислот в последовательности, y является 0,95 для 95%, 0,97 для 97% или 1,00 для 100%, а • является символом для оператора умножения, и где любой нецелочисленный результат xа и y округляется до ближайшего целого числа до вычитания его из xа.
«Выделенный» означает измененный «руками человека» из своего природного состояния, то есть, если это происходит в природе, измененный или удаленный из своей исходной окружающей среды, или оба варианта. Например, полинуклеотид или полипептид, естественным образом присутствующий в живом организме, не является «выделенным», но тот же самый полинуклеотид или полипептид, отделенный от сопутствующих в своем природном состоянии материалов, является «выделенным», включая, но не ограничиваясь этим, когда такой полинуклеотид или полипептид вводят обратно в клетку.
«Выделенными» или «по существу чистыми» нуклеиновой кислотой или полинуклеотидом (например, РНК, ДНК или смешанный полимер) являются те, которые по существу отделены от других клеточных компонентов, естественным образом сопровождающих нативный полинуклеотид в его природной клетке-хозяине, например, рибосомы, полимеразы и геномные последовательности, с которыми он ассоциирован в природе. Термин охватывает нуклеиновую кислоту или полинуклеотид, которые (1) были удалены из своей природной среды, (2) не ассоциированы с целым полинуклеотидом или его участком, в котором «выделенный полинуклеотид» встречается в природе, (3) функционально связаны с полинуклеотидом, с которым они не связаны в природе, или (4) не встречаются в природе. Термин «выделенный» или «по существу чистый» также может использоваться по отношению к рекомбинантной или клонированной выделенной ДНК, химически синтезированным аналогам полинуклеотидов или полинуклеотидным аналогам, биологически синтезированным в гетерологичных системах.
Однако термин «выделенный» не требует обязательно, чтобы таким образом описанные нуклеиновая кислота или полинуклеотид сами по себе были физически удалены из своего природного окружения. Например, эндогенная последовательность нуклеиновой кислоты в геноме организма считается «выделенной» в данном описании, если гетерологичная последовательность примыкает к эндогенной последовательности нуклеиновой кислоты таким образом, что изменяется экспрессия этой эндогенной последовательности нуклеиновой кислоты, например, увеличивается, уменьшается или прекращается. В этом контексте гетерологичной последовательностью является последовательность, которая в природе не примыкает к эндогенной последовательности нуклеиновой кислоты, вне зависимости от того, является ли сама гетерологичная последовательность эндогенной (полученной из той же клетки-хозяина или ее потомства) или экзогенной (полученной из другой клетки-хозяина или ее потомства). В качестве примера, промоторная последовательность может замещать (например, посредством гомологичной рекомбинации) нативный промотор гена в геноме клетки-хозяина таким образом, что этот ген имеет измененную картину экспрессии. Этот ген будет теперь являться «выделенным», поскольку он отделен по меньшей мере от некоторых последовательностей, которые фланкируют его в природе.
Нуклеиновая кислота также считается «выделенной», если она содержит любые модификации, которые естественным образом не возникают в соответствующей нуклеиновой кислоте в геноме. Например, эндогенная кодирующая последовательность считается «выделенной», если она содержит вставку, делецию или точечную мутацию, введенные искусственно, например, путем вмешательства человека. «Выделенная нуклеиновая кислота» также включает нуклеиновую кислоту, встроенную в хромосому клетки-хозяина в гетерологичном участке, и нуклеотидную конструкцию, присутствующую в виде эписомы. Более того, «выделенная нуклеиновая кислота» может быть по существу свободной от другого клеточного материала или по существу свободной от культуральной среды при получении рекомбинантными методами, или по существу свободной от химических предшественников или других химических веществ при получении химическим синтезом.
В контексте настоящего изобретения «последовательность нуклеиновой кислоты, кодирующая функциональный генный продукт» относится к любому участку кодирующей части гена. Последовательность нуклеиновой кислоты, кодирующая функциональный генный продукт, может быть участком фермента, который способен осуществлять по меньшей мере одну активность всего фермента или целого фермента.
В контексте настоящего изобретения «нуклеиновая кислота, необходимая для экспрессии по меньшей мере одного генного продукта» относится к последовательности нуклеиновой кислоты, которая кодирует любой участок гена и/или функционально связана с нуклеиновой кислотой, кодирующей генный продукт, но не включает обязательно кодирующую последовательность. В качестве примера, последовательность нуклеиновой кислоты, необходимая для экспрессии по меньшей мере одного генного продукта, включает, но не ограничивается ими, энхансеры, промоторы, регуляторные последовательности, стартовые кодоны, стоп-кодоны, последовательности полиаденилирования и/или кодирующие последовательности.
В контексте настоящего изобретения «протеолиз» или «генный продукт, отвечающий за протеолиз в клетке», относится к любому пептиду, полипептиду, белку и/или ферменту, или их участку, способному вызвать расщепление по меньшей мере одного пептида, полипептида и/или белка. Генный продукт, отвечающий за протеолиз, может непосредственно отвечать за расщепление (т.е. являться пептидазой) или может отвечать непрямым образом, являясь частью пути синтеза пептидазы. Примеры генных продуктов, которые ответственны за протеолиз в клетке, включают, но не ограничиваются ими, аспартильные протеазы, сериновые протеазы, секретируемые аспартильные протеазы, секретируемые сериновые протеазы, протеазы метилотрофных дрожжей, DPPIV-подобные эндопептидазы, металлоэндопептидазы, Prb1-подобные сериновые протеазы, сериновые протеазы Prb1 и CPY-подобные карбоксипептидазы. Также в это определение включены протеазы, которые могут секретироваться из клетки, но по-прежнему сохранять некоторую или всю протеолитическую активность, например, секретируемые сериновые протеазы. Секретируемая протеаза может отвечать за протеолиз внутри клетки и/или вне клетки.
В контексте настоящего изобретения «гликозилирование» или «генный продукт, отвечающий за гликозилирование в клетке» относится к любому пептиду, полипептиду, белку и/или ферменту, или их части, участвующему в добавлении по меньшей мере одного сахаридного звена к полипептиду или в удлинении по меньшей мере одной сахаридной цепи в клетке. Генный продукт, отвечающий за гликозилирование в клетке, может непосредственно отвечать за добавление сахарида к полипептиду в клетке, как, например, но не ограничиваясь этим, маннозилтрансферазы. Маннозилтранферазы могут переносить остаток от Dol-P-Man на сериновый и/или треониновый остатки в пептиде, полипептиде и/или белке, или могут переносить остаток маннозы с GPD-Man на сахарид, таким образом удлиняя сахаридную цепь. В альтернативном варианте генный продукт, отвечающий за гликозилирование, может быть частью пути гликозилирования и может непрямым образом отвечать за добавление полисахарида к полипептиду в клетке. Примеры генных продуктов, которые отвечают за гликозилирование в клетке, включают, но не ограничиваются ими, маннозилтранферазы.
«Полинуклеотид(ы)» в общем относится к любому полирибонуклеотиду или полидезоксирибонуклеотиду, который может представлять собой немодифицированную РНК или ДНК или модифицированную РНК или ДНК. «Полинуклеотид(ы)» включает, без ограничения, одно- и двухцепочечную ДНК, ДНК, которая представляет собой смесь одно- и двухцепочечных областей или одно-, двух- и трехцепочечных областей, одно- и двухцепочечную РНК и РНК, которая представляет собой смесь одно- и двухцепочечных областей, гибридные молекулы, содержащие ДНК и РНК, которые могут представлять собой одноцепочечные или, более типично, двухцепочечные или трехцепочечные области, или смесь одно- и двухцепочечных областей. Дополнительно, «полинуклеотид» в контексте настоящего изобретения относится к трехцепочечным областям, содержащим РНК или ДНК, или как РНК, так и ДНК. Цепи в таких областях могут принадлежать одной молекуле или разным молекулам. Области могут включать в себя целиком одну или несколько молекул, но обычно включают только область некоторых молекул. Одна из молекул трехспиральной области часто представляет собой олигонуклеотид. В контексте настоящего изобретения термин «полинуклеотид(ы)» включает также ДНК или РНК, описанные выше, которые содержат одно или несколько модифицированных оснований. Таким образом, ДНК или РНК со остовом, модифицированным для стабильности или по другим причинам, представляют собой «полинуклеотид(ы)» согласно тому, что подразумевает данный термин в настоящем документе. Более того, ДНК или РНК, содержащие необычные основания, такие как инозин, или модифицированные основания, такие как тритилированные основания (приведено только два примера), представляют собой полинуклеотиды согласно тому, что подразумевает данный термин в настоящем документе. Следует иметь в виду, что можно провести большое число модификаций ДНК и РНК, которые служат для различных полезных целей, известных специалистам в данной области техники. Термин «полинуклеотид(ы)» в контексте настоящего описания, охватывает такие химически, ферментативно или метаболически модифицированные формы полинуклеотидов, а также химические формы ДНК и РНК, характерные для вирусов и клеток, включая, например, простые и сложные клетки. «Полинуклеотиды» также охватывают короткие полинуклеотиды, часто называемые олигонуклеотидами.
«Полипептид(ы)» относится к любому пептиду или белку, содержащему две или несколько аминокислот, соединенных друг с другом пептидными связями или модифицированными пептидными связями. «Полипептид(ы)» относится как к коротким цепям, обычно называемым пептидами, олигопептидами и олигомерами, так и к длинным цепям, обычно называемым белками. Полипептиды могут содержать аминокислоты, отличающиеся от генетически кодируемых 20 аминокислот. «Полипептид(ы)» включает аминокислоты, модифицированные в результате любых природных процессов, таких как процессинг и другие посттрансляционные модификации, а также в результате химических методов модификации. Такие модификации хорошо описаны в базовых учебниках и в более подробных монографиях, а также в большом количестве исследовательской литературы, и они хорошо известны специалистам в данной области техники. Следует понимать, что один тип модификации может присутствовать в одинаковой или различной степени в нескольких участках в данном полипептиде. Также данный полипептид может содержать много типов модификаций. Модификации могут присутствовать в любом участке полипептида, включая пептидный остов, аминокислотные боковые цепи и амино- или карбоксильные концы. Модификации включают, например, ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гема, ковалентное присоединение нуклеотида или нуклеотидного производного, ковалентное присоединение липида или липидного производного, ковалентное присоединение фосфотидилинозитола, перекрестную сшивку, циклизацию, образование дисульфидной связи, деметилирование, образование ковалентных перекрестных сшивок, образование цистеина, образование пироглутамата, формилирование, гамма-карбоксилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитический процессинг, фосфорилирование, пренилирование, рацемизацию, гликозилирование, присоединение липидов, сульфирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и АДФ-рибозилирование, селеноилирование, сульфатирование, тРНК-опосредованное добавление аминокислот к белкам, такое как аргинилирование, и убиквитинилирование. См., например, PROTEINS - STRUCTURE AND MOLECULAR PROPERTIES, 2nd Ed., T. E. Creighton, W. H. Freeman and Company, New York (1993) и Wold, F., Posttranslational Protein Modifications: Perspectives and Prospects, pgs. 1-12 в POSTTRANSLATIONAL COVALENT MODIFICATION OF PROTEINS, B. C. Johnson, Ed., Academic Press, New York (1983); Seifter et al., Meth. Enzymol. 182: 626-646 (1990) и Rattan et al., Protein Synthesis: Posttranslational Modifications and Aging, Ann. N.Y. Acad. Sci. 663: 48-62 (1992). Полипептиды могут быть разветвленными или циклическими, с разветвлением или без него. Циклические, разветвленные и разветвленные циклические полипептиды могут быть результатом посттрансляционных природных процессов, а также могут быть изготовлены полностью синтетическими способами.
«Вариант» в соответствии с тем, как этот термин используется в данном документе, представляет собой полинуклеотид или полипептид, который соответственно отличается от эталонного полинуклеотида или полипептида, но сохраняет значимые свойства. Типичный вариант полинуклеотида отличается по нуклеотидной последовательности от другого, эталонного полинуклеотида. Изменения в нуклеотидной последовательности варианта могут изменять или могут не изменять аминокислотную последовательность полипептида, кодируемого эталонным полинуклеотидом. Нуклеотидные изменения могут привести к аминокислотным заменам, добавлениям, делециям, слияниям и укорачиваниям в полипептиде, кодируемом эталонной последовательностью, как описано ниже. Типичный вариант полипептида отличается по аминокислотной последовательности от другого, эталонного полипептида. Обычно различия ограничены, так что последовательности эталонного полипептида и варианта очень похожи в целом и во многих областях идентичны. Вариант и эталонный полипептид могут отличаться по аминокислотной последовательности одной или несколькими заменами, добавлениями, делециями в любой комбинации. Замещенный или вставленный аминокислотный остаток может кодироваться или может не может кодироваться генетическим кодом. Вариант полинуклеотида или полипептида может быть природным, таким как аллельный вариант, или это может быть вариант, который не встречается в природе. Настоящее изобретение также включает в варианты каждого из полипептидов по настоящему изобретению, то есть полипептиды, которые отличаются от эталонной последовательности консервативными аминокислотными заменами, в результате которых остаток заменен другим с похожими характеристиками. Как правило, такие замены происходят между Ala, Val, Leu и Ile; между Ser и Thr; между кислотными остатками Asp и Glu; между Asn и Gln; и между основными остатками Lys и Arg; или между ароматическими остатками Phe и Tyr. В частности, существуют варианты, в которых несколько, 5-10, 1-5, 1-3, 1-2 или 1 аминокислота замещены, удалены или добавлены в любой комбинации. Неприродные варианты полинуклеотидов и полипептидов могут быть получены методами мутагенеза или прямым синтезом. Варианты могут также включать, но не ограничиваются ими, полипептиды или их фрагменты, имеющие химическую модификацию одной или нескольких из боковых групп своих аминокислот. Химическая модификация включает, но не ограничиваются ими, добавление химических групп, создание новых связей и удаление химических групп. Модификации в боковых группах аминокислот включают, без ограничения, ацилирование ε-аминогруппы лизина, N-алкилирование аргинина, гистидина, лизина или алкилирование глутаминовой или аспарагиновой карбоксильных групп, и дезамидирование глутамина или аспарагина. Модификации концевой аминогруппы включают, без ограничения, дезамининирование, N-алкилирование низшими алкилами, N-диалкилирование низшими алкилами и N-ацильные модификации. Модификации концевой карбоксильной группы включают, без ограничения, амидирование, образование алкиламида с низшими алкилами, образование диалкиламида с низшими алкилами и образование сложных эфиров с низшими алкилами. Кроме того, одна или несколько боковых групп или концевых групп могут быть защищены защитными группами, обычно известными среднему специалисту в белковой химии.
В контексте настоящего изобретения «фрагмент» при использовании в отношении полипептида представляет собой полипептид, имеющий аминокислотную последовательность, которая является одинаковой с частью, но не со всей аминокислотной последовательностью полноразмерного природного полипептида. В контексте настоящего изобретения «фрагмент» при использовании в отношении полинуклеотида или последовательности нуклеиновой кислоты представляет собой полинуклеотид, кодирующий аминокислотную последовательность, которая является одинаковой с частью, но не со всей аминокислотной последовательностью полноразмерного природного полипептида. Фрагменты могут быть «одиночными» или входить в состав более крупного полипептида, в котором они образуют часть или область в виде одной непрерывной области в одном более крупном полипептиде. В качестве примера, фрагмент природного GLP-1 будет включать аминокислоты с 7-й по 36-ю из природных аминокислот с 1-й по 36-ю. Кроме того, фрагменты полипептида могут также представлять собой варианты природной частичной последовательности. Например, фрагмент GLP-1, содержащий аминокислоты 7-36 из природного GLP-1, также может представлять собой вариант, имеющий аминокислотные замены в данной частичной последовательности. В качестве другого примера, «фрагмент» может относиться к любому гетерологичному полипептиду или нуклеиновой кислоте, кодирующей указанный полипептид, описанный в данном документе, включая, но не ограничиваясь ими, Kex2р, Pdi1 и Ero1, где указанный фрагмент сохраняет по меньшей мере одну функциональную активность указанного полипептида или фермента дикого типа.
