Область техники, к которой относится изобретение
Настоящее изобретение относится к микробиологической промышленности, в частности к способу получения L-аминокислоты, принадлежащей к семейству глутамата, с использованием коринеформной бактерии, модифицированной таким образом, что экспрессия одного или нескольких генов бактериального оперона NCgl_2067-NCgl_2065 ослаблена.
Уровень техники
Традиционно L-аминокислоты в промышленном масштабе получают методом ферментации с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов. Обычно микроорганизмы модифицируют, для того чтобы увеличить продукцию L-аминокислот.
Описано множество методов увеличения продукции L-аминокислот, например, путем трансформации микроорганизма рекомбинантной ДНК (см., например, патент США 4,278,765). Другие методы основаны на повышении активности ферментов, вовлеченных в биосинтез аминокислот, и/или уменьшении чувствительности целевого фермента к ингибированию по типу обратной связи продуцируемой L-аминокислотой (см., например, патенты США 4,346,170; 5,661,012 и 6,040,160).
Описаны многие способы получения L-глутамина путем культивирования коринеформной бактерии. Например, в патенте ЕР 1229121 А2 описан способ получения L-глутамина путем культивирования коринеформной бактерии-продуцента L-глутамина, модифицированной таким образом, что ферментативная активность внутриклеточной глутаминсинтетазы повышена, и предпочтительно модифицированной далее таким образом, что повышена ферментативная активность внутриклеточной глутаматдегидрогеназы, с целью продукции и накопления L-глутамина в питательной среде и выделения L-глутамина из культуральной жидкости.
Известно, что транскрипция гена glnA из Bacillus subtilis, кодирующего глутаминсинтетазу, регулируется источником азота. Ген glnA находится в одном опероне с геном glnR. Ген glnR кодирует негативный регулятор, который напрямую контролирует экспрессию гена глутаминсинтетазы. Некоторые мутации в гене glnR, такие как большая делеция в пределах рамки считывая и мутация в стартовом кодоне, приводили к высокой конститутивности оперона, в то время как другие мутации вызывали низкую конститутивность (Schreier H.J. et al., J Mol Biol. 1989, 210(1):51-63).
Недавно в Streptococcus pneumoniae изучали регулирование метаболизма глутамина и глутамата под действием регуляторного белка GlnR (Kloosterman T.G. et al., J Biol Chem. 2006, 281(35):25097-25109). Используя метод ДНК-микрочипов (DNA microarray) при анализе дикого штамма S.pneumoniae D39 и его изогенного glnR-мутанта, который выращивался на среде, богатой азотом и содержащей глутамин, был определен ряд оперонов и генов, активированных в glnR-мутанте: glnA (кодирует глутаминсинтетазу GlnA), glnPQ (кодирует транспортер семейства ABC - аминокислота-связывающий белок/пермеазу) и gdhA (кодирует глутаматдегидрогеназу). Все эти гены зависимы от GlnR-оператора в их промоторной области. Также было найдено, что ген zwf, кодирующий глюкоза-6-фосфатдегидрогеназу и расположенный ниже и в той же ориентации, что и ген glnPQ, активирован.
В настоящее время нет данных, указывающих на присутствие предполагаемого гена glnR в геноме коринеформных бактерий. Также нет данных об ослаблении экспрессии одного или нескольких генов оперона NCgl_2067-NCgl_2065 с целью повышения продукции L-аминокислоты, принадлежащей к семейству глутамата.
Раскрытие изобретения
Целями настоящего изобретения являются повышение продуктивности штаммов-продуцентов L-аминокислоты семейства глутамата и предоставление способа получения L-аминокислоты семейства глутамата с использованием этих штаммов.
В геноме Corynebacterium glutamicum была обнаружена открытая рамка считывания (OPC) NCgl_2066, гомологичная гену glnR из S. pneumoniae. OPC NCgl_2066 расположена в опероне с двумя другими OPC - NCgl_2067 и NCgl_2065. Соответствующие OPC были названы как гены NCgl_2067, NCgl_2066 и NCgl_2065, a оперон - как NCgl_2067-NCgl_2065.
Вышеупомянутые цели настоящего изобретения были достигнуты благодаря обнаружению того факта, что ослабление экспрессии одного или нескольких генов бактериального оперона NCgl_2067-NCgl_2065 может приводить к увеличению продукции L-аминокислоты, принадлежащей к семейству глутамата, такой как L-глутаминовая кислота, L-глутамин, L-пролин, L-гистидин, L-аргинин, L-орнитин и L-цитруллин.
Целью настоящего изобретения является предоставление коринеформной бактерии, способной производить L-аминокислоту, принадлежащей к семейству глутамата, отличающейся тем, что указанная бактерия модифицирована таким образом, что экспрессия одного или нескольких генов оперона NCgl_2067-NCgl_2065 указанной бактерии ослаблена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия модифицирована таким образом, что экспрессия гена NCgl_2067 ослаблена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что экспрессия гена NCgl_2067 ослаблена путем инактивации гена NCgl_2067.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия модифицирована таким образом, что экспрессия гена NCgl_2066 ослаблена.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что экспрессия гена NCgl_2066 ослаблена путем инактивации гена NCgl_2066.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия принадлежит к роду Corynebacterium.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная бактерия принадлежит к роду Brevibacterium.
Также целью настоящего изобретения является предоставление описанной выше бактерии, отличающейся тем, что указанная L-аминокислота принадлежит к семейству глутамата и выбрана из группы, состоящей из L-глутаминовой кислоты, L-глутамина, L-пролина, L-гистидина, L-аргинина, L-орнитина и L-цитруллина.
Также целью настоящего изобретения является предоставление способа получения L-аминокислоты, принадлежащей к семейству глутамата, включающего:
- выращивание описанной выше бактерии в питательной среде; и
- выделение указанной L-аминокислоты из культуральной жидкости.
Также целью настоящего изобретения является предоставление описанного выше способа, отличающегося тем, что указанная L-аминокислота, принадлежащая к семейству глутамата, выбрана из группы, состоящей из L-глутаминовой кислоты, L-глутамина, L-пролина, L-гистидина, L-аргинина, L-орнитина и L-цитруллина.
Настоящее изобретение более подробно будет описано ниже со ссылкой на последующие, но не ограничивающие настоящее изобретение Примеры.
Фигуры и их краткое описание
На Фигуре 1 изображено выравнивание аминокислотных последовательностей белка GlnR из Streptococcus pneumoniae R6 и предполагаемого белка NCgl_2066 из Corynebacterium glutamicum ATCC13032.
