Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к областям биологии и медицины, более конкретно, к молекулярной и клеточной биологии, и относится к рекомбинантной инженерии и экспрессии микроорганизмами продуктов, таких как пептиды, полипептиды или белки. Более конкретно, настоящее изобретение относится к конструкциям полицистронной экспрессии или кассетам для экспрессии микроорганизмами таких продуктов и дополнительно к родственным векторам, трансформированным хозяевам, использованиям и применениям, таким как доставка, в особенности, доставка терапевтического средства, совместно экспрессируемых продуктов субъектам.
Предшествующий уровень техники настоящего изобретения
На сегодняшний день для различных биотехнологических областей применений было разработано большое количество систем экспрессии для рекомбинантных белков. Системы для экспрессии гетерологичных или гомологичных генов были определены у прокариот, дрожжей и грибов и у клеток млекопитающих.
Большинство полученных из дрожжей рекомбинантных белков были экспрессированы при помощи Saccharomyces cerevisiae в качестве системы-хозяина. Несмотря на это, для системы S. cerevisiae было выявлено несколько ограничений. Примерами являются выход продукта, который обычно является низким, и недостаточная секреция (множество белков S. cerevisiae не обнаруживаются в культуральной среде в свободном виде, но наоборот содержатся в периплазматическом пространстве или связаны с клеточной стенкой) (Dominguez et al. Int. Microbiol., 1998, vol. 1(2), 131-142). В результате ограничений выработки у дрожжей появился большой интерес к экспрессии белков в бактериях, которые легко выращивать в недорогостоящем бульоне, и их часто применяют для получения рекомбинантных белков. Среди прокариотических систем наиболее высокие уровни белков обычно получают при помощи рекомбинантной экспрессии у Escherichia coli (E. coli) (Jana & Deb. Appl. Microbiol. Biotechnol., 2005, vol. 67(3), 289-298). Тем не менее, у Е. coli наиболее часто применяемые стратегии получения являются внутриклеточными (в периплазме или цитоплазме) и, следовательно, предусматривают дорогостоящие и зачастую проблематичные последующие способы очистки.
Кисломолочные бактерии (LAB) приобретают повышенную важность в качестве хозяев для рекомбинантной экспрессии гетерологичных полипептидов in vitro (например, патент США №5559007), а также экспрессии in vivo или in situ и доставки антигенов и/или терапевтически необходимых полипептидов (например, WO 97/14806). Гетерологичные белки, выработанные у таких грамположительных бактериальных хозяев, могут быть легко секретированы в среду, таким образом, облегчая их очистку, а также их прямую доставку субъектам.
Большинство систем экспрессии могут очень хорошо управлять экспрессией одного отдельного белка (в качестве результата одной отдельной генной последовательности). Тем не менее, в некоторых случаях желательно иметь систему экспрессии, которая способна экспрессировать множество белков или полигенные белковые комплексы, например, in vitro экспрессия антител или белковых комплексов, а также in vivo или in situ экспрессия и доставка двух или более белков, которые оказывают синергический эффект при конкретном заболевании, или in vivo или in situ экспрессия и доставка антител или их функциональных (полигенных) фрагментов. В таких случаях, желательно иметь полимерные гены, которые кодируют необходимые белки или антитела под контролем одного промотора из-за необходимости тесной совместной регуляции полимерных генов.
Два наиболее широкоизвестных подхода получения рекомбинантных белковых комплексов заключаются в выполнении in vitro реконструкции отдельно экспрессированных и очищенных субъединиц или в осуществлении in vivo реконструкции путем совместной экспрессии субъединиц в соответствующем хозяине (Selleck & Tan, "Recombinant protein complex expression in E. coli, Curr. Protoc. Protein Sci., 2008, chapter 5:unit 5, 21). Несмотря на успешное применение реконструкции in vitro этот способ является трудоемким (каждая субъединица должна быть экспрессирована и очищена, а комплекс должен быть дополнительно очищен после реконструкции) и выходы реконструкции часто являются низкими. В отличие от этого, реконструкция in vivo посредством совместной экспрессии обеспечивает преимущества эффективности (только один цикл экспрессии и очистки) и потенциально более высокие выходы и качество необходимого комплекса (рефолдинг и сборка комплекса происходят в присутствии обеспечивающих сворачивание белка ферментов в клеточной среде) (Selleck & Tan 2008, ранее). Реконструкция in vivo была успешно выполнена путем совместного инфицирования бакуловирусом клеток насекомых, экспрессирующих отдельные субъединицы белка (Tirode et al. Mol. Cell, 1999, vol. 3(1), 87-95), и в бактериях из сложных плазмид (Johnston et al. Protein Expr. Purif., 2000, vol. 20(3), 435-443; McNally et al. Proc. Natl. Acad. Sci. USA, 1988, vol. 85(19), 7270-7273) или из специализированных полицистронных плазмид (Henricksen et al. J. Biol. Chem., 1994, vol. 269(15), 1112111132; Ishiai et al. J. Biol. Chem. 1996, vol. 271(34), 20868-20878; Li et al. Proc. Natl. Acad. Sci. USA, 1997, vol. 94(6), 2278-2283).
Основные системы полицистронной экспрессии для получения белковых комплексов в Е. coli были описаны в (Selleck & Tan 2008, ранее; Tan. Protein. Expr. Purif., 2001, vol. 21(1), 224-234; Tan et al. Protein Expr. Purif., 2005, vol. 40(2), 385-395). В таких системах использована концепция трансляционной кассеты, состоящей из кодирующего участка с соответствующими ИНИЦИИРУЮЩИМ и СТОП кодонами и с предшествующими сигналами инициации трансляции, такими как последовательность Шайна-Дальгарно (SD) и трансляционные энхансеры (Tan 2001, Tan et al. 2005, ранее). В транскрибированном в мРНК состоянии трансляционная кассета содержит необходимую и достаточную информацию для запуска механизма трансляции Е. coli и поддержания трансляции мРНК в необходимый полипептид (Selleck & Tan 2008, ранее).
Был описан вектор бицистронной экспрессии для интерлейкина-18 у Е. coli, однако, межгенный участок между двумя генами состоял из синтетического линкера и явно является гено-специфичным, поскольку экспрессия каспазы-4 была значительно выше экспрессии ICE. Smolke et al. ранее показали, что возможен дифференциальный контроль уровней белка, кодируемого двумя или более генами в опероне при помощи последовательностей синтетических межгенных участков (Smolke et al. Appl. Environ. Microbiol., 2000, vol. 66(12), 5399-5405; Smolke & Keasling. Biotechnol. Bioeng., 2002, vol. 80(7), 762-776). Тем не менее, в основе этого подхода лежат случайные комбинации, и он нуждается во введении синтетических последовательностей в клетку-хозяина для экспрессии.
В последние годы возникла потребность в новых и улучшенных системах выработки антител. Системы для экспрессии антител были определены у прокариот, дрожжей и грибов и у клеток млекопитающих. Несмотря на то, что одноцепочечные и однодоменные антитела легче получать от бактерий, полноразмерные антитела обычно характеризуются более высокими показателями сродства связывания и меньшим риском образования нейтрализующего антитела при введении.
Полноразмерные антитела можно получить из бактерий (Mazor et al. Nat. Biotechnol., 2007, vol. 25(5), 563-565; Simmons et al. J. Immunol. Methods, 2002, vol. 263(1-2), 133-147). В большинстве публикаций о рекомбинантной экспрессии у прокариот описано получение фрагментов антител, хотя фактически исключительно от Е. coli. Несмотря на то, что многие сконструированные LAB способны корректировать образование дисульфидной связи, в литературе содержится только ограниченное число примеров подобных антителу молекул, полученных от LAB (Kruger et al. Nature Biotechnology, 2002, vol. 20(7), 702-706; Beninati et al. Nature Biotechnology, 2000, vol. 18(10), 1060-1064; Chancey et al. J. Immunol, 2006, vol. 176(9), 5627-5636; Hultberg et al. BMC Biotechnol, 2007, vol. 7, 58; Yuvaraj et al. Mol. Nutr. Food. Res., 2008, vol. 52(8), 913-920). В таких публикациях описаны только одноцепочечные фрагменты антител, экспрессируемые у видов Lactobacillus, Lactococcus lactis и Streptococcus gordonii, но не полигенные, двухцепочечные фрагменты антител или полноразмерные антитела.
Системы полицистронной экспрессии могут быть очень важны для получения у прокариот эффективного синтеза и экспрессии сложных белков, таких как антитела. С момента одобрения FDA в 1986 году муромонаба-CD3, все еще одного из наиболее сильнодействующих иммунодепрессивных лекарственных средств, пригодных для контроля отторжения трансплантата (Hooks et al. Pharmacotherapy, 1991, vol. 11(1), 26-37), полноразмерные антитела и фрагменты антител становились все более важными и универсальными инструментальными средствами в медицине.
Несмотря на то, что из уровня техники известно несколько примеров систем полицистронной экспрессии в бактериальных клетках, эти примеры весьма ограничены, что подчеркивает необходимость в более эффективной системе введения и экспрессии полимерных генов. Соответственно, имеет место необходимость получения дополнительных последовательностей, которые можно благоприятно применять для экспрессии белков, предпочтительно экспрессии гетерологичного белка и еще более предпочтительно экспрессии множества гетерологичных белков.
В дополнение к изложенному выше, попытка получить более высокие количества рекомбинантного белка как для прямой доставки белка посредством рекомбинантных организмов, так и для массового получения белка и последующей очистки является технологически очень сложной. Существующий подход увеличения получения гетерологичных белков заключается в применении выбранных сильных промоторов (см., например, WO 2008/084115). В соответствии с настоящим подходом, осуществляют протеомный анализ для определения большинства избыточных эндогенных белков, экспрессируемых микроорганизмами. С помощью геномной последовательности можно определить и выделить соответствующие гены и промоторы. Такие сильные промоторы (например, промотор гена hIIA, PhIIA, Lactococcus lactis) можно расположить перед гетерологичным геном и, таким образом, может быть достигнут высокий уровень экспрессии. Тем не менее, уровень экспрессии, который вредит физиологии хозяина, может приводить к затруднению роста хозяина и давать в результате отрицательный отбор. Это, по сути, ограничивает наиболее высокую возможную экспрессию любого заданного гетерологичного белка в клетке-хозяине при экспрессии до определенного конкретного уровня. Это представляет собой особенно серьезное препятствие для разработки расположенных в хромосоме единиц экспрессии.
Проблему отрицательного отбора обычно решают путем обеспечения маркеров отбора. Фактически, положительный или отрицательный отбор, например путем обеспечения генов устойчивости к антибиотику, может предупредить потерю введенного гетерологичного гена. Альтернативно, или в дополнение к применению маркеров отбора, можно использовать индуцируемые системы экспрессии генов, которые предусматривают блокировку размножения хозяина и экспрессию гетерологичного белка, таким образом, предупреждая возможный отрицательный отбор в течение фазы размножения, когда гетерологичный ген не экспрессируется. В этом контексте, в документе ЕР 0569604 описана индуцируемая система экспрессии у Streptococcus thermophilus, у которой гетерологичный ген обязательно расположен на 5'-конце по отношению к гену LacZ. Таким образом, экспрессия гетерологичного гена не только является индуцируемой, но и дополнительно сохранение гетерологичного гена также отбирают путем выращивания бактерий в их естественной среде обитания, молоке с лактозой в качестве источника углерода, для чего необходима экспрессия гена LacZ.
Ясно, что описанные выше системы экспрессии гетерологичного гена ограничены в применении. Например, применение маркеров отбора, таких как гены устойчивости к антибиотику, не полностью подходят для применения в пищевой промышленности или в фармацевтических областях применения. Дополнительно, ограничение выращивания в естественной среде обитания или использования источника углерода из естественной среды обитания для роста значительно снижает адаптивность любой системы экспрессии гетерологичного гена. Также, применение индуцируемых систем, в сущности, зависит от условий роста хозяина, так что для обеспечения экспрессии гетерологичного белка необходимы определенные культуральные среды, к которым необходимо добавлять индуцирующий фактор.
Таким образом, в данной области техники также существует потребность в повышении экспрессии гетерологичных белков; и необходимы последовательности, системы клонирования и стратегии, которые могут обеспечивать высокие уровни экспрессии для получения достаточных количеств экспрессированных гетерологичных белков в промышленных и/или терапевтических установках, при этом, в то же время, являться адаптивными и широко применимыми в ряде различных условий. В таких установках также может быть особенно пригодно получение экспрессии множества белков, каждый из которых характеризуется собственной биологической активностью и терапевтическим эффектом.
Сущность настоящего изобретения
Аспекты и варианты осуществления по настоящему изобретению направлены на удовлетворение по меньшей мере некоторых, например, одной или нескольких, обсуждаемых выше потребностей в данной области техники.
Было неожиданно обнаружено, что грамположительные бактерии могут эффективно экспрессировать экзогенные или гетерологичные гены с единиц полицистронной экспрессии, также содержащих эндогенный(е) ген(ы) таких бактерий. Таким образом, грамположительные бактерии могут эффективно экспрессировать экзогенные или гетерологичные гены с единиц полицистронной экспрессии, если такие гены транскрипционно или трансляционно связаны с эндогенным(и) геном(генами) таких бактерий. Неожиданно было обнаружено, что транскрипционное и/или трансляционное связывание эндогенных генов и экзогенных генов в единицах полицистронной экспрессии дает в результате высокие уровни экспрессии экзогенных генов у грамположительных бактерий. В частности, было обнаружено, что уровни экспрессии экзогенных генов, транскрипционно и/или трансляционно связанных с эндогенными генами грамположительных бактерий, по меньшей мере сравнимы и преимущественно выше уровней экспрессии экзогенных генов, которые транскрипционно или трансляционно не связаны с эндогенными генами грамположительных бактерий.
Таким образом, в соответствии с одним аспектом настоящее изобретение относится к грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит один или несколько эндогенных генов и один или несколько экзогенных генов. Единицу полицистронной экспрессии, таким образом, также можно обозначить как содержащую эндогенный ген (например, без ограничения один эндогенный ген) и один или несколько экзогенных генов. Предпочтительно, единица полицистронной экспрессии последовательно содержит один или несколько эндогенных генов и один или несколько экзогенных генов. Такую единицу полицистронной экспрессии, таким образом, можно также обозначить как последовательно содержащую эндогенный ген (например, без ограничения один эндогенный ген) и один или несколько экзогенных генов. Единицу полицистронной экспрессии конфигурируют так, чтобы она затрагивала транскрипцию одного или нескольких эндогенных генов и одного или нескольких экзогенных генов в полицистронной мРНК. Следовательно, настоящую грамположительную бактерию можно, иным образом, обозначить как содержащую один или несколько эндогенных генов, с которыми транскрипционно или трансляционно связаны один или несколько экзогенных генов. Также, следовательно, настоящее изобретение относится к грамположительной бактерии, содержащей один или несколько эндогенных генов, с которыми транскрипционно и/или трансляционно связаны один или несколько экзогенных генов.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит ген, эндогенный по отношению к грамположительной бактерии, и один или несколько генов, экзогенных по отношению к грамположительной бактерии. Предпочтительно, единица полицистронной экспрессии последовательно содержит один или несколько эндогенных генов и один или несколько экзогенных генов. Следовательно, также настоящее изобретение относится к рекомбинантной нуклеиновой кислоте, содержащей единицу полицистронной экспрессии, содержащую один или несколько генов, эндогенных по отношению к грамположительной бактерии, с которой транскрипционно и/или трансляционно связаны один или несколько генов, экзогенных по отношению к грамположительной бактерии.
Предпочтительно, как подразумевают по всему настоящему описанию, указанный один или несколько экзогенных генов могут быть транскрипционно или трансляционно связаны с 3'-концом указанного одного или нескольких эндогенных генов. К удивлению было обнаружено, что такая конфигурация является благоприятной в отношении уровней экспрессии гетерологичного белка, сохранения и/или геномной стабильности единицы полицистронной экспрессии. Кроме того, было обнаружено, что дальнейшее расположение в геноме ниже по ходу транскрипции имеет меньшее значение или не имеет значения.
Транскрипция транскрипционно или трансляционно связанного одного или нескольких эндогенных генов и одного или нескольких экзогенных генов может надлежащим образом регулироваться или контролироваться промотором, способным обеспечивать транскрипцию у грамположительной бактерии, и предпочтительно может регулироваться или контролироваться эндогенным промотором указанной грамположительной бактерии. Следовательно, настоящее изобретение также относится к грамположительной бактерии, содержащей один или несколько эндогенных генов, расположенных в своем естественном хромосомном локусе, с которыми транскрипционно или трансляционно связаны один или несколько экзогенных генов. Предпочтительно, транскрипция таких транскрипционно или трансляционно связанных одного или нескольких эндогенных генов и одного или нескольких экзогенных генов, таким образом, контролируется или регулируется нативным промотором указанных одного или нескольких эндогенных генов. Соответственно, транскрипционное или трансляционное связывание можно осуществить в результате встраивания в хромосому одного или нескольких экзогенных генов в указанный локус, как, например, в результате встраивания в хромосому одного или нескольких экзогенных генов 3' от указанного одного или нескольких эндогенных генов в указанном локусе.
Таким образом, в соответствии с одним аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного одного или нескольких эндогенных генов, причем предпочтительно указанный один или несколько экзогенных генов являются наиболее 3' удаленными генами единицы полицистронной экспрессии.
К удивлению было обнаружено, что встраивание в хромосому экзогенного или гетерологичного гена (или нескольких гетерологичных генов), транскрипционно связанного 3' с нативным геном, который сам по себе может представлять собой полицистронный ген, такой как, например, оперон, дает на выходе стабильную единицу экспрессии, в которой, вопреки ожиданиям, отрицательный отбор по (одному или нескольким) экзогенному гену отсутствует или минимален.
Неожиданно было обнаружено, что описанные в настоящем документе преимущества проявляются все в большей степени, когда экспрессия единицы полицистронной экспрессии находится под воздействием определенных условий, в частности, определенных типов промоторов. Неожиданно было обнаружено, что описываемые в настоящем документе системы полицистронной экспрессии, в которых отрицательный отбор по гетерологичным белкам нельзя устранить традиционными мерами, такими как применение маркеров отбора, или применением индуцируемых систем, могут, тем не менее, стабильно сохраняться и экспрессироваться на высоких уровнях, таким образом являясь широко применимыми в ряде различных условий при отсутствии необходимости в средствах для отбора или индуцирующий факторах. Модули полицистронной экспрессии, которые описаны в настоящем документе, таким образом, позволяют применять не выбираемые эндогенные и/или экзогенные гены.
В соответствии с одним аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием конститутивного промотора.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием промотора гена центрального метаболизма.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием промотора гена "домашнего хозяйства".
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием промотора необходимого гена.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии не находится под влиянием индуцируемого генного промотора.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием промотора рибосомального гена.
В соответствии с другим аспектом настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием промотора гена гликолиза.
Также, как указано выше, в соответствии с предпочтительным вариантом осуществления описанные выше промоторы представляют собой промоторы эндогенных генов. Также, как дополнительно подробно описано ниже, предпочтительно используемые в настоящем изобретении промоторы представляют собой сильные промоторы. Предпочтительно без ограничения указанный эндогенный промотор можно выбрать из группы, состоящей из промоторов eno, usp45, gapB, pyk, rpmB и rplS. Очень предпочтительно, чтобы транскрипция трансляционно связанного эндогенного гена и одного или нескольких экзогенных генов могла регулироваться или контролироваться нативным промотором (одного из) указанного эндогенного гена.
Следует понимать, что характеристики описываемых в настоящем документе промоторов в соответствии с настоящим изобретением можно комбинировать. Таким образом, в соответствии с вариантами осуществления настоящее изобретение относится к рекомбинантной нуклеиновой кислоте или грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного эндогенного гена, причем экспрессия указанной единицы полицистронной экспрессии находится под влиянием, например, (эндогенного) конститутивного промотора гена "домашнего хозяйства", (эндогенного) конститутивного промотора гена центрального метаболизма, (эндогенного) конститутивного промотора необходимого гена, (эндогенного) конститутивного промотора рибосомального гена, (эндогенного) конститутивного промотора гена гликолиза, (эндогенного) промотора гена "домашнего хозяйства" и гена центрального метаболизма, (эндогенного) промотора необходимого гена и гена центрального метаболизма, (эндогенного) промотора необходимого гена, гена "домашнего хозяйства" и гена центрального метаболизма, (эндогенного) промотора необходимого гена и гена "домашнего хозяйства", (эндогенного) конститутивного промотора гена "домашнего хозяйства" и гена центрального метаболизма, (эндогенного) конститутивного промотора гена "домашнего хозяйства", необходимого гена и гена центрального метаболизма, (эндогенного) промотора необходимого гена и рибосомального гена, (эндогенного) промотора необходимого гена и гена гликолиза, (эндогенного) конститутивного промотора необходимого гена и рибосомального гена, (эндогенного) конститутивного промотора необходимого гена и гена гликолиза.
Предпочтительно, как указано по всему настоящему описанию, указанный один или несколько экзогенных генов могут быть транскрипционно или трансляционно связаны с 3'-концом указанного одного или нескольких эндогенных генов, причем один или несколько эндогенных генов присутствуют в своем естественном положении на бактериальной хромосоме. В соответствии с настоящей конфигурацией последовательность на 5'-конце одного или нескольких эндогенных генов (как минимум включающего промотор эндогенных генов) идентична последовательности из штамма дикого типа, и участок, следующий за 3'-концом одного или нескольких экзогенных генов, идентичен последовательности участка 3' от одного или нескольких эндогенных генов, как и в штамме дикого типа.
От экспрессии указанного терапевтического белка в конкретных выбранных микроорганизмах-хозяевах можно получить пользу для множества областей применений, связанных с экспрессией экзогенных белков, таких как, например, для доставки терапевтического белка посредством рекомбинантных организмов. Такие микроорганизмы можно выбрать на основании их колонизирующей способности, как, например, выбранные штаммы, происходящие из микробиоты человека или животных. Микроорганизмы также можно выбирать по их способности усиливать активность любого конкретного доставляемого терапевтического белка, например, по результату взаимодействия их клеточной стенки, поверхности клетки или внутриклеточного содержимого с иммунной системой хозяина, например, посредством взаимодействия с toll-подобными рецепторами, членами семейства Ig, комплементом, цитокинами и др. Конкретные микроорганизмы можно выбирать по их способности выживать в или на конкретных участках доставки с жесткими условиями, таких как внутриопухолевые, кожные, участки с высоким содержанием желчи, участки с низким рН и др. Упоминаемая в настоящем описании грамположительная бактерия предпочтительно может быть молочнокислой бактерией (LAB), более предпочтительно Lactococcus sp., еще более предпочтительно Lactococcus lactis или ее подвидом или штаммом. Альтернативно, указанная LAB предпочтительно может быть Enterococcus sp., более предпочтительно Enterococcus fecium или Enterococcus faecalis или ее подвидом или штаммом.
Во избежание латерального переноса гена в эндогенную микрофлору, экспрессия с включенной в хромосому единицей экспрессии высоко предпочтительна для применения у рекомбинантной микрофлоры в качестве инструментов доставки терапевтических белков в медицинских целях. Также, расположенные в хромосоме единицы экспрессии могут оказаться гораздо более стабильно наследуемыми в поколениях, так что расположенные в хромосоме единицы экспрессии могут быть желательной структурой для получения штаммов, применяемых при массовом получении белка. Как известно из уровня техники, вставку в хромосому осуществляют при помощи векторов типа нокин (KI), которые не реплицируются в зависимости от условий и которые содержат гетерологичный ген между фланкирующими участками, которые делают возможной гомологичную рекомбинацию. В соответствии с традиционным подходом (см., например, WO 2008/084115) KI-плазмиду строят в гомологичном хозяине (KI-плазмиду для L. lactis встраивают в L. lactis). Это особенно полезно для гетерологичной экспрессии, для которой необходима секреция белка, поскольку многие сигнальные последовательности секреции не подходят для применения у других хозяев. Применение сильных промоторов в конструкциях экспрессии, которые предназначены для размещения на бактериальной хромосоме, затрудняется экспрессией гетерологичного гена с промежуточных продуктов KI-плазмиды. Перед гетерологичным геном непосредственно идет сильный промотор, делая так, что экспрессия с KI-плазмиды, несмотря на то, что не предполагается и не необходима, по сути ограничивает применение наиболее сильных промоторов. Во многих случаях KI-плазмида имеет некоторое количество копий, которое является кратным числу хромосом у хозяина, делая так, что при встраивании экспрессия будет в несколько раз ниже. Таким образом, хромосомные единицы экспрессии будут по сути слабее, чем можно было бы достичь на наиболее высоком уровне. Этот недостаток преодолевают с помощью описанного в настоящем документе настоящего изобретения. В соответствии с настоящей методикой, гетерологичные гены будут расположены ниже и они будут транскрипционно и/или трансляционно связаны с (сильно экспрессирующимся) эндогенным геном на бактериальной хромосоме. Согласно такой стратегии присутствие эндогенного (сильного) промотора на KI-плазмиде не является необходимым. Точнее, выше гетерологичного гена расположен 3'-конец без промотора (сильно экспрессируемого) эндогенного гена. Этот тип KI-плазмиды является молчащим и не будет ограничивать применение сильных промоторов.
Транскрипционное или трансляционное связывание одного или нескольких экзогенных генов с одним или несколькими другими генами, как описано в настоящем документе, можно осуществить за счет межгенного участка, активного (т.е., функционального, эффективного) у грамположительной бактерии, предпочтительно за счет эндогенного межгенного участка грамположительной бактерии. Таким образом, дополнительный аспект относится к рекомбинантной нуклеиновой кислоте, содержащей межгенный участок, активный у грамположительной бактерии, предпочтительно эндогенный межгенный участок грамположительной бактерии, функционально связанный с геном, экзогенным по отношению к указанной грамположительной бактерии. Функциональная связь обеспечивает, чтобы транскрипт межгенного участка, присутствующий на мРНК вместе с транскриптом экзогенного гена, был способен предоставить сайт для инициации трансляции экзогенного гена у грамположительной бактерии. Предпочтительно, межгенный участок может быть расположен 5' от экзогенного гена. Нуклеиновая кислота может содержать два или более экзогенных генов в полицистронной структуре, причем каждому экзогенному гену предшествует межгенный участок. Межгенные участки могут быть одинаковыми или отличаться. Например, если межгенные участки отличаются, то они могут соответствовать межгенным участкам, полученным от различных генов одного вида или различных видов, или из одного гена разных видов. Такие нуклеиновые кислоты могут быть пригодны при построении единиц полицистронной экспрессии, содержащих один или несколько экзогенных генов, причем один или несколько других генов транскрипционно или трансляционно связаны с одним или несколькими экзогенными генами посредством межгенного участка. Например, такие нуклеиновые кислоты могут быть пригодны при построении описываемых в настоящем документе единиц полицистронной экспрессии, причем один или несколько эндогенных генов транскрипционно или трансляционно связаны с одним или несколькими экзогенными генами посредством межгенного участка. Предпочтительно первый цистрон единицы полицистронной экспрессии будет представлять собой сильно экспрессирующийся эндогенный ген.
Настоящие рекомбинантные нуклеиновые кислоты могут содержаться на репликоне. Таким образом, один аспект также относится к репликону или вектору, содержащему описываемую в настоящем документе нуклеиновую кислоту. Например, вектор может представлять собой прокариотический вектор экспрессии, предпочтительно прокариотический вектор полицистронной экспрессии. Разработка таких плазмидных систем экспрессии, тем не менее, может быть трудоемкой, поскольку комбинация определенных репликонов и сильных промоторов может быть нестабильной. Также может быть невозможна трансформация выбранных микроорганизмов рекомбинантными плазмидами и стабильное сохранение последней в микроорганизме по причине присутствия естественных плазмид, в частности это невозможно, если нельзя включить маркеры отбора по антибиотику в плазмиду экспрессии, как это может быть в случае применения для доставки терапевтического белка. Эту проблему можно обойти при помощи расположения гетерологичных генов ниже и транскрипционно или трансляционно связанных с (сильно экспрессирующимся) эндогенным геном на бактериальной хромосоме. Поскольку согласно такой стратегии переносимая плазмидой система экспрессии не является необходимой, стратегию можно применять в качестве обычного подхода для генетической инженерии любого типа выбранной микрофлоры. Единственную необходимую штаммоспецифичную информацию можно быстро установить с помощью методики, известной из уровня техники. Высокопроизводительное секвенирование в сочетании с протеомным анализом экспрессирующихся в большом количестве белков быстро даст нуклеотидную последовательность участков, кодирующих присутствующие в большом количестве белки. Таким образом, наиболее предпочтительно, описываемый в настоящем документе вектор можно сконфигурировать так, чтобы он затрагивал гомологичную рекомбинацию у грамположительной бактерии, к примеру, приводил к встраиванию в хромосому экзогенного гена(генов).
Дополнительно, настоящее изобретение относится к применению описываемых в настоящем документе рекомбинантной нуклеиновой кислоты или вектора для полицистронной экспрессии одного или нескольких экзогенных генов или для полицистронной экспрессии одного или нескольких эндогенных генов и одного или нескольких экзогенных генов у грамположительной бактерии. Также настоящее изобретение относится к грамположительной бактерии, содержащей (например, трансформированной с ее помощью) описываемые в настоящем документе рекомбинантную нуклеиновую кислоту или вектор, причем грамположительная бактерия способна к полицистронной экспрессии одного или нескольких экзогенных генов или к полицистронной экспрессии одного или нескольких эндогенных генов и одного или нескольких экзогенных генов. Дополнительно настоящее изобретение относится к способу воздействия на полицистронную экспрессию одного или нескольких экзогенных генов или воздействия на полицистронную экспрессию одного или нескольких эндогенных генов и одного или нескольких экзогенных генов у грамположительной бактерии, предусматривающему этап введения описываемых в настоящем документе рекомбинантной нуклеиновой кислоты или вектора в указанную грамположительную бактерию. Также настоящее изобретение относится к способу получения грамположительной бактерии, способной к полицистронной экспрессии одного или нескольких экзогенных генов или способной к полицистронной экспрессии одного или нескольких эндогенных генов и одного или нескольких экзогенных генов, предусматривающему этап введения описываемых в настоящем документе рекомбинантной нуклеиновой кислоты или вектора в указанную грамположительную бактерию.
Настоящее изобретение относится к экспрессии, предпочтительно сильной (высокой) экспрессии, отдельного экзогенного гена или множества (например, двух, трех или более) различных экзогенных генов у грамположительной бактерии. Указанный экзогенный ген или гены могут кодировать продукт или продукты экспрессии, такие как, предпочтительно, белок(белки), полипептид(полипептиды) и/или пептид (пептиды). К примеру и без ограничения, такой белок(белки), полипептид(полипептиды) и/или пептид(пептиды) могут охватывать антигены (например, для индукции иммунитета или иммунотолерантности), аллергены, невакциногенные терапевтические полипептиды (цитокины, факторы роста, факторы заживления ран…), антитела или их функциональные фрагменты (например, Fab-фрагменты), химерные белки, мультимерные белки и т.д. и любую их комбинацию.
Полицистронная организация может давать описываемые в настоящем документе единицы экспрессии, которые особенно подходят для экспрессии белков, содержащих две или более полипептидных цепей (например, мультимерных белков, белковых комплексов). Таким образом, два или более приведенных в настоящем документе экзогенных генов могут предпочтительно кодировать различные мономеры или субъединицы мультимерного белка, причем гены совместно транскрибируются в полицистронную мРНК, и с этой мРНК транслируются отдельные мономеры или субъединицы. Это может обеспечивать тесно регулируемую совместную экспрессию экзогенных генов, например, для достижения сбалансированной и оптимальной сборки отдельных мономеров или субъединиц в мультимерный белок.
Особенно предпочтительной иллюстрацией такого принципа является экспрессия антител и их функциональных фрагментов. Следовательно, два или более описываемых в настоящем документе экзогенных генов могут предпочтительно кодировать отдельные цепи антитела или его функционального фрагмента. Например, один экзогенный ген может кодировать легкую цепь (VL) антитела или ее функционального фрагмента, а другой экзогенный ген может кодировать тяжелую цепь (VH) антитела или его функционального фрагмента. Предпочтительно, функциональный фрагмент антитела может представлять собой Fab. В соответствии с конкретными неограничивающими вариантами осуществления указанный Fab может связываться цитокинами, рецепторами цитокинов, хемокинами или активирующими иммунный ответ/воспаление молекулами и/или ингибировать их биологическое действие. В соответствии с предпочтительным вариантом осуществления Fab может связываться с TNFα и/или ингибировать его биологическое действие, к примеру, без ограничения указанный Fab может представлять собой сА2 к TNF или CDP870 к TNF.
Экзогенные гены, кодирующие отдельные цепи антитела или его фрагмента, таким образом, транскрипционно или трансляционно связывают для полицистронной экспрессии у грамположительной бактерии. Предпочтительно, экзогенный ген, кодирующий VL или его функциональный фрагмент, можно транскрипционно или трансляционно связать с 3'-концом экзогенного гена, кодирующего VH или его функциональный фрагмент. Эта организация генов дает на выходе особенно эффективную экспрессию и сборку антитела или его функционального фрагмента.
Полицистронная организация может также давать описываемые в настоящем документе единицы экспрессии, особенно подходящие для совместной экспрессии продуктов, таких как белки, которые взаимодействуют с достижением синергического эффекта, например, синергического терапевтического или профилактического эффекта, например, при доставке in situ бактерией.
Другой аспект относится к описываемой в настоящем документе грамположительной бактерии, причем один или несколько экзогенных генов кодируют продукт или продукты, такие как белок(белки), полипептид(полипептиды) или пептид (пептиды), оказывающие терапевтический или профилактический эффект на субъекта. В частности, такая бактерия предусматривается для применения в качестве лекарственного средства, более конкретно, для применения при введении или доставке указанного продукта или продуктов субъекту, еще конкретнее для применения при лечении заболевания, которое может принести пользу в результате введения или доставки указанного продукта или продуктов. Таким образом, настоящее изобретение также относится к фармацевтической композиции, содержащей такую грамположительную бактерию.
Настоящее изобретение также относится к способу доставки продукта или продуктов, таких как белок(белки), полипептид(полипептиды) или пептид(пептиды) субъекту, предусматривающему введение описываемой в настоящем документе грамположительной бактерии субъекту, причем указанный продукт или продукты кодируют один или несколько экзогенных генов. Предпочтительно указанный продукт или продукты могут оказывать терапевтический или профилактический эффект на субъект.
Преимущественно, что касается доставки in situ настоящей грамположительной бактерии субъектам, то бактерия ближе сохраняет свой эндогенный характер в результате отсутствия введения или введения меньшего количества экзогенных или даже патогенных последовательностей наряду с последовательностями для экзогенных продуктов экспрессии. Таким образом, регуляторный статус GRAS или "общепризнан безопасным" поддерживают настолько, насколько это возможно, способствуя, таким образом, процессу получения клинического разрешения или допуска на рынок для применения сконструированных штаммов для человека и животных.
Дополнительные аспекты и варианты осуществления в соответствии с настоящим изобретением представлены ниже в пунктах (i)-(xxii).
(i) Грамположительная бактерия, содержащая единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного одного или нескольких эндогенных генов.
(ii) Рекомбинантная нуклеиновая кислота, содержащая единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит ген, эндогенный по отношению к грамположительной бактерии и одному или нескольким генам, экзогенным по отношению к грамположительной бактерии, транскрипционно связанным с 3'-концом указанного одного или нескольких эндогенных генов.
(iii) Грамположительная бактерия согласно (i) или рекомбинантная нуклеиновая кислота согласно (ii), причем указанный один или несколько экзогенных генов кодируют белок, полипептид и/или пептид, оказывающий терапевтический или профилактический эффект на субъекта, или антиген для индукции иммунитета или иммунотолерантности, невакциногенный терапевтически активный полипептид, антитело или его функциональный фрагмент, такой как Fab, химерный белок или мультимерный белок.
(iv) Грамположительная бактерия согласно (i) или рекомбинантная нуклеиновая кислота согласно (ii), причем один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид или пептид, причем продукт оказывает терапевтический или профилактический эффект на субъект, для применения в качестве лекарственного препарата, предпочтительно для применения при введении или доставке указанного продукта субъекту.
(v) Грамположительная бактерия согласно (i), (iii) или (iv) или рекомбинантная нуклеиновая кислота согласно (ii)-(iv), причем указанный один или несколько экзогенных генов являются наиболее 3' удаленным геном единицы полицистронной экспрессии.
(vi) Грамположительная бактерия согласно любому из (i), (iii), (iv) или (v) или рекомбинантная нуклеиновая кислота согласно любому из (ii)-(v), причем указанный эндогенный ген и указанный один или несколько экзогенных генов находятся под транскрипционным контролем промотора, эндогенного по отношению к грамположительной бактерии.
(vii) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (vi), причем указанный промотор является промотором необходимого гена, конститутивным промотором, промотором гена центрального метаболизма и/или промотором гена "домашнего хозяйства".
(viii) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (vi), причем указанный промотор является промотором рибосомального гена.
(ix) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (vi), причем указанный промотор является промотором гена гликолиза.
