Область применения
Изобретение относится к области биотехнологии и касается термостабильных ферментов с активностью липазы. Предлагается новая липаза из термоалкалофильной бактерии Thermosyntropha lipolytica и способ ее выделения.
Актуальность
Ферменты - биокатализаторы широко используются в различных отраслях промышленности, сельского хозяйства и медицины. Однако для многих задач необходимые ферменты отсутствуют. Причиной этого является то, что большинство известных ферментов не выдерживают «экстремальных» условий реакций, в которых они должны работать, - высокой температуры, низкой или, наоборот, высокой кислотности и пр. Источником таких ферментов могут являться микроорганизмы, обитающие в природных местообитаниях с требуемыми условиями - экстремофилы.
Липазы осуществляют реакции гидролиза и синтеза длинноцепочечных эфиров жирных кислот (Gupta et al., 2004; Sarda et al., 1958). Они широко используются в качестве биокатализаторов в различных биотехнологиях, в том числе в пищевой, целлюлозно-бумажной и текстильной промышленности, при проведении тонкого органического синтеза, производстве детергентов и др. (Jaeger et al., 1994; Pandey et al., 1999; Nagao et al., 2001; Kim et al., 2002). Перспективным направлением использования липаз является производство биодизеля из растительных масел (Bisen et al., 2010).
Основными коммерчески-значимыми характеристиками, которыми должны обладать липазы, являются термостабильность и устойчивость к щелочным условиям, а также к органическим растворителям и другим «экстремальным» условиям (Kulkumi and Gadre, 1999). Несмотря на продолжающийся поиск таких ферментов, было обнаружено лишь несколько липаз, сохраняющих активность при температурах 80-90°С, но их оптимум актиности достигался при нейтральных значения рН. Поэтому актуальной задачей, стоящей перед крупными исследовательскими центрами и ведущими биотехнологическими компаниями, является поиск новых липаз, обладающих высокой термостабильностью и активных в условиях щелочной среды.
Уровень техники
Липолитические ферменты катализируют гидролитическое расщепление сложных эфиров на спирты и кислоты, причем эстеразы расщепляют преимущественно короткие, а липазы - длинные цепи.
Липазы и эстеразы встречаются у всех живых организмов, но наибольшим потенциалом практического применения обладают ферменты из бактерий и грибов (Wu et al., 1996). Источником липолитических ферментов являются многим бациллы (Kim et al., 1994; Schmidt-Dannert et al., 1994; Rua et al., 1997). Большая часть ферментов активны при температурах не выше 60°С и нейтральных значениях pH, однако, липазы из Bacillus thermoleovorans и Rhizopus oryzae могут сохранять часть активности и при более высоких температурах и pH (Lee et al., 1999; Hiol et al., 2000).
Хотя некоторые липазы, выделенные из мезофильных организмов, обладают умеренной термостабильностью, естественным источником таких ферментов являются экстремофильные микроорганизмы, живущие при соответствующих физико-химических условиях (в первую очередь - температура и pH среды). Так, обладающие более высокой термостабильностью липазы были выделены из анаэробных гипертермофильных бактерий Thermoannaerobacter thermohydrosulfuricus и Caldanaerobacter subterraneus (Royter et al., 2009).
Thermosyntropha lipolytica - термофильная анаэробная бактерия, выделенная из щелочного озера Богория в Кении (Svetlitshnyi et al., 1996). Этот относящийся к порядку Clostridiales микроорганизм растет в широком диапазоне температур от 52 до 70°C при щелочных значениях pH (7.15-9.5). Т. lipolytica растет на органических субстратах, в том числе липидах, гидролиз которых осуществляется секретируемыми в окружающую среду липазами. Ранее из одного из штаммов Т. lipolytica, DSM 11003, были выделены и функционально охарактеризованы две липазы, проявлявшие максимум активности при температуре около 96°C и pH 9.5 (Salameh and Wiegel, 2007; Salameh and Wiegel, 2009). Ферменты были выделены из супернатанта культуры Т. lipolytica и очищены до гомогенного состояния, однако, кодирующие их гены не были идентифицированы (Salameh and Wiegel, 2007), что не позволяет получать рекомбинантные ферменты в практически-значимых количествах. Полные аминокислотные последовательности соответствующих белков не известны, были определены лишь короткие последовательности N-концевых районов (Salameh and Wiegel, 2007).
