Заявленное техническое решение (предлагаемый штамм) относится к области биотехнологии, а именно к микроорганизмам и ферментам. Может быть использовано в микробиологической промышленности и генетической инженерии.
Наличие дефекта по генам протеаз или дефицит таких генов у бактерий делает их (бактерии) универсальными продуцентами рекомбинантных белков, например, применяемых для производства лекарственных препаратов со стабильными свойствами.
Известен штамм бактерий Bacillus brevis 31-OK [1] с низкой протеолитической активностью, полученный путем спонтанной мутации после трансформации штамма Bacillus brevis HPD31 несущей ген гормона роста человека плазмидой pNU2ООhGH.
Недостатком [1] является зависимость продуктивности штамма по синтезу ферментов от состава питательной среды. Например, для приготовления питательной среды требуется специфическая питательная среда с обязательным добавлением полипептона PI в концентрации до 2%, отсутствие которого существенно снижает выход продукции - ферментов. Эта зависимость сужает перечень применяемых для культивирования штамма (Bacillus brevis 31-OK) питательных сред, что, в свою очередь, существенно ограничивает область применения этого штамма в качестве производителя рекомбинантных белков, например, в производстве лекарственных препаратов.
Известен полученный путем многократного УФ (ультрафиолетового) облучения дефектный по протеазам штамм бактерий Bacillus amyloliquefaciens A50-32 [2], обладающий 0,3% протеазной активностью (0,40 ед/мл).
Недостатком штамма бактерий Bacillus amyloliquefaciens A50-32 [2] является наличие нескольких неконтролируемых и непредсказуемых мутаций, например регуляторной мутации. Эти мутации оказывают плейотропный (множественный) эффект на синтез других экзоферментов. Из-за этого недостатка (неконтролируемых и непредсказуемых мутаций) продукты жизнедеятельности штамма (экзоферменты), например те, которые могут быть использованы для производства лекарственных препаратов, нестабильны по свойствам и действию, что существенно ограничивает область применения аналога.
Известен способ получения щелочной фосфатазы животного происхождения [3]. Недостатки [3]: (1) способ основан на использовании труднодоступного органического компонента хитозана, (2) применение этанола для осаждения щелочной фосфатазы приводит к удорожанию технологии получения фермента, что существенно ограничивает осуществление этого способа.
Известен способ получения щелочной фосфатазы животного происхождения [4]. Недостатком [4] является сложная технологическая цепочка получения фермента, которая приводит к увеличению времени и затрат на производство фермента этим способом.
Наиболее близким по существу заявляемым техническим решением (прототипом) является протеазодефицитный штамм бактерий Bacillus subtilis GX4924 [5] с направленной инактивировацией гена субтилизина (сокращенное название гена apr).
К недостаткам прототипа [5] относится наличие у прототипа гена другой протеазы (сокращенное название гена npr), выступающей «помехой», которая вызывает дополнительную деградацию рекомбинантных полипептидов, количественно ограничивающих синтез полезных экзоферментов, снижает количество и качество получаемого продукта, например, применяемого в фармацевтике для производства лекарственных препаратов в форме ферментативных добавок, например, для лечения недостаточности экзокринной функции поджелудочной железы. Кроме того, недостатком прототипа является использование сложной методологической базы (на основе профагов - то есть генома фага, интегрированного в хромосомную ДНК штамма бактерий Bacillus subtilis GX4924) для инактивации одного генетического элемента. Прототип также не обладает секреторной способностью в отношении фермента фосфатазы.
Целью заявленного технического решения является получение штамма-реципиента бактерий вида Bacillus pumilus с низкой протеолитической активностью для производства чистых гомологичных и рекомбинантных белков, повышение чистоты используемых в полезных целях продуктов жизнедеятельности бактерий, повышение его продуктивности как продуцента щелочной фосфатазы.
Цели достигают тем, что в качестве реципиента с низкой протеолитической активностью применяют штамм бактерий Bacillus pumilus 2A-5 ВКПМ В-10743, получаемый от исходного штамма бактерий Bacillus pumilus 3-19 путем инактивирования гена минорной сериновой протеазы (глутамилэндопептидазы) на хромосоме.