(x) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (vi), причем указанный промотор выбран из группы, состоящей из промотора eno, usp45, gap, pyk, rpmB и rplS указанной грамположительной бактерии.
(xi) Грамположительная бактерия согласно любому из (i) или (ii)-(x), причем эндогенный ген расположен в своем естественном локусе на хромосоме у грамположительной бактерии.
(xii) Грамположительная бактерия согласно (xi), причем эндогенный ген транскрипционно связан с одним или несколькими экзогенными генами посредством встраивания в хромосому одного или нескольких экзогенных генов в указанный локус, предпочтительно посредством встраивания в хромосому одного или нескольких экзогенных генов 3' от эндогенного гена в указанном локусе.
(xiii) Грамположительная бактерия согласно любому из (i) или (iii) до (xii) или рекомбинантная нуклеиновая кислота в соответствии с любым из от (ii) до (х), причем эндогенный ген и один или несколько экзогенных генов транскрипционно связаны посредством межгенного участка или участков, активных у грамположительной бактерии, причем предпочтительно межгенный участок или участки являются эндогенными по отношению к указанной грамположительной бактерии.
(xiv) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (xiii), причем указанный межгенный участок выбран из группы, состоящей из межгенных участков, предшествующих rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE и rplF.
(xv) Рекомбинантная нуклеиновая кислота, содержащая межгенный участок, активный у грамположительной бактерии, функционально связанный с геном, экзогенным по отношению к указанной грамположительной бактерии, причем предпочтительно межгенный участок является эндогенным межгенным участком грамположительной бактерии.
(xvi) Рекомбинантная нуклеиновая кислота согласно (xiv), причем указанный межгенный участок выбран из группы, состоящей из межгенных участков, предшествующих rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE и rplF.
(xvii) Грамположительная бактерия согласно любому из (i) или (iii)-(xiv) или рекомбинантная нуклеиновая кислота согласно любому из (ii)-(x) или (xiii)-(xvi), причем один экзогенный ген кодирует легкую цепь (VL) антитела или его функционального фрагмента, а другой экзогенный ген кодирует тяжелую цепь (VH) антитела или его функционального фрагмента, причем более предпочтительно функциональным фрагментом является Fab.
(xviii) Грамположительная бактерия или рекомбинантная нуклеиновая кислота согласно (xvii), причем экзогенный ген, кодирующий VL или ее функциональный фрагмент, транскрипционно связан с 3'-концом экзогенного гена, кодирующего VH или ее функциональный фрагмент.
(xix) Грамположительная бактерия согласно любому из (i), (iii)-(xiv), (xvii)-(xviii) или рекомбинантная нуклеиновая кислота согласно любому из (ii)-(x) или (xiii)-(xviii), причем грамположительная бактерия является молочнокислой бактерией, предпочтительно Lactococcus, Lactobacillus или Enterococcus, более предпочтительно Lactococcus lactis или Enterococcus faecium, или где грамположительной бактерией является представитель рода Bifidobacterium.
(xx) Фармацевтическая композиция, содержащая грамположительную бактерию согласно любому из (i), (iii)-(xiv) или (xvii)-(xix).
(xxi) Фармацевтическая композиция согласно (хх), причем указанный один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид или пептид, причем продукт оказывает терапевтический или профилактический эффект на субъект.
(xxii) Вектор, содержащий рекомбинантную нуклеиновую кислоту согласно любому из (ii)-(x) или (xiii)-(xix).
Приведенные выше и дополнительные аспекты и предпочтительные варианты осуществления по настоящему изобретению описаны в дальнейших разделах и в прилагаемой формуле изобретения. Объект прилагаемой формулы изобретения, таким образом, определенно включен в настоящее описание.
Краткое описание чертежей
Фигура 1: Окрашивание кумасси синим клеточных белков культуры конца лог-фазы штамма MG1363 Lactococcus lactis ssp. Cremoris. Заметные белковые бэнды обозначены 1-12.
Фигура 2: Изображение эталонной конструкции моноцистронной экспрессии (вверху, SAGX0090) и полицистронной (бицистронной, с двойным цистроном) конструкции в соответствии с вариантом осуществления по настоящему изобретению (внизу), причем ген X представляет собой эндогенный ген. Обе конструкции экспрессии предназначены для экспрессии β-глюкуронидазы с гена uidA Е. coli, который служит в настоящем описании в качестве иллюстративного экзогенного гена.
Фигура 3: Относительная активность β-глюкуронидазы (GUS) у эталонного хозяина (моноцистронный: PhILA»uidA, sAGX0090) и у хозяина, содержащего полицистронную (бицистронную) конструкцию в соответствии с вариантом осуществления по настоящему изобретению (эндогенный ген X»rpmD»uidA), организованный как на фигуре 2. Эндогенные гены X представляют собой, в настоящем примере, usp45, enoA, rplS, rpmB, pyk и gapB. В настоящем примере межгенный участок rpmD обеспечивает транскрипционную связь эндогенного и экзогенного гена. Экзогенный ген uidA E. coli кодирует β-глюкуронидазу. Все конструкции экспрессии встроены в бактериальную хромосому. Моноцистронная конструкция присутствует в локусе thyA, бицистронные конструкции встроены в естественном положении гена X. Из данных видно, что все бицистронные конструкции характеризуются b-галактозидазной активностью, превосходящей моноцистронную конструкцию PhILA»uidA.
Фигура 4: Количественное определение секреции проинсулина (ins) человека посредством Lactococcus lactis у эталонного хозяина (sAGX0122) и хозяев в соответствии с вариантом осуществления по настоящему изобретению (sAGX0121 и SAGX0164). (А) Схематическое представление модулей экспрессии ins. Штамм SAGX0122 несет конструкцию моноцистронной экспрессии, в котором промотор thyA управляет экспрессией слияния определяющей секрецию лидерной последовательности - проинсулина человека (SS::ins), встроенного в хромосоме MG1363 Lactococcus lactis в локусе thyA. Бицистронные конструкции экспрессии у SAGX0121 и SAGX0164 характеризуются транскрипционной связью эндогенного usp45 и enoA, соответственно, с SS::ins посредством межгенного участка rpmD. Такие конструкции расположены на хромосоме MG1363 Lactococcus lactis в естественных положениях генов usp45 и enoA, соответственно. (В) Уровни проинсулина, детектированные в супернатантах различных штаммов. Коды штаммов (sAGX0122, sAGX0121 и sAGX0164) указаны снизу столбцов, соответственно показывающих уровни проинсулина человека в супернатантах таких штаммов. Из данных видно, что штаммы, несущие обе бицистронных конструкции характеризуются уровнями проинсулина человека, превосходящими штамм, несущий моноцистронную конструкцию PthyA»ins.
Фигура 5: Экспрессия Fab сА2 к TNF у Lactococcus lactis. Гены, кодирующие фрагменты VLCL (L) и VHCH1 (Н) транскрипционно связывали посредством межгенных участков rpmD, rplB, rpsG, rpsE и rplN. Получали конструкции, у которых либо L, либо Н расположены в качестве первого гена бицистронной конструкции. Все конструкции экспрессии антител к TNF переносились плазмидой и находились под контролем промотора PthyA. Активность к TNF измеряли у супернатанов различных штаммов. Из данных видно, что имеет место более высокая активность к TNF у всех конструкций, где Н является первым геном бицистронной конструкции.
Фигура 6: Экспрессия Fab CDP870 к TNF у Lactococcus lactis. (А) слияния легкой и тяжелой цепей CDP870 с последовательностями, кодирующими определяющую секрецию лидерную последовательность usp45 (SS::CDP870 VLCL и SS::CDP870 VHCH1), включали в качестве второго и третьего цистрона ниже usp45 (sAGX0219, SAGX0220) в хромосоме MG1363 Lactococcus lactis. В sAGX0219 и sAGX0220 rpmD использовали для связывания генов SS::CD870 с usp45. Для предупреждения генетической нестабильности гены легкой и тяжелой цепей связывали посредством межгенного участка, предшествующего rplN. В sAGX0219 ген легкой цепи предшествовал гену тяжелой цепи, в то время как в SAGX0220 ген тяжелой цепи предшествовал гену легкой цепи. (В) Количественное определение активности к TNF человека в неочищенных культуральных супернатантах. Как тяжелая цепь, так и легкая цепи экспрессировались на высоком уровне конструкциями с двойным цистроном, что давало высокие уровни функционального Fab к TNF CDP870. Экспрессия CDP870 к TNF значимо повышалась, если тяжелую цепь располагали перед легкой цепью.
Фигура 7: Количественное определение секреции человеческого фактора "трилистника"-1 (hTFF1) посредством Lactococcus lactis у эталонного хозяина (sAGX0085) и хозяина в соответствии с вариантом осуществления по настоящему изобретению (sAGX0276). (А) Схематическое представление модулей экспрессии hTFF1. Штамм SAGX0085 несет конструкцию моноцистронной экспрессии, в которой промотор PhILA управляет экспрессией слияния определяющей секрецию лидерной последовательности - hTFF1 (SS::hTFF1), встроенной в хромосоме MG1363 Lactococcus lactis в локусе thyA. Конструкция бицистронной экспрессии в SAGX0276 характеризуется транскрипционной связью gapB с SS::hTFF1, посредством межгенного участка rpmD. Эта конструкция расположена на хромосоме MG1363 Lactococcus lactis в естественных положениях гена gapB. (В) Уровни hTFF1, детектированные в супернатантах различных штаммов. Коды штамма (sAGX0085 и SAGX0276) указаны снизу столбцов, соответственно показывающих уровни hTFF1 человека в супернатантах таких штаммов. Из данных видно, что SAGX0276, несущий бицистронную конструкцию, производит уровни hTFF1, превосходящие SAGX0085, который содержит моноцистронную конструкцию.
Фигура 8: Окрашивание кумасси синим клеточных белков культуры конца лог-фазы штамма LMG 15709 Enterococcus faecium. Заметные белковые бэнды обозначены 1-12.
Фигура 9: Изображение полицистронных (бицистронных, с двойным цистроном) конструкций в соответствии с вариантом осуществления по настоящему изобретению, причем ген X представляет собой эндогенный ген. Конструкции экспрессии предназначены для экспрессии β-глюкуронидазы с гена uidA Е. coli, служащего в настоящем описании в качестве иллюстративного экзогенного гена. Gap и eno являются иллюстративными "первыми" эндогенными генами.
Фигура 10: Относительная активность β-глюкуронидазы (GUS) у эталонного хозяина (моноцистронный: PhILA»uidA, sAGX0090) и у хозяина, содержащего полицистронную (бицистронную) конструкцию в соответствии с вариантом осуществления по настоящему изобретению (эндогенный ген X»rpmD»uidA), организованный как на фигуре 9. Эндогенные гены X представляют собой, в настоящем примере, gapB и eno. В настоящем примере межгенный участок rpmD обеспечивает транскрипционное связывание эндогенного и экзогенного гена. Экзогенный ген uidA E. coli кодирует β-глюкуронидазу. Все конструкции экспрессии встроены в бактериальную хромосому. Моноцистронная конструкция присутствует в локусе thyA, бицистронные конструкции встроены в естественном положении гена X. Из данных видно, что все бицистронные конструкции характеризуются β-галактозидазной активностью, превосходящей моноцистронную конструкцию PhILA»uidA.
Фигура 11: Количественное определение секреции интерлейкина-10 (hIL-10) человека посредством Enterococcus faecium у эталонного хозяина (sAGX0270) и хозяина в соответствии с вариантом осуществления по настоящему изобретению (sAGX0279). (А) Схематическое представление модулей экспрессии hIL-10. Конструкция бицистронной экспрессии у sAGX0279 характеризуется транскрипционной связью эндогенного gap с SS::hIL10 посредством межгенного участка rpmD. (В) Уровни hIL-10, детектированные в супернатантах различных штаммов.
Фигура 12: Количественное определение секреции интерлейкина-27 (hIL-27) человека посредством Enterococcus faecium у эталонного хозяина (sAGX0270) и хозяина в соответствии с вариантом осуществления по настоящему изобретению (sAGX0279). (А) Схематическое представление модулей экспрессии hIL-27. Конструкция бицистронной экспрессии у sAGX0317 характеризуется транскрипционной связью эндогенного gap с SS::hIL27 посредством межгенного участка rpmD. (В) Уровни hIL-27, детектированные в супернатантах различных штаммов.
Фигура 13: Экспрессия Fab CDP870 к TNF у Enterococcus faecium. (А) слияния легкой и тяжелой цепей CDP870 с последовательностями, кодирующими определяющую секрецию лидерную последовательность usp45 (SS::CDP870 VHCH1 и SS::CDP870 VLCL), включали в качестве второго и третьего цистрона ниже gap (sAGX0278). Для предупреждения генетической нестабильности гены легкой и тяжелой цепи связывали посредством межгенного участка, предшествующего rpmD от Lactococcus lactis (LL), в то время как rpmD от Enterococcus faecium (EF) использовали для связывания генов gap и тяжелой цепи. (В) Количественное определение активности к TNF человека в неочищенных культуральных супернатантах. Как тяжелая цепь, так и легкая цепи экспрессировались на высоком уровне конструкциями с двойным цистроном, что давало высокие уровни функционального Fab к TNF CDP870.
Фигура 14: Влияние производящих антитела к hTNF бактерий L.lactis (sAGX0220) на индуцированную hTNF токсичность и выработку цитокинов воспаления у A20IEC-KO мышей, (а) A20IEC-KО мышей (n=5 на группу) предварительно обрабатывали основой, SAGX0220 или MG1363 за 1 час до введения инъекцией 2 мкг (левый сегмент) и 6 мкг (правый сегмент) рекомбинантного hTNF и отслеживали температуру тела с течением времени. Одной группе A20IEC-KО мышей вводили инъекцией ремикейд перед введением инъекцией 6 мкг hTNF. (b) уровни МСР-1 в подвздошной кишке, проксимальном отделе толстой кишки и сыворотке крови через 5 часов после введения инъекцией 2 мкг hTNF. (с) уровни KС и IL-6 в гомогенатах подвздошной кишки через 5 часов после введения инъекцией 2 мкг hTNF. (d) уровни МСР-1 в подвздошной кишке, проксимальном отделе толстой кишки и сыворотке крови через 5 часов после введения инъекцией 6 мкг hTNF. (е) уровни KС и IL-6 в гомогенатах подвздошной кишки через 5 часов после введения инъекцией 6 мкг hTNF. Погрешности демонстрируют SEM. *,р<0,05.
Фигура 15: Выработка CDP870 в штаммах по варианту осуществления настоящего изобретения. (А) Тяжелая цепь и легкая цепь CDP870, встроенные в локус usp45, локус enoA или локус gapB. (В) Вестерн-блоттинг, показывающий экспрессию CDP870 в различных штаммах в соответствии с вариантом осуществления по настоящему изобретению. (С) и (D) Результаты ELISA-анализа, показывающие экспрессию CDP870 в различных штаммах в соответствии с вариантом осуществления по настоящему изобретению. (Е) Нейтрализирующая TNF активность различных штаммов в соответствии с вариантом осуществления по настоящему изобретению.
Фигура 16: Выживаемость мышей Tg1278 с индуцированным TNBS колитом после обработки штаммом в соответствии с вариантом осуществления по настоящему изобретению (секретирующим антитело к hTNF штаммом SAGX0309 L. lactis) по сравнению с мышами, обработанными штаммом дикого типа L. lactis, и мышами, обработанными симзией (Cimzia).
Фигура 17: Рост массы тела мышей Tg1278 с индуцированным TNBS колитом после обработки штаммом в соответствии с вариантом осуществления по настоящему изобретению (секретирующим антитело к hTNF штаммом sAGX0309 L. lactis) по сравнению с мышами, обработанными штаммом дикого типа L. lactis, и мышами, обработанными симзией. Верхний сегмент: абсолютная масса тела (г); нижний сегмент: масса тела по отношению к исходной массе тела (%).
Фигура 18: Гистологическая оценка ткани толстой кишки мышей Tg1278 с индуцированным TNBS колитом после обработки штаммом в соответствии с вариантом осуществления по настоящему изобретению (секретирующим антитело к hTNF штаммом sAGX0309 L. lactis) по сравнению с мышами, обработанными штаммом дикого типа L. lactis, и мышами, обработанными симзией. Средние значения указаны над столбцами. Степень выживаемости указана на группу.
Фигура 19: Секреция провоспалительных цитокинов у мышей Tg1278 с индуцированным TNBS колитом после обработки штаммом в соответствии с вариантом осуществления по настоящему изобретению (секретирующим антитело к hTNF штаммом sAGX0309 L. lactis) по сравнению со здоровыми мышами, мышами, обработанными штаммом дикого типа L. lactis, и мышами, обработанными симзией. На (А), (В) и (С) видны уровни miL6, mKC и mMCP1 в пг/мг в дистальных отделах толстого кишечника, соответственно.
Подробное раскрытие настоящего изобретения
Используемые в настоящем документе формы единственного числа включают формы как единственного, так и множественного числа, если контекст явно не диктует иное.
Используемые в настоящем документе термины "включающий", "включает" и "включающий в составе" являются синонимичными с "вмещающий", "вмещает" или "содержащий", "содержит", и являются включительными или неисчерпывающими, и не исключают дополнительных, неперечисленных членов, элементов и этапов способа. Будет понятно, что применяемые в данном документе термины "включающий", "включает" и "включающий в составе" включают термины "состоящий из", "состоит" и "состоит из", а также термины "по сути состоящий из", "по сути состоит" и "по сути состоит из".
Упоминание диапазонов числовых значений посредством крайних точек включает все числа и дробные части, относящиеся к соответствующим диапазонам, а также упомянутые крайние точки.
Применяемый в настоящем документе термин "приблизительно" или "примерно" в отношении измеряемого значения, такого как параметр, количество, продолжительность времени и т.п., понимают как охватывающий вариации +/-20% или менее, предпочтительно +/-10% или менее, более предпочтительно +/- 5% или менее, и еще более предпочтительно +/-1% или менее и от указанного значения, в такой мере такие вариации подходят для осуществления раскрываемого настоящего изобретения. Понятно, что значение к которому относится модификатор "приблизительно" или "примерно", само по себе также конкретно и предпочтительно раскрывается.
Несмотря на то, что термины "один или несколько" или "по меньшей мере один", такой как один или несколько или по меньшей мере один член(ы) из группы членов, сами по себе понятны, посредством дополнительного иллюстрирующего примера, термин охватывает, среди прочего, отсылку к любому из указанных членов, или любым двум или более из указанных членов, таких как, например, любые >3, >4, >5, >6 или >7 и т.д. из указанных членов и до всех указанных членов.
Все процитированные в настоящем описании справочные материалы включены в настоящее описание с помощью ссылки в их полных объемах. В частности, идеи, излагаемые во всех приведенных в настоящем документе справочных материалах, на которые конкретно делается ссылка, включены с помощью ссылки.
Если не указано иное, все применяемые при раскрытии настоящего изобретения термины, включая технические или научные термины, имеют значение, которое обычно понимается рядовым специалистом в данной области техники, к которой относится настоящее изобретение. В качестве дополнительной информации определения терминов включены для лучшего понимания идеи настоящего изобретения.
В последующих разделах различные аспекты настоящего изобретения определены подробнее. Определенный таким образом каждый аспект можно комбинировать с любым другим аспектом или аспектами, за исключением случаев, где явно указано противоположное. В частности, любой признак, указанный как являющийся предпочтительным или преимущественным, можно комбинировать с любым другим признаком или признаками, которые указаны как являющиеся предпочтительными или преимущественными.
Указание по всему настоящему описанию на "один вариант осуществления" или "вариант осуществления" означает, что конкретный признак, структура или характеристика, описанные в сочетании с вариантом осуществления, включены по меньшей мере в один вариант осуществления по настоящему изобретению. Таким образом, появления фраз "в соответствии с одним вариантом осуществления" или "в соответствии с вариантом осуществления" в различных местах по всему настоящему описанию не обязательно все указывают на один и тот же вариант осуществления, но могут. Кроме того, конкретные признаки, структуры или характеристики можно комбинировать любым подходящим способом, который будет очевиден специалисту в данной области техники из настоящего раскрытия, в соответствии с одним или несколькими вариантами осуществления. Кроме того, несмотря на то, что некоторые описываемые в настоящем документе варианты осуществления включают некоторые, но не другие признаки, включенные в другие варианты осуществления, комбинации признаков из различных вариантов осуществления понимают как входящие в объем настоящего изобретения, и образуют иные варианты осуществления, которые будут понятны специалистам в данной области техники. Например, в прилагаемой формуле изобретения любые из заявленных вариантов осуществления можно использовать в любой комбинации.
В приведенном далее подробном описании настоящего изобретения упоминаются прилагаемые чертежи, которые формируют его часть, и которые показаны лишь в качестве иллюстрации конкретных вариантов осуществления, в соответствии с которыми может быть осуществлено настоящее изобретение. Понятно, что могут быть использованы другие варианты осуществления и могут быть осуществлены структурные или логические изменения без отступления от объема настоящего изобретения. Последующее подробное описание, таким образом, не следует принимать во внимание в ограничивающем смысле, а объем настоящего изобретения определен прилагаемой формулы изобретения.
Стандартные справочные материалы, в которых изложены основные принципы методики с использованием рекомбинантной ДНК, включают: Molecular Cloning: А Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989; Current Protocols in Molecular Biology, ed. Ausubel et al., Greene Publishing and Wiley-lnterscience, New York, 1992 (с периодическими обнавлениями) ("Ausubel et al. 1992"); Innis et al., PCR Protocols: A Guide to Methods and Applications, Academic Press: San Diego, 1990. Основные принципы микробиологии изложены, например, в работе Davis, В.D. et al., Microbiology, 3rd edition, Harper & Row, publishers, Philadelphia, Pa. (1980).
Как упомянуто ранее, в соответствии с одним аспектом настоящее изобретение относится к грамположительной бактерии, содержащей эндогенный ген, с которым транскрипционно или трансляционно связаны один или несколько экзогенных генов. Предпочтительно, один или несколько экзогенных генов транскрипционно или трансляционно связаны ниже (т.е. на 3'-конце) эндогенного гена. Родственный аспект относится к грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит эндогенный ген и один или несколько экзогенных генов. Предпочтительно, единица полицистронной экспрессии последовательно содержит один или несколько эндогенных генов и один или несколько экзогенных генов. Следующий аспект относится к рекомбинантной нуклеиновой кислоте, содержащей единицу полицистронной экспрессии, содержащей ген, эндогенный по отношению к грамположительной бактерии, с которым транскрипционно или трансляционно связаны один или несколько генов, экзогенных по отношению к грамположительной бактерии. Предпочтительно, один или несколько экзогенных генов транскрипционно или трансляционно связаны ниже (т.е. на 3'-конце) эндогенного гена.
Предпочтительно, один или несколько экзогенных генов являются наиболее 3' удаленными генами единицы полицистронной экспрессии, т.е. один или несколько экзогенных генов являются последними или наиболее нижними генами единицы полицистронной экспрессии. Например, если эндогенный ген является моноцистронным, то один или несколько экзогенных генов расположены после или ниже (т.е. на 3'-конце) и транскрипционно связанны с открытой рамкой считывания гена. Аналогично, если эндогенный ген сам по себе является полицистронным, такой как оперон (часть оперона), то один или несколько экзогенных генов расположены после или ниже (т.е. на 3'-конце) последнего (т.е. наиболее низко или наиболее 3' удаленно) эндогенного гена эндогенного полицистронного гена.
Наиболее предпочтительно, упоминаемый по всему настоящему описанию эндогенный ген является моноцистронным. Таким образом, предпочтительно, эндогенный ген не формирует часть эндогенного оперона.
Предпочтительно, экспрессия описываемой в настоящем документе единицы полицистронной экспрессии находится под воздействием промотора, который может представлять собой один или несколько из приведенных далее или может характеризоваться одной или несколькими из следующих характеристик: конститутивные промоторы, промоторы гена центрального метаболизма, промоторы необходимого гена, сильные промоторы, промоторы гена "домашнего хозяйства", промоторы рибосомального гена, промоторы гена гликолиза. Наиболее предпочтительно, промотор является конститутивным промотором.
Применяемый в настоящем документе термин "грамположительная бактерия" имеет свое известное в данной области техники общепринятое значение. В качестве дополнительной информации, грамположительная бактерия может быть определена посредством окрашивания по Граму как удерживающая кристаллический фиолетовый краситель.
В соответствии с предпочтительным вариантом осуществления грамположительная бактерия согласно настоящему изобретению является непатогенной в том смысле, что она не наносит вред или не может привести к вредным эффектам при введении предполагаемому субъекту.
Предпочтительно, грамположительная бактерия согласно настоящему изобретению является молочнокислой бактерией (LAB), включая без ограничения роды Lactococcus, Lactobacillus, Leuconostoc, Pediococcus, Streptococcus, Aerococcus, Carnobacterium, Enterococcus, Oenococcus, Sporolactobacillus, Tetragenococcus, Vagococcus и Weisella. Более предпочтительно, LAB представляет собой виды Lactococcus, такие как без ограничения Lactococcus lactis, Lactococcus garvieae, Lactococcus piscium, Lactococcus plantarum и Lactococcus raffinolactis и любые их подвиды и штаммы. Наиболее предпочтительно, вид Lactococcus представляет собой Lactococcus lactis, и любой его подвид и штамм, такой как без ограничения Lactococcus lactis ssp. cremoris, Lactococcus lactis ssp. hordniae, Lactococcus lactis ssp. lactis, Lactococcus lactis ssp. bv. diacetylactis. В соответствии со следующим предпочтительным вариантом осуществления по настоящему изобретению Lactococcus lactis является Lactococcus lactis ssp. cremoris или Lactococcus lactis ssp. lactis, более предпочтительно Lactococcus lactis ssp.cremoris, и охватывает любые их штаммы, такие как, например, SK11 Lactococcus lactis ssp. cremoris, MG1363 Lactococcus lactis ssp. cremoris или IL1403 Lactococcus lactis ssp lactis. В соответствии с другим предпочтительным вариантом осуществления LAB является Enterococcus sp., предпочтительно Enterococcus faecalis, Enterococcus faecium и любыми их подвидами и штаммами, такими как без ограничения штамм LMG15709 Enterococcus faecium.
В соответствии с другим предпочтительным вариантом осуществления грамположительной бактерией согласно настоящему изобретению является представитель рода Bifidobacterium.
Bifidobacterium представляет собой род грамположительных, неподвижных, зачастую ветвящихся анаэробных бактерий. В контексте настоящего изобретения представители рода Bifidobacterium могут включать В. adolescentis, В. angulatum, В. animalis, В. asteroides, В. bifidum, В. bourn, В. breve, В. catenulatum, В. choerinum, В. coryneforme, В. cuniculi, В. denticolens, В. dentium, В. gallicum, В. gallinarum, В. indicum, В. infantis, В. inopinatum, В. lactis, В. longum, В. magnum, В. merycicum, В. minimum, В. pseudocatenulatum, В. pseudolongum, В. pullorum, В. ruminantium, В. saeculare, В. subtile, В. suis, В. thermacidophilum, В. thermophilum. Предпочтительно, представитель рода Bifidobacterium представляет собой В. adolescentis, В. bifidum, В. breve, В. infantis, В. longum. Следует понимать, что также включены все подвиды и штаммы представителей рода Bifidobacterium.
Применяемое в настоящем документе выражение "последовательно" в контексте эндогенных и экзогенных генов относится к 5'-3' порядку соответствующих генов в полинуклеиновой кислоте, векторе или хромосоме. Например, единица полицистронной экспрессии, последовательно содержащая один или несколько эндогенных генов и один или несколько экзогенных генов, относится к единице, в которой один или несколько эндогенных генов расположены выше одного или нескольких экзогенных генов. Следовательно, один или несколько экзогенных генов расположены после 3'-конца одного или нескольких эндогенных генов. Следует понимать, что описанное в настоящем документе последовательное связывание или упорядочение не обязательно предполагает непосредственное связывание эндогенного и экзогенного гена. Между эндогенным и экзогенным геном могут присутствовать дополнительные последовательности. Например, межгенный участок, определение которому в настоящем документе приведено далее, может присутствовать между (т.е. ниже или 3' от эндогенного гена и выше или 5' от экзогенного гена) последовательными эндогенными и экзогенными генами. Применяемые в настоящем документе термины "эндогенный ген", "эндогенный промотор", "эндогенный межгенный участок", "эндогенный сайт связывания рибосомы" относится к соответствующему гену, промотору, межгенному участку или сайту связывания рибосомы, которые являются естественными для грамположительной бактерии или могут быть обнаружены в природе у грамположительной бактерии. В связи с этим, термин эндогенный ген, промотор, межгенный участок или сайт связывания рибосомы охватывает ортологические гены, промоторы, межгенные участки и сайты связывания рибосом среди различных родов, видов, подвидов или штаммов грамположительных бактерий. В частности, ген, промотор, межгенный участок или сайт связывания рибосом, выделенный из одного рода, вида, подвида или штамма грамположительных бактерий, называют эндогенным для всех других родов, видов, подвидов или штаммов грамположительных бактерий независимо от возможных отличий последовательности полинуклеиновой кислоты при условии, что указанный другой род, вид, подвид или штамм грамположительных бактерий в природе также содержит такой ген, промотор, межгенный участок или сайт связывания рибосомы. Таким образом, такие дивергентные, но встречающиеся в природе последовательности гена, промотора, межгенного участка или сайта связывания рибосомы можно считать эндогенными. К примеру и без ограничения, кодирующий енолазу ген, enoА, который выделен из Lactococcus lactis ssp. lactis, также считают эндогенным по отношению к Lactococcus lactis ssp. cremoris.
Предпочтительно, тем не менее, "эндогенный" ген, промотор или межгенный участок указанного рода, вида, подвида или штамма рассматриваемой в настоящем документе грамположительной бактерии может означать ген, промотор или межгенный участок, который встречается в природе, т.е. он является естественным или собственным по отношению, соответственно, к тому же роду, виду, подвиду или штамму грамположительной бактерии. К примеру и без ограничения, кодирующий енолазу ген, enoA, который выделен из Lactococcus lactis ssp. lactis, предпочтительно можно считать "эндогенным" по отношению к Lactococcus lactis ssp. lactis, но не к Lactococcus lactis ssp. cremoris.
Применяемый в данном документе термин "экзогенный ген" относится к гену, который не является естественным по отношению к грамположительной бактерии или не может встречаться в природе у грамположительной бактерии. Термин экзогенный ген является синонимом с термином гетерологичный ген. Экзогенный ген может представлять собой ген полной длины или, в альтернативном случае, может представлять собой укороченный ген или фрагмент гена. К примеру, экзогенный ген может быть получен от вирусов, других прокариот, таких как грамотрицательная бактерия, или, альтернативно и предпочтительно, может быть получен от эукариот, таких как растения, животные, предпочтительно млекопитающие, наиболее предпочтительно человек. Альтернативно, экзогенный ген может быть полностью или частично синтетическим или искусственным в том смысле, что он полностью или в частичном виде не встречается в природе. В дополнение к этому, экзогенный ген может быть химерным в том смысле, что он может быть составлен из последовательностей, происходящих от различных видов, или комбинацией из встречающихся в природе, синтетических или искусственных последовательностей. Также охвачены химерные последовательности, состоящие из последовательностей грамположительных бактерий и последовательностей, экзогенных для грамположительных бактерий, таких как, например, последовательности, кодирующие химерные белки, состоящие из сигнальных пептидов секреции грамположительных бактерий и экзогенных белков.
Поскольку эукариотические гены в большинстве случаев наряду с экзонами содержат интроны, специалист в данной области техники поймет, что согласно настоящему изобретению любое упоминание экзогенного гена относится к безинтронной открытой рамке считывания такого гена, т.е. кодирующей белок последовательности такого гена. Термин "открытая рамка считывания" или ORF относится к последовательности кодирующих нуклеотидных триплетов, начиная с кодона инициации трансляции (например, ATG или GTG) и заканчивая кодоном терминации трансляции (например, ТАА, TAG или TGA), и причем последовательность кодирует отдельный полипептид.
Прокариотические гены, в частности, гены из грамположительных бактерий, не содержат интроны. Следовательно, кодирующая последовательность или открытая рамка считывания прокариотического гена соответствует последовательности кодирующих нуклеотидных триплетов, начиная с кодона инициации трансляции и заканчивая кодоном терминации трансляции, который расположен на прокариотическом геноме, в частности, бактериальной хромосоме.
Таким образом, в соответствии с одним аспектом настоящее изобретение относится к грамположительной бактерии, содержащей эндогенную открытую рамку считывания или кодирующую последовательность, с которой одна или несколько экзогенных открытых рамок считывания или кодирующие последовательности транскрипционно или трансляционно связаны.
Специалистам в данной области техники будет понятно, что, несмотря на то, что термин "ген" в целом может относиться к обнаруживаемому участку геномной последовательности, соответствующему единице наследования, который ассоциирован с транскрипционными и трансляционными регуляторными участками, такими как блок Прибнова, последовательность Шайн-Дальгарно, операторы, терминаторы, транскрибируемые участки и или другие функциональные участки последовательности, любая отсылка к термину "ген" в контексте описываемых в настоящем документе экзогенных последовательностей предпочтительно относится к кодирующей последовательности или открытой рамке считывания такого гена, если явно не указано иное. Любая отсылка к термину "ген" в контексте описываемых в настоящем документе эндогенных последовательностей может относиться к обнаруживаемому участку геномной последовательности, соответствующему единице наследования, который ассоциирован с регуляторными участками, транскрибируемыми участками и или другими функциональными участками последовательности, но альтернативно может также относиться к кодирующей последовательности или открытой рамке считывания такого гена.
Используемый в настоящем документе термин "трансляционно связанный" является синонимом с "трансляционно сцепленный" или "трансляционно соединенный". Такие термины, по сути, относятся к системам или единицам полицистронной экспрессии. Два или более генов, открытых рамок считывания или кодирующих последовательностей называют трансляционно связанными, когда общий(общие) регуляторный(регуляторные) элемент(элементы), такой(такие) как, в частности, общий промотор, влияет на транскрипцию указанных двух или более генов в одну мРНК, кодирующую указанные два или более генов, открытых рамок считывания или кодирующих последовательностей, которая затем может быть транслирована в две или более отдельных полипептидных последовательностей. Специалист в данной области техники поймет, что бактериальные опероны являются природными системами или единицами полицистронной экспрессии, в которых трансляционно или транскрипционно связаны два или более генов. Согласно настоящему изобретению транскрипционное связывание лежит в основе трансляционного связывания.
Таким образом, в соответствии с одним аспектом настоящее изобретение относится к грамположительной бактерии, содержащей эндогенный ген, с которым транскрипционно связаны один или несколько экзогенных генов, открытая рамка считывания или кодирующая последовательность. Предпочтительно, грамположительная бактерия последовательно содержит эндогенный ген, с которым транскрипционно связаны один или несколько экзогенных генов, открытая рамка считывания или кодирующая последовательность. Применяемый в настоящем документе термин "транскрипционно связанный" является синонимом с "транскрипционно сцепленный" и "транскрипционно соединенный". Такие термины обычно относятся к последовательностям полинуклеиновой кислоты, содержащим две или более открытых рамок считывания или кодирующих последовательностей, которые совместно транскрибируются как одна мРНК, которая может быть транслирована в два или более отдельных полипептидов.
В соответствии с другими аспектами настоящее изобретение относится к грамположительной бактерии или рекомбинантной нуклеиновой кислоте, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит эндогенный ген и один или несколько экзогенных генов, открытую рамку считывания или кодирующую последовательность.
Применяемый в настоящем документе термин "единица полицистронной экспрессии" или "система полицистронной экспрессии" относится к единице, в которой экспрессия двух или более генов регулируется посредством общих регуляторных механизмов, таких как промоторы, операторы и т.п. Применяемый в настоящем документе термин единица полицистронной экспрессии является синонимом с термином единица мультицистронной экспрессии. Примерами единиц полицистронной экспрессии являются без ограничения единицы бицистронной, трицистронной, тетрацистронной экспрессии. В рамках термина полицистронная охвачена любая мРНК, содержащая две или более, к примеру, 3, 4, 5, 6, 7, 8, 9, 10 или более, открытых рамок считывания или кодирующих участков, кодирующих отдельные продукты экспрессии, такие как белки, полипептиды и/или пептиды.
В соответствии с вариантом осуществления трансляционно или транскрипционно связанные один или несколько описываемых в настоящем документе эндогенных генов и один или несколько описываемых в настоящем документе экзогенных генов находятся под транскрипционным контролем промотора, который является эндогенным по отношению к грамположительной бактерии. В соответствии с другим вариантом осуществления описываемая в настоящем документе единица или система полицистронной экспрессии находится под транскрипционным контролем промотора, который является эндогенным по отношению к грамположительной бактерии.
Под "промотором" обычно понимают участок на молекуле нуклеиновой кислоты, предпочтительно молекуле ДНК, с которым связывается ДНК-полимераза и инициирует транскрипцию. Промотор предпочтительно, но не обязательно, расположен выше, т.е. 5', от последовательности, транскрипцию которой он контролирует.
