Область, к которой относится изобретение
Настоящее изобретение относится к клеткам, которые были генетически модифицированы в целях изменения их способности экспрессировать и/или продуцировать представляющие интерес белки. В частности, настоящее изобретение относится к модифицированным клеткам-хозяевам грамположительных микроорганизмов, таких как Bacillus sp., которые способны сверхэкспрессировать ymaH. Настоящее изобретение охватывает полинуклеотидные конструкции и экспрессионные векторы, содержащие полинуклеотидные последовательности, кодирующие YmaH, и модифицированные клетки-хозяева, содержащие указанные векторы. В частности, настоящее изобретение относится к композициям и к способам сверхэкспрессии YmaH для повышения уровня экспрессии и продуцирования представляющих интерес белков (например, протеаз) в Bacillus sp.
Предшествующий уровень техники
Генная инженерия позволяет модифицировать микроорганизмы, используемые в качестве биологических реакторов, клеточных «фабрик», а также для ферментации пищевых продуктов. В частности, микроорганизмы вида Bacillus продуцируют и секретируют большое количество ценных белков и метаболитов (Zukowski, «Production of commercially valuable products» In: Doi and McGlouglin (eds.) Biology of Bacilli: Applications to Industry, Butterworth-Heinemann, Stoneham. Mass pp 311-337 [1992]). Самыми широко используемыми в промышленности бациллами являются B. licheniformis, B. amyloliquefaciens и B. subtilis. Благодаря тому, что эти микроорганизмы имеют статус GRAS (generally recognized as safe) (обычно рассматриваемые как безопасные), штаммы этих видов Bacillus являются природными кандидатами на продуцирование белков, используемых в пищевой и фармацевтической промышленностях. Важными продуцируемыми ферментами являются α-амилазы, нейтральные протеазы и щелочные (или сериновые) протеазы. Однако несмотря на достижения в понимании продуцирования белков в клетках-хозяевах Bacillus, сохраняется потребность в усовершенствованных методах экспрессии и продуцирования этих белков микроорганизмами.
Сущность изобретения
Настоящее изобретение относится к клеткам, которые были генетически модифицированы в целях изменения их способности экспрессировать и/или продуцировать представляющие интерес белки. В частности, настоящее изобретение относится к модифицированным клеткам-хозяевам грамположительных микроорганизмов, таких как Bacillus sp., которые способны сверхэкспрессировать ymaH. Настоящее изобретение охватывает полинуклеотидные конструкции и экспрессионные векторы, содержащие полинуклеотидные последовательности, кодирующие YmaH, и модифицированные клетки-хозяева, содержащие указанные конструкции и векторы. В частности, настоящее изобретение относится к композициям и к способам сверхэкспрессии YmaH для повышения уровня экспрессии и продуцирования представляющих интерес белков (например, протеаз) в Bacillus sp.
В одном варианте изобретения настоящее изобретение относится к выделенному химерному полинуклеотиду, который содержит полинуклеотидную последовательность, определяющую промотор SigA, функционально связанный с полинуклеотидом, кодирующим белок YmaH.
В другом варианте настоящее изобретение относится к выделенному химерному полинуклеотиду, который содержит полинуклеотидную последовательность, определяющую промотор SigA, функционально связанный с полинуклеотидом, кодирующим белок YmaH, где указанный химерный полинуклеотид включает SEQ ID NO:2 или SEQ ID NO:3.
В другом варианте настоящее изобретение относится к вектору, содержащему полинуклеотидную конструкцию, содержащую полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH.
В другом варианте настоящее изобретение относится к вектору, содержащему полинуклеотидную конструкцию, содержащую полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где полинуклеотидная конструкция содержит SEQ ID NO:1, 2, 3 или 13.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, которая содержит вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать представляющий интерес белок.
В другом варианте настоящее изобретение относится к модифицированной клетке-хозяину Bacillus, выбранной из группы, состоящей из B. licheniformis, B subtilis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. coagulans, B. circulans, B. lautus, B. pumilus, B. thuringiensis, B. clausii и B. megaterium и содержащей вектор, который содержит полинуклеотидную конструкцию, содержащую полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать представляющий интерес белок.
В другом варианте настоящее изобретение относится к модифицированной клетке-хозяину Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать представляющий интерес белок, который является гомологичным или гетерологичным для модифицированной клетки.
В другом варианте настоящее изобретение относится к модифицированной клетке-хозяину Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать представляющий интерес белок и где экспрессия указанного представляющего интерес белка инициируется промотором aprE.
В другом варианте настоящее изобретение относится к модифицированной клетке-хозяину Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать представляющий интерес белок, выбранный из амилаз, протеаз, ксиланаз, липаз, лакказ, фенолоксидаз, оксидаз, кутиназ, целлюлаз, гемицеллюлаз, эстераз, пероксидаз, каталаз, глюкозооксидаз, фитаз, пектиназ, глюкозидаз, изомераз, трансфераз, киназ, фосфотаз, галактозидаз и хитиназ, гормонов, цитокинов, факторов роста, рецепторов, вакцин и антител.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать фермент.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать протеазу.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, содержащей вектор, содержащий полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где модифицированная клетка способна продуцировать по меньшей мере один субтилизин, выбранный из субтилизина 168, субтилизина BPN', субтилизина Карлсберга, субтилизина B. lentus, субтилизина B. clausii, субтилизина DY, субтилизина 147 и субтилизина 309 и их вариантов.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, продуцирующей протеазу, которая способна сверхэкспрессировать ymaH, где модифицированная клетка содержит мутацию по меньшей мере в одном гене, выбранном из degU, degQ, degS, sco4, spollE, degQ и degR.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus, продуцирующей протеазу, которая способна сверхэкспрессировать ymaH, где модифицированная клетка содержит мутацию deg(Hy)32.
В другом варианте настоящее изобретение относится к модифицированной клетке Bacillus subtilis, продуцирующей протеазу, способной сверхэкспрессировать ymaH, где модифицированная клетка содержит мутацию по меньшей мере в одном гене, выбранном из degU, degQ, degS, sco4, spollE, degQ и degR.
В другом варианте настоящее изобретение относится к способу получения модифицированной клетки Bacillus, включающему: трансформацию клетки-хозяина Bacillus вектором, содержащим полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где клетка-хозяин Bacillus способна экспрессировать представляющий интерес белок; и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии представляющего интерес белка.
В другом варианте настоящее изобретение относится к способу получения модифицированной клетки Bacillus, включающему: трансформацию клетки-хозяина Bacillus вектором, содержащим полинуклеотидную конструкцию, которая находится на реплицирующейся плазмиде и которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где клетка-хозяин Bacillus способна экспрессировать представляющий интерес белок; и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии представляющего интерес белка.
В другом варианте настоящее изобретение относится к способу получения модифицированной клетки Bacillus, включающему: трансформацию клетки-хозяина Bacillus вектором, содержащим полинуклеотидную конструкцию, которая интегрирована в геном модифицированной клетки и которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где клетка-хозяин Bacillus способна экспрессировать представляющий интерес белок; и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии представляющего интерес белка.
В другом варианте настоящее изобретение относится к способу получения модифицированной клетки Bacillus, включающему: трансформацию клетки-хозяина Bacillus вектором, содержащим полинуклеотидную конструкцию, которая содержит полинуклеотид, кодирующий белок YmaH, функционально связанный с полинуклеотидной последовательностью промотора sigA и/или sigH, где клетка-хозяин Bacillus способна экспрессировать по меньшей мере один субтилизин; и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии субтилизина.
В другом варианте настоящее изобретение относится к способу продуцирования представляющего интерес белка в модифицированной клетке Bacillus, включающему культивирование модифицированной клетки Bacillus, способной сверхэкспрессировать ymaH, и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии представляющего интерес белка. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу продуцирования представляющего интерес белка в модифицированной клетке Bacillus, включающему культивирование модифицированной клетки Bacillus, способной сверхэкспрессировать ymaH, выращивание модифицированной клетки в условиях роста, благоприятствующих экспрессии представляющего интерес белка, и выделение представляющего интерес белка. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу продуцирования представляющего интерес белка в модифицированной клетке Bacillus в течение меньшего периода времени, чем в соответствующем предшественнике клетке-хозяине, где указанный способ включает культивирование модифицированной клетки Bacillus, способной сверхэкспрессировать ymaH; и выращивание модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии представляющего интерес белка. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу продуцирования представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка инициируется промотором aprE и где указанный способ включает культивирование модифицированной клетки Bacillus, способной сверхэкспрессировать ymaH; и выращивание модифицированной клетки в условиях роста, благоприятствующих экспрессии представляющего интерес белка. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу усиления экспрессии представляющего интерес белка из Bacillus, включающему получение модифицированной клетки Bacillus с применением способа, который включает сверхэкспрессию ymaH в родительской клетке-хозяине Bacillus; выращивание полученной модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией того же самого представляющего интерес белка в родительской клетке-хозяине. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способам усиления экспрессии представляющего интерес белка из Bacillus, включающему получение модифицированной клетки Bacillus способом, который включает сверхэкспрессию ymaH в родительской клетке-хозяине Bacillus; выращивание полученной модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке усиливается по сравнению с экспрессией того же самого представляющего интерес белка в родительской клетке-хозяине и где сверхэкспрессия включает трансформацию родительской клетки-хозяина Bacillus полинуклеотидной конструкцией, содержащей полинуклеотид, кодирующий белок YmaH, где полинуклеотид функционально связан с полинуклеотидной последовательностью промотора sigA или sigH. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способам усиления экспрессии представляющего интерес белка из Bacillus, включающим получение модифицированной клетки Bacillus способом, включающим сверхэкспрессию ymaH в родительской клетке-хозяине Bacillus; выращивание модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке усиливается по сравнению с экспрессией того же самого представляющего интерес белка в родительской клетке-хозяине и где сверхэкспрессия включает трансформацию клетки-хозяина Bacillus полинуклеотидной конструкцией, содержащей последовательность, выбранную из SEQ ID NO: 1, 2, 3 и 13. В некоторых вариантах осуществления представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу усиления экспрессии представляющего интерес белка из Bacillus, включающему получение модифицированной клетки Bacillus способом, включающим сверхэкспрессию ymaH в клетке-хозяине Bacillus; выращивание полученной модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией того же самого представляющего интерес белка в клетке-хозяине Bacillus и где сверхэкспрессия включает трансформацию клетки-хозяина Bacillus полинуклеотидной конструкцией, которая находится на плазмиде или интегрирована в геном модифицированной клетки и содержит полинуклеотид, кодирующий белок YmaH, где полинуклеотид функционально связан с полинуклеотидной последовательностью промотора sigA или sigH. В некоторых вариантах осуществления представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу усиления экспрессии представляющего интерес белка из Bacillus, включающему получение модифицированной клетки Bacillus способом, включающим сверхэкспрессию ymaH в клетке-хозяине Bacillus дикого типа; выращивание модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией того же самого представляющего интерес белка в клетке-хозяине дикого типа и где сверхэкспрессия включает трансформацию клетки-хозяина Bacillus дикого типа полинуклеотидной конструкцией, которая содержит полинуклеотид, кодирующий белок YmaH, где полинуклеотид функционально связан с полинуклеотидной последовательностью промотора sigA или sigH. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
В другом варианте настоящее изобретение относится к способу усиления экспрессии представляющего интерес белка из Bacillus, включающему получение модифицированной клетки Bacillus способом, включающим сверхэкспрессию ymaH в измененной клетке-хозяине Bacillus; выращивание модифицированной клетки Bacillus в условиях, благоприятствующих клеточному росту, и экспрессию представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия представляющего интерес белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией того же самого представляющего интерес белка в измененной клетке-хозяине и где сверхэкспрессия включает трансформацию измененной клетки-хозяина Bacillus полинуклеотидной конструкцией, содержащей полинуклеотид, кодирующий белок YmaH, где полинуклеотид функционально связан с полинуклеотидной последовательностью промотора sigA или sigH. В некоторых вариантах осуществления, представляющим интерес белком является фермент, например субтилизин. В некоторых вариантах осуществления клеткой Bacillus является клетка Bacillus subtilis.
Краткое описание графического материала
На фигуре 1 (A-E) показана локализация праймеров, используемых для получения полинуклеотидных конструкций согласно некоторым вариантам осуществления настоящего изобретения. На панелях B-E указано положение праймеров, используемых для получения конструкций SigH, SigA1, SigA2 и SigA3, соответственно, по отношению к хромосомной последовательности Bacillus оперона miaA Bacillus subtilis (1865428-1867019 пар нуклеотидов штамма 168 Bacillus subtilis; NCBI рег. № NC000964), который показан на панели A. Пары праймеров P4 - P5 и P6 - P7 являются гибридными праймерами, которые на своем 5'-конце содержат «хвост» из пар оснований, которые гомологичны непосредственно амплифицированной последовательности и комплементарны друг другу. Комплементарные хвосты этих гибридных праймеров позволяют присоединить амплифицированную ДНК промотора Sigma A к амплифицированной YmaH-кодирующей ДНК с получением химерных полинуклеотидов, содержащих промоторную последовательность Sigma A, смежную с YmaH-кодирующей последовательностью, при этом большая часть miaA-кодирующей последовательности делетирована или вообще отсутствует.
На фигуре 2 показана полинуклеотидная последовательность части генома B. subtilis, которая включает последовательность, определяющую промотор sigA, присоединенный к концу последовательности, кодирующей белок YmaH. Эта последовательность схематично представлена на фигуре 1 панели А. Показано начало последовательности, кодирующей белок miaA, а полноразмерная miaA-кодирующая последовательность представлена жирным шрифтом; кроме того, показано начало последовательности, кодирующей белок YmaH, а полноразмерная YmaH-кодирующая последовательность представлена жирным шрифтом и подчеркнута.
На фигуре 3 (A-B), панели A, представлен график протеолитической активности субтилизина, продуцированного контрольными клетками-хозяевами Bacillus (42pBS) и модифицированными клетками-хозяевами Bacillus, которые сверхэкспрессируют ymaH (42SigA1 и 42SigH). На панели B проиллюстрирована активность субтилизина, продуцированного контрольными клетками-хозяевами Bacillus (41pBS) и модифицированными клетками-хозяевами Bacillus, которые сверхэкспрессируют ymaH (41SigH). Протеолитическая активность была определена как увеличение оптической плотности на 405 нм, обусловленное гидролизом и высвобождением п-нитроанилина. Уровень ферментативной активности является показателем эффективности сверхэкспрессии ymaH в отношении продуцирования субтилизина клетками-хозяевами Bacillus.
На фигуре 4 указан уровень продуцирования субтилизина контрольными клетками-хозяевами Bacillus 42pBS19 и модифицированными клетками-хозяевами Bacillus 42SigH и 42SigA1, которые сверхэкспрессируют ymaH.
Подробное описание изобретения
Настоящее изобретение относится к клеткам, которые были генетически модифицированы в целях изменения их способности экспрессировать и/или продуцировать представляющие интерес белки. В частности, настоящее изобретение относится к модифицированным клеткам-хозяевам, которые представляют собой грамположительные микроорганизмы, такие как Bacillus sp., способные сверхэкспрессировать ymaH. Настоящее изобретение относится к полинуклеотидным конструкциям и экспрессионным векторам, содержащим полинуклеотидные последовательности, кодирующие YmaH, и к модифицированным клеткам-хозяевам, содержащим указанные полинуклеотидные конструкции и экспрессионные векторы. В частности, настоящее изобретение относится к композициям и к способам сверхэкспрессии YmaH для повышения уровня экспрессии и продуцирования представляющих интерес белков (например, протеаз) в Bacillus sp.
Если это не оговорено особо, то настоящее изобретение осуществляют стандартными методами, известными специалистам и обычно применяемыми в молекулярной биологии, микробиологии, в очистке белков, в конструировании белков, в секвенировании белков и ДНК и в технике рекомбинантных ДНК. Эти методы известны специалистам и описаны во многих общих руководствах и в научной литературе. Все выше- и нижеупомянутые патенты, патентные заявки, статьи и публикации во всей своей полноте вводятся в настоящее описание посредством ссылки.
Если это не оговорено особо, то все технические и научные термины, используемые в настоящей заявке, имеют значения, в основном понятные среднему специалисту в области, к которой относится изобретение. Специалистам хорошо известны и доступны различные научные словари, в которых дается определение используемых в настоящем документе терминов. Хотя для осуществления настоящего изобретения, как в теории, так и на практике, могут быть применены любые методы и материалы, аналогичные или эквивалентные описанным в настоящем документе методам и материалам, однако предпочтительными являются некоторые методы и материалы, описанные в настоящей заявке. В соответствии с этим термины, описанные непосредственно ниже, более подробно и во всей своей полноте определены путем ссылки на описание настоящего изобретения. При этом следует отметить, что настоящее изобретение не ограничивается конкретно описанными в настоящем документе методами, протоколами и реагентами, которые могут варьироваться в зависимости от цели их применения.
В настоящем описании формы единственного числа включают формы множественного числа, если это явно не противоречит контексту изобретения. Если это не оговорено особо, то нуклеиновые кислоты записываются слева направо в направлении 5' → 3', а аминокислотные последовательности записываются слева направо в направлении от амино- до карбокси-конца, соответственно.
Все патенты, патентные заявки и другие публикации, включающие все цитируемые в настоящем документе последовательности, точно вводятся в настоящее описание посредством ссылки так, как если бы каждая отдельная публикация, патент или патентная заявка были конкретно и отдельно включены в данное описание посредством ссылки. Все документы, цитируемые в соответствующем разделе настоящей заявки, вводятся в настоящее описание посредством ссылки. Однако цитирование какого-либо документа не должно обязательно означать, что этот документ относится к прототипу настоящего изобретения.
В указанные числовые интервалы входят все входящие в него значения, включая его границы. При этом, предусматривается, что каждый максимальный числовой предел, указанный в настоящем описании, может включать каждый меньший числовой предел так, как если бы такой меньший числовой предел был точно определен в настоящей заявке. Каждый минимальный числовой предел, указанный в настоящем описании, может включать больший числовой предел так, как если бы такой больший числовой предел был точно определен в настоящей заявке. Каждый числовой интервал, указанный в описании настоящей заявки, включает каждый более узкий числовой интервал, который входит в такой более широкий числовой интервал так, как если бы все указанные более узкие числовые интервалы были точно определены в описании настоящей заявки.
Сущность изобретения не ограничивается различными приведенными в настоящем документе аспектами или вариантами осуществления изобретения, которые целиком могут быть включены в настоящее описание посредством ссылки. В соответствии с этим, как указывается выше, описанные ниже термины более подробно определены посредством ссылки на все описание в целом.
В настоящем документе термины «выделенный» и «очищенный» относятся к нуклеиновой кислоте или к аминокислоте (или к другому компоненту), которые были отделены по меньшей мере от одного компонента, с которым они связаны в своем природном окружении.
Термины «химерный полинуклеотид», «химерная полинуклеотидная конструкция» и «гетерологичная конструкция нуклеиновой кислоты» означают полинуклеотид, который состоит из частей различных генов, включая регуляторные элементы. Таким образом, в некоторых вариантах осуществления химерная полинуклеотидная конструкция включает белок-кодирующую область, функционально связанную с промотором, который не является нативным промотором. В некоторых вариантах осуществления химерный полинуклеотид означает полинуклеотидную последовательность, которая включает полинуклеотидную последовательность, определяющую промотор и функционально связанную с полинуклеотидной последовательностью, кодирующей белок. В некоторых вариантах осуществления, промотор и кодирующие полинуклеотиды являются смежными.
Термин «определяющий», если он употребляется при описании промотора, относится к полинуклеотидной последовательности, содержащей промоторные элементы, обеспечивающие транскрипцию.
В настоящем документе термин «промотор» означает последовательность нуклеиновой кислоты, которая инициирует/осуществляет транскрипцию расположенного ниже гена. Обычно такой промотор является подходящим для той клетки-хозяина, в которой осуществляется экспрессия гена. Промотор, вместе с другими последовательностями нуклеиновой кислоты, регулирующими транскрипцию и трансляцию (также называемыми «регуляторными последовательностями»), является необходимым для экспрессии данного гена. В общих чертах, последовательностями регуляции транскрипции и трансляции являются, но не ограничиваются ими, промоторные последовательности, сайты связывания с рибосомой, последовательности инициации и терминации транскрипции, последовательности инициации и терминации трансляции, вышерасположенные последовательности элементов, расположенных выше промотора (UP-элементы), и последовательности энхансеров или активаторов. В некоторых вариантах осуществления изобретения, промотор также включает транскрипционную лидерную последовательность.
В настоящем документе термины «промотор Sigma A» и «промотор SigA» означают полинуклеотидную последовательность, содержащую коровые промоторные последовательности, которые включают последовательности, непосредственно распознаваемые соответствующим фактором σА. Промотор SigA входит в последовательность, которая обычно расположена выше miaA-кодирующей области.
В настоящем документе термины «промотор Sigma H» и «промотор SigH» означают полинуклеотидную последовательность, содержащую коровые промоторные последовательности, которые включают последовательности, непосредственно распознаваемые соответствующим фактором σH. Промотор SigH входит в последовательность, которая обычно расположена выше ymaH-кодирующей области (Britton et al. J. Bacteriol. 184:4881-4890 [2002]). Коровый промотор включает промоторные последовательности, содержащие последовательности, непосредственно распознаваемые соответствующим фактором σ, и спейсерную последовательность, расположенную между последовательностями, непосредственно распознаваемыми соответствующим фактором σ.
В настоящем документе термин «промотор aprE» означает полинуклеотидную промоторную последовательность, которая обычно инициирует экспрессию субтилизина в B. subtilis (Ferrari et al., J Bacteriol. 170:289-295 [1988]). Что касается промотора aprE, то в настоящем документе термин «промотор aprE» означает промотор aprE дикого типа и его мутанты. В некоторых вариантах осуществления изобретения промотор aprE включает нуклеотидные последовательности, необходимые для регуляции транскрипции, осуществляемой под действием DegU, ScoC, AbrB и любого другого регулятора такого промотора, и/или транскрипционную лидерную последовательность AprE (Hambraeus et al., Microbiology 148:1795-1803 [2002]).
В некоторых альтернативных вариантах осуществления изобретения промотор aprE не включает все нуклеотидные последовательности, необходимые для регуляции транскрипции, осуществляемой под действием DegU, ScoC, AbrB и других регуляторов, и/или не включает транскрипционную лидерную последовательность aprE.
Термины «регуляторный сегмент», «регуляторная последовательность» и «последовательность регуляции экспрессии» означают полинуклеотидную последовательность ДНК, функционально связанную с полинуклеотидной последовательностью ДНК, которая кодирует аминокислотную последовательность полипептидной цепи, в результате чего осуществляется экспрессия кодируемой аминокислотной последовательности. Регуляторная последовательность может ингибировать, подавлять или стимулировать экспрессию функционально связанной полинуклеотидной последовательности, кодирующей аминокислоту. В некоторых осуществления изобретения регуляторная последовательность содержит промотор, функционально связанный с последовательностью ДНК, кодирующей регулятор транскрипции YmaH. В некоторых вариантах осуществления изобретения указанный промотор является гетерологичным по отношению к гену ymaH (например, указанным промотором является промотор, который непосредственно не участвует в инициации экспрессии белка YmaH). Так, например, в некоторых вариантах осуществления, указанным промотором является промотор Sigma A, функционально связанный с ДНК, кодирующей белок YmaH. В некоторых других вариантах осуществления указанным промотором является промотор, который непосредственно инициирует экспрессию белка YmaH и который функционально связан с ДНК, кодирующей YmaH, поскольку такая ДНК является природной для данных хозяев.
