ПЕРЕКРЕСТНАЯ ССЫЛКА НА ДРУГИЕ ЗАЯВКИ
Эта заявка испрашивается приоритет предварительной заявки на патент США № 61/486697, поданной 16 мая 2011, описание которой включено сюда посредством ссылки в его полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Это изобретение относится в общем к иммунологии, а конкретнее к применению метотрексата для уменьшения нежелательных иммунных реакций у пациентов.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
В настоящее время Управление по контролю качества пищевых продуктов и лекарственных средств США (FDA) разрешило свыше 130 являющихся белками терапевтических средств для клинического применения (Leader et al., Nat Rev Drug Discov, 7(1):21-39 (2008)). Эти терапевтические средства включают пептиды, рекомбинантные белки человека, вакцины на основе белков, препараты поликлональных антител, полученных из множества видов животных, и моноклональные антитела. В зависимости от гомологии своих последовательностей и конформационной гомологии с эндогенными аутоантигенами, состояния гликозилирования, уровня дозы, пути введения, местонахождения и связанных с процессом производства характеристик терапевтические белки могут индуцировать гуморальные иммунные ответы у пациента (Schellekens, Nat Rev Drug Discov, 1(6):457-62 (2002); Zinkernagel, Semin Immunol, 12(3):163-71 (2000), discussion 257-344; Thorland et al, Haemophilia, 5(2):101-5 (1999); Goodeve, Blood Coagul Fibrinolysis, 14 Suppl 1:S17-21 (2003)). Эти ответы, называемые образованиями антител против лекарственных средств (ADA), могут иногда влиять на безопасность для пациента и эффективность лекарственных средств.
В случае лизосомальной болезни накопления, болезни Помпе, ADA против рекомбинантной кислой альфа-глюкозидазы человека может развиться при ферментозаместительной терапии (ERT). У пациентов, которые не экспрессируют поддающиеся измерению количества эндогенного фермента, устойчивые уровни высокого титра антител коррелируют с ухудшением состояния здоровья пациентов (состояние CRIM у пациентов с болезнью Помпе) (Kishnani et al, Mol Genet Metab., 99(l):26-33 (2010); Hunley et al, Pediatrics, 114(4):e532-5 (2004); Amalfitano et al, Genet Med, 3(2):132-8 (2001)). Схожий подрыв безопасности и эффективности лекарственного средства был отмечен у пациентов с гемофилией, у которых развивается ADA против фактора IX (Thorland et al, Haemophilia, 5(2):101-5 (1999); Ewenstein et al, Blood, 89(3):1115-6 (1997)). В редких случаях ADA может также вызывать аутоиммунное заболевание, как и в случае рекомбинантного эритропоэтина человека (Schellekens, Clin Ther, 24(11):1720-40 (2002), discussion 1719; Locatelli et al, Perit Dial Int, 27 Suppl 2:S303-7 (2007)).
Ответы в виде ADA могут также возникать при использовании являющихся антителами терапевтических средств независимо от того, являются ли терапевтические средства нечеловеческого происхождения, гуманизированными или даже полностью человеческими. Иммуногенность терапевтических средств в виде и моноклональных, и поликлональных антител может оказывать влияние на безопасность для пациента и эффективность лекарственных средств. Антитела, которые образуются против терапевтических моноклональных антител, таких как инфликсимаб, адалимумаб, ритуксимаб и натализумаб, были связаны с уменьшением уровней в сыворотке и эффективности терапевтических антител (Bendtzen et al, Arthritis Rheum, 54(12):3782-9 (2006); Schmidt et al, Clin Immunol, 132(3):334-41 (2009); Bartelds et al, Ann Rheum Dis, 66(7):921-6 (2007); Baert et al, N Engl J Med, 348(7):601-8 (2003); Tahir et al, Rheumatology (Oxford), 44(4):561-2 (2005); Maini et al, Arthritis Rheum, 41(9):1552-63 (1998)). Аллергические реакции были связаны с антителами против инфликсимаба (Baert et al., N Engl J Med, 348(7):601-8 (2003)). Связанные с инфузиями реакции гиперчувствительности отмечались у небольшого процента пациентов с ремиттирующе-рецидивирующим рассеянным склерозом, подвергнутых лечению натализумабом (Phillips et al, Neurology, 67(9):1717-8 (2006)).
Алемтузумаб является другим терапевтическим антителом, которое может вызывать ADA у пациентов с ремиттирующе-рецидивирующим рассеянным склерозом (Coles et al, N Engl J Med, 2008, 359(17):1786-801 (2008)). Алемтузумаб является истощающим лимфоциты моноклональным антителом, которое взаимодействует с CD52, антигеном клеточной поверхности, представленным на иммуноцитах. Алемтузумаб находится на поздней стадии клинических испытаний в отношении лечения ремиттирующе-рецидивирующего рассеянного склероза, а также был оценен в отношении лечения ревматоидного артрита. Группа исследователей установила, что антитела против алемтузумаба образуются у 63% пациентов с ревматоидным артритом, подвергнутых лечению в исследовании с увеличением однократной дозы (Weinblatt et al., Arthritis Rheum, 1995. 38(11):1589-94 (1995)). В этом исследовании эффективность алемтузумаба, по-видимому, была изменена в результате наличия ADA (там же).
Применение терапевтического поликлонального антитела Thymoglobulin® также сопровождается вредным ADA у небольшой субпопуляции пациентов. Сывороточная болезнь, острая почечная недостаточность и сердечнососудистые реакции отмечались у подвергнутых лечению Thymoglobulin® реципиентов трансплантатов (Boothpur et al, Am J Kidney Dis., 55(1):141-3 (2009); Lundquist et al, Liver Transpl, 13(5):647-50 (2007); Busani et al, Minerva Anestesiol, 72(4):243-8 (2006); Tanriover et al, Transplantation, 80(2):279-81 (2005); Buchler et al., Clin Transplant, 17(6):539-45 (2003)).
Исследователи пытались найти пути для минимизации вредных эффектов ADA. Средства, которые были проверены в отношении их способности к индукции иммунотолерантности и уменьшению ADA при терапиях с использованием белков, включают, например, неистощающие антитела против CD4 (Cobbold et al, Semin Immunol, 2(6):377-87 (1990); Winsor-Hines et al, J Immunol, 173(7):4715-23 (2004)), не связывающиеся с клетками минимальные мутанты алемтузумаба (Gilliland et al., J Immunol, 162(6):3663-71 (1999); Somerfield et al, J Immunol, 185(l):763-8 (2010)), иммунодепрессивные терапии (Bennett et al, Blood, 106(10):3343-7 (2005); Dickson et al, J Clin Invest, 118(8):2868-76 (2008)), содержащие фосфатидилинозит липидные частицы, связывающиеся с фактором VIII (Peng et al, AAPS. J., 12(3):473-81 (2010)), и введение точно в печень рекомбинантной кислой альфа-глюкозидазы посредством инфицирования аденоассоциированным вирусом (Sun et al, Am J Human Genet, 81(5):1042-9 (2007); Sun et al, Mol Ther 18(2):353-60 (2009); Ziegler et al, Hum Gene Ther, 19(6):609-21 (2008)). Однако сохраняется потребность в разработке улучшенных способов уменьшения нежелательных гуморальных иммунных ответов при терапиях с использованием белков и в других ситуациях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящим изобретением обеспечивается способ индукции иммунологической толерантности у субъекта, нуждающегося в лечении терапевтическим средством. В этом способе субъекту вводят эффективное количество метотрексата в одном цикле, тем самым индуцируя иммунологическую толерантность к терапевтическому средству у субъекта.
Настоящим изобретением также обеспечивается способ ингибирования образований антител против терапевтического средства у субъекта, нуждающегося в лечении терапевтическим средством. В этом способе субъекту вводят эффективное количество метотрексата в одном цикле, тем самым ингибируя образования антител против терапевтического средства у субъекта.
Настоящим изобретением также обеспечивается способ облегчения инфузионной реакции на терапевтическое средство у субъекта, нуждающегося в лечении терапевтическим средством. В этом способе субъекту вводят эффективное количество метотрексата, тем самым облегчая инфузионную реакцию на терапевтический белок у субъекта.
Настоящим изобретением также обеспечивается способ ослабления вторичного аутоиммунитета у субъекта с аутоиммунным заболеванием, нуждающегося в лечении терапевтическим средством. В этом способе субъекту вводят эффективное количество метотрексата, тем самым ослабляя вторичный аутоиммунитет у субъекта.
Настоящим изобретением также обеспечивается способ увеличения эффективности терапевтического средства у субъекта, нуждающегося в лечении терапевтическим средством. В этом способе субъекту вводят эффективное количество метотрексата, тем самым увеличивая эффективность терапевтического белка у субъекта.
Настоящим изобретением также обеспечивается способ увеличения процента регуляторных T-клеток в Т-клеточной популяции у субъекта, подвергнутого лечению с использованием истощающей лимфоциты терапии, например, терапии с использованием алемтузумаба или терапии с использованием Thymoglobulin®. В этом способе субъекту вводят эффективное количество метотрексата, тем самым увеличивая указанный процент у указанного субъекта.
Настоящим изобретением также обеспечивается способ увеличения процента регуляторных В-клеток в В-клеточной популяции у субъекта, нуждающегося в лечении терапевтическим белком, таким как терапевтическое антитело. В этом способе субъекту вводят эффективное количество метотрексата, тем самым увеличивая указанный процент у указанного субъекта. В некоторых вариантах осуществления эффективное количество метотрексата вводят в одном цикле. В связанных вариантах осуществления настоящим изобретением обеспечивается способ увеличения экспрессирующих TGF-бета, IL-10 и/или FoxP3 B-клеток у субъекта, нуждающегося в лечении терапевтическим белком, включающий введение эффективного количества метотрексата в одном цикле.
Настоящим изобретением также обеспечивается способ продления фармакокинетики терапевтического средства у субъекта. В этом способе субъекту вводят метотрексат, до, или во время, или после введения терапевтического средства, в количестве, эффективном для продления фармакокинетики терапевтического средства.
Настоящим изобретением, кроме того, обеспечивается способ истощения лимфоцитов у являющегося человеком пациента, нуждающегося в этом. В этом способе пациента подвергают лечению истощающим лимфоциты средством, и ему вводят метотрексат, до, или во время, или после лечения истощающим лимфоциты средством, в количестве, эффективном для индукции у пациента иммунологической толерантности к истощающему лимфоциты средству или для ослабления вторичного аутоиммунитета. В некоторых вариантах осуществления эффективное количество метотрексата вводят в одном цикле. В некоторых вариантах осуществления истощающим лимфоциты средством может быть терапевтическое моноклональное антитело (например, алемтузумаб или ритуксимаб). В некоторых из этих вариантов осуществления пациентом может быть пациент с рассеянным склерозом (например, пациент с ремиттирующе-рецидивирующим рассеянным склерозом). В некоторых вариантах осуществления истощающим лимфоциты средством может быть терапевтическое поликлональное антитело (например, антитимоцитарное поликлональное антитело - глобулярный белок).
Настоящим изобретением также обеспечивается способ индукции иммунологической толерантности у субъекта, нуждающегося в трансплантации ткани. В этом способе субъекту вводят эффективное количество метотрексата, тем самым индуцируя иммунологическую толерантность к трансплантированной ткани у субъекта. В некоторых вариантах осуществления эффективное количество метотрексата вводят в одном цикле. В некоторых вариантах осуществления трансплантированной тканью является ткань почки или ткань сердца. В некоторых вариантах осуществления субъект также получает иммуномодулирующее средство (например, иммунодепрессант), такое как антитимоцитарное поликлональное антитело - глобулярный белок.
Настоящим изобретением также обеспечивается способ ингибирования T-клеточных реакций у субъекта, нуждающегося в терапевтическом средстве или трансплантации ткани. В этом способе субъекту вводят эффективное количество метотрексата до, одновременно с или после лечения субъекта терапевтическим средством или трансплантации ткани, тем самым ингибируя T-клеточные реакции у субъекта.
В некоторых вариантах осуществления способов настоящего изобретения терапевтическим средством является терапевтический белок.
В некоторых вариантах осуществления способов настоящего изобретения терапевтическим средством является терапевтическое антитело. Например, терапевтическим антителом может быть терапевтическое моноклональное антитело и/или истощающее лимфоциты средство (например, алемтузумаб), или терапевтическое поликлональное антитело (например, антитимоцитарное поликлональное антитело - глобулярный белок). В дополнительных вариантах осуществления, в которых терапевтическим антителом является алемтузумаб, субъектом может быть пациент с рассеянным склерозом. В дополнительных вариантах осуществления, в которых терапевтическим антителом является кроличье поликлональное антитимоцитарное антитело - глобулярный белок, субъектом может быть являющийся человеком пациент, который нуждается в трансплантации органа, страдает апластической анемией, и/или имеет или подвержен риску наличия гомологичной болезни. В других вариантах осуществления способов настоящего изобретения терапевтическим средством является фермент. Например, ферментом может быть альфа-галактозидаза A человека или кислая альфа-глюкозидаза человека.
В некоторых вариантах осуществления способов настоящего изобретения субъектом является человек.
В некоторых вариантах осуществления способов настоящего изобретения эффективное количество метотрексата может составлять от 0,1 мг/кг до 5 мг/кг. В некоторых вариантах осуществления один цикл метотрексата может состоять из 1 дня введения метотрексата или 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11 календарных дней введения метотрексата. В некоторых вариантах осуществления один цикл может назначаться между 48 часами до и 48 часами после начала терапевтического лечения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На каждой из фигур, описываемых ниже, звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05; **, p≤0,001; ***, p≤0,0001).
На фиг. 1A-B представлены образования антикроличьих IgG в ответ на один и множество курсов mATG, кроличьего поликлонального антитела - глобулярного белка против тимоцитов человека. Определяли средние титры антикроличьих IgG, и мышей, подвергнутых лечению только кроличьим IgG (rbIgG), использовали в качестве контроля. На фиг. 1A представлены ответы на один курс mATG в течение 8-недельного периода. На фиг. 1B представлены ответы на множество курсов mATG в течение 20-недельного периода. Стрелки указывают на моменты времени, в которые вводили mATG или rbIgG.
На фиг. 2 представлены средние титры алемтузумаб-специфических IgG после пяти ежемесячных лечений алемтузумабом. Стрелки указывают на моменты времени, в которые вводили алемтузумаб.
На фиг. 3 показано подавление образований IgG против mATG с помощью метотрексата (MTX) после одного курса mATG. Мышей подвергали лечению только mATG, только rbIgG, или mATG и метотрексатом.
На фиг. 4A-C представлены эффекты метотрексата на образования антикроличьих IgG на протяжении курса из пяти ежемесячных лечений mATG. Стрелки указывают на моменты времени, в которые вводили mATG или метотрексат. На фиг. 4A показано, что три цикла метотрексата значимо снижают средние титры антикроличьих IgG. На фиг. 4B показано, что один курс метотрексата значимо снижает средние титры антикроличьих IgG. На фиг. 4C сравниваются средние титры антикроличьих IgG у мышей, подвергнутых лечению только mATG, mATG и одним циклом метотрексата, или mATG и тремя циклами метотрексата. Один цикл метотрексата снижает титры антикроличьих IgG более значительно, чем три цикла метотрексата.
На фиг. 5 показано, что средние титры антикроличьих IgG являются уменьшенными после повторного введения mATG у мышей, подвергнутых лечению метотрексатом (с использованием одного или множества циклов метотрексата), по сравнению с мышами, которых подвергали лечению mATG, но не метотрексатом. Стрелки указывают на моменты времени, в которые вводили mATG или метотрексат.
На фиг. 6 показано, что средний титр антикроличьих IgG у мышей, подвергнутых лечению с использованием mATG и одного цикла метотрексата, в 100 раз меньше такового у мышей, подвергнутых лечению только mATG. Стрелки указывают на моменты времени, в которые вводили mATG или метотрексат.
На фиг. 7A-C показано, что метотрексат может снижать средние титры IgG против алемтузумаба у трансгенных по huCD52 мышей. Фиг. 7A: Титры антител против алемтузумаба ниже у мышей, подвергнутых лечению с использованием одного цикла метотрексата в дозе 1 мг/кг или 5 мг/кг, чем у мышей, подвергнутых или не подвергнутых лечению 0,5 мг/кг метотрексата. Мыши были подвергнуты лечению только алемтузумабом, или алемтузумабом и одним циклом метотрексата в дозе 0,5 мг/кг, 1 мг/кг или 5 мг/кг. Фиг. 7B: Дизайн исследования для исследования титров антител против алемтузумаба. Данные, относящиеся к титрам, получали через 24 часа после пятой дозы алемтузумаба, как указано звездочкой. Фиг. 7C: Метотрексат снижал титры антител против алемтузумаба у мышей, получающих алемтузумаб и метотрексат, по сравнению с мышами, получающими только алемтузумаб, через 24 часа после пятой дозы алемтузумаба.
На фиг. 8A-D представлено абсолютное количество клеток/мкл цельной крови для циркулирующих T-клеток у мышей, подвергнутых лечению с использованием пяти ежедневных доз 0,5 мг/кг алемтузумаба или забуференного фосфатом солевого раствора (PBS). На фиг. 8A показано, что количество T-клеток в целом (CD3+) было уменьшенным через две, три и четыре недели после лечения. На фиг. 8B показано, что количество В-клеток в целом (CD19+) было уменьшенным через три недели после лечения. На фиг. 8C показано, что количество T-клеток-хелперов (CD4+) было уменьшенным через две, три и четыре недели после лечения, а на фиг. 8D показано, что количество цитотоксических T-клеток (CD8+) было уменьшенным через две, три и четыре недели после лечения.
На фиг. 9A-B представлены ответы в виде образования алемтузумаб-специфических IgG. На фиг. 9A представлены ответы после трех циклов лечения алемтузумабом в дозе, равной 0,5 мг/кг, и одного цикла лечения метотрексатом в дозе, равной 0,5 мг/кг, 1 мг/кг или 2 мг/кг. Первый цикл лечения алемтузумабом состоял из пяти календарных дней введения доз, а каждый из второго и третьего циклов состоял из трех календарных дней введения доз. На фиг. 9B представлены титры IgG против алемтузумаба у каждого животного в группе на неделе 14.
На фиг. 10 показано, что один цикл метотрексата в дозе 5 мг/кг восстанавливает уровни циркулирующих mATG у мышей, подвергнутые лечению с использованием пяти ежемесячных доз mATG.
На фиг. 11 показано, что мыши, подвергнутые лечению с использованием пяти ежемесячных лечений mATG и одного цикла метотрексата, имеют увеличенное, mATG-опосредуемое истощение CD4+ и CD8+ T-клеток в крови после пятой дозы mATG. Эти данные являются объединенными из двух экспериментов.
На фиг. 12A-B показано, что мыши, подвергнутые лечению с использованием одного цикла метотрексата, демонстрируют увеличенные проценты регуляторных T-клеток (CD25+Foxp3+) после пятого ежемесячного лечения mATG. Мышей подвергали лечению с использованием пяти ежемесячных лечений только mATG, или пяти ежемесячных лечений mATG и одного цикла метотрексата, или пяти ежемесячных лечений mATG и трех циклов метотрексата. На фиг. 12A представлены уровни регуляторных T-клеток в селезенке. На фиг. 12B представлены уровни регуляторных T-клеток в крови.
На фиг. 13A показано, что титр антикроличьих IgG, превышающий 10000, может вносить помехи в фармакодинамику mATG. На фиг. 13B показано, что титр антикроличьих IgG, превышающий 100000, может мешать истощению CD4 и CD8 клеток. % от контроля до лечения относится к титрам CD4 и CD8 клеток относительно их соответствующих уровней до лечения mATG в процентах.
На фиг. 14 показано, что мыши, подвергнутые лечению 5 мг/кг метотрексата, демонстрируют увеличенное, опосредуемое алемтузумабом истощение циркулирующих CD3+ T-клеток и CD19+ B-клеток после пятой ежемесячной дозы алемтузумаба. Мышей подвергали лечению только алемтузумабом, или алемтузумабом и одним циклом метотрексата в течение первых трех дней исследования. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05; ***, p≤0,0001).
На фиг. 15A показано, что один цикл метотрексата может снижать титры специфических в отношении рекомбинантной кислой альфа-глюкозидазы человека (rhGAA) IgG на протяжении лечения, и даже через четыре недели после последнего лечения rhGAA, в 16-недельном исследовании. Стрелки указывают на моменты времени, в которые вводили rhGAA и метотрексат. Мышей подвергали лечению только rhGAA, или rhGAA и одним циклом метотрексата, или rhGAA и тремя циклами метотрексата. На фиг. 15B показано, что, в шестинедельном исследовании, один цикл метотрексата снижает титры rhGAA-специфических IgG. На фиг. 15C показано, что один цикл метотрексата снижает титры rhGAA-специфических IgG в 18-недельном исследовании.
На фиг. 16A показано, что метотрексат снижает титры антикроличьих IgG в модели аллогенной трансплантации сердца на мышах при введении с mATG, по сравнению с только mATG. На фиг. 16B показано, что метотрексат увеличивает уровни циркулирующих mATG в модели аллогенной трансплантации сердца на мышах. Мышей подвергали лечению солевым раствором, только mATG, или mATG и одним циклом метотрексата. mATG вводили в дозе 20 мг/кг в дни 0 и 4 исследования, в то время как 2 мг/кг метотрексата вводили в дни 0-6 исследования.
На фиг. 17 представлен график кривой выживаемости, полученной по методу Каплана-Мейера, который показывает, что комбинированное лечение с использованием mATG и метотрексата увеличивает продолжительность существования аллогенных сердец, трансплантированных мышам-реципиентам. Мышей оставляли без лечения, или подвергали лечению только 20 мг/кг mATG в дни 0 и 4 исследования, или лечению только 2 мг/кг метотрексата в дни 0-6 исследования, или лечению и mATG, и 2 мг/кг метотрексата в дни 0-6, или лечению и mATG, и 0,5 мг/кг метотрексата в дни 0-6 или дни 0-11.
На фиг. 18 показано, что мыши с трансплантированным аллогенным сердцем, подвергнутые лечению или только метотрексатом или метотрексатом в комбинации с mATG, демонстрируют уменьшение образований антител против аллотрансплантата. На фиг. 18A представлено связывание IgG реципиента с аллогенными фибробластами в день 21. На фиг. 18B представлены уровни аллоантител в день 21 у мышей, подвергнутых сингенным трансплантациям (Syn Tx) или аллогенным трансплантациям (Allo Tx) вместе с указанными лечениями. Представлено связывание IgG сыворотки отдельных мышей-реципиентов с аллогенными фибробластами, и оно представлено как отношение средней интенсивности флуоресценции (MFI) к неокрашенным фибробластам. На фиг. 18C представлены уровни аллоантител в день 21 у мышей, не подвергнутых лечению, подвергнутых лечению mATG, метотрексатом или mATG и метотрексатом. Уровни аллоантител были значимо ниже у мышей, подвергнутых лечению или mATG и метотрексатом (p=0,0008 и p<0,0001, соответственно).
На фиг. 19 показано, что количество B10 регуляторных B-клеток значимо увеличено после лечения метотрексатом. Мышей подвергали лечению только rhGAA или rhGAA и одним трехдневным циклом метотрексата или солевого раствора. Количества клеток подсчитывали в день 7 и день 8 исследования.
На фиг. 20 показано, что субпопуляции активированных В-клеток значимо увеличены в день 6 после лечения rhGAA и одним трехдневным циклом метотрексата, по сравнению с лечением только rhGAA. Абсолютные количества CD86+ B-клеток на стадии 2 иммунологической дифференциации, CD86+ В-клеток на стадии 3 иммунологической дифференциации, CD86+ фолликулярных B-клеток и CD86+ B-клеток краевой зоны подсчитывали в день 6 исследования. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05; **, p≤0,001).
На фиг. 21 показано, что в случае субпопуляций активированных B-клеток селезенки, таких как активированные B-клетки краевой зоны, активированные фолликулярные B-клетки и активированные В-клетки на стадии 3 иммунологической дифференциации, количества клеток остаются увеличенными даже после лечения rhGAA и метотрексатом, по сравнению с лечением только rhGAA. Стрелки отражают лечение rhGAA и метотрексатом. rhGAA вводили в день 1 и день 8. Метотрексат вводили в дни 1, 2 и 3, и дни 8, 9 и 10. Значимые различия представлены звездочками (*, p<0,05). Не представленные для дней 8 и 9 данные (указаны вертикальными пунктирными линиями).
На фиг. 22 показано, что популяции активированных T-клеток селезенки, такие как активированные Т-клетки-хелперы, активированные цитотоксические T-клетки и активированные регуляторные T-клетки оставались почти совершенно неизменными после лечения rhGAA и метотрексатом, по сравнению с лечением только rhGAA. Стрелки отражают лечение rhGAA и метотрексатом. rhGAA вводили в день 1 и день 8. Метотрексат вводили в дни 1, 2 и 3, и дни 8, 9 и 10. Значимые различия представлены звездочками (*, p<0,05). Не представленные для дней 8 и 9 данные (указаны вертикальными пунктирными линиями).
На фиг. 23 представлен дизайн исследования на протяжении шести месяцев для исследования лечения метотрексатом в комбинации с mATG. Сплошные стрелки отражают инъекции 5 мг/кг mATG, пунктирные стрелки отражают инъекции 5 мг/кг метотрексата, а точечные стрелки отражают конечные умерщвления.
На фиг. 24A-B показано, что один цикл метотрексата вместе с mATG обогащает B-клетки селезенки, по сравнению с только mATG или тремя циклами метотрексата. Фиг. 24A: активированные фолликулярные В-клетки; фиг. 24B: активированные B-клетки на стадии 3 иммунологической дифференциации.
На фиг. 25A-B представлены эффекты метотрексата на фармакодинамику алемтузумаба в крови. Мышей подвергали лечению 0,5 мг/кг алемтузумаба в течение пяти месяцев, или вместе с тремя еженедельными дозами 5 мг/кг/день метотрексата в связи с первым введением алемтузумаба или без них. Фиг. 25A: Метотрексат увеличивает истощение T-клеток в целом (CD3+), T-клеток-хелперов (CD4+) и регуляторных T-клеток (CD4+CD25+Foxp3+) с помощью алемтузумаба (AZM). Черные столбики отражают показатели у мышей, подвергнутых лечению только алемтузумабом, в то время как белые столбики отражают показатели у мышей, подвергнутых лечению алемтузумабом и метотрексатом. Фиг. 25B: Метотрексат увеличивает истощение B-клеток с помощью алемтузумаба. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05).
На фиг. 26 представлены эффекты метотрексата на фармакодинамику алемтузумаба в селезенке. Мышей подвергали лечению 0,5 мг/кг алемтузумаба в течение пяти месяцев, или вместе с тремя еженедельными дозами 5 мг/кг/день метотрексата в связи с первым введением алемтузумаба или без них. T-клетки истощены после пятого лечения алемтузумабом. CD8 CEN: CD8+ цетральные T-клетки памяти; CD8 EFF MEM: CD8+ эффекторные T-клетки памяти; CD4 CEN MEM: CD4+ центральные T-клетки памяти; CD4 EFF MEM: CD4+ эффекторные T-клетки памяти. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05).
На фиг. 27A-B представлены эффекты метотрексата на количества B-клеток в селезенке. Мышей подвергали лечению 0,5 мг/кг алемтузумаба в течение пяти месяцев, или вместе с тремя еженедельными дозами 5 мг/кг/день метотрексата в связи с первым введением алемтузумаба или без них. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05).
На фиг. 28A-B представлено истощение лимфоцитов селезенки через три дня после введения однократной дозы алемтузумаба. Фиг. 28A: T-клетки и B-клетки значимо истощены. Маленькие клетки отражают контрольных мышей, подвергнутых лечению PBS, а большие клетки отражают мышей, подвергнутых лечению алемтузумабом. Фиг. 28B: B-клетки (CD19+) истощены на 92% через 24 часа после лечения и остаются истощенными на 36% через три дня после лечения. Три графика отражают различные группы животных, кровоизвлечение у каждой из которой осуществляли в различные моменты времени. Звездочки указывают на показатели со статистически значимыми различиями (*р<0,05; **, p≤0,001; ***, p≤0,0001).
На фиг. 29A-B представлены эффекты метотрексата на уровни цитокинов. На фиг. 29A представлен дизайн шестимесячного исследования для определения уровней цитокинов в селезенке и лимфатических узлах. Звездочка указывает на то, что данные были получены через 24 часа после пятого лечения алемтузумабом. Стрелки указывают на моменты времени, в которые вводили алемтузумаб или метотрексат, или выполняли конечные умерщвления, как указано. На фиг. 29B представлены уровни активизирующего В-клетки фактора, относящегося к семейству TNF, (BAFF) у мышей, подвергнутых лечению только алемтузумабом или алемтузумабом и метотрексатом.
На фиг. 30A-B представлены уровни цитокинов после лечения алемтузумабом и метотрексатом. На фиг. 30A представлен дизайн четырехмесячного исследования. Звездочка указывает на то, что данные были получены через одну неделю после второго цикла метотрексата. Стрелки указывают на моменты времени, в которые вводили алемтузумаб или метотрексат, или выполняли конечные умерщвления, как указано. На фиг. 30В представлены уровни различных цитокинов у мышей, подвергнутых лечению только алемтузумабом (AZM) или с использованием 0,5 мг/кг, 1,0 мг/кг или 2,0 мг/кг метотрексата.
На фиг. 31 представлены эффекты метотрексата на титры антител против rhGAA у мышей с выключенным геном IL10 (IL10-/-) и C57BL/6. 20 мг/кг rhGAA вводили еженедельно в течение девяти недель мышам с выключенным геном IL10 и мышам дикого типа C57BL/6, вместе с 5 мг/кг/день метотрексата через 0, 24 и 48 часов после первых трех еженедельных лечений rhGAA или без него.
На фиг. 32 показаны некоторые, но не все, популяции В-клеток селезенки истощены через одну и/или две недели после лечения алемтузумабом. Столбики с мелкими штрихами отражают подвергнутых лечению забуференным фосфатом солевым раствором (PBS) трансгенных по huCD52 контрольных мышей; столбики с крупными штрихами отражают трансгенных по huCD52 мышей, подвергнутых лечению 0,5 мг/кг алемтузумаба в течение пяти календарных дней. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05; **, p≤0,001; ***, p≤0,0001).
На фиг. 33 представлен дизайн исследования для исследования эффектов метотрексата на популяции B-клеток в случае лечения алемтузумабом. Для патологической оценки использовали щитовидную железу и лимфатические узлы (LN). Стрелки указывают на моменты времени, в которые вводили алемтузумаб или метотрексат, или выполняли конечные умерщвления, как указано.
На фиг. 34 представлены эффекты только трех ежедневных доз 5 мг/кг/день метотрексата, только 0,5 мг/кг алемтузумаба и комбинации 0,5 мг/кг алемтузумаба и 5 мг/кг/день метотрексата на истощение популяций B-клеток. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05; **, p≤0,001; ***, p≤0,0001); ns, не значимые).
На фиг. 35 представлены эффекты метотрексата на истощение B-клеток после только пяти циклов лечения 0,5 мг/кг алемтузумаба или только трех ежедневных доз 5 мг/кг/день метотрексата или комбинации 0,5 мг/кг алемтузумаба и 5 мг/кг/день метотрексата. Звездочки указывают на показатели со статистически значимыми различиями (*, p<0,05).
На фиг. 36 показано, что уровни цитокинов MCP-1, IL-13, IL-6 и IL-12 являются уменьшенными у мышей, подвергнутых лечению с использованием одного цикла из трех дней 5 мг/кг метотрексата, вводимого в первый день лечения алемтузумабом. Данные получены через 24 часа после пятой дозы алемтузумаба, через четыре месяца после лечения метотрексатом.
На фиг. 37A-B представлены титры rhGAA на неделях 2, 6 и 12 у nu/nu бестимусных мышей. На фиг. 37A представлены титры rhGAA на неделях 2 и 6. Слева направо, показатели на неделе 2 у контрольных мышей, подвергнутых лечению солевым раствором, мышей, подвергнутых лечению rhGAA, и мышей, подвергнутых лечению rhGAA и метотрексатом, а затем показатели на неделе 6 у контрольных мышей, подвергнутых лечению солевым раствором, мышей, подвергнутых лечению rhGAA, и мышей, подвергнутых лечению rhGAA и метотрексатом. На фиг. 37B представлены показатели на неделе 12 у, слева направо, nu/nu мышей, подвергнутых лечению только rhGAA (Myo), nu/nu мышей, подвергнутых лечению метотрексатом и rhGAA, BL6 мышей, подвергнутых лечению только rhGAA, и BL6 мышей, подвергнутых лечению метотрексатом и rhGAA.
На фиг. 38A-B показано, что количество и процент экспрессирующих IL-10 B10 B-клеток увеличивается у мышей с индуцированной с помощью метотрексата толерантностью к rhGAA. B10 B-клетки, выделенные из животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, оценивали в отношении экспрессии белка IL-10 с помощью проточной цитометрии.
На фиг. 39A-B показано, что IL-10 экспрессируется и в активированных (CD86+), и в неактивированных (CD86-) B10 B-клетках. На фиг. 39A представлено полученное с использованием FACS (клеточного сортера с возбуждением флуоресценции) графическое изображение B10 B-клеток, окрашенных CD86, в то время как на фиг. 39B представлены количества CD86+IL10+ B10 B-клеток и CD86-IL10+ B10 B-клеток в ответ на лечение rhGAA или rhGAA и метотрексатом.
На фиг. 40A-B показано, что лечение метотрексатом вместе с rhGAA вызывает увеличение экспрессии в B 10 B-клетках TGF-бета. Вторая и третья панели фиг. 40A представляют собой полученные с использованием FACS графические изображения, на которых представлены B10 B-клетки, окрашенные TGF-бета и CD86, от животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом. На первой панели фиг. 40A представлено количество TGF-бета+ B10 B-клеток у животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом. На фиг. 40B представлены количества CD86+TGF-бета+ B10 B-клеток и CD8-TGF-бета+ B10 B-клеток.
На фиг. 41A показано, что B10 B-клетки, по-видимому, экспрессируют FoxP3 у животных, подвергнутых лечению rhGAA (фиг. 41A). На фиг. 41B показано, что количества FoxP3+ B-клеток увеличиваются при лечении и метотрексатом, и rhGAA. На фиг. 41C показано, что и активированные (CD86+), и неактивированные (CD86-) B10 B-клетки экспрессируют FoxP3.
На фиг. 42A-C показано, что фолликулярные В-клетки, В-клетки на стадии 2 иммунологической дифференциации и на стадии 3 иммунологической дифференциации (сверху вниз) экспрессируют IL-10, и что количества клеток субпопуляций экспрессирующих IL-10 B-клеток увеличиваются при лечении метотрексатом по сравнению с мышами, подвергнутыми лечению только rhGAA.
На фиг. 43A-C показано, что фолликулярные В-клетки, В-клетки на стадии 2 иммунологической дифференциации и на стадии 3 иммунологической дифференциации (сверху вниз) экспрессируют TGF-бета, и что количества клеток субпопуляций экспрессирующих TGF-бета В-клеток увеличиваются при лечении метотрексатом по сравнению с мышами, подвергнутыми лечению только rhGAA.
На фиг. 44A-C показано, что фолликулярные В-клетки, В-клетки на стадии 2 иммунологической дифференциации и на стадии 3 иммунологической дифференциации (сверху вниз) экспрессируют FoxP3, и что количества клеток субпопуляций экспрессирующих FoxP3 В-клеток увеличиваются при лечении метотрексатом по сравнению с мышами, подвергнутыми лечению только rhGAA.
На фиг. 45 представлены титры антител против rhGAA на неделе 6 у животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, в присутствии 5 мг/кг антитела против TGF-бета (1D11, Genzyme) или подобранного по изотипу контроля (13C4) или в его отсутствие. Титры антител определяли раз в две недели у четырех различных групп животных.
На фиг. 46A-C показано, что лечение 1D11 препятствовало индуцируемому метотрексатом увеличению количества B10 B-клеток, экспрессирующих TGF-бета, IL-10 или FoxP3. Селезенки получали от животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, которым также совместно вводили 1D11 или 13C4 через семь дней после одного лечения rhGAA или одного лечения rhGAA и метотрексатом. Клетки в каждой группе затем объединяли и культивировали в течение двух дней, а затем подсчитывали, используя проточную цитометрию.
На фиг. 47A-C показано, что лечение 1D11 препятствовало индуцируемому метотрексатом увеличению количества фолликулярных B-клеток, экспрессирующих TGF-бета или IL-10, хотя FoxP3+ фолликулярные B-клетки, по-видимому, не демонстрировали эффекты лечения 1D11. Селезенки получали от животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, которым также совместно вводили 1D11 или 13C4 через семь дней после одного лечения rhGAA или одного лечения rhGAA и метотрексатом. Клетки в каждой группе затем объединяли и культивировали в течение двух дней, а затем подсчитывали, используя проточную цитометрию.
На фиг. 48A-C показано, что, в случае В-клеток на стадии 2 иммунологической дифференциации, хотя лечение 1D11 мешало индуцируемому метотрексатом увеличению количества TGF-бета-экспрессирующих В-клеток на стадии 2 иммунологической дифференциации, эффекты не наблюдались по отношению к IL-10+ В-клеткам на стадии 2 иммунологической дифференциации. Селезенки получали от животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, которым также совместно вводили 1D11 или 13C4 через семь дней после одного лечения rhGAA или одного лечения rhGAA и метотрексатом. Клетки в каждой группе затем объединяли и культивировали в течение двух дней, а затем подсчитывали, используя проточную цитометрию.
На фиг. 49A-C показано, что в случае В-клеток на стадии 3 иммунологической дифференциации TGF-бета, IL-10 и FoxP3 поддаются детектированию, но нет явного эффекта лечения 1D11 на эти клетки. Селезенки получали от животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом, которым также совместно вводили 1D11 или 13C4 через семь дней после одного лечения rhGAA или одного лечения rhGAA и метотрексатом, и клетки подсчитывали, используя проточную цитометрию.
На фиг. 50A-C показано, что лечение 1D11 не влияет на основные уровни IL-10, TGF-бета и FoxP3 в фолликулярных клетках, В-клетках на стадии 2 иммунологической дифференциации и на стадии 3 иммунологической дифференциации.
Фиг. 51A является схемой, на которой показан перенос всех B-клеток селезенки от мыши с индуцированной толерантностью к hGAA (Myozyme® или «MYO») в не подвергнутую воздействию rhGAA мышь-реципиент. После переноса реципиентов (вместе с не подвергнутыми переносу контрольными животными, подвергнутыми лечению или rhGAA или rhGAA и метотрексатом) подвергали еженедельно лечению 20 мг/кг rhGAA.
На фиг. 51B представлено изучение титров, показывающее, что В-клетки в целом селезенки, выделенные от животных, подвергнутых лечению с использованием rhGAA и одного цикла метотрексата, могут переносить иммунологическую толерантность к rhGAA на не подвергнутых воздействию хозяев.
На фиг. 52 показаны количества клеток крови или селезенки нормальных мышей, подвергнутых лечению кроличьим IgG (rbIgG), только mATG, rbIgG и метотрексатом, или mATG и метотрексатом. Метотрексат не истощает CD4+, CD8+, регуляторные (CD4+CD25+FoxP3+) T-клетки или CD19+ B-клетки в целом у нормальных мышей.
На фиг. 53 показаны количества клеток крови или селезенки мышей с трансплантатами, подвергнутых лечению rbIgG, только mATG, rbIgG и метотрексатом, или mATG и метотрексатом, через 14 дней после трансплантации. Метотрексат не истощает CD4+, CD8+, регуляторные (CD4+CD25+FoxP3+) T-клетки или CD19+ B-клетки в целом у мышей с трансплантатами.
На фиг. 54 показано, что лечение метотрексатом вызывает статистически значимые увеличения IL-10 во множестве субпопуляций клеток, если судить по изменению средней интенсивности флуоресценции (MFI) этих белков у животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом.
На фиг. 55 показано, что лечение метотрексатом вызывает статистически значимые увеличения TGF-бета во множестве субпопуляций клеток, если судить по изменению средней интенсивности флуоресценции (MFI) этих белков у животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом.
На фиг. 56 показано, что лечение метотрексатом вызывает статистически значимые увеличения FoxP3 во множестве субпопуляций клеток, если судить по изменению средней интенсивности флуоресценции (MFI) этих белков у животных, подвергнутых лечению rhGAA или rhGAA и метотрексатом.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее изобретение основывается на неожиданном открытии авторов настоящего изобретения, что один цикл или короткий курс введения метотрексата уменьшает нежелательные иммунные ответы (такие как ответы в виде ADA и другие нежелательные опосредуемые T- и/или B-клетками иммунные ответы) у пациентов, получающих терапевтические белки, такие как заместительные ферменты или терапевтические антитела, и образования антител против трансплантата при трансплантации ткани. Это открытие привело к новым способам увеличения и безопасности, и эффективности терапий с использованием белков и трансплантации органов.
Конкретнее, исследования авторов настоящего изобретения показали, что один цикл метотрексата уменьшает ADA против терапевтических антител. Как детализировано ниже, один ряд исследований был выполнен с использованием мышиного варианта терапевтического поликлонального антитела, Thymoglobulin®. Thymoglobulin® является кроличьим поликлональным антителом -глобулярным белком против тимоцитов человека, используемым для иммуносупрессии в ситуациях трансплантации цельного органа, апластической анемии и для предупреждения гомологичной болезни. Было разработано кроличье поликлональное антитело - глобулярный белок против тимоцитов мыши (mATG). У него сохраняются характеристики, схожие с таковыми Thymoglobulin® (Ruzek, et al, Transplantation, 88(2):170-9 (2009)). Авторы настоящего изобретения показали, что один курс метотрексата может снизить титры IgG против mATG на >95%. В действительности, авторы настоящего изобретения неожиданно обнаружили, что один цикл метотрексата работает лучше в отношении уменьшения ADA, чем три цикла метотрексата. Кроме того, при этом уменьшении ADA сохраняются уровни циркулирующих mATG и увеличивается mATG-опосредованное истощение клеток и проценты регуляторных T-клеток после повторного введения дозы mATG. Другой ряд исследований был выполнен с использованием терапевтического моноклонального антитела, алемтузумаба. Авторы настоящего изобретения установили, что один курс метотрексата может сходно контролировать образования антител против алемтузумаба и увеличивать алемтузумаб-опосредованное истощение лимфоцитов.
Авторы настоящего изобретения обнаружили, что эта схема, предусматривающая один цикл метотрексата, может уменьшить ADA, когда терапевтическим белком является фермент. В двух из исследований авторов настоящего изобретения они показали, что один цикл метотрексата может эффективно контролировать образования антител против Myozyme® (рекомбинантной альглюкозидазы альфа человека или «rhGAA»), когда множество циклов, как первоначально полагали, требуется для уменьшения ADA.
Авторы настоящего изобретения обнаружили, что метотрексат также полезен при трансплантации органов. В ходе своих исследованияй авторы настоящего изобретения обнаружили, что один цикл метотрексата может контролировать образования антител против аллотрансплантата при аллотрансплантации сердца. При объединении с mATG продолжительность существования сердечного аллотрансплантата была значительно большей.
Исследования авторов настоящего изобретения показывают, что один цикл метотрексата, назначаемый в пределах первой недели терапии с использованием белка, может обеспечить долговременное уменьшение, составляющее более чем 95%, ADA в течение многих месяцев введения доз. Это уменьшение титра антител было долговременным, несмотря на отсутствие метотрексата на протяжении большей части исследований. Кроме того, авторы настоящего изобретения обнаружили, что один цикл метотрексата может контролировать реакции против аллотрансплантата (например, в модели аллогенной трансплантации сердца на мышах), демонстрируя контроль гуморальных иммунных ответов, направленных против множества антигенов одновременно.
Метотрексат классически известен как антагонист дигидрофолатредуктазы, который, как полагают, уничтожает пролиферирующие клетки в результате ингибирования метаболизма пуринов и препятствования de novo синтезу ДНК (Kremer, Arthritis Rheum, 50(5):1370-82 (2004)). Было нетрудно предположить, что метотрексат может просто уничтожать реактивные клетки посредством этого достаточно описанного механизма, но это представляется невероятным при использовании схемы, предусматривающей один цикл, которую открыли авторы настоящего изобретения. Метотрексат имеет короткий полупериод существования, и, невероятно, чтобы он находился в клетках или кровотоке достаточно долго, чтобы активно уничтожать клетки в течение трех-четырех месяцев после лечения (Walling, Invest New Drugs, 24(l):37-77 (2006); Slavikova et al, Neoplasma, 25(2):211-6 (1978)). Кроме того, авторы настоящего изобретения не находят доказательств истощения лимфоцитов после лечения метотрексатом. Наоборот, авторы настоящего изобретения неожиданно обнаружили, что схема, предусматривающая один цикл метотрексата, уменьшает нежелательные гуморальные иммунные ответы в результате индукции активного механизма иммунологического контроля, а не в результате беспорядочного истощения лимфоцитов.
Исследования авторов настоящего изобретения показывают, что один цикл метотрексата увеличивает количество B10 регуляторных B-клеток, а также активированных B-клеток краевой зоны, активированных фолликулярных B-клеток и активированных В-клеток на стадии 3 иммунологической дифференциации. Исследования авторов настоящего изобретения также показывают, что один цикл метотрексата увеличивает количество экспрессирующих IL-10, TGF-бета и FoxP3 B10 В-клеток, фолликулярных B-клеток, В-клеток на стадии 2 иммунологической дифференциации и «переходных» В-клеток, и что это увеличение опосредует TGF-бета. Эти исследования также показывают, что метотрексат увеличивает уровни экспрессии IL-10, TGF-бета и FoxP3. Увеличение популяций В-клеток селезенки является неожиданным, принимая во внимание современное понимание механизма действия метотрексата, и наводит на мысль, что в этом примере введения доз метотрексат работает уникальным, ранее неизвестным образом. Было установлено, что низкие, повторяющиеся дозы метотрексата у подвергнутых лечению инфликсимабом пациентов с ревматоидным артритом уменьшают образования антител против инфликсимаба; однако же, поскольку подвергание воздействию является повторяющимся, эта схема, более вероятно, приводит к постоянной иммунодепрессии, а не к индукции толерантности. Лечение только метотрексатом, как было установлено, уменьшает активность ревматоидного артрита при назначении раз в неделю в низких дозах. Недавняя публикация наводит на мысль, что при повторяющемся введении метотрексата в низкой дозе (через день) появляются аутоантиген-специфические регуляторные T-клетки, которые могут помочь обеспечить эффективность лечения метотрексатом при ревматоидном артрите (Xinqiang et al., Biomed Pharmacother, 64(7):463-471 (2010)). В отличие от этого, описанный здесь пример введения доз метотрексата является действительно уникальным, поскольку он включает короткий курс лечения метотрексатом, который может обеспечить длительный контроль желательных иммунологических реакций.
В общем, авторы настоящего изобретения установили уникальную схему введения доз метотрексата, которая может приводить к длительному контролю нескольких различных типов ответов в виде ADA и образований антител против аллотрансплантатов при трансплантации тканей, а также T-клеточных и B-клеточных реакций. Авторы настоящего изобретения установили, что иммунологическая толерантность, развивающаяся с помощью метотрексата, может быть передана от одного животного другому, например, посредством трансплантации B-клеток животного, у которого индуцирована толерантность, животному без толерантности. Данные авторов настоящего изобретения говорят о том, что метотрексат действует через уникальный механизм действия, который включает увеличение субпопуляций активированных B-клеток, которые могут представлять собой регуляторные B-клетки, действующие при супрессии иммунных ответов. Кроме того, метотрексат может также действовать через механизм увеличения регуляторных T-клеток.
Нежелательные иммунные ответы при биотерапии
Способы этого изобретения могут контролировать нежелательные иммунологические реакции (например, ответы в виде ADA и другие нежелательные опосредуемые T- и/или B-клетками иммунные ответы) в случае ряда биологических терапий (например, терапии с использованием биологического вещества, такого как белки, нуклеиновые кислоты, углеводы, липиды и метаболиты). Терапия с использованием белков относится к терапии, в которой терапевтическим средством является белковоподобное вещество, включая пептиды и белки. Белковыми терапевтическими средствами могут быть, например, ферменты, цитокины, факторы роста, иммуномодуляторы, тромболитики, антитела (в том числе поликлональные и моноклональные антитела), фрагменты антител или модифицированные антитела (например, Fab, F(ab')2, Fv, Fd, scFv, и dAb). Например, множество ферментозаместительных терапий было разработано для пациентов с некоторыми генетическими заболеваниями, включая Fabrazyme® (рекомбинантную альфа-галактозидазу человека) для болезни Фабри, Cerezyme® (имиглицеразу) для болезни Гоше, Aldurazyme® (ларонидазу) для мукополисахаридоза I (MPS I), и Myozyme® и Lumizyme® (альглюкозидазу альфа) для болезни Помпе. Примеры терапевтических антител включают Campath® (алемтузумаб), Thymoglobulin®, Avastin® (бевацизумаб), Lucentis® (ранибизумаб), Remicade® (инфликсимаб), Humira® (адалимумаб), Rituxan® (ритуксимаб), Tysabri® (натализумаб), Simulect® (базиликсимаб), Zenapax® (даклизумаб), OKT3® (муромонаб-CD3), Erbitux® (цетуксимаб), Mylotarg® (гемтузумаб), Herceptin® (трастузумаб) и Benlysta® (белимумаб). Примеры других терапевтических белков включают Enbrel® (этанерцент) и другие слитые белки.
Во многих случаях у пациента могут вызываться нежелательные иммунные ответы против терапевтического белка, приводя к переменным эффектам на результаты у пациента. Такие ответы имеют место, поскольку биологические терапевтические средства часто содержат последовательности и конформации, являющиеся несвойственными являющемуся человеком пациенту. Например, ADA препятствует терапевтической эффективности и/или увеличивает риски, относящиеся к безопасности. ADA может вызывать реакции гиперчувствительности, анафилаксию, сывороточную болезнь, болезнь иммунных комплексов, острую почечную недостаточность. За ADA у пациентов, получающих терапию с использованием белков, может следить клинический врач, используя твердо установившиеся способы, включая ELISA и иммуногистохимию.
В некоторых случаях «терапия с использованием белков», как здесь используется, относится к терапии с использованием вирусов, в которой вирусный вектор используется для доставки терапевтической нуклеиновой кислоты. Приводимые в качестве примера вирусы, используемые в таких терапиях, включают, но без ограничения, аденовирусы, аденоассоциированные вирусы и ретровирусы. Антитела могут образовываться против капсидных белков вируса, уменьшая эффективность и увеличивая относящиеся к безопасности риски таких терапий. Способы этого изобретения также применимы для контролирования нежелательных иммунологических реакций (например, ответов в виде ADA и других нежелательные опосредуемых T- и/или B-клетками иммунных ответов) при терапии с использованием вирусов.
Способы этого изобретения могут также использоваться для контролирования нежелательных иммунологических реакций при биотерапиях без использования белков. Приводимые в качестве примера терапии без использования белков включают, но без ограничения, терапии с использованием нуклеиновых кислот (например, антисмысловые терапии, терапии с использованием коротких интерферирующих РНК и терапии с использованием микроРНК).
Реакция против трансплантата при трансплантации
Способы этого изобретения можно также использовать для индукции иммунологической толерантности у пациента, которому трансплантируют ткань, например, ткань почки, печени, сердца и стволовые клетки. Отторжения вследствие реакций «хозяин против трансплантата» и «трансплантат против хозяина» часто происходят при трансплантации ткани, особенно алло- и ксенотрансплантации. Один цикл метотрексата может использоваться отдельно или вместе с другим иммуномодулятором (например, иммунодепрессантом, таким как Thymoglobulin®) для контролирования образования антител против трансплантата. Кроме того, комбинация Thymoglobulin® и метотрексата при трансплантации может работать с увеличением продолжительности существования трансплантата. Наконец, в том случае, когда Thymoglobulin® будет исследоваться в ситуациях хронического аутоиммунного заболевания, такого как ревматоидный артрит и рассеянный склероз, метотрексат может создать возможность для безопасного повторного лечения Thymoglobulin®, защищая пациента от образования значительных количеств антикроличьих антител (таких как IgG и/или IgM) и/или развития инфузионных реакций.
Контролирование нежелательных иммунологических реакций с помощью метотрексата
Авторы настоящего изобретения установили, что один, короткий цикл метотрексата может значительно уменьшить нежелательные иммунологические реакции, такие как ADA, у субъектов, получающих биологические терапевтические средства (например, терапевтические белки), и реакции против аллотрансплантата у пациентов, которым трансплантируют ткань. Уменьшение нежелательного ADA может не только повысить безопасность для пациентов, но также может увеличить эффективность терапевтического белка посредством улучшения фармакодинамики и/или фармакодинамики терапевтического белка.
Метотрексат, соединение в виде небольшой молекулы, использовался для лечения пациентов с тяжелой формой активного ревматоидного артрита, тяжелой формой псориаза и некоторыми типами рака, включая раки, которые возникают в тканях, которые образуются вокруг оплодотворенного яйца в матке, рак молочной железы, рак легкого, некоторые раки головы и шеи, некоторые типы лимфомы и лейкоз (рак, который возникает в лейкоцитах). Метотрексат лечит рак посредством замедления роста раковых клеток. Метотрексат лечит псориаз посредством замедления роста клеток кожи с остановкой образования чешуек. Метотрексат может лечить ревматоидный артрит посредством уменьшения активности иммунной системы.
Метотрексат был исследован с точки зрения контролирования ответов в виде ADA, индуцированных против α-галактозидазы A и α-глюкозидазы. Однако эти исследования были выполнены с использованием множества циклов лечения метотрексатом (Garman et al. Clin Exp Immunol, 137(3):496-502 (2004); Joseph et al, Clin Exp Immunol, 152(1):138-46 (2008); Mendelsohn et al, N Engl J Med, 360(2):194-5 (2009)), а не одного цикла метотрексата.
Исследования авторов настоящего изобретения неожиданно показывают, что одного цикла метотрексата хватает для значительного уменьшения ADA и антител против аллотрансплантата. Действительно, в исследованиях, включающих терапевтические антитела (например, mATG), авторы настоящего изобретения показывают, что лечение метотрексатом с использованием одного цикла является более эффективным в уменьшении ADA, чем лечение метотрексатом с использованием множества циклов. Предшествующие исследования с использованием метотрексата не свидетельствовали о том, что одного цикла метотрексата будет достаточно для уменьшения ADA, не говоря уже о том, что он будет более эффективным, чем лечение с использованием множества циклов. Метотрексат, как известно, является цитотоксическим. Поэтому в случае выдвижения гипотезы, что механизм уменьшения ADA с помощью метотрексата основан на этом свойстве, фактически можно было бы рассчитывать, что уменьшение числа циклов лечения будет уменьшать благотворные эффекты метотрексата. Кроме того, эти ранее опубликованные исследования не демонстрируют полезный эффект лечения метотрексатом на фармакокинетику, фармакодинамику, эффективность или безопасность, что неожиданно было продемонстрировано здесь с использованием одного курса метотрексата. Также в них не описывается, что метотрексат может уменьшать ADA при терапии с использованием антител.
