Предпосылки создания изобретения
Известно, что L-орнитин оказывает стимулирующее действие на функцию печени, и его часто применяют в качестве компонента в лекарственных средствах и продуктах питания для спортсменов.
В настоящее время L-орнитин получают с помощью различных методов. Одним из методов является получение посредством ферментации с использованием микроорганизмов. Другим методом является щелочной гидролиз аргинина, например, с применением гидроксида бария (CN 1594282 А). Еще одним методом является биотрансформация аргинина с помощью иммобилизованных микроорганизмов, обладающих аргиназной активностью (KR 589121 В1). В патентной литературе описан также метод получения L-орнитина из L-цитруллина (JP 42007767 В4).
В литературе описаны микроорганизмы, отличающиеся тем, что они высвобождают L-орнитин в культуральную среду. Примерами таких микроорганизмов могут служить бактерии p.Corynebacterium, Brevibacterium, Bacillus (JP 43010996 B4, JP 57041912 В), Escherichia (US 3668072 A), Providencia (JP 03195494) или Arthrobacter (US 3574061).
Продуцирующие L-орнитин микроорганизмы часто отличаются тем, что они являются ауксотрофами по аминокислотам L-аргинину или L-цитруллину (что описано для Brevibacterium, Bacillus, Corynebacterium в ЕР 392708 В1 и KR 161147 В1 и для Escherichia в US 366072 А). Кроме того, были описаны микроорганизмы, обладающие устойчивостью к 2-тиазол-аланину, сульфагуанидину или 2-фторпирувату (публикация выложенной заявки на японский патент №61-119194). В ЕР 0393708 В1 описаны продуценты L-орнитина, которые отличаются пониженной устойчивостью к орнитолу и микофеноловой кислоте. Указанные свойства могут присутствовать также в сочетании друг с другом.
Уровень высвобождения основных аминокислот, таких как L-лизин, L-аргинин и L-орнитин, посредством пассивной диффузии из клетки является очень низким (Bellmann и др., Microbiology, 147, 2001, сс.1765-1774). Это было подробно описано на примере лизина. Vrlijc с соавторами (Vrlijc и др., Journal of Bacteriology, 177(14), 1995, сс.4021-4027) изучили целый ряд мутантов Corynebacterium glutamicum, характеризующихся дефицитом экспорта. Для одного из мутантов было установлено, что внутриклеточная концентрация L-лизина составляла 174 мМ, в то время как его концентрация во внеклеточной среде составляла только 0,7 мМ.
В работах, опубликованных Vrlijc с соавторами (Molecular Microbiology, 22(5), 1996, сс.815-826 и Journal of Molecular Microbiology and Biotechnology, 1, 1999, cc. 327-336), и в ЕР 0868527 B1 представлены результаты идентификации и дано описание нового организма-экспортера, являющегося экспортером L-лизина (LysE). Выявленный нуль-мутант по гену LysE не обладал способностью транспортировать L-лизин из клетки. Полипептид, кодируемый геном lysE, состоит из 233 аминокислот или аминокислотных остатков и имеет последовательность, представленную в SEQ ID NO: 2. Было установлено, что в результате осуществления сверхэкспрессии гена lysE в продуценте лизина происходило увеличение высвобождения L-лизина.
Bellmann с соавторами (Microbiology, 147, 2001, сс.1765-1774) охарактеризовали более подробно экспортера LysE с точки зрения транспорта различных основных аминокислот в С.glutamicum. Авторы продемонстрировали, что транспортер обладает специфической способностью экспортировать аминокислоты L-лизин и L-аргинин из клетки. Кроме того, авторы изучили вопрос о том, может ли LysE экспортировать также и L-орнитин из клетки. Для этой цели, прежде всего, был создан ауксотрофный по L-аргинину штамм С.glutamicum, обозначенный как ATCC13032::argF.
Штамм культивировали в 50 мл (периодический процесс культивирования) минимальной среды, обозначенной как CGXII, которая содержала глюкозу в концентрации 40 г/л. После инкубации в течение 24 ч измерения показали, что концентрация L-орнитина составляла 60 мМ, что соответствовало 7,9 г/л. Внутриклеточная концентрация L-орнитина в клетках указанного штамма, измеренная после периода инкубации, составлявшего примерно 70 мин, составляла примерно 200 мМ. Для решения вопроса о том, обладает ли LysE способностью транспортировать также L-орнитин из клетки, осуществляли трансформацию штамма 13032::argF с помощью реплицирующейся плазмиды pEC71ysE. Это было сделано для того, чтобы придать штамму повышенную LysE-активность и тем самым придать ему способность с большей скоростью транспортировать L-орнитин в среду. Однако принятые меры не привели к увеличению скорости экспорта L-орнитина. Было установлено, что и контрольный штамм (13032::argF) и трансформант (13032::argF, несущий pEC71ysE) характеризовались одинаковой скоростью экспорта (0,6 нмоля·мин-1 (мг сухой массы)-1). На основе полученных результатов авторы сделали вывод о том, что L-орнитин не экспортируется экспортером LysE. Кроме того, они пришли к заключению о том, что в Corynebacterium glutamicum должна существовать другая еще неизвестная обеспечивающая экспорт L-орнитина функция (белок-экспортер) (Bellmann и др., 2001, с.1771, фиг.56 и с.1772, строки 21-28).
Был идентифицирован вариант LysE (см. SEQ ID NO:4) в штамме С.glutamicum R, который обличался от аминокислотной последовательности LysE- экспортера из штамма АТСС 13032, представленной в SEQ ID NO:2, удлиненным на три аминокислотных остатка N-концом. Аминокислотные остатки располагались в следующем порядке: метионин, валин, изолейцин (MVI). Этот LysE-полипептид из штамма R описан в ЕР 1266966 В1 в качестве варианта, который отличался от белка дикого типа с точки зрения формирования области петли, более конкретно, он не мог формировать указанную петлю, и вследствие этого обладал способностью более эффективно осуществлять экспорт L-лизина и L-аргинина.
Другой вариант LysE был описан Gunji и Yasueda (Journal of Biotechnology 127, 2006, cc. 1-13). Авторы изучали вопрос производства L-лизина облигатной метилотрофной бактерией, Methylophilus methylotrophus. Они трансформировали М. methylotrophus плазмидой, обозначенной как pSE, которая содержала ген lysE из штамма С.glutamicum АТСС13869, с целью повышения производства лизина М. methylotrophus. Однако авторы обнаружили, что им удалось стабильно интродуцировать в М. methylotrophus только мутантную форму гена lysE (lysE24). Открытая рамка считывания гена lysE была сдвинута в аллеле lysE24 вследствие инсерции остатка тимидина, это обусловливало то, что открытая рамка считывания заканчивалась после 432 пары оснований. Укороченная рамка считывания кодировала белок LysE, который был короче на 92 ак на С-конце по сравнению с белком LysE, присутствующим в штамме С.glutamicum АТСС 13869 дикого типа. Он состоял из 141 аминокислотного остатка. Кроме того, последние 6 С-концевых аминокислот укороченного белка (остатки 135-141) отличались от аминокислот аминокислотной последовательности LysE дикого типа. Штамм М. methylotrophus, несущий на плазмиде (pSE24) модифицированный аллель LysE, тестировали в отношении производства лизина. Для этой цели штамм анализировали в 0,3 л минимальной среды, обозначенной как SEIIc, в режиме периодического культивирования с подпиткой в течение 50 ч. Авторы установили, что трансформант продуцировал наряду с L-лизином (0,55 мМ) и L-аргинином (0,19 мМ) также в небольших количествах L-орнитин (0,07 мМ, что соответствовало концентрации 11,8 мг/л). Авторы пришли к выводу о том, что возможным объяснением обнаруженного образования L-орнитина может являться либо измененная специфичность мутантного транспортера в отношении субстрата, либо измененный внутриклеточный пул L-аргинина в штамме. В ЕР 1266966 В1 (на имя Gunji и Yasueda) описано положительное воздействие LysE24-TpancnopTepa на экскрецию L-лизина и L-аргинина.
Задача изобретения
В основу настоящего изобретения была положена задача разработать новый способ ферментативного получения L-орнитина.
Описание изобретения
Объектом изобретения является способ получения L-орнитина, отличающийся тем, что осуществляют следующие стадии, на которых:
а) осуществляют ферментацию высвобождающей L-орнитин бактерии, выбранной из группы, включающей бактерии p.p.Corynebacterium, Bacillus, Streptomyces, Arthrobacter и сем. Enterobacteriaceae, которая сверхэкспрессирует в среду полинуклеотид, кодирующий полипептид, который обладает активностью экспортера L-орнитина и имеет аминокислотную последовательность, идентичную по меньшей мере на (≥) 35%, ≥40%, ≥50%, ≥55%, ≥60%, ≥65%, ≥70%, ≥75%, ≥80%, ≥85%, ≥90%, ≥92%, ≥94%, ≥96%, ≥97%, (98%, ≥99% или 100%, предпочтительно на ≥70%, более предпочтительно на>90%, еще более предпочтительно на ≥96%, и наиболее предпочтительно на 100%, аминокислотной последовательности, представленной в SEQ ID NO:2,
б) накапливают L-орнитин в среде, получая в результате ферментативный бульон,
в) при этом для осуществления сверхэкспрессии не применяют плазмиду pEC71ysE, депонированную в DSM23239,
г) и при этом длина кодируемого полипептида составляет при необходимости от ≥146 до ≤286 аминокислот или аминокислотных остатков.
Предпочтительно диапазон длин выбирают из группы, включающей диапазоны, включающие от ≥171 до ≤286, от ≥196 до ≤261, от ≥203 до ≤258, от ≥218 до ≤243, от ≥228 до ≤236 и от ≥228 до ≤233 аминокислот или аминокислотных остатков.
Особенно предпочтительными диапазонами длин являются диапазоны от ≥203 до ≥258, от ≥218 до ≤243, от ≥228 до ≤236 и от ≥228 до ≤233, и наиболее предпочтительными диапазонами длин являются диапазоны от ≥228 до ≤236 и от ≥228 до ≤233.
Когда ниже в настоящем описании упоминается L-орнитин, то следует иметь в виду, что это понятие включает также его соли, такие, например, как моногидрохлорид L-орнитина или сульфат L-орнитина.
В способе, предлагаемом в изобретении, применяют бактерии, выбранные из группы, включающей бактерии p.p.Corynebacterium, Bacillus, Streptomyces, Arthrobacter и бактерии сем. Enter obacteriaceae.
В роде Corynebacterium, предпочтительными являются штаммы, выведенные из следующих видов:
Corynebacterium efficiens, например, типовой штамм DSM44549,
Corynebacterium glutamicum, например, типовой штамм АТСС13032 или штамм R, и
Corynebacterium ammoniagenes, например, штамм АТСС6871,
при этом наиболее предпочтительными являются штаммы, выведенные из Corynebacterium glutamicum.
Некоторые представители вида Corynebacterium glutamicum известны из существующего уровня техники под другими названиями. К ним относятся, например:
штамм АТСС13870, имеющий название Corynebacterium acetoacidophilum,
штамм DSM20137, имеющий название Corynebacterium lilium,
штамм АТСС 17965, имеющий название Corynebacterium melassecola,
штамм АТСС 14067, имеющий название Brevibacterium flavum,
штамм АТСС 13869, имеющий название Brevibacterium lactofermentum, и
штамм АТСС14020, имеющий название Brevibacterium divaricatum.
Для Corynebacterium glutamicum применяют также название «Micrococcus glutamicus». Некоторые представители вида Corynebacterium efficiens известны из существующего уровня техники также под названием Corynebacterium thermoaminogenes, например, штамм FERM BP-1539.
Среди бактерий p.Bacillus предпочтительными являются бактерии вида Bacillus subtilis.
Среди бактерий p.Arthrobacter предпочтительными являются бактерии вида Arthrobacter citreus.
Среди бактерий семейства Enterobacteriacae предпочтительными являются бактерии родов Escherichia, Erwinia, Providencia, Pantoea и Serratia. Наиболее предпочтительными являются бактерии p.Escherichia и p.Serratia. Наиболее предпочтительными являются: в p.Escherichia вид Escherichia coli, в p.Serratia вид Serratia marcescens, и в p.Providencia вид Providencia rettgeri.
Бактерии или штаммы (исходные штаммы), применяемые в способах обеспечения сверхэкспрессии экспортера L-орнитина, предпочтительно уже должны обладать способностью высвобождать L-орнитин в окружающую их питательную среду и накапливать его в ней. Для обозначения этого ниже в настоящем описании используют также выражение «продуцировать». Более конкретно, применяемые в способах обеспечения сверхэкспрессии штаммы должны обладать способностью концентрировать или накапливать L-орнитин в питательной среде до уровня, составляющего ≥0,1 г/л, ≥0,3 г/л, ≥1 г/л, ≥3 г/л, ≥10 г/л. Исходные штаммы предпочтительно представляют собой штаммы, которые были получены путем мутагенеза и селекции, с помощью технологий рекомбинантной ДНК или с помощью комбинации обоих методов.
