ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новому липолитическому ферменту, в частности к новому липолитическому ферменту и к кодирующим его нуклеотидным последовательностям. Настоящее изобретение также относится к способам получения нового липолитического фермента и к его применениям. Настоящее изобретение относится также к способам и применениям липолитического фермента.
ПРЕДШЕСТВУЮЩАЯ ОБЛАСТЬ ТЕХНИКИ
Полезное применение липолитических ферментов, действующих на гликолипиды, в изготовлении хлеба объясняют в EP 1193314. Объясняют, что обнаружили, что продукты частичного гидролиза лизогликолипидов обладают очень высокой эмульгирующей активностью. Однако, обнаружено, что ферменты, указанные в EP 1193314, обладают также значительной неселективной активностью в отношении триглицеридов, что приводит к чрезмерному повышению уровня свободной жирной кислоты.
Липолитический фермент из Fusarium oxysporum, обладающий фосфолипазной активностью, указан в EP 0869167. Этот липолитический фермент обладает высокой гидролизующей триацилглицериды (липазной) активностью. В настоящее время этот фермент выпускается Novozymes A/S (Denmark) как Lipopan F™.
В WO02/00852 описаны пять липазных ферментов и кодирующих их полинуклеотидов, выделенных из Fusarium venenatum, F. sulphureum, Aspergillus berkeleyanum, F. culmorum и F. solani. Все пять ферментов описаны как обладающие высокой гидролизующей триацилглицерин активностью, фосфолипазной и галактолипазной активностью.
Получены варианты липолитических ферментов, с конкретными аминокислотными заменами и слитые белки; некоторые из них обладают усиленной активностью в отношении полярных липидов по сравнению с исходными ферментами дикого типа. В WO01/39602 описан такой вариант, названный SP979, который является слитым белком липазы Thermomyces lanuginosus, и липаза Fusarium oxysporum описана в EP 0 869 167. Обнаружено, что этот вариант обладает значительно высокой степенью активности в отношении фосфолипидов и гликолипидов по сравнению с триглицеридами.
В WO02/094123 обнаружили, что можно достичь улучшенной функциональности выбором липолитических ферментов, активных в отношении полярных липидов (гликолипидов и фосфолипидов) в тесте, но, в основном, не активных в отношении триглицеридов или 1-моноглицеридов.
В находящейся одновременно на рассмотрении заявке PCT номер PCT/IDB2005/000875 описаны липолитические ферменты дикого типа, обладающие более высокой степенью активности в отношении полярных липидов по сравнению с триглицеридами. Однако в этом документе не указывают липолитических ферментов из видов Streptomyces, Thermobifida или Corynebacterium.
До настоящего изобретения не опубликовано липолитических ферментов из видов Streptomyces, обладающих активностью или значительной активностью в отношении гликолипидов. Аналогично, не опубликовано липолитических ферментов из видов Thermobifida или видов Corynebacterium, обладающих активностью или значительной активностью в отношении гликолипидов. Хотя липазы, т.е. гидролизующие триацилглицерин ферменты, выделены из видов Streptomyces (см., например, Vujaklija et al Arch Microbiol (2002) 178: 124-130), эти ферменты не были идентифицированы как обладающие гидролизующей гликолипид активностью.
АСПЕКТЫ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на основополагающем обнаружении липолитического фермента, обладающего значительной активностью для галактолипидов, из рода Streptomyces. В частности, липолитический фермент из рода Streptomyces обладает значительной гидролизующей галактолипид активностью и/или значительной ацилтрансферазной активностью для галактолипида, особенно при применении в способах и применениях по настоящему изобретению.
Кроме того, настоящее изобретение основано на основополагающем открытии, что липолитические ферменты из рода Thermobifida или Corynebacterium обладают значительной активностью в отношении галактолипида. В частности, липолитические ферменты из рода Thermobifida или Corynebacterium обладают значительной гидролизующей галактолипиды активностью и/или значительной ацилтрансферазной активностью в отношении галактолипида, особенно при применении в способах и применениях по настоящему изобретению.
В широком аспекте настоящее изобретение относится к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипиды и/или способному переносить ацильную группу по меньшей мере от гликолипида к одному или нескольким субстратам - акцепторам ацила, где фермент может быть получен, предпочтительно, получен из видов Streptomyces.
В следующем аспекте настоящее изобретение относится к липолитическому ферменту, способному гидролизовать по меньшей мере галактолипиды и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент кодирует нуклеиновая кислота, выбранная из группы, состоящей из:
a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, показанную на SEQ ID No:3;
b) нуклеиновой кислоты, родственной нуклеотидной последовательности SEQ ID No:3 вследствие вырожденности генетического кода; и
c) нуклеиновой кислоты, содержащей нуклеотидную последовательность, по меньшей мере на 70% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
Настоящее изобретение также относится к липолитическому ферменту, содержащему аминокислотную последовательность, как показано на SEQ ID No:4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
В другом аспекте настоящее изобретение относится к липолитическому ферменту, способному гидролизовать по меньшей мере галактолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент содержит аминокислотную последовательность, как показано на SEQ ED No. 4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
В следующем аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей липолитический фермент, содержащий аминокислотную последовательность, как показано на SEQ ID No:4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
SEQ ID No:3 показан на фигуре 3, и SEQ ID No:4 показан на фигуре 4.
Настоящее изобретение также относится к нуклеиновой кислоте, кодирующей липолитический фермент, где нуклеиновую кислоту выбирают из группы, состоящей из:
a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, показанную на SEQ ID No:3;
b) нуклеиновой кислоты, родственной нуклеотидной последовательности SEQ ID No:3 вследствие вырожденности генетического кода; и
c) нуклеиновой кислоты, содержащей нуклеотидную последовательность, по меньшей мере на 70% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
Настоящее изобретение также относится к применению липолитического фермента по настоящему изобретению на субстрате (предпочтительно, пищевом продукте) для получения лизогликолипида, например дигалактозилмоноглицерида (DGMG) или моногалактозилмоноглицерида (MGMG), посредством обработки гликолипида (например, дигалактозилдиглицерида (DGDG) или моногалактозилдиглицерида (MGDG)) липолитическим ферментом по настоящему изобретению для получения продукта частичного гидролиза, т.е. лизогликолипида.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению на субстрате (предпочтительно, пищевом продукте) для получения лизофосфолипида, например лизолецитина, посредством обработки фосфолипида (например, лецитина) ферментом по настоящему изобретению для получения продукта частичного гидролиза, т.е. лизофосфолипида.
В одном из общих аспектов настоящее изобретение относится к способу получения пищевого продукта, включающему смешивание липолитического фермента по настоящему изобретению с одним или несколькими ингредиентами пищевого продукта.
В другом общем аспекте настоящее изобретение относится к способу получения выпеченного продукта из теста, включающему смешивание липолитического фермента по настоящему изобретению с тестом.
В следующем аспекте настоящее изобретение относится к способу получения молочного продукта, включающему смешивание липолитического фермента по настоящему изобретению с одним или несколькими ингредиентами молочного продукта.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в производстве молочного продукта для уменьшения одного или нескольких из следующих неблагоприятных эффектов: неприятный запах и/или неприятный, и/или мыльный вкус.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе обработки яйца или продуктов на основе яиц для получения лизофосфолипидов.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе обработки яйца или продуктов на основе яиц для получения лизогликолипидов.
В другом аспекте настоящее изобретение относится к способу ферментативного рафинирования растительных или пищевых масел, включающему обработку пищевого или растительного масла липолитическим ферментом по настоящему изобретению так, чтобы гидролизовать большую часть полярных липидов (например, фосфолипид и/или гликолипид).
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе, включающем обработку фосфолипида так, чтобы гидролизовать жирные ацильные группы.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе уменьшения содержания фосфолипида в пищевом масле, включающем обработку масла липолитическим ферментом по настоящему изобретению так, чтобы гидролизовать большую часть фосфолипида, и отделение водной фазы, содержащей гидролизованный фосфолипид, от масла.
Изобретение также относится к способу получения липолитического фермента по настоящему изобретению, включающему трансформацию клетки-хозяина рекомбинантной нуклеиновой кислотой, содержащей нуклеотидную последовательность, кодирующую липолитический фермент, где клетка-хозяин способна экспрессировать нуклеотидную последовательность, кодирующую полипептид липолитического фермента, культивирование трансформированной клетки-хозяина в условиях, где нуклеиновая кислота экспрессируется, и сбор липолитического фермента.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в биоконверсии полярных липидов (предпочтительно, гликолипидов) для получения ценных продуктов, таких как сложные эфиры углеводорода, и/или сложные эфиры белка, и/или сложные эфиры субъединицы белка, и/или сложный эфир гидроксикислоты.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе ферментативного рафинирования растительного или пищевого масла, включающем обработку указанного пищевого или растительного масла указанным липолитическим ферментом так, чтобы гидролизовать большую часть полярных липидов.
В другом аспекте настоящее изобретение относится к применению липолитического фермента по настоящему изобретению в способе, включающем обработку фосфолипида так, чтобы гидролизовать жирные ацильные группы.
Настоящее изобретение также относится к иммобилизованному липолитическому ферменту по настоящему изобретению.
В другом аспекте настоящее изобретение относится к способу получения лизогликолипида, включающему обработку содержащего гликолипид субстрата по меньшей мере одним липолитическим ферментом для получения указанного лизогликолипида, где указанный липолитический фермент обладает гликолипазной активностью и где указанный липолитический фермент может быть получен из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida.
В другом аспекте настоящее изобретение относится к способу получения лизофосфолипида, включающему обработку содержащего фосфолипид субстрата по меньшей мере одним липолитическим ферментом для получения указанного лизофосфолипида, где указанный липолитический фермент обладает фосфолипазной активностью и где указанный липолитический фермент является получаемым из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida.
В другом аспекте настоящее изобретение относится к способу ферментативного рафинирования растительного или пищевого масла, включающему обработку указанного пищевого или растительного масла липолитическим ферментом, получаемым из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida, способным гидролизовать большую часть полярных липидов.
Настоящее изобретение далее относится к способу биоконверсии полярных липидов для получения ценных продуктов, включающему обработку указанных полярных липидов липолитическим ферментом, который может быть получен из одного их следующих родов: Streptomyces, Corynebacterium и Thermobifida, для получения указанных ценных продуктов, где указанный липолитический фермент способен гидролизовать указанные полярные липиды.
В другом аспекте настоящее изобретение относится к способу получения пищевого продукта, включающему смешивание по меньшей мере одного липолитического фермента с одним или несколькими ингредиентами пищевого продукта, где указанный липолитический фермент способен гидролизовать гликолипид и/или фосфолипид, присутствующий по меньшей мере в одном из указанных ингредиентов или в качестве него, и где указанный липолитический фермент можно получить из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida.
В другом аспекте настоящее изобретение относится к применению липолитического фермента на субстрате для получения лизофосфолипида, где указанный липолитический фермент обладает фосфолипазной активностью и где указанный липолитический фермент можно получить из одного из следующих: Streptomyces, Corynebacterium и Thermobifida.
Настоящее изобретение также относится к применению липолитического фермента, который может быть получен из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida, для ферментативного рафинирования растительного или пищевого масла так, чтобы гидролизовать большую часть полярных липидов.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, который может быть получен из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida по способу, включающему обработку фосфолипида так, чтобы гидролизовать жирные ацильные группы.
В другом аспекте настоящее изобретение относится к применению липолитического фермента для биоконверсии полярных липидов для получения ценных продуктов, где указанный липолитический фермент способен гидролизовать указанные полярные липиды и где указанные липолитические ферменты могут быть получены из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, получаемого из одного из следующих родов: Streptomyces, Corynebacterium и Thermobifida в получении пищевого продукта, где указанный липолитический фермент способен гидролизовать гликолипид и/или фосфолипид.
Аспекты настоящего изобретения представлены в формуле изобретения и в следующих пояснениях.
Другие аспекты, относящиеся к нуклеотидным последовательностям, которые можно использовать по настоящему изобретению, включают в себя: конструкцию, содержащую последовательности по настоящему изобретению; вектор, содержащий последовательности для применения по настоящему изобретению; плазмиду, содержащую последовательности для применения по настоящему изобретению; трансформированную клетку, содержащую последовательности для применения по настоящему изобретению; трансформированную ткань, содержащую последовательности для применения по настоящему изобретению; трансформированный орган, содержащий последовательности для применения по настоящему изобретению; трансформированного хозяина, содержащего последовательности для применения по настоящему изобретению; трансформированный организм, содержащий последовательности для применения по настоящему изобретению. Настоящее изобретение относится также к способам экспрессии нуклеотидной последовательности для применения по настоящему изобретению с использованием этих способов, таким как экспрессия в клетке-хозяине; включая способы переноса этой последовательности. Настоящее изобретение далее относится к способам выделения нуклеотидной последовательности, таким как выделение из клетки-хозяина.
Другие аспекты, относящиеся к аминокислотной последовательности для применения по настоящему изобретению, включают в себя: конструкцию, кодирующую аминокислотные последовательности для применения по настоящему изобретению; вектор, кодирующий аминокислотные последовательности для применения по настоящему изобретению; плазмиду, кодирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированную клетку, экспрессирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированную ткань, экспрессирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированный орган, экспрессирующий аминокислотные последовательности для применения по настоящему изобретению; трансформированного хозяина, экспрессирующего аминокислотные последовательности для применения по настоящему изобретению; трансформированный организм, экспрессирующий аминокислотные последовательности для применения по настоящему изобретению. Настоящее изобретение относится также к способам очистки аминокислотной последовательности для применения по настоящему изобретению с использованием этих способов, таким как экспрессия в клетке-хозяине; включая способы переноса этой последовательности, и затем очистка указанной последовательности.
Для простоты цитирования, эти и другие аспекты настоящего изобретения обсуждаются сейчас под соответствующими заголовками разделов. Однако объяснения в каждом разделе не обязательно ограничены каждым конкретным разделом.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Соответственно, липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемый любой из нуклеотидных последовательностей, показанных как SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидной последовательностью, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им.
Предпочтительно, липолитический фермент для использования в способах и применениях по настоящему изобретению представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент может быть получен, предпочтительно, получен из видов Streptomyces.
В одном из вариантов осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению предпочтительно представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент кодирует нуклеиновая кислота, выбранная из группы, состоящей из:
a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, показанную на SEQ ID No:3;
b) нуклеиновой кислоты, родственной нуклеотидной последовательности SEQ ID No:3 вследствие вырожденности генетического кода; и
c) нуклеиновой кислоты, содержащей нуклеотидную последовательность, по меньшей мере на 70% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
В одном из вариантов осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению предпочтительно представляет собой липолитический фермент, содержащий аминокислотную последовательность, как показано на SEQ ID No:4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению предпочтительно представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипид и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент содержит аминокислотную последовательность, как показано на SEQ ID No:4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
Предпочтительно, липолитический фермент для использования в способах и применениях по настоящему изобретению представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент может быть получен, предпочтительно, из видов Thermobifida, предпочтительно Thermobifida fusca.
Предпочтительно, липолитический фермент для использования в способах и применениях по настоящему изобретению представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент может быть получен, предпочтительно, из видов Corynebacterium, предпочтительно Corynebacterium efficiens.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий любую из аминокислотных последовательностей, показанных как SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую одной из нуклеотидных последовательностей, показанной как SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий любую из аминокислотных последовательностей, показанных как SEQ ID No:5, 7, 8, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для применений, описанных в настоящем описании.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий одну из аминокислотных последовательностей, показанных как SEQ ID No:5, 7 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для применений, описанных в настоящем описании.
Более предпочтительно, в одном из вариантов осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий аминокислотную последовательность, показанную как SEQ ID No:16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную ей.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий аминокислотную последовательность, показанную как SEQ ID No:12 или 14, или аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную ей.
В другом варианте осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, содержащий аминокислотную последовательность, показанную как SEQ ID No:8, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им.
В одном из вариантов осуществления липолитический фермент для использования в способах и применениях по настоящему изобретению может представлять собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент кодирует нуклеиновая кислота, выбранная из группы, состоящей из:
a) нуклеиновой кислоты, содержащей нуклеотидную последовательность, показанную на SEQ ID No:3;
b) нуклеиновой кислоты, родственной нуклеотидной последовательности SEQ ID No:3 вследствие вырожденности генетического кода; и
c) нуклеиновой кислоты, содержащей нуклеотидную последовательность, по меньшей мере на 70% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
В одном из вариантов осуществления липолитический фермент по настоящему изобретению может представлять собой липолитический фермент, который может быть получен, предпочтительно, из штаммов Streptomyces Ll30 или Ll31, депонированных Danisco A/S of Langebrogade 1, DK-1001 Copenhagen K, Denmark в рамках Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры в Национальной коллекции промышленных, пищевых и морских бактерий (NCIMB) 23 St Machar Street, Aberdeen Scotland, GB 25 июня 2004 под инвентарными номерами NCIMB 41226 и NCIMB 41227, соответственно.
Предпочтительно, липолитический фермент по настоящему изобретению действует по меньшей мере на гликолипид, такой как, например, дигалактозилдиглицерид (DGDG). Соответственно, липолитический фермент по настоящему изобретению может действовать также на один или несколько полярных липидных субстратов, таких как фосфолипид, например лецитин, например фосфатидилхолин.
Альтернативным способом выражения используемого в настоящем описании термина «способный гидролизовать гликолипиды», можно сказать, что липолитический фермент обладает гидролизующей гликолипид активностью.
Предпочтительно, липолитический фермент по настоящему изобретению гидролизует гликолипид, такой как, например, дигалактозилдиглицерид (DGDG), а также фосфолипид, такой как лецитин, например, фосфатидилхолин.
Предпочтительно липолитический фермент по настоящему изобретению действует на гликолипиды, такие как DGDG или MGDG.
В одном из аспектов липолитический фермент по настоящему изобретению гидролизует DGDG до DGMG и/или MGDG до MGMG.
В одном из аспектов липолитический фермент по настоящему изобретению гидролизует лецитин до лизолецитина.
В случае, когда липолитический фермент способен переносить ацильную группу по меньшей мере от гликолипида к донорному субстрату, полярный липидный субстрат можно обозначить в настоящем описании как «липидный ацильный донор».
В одном из вариантов осуществления фермент по настоящему изобретению, обладающий как фосфолипазной, так и/или гликолипазной активностью (обычно классифицируемым как E.C. 3.1.1.26; E.C. 3.1.1.4 или E.C. 3.1.1.32 согласно рекомендациям по номенклатуре ферментов (1992) Номенклатурного Комитета Международного союза по биохимии и молекулярной биологии), обладает также ацилтрансферазной активностью (обычно классифицируемой как E.C. 2.3.1.x), в соответствии с чем фермент способен переносить ацильную группу от липидного ацильного донора к одному или нескольким акцепторным субстратам, таким как один или несколько из следующих: стерин; станол; углеводород; белок; субъединица белка; глицерин.
Липид-ацилтрансферазы и их применения объясняют в находящейся в одновременном рассмотрении международной патентной заявке номер PCT/IB2004/000655. Этот документ приведен в настоящем описании в качестве ссылки. Однако липолитические ферменты из рода Streptomyces по настоящему изобретению не указаны в PCT/IB2004/000655.
В некоторых аспектах липолитический фермент для использования в способах и/или применениях по настоящему изобретению могут быть способными переносить ацильную группу от полярного липида (как определено в настоящем описании) к одному или нескольким из следующих субстратов - акцепторов ацила: стерину, станолу, углеводороду, белку, или его субъединицам, или глицерину.
В некоторых аспектах «акцептором ацила» по настоящему изобретению может быть любое соединение, содержащее гидроксигруппу (-OH), такое как, например, поливалентные спирты, включая глицерин; стерин; станолы; углеводороды; оксикислоты, включая фруктовые кислоты, лимонная кислота, винная кислота, молочная кислота и аскорбиновая кислота; белки или их субъединицы, такие как аминокислоты, гидролизаты белка и пептиды (частично гидролизованный белок); и их смеси и производные.
В некоторых аспектах «акцептор ацила» по настоящему изобретению может предпочтительно не являться водой.
В одном из вариантов осуществления акцептор ацила предпочтительно не является моноглицеридом и/или диглицеридом.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к стерину и/или к станолу.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к углеводороду.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к белку или к его субъединице. Соответственно, субъединица белка может представлять собой одно или несколько из следующего: аминокислота, гидролизат белка, пептид, дипептид, олигопептид, полипептид.
Соответственно в белке или субъединице белка акцептором ацила может быть одно или несколько из следующих составляющих белка или субъединицы белка: серин, треонин, тирозин или цистеин.
Когда субъединица белка представляет собой аминокислоту, соответственно, аминокислотой может быть любая подходящая аминокислота. Соответственно аминокислота может представлять собой одну или несколько аминокислот из числа, например, серина, треонина, тирозина или цистеина.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к глицерину.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к оксикислоте.
В одном из аспектов фермент предпочтительно способен переносить ацильную группу от липида к поливалентному спирту.
В одном из аспектов липолитический фермент может как являться способным переносить ацильную группу от липида к стерину и/или станолу, так и являться способным переносить ацильную группу от липида к одному или нескольким из следующего: углеводород, белок, субъединица белка, глицерин.
Термин “лецитин” в настоящем описании означает фосфатидилхолин, фосфатидилэтаноламин, фосфатидилинозитол, фосфатидилсерин и фосфатидилглицерин.
Для некоторых аспектов, липидный субстрат предпочтительно представляет собой по меньшей мере гликолипид, такой как, например, DGDG.
Для некоторых аспектов, липидный субстрат предпочтительно также может представлять собой фосфолипид, такой как лецитин, например фосфатидилхолин. Другие фосфолипидные субстраты по настоящему изобретению могут представлять собой один или несколько из N-ацил-фосфатидилэтаноламина (APE) или N-ацил-лизофосфатидилэтаноламина (ALPE).
Предпочтительно липидный субстрат представляет собой пищевой липид, то есть липидный компонент пищевого продукта.
Для некоторых аспектов, липолитический фермент по настоящему изобретению предпочтительно неспособен или по существу не способен действовать на триглицерид и/или 1-моноглицерид и/или 2-моноглицерид.
В одном из вариантов осуществления липолитический фермент по настоящему изобретению не обладает активностью или не обладает существенной активностью в отношении триглицерида и/или 1-моноглицеридов, и/или 2-моноглицеридов.
Соответственно, липидный субстрат или липидный ацильный донор может представлять собой один или несколько липидов, присутствующих в одном или нескольких из следующих субстратов: жиры, включая сало, жир и молочный жир; масла, включая масла, выделенные из или производные от пальмового масла, подсолнечного масла, соевого масла, сафлорового масла, хлопкового масла, масла земляного ореха, кукурузного масла, оливкового масла, арахисового масла, кокосового масла и рапсового масла. Лецитин из сои, рапсового семени или яичного желтка также представляет собой подходящий липидный субстрат. Липидный субстрат может представлять собой липид овса или материал на основе другого растения, содержащий галактолипиды.
В одном из аспектов липидный субстрат или липидный ацильный донор предпочтительно представляет собой лецитин (такой как фосфатидилхолин) яичного желтка.
Для некоторых аспектов по настоящему изобретению липиды можно выбирать из липидов с цепью жирной кислоты длиной от 8 до 22 атомов углерода.
Для некоторых аспектов по настоящему изобретению липид может быть выбран из липидов с длиной цепи жирной кислоты от 16 до 22 атомов углерода, более предпочтительно от 16 до 20 атомов углерода.
Для некоторых аспектов по настоящему изобретению липид можно выбирать из липидов с длиной цепи жирной кислоты не более чем 14 атомов углерода, соответственно из липидов с длиной цепи жирной кислоты от 4 до 14 атомов углерода, соответственно от 4 до 10 атомов углерода, соответственно от 4 до 8 атомов углерода.
Соответственно, липолитический фермент по настоящему изобретению демонстрирует по меньшей мере гликолипазную активность (E.C. 3.1.1.26). Соответственно липолитический фермент по настоящему изобретению может также демонстрировать активность фосфолипазы A2 (E.C. 3.1.1.4) и/или активность фосфолипазы A1 (E.C. 3.1.1.32).
Для некоторых аспектов липолитический фермент по настоящему изобретению может обладать только гликолипазной активностью (E.C. 3.1.1.26).
