Заявленное техническое решение (предлагаемый штамм) относится к области биотехнологии, а именно к микроорганизмам и ферментам. Может быть использовано в микробиологической промышленности и генетической инженерии.
Наличие дефекта по генам доминирующих протеаз или дефицит таких генов у бактерий делает их (бактерии) универсальными продуцентами рекомбинантных белков, например,- применяемых для производства лекарственных препаратов со стабильными свойствами.
Известен штамм бактерий Bacillus brevis 31-OK [1] с низкой протеолитической активностью, полученный путем спонтанной мутации после трансформации штамма Bacillus brevis HPD31 плазмидой pNU2OOhGH, несущей ген гормона роста человека. Недостатком [1] является зависимость продуктивности штамма по синтезу экзоферментов от состава питательной среды. Требуется специфическая питательная среда с обязательным добавлением, например, полипептона P1 - в концентрации до 2%, отсутствие которого существенно снижает выход продукции ферментов. Эта зависимость сужает перечень применяемых для культивирования штамма (Bacillus brevis 31-OK) питательных сред, что в свою очередь существенно ограничивает область применения этого штамма в качестве производителя рекомбинантных белков, например, в производстве лекарственных препаратов.
Известен полученный путем многократного УФ (ультрафиолетового) облучения дефектный по протеазам штамм бактерий Bacillus amyloliquefaciens A50-32 [2], обладающий 0,3% протеазной активностью. Недостатком штамма бактерий Bacillus amyloliquefaciens A50-32 [2] является наличие неконтролируемых и непредсказуемых мутаций, например регуляторной мутации. Эти мутации оказывают плейотропный (множественный) эффект на синтез экзоферментов. Из-за этого недостатка (неконтролируемых и непредсказуемых мутаций) продукты жизнедеятельности штамма (экзоферменты), например те, которые используют для производства лекарственных препаратов, нестабильны по свойствам и действию, что существенно ограничивает область применения аналога.
Наиболее близким по существу предлагаемому штамму (прототипом) является протеазо-дефицитный штамм бактерий Bacillus subtilis GX4924 [3] с направленной инактивацией гена субтилизина (сокращенное название гена - apr). Недостатком прототипа [3] является наличие «помехи» в геноме указанного штамма бактерий - гена другой мажорной нейтральной протеазы (сокращенное название гена - npr), доля которой составляет до 54% в общем протеолитическом пуле ферментов (продуктов жизнедеятельности прототипа). Наличие протеазы, выступающей помехой, вызывает дополнительную деградацию рекомбинантных полипептидов, количественно ограничивающих синтез полезных экзоферментов, снижает количество и качество получаемого продукта, например, применяемого в фармацевтике для производства лекарственных препаратов в форме ферментативных добавок, например, для лечения недостаточности экзокринной функции поджелудочной железы.