Является очевидным и не требует дополнительных объяснений, что бактерию, пригодную для применения в способах (подходах), предлагаемых в изобретении, можно получать также, осуществляя сначала сверхэкспрессию полинуклеотида, кодирующего полипептид, который обладает активностью экспортера L-орнитина и аминокислотная последовательность которого по меньшей мере на (≥) 35% идентична аминокислотной последовательности, представленной в SEQ ID NO:2, где длина кодируемых полипептидов при необходимости находится в указанных выше диапазонах длин, в штамме дикого типа, таком, например, как типовой штамм Corynebacterium glutamicum АТСС13032 или штамм АТСС14067, и затем с помощью дополнительных известных из существующего уровня техники генетических методов обеспечивать способность бактерии продуцировать L-орнитин. Осуществление только лишь трансформации штамма дикого типа, такого, например, как штамм АТСС13032, АТСС14067, АТСС13869 или АТСС17965, с помощью указанного полинуклеотида не представляет собой способ, предлагаемый в изобретении.
Примерами штаммов вида Corynebacterium glutamicum, которые обладают способностью высвобождать или продуцировать L-орнитин, являются:
штамм Brevibacterium lactofermentum FERM-BP 2344 и штамм Corynebacterium glutamicum FERM-BP 2345, описанные в US 5188947.
Примером штамма вида Arthrobacter citreus, который обладает способностью высвобождать или продуцировать L-орнитин, является:
штамм Arthrobacter citreus FERM-BP 2342, описанный в US 5188947.
Примером штамма вида Bacillus subtilis, который обладает способностью высвобождать или продуцировать L-орнитин, является:
штамм Bacillus subtilis BOR-32 (FERM-P 3647), описанный в JP 57041912.
Примером штамма вида Providencia rettgeri, который обладает способностью высвобождать или продуцировать L-орнитин, является:
штамм Providencia rettgeri ARGA6 (FERM P-l 1147), описанный в JP 03195494.
Примером штамма вида-Escherichia coli, который обладает способностью высвобождать или продуцировать L-орнитин, является:
штамм Escherichia coli В-9-19 (АТСС 21104), описанный в US 3668072.
Продуцирующие L-орнитин бактерии, как правило, являются ауксотрофами по таким аминокислотам, как L-цитруллин или L-аргинин. В альтернативном варианте можно рассматривать продуцирующие L-орнитин бактерии, которые являются брадитрофными по L-цитруллину или L-аргинину. Определения понятий «ауксотрофный» и «брадитрофный» даны, например, на с.9 в WO 01/09286. Брадитрофов называют в данной области также «просачивающимися» («протекающими») мутантами (т.е. мутантами с неполным выражением мутационного повреждения). Среди брадитрофных бактерий применяют прежде всего таких, в которых активность продуктов генов ArgF (орнитинкарбамоилтрансфераза), ArgG (аргининсукцинатсинтаза) или ArgH (аргининсукцинатлиаза) больше (>) нуля, но равна или меньше (≤) 10%, предпочтительно>нуля и ≤1% от указанной активности в бактерии дикого типа.
Из существующего уровня техники известны полинуклеотиды, которые обозначают как ген lysE и которые кодируют белки или полипептиды, обладающие активностью экспортера L-лизина. Указанные полипептиды сокращенно обозначают также как LysE.
Экспортер представляет собой белок, который локализован в клеточной мембране клетки и который транспортирует метаболит, например, L-лизин или L-орнитин, из цитоплазмы указанной клетки в окружающую среду. Если необходимая для этого энергия присутствует в форме аденозинтрифосфата (АТФ), то указанный процесс называют первичным активным транспортом или экспортом. Если указанная энергия поступает в форме ионного градиента, например, ионов натрия, то данный процесс называют вторичным активным транспортом или экспортом (Jeremy М. Berg, John L. Tymoczko и L. Stryer; Biochemie, 5-е изд, изд-во Spektrum Akademischer Verlag, Heidelberg, Germany, 2003, cc. 378-384). Instructions for determining L-ornitin export activity can be found in Bellmann et al. (Microbiology 2001; 147:1765-74).
При создании настоящего изобретения было установлено, что экспортеры лизина, являющиеся представителями p.Corynebacterium, предпочтительно Corynebacterium glutamicum, и p.Micrococcus, предпочтительно Micrococcus luteus, обладают помимо активности в отношении экспорта L-лизина также активностью, присущей экспортеру L-орнитина.
Согласно изобретению применяют гены, кодирующие полипептиды, которые обладают активностью в отношении экспорта L-орнитина и имеют аминокислотную последовательность, идентичную по меньшей мере на (≥) 35%, ≥40%, ≥50%, ≥55%, ≥60%, ≥65%, ≥70%, ≥75%, ≥80%, ≥85%, ≥90%, ≥92%, ≥94%, ≥96%, ≥97%, ≥98%, ≥99% или на 100%, предпочтительно>70%, более предпочтительно ≥90%, еще более предпочтительно ≥96%, и наиболее предпочтительно ≥100%, аминокислотной последовательности, представленной в SEQ ID NO:2, при этом длина кодируемого полипептида при необходимости находится в указанных выше диапазонах длин.
Примерами пригодных экспортеров L-орнитина являются экспортеры лизина или LysE-полипептиды штаммов Corynebacterium glutamicum АТСС 13032 (SEQ ID NO:2), Corynebacterium glutamicum R (SEQ ID NO:4), Corynebacterium glutamicum ATCC14067 (SEQ ID NO:5), Corynebacterium glutamicum ATCC13869 (SEQ ID NO:7), Corynebacterium efficiens YS-314 (SEQ ID NO:9), Corynebacterium diphteriae NCTC 13129 (SEQ ID NO:10), Corynebacterium striatum ATCC6940 (SEQ ID NO: 11), Corynebacterium aurimucosum ATCC700975 (SEQ ID NO:12), Corynebacterium matruchotii ATCC33806 (SEQ ID NO:13), Corynebacterium pseudogenitalium ATCC33035 (SEQ ID NO: 14), Corynebacterium accolens ATCC49725 (SEQ ID NO:15), Corynebacterium glucuronalyticum ATCC 51867 (SEQ ID NO:16), Micrococcus luteus NCTC2665 (SEQ ID NO:17), Corynebacterium tubuculostearicum SK141 (SEQ ID NO:18) и Corynebacterium matruchotii ATCC14266 (SEQ ID NO:19). Полипептиды, последовательности которых представлены в SEQ ID NO: 18 и SEQ ID NO:19, обозначают в данной области также как ArgO-полипептиды.
При создании настоящего изобретения была определена нуклеотидная последовательность генов lysE штаммов Corynebacterium glutamicum АТСС 14067 и Corynebacterium glutamicum АТСС13869 (SEQ ID NO:6 и SEQ ID NO:8). Аминокислотные последовательности LysE-полипептида штаммов Corynebacterium glutamicum АТСС 14067 и Corynebacterium glutamicum АТСС13869 представлены в SEQ ID NO:5 и SEQ ID NO:7. Они идентичны аминокислотной последовательности LysE штамма С.glutamicum АТСС13032, представленной в SEQ ID NO:2.
В таблице 1 приведены регистрационные номера LysE-полипептидов различных представителей p.Corynebacterium и p.Micrococcus luteus, взятые из баз данных Национального центра биотехнологической информации (NCBI, Бетесда, шт.Мэриленд, США). Кроме того, в таблице 1 даны ссылки на аминокислотные последовательности LysE-полипептида, представленные в перечне последовательностей. И, наконец, в таблице 1 указана длина (количество аминокислот) кодируемого LysE-полипептида.
На фиг.1 представлено множественное сравнение аминокислотных последовательностей LysE-полипептидов бактерий, перечисленных в таблице 1. Выравнивание аминокислотных последовательностей, представленных на фиг.1, осуществляли с помощью программы Clone Manager 9 Professional Edition (фирма Scientific & Educational Software, 600 Pinner Weald Way, Ste 202, Cary, шт.Северная Каролина, 27513, США). В качестве эталонной молекулы при осуществлении выравнивания использовали LysE-полипептид (LysE) штамма АТСС 13032. В качестве матрицы очков (матрицы баллов) была выбрана матрица «Blosum 62» (см. Jeremy М. Berg, John L. Tymoczko и L. Stryer; Biochemie, 5-e изд., изд-во Spektrum Akademischer Verlag, Heidelberg, Germany, cc. 194-197, 2003) was chosen.
При необходимости можно использовать также программы, известные из существующего уровня техники, такие, например, как программа ClustalX (Thompson J.D., Gibson T.J., Plewniak F., Jeanmougin F. и Higgins D.G., The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 25, 1997, cc. 4876-4882).
Аминокислотные остатки 4-236 LysE-полипептида штамма Corynebacterium glutamicum R (см. SEQ ID NO:4) соответствуют аминокислотной последовательности LysE штамма С.glutamicum АТСС 13032, представленной в SEQ ID NO:2. Полипептид штамма С.glutamicum R имеет на N-конце дополнительную последовательность, состоящую из трех аминокислотных остатков (метионин-валин-изолейцин). Указанные дополнительные остатки образуются в том случае, когда вместо стартового кодона гена lysE в штамме С.glutamicum АТСС 13032 (см. SEQ ID NO:1) используют стартовый кодон, расположенный на расстоянии 9 пар оснований против хода транскрипции относительно гена lysE.
Аминокислотная последовательность LysE-полипептида штамма С.efficiens YS-314 идентична на 71%, штамма С.diphteriae NCTC 13129 идентична 44%, штамма Corynebacterium striatum АТСС6940 идентична на 44%, штамма Corynebacterium aurimucosum АТСС700975 идентична на 42%, штамма Corynebacterium matruchotii АТСС33806 идентична на 43%, штамма Corynebacterium pseudogenitalium АТСС33035 идентична на 43%, штамма Corynebacterium accolens АТСС49725 идентична на 43%, штамма Corynebacterium glucuronalyticum АТСС 51867 идентична на 36%, штамма Micrococcus luteus NCTC2665 идентична на 40% аминокислотной последовательности LysE штамма С.glutamicum АТСС 13032, представленной в SEQ ID NO:2. Кроме того, аминокислотная последовательность ArgO-полипептида штамма С.tubuculostearicum SK141 идентична на 43% аминокислотной последовательности, представленной в SEQ ID NO:2. Кроме того, аминокислотная последовательность ArgO-полипептида штамма С.matruchotii АТСС14266 идентична на 44% аминокислотной последовательности, представленной в SEQ ID NO: 2. Проценты идентичности рассчитывали на основе глобального выравнивания последовательностей с помощью программы Clone Manager 9 с использованием задаваемой матрицы Blosum 62 (см. фиг.2).
Гены lysE, т.е. полинуклеотиды, которые кодируют полипептиды, обладающие активностью экспортера L-орнитина, можно выделять из организмов с помощью полимеразной цепной реакции (ПЦР) с использованием соответствующих праймеров. Соответствующие инструкции можно найти, в частности, в лабораторном руководстве «PCR» под ред. Newton и Graham, изд-во Spektrum Akademischer Verlag, Heidelberg, Germany, 1994, и в WO 2006/100211 на cc. 14-17.
Наиболее предпочтительно в способе, предлагаемом в изобретении, применяют гены, кодирующие полипептиды, которые обладают активностью в отношении экспорта L-орнитина и имеют аминокислотную последовательность, отличающуюся одним или несколькими характерными особенностями, выбранными из группы особенностей, которыми обладают следующие последовательности:
а) аминокислотная последовательность, представленная в SEQ ID NO:2 или SEQ ID NO:4,
б) аминокислотная последовательность, представленная в SEQ ID NO:2, которая имеет одну или несколько, вплоть до 25, 20, 15, 10, 5, 4, 3, 2 или 1, делецию(й) аминокислот,
в) аминокислотная последовательность, представленная в SEQ ID NO:2, которая имеет одну или несколько, вплоть до 25, 20, 15, 10, 5, 4, 3, 2 или 1, инсерцию(й) аминокислот, и
г) аминокислотная последовательность, представленная в SEQ ID NO: 2, которая имеет одну или несколько, вплоть до 140, 130, 120, 110, 100, 90, 80, 70, 60, 50, 40, 30, 25, 20, 15, 10, 5, 4, 3, 2 или 1, предпочтительно вплоть до 5, 4, 3, 2 или 1, замену/замен (замещение/замещений) аминокислот,
д) аминокислотная последовательность, представленная в SEQ ID NO:2, которая имеет одно или несколько, вплоть до 25, 20, 15, 10, 5, 4, 3, 2 или 1, предпочтительно вплоть до 5, 4, 3, 2 или 1, добавление(й) аминокислот на N-конце и/или на С-конце.
Предпочтительно применяют, если это необходимо, консервативные аминокислотные замены. В случае ароматических аминокислот консервативные замены представляют собой замены фенилаланина, триптофана и тирозина друг на друга. В случае гидрофобных аминокислот консервативные замены представляют собой замены лейцина, изолейцина и валина друг на друга. В случае полярных аминокислот консервативные замены представляют собой замены глутамина и аспарагина друг на друга. В случае основных аминокислот консервативные замены представляют собой замены аргинина, лизина и гистидина друг на друга. В случае кислых аминокислот консервативные замены представляют собой замены аспарагиновой кислоты и глутаминовой кислоты друг на друга. В случае аминокислот, содержащих гидроксильные группы, консервативные замены представляют собой замены серина и треонина друг на друга.