В соответствии с дополнительным вариантом осуществления трансляционно или транскрипционно связанные один или несколько описываемых в настоящем документе эндогенных генов и один или несколько описываемых в настоящем документе экзогенных генов находятся под транскрипционным контролем нативного промотора (одного из) указанных одного или нескольких эндогенных генов. В соответствии с другим вариантом осуществления описываемая в настоящем документе единица или система полицистронной экспрессии находится под транскрипционным контролем нативного промотора (одного из) указанного одного или нескольких эндогенных генов, содержащихся в указанной системе или единице полицистронной экспрессии. В соответствии с другим вариантом осуществления описываемая в настоящем документе единица или система полицистронной экспрессии функционально связана с эндогенным промотором грамположительным бактерии.
Применяемый в настоящем документе термин "функционально связанный" или "функциональная связь" является связью, в которой регуляторные ДНК-последовательности и ДНК-последовательность, которую необходимо экспрессировать, соединены таким образом, чтобы делать возможной экспрессию. Например, промотор называют функционально связанным с геном, открытой рамкой считывания или кодирующей последовательностью, если связь или соединение обеспечивает транскрипцию или влияет на транскрипцию указанного гена. В соответствии с дополнительным примером 5' и 3' ген, цистрон, открытую рамку считывания или кодирующую последовательность называют функционально связанной в единице полицистронной экспрессии, если связь или соединение обеспечивает транскрипцию или влияет на транскрипцию по меньшей мере 3' гена.
Например, последовательности ДНК, такие как, например, предпочтительно промотор и открытая рамка считывания, называют функционально связанными, если природа связи между последовательностями (1) не приводит к внесению мутации со сдвигом рамки, (2) не препятствуют возможности промотора управлять транскрипцией открытой рамки считывания или (3) не препятствует возможности открытой рамки транскрибироваться под действием последовательности промоторного участка.
В соответствии с иллюстративным вариантом осуществления промотор может быть расположен выше, т.е. 5', от открытой(открытых) рамки(рамок) считывания, с которой(которыми) он функционально связан.
Специалист в данной области техники поймет, что промотор может быть ассоциирован с дополнительными нативными регуляторными последовательностями или участками, например, операторами. Точная природа необходимых для экспрессии регуляторных участков может изменяться от организма к организму, но обычно должна включать промоторный участок, который у прокариот содержит как промотор (который управляет инициацией транскрипции РНК), так и последовательности ДНК, которые при транскрибции в РНК будут служить сигналом инициации синтеза белка. Такие участки в норме будут включать эти 5'-некодирующие последовательности, вовлеченные в инициацию транскрипции и трансляции, такие как блок Прибнова (см. ТАТА-блок), последовательность Шайн-Дальгарно и т.п.
В соответствии со следующим вариантом осуществления промотор является нативным промотором, наиболее 5' удаленным, т.е. наиболее верхним, от эндогенного гена в единице полицистронной экспрессии.
Применяемый в настоящем документе термин "конститутивный" в контексте промотора (или, как само собой разумеется, в отношении экспрессии эндогенного гена) относится к промотору, который обеспечивает непрерывную транскрипцию такого ассоциированного гена. В частности, транскрипция ассоциированного гена или генов под контролем такого промотора происходит независимо от любого индуцирующего фактора или другого регуляторного сигнала.
Применяемый в настоящем документе термин "ген «домашнего хозяйства»" или "промотор «домашнего хозяйства»" относится к гену или промотору гена, который необходим для поддержания основной клеточной функции. Несмотря на то, что некоторые гены "домашнего хозяйства" экспрессируются с относительно постоянными уровнями, другие гены "домашнего хозяйства" могут варьировать в зависимости от внешних или экспериментальных условий. Гены "домашнего хозяйства", например, могут быть вовлечены в метаболизм, экспрессию гена (к примеру, основной аппарат транскрипции), передачу сигналов, но также могут быть структурными генами.
Применяемый в настоящем документе термин "ген гликолиза" или "промотор гликолиза" относится к гену или промотору гена, вовлеченному в каскад реакций гликолиза, и включают промоторы генов, кодирующих ферменты гликолиза, в частности гексокиназу, изомеразу фосфоглюкозы, фосфофруктокиназу, фруктозобисфосфатальдолазу, триозафосфатизомеразу, глицеральдегидфосфатдегидрогеназу, фосфоглицераткиназу, фосфоглицератмутазу, енолазу и пируваткиназу.
Используемый в настоящем документе термин "рибосомальный ген" или "рибосомальный промотор" относится к гену или промотору рибосомального гена, в том числе к генам, кодирующим рибосомальные белки, а также к генам, транскибированным в рибосомальную РНК. Предпочтительно, он может относиться к гену или промотору рибосомального белка.
Используемый в настоящем документе термин "ген центрального метаболизма" или "промотор центрального метаболизма" или, в альтернативном случае, "ген основного метаболизма" или "промотор основного метаболизма" относится к гену или промотору гена, вовлеченному в каскад важных метаболических реакций, и включает гены, вовлеченные в гликолиз, пентозофосфатный путь и цикл трикарбоновых кислот (ТСА).
Используемый в настоящем документе термин "необходимый" в контексте гена (или, как само собой разумеющееся, в отношении промотора такого гена) относится к гену, отсутствие нативного продукта экспрессии которого является вредным, к примеру, в частности, летальным, для хозяина или альтернативно изменяет, ингибирует или мешает протеканию нормальных физиологических процессов или осуществлению нормальной функции, такой как, в частности, размножение или рост. Следует понимать, что применяемый в настоящем документе термин "необходимый" в контексте гена или промотора гена относится к конститутивно необходимому, который является противоположным необходимому в зависимости от условий. Например, гены лактозного оперона, такие как ген бета-галактозидазы, у нескольких грамположительных бактерий, в частности, кисломолочных бактерий, таких как Lactococcus sp., могут быть необходимыми при культивировании бактерий в среде, содержащей лактозу в качестве основного или единственного источника углерода, такие гены не являются необходимыми при культивировании бактерий в среде, содержащей альтернативные источники углерода. Таким образом, такие гены являются необходимыми только в зависимости от условий, но не конститутивно необходимыми, как подразумевается в настоящем документе.
В соответствии с предпочтительным вариантом осуществления описываемые в настоящем документе эндогенный промотор и/или эндогенный ген выбран из группы включающей промоторы и/или гены грамположительных бактерий, или состоящей из них, соответствующих приведенным далее промоторам и/или генам Lactococcus, более конкретно промоторам и/или генам штамма MG1363 Lactococcus lactis ssp. cremoris: 1) ДНК-зависимая РНК-полимераза, бета' субъединица / 160 кДа субъединица (rpoC), 2) ДНК-зависимая РНК-полимераза, бета субъединица / 140 кДа субъединица (rpb2 или rpoB), 3) ДНК-связывающий ферритин-подобный белок (защитник от окислительного повреждения) (dps), 4) пируваткианаза (руk), 5) глутамил- и глутаминил-тРНК синтетазы (glnS или gltX), 6) енолаза (eno), 7) глутаминсинтетаза (glnA) 8) транскрипционный регулятор НТН-типа (glnR), 9) Xaa-His дипептидаза (argE или pepV), 10) бета-субъединица АТФ-синтазы F0F1-типа (бета-субъединица АТФ-синтазы F1) (atpD), 11) 3-фосфоглицераткиназа (pgk), 12) глицеральдегид-3-фосфатдегидрогеназа / эритрозо-4-фосфатдегидрогеназа (gapA или gapB), 13) ацетаткиназа (ackA), 14) 3-оксоацил-(ацил-белок-носитель)-синтаза (fabB или fabF), 15) 3-оксоацил-(ацил-белок-носитель)-редуктаза (fabG), 16) ДНК-зависимая РНК-полимераза, альфа-субъединица / 40 кДа субъединица (rpoA), 17) Хаа-Pro-аминопептидаза (рерР), 18) фруктозо- / тогатозобифосфатальдолаза (tbp или fbaA), 19) рибосомальный белок S4 (rpsD), 20) супероксиддисмутаза (sodA), 21) рибосомальный белок S12 (rpsL) и рибосомальный белок S7 (rpsG), 22) рибосомальный белок L18 (rplR), и рибосомальный белок S5 (rpsE), и рибосомальный белок L30/L7E (rpmD), 23) S-рибозилгомоцистеинлиаза (luxS), 24) рибосомальный белок L19 (rplS), 25) рибосомальный белок S11 (rpsK), 26) рибосомальный белок L10 (rplJ), 27) рибосомальный белок L7/L12 (rplL), 28) ДНК-связывающий белок бактерильного нуклеоида / ДНК-связывающий белок HU (hup или hILA), 29) 50S рибосомальный белок L28 (rpmB), 30) целлобиоза-специфичный компонент IIB фосфотрансферазной системы (lace или ptcB), 31) альфа-субъединица АТФ-синтазы F0F1-типа (atpA), 32) система транспорта сахаров АВС-типа (АТФазный компонент) (maIK или msmK), 33) альфа-субъединица, компонент комплекса ацетоиндегидрогеназы Е1 (асоА или pdhA), 34) белок клеточного деления (difIVA или ftsA), 35) UDP-галактопираноза, мутаза (glf), 36) глутамил-аминопептидаза (frvX или pepA), 37) прогнозируемый родственный дегидрогеназе белок (mviM или IImg_0272), 38) рибосомальный белок S2 (rpsB), 39) инициирующий трансляцию фактор-3 (IF-3) (infC), 40) рибосомальный белок L4 (rplD), и рибосомальный белок L23 (rplW), и рибосомальный белок L2 (rplB), 41) ЕМАР-домен (ydjD), 42) транскрипционный фактор элонгации (greA), 43) протеазная субъединица АТФ-зависимой CIp протеазы (cIpP), 44) рибосомальный белок L15 (rplO), 45) рибосомальный белок L11 (rplK), 46) рибосомальный белок S8 (rpsH), 47) рибосомальный белок L21 (rplU), 48) рибосомальный белок S13 (rpsM), 49) рибосомальный белок S19 (rpsS), и рибосомальный белок L22 (rplU или rplV), и рибосомальный белок L16 (rplP), и рибосомальный белок L14 (rplN), 50) рибосомальный белок S10 (rpsJ), 51) ко-шаперонин GroES (Hsp10) (cpn10), 52) рибосомальный белок L24 (rplX), 53) гипотетический белок LACR_0137 (duf965) и 54) секретируемый 45 кДа белок (usp45). Предпочтительно, эндогенный промотор и/или эндогенный ген выбран из группы, включающей или состоящей из enoA, usp45, gapB, pyk, rpmB и rplS. Такие промоторы и их последовательности раскрыты например в WO 2008/084115, включенной в настоящий документ при помощи ссылки, например, в приведенной в ней таблице 1 и фигуре 1А-Н. В соответствии с вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, причем эндогенный ген и один или несколько экзогенных генов находятся под транскрипционным контролем промотора, эндогенного по отношению к грамположительной бактерии, предпочтительно эндогенного промотора, выбранного из группы, состоящей из промотора eno, usp45, gap, pyk, rpmB и rplS указанной грамположительной бактерии. В соответствии со следующим вариантом осуществления эндогенный ген расположен в своем естественном локусе на хромосоме у грамположительной бактерии.
В соответствии с предпочтительным вариантом осуществления указанный один или несколько экзогенных генов, открытые рамки считывания или кодирующие последовательности трансляционно или транскрипционно связаны с 3'-концом указанного одного или нескольких эндогенных генов, открытой рамки считывания или кодирующей последовательности. Таким образом, в соответствии с вариантом осуществления настоящее изобретение относится к грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит один или несколько 5' эндогенных генов и один или несколько 3' экзогенных генов. Предпочтительно, наиболее 5' удаленным геном единицы полицистронной экспрессии является эндогенный ген. К примеру и без ограничения, единица полицистронной экспрессии может содержать эндогенный ген от 5'-конца до 3'-конца, за которым следует один или несколько эндогенных генов, за которыми следует один или несколько экзогенных генов, или, по сути, состоять из них. Альтернативно и без ограничения единица полицистронной экспрессии может содержать эндогенный ген от 5'-конца до 3'-конца, за которым следует один или несколько экзогенных генов, или, по сути, состоять из них. Альтернативно, единица полицистронной экспрессии может содержать эндогенный ген от 5'-конца до 3'-конца, за которым следует один или несколько экзогенных генов, за которыми следует один или несколько эндогенных генов, или, по сути, состоять из них.
Трансляционно связаные или транскрипционно связанные один или несколько эндогенных генов и один или несколько экзогенных генов, или единица или система полицистронной экспрессии, которые описаны в настоящем документе, можно включить в состав репликона, который обеспечивает сохранение и/или воспроизводство и экспрессию эндогенных и экзогенных генов у грамположительных бактерий согласно описываемому в данном документе настоящему изобретению.
В соответствии с вариантом осуществления трансляционно связанные или транскрипционно связанные один или несколько эндогенных генов и один или несколько экзогенных генов, необязательно включающих (эндогенный) промотор, который описан в других разделах настоящего описания, или единица или система полицистронной экспрессии, которые описаны в настоящем документе, можно включить в состав вектора, предпочтительно вектора экспрессии, обеспечивающего экспрессию в грамположительных бактериях. Таким образом, настоящее изобретение также относится вектору, содержащему описываемую в настоящем документе рекомбинантную нуклеиновую кислоту.
Используемый в настоящем документе термин "вектор" относится к молекуле нуклеиновой кислоты, как правило, ДНК, в которую можно вставить и клонировать, т.е. размножить, фрагменты нуклеиновой кислоты. Следовательно, вектор, как правило, будет содержать один или несколько уникальных участков рестрикции и может быть способен к автономной репликации в определенном хозяине или организме-носителе так, чтобы можно было репродуцировать клонированную последовательность. Векторы могут включать без ограничения плазмиды, фагмиды, бактериофаги, полученные от бактериофагов векторы, РАС, ВАС, линейные нуклеиновые кислоты, например, линейную ДНК и т.д., сообразно обстоятельствам (см., например, Sambrook et al., 1989; Ausubel 1992).
Важные факторы при выборе конкретного вектора, например, плазмиды, включают, среди прочего, удобство, с которым клетки-реципиенты, которые содержат вектор, можно распознать и отобрать от клеток-реципиентов, которые не содержат вектор; число копий вектора, которое необходимо для конкретного хозяина; и то, необходима ли возможность использования вектора в роли челнока между клетками-хозяевами разных видов. Предпочтительные прокариотические векторы включают плазмиды, такие как векторы, способные к репликации в Е. coli (такие как, например, pBR322, ColE1, pSC101, pUC19 и т.д.). Такие плазмиды описаны, например, в работах Sambrook et al., 1989; Ausubel 1992. Особенно предпочтительными векторами могут быть векторы, которые способны к репликации в Е. coli (или в других грамотрицательных бактериях), а также в другой представляющей интерес клетке-хозяине, такой как грамположительная бактерия, молочнокислая бактерия, предпочтительно Lactococcus, более предпочтительно Lactococcus lactis (см., например, работу Kok et al. Appl. Environ. Microbiol., 1984, vol. 48(4), 726-31). Другими предпочтительными векторами могут быть векторы, которые способны к репликации и/или способны выступать в роли челнока между одной или несколькими грамположительными бактериями, но не в грамотрицательных бактериях. В соответствии с предпочтительным вариантом осуществления вектором является pT1NX, который описан в работе Steidler et al. Appl. Environ. Microbiol., 1995, vol. 61(4), 1627-1629, которая отдельно включена в настоящий документ с помощью ссылки.
В соответствии с другим вариантом осуществления трансляционно связаный или транскрипционно связанный один или несколько эндогенных генов и один или несколько экзогенных генов, или единица или система полицистронной экспрессии, которые описаны в настоящем документе, встроены в геноме или хромосоме грамположительных бактерий. Способы получения рекомбинантных грамположительных бактерий и случайной, а также гомологичной рекомбинации хорошо известны из уровня техники, а также векторы для влияния на рекомбинацию. В качестве дополнительной информации, такие способы и векторы, например, раскрыты в работе Steidler et al. (2003, Nature Biotechnology, 21:785-789), Law et al. (1995, J Bacteriol, 177(24): 7011-7018), Leenhouts et al. (1998, Methods in Cell Science, 20:35-50) и WO 2004/046346, которые включены в настоящий документ с помощью ссылки в их полном объеме. Предпочтительно, описываемую в настоящем документе единицу полицистронной экспрессии получают или вносят посредством сайт-направленного встраивания соответствующих последовательностей в бактериальную хромосому посредством гомологичной рекомбинации.
В соответствии с вариантом осуществления вектор рекомбинации содержит описываемый в других разделах настоящего описания эндогенный промотор и необязательно дополнительные регуляторные последовательности, а также описываемую в настоящем документе единицу полицистронной экспрессии. Предпочтительно, эндогенный промотор и единица полицистронной экспрессии являются функционально связанными. Гомологичную рекомбинацию можно осуществить в предопределенном локусе. Такая система является высокомодульной и предусматривает отдельный подбор и комбинирование промотора, регуляторных последовательностей, эндогенного гена и экзогенного гена, а также выбор сайта встраивания.
В соответствии с другим вариантом осуществления грамположительная бактерия согласно настоящему изобретению содержит описываемый в других разделах настоящего описания эндогенный промотор в своем естественном локусе, т.е. в своем естественном геномном контексте на бактериальной хромосоме, с которым функционально связана единица полицистронной экспрессии, содержащая один или несколько эндогенных генов, открытую рамку считывания или кодирующую последовательность и один или несколько экзогенных генов, открытую рамку считывания или кодирующую последовательность. Функциональную связь можно создать с помощью гомологичной рекомбинации между содержащим промотор локусом и вектором рекомбинации, содержащим единицу полицистронной экспрессии, фланкированную последовательностями, предназначенными для осуществления указанной гомологичной рекомбинации. Таким образом, в соответствии с вариантом осуществления настоящее изобретение относится к описываемой в настоящем документе грамположительной бактерии, причем эндогенный ген транскрипционно связан с одним или несколькими экзогенными генами посредством встраивания в хромосому одного или нескольких экзогенных генов в указанном локусе, предпочтительно посредством встраивания в хромосому одного или нескольких экзогенных генов 3' от эндогенного гена в указанном локусе.
Конструкция вектора может быть выбрана так, чтобы только открытая рамка считывания или кодирующие последовательности эндогенных и/или экзогенных генов встраивались в предполагаемый хромосомный локус. В этом случае под естественным геномным локусом промотора предусматривают регуляторные последовательности рядом с самим по себе промотором, которые влияют на транскрипцию и/или трансляцию, например, операторы, сайт инициации транскрипции, последовательность Шайн-Дальгарно, терминаторную последовательность и т.д. Альтернативно, такие последовательности могут быть представлены на векторе для рекомбинации, содержащем единицу полицистронной экспрессии. Во втором случае, в зависимости от потребностей, нативные регуляторные последовательности, ассоциированные с эндогенным промотором, могут быть удалены при гомологичной рекомбинации. Описываемые в настоящем документе системы являются модульными в отношении отдельного подбора эндогенного гена, экзогенного гена и, возможно, регуляторных последовательностей, но они предопределяют сайт встраивания в эндогенном локусе выбранного промотора.
В соответствии со следующим вариантом осуществления грамположительная бактерия согласно настоящему изобретению содержит эндогенный промотор, а также один или несколько эндогенных генов, оба из которых описаны в настоящем описании, в своем (их) естественном локусе, т.е. в своем (их) естественном геномном контексте на бактериальной хромосоме, с которым функционально связаны один или несколько экзогенных генов, открытая рамка считывания или кодирующая последовательность так, чтобы влиять на полицистронную экспрессию одного или нескольких эндогенных генов и одного или нескольких экзогенных генов. В такой системе эндогенный промотор и один или несколько эндогенных генов, а также регуляторные последовательности, влияющие на транскрипцию и трансляцию указанного одного или нескольких эндогенных генов, присутствуют в своем естественном локусе. Такая система максимально сохраняет естественный характер грамположительной бактерии.
Единица полицистронной экспрессии содержит по меньшей мере два/две гена, открытые рамки считывания или кодирующие последовательности. Для инициации трансляции всех генов каждый из таких генов обычно ассоциирован с последовательностями, влияющими на связывание с рибосомой, т.е. сайтами связывания рибосомы. У прокариот сайты связывания рибосом обозначают последовательностями (SD) Шайн-Дальгарно, которые имеют общую консенсусную последовательность 5'-AGGAGG-3'. Последовательность SD, в среднем, расположена приблизительно на 8 пар оснований выше (т.е. 5') кодона инициации трансляции или инициаторного кодона. В зависимости от расстояния (в количестве нуклеотидов) между стоп-кодоном 5' гена и инициаторным ко доном 3' гена, последовательности SD обычно могут быть расположены 1) в межгенном участке между обоими генами, если расстояние по меньшей мере равно размеру последовательности SD; 2) в межгенном участке между обоими генами, но перекрываясь со стоп-кодоном 5' гена в случае меньшего расстояния между 5' и 3' геном; или 3) 5' к стоп-кодону 5' гена, если, например, стоп-кодон 5' гена и инициаторный кодон 3' гена расположены очень близко или перекрываются.
В соответствии с вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, дополнительно содержащим одну или несколько последовательностей полинуклеиновой кислоты, содержащих сайт связывания рибосомы, сконфигурированный для влияния на трансляцию одного или нескольких экзогенных генов. В соответствии с другим вариантом осуществления настоящее изобретение относится к описываемой в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, дополнительно содержащим один или несколько сайтов связывания рибосомы, сконфигурированных для влияния на трансляцию одного или нескольких экзогенных генов. В соответствии со следующим вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, причем указанные один или несколько эндогенных генов и указанные один или несколько экзогенных генов транскрипционно или трансляционно связаны посредством сайта связывания рибосомы. В соответствии с еще одним вариантом осуществления настоящее изобретение относится к грамположительной бактерии или рекомбинантной нуклеиновой кислоте, содержащим описываемую в настоящем документе единицу полицистронной экспрессии, причем любой 5' ген связан с 3' геном посредством последовательности полинуклеиновой кислоты, содержащей сайт связывания рибосомы или (по сути) состоящей из него. В соответствии с предпочтительным вариантом осуществления указанный сайт связывания рибосомы является эндогенным по отношению к грамположительной бактерии. В соответствии со следующим предпочтительным вариантом осуществления указанный сайт связывания рибосомы включен в межгенный участок, предпочтительно межгенный участок оперона.
В соответствии с другим вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, дополнительно содержащим одну или несколько последовательностей полинуклеиновой кислоты, которые содержат межгенный участок, сконфигурированный для влияния на трансляцию одного или нескольких экзогенных генов. В соответствии с дополнительным вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, дополнительно содержащим один или несколько межгенных участков, сконфигурированных для влияния на трансляцию одного или нескольких экзогенных генов. В соответствии со следующим вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, причем указанные один или несколько эндогенных генов и указанные один или несколько экзогенных генов транскрипционно или трансляционно связаны посредством межгенного участка. В соответствии с еще одним вариантом осуществления настоящее изобретение относится к грамположительной бактерии или рекомбинантной нуклеиновой кислоте, содержащим описываемую в настоящем документе единицу полицистронной экспрессии, причем любой 5' ген связан с 3' геном посредством последовательности полинуклеиновой кислоты, содержащей межгенный участок или состоящей из него. В соответствии с предпочтительным вариантом осуществления указанный межгенный участок является эндогенным по отношению к грамположительной бактерии. В соответствии со следующим предпочтительным вариантом осуществления указанный межгенный участок является межгенным участком оперона.
Используемый в настоящем документе термин "межгенный участок" является синонимом с термином "межгенный линкер" или "межгенный спейсер". Межгенный участок определяют как последовательность полинуклеиновой кислоты между смежными (т.е. расположенными на одной последовательности полинуклеиновой кислоты) генами, открытыми рамками считывания, цистронами или кодирующими последовательностями. Разумеется, что межгенный участок может включать стоп-кодон 5' гена и/или инициаторный кодон 3' гена, которые связаны посредством указанного межгенного участка. Определяемый в настоящем документе, термин межгенный участок, в частности, относится к межгенным участкам между смежными генами в единице полицистронной экспрессии. Например, определяемый в настоящем документе межгенный участок можно обнаружить между смежными генами в опероне. Таким образом, в соответствии с вариантом осуществления определяемый в настоящем документе межгенный участок является межгенным участком оперона.
В соответствии с вариантом осуществления межгенный участок, линкер или спейсер выбраны из группы межгенных участков, содержащей межгенные участки предшествующие, т.е. 5', более конкретно непосредственно 5' к rplW, rplP, rpmD, rplB, rpsG, rpsE или rplN грамположительной бактерии, или состоящей из них. В соответствии с вариантом осуществления указанная грамположительная бактерия является молочнокислой бактерией, предпочтительно видом Lactococcus, более предпочтительно Lactococcus lactis и любым его подвидом или штаммом. В соответствии с вариантом осуществления указанный межгенный участок охватывает инициаторный кодон rplW, rplP, rpmD, rplB, rpsG, rpsE или rplN и/или стоп-кодон предшествующего, т.е. 5', гена. В соответствии с предпочтительным вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, причем эндогенный ген и один или несколько экзогенных генов транскрипционно связаны посредством межгенного участка или участками, активными у грамположительной бактерии, причем предпочтительно межгенный участок или участки являются эндогенными по отношению к указанной грамположительной бактерии, причем более предпочтительно эндогенный межгенный участок выбран из группы, состоящей из межгенных участков, предшествующих rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE и rplF.
Специалист в данной области техники поймет, что если межгенный участок охватывает 5' стоп-кодон и/или 3' инициаторный кодон, то эти соответствующие кодоны предпочтительно отсутствуют в генах, которые соединены указанными межгенными участками, во избежание двойных инициаторных кодонов и/или стоп-кодонов, что может повлиять на правильную инициацию и/или терминацию трансляции. Способы выявления межгенных участков известны из уровня техники. В качестве дополнительной информации, межгенные участки можно, например, выявить на основе прогноза оперонов и связанных с ними промоторов и открытых рамок считывания, для чего в данной области известно и доступно множественное программное обеспечение.
В соответствии со следующим вариантом осуществления последовательность указанного межгенного участка выбрана из группы, включающей, по сути состоящей или состоящей из любой из SEQ ID NO: 1-7:
SEQ ID NO: 1 TAATG,
SEQ ID NO: 2 TAATCCATG,
SEQ ID NO: 3 TAAGGAGGAAAAAATG,
SEQ ID NO: 4 TAATAGAGGAGGAAAATCGTG,
SEQ ID NO: 5 TAAGAAGGGAGATAAGTAAGAATG,
SEQ ID NO: 6 TAAGGAAAGGGGTAATTAAACATG,
SEQ ID NO: 7 TAAGCAAAACTAGGAGGAATATAGCATG.
В соответствии со следующим вариантом осуществления последовательность указанного межгенного участка выбрана из группы, включающей, по сути состоящей или состоящей из последовательностей, у которых наблюдают одно нарушение комплементарности или делецию или вставку нуклеотида относительно SEQ ID NO: 1 или SEQ ID NO: 2, последовательностей, у которых наблюдают одно, два или три нарушения комплементарности или делецию или вставку одного, двух или трех нуклеотидов относительно SEQ ID NO: 3 или SEQ ID NO: 4, и последовательностей, у которых наблюдают одно, два, три или четыре нарушения комплементарности или делецию или вставку одного, двух, трех или четырех нуклотидов относительно SEQ ID NO: 5, SEQ ID NO: 6 или SEQ ID NO: 7.
Все SEQ ID NO: 1-7 содержат 5' стоп-кодон и 3' инициаторный кодон. SEQ ID NO: 1-7 соответствуют межгенным участкам, предшетвующим, соответственно, rplW, rplP, rpmD, rplB, rpsG, rpsE и rplN у штамма MG1363 Lactococcus lactis ssp. cremoris (номер доступа в Genbank AM406671.1). Эти последовательности, среди прочих, идентичны соответствующим последовательностям штамма CV56 Lactococcus lactis ssp. lactis (номер доступа в Genbank СР002365.1), штамма NZ9000 Lactococcus lactis ssp. cremoris (номер доступа в Genbank СР002094.1), штамма KF147 Lactococcus lactis ssp. lactis (номер доступа в Genbank CP001834.1), штамма IL1403 Lactococcus lactis ssp. lactis (номер доступа в Genbank AE005176.1) и штамма SK11 Lactococcus lactis ssp. cremoris (номер доступа в Genbank CP000425.1).
В соответствии с другим вариантом осуществления межгенный участок, линкер или спейсер выбраны из группы межгенных участков, которая включает межгенные участки, предшествующие, т.е. 5'-, более конкретно непосредственно 5'-, rplP, rpmD, rplM, rpsE, rplE, или rplF грамположительной бактерии, или состоит из них. В соответствии с вариантом осуществления указанная грамположительная бактерия представляет собой молочнокислую бактерию, предпочтительно вид Enterococcus, более предпочтительно Enterococcus faecium, и любой его подвид или штамм. В соответствии с вариантом осуществления указанный межгенный участок охватывает инициаторный кодон rplP, rpmD, rplM, rpsE, rplE или rplF и/или стоп-кодон предшествующего, т.е. 5', гена. Специалист в данной области техники поймет, что если межгенный участок охватывает 5' стоп-кодон и/или 3' инициаторный кодон, то эти соответствующие кодоны предпочтительно отсутствуют в генах, которые соединены указанными межгенными участками, во избежание двойных инициаторных кодонов и/или стоп-кодонов, что может повлиять на правильную инициацию и/или терминацию трансляции. Способы выявления межгенных участков известы из уровня техники. Для дополнительно информации, межгенные участки можно, например, выявить на основе прогноза оперонов и связанных с ними промоторов и открытых рамок считывания, для чего в данной области известно и доступно множественное программное обеспечение.
В соответствии со следующим вариантом осуществления последовательность указанного межгенного участка выбрана из группы, включающей, по сути состоящей или состоящей из любой из SEQ ID NO: 8-13:
SEQ ID NO: 8 TAATC,
SEQ ID NO: 9 TAAG GAG G AC AAC AATA,
SEQ ID NO: 10 TAATAGGAGGGAATTTCA,
SEQ ID NO: 11 TTAGAAGAAGGAGGAATACCATTC,
SEQ ID NO: 12 TAAAAGTTTAAGGAAGGAGGGTCTTACTGA,
SEQ ID NO: 13 TAATCAAGTAGAATCTACAAGGAGGTGTCTTTAA.
В соответствии со следующим вариантом осуществления последовательность указанного межгенного участка выбрана из группы, включающей, по сути состоящей или состоящей из последовательностей, у которых наблюдают одно нарушение комплементарности или делецию или вставку нуклеотида относительно SEQ ID NO: 8, последовательностей, у которых наблюдают одно, два или три нарушения комплементарности, или делецию или вставку одного, двух или трех нуклеотидов относительно SEQ ID NO: 9 или SEQ ID NO: 10, и последовательностей, у которых наблюдают одно, два, три или четыре нарушения комплементарности или делецию или вставку одного, двух, трех или четырех нуклеотидов относительно SEQ ID NO: 11, SEQ ID NO: 12 или SEQ ID NO: 13. SEQ ID NO: 8-13 соответствуют межгенным участкам, предшествующим, соответственно, rplP, rpmD, rplM, rpsE, rplE и rpIF у штамма LMG15709 Enterococcus faecium.
В соответствии с вариантом осуществления описанные в настоящем документе межгенные участки, за исключением любого предшествующего стоп-кодона и за исключением любого последующего инициаторного кодона, могут содержать или состоять из более 1 нуклеотида, как, например, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более нуклеотидов, предпочтительно более 5 нуклеотидов, еще более предпочтительно 10 или более нуклеотидов. В соответствии с другим вариантом осуществления межгенный участок может содержать 1-50 нуклеотидов, как, например, 1-40, 1-30, 1-25, 1-20, 1-15 или 1-10, предпочтительно 5-50, 5-40, 5-30, 5-25, 5-20, 5-15 или 5-10 нуклеотидов, еще более предпочтительно 10-50, 10-40, 10-30, 10-25, 10-20 или 10-15 нуклеотидов.
Особенно предпочтительные варианты осуществления грамположительных бактерий, содержащих описанную в настоящем документе единицу полицистронной экспрессии, изложены в таблицах 1 и 2, причем указанная грамположительная бактерия содержит эндогенный промотор, на 3' у которого эндогенный ген связан с межгенным участком, изложенным в таблицах 1 и 2, на 3' у которого один или несколько экзогенных генов, открытая рамка считывания или кодирующая последовательность связаны с межгенным участком. В соответствии с предпочтительным вариантом осуществления каждый изложенный в таблицах 1 и 2 ген находится под транскрипционным контролем своего нативного промотора и необязательных регуляторных последовательностей. В соответствии с другим предпочтительным вариантом осуществления указанная единица полицистронной экспрессии встроена в бактериальную хромосому. В соответствии со следующим предпочтительным вариантом осуществления указанный эндогенный промотор и/или эндогенный ген присутствуют в своем естественном локусе в бактериальном геноме или хромосоме. Предпочтительно, инициаторный и стоп-кодоны, при наличии, заменяют, соответственно, инициаторный и стоп-кодоны указанного экзогенного гена и указанного эндогенного гена.
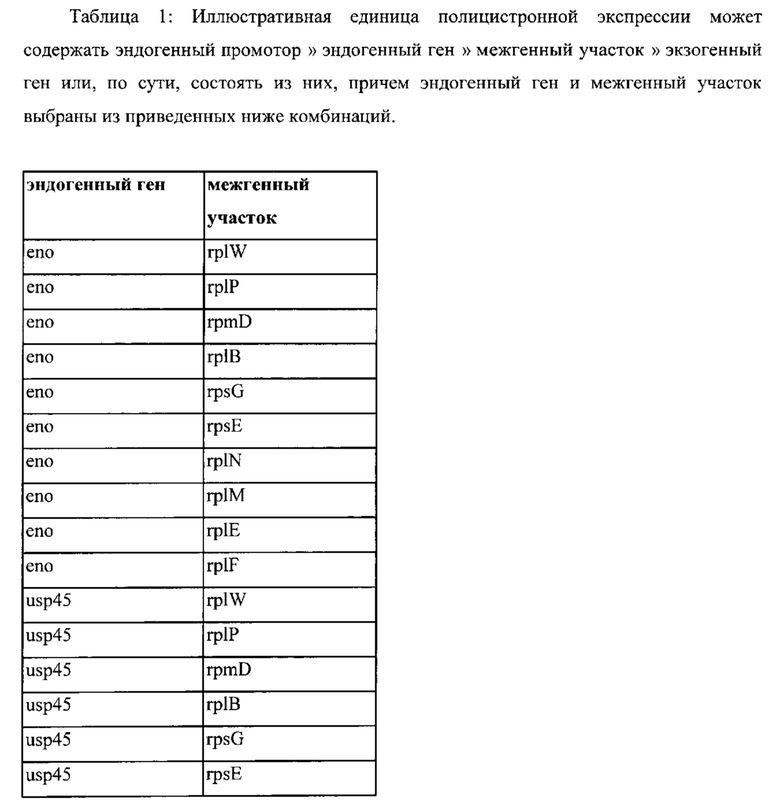
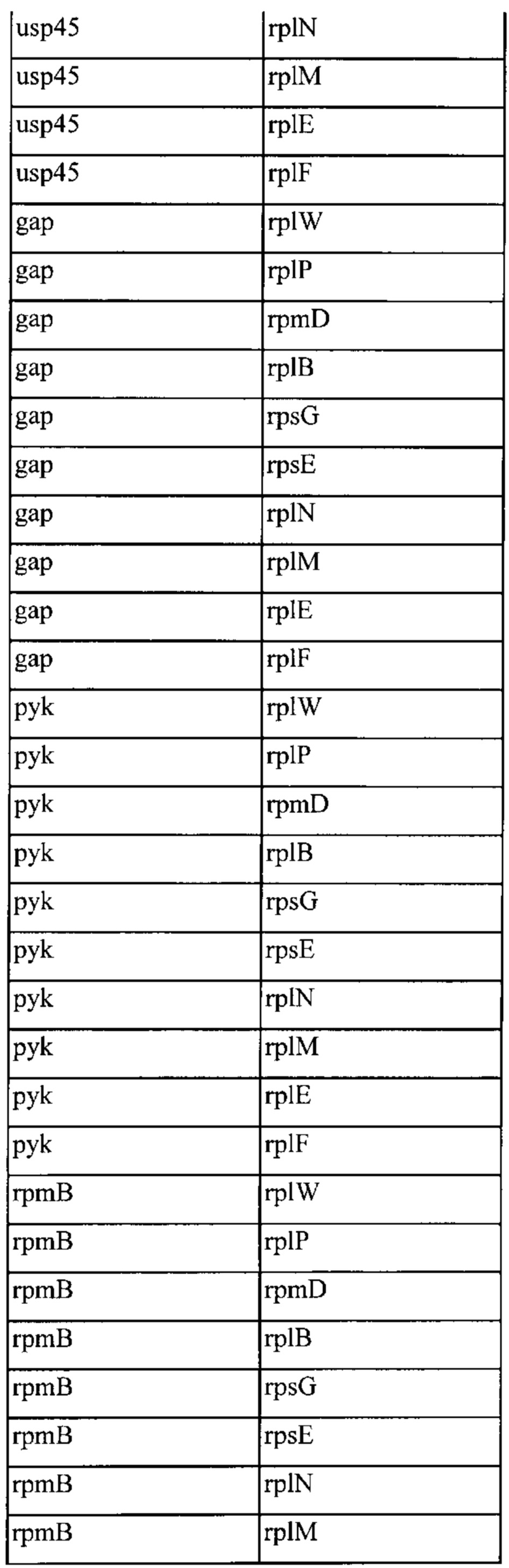

Предпочтительно, межгенные участки rplW, rplB, rpsG и rplN происходят от вида, подвида или штамма Lactococcus, предпочтительно Lactococcus lactis. Предпочтительно, межгенные участки rplP, rplM и rplE происходят от вида, подвида или штамма Enterococcus, предпочтительно Enterococcus faecalis или Enterococcus faecium. Предпочтительно, межгенные участки rplP, rpmD и rpsE происходят от вида, подвида или штамма Lactococcus, предпочтительно Lactococcus lactis, или от вида, подвида или штамма Enterococcus, предпочтительно Enterococcus faecalis или Enterococcus faecium.