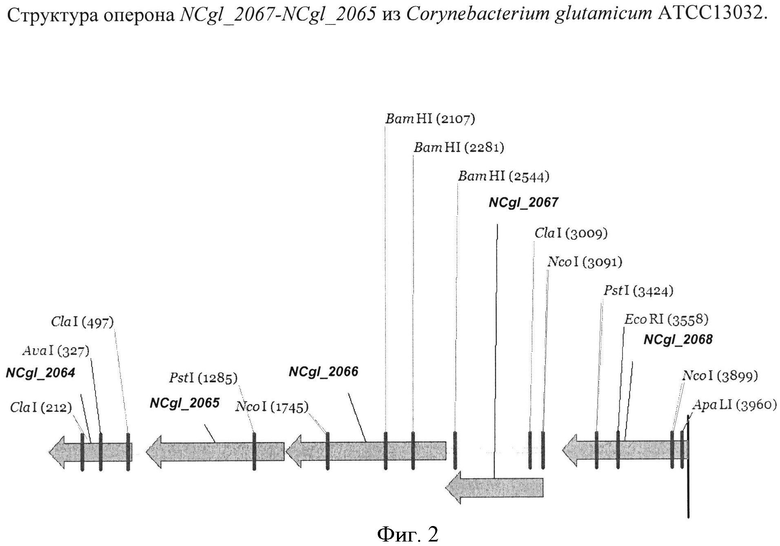
На Фигуре 2 изображена структура оперона NCgl_2067-NCgl_2065 из Corynebacterium glutamicum ATCC13032.
На Фигуре 3 изображена структура плазмиды pBS4S.
Наилучший способ осуществления настоящего изобретения
1. Бактерия согласно настоящему изобретению
Коринеформная бактерия, согласно настоящему изобретению, - бактерия, которая способна производить L-аминокислоту семейства глутамата, при этом бактерия модифицирована таким образом, что экспрессия оперона NCgl_2067-NCgl_2065 ослаблена.
Термин «бактерия-продуцент L-аминокислоты» может означать бактерию, обладающую способностью к продукции и секреции L-аминокислоты в питательную среду, когда бактерия выращивается в указанной питательной среде.
Термин «бактерия-продуцент L-аминокислоты» также может означать бактерию, которая способна к продукции и накоплению L-аминокислоты, принадлежащей к семейству глутамата, в ферментационной среде в больших количествах по сравнению с природным или родительским штаммом Е.coli, таким, как штамм Е.coli K-12, и также может означать, что бактерия способна накапливать в среде целевую L-аминокислоту в количестве, не менее чем 0.5 г/л или не менее чем 1.0 г/л в другом примере.
Термин «L-аминокислота, принадлежащая к семейству глутамата» может означать L-глутаминовую кислоту, L-глутамин, L-пролин, L-гистидин, L-аргинин, L-орнитин и L-цитруллин.
Термин «коринеформная бактерия» может обозначать обычную коринеформную бактерию в соответствии с классификацией, известной специалисту в области микробиологии, и бактерии, классифицируемые как принадлежащие к роду Brevibacterium, однако в настоящее время классифицируемые как принадлежащие к роду Corynebacterium (Liebl W et al., Int J Syst Bacteriol. 1991, 41(2):255-260), а также может обозначать бактерии рода Brevibacterium, которые в высокой степени родственны бактериям рода Corynebacterium. В качестве примеров подобных коринеформных бактерий можно привести, но не ограничиться этим, виды:
Corynebacterium acetoacidophilum
Corynebacterium acetoglutamicum
Corynebacterium alkanolyticum
Corynebacterium callunae
Corynebacterium glutamicum
Corynebacterium lilium
Corynebacterium melassecola
Corynebacterium thermoaminogenes (Corynebacterium efficiens)
Corynebacterium herculis
Brevibacterium divaricatum
Brevibacterium flavum
Brevibacterium immariophilum
Brevibacterium lactofermentum (Corynebacterium glutamicum)
Brevibacterium roseum
Brevibacterium saccharolyticum
Brevibacterium thiogenitalis
Brevibacterium ammoniagenes
Brevibacterium album
Brevibacterium cerinum
Microbacterium ammoniaphilum и т.д.
В качестве примеров коринеформной бактерии можно привести, но не ограничиться этим, штаммы С.acetoacidophilum ATCC 13870, С.acetoglutamicum ATCC 15806, С.alkanolyticum ATCC 21511, С.callunae ATCC 15991, С.glutamicum АТСС13020, АТСС 13032, АТСС 13060, АТСС 13869, С.lilium ATCC 15990, С.melassecola АТСС 17965, С.thermoaminogenes AJ12340 (FERM BP-1539), С.herculis АТСС 13868, B.divaricatum АТСС 14020, В.flavum АТСС 13826, АТСС 14067, AJ12418 (FERM ВР-2205), В.immariophilum АТСС 14068, В.lactofermentum (С.glutamicum) АТСС 13869, В. roseum АТСС 13825 В. saccharolyticum АТСС 14066, & thiogenitalis АТСС 19240, A ammoniagenes АТСС 6871, АТСС 6872, 5. album АТСС 15111, 5. cerinum АТСС 15112, Microbacterium ammoniaphilum АТСС 15354 и т.д.
Эти штаммы доступны в Американской коллекции типовых культур (АКТК, American Type Culture Collection, АТСС) по адресу: P.O. Box 1549, Манассас, VA 20108, США. Каждому штамму присвоен индивидуальный регистрационный номер, который содержится в каталоге АКТК. Штаммы могут быть заказаны согласно регистрационному номеру. Штамм AJ12340 был депонирован 27 октября 1989 года согласно условиям Будапештского Договора под инвентарным номером FERM BP-1539 в Национальном институте биологических наук и человеческих технологий. Агентство промышленной науки и технологии. Министерство международной торговли и промышленности (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry), в настоящее время называющийся Международный депозитарий организмов для целей патентования. Национальный институт современной промышленной науки и технологии (International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology), и находящийся по адресу: Тсукуба Централ 6, 1-1, Хигаши 1-Чоме, Тсукуба-ши, Ибараки-кен, 305-8566, Япония (Tsukuba Central 6, 1-1, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken 305-5466, Japan). Штамм AJ12418 был депонирован 5 января 1989 года согласно условиям Будапештского Договора под инвентарным номером FERM ВР-2205 в Национальном институте биологических наук и человеческих технологий, Агентство промышленной науки и технологии, Министерство международной торговли и промышленности.
Термин «бактерия, принадлежащая к роду Corynebacterium» может означать, что бактерия относится к роду Corynebacterium в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера бактерии, принадлежащей к роду Corynebacterium, можно отнести, но не ограничиться этим, бактерию Corynebacterium glutamicum.
Термин «бактерия, принадлежащая к роду Brevibacterium» может означать, что бактерия относится к роду Brevibacterium в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера бактерии, принадлежащей к роду Brevibacterium, можно отнести, но не ограничиться этим, бактерию Brevibacterium flavum.
Термин «бактерия, модифицированная таким образом, что экспрессия одного или нескольких генов в опероне NCgl_2067-NCgl_2065 ослаблена» может означать, что бактерия модифицирована таким образом, что модифицированная бактерия содержит пониженное количество белка(ов) NCgl_2067, NCgl_2066 и NCgl_2065 по сравнению с немодифицированной бактерией, или что модифицированная бактерия не способна синтезировать белок(ки) NCgl_2067, NCgl_2066 и NCgl_2065. Термин «бактерия, модифицированная таким образом, что экспрессия одного или нескольких генов в опероне NCgl_2067-NCgl_2065 ослаблена» также может означать, что бактерия модифицирована таким образом, что модифицированный(е) ген(ы) кодирует(ют) мутантный(е) белок(ки) NCgl_2067, NCgl_2066 и NCgl_2065 с пониженной активностью. Так как, предположительно, гены NCgl_2067, NCgl_2066 и NCgl_2065 образуют оперон, ослабление экспрессии одного из генов может приводить к противоположному эффекту, то есть противоположно влиять на экспрессию других генов оперона NCgl_2067-NCgl_2065.