Как здесь используется, схема, предусматривающая один цикл, относится к схеме лечения, или лечебному комплекту, состоящей из календарных или некалендарных дней, и начинается предпочтительно спустя не более пяти дней (например, не более трех дней) после введения дозы основного терапевтического белка или трансплантации. Если дозы основного терапевтического белка вводятся во множестве периодов, один цикл лечения метотрексатом предпочтительно не выходит за пределы первого периода введения дозы терапевтического белка. В качестве примера, в случае назначаемой раз в неделю, раз в месяц или раз год терапии с использованием белка один цикл метотрексата состоит из трех календарных дней приема метотрексата (например, перорально), начиная с дня 0, дня, когда пациенту в первый раз назначается основной терапевтический белок, или когда пациенту трансплантируют ткань. Затем пациент получает однократную дозу метотрексата в день 1 (спустя 24 часа) и в день 2 (спустя 48 часов). Один цикл метотрексата может также состоять, например, из 2, 3, 4, 5, 6, 7 или 8 последовательных ежедневных доз метотрексата, начиная с дня 0. Метотрексат может также назначаться в другие моменты времени, считающиеся подходящими, например, при контролировании вторичного аутоиммунитета, например, при истощающей лимфоциты терапии. Предпочтительно один цикл метотрексата не длится больше 8 дней. В некоторых вариантах осуществления один цикл метотрексата начинается между 48 часами до и 48 часами после начала основного терапевтического лечения (т.е. лечения с использованием биологического терапевтического средства). Например, один цикл метотрексата может начаться за 48 часов до, за 36 часов до, за 24 часа до, за 12 часов до начала основного терапевтического лечения, одновременно с ним, через 12 часов после, через 24 часа после, через 36 часов после или через 48 часов после начала основного терапевтического лечения.
Исследования авторов настоящего изобретения неожиданно показывают, что низкой дозы метотрексата хватает для контролирования нежелательных иммунологических реакций (например, ответов в виде ADA и других нежелательных опосредуемых T- и/или B-клетками иммунных ответов) при терапиях с использованием белков и трансплантации. Соответственно, в вариантах осуществления настоящего изобретения метотрексат можно назначать в более чем одном цикле, но в низкой общей дозе. Например, метотрексат можно назначать в двух или более (например, 3, 4, 5, 6 и т.д.) циклов, но с использованием общей кумулятивной дозы, составляющей не более чем 5 мг/кг пациента.
Доза метотрексата будет представлять собой количество метотрексата, эффективное в уменьшении нежелательных иммунологических реакций, таких как гуморальные или клеточные ответы, при назначении в одном цикле. Эффективное количество метотрексата у являющихся людьми пациентов может находиться в диапазоне от 0,05 мг/кг до 5 мг/кг. В некоторых вариантах осуществления эффективное количество составляет 0,1 мг/кг - 1,5 мг/кг. В некоторых вариантах осуществления эффективное количество составляет 0,12 мг/кг - 1,28 мг/кг. В некоторых вариантах осуществления эффективное количество составляет 0,12 мг/кг. В некоторых вариантах осуществления эффективное количество составляет 1,28 мг/кг. Рекомендуемая доза метотрексата может представлять минимальные риски безопасности, поскольку схема введения доз включает лишь короткий курс метотрексата на уровнях доз, которые больше соответствуют дозам для ревматоидного артрита, чем низким дозам для неопластических заболеваний. В исследования авторов настоящего изобретения проверенная общая доза метотрексата в каждом цикле для мышей составляла 14 или 15 мг/кг. 14 мг/кг метотрексата для мышей эквивалентны приблизительно 68 мг или 5,92 мг/м2 для среднего взрослого человека весом 60 кг. Пациенты с ревматоидным артритом могут получать вплоть до 25 мг метотрексата в неделю без страдания значительными токсичностями. Низкая доза метотрексата для неопластических заболеваний, как считают, составляет 30 мг/м2, значительно больше чем 5,92 мг/м2. Поэтому рекомендуемые выше дозы, в сочетании с кратковременным характером этой схемы введения метотрексата, будут, вероятно, хорошо переноситься взрослыми пациентами. Точная доза и схема введения метотрексата должны, конечно, устанавливаться клиническим врачом принимая во внимание физическое состояние пациента, его возраст, вес, пол, другие лекарственные средства, которые он/она принимает, и их известные побочные эффекты, и любые другие соответствующие факторы. За эффектом метотрексата на контролирование нежелательных гуморальных иммунных ответов у пациента можно следить с помощью хорошо известных способов, включающих клиническое обследование пациента, симптомов, тестов с использованием крови для исследования ADA и титров антител против аллотрансплантата, иммуногистохимических исследований (например, анализы отложения компонента комплемента C4 и другие твердофазные методы детектирования антител, такие как иммуноферментный твердофазный анализ (ELISA) и флуорометрические анализы на основе шариков). За эффектом можно также следить с помощью измерения уровней биомаркеров, таких как MCP-1, IL-13, IL-6 и IL-12, уровни которых, как было установлено авторами настоящего изобретения, снижаются при лечении метотрексатом, и В-клеток на стадии 2 иммунологической дифференциации, В-клеток на стадии 3 иммунологической дифференциации, фолликулярных B-клеток, B-клеток краевой зоны, B10 B-клеток и B1 B-клеток, количество которых, как было установлено авторами настоящего изобретения, увеличивается при лечении метотрексатом. Кроме того, TGF-бета, FoxP3, IL-5, IL-10, IL-15 и GM-CSF могут использоваться в качестве биомаркеров для слежения за эффектами метотрексата на нежелательные иммунные ответы по мере необходимости. Уровни биомаркеров могут также использоваться для слежения за эффектами метотрексата на реагирование T-клеток на терапевтическое средство (например, терапевтический белок). В качестве считываний данных для эффекта метотрексата на T-клеточные реакции можно также следить за биомаркерами активации Т-клеток, такими как IL-2, интерферон-γ и TNF-α.
Благодаря своей способности контролировать нежелательные иммунные ответы схема, предусматривающая один цикл метотрексата, этого изобретения может расширить применение многих терапевтических белков, многократные применения которых для конкретного пациента были ограничены в прошлом из-за забот о безопасности и эффективности. Например, сопутствующее применение метотрексата с Thymoglobulin® может расширить полезность Tymoglobulin® для других болезненных состояний, при которых желательно повторное введение дозы, таких как опосредуемые T-клетками аутоиммунные заболевания, включающие, но без ограничения, диабет, обыкновенную волчанку, склеродермию, ревматоидный артрит, псориаз и рассеянный склероз. Кроме того, эффект метотрексата может распространяться на эффективность и безопасность алемтузумаба, например, в случае таких аутоиммунных заболеваний, как рассеянный склероз, когда алемтузумаб обычно назначается в многократных ежегодных циклах, или в случае хронического B-клеточного лимфолейкоза, когда алемтузумаб назначают в 12-недельном цикле, начиная с дозы 3 мг/день (до тех пор, пока инфузионные реакции не станут равны или меньше степени 2), затем увеличивая до 10 мг/день (до тех пор, пока инфузионные реакции не станут равны или меньше степени 2) и, наконец, повышая до 30 мг/день (через день, 3 раза в неделю). Эти типы схем введения доз могут усиливать ингибиторные в отношении ADA реакции. Метотрексат может, таким образом, использоваться для контролирования ADA и любых других нежелательных иммунный ответов.
Улучшение истощающей лимфоциты терапии с помощью метотрексата
Приводимым в качестве примера применением этого изобретение является использование метотрексата для улучшения истощающей лимфоциты терапии, такой как терапия с использованием алемтузумаба для лечения рассеянного склероза (MS), такого как ремиттирующе-рецидивирующий MS. «Истощение лимфоцитов» представляет собой тип иммунодепрессии посредством уменьшения циркулирующих лимфоцитов, например, T-клеток и/или B-клеток, приводящее к лимфопении. Терапевтически, истощения лимфоцитов можно достичь с помощью терапевтического белка, такого как Thymoglobulin®, гуманизированного моноклонального антитела против CD52 CAMPATH-1H® (алемтузумаба) и ритуксимаба. Истощение лимфоцитов является желательным при лечении ряда аутоиммунных заболеваний, включая рассеянный склероз (Coles et al, Ann. Neurol. 46, 296-304 (1999); Coles et al, 2008), ревматоидный артрит, васкулит и обыкновенную волчанку.
Истощающая лимфоциты терапия может вызывать вторичный аутоиммунитет. Аутоиммунитет называют здесь «вторичным аутоиммунитетом», когда он возникает после проявления первого («основного») заболевания, например, «основного» аутоиммунного заболевания. Вторичный аутоиммунитет иногда возникает у пациентов с MS, имеющих, или имевших, лимфопению после, например, истощающей лимфоциты терапии. У некоторых индивидуумов вторичный аутоиммунитет возникает вскоре после истощающей лимфоциты терапии (например, лечения алемтузумабом). У других индивидуумов вторичный аутоиммунитет может не возникать впредь по истечения месяцев или лет после истощающей лимфоциты терапии; у некоторых из этих индивидуумов к тому времени, когда у них развивается вторичный аутоиммунитет, может произойти существенное восстановление лимфоцитов (общего числа лимфоцитов), так что они могут больше не иметь лимфопению. Истощение лимфоцитов может возникать в случае лечения терапевтическими антителами или терапевтическими средствами в виде небольших молекул.
Вторичный аутоиммунитет, возникающий у лимфопенических пациентов с MS, может быть любым типом аутоиммунного состояния, отличным от MS, включая, но без ограничения, тиреоидный аутоиммунитет (например, болезнь Грейвса), тромбоцитопеническую пурпуру, тромбоцитопению иммунного генеза (ITP), болезнь Гудпасчера, аутоиммунную нейропению, аутоиммунную гемолитическую анемию и аутоиммунную лимфопению. В некоторых вариантах осуществления вторичный аутоиммунитет опосредуется В-клетками, т.е. является B-клеточными реакциями, и аутоантитела непосредственно связаны с развитием заболевания и его патологией.
Методы диагностирования этих аутоиммунных заболеваний и слежения за ними, включая оценку симптомов и медицинское обследование, такое как анализ крови, хорошо известны квалифицированным в данной области техники специалистам. В настоящем изобретении предусматривается использование любых известных методов. Например, в качестве способа обнаружения признаков аутоиммунитета можно определить уровни аутоантител в жидкости организма пациента (например, крови). В частности, можно измерить антиядерные антитела, антитела против гладкой мышцы и антитела против митохондрий. В случае обнаружения антиядерных антител могут быть выполнены дополнительные анализы для измерения антител против двухцепочечной ДНК, антител против рибонуклеопротеинов и анти-La антител. Для обнаружения аутоиммунных заболеваний щитовидной железы можно измерить антитела против пероксидазы щитовидной железы (TPO) и антитела против рецептора стимулирующего гормона щитовидной железы (TSH); в случае обнаружения антител против TPO или рецептора TSH можно определить, затронута ли функция щитовидной железы, посредством измерения уровней свободного T3, свободного T4 и TSH. Антитела против тромбоцитов можно измерить для выявления аутоиммунной тромбоцитопении, а определение уровней тромбоцитов в крови может служить для определения того, является наличие антител против тромбоцитов причиной уменьшения количества тромбоцитов. См., также WO 2010/041149.
Схема, предусматривающая один цикл метотрексата, этого изобретения может использоваться для повышения безопасности и эффективности истощающей лимфоциты терапии в результате уменьшения ADA, а также минимизации вторичного аутоиммунитета. Без желания ограничиваться этой теорией, авторы настоящего изобретения полагают, что метотрексат может ослаблять вторичный аутоиммунитет посредством индукции толерантности к множеству аутоантигенов одновременно.
Кроме особо оговоренных случаев, все технические и научные термины, используемые здесь, имеют значение, одинаковое с тем, в котором они обычно понимаются специалистом со средним уровнем компетентности в данной области техники, к которой относится это изобретение. Ниже описываются примерные методы и материалы, хотя методы и материалы, схожие с методами и материалами, описанными здесь, или эквивалентные им, могут также использоваться при осуществлении на практике или проверке настоявшего изобретения. Все публикации или другие ссылочные документы, упоминаемые здесь, включены посредством ссылки в их полном объеме. В случае противоречия будет господствовать настоящее описание, включая определения. Хотя здесь приведен ряд документов, это приведение не является признанием, что любой из этих документов образует часть общего известного уровня техники. На протяжении всего этого описания и формулы изобретения слово «включать», или варианты, такие как «включает» или «включающий», как будет подразумеваться, предполагает включение заданного целого числа или группы целых чисел, а не исключение какого-либо целого числа или группы целых чисел. Материалы, методы и примеры являются лишь иллюстративными и, как предполагается, неограничивающими.
ПРИМЕРЫ
Дополнительные детали настоящего изобретения будут описаны в следующих неограничивающих примерах. Следует понимать, что эти примеры, хоть и указывающие на предпочтительные варианты осуществления настоящего изобретения, приведены лишь ради иллюстрации и не должны рассматриваться как ограничение прилагаемых вариантов осуществления. Исходя из описания настоящего изобретения и этих примеров, квалифицированный в данной области техники специалист может выяснить определенные характеристики этого изобретения и, не выходя за пределы его сущности и объема, может внести различные изменения и модификации в настоящее изобретение, чтобы адаптировать его к различным использованиям и условиям. Материалы и методы, используемые в этих демонстрационных примерах, описаны, как указано далее.
Мыши
Нормальных самок мышей C57BL/6 возрастом 6-12 недель использовали для in vivo исследований кроличьего поликлонального антитела - глобулярного белка против тимоцитов мыши (mATG) и получали из Jackson Laboratories (Bar Harbor, ME) или Taconic Laboratories (Hudson, NY). В исследованиях, относящихся к алемтузумабу, использовали трансгенных (Tg) по CD52 человека (huCD52) мышей возрастом 6-12 недель, которых получали из Charles River Laboratories/Genzyme Corp. Мышей размещали и содержали в соответствии с Руководством по содержанию и использованию лабораторных животных и в соответствии с аккредитацией Американской ассоциации по аккредитации содержания лабораторных животных I, и все протоколы с использованием животных, используемые в этих исследованиях, были одобрены Институциональным комитетом по содержанию и использованию животных.
Трансгенная по huCD52 мышь, использованная для неклинических фармакологических исследований, была создана Xenogen (Cranbury, NJ, США). Для создания этой мыши конструкцию в виде бакмиды, содержащую приблизительно 145 т.п.о. геномной ДНК из хромосомы 1 человека, неспецифическим образом интегрировали в геном CD-1 эмбриональных стволовых клеток мыши. На основании участка геномной ДНК человека, который содержала эта бакмида, конструкция включала всего 5 частичных и полных генов с неизвестной функцией помимо CD52 человека. 5 частичными или полными генными сегментами, содержащими в бакмиде, были следующие генные сегменты: ген CD52 человека, 3'-конец нового гена (DKFZP434L0117), ген SH3BGRL3 (подобного связывающемуся с SH3-доменом белку с высоким содержанием глютаминовой кислоты 3), ген для компаньона (SOC), ген AIM1L (подобного отсутствующему при меланоме 1) и ген белка «цинковые пальцы» 683. Были созданы три линии-основатели, и линия 107 была создана в Genzyme.
Введение антител
Поликлональные антитела mATG и rbIgG были приготовлены, как описано в Ruzek et al., Transplantation, 88(2):170-9 (2009), и вводились с помощью внутрибрюшинной инъекции в различных схемах в зависимости от эксперимента. Моноклональное антитело алемтузумаб вводили внутривенно или в виде однократной инъекции 0,5 мг/кг, или в трех- или пятидневном цикле введения 0,5 мг/кг/день.
Лечение Myozyme®
Рекомбинантную альглюкозидазу альфа человека (rhGAA, продаваемую Genzyme Corp. как Myozyme®) использовали в качестве приготовленного лекарственного продукта. Мышей подвергали еженедельному лечению 20 мг/кг rhGAA с помощью болюсной инъекции в хвостовую вену, если не изложено по-другому. Всех мышей подвергали профилактическому лечению с использованием 5-30 мг/кг дифенгидрамина (Baxter Healthcare Corporation, Deerfield, IL), вводимого внутрибрюшинно до введения rhGAA. Контрольных животных лечили с помощью внутривенного введения или стерильного 0,9% солевого раствора, или буфера для препарата rhGAA.
Лечение метотрексатом
Метотрексат (Каталожный № Calbiochem - 454125) вводили в дозе, составляющей 0,5, 1,0, 2,0 или 5 мг/кг, с помощью внутрибрюшинной инъекции в течение 1-3 циклов, где каждый цикл равен или трем, или шести, или семи календарным дням инъецирования в зависимости от эксперимента. В исследованиях, которые включали ежемесячное лечение mATG, метотрексат вводили внутрибрюшинно в дозе, составляющей 5 мг/кг, через 0, 24 и 48 часов после или первоначально лечения mATG, или трех первых лечений mATG. В исследованиях с использованием трансплантата, в которых введение доз mATG осуществляли в дни 0 и 4, метотрексат назначали ежедневно в дозе, равной 2 мг/кг, с дня 0 до дня 6, ежедневно в дозе, равной 0,5 мг/кг, с дня 0 до дня 6, или ежедневно в дозе, равной 0,5 мг/кг, с дня 0 до дня 11.
Лечение mATG
Поликлональное антитело mATG вводили в виде внутрибрюшинной инъекции 5 мг/кг каждые четыре недели или в виде двух доз, равных 20 мг/кг, назначаемых с интервалом в четыре дня в случае трансплантации, при этом первую дозу назначали в день трансплантации (день 0).
Приготовление клеток из различных тканей
Для приготовлений спленоцитов и клеток лимфатических узлов создавали суспензии отдельных клеток из полученных мышиных селезенок или паховых и кишечных лимфатических узлов посредством гомогенизации между предметными матированными стеклами в PBS, содержащем 2% FCS. Для приготовлений спленоцитов эритроциты лизировали посредством инкубации в течение 1-2 минуты с раствором для лизиса эритроцитов (BD Biosciences, San Diego, CA). Кровь получали с помощью ретроорбитального кровоизвлечения, и приготовления клеток осуществляли посредством лизиса эритроцитов с помощью раствора для лизиса эритроцитов (BD Biosciences) в течение 20-30 минут. В случае приготовлений из всех тканей живые клетки подсчитывали, используя автоматический счетчик ViCell (Beckman Coulter, Fullerton, CA). После выделения все клеточные препараты промывали PBS/2% FCS перед использованием в описанных ниже анализах.
Проточная цитометрия
Для оценки популяций клеток в различных тканях суспензии отдельных клеток тканей инкубировали с конъюгированными с флуорохромом антителами, которые включали антитела против CD4, CD8, CD25, CD44, CD62L мыши (все антитела от BD
Biosciences или eBioscience, San Diego, CA). Анализ внутриклеточной экспрессии Foxp3 выполняли в соответствии с протоколом производителя анти-Foxp3 (eBiosciences, San Diego, CA). После инкубации с антителами клетки промывали и анализировали с помощью проточной цитометрии (FACSCanto, программное обеспечение BD Biosciences и FCS Express, De Novo Software, Los Angeles, CA).
Оцененные популяции клеток были определены, как указано далее:
все CD4 T-клеток: CD4+CD8-,
все CD8 T-клетки: CD8+CD4-,
CD4 клетки, не подвергнутые воздействию антигенов: CD4+CD25-CD62L+CD44-,
CD4 клетки памяти: CD4+CD25-CD62L-CD44+,
не подвергнутые воздействию антигенов CD8 T-клетки: CD8+CD44-CD62L+,
CD8 клетки памяти: CD8+CD44+CD62L-,
регуляторные T-клетки: CD4+CD25+Foxp3+,
все B-клетки: CD19+,
B2/фолликулярные B-клетки: CD19+CD21средний уровеньCD23высокий уровень,
B1 B-клетки: CD19+CD43+CD11b+,
B-клетки на стадии 1 иммунологической дифференциации: CD19+CD93+CD23-IgMвысокий уровень,
B-клетки на стадии 2 иммунологической дифференциации: CD19+CD93+CD23+IgMвысокий уровень,
B-клетки на стадии 3 иммунологической дифференциации: CD19+CD93+CD23+IgMнизкий уровень,
B-клетки краевой зоны: CD19+CD21высокий уровеньCD23низкий уровень, и
B10 B-клетки: CD19+CD5+CD1d+.
С помощью in vitro исследований блокирования было определено, что вплоть до 100 мкг/мл mATG не мешают детектированию этих популяций.
ELISA IgG против mATG
Уровни IgG против mATG в мышиной сыворотке анализировали с помощью иммуноферментного твердофазного анализа (ELISA). Вкратце, 96-луночные планшеты (Corning Inc., Corning, NY, США) покрывали в течение ночи 1 мкг/мл кроличьего IgG в забуференном фосфатом солевом растворе (PBS). После блокирования с помощью буфера для блокирования Super Block Blocking Buffer (Thermo Scientific, Rockford, IL, США) последовательные разведения сыворотки добавляли в двух повторах в покрытые кроличьим IgG планшеты и инкубировали при 37°C в течение 1 ч. Планшеты промывали, и добавляли конъюгированное с пероксидазой хрена козье второе антитело против IgG мыши (Southern Biotechnology Associates, Birmingham, AL, США), и допускали инкубацию в течение 1 ч при 37°C. После конечной промывки добавляли субстрат 3,3',5,5'-тетраметилбензидин (BioFx, Owings Mills, MD, США), и допускали проявление цвета в течение 15 мин при комнатной температуре. Реакцию останавливали добавлением 1 н HCl, и значения оптической плотности считывали при 450/650 нм в считывающем устройстве для планшетов ELISA (Molecular Devices, Sunnyvale, CA, США). Конечные титры определяли как наибольшее разведение, которое дает в среднем значение, превышающее ОП=0,100, используя программное обеспечение Softmax (Molecular Devices, Sunnyvale, CA, США).
ELISA mATG-специфических IgG
Мышиную сыворотку анализировали с помощью ELISA. Вкратце, 96-луночные планшеты (Corning Inc., Corning, NY, США) покрывали в течение ночи 1 мкг/мл козьим антителом против Fc-фрагмента IgG кролика (Bethyl Laboratories, TX, США). После блокирования с помощью 0,5% BSA (высокой степени чистоты) стандартные контроли и образцы сыворотки разводили при необходимости и добавляли в двух повторах в лунки покрытых планшетов, и осуществляли инкубацию при 36-38°C в течение 1 часа с легким встряхиванием. Планшеты промывали, и добавляли конъюгированное с пероксидазой хрена козье антитело против Fc-фрагмента IgG кролика (Bethyl Laboratories, TX, США) в соответствующих случаях, и осуществляли инкубацию в течение 1 часа при 36-38°C с легким встряхиванием. После конечной промывки добавляли субстрат 3,3',5,5'-тетраметилбензидин (BioFx, Owings Mills, MD, США), и допускали проявление цвета в течение 15 мин при комнатной температуре. Реакцию останавливали добавлением 1 н HCl, и значения оптической плотности считывали при 450/650 нм в считывающем устройстве для планшетов ELISA (Molecular Devices, Sunnyvale, CA, США). Конечные концентрации интерполировали из стандартной кривой. Было заранее определено, что на измерение mATG-специфических IgG с помощью этого метода оказывают лишь умеренное влияние титры IgG против mATG, которые превышали 218000.
ELISA IgG против алемтузумаба
Кровоизвлечение у мышей осуществляли через 4-6 дней после лечения алемтузумабом, и специфические IgG против алемтузумаба измеряли с помощью ELISA. Вкратце, 96-луночные планшеты (Corning Inc., Corning, NY, США) покрывали в течение ночи 3 мкг/мл алемтузумаба в PBS (pH 7,2). После блокирования с помощью 0,1% BSA в PBS последовательные разведения сыворотки добавляли в двух повторах в покрытые алемтузумабом планшеты, и осуществляли инкубацию при 37°C в течение 1 ч. Планшеты промывали, и добавляли конъюгированное с пероксидазой хрена козье второе антитело против IgG мыши (Southern Biotechnology Associates, Birmingham, AL, США), и допускали инкубацию в течение 1 ч при 37°C. После конечной промывки добавляли субстрат TMB (BioFx, Owings Mills, MD, США), и допускали проявление цвета в течение 15 мин при комнатной температуре. Реакцию останавливали добавлением 1 н HCl, и значения оптической плотности считывали при 450/650 нм в считывающем устройстве для планшетов ELISA (Molecular Devices, Sunnyvale, CA, США). Конечные титры определяли как антилогарифм преобразованного посредством логарифмирования разведения образца, интерполируемого в значение оптической плотности, равное 0,2, используя программное обеспечение Excel (Microsoft, Redmond, WA, США).
ELISA IgG против rhGAA
Кровоизвлечение у мышей осуществляли через 4-6 дней после лечения rhGAA, и специфические IgG определяли с помощью ELISA. Вкратце, 96-луночные планшеты (Corning Inc., Corning, NY, США) покрывали в течение ночи 5 мкг/мл rhGAA в натрийацетатном буфере (pH 5,0). После блокирования с помощью 0,1% BSA в PBS последовательные разведения сыворотки добавляли в двух повторах в покрытые rhGAA планшеты, и осуществляли инкубацию при 37°C в течение 1 часа. Планшеты промывали, и добавляли конъюгированное с HRP козье второе антитело против IgG мыши (Southern Biotechnology Associates, Birmingham, AL, США), и допускали инкубацию в течение 1 ч при 37°C. После конечной промывки добавляли субстрат 3,3',5,5'-тетраметилбензидин (TMB, KPL, Gaithersburg, MD), и допускали проявление цвета в течение 15 мин при комнатной температуре. Реакцию останавливали добавлением 1 н HCl, и значения оптической плотности считывали при 450/650 нм в считывающем устройстве для планшетов ELISA (Molecular Devices, Sunnyvale, CA, США). Конечные титры определяли как обратную величину разведения сыворотки, которое дает в результате значение оптической плотности, равное 0,2, используя программное обеспечение Softmax (Molecular Devices, Sunnyvale, CA).
Ex vivo исследования
Мышам C57BL/6 (Jackson Laboratories) или E4GAAKO (Charles River) возрастом 8-12 недель назначали 5 мг/кг метотрексата (каталожный № Calbiochem - 454125) посредством внутрибрюшинной инъекции в течение 1-3 циклов, где 1 цикл равен 3 календарным дням инъецирования. 20 мг/кг Myozyme® (Genzyme Corporation) вводили посредством инъекции в хвостовую вену один раз или 2-6 доз еженедельно, начиная с первой дозы метотрексата. Животных умерщвляли еженедельно или ежедневно после инициирования лечения. Селезенку, кишечные и паховые лимфатические узлы собирали для анализа с помощью проточной цитометрии субпопуляций T- и B-клеток, а сыворотки получали для анализов ELISA. Селезенки подвергали обработке между предметными стеклами, и эритроциты (RBC) лизировали с помощью буфера для лизиса, купленного у BD Biosciences (каталожный № 555899) в соответствии с инструкциями производителя. Лимфатические узлы подвергали обработке между предметными стеклами и промывали забуференным фосфатом солевым раствором (PBS), содержащим 2% фетальной телячьей сыворотки (FCS). Клетки суспендировали в 200 мкл PBS, содержащего 4% фетальной телячьей сыворотки и 25 мкг/мл всех IgG мыши, и блокировали в течение 30 минут при 4°. Приблизительно 3 миллиона клеток селезенки и 1 миллион клеток лимфатических узлов подвергали окрашиванию различными смесями антител и анализировали с использованием высокопроизводительного пробоотборника (HTS) в проточном цитометре Becton Dickinson CANTOII. По крайней мере 100000 случаев - клеток захватывали в лимфоцит-дискриминационное окно. Смесь антимышиных антител состояла из PE-CD21/35 с каталожным № 552957, FITC- с каталожным № 553138, PE-CD138 с каталожным № 553714, PE-CD127 с каталожным № 552543, APC-Cy7-CD19 с каталожным № 557655, FITC-CD43 с каталожным № 553270, PE-Cy7-CD4 с каталожным № 552775, FITC-CD3e с каталожным № 553062, APC-CD11b с каталожным № 553312, PE-Cy7-IgM с каталожным № 552867, APC-Cy7-CD8 с каталожным № 557654, PE-CD273 (PD-L2) с каталожным № 557796, APC-CD138 с каталожным № 558626, PE-Cy7-CD11b с каталожным № 552850, PE-CD93 (для В-клеточной линии ранней дифференцировки) с каталожным № 558039, APC-CD69 с каталожным № 560689, Pe-Cy7-CD24 с каталожным № 560536, FITC-CD1d с каталожным № 553845, APC-CD5 с каталожным № 550035 и Percp-Cy5-7AAD с каталожным № 559925, все из которых приобретены у BD Pharmingen. Набор для внутриклеточного окрашивания FITC-FoxP3 был приобретен у eBioscience. Pacific Blue (PB)-CD25 с каталожным № 102022, PB-CD23 с каталожным № 101616 и PB-CD86 с каталожным № 105022 были приобретены у BioLegend. Анализ субпопуляций лимфоцитов выполняли с использованием программного обеспечения FCS express версии 3, предоставляемого De Novo Software. Проценты определяли с использованием опции пакетной обработки, и абсолютные количества рассчитывали в соответствии с полученными количествами клеток. Количества клеток селезенки и лимфатических узлов получали с использованием анализатора жизнеспособности Beckman Coulter Vi-cell XR в соответствии с инструкциями производителя.
In vitro анализ цитокинов
Мышам C57BL/6 (Jackson Laboratories) или E4GAAKO (Charles River) возрастом 8-12 недель назначали 5 мг/кг метотрексата (каталожный № Calbiochem - 454125) с помощью внутрибрюшинной инъекции в течение 1 цикла (3 последовательные ежедневные дозы), начиная с лечения 20 мг/кг rhGAA. В случае исследований с использованием 1D11 животных подвергали лечению с помощью внутрибрюшинных инъекций 5 мг/кг или 1D11, или 13C4 (Genzyme Corporation) 3 раза в неделю, через день, начиная с лечения rhGAA и метотрексатом. Животных умерщвляли в день 6 или 7 после инициации лечения rhGAA в зависимости от линии мышей. Селезенки приводили в состояние суспензии отдельных клеток и вносили в прибор RoboSep (STEMCELL technologies) в соответствии с инструкциями производителя, и подвергали В-клеточному негативному отбору. Очищенные B-клетки засевали в количестве 500000 клеток в каждую лунку в 96-луночные круглодонные планшеты (каталожный № Costar - 3799) и или инкубировали без стимуляции или со стимуляцией 10 мкг/мл LPS (каталожный № Sigma - L5014) в течение 48 часов при 37°C. Во все лунки вносили Monensin (каталожный № BD Bioscience - 554724) в соответствии с инструкциями производителя. Допускали инкубацию клеток в течение по крайней мере 4 часов при 37°C. Образцы переносили в лунки с V-образным дном (каталожный № USA Scientific - 651201) и подвергали центрифугированию при 1200 об./мин в течение 5 минут при 4°C. Клетки ресуспендировали в 200 мкл PBS, содержащего 4% фетальной телячьей сыворотки и 25 мкг/мл всех IgG мыши, и блокировали в течение 30 минут при 4°C. Планшеты снова подвергали центрифугированию, и осадок ресуспендировали в 90 мкл PBS/2% FCS с добавлением 10 мкл смеси антител, описанной выше, и инкубировали в течение 20 минут при 4°C с добавлением 5 мкл 7AAD на последние 10 минут процедуры окрашивания. Добавление 100 мкл буфера к образцам с последующим центрифугированием использовали в качестве промывки. Образцы могли ресуспендировать в буфере для анализа белка на поверхности и немедленного сбора данных или суспендировать в Fix/Perm (каталожный № eBioscience - 11-5773) для внутриклеточного окрашивания IL-10 (каталожный № BioLegend - 505008), TGF-бета (каталожный № BioLegend - 141404) и FoxP3 (каталожный № eBioscience - 11-5773-82) в соответствии с инструкциями производителя. Дополнительное окрашивание поверхности включало антитела против TGF-бета и Tim-1 (каталожный № BioLegend - 119506). Все образцы были получены и проанализированы, как описано выше.
Животные и модель аллотрансплантации сердца
Мышей C57BL/6 и BALB/c получали из Charles River (Kingston, NY или Raleigh, NC) и использовали в этих экспериментах в возрасте 8-13 недель. Аллогенных мышей-доноров C57BL/6 сначала анестезировали с помощью внутрибрюшинной инъекции Кетамина (Fort Dodge Animal Health/Pfizer, Fort Dodge, IA) и Ксилазина (Lloyd, Shenandoah, IA), и выполняли медианную стернотомию. Донорское сердце медленно перфузировали in situ с использованием 1 мл холодного гепаринизированного раствора лактата Рингера (Baxter Healthcare, Deerfield, IL) через нижнюю полую вену и аорту, прежде чем верхнюю полую вену и легочные вены лигировали и разделяли. Восходящую часть дуги аорты и легочную артерию затем рассекали, трансплантат отделяли от донора, и сердце хранили в холодном как лед солевом растворе до приживления трансплантата. Мышь-реципиента (Balb/c) анестезировали аналогичным образом и подготавливали, как описано выше для мыши-донора, за исключением того, что открывали брюшную полость. Используя хирургический микроскоп для обзора открытой брюшной полости изолировали брюшную аорту (AA) и нижнюю полую вену (IVC). Донорское сердце помещали в брюшную полость реципиента (вверх дном), и трансплантаты подвергали реваскуляризации с помощью анастомозов конец в бок между легочной артерией донора и нижней полой веной реципиента, а также аортой донора и брюшной аортой реципиента. После подтверждения гемостаза брюшную мышцу сшивали с помощью непрерывного 5-0 викрилового шва (Ethicon, Johnson & Johnson, Somerville, NJ), а кожу сшивали с помощью непрерывного 5-0 этилонового шва (Ethicon). Выполняли стандартную послеоперационную оценку боли и ее контролирование. Трансплантаты оценивали пальпацией 5-7 раз в неделю в течение первых 30 дней, а затем 3-4 раза в неделю до конца исследования.
Мышей C57B1/6 подвергали лечению одной 20 мг/кг внутривенной дозой rhGAA (Genzyme Corporation), 3 последовательными внутрибрюшинными дозами 5 мг/кг метотрексата (APP Pharmaceuticals, LLC) и 3 дозами через день 5 мг/кг или 1D11, или 13C4 (Genzyme Corporation). Лечение метотрексатом, 1D11 и 13C4 начиналось с инъекций rhGAA. Селезенки получали через 7 дней после начала лечения и подвергали обработке, как описано выше для B-клеток, культуры и анализа с использованием проточной цитометрии. Кроме того, данные, относящиеся к титрам rhGAA, получали при лечении животных, как описано выше, rhGAA, дозы которого вводили еженедельно в течение 12 недель, метотрексатом, дозы которого вводили в течение или 1, или 3 циклов, и 1D11 или 13C4, дозы которого вводили 3 раза в неделю через день в течение 12 недель. Образцы сыворотки получали каждые 2 недели для анализа ELISA.
Гистопатология и иммуногистохимия
Сердечные трансплантаты фиксировали в 10% нейтральном забуференном формалине, разрезали по продольной оси для обнажения правого и левого желудочков и выводного тракта, и обычным образом обрабатывали для заливки в парафин. Срезы делали толщиной 5 микрон и окрашивали гематоксилином и эозином (H&E) или трихромом по Массону. Последовательные срезы также подвергали иммуноокрашиванию, как описано ниже. Каждый скрашенный H&E срез подвергали качественной оценке в отношении различных признаков патологии отторжения трансплантата (например, васкулита, дегенерации миокарда и его некроза, миокардита), используя гистологическую разбаловку.
Иммуногистохимическое исследование выполняли, используя автоматизированную систему для иммуноокрашивания Bond-Max (Leica Microsystems Inc., Buffalo Grove, IL). Для выявления двойных: CD3 и Foxp3 - имммунопозитивных клеток тканевые срезы трансплантата подвергали двойному иммунокрашиванию с использованием антител против CD3 и Foxp3, используя набор Bond Polymer Refine Detection и набор Bond Polymer AP Red (Leica, Buffalo Grove, IL), следуя рекомендациям производителя. Вкратце, депарафинизированные срезы трансплантатов, залитых в парафин, подвергали термическому восстановлению эпитопов (25 мин при 99°C), инкубировали с бессыворотным раствором белков для блокирования (Dako, Carpentaria, CA), кроличьим моноклональным антителом против CD3 (Lab Vision/Neo Marker), конъюгированным с пероксидазой полимером, Peroxidase Block и реагентом для детектирования диаминобензидином, а затем с кроличьим антителом против Foxp3 мыши (eBioscience Inc., San Diego, CA), а затем с кроличьим антикрысиным антителом (Vector Laboratories, Inc., Burlingame, CA). Затем срезы инкубировали с Bond Polymer AP и смешанным красным реагентом для детектирования и, наконец, подвергали контрастному окрашиванию гематоксилином. В случае срезов в качестве отрицательных контролей первые антитела против CD3 и Foxp3 заменяли на IgG в целом кролика Chromepure (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) и IgG2a крысы (AbD Serotec, Raleigh, NC), соответственно.
Уровни аллоантител в сыворотке
Уровни аллоантител в сыворотке определяли посредством инкубации сыворотки от мышей с трансплантированным сердцем или нормальных мышей в разведениях 1:50 с линий трансформированных SV40 фибробластов C57BL/6 (SVB6) с последующим детектированием связавшихся с фибробластами антител (аллоантител), используя конъюгированное с FITC кроличье антитело против IgG мыши (Dako, Carpinteria, CA), и с помощью анализа с использованием проточной цитометрии. Средние геометрические интенсивности флуоресценции окрашенных антителами сыворотки фибробластов делили на таковые для окрашенных подобранным по изотипу контролем фибробластов для нормализации уровней аллоантител между экспериментами. Сывороточные антитела, связывающиеся с аллогенными фибробластами, были определенно аллоантителами, поскольку те же образцы сыворотки не связывались с линией трансформированных SV40 фибробластов BALB/c (SVBalb) (не представленные данные).
Модель адоптивного переноса на мышах
Мышей C57BL/6 получали из Jackson Laboratories и содержали в особых, лишенных патогенов условиях. Контрольным мышам назначали одну внутривенную инъекцию rhGAA в дозе, равной 20 мг/кг. Мышам с индуцируемой толерантностью назначали одну внутривенную инъекцию rhGAA в дозе, равной 20 мг/кг, помимо трех ежедневных последовательных внутрибрюшинных инъекций метотрексата в количестве 0,5 мг. Селезенки получали в день шесть после первоначальной инъекции rhGAA, из обеих донорских групп, и подвергали обработке с получением объединенных суспензий отдельных клеток. Клетки промывали и фильтровали через 0,22 мкм фильтр. Обогащение клеток в отношение B-клеток затем осуществляли, используя систему для разделения клеток StemCell Technologies RoboSep, и их ресуспендировали для создания возможности для 200 мкл внутривенной инъекции. Группы реципиентов с индуцируемой толерантностью и без нее получали высокую концентрацию клеток 10×106, или низкую концентрацию клеток 5×106 посредством внутривенных инъекций. Контрольным группам назначали только rhGAA, или rhGAA и метотрексат (один цикл из трех последовательных ежедневных инъекций MTX). Все группы получали ежедневные внутривенные инъекции 20 мг/кг rhGAA, и подвергались ретроорбитальному кровоизвлечению один раз в две недели в течение 16 недель, в пробирки для отделения сыворотки BD Vacutainer. Сыворотку отделяли и хранили при температуре ниже -20°C до использования для ELISA. Титры антител против rhGAA определяли, используя ELISA, считывали на SpectraMax M2 и рассчитывали, используя Softmax для экстраполяции значения титра. Файлы Softmax с необработанными данными и EXCEL электронные таблицы, содержащие контрольную информацию и информацию, относящуюся к титрам, хранили в сетевых серверах. Все графики и статистические данные были получены, используя программное обеспечение GraphPad Prism.
Пример 1: Антитела против лекарственных средств продуцируются в ответ на терапевтические антитела
Поликлональное антитело mATG связывается с множеством типом иммуноцитов, включая антигенпрезентирующие клетки. Данные авторов настоящего изобретения указывают на то, что один курс mATG (две дозы 25 мг/кг, назначаемые с интервалом в три дня, вводимые внутрибрюшинно) мог порождать титры IgG против mATG вплоть до 100000 (фиг. 1A) у мышей. После назначения в виде последовательных ежемесячных инъекций (5 мг/кг, каждые 4 недели), титры IgG против mATG дополнительно увеличивались вплоть до 5×106 после пяти ежемесячных инъекций (фиг. 1B). И в случае одного курса, и в случае ежемесячных инъекций кроличий IgG (rbIgG) использовали в качестве контроля.
Поскольку алемтузумаб не вступает в перекрестную реакцию с CD52 мыши, преклинические исследования с использованием алемтузумаба должны осуществляться на трансгенных по huCD52 мышах, у которых характер экспрессии трансгена схож с экспрессией CD52 у людей. Подобно mATG, внутривенное введение алемтузумаба (0,5 мг/кг) индуцировало значительные ответы в виде ADA у трансгенных по huCD52 мышей. Эти ответы увеличивались на протяжении первых четырех лечений, а затем уменьшались, так что после пятой дозы алемтузумаба трансгенные по huCD52 мыши больше не вырабатывали антитела против алемтузумаба (фиг. 2). Эта неотвечаемость говорит о том, что возникла естественная толерантность (Rosenberg et al, Blood, 93(6):2081-8 (1999)).
Эти данные указывает на то, что титры ADA против mATG у мышей C57BL/6 и титры ADA против алемтузумаба у трансгенных по huCD52 CD1 мышей были высокими (>100000). Этот высокий уровень иммуногенности может быть связан со способностью mATG и алемтузумаба к связыванию с антигенпрезентирующими клетками, тем самым увеличивая процессирование антигена и его презентацию для индукции иммунных ответов против них.
Пример 2: Метотрексат контролирует образования IgG против mATG при использовании одного цикла введения
Были представлены данные о повышенных титрах антител после лечения Thymoglobulin®, и были представлены описания случаев сывороточной болезни, острой почечной недостаточности и сердечнососудистых реакций у пациентов, подвергнутых лечению Thymoglobulin® (Boothpur et al., supra; Lundquist et al, Liver Transpl, 13(5):647-50 (2007); Busani et al, Minerva Anestesiol, 72(4):243-8 (2006); Tanriover et al, Transplantation, 80(2):279-81 (2005); Buchler et al., Clin Transplant, 17(6):539-45 (2003)). Для определения способности метотрексата к уменьшению образований антител против ATG, и, таким образом, к уменьшению беспокойств в отношении безопасности, трехдневная схема лечения метотрексатом, назначаемым только в виде одного цикла, была оценена в качестве способа контролирования образований IgG против mATG у мышей. Она отличается от схемы, которая была ранее опубликована, поскольку только один цикл метотрексат назначали вместе с mATG в отличие от по крайней мере трех циклов, которые назначали в случае ERT. Метотрексат (каталожный № Calbiochem - 454125), вводимый внутрибрюшинно в дозе, равной 5 мг/кг, в течение шести календарных дней, начиная с первого из двух введений mATG (25 мг/кг, с интервалом в 3 дня), мог подавлять образования IgG против mATG на протяжении по крайней мере восьми недель после лечения на 95% при сравнении площади под кривыми эффекта (фиг. 3).
Затем оценивали эффект метотрексата на титры антител против mATG после пяти ежемесячных инъекций mATG в расчете 5 мг/кг/инъекцию. Лечение в виде ежемесячных инъекций mATG осуществляли, поскольку репопуляция лимфоцитов является почти законченной через один месяц после лечения mATG (Ruzek, выше). Количественные определения образований антител против лекарственного средства затем проводили еженедельно на протяжении 20 недель ежемесячных лечений. Во время этого периода, несмотря на истощение CD4+ T-клеток с помощью mATG, титры антител достигали вплоть до 5 миллионов (фиг. 1B). Примечательно, что животные, которые получали неспецифический кроличий IgG на уровне дозы и с использованием схемы, одинаковой с таковыми для mATG, продемонстрировали небольшие образования антикроличьих IgG (фиг. 1B). Одним вероятным объяснением увеличенной иммуногенности mATG может быть специфическое связывание mATG с антигенпрезентирующими клетками (APC), такими как фолликулярные дендритные клетки, которые в присутствии комплемента могут значительно усилить B-клеточные реакции. Также были оценены две схемы лечения метотрексатом. В предшествующей работе с использованием ферментозаместительной терапии (ERT), три цикла метотрексата, назначаемые во время первых трех доз кислой альфа-глюкозидазы, обеспечили непрерывное уменьшение титра антител на протяжении по крайней мере восьми месяцев еженедельного введения доз ERT (Joseph et al, Clin Exp Immunol, 152(l):138-46 (2008)). Схожий курс метотрексата был оценен в случае mATG, когда 5 мг/кг метотрексата назначали в пределах 15 минут от введения mATG, а также через 24 и 48 часов после каждого из первых трех ежемесячных лечений mATG. Эта схема успешно уменьшала образования антител против mATG с титров, составляющих приблизительно 4 миллиона, до титров, составляющих 816000, приводя к составляющему 79% уменьшению при сравнении площади под кривыми эффекта (фиг. 4А).
Кроме того, эффект одного курса метотрексата, назначаемого вблизи только первого из пяти ежемесячных лечений mATG, на образования IgG против mATG определяли и сравнивали непосредственно со схемой, предусматривающей три цикла. Как ни удивительно, эта схема, предусматривающая один цикл, приводила к уменьшению титров IgG против mATG даже больше, чем схема, предусматривающая три цикла, до титра, составляющего приблизительно 50000 (фиг. 4B). При сравнении площади под кривыми эффекта схема, предусматривающая один цикл, приводила к уменьшению титров IgG против mATG на 98%, в то время как схема, предусматривающая три цикла, приводила к уменьшению титров на 69% (фиг. 4C). Увеличенные эффекты одного цикла в сравнение со схемой, предусматривающей три цикла, говорят о том, что увеличение подвергания воздействию метотрексата может в действительности противодействовать его эффектам индукции толерантности, возможно, в результате уничтожения клеток, которые опосредуют контроль титра антител.
Пример 3: Метотрексат запускает активный механизм иммунологической толерантности
Как продемонстрировано выше, очень короткий курс метотрексата может в значительной степени контролировать гуморальные иммунные ответы антител на протяжении множество циклов антигенного стимула. В этом случае коротким циклом был один цикл метотрексата, который вызывал длительные эффекты (и на титры антител, и на уровни цитокинов) в течение месяцев проверки. Этот долговременный контроль гуморального иммунного ответа наводит на мысль, что метотрексат может с успехом индуцировать иммунологическую толерантность. Метотрексат был пока оценен в случае пяти последовательных ежемесячных доз mATG. Для дальнейшего определения того, активировался ли механизм иммунологической толерантности, животным, которые сначала получили пять ежемесячных инъекций mATG, не предоставляли лечение в течение восьми недель. После этого периода покоя животным назначали конечную инъекцию mATG. Если механизм иммунологической толерантности был задействован, титры IgG против mATG не должны были значительно увеличиваться после шестого лечения mATG.
Данные авторов настоящего изобретения указывают на то, что животные, которым вводили дозы метотрексата, продемонстрировали увеличение титров IgG против mATG, как и ожидалось (фиг. 5). В отличие от этого, животные, которые получили только один цикл метотрексата с первой инъекцией mATG, не вырабатывали значимо более высокие титры IgG против mATG (фиг. 5). Схожая тенденция отмечалась в случае животных, подвергнутых лечению тремя циклами метотрексата, хотя эффект не был столь существенным (фиг. 5). При сравнении площади под кривыми эффекта один курс метотрексата снижал титры на 99%, в то время как схема, предусматривающая три цикла, снижала титры на 85%. Эти данные указывают на то, что метотрексат может поддерживать контроль ответов против «воскресшего» антигена, что говорит о том, что у мышей развилась толерантность к этому антигену.
Хотя у животных, подвергнутых лечению метотрексатом, вырабатывались поддающиеся измерению титры, которые увеличиваются с последующими лечениями mATG, в целом, уровни титров у подвергнутых лечению метотрексатом животных были сообразно в 100 раз ниже, чем таковые, отмечаемые у животных, подвергнутых лечению только mATG (фиг. 6). Более низкий уровень титров антител должен значительно уменьшить возможность связанных с безопасностью рисков и эффекты на эффективность. Хоть и без желания ограничиваться этой теорией, эти данные наводят на мысль, что был запущен активный механизм контроля, который может значительно уменьшить образования IgG против mATG, и он долго сохраняется после лечения метотрексатом.
Пример 4: Один цикл метотрексата может в значительной степени контролировать ответы против алемтузумаба
В случае ремиттирующе-рецидивирующего рассеянного склероза дозы алемтузумаба вводят в ежегодных циклах, и у пациентов может возникать ADA (Coles et al, N Engl J Med, 359(17):1786-801 (2008)). В то время как проверка иммуногенности и фармакокинетическое исследование проводятся в настоящее время во множестве исследований фазы III, неясно, будут ли антитела против алемтузумаба оказывать влияние на подвергание воздействию, эффективность и/или безопасность в субпопуляции пациентов. Соответственно, авторы настоящего изобретения определили, может ли один цикл метотрексата контролировать титры ADA после циклов в виде пяти ежемесячных инъекций алемтузумаба. Трансгенным по huCD52 мышам назначали (0,5 мг/кг) внутривенно ежемесячно в течение пяти календарных месяцев. Метотрексат назначали в дозе, равной 0,5, 1 или 5 мг/кг, за 15 минут до первой ежемесячной дозы алемтузумаба, а также через 24 и 48 часов после этой дозы. Доза в 1 мг/кг метотрексата давала некоторый эффект, поскольку она уменьшала ответы против алемтузумаб на 88% (фиг. 7A). Доза в 5 мг/кг метотрексата с успехом уменьшала образования IgG против алемтузумаб на 99% (фиг. 7A). Метотрексат, по-видимому, не оказывал эффект на естественную толерантность.
Второе исследование подтвердило вышеприведенные данные. Как указано выше, трагсгенных по huCD52 мышей подвергали лечению с использованием пяти ежемесячных доз 0,5 мг/кг алемтузумаба. Мышей также подвергали лечению с использованием трех ежедневных доз 5 мг/кг/день метотрексата в связи с первым введением алемтузумаба или без него (фиг. 7B). Образцы сыворотки получали на протяжении всего исследования для определения титров антител против алемтузумаба и подтверждения толерантности. Относящиеся к титрам данные получали через 24 часа после пятой ежемесячной дозы. Данные указывали на то, что метотрексат уменьшает титры антител против алемтузумаба (фиг. 7C).
Пример 5: Метотрексат может контролировать образования антител против алемтузумаба в случае клинически соответствующей схемы введения доз алемтузумаба
Был выполнен ряд экспериментов для определения, может ли метотрексат успешно контролировать ADA в случае клинически соответствующей схемы введения доз алемтузумаба у трансгенных по huCD52 мышей. В случае этой клинической схемы дозы алемтузумаба вводят в виде первоначального цикла из пяти ежедневных лечений в дозе 12 мг/день. Через двенадцать месяцев после первоначального цикла лечения пациентов назначают дополнительный цикл из трех ежедневных 12 мг-доз алемтузумаба. Во время второго цикла лечения уровни циркулирующих CD19+ B-клеток восстанавливались до значений на исходном уровне; однако уровни циркулирующих CD4+ T-клеток-хелперов и цитотоксических CD8+ T-клеток не полностью восстанавливались (Coles et al, N Engl J Med, 359(17):1786-801 (2008)).
Сначала кинетики истощения и репопуляции субпопуляций циркулирующих T- и B-клеток после пяти ежедневных лечений алемтузумабом исследовали у трансгенных по huCD52 мышей. В периферической крови трансгенных по huCD52 мышей, подвергнутых лечению алемтузумабом в течение пяти календарных дней в дозе 0,5 мг/кг (эквивалентной дозе для человека, равной 12 мг/кг), вводимой посредством внутривенной инъекции, CD3+ T-клетки в целом, CD4+ T-клетки-хелперы и цитотоксические CD8+ T-клетки не восстанавливались до уровней до лечения через четыре недели после лечения, в то время как количества CD19+ B-клеток возвращались к контрольным уровням (фиг. 8A-D).
Для имитирования состояния клеток, которое демонстрируют пациенты во время повторного лечения, алемтузумаб вновь вводили трансгенным по huCD52 мышам между 4 и 5 неделями после первого цикла. Поскольку первоначальный курс алемтузумаба представлял собой пятидневный цикл, метотрексат вводили за 15 минут до каждого ежедневного лечения алемтузумабом и в течение двух дней после. Максимальная кумулятивная доза метотрексата, назначаемая в этой схеме, составляет 14 мг/кг (2 мг/кг/день), что очень схоже с кумулятивной дозой, равной 15 мг/кг, когда метотрексат назначают в виде трехдневного курса в дозе 5 мг/кг/день. Авторы настоящего изобретения оценили эффекты введения 2, 1 и 0,5 мг/кг/день метотрексата на антитела против алемтузумаба на протяжении трех циклов лечения алемтузумабом. В дозе, равной 1 мг/кг, метотрексат, по-видимому, контролировал титры у 7 из 8 проверенных мышей и в целом уменьшал титры на 79% (фиг. 9B). В дозе, равной 2 мг/кг, метотрексат успешно уменьшал ADA на 98% при сравнении площади под кривыми эффекта (фиг. 9A).
Пример 6: Метотрексат может улучшить фармакокинетику и фармакодинамику mATG
ADA может вносить помехи в фармакокинетику и фармакодинамику терапевтических белков. Авторы настоящего изобретения оценили, вносят ли ответы в виде ADA против mATG помехи в фармакокинетику mATG. Мышей подвергали лечению в течение пяти месяцев с использованием ежемесячных инъекций или только mATG, или mATG вместе с одним циклом метотрексата. Метотрексат вводили в дозе, равной 5 мг/кг, ежедневно в течение трех дней. Кровь отбирали в различные моменты времени через месяц 1, месяц 3 и месяц 5 для исследования уровня циркулирующих mATG (фиг. 10).
В месяц 1 уровни циркулирующих mATG были схожими в обеих группах лечения, но в месяцы 3 и 5 только группа, подвергнутая лечению метотрексатом, имела поддающиеся измерению уровни циркулирующих mATG. Без введения метотрексата уровни mATG, определяемые после третьей и пятой доз mATG, были значительно ниже уровней, определяемых после первой дозы (фиг. 10). Предшествующие исследования показали, что метотрексат значительно уменьшает образования IgG против mATG. Таким образом, представляется, что антитела против mATG мешают подверганию воздействию mATG и вносят помехи в его фармакокинетику.