Например без ограничения, если единица полицистронной экспрессии включает два экзогенных гена, то структура, представленная в виде эндогенный промотор » эндогенный ген » межгенный участок » экзогенный ген » межгенный участок » экзогенный ген, может быть следующей: usp45 » usp45 » rpmD » экзогенный ген 1 » rplN » экзогенный ген 2; enoA » enoA » rpmD » экзогенный ген 1 » rplN » экзогенный ген 2; gapB » gapB » rpmD » экзогенный ген 1 » rplN » экзогенный ген 2. Например, такой порядок может, в частности, подходить для экспрессии тяжелых и легких цепей антител (предпочтительно в таком порядке), таких как излагаемые в настоящем документе антитела к TNFα.



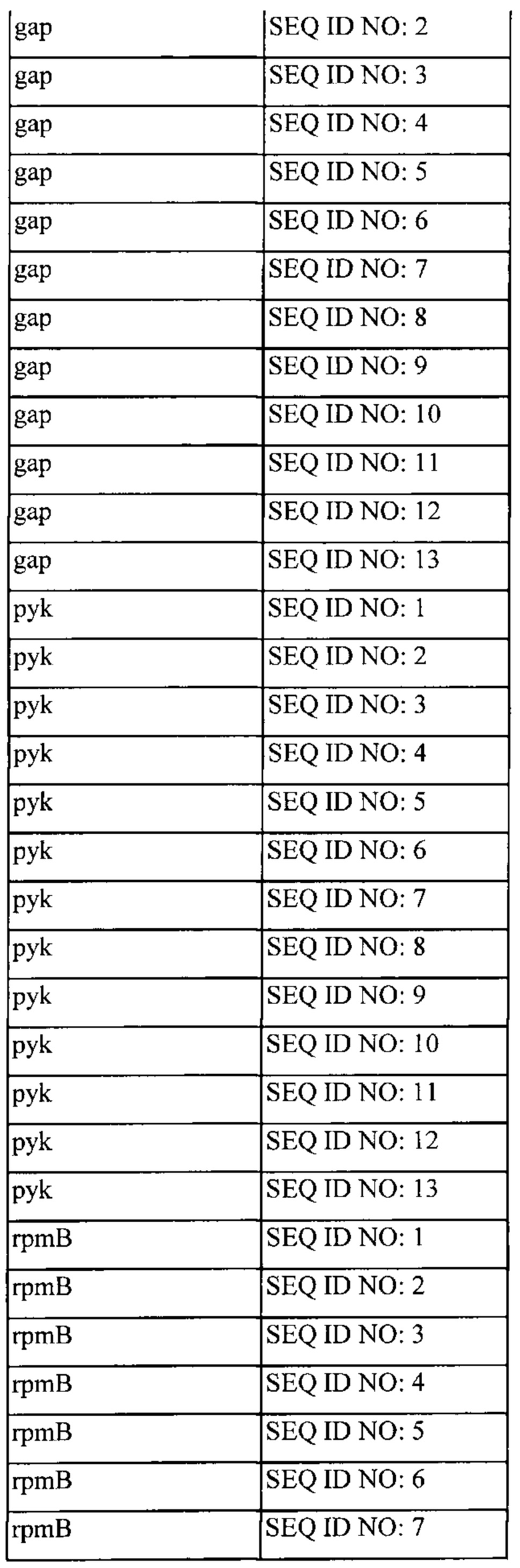
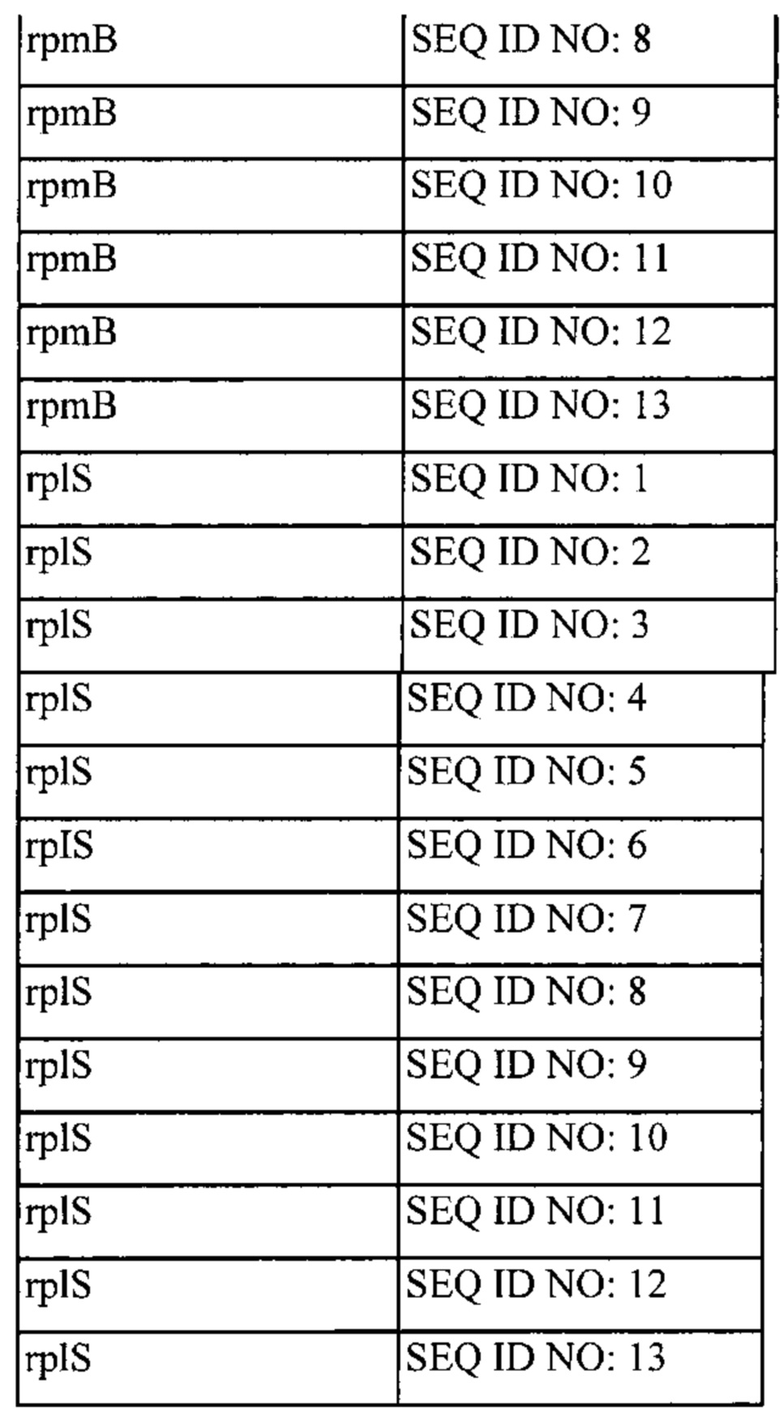
Предпочтительно, грамположительная бактерия с единицей полицистронной экспрессии, содержащей любую из SEQ ID NO: 1-7, является видом, подвидом или штаммом Lactococcus, предпочтительно Lactococcus lactis. Предпочтительно, грамположительная бактерия с единицей полицистронной экспрессии, содержащей любую из SEQ ID NO: 8-13, является видом, подвидом или штаммом Enterococcus, предпочтительно Enterococcus faecalis или Enterococcus faecium.
Специалист в данной области техники поймет, что экзогенные гены, открытые рамки считывания или кодирующие последовательности согласно настоящему изобретению могут быть связаны с дополнительными последовательностями, причем дополнительные последовательности выполняют конкретную задачу. Например, для повышения секреции продукта экзогенного гена ген можно связать с последовательностью нуклеиновой кислоты, кодирующий определяющий секрецию сигнальный пептид. В соответствии с особенно предпочтительным вариантом осуществления экзогенный ген, открытая рамка считывания или кодирующая последовательность согласно настоящему изобретению связаны на своем 5'-конце с последовательностью полинуклеиновой кислоты, кодирующий определяющий секрецию сигнал Usp45, предпочтительно происходящий от вида Lactococcus, более предпочтительно Lactococcus lactis и их подвидов и штаммов.
Как правило, определяющая секрецию сигнальная последовательность представляет собой сегмент длиной от приблизительно 16 до приблизительно 35 аминокислот, обычно содержащий гидрофобные аминокислоты, которые погружаются в липидную двухслойную мембрану и, таким образом, делают возможной секрецию сопровождающей белковой или пептидной последовательности из клетки-хозяина, и обычно она отщепляется от такого белка или пептида. Предпочтительно, определяющая секрецию сигнальная последовательность может проявлять подобную активность в клетке-хозяине, предназначенной для использования с нуклеиновой кислотой, содержащей указанную сигнальную последовательность.
Определяющие секрецию сигнальные последовательности, которые активны в подходящих клетках-хозяевах, известны из уровня техники; типичные сигнальные последовательности Lactococcus включают последовательности usp45 (см. патент US 5559007) и другие, см., например, работу Perez-Martinez et al. Mol. Gen. Genet, 1992, vol. 234, 401-11; Sibakov et al., Appl. Environ. Microbiol., 1991, vol. 57(2), 341-8. Предпочтительно, сигнальная последовательность расположена между промоторной последовательностью и ORF, т.е. сигнальная последовательность расположена 3' от промоторной последовательности и предшествует ORF представляющего интерес полипептида. В соответствии с предпочтительным вариантом осуществления сигнальная последовательность кодирует аминокислотную последовательность MKKKIISAILMSTVILSAAAPLSGVYA (usp45). Альтернативно, может быть использована мутантная сигнальная последовательность usp45 (usp45N), что в результате дает дополнительное контролируемое продуцирование и секрецию представляющего интерес полипептида. В частности, мутант содержит аспарагин (N) в положении 4 вместо лизина (K) или мутацию K4N. В соответствии с предпочтительным вариантом осуществления сигнальная последовательность кодирует аминокислотную последовательность MKKNIISAILMSTVILSAAA PLSGVYADTN.
Настоящее изобретение также относится к последовательности полинуклеиновой кислоты, содержащей описанную в настоящем документе единицу полицистронной экспрессии согласно настоящему изобретению. В частности, в одном аспекте настоящее изобретение относится к последовательности полинуклеиновой кислоты, содержащей описываемую в настоящем документе единицу полицистронной экспрессии согласно настоящему изобретению, причем указанная полицистронная единица содержит один или несколько генов, эндогенных по отношению к грамположительной бактерии, и один или несколько генов, открытых рамок считывания или кодирующих последовательностей, экзогенных по отношению к грамположительной бактерии, причем один или несколько эндогенных генов и один или несколько экзогенных генов трансляционно или транскрипционно связаны описываемым в настоящем документе способом. Предпочтительно, один или несколько эндогенных генов связаны с 5'-концом одного или нескольких экзогенных генов. Предпочтительно, один или несколько эндогенных генов и один или несколько экзогенных генов связаны посредством описанного в настоящем документе межгенного участка, предпочтительно межгенного участка перед rplW, rplP, rpmD, rplB, rpsG, rpsE, и rplN, как описано в других разделах настоящего документа, или межгенного участка, соответствующего любой из SEQ ID NO: 1-7, или межгенного участка перед rplP, rpmD, rplM, rpsE, rplE, или rplF, как описано в других разделах в настоящем документе, или межгенного участка, соответствующего любой из SEQ ID NO: 8-13, или родственных последовательностей, которые описаны выше. В соответствии с вариантом осуществления последовательность полинуклеиновой кислоты дополнительно содержит промотор, предпочтительно промотор, эндогенный по отношению к грамположительной бактерии. В соответствии с другим вариантом осуществления последовательность полинуклеиновой кислоты дополнительно содержит регуляторную последовательности, например оператор, терминатор и т.п. В соответствии с предпочтительным вариантом осуществления промотор является нативным промотором эндогенного гена.
В соответствии с дополнительным аспектом настоящее изобретение относится к репликону, содержащему описываемую в настоящем документе последовательность полинуклеиновой кислоты. Предпочтительно, указанный репликон представляет собой вектор, который описан в других разделах настоящего документа. В соответствии с вариантом осуществления указанный вектор подходит для экспрессии у прокариот.В соответствии с другим вариантом осуществления указанный вектор подходит для гомологичной рекомбинации у грамположительной бактерии.
В соответствии с другим аспектом настоящее изобретение относится к последовательности полинуклеиновой кислоты, содержащей сайт связывания рибосомы грамположительной бактерии и ген, открытую рамку считывания или кодирующую последовательность, экзогенные по отношению к указанной бактерии, причем сайт связывания рибосомы сконфигурирован с возможностью влиять на трансляцию экзогенного гена, открытой рамки считывания или кодирующей последовательности. В соответствии с вариантом осуществления последовательность полинуклеиновой кислоты содержит сайт связывания рибосомы грамположительной бактерии и ген, открытую рамку считывания или кодирующую последовательность, экзогенные по отношению к указанной бактерии, причем сайт связывания рибосомы соединен на 5'-конце экзогенного гена, открытой рамки считывания или кодирующей последовательности.
В соответствии с другим аспектом настоящее изобретение относится к последовательности полинуклеиновой кислоты, содержащей межгенный участок, предпочтительно межгенный участок оперона грамположительной бактерии, и ген, открытую рамку считывания или кодирующую последовательность, экзогенные по отношению к указанной бактерии, причем межгенный участок сконфигурирован с возможностью влиять на трансляцию экзогенного гена, открытой рамки считывания или кодирующей последовательности. В соответствии с вариантом осуществления последовательность полинуклеиновой кислоты содержит межгенный участок, предпочтительно межгенный участок оперона грамположительной бактерии, и ген, открытую рамку считывания или кодирующую последовательность, экзогенные по отношению к указанной бактерии, причем межгенный участок соединен на 5'-конце экзогенного гена, открытой рамки считывания или кодирующей последовательности. Предпочтительно, межгенный участок представляет собой межгенный участок, предшествующий rplW, rplP, rpmD, rplB, rpsG, rpsE или rplN, которые описаны в других разделах настоящего документа, или межгенный участок, соответствующий любой из SEQ ID NO: 1-7, или межгенный участок, предшествующий rplP, rpmD, rplM, rpsE, rplE, или rplF, как описано в других разделах в настоящем документе, или межгенный участок, соответствующий любой из SEQ ID NO: 8-13 или описанным ранее родственным последовательностям.
В соответствии с дополнительным аспектом настоящее изобретение относится к вектору для полицистронной экспрессии, содержащему межгенный участок, предшествующий rplW, rplP, rpmD, rplB, rpsG, rpsE или rplN, которые описаны в других разделах настоящего документа, или межгенный участок, соответствующий любой из SEQ ID NO: 1-7, или межгенный участок, предшествующий rplP, rpmD, rplM, rpsE, rplE или rplF, которые описаны в других разделах в настоящем документе, или межгенный участок, соответствующий любой из SEQ ID NO: 8-13 или описанным ранее родственным последовательностям. В соответствии с вариантом осуществления указанный вектор подходит для клонирования гена, открытой рамки считывания или кодирующей последовательности на 3'-конце указанного межгенного участка, предпочтительно гена, который является экзогенным по отношению к грамположительной бактерии. В соответствии с вариантом осуществления указанный вектор подходит для репликации у грамположительной бактерии. В соответствии со следующим вариантом осуществления указанный вектор подходит для влияния на гомологичную рекомбинацию у грамположительной бактерии, в частности, для встраивания в хромосому указанного межгенного участка и гена, открытой рамки считывания или кодирующей последовательности на 3'-конце указанного межгенного участка. В соответствии с вариантом осуществления указанный вектор дополнительно содержит один или несколько промоторов, предпочтительно промотор грамположительной бактерии. В соответствии со следующим вариантом осуществления указанный вектор дополнительно содержит регуляторные последовательности, например оператор, терминатор и тому подобные. В соответствии с еще одним вариантом осуществления указанный вектор дополнительно содержит один или несколько маркеров для отбора, такие как гены устойчивости к антибиотику.
В соответствии с другим аспектом настоящее изобретение относится к способу экзогенной экспрессии гена у грамположительной бактерии, предусматривающему этап трансформации указанной грамположительной бактерии вектором, содержащим экзогенный ген, открытую рамку считывания или кодирующую последовательность, необязательно дополнительно содержащим описываемый в настоящем документе (эндогенный) промотор.
В соответствии с дополнительным аспектом настоящее изобретение относится к применению последовательности полинуклеиновой кислоты, содержащей описываемый в настоящем документе межгенный участок грамположительной бактерии для полицистронной экспрессии одного или нескольких генов, открытых рамок считывания или кодирующих последовательностей, экзогенных по отношению к указанной грамположительной бактерии. В соответствии с вариантом осуществления настоящее изобретение относится к применению последовательности полинуклеиновой кислоты, содержащей описываемый в настоящем документе межгенный участок грамположительной бактерии, для полицистронной экспрессии одного или нескольких генов, открытых рамок считывания или кодирующих последовательностей, экзогенных по отношению к указанной грамположительной бактерии, и один или несколько генов, открытых рамок считывания или кодирующих последовательностей, эндогенных по отношению к указанной грамположительной бактерии. В соответствии с вариантом осуществления указанный один или несколько генов, экзогенных по отношению к указанной грамположительной бактерии, связаны с 3'-концом указанного эндогенного гена. Предпочтительно, межгенный участок представляет собой межгенный участок, предшествующий rplW, rplP, rpmD, rplB, rpsG, rpsE или rplN, которые описаны в других разделах настоящего документа, или межгенный участок, соответсвующий любой из SEQ ID NO: 1-7, или межгенный участок, предшествующий rplP, rpmD, rplM, rpsE, rplE или rplF, которые описаны в других разделах настоящего документа, или межгенный участок, соответствующий любой из SEQ ID NO: 8-13 или описанным ранее родственным последовательностям.
В соответствии с другим аспектом настоящее изобретение относится к способу экспрессии одного или нескольких экзогенных белков у грамположительной бактерии, предусматривающему этап введения описываемых в настоящем документе последовательности полинуклеиновой кислоты, кодирующей указанный один или несколько экзогенных белков, или вектора в указанную грамположительную бактерию так, чтобы они транскрибировались в полицистронную мРНК.
В соответствии с еще одним аспектом настоящее изобретение относится к способу получения грамположительной бактерии, способной экспрессировать один или несколько экзогенных белков, предусматривающему этап введения описываемых в настоящем документе последовательности полинуклеиновой кислоты, кодирующий один или несколько экзогенныъ белков, или вектора в указанную грамположительную бактерию так, чтобы они транскрибировались в полицистронную мРНК.
Согласно настоящему изобретению один или несколько экзогенных генов, открытых рамок считывания или кодирующих последовательностей могут быть любого вида или происхождения. В соответствии с вариантом осуществления один или несколько экзогенных генов кодируют белок, полипептид и/или пептид, предпочтительно белок, полипептид и/или пептид с терапевтическим или профилактическим эффектом у субъекта, или предпочтительно антиген, такой как антиген для стимуляции иммунитета или иммунотолерантности, невакциногенный терапевтически активный полипептид, антитело или его функциональный фрагмент, такой как Fab, химерный белок или мультимерный белок. В соответствии с предпочтительным вариантом осуществления один или несколько экзогенных генов кодируют антитело или функциональный фрагмент антитела. В контексте настоящего документа термин "функциональный" относится к фрагменту антитела, который все еще может проявлять свою предусмотренную функцию, т.е. связывание антигена. Используемый в настоящем документе термин антитело включает без ограничения традиционные антитела, химерные антитела, dAb, биспецифическое антитело, триспецифическое антитело, полиспецифическое антитело, бивалентное антитело, тривалентное антитело, поливалентное антитело, VHH, нанотело, Fab, Fab', F(ab')2 scFv, Fv, dAb, Fd, диатело, триатело, одноцепочечное антитело, однодоменное антитело, отдельный вариабельный домен антитела.
В настоящем контексте термин "антитело" применяют для описания иммуноглобулина либо природного, либо частично или полностью сконструированного. Поскольку антитело можно модифицировать рядом способов, термин "антитело" следует интерпретировать как охватывающий любую специфично связывающую молекулу или вещество со связывающим доменом с необходимой специфичностью связывания в отношении другого члена пары молекул, т.е. целевой молекулы, определение которой дано ранее. Таким образом, этот термин охватывает фрагменты, производные, функциональные эквиваленты антител и гомологи антител, а также одноцепочечные антитела, бифункциональные антитела, бивалентные антитела, VHH, нанотела, Fab, Fab', F(ab')2, scFv, Fv, dAb, Fd, диатела, триотела и верблюжьи антитела, в том числе любой полипептид, содержащий связывающий домен иммуноглобулина, независимо от того, является ли оно природным или частично или полностью сконструированным. Таким образом, включены химерные молекулы, содержащие связывающий домен иммуноглобулина, или слитый с другим полипептидом эквивалент.Термин также охватывает любой полипептид или белок со связывающим доменом, который представляет собой связывающий домен антитела, например миметики антител, или гомологичен ему. Примерами антител являются изотипы иммуноглобулина и их изотипические подклассы, в том числе IgG (IgG1, IgG2a, IgG2b, IgG3, IgG4), IgA, IgD, IgM и IgE. Специалист в данной области техники, следовательно, поймет, что настоящее изобретение также относится к фрагментам антител, содержащим антиген-связывающий домен, таким как VHH, нанотела Fab, scFv, Fv, dAb, Fd, диатела и триотела. В соответствии с вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или рекомбинантной нуклеиновой кислоте, причем один экзогенный ген кодирует легкую цепь (VL) антитела или ее функциональный фрагмент, а другой экзогенный ген кодирует тяжелую цепь (VH) антитела или ее функциональный фрагмент, причем более предпочтительно функциональный фрагмент представляет собой Fab. В соответствии с вариантом осуществления экзогенный ген, кодирующий VL или его функциональный фрагмент, транскрипционно связан с 3'-концом экзогенного гена, кодирующего VH или ее функциональный фрагмент.
В соответствии с вариантом осуществления описываемое в настоящем документе антитело по меньшей мере частично или полностью блокирует, ингибирует или нейтрализует биологическую активность целевой молекулы, такой как цитокин или хемокин. В контексте настоящего документа экспрессия "нейтрализует" или "нейтрализация" означает ингибирование или снижение биологической активности цитокина, измеренной in vivo или in vitro при помощи известных из уровня техники способов, таких как, например, описанных в примерах. В частности, ингибирование или снижение можно измерить путем определения колитного балла или путем определения целевой молекулы в образце ткани или крови. В контексте настоящего документа экспрессия "нейтрализует" или "нейтрализация" означает ингибирование или снижение биологической активности цитакина, измеренной in vivo или in vitro, по меньшей мере на 10% или более предпочтительно по меньшей мере на 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% и еще предпочтительнее на 100%.
Предпочтительно, указанные связывающие молекулы связываются с цитокинами, выбранными из перечня из IL-1β, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-12 (или его субъединицы IL-12p35 и IL12p40), IL-13, IL-15, IL-16, IL-17, IL-18, IL-21, IL-23 (или его субъединицы IL-23p19), IL-27, IL-32 (и его сплайс-варианты), IFN (α, β, γ) и TNFα, и ингибируют биологическое действие. Предпочтительно, указанные связывающие молекулы представляют собой растворимые рецепторы цитокинов, такие как gp130, или они связываются с рецепторами указанных цитокинов, например IL-2R (CD25, CD122, CD132), IL-12R (бета 1, бета 2), IL15R, IL-17R, IL-23R или IL-6R, без запуска воспалительного сигнала. Предпочтительно, указанные связывающие молекулы нейтрализуют хемокины, выбранные из перечня из MIF, MIP-1α, МСР-1, RANTES и эотаксина. Предпочтительно, указанные связывающие молекулы осуществляют блокировку активации иммунной системы посредством связывания с костимулирующими молекулами из перечня из CD3/CD28, HVEM, В7.1/В7.2, CD40/CD40L(CD154), ICOS/ICOSL, OX40/X40L, CD27/CD27L(CD70), CD30/CD30L(CD153) и 41BB/41BBL. Предпочтительно, указанные связывающие молекулы осуществляют блокировку воспаления посредством связывания с адгезивными молекулами из перечня I-CAM1, интегрина α4 и интегрина α4β7. Предпочтительно, указанные связывающие молекулы обладают костимулирующим и агонистическим эффектом на CD3, CTLA4 и/или PD1. Предпочтительно, указанные связывающие молекулы нейтрализуют активность Т-клеток или В-клеток путем целенаправленного воздействия на CD25, CD20, CD52, CD95, BAFF, APRIL и/или IgE. Предпочтительно, указанные связывающие молекулы осуществляют блокировку воспаления путем связывания с ферментами из семейства ММР. Предпочтительно, указанные связывающие молекулы оказывают анти-ангиогенный эффект, такой как нейтрализация активности αvβ3/α5β1 и IL-8. В соответствии со следующим предпочтительным вариантом осуществления указанная связывающая молекула может нейтрализовать биологический эффект TNFα, IL-12, IFNγ, IL-23 или IL-17. Предпочтительно, указанная связывающая молекула выбрана из группы, состоящей из
- антитела к TNFα, фрагмента антитела к TNFα, отдельного вариабельного домена антитела к TNFα, растворимого рецептора для TNF или доминантного отрицательного варианта TNFα;
- антитела к IL-12, фрагмента антитела к IL-12, отдельного вариабельного домена антитела к IL-12, растворимого рецептора для IL-12, доминантного отрицательного варианта IL-12 или dAb к IL-12;
- антитела к IL-12p35, фрагмента антитела к IL-12p35, отдельного вариабельного домена антитела к IL-12p35, растворимого рецептора для IL-12p35, доминантного отрицательного варианта IL-12p35 или dAb к IL-12p35;
- антитела к IL-12p40, фрагмента антитела к IL-12p40, отдельного вариабельного домена антитела к IL-12p40, растворимого рецептора для IL-12p40, доминантного отрицательного варианта IL-12p40 или dAb к IL-12p40;
- антитела к IL-23, фрагмента антитела к IL-23, отдельного вариабельного домена антитела к IL-23, растворимого рецептора для IL-23, доминантного отрицательного варианта IL-23 или dAb к IL-23;
- антитела к IL-23p19, фрагмента антитела к IL-23p19, отдельного вариабельного домена антитела к IL-23p19, растворимого рецептора для IIL-23p19, доминантного отрицательного варианта IL-23p19 или dAb к IL-23p19;
- антитела к IFNγ, фрагмента антитела к IFNγ отдельного вариабельного домена антитела к IFNγ, растворимого рецептора для IFNγ или доминантного отрицательного варианта IFNγ;
- антитела к IL-17, фрагмента антитела к IL-17, отдельного вариабельного домена антитела к IL-17, растворимого рецептора для IL-17, доминантного отрицательного варианта IL-17 или dAb к IL-17;
- антитела к МСР-1, фрагмента антитела к МСР-1, отдельного вариабельного домена антитела к МСР-1, растворимого рецептора для МСР-1, доминантного отрицательного варианта МСР-1 или dAb к МСР-1.
В соответствии с предпочтительным вариантом осуществления указанное антитело представляет собой Fab-фрагмент (антиген-связывающий фрагмент). Fab-фрагменты хорошо известны из уровня техники. В качестве дополнительной информации, Fab-фрагмент представляет собой участок антитела, который связывается с антигенами. Он состоит из одного константного и одного вариабельного домена каждой из тяжелой и легкой цепи.
В соответствии с вариантом осуществления Fab представляет собой Fab сА2 к TNF (полинуклеотидная и полипептидная последовательности варибального домена тяжелой цепи и легкой цепи которого раскрыты в US 6790444 в виде SEQ ID NO: 4 и 5 (тяжелая цепь) и SEQ ID NO: 2 и 3 (легкая цепь), соответственно) или Fab CDP870 к TNF (полинуклеотидная и полипептидная последовательности тяжелой цепи и легкой цепи которого раскрыты в WO 01/94585 в виде SEQ ID NO: 114 и 115 (тяжелая цепь) и SEQ ID NO: 112 и 113 (легкая цепь), соответственно).
Специалист в данной области техники поймет, что антитела, которые являются функциональными фрагментами антител, и, в частности, Fab-фрагментами, состоят из различных отдельных полипептидов, которые могут ковалентно соединяться дисульфидными мостиками. В частности, тяжелая цепь и легкая цепь кодируются различными отдельными кодирующими последовательностями.
Соответственно, кодирующие участки каждой из тяжелой и легкой цепей могут быть включены в описываемую в настоящем документе единицу полицистронной экспрессии. Последовательности полинуклеиновой кислоты, кодирующие тяжелую и легкую цепи могут, быть включены в различные единицы полицистронной экспрессии. Предпочтительно, последовательности полинуклеиновой кислоты, кодирующие тяжелую и легкие цепи, включены в одну единицу полицистронной экспрессии. Таким образом, в соответствии с вариантом осуществления настоящее изобретение относится к описываемой в настоящем документе грамположительной бактерии, содержащей один или несколько эндогенных генов, одну или несколько последовательностей полинуклеиновой кислоты, кодирующих тяжелую цепь антитела, или ее фрагмент, предпочтительно функциональный фрагмент, и одну или несколько последовательностей полинуклеиновой кислоты, кодирующих легкую цепь антитела, или ее фрагмент, предпочтительно функциональный фрагмент, которые трансляционно или транскрипционно связаны. В соответствии с другим вариантом осуществления настоящее изобретение относится к грамположительной бактерии, содержащей единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит один или несколько эндогенных генов, одну или несколько последовательностей полинуклеиновой кислоты, кодирующих тяжелую цепь антитела, или ее фрагмент, предпочтительно функциональный фрагмент, и одну или несколько последовательностей полинуклеиновой кислоты, кодирующих легкую цепь антитела, или ее фрагмент, предпочтительно функциональный фрагмент.В соответствии с еще одним вариантом осуществления кодирующая легкую цепь последовательность полинуклеиновой кислоты транскрипционно или трансляционно связана с 3'-концом кодирующей тяжелую цепь последовательности полинуклеиновой кислоты. Предпочтительно, такая связь дополнительно повышает экспрессию как тяжелой, так и легкой цепей.
Настоящее изобретение также относится к применению описываемых в настоящем документе грамположительных бактерий согласно настоящему изобретению для терапии. Настоящее изобретение дополнительно относится к фармацевтической композиции, содержащей описываемую в настоящем документе грамположительную бактерию согласно настоящему изобретению.
Таким образом, в соответствии с одним аспектом настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или фармацевтической композиции, содержащей грамположительную бактерию согласно настоящему изобретению, для применения в качестве лекарственного препарата. В соответствии с другим аспектом настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или фармацевтической композиции, содержащей грамположительную бактерию согласно настоящему изобретению, для применения при терапии или лечении. В соответствии со следующим аспектом настоящее изобретение относится к применению описываемых в настоящем документе грамположительной бактерии или фармацевтической композиции, содержащей грамположительную бактерию согласно настоящему изобретению, для получения лекарственного препарата. В соответствии с еще одним аспектом настоящее изобретение относится к способу лечения, предусматривающему введение описываемых в настоящем документе грамположительной бактерии или фармацевтической композиции, содержащей грамположительную бактерию согласно настоящему изобретению. В соответствии с вариантом осуществления настоящее изобретение относится к описываемым в настоящем документе грамположительной бактерии или фармацевтической композиции, содержащей грамположительную бактерию, причем один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид или пептид, причем продукт оказывает терапевтический или профилактический эффект на субъект, предпочтительно для применения в качестве лекарственного препарата, предпочтительно для применения при введении или доставки указанного продукта субъекту.
В соответствии с родственным аспектом настоящее изобретение относится к способу доставки полипептида, кодируемого одним или несколькими экзогенными генами, открытой рамкой считывания или кодирующей последовательностью, которые содержаться в грамположительной бактерии согласно настоящему изобретению, нуждающемуся в этом человеку или животному, предусмающему введение указанному человеку или животному терапевтически эффективного количество описываемых в настоящем документе грамположительных бактерий согласно настоящему изобретению. Животное предпочтительно может быть млекопитающим, таким как, например, домашние животные, сельскохозяйственные животные, животные в зоопарке, спортивные животные, домашние питомцы и экспериментальные животные, такие как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, крупный рогатый скот, коровы; приматы, такие как человекообразные обезьяны, мартышки, орангутанги и шимпанзе; псовые, такие как собаки и волки; кошачьи, такие как кошки, львы и тигры; непарнокопытные, такие как лошади, ослы и зебры; домашние животные, такие как коровы, свиньи и овцы; копытные, таки как олень и жирафы; грызуны, такие как мыши, крысы, хомяки и морские свинки, и т.д.
Применяемые в настоящем документе термины "лечить" или "лечение" относится как к терапевтическому лечению, так и к профилактическим или предупредительным мерам, причем объект должен предупреждать или замедлять (ослаблять) нежелательное физиологическое изменение или нарушение. Выражение "нуждающиеся в лечении человек или животное" включает таких, которые получат пользу от лечения указанного состояния.
Выражение "терапевтически эффективное количество" относится к количеству терапевтического вещества или композиции, которое эффективно для лечения заболевания или нарушения у субъекта, например, человека или животного, т.е. для получения необходимого локального или системного эффекта и действия. К примеру, терапевтически эффективное количество бактерий может включать по меньшей мере 1 бактерию, или по меньшей мере 10 бактерий, или по меньшей мере 102 бактерий, или по меньшей мере 103 бактерий, или по меньшей мере 104 бактерий, или по меньшей мере 105 бактерий, или по меньшей мере 106 бактерий, или по меньшей мере 107 бактерий, или по меньшей мере 108 бактерий, или по меньшей мере 109, или по меньшей мере 1010, или по меньшей мере 1011, или по меньшей мере 1012, или по меньшей мере 1013, или по меньшей мере 1014, или по меньшей мере 1015 или более грамположительных бактерий, например, в однократной или повторной дозе.
Грамположительные бактерии согласно настоящему изобретению можно вводить отдельно или в комбинации с одним или несколькими активными соединениями. Последние можно вводить до, после или одновременно с введением грамположительных бактерий.
Существует ряд раскрытий в данной области, касающихся доставки антигенов и/или терапевтически активных полипептидов, и следует понимать, что такие раскрытия можно дополнительно преимущественно модифицировать с помощью грамположительных бактерий согласно настоящему изобретению. К примеру и без ограничения, бактериальную доставку интерлейкинов, в частности IL-10, для лечения колита (например, WO 00/23471), IL-27 для модуляции воспалительного ответа (WO 2004/069177), доставку антигенов в качестве вакцин (например, WO 97/14806), доставку GLP-2 и родственных аналогов можно применять для лечения болезни укороченного кишечника, болезни Крона, остеопороза и в качестве вспомогательной терапии в ходе химиотерапии злокачественной опухоли и т.д. Кроме того, бактериальную доставку пептиды фактора "трилистника" можно применять для лечения заболеваний пищеварительного тракта (например WO 01/02570). В частности, применение белков или пептидов фактора "трилистника" для лечения нарушений и повреждений пищеварительного тракта, в том числе рта, пищевода, желудка и толстого и тонкого кишечника, а также для защиты и лечения тканей, которые лежат за пределами пищеварительного тракта, описаны в международных заявках WO 97/38712 и WO 92/14837. Эти белки можно применять либо для лечения поражений в этих областях, либо для ингибирования формирования поражений. Эти поражения могут быть обусловлены: лучевой терапией или химиотерапией для лечения злокачественной опухоли, любым другим лекарственным средством, в том числе спиртом, который повреждает пищеварительный тракт, случайным воздействием излучения или щелочным веществом, инфекцией, нарушением пищеварения, в том числе без ограничения мукозитом слизистой оболочки полости рта, мукозитом слизистой оболочки кишечника, эзофагитом, проктитом, неязвенной диспепсией, гастритом, пептической язвой или язвой двенадцатиперстной кишки, злокачественной опухолью желудка, злокачественной опухолью толстой кишки, лимфомой лимфоидной ткани слизистых оболочек, синдромом Менетрие, связанным с гастроэзофагальным рефлюксом заболеванием, болезнью Крона, язвенным колитом и острым колитом химического, бактериального или неясного происхождения. Пептиды фактора "трилистника" особенно пригодны для лечения острого колита, мукозита слизистой оболочки полости рта, мукозита слизистой оболочки кишечника, эзофагита, проктита. Предусмотрены дополнительные терапевтические применения с использованием промоторов и клеток-хозяев согласно настоящему изобретению.