Заявленное техническое решение поясняется Фиг.1., 2., 3 и таблицей, на которых представлены:
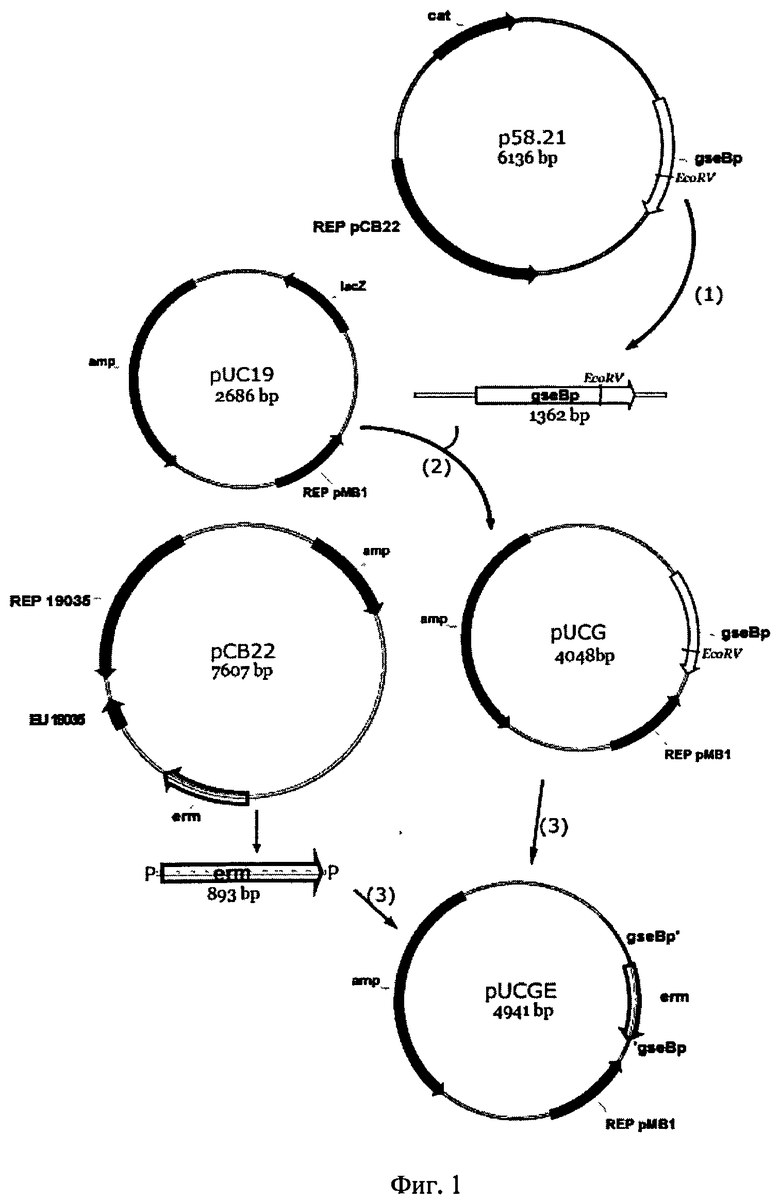
На Фиг.1. представлена схема поэтапного создания плазмиды (pUCGE), содержащей рекомбинирующую конструкцию для инактивации гена gseBp в геноме Bacillus pumilus 3-19.
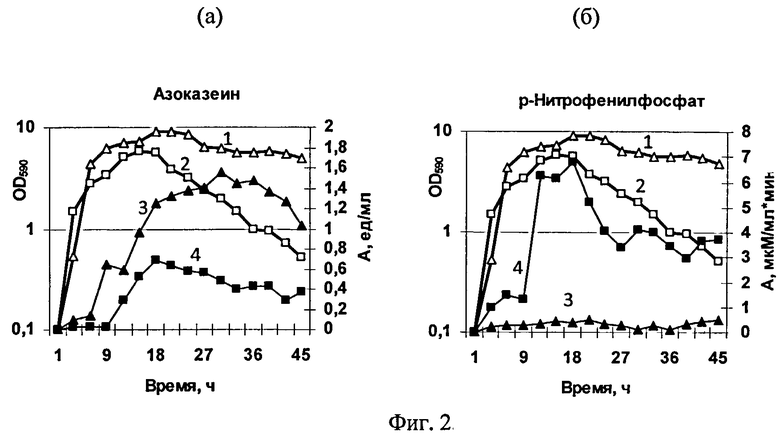
На Фиг.2. представлена динамика роста (1/2) и активность протеолитических ферментов (3/4) штаммов бактерий Bacillus pumilus: исходный стрептомицинустойчивый 3-19 (треугольники); предлагаемый штамм 2А-5 с генотипом gseBp::erm (квадраты).