Для некоторых аспектов липолитический фермент по настоящему изобретению представляет собой галактолипазу (E.C. 3.1.1.26). Однако тот факт, что фермент определяют как галактолипазу, не мешает ему обладать другими побочными активностями, такими как, например, активность в отношении других полярных липидов.
В настоящем описании термины «гликолипазная активность» и «галактолипазная активность» используют взаимозаменяемо.
Соответственно, для некоторых аспектов липолитический фермент по настоящему изобретению может быть способным переносить ацильную группу от гликолипида и/или фосфолипида к одному или нескольким акцепторным субстратам.
Соответственно, акцепторный субстрат может представлять собой один или несколько из следующих субстратов: стерин, станол, углеводород, белок, глицерин.
Термин «полярные липиды» в настоящем описании означает фосфолипиды и/или гликолипиды. В некоторых аспектах термин полярные липиды предпочтительно обозначает по меньшей мере гликолипиды.
Гликолипазную активность; фосфолипазную активность и/или триацилглицеринлипазную активность фермента можно определить с применением анализов, представленных ниже.
Определение галактолипазной активности (анализ гликолипазной активности (GLU-7)):
Субстрат
0,6% дигалактозилдиглицерид (Sigma D 4651), 0,4% Тритон-X 100 (Sigma X-100) и 5 мМ CaCl2 растворяли в буфере 0,05 М HEPES, pH 7.
Процедура анализа:
400 мкл субстрата добавляли в 1,5 мл пробирку Эппендорф и помещали в термосмеситель Эппендорф при 37°C на 5 минут. Во время t=0 мин добавляли 50 мкл раствора фермента. Также анализировали контроль с водой вместо фермента. Образец перемешивали при 10×100 об/мин в термосмесителе Эппендорф при 37°C в течение 10 минут. Во время t=10 мин пробирку Эппендорф помещали в другой термосмеситель при 99°C на 10 минут, чтобы остановить реакцию.
Свободную жирную кислоту в образцах анализировали с применением набора NEFA C от WAKO GmbH.
Ферментативную активность GLU при pH 7 вычисляли как микромоль жирной кислоты, полученных в минуту при условиях анализа.
Определение фосфолипазной активности (анализ фосфолипазной активности (PLU-7)):
Субстрат
0,6% растительный L-α-фосфатидилхолин 95% (Avanti #441601), 0,4% Тритон-X 100 (Sigma X-100) и 5 мМ CaCl2 диспергировали в буфере 0,05 М HEPES pH 7.
Процедура анализа: 400 мкл субстрата добавляли в 1,5 мл пробирку Эппендорф и помещали в термосмеситель Эппендорф при 37°C на 5 минут. Во время t= 0 мин добавляли 50 мкл раствора фермента. Также анализировали контроль с водой вместо фермента. Образец перемешивали при 10×100 об/мин в термосмесителе Эппендорф при 37°C в течение 10 минут. Во время t=10 мин пробирку Эппендорф помещали в другой термосмеситель при 99°C на 10 минут, чтобы остановить реакцию.
Свободную жирную кислоту в образцах анализировали с применением набора NEFA C от WAKO GmbH.
Ферментативную активность PLU-7 при pH 7 вычисляли как микромоль жирной кислоты, полученных в минуту при условиях анализа.
Определение триацилглицеридлипазной активности: анализ, основанный на триглицериде (трибутирине) в качестве субстрата (LIPU):
Липазную активность, основанную на трибутирине, измеряли по Food Chemical Codex, Forth Edition, National Academy Press, 1996, p.803. С модификацией, что образец растворяют в деионизованной воде вместо глицинового буфера, и установленное заданное значение pH составляет 5,5 вместо 7.
1 LIPU определяют как количество фермента, которое может высвобождать 1 микромоль масляной кислоты в мин в услових анализа.
В одном из вариантов осуществления, липолитический фермент по настоящему изобретению предпочтительно представляет собой липолитический фермент дикого типа.
Термины «природный» и «дикого типа» в настоящем описании означают существующий в природе фермент. То есть это фермент, экспрессированный с эндогенного генетического кода и выделенный из его эндогенного организма-хозяина, и/или гетерологично полученный фермент, не являющийся мутированным (т.е., не содержащий делеций, вставок или замен аминокислот) по сравнению с эндогенно полученной зрелой последовательностью (после событий ко- и посттрансляционного расщепления). Природные и дикого типа белки по настоящему изобретению могут являться кодированными полинуклеотидами с оптимизированными кодонами для гетерологичной экспрессии и могут содержать также не эндогенные сигнальные пептиды, выбранные для экспрессии в данном хозяине.
Термин «вариант» в настоящем описании означает белок, экспрессированный с не эндогенного генетического кода, что приводит к одному или нескольким изменениям аминокислот (т.е. делециям, вставкам или заменам аминокислот) по сравнению с природной или дикого типа последовательностью в последовательности зрелого белка.
Липолитический фермент по настоящему изобретению предпочтительно является получаемым (соответственно, может быть получен) из бактерии.
Липолитический фермент по настоящему изобретению предпочтительно может быть получен (предпочтительно, получен) из видов Streptomyces. Липолитический фермент по настоящему изобретению предпочтительно может быть получен (предпочтительно, получен) из Streptomyces штамма L131 или Streptomyces штамма L130.
Липолитический фермент по настоящему изобретению предпочтительно содержит аминокислотную последовательность, по меньшей мере на 70%, предпочтительно по меньшей мере на 75%, предпочтительно по меньшей мере на 80%, предпочтительно по меньшей мере на 90%, предпочтительно по меньшей мере на 95%, предпочтительно по меньшей мере на 98%, предпочтительно по меньшей мере на 99% идентичную аминокислотной последовательности, показанной как SEQ ID No:4.
Нуклеиновая кислота, кодирующая липолитический фермент по настоящему изобретению, предпочтительно содержит нуклеотидную последовательность, по меньшей мере на 75%, предпочтительно по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, предпочтительно по меньшей мере на 90%, предпочтительно по меньшей мере на 95%, предпочтительно по меньшей мере на 98%, предпочтительно по меньшей мере на 99% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
В одном из вариантов осуществления оптимальный pH для фермента для галактолипидного субстрата составляет, соответственно, приблизительно 6-8, предпочтительно приблизительно 6,5 - 7,5, более предпочтительно приблизительно 7.
Соответственно, липолитический фермент по настоящему изобретению может не ингибироваться или по существу не ингибироваться ингибиторами липаз, присутствующими в пшеничной муке. Термин «по существу не ингибированный» в настоящем описании означает, что фермент является менее чувствительным к ингибиторам липазы, присутствующим в пшеничной муке, по сравнению с эквивалентной дозой (PLU) LipopanF™ (Novozymes A/S, Denmark), на основании обычного фосфолипазного анализа (PLU-7), определенного в настоящем описании.
Соответственно, липолитический фермент по настоящему изобретению способен гидолизовать по меньшей мере 10% сложного диэфира галактолипида в субстрате (т.е., в пищевом продукте, например тесте) до сложного моноэфира. Предпочтительно, фермент способен гидролизовать по меньшей мере 20%, более предпочтительно по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% сложного диэфира галактолипида до сложного моноэфира. Соответственно, сложный диэфир галактолипида может представлять собой один или несколько из MGDG или DGDG, а сложный моноэфир может представлять собой один или несколько из MGMG или DGMG, соответственно.
Соответственно, липолитический фермент по настоящему изобретению можно выделить из среды для ферментации Streptomyces штамма L131 или Streptomyces штамма L130.
Соответственно, фермент можно очистить жидкостной хроматографией.
Аминокислотную последовательность очищенного липолитического фермента можно определить расщеплением по Эдману, анализом LC-MS и MALDI-TOF.
Соответственно, фермент, как определено в настоящем описании, может катализировать одну или несколько из следующих реакций: переэтерификацию, трансэтерификацию, алкоголиз, гидролиз.
Термин «переэтерификация» относится к катализируемому ферментом переносу ацильных групп между липидным донором и липидным акцептором, где липидный донор не является свободной ацильной группой.
Термин «трансэтерификация» в настоящем описании означает катализируемый ферментом перенос ацильной группы от липидного донора (отличного от свободной жирной кислоты) к акцептору ацила (отличного от воды).
В настоящем описании термин «алкоголиз» относится к ферментативному расщеплению ковалентной связи производного кислоты посредством реакции со спиртом ROH, так что один из продуктов соединяется с H спирта, а другой продукт соединяется с OR-группой спирта.
В настоящем описании термин «спирт» относится к алкильному соединению, содержащему гидроксильную группу.
В настоящем описании термин «гидролиз» относится к катализируемому ферментом переносу ацильной группы от липида к OH-группе молекулы воды. Перенос ацила в результате гидролиза требует расщепления молекулы воды.
Термин «пищевой продукт» в настоящем описании относится к веществу, пригодному для употребления человеком и/или животным.
Соответственно, термин «пищевой продукт» в настоящем описании может означать пищевой продукт в форме, готовой к употреблению. Однако альтернативно или дополнительно, термин пищевой продукт в настоящем описании может означать одно или несколько питательных веществ, которые используют для получения пищевого продукта. Только в качестве примера, термин пищевой продукт охватывает как хлебобулочные изделия, полученные из теста, так и тесто, используемое для получения указанных хлебобулочных изделий.
В предпочтительном аспекте настоящее изобретение относится к пищевому продукту, как определено выше, где пищевой продукт выбирают из одного или нескольких следующих: яйца, продукты на основе яиц, включая в качестве неограничивающих примеров майонез, заправки для салата, соусы, мороженое, яичный порошок, модифицированный яичный желток и полученные из него продукты; хлебобулочные изделия, включая хлеб, торты, продукты из сдобного теста, слоеное тесто, жидкое взбитое тесто, сдобу, пончики, печенье, сухое печенье и булочки; кондитерские изделия, включая шоколад, леденцы, карамель, халву, гумми, включая не содержащие сахара и подслащенные сахаром гумми, надувную жевательную резинку, мягкую жевательную резинку, жевательную резинку и пудинги; замороженные продукты, включая фруктовое мороженое, предпочтительно замороженные молочные продукты, включая пломбир и молочное мороженое; молочные продукты, включая сыр, масло, молоко, сливки для кофе, взбитые сливки, яичный крем, молочные напитки и йогурты; муссы, взбитые растительные сливки, мясные продукты, включая готовые мясные блюда; пищевые масла и жиры, газированные и негазированные взбитые продукты, эмульсии типа масло в воде, эмульсии типа вода в масле, маргарин, кулинарные жиры и пасты, включая низкожирные и очень низкожирные пасты; заправки, майонез, подливы, соусы на основе сливок, супы на основе сливок, напитки, эмульсии специй и соусы.
Соответственно, пищевой продукт по настоящему изобретению может представлять собой "высококачественные продукты", включая торты, пирожные, кондитерские изделия, шоколад, помадку и подобные.
В одном из аспектов пищевой продукт по настоящему изобретению может представлять собой тестовую заготовку или выпеченный продукт, такой как хлеб, жареный продукт, закуску, торты, пироги, шоколадные кексы, сухое печенье, лапшу, легкие закуски, такие как печенье, печенье из муки грубого помола, крендели, картофельные чипсы и макаронные изделия.
В другом аспекте пищевой продукт по настоящему изобретению может представлять собой пищевой продукт из растений, такой как мука, готовые смеси, масла, жиры, какао-масло, забеливатель для кофе, заправки для салата, маргарин, пасты, арахисовое масло, кулинарные жиры, пломбир, кулинарные масла.
В другом аспекте пищевой продукт по настоящему изобретению может представлять собой молочный продукт, включая масло, молоко, сливки, сыр, такой как натуральный, плавленый и искусственный сыры во множестве форм (включая куски, брусок, ломтики или тертый), сливочный сыр, пломбир, замороженные десерты, йогурт, йогуртовые напитки, молочный жир, безводный молочный жир, другие молочные продукты. Фермент по настоящему изобретению может улучшать стабильность жира в молочных продуктах.
Особенно предпочтительно применять фермент по настоящему изобретению в сыре. Так, липолитический фермент по настоящему изобретению можно использовать для получения сыра. Липолитический фермент катализирует гидролиз фосфолипидов в молоке, что приводит к увеличенному выходу сыра. Предпочтительно липолитический фермент по настоящему изобретению можно добавлять в молоко (обозначенное как молоко для сыроделия) перед процессом получения сыра или во время него.
В другом аспекте пищевой продукт по настоящему изобретению может представлять собой пищевой продукт, содержащий животные ингредиенты, такой как готовые мясные блюда, кулинарные масла, кулинарные жиры.
В дополнительном аспекте пищевой продукт по настоящему изобретению может представлять собой напиток фруктовый, из смеси фруктов, овощной или вино. В некоторых случаях напиток может содержать до 20 г/л добавленных фитостеринов.
В другом аспекте пищевой продукт по настоящему изобретению может представлять собой корм для животных. Корм для животных можно обогащать фитостерином и/или фитостанолами, предпочтительно бета-ситостерин/станолом. Соответственно, корм для животных может представлять собой корм для птиц. Когда пищевой продукт представляет собой корм для птиц, настоящее изобретение можно использовать для понижения содержания холестерина в яйцах, полученных от домашней птицы, которую кормят пищевым продуктом по настоящему изобретению.
В одном из аспектов пищевой продукт предпочтительно выбирают из одного или нескольких из следующего: яйца, продукты на основе яиц, включая майонез, заправки для салата, соусы, пломбир, яичный порошок, модифицированный яичный желток и полученные из него продукты.
Пищевой продукт по настоящему изобретению предпочтительно представляет собой содержащий воду пищевой продукт. Соответственно, пищевой продукт может содержать 10-98% воды, соответственно 14-98%, соответственно 18-98% воды, соответственно 20-98%, соответственно 40-98%, соответственно 50-98%, соответственно 70-98%, соответственно 75-98%.
Для некоторых аспектов пищевой продукт по настоящему изобретению может не являться чистым полученным из растений маслом, таким как, например, оливковое масло, подсолнечное масло, арахисовое масло, рапсовое масло. Во избежание недоразумений, в некоторых аспектах по настоящему изобретению пищевой продукт по настоящему изобретению может содержать масло, но пищевой продукт не состоит главным образом из масла или смесей масла. Для некоторых аспектов пищевой продукт предпочтительно содержит менее чем 95% липидов, предпочтительно менее чем 90% липидов, предпочтительно менее чем 85%, предпочтительно менее чем 80% липидов. Таким образом, для некоторых аспектов по настоящему изобретению масло может являться компонентом пищевого продукта, но предпочтительно пищевой продукт не является маслом по существу.
Преимущества применения липолитического фермента, способного переносить ацильную группу, в качестве пищевой добавки объясняют в патентных заявках WO2004/064987, WO2004/064537, PCT/IB2004/004374 и GB0513859.9, приведенных в настоящем описании в качестве ссылки.
Образование свободных жирных кислот может являться вредным для пищевых продуктов. Свободные жирные кислоты связаны с неприятными запахами и/или неприятными привкусами пищевых продуктов, а также и с другими неблагоприятными эффектами, включая мыльный вкус молочных продуктов, таких, например, как сыр. Соответственно, в некоторых вариантах осуществления настоящего изобретения липолитический фермент способен переносить жирную кислоту от липида к рецептору ацила, например стерину и/или станолу. Следовательно, общий уровень свободных жирных кислот в пищевом продукте не увеличивается или увеличивается только в незначительной степени. Таким образом, липолитический фермент, способный переносить ацильную группу по настоящему изобретению, может предоставлять один или несколько из следующих неожиданных технических эффектов в производстве сыра: уменьшение эффекта обезжиривания в сыре; увеличение выхода сыра; улучшение вкуса; уменьшению аномального запаха; уменьшению «мыльного» вкуса.
Использование липолитического фермента, описанного в настоящем описании, способного переносить ацильную группу к углеводороду, а также к стерину и/или станолу, особенно благоприятно для пищевых продуктов, содержащих яйца. В частности, присутствие сахаров, особенно глюкозы, в яйцах и яичных продуктах часто представляют неблагоприятным. Яичный желток может содержать вплоть до 1% глюкозы. По настоящему изобретению этот нежелательный сахар можно легко удалить «этерификацией» сахара для получения сложного эфира сахара.
Присутствие диглицеридов в пищевых маслах является неблагоприятным. В частности, диглицериды в пищевых маслах (в частности, в пальмовом масле) могут привести к низкому качеству масла. Соответственно, в некоторых вариантах осуществления настоящего изобретения липолитический фермент, описанный в настоящем описании, способен переносить жирную кислоту от липида к акцептору ацила, что уменьшает уровень диглицеридов в масле без увеличения или существенного увеличения уровня свободных жирных кислот.
Липолитический фермент, указанный в настоящем описании, способен гидролизовать большую часть фосфолипидов в пищевом или растительном масле. Это очень благоприятно для ферментативного рафинирования растительных или пищевых масел. Соответственно, в некоторых вариантах осуществления настоящего изобретения липолитический фермент может являться способным переносить жирную кислоту от липида к акцептору ацила. Следовательно, общий уровень свободных жирных кислот в масле преимущественно не увеличивается или увеличивается до незначительной степени. Образование свободных жирных кислот в пищевом масле может являться вредным. Предпочтительно, способ по настоящему изобретению приводит к рафинированию пищевого масла, где накопление свободных жирных кислот уменьшено и/или прекращено.
Формула настоящего изобретения построена так, чтобы включать каждый из пищевых продуктов, перечисленных выше.
Для некоторых применений, приведенных в настоящем описании, особенно применений в качестве пищевой добавки, таких как применения в хлебопечении, липолитический фермент по настоящему изобретению можно использовать с одним или несколькими общепринятыми эмульгаторами, включая, например, моноглицериды, сложные эфиры диацетилвинной кислоты с моно- и диглицеридами жирных кислот, сложные эфиры сахара, стеароиллактилат натрия (SSL) и лецитины.
Дополнительно или альтернативно, фермент по настоящему изобретению можно использовать с одним или несколькими подходящими пищевыми ферментами. Таким образом, в объем настоящего изобретения попадает то, что, в дополнение к липолитическому ферменту по настоящему изобретению, по меньшей мере один дополнительный фермент можно добавлять к выпеченному продукту и/или тесту. Такие дополнительные ферменты включают в себя расщепляющие крахмал ферменты, такие как эндо- или экзоамилазы, пуллуланазы, деветвящие ферменты, гемицеллюлазы, включая ксиланазы, целлюлазы, оксидоредуктазы, например глюкозооксидаза, пиранозооксидаза, сульфгидрилоксидаза или карбогидрат-оксидаза, такая как окисляющая мальтозу, например, гексоза-оксидаза (HOX), липазы, фосфолипазы и гексоза-оксидаза, протеазы и ацилтрансферазы (такие как описанные, например, в PCT/IB2004/000575).
Настоящее изобретение относится к композициям пищевых ферментов, включая композиции для улучшения хлеба и/или теста, содержащие фермент по настоящему изобретению и, необязательно, дополнительно содержащие другой фермент, такой как один или несколько подходящих пищевых ферментов, включая расщепляющие крахмал ферменты, такие как эндо- или экзоамилазы, пуллуланазы, деветвящие ферменты, гемицеллюлазы, включая ксиланазы, целлюлазы, оксидоредуктазы, например глюкозооксидазу, пиранозооксидазу, сульфгидрилоксидазу или карбогидрат-оксидазу, такую как окисляющая мальтозу, например гексоза-оксидаза (HOX), липазы, фосфолипазы и гексоза-оксидаза, протеазы и ацилтрансферазы (такие, как описано, например, в PCT/IB2004/000575).
Для некоторых применений, описанных в настоящем описании, особенно для применений в качестве пищевой добавки, таких как применения в хлебопечении, липолитический фермент по настоящему изобретению можно добавлять в комбинации или последовательно с одним или несколькими субстратами фермента. Только в качестве примера, липолитический фермент по настоящему изобретению можно добавлять вместе с одним или несколькими полярными липидными субстратами и/или одним или несколькими субстратами - акцепторами ацила.
Для некоторых применений, упомянутых в настоящем описании, особенно применений в качестве пищевой добавки, липолитический фермент по настоящему изобретению можно использовать с одной или несколькими гидроксикислотами, включая, например, винную кислоту, лимонную кислоту, молочную кислоту, янтарную кислоту или аскорбиновую кислоту.
Термин «улучшенные свойства» в настоящем описании означает любое свойство, которое можно улучшить воздействием липолитического фермента по настоящему изобретению. В частности, применение липолитического фермента по настоящему изобретению приводит к одной или нескольким из следующих характеристик: увеличенный объем выпеченного продукта; улучшенная структура мякиша выпеченного продукта; препятствующие зачерствению свойства выпеченного продукта; увеличенная прочность, увеличенная стабильность, уменьшенная слипаемость и/или улучшенная механическая обрабатываемость теста.
Улучшенные свойства оценивают по сравнению с тестом и/или выпеченным продуктом, полученным без добавления липолитического фермента по настоящему изобретению.
Термин «выпеченный продукт» в настоящем описании включает в себя продукт, полученный из теста. Примеры выпеченных продуктов (либо белого, светлого или темного типа), которые можно преимущественно получать по настоящему изобретению, включают в себя один или несколько из следующих продуктов: хлеб (включая белый, из муки цельного зерна и ржаной хлеб), обычно в форме буханки или батонов, паровых булочек, французского хлеба типа багета, питы, тако, кукурузной тортильи, пшеничной тортильи, пирогов, блинов, бисквитов, хрустящего хлеба, пасты, лапши и подобное.
Тесто по настоящему изобретению может представлять собой кислое тесто или тесто, подвергаемое разрыхлению. Тесто можно разрыхлять различными способами, такими как добавление бикарбоната натрия или подобного, или добавлением подходящей культуры дрожжей, такой как культура Saccharomyces cerevisiae (пекарские дрожжи).
Настоящее изобретение далее относится к применению липолитического фермента по настоящему изобретению для получения теста для макаронных изделий, предпочтительно полученного из макаронной муки или муки сравнимого качества.
Липолитический фермент по настоящему изобретению пригоден для ферментативного рафинирования растительных или пищевых масел. В переработке растительного или пищевого масла пищевое или растительное масло обрабатывают липолитическим ферментом по настоящему изобретению так, чтобы гидролизовать большую часть полярных липидов (например, фосфолипид). Предпочтительно, гидролизуют жирные ацильные группы из полярных липидов. Процесс рафинирования обычно приводит к уменьшению содержания полярных липидов, особенно фосфолипидов, в пищевом масле из-за гидролиза большей части (т.е. более чем 50%) полярного липида, например фосфолипида. Обычно водную фазу, содержащую гидролизованный полярный липид (например, фосфолипид), отделяют от масла. Соответственно, пищевое или растительное масло может исходно (перед обработкой ферментом по настоящему изобретению) обладать содержанием фосфора 50-250 промилле.
В одном из вариантов осуществления настоящее изобретение относится к применению липолитического фермента по настоящему изобретению для биоконверсии полярных липидов (предпочтительно гликолипидов) для получения ценных продуктов, таких как сложные эфиры углеводорода и/или сложные эфиры белка, и/или сложные эфиры субъединицы белка, и/или сложный эфир гидроксикислоты. Применение липолитического фермента, в частности липолитического фермента, способного переносить ацильные группы от полярного липидного субстрата (предпочтительно, гликолипида) к акцептору ацила, для биоконверсии полярных липидов и его преимущества подробно описаны в PCT/IB2004/004374, содержание которой включено в настоящем описании в качестве ссылки.
В одном из вариантов осуществления липолитический фермент для применения в способах по настоящему изобретению может быть иммобилизованным. В случае, когда фермент иммобилизован, смесь, содержащую донор ацила, не обязательно, акцептор ацила и, не обязательно, воду, можно пропускать через колонку, например, содержащую иммобилизованный фермент. Благодаря иммобилизации фермента, возможно легко использовать его повторно.
Соответственно, иммобилизованный фермент можно использовать в проточном реакторе или в периодическом реакторе, содержащем реакционную смесь, содержащую липидный донор ацила и, не обязательно, акцептор ацила, растворенные в воде. Когда присутствует акцептор ацила, донор и акцептор присутствуют в двухфазной системе или в эмульсии. Реакционную смесь можно, не обязательно, перемешивать или диспергировать с помощью ультразвука. Как только реакция достигнет равновесия, например, реакционную смесь и иммобилизованный фермент можно разделить. Соответственно, продукт реакции можно фракционировать, например, хроматографией гидрофобного взаимодействия, кристаллизацией или перегонкой в глубоком вакууме.
