Область изобретения
Изобретение относится к новой бактерии, продуцирующей L-глутаниновую кислоту, и к способу получения L-глутаминовой кислоты методом ферментации с использованием указанной бактерии. L-глутаминовая кислота является важной аминокислотой, входящей в состав пищи, лекарственных средств и т.п.
Предпосылки изобретения
До настоящего времени L-глутаминовую кислоту получали методом ферментации с использованием, главным образом, так называемых коринеформных бактерий, продуцирующих L-глутаминовую кислоту и принадлежащих к роду Breuibacterium, Corynebacterium или Microbacterium, или их вариантов ("Amino Acid Fermentation", Gakkai Shuppan Center, pp. 195-215, 1986). Известные способы получения L-глутаминовой кислоты путем ферментации с использованием других штаммов включают способ, в котором используется микроорганизм, относящийся к роду Bacillus, Streptomyces или Penicillium (патент США 3220929); способ, в котором используется микроорганизм, относящийся к роду Pseudomonas, Arthrobacter, Serratia или Candida и т.п.(патент США 3563857); способ, в котором используется микроорганизм, относящийся к роду Bacillus, Pseudomonas, Serratia или Aerobacter aerogenes (называемый в настоящее время Enterobacter aerogenes) (публикация прошедшей экспертизу заявки на патент Японии 32-9393); способ, в котором используется мутантный штамм Escherichia coli (выложенная заявка на патент Японии 5-244970) и т.п.
Производство L-глутаминовой кислоты было значительно увеличено путем выведения вышеуказанных микроорганизмов или путем усовершенствования способов получения. Необходимость в разработке менее дорогостоящего и более эффективного способа получения L-глутаминовой кислоты, который удовлетворял бы растущий спрос на L-глутаминовую кислоту, пока еще остается актуальной.
С учетом вышеуказанной ситуации авторами настоящего изобретения было проведено широкое исследование и изучение микроорганизмов, являющихся продуцентами L-глутаминовой кислоты. В результате было обнаружено, что микроорганизм, обладающий высокой продуктивностью L-глутаминовой кислоты, может быть получен путем увеличения активности фермента, который катализирует реакцию биосинтеза L-глутаминовой кислоты (цитратсинтазы, фосфоенолпируваткарбоксилазы, глутаматдегидрогеназы) микроорганизмом, относящимся к роду Enterobacter, Serratia, Klebsiella или Erwinia (выложенные заявки на патент Японии 10-224909 и 10-297129).
Авторами настоящего изобретения также было обнаружено, что микроорганизм, обладающий высокой продуктивностью L-глутаминовой кислоты, может быть получен путем увеличения активности каждого из этих ферментов посредством введения гена, кодирующего цитратсинтазу (далее иногда сокращенно называемую "CS") и фосфоенолпируваткарбоксилазу бактерий рода Escherichia, в восприимчивый к валину штамм, относящийся к роду Escherichia (WO 97/08294).
Между тем, сообщалось, что введение гена (гена CS), кодирующего цитратсинтазу Escherichia coli или Corynebacterium glutamicum, является эффективным способом увеличения продуктивности L-глутаминовой кислоты бактериями Corynebacterium или Brevibacterium (публикация прошедшей экспертизу заявки на патент Японии 7-121228). Когда в качестве хозяина использовали коринеформную бактерию, то введение гена CS из Corynebacterium glutamicum того же самого вида, что и хозяин, давало несколько больший эффект, чем введение гена CS из Escherichia coli, но заметной разницы между ними не отмечалось.
Как указывалось выше, известно, что ген CS вводят в различные микроорганизмы для увеличения продуктивности L-глутаминовой кислоты. Однако пока еще не известен пример, когда ген CS из коринеформной бактерии вводили бы в микроорганизм, относящийся к энтеробактериям, такой как бактерия, относящаяся к роду Escherichia.
Краткое описание изобретения
Целью настоящего изобретения является обнаружение новой бактерии, продуцирующей L-глутаминовую кислоту и обладающей продуктивностью L-глутаминовой кислоты, для разработки дешевого и эффективного способа продуцирования L-глутаминовой кислоты.
Авторами настоящего изобретения были выведены энтеробактерии путем введения в них гена, улучшающего их продуктивность L-глутаминовой кислоты. В основном считается, что лучший эффект достигается при введении эндогенного гена хозяина или гена микроорганизма, который является родственным этому хозяину, чем при введении гетерологичного гена, в том случае если данный хозяин содержит целевой ген для селективного выведения микроорганизма путем амплификации генов. Однако авторами настоящего изобретения было обнаружено, что в случае энтеробактерий гораздо более эффективным методом повышения продуктивности L-глутаминовой кислоты данным микроорганизмом является введение гена CS, происходящего из коринеформной бактерии, чем введение гена CS, происходящего от микроорганизма того же самого вида, что и энтеробактерия. И на основе этого обнаруженного факта было разработано настоящее изобретение.
Таким образом, настоящее изобретение относится:
(1) к микроорганизму, относящемуся к энтеробактериям и обладающему способностью продуцировать L-глутаминовую кислоту, в который был введен ген цитратсинтазы, происходящий из коринеформной бактерии;
(2) к микроорганизму, указанному в (1), где этой коринеформной бактерией является Brevibacterium lactofermentum;
(3) к микроорганизму, указанному в (1) или (2), где этот микроорганизм относится к энтеробактериям, которые относятся к роду Enterobacter или Klebsiella;
(4) к микроорганизму, указанному в (3), где указанная бактерия относится к Enterobacter agglomerans или Klebsiella planticola; и
(5) к способу получения L-глутаминовой кислоты, предусматривающему стадии культивирования микроорганизма, указанного в любом из (1)-(4), в жидкой среде с продуцированном и накоплением в этой среде L-глутаминовой кислоты и сбором L-глутаминовой кислоты из этой среды.
