ССЫЛКА НА СВЯЗАННЫЕ ЗАЯВКИ
Сделана ссылка на следующие связанные заявки: заявка США № 09/750990, поданная 20 июля 1999 г.; заявка США № 10/409391; заявка США № 60/489441, поданная 23 июля 2003 г.; заявка Великобритании № GB 0330016,7, поданная 24 декабря 2003 г. и международная патентная заявка № PCT/IB2004/000655, поданная 15 января 2004 г. Каждая из этих заявок и каждый из приводимых в этих заявках документов ("цитируемые в заявке документы") и каждый документ, на который ссылались или который цитировали в документах ссылочной заявки, в тексте или при выполнении этих заявок, а также во всех подтверждениях патентоспособности, поданных в процессе ее ведения, таким образом, приведен здесь в качестве ссылки. Также в данном тексте процитированы различные документы ("здесь ссылочные документы"). Каждый из ссылочных документов и каждый документ, который процитирован или на который сделана ссылка в ссылочных документах, таким образом, приведен здесь в качестве ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам получения вариантов ферментов. Настоящее изобретение дополнительно относится к новым вариантам ферментов и к применению этих новых вариантов ферментов.
УРОВЕНЬ ТЕХНИКИ
В последнее временя были найдены ферменты липид:холестеринацилтрансферазы (см., например, Buckley - Biochemistry 1983, 22, 5490-5493). В частности, были обнаружены глицерофосфолипид:холестеринацилтрансферазы (GCAT), которые, подобно лецитин:холестеринацилтрансферазам (LCAT) растений и/или млекопитающих, катализируют перенос жирной кислоты между фосфатидилхолином и холестерином.
Upton и Buckley (TIBS 20, May 1995, p178-179), а также Brumlik и Buckley (J. of Bacteriology Apr. 1996, p2060-2064) указывают, что липаза/ацилтрансфераза из Aeromonas hydrophila способна осуществлять перенос ацила на спиртовые рецепторы в водной среде.
Для ацетилтрансферазы A. hydrophila были идентифицированы предполагаемые домен связывания субстрата и активный центр (см., например, Thornton et al 1988 Biochem. et Biophys. Acta. 959, 153-159 and Hilton & Buckley 1991 J. Biol. Chem. 266, 997-1000).
Buckley et al. (J. Bacteriol 1996, 178 (7) 2060-4) указали, что Ser16, Asp116 и His291 являются необходимыми аминокислотами, которые необходимы для поддержания ферментативной активности.
Robertson et al. (J. Biol. Chem. 1994, 269, 2146-50) указали на некоторые конкретные мутации ацилтрансферазы A. hydrophila, а именно Y226F, Y230F, Y30F, F13S, S18G, S18V, ни к одной из которых не относится настоящее изобретение.
КРАТКОЕ ИЗЛОЖЕНИЕ АСПЕКТОВ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на открытии конкретных вариантов содержащего GDSX фермента липидацилтрансферазы, которые обладают повышенной трансферазной активностью по сравнению с исходным ферментом. В частности, варианты по настоящему изобретению обладают повышенной по сравнению с исходным ферментом трансферазной активностью, используя в качестве донора ацила галактолипид. Эти липидацилтрансферазы в настоящем описании обозначены как гликолипидацилтрансферазы. Варианты по настоящему изобретению дополнительно могут обладать повышенным отношением трансферазной активности, используя галактолипиды в качестве доноров ацила к фосфолипидтрансферазной активности (отношение GL:PL), и/или повышенным отношением по сравнению с исходным ферментом отношением трансферазной активности, используя в качестве донора ацила галактолипиды, к галактолипидгидролизной активности (отношение GLt:GLh) по сравнению с отношением исходного фермента.
В первом аспекте настоящее изобретение относится к способу получения варианта фермента гликолипидацилтрансферазы, предусматривающему: (a) отбор исходного фермента, представляющего собой фермент липидацилтрансферазу, отличающуюся тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S; (b) модификацию одной или нескольких аминокислот с получением варианта липидацилтрансферазы; (c) тестирование варианта липидацилтрансферазы на активность в отношении галактолипидного субстрата и, необязательно, фосфолипидного субстрата и/или, необязательно, триглицеридного субстрата; (d) отбор варианта фермента с повышенной активностью в отношении галактолипидов по сравнению с исходным ферментом; и, необязательно, (e) получение большого количества варианта фермента.
В другом аспекте настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, отличающегося тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S, и где вариант фермента по сравнению с исходной последовательностью содержит одну или несколько модификаций аминокислот в любых, одном или нескольких, аминокислотных остатках, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7 (определенных здесь ниже).
В дополнительном аспекте настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, отличающемуся тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S, и где вариант фермента по сравнению с исходной последовательностью содержит одну или несколько модификаций аминокислот в любых, одном или нескольких, аминокислотных остатках, подробно описанных в наборе 2, или наборе 4, или наборе 6, или наборе 7 (определенных здесь ниже), идентифицируемых указанной исходной последовательностью, структурно сравненной с определенной здесь структурной моделью P10480, которая предпочтительно получена посредством структурного сравнения координат кристаллической структуры P10480 с 1IVN.PDB и/или 1DEO.PDB, как указано здесь.
Дополнительно настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, отличающемуся тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S, и где вариант фермента содержит одну или несколько модификаций аминокислот по сравнению с исходной последовательностью в любых, одной или нескольких, аминокислотных остатках указывали в наборе 2, идентифицируемых при сравнении исходной последовательности с консенсусной последовательностью pfam (SEQ ID NO:1) и модифицируемых по структурной модели P10480 для обеспечения наибольшего перекрывания (см. фиг.55), как указано.
В дополнительном аспекте настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, где вариант фермента содержит аминокислотную последовательность, которая приведена в SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45, за исключением одной или нескольких модификаций аминокислот в любых, одном или нескольких, аминокислотных остатках, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7 (определенных здесь ниже), идентифицируемых сравнением последовательности с SEQ ID NO:2.
В дополнительном аспекте настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, где вариант фермента содержит аминокислотную последовательность, которая приведена в SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45, за исключением одной или нескольких модификаций аминокислот в любых, одном или нескольких, аминокислотных остатках, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7, идентифицируемых указанной исходной последовательностью, структурно сравненной с определенной здесь структурной моделью P10480, предпочтительно полученной посредством структурного сравнения координат кристаллической структуры P10480 с 1IVN.PDB и/или 1DEO.PDB, как указано здесь.
В дополнительном аспекте настоящее изобретение относится к варианту фермента гликолипидацилтрансферазы, где вариант фермента содержит аминокислотную последовательность, которая приведена в SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45, за исключением одной или нескольких модификаций аминокислот в любых, одном или нескольких, аминокислотных остатках указывали в наборе 2, идентифицируемых при сравнении исходной последовательности с консенсусной последовательностью pfam (SEQ ID NO:1) и модифицируемых по структурной модели P10480 для обеспечения наибольшего перекрывания (см. фиг.55), как указано.
Дополнительно настоящее изобретение относится к применению варианта гликолиполитического фермента по настоящему изобретению или полученного способом по настоящему изобретению для производства субстрата (предпочтительно, пищевого продукта) для получения лизогликолипида, например, дигалактозилмоноглицерида (DGMG) или моногалактозилмоноглицерида (MGMG), посредством обработки гликолипида (например, дигалактозилдиглицерида (DGDG) или моногалактозилдиглицерида (MGDG)) вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению с получением продукта частичного гидролиза, т.е. лизогликолипида.
В дополнительном аспекте настоящее изобретение относится к применению варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению для производства субстрата (предпочтительно, пищевого продукта) для получения лизофосфолипида, например, лизолецитина, посредством обработки фосфолипида (например, лецитина) вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению с получением продукта частичного гидролиза, т.е. лизофосфолипида.
В одном из аспектов настоящее изобретение относится к способу получения пищевого продукта, где способ предусматривает добавление варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению к одному или нескольким ингредиентам пищевых продуктов.
В другом аспекте настоящее изобретение относится к способу получения выпеченного из теста продукта, где способ предусматривает добавление варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению в тесто.
В другом аспекте настоящее изобретение относится к применению варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению в производстве яичного продукта для получения лизофосфолипидов.
В другом аспекте предоставлен способ обработки яиц или яичных продуктов, предусматривающий добавление варианта липолитического фермента по настоящему изобретению в яйцо или яичный продукт для получения лизофосфолипида.
Варианты изобретения можно применять в способе получения полуфабрикатов пищевых продуктов, таких как макаронные изделия быстрого приготовления по аналогии с WO 02/065854.
Настоящее изобретение относится к применению варианта липидацилтрансферазы по настоящему изобретению для получения предпочтительного технического результата или сочетания технических результатов, например, в пищевом продукте (такие как технические результаты, перечисленные здесь в разделе "Технические результаты").
В дополнительном аспекте настоящее изобретение относится к способу ферментативного рафинирования растительных или пищевых масел, предусматривающему обработку пищевого или растительного масла вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению так, чтобы гидролизовать большую часть полярных липидов (например, фосфолипиды и/или гликолипиды).
В другом аспекте настоящее изобретение относится к способу, предусматривающему обработку фосфолипида с тем, чтобы гидролизовать жирные ацильные группы, где способ предусматривает смешивание указанных фосфолипидов с вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению.
В другом аспекте настоящее изобретение относится к способу уменьшения содержания фосфолипидов в пищевом масле, предусматривающему обработку масла вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению с тем, чтобы гидролизовать большую часть фосфолипидов, и отделение содержащей гидролизованный фосфолипид водной фазы от масла.
Также предоставлен способ получения варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению, где способ предусматривает трансформацию клетки-хозяина рекомбинантной нуклеиновой кислотой, содержащей нуклеотидную последовательность, кодирующую указанный вариант липолитического фермента, где клетка-хозяин способна к экспрессии нуклеотидной последовательности, кодирующей полипептид липолитического фермента, культивирование трансформированной клетки-хозяина в условиях, при которых возможна экспрессия нуклеиновой кислоты и получение варианта липолитического фермента.
В дополнительном аспекте настоящее изобретение относится к применению варианта липолитического фермента по настоящему изобретению или полученного способом по настоящему изобретению в биоконверсии полярных липидов (предпочтительно гликолипидов) для получения высокоценных продуктов, таких как сложные эфиры углеводов и/или сложные эфиры белков и/или сложные эфиры белковых субъединиц и/или сложные эфиры оксикислот.
Способ биоконверсии полярных липидов (предпочтительно, гликолипидов) в высокоценные продукты включает в себя смешивание указанного полярного липида с вариантом липолитического фермента по настоящему изобретению или полученным способом по настоящему изобретению.
Также настоящее изобретение дополнительно относится к иммобилизованному варианту липолитического фермента по настоящему изобретению или полученному способом по настоящему изобретению.
Аспекты настоящего изобретения предоставлены в формуле изобретения и в следующих ниже комментариях.
Другие аспекты, относящиеся к нуклеотидным последовательностям, которые можно использовать по настоящему изобретению включают в себя: конструкцию, содержащую последовательности по настоящему изобретению; вектор, содержащий последовательности для применения по настоящему изобретению; плазмиду, содержащую последовательности для применения по настоящему изобретению; трансформированную клетку, содержащую последовательности для применения по настоящему изобретению; трансформированную ткань, содержащую последовательности для применения по настоящему изобретению; трансформированный орган, содержащий последовательности для применения по настоящему изобретению; трансформированного хозяина, содержащего последовательности для применения по настоящему изобретению; трансформированный организм, содержащий последовательности для применения по настоящему изобретению. Настоящее изобретение также относится к способам экспрессии нуклеотидной последовательности для применения по настоящему изобретению с ее использованием, таким как экспрессия в клетке-хозяине, включая сюда способы для ее переноса. Настоящее изобретение дополнительно относится к способам выделения нуклеотидной последовательности, таким как выделение из клетки-хозяина.
Другие аспекты, относящиеся к аминокислотной последовательности для применения по настоящему изобретению включают в себя: конструкцию, кодирующую аминокислотные последовательности для применения по настоящему изобретению; вектор, кодирующий аминокислотные последовательности для применения по настоящему изобретению; плазмиду, кодирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированную клетку, экспрессирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированную ткань, экспрессирующую аминокислотные последовательности для применения по настоящему изобретению; трансформированный орган, экспрессирующий аминокислотные последовательности для применения по настоящему изобретению; трансформированного хозяина, экспрессирующего аминокислотные последовательности для применения по настоящему изобретению; трансформированный организм, экспрессирующий аминокислотные последовательности для применения по настоящему изобретению. Настоящее изобретение также относится к способам очистки аминокислотной последовательности для применения по настоящему изобретению с ее использованием, таким как экспрессия в клетке-хозяине; включая сюда способы ее переноса, а затем очистки указанной последовательности.
Для удобства использования, эти и дополнительные аспекты настоящего изобретения будут обсуждены под соответствующими заголовками разделов. Однако информация каждого раздела не обязательно ограничена каждым конкретным разделом.
ОПРЕДЕЛЕНИЕ НАБОРОВ
Набор аминокислот 1:
Набор аминокислот 1 (необходимо отметить, что они представляют собой аминокислоты в 1IVN - фиг.57 и фиг.58).
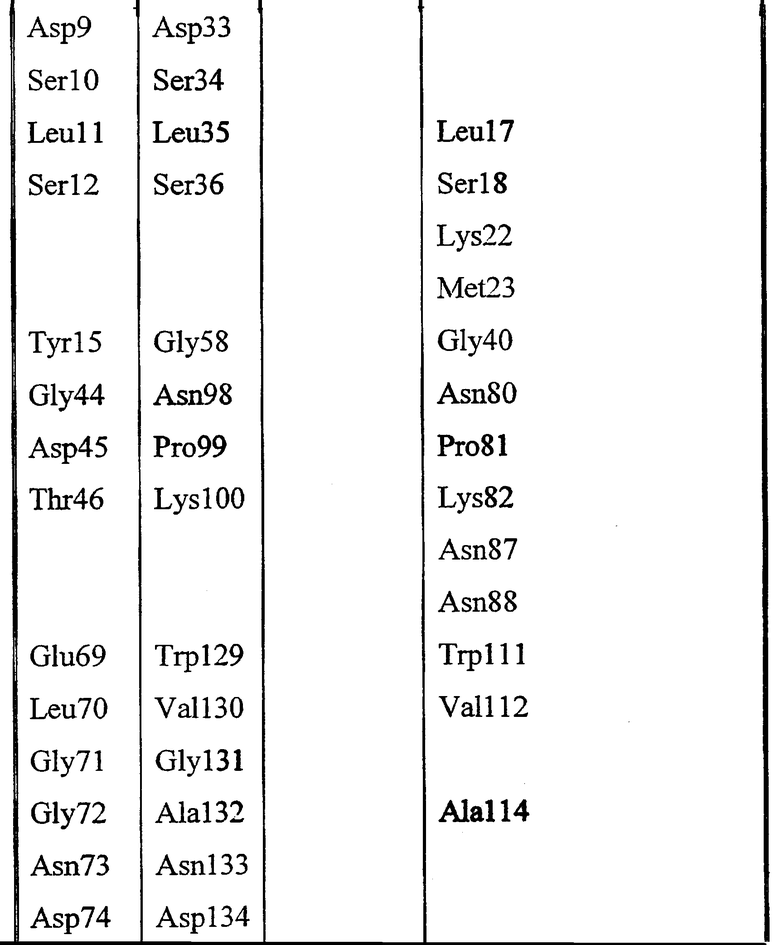
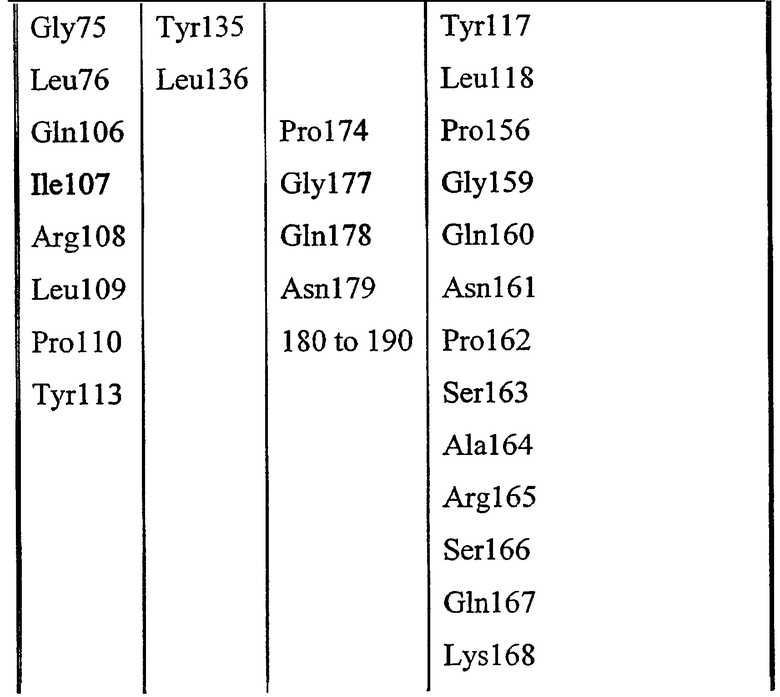
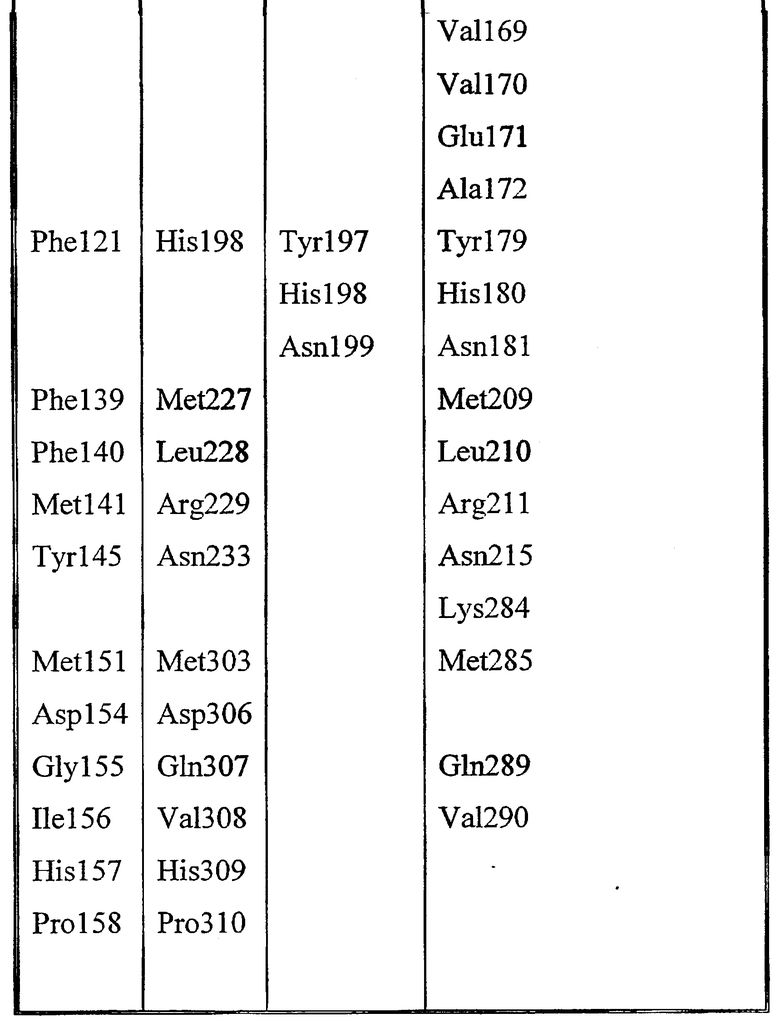
Gly8, Asp9, Ser10, Leu11, Ser12, Tyr15, Gly44, Asp45, Thr46, Glu69, Leu70, Gly71, Gly72, Asn73, Asp74, Gly75, Leu76, Gln106, Ile107, Arg108, Leu109, Pro110, Tyr113, Phe121, Phe139, Phe140, Met141, Tyr145, Met151, Asp154, His157, Gly155, Ile156, Pro158.
Высококонсервативные мотивы, такие как GDSX, и входящие в состав каталитического центра остатки исключены из набора 1 (подчеркнутые остатки). Во избежание неопределенности, набор 1 определен аминокислотными остатками в пределах 10 Å от центрального углеродного атома глицерина в активном центре модели 1IVN.
Набор аминокислот 2:
Набор аминокислот 2 (необходимо отметить, что нумерация аминокислот относится к аминокислотам в зрелой последовательности P10480).
Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289 и Val290.
Таблица выбранных остатков в наборе 1 по сравнению с набором 2:

Набор аминокислот 3:
Набор аминокислот 3 идентичен набору 2, но относится к кодирующей последовательности Aeromonas salmonicida (SEQ ID NO:28), т.е. номера аминокислотных остатков в наборе 3 на 18 больше, т.к. это отражает разницу между нумерацией аминокислот в зрелом белке (SEQ ID NO:2) по сравнению с белком, включающем в себя сигнальную последовательность (SEQ ID NO:28).
Зрелые белки Aeromonas salmonicida GDSX (SEQ ID NO:28) и Aeromonas hydrophila GDSX (SEQ ID NO:26) отличаются пятью аминокислотами. Они представляют собой Thr3Ser, Gln182Lys, Glu309Ala, Ser310Asn, Gly318-, где остаток salmonicida приведен первым, а остаток hydrophila приведен последним (фиг.59). Белок hydrophila состоит в длину только из 317 аминокислот и у него отсутствует остаток в положении 318. Aeromonas salmonicidae GDSX обладает значительно большей активностью в отношении полярных липидов, таких как галактолипидные субстраты, чем белок Aeromonas hydrophila. Сканирование участков проводилось на всех пяти позициях аминокислот.
Набор аминокислот 4:
Набор аминокислот 4 представляет собой S3, Q182, E309, S310, и -318.
Набор аминокислот 5:
F13S, D15N, S18G, S18V, Y30F, D116N, D116E, D157N, Y226F, D228N, Y230F.
Набор аминокислот 6:
Набор аминокислот 6 представляет собой Ser3, Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Gln182, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289, Val290, Glu309, Ser310, -318.
Нумерация аминокислот в наборе 6 относится к аминокислотным остаткам в P10480 (SEQ ID NO:2) - соответствующие аминокислоты в других основных цепях последовательностей можно определить посредством гомологичного сравнения и/или структурного сравнение с P10480 и/или 1IVN.
Набор аминокислот 7:
Набор аминокислот 7 представляет собой Ser3, Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Gln182, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289, Val290, Glu309, Ser310, -318, Y30X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Y226X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Y230X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), S18X (где X выбран из A, C, D, E, F, H, I, K, L, M, N, P, Q, R, T, W или Y), D157X (где X выбран из A, C, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y).