Наличие гена(ов) оперона NCgl_2067-NCgl_2065 в бактериальной хромосоме может быть определено при помощи известных методов, например, метода полимеразной цепной реакции (ПЦР), блоттинга по Саузерну и подобных. Уровень экспрессии гена(ов) может быть определен путем измерения уровня транскрибированной мРНК при помощи многих известных методов, например, метода Нозерн-блоттинга, количественной ОТ-ПЦР и подобных. Количество и молекулярную массу белков, кодируемых генами оперона NCgl_2067-NCgl_2065, можно определить известными методами, включая электрофорез в SDS-ПААГ с последующим иммуноблоттингом (Вестерн-блоттинг) и подобными.
Термин «инактивация одного или нескольких генов в опероне NCgl_2067-NCgl_2065» может означать, что модифицированный(е) ген(ы) кодирует(ют) полностью неактивный(е) белки(ок). Возможно также, что естественная экспрессия модифицированного участка ДНК невозможна из-за делеции части гена, сдвига рамки считывания, введения миссенс/нонсенс мутации(-ий) или модификации примыкающих к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промоторы, энхансеры, аттенюаторы и т.д.
Ген NCgl_2067 (синонимы - CG2358, Cgl2147) кодирует гипотетический белок с неизвестной функцией. Ген NCgl_2067 из С.glutamicum (нуклеотиды, комплементарные нуклеотидам в положении с 2,270,870 по 2,270,256, инвентарный номер в GenBank NC 003450; gi: 58036263) расположен между генами NCgl_2066 и HeS на хромосоме штамма С.glutamicum ATCC 13032. Нуклеотидная последовательность гена NCgl_2067 и аминокислотная последовательность белка NCgl_2067, кодируемого геном NCgl_2067, приведены в SEQ ID NO:1 и SEQ ID NO:2 соответственно.
Ген NCgl_2066 (синонимы - CG2357, Cgl2146) кодирует гипотетический белок с неизвестной функцией. Ген NCgl_2066 из С.glutamicum (нуклеотиды, комплементарные нуклеотидам в положении с 2,270,262 по 2,269,258, инвентарный номер в GenBank NC_003450; gi: 58036263) расположен между генами NCgl_2067 и NCgl_2065 на хромосоме штамма С.glutamicum ATCC 13032, при этом оба гена ориентированы в том же направлении, что и ген NCgl_2066, и, вероятно, образуют оперон. Нуклеотидная последовательность гена NCgl_2066 и аминокислотная последовательность белка NCgl_2066, кодируемого геном NCgl_2066, приведены в SEQ ID NO:3 и SEQ ID NO:4 соответственно.
Ген NCgl_2065 (синонимы - CG2356, Cgl2145) кодирует гипотетический белок с неизвестной функцией. Ген NCgl_2065 из С.glutamicum (нуклеотиды, комплементарные нуклеотидам в положении с 2,269,246 по 2,268,386, инвентарный номер в GenBank NC_003450; gi: 58036263) расположен между генами NCgl_2066 и NCgl_2064 на хромосоме штамма С.glutamicum ATCC 13032. Нуклеотидная последовательность гена NCgl_2065 и аминокислотная последовательность белка NCgl_2065, кодируемого геном NCgl_2065, приведены в SEQ ID NO:5 и SEQ ID NO:6 соответственно.
Ввиду того, что могут быть некоторые различия в последовательностях ДНК между родами и штаммами бактерий семейства Enterobacteriaceae, нуклеотидные последовательности инактивируемых генов оперона NCgl_2067-NCgl_2065 не ограничиваются нуклеотидными последовательностями генов, приведенных в SEQ ID NO:1, SEQ ID NO:3 и SEQ ID NO:5, однако могут включать гены, гомологичные нуклеотидным последовательностям SEQ ID NO:I, SEQ ID NO:3 и SEQ ID NO:5, кодирующие варианты белков NCgl_2067, NCgl_2066 и NCgl_2065.
Термин «вариант белка», используемый в настоящем изобретении, может означать белок, который содержит изменения в последовательности, которыми могут быть делении (удаления), вставки, добавления или замены аминокислотных остатков, при условии, что активность такого белка такая же, как и у белков NCgl_2067, NCgl_2066 и NCgl_2065. Количество изменений в вариантах белков зависит от вида или положения аминокислотного остатка в третичной структуре белка. Оно может быть от 1 до 30, в другом примере от 1 до 15 и еще в другом примере от 1 до 5 в SEQ ID NO:2, SEQ ID NO:4 и SEQ ID NO:6. В вариантах белков подобные изменения могут быть в областях, не критичных для функции белка, так как некоторые аминокислоты в высокой степени гомологичны другим, и поэтому третичная структура или активность белка не затрагивается ввиду этих изменений.
Следовательно, варианты белков, кодируемые генами оперона NCgl_2067-NCgl_2065, могут иметь гомологию, не менее чем 80%, или в другом примере не менее чем 90%, или в другом примере не менее чем 95% по отношению к полной аминокислотной последовательности, приведенной в SEQ ID NO:2, SEQ ID NO:4 и SEQ ID NO:6, при условии, что активность или функция соответствующего белка NCgl_2067, NCgl_2066 и NCgl_2065 сохранена до инактивации генов оперона NCgl_2067-NCgl_2065.
Гомология между двумя аминокислотными последовательностями может быть определена с использованием известных методов, например, компьютерной программы BLAST 2.0, которая считает три параметра: число аминокислотных остатков, идентичность и сходство.
Кроме того, гены оперона NCgl_2067-NCgl_2065 могут быть вариантами, если они гибридизуются в жестких условиях с нуклеотидными последовательностями, комплементарными нуклеотидным последовательностям, приведенным в Перечне последовательностей под номерами SEQ ID NO:1, SEQ ID NO:3 и SEQ ID NO:5, или зондом который может быть синтезирован на основе нуклеотидной последовательности при условии, что до инактивации она кодирует функциональный белок NCgl_2067, NCgl_2066 или NCgl_2065.
Под «жесткими условиями» понимаются такие условия, в которых образуются специфические гибриды, например, гибриды, имеющие гомологию, не менее чем 60%, или в другом примере не менее чем 70%, или в другом примере не менее чем 80%, или в другом примере не менее чем 90%, или в другом примере не менее чем 95%, и в которых не образуются неспецифические гибриды, например, гибриды, имеющие гомологию меньшую, чем указанно выше. Практическим примером жестких условий является однократная отмывка, предпочтительно двух- или трехкратная, при концентрации солей 1×SSC, 0.1% SDS, или в другом примере 0.1×SSC, 0.1% SDS, при 60°С. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для нейлоновой мембраны Hybond™ N+ (Amersham) при строгих условиях составляет 15 минут. Предпочтительна двух - трехкратная отмывка. Длина зонда может быть выбрана в зависимости от условий гибридизации, в данном конкретном случае она может быть около 100-1000 п.н.