Кроме того, можно применять полинуклеотиды, которые гибридизуются в строгих условиях с нуклеотидной последовательностью, комплементарной последовательности, которая представлена в SEQ ID NO: 1, предпочтительно с кодирующей областью последовательности, представленной в SEQ ID NO: 1, и которые кодируют полипептид, обладающий активностью в отношении экспорта L-орнитина, где аминокислотная последовательность кодируемого белка идентична по меньшей мере на ≥70% аминокислотной последовательности, представленной в SEQ ID NO: 2, и длина кодируемого полипептида при необходимости находится в указанных выше диапазонах длин.
Специалист в данной области может найти инструкции касательно гибридизации нуклеиновых кислот и полинуклеотидов соответственно среди прочего в руководстве «The DIG System Users Guide for Filter Hybridization)) фирмы Boehringer Mannheim GmbH (Маннгейм, Германия, 1993) и у Liebl и др., International Journal of Systematic Bacteriology 41, 1991, cc. 255-260. Если гибридизацию осуществляют в строгих условиях, то это означает, что образуются только такие гибриды, в которых зонд, т.е. полинуклеотид, который содержит нуклеотидную последовательность, комплементарную последовательности, представленной в SEQ ID NO:1, предпочтительно кодирующей области последовательности, представленной в SEQ ID NO:1, и последовательность-мишень, т.е. полинуклеотиды, которые обрабатывают или идентифицируют с помощью рассматриваемого зонда, идентичны по меньшей мере на 70%. Известно, что на строгость гибридизации, включая стадии отмывки, можно оказывать воздействие или задавать ее посредством вариации состава буфера, температуры и концентрации соли. Реакцию гибридизации, как правило, осуществляют в условиях относительно низкой строгости по сравнению со стадиями отмывки (руководство Hybaid Hybridisation Guide, фирма Hybaid Limited, Теддингтон, Великобритания, 1996).
Например, для реакции гибридизации можно применять 5х SSC-буфер при температуре примерно 50-68°C. В этом случае зонды могут гибридизоваться также с полинуклеотидами, идентичными менее чем на 70% нуклеотидной последовательности применяемого зонда. Такие гибриды менее стабильны и их удаляют путем отмывки в строгих условиях. Это можно осуществлять, например, снижая концентрацию соли до 2х SSC или lx SSC и затем, при необходимости, до 0,5х SSC (руководство The DIG System User's Guide for Filter Hybridisation), фирма Boehringer Mannheim, Маннгейм, Германия, 1995), и устанавливая температуру на уровне примерно 50-68°C, примерно 52-68°C, примерно 54-68°C, примерно 56-68°C, примерно 58-68°C, примерно 60-68°C, примерно 62-68°C, примерно 64-68°C, примерно 66-68°C. Предпочтительными диапазонами температур являются примерно 64-68°C или примерно 66-68°C. Необязательно можно снижать концентрацию соли до концентрации, соответствующей 0,2х SSC или 0,1х SSC. SSC-буфер необязательно содержит додецилсульфат натрия (ДСН) в концентрации 0,1%. Путем постепенного ступенчатого (примерно на 1-2°C) повышения температуры гибридизации от 50°C до 68°C, можно выделять полинуклеотидные фрагменты, идентичные по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 92%, по меньшей мере на 94%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99%, при необходимости на 100%, последовательности применяемого зонда или последовательности, комплементарной последовательности применяемого зонда, и которая кодирует полипептид, обладающий активностью в отношении экспорта L-орнитина. Дополнительные инструкции касательно гибридизации содержатся в продуктах, поступающих в продажу в форме «наборов» (например, набор DIG Easy Hyb фирмы Roche Diagnostics GmbH, Маннгейм, Германия, каталожный номер 1603558).
Согласно изобретению осуществляют сверхэкспрессию полинуклеотида, который кодирует белок, обладающий активностью в отношении экспорта L-орнитина, в бактерии или исходном или родительском штамме, продуцирующем L-орнитин, где аминокислотная последовательность кодируемого белка идентична на ≥35% аминокислотной последовательности, представленной в SEQ ID NO: 2, и длина кодируемого полипептида при необходимости находится в указанных выше диапазонах.
В целом, сверхэкспрессия означает повышение внутриклеточной концентрации или активности рибонуклеиновой кислоты, белка (полипептида) или фермента по сравнению с исходным штаммом (родительским штаммом) или штаммом дикого типа, если последний применяют в качестве исходного штамма. Исходный штамм (родительский штамм) представляет собой штамм, который подвергали манипуляции, приводящей к сверхэкспрессии.
Понятия «белок» и «полипептид» рассматриваются как взаимозаменяемые.
Для обеспечения сверхэкспрессии предпочтительно применяют методы рекомбинантной сверхэкспрессии. Они включают любые методы, в которых микроорганизм создают с использованием ДНК-молекулы, полученной in vitro. Примерами таких ДНК-молекул являются промоторы, кассеты экспрессии, гены, аллели, кодирующие области и т.д. Их можно переносить в требуемый микроорганизм с помощью методов трансформации, конъюгации, трансдукции или других подобных методов.
Манипуляции, осуществляемые для обеспечения сверхэкспрессии, повышают активность или концентрацию соответствующего полипептида, как правило, по меньшей мере на 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500%, предпочтительно вплоть до 1000%, 2000%, 4000%, 10000% или 20000%, по отношению к уровню активности или концентрации рассматриваемого полипептида в штамме до осуществления манипуляции, приводящей к сверхэкспрессии.
Когда применяют штаммы вида Corynebacterium glutamicum, то при необходимости в качестве пригодного эталона (референс-точки) при оценке сверхэкспрессии используют активность в отношении экспорта L-орнитина в штамме АТСС13032 или АТСС14067, или АТСС13869, или АТСС17965. Когда применяют штаммы на основе штамма АТСС13032 или выведенные из него штаммы, то пригодным эталоном является указанный штамм АТСС 13032. Соответствующим примером является штамм, полученный при создании настоящего изобретения, а именно, ATCC13032_Delta_argFRGH/pVWExl_lysE, созданный на основе штамма АТСС 13032. Когда применяют штаммы на основе штамма АТСС 14067 или выведенные из него штаммы, то пригодным эталоном является указанный штамм АТСС 14067. Когда применяют штаммы на основе штамма АТСС 13869 или выведенные из него штаммы, то пригодным эталоном является указанный штамм АТСС13869. Соответствующим образом выбирают другие пригодные эталоны.
Когда применяют штаммы вида Escherichia coli, предпочтительно штамм Escherichia coli К12, то при необходимости в качестве пригодного эталона при оценке сверхэкспрессии используют активность в отношении экспорта L-орнитина в штамме MG1655.
Для достижения сверхэкспрессии применяют многочисленные методы, известные из существующего уровня техники.
Они включают увеличение количества копий и модификацию нуклеотидных последовательностей, регулирующих или контролирующих экспрессию гена. Транскрипцию гена контролируют среди прочего с помощью промотора и необязательно с помощью белков, которые подавляют (белки-репрессоры) или стимулируют (белки-активаторы) транскрипцию. Трансляцию образовавшейся РНК контролируют среди прочего с помощью сайта связывания рибосом и стартового кодона. Полинуклеотиды или ДНК-молекулы, которые включают промотор и сайт связывания рибосом, и необязательно стартовый кодон, называют также кассетой экспрессии.
Указанные методы включают также применение вариантов полипептидов или ферментов, обладающих повышенной каталитической активностью.
Количество копий можно увеличивать с использованием плазмид, которые реплицируются в цитоплазме бактерии. Для этой цели можно применять целый ряд плазмид, известных из существующего уровня техники, которые пригодны для отличающихся большим разнообразием групп микроорганизмов, указанные плазмиды можно использовать для достижения требуемого увеличения количества копий гена. Плазмиды, которые можно применять для бактерий р. Escherichia, описаны, например, в руководстве по молекулярной биологии Labfax (под ред. Т.А. Brown, изд-во Bios Scientific, Oxford, UK, 1991). Плазмиды, которые можно применять для бактерий p.Corynebacterium, описаны, например, у Tauch и др., Journal of Biotechnology, 104 (1-3), 2003, сс.27-40 или у Stansen и др., Applied and Environmental Microbiology 71, 2005, сс.5920-5928.
Подход, основанный на применении плазмиды pEC71ysE, депонированной под регистрационным номером DSM 23239, с целью увеличения количества копий в штаммах Corynebacterium glutamicum исключено из числа подходов, предназначенных для осуществления настоящего изобретения. Нуклеотидная последовательность плазмиды pEC7lysE была определена и она представлена в SEQ ID NO:29.
Количество копий можно дополнительно увеличивать по меньшей мере на одну (1) копию путем интродукции дополнительных копий в хромосому бактерии. Соответствующие методы, которые можно применять для бактерий р. Corynebacterium, предпочтительно Corynebacterium glutamicum, описаны, например, в международных заявках на патент WO 03/014330, WO 03/040373 и WO 04/069996. В WO 03/014330 описаны методы тандемного удвоения генов в нативном локусе генов. В WO 03/040373 описаны методы встраивания второй или третьей копии гена в другие локусы генов, при этом конкретный локус генов не является существенным для роста или производства конкретной аминокислоты, которой в случае настоящего изобретения является L-орнитин. Примерами локусов генов, пригодных для включения второй или следующей копии гена lysE согласно способу, предлагаемому в изобретении, являются гены odh, sucA, dapA, dapB, ddh, lysA, argR, argF, argG и argH. В WO 04/069996 (см. таблицы 12 и 13) описаны межгенные области и гены С.glutamicum, кодирующие фаги и компоненты фагов, которые пригодны для встраивания дополнительных копий гена lysE.
Примерами методов, которые можно применять для бактерий р. Escherichia, являются встраивание копии гена в сайт att фага (Yu и Court, Gene 223, 1988, сс.77-81), амплификация хромосомального гена с помощью фага Mu, описанная в ЕР 0332448, или методы замены гена с помощью условно реплицирующейся плазмиды, описанные у Hamilton и др., Journal of Bacteriology 174, 1989, сс.4617-4622 или у Link и др., Journal of Bacteriology 179, 1997, сс.6228-6237).
Экспрессию гена можно дополнительно повышать путем применения сильного промотора, функционально связанного с предназначенным для экспрессии геном. Предпочтительно следует применять промотор, более сильный, чем встречающийся в естественных условиях промотор, т.е. промотор, присутствующий в штамме дикого типа или в родительском штамме. Для этой цели можно применять целый ряд методов, известных из существующего уровня техники.
Промоторы и системы экспрессии, которые можно применять для представителей p.Corynebacterium, описаны среди прочего в следующих патентах и заявках на патент: ЕР 0629699 А2, US 2007/0259408 А1 (промотор gap), WO 2006/069711, ЕР 1881076 Al, WO 2008/088158, WO 2009/025470 (промотор butA, промотор рук), US 6861246 (варианты МС20 и МА16 промотора dap А) и ЕР 1918378 А1 (промотор sod), а также обзорах, таких как «Handbook of Corynebacterium glutamicum» (под ред. Lothar Eggeling и Michael Bott, изд-во CRC Press, Boca Raton, US, 2005), или в книге «Corynebacteria, Genomics and Molecular Biology» (под ред. Andreas Burkovski, изд-во Caister Academic Press, Norfolk, UK, 2008). Примеры промоторов, которые обеспечивают контролируемую, т.е. индуцибельную или подавляемую, экспрессию, описаны, например, у Tsuchiya и Morinaga, Bio/Technology 6, 1988, сс.428-430.
Промоторы, которые можно применять для представителей p.Escherichia, известны уже в течение длительного периода времени. К ним относятся, среди прочего, классические промоторы, такие как промотор lac, промотор trp, гибридные промоторы tac и trc, промоторы PL и PR фага λ. Можно применять также промоторы фага Т7, промоторы gear-box, промотор nar или промоторы генов rrsG, rnpB, csrA, csrB, ompA, fusA, pepQ, rplX или rpsG. Контролируемую экспрессию можно обеспечивать, например, с помощью системы cI857-PR или CI857-PL фага λ (Gotting и др., BioTechniques 24, 1988, сс.362-366). Соответствующие обзоры приведены у Makrides, Microbiological Reviews 60(3), 1996, сс.512-538 или в руководстве «Escherichia coli and Salmonella, Cellular and Molecular Biology» (главный редактор F.C.Neidhardt, изд-во ASM Press, Washington, US, 1996).
Такие промоторы или кассеты экспрессии, как правило, помещают на расстоянии от 1 до 1000, предпочтительно от 1 до 500, нуклеотидов против хода транскрипции от первого нуклеотида стартового кодона кодирующей области гена. Расстояние, равное 1, означает, что промотор или кассета экспрессии расположен/расположена непосредственно перед первым основанием стартового кодона кодирующей области.
Для повышения экспрессии гена lysE в С.glutamicum пригодные промоторы, такие, например, как промотор sod С.glutamicum (см. SEQ ID NO:1, представленную в ЕР 1918378 А1) или промотор gap С.glutamicum (см. SEQ ID NO: 3, представленную в US 2007/0259408), предпочтительно встраивают между положениями 930 и 990 SEQ ID NO:1.