Нумерация аминокислот в наборе 7 относится к аминокислотным остаткам в P10480 (SEQ ID NO:2) - соответствующие аминокислоты в других основных цепях последовательностей можно определить посредством гомологичного сравнения и/или структурного сравнение с P10480 и/или 1IVN.
ПОДРОБНОЕ ИЗЛОЖЕНИЕ АСПЕКТОВ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Предпочтительно, исходный фермент липидацилтрансфераза содержит любую из следующих ниже аминокислотных последовательностей: SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45 или аминокислотную последовательность, обладающую 75% или более идентичностью с любой из последовательностей, представленных в SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45.
Таким образом, исходный фермент липидацилтрансфераза по настоящему изобретению содержит аминокислотную последовательность, которая по меньшей мере, на 80%, предпочтительно, по меньшей мере, на 85%, более предпочтительно, по меньшей мере, на 90%, более предпочтительно, по меньшей мере, на 95%, больше чем, по меньшей мере, на 98% гомологична любой последовательности, представленной как SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45.
Таким образом, исходный фермент липидацилтрансфераза может кодировать любая из следующих ниже нуклеотидных последовательностей: SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:35, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44 или SEQ ID NO:46 или нуклеотидная последовательность, которая по меньшей мере, на 75% или более идентична любой из последовательностей, представленных как SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:35, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44 или SEQ ID NO:46.
Таким образом, нуклеотидная последовательность может быть на 80% или более, предпочтительно, на 90% или более, более предпочтительно, на 95% или более, даже более предпочтительно, на 98% или более, идентична любой из последовательностей, представленных как SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:35, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44 или SEQ ID NO:46.
Предпочтительно, исходный фермент модифицирован по одному или нескольким из аминокислотных остатков, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7 при сравнении с эталонной последовательностью (SEQ ID NO:2) или структурном сравнении по структурной модели P10480, или сравненных по консенсусной последовательности pfam и модифицированных по структурной модели P10480.
Таким образом, вариант фермента по сравнению с исходным ферментом может обладать повышенным отношением активности в отношении галактолипидов к активности в отношении либо фосфолипидов, либо/и триглицеридов.
Таким образом, способ по настоящему изобретению может предусматривать тестирование варианта липидацилтрансферазы на:
(i) трансферазную активность в отношении галактолипидного субстрата, и
(ii) трансферазную активность в отношении фосфолипидного субстрата; и
отбор варианта фермента, который по сравнению с исходным ферментом обладает повышенным отношением трансферазной активности в отношении галактолипидов к трансферазной активности в отношении фосфолипидов.
Таким образом, отношение трансферазной активности варианта фермента по настоящему изобретению в отношении галактолипидов к трансферазной активности в отношении фосфолипидов может представлять собой, по меньшей мере, 1, по меньшей мере, 2, по меньшей мере, 3, по меньшей мере, 4 или, по меньшей мере, 5.
Таким образом, способ по настоящему изобретению может предусматривать тестирование варианта липидацилтрансферазы на:
(a) трансферазную активность в отношении галактолипидного субстрата, и
(b) гидролитическую активность в отношении галактолипидного субстрата; и
отбор варианта фермента с повышенным отношением трансферазной активности в отношении галактолипидов к его гидролитической активности в отношении гликолипидов по сравнению с отношением исходного фермента.
Таким образом, отношение трансферазной активности для галактолипидов к гидролитической активности в отношении галактолипидов может составлять более 1, по меньшей мере, 1,5, по меньшей мере, 2, по меньшей мере, 4 или, по меньшей мере, 5.
Анализ для определения трансферазной и гидролитической активности в отношении галактолипидов и/или фосфолипидов указан, например, в примере 8.
Термин "повышенная активность в отношении галактолипидов" означает фермент, который по сравнению с исходным ферментом обладает повышенной (т.е. большей) трансферазной активностью, если липидным донором ацила является галактолипид, (галактолипидтрансферазная активность), и/или по сравнению с исходным ферментом обладает повышенным отношением галактолипидтрансферазной активности к фосфолипидтрансферазной активности (отношение GLt:PLt), и/или по сравнению с исходным ферментом обладает повышенным отношением галактолипидтрансферазной активности к галактолипидгидролазной активности (отношение GLt:GLh).
Таким образом, вариант фермента по сравнению с исходным ферментом может обладать повышенной галактолипидтрансферазной активностью и либо такой же, либо меньшей галактолипидгидролитической активностью. Другими словами, таким образом, вариант фермента может обладать большей галактолипидтрансферазной активностью по сравнению с его галактолипидгидролитической активностью по сравнению с исходным ферментом. Таким образом, вариант фермента может предпочтительно переносить ацильную группу с галактолипида на акцептор ацила вместо простого гидролиза галактолипида.
В одном из вариантов осуществления фермент по настоящему изобретению может обладать повышенной трансферазной активностью в отношении фосфолипидов (т.е. повышенной фосфолипидтрансферазной активностью) по сравнению с исходным ферментом. Эта повышенная фосфолипидтрансферазная активность может быть независимой от повышенной активности в отношении галактолипидов. Однако, таким образом, вариант фермента может обладать повышенной галактолипидтрансферазной активностью и повышенной фосфолипидтрансферазной активностью.
В одном из вариантов осуществления настоящее изобретение относится к варианту фермента липидацилтрансферазы, отличающемуся тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S, где вариант обладает повышенной активностью в отношении фосфолипидов, предпочтительно, повышенной фосфолипидтрансферазной активностью, по сравнению с исходным ферментом, и где вариант фермента содержит одну или несколько модификаций аминокислот по сравнению с исходной последовательностью в любых, одном или нескольких, аминокислотных остатках, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7.
Как применяют здесь, термин "модифицирование" означает добавление, замену и/или удаление. Предпочтительно термин "модифицирование" означает "замену".
Во избежание неопределенности, когда в исходном ферменте замещена аминокислота, она предпочтительно замещена аминокислотой, отличающейся от аминокислоты, первоначально обнаруженной в исходном ферменте в этом положении, таким образом, чтобы получить вариант фермента. Другими словами, термин "замена" не предназначен для обозначения замещения аминокислоты той же аминокислотой.
Предпочтительно, исходный фермент представляет собой фермент, содержащий аминокислотную последовательность, представленную как SEQ ID NO:2 и/или SEQ ID NO:28.
Предпочтительно, вариант фермента представляет собой фермент, содержащий аминокислотную последовательность, где данная аминокислотная последовательность представлена как SEQ ID NO:2, за исключением одной или нескольких модификаций аминокислот в любых одном или нескольких аминокислотных остатках, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7.
В одном из вариантов осуществления предпочтительный вариант фермента содержит по сравнению с исходной последовательностью одну или несколько модификаций аминокислот, по меньшей мере, в одном из аминокислотных остатков, определенных в наборе 4.
Таким образом, вариант фермента по сравнению с исходным ферментом содержит одну или несколько из следующих ниже модификаций аминокислот:
S3E, A, G, K, M, Y, R, P, N, T или G
E309Q, R или A, предпочтительно Q или R
-318Y, H, S или Y, предпочтительно Y.
Предпочтительно, X из мотива GDSX представляет собой L. Таким образом, предпочтительно исходный фермент содержит аминокислотный мотив GDSL.
Предпочтительно способ получения варианта фермента липидацилтрансферазы дополнительно предусматривает одну или несколько из следующих ниже стадий:
1) картирование структурной гомологии или
2) сравнение по гомологии последовательностей.
Таким образом, картирование структурной гомологии может предусматривать одну или несколько из следующих ниже стадий:
i) сравнение исходной последовательности со структурной моделью (1IVN.PDB), показанной на фиг.52;
ii) выбор одного или нескольких аминокислотных остатков в пределах сферы размером 10 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53) (такие как один или несколько из аминокислотных остатков, определенных в наборе 1 или наборе 2); и
iii) модификацию указанной исходной последовательности одной или нескольких аминокислот, выбранных в соответствии со стадией (ii).
В одном из вариантов осуществления выбранный аминокислотный остаток может находиться в пределах сферы размером 9, предпочтительно, в пределах 8, 7, 6, 5, 4, или 3 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53).
Таким образом, картирование структурной гомологии может предусматривать одну или несколько из следующих ниже стадий:
i) сравнение исходной последовательности со структурной моделью (1IVN.PDB), показанной на фиг.52;
ii) выбор одой или нескольких аминокислот в пределах сферы размером 10 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53) (таких как один или несколько из аминокислотных остатков, определенных в наборе 1 или наборе 2);
iii) определение того, являются ли один или несколько аминокислотных остатков, выбранных в соответствии со стадией (ii) высоко консервативными (особенно являются ли они остатками активного центра, и/или частью мотива GDSX, и/или частью мотива GANDY); и
iv) модификацию указанной исходной последовательности одной или нескольких аминокислот, выбранных в соответствии со стадией (ii), за исключением консервативных областей, идентифицированных в соответствии со стадией (iii).
В одном из вариантов осуществления выбранный аминокислотный остаток может находиться в пределах сферы размером 9, предпочтительно, в пределах 8, 7, 6, 5, 4, или 3 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53).
Альтернативно описанному выше картированию структурной гомологии или совместно с ним, картирование структурной гомологии можно проводить посредством выбора конкретных областей петель (LR) или промежуточных областей (IVR), полученных из сравнения с pfam (сравнение 2, фиг.56), перекрывающихся с моделью P10480 и 1IVN. Области петель (LR) или промежуточные области (IVR) определены ниже в таблице:

В некоторых вариантах осуществления настоящего изобретения вариант фермента ацилтрансферазы содержит не только модификации аминокислот по одной или нескольким из аминокислот, определенных в любом из наборов 1-4 и 6-7, но также содержит по меньшей мере одну модификацию аминокислоты, одной или нескольких, определенных выше промежуточных областей (IVR1-6) (предпочтительно, в одной или нескольких из IVR 3, 5 и 6, более предпочтительно, в IVR5 или IVR 6) и/или в одной или нескольких из определенных выше областей петель (LR1-5) (предпочтительно, в одной или нескольких из LR1, LR2 или LR5, более предпочтительно, в LR5).
В одном из вариантов осуществления, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые определены не только в одном или нескольких наборах 2, 4, 6 и 7, но также находятся в одной или нескольких IVR 1-6 (предпочтительно, в пределах IVR 3, 5 или 6, более предпочтительно, в пределах IVR 5 или IVR 6) или в одной или нескольких из LR 1-5 (предпочтительно, в пределах LR1, LR2 или LR5, более предпочтительно, в пределах LR5).
Таким образом, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые находятся не только в наборе 1 или 2, но также расположены в пределах IVR 3.
Таким образом, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые находятся не только в наборе 1 или 2, но также расположены в пределах IVR 5.
Таким образом, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые находятся не только в наборе 1 или 2, но также расположены в пределах IVR 6.
Таким образом, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые находятся не только в наборе 1 или 2, но также расположены в пределах LR 1.
Таким образом, вариант ацилтрансферазы по настоящему изобретению или полученный способом по настоящему изобретению может содержать одну или несколько модификаций аминокислот, которые находятся не только в наборе 1 или 2, но также расположены в пределах LR 2.
Подобным образом, в некоторых вариантах осуществления настоящего изобретения вариант фермента ацилтрансферазы не только содержит модификацию аминокислоты в одном или нескольких аминокислотных остатках, находящихся в пределах сферы размером 10, предпочтительно, в пределах 9, 8, 7, 6, 5, 4, или 3 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53), но также содержит по меньшей мере одну модификацию аминокислоты в одной или нескольких из определенных выше промежуточных областях (IVR1-6) (предпочтительно, в одной или нескольких из IVR 3, 5 и 6, более предпочтительно, в IVR 5 или IVR 6) и/или в одной или нескольких из определенных выше областей петель (LR1-5) (предпочтительно, в одной или нескольких из LR1, LR2 или LR5, более предпочтительно, в LR5).
В одном из вариантов осуществления предпочтительная модификация аминокислоты расположена в одном или нескольких аминокислотных остатках, находящихся в пределах сферы размером 10 Å, а также в пределах LR5.
Таким образом, картирование структурной гомологии может предусматривать одну или несколько из следующих ниже стадий:
i) сравнение исходной последовательности со структурной моделью (1IVN.PDB), показанной на фиг.52;
ii) выбор одного или нескольких аминокислотных остатков в пределах сферы 10 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53) (такие как один или несколько аминокислотных остатков, определенных в наборе 1 или наборе 2); и/или выбор одного или нескольких аминокислотных остатков в пределах IVR1-6 (предпочтительно, в пределах IVR 3, 5 или 6, более предпочтительно, в пределах IVR 5 или IVR6); и/или выбор одного или нескольких аминокислотных остатков в пределах LR1-5 (предпочтительно, в пределах LR1, LR2 или LR5, более предпочтительно, в пределах LR5); и
iii) модификацию в указанной исходной последовательности одной или нескольких аминокислот, выбранных в соответствии со стадией (ii).
В одном из вариантов осуществления выбранный аминокислотный остаток может находиться в пределах сферы размером 9, предпочтительно, в пределах 8, 7, 6, 5, 4, или 3 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53).
Таким образом, картирование структурной гомологии может предусматривать одну или несколько из следующих ниже стадий:
i) сравнение исходной последовательности со структурной моделью (1IVN.PDB), показанной на фиг.52;
ii) выбор одой или нескольких аминокислот в пределах сферы 10 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53) (таких как один или несколько из аминокислотных остатков, определенных в наборе 1 или наборе 2); и/или выбор одного или нескольких аминокислотных остатков в пределах IVR1-6 (предпочтительно, в пределах IVR 3, 5 или 6, более предпочтительно, в пределах IVR 5 или IVR 6); и/или выбор одного или нескольких аминокислотных остатков в пределах LR1-5 (предпочтительно, в пределах LR1, LR2 или LR5, более предпочтительно, в пределах LR5);
iii) определение того, являются ли один или несколько аминокислотных остатков, выбранных в соответствии со стадией (ii) высококонсервативными (особенно являются ли они остатками активного центра и/или частью мотива GDSX, и/или частью мотива GANDY); и
модификацию в указанной исходной последовательности одной или нескольких аминокислот, выбранных в соответствии со стадией (ii), за исключением консервативных областей, идентифицированных в соответствии со стадией (iii).
Таким образом, одна или несколько аминокислот, выбранных в подробно описанных выше способах расположен не только в пределах сферы размером 10 Å с центром в центральном углеродном атоме молекулы глицерина в активном центре (см. фиг.53) (такие как один или несколько аминокислотных остатков, определенных в наборе 1 или наборе 2), но также расположены в пределах одной или нескольких из IVR 1-6 (предпочтительно, в пределах IVR 3, 5 или 6, более предпочтительно, в пределах IVR 5 или IVR 6) или в пределах одной или нескольких из LR 1-5 (предпочтительно, в пределах LR1, LR2 или LR5, более предпочтительно, в пределах LR5).
В одном из вариантов осуществления предпочтительные одна или несколько модификаций аминокислот находятся в пределах LR5. В том случае когда модификация(и) расположена в пределах LR5, модификация не является одной из тех, которые определенны в наборе 5. Таким образом, одна или несколько модификаций аминокислот не только попадают в область, определенную посредством LR5, но также состоят из аминокислот в пределах одной или нескольких из набора 2, набора 4, набора 6 или набора 7.
Таким образом, сравнение по гомологии последовательностей может предусматривать одну или несколько из следующих ниже стадий:
i) выбор первой исходной липидацилтрансферазы;
ii) идентификацию второй родственной липидацилтрансферазы с желаемой активностью;
iii) сравнение указанной первой исходной липидацилтрансферазы и второй родственной липидацилтрансферазы;
iv) идентификацию аминокислотных остатков, отличающихся у двух последовательностей; и
v) модификацию в указанной исходной липидацилтрансферазе одного или нескольких из аминокислотных остатков, идентифицированных в соответствии со стадией (iv).
Таким образом, сравнение по гомологии последовательностей может предусматривать одну или несколько из следующих ниже стадий:
i) выбор первой исходной липидацилтрансферазы;
ii) идентификацию второй родственной липидацилтрансферазы с желаемой активностью;
iii) сравнение указанной первой исходной липидацилтрансферазы и второй родственной липидацилтрансферазы;
iv) идентификацию аминокислотных остатков, отличающихся у двух последовательностей; и
v) определение того, являются ли один или несколько аминокислотных остатка, выбранных в соответствии со стадией (iv) высококонсервативными (особенно являются ли они остатками активного центра, и/или частью мотива GDSX, и/или частью мотива GANDY); и
vi) модификацию в указанной исходной последовательности одного или нескольких из аминокислотных остатков, идентифицированных в соответствии со стадией (iv) за исключением консервативных областей, идентифицированных в соответствии со стадией (v).
Таким образом, указанная первая исходная липидацилтрансфераза может содержать любую из следующих ниже аминокислотных последовательностей: SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45.
Таким образом, указанная вторая родственная липидацилтрансфераза может содержать любую из следующих ниже аминокислотных последовательностей: SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43 или SEQ ID NO:45.
Вариант фермента по сравнению с исходным ферментом должен содержать по меньшей мере одну модификацию аминокислоты. В некоторых вариантах осуществления, вариант фермента по сравнению с исходным ферментом может содержать, по меньшей мере 2, предпочтительно, по меньшей мере 3, предпочтительно, по меньшей мере 4, предпочтительно, по меньшей мере 5, предпочтительно, по меньшей мере 6, предпочтительно, по меньшей мере 7, предпочтительно, по меньшей мере 8, предпочтительно, по меньшей мере 9, предпочтительно, по меньшей мере 10 модификаций аминокислот.
Таким образом, способы по настоящему изобретению могут предусматривать дополнительную стадию получения варианта фермента в композицию фермента и/или композицию пищевого продукта, такую как улучшенная хлебобулочная композиция.
Для сравнения последовательности полипептида GDSX (исходная последовательность) с SEQ ID NO:2 (P01480), можно применять сравнение последовательностей, такое как парное сравнение (http://www.ebi.ac.uk/emboss/align/index.html). Таким образом, можно определить и модифицировать эквивалентные аминокислоты в альтернативных исходных полипептидах GDSX, которые соответствуют одной или нескольким из аминокислот, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7 в отношении SEQ ID NO:2. Как легко поймут специалисты, при применении парного сравнения emboss, как правило, достаточно стандартных установок. Соответствующие остатки можно идентифицировать с применением "needle" для получения сравнения, охватывающего всю длину обоих последовательностей. Однако также возможно найти лучшую область сходства между двумя последовательностями с применением "water".
Альтернативно, особенно в случаях, когда исходные полипептиды GDSX обладают низкой гомологией с SEQ ID NO:2, соответствующие аминокислоты в альтернативных исходных полипептидах GDSX, которые соответствуют одной или нескольким из аминокислот, определенных в наборе 2, наборе 4, наборе 6 или наборе 7 в отношении SEQ ID NO:2, можно определить посредством структурного сравнения со структурной моделью P10480, полученного посредством сравнения полученной из P10480 структурной модели со структурными координатами 1IVN.PDB и 1DEO.PDB с применением "Deep View Swiss-PDB viewer" (полученного с www.expasy.org/spdbv/) (фиг.53 и пример 1). Эквивалентные остатки идентифицируют как остатки, перекрывающиеся или находящиеся в непосредственной близости с остатками в полученной структурной модели P10480, как проиллюстрировано в таблице, сравнивающей набор 1 и набор 2 (см. раздел, озаглавленный "Определение наборов", здесь выше). Таким образом другие полипептиды GDSX можно сравнивать по отношению к координатам кристалла 1IVN.PBD, и определять эквивалентные набору 1 остатки.
Альтернативно, особенно в случаях, когда исходный полипептид GDSX обладает низкой гомологией с SEQ ID NO:2, эквивалентные аминокислоты в альтернативных исходных полипептидах GDSX, которые соответствуют одной или нескольким из аминокислот, определенных в наборе 2, или наборе 4, или наборе 6, или наборе 7 в отношении SEQ ID NO:2, можно определить из сравнения, полученного на основании базы данных PFAM (консенсус PFAM), модифицированного на основании структурного сравнения, как показано в Выравнивании1 (фиг.55). Модификация на основе структурных моделей может являться необходимой для незначительного сдвига сравнения для обеспечения наибольшего перекрывания. Сравнение1 (фиг.55) предоставляет руководство для этого.
Вариант фермента по настоящему изобретению предпочтительно не содержит одну или несколько из модификаций аминокислот, определенных в наборе 5.
Таким образом, вариант фермента можно получить с применением сайт-специфического мутагенеза.
Альтернативно, мутации можно вводить случайным образом, например, с применением коммерческого набора, такого как набор GeneMorph PCR mutagenesis kit из Stratagene, или набор Diversify PCR random mutagenesis kit из Clontech. К способам оптимизации мутагенеза, основанного на ПЦР, который также можно сочетать с применением мутагенных аналогов ДНК, таких, как описаны в EP 0 866 796, относится EP 0 583 265. Для получения вариантов липидацилтрансфераз с предпочтительными характеристиками подходят технологии подверженной ошибкам ПЦР. К молекулярному преобразованию липаз относится WO 0206457.
Третий способ для получения новых последовательностей представляет собой фрагментирование неидентичных нуклеотидных последовательностей, или посредством применения ряда рестрикционных ферментов или такого фермента, как ДНКаза I, и повторную сборку полноразмерных нуклеотидных последовательностей, кодирующих функциональные белки (далее здесь называют "перестановка"). Альтернативно можно применять одну или несколько неидентичных нуклеотидных последовательностей и вносить мутации в течение повторной сборки полноразмерной нуклеотидной последовательности. Для получения вариантов липидацилтрансфераз с предпочтительными характеристиками пригодны технологии перестановки ДНК и перестановки семейств. Пригодные способы для проведения "перестановки" можно найти в EP 0 752 008, EP 1 138 763, EP 1 103 606. Перестановку также можно сочетать с другими формами мутагенеза ДНК, как описано в патенте США 6180406 и WO 01/34835.
Таким образом, в нуклеотидной последовательности можно получать множество участков направленных или случайных мутаций, или in vivo, или in vitro, и далее различными способами можно исследовать кодируемый вариант полипептида на улучшенную функциональность.
В качестве неограничивающего примера, кроме того, мутантные или природные варианты полинуклеотидной последовательности для получения новых вариантов можно рекомбинировать или с диким типом или с другими мутантными или природными вариантами. Такие новые варианты также можно исследовать на улучшенную функциональность кодируемого полипептида.
Для локализованного случайного мутагенеза и/или перестановки предпочтительно можно выбирать следующие области: IVR3, IVR5, IVR6, LR1, LR2, и/или LR5, наиболее предпочтительно, LR5.