Краткое описание чертежей
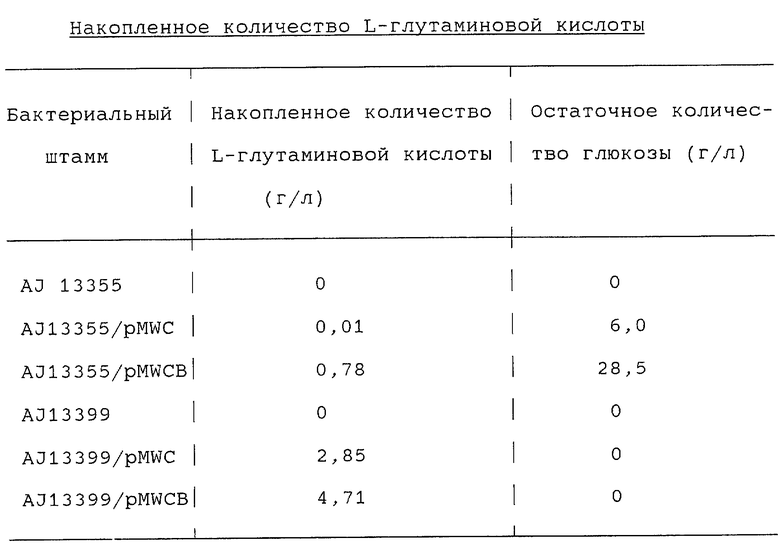
На фиг. 1 показана схема конструирования плазмиды pMWCB, содержащей ген gltA, и
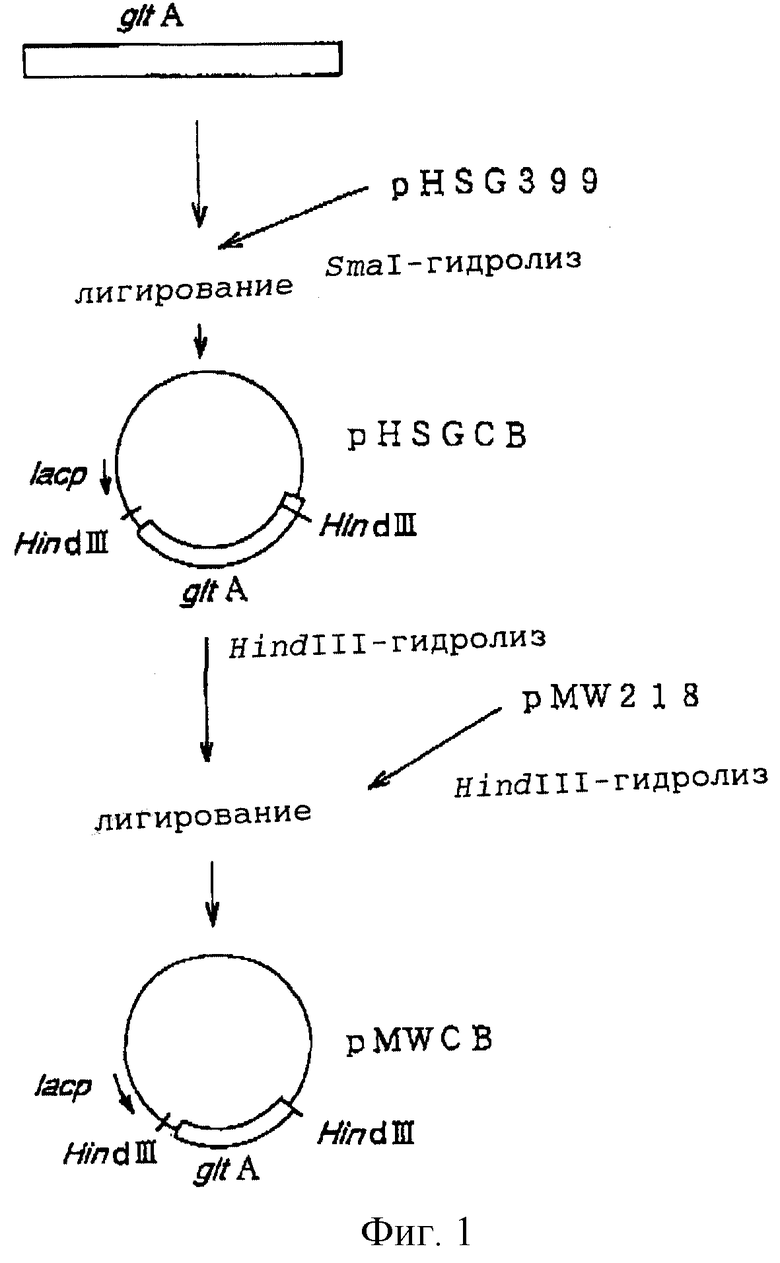
на фиг. 2 показана схема конструирования плазмиды pMWC, содержащей ген gltA.
Подробное описание изобретения
Ниже приводится подробное описание изобретения.
Микроорганизм настоящего изобретения
Микроорганизм настоящего изобретения, относящийся к энтеробактериям, не ограничивается конкретно принадлежностью к энтеробактериям, которым может быть сообщено или у которых может быть повышено продуцирование L-глутаминовой кислоты благодаря введению гена CS коринеформной бактерии. Примером этого микроорганизма является бактерия, относящаяся к роду Enterobacter, Klebsiella, Serratia, Erwinia или Escherichia. Из этих бактерий предпочтительными являются бактерии, относящиеся к роду Enterobacter или Klebsiella. Показательные примеры этих бактерий описаны ниже, но микроорганизм настоящего изобретения не ограничивается этими примерами.