Дополнительно неограничивающие примеры типов заболеваний, которые можно лечить у людей или животных путем доставки терапевтических полипептидов согласно настоящему изобретению, включают без ограничения, например, воспалительные заболевания кишечника, в том числе болезнь Крона и язвенный колит (которые можно лечить с помощью, например, IL-Ira, IL-10, IL-27 или пептидов фактора "трилистника"); аутоиммунные заболевания, в том числе без ограничения псориаз, ревматоидный артрит, красная волчанка (которую можно лечить с помощью, например, IL-Ira, IL-27, IL-10 или соответствующего аутоантигена); аллергические заболевания, в том числе без ограничения астму, пишевые аллергии, (которые можно лечить с помощью соответствующего аллергена); глютеновую болезнь (которую можно с помощью глютеновых аллергенов); неврологические нарушения, в том числе без ограничения болезнь Альцгеймера, болезнь Паркинсона и амиотрофический боковой склероз (который можно лечить с помощью, например, нейротропного фактора головного мозга и цилиарного нейротрофического фактора); злокачественную опухоль (которую можно лечить с помощью, например, IL-1, колониестимулирующих факторов или интерферона- W); остеопороз (который можно лечить с помощью, например, трансформирующего фактора роста f3); диабет (который можно лечить с помощью, например, инсулина); сердечно-сосудистое заболевание (которое можно лечить с помощью, например, тканевого активатора плазминогена); атеросклероз (который можно лечить с помощью, например, цитокинов и антагонистов цитокинов); гемофилию (которую можно лечить с помощью, например, факторов свертывания крови); дегенеративное заболевание печени (которое можно лечить с помощью, например, фактора роста гепатоцитов или интерферона а); заболевания легких, такие как муковисцидоз (который можно лечить с помощью, например, альфа-антитрипсина); ожирение; инфекции патогенами, например, вирусные или бактериальные инфекции (которые можно лечить с помощью любого количества вышеупомянутых композиций или антигенов) и т.д.
Грамположительные бактерии согласно настоящему изобретению также можно применять для лечения инфекционных заболеваний. В соответствии с вариантом осуществления можно получить пассивную иммунизацию от связанного с Clostridium заболевания, предпочтительно связанного с Clostridium dificile заболевания (CDAD), нейтрализующими токсин антителами, которые локально продуцируются и секретируются грамположительной бактерией согласно настоящему изобретению. Предпочтительно, указанная грамположительная бактерия представляет собой Lactococcus sp., более предпочтительно Lactococcus lactis или ее подвид или штамм.
CDAD опосредуется двумя экзотоксинами, токсином А (энтеротоксином; см., например, Genbank NC_009089.1, участок: 795843..803975 для ДНК-последовательности или YP_001087137.1 для последовательности белка) и токсином В (цитотоксином; см., например, Genbank NC_009089.1, участок: 787393..794493 для ДНК-последовательности или YP_001087135.1 для последовательности белка). Оба они являются высокомолекулярными белками, которые связываются с поверхностью клеток кишечного эпителия, где они интернализируются и катализируют гликозилирование цитоплазматических белков rho, что приводит к гибели клеток, воспалению и диарее. Также они были вовлечены в стимуляцию вирулентности, колонизации С. difficile, активацию и хемотаксис нейтрофилов. Сама по себе бактерия не является инвазивной и не вызывает повреждение тканей. В результате нейтрализации антителами токсинов С. difficile патогенный механизм патогена блокируется, его способность размножаться в кишечнике может снизиться, а влияние на экологию микроорганизмов может быть сведено к минимому, что делает возможным восстановление нормальной микрофлоры. Преимущество такого подхода в медицинском плане может включать более быстрое восстановление, меньшее количество случаев рецидива и ослабление давления отбора по устойчивости к антибиотику у нормальной микрофлоры кишечника.
Таким образом, в соответствии с вариантом осуществления настоящее изобретение относится к описываемой в настоящем документе грамположительной бактерии, у которой единица полицистронной экспрессии охватывает антитело или его фрагмент, предпочтительно Fab, который описан в других разделах настоящего документа, направленный против токсина А и/или токсина В Clostridium. Наиболее предпочтительно, указанное антитело или его фрагмент представляет собой нейтрализующее антитело. В соответствии со следующим вариантом осуществления настоящее изобретение относится к грамположительной бактерии, предпочтительно Lactococcus sp., такой как Lactococcus lactis, или Enterococcus sp., такой как Enterococcus faecalis или Enterococcus faecium, содержащей единицу полицистронной экспрессии, предпочтительно встроенную в бактериальную хромосому, причем указанная единица полицистронной экспрессии содержит эндогенный ген, предпочтительно выбранный из группы, состоящей из eno, usp45, gap, pyk, rpmB и rplS, предпочтительно происходящих от Lactococcus sp. или Enterococcus sp., и один или несколько экзогенных генов, кодирующих нейтрализующее антитело или фрагмент антитела, предпочтительно Fab, к токсину А и/или токсину В Clostridium, предпочтительно Clostridium dificile, причем указанная единица полицистронной экспрессии предпочтительно встроена в хромосому в естественном локусе указанного эндогенного гена, а указанный ген антитела (фрагмента) к токсину А и/или токсину В предпочтительно транскрипционно связан с 3'-концом указанного эндогенного гена, причем указанная транскрипционная связь предпочтительно находится под влиянием межгенного участка, предпочтительно выбранного из группы, состоящей из межгенных участков, предшествующих rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE и rplF у грамположительной бактерии, предпочтительно Lactococcus sp. или Enterococcus sp. Описываемые в настоящем документе антитела к токсину А и токсину В Clostridium известны из уровня техники (см., например, работу Leung et al., J Pediatr 1991; 118(4 Pt 1):633-637; Wilcox. J Antimicrob Chemother 2004; 53(5):882-884; Sougioultzis et al., Gastroenterology 2005; 128(3):764-770; Kyne et al., N Engl J Med 2000; 342(6):390-397; Lowy et al., N Engl J Med; 362(3): 197-205). Информация как об антителах, так и их фрагментах может быть расположена на различных единицах полицистронной экспрессии у одной или разных грамположительных бактерий, но предпочтительно она расположена на одной единице полицистронной экспрессии. Настоящее изобретение дополнительно относится к способу предупреждения и/или лечения CDAD, предусматривающему введение такой грамположительной бактерии.
Специалист в данной области поймет, что упомянутые отдельно в настоящем документе заболевания являются всего лишь иллюстративными, и их упоминание никоим образом не предназначено для ограничения применения реагентов, к которым относится настоящее изобретение, например, промоторов, нуклеиновых кислот, векторов и клеток-хозяев по настоящему изобретению, в отношении таких конкретных заболеваний. Наоборот, специалист в данной области техники поймет, что реагенты по настоящему изобретению можно применять для экспрессии, в принципе, любых представляющих интерес продуктов экспрессии, предпочтительно полипептидов, которые могут иметь терапевтическое значение не только в упомянутых случаях, но также при различных других заболеваниях или состояниях у людей и животных. Следовательно, после того, как подходящий продукт экспрессии, предпочтительно полипептид, например, антиген, антитело (фрагмент) и/или невакциногенный терапевтически активный полипептид, был выбран или выявлен для определенной болезни, специалист в данной области сможет осуществить его экспрессию, выделение и/или доставку с применением реагентов по настоящему изобретению.
Настоящее изобретение также относится к лечению заболеваний у других животных, в том числе собак, лошадей, кошек и птиц. Заболевания у собак включают без ограничения собачью чуму (парамиксовирус), собачий гепатит (аденовирус Cav-1), кашель псарен или ларинготрахеит (аденовирус Cav-2), инфекционный энтерит собак (коронавирус) и геморрагический энтерит (парвовирус).
Заболевания у кошек включают без ограничения вирусных ринотрахеит (герпесвирус), калицивирусное заболевание кошачьих (калицивирус), инфекционный перитонит кошачьих (парвовирус) и лейкоз кошачьих (вирус лейкоза кошачьих). Также предусмотрено, что с помощью способов и композиций согласно настоящему изобретению лечению поддаются другие вирусные заболевания у лошадей и птиц. С этой целью применение экспрессирующих рекомбинантные интерфероны микроорганизмов будет особенно предпочтительным.
В контексте настоящего документа, фармацевтическая композиция предпочтительно включает терапевтически эффективное количество грамположительных бактерий согласно настоящему изобретению и фармацевтически приемлемый носитель, т.е. один или несколько фармацевтически приемлемых веществ-носителей и/или добавок, например, буферов, носителей, наполнителей, стабилизаторов и т.д.
Применяемый в настоящем документе термин "фармацевтически приемлемый" согласуются с термином в данной области и означает совместимый с другими ингредиентами фармацевтической композиции и не оказывающий вредного действия на принимающего его пациента.
Грамположительные бактерии согласно настоящему изобретению можно суспендировать в фармацевтическом составе для введения человеку или животному с подлежащим лечению заболеванием. Такие фармацевтические составы включают без ограничения живые грамположительные бактерии и подходящую для введения среду. Грамположительные бактерии можно лиофилизировать в присутствии традиционных наполнителей, таких как лактоза, другие сахара, стеарат, карбонат и/или сульфат щелочного и/или щелочноземельного металла, (например, стеарат магния, карбонат натрия и сульфат натрия), каолин, кремнезем, придающие вкус и запах добавки. Лиофилизированные таким способом грамположительные бактерии можно получить в форме капсул, таблеток, гранулятов и порошков (например, порошка для полоскания рта), каждый из которых можно вводить пероральным путем. Альтернативно, некоторые грамположительные бактерии можно получить в виде водных суспензий в подходящих средах, или же лиофилизированные бактерии можно суспендировать в подходящей среде непосредственно перед применением, причем такая среда включает указанные в настоящем документе наполнители и другие наполнители, такие как глюкоза, глицин и сахаринат натрия.
Для перорального применения можно составить резистентные к действию желудочного сока пероральные лекарственные формы, причем лекарственные формы могут также включать соединения, обеспечивающие контролируемое высвобождение грамположительных бактерий, и, таким образом, обеспечивать контролируемое высвобождение необходимого закодированного в них белка. Например, пероральная лекарственная форма (в том числе капсулы, таблетки, пеллеты, грануляты, порошки) можно покрыть тонким слоем наполнителя (обычно полимеров, производных целлюлозы и/или липофильных материалов), которые резистентны к растворению или разрушению в желудке, но не в кишечнике, что, таким образом, делает возможным прохождение через желудок с целью распада, растворения и всасывания в кишечнике.
Пероральную лекарственную форму можно разработать так, чтобы было возможно медленное высвобождения грамположительных бактерий и произведенных экзогенных белков, например, в виде таблеток или капсул с контролируемым высвобождением, замедленным высвобождением, пролонгированным высвобождением, пролонгированным действием. Такие лекарственные формы обычно содержат традиционные и хорошо известные наполнители, такие как липофильные, полимерные, целлюлозные, нерастворимые, разбухаемые наполнители. Составы с контролируемым высвобождением также можно применять для любых других мест доставки, в том числе доставки в кишечник, толстую кишку, биоадгезивной или подъязычной доставки (т.е., доставка к слизистой пародонта) и бронхиальной доставки. Если композиции согласно настоящему изобретению необходимо ввести ректально или вагинально, то фармацевтические составы могут включать мази, суппозитории и кремы. В таком случае грамположительные бактерии суспендируют в смеси традиционных наполнителей, которые также включают липиды. Каждый из вышеупомянутых составов хорошо известны из уровня техники и описаны, например, в следующих справочных материалах: Hansel et al., Pharmaceutical dosage forms and drug delivery systems, 5th edition, William and Wilkins, 1990; Chien 1992, Novel drug delivery system, 2nd edition, M. Dekker; Prescott et al. (1989, Novel drug delivery, J.Wiley & Sons); Cazzaniga et al, (1994, Oral delayed release system for colonic specific delivery, Int. J. Pharm.i08:7'.
Предпочтительно, для ректального введения можно применять составы для введения посредством клизмы. Термин "клизма" применяют для охватывания жидких препаратов, предназначенных для ректального применения. Обычно клизму поставляют в емкостях с однократной дозой, и она содержит одно или несколько активных веществ, растворенных или диспергированных в воде, глицерине или макроголях или других подходящих растворителях.
Таким образом, согласно настоящему изобретению в соответствии с предпочтительным вариантом осуществления описываемые в настоящем документе грамположительные бактерии согласно настоящему изобретению, кодирующие необходимый экзогенный ген, можно вводить животному или человеку через слизистую, например, пероральным, назальным, ректальным, вагинальным или бронхиальным путем, с помощью любых известных из уровня техники составов, которые применимы для конкретного пути. Дозировки грамположительных бактерий для введения будут варьировать в зависимости от множества факторов, в том числе типа бактерий и кодируемых ими гена, типа и тяжести подлежащего лечению заболевания и пути введения, который необходимо использовать.
Таким образом, точные дозировки невозможно определить для всех без исключения вариантов осуществления согласно настоящему изобретению, но они будут вполне очевидны специалистам в данной области, взявшим на вооружение настоящее изобретение. Дозировку можно, так или иначе, определить в каждом конкретном случае путем измерения уровней концентраций рекомбинантного белка в сыворотке крови после введения предопределенного количества клеток с помощью таких хорошо известных способов, которые известны под названием ELISA или Biacore (см. примеры). Анализ кинетического профиля и времени полужизни доставленного рекомбинантного белка дает достаточную информацию для того, чтобы сделать возможным определение диапазона эффективных доз трансформированных клеток-хозяев.
В соответствии с вариантом осуществления, если описываемые в настоящем документе грамположительные бактерии согласно настоящему изобретению экспрессируют антиген, то настоящее изобретение, таким образом, может также относиться к вакцине.
Термин "вацина" указывает на фармацевтически приемлемую композицию, которая при введении в эффективном количестве субъекту-человеку может индуцировать выработку антител на иммуноген, содержащийся в вакцине, и/или формирует защитный иммунитет у субъекта.
Вакцина согласно настоящему изобретению будет содержать описываемые в настоящем документе грамположительные бактерии согласно настоящему изобретению и необязательно дополнительно наполнитель. Такие вакцины также могут содержать адъювант, т.е. соединение или композицию, которая усиливает иммунный ответ на антиген. Адъюванты включают без ограничения полный адъювант Фрейнда, неполный адъювант Фрейнда, сапонин, минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюрониловые многоатомные спирты, полианионы, пептиды, эмульсии на основе масла или углеводорода и потенциально полезные фармацевтически приемлемые адъюванты для человека, такие как BCG (бацилла Кальмета-Герена) и Corynebacterium parvum.
Таким образом, становится очевидным, что настоящее изобретение относится к биомаркерам, применениям и способам, которые обеспечивают существенные преимущества при постановке диагноза, предопределении, погнозировании и/или наблюдении за лечением причиняющих ущерб нарушений. Несмотря на то, что настоящее изобретение было описано в связи с его конкретными вариантами осуществления, понятно, что в свете вышеприведенного описания специалистам в данной области будут очевидны многие альтернативы, модификации и варианты. Соответственно, подразумевается, что оно охватывает все такие альтернативы, модификации и варианты, которые соответствуют идее и широкому объему прилагаемой формулы изобретения.
Аспекты и варианты осуществления согласно настоящему изобретению дополнительно подкреплены приведенными далее неограничивающими примерами.
Примеры
Пример 1: Выбор межгенных участков из генома Lactococcus lactis
Клеточные белки культуры конца лог-фазы штамма MG1363 Lactococcus lactis ssp. cremoris визуализировали на геле белкового фореза с окрашиванием красителем кумасси синий, как показано на фигуре 1. Дорожки А и В содержали 29 мкг и 58 мкг белков MG1363, соответственно. Из геля выделяли 12 бэндов определенных белков из дорожки А. Выделяли белки и идентифицировали межгенные участки путем:
1) идентификации экспрессирующихся в большом количестве белков в фрагментах с помощью частичного секвенирования пептидов (MALDI-TOF/TOF) и поиска по базам данных с применением объединенных масс пептидов и информации о последовательности;
2) идентификации с использованием хромосомной последовательности штамма MG1363 Lactococcus lactis ssp. cremoris (Wegmann et al) генов, кодирующих экспрессирующиеся в большом количестве белки (1), которые присутствуют в опероне, но не в "первом гене";
3) идентификации межгенных участков, которые предшествуют этим экспрессирующимся в большом количестве генам.
В таблице 3, таким образом, приведены межгенные участки. Подчеркнутые последовательности представляют собой сайты связывания рибосомы.
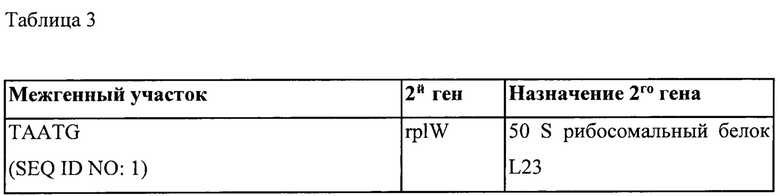

Пример 2: Выбор сайтов для бицистронной экспрессии
В таблице 4 приведены целевые промоторы, идентифицированные в примере 1 как обеспечивающие высокий уровень экспрессии. Такие промоторы можно использовать в качестве целевых сайтов для полицистронной экспрессии экзогенных генов.
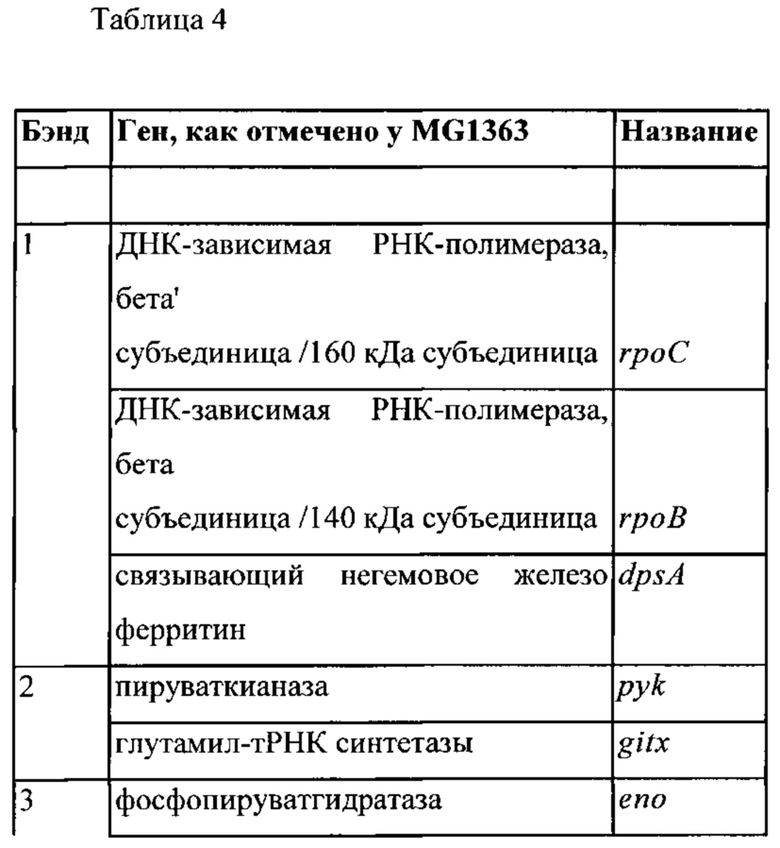
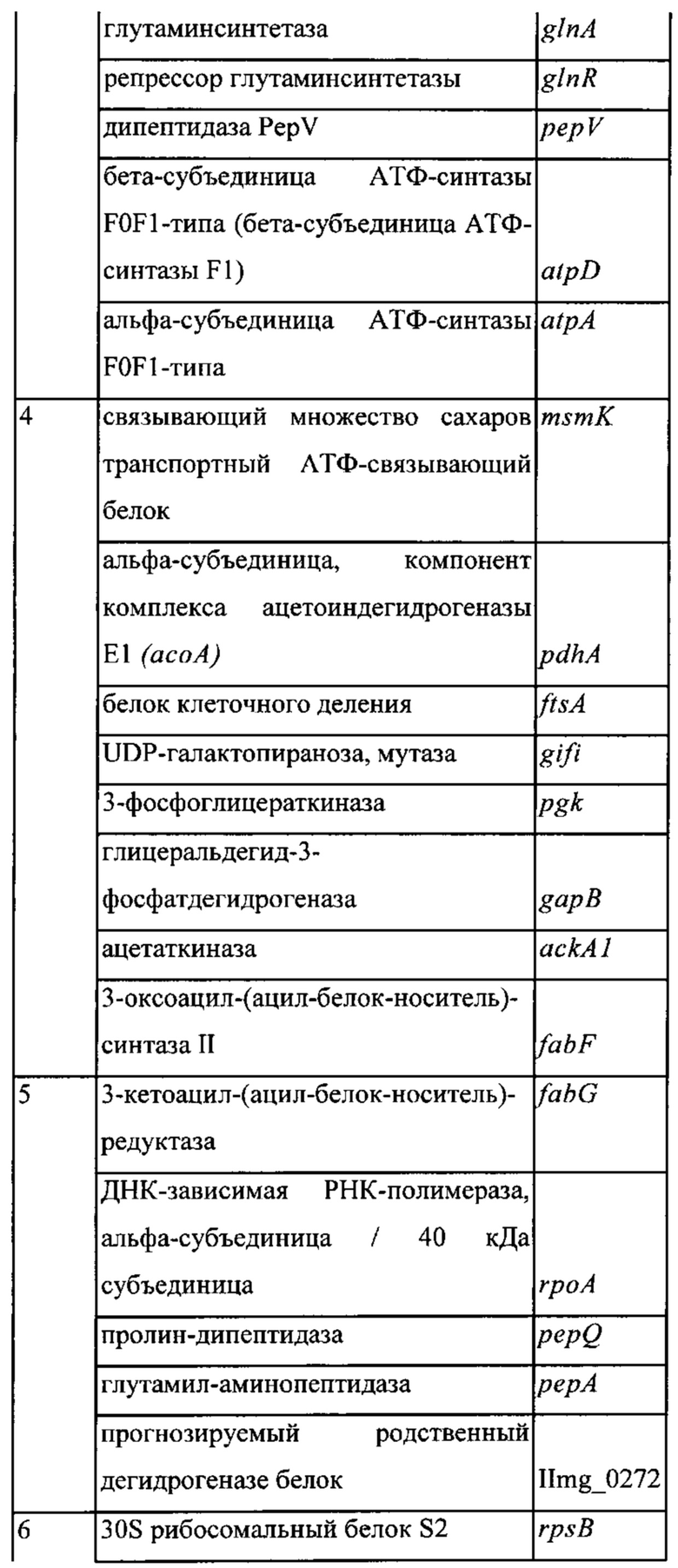


Ген β-глюкуронидазы (uidA) из Е. coli вводили в качестве гена-репортера в MG1363 Lactococcus lactis. β-глюкуронидаза, продукт гена uidA, катализирует расщепление широкого спектра β-глюкуронидов, которые коммерчески доступны в качестве субтратов для гистохимических и спектрофотометрических исследований. Штамм sAGX0090 имеет кассету экспрессии PhIIA»uidA в локусе thyA (фигура 2). Этот промотор также использовали в штаммах sAGX0037 и SAGX0085.
Как изображено на фигуре 2, конструкции с двойными цистронами получали в результате вставки uidA в хромосоме MG1363 Lactococcus lactis на 3'-конце нескольких эндогенных генов (ген X). Таким образом, rpmD использовали в качестве межгенной последовательности между представляющими интерес эндогенными генами из таблицы 4 и uidA для определения сайтов, которые в результате дают более высокую β-глюкуронидазную (GUS) активность по сравнению с SAGX0090.
Культуры Lactococcus lactis выращивали в течение 16 часов при температуре 30°С в GM17, при необходимости дополненной тимидином. Клетки 1 мл культуры промывали и ресуспендировали в 1 мл деминерализованной воды. Клетки разрушали при помощи лизирующей матрицы В от MP Biomedicals и устройства Fastprep-24 со скоростью 6 м/с в течение 40 секунд. Пробирки центрифугировали и получали серию разведений клеточного супернатанта. GUS-активность измеряли путем добавления п-нитрофенилового субстрата и β-меркаптоэтанола, что давало раствор желтого цвета при наличии β-глюкуронидазы. GUS-активность измеряли при 405 нм и выражали относительно эталонного штамма SAGX0090. Все штаммы обрабатывали параллельно.
На фигуре 3 показана относительная GUS-активность у конструкций с двойными цистронами ген X»rpmD»uidA. GUS-активность выражали относительно эталонного штамма SAGX0090, который нес кассету экспрессии PhIIA»uidA, и указывали ее на оси Y. Было обнаружено, что GUS-активность у всех штаммов с двойными цистронами была выше, чем у эталонного штамма. В частности, было обнаружено, что GUS-активность у sAGX0168 (enoA»rpmD»uidA) и sAGX0222 (gapB»rpmD»uidA) была, соответственно, в 6,89 и 2099 раз выше по сравнению с SAGX0090.
Эти результаты явно подтверждали, что бицистронная экспрессия обеспечивала повышенные уровни экспрессии белка в широком спектре параметров.
Пример 3: Бицистронная экспрессия проинсулина человека
Определяющую секрецию лидерную последовательность usp45 (SS) сливали с проинсулином человека (ins) с получением секретирующейся последовательности проинсулина (SS::ins). Кассету экспрессии [SS::ins] либо встраивали в хромосому MG1363 Lactococcus lactis в локусе thyA и непосредственно экспрессировали с PthyA (sAGX0121), либо вставляли, наряду с межгенным участком rpmD, предшествующим SS::ins, в качестве второго цистрона ниже usp45 (sAGX0121) или enoA (sAGX0164) (фигура 4а). Способность секретироваться у инсулина количественно определяли с помощью ELISA для оценки бицистронной экспрессии груза по сравнению с экспрессией под управлением PthyA в локусе thyA.
Попытки построить плазмиду для встраивания PhIIA»SS::Ins терпели неудачу.
Штаммы инокулировали из отдельной колонии в 2 мл GM17, по необходимости дополненной 200 мкМ тимидином, и выращивали в течение 16 часов при температуре 30°С. Для количественного определения секреции проинсулина эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17 и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9 и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9 содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Количество секретированного проинсулина человека в культуральном суперанатанте количественно оценивали с помощью ELISA-набора, предоставленного компанией Mercodia. Все штаммы обрабатывали параллельно.
На фигуре 4b представлены результаты количественного определения секреции проинсулина человека штаммами sAGX0122, sAGX0121 и sAGX0164 Lactococcus lactis. Количество секретированного проинсулина выражали в нг/мл и указывали на оси Y. Из фигуры явно видно, что штаммы, содержавшие бицистронную кассету экспрессии, имели значимо более высокую экспрессию груза, чем эталонный штамм. В частности, секреция инсулина была наиболее высокой в случае, когда SS::ins связывали посредством rpmD с enoA.
Пример 4: Бицистронная экспрессия Fab сА2
Получали конструкции с двойными цистронами экспрессии с тяжелой цепью и легкой цепью с Fab сА2 к hTNF. Все единицы экспрессии конструировали под управлением промотора thyA и размещали на плазмидах. Все они несли гены легкой цепи, VLCL (L) и Fab-фрагмента тяжелой цепи, VHCH1 (Н), полученные от моноклонального антитела инфликсимаб сА2. L»H и H»L конфигурации соединены межгенными участками, предшествующими rpmD, rplB, rpsG, rpsE или rplN. Все конструкции переносились плазмидой.
Из фигуры 5 видно, что как тяжелая цепь, так и легкие цепи имели высокий уровень экспрессии у конструкций с двойными цистронами, что давало высокие уровни функционального Fab сА2 к TNF. Из фигуры 5 дополнительно видно, что экспрессия сА2 к TNF понижалась, если тяжелую цепь располагали перед легкой цепью, независимо от межгенного участка.
Для количественного определения секреции элементов антитела к hTNF штаммы инокулировали из отдельной колонии в 2 мл GM17 и выращивали в течение 16 часов при температуре 30°С. Эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17 и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9 и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9 содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Необработанные супернатанты из несущих отдельные конструкции штаммов готовили параллельно и анализировали на наличие активности в отношении TNF. Это осуществляли с помощью прямого ELISA с применением TNF человека в качестве белка захвата. VLCL-части детектировали с помощью антисыворотки кролика к IgG человека и проявляли с помощью конъюгированных с щелочной фосфатазой антител к антисыворотке кролика. Фосфатазную активность измеряли колориметрическим анализов и считывали как OD402. Все штаммы обрабатывали параллельно.
Пример 5: Бицистронная экспрессия CDP870
Получали конструкции с двойными цистронами экспрессии с тяжелой цепью и легкой цепью с Fab CDP870 к TNF. Все единицы экспрессии располагали на бактериальной хромосоме.
Фигура 6а: слияния легкой и тяжелой цепей CDP870 с последовательностями, кодирующими определяющую секрецию лидерную последовательность usp45 (SS::CDP870 VHCH1 и SS::CDP870 VLCL), включали в качестве второго и третьего цистрона ниже usp45 (sAGX0219, sAGX0220). В SAGX0219 и sAGX0220 rpmD использовали для связывания генов SS::CD870 с usp45. Для предупреждения генетической нестабильности гены легкой и тяжелой цепей связывали посредством межгенного участка, предшествующего rplN. В sAGX0219, ген легкой цепи предшествовал гену тяжелой цепи, в то время как в sAGX0220, ген тяжелой цепи предшествовал гену легкой цепи.
Для количественного определения секреции элементов антитела к hTNF штаммы инокулировали из отдельной колонии в 2 мл GM17 и выращивали в течение 16 часов при температуре 30°С. Эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17 и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9 и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9 содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Необработанные супернатанты из несущих отдельные конструкции штаммов готовили параллельно и анализировали на наличие активности в отношении TNF. Это осуществляли с помощью прямого ELISA с применением TNF человека в качестве белка захвата с ремикейдом в качестве эталонного стандарта. VLCL-части детектировали с помощью антисыворотки кролика к IgG человека и проявляли с помощью конъюгированных с щелочной фосфатазой антител к антисыворотке кролика. Фосфатазную активность измеряли колориметрическим анализов и считывали как OD402. Все штаммы обрабатывали параллельно.
Из фигуры 6b видно, что как тяжелая цепь, так и легкие цепи имели высокий уровень экспрессии у конструкций с двойными цистронами, что давало высокие уровни функционального Fab CDP870 к TNF. Из фигуры 6b дополнительно видно, что экспрессия CDP870 к TNF значимо повышалась, если тяжелую цепь располагали перед легкой цепью.
Пример 6: Бицистронная экспрессия человеческого фактора "трилистника" 1 (hTFF1)
Получали конструкции экспрессии с последовательностью, кодирующей определяющую секрецию лидерную последовательность usp45, слитой с hTFF1 (SS::hTFF1). Все единицы экспрессии располагали на бактериальной хромосоме. Невозможно построить плазмиды для встраивания для моноцистронной экспрессии hTFF1 с использованием промоторов сильнее PhIIA.
Фигура 7а: Последовательность, кодирующую определяющую секрецию лидерную последовательность usp45 (SS), сливали с hTFF1 с получением секретирующейся последовательности hTFF1 (SS::hTFF1). Кассету экспрессии SS::hTFF1 либо встраивали в хромосому MG1363 Lactococcus lactis в локусе thyA и непосредственно экспрессировали с PhIIA (sAGX0085), либо вставляли, наряду с межгенным участком rpmD, предшествующим SS::hTFF1, в качестве второго цистрона ниже gapB (SAGX0276).
Фигура 7b: Способность секретироваться у hTFF1 количественно определяли с помощью ELISA для оценки бицистронной экспрессии груза по сравнению с экспрессией под управлением PhIIA в локусе thyA.
Для количественного определения секреции элементов антитела к hTFF1 штаммы инокулировали из отдельной колонии в 2 мл GM17 и выращивали в течение 16 часов при температуре 30°С. Эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17 и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9 и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9 содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. На этой стадии у всех культур определяли колониеобразующие единицы (CFU). Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Необработанные супернатанты из несущих отдельные конструкции штаммов готовили параллельно и анализировали с помощью ELISA с использованием очищенного hTFF1 в качестве эталонного стандарта. Количество секретированного hTFF1 выражали как нг/мл и нг/109 CFU. Все штаммы обрабатывали параллельно.
Из фигуры явно видно, что штамм (sAGX0276), содержавший бицистронную кассету экспрессии, имел значимо более высокую экспрессию груза, чем эталонный штамм (sAGX0085). Величина экспрессии секретированного hTFF1 была существенно повышена (>5 раз на мл; >12 раз на CFU), если hTFF1 связывали посредством rpmD с gapB.
Пример 7: Селекция межгенных участков из генома Enterococcus faecium
Клеточные белки культуры конца лог-фазы штамма LMG 15709 Enterococcus faecium (Enterococcus faecium [Orla-Jensen 1919] Schleifer and Kilpper-Balz 1984 VP; LMG 15709; ATCC 6057; DSM 2146; NCIMB 8842) визуализировали на геле белкового фореза с окрашиванием красителем кумасси синий, как показано на фигуре 8. Дорожки А, В и С содержали клеточные белки лизированного клеточного эквивалента 284 мкл, 142 мкл и 56,8 мкл культуры конца лог-фазы LMG 15709 Entercococcus faecium, соответственно. Из геля выделяли 12 бэндов определенных белков из дорожки С. Выделяли белки и идентифицировали межгенные участки путем:
1) идентификации экспрессирующихся в большом количестве белков в фрагментах с помощью частичного секвенирования пептидов (MALDI-TOF/TOF) и поиска по базам данных с применением объединенных масс пептидов и информации о последовательности;
2) идентификации с помощью хромосомной последовательности РС4.1 Enterococcus faecium (найденной в банке данных NCBI Genome, номер доступа в Genbank ADMM01000000) генов, кодирующих экспрессирующиеся в большом количестве белки (1), которые присутствуют в опероне, но не в качестве "первого гена";
3) идентификации межгенных участков, которые предшествуют этим экспрессирующимся в большом количестве генам (таблица 5).
В таблице 5 приведены идентифицированные межгенные участки у LMG15709 Enterococcus faecium. Подчеркнутые последовательности представляют собой сайты связывания рибосомы.

Пример 8: Выбор сайтов в геноме Enterococcus faecium для бицистронной экспрессии
В таблице 6 приведены гены с высоким уровнем экспрессии Enterococcus faecium, идентифицированные в примере 7 как обеспечивающие высокий уровень экспрессии. Управляющие этими генами промоторы можно использовать в качестве целевых сайтов для полицистронной экспрессии экзогенных генов. Эти эндогенные гены можно дополнительно использовать в модуле полицистронной экспрессии в качестве первого гена, транскрипционно или трансляционно связанного посредством межгенного участка с находящимися ниже экзогенными генами.