Когда применяют кассеты экспрессии, содержащие промотор и сайт связывания рибосом (RBS), такие, например, как экспрессионная конструкция гена sod С.glutamicum (см. SEQ ID NO:2, представленную в ЕР 1918378 А1) или экспрессионная конструкция гена gap С.glutamicum, описанный в US 2007/0259408 и представленный в SEQ ID NO:28 (обозначен в указанном документе как PgapRBS), их встраивают, например, в случае С.glutamicum предпочтительно между положениями 930 и 1001, наиболее предпочтительно между положениями 1000 и 1001 SEQ ID NO:1. Примером пригодного сайта связывания рибосом в такой кассете экспрессии является нуклеотидная последовательность 5'-agaaaggagg-3', охарактеризованная Amador (Microbiology 145, 1999, сс.915-924).
Можно также помещать несколько промоторов против хода транскрипции относительно целевого гена или функционально связывать их с геном, подлежащим экспрессии, и тем самым достигать повышения уровня экспрессии. Этот подход описан, например, в WO 2006/069711.
Структура промоторов из Corynebacterium glutamicum и Escherichia coli хорошо известна. Поэтому можно повышать силу промотора путем модификации его последовательности с помощью одной замены или нескольких замен, и/или одной инсерции или нескольких инсерций, и/или одной делеции или нескольких делеций нуклеотидов. Примеры такого подхода описаны среди прочего в «Herder Lexikon der Biologie» [Herder's Encyclopaedia of Biology], изд-во Spektrum Akademischer Verlag, Heidelberg, Germany, 1994.
Таким образом, пригодный для достижения сверхэкспрессии гена lysE подход заключается в осуществлении модификации или мутации промотора указанного гена lysE.
Структура сайтов связывания рибосомы Corynebacterium glutamicum и Escherichia coli также хорошо известна, она описана, например, у Amador, Microbiology 145, 1999, сс.915-924, и в руководствах и учебниках по генетике, таких, например, как «Gene und КЛопе» [Genes and Clones], под ред. Winnacker, изд-во Verlag Chemie, Weinheim, Germany, 1990 или «Molecular Genetics of Bacteria», Dale и Park, изд-во Wiley and Sons Ltd., Chichester, UK, 2004. Характеризующиеся высоким уровнем экспрессии гены, т.е. наиболее важные структурные гены в организме, имеют эффективный сайт связывания рибосомы (Amador, Microbiology 145, 1999, сс.915-924), это означает, что он обладает большим сходством с консенсусной последовательностью или соответствует ей. В литературе продемонстрировано, что характеризующиеся высоким уровнем экспрессии гены имеют сильный сайт связывания рибосомы (Karlin и Mrazek, Journal of Bacteriology; 182(18), 2000, сс.5238-5250). Следовательно, эффективную трансляцию гена или мРНК можно обеспечивать путем регулирования сайта связывания рибосомы.
Эффективность трансляции можно повышать также путем регулирования наиболее часто встречающихся кодонов в генах, подлежащих экспрессии (см., например, Najafabiad и др., Nucleic Acids Research, 37 (21), 2009, сс.7014-7023).
Сверхэкспрессию можно обеспечивать также путем повышения уровня экспрессии белков-активаторов или путем снижения или «выключения» экспрессии белков-репрессоров.
Белок LysG, являющийся активатором экспрессии lysE, был описан у Bellmann и др., Microbiology; 147, 2001, сс.1765-1774, и он был обозначен указанными авторами как «положительный регулятор». Аминокислотная последовательность LysG штамма Corynebacterium glutamicum АТСС13032 представлена в SEQ ID NO: 30. При глобальном выравнивании последовательностей было установлено, что аминокислотная последовательность полипептида LysG штамма Corynebacterium diphteriae NCTC13129 идентична на 62%, аминокислотная последовательность полипептида LysG штамма Corynebacterium efficiens YS-314 идентична на 81%, а аминокислотная последовательность полипептида LysG штамма Corynebacterium glutamicum R идентична на 94% последовательности, представленной в SEQ ID NO:30.
Касательно белков-активаторов предпочтительным является полипептид, аминокислотная последовательность которого идентична на>(по меньшей мере) 55%, предпочтительно на>80%, более предпочтительно на ≥90%, ≥92% или на ≥94%, veine более предпочтительно на ≥99%, и наиболее предпочтительно на 100%, аминокислотной последовательности, представленной в SEQ ID NO:30.
Указанные выше подходы, направленные на достижение сверхэкспрессии, которые предпочтительно выбирают из группы, включающей увеличение количества копий, применение сильного промотора, осуществление мутации промотора, применение соответствующей кассеты экспрессии и осуществление сверхэкспрессии белка-активатора, можно объединять друг с другом соответствующим образом. Так, можно, например, объединять применение пригодного промотора с увеличением количества копий, или осуществлять сверхэкспрессию белка-активатора в сочетании с применением пригодного промотора или пригодной кассеты экспрессии.
Можно также помимо осуществления манипуляций, связанных с полинуклеотидом, кодирующим белок, который обладает активностью в отношении экспорта L-орнитина, ослаблять определенные гены биосинтеза.
Так, для повышения производства L-орнитина можно, если это является целесообразным, дополнительно ослаблять один или несколько генов, выбранных из группы, включающей:
а) ген odhA, кодирующий субъединицу Е1 альфа-кетоглутарат-дегидрогеназы (КФ 1.2.4.2),
б) ген sucA, кодирующий дигидролипоамид-сукцинилтрансферазу (КФ 2.3.1.61),
в) ген dapA, кодирующий дигидродипиколинатсинтазу (DapA, КФ 4.2.1.52),
г) ген dapB, кодирующий дигидродипиколинатсинтазу (DapB, КФ 1.3.1.26),
д) ген ddh, кодирующий мезо-диаминопимелатдегидрогеназу (Ddh, КФ 1.4.1.16),
е) ген lysA, кодирующий диаминопимелатдекарбоксилазу (LysA, КФ 4.1.1.20),
ж) ген argR, кодирующий любой/конкретный репрессор (ArgR) биосинтеза L-аргинина,
з) ген argF, кодирующий орнитинкарбомоилтрансферазу (ArgF, КФ 2.1.3.3),
и) ген argG, кодирующий аргининсукцинатсинтазу (ArgG, КФ 6.3.4.5), к) ген argH, кодирующий аргининсукцинатлиазу (ASAL) (ArgH, КФ 4.3.2.1),
л) ген lysC, кодирующий аспартаткиназу (LysC, КФ 2.7.2.4), и м) ген asd, кодирующий аспартат-полуальдегид-дегидрогеназу (Asd, КФ 1.2.1.11).
Предпочтительно ослабляют один или несколько генов, выбранных из группы, включающей lysA, odhA, argR, argF, argG и argH. Более предпочтительно ослабляют один или несколько генов, выбранных из группы, включающей lysA, odhA и argF. Наиболее предпочтительно ослабляют ген(ы) lysA и/или argF.
В контексте настоящего изобретения понятие «ослабление» обозначает снижение или «выключение» внутриклеточной активности одного или нескольких ферментов (белков) в бактерии, которые кодируются соответствующей ДНК, путем применения, например, слабого промотора или гена, или аллеля, который кодирует соответствующий фермент, обладающий низкой активностью, или путем инактивации соответствующего гена или фермента (белка), и необязательно путем применения комбинации указанных подходов.
Обзор известных промоторов различной силы в Corynebacterium glutamicum, приведен у Patek и др., Journal of Biotechnology 104, 2003, сс.311-323. Другие слабые промоторы описаны в сообщении 512057, опубликованном в журнале Research Disclosure, декабрь 2006 г., сс.1616-1618.
Мутации, которые можно рассматривать для достижения ослабления, представляют собой транзиции, трансверсии, инсерции и делеции по меньшей мере одной/одного (1) пары оснований или нуклеотида в кодирующей области представляющего интерес гена. В зависимости от воздействия, которое оказывает обусловленная мутацией аминокислотная замена на активность белка или фермента, мутации обозначают как миссенс-мутации или нонсенс-мутации.
Миссенс-мутация приводит к замене рассматриваемой аминокислоты в белке на другую, при этом замена, как правило, представляет собой неконсервативную аминокислотную замену. Такая замена ослабляет функциональную способность или активность белка и снижает их уровень на величину, составляющую от ≥0 до 75%, от ≥0 до 50%, от ≥0 до 25%, от ≥0 до 10% или от ≥0 до 5%.
Нонсенс-мутации приводят к появлению стоп-кодона в кодирующей области гена и, следовательно, к преждевременному прекращению трансляции и затем к ее «выключению». Инсерции или делеции по меньшей мере одной пары оснований в гене приводят к мутациям со сдвигом рамки считывания, в результате чего происходит встраивание неправильных аминокислот или преждевременное прекращение трансляции. Если мутация приводит к появлению стоп-кодона в кодирующей области, то это также вызывает преждевременное прекращение трансляции. Нонсенс-мутации предпочтительно создают в 5'-концевой части кодирующей области, которая кодирует N-конец полипептида. Если общую длину полипептида (которую представляют в виде количества химически связанных L-аминокислот) принять за 100%, то (в контексте настоящего изобретения) N-конец полипептида включает часть аминокислотной последовательности, которая, если считать, начиная с первой (стартовой) аминокислоты, т.е. L-формил-метионина, по ходу транскрипции, содержит 80% расположенных по ходу транскрипции L-аминокислот.
Методы мутагенеза in vivo описаны, например, в руководстве «Manual of Methods for General Bacteriology» под ред. Gerhard и др., изд-во American Society for Microbiology, Washington, DC, USA, 1981 или у Tosaka и др., Agricultural and Biological Chemistry 42(4), 1978, cc. 745-752, или у Konicek и др., Folia Microbiologica 33, 1988, cc. 337-343.
Пригодными методами мутагенеза in vitro являются, среди прочего, обработка гидроксиламином согласно методу Миллера (Miller J.H., A Short Course in Bacterial Genetics. A Laboratory Manual and Handbook for Escherichia coli and OxyRated Bacteria, изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1992), применение мутагенных олигонуклеотидов (Т.A. Brown, Gentechnologie fur Einsteiger [Genetic Engineering for Beginners], изд-во Spektrum Akademischer Verlag, Heidelberg, 1993 и R.M. Horton, PCR-Mediated Recombination and Mutagenesis, Molecular Biotechnology 3, 1995, cc. 93-99), и применение полимеразной цепной реакции с использованием ДНК-полимеразы с высокой частотой ошибок. Примером такой ДНК-полимеразы является ДНК-полимераза Mutazyme (набор для ПЦР-мутагенеза GeneMorph, №600550) фирмы Stratagene (Ла-Джолла, шт.Калифорния, США).
Дополнительные инструкции и обзоры методов создания мутаций in vivo или in vitro можно почерпнуть из существующего уровня техники, а также в известных учебниках по генетике и молекулярной биологии, таких, например, как учебник Knipper, «Molekulare Genetik», 6-е изд., изд-во Georg Thieme Verlag, Stuttgart, Germany, 1995, учебник Winnacker, «Gene and Klone», изд-во VCH Verlagsgesellschaft, Weinheim, Germany, 1990, или учебник Hagemann, «Allgemeine Genetik» [General Genetics], изд-во Gustav Fischer Verlag, Stuttgart, 1986.
С помощью известных методов замены гена или аллеля, основы которых описаны у Schwarzer and Puhler, Bio/Technology 9, 1991, сс.84-87, можно переносить мутации, созданные in vitro, или полинуклеотид, содержащий требуемую мутацию, в хромосому. Schafer с соавторами (Gene 145, 1994, сс.69-73) применяли указанный метод для осуществления делеции в опероне hom-thrB С.glutamicum. Nakagawa с соавторами (ЕР 1108790) и Ohnishi с соавторами (Applied Microbiology and Biotechnology 58(2), 2001, сс.217-223) применяли указанный метод для включения различных мутаций, созданных в выделенных аллелях, в хромосому С.glutamicum.
Один из методов направленного снижения уровня экспрессии гена заключается в том, что ген, предназначенный для ослабления, помещают под контроль промотора, который можно индуцировать путем добавления в определенных количествах ИПТГ (изопропил-β-D-тиогалактопиранозид), такого, например, как промотор trc или промотор tac. Для этой цели можно применять, например, такие векторы, как экспрессионный вектор рХК99Е для Escherichia coli (WO 0226787; депонированный согласно Будапештскому договору 31 июля 2001 г в DH5alpha/pXK99E под регистрационным номером DSM14440 в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ, Брауншвейг, Германия)), рЕКЕх2 (регистрационный номер AY585307 в NCBI) или pVWEx2 (Wendisch, Ph.D.-диссертация, Berichte des Forschungszentrums Jiilich, Jul-3397, ISSN 0994-2952, Jttlich, Germany, 1997), которые позволяют осуществлять зависящую от ИПТГ экспрессию клонированного гена в Corynebacterium glutamicum.
Указанный метод был применен, например, в заявке на патент WO 02266787 для осуществления регулируемой экспрессии гена deaD с помощью встраивания вектора pXK99EdeaD в геном Corynebacterium glutamicum, и Simic с соавторами (Applied and Environmental Microbiology 68, 2002, сс.3321-3327) для осуществления регулируемой экспрессии гена glyA с помощью встраивания вектора pK18mobglyA' в Corynebacterium glutamicum.