В настоящем документе термины «полинуклеотид» и «нуклеиновая кислота» являются взаимозаменяемыми и означают полимерную форму нуклеотидов любой длины. Эти термины включают, но не ограничиваются ими, одноцепочечную ДНК, двухцепочечную ДНК, геномную ДНК, кДНК, или полимер, содержащий пуриновые и пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания. Неограничивающими примерами полинуклеотидов являются гены, генные фрагменты, хромосомные фрагменты, EST, экзоны, интроны, мРНК, тРНК, рРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК, имеющая любую последовательность, выделенная РНК, имеющая любую последовательность, нуклеиновокислотные зонды и праймеры. Следует отметить, что из-за вырожденности генетического кода может продуцироваться множество нуклеотидных последовательностей, кодирующих данный белок.
В настоящем документе термин «ген» означает хромосомный сегмент ДНК, участвующий в продуцировании полипептидной цепи, которая может включать, а может и не включать, области, расположенные до и после кодирующих областей (например, 5'-нетранслируемые (5'-UTR) или лидерные последовательности и 3'-нетранслируемые (3'-UTR) трейлерные последовательности, а также промежуточные последовательности (интроны), расположенные между отдельными кодирующими сегментами (экзонами)). В некоторых вариантах осуществления изобретения ген кодирует коммерчески доступные и важные с промышленной точки зрения белки или пептиды, такие как ферменты, включая, но не ограничиваясь ими, протеазы, целлюлазы, карбогидразы, такие как амилазы и глюкоамилазы, целлюлазы, оксидазы, изомеразы, трансферазы, киназы, фосфатазы и липазы. В некоторых других вариантах осуществления изобретения указанный ген кодирует белки, кодируемые опероном, в котором присутствует miaA (например, miaA или ymaH). Однако следует отметить, что настоящее изобретение не ограничивается каким-либо конкретным ферментом или белком. В некоторых других вариантах осуществления, указанный ген кодирует другие белки или пептиды, такие как факторы роста, цитокины, лиганды, рецепторы и ингибиторы, а также вакцины и антитела. В некоторых вариантах осуществления представляющим интерес геном является природный ген, а в других вариантах осуществления таким геном является мутированный ген или синтетический ген.
В настоящем документе термин «синтетический» относится к полинуклеотидной молекуле, продуцируемой in vitro методами химического или ферментативного синтеза. Этот термин включает, но не ограничивается ими, варианты нуклеиновых кислот, полученные с использованием оптимальных кодонов, встречающихся в организмах-хозяевах, таких как, но не ограничивающихся ими, Bacillus sp.
В настоящем документе термин «полимеразная цепная реакция» («ПЦР») относится к методам, описанным в патентах США №№ 4683195, 4683202 и 4965188, которые вводятся в настоящее описание посредством ссылки, где указанные методы включают увеличение концентрации сегмента последовательности-мишени в смеси геномной ДНК без проведения клонирования или очистки. Такой способ амплификации последовательности-мишени состоит из введения большого избытка двух олигонуклеотидных праймеров в смесь ДНК, содержащую нужную последовательность-мишень, с последующим проведением определенного ряда термоциклов в присутствии ДНК-полимеразы. Два праймера являются комплементарными их соответствующим цепям двухцепочечной последовательности-мишени. Для осуществления амплификации смесь денатурируют, а затем праймеры гибридизуют с комплементарными им последовательностями в молекуле-мишени. После отжига праймеры удлиняют под действием полимеразы, в результате чего образуется новая пара комплементарных цепей. Стадии денатурации, отжига праймеров и удлинения под действием полимеразы могут быть проведены многократно (то есть стадии денатурации, отжига и удлинения составляют один «цикл», а могут составлять и множество «циклов») с получением высокой концентрации амплифицированного сегмента нужной последовательности-мишени. Длину амплифицированного сегмента нужной последовательности-мишени определяют по положениям праймеров по отношению друг к другу, а поэтому такая длина является контролируемым параметром. Поскольку данный процесс носит повторяющийся характер, то такой метод называется «полимеразной цепной реакцией» (далее обозначаемой «ПЦР»). Так как нужные амплифицированные сегменты последовательности-мишени становятся преобладающими последовательностями (с точки зрения их концентрации) в данной смеси, то их принято называть «ПЦР-амплифицированными».
В настоящем документе термин «реагенты для амплификации» означает реагенты (дезоксирибонуклеотид-трифосфаты, буфер и т.п.), необходимые для амплификации, за исключением праймеров, нуклеиновокислотной матрицы и фермента для амплификации. Обычно реагенты для амплификации, вместе с другими компонентами реакционной смеси, помещают в реакционный сосуд и хранят в таком сосуде (в тест-пробирке, микролунке и т.п.).
С помощью ПЦР можно амплифицировать одну копию специфической последовательности-мишени в геномной ДНК до уровня, который может быть детектирован с применением нескольких различных методов (например, гибридизации с меченым зондом; введения биотинилированных праймеров с последующим детектированием конъюгата «авидин-фермент»; и введения 32P-меченных дезоксинуклеотид-трифосфатов, таких как dCTP или dATP, в амплифицированный сегмент). Помимо геномной ДНК любая олигонуклеотидная или полинуклеотидная последовательность может быть амплифицирована с использованием соответствующего набора молекул-праймеров. В частности, амплифицированные сегменты, полученные с помощью ПЦР, сами являются матрицами, которые могут быть использованы для последующих ПЦР-амплификаций.
В настоящем документе термины «ПЦР-продукт», «ПЦР-фрагмент» и «продукт амплификации» означают смесь соединений, полученную после проведения двух или более циклов ПЦР-стадий денатурации, отжига и удлинения. Эти термины включают случаи, когда происходит амплификация одного или нескольких сегментов одной или нескольких последовательностей-мишеней.
В настоящем документе термины «эндонуклеазы рестрикции» и «ферменты рестрикции» означают бактериальные ферменты, каждый из которых разрезает двухцепочечную ДНК в конкретной нуклеотидной последовательности или возле этой последовательности.
В настоящем документе термин «рекомбинантный» относится к полинуклеотиду или полипептиду, который обычно не присутствует в данной клетке-хозяине. В некоторых вариантах изобретения рекомбинантные молекулы содержат две или более природных последовательности, которые функционально связаны друг с другом, так, как это обычно не встречается в природе.
Нуклеиновая кислота является «функционально связанной», если она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Так, например, полинуклеотидная промоторная последовательность функционально связана с полинуклеотидом, кодирующим полипептид, если он влияет на транскрипцию последовательности. В некоторых других вариантах осуществления изобретения сайт связывания с рибосомой является функционально связанным с кодирующей последовательностью, если его локализация облегчает трансляцию. В некоторых вариантах изобретения термин «функционально связанный» означает, что связанные друг с другом полинуклеотидные последовательности являются смежными. Связывание осуществляется лигированием в подходящих рестрикционных сайтах. Если такие сайты отсутствуют, то в соответствии со стандартной практикой могут быть использованы синтетические олигонуклеотидные адаптеры или линкеры.
В настоящем документе термин «гомологичные гены» означает гены, которые отличаются друг от друга, но обычно являются родственными молекулами, которые соответствуют друг другу и либо являются идентичными, либо имеют очень высокую степень сходства. Этот термин охватывает гены, которые были разделены в процессе видообразования (то есть образования новых видов) (например, ортологичные гены), а также гены, которые были разделены в процессе генетической дупликации (например, паралогичные гены).
В настоящем документе термин «ортолог» и «ортологичные гены» означает гены, присутствующие у различных видов, которые эволюционировали от общего гена-предка (то есть гомологичного гена) в процессе видообразования. Обычно ортологи сохраняют свою функцию в процессе эволюции. Идентификация ортологов применяется для точного предсказания функции гена в только что секвенированных геномах.
В настоящем документе термин «паралог» и «паралогичные гены» означает гены, которые стали родственными в результате дупликации в геноме. Ортологи сохраняют свои функции в процессе эволюции, а паралоги приобретают новые функции, хотя некоторые функции паралогов часто соответствуют их исходным функциям. Примерами генов-паралогов являются, но не ограничиваются ими, гены, кодирующие трипсин, химотрипсин, эластазу и тромбин, и все они являются сериновыми протеиназами и в совокупности присутствуют у одного и того же вида.
В настоящем документе термин «гомология» означает сходство или идентичность последовательностей, а предпочтительно - идентичность. Гомологию определяют стандартными методами, известными специалистам (см., например, в публикации Smith and Waterman, Adv Appl Math, 2:482, 1981; Needleman and Wunsch, J. Mol. Biol., 48:443, 1970; Pearson and Lipman, Proc. Natl. Acad. Sci. USA, 85:2444, 1988; и программы, такие как GAP, BESTFIT, FASTA и TFASTA, входящие в пакет программного обеспечения Wisconsin Genetics Software Package, (Genetics Computer Group, Madison, WI); и Devereux et al, Nucl. Acid. Res., 12:387-395, 1984).
В настоящем документе термин «аналогичная последовательность» означает последовательность, в которой функция гена по существу аналогична функции гена, сконструированного на основе предпочтительного штамма Bacillus subtilis (то есть Bacillus subtilis 168). Кроме того, аналогичными генами являются гены, последовательности которых по меньшей мере примерно на 60%, примерно на 65%, примерно на 70%, примерно на 75%, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 97%, примерно на 98%, примерно на 99% или примерно на 100% идентичны последовательности гена штамма Bacillus subtilis 168. Альтернативно, аналогичные последовательности при их выравнивании имеют 70-100%-ное соответствие с генами, присутствующими в области генома Bacillus subtilis 168 и/или имеют по меньшей мере 5-10 генов, обнаруженных в области их выравнивания с генами геномной области B. subtilis 168. В дополнительных вариантах изобретения, указанная последовательность обладает более чем одним из вышеуказанных свойств. Аналогичные последовательности определяют известными методами выравнивания последовательностей. Наиболее распространенным методом выравнивания является BLAST, хотя, как указывается выше и ниже, для выравнивания последовательностей могут быть также применены и другие методы.
Одним из примеров подходящего алгоритма является алгоритм PILEUP. Алгоритм PILEUP позволяет осуществлять выравнивание множества последовательностей из группы родственных последовательностей путем последовательного попарного выравнивания. Для осуществления выравнивания может быть также построена древовидная схема, иллюстрирующая кластерные взаимосвязи. В программе PILEUP используется упрощенный метод последовательного выравнивания, описанный Feng и Doolittle (Feng and Doolittle, J. Mol. Evol., 35:351-360, 1987). Этот метод аналогичен методу, описанному Higgins и Sharp (Higgins and Sharp, CABIOS 5:151-153, 1989). Подходящими параметрами PILEUP являются: «цена» пробела 3,00 по умолчанию, «цена» длины пробела 0,10 по умолчанию и взвешенные значения концевых пробелов.
Другим примером подходящего алгоритма является алгоритм BLAST, описанный Altschul et al., (Altschul et al, J. Mol. Biol, 215:403-410, [1990]; и Karlin et al., Proc. Natl. Acad. Sci. USA, 90:5873-5787, [1993]). Особенно подходящей программой BLAST является программа WU-BLAST-2 (см., Altschul et al., Meth. Enzymol., 266:460-480, [1996]). В программе WU-BLAST-2 используется несколько параметров поиска, большинство из которых принимаются как величины по умолчанию. Корректируемые параметры установлены со следующими величинами по умолчанию: охват с перекрыванием = 1, перекрывающаяся фракция = 0,125, предельная длина слова (T) = 11. Параметрами HSP S и HSP S2 являются динамические величины, и эти параметры устанавливаются самой программой в зависимости от состава конкретной последовательности и состава конкретной базы данных, в которой осуществляется поиск представляющей интерес последовательности. Однако для повышения чувствительности эти величины могут быть скорректированы. Процент (%) идентичности аминокислотных последовательностей определяют по числу соответствующих идентичных остатков, деленному на общее число остатков «более длинной» последовательности в выравниваемой области. «Более длинной» последовательностью является последовательность, имеющая наиболее ценные остатки в выравниваемой области (пробелы, вводимые с помощью программы WU-Blast-2 для максимизации «веса» выравнивания, не учитываются).
Таким образом, «процент (%) идентичности последовательностей нуклеиновой кислоты» определяют как процент нуклеотидных остатков в последовательности-кандидате, которые идентичны нуклеотидным остаткам исходной последовательности (то есть, представляющей интерес последовательности). В предпочтительном методе используется модуль BLASTN программы WU-BLAST-2, установленный по умолчанию, где перекрывание и перекрывающаяся фракция имеют значения 1 и 0,125, соответственно.
В некоторых вариантах осуществления изобретения выравнивание включает введение пробелов в выравниваемые последовательности. Кроме того, для последовательностей, содержащих большее число или меньшее число нуклеотидов по сравнению с последовательностями нуклеиновой кислоты-кандидата, процент гомологии определяют исходя из числа гомологичных последовательностей по отношению к общему числу нуклеотидов. Так, например, гомология последовательностей, которые являются более короткими, чем последовательности, идентифицированные в настоящей заявке и обсуждаемые ниже, определяют с использованием числа нуклеотидов в более короткой последовательности.
В настоящем документе термины «полинуклеотидная конструкция», «экспрессионный кластер» и «экспрессионный вектор» означают конструкцию ДНК, содержащую последовательность ДНК, которая функционально связана с подходящей регуляторной последовательностью, способной осуществлять экспрессию ДНК в подходящем хозяине. Такими регуляторными последовательностями являются промотор, инициирующий транскрипцию; необязательная последовательность-оператор, регулирующая такую транскрипцию; последовательность, кодирующая подходящие сайты связывания с рибосомой, присутствующие в мРНК; и последовательности, регулирующие терминацию транскрипции и трансляции. Обычно полинуклеотидная конструкция включает область регуляции транскрипции (например, промотор), функционально связанную с белок-кодирующей областью. В некоторых вариантах осуществления изобретения полинуклеотидная конструкция содержит белок-кодирующую область, которая функционально связана с нативным промотором (то есть, с промотором, который является смежным с природной кодирующей последовательностью). Так, например, полинуклеотидная конструкция согласно изобретению содержит промотор sigH и ymaH-кодирующую последовательность. В других вариантах изобретения, полинуклеотидная конструкция содержит белок-кодирующую область, которая функционально связана с промотором, который, по своей природе, не является смежным с кодирующей последовательностью (то есть, указанная полинуклеотидная конструкция содержит химерный полипептид, в котором промотор находится в положении, не соответствующем его природному положению по отношению к кодирующей последовательности). Так, например, промотор sigA функционально связан с ymaH-кодирующей последовательностью. В других вариантах изобретения, полинуклеотидная последовательность содержит более, чем один промотор и более, чем одну белок-кодирующую область (например, полинуклеотидная конструкция содержит полицистронную последовательность, включающую промоторы и кодирующие области, присутствующие в опероне). В некоторых других вариантах изобретения полинуклеотидная конструкция или экспрессионный кластер содержит селективный маркер (например, маркер резистентности к антибиотику, такой как ген, кодирующий хлорамфениколацетил-трансферазу), который, в случае присутствия соответствующего антибиотика, позволяет амплифицировать полинуклеотидную конструкцию в геноме клетки-хозяина. Полинуклеотидная конструкция может быть введена в плазмиду, геном, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. В некоторых вариантах изобретения указанным вектором является плазмида, фаговая частица или просто потенциальная геномная вставка. В некоторых других вариантах изобретения вектор, после его трансформации в клетку-хозяина, реплицируется и функционирует независимо от генома-хозяина, или, в некоторых альтернативных случаях, интегрируется в свой собственный геном. В настоящем документе термины «плазмида», «экспрессионная плазмида» и «вектор» часто являются взаимозаменяемыми, поскольку плазмидой является наиболее распространенная форма вектора, обычно используемого в настоящее время. Однако настоящее изобретение включает и другие формы экспрессионных векторов, которые имеют эквивалентные функции и которые либо являются известными специалистам, либо будут известны в будущем. «Векторами» являются клонирующие векторы, экспрессионные векторы, челночные векторы, плазмиды, фаговые или вирусные частицы, конструкции ДНК, кластеры и т.п.
В настоящем документе термин «плазмида» означает кольцевую двухцепочечную (дц) ДНК-конструкцию, используемую в качестве клонирующего вектора, которая образует внехромосомный самореплицирующийся генетический элемент во многих бактериях и в некоторых эукариотах. В некоторых вариантах изобретения плазмиды вводят в геном клетки-хозяина. Термин «плазмида» включает многокопийные плазмиды, которые могут интегрироваться в геном клетки-хозяина посредством гомологичной рекомбинации.
В настоящем документе термины «трансформируемый» и «стабильно трансформируемый» относятся к клетке, которая имеет не-нативную (гетерологичную) полинуклеотидную последовательность, интегрированную в ее геном, или существующую в виде эписомной плазмиды, сохраняющейся по меньшей мере в двух или более генерациях. В настоящем документе термин «экспрессия» означает процесс, посредством которого полинуклеотид транскрибируется, и полученный транскрипт транслируется с образованием полипептида. Такой процесс включает транскрипцию и трансляцию.
В настоящем документе термин «сверхэкспрессия» означает процесс, посредством которого ген, содержащий последовательность, кодирующую полипептид, искусственно экспрессируется в модифицированной клетке на уровне экспрессии кодируемого полипептида, превышающего уровень экспрессии того же полипептида в предшественнике клетки-хозяина. Таким образом, хотя этот термин обычно употребляется по отношению к гену, однако термин «сверхэкспрессия» может также употребляться по отношению к белку, и в этом случае, он означает повышенный уровень белка, образующегося в результате сверхэкспрессии гена, кодирующего этот белок. В некоторых вариантах осуществления изобретения сверхэкспрессия гена, кодирующего белок, достигается путем увеличения числа копий гена, кодирующего белок. В других вариантах изобретения сверхэкспрессия гена, кодирующего белок, достигается путем увеличения силы связывания промоторной области и/или сайта связывания с рибосомой, что приводит к повышению уровня транскрипции и/или трансляции гена, кодирующего белок. В других вариантах изобретения сверхэкспрессия может достигаться путем увеличения числа копий гена и силы связывания промоторной области и/или сайта связывания с рибосомой. В некоторых вариантах изобретения сверхэкспрессия гена, кодирующего белок, происходит в результате экспрессии по меньшей мере одной копии соответствующего кодирующего полинуклеотида, присутствующего на многокопийной плазмиде, которая была введена в клетку-хозяина. В других вариантах изобретения сверхэкспрессия гена, кодирующего белок, происходит в результате экспрессии двух или более копий соответствующего кодирующего полинуклеотида, интегрированного в геном клетки-хозяина.
Термин «клетка-хозяин» означает подходящую клетку среди тех клеток, которые служат в качестве хозяев для экспрессионного вектора, содержащего ДНК согласно изобретению. Подходящей клеткой-хозяином может быть природная клетка-хозяин или клетка-хозяин дикого типа либо такой клеткой может быть модифицированная клетка-хозяин. В одном из вариантов изобретения указанной клеткой-хозяином является грамположительный микроорганизм. В некоторых предпочтительных вариантах изобретения, указанный термин означает клетки, принадлежащие к роду Bacillus.
В настоящем документе термин «клетка Bacillus» включает все члены этого рода, известные специалистам, включая, но не ограничиваясь ими, B. subtilis, B. licheniformis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. clausii, B. halodurans, B. megaterium, B. coagulans, B. circulans, B. lautus и B. thuringiensis. Следует отметить, что бактерии рода Bacillus постоянно подвергаются таксономической реорганизации. Таким образом, предусматривается, что микроорганизмами такого рода являются виды, которые были переклассифицированы, включая, но не ограничиваясь ими, такие микроорганизмы как B. stearothermophilus, который в настоящее время называется «Geobacillus stearothermophilus». Продуцирование резистентных эндоспор в присутствии кислорода рассматривается как признак, определяющий род Bacillus, хотя такой признак также является характерным для недавно идентифицированных бактерий, таких как Alicyclobacillus, Amphibacillus, Aneurinibacillus, Anoxybacillus, Brevibacillus, Filobacillus, Gracilibacillus, Halobacillus, Paenibacillus, Salibacillus, Thermobacillus, Ureibacillus и Virgibacillus.
Термин «клетка-хозяин дикого типа» означает клетку-хозяина, которая не была генетически модифицирована рекомбинантными методами.
В настоящем документе термин «дикого типа» относится к гену или генному продукту, который имеет свойства гена или генного продукта, выделенного из природного источника. Обычно ген дикого типа представляет собой ген, который наиболее часто встречается в определенной популяции клеток, а поэтому его иногда называют «нормальным геном» или «геном дикого типа». В настоящем документе термины «последовательность дикого типа» и «ген дикого типа» являются взаимозаменяемыми и означают последовательность, которая является нативной или обычно присутствует в клетке-хозяине. Последовательность дикого типа может кодировать гомологичный или гетерологичный белок.
В настоящем документе термины «измененная клетка-хозяин», «измененная клетка» и «измененный штамм» означают генетически сконструированную клетку-хозяина (микроорганизм), в которой представляющий интерес белок экспрессируется и/или продуцируется на уровне экспрессии или продуцирования, превышающем уровень экспрессии и/или продуцирования того же самого представляющего интерес белка при культивировании неизмененной клетки-хозяина или клетки-хозяина дикого типа, по существу в тех же самых условиях роста. В некоторых вариантах изобретения измененной клеткой-хозяином является клетка, продуцирующая рекомбинантную протеазу.
В настоящем документе термины «модифицированная клетка» и «модифицированная клетка-хозяин» означают клетку-хозяина дикого типа или измененную клетку-хозяина, которые были генетически сконструированы так, чтобы они сверхэкспрессировали ген, кодирующий белок YmaH. В некоторых вариантах изобретения модифицированной клеткой-хозяином является клетка-хозяин, продуцирующая рекомбинантную протеазу. В некоторых вариантах изобретения модифицированная клетка-хозяин обладает способностью экспрессировать и/или продуцировать представляющий интерес белок на более высоком уровне, чем клетка-хозяин дикого типа или измененная родительская клетка-хозяин.
В настоящем документе термин «родительская клетка» или «клетка-предшественник» означает клетку, от которой происходит модифицированная клетка-хозяин, причем родительской клеткой или клеткой-предшественником может быть клетка дикого типа или измененная клетка. В некоторых вариантах осуществления изобретения измененная родительская клетка обладает способностью экспрессировать и/или продуцировать представляющий интерес белок на более высоких уровнях, чем неизмененная родительская клетка/клетка-предшественник или родительская клетка/клетка-предшественник дикого типа.
Термин «вводимый», в настоящем документе в отношении введения последовательности нуклеиновой кислоты в клетку, относится к любому методу, подходящему для переноса последовательности нуклеиновой кислоты в клетку. Такими методами введения являются, но не ограничиваются ими, слияние протопластов, трансфекция, трансформация, конъюгирование и трансдукция (см., например, Ferrari et al., "Genetics," in Hardwood et al., (eds.), Bacillus , Plenum Publishing Corp., pages 57-72, [1989]).
В настоящем документе термины «трансформированный» и «стабильно трансформированный» относятся к клетке, которая имеет ненативную (гетерологичную) полинуклеотидную последовательность, интегрированную в ее геном, или гетерологичную полинуклеотидную последовательность, присутствующую в виде эписомной плазмиды, которая сохраняется по меньшей мере в двух последующих генерациях.
В настоящем документе термины «трансформирующая ДНК/трансформирующий полинуклеотид», «трансформирующая последовательность» и «ДНК-конструкция/полинуклеотидная конструкция» означают ДНК, используемую для введения последовательностей в клетку-хозяин или в организм-хозяин. «Трансформирующая ДНК» представляет собой ДНК, используемую для введения последовательностей в клетку-хозяин или в организм-хозяин. ДНК может быть получена in vitro с помощью ПЦР или любыми другими подходящими методами. В некоторых предпочтительных вариантах осуществления изобретения трансформирующая ДНК включает встраивающуюся последовательность, а в других предпочтительных вариантах осуществления изобретения, она также включает встраивающуюся последовательность, фланкированную гомологичными боксами. В еще одном варианте осуществления изобретения трансформирующая ДНК включает другие негомологичные последовательности, присоединенные по концам (то есть «лишние последовательности» или фланги). В некоторых вариантах изобретения концы замыкают так, чтобы трансформирующая ДНК образовывала замкнутый цикл, такой как, например, вставка в вектор.