Сущность заявленного технического решения заключается в следующем: штамм бактерий Bacillus pumilus 2A-5, полученный путем инактивации гена глутамилэндопептидазы в геноме Bacillus pumilus 3-19, используют в качестве реципиента с низкой протеолитической активностью; штамм бактерий применяют для производства гомологичных белков Bacillus pumilus; штамм бактерий применяют для производства гетерологичных белков рода Bacillus; штамм бактерий применяют для производства фосфатазы.
Предлагаемый штамм Bacillus pumilus 2A-5 получают описанным далее путем.
С применением известных компьютерных программ, например программного обеспечения Oligo Calc [7], известным путем [8] конструируют олигонулеотидные праймеры с использованием общедоступной базы данных NCBI (National Center for Biotechnology Information - Национальный центр биотехнологической информации) [9]. Последовательности гена глутамилэндопептидазы (gseBp) и гена резистентности к антибиотику эритромицину (erm) взяты из общедоступной базы данных NCBI. Последовательности праймеров приведены в Таблице.
Дальнейшие действия показаны на Фиг.1.
(1) С плазмиды р58.21 известным методом полимеразно-цепной реакции (далее по тексту ПЦР), с использованием праймеров Glu pd/Glu stop амплифицируют ген протеазы gseBp.
(2) Полученный продукт амплификации по сайтам рестрикции BamHI и HindIII клонируют в вектор pUC19 и получают плазмиду pUCG.
(3) Поскольку ген gseBp несет уникальный (единственный) сайт рестрикции EcoRV, его используют для клонирования гена erm. Для этого проводят амплификацию гена erm с плазмиды рСВ22 с использованием праймеров Em 1/Em 2. Затем с помощью Т4-полинуклеотидкиназы, например, фирмы «Fermentas», Литва, присоединяют фосфатную группу к 5'-концам амплификата (erm) и впоследствии клонируют его (Р~erm~Р) в плазмиду pUCG. Вектор pUCG предварительно лианеризуют по сайту EcoRV, например, с использованием фермента фирмы «Сибэнзим», Москва. В результате получают плазмиду pUCGE.
Заключительным этапом по получению протеазодефицитного штамма бактерий Bacillus pumilus является трансформация клеток бактерий Bacillus pumilus 3-19 полученной плазмидой pUCGE с происходящей вследствие этого гомологичной рекомбинацией участков ДНК. Трансформацию проводят по методу [10], при этом используют синтетические минимальные питательные среды Спецайзена I/II. Солевая основа среды Спецайзена (%): К2НРО4×3H2O - 18,34; KH2PO4 (безводный) - 6,0; (NH4)2SO4 - 2,0; цитратNa - 1,2. Отдельно готовят растворы: глюкоза - 40%; казаминовые кислоты - 200 мг/мл; Mg2SO4 - 0,2 г/мл. Среда Спецайзена I: на 100 мл H2O: глюкоза - 2 мл; казаминовые кислоты - 2 мл; дрожжевой экстракт - 2 мл; Mg2SO4 - 0,1 мл; солевая основа - 10 мл. Среда Спецайзена II: на 100 мл H2O: глюкоза - 2 мл; казаминовые кислоты - 1 мл; Mg2SO4 - 8 мкл; солевая основа - 10 мл.