Настоящее изобретение касается выделения и характеристики новой липазы из термоалкалофильной бактерии Т. lipolytica.
Раскрытие изобретения
Анализ определенной нуклеотидной последовательности генома бактерии Т. lipolytica выявил наличие в геноме этого организма как минимум 7 генов, кодирующих ферменты, относящиеся к классу липаз и эстераз. Аминокислотные последовательности двух липаз содержали N-концевые сигнальные последовательности, которые могут обеспечивать их секрецию из клетки в окружающую среду. Один из этих генов, TSLip1, имеющий нуклеотидную последовательность SEQ ID NO1, кодирует белок, состоящий из 478 а.о., включающий N-концевую сигнальную последовательность длиной 28 а.о.
Методом культивирования рекомбинантного штамма Escherichia coli Rosetta-gami (DE3), трансформированного вектором pQE80L (Qiagen), содержащим фрагмент (с 85 по 1437 нуклеотид) последовательности SEQ ID NO 1, кодирующий «зрелый» белок без сигнального пептида, была выделена и очищена с помощью металло-хелатной хроматографии новая рекомбинантная термостабильная липаза TSLip1, содержащая аминокислотную последовательность SEQ ID NO2. Функциональная характеристика фермента показала, что рекомбинантная липаза проявляла гидролитическую активность в отношении паранитрофениловых эфиров с различной длиной цепи, а также триглицеридов, в том числе растительных масел. Оптимальные условия реакции достигались при температуре 70-80°С и рН 8. Фермент сохраняет более 80% активности в присутствии 10% метанола. Таким образом, новая термостабильная липаза может быть перспективным биокатализатором для органического синтеза, применения в пищевой промышленности, производства детергентов и получения биодизеля.
Краткое описание рисунков
Фиг.1. Аминокислотная последовательность белкового продукта гена TSLip1. Последовательность N-концевого сигнального пептида подчеркнута, консервативный мотив [LIV]-X-[LIVFY]-[LIVMST]-G-[HYWV]-S-XG-[GSTAC], присутствующий в липазах и эстеразах, выделен серым.
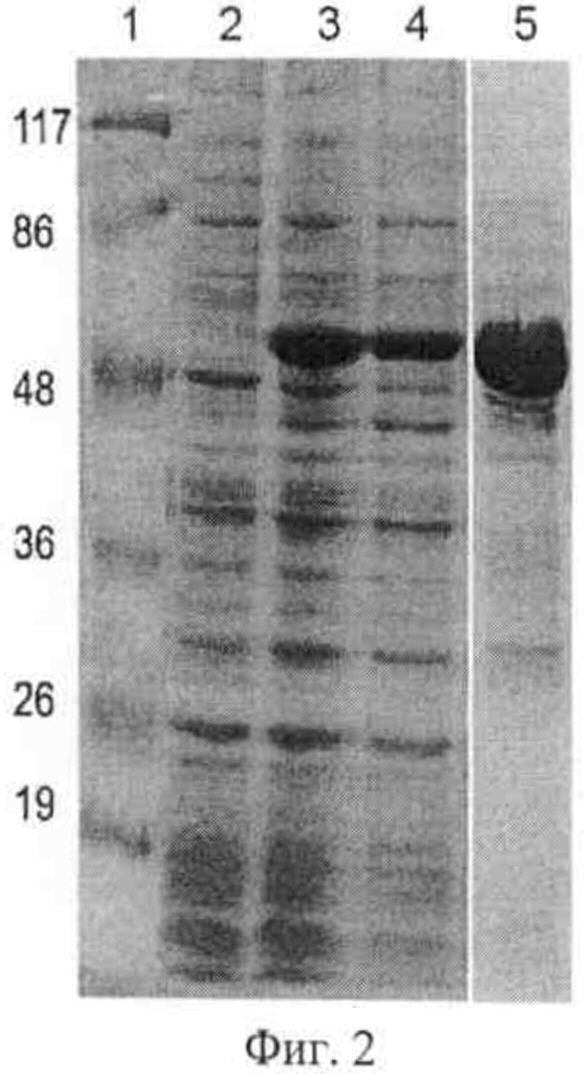
Фиг.2. Экспрессия TSLip1 в клетках Е.coli.