Примерами микроорганизмов, относящихся к роду Enterobacter, которые могут быть использованы в настоящем изобретении, являются нижеследующие микроорганизмы:
Enterobacter agglomerans
Enterobacter aerogenes
Enterobacter amnigenus
Enterobacter asburiae
Enterobacter cloacae
Enterobacter dissolvens
Enterobacter gergoviae
Enterobacter hormaechei
Enterobacter intermedius
Enterobacter nimipressuralis
Enterobacter sakazakii
Enterobacter taylorae
В качестве более предпочтительных микроорганизмов могут быть упомянуты нижеследующие бактериальные штаммы:
Enterohacter agglomerans AJ13355
Serratia liquefacience ATCC 14460.
Enterobacter agglomerans. Штамм АJ13355 был депонирован National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry 19 февраля 1998 года под номером доступа FERM Р-16644, а затем в соответствии с Будапештским договором он был переведен на международное депонирование 11 января 1999 года под номером доступа FERM BP-16644. Штаммы Enterobacter agglomerans ATCC 12287 и Serratia liquefacience ATCC 14460 могут быть востребованы из ATCC.
Enterobacter agglomerans. Штамм АJ13355 представляет собой штамм, выделенный из почвы в Iwatashi, Shizuoka, Япония.
Штамм АJ13355 имеет следующие физиологические свойства:
(1) Окрашивание по Граму: отрицателен
(2) Аэрофобность: факультативный анаэроб
(3) Каталаза: положителен
(4) Оксидаза: отрицателен
(5) Восстановление нитрата: отрицателен
(6) Реакция Фогеса-Проскауэра: положителен
(7) Тест на метиловый красный: отрицателен
(8) Уреаза: отрицателен
(9) Продуцирование индола: положителен
(10) Подвижность: присутствует
(11) Продуцирование сероводорода в культуральной среде TSI: слегка активен
(12) β-Галактозидаза: положителен
(13) Усвояемость сахара:
Арабиноза: положителен
Сахароза: положителен
Лактоза: положителен
Ксилоза: положителен
Сорбит: положителен
Инозит: положителен
Трегалоза: положителен
Мальтоза: положителен
Мелибиоза: положителен
Адонит: отрицателен
Раффиноза: положителен
Салицин: отрицателен
(14) Усвояемость глицерозы: положителен
(15) Усвояемость органических кислот:
Лимонная кислота: положителен
Винная кислота: отрицателен
Глюконовая кислота: положителен
Уксусная кислота: положителен
Малоновая кислота: отрицателен
(16) Аргининдегидратаза: отрицателен
(17) Орнитиндекарбоксилаза: отрицателен
(18) Лизиндекарбоксилаза: отрицателен
(19) Фенилаланиндезаминаза: отрицателен
(20) Образование пигмента: желтый
(21) Способность к разжижению желатина: положителен
(22) Рост при рН: плохой рост при рН 4; хороший рост при рН 4,5-7
(23) Температура роста: хороший рост при 25oС, хороший рост при 30oС, хороший рост при 37oС, возможен рост при 42oС, рост отсутствует при 45oС.
Исходя из указанных бактериологических свойств штамм AJ13355 определен как Enterobacter agglomerans.
Примерами микроорганизмов, относящихся к роду Klebsiella, которые могут быть использованы в настоящем изобретении, перечислены ниже.
Klebsiella planticola
Klebsiella terrigena
Примером более предпочтительного микроорганизма является Klebsiella planticola АJ13399.
Штамм Klebsiella planticola АJ13399 был депонирован National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry 19 февраля 1998 года под номером доступа FERM Р-16646, а затем в соответствии с Будапештским договором он был переведен на международное депонирование 11 января 1999 г. под номером доступа FERM BP-6616.
Klebsiella planticola АJ13399 представляет собой штамм, выделенный из почвы в Sapporoshi, Хоккайдо Япония.
Штамм АJ13399 имеет следующие физиологические свойства
(1) Морфология клетки: палочка
(2) Подвижность: отсутствует
(3) Образование спор: отсутствует
(4) Морфология колоний на питательном агаре с LabM: кольцевая гладкая поверхность кремового цвета, ровная, выпуклая и блестящая
(5) OF-тест на глюкозу: положителен на сбраживаемость
(6) Окрашивание по Граму: отрицателен
(7) Аэрофобность: факультативный анаэроб
(8) Каталаза: положителен
(9) Оксидаза: отрицателен
(10) Уреаза: положителен
(11) Цитохромоксидаза: отрицателен
(12) β-Галактозидаза: положителен
(13) Аргининдегидратаза: отрицателен
(14) Орнитиндекарбоксилаза: отрицателен
(15) Лизиндекарбоксилаза: положителен
(16) Триптофандезаминаза: отрицателен
(17) Реакция Фогеса-Проскауэра: положителен
(18) Продуцирование индола: положителен
(19) Продуцирование сероводорода в культуральной среде TSI: отрицателен
(20) Усвояемость лимонной кислоты: положителен
(21) Усвояемость м-гидроксибензойной кислоты: отрицателен
(22) Способность к разжижению желатина: отрицателен
(23) Продуцирование кислоты из сахара:
Глюкоза: положителен
Маннит: положителен
Рамноза: положителен
Арабиноза: положителен
Сахароза: положителен
Сорбит: положителен
Инозит: положителен
Мелибиоза: положителен
Амигдалин: положителен
Адонит-пептон-вода: положителен
Целлобиоза-пептон-вода: положителен
Дульцин-пептон-вода: отрицателен
Раффиноза-пептон-вода: положителен
(24) Температура роста: хороший рост при 37oС, рост отсутствует при 45oС.