Суспензию компетентных клеток Bacillus pumilus 3-19, содержащих плазмиду pUCGE, рассевают на чашки с агаризованной средой, содержащей антибиотик эритромицин, например, в концентрации 20 мкг/мл. После инкубации в течении от 24 до 48 часов при плюс 37°С отбирают, например, три растущих на среде с эритромицином независимых клона. Проверку наличия или отсутствия гена протеазы выполняют с применением ПЦР. Разрушение функционального гена протеазы gseBp по результату ПЦР (достижение цели изобретения) подтверждают наличием вставки гена erm. Выбранные колонии модифицированного штамма бактерий именуют как Bacillus pumilus 2A-5 (т.е. предлагаемый штамм имеет генотип gseBp::erm). Далее проводят определение протеолитической активности нокаутированного по гену gseBp штамма бактерий на питательной среде, например, содержащей 2% молочного агара. Наличие протеолитической активности при этом определяют по появлению области просветления на молочном агаре (зоны расщепления белков молока). У выбранных клонов, например, трех колоний, которые не имеют зон просветления на молочной среде, определяют протеолитическую активность с использованием специфического хромогенного субстрата для фермента глутамилэндопептидазы Z-Glu-pNa, как описано в [11].
Из отобранных на молочном агаре клонов с отсутствием зон протеолиза селекционируют клоны, не проявляющие активность в отношении Z-Glu-pNa субстрата.
В результате селекционирования получают предлагаемый штамм, свойства которого приводятся далее.
Описание предлагаемого штамма
Штамм бактерий Bacillus pumilus 2A-5 депонирован во Всероссийской коллекции промышленных микроорганизмов ФГУП ГосНИИГенетика под регистрационным номером ВКПМ В-10743. Предлагаемый штамм является производным от штамма бактерий Bacillus pumilus 3-19 и содержит гены протеаз, например субтилизиноподобной протеазы, адамализиноподобной металлопротеазы.
Генетические особенности штамма
В геноме предлагаемого штамма бактерий Bacillus pumilus 2A-5 методом гомологичной рекомбинации ген глутамилэндопептидазы разрушен путем встраивания кодирующей области гена эритромицина (gseBp::erm).
Культурально-морфологические особенности штамма
Предлагаемый штамм Bacillus pumilus 2A-5 представляет собой аэробные грамположительные бактерии. Клетки палочковидные, прямые или изогнутые, располагаются скоплениями или обособленно. Размеры клеток односуточный культуры, выращенной на агарозной среде, составляют 13…15 мкм. Свободные споры появляются на сороковой час роста (не более 5%), далее их (спор) количество нарастает и достигает максимального значения (до 80%) на 76-80-й час культивирования. При росте на агаризованной среде Лурия-Бертани (далее по тексту LB) колонии имеют белую окраску, более шероховатую поверхность и неровные края по сравнению с более гладкими и ровными колониями исходного штамма бактерий Bacillus pumilus 3-19, пигмента не образуют.
Физиолого-биохимические свойства
Оптимальная температура роста: +37°С. Среда LB (г/л): триптон - 1,0; дрожжевой экстракт - 0,5; Nad - 0,5; рН 8.5. Для создания твердой питательной среды дополнительно вносят 2% агар. Штамм устойчив к антибиотикам, например стрептомицину в концентрации 500 мкг/мл, эритромицину в концентрации 20 мкг/мл.
Протеолитическая активность предлагаемого штамма
Активность предлагаемого штамма определяют различными способами.
Определение активности предлагаемого штамма с применением азоказеина
Азоказеин - неспецифический хромогенный растворимый белковый субстрат, который расщепляется большинством внеклеточных протеаз. Несмотря на то что в предлагаемом штамме бактерий Bacillus pumilus 2A-5 инактивирован ген минорного фермента глутамилэндопептидазы (GseBp) и клетки должны сохранять способность к расщеплению предпочтительного белкового субстрата (азоказеина), уровень активности штамма по гидролизу азоказеина снижается в 3.5 раза (Фиг.2 а). Протеолитическая активность предлагаемого штамма бактерий Bacillus pumilus 2A-5 появляется после 10 ч культивирования и достигает в среднем 40-50% от активности исходного штамма бактерий (Bacillus pumilus 3-19). Такой полярный эффект, вызванный инактивацией гена gseBp, связан с участием фермента глутамилэндопептидазы во множественном процессинге (созревании) активных форм различных протеаз Bacillus pumilus. Протеолитическую активность по расщеплению азоказеина определяют, как описано в [12]. Методика: к 100 мкл субстрата (раствор азоказеина («Sigma», США) в концентрации 10 мг/мл в 0.05 М Трис-HCl буфере, рН 7.3) добавляют 50 мкл культуральной жидкости (далее по тексту КЖ) и инкубируют при +37°С в течение 1 ч. Реакционную смесь осаждают 200 мкл 10% трихлоруксусной кислотой (далее по тексту ТХУ) и выдерживают 10 мин на ледяной бане. Далее пробы центрифугируют 5 мин при 13 тыс. об/мин, отбирают 250 мкл супернатанта и добавляют 50 мкл 4 н. NaOH. Контрольным является образец, в котором изначально к субстрату добавляют ТХУ, выдерживают вместе с опытными пробами в течение 1 ч при +37°С, а затем добавляют 50 мкл КЖ. Против данного контроля проводят измерение оптической плотности при длине волны излучения λ=450 нм, например, на спектрофотометре SmartSpec Plus (BioRad, США). За единицу активности принимают количество фермента, гидролизующего в условиях эксперимента 1 мкг субстрата за 1 мин.