Приведены результаты электрофоретического анализа белковых препаратов:
1 - маркер молекулярной массы, размеры указаны в кД,
2 - суммарный белковый препарат, выделенный из клеток штамма E.coli Rosseta-gami (DE3), содержащего плазмиду pQE80-TSLip1, до индукции синтеза TSLip1,
3 - то же, что на дорожке 2, но через 3 часа после индукции синтеза TSLip1 внесением в среду 1 мМ Изопропил-бета-D-тиогалактозид (ИПТГ),
4 - фракция растворимых белков, через 3 часа после индукции синтеза TSLip1 внесением в среду 1 мМ ИПТГ,
5 - очищенный препарат рекомбинантной липазы TSLip1.
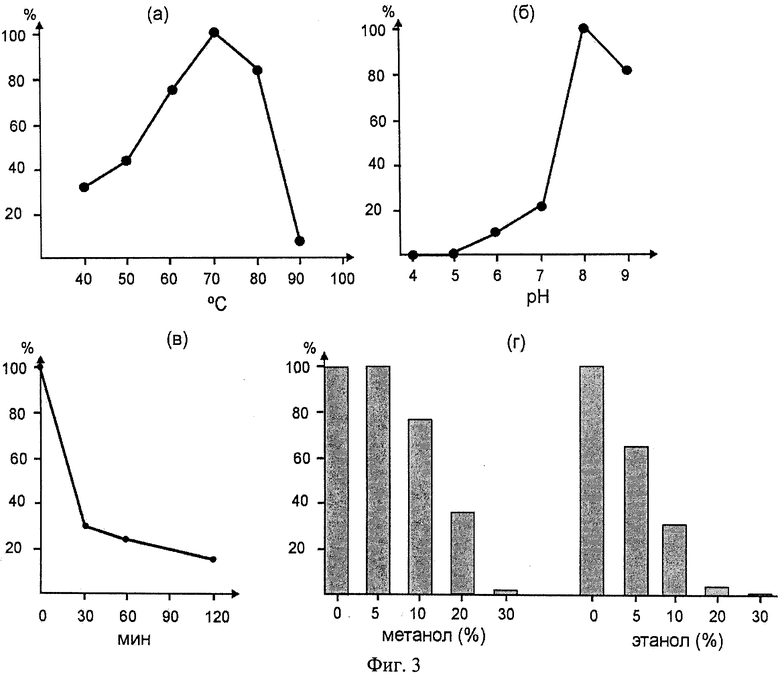
Фиг.3. Гидролитическая активность TSLip1 в отношении 4-нитрофениловых эфиров
(а) Зависимость активности TSLip1 от температуры. В качестве субстрата использовалии 4-нитрофенил бутират. Реакцию проводили в течение 2.5 минут в реакционном буфере с рН 7.0 при указанной температуре.
(б) Зависимость активности TSLip1 от рН буфера. В качестве субстрата использовали 4-нитрофенил бутират. Реакцию проводили в течение 3 минут в реакционном буфере с указанным рН при температуре 60°С.
(в) Термостабильность TSLip1 при 80°С. Фермент инкубировали в реакционном буфере с рН 7.0 при 80°С в течение указанного времени, затем проводили реакцию гидролиза с 4-нитрофенил бутиратом при 60°С в течение 3 мин.