Иммобилизованнцую липид-ацилтрансферазу можно получить способами иммобилизации, известными в данной области. Существуют многочислелнные способы получения иммобилизованных ферментов, которые будут очевидны специалистам в данной области (например, способы, на которые ссылаются в EP 0 746 608; или Balcao V.M. et al Enzyme Microb Technol. 1996 May 1; 18(6):392-416; или Retz et al Chem Phys Lipids 1998 June:93(l-2): 3-14; Bornscheuer et al Trends Biotechnol. 2002 Oct; 20(10):433-7; Plou et al Biotechnology 92 (2002) 55-66; Warmuth et al 1992 Bio Forum 9, 282-283; Ferrer et al 2000 J. Chem Technol. Biotechnol. 75, 1-8; или Christensen et al 1998 Nachwachsende Rohstoff 10, 98-105; Petersen and Christenen 2000 Applied Biocatalysis Harwood Academic Publishers, Amsterdam (каждая из которых приведена в настоящем описании в качестве ссылки).
Способы, которые можно использовать в настоящем описании, включают в себя, например, ковалентное присоединение к Eupergit C, адсорбцию на полипропилене и грануляцию диоксидом кремния.
ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ ПО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
Липолитический фермент для применения по настоящему изобретению и/или в способах, описанных в настоящем описании, предпочтительно представляет собой липолитический фермент, способный гидролизовать по меньшей мере галактолипиды и/или способный переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким субстратам - акцепторам ацила, где фермент кодирует нуклеиновая кислота, выбранная из группы, состоящей из:
d) нуклеиновой кислоты, содержащей нуклеотидную последовательность, показанную на SEQ ID No:3;
e) нуклеиновой кислоты, родственной нуклеотидной последовательности SEQ ID No:3 вследствие вырожденности генетического кода; и
f) нуклеиновой кислоты, содержащей нуклеотидную последовательность, по меньшей мере на 70% идентичную нуклеотидной последовательности, показанной на SEQ ID No:3.
Предпочтительно, липолитический фермент, используемый по настоящему изобретению и/или в способах, описанных в настоящем описании, представляет собой липолитический фермент, содержащий аминокислотную последовательность, как показано на SEQ ID No:4, или аминокислотную последовательность, по меньшей мере на 60% идентичную ей.
Однако липолитический фермент для применения по настоящему изобретению и/или в способах по настоящему изобретению может представлять собой любой липолитический фермент, который может быть получен из видов Streptomyces, способный гидролизовать по меньшей мере галактолипид и/или способный переносить ацильную группу от галактолипида к одному или нескольким субстратам - акцепторам ацила.
Подходящие липолитические ферменты, обладающие галактолипазной активностью для применения по настоящему изобретению и/или в способах по настоящему изобретению, могут содержать любую из следующих аминокислотных последовательностей и/или могут кодироваться следующими нуклеотидными последовательностями:













Таким образом, в другом аспекте настоящее изобретение относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, в пищевом продукте для получения лизогликолипида, например дигалактозилмоноглицерида (DGMG) или моногалактозилмоноглицерида (MGMG), посредством обработки гликолипида (например, дигалактозилдиглицерида (DGDG) или моногалактозилдиглицерида (MGDG)) липолитическим ферментом по настоящему изобретению с получением продукта частичного гидролиза, т.е. лизогликолипида.
В другом аспекте настоящее изобретение также относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, в пищевом продукте для получения лизофосфолипида, например лизолецитина, посредством обработки фосфолипида (например, лецитина) ферментом для получения продукта частичного гидролиза, т.е. лизофосфолипида.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для яйца или продукта на основе яиц для гидролиза фосфолипидов и/или гликолипидов.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для субстрата (предпочтительно, пищевого продукта) для гидролиза жирных ацильных групп.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для пищевого масла для уменьшения содержания фосфолипида.
В другом аспекте настоящее изобретение относится к применению липолитического фермента, содержащего любую из аминокислотных последовательностей, показанных на SEQ ID No:4, 5, 7, 8, 12, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или кодируемую любой из нуклеотидных последовательностей, показанных на SEQ ID No:3, 6, 9, 13, 15 или 17, или нуклеотидную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для субстрата (предпочтительно, смеси для биоконверсии, содержащую полярные липиды (предпочтительно, гликолипиды)) для получения ценных продуктов, таких как сложные эфиры углеводорода, и/или сложные эфиры белка, и/или сложные эфиры субъединицы белка, и/или сложный эфир оксикислоты.
В предпочтительном аспекте настоящее изобретение относится к липолитическому ферменту, содержащему любую из аминокислотных последовательностей, показанных на SEQ ID No:8, 14 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, для применений, описанных в настоящем описании.
Более предпочтительно настоящее изобретение относится к применению липолитического фермента, содержащего аминокислотную последовательность, показанную на SEQ ID No:16, или аминокислотную последовательность по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную ей.
В широком аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент может быть получен, предпочтительно, из видов Thermobifida, предпочтительно T.fusca.
В другом широком аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент может быть получен, предпочтительно, из видов Corynebacterium, предпочтительно C. efficiens.
В другом широком аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент может быть получен, предпочтительно, из Streptomyces avermitilis.
В другом аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент содержит SEQ ID No:5, 7, 8, 12 или 16, или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную им, или фермент является кодированным любой из нуклеотидных последовательностей, показанных на SEQ ID No:6, 9, 13 или 17, или нуклеотидной последовательностью, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичной им.
В другом аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент содержит SEQ ID No:14 или аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную ей, или фермент является кодированным любой из нуклеотидных последовательностей, показанных на SEQ ID No:15, или нуклеотидной последовательностью, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичной ей.
В другом аспекте настоящее изобретение может относиться к липолитическому ферменту, способному гидролизовать по меньшей мере гликолипид и/или способному переносить ацильную группу по меньшей мере от галактолипида к одному или нескольким акцепторам ацила, где фермент содержит SEQ ID No:16 или аминокислотную последовательность, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичную ей, или фермент является кодированным любой из нуклеотидных последовательностей, показанных на SEQ ID No:17, или нуклеотидной последовательностью, по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98% идентичной ей.
В одном из вариантов осуществления настоящего изобретения виды Streptomyces, из которых можно получить (или получен) липолитический фермент, предпочтительно не являются Streptomyces rimosus.
В одном из вариантов осуществления настоящего изобретения виды Streptomyces, из которых можно получить (или получен) липолитический фермент, предпочтительно не являются Streptomyces coelicolor.
ПРЕИМУЩЕСТВА
Одним из преимуществ настоящего изобретения является то, что липолитический фермент обладает значительной гидролизирующей гликолипиды активностью. Это оказалось неожиданным для липолитического фермента из видов Streptomyces. Кроме того, это оказалось неожиданным для липолитического фермента из видов Thermobifida и Corynebacterium.
Дополнительным преимуществом настоящего изобретения является то, что липолитический фермент не обладает или по существу не обладает значительной гидролизирующей триацилглицерины активностью.
ВЫДЕЛЕННАЯ
В одном из аспектов последовательность предпочтительно присутствует в выделенной форме. Термин «выделенная» означает, что последовательность является по существу свободной по меньшей мере от одного из других компонентов, с которыми последовательность естественно связана в природе, и как обнаружено в природе.
ОЧИЩЕННАЯ
В одном из аспектов последовательность предпочтительно присутствует в очищенной форме. Термин «очищенная» означает, что последовательность присутствует в относительно чистом состоянии - например, по меньшей мере 90% чистоты, или по меньшей мере приблизительно 95% чистоты, или по меньшей мере приблизительно 98% чистоты.
НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
Объем настоящего изобретения входят нуклеотидные последовательности, кодирующие ферменты, обладающие конкретными свойствами, как определено в настоящем описании.
Термин «нуклеотидная последовательность» в настоящем описании относится к олигонуклеотидной последовательности или полинуклеотидной последовательности и ее вариантам, гомологам и производным (таким как ее части). Нуклеотидная последовательность может быть геномного, синтетического или рекомбинантного происхождения, может быть двухцепочечной или одноцепочечной, представляющей либо смысловую, либо антисмысловую цепь.
Термин «нуклеотидная последовательность» по отношению к настоящему изобретению включает в себя геномную ДНК, кДНК, синтетическую ДНК и РНК. Предпочтительно он означает ДНК, более предпочтительно кодирующую последовательность кДНК по настоящему изобретению.
В предпочтительном варианте осуществления нуклеотидная последовательность, когда относится к настоящему изобретению и когда охвачена объемом настоящего изобретения по существу, не включает природную нуклеотидную последовательность по настоящему изобретению, когда она присутствует в ее природном окружении и когда она связана с естественно ассоциированной с ней последовательностью(последовательностями), которая также присутствует/присутствуют в их природном окружении. Для упрощения ссылки, авторы настоящего изобретения будут обозначать этот предпочтительный вариант осуществления «неприродной нуклеотидной последовательностью». В этом смысле, термин «природная нуклеотидная последовательность» означает полную нуклеотидную последовательность, присутствующую в ее природном окружении и функционально связанную с полным промотором, с которым она ассоциирована в природе, где промотор присутсвует также в его природном окружении. Однако аминокислотную последовательность, входящую в объем настоящего изобретения, можно выделить и/или очистить после экспрессии нуклеотидной последовательности в ее природном организме. Предпочтительно, однако, чтобы аминокислотную последовательность, входящую в объем настоящего изобретения, можно было экспрессировать посредством нуклеотидной последовательности в ее природном организме, но чтобы нуклеотидная последовательность находилась не под контролем промотора, с которым она естественно ассоциирована в организме.
ПОЛУЧЕНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ
Обычно нуклеотидную последовательность, входящую в объем настоящего изобретения, получают с применением способов рекомбинантной ДНК (т.е. рекомбинантной ДНК). Однако в альтернативном варианте осуществления настоящего изобретения нуклеотидную последовательность можно синтезировать, полностью или частично, с применением химических способов, хорошо известных в данной области (смотри Caruthers MH et al, (1980) Nuc Acids Res Symp Ser 215-23 и Horn T et al, (1980) Nuc Acids Res Symp Ser 225-232).
Нуклеотидную последовательность, кодирующую фермент с конкретными свойствами, как определено в настоящем описании, можно идентифицировать и/или выделить, и/или очистить из любой клетки или организма, продуцирующих указанный фермент. В данной области хорошо известны различные способы для идентификации и/или выделения, и/или очистки нуклеотидных последовательностей. В качестве примера, можно использовать способ амплификации PCR для получения большего количества последовательности после того, как подходящую последовательность идентифицировали, и/или выделили, и/или очистили.
В качестве дополнительного примера, можно сконструировать библиотеку геномной ДНК и/или кДНК с применением хромосомной ДНК или матричной РНК из организма, продуцирующего фермент. Если известна аминокислотная последовательность фермента или часть аминокислотной последовательности фермента, можно синтезировать меченые олигонуклеотидные пробы и применять для идентификации кодирующих фермент клонов из полученной из организма геномной библиотеки. Альтернативно, для идентификации кодирующих фермент клонов можно использовать меченые олигонуклеотидные зонды, содержащие последовательности, гомологичные гену другого известного фермента. В последнем случае используют условия гибридизации и отмывки пониженной строгости.
Альтернативно, кодирующие фермент клоны можно идентифицировать вставкой фрагментов геномной ДНК в экспрессирующий вектор, такой как плазмида, трансформацией отрицательных по ферменту бактерий полученной библиотекой геномной ДНК и затем посевом трансформированных бактерий на чашки с агаром, содержащие субстрат для фермента (например, мальтозу для продуцирующих фермент глюкозидазу (мальтазу)), таким обоазом позволяя идентификацию клонов, экспрессирующих фермент.
В качестве дополнительной альтернативы, нуклеотидную последовательность, кодирующую фермент, можно получать синтетически установившимися общепринятыми способами, например фосфорамидатным способом, описанном Beucage S.L. et al, (1981) Tetrahedron Letters 22, p.1859-1869, или способом, описанным Matthes et al, (1984) EMBO J.3, p 801-805. По фосфорамидатному способу олигонуклеотиды синтезируют, например, в автоматическом синтезаторе ДНК, очищают, отжигают, лигируют и клонируют в подходящих векторах.
Нуклеотидная последовательность может быть смешанного геномного и синтетического происхождения, смешанного синтетического и кДНК происхождения или смешанного геномного и кДНК происхождения, полученной лигированием фрагментов синтетического, геномного или кДНК происхождения (в соответствующих случаях) по общепринятым способам. Каждый лигированный фрагмент соответствует различным частям полной нуклеотидной последовательности. Последовательность ДНК можно получить также полимеразной цепной реакцией (PCR) с применением спецфических праймеров, например, как описано в US 4683202 или в Saiki R K et al, (Science (1988) 239, p.487-491).
Вследствие вырожденности генетического кода можно легко получить нуклеотидные последовательности, в которых применение триплетных кодонов для некоторых или всех аминокислот, кодируемых исходной нуклеотидной последовательностью, изменено, таким образом получая нуклеотидную последовательность с низкой гомологией с исходной нуклеотидной последовательностью, но кодирующую такую же аминокислотную последовательность, какую кодирует исходная нуклеотидную последовательность, или ее вариант. Например, для большинства аминокислот существует вырожденность генетического кода для третьего положения триплетного кодона (неоднозначное положение) (для ссылки см. Stryer, Lubert, Biochemistry, Third Edition, Freeman Press, ISBN 0-7167-1920-7), таким образом, нуклеотидная последовательность, в которой все триплетные кодоны являются «неоднозначными» в третьем положении, будет являться приблизительно на 66% идентичной исходной нуклеотидной последовательности, однако, измененная нуклеотидная последовательность будет кодировать такую же первичную аминокислотную последовательность, или ее вариант как исходная нуклеотидная последовательность.
Таким образом, настоящее изобретение далее относится к любой нуклеотидной последовательности с использованием альтернативного триплетного кодона по меньшей мере для одной аминокислоты, кодируемой триплетным кодоном, но кодирующей такую же или вариант последовательности полипептида, как последовательность полипептида, кодированная исходной нуклеотидной последовательностью.
Кроме того, конкретные организмы обычно обладают смещением предпочтения относительно того, какие триплетные кодоны используют для кодирования аминокислот. Таблицы предпочтительного использования кодонов широко доступны и их можно использовать для получения оптимизированных по кодонам генов. Такие способы оптимизации кодонов используют в общепринятой практике для оптимизации экспрессии трансгенов в гетерологичном хозяине.
МОЛЕКУЛЯРНАЯ ЭВОЛЮЦИЯ
После выделения кодирующей фермент нуклеотидной последовательности или идентификации предполагаемой кодирующей фермент нуклеотидной последовательности может быть желательно модифицировать выбранную нуклеотидную последовательность, например, может быть желательно внести в последовательность мутации, чтобы получить фермент по настоящему изобретению.
Мутации можно вводить с применением синтетических олигонуклеотидов. Эти олигонуклеотиды содержат нуклеотидные последовательности, фланкирующие желательные участки мутации.
Подходящий способ описан в Morinaga et al (Biotechnology (1984) 2, p.646-649). Другой способ введения мутаций в кодирующие фермент нуклеотидные последовательности описан в Nelson and Long (Analytical Biochemistry (1989), 180, p.147-151).
Вместо сайт-специфического мутагенеза, такого, как описан выше, можно вводить мутации случайным образом, например, с использованием коммерческого набора, такого, как набор для мутагенеза GeneMorph PCR от Stratagene, или набор для случайного мутагенеза Diversify PCR от Clontech. EP 0583265 относится к способам мутагенеза, основанным на оптимизированной PCR, которые можно сочетать также с использованием мутагенных аналогов ДНК, таких как описанные в EP 0866796. Способы ошибочно-направленной PCR пригодны для получения вариантов липолитических ферментов с предпочтительными характеристиками. WO 0206457 относится к молекулярной эволюции липаз.
Третьим способом получения новых последовательностей является фрагментация не идентичных нуклеотидных последовательностей, или с использованием любого числа рестрикционных ферментов, или таким ферментом, как ДНКаза I, и повторная сборка полных нуклеотидных последовательностей, кодирующих функциональные белки. Альтернативно, можно использовать одну или множество не идентичных нуклеотидных последовательностей и вводить мутации во время повторной сборки полной нуклеотидной последовательности. Способы перетасовки ДНК и способы родственной перетасовки пригодны для получения вариантов липид-ацилтрансфераз с предпочтительными характеристиками. Подходящие способы для проведения «перетасовки» можно найти в EP0 752 008, EP1 138 763, EP1 103 606. Перетасовку можно сочетать также с другими формами мутагенеза ДНК, как описано в US 6180406 и WO 01/34835.
Таким образом, возможны получение многочисленных участков направленных или случайных мутаций в нуклеотидной последовательности in vivo или in vitro и последующий скрининг различными способами по улучшенной функциональности кодируемого полипептида. С использованием способов рекомбинации in silico и экзо-опосредованных способов (см. WO 00/58517, US 6344328, US 6361974), например, можно проводить молекулярную эволюцию, где полученный вариант сохраняет очень низкую гомологию с известными ферментами или белками. Эти варианты, полученные таким образом, могут обладать значительной структурной аналогией с известными липолитическими ферментами, но обладать очень низкой гомологией аминокислотной последовательности.
Кроме того, в качестве неограничивающего примера, мутантные или природные варианты полинуклеотидной последовательности можно подвергать рекомбинации либо с диким типом, либо с другими мутантными или природными вариантами для получения новых вариантов. Такие новые варианты также можно скринировать по улучшенной функциональности кодируемого полипептида.
Применение вышеупомянутых и подобных способов молекулярной эволюции позволяет идентификацию и отбор вариантов ферментов по настоящему изобретению, обладающих предпочтительными характеристиками, без каких-либо предварительных знаний о структуре или функции белка, и позволяет получение непредсказуемых, но выгодных мутаций или вариантов. Существует множество примеров применения молекулярной эволюции в области оптимизации и изменения ферментативной активности, такие примеры включают, но без ограничения одно или несколько из следующего: оптимизированная экспрессия и/или активность в клетке-хозяине или in vitro, увеличенная ферментативная активность, измененная специфичность субстрата и/или продукта, увеличенная или уменьшенная ферментативная или структурная стабильность, измененная ферментативная активность/специфичность в предпочтительных условиях внешней среды, например, температура, pH, субстрат.
Специалисту в данной области будет очевидно, что с использованием способов молекулярной эволюции можно изменять фермент для улучшения функциональности фермента.
Соответственно, липолитический фермент, используемый по настоящему изобретению, может представляет собой вариант, т.е. может содержать по меньшей мере одну замену, делецию или вставку аминокислоты по сравнению с исходным ферментом. Вариант фермента сохраняет по меньшей мере 20%, 30%, 40%, 50 %, 60%, 70%, 80%, 90%, 95%, 97%, 99% гомологию с исходным ферментом. Подходящие исходные ферменты могут включать в себя любой фермент с активностью эстеразы или липазы. Предпочтительно, исходный фермент выравнивают с консенсусной последовательностью pfam00657.
В предпочтительном варианте осуществления вариант липолитического фермента сохраняет или содержит по меньшей мере один или несколько аминокислотных остатков консенсусной последовательности pfam00657, обнаруженных в блоках GDSx, GANDY и HPT.
В ферменты, такие как липазы, с отсутствием или низкой галактолипазной и/или фосфолипазной активностью в водном окружении можно вводить мутации с применением способов молекулярной эволюции для введения или улучшения галактолипазной и/или фосфолипазной активности, таким образом получая липолитический фермент с существенной галактолипазной и/или фосфолипазной активностью, пригодный для использования в композициях и способах по настоящему изобретению.
Соответственно, вариант фермента может не обладать активностью в отношении триглицеридов и/или моноглицеридов, и/или диглицеридов.
Альтернативно, вариант фермента для применения по настоящему изобретению может обладать увеличенной активностью в отношении триглицеридов и/или может также обладать увеличенной активностью в отношении одного или нескольких из полярных липидов, фосфолипидов, лецитина, фосфатидилхолина, гликолипидов, дигалактозилмоноглицерида, моногалактозилмоноглицерида.
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
Объем настоящего изобретения охватывает также аминокислотные последовательности ферментов, обладающих специфическими свойствами, как определено в настоящем описании.
В настоящем описании термин «аминокислотная последовательность» является синонимом термина «полипептид» и/или термина «белок». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «пептид». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «фермент».
Аминокислотную последовательность можно получить/выделить из подходящего источника, или ее можно получить синтетически, или ее можно получить с применением способов рекомбинантной ДНК.
Ферменты, относящиеся к настоящему изобретению, можно использовать вместе с другими ферментами. Таким образом, настоящее изобретение относится также к комбинации ферментов, где комбинация содержит фермент по настоящему изобретению и другой фермент, который может представлять собой другой фермент по настоящему изобретению. Этот аспект обсуждают в последующем разделе.
Предпочтительно, аминокислотная последовательность, когда относится к настоящему изобретению и по существу входит в его объем, представляет собой не природный фермент. В этом отношении, термин «природный фермент» означает полный фермент, присутствующий в его природном окружении и экспрессирующийся с его природной нуклеотидной последовательностью.
ИДЕНТИЧНОСТЬ/ГОМОЛОГИЯ
Настоящее изобретение относится также к применению гомологов любой аминокислотной последовательности фермента или любой нуклеотидной последовательности, кодирующей такой фермент.
Термин «гомолог» в настоящем описании обозначает объект, обладающий определенной гомологией с аминокислотными последовательностями и нуклеотидными последовательностями. Термин «гомология» в настоящем описании можно приравнять к «идентичности». Эти термины в настоящем описании являются взаимозаменяемыми.
В настоящем контексте за гомологичную аминокислотную последовательность принимают содержащую аминокислотную последовательность, которая может быть по меньшей мере на 87 или 90% идентичной, предпочтительно по меньшей мере на 95, 96, 97, 98 или 99% идентичной последовательности. Обычно гомологи содержат такие же активные участки и т.д. - например, как рассматриваемая аминокислотная последовательность. Хотя гомологию можно рассматривать также в переводе на сходство (т.е. аминокислотные остатки, обладающие сходными химическими свойствами/функциями), в контексте настоящего изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательности.
В настоящем контексте за гомологичную нуклеотидную последовательность принимают нуклеотидную последовательность, которая может быть по меньшей мере на 85 или 90% идентичной, предпочтительно по меньшей мере на 95, 96, 97, 98 или 99% идентичной нуклеотидной последовательности, кодирующей фермент по настоящему изобретению (рассматриваемой последовательности). Обычно гомологи содержат одинаковые последовательности, которые кодируют активные участки и т.д., как рассматриваемая последовательность. Хотя гомологию можно рассматривать также с точки зрения сходства (т.е. аминокислотные остатки, обладающие сходными химическими свойствами/функциями), в контексте настоящего изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательности.
Для аминокислотных последовательностей и нуклеотидных последовательностей, сравнения гомологии можно проводить визуально, или более обычно, с помощью легко доступных программ для сравнения последовательности. Эти коммерчески доступные компьютерные программы могут вычислять % гомологии между двумя или более последовательностями.
% гомологии можно вычислять для смежных последовательностей, т.е. где одну последовательность выравнивают с другой последовательностью и каждую аминокислоту в последовательности непосредственно сравнивают с соответствующей аминокислотой в другой последовательности, один остаток за один раз. Это называют выравниванием «без пропусков». Обычно такие выравнивания без пропусков проводят только для относительно небольшого числа остатков.
Хотя это представляет собой очень простой и последовательный способ, он не может учитывать того, что, например, в паре последовательностей в остальном идентичной, одна вставка или делеция будет вызывать выпадение из выравнивания следующих аминокислотных остатков, таким образом потенциально приводя к большому уменьшению % гомологии при проведении общего выравнивания. Поэтому большинство способов сравнения последовательностей разрабатывают для получения оптимальных выравниваний, которые учитывают возможные вставки и делеции без чрезмерных штрафов для общего показателя гомологии. Этого достигают вставкой «пропусков» в выравнивание последовательности, чтобы попытаться максимизировать локальную гомологию.