В контексте настоящего изобретения «конъюгат» или «конъюгированный» относится к двум молекулам, которые связаны друг с другом. Например, первый полипептид может быть ковалентно или нековалентно связан со вторым полипептидом. Первый полипептид может быть ковалентно связан с помощью химического линкера или может быть генетически слит со вторым полипептидом, когда первый и второй полипептиды имеют общий полипептидный остов. Рекомбинантные полипептиды, экспрессированные в клетках-хозяевах по настоящему изобретению, могут содержать по меньшей мере один терапевтический полипептид, конъюгированный с сывороточным альбумином человека. Другие конъюгаты также включают, но не ограничиваются ими, по меньшей мере один терапевтический полипептид, конъюгированный с трансферрином, одноцепочечным вариабельным доменом и/или по меньшей мере одной Fc-областью антитела. Конъюгаты могут содержать или не содержать линкер.
В контексте изобретения «тандемно ориентированные» относится к двум или нескольким полипептидам, которые примыкают друг к другу в виде части одной молекулы. Они могут быть связаны ковалентно или нековалентно. Два или несколько тандемно ориентированных полипептидов могут входить в состав одного полипептидного остова. Тандемно ориентированные полипептиды могут иметь прямую или обратную ориентацию и/или могут быть разделены другими аминокислотными последовательностями.
В контексте настоящего изобретения «альбиглютид» относится к рекомбинантному слитому белку, состоящему из 2 копий 30-аминокислотной последовательности модифицированного глюкагон-подобного пептида 1 человека (GLP-1, фрагмент 7-36(A8G)), генетически слитому с сохранением рамки считывания с рекомбинантным сывороточным альбумином человека. Аминокислотная последовательность альбиглютида приведена ниже как SEQ ID NO:1.

«Рекомбинантные экспрессионные системы» относятся к экспрессионным системам или их участкам, или к полинуклеотидам по изобретению, введенным, трансфицированным или трансформированным в клетку-хозяина или лизат клеток-хозяев, для получения полинуклеотидов и полипептидов по настоящему изобретению.
В контексте настоящего изобретения «белок, слитый с альбумином» включает по меньшей мере фрагмент или вариант терапевтического полипептида и по меньшей мере фрагмент или вариант сывороточного альбумина человека, которые связаны друг с другом, предпочтительно, путем генетического слияния.
Полипептиды, имеющие активность GLP-1, могут содержать по меньшей мере один фрагмент и/или вариант GLP-1 человека. Два природных фрагмента GLP-1 человека представлены в SEQ ID NO: 2.
 ,
,
где Хаа в позиции 37 является Gly (далее обозначается как «GLP-1(7-37)») или -NH2 (далее обозначается как «GLP-1(7-36)»). Фрагменты GLP-1 могут включать, но не ограничиваются ими, молекулы GLP-1, содержащие или, в альтернативном варианте, состоящие из аминокислот 7-36 GLP-1 человека (GLP-1(7-36)). Варианты GLP-1 или их фрагменты могут включать, но не ограничиваются ими, одну, две, три, четыре, пять или более аминокислотных замен в GLP-1 дикого типа или в природных фрагментах GLP-1, представленного в SEQ ID NO:2. Варианты GLP-1 или фрагменты GLP-1 могут включать, но не ограничиваются ими, замены остатка аланина, аналогичного аланину 8 в GLP-1 дикого типа, где аланин заменен на глицин (далее обозначается как «A8G») (см., например, мутанты, описанные в патенте США № 5545618, полное содержание которого включено в настоящий документ путем ссылки).
В контексте настоящего изобретения «KEX2» относится к гену, кодирующему белок, называемый «протеазой Killer Expression» или «Kex2p», также называемый в данном описании «kexp». Kex2p является кальций-зависимой сериновой протеазой, участвующей в процессинге пробелков. Эта протеаза расщепляет полипептиды по карбоксильному концу узнаваемых последовательностей Arg-Arg/X и Lys-Arg/X. Другие виды активности Kex2p включают, но не ограничиваются ими, гидролазную активность, способность к связыванию ионов металлов, активность эндопептидаз серинового типа, пептидазную активность и активность пептидаз серинового типа. Другие названия KEX включают в себя: Pcsk2, Pcsk4, kpc-1. Ген эндопептидазы (KEX2) из Saccharomyces cerevisiae имеет ID NO 855483 в GenBank и кодирует белковую последовательности NCBI Ref Seq NP:014161.1. Ген KEX2 является консервативным в плодовой мушке, S. cerevisiae, K. lactis, E. gossypii, S. pombe, M. oryzae и N. crassa. Вариант KEX2 может представлять собой полинуклеотид, который имеет по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% идентичности по последовательности с геном (KEX2) из Saccharomyces cerevisia или кодирует белок, имеющий аминокислотную последовательность по меньшей мере с 85%, 90%, 95%, 96%, 97%, 98%, 99% идентичности по последовательности с Kex2p из Saccharomyces cerevisiae. Функциональный фрагмент и/или вариант Kex2p сохранял бы по меньшей мере одну функцию Kex2p, включая, но не ограничиваясь ими, способность расщеплять полипептиды по карбоксильному концу узнаваемых последовательностей Arg-Arg/X и Lys-Arg/X. Последовательность гена KEX2 (SEQ ID NO:3) и соответствующая аминокислотная последовательность Kex2p (SEQ ID NO:4) из S. cerevisiae приведены ниже:

ПРОИСХОЖДЕНИЕ: S. cerevisiae

В контексте настоящего изобретения «PDI» или «PDI1» относится к гену, кодирующему «pdi» или «Pdi1p», также известному как «протеин-дисульфидизомераза», которая представляет собой фермент в эндоплазматическом ретикулуме у эукариот, катализирующий образование и разрушение дисульфидных связей между остатками цистеина в белках при их сворачивании (Wilkinson B, Gilbert HF (June 2004). "Protein disulfide isomerase". Biochimica et Biophysica Acta 1699 (1-2): 35-44 и Gruber CW, Cemazar M, Heras B, Martin JL, Craik DJ (August 2006). "Protein disulfide isomerase: the structure of oxidative folding". Trends in Biochemical Sciences 31 (8): 455-64)) и может выступать в качестве шаперона белков (Wang, CC and Tsou, CL FASEB J. 1993 Dec; 7(15): 1515-7). Протеин-дисульфидизомераза является многофункциональным белком, присутствующим в просвете эндоплазматического ретикулума и необходимым для образования дисульфидных связей в секреторных белках и белках клеточной поверхности и ликвидации неприродных дисульфидных связей; она образует комплекс с Mnl1p, которая имеет экзоманнозидазную активность для процессинга Man8GlcNAc2-олигосахаридов, связанных с несвернутыми белками до Man7GlcNAc2, который усиливает деградацию несвернутого белка. Pdi1 также имеет окислительно-редуктазную активность. Pdi1p из S. cerevisiae кодируется последовательностью с ID NO 850314 в базе данных GenBank. Функциональный фрагмент и/или вариант pdi может представлять собой полипептид, который имеет по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% идентичности по последовательности с аминокислотной последовательностью NCBI Ref Seq NP_009887, кодируемой полинуклеотидом, имеющим по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% идентичности по последовательности с GenBank ID NO: 850314 и сохраняющим по меньшей мере одну функцию PDI, включая, но не ограничиваясь этим, изомеразную активность. Функциональная активность Pdi1 включает, но не ограничивается этим, катализ образования и/или разрушения дисульфидных связей, способствуя правильной укладке неправильно свернутых белков. Другие названия PDI включают: PDI1, PDIA2, Pdia3, P4HB, PADI1, Padi2, EUG1, NCU09223, SOAC1F5.02 и ген AGOS_AFR718W. Ген PDI1 является консервативным в человеке, макаке-резус, собаке, корове, мыши, крысе, курице, данио рерио, плодовой мушке, комаре, C. elegans, S. cerevisiae, K. lactis, E. gossypii, S. pombe, M. oryzae, N. crassa, A. thaliana и рисе. Последовательность гена PDI (SEQ ID NO:5) и соответствующая аминокислотная последовательность Pdi1 (SEQ ID NO:6) из S. cerevisiae приведены ниже:


PDI постоянно присутствует в просвете эндоплазматического ретикулума клеток. Совокупность данных о клеточном распределении фермента, его субклеточное расположение и его свойства в процессе развития показывают, что он участвует в биосинтезе белка и секреторном пути (Freedman, 1984, Trends Biochem. Sci. 9, pp.438-41), и это подтверждается прямыми исследованиями перекрестной сшивки in situ (Roth and Pierce, 1987, Biochemistry, 26, pp.4179-82). Тот факт, что микросомальные мембраны, дефицитные по PDI, имеют нарушения в образовании дисульфидных связей в белке (Bulleid and Freedman, 1988, Nature, 335, pp.649-51), позволяет предположить, что фермент функционирует в качестве катализатора образования природных дисульфидных связей в ходе биосинтеза секреторных белков и белков клеточной поверхности. Эта роль согласуется с тем, что известно о каталитических свойствах фермента in vitro: он катализирует тиол-дисульфидные реакции обмена, приводя к образованию, разрушению или изомеризации сети дисульфидных связей в белке, и может катализировать укладку белка и образование нативных дисульфидных связей у широкого круга восстановленных, несвернутых белковых субстратов (Freedman et al., 1989, Biochem. Soc. Symp., 55, pp.167-192). Последовательность ДНК и аминокислотная последовательность фермента известна для нескольких видов (Scherens, B. et al., 1991, Yeast, 7, pp. 185-193; Farquhar, R., et al., 1991, Gene, 108, pp. 81-89), и увеличивается количество информации в отношении механизма действия фермента, очищенного до гомогенности из печени млекопитающих (Creighton et al., 1980, J. Mol. Biol., 142, pp.43-62; Freedman et al., 1988, Biochem. Soc. Trans., 16, pp.96-9; Gilbert, 1989, Biochemistry 28, pp.7298-7305; Lundstrom and Holmgren, 1990, J. Biol. Chem., 265, pp.9114-9120; Hawkins and Freedman, 1990, Biochem. J., 275, pp.335-339). Из многих белковых факторов, известных на сегодняшний день в качестве медиаторов укладки, сборки и переноса в клетке белков (Rothman, 1989, Cell 59, pp.591-601), PDI необычен тем, что имеет хорошо определенную каталитическую активность.
PDI легко выделяется из тканей млекопитающих, и гомогенный фермент представляет собой гомодимер (2×57 кДа) с характерной кислой pI (4,0-4,5) (Hillson et al., 1984, Methods Enzymol., 107, pp.281-292). Фермент также был выделен из пшеницы и из водорослей Chlamydomonas reinhardii (Kaska et al., 1990 Biochem. J. 268, pp.63-68). Его активность была обнаружена в обширном ряде источников, и согласно предварительным данным активность PDI присутствует в S. cerevisiae (Williams et al., 1968, FEBS Letts., 2, pp.133-135). Недавно были опубликованы полные аминокислотные последовательности ряда PDI, в основном выведенные из клонированных кДНК-последовательностей, они включают PDI из крысы (Edman et al., 1985, Nature, 317, pp.267-270), быка (Yamauchi et al., 1987, Biochem. Biophys. ReS. Comm., 146, pp.1485-1492), человека (Pihlajaniemi et al., 1987, EMBO J., 6, pp.643-9), дрожжей (Scherens, B., et al., выше; Farquhar, R. et al., выше) и курицы (Parkkonen et al., 1988, Biochem. J., 256, pp.1005-1011). Белки из этих видов позвоночных показывают высокую степень консервативности последовательностей между собой, и все имеют несколько общих признаков, впервые замеченных в последовательности крысиной PDI (Edman et al., 1985 выше).
Последовательности, соответствующие или близкородственные PDI, были идентифицированы в работе, направленной на анализ функций, отличающихся от функции образования дисульфидных связей. Например, есть явное доказательство того, что PDI действует в качестве β-субъединицы тетрамерного αβ-фермента пролил-4-гидроксилазы, которая катализирует главную посттрансляционную модификацию формирующихся или новосинтезированных проколлагеновых полипептидов в эндоплазматическом ретикулуме (Pihlajaniemi et al., 1987, выше; Koivu et al., 1987, J. Biol. Chem., 262, pp.6447-49)). Существует также доказательство того, что PDI участвует в системе котрансляционного N-гликозилирования (Geetha-Habib et al., 1988, Cell, 4, pp.63-68), и недавно было высказано предположение, что фермент участвует в комплексе, который переносит триглицерид с формирующихся секреторных липопротеинов (Wetterau at al., 1990, J. Biol. Chem., 265, pp.9800-7). Поэтому PDI может иметь множество функций в ко- и посттрансляционных модификациях секреторных белков (Freedman, 1989, Cell, 57, pp.1069-72).
Повышение активности Pdi1 в экспрессионных системах на основе клеток бактерий, дрожжей и насекомых может привести к повышению секреции рекомбинантных белков, содержащих дисульфидные связи. Альбиглютид (ALB), аминокислотная последовательность которого показана в SEQ ID NO:1, состоит из устойчивого к DPP-4 димера GLP-1, слитого с альбумином человека. Белок содержит 8 дисульфидных связей. Вполне возможно, что сверхэкспрессия Pdi1 может улучшить правильное сворачивание и секрецию SEQ ID NO:1 в клетках-хозяевах и/или из них.
ERO1 представляет собой ген, кодирующий оксидоредуктин 1 эндоплазматического ретикулума (Ero1), который является оксидоредуктазным ферментом, катализирующим образование и изомеризацию дисульфидных связей в белках в эндоплазматическом ретикулуме (ER) эукариот. (Frand AR, Cuozzo JW, Kaiser CA (2000). "Pathways for protein disulphide bond formation". Trends Cell Biol. 10 (5): 203-10 и Frand AR, Kaiser CA (2000). "Two pairs of conserved cysteines are required for the oxidative activity of Ero1p in protein disulfide bond formation in the endoplasmic reticulum". Mol. Biol. Cell 11 (9): 2833-43). ERO1 из S. cerevisiae имеет следующие номера в базах данных: NCBI Gene ID NO 854909 и NCBI Protein Ref Seq NP_013576. Другие названия для ERO1 включают, но не ограничивается ими: ERO1L, ERO1LB, Ero1a, Ero1b, ero-1, NCU02074. Ген ERO1 консервативен у человека, шимпанзе, макака-резуса, собаки, коровы, мыши, крысы, курицы, данио рерио, C. elegans, S. cerevisiae, K. lactis, E. gossypii, S. pombe, M. oryzae, N. crassa, A. thaliana и в рисе. Виды активности ERO1 включают, но не ограничиваются ими, связывание флавин-аденинового динуклеотида, оксидоредуктазную активность, протеин-дисульфидизомеразную активность и тиолоксидазную активность. Функциональным фрагментом и/или вариантом Ero1 был бы полипептид, который сохраняет по меньшей мере одну функциональную активность Ero1 дикого типа.