(г) Зависимость активности TSLip1 от содержания метанола и этанола (%) в реакционной смеси. В качестве субстрата использовали 4-нитрофенил бутират. Реакцию проводили в течение 3 минут в реакционном буфере с рН 7.0 при температуре 70°С.
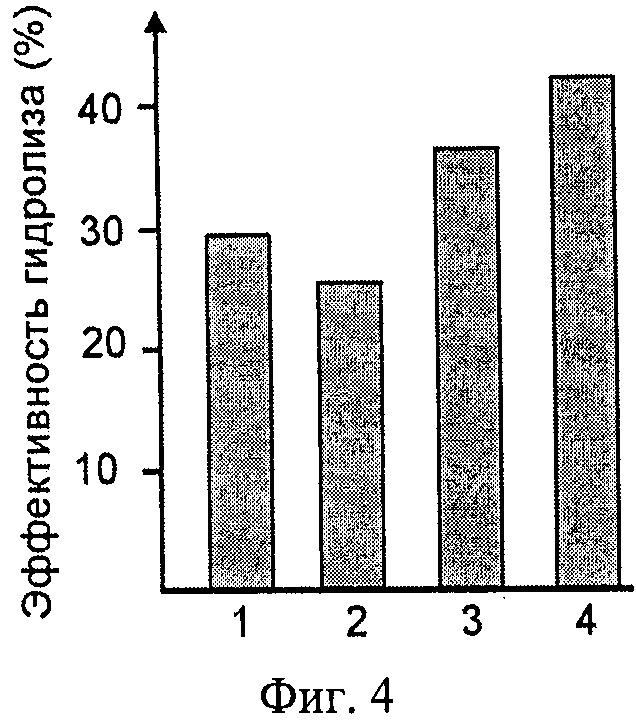
Фиг.4. Гидролиз растительных масел липазой TSLip1. Показан процент гидролиза соевого (1), оливкового (2), кукурузного (3) и подсолнечного (4) масел. Реакцию проводили в течение 2 часов при температуре 50°С.
Осуществление изобретения
Пример 1. Идентификация гена липазы в геноме Т. lipolytica.
Термоалкалофильная бактерия Т. lipolytica (Svetlitshnyi et al., 1996) была выделена из щелочного озера Богория в Кении. Определена нуклеотидная последовательность генома этого микроорганизМа (штамма из нашей коллекции, отличного от использованного в работе Salameh and Wiegel, 2007), в результате чего были идентифицированы белок-кодирующие гены (Гумеров и др. 2012). Сравнение аминокислотных последовательностей предсказанных белков Т. lipolytica с представленными в GenBank аминокислотными последовательностями выявило наличие в геноме Т. lipolytica как минимум трех генов ферментов, относящихся к классу липаз и эстераз, аминокислотные последовательности двух белков содержали N-концевые сигнальные последовательности, которые могут обеспечивать их секрецию из клетки.
Ген TSLip1, имеющий нуклеотидную последовательность SEQ ID NO1, кодирует полипептид длиной 478 а.о., включающий N-концевую сигнальную последовательность длиной 28 а.о. (Фиг.1). Анализ аминокислотной последовательности предсказанного белкового продукта этого гена выявил наличие консервативного мотива [LIV]-X-[LIVFY]-[LIVMST]-G-[HYWV]-S-XG-[GSTAC], входящего в активный центр эстераз и липаз (Blow, 1990). В С-концевой области белка отсутствовали трансмембранные домены, которые могут обеспечивать "заякоривание" фермента на поверхности клетки, что вероятно свидетельствует о том, что липаза секретируется в среду, а не остается связанной с клетками. В результате поиска сходных последовательностей в GenBank была обнаружено, что ближайшими гомологами идентифицированной липазы являются липазы бактерий типа Firmicutes, - Clostridium tetani, Clostridium botulinum и Aneurinibacillus thermoaerophilus (50-55% идентичности). "Зрелый" белок без N-концевой сигнальной последовательности имеет аминокислотную последовательность SEQ ID NO2. Отметим, что по аминокислотной последовательности продукт гена TSLip1 отличается от липаз из другого штамма Т. lipolytica, описанных в работе (Salameh and Wiegel, 2007).