Определение активности щелочной фосфатазы (фосфогидролазы) предлагаемого штамма
Для исходного штамма бактерий Bacillus pumilus 3-19 ранее показана способность секретировать в среду щелочную фосфогидролазу при выращивании культуры на среде с низким содержанием фосфата [13]. Фосфатаза Bacillus pumilus 3-19 обладает широкой субстратной специфичностью и способна мобилизировать неорганический фосфат из природных субстратов. В неиндуцированных условиях исходный штамм бактерий Bacillus pumilus 3-19 показал отсутствие фосфатазой активности на всех фазах развития культуры, поскольку в состав питательной среды (LB) входит триптон, обогащенный фосфатами. В то же время для предлагаемого штамма бактерий Bacillus pumilus 2A-5 с инактивированным геном gseBp наблюдали повышение активности фосфатазы в среднем в 10 раз на всем протяжении культивирования (Фиг.2б). Таким образом, инактивация гена gseBp привела к активации фермента фосфатазы, что может свидетельствовать об инактивации фосфатазы у дикого штамма глутамилэндопептидазой и является существенным обстоятельством при изыскании продуцентов фосфатазы. Фосфомоноэстеразную активность [14] определяют по действию фермента (или КЖ) на р-нитрофенилфосфат (p-NPP) («Serva», Германия). Субстрат растворяют в 0.2 М Трис-HCl буфере, рН 8.5, содержащем 0.05 М MgCl2 Реакционную смесь, содержащую 0,2 мл субстрата (2 мг/мл) и 0,2 мл раствора фермента, инкубируют в течение одного часа при +37°С до появления желтой окраски. Реакцию останавливают добавлением 0,8 мл 0.4 н. NaOH и пробы центрифугируют при 13000 об/мин в течение 3 мин. В надосадочной жидкости измеряют оптическую плотность на спекрофотометре, например SmartSpec Plus (BioRad, США) при длине волны излучения λ=410 нм. Параллельно с опытными пробами инкубируют контрольную пробу, исключающую поглощение самого субстрата и фермента, в которую фермент добавляют после осаждения 0.4 н. NaOH. За единицу активности фосфатазы принимают количество фермента, которое вызывает увеличение оптической плотности на одну спекрофотометрическую единицу при λ=410 нм в пересчете на 1 мл ферментного раствора за 1 час инкубации. Активность (Д) рассчитывали по формуле:
А=(A410-ΣА410контр.)×3×a×b×t,
где A - активность, выраженная в условных единицах;
A410 - плотность опытной пробы;
ΣА410контр. - сумма плотностей контрольных проб;
3 (три) - коэффициент разведения при остановке реакции раствором щелочи;
а - пересчет на 1 мл фермента;
b - пересчет активности фермента с учетом разведения;
t - время проявления активности фермента (для пересчета активности на 1 час).
Применение предлагаемого штамма бактерий Bacillus pumilus 2A-5
Предлагаемый штамм применяют, например, следующим путем для производства и синтеза рекомбинантных гомо- и гетерологичных белков р. Bacillus, используемых для создания лекарственных препаратов. Предлагаемый штамм лишен способности продуцировать минорную протеазу - глутамилэндопептидазу GseBp и его общая протеолитическая активность достигает ~40% от активности исходного штамма. Производство протеаз возможно благодаря локализации желаемых генов на плазмидах или на хромосоме предлагаемого штамма бактерий. При этом предлагаемый штамм бактерий Bacillus pumilus 2A-5 позволяет эффективно отбирать клоны, содержащие конструкции с генами протеаз благодаря интегрированному в хромосому селекционному маркеру - резистентности к антибиотику эритромицину.