Поскольку антитела против mATG, как представляется, значительно снижают уровни циркулирующих mATG после повторного введения дозы, можно ожидать, что на фармакодинамику mATG после повторного введения дозы также оказывается отрицательное влияние. Истощение лимфоцитов в крови, селезенке и лимфатических узлах было оценено после пятого ежемесячного лечения mATG, когда титры были на самых высоких уровнях. Как описано выше, животным, подвергнутым лечению метотрексатом, был назначен только один цикл лечения.
Уровни циркулирующих CD4+ и CD8+ T-клеток были неизменными после пятого лечения mATG у животных, подвергнутых лечению mATG, но не метотрексатом. Однако у животных, подвергнутых лечению с использованием mATG и одного цикла метотрексата, циркулирующие CD4+ и CD8+ T-клетки были значимо истощены (фиг. 11). Этот эффект также отмечался в селезенке и лимфатических узлах. Лечение метотрексатом увеличивало способность mATG к увеличению процента регуляторных T-клеток в селезенке и крови (фиг. 12A-B). Схожий эффект отмечался в крови и лимфатических узлах. Увеличенное присутствие регуляторных T-клеток после лечения mATG и Thymoglobulin®, как было постулировано, способствует эффективности этого терапевтического средства (Ruzek et al., Blood, 111(3):1726-34 (2008)). Способность метотрексата к содействию в сохранении этого эффекта после последовательных курсов mATG является потенциальным добавленным эффектом значительного уменьшения титров антикроличьих IgG.
До сих пор, исследования фармакокинетики и эффективности говорят о том, что антитела против mATG мешают подверганию воздействию и эффективности mATG. Прямое сравнение между титром IgG против mATG и поверганием воздействию mATG показывает, что когда конечные титры превышают 10000, уровень циркулирующих mATG значительно снижен (фиг. 13a). Более того, когда титры IgG против mATG превышают 100000, ингибируется mATG-опосредуемое истощение клеток (фиг. 13b и 13c). Квадрат линейного коэффициента корреляции R2 между титром и истощением клеток >0,7.
Пример 7: Метотрексат может улучшить фармакодинамику алемтузумаба
Метотрексат не только улучшает фармакодинамику mATG, но также восстанавливает опосредуемое алемтузумабом истощение циркулирующих T- и B-клеток, когда ответы против алемтузумаба, по-видимому, нейтрализуют некоторую степень истощающей активности. В исследованиях, описанных в этом примере, пять ежемесячных инъекций алемтузумаба назначали трансгенным по huCD52 мышам вместе с метотрексатом или без него. 5 мг/кг метотрексата вводили ежедневно в первые три дня исследования в течение 6 месяцев. Кровь отбирали у животных обеих групп лечения за два дня до пятой дозы и через один день после пятой дозы алемтузумаба. Популяции клеток оценивали с помощью проточной цитометрии, как описано в примере 6.
Данные авторов настоящего изобретения указывают на то, что пятая ежемесячная доза алемтузумаба, как представлялось, больше не истощает T-клетки, поскольку абсолютное количество циркулирующих T-клеток казалось схожим до и после лечения (фиг. 14 (каждый момент времени представляет отличную группу животных)). В отличие от этого, метотрексат, как представлялось, восстановил способность алемтузумаба к истощению T-клеток (Р=0,012). При сравнении абсолютных количеств циркулирующих T-клеток у животных, подвергнутых лечению только алемтузумабом, и у животных, подвергнутых лечению и алемтузумабом, и метотрексатом или за два дня до, или через один день после введения дозы алемтузумаба, количества циркулирующих T-клеток были значимо меньшими у животных, подвергнутых лечению метотрексатом (Р=0,034 и 0,02; фиг. 14). Так же лечение метотрексатом, как представлялось, увеличивает истощение B-клеток с помощью алемтузумаба (P=1,2×l0-5, P=0,02, соответственно), и количество циркулирующих B-клеток уменьшалось дополнительно у животных, подвергнутых лечению метотрексатом, через день после лечения (фиг. 14).
Пример 8: Один цикл метотрексата может в значительной степени контролировать образования антител против rhGAA
Авторы настоящего изобретения также изучили эффект метотрексата при ферментозаместительной терапии с использованием rhGAA. В этом исследовании животным инъецировали еженедельно rhGAA в течение двенадцати календарных недель, затем оставляли в покое на четыре недели, а затем вновь вводили rhGAA на неделе 16. Животным также назначали один цикл из трех последовательных ежедневных доз 5 мг/кг метотрексата на неделе 1, или назначали один цикл на каждой из недель 1, 2 и 3 (всего три цикла). Титры rhGAA-специфических IgG определяли у животных на неделе 0 (до какого-либо лечения), 6, 8, 12, 16, 18 и 20. Данные авторов настоящего изобретения указывают на то, что один цикл метотрексата контролировал ответы против rhGAA на протяжении по крайней мере 20 недель, также как схема, предусматривающая три цикла (фиг. 15).
Были также проведены исследования на мышах Nu/Nu с дефицитом Т-клеток для установления роли T-клеток в выработке титров антител против rhGAA. В этих экспериментах авторы настоящего изобретения неоднократно отмечали, что у этих мышей с дефицитом Т-клеток не развивается или развивается незначительный ответ в виде ADA против rhGAA (фиг. 37A-B). Эти данные подкрепляют представление о том, что T-клетки вносят вклад в титры антител против rhGAA. Таким образом, в то время как метотрексат может контролировать ADA против rhGAA, он также, вероятно, оказывает воздействие на T-клеточные реакции на rhGAA.
Пример 9: Метотрексат увеличивает продолжительность существования, опосредуемого mATG, аллогенных трансплантатов сердца
Помимо установления того, может ли метотрексат увеличивать эффективность mATG у нормальных мышей, авторы настоящего изобретения исследовали, может ли метотрексат увеличить действие mATG в условиях трансплантации. Поскольку Thymoglobulin® используется клинически в качестве индукционной терапии для увеличения продолжительности существования трансплантата, авторы настоящего изобретения установили, может ли добавление метотрексата увеличить эффективность mATG в модели аллогенной трансплантации сердца на мышах. 20 мг/кг mATG вводили в дни 0 и 4, в то время как 2 мг/кг метотрексата вводили в виде одного цикла лечения в дни 0-6.
Кроме того, авторы настоящего изобретения исследовали дозы метотрексата, которые меньше в 4 раза, (0,5 мг/кг), назначаемые в соответствии с той же схемой или с использованием удлиненной схемы из 12 календарных дней. Группы мышей или не получали лечение (контроль в виде солевого раствора), получали только mATG или комбинацию mATG и схем меторексата. Подобно исследованиям на нормальных мышах, метотрексат, вводимый вместе с mATG, уменьшал титры антител против лекарственного средства - mATG независимо от используемой схемы (фиг. 16A и таблица 1). Кроме того, совпадающим с уменьшением титров антител было отмечаемое увеличение подвергания воздействию mATG при этой трансплантации (фиг. 16). В связи с вероятно адъювантным эффектом в условиях сильной, совпадающей иммунной реакции против трансплантированной ткани, титры антител против лекарственного средства увеличивались даже быстрее, чем в случае нормальных мышей и приводили к уровням mATG, почти не поддающимся измерению в пределах 7 дней от первого введения mATG (фиг. 16B). В противоположность этому, к семи дням после трансплантации, титры антител против IgG кролика были значительно ниже у мышей, подвергнутых лечению метотрексатом, а уровни циркулирующих mATG были значительно выше у этих мышей. Это подчеркивает, что в условиях продолжающейся воспалительной реакции образования антител против лекарственного средства могут ускоряться и, возможно, оказывают даже большее влияние на фармакодинамику и эффективность. Важно отметить, что даже в этих условиях метотрексат оказывал сильный ингибиторный эффект на антитела против лекарственного средства - mATG и увеличивал подвергание воздействию mATG. Однако, поскольку уровни циркулирующих mATG были все еще низкими или не поддающимися измерению к дню 21 при использовании комбинированного лечения mATG и метотрексатом, дополнительные механизмы толерантности, вероятно, играют роль, принимая во внимание составляющую >100 дней продолжительность существования трансплантата. Эти исследования демонстрируют поразительный синергизм между лечением mATG и лечением метотрексатом в отношении продолжительности существования аллогенных трансплантатов и указывают на уровень уменьшения антител против лекарственного средства - mATG и увеличения подвергания воздействию mATG, схожий с таковым, который отмечался у нормальных мышей.
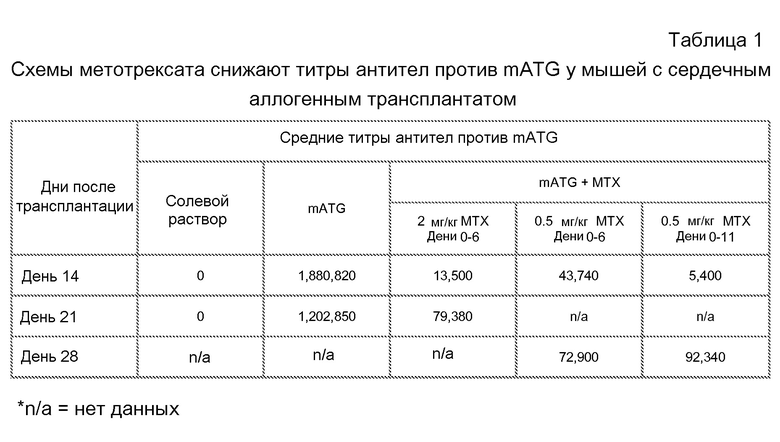
Дополнительные данные подтвердили, что комбинированное лечение с использованием mATG и метотрексата значительно увеличивало продолжительность существования аллотрансплантатов сердца помимо уменьшения реакций против аллотрансплантатов (фиг. 17 и 18). В самом деле, в то время как оба лечения только mATG или только метотрексатом обеспечивали ограниченный эффект в виде среднего увеличения продолжительности существования до 15 или 20 дней, соответственно, совместное назначение mATG и любой из схем метотрексата, которые оценивались, продемонстрировало существенный эффект на продолжительность существования сердечного трансплантата, при этом у большей части мышей трансплантаты сохранялись в течение вплоть до более 100 дней (фиг. 17). Поскольку существование сердечного трансплантата продолжается дольше после ранних, коротких индукционных лечений с использованием mATG и метотрексата, эта схема, по-видимому, является толерогенной, а не иммунодепрессивной. Этот эффект присущ только комбинации mATG и метотрексата, поскольку другие иммунодепрессанты (например, микофенолат мофетил, дексаметазон, рапамицин и циклофосфамид) не увеличивали значительно продолжительность существования трансплантата при совместном введении с mATG. Примечательно, что лечение только метотрексатом было способно к значительному уменьшению антител против аллотрансплантата, что служит дополнительным доказательством эффектов метотрексата на контролирование гуморальных иммунных ответов вообще (фиг. 18). В этом сценарии, метотрексат, по-видимому, способен контролировать образования антител против множества антигенов сердечного аллотрансплантата одновременно. Дальнейшее уменьшение было вызвано при объединении mATG с лечением метотрексатом (фиг. 18C).
Пример 10: Механизм индуцируемой метотрексатом толерантности отличен от известного и общепринятого в настоящей время действия метотрексата
Схема введения доз метотрексата, описанная в приведенных выше примерах, как представляется, запускает механизм, который отличен от ранее описанного механизма. Метотрексат является антагонистом фолата, который, как полагают, опосредует свои эффекты подавления в результате индукции гибели пролиферирующих клеток. Представленные выше данные, относящиеся к mATG, указывают на то, что гуморальные иммунные ответы индуцируются, но остаются значительно уменьшенными при использовании каждого последующего лечения mATG у животных, подвергнутых лечению метотрексатом. Эти данные наводят на мысль, что B-клеточные реакции активно контролируются. Без желания ограничиваться этой теорией, авторы настоящего изобретения выдвигают гипотезу, что метотрексат может индуцировать популяцию(и) регуляторных клеток, которая может контролировать эти реакции, когда они возникают. Авторы настоящего изобретения исследовали эффект лечения метотрексатом на ряд субпопуляций B- и T-клеток селезенки у животных, подвергнутых лечению Myozyme® и метотрексатом, по сравнению с животными, подвергнутыми лечению только Myozyme®. Авторы настоящего изобретения отметили значительные увеличения популяции B10 регуляторных B-клеток через семь и восемь дней после лечения Myozyme® (через четыре и пять дней после лечения метотрексатом; фиг. 19).
Кроме того, ряд субпопуляций активированных B-клеток был значимо увеличен после лечения метотрексатом (фиг. 20). Эти популяции включали активированные B-клетки краевой зоны, активированные фолликулярные B-клетки и активированные B-клетки на стадии 2 и 3 иммунологической дифференциации. Популяции клеток были определены, как указано далее: B2/фолликулярные B-клетки: CD19+CD21средний уровеньCD23высокий уровень, B-клетки на стадии 2 иммунологической дифференциации: CD19+CD93+CD23+IgMвысокий уровень, и B-клетки на стадии 3 иммунологической дифференциации: CD19+CD93+CD23+IgMнизкий уровень, B-клетки краевой зоны: CD19+CD21высокий уровеньCD23низкий уровень. Ежедневная оценка популяций клеток селезенки у животных, которым назначали два цикла Myozyme® и метотрексата, причем Myozyme® назначали в дни 1 и 8, а 5 мг/кг метотрексата назначали в дни 1-3 и 8-10, показала, что эти популяции активированных B-клеток остаются увеличенными (фиг. 21). Этот результат является неожиданным, поскольку ожидаемой реакцией после лечения метотрексатом была бы гибель активированных, пролиферирующих клеток. В противоположность этому, популяции активированных Т-клеток-хелперов, цитотоксических T-клеток и регуляторных T-клеток оставались почти совершенно неизменными (фиг. 22). T-клетки-хелперы были определены как CD4+, цитотоксические T-клетки были определены как CD8+, а регуляторные T-клетки были определены как CD4+CD25+ и FoxP3+. Эти данные наводят на мысль, что увеличенные популяции B-клеток могут оказывать помощь в опосредовании
метотрексат-индуцируемой иммунологической толерантности.
Пример 11: Метотрексат увеличивает некоторые популяции B-клеток в комбинации с mATG
В вышеприведенных примерах, после каждого лечения mATG, титры ADA и у мышей, подвергнутых лечению только mATG, и у мышей, подвергнутых лечению mATG и метотрексатом, увеличивались, что наводит на мысль, что обе группы животных содержат B-клетки, которые способны вносить вклад в гуморальный иммунный ответ (фиг. 6). В таком случае можно выдвинуть по крайней мере две гипотезы. Первая гипотеза заключается в том, что метотрексат может уничтожать значительное большинство B-клеток, способных отвечать на Myozyme®, и что небольшое количество, которое остается, ответственно за этот небольшой ответ. Хоть это и является возможным, данные проточной цитометрии, полученные во множестве экспериментов, не указывали на уменьшение популяций B-клеток после лечения Myozyme® и метотрексатом. Вторая гипотеза заключается в том, что популяции B-клеток, которые могут отвечать на Myozyme®, не уничтожаются в результате этого короткого курса метотрексата, а остаются и контролируются регуляторными клетками после каждого подвергания воздействию Myozyme®. До сих пор, данные, относящиеся к фенотипам, описывают увеличение субпопуляций B-клеток после лечения метотрексатом и Myozyme®. Фенотип этих B-клеток кажется схожим с субпопуляциями регуляторных B-клеток, которые были описаны в исследованиях животных и у пациентов, толерантных к трансплантату. Поэтому авторы настоящего исследования попытались проверить эту вторую гипотезу в случае различных лечений.
Животных подвергали ежемесячно лечению mATG в течение пяти месяцев, и они получали или один цикл метотрексата в дозе 5 мг/кг в первые три дня исследования, три цикла метотрексата в дозе 5 мг/кг или не получали метотрексат (фиг. 23). Затем определяли различия в популяциях клеток в этих трех группах лечения посредством сравнения популяций у животных за один день до пятой дозы mATG и через два дня после пятой дозы mATG.
Неожиданно, через пять месяцев после лечения с использованием одного цикла метотрексата, отмечались различия между мышами, которые получили один цикл метотрексата и mATG, и мышами, которые получили или только mATG, или mATG с тремя циклами метотрексата. Двумя популяциями клеток, которые неожиданно продемонстрировали эффекты, были активированные фолликулярные B-клетки и активированные B-клетки на стадии 3 иммунологической дифференциации (фиг. 24A-B, соответственно). У мышей, подвергнутых лечению или только mATG, или в комбинации с тремя циклами метотрексата, отмечались уменьшения абсолютного количества клеток этих популяций клеток. Однако у мышей, которые получили только один цикл метотрексата, не отмечались статистически значимые уменьшения этих популяций, что наводит на мысль, что эта схема введения доз метотрексата индуцировала некоторое обогащение этих популяций. Примечательно, что обе из этих популяций клеток, как было установлено, также являются обогащенными непосредственно после лечения метотрексатом в комбинации с Myozyme® (фиг. 21). Схожие субпопуляции также были идентифицированы у пациентов, толерантных к трансплантату. Возможно, что один цикл лечения метотрексатом может индуцировать эти субпопуляции B-клеток, которые, после подвергания воздействию антигена, становятся активированными и подавляющими.
Пример 12: Метотрексат увеличивает некоторые популяции B-клеток в комбинации с алемтузумабом
Как описано в примере 4, трансгенных по huCD52 мышей подвергали лечению с использованием однократных ежемесячных доз, составляющих 0,5 мг/кг алемтузумаба, в течение пяти месяцев, или с использованием трех ежедневных доз 5 мг/кг/день метотрексата в связи с первым введением алемтузумаба или без такого использования. Популяции клеток оценивали в крови и селезенке мышей за 2 дня до и через 1, 7 и/или 28 дней после пятой дозы алемтузумаба с помощью проточной цитометрии.
В крови фармакодинамический эффект алемтузумаба был усилен у животных, подвергнутых лечению метотрексатом, через 24 часа после пятой дозы алемтузумаба. Статистически значимое истощение клеток отмечалось в субпопуляциях T-клеток и B-клеток через один день после пятой дозы, в соответствии с предыдущими данными, указывающими на то, что титры антител против алемтузумаба ниже у этих животных и не будут, вероятно, препятствовать опосредуемому алемтузумабом истощению. В противоположность этому, мыши, подвергнутые лечению только алемтузумабом, могут иметь больше алемтузумаб-нейтрализующих антител, которые вносят помехи в фармакодинамику алемтузумаба. Как показано на фиг. 25A, не отмечалось значимое истощение субпопуляций T-клеток у мышей, подвергнутых лечению алемтузумабом, но мыши, подвергнутые лечению алемтузумабом и метотрексатом, демонстрируют значимое, опосредуемое алемтузумабом истощение T-клеток в целом, T-клеток-хелперов и регуляторных T-клеток через один день после введения дозы алемтузумаба. Схожие данные были получены в случае субпопуляций циркулирующих B-клеток (фиг. 25B), хотя тенденции к уменьшению количества клеток также отмечались у животных, подвергнутых лечению только алемтузумабом. Как и в случае популяций циркулирующих T-клеток, популяции T-клеток селезенки были значимо истощены у животных, подвергнутых лечению метотрексатом, через день после пятого лечения алемтузумабом (фиг. 26A-B).
В противоположность истощению T-клеток у мышей, подвергнутых лечению алемтузумабом и метотрексатом, все популяции B-клеток селезенки, которые были проанализированы, за исключением фолликулярных В-клеток, не были значимо истощены (фиг. 27). Эта оценка включала популяцию регуляторных B-клеток, B10 B-клеток. Без желания ограничиться этой теорией, авторы настоящего изобретения выдвигают гипотезу, что метотрексат может обогащать некоторые или все из этих популяций B-клеток, что противостоит их истощению, опосредуемому алемтузумабом. Это обогащение, как полагают, не происходит в быстрой, жидкой среде крови, поскольку иммуноциты не дифференцируются в крови. Вместо этого, иммунные ответы возникают в селезенке и другой периферической лимфоидной ткани, где межклеточные взаимодействия и инициируемые цитокинами/хемокинами реакции могут происходить в разнообразных нишах ткани. Этим могут объясняться дифференциальные эффекты опосредуемого алемтузумабом истощения B-клеток, отмечаемые в крови и селезенке. Эти данные схожи с данными, полученными при использовании mATG, согласно которым B-клетки селезенки, как представлялось, являются обогащенными у мышей, подвергнутых лечению с использованием одного цикла метотрексата в комбинации с mATG (фиг. 24A-B). Действительно, во всех из описанных здесь исследований, обогащение после лечения метотрексатом отмечалось при оценке специфических популяций активированных клеток. Однако, при оценке общих количеств клеток популяции, которая включает как активированные, так и неактивированные клетки, такие как фолликулярные B-клетки в целом, значимое обогащение у животных, подвергнутых лечению метотрексатом, не отмечалось.
В отличие от популяций клеток селезенки через 24 часа после лечения алемтузумабом, через три дня после лечения с использованием одной дозы алемтузумаба (но не метотрексата) субпопуляции клеток селезенки были значимо истощены (фиг. 28A). Возможно, что через 24 часа истощение B-клеток может быть больше у животных, подвергнутых лечению алемтузумабом, чем через три дня лечения алемтузумабом, поскольку репопуляция B-клеток могла начаться к отметке в три дня. Это было продемонстрировано в нескольких исследованиях, в которых оценивались эти популяции в периферической крови. Истощение отмечалось уже через три часа после введения дозы в периферической крови. К трем дням после лечения, пулы циркулирующих B-клеток были все еще значимо меньше у мышей, подвергнутых лечению алемтузумабом, чем у контрольных мышей, подвергнутых лечению забуференным фосфатом солевым раствором (PBS), хотя процент истощения не был столь высок как через 24 часа. Процент истощения через 24 часа составлял 92%, тогда как истощение через три дня составляло 36% (фиг. 28B). Восстановление B-клеток, по-видимому, происходит быстро после однократной дозы.
В заключение, по-видимому, через пять месяцев после получения животными метотрексата и непосредственно после подвергания воздействию антигена метотрексат, возможно, обогатил популяции B-клеток, которые потенциально оказывают помощь в опосредовании индукции толерантности. Популяции, которые кажутся обогащенными, схожи с теми, которые увеличиваются непосредственно после лечения метотрексатом (фиг. 21). В своей совокупности, это говорит о том, что метотрексат может индуцировать обогащение, которое делает возможным активный контроль иммунных ответов во время подвергания воздействию антигена, даже спустя много времени от лечения метотрексатом.
Пример 13: Эффекты метотрексата на уровни цитокинов
Цитокины играют двоякую роль в B-клеточных реакциях. Например, активирующие B-клетки цитокины IL-6 и BAFF также необходимы для дифференциации B-клеток. Поскольку метотрексат, как представляется, увеличивает популяции B-клеток, можно ожидать увеличения этих цитокинов. Однако IL-6 является также провоспалительным, и, следовательно, повышенные уровни могут препятствовать индуцируемым метотрексатом эффектам. Подобно IL-6, IL-10 участвует в дифференциации B-клеток в плазмоциты, а также в иммуносупрессии.
Данные, относящиеся к BAFF, были получены, исходя из образцов сыворотки, отобранных через 24 часа после 5-ой дозы алемтузумаба (фиг. 29A). Они являются продолжением относящихся к клеткам данных из этого исследования, представленного выше. В этот момент времени не отмечалось различие в уровнях BAFF (фиг. 29B).

Уровни цитокинов определяли через одну неделю после второго цикла алемтузумаба (фиг. 29B). Как правило, через одну неделю после второго цикла алемтузумаба уровни цитокинов казались низкими. В этот момент времени отмечалось статистически значимое увеличение уровней TNF-альфа у животных, подвергнутых лечению 2 мг/кг метотрексата. Это увеличение может служить отражением изменения, которое связано с индуцируемой метотрексатом толерантностью, или которое является признаком воспалительной реакции. Также отмечались такие тенденции в отношении других цитокинов, как видимые увеличения IL-6 и потенциально небольшие уменьшения IL-7 (фиг. 30).
Регуляторные B-клетки были связаны с секрецией IL-10. Одним способом определения, играют ли секретирующие IL-10 регуляторные B-клетки роль в индуцируемой метотрексатом толерантности, является установление того, может ли метотрексат контролировать гуморальные ответы у животных с дефицитом IL-10. Этот тип определения может быть сложной задачей, поскольку, как отмечено выше, IL-10 необходим для дифференциации в плазмоциты и, следовательно, гуморальные ответы могут быть меньшими у этих животных. Учитывая это предостережение, авторы настоящего изобретения отметили интересную тенденцию, говорящую о том, что IL-10 может играть роль в индуцируемой метотрексатом толерантности.
В этом исследовании животные получали 20 мг/кг Myozyme®, вводимого внутривенно еженедельно в течение девяти недель. Три цикла метотрексата назначали в дозе 5 мг/кг/день через 0, 24 и 48 часов после первых трех еженедельных лечений Myozyme®. Титры антител против Myozyme® определяли на неделях 4, 6 и 9 (фиг. 31). При сравнении средних значений титров у мышей с выключенным геном IL-10, подвергнутых лечению rhGAA и rhGAA/метотрексатом, значимое уменьшение титра не отмечалось, хотя незначительная тенденция была отмечена на неделе 9. Как и ожидалось, титры антител не были столь высокими у животных с выключенным геном IL-10, как у животных дикого типа C57BL/6. Ответы против rhGAA у животных дикого типа C57BL/6, подвергнутых лечению rhGAA и метотрексатом, были уменьшенными на неделях 4, 6 и 9. В противоположность этому, титры антител против rhGAA на неделях 4 и 6 не были уменьшенными у животных с выключенным геном IL-10, которые были подвергнуты лечению rhGAA и метотрексатом. На неделе 9 отмечалось небольшое уменьшение, которое может указывать на отсроченную индукцию толерантности у мышей с дефицитом IL-10. Это, вероятно, находится в соответствии с сообщениями о том, что IL-10 не является лишь подавляющим цитокином, секретируемым регуляторными B-клетками (Sagoo et al, J. Clin. Investigation; 120(6):1848-1861 (2010)). TGF-бета также был связан с реакцией регуляторных B-клеток. Если существует такая отсрочка, другие цитокины, такие как TGF-бета, могут быть способными оказать помощь в опосредовании эффекта индукции толерантности с помощью метотрексата. Эти данные до сих пор говорят о том, что IL-10 может играть роль в индуцируемой метотрексатом толерантности.
Пример 14: Роль метотрексата в лечении связанного с алемтумузабом вторичного аутоиммунитета
У подвергнутых лечению алемтузумабом пациентов с рассеянным склерозом может развиваться вторичный аутоиммунитет. Самыми часто встречающимися аутоиммунными нарушениями, которые развиваются после лечения алемтузумабом, являются те, которые относятся к тиреоидному аутоиммунитету. Кроме того, тромбоцитопеническая пурпура иммунного генеза и синдром Гудпасчера также отмечались у пациентов с рассеянным склерозом, которые были подвергнуты лечению алемтузумабом. Все три типа аутоиммунитета являются опосредованными B-клетками, поскольку B-клеточные реакции и аутоантитела непосредственно связаны с развитием заболевания и его патологией. Связь лечения алемтузумабом с развитием этих вторичных заболеваний недостаточно понята.
После лечения алемтузумабом T-клетки и В-клетки истощены. Большой процент этих истощенных T- и B-клеток является, вероятно, аутореактивными клетками, которые взаимодействуют с антигенами, экспрессируемыми в центральной нервной системе. В результате, их последующее истощение с помощью алемтузумаба, как полагают, вносит вклад в терапевтический эффект терапии с использованием этого моноклонального антитела. Пациенты, страдающие аутоиммунным заболеванием, как было описано, обладают аутореактивностями (т.е. содержат аутореактивные B-клетки и аутореактивные аутоантитела) с множеством антигенов, которые связаны с множеством аутоиммунных заболеваний. Все из факторов окружающей среды, физиологических и генетических факторов вносят вклад в определение того, последует ли по всей вероятности аутоиммунное заболевание, и влияние на то, которое аутоиммунное заболевание будет представлено у пациента. Нередко, когда у пациентов, которые страдают одним типом аутоиммунного заболевания, также развивается другой тип.
В случае алемтузумаба одна гипотеза заключается в том, что после лечения алемтузумабом в ситуации истощения лимфоцитов допускается развитие врожденных аутореактивностей, которые не были столь заметными, как те, которые связаны с рассеянным склерозом. Люди, у которых развивается аутоиммунное заболевание, обычно обладают аутореактивностями с рядом различных антигенов и, следовательно, предрасположенностью к развитию других аутоиммунитетов (т.е. «врожденной аутореактивностью»). Подтверждением этой гипотезы служат данные, полученные на трансгенных по huCD52 мышах, которые указывают на то, что алемтузумаб не истощает в равной степени все популяции B-клеток (фиг. 27). На самом деле, существует, по-видимому, дисбаланс в популяции B-клеток, который поддерживает представительство аутореактивных В-клеток с низкой аффинностью, особенно В-клеток краевой зоны. Важно отметить, что В-клетки краевой зоны были связаны с тиреоидным аутоиммунитетом (Segundo et al, Thyroid, 11(6):525-530 (2001). Кроме того, субпопуляция регуляторных B-клеток, B10 B-клетки, которые, как было установлено, помогают подавить аутоиммунитет в модели множественного склероза на мышах, EAE (Matsushita et al, J. Clin. Investigation, 118:3420-3430 (2008)), и, которые как было установлено, существуют у людей (Iwata et al, Blood, 117:530-541 (2011)), является истощенной в течение больших периодов времени, чем B-клетки краевой зоны. Короткий курс лечения метотрексатом во время первого цикла алемтузумаба может оказывать помощь в увеличении представительства регуляторных B10 B-клеток и в восстановлении баланса B-клеток, так что B-клетки краевой зоны представлены в более равной степени в популяции В-клеток после лечения алемтузумабом, и/или дифференциации B-клеток краевой зоны в регуляторные B-клетки краевой зоны (CD1d+ B-клетки краевой зоны).
Популяции В-клеток селезенки исследовали для определения эффектов алемтузумаба на истощение В-клеток (фиг. 32). 0,5 мг/кг алемтузумаба вводили внутривенно в течение пяти календарных дней трансгенным по huCD52 мышам. В частности, были исследованы популяции фолликулярных B-клеток (которые обычно не являются аутореактивными), B1 B-клеток и B-клеток краевой зоны (и те, и другие являются аутореактивными), B10 B-клеток (которые являются регуляторными), и «переходных» В-клеток и В-клеток краевой зоны (которые, как полагают, способны к дифференциации в регуляторные B-клетки).
В то время как фолликулярные, B1 и регуляторные B-клетки были истощены через одну и/или две недели после лечения алемтузумабом, B-клетки краевой зоны и В-клетки на стадии 1 иммунологической дифференциации (T1) и на стадии 2 иммунологической дифференциации (T2) не были истощены в какой-либо из моментов времени (фиг. 32). В-клетки на стадии 3 иммунологической дифференциации (T3) были только истощены спустя одну неделю лечения, и количество регуляторных B-клеток в целом кажется меньше у мышей, подвергнутых лечению алемтузумабом, чем у животных, подвергнутых лечению контролем, хотя статистическая значимость не отмечалась на неделях 2 и 4.
Для исследования эффектов метотрексата на популяции B-клеток в случае лечения алемтузумабом было выполнено второе исследование, как указано в таблице 3 и на фиг. 33. Неожиданно, что комбинированное лечение с использованием метотрексата и алемтузумаба может создать возможность для более сильного истощения В-клеток краевой зоны вскоре после лечения алемтузумабом, чем истощение, наблюдаемое при использовании только алемтузумаба (фиг. 34), тем самым поддерживая сбалансированную иммунологическую среду с помощью соответствующего уровня встречающихся в природе аутореактивных B-клеток с низкой аффинностью, необходимых для надлежащей ранней реакции на инфекцию.
В это исследование была включена группа мышей, подвергнутых лечению только алемтузумабом, и было обнаружено, что мыши, подвергнутые лечению только метотрексатом в этой схеме, предусматривающей короткий цикл, проявляют эффекты на клетки в отсутствие антигенной стимуляции, которые могут быть отличными от эффектов, отмечаемых у контрольных мышей, подвергнутых лечению только метотрексатом (фиг. 34). Данные, полученные в этом исследовании, в котором популяции B-клеток селезенки оценивались через два дня после последнего дня лечения метотрексатом, говорят о том, что метотрексат может усиливать истощение В-клеток краевой зоны. Это служит подтверждением гипотезы, что метотрексат, в случае доставки вместе с алемтузумабом, может приводить к клеточной ситуации, в которой существует соответственно баланс между встречающимися в природе B-клеточными субпопуляциями и субпопуляциями неаутореактивных B-клеток.
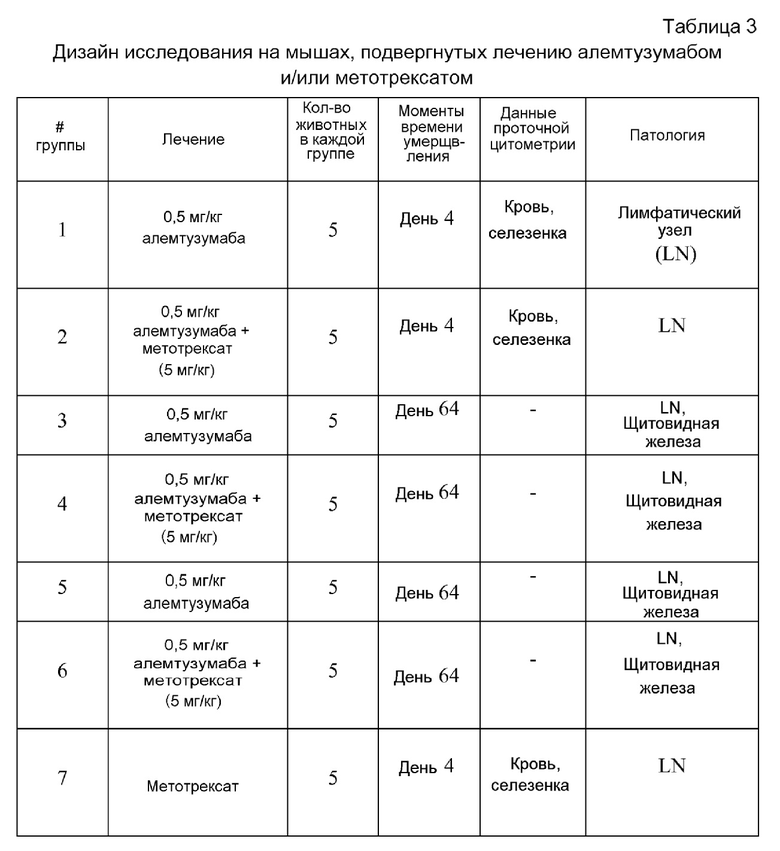
На основе ежедневной оценки популяций клеток, которые следовали непосредственно после лечения метотрексатом вместе с Myozyme®, авторы настоящего изобретения выдвигают гипотезу, что в случае алемтузумаба метотрексат будет приводить к обогащению популяций B10 B-клеток и популяций других потенциально регуляторных B-клеток не раньше чем через 5-6 дней после лечения метотрексатом. Авторы настоящего изобретения еще не наблюдали обогащение таких популяций через два дня после лечения метотрексатом, что находится в соответствии с этой гипотезой. В отличие от этого, эффекты, по-видимому, являются отличными через большие периоды времени после лечения метотрексатом, как видно в случае лечения mATG и алемтузумабом. В обоих из этих сценариев, через пять месяцев после лечения метотрексатом субпопуляции B-клеток казались обогащенными у мышей, подвергнутых лечению метотрексатом, через один и два дня после введения доз mATG и алемтузумаба (см. фиг. 35 для данных, относящихся к алемтузумабу). Примечательно, что B-клетки краевой зоны в этот момент времени также казались увеличенными (хотя это увеличение не является статистически значимым).
Метотрексат индуцирует толерантность к терапевтическим белкам и антигенам трансплантированной ткани сердца, тем самым аннулируя B-клеточные иммунные реакции, которые относятся к продукции и секреции ADA и антител против аллотрансплантата. В связи с этим авторы настоящего изобретения выдвигают гипотезу, что метотрексат может не только оказывать помощь в контролировании клеточной ситуации после лечения алемтузумабом, но что он также может индуцировать толерантность к собственным белкам и ослаблять B-клеточные иммунные реакции, которые относятся к выработке аутоантител, которые вносят вклад в развитие и патологию опосредуемых B-клетками аутоиммунных заболеваний.
Для проверки этой гипотезы животным ежемесячно вводили дозы алемтузумаба в течение пяти месяцев. Метотрексат назначали группе животных в дозе 5 мг/кг в течение трех календарных дней после только первого лечения алемтузумабом. За два дня до и через один день после лечения с использованием пятой дозы алемтузумаба, животных умерщвляли для проведения анализа. Уровни цитокинов в сыворотке определяли до и после лечения алемтузумабом. Как ни удивительно, эти результаты говорят о том, что уровни провоспалительных цитокинов MCP-1, IL-13, IL-6, и IL-12 были уменьшенными у животных, подвергнутых лечению метотрексатом, через 24 часа после пятой дозы алемтузумаба, через пять месяцев после введения какой-либо дозы метотрексата (фиг. 36). Эти цитокины не только стимулируют B-клеточные реакции и рекрутмент иммуноцитов, но они также могут играть роль в реакциях гиперчувствительности. Эти данные говорят о том, что метотрексат может оказывать помощь в ограничении инфузионных реакций.
Пример 15: Метотрексат индуцирует иммунологическую толерантность посредством специфической индукции популяций регуляторных B-клеток
Данные авторов настоящего изобретения неожиданно указывают на то, что метотрексат индуцирует иммунологическую толерантность не с помощью ожидаемого способа уничтожения пролиферирующих клеток, как предполагалось другими (Messinger et al, Genetics in Medicine 14:135-142 (2012) и Lacana et al, Am J Med Genet Part C Semin Med Genet 160C:30-39 (2012)), а посредством специфической индукции популяций регуляторных B-клеток, которые экспрессируют TGF-бета, IL-10 и FoxP3. B-клетки от мышей с индуцированной с помощью одного цикла метотрексата толерантностью к Myozyme®, как представлялось, переносят иммунологическую толерантность на не подвергшихся воздействию животных. Кроме того, и IL-10, и TGF-бета, как представлялось, необходимы для индуцируемой метотрексатом иммунологической толерантности. Также представлялось, что в некоторых субпопуляциях клеток метотрексат индуцирует TGF-бета, который в свою очередь индуцирует IL-10 и FoxP3. Этот механизм является новым и неожиданным, и подвергает сомнению ценность современного клинического протокола индукции иммунологической толерантности, который включает комбинированное лечение с использованием трех циклов метотрексата, Rituximab® (истощающего B-клетки средства) и необязательно внутривенно вводимого иммуноглобулина (IVIG) (Messinger et al., выше). Хотя это комбинированное лечение кажется успешным, данные авторов настоящего изобретения говорят о том, что 1) один цикл метотрексата может потенциально привести даже к более низким титрам ADA, чем те, которые отмечаются в настоящее время, и 2) в случае введения слишком большой дозы метотрексата и ритуксимаба иммунологическая толерантность может не сохраняться. Первоначальные дозы ритуксимаба могут не быть слишком вредными, поскольку опосредуемое ритуксимабом истощение B-клеток, как полагают, не является всеобъемлющим истощением всех B-клеток в крови и тканях. Как видно из исследований, описанных выше, хотя алемтузумаб активно истощает B10 B-клетки, лечение метотрексатом все еще может иметь доступ к тем клеткам, которые, по-видимому, оказывают помощь в сохранении толерантности к алемтузумабу в течение многих месяцев после первоначального цикла метотрексата. Кроме того, за лечением ритуксимабом быстро следует увеличенное представительство «переходных» В-клеток, на которые, как здесь показано, по-видимому, оказывает влияние метотрексат для индукции и опосредования иммунологической толерантности. Поскольку эти относящиеся к механизмам действия данные являются парадоксальными и неожиданными, один цикл метотрексата в низкой дозе является неожиданным и эффективным способом индукции иммунологической толерантности к, среди прочего, истощающим лимфоциты терапевтическим белкам.
Как описано выше, несколько субпопуляций B-клеток значимо увеличивались в процентном и/или количественном отношении вскоре после совместного введения метотрексата с терапевтическим белком. Кроме того, эти субпопуляции кажутся увеличенными у мышей с индуцированной метотрексатом толерантностью в течение долгого времени после лечения с использованием одного цикла метотрексата (см., например, фиг. 24, 27 и 35). Вместе, эти данные говорят о том, что эти популяции клеток могут активно опосредовать и индукцию, и сохранение индукции иммунологической толерантности. Для дальнейшего подтверждения этой гипотезы авторы настоящего изобретения исследовали, экспрессируют ли эти популяции клеток цитокины и другие белки, часто связанные с иммунорегуляцией.
Одним типом клеток, связанным с иммунорегуляцией, являются B10 B-клетки. B10 B-клетки и у человека, и у мыши характеризуются экспрессией в них IL-10 (Matsushita et al., J. Clin. Invest. 118:3420-3430 (2008), Iwata et al, Blood 117:530-541 (2011)) и могут только подавлять иммунные ответы у IL-10-компетентных мышей. B10 B-клетки были увеличены у мышей с индуцированной метотрексатом толерантностью (фиг. 19), а животные с выключенным геном IL-10 были неотвечающими на индуцируемую метотрексатом иммунологическую толерантность (фиг. 31). B10 B-клетки, выделенные от животных, подвергнутых лечению Myozyme® или Myozyme® и метотрексатом, оценивали в отношении экспрессии белка IL-10 с помощью проточной цитометрии. Хотя IL-10 экспрессировался в B10 клетках от обеих групп лечения, количество B10 B-клеток, экспрессирующих IL-10, было увеличенным у животных, подвергнутых лечению Myozyme® и метотрексатом (фиг. 38). IL-10 экспрессировался и в активированных, CD86+, и в неактивированных, CD86- B10 B-клетках после двух дней культивирования (фиг. 39). Предыдущие исследования, как представляется, указывают на то, что экспрессия IL-10 определяется только после in vitro стимуляции клеток такими стимулами, как PMA/Иономицин или LPS (Carter et al, J Immunol 186:5569-5579 (2011); Yanaba et al, J Immunol 182:7459-7472 (2009)). Неожиданно, в исследованиях авторов настоящего изобретения, какая-либо стимуляция культивируемых В-клеток селезенки или манипуляции ими не требовалась, чтобы сделать возможным измерение IL-10, что снова напрямую говорит о том, что эти B10 B-клетки экспрессируют IL-10 in vivo. Представленные здесь данные были получены в не подвергнутых стимуляции культурах. Кроме того, авторы настоящего изобретения полагают, что они впервые продемонстрировали, что метотрексат может специфически увеличить популяции клеток, которые экспресссируют IL-10. Кроме того, эти популяции клеток, по-видимому, экспрессируют больше IL-10, когда они выделены от животных, подвергнутых лечению Myozyme® и метотрексатом, чем когда они выделены от животных, подвергнутых лечению только Myozyme® (фиг. 54).
Экспрессию TGF-бета связывают с иммунорегуляцией в регуляторных T-клетках и часто связывают с экспрессией IL-10 в регуляторных T-клетках. Кроме того, некоторые сообщения, как представляется, указывают на то, что регуляторные B-клетки могут экспрессировать TGF-бета. Хотя никогда не сообщалось, что B10 B-клетки экспрессируют TGF-бета, авторы настоящего изобретения решили определить экспрессию TGF-бета в B10 B-клетках мышей, подвергнутых лечению только Myozyme® или Myozyme® и метотрексатом, используя проточную цитометрию. Неожиданно авторы настоящего изобретения обнаружили, что B10 B-клетки экспрессируют TGF-бета, и что количество TGF-бета-экспрессирующих клеток увеличено у животных с индуцированной метотрексатом толерантностью (фиг. 40A). Кроме того, в этих культивируемых клетках TGF-бета экспрессируется и в активированных (CD86+), и в неактивированных (CD86-) B10 B-клетках (фиг. 40B). То, что лечение метотрексатом увеличивает количество клеток, которые экспрессируют TGF-бета, является дополнительными новыми данными наблюдений. Кроме того, метотрексат увеличивает уровень экспрессии TGF-бета (фиг. 55).
FoxP3 является другим белком, связанным с иммунорегуляцией. FoxP3 является маркером регуляторных T-клеток. Не сообщалось, что FoxP3 экспрессируется в B10 B-клеткам у мышей. Авторы настоящего изобретения исследовали экспрессию FoxP3 в B10 B-клетках при наличии индуцированной метотрексатом иммунологической толерантности и в ее отсутствие, используя проточную цитометрию. B10 B-клетки, по-видимому, экспрессируют FoxP3, как видно у животных, подвергнутых лечению только Myozyme® (фиг. 41A). Количества FoxP3+ B-клеток, как представляется, увеличивается при лечении и метотрексатом, и Myozyme® (фиг. 41B). Кроме того, культивируемые и активированные (CD86+), и неактивированные (CD86-) B10 B-клетки, как представляется, экспрессируют FoxP3 (фиг. 41). Это было первым сообщением о том, что B10 B-клетки экспрессируют FoxP3. Авторы настоящего изобретения обнаружили, что FoxP3 экспрессируется и в активированных (CD86+), и в неактивированных (CD86-) B10 B-клетках, и что экспрессия FoxP3 увеличивается при лечении метотрексатом (фиг. 56).
Поскольку дополнительные типы B-клеток значимо увеличены при индуцированной одним циклом метотрексата иммунологической толерантности, авторы настоящего изобретения исследовали, экспрессируют ли некоторые из этих типов клеток IL-10, TGF-бета и FoxP3. Было установлено, что В-клетки на стадии 2 иммунологической дифференциации, на стадии 3 иммунологической дифференциации и фолликулярные B-клетки экспрессируют IL-10 (фиг. 42), TGF-бета (фиг. 43) и FoxP3 (фиг. 44), что было новым и неожиданным для каждой из этих субпопуляций B-клеток. Как было отмечено в случае B10 клеток, абсолютные количества клеток субпопуляций IL-10, TGF-бета и FoxP3 B-клеток были увеличенными у мышей, подвергнутых лечению метотрексатом (фиг. 42-44B и C), по сравнению с мышами, подвергнутыми лечению только Myozyme® (фиг. 42-44A и C). Лечение метотрексатом также вызывало статистически значимые увеличения IL-10, TGF-бета и FoxP3 во множестве субпопуляцией клеток, если судить по изменению средней интенсивности флуоресценции этих белков у животных, подвергнутых лечению только Myozyme® или Myozyme® и метотрексатом (фиг. 54-56).
Для дальнейшего исследования множества популяций TGF-бета-экспрессирующих B-клеток, обогащенных в результате лечения метотрексатом и Myozyme®, авторы настоящего изобретения затем рассчитывали определить, требовался ли TGF-бета для индуцируемой метотрексатом иммунологической толерантности. Животных подвергали лечению Myozyme® или Myozyme® и метотрексатом вместе с 5 мг/кг антитела против TGF-бета (1D11, Genzyme) или подобранного по изотипу контроля (13C4), вводимого три раза в неделю посредством внутрибрюшинной инъекции на протяжении всего исследования, или без него. Титры антител определяли раз в две недели у четырех различных групп животных. Если TGF-бета требовался для индуцируемой метотрексатом иммунологической толерантности, то ожидалось бы, что животные, подвергнутые лечению антителом против TGF-бета, не должны демонстрировать уменьшенные титры антител против Myozyme®. Титры на неделе 6 представлены на фиг. 45 и говорят о том, что TGF-бета может быть необходим для индуцируемой метотрексатом иммунологической толерантности. Поскольку неделя 6 является ранним моментом времени, только некоторые из животных имели время для выработки антител против Myozyme®. Важно отметить, что в этот момент времени титры кажутся схожими с теми, которые представлены на фиг. 15C на неделе 6, когда два животных среди животных, подвергнутых лечению только rhGAA, и подвергнутые лечению rhGAA и метотрексатом, и 1D11 животные продемонстрировали высокие титры. По сравнению с этим, ни одно из животных, подвергнутых лечению rhGAA и метотрексатом или rhGAA и метотрексатом и 13C4, не продемонстрировало высокие титры.
Кроме того, селезенки были выделены от животных, подвергнутых лечению Myozyme® или Myozyme® и метотрексатом, которым также одновременно вводили 1D11 или 13C4 через день дней после одного лечения Myozyme® или одного лечения Myozyme® и метотрексатом. В этот момент времени, В-клетки на стадии 2 иммунологической дифференциации, на стадии 3 иммунологической дифференциации, В10 и фолликулярные B-клетки, которые экспрессировали IL-10, TGF-бета и FoxP3, были увеличенными у животных, подвергнутых лечению Myozyme® и метотрексатом. Клетки в каждой группе затем объединяли и культивировали в течение двух дней, а затем оценивали с помощью проточной цитометрии для определения, препятствует ли лечение антителом против TGF-бета - 1D11 увеличению количества клеток, которые экспрессируют TGF-бета, IL-10 и FoxP3, по сравнению с подобранным по изотипу контрольным антителом (13C4).
Довольно неожиданно, лечение 1D11 препятствовало не только индуцируемому метотрексатом увеличению количества клеток, экспрессирующих TGF-бета, но также увеличению некоторых субпопуляций, экспрессирующих IL-10 и FoxP3. Это было верно в частности для B10 B-клеток (фиг. 46) и фолликулярных B-клеток (фиг. 47), хотя FoxP3+ фолликулярные B-клетки, по-видимому, не демонстрировали эффекты лечения 1D11. В случае В-клеток на стадии 2 иммунологической дифференциации, хотя лечение 1D11 мешало увеличению количества TGF-бета-экспрессирующих В-клеток на стадии 2 иммунологической дифференциации, эффекты не наблюдались по отношению к IL-10+ В-клеткам на стадии 2 иммунологической дифференциации (включая активированные В-клетки на стадии 2 иммунологической дифференциации; фиг. 48). Кроме того, эффекты в FoxP3+ В-клетках на стадии 2 иммунологической дифференциации, оказываемые 1D11, не казались явными в этой небольшой группе лечения, если не посмотреть на активированные, CD86+ В-клетки на стадии 2 иммунологической дифференциации. Важно отметить, что эти данные говорят о том, что индуцируемый метотрексатом TGF-бета связан с IL-10 и FoxP3 только в некоторых типах клеток. В случае В-клеток на стадии 3 иммунологической дифференциации, даже хотя TGF-бета поддается детектированию в субпопуляции клеток на стадии 3 иммунологической дифференциации (фиг. 49), нет явного эффекта лечения 1D11 на эти клетки (P<0,05; фиг. 49). Примечательно, в случае активированных CD86+ В-клеток на стадии 3 иммунологической дифференциации, по-видимому, существуют большие количества IL-10+ В-клеток на стадии 3 иммунологической дифференциации у подвергнутых лечению 1D11 мышей. Это может говорить о том, что эта популяция могла увеличиваться в попытке помочь компенсировать уменьшения количеств других типов клеток вследствие лечения 1D11. Также примечательно, что лечение 1D11 не влияло на основные уровни IL-10, TGF-бета и FoxP3 в этих клетках (фиг. 50A-C, соответственно), но оно, как представлялось, влияет на эффекты метотрексата на клетки, которые экспрессируют эти цитокины.
В заключение, вмешательство в TGF-бета посредством инъекции антитела против TGF-бета во время индуцируемой метотрексатом иммунологической толерантности уменьшает количество клеток, которые экспрессируют TGF-бета, по сравнению с животными, подвергнутыми лечению подобранным по изотипу контролем. Кроме того, лечение 1D11 подавляет типичные увеличения, отмечаемые за счет индуцируемой метотрексатом толерантности, экспрессирующих IL-10 и FoxP3 B10 B-клеток. Лечение 1D11, по-видимому, влияет на эффекты метотрексата на TGF-бета, IL-10 и/или FoxP3 в некоторых типах B-клеток, но не во всех типах B-клеток. Эти данные наблюдений являются неожиданными и говорят о том, что метотрексат индуцирует TGF-бета, который в свою очередь индуцирует IL-10 и возможно FoxP3 в некоторых клетках, таких как B10 B-клетки. Хотя о связи TGF-бета с IL-10 и FoxP3, по-видимому, сообщалось раньше, она не была установлена в этих типах клеток. Кроме того, метотрексат не был одновременно связан с этим сложным каскадом передачи сигналов.
До сих пор, авторы настоящего изобретения показали, что некоторые B-клетки значимо увеличены за счет индуцируемой метотрексатом толерантности, и что эти B-клетки экспрессируют белки, связанные с иммунорегуляцией (супрессией). Для более прямой оценки, опосредуют ли сами B-клетки индуцируемую метотрексатом толерантность, авторы настоящего изобретения выполнили эксперимент адаптивного переноса, в котором определялось, могут ли B-клетки селезенки в целом от мышей с индуцированной метотрексатом толерантностью переносить иммунологическую толерантность на не подвергнутых воздействию мышей. Авторы настоящего изобретения выполнили этот эксперимент, используя Myozyme® и один цикл лечения метотрексатом. Авторы настоящего изобретения выделили селезенки из животных, подвергнутых лечению только Myozyme® или Myozyme® и метотрексатом, через 7 дней после одного лечения Myozyme® или Myozyme® и метотрексатом (когда отмеченные выше субпопуляции B-клеток были увеличены под действием метотрексата), а затем очистили все из B-клеток селезенок. Эти клетки затем переносили в мышей-реципиентов, не подвергнутых воздействию Myozyme® (фиг. 51A). После переноса реципиентов (вместе с не подвергнутыми переносу контрольными животными, которых лечили Myozyme® или Myozyme® и метотрексатом) подвергали еженедельно лечению 20 мг/кг Myozyme®. Кровь отбирали каждые две недели для определения титров антител против Myozyme®. В момент получения селезенок субпопуляцию B-клеток от каждой группы доноров оценивали с помощью проточной цитометрии для подтверждения ожидаемых увеличений TGF-бета+, IL-10+ и/или FoxP3+ В-клеток на стадии 2 иммунологической дифференциации, на стадии 3 иммунологической дифференциации, B10 и фолликулярных B-клеток. Было подтверждено, что группы доноров имеют ожидаемый фенотип. Анализ титров говорил о том, что В-клетки в целом селезенки, выделенные из животных, подвергнутых лечению с использованием Myozyme® и одного цикла метотрексата, могут переносить иммунологическую толерантность к Myozyme® на не подвергнутых воздействию хозяев (фиг. 51B). Это служит подтверждением того, что B-клетки могут опосредовать индуцируемую метотрексатом иммунологическую толерантность, и впервые было описано, что B-клетки обогащаются (а не уничтожаются) под действием метотрексата, чтобы помочь в опосредовании противовоспалительных эффектов. Эти данные также напрямую подвергают сомнению сопутствующее применение истощающего B-клетки средства с метотрексатом для индукции иммунологической толерантности, которое в настоящее время осуществляют в отношении пациентов (Messinger et al, выше и Lacana et al., выше).
Пример 16: Метотрексат уменьшает степень патологии, относящейся к трансплантату.