Однако в этих более сложных способах назначают «штрафы за пропуски» для каждого пропуска, встретившегося в выравнивании, так что для одинакового числа идентичных аминокислот, выравнивание последовательности с наименее возможным числом пропусков - отражающее более сильное родство между двумя сравниваемыми последовательностями - будет достигать более высокого показателя гомологии, чем выравнивание со многими пропусками. Обычно используют «аффинную стоимость пропусков», где начисляют относительно высокую стоимость за наличие пропуска и меньший штраф за каждый последующий остаток в пропуске. Это представляет собой наиболее широко применяемую систему оценки пропусков. Конечно, с высокими штрафами за пропуски получают оптимизированные выравнивания с меньшими пропусками. Большинство программ выравнивания позволяют модифицировать штрафы за пропуски. Однако при использовании такого программного обеспечения для сравнений последовательностей предпочтительно применять значения по умолчанию. Например, при использовании пакета GCG Wisconsin Bestfit штраф за пропуск для аминокислотных последовательностей по умолчанию составляет -12 для пропуска и -4 для каждого продолжения.
Таким образом, вычисление максимального % гомологии сначала требует получения оптимального выравнивания, принимая во внимание штрафы за пропуски. Подходящей компьютерной программой для проведения такого выравнивания является пакет GCG Wisconsin Bestfit (Devereux et al 1984 Nuc. Acids Research 12, p.387). Другое программное обеспечение, которое может проводить сравнения последовательностей, включает в себя в качестве неограничивающих примеров пакет BLAST (см. Ausubel et al, 1999 Short Protocols in Molecular Biology, 4th Ed - Chapter 18), FASTA (Altschul et al, 1990 J. MoI. Biol. 403-410) и набор инструментов для сравнения GENEWORKS. Как BLAST, так и FASTA доступны для автономного и интерактивного поиска (смотри Ausubel et al, 1999, Short Protocols in Molecular Biology, pages 7-58, to 7-60).
Однако для некоторых применений предпочтительно использовать программу GCG Bestfit. Новый инструмент, названный BLAST 2 Sequences, также доступен для сравнения белковой и нуклеотидной последовательности (см. FEMS Microbiol Lett 1999 174(2): 247-50; и FEMS Microbiol Lett 1999 177(1): 187-8).
Хотя конечный % гомологии можно измерить с точки зрения идентичности, сам процесс выравнивания обычно не основан на парном сравнении «все или ничего». Вместо этого обычно используют матрицу баллов, назначающую баллы для каждого парного сравнения на основе химического сходства или эволюционного расстояния. Примером такой общераспространенной матрицы является матрица BLOSUM62 - матрица по умолчанию для набора программ BLAST. Программы GCG Wisconsin обычно используют общедоступные значения по умолчанию или пользовательскую таблицу сравнения символов, если поставляется (дополнительные подробности смотри в руководстве пользователя). Для некоторых применений предпочтительно использовать общедоступные значения по умолчанию для пакета GCG, или, в случае другого программного обеспечения, матрицу по умолчанию, такую как BLOSUM62.
Альтернативно, процентные гомологии можно вычислить с использованием возможности множественного выравнивания в DNASIS™ (Hitachi Software), основанной на алгоритме, аналогичном CLUSTAL (Higgins DG & Sharp PM (1988), Gene 73(1), 237-244).
После получения оптимального выравнивания программным обеспечением возможно вычислить % гомологии, предпочтительно % идентичности последовательности. Обычно программное обеспечение производит это как часть сравнения последовательности и выводит численный результат.
В предпочтительном аспекте настоящего изобретения используют следующее программное обеспечение и установочные параметры для вычисления процентной гомологии/идентичности последовательности. Для последовательностей аминокислот процент идентичных (гомологичных) или «положительных» для каждой возможной пары аминокислотных последовательностей вычисляли посредством AlignX VectorNTI (Vector NTI Advance 9.1 от Invitrogen Corporation, Carlsbad, California, USA.). Установочные параметры представляют собой параметры по умолчанию (штраф за внесение пропуска - 10, штраф за продолжение пропуска 0,1).
Для последовательностей нуклеиновой кислоты процент идентичных (гомологичных) или «положительных» для каждой возможной пары последовательностей нуклеиновых кислот вычисляли посредством программы AlignX Vector NTI от Informax Inc. (USA). Установочные параметры представляют собой параметры по умолчанию, которые для ДНК составляют: штраф за внесение пропуска: 15 и штраф за продолжение пропуска: 6,66 (такие же установочные параметры для множественного выравнивания).
Предпочтительно, аминокислотную идентичность (гомологию) вычисляют на протяжении полноразмерной аминокислотной последовательности (например, SEQ ID 4, 5, 7, 8, 10, 12 и 14) или для нуклеиновой кислоты соответствующего полинуклеотида, кодирующего соответствующую полноразмерную аминокислотную последовательность. Идентичность (гомологию) аминокислот или нуклеиновой кислоты можно, предпочтительно, вычислить посредством сравнения гомологии/идентичности на протяжении зрелой полипептидной последовательности, т.е. полипептидной последовательности, являющейся ко- или посттрансляционно процессированной, например, посредством отщепления N-концевого сигнального пептида, или явления C-концевого отщепления.
Последовательности могут содержать также делеции, вставки или замены аминокислотных остатков, дающие молчащие замены, приводящие к функционально эквивалентному веществу. Можно производить преднамеренные замены аминокислот на основе сходства свойств аминокислот (таких как полярность, заряд, растворимость, гидрофобность, гидрофильность, и/или амфипатическая природа остатков), и таким образом это может быть использовано для группировки аминокислоты в функциональные группы. Аминокислоты можно группировать на основе свойств только их боковых цепей. Однако более полезно включать также данные по мутациям. Выведенные таким образом группы аминокислот, по-видимому, являются консервативными по структурным причинам. Эти группы могут быть описаны в форме диаграммы Венна (Livingstone CD. and Barton GJ. (1993) "Protein sequence alignments: a strategy for the hierarchical analysis of residue conservation" Comput.Appl Biosci. 9: 745-756)(Taylor W.R. (1986) "The classification of amino acid conservation" J.Theor.Biol. 119; 205-218). Консервативные замены можно осуществлять, например, согласно таблице ниже, описывающей общепринятую диаграмму Венна, группирующую аминокислоты.
Настоящее изобретение относится также к гомологичным заменам (как замену, так и замещение используют в настоящем описании для обозначения обмена существующего аминокислотного остатка на альтернативный остаток), которые могут происходить, т.е. заменам подобного-на-подобный, таким как основного на основной, кислого на кислый, полярного на полярный и т.д. Могут происходить также негомологичные замены, т.е. из одного класса остатков на другой или, альтернативно, с вовлечением неприродных аминокислот, таких как орнитин (обозначенный ниже как Z), диаминомасляная кислота-орнитин (обозначенная ниже как B), норлейцин-орнитин (обозначенный ниже как O), пирилаланин, тиенилаланин, нафтилаланин и фенилглицин.
Можно также осуществлять замены неприродными аминокислотами.
Варианты аминокислотных последовательностей могут содержать подходящие спейсерные группы, которые можно вставлять между любыми двумя аминокислотными остатками в последовательности, включая алкильные группы, такие как метильные, этильные или пропильные группы в дополнение к аминокислотным спейсерам, таким как остатки глицина или β-аланина. Дополнительная форма варианта включает в себя присутствие одного или нескольких аминокислотных остатков в пептоидной форме и будет хорошо понятна специалистам в данной области. Чтобы избежать неопределенности, «пептоидную форму» используют для обозначения вариантов аминокислотных остатков, где замещающая группа α-углерода находится на атоме азота остатка, а не на атоме α-углерода. Способы получения пептидов в пептоидной форме известны в данной области, например Simon RJ et al, PNAS (1992) 89(20), 9367-9371 и Horwell DC, Trends Biotechnol. (1995) 13(4), 132-134.
Нуклеотидные последовательности для применения по настоящему изобретению могут включать в себя синтетические или модифицированные нуклеотиды. Ряд различных типов модификации олигонуклеотидов известен в данной области. Они включают в себя метилфосфонатный и фосфоротиоатный остовы и/или добавление акридиновых или полилизиновых цепей на 3' и/или 5'-концах молекулы. Понятно, что для целей настоящего изобретения описанные в настоящем описании нуклеотидные последовательности можно модифицировать любым способом, доступным в данной области. Такие модификации могут быть выполнены для усиления активности или времени жизни in vivo нуклеотидных последовательностей по настоящему изобретению.
Настоящее изобретение относится также к применению нуклеотидных последовательностей, комплементарных представленным в настоящем описании последовательностям или любому их производному, фрагменту или его производному. Если последовательность является комплементарной их фрагменту, эту последовательность можно применять в качестве пробы для идентификации сходных кодирующих последовательностей в других организмах и т.д.
Полинуклеотиды, не являющиеся на 100% гомологичными последовательностям по настоящему изобретению, но попадающие в объем изобретения, можно получить рядом способов. Другие варианты описанных в настоящем описании последовательностей можно получить, например, анализом зондами библиотек ДНК, полученных от ряда индивидуумов, например, индивидуумов из различных популяций. Кроме того, можно получить другие гомологи, и такие гомологи и их фрагменты, как правило, будут способны избирательно гибридизоваться с последовательностями, показанными в списке последовательностей в настоящем описании. Такие последовательности можно получить анализом зондами библиотек кДНК или геномной ДНК из других видов и анализом данных библиотек зондами, содержащими всю или часть любой последовательности из прилагаемого списка последовательностей, в условиях от средней до высокой строгости. Сходные соображения используют для получения видовых гомологов и аллельных вариантов полипептидных или нуклеотидных последовательностей по изобретению.
Варианты и видовые/штаммовые гомологи можно получить также с использованием вырожденной PCR, для которой используют праймеры, сконструированные нацеленными на последовательности в вариантах и гомологах, кодирующие аминокислотные последовательности, консервативные в последовательностях по настоящему изобретению. Консервативные последовательности можно предсказать, например, выравниванием аминокислотных последовательностей из нескольких вариантов/гомологов. Выравнивания последовательностей можно проводить с применением компьютерного программного обеспечения, известного в данной области. Например, широко распространенной является программа GCG Wisconsin PileUp.
Праймеры, использумые для PCR, содержат одно или несколько вырожденных положений и их можно использовать в условиях более низкой строгости, чем используемые для клонирования последовательностей с единственными последовательностями праймеров против известных последовательностей.
Альтернативно, такие полинуклеотиды могут быть получены сайт-специфическим мутагенезом охарактеризованных последовательностей. Это может быть применимо, например, если требуются молчащие изменения последовательности кодонов для оптимизации кодонных предпочтений для конкретной клетки-хозяина, в которой экспрессируют полинуклеотидные последовательности. Другие изменения последовательности могут являться желательными, для введения участков распознавания рестрикционных ферментов или для изменения свойства или функции кодируемых полинуклеотидами полипептидов.
Полинуклеотиды (нуклеотидные последовательности) по изобретению можно использовать для получения праймера, например PCR-праймера, праймера для альтернативной реакции амплификации, зонда, например, меченого обнаруживаемой меткой общепринятыми способами с использованием радиоактивных или нерадиоактивных меток, или полинуклеотиды можно клонировать в векторы. Такие праймеры, зонды и другие фрагменты могут составлять по меньшей мере 15, предпочтительно по меньшей мере 20, например, по меньшей мере 25, 30 или 40 нуклеотидов в длину, и также охвачены термином полинуклеотиды по изобретению.
Полинуклеотиды, такие как ДНК-полинуклеотиды и зонды по настоящему изобретению, можно получать рекомбинантно, синтетически или любыми способами, доступными специалистам в данной области. Их можно также клонировать общепринятыми способами.
Как правило, праймеры получают синтетическими способами, включающими пошаговое изготовление желаемой последовательности нуклеиновой кислоты по одному нуклеотиду за один раз. Способы для осуществления этого с использованием автоматизированных способов легко доступны в данной области.
Более длинные полинуклеотиды, как правило, получают с использованием рекомбинантных способов, например, с использованием способов клонирования PCR (полимеразная цепная реакция). Можно сконструировать праймеры, содержащие подходящие участки узнавания рестрикционных ферментов, так что амплифицированную ДНК можно клонировать в подходящий клонирующий вектор.
БИОЛОГИЧЕСКИ АКТИВНЫЙ
Предпочтительно, варианты последовательностей и т.д. являются по меньшей мере настолько биологически активными, как последовательности, представленные в настоящем описании.
В настоящем описании, «биологически активный» относится к последовательности, обладающей сходной структурной функцией (но не обязательно в той же самой степени) и/или сходной регуляторной функцией (но не обязательно в той же самой степени), и/или сходной биохимической функцией (но не обязательно в той же самой степени), как встречающаяся в природе последовательность.
ГИБРИДИЗАЦИЯ
Настоящее изобретение относится также к последовательностям, комплементарным последовательностям нуклеиновой кислоты по настоящему изобретению или последовательностям, способным гибридизоваться или с последовательностями по настоящему изобретению, или с последовательностями, комплементарными им.
Термин «гибридизация» в настоящем описании включает в себя «процесс, при котором цепь нуклеиновой кислоты соединяется с комплементарной цепью посредством спаривания оснований», а также процесс амплификации, как проводят в способах полимеразной цепной реакции (PCR).
Настоящее изобретение относится также к применению нуклеотидных последовательностей, способных гибридизоваться с последовательностями, комплементарными последовательностям, представленным в настоящем описании, или любым их производным, фрагментом или его производным.
Термин «вариант» также охватывает последовательности, комплементарные последовательностям, способным гибридизоваться с нуклеотидными последовательностями, представленными в настоящем описании.
Предпочтительно, термин «вариант» также охватывает последовательности, комплементарные последовательностям, способным гибридизоваться в строгих условиях (например, 50°C и 0,2×SSC {l×SSC = 0,15 М NaCl, 0,015 М Na3цитрат, pH 7,0}) с нуклеотидными последовательностями, представленными в настоящем описании.
Более предпочтительно, термин «вариант» охватывает последовательности, комплементарные последовательностям, способным гибридизоваться в высоко строгих условиях (например, 65°C и 0,1×SSC {1×SSC = 0,15 М NaCl, 0,015 М Na3цитрат, pH 7,0}) с нуклеотидными последовательностями, представленными в настоящем описании.
Настоящее изобретение относится также к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями по настоящему изобретению (включая последовательности, комплементарные последовательностям, представленным в настоящем описании).
Настоящее изобретение относится также к нуклеотидным последовательностям, комплементарным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями по настоящему изобретению (включая последовательности, комплементарные представленным в настоящем описании).
В объем настоящего изобретения включены также полинуклеотидные последовательности, способные гибридизоваться с нуклеотидными последовательностями, представленными в настоящем описании, в условиях от средней до максимальной строгости.
В предпочтительном аспекте настоящее изобретение относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидной последовательностью по настоящему изобретению или комплементарной ей, в строгих условиях (например, 50°C и 0,2×SSC).
В более предпочтительном аспекте настоящее изобретение относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидной последовательностью по настоящему изобретению или комплементарной ей, в высоко строгих условиях (например, 65°C и 0,1×SSC).
РЕКОМБИНАНТНЫЙ
В одном из аспектов последовательность для применения по настоящему изобретению представляет собой рекомбинантную последовательность, т.е. последовательность, полученную с использованием способов рекомбинантной ДНК.
Эти способы рекомбинантной ДНК находятся в компетенции обычных специалистов в данной области. Такие способы описаны в литературе, например, J. Sambrook, E.F. Fritsch, and T. Maniatis, 1989, Molecular Cloning: A Laboratory Manual, Second Edition, Books 1-3, Cold Spring Harbor Laboratory Press.
СИНТЕТИЧЕСКИЙ
В одном из аспектов последовательность для применения по настоящему изобретению представляет собой синтетическую последовательность, т.е. последовательность, полученную химическим или ферментативным синтезом in vitro. Она включает в себя в качестве неограничивающих примеров последовательность, полученную с оптимальным использованием кодонов для организма-хозяина - такого как метилотрофные дрожжи Pichia и Hansenula.
ЭКСПРЕССИЯ ФЕРМЕНТОВ
Нуклеотидную последовательность для применения по настоящему изобретению можно встраивать в рекомбинантный реплицирующийся вектор. Вектор можно использовать для репликации и экспрессии нуклеотидной последовательности, в форме фермента в подходящей клетке-хозяине и/или из нее.
Экспрессию можно контролировать с использованием контрольных последовательностей, например регуляторных последовательностей.
Фермент, продуцируемый рекомбинантной клеткой-хозяином посредством экспрессии нуклеотидной последовательности, может секретироваться или может содержаться внутри клетки в зависимости от последовательности и/или используемого вектора. Кодирующие последовательности можно конструировать с сигнальными последовательностями с прямой секрецией вещества, кодируемого последовательностями, через конкретную мембрану прокариотической или эукариотической клетки.
ЭКСПРЕССИРУЮЩИЙ ВЕКТОР
Термин «экспрессирующий вектор» означает конструкцию, способную экспрессироваться in vivo или in vitro.
Предпочтительно, экспрессирующий вектор встроен в геном подходящего организма-хозяина. Термин «встроенный» предпочтительно охватывает стабильное встраивание в геном.
Нуклеотидная последовательность по настоящему изобретению может присутствовать в векторе, в котором нуклеотидная последовательность функционально связана с регуляторными последовательностями, способными обеспечивать экспрессию нуклеотидной последовательности подходящим организмом-хозяином.
Векторы для применения по настоящему изобретению можно трансформировать в подходящую клетку-хозяина, как описано ниже, для обеспечения экспрессии полипептида по настоящему изобретению.
Выбор вектора, например, плазмидного, космидного или фагового вектора, часто зависит от клетки-хозяина, в которую он подлежит введению.
Векторы для применения по настоящему изобретению могут содержать один или несколько генов селективных маркеров - таких как ген, придающий устойчивость к антибиотику, например устойчивость к ампициллину, канамицину, хлорамфениколу или тетрациклину. Альтернативно, селекцию можно осуществлять котрансформацией (как описано в WO91/17243).
Векторы можно использовать in vitro, например, для получения РНК, или использовать для трансфекции, трансформации, трансдукции или инфекции клетки-хозяина.
Таким образом, в другом варианте осуществления, изобретение относится к способу получения нуклеотидных последовательностей по настоящему изобретению введением нуклеотидной последовательности по настоящему изобретению в реплицируемый вектор, введением вектора в совместимую клетку-хозяина и выращиванием клетки-хозяина в условиях, приводящих к репликации вектора.
Вектор может дополнительно содержать нуклеотидную последовательность, позволяющую вектору реплицироваться в рассматриваемой клетке-хозяине. Примерами таких последовательностей являются точки начала репликации плазмид pUC19, pACYC177, pUB110, pE194, pAMB1 и pIJ702.
РЕГУЛЯТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
В некоторых применениях нуклеотидная последовательность для использования по настоящему изобретению является функционально связанной с регуляторной последовательностью, способной обеспечивать экспрессию нуклеотидной последовательности, такую как в выбранной клетке-хозяине. В качестве примера, настоящее изобретение относится к вектору, содержащему нуклеотидную последовательность по настоящему изобретению, функционально связанную с такой регуляторной последовательностью, т.е. вектор является экспрессирующим вектором.
Термин «функционально связанный» относится к непосредственному соседству, где описанные компоненты находятся в связи, позволяющей им функционировать предназначенным для них способом. Регуляторная последовательность, «функционально связанная» с кодирующей последовательностью, лигирована таким образом, что экспрессии кодирующей последовательности достигают в условиях, согласующихся с контрольными последовательностями.
Термин «регуляторные последовательности» включает в себя промоторы, энхансеры и другие сигналы регуляции экспрессии.
Термин «промотор» применяют в обычном в данной области смысле, например, участок связывания РНК-полимеразы.
Усиленной экспрессии нуклеотидной последовательности, кодирующей фермент по настоящему изобретению, можно также достичь выбором гетерологичных регуляторных областей, например областей промотора, секреторного лидера и терминатора.
Предпочтительно, нуклеотидная последовательность по настоящему изобретению является функционально связанной по меньшей мере с промотором.
Примеры подходящих промоторов для управления транскрипцией нуклеотидной последовательности в бактериальном, грибковом или дрожжевом хозяине хорошо известны в данной области.
КОНСТРУКЦИИ
Термин «конструкция» - который является синонимом таким терминам, как «конъюгат», «кассета» и «гибрид», включает в себя нуклеотидную последовательность для применения по настоящему изобретению, непосредственно или опосредованно присоединенную к промотору.
Примером опосредованного присоединения является предоставление подходящей спейсерной группы, такой как последовательность интрона, такого как Sh1-интрон или интрон ADH, находящейся между промотором и нуклеотидной последовательностью по настоящему изобретению. То же самое справедливо для термина «слитый» по отношению к настоящему изобретению, который включает в себя непосредственное или опосредованное присоединение. В некоторых случаях термины не охватывают природное сочетание нуклеотидной последовательности, кодирующей белок, обычно связанной с промотором дикого типа, когда они оба присутствуют в их природном окружении.
Конструкция может даже содержать или экспрессировать маркер, позволяющий селекцию генетической конструкции.
Для некоторых применений конструкция по настоящему изобретению предпочтительно содержит по меньшей мере нуклеотидную последовательность по настоящему изобретению, функционально связанную с промотором.
КЛЕТКА-ХОЗЯИН
Термин «клетка-хозяин» по отношению к настоящему изобретению включает в себя любую клетку, содержащую или нуклеотидную последовательность, или экспрессирующий вектор, как описано выше, которую используют для рекомбинантной продукции фермента, обладающего конкретными свойствами, как определено в настоящем описании.
Таким образом, в другом варианте осуществления настоящее изобретение относится к клеткам-хозяевам, трансформированным или трансфицированным нуклеотидной последовательностью, экспрессирующей фермент по настоящему изобретению. Клетки выбирают по принципу совместимостии с указанным вектором, и они могут, например, быть прокариотическими (например, бактериальными), грибковыми, дрожжевыми или растительными клетками. Предпочтительно, клетки-хозяева не являются человеческими клетками.
Примерами подходящих бактериальных организмов-хозяев являются грамположительные или грамотрицательные виды бактерий.
В зависимости от природы нуклеотидной последовательности, кодирующей фермент по настоящему изобретению, и/или желательности дальнейшего процессинга экспрессированного белка, могут быть предпочтительны эукариотические хозяева, такие как дрожжи или другие грибы. Как правило, дрожжевые клетки предпочтительнее, чем другие клетки грибов, так как с ними легче манипулировать.
Однако некоторые белки или плохо секретируются из дрожжевых клеток, или в некоторых случаях не процессированы надлежащим образом (например, гипергликозилирование в дрожжах). В этих случаях следует выбирать другой грибковый организм-хозяин.
Использование подходящих клеток-хозяев - таких как дрожжевые, грибковые или растительные клетки-хозяева - может обеспечивать посттрансляционные модификации (например, миристоилирование, гликозилирование, укорочение, липидизацию и фосфорилирование тирозина, серина или треонина), что может требоваться для придания оптимальной биологической активности продуктов рекомбинантной экспрессии по настоящему изобретению.
Клетка-хозяин может представлять собой дефектный по протеазе или лишенный протеазы штамм.
Генотип клетки-хозяина можно модифицировать для улучшения экспрессии.
Примеры модификаций клетки-хозяина включают в себя дефектность по протеазе, дополнение редких тРНК и модификацию восстановительного потенциала в цитоплазме для усиления формирования дисульфидной связи.
Например, клетка-хозяин E. coli может сверхэкспрессировать редкие тРНК для улучшения экспрессии гетерологичных белков, как проиллюстрировано/описано в Kane (Curr Opin Biotechnol (1995), 6, 494-500 "Effects of rare codon clusters on high-level expression of heterologous proteins in E.coli"). Клетка-хозяин может являться дефектной по ряду восстанавливающих ферментов, содействующих формированию стабильных дисульфидных связей, как проиллюстрировано/описано в Bessette (Proc Natl Acad Sci USA (1999), 96, 13703-13708 " Efficient folding of proteins with multiple disulphide bonds in the Escherichia coli cytoplasm").
