Данное изобретение относится к способу выделения плюрипотентных/мультипотентных стволовых клеток, происходящих из пуповинной крови, где способ характеризуется культивированием моноцитов, выделенных из пуповинной крови, в сосуде для культивирования, содержащем фибронектин, и последующим сбором стволовых клеток из культуры, с выделением, таким образом, происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток; и к клеточному терапевтическому средству, содержащему происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки или дифференцированные из них клетки. Данное изобретение также относится к новой среде для культивирования стволовых клеток, способу культивирования стволовых клеток, где способ характеризуется культивированием и пролиферацией стволовых клеток в среде для культивирования, и способу увеличения «стволовости» стволовых клеток, которая характеризуется сферичной или трехмерной культурой стволовых клеток.
Предшествующий уровень техники
Стволовые клетки, характеризующиеся тем, что являются самообновляющимися, подвергаются дифференцировке и являются иммортализованными, были предложены для решения проблем регенеративной медицины и тканевой реплантации, таким образом, их можно использовать для лечения различных дегенеративных заболеваний, так же как для предоставления возможности глубокого понимания клеточной биологии. Стволовые клетки взрослых, получаемые из различных тканей, являются более предпочтительными, чем эмбриональные стволовые клетки, вследствие того, что стволовые клетки взрослых можно получать из неограниченного числа источников, и этические соображения для использования человеческих эмбрионов в качестве источника клеток утрачивают свое значение. Кроме того, стволовые клетки, выделенные из пуповинной крови, обладают преимуществами по сравнению с другими стволовыми клетками взрослых в том отношении, что доноры пуповинной крови не подвергаются травмированию, в отличие от доноров костного мозга или жировой ткани.
Происходящие из пуповинной крови мезенхимальные стволовые клетки успешно вводили in vitro в различные виды клеток, включающие нервные клетки, гепатоциты, остеоциты и т.д. (Sun, W. et al., Stem cells, 23:931, 2005; Hong SH. et al., Biochem. Biophys. Res. Commun., 30:1153, 2005; Hutson EL. et al., Tissue Engineering, 11:1407, 2005). Также сообщали об успешности in vivo трансплантации происходящих из пуповинной крови мезенхимальных стволовых клеток при травмах, сахарном диабете и инфаркте миокарда (Nonome, K. et al., Am, J. Physiol. Gastrointest. Liver Physiol., 289:1091, 2005; Yoshida, S. et al., Stem cells, 23:1409, 2005; Kim Bo. et al., 25 Circulation, 112:96, 2005). Вследствие низкой вероятности заражения инфекционными заболеваниями и низкой вероятности возникновения реакции «трансплантат против хозяина», трансплантацию происходящих из пуповинной крови мезенхимальных стволовых клеток все чаще применяют в отношении как пациентов детского возраста, так и взрослых пациентов (Claudio G. B. et al., Annual Review of Medicine, 57:403, 2006). Несмотря на то что трансплантацию пуповинной крови применяли в качестве терапии некоторых заболеваний, а конкретно - заболеваний, связанных с дефектом в системе кроветворения (Grewal, SS. et al., Blood, 103:1147, 2004; Knutsen, AP. et al., Journal pediatrics, 142:519, 2003; Ooi, J. et al., Blood, 103:489, 2004; Sanz GF. et al., Blood, 103:489, 2004), количество исследований клинического применения происходящих из пуповинной крови мезенхимальных стволовых клеток все еще ограничено. Например, существуют сообщения, что стволовые клетки используют для не полностью, но частично успешного лечения женщин с поражениями спинного мозга и пациентов с болезнью Бюргера (Kim, SW. et al., Stem cells, 2006; Kang, KS. et al., Cytotherapy, 7:368, 2005). Однако механизм развития или роста и пролиферации клеток все еще не известен.
Выделить мезенхимальные стволовые клетки из пуповинной крови трудно. Например, если способ, используемый для выделения происходящих из костного мозга мезенхимальных стволовых клеток, применяют для пуповинной крови, степень выделения остается на уровне приблизительно 20%. До 50% мезенхимальных стволовых клеток можно выделить из свежей крови, которую отобрали за 5 часов до выделения. Однако степень выделения снижается до 20% или менее, если кровь отобрали более чем за 5 часов до выполнения выделения, и клетки, хотя и выделены, но плохо пролиферируют.
Описание
Техническая проблема
В результате предпринятого авторами изобретения интенсивного и всестороннего исследования в области выделения и массовой пролиферации стволовых клеток из пуповинной крови, приведшего к настоящему изобретению, получили данные о том, что при культивировании в присутствии фибронектина, моноциты, выделенные из пуповинной крови человека, активно пролиферируют и образуют колонии, демонстрирующие веретенообразную морфологию, и даже после многочисленных пассажей сохраняют свойства, присущие стволовым клеткам. Таким образом, на основе полученных данных разработан эффективный способ выделения происходящих из пуповинной крови негематопоэтических плюрипотентных/мультипотентных стволовых клеток и мезенхимальных стволовых клеток и способ их массовой пролиферации.
Таким образом, задачей настоящего изобретения является разработка способа выделения происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, содержащих культивируемые в присутствии фибронектина моноциты, выделенные из пуповинной крови и выделение стволовых клеток из культуры.
Другой задачей настоящего изобретения является предоставление происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, выделенных посредством способа выделения.
Дополнительной задачей настоящего изобретения является предоставление клеточного терапевтического средства, содержащего происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки или дифференцированные из них клетки.
Еще одной дополнительной задачей настоящего изобретения является разработка новой среды для культивирования стволовых клеток.
Другой задачей настоящего изобретения является разработка способа культивирования стволовых клеток с использованием среды для культивирования.
Это способ увеличения «стволовости» стволовых клеток с использованием сферичной или трехмерной культуры стволовых клеток.
Техническое решение
В соответствии с аспектом изобретения, изобретение относится к способу выделения происходящих из пуповинной крови стволовых клеток, где способ характеризуется культивированием выделенных из пуповинной крови моноцитов в сосуде для культивирования, содержащем фибронектин, и получение клеток из культуры.
В одном из вариантов осуществления стадия получения стволовых клеток из культуры включает использование иммунологического свойства стволовых клеток для отделения стволовых клеток.
Выделение моноцитов из пуповинной крови можно осуществлять с использованием общепринятого способа. После смешивания пуповинной крови с Hetasep для удаления эритроцитов, выделяют моноциты, используя Фиколл-пак. В настоящем документе, Hetasep предпочтительно используют в количестве 0,5~2 мл на 5 мл Hetasep.
Для увеличения выхода, при выделении моноцитов из пуповинной крови, используемая в настоящем изобретении пуповинная кровь предпочтительно представляет собой такую пуповинную кровь, которую предпочтительно получают непосредственно после родов, хранят при комнатной температуре в течение 12~48 часов после получения или хранят при 3~5°C в течение 6~72 часов после получения.
Выделение стволовых клеток из происходящих из пуповинной крови моноцитов характеризуется использованием фибронектина. Как применяют в настоящем документе, термин «сосуд для культивирования, содержащий фибронектин», предназначен для обозначения условия, при котором моноциты могут подвергаться взаимодействию с фибронектином. Например, фибронектин можно наслаивать на сосуд для культивирования или фибронектин может содержаться в среде для культивирования в форме сфер или трехмерных структур. В одном из вариантов осуществления настоящего изобретения, в случае, если сосуд для культивирования покрывают фибронектином, фибронектин может содержаться из расчета удельного содержания от 0,1 до 1 мг/мл.
Фибронектин, пригодный в настоящем изобретении, может быть животного происхождения, без ограничения, и предпочтительно человеческого происхождения. Также, фибронектин можно получать искусственным синтезом (например, химический синтез, синтез с использованием пептидного синтезатора и т.д.) или биосинтезом (например, технология рекомбинантных ДНК, культура фибробластов и т.д.) или можно выделять из плазмы крови животных, включая людей, из внеклеточных матриксов. Фибронектин может представлять собой фрагмент или пептидную последовательность фибронектина или может содержать фрагмент или пептид.
Никаких конкретных ограничений не предъявляют к среде при культивировании моноцитов в сосуде для культивирования, содержащем фибронектин. В качестве основной среды для культивирования моноцитов предпочтительно используют SNU-1 или EGM-2.
Среда SNU-1 содержит следующую композицию (Таблица 1).
В настоящем изобретении, основную среду предпочтительно обогащают FGF-B (Фактор роста фибробластов), аскорбиновой кислотой, EGF (эпидермальный фактор роста), гидрокортизоном, IGF-1 (инсулиноподобный фактор роста-1) или VEGF (Фактор роста эндотелия сосудов) и гепарином и необязательно GA-1000 (Гентамицина сульфат, Амфотерицин-B), если необходимо.
Более предпочтительно, основную среду обогащают 20% эмбриональной телячьей сывороткой (FBS), bFGF (Фактор роста фибробластов) 1~40 нг/мл, аскорбиновой кислотой 0,1~5,0 мкг/мл, EGF (эпидермальный фактор роста) 1~40 нг/мл, гидрокортизоном 0,1~1 мкг/мл, IGF-I (инсулиноподобный фактор роста-1) 1~40 нг/мл или VEGF (фактор роста эндотелия сосудов) 1~5 нг/мл и гепарином 20~25 мкг/мл и необязательно GA-1000 (Гентамицина сульфат, Амфотерицин-B), если необходимо.
После культивирования в течение трех суток, моноциты, которые остались в суспензии, удаляли, и подвергали культивированию только прикрепленные клетки. Среди моноцитов, прикрепленных к сосуду, пролиферируют только стволовые клетки. Через 12-20 суток после выделения можно наблюдать быструю пролиферацию стволовых клеток. В связи с этим, предпочтительно заменять среду свежей каждые двое или трое суток.
Для получения из культуры происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток можно использовать FACS с использованием проточного цитометра с функцией сортировки (Int. Immunol., 10(3):275, 1998), магнитные шарики или способ пэннинга, основанный на специфичности антител к мезенхимальным стволовым клеткам (J. Immunol., 141(8):2797, 1998). Для получения плюрипотентных/мультипотентных стволовых клеток из массовой культуры можно использовать колонку, на которой по отдельности или в комбинации иммобилизованы антитела, специфичные для молекул, экспрессированных на клеточной поверхности (далее в настоящем документе, обозначаемые как «поверхностный антиген»).