Авторы настоящего изобретения также получили дополнительные данные в случае мышиного антитимоцитарного глобулярного белка, которые указывают на то, что метотрексат не истощает CD4+, CD8+, регуляторные (CD4+CD25+FoxP3+) T-клетки и CD19+ B-клетки в целом у нормальных животных (фиг. 52) и у животных с трансплантатом (фиг. 53). При сравнении нормальных животных, подвергнутых лечению неспецифическим кроличьим IgG отдельно или в комбинации с метотрексатом, не было значимых изменений популяций клеток в крови и селезенке (фиг. 52). Кроме того, при сравнении тех популяций, которые были выделены из животных, подвергнутых лечению только mATG или mATG и метотрексатом, не было увеличенного истощения. Точнее, авторы настоящего изобретения только отмечали удлинение опосредуемых mATG эффектов на CD4+, CD8+ и регуляторные T-клетки, которое, весьма правдоподобно, обусловлено увеличением подвергания воздействию mATG под действием метотрексата (фиг. 52). Кроме того, только лечение метотрексатом у животных, подвергаемых аллогенной трансплантации, не истощало эти конкретные субпопуляции клеток (фиг. 53). Меньшие количества CD4+ и CD8+ T-клеток у животных, подвергнутых лечению mATG и метотрексатом, по сравнению с животными, подвергнутыми лечению только mATG, могут объясняться пролонгированным эффектом mATG при совместном лечении метотрексатом. Важно отметить, что метотрексат не вызывал уменьшений B-клеток, как ожидалось бы, если бы метотрексат уничтожал активированные B-клетки для уменьшения гуморальных иммунных ответов или у нормальных, или у подвергнутых трансплантации животных (фиг. 52 и 53). Эти данные говорят о том, что эффекты метотрексата на антитела против лекарственного средства не могут опосредоваться индуцируемым антифолатной активностью истощением активированных B-клеток. Как и ожидалось, лечение mATG также не оказывало воздействие на общее количество В-клеток в этой модели гетеротопной аллотрансплантации сердца. В общем, комбинированное лечение с использованием mATG и метотрексата уменьшало тяжесть патологии отторжения трансплантата и сопровождалось уменьшением T-клеточного инфильтрата в трансплантате, но без увеличений клеток с фенотипом регуляторных T-клеток. Это находится в соответствии с результатами, полученными в случае Myozyme® и алемтузумаба.
Для определения гистологических изменений в трансплантатах с длительной продолжительностью существования, а также понимания механизма действия комбинации mATG и метотрексата на выживание трансплантата, трансплантаты сердца были собраны и оценены в отношении патологии, а также клеточного состава. В частности, поскольку регуляторные T-клетки были связаны с длительным выживанием трансплантата при трансплантации, индуцируются под действием Thymoglobulin® и mATG и, как было продемонстрировано, ответственны за отсроченное отторжение трансплантата после лечения mATG, были оценены CD3+Foxp3+ клетки, которые имеют фенотип, соответствующий регуляторным T-клеткам. А именно, трансплантаты сердца получали из группы, подвергнутой лечению комбинацией mATG и метотрексата, и не подвергнутой лечению сингенной группы через по крайней мере 100 дней после трансплантации. Трансплантаты от не подвергнутых лечению мышей и мышей, подвергнутых лечению только mATG или только метотрексатом, были