В одном из вариантов осуществления клетка-хозяин представляет собой бактерии, предпочтительно грамположительные бактерии, предпочтительно клетку-хозяина выбирают из Actinobacteria, таких как Biofidobacteria и Aeromonas, особенно предпочтительно Aeromonas salmonicida. Еще более предпочтительными являются Actinomicetales, такие как Corynebacteria, в частности Corynebacterium glutamicum и Nocardia. Особенно предпочтительными являются Streptomycetaceae, такие как Streptomyces, особенно S.lividans.
Для экспрессии гена галактолипазы можно использовать микробного хозяина, например Eubacteria, Archea; или Fungi, включая дрожжи. Предпочтительными являются Eubacteria, например, Firmicutes (грамположительные бактерии с низким GC), такие как Bacillus subtilis и другие виды бацилл, молочнокислые бактерии, такие как виды родов Lactobacillus и Lactococcus.
Также предпочтительными являются грамотрицательные Proteobacteria, в частности, Gammaproteobacteria, такие как виды-хозяева, принадлежащие к родам Pseudomonas, Xanthomonas, Citrobacter и Escherichia, особенно Escherichia coli.
В другом варианте осуществления клетка-хозяин того же рода, что и виды природного хозяина, т.е. рекомбинантный ген введен повторно и экспрессирован в видах того же самого рода, как виды, из которых выделен рекомбинантный ген.
В другом варианте осуществления клетка-хозяин представляет собой виды природного хозяина, т.е. рекомбинантный ген введен повторно и экспрессирован в тех же видах, из которых выделен рекомбинантный ген.
ОРГАНИЗМ
Термин «организм» по отношению к настоящему изобретению включает в себя любой организм, который может содержать нуклеотидную последовательность, кодирующую фермент по настоящему изобретению, и/или полученные из нее продукты, и/или где промотор может позволять экспрессию нуклеотидной последовательности по настоящему изобретению, когда присутствует в организме.
Подходящие организмы могут включать в себя прокариоты, гриб, дрожжи или растение.
Термин «трансгенный организм» по отношению к настоящему изобретению включает в себя любой организм, содержащий нуклеотидную последовательность, кодирующую фермент по настоящему изобретению, и/или полученные из нее продукты, и/или где промотор может позволять экспрессию нуклеотидной последовательности по настоящему изобретению в организме. Предпочтительно, нуклеотидную последовательность встраивают в геном организма.
Термин «трансгенный организм» не охватывает природные кодирующие нуклеотидные последовательности в их природном окружении, когда они находятся под контролем их природного промотора, который также присутствует в его природном окружении.
Следовательно, трансгенный организм по настоящему изобретению включает в себя организм, содержащий нуклеотидную последовательность, кодирующую фермент по настоящему изобретению, конструкции по настоящему изобретению, векторы по настоящему изобретению, плазмиды по настоящему изобретению, клетки по настоящему изобретению, ткани по настоящему изобретению или их продукты, или их комбинацию.
Например, трансгенный организм может содержать также нуклеотидную последовательность, кодирующую фермент по настоящему изобретению под контролем гетерологичного промотора.
ТРАНСФОРМАЦИЯ КЛЕТОК-ХОЗЯЕВ/ОРГАНИЗМА
Как указано ранее, организм-хозяин может представлять собой прокариотический или эукариотический организм. Примеры подходящих прокариотических хозяев включают в себя E. coli и Bacillus subtilis.
Способы трансформации прокариотических хозяев хорошо задокументированы в данной области, например, см. Sambrook et al (Molecular Cloning: A Laboratory Manual, 2nd edition, 1989, Cold Spring Harbor Laboratory Press). Если используют прокариотического хозяина, тогда нуклеотидная последовательность может нуждаться в соответствующей модификации перед трансформацией, такой как удаление интронов.
Клетки мицелиальных грибов можно трансформировать с использованием различных способов, известных в данной области, таких как способ, предусматривающий формирование протопласта и трансформацию протопластов с последующей регенерацией клеточной стенки известным способом. Применение Aspergillus в качестве микроорганизма-хозяина описано в EP 0238 023.
Другой организм-хозяин может представлять собой растение. Обзор основных способов, используемых для трансформации растений, можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant Mol Biol [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994 17-27). Дополнительные объяснения трансформации растений можно найти в EP-A-0449375.
Общие объяснения трансформации грибов, дрожжей и растений представлены в следующих разделах.
ТРАНСФОРМИРОВАННЫЕ ГРИБЫ
Организм-хозяин может представлять собой гриб - такой как мицелиальный гриб. Примеры таких подходящих хозяев включают в себя любой организм, принадлежащий к родам Thermomyces, Acremonium, Aspergillus, Penicillium, Mucor, Neurospora, Trichoderma и подобным.
Обзор описаний трансформации мицелиальных грибов дан в US-A-5741665, где заявлено, что общепринятые способы трансформации мицелиальных грибов и культивирования грибов хорошо известны в данной области. Обширный обзор методов в применении к N. crassa обнаружен, например, у Davis and de Serres, Methods Enzymol (1971) 17A: 79-143.
Дополнительный обзор описаний трансформации мицелиальных грибов дан в US-A-5674707.
В одном из аспектов организм-хозяин может быть из рода Aspergillus, такой как Aspergillus niger.
Трансгенный Aspergillus по настоящему изобретению можно также получить, например, по следующим объяснениям Turner G. 1994 (Vectors for genetic manipulation. In: Martinelli S.D., Kinghorn J.R.( Editors) Aspergillus: 50 years on. Progress in industrial microbiology vol 29. Elsevier Amsterdam 1994. p.641-666).
Обзор экспрессии генов в мицелиальных грибах дан в Punt et al. (2002) Trends Biotechnol 2002 May;20(5):200-6, Archer & Peberdy Crit Rev Biotechnol (1997) 17(4):273-306.
ТРАНСФОРМИРОВАННЫЕ ДРОЖЖИ
В другом варианте осуществления трансгенный организм может представлять собой дрожжи.
Обзор принципов экспрессии гетерологичных генов в дрожжах дан, например, в Methods Mol Biol (1995), 49:341-54 и Curr Opin Biotechnol (1997) Oct;8(5):554-60.
В данном отношении дрожжи - такие как виды Saccharomyces cereviseae или Pichia pastoris (см. FEMS Microbiol Rev (2000 24(l):45-66), можно использовать в качестве средства экспрессии гетерологичного гена.
Обзор принципов экспрессии гетерологичного гена в Saccharomyces cerevisiae и секреции продуктов гена дан в E Hinchcliffe E Kenny (1993, "Yeast as a vehicle for the expression of heterologous genes", Yeasts, Vol 5, Anthony H Rose and J Stuart Harrison, eds, 2nd edition, Academic Press Ltd.).
Для трансформации дрожжей разработано несколько протоколов трансформации. Например, можно получить трансгенные Saccharomyces по настоящему изобретению, следуя методикам Hinnen et al. (1978, Proceedings of the National Academy of Sciences of the USA 75, 1929); Beggs, J D (1978, Nature, London, 275, 104); и Ito, H et al. (1983, J Bacteriology 153, 163-168).
Трансформированные клетки дрожжей можно отбирать с использованием различных селективных маркеров, таких как ауксотрофные маркеры, доминантные маркеры устойчивости к антибиотикам.
ТРАНСФОРМИРОВАННЫЕ РАСТЕНИЯ/КЛЕТКИ РАСТЕНИЙ
Организм-хозяин, подходящий для целей настоящего изобретения, может представлять собой растение. Обзор общих способов можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant MoI Biol [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994, 17-27).
КУЛЬТИВИРОВАНИЕ И ПРОДУКЦИЯ
Клетки-хозяева, трансформированные нуклеотидной последовательностью по настоящему изобретению, можно культивировать в условиях, способствующих продукции кодированного фермента и облегчающих выделение фермента из клеток и/или культуральной среды.
Среда, используемая для культивирования клеток, может представлять собой любую известную среду, пригодную для роста рассматриваемой клетки-хозяина и осуществления экспрессии фермента.
Белок, продуцируемый рекомбинантной клеткой, может экспонироваться на поверхности клетки.
Фермент может секретироваться из клеток-хозяев и его можно удобно выделять из культуральной среды с использованием хорошо известных способов.
СЕКРЕЦИЯ
Часто желательно, чтобы фермент секретировался из экспрессирующего хозяина в культуральную среду, из которой фермент можно легко выделить. По настоящему изобретению последовательность секреторного лидера может быть выбрана на основе желаемого экспрессирующего хозяина. В контексте настоящего изобретения можно также использовать гибридные сигнальные последовательности.
Конкретными примерами последовательностей гетерологичных секреторных лидеров являются происходящие из гена амилоглюкозидазы (AG) грибов (glaA - как 18, так и 24 аминокислотные варианты, например, из Aspergillus), гена a-фактора (дрожжей, например, Saccharomyces, Kluyveromyces и Hansenula) или ген α-амилазы (Bacillus).
В качестве примера, обзор секреции гетерологичных белков в E. coli дан в Methods Enzymol (1990) 182:132-43.
ДЕТЕКЦИЯ
Множество протоколов для детекции и измерения экспрессии аминокислотной последовательности известны в данной области. Примеры включают в себя твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) и флюоресцентно-активированную сортировку клеток (FACS).
Широкое множество меток и способов конъюгации известно специалистам в данной области и их можно использовать в различных анализах нуклеиновых кислот и аминокислот.
Ряд компаний, таких как Pharmacia Biotech (Piscataway, NJ), Promega (Madison, WI), и US Biochemical Corp (Cleveland, OH) поставляют коммерческие наборы и протоколы для этих способов.
Подходящие репортерные молекулы или метки включают в себя радиоизотопы, ферменты, флуоресцентные, хемилюминесцентные или хромогенные вещества, а так же субстраты, кофакторы, ингибиторы, магнитные частицы и подобные. Патенты, в которых описано объясняющие применение таких меток, включают в себя US-A-3817837; US-A-3850752; US-A-3939350; US-A-3996345; US-A-4277437; US-A-4275149 и US-A-4366241.
Также можно получать рекомбинантные иммуноглобулины, как показано в US-A-4816567.
СЛИТЫЕ БЕЛКИ
Аминокислотная последовательность для применения по настоящему изобретению может быть получена в виде слитого белка, например, для облегчения выделения и очистки. Примеры партнеров для слитого белка включают в себя глутатион-S-трансферазу (GST), 6xHis, GAL4 (ДНК-связывающий и активирующий транскрипцию домены) и (β-галактозидазу). Может быть удобно также включать участок протеолитического расщепления между партнером для слитого белка и представляющей интерес последовательностью белка, чтобы позволить удаление последовательностей слитого белка.
Предпочтительно, слитый белок не мешает активности последовательности белка.
Обзор экспрессии систем слитых генов в E. coli дан в Curr Opin Biotechnol (1995) 6(5):501-6.
В другом варианте осуществления изобретения, аминокислотную последовательность можно лигировать с гетерологичной последовательностью для кодирования слитого белка. Например, для скрининга пептидных библиотек по средствам, способным влиять на активность вещества, может быть полезно кодировать химерную молекулу, экспрессирующую гетерологичный эпитоп, распознаваемый коммерчески доступным антителом.
КРУПНОМАСШТАБНОЕ ПРИМЕНЕНИЕ
В одном из предпочтительных вариантов осуществления аминокислотную последовательность используют для крупномасштабных применений.
Предпочтительно, аминокислотную последовательность получают в количестве от 1 г на литр до приблизительно 2 г на литр общего объема культуры после культивирования организма-хозяина.
Предпочтительно, аминокислотную последовательность получают в количестве от 100 мг на литр до приблизительно 900 мг на литр общего объема культуры после культивирования организма-хозяина.
Предпочтительно, аминокислотную последовательность получают в количестве от 250 мг на литр до приблизительно 500 мг на литр общего объема культуры после культивирования организма-хозяина.
ПИЩЕВОЙ ПРОДУКТ
Композицию по настоящему изобретению можно применять в качестве - или для получения - пищевого продукта. В настоящем описании термин «пищевой продукт» применен в широком смысле и включает пищевой продукт для людей, а также пищевой продукт для животных (т.е. корм). В предпочтительном аспекте пищевой продукт служит для употребления человеком.
Пищевой продукт может присутствовать в форме раствора или в твердом виде - в зависимости от применения и/или способа применения и/или способа введения.
ИНГРЕДИЕНТ ПИЩЕВОГО ПРОДУКТА
Композицию по настоящему изобретению можно использовать в качестве ингредиента пищевого продукта.
В настоящем описании, термин «ингредиент пищевого продукта» включает состав, который можно добавлять к функциональным пище или пищевым продуктам, и включает составы, низкий уровень которых можно использовать в широком множестве продуктов, требующих, например, окисления или эмульгирования.
Компонент пищевого продукта может присутствовать в форме раствора или в твердом виде - в зависимости от применения и/или способа применения, и/или способа введения.
ПРОДУКТЫ ПИТАНИЯ
Композиции по настоящему изобретению можно использовать для получения продуктов питания, таких как один или несколько из: кондитерских изделий, молочных продуктов, мясных продуктов, продуктов из птиц, рыбопродуктов и хлебобулочных изделий.
Настоящее изобретение относится также к способу получения пищевого продукта или ингредиента пищевого продукта, включающему смешивание липолитического фермента по настоящему изобретению с другим ингредиентом пищевого продукта.
Дополнительные предпочтительные аспекты представлены в сопровождающей формуле изобретения и в следующих фигурах и примерах.
На фигуре 1 показан фрагмент PCR SEQ ID No:1, представляющий собой часть не кодирующего фермент полинуклеотида; эта последовательность представляет собой ген рибосомальной 16S РНК, широко используемый для таксономических сравнений;
На фигуре 2 показан фрагмент PCR SEQ ID No:2, представляющий собой часть не кодирующего фермент полинуклеотида; эта последовательность представляет собой ген рибосомальной 16S РНК, широко используемый для таксономических сравнений;
На фигуре 3 показан полинуклеотид, кодирующий липолитический фермент по настоящему изобретению (SEQ ID No:3);
На фигуре 4 показана аминокислотная последовательность липолитического фермента по настоящему изобретению (SEQ ID No:4);
На фигуре 5 показана структура экспрессирующих липолитический фермент векторов pGTK(L131) и pET11(131-51);
На фигуре 6 показан график эффекта липолитического фермента из Streptomyces sp. L130 на дигалактозилдиглицерид в тесте;
На фигуре 7 в графической форме показан эффект липолитического фермента из Streptomyces sp. L131 на дигалактозилдиглицерид в тесте;
На фигуре 8 в графической форме показан эффект липолитического фермента из Streptomyces на триглицерид в тесте;
На фигуре 9 показан профиль pH липолитического фермента, полученного из Streptomyces sp. L131 для галактолипидного субстрата;
На фигуре 10 показана пластина TLC липидов, выделенных из теста, обработанного липолитическим ферментом из Streptomyces, экспрессированным в E.coli, обозначенным #236; дорожка 1=контроль; дорожка 2=#236, 0,225 PLU-7/г муки; дорожка 3=#236, 0,45 PLU-7/ г муки; дорожка 4=#236, 0,675 PLU-7/г муки; дорожка 5= контрольное вещество DGDG.
На фигуре 11 показана конструкция экспрессирующего вектора pRX487 из pUC18(L131R) и pIJ48;
На фигуре 12 в графической форме показано влияние температуры на стабильность липолитического фермента из Streptomyces sp L131;
На фигуре 13 в графической форме показаны субстратные специфичности галактолипаз из Streptomyces sp L131, Streptomyces avermitillis, Corynebacterium efficiens и Thermobifida fusca;
На фигуре 14 показана структура экспрессирующего вектора pCB5(TF) для экспрессии липазы Thermobifida fusca в C. glutamicum;
На фигуре 15 показано выравнивание последовательности L131 и гомологов S. avermitilis и T. fusca;
На фигуре 16 показана пластина HPTLC продуктов реакции обработки ферментом образцов неочищенного соевого масла. Дорожка 1 = контроль, дорожка 2 = 99% неочищенного масла и 1% 10%-го K371 в воде, дорожка 3 = 98% неочищенного масла и 2% 10%-го K371 в воде, дорожка 4 = 97% неочищенного масла и 3% K371 10%-го K371 в воде, дорожка 5 = 99,7% неочищенного масла и 0,3% 1%-й лецитазы Ultra™ #3108 в воде, дорожка 6 = 99% неочищенного масла, 0,3% 1%-й лецитазы Ultra™ #3108 в воде и 0,7% воды. В качестве контроля анализировали фосфатидилхолин (PC); и
На фигуре 17 показана пластина HPTLC продуктов реакции обработки ферментом образцов неочищенного соевого масла. Дорожка 1 = контроль, дорожка 2 = 99% неочищенного масла и 1% 10%-го K371 в воде, дорожка 3 = 98% неочищенного масла и 2% 10%-го K371 в воде, дорожка 4 = 97% неочищенного масла и 3% 10%-го K371 в воде, дорожка 5 = 99,7% неочищенного масла и 0,3% 1%-й лецитазы Ultra™ #3108 в воде, дорожка 6 = 99% неочищенного масла, 0,3% 1%-й лецитазы Ultra™ #3108 в воде и 0,7% воды, вместе с контрольными линиями сложного эфира холестерина, моноглицерида, диглицерида, триглицерида и растительного стерина.
Пример 1: Идентификация бактериального штамма, продуцирующего галактолипазу
Два штамма микроорганизма со сходным фенотипом, обозначенные L130 и L131, выделили из почвы, собранной в Южной Финляндии. Гены 16S РНК данных двух штаммов амплифицировали обычным PCR с применением олигонуклеотидных праймера 536f (CAGCMGCCGCGGTAATWC) и 1392r-праймера (ACGGGCGGTGTGTRC). Полученные фрагменты PCR частично секвенировали. SEQ ID No:1 и 2 представляют собой не кодирующие фермент полинуклеотиды. Эти последовательности представляют собой гены рибосомальной 16S RNA, широко применяемые для таксономических сравнений. Обнаружили, что SEQ ID No:1 и SEQ ID No:2 обладают большим сходством. Затем последовательности сравнивали с последовательностями 16S РНК в GenBank. Для обоих изолятов наибольшую гомологию (97%) обнаружили с последовательностью гена 16S РНК из Streptomyces thermosacchari. Таким образом, штаммы обозначили Streptomyces sp. L130 и Streptomyces sp. L131.
Пример 2: Получение образцов липолитического фермента (галактолипазы) из штаммов Streptomyces sp. L130 и L131
0,5 л среды LB инокулировали Streptomyces L130 и культивировали в ротационном шейкере при 200 об/мин и 30°C в течение 2 суток. Полученную культуру использовали в качестве инокулята для 10 л ферментера, содержащего такую же среду. Культивирование продолжали в течение 3 суток при 30°C, скорости перемешивания 600 об/мин и аэрации 0,5 об/об. Среду для ферментации осветляли центрифугированием (15 мин при 5000 об/мин) и добавляли Тритон X-100 до конечной концентрации 0,1%. Раствор концентрировали до 300 мл с применением камеры для ультрафильтрации Vivaflow 200 (Vivascience AG, Hannover, Germany). Концентрат диализовали против 10 л буфера 20 мМ Трис HCl, pH 7, содержащего 2 мМ CaCl2 и 2 мМ MgCl2 с последующим диализом против 0,5 л мл 85% глицерина. Полученные препараты содержали 90 ед. галактолипазной активности, анализируемой, как определено выше (GLU-7).
Штамм Streptomyces L131 культивировали в таких же условиях и его культуральную жидкость концентрировали таким же способом. Полученные препараты содержали галактолипазы 70 ед. активности.
Пример 3 - Эксперименты по выпечке
Галактолипазы из бактериальных изолятов L130 и L131 демонстрируют высокую активность в отношении полярных липидных субстратов, галактолипидов (DGDG) и фосфолипидов (активность галактолипазы и фосфолипазы), эквивалентную активности липазы Fusarium oxysporum (Lipopan F™ Novozymes A/S Denmark): однако, было обнаружено, что галактолипазы из бактериальных изолятов L130 и L131 (т.е. липолитический фермент по настоящему изобретению) по существу не обладали активностью в отношении триглицеридов. Это резко отличается от активности липазы Fusarium oxysporum - LipopanF™.
Липолитические ферменты из бактериальных изолятов L130 и L131 получали, как описано в примере 2, и анализировали для характеризации их активности в отношении гликолипидов, фосфолипидов и триглицеридов, как в обычных условиях, так и в тесте.
Эксперименты по выпечки в малом масштабе и модельная система теста. Оба фермента являлись очень активными в отношении галактолипидов в муке.
Материалы и методы.
Получали по три образца каждого фермента, как в примере 3. Каждый образец обозначали, как показано в таблице 1:
Фосфолипазную и галактолипазную активность ферментов оценивали, используя анализ фосфолипазной активности (PLU-7) и анализ галактолипазной активности (GLU-7), приведенные выше.
Эксперимент с жидким тестом
0,8 грамм пшеничной муки взвешивали в 12 мл центрифужной пробирке с крышкой. Добавляли 1,5 мл воды, содержащей фермент. Образец перемешивали на Whirley и помещали в термошкаф при 30°С на 60 минут. Добавляли 6 мл n-бутанола:этанола 9:1 и образец опять перемешивали, пока мука окончательно не распределялась в растворителе. Затем образцы помещали на водяную баню при 95°C на 10 минут. Затем снова перемешивали и помешали во вращающее устройство при 45 об/мин на 45 минут.
Затем образец центрифугировали при 2000 g в течение 10 минут. И 2 мл супернатанта переносили в 10 мл малый стакан. Растворитель выпаривали при 70°C под парами азота.
Выделенные липиды анализировали посредством GLC.
Газовая хроматография
Капиллярный газовый хроматограф Perkin Elmer 8420, оборудованный кварцевой капиллярной колонкой WCOT 12,5 м × 0,25 мм ID × 0,1 мкм 5% фенилметилсиликона (CP Sil 8 CB от Crompack).
Носитель: Гелий.
Ввод пробы: 1,5 мкл с разделением.
Детектор: FID. 385°C.
Получение образца: Липиды, экстрагированные из 0,2 грамм муки, растворяли в 2 мл гептана:пиридина 2:1, содержащего внутренний контроль гептадекан, 2 мг/мл. 500 мкл образца переносили в завальцованный флакон. Добавляли 100 мкл MSTFA (N-метил-N-(триметилсилил)трифторацетамида) и реакционную смесь инкубировали 15 минут при 90°C.
Вычисление: Определяли факторы отклика для моноглицеридов, диглицеридов, триглицеридов, свободных жирных кислот и галактолипидов по контрольным смесям данных компонентов. На основании данных факторов отклика вычисляли уровень липидов в тесте.
Результаты
Анализировали фосфолипазную и галактолипазную активность в образцах фермента из Streptomyces, результаты показаны в таблице 2. Вычисляли также отношение активности PLU-7/GLU-7. Среднее отношение для образцов составляло 1,4, хотя с некоторыми отклонениями в некоторых образцах, что можно объяснить аналитическими отклонениями.
Таблица 2
Эксперимент с тестом
Активность фермента в отношении липидов пшеницы тестировали в эксперименте с жидким тестом, как упомянуто в материалах и методах. Выделенные липиды из теста анализировали посредством GLC, как показано в таблице 3.
Анализ GLC липидов теста (% на основе веса муки). FFA = свободные жирные кислоты. MGMG = моногалактозилмоноглицерид. DGMG = дигалактозилдиглицерид. MGDG = моногалактозилдиглицерид. DGDG = дигалактозилдиглицерид. TRI = триглицерид.
Результаты из таблицы 3 и таблицы 4 подтверждают, что ферменты, выделенные из супернатанта от ферментации Streptomyces sp L130 и L131, являются очень активными в отношении галактолипидов в тесте. Сложные диэфиры DGDG и MGDG гидролизованы до соответствующих сложных моноэфиров DGMG и MGMG. Результаты проиллюстрированы также графически на фигурах 6 и 7. Данные результаты подтверждают, что оба фермента являются очень активными в низкой дозе 0-0,2 единиц/г муки, и получено соответствующее количество сложного моноэфира. В более высокой дозе 0,4-1 единиц/г муки DGDG расщепляется дальше, однако обнаружен также некоторый гидролиз сложных моноэфиров. Это может указывать на то, что ферменты не являются специфичными к положению жирной кислоты в молекуле галактолипида.