Экспрессия генов оперона NCgl_2067-NCgl_2065 может быть ослаблена путем введения в ген хромосомы такой мутации, что внутриклеточная активность кодируемого геном белка снижается по сравнению с немодифицированным штаммом. Мутации, результатом которых является ослабление экспрессии гена, включают замену одного или более оснований для аминокислотной замены в кодируемом геном белке («миссенс»-мутация), введение стоп-кодона («нонсенс»-мутация), делецию одного или более оснований для сдвига рамки считывания, вставку гена устойчивости к антибиотику или делецию гена или его части (Qiu, Z et al., J. Biol. Chem. 1997, 272:8611-8617; Kwon, DH et al, J. Antimicrob. Chemother. 2000, 46:793-796). Экспрессия генов оперона NCgl_2067-NCgl_2065 также может быть ослаблена путем модификации экспрессии регуляторных последовательностей, таких как промотор, последовательность Шайна-Дальгарно (Shine-Dalgarno) и т.д. (заявка РСТ WO95/34672; Carrier, ТА et al., Biotechnol. Prog. 1999, 15:58-64).
Например, для введения мутаций путем генной рекомбинации могут применяться следующие методы. Конструируется мутантный ген, кодирующий мутантный белок со сниженной активностью, и бактерия для ее модификации трансформируется фрагментом ДНК, содержащим мутантный ген. Затем нативный ген на хромосоме замещается путем гомологичной рекомбинации мутантным геном, и полученный штамм отбирается. Замещение гена с использованием гомологичной рекомбинации может быть проведено с использованием линейной ДНК - методом, известным как "Red-зависимая интеграция" или "интеграция посредством Red-системы" (Datsenko, K.A., Wanner, B.L., Proc.Natl.Acad.Sci.USA 2000, 97(12):6640-6645), - или с использованием плазмиды, репликация которой чувствительна к температуре (патент США 6,303,383 или патентная заявка Японии JP 05-007491A). Кроме того, введение сайт-специфической мутации путем замещения гена с использованием вышеупомянутой гомологичной рекомбинации может также быть осуществлено с использованием плазмиды с пониженной способностью к репликации в клетке хозяина.
Экспрессия гена также может быть ослаблена вставкой транспозона или IS-фактора в кодирующую область гена (патент США 5,175,107) или традиционными методами, такими как мутагенез с использованием УФ-излучения или обработка нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин). Инактивация гена также может быть осуществлена такими методами, как сайт-направленный мутагенез, разрушение гена с использованием гомологичной рекомбинации или/и мутагенез за счет вставки-делеции (Yu, D. et al., Proc. Natl. Acad. Sci. USA 2000, 97(12): 5978-5983 и Datsenko, K.A. and Wanner, B.L., Proc. Natl. Acad. Sci. USA 2000, 97(12): 6640-6645), также называемого "Red-зависимая интеграция".
Методы приготовления плазмидной ДНК, рестрикции и лигирования ДНК, трансформации, выбора нуклеотидов в качестве праймера и т.п. могут быть обычными методами, известными специалисту в этой области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия-продуцент L-аминокислоты
В соответствии с настоящим описанием изобретения в качестве бактерии может быть использована бактерия, модифицированная таким образом, что экспрессия генов оперона NCgl_2067-NCgl_2065 в указанной бактерии ослаблена, и способная к продукции L-аминокислоты семейства глутамата.
Указанная бактерия в соответствии с настоящим описанием изобретения может быть получена путем ослабления экспрессии одного или нескольких генов оперона NCgl_2067-NCgl_2065 в бактерии, уже обладающей способностью к продукции L-аминокислоты. С другой стороны, указанная бактерия может быть получена путем придания бактерии, в которой экспрессия одного или нескольких генов оперона NCgl_2067-NCgl_2065 уже ослаблена, способности продуцировать L-аминокислоту.
Бактерия-продуцент L-глутаминовой кислоты
Примеры родительских штаммов для получения бактерии-продуцента L-глутаминовой кислоты согласно настоящему изобретению могут включать, но не ограничиваться этим, штаммы коринеформной бактерии, такие как С.glutamicum ATCC 13032, ATCC 31833, С.acetoacidophilum ATCC 13870, С.herculis ATCC 13868, С.lilium ATCC 15990, В.divaricatum ATCC 14020, В.flavum ATCC 14067, В.imaliophilum ATCC 14068, В.lactofermentum ATCC 13869, В.thiogenitalis ATCC 19240 и т.д.
Примеры родительских штаммов, которые могут быть использованы для получения бактерий-продуцентов L-глутаминовой кислоты согласно настоящему изобретению, также включают штаммы, в которых усилена экспрессия гена(ов), кодирующего(их) фермент(ы), катализирующий(е) реакцию(и) пути биосинтеза L-глутаминовой кислоты. В качестве примеров подобных генов можно привести, но не ограничиться ими, гены, кодирующие глутаматдегидрогеназу, глутаминсинтазау, глутаматсинтетазу, изоцитратдегидрогеназу, аконитатгидратазу, цитратсинтазу, фосфоенолпируваткарбоксилазу, пируваткарбоксилазу, пируватдегидрогеназу, пируваткиназу, фосфоенолпируватсинтазу, энолазу, фосфоглицеромутазу, фосфоглицераткиназу, глицеральдегид-3-фосфатдегидрогеназу, триозофосфатизомеразу, фруктозобисфосфатальдолазу, фосфофруктокиназу и клюкозофосфатизомеразу.
Усиление экспрессии гена(ов) может быть достигнуто за счет увеличения числа копий гена(ов) путем введения гена(ов) в вектор, способный функционировать в коринеформной бактерии.
Усиление экспрессии гена(ов) также может быть достигнуто путем введения нескольких копий гена в хромосомную ДНК бактерии, например, методом гомологичной рекомбинации, Mu интеграции и т.п. В частности, увеличение числа копий гена в бактериальной ДНК методом гомологичной рекомбинации может быть достигнуто при использовании в качестве целевых последовательностей такие последовательности, которые присутствуют в хромосоме во множестве копий. Экспрессия гена(ов) также может быть усилена путем подстановки фрагмента ДНК под контроль сильного промотора.
С другой стороны, действие промотора может быть усилено, например, введением в промотор мутации, ведущей к увеличению уровня транскрипции локализованного за промотором гена. Кроме того, известно, что замена нескольких нуклеотидов в области между сайтом связывания рибосомы (ribosome binding site - RBS) и стартовым кодоном, особенно в последовательности непосредственно перед стартовым кодоном, существенно влияет на трансляционную способность мРНК. Например, обнаружен 20-кратный разброс в уровне экспрессии в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold et al., Annu. Rev. Microbiol, 1981, 35:365-403; Hui et al., EMBO J., 1984, 3:623-629).