взяты после отторжения трансплантатов для сравнения. Срезы тканей, окрашенные H&E или трихромом по Массону или подвергнутые иммуноокрашиванию антителами против CD3 и Foxp3, были микроскопически исследованы в отношении гистологических изменений, указывающих на отторжение трансплантата, например, миокардита легкой-средней степени тяжести, дегенерации миокарда и его некроза, васкулопатии в сердечном аллотрансплантате (CAV) и T-клеточной инфильтрации. В день 100 в аллотрансплантатах от животных, подвергнутых комбинированному лечению с использованием mATG и метотрексата, выявлены незначительные-легкие патологические изменения в виде CAV и отсутствие и незначительная дегенерация миокарда и миокардит. Гистологические изменения, говорящие об отторжении трансплантата, не были явными в сингенных трансплантатах в этот поздний момент времени (фиг. 54). В аллотрансплантатах из группы, подвергнутой комбинированному лечению, выявлена умеренная T-клеточная инфильтрация в миокарде с небольшим количеством клеток, инфильтрирующих кровеносные сосуды миокарда. Сингенные трансплантаты содержали редкие T-клетки в миокарде. Кластеры T-клеток с редкими имммунопозитивными и по CD3, и по Foxp3 клетками присутствовали в эпикарде и сингенных трансплантатов, и подвергнутых комбинированному лечению аллотрансплантатов. Таким образом, трансплантаты с длительным выживанием продемонстрировали минимальные признаки отторжения трансплантата, которые коррелируют с уменьшенным воспалением.
Поскольку эффекты комбинированного лечения с использованием mATG и метотрексат, вероятно, более активно происходили ближе ко времени трансплантации, авторы настоящего изобретения оценили патологию и охарактеризовали клеточный инфильтрат в трансплантированных сердечных аллотрансплантатах в дни 7 (для не подвергнутых лечению мышей) или 14 (для всех групп лечения) после трансплантации. Не подвергнутые лечению аллотрансплантаты продемонстрировали патологию отторжения трансплантата, включая миокардит, дегенерацию миокарда и его некроз и в эпикардиальных, и в интрамиокардиальных ветвях коронарных артерий. В отличие от этого, в аллотрансплантатах, выделенных от животных, подвергнутых лечению и mATG, и метотрексатом, выявлена менее тяжелая CAV. Это отмечалось по сравнению с аллотрансплантатами от не подвергнутых лечению животных, а также от животных, подвергнутых лечению или mATG, или метотрексатом (фиг. 58). Как и ожидалось, в сингенных трансплантатах от не подвергнутых лечению мышей не была выявлена или была выявлена незначительная патология в эти моменты времени. CD3+ T-клеточная инфильтрация наблюдалась в миокарде и эпикарде в аллотрансплантатах от не подвергнутых лечению мышей и мышей, подвергнутых лечению только mATG или метотрексатом. Небольшое количество CD3+ T-клеток также присутствовало в воспалительном клеточном инфильтрате, связанном с патологическими изменениями CAV в этих трансплантатах. В отличие от этого, аллотрансплантаты от животных, подвергнутых лечению комбинацией mATG и метотрексата, продемонстрировали значительную меньшую CD3+ T-клеточную инфильтрацию в миокарде и лишь минимальную CD3+ T-клеточную инфильтрацию в эпикарде. Сингенные трансплантаты сердца продемонстрировали минимальную CD3+ T-клеточную инфильтрацию только в эпикарде. Небольшой процент T-клеток в воспалительных клеточных инфильтратах в эпикарде, как представлялось, имел фенотип регуляторных T-клеток, на что указывала иммунореактивность и с CD3, и с Foxp3, но эта встречаемость не казалась выше в воспалительных инфильтратах, чем в других группах. Следовательно, уменьшенная степень патологии также отмечалась вскоре после лечения mATG и метотрексатом и сопровождалась как уменьшенной, так и ограниченной эпикардом T-клеточной инфильтрацией.