Заявленное техническое решение поясняется Фиг.1 - 3 и таблицей, на которых представлены:
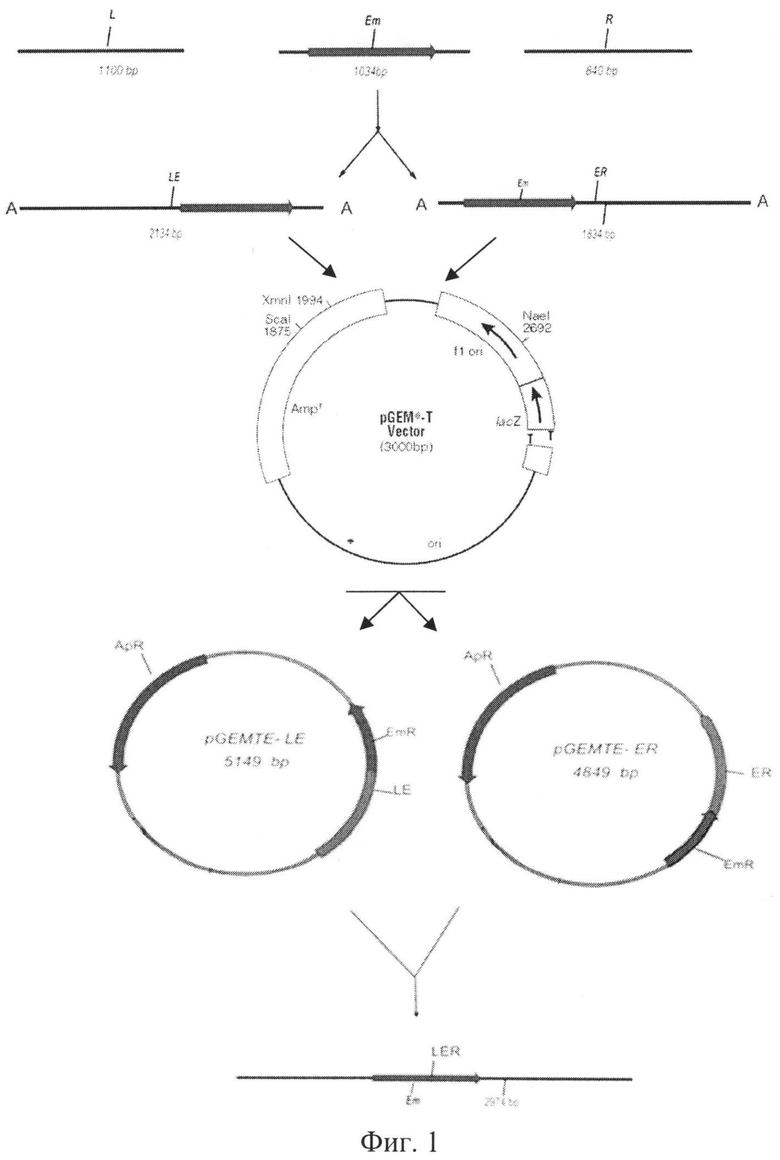
На Фиг.1 представлена схема поэтапного создания тройной рекомбинирующей конструкции, состоящей из левой (L), правой (R) фланкирующей областей гена aprBp Bacillus pumilus 3-19 и гена резистентности к антибиотику эритромицину erm (E) с использованием плазмиды pGEM-T.
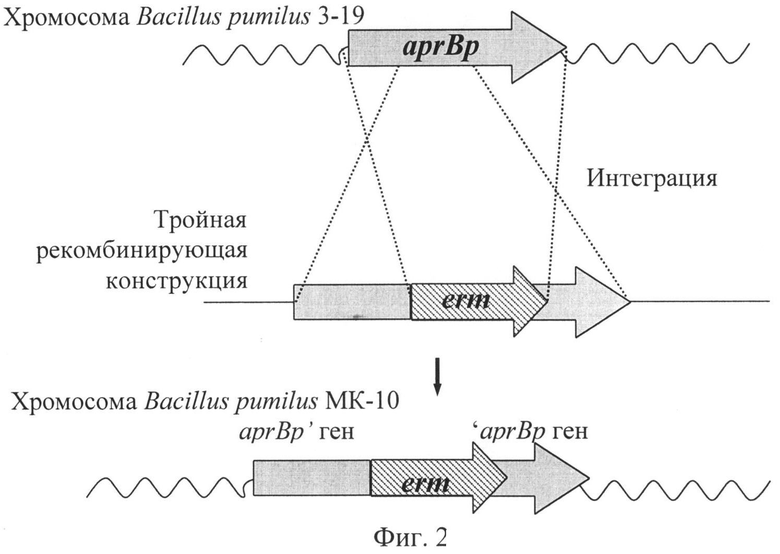
На Фиг.2. представлена схема интеграции тройной рекомбинирующей конструкции, инактивирующей ген aprBp в геном Bacillus pumilus 3-19. Обозначения: aprBp - ген субтилизиноподобной протеазы, erm - ген резистентности к антибиотику эритромицину, волнистой линией обозначены хромосома Bacillus pumilus 3-19 и хромосома Bacillus pumilus MK-10, пунктирной линией обозначен путь интеграции. На Фиг.3. представлены колонии исходного штамма Bacillus pumilus 3-19 (контроль) и нокаутированного/протеазо-негативного штамма Bacillus pumilus MK-10 (опыт) на 2% молочном агаре.