Для получения библиотек вариантов можно применять микробных эукариотических или прокариотических экспрессирующих хозяев. Для обеспечения одинаковой экспрессии в пределах библиотеки вариантов предпочтительными могут быть системы с низкой копийностью, предпочтительно с одним фактом хромосомной экспрессии. Также предпочтительны экспрессирующие системы с высокой частотой трансформации, особенно для экспрессии больших библиотек вариантов (> 1000 колоний), таких как библиотеки, полученные с применением технологий случайного мутагенеза и/или перестановки.
Пригодные способы для применения эукариотического экспрессирующего хозяина, т.е. дрожжей, для получения ферментов описаны в EP 1131416. Микробные эукариотические экспрессирующие хозяева, такие как дрожжи, могут быть предпочтительны для экспрессии библиотек вариантов, полученных с применением исходного гена эукариотической ацилтрансферазы.
Пригодные способы с применением в качестве экспрессирующего хозяина для получения ферментов Bacillus, т.е. Bacillus subtilis описаны в WO 02/14490. Микробные прокариотические экспрессирующие хозяева, такие как Bacillus, могут быть предпочтительны для экспрессии библиотек вариантов, полученных с применением исходного гена прокариотической ацилтрансферазы, например, эталонной последовательности P10480 (SEQ ID NO:2).
Таким образом, вариант липидацилтрансферазы по настоящему изобретению сохраняет по меньшей мере 70%, предпочтительно, по меньшей мере 80%, предпочтительно, по меньшей мере 90%, предпочтительно, по меньшей мере 95%, предпочтительно, по меньшей мере 97%, предпочтительно, по меньшей мере 99% гомологии с исходным ферментом.
Пригодные исходные ферменты могут включать в себя любой фермент с эстеразной или липазной активностью.
Предпочтительно, исходный фермент сравнен с консенсусной последовательностью pfam00657.
В предпочтительном варианте осуществления вариант фермента липидацилтрансферазы сохраняет или включает в себя по меньшей мере один или несколько аминокислотных остатков консенсусной последовательности pfam00657, находящихся в участках GDSX, GANDY и HPT.
Ферменты, такие как липазы, без или с низкой липидацилтрансферазной активностью в водной среде для введения или усиления трансферазной активности можно подвергнуть мутированию с применением средств молекулярного преобразования, таким образом получая вариант фермента липидацилтрансферазы со значительной трансферазной активностью, пригодный для применения в композициях и способах по настоящему изобретению.
Таким образом, липидацилтрансфераза для применения по изобретению может представлять собой вариант с повышенной ферментативной активностью в отношении полярных липидов, предпочтительно гликолипидов, по сравнению с исходным ферментом. Предпочтительно, такие варианты также обладают низкой или не обладают активностью в отношении полярных лизолипидов. Повышенная активность в отношении полярных липидов, предпочтительно гликолипидов, может являться результатом гидролиза и/или трансферазной активности или их сочетания.
Вариант липидацилтрансферазы для применения по изобретению может обладать уменьшенной активностью в отношении триглицеридов, и/или моноглицеридов, и/или диглицеридов по сравнению с исходным ферментом.
Таким образом, у варианта фермента может отсутствовать активность в отношении триглицеридов, и/или моноглицеридов, и/или диглицеридов. Низкая активность в отношении триглицеридов предпочтительна в вариантах белков, которые нужно использовать для применений в хлебопечении для обработки яиц или яичных продуктов и/или для рафинирования масел.
Таким образом, в одном из вариантов осуществления вариант фермента может обладать высокой активностью в отношении диглицеридов и у него может отсутствовать или быть низкой активность в отношении триглицеридов.
При обозначении здесь конкретных аминокислотных остатков нумерация представляет собой нумерацию, полученную при сравнении варианта последовательности с эталонной последовательностью, представленной как SEQ ID NO:2.
В одном из аспектов вариант фермента предпочтительно содержит одну или несколько из следующих ниже аминокислотных замен:
S3A, С, D, E, F, G, H, I, К, L, M, N, P, Q, R, T, V, W или Y; и/или
L17A, C, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W или Y; и/или
S18A, C, D, E, F, H, I, K, L, M, N, P, Q, R, T, W или Y; и/или
K22A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y; и/или
M23A, C, D, E, F, G, H, I, K, L, N, P, Q, R, S, T, V, W или Y; и/или
Y30A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; и/или
G40A, C, D, E, F, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
N80A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
P81A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; и/или
K82A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y; и/или
N87A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
N88A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
W111A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
V112A, С, D, E, F, G, H, I, К, L, M, N, P, Q, R, S, T, W или Y; и/или
A114C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
Y117A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; и/или
L118A, С, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W или Y; и/или
P156A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; и/или
D157A, C, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
G159A, C, D, E, F, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
Q160A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y; и/или
N161A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
P162A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; и/или
S163A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y; и/или
A164C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
R165A, C, D, E, F, G, H, I, K, L, M, N, P, Q, S, T, V, W или Y; и/или
S166A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y; и/или
Q167A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y; и/илиK168A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y; и/или
V169A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y; и/или
V170A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y; и/или
E171A, C, D, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
A172C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
Y179A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; и/или
H180A, C, D, E, F, G, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
N181A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; и/или
Q182A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y, предпочтительно K; и/или
M209A, C, D, E, F, G, H, I, K, L, N, P, Q, R, S, T, V, W или Y; и/или
L210 A, C, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W или Y; и/или
R211 A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
N215 A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
Y226A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; и/или
Y230A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; и/или
K284A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y; и/или
M285A, C, D, E, F, G, H, I, K, L, N, P, Q, R, S, T, V, W или Y; и/или
Q289A, С, D, E, F, G, H, I, К, L, M, N, P, R, S, T, V, W или Y; и/или
V290A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y; и/или
E309A, C, D, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; и/или
S310A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y.
В дополнение или альтернативно к этому, могут присутствовать одно или несколько расширений на C-конце. Предпочтительно дополнительное расширение на C-конце состоит из одной или нескольких алифатических аминокислот, предпочтительно, неполярных аминокислот, более предпочтительно, из I, L, V или G. Таким образом, настоящее изобретение дополнительно относится к варианту фермента, включающему в себя одно или несколько из следующих дополнений на C-конце: 318I, 318L, 318V, 318G.
В том случае когда остатки в исходной основной цепи отличаются от остатков в P10480 (SEQ ID NO:2), как определено посредством гомологичного сравнения и/или структурного сравнения с P10480 и/или 1IVN, может быть желательным заменить остатки, выравнивающиеся с любым одним или несколькими из следующих ниже остатков в P10480 (SEQ ID NO:2): Ser3, Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Gln182, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289, Val290, Glu309 или Ser310, на найденный в P10480 остаток, соответственно.
Найдено, что следующие ниже остатки дикого типа P10480 предпочтительны для сохранения хорошей активности, особенно хорошей трансферазной активности для галактолипида: L17, W111, R221, S3, G40, N88, K22, Y117, L118, N181, M209, M285, E309, M23. Таким образом, вариант фермента предпочтительно содержит в любом одном или нескольких из этих участков найденный в P10480 аминокислотный остаток.
Варианты ферментов с повышенной гидролитической активностью в отношении полярного липида также могут обладать повышенной трансферазной активностью для полярного липида.
Варианты ферментов с повышенной гидролитической активностью в отношении фосфолипида, такого как фосфатидилхолин (PC) также могут обладать повышенной трансферазной активностью для фосфолипида.
Варианты ферментов с повышенной гидролитической активностью в отношении галактолипида, такого как DGDG, также могут обладать повышенной трансферазной активностью для галактолипида.
Варианты ферментов с повышенной трансферазной активностью для фосфолипида, такого как фосфатидилхолин (PC), также могут обладать повышенной гидролитической активностью в отношении фосфолипида.
Варианты ферментов с повышенной трансферазной активностью для галактолипида, такого как DGDG, также могут обладать повышенной гидролитической активностью в отношении галактолипида.
Варианты ферментов с повышенной трансферазной активностью для полярного липида также могут обладать повышенной гидролитической активностью в отношении полярного липида.
Таким образом, в связывание субстрата могут быть вовлечены один или несколько из следующих ниже участков: Leu17; Ala114; Tyr179; His180; Asn181; Met209; Leu210; Arg211; Asn215; Lys284; Met285; Gln289; Val290.
Вариант фермента по настоящему изобретению по сравнению с исходным ферментом может обладать одной или несколькими из следующих ниже функциональных характеристик:
1) повышенной относительной трансферазной активностью в отношении галактолипида (DG) по сравнению с PC, подсчитанной как % TDG/TPC (как проиллюстрировано в примере 8);
2) повышенной абсолютной трансферазной активностью в отношении галактолипида (DG) (как проиллюстрировано в примере 8);
3) повышенной трансферазной активностью с применением галактолипида в качестве донора (TDG) по отношению к гидролитической активности HDG у галактолипида (DG) (как проиллюстрировано в примере 8);
4) повышенной абсолютной трансферазной активностью в отношении PC (как проиллюстрировано в примере 8);
где DG представляет собой галактолипид (например, DGDG) (и также может быть обозначен здесь как GL), а PC представляет собой фосфолипид (например, лецитин). Варианты с повышенной активностью в отношении галактолипида включают в себя варианты в пределах категорий 1), 2) и 3), указанных выше. Варианты с повышенной активностью в отношении галактолипидов также могут обладать повышенной активностью у фосфолипидов (как для категории 4, выше).
1. К варианту фермента с повышенной относительной трансферазной активностью в отношении DG по сравнению с PC, подсчитанной как %T DG /T PC , может приводить модификация по одному или нескольким из следующих ниже остатков:
-318, N215, L210, S310, Е309, Н180, N80, V112, Y30X (где X выбран из А, С, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), V290, Q289, K22, G40, Y179, M209, L211, K22, P81, N87, Y117, N181, Y230X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Q182.
Как правило, предпочтительными могут являться одна или несколько из следующих ниже замен:
S3A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y, предпочтительно M, R, N, G, T, Q, P, Y, S, L, E, W, наиболее предпочтительно Q
K22A, E, C, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно A, C, E или R
Y30A, C, D, H, K, M, N, P, Q, R, T, V, W, G, I, L, S, M, A, R или E, предпочтительно H, T, W, N, D, C, Q, G, I, L, S, M, A, R или E
G40L, N, T, V или A
N80N, R, D, A, C, E, F, G, H, I, K, L, M, P, Q, S, T, V, W или Y, предпочтительно H, I, Y, C, Q, M, S, W, L, N, R, D или F
P81A, C, D, E, F, G H, I, K, L, M, N, Q, R, S, T, V, W или Y, предпочтительно I, M, F, G, V, Y, D, C или A
K82A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно H, K, S или R
N87A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y, предпочтительно I, Y, M, T, Q, S, W, F, V или P
N88A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y, предпочтительно C, V, A или F
V112C
Y117A, С, D, E, F, H, T, G, I, K, L, M, N, P, Q, R, S, V или W, предпочтительно A, N, E, H, T, I, F, C, P или S
L118A, C, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W или Y, предпочтительно F
V112A, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y, предпочтительно I, M, F, Y, N, E, T, Q, H или P
Y179A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V или W, предпочтительно F, C, H, I, L, M, P, V или W
H180K, Q, A, C, D, E, F, G, I, L, M, P, R, S, T, V, W или Y, предпочтительно M, F, С, К или Q
N181A или V
Q182A, C, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W, или Y, предпочтительно K
M209L, K, M, A, C, D, E, F, G, H, I, N, P, Q, R, S, T, V, W, или Y, предпочтительно I, F, T, D, C, H, L, K, M или P
L210 G, I, H, E, M, S, W, V, A, R, N, D, Q, T, C, F, K, P или Y, предпочтительно G, I, H, E, M, S, W, V, A, R, N, D, Q, Т, Y или F
R211G, Q, K, D, A, C, E, F, H, I, L, M, N, P, R, S, T, V, W или Y, предпочтительно G, Q, K, D, H, I, M, F, P, S, Y, N, C, L или W
N215A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно I, F, P, T, W, H или A
Y230A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W, предпочтительно I, T, G, D, R, E, V, M или S, наиболее предпочтительно I, D, R или E
Q289A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y, предпочтительно F, W, H, I, Y, L, D, C, K, V, E, G, R, N или P, более предпочтительно R, T, D, K, N или P
V290A, C, D, E, H, F, G, I, K, L, M, N, P, Q, R, S, T, W или Y;
E309S, Q, R, A, C, D, F, G, H, I, K, L, M, N, P, T, V, W или Y, предпочтительно F, W, N, H, I, M, S, Q, R, A или Y
S310A, P, T, H, M, K, G, C, D, E, F, I, L, N, Q, R, V, W или Y, предпочтительно F, Y, C, L, K, A, P, T, H, M, K или G
-318 A, C, D, E, F, G, I, K, L, M, N, P, Q, R, T, V, W, Y, H или S
Предпочтительно, к варианту фермента с повышенной относительной трансферазной активностью в отношении DG по сравнению с PC, подсчитанной как %TDG/TPC, может приводить одна или несколько из следующих ниже модификаций:
S3A, С, D, E, F, G, H, I, К, L, M, N, P, Q, R, T, V, W или Y, предпочтительно M, R, N, G, T, Q, P, Y, S, L, E, W, наиболее предпочтительно Q
G40L, N, T, V или A
K82A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно H, K, S или R
N88A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y, предпочтительно C, V, A или F
Y230A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W, предпочтительно I, T, G, D, R, E, V, M или S, наиболее предпочтительно D, R или E
Q289A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y, предпочтительно F, W, H, I, Y, L, D, C, K, V, E, G или P, более предпочтительно R, T, D, K или P
К варианту фермента с повышенной относительной трансферазной активностью в отношении DG по сравнению с PC, подсчитанной как %TDG/TPC, приводит модификация одной или нескольких из следующих ниже модификаций:
-318Y, H или S
N215H
L210G, I, H, E, M, S, W, V, A, R, N, D, Q или T
S310A, P, T, H, M, K или G
E309S, Q, A или R
H180K, T или Q
N80N, R или D
V112C
Y30G, I, L, S, M, A, R или E, более предпочтительно Y30M, A или R
V290R, E, H или A
Q289R, T, D или N
K22E
G40L
Y179V или R
M209L, K или M
L211G, Q, K или D
Y230V
G40Q, L или V
N88W
N87R или D
Для некоторых вариантов осуществления также могут быть пригодными следующие ниже замены:
K22A или C
P81G
N87M
Y117A, N, E, H или Т
N181A или V
Y230I
V290H
N87R, D, E или М
Q182T
Предпочтительно, остатки, модифицированные для увеличения отношения галактолипидтрансферазной активности к фосфолипидтрансферазной активности, представляют собой один или несколько из следующих ниже: -318, N215, L210, E309, H180, N80.
Как правило, предпочтительны одна или несколько из следующих ниже замен:
-318 Y, H или S, наиболее предпочтительно Y
N215H
L210D, Q или Т
E309Q или R
H180K или Q
N80N, R или D
2. К варианту с повышенной абсолютной трансферазной активностью в отношении DG может приводить модификация по одному или нескольким из следующих ниже остатков:
-318, N215, L210, S310, Е309, Н180, N80, V112, Y30X (где X выбран из A, С, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), V290, Q 289, K22, G40, Y179, M209, L211, K22, P81, N87, Y117, N181, Y230X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), V290, N87, Q182, S3, S310, K82, A309.
В частности, к варианту с повышенной абсолютной трансферазной активностью в отношении DG может приводить одна или несколько из следующих ниже модификаций:
-318Y, H, S, A, C, D, E, F, G, I, K, L, M, N, P, Q, R, T, V или W, предпочтительно Y, H, S или I
N215A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; предпочтительно H, I, F, P, T, W или A, наиболее предпочтительно H, S, L, R, Y
L210G, I, H, E, M, S, W, V, A, R, N, D, Q, T, C, F, K, P или Y, предпочтительно D, Q, T, Y или F
S310A, P, T, H, M, K, G, C, D, E, F, I, L, N, Q, R, V, W или Y, предпочтительно F, Y, C, L, K или P,
E309S, Q, R, A, C, D, F, G, H, I, K, L, M, N, P, S, T, V, W или Y; предпочтительно S, Q, R, F, W, N, H, I, M или Y, наиболее предпочтительно S, Q, R, N, P или A
H180A, C, D, E, F, G, I, K, Q, L, M, P, R, S, T, V, W или Y; предпочтительно K, Q, M, F или C, наиболее предпочтительно Т, К или Q
N181A или V
N80N, R, D, A, C, E, F, G, H, I, K, L, M, P, Q, S, T, V, W или Y, предпочтительно H, I, Y, C, Q, M, S, W, L, N, R, D или F, наиболее предпочтительно N, R, D, P, V, A или G
V112A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y; предпочтительно I, M, F, Y, N, E, T, Q, H или P
Y30G, I, L, S, A, E, C, D, H, K, M, N, P, Q, R, T, V или W, предпочтительно H, T, W, N, D, C или Q
V290A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, W или Y;
Q289A, С, D, E, F, G, H, I, К, L, M, N, P, R, S, T, V, W или Y; предпочтительно R, E, G, P, N или R
K22A, C, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно С
Y179A, С, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; предпочтительно F, C, H, I, L, M, P или W, более предпочтительно E, R, N, V, K, S
M209A, C, D, E, F, G, H, I, L, K, M, N, P, Q, R, S, T, V, W или Y; предпочтительно R, N, Y, E или V
R211A, C, E, F, G. H, I, L, M. N, P, Q, K, D, R, S, T, V, W или Y, предпочтительно H, I, M, F, P, S, Y, N, C, L или W, наиболее предпочтительно R
S310 C, D, E, F, I, L, N, Q, R, V, W или Y, предпочтительно F, Y, C, L, K или P
S3A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y, предпочтительно M, R, N, A, G, T, Q, P, Y или S наиболее предпочтительно Q или N
K82A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y, предпочтительно H, K, S, E или R
P81A, C, D, E, F, G, H, I, K. L, M, N, Q, R, S, T, V, W или Y; предпочтительно I, M, F, V, Y, D, C или A
N87A, C, F, G, H, I, K, L, M, P, Q, R, D, E, S, T, V, W или Y; предпочтительно L, G или A
Y117A, N, E, H, T, C, D, F, G, I, K, L, M, P, Q, R, S, V или W; предпочтительно I, F, C, P или S
N87A, C, F, G, H, I, K, L, P, Q, S, T, V, W или Y; предпочтительно I, Y, T, Q, S, W, F, V или P
Q182A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y, предпочтительно D или K
Y230A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W, предпочтительно W, H, Q, L, P или C, наиболее предпочтительно T или G
D157A, C, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y, предпочтительно C
G40L
Y226I
Как правило, предпочтительны одна или несколько из следующих ниже замен:
-318Y, H или S
N215H
L210G, I, H, E, M, S, W, V, A, R, N, D, Q или T
S310A, P, T, H, M, K или G
E309S, Q или R
H180K или Q
N80N, R или D
V112C
Y30G, I, L, S, M, A, R или Е, более предпочтительно Y30M, A или R
V290R, E, H или А
Q289R или N
K22E
Q40L
Y179V
M209L, K или М
L211G, Q, K или D
Для некоторых вариантов осуществления также могут быть пригодными следующие ниже замены:
K22A или C
P81G
N87 M
Y117A, N, E, H или Т
N181A или V
Y230 I
V290H
N87R, D, E или М
Q182T
Предпочтительно, остатки, модифицированные для увеличения трансферазной активности для галактолипидного субстрата (DGDG), представляют собой один или несколько из следующих ниже: -318, N215, L210, E309, H180, N80.
Как правило, предпочтительны одна или несколько из следующих ниже замен:
-318Y, H или S, наиболее предпочтительно Y
N215H
L210D, Q или Т
E309Q или R
H180K или Q
N80N, R или D
3. К варианту фермента с повышенной трансферазной активностью T DG по отношению к гидролитической активности H DG в отношении DG может приводить модификация в одном или нескольких из следующих ниже остатков:
Y230, S310, H180, Q289, G40, N88, Y179, N215, L210, N80, Y30X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), N87, M209, R211, S18X (где X конкретно выбран из A, C, D, E, F, H, I, K, L, M, N, P, Q, R, T, W или Y).
Предпочтительно, к варианту фермента с повышенной трансферазной активностью TDG по отношению к гидролитической активности HDG в отношении DG может приводить одна или несколько из следующих ниже модификаций:
Y230A, С, D, E, G, H, I, К, L, M, N, P, Q, R, S, Т или W, предпочтительно W, H, Q, L, P или С
S310A, С, D, E, F, G, H, I, K, L, M, N, Q, R, T, V, W или Y, предпочтительно F, Y, C, L, К или P
Y179A, C, D, E, F, G, H, I, K, L, M, N, P, Q, S, T, V, или W, предпочтительно F, C, H, I, L, M, P или W
H180A, C, D, E, F, G, I, K, L, M, P, Q, R, S, V, W или Y, предпочтительно M, F или C
Q289A, C, E, F, G, H, I, K, L, M, N, P, R, S, V, W или Y; предпочтительно F, W, H, I, Y, L, D, C, K, V, E, G или P
G40A, C, D, E, F, H, I, K, M, N, P, R, S, T, W или Y; предпочтительно I, P, W или Y
N88A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V или Y; предпочтительно I или H
N87A, C, E, F, G, H, I, K, L, M, P, Q, S, T, V, W или Y; предпочтительно I, Y, T, Q, S, W, F, V или P
Как правило, предпочтительны одна или несколько из следующих ниже замен (такие варианты ферментов могут обладать сниженной гидролитической активностью (галактолипид и/или фосфолипиды) и/или повышенной трансферазной активностью для галактолипида):
Y179 E, R, N или Q
N215G
L210D, H, R, E, A, Q, P, N, K, G, R, T, W, I, V или S
N80G
Y30L
N87G
Как правило, предпочтительны одна или несколько из следующих ниже замен (такие варианты ферментов могут обладать сниженной гидролитической активностью (галактолипид и/или фосфолипиды) при сохранении значительной трансферазной активности для галактолипида:
Y179 E, R, N, Q
N215 G
L210 D, H, R, E, A, Q, P, N, K, G, R, T, W, I, V и S
N80 G
Y30 L
N87 G
H180 I, T
M209 Y
R211 D, T и G
S18 G, M и T
G40 R и M
N88 W
N87 C, D, R, E и G
4. К варианту фермента с повышенной абсолютной трансферазной активностью в отношении фосфолипида может приводить модификация одного или нескольких из следующих ниже остатков:
S3, D157, S310, Е309, Y179, N215, К22, Q289, М23, Н180, М209, L210, R211, Р81, V112, N80, L82, N88; N87
Конкретные модификации, которые могут обеспечить вариант фермента с улучшенной трансферазной активностью для фосфолипида, можно выбрать из одной или нескольких из следующих ниже:
S3A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y; предпочтительно N, E, K, R, A, P или M, наиболее предпочтительно S3A
D157A, C, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V, W или Y; предпочтительно D157S, R, E, N, G, T, V, Q, K или C
S310A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y; предпочтительно S310T
-318 E
E309A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, T, V, W или Y; предпочтительно Е309 R, E, L, R или A
Y179A, C, D, E, F, G, H, I, K, L, M, N, P, Q, R, S, T, V или W; предпочтительно Y179 D, T, E, R, N, V, K, Q или S, более предпочтительно E, R, N, V, K или Q
N215A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; предпочтительно N215 S, L, R или Y
K22A, C, D, E, F, G, H, I, L, M, N, P, Q, R, S, T, V, W или Y; предпочтительно К22 E, R, C или A
Q289A, C, D, E, F, G, H, I, K, L, M, N, P, R, S, T, V, W или Y; предпочтительно Q289 R, E, G, P или N
M23A, C, D, E, F, G, H, I, K, L, N, P, Q, R, S, T, V, W или Y; предпочтительно М23 K, Q, L, G, T или S
H180A, C, D, E, F, G, I, K, L, M, P, Q, R, S, T, V, W или Y; предпочтительно H180 Q, R или K
М209 A, C, D, E, F, G, H, I, K, L, N, P, Q, R, S, T, V, W или Y; предпочтительно М209 Q, S, R, A, N, Y, E, V или L
L210A, C, D, E, F, G, H, I, K, M, N, P, Q, R, S, T, V, W или Y; предпочтительно L210 R, A, V, S, T, I, W или M
R211A, C, D, E, F, G, H, I, K, L, M, N, P, Q, S, T, V, W или Y; предпочтительно R211T
P81A, C, D, E, F, G, H, I, K, L, M, N, Q, R, S, T, V, W или Y; предпочтительно P81G
V112A, С, D, E, F, G, H, I, К, L, M, N, P, Q, R, S, T, W или Y; предпочтительно V112C
N80A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; предпочтительно N80 R, G, N, D, P, T, E, V, A или G
L82A, C, D, E, F, G, H, I, M, N, P, Q, R, S, T, V, W или Y; предпочтительно L82N, S или E
N88A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; предпочтительно N88C
N87A, C, D, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y; предпочтительно N87M или G
К варианту фермента с повышенной абсолютной трансферазной активностью в отношении фосфолипида приводит модификация одного или нескольких из следующих ниже остатков:
S3 N, R, A, G
M23 K, Q, L, G, T, S
H180 R
L82 G
Y179 E, R, N, V, K или Q
E309 R, S, L или A
5. Остатки, модификация которых приводит к повышенной трансферазной активности для галактолипидного субстрата (DGDG) и увеличению отношения галактолипидтрансферазной активности к фосфолипидтрансферазной активности, включают в себя один или несколько из: -318, N215, L210, S310, E309, H180, N80, V112, Y30X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), V290, Q 289, K22, G40, Y179, M209, L211, K22, P81, N87, Y117, N181, Y230X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Q182.