Кроме того, возможно введение нуклеотидной замены в область промотора гена на бактериальной хромосоме, результатом чего является усиление функции промотора. Изменение последовательности, контролирующей экспрессию, можно осуществить, например, таким же способом, что и замена гена с использованием чувствительной к температуре плазмиды, как раскрыто в международной заявке WO 00/18935 и японской заявке JP 1-215280 А.
Примеры микроорганизмов, модифицированных таким образом, что экспрессия гена цитратсинтазы, гена фосфоенолпируваткарбоксилазы и/или гена глутаматдегидрогеназы усилена, могут включать микроорганизмы, описанные в патентах JP2001-333769A (ЕР1078989А), JP2000-106869A (ЕР955368А), JP2000-189169А (ЕР952221А) и JP2001-333769A (ЕР1078989А). В качестве примеров методов, направленных на придание коринеформным бактериям способность продуцировать L-глутаминовую кислоту, можно привести усиление экспрессии гена fasR и повышение чувствительности к поверхностно-активным веществам (WO2007024010 А1). Примером модифицированной бактерии-продуцента L-глутаминовой кислоты может служить коринеформная бактерия с усиленной экспрессией гена yggB (JP2007-097573 А).
Примеры родительских штаммов, которые могут использоваться для получения бактерий, продуцирующих L-глутаминовую кислоту, также включают штаммы, в которых снижена активность гена(ов), кодирующего(их) фермент(ы), катализирующий(е) реакцию(и) синтеза отличных от L-глутаминовой кислоты соединений, ответвляющихся от основного пути биосинтеза L-глутаминовой кислоты. Примеры ферментов, кодируемые подобными генами, включают изоцитратлиазу, α-кетоглутаратдегидрогеназу, фосфотрансацетилазу, ацетаткиназу, ацетогидроксатсинтазу, ацетолактатсинтазу, формилацетилтрансферазу, лактатдегидрогеназу и глутаматдекарбоксилазу. Для ослабления экспрессии генов, кодирующих подобные белки, могут применяться описанные выше методы.
Примерами коринеформной бактерии-продуцента L-глутаминовой кислоты могут быть штаммы, к которых понижена активность α-кетоглутаратдегидрогеназы (JP7-834672А и JP06-237779), неактивен ген gluX (US 2010279363 А1), ослаблена экспрессия генов sucC и/или sucD (EP1103611 А1) и т.д.
Специфические примеры штаммов, обладающих способностью к продукции L-глутаминовой кислоты, также включают штаммы, устойчивые к бензопирону или нафтохинону (JP56-1889A), монофторуксусной кислоте (JP 50-113209A), аденину и тимину (JP57-065198), α-кетомалоновой кислоте (JP57-2689A), гуанидину (JP56-35981А), дауномицину (JP58-158192A), а также штаммы, устойчивые к пенициллину (JP04-88994A).
Специфические примеры штаммов, обладающих способностью к продукции L-глутаминовой кислоты, также включают следующие штаммы: В.flavum AB949 (PERM BP-2632) (JP 50-113209A), AJ11355 (FERM P-5007) (JP56-1889A), AJ11217 (PERM P-4318) (JP57-2689A), AJ11564 (PERM P-5472) (JP56-140895A), AJ11439 (PERM P-5136) (JP56-35981A), В.lactofermentum AJ11426 (PERM P5123) (JP56-048890A), AJ11796 (PERM P6402) (JP58-158192A), С.glutamicum AJ11628 (PERM P-5736) (JP 57-065198A), AJ11440 (PERM P5137) (JP56-048890A), H7684 (PERM BP-3004; JP04-88994A), AJ11355 (PERM P-5020) (JP56-1889A), AJ11218 (PERM P-4319) (JP57-2689A).
Бактерия-продуцент L-глутамина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-глутамина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как В.flavum AJ11573 (PERM P-5492) (JP56-161495A) AJ11576 (PERM BP-10381) (JP56-161495A) AJ12212 (PERM P-8123) (JP61-202694A), AJ12418 (PERM BP-2205) (JP02-186994A), DH18 (PERM P-11116) (JP03-232497A), С.glutamicum AJ11574 (PERM P-5493) (JP56-151495А), С.melassecola DH344 (PERM P-11117) (JP3-232497A).
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-глутамина, также включают в себя, но не ограничиваются ими, штаммы, в которых усилена экспрессия одного или нескольких генов, кодирующих ферменты биосинтеза L-глутамина. Примеры таких ферментов включают глутаминсинтетазу и глутаматдегидрогеназу (US7262035 В2, ЕР 1229121 Bl) и т.п.
Примеры других родительских штаммов, которые могут использоваться для получения бактерий, продуцирующих L-глутамин, также включают штаммы, в которых снижена экспрессия гена(ов), кодирующего(их) фермент(ы), катализирующий(е) реакцию(и) образования отличных от L-глутамина соединений, ответвляющихся от основного пути биосинтеза L-глутамина. Для ослабления экспрессии генов, кодирующих подобные белки, могут применяться описанные выше методы. В качестве родительских штаммов-продуцентов L-глутамина может использоваться штамм с пониженной активностью глутаминазы (ЕР 1424397 B1) или штамм, в котором оперон gluABCD системы транспорта глутамина подвержен пониженной регуляции или удален (WO2008026698 А1), и т.п.
Специфические примеры штаммов, обладающих способностью к продукции L-глутамина, также включают штаммы, устойчивые к аналогам L-аминокислот: штаммы, устойчивые к 6-диазо-5-оксонорлейцин (JP-A-3-232497), пуриновым аналогам и/или метионинсульфоксиду (JP-A-61-202694), α-кетомалоновой кислоте (JP-A-56-151495) и т.д.
Бактерия-продуцент L-гистидина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-гистидина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как С.glutamicum FERM ВР-485 и FERM ВР-486 (Патент США No. 4,495,283), С.acetoacidophilum AJ 1562 и Microbacterium flavum (С.flavescens) ATCC 10340 (JP 47002549 В), С.glutamicum AJ 12092 (FERM P-7273) и AJ 12426 (FERM BP-2213), B.flavum AJ 3420 (FERM P-2316) и В.flavum AJ 12425 (FERM BP-2212) (Патент США No. 5,294,547), С.glutamicum (В.flavum) ATCC 21604 (VKPM B-1080) (Патент США No. 3,713,977), С.glutamicum ("Micrococcus glutamicus") ATCC 13761 (NCIB 10334, VKPM B-4382) (Патент США No. 3,220,929), С.glutamicum ATCC 21339 (VKPM B-1003) (Патент США No. 3,676,301).
Специфические примеры штаммов, обладающих способностью к продукции L-гистидина, также включают штаммы, принадлежащие к коринеформной бактерии, такие как В.flavum FERM-P 2317, В.lactofermentum FERM-P 1565 (AJ 3386), С.glutamicum ATCC 14297, В.lactofermentum ATCC 21086, С.acetoacidophilum ATCC 21407, В.flavum ATCC 21406 (AJ 3225) и т.д.