Целью предлагаемого изобретения является получение штамма-реципиента бактерий вида Bacillus pumilus с низкой протеолитической активностью для производства чистых гомологичных и рекомбинантных белков, повышение чистоты используемых в полезных целях продуктов жизнедеятельности бактерий.
Цели достигают тем, что в качестве реципиента с низкой протеолитической активностью применяют штамм бактерий Bacillus pumilus MK-10 ВКПМ В-10742, получаемый от исходного штамма бактерий Bacillus pumilus 3-19 путем инактивирования гена мажорной сериновой протеазы на хромосоме. Штамм бактерий Bacillus pumilus MK-10 ВКПМ В-10742 используют в качестве реципиента с низкой протеолитической активностью. Штамм бактерий применяют для производства гомологичных белков Bacillus pumilus. Штамм бактерий применяют для производства гетерологичных белков рода Bacillus.
Предлагаемый штамм Bacillus pumilus MK-10 получают, например, описанным далее путем. С применением известных компьютерных программ, например программного обеспечения Oligo Calc [4], известным путем [5] конструируют олигонуклеотидные праймеры с использованием общедоступной базы данных NCBI (National Center for Biotechnology Information - Национальный центр биотехнологической информации) [6]. Последовательности гена субтилизиноподобной протеазы (aprBp) и гена резистентности к антибиотику эритромицину (erm) используют, например, из общедоступной базы данных NCBI. Последовательности праймеров приведены в таблице.
Дальнейшие действия показаны на Фиг.1. Методом полимеразно-цепной реакции (далее по тексту - ПЦР) проводят амплификацию левого (L) и правого (R) фланкирующих участков гена aprBp с геномной ДНК штамма бактерий Bacillus pumilus 3-19 при использовании олигонулеотидных пар Sub left up/Sub left low и Sub right up/Sub right low соответственно. ПЦР для получения гена erm проводят с плазмиды pCS9 с использованием праймеров Em up/Em low. В результате нарабатывают амплификаты одинарных конструкций (с длиной, например, ~1 kb или одной килобазы).
Затем амплификаты одинарных конструкций используют как сырье и путем ПЦР получают двойные конструкции: левый фланкирующий регион aprBp+erm (LE) и правый фланкирующий регион aprBp+erm (ER). При этом в ПЦР используют олигонуклеотидные пары Sub left up/Em low (LE) и Em up/Sub right low (ER) соответственно. Продукты амплификации (двойные конструкции), электрофоретически соответствующие ожидаемой длине (продуктов амплификации) в ~2 kb, выделяют из агарозного геля, например - с использованием набора реактивов фирмы Fermentas (Литва).
Полученные двойные конструкции (LE и ER) клонируют в плазмиду pGEM-T Easy (Promega, США) путем лигирования по липким Т-концам плазмиды и А-концам амплификатов. Лигазной смесью трансформируют известный штамм бактерий Escherichia coli XL-1 Blue. Трансформацию проводят, например, по известной методике [7]. Суспензию трансформированных клеток высевают на чашки с агаризованной средой, содержащей 5-бромо-4-хлоро-3-индоил-бета-D-галактопиранозид, например, в концентрации 40 mg/ml, изопропил-β-D-1-тиогалактопиранозид, например, в концентрации 32 mg/ml и ампициллин, например, в концентрации 20 mg/ml. Используют, например, реактивы фирмы Fermentas (Литва). После инкубации трансформантов в течение 24 часов при плюс 37°С на чашках образуются белые (содержащие LE и ER клонированные конструкции) и синие (не содержащие конструкций) колонии бактериальных клеток.