Как правило, предпочтительны одна или несколько из следующих ниже замен:
-318 Y, H или S
N215H
L210G, I, H, E, M, S, W, V, A, R, N, D, Q или T
S310A, P, T, H, M, K или G
E309S, A, Q или R
H180K или Q
N80N, R или D
V112C
Y30G, I, L, S, M, A, R или E, более предпочтительно Y30M, A или R
V290R, E, H или А
Q289R или N
K22E
G40L
Y179V
M209L, K или М
K211G, Q, K или D
Для некоторых вариантов осуществления также могут быть пригодными следующие ниже замены:
K22A или С
P81G
N87 M
Y117A, N, E, H или T
N181A или V
Y230I
V290H
N87R, D, E или М
Q182T
Предпочтительно, остатки, модифицированные для увеличения трансферазной активности для галактолипидного субстрата (DGDG) и/или увеличения отношения галактолипидтрансферазной активности к фосфолипидтрансферазной активности представляют собой один или несколько из следующих ниже: -318, N215, L210, E309, H180, N80.
Как правило, предпочтительны одна или несколько из следующих ниже замен:
-318 Y, H или S, более предпочтительно Y
N215H
L210D, Q или T
E309Q или R
H180K или Q
N80N, R или D
6. Выявлено, что для сохранения хорошей активности, особенно хорошей трансферазной активности для галактолипида предпочтительны следующие остатки дикого типа P10480:
W111, R211, N181, S3, L17, G40, N88, Y117, L118, N181, K22, M209, M285, M23.
Предпочтительно, данные остатки в варианте фермента сохранены.
При получении варианта GDSX-ацилтрансферазы для повышенной трансферазной активности в отношении галактолипидных субстратов, где исходный фермент несет остаток, соответствующий остаткам последовательности P10480 в положениях W111, R211, N181, S3, L17, G40, N88, Y117, L118, N181, K22, M209, M285, M23, отличный от остатка найденного в P10480, вариант может предпочтительно в соответствующем положении содержать замену для включения в него аминокислотного остатка, найденного в последовательности P10480.
L17 предпочтительно представляет собой гидрофобный аминокислотный остаток.
7. Повышенной трансферазной активностью для галактолипидного субстрата (DGDG) и/или увеличенным отношением галактолипидтрансферазной активности к фосфолипидтрансферазной активности могут обладать следующие ниже сочетания:
N215H и -318Y
N215H и L210D, Q или T
-318Y и L210D, Q или Т
N215H и -318Y и L210D, Q или Т
Также указанные выше сочетания необязательно могут включать в себя добавление аминокислоты на C-конце, такое как -318Y, H или S, или, предпочтительно, -318Y.
Также указанные выше сочетания необязательно могут включать в себя следующую ниже модификацию:
Таким образом, повышенной трансферазной активностью для галактолипидного субстрата (DGDG) и/или увеличенным отношением галактолипидтрансферазной активности к фосфолипидтрансферазной активности может обладать одно или несколько из следующих ниже сочетаний:
Е309А, Q или R
N215H и -318Y, H или S, предпочтительно Y
L210D, Q или T и -318Y, H или S, предпочтительно Y
N215H и Е309А, Q или R
L210D, Q или Т и Е309А, Q или R
-318Y и Е309А, Q или R
Также указанные выше сочетания необязательно могут включать в себя замену в положении Q182, предпочтительно, Q182K.
Повышенной трансферазной активностью для галактолипидного субстрата (DGDG) и/или увеличенным отношением галактолипидтрансферазной активности к фосфолипидтрансферазной активности, и/или повышенным отношением галактолипидтрансферазной активности к гидролитической активности могут обладать следующие ниже сочетания:
N215H и N80G
-318Y и N80G
L210D или Q и N80G
N215H и N88N
-318Y и N88N
L210D или Q и N88N
N215H и Y30L
-318Y и Y30L
L210D или Q и Y30L
N215H и N87G
-318Y и N87G
L210D или Q и N87G
N215H и Y179E, R, N или Q
-318Y и Y179E, R, N или Q
L210D или Q и Y179E, R, N или Q
Как указано выше, при обозначении здесь конкретных аминокислотных остатков нумерация представляет собой нумерацию, полученную из сравнения варианта последовательности с эталонной последовательностью, представленной как SEQ ID NO:2.
Во избежание неопределенности, когда конкретная аминокислота указана в конкретном участке, например L118, например, это относится к конкретной аминокислоте остатка номер 118 в SEQ ID NO:2. Однако аминокислотный остаток в участке 118 в другом исходном ферменте может быть отличным от лейцина.
Таким образом, когда указано заменить аминокислоту в остатке 118, хотя указание может быть сделано в отношении L118, специалист в данной области легко поймет, что когда исходный фермент отличен от исходного фермента, представленного в SEQ ID NO:2, замещаемая аминокислота может не быть лейцином. Следовательно, возможно, что при замене в исходном ферменте аминокислотной последовательности, которая не является ферментом с аминокислотной последовательностью, представленной как SEQ ID NO:2, новая (замещающая) аминокислота может являться такой же, как указана в SEQ ID NO:2. Например, это может происходить в том случае, когда аминокислота в указанном остатке 118 не является лейцином и, следовательно, отлична от аминокислоты в остатке 118 в SEQ ID NO:2. Другими словами, в остатке 118, например, если в исходном ферменте в данном положении находится отличная от лейцина аминокислота, данную аминокислоту по настоящему изобретению можно заменить на лейцин.
Как применяют здесь, термин "липидацилтрансфераза" означает фермент, обладающий ацилтрансферазной активностью (как правило классифицируемой в соответствии с рекомендациями по номенклатуре ферментов (1992) Nomenclature Committee of the International Union of Biochemistry и Molecular Biology как E.C. 2.3.1.x), вследствие чего фермент способен к переносу ацильной группы с липида на один или несколько акцепторных субстратов, таких как один или несколько из следующих ниже: стерин; станол; углевод; белок; белковая субъединица; глицерин.
Предпочтительно липидацилтрансфераза способна к переносу ацильной группы с липида, по меньшей мере, на стерин и/или станол, например, на холестерин.
Предпочтительно, вариант липидацилтрансферазы по настоящему изобретению и/или для применения в способах и/или применениях настоящего изобретения способен к переносу ацильной группы с липида (как определено здесь) на один или несколько из следующих ниже субстратов-акцепторов ацила: стерин, станол, углевод, белок или его субъединицы или глицерин.
Для некоторых аспектов "акцептор ацила" по настоящему изобретению может представлять собой любое соединение, включающее в себя гидроксильную группу (-OH), такое как, например, многоатомные спирты, включающие в себя глицерин; стерин; станолы; углеводы; гидроксикислоты, включающие в себя фруктовые кислоты, лимонную кислоту, винную кислоту, молочную кислоту и аскорбиновую кислоту; белки или их субъединицы, такие как, например, аминокислоты, гидролизаты белков и пептиды (частично гидролизованный белок); и их смеси и производные. Предпочтительно, "акцептор ацила" по настоящему изобретению не является водой.
В одном из вариантов осуществления, акцептор ацила предпочтительно не является моноглицеридом и/или диглицеридом.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на стерин и/или станол.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на углевод.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на белок или его субъединицы. Таким образом, белковая субъединица может представлять собой одну или несколько из следующих ниже: аминокислоту, гидролизат белка, пептид, дипептид, олигопептид, полипептид.
Таким образом, в белке или белковой субъединице акцептор ацила может представлять собой одно или несколько из следующих ниже составляющих белка или белковой субъединицы: серин, треонин, тирозин, или цистеин.
Когда белковая субъединица представляет собой аминокислоту, тогда аминокислота может представлять собой любую подходящую аминокислоту. Таким образом, аминокислота может представлять собой, например, одну или несколько из серина, треонина, тирозина или цистеина.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на глицерин.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на гидроксикислоту.
В одном из аспектов вариант фермента предпочтительно способен к переносу ацильной группы с липида на многоатомный спирт.
В одном из аспектов вариант липидацилтрансферазы также может быть способным к переносу ацильной группы с липида на стерин и/или станол, дополнительно быть способным к переносу ацильной группы с липида на один или несколько из следующих ниже: углевод, белок, белковая субъединица, глицерин.
Предпочтительно, липидный субстрат на который действует вариант липидацилтрансферазы по настоящему изобретению представляет собой один или несколько из следующих ниже липидов: фосфолипид, такой как лецитин, например, фосфатидилхолин, триацилглицерид, кардиолипин, диглицерид или гликолипид, например, такой как, дигалактозилдиглицерид (DGDG) или моногалактозилдиглицерид (MGDG). Более предпочтительно вариант фермента по настоящему изобретению действует на один или оба из DGDG и MGDG. Предпочтительно, вариант фермента по настоящему изобретению не обладает (или обладает только ограниченной) активностью в отношении дигалактозилмоноглицерида (DGMG) и моногалактозилмоноглицерида (MGMG). Таким образом, предпочтительно, липидный субстрат не является одним или обоими из DGMG или MGMG. Данный липидный субстрат может быть обозначен здесь как "липидный донор ацила". Как применяют здесь, термин лецитин включает в себя фосфатидилхолин, фосфатидилэтаноламин, фосфатидилинозитол, фосфатидилсерин и фосфатидилглицерин.
Как применяют здесь, термин "галактолипид" означает один или несколько из DGDG или DGMG.
Как применяют здесь, термин "фосфолипид" обозначает лецитин, включающий в себя фосфатидилхолин.
Как применяют здесь, термин "полярный липид" обозначает фосфолипиды и/или галактолипид, предпочтительно фосфолипиды и галактолипид.
Для некоторых аспектов, липидный субстрат, на который действует вариант липидацилтрансферазы, предпочтительно представляет собой фосфолипид, такой как лецитин, например, фосфатидилхолин.
Для некоторых аспектов, липидный субстрат предпочтительно представляет собой гликолипид, например, такой как DGDG или MGDG.
Предпочтительно липидный субстрат представляет собой пищевой липид, то есть, липидный компонент пищевого продукта.
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно неспособен или по существу не способен, действовать на триглицерид и/или на 1-моноглицерид и/или 2-моноглицерид.
Таким образом, липидный субстрат или липидный донор ацила может представлять собой один или несколько липидов присутствующих в одном или нескольких из следующих ниже субстратов: жиры, включающие в себя свиной, топленый жир или жир масла; масла, включающие в себя масла, экстрагированные или полученные из пальмового масла, подсолнечного масла, масла из соевых бобов, сафлоровое масло, масло из семян хлопка, масло из земляного ореха, кукурузное масло, оливковое масло, арахисовое масло, кокосовое масло и масло из рапсового семени. Лецитин из сои, рапсового семени или яичного желтка также представляет собой пригодный липидный субстрат. Липидный субстрат может представлять собой липид овса или другое вещество из растений, содержащее галактолипиды.
В одном из аспектов липидный донор ацила предпочтительно представляет собой лецитин (такой как фосфатидилхолин) в яичном желтке.
Для некоторых аспектов настоящего изобретения липид можно выбрать из липидов с длиной цепей жирных кислот от 8 до 22 атомов углерода.
Для некоторых аспектов настоящего изобретения, липид можно выбрать из липидов с длиной цепей жирных кислот от 16 до 22 атомов углерода, более предпочтительно, от 16 до 20 атомов углерода.
Для некоторых аспектов настоящего изобретения, липид можно выбрать из липидов с длиной цепей жирных кислот не более чем 14 атомов углерода, соответственно, из липидов с длиной цепей жирных кислот от 4 до 14 атомов углерода, соответственно, от 4 до 10 атомов углерода, соответственно, от 4 до 8 атомов углерода.
Таким образом, вариант липидацилтрансферазы по настоящему изобретению может проявлять один или несколько из следующих ниже видов липазной активности: гликолипазная активность (E.C. 3.1.1.26), триацилглицероллипазная активность (E.C. 3.1.1.3), активность фосфолипазы A2 (E.C. 3.1.1.4) или активность фосфолипазы A1 (E.C. 3.1.1.32). Как применяют здесь, термин "гликолипазная активность" включает в себя "галактолипазную активность".
Таким образом, вариант липидацилтрансферазы по настоящему изобретению может обладать, по меньшей мере, одним или несколькими из следующих ниже видов активности: гликолипазная активность (E.C. 3.1.1.26) и/или активность фосфолипазы A1 (E.C. 3.1.1.32) и/или активность фосфолипазы A2 (E.C. 3.1.1.4).
Для некоторых аспектов, вариант липидацилтрансферазы по настоящему изобретению может обладать, по меньшей мере, гликолипазной активностью (E.C. 3.1.1.26).
Таким образом, для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению может быть способным к переносу ацильной группы с гликолипида и/или фосфолипида на один или несколько из следующих ниже акцепторных субстратов: стерин, станол, углевод, белок, глицерин.
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно способен к переносу ацильной группы с гликолипида и/или фосфолипида на стерин и/или станол с формированием, по меньшей мере, сложного эфира стерина и/или сложного эфира станола.
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно способен к переносу ацильной группы с гликолипида и/или фосфолипида на углевод с формированием, по меньшей мере, сложного эфира углевода.
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно способен к переносу ацильной группы с гликолипида и/или фосфолипида на белок с формированием, по меньшей мере, сложного эфира белка (или конденсата белка с жирной кислотой).
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно способен к переносу ацильной группы с гликолипида и/или фосфолипида на глицерин с формированием, по меньшей мере, диглицерида и/или моноглицерида.
Для некоторых аспектов вариант липидацилтрансферазы по настоящему изобретению предпочтительно не проявляет триацилглицеринлипазной активности (E.C. 3.1.1.3).
В некоторых аспектах вариант липидацилтрансферазы может быть способен к переносу ацильной группы с липида на стерин и/или станол. Таким образом, в одном из вариантов осуществления "акцептор ацила" по настоящему изобретению может представлять собой или стерин, или станол, или сочетание стерина и станол.
Таким образом, в одном из вариантов осуществления стерин и/или станол может содержать одну или несколько из следующих ниже структурных особенностей:
i) 3-бета гидроксильная группа или 3-альфа гидроксильная группа; и/или
ii) циклы A:B в цис-положении или циклы A:B в транс-положении или C5-C6 является ненасыщенной.
Пригодные стериновые акцепторы ацила включают в себя холестерин и фитостерины, например, альфа-ситостерин, бета-ситостерин, стигмастерин, эргостерин, кампастерин, 5,6-дигидростерин, брассикастерин, альфа-спинастерин, бета-спинастерин, гамма-спинастерин, дельта-спинастерин, фукостерин, димостерин, аскостерин, серебистерин, эпистерин, анастерин, гипостерин, хондрилластерин, десмостерин, халиностерин, пролиферастерин, клионастерин, гликозиды стерина и другие природные или синтетические изомерные формы и производные.
Таким образом, в одном из аспектов настоящего изобретения в качестве акцептора ацила может действовать более чем один стерин и/или станол, таким образом, в качестве акцептора ацила может действовать более чем два стерина и/или станола. Другими словами, таким образом, в одном из аспектов настоящего изобретения может быть получен более чем один сложный эфир стерина и/или сложный эфир станола. Таким образом, когда акцептором ацила является холестерин, в качестве акцепторов ацила также могут действовать один или несколько дополнительных стеринов или один или несколько станолов. Таким образом, в одном из аспектов, настоящее изобретение относится к способу получения in situ эфира холестерина в сочетании, по меньшей мере, с одним сложным эфиром стерина или сложным эфиром станола. Другими словами, для некоторых аспектов настоящего изобретения липидацилтрансфераза может переносить ацильную группу с липида на холестерин и, по меньшей мере, один дополнительный стерин и/или, по меньшей мере, один станол.
В одном из аспектов стериновый акцептор ацила предпочтительно представляет собой один или несколько из следующих ниже: альфа-ситостерин, бета-ситостерин, стигмастерин, эргостерин и кампестерин.
В одном из аспектов стериновый акцептор ацила предпочтительно представляет собой холестерин. В том случае когда акцептором ацила для варианта липидацилтрансферазы является холестерин, количество свободного холестерина в пищевом продукте по сравнению с пищевым продуктом до воздействия варианта липидацилтрансферазы и/или по сравнению с эквивалентным пищевым продуктом, не обработанным вариантом липидацилтрансферазы, уменьшено.
Пригодные станоловые акцепторы ацила включают в себя фитостанолы, например, бета-ситостанол или ss-ситостанол.
В одном из аспектов стериновый и/или станоловый акцептор ацила предпочтительно представляют собой стерин и/или станол, отличный от холестерина.
В некоторых аспектах, пищевой продукт, полученный по настоящему изобретению, можно применять для уменьшения холестерина в сыворотке крови и/или для уменьшения липопротеинов низкой плотности. Холестерин сыворотки крови и липопротеины низкой плотности у людей ассоциированы с определенными болезнями, например, такими как атеросклероз и/или заболевание сердца. Таким образом, предусмотрено, что пищевые продукты, полученные по настоящему изобретению, можно применять для уменьшения риска таких заболеваний.
Таким образом, в одном из аспектов настоящее изобретение относится к применению пищевого продукта по настоящему изобретению для применения в лечении и/или профилактике атеросклероза и/или заболевания сердца. Таким образом, в одном из аспектов пищевой продукт можно рассматривать как нутрицевтик.
В дополнительном аспекте настоящее изобретение относится к лекарственному средству, включающему в себя пищевой продукт по настоящему изобретению.
В дополнительном аспекте настоящее изобретение относится к способу лечения и/или профилактики заболевания у человека или животного, где способ включает в себя введение пациенту эффективного количества пищевого продукта по настоящему изобретению.
Таким образом, стериновый и/или станоловый "акцептор ацила" может находиться в пищевом продукте естественным образом. Альтернативно, стерин и/или станол можно добавлять в пищевой продукт. В том случае, когда стерин и/или станол добавляют к пищевому продукту, стерин и/или станол можно добавлять перед добавлением липидацилтрансферазы по настоящему изобретению, одновременно с ним, и/или после него. Таким образом, настоящее изобретение может включать в себя добавление экзогенных стеринов/станолов, особенно фитостеринов/фитостанолов, в пищевой продукт до добавления варианта фермента по настоящему изобретению или одновременно с ним.
Для некоторых аспектов один или несколько стеринов находящихся в пищевом продукте можно конвертировать в один или несколько станолов до или в то же время, когда по настоящему изобретению добавляют вариант липидацилтрансферазы. Для преобразования стеринов в станолы можно применять любой пригодный способ. Например, преобразование можно осуществлять, например, посредством химической гидрогенизации. Преобразование можно проводить до добавления варианта липидацилтрансферазы по настоящему изобретению или одновременно с добавлением варианта липидацилтрансферазы по настоящему изобретению. Таким образом, ферменты для преобразования стерина в станолы указаны в WO 00/061771.
Таким образом, настоящее изобретение можно применять для получения в пищевом продукте сложных эфиров фитостанолов in situ. Сложные эфиры фитостанолов обладают повышенной растворимостью в липидных мембранах, биодоступностью и полезны для здоровья (см., например, WO 92/99640).
В некоторых вариантах осуществления настоящего изобретения сложный эфир станола и/или сложный эфир стерина может представлять собой ароматизатор и/или структурообразователь. В таких случаях настоящее изобретение относится к получению ароматизаторов и/или структурообразователей in situ.
В одном из вариантов осуществления, настоящее изобретение относится к способу получения сложного эфир стерина и/или сложного эфира станола из растений и лизолецитина в пищевом масле (таком как растительное масло, например, таком как масло из бобов сои) без образования свободных жирных кислот обработкой масла вариантом фермента по настоящему изобретению. В таких случаях полученный таким образом лизолецитин можно удалять с применением способа рафинирования. Можно использовать любой способ рафинирования, такой как один или несколько из известных способов рафинирования. Если необходимо, любые свободные жирные кислоты можно удалять посредством дезодорирования. Необходимо отметить, что никакой полученный в масле сложный эфир станола/стерина способом дезодорирования не удаляется. Таким образом, полученное пищевое масло содержит сложные эфиры стеринов и/или сложные эфиры станолов, которые могут обладать полезными питательными и/или нутрицевтическими эффектами, такими как уменьшение уровней холестерина в крови.