Пример 2. Получение штамма Е. coli - продуцента рекомбинантной липазы, выделение и очистка фермента.
Праймеры LipF_BamHI (5'-ATGGGATCCCAAGAAATTTCGAGTAACAA-3') и LipR_SalI (5'-AAGGTCGACTTCTACTCTTCAGCAATAT-3') были использованы для ПЦР-амплификации фрагмента гена TSLip1 (с 85 по 1437 нуклеотид в последовательности SEQ ID NO 1), соответствующего «зрелому» белку без N-концевой сигнальной последовательности, в качестве матрицы использовали геномную ДНК Т. lipolytica. Полученный фрагмент обрабатывали рестриктазами BamHI и SalI и клонировали по сайтам BamHI и SalI в экспрессионном векторе pQE80L (Qiagen), позволяющем экспрессировать целевой белок, слитый с 6 аминокислотными остатками гистидина на N-конце. В результате был получен экспрессионный вектор pQE80-TSLip1, обеспечивающий продукцию рекомбинантного фермента, TSLip1, содержащего N-концевой шести-гистидиновый "таг" и последовательность «зрелой» липазы.
Для продукции рекомбинантной липазы TSLip1 экспрессионным вектором pQE80-TSLip1 трансформировали штамм Е. coli Rosetta-gami (DE3). Указанный штамм, содержащий pQE80-TSLip1, выращивали в среде LB с добавлением ампициллина (100 мг/л) на шейкере при 37°C до середины логарифмической фазы роста (OD600~0.6), затем индуцировали синтез рекомбинантного белка, внося изопропил-β-D-тиогалактопиранозид (ИПТГ) до 1 мМ, и продолжали выращивать культуру в течение 20 ч при 37°C.
Клетки штамма-продуцента собирали центрифугированием, выделение и очистку рекомбипантной липазы TSLip1 осуществляли методом металл-афинной хромотографии с помощью набора Spin-NTA KIT (Qiagen, Германия). Препарат диализовали с помощью колонок Illustra NAP-10 (GE Healthcare) и хранили при -20°C в буфере, содержащем 20 мМ Трис-HCl, pH 8.0, 100 mM NaCl, 5 тМ 2-МЕ, и 40% глицерин.
Гомогенность препарата TSLip1 и соответствие молекулярного веса рекомбинантного белка расчетному значению 51.1 кД была подтверждена при помощи SDS-PAGE (Фиг.2).
Пример 3. Зависимость гидролитической активности TSLip1 от температуры, pH и присутствия органических растворителей.
Для определения гидролитической активности TSLip1 в зависимости от температуры, pH и присутствия органических растворителей в реакционной среде, в качестве субстрата использовали 4-нитрофенил бутират (pNB, Sigma N9876). Субстрат разводили в ацетонитриле до концентрации 100 мМ. Реакционная смесь содержала 5 мкМ CaCl2, 100 мМ Hepes (pH 7.5), 1.25 мкМ pNB и 0.02 мкг фермента в объеме 1 мл. Реакцию останавливали добавлением 0.5 объема этанола. Количество освобожденного в результате гидролиза паранитрофенола определяли спектрофотометрически при длине волны 405 нм.
Определение зависимости активности TSLip1 от температуры и pH проводили, определяя эффективность гидролиза 4-нитрофенил бутирата. Оптимальная температура реакции составляла 70°C, не менее 80% активности липаза проявляла при 60°C и 80°C (Фиг.3а). При более высоких температурах активность фермента резко снижалась. Зависимость активности липазы от pH (Фиг.3б) определяли в 0.1М ацетатном (для pH 3-5), 0.1М фосфатном (pH 6.0) или в 0.1М трис (pH 6-9) буферах, проводя реакцию гидролиза 4-нитрофеиил бутирата при температуре 70°C. Фермент не проявлял активности при pH ниже 5.0, максимальная активность наблюдалась при pH 8, при pH 9 активность незначительно снижалась. Нестабильность субстрата в щелочных условиях не позволила провести измерения активности липазы при более высоких значениях pH.