В настоящем документе термин «нуклеотидная последовательность, кодирующая селективный маркер» означает нуклеотидную последовательность, которая обладает способностью экспрессироваться в клетках-хозяевах, где экспрессия селективного маркера сообщает клеткам, содержащим экспрессируемый ген, способность расти в присутствии соответствующего селективного агента или в отсутствии основного питательного вещества.
В настоящем документе термин «селектируемый маркер» означает ген, способный экспрессироваться в клетках-хозяевах, что облегчает отбор этих хозяев, содержащих данный вектор. Примерами таких селектируемых маркеров являются, но не ограничиваются ими, противомикробные средства (например, канамицин, эритромицин, актиномицин, хлорамфеникол и тетрациклин). Таким образом, термин «селектируемый маркер» означает гены, которые указывают на поглощение данной клеткой-хозяином встраивающейся и представляющей интерес ДНК или на осуществление некоторых других реакций. Обычно селектируемые маркеры представляют собой гены, которые сообщают клеткам-хозяевам резистентность к микробам или метаболическое преимущество, что позволяет дифференцировать клетки, содержащие экзогенную ДНК, от клеток, которые не приобретали какой-либо экзогенной последовательности в процессе трансформации. Термин «эндогенный селектируемый маркер» означает маркер, локализованный на геноме трансформируемого микроорганизма. Эндогенный селектируемый маркер кодирует ген, который отличается от гена селектируемого маркера, присутствующего на трансформирующей ДНК-конструкции.
В настоящем документе термины «амплификация» и «амплификация гена» означают процесс, благодаря которому конкретные последовательности ДНК непропорционально реплицируются, в результате чего амплифицированный ген присутствует в геноме с большим числом копий, чем в исходном состоянии. В некоторых вариантах изобретения отбор клеток путем их культивирования в присутствии лекарственного средства (например, ингибитора ингибируемого фермента) приводит к амплификации любого эндогенного гена, кодирующего генный продукт, необходимый для роста клеток в присутствии лекарственного средства, или к амплификации экзогенных (то есть встраиваемых) последовательностей, кодирующих этот генный продукт, или к той и другой амплификации. Отбор клеток путем культивирования в присутствии лекарственного средства (например, ингибитора ингибируемого фермента) может приводить к амплификации любого эндогенного гена, кодирующего генный продукт, необходимый для роста клеток в присутствии лекарственного средства, или к амплификации экзогенных (то есть, встраиваемых) последовательностей, кодирующих этот генный продукт, или к той и другой амплификации.
В настоящем документе термин «полипептид» означает соединение, состоящее из аминокислотных остатков, связанных пептидными связями. В некоторых вариантах изобретения в настоящем документе термин «белок» является синонимом термину «полипептид». В некоторых альтернативных вариантах изобретения этот термин означает комплекс из двух или более полипептидов. Таким образом, в настоящем документе термины «белок» и «полипептид» являются взаимозаменяемыми.
В настоящем документе термины «белок YmaH» и «белок Hfq» являются взаимозаменяемыми и означают белок, который усиливает экспрессию представляющего интерес белка. В контексте настоящего описания «белок YmaH» означает белок YmaH дикого типа и его варианты, включая ортологи.
В настоящем документе термин «вариант» означает белок, образующийся от белка-предшественника (например, белка YmaH B. subtilis) в результате присоединения одной или нескольких аминокислот к С-концу или к N-концу или к обоим концам, замены одной или нескольких аминокислот в одном или в нескольких различных сайтах аминокислотной последовательности, делеции одной или нескольких аминокислот у любого конца белка или у обоих концов белка, или в одном или нескольких сайтах аминокислотной последовательности, и/или инсерции одной или нескольких аминокислот в одном или нескольких сайтов аминокислотной последовательности. Термин «белок YmaH B. subtilis» означает белок YmaH B. subtilis, модифицированный как описано ниже. Продуцирование варианта белка YmaH B. subtilis предпочтительно достигается путем модификации последовательности ДНК, кодирующей нативный белок, переноса такой последовательности ДНК в подходящего хозяина и экспрессии модифицированной последовательности ДНК с образованием дериватизированного фермента. Вариантами белков YmaH B. subtilis являются пептиды, имеющие аминокислотные последовательности, которые отличаются от аминокислотной последовательности фермента-предшественника, где указанный вариант белка YmaH B. subtilis сохраняет способность усиливать продуцирование представляющего интерес белка в клетках B. subtilis, в которых сверхэкспрессируется белок YmaH. Активность данного варианта может быть повышенной или пониженной по отношению к фактору секреции предшественника. При этом считается, что варианты согласно изобретению могут происходить от ДНК-фрагмента, кодирующего вариант белка YmaH B. subtilis, причем экспрессированный вариант белка YmaH B. subtilis сохраняет свою функциональную активность.
Термины «представляющий интерес белок» и «представляющий интерес полипептид» означает белок/полипептид, продуцируемый клеткой-хозяином. Обычно представляющими интерес белками являются нужные белки, имеющие коммерческую ценность. Представляющий интерес белок может быть гомологичным или гетерологичным по отношению к хозяину. В некоторых вариантах осуществления изобретения представляющим интерес белком является секретированный полипептид, а в частности, фермент, включая, но не ограничиваясь ими, амилолитические ферменты, протеолитические ферменты, целлюлитические ферменты, оксидоредуктазные ферменты и ферменты, разрушающие стенки растений. В других вариантах осуществления изобретения такими ферментами являются, но не ограничиваются ими, амилазы, протеазы, ксиланазы, липазы, лакказы, фенолоксидазы, оксидазы, кутиназы, целлюлазы, гемицеллюлазы, эстеразы, пероксидазы, каталазы, глюкозо-оксидазы, фитазы, пектиназы, глюкозидазы, изомеразы, трансферазы, галактозидазы и хитиназы. В других вариантах изобретения экспрессируемым полипептидом являются гормон, цитокин, фактор роста, рецептор, вакцина, антитело или т.п. Следует отметить, что настоящее изобретение не ограничивается какими-либо конкретными белками/полипептидами, и в некоторых наиболее предпочтительных вариантах изобретения, представляющим интерес экспрессируемым белком является протеаза.
В настоящем документе термин «гетерологичный белок» означает белок или полипептид, который обычно не присутствует в клетке-хозяине. Примерами гетерологичных белков являются ферменты, такие как гидролазы, включая протеазы, целлюлазы, амилазы, другие карбогидразы и липазы; изомеразы, такие как рацемазы, эпимеразы, таутомеразы или мутазы; трансферазы, киназы фосфотаз. В некоторых вариантах изобретения указанными белками являются терапевтически ценные белки или пептиды, включая, но не ограничиваясь ими, факторы роста, цитокины, лиганды, рецепторы и ингибиторы, а также вакцины и антитела. В некоторых альтернативных вариантах осуществления изобретения указанным белком являются коммерчески доступный и представляющий промышленный интерес белок или пептид (например, протеазы, карбогидразы, такие как амилазы и глюкоамилазы, целлюлазы, оксидазы и липазы). В некоторых вариантах изобретения генами, кодирующими белки, являются природные гены, а в других вариантах изобретения используются мутированные и/или синтезированные гены. В некоторых вариантах изобретения генами, кодирующими белки, являются природные гены, а в других вариантах изобретения используются мутированные и/или синтезированные гены.
В настоящем документе термин «гомологичный белок» означает нативный или природный белок или полипептид либо белок или полипептид, присутствующий в клетке-хозяине. Настоящее изобретение охватывает клетки-хозяева, продуцирующие гомологичный белок и полученные методами рекомбинантных ДНК. В альтернативных вариантах осуществления изобретения гомологичным белком является нативный белок, продуцируемый другими микроорганизмами, включая, но не ограничиваясь ими, E. coli. Настоящее изобретение охватывает клетки-хозяева, продуцирующие гомологичный белок в результате применения техники рекомбинантных ДНК. Настоящее изобретение также охватывает клетки-хозяева, в которых имеются одна или несколько делеций или один или несколько разрывов в нуклеиновых кислотах, кодирующих природный(е) гомологичный(е) белок(белки) (например, протеазу), и в которых присутствует нуклеиновая кислота, кодирующая повторяющийся(иеся) гомологичный(е) белок(белки) в рекомбинантной форме (то есть в экспрессионном кластере). В других вариантах изобретения клетка-хозяин продуцирует гомологичный белок.
В настоящем документе термины «протеаза» и «протеолитическая активность» относятся к белку или пептиду, обладающему способностью гидролизовать пептиды или субстраты, имеющие пептидные связи. Для определения протеолитической активности существует множество хорошо известных методов (Kalisz, "Microbial Proteinases," In: Fiechter (ed.), Advances in Biochemical Engineering/Biotechnology, [1988]). Так, например, протеолитическая активность может быть определена путем проведения сравнительных анализов, которые позволяют проанализировать соответствующую способность протеазы гидролизовать коммерчески доступный субстрат. Репрезентативными субстратами, которые могут быть использованы в таком анализе на протеазную или протеолитическую активность, являются, но не ограничиваются ими, диметилказеин (Sigma C-9801), бычий коллаген (Sigma C-9879), бычий эластин (Sigma E-1625) и бычий кератин (ICN Biomedical 902111). Колориметрические анализы, в которых используются такие субстраты, хорошо известны специалистам (см., например, заявку WO 99/34011 и патент США № 6376450, которые вводятся в настоящее изобретение посредством ссылки). Анализ AAPF (см., например, Del Mar et al., Anal Biochem, 99:316-320, [1979]) также применяется для определения уровня продуцирования зрелой протеазы. В этом анализе измеряют скорость высвобождения п-нитроанилина, поскольку указанный фермент гидролизует растворимый синтетический субстрат, то есть сукцинил-аланин-аланин-пролин-фенилаланин-п-нитроанилид (sAAPF-pNA). Скорость продуцирования желтой окраски после реакции гидролиза измеряют на спектрофотометре на длине волны 410 нм, и такая скорость пропорциональна концентрации активного фермента.
В настоящем документе термин «активность» означает биологическую активность, ассоциированную с конкретным белком, такую как протеолитическая активность, ассоциированная с протеазой. Термин «биологическая активность» означает любую активность, которой обычно, как предполагается специалистом, обладает данный белок.
Термин «продуцирование», если он относится к представляющему интерес белку, охватывает стадии процессинга при продуцировании полипептидов, включая удаление про-области, что обычно приводит к образованию активной зрелой формы полипептида, которая, как известно, образуется в процессе созревания. В некоторых вариантах изобретения продуцирование полипептида включает удаление сигнального пептида, что, как известно, происходит в процессе секреции белка (см, например, Wang et al., Biochemistry 37:3165-3171 (1998); и Power et al., Proc. Natl. Acad. Sci. USA 83:3096-3100 [1986]). В некоторых вариантах осуществления изобретения экспрессированный белок присутствует во внутриклеточной среде, в которой он экспрессируется, а в других вариантах осуществления изобретения экспрессированный белок секретируется во внеклеточное пространство. Таким образом, В некоторых вариантах изобретения продуцирование представляющего интерес белка включает осуществления экспрессии белка в клетках и его секрецию во внеклеточную среду. Так, например, продуцирование протеазы охватывает две стадии процессинга полноразмерной протеазы, включая: 1) удаление сигнального пептида, которое, как известно, происходит во время секреции белка и 2) удаление про-области, которое, как известно, приводит к образованию активной зрелой формы фермента и которое, как известно, происходит в процессе созревания (Wang et al., Biochemistry 37:3165-3171 (1998); Power et al., Proc Natl Acad Sci USA 83:3096-3100 [1986]).
В настоящем документе термин «ранняя экспрессия и/или раннее продуцирование» означает, что экспрессии и/или продуцирование представляющего интерес белка в клетке-хозяине происходит раньше, чем это обычно наблюдается в предшественнике/хозяине-родителе. В некоторых вариантах изобретения «ранняя экспрессия и/или раннее продуцирование» представляющего интерес белка в хозяине, в котором наблюдается сверхэкспрессия YmaH, происходит раньше, чем в хозяине, в котором не происходит сверхэкспрессии ymaH.
В настоящем документе термин «усиление» относится к повышенному уровню продуцирования представляющих интерес белков. В своих предпочтительных вариантах, настоящее изобретение относится к усилению (то есть к повышению уровня) продуцирования представляющего интерес белка в модифицированном хозяине. В этих вариантах изобретения термин «усиление» продуцирования означает повышение уровня продуцирования по сравнению с нормальными уровнями продуцирования немодифицированным хозяином дикого типа или модифицированным хозяином-родителем (например, клетками дикого типа или модифицированными клетками, которые не сверхэкспрессируют активатор транскрипции, такой как YmaH).
Полипептиды YmaH и полинуклеотидные конструкции, кодирующие эти полипептиды
В некоторых вариантах осуществления настоящее изобретение относится к полинуклеотидным конструкциям, содержащим промотор и полинуклеотидную последовательность, кодирующую белок YmaH. YmaH B. subtilis, также известный как HFQ_BACSU, представляет собой РНК-связывающий белок, являющийся членом Hfq-семейства РНК-связывающих белков (Sauter et al., Nucleic Acid Res 31:4091-4098, [2003]). Белок YmaH кодируется в Bacillus subtilis геном ymaH, который представляет собой ортолог гена hfq E.coli (Silvaggi et al., J Bacteriol. 187(19): 6641-6650, [2005]). YmaH представляет собой часто встречающийся и повсеместно распространенный РНК-связывающий белок, который действует как плейотропный регулятор метаболизма РНК у прокариотов и является необходимым для стабилизации некоторых транскриптов и разложения других компонентов. YmaH связывается преимущественно с неструктурированными A/U-богатыми последовательностями РНК и, по своей последовательности и структуре, аналогичен эукариотическим белкам Sm. Также известно, что YmaH связывается с небольшими молекулами РНК, называемыми риборегуляторами, которые повышают стабильность или эффективность трансляции РНК-транскриптов.
Настоящее изобретение относится к способам и композициям, применяемым для осуществления сверхэкспрессии ymaH, где указанные способы и композиции способствуют повышению уровня продуцирования представляющего интерес белка в клетках-хозяевах, которые были модифицированы так, чтобы они сверхэкспрессировали ymaH. Кроме того, как указывается в описании настоящей заявки, сверхэкспрессия ymaH способствует повышению уровня продуцирования протеазы в клетках-хозяевах Bacillus. Сверхэкспрессия ymaH может быть достигнута различными методами, включая повышение уровня транскрипции и/или трансляции YmaH-кодирующего полинуклеотида. Так, например, на уровне транскрипции, сверхэкспрессия ymaH может быть достигнута путем увеличения числа полинуклеотидных последовательностей, кодирующих ymaH в клетках-хозяевах и/или увеличения силы связывания промотора ymaH для повышения активности когнатной РНК-полимеразы. На уровне трансляции сверхэкспрессия ymaH может быть достигнута путем повышения трансляционной активности посредством мутации сайта связывания с рибосомой (RBS) в целях повышения аффинности рибосом к RBS. Для специалиста в данной области очевидно, что сверхэкспрессия ymaH может быть достигнута путем увеличения числа копий гена ymaH, взятого отдельно или в комбинации с другими возможными модификациями, введенными в ген ymaH в целях достижения сверхэкспрессии ymaH.
Настоящее изобретение относится к композициям, содержащим полинуклеотидные конструкции, векторы и клетки-хозяева, способные сверхэкспрессировать ymaH. Настоящее изобретение также относится к способам применения композиций согласно изобретению в целях сверхэкспрессии представляющего интерес белка. Полинуклеотидные конструкции согласно изобретению содержат полинуклеотидые последовательности, кодирующие белок YmaH и промотор SigA и/или SigH.
В одном из осуществлений настоящее изобретение относится к сверхэкспрессии ymaH путем увеличения числа полинуклеотидных последовательностей, кодирующих ymaH. Так, например, настоящее изобретение относится к полинуклеотидным конструкциям, содержащим полинуклеотидую последовательность, кодирующую ymaH, функционально связанный с промотором ymaH. Промотором ymaH может быть любой промотор, инициирующий экспрессию ymaH (например, промотор SigA и/или SigH), и любая последовательность нуклеиновой кислоты, которая обладает транскрипционной активностью в выбранной клетке-хозяине, а также мутантные, усеченные и гибридные промоторы, которые могут быть получены от генов, кодирующих внеклеточные или внутриклеточные полипептиды, которые являются либо гомологичными, либо гетерологичными по отношению к клеткам-хозяевам. Промоторная последовательность может быть нативной или чужеродной для клеток-хозяев.
В некоторых вариантах осуществления изобретения промоторная последовательность может быть получена от бактериального источника. В некоторых вариантах осуществления изобретения промоторная последовательность может быть получена от грамположительных бактерий, таких как штамм Bacillus (например, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis, или Bacillus thuringiensis); или штамм Streptomyces (например, Streptomyces lividans или Streptomyces murinus); или от грамотрицательных бактерий (например, E. coli или Pseudomonas sp.).
Транскрипция ymaH обычно запускается двумя промоторами: промотором SigA, который расположен выше miaA-кодирующей области, и промотором SigH, который расположен выше сразу за ymaH-кодирующей областью в опероне miaA B. subtilis. В некоторых своих вариантах настоящее изобретение относится к полинуклеотидным конструкциям, содержащим полинуклеотидную последовательность, кодирующую YmaH и промотор SigA (например, SEQ ID NO:2 и 3). В SEQ ID NO:2 и 3 представлены варианты, в которых ymaH-кодирующая последовательность является смежной с последовательностью промотора SigA и образует химерную полинуклеотидную конструкцию. В некоторых предпочтительных вариантах изобретения химерные полинуклеотидные конструкции содержат промоторную последовательность, которая по своей природе не является смежной с ymaH-кодирующей последовательностью. Так, например, в SEQ ID NOS:2 и 3 представлены химерные конструкции SigA1 и SigA2 соответственно, где каждая из этих конструкций содержит промотор SigA, функционально связанный с полинуклеотидной последовательностью, кодирующей ymaH.
GCGCCGAATTCTCATACCCTGAAAGGAAAGACAAGGGAAATTGTCGGCAATGAGCCGCTC GGCAGGTAGAAGGATGTTTACCGATGCAAAAAAAGGGCAAAATGGATAGGTGGTTGTCCA TGTTGAATGCTATAATGGGGGAGATTTATAAAAGAGAGTGATACATATTGAATAATACGAAG CAGCCCCACACATATAGCAGGAAAACTCGAACTTTAATCGAAACTGTATGATATAGAGAAT CAAGGAGGACGAAACATGAAACCGATTAATATTCAGGATCAGTTTTTGAATCAAATCCGGA AAGAAAATACGTATGTCACTGTTTTTTTGCTGAACGGCTTTCAGTTGCGGGGCCAGGTGAA AGGCTTTGATAACTTTACCGTATTGTTGGAATCGGAAGGTAAGCAGCAGCTTATATATAAAC ATGCGATCTCAACGTTTGCGCCGCAAAAAAACGTCCAGCTTGAACTCGAATAGATCAAAAA ATGCCATGTCAAGACATGAGGAAAGGCTGTCGGGGGTTCCCGGCGGCCATTTTTAACATG AATCCACTTTTGCTCCAAGCTTTTTGTGTAAGCTGACCATGCCAAGGCACGGTCTTTTTTTA TGAGGGATCCGGTGCC (SEQ ID NO:2)
GCGCCGAATTCTCATACCCTGAAAGGAAAGACAAGGGAAATTGTCGGCAATGAGCCGCTC
GGCAGGTAGAAGGATGTTTACCGATGCAAAAAAAGGGCAAAATGGATAGGTGGTTGTCCA TGTTGAATGCTATAATGGGGGAGATTTATAAAAGAGAGTGCTCGAACTTTAATCGAAACTG TATGATATAGAGAATCAAGGAGGACGAAACATGAAACCGATTAATATTCAGGATCAGTTTTT
GAATCAAATCCGGAAAGAAAATACGTATGTCACTGTTTTTTTGCTGAACGGCTTTCAGTTGC
GGGGCCAGGTGAAAGGCTTTGATAACTTTACCGTATTGTTGGAATCGGAAGGTAAGCAGC
AGCTTATATATAAACATGCGATCTCAACGTTTGCGCCGCAAAAAAACGTCCAGCTTGAACT CGAATAGATCAAAAAATGCCATGTCAAGACATGAGGAAAGGCTGTCGGGGGTTCCCGGCG GCCATTTTTAACATGAATCCACTTTTGCTCCAAGCTTTTTGTGTAAGCTGACCATGCCAAGG CACGGTCTTTTTTTATGAGGGATCCGGTGCC (SEQ ID NO:3)
В другом варианте осуществления настоящее изобретение относится к полинуклеотидным конструкциям, содержащим полинуклеотидную последовательность, которая кодирует YmaH и промотор SigA (например, SigH-конструкцию SEQ ID NO:1, указанную ниже). В SEQ ID NO:1 также представлена полинуклеотидная конструкция, содержащая ymaH-кодирующую последовательность, которая по своей природе является смежной с промотором SigH.
GGCACCGAATTCGACGTGGTTTCGCAACAAAATGCAGGTCACATGGTTCGATATGACACC GCCTGTTGATATGGAGCTGAAAAAAAAGGAAATTTTCACACATATAGCAGGAAAACTCGAA CTTTAATCGAAACTGTATGATATAGAGAATCAAGGAGGACGAAACATGAAACCGATTAATAT TCAGGATCAGTTTTTGAATCAAATCCGGAAAGAAAATACGTATGTCACTGTTTTTTTGCTGA ACGGCTTTCAGTTGCGGGGCCAGGTGAAAGGCTTTGATAACTTTACCGTATTGTTGGAATC GGAAGGTAAGCAGCAGCTTATATATAAACATGCGATCTCAACGTTTGCGCCGCAAAAAAAC GTCCAGCTTGAACTCGAATAGATCAAAAAATGCCATGTCAAGACATGAGGAAAGGCTGTCG GGGGTTCCCGGCGGCCATTTTTAACATGAATCCACTTTTGCTCCAAGCTTTTTGTGTAAGC TGACCATGCCAAGGCACGGTCTTTTTTTATGAGGGATCCGGAGCC (SEQ ID NO: 1)
В другом варианте осуществления настоящее изобретение относится к полинуклеотидным конструкциям, содержащим полинуклеотидную YmaH-кодирующую последовательность и промоторы SigA и SigH (например, конструкции SigA3 SEQ ID NO: 13, представленной ниже).