Другим методом специфического снижения уровня экспрессии гена является антисмысловой метод, в котором применяют введение в клетки-мишени коротких олигодезоксинуклеотидов или векторов для синтеза более длинной антисмысловой РНК. После этого антисмысловая РНК может связываться с комплементарными участками специфических мРНК и уменьшать их стабильность или блокировать способность к трансляции. Специалист в данной области может найти пример такого применения у Srivastava и др., Applied Environmental Microbiology, 66 (10), октябрь 2000 г., сс.4366-4371.
Скорость удлинения зависит от частоты встречаемости кодонов. Экспрессию гена можно ослаблять путем применения кодонов для тРНК, которые редко встречаются в родительском штамме. Этот подход подробно описан в WO 2008049781 и WO 2009133063. Например, заменяя стартовый кодон ATG на менее часто встречающиеся кодоны GTG или TTG можно ослаблять трансляцию, поскольку кодон AUG в два-три раза более эффективен, чем кодоны GUG и UUG (см., например, Khudyakov и др., FEBS Letters 232(2), 1988, сс.369-371; Reddy и др., Proceedings of the National Academy of Sciences of the USA 82(17), 1985, cc. 5656-5660).
Можно также в дополнение к манипуляциям, которым подвергают полинуклеотид, кодирующий белок, обладающий активностью в отношении экспорта L-орнитина, усиливать определенные гены биосинтеза.
Так, для увеличения производства L-орнитина может оказаться целесообразным при необходимости дополнительно повышать ферментативную активность одного или нескольких белков, выбранных из группы, включающей:
а) глутаматдегидрогеназу (КФ 1.4.1.3), кодируемую геном gdh,
б) глутамат-N-ацетилтрансферазу (КФ 2.3.1.35 и КФ 2.3.1.1), кодируемую геном argJ,
в) ацетилглутаматкиназу (КФ 2.7.2.8), кодируемую геном argB,
г) N-ацетил-гамма-глутамил-фосфатредуктазу (КФ 1.2.1.38), кодируемую геном argC,
д) ацетилорнитинаминотрансферазу (КФ 2.6.1.11), кодируемую геном argD,
е) специфический в отношении глюкозы компонент EIIB (PtsG) (КФ 2.7.1.69) системы поглощения глюкозы, кодируемый геном ptsG,
ж) специфический в отношении сахарозы компонент EIIB (PtsS) (КФ 2.7.1.69) системы поглощения сахарозы, кодируемый геном ptsS,
з) глюкозо-6-фосфат-1-дегидрогеназу (КФ 1.1.1.49), кодируемую геном zwf,
и) глюкозо-6-фосфатизомеразу (КФ 5.3.1.9), кодируемую геном pgi,
к) фосфофруктокиназу (КФ 2.7.1.11), кодируемую геном pfkA,
л) фруктозобифосфатальдолазу (КФ 4.1.2.13), кодируемую геном fda,
м) глицеральдегид-3-фосфатдегидрогеназу (КФ 1.2.1.59), кодируемую геном gap,
н) фосфоглицераткиназу (КФ 2.7.2.3), кодируемую геном pgk,
о) пируваткиназу (КФ 2.7.1.40), кодируемую геном рук,
п) Е1-субъединицу пируватдегидрогеназы (КФ 1.2.4.1), кодируемую геном aceE,
р) фосфоенолпируваткарбоксилазу (КФ 4.1.1.31), кодируемую геном ррс,
с) пируваткарбоксилазу (КФ 6.4.1.1), кодируемую геном рус,
т) аконитазу (КФ 4.2.1.3), кодируемую геном acn, и
у) изоцитратдегидрогеназу (КФ 1.1.1.42), кодируемую геном icd.
Понятие «усиление» («повышение») включает осуществление мер, обеспечивающих сверхэкспрессию, и применение вариантов, обладающих увеличенной каталитической активностью по сравнению с белком дикого типа.
Наиболее предпочтительно усиливают один или несколько ферментов, выбранных из группы, включающей глутаматдегидрогеназу, глутамат-N-ацетилтрансферазу и ацетилглутаматкиназу.
Дополнительные перечисленные манипуляции, направленные на ослабление, можно объединять с дополнительными манипуляциями, направленными на усиление.
Инструкции по применению ДНК, расщеплению и лигированию ДНК, осуществлению трансформации и отбору трансформантов можно найти среди прочего в известном руководстве Sambrook и др., «Molecular Cloning: А Laboratory Manual», 2-е изд., изд-во Cold Spring Harbor Laboratory Press, 1989.
Уровень экспрессии или сверхэкспрессии можно оценивать путем измерения количества или концентрации мРНК, транскрибируемой с гена, путем определения количества или концентрации полипептида и путем определения уровня ферментативной активности.
Количество мРНК можно определять, среди прочего, с помощью методов «Нозерн-блоттинга» и количественной ОТ-ПЦР. При проведении количественной ОТ-ПЦР перед полимеразной цепной реакцией осуществляют обратную транскрипцию. Для этой цели можно применять, например, систему LightCycler™ фирмы Roche Diagnostics (фирма Boehringer Mannheim GmbH, фирма Roche Molecular Biochemicals, Маннгейм, Германия), как описано у Jungwirth и др., FEMS Microbiology Letters 281, 2008, сс.190-197. Концентрацию белка можно определять путем фракционирования белка методом 1-й 2-мерного гель-электрофореза и последующей идентификации концентрации белка в геле оптическими методами с использованием соответствующего программного обеспечения для осуществления анализа. В качестве общего метода приготовления содержащих белок гелей в случае применения коринеформных бактерий и идентификации белков можно применять процедуру, описанную у Hermann и др., Electrophoresis, 22, 2001, сс.1712-1723. Концентрацию белка можно определять также с помощью гибридизации по методу Вестерн-блоттинга с использованием антитела, специфического в отношении предназначенного для выявления белка (Sambrook и др., Molecular cloning: a laboratory manual, 2-е изд., изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989), и последующего анализа оптическими методами с использованием соответствующего программного обеспечения, предназначенного для определения концентрации (Lohaus и Meyer, Biospektrum 5, 1998, сс.32-39; Lottspeich, Angewandte Chemie 321, 1999, сс.2630-2647).
Для получения L-орнитина сконструированные бактерии можно культивировать непрерывно (как описано, например, в WO 05/021772) или с перерывами с помощью периодического процесса (культивирование партий) или периодического процесса с подпиткой или периодического процесса с повторной подпиткой (например, как описано в US 6562601). Обзор общих принципов, на которых основаны известные методы культивирования, можно найти в учебнике Chmiel, Bioprozesstecknik [Bioprocess Technology], 1. Einfuhrung in die Bioverfahrenstechnik [Introduction to Bioprocess Engineering], изд-во Gustav Fischer Verlag, Stuttgart, 1991, или в учебнике Storhas, Bioreaktoren und periphere Einrichtungen [Bioreactors and Peripheral Equipment], изд-во Vieweg Verlag, Braunschweig/Wiesbaden, Germany, 1994.
Предназначенная для применения культуральная среда или ферментационная среда должна удовлетворять соответствующим образом требованиям, зависящим от конкретных штаммов. В руководстве «Manual of Methods for General Bacteriology» Американского общества бактериологов (Washington D.C., USA, 1981) приведено описание культуральных сред для различных микроорганизмов. Понятия «среда для роста», «культуральная среда» и «ферментационная среда» или среда используются взаимозаменяемо.
В качестве источника углерода можно использовать сахара и углеводы, такие, например, как глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, содержащие сахарозу растворы, полученные в результате обработки сахарной свеклы или сахарного тростника, крахмал, гидролизат крахмала и целлюлозу, масла и жиры, такие, например, как соевое масло, подсолнечное масло, арахисовое масло и кокосовый жир, жирные кислоты, такие, например, как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие, например, как глицерин, метанол и этанол, и органические кислоты, такие, например, как уксусная кислота или молочная кислота.
Среди Сахаров предпочтительными являются глюкоза, фруктоза, сахароза, смеси глюкозы и фруктозы и смеси глюкозы, фруктозы и сахарозы. Наиболее предпочтительной является, если это возможно, сахароза.
Среди спиртов предпочтительным является глицерин.
В качестве источника азота можно использовать содержащие азот органические соединения, такие как пептоны, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный экстракт, соевая мука и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота можно применять индивидуально или в виде смеси.
В качестве источника фосфора можно использовать фосфорную кислоту, первичный кислый фосфат калия или вторичный кислый фосфат калия, или соответствующие содержащие натрий соли.
Кроме того, культуральная среда может содержать соли, например, в форме хлоридов или сульфатов металлов, таких, например, как натрий, калий, магний, кальций и железо, такие, например, как сульфат магния или сульфат железа, которые необходимы для роста. И, наконец, помимо указанных выше субстанций можно применять незаменимые факторы роста, такие как аминокислоты, например, гомосерин, и витамины, например, тиамин, биотин или пантотеновую кислоту.
Указанные исходные продукты можно добавлять в культуру в виде одной партии или осуществлять соответствующим образом подпитку в процессе культивирования.
Значение pH культуры можно контролировать соответствующим образом путем применения оснований, таких как гидроксид натрия, гидроксид калия, аммиак или водный раствор аммиака, или кислот, таких как фосфорная кислота или серная кислота. Как правило, pH доводят до значения, составляющего от 6,0 до 8,5, предпочтительно от 6,5 до 8. Для контроля пенообразования можно применять противовспенивающие средства, такие, например, как эфиры жирной кислоты и полигликоля. Для поддержания стабильности плазмид можно добавлять в среду пригодные обладающие избирательным действием субстанции, такие, например, как антибиотики. Ферментацию предпочтительно осуществляют в аэробных условиях. Для поддержания таких условий в культуру вводят кислород или кислородсодержащие газовые смеси, такие, например, как воздух. Можно применять также жидкости, обогащенные пероксидом водорода. При необходимости ферментацию осуществляют при избыточном давлении, например, при избыточном давлении, составляющем от 0,03 до 0,2 МПа. Температура культуры обычно составляет от 20 до 45°C и предпочтительно от 25 до 40°C, наиболее предпочтительно от 30 до 37°C. В случае периодических процессов культивирование продолжают до тех пор, пока не образуется достаточное для выделения количество требуемого L-орнитина. Как правило, для достижения этой цели требуется от 10 до 160 ч. В случае непрерывных процессов может потребоваться осуществлять культивирование в течение более длительных промежутков времени. В результате активности бактерий происходит обогащение или повышение концентрации (накопление) L-орнитина в ферментационной среде.
Примеры пригодных ферментационных сред описаны, среди прочего, в патентах JP 43010996 В4 (для В. subtilis), US 3668072 А (для Е. coli) и JP 57041912 В (для B.flavum).
Объем ферментационной среды в процессе, предлагаемом в изобретении, может составлять при необходимости ≥0,5 л, ≥1 л, ≥5 л, ≥10 л, ≥50 л, ≥100 л, ≥500 л, ≥1000, предпочтительно ≥1 л, более предпочтительно ≥10 л, еще более предпочтительно ≥100 л и наиболее предпочтительно ≥1000 л.
Для определения концентрации L-орнитина в один момент времени или несколько моментов времени в процессе ферментации можно анализировать его содержание путем выделения L-аминокислот с помощью ионообменной хроматографии, предпочтительно катионообменной хроматографии, с последующей постколоночной дериватизацией с использованием нингидрина согласно методу, описанному у Spademan и др., Analytical Chemistry 30, 1958, сс.1190-1206. Для постколоночной дериватизации можно применять орто-фталдиальдегид вместо нингидрина. Обзор публикаций, в которых описана ионообменная хроматография, приведен у Pickering, LC.GC, Magazine of Chromatographic Science, 7(6), 1989, сс.484-487.
Можно осуществлять также предколоночную дериватизацию, например, с использованием орто-фталдиальдегида или фенилизотиоцианата, и осуществлять фракционирование полученных производных аминокислот с помощью обращенно-фазовой хроматографии (ОФ), предпочтительно в форме жидкостной хроматографии высокого разрешения (ЖХВР). Такой метод описан, например, у Lindroth и др., Analytical Chemistry 51, 1979, сс.1167-1174. Обнаружение осуществляют фотометрическим методом (абсорбция, флуоресценция).
Обзор методов анализа аминокислот представлен среди прочего в учебнике Lottspeich и Zorbas, «Bioanalytik», изд-во Spektrum Akademischer Verlag, Heidelberg, Germany 1998.
Производительность процессов или ферментационных процессов, предлагаемых в изобретении, касательно одного или нескольких параметров, выбранных из группы, включающей концентрацию L-орнитина (количество образовавшегося L-орнитина на единицу объема), выход L-орнитина (количество образовавшегося L-орнитина на единицу поглощенного источника углерода), образование L-орнитина (количество образовавшегося L-орнитина на единицу объема в единицу времени) и удельное образование L-орнитина (количество образовавшегося L-орнитина на единицу сухого клеточного вещества или сухой биомассы в единицу времени, или количество образовавшегося L-орнитина на единицу клеточного белка в единицу времени), или других параметров процесса и их комбинаций, повышают по меньшей мере на 0,5%, по меньшей мере на 1%, по меньшей мере на 1,5% или по меньшей мере на 2%, по сравнению с процессами или ферментационными процессами с применением бактерий, в которых не происходит сверхэкспрессия белка, обладающего активностью в отношении экспорта L-орнитина, или которые не подвергали манипуляциям с целью обеспечения сверхэкспрессии.