Для характеристики термостабильности TSLip1 препарат фермента инкубировали в реакционном буфере при 80°C различное время, затем охлаждали до 60°C, вносили 4-нитрофенил бутирате и проводили реакцию при 60°C в стандартном реакционном буфере. Быстрое снижение активности примерно в 3 раза наблюдалось в течение первых 30 мин инкубации при 80°C, в дальнейшем активность снижалась намного медленнее (время 50%-ной инактивации фермента - около 90 мин, Фиг.3в). Быстрая первоначальная инактивация фермента при 80°C может объяснять наблюдавшееся резкое снижение активности при температурах 80°C и выше, реальный температурный оптимум активности фермента может быть выше 70°C.
Поскольку одним из перспективных направлений практического использования липаз является получение биодизеля из растительных масел, т.е. конверсия триглицеридов растительных масел в эфиры жирных кислот в реакции трансэстерификации с помощью спиртов, обычно, этанола или метанола, протестировали устойчивость фермента к присутствию этих спиртов в реакционной смеси. Для этого реакции гидролиза 4-нитрофенил бутирата проводили в присутствии этанола или метанола. Полученные результаты (Фиг.3г) показывают, что метанол в концентрации до 10% практически не оказывает ингибирующего воздействия, полное ингибирование активности липазы наблюдается лишь в присутствии 30% метанола. Этанол вызывал более сильное ингибирование активности TSLip1, наблюдавшееся уже при концентрации 5%, внесение этанола до 20% практически полностью ингибировало реакцию.
Пример 5. Гидролиз растительных масел липазой TSLip1.
В ходе экспериментов по культивированию штамма-продуцента TSLip1 на чашках Петри с добавлением трибутирина в условиях индукции экспрессии липазы было обнаружено образование светлого «гало» вокруг колоний, указывающего на специфический гидролиз субстрата. В качестве «природных» триглицеридов для функциональной характеристики TSLip1 использовали растительные масла - оливковое, соевое, кукурузное и подсолнечное. Активность липазы измеряли методом титрометрии.
Реакционную смесь, содержащую 100 мкл масла и 100 мкл 0.1 М фосфатного буфера (pH 7.0), прогревали при температуре проведения реакции в течение 10 мин, затем вносили 10 мкг фермента (0.01% вес/объем) и инкубировали в течение 2 ч при периодическом перемешивании. По завершении гидролиза к пробе добавляли 10 мл смеси ацетона с этанолом (50:50) для растворения масла и остановки реакции. Затем вносили 11 мкл индикатора фенолфталеина (1% в 95% этаноле) и проводили титрование образовавшихся свободных жирных кислот 0.1 М КОН.
Процент гидролиза определяли как процент образовавшихся в образце свободных жирных кислот от теоретически возможного максимального количества, определенный по формуле (Rooney and Weatherley, 2001): % гидролиза =((V*М*MW)/(Wt*f))*100, где V - объем, взятой на реакцию КОН, М - молярность КОН (0.1 М), MW - средняя молекулярная масса жирных кислот (гр/моль), Wt - вес реакционной смеси, f - доля масла в начале реакции. Полученные результаты (Фиг.4) показывают, что TSLip1 осуществляет гидролиз всех использованных растительных масел, причем максимальная активность наблюдалась в отношении подсолнечного и кукурузного масел.
СПИСОК ЛИТЕРАТУРЫ
1. Гумеров В.М., Марданов А.В., Колосов П.М., Равин Н.В. (2012) Выделение и характеристика липазы из термоалкалофильной бактерии Thermosyntropha lipolytica. Прикладная биохимия и микробиология, т.48, №4, с.376-382.