TCATACCCTGAAAGGAAAGACAAGGGAAATTGTCGGCAATGAGCCGCTCGGCAGGTAGAA GGATGTTTACCGATGCAAAAAAAGGGCAAAATGGATAGGTGGTTGTCCATGTTGAATGCTA TAATGGGGGAGATTTATAAAAGAGAGTGATACATATTGAATAATACGAAGCAGCCCGTTGT CATTTTAGTCGGACCGACGGCAGTGGGGAAAACCAATTTAAGTATTCAGCTAGCCAAATCC TTAAACGCGGAAATTATCAGCGGAGATTCGATGCAGATTTATAAAGGGATGGATATTGGAA CAGCTAAAATTACCGAACAGGAGATGGAGGGAGTGCCCCATCATCTGATTGACATTTTAGA TCCCCAAGACTCTTTCTCTACTGCCGATTATCAAAGCTTAGTAAGAAATAAAATCAGCGAGA TTGCAAATAGAGGAAAGCTTCCGATGATTGACGGCGGTACAGGGCTTTATATACAATCTGA GCTTTACGATTATACATTTACGGAAGAGGCAAATGATCCCGTGTTTCGAGAGAGCATGCAA ATGGCTGCTGAGCGGGAAGGCGCTGACTTTCTTCATGCCAAACTTGCTGCAGCAGATCCC GAGGCAGCAGCTGCGATTCATCCGAATAATACAAGAAGAGTCATTCGCGCACTGGAAATTT TACATACGTCCGGAAAAACGATGTCCCAGCATTTGAAGGAACAAAAACGAGAACTTCTGTA CAATGCAGTGTTAATTGGCCTGACAATGGATAGAGACACGCTTTACGAAAGAATTAATCAG CGGGTCGATTTGATGATGCAGTCAGGCCTTCTTCCGGAAGTGAAACGCTTATACGACAAG AACGTGAGAGACTGTCAATCAATACAGGCGATAGGCTATAAAGAGCTGTATGCATATTTTG ACGGTTTTGTGACACTTTCCGATGCTGTCGAACAGCTAAAGCAGAACTCGAGGCGGTATG CGAAACGCCAGCTGACGTGGTTTCGCAACAAAATGCAGGTCACATGGTTCGATATGACAC CGCCTGTTGATATGGAGCTGAAAAAAAAGGAAATTTTCACACATATAGCAGGAAAACTCGA ACTTTAATCGAAACTGTATGATATAGAGAATCAAGGAGGACGAAACATGAAACCGATTAATA TTCAGGATCAGTTTTTGAATCAAATCCGGAAAGAAAATACGTATGTCACTGTTTTTTTGCTG AACGGCTTTCAGTTGCGGGGCCAGGTGAAAGGCTTTGATAACTTTACCGTATTGTTGGAAT CGGAAGGTAAGCAGCAGCTTATATATAAACATGCGATCTCAACGTTTGCGCCGCAAAAAAA CGTCCAGCTTGAACTCGAATAGATCAAAAAATGCCATGTCAAGACATGAGGAAAGGCTGTC GGGGGTTCCCGGCGGCCATTTTTAACATGAATCCACTTTTGCTCCAAGCTTTTTGTGTAAG CTGACCATGCCAAGGCACGGTCTTTTTTTATGAG (SEQ ID NO: 13)
Примерами подходящих промоторов для регуляции экспрессии гена ymaH являются промоторы SigA и SigH, происходящие от оперона B. subtilis, который охватывает ген, кодирующий miaA. Так, например, в одном из своих вариантов, настоящее изобретение относится к полинуклеотидной последовательности, определяющей промотор SigA (SEQ ID NO: 14, представленной ниже).
TCATACCCTGAAAGGAAAGACAAGGGAAATTGTCGGCAATGAGCCGCTCGGCAGGTAGAA GGATGTTTACCGATGCAAAAAAAGGGCAAAATGGATAGGTGGTTGTCCATGTTGAATGCTA TAATGGGGGAGATTTATAAAAGAGAGTGATACATA (SEQ ID NO:14)
В другом варианте настоящее изобретение относится к полинуклеотидной последовательности, определяющей промотор SigH (SEQ ID NO: 16, представленной ниже).
AAAGGAAATTTTCACACATATAGCAGGAAAACTCGAACTTTAATCGAAACTGTATGATATAG
AGAATCAAGGAGGACGAAAC (SEQ ID NO:16)
Другими примерами промоторов, которые могут быть использованы для экспрессии гена ymaH, являются промоторы Sigma A, распознаваемые фактором σА, включая промотор гена агаразы Streptomyces coelicolor (dagA), промотор гена щелочной протеазы Bacillus lentus (aprH), промотор гена щелочной протеазы Bacillus licheniformis (ген субтилизина Карлсберга), промотор гена левансахаразы Bacillus subtilis (sacB), промотор гена альфа-амилазы Bacillus subtilis (amyE), промотор гена альфа-амилазы Bacillus licheniformis (amyL), промотор гена мальтогенной амилазы Bacillus stearothermophilus (amyM), и промотор гена альфа-амилазы Bacillus amtyloliquefacietis (amyQ). Примерами промоторов, которые могут быть использованы для экспрессии гена ymaH, являются промоторы Sigma H, распознаваемые факторами σH, включая spoOA, spoOF, spoVG и citG (см., Helmann, J. D. and C. P. Moran. 2002. RNA polymerase and sigma factors, pp.289-312 in A. L. Sonenshein, J. A. Hoch and R. Losick (ed), Bacillus subtilis and its closest relatives: from genes to cells. American Society for Microbiology, Washington, D.C.).
В некоторых вариантах осуществления настоящее изобретение относится к применению консенсусных промоторов SigA и/или SigH. Конструирование консенсусного промотора может быть осуществлено методом сайт-направленного мутагенеза с получением промотора, конформация которого более точно соответствует конформации известных консенсусных последовательностей для "-10" и "-35"-областей вегетативных промоторов «типа SigmaA» Bacillus subtilis (Voskuil et at., Mol. Microbiol 17: 271 279 [1995]). В других вариантах изобретения консенсусный промотор конструируют методом сайт-направленного мутагенеза с получением промотора, конформация которого более точно соответствует конформации известных консенсусных последовательностей для "-10" и "-35"-областей вегетативных промоторов «типа Sigma Н» Bacillus subtilis (см., публикации Helman и Moran in Bacillus subtilis и наиболее близкородственные публикации, Ch.21, pg289-312; Sonenshein et al (2002 ASM Press, Washington, D.C.). Консенсусной последовательностью для "-35"-области промотора типа sigma A является TTGaca, для "-10"-области - tgnTATaat, а консенсусной последовательностью для «-35»-области промотора sigma H является последовательность RnAGGAwWW, и для «-10»-области - RnnGAAT. Заглавными буквами обозначены высококонсервативные положения; строчными буками обозначены менее консервативные положения; аббревиатура R может означать A или G, а W может означать A или T. Консенсусная последовательность промотора может быть получена для любого промотора, который может функционировать в клетке-хозяине Bacillus.
В некоторых вариантах осуществления изобретения промотор SigA, который охватывает SEQ ID NO: 14, определяется полинуклеотидной последовательностью, которая обычно расположена выше miaA-кодирующей последовательностью (SEQ ID NO: 15, представленной ниже), а промотор SigH, который охватывает SEQ ID NO: 16, определяется полинуклеотидной последовательностью, которая обычно расположена выше ymaH-кодирующей области (SEQ ID NO:17, представленной ниже).
TTGAATAATACGAAGCAGCCCGTTGTCATTTTAGTCGGACCGACGGCAGTGGGGAAAACC AATTTAAGTATTCAGCTAGCCAAATCCTTAAACGCGGAAATTATCAGCGGAGATTCGATGC AGATTTATAAAGGGATGGATATTGGAACAGCTAAAATTACCGAACAGGAGATGGAGGGAGT GCCCCATCATCTGATTGACATTTTAGATCCCCAAGACTCTTTCTCTACTGCCGATTATCAAA GCTTAGTAAGAAATAAAATCAGCGAGATTGCAAATAGAGGAAAGCTTCCGATGATTGACGG CGGTACAGGGCTTTATATACAATCTGAGCTTTACGATTATACATTTACGGAAGAGGCAAAT GATCCCGTGTTTCGAGAGAGCATGCAAATGGCTGCTGAGCGGGAAGGCGCTGACTTTCTT CATGCCAAACTTGCTGCAGCAGATCCCGAGGCAGCAGCTGCGATTCATCCGAATAATACA AGAAGAGTCATTCGCGCACTGGAAATTTTACATACGTCCGGAAAAACGATGTCCCAGCATT TGAAGGAACAAAAACGAGAACTTCTGTACAATGCAGTGTTAATTGGCCTGACAATGGATAG AGACACGCTTTACGAAAGAATTAATCAGCGGGTCGATTTGATGATGCAGTCAGGCCTTCTT CCGGAAGTGAAACGCTTATACGACAAGAACGTGAGAGACTGTCAATCAATACAGGCGATA GGCTATAAAGAGCTGTATGCATATTTTGACGGTTTTGTGACACTTTCCGATGCTGTCGAAC AGCTAAAGCAGAACTCGAGGCGGTATGCGAAACGCCAGCTGACGTGGTTTCGCAACAAAA TGCAGGTCACATGGTTCGATATGACACCGCCTGTTGATATGGAGCTGAAAAAAAAGGAAAT TTTCACACATATAGCAGGAAAACTCGAACTTTAA (SEQ ID NO: 15)
ATGAAACCGATTAATATTCAGGATCAGTTTTTGAATCAAATCCGGAAAGAAAATACGTATGT CACTGTTTTTTTGCTGAACGGCTTTCAGTTGCGGGGCCAGGTGAAAGGCTTTGATAACTTT ACCGTATTGTTGGAATCGGAAGGTAAGCAGCAGCTTATATATAAACATGCGATCTCAACGT TTGCGCCGCAAAAAAACGTCCAGCTTGAACTCGAATAG (SEQ ID NO: 17)
Настоящее изобретение также охватывает промоторные последовательности, которые были мутированы для повышения активности промотора по сравнению с активностью соответствующего промотора дикого типа, и тем самым для сверхэкспрессии белка YmaH. Таким образом, следует отметить, что в конструкциях согласно изобретению могут быть использованы варианты последовательностей, определяющих промоторы SigA и SigH. Методы конструирования вариантов промоторов в Bacillus sp. хорошо известны специалистам (см., например, Helmann et al., 2002. RNA polymerase and sigma factors, pp289-312 In A. L. Sonenshein, J. A. Hoch and R. Losick (ed), Bacillus subtilis and its closest relatives: from genes to cells. American Society for Microbiology, Washington, D.C.). При этом предусматривается, что настоящее изобретение не ограничивается какими-либо конкретными промоторами, поскольку известно, что в настоящем изобретении может быть использован любой подходящий промотор. Тем не менее, В некоторых вариантах изобретения указанным промотором является промотор sigH B. subtilis, а в других вариантах изобретения указанным промотором является промотор sigA B. subtilis. В других вариантах изобретения промоторы sigH и sigA служат для осуществления сверхэкспрессии белка YmaH.
В некоторых вариантах осуществления изобретения полинуклеотидные конструкции согласно изобретению также содержит необходимый сайт связывания с рибосомой, обеспечивающий оптимальную трансляцию РНК-транскрипта ymaH. В некоторых вариантах изобретения полинуклеотидная конструкция содержит последовательность сайта связывания с рибосомой (RBS) гена miaA (AAGAGAG; SEQ ID NO:21), а в других вариантах изобретения полинуклеотидная конструкция содержит последовательность RBS гена ymaH (GGAGG; SEQ ID NO:22). В других вариантах изобретения полинуклеотидная конструкция содержит последовательности сайта связывания с рибосомой генов miaA и ymaH. В некоторых своих вариантах настоящее изобретение относится к конструкциям, имеющим промотор и последовательности сайта связывания с рибосомой, расположенные выше ymaH-кодирующей последовательности. Настоящее изобретение не ограничивается описанными в настоящем документе последовательностями сайта связывания с рибосомой, поскольку настоящее изобретение также охватывает любые подходящие последовательности сайта связывания с рибосомой, которые были мутированы для повышения уровня экспрессии гена ymaH. Методы получения мутированных последовательностей сайта связывания с рибосомой, повышающих уровень экспрессии гена в Bacillus, известны специалистам. Так, например, Бэндом (Band) и Хеннером (Henner) было успешно осуществлено повышение уровня экспрессии интерферона в B. subtilis путем модификации RBS с получением пары оснований, тесно связанной с рРНК 16S (Band, L. and D. J. Henner, DNA 3:17-21 [1984]).
Природный белок YmaH Bacillus subtilis представляет собой белок из 73 аминокислот (SEQ ID NO:4), кодируемый полинуклеотидом из 219 пар нуклеотидов (222, включая стоп-кодон) (EMBL Primary Accession Number Z99113; SEQ ID NO:17).
MKPINIQDQFLNQIRKENTYVTVFLLNGFQLRGQVKGFDNFTVLLESEGKQQLIYKHAISTFAPQKNVQLELE (SEQ ID NO:4)
Таким образом в некоторых вариантах изобретения последовательность полинуклеотидной конструкции, кодирующей YmaH, представляет собой природную полинуклеотидную последовательность, присутствующую в геноме штамма 168 Bacillus subtilis дикого типа (SEQ ID NO:4). Настоящее изобретение также охватывает варианты белков YmaH, включая варианты белков YmaH, полученные из белков дикого типа в результате делеции (то есть усечения), добавления или замены одной или нескольких аминокислот в одном или нескольких положениях нативного белка. Методы введения таких делеций, добавлений и замен, в основном известны специалистам. Так, например, варианты аминокислотной последовательности полипептида могут быть получены путем введения мутаций в клонированную последовательность ДНК, кодирующую представляющий интерес нативный белок. Методы мутагенеза и модификаций нуклеотидной последовательности хорошо известны специалистам (см., например, публикации Kunkel (1985) Proc. Natl. Acad. Sci. USA 82:488 492; Kunkel et al. (1987) Methods Enzymol. 154:367 382; патент США 4873192; и цитируемые в них работы, которые вводятся в настоящее описание посредством ссылки). При конструировании вариантов представляющих интерес белков модификации в нуклеотидных последовательностях, кодирующих указанные варианты, должны быть сделаны так, чтобы указанные варианты сохраняли желаемую активность. Как известно специалистам, из-за вырожденности генетического кода, белок YmaH кодируется различными модифицированными полинуклеотидами. В некоторых других своих вариантах настоящее изобретение относится к полинуклеотидам, содержащим нуклеотидную последовательность, которая по меньшей мере примерно на 70%, по меньшей мере примерно на 75%, по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 92%, по меньшей мере примерно на 95%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% идентична полинуклеотидной последовательности SEQ ID NO: 17.
В других вариантах осуществления изобретения полинуклеотидные конструкции согласно изобретению содержат YmaH-кодирующие последовательности, аналогичные YmaH-кодирующей последовательности штамма 168 Bacillus subtilis. Геном этого штамма, составляющий один 4215 т.п.н.-геном, был хорошо охарактеризован (см., Kunst et al., Nature 390:249-256 [1997]; и Henner et al., Microbiol. Rev., 44:57-82 [1980]). В некоторых вариантах изобретения полинуклеотидные конструкции согласно изобретению содержат полинуклеотидные последовательности, кодирующие белок YmaH, аминокислотная последовательность которого по меньшей мере примерно на 65%, по меньшей мере примерно на 70%, по меньшей мере примерно на 75%, по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 92%, по меньшей мере примерно на 95%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% и по меньшей мере примерно на 99% идентична аминокислотной последовательности белка YmaH дикого типа, где указанный белок обладает сравнимой или повышенной способностью усиливать продуцирование представляющего интерес белка в клетках-хозяевах по сравнению с полипептидом дикого типа (SEQ ID NO:4) и сохраняет способность повышать уровень экспрессии представляющего интерес белка в клетках-хозяевах. В других своих вариантах настоящее изобретение относится к полинуклеотидным конструкциям, содержащим полинуклеотидные последовательности, которые являются гомологичными, ортологичными или паралогичными генам, имеющим последовательность Bacillus дикого типа SEQ ID NO:17, и которые сохраняют способность повышать уровень продуцирования представляющего интерес белка.
Настоящее изобретение также охватывает полинуклеотидные конструкции, содержащие последовательности, кодирующие белки YmaH, которые имеют структурное и/или функциональное сходство. В некоторых вариантах осуществления изобретения эти белки происходят от микроорганизмов различного рода и/или вида, включая микроорганизмы различных классов (например, бактериальный белок и белок грибов). В некоторых вариантах изобретения эти белки происходят от микроорганизмов различного рода и/или вида. В дополнительных вариантах изобретения родственные белки происходят от одного и того же вида. Действительно, предусматривается, что настоящее изобретение не ограничивается родственными белками, происходящими от какого-либо (каких-либо) конкретного(ых) источника(ов). Кроме того, термин «родственные белки” охватывает гомологи с третичной структурой и гомологи с первичной последовательностью (например, YmaH согласно изобретению). Так, например, настоящее изобретение охватывает указанные гомологи, включая, но не ограничиваясь ими, белки YmaH, такие как YmaH E. coli, (HFQ_ECOLI), Shighella flexneri (HFQ_SHIFL), Salmonella typhimurium (HFQ_SALTY), Yersinia enterocolitica (HFQ_YEREN), Yersinia pestis (HFQ_YERPE), Erwinia carotovora (HFQ_ERWCA), Haemophilus influenzae (HFQ_HAEIN), Pasteurella multocida (HFQ_PASMU), Vibrio cholerae (HFQ_VIBCH), Pseudomonas aeruginosa (HFQ_PSEAE), Xanthomonas axonopodis (HFQ_XANAC), Xanthomonas campestris (HFQ_XANCP), Xylella fastidiosa (GSQ_XYLFA), Neisseria meningitidis (HFQ_NEIMA), Ralstonia solanacearum (HFQ_RALSO), Agrobacterium tumefaciens (HFQ_AGRTS), Brucella melitensis (HFQ_BRUME), Rhizobium loti (HFQ_RHILO), Azorhizobium caulinodans (HFQ_AZOCA), Caulobacter crescentus (HFQ_CAUCR), Aquifex melitensis (HFQ_AQUAE), Thermotoga maritime (HFQ_THEMA), Clostridium acetobutylicum (HFQ_CLOAB), Clostridium perfringens (HFQ_CLQPE), Bacillus halodurans (HFQ_BACHD), Bacillus subtilis (HFQ_BACSU), Thermoanaerobacter tengcongensis (HFQ_THETN), S. aureaus (Q99UG9) и M. jannasci (Q58830) (Sauter et al., Nucleic Acids Res. 31:4091-4098 [2003]).
Родственные белки (и их производные) включают варианты белков YmaH. В некоторых предпочтительных вариантах осуществления изобретения, варианты белков отличаются от родительского белка и друг от друга небольшим числом аминокислотных остатков. Число отличающихся аминокислотных остатков может составлять 1 или более, а предпочтительно примерно 1, 2, 3, 4, 5, 10, 15, 20, 30, 40, 50 или более аминокислотных остатков. В некоторых предпочтительных вариантах изобретения число различных аминокислот в этих вариантах составляет примерно от 1 до 10. В некоторых особенно предпочтительных вариантах осуществления изобретения родственные белки, а в частности, варианты белков имеют аминокислотные последовательности, идентичные по меньшей мере примерно на 70%, примерно на 75%, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 97%, примерно на 98% или примерно на 99%. Для получения вариантов белков YmaH согласно изобретению могут быть применены различные методы, известные специалистам, включая, но не ограничиваясь ими, сайт-насыщающий мутагенез, сканирующий мутагенез, инсерционный мутагенез, неспецифический мутагенез, сайт-направленный мутагенез и прямая эволюция, а также различные другие рекомбинаторные методы.
Характеризацию белков дикого типа и мутантных белков осуществляют любыми подходящими методами, предпочтительно исходя из оценки представляющих интерес свойств. Так, например, предусматривается, что могут быть использованы белки YmaH, способные повышать уровень продуцирования представляющего интерес белка.
В некоторых вариантах изобретения рекомбинантные полинуклеотиды согласно изобретению включают полинуклеотидные последовательности, которые могут быть оптимизированы по кодонам для экспрессии белка YmaH в используемой клетке-хозяине. Поскольку таблицы кодонов, в которых указана частота встречаемости для каждого кодона, хорошо известны специалистам (см., например, Nakamura et al., Nucl. Acids Res., 28:292 [2000]) или могут быть легко получены, то могут быть легко выбраны такие нуклеиновые кислоты, которые дают соответствующую аминокислотную последовательность экспрессируемого белка. В некоторых вариантах изобретения оптимизированные по кодонам последовательности содержат полинуклеотиды, кодирующие белок YmaH, который по меньшей мере примерно на 70% идентичен последовательности SEQ ID NO:4.
Векторы, содержащие ymaH
Настоящее изобретение относится к векторам, содержащим полинуклеотидные конструкции согласно изобретению. Указанные векторы вводят в клетку-хозяина для осуществления сверхэкспрессии белка YmaH.
В некоторых вариантах изобретения сверхэкспрессия полипептида происходит в результате экспрессии одной или нескольких копий соответствующего YmaH-кодирующего полинуклеотида, присутствующего на многокопийной/реплицирующейся плазмиде, которая была введена в клетку-хозяина. Таким образом, в некоторых своих вариантах настоящее изобретение относится к вектору, содержащему полинуклеотидную конструкцию, введенную в данный вектор. В некоторых вариантах изобретения указанным вектором является многокопийный/реплицирующийся плазмидный вектор, образующий внехромосомный самореплицирующийся генетический элемент, который сверхэкспрессирует YmaH в клетке-хозяине. Обычно, таким вектором является плазмидный вектор, несущий селектируемый маркерный ген, который позволяет легко осуществлять отбор клеток-хозяев, содержащих данную плазмиду. Векторами, которые автономно реплицируются в клетке-хозяине, являются векторы, содержащие ориджин репликации, что позволяет данному вектору автономно реплицироваться в клетке Bacillus. Примерами бактериальных ориджинов репликации являются ориджины репликации плазмид pBR322, pUC19, pACYC177 и pACYC184, обеспечивающие репликацию в E.coli, и плазмид pUB110, pC194, pE194, pTA1060 и pAMβ1, обеспечивающие репликацию в Bacillus. Такой ориджин репликации может иметь мутацию, сообразную данной плазмиде термочувствительность в клетке Bacillus (см., например, Ehrlich, Proceedings of the National Academy of Sciences USA 75:1433 [1978]).
Как указывалось выше, в некоторых вариантах изобретения полинуклеотид, кодирующий белок YmaH, вводят в клетку-хозяина посредством экспрессии вектора, способного реплицироваться в клетке-хозяине. Подходящие плазмиды, способные реплицироваться и интегрироваться в Bacillus, известны специалистам (см., например, публикацию Harwood и Cutting (eds), Molecular Biological Methods for Bacillus , John Wiley & Sons, [1990], а в частности главу 3; подходящими реплицирующимися плазмидами для B. subtilis являются плазмиды, перечисленные на стр. 92).
В некоторых вариантах осуществления изобретения сверхэкспрессия полипептида YmaH происходит в результате экспрессии по меньшей мере одной копии YmaH-кодирующего полинуклеотида, интегрированного в геном клетки-хозяина. Таким образом, в некоторых вариантах осуществления изобретения, если вектор введен в клетку-хозяина, то он интегрируется в ее геном и реплицируется вместе с геномом, в который он был интегрирован. При этом могут быть интегрированы множество копий гена ymaH в нескольких положениях генома клетки-хозяина. Альтернативно, амплифицируемый экспрессионный кластер, несущий последовательность, кодирующую YmaH, и селективный маркер (например, маркер резистентности к противомикробному средству, такой как ген, кодирующий хлорамфеникол-ацетилтрансферазу), может быть интегрирован в геном посредством одного события кроссинговера, а затем амплифицирован путем стимуляции трансформированной клетки-хозяина увеличивающимися концентрациями соответствующего противомикробного средства (например, хлорамфеникола).
В других вариантах осуществления настоящее изобретение относится к полинуклеотидной конструкции, включенной в вектор для интеграции. В некоторых вариантах изобретения полинуклеотидные конструкции согласно изобретению, которые вводят в вектор для интеграции, доставляют в хромосомные последовательности клеток-хозяев Bacillus в целях создания модифицированных клеток-хозяев, содержащих стабильные тандемные агрегаты из множества копий вектора. Полинуклеотидная конструкция, которую включают в вектор для интеграции, обычно содержит селектируемый маркерный ген, который сообщает клеткам резистентность к противомикробному средству и обеспечивает амплификацию интегрированной конструкции ymaH. При этом может происходить тандемная интеграция в один сайт, а также однокопийная и двухсайтовая интеграция. Полинуклеотидная конструкция, независимо от того, включена ли она в вектор или применяется отдельно без плазмидной ДНК, может быть использована для трансформации клеток-хозяев любым подходящим методом, известным специалистам.
Методы введения ДНК в клетки Bacillus, проводимые с использованием плазмидных конструкций и путем переноса плазмид в бактериальные клетки-хозяева, хорошо известны специалистам. В некоторых вариантах изобретения плазмиды выделяют из E.coli и переносят в Bacillus. Однако метод встраивания в микроорганизмы, такие как E.coli, не имеет большого значения, a в некоторых вариантах изобретения ДНК-конструкцию или ДНК-вектор непосредственно вводят в клетку-хозяина Bacillus.