Посредством ферментации получают ферментационный бульон, который содержит требуемый L-орнитин.
Затем продукт, содержащий L-орнитин, запасают или получают, или выделяют в жидкой или твердой форме.
Ферментационный бульон представляет собой ферментационную среду или среду для роста, в которой осуществляли культивирование микроорганизма в течение определенного периода времени и при определенной температуре. Ферментационная среда или среды, применяемая/применяемые при ферментации, содержит/содержат все субстанции или компоненты, обеспечивающие производство L-орнитина, и, как правило, размножение и жизнеспособность микроорганизма.
После завершения ферментации полученный ферментационный бульон содержит соответственно:
а) бактериальную биомассу (клеточную массу), образовавшуюся в результате размножения бактериальных клеток,
б) L-орнитин, образовавшийся в результате ферментации,
в) органические побочные продукты, образовавшиеся в процессе ферментации, и
г) компоненты применявшейся ферментационной среды или исходных продуктов, например, витамины, такие как биотин, или соли, такие как сульфат магния, которые не были поглощены в процессе ферментации.
К органическим побочным продуктам относятся субстанции, которые были произведены бактериями, применяемыми для ферментации, помимо L-орнитина, и которые необязательно были высвобождены. Они включают также сахара, такие, например, как трегалоза.
Ферментационный бульон удаляют из резервуара для культивирования или ферментационного бака, необязательно собирают и используют для получения содержащего L-орнитин продукта в жидкой или твердой форме. Для обозначения этой операции применяют также выражение «выделяют содержащий L-орнитин продукт». В простейшем случае выделенный продукт представляет собой сам содержащий L-орнитин ферментационный бульон, который был удален из ферментационного бака.
Концентрацию или очистку L-орнитина из ферментационного бульона осуществляют с помощью одной или нескольких процедур, выбранных из группы, включающей:
а) удаление от частичного (от >0 до <80%) до полного (100%) или практически полное удаление (≥80%, ≥90%, ≥95%, ≥96%, ≥97%, ≥98% или от ≥99 до <100%) воды,
б) удаление от частичного (от>0 до<80%) до полного (100%) или практически полное удаление (≥80%, ≥90%, ≥95%, ≥96%, ≥97%, ≥98% или от ≥99 до<100%) биомассы, которую необязательно инактивируют перед осуществлением удаления,
в) удаление от частичного (от >0 до <80%) до полного (100%) или практически полное удаление (≥80%, (90%, ≥95%, ≥96%, ≥97%, ≥98% или от ≥99 до <100%) органических побочных продуктов, образовавшихся в процессе ферментации, и
г) удаление от частичного (от >0 до <80%) до полного (100%) или практически полное удаление (≥80%, ≥90%, ≥95%, ≥96%, ≥97%, ≥98% или от ≥99 до <100%) компонентов применявшейся ферментационной среды или исходных продуктов, которые не были поглощены при ферментации. Таким путем выделяют продукты, имеющие требуемое содержание L-орнитина.
Удаление от частичного (от>0 до<80%) до полного (100%) или практически полное удаление (≥80%, ≥90%, ≥95%, ≥96%, ≥97%, ≥98% или от ≥99 до<100%) воды (процедура а)) называют также сушкой.
В одном из вариантов осуществления способа, полное или практически полное удаление воды, биомассы, органических побочных продуктов и непоглощенных компонентов ферментационной среды приводит к получению содержащего L-орнитин продукта в чистой форме (≥80 мас.% или ≥90 мас.%) или высокоочищенной форме (≥95 мас.%, ≥97 мас.% или ≥99 мас.%). Из существующего уровня техники известны многочисленные технические инструкции касательно осуществления процедур, указанных в подпунктах а), б), в) или г).
В случае аминокислоты L-орнитина или ее солей из существующего уровня техники известны в основном три различных продукта.
К одной из групп относится L-орнитин-HCL, в этом случае L-орнитин очищают из ферментационного раствора после удаления клеток с помощью ионообменника и затем кристаллизуют посредством кристаллизации в виде монохлорида L-орнитина и перекристаллизации в виде монохлорида L-орнитина (US 2988489). Получаемый в этом случае L-орнитирн-HCL имеет степень чистоты более чем 90%, предпочтительно более чем 95%, еще более предпочтительно более чем 98%, и наиболее предпочтительно более чем 99%.
Другой процесс описан в заявке на патент ЕР 1995322. В нем содержащий биомассу ферментационный бульон вносят наверх слабокислого ионообменника с диаметром частиц>300 мкм и на этой стадии производят очистку L-орнитина. Выбирая соответствующий диаметр частиц, предупреждают блокирование смолы биомассой. Эффективность удаления клеток составляла 99%.
Затем очищенный L-орнитин можно использовать для получения различных солей L-орнитина, таких, например, как моно- или ди-L-орнитин-α-кетоглутарат, L-орнитин-L-аспартат и т.д.
В ЕР 0477991 описан, например, процесс получения L-орнитин-L-аспартата. Согласно этому процессу добавляют к водному раствору L-орнитина и L-аспартата водорастворимый растворитель до получения раствора, насыщенного по меньшей мере на 90%, или перенасыщенного раствора. Полученный раствор выдерживают при температуре дефлегмации до завершения образования кристаллов. Затем продолжают добавлять при температуре дефлегмации смешивающийся с водой растворитель до формирования кристаллов соли. Кристаллы можно удалять, например, путем центрифугирования, и затем сушить в вакууме. Чистота продукта, как правило, составляет более 98,5%.
В JP 46003194 описан процесс получения L-орнитин-L-кетоглутарата. Согласно этому процессу орнитин L-орнитина-HCL превращают в свободное основание путем адсорбции с использованием кислого ионообменника и элюции с использованием водного раствора аммиака, добавления α-кетоглутарата и выпаривания раствора в вакууме до кристаллизации продукта.
Плазмиду pEC71ysE депонировали в форме штамма Escherichia coli DH5alpha/pEC71ysE (DM2204) в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (DSMZ, Брауншвейг, Германия) под регистрационным номером DSM 23239 15 января 2010 г.
Примеры
Пример 1
Клонирование и секвенирование гена lysE из штамма Corynebacterium glutamicum АТСС 13032
Ген lysE из штамма АТСС 13032 клонировали в бифункциональном (Е. coli/С.glutamicum) и экспрессионном векторе pVWExl (Peters-Wendisch и др., J.Mol. Microbiol. Biotechnol., 3(2), 2001, сс.295-300).
Процесс клонирования состоял из двух стадий. Сначала с помощью полимеразной цепной реакции (ПЦР) амплифицировали ген из штамма Corynebacterium glutamicum АТСС13032 с использованием представленных ниже олигонуклеотидных праймеров, созданных на основе последовательности, которая представлена в SEQ ID NO:1. Указанные олигонуклеотиды включали дополнительные сайты рестрикции на своем 5'-конце (подчеркнуты: EcoRV для
lysE_l.p и Avrll или Sspl для lysE_2.p).
lysE_l.р: 5' - [TCGATATCATGGAAATCTTCATTACAGG]-3'
(см. SEQ ID NO:22);
lysE_2.p:5,-[TGCCTAGGTCAATATTTGGGCGAAGGCCACCGl-3, (см. SEQ ID NO:23).
Реакцию ПЦР осуществляли в присутствии 200 мкМ дезоксинуклеозидтрифосфатов (дАТФ, дЦТФ, дГТФ, дТТФ), 0,5 мкМ каждого соответствующего олигонуклеотида, 100 нг хромосомальной ДНК штамма Corynebacterium glutamicum АТСС13032, 1/5 объема 5-кратного реакционного буфера HF и 0,02 ед./мкл ДНК-полимеразы Phusion® Hot Start (фирма Biozym Scientific GmbH, D-31840 Hess, Олдендорф) в термоячейке (Mastercycler, фирма Eppendorf AG, Гамбург) в следующих условиях: 98°C в течение 1 мин; 30 циклов х (98°C, 20 с; 63°C, 20 с; 72°C, 40 с); 72°C в течение 6 мин.
Полученный в результате ПЦР фрагмент lysE, состоящий из 761 пары оснований (см. SEQ ID NO: 3), клонировали в pVWExl согласно описанному ниже методу:
Получение вектора: 1 мкг плазмидной ДНК pVWExl расщепляли в фермент-специфической буферной системе, содержащей 10 единиц фермента PstI, посредством инкубации в течение 1 ч при 37°С.Сразу после этого полученную в результате расщепления смесь обрабатывали с использованием набора Quick Blunting (фирма New England Biolabs GmbH, Франкфурт-на Майне) согласно инструкциям производителя и затем очищали с использованием набора для очистки QiaExII (фирма Qiagen AG, Гильден, Германия) согласно инструкциям производителя. Затем предварительно обработанный таким путем вектор расщепляли с помощью 10 единиц Xbal в вектор-специфической буферной системе в течение 1 ч при 37°C и после этого снова производили очистку с использованием набора для очистки QiaExII.
Получение вставки: ПЦР-фрагмент lysE расщепляли с помощью 10 единиц каждого из ферментов Avrll и EcoRV и затем очищали с использованием набора для очистки QiaExII согласно инструкциям производителя.
Лигирование: вектор и вставку смешивали в молярном соотношении 1:5 и лигировали с использованием ДНК-лигазы Т4 при 16°C в течение 1 ч. Химически компетентные клетки штамма Е. coli DH5alpha (Subcloning Efficiency™, Invitrogen GmbH, Карлсруэ, Германия) трансформировали с использованием 3 мкл смеси для лигирования.
Идентификацию трансформантов осуществляли на основе их устойчивости к канамицину на пластинах с LB-агаром, содержащем 50 мкг/мл сульфата канамицина. Плазмидную ДНК выделяли из 4 указанных трансформантов и плазмиды анализировали с помощью рестрикционного анализа в отношении присутствия вставки, представляющей собой фрагмент длиной 0,75 т.п.н. Рекомбинантную плазмиду, полученную таким путем, обозначили как pVWExl_lysE.
Нуклеотидную последовательность фрагмента длиной 0,75 т.п.н., присутствующего в плазмиде pVWExl-lysE, определяли методом терминации дидезокси-цепи, разработанному Sanger и др., Proceedings of the National Academy of Sciences of the United States of America, 74, 1977, cc. 5463-5467. Для этой цели осуществляли секвенирование полной вставки плазмиды pVWExl_lysE с использованием олигонуклеотидных праймеров pVW_l.p (5'-TGA GCG GAT ААС ААТ ТТС АСА С-3') и pVW_2.p (5'-CGA CGG CCA GTG AAT TCG AG-3') на фирме Eurofins MWG Operon GmbH (Эберсберг, Германия).
Полученную нуклеотидную последовательность анализировали с помощью программы Clone Manager 9, и она представлена в SEQ ID NO: 20.
Пример 2
Конструирование вектора pK18mobsacB_DargFRGH для осуществления делеции области argFRGH в Corynebacterium glutamicum
Для указанной цели сначала выделяли хромосомальную ДНК из штамма С.glutamicum АТСС13032 согласно методу, описанному у Tauch и др., Plasmid 33, 1995, сс.168-179. Для получения содержащей делецию argFRGH конструкции были выбраны представленные ниже олигонуклеотиды на основе последовательности гена argFRGH С.glutamicum. Указанную содержащую делецию конструкцию создавали с помощью полимеразной цепной реакции (ПЦР), более конкретно, с помощью метода Gene SOEing (сплайсинг генов путем удлинения с перекрыванием (Gene Splicing by Overlap Extension), Horton,
Molecular Biotechnology 3, 1995, cc. 93-98).
argFRGH_dl:
5'-GGT GGT GCT AGC CCG GCG ATT TCT TTG С AC AT-3'
(cm. SEQ ID NO:24)
argFRGH_d2:
5'-AAT GCT TAT CGA CGT ACC CCC CTG TGG TTG TGA AGT CAT A-3'
(см. SEQ ID NO:25)
argFRGH_d3:
5'-GGG GTA CGT CGA TAA GCA TT-3'
(cm. SEQ ID NO: 26)
argFRGH_d4:
5'-GGT GGT ATG CAT GGT GAT GGT TCC GAA TGT TG-3' (см. SEQ ID NO: 27).
Указанные олигонуклеотидные праймеры получали от фирмы Eurofins MWG Operon GmbH (Эберсберг, Германия). ПЦР осуществляли с использованием ДНК-полимеразы Phusion® Hot Start (фирма Biozym Scientific GmbH, D-31840 Hess, Олдендорф) в термоячейке (Mastercycler, фирма Eppendorf AG, Гамбург).
Праймер argFRGH_d2 состоял из двух областей. Одна часть нуклеотидной последовательности комплементарна области, простирающейся от 1-й пары оснований, находящейся против хода транскрипции, до 19-й пары оснований, находящейся по ходу транскрипции, относительно стартового кодона гена argF. Другая часть нуклеотидной последовательности комплементарна области, простирающейся от нуклеотида 1419 гена argH до нуклеотида 5, расположенного по ходу транскрипции относительно гена argH.