Пригодные масла, для которых можно проводить данный способ, представляют собой масла, среди прочего включающие в себя лецитин и стерин/станол. Таким образом, масло после обработки представляет собой неочищенное масло. Таким образом, пригодное в пищу масло может представлять собой одно или несколько из следующих ниже: масло ростков злаков, масло семян хлопка, масло льняного семени, пальмовое масло, арахисовое масло, масло семян рапса, кунжутное масло, соевое масло, подсолнечное масло и масло ростков пшеницы.
Для некоторых аспектов настоящего изобретения, вариант липидацилтрансферазы по настоящему изобретению может использовать в качестве акцептора ацила углевод. Углеводный акцептор ацила может представлять собой один или несколько из следующих ниже: моносахарид, дисахарид, олигосахарид или полисахарид. Предпочтительно, углевод представляет собой один или несколько из следующих ниже: глюкоза, фруктоза, ангидрофруктоза, мальтоза, лактоза, сахароза, галактоза, ксилоза, ксилоолигосахариды, арабиноза, мальтоолигосахариды, тагатоза, микротецин, аскопирон P, аскопирон T, кортальцерон.
Таким образом, углеводный "акцептор ацила" может находиться в пищевых продуктах от природы. Альтернативно, углевод можно добавлять в пищевой продукт. В том случае, когда углевод добавляют в пищевой продукт, углевод можно добавлять перед добавлением варианта липидацилтрансферазы по настоящему изобретению, вместе с ним и/или после него.
Сложные эфиры углеводов могут функционировать в пищевых продуктах как полноценные эмульгаторы. Таким образом, в том случае когда фермент действует для переноса ацильной группы на сахар, изобретение относится к получению второго эмульгатора в пищевом продукте in situ.
В некоторых вариантах осуществления, вариант липидацилтрансферазы может использовать в качестве акцептора ацила и стерин и/или станол, и углевод.
Применение варианта липидацилтрансферазы, способной переносить ацильную группу на углевод, а также на стерин и/или станол, особенно предпочтительно для пищевых продуктов, содержащих яйца. В частности, присутствие сахаров, в частности глюкозы, в яйцах и яичных продуктах часто рассматривают как неблагоприятное. Яичный желток может содержать до 1% глюкозы. Как правило, яичные продукты или продукты, основанные на яичных продуктах, для удаления части или всей этой глюкозы можно обрабатывать оксидазой глюкозы. Однако по настоящему изобретению этот нежелательный сахар можно легко удалить посредством "этерификации" сахара с формированием сложного эфира сахара.
Для некоторых аспектов настоящего изобретения вариант липидацилтрансферазы по настоящему изобретению в качестве акцептора ацила может использовать белок. Таким образом, белок может представлять собой один несколько из белков, находящихся в пищевом продукте, например, в молочном продукте и/или мясном продукте. В качестве неограничивающих примеров пригодные белки могут представлять собой белки, находящиеся в твороге или сыворотке, такие как лактоглобулин. Другие пригодные белки включают в себя яичный овальбумин, глиадин, глютенин, пуроиндолин, переносящие липиды белки из злаков и миозин из мяса.
Предпочтительно, исходный фермент липидацилтрансфераза по настоящему изобретению может отличаться по следующим критериям:
(i) фермент обладает ацилтрансферазной активностью, которую можно определить как активность по переносу сложного эфира, где ацильная часть исходной эфирной связи липидного донора ацила переносится на акцептор ацила с формированием нового сложного эфира; и
(ii) фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S.
Предпочтительно, X из мотива GDSX представляет собой L. Таким образом, предпочтительно фермент по настоящему изобретению включает в себя аминокислотный мотив GDSL.
Мотив GDSX состоит из четырех консервативных аминокислот. Предпочтительно серин в мотиве представляет собой каталитический серин фермента липидацилтрансферазы. Таким образом, серин мотива GDSX может находиться в положении, соответствующем Ser-16 в липолитическом ферменте Aeromonas hydrophila, как указано в Brumlik и Buckley (Journal of Bacteriology Apr. 1996, Vol. 178, No. 7, p. 2060-2064).
Для определения того, есть ли в белке мотив GDSX по настоящему изобретению, предпочтительно последовательность сравнивают с профилями скрытой марковской модели (профили HMM) базы данных pfam.
Pfam представляет собой базу данных семейств белковых доменов. Pfam содержит выверенные сравнения множества последовательностей для каждого семейства, а также профиль скрытых марковских моделей (профиль HMM) для идентификации этих доменов в новых последовательностях. Введение в pfam можно найти у Bateman A et al. (2002) Nucleic Acids Res. 30; 276-280. Скрытые марковские модели применяют в ряде баз данных, стремящихся классифицировать белки, для обзора см. Bateman A and Haft DH (2002) Brief Bioinform 3; 236-245.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve& db=PubMed&list_uids=12230032&dopt=Abstract
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve& db=PubMed&list_uids=11752314&dopt=Abstract
Для подробного разъяснения скрытых марковских моделей и того, как их применяют в базе данных pfam см. Durbin R, Eddy S, and Krogh A (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4. Пакет программного обеспечения Hammer можно получить из Washington University, St Louis, USA.
Альтернативно мотив GDSX можно идентифицировать с применением пакета программного обеспечения Hammer, инструкции предоставлены в Durbin R, Eddy S, и Krogh A (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4 и ссылках в ней, и профиль HMMER2, предоставленный в пределах данной спецификации.
К базе данных PFAM можно получить доступ, например, через несколько серверов, в настоящее время расположенных на следующих ниже веб-сайтах:
http://www.sanger.ac.uk/Software/pfam/index.shtml
http://pfam.wustl.edu/
http://pfam.jouy.inra.fr/
http://pfam.cgb.ki.se/
База данных предлагает средство поиска, где можно вводить белковую последовательность. Затем, с применением параметров по умолчанию базы данных, белковую последовательность анализируют на присутствие доменов pfam. Домен GDSX является известным в базе данных доменом и по существу распознается его присутствие в любой введенной в запросе последовательности. База данных возвращает сравнение консенсусной последовательности pfam00657 с вводимой в запросе последовательностью.
Множественное сравнение, включающее в себя Aeromonas salmonicida или Aeromonas hydrophila можно получить:
a) вручную
получение сравнения представляющего интерес белка с консенсусной последовательностью pfam00657 и получение сравнения P10480 с консенсусной последовательностью pfam00657 по описанной выше процедуре;
или
b) с помощью базы данных
После идентификации консенсусной последовательности pfam00657 база данных предоставляет возможность показать сравнение вводимой в запросе последовательности по выбранному сравнению консенсусной последовательности pfam00657. P10480 является частью этого выбранного сравнения и обозначается как GCAT_AERHY. И вводимая в запросе последовательность и P10480 отображаются в одном и том же окне.
Эталонная последовательность Aeromonas hydrophila.
Остатки липазы Aeromonas hydrophila GDSX пронумерованы в файле P10480 в NCBI, номера в данном тексте относятся к номерам, присвоенным в данном файле, который в настоящем изобретении используют для определения конкретных аминокислотных остатков, которые в предпочтительном варианте осуществления присутствуют в ферментах липидацилтрансферазы по изобретению.
Проводили сравнение pfam (фиг.33 и фиг.34).
Можно распознать следующие ниже консервативные остатки, и в предпочтительном варианте осуществления они могут присутствовать в вариантах ферментов для применения в композициях и способах по изобретению:

где 'hid' означает гидрофобный остаток, выбранный из Met, Ile, Leu, Val, Ala, Gly, Cys, His, Lys, Trp, Tyr, Phe.
Предпочтительно исходный фермент липидацилтрансфераза и/или его вариант для применения в композициях/способах по изобретению можно сравнивать с применением консенсусной последовательности pfam00657.
Предпочтительно, положительное совпадение с профилем скрытой марковской модели (профиль HMM) семейства доменов pfam00657 указывает на присутствие домена GDSL или GDSX по настоящему изобретению.
Предпочтительно при сравнении с консенсусной последовательностью pfam00657 исходная липидацилтрансфераза и/или ее вариант для применения в композициях/способах по изобретению обладает, по меньшей мере, одним, предпочтительно более чем одним, предпочтительно - более чем двумя, из следующих ниже, участок GDSX, участок GANDY, участок HPT. Таким образом, исходная липидацилтрансфераза и/или ее вариант может обладать участком GDSX и участком GANDY. Альтернативно, исходный фермент и/или его вариант может обладать участком GDSX и участком HPT. Предпочтительно исходный фермент и/или его вариант содержит, по меньшей мере, участок GDSX.
Предпочтительно, при сравнении с консенсусной последовательностью pfam00657 исходный фермент и/или его вариант для применения в композициях/способах по изобретению по сравнению с эталонной полипептидной последовательностью A.hydrophilia обладает, по меньшей мере, одним, предпочтительно, более чем одним, предпочтительно, более чем двумя, предпочтительно, более чем тремя, предпочтительно, более чем четырьмя, предпочтительно, более чем пятью, предпочтительно, более чем шестью, предпочтительно, более чем семью, предпочтительно, более чем восемью, предпочтительно, более чем девятью, предпочтительно, более чем десятью, предпочтительно, более чем одиннадцатью, предпочтительно, более чем двенадцатью, предпочтительно, более чем тринадцатью, предпочтительно, более чем четырнадцатью, из следующих ниже аминокислотных остатков, т.е. SEQ ID NO:26: 28hid, 29hid, 30hid, 31hid, 32gly, 33Asp, 34Ser, 35hid, 130hid, 131Gly, 132Hid, 133Asn, 134Asp, 135hid, 309His.
Домен GDSX pfam00657 является уникальным идентификатором, отличающим обладающие данным доменом белки от других ферментов.
Консенсусная последовательность pfam00657 представлена на фигуре 1 как SEQ ID NO:1. Она получена из идентификации семейства pfam 00657, база данных версии 6, которое здесь также может быть обозначено как pfam00657.6.
Консенсусная последовательность может быть обновлена при использовании последующих выпусков базы данных pfam.
Например, на фиг.33 и 34 показано сравнение pfam семейства 00657, из базы данных версии 11, которое здесь также может быть обозначено как pfam00657.11.
Наличие участков GDSX, GANDY и HPT выявлено в семействе pfam 00657 с применением обоих выпусков базы данных. Для идентификации семейства pfam 00657 можно применять последующие выпуски базы данных pfam.
Предпочтительно, исходный фермент липидацилтрансферазу по настоящему изобретению можно охарактеризовать с применением следующих критериев:
(i) фермент обладает ацилтрансферазной активностью, которую можно определить как активность по переносу сложного эфира, где ацильная часть исходной эфирной связи липидного донора ацила переносится на акцептор ацила с формированием нового сложного эфира; и
(ii) фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S.
(iii) фермент содержит His-309 или содержит гистидиновый остаток в положении, соответствующем His-309 в липолитическом ферменте Aeromonas hydrophila, показанном на фиг.2 (SEQ ID NO:2 или SEQ ID NO:26).
Предпочтительно, аминокислотный остаток мотива GDSX представляет собой L.
В SEQ ID NO:26 первые 18 аминокислотных остатков формируют сигнальную последовательность. His-309 полноразмерной последовательности, являющейся белком, включающим в себя сигнальную последовательность, соответствует His-291 зрелой части белка, т.е. последовательности без сигнальной последовательности.
Предпочтительно, исходный фермент липидацилтрансфераза по настоящему изобретению содержит следующую каталитическую триаду: Ser-16, Asp-116 и His-291 или содержит остаток серина, остаток аспарагиновой кислоты и остаток гистидина, соответственно, в положениях, соответствующих Ser-16, Asp-116 и His-291 в липолитическом ферменте Aeromonas hydrophila, показанном на фиг.2 (SEQ ID NO:2) или в положениях, соответствующих Ser-34, Asp-134 и His-309 полноразмерной последовательности, показанной на фиг.28 (SEQ ID NO:26). Как указано выше, в последовательности, приведенной в SEQ ID NO:26, первые 18 аминокислотных остатков формируют сигнальную последовательность. Ser-34, Asp-134 и His-309 полноразмерной последовательности, являющейся белком, включающим в себя сигнальную последовательность, соответствуют Ser-16, Asp-116 и His-291 зрелой части белка, т.е. последовательности без сигнальной последовательности. В консенсусной последовательности pfam00657, как представлено на фиг.1 (SEQ ID NO:1), остатки активного центра соответствуют Ser-7, Asp-157 и His-348.
Предпочтительно, исходный фермент липидацилтрансферазу по настоящему изобретению можно охарактеризовать с применением следующих критериев:
(i) фермент обладает ацилтрансферазной активностью, которую можно определить как активность по переносу сложного эфира, где ацильная часть исходной эфирной связи первого липидного донора ацила переносится на акцептор ацила с формированием нового сложного эфира; и
(ii) фермент содержит, по меньшей мере, Gly-14, Asp-15, Ser-16, Asp-116 и His-191 в положениях, соответствующих ферменту Aeromonas hydrophila на фиг.2 (SEQ ID NO:2), эквивалентных положениям Gly-32, Asp-33, Ser-34, Asp-134 и His-309, соответственно, на фиг.28 (SEQ ID NO:26).
Таким образом, исходный фермент липидацилтрансферазу по настоящему изобретению может быть получен, предпочтительно получен из организмов одного или нескольких из следующих ниже родов: Aeromonas, Corynebacterium, Novosphingobium, Termobifida, Streptomyces, Saccharomyces, Lactococcus, Mycobacterium, Streptococcus, Lactobacillus, Desulfitobacterium, Bacillus, Campylobacter, Vibrionaceae, Xylella, Sulfolobus, Aspergillus, Schizosaccharomyces, Listeria, Neisseria, Mesorhizobium, Ralstonia, Xanthomonas и Candida.
Таким образом, исходный фермент липидацилтрансфераза по настоящему изобретению может быть получен, предпочтительно получен, из одного или нескольких из следующих ниже организмов: Aeromonas hydrophila, Aeromonas salmonicida, Streptomyces coelicolor, Streptomyces rimosus, Mycobacterium, Streptococcus pyogenes, Lactococcus lactis, Streptococcus pyogenes, Streptococcus thermophilus, Lactobacillus helveticus, Desulfitobacterium dehalogenans, виды Bacillus, Campylobacter jejuni, Vibrionaceae, Xylella fastidiosa, Sulfolobus solfataricus, Saccharomyces cerevisiae, Aspergillus terreus, Schizosaccharomyces pombe, Listeria innocua, Listeria monocytogenes, Neisseria meningitidis, Mesorhizobium loti, Ralstonia solanacearum, Xanthomonas campestris, Xanthomonas axonopodis, Corynebacterium efficens, Novosphingobium aromaticivorans, Termobifida fusca и Candida parapsilosis.
В одном из аспектов исходный фермент липидацилтрансфераза по настоящему изобретению предпочтительно может быть получен, предпочтительно получен, из одного или нескольких из Aeromonas hydrophila или Aeromonas salmonicida.
В одном из аспектов, исходная липидацилтрансфераза по настоящему изобретению может представлять собой лецитин:холестеринацилтрансферазу (LCAT) или ее вариант (например, вариант может представлять собой молекулярное преобразование).
В данной области известны подходящие LCAT и их можно получить, например, из одного или нескольких из следующих ниже организмов: млекопитающие, крыса, мышь, куры, Drosophila melanogaster, растения, включающие в себя Arabidopsis и Oryza sativa, нематоды, грибы и дрожжи.
Предпочтительно, при выполнении способа по настоящему изобретению продукт (т.е. пищевой продукт) получают без увеличения или существенного увеличения свободных жирных кислот в пищевом продукте.
Как применяют здесь, термин "трансфераза" является взаимозаменяемым с термином "липидацилтрансфераза".
Как применяют здесь термин "галактолипидтрансферазная активность" обозначает способность фермента катализировать перенос ацильной группы с галактолипидного донора на акцепторную молекулу (отличную от воды), например, такую как глицерин.
Подобным образом, как применяют здесь термин "фосфолипидтрансферазная активность" обозначает способность фермента катализировать перенос ацильной группы с фосфолипидного донора на акцепторную молекулу (отличную от воды), например, такую как глицерин.
Как применяют здесь, термин "повышенное отношение галактолипазной трансферазной активности к фосфолипидтрансферазной активности" обозначает вариант фермента, который по сравнению с исходным ферментом способен к катализу в качестве галактолипидтрансферазы в большей степени по сравнению с катализом в качестве фосфолипидтрансферазы. Это может означать, что и галактолипидтрансферазная активность, и фосфолипидтрансферазная активность увеличены по сравнению с исходным ферментом или что галактолипидтрансферазная активность увеличена, тогда как фосфолипидтрансферазная активность уменьшена по сравнению с исходным ферментом. Важным является конечное отношением между двумя этими видами активности.
Таким образом, как определено здесь липидацилтрансфераза катализирует одну или несколько из следующих ниже реакций: интерэстерификация, трансэстерификация, алкоголиз, гидролиз.
Термин "интерэстерификация" относится к ферментативно катализируемому переносу ацильных групп между липидным донором и липидным акцептором, где липидный донор не является свободной ацильной группой.
Как применяют здесь, термин "трансэстерификация" обозначает ферментативно катализируемый перенос ацильной группы с липидного донора (отличного от свободной жирной кислоты) на акцептор ацила (отличного от воды).
Как применяют здесь, термин "алкоголиз" относится к ферментативному расщеплению ковалентной связи производного кислоты посредством реакции со спиртом ROH так, что один из продуктов соединяется с H спирта, а другой продукт соединяется с группой OR спирта.
Как применяют здесь, термин "спирт" относится к алкильному соединению, содержащему гидроксильную группу.
Как применяют здесь, термин "гидролиз" относится к ферментативно катализируемому переносу ацильной группы с липида на OH-группу молекулы воды. Перенос ацила, происходящий вследствие гидролиза, требует разделения молекулы воды.
Термин "галактолипидгидролитическая активность" как применяют здесь обозначает способность фермента катализировать гидролиз галактолипида посредством переноса ацильной группы с галактолипида на OH-группу молекулы воды.
Подобным образом, как применяют здесь, термин "фосфолипидгидролитическая активность" обозначает способность фермента катализировать гидролиз фосфолипида посредством переноса ацильной группы с фосфолипида на OH-группу молекулы воды.
Как применяют здесь, термин "повышенное отношение галактолипазной трансферазной активности к активности гидролиза галактолипидов" обозначает вариант фермента, который по сравнению с исходным ферментом способен к катализу в качестве галактолипидтрансферазы в большей степени, чем к катализу гидролиза галактолипидов. Это может означать, что и галактолипидтрансферазная активность, и активность гидролиза галактолипидов увеличены по сравнению с исходным ферментом, или что галактолипидтрансферазная активность увеличена, тогда как активность гидролиза галактолипидов уменьшена по сравнению с исходным ферментом. Важным является конечное отношением между двумя этими видами активности.
Как применяют здесь, термин "без увеличения или без существенного увеличения свободных жирных кислот" обозначает, что предпочтительно липидацилтрансфераза по настоящему изобретению обладает 100% трансферазной активностью (т.е. переносит 100% ацильных групп с донора ацила на акцептор ацила, без гидролитической активности); однако фермент может переносить на акцептор ацила менее чем 100% ацильных групп, присутствующих в липидном доноре ацила. В таком случае, ацилтрансферазная активность предпочтительно составляет, по меньшей мере 5%, более предпочтительно, по меньшей мере 10%, более предпочтительно, по меньшей мере 20%, более предпочтительно, по меньшей мере 30%, более предпочтительно, по меньшей мере 40%, более предпочтительно, 50%, более предпочтительно, по меньшей мере 60%, более предпочтительно, по меньшей мере 70%, более предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, а более предпочтительно, по меньшей мере 98% общей ферментативной активности. % Трансферазной активности (т.е. трансферазная активность как процент общей ферментативной активности) можно определить посредством следующего протокола:
Протокол для определения % ацилтрансферазной активности:
Пищевой продукт к которому добавляли липидацилтрансферазу по настоящему изобретению после ферментативной реакции можно экстрагировать посредством CHCl3:CH3OH 2:1 и органическую фазу, содержащую липидное вещество выделяют и анализируют посредством GLC следуя подробно описанной здесь ниже процедуре. Из анализа GLC (и, если необходимо, анализа ВЭЖХ) определяют количество свободных жирных кислот и одного или нескольких из сложных эфиров стерина/станола; сложных эфиров углеводов, сложных эфиров белков; диглицеридов или моноглицеридов. Контрольный пищевой продукт, к которому не добавляли фермент по настоящему изобретению, анализируют тем же способом.
Подсчет:
Из результатов GLC (и, необязательно, анализа ВЭЖХ) можно подсчитать увеличение содержания свободных жирных кислот и сложных эфиров стерина/станола, и/или сложных эфиров углеводов, и/или сложных эфиров белков, и/или диглицеридов, и/или моноглицеридов:
Δ % жирной кислоты = % жирной кислоты (фермент) - % жирной кислоты (контроль); Mv жирной кислоты = средняя молекулярная масса жирных кислот;
A = Δ % сложного эфира стерина/Mv сложного эфира стерина (где Δ % сложного эфира стерина = % сложного эфира стерина/станола (фермент) - % сложного эфира стерина/станола (контроль), а Mv сложного эфира стерина = средняя молекулярная масса сложных эфиров стеринов/станолов) - применимо, когда акцептор ацила представляет собой стерин и/или станол;
B = Δ % сложного эфира углевода/Mv сложного эфира углевода (где Δ % сложного эфира углевода = % сложного эфира углевода (фермент) - % сложного эфира углевода (контроль), а Mv сложного эфира углевода = средняя молекулярная масса сложного эфира углевода) - применимо, когда акцептор ацила представляет собой углевод;
C = Δ % сложного эфира белка/Mv сложного эфира белка (где Δ % сложного эфира белка = % сложного эфира белка (фермент) - % сложного эфира белка (контроль), а Mv сложного эфира белка = средняя молекулярная масса сложного эфира белка) - применимо, когда акцептор ацила представляет собой белок; и
D = абсолютное значение диглицерида и/или моноглицерида/Mv ди/моноглицерида (где Δ % диглицерида и/или моноглицерида = % диглицерида и/или моноглицерида (фермент) - % диглицерида и/или моноглицерида (контроль), а Mv ди/моноглицерида = средняя молекулярная масса диглицерида и/или моноглицерида) - применимо, когда акцептор ацила представляет собой глицерин.
Трансферазную активность подсчитывают как процент общей ферментативной активности:
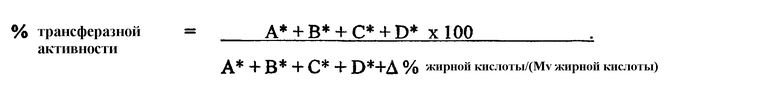
* - удалить при необходимости.