Таким образом, предлагаемый штамм Bacillus pumilus 2A-5 обеспечивает возможность его основного использования в качестве реципиента для экспрессии гомологичных генов из бактерий рода Bacillus pumilus. Данное свойство связано с отсутствием вариаций последовательностей Shine Dalgamo (Шайна Дальгарно) в промоторах генов, выделенных из штаммов Bacillus pumilus. Поэтому предлагаемый штамм является наиболее эффективным при производстве промышленно-важных ферментов, например субтилизиноподобной протеазы Bacillus pumilus, имеющей потенциал в создании тромболитических лекарственных препаратов [15]. То есть предлагаемый штамм позволяет избавиться от присущих прототипу возможных изменений эффективности трансляции белков и тем самым добиться стабильности свойств продуктов, получаемых как результат жизнедеятельности этих микроорганизмов и реализуемых в биотехнологиях производства препаратов, например, применяемых в медицине, ветеринарии, сельскохозяйственном производстве и производстве продуктов питания.
Предлагаемое изобретение представляет собой штамм Bacillus pumilus 2A-5, который авторы используют в качестве хозяина для экспрессии гомологичных генов Bacillus pumilus, например, нижеописанным методом.
1. Клетки бактерий предлагаемого штамма Bacillus pumilus 2A-5 трансформируют плазмидой, содержащей ген рекомбинантного белка (например, гена субтилизиноподобной протеазы aprBp штамма Bacillus pumilus 3-19) под контролем индуцибельного или конститутивного промотора.
2. Полученные трансформированные клетки подращивают на питательной среде, например LB, при температуре инкубации +37°С. Индукцию синтеза белка осуществляют в соответствии с выбранным промотором.
3. Пробы для анализа белков отбирают из КЖ путем отделения супернатанта от клеточной массы, например через 1 ч и 16 ч после индукции.
4. Внеклеточные белки в пробе (КЖ) характеризуют с помощью электрофореза в полиакриламидном геле по Лэммли [16] и определением активности.
5. Рекомбинантные белки далее очищают различными методами, например с помощью аффинной хроматографии, гель-фильтрации, ионно-обменной хроматографии, высаливания.
Таким образом, предлагаемый штамм бактерий Bacillus pumilus 2A-5 позволяет повысить уровень синтеза и качество целевого белка за счет снижения его (белка) деградации. Также предлагаемый штамм применяют для экспрессии гетерологичных генов р. Bacillus по приведенному выше методу.
Другое применение предлагаемого штамма бактерий Bacillus pumilus 2A-5 выполняют для производства щелочной бактериальной фосфогидролазы, полученной по упрощенной методике. То есть только инактивацией минорного протеолитического фермента (GseBp) в исходном штамме Bacillus pumilus 3-19 удалось повысить выход фосфатазы. Подобный эффект описан, в частности, для клеток Bacillus subtilis, где при делегировании гена определенной протеазы повышается уровень других ферментов [17]. Фосфатаза имеет потенциал использования в генной инженерии как фермент, гидролизующий нуклеиновые кислоты (РНК и ДНК) до нуклеозидов, т.е. востребована, в первую очередь, для получения химических реактивов в научно-исследовательской работе. Фосфатазы применяют также для диагностики вирусных, бактериальных и паразитарных заболеваний. Таким образом, получение предлагаемого штамма по описанному пути является простым и легким способом повышения выхода фермента - щелочной бактериальной фосфогидролазы. Использование предлагаемого штамма бактерий Bacillus pumilus 2A-5 для выделения и очистки щелочной фосфатазы не требует больших затрат материалов, сырья, рабочего и технологического времени.