С помощью полимеразной цепной реакции (ПЦР) с использованием праймеров argFRGH_l и argFRGH_2 можно амплифицировать фрагмент ДНК длиной 543 пары оснований, а с использованием праймеров argFRGH_3 и argFRGH_4 можно амплифицировать фрагмент ДНК длиной 513 пар оснований. Ампликоны получали с помощью ПЦР, анализировали методом электрофореза в 0,8%-ном агарозном геле, выделяли из агарозного геля с использованием набора для очистки ПЦР-продуктов High Pure PCR Product Purification Kit (номер продукта 1732676, фирма Roche Diagnostics GmbH, Маннгейм, Германия), и применяли в качестве матрицы для следующей ПЦР с использованием праймеров argFRGH_l и argFRGH_4. Таким путем создавали производное длиной 1036 пар оснований, имеющее делецию DargFRGH (см. также SEQ ID NO: 21). Указанная конструкция включала 477 пар оснований 3'-концевой области гена argD, 19 пар оснований 5'-концевой области гена argF, 15 пар оснований 3'-концевой области гена argH и 420 пар оснований 5'-концевой области рамки считывания cgl589. Амплифицированный таким путем продукт анализировали методом электрофореза в 0,8%-ном агарозном геле.
ПЦР-продукт DargFRGH длиной 1,04 т.п.н. (SEQ ID NO:21) полностью расщепляли ферментами Ndel и Nsil. Затем фрагмент очищали с помощью набора для очистки ПЦР-продукта (фирма Qiagen, Гильден, Германия). Имеющее делецию DargFRGH производное, предварительно обработанное таким путем, применяли в сочетании с мобилизуемым клонирующим вектором pK18mobsacB (Schafer и др., Gene 14, 1994, сс.69-73) для лигирования. Указанный клонирующий вектор предварительно полностью расщепляли с помощью рестриктаз Xbal и Pstl. В результате получали концы ДНК, совместимые с концами вставки, созданной путем расщепления с помощью Ndel и Nsil. Полученный таким путем вектор смешивали с фрагментом DargFRGH в молярном соотношении 1:5 и осуществляли лигирование с использованием ДНК-лигазы Т4 (фирма Amersham-Pharmacia, Фрейбург, Германия) при 16°С в течение 1 ч. Химически компетентные клетки штамма Е. coli DH5alpha (Subcloning Efficiency™, фирма Invitrogen GmbH, Карлсруэ, Германия) трансформировали с использованием 3 мкл смеси для лигирования. Трансформанты идентифицировали на основе их устойчивости к канамицину на пластинах с LB-агаром, содержащих сульфат канамицина в концентрации 50 мкг/мл. Плазмидную ДНК выделяли из 4 трансформантов (набор QIAprep Spin Miniprep фирмы Qiagen (Гильден)), и плазмиды анализировали с помощью рестрикционного анализа в отношении присутствия вставки, представляющей собой фрагмент длиной 1,04 т.п.н. Полученную таким путем рекомбинантную плазмиду обозначили как pK18mobsacB_DargFRGH. Штамм был обозначен как £.co//_DH5alpha/pK18mobsacB_DargFRGH.
Нуклеотидную последовательность фрагмента длиной 1,04 т.п.н. (SEQ ID NO: 21) в плазмиде pK18mobsacB_DargFRGH определяли методом терминации дидезокси-цепи, разработанным Sanger и др., Proceedings of the National Academy of Sciences of the United States of America, 74, 1977, cc. 5463-5467. Для этой цели секвенировали полную вставку плазмиды pK18mobsacB_DargFRGH с использованием праймеров М13 uni (-21) (5'-TGT AAA ACG ACG GCC AGT-3') и M13 rev (-49) (5'-GAG CGG ATA АСА ATT TCA С AC AGG-3') на фирме Eurofins MWG Operon (Эберсберг, Германия) и тем самым проверяли ее правильность.
Пример 3
Получение штамма Corynebacterium zlutamicum АТСС 13032 DargFRGH Вектор, указанный в примере 2, а именно pK18mobsacB_DargFRGH, встраивали путем конъюгации согласно протоколу, разработанному Schafer и др., Journal of Microbiology 172, 1990, сс.1663-1666, в штамм Corynebacterium glutamicum АТСС 13032. Для этой цели вектором предварительно трансформировали штамм Е. coli SI7-1 (Simon и др., Biotechnology 1, сс.784-791). Вектор, присутствующий в S17-1, анализировали в отношении его идентичности аналогично тому, как это осуществляли при обнаружении в штамме Е. coli DH5alpha (см. пример 2).
Векторы pK18mobsacB и pK18mobsacB_DargFRGH не могут самореплицироваться в штамме С.glutamicum АТСС13032 и остаются в клетке только в том случае, когда они интегрированы в хромосому посредством рекомбинации. Клоны с интегрированной pK18mobsacB_DargFRGH отбирали путем высевания конъюгированной смеси на LB-агар (Sambrook и др., Molecular Cloning: A Laboratory Manual, 2-е изд., изд-во Cold Spring Harbor, New York, 1989), дополненный 15 мг/мл канамицина и 50 мг/мл налидиксовой кислоты. Полученные клоны стряхивали на LB-агаровые пластины, содержащие 25 мг/мл канамицина и инкубировали при 33°С в течение 16 ч. Мутанты, из которых плазмида была вырезана в результате второго этапа рекомбинации, отбирали путем культивирования клонов в жидкой LB-среде без осуществления селекции в течение 20 ч, затем их стряхивали на LB-агар, содержащий 10% сахарозы, после чего инкубировали в течение 24 ч.
Плазмида pK18mobsacB_DargFRGH содержала аналогично исходной плазмиде pK18mobsacB помимо гена, обусловливающего устойчивость к канамицину, копию гена sacB, кодирующего левансахарозу Bacillus subtilis. Индуцируемая сахарозой экспрессия приводила к образованию левансахарозы, которая катализирует синтез продукта левана, токсичного для С.glutamicum. Вследствие этого только те клоны, из которых вырезана интегрированная плазмида pK18mobsacB_DargFRGH, снова приобретали способность расти на LB-arape, содержащем сахарозу. Вырезание может заключаться в вырезании плазмиды вместе с полной хромосомальной копией argFRGH или с неполной копией, имеющей внутреннюю делецию argFRGH.
Примерно 40-50 колоний подвергали тестированию в отношении фенотипа «рост в присутствии сахарозы» и «отсутствие роста в присутствии канамицина». Для доказательства того, что подвергнутый делеции аллель argFRGH сохранился в хромосоме, примерно 20 колоний, имеющих фенотип «рост в присутствии сахарозы» и «отсутствие роста в присутствии канамицина», анализировали с помощью полимеразной цепной реакции согласно стандартному ПЦР-методу (см. Innis и др., PCR Protocols. A Guide to Methods and Applications, изд-во Academic Press, 1990). Для этого амплифицировали из хромосомальной ДНК колоний ДНК-фрагмент, содержащий области, окружающие удаленную в результате делеции argFRGH-область. Для ПЦР были выбраны следующие олигонуклеотидные праймеры: argFRGH_dl (SEQ ID NO:24):
5'-GGT GGT GCT AGC CCG GCG ATT TCT TTG С AC AT-3' argFRGH_d4 (SEQ ID NO:27):
5'-GGT GGT ATG CAT GGT GAT GGT TCC GAA TGT TG-3'.
В контрольных клонах, содержащих полный локус argFRGH, праймеры позволяли амплифицировать ДНК-фрагмент длиной примерно 5,35 т.п.н. В клонах, в которых локус argFRGH был удален путем делеции, в результате амплификации получали ДНК-фрагменты длиной примерно 1,04 т.п.н.
Амплифицированные ДНК-фрагменты идентифицировали с помощью электрофореза в 0,8%-ном агарозном геле. С помощью этого метода было продемонстрировано, что штамм имел хромосому, из которой был удален путем делеции аллель argFRGH. Указанный штамм обозначили как штамм Corynebacterium glutamicum Delta_argFRGH.
Пример 4
Экспрессия гена lysE в штамме Corynebacterium glutamicum АТСС 13032 Delta argFRGH
Плазмиду pVWExl_LysE и «пустую» плазмиду pVWExl встраивали путем электропорации (Haynes и др., FEMS Microbiology Letters, 61, 1989, сс.329-334) в продуцирующий L-орнитин штамм АТСС 13032_Delta_argFGH. Трансформанты идентифицировали на основе их устойчивости к канамицину на пластинах с Caso-агаром, содержащем 25 мкг/мл канамицина. Затем 5 индивидуальных клонов анализировали в отношении правильности трансформированной плазмиды. Для этой цели выделяли плазмидную ДНК (набор для выделения плазмид, фирма Qiagen) и полученную ДНК анализировали с помощью рестрикционного анализа в отношении правильности схемы расщепления. Таким путем получали штаммы С.glutamicum АТСС 13032_Delta_argFRGH/pVWExl_lysE и АТСС 13032_Delta_argFRGH/pVWExl.
Пример 5
Получение L-орнитина с использованием Corynebacterium slutamicum
Для изучения способности продуцировать L-орнитин осуществляли предварительное культивирование в каждом случае трех клонов штамма АТСС 13032_Delta_argFRGH/pVWExl_lysE и трех клонов штамма АТСС 13032_Delta_argFRGH/pVWExl в каждом случае в 10 мл тест-среды при 33°C в течение 16 ч. Для анализа продуктивности в каждом случае инокулировали 10 мл тест-среды полученной предварительной культурой таким образом, чтобы величина ОПбоо (оптическая плотность при 600 нм) в начале эксперимента составляла 0,1. Каждый клон тестировали в трех встряхиваемых колбах, таким образом, в соответствующий момент времени сбора для каждого штамма анализировали содержимое в общей сложности девяти встряхиваемых колб. Тест-среда была идентичной среде CgXII, описанной у Keilhauer и др., Journal of Bacteriology, 175, 1993, сс.5593-5603, но дополнительно содержала 7,5 г/л дрожжевого экстракта (фирма Difco), 25 мкг/мл канамицина, 1 мМ ИПТГ (изопропил-бета-D-тиогалактопиранозид) и 40 г/л сахарозы вместо глюкозы. Для наглядности состав тест-среды обобщен в приведенной ниже таблице 2.
Культивирование осуществляли во встряхиваемых колбах вместимостью 100 мл при 33°C и 200 об/мин. Отклонение шейкера составляло 5 см. Через 24 и 48 ч собирали по три культуры клона. Для этой цели из культур брали образцы и определяли оптическую плотность, содержание сахарозы и содержание L-орнитина. Для определения содержания сахарозы и содержания L-орнитина клетки удаляли путем кратковременного центрифугирования (настольная центрифуга типа 5415D (фирма Eppendorf) при 13000 об/мин в течение 10 мин при комнатной температуре).
Оптическую плотность определяли при длине волны 660 нм с помощью фотометра для титрационных микропланшетов GENios (фирма Тесап, Ридинг, Великобритания). Образцы разводили в соотношении 1:100 деминерализованной водой перед осуществлением измерения.
Содержание сахарозы определяли с помощью системы анализа (каталожный №10716251035) фирмы R-Biopharm AG (Дармштадт, Германия). При использовании указанного подхода осуществляют инверсию сахарозы и образовавшуюся глюкозу выявляют с помощью анализа с использованием совместно иммобилизованных ферментов (гексокиназа/глюкозо-6-фосфатдегидрогеназа) по образованию НАД·Н.
Количественное определение концентрации аминокислоты вне клетки, а именно в супернатанте культуры, осуществляли с помощью ЖХВР с обращенной фазой (Lindroth и др., Analytical Chemistry 51, 1979, сс.1167-1174) с использованием устройства для ЖХВР серии HP 1100 (фирма Hewlett-Packard, Вальдбронн, Германия) с присоединенным флуоресцентным детектором (G1321 А); управление системой и анализ данных осуществляли с помощью станции HP ChemStation (фирма Hewlett-Packard). 1 мкл раствора аминокислоты, подлежащего анализу, смешивали в автоматическом устройстве для предколоночной дериватизации с 20 мкл готового для применения реагента орто-фталальдегид/2-меркаптоэтанол (фирма Pierce Europe BV, Оуд-Бейерланд, Нидерланды). Образовавшиеся флуоресцентные тио-замещенные изоиндолы (Jones и др., Journal of Chromatography, 266, 1983, сс.471-482) подвергали фракционированию на комбинации предколонки (40×4 мм Hypersil ODS 5) и основной колонки (Hypersil ODS 5, обе колонки получали от фирмы CS-Chromatographie Service GmbH, Лангервеэ, Германия) с использованием градиентной программы с возрастанием неполярной фазы (метанол). В качестве полярного элюента применяли ацетат натрия (0,1М; pH 7,2); скорость потока составляла 0,8 мл/мин. Флуоресценцию дериватизированных аминокислот обнаруживали при длине волны возбуждения 230 нм и длине волны испускания 450 нм. Концентрации L-орнитина и/или гидрохлорида L-орнитина рассчитывали на основе сравнения с внешним стандартом и L-аспарагином, служащим в качестве дополнительного внутреннего стандарта.
Молекулярная масса гидрохлорида L-орнитина составляет 168,6 г × моль-1, молекулярная масса L-орнитина составляет 132,1 г × моль-1.
Выход рассчитывали путем деления количества образовавшегося L-орнитина (измеренного по количеству гидрохлорида L-орнитина) на количество поглощенной сахарозы.
Результаты представлены в таблице 3.