Исходя из указанных бактериологических свойств было определено, что микроорганизм АJ13399 является Klibsiella planticola.
Примеры микроорганизма, относящегося к роду Serratia, который может быть использован в настоящем изобретении, перечислены ниже:
Serratia liquefacience
Serratia entomophila
Serratia ficaria
Serratia fonticola
Serratia grimesii
Serratia proteamaculans
Serratia odorifera
Serratia plymuthica
Serratia rubidaea
В качестве более предпочтительного примера может быть упомянут Serratia liquefacience ATCC 14460. Serratia liquefacience ATCC 14460 может быть востребован из ATCC.
Примеры микроорганизма, относящегося к роду Erwinia, который может быть использован в настоящем изобретении, перечислены ниже.
Erwinia herbicola (в настоящее время классифицирован как Pantoea aggiomerans)
Erwinia ananas
Erwinia cacticida
Erwinia chrysanthemi
Erwinia mallotivora
Erwinia persicinus
Erwinia psidii
Erwinia quercina
Erwinia rhapontici
Erwinia rubrifaciens
Erwinia salicis
Erwinia uredovora
Более предпочтительным является штамм Erwinia herbicola IAM1595 (Pantoea agglomerans AJ12666). Штамм Erwinia herbicola IAM1595 может быть востребован из Institute of Molecular and Cellular Biosciences, the University of Tokyo. Следует отметить, что в "Определителе бактерий Берджи, девятое издание" бактерия Erwinia herbicola не описана, и микроорганизмы, которые были классифицированы как Erwinia herbicola, классифицированы как Pantoea agglomerans. Микроорганизмы, которые были классифицированы как Erwinia herbicola, в настоящее время были классифицированы как Pantoea agglomerans. Таким образом, микроорганизмы, относящиеся к роду Erwinia, и микроорганизмы, относящиеся к роду Pantoea, являются близкородственными. Поэтому любые микроорганизмы, относящиеся к роду Pantoea, могут быть использованы аналогично микроорганизмам, относящимся к роду Erwinia. Такими микроорганизмами являются Pantoea agglomerans и Pantoea dispersa. Erwinia herbicola IAM1595 получил название Pantoea agglomerans AJ2666 и был депонирован 25 февраля 1999 года National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry, в рамках международного депонирования в соответствии с Будапештским договором и получил номер доступа FERM ВР-6660.
Примерами микроорганизма, относящегося к роду Escherichia, который может быть использован в настоящем изобретении, является Escherichia coli.
Более предпочтительными Escherichia coli, обладающими резистентностью к валину, являются, например, следующие штаммы:
Escherichia coli K-12 (АТСС 10798)
Escherichia coli В (АТСС 11303)
Escherichia coli W (АТСС 9637).
Escherichia coli K-12 (ATCC 10798), Escherichia coli В (АТСС 11303) и Escherichia coli W (ATCC 9637) могут быть востребованы из АТСС.
Следует отметить, что метаболизм сахаров бактериями, относящимися к роду Enterobacter, Klebsiella, Serratia, Erwinia и Escherichia, такими как бактерии, упомянутые выше, происходит по пути Эмбдена-Мейерхофа, и продуцированная в результате этого пировиноградная кислота окисляется в цикле трикарбоновых кислот как в аэробных условиях. Биосинтез L-глутаминовой кислоты происходит исходя из α-кетоглутаровой кислоты, являющейся промежуточным соединением цикла трикарбоновых кислот, под действием фермента GDH или глутаминсинтетазы/глутаматсинтазы. Таким образом, эти микроорганизмы имеют один и тот же путь биосинтеза L-глутаминовой кислоты, и вышеупомянутые микроорганизмы входят в один из аспектов настоящего изобретения. Поэтому микроорганизмы, относящиеся к кишечным бактериям, отличающимся от видов и штаммов, упомянутых выше, также входят в объем настоящего изобретения.
Микроорганизмом настоящего изобретения является микроорганизм, относящийся к кишечным бактериям и обладающий способностью продуцировать L-глутаминовую кислоту. Выражение "обладающий способностью продуцировать L-глутаминовую кислоту", используемое в настоящем описании, означает способность этого микроорганизма накапливать L-глутаминовую кислоту в культуральной среде в процессе культивирования. Такой способностью продуцировать L-глутаминовую кислоту может обладать либо штамм дикого типа как присущим ему свойством, либо штамм, наделенный этим свойством, либо штамм, у которого это свойство было усилено путем селективного разведения. Может быть также использован микроорганизм, который может быть наделен способностью продуцировать L-глутаминовую кислоту благодаря введению в него гена gltA. Микроорганизмами, относящимися к кишечным бактериям и обладающими способностью к продуцированию L-глутаминовой кислоты, являются, например, такие микроорганизмы, которые обладают повышенной активностью одного или нескольких ферментов, катализирующих одну или несколько реакций биосинтеза L-глутаминовой кислоты, и такие микроорганизмы, которые обладают пониженной активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой, или вообще не обладают такой активностью. Другими такими микроорганизмами являются микроорганизмы, которые обладают повышенной активностью одного или нескольких ферментов, катализирующих одну или несколько реакций биосинтеза L-глутаминовой кислоты, и которые обладают пониженной активностью фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-глутаминовой кислоты и продуцирующую соединение, не являющееся L-глутаминовой кислотой, или вообще не обладает такой активностью.