Специфические примеры штаммов, обладающих способностью к продукции L-гистидина, также включают штаммы, принадлежащие к роду Brevibacterium, такие как В.flavum AJ11846, устойчивый к антагонистам тиамина (2-тиазолаланину, триазолкарбоксамиду и кобаламину) (JP 02018838 В), и В.falvum AJ 3579, требующий треонин, пролин, шикимовую кислоту, ксантин или гуанин и устойчивый к 2-тиазолаланину (JP 51024594).
Бактерия-продуцент L-пролина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-пролина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как В.lactofermentum AJ 11225 (FERM Р-4370) (японская Патентная заявка No. 60-87788), В.flavum AJ11512 (FERM Р-5332), AJ11513 (FERM P-5333), AJ11514 (FERM P-5334), С.glutamicum AJ11522 (FERM P-5342), AJ11523 (FERM P-5343) (японская Патентная заявка No. 62-36679) и т.д.
Примеры родительских штаммов, которые могут использоваться для получения бактерий, продуцирующих L-пролин, также включают штаммы, в которых повышена экспрессия гена(ов), кодирующего(их) фермент(ы), катализирующий(е) реакцию(и) пути биосинтеза L-аргинина. Примеры ферментов пути биосинтеза L-пролина включают глутаматкиназу, γ-глутамилфосфатредуктазу и пирролин-5-карбоксилатредуктазу (Ankri S. et al., J Bacteriol. 1996, 178(15):4412-4419). Также могут использоваться бактерии-продуценты L-пролина, в которых снижена активность ферментов, катализирующих реакции образования отличных от L-пролина соединений, ответвляющихся от основного пути биосинтеза L-пролина. Способность бактерий продуцировать L-пролин может быть сообщена путем уменьшения орнитинаминотрансферазной активности (Ankri S. et al., J Bacteriol. 1996, 178(15):4412-4419).
Бактерия-продуцент L-аргинина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-аргинина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как С.glutamicum AJ12092 и В.flavum AJ11169 (ЕР 378223 B1, JP 2817155 В2, US 5284757 А), В.flavum AJ12144 (FERM Р-7642) и С.glutamicum AJ12145 (PERM P-7643) (японская Патентная заявка No. 5-27388), В.flavum ATCC 21493 и С.glutamicum ATCC 21659 (японская Патентная заявка No. 5-3793), В.flavum FERM-P 4948, С.glutamicum FERM-P 7274 (AJ12093), В.flavum NRRL 12235 (AJ11337), С.acetoglutamicum NRRL 12239 (AJ11342).
Известно, что способность штаммов продуцировать L-аргинин может быть существенно повышена путем использования мутантной N-ацетилглутаматкиназы, устойчивой к обратному ингибированию L-аргинином (WO 2006035831 A1).
Специфические примеры штаммов, обладающих способностью к продукции L-аргинина, включают штаммы, принадлежащие к роду Brevibacterium или к роду Corynebacterium, устойчивые аргинингидроксамату, антагонистам аргинина, аргининолу, сульфамидам, цистеину или их аналогам (ЕР 336387 B1, JP 2578468 В2, US 5034319 А), гуанидину, содержащему алифатические заместители и их производные (JP 2817155 В2, ЕР 0378223 B1), и т.д.
Способность коринеформных бактерий продуцировать L-аргинин может быть повышена путем придания устойчивости к серосодержащим веществам, аргининолу, 8-азагуанину или α-амино-β-гидроксивалериановой кислоте и т.д.
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-аргинина, также включают штаммы, ауксотрофные по таким аминокислотам, как L-гистидин, L-пролин, L-треонин, L-триптофан, L-лизин и т.д.
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-аргинина, также включают штаммы, ауксотрофные по цитруллину или аргинину (японская Патентная заявка No. 43-8712), варианты ауксотрофов по аргинину (японская Патентная заявка No. 53-24096), варианты рода Corynebacterium, имеющие устойчивость к 2-тиазолаланину, сульфагуанидину или 2-фторопировиноградной кислоте (японская Патентная заявка No. 61-119194) и т.д.
Примеры родительских штаммов, которые могут использоваться для получения бактерий, продуцирующих L-аргинин, также включают штаммы, в которых повышена экспрессия гена(ов), кодирующего(их) фермент(ы), катализирующий(е) реакцию(и) пути биосинтеза L-аргинина. Примеры ферментов, которые катализируют реакции пути биосинтеза L-аргинина, включают N-ацетилглутамилфосфатредуктазу (argC), орнитинацетилтрансферазу (argJ), N-ацтилглутаматкиназу (argB), ацетилорнитинтрансаминазу (argD), орнитинкарбамоилтрансферазу (argF), аргининосукцинатсинтетазу (argG), аргининосукцинатлиазу (argH) и карбамоилфосфатсинтетазу (carAB).
Способность бактерии продуцировать L-аргинин может быть повышена путем инактивации аргининового репрессора, кодируемого геном argR. Способы инактивации гена argR описаны выше.
Бактерия-продуцент L-орнитина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-орнитина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как В.lactofermentum PERM BP-2344, FERM-P 5936 (AJ11678) и С.glutamicum FERM BP-2345 (JP 2817185 (В2), ЕР 393708 В1), С.glutamicum FERM-P 5644 (AJ11589), АТСС 13232 и т.п.
Так как L-орнитин является промежуточным соединением пути биосинтеза L-аргинина, родительские штаммы, которые могут быть использованы для получения бактерии-продуцента L-орнитина, могут быть получены из любой бактерии-продуцента L-аргинина, например, путем инактивации орнитинкарбамоилтрансферазы, кодируемой как геном argF, так и геном argI. Способы инактивации орнитинкарбамоилтрансферазы описаны выше.
Бактерия-продуцент L-цитруллина
Примеры родительских штаммов, которые могут быть использованы для получения бактерии-продуцента L-цитруллина, включают в себя, но не ограничиваются ими, штаммы, принадлежащие к коринеформной бактерии, такие как С.glutamicum FERM-P 5643 (AJ11588), B.flavum FERM-P 1645 (AJ 3408) и т.д.
Так как L-цитруллин является промежуточным соединением пути биосинтеза L-аргинина, родительские штаммы, которые могут быть использованы для получения бактерии-продуцента L-цитруллина, могут быть получены из любой бактерии-продуцента L-аргинина, например, путем инактивации аргининосукцинатсинтазы, кодируемой как геном argG. Способы инактивации аргининосукцинатсинтазы описаны выше.
2. Способ
Способом согласно настоящему изобретению является способ получения L-аминокислоты, включающий стадии выращивания бактерии в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде и выделения L-аминокислоты из культуральной жидкости.
Выращивание, выделение и очистка L-аминокислоты от культуральной или подобной ей жидкости могут быть осуществлены способами, подобными традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза или сахароза и другие органические кислоты. В зависимости от типа ассимиляции используемого микроорганизма может также использоваться спирт, включая этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Выращивание может осуществляться в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией при температуре в пределах от 20 до 40°С, или в пределах от 30 до 38°С в другом примере. Кислотность среды (рН) поддерживают в пределах от 5 до 9 или от 6.5 до 7.2 в другом примере. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно, выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной среде.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и/или кристаллизации.