«Оксидоредуктин эндоплазматического ретикулума» или «ERO» представляет собой оксидоредуктазный фермент, который катализирует образование и изомеризацию дисульфидных связей в белках в эндоплазматическом ретикулуме эукариотических клеток. Образование дисульфидных связей является окислительным процессом. После того как протеин-дисульфидизомераза (PDI) катализирует образование дисульфидных связей в формирующемся полипептиде, PDI становится восстановленной в ходе реакции тиол-дисульфидного обмена. ERO необходим для окисления PDI. В S. cerevisiae окисидоредуктин эндоплазматического ретикулума кодируется ERO1.
Последовательность гена ERO1 (SEQ ID NO:7) и соответствующая аминокислотная последовательность Ero1 (SEQ ID NO:8) из S. cerevisiae показаны ниже:


«Микроорганизм(ы)» означает (i) прокариоты, включающие, но не ограниченные ими, член рода Streptococcus, Staphylococcus, Bordetella, Corynebacterium, Mycobacterium, Neisseria, Haemophilus, Actinomycetes, Streptomycetes, Nocardia, Enterobacter, Yersinia, Fancisella, Pasturella, Moraxella, Acinetobacter, Erysipelothrix, Branhamella, Actinobacillus, Streptobacillus, Listeria, Calymmatobacterium, Brucella, Bacillus, Clostridium, Treponema, Escherichia, Salmonella, Kleibsiella, Vibrio, Proteus, Erwinia, Borrelia, Leptospira, Spirillum, Campylobacter, Shigella, Legionella, Pseudomonas, Aeromonas, Rickettsia, Chlamydia, Borrelia и Mycoplasma, и дополнительно включающие, но не ограниченные ими, член вида или группы - группы A Streptococcus, группы B Streptococcus, группы C Streptococcus, группы D Streptococcus, группы G Streptococcus, Streptococcus pneumoniae, Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus faecalis, Streptococcus faecium, Streptococcus durans, Neisseria gonorrheae, Neisseria meningitidis, Staphylococcus aureus, Staphylococcus epidermidis, Corynebacterium diptheriae, Gardnerella vaginalis, Mycobacterium tuberculosis, Mycobacterium bovis, Mycobacterium ulcerans, Mycobacterium leprae, Actinomyctes israelii, Listeria monocytogenes, Bordetella pertusis, Bordatella parapertusis, Bordetella bronchiseptica, Escherichia coli, Shigella dysenteriae, Haemophilus influenzae, Haemophilus aegyptius, Haemophilus parainfluenzae, Haemophilus ducreyi, Bordetella, Salmonella typhi, Citrobacter freundii, Proteus mirabilis, Proteus vulgaris, Yersinia pestis, Kleibsiella pneumoniae, Serratia marcessens, Serratia liquefaciens, Vibrio cholera, Shigella dysenterii, Shigella flexneri, Pseudomonas aeruginosa, Franscisella tularensis, Brucella abortis, Bacillus anthracis, Bacillus cereus, Clostridium perfringens, Clostridium tetani, Clostridium botulinum, Treponema pallidum, Rickettsia rickettsii и Chlamydia trachomitis, (ii) археи, включающие, но не ограниченные ими, Archaebacter, и (iii) одноклеточные или нитевидные эукариоты, включающие, но не ограниченные ими, протозои, грибы, член рода Saccharomyces, Kluveromyces или Candida и член вида Saccharomyces ceriviseae, Kluveromyces lactis или Candida albicans.
«Бактерия(и)» («бактериальный») означает (i) прокариоты, включающие, но не ограниченные ими, член рода Streptococcus, Staphylococcus, Bordetella, Corynebacterium, Mycobacterium, Neisseria, Haemophilus, Actinomycetes, Streptomycetes, Nocardia, Enterobacter, Yersinia, Fancisella, Pasturella, Moraxella, Acinetobacter, Erysipelothrix, Branhamella, Actinobacillus, Streptobacillus, Listeria, Calymmatobacterium, Brucella, Bacillus, Clostridium, Treponema, Escherichia, Salmonella, Kleibsiella, Vibrio, Proteus, Erwinia, Borrelia, Leptospira, Spirillum, Campylobacter, Shigella, Legionella, Pseudomonas, Aeromonas, Rickettsia, Chlamydia, Borrelia и Mycoplasma, и дополнительно включающие, но не ограниченные ими, член вида или группы - группы A Streptococcus, группы B Streptococcus, группы C Streptococcus, группы D Streptococcus, группы G Streptococcus, Streptococcus pneumoniae, Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus faecalis, Streptococcus faecium, Streptococcus durans, Neisseria gonorrheae, Neisseria meningitidis, Staphylococcus aureus, Staphylococcus epidermidis, Corynebacterium diptheriae, Gardnerella vaginalis, Mycobacterium tuberculosis, Mycobacterium bovis, Mycobacterium ulcerans, Mycobacterium leprae, Actinomyctes israelii, Listeria monocytogenes, Bordetella pertusis, Bordatella parapertusis, Bordetella bronchiseptica, Escherichia coli, Shigella dysenteriae, Haemophilus influenzae, Haemophilus aegyptius, Haemophilus parainfluenzae, Haemophilus ducreyi, Bordetella, Salmonella typhi, Citrobacter freundii, Proteus mirabilis, Proteus vulgaris, Yersinia pestis, Kleibsiella pneumoniae, Serratia marcessens, Serratia liquefaciens, Vibrio cholera, Shigella dysenterii, Shigella flexneri, Pseudomonas aeruginosa, Franscisella tularensis, Brucella abortis, Bacillus anthracis, Bacillus cereus, Clostridium perfringens, Clostridium tetani, Clostridium botulinum, Treponema pallidum, Rickettsia rickettsii и Chlamydia trachomitis, и (ii) археи, включающие, но не ограниченные ими, Archaebacter.
В контексте настоящего изобретения «гетерологичная последовательность нуклеиновой кислоты» относится к последовательности нуклеиновой кислоты, которая встроена, трансформирована или трансфицирована в клетку-хозяина или микроорганизм, представляющий интерес. Гетерологичная последовательность нуклеиновой кислоты может представлять собой кодирующую последовательность для всего полипептида или его части, и/или она может содержать некодирующие регуляторные элементы, такие как промотор, энхансер, участок связывания рибосомы или область полиаденилирования. Гетерологичная последовательность нуклеиновой кислоты может представлять собой последовательность нуклеиновой кислоты, которая в природе не встречается в клетке-хозяине, такую как последовательность нуклеиновой кислоты, которая кодирует полипептид из другого организма, рода или вида, нежели клетка-хозяин. В альтернативном варианте гетерологичная последовательность нуклеиновой кислоты может являться нативной для генома клетки-хозяина, но может быть встроена, трансформирована или трансфицирована в клетку-хозяина для увеличения функции нативной последовательности нуклеиновой кислоты или экспрессии полипептида, кодируемого указанной последовательностью нуклеиновой кислоты. Например, S. cerevisiae дикого типа могут содержать нуклеотидные последовательности, которые кодируют Kex2p дикого типа, а гетерологичная нуклеиновая кислота, кодирующая Kex2p дикого типа, может быть трансформирована в указанные S. cerevisiae для увеличения продукции Kex2p клеткой-хозяином. Аналогичным образом, S. cerevisiae дикого типа могут содержать нуклеотидные последовательности, которые кодируют Kex2p дикого типа, а гетерологичная нуклеиновая кислота, кодирующая Kex2p из другого организма, может быть встроена в указанные S. cerevisiae для увеличения продукции Kex2p клеткой-хозяином. Также предметом настоящего изобретения являются клетки-хозяева, которые содержат гетерологичные последовательности нуклеиновых кислот, которые являются вариантами и/или фрагментами нуклеиновых кислот дикого типа из того же биологического вида клетки-хозяина.
В контексте настоящего изобретения «рекомбинантный полипептид(ы)» и грамматические варианты этого выражения относятся к полипептиду, который в природе не синтезируется в трансформированной клетке-хозяине или представляющем интерес микроорганизме, и вводится в клетку-хозяина или микроорганизм с помощью рекомбинантной ДНК. Например, S. cerevisiae может действовать в качестве клетки-хозяина для экспрессии сывороточного альбумина человека, который не встречается в нетрансформированных или нетрансфицированных S. cerevisiae . Рекомбинантные полипептиды могут включать полипептиды, которые были модифицированы для облегчения выделения.
В контексте настоящего изобретения «аффинный тег» относится к любому фрагменту, связанному с молекулой, который может придать указанной молекуле селективное сродство к другому веществу или молекуле. Например, аффинный тег может использоваться для облегчения очистки молекулы, обеспечивая молекулу с селективной аффинностью к материалу-наполнителю колонки. Неограничивающим примером аффинного тега является His-тег.
Нуклеиновая кислота является «функционально связанной», когда она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК предшествующей последовательности или секреторного лидера функционально связана с ДНК полипептида, если она экспрессируется в виде белка-предшественника, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. В общем, «функционально связанный» означает, что последовательности ДНК, будучи связанными, являются непрерывными, а в случае секреторного лидера - непрерывными и в одной рамке считывания. Однако энхансеры не должны быть непрерывными. Связывание осуществляют лигированием в удобных сайтах рестрикции. Если такие сайты не существуют, то используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с обычной практикой.
Используемый в данном документе термин «сбор» клеток относится к сбору клеток из клеточной культуры. В процессе сбора клетки могут быть сконцентрированы для отделения их от культурального бульона, например, путем центрифугирования или фильтрации. Сбор клеток может дополнительно включать стадию лизиса клеток для получения внутриклеточного материала, такого как, но не ограниченного ими, полипептиды и полинуклеотиды. Опытный специалист в данной области должен понимать, что некоторый клеточный материал, включающий, но не ограниченный ими, гетерологично экспрессированные полипептиды, может высвобождаться из клеток в процессе культивирования. Поэтому представляющий интерес продукт (например, рекомбинантный экспрессированный полипептид) может оставаться в культуральном бульоне после сбора клеток.
Также предложены способы, в которых рекомбинантная ДНК-конструкция кодирует селективный маркер. Такой селективный маркер обеспечивает либо положительную, либо отрицательную селекцию. Также предложены способы, включающие экспрессию указанного селективного маркера и сравнение количества селективного маркера, продуцируемого по меньшей мере одной первой трансформированной клеткой на стадии отбора, с количеством селективного маркера, продуцируемого по меньшей мере одной второй трансформированной клеткой на стадии отбора, где первая и вторая трансформированные клетки продуцируют один и тот же селективный маркер. Как известно в данной области, селективные маркеры включают, но не ограничиваются ими, дигидрофолатредуктазу (DHFR), β-галактозидазу, флуоресцентный белок, секретируемую форму плацентарной щелочной фосфатазы человека, бета-глюкуронидазу, дрожжевые селективные маркеры LEU2 и URA3, гены резистентности к апоптозу и антисмысловые олигонуклеотиды, а также гены устойчивости к антибиотикам, придающие способность к росту в присутствии антибиотиков, включая неомицин (neo), канамицин, генетицин, гигромицин, пуромицин, зеоцин, бластицидин, нурсеотрицин, биалафос, флеомицин и ампициллин. Как еще известно в данной области, клетки могут быть отсортированы различными средствами, включающими, но не ограниченными ими, визуальный осмотр или клеточный сортер, такой как BD FACS Aria, с помощью которых можно обнаружить экспрессию селективного маркера.
Термин «дикий тип», известный в данной области, относится к клетке-хозяину, или полипептиду или полинуклеотидной последовательности, которые присутствуют в природной популяции без генетической модификации. Например, «клетка-хозяин дикого типа» относится к немодифицированному штамму клетки-хозяина до любой генетической модификации, проведенной или возникшей в геноме клетки-хозяина.
В контексте настоящего изобретения «титр» или «итоговый титр» относится к концентрации продукта (например, рекомбинантно экспрессированного полипептида) в растворе (например, культуральном бульоне, лизате клеток или буфере) и обычно выражается в мг/л или г/л. Повышение итогового титра может относиться к абсолютному или относительному увеличению концентрации продукта, полученного при двух определенных наборах условий.
«Инкретиновый гормон» в контексте настоящего изобретения означает любой гормон, который усиливает секрецию инсулина или иным образом повышает уровень инсулина. Одним из примеров инкретиновых гормонов является GLP-1. Инкретин GLP-1 секретируется кишечными L-клетками в ответ на прием пищи. У здоровых людей GLP-1 играет важную роль в регуляции уровня глюкозы в крови после еды, стимулируя глюкозозависимую секрецию инсулина поджелудочной железой, приводящую к увеличению поглощения глюкозы на периферии. GLP-1 также подавляет секрецию глюкагона, что приводит к снижению печеночной продукции глюкозы. В дополнение к этому, GLP-1 задерживает время опорожнения желудка и замедляет моторику тонкой кишки, задерживая абсорбцию пищи. GLP-1 способствует пролонгации нормального функционирования бета-клеток, стимулируя транскрипцию генов, вовлеченных в глюкозозависимую секрецию инсулина и стимулируя неогенез бета-клеток (Meier, et al. Biodrugs 2003; 17 (2): 93-102).
«Активность GLP-1» в контексте настоящего изобретения означает одну или несколько из видов активности природного GLP-1 человека, включающих, но не ограниченных ими, снижение содержания глюкозы в крови и/или в плазме, стимуляцию глюкозозависимой секреции инсулина или иное повышение уровня инсулина, подавление секреции глюкагона, снижение уровня фруктозамина, увеличение доставки глюкозы и метаболизма в головном мозге, задерживание опорожнения желудка а также пролонгацию нормального функционирования бета-клеток и/или неогенез. Любой из этих видов активности и иная активность, ассоциированная с активностью GLP-1, могут быть вызваны непосредственно или косвенно композицией, имеющей активность GLP-1 или содержащей агонист GLP-1. Например, композиция, имеющая активность GLP-1, может прямо или косвенно стимулировать глюкозозависимую секрецию инсулина, в то время как стимуляция продукции инсулина может косвенно снижать уровень глюкозы в плазме млекопитающего.
«Инкретиновый миметик» в контексте настоящего изобретения представляет собой соединение, способное усиливать секрецию инсулина или иным образом повышать уровень инсулина. Инкретиновый миметик может быть способен стимулировать секрецию инсулина, увеличивать неогенез бета-клеток, ингибировать апоптоз бета-клеток, ингибировать секрецию глюкагона, задерживать опорожнение желудка и вызывать чувство насыщения у млекопитающего. Инкретиновый миметик может включать, но не ограничен ими, любой полипептид, который имеет активность GLP-1, включая, но не ограничиваясь ими, эксендин 3 и эксендин 4, в том числе любые их фрагменты и/или варианты и/или конъюгаты.
«Доменное антитело» или «dAb» может рассматриваться, как то же самое, что и «одиночный вариабельный домен», который способен связываться с антигеном. Одиночный вариабельный домен может являться вариабельным доменом антитела человека, но также включает в себя одиночные вариабельные домены антител из других видов, таких как грызуны (например, как описанные в WO 00/29004), акулы-няньки и VHH-доменные антитела представителей семейства верблюдовых. VHH верблюдовых представляют собой одиночные вариабельные домены иммуноглобулинов, выделяемых из видов, включающих верблюда, ламу, альпаку, дромадера и гуанако, которые продуцируют тяжелые цепи антител, по своей природе лишенные легких цепей. Такие VHH-домены могут быть гуманизированы в соответствии со стандартными методиками, доступными в данной области, и такие домены считаются «доменными антителами». В контексте настоящего изобретения VH включает VHH-домены верблюдовых.