"Коринеформными бактериями", которые могут быть источником для гена gltA, вводимого в кишечную бактерию, являются бактерии, которые были ранее классифицированы как род Brevibacterium, но в настоящее время они были объединены в род Corynebacterium (Int. 3. Syst. Bacteriol., 41, 255 (1981)), и включают бактерии, относящиеся к роду Brevibacterium, который является близкородственным роду Corynebacterium. Примерами таких коринеформных бактерий, продуцирующих L-глутаминовую кислоту, являются следующие бактерии:
Corynebacterium acetoacidophilum
Corynebacterium acetoglutamicum
Corynebacterium alkanolyticum
Corynebacterium callunae
Corynebacterium glutamicuin
Corynebacterium lilium (Corynebacterium glutamicum)
Corynebacterium melassecola
Corynebacterium thermoaminogenes
Corynebacterium herculis
Breuibacterium divaricatum (Corynebacterium glutamicum)
Brevibacterium flavum (Corynebacterium glutamicum)
Brevibacterium immariophilum
Brevibacterium lactofermentum (Corynebacterium glutamicum)
Brevibacterium roseum
Brevibacterium saccharolyticum
Brevibacterium thiogenitalis
Brevibacterium album
Brevibacterium cerinum
Microbacterium ammoniaphilum
Конкретными примерами нижеследующих штаммов этих бактерий являются штаммы:
Corynebacterium acetoacidophilum ATCC 13870
Corynebacterium acetoglutamicum ATCC 15806
Corynebacterium alkanolyticum ATCC 21511
Corynebacterium callunae ATCC 15991
Corynebacterium glutamicum ATCC 13020, 13032, 13060
Corynebacterium lilium (Corynebacterium glutamicum) ATCC 15990
Corynebacterium melassecola ATCC 17965
Corynebacterium thermoaminogenes АJ12340 (FERM BP-1539)
Corynebacterium herculis ATCC 13868
Brevibacterium divaricatum (Corynebacterium glutamicum) ATCC 14020
Brevibacterium flavum (Corynebacterium glutamicum) ATCC 13826, ATCC 14067
Brevibacterium immariophilum ATCC 14068
Brevibacterium lactofermentum (Corynebacterium glutamicum) ATCC 13665, ATCC 13869
Brevibacteriun roseum ATCC 13825
Brevibacterium saccharolyticum ATCC 14066
Brevibacterium thiogenitalis ATCC 19240
Brevibacterium album ATCC 15111
Brevibacterium cerinum ATCC 15112
Microbacteriuni ammoniaphilum ATCC 15354.
Ген gltА, происходящий от коринеформной бактерии, может быть получен путем выделения ДНК-фрагмента, комплементирующего ауксотрофию бактерии, у которой отсутствует CS-активность, такой как мутант коринеформной бактерии, и выделенного из хромосомной ДНК коринеформной бактерии. Известна нуклеотидная последовательность гена qltA коринеформной бактерии (Microbiology, 140, 1817-1828, 1994). Поэтому ген gltA может быть получен методом ПЦР с использованием хромосомной ДНК в качестве матрицы и праймеров, синтезированных на основе этой нуклеотидной последовательности. Примерами праймеров являются олигонуклеотиды, имеющие нуклеотидную последовательность, показанную в SEQ ID 1 и 2.
Для введения гена CS коринеформной бактерии в микроорганизм, относящийся к энтеробактериям, ген CS может быть клонирован в соответствующую плазмиду, и вышеуказанный исходный родительский штамм, который служит в качестве хозяина, может быть трансформирован полученной рекомбинантной плазмидой. Это может приводить к увеличению числа копий гена CS (обозначаемого ниже "геном gltA") в клетке трансформанта, в результате чего активность CS может быть увеличена.
Тип используемой плазмиды не имеет конкретных ограничений при условии, что эта плазмида способна автономно реплицироваться в микроорганизме, относящемся к кишечным бактериям, и примерами такой плазмиды являются pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218 и т.п. Могут быть также использованы другие плазмиды и фаговые ДНК-векторы.
Введение гена gltA может быть осуществлено путем получения гена gltA, присутствующего на хромосомной ДНК вышеупомянутого родительского штамма, который служит в качестве хозяина, предпочтительно, во множестве копий. Для введения гена gltA в множестве копий в хромосомную ДНК микроорганизма, относящегося к роду энтеробактерий, может быть использована последовательность, присутствующая на хромосомной ДНК в множестве копий, такая как повторяющаяся ДНК или инвертированный повтор, присутствующий в концевой области мобильного генетического элемента. Альтернативно, ген gltA может быть также введен в хромосомную ДНК во множестве копий путем встраивания этого гена в транспозон и путем транспозиции этого транспозона. Это позволяет увеличить число копий гена gltA в клетке трансформанта, в результате чего увеличивается активность CS.
Трансформация может быть осуществлена, например, методом D.M.Morrison (Methods in Enzymology, 68, 326 (1979)); методом, в котором для повышения проницаемости клеток-реципиентов для ДНК эти клетки обрабатывают хлоридом кальция (Mandel, М. & Higa, A., J-Mol.Biol., 53, 159 (1970)).
Ген gltA, предназначенный для введения, вместо присущего ему собственного промотора, может иметь промотор, подходящий для клетки микроорганизма, относящегося к энтеробактерий, такой как lac, tpr или РL.
Такие методы, как клонирование гена, гидролиз или лигирование ДНК, и метод трансформации описан в руководстве Molecular Cloning, 2nd edition. Cold Spring Harbor Press (1989).