2. Hiol A., Jonzo M.D., Rugani N., Druet D. Sarda L., Comeau L.C. (2000) Purification and characterization of an extracellular lipase from a thermophilic Rhizopus oryzae strain isolated from palm fruit. Enzyme Microb. Technol. 26, 421-430.
3. Bisen P.S., Sanodiya B.S., Thakur G.S., Baghel R.K., Prasad G.B. (2010) Biodiesel production with special emphasis on lipase-catalyzed transesterification. Biotechnol. Lett. 32(8), 1019-1030.
4. Blow D. (1990) Enzymology. More of the catalytic triad. Nature. 343(6260), 694-695.
5. Lee D., Koh Y., Kirn K., Kirn В., Choi H., Kim D., Suhartono M.T., Pyun Y. (1999) Isolation and characterisation of thermophilic lipase from Bacillus thermoleovorans ID-1. FEMS Microbiol. Lett. 179, 393-400.
6. Gupta R., Gupta N., Rathi P. (2004) Bacterial lipases: an overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 64(6), 763-781.
7. Jaeger K.E., Ransac S., Dijkstra B.W., Colson C., van Heuvel М., Misset O. (1994) Bacterial lipases. FEMS Microbiol. Rev. 15, 29-63.
8. Kim H., Sung М., Kim М., Oh T. (1994) Occurrence of thermostable lipase in thermophilic Bacillus sp. strain 398. Biosci. Biotech. Biochem. 58, 961-962.
9. Kim I., Kim H., Lee K., Chung S., Ко S. (2002) Lipase-catalyzed acidolysis of perilla oil with caprylic acid to produce structured lipids. JAOCS 79, 363-367.
10. Kulkumi N., Gadre R.V. (1999) A novel alkaline, thermostable, protease free lipase from Pseudomonas sp. Biotechnol. Lett. 21, 897-899.
11. Rua M.L., Schmidt-Dannert C., Wahl S., Sprauer A., Schmid R.D. (1997) Thermoalkalophilic lipase of Bacillus thermocatenulatus. Large scale production, purification and properties: aggregation behavior and its effect on activity. J. Biotechnol. 56, 89-102.
12. Nagao Т., Shimada Y., Sugihara A., Murata A., Komemushi S., Tominaga Y. (2001). Use of thermostable Fusarium heterosporum lipase for production of structured lipid containing oleic and palmitic acids in organic solvent- free system. JAOCS. 78, 167-172.
13. Pandey A., Benjamin S., Soccol C.R., Nigam P., KriegerN., Soccol V.T. (1999) The realm of microbial Upases in biotechnology: a review. Biotechnol. Appl. Biochem. 29, 119-131.
14. Rooney D., Weatherley L.R. (2001) The effect of reaction conditions upon lipase catalysed hydrolysis of high oleate sunflower oil in a stirred liquid-liquid reactor. Process Biochem. 36(1), 947-953.
15. Royter M., Schmidt M., Elend С., Hobenreich H., Schafer Т., Bomscheuer U.T., Antranikian G. (2009) Thermostable Upases from the extremophilic anaerobic bacteria Thermoannaerobacterthermohydrosulfuricus SOLI and Caldanaerobacter subterraneus subsp. tengcongensis. Extremophiles. 2009, 13, 769-783.
16. Salameh M.A., Wiegel J. (2007) Purification and characterization of two highly thermophilic alkaline lipases from Thermosyntropha lipolytica. Appl Environ Microbiol. 73(23), 7725-7731.
17. Salameh M.A., Wiegel J. (2009) Synthesis of fatty acid esters and diacylglycerols at elevated temperatures by alkalithermophilic lipases from Thermosyntropha lipolytica. J. Ind. Microbiol. Biotechnol. 36(10), 1281-1287.
18. Sarda L., Desnuelle P. (1958) Actions of pancreatic lipase on esters in emulsions. Biochim. Biophys. Acta. 30(3), 513-521.