Аминокислоты, соответствующие терминам "неполярная", "полярная - незаряженная", "полярная - заряженная" приведены в таблице ниже, как аминокислоты, соответствующие терминам "алифатическая" и "ароматическая". Термин "полярная" относится и к "полярным - незаряженным", и к "полярным - заряженным" аминокислотам.
Анализ GLC
Хроматограф Perkin Elmer Autosystem 9000 Capillary Gas Chromatograph, снаряженный колонкой WCOT из расплавленного диоксида кремния 12,5 м × 0,25 мм ID × 0,1 мк толщины пленки 5% фенилметилсиликона (CP Sil 8 CB from Chrompack).
Газ-носитель: гелий.
Инжектор: холодная раздельная инъекция PSSI (исходная температура 50°C с нагревом до 385°C), объем 1,0 мкл
Детектор FID: 395°C
Получение образца: 30 мг образца растворяли в 9 мл гептана:пиридина, 2:1, содержащем внутренний стандарт гептадекан, 0,5 мг/мл. 300 мкл раствора образца переносили в изогнутый флакон, добавляли 300 мкл MSTFA (N-метил-N-триметилсилилтрифторацетамид) и проводили реакцию в течение 20 минут при 60°C.
Подсчет: факторы отклика для моно-ди-триглицеридов и свободной жирной кислоты определяли на основании стандарта 2 (моно-ди-триглицерид), для холестерина, холестерилпальмитата и холестерилстеарата факторы отклика определяли на основании чистого эталонного вещества (навеска чистого вещества составляла 10 мг).
ТЕХНИЧЕСКИЕ РЕЗУЛЬТАТЫ
Настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в яичных продуктах, особенно в майонезе: повышенная термоустойчивость в течение пастеризации; улучшенные органолептические свойства, улучшенная консистенция.
Варианты ферментов с повышенной фосфолипидтрансферазной активностью, особенно с повышенной трансферазной активностью между фосфолипидом и стерином и/или станолом, таким как холестерин, могут быть особенно пригодны в способах получения лизофосфолипидов и/или для ферментативного рафинирования прищевых масел и/или для получения яичных продуктов с улучшенными свойствами эмульсификации и/или пользой для здоровья.
Для применения в способах ферментативного рафинирования предпочтительны варианты с повышенной абсолютной фосфолипидтрансферазной активностью для стерина.
Таким образом, настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в яйцах или яичных продуктах: повышенная стабильность эмульсии; термостабильность эмульсии; улучшенный вкус; сниженный неприятный запах; улучшенная способность к загущению, улучшенная консистенция.
Настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в продуктах из теста и/или выпеченных продуктах: повышенный удельный объем или продуктов из теста или выпеченных продуктов (например, хлеба и/или пирога); повышенная стабильность теста; улучшенное формирование корки (например, более тонкая и/или хрустящая корка хлеба), улучшенное формирование мякоти (например, более гомогенное распределение мякоти и/или более тонкая структура мякоти и/или более мягкая мякоть); улучшенный внешний вид (например, гладкая поверхность без вздутий или отверстий или по существу без вздутий или отверстий); уменьшенное очерствение; улучшенная мягкость; улучшенный запах; улучшенный вкус.
Таким образом, настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в пищевом продукте: улучшенный внешний вид, улучшенное ощущение во рту, повышенная стабильность, в частности повышенная термостабильность, улучшенный вкус, повышенная мягкость, улучшенная упругость, улучшенная эмульсификация.
Таким образом, настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в молочных продуктах, например, таких как мороженное: улучшенное разжевывание (предпочтительно сметанообразное ощущение во рту); улучшенный вкус; улучшенное плавление.
Конкретные технические результаты, ассоциированные с применением липидацилтрансферазы, как определено здесь для получения пищевых продуктов перечислены в таблице ниже:
Таким образом, настоящее изобретение может обеспечивать один или несколько из следующих ниже неожиданных технических результатов в сыре: уменьшение эффекта обезжиривания в сыре; увеличение образования сыра; улучшение вкуса; уменьшенный неприятный запах; сниженный "мыльный" вкус.
В одном из аспектов, настоящее изобретение частично основано на варианте осуществления, позволяющем получать продукты питания, такие как сыр, которые можно улучшить применением липидацилтрансферазы. Кроме того, или альтернативно, можно улучшить вкус, структуру, устойчивость к окислению и/или срок хранения продукта питания. Кроме того или альтернативно, продукт питания может обладать сниженным уровнем холестерина или повышенным содержанием сложных эфиров фитостеринов/станолов.
Настоящее изобретение в одном из аспектов может относится к композиции пищевой добавки, содержащей липидацилтрансферазу, как определено здесь.
Настоящее изобретение в другом аспекте может относится к косметической композиции, содержащей липидацилтрансферазу, как определено здесь.
Кроме того, настоящее изобретение может относится к применению ацилтрансферазы, как определено здесь, для получения косметической композиции.
ПРЕИМУЩЕСТВА
Варианты трансфераз по настоящему изобретению по сравнению с исходным ферментом обладают одним или несколькими из следующих ниже преимущественных свойств:
i) повышенной активностью для полярных липидов и/или повышенной активностью для полярных липидов по сравнению с триглицеридами.
ii) повышенной активностью в отношении галактолипидов (гликолипиды), таких как один или несколько из дигалактозилдиглицерида (DGDG) и/или моногалактозилдиглицерида (MGDG).
iii) повышенным отношением активности в отношении галактолипидов (гликолипиды) к активности в отношении фосфолипидов и/или триглицеридов.
Предпочтительно варианты трансфераз по изобретению обладают повышенной активностью в отношении дигалактозилдиглицерида (DGDG) и/или моногалактозилдиглицерида (MGDG).
Предпочтительно варианты трансфераз по настоящему изобретению обладают повышенной активностью в отношении DGDG и/или MGDG и уменьшенной активностью для DGMG и/или MGMG.
Варианты трансфераз по изобретению также могут обладать повышенной активностью в отношении триглицеридов.
Варианты трансфераз по изобретению также могут обладать повышенной активностью в отношении фосфолипидов, таких как лецитин, включая сюда фосфатидилхолин.
Варианты трансфераз по настоящему изобретению могут обладать уменьшенной активностью в отношении триглицеридов, и/или моноглицеридов, и/или диглицеридов.
Термин полярный липид относится к полярным липидам, обычно находящимся в тесте, предпочтительно галактолипидам и фосфолипидам.
При применении в получении теста или выпеченного из теста продукта вариант трансферазы по изобретению может приводить к одному или нескольким из следующих ниже неожиданных технических результатов в тесте и/или выпеченных из теста продуктах: повышенный удельный объем теста или выпеченных из теста продуктов (например, хлеб и/или пирог); повышенная стабильность теста; улучшенные свойства корки (например, более тонкая и/или хрустящая хлебная корка), улучшенные свойства мякоти (например, более гомогенное распределение мякоти и/или более тонкая структура мякоти и/или боле мягкая мякоть); улучшенный внешний вид (например, гладкая поверхность без вздутий или отверстий или по существу без вздутий или отверстий); уменьшенное очерствение; улучшенная мягкость; улучшенный запах; улучшенный вкус.
ВЫДЕЛЕННЫЙ
В одном из аспектов, полипептид или белок для применения по настоящему изобретению предпочтительно находится в выделенной форме. Термин "выделенный" означает, что последовательность, по меньшей мере, по существу свободна, по меньшей мере, от одного другого компонента, с которым последовательность самопроизвольно ассоциирована в природе и как находят в природе.
ОЧИЩЕННЫЙ
В одном из аспектов, полипептид или белок для применения по настоящему изобретению предпочтительно находятся в очищенной форме. Термин "очищенный" означает, что последовательность находится в относительно чистом состоянии, например по меньшей мере приблизительно 51% чистоты, или, по меньшей мере приблизительно 75%, или, по меньшей мере приблизительно 80%, или, по меньшей мере приблизительно 90% чистоты, или, по меньшей мере приблизительно 95% чистоты или, по меньшей мере приблизительно 98% чистоты.
КЛОНИРОВАНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩЕЙ ПОЛИПЕПТИД ПО НАСТОЯЩЕМУ ИЗОБРЕТЕНИЮ
Нуклеотидную последовательность, кодирующую полипептид, обладающий конкретными свойствами, как определено здесь, или полипептид, пригодный для модификации, можно выделять из любой клетки или организма, продуцирующих указанный полипептид. В данной области хорошо известны различные способы выделения нуклеотидных последовательностей.
Например, можно сконструировать библиотеку геномной ДНК и/или кДНК с применением хромосомной ДНК или матричной РНК из организма, продуцирующего полипептид. Если аминокислотная последовательность полипептида известна, можно синтезировать меченные олигонуклеотидные зонды и применять для идентификации кодирующих полипептид клонов из геномной библиотеки, полученной из организма. Альтернативно, для идентификации кодирующих полипептид клонов можно применять меченный олигонуклеотидный зонд, содержащий последовательности, гомологичные другому известному гену полипептида. В последнем случае, применяют условия гибридизации и отмывки с меньшей строгостью.
Альтернативно, кодирующие полипептид клоны можно идентифицировать посредством вставки фрагментов геномной ДНК в экспрессирующий вектор, такой как плазмида, трансформации отрицательных по ферменту бактерий полученной библиотекой геномной ДНК и затем нанесения трансформированных бактерий на агар, содержащий фермент, ингибируемый полипептидом, обеспечивая таким образом идентификацию клонов, экспрессирующих полипептид.
В еще одной альтернативе кодирующую полипептид нуклеотидную последовательность можно получать синтетическим способом посредством общепризнанных стандартных способов, например, фосфорамидатный способ, описанный Beucage S.L. et al. (1981) Tetrahedron Letters 22, p 1859-1869, или способ, описанный у Matthes et al. (1984) EMBO J. 3, p 801-805. В фосфорамидатном способе олигонуклеотиды синтезируют, например, в автоматическом синтезаторе ДНК, очищают, отжигают, лигируют и клонируют в подходящие векторы.
Нуклеотидная последовательность может быть смешанного геномного и синтетического происхождения, смешанного синтетического происхождения и происходящей из кДНК или смешанного геномного происхождения и происходящей из кДНК, полученной посредством лигирования фрагментов синтетического, геномного происхождения или происходящих из кДНК (соответственно) стандартными способами. Каждый лигированный фрагмент соответствует различным частям полной нуклеотидной последовательности. Последовательность ДНК также можно получать посредством полимеразной цепной реакции (ПЦР) с применением специфических праймеров, например, как описано в патенте США 4683202 или у Saiki R K et al. (Science (1988) 239, pp 487-491).
НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
Настоящее изобретение также относится к нуклеотидным последовательностям, кодирующим полипептиды с конкретными свойствами, как определено здесь. Как применяют здесь, термин "нуклеотидная последовательность" относится к олигонуклеотидной последовательности или полинуклеотидной последовательности и к их варианту, гомологу, фрагментам и производным (таким как их части). Нуклеотидная последовательность может быть геномного, или синтетического, или рекомбинантного происхождения, и может быть двухцепочечной или одноцепочечной, представляя собой смысловую или антисмысловую цепь.
Термин "нуклеотидная последовательность" относительно настоящего изобретения включает в себя геномную ДНК, кДНК, синтетическую ДНК и РНК. Предпочтительно он означает ДНК, более предпочтительно, кДНК для кодирующей последовательности.
В предпочтительном варианте осуществления, нуклеотидная последовательность, непосредственно кодирующая полипептид с конкретными свойствами, как определено здесь, не покрывает природную нуклеотидную последовательность в ее природном окружении, когда она связана с ассоциированной с ней в природе последовательностью(ями), которая(ые) также находится(ятся) в ее(их) природном окружении. Для простоты ссылки авторы предлагают называть данный предпочтительный вариант осуществления "неприродная нуклеотидная последовательность". В данном отношении, термин "природная нуклеотидная последовательность" означает полную нуклеотидную последовательность, находящуюся в ее природном окружении и функционально связанную с полным промотором, с которым она ассоциирована в природе, где этот промотор также находится в его природном окружении. Таким образом, полипептид по настоящему изобретению можно экспрессировать посредством нуклеотидной последовательности в его естественном организме, но где нуклеотидная последовательность находится не под контролем того промотора, с которым она в природе ассоциирована в данном организме.
Предпочтительно полипептид не является природным полипептидом. В этом отношении, термин "природный полипептид" означает полноразмерный полипептид, находящийся в его природном окружении, и когда он экспрессирован с его природной нуклеотидной последовательности.
Как правило, нуклеотидную последовательность, кодирующую полипептиды с конкретными свойствами, как определено здесь, получают с применением способов рекомбинантной ДНК (т.е. рекомбинантная ДНК). Однако в альтернативном варианте осуществления изобретения нуклеотидную последовательность целиком или полностью можно синтезировать с применением химических способов, хорошо известных в данной области (см. Caruthers MH et al. (1980) Nuc Acids Res Symp Ser 215-23 и Horn T et al. (1980) Nuc Acids Res Symp Ser 225-232).
МОЛЕКУЛЯРНОЕ ПРЕОБРАЗОВАНИЕ
После выделения кодирующей фермент нуклеотидной последовательности, или после идентификации предполагаемой кодирующей фермент нуклеотидной последовательности, выбранную нуклеотидную последовательность может быть желательным модифицировать, например, может быть желательным подвергнуть последовательность мутированию для получения фермента по настоящему изобретению.
Мутации можно вносить с применением синтетических олигонуклеотидов. Эти олигонуклеотиды содержат нуклеотидные последовательности, фланкирующие желательные участки мутации.
Подходящий способ описан у Morinaga et al. (Biotechnology (1984) 2, p646-649). Другой способ внесения мутаций в кодирующие фермент нуклеотидные последовательности описан у Nelson и Long (Analytical Biochemistry (1989), 180, p147-151).
Вместо сайт-специфического мутагенеза, такого, как описан выше, мутации можно вносить случайным образом, например, с применением коммерческого набора, такого как набор GeneMorph PCR mutagenesis kit из Stratagene, или набор Diversify PCR random mutagenesis kit из Clontech. К способам оптимизации основанного на ПЦР мутагенеза, который также можно сочетать с применением мутагенных аналогов ДНК, таких как описанные в EP0 866 796, относится EP0 583 265. Для получения вариантов липидацилтрансфераз с предпочтительными характеристиками подходят технологии подверженной ошибкам ПЦР. К молекулярному преобразованию липаз относится WO 0206457.
Третий способ получения новых последовательностей состоит во фрагментировании неидентичных нуклеотидных последовательностей, или с применением любого количества рестрикционных ферментов или такого фермента, как ДНКаза I, и повторной сборке полноразмерных нуклеотидных последовательностей, кодирующих функциональные белки. Альтернативно можно применять одну или несколько неидентичных нуклеотидных последовательностей и вносить мутации в течении повторной сборки полноразмерной нуклеотидной последовательности. Для получения вариантов липидацилтрансфераз с предпочтительными характеристиками пригодны технологии перестановки ДНК и перестановки семейств. Пригодные способы для проведения "перестановки" можно найти в EP0 752 008, EP1 138 763, EP1 103 606. Перестановку также можно сочетать с другими формами мутагенеза ДНК, как описано в патенте США 6180406 и WO 01/34835.
Таким образом, в нуклеотидной последовательности можно получать множество участков направленных или случайных мутаций, или in vivo, или in vitro, и далее различными способами исследовать кодируемый вариант полипептида на улучшенную функциональность. Например, с применением рекомбинантных способов in silico и опосредованных exo (см. WO 00/58517, патент США № 6344328, патент США № 6361974) молекулярное преобразование можно проводить, когда полученный вариант обладает очень низкой гомологией с известными ферментами или белками. Такие варианты, полученные таким образом, могут обладать значительной структурной аналогией с известными трансферазными ферментами, но обладать очень низкой гомологией аминокислотных последовательностей.
Кроме того, в качестве неограничивающего примера, для получения новых вариантов мутантные или природные варианты полинуклеотидной последовательности можно рекомбинировать с диким типом или с другими мутантными или природными вариантами. Такие новые варианты также можно скринировать на улучшенную функциональность кодируемого полипептида.
Применения указанного выше и сходные способы молекулярного преобразования позволяют идентифицировать и выбрать варианты ферментов по настоящему изобретению, обладающие предпочтительными характеристиками, без любого предварительного знания о структуре или функции белка, и позволяют получать непредсказуемые, но выгодные мутации или варианты. Существует множество примеров применения молекулярного преобразования в данной области для оптимизации или изменения ферментативной активности, такие примеры включают в себя в качестве неограничивающих примеров один или несколько из следующих ниже: оптимизированная экспрессия и/или активность в клетке-хозяине или in vitro, повышенная ферментативная активность, измененная субстратная специфичность и/или специфичность в отношении продукта, повышенная или уменьшенная ферментативная или структурная стабильность, измененная ферментативная активность/специфичность в предпочтительных условиях окружающей среды, например, температура, pH, субстрат.
Как понятно специалистам в данной области, с применением средств молекулярного преобразования, фермент можно изменять для улучшения функциональности фермента.
Таким образом, применяемая по изобретению липидацилтрансфераза может представлять собой вариант, т.е. по сравнению с исходным ферментом может содержать по меньшей мере одну замену, делецию или добавление аминокислоты. Варианты белков сохраняют, по меньшей мере 25%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% гомологии с исходным ферментом. Пригодные исходные ферменты могут включать в себя любой фермент с эстеразной или липазной активностью. Предпочтительно, исходный фермент сравнен с консенсусной последовательностью pfam00657.
В предпочтительном варианте осуществления вариант фермента липидацилтрансферазы сохраняет или содержит, по меньшей мере, одну или несколько, аминокислотных остатков консенсусной последовательности pfam00657, находящихся в участках GDS, GANDY и HPT.
Ферменты, такие как липазы с отсутствием или низкой липидацилтрансферазной активностью в водной среде, можно подвергать мутированию с применением средств молекулярного преобразования для введения или усиления трансферазной активности, таким образом, получая фермент липидацилтрансферазу со значительной трансферазной активностью, пригодный для применения в композициях и способах по настоящему изобретению.
Таким образом, липидацилтрансфераза для применения по изобретению может представлять собой вариант с повышенной ферментативной активностью в отношении полярных липидов, предпочтительно фосфолипидов и/или гликолипидов, по сравнению с исходным ферментом. Предпочтительно, такие варианты также обладают низкой активностью или у них нет активности в отношении полярные лизолипидов. Повышенная активность в отношении полярных липидов, фосфолипидов и/или гликолипидов может быть результатом гидролитической и/или трансферазной активности или сочетания обоих.
Вариант липидацилтрансферазы для применения по изобретению по сравнению с исходным ферментом может обладать уменьшенной активностью в отношении триглицеридов, и/или моноглицеридов, и/или диглицеридов.
Таким образом, вариант фермента может не обладать активностью в отношении триглицеридов, и/или моноглицеридов, и/или диглицеридов.
Альтернативно, вариант фермента для применения по изобретению может обладать повышенной активностью в отношении триглицеридов и/или также может обладать повышенной активностью в отношении одного или несколько из следующих ниже, полярных липидов, фосфолипидов, лецитина, фосфатидилхолина, гликолипидов, дигалактозилмоноглицерида, моногалактозилмоноглицерида.
Варианты липидацилтрансфераз известны, и один или несколько из таких вариантов может быть пригоден для применения в способах и применениях по настоящему изобретению и/или в композициях фермента по настоящему изобретению. В качестве неограничивающих примеров, по настоящему изобретению можно применять варианты липидацилтрансфераз, описанные в следующих ниже ссылках: Hilton & Buckley J Biol. Chem. 1991 Jan 15: 266 (2): 997-1000; Robertson et al. J. Biol. Chem. 1994 Jan 21; 269 (3): 2146-50; Brumlik et al. J. Bacteriol 1996 Apr; 178 (7): 2060-4; Peelman et al. Protein Sci. 1998 Mar; 7 (3):587-99.
АМИНОКИСЛОТНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
Настоящее изобретение также относится к аминокислотным последовательностям полипептидов с конкретными свойствами, как определено здесь.
Как применяют здесь, термин "аминокислотная последовательность" является синонимом с термином "полипептид" и/или с термином "белок". В некоторых случаях термин "аминокислотная последовательность" является синонимом с термином "пептид".
Аминокислотную последовательность можно получить/выделить из природного источника или ее можно получить синтетическим способом или ее можно получить с применением технологий рекомбинантных ДНК.
Таким образом, аминокислотные последовательности можно получать из указанных здесь выделенных полипептидов посредством стандартных способов.
Один из пригодных способов определения аминокислотных последовательностей из выделенных полипептидов представляет собой следующее:
Очищенный полипептид можно высушивать замораживанием, и 100 мкг высушенного замораживанием вещества можно растворять в 50 мкл смеси 8 М мочевины и 0,4 М кислого углекислого аммония, pH 8,4. Растворенный белок можно денатурировать и восстанавливать в течение 15 минут при 50°C с последующим нанесением азота и добавлением 5 мкл 45 мМ дитиотреитола. После охлаждения до комнатной температуры, можно добавлять 5 мкл 100 мМ йодацетамида для дериватизации остатков цистеина в течение 15 минут при комнатной температуре в темноте в азоте.
К указанной выше реакционной смеси можно добавлять 135 мкл воды и 5 мкг эндопротеиназы Lys-C в 5 мкл воды и в течение 24 часов можно проводить расщепление при 37°C в азоте.
Полученные пептиды можно разделять посредством ВЭЖХ с обратной фазой на колонке VYDAC C18 (0,46×l5см; 10мкм; The Separation Group, California, USA) с применением растворителя A: 0,1% TFA в воде и растворителя B: 0,1% TFA в ацетонитриле. До N-концевого определения последовательности выбранные пептиды можно повторно хроматографировать на колонке Develosil C18 с применением той же системы растворителей. Определение последовательности можно проводить с применением секвенатора Applied Biosystems 476A с применением быстрых циклов пульсирующей жидкости по инструкциям производителя (Applied Biosystems, California, USA).
ИДЕНТИЧНОСТЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ИЛИ ГОМОЛОГИЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящее изобретение также относится к применению последовательностей с определенной степенью идентичности последовательности или гомологии последовательности с аминокислотной последовательностью(ями) полипептида, обладающего конкретными определенными здесь свойствами, или с любой нуклеотидной последовательностью, кодирующей такой полипептид (далее обозначаемых здесь как "гомологичная последовательность(и)"). Здесь термин "гомолог" означает объект, обладающий определенной гомологией с аминокислотными последовательностями по изобретению и нуклеотидными последовательностями по изобретению. Здесь термин "гомология" можно приравнять к "идентичности".
Гомологичная аминокислотная последовательность и/или нуклеотидная последовательность должна предоставлять и/или кодировать полипептид, сохраняющий функциональную активность и/или усиливающий активность фермента.