Примеры осуществления настоящего изобретения
Пример 1: Конструирование штамма с пониженной экспрессией гена
NCgl_2067
1-1. Удаление гена NCgl_2067
Штамм с удаленным геном NCgl_2067 был сконструирован с помощью нерепликативной плазмиды pSB4S (Фиг.3; конструирование плазмиды описано в патенте США No. 7,794,989). Была выполнена внутрирамочная делеция. Для этого была проведена ПЦР, позволяющая получать ДНК-фрагменты с перекрывающимися участками. Сначала был получен ДНК-фрагмент, используя олигонуклеотидные праймеры NC2067F (SEQ ID NO:7) и NC2067R1 (SEQ ID NO:8) и хромосомальную DNA В.flavum В-6642 в качестве матрицы, как описано в патенте РФ 2084520 С2). Затем методом ПЦР был получен ДНК-фрагмент, используя олигонуклеотидные праймеры NC2067F1 (SEQ ID NO:9) и NC2067R (SEQ ID NO:10) и хромосомальную DNA В.flavum В-6642 в качестве матрицы. После этого оба полученных ДНК-фрагмента использовали как матрицу в первом цикле ПЦР с олигонуклеотидными праймерами NC2067-SmF (SEQ ID NO:11) и NC2067-SmR (SEQ ID NO:12). Температурные условия были следующие: начальная денатурация ДНК в течение 30 сек при 94°С; затем 25 циклов: денатурация при 94°С в течение 30 сек, отжиг при 58°С в течение 30 сек и элонгация при 72°С в течение 60 сек; конечная элонгация в течение 2 мин при 72°С. Полученный ПЦР-продукт очищали, обрабатывали эндонуклеазой SmaI и лидировали в обработанную той же эндонуклеазой плазмиду pBS4S, получив, таким образом, плазмиду pBS4SΔ2067.
Электрокомпетентные клетки готовили следующим образом: штамм В.flavum В-6642 выращивали в течение ночи при 30°С на среде CM2GxYE, содержащей пептон (10 г/л), дрожжевой экстракт (20 г/л), NaCl (5 г/л) и глюкозу (5 г/л), рН 7.0. Полученную культуру разводили в 20 раз в свежей среде CM2GxYE. Клетки выращивали с аэрацией при 30°С до оптической плотности бактериальной культуры OD600≈0.6, затем добавляли ампициллин (10 мг/л) и выращивали клетки еще 1 час. Полученные клетки промывали три раза ледяной деионизированной водой. Для электропорации использовали 50 мкл клеток и «5 мкг ПЦР-продукта pBS4SΔ2067, применяя следующие условия: напряжение 2 kV, расстояние между электродами 0,1 см. Чтобы отобрать KmR - рекомбинанты, клетки после электропорации инкубировали в 1 мл CM2GxYE-среды в течение 2 часов при 30°С при встряхивании, затем пересевали на чашки с 2LA-агаром, содержащим пептон (10 г/л), дрожжевой экстракт (10 г/л), NaCl (5 г/л), агар (20 г/л) и канамицин (25 мкг/мл), рН 7.0, и выращивали при 30°С в течение 48 ч. Полученные рекомбинанты были неспособны расти на среде 2LA, содержащей 10% сахарозу.
1-2. Проверка удаления гена NCgl_2067 методом ПЦР
Удаление гена NCgl_2067 в мутантах проводили методом ПЦР. Использовали локус-специфичные олигонуклеотидные праймеры P1 (SEQ ID NO:13) и Р2 (SEQ ID NO:14). ПЦР-продукт, полученный при использовании в качестве матрицы ДНК родительского штамма В.flavum B-6642, имел длину ~1.35 т.п.н., тогда как ПЦР-продукт, полученный при использовании в качестве матрицы ДНК мутантного штамма, имел длину ~0.78 т.п.н. Удаление гена NCgl_2067 также было подтверждено методом секвенирования соответствующего фрагмента хромосомной ДНК. Полученный мутантный штамм был назван В.flavum B-6642ΔNCgl_2067.
Пример 2: Конструирование штамма с пониженной экспрессией гена
NCgl_2066
1-3. Удаление гена NCgl_2066
Штамм с удаленным геном NCgl_2066 был сконструирован, как описано в Примере 1. Была выполнена внутрирамочная делеция. Для этого была проведена ПЦР, позволяющая получать ДНК-фрагменты с перекрывающимися участками. Сначала был получен ДНК-фрагмент, используя олигонуклеотидные праймеры NC2066F (SEQ ID NO:15) и NC2066R1 (SEQ ID NO:16) и хромосомальную DNA В.flavum B-6642 в качестве матрицы, как описано патенте РФ 2084520 С2). Затем, методом ПЦР был получен ДНК-фрагмент, используя олигонуклеотидные праймеры NC2066F1 (SEQ ID NO:17) и NC2066R (SEQ ID NO:18) и хромосомальную DNA В.flavum B-6642 в качестве матрицы. После этого оба полученных ДНК-фрагмента использовали как матрицу в первом цикле ПЦР с олигонуклеотидными праймерами NC2066-SmF (SEQ ID NO:19) и NC2066-SmR (SEQ ID NO:20). Температурные условия были, как описано в Примере 1. Полученный ПЦР-продукт очищали, обрабатывали эндонуклеазой SmaI и лидировали в обработанную той же эндонуклеазой плазмиду pBS4S, получив, таким образом, плазмиду pBS4SΔ2066.
Электрокомпетентные клетки готовили следующим образом: штамм В.flavum В-6642 выращивали в течение ночи при 30°С на среде CM2GxYE, содержащей пептон (10 г/л), дрожжевой экстракт (20 г/л), NaCl (5 г/л) и глюкозу (5 г/л), рН 7.0. Полученную культуру разводили в 20 раз в свежей среде CM2GxYE. Клетки выращивали с аэрацией при 30°С до оптической плотности бактериальной культуры OD600≈0.6, затем добавляли ампициллин (10 мг/л) и выращивали клетки еще 1 час. Полученные клетки промывали три раза ледяной деионизированной водой. Для электропорации использовали 50 мкл клеток и ≈5 мкг ПЦР-продукта pBS4SΔ2066, применяя следующие условия: напряжение 2 kV, расстояние между электродами 0,1 см. Чтобы отобрать KmR - рекомбинанты, клетки после электропорации инкубировали в 1 мл CM2GxYE-среды в течение 2 часов при 30°С при встряхивании, затем пересевали на чашки с 2bA-агаром, содержащим пептон (10 г/л), дрожжевой экстракт (10 г/л), NaCl (5 г/л), агар (20 г/л) и канамицин (25 мкг/мл), рН 7.0, и выращивали при 30°С в течение 48 ч. Полученные рекомбинанты были неспособны расти на среде 2LA, содержащей 10% сахарозу.