Фраза «одиночный вариабельный домен» относится к вариабельному домену антигенсвязывающего белка (например, VH, VHH, VL), который специфично связывает антиген или эпитоп независимо от других вариабельных области или домена.
Термин «антигенсвязывающий белок», используемый в данном документе, относится к антителам, фрагментам антител и другим белковым конструкциям, таким как домены, но не ограниченные ими, вариабельные домены и доменные антитела, которые способны связываться с антигеном.
В контексте настоящего изобретения «сниженное количество» и грамматические варианты в отношении фермента или его фрагмента, или ферментативной активности, сравниваемых в генетически модифицированной клетке-хозяине, относится к генетически модифицированной клетке-хозяину, которая производит меньший уровень по меньшей мере одного фермента или имеет меньший уровень по меньшей мере одного вида ферментативной активности по сравнению с генетически немодифицированной клеткой-хозяином. Как правило, сравнение ферментативной активности, осуществляемой генетически модифицированной клеткой-хозяином, проводится со штаммом дикого типа того же биологического вида. Однако сравнение может также проводиться между генетически модифицированной клеткой-хозяином и клеткой-хозяином дикого типа того же рода, но другого вида или штамма, или с другим генетически модифицированным штаммом. Снижение по меньшей мере количества одного фермента или ферментативной активности также включает в себя полное удаление по меньшей мере одного фермента или ферментативной активности, когда никакое количество из по меньшей мере одного фермента не производится в генетически модифицированной клетке-хозяине, и/или никакое количество из по меньшей мере одного фермента не является функциональным или не имеет активность. Также в это определение входит снижение уровня по меньшей мере одной ферментативной активности. То есть ферменты, которые имеют более одного вида активности, могут сохранять уровень первого вида активности, тогда как уровень второго вида активности этого же фермента снижается.
В контексте настоящего изобретения «повышенное количество» и грамматические варианты в отношении фермента или его фрагмента, или ферментативной активности в генетически модифицированной клетке-хозяине относится к генетически модифицированной клетке-хозяину, которая производит больший уровень по меньшей мере одного фермента или имеет больший уровень по меньшей мере одного вида активности фермента по сравнению с генетически немодифицированной клеткой-хозяином. Как правило, сравнение ферментативной активности, осуществляемой генетически модифицированной клеткой-хозяином, проводится со штаммом дикого типа того же биологического вида. Однако сравнение может также проводиться между генетически модифицированной клеткой-хозяином и клеткой-хозяином дикого типа того же рода, но другого вида или штамма, или с другим генетически модифицированным штаммом. Также в это определение входит повышение уровня по меньшей мере одной ферментативной активности. То есть, ферменты, которые имеют более одного вида активности, могут сохранять уровень первого вида активности, а второй вид активности того же фермента увеличивается. Дополнительно этот термин включает в себя повышение ферментативной активности вне зависимости от количества фермента, продуцируемого клеткой-хозяином. Например, генетически модифицированные клетки-хозяева могут продуцировать одинаковое или аналогичное количество фермента или его фрагмента и/или варианта, которое продуцируется клеткой-хозяином дикого типа, измеряемое по массе или по количеству, но может наблюдаться заметное повышение уровня по меньшей мере одной функциональной активности указанного фермента по сравнению с диким типом.
Используемые в данном документе термины «жесткие условия» и «жесткие условия гибридизации» означают, что гибридизация будет происходить только тогда, если существует по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 95% идентичности между последовательностями. Примером жестких условий гибридизации является инкубация в течение ночи при 42°С в растворе, содержащем 50% формамида, 5xSSC (150 мМ NaCl, 15 мМ тринатрий цитрат), 50 мМ фосфат натрия (рН 7,6), 5× раствор Денхардта, 10% декстрансульфата и 20 мкг/мл денатурированной, фрагментированной ДНК спермы лосося, с последующей отмывкой фильтров в 0,1х SSC при температуре приблизительно 65°С. Условия гибридизации и отмывки хорошо известны и приведены в руководстве Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989), в частности, в 11-й главе в нем, раскрытие которого полностью включено в данное описание путем ссылки.
В контексте настоящего изобретения «генетическая модификация» или «генетически модифицированный» относится к любым подавлению, замене, делеции и/или вставке одного или нескольких оснований или фрагментов клеточной ДНК-последовательности (последовательностей). Такая генетическая модификация может быть выполнена in vitro (непосредственно на выделенной ДНК) или in situ, например, с помощью методов генной инженерии или путем воздействия на клетки мутагенным агентом. Мутагенные агенты включают, например, физические агенты, такие как высокоэнергетическое излучение (рентгеновское излучение, γ-излучение, УФ-излучение и т.п.), или химические агенты, способные вступать в реакции с различными функциональными группами ДНК, такие как алкилирующие агенты (EMS, NQO и т.д.), биалкилирующие агенты, интеркалирующие агенты и т.д. Генетические модификации могут быть также получены с помощью нарушения целостности генов, например, в соответствии со способом, описанным Rothstein et al. (Meth. Enzymol. 194:281-301(1991)). Согласно этому способу, часть гена или весь ген замещают посредством гомологичной рекомбинации на in vitro модифицированную версию. Генетические модификации также могут быть выполнены с помощью любой мутирующей вставки в ДНК-последовательности, например, с помощью транспозонов, фагов и т.д. Также в контексте настоящего изобретения «генетически модифицированный» может относиться к гену, кодирующему полипептид, или к полипептиду, имеющему по меньшей мере одну делецию, замену или подавление нуклеиновой кислоты или аминокислоты, соответственно. Например, полипептид, в котором по меньшей мере одна аминокислота замещена относительно формы дикого типа, будет считаться генетически модифицированным.
Генетическая модификация может быть обратимой или может быть ослаблена в результате действия клеточного механизма. В альтернативном варианте мутации могут представлять собой необратимые и/или не «текущие». «Текущие мутации» включают мутации, которые приводят к частичной, а не полной инактивации функции дикого типа.
Генетические модификации, которые несут клетки-хозяева по изобретению, могут находиться в кодирующей области последовательности ДНК клетки и/или в области, влияющей на экспрессию гена. Поэтому модификации по изобретению обычно будут влиять на генный продукт либо на регулирование или увеличение генного продукта в случае белков и/или ферментов, участвующих в протеолизе и/или гликозилировании. Сниженная способность клеток по изобретению к протеолитическому расщеплению и/или гликозилированию гетерологично экспрессированного полипептида может быть обусловлена структурными и/или конформационными изменениями, являться следствием продукции одного или нескольких ферментов, имеющих измененные биологические свойства, следствием отсутствия продукции указанных одного или нескольких ферментов или следствием продукции одного или нескольких ферментов на низком уровне.
Генетические модификации по изобретению также влияют на генный продукт либо на регулирование или увеличение генного продукта в отношении белков и/или ферментов, участвующих в любом из видов функциональной активности Kex2p, Pdi1 и Ero1, описанных в данном документе. Повышенная способность клеток по настоящему изобретению к правильной укладке и секреции рекомбинантно экспрессируемых полипептидов может быть связана с ферментами, участвующими в этих процессах, которые имеют измененные биологические свойства или продуцируются на высоком уровне.
В одном аспекте настоящего изобретения предложены генетически модифицированные клетки-хозяева, содержащие по меньшей мере один выделенный полинуклеотид, кодирующий Killer Expression (Kex) протеазу (Kex2p) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность протеазы Kex2p, и по меньшей мере один выделенный полинуклеотид, кодирующий протеин-дисульфидизомеразу (PDI) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Pdi1. Генетически модифицированные клетки-хозяева по настоящему изобретению включают генетически модифицированные клетки-хозяева, содержащие по меньшей мере один выделенный полинуклеотид, кодирующий оксидоредуктин эндоплазматического ретикулума (Ero1) или его фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность ERO.
Генетически модифицированные клетки-хозяева по настоящему изобретению также включают генетически модифицированные клетки-хозяева, которые экспрессируют или сверхэкспрессирует по меньшей мере один генный продукт по меньшей мере одного выделенного полинуклеотида, кодирующего белок и/или его вариант, который имеет по меньшей мере одну функциональную активность указанного белка, выбранного из Kex2p, Pdi1 или Ero1, когда указанную генетически модифицированную клетку-хозяина выращивают в культуре, по сравнению со второй клеткой-хозяином, где указанная вторая клетка-хозяин не экспрессирует или не сверхэкспрессирует по меньшей мере один генный продукт, выбранный из KEX, PDI и ERO. Также в настоящее изобретение включены генетически модифицированные клетки-хозяева, которые сверхэкспрессируют по меньшей мере два белка или их фрагмента и/или варианта, которые имеют по меньшей мере одну функциональную активность указанного белка, выбранного из Kex2p, Pdi1 или Ero1, когда указанную генетически модифицированную клетку-хозяина выращивают в культуре, по сравнению со второй клеткой-хозяином, где указанная вторая клетка-хозяин принадлежит к тому же биологическому виду и выращивается при тех же условиях культивирования, но не сверхэкспрессирует по меньшей мере два генных продукта, выбранных из KEX, PDI и ERO. В некоторых случаях вторая клетка-хозяин может иметь генетическую модификацию, но не имеет генетические модификации, которые позволяют ей экспрессировать или сверхэкспрессировать по меньшей мере один генный продукт по меньшей мере одного выделенного полинуклеотида, кодирующего белок и/или его вариант, который имеет по меньшей мере одну функциональную активность указанного белка, выбранного из Kex2p, Pdi1 или Ero1. В некоторых случаях вторая клетка-хозяин может являться клеткой дикого типа (то есть без генетических модификаций) того же биологического вида, что и модифицированная клетка-хозяин. В некоторых случаях вторая клетка-хозяин может содержать все те же генетические модификации, что и генетически модифицированная клетка-хозяин, за исключением содержания нуклеиновой кислоты, кодирующей белок и/или его вариант, который имеет по меньшей мере одну функциональную активность указанного белка, выбранного из Kex2p, Pdi1 или Ero1. Также в настоящем изобретении рассматриваются клетки-хозяева, которые генетически модифицированы для усиления экспрессии эндогенных полипептидов, включающих, но не ограниченных ими, Kex2p, Pdi1 и Ero1, с генов, уже содержащихся в клетке-хозяине.
В другом аспекте настоящего изобретения предложены генетически модифицированные клетки-хозяева, которые дополнительно содержат по меньшей мере одну из следующих генетических модификаций: нокаут протеазы рер4, сниженную активность ubc4 и/или ubc5 по сравнению с клеткой-хозяином дикого типа, нокаут yps1, нокаут hsp150 и нокаут pmt1. Было найдено, что эти генетические модификации увеличивают секрецию рекомбинантного сывороточного альбумина человека и снижают нежелательные посттрансляционные модификации.
Штаммы дрожжей, используемые для продукции слитых с альбумином белков, включают, но не ограничиваются ими, D88, DXY1 и BXP10. D88 [leu2-3, leu2-122, can1, pra1, ubc4] является производным родительского штамма AH22his.sup+ (также известного как DB1; см., например, Sleep et al. Biotechnology 8:42-46 (1990)). Штамм содержит мутацию leu2, которая позволяет ауксотрофную селекцию плазмид на основе плазмиды 2-micron, содержащей ген leu2. D88 также способен к дерепрессии PRB1 при избытке глюкозы. Промотор PRB1 обычно регулируется на двух уровнях, которые контролируют уровень глюкозы и стадии роста. Промотор активируется в дрожжах дикого типа при истощении глюкозы и вхождении в стационарную фазу. Штамм D88 репрессируется глюкозой, но сохраняет способность к индукции после вхождения в стационарную фазу. Ген PRA1 кодирует дрожжевую вакуолярную протеазу, эндопротеазу YscA, которая локализуется в ER. Ген UBC4 относится к пути убиквитинирования и участвует в мечении короткоживущих и аномальных белков для убиквитин-зависимой деградации. Было найдено, что выделение этой мутации ubc4 увеличивает число копий экспрессионной плазмиды в клетке и вызывает повышение уровня экспрессии целевого белка, экспрессируемого с плазмиды (см., например, международную публикацию № WO99/00504, таким образом полностью включенную в данный документ путем ссылки).
DXY1, производное D88, имеет следующий генотип: [leu2-3, leu2-122, can1, pra1, ubc4, ura3:yap3]. В дополнение к мутациям, выделенным в D88, этот штамм также имеет нокаут по протеазе YAP3. Эта протеаза расщепляет главным образом по двум основным остаткам (RR, RK, KR, KК), но может также усиливать расщепление по одиночным основным остаткам в белках. Выделение этой мутации yap3 привело к более высокому уровню продукции полноразмерного HSA (см., например, патент США № 5965386 и Kerry-Williams et al., Yeast 14:161-169 (1998), таким образом полностью включенные в данный документ путем ссылки).
BXP10 имеет следующий генотип: leu2-3, leu2-122, can1, pra1, ubc4, ura3, yap3::URA3, lys2, hsp150::LYS2, pmt1::URA3. В дополнение к мутациям, выделенным в DXY1, этот штамм также имеет нокаут по генам PMT1 и HSP150. Ген PMT1 является членом эволюционно консервативного семейства протеин-О-маннозилтрансфераз, переносящих долихолфосфат-D-маннозу (Pmt). Трансмембранная топология Pmt1p предполагает, что она является встроенным в мембрану белком эндоплазматического ретикулума с ролью в O-связанном гликозилировании. Эта мутация служит для уменьшения/устранения O-связанного гликозилирования слитых с HSA белков (см., например, международную публикацию № WO00/44772, таким образом полностью включенную в данный документ путем ссылки). Исследования показали, что белок Hsp150 неэффективно отделяется от rHA с помощью ионообменной хроматографии. Мутация в гене HSP150 удаляет потенциальную примесь, которая оказалась трудной для удаления стандартными методами очистки. См., например, патент США № 5783423, таким образом полностью включенный в данный документ путем ссылки.
Генетически модифицированные клетки-хозяева по настоящему изобретению, включают, но не ограничиваются ими, клетки грибов, дрожжевые клетки и клетки млекопитающих. Генетически модифицированные клетки-хозяева по настоящему изобретению включают, но не ограничиваются ими: Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces и Yarrowia. Генетически модифицированные клетки-хозяева по настоящему изобретению также включают в себя, но не ограничиваются ими, S. cerevisiae .
Генетически модифицированные клетки-хозяева по настоящему изобретению могут дополнительно содержать по меньшей мере один полинуклеотид, кодирующий рекомбинантный полипептид. Полинуклеотид, способный экспрессировать по меньшей мере один гетерологичный полипептид, включает, но не ограничивается ими, векторы, ДНК, трансформированную в геном клетки-хозяина, вирус или часть вируса, и/или плазмиды. Полинуклеотид, способный экспрессировать гетерологичный полипептид, может быть трансформирован в геном клетки-хозяина и/или может быть частью экспрессионного вектора и/или эписомальной экспрессионной системы.
В некоторых аспектах настоящего изобретения нуклеиновая кислота, кодирующая рекомбинантный полипептид, содержится в плазмиде. В других аспектах нуклеиновой кислотой, кодирующей рекомбинантный полипептид, трансформируют геном клетки-хозяина по настоящему изобретению.