Проточную цитометрию с сортировкой клеток можно выполнять посредством электростатической зарядки капель или «улавливания» клеток. В обоих процессах антитела, специфично распознающие поверхностный антиген, метят флуоресцентной меткой, после чего измеряют интенсивность флуоресценции меченого конъюгата антитело-антиген и сигнал переводят в электрические сигналы для определения уровня экспрессии антигена на поверхности клеток. Дополнительно, различные виды флуорофоров можно использовать в комбинации для разделения клеток, экспрессирующих различные поверхностные антигены. Среди флуорофоров представлены FITC (флуоресцеинизотиоцианат), PE (фикоэритрин), APC (алло-фикоцианин), TR (TexasRed), Cy3, CyChrome, Red613, Red670, TRI-Color и QuantumRed.
В FACS с использованием проточного цитометра, стволовые клетки, собранные из культуры, например, центрифугированием, могут подвергаться прямому иммуноокрашиванию с антителом или пролиферировать в подходящей среде перед иммуноокрашиванием с антителом. Для иммуноокрашивания образец клетки-мишени смешивают с первичным антителом, специфичным для поверхностного антигена, и инкубируют в течение 0,5-1 часа на льду. Если первичное антитело помечено флуорофором, клеточный образец промывают и разделяют в проточном цитометре. Если первичное антитело не помечено флуорофором, клеточный образец, обработанный первичным антителом, промывают и смешивают с флуоресцентно меченным вторичным антителом, которое может связываться с первичным антителом. Затем иммуноокрашенные клетки вновь инкубируют в течение от 0,5 до 1 часа на льду и промывают перед разделением посредством проточного цитометра.
Происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки, выделенные по настоящему изобретению, обладают по меньшей мере одним из следующих свойств:
(a) демонстрируют положительную иммунологическую характеристику по отношению к факторам транскрипции c-myc и ZNF281;
(b) прикрепляются к поверхности покрывающего внеклеточного матрикса и через 5-30 суток после прикрепления образуют клеточные колонии веретенообразной или сферической формы;
(c) демонстрируют CPDL (кумулятивный уровень удвоения популяций) от 30 до 45;
(d) демонстрирует негативную иммунологическую характеристику в отношении CD14, CD31, CD34, CD45 и HLA-DR;
(e) способность к дифференцировке в мезодермальные, эндодермальные и эктодермальные клетки;
(f) секретируют по меньшей мере один цитокин или хемокин, выбранный из группы, состоящей из TIMP-2, TGF-β, RANTES CINC-3, ЭОТАКСИН, GM-CSF, IFN-γ, IL-1b, IL-3, IL-6, IL-8, IL-10, IL12p40, IL13, IL-16, IP-10, лептин, MCP-2, MIG, MIP-3a, b-NGFm, sTNFRI, и PFGF-bb.
Факт того, что полученные из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению остаются недифференцированными, очевиден из наличия экспрессии Oct-4, Sox-2, Rex-1, c-myc и ZNF281 в стволовых клетках.
Кроме того, демонстрирующие CPDL (кумулятивный уровень удвоения популяций) от 30 до 45, полученные из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению являются высоко пролиферативными. Анализ кариотипа показал, что клетки по настоящему изобретению активно пролиферируют, но обладают нормальной хромосомной структурой.
Происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению являются иммунологически негативными в отношении CD14, CD31, CD34, CD45 и HLA-DR, каждый из которых известен как маркер гематопоэтических стволовых клеток или связанный с иммунным отторжением маркер. Вследствие отсутствия подобных маркеров гемопоэза или маркеров, связанных с иммунным отторжением, полученные из пуповинной крови стволовые клетки по настоящему изобретению можно трансплантировать с минимальной васкуляризацией и иммунным отторжением и таким образом можно эффективно использовать в аллогенной трансплантации.
Происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению также могут дифференцировать в гепатоциты из клеток, родственных мезодермальным клеткам, нейронам и клеткам сетчатой оболочки глаза из эктодермы, а также из мезодермы, в остеобласты, хондроциты и адипоциты. Таким образом, происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению можно использовать для лечения различных заболеваний.
Дополнительно, происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению могут секретировать различные цитокины или хемокины, включающие TIMP-2, TGF-β, RANTES CINC-3, ЭОТАКСИН, GM-CSF, IFN-γ, IL-1b, IL-3, IL-6, IL-8, IL-10, IL12p40, IL13, IL-16, IP-10, Лептин, MCP-2, MIG, MIP-3a, b-NGFm, sTNFRI и PFGF-bb. Происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению, обладающие способностью к секреции таких цитокинов или хемокинов, можно использовать для лечения различных заболеваний.
Новая особенность происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению происходит из того, что эти клетки обладают перечисленными выше характеристиками.
Как описано выше, происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению могут дифференцироваться в различные типы клеток, включающие остеобласты, хондроциты, адипоциты, гепатоциты и нейроны, и таким образом находить применения в терапии различных соответствующих заболеваний. Таким образом, настоящее изобретение относится к средству клеточной терапии, включающему происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению или дифференцированные из них клетки. Средство клеточной терапии по настоящему изобретению можно применять для лечения различных заболеваний, включающих, например, заболевания нервной системы (например, дегенеративные заболевания нервной системы), остеоартрит (например, дегенеративный артрит, ревматоидный артрит), остеопороз (например, остеопороз), заболевания печени (например, цирроз печени), и сердечно-сосудистые заболевания.
Предпочтительно, средство клеточной терапии по настоящему изобретению содержит по меньшей мере один разбавитель, который может защищать и поддерживать жизнеспособность клеток. В качестве разбавителей можно использовать буферные растворы, такие как физиологический раствор, PBS (фосфатно-солевой буфер), HBSS (уравновешенный солевой раствор Хэнка) и компоненты плазмы или сыворотки.
В другом аспекте настоящее изобретение относится к среде для культивирования новых стволовых клеток. Среда основана на среде EGM-2 или SNU-1 и, если необходимо, обогащена 20% эмбриональной телячьей сывороткой (FBS), bFGF (фактор роста фибробластов) 1~40 нг/мл, аскорбиновой кислотой 0,1-5,0 мкг/мл, EGF (эпидермальный фактор роста) 1~40 нг/мл, гидрокортизоном 0,1~1 мкг/мл, IGF-I (инсулиноподобный фактор роста-1) 1~40 нг/мл или VEGF (фактор роста эндотелия сосудов) 1~5 нг/мл и гепарином 20~25 мкг/мл, и необязательно GA-1000 (гентамицина сульфат, амфотерицин-B).
Среда для культивирования стволовых клеток является новой и используется в способе выделения происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению. Дополнительно, среда для культивирования по настоящему изобретению является пригодной для пролиферации всех стволовых клеток взрослых, включая происходящие из пуповинной крови стволовые клетки, и применима для культивирования стволовых клеток взрослых.
Согласно дополнительному аспекту настоящее изобретение относится к способу культивирования стволовых клеток, где способ включает культивирование и пролиферацию стволовых клеток в среде по настоящему изобретению. Предпочтительно, стволовые клетки могут представлять собой стволовые клетки взрослых.
В одном из вариантов осуществления среду для культивирования стволовых клеток в соответствии с настоящим изобретением можно использовать для культивирования происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению. При культивировании в среде по настоящему изобретению, полученные из пуповинной крови плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению можно подвергать пассированию предпочтительно через 3-5 суток после образования клеточных колоний веретенообразной формы. Культивирование предпочтительно осуществляют в условиях 5% CO2 и его можно проводить в течение 5~30 суток, но настоящее изобретение не ограничено этими условиями.
В соответствии с еще одним аспектом, настоящее изобретение относится к способу увеличения «стволовости» стволовых клеток, для которого характерно использование сферической культуры или трехмерной культуры стволовых клеток, таким образом, культивируют стволовые клетки. Для трехмерной культуры предпочтительно используют MEF (эмбриональные клетки-фибробласты мыши). Дополнительно, стволовые клетки предпочтительно могут представлять собой взрослые стволовые клетки.
Как применяют в настоящем документе, термин «увеличение стволовости» обозначает образование колонии, подобной колонии эмбриональной стволовой клетки, или экспрессию на высоком уровне факторов транскрипции, таких как Oct4, Sox2 и т.д.
Полезные эффекты
При культивировании в присутствии фибронектина, как подробно описано выше, полученные из пуповинной крови плюрипотентные/мультипотентные стволовые клетки человека по настоящему изобретению активно пролиферируют без дифференцировки в течение продолжительного периода времени, по сравнению с общепринятыми стволовыми клетками взрослых. Кроме того, обладающие способностью к дифференцировки в различные типы клеток, такие как хондроциты, остеобласты и адипоциты, плюрипотентные/мультипотентные стволовые клетки по настоящему изобретению можно эффективно использовать для лечения традиционно неизлечимых заболеваний, а также заболеваний нервной системы, сердечно-сосудистых заболеваний и заболеваний костной системы.