Подходящие методы введения полинуклеотидных последовательностей в клетки Bacillus хорошо известны специалистам (см., например, Ferrari et al., "Genetics," in Harwood et al. (ed.), Bacillus, Plenum Publishing Corp. [1989], pages 57-72; Saunders et al., J. Bacteriol., 157:718-726 [1984]; Hoch et al., J. Bacteriol., 93:1925 -1937 [1967]; Mann et al., Current Microbiol., 13:131-135 [1986]; and Holubova, Folia Microbiol., 30:97 [1985]; Chang et al., Mol. Gen. Genet, 168:11-115 [1979]; Vorobjeva et al., FEMS Microbiol. Lett., 7:261-263 [1980]; Smith et al., Appl. Env. Microbiol., 51:634 [1986]; Fisher et al., Arch. Microbiol., 139:213-217 [1981]; и McDonald, J. Gen. Microbiol., 130:203 [1984]). Действительно, такие методы как трансформация, включая трансформацию и интеграцию протопластов, а также трансдукция и слияние протопластов, являются известными и могут быть использованы в настоящем изобретении. Методы трансформации являются особенно предпочтительными для введения ДНК-конструкции, полученной в соответствии с настоящим изобретением, в клетку-хозяина.
В некоторых вариантах осуществления изобретения трансформацию клеток-хозяев, помимо общеизвестных методов, осуществляют непосредственно (то есть перед введением ДНК-конструкции в клетку-хозяина, эти клетки не подвергают промежуточной амплификации или каким-либо другим процессам). Известными физическими и химическими методами, которые обычно применяются специалистами для введения ДНК в клетку-хозяина, является введение ДНК-конструкции в клетку-хозяина, что позволяет не прибегать к введению этой конструкции в плазмиду или вектор. Такими методами являются, но не ограничиваются ими, электропорация, введение «оголенной» ДНК или липосом и т.п. В дополнительных вариантах изобретения ДНК-конструкции вводят вместе с плазмидой, не встраивая эти конструкции в плазмиду. В других вариантах осуществления изобретения селективный маркер удаляют из модифицированного штамма Bacillus методами, известными специалистам (см., Stahl et al., J. Bacteriol., 158:411-418 [1984]; и Palmeros et al., Gene 247:255 -264 [2000]).
Известными методами, которые применяются специалистами для трансформации Bacillus, являются такие методы, как трансформация с сохранением плазмидного маркера, которая включает поглощение донорной плазмиды компетентными клетками, несущими частично гомологичную плазмиду-резидента (Contente et al., Plasmid 2:555-571 [1979]; Haima et al., Mol. Gen. Genet, 223:185-191 [1990]; Weinrauch et al., J. Bacteriol., 154:1077-1087 [1983]; и Weinrauch et al., J. Bacteriol., 169:1205-1211 [1987]). Этот метод основан на рекомбинации между встраивающейся донорной плазмидой и гомологичной областью «хелперной» плазмиды-резидента по механизму имитации трансформации хромосомы.
Другие методы трансформации, включая трансформацию протопластов, хорошо известны специалистам (см., например, Chang and Cohen, Mol. Gen. Genet, 168:111-115 [1979]; Vorobjeva et al., FEMS Microbiol. Lett., 7:261-263 [1980]; Smith et al., Appl. Env. Microbiol.,51:634 [1986]; Fisher et al., Arch. Microbiol., 139:213-217 [1981]; McDonald [1984] J. Gen. Microbiol., 130:203 [1984]; и Bakhiet et al., 49:577 [1985]). Кроме того, в публикации Mann et al., (Mann et al., Curr. Microbiol., 13:131-135 [1986]) описана трансформация протопластов Bacillus, а в публикации Holubova (Holubova, Microbiol., 30:97 [1985]) описаны методы введения ДНК в протопласты с использованием ДНК-содержащих липосом. В некоторых вариантах осуществления изобретения для того, чтобы узнать, присутствует ли представляющий интерес ген в клетке-хозяине, используют маркерные гены. В некоторых вариантах осуществления изобретения полинуклеотидная последовательность ymaH, содержащаяся в векторе согласно изобретению, кодирует белок YmaH, имеющий SEQ ID NO:4, или его варианты.
В других вариантах осуществления изобретения помимо этих методов, осуществляют прямую трансформацию клеток-хозяев. При «прямой трансформации», перед введением модифицированного полинуклеотида в клетку-хозяина (то есть, Bacillus), эти клетки не подвергают промежуточной стадии амплификации или каким-либо другим процессам. Известными физическими и химическими методами, которые обычно применяются специалистами для введения модифицированного полинуклеотида в клетку-хозяина, является введение модифицированного полинуклеотида в клетку-хозяина, что позволяет не прибегать к введению этого полинуклеотида в плазмиду или вектор. Такими методами введения ДНК в клетки являются, но не ограничиваются ими, использование компетентных клеток, а также применение «искусственных методов», таких как преципитация хлоридом кальция, электропорация и т.п. Таким образом, в соответствии с настоящим изобретением используется «оголенная» ДНК, липосомы и т.п. В других вариантах осуществления изобретения модифицированные полинуклеотиды вводят вместе с плазмидой, не встраивая эти полинуклеотиды в плазмиду. В некоторых вариантах осуществления настоящее изобретение относится к вектору, содержащему полинуклеотид, кодирующий белок YmaH и функционально связанный с промотором sigA (например, SEQ ID NO:2 и 3). В других вариантах изобретения вектор содержит YmaH-кодирующий полинуклеотид, функционально связанный с промотором sigH (например, SEQ ID NO:1). В других вариантах осуществления изобретения указанный вектор содержит полинуклеотидную конструкцию, которая содержит YmaH-кодирующую последовательность, промотор sigA и промотор sigH (например, SEQ ID NO: 13).
В некоторых вариантах осуществления изобретения ymaH сверхэкспрессируется в неинтегрирующемся векторе. В некоторых вариантах изобретения ymaH сверхэкспрессируется в клетке-хозяине, в которой один или несколько хромосомных генов были модифицированы (например, degU) и/или делетированы (например, nprE) из генома Bacillus. В некоторых вариантах изобретения одна или несколько природных хромосомных областей были модифицированы и/или делетированы из соответствующего генома хозяина Bacillus дикого типа. В некоторых своих предпочтительных вариантах настоящее изобретение относится к способам и композициям, применяемым для повышения уровня экспрессии и/или секреции по меньшей мере одного представляющего интерес белка в Bacillus.
Клетки-хозяева, содержащие ymaH
Настоящее изобретение относится к модифицированным клеткам-хозяевам, которые были генетически модифицированы в целях сверхэкспрессии ymaH и которые обладают повышенной способностью продуцировать представляющие интерес белки. В частности, настоящее изобретение относится к модифицированным клеткам-хозяевам грамположительных микроорганизмов, таких как Bacillus sp., которые сверхэкспрессируют ymaH. В некоторых вариантах осуществления изобретения, ymaH сверхэкспрессируется в микроорганизмах дикого типа, а в других вариантах осуществления изобретения YmaH сверхэкспрессируется в модифицированных клетках-хозяевах. В некоторых вариантах осуществления изобретения модифицированная клетка-хозяин обладает способностью продуцировать представляющий интерес белок на более высоком уровне, чем его предшественник дикого типа. В некоторых особенно предпочтительных вариантах осуществления изобретения сверхэкспрессия YmaH в сверхпродуцирующих модифицированных хозяевах-родителях также приводит к повышению уровня продуцирования представляющего интерес белка. В некоторых вариантах осуществления изобретения сверхэкспрессия YmaH в модифицированном хозяине-родителе индуцирует продуцирование представляющего интерес белка за меньший период времени, чем это происходит в соответствующем немодифицированном хозяине-родителе. Сверхэкспрессия ymaH в клетке-хозяине достигается с использованием векторов и конструкций согласно изобретению, описанных в настоящей заявке. Таким образом, в некоторых вариантах осуществления настоящее изобретение относится к модифицированной клетке-хозяину, которую получают путем трансформации клетки-хозяина дикого типа или модифицированной клетки-хозяина вектором, содержащим YmaH-кодирующую последовательность, функционально связанную с промотором sigA и/или sigH. В частности, модифицированные клетки-хозяева согласно изобретению обладают способностью продуцировать представляющий интерес белок, а в некоторых вариантах осуществления изобретения модифицированные клетки-хозяева содержат полинуклеотидные конструкции, кодирующие YmaH (например, SEQ ID NO:1, 2, 3 или 13).
В некоторых своих вариантах настоящее изобретение относится к способам сверхэкспрессии ymaH в клетках-хозяевах для повышения уровня продуцирования представляющего интерес белка. Представляющий интерес белок может быть либо гомологичным, либо гетерологичным по отношению к хозяину. В некоторых вариантах осуществления изобретения представляющим интерес белком является секретированный полипептид, а в частности фермент, включая, но не ограничиваясь ими, амилолитические ферменты, протеолитические ферменты, целлюлитические ферменты, оксидоредуктазные ферменты и ферменты, разрушающие стенки растений. В других вариантах изобретения такими ферментами являются, но не ограничиваются ими, амилазы, протеазы, ксиланазы, липазы, лакказы, фенолоксидазы, оксидазы, кутиназы, целлюлазы, гемицеллюлазы, эстеразы, пероксидазы, каталазы, глюкозо-оксидазы, фитазы, пектиназы, глюкозидазы, изомеразы, трансферазы, киназы фосфотаз, галактозидазы и хитиназы. В других вариантах осуществления изобретения представляющим интерес белком является гормон, цитокин, фактор роста, рецептор, вакцина, антитело или т.п. Следует отметить, что настоящее изобретение не ограничивается какими-либо конкретными белками, а в некоторых вариантах осуществления изобретения представляющим интерес белком является протеаза.
В некоторых вариантах осуществления изобретения клетками-хозяевами являются Bacillus sp., Streptomyces sp., Escherichia sp. или Aspergillus sp. В других вариантах изобретения представляющими интерес белками являются протеазы, продуцируемые клетками-хозяевами рода Bacillus (см., например, патент США № 5264366, патент США № 4760025 и RE 346060). В некоторых вариантах изобретения представляющим интерес штаммом Bacillus является алкалофильный микроорганизм Bacillus. Специалистам известно множество алкалофильных штаммов Bacillus (см., например, патент США № 5217878; и Aunstrup et al., Proc IV IFS: Ferment. Tech. Today, 299-305 [1972]). Другим типом представляющего особый интерес штамма Bacillus является клетка промышленного штамма Bacillus. Примерами промышленных штаммов Bacillus являются, но не ограничиваются ими, B. licheniformis, B. lentus, B. subtilis, B. clausii и B. amyloliquefaciens. В дополнительных вариантах изобретения штамм-хозяин Bacillus выбирают из группы, состоящей из B. licheniformis, B subtilis, B. lentus, B. brevis, B. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. coagulans, B. circulans, B. lautus, B. pumilus, B. thuringiensis и B. megaterium, а также других микроорганизмов рода Bacillus. В предпочтительных вариантах изобретения используются клетки B. subtilis.
В некоторых вариантах осуществления изобретения промышленные штаммы-хозяева выбирают из группы, состоящей из не-рекомбинантных штаммов Bacillus sp., мутантов природного штамма Bacillus и рекомбинантных штаммов-хозяев Bacillus. Предпочтительным штаммом-хозяином является рекомбинантный штамм, где полинуклеотид, кодирующий представляющий интерес полипептид, был предварительно введен в клетку-хозяина. Другим предпочтительным штаммом-хозяином является штамм-хозяин Bacillus subtilis, а в частности рекомбинантный штамм-хозяин Bacillus subtilis. Многие штаммы B. subtilis известны специалистам и являются подходящими для их использования в настоящем изобретении (см., например, 1A6 (ATCC 39085), 168 (1A01), SB19, W23, Ts85, B637, PB1753-PB1758, PB3360, JH642, 1A243 (ATCC 39087), ATCC 21332, ATCC 6051, MI 13, DE100 (ATCC 39094), GX4931, PBT 110 и штамм PEP 211; Hoch et al., Genetics, 73:215-228 [1973]; патент США № 4450235; патент США № 4302544; EP 0134048). B. subtilis широко применяется специалистами в качестве хозяина для осуществления экспрессии (см. Palva et al., Gene, 19:81-87 [1982]; Fahnestock and Fischer, J. Bacteriol., 165:796-804 [1986]; и Wang et al, Gene 69:39-47 [1988]).
Клетками-хозяевами, представляющими особый интерес, являются клетки промышленных штаммов Bacillus, продуцирующих протеазу. Высокая эффективность продуцирования протеазы этими штаммами может быть еще больше увеличена с использованием модифицированных штаммов Bacillus, полученных в соответствии с настоящим изобретением. Особенно предпочтительными хозяевами, подходящими для экспрессии, являются промышленные штаммы Bacillus, продуцирующие протеазу. В некоторых вариантах осуществления изобретения применение этих штаммов согласно изобретению также позволяет еще больше повышать уровень продуцирования протеазы. Как указывалось выше, существует два основных типа протеаз, обычно секретируемых Bacillus sp., а именно нейтральная протеаза (или «металлопротеазы») и щелочная протеаза (или «сериновая» протеаза). Кроме того, как указывалось выше, субтилизин представляет собой предпочтительную сериновую протеазу, используемую в настоящем изобретении. Были идентифицированы и секвенированы субтилизины Bacillus широкого ряда, например субтилизин 168, субтилизин BPN', субтилизин Карлсберга, субтилизин DY, субтилизин 147, субтилизин 309 (см., например, EP 414279 B; WO 89/06279; и Stahl et al., J. Bacterid., 159:811-818 [1984]), субтилизин B. lentus и субтилизин B. clausii (J. C. van der Laan, G. Gerritse, L. J. Mulleners, R. A. van der Hoek and W. J. Quax. Appl Environ Microbiol. 57: 901-909 [1991]).
В некоторых вариантах осуществления настоящего изобретения штаммы-хозяева Bacillus продуцируют мутантные протеазы (например, варианты). В литературе приводится множество примеров модифицированных и стандартных протеаз (см., например, WO 99/20770; WO 99/20726; WO 99/20769; WO 89/06279; RE 34606; патент США № 4914031; патент США № 4980288; патент США № 5208158; патент США № 5310675; патент США № 5336611; патент США № 5399283; патент США № 5441882; патент США № 5482849; патент США № 5631217; патент США № 5665587; патент США № 5700676; патент США № 5741694; патент США № 5858757; патент США № 5880080; патент США № 6197567; и патент США № 6218165.
В некоторых вариантах осуществления изобретения, экспрессия представляющего интерес белка в клетках-хозяевах инициируется промотором aprE гена aprE, из которого обычно транскрибируется субтилизин B. subtilis. Ген aprE транскрибируется фактором sigma A (σА), а его экспрессия в значительной степени регулируется несколькими регуляторами, такими как: DegU/DegS, AbrB, Hpr и SinR (Valle and Ferrari (1989) In: Smith I, Slepecky RA, Setlow P (eds) Regulation of Procaryotic Development. American Society for Microbiology. Washington, DC pp 131-146), при этом был идентифицирован консенсусный промтор Sigma A: TGGGTCTTGACAAATATTATTCCATCTATTACAATAAATTCACAGA (SEQ ID NO:23; US 2003014846; Helman et al., 1995, Nucleic Acid Research, Vol. 24, pp. 2351-2360). В некоторых вариантах осуществления изобретения клетка-хозяин содержит промотор aprE, который представляет собой промотор aprE дикого типа: TGGGTCTACTAAAATATTATTCCATCTATTACAATAAATTCACAGA (SEQ ID NO:24; публикация заявки на патент США № 20030148461).
В других вариантах осуществления изобретения экспрессия представляющего интерес белка клетками-хозяевами инициируется мутантными промоторами aprE B. subtilis. В некоторых вариантах осуществления, настоящее изобретение относится к клеткам-хозяевам Bacillus, содержащим мутантный промотор aprE, функционально связанный с полинуклеотидной последовательностью, кодирующей представляющий интерес белок. Таким образом, настоящее изобретение охватывает клетки-хозяева, экспрессирующие представляющий интерес белок из мутантного промотора aprE. Примером мутантного промотора aprE является мутантный промотор aprE, имеющий последовательность TGGGTCTTGACAAATATTATTCCATCTATTACAATAAATTCACAGA (SEQ ID NO: 25), описанную в публикации заявки на патент США № 20030148461. Любой из описанных в настоящем документе представляющих интерес белков (например субтилизины Bacillus) может транскрибироваться из промотора aprE. В некоторых своих вариантах настоящее изобретение относится к модифицированной клетке-хозяину Bacillus, способной экспрессировать представляющий интерес белок из промотора aprE. В некоторых вариантах изобретения модифицированной клеткой-хозяином является модифицированная клетка-хозяин B. subtilis, способная экспрессировать протеазу под действием промотора aprE. В некоторых вариантах изобретения промотор aprE включает регуляторные элементы промотора aprE и/или транскрипционную лидерную последовательность aprE, а в других вариантах изобретения промотор aprE не включает регуляторные элементы промотора aprE и/или транскрипционную лидерную последовательность aprE.
Помимо промотора aprE настоящее изобретение также охватывает композиции и способы, применяемые для экспрессии представляющего интерес белка клеткой-хозяином, где экспрессия гена, кодирующего представляющий интерес белок, инициируется любым промотором, подходящим для инициации транскрипции представляющего интерес гена, при условии, что указанный промотор содержит транскрипционную лидерную последовательность гена aprE.
В другом варианте осуществления изобретения хозяином Bacillus является Bacillus sp., который содержит мутацию или делецию по меньшей мере в одном из генов degU, degS, degR и/или degQ. Предпочтительной мутацией является мутация в гене degU, а более предпочтительной мутацией является мутация degU(Hy)32 (см. Msadek et al., J. Bacterid., 172:824-834 [1990]; и Olmos et al., Mol. Gen. Genet., 253:562-567 [1997]). Наиболее предпочтительным штаммом-хозяином является бактерия Bacillus subtilis, несущая мутацию degU(Hy) 32. В другом варианте осуществления изобретения хозяин Bacillus содержит мутацию или делецию в scoC4 (см. Caldwell et al., J. Bacteriol., 183:7329-7340 [2001]); и spollE (See, Arigoni et al., Mol. Microbiol., 31:1407-1415 [1999]). В некоторых вариантах изобретения такие мутации являются одиночными, а в других вариантах изобретения присутствуют комбинации таких мутаций. В некоторых вариантах осуществления изобретения модифицированную Bacillus согласно изобретению получают из штамма-хозяина Bacillus, который уже содержит мутацию в одном или нескольких вышеупомянутых генах. В некоторых вариантах изобретения модифицированного хозяина Bacillus, обладающего повышенной способностью продуцировать представляющий интерес белок, выбирают в качестве клетки-хозяина согласно изобретению. В некоторых вариантах изобретения модифицированная клетка-хозяин Bacillus обладает повышенной способностью продуцировать протеазу.
Методы культивирования
Настоящее изобретение относится к способам продуцирования представляющего интерес белка в модифицированной клетке Bacillus, способной сверхэкспрессировать ymaH, путем культивирования модифицированной клетки, обладающей способностью продуцировать представляющий интерес белок, и выращивания клетки в условиях роста, подходящих для экспрессии представляющего интерес белка. В некоторых вариантах изобретения клетки-хозяева и модифицированные клетки-хозяева согласно изобретению культивируют в стандартных питательных средах. Подходящие конкретные условия культивирования, такие как температура, рН и т.п., известны специалистам в данной области. Дополнительные предпочтительные условия культивирования хорошо известны специалистам и описаны в различных общих публикациях.
В некоторых вариантах осуществления изобретения представляющий интерес белок, продуцируемый модифицированной клеткой-хозяином, присутствует во внутриклеточной среде клетки-хозяина, а в других вариантах изобретения представляющий интерес белок, продуцируемый клеткой-хозяином, секретируется во внеклеточное пространство (то есть в культуральную среду). Таким образом, в некоторых вариантах осуществления изобретения представляющий интерес белок может быть выделен из внутриклеточной среды, в которой он экспрессируется, путем лизиса клетки-хозяина и выделения представляющего интерес белка методами, известными специалистам. В других вариантах осуществления изобретения модифицированные клетки-хозяева культивируют в условиях, подходящих для экспрессии и выделения представляющего интерес белка из клеточной культуры. Представляющий интерес белок, продуцированный модифицированной клеткой-хозяином, сверхэкспрессирующей ymaH согласно изобретению, секретируется в культуральную среду. В некоторых вариантах изобретения представляющий интерес белок (например, протеаза), продуцируемый клетками, выделяют из культуральной среды стандартными методами, включая, но не ограничиваясь ими, выделение клеток-хозяев из среды путем центрифугирования или фильтрации, осаждение белковых компонентов супернатанта или фильтрата с использованием соли (например, сульфата аммония), хроматографическую очистку (например, очистка с помощью ионообменной хроматографии, гель-фильтрации, аффинной хроматографии и т.п.). Таким образом, в настоящем изобретении для выделения протеазы (протеаз) согласно изобретению может быть применен любой метод. Действительно, предусматривается, что настоящее изобретение не ограничивается каким-либо конкретным методом очистки.
В некоторых вариантах осуществления изобретения используются и другие рекомбинантные конструкции, позволяющие присоединять гетерологичные или гомологичные полинуклеотидные последовательности, кодирующие представляющие интерес белки, к нуклеотидной последовательности, кодирующей полипептидный домен, облегчающий очистку растворимых белков (Kroll D.J. et al., DNA Cell Biol 12:441-53 [1993]). Такими облегчающими очистку доменами являются, но не ограничиваются ими, пептиды, образующие хелатные комплексы с металлами, такие как модули «гистидин-триптофан», которые позволяют проводить очистку на иммобилизованных металлах (Porath, Protein Expr Purif 3:263-281 [1992]), домены белка A, которые позволяют проводить очистку на иммобилизованном иммуноглобулине, и домен, используемый в системе удлинения/аффинной очистки FLAGS (Immunex Corp, Seattle WA). Для облегчения очистки, между доменом для очистки и гетерологичным белком может быть также включена отщепляемая линкерная последовательность, такая как фактор XA или энтерокиназа (Invitrogen, San Diego CA).
В некоторых вариантах осуществления изобретения трансформированные клетки-хозяева согласно изобретению культивируют в подходящей питательной среде в условиях, благоприятствующих экспрессии представляющего интерес белка (например, протеазы), с последующим выделением полученной протеазы из культуры. Среда, используемая для культивирования клеток, включает любую стандартную среду, подходящую для культивирования клеток-хозяев, такую как минимальная или комплексная среда, содержащая соответствующие добавки. Подходящие среды поставляются коммерческими фирмами-поставщиками либо они могут быть получены в соответствии с опубликованными протоколами (например, как описано в каталогах Американской коллекции типовых культур). В некоторых вариантах осуществления изобретения клетки-хозяева культивируют в условиях ферментации в периодической культуре, в периодической культуре с подпиткой или в непрерывной культуре. Классические методы ферментации в периодической культуре осуществляют в закрытой системе, где культуральную среду приготавливают до начала проведения ферментации, а затем среду инокулируют желательным(и) микроорганизмом(ами), и такая ферментация не требует добавления в среду каких-либо компонентов. В некоторых случаях, при проведении ферментации в периодической культуре, в культуральной среде изменяют рН и содержание кислорода, а содержание источника углерода остается неизменным. Содержание метаболитов и клеточной биомассы в системе ферментации в периодической культуре постоянно изменяют вплоть до прекращения ферментации. В системе ферментации в периодической культуре рост клеток обычно проходит через статическую лаг-фазу до фазы значительного логарифмического роста и, наконец, до стационарной фазы, при которой скорость роста снижается или такой рост вообще прекращается. Необработанные клетки в стационарной фазе в конечном счете погибают. Вообще говоря, наибольшее количество белка продуцируется клетками в логарифмической фазе.