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ВОЛЧАНКИ | 2010 |
|
RU2607022C2 |
| БИОМАРКЕРЫ ИММУННОЙ ТОЛЕРАНТНОСТИ, ИНДУЦИРОВАННОЙ МЕТОТРЕКСАТОМ | 2018 |
|
RU2795201C2 |
| ИНГИБИТОРЫ Т-КЛЕТОЧНОЙ АКТИВАЦИИ | 2012 |
|
RU2657440C2 |
| КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ РАССЕЯННОГО СКЛЕРОЗА | 2008 |
|
RU2492234C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-ТФР-БЕТА-АНТИТЕЛА | 2005 |
|
RU2386638C2 |
| КОМПОЗИЦИЯ ДЛЯ ИНДУЦИРОВАНИЯ СПЕЦИФИЧЕСКОЙ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ | 2010 |
|
RU2782223C2 |
| АНТИ-ICOS АНТИТЕЛА | 2017 |
|
RU2764548C2 |
| АНТАГОНИСТ CD40L И ПУТИ ЕГО ПРИМЕНЕНИЯ | 2019 |
|
RU2812919C2 |
| ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ АГОНИСТА TLR И КОМБИНИРОВАННАЯ ТЕРАПИЯ | 2011 |
|
RU2603467C2 |
| КОМБИНИРОВАННАЯ ПРОТИВООПУХОЛЕВАЯ ТЕРАПИЯ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ НЕЕ | 2005 |
|
RU2396960C2 |
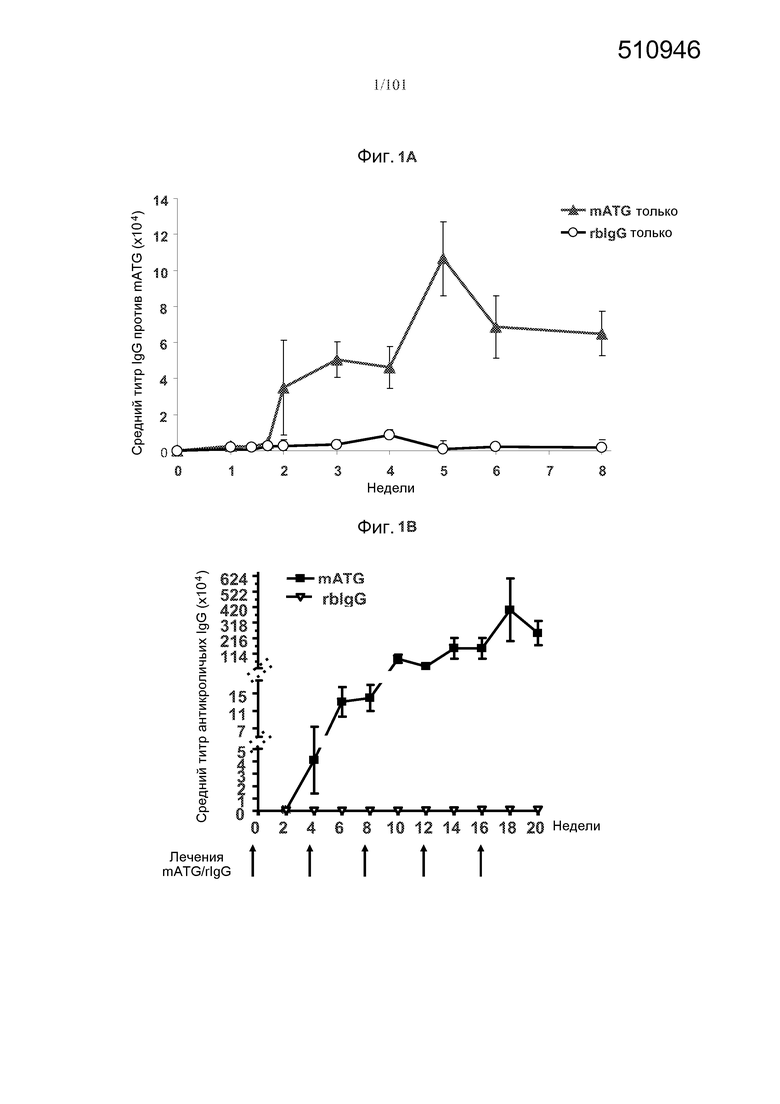
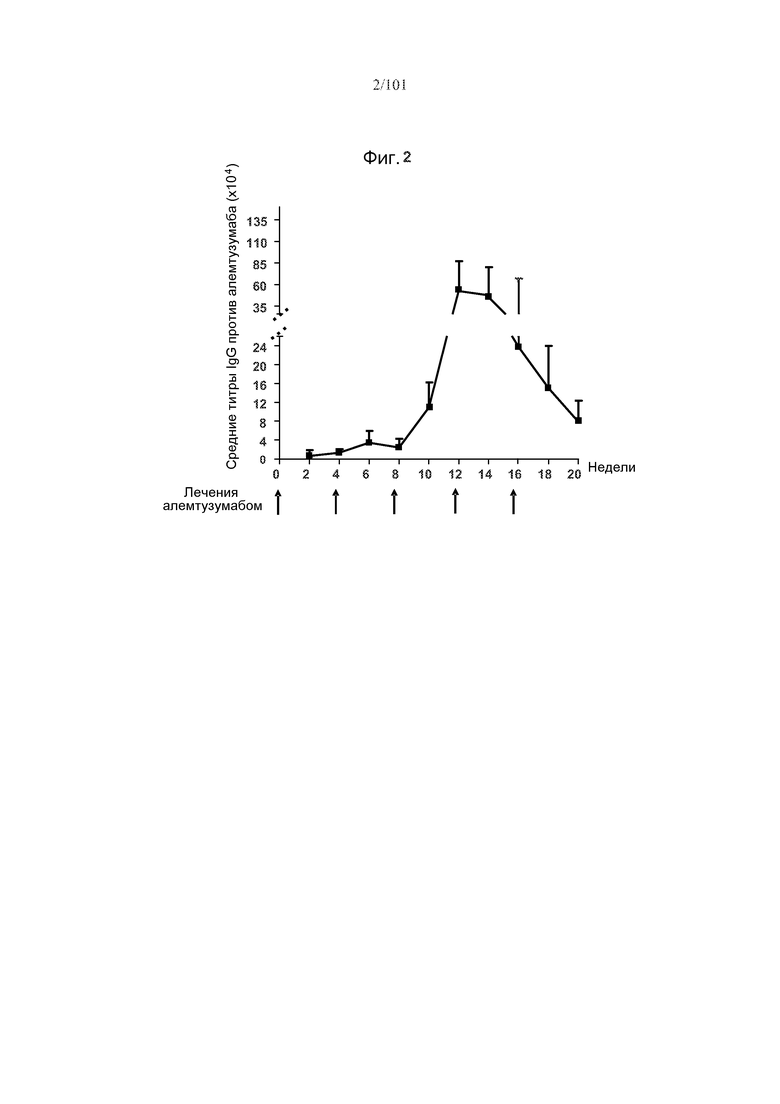
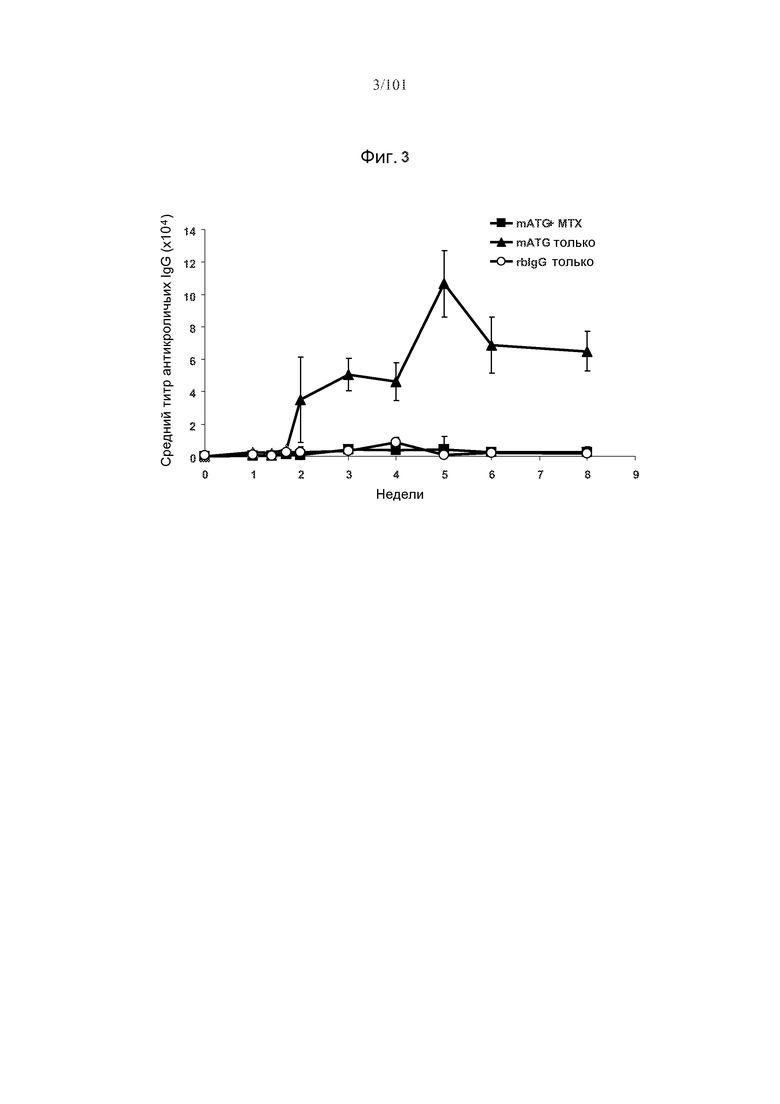
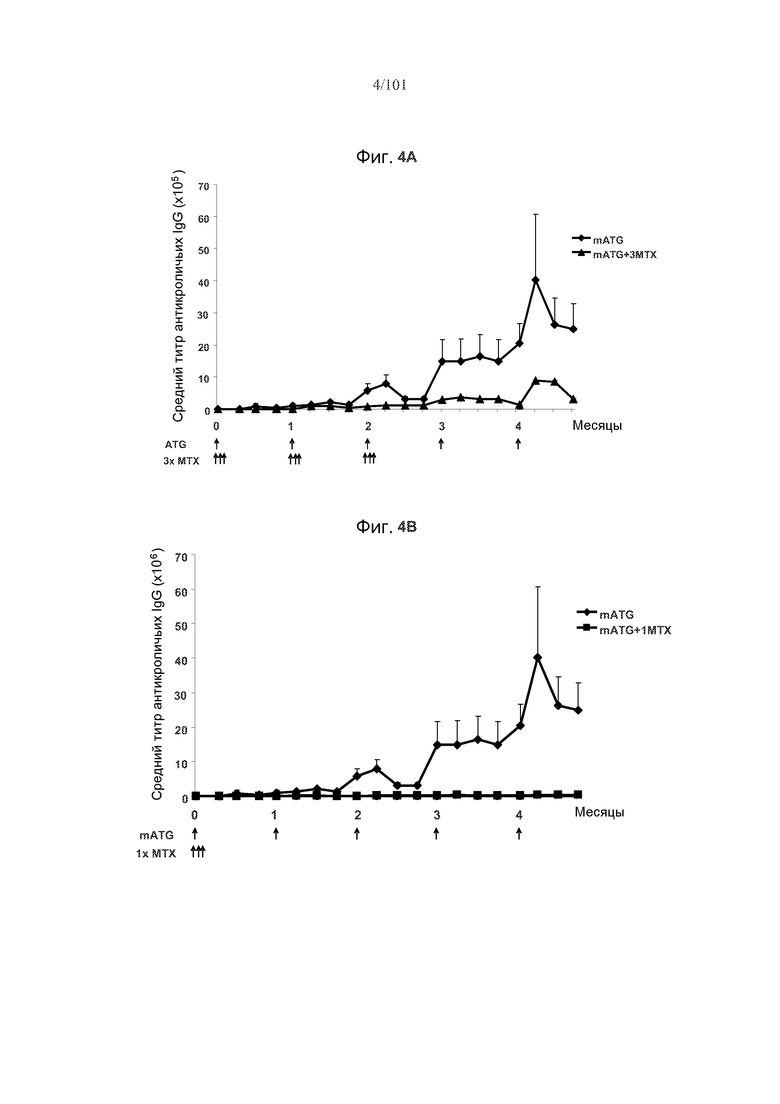
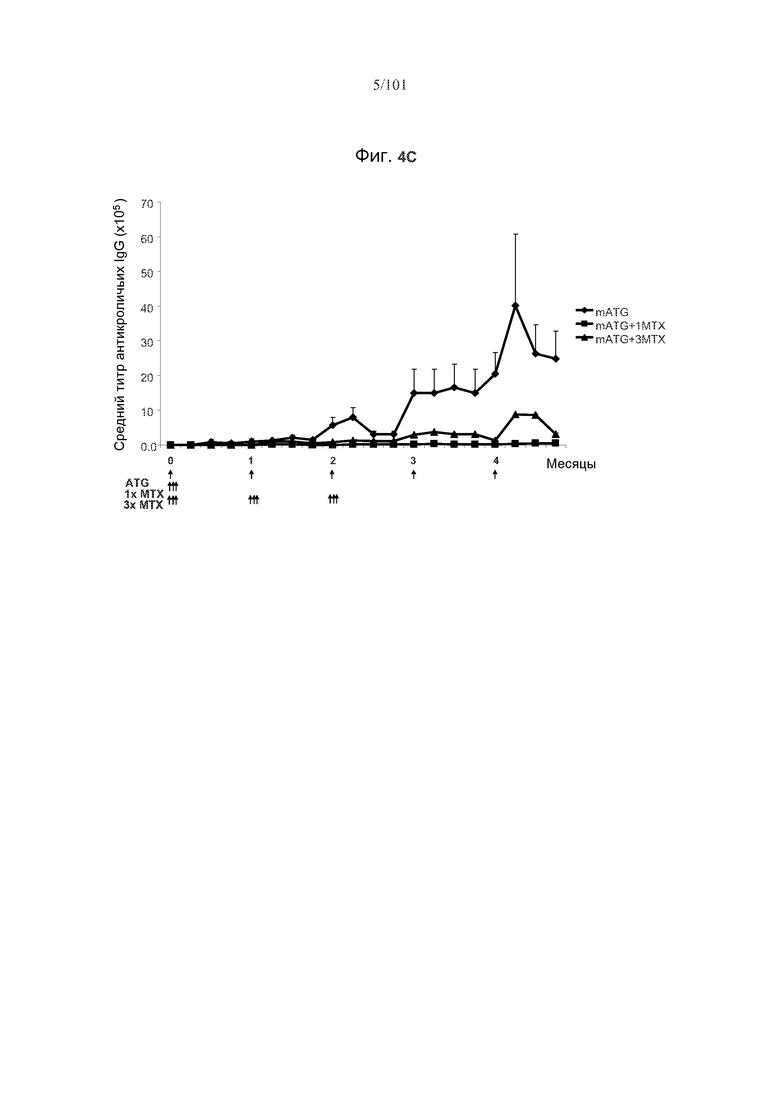
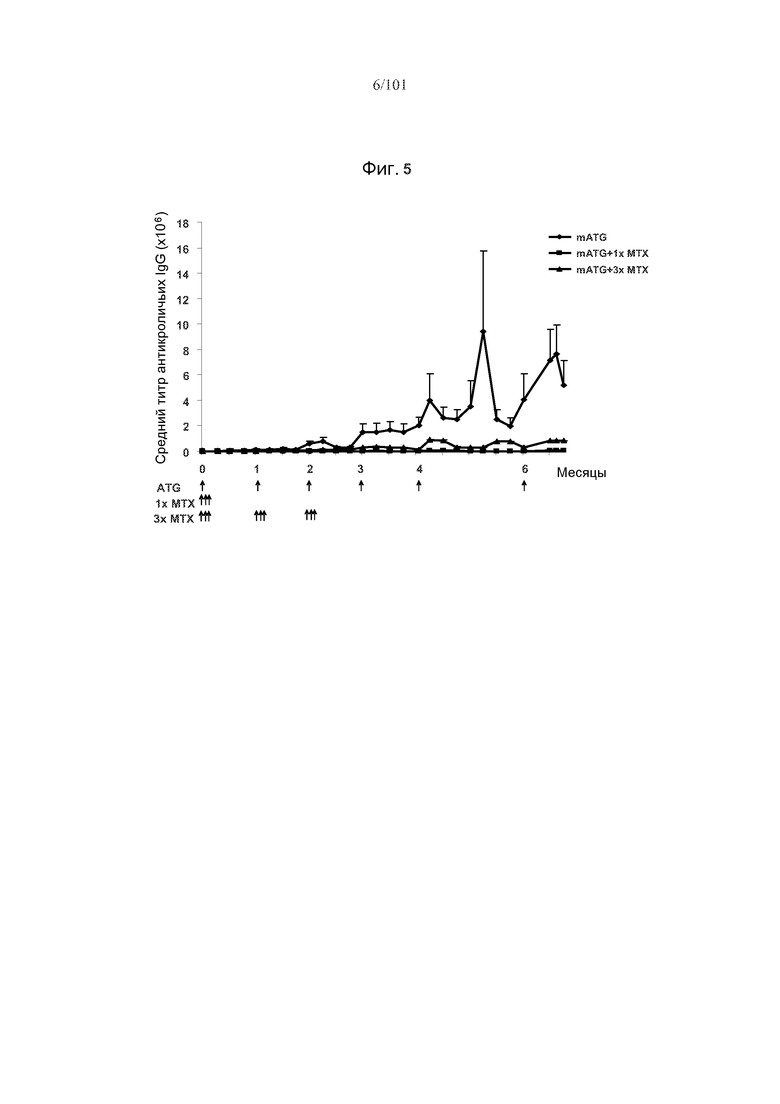
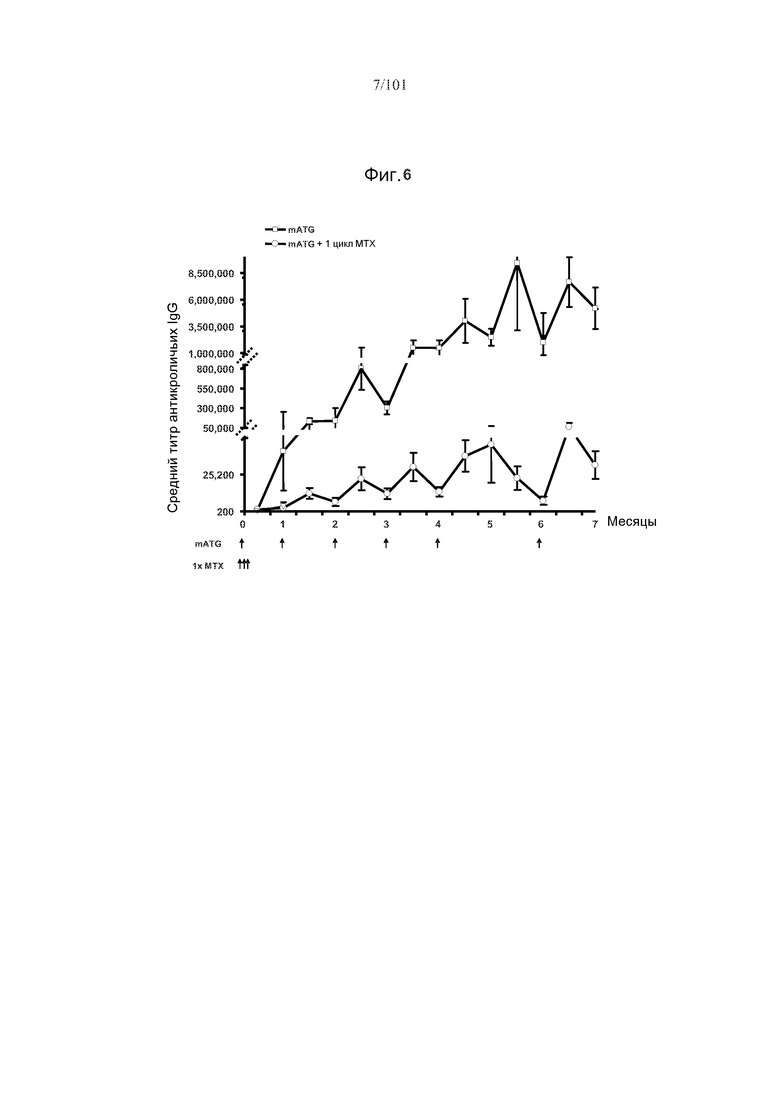
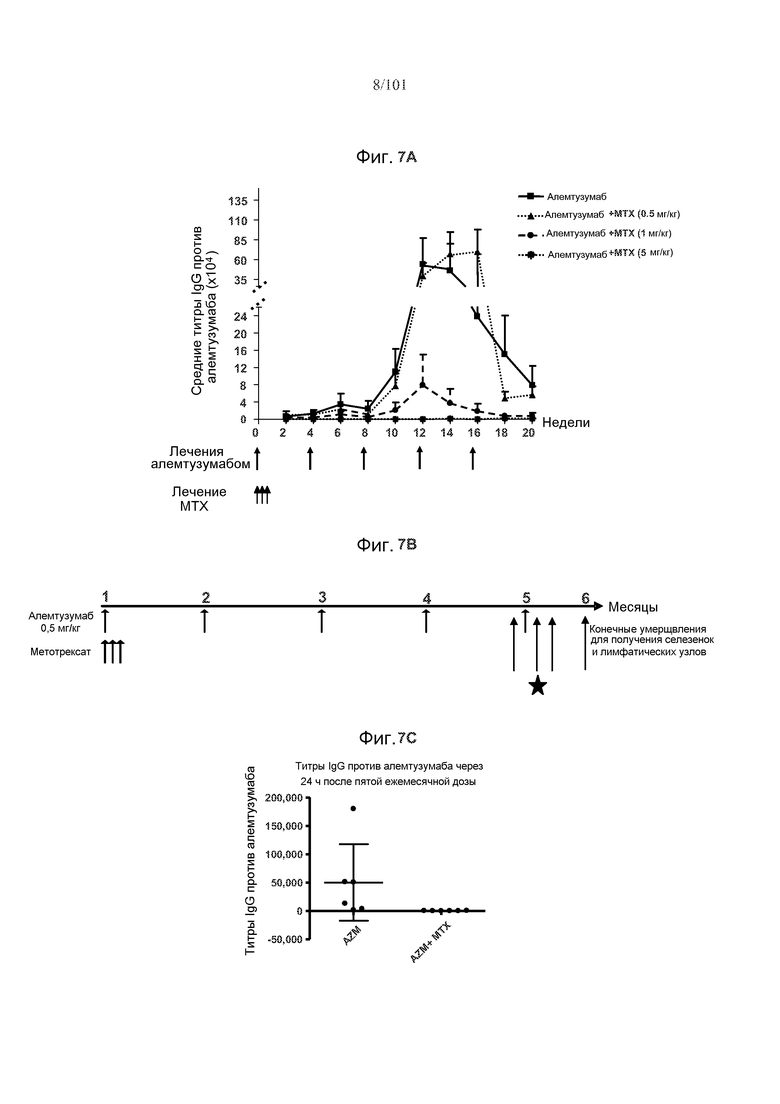
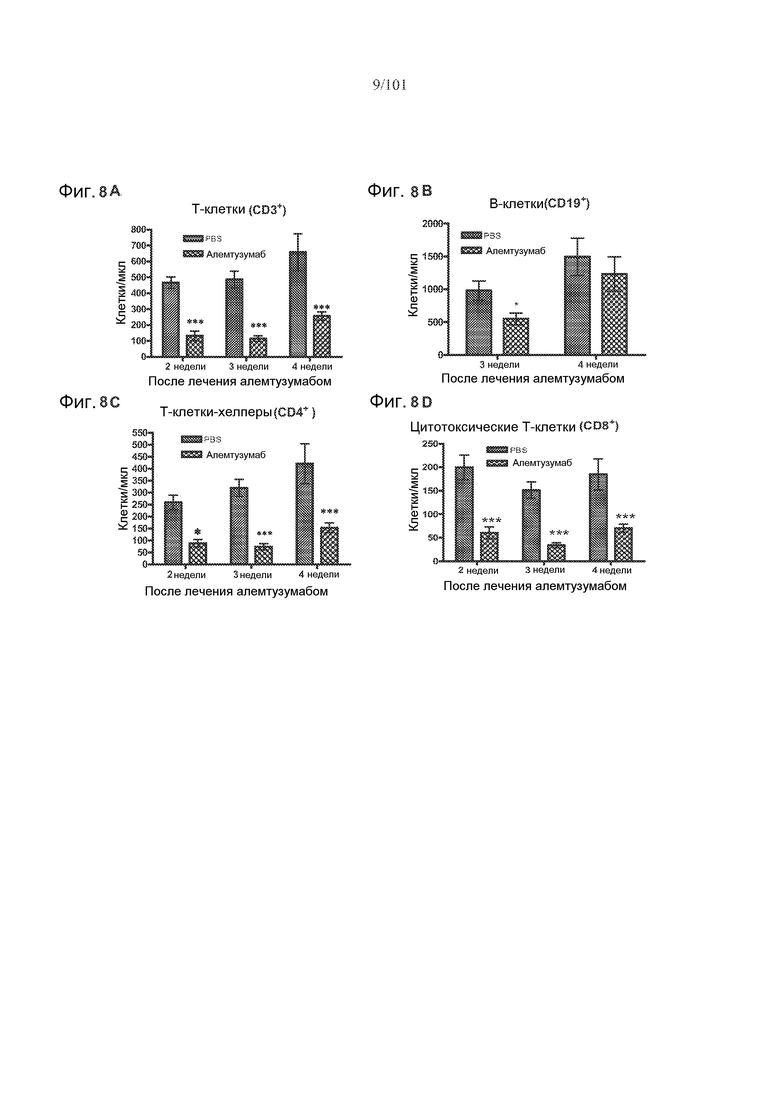
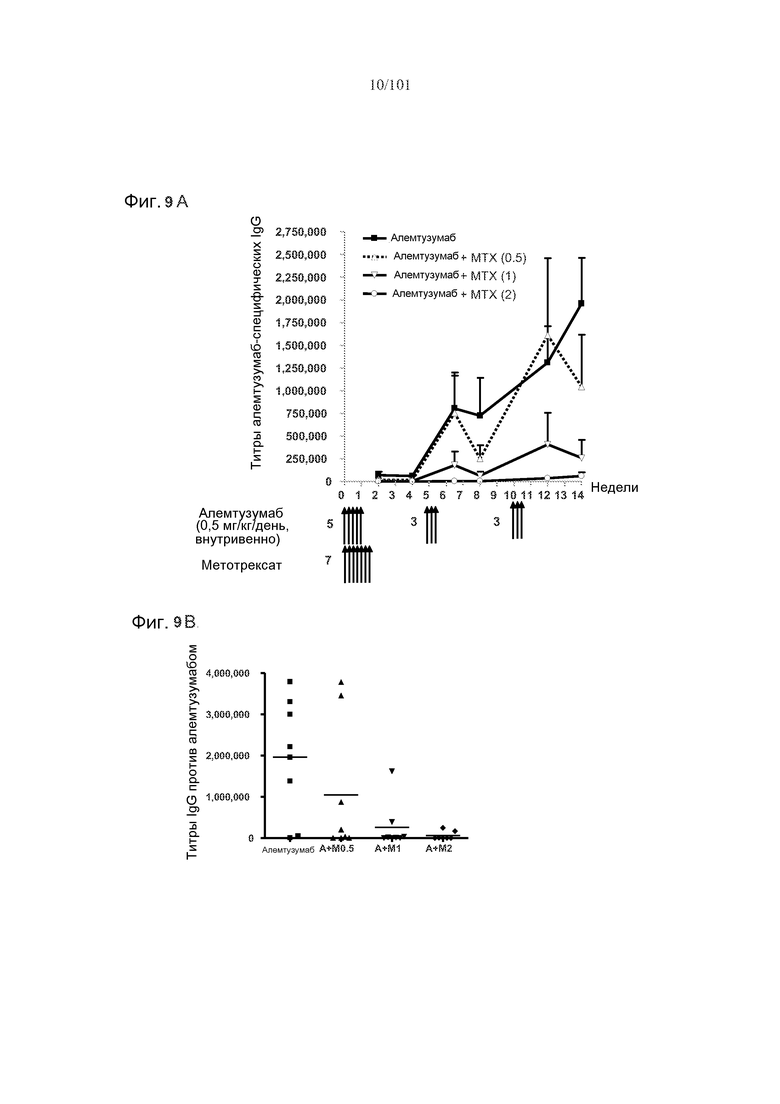
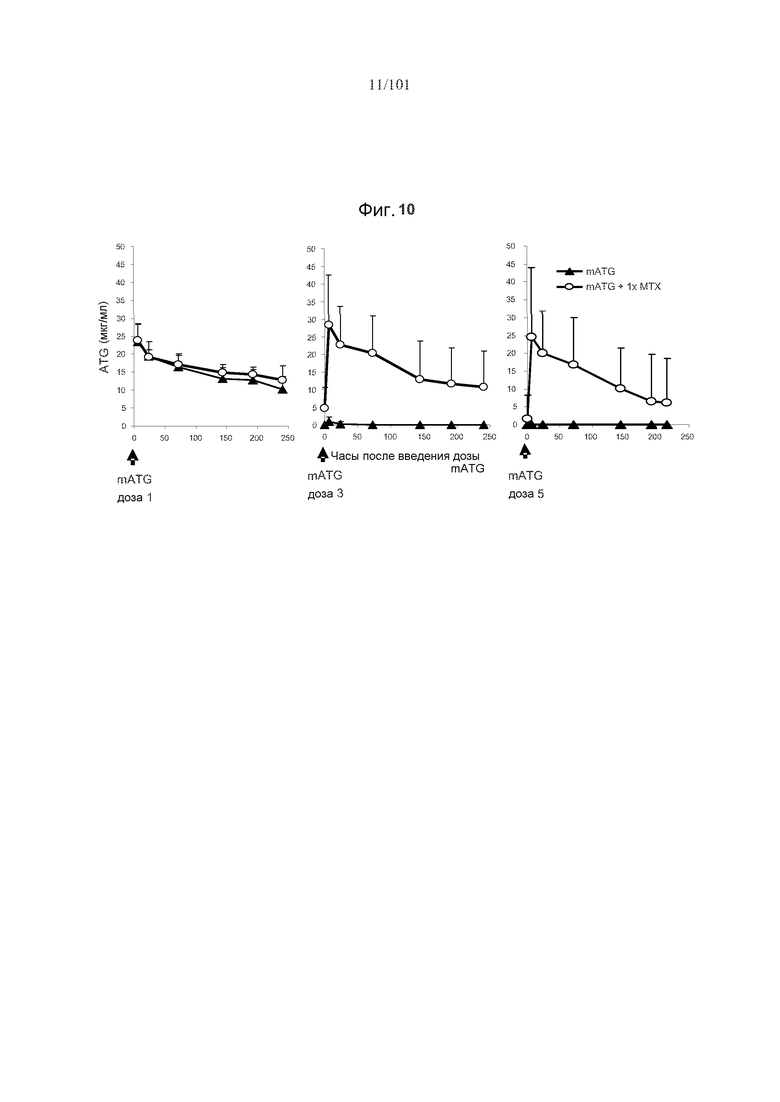
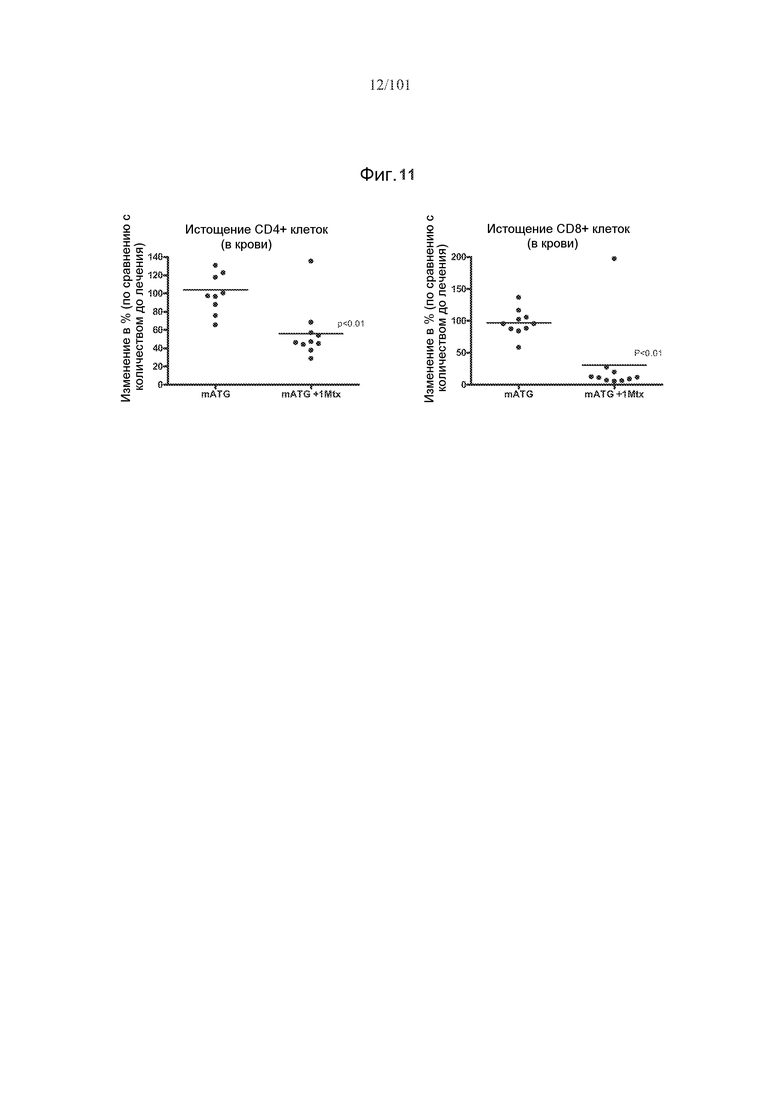
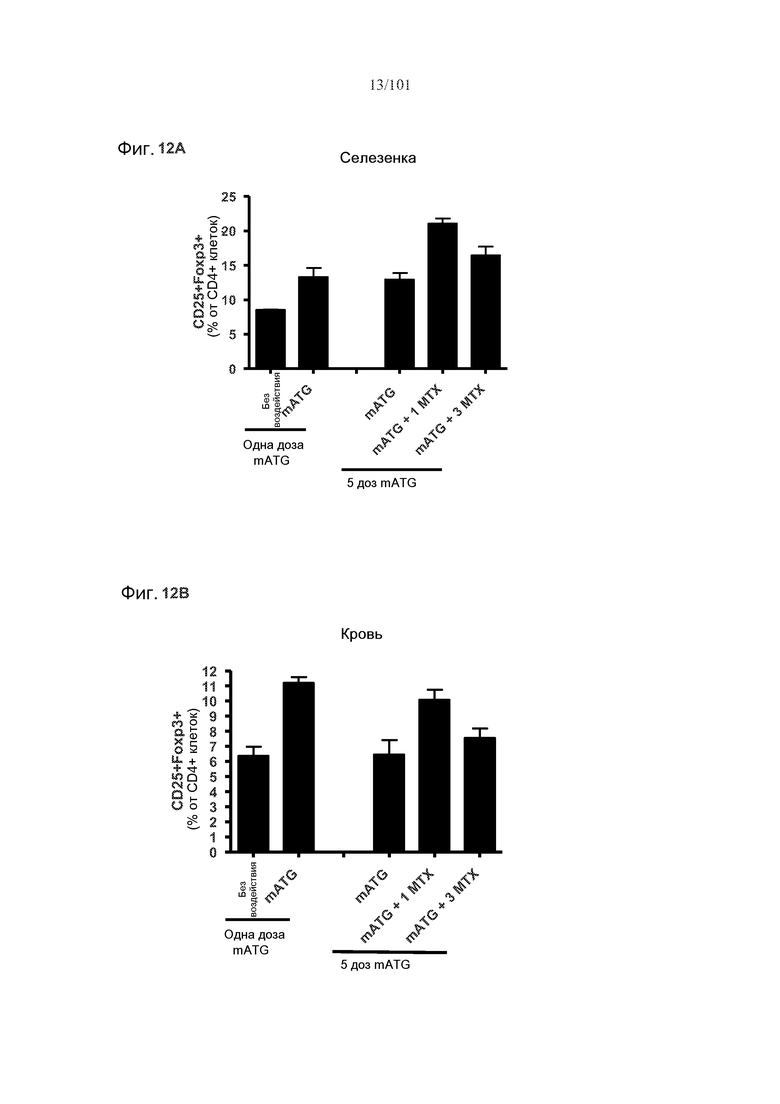
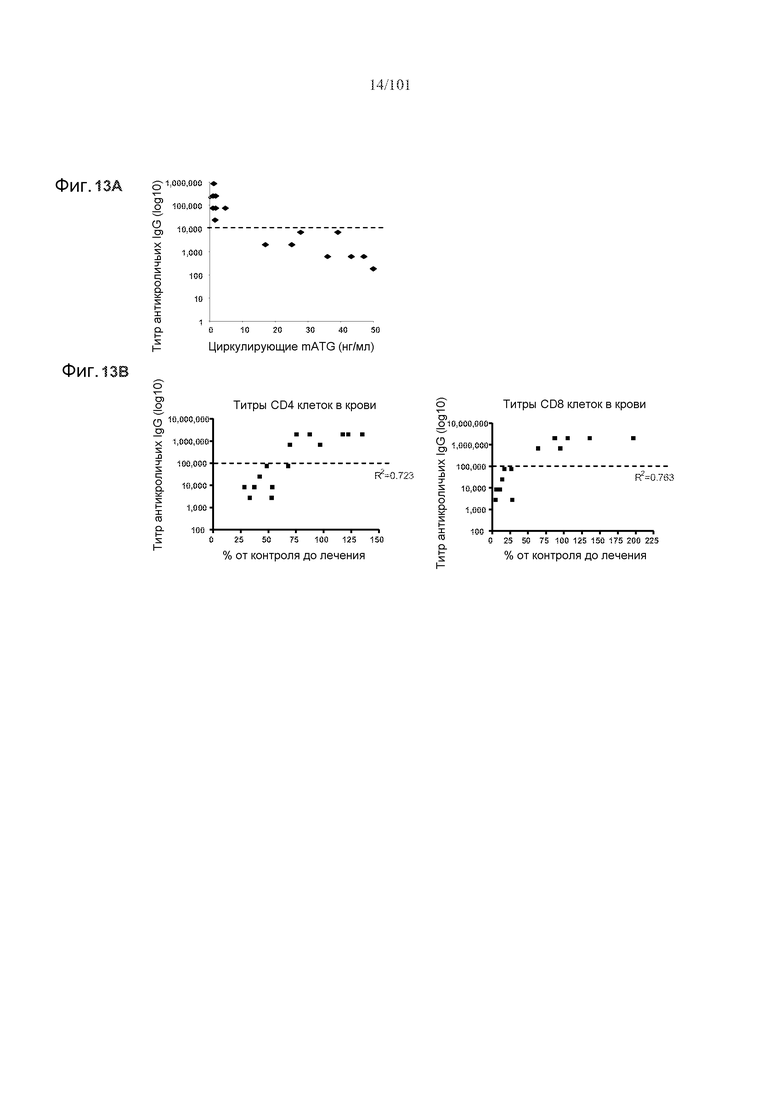
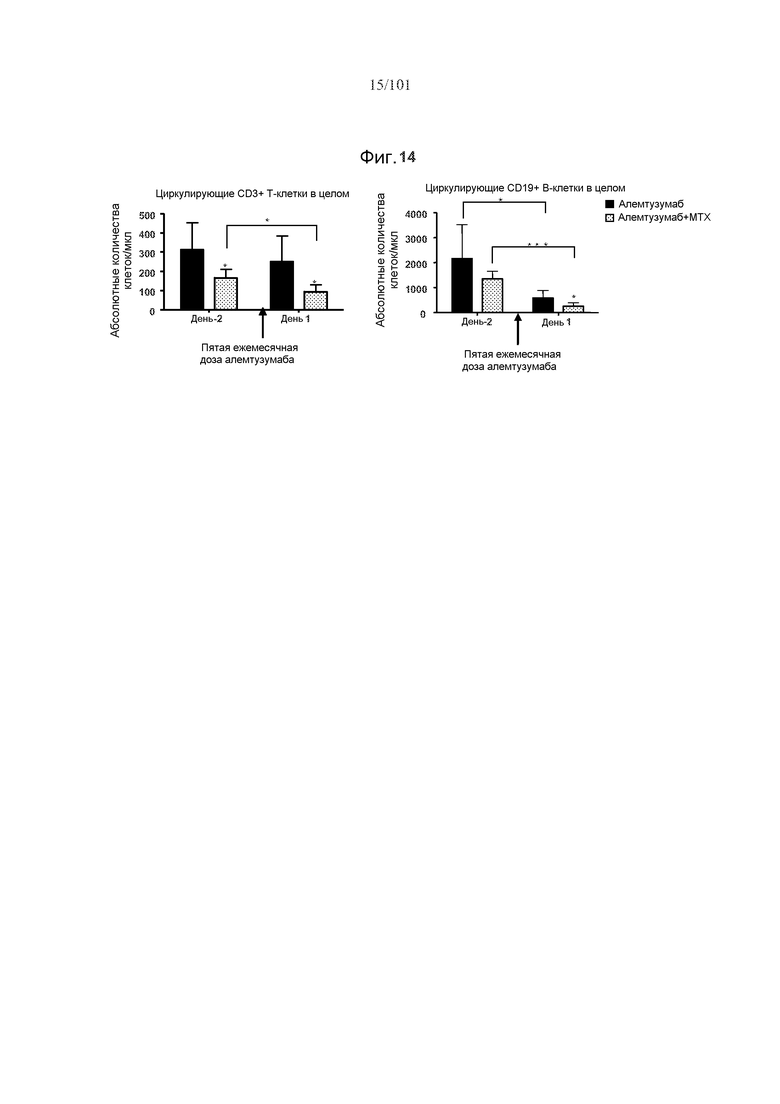
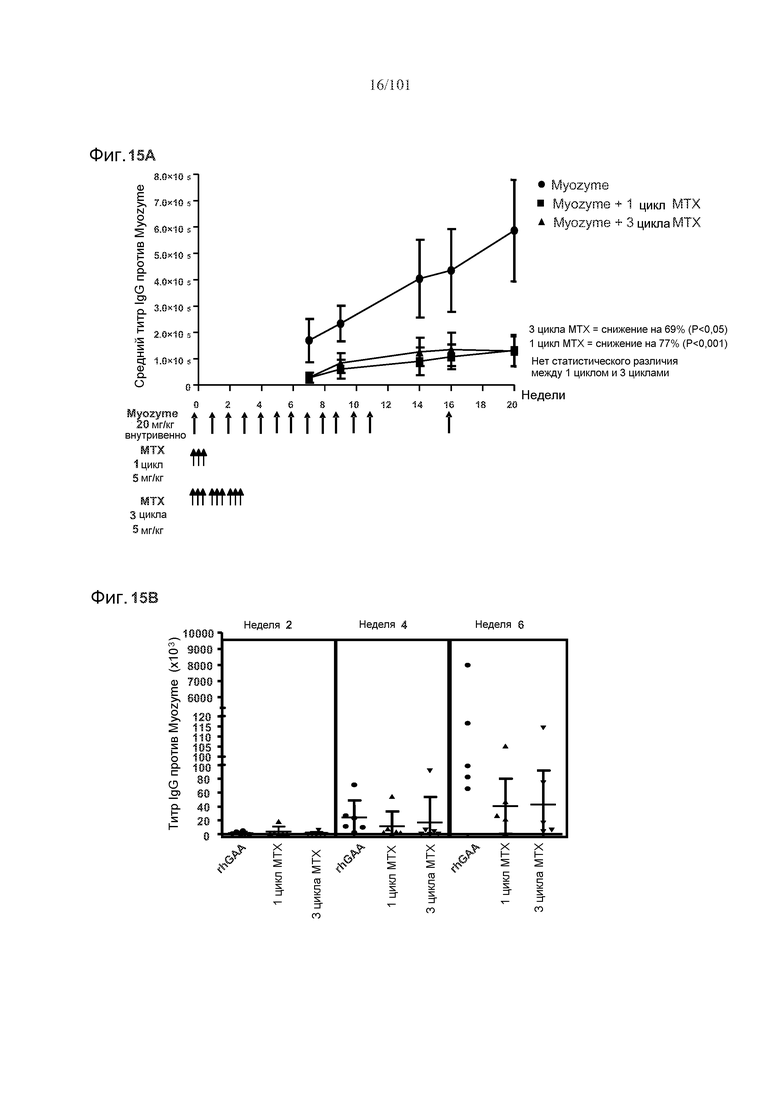
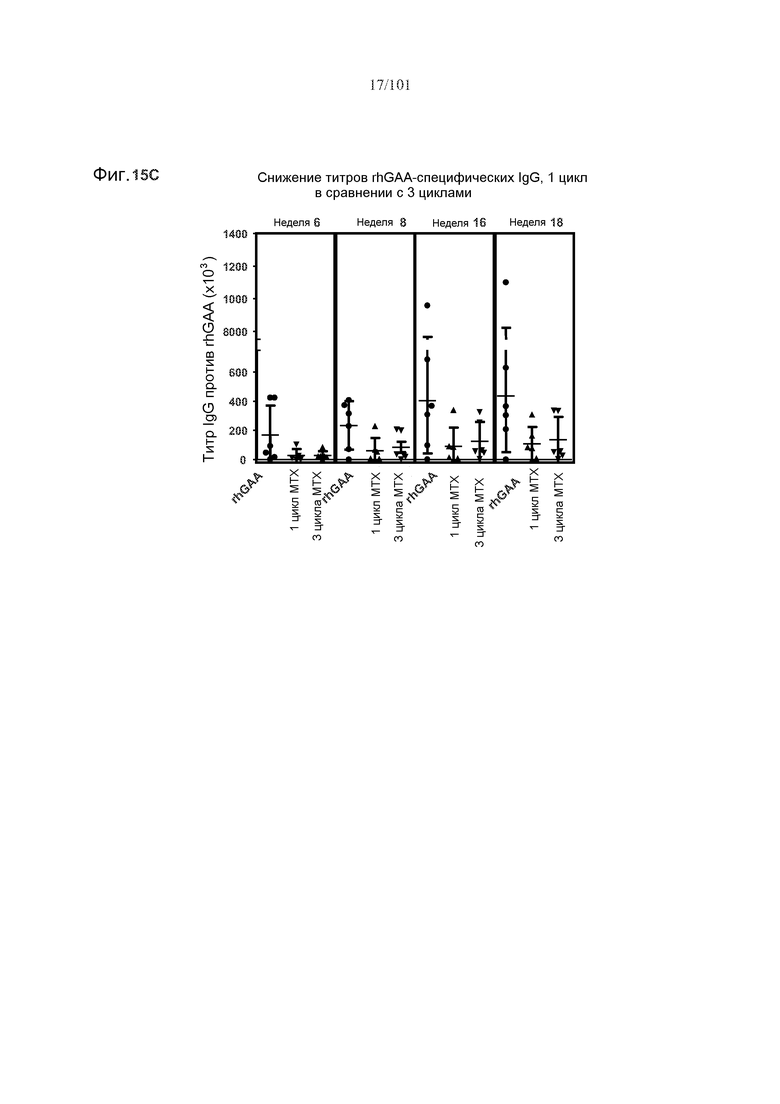
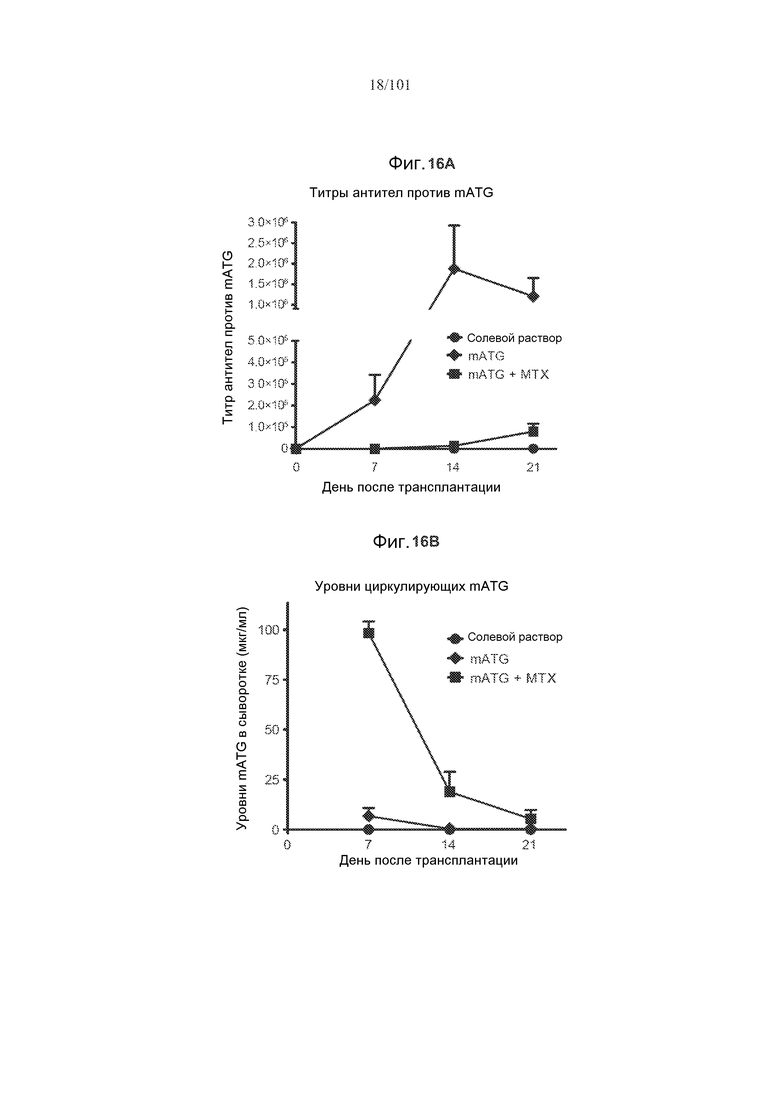
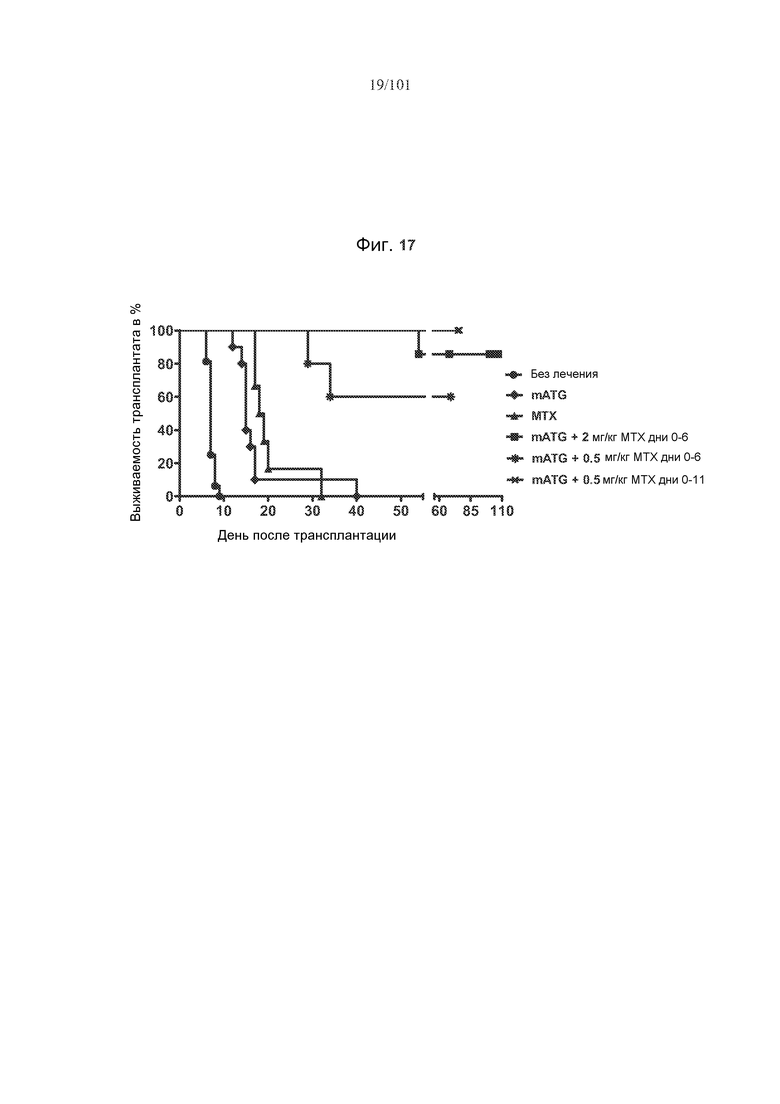
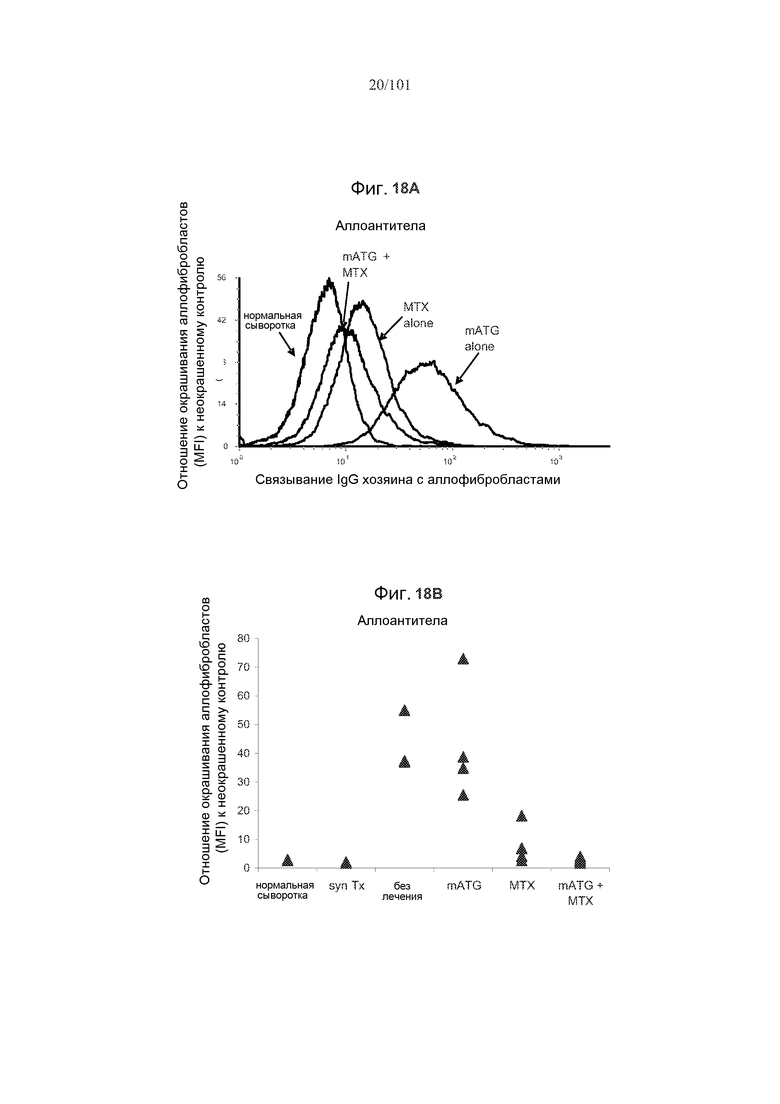
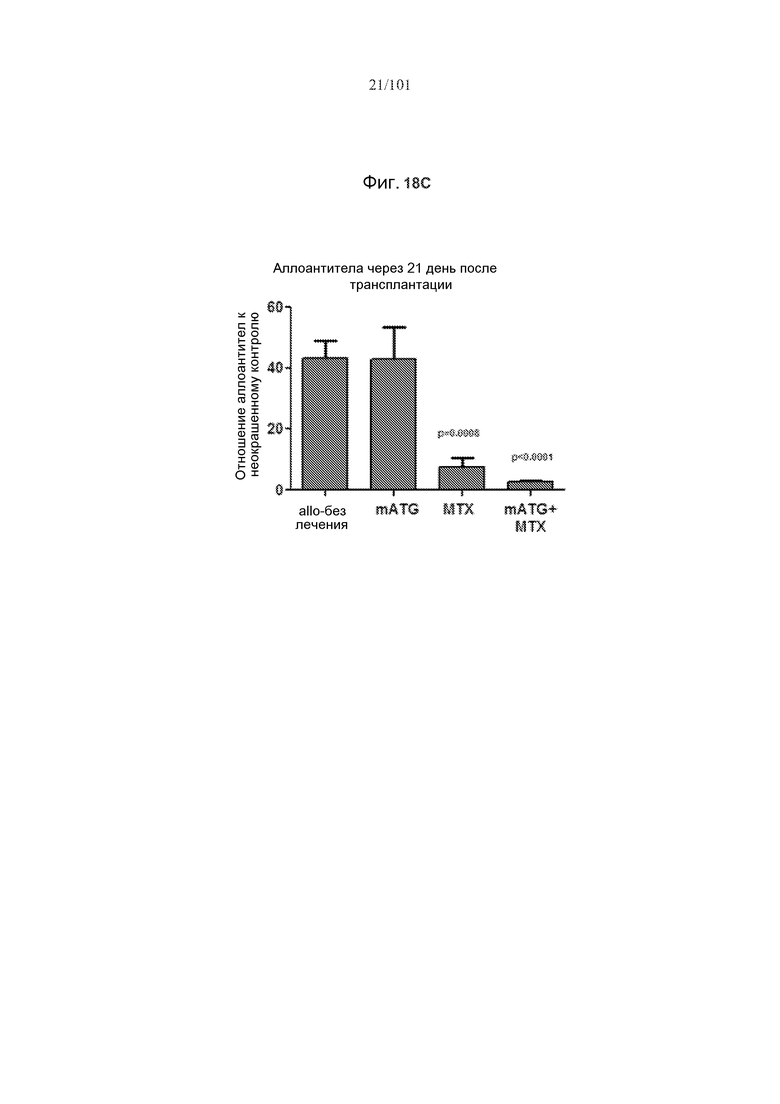
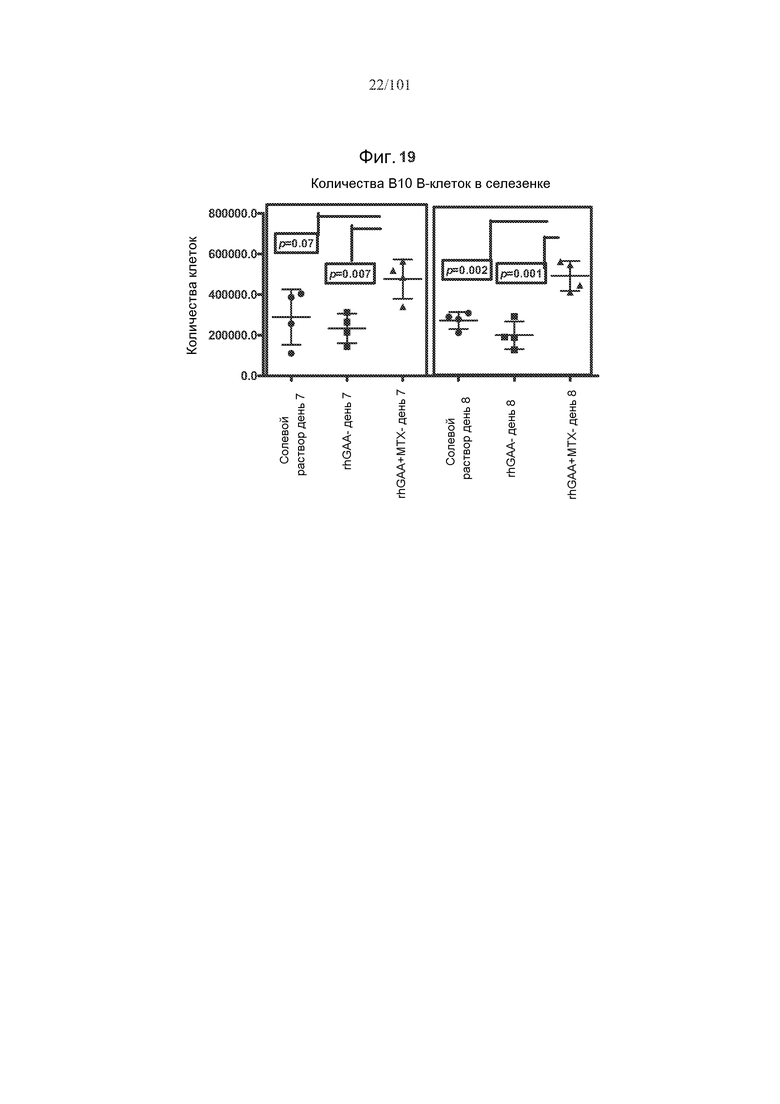
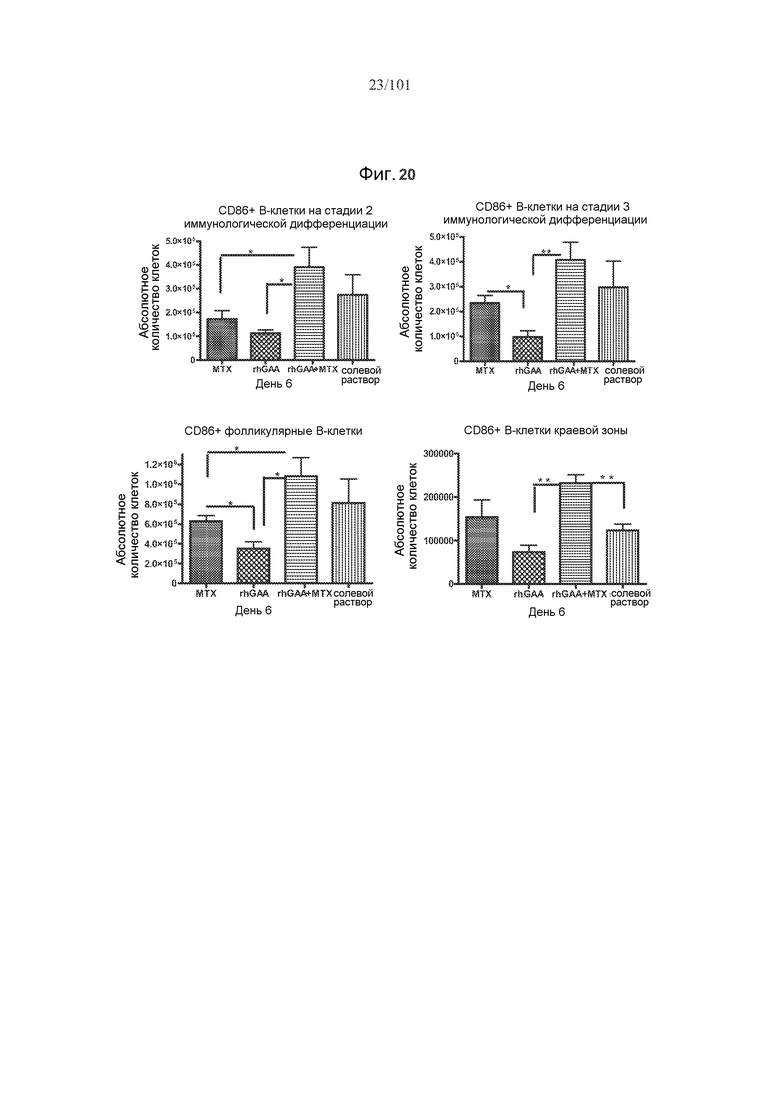
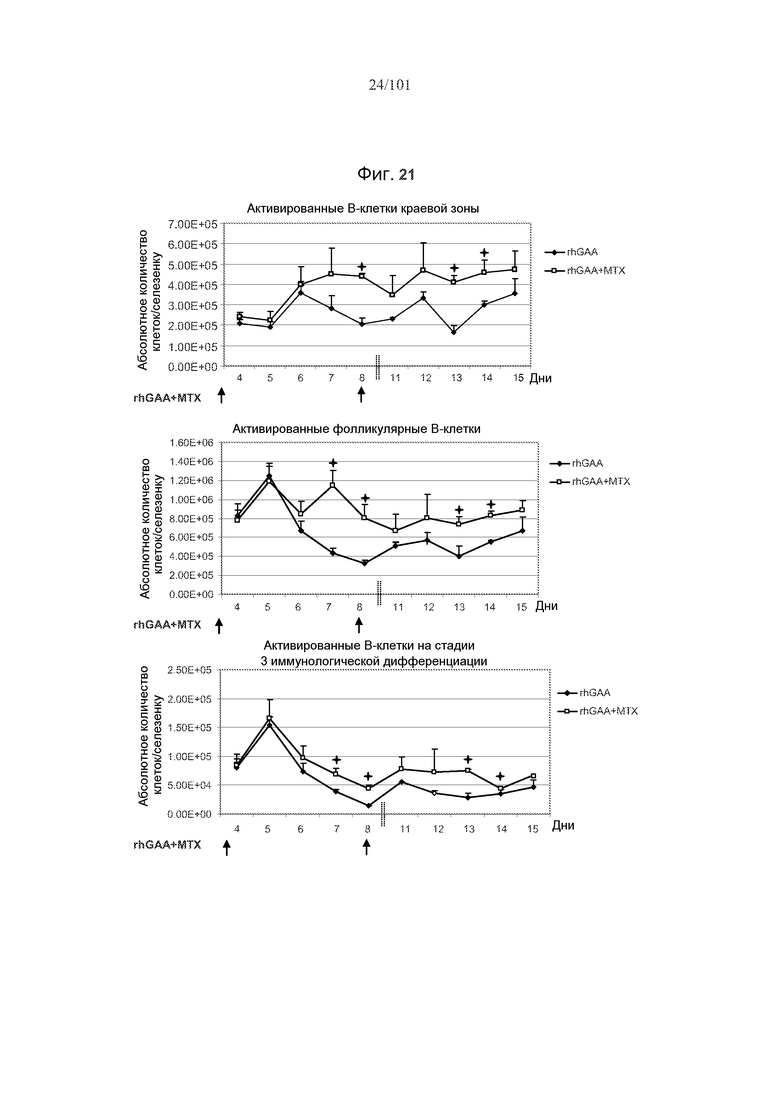
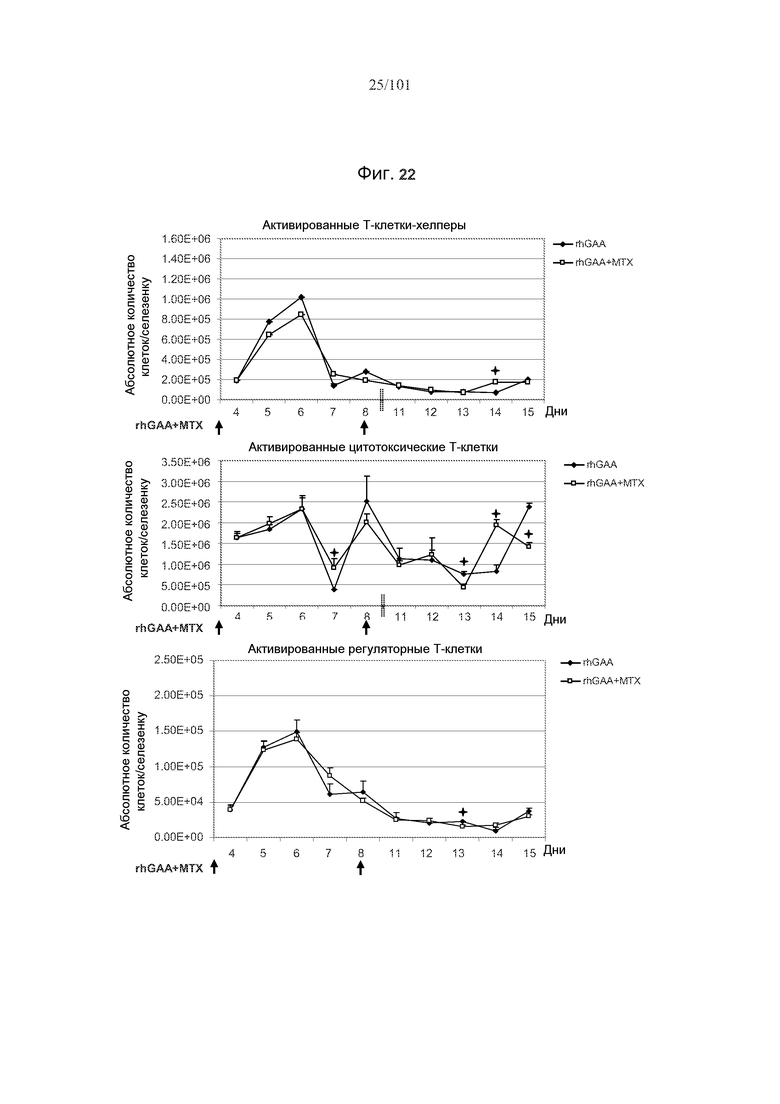

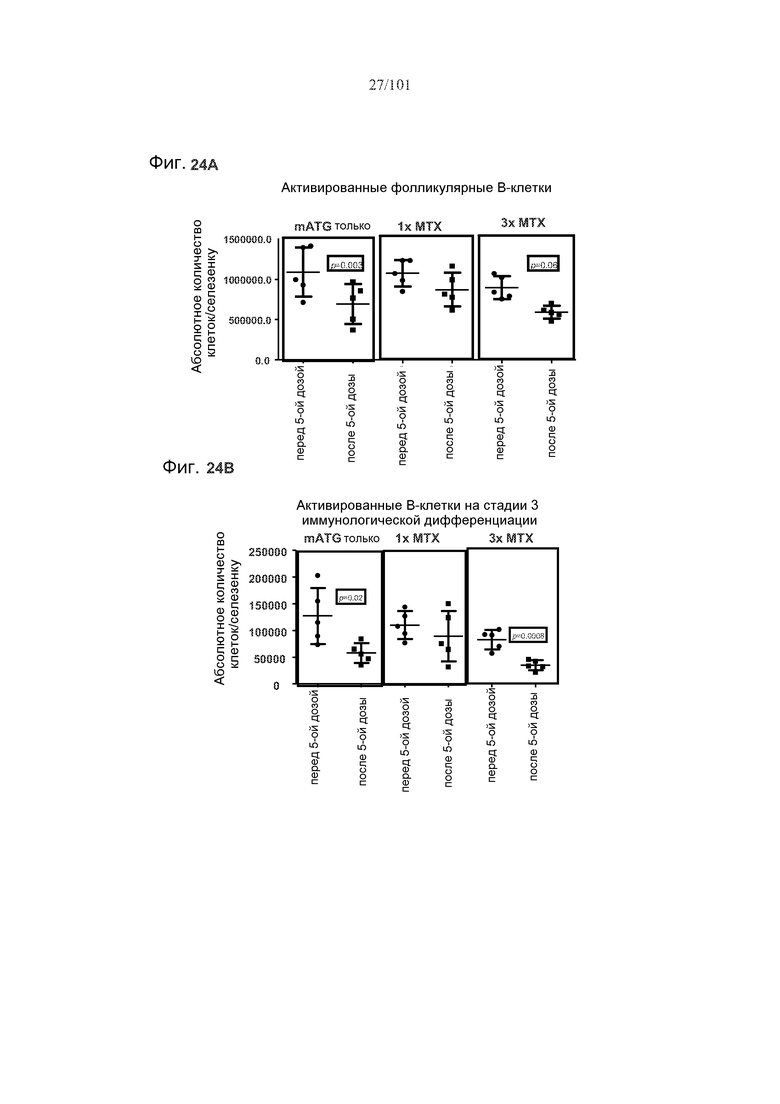
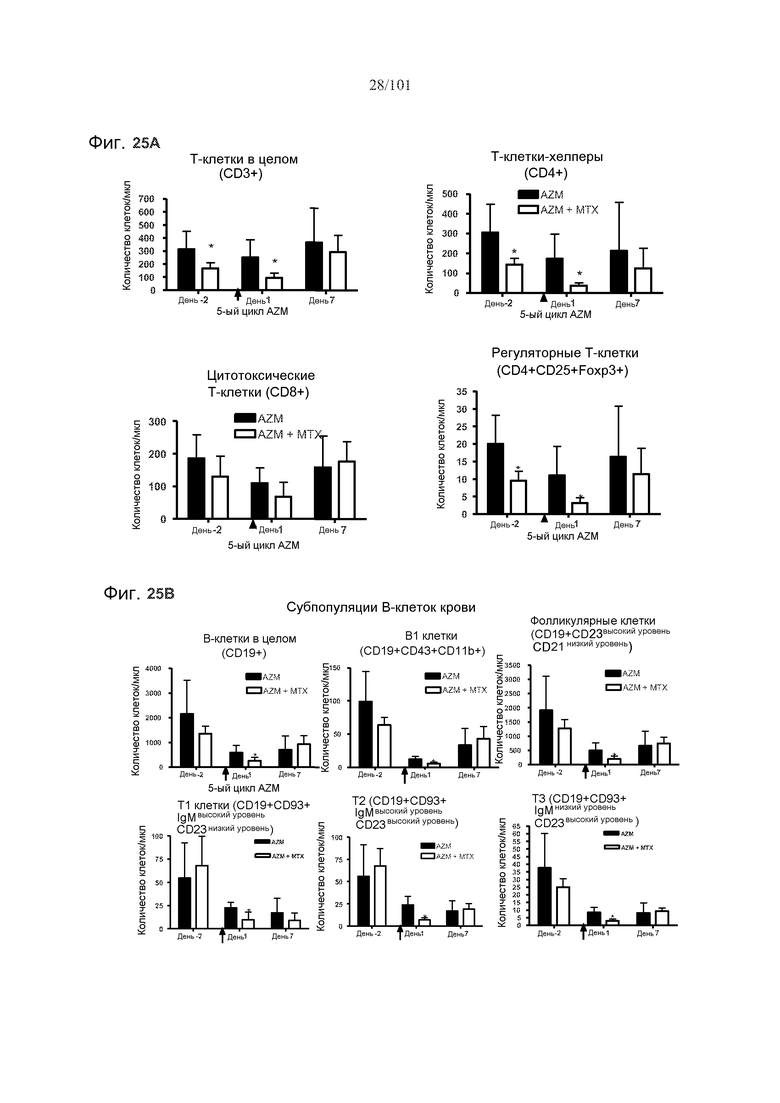
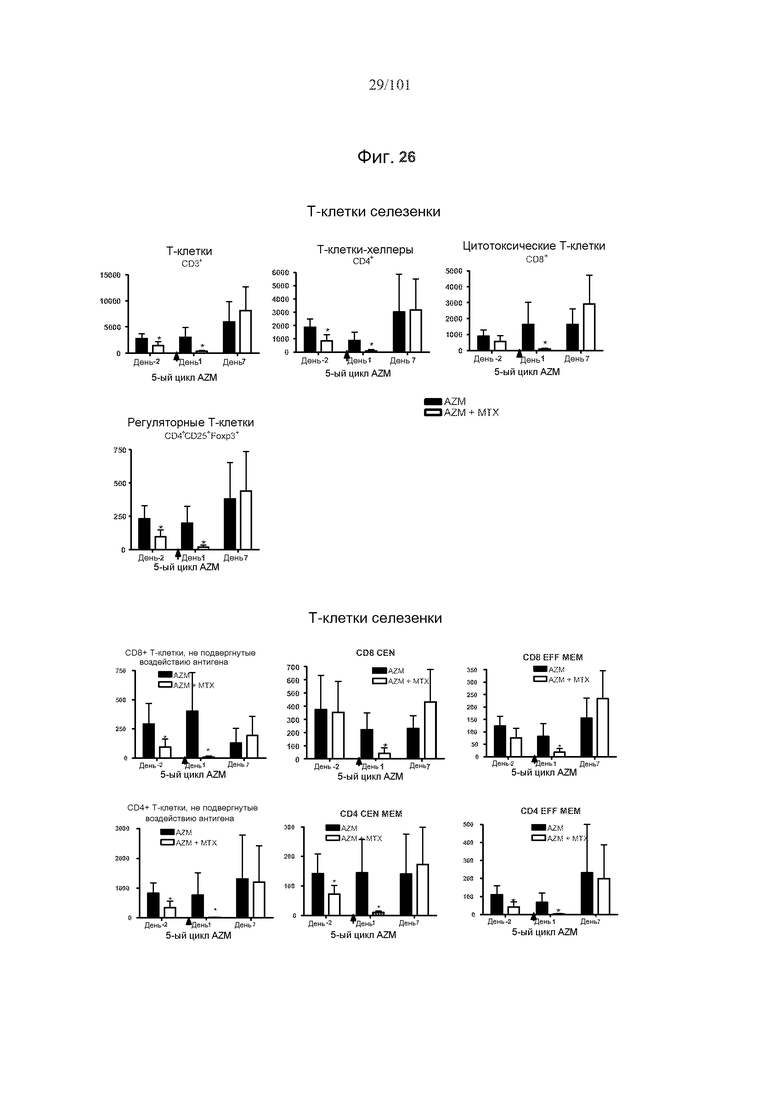
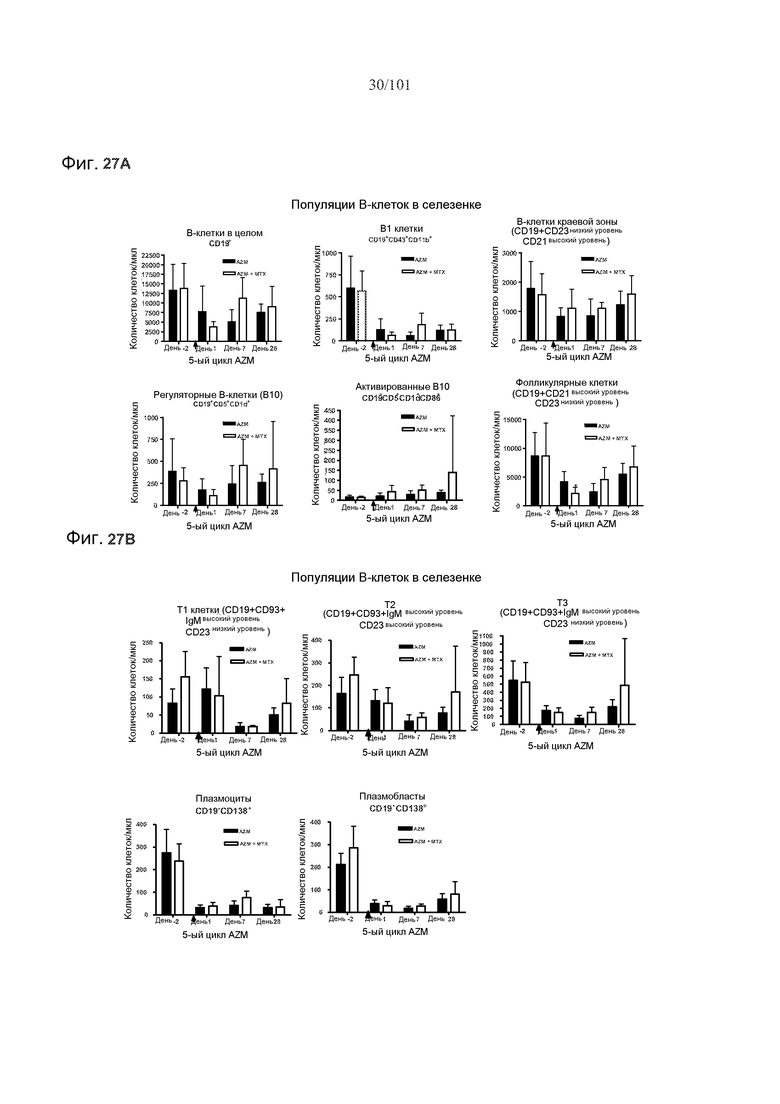
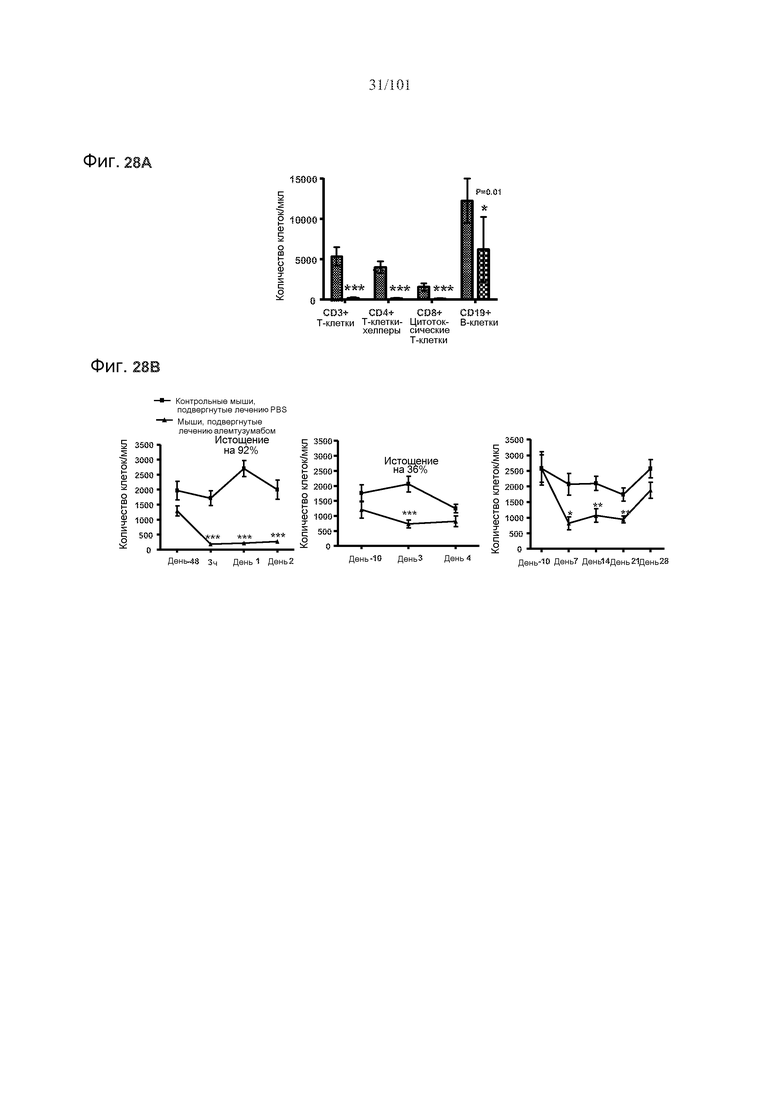
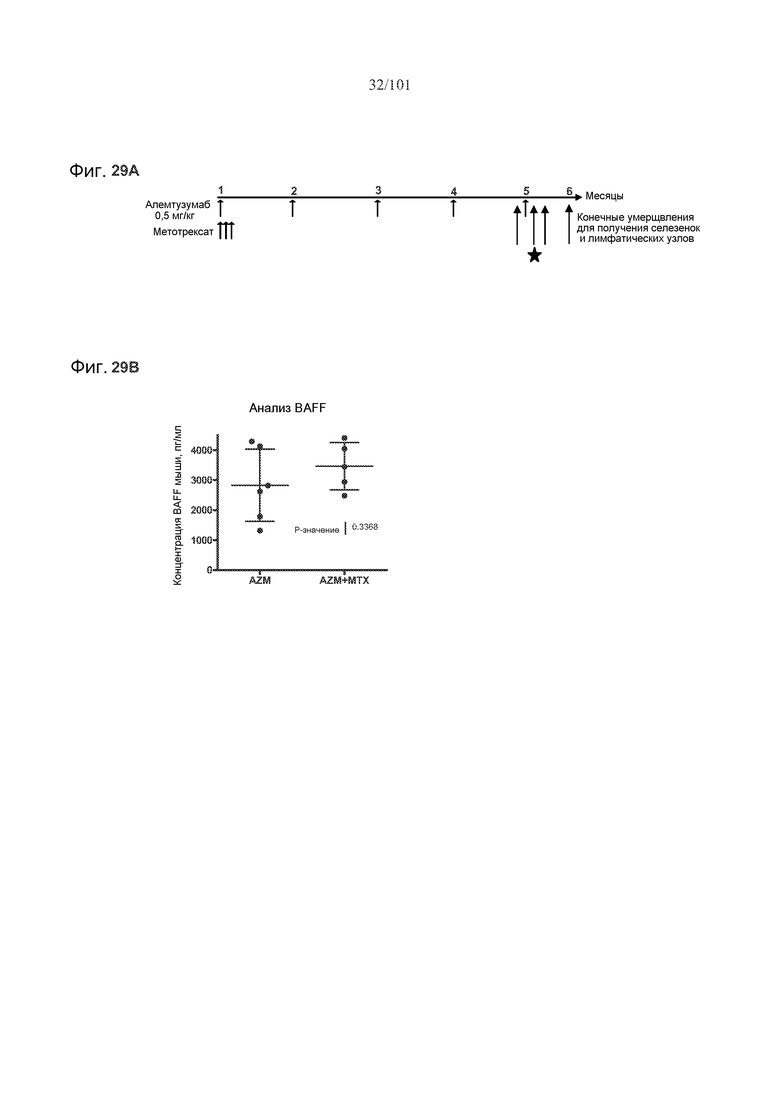

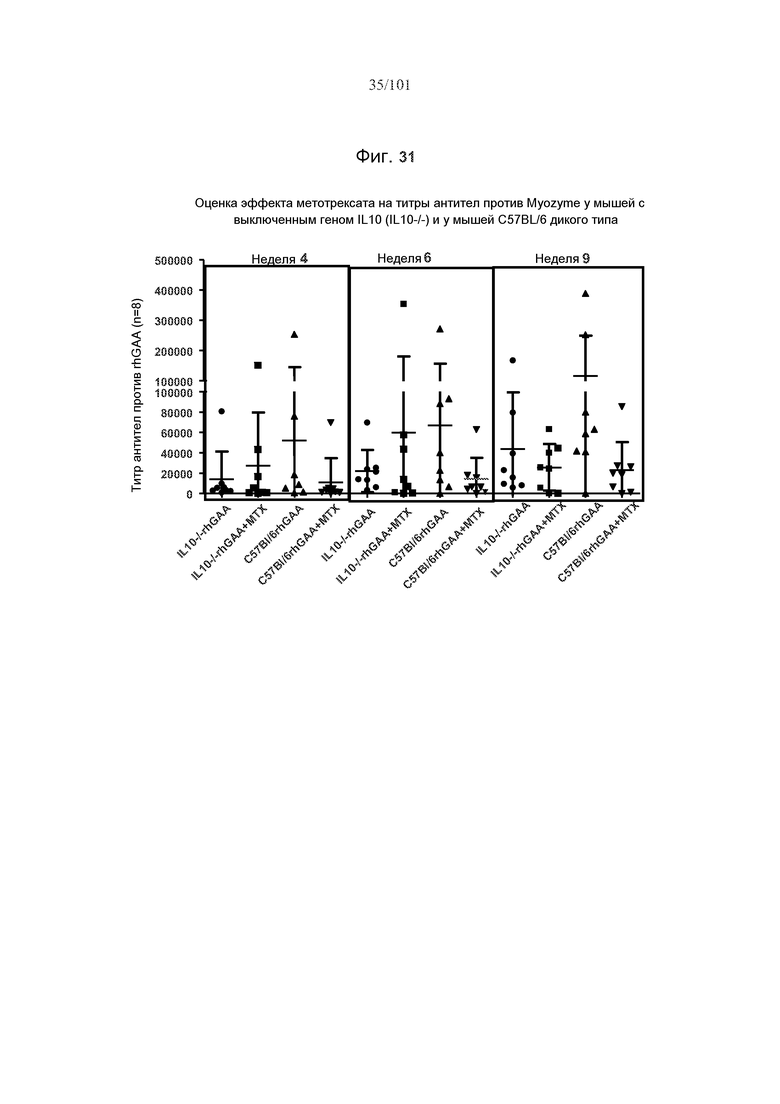
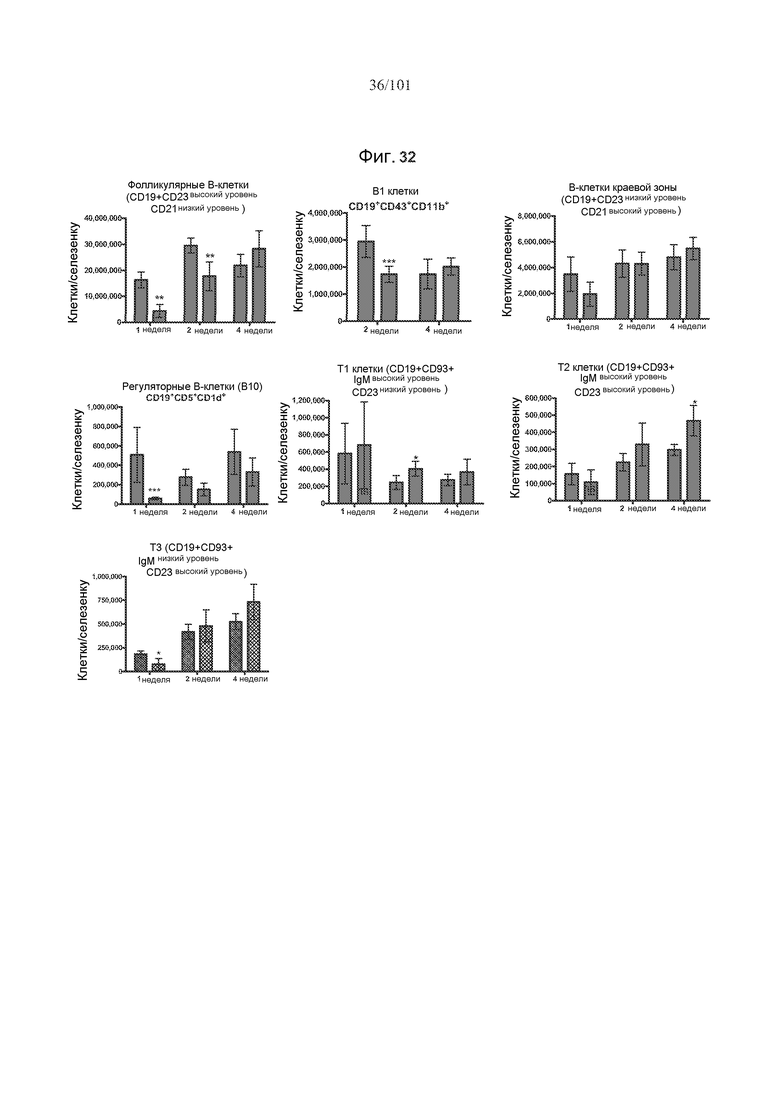

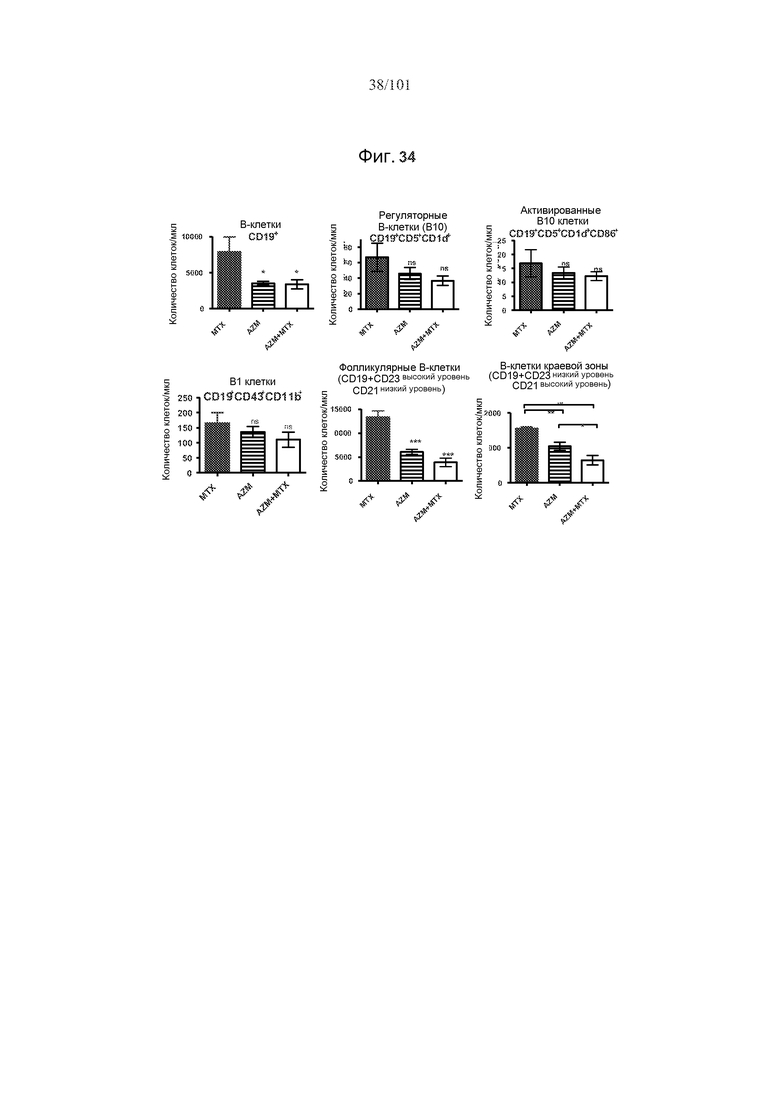
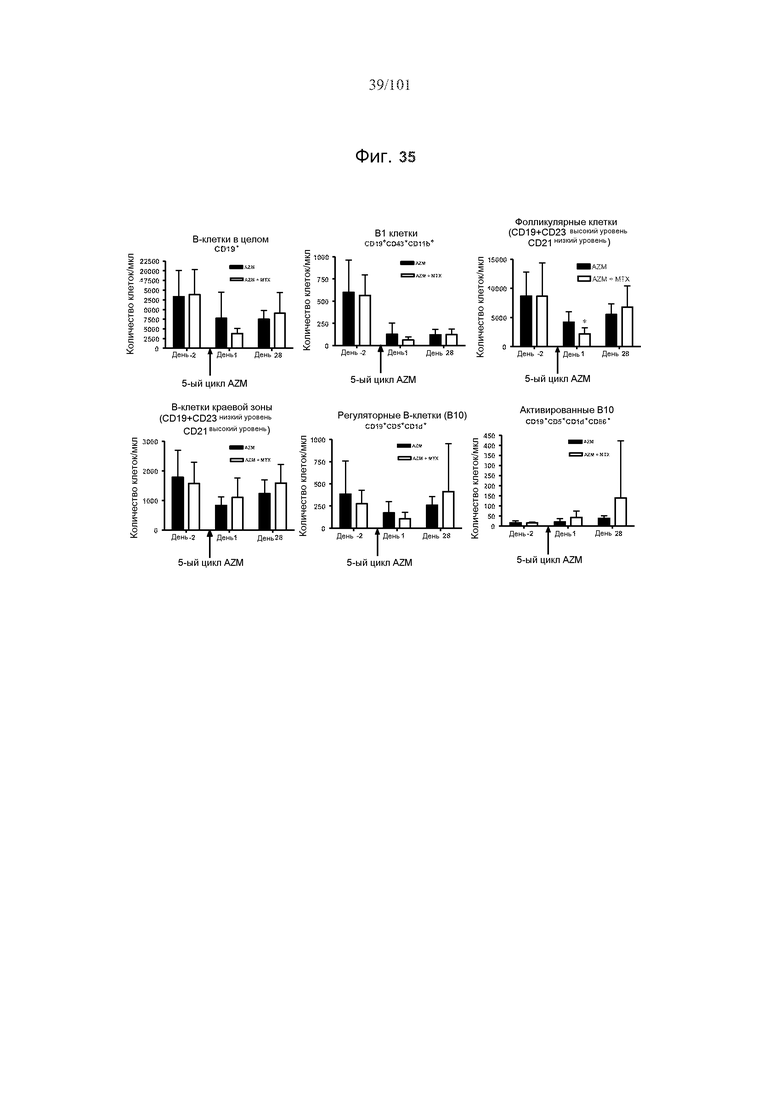
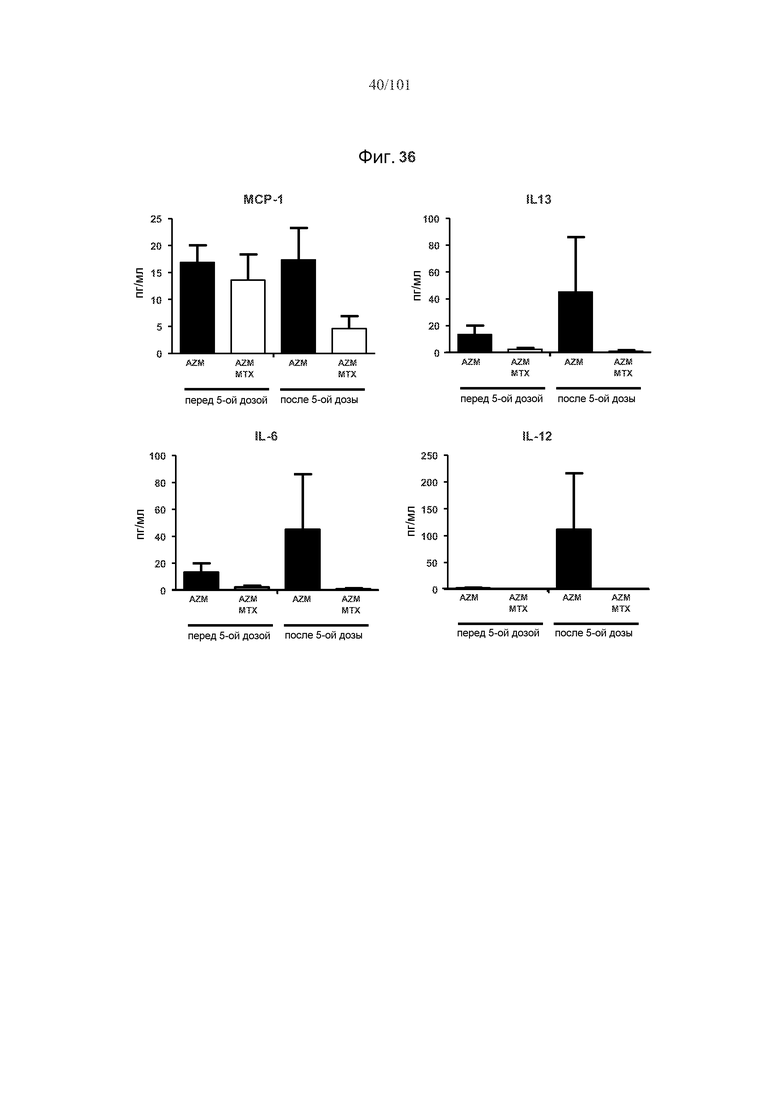
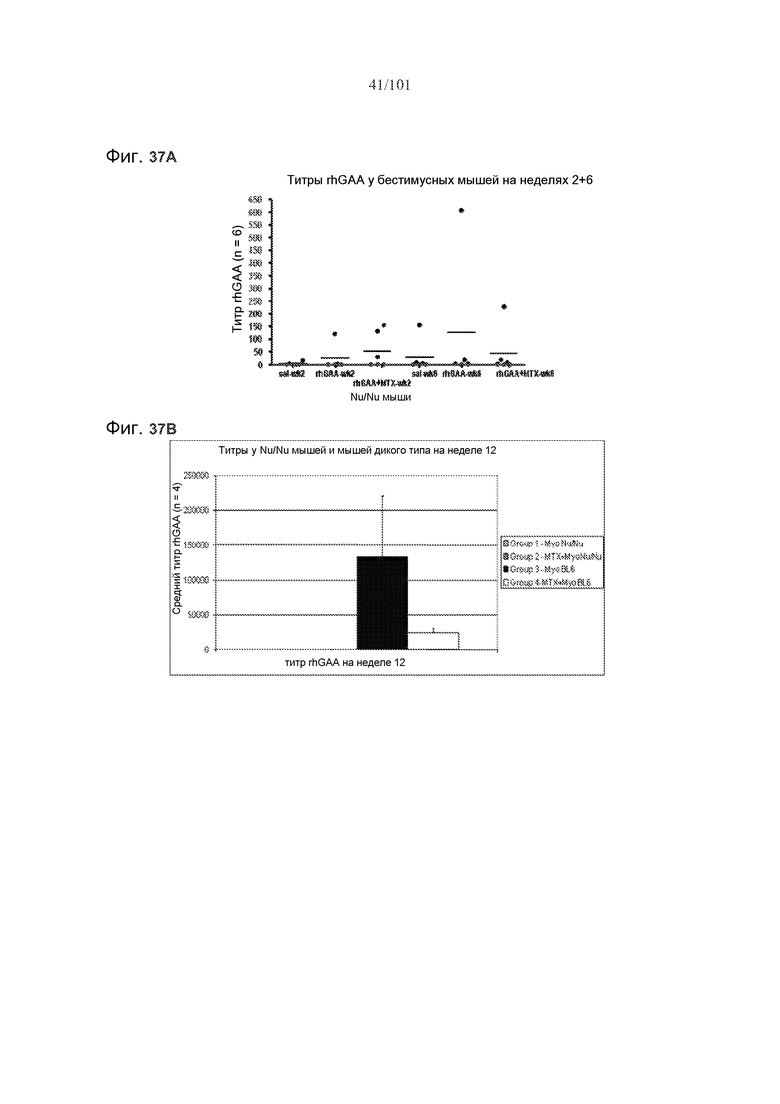
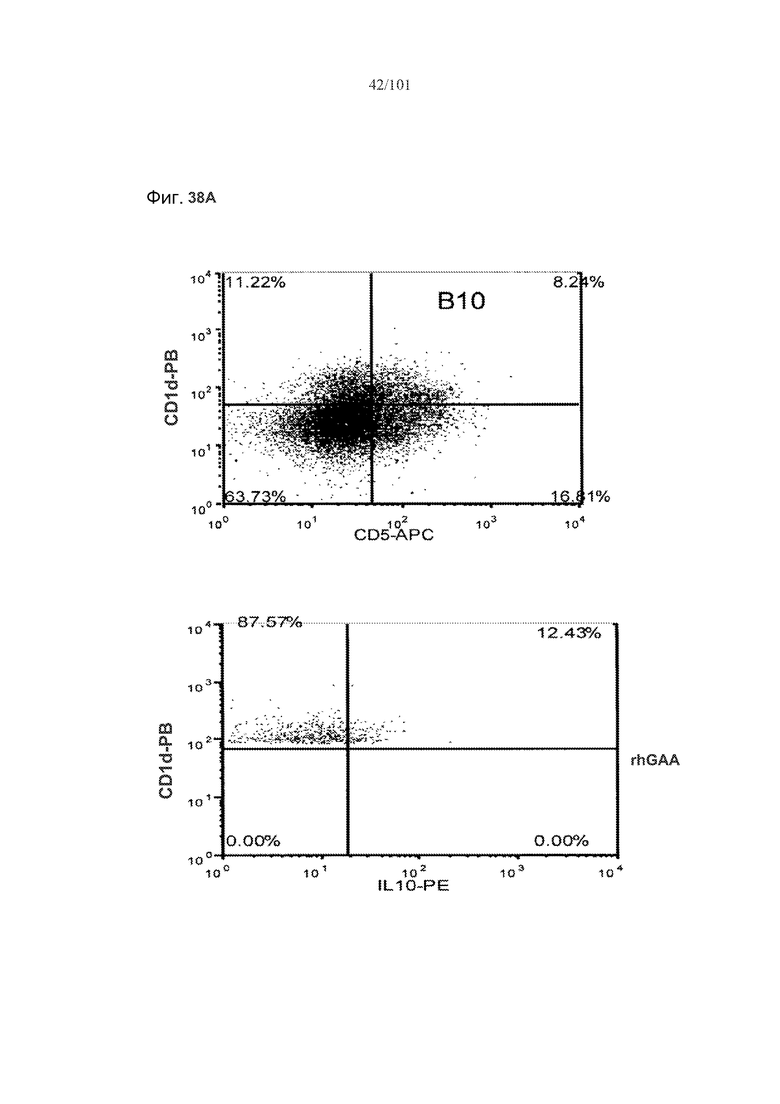
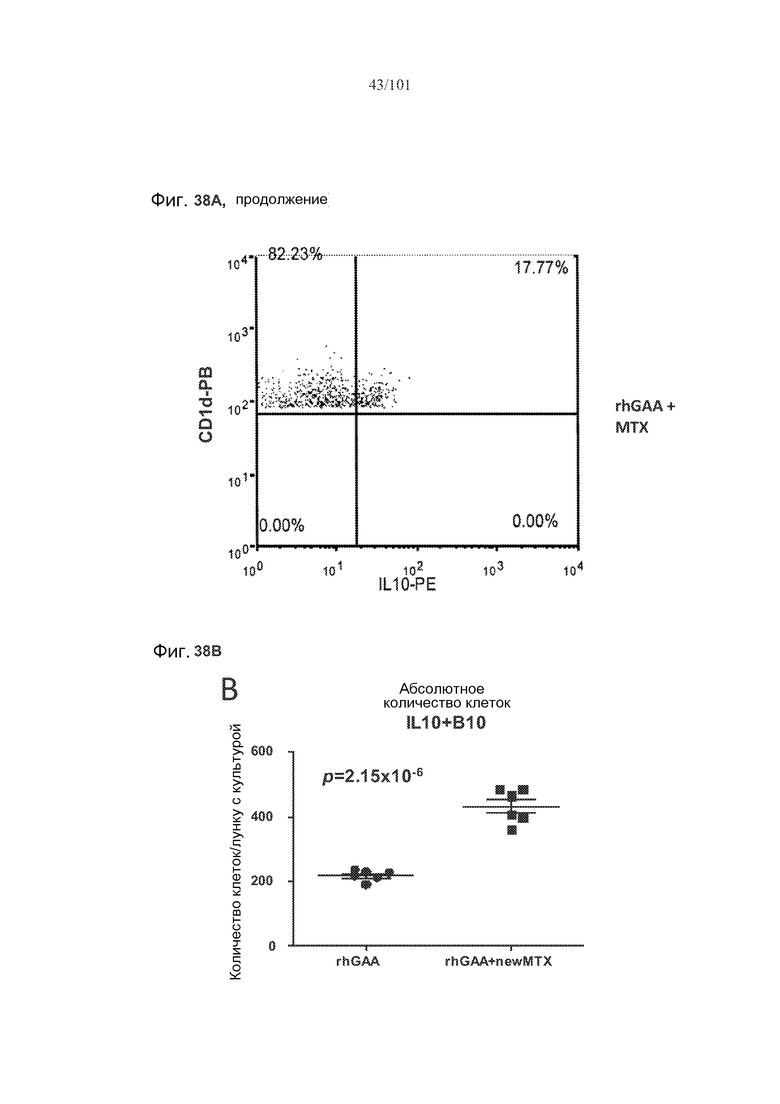
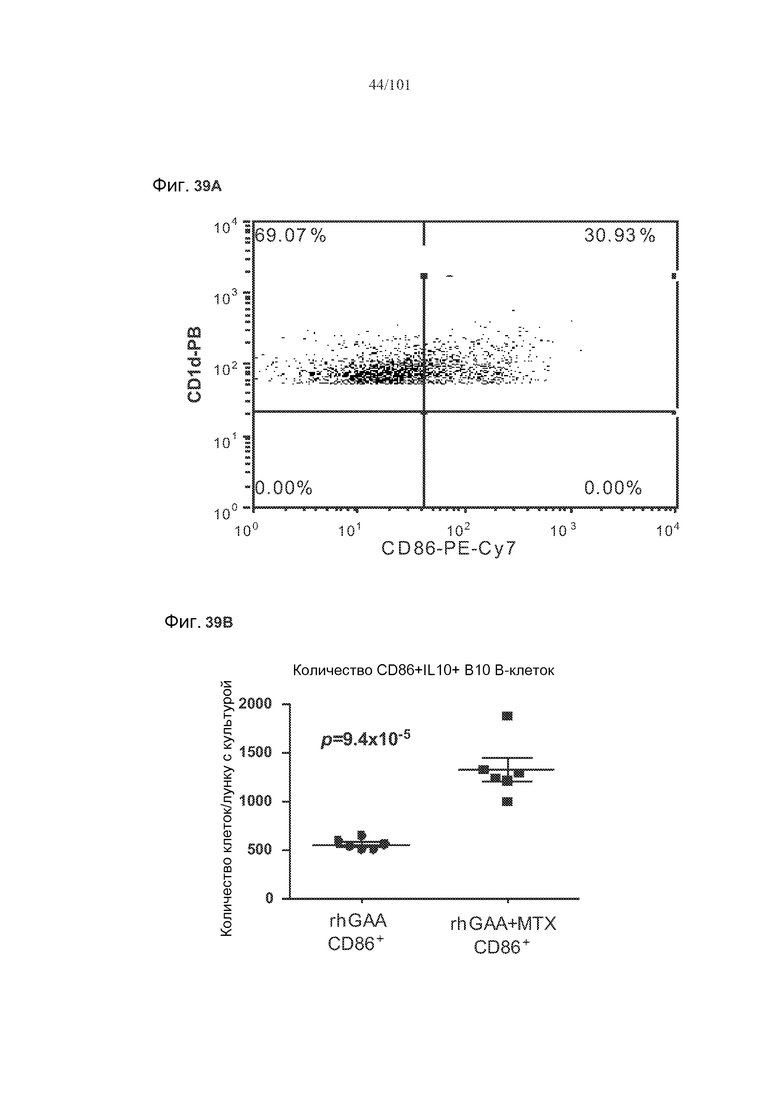
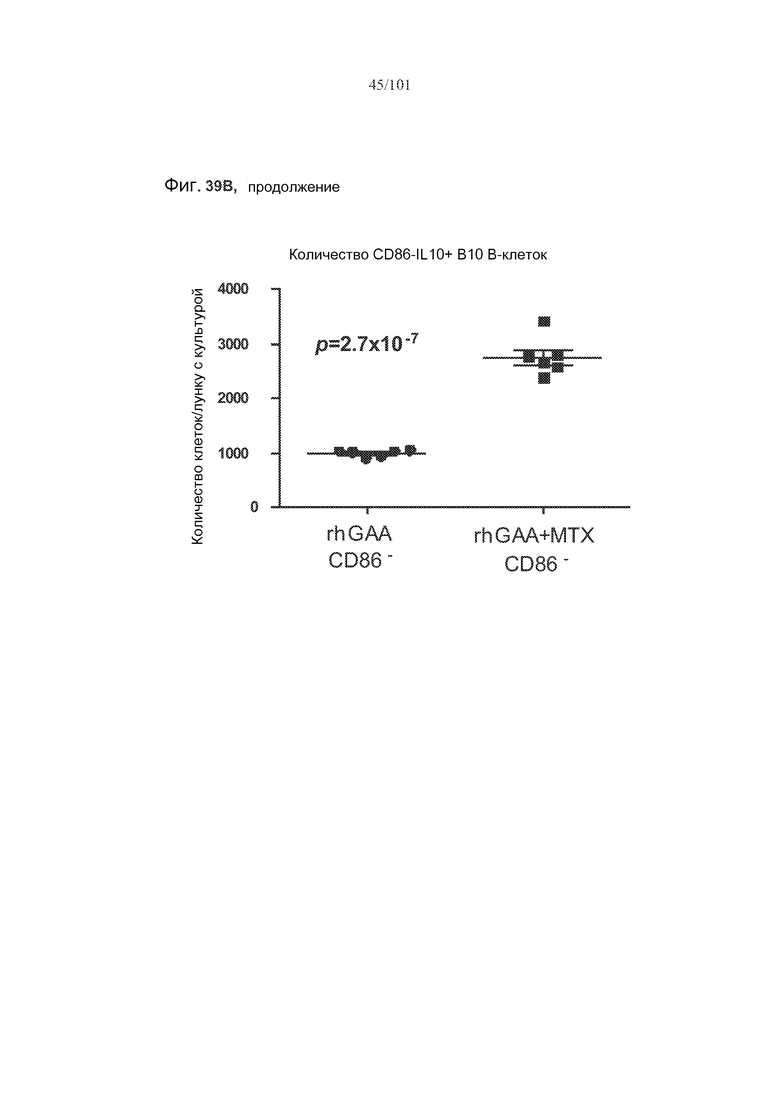
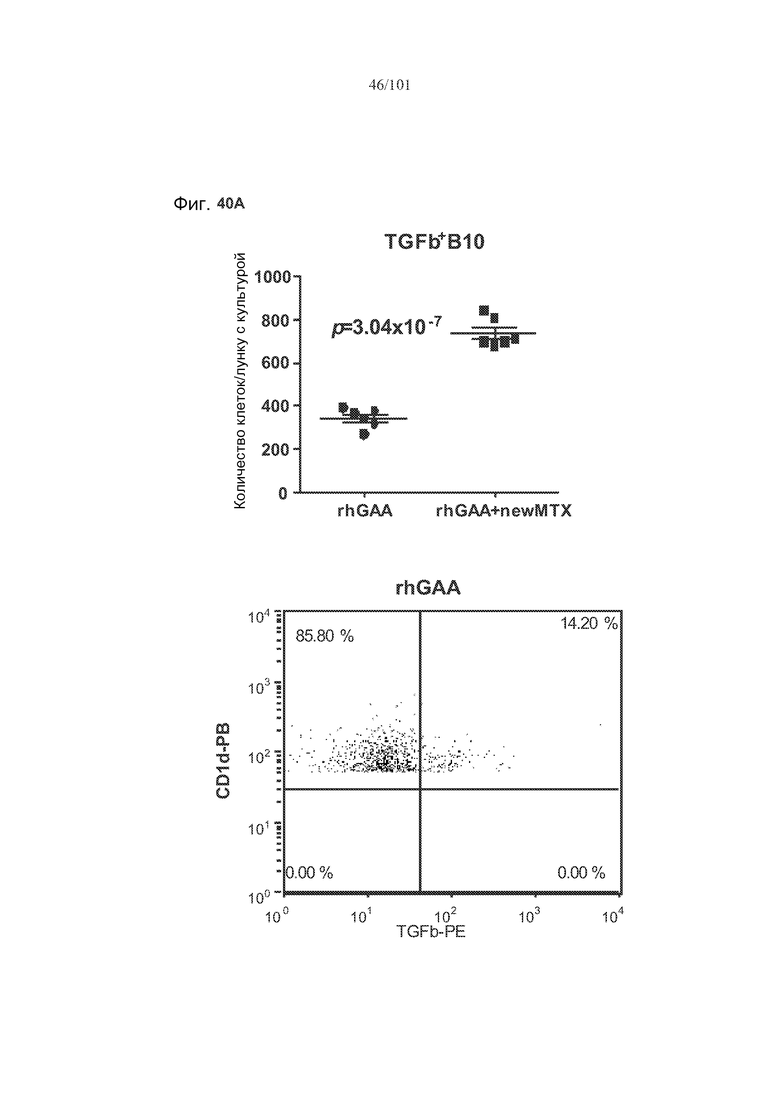
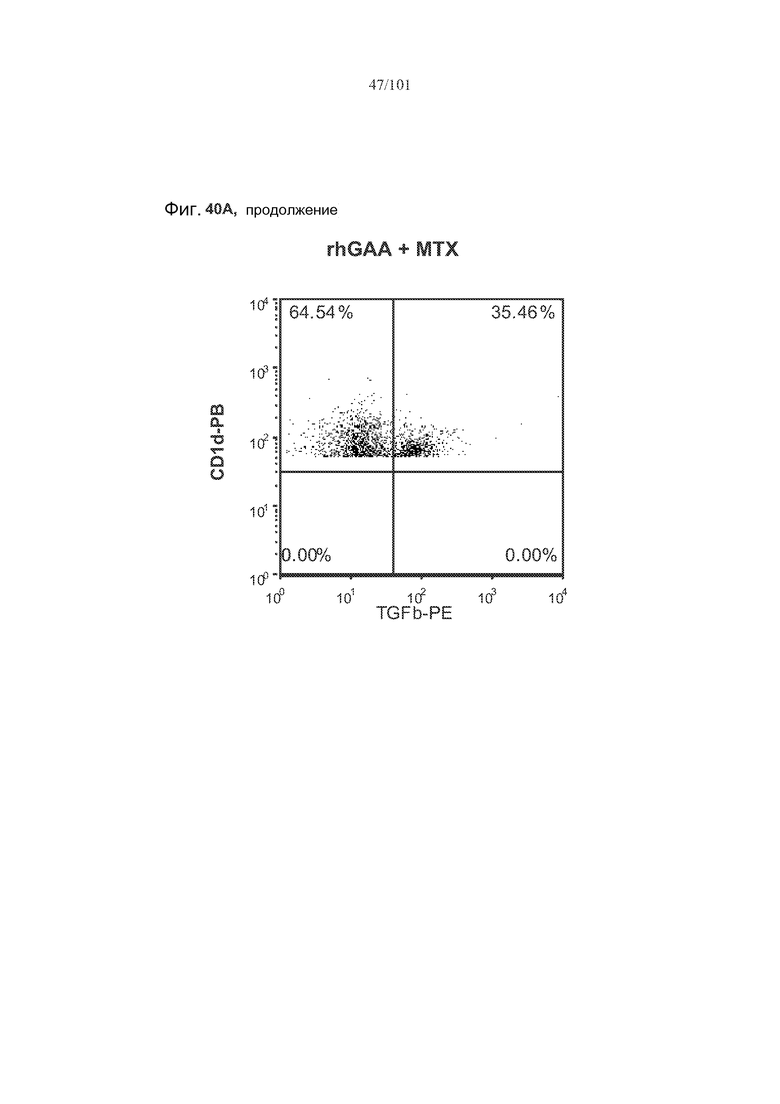
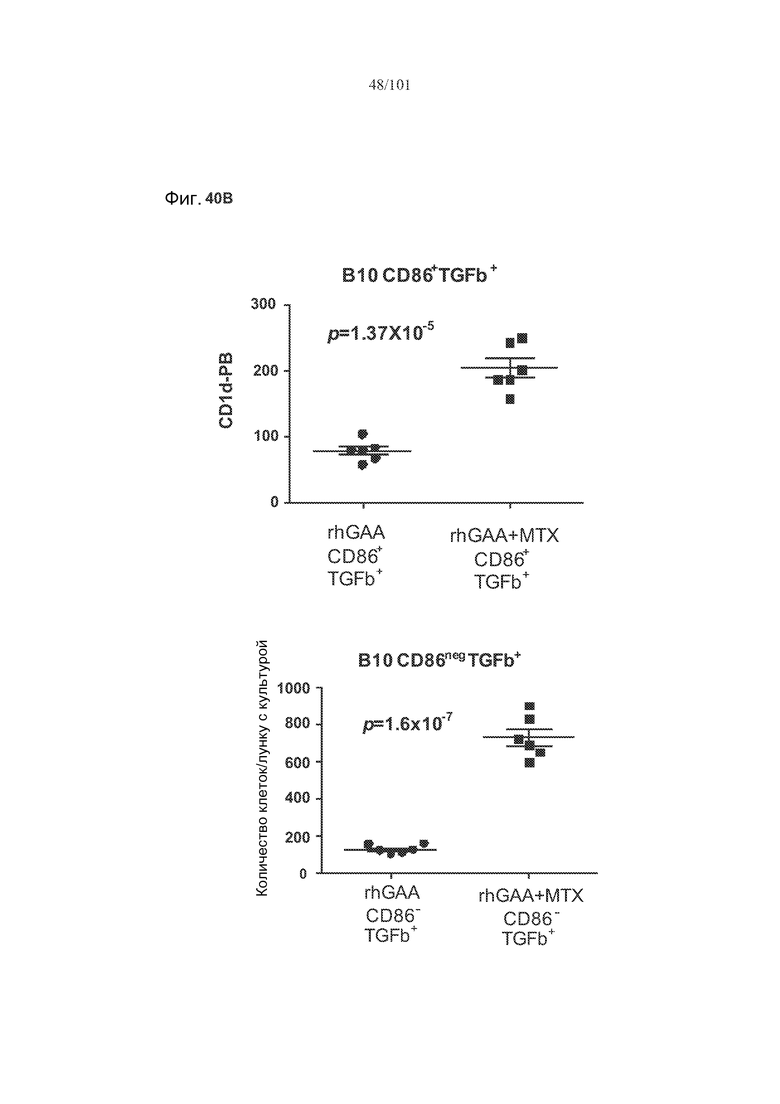
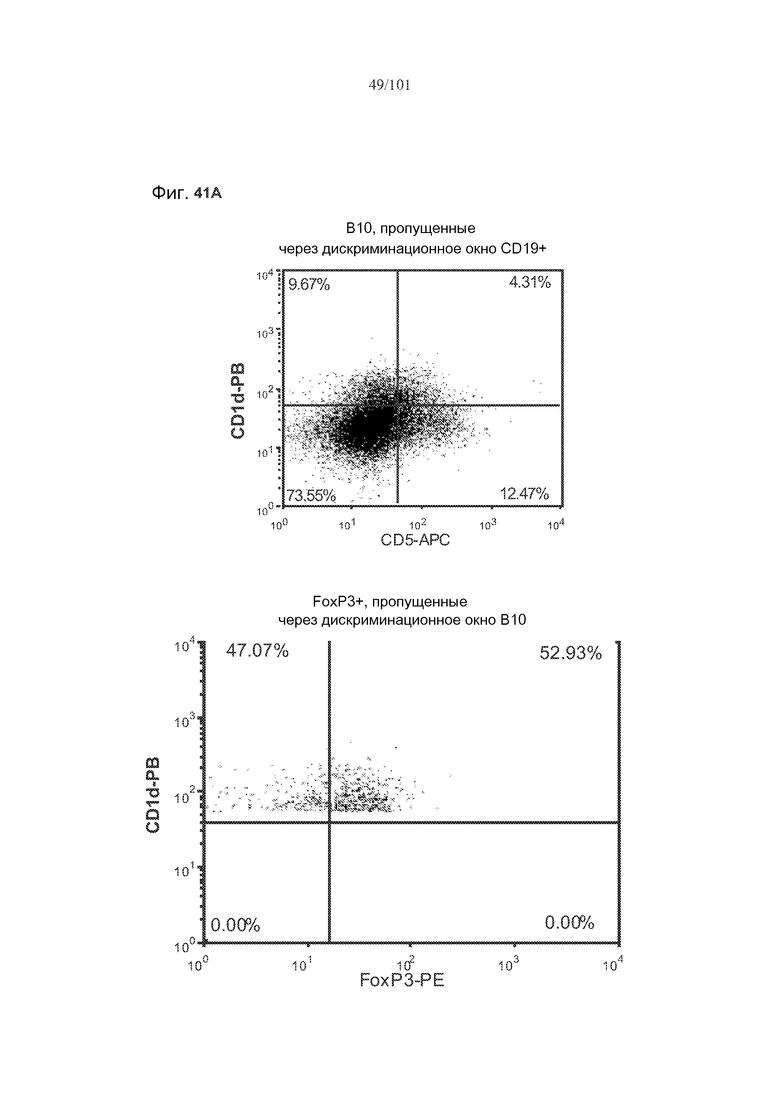
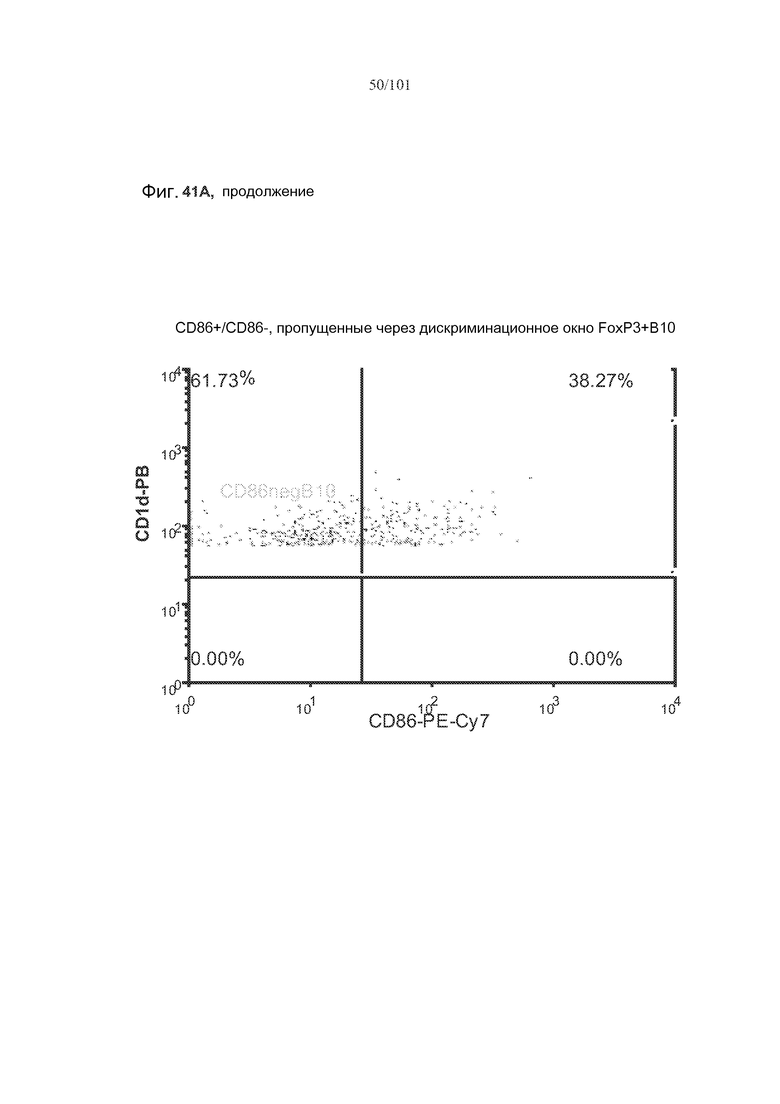