1-4. Проверка удаления гена NCgl_2066 методом ПЦР
Удаление гена NCgl_2066 в мутантах проводили методом ПЦР. Использовали локус-специфичные олигонуклеотидные праймеры NC2066F (SEQ ID NO:15) и NC2066R (SEQ ID NO:18). ПЦР-продукт, полученный при использовании в качестве матрицы ДНК родительского штамма В.flavum B-6642, имел длину ~2.12 т.п.н., тогда как ПЦР-продукт, полученный при использовании в качестве матрицы ДНК мутантного штамма, имел длину ~1.15 т.п.н. Удаление гена NCgl_2066 также было подтверждено методом секвенирования соответствующего фрагмента хромосомной ДНК. Полученный мутантный штамм был назван В.flavum B-6642ΔNCgl_2066.
Пример 3: Продукция L-глутаминовой кислоты и L-глутамина штаммом
В.flavum B-6642ΔNCgl_2067
Штаммы В.flavum B-6642ΔNCgl_2067 и В-6642 выращивали при 30°С в течение 18-24 ч на чашках с 2LA-агаром. Затем одна петля клеток каждой культуры была перенесена в 2 мл питательной среды в пробирках 20×200 мм. Питательная среда содержала: (NH4)2SO4 (70 г/л), KH2PO4 (2,5 г/л), тиамин (350 мг/л), биотип (100 мг/л), MgSO4 7H2O (0.4 г/л), мамено (mameno, гидролизат соевых бобов) (5.7 мг/л), сахароза (120 г/л) и СаСО3 (50 г/л), рН 7. Клетки выращивали при 30°С в течение 48 ч на роторной качалке.
После выращивания количество L-глутаминовой кислоты и L-глутамина, накопленного в среде, определяли методом тонкослойной хроматографии (ТСХ) на пластине с силикагелем с использованием следующей подвижной фазы: бутанол: уксусная кислота: вода=4:1:1 (об.). Для визуализации использовали 1% раствор нингидрина в ацетоне. Пятна, содержащие L-глутаминовую кислоту и L-глутамин, вырезали и L-глутаминовую кислоту и L-глутамин элюировали 0.5% водным раствором CdCl2. Количества L-глутаминовой кислоты и L-глутамина определяли спектрофотометрически при 540 нм.
Результаты двух независимых пробирочных ферментации представлены в Таблице 1. Как видно из Таблицы 1, штамм B-6642ΔNCgl_2067 с ослабленной экспрессией гена NCgl_2067 продуцировал большее количество L-глутаминовой кислоты и L-глутамина по сравнению с родительским штаммом В-6642.
Пример 4: Продукция L-глутаминовой кислоты и L-глутамина штаммом
В.flavum B-6642ΔNCgl_2066
Штаммы В.flavum B-6642ΔNCgl_2066 и В-6642 выращивали при 30°С в течение 18-24 ч на чашках с 2LA-агаром. Затем одна петля клеток каждой культуры была перенесена в 2 мл питательной среды в пробирках 20×200 мм. Питательная среда содержала: (NH4)2SO4 (70 г/л), KH2PO4 (2,5 г/л), тиамин (350 мг/л), биотин (100 мг/л), MgSO4 7H2O (0.4 г/л), мамено (mameno, гидролизат соевых бобов) (5.7 мг/л), сахароза (120 г/л) и СаСО3 (50 г/л), рН 7. Клетки выращивали при 30°С в течение 48 ч на роторной качалке.
После выращивания количество L-глутаминовой кислоты и L-глутамина, накопленного в среде, определяли методом тонкослойной хроматографии (ТСХ) на пластине с силикагелем с использованием следующей подвижной фазы: бутанол: уксусная кислота: вода=4:1:1 (об.). Для визуализации использовали 1% раствор нингидрина в ацетоне. Пятна, содержащие L-глутаминовую кислоту и L-глутамин, вырезали и L-глутаминовую кислоту и L-глутамин элюировали 0.5% водным раствором CdCl2. Количества L-глутаминовой кислоты и L-глутамина определяли спектрофотометрически при 540 нм.
Результаты двух независимых пробирочных ферментации представлены в Таблице 2. Как видно из Таблицы 2, штамм B-6642ΔNCgl_2066 с ослабленной экспрессией гена NCgl_2066 продуцировал большее количество L-глутаминовой кислоты и L-глутамина по сравнению с родительским штаммом В-6642.
Хотя указанное изобретение описано в деталях со ссылкой на Наилучшие способы осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения. Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.














Изобретение относится к биотехнологии и представляет собой коринеформную бактерию-продуцент L-аминокислоты семейства глутамата. Причем указанная бактерия модифицирована таким образом, что один или несколько генов, выбранных из NCgl_2067 и NCgl_2066, оперона NCgl_2067-NCgl_2065 инактивирован(ы). Изобретение относится также к способу получения L-аминокислоты семейства глутамата с использованием указанной бактерии. Изобретение позволяет получать L-аминокислоты семейства глутамата с высокой степенью эффективности. 2 н. и 8 з.п. ф-лы, 3 ил., 2 табл., 4 пр.
1. Коринеформная бактерия-продуцент L-аминокислоты семейства глутамата, модифицированная таким образом, что один или несколько генов, выбранных из NCgl_2067 и NCgl_2066, оперона NCgl_2067-NCgl_2065 указанной бактерии инактивирован(ы).
2. Бактерия по п.1, отличающаяся тем, что указанная бактерия модифицирована таким образом, что ген NCgl_2067 инактивирован.
3. Бактерия по п.2, отличающаяся тем, что указанный ген NCgl_2067 инактивирован за счет делеции указанного гена NCgl_2067.
4. Бактерия по п.1, отличающаяся тем, что указанная бактерия модифицирована таким образом, что ген NCgl_2066 инактивирован.
5. Бактерия по п.4, отличающаяся тем, что указанный ген NCgl_2066 инактивирован за счет делеции указанного гена NCgl_2066.
6. Бактерия по п.1, отличающаяся тем, что указанная бактерия принадлежит к роду Corynebacterium.
7. Бактерия по п.1, отличающаяся тем, что указанная бактерия принадлежит к роду Brevibacterium.
8. Бактерия-продуцент L-аминокислоты по любому из пп.1-7, отличающаяся тем, что указанная L-аминокислота выбрана из группы, состоящей из L-глутаминовой кислоты, L-глутамина, L-пролина, L-аргинина, L-орнитина и L-цитруллина.
9. Способ получения L-аминокислоты, принадлежащей к семейству глутамата, включающий:
- выращивание бактерии по любому из пп.1-8 в питательной среде; и
- выделение указанной L-аминокислоты из культуральной жидкости.
10. Способ по п.9, отличающийся тем, что указанная L-аминокислота выбрана из группы, состоящей из L-глутаминовой кислоты, L-глутамина, L-пролина, L-аргинина, L-орнитина и L-цитруллина.
| RU 2009102012 А, 27.07.2010 | |||
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ АМИНОКИСЛОТ СЕМЕЙСТВА АСПАРТАТОВ И/ИЛИ ГЛУТАМАТОВ И ИСПОЛЬЗУЕМЫЕ В СПОСОБЕ СРЕДСТВА | 1998 |
|
RU2231552C2 |