Описание чертежей
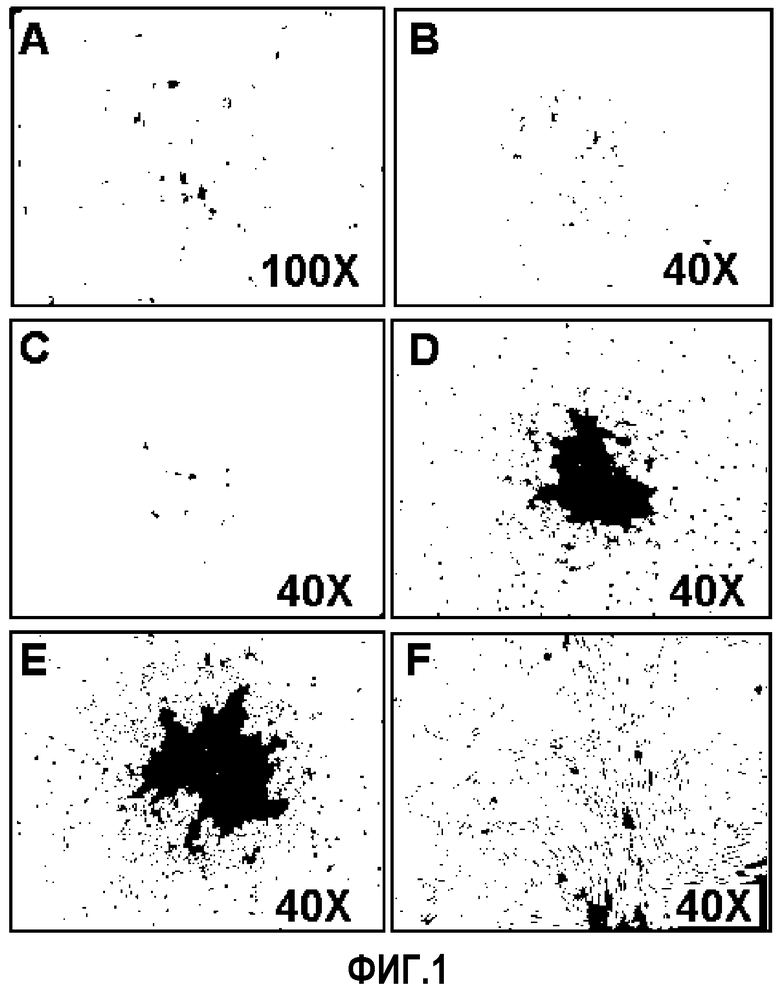
На ФИГ.1 представлены фотографии, демонстрирующие колонии, образованные после того, как моноциты, выделенные из пуповинной крови человека, культивировали в течение 14, 15, 16, 17 и 18 суток (A, B, C, D и E соответственно) и клетки после 3 пассажа (F);
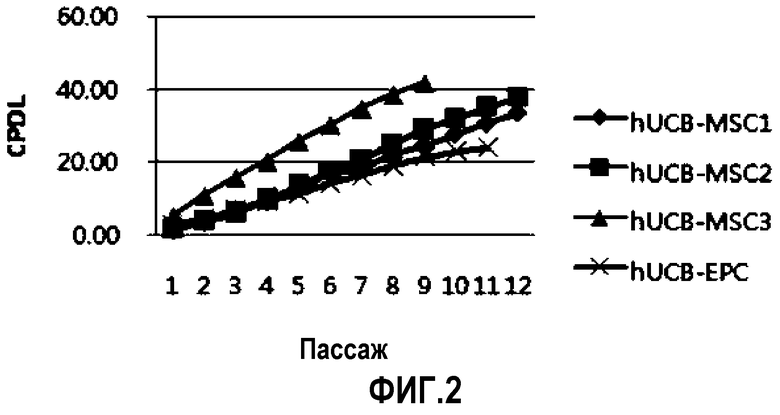
На ФИГ.2 представлен график, на котором изображен кумулятивный клеточный рост в зависимости от времени.
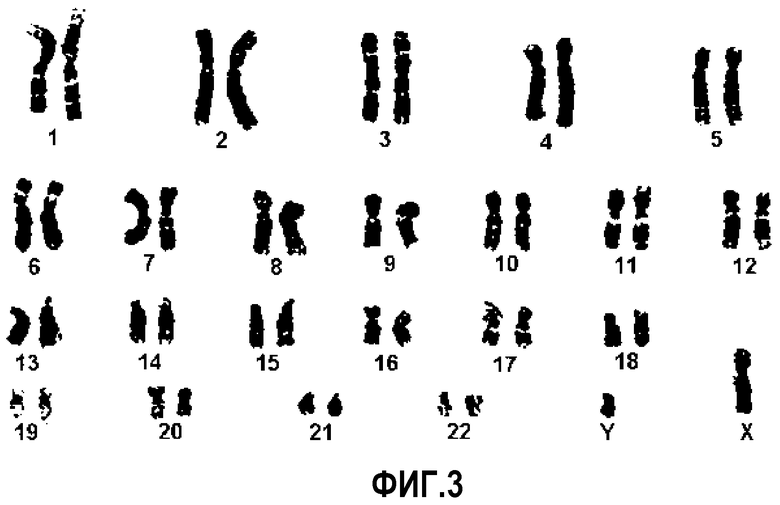
На ФИГ.3 представлены результаты кариотипического анализа после культивирования происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в течение длительного периода времени.
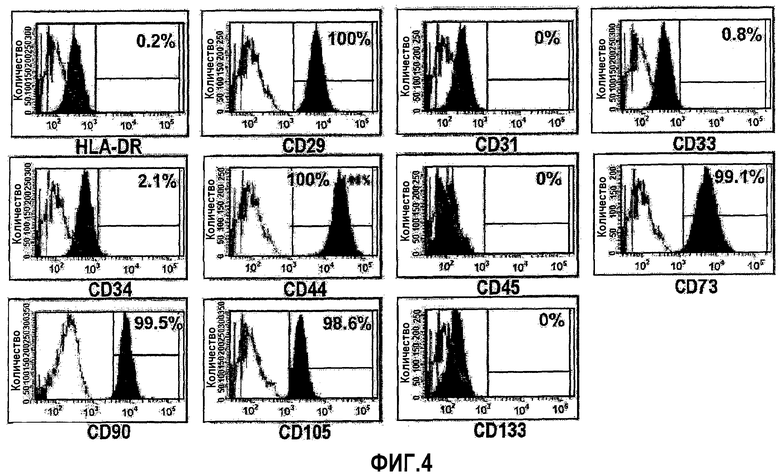
На ФИГ.4 представлены цитограммы проточного цитометра, демонстрирующие профили экспрессии различных маркеров на происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению.
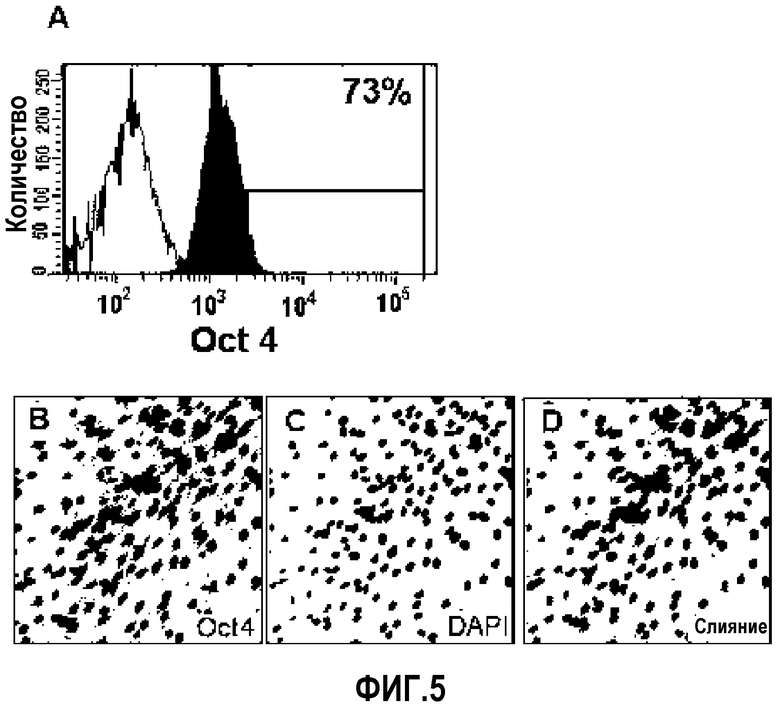
На ФИГ.5 представлены профили экспрессии различных маркеров недифференцированных клеток на происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению, как измерено посредством проточной цитометрии и иммуноокрашивания (A: цитограмма проточной цитометрии клеток, экспрессирующих Oct4, B: изображение подвергнутого иммуноокрашиванию Oct4, C: изображение окрашенного ядра в клетках, экспрессирующих Oct4, D: совмещенное изображение экспрессии Oct4 и окрашивания ядер).
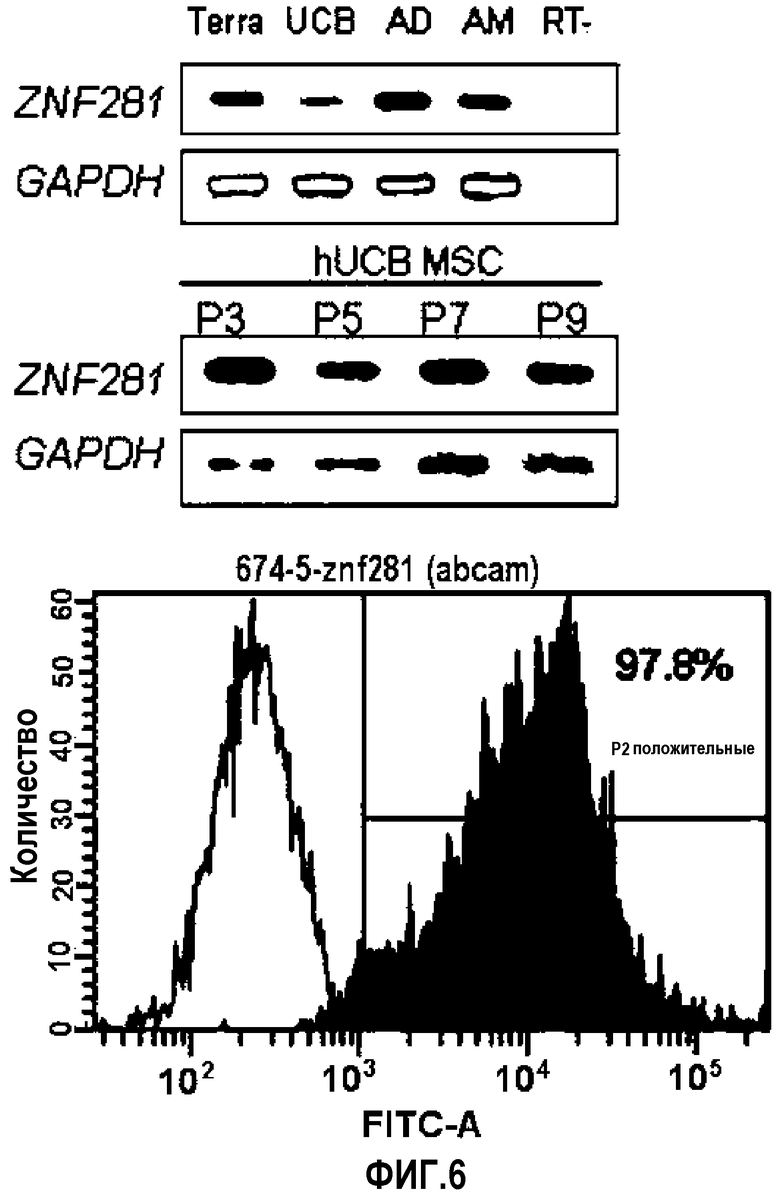
На ФИГ.6 представлена экспрессия ZNF281 в terra-1, hUCB-MSC, AD-MSC и AM (верхняя вставка) и в hUCB-MSC после 3~9 пассажей (нижняя вставка), как проанализировано посредством FACS.