Вариантом стандартной системы периодической ферментации является система периодической ферментации с подпиткой. В этой системе питательные вещества (например, источник углерода, источник азота, O2 и обычно другие питательные вещества) добавляют только в том случае, когда их концентрация в культуре падает ниже порогового уровня. Системы периодической ферментации с подпиткой являются подходящими в том случае, когда подавление катаболитов может ингибировать метаболизм клеток и в случае, когда желательно ограничить количество питательных веществ в среде. Определение фактической концентрации питательных веществ в системах периодической ферментации с подпиткой осуществляют исходя из изменения определяемых факторов, таких как pH, растворенный кислород и парциальное давление отработанных газов, таких как CO2. Ферментация в периодической культуре и ферментация в периодической культуре с подпиткой являются стандартными методами, хорошо известными специалистам.
Непрерывная ферментация представляет собой открытую систему, в которой определенную культуральную среду непрерывно добавляют в биореактор, и такое же количество кондиционированной среды одновременно удаляют для переработки. При непрерывной ферментации культуру постоянно поддерживают при высокой плотности, где клетки находятся, главным образом, в логарифмической фазе роста.
Непрерывная ферментация позволяет модулировать один фактор или любое число факторов, влияющих на рост клеток и/или концентрацию конечного продукта. Так, например, в некоторых вариантах осуществления изобретения ограничение питательных веществ, таких как источник углерода или источник азота, поддерживается на постоянном уровне, а все другие параметры можно изменять. В других системах ряд факторов, влияющих на рост клеток, постоянно изменяется, а концентрация клеток, определяемая по мутности среды, остается постоянной. В непрерывных системах клетки следует поддерживать в стационарной фазе роста. Таким образом, потеря клеток, происходящая в результате оттока среды, может быть компенсирована за счет скорости роста клеток при ферментации. Методы модуляции питательных веществ и факторов роста при непрерывной ферментации, а также методы максимизации скорости образования продукта известны специалистам в данной области и могут применяться для продуцирования представляющего интерес белка (например, протеазы) способами согласно изобретению.
Как указывалось выше, сверхэкспрессия ymaH в клетке-хозяине приводит к повышению уровня продуцирования представляющего интерес белка по сравнению с уровнем продуцирования того же самого белка в соответствующей клетке-хозяине дикого типа или модифицированной родительской клетке-хозяине. В некоторых вариантах осуществления изобретения сверхэкспрессия ymaH в клетке-хозяине Bacillus приводит к повышению уровня продуцирования представляющего интерес белка, которое превышает уровень продуцирования, достигаемый в соответствующей клетке, которая не сверхэкспрессирует ymaH. В некоторых вариантах осуществления настоящее изобретение относится к клеткам-хозяевам дикого типа или рекомбинантным (модифицированным) клеткам-хозяевам Bacillus, которые сверхэкспрессируют ymaH. В некоторых вариантах изобретения указанная рекомбинантная клетка-хозяин Bacillus была модифицирована в целях сообщения ей способности продуцировать более высокие уровни протеазы, чем в немодифицированной родительской клетке/клетке-предшедственнике Bacillus при культивировании в тех же самых условиях роста.
Настоящее изобретение также включает способы продуцирования представляющего интерес белка в модифицированной клетке, которая сверхэкспрессирует ymaH, в течение меньшего периода времени, чем в клетке-предшественнике. Так, например, модифицированные клетки-хозяева согласно изобретению обладают способностью продуцировать представляющий интерес белок на более высоком уровне и в течение меньшего периода времени, чем соответствующая немодифицированная клетка-предшественник. Таким образом, в некоторых своих вариантах настоящее изобретение относится к способам продуцирования представляющего интерес белка (например, протеазы) на более высоком уровне, чем родительская клетка-хозяин, и за период времени, составляющий 1/6 периода времени, за который клетка-предшественник будет экспрессировать такой белок на максимальном уровне. В других вариантах осуществления изобретения модифицированный хозяин продуцирует представляющий интерес белок за период времени, составляющий примерно 1/5, примерно 1/4, примерно 1/3 или примерно 1/2 от времени, за которое клетка-предшественник будет экспрессировать такой белок на максимальном уровне.
Измерение уровня продуцирования/активности
В некоторых своих вариантах настоящее изобретение относится к способам повышения уровня экспрессии представляющего интерес белка клеткой-хозяином Bacillus путем получения модифицированного хозяина Bacillus, который сверхэкспрессирует ymaH, культивирования модифицированного хозяина Bacillus в подходящих условиях роста и продуцирования клеткой-хозяином представляющего интерес белка на более высоком уровне по сравнению с клеткой-предшественником. В некоторых вариантах изобретения модифицированную клетку Bacillus получают из клетки Bacillus дикого типа. В других вариантах осуществления изобретения модифицированную клетку Bacillus получают из модифицированной клетки-хозяина. Уровень продуцирования представляющего интерес белка клеткой-хозяином может быть определен как функция активности продуцированного белка. В некоторых вариантах изобретения в способе усиления экспрессии представляющего интерес белка используют модифицированную клетку-хозяина Bacillus, которая была трансформирована полинуклеотидной конструкцией, кодирующей YmaH и функционально связанной с промотором sigA и/или sigH. В некоторых вариантах изобретения полинуклеотидные конструкции содержат последовательность, выбранную из SEQ ID NO: 1, 2, 3, и 13. В некоторых вариантах изобретения полинуклеотидная конструкция присутствует на плазмиде, которая реплицируется в клетках Bacillus, а в других вариантах изобретения полинуклеотидная конструкция интегрируется в геном модифицированной клетки Bacillus. Как обсуждалось выше, модифицированная клетка Bacillus способна продуцировать представляющий интерес белок, включая, но не ограничиваясь ими, белок, выбранный из амилолитических ферментов, протеолитических ферментов, целлюлитических ферментов, оксидоредуктазных ферментов и ферментов, разрушающих стенки растений. В других вариантах изобретения такими ферментами являются, но не ограничиваются ими, амилазы, протеазы, ксиланазы, липазы, лакказы, фенолоксидазы, оксидазы, кутиназы, целлюлазы, гемицеллюлазы, эстеразы, пероксидазы, каталазы, глюкозо-оксидазы, фитазы, пектиназы, глюкозидазы, изомеразы, трансферазы, каназы фосфатаз, галактозидазы и хитиназы. В других вариантах изобретения представляющим интерес белком является гормон, цитокин, фактор роста, рецептор, вакцина, антитело или т.п. При этом предусматривается, что настоящее изобретение не ограничивается какими-либо конкретными белками, а в некоторых особенно предпочтительных вариантах изобретения представляющим интерес белком является протеаза.
Существуют различные анализы, известные среднему специалисту в этой области и применяемые для детектирования и измерения активности представляющих интерес белков, продуцируемых клетками-хозяевами согласно изобретению. В частности, эти анализы являются подходящими для измерения протеазной активности, которое осуществляют исходя из уровня высвобождения растворимых в кислоте пептидов из казеина или гемоглобина, и такую активность определяют путем измерения оптической плотности на 405 нм или колориметрическим методом Фолина (см., например, Bergmeyer et al., "Methods of Enzymatic Analysis" vol. 5, Peptidases, Proteinases and their Inhibitors, Verlag Chemie, Weinheim [1984]). Могут быть проведены и некоторые другие анализы, включающие солюбилизацию хромогенных субстратов (см., например, Ward, "Proteinases," in Fogarty (ed.)., Microbial Enzymes and Biotechnology, Applied Science, London, [1983], pp 251-317). Другими репрезентативными анализами являются, но не ограничиваются ими, анализ, проводимый с использованием сукцинил-Ala-Ala-Pro-Phe-пара-нитроанилида (SAAPFpNA) и натриевой соли сульфоната 2,4,6-тринитробензола (TNBS-анализ). Подходящие методы можно найти в различных других публикациях, известных специалистам (см., например, Wells et al., Nucleic Acids Res. 11:7911-7925 [1983]; Christianson et al., Anal. Biochem., 223:119-129 [1994]; и Hsia et al., Anal Biochem., 242:221-227 [1999]). При этом, не следует считать, что настоящее изобретение ограничивается какими-либо конкретными аналитическими методами.
Другими средствами определения уровней продуцирования представляющего интерес белка в клетках-хозяевах и детектирования экспрессируемых белков являются проведение иммуноанализов с использованием поликлональных или моноклональных антител, специфичных к указанному белку. Примерами таких анализов являются твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (РИА), флуоресцентный иммуноанализ (ФИА) и клеточный сортинг с активацией флуоресценции (FACS). Однако известны и другие методы, которые могут быть применены для оценки представляющего интерес белка (см., например, Hampton et al., Serological Methods, A Laboratory Manual. APS Press, St. Paul, MN [1990]; и Maddox et al., J. Exp. Med., 158:1211 [1983]). В некоторых предпочтительных вариантах осуществления изобретения уровень продуцирования представляющего интерес белка в клетках-хозяевах согласно изобретению превышает уровень продуцирования белка в соответствующем хозяине дикого типа или в модифицированном хозяине. Как известно специалистам, клетки Bacillus, продуцированные в соответствии с настоящим изобретением, поддерживают и культивируют в условиях, подходящих для экспрессии и выделения представляющего интерес полипептида из клеточной культуры. При этом следует отметить, что настоящее изобретение не ограничивается какими-либо конкретными аналитическими методами.
Уровень повышения продуцирования протеазы модифицированными клетками-хозяевами согласно изобретению по сравнению с уровнем продуцирования той же самой протеазы клетками-хозяевами дикого типа или модифицированными родительскими клетками-хозяевами может быть определен как отношение активностей. Отношение активностей может быть выражено как отношение ферментативной активности представляющего интерес белка, продуцируемого клеткой-хозяином, которая сверхэкспрессирует ymaH, к ферментативной активности представляющего интерес белка, продуцируемого соответствующей клеткой-хозяином, которая не сверхэкспрессирует ymaH. Отношение, равное или превышающее 1, означает, что представляющий интерес белок, продуцируемый клеткой-хозяином, которая сверхэкспрессирует ymaH, продуцируется на уровне, равном или превышающим уровень, на котором продуцируется тот же самый представляющий интерес белок в соответствующей клетке-хозяине, которая не сверхэкспрессирует ymaH. Так, например, отношение активностей 1,5 означает, что уровень представляющего интерес белка, продуцируемого клеткой-хозяином, которая сверхэкспрессирует ymaH, в 1,5 раз превышает уровень продуцирования того же самого представляющего интерес белка в соответствующей клетке-хозяине, которая не сверхэкспрессирует ymaH при культивировании в тех же самых условиях (то есть клетки-хозяева, экспрессирующие ymaH, продуцируют представляющий интерес белок на 50% больше, чем соответствующая клетка-хозяин, которая не сверхэкспрессирует ymaH). В некоторых вариантах осуществления изобретения отношение активностей составляет по меньшей мере 1, по меньшей мере примерно 1,05, по меньшей мере примерно 1,1, по меньшей мере примерно 1,2, по меньшей мере примерно 1,3, по меньшей мере примерно 1,4, по меньшей мере примерно 1,5, по меньшей мере примерно 1,6, по меньшей мере примерно 1,7, по меньшей мере примерно 1,8, по меньшей мере примерно 1,9, или по меньшей мере примерно 2. В других вариантах осуществления изобретения, отношение активностей составляет по меньшей мере примерно 2,1, по меньшей мере примерно 2,2, по меньшей мере примерно 2,3, по меньшей мере примерно 2,4, по меньшей мере примерно 2,5, по меньшей мере примерно 2,6, по меньшей мере примерно 2,7, по меньшей мере примерно 2,8, по меньшей мере примерно 2,9 или по меньшей мере примерно 3. В других вариантах изобретения отношение активностей составляет по меньшей мере примерно 3,5, по меньшей мере примерно 4,0 или по меньшей мере примерно 5. В некоторых вариантах изобретения желаемое отношение равно 1 или выше.
Альтернативно, уровень продуцирования протеазы в модифицированной клетке-хозяине согласно изобретению может быть выражен как процент увеличения уровня продуцирования по сравнению с уровнем продуцирования в родительской клетке-хозяине. В способах согласно изобретению модифицированная клетка Bacillus продуцирует полипептид на уровне, который предпочтительно по меньшей мере примерно на 25%, более предпочтительно по меньшей мере примерно на 50%, более предпочтительно по меньшей мере примерно на 75%, более предпочтительно по меньшей мере примерно на 100%, еще более предпочтительно по меньшей мере примерно на 200%, наиболее предпочтительно по меньшей мере примерно на 300% и особенно предпочтительно по меньшей мере примерно на 400% превышает уровень продуцирования полипептида в хозяине дикого типа или в модифицированном хозяине-родителе. Таким образом, в некоторых вариантах изобретения уровень продуцирования представляющего интерес белка в клетке-хозяине, которая сверхэкспрессирует ymaH, повышается по меньшей мере примерно на 0,5%, примерно на 1,0%, примерно на 1,5%, примерно на 2,0%, примерно на 2,5%, примерно на 3,0%, примерно на 4,0%, примерно на 5,0%, примерно на 8,0%, примерно на 10%, примерно на 15%, примерно на 20%, примерно на 25%, примерно на 30%, примерно на 40%, примерно на 50%, по меньшей мере примерно на 60%, по меньшей мере примерно на 70%, по меньшей мере примерно на 80%, по меньшей мере примерно на 90%, по меньшей мере примерно на 100% или более по сравнению с уровнем продуцирования того же самого представляющего интерес белка в клетке-хозяине, которая не сверхэкспрессирует ymaH. В других вариантах изобретения уровень продуцирования представляющего интерес белка в клетке-хозяине, которая сверхэкспрессирует ymaH, повышается по меньшей мере примерно на 110%, примерно на 120%, примерно на 130%, примерно на 140%, примерно на 150%, примерно на 160%, примерно на 170%, примерно на 180%, примерно на 190%, и по меньшей мере примерно на 200% или более по сравнению с уровнем продуцирования того же самого представляющего интерес белка в клетке-хозяине, которая не сверхэкспрессирует ymaH.
Для дополнительной иллюстрации настоящего изобретения и его преимуществ ниже приводятся конкретные примеры, которые позволяют лучше понять настоящее изобретение и не должны рассматриваться как ограничение его объема.
Экспериментальная часть
В целях иллюстрации некоторых предпочтительных вариантов и аспектов настоящего изобретения ниже приводятся примеры, которые не должны рассматриваться как ограничение объема изобретения.
В описании экспериментальной части, которое приводится ниже, используются следующие сокращения: м.д. (миллионные доли); M (молярность); мМ (миллимоли); мкМ (микромоли); нМ (наномоли); моль (моли); ммоль (миллимоли); мкмоль (микромоли); нмоль (наномоли); г (граммы); мг (миллиграммы); мкг (микрограммы); пг (пикограммы); л (литры); мл (миллилитры); мкл (микролитры); см (сантиметры); мм (миллиметры); мкм (микрометры); нм (нанометры); ед. (единицы); В (вольт); MW (молекулярная масса); с (секунды); мин (минута/минуты); ч (час/часы); °C (градусы Цельсия); дост.кол. (достаточное количество); ND (не проводили); NA (не применяется); об/мин (оборотов в минуту); H2O (вода); dH2O (деионизованная вода); HCl (соляная кислота); а.к. (аминокислота); п.н. (пар нуклеотидов); т.п.н. (тысяча пар оснований); кД (килодальтон); кДНК (копийная или комплементарная ДНК); ДНК (дезоксирибонуклеиновая кислота); оцДНК (одноцепочечная ДНК); дцДНК (двухцепочечная ДНК); dNTP (дезоксирибонуклеотид-трифосфат); РНК (рибонуклеиновая кислота); MgCl2 (хлорид магния); NaCl (хлорид натрия); Cm (хлорамфеникол); масс./об. (масса к объему); об./об. (объем/объем); g (вес); OD (оптическая плотность); забуференный фосфатом раствор Дульбекко (DPBS); OD280 (оптическая плотность на 280 нм); OD600 (оптическая плотность на 600 нм); A405 (оптическая плотность на 405 нм); PAGE (электрофорез в полиакриламидном геле); PBS (забуференный фосфатом физиологический раствор [150 мМ NaCl, 10 мМ фосфатно-натриевый буфер, pH 7,2]); PBST (PBS + 0,25% Твин®-20); ПЭГ (полиэтиленгликоль); ПЦР (полимеразная цепная реакция); ДСН (додецилсульфат натрия); трис (трис(гидроксиметил)аминометан); HEPES (N-[2-гидроксиэтил]пиперазин-N-[2-этансульфоновая кислота]); HBS (HEPES-забуференный физиологический раствор); ДСН (додецилсульфат натрия); bME, BME и βME (бета-меркаптоэтанол или 2-меркаптоэтанол); трис-HCl (гидрохлорид трис[гидроксиметил]аминометана); трицин (N-[трис-(гидроксиметил)метил]глицин); ДМСО (диметилсульфоксид); Taq (ДНК-полимераза Thermus aquaticus); фрагмент Кленова (крупный фрагмент ДНК-полимеразы I (Кленова)); об/мин. (оборотов в минуту); EGTA (этиленгликоль-бис(β-аминоэтиловый эфир)-N,N,N', N'-тетрауксусная кислота); EDTA (этилендиаминтетрауксусная кислота); bla (ген β-лактамазы или ген резистентности к ампициллину); DNA2.0 (DNA2,0, Menlo Park, CA); OXOID (Oxoid, Basingstoke, Hampshire, UK); Corning (Corning Life Sciences, Corning, NY); ATCC (Американская коллекция типовых культур, Rockville, MD); Sequetech (Sequetech Corporation, Mountainview, CA); Gibco/BRL (Gibco/BRL, Grand Island, NY); Sigma (Sigma Chemical Co., St. Louis, MO); Pharmacia (Pharmacia Biotech, Pisacataway, NJ); NCBI (Национальный центр биотехнологической информации); Applied Biosystems (Applied Biosystems, Foster City, CA); Clontech (CLONTECH Laboratories, Palo Alto, CA); Operon Technologies (Operon Technologies, Inc., Alameda, CA); Bachem (Bachem Bioscience, Inc., King of Prussia, PA); Difco (Difco Laboratories, Detroit, Ml); GIBCO BRL или Gibco BRL (Life Technologies, Inc., Gaithersburg, MD); Millipore (Millipore, Billerica, MA); Bio-Rad (Bio-Rad, Hercules, CA); Invitrogen (Invitrogen Corp., San Diego, CA); NEB (New England Biolabs, Beverly, MA); Sigma (Sigma Chemical Co., St. Louis, MO); Pierce (Pierce Biotechnology, Rockford, IL); Takara (Takara Bio Inc. Otsu, Japan); Roche (Hoffmann-La Roche, Basel, Switzerland); EM Science (EM Science, Gibbstown, NJ); Qiagen (Qiagen, Inc., Valencia, CA); Molecular Devices (Molecular Devices, Corp., Sunnyvale, CA); R&D Systems (R&D Systems, Minneapolis, MN); Stratagene (Stratagene Cloning Systems, La JoIIa, CA); и Microsoft (Microsoft, Inc., Redmond, WA).
Пример 1
Получение полинуклеотидных конструкций SigA и SigH
Были получены полинуклеотидные конструкции SigH, SigA1 и SigA2, которые сверхэкспрессируют ymaH в клетках-хозяевах B. subtilis.
Были сконструированы ПЦР-праймеры, которые являются гомологичными геному Bacillus subtilis (фигура 1А) и содержат рестрикционный сайт из 6 пар нуклеотидов, расположенный на расстоянии 6 пар нуклеотидов от 5'-конца праймера. Эти праймеры были сконструированы для создания уникальных рестрикционных сайтов, расположенных выше и ниже от концов данной конструкции. Основной источник геномной последовательности (Kunst et al., Nature 390:249-256 [1997]), сайт локализации гена и информация о старт- и стоп-кодонах были получены из базы данных NCBI: полный геном Bacillus subtilis subsp. subtilis штамм 168, или из сервера SubtiList World Wide Web Server, известных специалистам (Moser, I. 1998. FEBS Lett. 430(1-2):28-36). В базе данных NCBI, ACC No NC000964, рассматриваемая последовательность дана как SEQ ID NO: 13 с координатами 1865428-18670191.
Была получена SigH-конструкция (SEQ ID NO:1), содержащая полинуклеотидную последовательность, включающую промотор Sigma H и смежную последовательность, кодирующую белок YmaH. Промотор Sigma H обычно расположен в полинуклеотидной последовательности, содержащей ген miaA, локализованный возле 3'-конца и непосредственно выше гена ymaH. Всю последовательность промотора Sigma H и смежную кодирующую последовательность ymaH амплифицировали с помощью ПЦР с использованием прямого праймера P1: GGCACCGAATTCGACGTGGTTTCGCAACAAAATGCAG (SEQ ID NO:5; в положениях 987-1011 SEQ ID NO:13), с рестрикционным EcoRI-сайтом, присоединенным к 5'-концу, и обратного праймера Р2: GGCACCGGATCCCTCATAAAAAAAGACCGTGCCTTGG (SEQ ID NO:6, в положениях 1472-1496 SEQ ID NO:13), с рестрикционным BamHI-сайтом (фигура 1B).
SigA1- и SigA2-конструкции получали в три стадии путем: 1) амплификации отдельных фрагментов хромосомной ДНК B. subtilis, 2) очистки и сборки фрагментов и 3) амплификации собранного продукта с помощью ПЦР.
SigA1-конструкцию (SEQ ID NO:2) получали с использованием двух наборов праймеров (фигура 1C). Первый набор праймеров: прямой праймер P3: GCGCCGAATTCTCATACCCTGAAAGGAAAGACAAGG (SEQ ID NO: 7), локализованный у 5'-конца SEQ ID NO: 13; и обратный праймер P4: TTCGAGTTTTCCTGCTATATGTGTGGGGCTGCTTCGTATTATTCAATATG (SEQ ID NO:8), локализованный в положениях от 153 до 177 п.н. на SEQ ID NO 13, был использован для амплификации первого фрагмента, содержащего промотор SigA, сайт связывания с рибосомой, старт-кодон и первые несколько кодонов гена miaA. Второй набор праймеров: прямой праймер P5: CATATTGAATAATACGAAGCAGCCCCACACATATAGCAGGAAAACTCGAA (SEQ ID NO:9), локализованный в положениях от 1071 до 1095 п.н. на SEQ ID NO: 13; и обратный праймер P2 (SEQ ID NO:6) был использован для амплификации второго фрагмента, содержащего последовательность ДНК, кодирующую белок YmaH. Обратный праймер P4 и прямой праймер P5 представляют собой гибридные праймеры, которые были сконструированы так, чтобы они содержали «хвосты», комплементарные друг другу, но не гомологичные последовательности, которая была амплифицирована в целях элиминации встроенной кодирующей последовательности miaA. Затем два фрагмента отжигали, в результате чего полученная SigA1-конструкция содержала промотор SigA, сайт связывания с рибосомой и сайт инициации транскрипции гена miaA. SigA1-конструкцию амплифицировали с использованием прямого праймера P3 (SEQ ID NO:7) и обратного праймера P2 (SEQ ID NO:6), которые содержали рестрикционный сайт EcoRI и рестрикционный сайт BamHl соответственно, а затем лигировали с полилинкером реплицирующейся плазмиды pBS19. Полинуклеотидная последовательность pBS19 представлена ниже (SEQ ID NO:12). Плазмида pBS19 может реплицироваться в E.coli и B. subtilis и содержит маркерный ген для отбора на резистентность к хлорамфениколу.