Как известно в данной области, ДНК может быть трансформирована в клетку-хозяина несколькими различными способами. В дрожжах может использоваться любой удобный способ переноса ДНК, например, электропорация, метод с использованием хлорида лития, метод с ацетатом лития или метод сферопластов. Для получения стабильного штамма, подходящего для ферментации с высокой плотностью клеток, желательно интегрировать (встроить) ДНК в хромосому хозяина. Интеграцию проводят посредством гомологичной рекомбинации с использованием способов, известных в данной области техники. Например, ДНК, способная экспрессировать по меньшей мере один гетерологичный белок, может быть снабжена фланкирующими последовательностями, гомологичными последовательностями организма-хозяина. Таким образом, интеграция происходит в определенном участке в геноме хозяина без нарушения желательных или необходимых генов. Дополнительно и в качестве альтернативы, ДНК, способную экспрессировать по меньшей мере один гетерологичный белок, встраивают в нежелательный ген в хромосоме хозяина, вызывая разрушение или удаление гена или экспрессию продукта этого гена.
Повышенная экспрессия или сверхэкспрессия генного продукта может быть достигнута путем интеграции дополнительных копий ДНК, способной экспрессировать генный продукт, в хромосому хозяина. Дополнительно и в качестве альтернативы, ДНК, кодирующая генный продукт, может быть функционально связана с сильным промотором, и вся экспрессионная кассета может быть встроена в хромосому хозяина в определенном участке. Например, интеграция ДНК, кодирующей Kex2p, Pdi1 или Ero1 и функционально связанной с сильным промотором (например, PGK1-промотором), в участок NTS2-2 позволяет сверхэкспрессировать соответствующий генный продукт. В других вариантах осуществления изобретения ДНК может быть введена в организм хозяина посредством хромосомы, плазмиды, ретровирусного вектора или случайной интеграции в геном хозяина.
Генетически модифицированные клетки-хозяева по настоящему изобретению включают генетически модифицированные клетки-хозяева, в которых по меньшей мере один выделенный полинуклеотид, кодирующий протеазу kex или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Kex2p, функционально связан по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, МЕТ25, CUP1, PHО5, tetО-CYC1, CaMV, HXT6, HXT7 и ARE. Подходящим промотором является PGK1. Генетически модифицированные клетки-хозяева по настоящему изобретению также включают генетически модифицированные клетки-хозяева, в которых по меньшей мере один выделенный полинуклеотид, кодирующий PDI или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Pdi1, функционально связан по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, МЕТ25, CUP1, PHО5, tetО-CYC1, CaMV, HXT6, HXT7 и ARE. Подходящим промотором является PGK1. Дополнительно, генетически модифицированные клетки-хозяева по настоящему изобретению включают генетически модифицированные клетки-хозяева, в которых по меньшей мере один выделенный полинуклеотид, кодирующий ERO или его фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность ERO, функционально связан по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, МЕТ25, CUP1, PHО5, tetО-CYC1, CaMV, HXT6, HXT7 и ARE. Подходящим промотором является PGK1.
В другом аспекте рекомбинантный полипептид, экспрессируемый в генетически модифицированных клетках-хозяевах по настоящему изобретению, имеет по меньшей мере одну дисульфидную связь. В некоторых аспектах рекомбинантный полипептид является слитым с альбумином белком. В некоторых аспектах рекомбинантный полипептид содержит по меньшей мере один конъюгированный с альбумином терапевтический полипептид, имеющий активность GLP-1.
В некоторых аспектах по меньшей мере один фрагмент и вариант GLP-1 содержит GLP-1(7-36(A8G)) и генетически слит с сывороточным альбумином человека. В дополнительном варианте осуществления изобретения полипептиды по изобретению содержат одну, две, три, четыре, пять или большее число тандемно ориентированных молекул GLP-1 и/или его фрагментов и/или вариантов, слитых с N- или С-концом человеческого сывороточного альбумина или его варианта. Другие варианты осуществления включают такие A8G-полипептиды, слитые с N- или С-концом альбумина или его варианта. Пример двух тандемно ориентированных GLP-1(7-36)(A8G)-фрагментов и/или вариантов, слитых с N-концом сывороточного альбумина человека, включает SEQ ID NO:1, представленную на фиг. 3. В другом аспекте по меньшей мере один фрагмент и вариант GLP-1 содержит по меньшей мере два GLP-1(7-36(A8G)) тандемно и генетически слитых с сывороточным альбумином человека. По меньшей мере два GLP-1(7-36(A8G)) могут быть генетически слиты с N-концом сывороточного альбумина человека. По меньшей мере один полипептид, имеющий активность GLP-1, может содержать последовательность SEQ ID NO:1.
Варианты GLP-1(7-37) могут быть обозначены, например, как Glu22-GLP-1(7-37)OH, который обозначает вариант GLP-1, в котором глицин, обычно присутствующий в позиции 22 в GLP-1(7-37)ОН, заменен на глутаминовую кислоту; Val8-Glu22-GLP-1(7-37)ОН обозначает соединение GLP-1, в котором аланин, обычно присутствующий в позиции 8, и глицин, обычно присутствующий в позиции 22 в GLP-1(7-37)ОН, заменены на валин и глутаминовую кислоту, соответственно. Примеры вариантов GLP-1 включают, но не ограничиваются ими:

Варианты GLP-1 также могут включать в себя, но не ограничиваются ими, GLP-1 или фрагменты GLP-1, имеющие химическую модификацию одной или нескольких боковых групп своих аминокислот. Химическая модификация включает в себя, но не ограничивается ими, добавление химических групп, создание новых связей и удаление химических групп. Модификации боковых групп аминокислот включают, без ограничения, ацилирование ε-аминогруппы лизина, N-алкилирование аргинина, гистидина или лизина, алкилирование карбоксильных групп глутаминовой или аспарагиновой кислот и дезамидирование глутамина или аспарагина. Модификации концевой аминогруппы включают, без ограничения, дезамининирование, N-алкилирование низшими алкилами, N-диалкилирование низшими алкилами и N-ацильные модификации. Модификации концевой карбоксильной группы включают, без ограничения, амидирование, образование алкиламида с низшими алкилами, образование диалкиламида с низшими алкилами и образование сложных эфиров с низшими алкилами. Кроме того, одна или несколько боковых или концевых групп могут быть защищены защитными группами, обычно известными среднему специалисту в белковой химии.
Фрагменты или варианты GLP-1 могут также включать полипептиды, в которых одна или несколько аминокислот были добавлены на N- и/или С-конце GLP-1(7-37)OH указанного фрагмента или варианта. Аминокислоты в GLP-1, в котором аминокислоты были добавлены на N- или C-конце, обозначены таким же номером, что и соответствующие аминокислоты в GLP-1(7-37)ОН. Например, N-концевая аминокислота в соединении GLP-1, полученном добавлением двух аминокислот к N-концу GLP-1(7-37)ОН, находится в позиции 5; а С-концевая аминокислота в соединении GLP-1, полученном добавлением одной аминокислоты на C-конце GLP-1(7-37)OH, находится в позиции 38. Таким образом, позиция 12 занята фенилаланином, а позиция 22 занята глицином, в обоих этих соединениях GLP-1, как и в GLP-1(7-37)OH. Аминокислоты 1-6 из GLP-1 с аминокислотами, добавленными на N-конце, могут быть такими же или представлять собой консервативную замену аминокислот в соответствующих позициях в GLP-1(1-37)ОН. Аминокислоты 38-45 из GLP-1 с аминокислотами, добавленными на С-конце, могут быть такими же или представлять собой консервативную замену аминокислот в соответствующих позициях глюкагона или эксендина-4.
В другом аспекте по меньшей мере один полипептид, имеющий активность GLP-1, содержит по меньшей мере один фрагмент и/или вариант GLP-1 человека, слитый с сывороточным альбумином человека. В другом аспекте по меньшей мере один фрагмент и вариант GLP-1 содержит GLP-1(7-36(A8G)). По меньшей мере один фрагмент и вариант GLP-1 является генетически слитым с сывороточным альбумином человека. В другом аспекте рекомбинантный полипептид по настоящему изобретению содержит по меньшей мере два GLP-1(7-36(A8G)) тандемно и генетически слитых с сывороточным альбумином человека. Два GLP-1(7-36(A8G)) являются генетически слитыми с N-концом сывороточного альбумина человека. В некоторых примерах рекомбинантный полипептид содержит последовательность SEQ ID NO:1.
В одном варианте осуществления настоящего изобретения рекомбинантный полипептид содержит полипептид, имеющий 99% идентичности по последовательности с полипептидом, приведенным в SEQ ID NO:1, или с полипептидом, имеющим аминокислотную последовательность SEQ ID NO:1, которая укорочена с C- и/или с N-конца. В одном аспекте рекомбинантный полипептид имеет активность GLP-1. В одном аспекте полипептид укорачивается с N-конца на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 аминокислот по сравнению с SEQ ID NO:1 или с полипептидом, имеющим 99% идентичности по последовательности с SEQ ID NO:1 по всей длине. В одном аспекте рекомбинантный полипептид укорачивается с C-конца на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 аминокислот по сравнению с SEQ ID NO:1 или с полипептидом, имеющим 99% идентичности по последовательности с SEQ ID NO:1 по всей длине.
В еще одном варианте осуществления изобретения по меньшей мере один рекомбинантный полипептид, экспрессируемый в клетках-хозяевах по изобретению, содержит одну или несколько из следующих молекул: по меньшей мере один антигенсвязывающий белок, по меньшей мере один единичный вариабельный домен и/или по меньшей мере однодоменное антитело. Полипептиды, содержащие по меньшей мере один антигенсвязывающий домен, могут также содержать по меньшей мере один полипептидный и/или пептидный агонист и/или антагонист рецептора. В некоторых случаях полипептидным агонистом может являться агонист рецептора GLP-1. Как известно в данной области, в одной клетке может экспрессироваться более одного рекомбинантного полипептида. В качестве примера, рекомбинантный полипептид, имеющий активность GLP-1, может экспрессироваться в той же клетке, что и антигенсвязывающий белок. Полипептид, имеющий активность GLP-1, может экспрессироваться с того же полинуклеотида, что и антигенсвязывающий белок, функционально связанного с нуклеотидными последовательностями, необходимыми для экспрессии. В качестве альтернативы и в качестве примера, полипептид, имеющий активность GLP-1, может экспрессироваться независимо от второго рекомбинантного полипептида, такого как антигенсвязывающий белок, или с той же эписомальной ДНК или с генома, но функционально связанными с разными полинуклеотидными последовательностями, необходимыми для экспрессии, или с ДНК-последовательностей, находящихся в отдельных векторах.
Также предложены генетически модифицированные клетки-хозяева, содержащие по меньшей мере один выделенный полипептид, кодирующий Killer Expression (KEX) протеазу (Kex2p) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Kex2p, по меньшей мере один выделенный полипептид, кодирующий протеин-дисульфидизомеразу (Pdi1) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность PDI, и по меньшей мере одну последовательность гетерологичной нуклеиновой кислоты, кодирующую оксидоредуктин эндоплазматического ретикулума (Ero1) или его фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность ERO. Генетически модифицированные клетки-хозяина по настоящему изобретению содержат по меньшей мере одну нуклеиновую кислоту, кодирующую рекомбинантный полипептид. В другом аспекте настоящего изобретения генетически модифицированные клетки-хозяива увеличивают экспрессию указанного рекомбинантного полипептида при выращивании в культуре по сравнению с клеткой-хозяином, относящейся к тому же виду и имеющей такие же генетические модификации, но которая не содержит по меньшей мере одну выделенную полинуклеотидную последовательность, кодирующую Killer Expression (KEX) протеазу Kex2p или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность KEX, по меньшей мере один выделенный полинуклеотид, кодирующий протеин-дисульфидизомеразу (Pdi1) или ее фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Pdi1, и по меньшей мере один выделенный полинуклеотид, кодирующий оксидоредуктин эндоплазматического ретикулума (Ero1) или его фрагмент и/или вариант, который имеет по меньшей мере одну функциональную активность Ero1. В некоторых случаях генетически модифицированными клетками-хозяевами являются S. cerevisiae .
В другом аспекте рекомбинантный полипептид, экспрессируемый в генетически модифицированной клетке-хозяине по настоящему изобретению, содержит аминокислотную последовательность, имеющую по меньшей мере 95%, 96%, 97%, 98%, 99% или 100% идентичности по последовательности с SEQ ID NO:1.
В другом аспекте предложены способы получения рекомбинантного полипептида, включающие культивирование генетически модифицированной клетки-хозяина по настоящему изобретению. В других аспектах способы дополнительно включают выделение указанного рекомбинантного полипептида из культуральной среды. В других аспектах предложен рекомбинантный полипептид, изготовленный указанными способами. В другом аспекте рекомбинантный полипептид, изготовленный способами по настоящему изобретению, содержит аминокислотную последовательность, имеющую 99% идентичности по последовательности с SEQ ID NO:1. В других аспектах рекомбинантный полипептид содержит аминокислотную последовательность, представленную в SEQ ID NO:1. В другом аспекте рекомбинантный полипептид содержит лидерную последовательность. В одном аспекте лидерная последовательность представляет собой модифицированную лидерную последовательность KEX, содержащую аминокислотную последовательность, представленную в SEQ ID NO:10. Как известно в данной области, клетка-хозяин и условия роста могут влиять на конечный продукт при рекомбинантной продукции белка клеткой-хозяином. Например, посттрансляционные модификации могут зависеть от типа клетки-хозяина и условий роста. Эти посттрансляционные модификации, включающие, но не ограниченные ими, гликозилирование и метилирование рекомбинантного белка, могут влиять на такие аспекты (но не ограниченные ими), как укладка белка и активность или удельная активность рекомбинантного белка, продуцируемого указанной клеткой-хозяином.
В еще одном аспекте настоящего изобретения предложена фармацевтическая композиция, содержащая рекомбинантный полипептид, изготовленный способами по настоящему изобретению. Также предложены способы лечения пациента, нуждающегося в этом, включающие введение терапевтически эффективного количества указанной фармацевтической композиции. В некоторых случаях пациент имеет заболевание или состояние, выбранное из диабета I типа, диабета II типа, нарушения толерантности к глюкозе, гипергликемии, болезни Альцгеймера, ожирения, сердечно-сосудистого заболевания, застойной сердечной недостаточности и ретинопатии.
В контексте настоящего изобретения «терапевтический полипептид» относится к белкам, полипептидам, антителам, пептидам или их фрагментам или вариантам, имеющим один или несколько видов терапевтической и/или биологической активности, и, в частности, по меньшей мере, одну биологическую активность, которая полезна для лечения, профилактики или ослабления заболевания. Терапевтические полипептиды, охватываемые настоящим изобретением, включают, но не ограничиваются ими, белки, полипептиды, пептиды, антитела и биологически активные соединения. (Термины пептиды, белки и полипептиды используются в настоящем описании взаимозаменяемо). Неполный перечень видов биологической активности, которыми может обладать терапевтический полипептид, включает любой из видов активности GLP-1, описанных в данном документе, усиление иммунного ответа, стимуляцию ангиогенеза, ингибирование ангиогенеза, регуляцию эндокринной функции, регуляцию гематопоэтических функций, стимуляцию роста нервов, усиление иммунного ответа или ингибирование иммунного ответа.