Отбирают белые колонии бактерий, с которых проводят ПЦР-скрининг с фланкирующих праймеров (Т7, SP6) на наличие вставки в плазмиде pGEM-T Easy длиной (продукта амплификации) в ~2 kb. Из положительных (содержащих продукт амплификации) колоний бактерий путем щелочного лизиса выделяют плазмиды pGEMTE-LE и pGEMTE-ER со вставками LE и ER соответственно.
Проводят реакцию амплификации и получают тройную рекомбинирующую конструкцию (LER). Для этого используют плазмиды pGEMTE-LE и pGEMTE-ER в качестве матрицы и праймеры Sub left up/Sub right low. Продукт реакции амплификации, соответствующий электрофоретически ожидаемой длине в ~3 kb, очищают из агарозного геля, например, с использованием набора фирмы Fermentas (Литва).
Заключительным этапом по получению предлагаемого протеазо-дефицитного штамма бактерий Bacillus pumilus является трансформация клеток бактерий Bacillus pumilus 3-19 полученной тройной рекомбинирующей конструкцией. Трансформацию проводят по методу [8], при этом используют синтетические минимальные среды Спецайзена 1/11. Солевая основа среды Спецайзена (%): К2НРО4×3Н2О - 18,34; KH2PO4 (безводный) - 6,0; (NH4)2SO4 - 2,0; цитрат - Na - 1.2. Отдельно готовят растворы: глюкоза - 40%; казаминовые кислоты - 200 mg/ml; Mg2SO4 - 0,2 g/ml. Среда Спецайзена I: на 100 ml H2O: глюкоза - 2 ml; казаминовые кислоты - 2 ml; дрожжевой экстракт - 2 ml; Mg2SO4 - 0,1 ml; солевая основа - 10 ml. Среда Спецайзена II: на 100 ml H2O: глюкоза - 2 ml; казаминовые кислоты - 1 ml; Mg2SO4 - 8 µ1; солевая основа - 10 ml).
Суспензию содержащих тройную рекомбинирующую конструкцию компетентных клеток Bacillus pumilus 3-19 рассевают на чашки с агаризованной средой с антибиотиком эритромицином, например, в концентрации 20 Hg/ml. После инкубации в течение от 24 до 48 часов при +37°С отбирают, например, три независимых клона, растущих на среде с эритромицином. Дополнительную проверку наличия (или отсутствия) гена протеазы выполняют с применением ПЦР. Выявленное в результате ПЦР наличие вставки гена erm (внутри гена протеазы aprBp) подтверждает разрушение функционального гена, т.е. достижение цели изобретения - получение имеющих генотип aprBp::.erm модифицированного штамма бактерий, именуемых как Bacillus pumilus MK-10.
Описание предлагаемого штамма
Штамм бактерий Bacillus pumilus MK-10 депонирован во Всероссийской коллекции промышленных микроорганизмов ФГУП ГосНИИГенетика под регистрационным номером ВКПМ В-10742. Предлагаемый штамм является производным от штамма Bacillus pumilus 3-19 и содержит гены минорных протеаз, например глутамилэндопептидазы, адамализиноподобной металлопротеазы.
Генетические особенности штамма
Предлагаемый штамм бактерий Bacillus pumilus MK-10 несет ген резистентности к антибиотику эритромицину, фланкированный участками гена субтилизиноподобной протеазы Bacillus pumilus 3-19. Ген aprBp инактивирован посредством гомологичной рекомбинации (Фиг.2).
Культурально-морфологические особенности штамма
Предлагаемый штамм бактерий Bacillus pumilus МК10 - аэробные, грамположительные бактерии. Клетки палочковидные, прямые или изогнутые, располагаются скоплениями или обособленно. Размер выращенных на агаризованной среде клеток односуточной культуры составляет 14-15 мкм. Свободные споры появляются на сороковые часы роста (не более 5%), далее их количество нарастает и достигает максимального значения (до 80%) в поздней стационарной фазе роста. При росте на агаризованной среде Лурия-Бертани (далее по тексту - LB) колонии имеют более шероховатую поверхность и неровные края по сравнению с более гладкими и ровными колониями исходного штамма Bacillus pumilus 3-19.