GAATTCGAGCTCGGTACCCGGGGATCCTCTAGAGTCGACCTGCAGGCATGCAAGCTTGG
CGATCCTGCCTCGCGCGTTTCGGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCG
GAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGGAGCAGACAAGCCCGTCAGGGCGC
GTCAGCGGGTGTTGGCGGGTGTCGGGGCGCAGCCATGACCCAGTCACGTAGCGATAGC
GGAGTGTATACTGGCTTAACTATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCATAT
GCGGTGTGAAATACCGCACAGATGCGTAAGGAGAAAATACCGCATCAGGCGCTCTTCCGC
TTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAGCGGTATCAGCTCA
CTCAAAGGCGGTAATACGGTTATCCACAGAATCAGGGGATAACGCAGGAAAGAACATGTG
AGCAAAAGGCCAGCAAAAGGCCAGGAACCGTAAAAAGGCCGCGTTGCTGGCGTTTTTCCA
TAGGCTCCGCCCCCCTGACGAGCATCACAAAAATCGACGCTCAAGTCAGAGGTGGCGAAA
CCCGACAGGACTATAAAGATACCAGGCGTTTCCCCCTGGAAGCTCCCTCGTGCGCTCTCC TGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGCCTTTCTCCCTTCGGGAAGCGTGGC
GCTTTCTCATAGCTCACGCTGTAGGTATCTCAGTTCGGTGTAGGTCGTTCGCTCCAAGCTG
GGCTGTGTGCACGAACCCCCCGTTCAGCCCGACCGCTGCGCCTTATCCGGTAACTATCGT
CTTGAGTCCAACCCGGTAAGACACGACTTATCGCCACTGGCAGCAGCCACTGGTAACAGG
ATTAGCAGAGCGAGGTATGTAGGCGGTGCTACAGAGTTCTTGAAGTGGTGGCCTAACTAC
GGCTACACTAGAAGGACAGTATTTGGTATCTGCGCTCTGCTGAAGCCAGTTACCTTCGGAA
AAAGAGTTGGTAGCTCTTGATCCGGCAAACAAACCACCGCTGGTAGCGGTGGTTTTTTTGT
TTGCAAGCAGCAGATTACGCGCAGAAAAAAAGGATCTCAAGAAGATCCTTTGATCTTTTCT
ACGGGGTCTGACGCTCAGTGGAACGAAAACTCACGTTAAGGGATTTTGGTCATGAGATTAT
CAAAAAGGATCTGGAGCTGTAATATAAAAACCTTCTTCAACTAACGGGGCAGGTTAGTGAC
ATTAGAAAACCGACTGTAAAAAGTACAGTCGGCATTATCTCATATTATAAAAGCCAGTCATT
AGGCCTATCTGACAATTCCTGAATAGAGTTCATAAACAATCCTGCATGATAACCATCACAAA
CAGAATGATGTACCTGTAAAGATAGCGGTAAATATATTGAATTACCTTTATTAATGAATTTTC
CTGCTGTAATAATGGGTAGAAGGTAATTACTATTATTATTGATATTTAAGTTAAACCCAGTAA
ATGAAGTCCATGGAATAATAGAAAGAGAAAAAGCATTTTCAGGTATAGGTGTTTTGGGAAA
CAATTTCCCCGAACCATTATATTTCTCTACATCAGAAAGGTATAAATCATAAAACTCTTTGAA
GTCATTCTTTACAGGAGTCCAAATACCAGAGAATGTTTTAGATACACCATCAAAAATTGTAT
AAAGTGGCTCTAACTTATCCCAATAACCTAACTCTCCGTCGCTATTGTAACCAGTTCTAAAA
GCTGTATTTGAGTTTATCACCCTTGTCACTAAGAAAATAAATGCAGGGTAAAATTTATATCC
TTCTTGTTTTATGTTTCGGTATAAAACACTAATATCAATTTCTGTGGTTATACTAAAAGTCGT
TTGTTGGTTCAAATAATGATTAAATATCTCTTTTCTCTTCCAATTGTCTAAATCAATTTTATTA
AAGTTCATTTGATATGCCTCCTAAATTTTTATCTAAAGTGAATTTAGGAGGCTTACTTGTCTG
CTTTCTTCATTAGAATCAATCCTTTTTTAAAAGTCAATATTACTGTAACATAAATATATATTTT
AAAAATATCCCACTTTATCCAATTTTCGTTTGTTGAACTAATGGGTGCTTTAGTTGAAGAATA
AAAGACCACATTAAAAAATGTGGTCTTTTGTGTTTTTTTAAAGGATTTGAGCGTAGCGAAAA
ATCCTTTTCTTTCTTATCTTGATAATAAGGGTAACTATTGCCGGTTGTCCATTCATGGCTGA
ACTCTGCTTCCTCTGTTGACATGACACACATCATCTCAATATCCGAATAGGGCCCATCAGT
CTGACGACCAAGAGAGCCATAAACACCAATAGCCTTAACATCATCCCCATATTTATCCAATA
TTCGTTCCTTAATTTCATGAACAATCTTCATTCTTTCTTCTCTAGTCATTATTATTGGTCCATT
CACTATTCTCATTCCCTTTTCAGATAATTTTAGATTTGCTTTTCTAAATAAGAATATTTGGAG
AGCACCGTTCTTATTCAGCTATTAATAACTCGTCTTCCTAAGCATCCTTCAATCCTTTTAATA
ACAATTATAGCATCTAATCTTCAACAAACTGGCCCGTTTGTTGAACTACTCTTTAATAAAATA
ATTTTTCCGTTCCCAATTCCACATTGCAATAATAGAAAATCCATCTTCATCGGCTTTTTCGTC
ATCATCTGTATGAATCAAATCGCCTTCTTCTGTGTCATCAAGGTTTAATTTTTTATGTATTTC
TTTTAACAAACCACCATAGGAGATTAACCTTTTACGGTGTAAACCTTCCTCCAAATCAGACA
AACGTTTCAAATTCTTTTCTTCATCATCGGTCATAAAATCCGTATCCTTTACAGGATATTTTG
CAGTTTCGTCAATTGCCGATTGTATATCCGATTTATATTTATTTTTCGGTCGAATCATTTGAA CTTTTACATTTGGATCATAGTCTAATTTCATTGCCTTTTTCCAAAATTGAATCCATTGTTTTTG
ATTCACGTAGTTTTCTGTATTCTTAAAATAAGTTGGTTCCACACATACCAATACATGCATGT
GCTGATTATAAGAATTATCTTTATTATTTATTGTCACTTCCGTTGCACGCATAAAACCAACAA
GATTTTTATTAATTTTTTTATATTGCATCATTCGGCGAAATCCTTGAGCCATATCTGACAAAC
TCTTATTTAATTCTTCGCCATCATAAACATTTTTAACTGTTAATGTGAGAAACAACCAACGAA
CTGTTGGCTTTTGTTTAATAACTTCAGCAACAACCTTTTGTGACTGAATGCCATGTTTCATT
GCTCTCCTCCAGTTGCACATTGGACAAAGCCTGGATTTACAAAACCACACTCGATACAACT
TTCTTTCGCCTGTTTCACGATTTTGTTTATACTCTAATATTTCAGCACAATCTTTTACTCTTTC
AGCCTTTTTAAATTCAAGAATATGCAGAAGTTCAAAGTAATCAACATTAGCGATTTTCTTTTC
TCTCCATGGTCTCACTTTTCCACTTTTTGTCTTGTCCACTAAAACCCTTGATTTTTCATCTGA
ATAAATGCTACTATTAGGACACATAATATTAAAAGAAACCCCCATCTATTTAGTTATTTGTTT
AGTCACTTATAACTTTAACAGATGGGGTTTTTCTGTGCAACCAATTTTAAGGGTTTTCAATA
CTTTAAAACACATACATACCAACACTTCAACGCACCTTTCAGCAACTAAAATAAAAATGACG
TTATTTCTATATGTATCAAGATAAGAAAGAACAAGTTCAAAACCATCAAAAAAAGACACCTTT
TCAGGTGCTTTTTTTATTTTATAAACTCATTCCCTGATCTCGACTTCGTTCTTTTTTTACCTCT
CGGTTATGAGTTAGTTCAAATTCGTTCTTTTTAGGTTCTAAATCGTGTTTTTCTTGGAATTGT
GCTGTTTTATCCTTTACCTTGTCTACAAACCCCTTAAAAACGTTTTTAAAGGCTTTTAAGCC
GTCTGTACGTTCCTTAAG (SEQ ID NO: 12)
SigA2-конструкция (SEQ ID NO:3) была получена методом, описанным для получения SigA1-конструкции, с использованием нижеследующих праймеров (фигура 1D). Первый фрагмент, содержащий промотор SigA, амплифицировали с использованием прямого праймера P3 (SEQ ID NO:7) и обратного гибридного праймера P7: CATACAGTTTCGATTAAAGTTCGAGCACTCTCTTTTATAAATCTCCCCCA (SEQ ID NO:11), локализованного в положениях от 125 до 149 п.н. на SEQ ID NO:13. Второй фрагмент, содержащий последовательность ДНК, кодирующую белок YmaH, амплифицировали с использованием прямого гибридного праймера P6: TGGGGGAGATTTATAAAAGAGAGTGCTCGAACTTTAATCGAAACTGTATG (SEQ ID NO: 10), локализованного в положениях от 1090 до 1114 п.н. на SEQ ID NO:13, и обратного праймера P2 (SEQ ID NO:6). Затем два фрагмента отжигали, в результате чего полученная SigA2-конструкция содержала промотор SigA, сайт связывания с рибосомой и сайт инициации транскрипции гена ymaH.
Настоящее изобретение также включает четвертую SigA-конструкцию (SigA3; SEQ ID NO: 13; фигура 1E), которая была получена путем амплификации miaA ymaH в области хромосомной ДНК Bacillus, которая включает промотор SigA, область, кодирующую белок MiaA, промотор SigH-YmaH и область, кодирующую белок YmaH.
SigA3-конструкция была получена с использованием прямого праймера GCGCGCGAATTCAGGGAAATTGTCGGCAATGAGCCGCTCGGC (SEQ ID NO: 18) и обратного праймера GCGCGCCATGGCTGATTCGTCTCAGTTCTGCTTCACTTTCA (SEQ ID NO: 19). SEQ ID NO:13 имеет рестрикционный сайт EcoRI у 5'-конца фрагмента, а SEQ ID NO:19 имеет NcoI-сайт на другом конце. Это позволяет клонировать фрагмент в вектор pBN3, представленный ниже как SEQ ID NO:20.
GACGTCTAAGAAACCATTATTATCATGACATTAACCTATAAAAATAGGCGTATCACGAGGCC
CTTTCGTCTTCAAGAATTAATTCTCATGTTTGACAGCTTATCATCGATAAGCTTGCATGCCT
GCAGGTCGACTCTAGAGGATCCCCGGGTACCGAGCTCGAATTCCTTAAGGAACGTACAGA
CGGCTTAAAAGCCTTTAAAAACGTTTTTAAGGGGTTTGTAGACAAGGTAAAGGATAAAACA
GCACAATTCCAAGAAAAACACGATTTAGAACCTAAAAAGAACGAATTTGAACTAACTCATAA
CCGAGAGGTAAAAAAAGAACGAAGTCGAGATCAGGGAATGAGTTTATAAAATAAAAAAAGC
ACCTGAAAAGGTGTCTTTTTTTGATGGTTTTGAACTTGTTCTTTCTTATCTTGATACATATAG
AAATAACGTCATTTTTATTTTAGTTGCTGAAAGGTGCGTTGAAGTGTTGGTATGTATGTGTT
TTAAAGTATTGAAAACCCTTAAAATTGGTTGCACAGAAAAACCCCATCTGTTAAAGTTATAA
GTGACTAAACAAATAACTAAATAGATGGGGGTTTCTTTTAATATTATGTGTCCTAATAGTAG
CATTTATTCAGATGAAAAATCAAGGGTTTTAGTGGACAAGACAAAAAGTGGAAAAGTGAGA
CCATGGAGAGAAAAGAAAATCGCTAATGTTGATTACTTTGAACTTCTGCATATTCTTGAATT
TAAAAAGGCTGAAAGAGTAAAAGATTGTGCTGAAATATTAGAGTATAAACAAAATCGTGAAA
CAGGCGAAAGAAAGTTGTATCGAGTGTGGTTTTGTAAATCCAGGCTTTGTCCAATGTGCAA
CTGGAGGAGAGCAATGAAACATGGCATTCAGTCACAAAAGGTTGTTGCTGAAGTTATTAAA
CAAAAGCCAACAGTTCGTTGGTTGTTTCTCACATTAACAGTTAAAAATGTTTATGATGGCGA
AGAATTAAATAAGAGTTTGTCAGATATGGCTCAAGGATTTCGCCGAATGATGCAATATAAAA
AAATTAATAAAAATCTTGTTGGTTTTATGCGTGCAACGGAAGTGACAATAAATAATAAAGAT
AATTCTTATAATCAGCACATGCATGTATTGGTATGTGTGGAACCAACTTATTTTAAGAATAC
AGAAAACTACGTGAATCAAAAACAATGGATTCAATTTTGGAAAAAGGCAATGAAATTAGACT
ATGATCCAAATGTAAAAGTTCAAATGATTCGACCGAAAAATAAATATAAATCGGATATACAA
TCGGCAATTGACGAAACTGCAAAATATCCTGTAAAGGATACGGATTTTATGACCGATGATG
AAGAAAAGAATTTGAAACGTTTGTCTGATTTGGAGGAAGGTTTACACCGTAAAAGGTTAATC
TCCTATGGTGGTTTGTTAAAAGAAATACATAAAAAATTAAACCTTGATGACACAGAAGAAGG
CGATTTGATTCATACAGATGATGACGAAAAAGCCGATGAAGATGGATTTTCTATTATTGCAA
TGTGGAATTGGGAACGGAAAAATTATTTTATTAAAGAGTAGTTCAACAAACGGGCCAGTTT
GTTGAAGATTAGATGCTATAATTGTTATTAAAAGGATTGAAGGATGCTTAGGAAGACGAGTT
ATTAATAGCTGAATAAGAACGGTGCTCTCCAAATATTCTTATTTAGAAAAGCAAATCTAAAAT
TATCTGAAAAGGGAATGAGAATAGTGAATGGACCAATAATAATGACTAGAGAAGAAAGAAT
GAAGATTGTTCATGAAATTAAGGAACGAATATTGGATAAATATGGGGATGATGTTAAGGCT
ATTGGTGTTTATGGCTCTCTTGGTCGTCAGACTGATGGGCCCTATTCGGATATTGAGATGA
TGTGTGTCATGTCAACAGAGGAAGCAGAGTTCAGCCATGAATGGACAACCGGTGAGTGGA
AGGTGGAAGTGAATTTTGATAGCGAAGAGATTCTACTAGATTATGCATCTCAGGTGGAATC AGATTGGCCGCTTACACATGGTCAATTTTTCTCTATTTTGCCGATTTATGATTCAGGTGGAT
ACTTAGAGAAAGTGTATCAAACTGCTAAATCGGTAGAAGCCCAAACGTTCCACGATGCGAT
TTGTGCCCTTATCGTAGAAGAGCTGTTTGAATATGCAGGCAAATGGCGTAATATTCGTGTG
CAAGGACCGACAACATTTCTACCATCCTTGACTGTACAGGTAGCAATGGCAGGTGCCATGT
TGATTGGTCTGCATCATCGCATCTGTTATACGACGAGCGCTTCGGTCTTAACTGAAGCAGT
TAAGCAATCAGATCTTCCTTCAGGTTATGACCATCTGTGCCAGTTCGTAATGTCTGGTCAA
CTTTCCGACTCTGAGAAACTTCTGGAATCGCTAGAGAATTTCTGGAATGGGATTCAGGAGT
GGACAGAACGACACGGATATATAGTGGATGTGTCAAAACGCATACCATTTTGAACGATGAC
CTCTAATAATTGTTAATCATGTTGGTTACCTGCCTCGCGCGTTTCGGTGATGACGGTGAAA
ACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGG
AGCAGACAAGCCCGTCAGGGCGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCGCAGCCA
TGACCCAGTCACGTAGCGATAGCGGAGTGTATACTGGCTTAACTATGCGGCATCAGAGCA
GATTGTACTGAGAGTGCACCATATGCGGTGTGAAATACCGCACAGATGCGTAAGGAGAAA
ATACCGCATCAGGCGCTCTTCCGCTTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCG
GCTGCGGCGAGCGGTATCAGCTCACTCAAAGGCGGTAATACGGTTATCCACAGAATCAGG
GGATAACGCAGGAAAGAACATGTGAGCAAAAGGCCAGCAAAAGGCCAGGAACCGTAAAAA
GGCCGCGTTGCTGGCGTTTTTCCATAGGCTCCGCCCCCCTGACGAGCATCACAAAAATCG
ACGCTCAAGTCAGAGGTGGCGAAACCCGACAGGACTATAAAGATACCAGGCGTTTCCCCC
TGGAAGCTCCCTCGTGCGCTCTCCTGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGC
CTTTCTCCCTTCGGGAAGCGTGGCGCTTTCTCATAGCTCACGCTGTAGGTATCTCAGTTCG
GTGTAGGTCGTTCGCTCCAAGCTGGGCTGTGTGCACGAACCCCCCGTTCAGCCCGACCG
CTGCGCCTTATCCGGTAACTATCGTCTTGAGTCCAACCCGGTAAGACACGACTTATCGCCA
CTGGCAGCAGCCACTGGTAACAGGATTAGCAGAGCGAGGTATGTAGGCGGTGCTACAGA
GTTCTTGAAGTGGTGGCCTAACTACGGCTACACTAGAAGGACAGTATTTGGTATCTGCGCT
CTGCTGAAGCCAGTTACCTTCGGAAAAAGAGTTGGTAGCTCTTGATCCGGCAAACAAACCA
CCGCTGGTAGCGGTGGTTTTTTTGTTTGCAAGCAGCAGATTACGCGCAGAAAAAAAGGAT
CTCAAGAAGATCCTTTGATCTTTTCTACGGGGTCTGACGCTCAGTGGAACGAAAACTCACG
TTAAGGGATTTTGGTCATGAGATTATCAAAAAGGATCTTCACCTAGATCCTTTTAAATTAAAA
ATGAAGTTTTAAATCAATCTAAAGTATATATGAGTAAACTTGGTCTGACAGTTACCAATGCTT
AATCAGTGAGGCACCTATCTCAGCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTC
CCCGTCGTGTAGATAACTACGATACGGGAGGGCTTACCATCTGGCCCCAGTGCTGCAATG
ATACCGCGAGACCCACGCTCACCGGCTCCAGATTTATCAGCAATAAACCAGCCAGCCGGA
AGGGCCGAGCGCAGAAGTGGTCCTGCAACTTTATCCGCCTCCATCCAGTCTATTAATTGTT
GCCGGGAAGCTAGAGTAAGTAGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTG
CTACAGGCATCGTGGTGTCACGCTCGTCGTTTGGTATGGCTTCATTCAGCTCCGGTTCCC
AACGATCAAGGCGAGTTACATGATCCCCCATGTTGTGCAAAAAAGCGGTTAGCTCCTTCG
GTCCTCCGATCGTTGTCAGAAGTAAGTTGGCCGCAGTGTTATCACTCATGGTTATGGCAGC ACTGCATAATTCTCTTACTGTCATGCCATCCGTAAGATGCTTTTCTGTGACTGGTGAGTACT
CAACCAAGTCATTCTGAGAATAGTGTATGCGGCGACCGAGTTGCTCTTGCCCGGCGTCAA
TACGGGATAATACCGCGCCACATAGCAGAACTTTAAAAGTGCTCATCATTGGAAAACGTTC
TTCGGGGCGAAAACTCTCAAGGATCTTACCGCTGTTGAGATCCAGTTCGATGTAACCCACT
CGTGCACCCAACTGATCTTCAGCATCTTTTACTTTCACCAGCGTTTCTGGGTGAGCAAAAA
CAGGAAGGCAAAATGCCGCAAAAAAGGGAATAAGGGCGACACGGAAATGTTGAATACTCA
TACTCTTCCTTTTTCAATATTATTGAAGCATTTATCAGGGTTATTGTCTCATGAGCGGATACA
TATTTGAATGTATTTAGAAAAATAAACAAATAGGGGTTCCGCGCACATTTCCCCGAAAAGTG
CCACCT (SEQ ID NO:20)
Все ПЦР-реакции проводили в объеме 50 мкл, содержащих 1-2 мкл ДНК, или в смеси для ресуспендирования колоний, 5 мкл 10X буфера Pfu Ultra (Stratagene), 1 мкл 10 мМ dNTP-смеси (Roche), 0,5 мкл 0,2 мкМ праймеров, 1 мкл полимеразы Pfu Ultra High Fidelity, а затем добавляли воду до общего объема 50 мкл. ПЦР проводили в следующих условиях: 95°С, 2 минуты, 30 циклов: 95°С, 30 секунд, 62°С, 30 секунд, 72°С, 1 минута, а затем 1 цикл: 72°С, 10 минут.
Полученные ПЦР-фрагменты подвергали гель-очистке с использованием набора для гель-очистки Qiagen в соответствии с инструкциями производителей.
Гибридные конструкции получали путем отжига 0,25 мкл аликвот очищенных ПЦР-фрагментов, которые были смешаны и добавлены в свежеприготовленную ПЦР-смесь, полученную в соответствии с вышеописанным протоколом с использованием праймеров Р3 и P2. Общий объем ПЦР-смеси составлял 50 мкл. Условия проведения ПЦР были аналогичны вышеописанным условиям, за исключением того, что температуру отжига корректировали в соответствии с Tm праймеров.
Нужные SigH-, SigA1-, и SigA2-конструкции лигировали в плазмиды pBS19, которые были гидролизованы ферментами EcoRI и BamHI с получением векторов, экспрессирующих SigA и SigH и используемых для трансформации клеток-хозяев как описано в примере 2.
Смесь для трансформации высевали на планшеты, содержащие среду LB + 1,6% сепарированного молока + 5 мкг/мл хлорамфеникола. На следующий день колонии, образующие гало, собирали и высевали для получения моноколоний. Очистку колоний проводили два раза. Пять отдельных клонов анализировали путем секвенирования области промотора aprE. Все эти клоны имели консенсусную последовательность в -35-области промотора aprE.
Пример 2
Трансформация клеток-хозяев и экспрессия протеазы aprE
Пять микролитров лигированной смеси, содержащей конструкции SigA или SigH, использовали для трансформации клеток Top10 E.coli (Invitrogen) путем электропорации. Трансформированные клетки высевали на планшеты с агаром LB, содержащие 5 м.д./мл хлорамфеникола (Cm), и колонии оставляли на ночь для культивирования при 37°С. Отдельные колонии собирали и переносили в пробирки, содержащие 5 мл LB + 5 м.д./мл Cm. Культуры выращивали в течение ночи при 37°С со встряхиванием при 250 об/мин. Плазмидную ДНК получали из культур E.coli, и часть плазмидного ДНК-препарата секвенировали (Sequetech). Автоматизированный анализ последовательности осуществляли с использованием компьютерных программ Phrep, Phrap, Consed, Custal W.