Как проиллюстрировано на фигуре 8, активность ферментов в отношении триглицерида почти отсутствует. Из этого заключают, что тестируемые ферменты не оказывают существенного воздействия на триглицерид. Это согласуется также с некоторыми экспериментами, проведенными на трибутирине в качестве субстрата, где не обнаружено активности.
РЕЗУЛЬТАТЫ
Липолитический фермент выделяли из супернатанта от ферментации видов Streptomyces.
Обнаружено, что липолитический фермент обладает как фосфолипазной, так и галактолипазной активностью, но по существу отсутствует активность в отношении триглицеридов. Отношение фосфолипазная: галактолипазная активность для тестированных образцов составляло приблизительно 1,4.
Эксперименты с жидким тестом подтверждают, что ферменты являются активными в отношении галактолипидов в муке. Ферменты являлись активными в тесте в очень низкой дозе 0-0,2 единицы/г муки. Чтобы получить такое же действие на галактолипиды, коммерческие фосфолипазы, подобные Lipopan F™ (Novozymes A/S, Denmark), необходимо дозировать в 3-4 раза более высокой дозе. Эксперименты с жидким тестом подтверждают также, что ферменты из видов Streptomyces не обладают измеряемой активностью в отношении триглицеридов.
Пример 4: Клонирование гена липолитического фермента из Streptomyces sp. L131
Хромосомную ДНК выделяли из Streptomyces sp. L131 с применением модификации общепринятого способа. Бактерии растили в ротационном шейкере на среде LB при 30°C и высокой аэрации (100 мл среды в 0,5 л колбе с отражателем, 200 об/мин) до ранней стационарной фазы. Клетки из 500 мл бактериальной культуры собирали центрифугированием и промывали один раз буфером для лизиса (550 мМ глюкоза, 100 мМ Трис, 2 мМ EDTA, pH 8,0).
Осадок клеток ресуспендировали в 10 мл буфера для лизиса и добавляли лизоцим до 1 мг/мл. Клетки инкубировали при 37°C по меньшей мере 15 мин. За протеканием расщепления лизоцимом следили посредством перенесения аликвот бактериальной суспензии в раствор 1% SDS и измерением абсорбции полученной смеси при 600 нм. Количество лизоцима и время инкубации регулировали так, чтобы по меньшей мере 70-90% всех клеток являлось лизированными, что доказывали уменьшением A600. В данный момент времени к бактериальной суспензии добавляли SDS до 1% и протеиназу K до 0,1 мг/мл. Суспензию инкубировали при 56°C в течение 30 мин с последующей экстракцией фенолом и хлороформом. После экстракции хлороформом ДНК осаждали ацетатом натрия (конечная концентрация 0,5 М) и изопропанолом (0,6 об/об), и осадок ДНК промывали 70% этанолом, сушили в вакууме и растворяли в буфере TE (10 мМ Трис, 1 мМ EDTA), содержащем РНКазу A (0,01 мг/мл).
ДНК частично расщепляли рестрикционной эндонуклеазой Sau3A и гидролизаты фракционировали в 0,8% агарозном геле. Фракцию 3-10 т.п.н. после Sau3A выделяли из агарозных гелей электроэлюированием. Полученный препарат ДНК применяли для конструирования библиотеки генов, используя набор для клонирования ZAP Express/Predigested Vector/Gigapack (продукт #239615) от Stratagene (LaJolla, USA). Лигирование, упаковку, амплификацию библиотеки и ее перевод в фагмиду проводили согласно протоколам, предоставленным Stratagene. Плазмиды из полученной библиотеки генов скринировали на индикаторных чашках, полученных следующим образом. 80 мл стерильного LB-агара, содержащего 25 мг/л канамицин, помещали в каждую 15 см чашку Петри и давали возможность затвердеть. Затем добавляли 10 мл слой верхнего агара, содержащего 0,5% DGDG и 0,0005% сафранина O. Библиотеку генов рассевали с плотностью приблизительно 5000 колоний на 15 см чашку. Чашки инкубировали при 37°C в течение 24 час с последующей инкубацией в течение четырех суток при комнатной температуре. Из библиотеки отбирали клон, формирующий красное гало на индикаторной чашке, и очищали клонированием на новой индикаторной чашке.
Плазмиду, выделенную из данного клона (названную pBK(L131)), применяли для повторной трансформации E. coli штамма XL1-Blue MRF' до устойчивости к канамицину. Все такие трансформанты демонстрировали положительный по галактолипазе фенотип. pBK(L131) содержал вставку приблизительно 7,5 т.п.н. Данную вставку секвенировали. Было обнаружено, что одна из секвенированных областей (SEQ ID No:3) содержит открытую рамку считывания, кодирующую белок (SEQ ID No:4), демонстрирующий гомологию с известной липазой из Streptomyces rimosus. Известно только, что данная липаза, член так называемого семейства GDSX липаз/эстераз/ацилтрансфераз, способна гидролизовать нейтральные липиды и искусственные субстраты липазы.
Серии делеций и субклонов исходной вставки сконструировали и тестировали на галактолипазную активность. Было обнаружено, что делеционное производное, содержащее фрагмент 3 т.п.н. EcoRI-SacI исходной вставки, еще сохраняет полную активность DGDG-азы. Эти данные хорошо коррелируют с результатами по частичной ДНК. Одна область показала гомологию с известными липазами. Данную область затем секвенировали полностью. Сравнением данной последовательности с GenBank показали, что наиболее близким биохимически охарактеризованным гомологом (58,5%) галактолипазы L131 является липаза S. rimosus, идентифицированная как липид-ацилтрансфераза в WO04/064987 и WO04/064537.
Экспрессия галактолипазы L131 в E. coli
Для экспрессии галактолипазы L131 в E. coli применяли стандартную систему pET-систему, в которой ген находится под контролем промотора фага T7.
Экспрессия галактолипазы L131 в Streptomyces lividans
Челночный вектор pRX487-5 (фигура 11) (полученный из pIJ4987: Kieser T. et al Practical Streptomyces genetics. The John Innes Foundation, Crowes, Norwich, England (2000)), применяемый для экспрессии галактолипазы L131 в S. lividans, сочетает плазмиду E. coli pUC18 и плазмиду S. lividans IJ487. В pRX487-5 lac-промотор pUC18 расположен выше лишенного промотора гена канамицин-фосфотрансферазы pIJ487. Действительно, плазмида трансформирует E. coli до устойчивости не только к ампициллину, но также по меньшей мере к низкому уровню (5 мг/л) канамицина. Вектор содержит немодифицированный EcoRI-XbaI фрагмент хромосомной ДНК S. thermosacchari, содержащий полную кодирующую последовательность гена галактолипазы, приблизительно 160 п.н. некодирующей последовательности выше и 420 п.н. некодирующей последовательности ниже. Все трансформанты демонстрировали сходные уровни галактолипазной активности, как оценивали по формированию гала на индикаторных чашках. Аналогично, когда S. lividans, несущий pRX487-5, культивировали в масштабе 10 л, и полученную культуру клонировали на индикаторных чашках, для всех клонов, по-видимому, получали равные уровни галактолипазной активности. В культурах в небольших встряхиваемых колбах, инокулированных вегетативными клетками непосредственно с чашек, для трансформантов обычно получали приблизительно 10-20 миллиединиц/мл галактолипазной активности после 3 суток культивирования. Когда культуры во встряхиваемых колбах инокулировали спорами рекомбинантного S. lividans, измеряли более высокие активности галактолипазы (приблизительно 30 миллиединиц/мл), что коррелирует с более высоким накоплением биомассы. В эксперименте, где S. lividans, несущий pRX487-5, растили в ферментере в условиях высокой аэрации и режима периодической подпитки (подробности смотри в материалах и методах), накопление биомассы достигало 170 г/л (масса во влажном состоянии). Соответственно, обнаруживали намного более высокую галактолипазную активность - приблизительно 1 ед./мл.
Биохимические свойства L131.
Тестировали некоторые биохимические свойства L131. Было обнаружено, что оптимальный pH для фермента составляет приблизительно 6,5-7,5 (фигура 9). Фермент демонстрировал максимальную активность в отношении DGDG при температуре приблизительно 50°C. Выше данной температуры происходила инактивация, однако не резко, и после 20 мин инкубации при 90°C обнаруживали приблизительно 10% остаточной активности (фигура 12).
Пример 5: Экспрессия гена липолитического фермента Streptomyces L131 по настоящему изобретению в E. coli
Открытую рамку считывания pBK(L131), кодирующую предполагаемый липолитический фермент по настоящему изобретению, амплифицировали PCR с применением праймеров oL131-5 (GGTGAATTCATGAGATTGACCCGATCCCTGTCGG, смысловой праймер) и oL131-3 (ACTTCTAGAGCGGCGCCACCGTGACGTACA, антисмысловой праймер). Амплифицированный фрагмент ДНК расщепляли EcoRI и XbaI и клонировали в челночный вектор B. subtilis - E. coli pGTK44. Этот вектор конструировали заменой фрагмента SalI-EcoRI плазмиды pGTK44 (Povelainen et al., Biochem J. 371, 191-197 (2003)), содержащего промотор degQ36, фрагментом EcoRI-Sall pGT44 (Kerovuo J. et al. Biotechnology Letters 22, 1311-1317 (2000)).
Галактолипазную активность в E. coli, трансформированной полученной плазмидой pGTK44(L131) (фигура 5), детектировали с применением индикаторных чашек. Контрольные трансформанты (содержащие родительскую плазмиду pGTK44) являлись отрицательными по галактолипазе. Таким образом, последовательность белка, представленная на SEQ ID No:4, действительно обладала галактолипазной активностью. С той же парой праймеров амплифицировали фрагмент такого же размера (по электрофорезу в агарозном геле) с хромосомной ДНК Streptomyces sp. L130, подтверждая более ранние наблюдения о близком сходстве двух выделенных штаммов и их генов галактолипазы.
Для экспрессии в E.coli под контролем промотора фага T7, выведенную кодирующую область галактолипазы амплифицировали PCR с применением в качестве матрицы хромосомной ДНК Streptomyces sp. L131 и двух олигонуклеотидных праймеров (oL131-51 GGTCATGCTAGCATGAGATTGACCCGATCCCTGTCGG и oL131-31 GCATGGATCCGCGGCGCCACCGTGACGTACA). Продукт PCR расщепляли NheI и BamHI и лигировали в вектор pETlla (Novagen, USA), расщепленный такими же рестрикционными эндонуклеазами. Смесь после лигирования применяли для трансформации E.coli штамма XL-Blue1 MRF' и выделили 12 различных плазмидных клонов с картинами рестрикции, соответствующими структуре pET11(131-51) (фиг. 4). Каждый плазмидный клон применяли для отдельной трансформации E. coli штамма BL21(DE3), полученные трансформанты выращивали на слое LB-ампициллина, содержащего индикатор галактолипазной активности (пример 4). Большинство клонов действительно экспрессировали активную галактолипазу. Один из клонов (pETll(131-51)-12) выбрали в качестве источника рекомбинантной галактолипазы для последующей характеризации.
Анализировали фермент, экспрессированный в E. coli (обозначенный #236), и при анализе с применением описанных выше анализа GLU-7 и анализа PLU-7 обнаружили, что он обладает: 0,33 GLU/мл и 0,36 PLU/мл.
В жидкой культуре E. coli BL21(DE3) экспрессировала приблизительно 2 миллиединиц/мл галактолипазной активности после 40 час культивирования в LB-ампициллиновой среде (37°C, покачивание 200 об/мин). По существу всю активность обнаружили в культуральной среде. Не обнаружили галактолипазной активности в E.coli BL21(DE3), трансформированной pET11a (Novagen, USA) и культивированной в таких же условиях.
Приблизительно четыре литра содержащей галактолипазу культуральной среды концентрировали в роторном испарителе до приблизительно 300 мл и диализовали против 15 л буфера 20 мМ Трис HCl, pH 7, содержащего 2 мМ CaCl2 и 2 мМ MgCl2. Диализованный материал снова концентрировали в роторном испарителе до приблизительно 30 мл и снова диализовали против 2 л 50% глицерина. Полученный препарат (18 мл) содержал приблизительно 100 миллиединиц/мл галактолипазной активности.
Фермент, экспрессированный в E. coli (обозначенный #236), тестировали также в тесте. Как видно из фигуры 10, показывающей пластину TLC, в тесте обнаружили высокую активность в отношении галактолипидов.
Пример 6: Экспрессия гена липолитического фермента из Streptomyces sp. L131 по настоящему изобретению в различных хозяевах
Конструкция вектора pGTK44(L131) изложена в примере 5. Помимо E. coli, этот вектор можно применять для получения липолитического фермента Streptomyces L131 по настоящему изобретению в Bacillus. Применение данного вектора является только одним из многих возможных способов экспрессии липолитического фермента L131 по настоящему изобретению в Bacillus. Например, промотор pst, применяемый в pGTK44(L131), можно заменить любым другим сильным конститутивным или регулируемым промотором, активным в Bacillus. Множество таких промоторов известно в данной области. Например, промотор degQ36 (Yang M et al. J. Bacteriol. 166, 113-119 (1986)), промотор cdd, известный также как p43 (Wang PZ, Doi RH. J. Biol. Chem. 259, 8619-8625 (1984), промоторы амилазы или нейтральной протеазы и т.д. Помимо pGTK44(L131) и других векторов для Bacillus на основе репликона pTZ12 (Aoki T. et al., MoI. Gen. Genet 208, 348-352 (1987)), можно применять любой другой плазмидный вектор (например, pUB110, Gryczan TJ et al. J. Bacteriol. 134, 318-29 (1978) и его производные).
Другими предпочтительными хозяевами для экспрессии гена липолитического фермента Streptomyces L131 по настоящему изобретению являются грамположительные бактерии с высоким GC, в частности, Streptomyces, (например, S. lividans, S. coelicolor, S. griseus, S. natalensis, S. rubiginosus, S. olivaceus, S. olivochromogenes, S. violaceoruber). В такие хозяева ген липолитического фермента по настоящему изобретению можно вводить под его собственным промотором в мультикопийном векторе (например, с применением производных pIJ110, таких как pIJ486, Ward et al. MoI. Gen. Genet. 203, 468-478 (1986)), или помещать под контроль сильного промотора Streptomyces, например, ermE* (Schmitt-John T, Engels JW. Appl. Microbiol. Biotechnol. 36, 493-498 (1992)), или индуцируемого тиострептоном промотора tip A (Kieser T et al. in Practical Streptomyces Genetics, p. 386, The John Innes Foundation, Norwich UK (2000)).
Помимо прокариотических хозяев ген липолитического фермента L131 можно экспрессировать в одном из множества подходящих видов грибов. В частности, подходят дрожжи, такие как Saccharomyces cerevisiae, Schizosaccharomyces pombe, Pichia pastoris, Hansenula polymorpha. В дрожжах ген липолитического фермента можно помещать под контроль любого из известных сильных дрожжевых промоторов, таких как гликолитические промоторы (PGK, GDH, ENO и т.д.), индуцируемые фосфатным голоданием промоторы, такие как PHO5, промоторы метаболизма этанола/метанола, такие как промотор ADH1 в S. cerevisiae или индуцируемые метанолом промоторы в H. polymorpha или P. pastoris.
При экспрессии гена липолитического фермента в любом хозяине будет являться преимущественным конструирование синтетического или полусинтетического гена, кодирующего последовательность SEQ ID No:4. Подобным образом, можно сконструировать частично или полностью синтетические гены на основе последовательностей, доступных по поискам гомологии in silico, как объясняют в примере 4. Данные последовательности могут содержать ряд полезных свойств, отсутствующих у генов липолитических ферментов дикого типа. Например, можно корректировать смещение предпочтения кодонов, для лучшего соответствия кодонным предпочтениям экспрессирующих хозяев. Одним из частных случаев коррекции смещения, применимым для всех хозяев, является перевод инициирующего кодона GTG из SEQ ID No:3 в ATG. Другой обычной модификацией, очевидной для специалиста в данной области, является замена природной сигнальной последовательности липолитического фермента L131 Streptomyces на сигнальную последовательность, природную для выбранного экспрессирующего хозяина, или известную, как функционирующую в нем.
Предыдущие примеры применимых экспрессирующих систем для липолитического фермента L131 направлены на применение плазмидных векторов для введения гена липолитического фермента в экспрессирующего хозяина. Это действительно является предпочтительным способом осуществления настоящего изобретения. Однако альтернативный способ встраивания экспрессирующей кассеты (включая промотор, кодирующую область гена липолитического фермента и, не обязательно, терминатор транскрипции) в хромосому также осуществим. В особенности эффективным будет мультикопийное встраивание экспрессирующей кассеты в хромосому хозяина.
Рекомбинантные хозяева, экспрессирующие ген липолитического фермента, могут, преимущественно, являться мутантными для уменьшения уровня протеазной активности в культуральной среде. Культивирование любого из таких рекомбинантных хозяев можно проводить в присутствии соединений, стабилизирующих фермент. Такие соединения могут представлять собой различные белки (например, казеин, пептон или сывороточный альбумин) или различные липиды, лизолипиды или детергенты (например, галактолипиды, моно- и диацилглицерины или Тритон X-100).
Пример 7: Ацилтрансферазная активность липолитического фермента Streptomyces L131 и его производных
Некоторые липазы могут также обладать ацилтрансферазной активностью. В частности, некоторые члены семейства GDSX, например, ацилтрансфераза Aeromonas hydrophila (P 10480) (описанная в находящейся в одновременном рассмотрении международной патентной заявке No PCT/IB2004/000655) обладает высокой ацилтрансферазной активностью. Таким образом, можно предсказать, что липолитический фермент Streptomyces L131 также равным образом обладает ацилтрансферазной активностью. Данную активность можно далее усилить посредством случайного мутагенеза/направленной эволюции. Кроме того, так как ацилтрансфераза A. hydrophila и липолитический фермент Streptomyces L131 разделяют одну и ту же укладку белка, возможна комбинация субстратной специфичности липолитического фермента Streptomyces L131 с высокой трансферазной эффективностью фермента Aeromonas. Этого сочетания можно достичь посредством известных способов направленного мутагенеза/белкового дизайна или перетасовкой генов.
Пример 8: Идентификация альтернативных липолитических ферментов из других видов Streptomyces
Семейство эстераз GDSX (Upton C, Buckley JT. Trends Biochem. Sic. 20, 178-179 (1995), pfam00657.11) представляет собой группу эстераз/липаз/ацилтрансфераз, разделяющих специфический мотив последовательности около серина активного участка (GDSX, где X представляет собой остаток гидрофобной аминокислоты). Эта группа ферментов известна также как семейство липаз II (Arpigny JL, Jaeger K-E. Biochem. J. 343, 177-183 (1999)). Хотя данное семейство включает в себя различные типы эстераз, липаз и ацилтрансфераз, липолитический фермент по настоящему изобретению представляет собой GDSX-фермент.
Таким образом, указанные в настоящем изобретении последовательности липолитического фермента Streptomyces sp. L131 (галактолипазы) можно применять in silico для идентификации других галактолипаз из других видов Streptomyces.
Чтобы определить, обладает ли белок мотивом GDSX по настоящему изобретению, последовательность предпочтительно сравнивают с профилями скрытой марковской модели (профили HMM) базы данных pfam.
Pfam представляет собой базу данных по семействам доменов белка. Pfam содержит упорядоченные множественные выравнивания последовательностей для каждого семейства, а так же профиль скрытых марковских моделей (профиль HMM) для того, чтобы идентифицировать эти области в новых последовательностях. Введение в Pfam можно найти в Bateman A et al. (2002) Nucleic Acids Res. 30; 276-280. Скрытые марковские модели применяют в ряде баз данных, предназначенных для классификации белков, обзор смотри в Bateman A and Haft DH (2002) Brief Bioinform 3; 236-245.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=12230032&dopt=Abstract
http://www.ncbi.nlm.nih.gov/entrez/querv.fcgi?cmd=Retrieve&db=PubMed&list_uids=11752314&dopt=Abstract
Подробное объяснение скрытых марковских моделей и их применения в базе данных Pfam смотри в Durbin R, Eddy S, and Krogh A (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4. Пакет программного обеспечения Hammer можно получить из Washington University, St Louis, USA.
Альтернативно, мотив GDSX можно идентифицировать с применением пакета программного обеспечения Hammer, инструкции предоставлены в Durbin R, Eddy S, and Krogh A (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4 и в ссылках там, и профиль предоставлен в данном описании.
Доступ к базе данных PFAM можно получить, например, через несколько серверов, в настоящее время локализованных на следующих веб-узлах.
http://www.sanger.ac.uk/Software/Pfam/mdex.shtml
http://pfam.wustl.edu/
http://pfam.iouy.inra.fr/
http://pfam.cgb.ki.se/
База данных предлагает средство поиска, где можно ввести последовательность белка. Затем с применением параметров по умолчанию базу данных последовательности белка анализируют на присутствие доменов Pfam. Домен GDSX является доменом, зафиксированным в базе данных, и таким образом будет распознано его присутствие в любой запрашиваемой последовательности. База данных будет возвращать выравнивание консенсусной последовательности Pfam00657 с запрашиваемой последовательностью.
Предпочтительно при выравнивании с консенсусной последовательностью Pfam00657 липолитический фермент для использования в композициях/способах по изобретению обладает по меньшей мере одним, предпочтительно более, чем одним, предпочтительно более, чем двумя, из следующего, блок GDSx, блок GANDY, блок HPT. Соответственно, липолитический фермент может обладать блоком GDSx и блоком GANDY. Альтернативно, фермент может обладать блоком GDSx и блоком HPT. Предпочтительно, фермент содержит по меньшей мере блок GDSx.
Домен pfam00657 GDSX представляет собой уникальный идентификатор, отделяющий белки, обладающие данным доменом, от других ферментов.
В дополнение или в качестве альтернативы этому, альтернативные липолитические ферменты из других видов Streptomyces можно идентифицировать проведением сравнения идентичности последовательности и/или гибридизации с одним или несколькими фрагментами PCR последовательностей, показанных как SEQ ID No:1 или SEQ ID No:2. Соответственно, сравнения можно проводить с фрагментами, содержащими более 15 нуклеотидов из SEQ ED No 1 или SEQ ID No:2, предпочтительно, с фрагментами, содержащими более 20 нуклеотидов из SEQ ED No 1 или SEQ ID No:2. Соответственно, можно применять полные последовательности, показанные как SEQ ID No:1 или SEQ ID No:2. Предпочтительно, гибридизацию проводят в условиях высокой или очень высокой строгости. Нуклеотидные последовательности, по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичные SEQ ID No:1 или SEQ ID No:2, указывают на штаммы Streptomyces, которые могут являться источниками липолитического фермента, т.е. галактолипазы по настоящему изобретению.
Пример 9: Идентификация галактолипаз для использования в способах и применениях по настоящему изобретению
Как указано выше, последовательность нового Streptomyces thermosacchari L131 открывает возможность для идентификации in silico новых галактолипаз семейства II. В этом отношении одной из конкретных областей, которая может представлять особенный интерес, является мотив GDSX.
Мотив GDSX содержит четыре консервативные аминокислоты. Предпочтительно, серин в мотиве представляет собой каталитический серин фермента липид-ацилтрансферазы. Соответственно, серин в мотиве GDSX может присутствовать в положении, соответствующем Ser-16 в липолитическом ферменте Aeromonas hydrophila, описанном в Brumlik & Buckley (Journal of Bacteriology Apr. 1996, Vol. 178, No. 7, p.2060-2064).
Чтобы определить, обладает ли белок мотивом GDSX, последовательность предпочтительно сравнивают с профилями скрытой марковской модели (профилями HMM) базы данных pfam. Как указано в примере 8, pfam представляет собой базу данных семейств доменов белков. Таким образом, базу данных pfam можно применять также для идентификации подходящих ферментов из родов, отличных от Streptomyces.
Альтернативно, мотив GDSX можно идентифицировать с применением пакета программного обеспечения Hammer, инструкции предоставлены в Durbin R, Eddy S, and Krogh A (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4 и ссылках там, и профиль HMMER2 предоставлен в данном описании.
Предпочтительно, липолитический фермент по настоящему изобретению содержит мотив GDSX.