В микроорганизме настоящего изобретения помимо введения гена gltA коринеформной бактерии может быть усилена активность фермента, который катализирует биосинтез L-глутаминовой кислоты и который не является CS. Иллюстрирующими примерами фермента, катализирующего биосинтез L-глутаминовой кислоты, являются глутаматдегидрогеназа (GDH), глутаминсинтетаза, глутаматсинтаза, изоцитратдегидрогеназа, аконитатгидратаза, фосфоенолпируваткарбоксилаза (РЕРС), пируватдегидрогеназа, пируваткиназа, енолаза, фосфоглицеромутаза, фосфоглицераткиназа, глицеральдегид-3-фосфатдегидрогеназа, триозофосфатизомераза, фруктозобифосфатальдолаза, фосфофруктокиназа, глюкозофосфатизомераза и т.п.
Активность фермента, который катализирует реакцию продуцирования соединения, отличающегося от L-глутаминовой кислоты, путем отхода от пути биосинтеза L-глутаминовой кислоты, может быть уменьшена или исключена. Показательными примерами фермента, который катализирует реакцию продуцирования соединения, отличающегося от L-глутаминовой кислоты, посредством отхода от пути биосинтеза L-глутаминовой кислоты, являются α-кетоглутаратдегидрогеназа (αKGDH), изоцитратлиаза, фосфатацетилтрансфераза, ацетаткиназа, ацетогидроксиматсинтаза, ацетолактатсинтаза, формиатацетилтрансфераза, лактатдегидрогеназа, глутаматдекарбоксилаза, 1-пирролиндегидрогеназа и т.п. Из этих ферментов предпочтительным является αKGDH.
Каждый из генов, кодирующих РЕРС и GDH, может быть получен из хромосомной ДНК любого из вышеупомянутых микроорганизмов путем выделения ДНК-фрагмента, комплементирующего ауксотрофию штамма-варианта, у которого отсутствует РЕРС- и GDH-активность. Альтернативно, поскольку нуклеотидные последовательности этих генов в бактериях рода Escherichia или Corynebacterium были уже выявлены (Biochemistry, 22, 5243-5249 (1983); J.Biochem. 95, 909-916 (1984); Gene, 27, 193-199 (1984); Mol.Gen.Genet. 218, 330-339 (1989) и Molecular Microbiology, 6, 317-326 (1992)), то эти гены могут быть получены посредством PCR с использованием праймера, синтезированного на основе каждой из выявленных нуклеотидных последовательностей и хромосомной ДНК в качестве матрицы.
Для достижения такого снижения или отсутствия вышеупомянутой ферментативной активности в микроорганизме, относящемся к кишечным бактериям, в ген, кодирующий этот фермент, с помощью стандартной техники мутагенеза или техники генной инженерии может быть введена мутация, приводящая к снижению или отсутствию данной ферментативной активности.
Примерами техники мутагенеза могут служить, например, метод с использованием облучения рентгеновским или ультрафиолетовым излучением; метод с использованием обработки мутагенным агентом, таким как N-метил-N-нитро-N-нитрозогуанидин, и т.п. Область гена, в которую вводят мутацию, может быть кодирующей областью белка-фермента, или областью, регулирующей экспрессию, такой как промотор.
Примерами техники генной инженерии являются, например, генетическая рекомбинация, генетическая трансдукция, слияние клеток и т.п. Так, например, для продуцирования функционально инактивированного гена (дефектного гена) в нужный ген встраивают ген резистентности к лекарственному средству. Затем этот ген делеционного типа вводят в клетку микроорганизма, относящегося к роду кишечных бактерий, и целевой ген на хромосоме заменяется геном делеционного типа посредством гомологичной рекомбинации (разрушение гена).
Для того чтобы определить, имеет ли данный микроорганизм пониженную активность целевого фермента, или эта активность отсутствует, или для того, чтобы определить степень снижения ферментативной активности, может быть измерена ферментативная активность экстракта бактериальных клеток или очищенной фракции штамма-кандидата с последующим сравнением этой активности с активностью штамма дикого типа или родительского штамма. Так, например, активность фермента αKGDH может быть измерена методом Reed и др. (L.Д.Reed & В.В. Mukherjee. Methods in Enzymology 1969, 13, р. 55-61).
Для некоторых ферментов нужный мутант может быть выбран из фенотипа этого мутанта. Так, например, мутант, в котором отсутствует (αKGDH-активность или эта активность является пониженной, не может расти или имеет значительно меньшую скорость роста на минимальной среде, содержащей глюкозу, или на минимальной среде, содержащей уксусную кислоту или L-глутаминовую кислоту в качестве единственного источника углерода. Однако даже при тех же самых условиях может наблюдаться нормальный рост, если в минимальную среду, содержащую глюкозу, добавить янтарную кислоту или L-лизин, L-метионин и диаминопимелиновую кислоту. С использованием этого факта в качестве критерия можно также провести скрининг на мутант, в котором отсутствует αKGDH-активность, либо эта активность является пониженной.
Способ продуцирования штамма Brevibacterium lactofermentum, не содержащего гена αKGDH, основанный на гомогенной рекомбинации, подробно описан в WО 95/34672, и аналогичный способ может быть использован для продуцирования микроорганизмов, относящихся к кишечным бактериям.
Примерами мутантного штамма с отсутствием или с пониженной (αGDH-активностью, полученного, как описано выше, является штамм Enterobacter agglomerans АJ13356 и Klebsiella planticola АJ13410. Штаммы Enterobacter agglomerans АJ13356 и Klebsiella planticola АJ13410 были депонированы 19 февраля 1998 г. National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry под номерами доступа FERM P-16645 и FERM Р-16647, а затем в соответствии с Будапештским договором, они были переведены на международное депонирование 11 января 1999 г. под номерами доступа FERM BP-6615 и FERM BP-6617 соответственно.
L-глутаминовая кислота может быть получена и накоплена в жидкой культуральной среде путем культивирования в этой среде микроорганизма, который относится к кишечным бактериям и в который был введен ген gltA, происходящий от коринеформной бактерии.