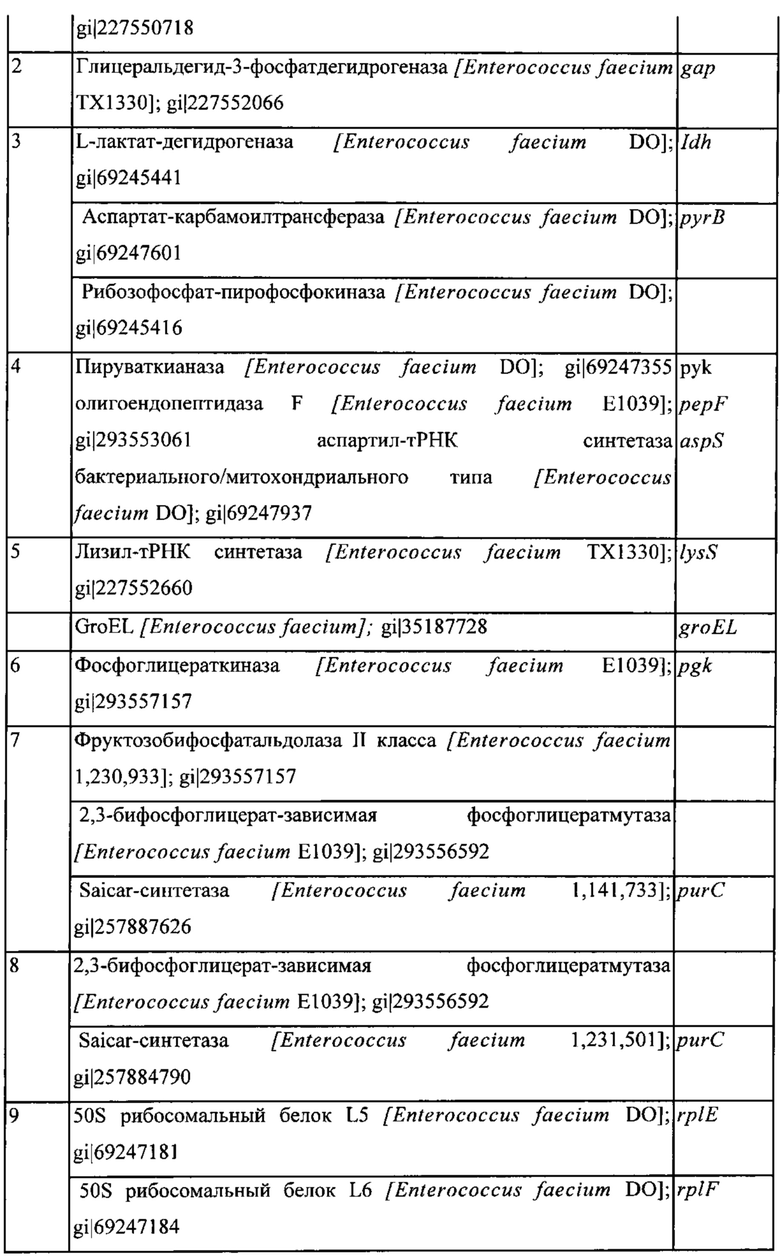
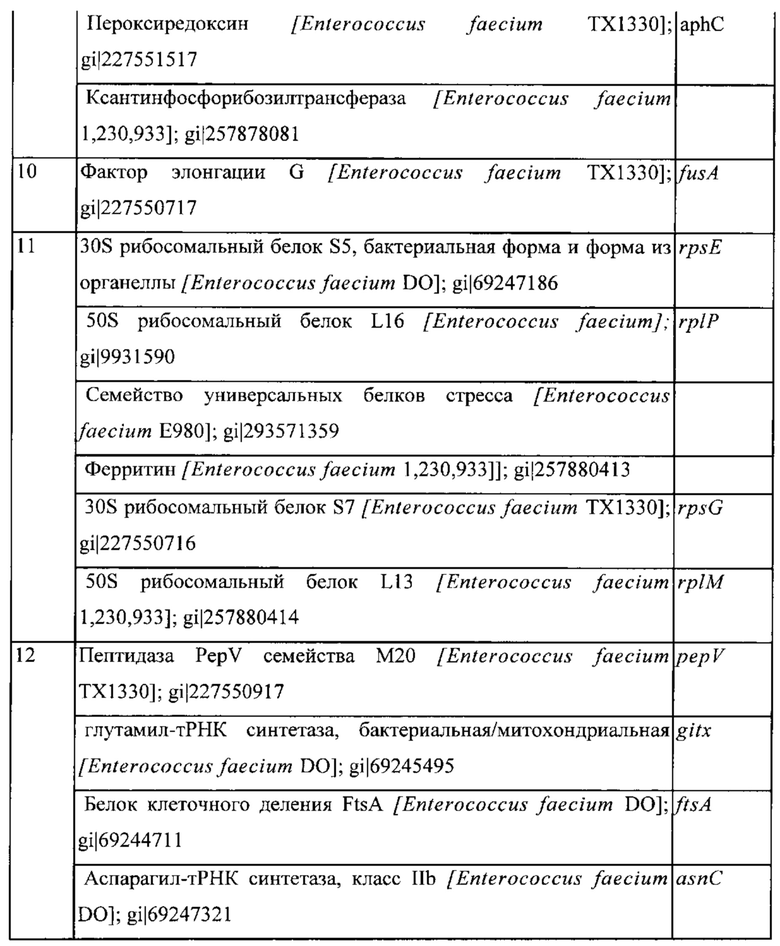
Таким способом ген β-глюкуронидазы (uidA) из Е. coli вводили в качестве гена-репортера в LMG15709 Enterococcus faecium. β-глюкуронидаза, продукт гена uidA, катализирует расщепление широкого спектра β-глюкуронидов, которые коммерчески доступны в качестве субтратов для гистохимических и спектрофотометрических исследований. Как изображено на фигуре 9, конструкции с двойным цистроном получали путем вставки uidA в хромосому LMG15709 Enterococcus faecium на 3'-конце нескольких эндогенных генов (ген X, в настоящем примере gap и епо; фигура 9). Таким образом, rpmD Enterococcus faecium использовали в качестве межгенного участка между представляющими интерес эндогенными генами из таблицы 6 и uidA для определения сайтов, которые в результате дают наиболее высокую β-глюкуронидазную (GUS) активность (фигура 10).
Культуры Enterococcus faecium выращивали в течение 16 часов при температуре 30°С в GM17, дополненной тимидином. Клетки 1 мл культуры промывали и ресуспендировали в 1 мл деминерализованной воды. Клетки разрушали при помощи лизирующей матрицы В от MP Biomedicals и устройства Fastprep-24 со скоростью 6 м/с в течение 40 секунд. Пробирки центрифугировали и получали серию разведений клеточного супернатанта. GUS-активность измеряли путем добавления п-нитрофенилового субстрата и β-меркаптоэтанола, что давало раствор желтого цвета при наличии β-глюкуронидазы. GUS-активность измеряли при 405 нм и выражали относительно эталонного штамма sAGX0090 Lactococcus lactis. Все штаммы обрабатывали параллельно.
На фигуре 10 показана относительная GUS-активность у конструкций с двойными цистронами ген X»rpmD»uidA. GUS-активность выражали относительно эталонного штамма sAGX0090, который нес кассету экспрессии PhIIA»uidA, и указывали ее на оси Y. Было обнаружено, что GUS-активность у всех штаммов с двойными цистронами была выше, чем у эталонного штамма.
В частности, было обнаружено, что GUS-активность у sAGX0270 (gap»rpmD»uidA) и sAGX0271 (eno»rpmD»uidA) была, соответственно, в 30,6 и 26,9 раз выше по сравнению с SAGX0090.
Эти результаты явно подтверждали, что бицистронная экспрессия обеспечивала повышенные уровни экспрессии белка в широком спектре параметров.
Пример 9: Бицистронная экспрессия интерлейкина-10 человека (hIL10) Enterococcus faecium
Кодирующую ДНК-последовательность определяющей секрецию лидерной последовательности usp45 Lactococcus lactis (SS) сливали в рамке с последовательностью ДНК зрелого hIL10 с получением секреции hIL10. Кассету экспрессии [SS::hIL10] вставляли, наряду с межгенным участком rpmD Enterococcus faecium, предшествующим SS::hIL10, в качестве второго цистрона ниже gap (sAGX0279; фигура 11a). Способность секретироваться hIL10 количественно оценивали с помощью ELISA для оценки бицистронной экспрессии груза у Enterococcus faecium. sAGX0270 Enterococcus faecium служил в качестве отрицательного контроля.
Штаммы инокулировали из отдельной колонии в 10 мл GM17, по необходимости дополненной 200 мкМ тимидином (GM17T), и выращивали в течение 16 часов при температуре 30°С. Для количественного определения секреции hIL10 эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17T и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9 и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9Т содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Количество секретированного hIL10 человека в культуральном суперанатанте количественно оценивали с помощью набора для проведения ELISA по типу сэндвича hIL10. Все штаммы обрабатывали параллельно.
На фигуре 11b видны результаты количественного определения секреции hIL10 человека штаммами SAGX0270 и sAGX0279 Enterococcus faecium. Количество секретированного hIL10 выражали в виде нг/109 клеток CFU через 3 часа и указывали на оси Y. Из фигуры явно видно, что содержавшие кассету бицистронной экспрессии штаммы Enterococcus faecium были способны секретировать существенные количества белка-груза hIL10.
Пример 10: Бицистронная экспрессия интерлейкина-27 человека (hIL27) Enterococcus faecium
Кодирующую ДНК-последовательность определяющей секрецию лидерной последовательности usp45 Lactococcus lactis (SS) сливали в рамке с последовательностью ДНК зрелого hIL27 с получением секреции hIL27. Кассету экспрессии [SS::hIL27] вставляли, наряду с межгенным участком rpmD Enterococcus faecium, предшествующим SS::hIL27, в качестве второго цистрона ниже gap (sAGX0317; фигура 12а). Способность секретироваться hIL27 количественно оценивали с помощью ELISA для оценки бицистронной экспрессии hIL27. sAGX0270 Enterococcus faecium служил в качестве отрицательного контроля.
Штаммы инокулировали из отдельной колонии в 10 мл GM17, дополненной 200 мкМ тимидина (GM17T), и выращивали в течение 16 часов при температуре 30°С. Для количественного определения секреции hIL27 эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17T и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВМ9Т и культивировали в течение еще 3 часов при температуре 30°С. ВМ9Т содержала соли М9, 0,5% казитона, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Количество секретированного hIL27 человека в культуральном суперанатанте количественно оценивали с помощью набора для проведения ELISA по типу сэндвича hIL27 (R&D systems). Все штаммы обрабатывали параллельно.
На фигуре 12b видны результаты количественного определения секреции hIL27 человека штаммами sAGX0270 и sAGX0317 Enterococcus faecium. Количество секретированного hIL27 выражали в виде нг/109 клеток CFU через 3 часа и указывали на оси Y. Из фигуры явно видно, что штамм sAGX0317 Enterococcus faecium, содержавший кассету бицистронной экспрессии, расположенную ниже гена gap Enterococcus faecium, мог эффективно секретировать существенные количества экзогенного белка hIL27.
Пример 11: Бицистронная экспрессия Fab CDP870 Enterococcus faecium
Получали конструкции с двойными цистронами экспрессии с генами тяжелой цепи и легкой цепи с Fab CDP870. Все единицы экспрессии располагали на бактериальной хромосоме.
Фигура 13а: продукты слияния Fab тяжелой и легкой цепей CDP870 с последовательностями [SS::CDP870 VHCH1] и [SS::CDP870 VLCL], кодирующими определяющую секрецию лидерную последовательность (SS) usp45 штамма MG1363 Lactococcus lactis, вставляли в качестве второго и третьего цистрона ниже гена gap (sAGX0278) LMG15709 Enterococcus faecium. У sAGX0278 межгенный участок rpmD Enterococcus faecium использовали для связывания кассет экспрессии SS::CD870 с gap. Для предупреждения генетической нестабильности гены тяжелой и легкой цепи связывали посредством межгенного участка, предшествующего rpmD Lactococcus lactis, а в сигнальных последовательностях секреции usp45 двух кассет экспрессии SS::CD870 использовали различную частоту использования кодона. У sAGX0278 кассета экспрессии тяжелой цепи предшествовала кассете экспрессии легкой цепи.
Для количественного определения секреции fab к CDP870 штаммы инокулировали из отдельной колонии в 10 мл GM17T и выращивали в течение 16 часов при температуре 30°С. Эти насыщенные полученные за ночь культуры разводили 1/25 в 5 мл свежей среды GM17T и выращивали в течение 4 часов при температуре 30°С. Клетки собирали центрифугированием при 3220 × g в течение 10 минут, ресупендировали в равном количестве среды ВАМ9Т и культивировали в течение еще 3 часов при температуре 30°С. ВАМ9Т содержала соли М9, 0,5% аминоплазмаля, 0,5% глюкозы, 25 мМ NaHCO3, 25 мМ Na2CO3, 2 мМ MgSO4, 0,1 мМ CaCl2 и 200 мкМ тимидина. Клетки и культуральные супернатанты разделяли с помощью центрифугирования при 3220 × g в течение 10 минут. Необработанные супернатанты из несущих отдельные конструкции штаммов готовили параллельно и анализировали на наличие TNF-связывающей активности. Это осуществляли с помощью прямого ELISA с применением TNF человека в качестве белка захвата с симзией (Cimzia®) (fab CDP870, связанный с PEG) в качестве эталонного стандарта. Fab CDP870 детектировали с помощью антисыворотки козы к Fab человека и проявляли с помощью конъюгированных с HRP антител осла к антисыворотке IgG(H+L) козы. HRP-активность визуализировали с помощью субстрата ТМВ. Реакцию останавливали через 30 минут путем добавления HCl. Поглощение измеряли при 450 нм с 595 нм в качестве длины волны сравнения. Все штаммы обрабатывали параллельно.
Из фигуры 13b видно, что как тяжелая цепь, так и легкие цепи экспрессировались конструкциями с множественными цистронами у Enterococcus faecium, что приводило к секреции функционального Fab CDP870 к TNF.
Пример 12: Бактерии L. lactis, вырабатывающие антитела к hTNF (согласно настоящему изобретению), защищают от hTNF-индуцированного повреждения кишечника у A20IEC-KO мышей
Получение мышей с тканеспецифичным дефицитом по А20
Мышей с зависящим от условий нокаутом A20/tnfaip3, у которых экзоны IV и V гена tnfaip3 фланкированы двумя сайтами LoxP, получали как описано (Piguet et al., 1999, Lab Invest 79, 495-500). Мышей с аллелем А20, фланкированным LoxP-сайтами, скрещивали с трансгенными мышами Villin-Cre с получением IEC-специфичных нокаутированных по А20 мышей (A20IEC-KO) (Madison et al., 2002, J Biol Chem 277, 33275-33283). Эксперименты проводили на мышах, подвергнутых возвратному скрещиванию на генетический фон C57BL/6 в течение по меньшей мере четырех поколений.
Токсичность TNF in vivo
Мышам интраперитонеально вводили различные дозы hTNF (50, 10, 8, 6, 4 и 2 мкг hTNF/20 г массы тела). Полученный от Е. coli рекомбинантный hTNF имел удельную активность 6,8×107 МЕ/мг. Получали TNF человека и очищали до гомогенного состояния в своей лаборатории, и уровни эндотоксина не превышали 1 нг/мг белка. Введение hTNF A20IEC-KO мышам вызывало несколько патологических и иммунологических изменений, которые указывали на повреждение кишечника. Низкая доза hTNF не вызывала тяжелые системные эффекты и летальный исход, но вызывала экспрессию провоспалительных цитокинов и хемокинов, которую можно было измерить как в сыворотке, так и в гомогенатах ткани кишечника. Мышей подвергали эвтаназии спустя 4 или 5 часов для гистологического анализа. Для терапевтических исследований A20IEC-KО мыши получали продуцирующие антитела к TNF бактерии L. lactis (согласно настоящему изобретению) путем 5-кратного искусственного кормления через рот (5× 1010 CFU с 30-минутным интервалом) перед интраперитонеальной инъекцией hTNF (2 мкг и 6 мкг hTNF/20 г массы тела). Контрольные группы обрабатывали либо исходным штаммом MG1363 L. lactis, либо бактериальной средой ВМ9Т (основа). Кроме того, группе положительного контроля производили однократное введение ремикейда (30 мг/кг). Температуру организма отслеживали каждый час.
Получение препарата образца ткани
Свежевыделенные сегменты толстой кишки и подвздошной кишки промывали PBS для удаления калового содержимого, а затем промывали формалином (4% формальдегид в фосфатно-солевом буферном растворе (PBS)) и фиксировали путем инкубации в течение ночи в 10-кратном избытке формалина при температуре 4°С. Формалин удаляли и сегменты кишечника дважды отмывали PBS перед заливкой парафиновым воском с применением стандартных способов.
Гистология
Отрезали срезы тканей толщиной 4 мкм и окрашивали гематоксилином/эозином с применением стандартным методик. Для объединенных окрашиваний красителями альциановый синий (АВ) и PAS депарафинизированные срезы увлажняли дистиллированной водой и инкубировали в альциановом синем в течение 20 минут. Затем срезы инкубировали в 1% перйодной кислоте в течение 10 минут с последующей инкубацией в реактиве Шиффа в течение 10 минут. Ядра докрашивали гематоксилином Майера в течение 30 секунд. Детектирование щелочной фосфатазы осуществляли путем инкубации депарафинизированных и увлажненных срезов ткани в растворе NBT/BCIP в темноте (70 мкл NBT+70 мкл BCIP+4860 мкл буфера A, NBT=1,5% раствор нитросинего тетразолия хлорида в 70% диметилформамиде, BCIP=1% 5-бром-4-хлор-3-индолилфосфатаза в 100% диметилформамиде, буфер А=0,1 М TrisHCl, 0,1 М NaCl, 0,05 М MgCl2, рН 9,5). Для иммунохимического исследования срезы депарафинизировали и инкубировали в растворе для демаскировки антигена Dako, и кипятили в течение 20 минут в устройстве для термообработки клеток Пика, и охлаждали в течение 2,5 часов. Активность эндогенной пероксидазы блокировали путем погружения микроскопических препаратов в буфер для блокировки пероксидазы (0,040 М лимонная кислота, 0,121 М двузамещенный фосфорнокислый натрий, 0,030 М азид натрия, 1,5% перекись водорода) на 15 минут при комнатной температуре. Добавляли буфер для блокировки (1% бычий сывороточный альбумин в PBS) к микроскопическим препаратам на 30 минут при комнатной температуре. Добавляли первые антитела (антитела к лизозиму кролика; 1:1750 разведение - Dako; антитела к муцину-2 кролика; 1:500 22) в буфер для блокировки и срезы тканей инкубировали в течение ночи. Добавляли второе антитело (полимерное меченное пероксидазой хрена антитело к антителу кролика, Envision) на 30 минут при комнатной температуре. Пероксидазу детектировали путем добавления субстрата диаминомасляная кислота (DAB) на 10 минут при комнатной температуре и докрашивали ядра гематоксилином Майера в течение 2 минут.Микроскопическое измерение радиусов гранул клеток Панета осуществляли с помощью программного обеспечения Leica Image manager 500.
Количественное определение цитокинов
Цитокины и хемокины в сыворотке крови и гомогенатах ткани количественно оценивали с помощью наборов Cytometric Bead Array (СВА) (BD Biosciences) на FACS-цитометре Calibur, оснащенном программным обесепечением CellQuest Pro и СВА (BD Biosciences).
Количественная ПЦР в реальном времени
Сегменты подвздошной кишки длиной 5 см выделяли свежими и промывали PBS с удалением калового содержимого. Один конец зашивали и сегменты заполнили РНК-лизирующим буфером (набор для выделения РНК Aurum Total RNA Mini, Bio-Rad Laboratories) и инкубировали на льду в течение 5 минут.РНК очищали из раствора лизата с помощью набора Aurum Total RNA Mini (Bio-Rad Laboratories). Синтез кДНК осуществляли с помощью набора для синтеза кДНК iScript (Bio-Rad Laboratories) в соответствии с инструкциями производителя. 10 нг кДНК использовали для количественной ПЦР в общем объеме 10 мкл с помощью мастер-микса LightCycler 480 SYBR Green I Master Mix (Roche) и специфическими праймерами на LightCycler 480 (Roche). Реакции ПЦР в реальном времени проводили в трех повторах. Использовали следующие специфические для мыши праймеры: убиквинтиновый прямой, 5'-AGGTC АААС AGG AAGAC AGACGTA-3' (SEQ ID NO: 14); и убиквинтиновый обратный, 5'-TCACACCCAAGAACAAGCACA-3' (SEQ ID NO: 15). Лизозима-Р, прямой, 5'- GCCAAGGTCTAACAATCGTTGTGAGTTG-3' (SEQ ID NO: 16); лизозима-P, обратный, 5'-CAGTCAGCCAGCTTGACACCACG-3' (SEQ ID NO: 17); криптидина-1, прямой, 5'-TCAAGAGGCTGCAAAGGAAGAGAAC-3' (SEQ ID NO: 18); криптидина-1, обратный, 5'-TGGTCTCCATGTTCAGCGACAGC-3' (SEQ ID NO: 19).
Статистические анализы
Результаты выражали как среднее ± SEM. Статистическую значимость между экспериментальными группами оценивали с помощью непарного t-критерия Стьюдента для двух выборок.
Характеристика продуцирующих антитела к hTNF бактерий L. lactis в отношении защиты от вызванных hTNF патологических и иммунологических изменений у A20IEC-KO мышей
У обеих контрольных групп, которым ввели 2 мкг hTNF, наблюдали более крутое падение температуры организма по сравнению с A20IEC-KO мышами, обработанными бактериями L. lactis, продуцирующими антитела к hTNF (фиг. 14а, левый сегмент). Защитный эффект обработки sAGX0220, наблюдаемый у мышей, обработанных посредством 2 мкг hTNF, нельзя больше было наблюдать при введении мышам более высокой концентрации hTNF (6 мкг), в то время как обработка ремикейдом явно оказывала защитный эффект (фиг. 14а, правый сегмент). Предварительная обработка sAGX0220 значимо снижала уровни МСР-1, KС и IL-6 в гомогенатах подвздошной и толстой кишки и в сыворотке крови мышей, обработанных любыми 2 мкг hTNF, по сравнению с мышами, обработанными основами (фиг. 14b, с). Тем не менее, имело место сравнимое уменьшение у мышей, обработанных парентеральным штаммом MG1363 L. lactis, что указывало на то, что введение бактерий само по себе могло легко оказать пробиотический положительный эффект. У мышей, которым повторно вводили 6 мкг hTNF, наблюдали подобное уменьшение уровней цитокинов и хемокинов в сыворотке крови и ткани при предварительной обработке sAGX0220 до такого же уровня, как оказывал ремикейд (фиг. 14d, e). При таких условиях, тем не менее, защитный эффект исходного штамма MG1363 в основном терялся (фиг. 14d, e).
Пример 13: Пассивная иммунизация от ассоциированного с Clostridium difficile заболевания токсин-нейтрализующими антителами, которые локально вырабатываются и секретируются Lactococcus lactis.
а) Синтез гена VLCL и VHCH1 mAb, нейтрализующего токсин А С. difficile и токсин В С. difficile
De novo синтез генов (оптимизированный по частоте использования кодона для L. lactis) осуществляли на основании информации об аминокислотной последовательности VH и VL в соответствии с общепринятым способом (Stemmer et al., Gene 1995; 164(1):49-53). Такой способ предусматривает применение синтетических олигонуклеотидов из 40-мономеров, которые охватывают всю кодирующую, а также некодирующую цепь таким образом, что каждый олигонуклеотид из кодирующей цепи будет на 100% комплементарен, соответственно, последним и первым 20 парам оснований двух последовательных олигонуклеотидов в некодирующей цепи, и наоборот.
На 5'-конце генов VLCL и VHCH1 добавляли генетическую информацию, которая кодирует определяющую секрецию лидерную последовательность L. lactis неустановленного секретирующегося 45-кДа белка (Usp45).
Для обеспечения скоординированной экспрессии эти синтетические гены VLCL и VHCH1 размещали в тандеме и соединяли межгенным участком rpmD. Это в результате приводило к образованию функциональных оперонов VLCL » VHCH1. Также строили варианты описанных выше синтетических генов, которые также кодировали С-концевые пептидные метки Е и FLAG. Это обеспечивало визуализацию полноразмерных и/или потенциальных продуктов распада и давало возможность подтверждения сборки легкой цепи и тяжелой цепи и связывания токсина.
У полученных генных конструкций проверяли последовательность.
b) Создание штаммов L. lactis, секретирующих нейтрализующее токсин А/В Fab: встраивание в локус Usp45
Эта задача состояла из следующих этапов:
1. Создание встраиваемых векторов для встраивания ниже локуса usp45
- VLCL » VHCH1 Mab, нейтрализующего токсин А С. difficile,
- VLCL » VHCH1 Mab, нейтрализующего токсин В С.difficile,
- меченных Е и FLAG на 3'-конце описанных выше вариантов.
Синтетические опероны VLCL » VHCH1 фланкировали на 5'-конце посредством 1 т.о. 3'-конца гена usp45 L. lactis, за которым следовал межгенный участок rplN. Опероны VLCL » VHCH1 фланкировали на 3'-конце фрагментом длиной 1 т.о. от 3' и ниже фланкируюшего участка гена usp45 L. lactis. Получали конструкции путем отжига ДНК в перекрывающемся ПЦР. У полученных плазмид подтверждали последовательности.
2. Вышеописанные полученные встраиваемые плазмиды получали для встраивания посредством трансляционного связывания с геном usp45 MG1363 L. lactis. Встраивание осуществляли с помощью двойной гомологичной рекомбинации как на 5', так и на 3' фланкирующих участках (соответствующих участкам длиной 1 т.о., фланкирующим 3'-конец гена usp4S) и подтверждали с помощью ПЦР и секвенирования ДНК.
c) Создание штаммов L. lactis, секретирующих нейтрализующее токсин А/В Fab: встраивание в локус enoA
Эта задача состояла из следующих этапов:
1. Создание встраиваемых векторов для встраивания ниже локуса enoA
- VLCL » VHCH1 Mab, нейтрализующего токсин А С. difficile,
- VLCL » VHCH1 Mab, нейтрализующего токсин В С. difficile,
- меченных Е и FLAG на 3'-конце описанных выше вариантов.
Синтетические опероны VLCL » VHCH1 из задания 7 фланкировали на 5'-конце посредством 1 т.о. 3'-конца гена enoA L. lactis, за которым следовал межгенный участок rplN. Опероны VLCL » VHCH1 фланкировали на 3'-конце фрагментом длиной 1 т.о. от 3' и ниже фланкируюшего участка гена enoA L. lactis. Получали конструкции путем отжига ДНК в перекрывающемся ПЦР. У полученных плазмид подтверждали последовательности.
2. Вышеописанные полученные встраиваемые плазмиды получали для встраивания посредством трансляционного связывания с геном enoA MG1363 L. lactis. Встраивание осуществляли с помощью двойной гомологичной рекомбинации как на 5', так и на 3' фланкирующих участках (соответствующих участкам длиной 1 т.о., фланкирующим 3'-конец гена enoA) и подтверждали с помощью ПЦР и секвенирования ДНК.
d) Обоснование совместимой модели CDAD TopAct™ на хомяках
Модель CDAD на хомяках является общепринятой моделью исследования вызванных токсином, ассоциированных с антибиотиком диареи и колита. Наиболее широко исследованной моделью инфекции С. difficile у хомяков является модель первичного заражения. Вкратце, хомяков предварительно орогастрально обрабатывали клиндамицином (10-30 мг/кг) за 24 часа до введения спор С. difficile для нарушения нормальной флоры толстой кишки у хомяков. Споры С. difficile (например, 100 спор штамма 630 или В1; см. работу Goulding et al., Infect Immun 2009; 77(12):5478-5485) вводили орогастрально и наблюдали за хомяками (модель первичного заражения CDAD). Как правило, 100% хомяков погибали от заболевания спустя от 36 до 72 часов после введения спор. Перед гибелью симптомы заболевания включали диарею и потерю массы. В целом, симптомы были намного тяжелее симптомов, наблюдаемых у людей, но модель на хомяках соответствует терапевтическим приемам, используемым при клиническом заболевании, таким как лечение ванкомицином, и поэтому ее широко используют при исследовании CDAD.
Для моделирования контроля над рецидивом CDAD использовали модифицированную модель первичного заболевания на хомяках. Ванкомицин защищает хомяков от вызываемого С. difficile заболевания так же, как он действует у людей. При прерывании лечения ванкомицином у хомяков начинается рецидив тяжелого заболевания, но степень обострения варьирует.Вкратце, хомякам давали однократную дозу клиндамицина с последующим орогастральным введением спор штамма В1 С. difficile 1 день спустя. Лечение ванкомицином начинали в день заражения спорами или через 24 часа и продолжали ежедневно в течение двух-четырех последующих дней (модель рецидива CDAD). Такой протокол можно дополнительно оптимизировать для обеспечения рецидива после терапии ванкомицином.
Для оценки полезного эффекта нейтрализующего токсин Fab, доставляемого в кишечник при помощи L. lactis, при предупреждении смертности в модели первичного заражения и/или модели рецидива инфекции у хомяка, важно задокументировать воздействие клиндамицина и ванкомицина на рост и способность вырабатывать Fab у L. lactis. Поэтому осуществляли исследования in vitro/in vivo для определения жизнеспособности и метаболической активности у секретирующих Fab штаммов L. lactis.
Оценка in vitro: выработку Fab (посредством ELISA) и рост (посредством культивирования) у дополненных клиндамицином/ванкомицином культур L. lactis сравнивали с не содержащей антибиотик культурой и с культурой, дополненной хлорамфениколом (Cm) в концентрации 5 мкг/мл. В такой концентрации Cm является известным ингибитором синтеза белка и роста, к которому чувствительна L. lactis.
Оценка in vivo: выработку Fab (посредством ELISA) и жизеспособность (посредством культивирования) определяли в тонком/толстом кишечнике после искусственного кормления хомяков и сопутствующего лечения различными дозами клиндамицина/ванкомицина.
Эти оценки позволили разработать и адаптировать модель CDAD (заражения и рецидива) у хомяков, которая хорошо подходила для оценки системы доставки L. lactis в отношении ее предупреждающих и лечебных эффектов: она заключается в применении (более низких) концентраций клиндамицина (и ванкомицина) и/или другой смеси антибиотиков, к которой наблюдали восприимчивость у инфекции С. difficile, без отрицательного эффекта на жизнеспособность и метаболическую активность L. lactis.
е) Проверка модели CDAD у хомяков
В модели первичного заражения хомяков сирийским золотым хомякам (70-80 г) давали различные пероральные дозы (ежедневно, два раза в день, три раза в день) выбранной секретирующей антитела к токсину А/В L. lactis (отдельно антитело к токсину А и антитело к токсину В и комбинированные антитела) или 1 мл mAb к токсину А/В (в качестве положительного контроля) интраперитонеально в течение 4 дней, начиная за 3 дня до введения спор С. difficile. Клиндамицин (дозу или другую смесь антибиотиков, как указано ранее) вводили орогастрально за 24 часа до заражения спорами С. difficile с применением стандартной иглы для кормления небольших животных. За животными наблюдали в отношении заболеваемости и смертности, ткани кишечника оценивали в отношении гистологического и макроскопического повреждения и определяли выработку токсина С difficile в содержимом просвета и фекалиях.
В модели рецидива у хомяков сирийским золотым хомякам (70-80 г) давали клиндамицин (дозу или другую смесь антибиотиков, как указано ранее) орогастрально и через 24 часа орогастрально заражали спорами В1 С. difficile. Во время введения спор или через 24 часа начинали орогастральное лечение ванкомицином (обозначенной выше дозой) и продолжали ежедневно в течение всего 2-4 дней. Начиная с 1, 2, 3, 4 или 5 дней после лечения ванкомицином вводили различные пероральные дозы (один раз в день, два раза в день, три раза в день) выбранной секретирующей антитела к токсину А/В L. lactis (отдельно антитело к токсину А и антитело к токсину В и комбинированные антитела) или 1 мл mAb к токсину А/В (в качестве положительного контроля интраперитонеально) интраперитонеально в течение всего 5-10 дней. За животными наблюдали в отношении заболеваемости и смертности, ткани кишечника оценивали в отношении гистологического и макроскопического повреждения и определяли выработку токсина С difficile в содержимом просвета и фекалиях.
Можно сделать вывод, что грамположительные бактерии согласно настоящему изобретению можно эффективно применять для иммунизации от CDAD. В частности, грамположительные бактерии согласно настоящему изобретению могут предупреждать возникновение CDAD, предупреждать рецидив CDAD, а также лечить CDAD.
Пример 14: Порошок для полоскания рта для растворения
Лекарственное вещество (DS) состава для полоскания рта представляет собой гомогенный лиофилизированный порошок сконструированного штамма согласно настоящему изобретению, и он смешан с криопротекторами (декстрином, сорбитом и глутаматом натрия).
Способ получения лекарственного вещества предусматривает следующие последовательные этапы: ферментация, концентрирование биомассы (путем диафильтрации или центрифугирования), составление с криопротекторами, наполнение в подходящие лотки и крупномасштабная лиофилизация. Осуществляли гомогенизацию и просеивание лиофилизированного осадка с получением гомогенного порошка (лекарственного вещества), подходящего для смешивания с наполнителем, и наполнения в необходимую фармацевтическую лекарственную форму.
Порошок лекарственного продукта (DP) для полоскания рта для растворения состоит из высушенных сублимацией бактерий L. lactis, смешанных с маннитом в качестве наполнителя и представленных в виде (спрессованного) порошка. Клинический состав представляет собой средство для перерального местного применения в форме ополаскивателя для рта. Такую суспензию для полоскания рта готовили путем растворения DP в выбранном растворе.
Способ получения порошка для полоскания рта для растворения предусматривает серию последовательных этапов:
- смешивание массового DS с маннитом,
- спрессовывание 500 мг порошковой смеси DP в 500 мг диспергируемые порошковые брикеты,
- наполнение спрессованным порошком стеклянных флаконов,
- закрытие флаконов защищенными от детей крышками с резьбой с контролем первого вскрытия и
- упаковка флаконов в алюминиевые (Alu) пакеты.
Пример 15: Бицистронная экспрессия CDP870
Получали конструкции с двойными цистронами экспрессии с тяжелой цепью и легкой цепью с Fab CDP870 к TNF. Все единицы экспрессии располагали на бактериальной хромосоме.
Фигура 15 А, схематический общий вид единиц экспрессии CDP870 к TNF у различных штаммов: слияния легкой и тяжелой цепей CDP870 с последовательностями, кодирующими определяющую секрецию лидерную последовательность usp45 (SS::CDP870 VHCH1 и SS::CDP870 VLCL), включали в качестве второго и третьего цистрона ниже usp45 (sAGX0309, SAGX0319), enoA (sAGX0275) и gapB (sAGX0323, SAGX0326). В этих штаммах rpmD использовали для соединения генов SS::CD870 с usp45, enoA или gapB, соответственно. Для предупреждения генетической нестабильности гены легкой и тяжелой цепей связывали посредством межгенного участка, предшествующего rplN. Анализировали и документировали 4 идентичных клона sAGX0326 (клон 1-4) из фигуры 15 В и С. В ходе экспериментов штаммы обрабатывали параллельно.
Для визуализации и количественного определения секреции CDP870 к hTNF штаммы инокулировали из отдельной колонии в 10 мл GM17T (Difco™M17, BD, Sparks, MD, +0,5% глюкозы + 200 мкМ тимидина) и выращивали в течение 16 часов при температуре 30°С. Бактерии из этих насыщенных полученных за ночь культур собирали путем центрифугирования при 3220 × g в течение 10 минут, и ресуспендировали в 10 мл свежей среды GM17T, и выращивали в течение 2 часов при температуре 30°С. Бактерии и неочищенные культуральные супернатанты разделяли путем центрифугирования при 3220 × g в течение 10 минут. Неочищенные супернатанты из всех штаммов получали параллельно и разделяли по штамму для анализа (фигура 15 В и С).
Общий белок из 5 мл объемов неочищенных культуральных супернатантов экстрагировали фенолом, осаждали этанолом и ресуспендировали в буфере для разведения образцов для SDS-PAGE.
Эквиваленты 1 мл неочищенных культуральных супернатантов анализировали с помощью вестерн-блоттинга с применением антител козы к Fab человека в качестве первой антисыворотки и проявляли при помощи антител кролика к АР козы и окрашивания NBT/BCIP (фигура 15 В; штаммы указаны справа от соответствующих дорожек).
Необработанные супернатанты из несущих отдельные конструкции штаммов анализировали на наличие hTNF-связывающей активности. Это осуществляли с помощью прямого ELISA с применением hTNF в качестве белка захвата с симзией в качестве эталонного стандарта. VLCL-части детектировали с помощью антисыворотки козы к IgG человека и проявляли с помощью конъюгированных с пероксидазой хрена (HRP) антител к антисыворотке козы. HRP-активность измеряли с помощью колориметрического анализа. Данные представлены на фигуре 15 С, штаммы указаны под соответствующими столбцами.
Из фигуры 15 (В и С) видно, что как тяжелая цепь, так и легкие цепи имели высокий уровень экспрессии у конструкций с двойными цистронами, что давало высокие уровни функционального Fab CDP870 к TNF. Из фигуры 15 (В и С) видно, что экспрессия CDP870 к TNF слегка снижалась при вставке генов тяжелой и легкой цепей в качестве второго и третьего цистрона ниже enoA по сравнению со вставкой ниже usp45. Из фигуры 15 (В и С) дополнительно видно, что экспрессия CDP870 к TNF существенно снижалась при вставке генов тяжелой и легкой цепей в качестве второго и третьего цистрона ниже gapB по сравнению со вставкой ниже usp45 или enoA.
Для определения удельной hTNF-нейтрализующей способности (биологической активности на количество TNF-связывающего белка) штаммы инокулировали из отдельной колонии в 5 мл GM17T и выращивали в течение 16 часов при температуре 30°С. Бактерии из этих насыщенных полученных за ночь культур собирали путем центрифугирования при 3220 × g в течение 5 минут, и ресуспендировали в 10 мл среды ВМ9Т, и выращивали в течение 2 часов при температуре 30°С. Бактерии и неочищенные культуральные супернатанты разделяли путем центрифугирования при 3220 × g в течение 10 минут. Неочищенные супернатанты из штаммов, несущих отдельные конструкции, получали параллельно и разделяли по штамму для анализа (фигура 15 D и Е).
Необработанные супернатанты из несущих отдельные конструкции штаммов анализировали на наличие TNF-связывающей активности. Это осуществляли с помощью прямого ELISA с применением TNF человека в качестве белка захвата с симзией в качестве эталонного стандарта. VLCL-части детектировали с помощью антисыворотки козы к IgG человека и проявляли с помощью конъюгированных с пероксидазой хрена (HRP) антител к антисыворотке козы. HRP-активность измеряли с помощью колориметрического анализа. Все штаммы обрабатывали параллельно. Данные представлены на фигуре 15 D, штаммы указаны под соответствующими столбцами.