Плазмиду, несущую конструкцию, расположенную справа от каждого из трех экспрессионных векторов, использовали для трансформации клеток-хозяев B. subtilis. Экспрессионные векторы, содержащие конструкции SigH (SEQ ID NO:1) и SigA1 (SEQ ID NO:2) и SigA2 (SEQ ID NO:3), которые были обозначены pBS19 ymaH-H и pBS19 ymaH-A1 и pBS19 ymaH-A2, переносили в штаммы BG2941 и BG2942 B. Subtilis, как описано ниже. Два микролитра плазмидной ДНК, несущей соответствующие конструкции, использовали для трансформации 100 мкл клеток штамма BG 2941 B. subtilis (ΔnprE, amyE::PxylRA-comK-phleoR) и штамма BG2942 B. subtilis (ΔnprE, degU(Hy)32, amyE::PxylRA-comK-phleoR). Трансформанты BG2941 и BG2942, несущие конструкции SigH, обозначали 41SigH и 42SigH соответственно, а трансформанты BG2941 и BG2942, несущие конструкции SigA1, обозначали 41SigA1 и 42SigA1 соответственно. Некоторые клетки-хозяева BG2941 и BG2942 были также трансформированы контрольной плазмидой pBS19 и обозначены 41pBS19 и 42pBS19. Клетки-хозяева BG2941 и BG2942 несут делецию гена nprE, что приводит к элиминации большей части фоновой протеолитической активности, не относящейся к aprE, и тем самым облегчает определение уровня продуцирования щелочной протеазы (aprE). Клетки-хозяева BG2941 и 2942 также несут кластер amyE::PxylRA-comK-phleoR, что позволяет получать компетентные клетки путем индуцирования роста культуры в присутствии ксилозы (Hahn et al., Mol Microbiol. 18:755-67 [1995]). Клетки-хозяева 2942 также несут мутацию в гене degU (мутация degU(Hy)32), которая сама по себе в несколько раз повышает уровень субтилизина, секретируемого клетками-хозяевами, по сравнению с уровнем субтилизина, секретируемого клетками-хозяевами, которые не несут мутацию degU(Hy) (Msadek et al. J Bacteriol, 172:824-834 [1990]).
Эффект сверхэкспрессии YmaH в клетках-хозяевах Bacillus качественно и количественно оценивали в анализах, описанных в примере 3.
Пример 3
Влияние сверхэкспрессии ymaH на продуцирование протеазы
Анализ с использованием казеина: - Влияние сверхэкспрессии ymaH на продуцирование протеазы в клетках-хозяевах Bacillus сначала оценивали с помощью количественного анализа, в котором сравнивали размер гало, продуцируемого колониями, выращенными на агаровых планшетах, содержащих казеин в виде сепарированного молока. Поскольку фермент протеаза секретируется клетками Bacillus, то он разлагает казеин, содержащийся в сепарированном молоке, и образует зоны прозрачности или гало вокруг растущей колонии. Клетки-хозяева, которые имеют инактивированную протеазу, будут образовывать небольшое гало или вообще не будут образовывать гало вокруг колоний. Таким образом, по размеру гало можно проводить качественную оценку количества протеазы, продуцируемой секретирующей ее колонией (Wells, T. A. et al. Nucleic Acids Res., 11, 7911-7925: [1983]).
Клетки-хозяева BG2941 и BG2942 B. subtilis, трансформированные SigH- или SigA1-экспрессирующими векторами, высевали на планшеты с агаром LB, содержащим 1,6% сепарированного молока и 5 м.д./мл Cm, и инкубировали в течение ночи при 37°С. На следующий день колонии от некоторых трансформантов образовывали отдельные моноколонии на планшетах с агаром LB, содержащих 5 м.д./мл Cm, после чего планшеты инкубировали в течение ночи при 37°С. Изоляты моноколоний собирали и наносили пятнами на планшеты того же типа и снова инкубировали при 37°С в течение ночи.
Самые крупные гало были продуцированы клетками-хозяевами 42SigH, несущими мутацию degU(Hy)32 и конструкцию SigH, способную сверхэкспрессировать ymaH. В частности, размер гало клеток 42SigH указывает на то, что сверхэкспрессия ymaH также приводит к повышению уровня продуцирования субтилизина в клетках-хозяевах, которые уже продуцировали данный фермент на уровне, превышающем уровень продуцирования этого фермента в клетках дикого типа, то есть клетки 42SigH продуцируют гало, размер которого превышает размер гало, продуцируемого клетками 42pBS19, несущими мутацию degU(Hy), но не несущими конструкцию, которая способна сверхэкспрессировать ymaH, и эти клетки в свою очередь продуцируют гало, размер которого превышает размер гало, продуцируемого клетками 41pBS19, не несущими мутацию degU(Hy)32 и не несущими конструкцию, которая способна сверхэкспрессировать ymaH. Размер гало, продуцируемого клетками 42SigH, также превышает размер гало, продуцируемого клетками 41 SigH, не несущими мутацию degU(Hy), но несущими конструкцию SigH, способную сверхэкспрессировать ymaH.
AAPF-анализ - продуцирование субтилизина трансформированными клетками-хозяевами Bacillus 42SigH, 42SigA1, 41SigA2, которые сверхэкспрессируют ymaH, и их соответствующими контрольными клетками 42pBS19 и 41pBS19, количественно оценивали как функцию активности секретированной протеазы aprE. Протеолитическую активность секретированной протеазы определяли как скорость гидролиза субстрата сукцинил-L-Ala-L-Ala-L-Pro-L-Phe-п-нитроанилида (AAPF от Sigma Chemical Co). В этом анализе измеряли уровень продуцирования протеазы как величину оптической плотности на 405 нм/мин, полученную после гидролиза и высвобождения п-нитроанилида (Estell et al., J. Biol. Chem., 260:6518-6521 [1985]). Измерения проводили с использованием компьютерной программы Sofmax Pro, в конкретных условиях, таких как: тип реакции: кинетическая; восстановление: точки Vmax (наилучшее считывание в 15 из 28 точек); Lm1: 405 нм; время: 5 минут; и интервал: 11 секунд.
Жидкие культуры контрольных клеток-хозяев 41pBS19 и 42pBS19 B. subtilis, и клеток-хозяев, сверхэкспрессирующих ymaH, получали путем инокуляции 5 мл среды LB, содержащей 5 м.д./мл хлорамфеникола (Cm), моноколониями трансформированных клеток 41 SigH и 42SigA1 и 42SigH и культивирования клеток со встряхиванием при 37°С до тех пор, пока клетки не достигали средней фазы логарифмического роста. Каждую культуру разводили 1:100 свежей комплексной средой, содержащей 5 м.д./мл Cm, и оставляли для культивирования при 37°С со встряхиванием при 250 об/мин. Образцы культур брали через периоды времени, указанные на фигурах. Образцы центрифугировали, и супернатанты анализировали на продуцирование субтилизина.
Десять микролитров каждого супернатанта культур B. subtilis разводили 100 микролитрами трис-буфера, содержащего 10 мМ триса + 0,005% твина®-80, pH 8,6; и 25 мкл 100 мг/мл AAPF. Затем определяли активность каждой из этих протеаз, и результаты влияния сверхэкспрессии YmaH на продуцирование протеазы представлены на фигурах 3A-B и на фигуре 4.
На фигурах 3A и 3B показано, что сверхэкспрессия ymaH в клетках-хозяевах Bacillus, независимо от присутствия (42SigA и 42SigH; фигура 3A) или отсутствия (41 SigH; фигура 3B) мутации degU(Hy), приводит к повышению уровня продуцирования субтилизина aprE, который в несколько раз превышает уровень, продуцируемый соответствующими контрольными клетками 41pBS19 и 42pBS19. Кроме того, клетки, которые сверхэкспрессируют ymaH, продуцируют субтилизин на уровне, превышающем уровень продуцирования субтилизина клетками, которые не сверхэкспрессируют ymaH. Так, например, на фигуре 3A показано, что клетки 42sigH за 20 часов культивирования продуцируют почти такое же количество субтилизина, которое продуцируют родительские контрольные клетки за 48 часов. Аналогичным образом, на фигуре 3B показано, что клетки 41sigH за 25 часов продуцируют большее количество субтилизина, чем контрольные клетки 41pBS за 48 часов. На графике, представленном на фигуре 4, показано, что клетки, которые экспрессируют ymaH под действием промотора SigH (42SigH), продуцируют субтилизин на уровне, превышающем уровень субтилизина, продуцируемого клетками, в которых экспрессия ymaH индуцируется промотором sigma (42SigA). На фигуре 4 также показано, что сверхэкспрессия ymaH, независимо от того, индуцируется ли она промотором SigH или SigA, приводит к повышению уровня продуцирования субтилизина уже по меньшей мере через один час после начала культивирования клеток.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ АМПЛИФИЦИРОВАНИЯ ЛОКУСОВ В БАКТЕРИАЛЬНОЙ КЛЕТКЕ | 2009 |
|
RU2577985C2 |
| СИГНАЛЬНЫЕ ПЕПТИДЫ ТАТ ДЛЯ ПРОДУКЦИИ БЕЛКОВ В ПРОКАРИОТАХ | 2008 |
|
RU2487937C2 |
| Способы и композиции, содержащие варианты сериновой протеазы | 2012 |
|
RU2663114C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |
| ПРОТЕАЗА Streptomyces | 2008 |
|
RU2486244C2 |
| ВАРИАНТЫ ПРОТЕАЗ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2790443C2 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| ПРИМЕНЕНИЕ ВАРИАНТОВ ПРОТЕАЗЫ | 2010 |
|
RU2639534C2 |
| CПОСОБ МЫТЬЯ ПОСУДЫ | 2007 |
|
RU2509152C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |









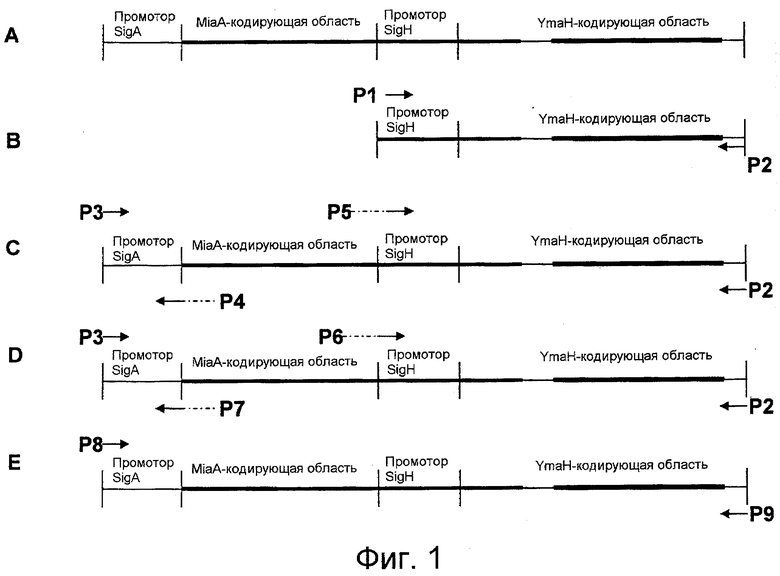

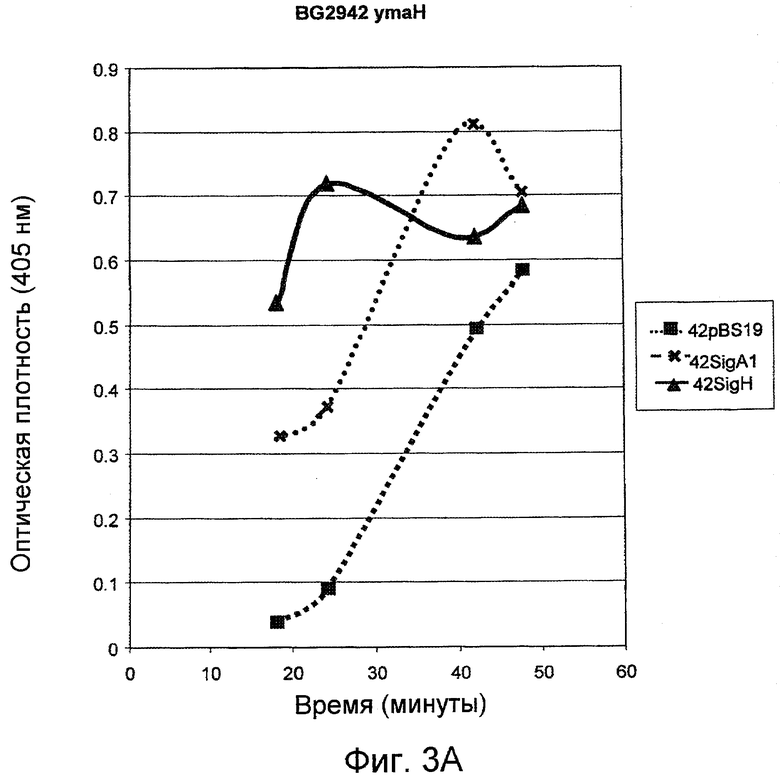
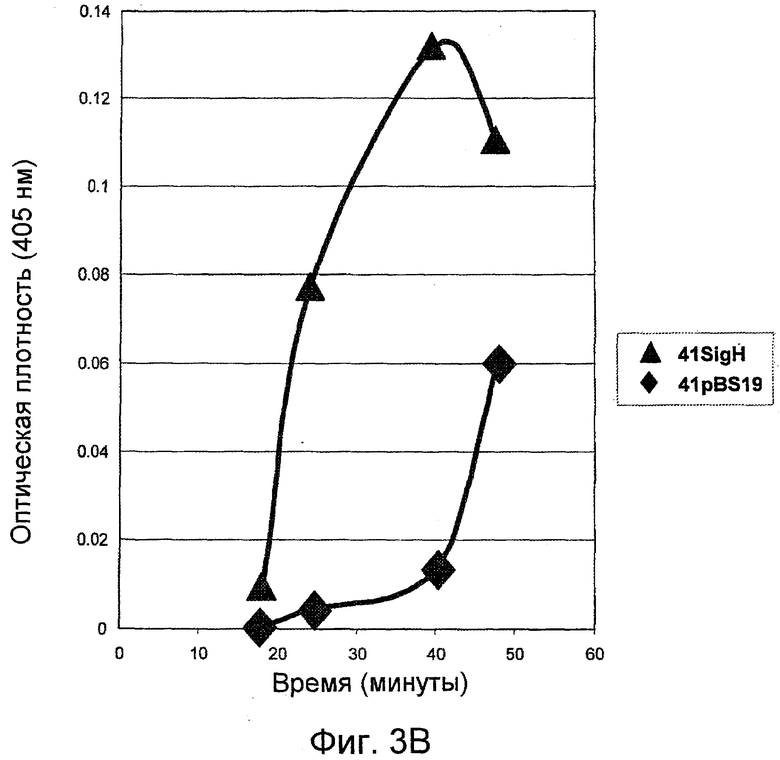
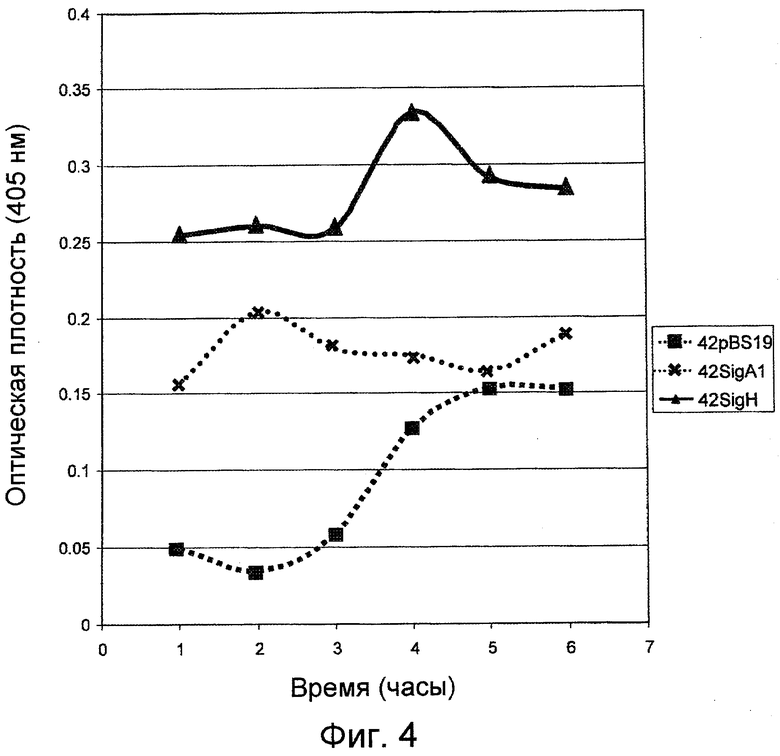
Изобретение относится к области биотехнологии. Представлен выделенный химерный полинуклеотид для усиления продуцирования представляющего интерес гетерологичного белка, содержащий полинуклеотидную последовательность промотора SigA или SigH, функционально связанную с полинуклеотидом, кодирующим белок YmaH, причем химерный полинуклеотид содержит последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 1, 2, 3 или 13. Также описаны вектор экспрессии, содержащий указанную нуклеотидную конструкцию, и клетка-хозяин Bacillus для продукции представляющего интерес гетерологичного белка, содержащая указанный вектор. Предложен способ получения модифицированной клетки Bacillus, включающий трансформацию клетки-хозяина Bacillus-продуцента представляющего интерес гетерологичного белка указанным вектором; и выращивание указанной модифицированной клетки в оптимальных условиях. Описан способ получения представляющего интерес белка в модифицированной клетке Bacillus, где способ включает культивирование указанной клетки-хозяина; и выращивание указанной модифицированной клетки Bacillus в оптимальных условиях. Кроме того, раскрыт способ усиления экспрессии представляющего интерес гетерологичного белка из Bacillus, включающий получение указанной модифицированной клетки Bacillus; выращивание модифицированной клетки Bacillus в оптимальных условиях; и экспрессию указанного представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия указанного представляющего интерес гетерологичного белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией указанного представляющего интерес белка в указанной родительской клетке-хозяине Bacillus. Изобретение позволяет повысить выход целевого белка за счет сверхэкспрессии белка YmaH. 7 н. и 23 з.п. ф-лы, 4 ил., 3 пр.
1. Выделенный химерный полинуклеотид для усиления продуцирования представляющего интерес гетерологичного белка, содержащий полинуклеотидную последовательность промотора SigA или SigH, функционально связанную с полинуклеотидом, кодирующим белок YmaH, причем химерный полинуклеотид содержит последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 1, 2, 3 или 13.
2. Выделенный химерный полинуклеотид по п.1, включающий SEQ ID NO: 1, 2, 3 или 13.
3. Вектор экспрессии химерного полинуклеотида для повышения продуцирования представляющего интерес гетерологичного белка, содержащий химерный полинуклеотид по п.1.
4. Вектор по п.3, где указанный химерный полинуклеотид содержит SEQ ID NO: 1, 2, 3 или 13.
5. Модифицированная клетка-хозяин Bacillus, содержащая вектор по п.3, где указанная модифицированная клетка-хозяин Bacillus способна продуцировать представляющий интерес гетерологичный белок.
6. Модифицированная клетка по п.5, где клетка Bacillus выбрана из группы, состоящей из В. licheniformis, В subtilis, В. lentus, В. brevis, В. stearothermophilus, B. alkalophilus, B. amyloliquefaciens, B. coagulans, В. circulans , В. lautus , B. pumilus, B. thuringiensis, B. clausii и B. megaterium.
7. Модифицированная клетка-хозяин по п.5, где представляющий интерес белок является гомологичным или гетерологичным по отношению к указанной модифицированной клетке-хозяину.
8. Модифицированная клетка-хозяин по п.5, где промотор aprE инициирует экспрессию указанного представляющего интерес белка.
9. Клетка-хозяин по п.5, где указанный представляющий интерес белок выбран из группы, состоящей из амилаз, протеаз, ксиланаз, липаз, лакказ, фенолоксидаз, оксидаз, кутиназ, целлюлаз, гемицеллюлаз, эстераз, пероксидаз, каталаз, глюкозооксидаз, фитаз, пектиназ, глюкозидаз, изомераз, трансфераз, киназ, фосфотаз, галактозидаз и хитиназ, гормонов, цитокинов, факторов роста, рецепторов, вакцин и антител.
10. Клетка-хозяин по п.5, где указанным представляющим интерес белком является фермент.
11. Клетка-хозяин по п.10, где указанным ферментом является протеаза.
12. Клетка-хозяин по п.11, где указанной протеазой является субтилизин, выбранный из группы, состоящей из субтилизина 168, субтилизина BPN′, субтилизина Карлсберга, субтилизина B. lentus, субтилизина B. clausii, субтилизина DY, субтилизина 147, субтилизина 309 и их вариантов.
13. Модифицированная клетка Bacillus, содержащая вектор по п.3, которая является клеткой, продуцирующей протеазы, способной сверхэкспрессировать YmaH, где модифицированная клетка содержит мутацию по меньшей мере в одном гене, выбранном из degU, degQ, degS, sco4, spollE, degQ и degR, где мутация усиливает способность продуцировать протеазу.
14. Модифицированная клетка по п.13, где указанной мутацией является deg(Hy)32.
15. Модифицированная клетка по п.13, где указанной клеткой Bacillus является клетка В. subtilis.
16. Способ получения модифицированной клетки Bacillus, включающий:
а) трансформацию клетки-хозяина Bacillus вектором по п.3, где клетка Bacillus способна экспрессировать представляющий интерес гетерологичный белок; и
b) выращивание указанной модифицированной клетки в условиях роста, благоприятствующих экспрессии указанного представляющего интерес гетерологичного белка.
17. Способ по п.16, где указанный вектор, содержащий указанную полинуклеотидную конструкцию, находится на реплицирующейся плазмиде.
18. Способ по п.16, где указанная конструкция интегрирована в геном модифицированной клетки.
19. Способ по п.16, где указанным представляющим интерес белком является субтилизин.
20. Способ получения представляющего интерес белка в модифицированной клетке Bacillus, способной продуцировать указанный представляющий интерес гетерологичный белок, где указанный способ включает:
а) культивирование модифицированной клетки-хозяина по п.5, где модифицированная клетка способна сверхэкспрессировать YmaH; и
b) выращивание указанной модифицированной клетки Bacillus в условиях роста, благоприятствующих экспрессии указанного представляющего интерес гетерологичного белка.
21. Способ по п.20, дополнительно включающий стадию выделения указанного представляющего интерес белка.
22. Способ по п.20, где указанный представляющий интерес белок продуцируется за меньший период времени, чем период времени, за который указанный белок продуцируется соответствующей клеткой-хозяином-предшественником.
23. Способ по п.20, где промотор aprE инициирует экспрессию указанного представляющего интерес белка.
24. Способ по п.20, где указанным представляющим интерес белком является фермент.
25. Способ усиления экспрессии представляющего интерес гетерологичного белка из Bacillus, включающий:
а) получение модифицированной клетки Bacillus, содержащей сверхэкспрессирующийся YmaH, путем трансформации родительской клетки-хозяина Bacillus химерным полинуклеотидом по п.1;
b) выращивание модифицированной клетки Bacillus в условиях,
благоприятствующих клеточному росту, и
c) экспрессию указанного представляющего интерес белка в модифицированной клетке Bacillus, где экспрессия указанного представляющего интерес гетерологичного белка в модифицированной клетке Bacillus усиливается по сравнению с экспрессией указанного представляющего интерес белка в указанной родительской клетке-хозяине Bacillus.
26. Способ по п.25, где указанный химерный полинуклеотид содержит полинуклеотидную последовательность, выбранную из SEQ ID NOS: 1, 2, 3 и 13.
27. Способ по п.25, где указанный химерный полинуклеотид находится в плазмиде или интегрирован в геном модифицированной клетки.
28. Способ по п.25, где указанной клеткой-хозяином является клетка дикого типа.
29. Способ по п.25, где указанной клеткой-хозяином является измененная клетка-хозяин.
30. Способ по п.25, где указанным представляющим интерес белком является фермент.
| ROBERT A | |||
| BRITTON, et al | |||
| "Genome-Wide Analysis of the Stationary-Phase Sigma Factor (Sigma-H) Regulon of Bacillus subtilis", Journal of bacteriology, Sept | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Переносная печь-плита | 1920 |
|
SU184A1 |
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| Печь для низких температур | 1926 |
|
SU4881A1 |
| JESSICA M | |||
| SILVAGGI et al "Small Untranslated RNA Antitoxin in Bacillus subtilis", JOURNAL OF BACTERIOLOGY, Oct | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Индукционная катушка | 1920 |
|
SU187A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| УСТРОЙСТВО ДЛЯ ИЗМЕРЕНИЯ КОЛИЧЕСТВА ПОТРЕБЛЯЕМОГО ТЕПЛА В НАГРЕВАТЕЛЬНЫХ УСТАНОВКАХ | 1926 |
|
SU6641A1 |