Физиолого-биохимические свойства
Среда для выращивания, например LB: триптон - 10 g/l, дрожжевой экстракт - 5 g/l, NaCl - 10 g/l, pH 8.5 с антибиотиком эритромицином, например, в конечной концентрации 20 µg/ml. Для создания твердой питательной среды дополнительно вносят 2% агар. Оптимальная температура роста +37°С.
Протеолитическая активность предлагаемого штамма
Активность штамма определяют, например, следующим способом.
Определение протеолитической активности протеазо-дефицитного штамма Bacillus pumilus MK-10 проводят на среде, содержащей 2% молочного агара. Наличие протеолитической активности при этом определяют по появлению области просветления на молочном агаре (зоны расщепления белков молока). В результате селекции (отбора), например, трех колоний, которые не имеют зон просветления на молочной среде, получают предлагаемый штамм.
Выраженные зоны протеолиза на молочном агаре (2% агара и 1% молоко) наблюдают только у исходного штамма бактерий Bacillus pumilus 3-19. У предлагаемого штамма Bacillus pumilus MK-10 просветление менее выражено по сравнению с исходным штаммом Bacillus pumilus 3-19 (Фиг.3), что указывает на инактивацию гена секретируемой субтилизиноподобной протеазы. Наличие остаточных зон лизиса объясняется присутствием активности других секретируемых протеаз, например металлопротеазы. Таким образом, предлагаемый штамм бактерий Bacillus pumilus MK-10 обладает низкой протеолитической активностью.
Пример применения предлагаемого штамма бактерий Bacillus pumilus MK-10 приведен далее.
ПРИМЕР. Использование предлагаемого штамма для производства и синтеза рекомбинантных гомо- и гетерологичных белков р.Bacillus, применяемых для создания лекарственных препаратов.
Отличительным свойством исходного штамма Bacillus pumilus 3-19, из которого получен предлагаемый штамм, является наличие в хромосоме гена одной мажорной сериновой субтилизиноподобной протеазы. То есть у предлагаемого штамма, в отличие от прототипа, отсутствует вторая мажорная секретируемая протеаза, наличие которой приводило бы к деградации рекомбинантных белков и снижало эффективность (уровень синтеза и качество целевого белка) получаемой продукции. Инактивация гена единственного мажорного фермента (субтилизиноподобной протеазы) позволяет получить штамм Bacillus pumilus MK-10 с низкой, по сравнению с прототипом, протеолитической активностью. В результате общая протеолитическая активность штамма снижена в 2,5 раза по сравнению с активностью исходного штамма. Таким образом, предлагаемый штамм Bacillus pumilus MK-10 обеспечивает его использование в качестве реципиента для экспрессии гомологичных генов из бактерий рода Bacillus pumilus. Данное свойство связано с отсутствием вариаций последовательностей Shine Dalgarno (Шайна Дальгарно) в промоторах генов, выделенных из штаммов Bacillus pumilus. Поэтому предлагаемый штамм является наиболее эффективным при производстве промышленно важных ферментов, например, субтилизиноподобной протеазы Bacillus pumilus, имеющей потенциал в создании тромболитических лекарственных препаратов [9]. То есть предлагаемый штамм позволяет избавиться от присущих прототипу возможных изменений эффективности трансляции белков и тем самым добиться стабильности свойств продуктов, получаемых как результат жизнедеятельности этих микроорганизмов и реализуемых в биотехнологиях производства препаратов, например, применяемых в медицине, ветеринарии, сельскохозяйственном производстве и производстве продуктов питания.
Предлагаемый штамм в качестве хозяина применяют, например, нижеописанным методом для экспрессии гомологичных генов Bacillus pumilus.