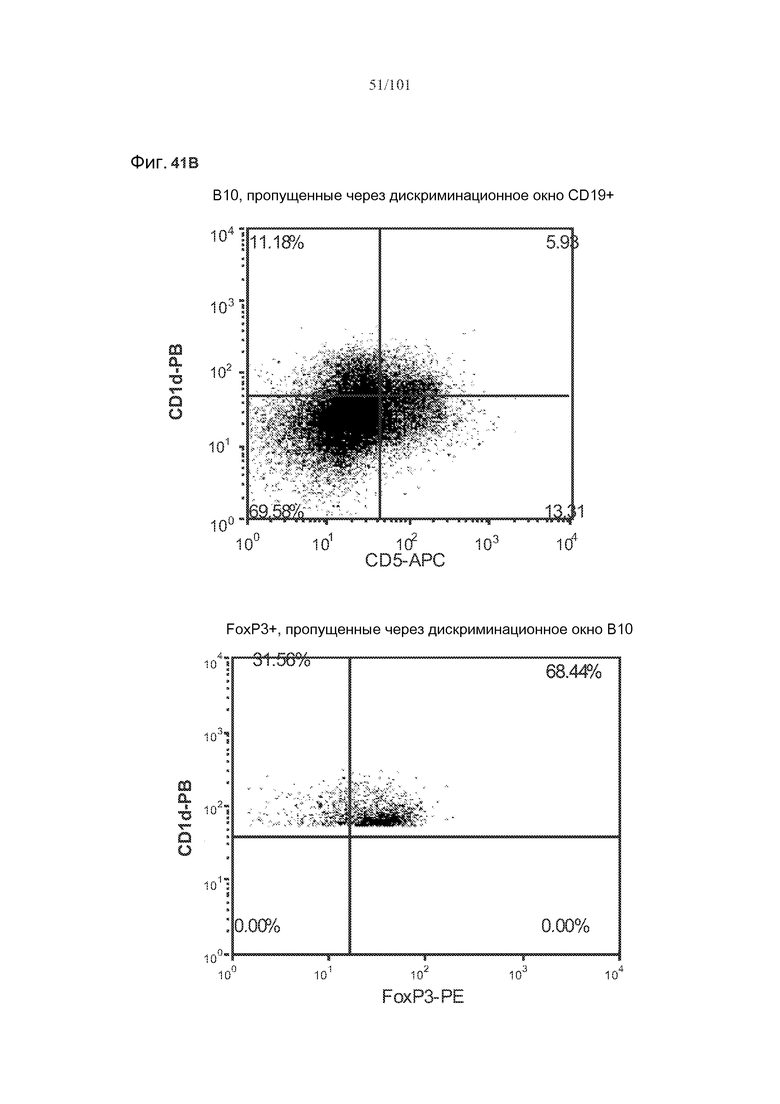
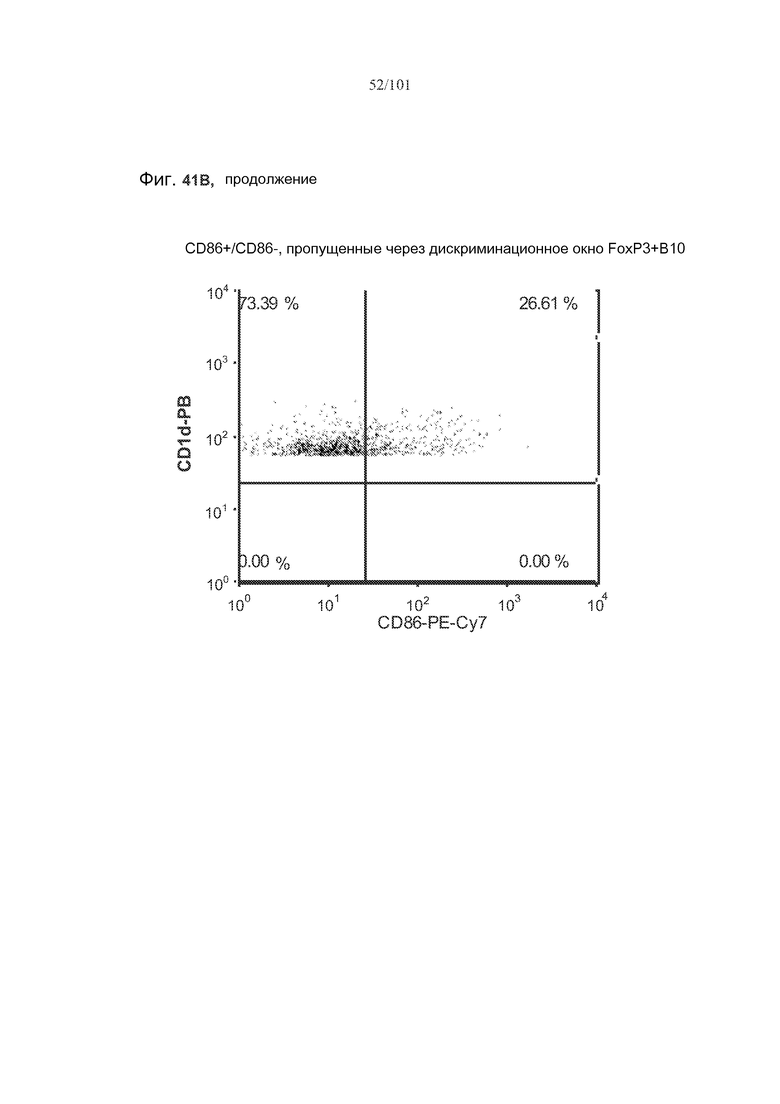
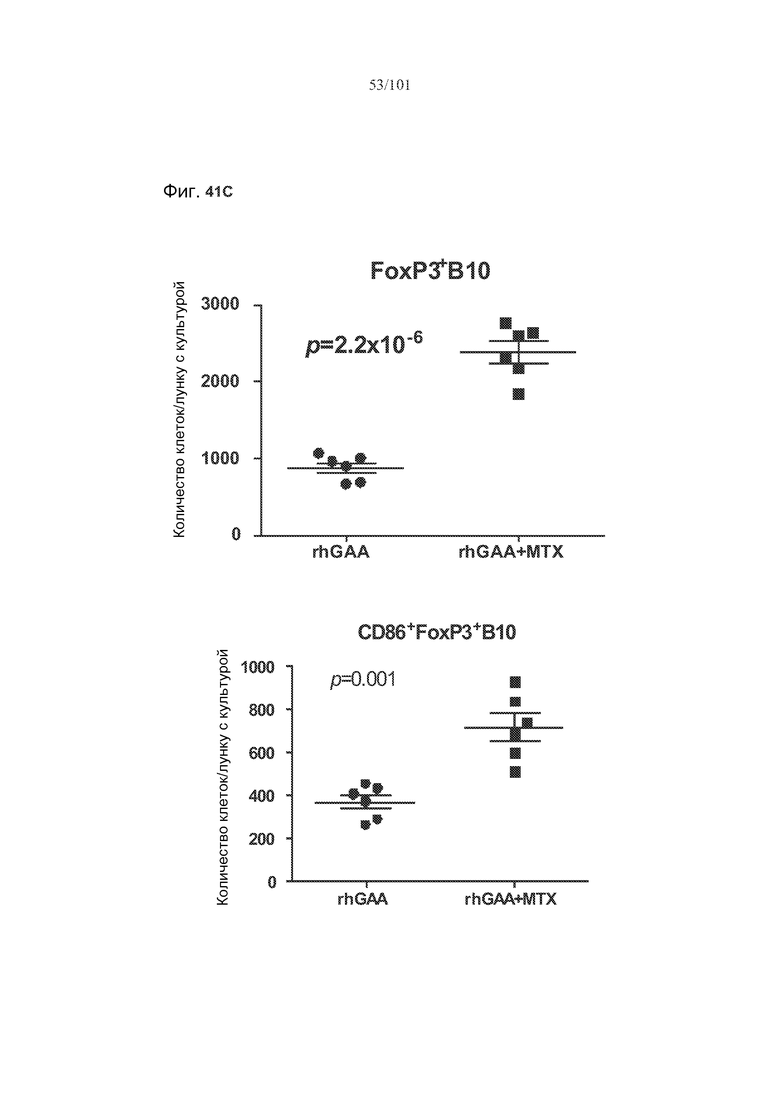
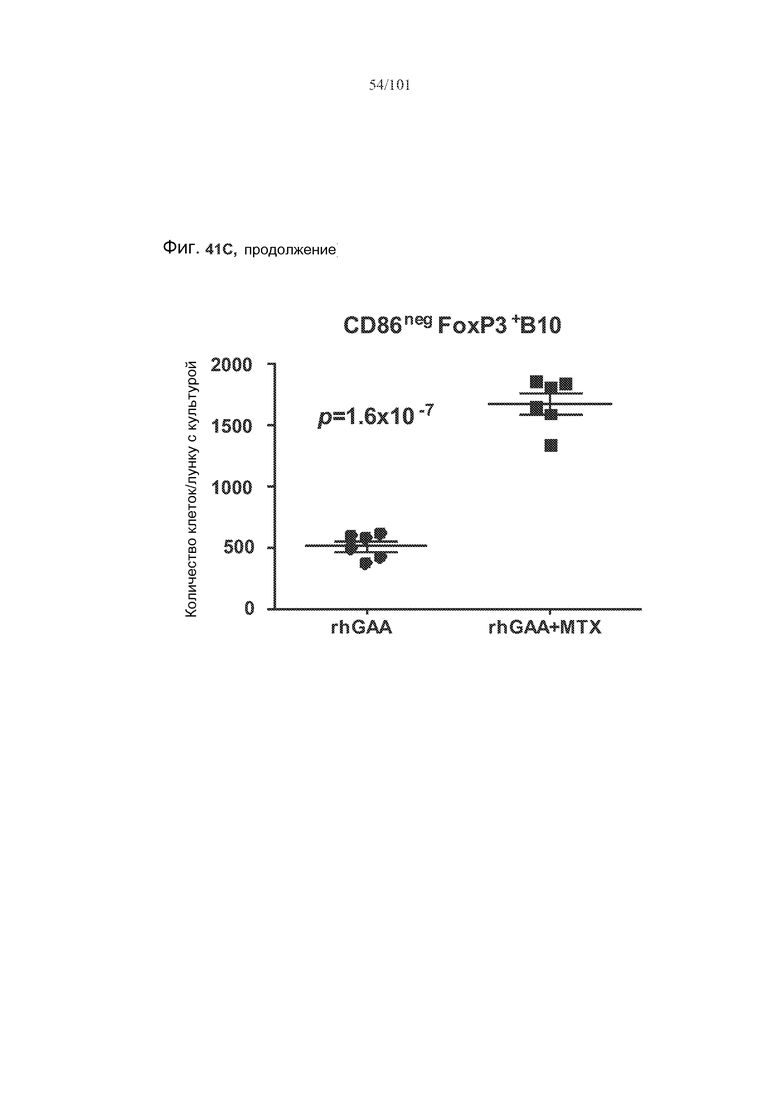
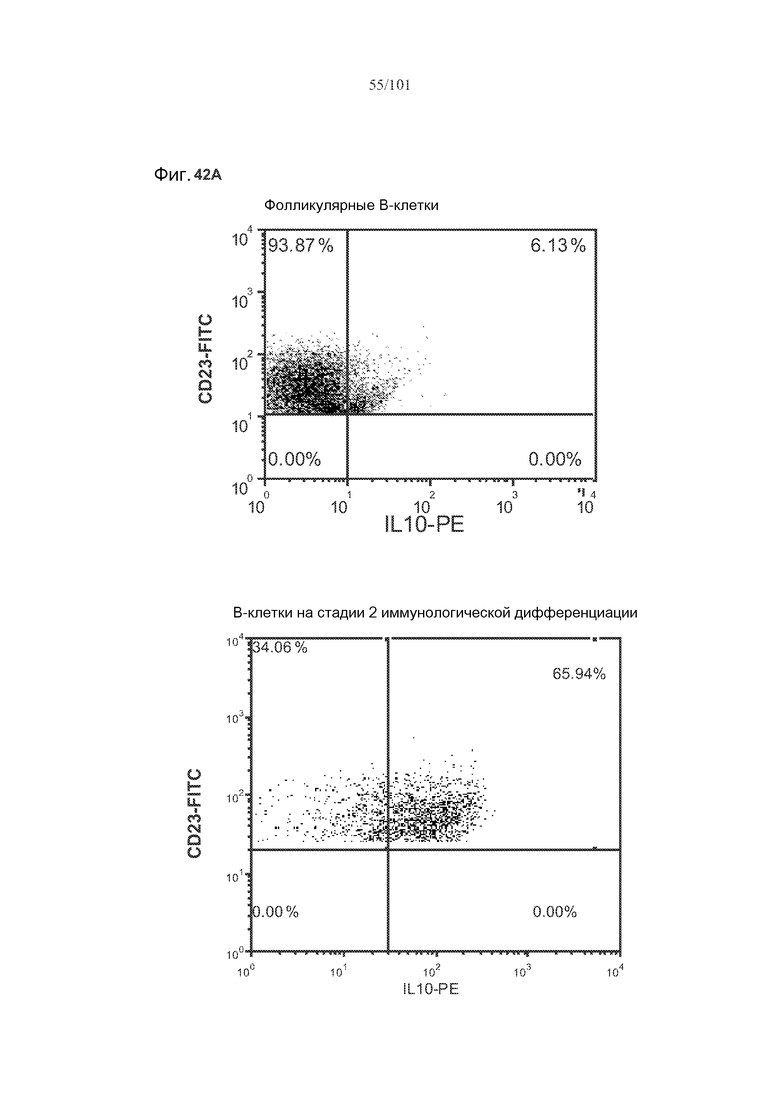
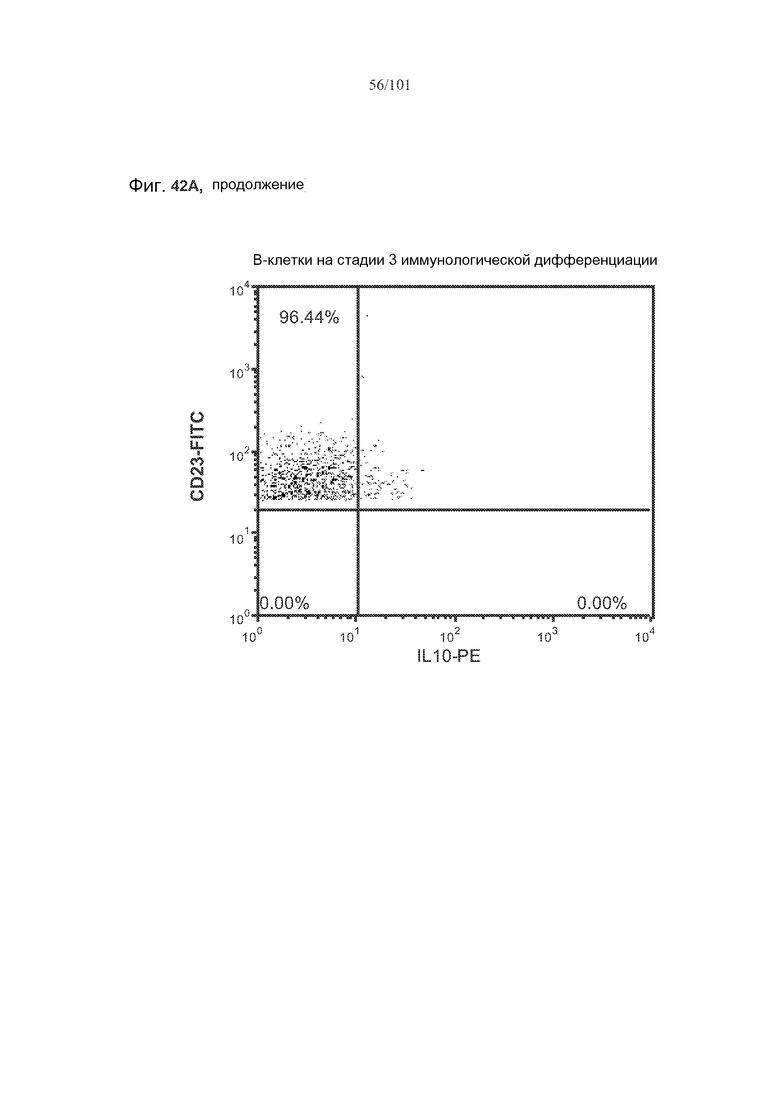
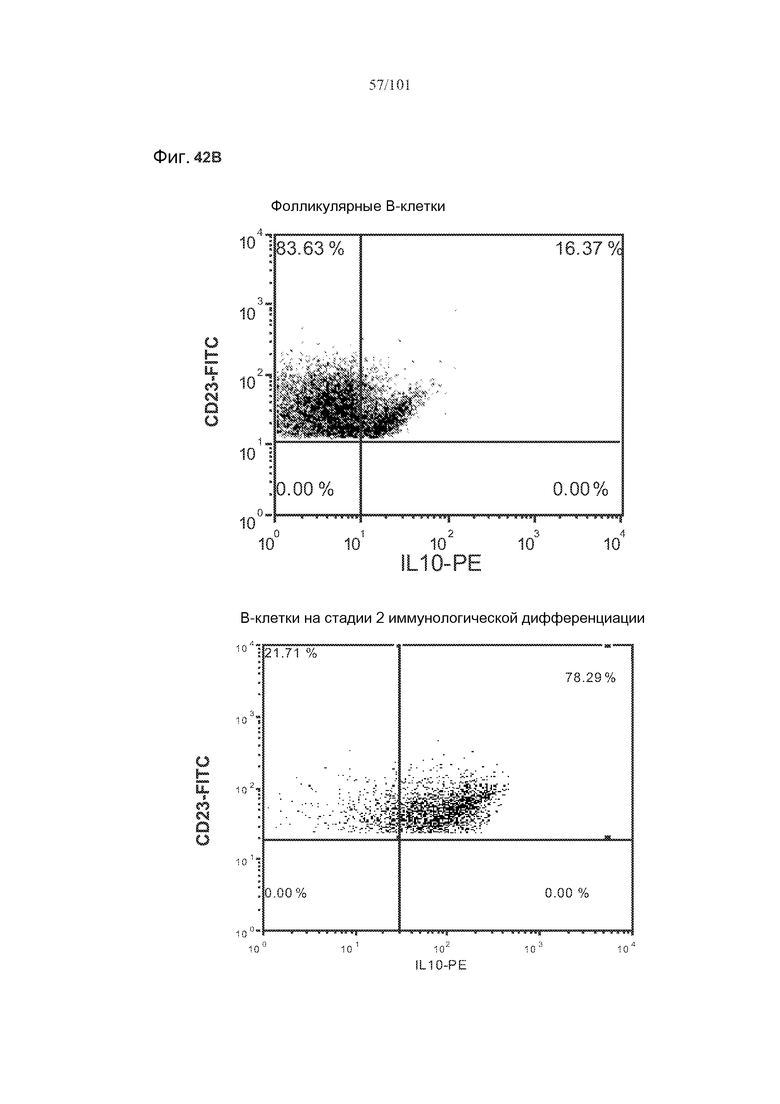
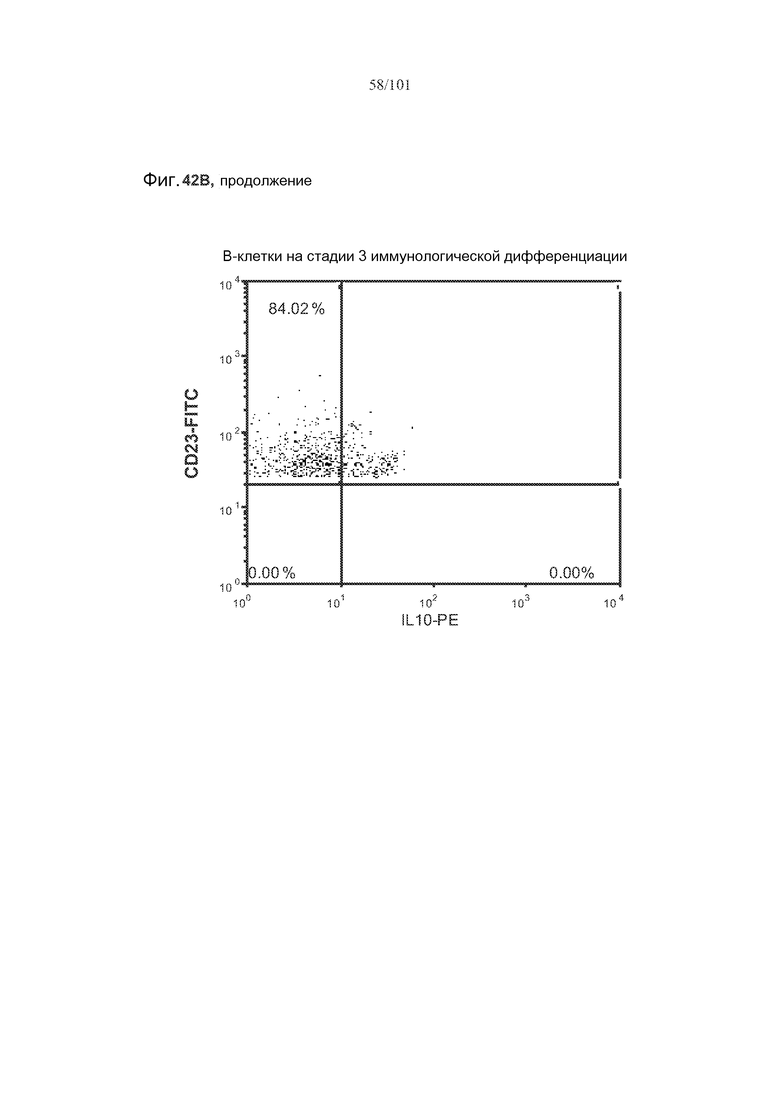
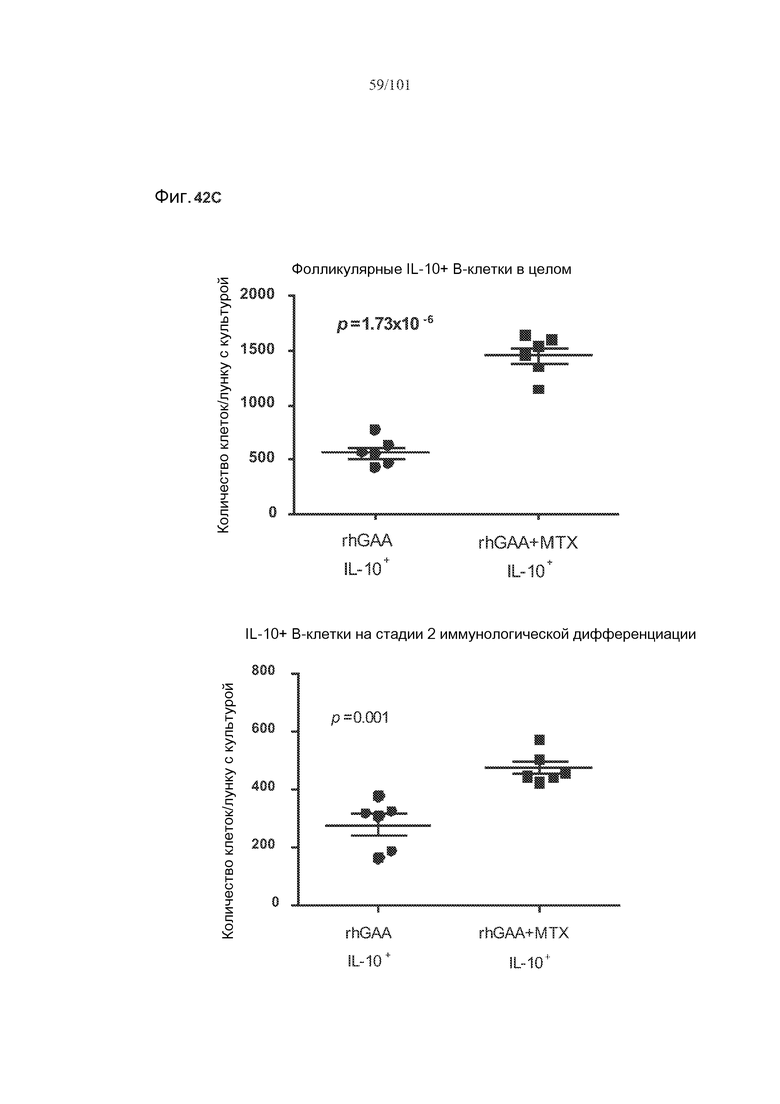
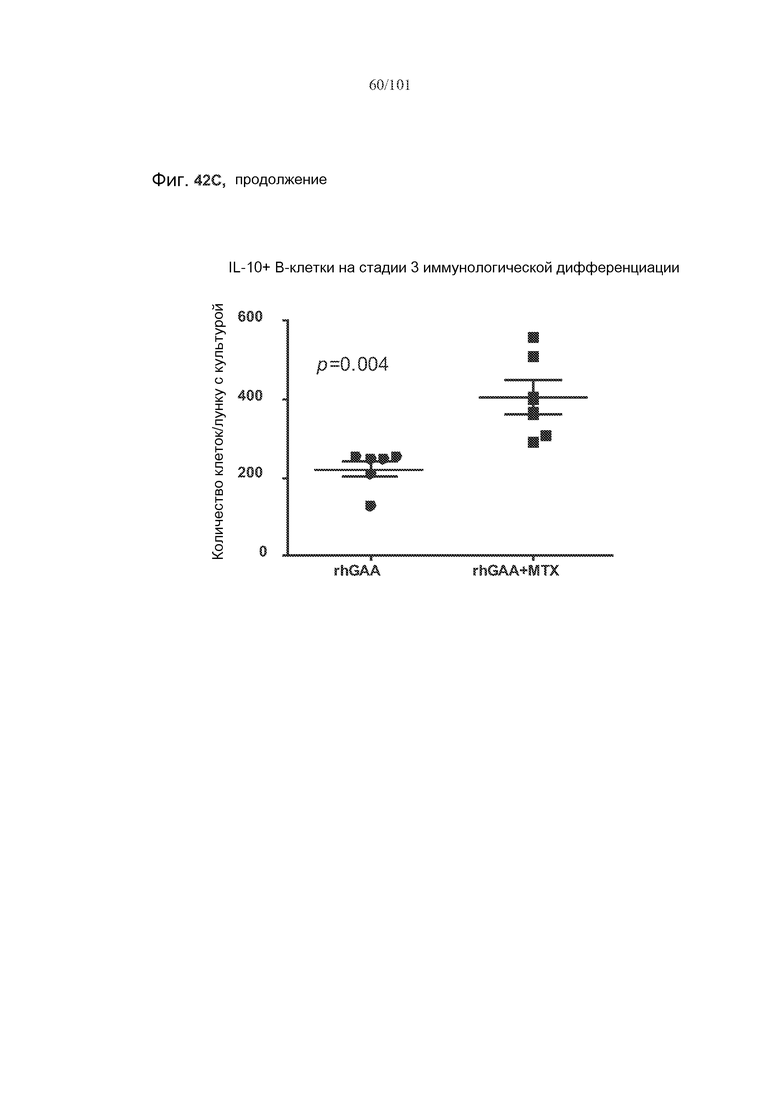
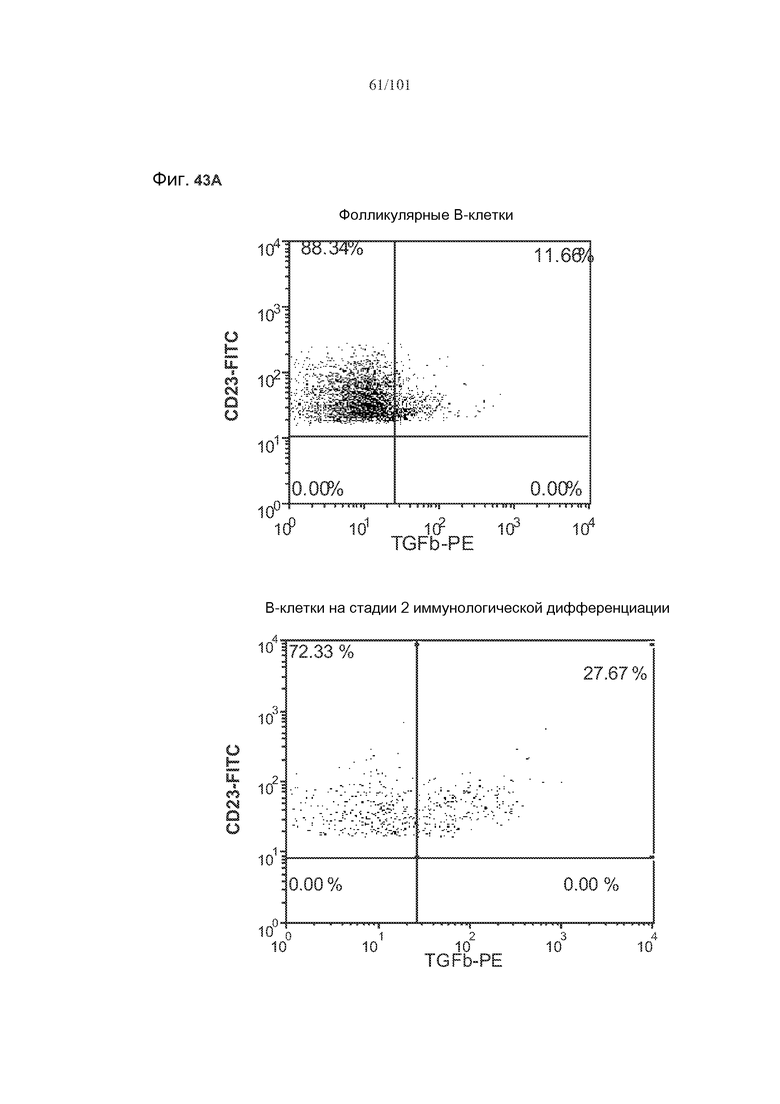
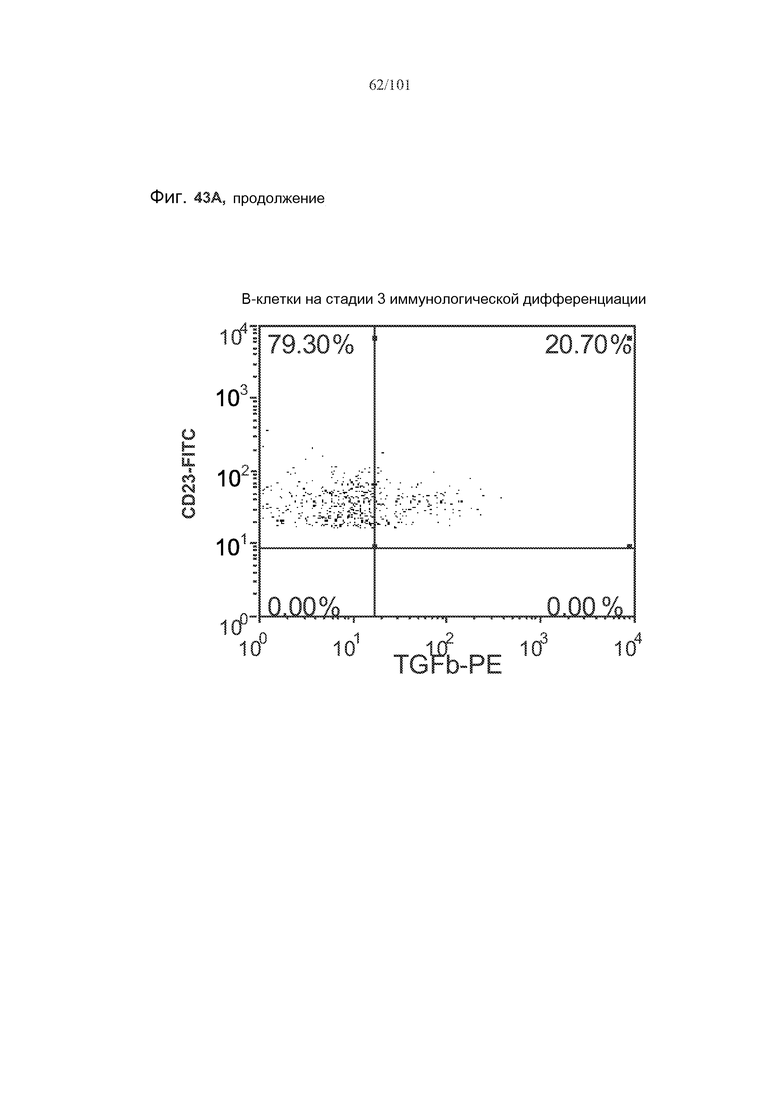
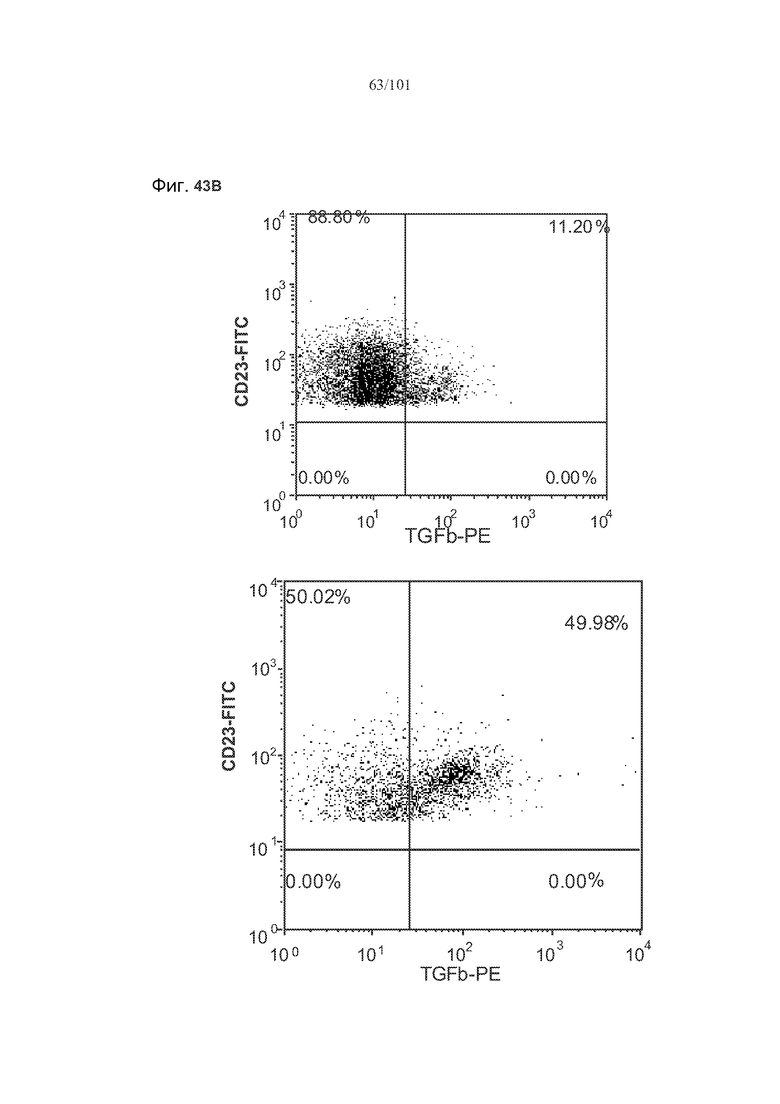
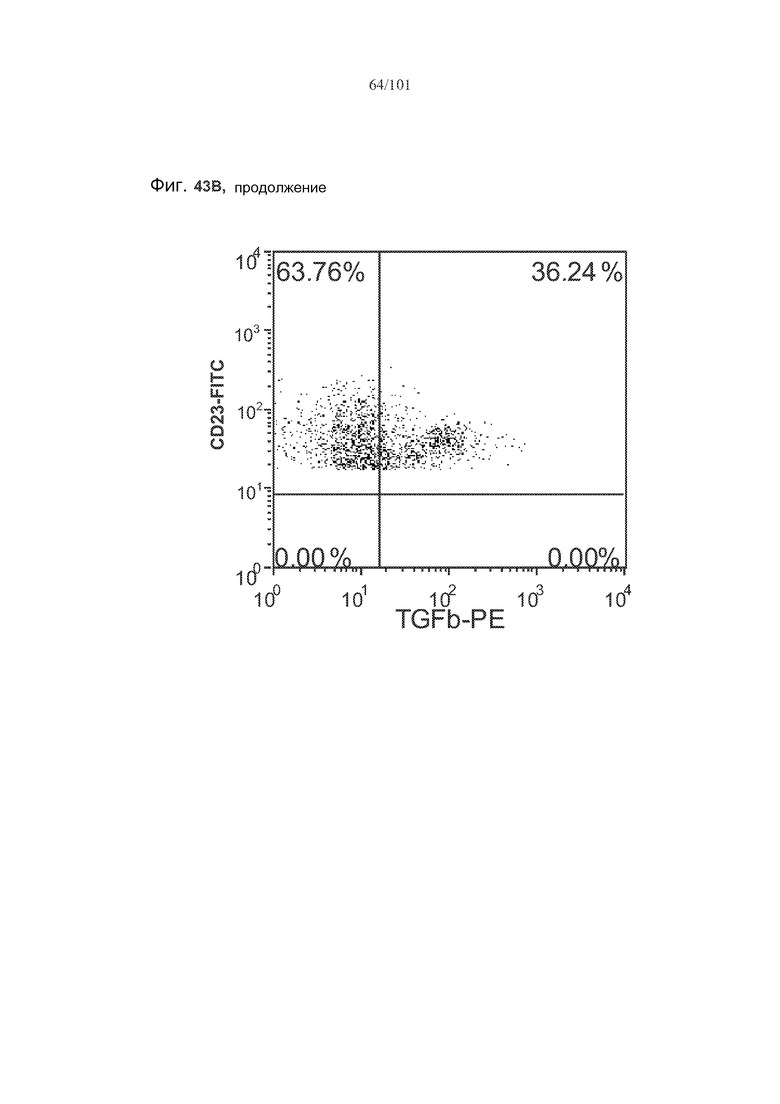
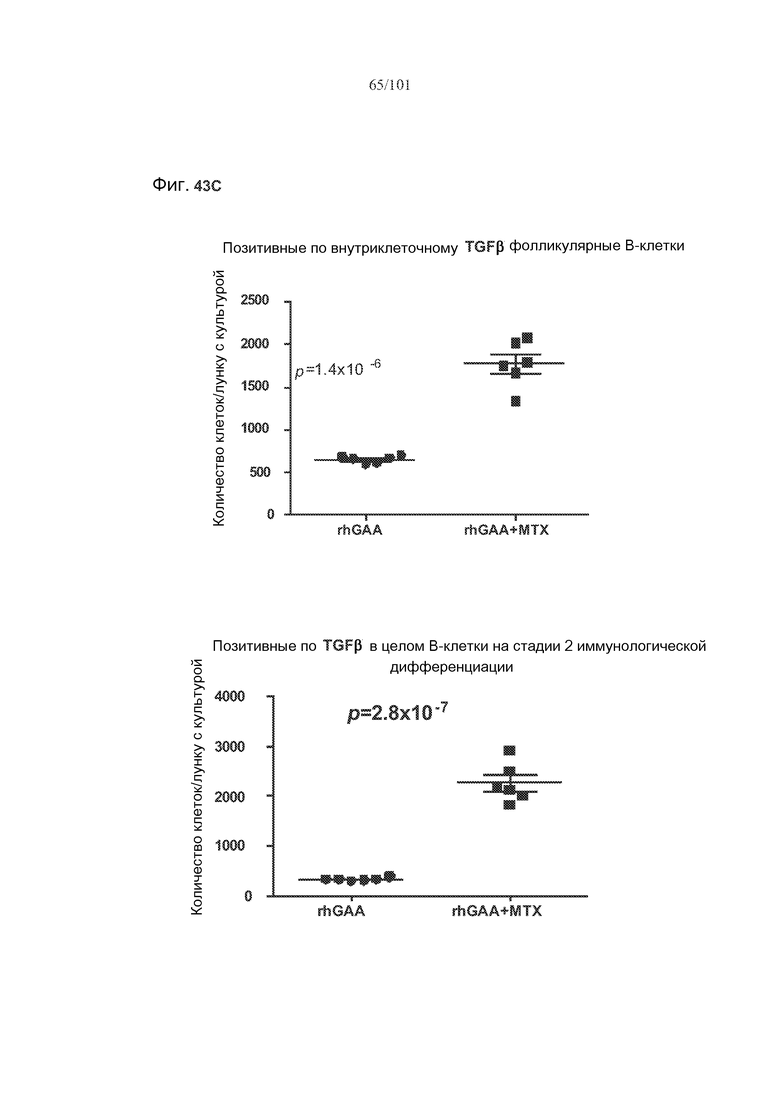
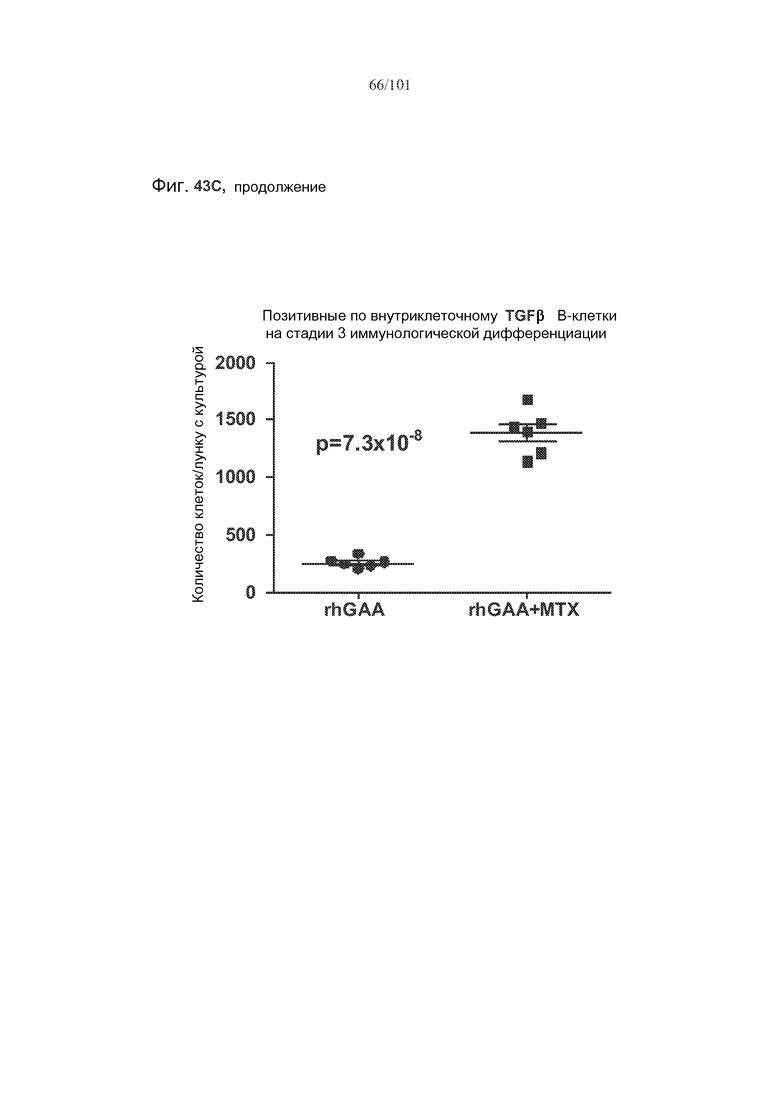
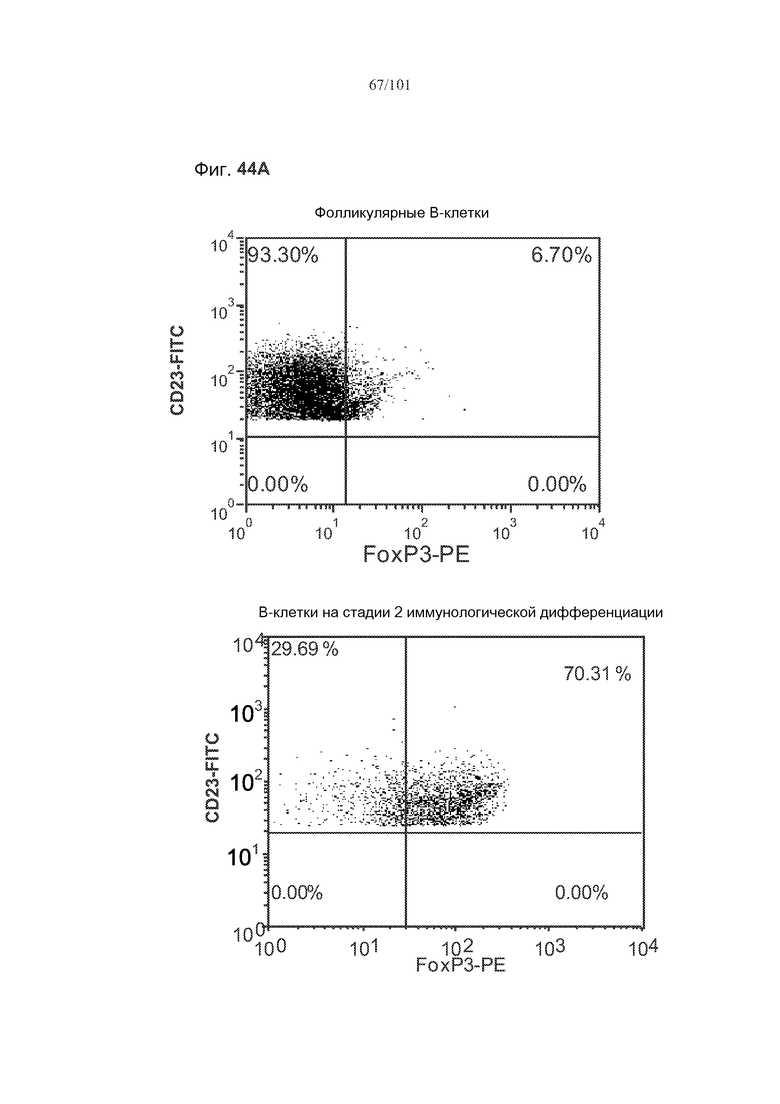
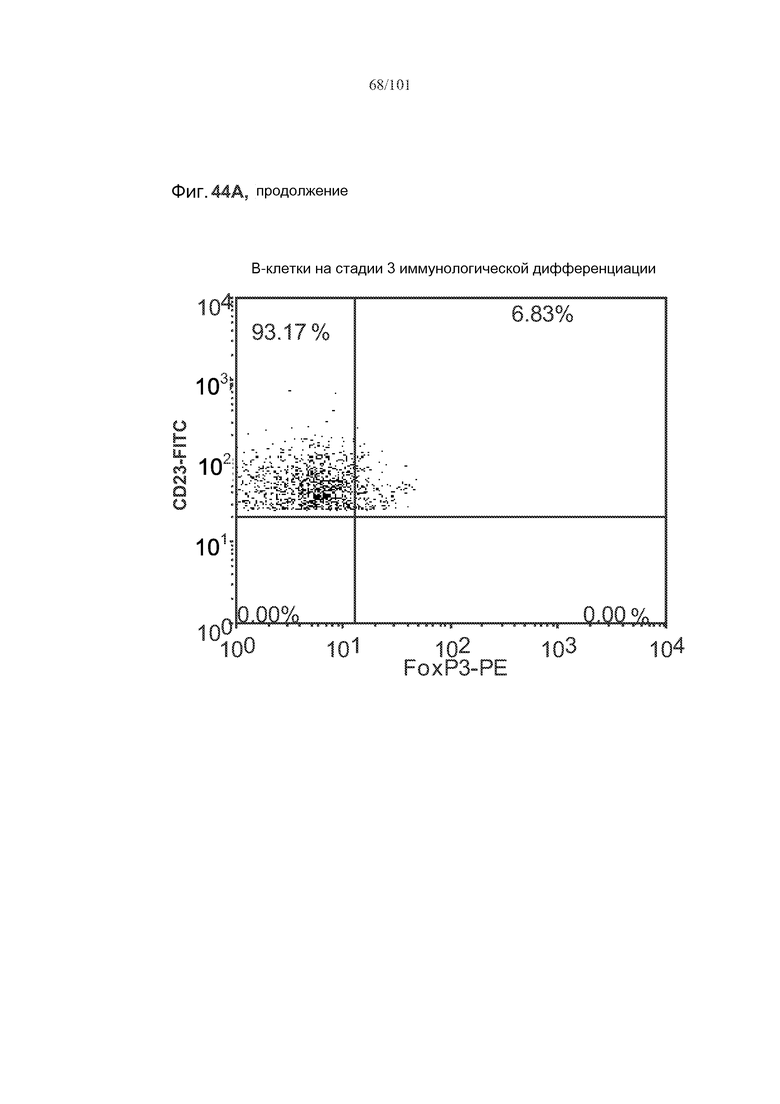
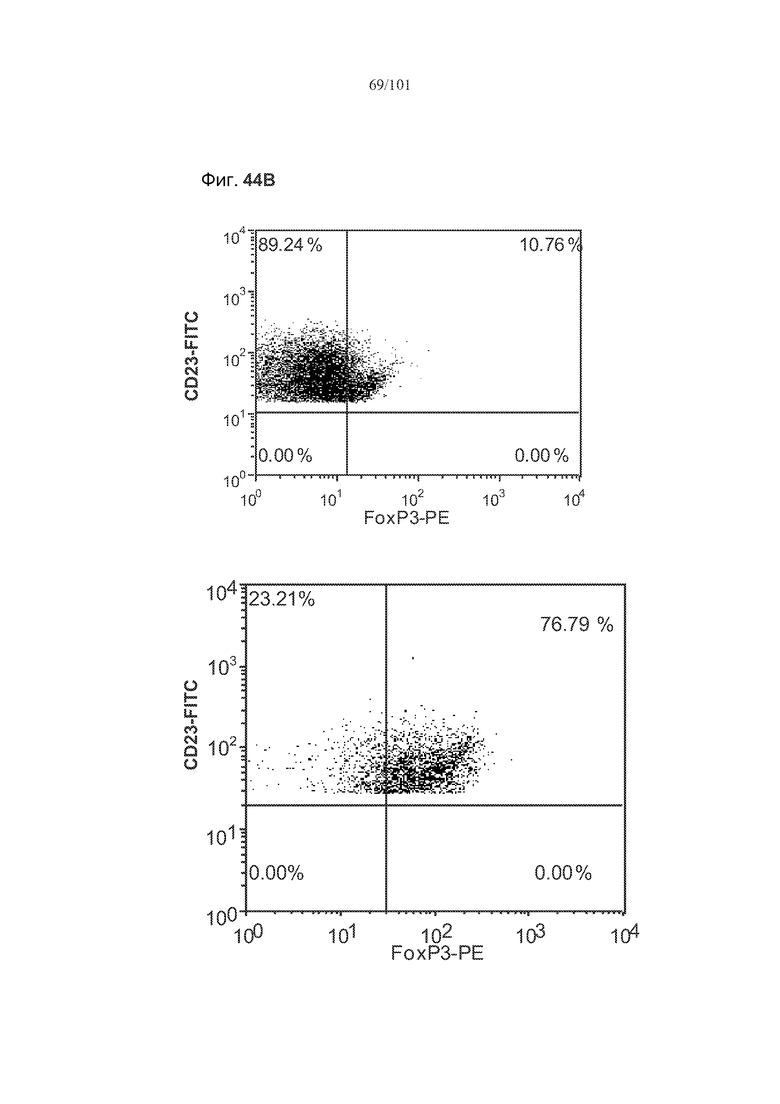
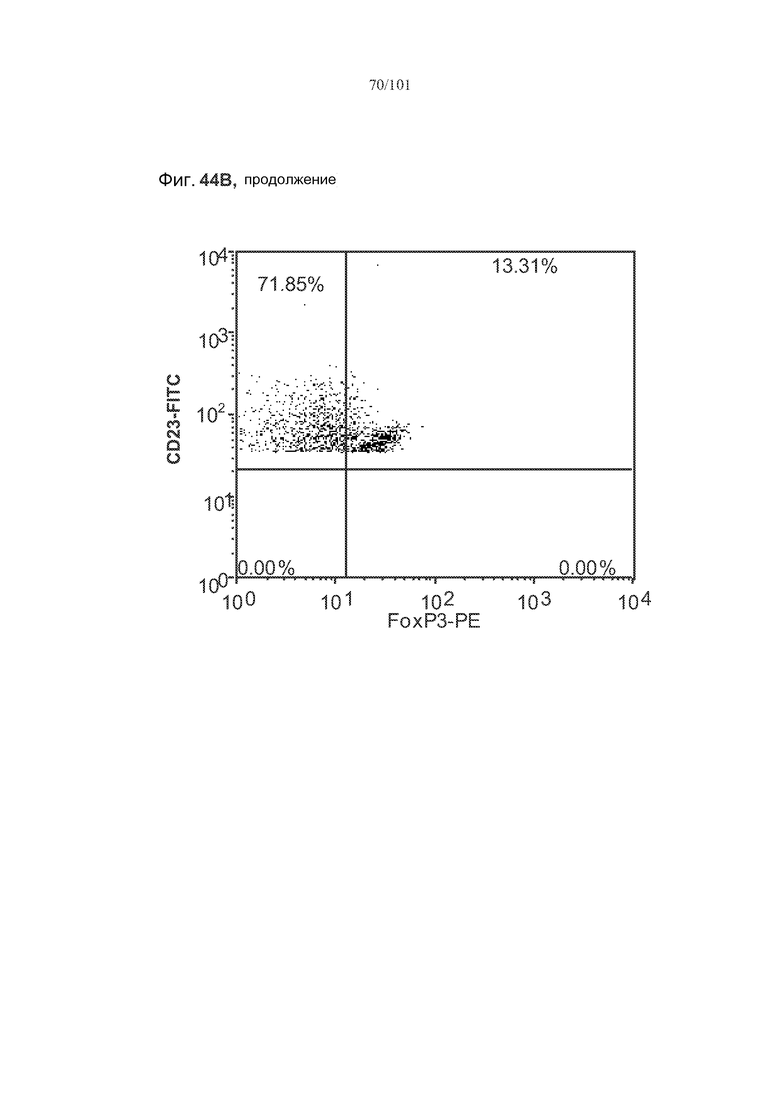
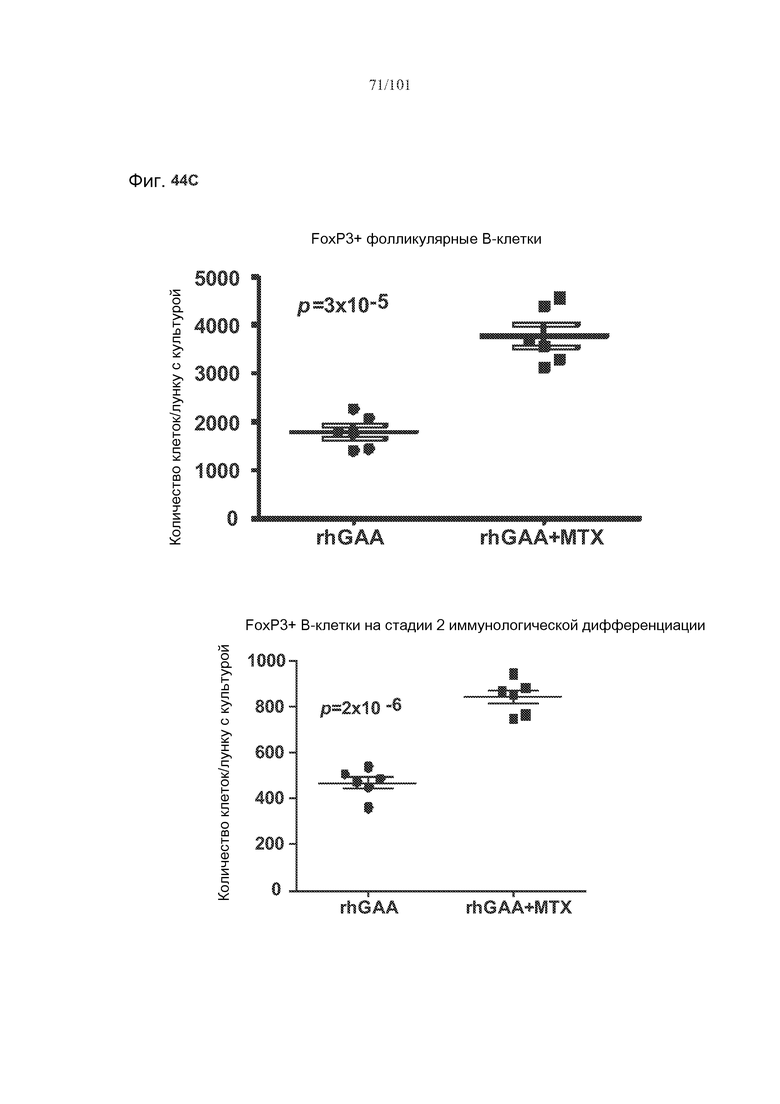
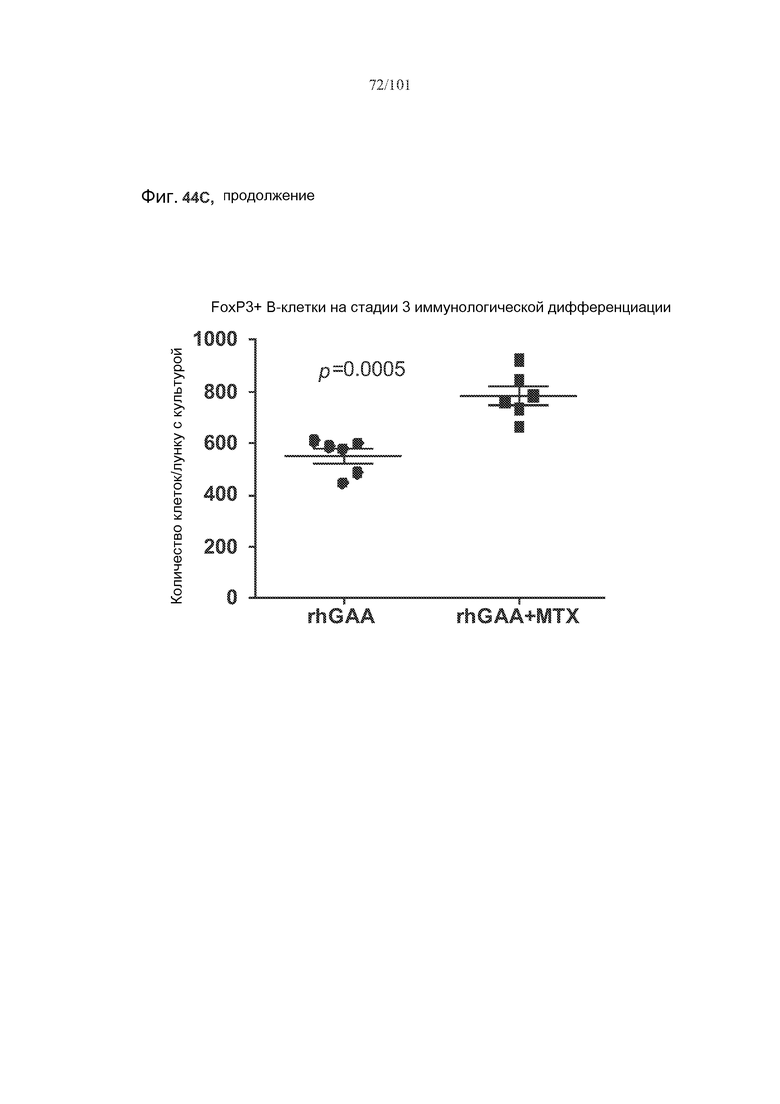
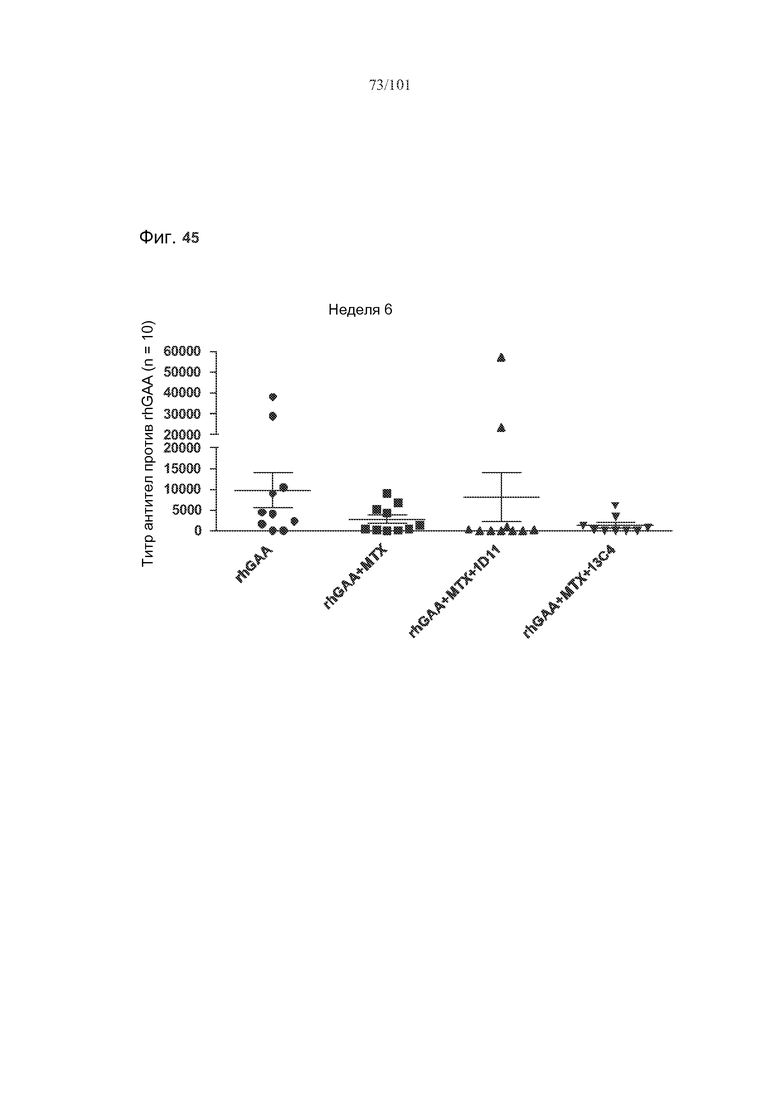
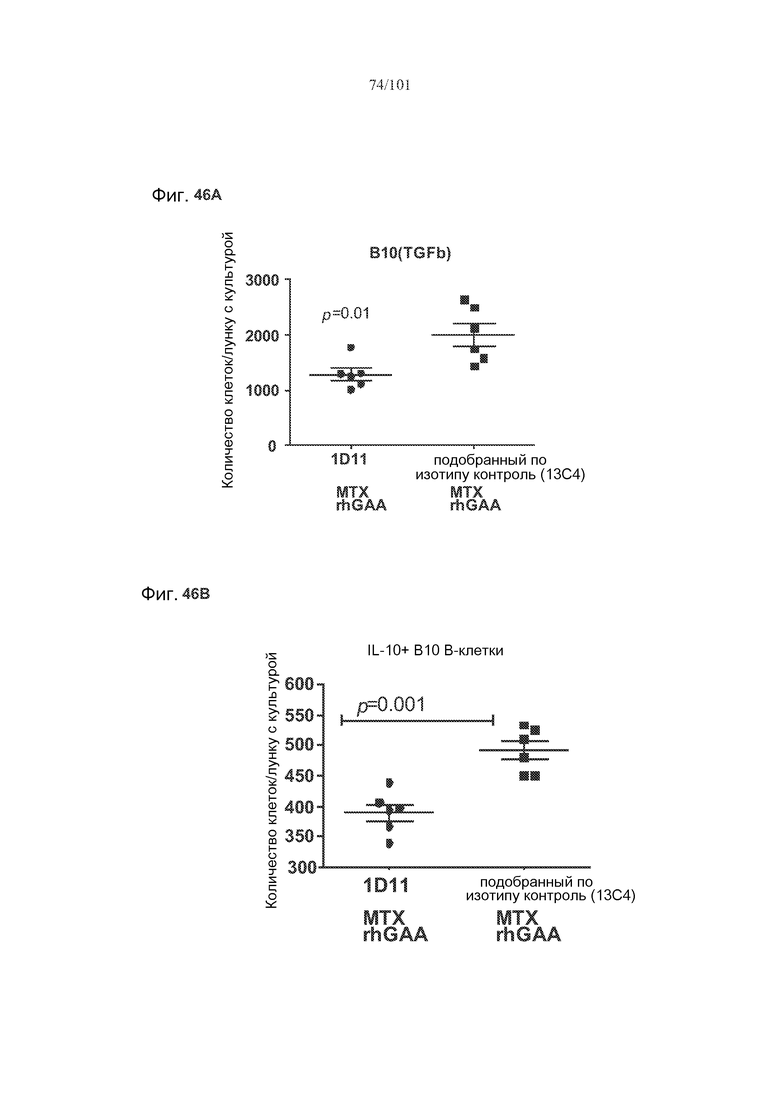
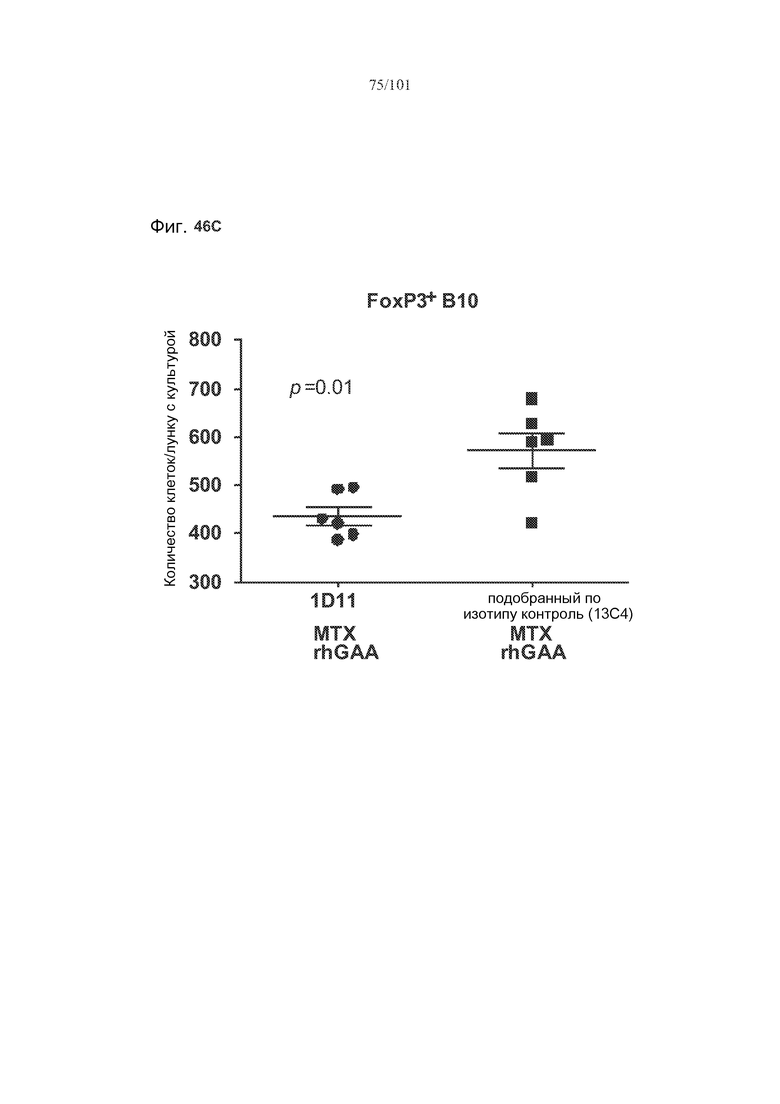
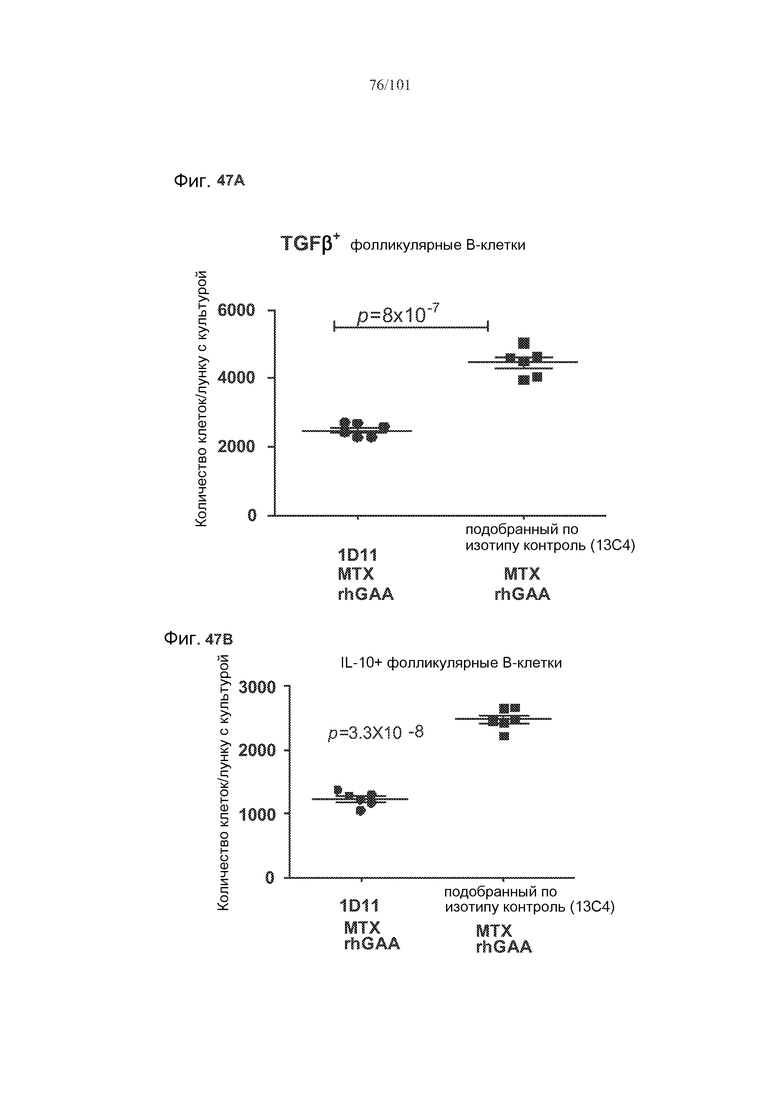
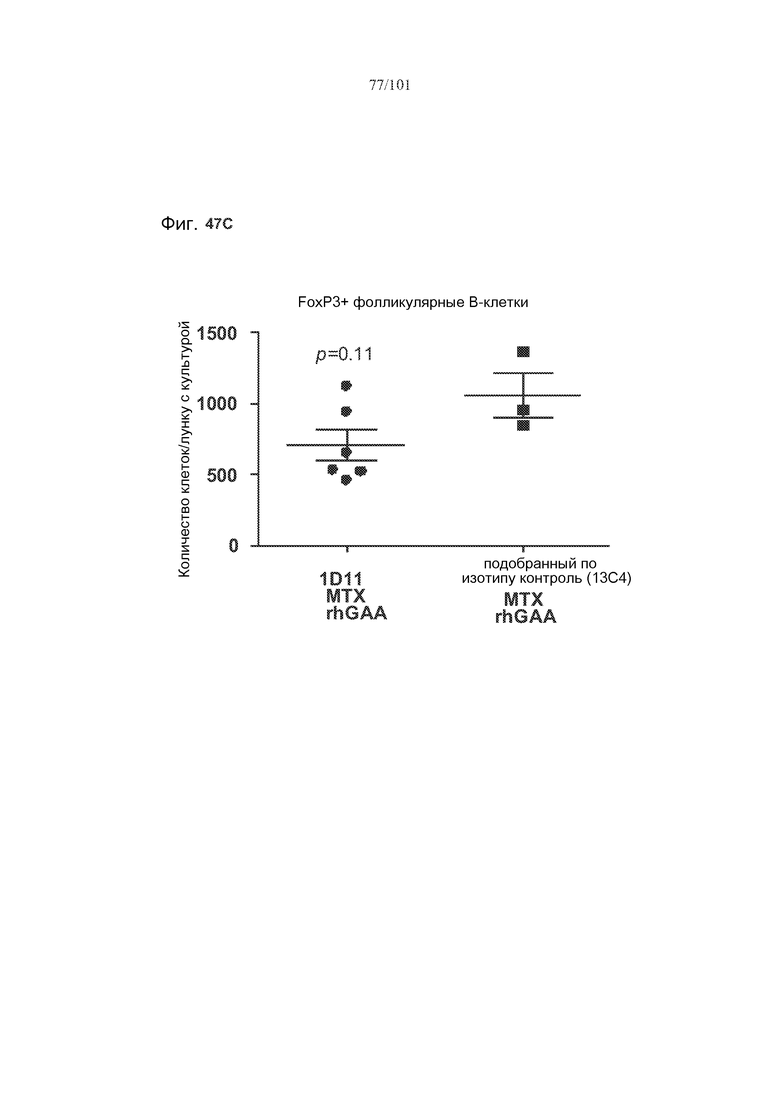
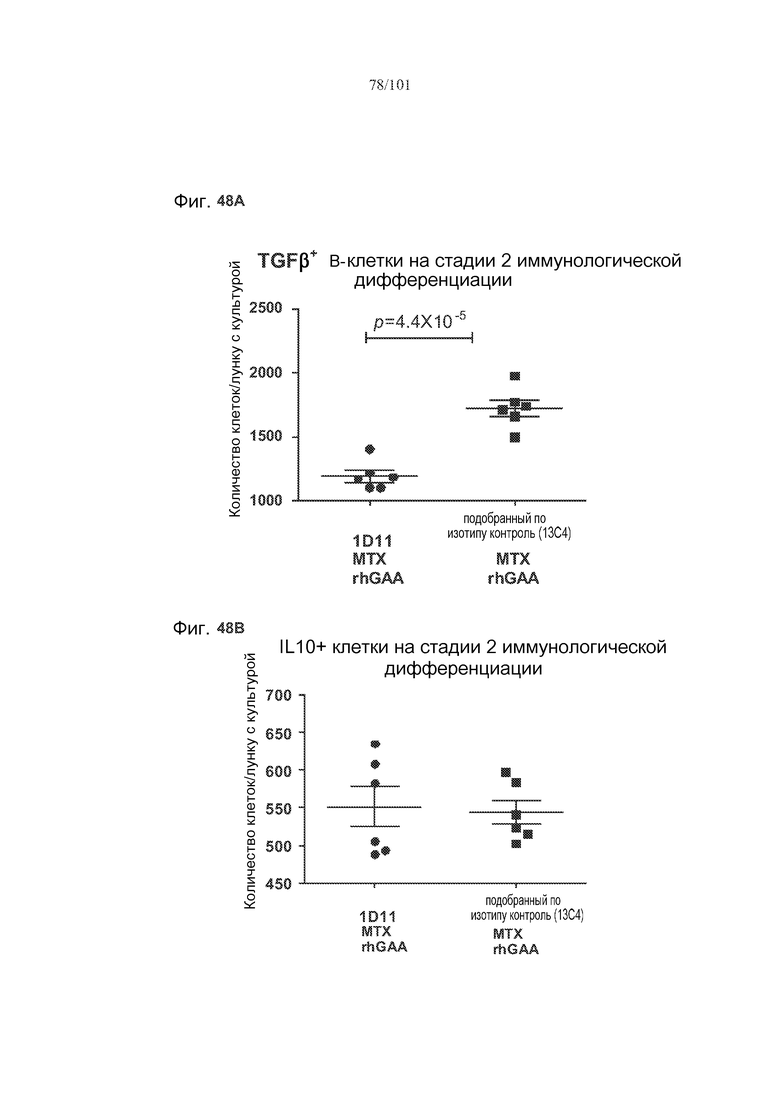
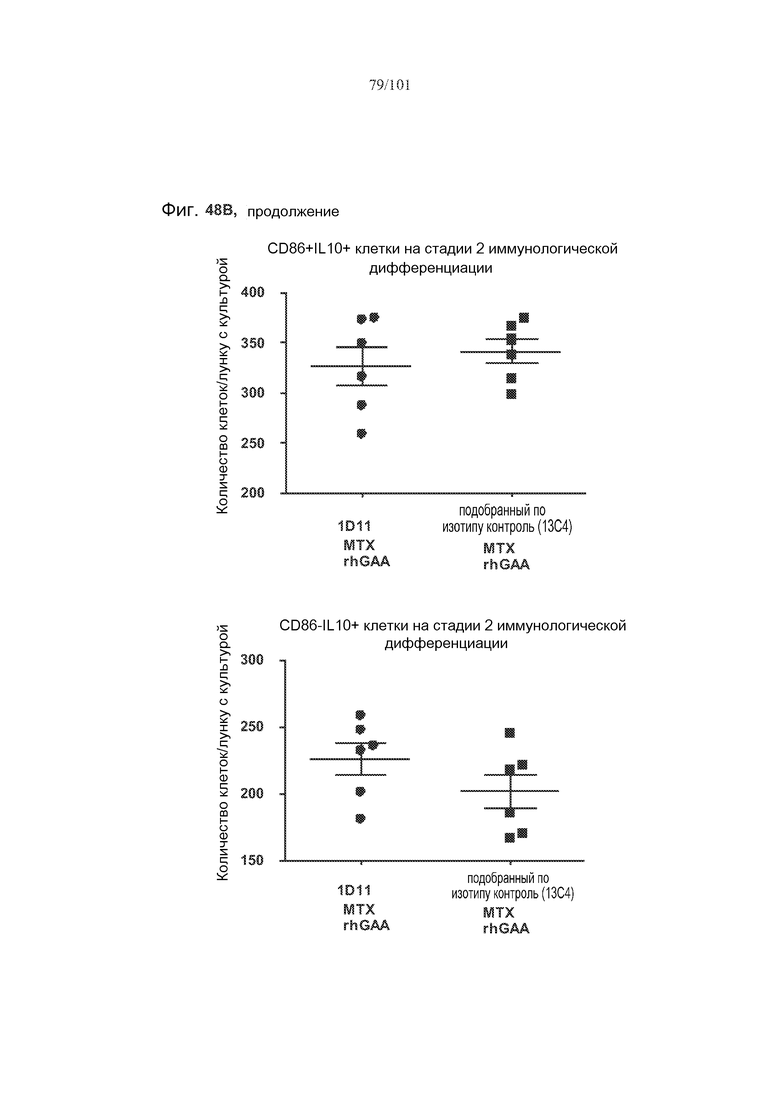
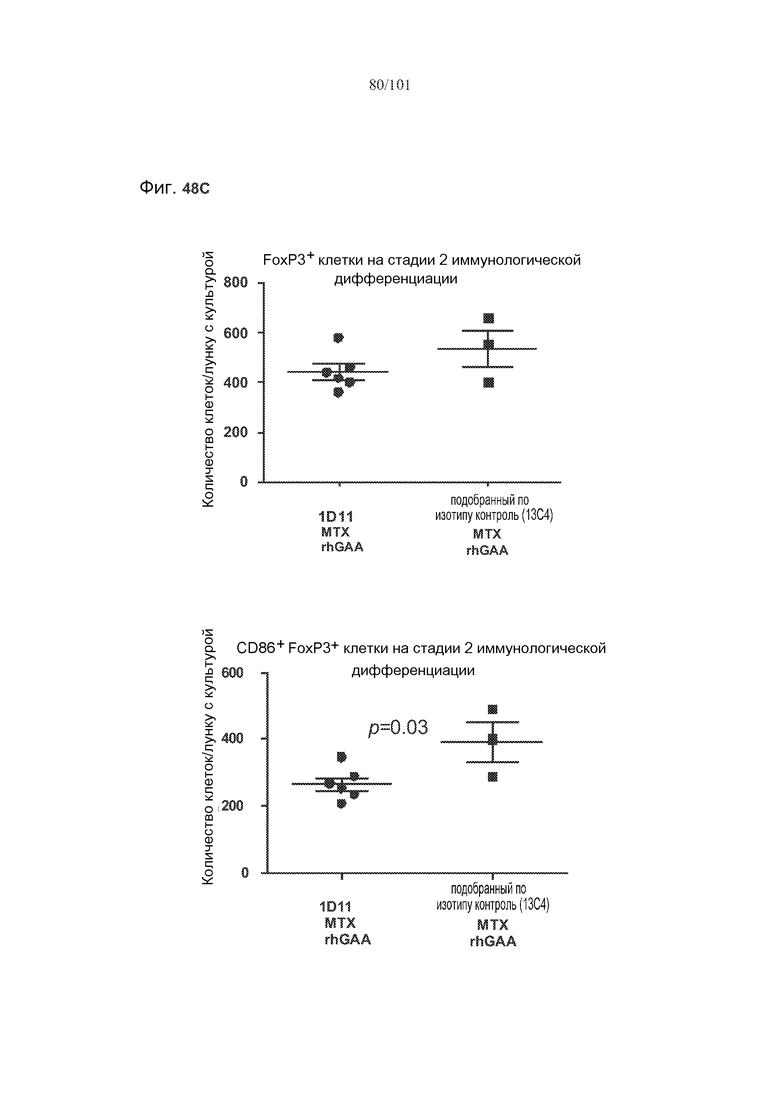
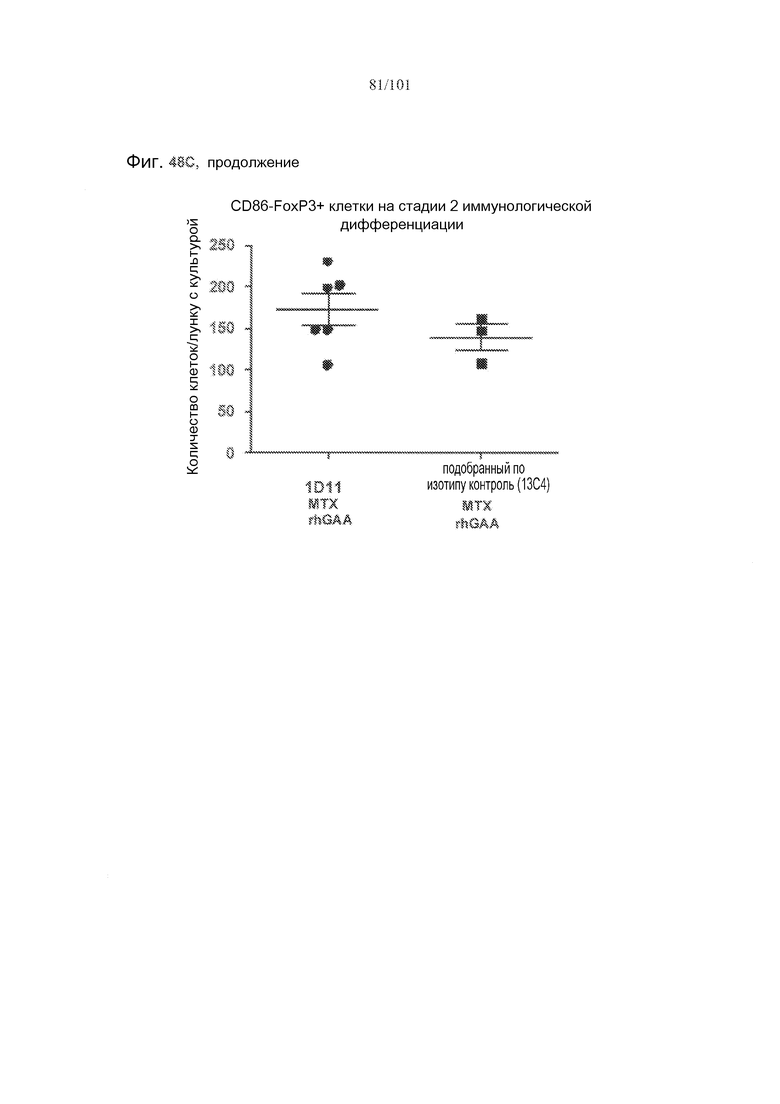
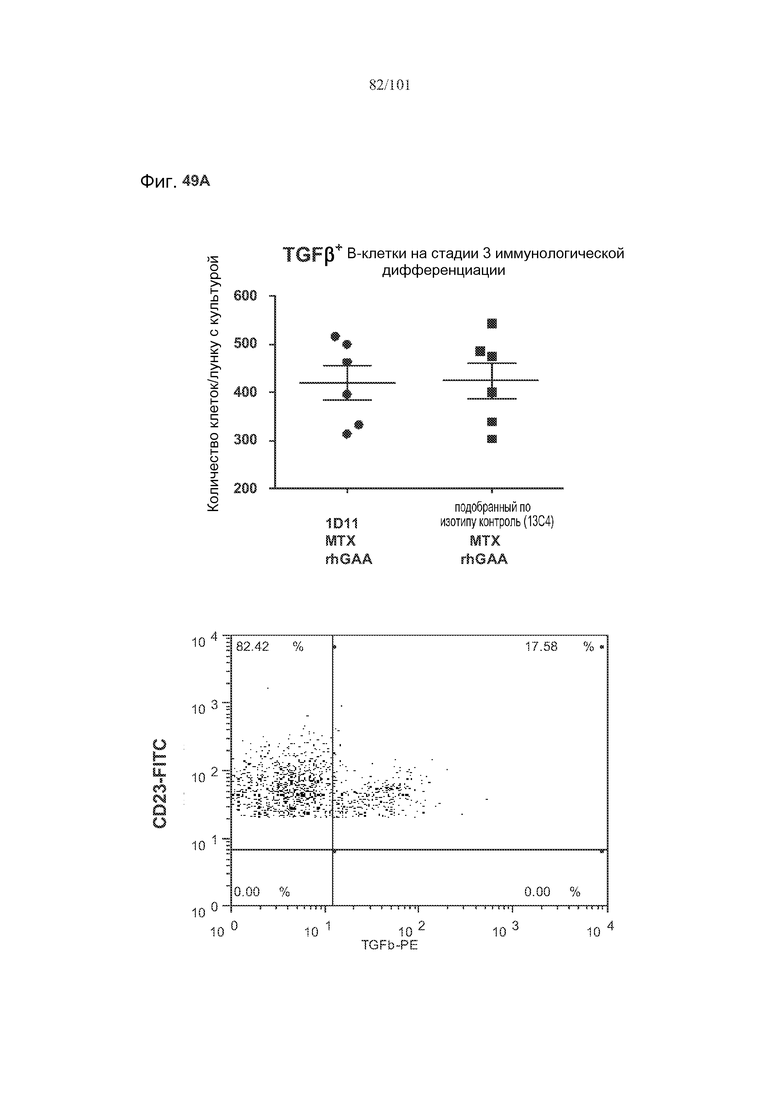
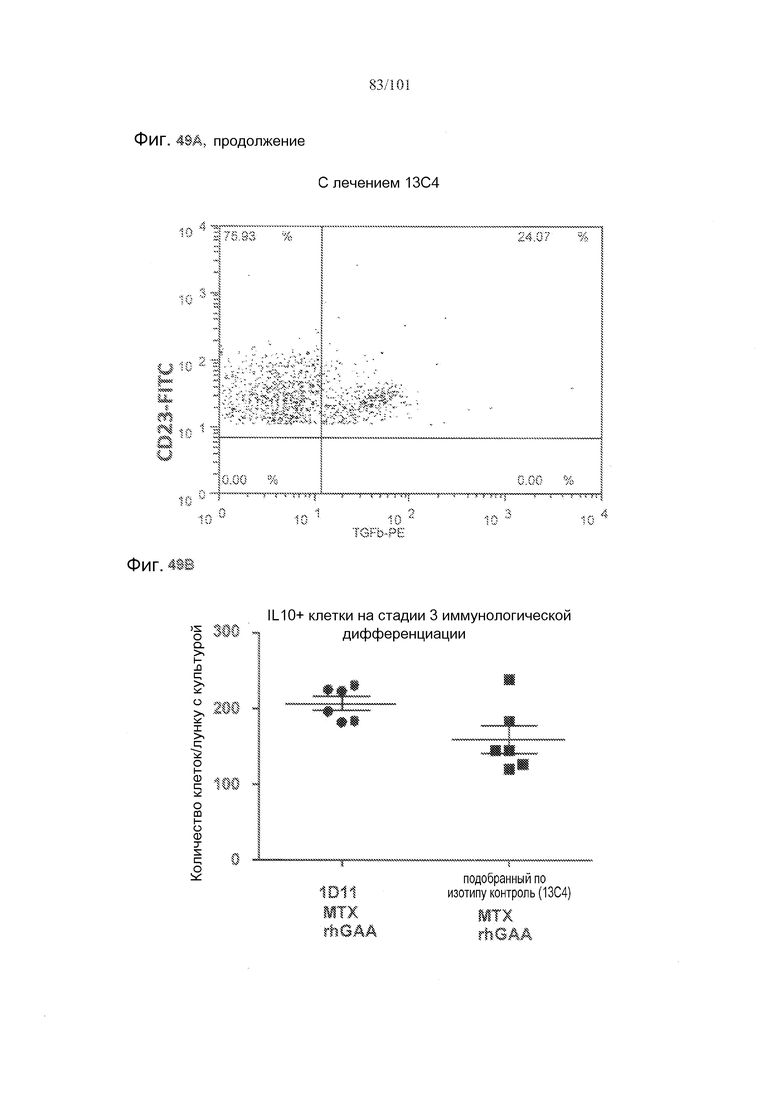
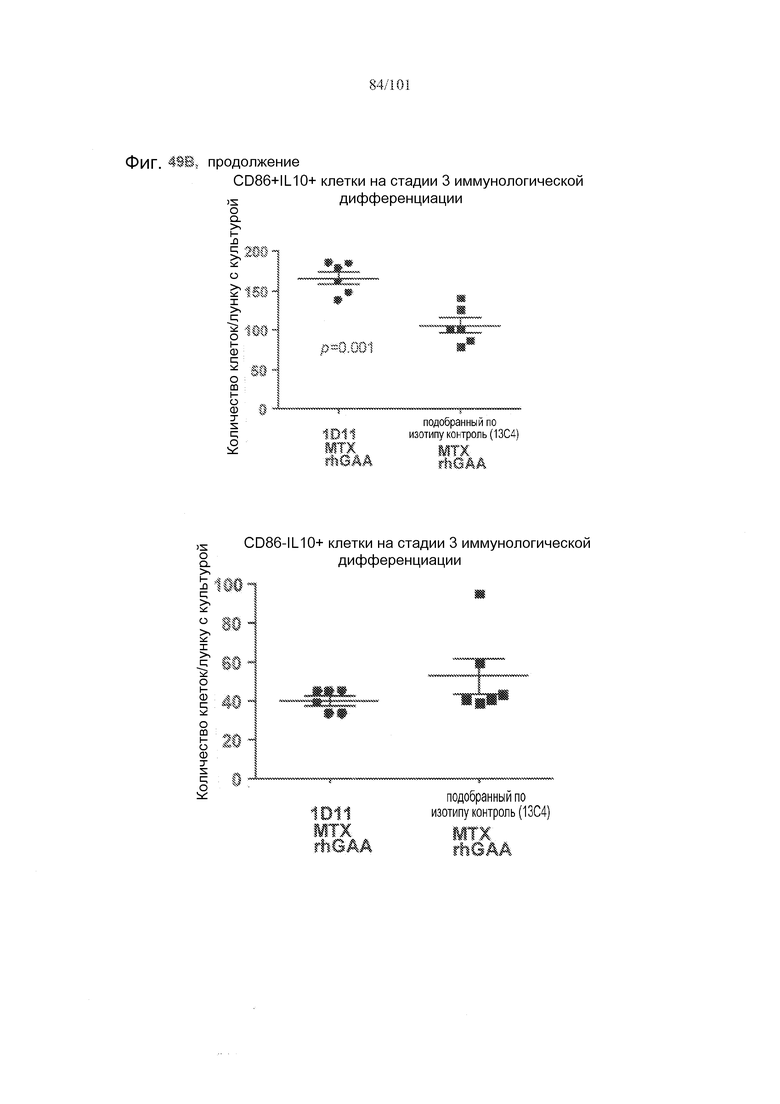
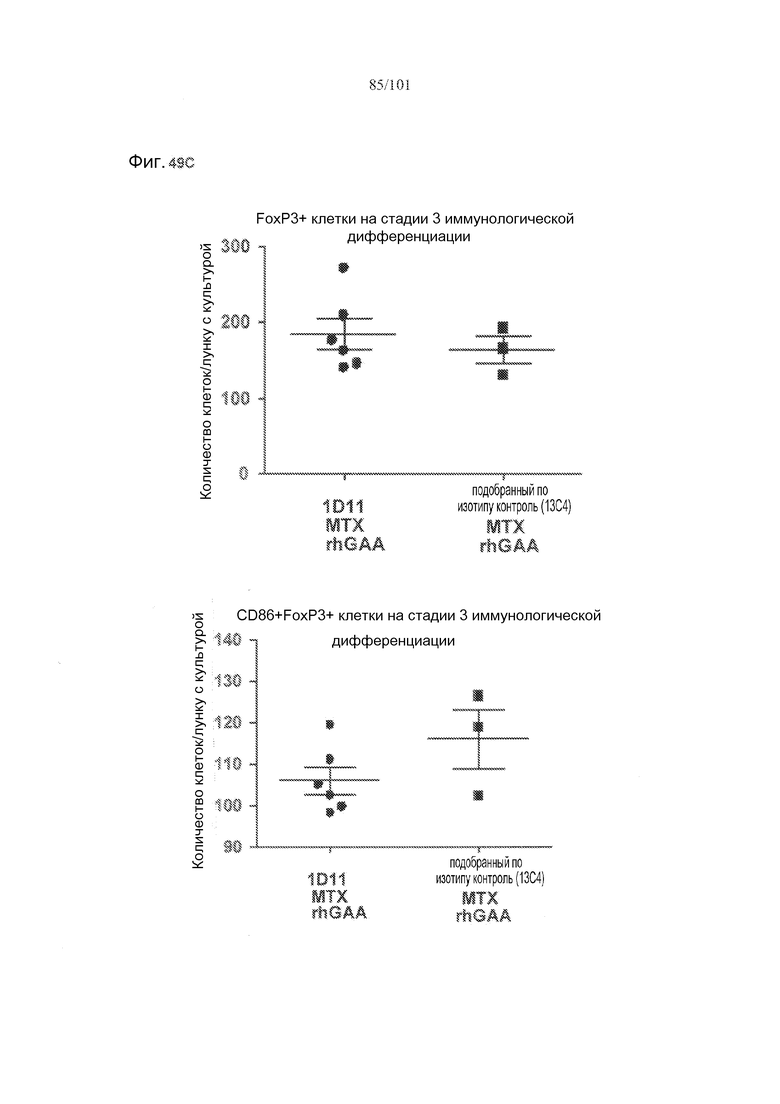
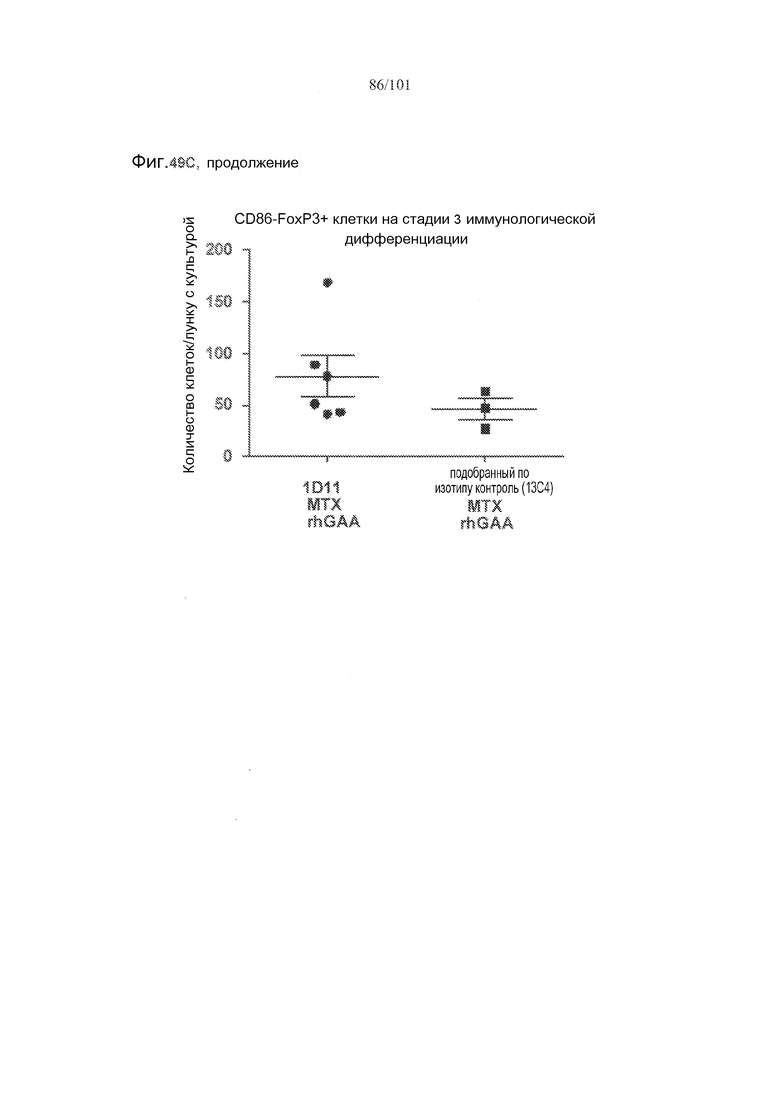
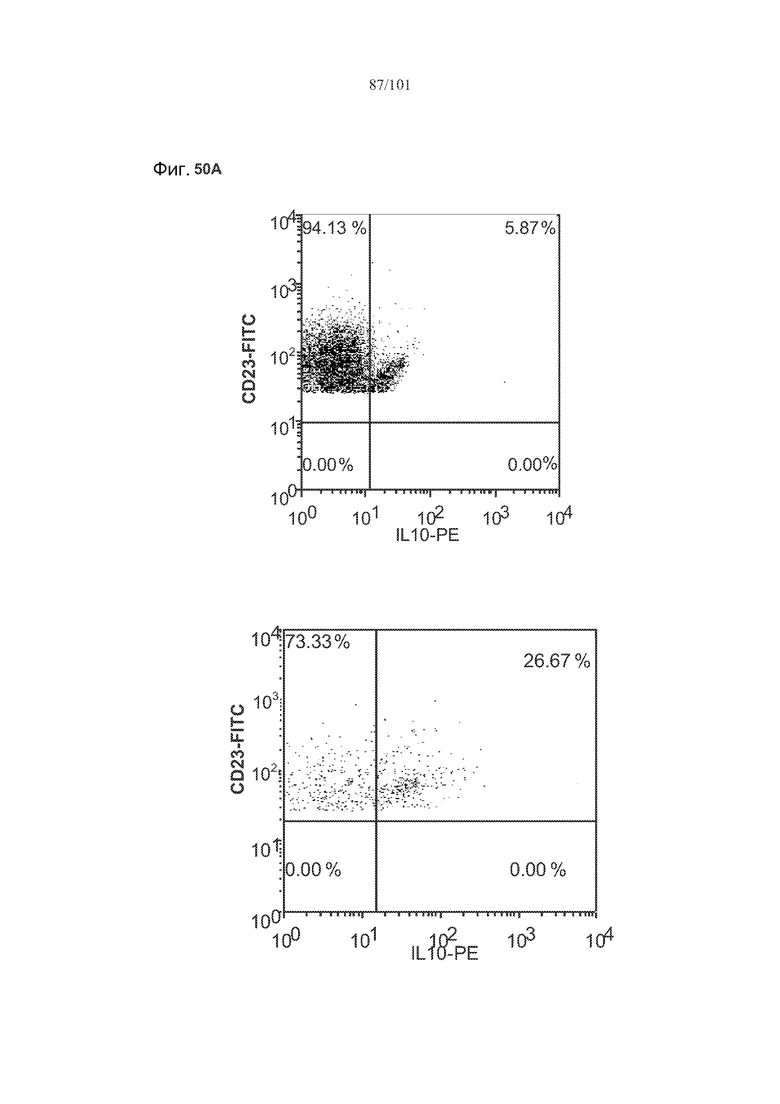
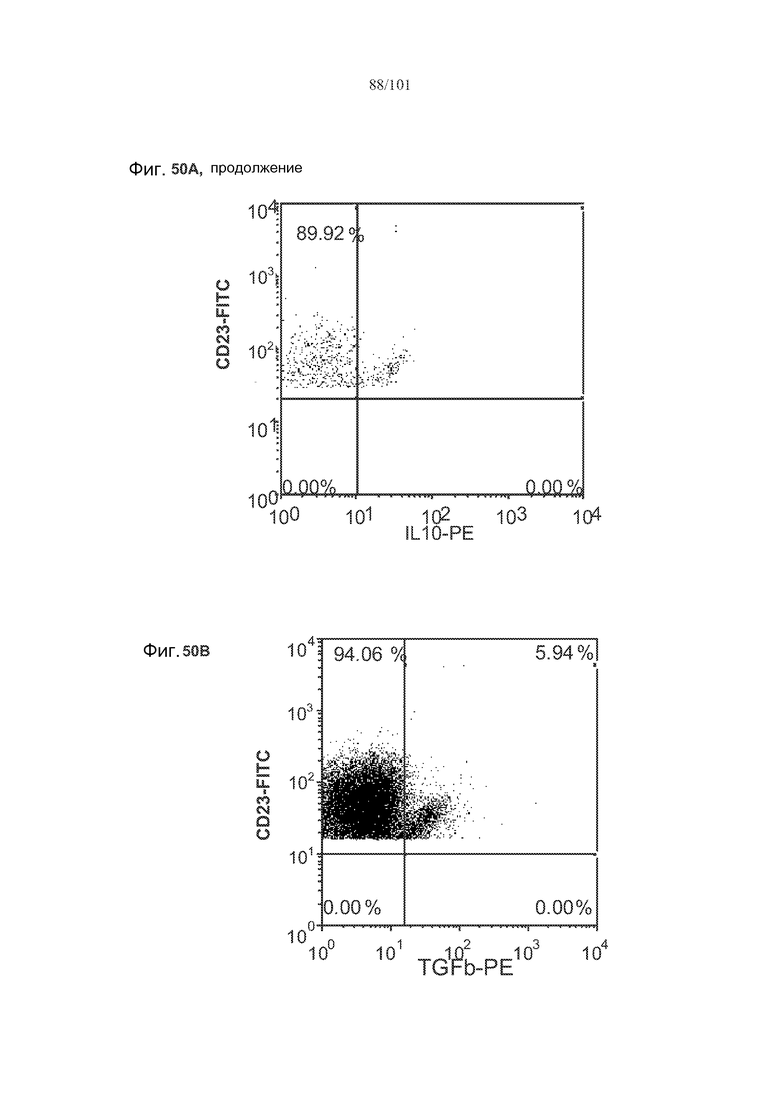
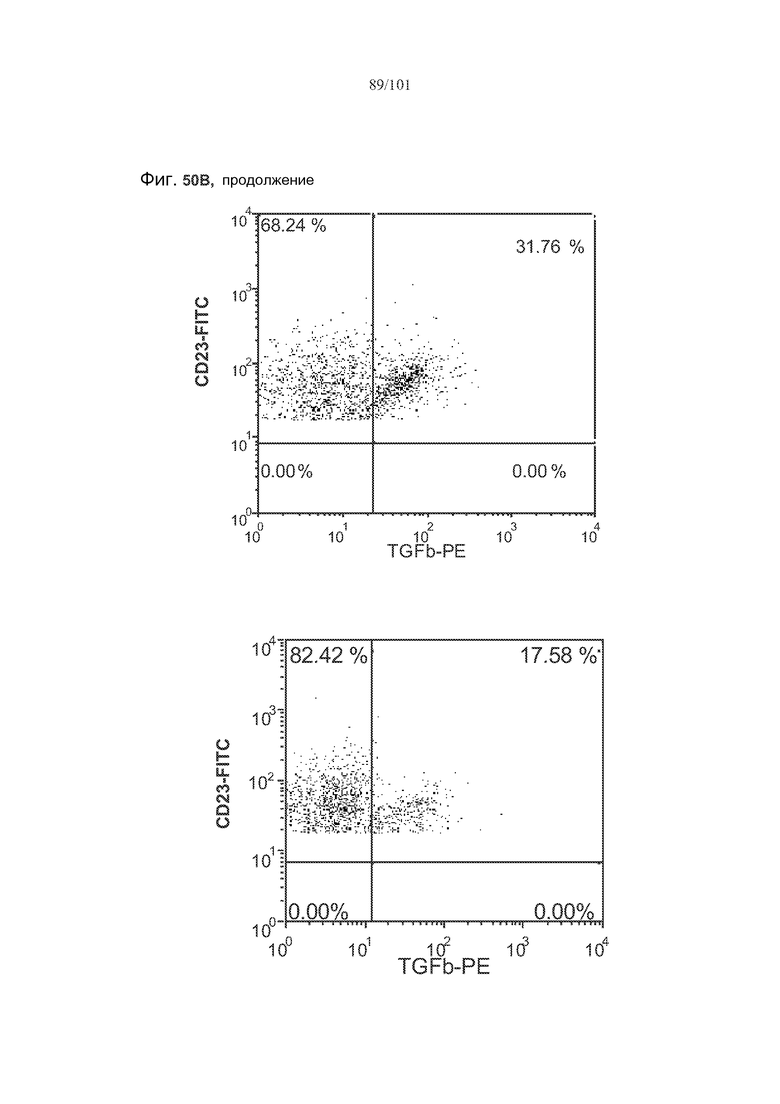
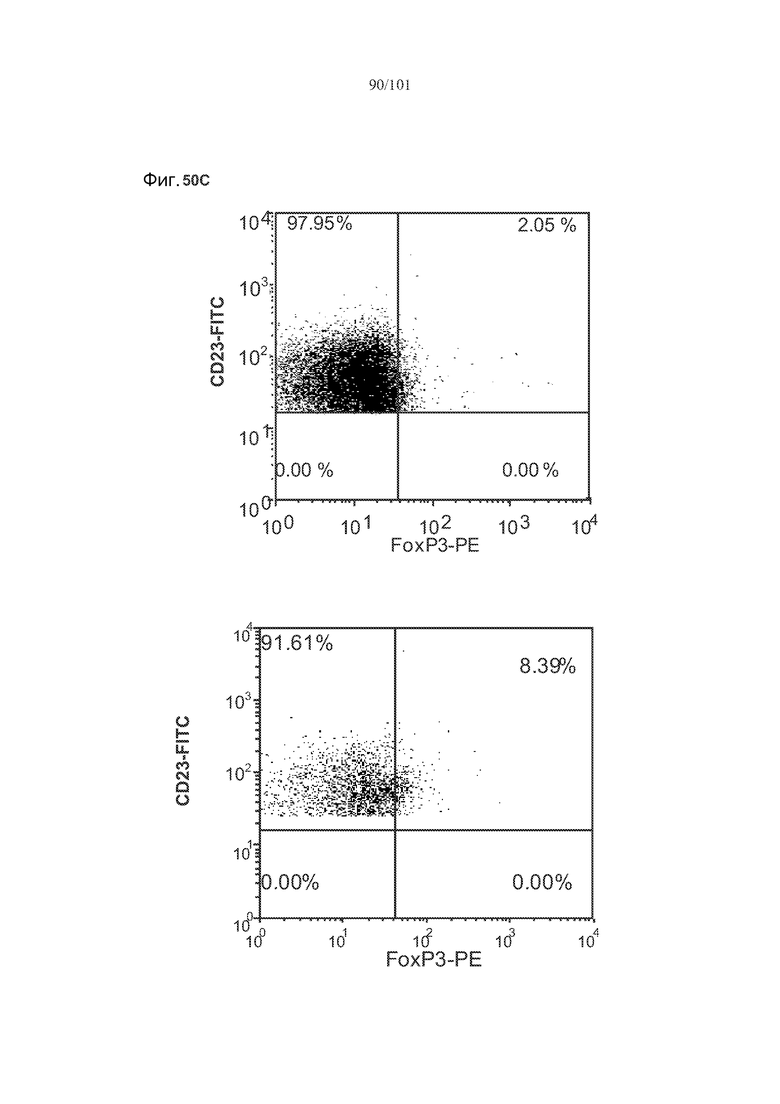
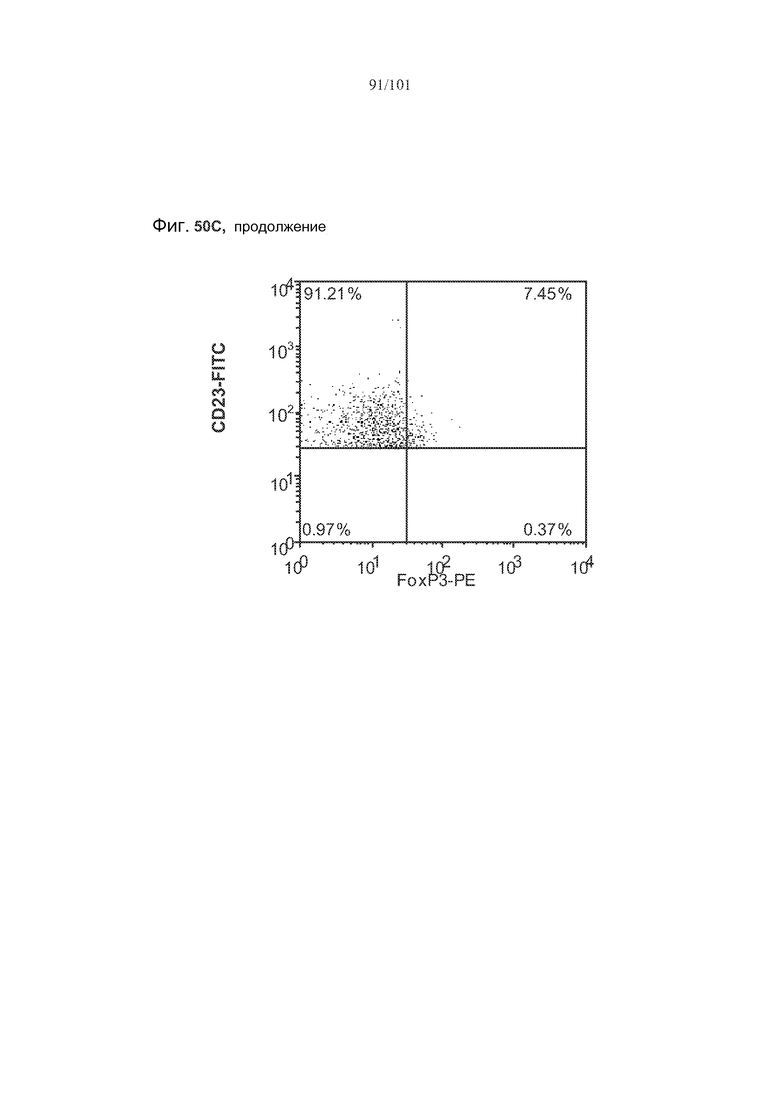
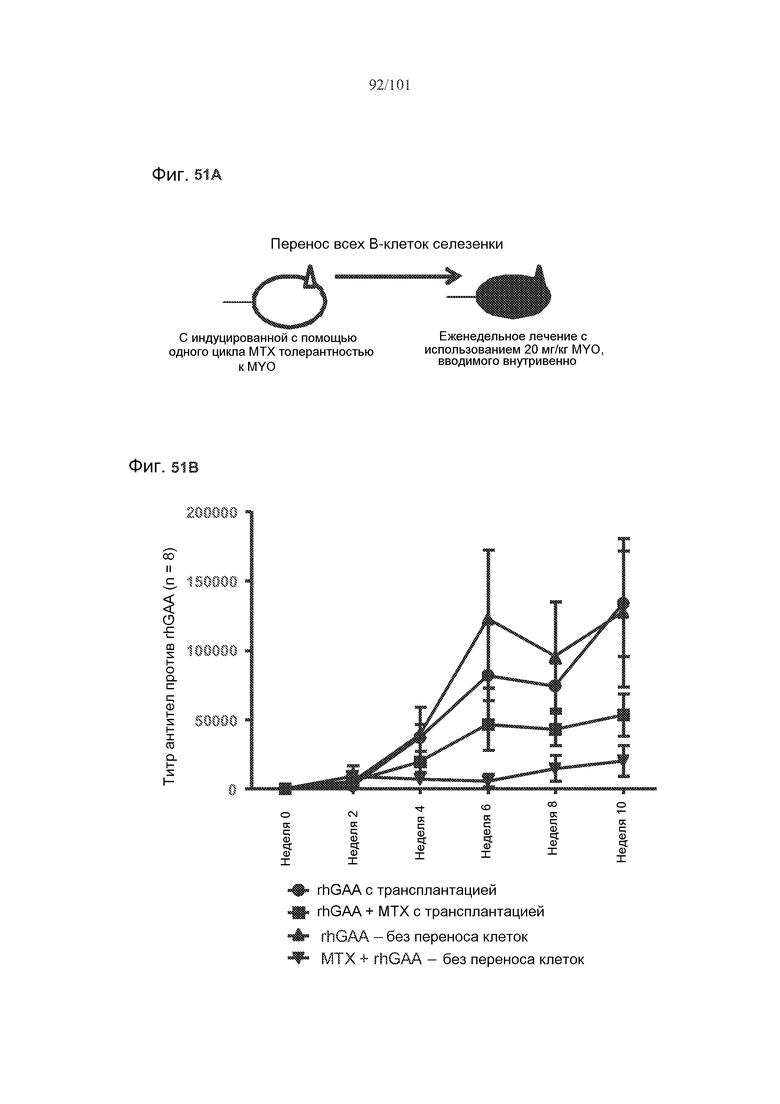
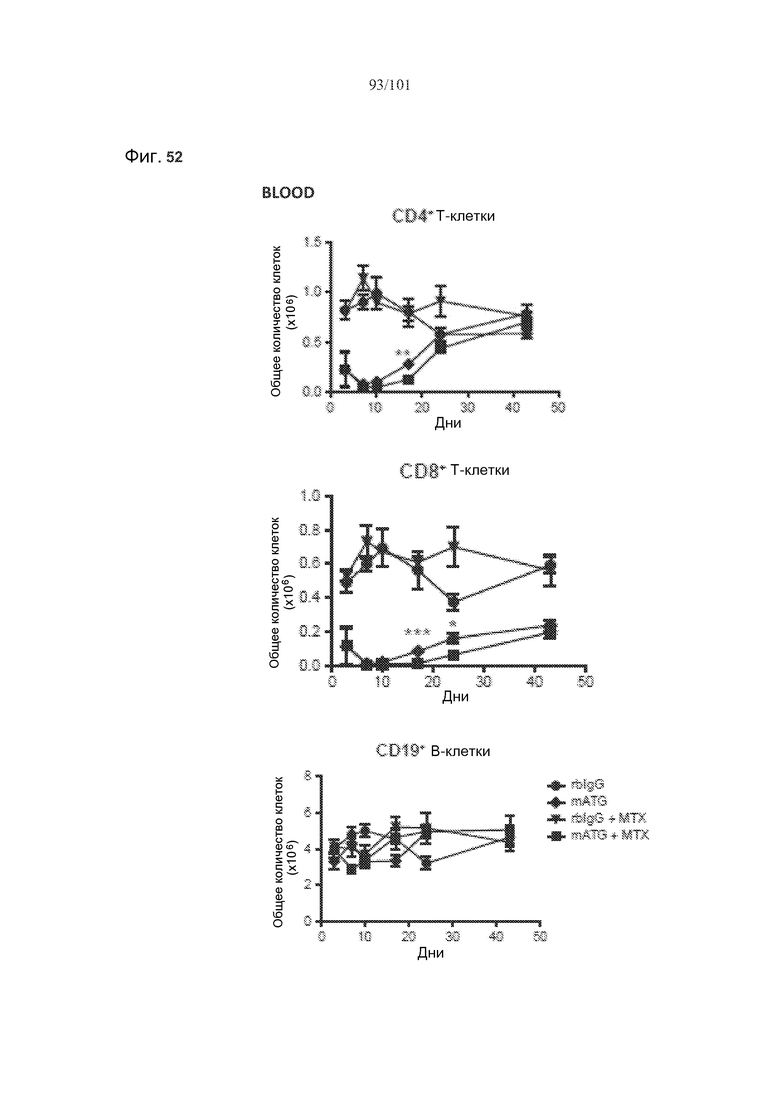
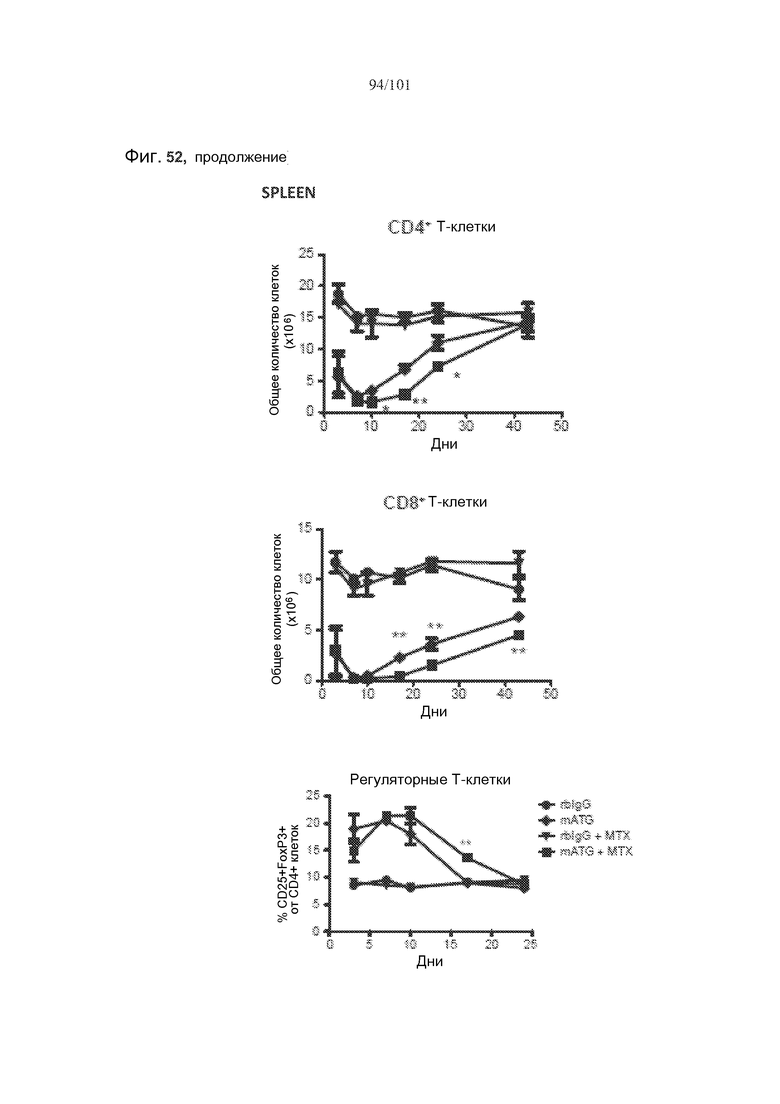
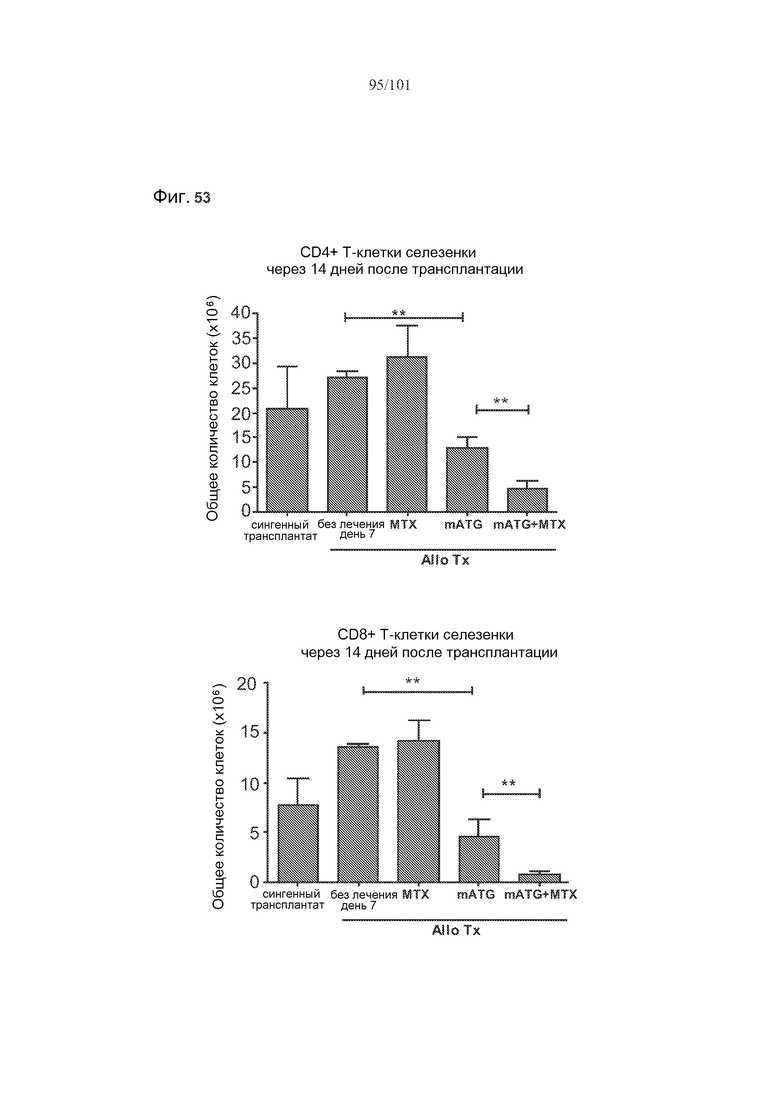
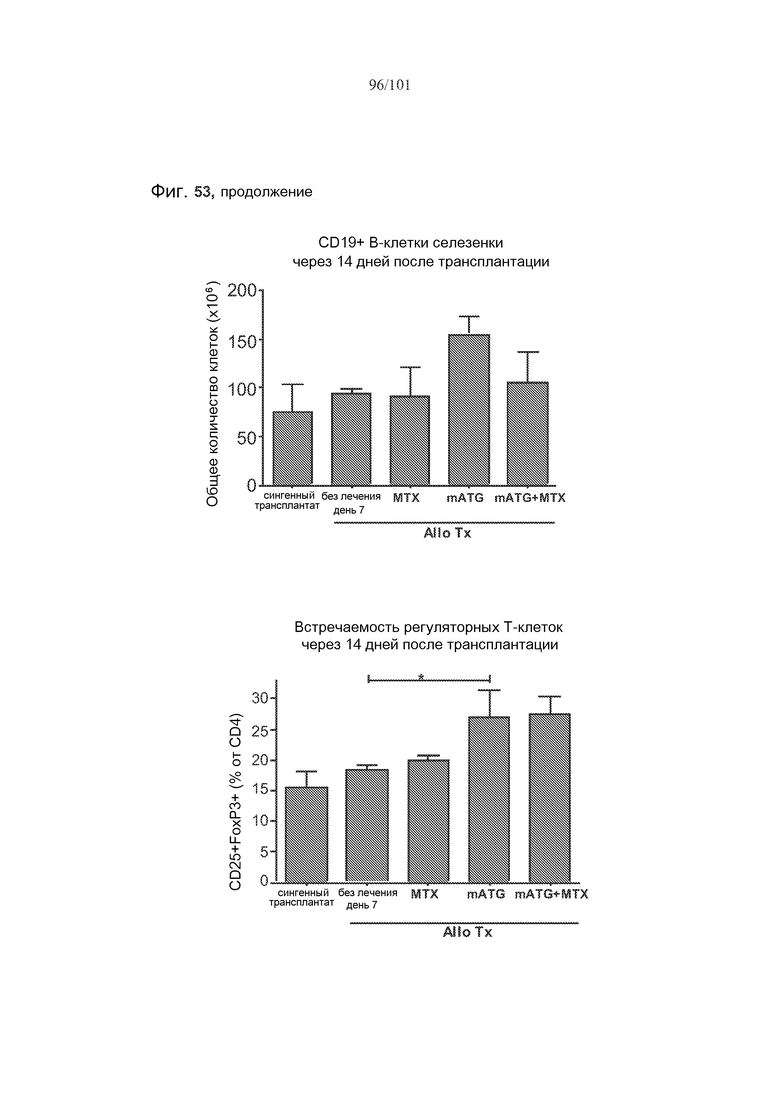
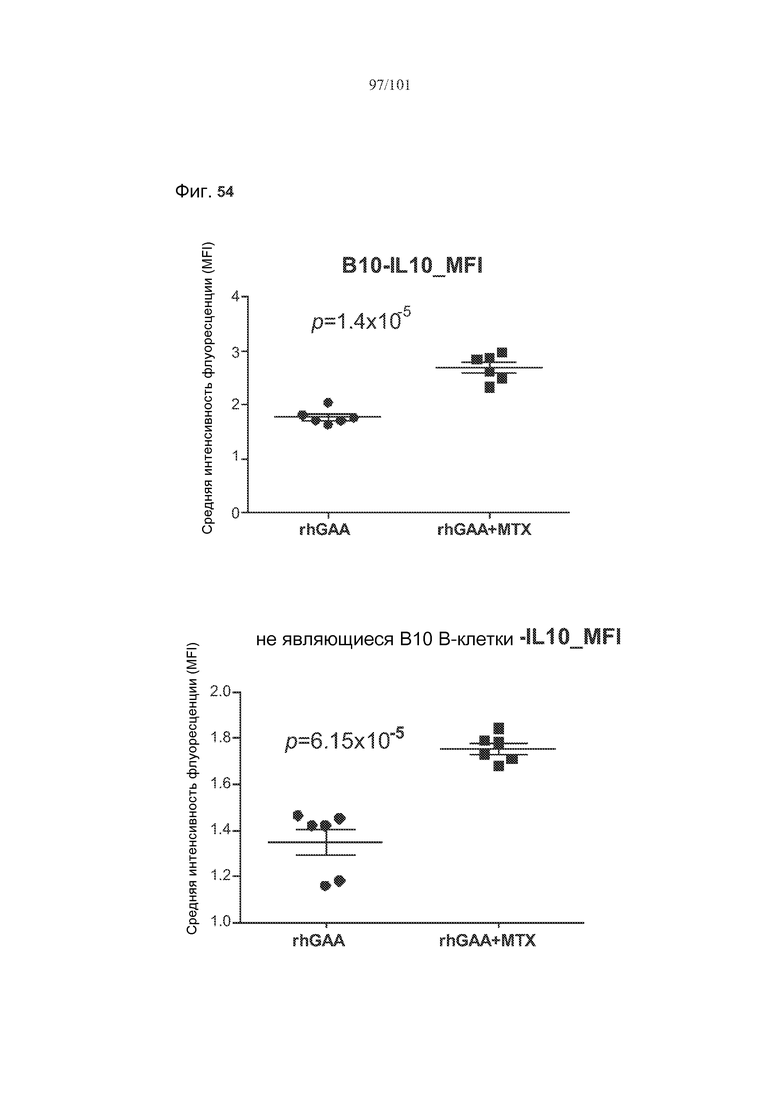
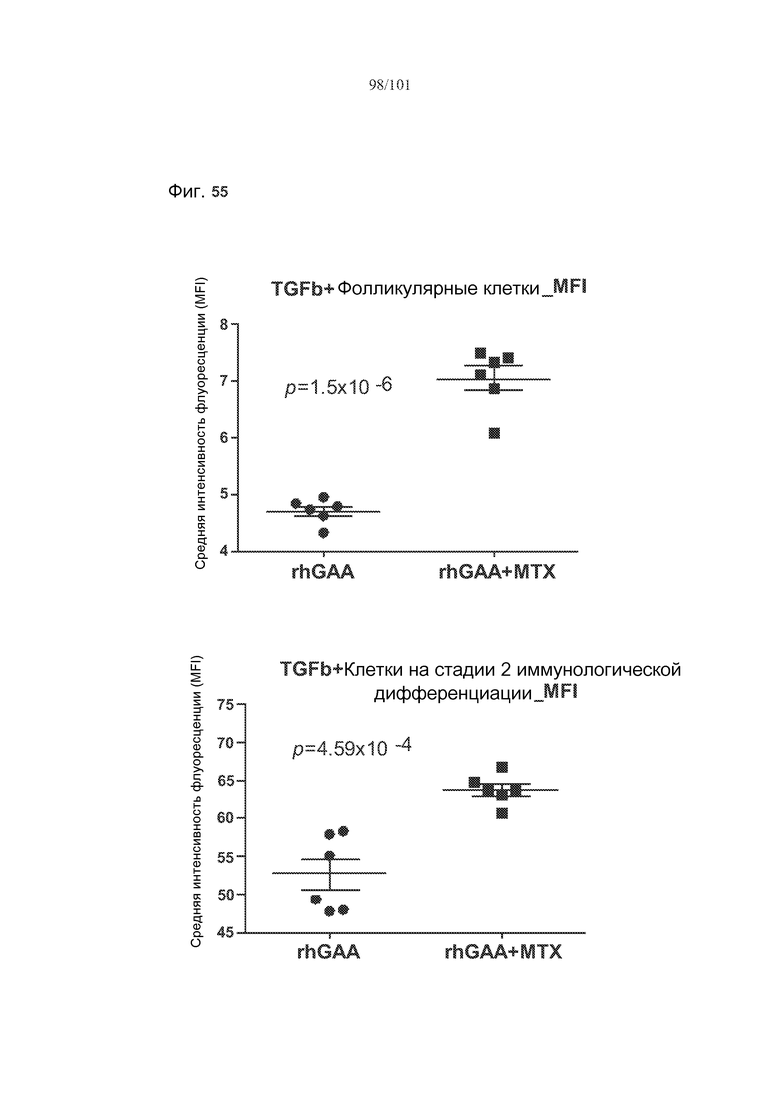
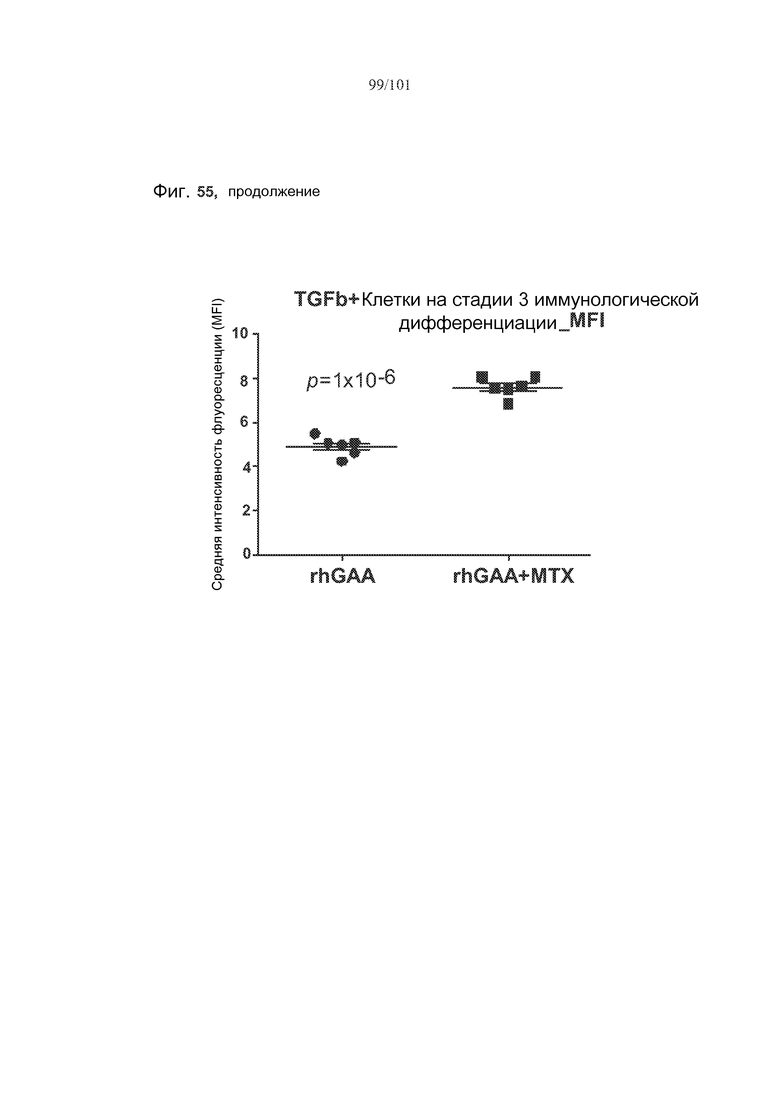
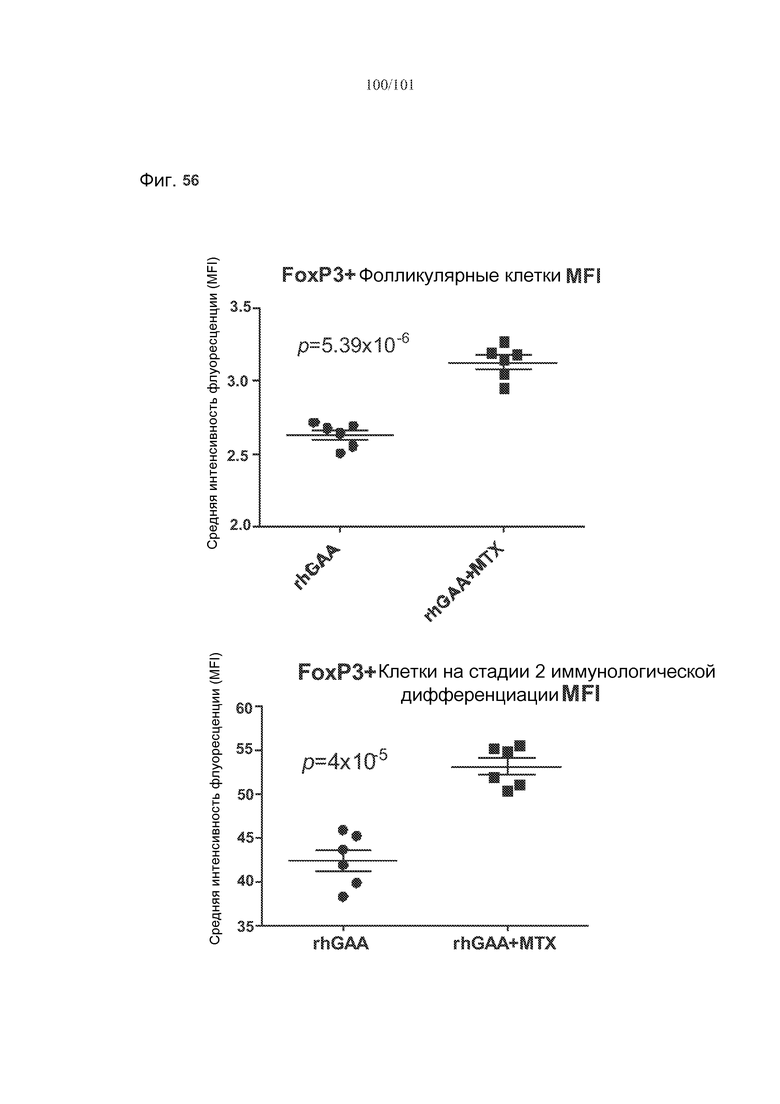
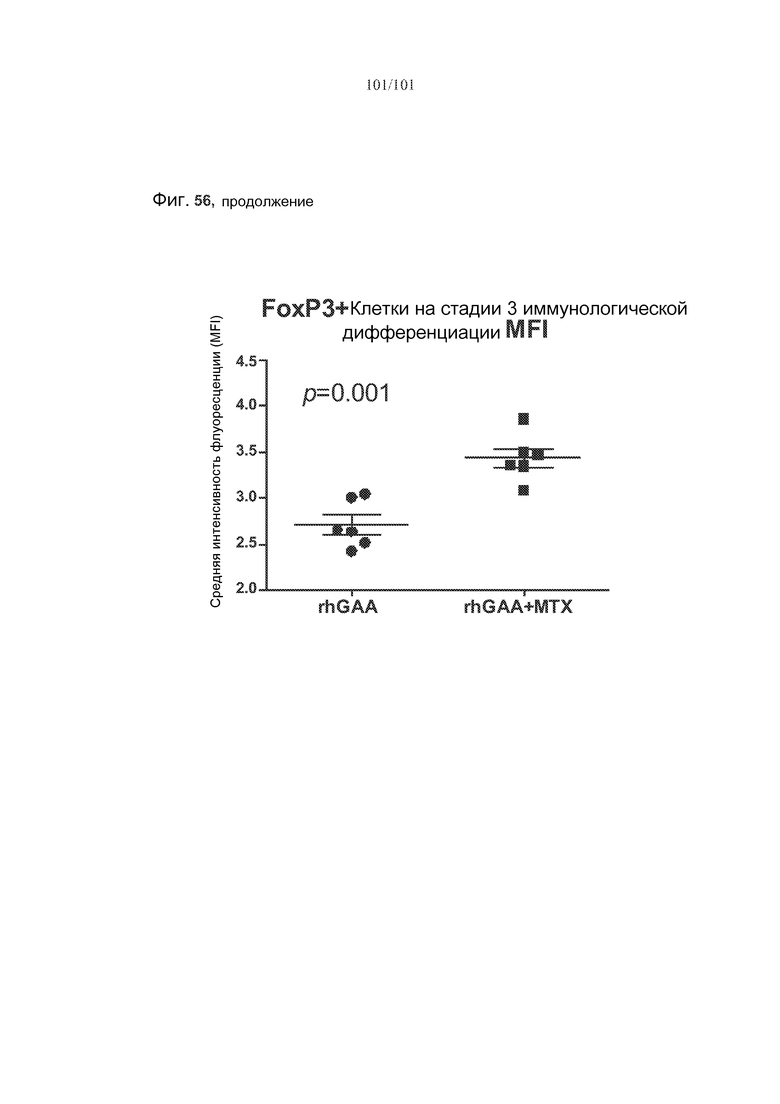
Изобретение относится к медицине, в частности онкологии, и может быть использовано для увеличения эффективности терапевтического белка у субъекта. Для этого способ включает введение субъекту эффективного количества метотрексата, где метотрексат вводят индивидууму в эффективном количестве в одном цикле, где один цикл метотрексата состоит из 1 дня введения метотрексата или 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11 последовательных дней приема метотрексата. Метотрексат вводят до, или во время, или после введения терапевтического белка, где терапевтический белок выбран из группы, состоящей из кроличьего поликлонального антитела - глобулярного белка против тимоцитов мыши (mATG), алемтузумаба и альглюкозидазы альфа человека (rhGAA), и где способ ингибирует реакцию антитела на терапевтический белок, тем самым увеличивая эффективность терапевтического белка у субъекта. Использование данного способа позволяет при лечении одним циклом метотрексата снизить нежелательные ответы антител против лекарственных антител (ADA). 14 з.п. ф-лы, 16 пр., 3 табл., 56 ил.