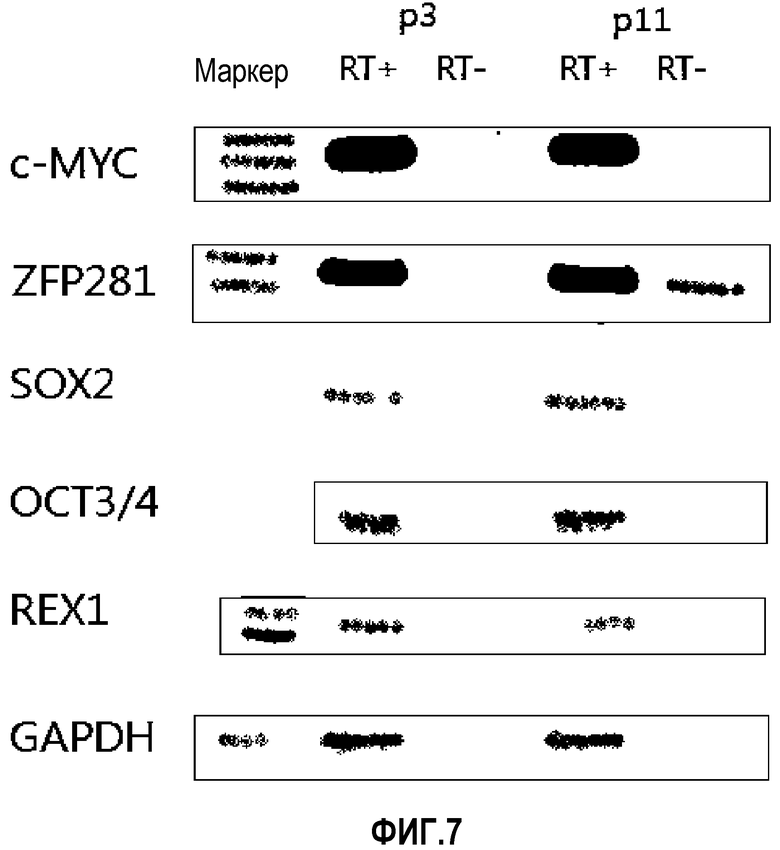
На ФИГ.7 представлена экспрессия ZNF281, Oct4, Sox2, c-myc и Rex-1, каждый из которых важен для поддержания недифференцированного состояния, у происходящих из пуповинной крови человека плюрипотентных/мультипотентных стволовых клеток по настоящему изобретению, как измерено посредством ОТ-ПЦР.
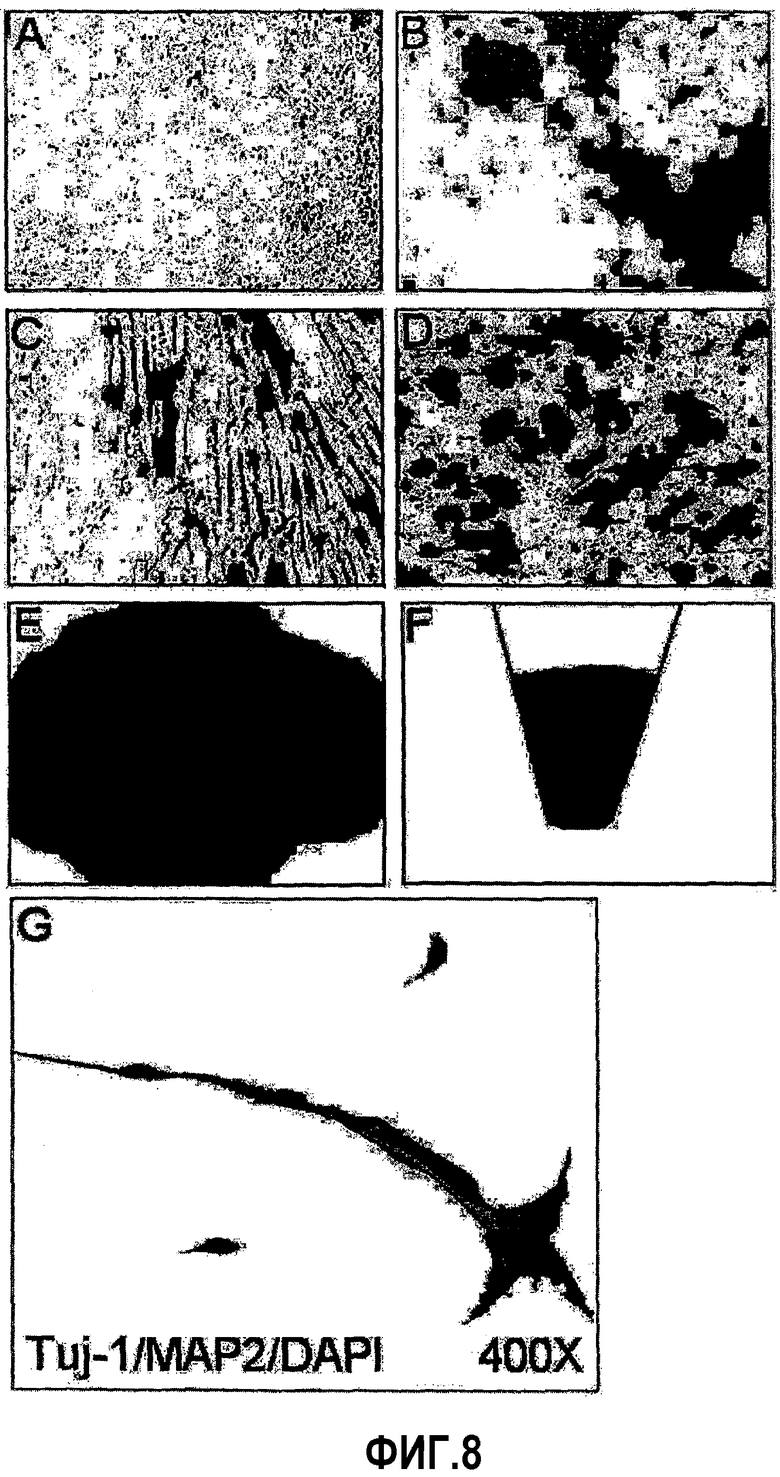
На ФИГ.8 представлены фотографии, показывающие дифференциацию происходящих из пуповинной крови человека плюрипотентных/мультипотентных стволовых клеток в остеобласты, адипоциты, хондроциты и нейроны (A: индуцированный к неостеогенной дифференцировке контроль после окрашивания ализариновым красным S, B: индуцированные к остеогенной дифференцировке клетки после окрашивания ализариновым красным S, C: индуцированный к неадипогенной дифференцировке контроль после окрашивания масляным красным O, D: индуцированные к адипогенной дифференцировке клетки после окрашивания масляным красным O, E: индуцированные к хондрогенной дифференцировке клетки после окрашивания толуидиновым синим, F: сгусток индуцированных к хондрогенной дифференцировке клеток, G: клетки, подвергнутые иммуноокрашиванию на относящиеся к нервной системе маркеры Tuj-1 и MAP2 после нейрогенной дифференцировки).
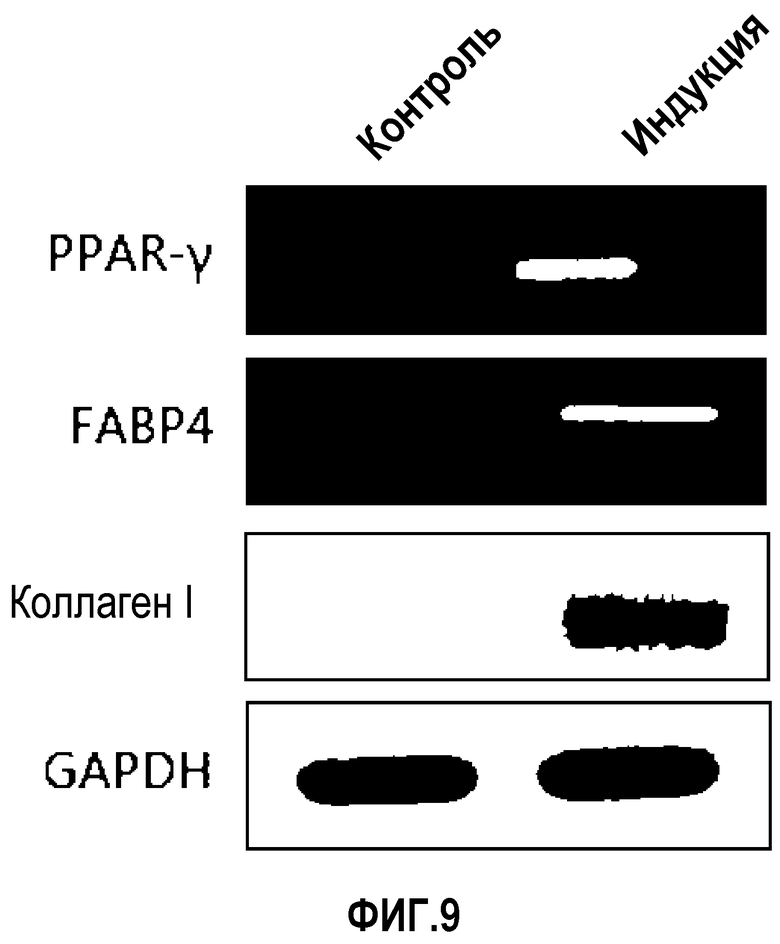
На ФИГ.9 представлены уровни РНК в происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клетках после индукции остеогенной дифференцировки и адипогенной дифференцировки, как измерено посредством RT-ПЦР (PPAR-γ и FABP-4 представляют собой маркеры адипогенной дифференцировки, и коллаген 1 типа представляет собой маркер остеогенной дифференцировки. PPAR-γ: γ-рецептор, активирующий пролиферацию пероксисом, FABP-4: белок-4, связывающий жирные кислоты, GAPDH: глицеральдегид-3-фосфатдегидрогеназа).