В контексте настоящего изобретения «пациент» представляет собой животное, предпочтительно млекопитающее и наиболее предпочтительно человека, имеющего заболевание, патологическое состояние или расстройство.
В контексте настоящего изобретения «терапевтически эффективное количество» относится к количеству, которое эффективно для лечения, профилактики или облегчения заболевания, патологического состояния или расстройства. Количество фармацевтической композиции по изобретению, которое будет эффективно для лечения, ингибирования и предупреждения заболевания или расстройства, связанного с аберрантной экспрессией и/или активностью терапевтического полипептида, может быть определено стандартными клиническими методами. Дополнительно (но необязательно) для определения оптимальных диапазонов доз можно использовать методы анализа in vitro. Точная доза, используемая в композиции, также будет зависеть от пути введения и тяжести заболевания или расстройства, и должна определяться лечащим врачом и в зависимости от состояния каждого пациента. Эффективные дозы могут быть экстраполированы из кривых «доза-ответ», полученных в тестовых системах in vitro и на животных моделях.
В контексте настоящего изобретения «фармацевтическая композиция» включает терапевтический полипептид и фармацевтически приемлемый носитель. В конкретном варианте осуществления термин «фармацевтически приемлемый» означает одобренный регулирующим ведомством федерального правительства или правительства штата или указанный в Фармакопее США или другой общепризнанной фармакопее для применения на животных и, более конкретно, у людей. Термин «носитель» означает разбавитель, адъювант, наполнитель или носитель, с которым вводится терапевтическое соединение. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая получаемые из нефти или животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем при внутривенном введении фармацевтической композиции. Солевые растворы и водные растворы декстрозы и глицерина также могут использоваться в качестве жидких носителей, в частности, для растворов для инъекций. Подходящие фармацевтические наполнители включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и т.п. Композиция, если желательно, может также содержать незначительное количество смачивающих или эмульгирующих агентов или рН-буферных агентов. Эти композиции могут иметь форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, препаратов с замедленным высвобождением и т.п. Композиция может быть составлена в виде суппозитория с традиционными связывающими агентами и носителями, такими как триглицериды. Пероральные составы могут включать стандартные носители, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлоза, карбонат магния и т.п. Примеры подходящих фармацевтических носителей описаны в "Remington's Pharmaceutical Sciences", автор E. W. Martin. Такие композиции будут содержать терапевтически эффективное количество соединения или терапевтического полипептида, предпочтительно в очищенной форме вместе с подходящим количеством носителя, чтобы обеспечить форму для надлежащего введения пациенту. Состав должен соответствовать способу введения.
Как известно в данной области, «выращивание в культуре» и грамматические варианты этой фразы относятся к инокуляции питательной среды клетками-хозяевами и к инкубации культуры клеток, как правило, в условиях, оптимальных или стандартных для роста конкретной клетки-хозяина, позволяющих клеткам расти и/или делиться. Как известно в данной области, ферментативная активность одного или нескольких ферментов, продуцируемых клетками-хозяевами в культуре, может зависеть от условий роста культуры. Например, протеолитическая активность протеазы, продуцируемой клеткой-хозяином в культуре, может быть снижена путем изменения одного или нескольких из следующих условий: рН, содержания растворенного кислорода, температуры, осмолярности, одного или нескольких компонентов среды, специфических протеазных ингибиторов, времени и/или скорости роста, концентрации клеток, продолжительности культивирования и/или скорости подачи глюкозы (например, подачи партиями). Добавление сложных белковых гидролизатов в культуру может быть особенно эффективно для ингибирования протеолиза. Более того, условия могут быть изменены в одной или нескольких определенных временных точках во время культивирования для максимизации эффекта. Аналогичным образом, гликозилирование белков, продуцируемых в культуре, может зависеть от аналогичных факторов. Таким образом, условия роста для снижения или повышения ферментативной активности клетки-хозяина (например, протеолитической или гликозилирующей активности) в культуре могут быть оптимизированы путем корректировки одного или нескольких из неограничивающих факторов, перечисленных выше.
Так же, как известно в данной области, продукция гетерологичного и/или рекомбинантного белка в клетке-хозяине может быть увеличена путем регуляции множества тех же факторов, что указаны выше. Кроме того, добавление факторов, увеличивающих число копий вектора, включая, но не ограничиваясь этим, добавление рапамицина в ростовую среду, также может увеличить продукцию. Другие факторы, которые могут увеличить продукцию, включают, но не ограничиваются ими, совместную экспрессию одного или нескольких шаперонных белков, таких как протеин- дисульфидизомераза (PDI). Кроме того, гемоглобин (HB) может совместно экспрессироваться по меньшей мере с одним гетерологичным полипептидом в клетке-хозяине, чтобы повысить доступность кислорода для окислительного метаболизма, таким образом, увеличивая продукцию полипептида.
В другом аспекте рекомбинантный полипептид, экспрессируемый в генетически модифицированных клетках-хозяевах по настоящему изобретению, содержит лидерную последовательность. В некоторых аспектах лидерная последовательность представляет собой лидерную последовательность KEX2 или модифицированную лидерную последовательность KEX2.
Лидерная последовательность КЕХ дикого типа представлена ниже как SEQ ID NO:9.

В некоторых случаях используется модифицированная лидерная последовательность КЕХ, представленная как SEQ ID NO:10.

В некоторых случаях лидерная последовательность КЕХ может иметь 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности по последовательности с SEQ ID NO:9 по всей длине последовательности.
Гетерологичные или рекомбинантные белки, которые секретируются из клетки-хозяина в процессе продукции, могут содержать лидерную последовательность, которая облегчает секрецию. Лидерные последовательности могут быть модифицированы для улучшения секреции и, следовательно, общей продукции и выделения экспрессированного гетерологичного белка - например, различные лидерные последовательности из различных секретируемых белков могут быть функционально связаны с гетерологичным белком и оценены по усилению экспрессии. Кроме того, данная лидерная последовательность может быть модифицирована с помощью сайт-направленного мутагенеза или с использованием комбинаторной библиотеки для определения улучшенного варианта лидерной последовательности. Для увеличения уровня экспрессии гетерологичного белка могут быть созданы химерные лидерные последовательности, содержащие области из двух или нескольких лидерных пептидов.
Примеры
В следующих примерах проиллюстрированы различные неограничивающие аспекты данного изобретения.
Пример 1 - штамм S. cerevisiae, сверхэкспрессирующий KEX2, PDI1 и ERO1
Штамм S. ceresiviae, сверхэкспрессирующий KEX2, PDI1 и ERO1, был сконструирован с использованием дрожжевой экспрессионной системы (S. cerevisiae BXP10), созданной в компании Delta Biotechnology Ltd. (Delta) в Ноттингеме, Соединенное королевство. BXP10 происходит от штамма AH22 S. cerevisiae, полученного из ATCC, который является производным s288c. Создание BXP10 включало серию процедур случайного мутагенеза и направленного выключения генов для увеличения способности к секреции рекомбинантного сывороточного альбумина человека (rHSA) и снижения нежелательных посттрансляционных модификаций.
На фигуре 1 показано создание BXP10-KEX2-PDI1-ERO1, штамма, который сверхэкспрессирует KEX2, PDI1 и ERO1. Экспрессионные кассеты, содержащие КЕX2-KanMX, функционально связанный с PGK1-промотором, PDI-HphMX, функционально связанный с PGK1-промотором, или ERO-BsdMX, функционально связанный с PGK1-промотором, были последовательно встроены в локусы NTS2-2 (нетранскрибируемой спейсерной области в повторах рДНК) штамма BXP10, так что были созданы следующие штаммы: BXP10-KEX2 (штамм, сверхэкспрессирующий KEX2), BXP10-KEX2-PDI1 (штамм, сверхэкспрессирующий KEX2 и PDI1) и BXP10-KEX2-PDI1-ERO1 (штамм, сверхэкспрессирующий KEX2, PDI1 и ERO1).
Для создания кассеты для экспрессии KEX2, открытая рамка считывания (ORF) KEX2, промотор гена PGK1 (PPGK1) и последовательность терминации транскрипции гена ADH1 (TADH1) были амплифицированы индивидуально с помощью ПЦР с геномной ДНК ВРХ10 и собраны после с использованием другой ПЦР-реакции («сшивающей» или «сливающей» ПЦР). Собранный фрагмент PPGK1-KEX2-TADH1 был клонирован в pRS314KanMX для создания pRS314KanMXpPGK1-КЕХ2. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали ВХР10 с помощью электропорации и высевали на чашки, содержащие G418. Клоны, резистентные к G418, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.
Плазмида pRS314HphMXpPGK1-PDI1, которая несет кассету для экспрессии PDI1, была сконструирована замещением областей ORF KEX2 и KanMX в pRS314KanMXpPGK1-КЕХ2 на амплифицированные с помощью ПЦР ORF PDI1 и маркер резистентности к гигромицину B, HphMX. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали штаммы ВХР10-KEX2 с помощью электропорации и высевали на чашки, содержащие гигромицин В. Клоны, резистентные к гигромицину В, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.
ORF ERO1 была амплифицирована с геномной ДНК BXP10 с помощью ПЦР и далее клонирована в pRS314pPGK1BsdMX для создания pRS314BsdMXpPGK1-ERO1. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали штамм-хозяин ВXP10-KEX2-PDI1 с помощью электропорации и высевали на чашки, содержащие бластицидин S. Клоны, резистентные к бластицидину S, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.
На фигуре 2 показан анализ методом Саузерн-блота, подтверждающий встраивание KEX2 и PDI1 в локусы NTS2-2 штамма BXP10, для создания штаммов BXP10-KEX2 и BXP10-KEX2-PDI1. В случае КЕХ2, полоса 2,6 т.п.о. соответствует эндогенной копии КЕХ2, а полоса 1,6 т.п.о. соответствует успешно встроенной копии. Для PDI1 полоса 1,3 т.п.о. соответствует эндогенной копии PDI1, а полоса 1,7 т.п.о. соответствует успешно встроенной копии.
На фигуре 3 показан анализ PDI1 и KEX2 методом Вестерн-блота, показывающий сверхэкспрессию PDI1 и KEX2 в клонах BXP10-KEX2-PDI1. Из пяти клонов BXР10-KEX2-PDI1, клон #2 имел наибольшую экспрессию как PDI1, так и KEX2 и поэтому был выбран в качестве штамма-хозяина для создания BXP10-KEX2-PDI1-ERO1.
Клетки-хозяева затем были трансформированы плазмидой pCID3610, которая содержит рекомбинантный слитый белок («белок pCID3610»), состоящий из двух копий глюкагон-подобного пептида 1 человека (GLP-1, фрагмент 7-36(A8G)) и рекомбинантного альбумина человека (rHA). Каждая последовательность GLP-1 была модифицирована глицином, замещающим природный аланин в позиции 8 для придания резистентности к протеолизу. Вторая последовательность GLP-1 действует в качестве пептидного спейсера между первой последовательностью GLP-1 и rHA. Белок pCID3610 представляет собой негликозилированный белок, состоящий из 645 аминокислот и имеющий молекулярный вес 72970,4 Да. PCID3610 подробно описан в патенте США № 7569384, полное содержание которого включено в данный документ.
Плазмида pCID3610 была сконструирована в компании Human Genome Sciences, находящейся в городе Роквилл, штат Мэриленд, с использованием экспрессионного вектора на основе pSAC35. pSAC35 содержит ген LEU2 из S. cerevisiae в качестве селективного маркера, который восполняет лейциновую ауксотрофность в BXP10. pSAC35 также содержит сильный дрожжевой промотор (PRB1), уникальный сайт для клонирования (NotI) и последовательность из pUC9-плазмиды для E. coli, позволяющую клонирование и размножение в E. coli. Кроме того, pSAC35 является дезинтегративным вектором, и при трансформации в дрожжи принадлежащие pUC9 последовательности вырезаются путем сайт-специфичной рекомбинации. Это вырезание выполняется с использованием распознаваемых FLP участков (FRT) и экспрессии дрожжевой рекомбиназы FLP («flip») с плазмиды 2-micron. Другие сегменты в pSAC35 включают области REP1 и REP2 D-гена. Гены REP1 и REP2 кодируют продукты, которые помогают регулировать число копий плазмиды и также участвуют в сегрегации плазмиды в ходе деления клетки. Продукт D-гена повышает экспрессию FLP путем снятия репрессии, вызываемой REP1 и REP2. pCID3610 подробно описана в патенте США № 7569384, полное содержание которого включено в данный документ.
Полноразмерная кДНК альбумина человека (НА) была выделена из библиотеки кДНК человека и клонирована в плазмиду pAT153ALB в лаборатории доктора F.E. Baralle в Оксфордском университете Соединенного Королевства. pAT153ALB была впоследствии модифицирована компанией Delta путем введения новых рестрикционных сайтов для облегчения клонирования в pSAC35.
Экспрессионный вектор, плазмида pCID3610, был сконструирован путем введения слитого гена GLP-1-rHSA, созданного, как указано ниже. Во-первых, были получены синтетические гены, кодирующие лидерный пептид и зрелый вариант GLP-1, имеющий одну замену A на G в позиции 2 зрелого пептида. Вариант пептида GLP-1 был обратно транслирован с использованием оптимальных кодонов для дрожжей, и посредством ПЦР были синтезированы тандемные копии с использованием перекрывающихся олигонуклеотидов. Этот синтетический ген использовали в качестве матрицы во втором раунде ПЦР для добавления 5'- и 3'-сайтов рестрикции, позволяющих его клонирование в 5'-конец гена rHSA. Наконец, последовательность, кодирующую сигнальный пептид, лигировали в 5'-конец конструкции GLP-1. Полученный фрагмент лигировали в pSAC35 в уникальный сайт NotI и трансформировали в DH5α, получая в результате экспрессионный вектор pCID3610. Была подтверждена нуклеотидная последовательность pCID3610. После чего pCID3610 повторно трансформировали в DH5α для дальнейшей амплификации и выделения плазмидной ДНК.
Для создания штамма BXP10-KEX2-PDI1-ERO1, экспрессирующего pCID3610, плазмидой pCID3610 трансформировали BXP10-KEX2-PDI1-ERO1 с помощью электропорации, клетки высевали на чашки с ESFM2-агарозой, и колонии Leu+ отбирали после четырех дней инкубации при 30°C. Двенадцать колоний трансформантов дополнительно перештириховывали на чашки к ESFM2-агарозой для получения одиночных колоний. Одной колонией из каждого штриха заражали ESFM2-среду для скрининга с использованием 24-луночных культуральных планшетов с глубокими лунками.
После 3 дней инкубации с качанием супернатанты от каждой из 12 клональных культур анализировали электрофорезом в ДСН-ПААГ. На фигуре 4 показаны результаты электрофореза в ДСН-ПААГ образцов 12 супернатантов. Затем 4 из 12 клонов (клоны #2, #8, #10 и #12) были отобраны для дальнейших ферментационных тестов (отобранные клоны показаны стрелками на фигуре 4). Вследствие различной степени испарения среды из каждой лунки в культуральном планшете, четыре клона были выбраны с учетом конечного объема каждой культуры в конце роста, измерения OD клеточной культуры и интенсивности полосы на геле.