19. Schmidt-Dannert С., Sztajer H., Stocklein W., Menge U., Schmid R.D. (1994) Screening, purification and properties of a thermophilic lipase from Bacillus thermocatelnulatus. Biochem. Biophys. Acta. 1214, 43-53.
20. Svetlitshnyi V., Rainey F., Wiegel J. (1996) Thermosyntropha lipolytica gen. nov., sp.nov., a lipolytic, anaerobic, alkalitolerant, thermophilic bacterium utilizing short- and long-chain fatty acids in syntrophic coculture with a methanogenic archaeum. Int J Syst Bacteriol. 46(4): 1131-1137.
21. Wu X.Y, Jaaskelainen S., Linko Y.Y. (1996) An investigation of crude lipase for hydrolysis, esterification, and transesterification. Enzyme Microb. Technol. 19, 226-231.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИИ Escerichia coli XL1-blue/pQS-G3, ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ ЛИПАЗЫ БАКТЕРИИ Geobacillus stearothermophilus G3 | 2013 |
|
RU2540873C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pEstPc, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5, И ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5 | 2011 |
|
RU2478708C1 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОЙ БЕТА-ГАЛАКТОЗИДАЗЫ AsBgl 1390 ИЗ АРХЕИ SACCHAROVORANS В КАЧЕСТВЕ БЕТА-ГЛЮКОЗИДАЗЫ, БЕТА-КСИЛОЗИДАЗЫ И БЕТА-МАННОЗИДАЗЫ | 2012 |
|
RU2523581C2 |
| Рекомбинантная плазмида pBU-LipA, обеспечивающая синтез белка липазы А штамма Bacillus natto IAN | 2023 |
|
RU2808501C1 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ BACILLUS LICHENIFORMIS - ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ ЛИПАЗЫ | 2012 |
|
RU2500812C1 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ГИДРОЛАЗЫ, КОДИРУЮЩИЕ ИХ НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2009 |
|
RU2525675C2 |




Изобретение относится к области биотехнологии и представляет собой новую термостабильную липазу из термоалкалофильной бактерии Thermosyntropha lipolytica. Ген, кодирующий липазу, был идентифицирован в геноме Thermosyntropha lipolytica, соответствующий рекомбинантный фермент, TSLip1, был экспрессирован в Escherichia coli, очищен и охарактеризован. Установлено, что TSLip1 проявляет гидролитическую активность в отношении паранитрофенил бутирата, а также триглицеридов, в том числе растительных масел. Оптимальные условия реакции достигались при температуре 70-80°С и рН 8. Фермент устойчив к органическим растворителям и сохраняет более 80% активности в присутствии 10% метанола. Изобретение позволяет получить новую липазу с высокой термостабильностью, активную в условиях щелочной среды. 2 н.п.ф-лы, 4 ил., 5 пр.
1. Рекомбинантная термостабильная липаза TSLip1 из термоалкалофильной бактерии Thermosyntropha lipolytica, проявляющая активность в реакциях гидролиза 4-нитрофенил бутирата и триглицероидов, в том числе растительных масел, при температурах до 80°C в условиях щелочной среды, содержащая аминокислотную последовательность SEQ ID NO:2, полученная из рекомбинантного штамма Escherichia coli.
1. Нуклеиновая кислота, кодирующая термостабильную липазу из термоалкалофильной бактерии Thermosyntropha lipolytica, имеющая нуклеотидную последовательность SEQ ID NO:1.
| SVETLITSHNYI et | |||
| al | |||
| Thermosyntropha lipolytica gen | |||
| nov., sp | |||
| nov., a lipolytic, anaerobic, alkalitolerant, thermophilic bacterium utilizing short- and long-chain fatty acids in syntrophic coculture with a methanogenic archaeum, J Syst Bacteriol | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| SALAMEH MA et | |||
| al | |||
| Purification and characterization of two highly | |||