Таким образом, данное в заявке описание в основном раскрывает два специфичных выполнения/применения изобретения. Одно из них - это наработка (гомологичных или гетерологичных) внеклеточных белков в протеазо-негативном предлагаемом штамме бактерий Bacillus pumilus 2A-5 на основе плазмид или путем генного обмена на хромосоме. Такие ферменты (например амилазы, липазы, протеазы и др.) востребованы в детергентной промышленности, медицине, фармакологии. Второе - получение щелочной фосфатазы Bacillus pumilus с повышенной активностью. Активность фермента выше в 4-6 раз по сравнению с исходным штаммом.
Заявляемое техническое решение - предлагаемый штамм бактерий Bacillus pumilus 2A-5 удовлетворяет критерию «новизны», так как при определении уровня аналогичных описанных штаммов не обнаружен штамм бактерий, которому присущи признаки, идентичные (то есть совпадающие по исполняемой им функции и форме выполнения этих признаков) всем признакам, перечисленным в формуле изобретения. Предлагаемый штамм имеет изобретательский уровень, поскольку не выявлены технические решения, имеющие признаки, совпадающие с отличительными признаками данного изобретения, и не установлена известность влияния отличительных признаков на указанный технический результат. Таким образом, описание и характеристика предлагаемого штамма бактерий Bacillus pumilus 2A-5 предполагает его применение в биотехнологии получения и разработки рекомбинантных белков, фосфатазы и способствует выявлению регуляторных свойств инактивированного фермента GseBp, что вносит свой вклад в фундаментальную молекулярную биологию.
Источники информации
1. Kajino Т., Kato К., Miyazaki С., Asami О., Hirai M., Yamada Y., Udaka S. Isolation of a protease-deficient mutant of Bacillus brevis and efficient secretion of a fungal protein disulfide isomerase by the mutant / J. Biosci Bioeng. 1999. 87(1): 37-42.
2. Plotnikova Т.О., Selivanova G.N., Iomantas lu.V., Kozlov Iul. Isolation and analysis of protease-deficient mutants of Bacillus amyloliquefaciens I Genetika. - 1992. 28(5): 66-72.
3. Авторское свидетельство СССР SU 1738108 «Способ получения щелочной фосфатазы», Заявка: 4886245/13, 28.11.1990. http://wwwl.fips.ru
4. Патент России RU 2016899 «Способ получения щелочной фосфатазы», Заявка: 4946819/13, 19.06.1991. http://wwwl.fips.ru
5. Fahnestock S.R., Fisher K.E. Protease-deficient Bacillus subtilis host strains for production of Staphylococcal protein A / Appl. Environ. Microbiol. 1987. 53(2): 379-84.
6. J. Sambrook, D. W. Russell Molecular cloning. A laboratory manual. Third edition. Cold Spring Harbor Laboratory Press, New York. 2001.
7. http://www.basic.northwestern.edu/biotools/oligocalc.html
8. Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии. Молекулярное клонирование // Москва «Мир». 1984.
9. http://www.ncbi.nlm.nih.gov/
10. Harwood, С.R., S.M.Cutting. 1990. Molecular biological methods for Bacillus. John Wiley & Sons, Chichester, England.
11. Houmard J. // Eur. J. Biochem. - 1967. - V.68. - P.621-628.
12. Charney J., Tomarelli R.M. A colorimetric method for the determination of the proteolytic activity of duodenal juice // J Biol Chem. 1947. 171(2): 501-505.
13. Шарипова М.Р., Балабан Н.П., Лещинаская И.Б. Продукция внеклеточной щелочной фосфатазы антибиотикоустойчивыми штаммами Bacillus intermedius // Микробиология. 1994. Т.63. Вып.1. С.52-58.
14. Лещинская И.Б., Балабан Н.П., Капранова М.Н., Голубенко И.А. Методы определения активности нуклеаз и родственных ферментов [Текст] // Современные методы изучения нуклеиновых кислот и нуклеаз микроорганизмов, Издательство КГУ. - Казань. - 1980. - С.53-60.
15. Ицкович Е.Л., Лютова Л.И., Балабан Н.П., Марданова A.M., Шакиров Е.В., Шарипова М.Р., Лещинская И.Б., Руденская Г.Н. Тромболитические и антикоагулянтные свойства тиолзависимой сериновой протеиназы Bacillus intermedius 3-19 / Вопросы медицинской химии, 1999. №5.