Четыре отобранных клона затем выращивали в мини-биореакторах DASGIP с помощью программы ферментации, а полученные итоговый титр и качество белка сравнивали полученными при экспрессии pCID3610 в BXP10, штамме-хозяине, который не сверхэкспрессирует KEX2, PDI1 и ERO1. На фигуре 5 показан анализ итогового титра и качества белка, получаемого в процессе ферментации. Качество белка измеряется по процентному содержанию белкового продукта, имеющего дополнительные 6 аминокислот (6-AA) на N-конце из-за неэффективного расщепления лидерной последовательности. Клоны #2 и #8 показали значительное увеличение концентрации белка pCID3610 по сравнению с BXP10, экспрессирующим pCID3610, который синтезирует только до 1,6 г/л белка pCID3610 при тех же условиях ферментации (данные для BXP10, экспрессирующего pCID3610, не показаны на фиг.5). Кроме того, содержание 6-AA во всех клонах (<1% белкового продукта, имеющего дополнительные 6 аминокислот) было значительно снижено по сравнению с содержанием 6-AA в BXP10, экспрессирующем белок pCID3610, который содержал 4-7% белкового продукта, имеющего дополнительные 6-AA (данные для BXP10, экспрессирующего pCID3610, не показаны на фиг.5). Эти результаты показывают, что сверхэкспрессия KEX2, PDI1 и ERO1 значительно улучшала штамм-хозяин (BXP10) в отношении получения белка pCID3610 с более лучшим качеством.
Хотя итоговый титр и уровень 6-АА у клона #2 и клона #8 были сравнимы, клон #8 был выбран в качестве основного клона, поскольку он давал немного более лучшие результаты при ферментации. Для подтверждения улучшенных итогового титра и качества белка для pCID3610, клон #8 ферментировали в 15-литровом ферментере. Средний итоговый титр для четырех партий составлял 2,5 г/л, что составляло 40-50%-ое увеличение итогового титра при использовании BXP10, экспрессирующего pCID3610.
Были приготовлены два отдельных замороженных законсервированных запаса клона #8 ВXP10-KEX2-PDI1-ERO1. Первый замороженный законсервированный запас (ампулы «Исследовательского клеточного банка») был приготовлен путем выращивания клона #8 в 200 мл среды ESFM2, которая содержала все три антибиотика (G418, гигромицин и бластицидин). Когда клеточная культура достигла OD600 ~3,0, клетки были собраны, промыты, ресуспендированы и разделены на аликвоты для получения замороженного запаса, содержащего 20% трегалозы.
Клетки из ампулы Исследовательского клеточного банка затем размораживали и выращивали на среде ESFM2 при 30°C и 250 об./мин. Плотность культуры контролировали путем измерения OD600 культуры. На фигуре 6 показаны кривая роста клеток. При OD600, приблизительно равной 2,54, клетки собирали, промывали и ресуспендировали для получения второго замороженного законсервированного запаса («Предварительного главного клеточного банка»). На фигуре 7 показана кривая роста клеток из Предварительного главного клеточного банка.
Стабильность клона #8 ВXP10-KEX2-PDI1-ERO1 затем проверяли старением клеток in vitro и получением в 15-литровом объеме. Кратко, клетки клона #8 из Предварительного главного клеточного банка пассировали через семь последовательных стадий выращивания в колбах при встряхивании, что соответствует приблизительно 51 поколению клеток. Затем клетками засевали 15-литровые ферментеры и выращивали в программе ферментации. Этот процесс ферментации добавлял еще 14 поколений. Итоговый титр супернатанта достигал 5,3 г/л с уровнем 6-АА менее 1,5%. Эти данные указывают на то, что клон #8 BXP10-KEX2-PDI1-ERO1 стабильно продуцирует белок pCID3610 после примерно 65 поколений.
Для создания кассеты для экспрессии KEX2, промотор гена PGK1 (PPGK1), ORF KEX2 и последовательность терминации транскрипции гена ADH1 (TADH1) были амплифицированы индивидуально с помощью ПЦР с геномной ДНК ВРХ10 и собраны после с использованием другой ПЦР-реакции («сшивающей» или «сливающей» ПЦР). Собранный фрагмент PPGK1-KEX2-TADH1 был клонирован в pRS314KanMX для создания pRS314KanMXpPGK1-КЕХ2. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали ВХР10 с помощью электропорации и высевали на чашки, содержащие G418. Клоны, резистентные к G418, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.
ORF PDI1 амплифицировали на геномной ДНК из ВХР10. Этот ДНК-фрагмент и маркер резистентности к гигромицину B, HphMX, использовали для замещения областей ORF KEX2 и KanMX в pRS314KanMXpPGK1-КЕХ2, что давало плазмиду pRS314HphMXpPGK1-PDI1. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали штаммы-хозяева ВХР10-KEX2 с помощью электропорации и высевали на чашки, содержащие гигромицин В. Клоны, резистентные к гигромицину В, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.
Аналогичным образом, ORF ERO1 была амплифицирована с геномной ДНК BXP10 с помощью ПЦР и далее клонирована в pRS314pPGK1BsdMX для создания pRS314BsdMXpPGK1-ERO1. Эту плазмиду использовали в качестве матрицы в финальном раунде ПЦР для добавления 5'- и 3'-фланкирующих последовательностей (105 п.о. и 101 п.о., соответственно), которые являются гомологичными участкам интеграции NTS2-2. Полученным фрагментом ДНК трансформировали штамм-хозяин ВXP10-KEX2-PDI1 с помощью электропорации и высевали на чашки, содержащие бластицидин S. Клоны, резистентные к бластицидину S, дополнительно подтверждали, как положительные по сайт-специфичной интеграции, с помощью ПЦР на колониях.










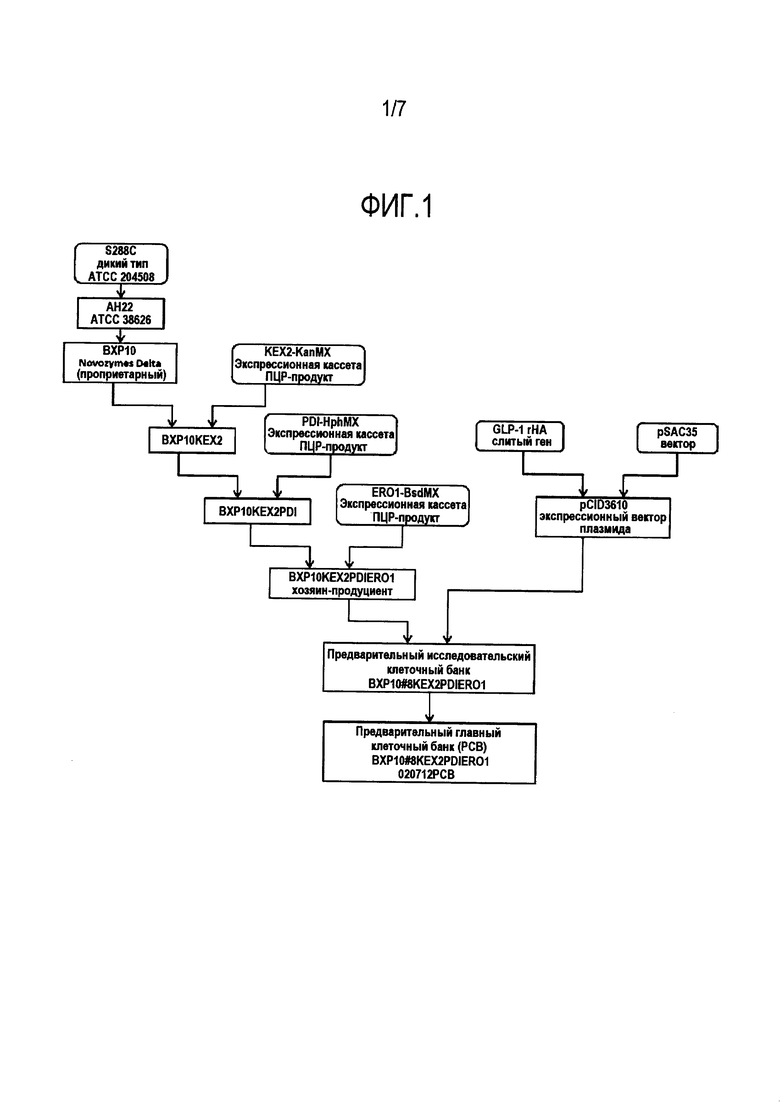
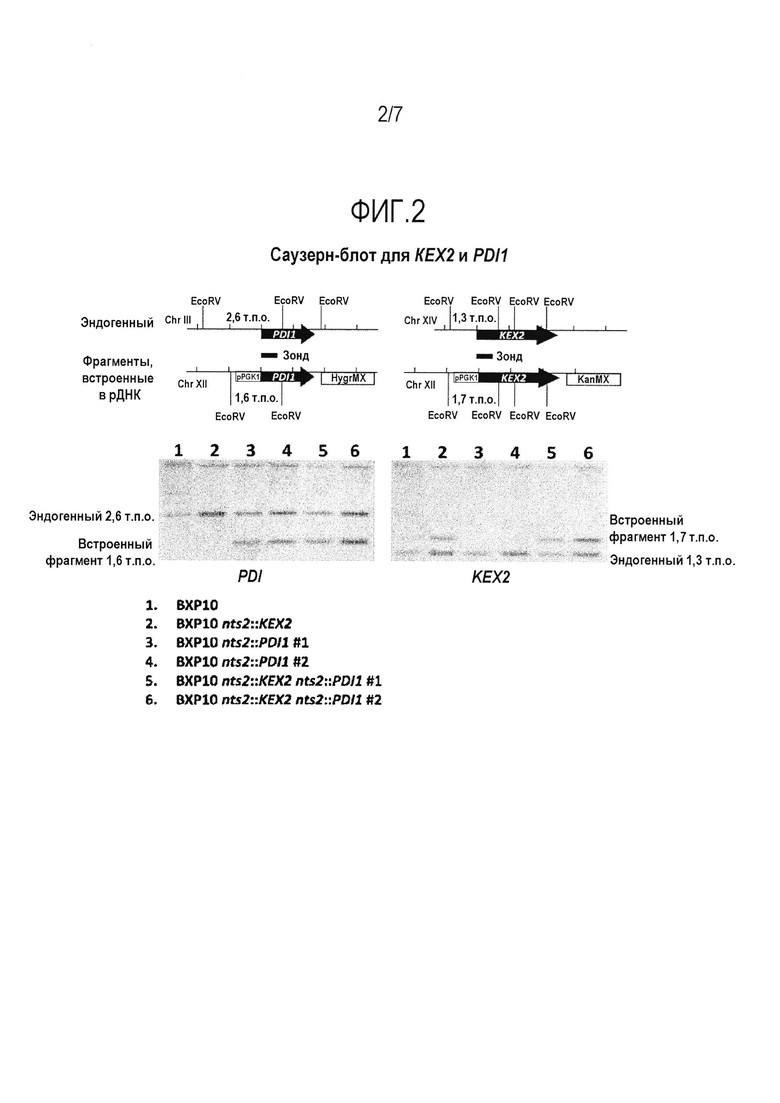
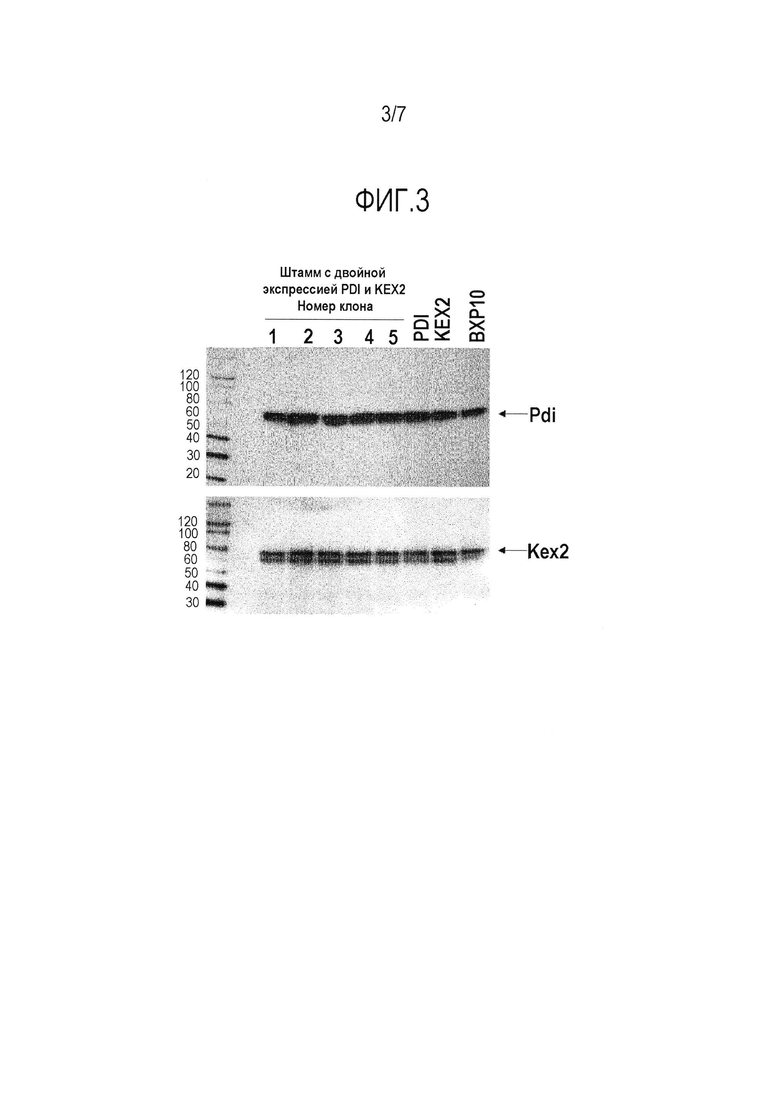

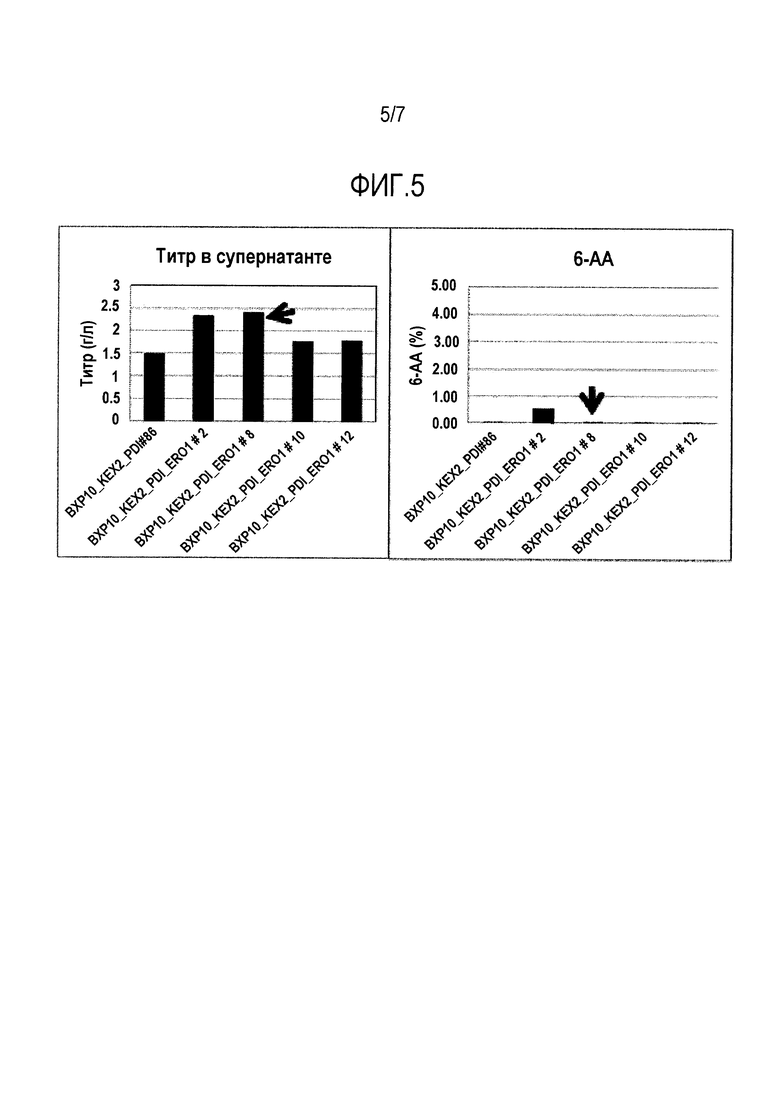
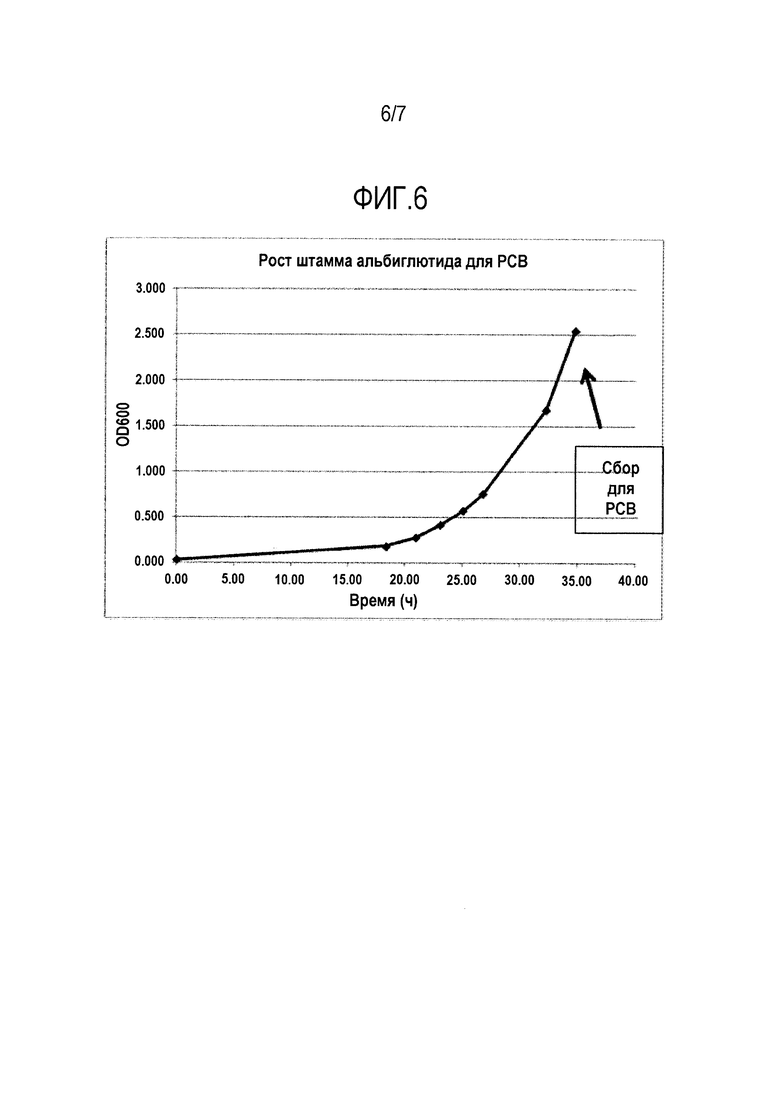
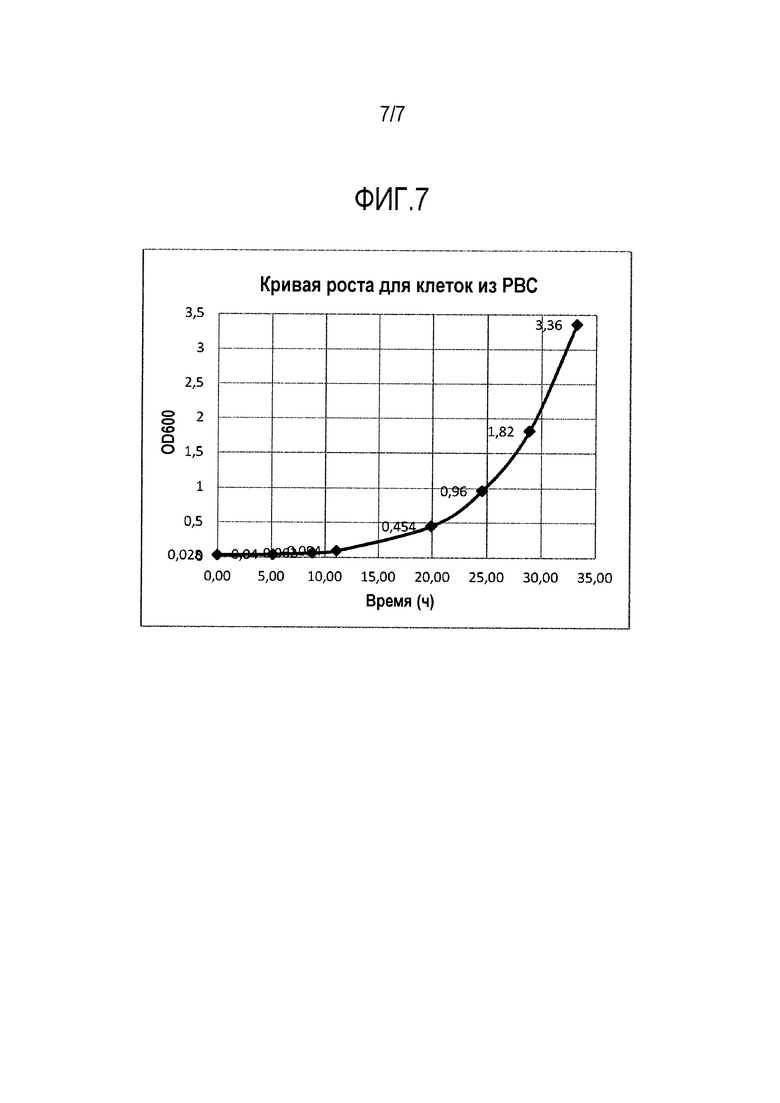
Группа изобретений относится к области биотехнологии. Предложена клетка-хозяин для получения рекомбинантного полипептида, содержащая гетерологичную последовательность нуклеиновой кислоты, кодирующую протеазу Killer Expression (Kех2р), гетерологичную последовательность нуклеиновой кислоты, кодирующую протеин-дисульфидизомеразу (Pdi1), и нуклеиновую кислоту, кодирующую рекомбинантный полипептид. Предложен способ получения рекомбинантного полипептида, включающий культивирование указанной клетки-хозяина. Предложена клетка-хозяин для получения рекомбинантного полипептида, содержащая гетерологичную последовательность нуклеиновой кислоты, кодирующую протеазу Killer Expression (Kex2p), гетерологичную последовательность нуклеиновой кислоты, кодирующую протеин-дисульфидизомеразу (Pdi1), гетерологичную последовательность нуклеиновой кислоты, кодирующую оксидоредуктин эндоплазматического ретикулума (Ero1), и нуклеиновую кислоту, кодирующую рекомбинантный полипептид. Группа изобретений позволяет повысить продукцию рекомбинантного полипептида. 3 н. и 28 з.п. ф-лы, 7 ил., 1 пр.
1. Клетка-хозяин для получения рекомбинантного полипептида, где указанная клетка-хозяин содержит по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеазу Killer Expression (Kех2р) или ее функциональный фрагмент и/или вариант, по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеин-дисульфидизомеразу (Pdi1) или ее функциональный фрагмент и/или вариант, и по меньшей мере одну нуклеиновую кислоту, кодирующую рекомбинантный полипептид.