Таблица 3: Образование L-орнитина после инкубации в течение 24 ч (таблица 3А) и 48 ч (таблица ЗБ). Сокращения: символ * обозначает АТСС 13032_Delta_argFRGH; Orn-HCl обозначает гидрохлорид L-орнитина.
Пример 6: Секвенирование и депонирование плазмиды pEC71ysE
Плазмиду pEC71ysE получали в форме водного раствора от д-ра Lothar Eggeling (фирма Forschungszentrum Julich GmbH, D-52425, Юлих), автора-корреспондента публикации Bellmann и др., Microbiology, 147, 2001, сс.1765-1774.
Аликвоту полученного раствора ДНК использовали для трансформации компетентных клеток штамма DH5alpha Escherichia coli (Subcloning Efficiency™, генотип: F-<P80/acZΔM15 Δ{lacZY A-argF) U169 recAl endAl hsdR17 (rK-, mk+) phoA supE44λ-thi-1 gyrA96 relAl), полученного от фирмы Invitrogen GmbH (Пейсли, Великобритания) согласно инструкциям производителя. Отбор трансформантов осуществляли на Луриа-Бертани-агаре, дополненном 50 мкг/мл канамицина.
Один трансформант, обозначенный как штамм Escherichia coli DH5alpha/pEC71ysE(DM2204), был депонирован в соответствии с Будапештским договором в Немецкой коллекции микроорганизмов и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Брауншвейг, Германия) 15 января 2010 г.под регистрационным номером DSM 23239.
Последовательность плазмиды pEC71ysE из штамма DSM 23239 была полностью секвенирована в соответствии с договором на проведение секвенирования (Walking Service) на фирме Eurofins MWG Operon GmbH (Мартинсрид, Германия). Последовательность pEC71ysE представлена в SEQ ID NO:29.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ СЕРУСОДЕРЖАЩИХ АМИНОКИСЛОТ | 2012 |
|
RU2651511C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА С ИСПОЛЬЗОВАНИЕМ КОРИНЕФОРМНОЙ БАКТЕРИИ | 2011 |
|
RU2496867C2 |
| ПОЛИНУКЛЕОТИД, СОДЕРЖАЩИЙ НУКЛЕОТИДНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩУЮ КОМПОНЕНТ Н СИСТЕМЫ ФОСФОТРАНСФЕРАЗЫ, ПЛАЗМИДНЫЙ ВЕКТОР, СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ, ШТАММ БАКТЕРИЙ И ПОЛИНУКЛЕОТИД, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПОЛУЧЕНИЯ ПОЛИНУКЛЕОТИДОВ, КОДИРУЮЩИХ ПРОДУКТ ГЕНА ptsH | 2001 |
|
RU2268938C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 2000 |
|
RU2264461C2 |
| Микроорганизмы, продуцирующие путресцин, и способ получения путресцина с использованием этих микроорганизмов | 2014 |
|
RU2665825C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ УЛУЧШЕННЫХ ШТАММОВ СЕМЕЙСТВА Enterobakteriaceae | 2013 |
|
RU2672614C2 |
| НОВЫЕ КОДИРУЮЩИЕ НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ ГЕНА ZWAL | 2000 |
|
RU2260048C2 |
| МИКРООРГАНИЗМ, СОДЕРЖАЩИЙ ВАРИАНТ LysE, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2805078C1 |
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ ЛИЗИН, И СПОСОБ ПРОДУЦИРОВАНИЯ ЛИЗИНА | 2021 |
|
RU2797843C9 |
| РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ, ОБЛАДАЮЩИЙ ПОВЫШЕННОЙ СПОСОБНОСТЬЮ ПРОДУЦИРОВАТЬ ПУТРЕСЦИН, И СПОСОБ ПОЛУЧЕНИЯ ПУТРЕСЦИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2013 |
|
RU2603089C2 |







































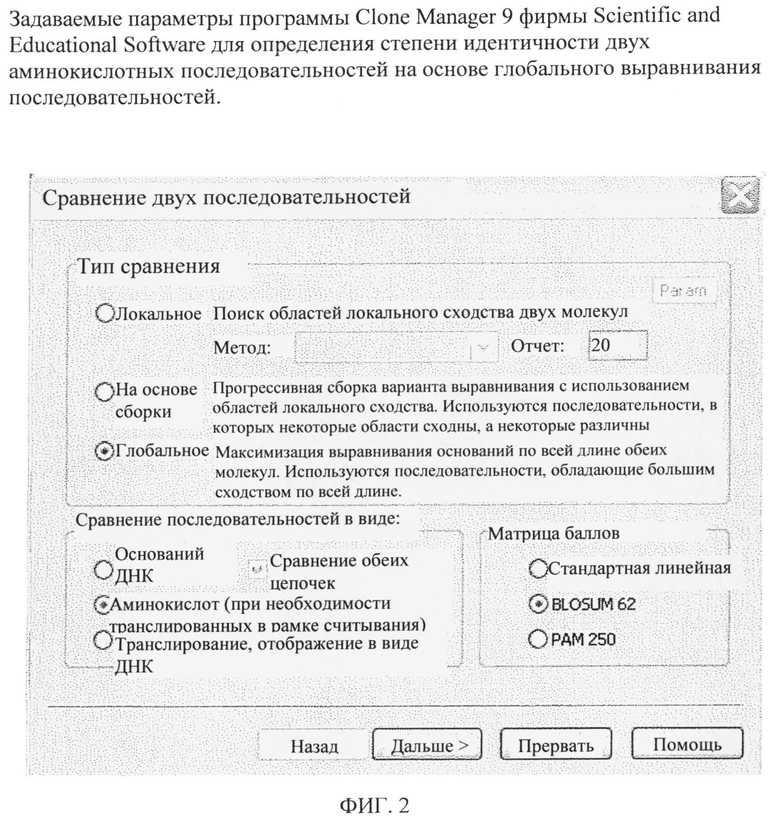
Изобретение относится к области биохимии. Предложен способ получения L-орнитина. Способ включает: а) осуществление ферментации высвобождающей в среду L-орнитин бактерии, в которой происходит сверхэкспрессия полинуклеотида, кодирующего полипептид, который обладает активностью экспортера L-орнитина и аминокислотная последовательность которого идентична аминокислотной последовательности, представленной в SEQ ID NO: 2; б) накопление L-орнитина в среде, получая в результате ферментационный бульон. Способ осуществляется при условии, что для осуществления сверхэкспрессии не применяют плазмиду pEC7lysE, депонированную под регистрационным номером DSM23239. Изобретение позволяет повысить производительность ферментационного процесса получения L-орнитина. 9 з.п. ф-лы, 2 ил., 4 табл., 6 пр.
1. Способ получения L-орнитина, отличающийся тем, что осуществляют следующие стадии, на которых:
а) осуществляют ферментацию бактерии, высвобождающей в среду L-орнитин, которая выбрана из группы, включающей бактерии p.p. Corynebacterium, Bacillus, Streptomyces, Arthrobacter и сем. Enterobacteriaceae, в которой происходит сверхэкспрессия полинуклеотида, кодирующего полипептид, который обладает активностью экспортера L-орнитина и аминокислотная последовательность которого идентична аминокислотной последовательности, представленной в SEQ ID NO: 2, или аминокислотная последовательность которого может быть получена из последовательности, представленной в SEQ ID NO: 2, путем осуществления в общей сложности максимум 25 делеций, инсерций, замен или добавлений аминокислот на N- или соответственно на С-конце,
б) накапливают L-орнитин в среде, получая в результате ферментационный бульон, и
при условии, что для осуществления сверхэкспрессии не применяют плазмиду pEC7lysE, депонированную под регистрационным номером DSM23239.
2. Способ по п. 1, отличающийся тем, что при применении Corynebacterium glutamicum в результате сверхэкспрессии повышают уровень активности в отношении экспорта L-орнитина по меньшей мере на 10% по сравнению с активностью штамма ATCC13032, или ATCC14067, или ATCC13869.
3. Способ по пп. 1-2, отличающийся тем, что сверхэкспрессию обеспечивают с помощью одной или нескольких мер, выбранных из группы, включающей:
а) увеличение количества копий,
б) применение сильного промотора, и
в) осуществление мутации промотора, и
г) сверхэкспрессию белка-активатора.
4. Способ по п. 1, отличающийся тем, что бактерия представляет собой бактерию р. Corynebacterium, предпочтительно Corynebacterium glutamicum.
5. Способ по п. 2, отличающийся тем, что бактерия представляет собой бактерию р. Corynebacterium, предпочтительно Corynebacterium glutamicum.
6. Способ по п. 3, отличающийся тем, что бактерия представляет собой бактерию р. Corynebacterium, предпочтительно Corynebacterium glutamicum.
7. Способ по п. 1, отличающийся тем, что дополнительно ослабляют один или несколько генов, выбранных из группы, включающей:
а) ген odhA, кодирующий субъединицу E1 альфа-кетоглутаратдегидрогеназы (КФ 1.2.4.2),
б) ген sucA, кодирующий дигидролипоамид-сукцинилтрансферазу (КФ 2.3.1.61),
в) ген dapA, кодирующий дигидродипиколинатсинтазу (DapA, КФ 4.2.1.52),
г) ген dapB, кодирующий дигидродипиколинатсинтазу (DapB, КФ 1.3.1.26),
д) ген ddh, кодирующий мезо-диаминопимелатдегидрогеназу (Ddh, КФ 1.4.1.16),
е) ген lysA, кодирующий диаминопимелатдекарбоксилазу (LysA, КФ 4.1.1.20),
ж) ген argR, кодирующий любой/конкретный репрессор (ArgR) биосинтеза L-аргинина,
з) ген argF, кодирующий орнитинкарбомоилтрансферазу (ArgF, КФ 2.1.3.3),
и) ген argG, кодирующий аргининсукцинатсинтазу (ArgG, КФ 6.3.4.5),
к) ген argH, кодирующий аргининсукцинатлиазу (ASAL) (ArgH, КФ 4.3.2.1),
л) ген lysC, кодирующий аспартаткиназу (LysC, КФ 2.7.2.4), и
м) ген asd, кодирующий аспартат-полуальдегид-дегидрогеназу (Asd, КФ 1.2.1.11).
8. Способ п. 1, отличающийся тем, что дополнительно усиливают один или несколько генов, выбранных из группы, включающей:
а) глутаматдегидрогеназу (КФ 1.4.1.3), кодируемую геном gdh,
б) глутамат-N-ацетилтрансферазу (КФ 2.3.1.35 и КФ 2.3.1.1), кодируемую геном argJ,
в) ацетилглутаматкиназу (КФ 2.7.2.8), кодируемую геном argB,
г) N-ацетил-гамма-глутамил-фосфатредуктазу (КФ 1.2.1.38), кодируемую геном argC,
д) ацетилорнитинаминотрансферазу (КФ 2.6.1.11), кодируемую геном argD,
е) специфический в отношении глюкозы компонент EIIB (PtsG) (КФ 2.7.1.69) системы поглощения глюкозы, кодируемый геном ptsG,
ж) специфический в отношении сахарозы компонент EIIB (PtsS) (КФ 2.7.1.69) системы поглощения сахарозы, кодируемый геном ptsS,
з) глюкозо-6-фосфат-1-дегидрогеназу (КФ 1.1.1.49), кодируемую геном zwf,
и) глюкозо-6-фосфатизомеразу (КФ 5.3.1.9), кодируемую геном pgi,
к) фосфофруктокиназу (КФ 2.7.1.11), кодируемую геном pfkA,
л) фруктозобифосфатальдолазу (КФ 4.1.2.13), кодируемую геном fda,
м) глицеральдегид-3-фосфатдегидрогеназу (КФ 1.2.1.59), кодируемую геном gap,
н) фосфоглицераткиназу (КФ 2.7.2.3), кодируемую геном pgk,
о) пируваткиназу (КФ 2.7.1.40), кодируемую геном pyk,
п) Е1-субъединицу пируватдегидрогеназы (КФ 1.2.4.1), кодируемую геном aceE,
р) фосфоенолпируваткарбоксилазу (КФ 4.1.1.31), кодируемую геном ppc,
с) пируваткарбоксилазу (КФ 6.4.1.1), кодируемую геном pyc,
т) аконитазу (КФ 4.2.1.3), кодируемую геном acn, и
у) изоцитратдегидрогеназу (КФ 1.1.1.42), кодируемую геном icd.
9. Способ по п. 1, отличающийся тем, что процесс ферментации выбирают из группы, включающей периодический процесс, периодический процесс с подпиткой, периодический процесс с повторной подпиткой и непрерывный процесс.
10. Способ по п. 1, отличающийся тем, что выделяют L-орнитин или жидкий или твердый содержащий L-орнитин продукт из содержащего L-орнитин ферментационного бульона.
| Y | |||
| GUNJI et al., Enhancement of L-lysine production in methylotroph Methylophilus methylotrophus by introducing a mutant LysE exporter, Journal of Biotechnology Vol | |||
| Способ получения морфия из опия | 1922 |
|
SU127A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| M | |||
| VRLJIC et al., A new type of transporter with a new type of cellular function: L-lysine export from Corynebacterium glutamicum, Molecular Microbiology | |||