При сравнении с консенсусной последовательностью Pfam00657 (как описано в WO04/064987) и/или последовательностью L131, описанной в настоящем описании (SEQ ID No:4)
i) Фермент галактолипаза/липид-ацилтрансфераза по изобретению или для применения в способах по изобретению предпочтительно обладает мотивом GDSx, более предпочтительно мотивом GDSY;
и/или
ii) фермент галактолипаза/ липид-ацилтрансфераза по изобретению или для применения в способах по изобретению, предпочтительно обладает блоком GANDY, более предпочтительно блоком GANDY, содержащим амино GGNDx, более предпочтительно GGNDA или GGNDL;
и/или
iii) фермент по изобретению или для применения в способах по изобретению, предпочтительно обладает блоком http;
и предпочтительно
iv) фермент галактолипаза/липид-ацилтрансфераза по изобретению или для применения в способах по изобретению, предпочтительно обладает мотивом GDSY и блоком GANDY, содержащим амино GGNDx, предпочтительно GGNDA или GGNDL, и блоком HTP (консервативный гистидин).
В этом отношении авторы настоящего изобретения идентифицировали последовательность, гомологичную Streptomyces L131, которая не содержит мотив GDSX: а именно Novosphingobium aromaticivorans (NAL)
Novosphingobium\aromaticivorans\GDSx 284 ак


Этот фермент содержит последовательность «GSSF» в отличие от GDSX.
При тестировании обнаружено, что данный фермент не обладает гликолипазной активностью по настоящему изобретению.
Следовательно, мотив GDSx может являться важным при попытках идентификации других пригодных галактолипаз.
А именно, известно, что фермент из S. rimosus, который очистили и охарактеризовали биохимически, показывающий приблизительно 56% гомологии последовательности с Streptomyces L131 (Abramic M., et al. (1999); Vujaklija D. et al. (2002)), гидролизует нейтральные липиды, такие как триолеин, или сложные нитрофениловые эфиры жирных кислот. Фермент из S. rimosus может также гидролизовать галактолипазу по настоящему изобретению. Подобным образом, два других вида Streptomyces, для которых доступны данные геномной последовательности - S. coelicolor A2(3) и S. Avermitilis, могут содержать ферменты, обладающие галактолипазной активностью, например, (NP_625998 и NP_827753) в настоящее время аннотированы в GenBank как "предполагаемые секретируемые гидролазы".
С применением подобного подхода можно идентифицировать много других применимых гомологов галактолипазы Streptomyces L131. Подходящие ферменты галактолипазы/липид-ацилтрансферазы для применения в способах по изобретению можно идентифицировать выравниванием последовательности L131 с применением алгоритма попарного выравнивания Align X, Clustal W из VectorNTI, с применением параметров по умолчанию.
Альтернативно, подходящую галактолипазу для применения в способах по изобретению можно идентифицировать выравниванием консенсусной последовательности pfam Pfam00657 (как описано в WO04/064987).
На фигуре 15 показано выравнивание последовательности L131 и гомологов из S.avermitilis и T.fusca. Фигура 15 иллюстрирует консервативность мотива GDSx (GDSY в L131 и S.avermitilis и T.fusca), бокса GANDY, который представляет собой или GGNDA, или GGNDL, и блока HPT (считающегося консервативным каталитическим гистидином). Данные три консервативные блока выделены на фигуре 15.
При выравнивании или с консенсусной последовательностью Pfam00657 (как описано в WO04/064987) и/или с последовательностью L131, описанной в настоящем описании (SEQ ID No:4), возможно идентифицировать три консервативных области, блок GDSx, блок GANDY и блок HTP (дополнительные подробности смотри в WO04/064987).
Пример 10: Клонирование генов и конструирование экспрессирующих векторов
Для выделения генов, гомологичных гену галактолипазы S. thermosacchari L131, применяли Corynebacterium efficiens DSM 44549, Thermobifida fusca DSM 43792 и Streptomyces avermitilis DSM46492.
Штаммы согласно номеру DSM депонированы и публично доступны в Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM).
Штаммы Escherichia coli XL-Blue MRF', BL21(DE3) (Novagen) и S17-1 (Simon R. et ah, 1983), Bacillus subtilis BD 170, штамм Streptomyces lividans 1326 (John Lines Centre), Corynebacterium glutamicum DSM20300 применяли в качестве хозяев для гетерологичной экспрессии. Штамм Aeromonas salmonicida (DSM 12609) также применяли в качестве экспрессионного хозяина.
S. thermosacchari L131, Citrobacter freundii P3-42 и Enterobacter nimipressuralis P 1-60 выделили в лаборатории авторов настоящего изобретения из природного окружения и таксономически идентифицировали секвенированием гена 16S рРНК.
В данном исследовании применяли следующую культуральную среду. LB (5 г/л дрожжевого экстракта, 10 г/л триптона, 10 г/л NaCl, pH 7,0), 2xYT (10 г/л NaCl, 10 г/л дрожжевого экстракта, 16 г/л триптона) применяли для культивирования E. coli и других грамотрицательных бактерий. Питательный бульон (3 г/л говяжьего экстракта, 5 г/л пептона, pH 7,0) применяли для выращивания C. efficiens и N. aromaticivorans, YM-среду (3 г/л дрожжевого экстракта, 3 г/л солодового экстракта, 5 г/л пептона, 10 г/л декстрозы, pH 7,0) применяли для культивирования S. avermitilis, среду 65 (4 г/л глюкозы, 4 г/л триптона, 10 г/л солодового экстракта, 2 г/л CaCO3, pH 7,2) применяли для T. fusca.
Выделение ДНК
Для выделения плазмид применяли обычный способ щелочного лизиса в сочетании со способом очистки на колонке Qiagen. Единственное исключение составляло препаративное выделение плазмидной ДНК из Streptomyces. В данном случае на конечной стадии очистки применяли равновесное центрифугирование в градиенте CsCl.
Способы введения ДНК в микробные штаммы.
Штаммы как E. coli, так и C. glutamicum трансформировали электропорацией с применением кювет 1 мм и следующих установок параметров электропорации: 1800 V, 25°F, 200 мкл. B. subtilis BDl трансформировали 70 «парижским» способом, основанным на естественной компетенции (Harwood CR. and Cutting S.M., 1990). Streptomyces lividans трансформировали способом протопластов (Kieser T. et al., 2000). ДНК в A. salmonicida вводили посредством конъюгации с E. coli с применением способа скрещивания на фильтрах Harayama et al. (1980).
Конструирование устойчивого к рифампицину мутанта A salmonicida.
Приблизительно 108 клеток из ночной культуры A. salmonicida DSM12609 высевали на серии чашек с LB-агаром, содержащих 5-30 мг/л рифампицина. Чашки облучали коротковолновым УФ-светом с использованием устройства SpectroLinker XL-1500 (Spectronics Corp. USA). Доза облучения составляла 4-6 J/м2. Чашки инкубировали при 30°C в течение 2 суток. Несколько колоний, выращенных на 30 мг/л рифампицина, отбирали и дополнительно тестировали на 50 мг/л рифампицина. Один из клонов, устойчивых к 50 мг/л рифампицина (обозначенный Rl), выбирали для последующей работы.
Конструирование экспрессирующий векторов E. coli для гомологов галактолипазы L131
Ген липазы из Streptomyces avermitilis амплифицировали PCR с применением хромосомной ДНК в качестве матрицы и двух олигонуклеотидных праймеров oSAL-5 (GGG AATTCCATATGAGACGTTCCCGAATTACG) и oSAL-3 (GCATGGATCCGG TGACCTGTGCGACGG). Для амплификации генов липазы из Thermobifida fusca и Corynebacterium efficiens применяли олигонуклеотидные праймеры oTFL-5 (GGGAATTCCATATGGGCAGCGGACCACGTG) и oTFL-3 (GCATGGATCCGACACGCACGGCTCAACG), OCEL-5 (GGGAATTCC ATATGAGGACAACGGTCATCG) и oCEL-3 (GCATGGATCCGGCATCGGGCTCATCC), соответственно. Продукты PCR расщепляли NdeI и BamHI и лигировали с вектором pETlla (Novagen, USA), расщепленным такими же рестрикционными эндонуклеазами.
Экспрессирующий галактолипазу L131 вектор для S. lividans конструировали следующим образом. Плазмиду pUC18(L131RX), содержащую 1,37 т.п.н. EcoRI-XbaI фрагмент исходного клонированного фрагмента ДНК, несущего ген липазы L131 (pBK(L131)), расщепляли EcoRI и лигировали с расщепленной EcoRI pIJ487 (Kieser et al., 2000). Данное лигирование приводило к получению двух рекомбинантных плазмид, отличающихся относительной ориентацией pIJ487 и pUC18(L131RX). Для последующей работы на основании рестрикционного анализа выбрали вариант, где промотор lac pUC18 фланкирует лишенный промотора ген neo R pIJ487. Данную конструкцию обозначили pRX487-5 (фигура 11). Помимо устойчивости к ампициллину данная плазмида также придает E. coli устойчивость по меньшей мере к 3 мг/л канамицина. Протопласты S. lividans 1326 трансформировали 0,1-10 мкг pRX487-5 до устойчивости к тиострептону (1,2 мг/л) и канамицину (5 мг/л). Данные трансформанты продуцировали активную галактолипазу, как показывали анализом на индикаторной чашке DGDG-сафранин. Трансформанты высевали на чашки SM (Kieser et al., 2000), дополненные 5 мг/мл канамицина и давали возможность споруляции. Полученные споры применяли для инокуляции культур во встряхиваемой колбе и в ферментере.
Конструирование экспрессирующих векторов для Corynebacterium glutamicum
Все применяемые в данной работе экспрессирующие векторы основаны на плазмиде pCB5, представляющей собой челночный вектор, несущий репликон C. glutamicum из плазмиды pSRl (Yoshihama et al., 1985) и репликон ColE1 из E. coli. Промотор, применяемый в данном векторе, получен из гена cop1, кодирующего главный секретируемый белок C. glutamicum - PS1. Ферменты экспрессировали с их природных генов, включая немодифицированные сигнальные пептиды, например, T.fusca (фигура 14).
УСЛОВИЯ ФЕРМЕНТАЦИИ
Ферментация продуцирующих липазу штаммов Streptomyces
Продуцирующие липазу рекомбинантные штаммы S. lividans растили во встряхиваемой колбе в среде, содержащей (на литр) 10 г пептона, 5 г дрожжевого экстракта, 2 г K2HPO4 и 10 г глюкозы (pH 7,0), дополненной соответствующими антибиотиками: тиострептон применяли при 1,2 мг/л, канамицин при 20 мг/л, хлорамфеникол при 1,5 мг/л и эритромицин при 1,5 мг/л. Суспензии спор, полученные выращиванием трансформантов на чашках SM, применяли для начала культивирований.
Для ферментаций с подпиткой применяли ферментер Braun Biostat E (10 л). Исходная среда (7 л) содержала (на литр): 20 г пептона, 10 г дрожжевого экстракта, 20 г глюкозы и соответствующие антибиотики, как описано выше (за исключением тиострептона, который не применяли в 10 л культурах). Культивирование проводили при 30°C, постоянной аэрации 10 л/мин и скорости перемешивания 600 об./мин. Инокулят (2 x 250 мл на ферментацию) растили в 2 л колбе Эрленмейера, как описано в предыдущем разделе. Ферментацию проводили в периодическом режиме в течение 18-20 час, времени, после которого культуру в ферментере подпитывали раствором, содержащим 30 % глюкозы и 12,5 % пептона со скоростью 0,5 мл/мин. Образцы (30 мл) культуры отбирали асептически дважды в сутки.
Ферментация рекомбинантных штаммов C. glutamicum
Культуры C. glutamicum во встряхиваемой колбе выращивали в LB, содержащей 50 мг/л канамицина при 30°C и скорости перемешивания 200 об./мин.
Ферментация рекомбинантных штаммов A salmonicida
Рекомбинантные штаммы A. salmonicida культивировали во встряхиваемых колбах в среде 2xYT, дополненной стрептомицином и канамицином (при 25 мг/л). Чтобы индуцировать промотор tac к питательной среде добавляли IPTG (1-5 мМ) или лактозу (1-10%).
Два набора условий для продукции рекомбинантной ацилтрансферазы в A. salmonicida тестировали в масштабе ферментера. В первом эксперименте исходная среда (7 л) представляла собой 2xYT, дополненную 2% глюкозой, 50 мг/л канамицина и 50 мг/л стрептомицина, а раствор для подпитки (3 л) содержал 25% глюкозы, 10% триптона и 5% дрожжевого экстракта, 100 мг/л как канамицина, так и стрептомицина. Культивирование проводили при аэрации 10 л/мин, скорости перемешивания 600 об./мин и 28°C. pH доводили до 7,5 25% NH3 и 10% фосфорной кислотой. 0,5 л ночной культуры A. salmonicida инокулировали в ферментер и выращивали в режиме с подпиткой в течение 24 час. На этой стадии добавляли IPTG до 5 мМ и начинали подпитку при скорости 50 мл/час.
Во втором эксперименте исходную среду модифицировали заменой глюкозы лактозой. Раствор для подпитки представлял собой 2 л 20% лактозы. Температуру ферментации увеличивали до 30°C и pH культуральной среды уменьшали до 7,0. Инокуляцию проводили как в первом эксперименте и подпитку (100 мл/час) начинали после 20 час культивирования в исходной среде.
АНАЛИЗЫ ФЕРМЕНТОВ
Способ скрининга на чашке с сафранином
В скрининге на чашке с сафранином нижний слой содержал культуральную среду + добавки, 1,5% агарозу и 0,002% сафранин (0,2% маточный раствор в воде, стерильно профильтрованный) и верхний слой - 0,7% агарозу, 1% DGDG и 0,002% сафранин.
Определение галактолипазной активности (анализ гликолипазной активности (GLU-7)):
Субстрат:
0,6% дигалактозилдиглицерид (Sigma D 4651), 0,4% Тритон-X 100 (Sigma X-100) и 5 мМ CaCl2 растворяли в 0,05 М буфере HEPES, pH 7.
Процедура анализа:
400 мкл субстрата добавляли в 1,5 мл пробирку Эппендорф и помещали в термосмеситель Эппендорф при 37°C на 5 минут. Во время t=0 мин добавляли 50 мкл раствора фермента. Также анализировали контроль с водой вместо фермента. Образец перемешивали при 10×100 об/мин в термосмесителе Эппендорф при 37°C в течение 10 минут. Во время t=10 мин пробирку Эппендорф помещали в другой термосмеситель при 99°C на 10 минут, чтобы остановить реакцию.
Свободную жирную кислоту в образцах анализировали с применением набора NEFA C от WAKO GmbH. Ферментативную активность GLU при pH 7 вычисляли как микромоль жирной кислоты, полученных в минуту при условиях анализа.
Определение фосфолипазной активности (анализ фосфолипазной активности (PLU-7)):
Субстрат
0,6% растительный L-α-фосфатидилхолин 95% (Avanti #441601), 0,4% Тритон-X 100 (Sigma X-100) и 5 мМ CaCl2 диспергировали в буфере 0,05 М HEPES pH 7.
Процедура анализа: 400 мкл субстрата добавляли в 1,5 мл пробирку Эппендорф и помещали в термосмеситель Эппендорф при 37°C на 5 минут. Во время t=0 мин добавляли 50 мкл раствора фермента. Также анализировали контроль с водой вместо фермента. Образец перемешивали при 10×100 об/мин в термосмесителе Эппендорф при 37°C в течение 10 минут. Во время t=10 мин пробирку Эппендорф помещали в другой термосмеситель при 99°C на 10 минут, чтобы остановить реакцию.
Свободную жирную кислоту в образцах анализировали с применением набора NEFA C от WAKO GmbH.
Ферментативную активность PLU-7 при pH 7 вычисляли как микромоль жирной кислоты, полученной за минуту в условиях анализа.
Спектрофотометрический анализ с p-нитрофенилпальмитатом (pNPP)
Липазную активность измеряли спектрофотометрическим анализом при 30°C с pNPP в качестве субстрата с применением буфера 50 мМ Трис-малеат (pH 6,5) с 0,4 % Тритона X-100 и 0,1 % гуммиарабика. Получали маточный раствор субстрата (100 мМ) в диоксане. Измерение кинетики начинали добавлением фермента к реакционной смеси. Чтобы оценить начальную гидролитическую активность, следили за увеличением абсорбции при 410 нм каждые 20 с в течение 20 мин на планшет-ридере Spectramax. Одну единицу липазной активности определяли как количество фермента, высвобождающего 1 мкмоль p-нитрофенола в мин. Активность для других сложных эфиров p-NP измеряли таким же способом с применением 1 мМ каждого субстрата. (Abramic M. et al. (1999))
Определение влияния pH и температуры на липазную активность
Для определения влияния pH на ферментативную активность, ее измеряли в диапазоне pH 2-10 с применением анализа галактолипазной активности за исключением того, что буферы, применяемые в эксперименте, являлись следующими: pH 2-3,5 Глицин-HCl; pH 4-5 NaOAc; pH 5,5-7,5 Трис-малеат; pH 7,5-9 Трис-HCl; pH 10 CAPS.
Влияние температуры на стабильность галактолипазы определяли инкубацией аликвот фермента в течение 20 мин при различных температурах (22°C-90°C) с последующей инкубацией во льду в течение 60 мин. Остаточную активность анализировали анализом галактолипазной активности.
Для детекции оптимальной температуры для галактолипазной активности обычную смесь для анализа уравновешивали при требуемой температуре (диапазон 20°C-70°C) и добавляли 2 или 4 мкл фермента для запуска реакции. Активность анализировали анализом галактолипазной активности, но с применением более короткого периода времени (20 мин).
Пример 11: Характеризация галактолипаз-кандидатов из анализа биоразнообразия
Последовательность галактолипазы Streptomyces thermosacchari L131 открывает возможность для идентификации in silico нового семейства галактолипаз II.
Можно идентифицировать много других применимых гомологов галактолипазы Streptomyces L131, например, «гипотетический белок» из Thermobifida fusca (ZP_00058717) и «гипотетический белок» из Corynebacterium efficiens (NP_738716).
Авторы настоящего изобретения клонировали и экспрессировали 3 гомолога галактолипазы Streptomyces L131: гены Streptomyces avermitilis (SAL), Thermobifida fusca (TFL) и Corynebacterium efficiens (CEL). Все гены экспрессировали в E. coli с применением экспрессирующей системы pET. Сначала рекомбинантные штаммы E. coli анализировали с применением DGDG-индикаторных чашек с сафранином и обнаружили, что ферменты S. avermitilis, T. fusca и C. efficiens обладают галактолипазной активностью.
Ферменты, демонстрирующие галактолипазную активность, исследовали далее. Изучали субстратные специфичности данных галактолипаз-кандидатов (фигура 13). Тестировали активность ферментов-кандидатов в отношении DGDG, лецитина, оливкового масла, нитрофенил-бутирата, нитрофенил-деканоата (NP-D) и нитрофенил-пальмитата. Обнаружили, что ферменты обладали очень разными профилями субстратной специфичности. Ацилтрансферазную активность тестировали в анализе с применением NP-D в качестве субстрата и количественного определения как высвобождения нитрофенола, так и свободных жирных кислот набором NEFA. Предварительные данные позволяют предполагать, что по меньшей мере фермент из Thermobifida fusca обладает трансферазной активностью в отношении глицерина и глюкозы.
Тестировали термостабильность галактолипаз-кандидатов. Обнаружили, что фермент Corynebacterium efficiens является наиболее термостабильным, тогда как фермент Streptomyces avermitilis является наиболее термочувствительным.
Пример 12: Испытание рафинирования для Streptomyces thermosacchari L131
Фосфолипазу из Streptomyces thermosacchari L131 тестировали на неочищенном соевом масле.
Материалы и методы
K371: Фермент Streptomyces thermosacchari L131, экспрессированный в S. lividans, лиофилизировали на крахмале.
(Активность: 108 PLU-7/г).
Лецитаза Ultra (#3108) от Novozymes, Denmark
Сложный эфир холестерина, Fluka 26950
Растительный стерин: Generol 122 N от Henkel, Germany
Неочищенное соевое масло от The Solae Company, Aarhus Denmark
Лецитин: растительный L-α-фосфатидилхолин 95% (Avanti #441601)
Фосфолипазная активность
Анализ фосфолипазы аналогичен, используемому в примере 10.
HPTLC
Аппликатор: Automatic TLC Sampler 4, CAMAG
Пластина HPTLC: 20 × 10 см, Merck no. 1.05641. Активировали 30 минут при 160°C перед применением. Нанесение: 1 мкл 8% раствора масла в буфере наносили на пластину HPTLC с применением автоматического аппликатора TLC.
Буфер для прогона 4: смесь хлороформ:метанол:вода 75:25:4
Буфер для прогона 5: смесь P-эфир:метил-трет-бутилкетон:уксусная кислота 70:30:1
Время нанесения/элюирования:
Буфер для прогона 4: 20 мин
Буфер для прогона 5: 10 мин
Проявление TLC
Пластину сушили в печи в течение 10 минут при 160°C, охлаждали и погружали в 6% ацетат меди в 16% H3PO4. Сушили дополнительные 10 минут при 160°C и сразу оценивали.
Эксперимент по рафинированию
Для исследования рафинирования применяли Streptomyces thermosacchari L131 (K371) в составах, указанных в таблице 4.
Образцы помещали при 40°C на 18 часов с перемешиванием, после чего образец собирали для анализа HPTLC посредством растворения образца в смеси хлороформ:метанол 2:l
Рафинирование неочищенного соевого масла Streptomyces thermosacchari L131 и лецитазой Ultra™
Результаты анализа HPTLC показаны на фигуре 16 и фигуре 17.
На фигуре 16 показана пластина TLC (буфер 4) продуктов реакции обработки ферментом образцов неочищенного соевого масла согласно таблице 4. Как упомянуто, анализировали также фосфатидилхолин (PC). Указаны также фосфатидилэтаноламин (PE) и лизофосфатидилхолин (LPC).
Результаты TLC на фигуре 16 ясно показывают, что фосфатидилхолин полностью удален посредством добавления к маслу Streptomyces thermosacchari L131. Только наименьшая доза (дорожка 2) не полностью гидролизовала фосфолипиды. Лецитаза Ultra™ также гидролизовала фосфолипиды в масле при наличии 5% воды (дорожка 6), но без добавления дополнительной воды (дорожка 5) только часть фосфолипидов являлась гидролизованной.
На фигуре 17 показана TLC (буфер 5) продуктов реакции обработки ферментом образцов неочищенного соевого масла согласно таблице 4. Как указано, сложный эфир холестерина, моноглицерид, диглицерид, триглицерид и растительный стерин. Указана также свободная жирная кислота (FFA).
Результаты, показанные на фигуре 17, показывают, что гидролиз фосфолипидов совпадает с формированием свободной жирной кислоты.
Заключение.
Результаты подтверждают, что Streptomyces thermosacchari L131 эффективно гидролизует фосфолипиды в неочищенном соевом масле и является пригодным альтернативным ферментом для рафинирования растительных масел.
Все публикации, приведенные в описании выше, включены в настоящее описание в качестве ссылки. Различные модификации и варианты описанных способов и систем по настоящему изобретению будут очевидны специалистам в данной области без отклонения от объема и сущности настоящего изобретения. Хотя настоящее изобретение описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что изобретение согласно формуле изобретения не должно быть каким-либо образом ограничено данными конкретными вариантами осуществления. Действительно, различные модификации описанных способов для осуществления изобретения, очевидные для специалистов в области биохимии и биотехнологии, предназначены для включения в объем следующей формулы изобретения.
Список литературы
Abramic M., Lescic I., Korica Т., Vitale L., Saenger W., Pigac J. Purification and properties of extracellular lipase from Streptomyces rimosus. Enzyme and Microbial Technology 25 522-529 (1999).
Harayama S., Masataka Т., lino T. High-frequency mobilisation of the chromosome of Escherichia coli by a mutant of plasmid RP4 temperature sensitive for maintenance. Mol. Gen. Genet. 180, 47-56 (1980).
Harwood C.R. and Cutting S.M. Molecular biological methods for Bacillus. John Wiley & Sons Ltd., West Sussex, England (1990).
Kieser Т., Bibb M J., Buttner M.J., Chater K.F., Hopwood D.A. Practical Streptomyces genetics. The John Innes Foundation, Crowes, Norwich, England (2000).