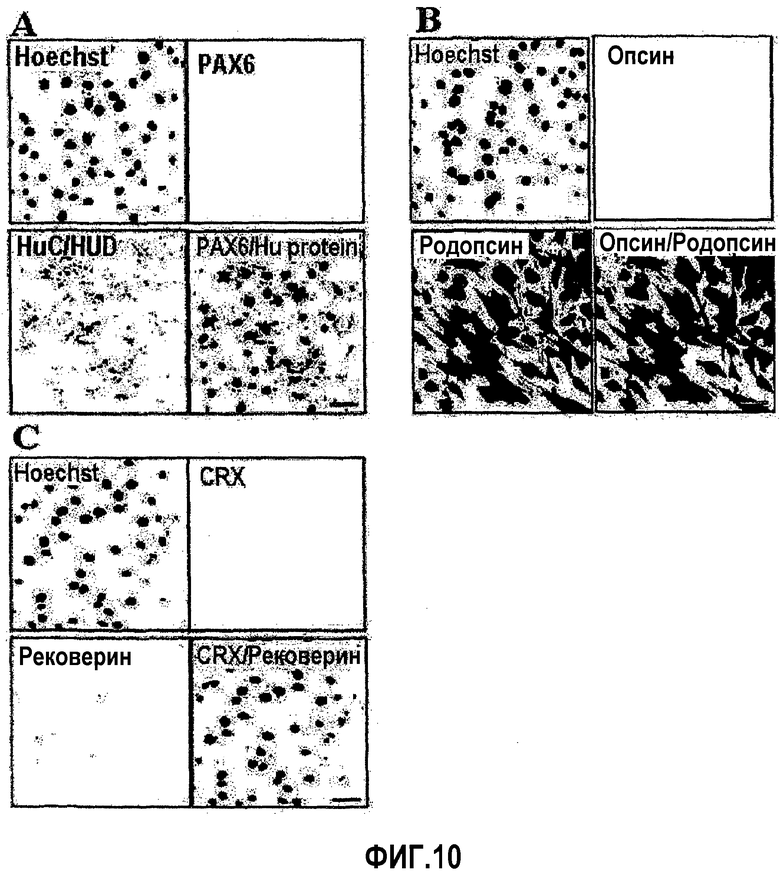
На ФИГ.10 представлены профили экспрессии связанных с сетчаткой глаза белков в происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клетках по настоящему изобретению.
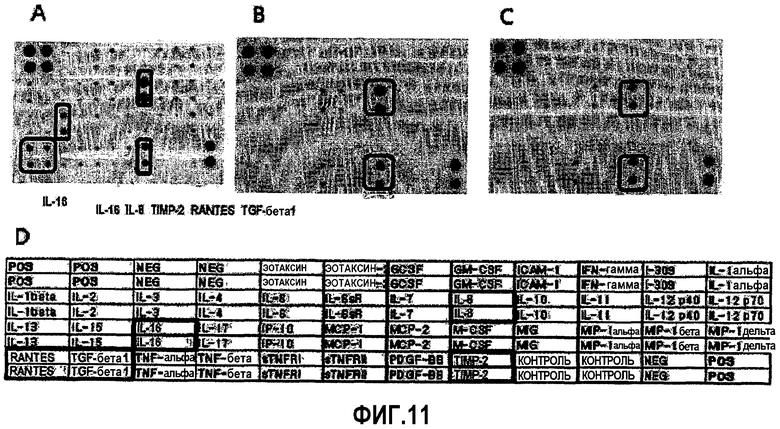
На ФИГ.11 представлена секреция различных цитокинов в средах для культивирования происходящих из пуповинной крови человека плюрипотентных/мультипотентных стволовых клеток, как проанализировано посредством микроматрицы антител (A: hUCB-MSC1, B: hUCB-MSC2, C: hUCB-MSC3, D: расположение антител, POS: положительный контроль, NEG: отрицательный контроль, GCSF: гранулоцитарный колониестимулирующий фактор, GM-CSF: гранулоцитарно-моноцитарный колониестимулирующий фактор, ICAM-1: внутриклеточная молекула адгезии, IFN-γ: интерферон-γ, IL: интерлейкин, MCP: моноцитарный белок-хемоаттрактант, M-CSF: моноцитарный колониестимулирующий фактор, MIG: монокин, индуцированный интерфероном-гамма, MIP: макрофагальный белок воспаления, RANTES: регулируемый при активации, экспрессируемый и секретируемый нормальными T-клетками, TGF-β: трансформирующий фактор роста-β, TNF: Фактор некроза опухоли, sTNFR: растворимый рецептор фактора некроза опухоли, PDGF-BB: полученный из тромбоцитов фактор роста-BB, TIMP2: тканевой ингибитор металлопротеиназ-2).
На ФИГ.12 и 13 представлены фотографии, демонстрирующие трехмерные культуры происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, с использованием способа сферической культуры и клетки STO и профили экспрессии факторов транскрипции Oct4 и Sox2, как измерено посредством ОТ-ПЦР (фиг.12: трехмерная культура происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток посредством способа сферической культуры, ФИГ.13: трехмерная культура происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток на клетках STO).
Способ изобретения
Лучшего понимания сущности изобретения можно достичь посредством следующих примеров, которые представлены для иллюстрации, но которые не следует рассматривать как ограничивающие настоящее изобретение.
ПРИМЕР 1: Культивирование, пролиферация и кариотипический анализ плюрипотентных/мультипотентных стволовых клеток, происходящих из пуповинной крови
Образцы UCB доношенных детей (n=20) отбирали непосредственно после получения от матерей информированного согласия на проведение отбора. Образцы UCB доношенных детей смешивали с HetaSep (Stem cells Technologies INC, Vancouver, BC) для уменьшения количества эритроцитов. Далее получали моноциты посредством общепринятого центрифугирования в градиенте плотности Фиколла. Моноциты высевали из расчета плотности 1×105~8 клеток/лунку на 6-луночные планшеты, покрытые 0,1 мг/мл - 1 мг/мл фибронектина и сохраняли в SNU-1 или EGM-2 (Lonza), обогащенными EGM-2 SingleQuots, содержащим 20% FBS. EGM-2 SingQouts содержал гепарин, аскорбиновую кислоту, rhEGF, гидрокортизон, VEGF, rhFGF-B, R3-IGF-1 и GA-1000. Моноциты, которые через трое суток культивирования все еще оставались неприкрепленными, удаляли, кроме того, среду заменяли свежей каждые двое или трое суток.
Прикрепленные клетки, как наблюдали, образуют колонии, демонстрирующие веретенообразную морфологию через 5~30 суток после культивирования в условиях с 5% CO2 (фиг.1A). Образовавшись, колонии быстро пролиферировали. Клетки суспендировали в 0,125% Трипсин-ЭДТА через 3~7 суток после образования колоний и переносили в новые планшеты, где продолжали поддерживание культуры клеток (ФИГ.1B, 1C, 1D, 1E и 1F). На ФИГ.1 представлены фотографии, демонстрирующие рост культивируемых клеток. Как видно из ФИГ.1, клеточная колония увеличивается с течением времени. Кроме того даже после пассажа морфология клеток оставалась одинаковой (ФИГ.1. выделение и пролиферация происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток. Колонии, образованные после того, как моноциты культивировали в течение 14 суток (A), 15 суток (B), 16 суток (C), 17 суток (D) и 18 суток (E) и клетки после 3 пассажа (F)).
Так как культивирование клеток продолжалось, то анализировали способность к пролиферации выделенных плюрипотентных/мультипотентных стволовых клеток, происходящих из пуповинной крови, измеряя CPDL (кумулятивный уровень удвоения популяций) (Cristofalo et al., Proc.) Natl. Acad. Sci., USA 95, 1998). Клетки размножали делением надвое. Таким образом, уровень роста клеток можно определять периодом времени, которое необходимо одной клетке для разделения надвое, называемое временем удвоения. CDPL 10 обозначает 10 делений одной клетки, что приводит в результате к пролиферации одной клетки до приблизительно 1000 клеток. Одной из самых важных проблем традиционных происходящих из пуповинной крови стволовых клеток является то, что они имеют существенно более низкую способность к пролиферации по сравнению с мезенхимальными стволовыми клетками, полученными из жировой ткани или из костного мозга. Так как при клиническом применении требуется большое количество стволовых клеток, способность к пролиферации у стволовых клеток является важным параметром. Измерение проводили, как указано далее. Во-первых, происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки трех типов выделяли из различных образцов пуповинной крови и высевали на 100 планшетов из расчета плотности 2×105 клеток и подвергали пассажам через равные промежутки времени от трех до четырех суток, так как клетки подлежали подсчету с использованием цитометра. Подсчет клеток проводили до прекращения роста клеток. На основе измерений количества клеток получали значения CPDL, согласно следующей математической формуле 1
[Математическая формула 1]
Nн/NI=2X или [log(NH)-log(NI)]/log(2)=X
где NI обозначает количество клеток в первоначальной культуре; NH обозначает количество клеток в условии насыщения при пассаже.
Значения CPDL рассчитывали, пока продолжали поддерживать культуру клеток. В качестве контроля для сравнения использовали hUCB-EPC (эндотелиальные клетки-предшественники, полученные из пуповинной крови человека). Результаты представлены на ФИГ.2. Как можно видеть из графика на ФИГ.2, для hUCB-EPC значение CPDL составляет приблизительно 20 в течение двух месяцев, в то время как значения CPDL всех происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток из трех различных образцов, как наблюдали, варьирует между 40 и 45. Эти численные значения показывают, что клетки могут теоретически расти до значения 1012 в одной колонии.
В итоге, клетки со способностью к быстрой пролиферации могут стать злокачественными, демонстрирующими бурный рост. Став злокачественными, клетки, по-видимому, пролиферируют независимо от различных регуляторных сигналов в организме, и таким образом не могут быть использованы в качестве средства клеточной терапии и являются несовместимыми с целью исследования. Таким образом, необходимо определять, является ли нормальным хромосомный набор плюрипотентных/мультипотентных стволовых клеток, происходящих из пуповинной крови и выделенных по настоящему изобретению. В связи с этим, для исследования хромосом клеток проводили кариотипический анализ. Как можно видеть на ФИГ.3, обнаружили, что даже после 10 пассажа клетки обладают нормальной хромосомной структурой.