Этой средой может быть обычная питательная среда, содержащая источник углерода, источник азота и неорганические соли, а также, если это необходимо, органические питательные вещества, такие как аминокислоты, витамины, и т. п. Такой средой может быть синтетическая среда или природная среда. Для культуральной среды могут быть использованы любые углеродные и азотные источники при условии, что они могут быть утилизованы культивируемым микроорганизмом.
Источником углерода может быть сахарид, такой как глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизаты крахмала, мелассы и т.п. Кроме того, может быть также использована органическая кислота, такая как уксусная кислота и лимонная кислота, отдельно или в комбинации с другими углеродными источниками.
Источником азота может быть аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония и ацетат аммония, а также нитраты и т.п.
В качестве органических микроэлементов используются аминокислоты, витамины, жирные кислоты, нуклеиновые кислоты, материалы, содержащие эти вещества, такие как пептон, казаминокислота, дрожжевой экстракт и продукты разложения соевого белка и т.п., а в случае ауксотрофного варианта, которому для его роста требуется аминокислота или т.п., эти микроэлементы необходимы для восполнения требуемых питательных веществ.
В качестве неорганической соли используются фосфаты, соли магния, соли кальция, соли железа, соли марганца и т.п.
Что касается условий культивирования, то это культивирование может быть осуществлено в аэробных условиях при температуре 20-42oС и при рН 4-8. Для аккумуляции значительного количества L-глутаминовой кислоты в жидкой культуральной среде культивирование может продолжаться от 10 часов до 4 дней.
После завершения культивирования L-глутаминовая кислота, аккумулированная в культуральной среде, может быть собрана известным методом. Так например, она может быть выделена методом, предусматривающим концентрирование среды после удаления клеток с кристаллизацией продукта; методом ионообменной хроматографии или т.п.
Поскольку в соответствии с настоящим изобретением микроорганизм, относящийся к энтеробактериям, может быть наделен способностью к эффективному продуцированию L-глутаминовой кислоты, то предполагается, что данным микроорганизмам может быть сообщена более высокая продуктивность путем стандартной техники размножения коринеформных бактерий, продуцирующих L-глутаминовую кислоту. Ожидается, что исследования условий культивирования и т.п. позволят разработать дешевый и эффективный способ продуцирования L-глутаминовой кислоты.
Наилучший вариант осуществления настоящего изобретения
Более подробно настоящее изобретение описано ниже со ссылками на следующие примеры.
(1) Конструирование плазмиды, содержащей ген gltA
Плазмиду, содержащую ген gltA, происходящий от Brevibacterium lactofermentum, конструировали следующим образом. ПЦР проводили с использованием праймеров, имеющих нуклеотидные последовательности, представленные в SEQ ID 1 и 2, и выбранные исходя из нуклеотидной последовательности гена gltA Corynebacterium glutamicum (Microbiology, 1994, 140, 1817-1828) и хромосомной ДНК Brevibacterium lactofermentum ATCC 13869, используемой в качестве матрицы для получения фрагмента гена gltA размером около 3 т.п.о. Этот фрагмент встраивали в плазмиду pHSG399 (закупленную у фирмы Такаrа Shuzo), гидролизованную ферментом SmaI, и получали плазмиду pHSGCB (фиг.1). Затем плазмиду pHSGCB гидролизовали ферментом HindIII и вырезанный фрагмент гена gltA размером около 3 т.п.о. встраивали в плазмиду pMW218 (закупленную у фирмы Nippon Gene), гидролизованную ферментом HindIII, в результате чего получали плазмиду pMWCB (фиг. 1). Экспрессию гена gltA в полученной плазмиде pMWCB подтверждали путем определения ферментативной активности в штамме Enterobacter agglomerans АJ13355.
(2) Конструирование плазмиды, имеющей ген gltA, происходящий от E.coli.
В качестве контроля плазмиду, содержащую ген gltA, происходящий от Escherichia coli, конструировали следующим образом. Плазмиду pTWVC, содержащую ген gltA, происходящий от Escherichia coli (WО 97/08294), гидролизовали ферментами HindIII и EcoRI, и полученный ДНК-фрагмент, содержащий ген gltA, очищали и собирали, а затем вводили в HindIII-ЕсоR1-сайт плазмиды pMW219 с получением плазмиды pMWC (фиг.2). Экспрессию гена gltA полученной плазмидой pMWC подтверждали путем определения ферментативной активности и комплементации ауксотрофного штамма E.coli, не содержащего гена gltA.
(3) Введение гена gltA в Enterobacter agglomerans или Klebsiella planticola и оценка продуцируемости L-глутаминовой кислоты.