16. Laemmli, U. К. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4 [Text] / U.K.Laemmli // Nature. - 1970. - V.227. - P.680-685.
17. Westers L., Westers H., Zanen G., Antelmann H., Hecker M., Noone D., Devine K.M., van Diji J.M., Quax W.J. Genetic or chemical protease inhibition causes significant changes in the Bacillus subtilis exoproteome / Proteomics. - 2008. - V.8. - No.13. - P.2704-2713.
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ БАКТЕРИЙ BACILLUS PUMILUS МК-10 С НИЗКОЙ ПРОТЕОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2510821C1 |
| СПОСОБ ЭКСПРЕССИИ | 2012 |
|
RU2642324C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pPHOFus, КОДИРУЮЩАЯ СУБСТРАТ ЛЕТАЛЬНОГО ФАКТОРА СИБИРСКОЙ ЯЗВЫ, СЛИТЫЙ СО ЩЕЛОЧНОЙ ФОСФАТАЗОЙ Escherichia coli, И ШТАММ Escherichia coli BL-PHOFus, ПРОДУЦИРУЮЩИЙ БЕЛОК - СУБСТРАТ ЛЕТАЛЬНОГО ФАКТОРА СИБИРСКОЙ ЯЗВЫ В СОСТАВЕ ЩЕЛОЧНОЙ ФОСФАТАЗЫ Escherichia coli | 2009 |
|
RU2416637C1 |
| СПОСОБ ПОЛУЧЕНИЯ СЕРИНОВОЙ ПРОТЕАЗЫ, ШТАММ ЩЕЛОЧЕФИЛЬНЫХ BACILLUS-ПРОДУЦЕНТ СЕРИНОВОЙ ПРОТЕАЗЫ | 1988 |
|
RU2023723C1 |
| Лабораторный способ биосинтеза бациллярной рибонуклеазы (варианты) | 2024 |
|
RU2835675C1 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОГО ШТАММА BACILLUS SUBTILIS ТВ-06 В КАЧЕСТВЕ ПРОДУЦЕНТА ГЛУТАМИЛЭНДОПЕПТИДАЗЫ, ВЕЩЕСТВА С ТРОМБОЛИТИЧЕСКИМИ И АНТИКОАГУЛЯНТНЫМИ СВОЙСТВАМИ | 2015 |
|
RU2611378C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПУРИНОВЫХ РИБОНУКЛЕОЗИДОВ И РИБОНУКЛЕОТИДОВ | 2008 |
|
RU2422510C2 |
| УЛУЧШЕННАЯ ПРОДУКЦИЯ БЕЛКА В BACILLUS | 2008 |
|
RU2515112C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |
Изобретение относится к области биотехнологии и представляет штамм - реципиент Bacillus pumilus 2A-5 ВКПМ В-10743 с низкой протеолитической активностью. Изобретение позволяет расширить ассортимент штаммов с низкой протеолитической активностью для производства чистых гомологичных и рекомбинантных белков, а также с повышенной продуктивностью в качестве продуцента щелочной фосфатазы. 3 з.п. ф-лы, 2 ил., 1 табл.
1. Штамм бактерий Bacillus pumilus 2A-5 ВКПМ В-10743 - реципиент с низкой протеолитической активностью, полученный путем инактивации гена глутамилэндопептидазы в геноме Bacillus pumilus 3-19.
2. Штамм бактерий по п.1 применяют для производства гомологичных белков Bacillus pumilus.
3. Штамм бактерий по п.1 применяют для производства гетерологичных белков рода Bacillus.
4. Штамм бактерий по п.1 применяют для производства фосфатазы.
| FAHNESTOCK S.R | |||
| et.al | |||
| Protease-deficient Bacillus subtilis host strains for production of Staphylococcal protein A., Appl Environ Microbiol | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| RU 2060276 C1, 20.05.1996 | |||
| WO8601825 A1, 27.03.1986 | |||
| ШТАММ БАКТЕРИИ DELCYA MARINA - ПРОДУЦЕНТ ЩЕЛОЧНОЙ ФОСФАТАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ЩЕЛОЧНОЙ ФОСФАТАЗЫ | 1994 |
|
RU2077577C1 |
| . | |||