Необработанные супернатанты из несущих отдельные конструкции штаммов анализировали на наличие TNF-нейтрализующей активности. Это осуществляли путем инкубирования восприимчивых к hTNF WEHI-клеток с TNF человека и добавления антител к TNF. Антитела к TNF будут захватывать hTNF и будут защищать WEHI-клетки от гибели клеток. Серию из 14 разведений неочищенных супернатантов, а также эталонный стандарт (симзию в количестве 63 нг/мл) добавляли к культурам клеток, подвергнутых воздействию hTNF. Определяли влияние на гибель клеток. Данные представлены на фигуре 15 Е, штаммы указаны под соответствующими столбцами.
Из фигуры 15 D видно, что как тяжелая цепь, так и легкие цепи имели высокий уровень экспрессии у конструкций с двойными цистронами, что давало высокие уровни функционального Fab CDP870 к TNF. Из фигуры 15 D также видно, что экспрессия CDP870 к TNF существенно снижалась при вставке генов тяжелой и легкой цепей в качестве второго и третьего цистрона ниже gapB по сравнению со вставкой ниже usp45.
Из фигуры 15 D и Е видно, что удельная TNF-нейтрализующая способность (биологическая активность на количество TNF-связывающего белка) антител CDP870 к TNF у неочищенных культуральных супернатантов штаммов SAGX0323 и SAGX0326 идентична таковой у симзии.
Пример 16: Эффективность L. lactis, секретирующих Fab-фрагмент к hTNFα
Экспериментальная модель основана на Tg1278TNFko мыши, трансгенной мыши с нормально регулируемой экспрессией TNF человека в отсутствии мышиного TNF (Keffer et al. EMBO. J 10, 4025-4031, 1991). Колит индуцировали путем заражения ректальным введением 4% TNBS в 40% этаноле после одной подкожной предварительной сенсибилизации. Вкратце, мышей сенсибилизировали за 7 дней (день -7) перед внутриректальным заражением путем нанесения 1 объема 5% TNBS + 4 объема 4:1 ацетон:оливковое масло на побритую 1,5×1,5 см зону кожи на спине. В день заражения (день 0) мышей сначала наркотизировали кетамином/ксилазином, после этого 100 мкл 4% TNBS/40% EtOH вводили ректально при помощи гибкого катетера, введенного в прямую кишку на 4 см. Для обеспечения равномерного распределения клизмы в толстой кишке мышей удерживали в вертикальном положении в течение 30 секунд сразу после ректального заражения.
Лечение начинали за 1 день до ректального заражения TNBS (день -1) и продолжали в течение еще 4 дней (день +3). Три группы мышей получали один раз в день внутрижелудочные засевы 1010 CFU MG1363 (отрицательный контроль), 1010 CFU SAGX0309 или 10 мкг симзии (положительный контроль). Начиная со дня 0 и затем ежедневно мышей отслеживали в отношении массы тела, заболеваемости и выживания. На день +3 мышей умерщвляли и собирали образцы толстой кишки и сыворотки крови для гистологического анализа (толстой кишки) и анализа цитокинов (толстой кишки и сыворотки крови).
Лечение штаммом в соответствии с вариантом осуществления настоящего изобретения (секретирующим антитела к hTNF штаммом sAGX0309 L. lactis) приводило к повышению выживаемости (фигура 16 и таблица 7) по сравнению со штаммом MG1363 L. lactis дикого типа и, к удивлению, еще более высокому проценту выживаемости, чем мышей, которых лечили симзией.
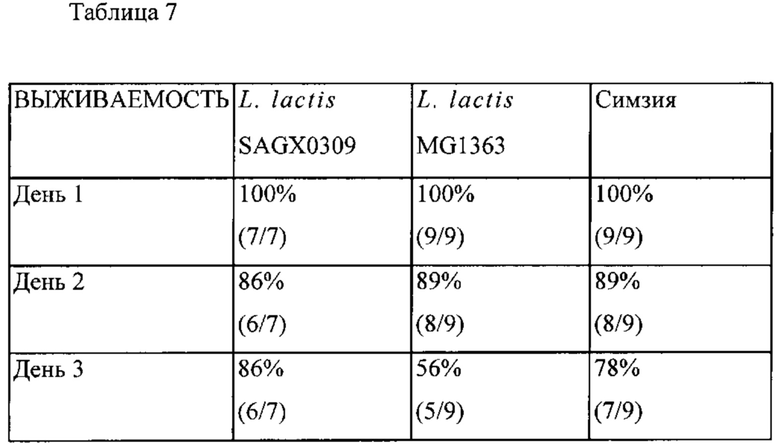
Массу тела мышей также отслеживали в ходе лечения, и результаты изображены на фигуре 17. Из фигуры 17 видно, что потеря массы снижалась после лечения штаммом в соответствии с вариантом осуществления настоящего изобретения по сравнению с лечением симзией.
Также анализировали гистологическое состояние толстой кишки и гистологическую оценку ставили в соответствии с таблицей 8. Результаты показаны на фигуре 18. Из фигуры 18 видно, что значимое улучшение гистологической оценки и, следовательно, уменьшенная патология колита, видна после лечения штаммом в соответствии с вариантом осуществления настоящего изобретения.
Наконец, из фигуры 19 видно, что лечение штаммом в соответствии с вариантом осуществления по настоящему изобретению приводило к подавлению секреции провоспалительных цитокинов в толстой кишке.
В частных воплощениях настоящее изобретение может относится к следующим вариантам осуществления:
Вариант 1. Грамположительная бактерия, содержащая единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный ген и один или несколько экзогенных генов, транскрипционно связанных с 3'-концом указанного одного или нескольких эндогенных генов, причем указанный один или несколько экзогенных генов являются наиболее 3' удаленными генами единицы полицистронной экспрессии.
Вариант 2. Рекомбинантная нуклеиновая кислота, содержащая единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии последовательно содержит эндогенный по отношению к грамположительной бактерии ген и один или несколько экзогенных по отношению к грамположительной бактерии генов, транскрипционно связанных с 3'-концом указанного одного или нескольких эндогенных генов, причем указанные один или несколько экзогенных генов являются наиболее 3' удаленными генами единицы полицистронной экспрессии.
Вариант 3. Грамположительная бактерия по варианту 1 или рекомбинантная нуклеиновая кислота по варианту 2, в которой указанный один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид и/или пептид, оказывающий терапевтический или профилактический эффект на субъекта, такой как антиген для индукции иммунитета или иммунотолерантности, невакциногенный терапевтически активный полипептид, антитело или его функциональный фрагмент, такой как Fab, химерный белок или мультимерный белок.
Вариант 4. Грамположительная бактерия по варианту 1 или рекомбинантная нуклеиновая кислота по варианту 2, в которой один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид и/или пептид, оказывающий терапевтический или профилактический эффект на субъект, такой как антиген для индукции иммунитета или иммунотолерантности, невакциногенный терапевтически активный полипептид, антитело или его функциональный фрагмент, такой как Fab, химерный белок или мультимерный белок, для применения в качестве лекарственного средства, предпочтительно для применения при введении или доставке указанного продукта субъекту.
Вариант 5. Грамположительная бактерия по любому из вариантов 1, 3 или 4 или рекомбинантная нуклеиновая кислота по любому из вариантов 2-4, в которой указанный эндогенный ген и указанный один или несколько экзогенных генов находятся под транскрипционным контролем промотора, эндогенного по отношению к грамположительной бактерии.
Вариант 6. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 5, в которой указанный промотор является промотором необходимого гена, конститутивным промотором, промотором гена центрального метаболизма и/или промотором гена "домашнего хозяйства".
Вариант 7. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 5, в которой указанный промотор является промотором рибосомального гена.
Вариант 8. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 5, в которой указанный промотор является промотором гена гликолиза.
Вариант 9. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 5, в которой указанный промотор выбран из группы, состоящей из промотора eno, usp45, gap, pyk, rpmB и rpIS указанной грамположительной бактерии.
Вариант 10. Грамположительная бактерия по любому из вариантов 1 или 3-9, в которой эндогенный ген расположен в своем естественном локусе на хромосоме у грамположительной бактерии.
Вариант 11. Грамположительная бактерия по варианту 10, в которой эндогенный ген транскрипционно связан с одним или несколькими экзогенными генами посредством встраивания в хромосому одного или нескольких экзогенных генов в указанный локус, предпочтительно посредством встраивания в хромосому одного или нескольких экзогенных генов 3' от эндогенного гена в указанном локусе.
Вариант 12. Грамположительная бактерия по любому одному из варианту 1 или вариантам 3-11 или рекомбинантная нуклеиновая кислота по любому из вариантов 2-9, в которой эндогенный ген и один или несколько экзогенных генов транскрипционно связаны посредством межгенного участка или участков, активных у грамположительной бактерии, причем предпочтительно межгенный участок или участки являются эндогенными по отношению к указанной грамположительной бактерии.
Вариант 13. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 12, в которой указанный межгенный участок выбран из группы, состоящей из межгенных участков, предшествующих rpIW, rpIP, rpmD, rpIB, rpsG, rpsE, rpIN, rpIM, rpIE и rpIF.
Вариант 14. Рекомбинантная нуклеиновая кислота, содержащая межгенный участок, активный у грамположительной бактерии, функционально связанный с геном, экзогенным по отношению к указанной грамположительной бактерии, причем предпочтительно межгенный участок является эндогенным межгенным участком грамположительной бактерии.
Вариант 15. Рекомбинантная нуклеиновая кислота по варианту 14, в которой указанный межгенный участок выбран из группы, состоящей из межгенных участков, предшествующих rpIW, rpIP, rpmD, rpIB, rpsG, rpsE, rpIN, rpIM, rpIE и rpIF.
Вариант 16. Грамположительная бактерия по любому из вариантов 1 или 3-13 или рекомбинантная нуклеиновая кислота по любому из вариантов 2-9 или вариантов 12-15, в которой один экзогенный ген кодирует легкую цепь (VL) антитела или ее функциональный фрагмент, а другой экзогенный ген кодирует тяжелую цепь (VH) антитела или ее функциональный фрагмент, причем более предпочтительно функциональным фрагментом является Fab.
Вариант 17. Грамположительная бактерия или рекомбинантная нуклеиновая кислота по варианту 16, в которой экзогенный ген, кодирующий VL или ее функциональный фрагмент, транскрипционно связан с 3'-концом экзогенного гена, кодирующего VH или ее функциональный фрагмент.
Вариант 18. Грамположительная бактерия по любому из вариантов 1,3-13, 16 или 17 или рекомбинантная нуклеиновая кислота по любому из вариантов 2-9 или вариантов 12-17, где грамположительная бактерия является молочнокислой бактерией, предпочтительно Lactococcus, Lactobacillus или Enterococcus, более предпочтительно Lactococcus lactis или Enterococcus faecium, или где грамположительной бактерией является представитель рода Bifidobacterium.
Вариант 19. Фармацевтическая композиция, содержащая грамположительную бактерию по любому из вариантов 1,3-13 или 16-18.
Вариант 20. Фармацевтическая композиция по варианту 19, где указанный один или несколько экзогенных генов кодируют продукт, такой как белок, полипептид или пептид, причем продукт оказывает терапевтический или профилактический эффект на субъект.
Вариант 21. Вектор, содержащий рекомбинантную нуклеиновую кислоту по любому из вариантов 2-9 или вариантов 12-18.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Actogenix N.V.
<120> Polycistronic expression system for bacteria
<130> AGX-030-PCT
<150> 11168495.7
<151> 2011-06-01
<150> 11173588.2
<151> 2011-07-12
<160> 19
<170> PatentIn version 3.3
<210> 1
<211> 5
<212> DNA
<213> Lactococcus lactis
<400> 1
taatg 5
<210> 2
<211> 9
<212> DNA
<213> Lactococcus lactis
<400> 2
taatccatg 9
<210> 3
<211> 16
<212> DNA
<213> Lactococcus lactis
<400> 3
taaggaggaa aaaatg 16
<210> 4
<211> 21
<212> DNA
<213> Lactococcus lactis
<400> 4
taatagagga ggaaaatcgt g 21
<210> 5
<211> 24
<212> DNA
<213> Lactococcus lactis
<400> 5
taagaaggga gataagtaag aatg 24
<210> 6
<211> 24
<212> DNA
<213> Lactococcus lactis
<400> 6
taaggaaagg ggtaattaaa catg 24
<210> 7
<211> 28
<212> DNA
<213> Lactococcus lactis
<400> 7
taagcaaaac taggaggaat atagcatg 28
<210> 8
<211> 5
<212> DNA
<213> Enterococcus faecium
<400> 8
taatc 5
<210> 9
<211> 17
<212> DNA
<213> Enterococcus faecium
<400> 9
taaggaggac aacaata 17
<210> 10
<211> 18
<212> DNA
<213> Enterococcus faecium
<400> 10
taataggagg gaatttca 18
<210> 11
<211> 24
<212> DNA
<213> Enterococcus faecium
<400> 11
ttagaagaag gaggaatacc attc 24
<210> 12
<211> 30
<212> DNA
<213> Enterococcus faecium
<400> 12
taaaagttta aggaaggagg gtcttactga 30
<210> 13
<211> 34
<212> DNA
<213> Enterococcus faecium
<400> 13
taatcaagta gaatctacaa ggaggtgtct ttaa 34
<210> 14
<211> 24
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 14
aggtcaaaca ggaagacaga cgta 24
<210> 15
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 15
tcacacccaa gaacaagcac a 21
<210> 16
<211> 28
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 16
gccaaggtct aacaatcgtt gtgagttg 28
<210> 17
<211> 23
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 17
cagtcagcca gcttgacacc acg 23
<210> 18
<211> 25
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 18
tcaagaggct gcaaaggaag agaac 25