1. Клетки бактерий предлагаемого штамма Bacillus pumilus МК-10 трансформируют плазмидой, содержащей ген рекомбинантного белка (например, гена субтилизиноподобной протеазы aprBp штамма Bacillus pumilus 3-19) под контролем индуцибельного или конститутивного промотора. Полученные трансформированные клетки подращивают на питательной среде, например LB, при температуре инкубации +37°С. Индукцию синтеза белка осуществляют в соответствии с выбранным промотором.
2. Пробы для анализа белков отбирают из КЖ путем отделения супернатанта от клеточной массы, например, через 1 ч и 16 ч после индукции.
3. Внеклеточные белки в пробе (культуральной жидкости или КЖ) характеризуют с помощью электрофореза в полиакриламидном геле по Лэммли [10].
4. Рекомбинантные белки далее очищают различными методами, например с помощью аффинной хроматографии, гель-фильтрации, ионно-обменной хроматографии, высаливания.
Таким образом, пример показывает, что предлагаемый штамм бактерий Bacillus pumilus МК-10 за счет снижения деградации белкового продукта позволяет получать рекомбинантные белки, например, для применения в медицине, ветеринарии, в сельском хозяйстве для растениеводства.
Кроме того, предлагаемый штамм применяют для экспрессии гетерологичных генов р.Bacillus по приведенному выше методу.
Пример применения предлагаемого штамма бактерий показывает его полезность в биотехнологии при промышленном производстве рекомбинантных белков. Использование предлагаемого штамма также способствует выявлению регуляторных свойств инактивированного фермента AprBp, что вносит свой вклад в фундаментальную молекулярную биологию. Предлагаемый штамм имеет изобретательский уровень, поскольку в опубликованной литературе не выявлены протеазо-дефицитные штаммы вида Bacillus pumilus.
ИСПОЛЬЗОВАННЫЕ ИСТОЧНИКИ
1. Kajino Т., Kato К., Miyazaki С., Asami О., Hirai M., Yamada Y., Udaka S. Isolation of a protease-deficient mutant of Bacillus brevis and efficient secretion of a fungal protein disulfide isomerase by the mutant / J Biosci Bioeng. 1999. 87(1): 37-42.
2. Plotnikova Т.О., Selivanova G.N., Iomantas Iu.V., Kozlov IuI. Isolation and analysis of protease-deficient mutants of Bacillus amyloliquefaciens I Genetika. - 1992. 28(5):66-72.
3. Fahnestock S.R., Fisher K.E. Protease-deficient Bacillus subtilis host strains for production of Staphylococcal protein A / Appl. Environ. Microbiol. 1987. 53(2):379-84.
4. http://www.basic.northwestern.edu/biotools/oligocalc.html
5. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии. Молекулярное клонирование // - M.: «Мир», 1984.
6. http://www.ncbi.nlm.nih.gov/
7. Sambrook J. Molecular cloning: a laboratory manual / J.Sambrook, E.F.Fritsch, T.Maniatis. - 2nd ed. NY: Cold Spring Harbor Laboratory Press, 1989.
8. Harwood, С.R., S.M.Cutting. 1990. Molecular biological methods for Bacillus. John Wiley & Sons, Chichester, England.
9. Ицкович Е.Л., Лютова Л.И., Балабан Н.П., Марданова A.M., Шакиров Е.В., Шарипова М.Р., Лещинская И.Б., Руденская Г.Н. Тромболитические и антикоагулянтные свойства тиолзависимой сериновой протеиназы Bacillus intermedms 3-19 / Вопросы медицинской химии, 1999, №5.