В настоящем контексте гомологичная последовательность взята так, чтобы содержать аминокислотную последовательность, которая может быть, по меньшей мере на 75, 85 или 90% идентична, предпочтительно, по меньшей мере на 95 или 98% идентична последовательности по изобретению. Как правило, гомологи содержат те же активные центры и т.д., что и аминокислотная последовательность по изобретению. Хотя гомологию также можно рассматривать с точки зрения сходства (т.е. аминокислотные остатки со сходными химическими свойствами/функциями), в контексте настоящего изобретения предпочтительно выражать гомологию в терминах идентичности последовательностей.
В настоящем контексте, гомологичная последовательность взята так, чтобы содержать нуклеотидную последовательность, которая может быть, по меньшей мере на 75, 85 или 90% идентична, предпочтительно, по меньшей мере на 95 или 98% идентична нуклеотидной последовательности, кодирующей полипептид по настоящему изобретению (последовательность по изобретению). Как правило, гомологи будут содержать те же последовательности, которые кодируют активные центры и т.д., как и последовательность по изобретению. Хотя гомологию также можно рассматривать с точки зрения сходства (т.е. аминокислотные остатки со сходными химическими свойствами/функциями), в контексте настоящего изобретения предпочтительно выражать гомологию в терминах идентичности последовательностей.
Сравнения гомологии можно проводить на глаз или, как правило, с помощью легко доступных программ сравнения последовательностей. Данные коммерчески доступные компьютерные программы могут рассчитывать % гомологии двух или более последовательностей.
% гомологии можно рассчитать у непрерывных последовательностей, т.е. одна последовательность сравнена с другой последовательностью и каждая аминокислота в одной последовательности непосредственно сравнивается с соответствующей аминокислотой в другой последовательности, один остаток за раз. Это называется сравнение "без пропусков". Как правило, такие сравнения без пропусков проводят только для относительно короткого количества остатков.
Хотя это является очень простым и логичным способом, он не может принимать в расчет, например, то, что в идентичных во всем остальном парах последовательностей инсерция или делеция будет вызывать сдвиг следующих аминокислотных остатков из сравнения, таким образом потенциально приводя к большому уменьшению % гомологии при проведении глобального сравнения. Следовательно, большинство способов сравнения последовательностей разработаны так, чтобы получать оптимальные сравнения, принимающие в расчет возможные инсерции и делеции без чрезмерного ухудшения показателя общей гомологии. Этого достигают посредством вставки в сравнение последовательностей "пробелов", чтобы попытаться максимизировать локальную гомологию.
Однако в этих более сложных способах устанавливают "штрафы за пробелы" для каждого пробела, находящегося в сравнении так, что для одного и того же количества идентичных аминокислот сравнение последовательностей с как можно меньшим числом пробелов насколько возможно - отражая большую связанность между двух сравниваемых последовательностей - будет обеспечивать больший показатель, чем с многими пробелами. Применяют, как правило, "цены однородных пробелов", устанавливающие относительно высокую цену за существование пробела и меньший штраф за каждый последующий остаток в пробеле. Это является наиболее часто используемой системой оценки пробелов. Конечно, высокие штрафы за пробел дадут оптимизированные сравнения с меньшим количеством пробелов. Большинство программ сравнения позволяют модифицировать штрафы за пробел. Однако при использовании такого программного обеспечения для сравнений последовательностей предпочтительно использовать значения по умолчанию. Например, при применении пакета GCG Wisconsin Bestfit значение штрафа за пробел для аминокислотных последовательностей по умолчанию составляет -12 за пробел и -4 за каждое продолжение.
Таким образом, для подсчета максимума % гомологии во-первых необходимо получение оптимального сравнения, принимая в расчет штрафы за пробелы. Подходящая компьютерная программа для проведения такого сравнения представляет собой пакет GCG Wisconsin Bestfit (Devereux et al. 1984 Nuc. Acids Research 12 p387). Примеры другого программного обеспечения, способного проводить сравнения последовательностей, включают в себя в качестве неограничивающих примеров пакет BLAST (см. Ausubel et al 1999 Short Protocols in Molecular Biology, 4th Ed - Chapter 18), FASTA (Altschul et al. 1990 J. Mol. Biol. 403-410) и набор средств сравнения GENEWORKS. И BLAST, и FASTA доступны для поиска в отключенном и подключенном к сети состоянии (см. Ausubel et al. 1999, страницы с 7-58 до 7-60). Однако для некоторых приложений предпочтительно применять программу GCG Bestfit. Также доступно новое средство для сравнения белка и нуклеотидной последовательности, называемое BLAST 2 Sequences (см. FEMS Microbiol Lett 1999 174 (2): 247-50; FEMS Microbiol Lett 1999 177(1): 187-8 и tatiana@ncbi.nlm.nih.gov).
Хотя конечный % гомологи можно измерять в терминах идентичности, сам процесс сравнения, как правило, не основывается на категорическом сравнении пар. Вместо этого, как правило, используют матрицу оценки взвешенного подобия, назначающую оценку каждому парному сравнению на основе химического сходства или эволюционного расстояния. Примером такой широко применяемой матрицы является матрица BLOSUM62 - матрица по умолчанию для набора программ BLAST. В программах GCG Wisconsin, как правило, применяют или общедоступные значения по умолчанию, или пользовательскую таблицу сравнения символов, если снабжено (см. руководство пользователя для дальнейших подробностей). Для некоторых применений предпочтительно применять общедоступные значения по умолчанию для пакета GCG или, в случае другого программного обеспечения, матрицу по умолчанию, такую как BLOSUM62.
Альтернативно, процент гомологии можно подсчитать с применением возможности множественного сравнения в DNASIS™ (Hitachi Software), на основе алгоритма, аналогичного CLUSTAL (Higgins DG & Sharp PM (1988), Gene 73(1), 237-244).
После получения с применением программного обеспечения оптимального сравнения можно подсчитать % гомологии, предпочтительно % идентичности последовательностей. Программное обеспечение, как правило, делает это как часть сравнения последовательностей и выдает численный результат.
Последовательности также могут обладать делециями, инсерциями или заменами аминокислотных остатков, дающие молчащее изменение и приводящие к функционально эквивалентному веществу. На основании сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков можно делать продуманные аминокислотные замены при условии сохранения вторичной активности связывания вещества. Например, отрицательно заряженные аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают в себя лизин и аргинин; а аминокислоты с незаряженными полярными концевыми группами, обладающие сходными значениями гидрофильности, включают в себя лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин.
Могут быть сделаны консервативные замены, например, такие, как показаны в таблице ниже. Аминокислоты в одной ячейке во второй колонке и предпочтительно в одной и той же строке третьей колонки могут замещать друг друга.
Настоящее изобретение также относится к гомологичной замене (и замену, и замещение здесь используют для обозначения обмена существующего аминокислотного остатка на альтернативный остаток), которую можно провести, т.е. сходную замену, такую как основную на основную, кислая на кислую, полярная на полярную и т.д. Также можно производить негомологичную замену, т.е. с одного класса остатков на другой, или, альтернативно, включающую в себя введение неприродных аминокислот, таких как орнитин (далее здесь обозначаемый как Z), диаминомасляная кислота орнитин (далее здесь обозначаемый как B), норлейцин орнитин (далее здесь обозначаемый как O), пириилаланин, тиенилаланин, нафтилаланин и фенилглицин.
Замещения также можно производить неприродными аминокислотами.
Варианты аминокислотных последовательностей в дополнение к аминокислотным спейсерам, таким как остатки глицина или β-аланина, могут включать в себя подходящие спейсерные группы, которые можно вставлять между любыми двумя аминокислотными остатками в последовательности и которые включают в себя алкильные группы, такие как метильная, этильная или пропильная группы. Дополнительная форма варьирования, включающая в себя присутствие одного или нескольких аминокислотных остатков в пептоидной форме, будет хорошо понятна специалистам в данной области. Во избежание неопределенности, "пептоидную форму" применяют для обозначения вариантов аминокислотных остатков, где замещающая группа α-углерода находится на атоме азота остатка, а не на α-углероде. Способ получения пептидов в пептоидной форме известен в данной области, например, Simon RJ et al., PNAS (1992) 89 (20), 9367-9371 и Horwell DC, Trends Biotechnol. (1995) 13 (4), 132-134.
Нуклеотидные последовательности для применения по настоящему изобретению или кодирующие полипептид с конкретными определенными здесь свойствами могут содержать внутри себя синтетические или модифицированные нуклеотиды. В данной области известен ряд различных типов модификаций олигонуклеотидов. Они включают в себя метилфосфонатные и фосфоротиоатные основные цепи и/или добавление акридиновых или полилизиновых цепей на 3'- и/или 5'-концы молекулы. Для целей настоящего изобретения необходимо понимать, что описанные здесь нуклеотидные последовательности можно модифицировать любым доступным в данной области способом. Такие модификации можно проводить для увеличения активности или времени жизни нуклеотидных последовательностей in vivo.
Настоящее изобретение также относится к применению нуклеотидных последовательностей, комплементарных описанным здесь последовательностям, или любого производного, фрагмента или его производного. Если последовательность комплементарна их фрагментам, тогда эту последовательность можно применять в качестве зонда для идентификации сходных кодирующих последовательностей в других организмах и т.д.
Полинуклеотиды, не на 100% гомологичные последовательностям по настоящему изобретению, но находящиеся в пределах объема изобретения, можно получать рядом способов. Другие варианты описанных здесь последовательностей можно получать, например, зондированием библиотек ДНК, полученных из ряда индивидуумов, например, индивидуумов из различных популяций. Кроме того, можно получать другие вирусные/бактериальные или клеточные гомологи, особенно клеточные гомологи, находящиеся в клетках млекопитающих (например, крысиные, мышиные, бычьи клетки и клетки приматов) и такие гомологи и их фрагменты, в большинстве случаев, способны к селективной гибридизации с последовательностями, представленными в перечисленных здесь последовательностях. Такие последовательности можно получать зондированием библиотек кДНК, полученных из других видов животных, или библиотек их геномных ДНК и зондированием таких библиотек зондами, содержащими всю или часть любой последовательности в приложенных списках последовательностей в условиях от средней до высокой строгости. Подобные рассуждения применяют для получения видовых гомологов и аллельных вариантов полипептида или нуклеотидных последовательностей по изобретению.
Варианты и штаммовые/видовые гомологи также можно получить с применением вырожденной ПЦР, в которой применяют праймеры, разработанные для соответствия последовательностям в вариантах и гомологах, кодирующих консервативные аминокислотные последовательности в последовательности по настоящему изобретению. Консервативные последовательности можно предсказывать, например, посредством сравнения аминокислотных последовательностей различных вариантов/гомологов. Сравнения последовательностей можно проводить с применением известного в данной области компьютерного программного обеспечения. Например, широко применяют программу GCG Wisconsin PileUp.
Применяемые в вырожденной ПЦР праймеры содержат одно или несколько вырожденных положений и их применяют в условиях строгости, меньшей, чем условия строгости, применяемые для клонирования последовательностей при использовании праймеров с одной последовательностью для известных последовательностей.
Альтернативно, такие полинуклеотиды можно получать посредством сайт-специфического мутагенеза охарактеризованных последовательностей. Это может быть пригодным, например, когда необходимы молчащие изменения последовательности кодонов для оптимизации предпочтительных кодонов для конкретной клетки-хозяина, в которой экспрессируются полинуклеотидные последовательности. Для введения участков распознавания рестрикционных полипептидов или для изменения свойства или функционирования кодируемых полинуклеотидами полипептидов могут быть желательными другие изменения последовательностей.
Полинуклеотиды (нуклеотидные последовательности) по изобретению можно применять для получения праймера, например праймера для ПЦР, праймера для альтернативной реакции амплификации, зонда, например, меченого выявляемой меткой традиционными способами с применением радиоактивных или нерадиоактивных меток, или полинуклеотиды можно клонировать в векторы. Такие праймеры, зонды и другие фрагменты составляют, по меньшей мере 15, предпочтительно, по меньшей мере 20, например, по меньшей мере, 25, 30 или 40 нуклеотидов в длину, и также обозначены термином полинуклеотиды по изобретению, как применяют здесь.
Полинуклеотиды, такие как полинуклеотиды и зонды ДНК по изобретению, можно получать рекомбинантным способом, синтетически или любыми доступными специалистам в данной области способами. Их также можно клонировать посредством стандартных способов.
В большинстве случаев, праймеры получают синтетическими способами, включающими в себя пошаговое получение желательной последовательности нуклеиновой кислоты по одному нуклеотиду за один раз. Способы выполнения этого с применением автоматических способов легко доступны в данной области.
Более длинные полинуклеотиды, как правило, получают с применением рекомбинантных способов, например с применением способов клонирования посредством ПЦР (полимеразная цепная реакция). Это включает в себя получение пары праймеров (например, приблизительно от 15 до 30 нуклеотидов), фланкирующих область направленной к липиду последовательности, которую желательно клонировать, приведение праймеров в контакт с мРНК или кДНК, полученных из животной или человеческой клетки, проведение полимеразной цепной реакции в условиях, в которых проходит амплификация желательной области, выделение амплифицированного фрагмента (например, посредством очистки реакционной смеси на агарозном геле) и восстановление амплифицированной ДНК. Праймеры можно разработать так, чтобы они содержали подходящие участки распознавания рестрикционными ферментами, так чтобы амплифицированную ДНК можно было клонировать в подходящий клонирующий вектор.
ГИБРИДИЗАЦИЯ
Настоящее изобретение также относится к последовательностям, комплементарным последовательностям по настоящему изобретению, или последовательностям, способным гибридизоваться или с последовательностями по настоящему изобретению или с последовательностями, комплементарными им.
Как применяют здесь, термин "гибридизация" должен включать в себя "процесс, посредством которого цепь нуклеиновой кислоты связывается с комплементарной цепью посредством спаривания оснований", а также процесс амплификации, как осуществляют в технологиях полимеразной цепной реакции (ПЦР).
Настоящее изобретение также относится к применению нуклеотидных последовательностей, способных к гибридизации с последовательностями, комплементарными с описанными здесь последовательностями по настоящему изобретению, или любых производных, фрагментов или их производных.
Настоящее изобретение также относится к последовательностям, которые комплементарны к последовательностям, способным гибридизоваться с описанными здесь нуклеотидными последовательностями.
Условия гибридизации основаны на температуре плавления (Tm) комплекса связанных нуклеотидов, как указано у Berger и Kimmel (1987, Guide to Molecular Cloning Techniques, Methods in Enzymology, Vol. 152, Academic Press, San Diego CA), и обладают определенной "строгостью", как указано ниже.
Максимальная строгость, как правило, имеет место приблизительно при Tm-5°C (5°C ниже Tm зонда); высокая строгость, приблизительно при температурах от 5°C до 10°C ниже Tm; промежуточная строгость, приблизительно при температурах от 10°C до 20°C ниже Tm; и низкая строгость, приблизительно при температурах от 20°C до 25°C ниже Tm. Как будет понятно специалистам в данной области, гибридизацию с максимальной строгостью можно применять для идентификации или выявления идентичных нуклеотидных последовательностей, тогда как гибридизацию с промежуточной строгостью (или низкой) можно применять для идентификации или выявления сходных или родственных полинуклеотидных последовательностей.
Предпочтительно, настоящее изобретение относится к последовательностям, которые комплементарны к последовательностям, способным гибридизоваться в условиях с высокой строгостью или в условиях с промежуточной строгостью с нуклеотидными последовательностями, кодирующими полипептиды с конкретными свойствами, как определено здесь.
Более предпочтительно, настоящее изобретение относится к последовательностям, комплементарным к последовательностям, способным к гибридизация в условиях с высокой строгостью (например, 65°C и 0,1 × SSC {1 × SSC = 0,15 М NaCl, 0,015 М цитрат Na pH 7,0}) с нуклеотидными последовательностями, кодирующими полипептиды с конкретными свойствами, как определено здесь.
Настоящее изобретение также относится к нуклеотидным последовательностям, способным гибридизоваться с описанными здесь нуклеотидными последовательностями (включая сюда последовательности, комплементарные к описанным здесь последовательностям).
Настоящее изобретение также относится к нуклеотидным последовательностям, которые комплементарны к последовательностям, способным гибридизоваться с описанными здесь нуклеотидными последовательностями (включая сюда последовательности комплементарные к описанным здесь последовательностям).
Также в объем настоящего изобретения включены полинуклеотидные последовательности, способные гибридизоваться с описанными здесь нуклеотидными последовательностями в условиях строгости от промежуточной до максимальной.
В предпочтительном аспекте настоящее изобретение относится к нуклеотидным последовательностям, способным гибридизоваться с описанными здесь нуклеотидными последовательностями или с комплементарными им последовательностями в строгих условиях (например, 50°C и 0,2 × SSC).
В более предпочтительном аспекте настоящее изобретение относится к нуклеотидным последовательностям, способным гибридизоваться с описанными здесь нуклеотидными последовательностями или комплементарными им последовательностями в условиях с высокой строгостью (например, 65°C и 0,1 × SSC).
ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ
Нуклеотидную последовательность для применения по настоящему изобретению или кодирующую полипептид с конкретными свойствами, как определено здесь, можно вставить в рекомбинантный реплицирующийся вектор. Вектор можно применять для репликации и экспрессии нуклеотидной последовательности, в форму полипептида в совместимой клетке-хозяине и/или из нее. Экспрессию можно контролировать с применением контрольных последовательностей, включающих в себя промоторы/энхансеры и другие сигналы регуляции экспрессии. Можно применять прокариотические промоторы и промоторы, функционирующие в эукариотических клетках. Можно применять тканеспецифические или специфичные к стимулам промоторы. Также можно применять химерные промоторы, включающие в себя элементы последовательностей из двух или большего числа различных промоторов, описанных выше.
Полипептид, продуцируемый рекомбинантной клеткой-хозяином при экспрессии нуклеотидной последовательности в зависимости от применяемых последовательности и/или вектора, может быть секретируемым или может содержаться внутриклеточно. Кодирующие последовательности можно разработать так, чтобы они содержали сигнальные последовательности, направляющие секрецию кодирующих вещество последовательностей за конкретную прокариотическую или эукариотическую клеточную мембрану.
ЭКСПРЕССИРУЮЩИЙ ВЕКТОР
Термин "экспрессирующий вектор" означает конструкцию, способную к экспрессии in vivo или in vitro.
Предпочтительно, экспрессирующий вектор включен в геном организма. Термин "включен" предпочтительно относится к стабильному включению в геном.
Нуклеотидная последовательность по настоящему изобретению или кодирующая полипептид с конкретными свойствами, как определено здесь, может находиться в векторе, в котором нуклеотидная последовательность функционально связана с регуляторными последовательностями так, что регуляторные последовательности способны обеспечивать экспрессию нуклеотидной последовательности подходящим организмом-хозяином, т.е. вектор представляет собой экспрессирующий вектор.
Векторы по настоящему изобретению можно трансформировать в подходящую клетку-хозяина, как описано ниже, для обеспечения экспрессии полипептида с конкретными свойствами, как определено здесь.
Выбор вектора, например, плазмидного, космидного, вирусного или фагового вектора, часто зависит от клетки-хозяина, в которую его вводят.
Векторы могут содержать один или несколько селективных маркерных генов, таких как ген, обеспечивающий устойчивость к антибиотику, например, устойчивость к ампициллину, канамицину, хлорамфениколу или тетрациклину. Альтернативно, выбор можно проводить посредством совместной трансформации (как описано в WO 91/17243).
Векторы можно применять in vitro, например, для получения РНК или применять для трансфекции или трансформации клетки-хозяина.
Таким образом, в дополнительном варианте осуществления, изобретение относится к способу получения нуклеотидных последовательностей по настоящему изобретению или нуклеотидных последовательностей, кодирующих полипептиды с конкретными свойствами, как определено здесь, посредством введения нуклеотидной последовательности в реплицируемый вектор, введения вектора в совместимую клетку-хозяина и выращивания клетки-хозяина в условиях, в которых осуществляется репликация вектора.
Вектор может дополнительно содержать нуклеотидную последовательность, позволяющую вектору реплицироваться в данной клетке-хозяине. Примеры таких последовательностей представляют собой точки начала репликации плазмид pUC19, pACYC177, pUB110, pE194, pAMB1 и pIJ702.
РЕГУЛЯТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
В определенных применениях нуклеотидную последовательность для применения по настоящему изобретению или нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, можно функционально связывать с регуляторной последовательностью, способной к обеспечению экспрессии нуклеотидной последовательности, например, посредством выбранной клетки-хозяина. В качестве примера настоящее изобретение относится к вектору, включающему в себя нуклеотидную последовательность по настоящему изобретению, функционально связанную с такой регуляторной последовательностью, т.е. вектор представляет собой экспрессирующий вектор.
Термин "функционально связанный" относится к смежному положению, где описанные компоненты находятся в связи, позволяющей им функционировать предназначенным для них образом. Регуляторная последовательность, "функционально связанная" с кодирующей последовательностью, лигирована таким образом, что в условиях, соответствующих контрольной последовательности, достигается экспрессия кодирующей последовательности.
Термин "регуляторные последовательности" включает в себя промоторы и энхансеры и другие регулирующие экспрессию сигналы.
Термин "промотор" применяют в обычном для данной области смысле, например, участок связывания РНК-полимеразы.
Повышенной экспрессии нуклеотидной последовательности, кодирующей фермент с конкретными свойствами, как определено здесь, также можно достичь посредством выбора гетерологичных регуляторных областей, например, промотора, областей секреторного лидера и терминатора.
Предпочтительно, нуклеотидная последовательность по настоящему изобретению может быть функционально связана, по меньшей мере, с промотором.
Примеры подходящих промоторов для направления транскрипции нуклеотидной последовательности в бактериальном, грибном или дрожжевом хозяине хорошо известны в данной области.
КОНСТРУКЦИИ
Термин "конструкция", являющийся синонимом с такими терминами, как "конъюгат", "кассета" и "гибрид", включает в себя нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, для применения по настоящему изобретению непосредственно или опосредованно присоединенную к промотору. Пример непрямого присоединения представляет собой предоставление подходящей спейсерной группы, такой как интронная последовательность, такая как интрон Sh1 или интрон ADH, находящейся между промотором и нуклеотидной последовательностью по настоящему изобретению. Тоже самое верно для термина "гибридный" в отношении настоящего изобретения, включающего в себя прямое или непрямое соединение. В некоторых случаях термины не подразумевают природное сочетание кодирующей белок нуклеотидной последовательности, обычным способом ассоциированной с генным промотором дикого типа и когда они оба находятся в их природном окружении.
Конструкция может даже содержать или экспрессировать маркер, позволяющий отбор генетической конструкции.
Для некоторых применений конструкция предпочтительно содержит, по меньшей мере, нуклеотидную последовательность по настоящему изобретению или нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, функционально связанный с промотором.
КЛЕТКИ-ХОЗЯЕВА
Термин "клетка-хозяин" в отношении настоящего изобретения включает в себя клетку, содержащую или нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, или экспрессирующий вектор, как описано выше, и которую применяют в рекомбинантном получении полипептида с конкретными свойствами, как определено здесь.