Vujaklija D., Schroder W., Abramic M., Zou P., Lescic L, Franke P., Pigac J. A novel streptomycete lipase: cloning, sequencing and high-level expression of the Streptomyces rimosus GDS(L)-lipase gene. Arch. Microbiol. 178,124-130 (2002).
Yoshihama M, Higashiro K, Rao EA, Akedo M, Shanabruch WG, Follettie MT, Walker GC, Sinskey AJ. Cloning vector system for Corynebacterium glutamicum. J Bacteriol. 162 (2):591-597 (1985).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| ВАРИАНТЫ ГЛИКОЛИПИДАЦИЛТРАНСФЕРАЗЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2004 |
|
RU2377300C2 |
| БЕЛКИ | 2004 |
|
RU2518345C2 |
| БЕЛОК | 2005 |
|
RU2380414C2 |
| СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА УГЛЕРОДА, СЛОЖНОГО ЭФИРА БЕЛКА, СЛОЖНОГО ЭФИРА БЕЛКОВОЙ СУБЪЕДИНИЦЫ ИЛИ СЛОЖНОГО ЭФИРА ГИДРОКСИКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЛИПИДАЦИЛТРАНСФЕРАЗЫ | 2004 |
|
RU2371474C2 |
| СПОСОБ | 2004 |
|
RU2376868C2 |
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |
| ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ ДЛЯ ПРИМЕНЕНИЯ В ХЛЕБОПЕЧЕНИИ | 2018 |
|
RU2763469C2 |
| СПОСОБЫ ПРИМЕНЕНИЯ ТЕРМОСТАБИЛЬНЫХ СЕРИНОВЫХ ПРОТЕАЗ | 2017 |
|
RU2771261C2 |

























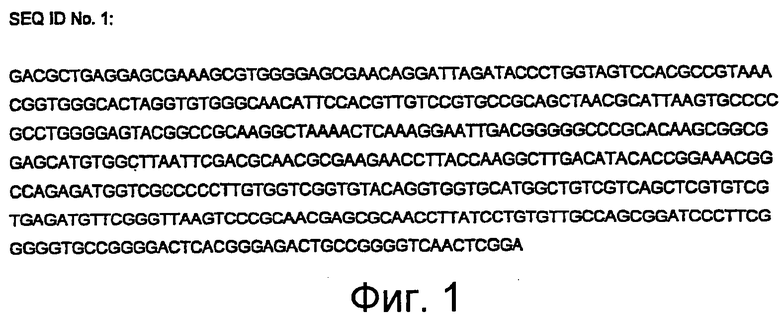
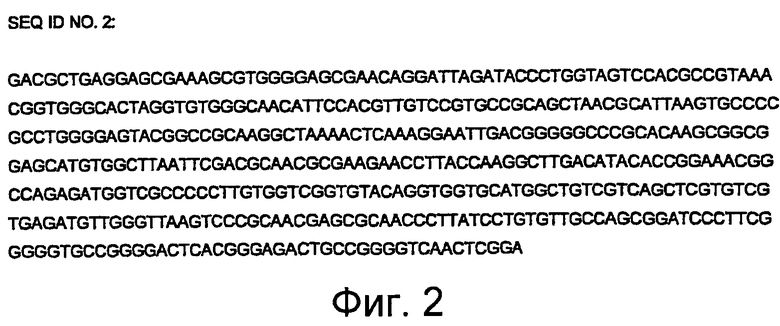


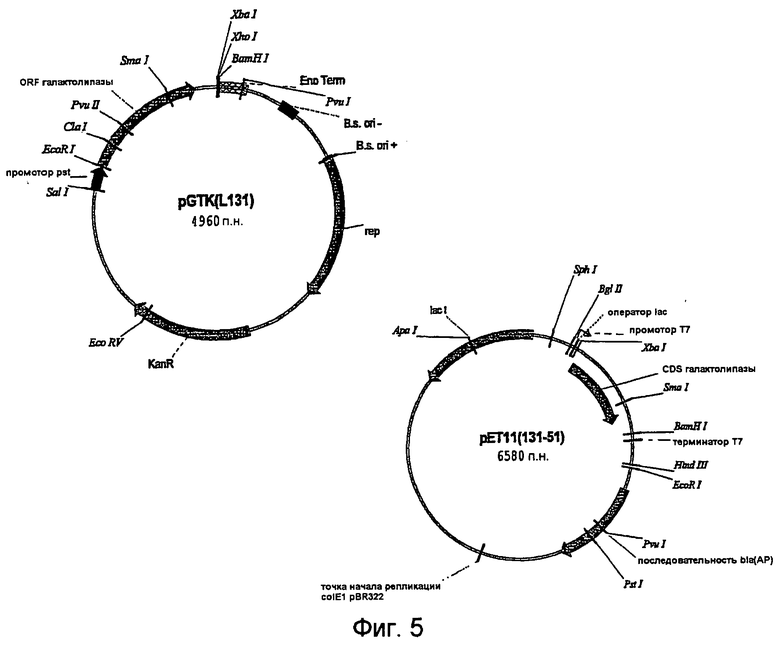
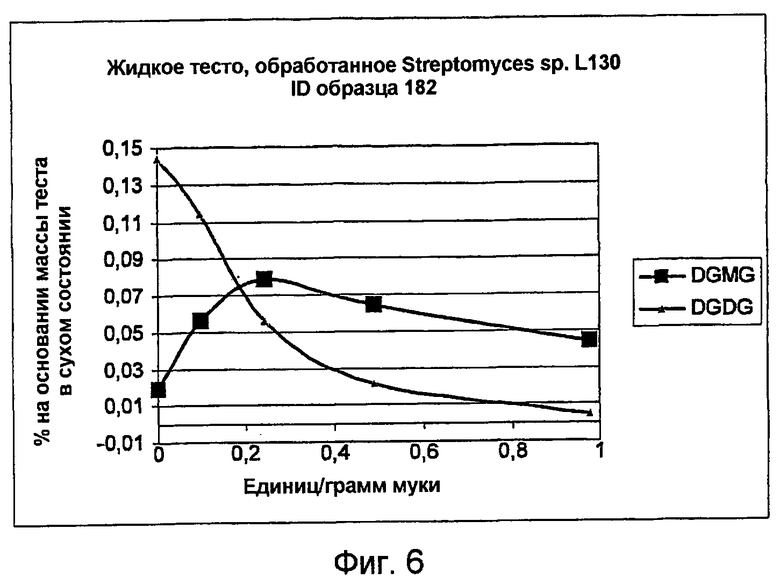
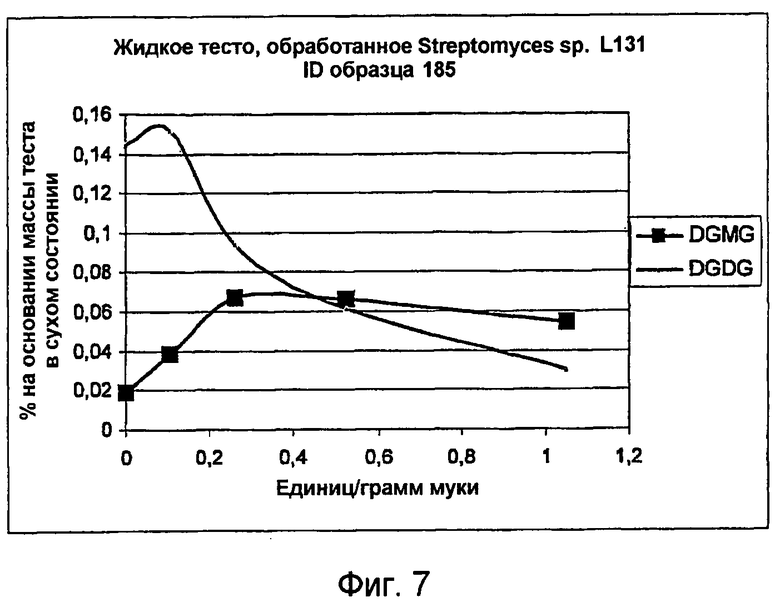
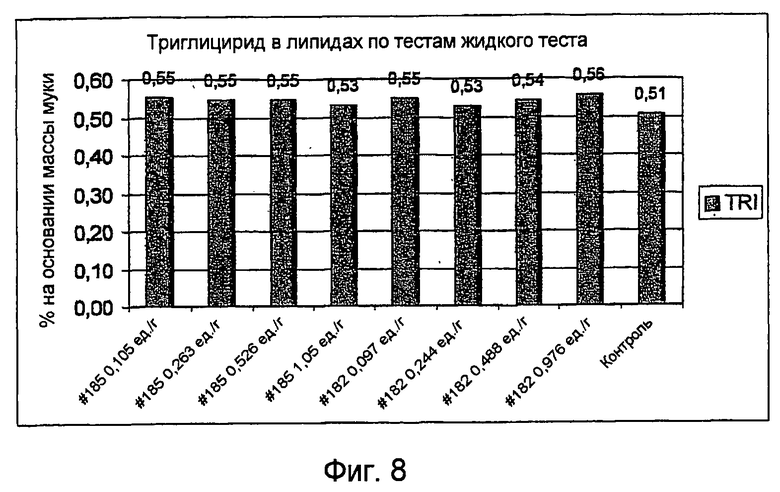
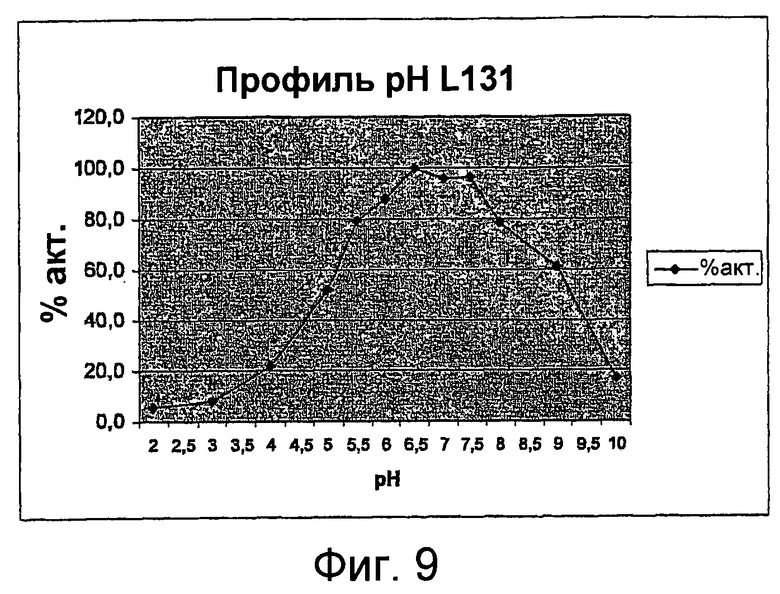

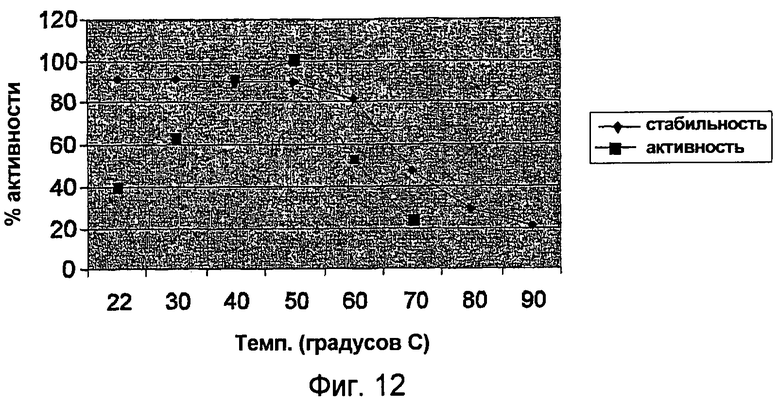
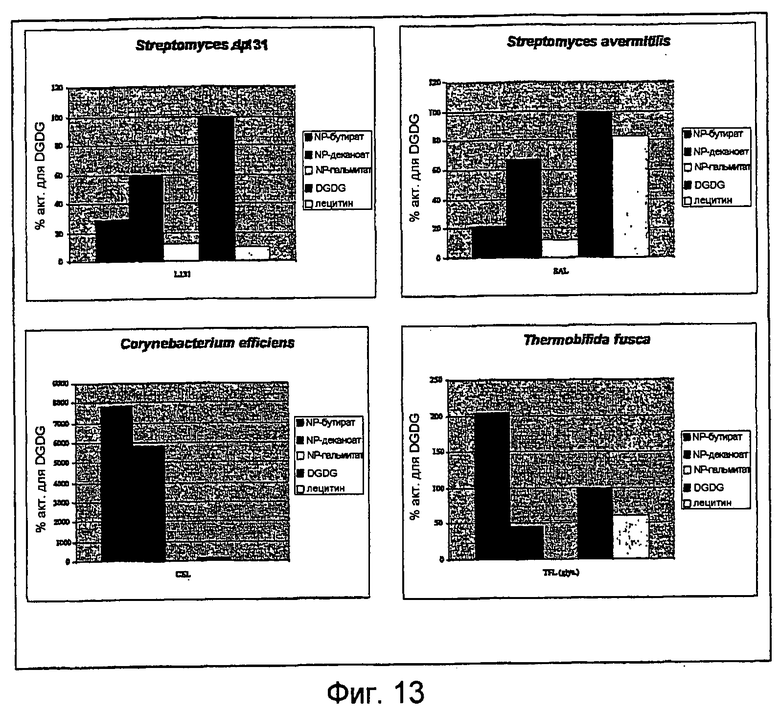
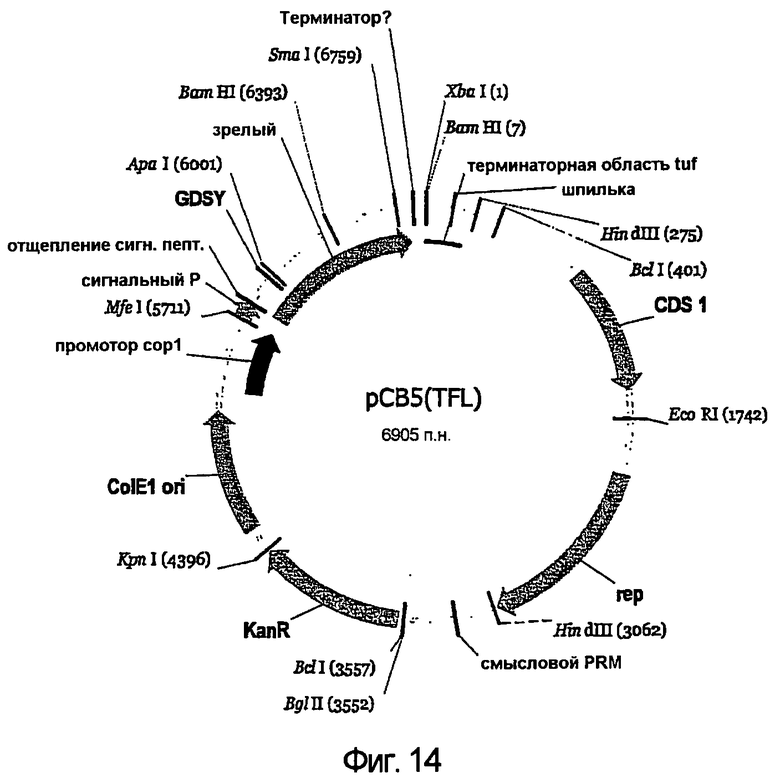


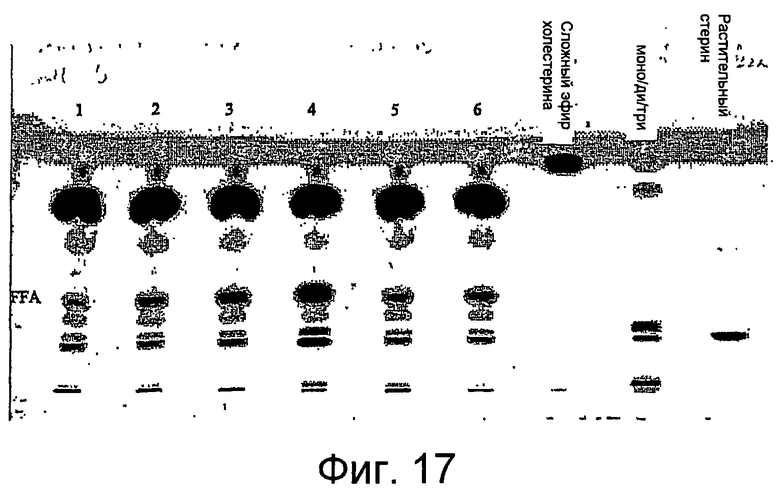
Изобретение относится к биохимии и представляет собой способ получения лизогликолипида, включающий обработку субстрата, содержащего гликолипид, по меньшей мере одним липолитическим ферментом для получения указанного лизогликолипида, где указанный липолитический фермент обладает гликолипазной активностью и где указанный липолитический фермент получают из рода Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им. Указанный липолитический фермент используется также для получения лизофосфолипида, для рафинирования растительного или пищевого масла, для конверсии полярных липидов, а также для получения пищевого продукта. Изобретение позволяет расширить арсенал липолитических ферментов. 11 н. и 17 з.п. ф-лы, 17 ил., 4 табл., 12 пр.
1. Способ получения лизогликолипида, включающий обработку субстрата, содержащего гликолипид, по меньшей мере одним липолитическим ферментом для получения указанного лизогликолипида, где указанный липолитический фермент обладает гликолипазной активностью и где указанный липолитический фермент получают из рода Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
2. Способ получения лизофосфолипида, включающий обработку субстрата, содержащего фосфолипид, по меньшей мере одним липолитическим ферментом для получения указанного лизофосфолипида, где указанный липолитический фермент обладает фосфолипазной активностью и где указанный липолитический фермент получают из рода Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
3. Способ ферментативного рафинирования растительного или пищевого масла, включающий обработку указанного пищевого или растительного масла липолитическим ферментом, который получают из видов Thermobifida, способным гидролизовать большую часть полярных липидов, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
4. Способ биоконверсии полярных липидов для получения ценных продуктов, включающий обработку указанных полярных липидов липолитическим ферментом, который получают из рода Thermobifida, с получением высокоценных продуктов, где указанный липолитический фермент способен гидролизовать указанные полярные липиды, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
5. Способ по п.4, где указанные ценные продукты представляют собой один или несколько следующих продуктов: сложный эфир углеводорода, сложный эфир белка, сложный эфир субъединицы белка и сложный эфир оксикислоты.
6. Способ получения пищевого продукта, включающий смешивание по меньшей мере одного липолитического фермента с одним или несколькими ингредиентами пищевого продукта, где указанный липолитический фермент способен гидролизовать гликолипид и/или фосфолипид, присутствующий в или представляющий собой по меньшей мере один указанный ингредиент и где указанный липолитический фермент получают из рода Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
7. Способ по любому из пп.1-6, где указанный липолитический фермент способен переносить ацильную группу от гликолипида к одному или нескольким субстратам - акцепторам ацила.
8. Способ по любому из пп.1-6, где указанный липолитический фермент содержит аминокислотную последовательность SEQ ID NO:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична указанной последовательности, или содержит нуклеотидную последовательность SEQ ID NO:6 или 17 или нуклеотидную последовательность, которая по меньшей мере на 70% идентична указанной последовательности.
9. Способ по п.8, где указанный липолитический фермент содержит аминокислотную последовательность SEQ ID NO:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична указанной последовательности.
10. Способ по п.6, где указанный пищевой продукт выбран из одного или нескольких следующих продуктов:
яиц, продуктов на основе яиц, включая в качестве неограничивающих примеров майонез, заправки для салата, соусы, мороженое, яичный порошок, модифицированный яичный желток и полученные из него продукты;
хлебобулочных изделий, включая хлеб, торты, продукты из сдобного теста, слоеное тесто, жидкое взбитое тесто, сдобу, пончики, печенье, сухое печенье и булочки;
кондитерских изделий, включая шоколад, леденцы, карамель, халву, резинку, включая не содержащую сахара и подслащенную сахаром резинку, надувную жевательную резинку, мягкую надувную жевательную резинку, жевательную резинку и пудинги;
замороженных продуктов, включая фруктовое мороженое, предпочтительно замороженные молочные продукты, включая пломбир и молочное мороженое;
молочных продуктов, включая сыр, масло, молоко, сливки для кофе, взбитые сливки, яичный крем, молочные напитки и йогурты;
муссов, взбитых растительных сливок, мясных продуктов, включая готовые мясные блюда;
пищевых масел и жиров, газированных и негазированных взбитых продуктов, эмульсий типа масло в воде, эмульсий типа вода в масле, маргарина, кулинарных жиров и паст, включая низкожирные и очень низкожирные пасты;
заправок, майонеза, подлив, соусов на основе сливок, супов на основе сливок, напитков, эмульсий специй и соусов.
11. Способ по п.10, где указанный пищевой продукт представляет собой выпеченный продукт и по меньшей мере один из указанных ингредиентов представляет собой тесто.
12. Способ по п.10, где указанный пищевой продукт представляет собой яйцо или продукт на основе яиц.
13. Способ по п.10, где указанный пищевой продукт представляет собой молочный продукт.
14. Применение липолитического фермента на субстрате для получения лизогликолипида, где указанный липолитический фермент обладает гликолипазной активностью и где указанный липолитический фермент получают из Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
15. Применение липолитического фермента на субстрате для получения лизофосфолипида, где указанный липолитический фермент обладает фосфолипазной активностью и где указанный липолитический фермент получают из Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
16. Применение липолитического фермента, полученного из Thermobifida, для ферментативного рафинирования растительного или пищевого масла для гидролиза большей части полярных липидов, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
17. Применение липолитического фермента, полученного из Thermobifida, в процессе, включающем обработку фосфолипида с гидролизом ацильных групп жирных кислот, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
18. Применение липолитического фермента в биоконверсии полярных липидов для получения ценных продуктов, где указанный липолитический фермент способен гидролизовать указанные полярные липиды и где указанный липолитический фермент получен из рода Thermobifida, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
19. Применение по п.18, где указанные ценные продукты представляют собой один или несколько из следующих продуктов: сложный эфир углеводорода, сложный эфир белка, сложный эфир субъединицы белка и сложный эфир оксикислоты.
20. Применение липолитического фермента, полученного из рода Thermobifida для получения пищевого продукта, где указанный липолитический фермент способен гидролизовать гликолипид и/или фосфолипид, где липолитический фермент содержит любую аминокислотную последовательность SEQ ID N0:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична им, или кодируется любой нуклеотидной последовательностью SEQ ID N0:6 или 17 или нуклеотидной последовательностью, которая по меньшей мере на 70% идентична им.
21. Применение по пп.14-20, где указанный липолитический фермент способен переносить ацильную группу от гликолипида на один или несколько субстратов - акцепторов ацила.
22. Применение по любому из пп.14-20, где указанный липолитический фермент содержит аминокислотную последовательность SEQ ID NO:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична указанной последовательности, или содержит нуклеотидную последовательность SEQ ID NO:6 или 17 или нуклеотидную последовательность, которая по меньшей мере на 70% идентична указанной последовательности.
23. Применение по п.22, где указанный липолитический фермент содержит аминокислотную последовательность SEQ ID NO:5, 7 или 16 или аминокислотную последовательность, которая по меньшей мере на 70% идентична указанной последовательности.
24. Применение по п.14, где указанный лизогликолипид представляет собой DGMG или MGMG.
25. Применение по п.20, где указанный пищевой продукт представляет собой молочный продукт.
26. Применение по п.25, где указанный пищевой продукт представляет собой яйцо или продукт на основе яиц и где указанный липолитический фермент способен переносить ацильную группу на один или несколько субстратов - акцепторов ацила для уменьшения одного или нескольких из следующих неблагоприятных эффектов: неприятный запах и/или неприятный и/или мыльный вкусы.
27. Применение по п.20, где указанный пищевой продукт представляет собой выпеченный продукт.
28. Применение по п.14, где указанный субстрат представляет собой пищевое масло.
| Устройство к горизонтально ковочным машинам для нагревания и перемещения заготовок в ручьях штампа | 1939 |
|
SU58717A1 |
| US 20030144165 А1, 31.07.2003 | |||
| RU 2001117497 A, 27.07.2003 | |||