ПРИМЕР 2: Анализ поверхностного антигена происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток
Для анализа характеристик клеток, суспендированных в среде, осуществляли способ проточной цитометрии. Для фенотипирования поверхностных клеточных антигенов клетки, собранные после 3~4 пассажей, окрашивали антителом, конъюгированным с флуоресцеинизотиоцианатом (FITC) или фикоэритрином (PE), и анализировали FACSAria (Becton Dickinson, NY).
Для анализа характеристик происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток (в частности, плюрипотентная/мультипотентная стволовая клетка), выделенных в настоящем изобретении, использовали поверхностные антигены, которые включали CD10 (маркер T-клеток), CD14 (маркер моноцитов), CD24 (маркер эпителиальных клеток), CD29 (маркер моноцитов), CD31 (маркер эндотелиальных клеток), CD34 (маркер гематопоэтических стволовых клеток), CD44 (маркер мезенхимальных стволовых клеток), CD45 (маркер негематопоэтических стволовых клеток), CD51/61 (маркер остеокластов), CD73 (маркер мезенхимальных стволовых клеток), CD90 (маркер мезенхимальных стволовых клеток), CD105 (маркер мезенхимальных стволовых клеток), CD133 (маркер гематопоэтических стволовых клеток) и HLA-DR (маркер иммунного отторжения) и анализировали с использованием проточного цитометра. Результаты суммированы в таблице 2 и графически представлены на ФИГ.4.
ПРИМЕР 3: Профиль экспрессии ZNF281 и основного фактора транскрипции в происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клетках
ZNF281 (белок 281 «цинковые пальцы») является одним из основных факторов транскрипции ESC (Wang J et al. (2006) Nature 444, 364-368). ZNF281, также называемый ZBP-99, содержит четыре Kruppel-подобных «цинковых пальца», которые в совокупности разделяют 91% сходности аминокислотной последовательности и 79% идентичности последовательности с Kruppel-подобными «цинковыми пальцами», найденными в ZBP-89. Кроме того, существуют высококонсервативные аминокислотные последовательности в карбоксиконцевых сегментах двух генов. Предполагаемая открытая рамка считывания кДНК ZNF281 кодирует белок массой 99 кДа. EMSA (анализ сдвига электрофоретической подвижности) показал, что белок ZNF281 специфически связывается с богатыми GC последовательностями промоторными элементами генов GASTRIN и ОРНИТИН-ДЕКАРБОКСИЛАЗЫ (Law DJ et al. (1999) Biochem Biophys Res Commun 262, 113-120; Lisowsky T et al. (1999) FEBS Lett 453, 369-374). ZNF281 идентифицировали как белок, связанный с c-MYC посредством техники масс-спектрометрической многомерной белковой идентификации и тандемной аффинной очисткой (Koch HB et al. (2007) Cell Cycle 6, 205-217.).
Как известно, гены Oct3/4, такие как семейство факторов транскрипции POU, не присутствуют в дифференцированных тканях, но экпрессируются, в частности, в недифференцированных стволовых клетках, которые обладают высокой пролиферативной способностью (Tai M-H. et al., Carcinogenesis 26:4 95, 2005; Tondreau T. et al., Stem cells, 23:1105, 2005). Таким образом, Oct3/4 в основном используют в качестве маркеров для эмбриональных стволовых клеток и также в качестве маркеров, указывающих на недифференцированные клетки. Клеточные колонии окрашивали, используя Oct4 как маркер «стволовости». В результате было обнаружено, что многие клетки имели Oct4, окрашиваемый вокруг ядра. Проточная цитометрия также показала экспрессию Oct4 в клетках.
Для окрашивания внутриклеточных белков, клетки фиксировали при 4°C в течение ночи с 4% формальдегидом и увеличивали проницаемость мембран в течение 10 мин с 0,1% Triton X-100 (Sigma-Aldrich). Предметные стекла и планшеты инкубировали в течение одного часа с первичным антителом мыши против Oct4 человека (1:200), промывали PBS (фосфатно-солевой буфер; Gibco), подвергали иммуноокрашиванию в течение одного часа с конъюгированным с Alexa 594 вторичным антителом козы к IgG мыши (Invitrogen), в то время как ядро окрашивали DAPI.
Как видно из Фиг.5 и 6, показано, что множество происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток экспрессируют Oct4 (фиг.5A: цитограмма проточной цитометрии клеток, экспрессирующих Oct4, B: изображение иммуноокрашенного Oct4, C: изображение окрашенного ядра клеток, экспрессирующих Oct4, D: совмещенное изображение экспрессии Oct4 и окрашивания ядер).
Гены, такие как Oct4, тесно связаны со «стволовостью» стволовых клеток. По существу, недавно были предприняты попытки индуцировать наличие у взрослых стволовых клеток плюрипотентности, схожей с плюрипотентностью эмбриональных стволовых клеток, придавая им способность к сверхэкспрессии генов, таких как Oct4, Sox2 и т.п. (Takahashi et al, Cell, 131(5), 861-872, 2007). Таким образом, экспрессия этих генов в стволовых клетках является очень важной для сохранения стволовых клеток в недифференцированном состоянии без потери «стволовости». Таким образом, для исследования профилей экспрессии ZNF281, Oct-4 и Sox2 в происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клетках, изолированных по способу настоящего изобретения, проводили ОТ-ПЦР.
Для достижения этой цели конструировали праймеры, как представлено в таблице 3.
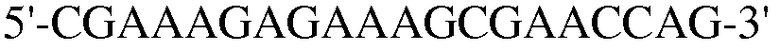
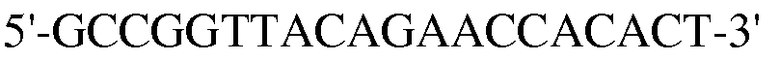
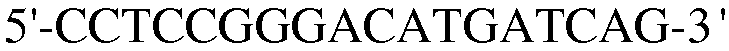
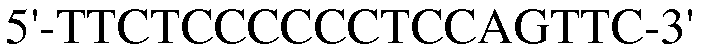
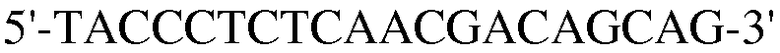
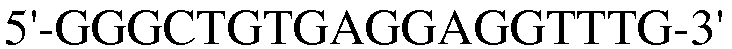
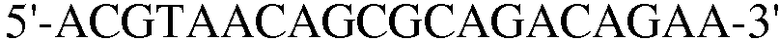
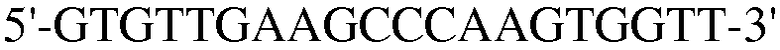
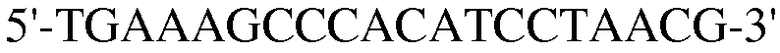
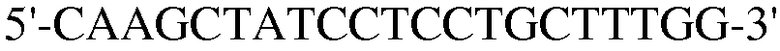
Как видно на ФИГ.7, Oct-4, Sox2, c-myc, ZNF281 и REX-1 экспрессируются, демонстрируя, что полученные из пуповинной крови стволовые клетки по настоящему изобретению обладают плюрипотентностью.
ПРИМЕР 4: Дифференцировка происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в остеобласты
Клетки индуцировали к дифференцировке в остеобласты. В связи с этим, клеткам позволяли прикрепиться к культуральному планшету и инкубировали до смыкания монослоя приблизительно 70-80%. Затем среду для культивирования замещали средой, индуцирующей остеогенную дифференцировку. Среду, индуцирующую остеогенную дифференцировку, получали посредством обогащения среды DMEM с низким содержанием глюкозы 10% FBS и 10 мМ бета-глицерофосфатом (Sigma-Aldrich), 0,1 мкМ D экзаметазоном (Sigma-Aldrich) и 50 мкМ аскорбатом (Sigma-Aldrich). Среду заменяли свежей каждые трое суток, пока в течение приблизительно двух недель индуцировали дифференцировку.
Через две недели кальциевую минерализацию, связанную с остеогенной дифференцировкой, исследовали посредством окрашивания с ализариновым красным S. Окрашивание выполняли следующим образом. После удаления среды, клетки дважды промывали дистиллированной водой и фиксировали при 4°C в течение одного часа в холодном 70% EtOH. Затем клетки вновь дважды промывали дистиллированной водой и окрашивали при комнатной температуре в течение 10 мин с 40 мМ ализаринового красного S, с последующим пятикратным промыванием дистиллированной водой.
Как видно на ФИГ.8A и 8B, на ФИГ.8A посредством окрашивания с ализариновым красным S в отсутствие дифференцировки кальция не обнаружили, в то время как на ФИГ.8B кальций окрашивался красным красителем при индукции клеток к дифференцировке в остеобласты, что показывает, что дифференцированные в остеобласты происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки высвобождают кальций.
ПРИМЕР 5: Дифференцировка происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в адипоциты
Клетки индуцировали к дифференцировке в адипоциты. В связи с этим, клеткам позволяли прикрепляться к культуральным планшетам и культивировали до смыкания монослоя приблизительно 70-80%. Затем среду заменяли средой, индуцирующей дифференцировку в адипоциты. Среду, индуцирующую дифференцировку в адипоциты, получали посредством обогащения среды DMEM с низким содержанием глюкозы 10% FBS и 1 мкМ дексаметазоном, 10 мкг/мл инсулина (Sigma-Aldrich), 0,5 мМ 3-изобутил-1-метилксантин (Sigma-Aldrich) и 0,2 мМ индометацином (Sigma-Aldrich). Среду для дифференцировки заменяли свежей один раз в трое суток, пока в течение приблизительно 2-3 недель индуцировали дифференцировку.
Через 2~3 недели, дифференцировку в адипоциты исследовали посредством окрашивания масляным красным О. Для этой цели удаляли среду и клетки промывали в PBS и инкубировали при комнатной температуре в течение 5 мин в 10% формалине. Формалин заменяли на тот же объем свежего формалина, с последующей фиксацией клеток при комнатной температуре в течение по меньшей мере одного часа. После удаления формалина клетки промывали 60% изопропанолом. Перед окрашиванием клеток при комнатной температуре в течение 10 мин масляным красным О спирт полностью выпаривали. Сразу после окончания времени окрашивания краситель удаляли, клетки промывали дистиллированной водой.