<210> 19
<211> 23
<212> DNA
<213> Artificial
<220>
<223> Primer
<400> 19
tggtctccat gttcagcgac agc 23
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СИСТЕМА ПОЛИЦИСТРОННОЙ ЭКСПРЕССИИ ДЛЯ БАКТЕРИЙ | 2012 |
|
RU2644346C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ БАКТЕРИИ, СТАБИЛЬНО ЭКСПРЕССИРУЮЩИЕ ИЛ-10 И ИНСУЛИН | 2017 |
|
RU2768027C2 |
| МУКОАДГЕЗИВНЫЙ МИКРООРГАНИЗМ | 2017 |
|
RU2762940C2 |
| ПРОМОТОРЫ Lactococcus И ИХ ПРИМЕНЕНИЕ: | 2008 |
|
RU2495129C2 |
| МОДИФИЦИРОВАННЫЕ ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2773435C2 |
| МОДИФИЦИРОВАННЫЕ ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2653757C2 |
| РЕКОМБИНАНТНЫЕ ПРОБИОТИЧЕСКИЕ БАКТЕРИИ | 2015 |
|
RU2723324C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ДИАБЕТА 1 ТИПА | 2017 |
|
RU2760997C2 |
| ХЛОРИД-ИНДУЦИРУЕМАЯ ПРОКАРИОТИЧЕСКАЯ СИСТЕМА ЭКСПРЕССИИ | 2019 |
|
RU2812766C2 |
| МИКРООРГАНИЗМ, ЭКСПРЕССИРУЮЩИЙ КСИЛОЗОИЗОМЕРАЗУ | 2009 |
|
RU2553537C2 |
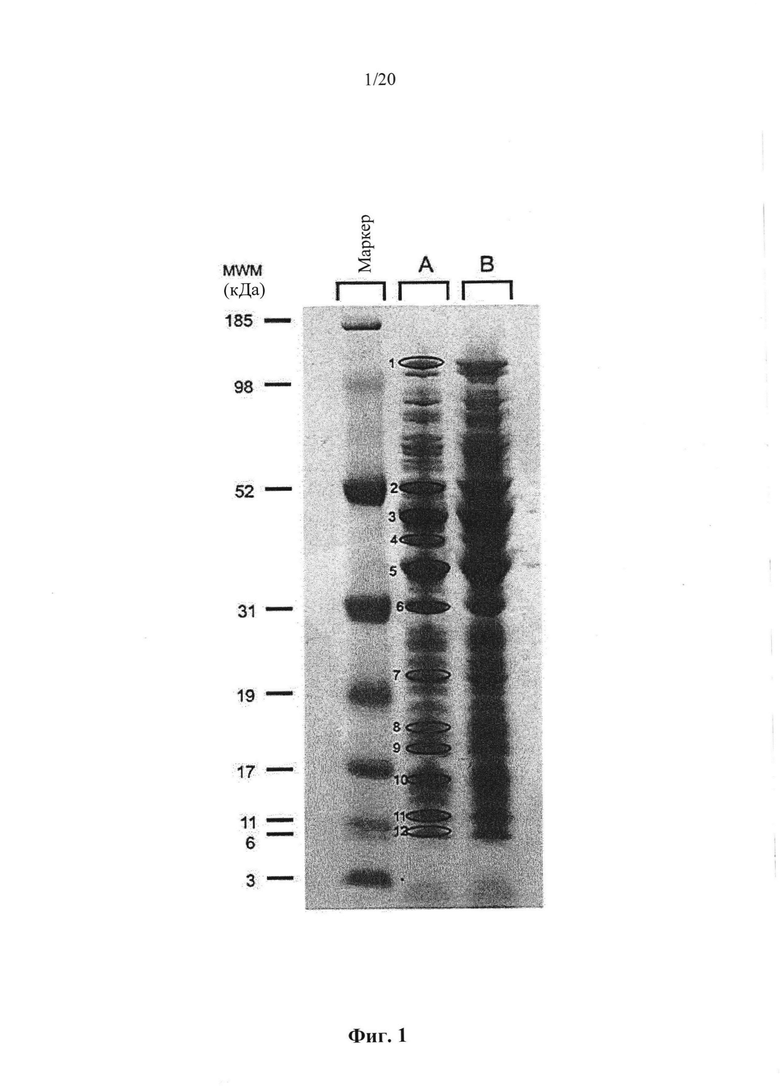
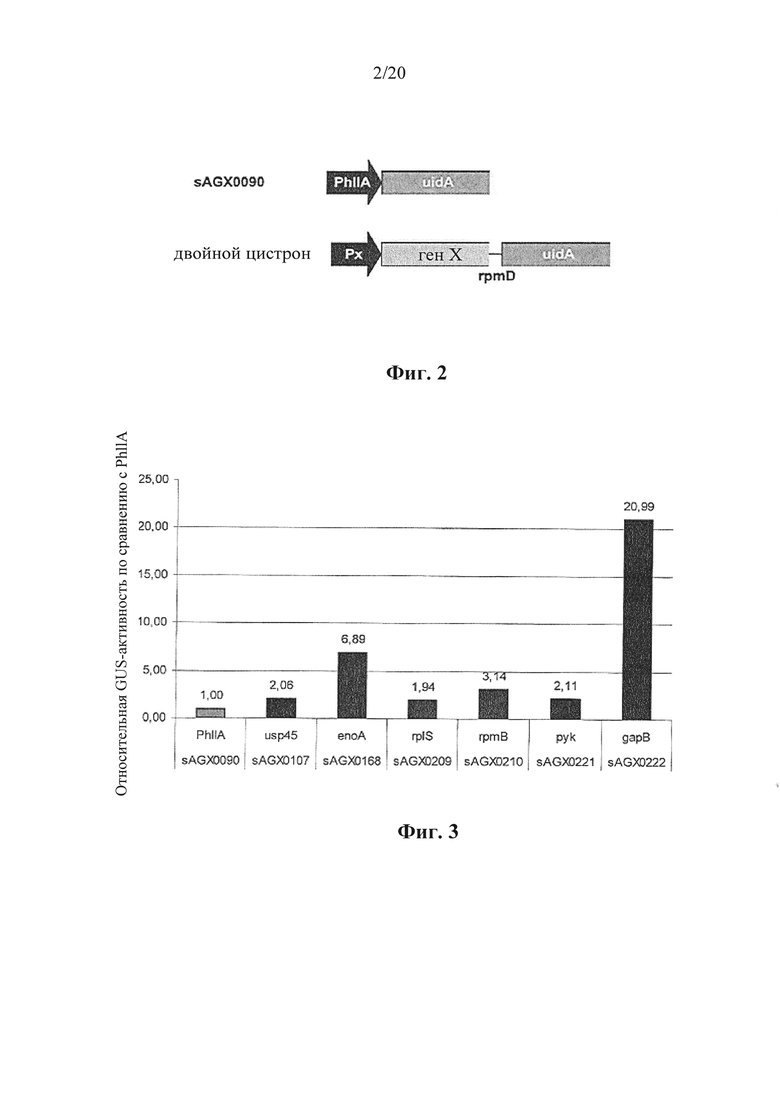
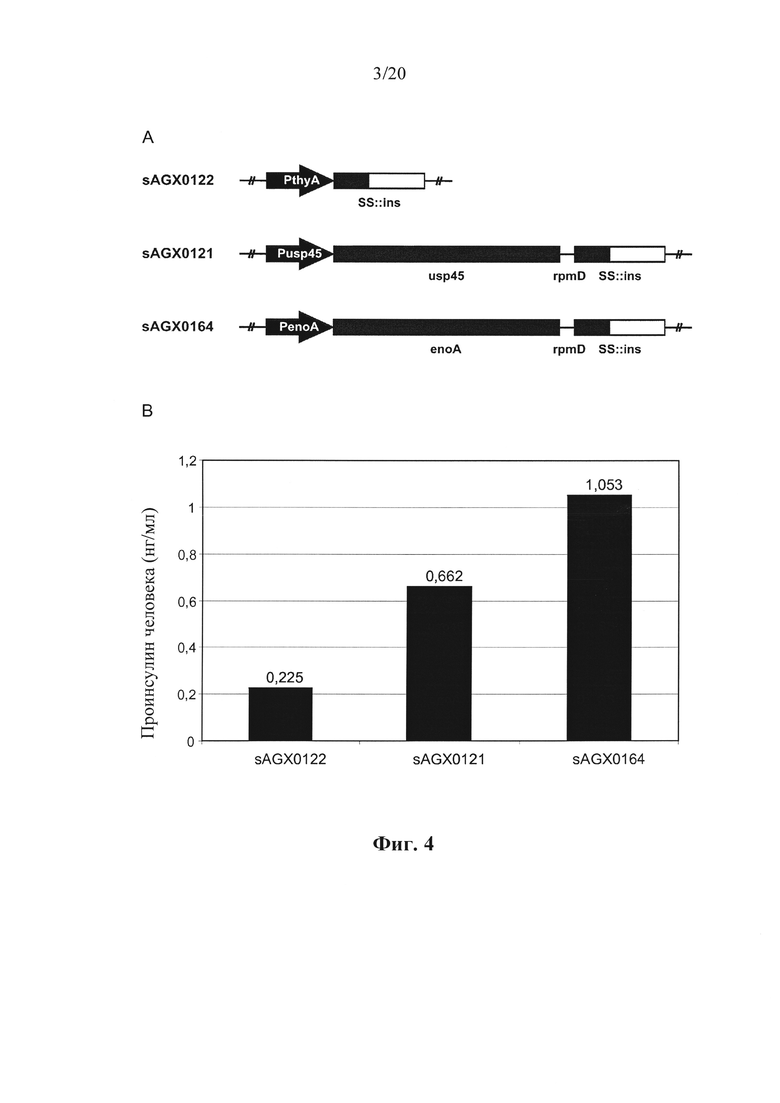
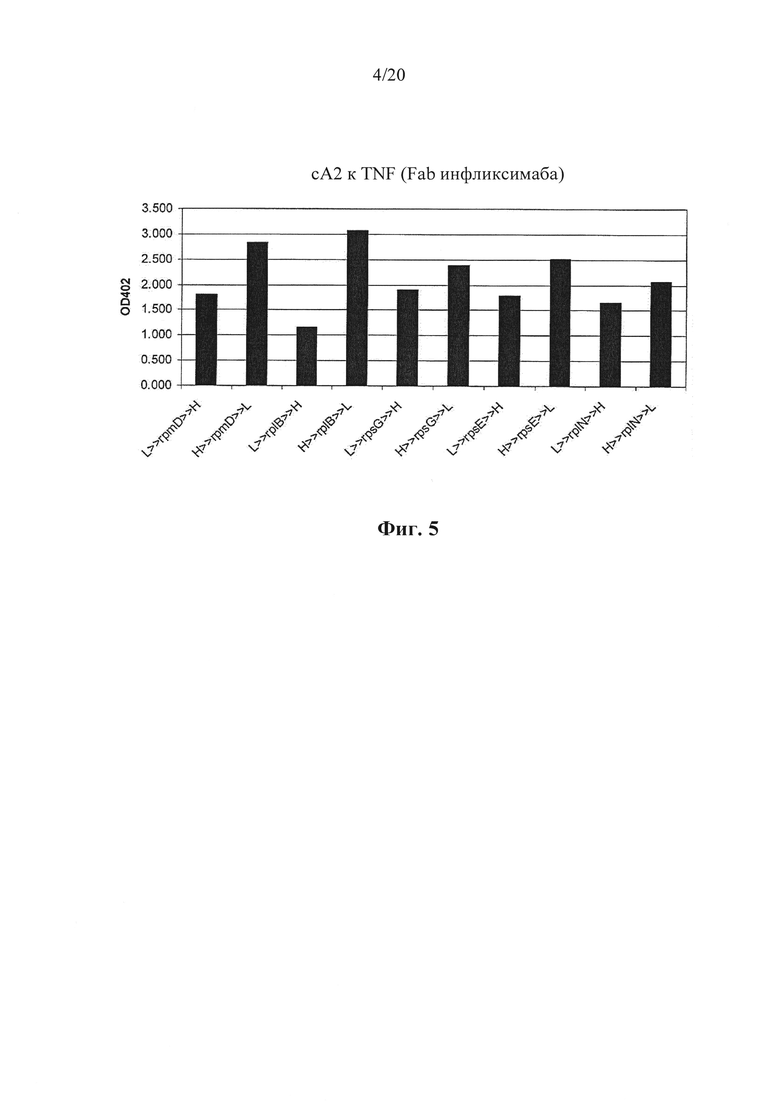

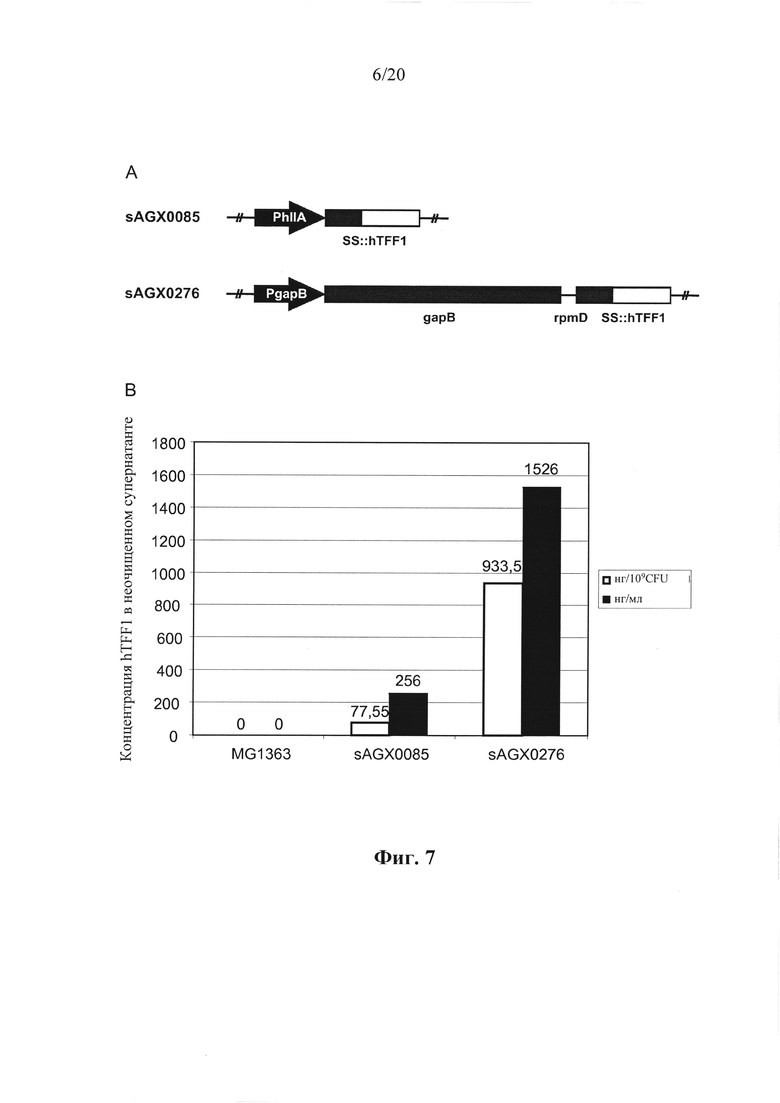
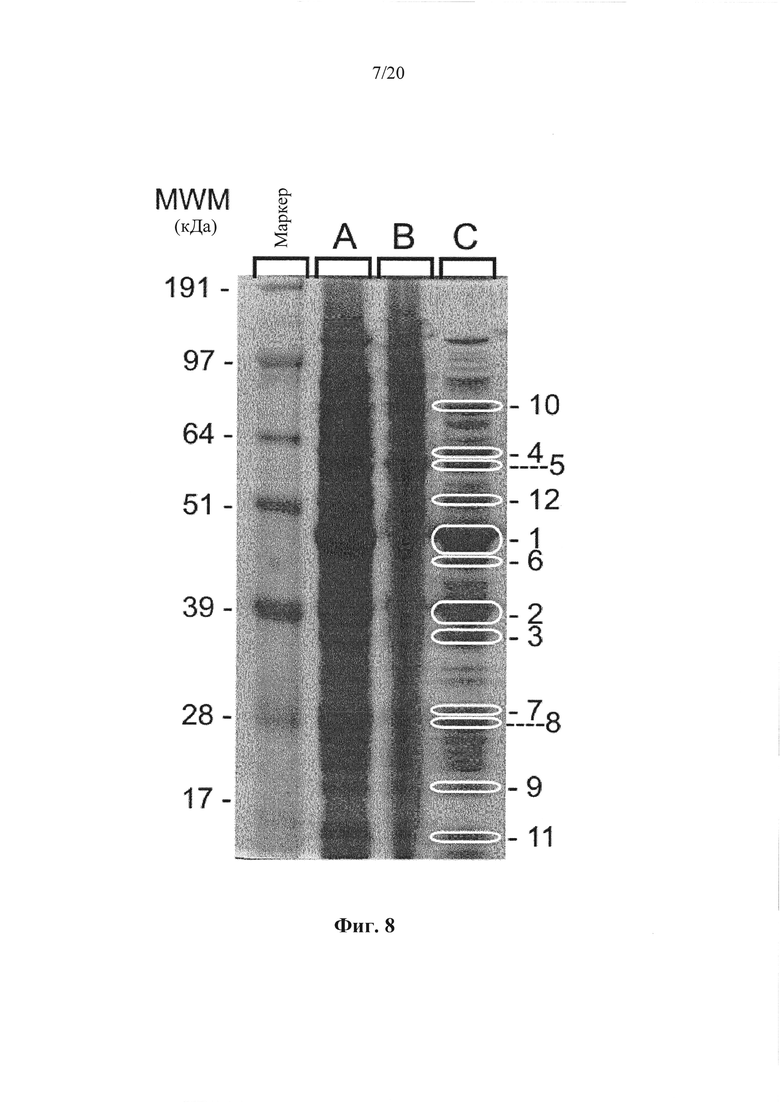
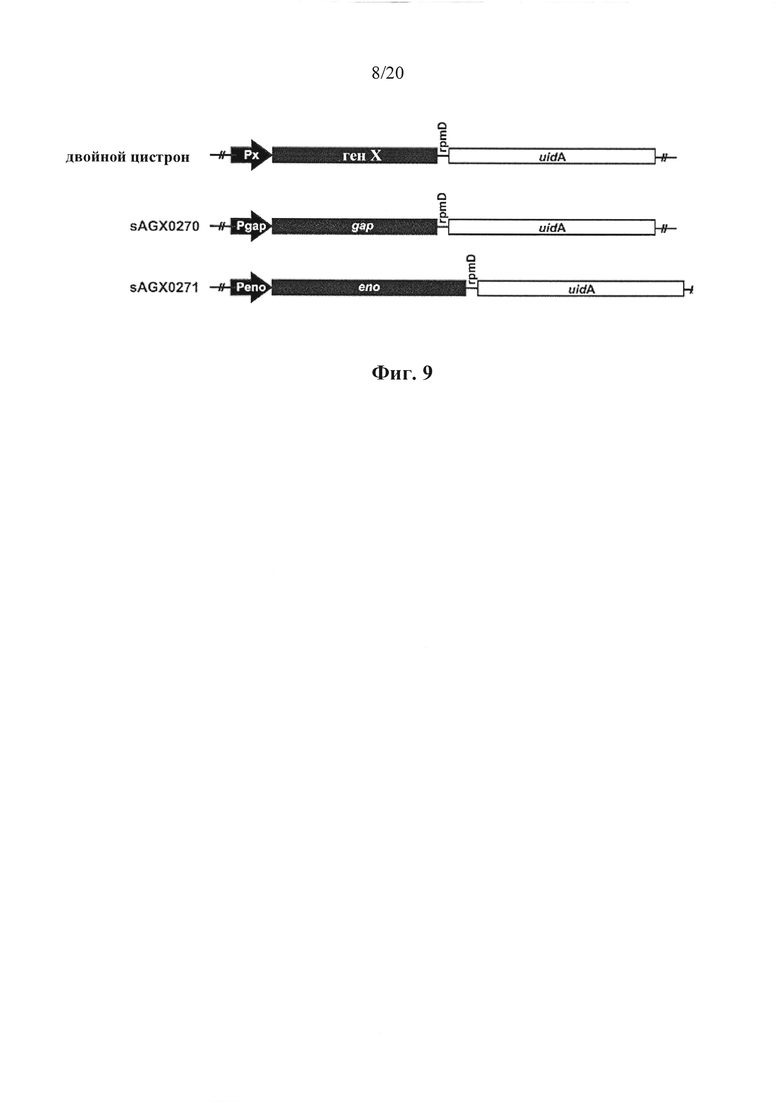

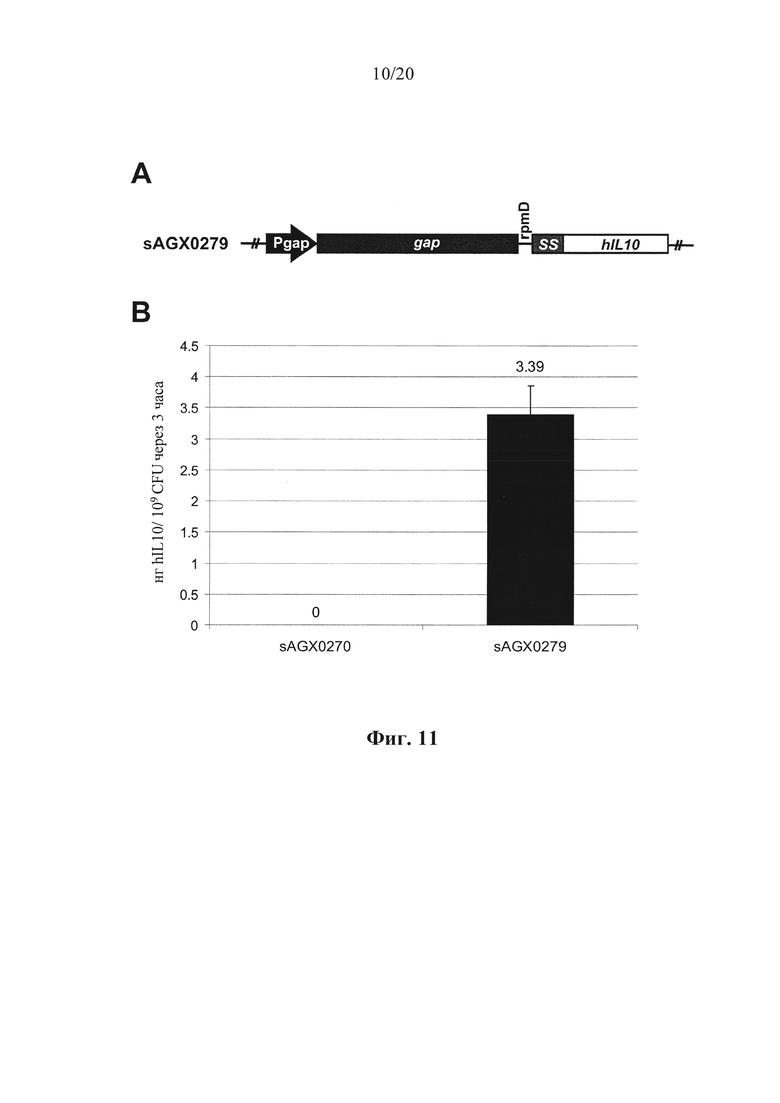
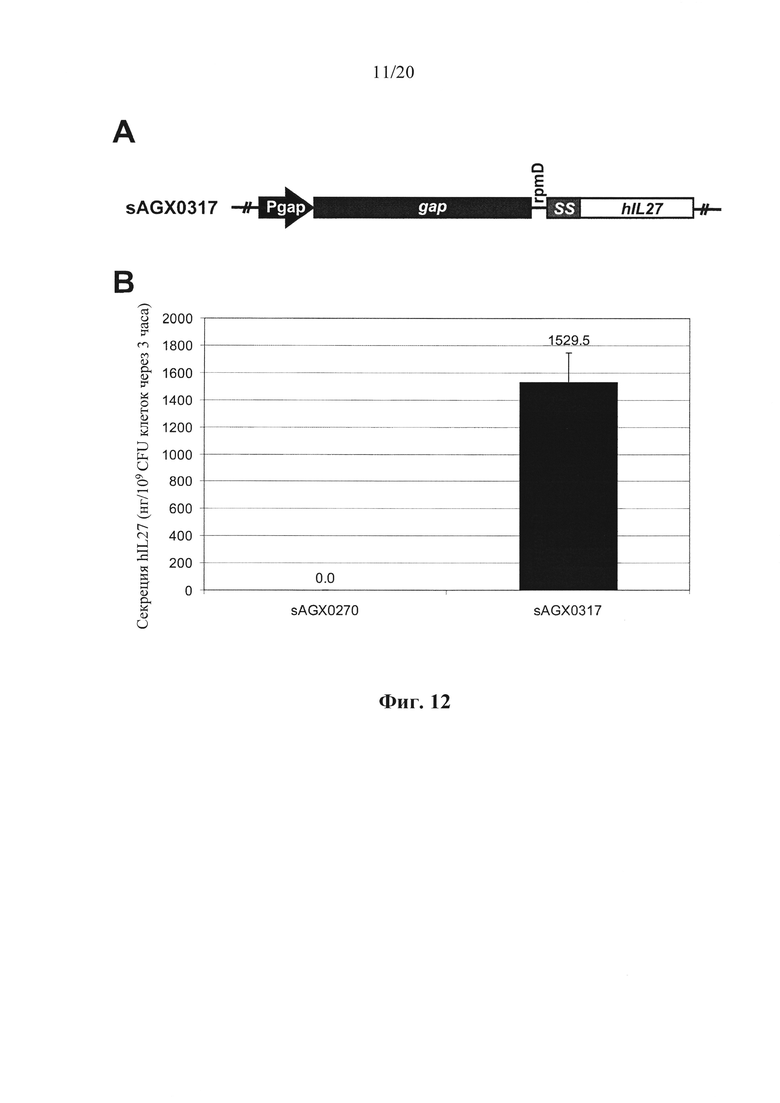
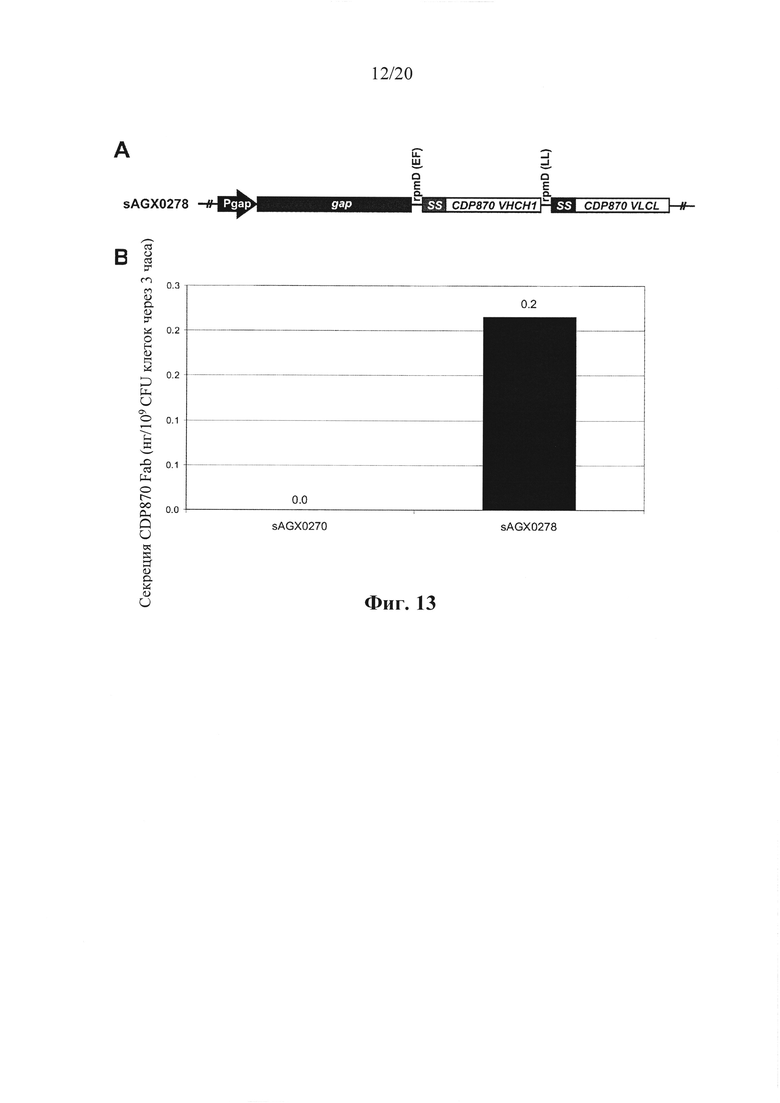
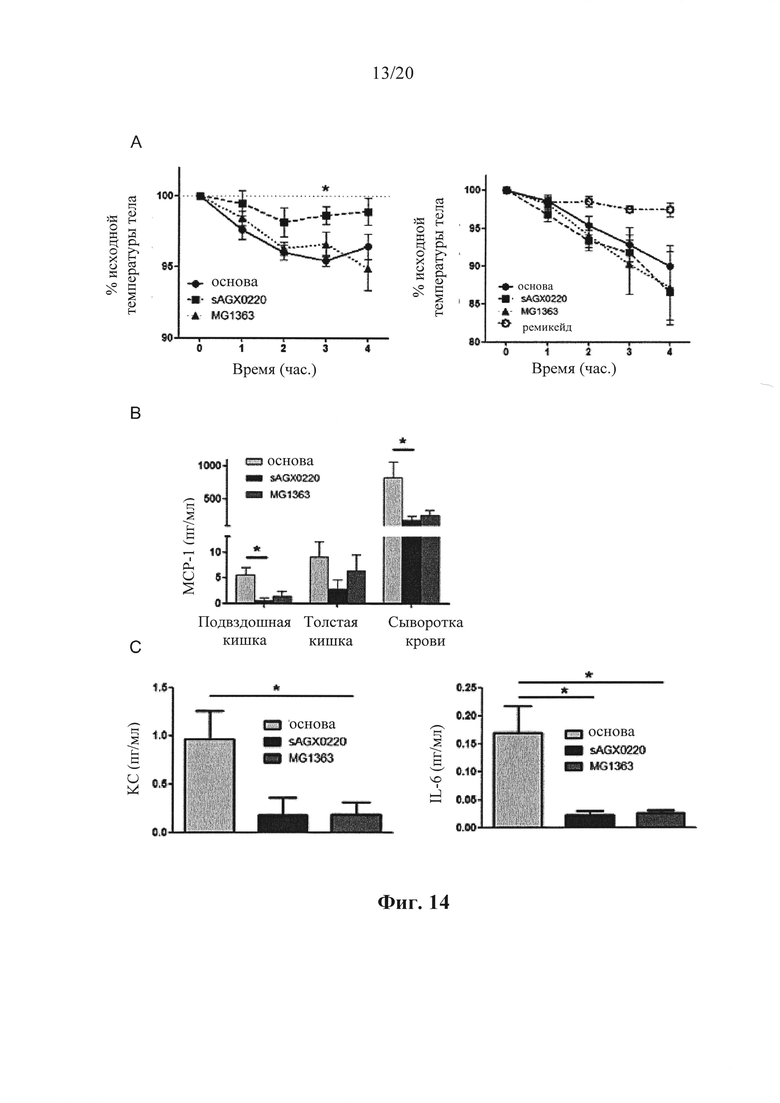
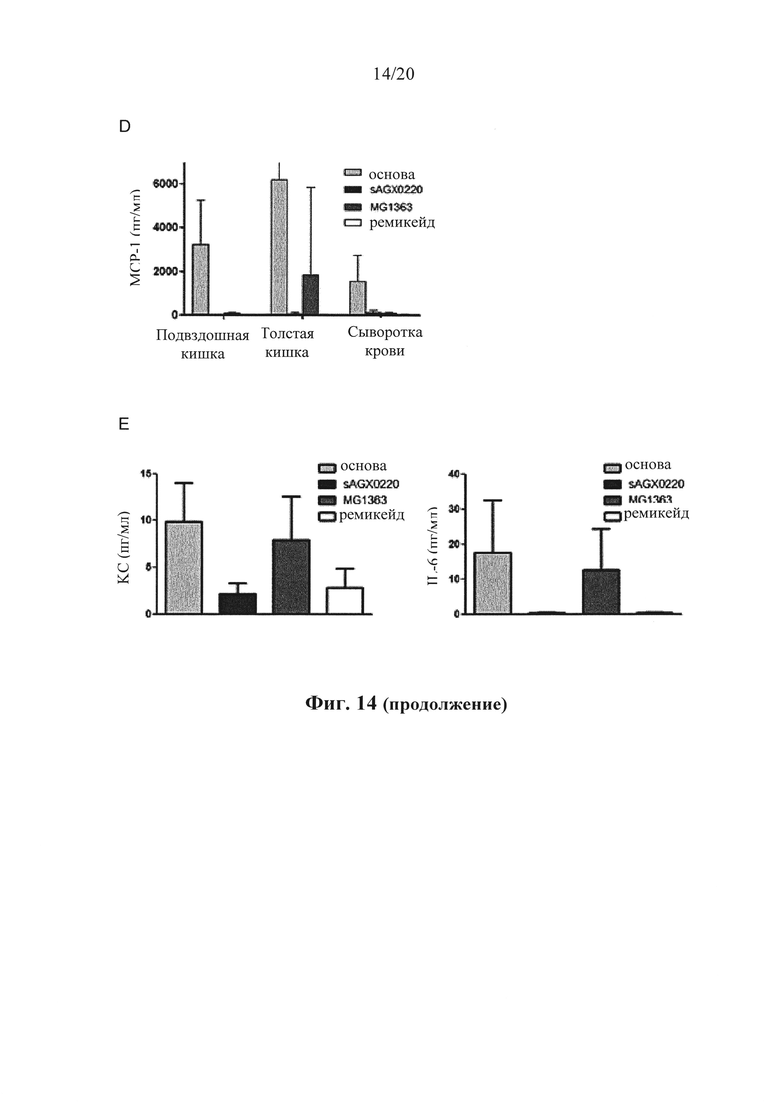
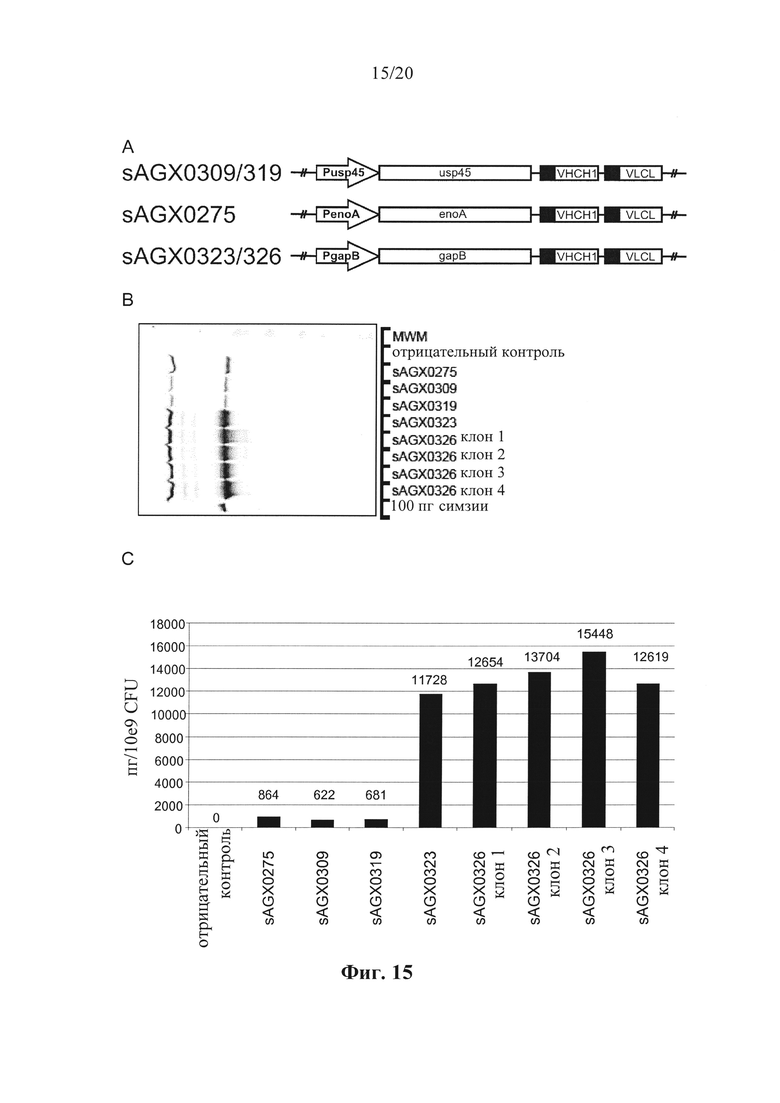
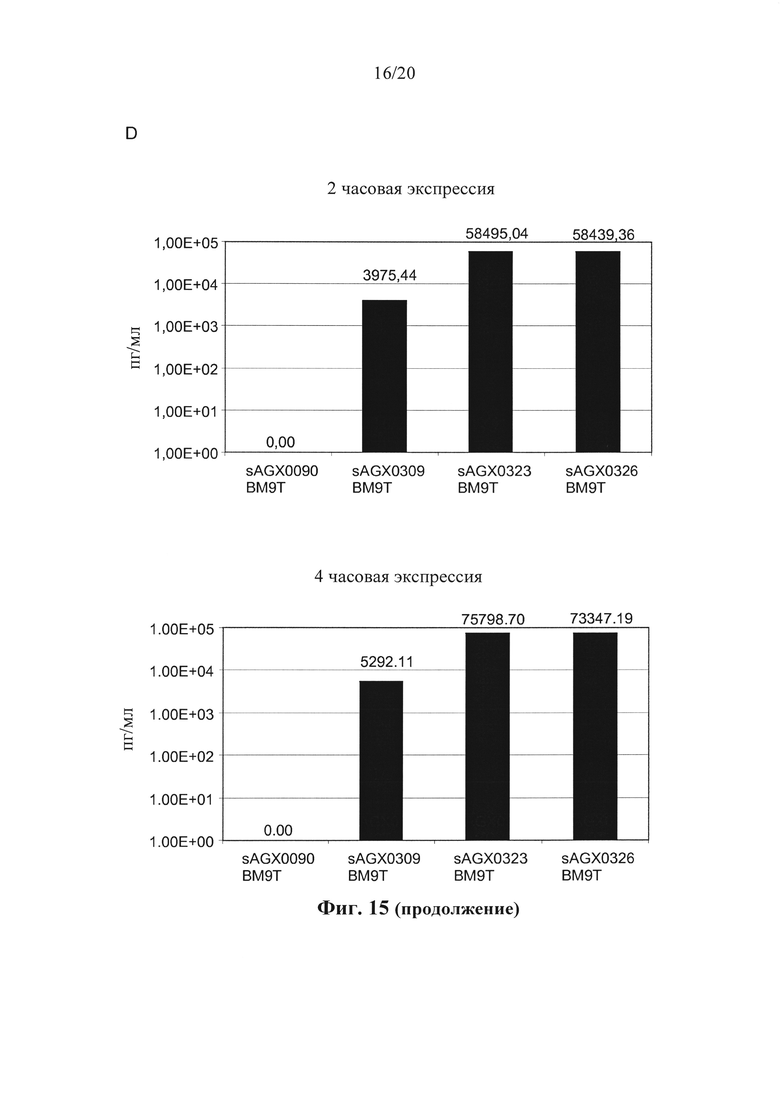
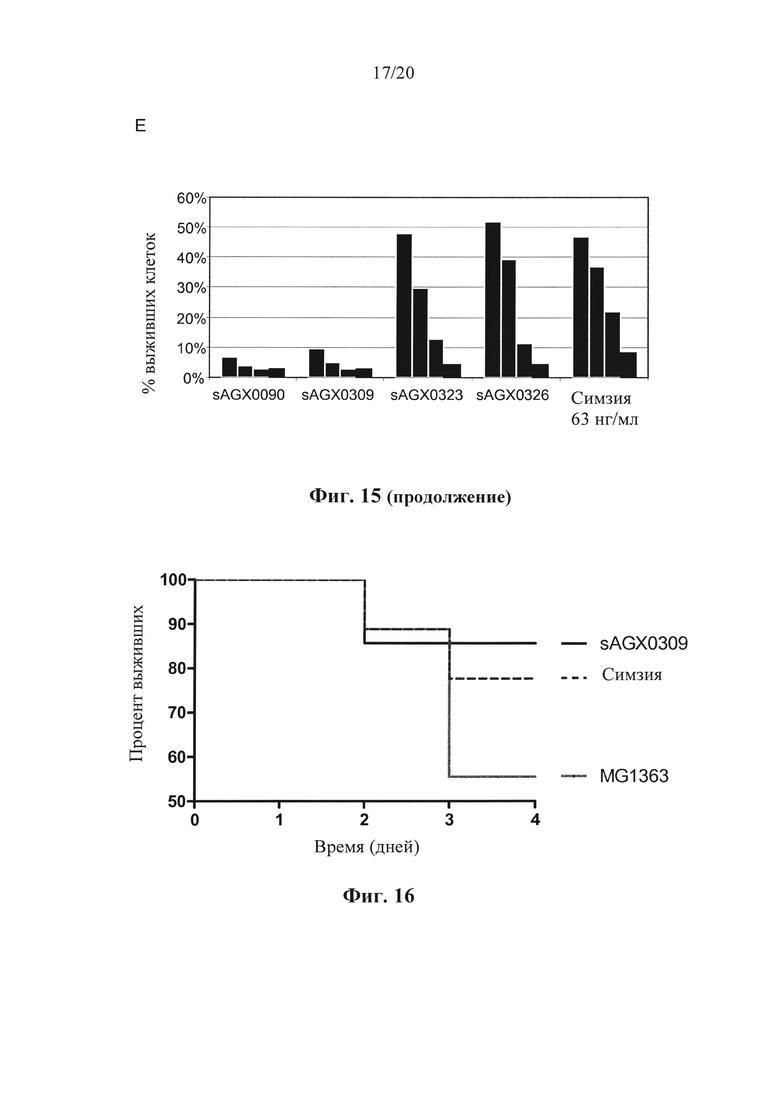
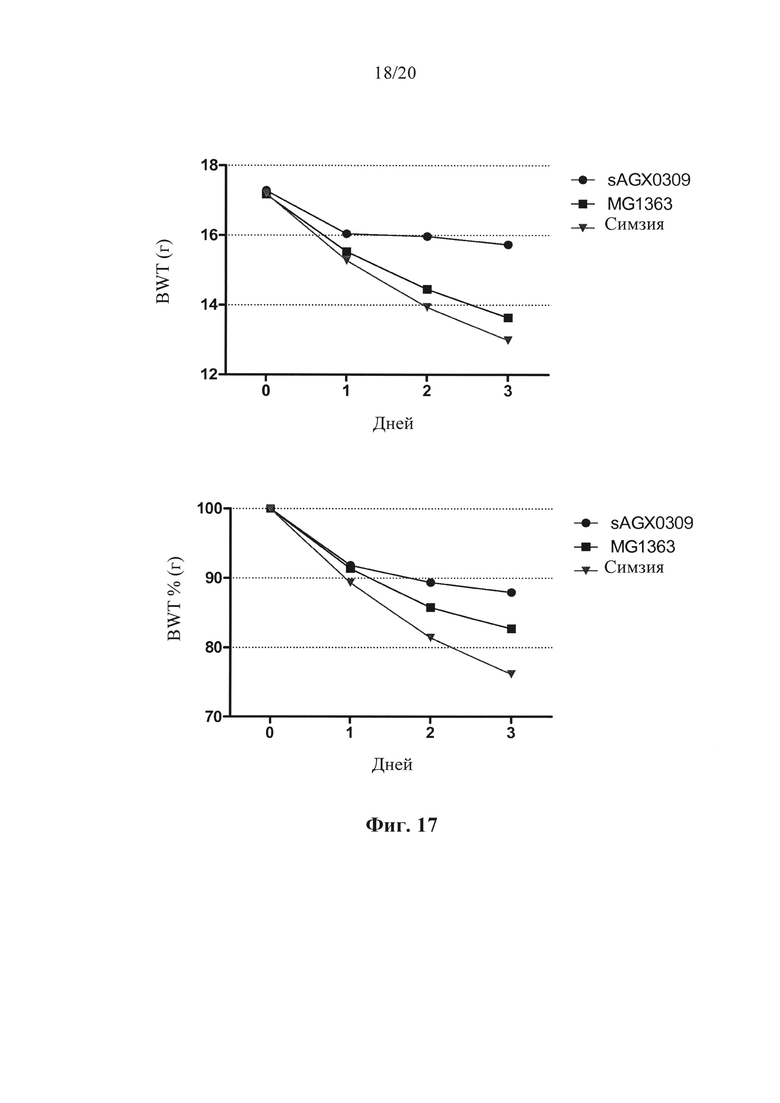
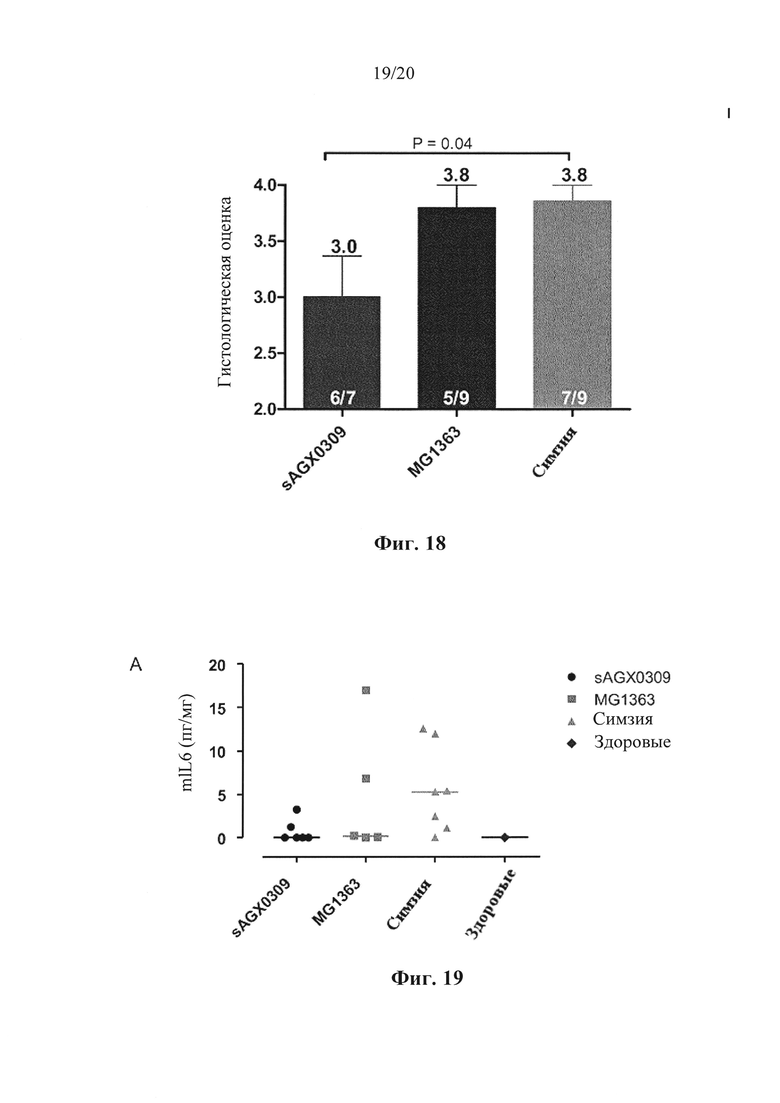
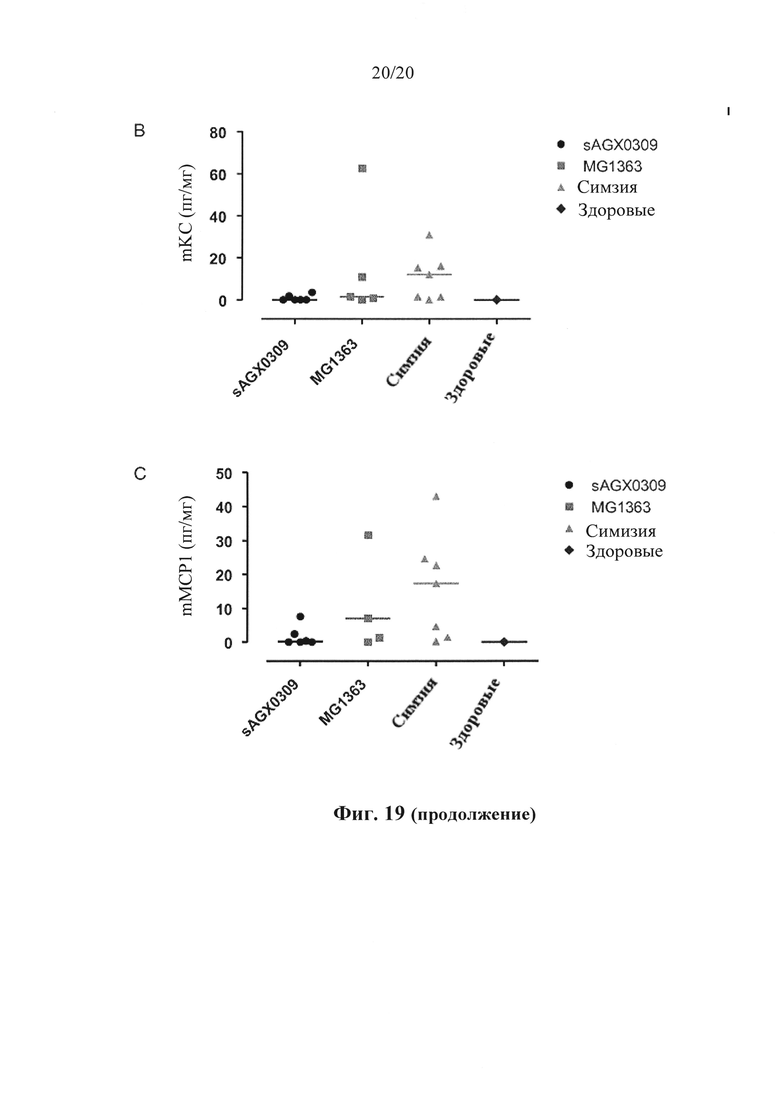
Настоящие изобретения относятся к полицистронной экспрессии у грамположительной бактерии и касаются молочнокислой бактерии для экспрессии одного или нескольких экзогенных генов, фармацевтической композиции, содержащей такую молочнокислую бактерию, нуклеиновой кислоты и вектора. Предложена молочнокислая бактерия, содержащая единицу полицистронной экспрессии. При этом указанная единица полицистронной экспрессии содержит в направлении от 5' к 3': (i) эндогенный промотор, представляющий собой промотор еnо (Реnо), промотор usp45 (Pusp45) или промотор gap В (PgapB); (ii) эндогенный ген; и (iii) один или несколько экзогенных генов, расположенные 3' по отношению к указанному эндогенному гену. При этом указанные эндогенный ген и один или несколько экзогенных генов являются транскрипционно связанными с по меньшей мере одним эндогенным межгенным участком. Изобретения обеспечивают высокие уровни экспрессии экзогенных генов у грамположительных бактерий. 5 н. и 32 з.п. ф-лы, 19 ил., 8 табл., 16 пр.
1. Молочнокислая бактерия для экспрессии одного или нескольких экзогенных генов, содержащая единицу полицистронной экспрессии, при этом указанная единица полицистронной экспрессии содержит в направлении от 5' к 3':
(i) эндогенный промотор, представляющий собой промотор епо (Репо), промотор usp45 (Pusp45) или промотор gap В (PgapB);
(ii) эндогенный ген; и
(iii) один или несколько экзогенных генов, расположенные 3' по отношению к указанному эндогенному гену;
при этом указанные эндогенный ген и один или несколько экзогенных генов являются транскрипционно связанными с по меньшей мере одним эндогенным межгенным участком, где указанный эндогенный межгенный участок включает нуклеотидные последовательности между (а) инициаторным кодоном rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE или rplF и (b) стоп-кодоном предшествующего 5' гена.
2. Молочнокислая бактерия по п.1, при этом указанный эндогенный межгенный участок имеет нуклеотидную последовательность, включающую любой из SEQ ID NO: 1-13.
3. Молочнокислая бактерия по п.1 или 2, при этом молочнокислая бактерия представляет собой Lactococcus, Lactobacillus или Enterococcus.
4. Молочнокислая бактерия по любому из пп. 1-3, при этом молочнокислая бактерия представляет собой Lactococcus lactis.
5. Молочнокислая бактерия по любому из пп. 1-4, в которой указанный промотор и указанный эндогенный ген расположены в своем естественном локусе на хромосоме у молочнокислой бактерии, и при этом указанный эндогенный ген транскрипционно связан с одним или несколькими экзогенными генами посредством встраивания в хромосому одного или нескольких экзогенных генов в указанный локус, предпочтительно посредством встраивания в хромосому одного или нескольких экзогенных генов 3' от указанного локуса.
6. Молочнокислая бактерия по любому из пп. 1-5, в которой указанный эндогенный межгенный участок включает нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD.
7. Молочнокислая бактерия по любому из пп. 1-6, в которой указанный межгенный участок содержит нуклеотидную последовательность SEQ ID NO: 3.
8. Молочнокислая бактерия по любому из пп. 1-7, в которой указанный один или несколько экзогенных генов кодируют один или несколько полипептидов.
9. Молочнокислая бактерия по п. 8, в которой указанный один или несколько экзогенных генов трансляционно связаны с нуклеиновой кислотой, кодирующей определяющий секрецию сигнальный пептид.
10. Молочнокислая бактерия по п. 9, в которой указанный определяющий секрецию сигнальный пептид представляет собой кодирующий определяющий секрецию сигнал Usp45 Lactococcus.
11. Молочнокислая бактерия по любому из пп. 8-10, в которой указанный один или несколько полипептидов имеют терапевтический или профилактический эффект на субъект.
12. Молочнокислая бактерия по любому из пп. 8-11, в которой указанный один или несколько полипептидов выбраны из группы, состоящей из:
(i) интерлейкина-10 (IL-10);
(ii) фактора «трилистника»-1 (TFF1);
(iii) проинсулина (ins); и/или
(iv) интерлейкина-27 (IL-27).
13. Молочнокислая бактерия по п. 11 или 12, в которой указанная единица полицистронной экспрессии выбрана из:
(i) gapB, слитый с SS:: TFF1 через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD;
(ii) usp45, слитый с SS::ins через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD; и/или
(iii) enoA, слитый с SS::ins через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD;
при этом SS:: представляет собой трансляционно слитый определяющий секрецию сигнал.
14. Молочнокислая бактерия по п. 13, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' PgapB, gapB нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::TFF1.
15. Молочнокислая бактерия по п. 13, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' Pusp45, usp45 нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::ins.
16. Молочнокислая бактерия по п. 13, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' РеnоА, еnоА нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::ins.
17. Молочнокислая бактерия по любому из пп. 13-16, в которой указанная единица полицистронной экспрессии расположена в локусе, эндогенном для указанного эндогенного промотора.
18. Молочнокислая бактерия по любому из пп. 8-17 для применения при введении или доставке указанного одного или нескольких полипептидов субъекту.
19. Фармацевтическая композиция для применения при введении или доставке одного или нескольких полипептидов, оказывающих терапевтический или профилактический эффект на субъекта, содержащая терапевтически эффективное количество молочнокислой бактерии по любому из пп. 11-17 и фармацевтически приемлемый носитель.
20. Рекомбинантная нуклеиновая кислота для экспрессии одного или нескольких экзогенных генов, содержащая единицу полицистронной экспрессии, причем указанная единица полицистронной экспрессии содержит в направлении от 5' к 3':
(i) промотор, эндогенный по отношению к указанной молочнокислой бактерии, представляющий собой промотор еnо (Реnо), промотор usp45 (Pusp45) или промотора gapB (PgapB);
(ii) эндогенный ген, который является эндогенным по отношению к указанной молочнокислой бактерии; и
(iii) один или несколько экзогенных генов, которые являются экзогенными по отношению к указанной молочнокислой бактерии, причем указанные один или несколько экзогенных генов расположены 3' к указанному эндогенному гену;
при этом указанные эндогенный ген и один или несколько экзогенных генов являются транскрипционно связанными с по меньшей мере одним межгенным участком, эндогенным к указанной молочнокислой бактерии, и где указанный межгенный участок, включающий нуклеотидные последовательности между (а) инициаторным кодоном rplW, rplP, rpmD, rplB, rpsG, rpsE, rplN, rplM, rplE или rplF и (b) стоп-кодоном предшествующего 5' гена.
21. Рекомбинантная нуклеиновая кислота по п. 20, при этом указанный эндогенный межгенный участок имеет нуклеотидную последовательность, включающую любой из SEQ ID NO: 1-13.
22. Рекомбинантная нуклеиновая кислота по п. 20 или 21, при этом молочнокислая бактерия выбрана из группы, состоящей из Lactococcus, Lactobacillus или Enterococcus.
23. Рекомбинантная нуклеиновая кислота по любому из пп. 20-22, при этом молочнокислая бактерия представляет собой Lactococcus lactis.
24. Рекомбинантная нуклеиновая кислота по любому из пп. 20-23, в которой указанный эндогенный ген и указанные один или несколько экзогенных генов находятся под транскрипционным контролем нативного промотора указанного эндогенного гена.
25. Рекомбинантная нуклеиновая кислота по любому из пп. 20-24, в которой указанный межгенный участок включает нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD.
26. Рекомбинантная нуклеиновая кислота по любому из пп. 20-25, в которой указанный межгенный участок содержит нуклеотидную последовательность SEQ ID NO: 3.
27. Рекомбинантная нуклеиновая кислота по любому из пп. 20-26, в которой указанный один или несколько экзогенных генов кодируют один или несколько полипептидов.
28. Рекомбинантная нуклеиновая кислота по п. 27, в которой указанный один или несколько экзогенных генов трансляционно связаны с нуклеиновой кислотой, кодирующей определяющий секрецию сигнальный пептид.
29. Рекомбинантная нуклеиновая кислота по п. 28, в которой указанный определяющий секрецию сигнальный пептид представляет собой кодирующий определяющий секрецию сигнал Usp45 Lactococcus.
30. Рекомбинантная нуклеиновая кислота по любому из пп. 27-29, в которой указанный один или несколько полипептидов имеют терапевтический или профилактический эффект на субъект.
31. Рекомбинантная нуклеиновая кислота по любому из пп. 27-30, в которой указанный один или несколько полипептидов выбраны из:
(i) интерлейкина-10 (IL-10);
(ii) фактора «трилистника»-1 (TFF1);
(iii) проинсулина (ins); и/или
(iv) интерлейкина-27 (IL-27).
32. Рекомбинантная нуклеиновая кислота по любому из пп. 19-29, в которой указанная единица полицистронной экспрессии содержит:
(i) gapB, слитый с SS:: TFF1 через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD;
(ii) usp45, слитый с SS::ins через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD; или
(iii) enoA, слитый с SS::ins через нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD;
при этом SS:: представляет собой трансляционно слитый определяющий секрецию сигнал.
33. Рекомбинантная нуклеиновая кислота по п. 32, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' PgapB, gapB нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::TFFJ.
34. Рекомбинантная нуклеиновая кислота по п. 32, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' Pusp45, usp45 нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::ins.
35. Рекомбинантная нуклеиновая кислота по п. 32, в которой указанная единица полицистронной экспрессии содержит в направлении от 5' к 3' РеnоА, еnоА, нуклеотидные последовательности между (а) инициаторным кодоном rpmD и (b) стоп-кодоном предшествующего 5' гена к rpmD и SS::ins.
36. Вектор для экспрессии одного или нескольких экзогенных генов, содержащий рекомбинантную нуклеиновую кислоту по любому из пунктов 20-35.
37. Молочнокислая бактерия для экспрессии одного или нескольких экзогенных генов, содержащая рекомбинантную нуклеиновую кислоту по любому из пунктов 20-35 или вектор по п. 36.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| MACHIELSEN R | |||
| ET AL | |||
| Indigenous and environmental modulation of frequencies of mutation in Lactobacillus plantarum | |||
| Appl Environ Microbiol | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| KIM E.B | |||
| ET AL | |||
| Cloning and Characterization of a Novel tuf Promoter from Lactococcus lactis subsp | |||
| Резервуар для керосиновых ламп типа "Примус" | 1924 |
|
SU1403A1 |
| Curr | |||