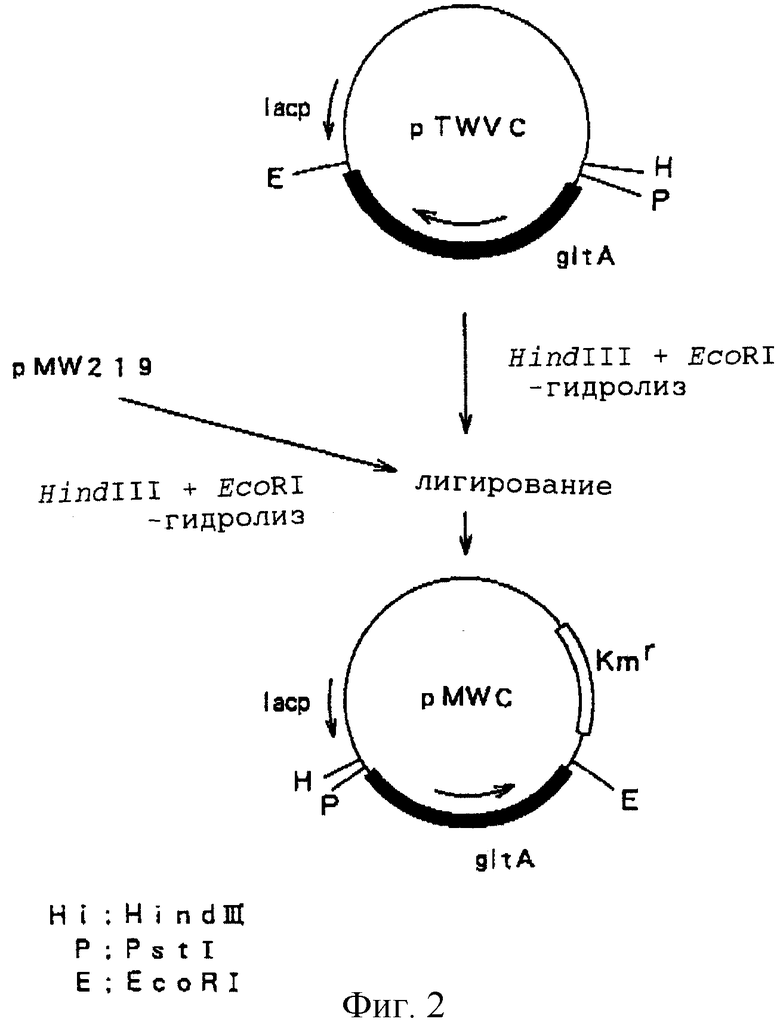
Штаммы Enterobacter agglomerans АJ13355 и Klebsiella planticola AJ13399 трансформировали плазмидами pMWC или pMWCB. Каждый из полученных трансформантов AJ13355/pMWC, AJ13355/рMWCB, AJ13399/pMWC и AJ13399/pMWCB и родительские штаммы инокулировали в колбе объемом 500 мл, содержащей 20 мл культуральной среды, включающей 40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л дигидрофосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л гептагидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия, 2 г/л дрожжевого экстракта и 30 г/л карбоната кальция, и культивировали, встряхивая, при 37oС в течение 15 часов. После завершения культивирования измеряли количество L-глутаминовой кислоты, накопленной в культуральной среде, и количество остаточной глюкозы. Результаты представлены в таблице.
Продуктивность L-глутаминовой кислоты наблюдали как в штамме Enterobacter agglomerans АJ13355, так и в штамме Kcebsiella planticola АJ13399, в которые был введен ген gltA. Аккумуляция L-глутаминовой кислоты была более заметной в том случае, когда был введен ген gltA, происходящий от Brevibacterium lactofermentum, чем в том случае, когда был введен ген gltA, происходящий от Escherichia coli. Большие остаточные количества глюкозы наблюдались в случае использования штамма AJ13355/pMWCB, который не поглощал глюкозу в вышеуказанных условиях. Предполагается, что при культивировании до тех пор, пока не будет израсходована вся глюкоза, может быть аккумулировано 1,5-2 г/л L-глутаминовой кислоты.
При этом какого-либо заметного отличия в числе копий плазмид между штаммами AJ13355/pMWC и AJ13355/pMWCB и между штаммами AJ13399/pMWC и AJ13399/pMWCB не наблюдалось.
Список последовательностей
<110> Ajinomoto Co., Inc.
<120> Бактерия, продуцирующая L-глутаминовую кислоту, и способ получения L-глутаминовой кислоты
<130> ОР892
<141> 1999
<150> JP 10-297350
<151> 1999-10-19
<160> 2
<170> Patentin Ver. 2,0
<210> 1
<211> 30
<212> ДНК
<213> Синтетическая последовательность
<220>
<223> Описание синтетической последовательности: праймер
<400> 1
gtcgacaata gccygaatct gttctggtcg 30
<210> 2
<211> 30
<212> ДНК
<213> Синтетическая последовательность
<220>
<223> Описание синтетической последовательности: праймер
<400> 2
aagcttatcg acgctcccct ccccaccgtt 30г
Изобретение относится к способу получения L-глутаминовой кислоты. Культивируют в среде энтеробактерии рода Enterobacter, Klebsiella, Serratia, Erwinia, Pantoea или Escherichia. В энтеробактерии вводят ген цитратсинтазы, происходящий от коринеформной бактерии. Происходит продуцирование и накопление в этой среде L-глутаминовой кислоты. Затем L-глутаминовую кислоту извлекают из среды. Преимущественно используют Enterobacter agglomerans или Klebsiella planticola. Для выделения гена цитратсинтазы используют преимущественно Brevibacterium lacto-fermentum. Изобретение позволяет получать L-глутаминовую кислоты более дешевым и эффективным способом. 2 з.п.ф-лы, 1 табл., 2 ил.
| US 5573945 А, 12.11.1996 | |||
| US 5378616 А, 03.01.1995 | |||
| US 4393135 А, 12.07.1983 | |||
| Способ получения -глютаминовой кислоты и ее производных | 1971 |
|
SU528338A1 |
| 0 |
|
SU294321A1 | |
| 0 |
|
SU285631A1 | |
| SU 229311 А, 18.11.1969 | |||
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ ГЛУТАМИНОВОЙ КИСЛОТЫ | 0 |
|
SU255162A1 |
| ШТАММ БАКТЕРИЙ CORYNEBACTERIUM GLUTAMICUM В-7198-ПРОДУЦЕНТ L-ГЛЮТАМИНОВОЙ КИСЛОТЫ | 1996 |
|
RU2107723C1 |