Адипоциты окрашиваются в красный цвет при обработке масляным красным, так как краситель окрашивает липидные капли в красный цвет. Как можно видеть на ФИГ.8C и 8D, на ФИГ.8C не обнаружили ни липидных капель, ни окрашивание в красный цвет в отсутствие дифференцировки в адипоциты, в то время как на ФИГ.8D, на котором показаны результаты по индуцированию дифференцировки в адипоциты, наблюдали клетки, которые содержат множество липидных капель и, таким образом, красное окрашивание).
ПРИМЕР 6: Дифференцировка происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в хондроциты
Для индукции дифференцировки клеток в хондроциты, клетки в течение трех недель обрабатывали средой для дифференцировки в хондроциты, содержащей rTGF-бета 3 (PT-3003) от Lonza, кроме того, среду заменяли свежей два раза в неделю. Процесс хондрогенеза оценивали раз в неделю.
Для исследования хондрогенной дифференцировки выполняли окрашивание толуидиновым синим. Клетки фиксировали в течение 10 часов 4% формальдегидом и затем дополнительно в течение 10 часов пикриновой кислотой. После изготовления криосрезов, клетки окрашивали в течение 3 мин толуидиновым синим и докрашивали в течение 3 сек гематоксилином.
Группа хондроцитов не разрушается, но сохраняет постоянную морфологию и окрашивается в синий цвет при обработке толуидиновым синим. Как можно видеть на ФИГ.8E и 8F, обнаружили, что клетки, подверженные хондрогенной дифференцировке, окрашивались в синий цвет и сохраняли свою морфологию.
ПРИМЕР 7: Изменение в уровне экспрессии генов после дифференцировки происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в остеобласты и адипоциты
У стволовых клеток наблюдали значительное изменение в профиле экспрессии генов, во время прохождения процесса дифференцировки. Как правило, уровень экспрессии PPAR-γ (рецептор γ, активирующий пролиферацию пероксисом) или FABP4 (белок 4, связывающий жирные кислоты) увеличивается при дифференцировке стволовых клеток в адипоциты, в то время как уровень экспрессии коллагена 1 типа увеличивается при дифференцировке стволовых клеток в остеобласты (Mat hews et al., J Am Acad Dermatol, 56(3), 472-492, 2007; Cho et al., J. Cell. Biochem., 96, 533-542, 2005). Таким образом, после индукции дифференцировки уровень экспрессии гена, который экспрессируется в клетках определенного типа, напрямую показывает, дифференцированы ли стволовые клетки в клетки определенного вида. Эксперименты выполняли, как указано далее.
Через две или три недели после дифференцировки в остеобласты и адипоциты, из клеток выделяли суммарную РНК, используя Trizol (Invitrogen). Синтезировали кДНК, используя AccuPower RT Premix (Bioneer), с последующим ПЦР с использованием Maxime PCRPreMix Kit (Intronbio).
Для этой цели сконструировали праймеры, как представлено в таблице 4 ниже.
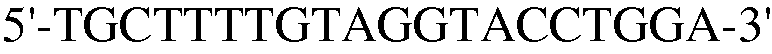
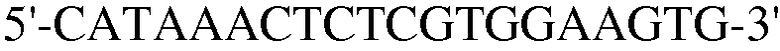
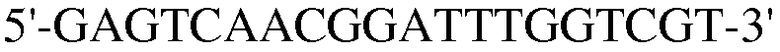
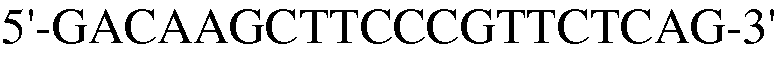
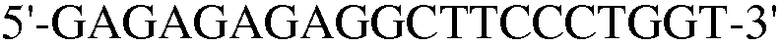

Как ясно из данных, представленных на ФИГ.9, уровень экспрессии PPAR-γ и FABP4, которые представляют собой маркеры дифференцировки в адипоциты, значительно повышены в случае, если клетки индуцировали к дифференцировке в адипоциты. Также, уровень экспрессии коллагена 1, маркера остеогенной дифференцировки, значительно повышался в клетках, проходящих процесс дифференцировки. При этом детектировали одинаковый уровень нанесенного для контроля GAPDH как в клетках, которые прошли через дифференцировку, так и в клетках, которые не прошли дифференцировку, что подтверждает обоснованность эксперимента.
ПРИМЕР 8: Дифференцировка происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток в нейроны
Клетки индуцировали к дифференцировке в нейроны.
Во-первых, клетки подвергали предварительной инкубации в течение 24 часов в DMEM, обогащенной 5% FBS и 10 нг/мл bFGF (основной фактор роста фибробластов). После чего клетки обрабатывали в течение 24 часов DMEM, содержащей 1% ДМСО, 100 мкМ BHA, 0,5 мМ VPA, 10 мМ KCl и 10 нг/мл NGF и B27 для индукции дифференцировки в нейроны. Для исследования дифференцировки в нейроны, клетки фиксировали 4% параформальдегидом и подвергали иммуноокрашиванию на маркеры нейронов Tuj-1, MAP-2, GFAP и нейрофиламент-160. В результате обнаружили, что экспрессируются четыре маркера (фиг.8G).
ПРИМЕР 9: Экспрессия специфичных для сетчатки глаза белков в происходящих из пуповинной крови мезенхимальных стволовых клетках
С использованием иммунофлуоресценции исследовали характеристики происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, ассоциированных с сетчаткой глаза.
Исследовали профили экспрессии в клетках специфичных для сетчатки глаза белков.
На ФИГ.10 представлены фотографии, демонстрирующие профили экспрессии специфичных для сетчатки глаза белков, как измерено посредством иммунофлуоресценции. Профили экспрессии PAX6 и белка Hu представлены на ФИГ.10A. PAX6 известен как маркер клеток-предшественников сетчатки глаза, белок Hu специфично экспрессируется в ганглиоцитах и амакринных клетках, которые являются составными компонентами сетчатки глаза. Они не обнаружены в контрольной культуре.
На ФИГ.10B детектируется опсин и родопсин. Опсин специфически экспрессируется в колбочковидных клетках, в то время как родопсин является специфичным для палочковидных клеток. В контрольной культуре опсин не детектировали, но наблюдали экспрессию родопсина.
На ФИГ.10C представлены профили экспрессии CRX и рековерина. CRX известен как пан-фоторецепторный маркер, а рековерин известен как фоторецепторный маркер. Эти маркеры не детектируют в контрольной культуре (увеличение ×400, измерительная линейка = 50 мкм).
ПРИМЕР 10: Анализ цитокинов, высвобождающихся из происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток
Терапевтический потенциал стволовых клеток является, в значительной степени, связанным с двумя эффектами: во-первых, с прямой дифференцировкой стволовых клеток в поврежденные клетки; и во-вторых, способностью секретировать различные цитокины или факторы роста, индуцирующие положительные изменения, которые приводят к терапевтическому воздействию на уже существующие клетки. Известно, что стволовые клетки, как правило, секретируют различные цитокины или факторы роста, демонстрируя так называемые паракринные эффекты (Kim et al. Cytokine. 2005). Для исследования профилей секреции происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, выделенных по настоящему изобретению, использовали микрочип с антителами к цитокинам человека (RaybioTech. Norc ross, USA).
Во-первых, клетки стабилизировали в течение 24 часов в среде без FBS и добавок, после чего среду в количестве 1 мл отбирали через равные двухчасовые интервалы. Каждый из образцов среды отбирали в количестве 100 мкл для получения пула образцов, в котором количественно анализировали уровень белков, с последующим исследованием на микрочипах.
Как показано на ФИГ.11, для всех происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток, выделенных из трех различных образцов, обнаружили наличие секреции IL-8 и TIMP-2. Кроме того, также наблюдали секрецию различных цитокинов, включая TGF-β, RANTES, CINC-3, ЭОТАКСИН, GM-CSF, IFN-γ, IL-1b, IL-3, IL-6, IL-10, IL12p40, IL13, IL-16, IP-10, Лептин, MCP-2, MIG, MIP-3a, b-NGFm, sTNFRI и PFGF-bb (на фиг.11 представлены результаты анализа микрочипа с hUCB-MSC1(A), hUCB-MSC2 (B) и hUCB-MSC3 (C) и распределение антител (D)).
ПРИМЕР 11: 3-мерная культура происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток
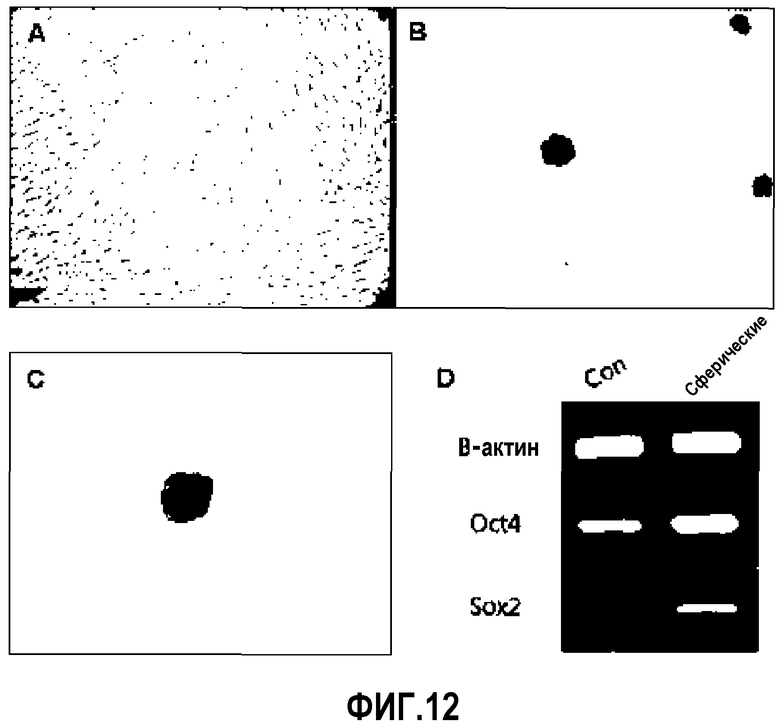
Как правило, взрослые стволовые клетки растут, образуя монослой в культуральном планшете. Однако эксперименты показали, что «стволовость» взрослых стволовых клеток увеличивалась в случае, когда взрослым стволовым клеткам позволяли расти не в 2-мерной структуре, а в виде сферы в 3-мерной структуре. В связи с этим, во-первых, культуральные планшеты покрывали 0,7% агарозой толщиной 5 мм или более, для того чтобы клетки не могли прикрепляться к дну планшета, а могли образовывать сферу. Клетки высевали из расчета плотности менее чем 2000 клеток/см2 для предотвращения адгезии между отдельными клетками. Образованные таким образом сферы отделяли от одиночных клеток, используя 40 мкм сито. Как можно видеть на ФИГ.12, стволовые клетки не подвергаются гибели, а при культивировании в системе для сферической культуры образуют сферы, сохраняющие характеристики стволовых клеток. Как представлено на ФИГ.12A-D, наблюдали, что уровни экспрессии эмбриональных маркеров, таких как OCT4, SOX2 и т.п. в клетках, которые поддерживали в сферической культуре, являлись значительно более высокими по сравнению с уровнями экспрессии эмбриональных маркеров в клетках, которые поддерживали в виде монослоя.