1. Способ увеличения эффективности терапевтического белка у субъекта, нуждающегося в лечении терапевтическим белком, включающий введение субъекту эффективного количества метотрексата, где метотрексат вводят индивидууму в эффективном количестве в одном цикле, где один цикл метотрексата состоит из 1 дня введения метотрексата или 2, 3, 4, 5, 6, 7, 8, 9, 10 или 11 последовательных дней приема метотрексата, где метотрексат вводят до, или во время, или после введения терапевтического белка, где терапевтический белок выбран из группы, состоящей из кроличьего поликлонального антитела - глобулярного белка против тимоцитов мыши (mATG), алемтузумаба и альглюкозидазы альфа человека (rhGAA), и где способ ингибирует реакцию антитела на терапевтический белок, тем самым увеличивая эффективность терапевтического белка у субъекта.
2. Способ по п. 1, где один цикл метотрексата вводят между 48 ч до и 48 ч после начала лечения терапевтическим белком.
3. Способ по п. 1, где эффективное количество метотрексата составляет от 0,1 мг/кг до 5 мг/кг.
4. Способ по п. 1, который включает:
a. индукцию иммунологической толерантности к терапевтическому белку у субъекта;
b. ингибирование образований антител против терапевтического белка у субъекта;
c. облегчение инфузионной реакции на терапевтический белок у субъекта;
d. ослабление вторичного аутоиммунитета у субъекта;
e. увеличение процента регуляторных T-клеток в Т-клеточной популяции у субъекта;
f. увеличение процента регуляторных В-клеток в В-клеточной популяции у субъекта; или
g. ингибирование Т-клеточных реакций у субъекта;
h. продление фармакокинетики терапевтического белка; или
i. истощение лимфоцитов.
5. Способ по любому из пп. 1-4, где субъектом является человек.
6. Способ по п. 1, где терапевтический белок представляет собой алемтузумаб.
7. Способ по п. 1, где терапевтический белок представляет собой кислую альфа-глюкозидазу человека.
8. Способ по любому из пп. 1-4, где субъект дополнительно получает иммуномодулирующее средство.
9. Способ по п. 8, где иммуномодулирующее средство представляет собой иммунодепрессант.
10. Способ по п. 1, где субъект имеет рассеянный склероз.
11. Способ по п. 10, где субъект имеет ремиттирующе-рецидивирующий рассеянный склероз.
12. Способ по п. 1, где субъект нуждается в трансплантации ткани или органа.
13. Способ по п. 12, где ткань или орган для трансплантации представляют собой почечную ткань или сердечную ткань.
14. Способ по п. 1, где субъект имеет апластическую анемию или имеет риск заболевания «трансплантат против хозяина».
15. Способ по п. 1, где у субъекта болезнь Помпе.
| JOSEPH A | |||
| et al | |||
| Immune tolerance induction to enzyme-replacement therapy by co-administration of short-term, low-dose methotrexate in a murine Pompe disease model // Clin Exp Immunol | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| GARMAN R.D | |||
| et al | |||
| Methotrexate reduces antibody responses to recombinant human α-galactosidase A therapy in a mouse model of Fabry disease // Clin Exp Immunol | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| SAKUMA Y | |||
| et al | |||
| Short-course methotrexate and long-term acceptance of fully allogeneic rat cardiac grafts: a possible mechanismof tolerance // Transpl Immunol | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| US 2009155207 A1, 18.06.2009. | |||