2. Клетка-хозяин по п. 1, дополнительно содержащая по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую оксидоредуктин эндоплазматического ретикулума (Ero1) или его функциональный фрагмент и/или вариант.
3. Клетка-хозяин по п. 1 или 2, где указанная клетка-хозяин экспрессирует или сверхэкспрессирует по меньшей мере один генный продукт указанной по меньшей мере одной гетерологичной последовательности нуклеиновой кислоты, кодирующей белок или его функциональный фрагмент и/или вариант, выбранный из Kex2p, Pdi1 и Ero1, когда указанную клетку-хозяина выращивают в культуре.
4. Клетка-хозяин по п. 1, где указанная клетка-хозяин представляет собой клетку гриба.
5. Клетка-хозяин по п. 4, где указанной клеткой гриба является дрожжевая клетка.
6. Клетка-хозяин по п. 5, где род указанной дрожжевой клетки выбирают из группы, состоящей из Saccharomyces, Kluyveromyces, Candida, Pichia, Schizosaccharomyces, Hansenula, Kloeckera, Schwanniomyces и Yarrowia.
7. Клетка-хозяин по п. 6, где указанная клетка-хозяин представляет S. cerevisiae.
8. Клетка-хозяин по п. 1, где указанная клетка-хозяин представляет собой клетку млекопитающего.
9. Клетка-хозяин по п. 1 или 2, где указанная по меньшей мере одна гетерологичная последовательность нуклеиновой кислоты, кодирующая Кех2р или ее функциональный фрагмент и/или вариант, функционально связана по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, МЕТ25, CUP1, РН05, tetO-CYC1, CaMV, НХТ6, HXT7 и ARE.
10. Клетка-хозяин по п. 1 или 2, где указанная по меньшей мере одна гетерологичная последовательность нуклеиновой кислоты, кодирующая Pdi1, функционально связана по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, МЕТ25, CUP1, PHO5, tetO-CYC1, CaMV, HXT6, HXT7 и ARE.
11. Клетка-хозяин по п. 2, где указанная по меньшей мере одна гетерологичная последовательность нуклеиновой кислоты, кодирующая Ero1, функционально связана по меньшей мере с одним промотором, выбранным из группы: TEF1, PRB1, ADH1, ADH2, PYK1, PGK1, ENO, GAL1.10.7, GALS, MET25, CUP1, PHO5, tetO-CYC1, CaMV, HXT6, HXT7 и ARE.
12. Клетка-хозяин по п. 1 или 2, где указанная клетка-хозяин дополнительно содержит по меньшей мере одну из следующих генетических модификаций: нокаут протеазы рер4, сниженная активность ubc4 и/или ubc5 по сравнению с клеткой-хозяином дикого типа, нокаут yps1, нокаут hsp150 и нокаут pmt1.
13. Клетка-хозяин по п. 1, где указанная по меньшей мере одна нуклеиновая кислота, кодирующая рекомбинантный полипептид, содержится в плазмиде.
14. Клетка-хозяин по п. 1, где указанная по меньшей мере одна нуклеиновая кислота, кодирующая рекомбинантный полипептид, встроена в геном указанной клетки-хозяина.
15. Клетка-хозяин по пп. 13, 14, где указанный рекомбинантный полипептид имеет по меньшей мере одну дисульфидную связь.
16. Клетка-хозяин по пп. 13, 14, где указанный рекомбинантный полипептид является белком, слитым с альбумином.
17. Клетка-хозяин по пп. 13, 14, где указанный рекомбинантный полипептид содержит по меньшей мере один терапевтический полипептид, имеющий активность GLP-1, генетически слитый с альбумином.
18. Клетка-хозяин по пп. 13, 14, где указанный рекомбинантный полипептид содержит полипептид, имеющий по меньшей мере 95% идентичности по последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 1.
19. Клетка-хозяин по пп. 13, 14, где рекомбинантный полипептид содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1.
20. Клетка-хозяин по пп. 13, 14, где указанный рекомбинантный полипептид содержит лидерную последовательность.
21. Клетка-хозяин по п. 20, где указанная лидерная последовательность является лидерной последовательностью KEX2 или модифицированной лидерной последовательностью KEX2.
22. Клетка-хозяин по п. 1, где рекомбинантный белок содержит модифицированную лидерную последовательность, приведенную в SEQ ID NO: 10.
23. Способ получения рекомбинантного полипептида, включающий культивирование клетки-хозяина по пп. 1-22.
24. Способ по п. 23, дополнительно включающий выделение указанного рекомбинантного полипептида из культуральной среды.
25. Клетка-хозяин для получения рекомбинантного полипептида, содержащая по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеазу Killer Expression (Kex2p) или ее функциональный фрагмент и/или вариант, по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеин-дисульфидизомеразу (Pdi1) или ее функциональный фрагмент и/или вариант, по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую оксидоредуктин эндоплазматического ретикулума (Ero1) или его функциональный фрагмент и/или вариант, и по меньшей мере одну нуклеиновую кислоту, кодирующую рекомбинантный полипептид.
26. Клетка-хозяин по п. 25, где указанная Kex2p или ее функциональный фрагмент и/или вариант представляет собой функциональный генный продукт SEQ ID NO: 3.
27. Клетка-хозяин по п. 25, где указанная Pdi1 или ее функциональный фрагмент и/или вариант представляет собой функциональный генный продукт SEQ ID NO: 5.
28. Клетка-хозяин по п. 25, где указанный Ero1 или его функциональный фрагмент и/или вариант представляет собой функциональный генный продукт SEQ ID NO: 7.
29. Клетка-хозяин по любому одному из пп. 25-28, где указанная клетка-хозяин представляет собой S. cerevisiae.
30. Клетка-хозяин по п. 25, где указанный рекомбинантный полипептид содержит полипептид, имеющий по меньшей мере 99% идентичности по последовательности с аминокислотной последовательностью, приведенной в SEQ ID NO: 1.
31. Клетка-хозяин по п. 30, где указанная клетка-хозяин увеличивает итоговый титр указанного рекомбинантного полипептида при выращивании в культуре по сравнению с клеткой-хозяином, относящейся к тому же виду и имеющей такие же генетические модификации, но которая не содержит по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеазу Killer Expression (Kex2p) или ее функциональный фрагмент и/или вариант, по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую протеин-дисульфидизомеразу (Pdi1) или ее функциональный фрагмент и/или вариант, и по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты, кодирующую оксидоредуктин эндоплазматического ретикулума (Ero1) или его функциональный фрагмент и/или вариант.
| US 20060046253 A1, 02.03.2006 | |||
| US 8034607 B2, 11.10.2011 | |||
| US 2009123452 A1, 14.05.2009 | |||
| US 2011014651 A1, 20.01.2011 | |||
| US 6361964 B1, 26.03.2002 | |||
| Электрический прожектор для экипажей с несколькими лампами | 1926 |
|
SU13134A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| СПОСОБ ЭЛЕКТРОЛИТИЧЕСКОГО ХРОМИРОВАНИЯ | 1929 |
|
SU17967A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| ПРИБОР ДЛЯ РЫТЬЯ КАНАВ ИЛИ ОЧИСТКИ СНЕГА С ЖЕЛЕЗНОДОРОЖНЫХ ПУТЕЙ | 1924 |
|
SU3103A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| US 8202837 B2, 19.06.2012 (см | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА, ШТАММ Saccharomyces cerevisiae - ПРОДУЦЕНТ ЗРЕЛОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ) | 2010 |
|
RU2427645C1 |