10. Laemmli, U.К. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4 [Text] / U.K.Laemmli //Nature. - 1970. - V.227. - P.680-685.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ BACILLUS PUMILUS 2A-5 С НИЗКОЙ ПРОТЕОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ, ПОВЫШЕННОЙ АКТИВНОСТЬЮ ФОСФАТАЗЫ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2012 |
|
RU2511416C1 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОГО ШТАММА BACILLUS SUBTILIS EO-11 ВКПМ В-11978 В КАЧЕСТВЕ ПРОДУЦЕНТА СУБТИЛИЗИНОПОДОБНОЙ ПРОТЕИНАЗЫ, ВЕЩЕСТВА С ТРОМБОЛИТИЧЕСКИМИ И АНТИКОАГУЛЯНТНЫМИ СВОЙСТВАМИ | 2015 |
|
RU2622006C2 |
| СПОСОБ ЭКСПРЕССИИ | 2012 |
|
RU2642324C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ ДЕЛЬТА-ЭНДОТОКСИНА Cry IIIA, И ШТАММ BACILLUS THURINGIENSIS SSP. KURSTAKI, ПОЛУЧЕННЫЙ НА ОСНОВЕ РЕКОМБИНАНТНОЙ ПЛАЗМИДНОЙ ДНК | 2004 |
|
RU2278161C1 |
| Способ получения аттенуированного бесплазмидного штамма F.tularensis 15 CMSA, синтезирующего микобактериальный антиген супероксиддисмутазу А | 2019 |
|
RU2745161C1 |
| ЛИТИЧЕСКАЯ ПРОТЕАЗА AlpA БАКТЕРИИ LYSOBACTER SP. XL1, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ЛИТИЧЕСКУЮ ПРОТЕАЗУ AlpA БАКТЕРИИ LYSOBACTER SP. XL1, И СПОСОБ ПОЛУЧЕНИЯ ЛИТИЧЕСКОЙ ПРОТЕАЗЫ AlpA БАКТЕРИИ LYSOBACTER SP. XL1 | 2009 |
|
RU2407782C2 |
| ИНТЕГРАТИВНАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pWpr-le | 2012 |
|
RU2539768C2 |
| ПЛАЗМИДА pNAN5, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ L-АСПАРАГИНАЗЫ ЕсА2, ШТАММ BACILLUS CEREUS 1576-pNAN5 - ПРОМЫШЛЕННЫЙ ШТАММ-ПРОДУЦЕНТ РЕКОМБИНАНТНОЙ L-АСПАРАГИНАЗЫ ЕсА2 И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2006 |
|
RU2313575C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНКРTV241, СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНОЙ ПЛАЗМИДНОЙ ДНКРTV241, СПОСОБ КОНСТРУИРОВАНИЯ ПЛАЗМИДЫ, СОДЕРЖАЩЕЙ ГЕН CRY III A В СОСТАВЕ ТРАНСПОЗОНА TN917CAT, ШТАММ БАКТЕРИЙ BACILLUS THURINGIENSIS SUBSP. THURINGIENSIS, АКТИВНЫЙ ПРОТИВ КОЛОРАДСКОГО ЖУКА | 1995 |
|
RU2125091C1 |
| УЛУЧШЕННАЯ ПРОДУКЦИЯ БЕЛКА В BACILLUS | 2008 |
|
RU2515112C2 |

Изобретение относится к области биотехнологии, а именно к микроорганизмам и ферментам, и представляет собой штамм бактерий Bacillus pumilus МК-10 ВКПМ В-10742. данный штамм получают от исходного штамма бактерий Bacillus pumilus 3-19 путем инактивирования гена мажорной сериновой протеазы на хромосоме. Изобретение позволяет расширить ассортимент штаммов бактерий, используемых в качестве реципиента с низкой протеолитической активностью при производстве гетерологичных белков. 2 з.п.ф-лы, 3 ил., 1 табл., 1 пр.
1. Штамм бактерий Bacillus pumilus MK-10 ВКПМ В-10742 - реципиент с низкой протеолитической активностью.
2. Штамм бактерий по п.1 применяют для производства гомологичных белков Bacillus pumilus.
3. Штамм бактерий по п.1 применяют для производства гетерологичных белков рода Bacillus.
| FAHNESTOCK S.R | |||
| et.al | |||
| Protease-deficient Bacillus subtilis host strains for production of Staphylococcal protein A., Appl Environ Microbiol | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| RU 2060276 C1, 20.05.1996 | |||
| WO8601825 A1, 27.03.1986 | |||
| . | |||