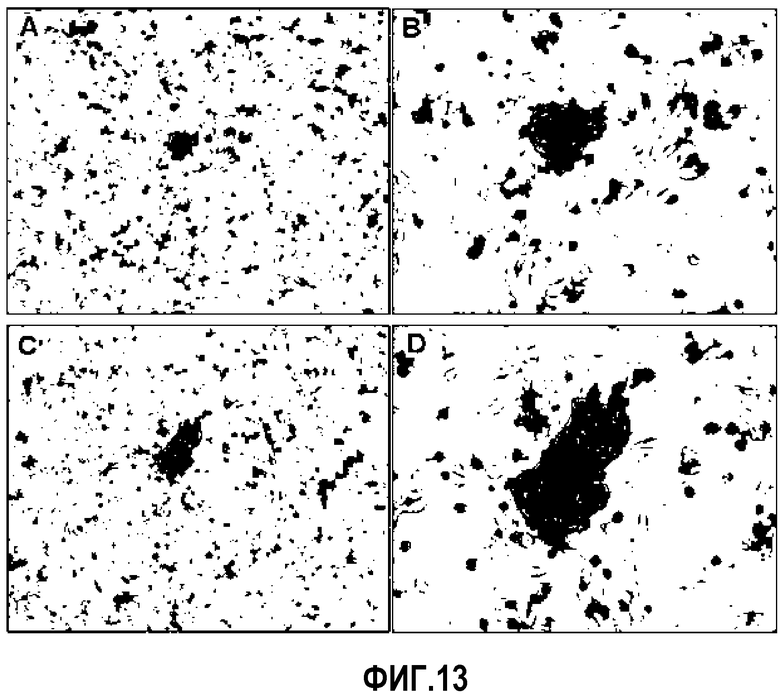
Эмбриональные стволовые клетки, как правило, культивируют на слое клеток - эмбриональных фибробластов мыши, так как кемокины, такие как LIF, из клеток-эмбриональных фибробластов мыши участвуют в поддержании морфологии клеток ES и предотвращают дифференцировку клеток ES. При культивировании на слое клеток - эмбриональных фибробластов мыши, происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки не росли в плоской форме, подобно типичным взрослым стволовым клеткам, а формировали 3-мерные колонии. Затем пролиферацию подавляли посредством обработки 0,1 мг/мл митомицина C, клетки STO высевали из расчета плотности 2×105 клетки/мл на покрытые 0,1% желатином планшеты и инкубировали в течение 24 часов, с последующим высеванием происходящих из пуповинной крови плюрипотентных/мультипотентных стволовых клеток на слой клеток STO. Как представлено на ФИГ.13, на клетках STO для клеток наблюдали картину пролиферации, которая была похожа, с течением времени, на картину пролиферации эмбриональной стволовой клетки.
Промышленная применимость
По настоящему изобретению, при культивировании в присутствии фибронектина происходящие из пуповинной крови плюрипотентные/мультипотентные стволовые клетки человека активно пролиферируют без дифференцировки в течение более долгого периода времени, по сравнению с традиционными взрослыми стволовыми клетками. Кроме того, по настоящему изобретению обладающие способностью к дифференцировке в различные виды клеток, такие как хондроциты, остеобласты и адипоциты, плюрипотентные/мультипотентные стволовые клетки можно эффективно применять для лечения некурабельного заболевания, а также заболевания нервной системы, сердечно-сосудистой системы и заболевания костной системы.




Изобретение относится к области молекулярной биологии, биотехнологии и медицины. Предложен способ выделения происходящих из пуповинной крови стволовых клеток, экспрессирующих ZNF281, с культивированием в сосуде, содержащем фибронектин, и последующим получением стволовых клеток из культуры, а также среда для культивирования стволовых клеток, способ культивирования стволовых клеток, клеточное терапевтическое средство, и способ увеличения «стволовости» стволовых клеток, который характеризуется наличием сферической культуры или трехмерной культуры стволовых клеток. Изобретение может быть использовано в медицине для целей трансплантологии. 6 н. и 9 з.п. ф-лы, 13 ил., 4 табл., 11 пр.
1. Способ выделения происходящих из пуповинной крови стволовых клеток, экспрессирующих ZNF281, из пуповинной крови, где способ включает культивирование выделенных из пуповинной крови моноцитов в культуральном сосуде, содержащем фибронектин, и получение из культуры стволовых клеток, где моноциты культивируют в среде EGM-2 или SNU-1, обогащенной эмбриональной телячьей сывороткой (FBS), 1-40 нг/мл bFGF, аскорбиновой кислотой 0,1-5,0 мкг/мл, 1-40 нг/мл EGF, гидрокортизоном 0,1-1 мкг/мл, 1-40 нг/мл IGF-I или 1-5 нг/мл VEGF и гепарином 20-25 мкг/мл.
2. Способ по п.1, где моноциты выделяют, смешивая пуповинную кровь с Hetasep для удаления эритроцитов, и затем используют Фиколл-пак.
3. Способ получения происходящих из пуповинной крови стволовых клеток, экспрессирующих ZNF281, включающий культивирование моноцитов, выделенных из пуповинной крови, в культуральном сосуде, содержащем фибронектин, и выделение происходящих из пуповинной крови стволовых клеток, экспрессирующих ZNF281, из культуры, где моноциты культивируют в среде EGM-2 или SNU-1, обогащенной эмбриональной телячьей сывороткой (FBS), 1-40 нг/мл bFGF, аскорбиновой кислотой 0,1-5,0 мкг/мл, 1-40 нг/мл EGF, гидрокортизоном 0,1-1 мкг/мл, 1-40 нг/мл IGF-I или 1-5 нг/мл VEGF и гепарином 20-25 мкг/мл.
4. Способ по п.1, где стволовые клетки получают из клеточных колоний, образованных после 5-30 суток после начала культивирования.
5. Способ по п.1, где фибронектин наслаивают на культуральный сосуд или где фибронектин содержится в среде для культивирования в форме сфер или трехмерных структур.
6. Способ по п.5, где культуральный сосуд покрыт фибронектином, фибронектин содержится из расчета плотности от 0,1 до 1 мг/мл.
7. Способ по п.3, где клетки, экспрессирующие ZNF281, культивируют в сферической культуре или трехмерной культуре.
8. Способ по п.1, где стадия получения стволовых клеток из культуры включает использование иммунологических свойств стволовых клеток для разделения стволовых клеток.
9. Происходящие из пуповинной крови стволовые клетки, экспрессирующие ZNF281, где стволовые клетки обладают по меньшей мере одним из следующих свойств:
(а) демонстрируют положительные иммунологические характеристики по отношению к фактору транскрипции c-myc;
(b) прикрепляются к поверхности, покрытой внеклеточным матриксом, и образуют через 5-30 суток после прикрепления клеточные колонии веретенообразной или сферической формы;
(с) демонстрируют CPDL (кумулятивный уровень удвоения популяций) со значением от 30 до 45;
(d) демонстрируют негативную иммунологическую характеристику по отношению CD14, CD31, CD34, CD45 и HLA-DR;
(е) способны дифференцироваться в мезодермальные, эндодермальные и эктодермальные клетки; и
(f) секретируют по меньшей мере один цитокин или хемокин, выбранный из группы, состоящей из TIMP-2, TGF-β, RANTES CINC-3, ЭОТАКСИНА, GM-CSF, IFN-γ, IL-1b, IL-3, IL-6, IL-8, IL-10, IL12p40, IL-13, IL-16, IP-10, Лептина, MCP-2, MIG, MIP-3a, b-NGFm, sTNFRI и PFGF-bb.
10. Происходящие из пуповинной крови стволовые клетки по п.9, которые представляют собой плюрипотентные или мультипотентные стволовые клетки.
11. Клеточное терапевтическое средство, содержащее происходящие из пуповинной крови стволовые клетки, экспрессирующие ZNF281, по п.9 или дифференцированные из них клетки.
12. Среда для культивирования выделенных из пуповинной крови стволовых клеток, экспрессирующих ZNF281, включающая среду EGM-2 или SNU-1, обогащенную эмбриональной телячьей сывороткой (FBS), 1-40 нг/мл bFGF, аскорбиновой кислотой 0,1-5,0 мкг/мл, 1-40 нг/мл EGF, гидрокортизоном 0,1-1 мкг/мл, 1-40 нг/мл IGF-I или 1-5 нг/мл VEGF и гепарином 20-25 мкг/мл.
13. Среда для культивирования по п.12, где стволовые клетки представляют собой взрослые стволовые клетки.
14. Способ культивирования выделенных из пуповинной крови стволовых клеток, экспрессирующих ZNF281, где способ включает использование среды для культивирования по п.12 или п.13.
15. Способ увеличения «стволовости» выделенных из пуповинной крови стволовых клеток, экспрессирующих ZNF281, включающий культивирование происходящих из пуповинной крови стволовых клеток, экспрессирующих ZNF281, в сферической культуре или трехмерной культуре.
| US 20040005559 A1, 08.01.2004 | |||
| US 20030224411A1, 04.12.2003 | |||
| US 20040136967 A1, 15.07.2004 | |||
| Эмалевый шликер для грунтового покрытия | 1988 |
|
SU1671627A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЯДРОСОДЕРЖАЩИХ КЛЕТОК ИЗ ПУПОВИННОЙ КРОВИ | 2007 |
|
RU2343928C1 |