Таким образом, дополнительный вариант осуществления настоящего изобретения относится к клеткам-хозяевам, трансформированным или трансфицированным нуклеотидной последовательностью по настоящему изобретению или нуклеотидной последовательностью, экспрессирующей полипептид с конкретными свойствами, как определено здесь. Клетки следует выбирать так, чтобы они сочетались с указанным вектором и они могут быть, например, прокариотическими (например, бактериальными), грибными, дрожжевыми или растительными клетками. Предпочтительно, клетки-хозяева не являются человеческими клетками.
Примеры подходящих бактериальных хозяйских организмов представляют собой грамотрицательные бактерии и грамположительные бактерии.
В зависимости от природы нуклеотидной последовательности, кодирующей полипептид с конкретными свойствами, как определено здесь, и/или желательности дальнейшего процессинга экспрессируемого белка, предпочтительными могут являться эукариотические хозяева, такие как дрожжи или другие грибы. В большинстве случаев дрожжевые клетки предпочтительнее клеток грибов, так как ими проще манипулировать. Однако некоторые белки или плохо секретируются из дрожжевой клетки, или, в некоторых случаях, не подвергаются правильному процессингу (например, гипергликозилирование в дрожжах). В этих случаях следует выбирать другой грибной хозяйский организм.
Применение подходящих клеток-хозяев, таких как дрожжи, грибы и растительные клетки-хозяева, может обеспечить посттрансляционные модификации (например, миристоилирование, гликозилирование, укорочение, липидизация и фосфорилирование по тирозину, серину или треонину), как может быть необходимо для обеспечения рекомбинантным продуктам экспрессии по настоящему изобретению оптимальной биологической активности.
Клетка-хозяин может быть с дефицитом по протеазе или минус-штаммом по протеазе.
ОРГАНИЗМ
Термин "организм" по отношению к настоящему изобретению включает в себя любой организм, способный содержать нуклеотидную последовательность по настоящему изобретению или нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, и/или продукты, полученные из него.
Пригодные организмы могут включать в себя прокариоты, грибы, дрожжи или растения.
Термин "трансгенный организм" в отношении настоящего изобретения включает в себя любой организм, который содержит нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, и/или получаемые из него продукты, и/или где промотор может обеспечивать экспрессию нуклеотидной последовательности, кодирующей полипептид с конкретными свойствами, как определено здесь, в организме. Предпочтительно нуклеотидная последовательность включена в геном организма.
Термин "трансгенный организм" не подразумевает природного нуклеотида, кодирующего последовательности в их природном окружении, когда они находятся под контролем их природного промотора, который также находится в природном окружении.
Таким образом, трансгенный организм по настоящему изобретению включает в себя организм, содержащий любую из нуклеотидных последовательностей, кодирующих полипептид с конкретными свойствами, как определено здесь, конструкции, как определено здесь, векторы, как определено здесь, плазмиды, как определено здесь, клетки, как определено здесь, или их продукты или их сочетания. Например, трансгенный организм также может содержать нуклеотидную последовательность, кодирующую полипептид с конкретными свойствами, как определено здесь, под контролем гетерологичного промотора.
ТРАНСФОРМАЦИЯ ХОЗЯЙСКИХ КЛЕТОК/ОРГАНИЗМА
Как указано ранее, организм-хозяин может представлять собой прокариотический или эукариотический организм. Примеры подходящих прокариотических хозяев включают в себя E. coli и Bacillus subtilis.
Руководства о трансформации прокариотических хозяев хорошо документированы в данной области, например, см. Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd edition, 1989, Cold Spring Harbor Laboratory Press). Если используют прокариотического хозяина, тогда нуклеотидную последовательность перед трансформацией может быть необходимо подходящим образом модифицировать, например, посредством удаления интронов.
В другом варианте осуществления трансгенный организм может представлять собой дрожжи.
Клетки нитчатых грибов можно трансформировать с применением различных известных в данной области способов, таких как способ, включающий в себя формирование протопластов и трансформацию протопластов с последующей регенерацией клеточной стенки известным образом. В EP0 238 023 в качестве микроорганизма-хозяина описано применение Aspergillus.
Другой организм-хозяин может представлять собой растение. Обзор общих способов, применяемых для трансформации растений, можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant Mol Biol [1991] 42: 205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994 17-27). Дополнительные способы трансформации растений можно найти в EP-A-0449375.
Общие руководства о трансформации грибов, дрожжей и растений представлены в следующих ниже разделах.
ТРАНСФОРМИРОВАННЫЕ БАКТЕРИИ
Организм-хозяин может представлять собой бактерию, такую как Streptomyces, Bacillus subtilis или E. coli. Пригодные способы гетерологической экспрессии в E. coli описаны в WO04/064537. Пригодные способы гетерологической экспрессии в Bacillus описаны в WO02/214490. Примеры пригодных бактериальных организмов-хозяев представляют собой грамположительные виды бактерий, такие как Bacillaceae, включающие в себя Bacillus subtilis, Bacillus licheniformis, Bacillus lentus, Bacillus brevis, Bacillus stearothermophilus, Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus coagulans, Bacillus lautus, Bacillus megaterium и Bacillus thuringiensis, виды Streptomyces, такие как Streptomyces murinus, бактериальные виды, образующие молочную кислоту, включающие в себя виды Lactococcus, такие как Lactococcus lactis, виды Lactobacillus, включающие в себя Lactobacillus reuteri, виды Leuconostoc, виды Pediococcus и виды Streptococcus. Альтернативно, в качестве организма-хозяина можно выбрать штаммы грамотрицательных видов бактерий, принадлежащих к Enterobacteriaceae, включающие в себя E. coli, или к Pseudomonadaceae.
ТРАНСФОРМИРОВАННЫЕ ГРИБЫ
Организм-хозяин может представлять собой гриб, такой как нитчатый гриб. Примеры таких пригодных хозяев включают в себя любого представителя, принадлежащего родам Thermomyces, Acremonium, Aspergillus, Penicillium, Mucor, Neurospora, Trichoderma и т.п.
Руководства по трансформации нитчатых грибов рассмотрены в US-A-5741665, который указывает, что стандартные способы трансформации нитчатых грибов и культивирования грибов хорошо известны в данной области. Обширный обзор способов в отношении N. crassa находится, например, у Davis и de Serres, Methods Enzymol (1971) 17A: 79-143.
Дополнительные руководства по трансформации нитчатых грибов рассмотрены в US-A-5674707. В одном из аспектов, организм-хозяин может быть из рода Aspergillus, такой как Aspergillus niger.
Трансгенный Aspergillus по настоящему изобретению также можно получать, например, по указаниям Turner G. 1994 (Vectors for genetic manipulation. In: Martinelli S. D., Kinghorn J. R. (Editors) Aspergillus: 50 years on. Progress in industrial microbiology vol 29. Elsevier Amsterdam 1994. pp. 641-666).
Экспрессия генов в нитчатых грибах рассмотрена у Punt et al. (2002) Trends Biotechnol 2002 May; 20 (5): 200-6, Archer & Peberdy Crit Rev Biotechnol (1997) 17 (4): 273-306.
ТРАНСФОРМИРОВАННЫЕ ДРОЖЖИ
В другом варианте осуществления трансгенный организм может представлять собой дрожжи.
Обзор принципов гетерологической экспрессии генов в дрожжах предоставлен, например, в Methods Mol Biol (1995), 49:341-54, и Curr Opin Biotechnol (1997) Oct; 8 (5):554-60.
В данном отношении в качестве средства для гетерологической генной экспрессии можно применять дрожжи, такие как виды Saccharomyces cerevisi или Pichia pastoris (см. FEMS Microbiol Rev (2000 24(1): 45-66).
Обзор принципов гетерологической экспрессии генов в Saccharomyces cerevisiae и секреции продуктов генов предоставлен в E Hinchcliffe E Kenny (1993, "Yeast as a vehicle for the expression of heterologous genes", Yeasts, Vol 5, Anthony H Rose и J Stuart Harrison, eds, 2nd edition, Academic Press Ltd.).
Для трансформации дрожжей разработано несколько протоколов трансформации. Например, трансгенные Saccharomyces по настоящему изобретению можно получить по указаниям Hinnen et al., (1978, Proceedings of the National Academy of Sciences of the USA 75, 1929); Beggs, J D (1978, Nature, London, 275, 104) и Ito, H et al (1983, J Bacteriology 153, 163-168).
Трансформированные дрожжевые клетки можно отобрать с применением различных селективных маркеров, таких как ауксотрофные маркеры, маркеры доминантной устойчивости к антибиотикам.
Подходящий дрожжевой организм-хозяин можно выбрать из биологически релевантных видов дрожжей, в качестве неограничивающих примеров, таких как виды дрожжей, выбранных из видов Pichia, видов Hansenula, Kluyveromyces, видов Yarrowinia, видов Saccharomyces, включающих в себя S. cerevisiae или, видов Schizosaccharomyce, включающих в себя Schizosaccharomyce pombe.
В качестве организма-хозяина можно применять штамм вида метилотрофных дрожжей Pichia pastoris.
В одном из вариантов осуществления организм-хозяин может представлять собой виды Hansenula, такие как H. polymorpha (как описано в WO01/39544).
ТРАНСФОРМИРОВАННЫЕ РАСТЕНИЯ/КЛЕТКИ РАСТЕНИЙ
Организм-хозяин, пригодный для настоящего изобретения, может представлять собой растение. Обзор общих способов можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant Mol Biol [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994 17-27), или в WO01/16308. Например, трансгенное растение может продуцировать повышенные уровни сложных эфиров фитостеринов и сложных эфиров фитостанолов.
Таким образом, настоящее изобретение также относится к способу получения трансгенного растения с повышенными уровнями сложных эфиров фитостеринов и сложных эфиров фитостанолов, включающему в себя стадии трансформации растительной клетки липидацилтрансферазой, как определено здесь (в частности, экспрессирующим вектором или конструкцией, включающих в себя липидацилтрансферазу, как определено здесь), и выращивания растения из трансформированной растительной клетки.
СЕКРЕЦИЯ
Часто желательно, чтобы полипептид из экспрессирующего хозяина секретировался в культуральную среду, откуда фермент можно легче восстанавливать. По настоящему изобретению в зависимости от желательного экспрессирующего хозяина можно выбрать секреторную лидерную последовательность. Также в контексте настоящего изобретения можно применять гибридные сигнальные последовательности.
Обычные примеры гетерологических секреторных лидерных последовательностей представляют собой последовательности гена амилоглюкозиды (AG) грибов (glaA - версии и из 18, и из 24 аминокислот, например, из Aspergillus), гена a-фактора (дрожжи например, Saccharomyces, Kluyveromyces и Hansenula) или гена α-амилазы (Bacillus).
ДЕТЕКЦИЯ
В данной области известно множество протоколов для детекции и измерения экспрессии аминокислотной последовательности. Примеры включают в себя твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) и активируемую флуоресценцией сортировку клеток (FACS).
Специалистам в данной области известно большое множество меток и способов конъюгации и их можно применять во множестве анализов нуклеиновых кислот и аминокислот.
Ряд компаний, таких как Pharmacia Biotech (Piscataway, NJ), Promega (Madison, WI), и US Biochemical Corp (Cleveland, OH) поставляют коммерческие наборы и протоколы для этих процедур.
Подходящие репортерные молекулы или метки включают в себя радионуклиды, ферменты, флуоресцентные, хемилюминесцентные или хромогенные средства, а также субстраты, кофакторы, ингибиторы, магнитные частицы и т.п. Патенты, в которых предоставлены руководства по применению таких меток, включают в себя US-A-3817837; US-A-3850752; US-A-3939350; US-A-3996345; US-A-4277437; US-A-4275149 и US-A-4366241.
Также можно получать рекомбинантные иммуноглобулины, как указано в US-A-4816567.
ГИБРИДНЫЕ БЕЛКИ
Полипептид с конкретными свойствами, как определено здесь, можно получать в качестве гибридного белка, например, для облегчения его выделения и очистки. Примеры партнеров для гибридного белка включают в себя глутатион-S-трансферазу (GST), 6×His, GAL4 (связывающие ДНК и/или активирующие транскрипцию домены) и β-галактозидазу. Также может быть удобно вставлять между партнером по гибридному белку и представляющей интерес белковой последовательностью участок протеолитического расщепления для обеспечения удаления гибридной белковой последовательности. Предпочтительно гибридный белок не препятствует активности белковой последовательности.
Экспрессирующие системы слитых генов у E. coli рассмотрены в Curr. Opin. Biotechnol. (1995) 6 (5):501-6.
В другом варианте осуществления изобретения аминокислотную последовательность полипептида с конкретными свойствами, как определено здесь, можно лигировать с гетерологичной последовательностью для кодирования гибридного белка. Например, для скрининга пептидных библиотек на средства, способные воздействовать на активность вещества, может быть пригодным кодировать химерные вещества, экспрессирующие гетерологичный эпитоп, узнающийся коммерчески доступным антителом.
В настоящий момент изобретение будет описано посредством неограничивающих примеров со ссылкой к следующим ниже фигурам и примерам.
На фиг.1 показана консенсусная последовательность pfam00657 из базы данных версии 6 (SEQ ID NO:1);
На фиг.2 показана аминокислотная последовательность (SEQ ID NO:2), полученная из организма Aeromonas hydrophila (P10480; GI:121051). Данная аминокислотная последовательность представляет собой эталонный фермент, который может быть исходным ферментом по настоящему изобретению;
На фиг.3 показана аминокислотная последовательность (SEQ ID NO:3), полученная из организма Aeromonas salmonicida (AAG098404; GI:9964017);
На фиг.4 показана аминокислотная последовательность (SEQ ID NO:4), полученная из организма Streptomyces coelicolor A3(2) (инвентарный номер Genbank NP_631558);
На фиг.5 показана аминокислотная последовательность (SEQ ID NO:5), полученная из организма Streptomyces coelicolor A3(2) (инвентарный номер Genbank: CAC42140);
На фиг.6 показана аминокислотная последовательность (SEQ ID NO:6), полученная из организма Saccharomyces cerevisiae (инвентарный номер Genbank P41734);
На фиг.7 показано сравнение выбранных последовательностей с консенсусной последовательностью pfam00657;
На фиг.8 показано парное сравнение SEQ ID NO:3 с SEQ ID NO:2, демонстрирующее 93% идентичности аминокислотной последовательности. Сигнальная последовательность подчеркнута + отмеченные различия. Мотив GDSX, содержащий серин 16 активного центра, и аспарагиновая кислота 116, и гистидин 291 активных центров выделены (см. затененные области). Номера после аминокислот представлены за минусом сигнальной последовательности;
На фиг.9 показана нуклеотидная последовательность (SEQ ID NO:7), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Aeromonas hydrophila;
На фиг.10 показана нуклеотидная последовательность (SEQ ID NO:8), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Aeromonas salmonicida;
На фиг.11 показана нуклеотидная последовательность (SEQ ID NO:9), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Streptomyces coelicolor A3(2) (инвентарный номер Genbank NC_003888.1:8327480..8328367);
На фиг.12 показана нуклеотидная последовательность (SEQ ID NO:10), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Streptomyces coelicolor A3(2) (инвентарный номер Genbank AL939131.1:265480..266367);
На фиг.13 показана нуклеотидная последовательность (SEQ ID NO:11), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Saccharomyces cerevisiae (инвентарный номер Genbank Z75034);
На фиг.14 показана аминокислотная последовательность (SEQ ID NO:12), полученная из организма Ralstonia (инвентарный номер Genbank: AL646052);
На фиг.15 показана нуклеотидная последовательность (SEQ ID NO:13), кодирующая липидацилтрансферазу по настоящему изобретению, полученную из организма Ralstonia;
На фиг.16 показана SEQ ID NO:14. Консервативный гипотетический белок с кодом доступа NCBI белка Scoe1 CAB39707.1 GI:4539178 [Streptomyces coelicolor A3(2)];
На фиг.17 показана нуклеотидная последовательность, представленная как SEQ ID NO:15, кодирующая консервативный гипотетический белок с кодом доступа NCBI белка CAB39707.1 GI:4539178 [Streptomyces coelicolor A3(2)];
На фиг.18 показана аминокислотная последовательность, представленная как SEQ ID NO:16. Консервативный гипотетический белок с кодом доступа NCBI белка Scoe2 CAC01477.1 GI:9716139 [Streptomyces coelicolor A3(2)];
На фиг.19 показана нуклеотидная последовательность, представленная как SEQ ID NO:17, кодирующая консервативный гипотетический белок с кодом доступа NCBI белка Scoe2 CAC01477.1 GI:9716139 [Streptomyces coelicolor A3(2)];
На фиг.20 показана аминокислотная последовательность (SEQ ID NO:18) предполагаемого секретируемого белка с кодом доступа NCBI белка Scoe3 CAB88833.1 GI:7635996 [Streptomyces coelicolor A3(2)];
На фиг.21 показана нуклеотидная последовательность, представленная как SEQ ID NO:19, кодирующая предполагаемый секретируемый белок с кодом доступа NCBI белка Scoe3 CAB88833.1 GI:7635996 [Streptomyces coelicolor A3(2)];
На фиг.22 показана аминокислотная последовательность (SEQ ID NO:20) предполагаемого секретируемого белка с кодом доступа NCBI белка Scoe4 CAB89450.1 GI:7672261 [Streptomyces coelicolor A3(2)];
На фиг.23 показана нуклеотидная последовательность, представленная как SEQ ID NO:21, кодирующая предполагаемый секретируемый белок с кодом доступа NCBI белка Scoe4 CAB89450.1 GI:7672261. [Streptomyces coelicolor A3(2)];
На фиг.24 показана аминокислотная последовательность (SEQ ID NO:22) предполагаемого липобелка с кодом доступа NCBI белка Scoe5 CAB62724.1 GI:6562793 [Streptomyces coelicolor A3(2)];
На фиг.25 показана нуклеотидная последовательность, представленная как SEQ ID NO:23, кодирующая предполагаемый липобелок с кодом доступа NCBI белка Scoe5 CAB62724.1 GI:6562793 [Streptomyces coelicolor A3(2)];
На фиг.26 показана аминокислотная последовательность (SEQ ID NO:24) GDSL-липазы с кодом доступа NCBI белка Srim1 AAK84028.1 GI:15082088 [Streptomyces rimosus];
На фиг.27 показана нуклеотидная последовательность, представленная как SEQ ID NO:25, кодирующая GDSL-липазу с кодом доступа NCBI белка Srim1 AAK84028.1 GI:15082088 [Streptomyces rimosus];
На фиг.28 показана аминокислотная последовательность (SEQ ID NO:26) представляющая собой липидацилтрансферазу из Aeromonas hydrophila (ATCC № 7965);
На фиг.29 показана нуклеотидная последовательность (SEQ ID NO:27), кодирующая липидацилтрансферазу из Aeromonas hydrophila (ATCC № 7965);
На фиг.30 показана аминокислотная последовательность (SEQ ID NO:28) липидацилтрансферазы из Aeromonas salmonicida подвида Salmonicida (ATCC № 14174);
На фиг.31 показана нуклеотидная последовательность (SEQ ID NO:29), кодирующая липидацилтрансферазу из Aeromonas salmonicida подвида Salmonicida (ATCC № 14174);
На фиг.32 показано, что гомологи генов Aeromonas можно идентифицировать с применением основного средства поиска локального сравнение сервиса в National Center for Biotechnology Information, NIH, MD, USA и полных базах данных генома. При поиске в базе данных применяли мотив GDSX и идентифицировали ряд последовательностей/генов, потенциально кодирующих ферменты с липолитической активностью. Гены идентифицированы в родах Streptomyces, Xanthomonas и Ralstonia. В качестве примера ниже Ralstonia solanacearum сравнивали по гену Aeromonas salmonicida (satA). Парное сравнение показало 23% идентичности. Серин активного центра находится на аминоконце и можно идентифицировать входящие в состав каталитического центра остатки гистидина и аспарагиновой кислоты;
На фиг.33 показаны консенсусная последовательность pfam00657.11 [семейство 00657, база данных версии 11] (ниже здесь обозначаемая как консенсус pfam) и сравнение различных последовательностей с консенсусной последовательностью pfam. Стрелки указывают остатки активного центра, подчеркнутые участки указывают три участка гомологии, указанных у [Upton C and Buckley JT (1995) Trends Biochem Sci 20; 179-179]. Заглавные буквы в консенсусе pfam указывают консервативные у многих представителей семейства остатки. Символ - указывает положение, где по скрытой марковской модели консенсуса pfam ожидали найти остаток, но которого нет, так что вставляют пробел. Символ . указывает остаток без соответствующего остатка в консенсусе pfam. Последовательности представляют собой аминокислотные последовательности, представленные на фиг.16, 18, 20, 22, 24, 26, 28 и 30.
На фиг.34 показаны консенсусная последовательность pfam00657.11 [семейство 00657, база данных версии 11] (ниже здесь обозначаемая как консенсус pfam) и сравнение различных последовательностей с консенсусной последовательностью pfam. Стрелки указывают остатки активного центра, подчеркнутые участки указывают три участка гомологи указанных у [Upton C and Buckley JT (1995) Trends Biochem Sci 20; 179-179]. Заглавные буквы в консенсусе pfam указывают консервативные у многих представителей семейства остатки. Символ - указывает положение, где под скрытой марковской модели консенсуса pfam ожидали найти остаток, но которого нет, так что вставляют пробел. Символ . указывает остаток без соответствующего остатка в консенсусе pfam. Последовательности представляют собой аминокислотные последовательности, представленные на фиг.2, 16, 18, 20, 26, 28 и 30. Выявлено, что все эти белки активны в отношении липидных субстратов.
На фиг.35 показана аминокислотная последовательность (SEQ ID NO:30) гибридной конструкции, применяемой для мутагенеза гена липидацилтрансферазы Aeromonas hydrophila в примере 7. Подчеркнутые аминокислоты представляют собой сигнальный пептид ксиланазы;
На фиг.36 показана нуклеотидная последовательность (SEQ ID NO:31), кодирующая фермент липидацилтрансферазу Aeromonas hydrophila, включающий в себя сигнальный пептид ксиланазы;
На фиг.37 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из Streptomyces (SEQ ID NO:32);
На фиг.38 показана полипептидная последовательность фермента липидацилтрансферазы из Streptomyces (SEQ ID NO:33);
На фиг.39 показана полипептидная последовательность фермента липидацилтрансферазы из Termobifida (SEQ ID NO:34);
На фиг.40 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из Termobifida (SEQ ID NO:35);
На фиг.41 показана полипептидная последовательность фермента липидацилтрансферазы из Termobifida (SEQ ID NO:36);
На фиг.42 показан полипептид фермента липидацилтрансферазы из 300 а.к. Corynebacterium\effciens\GDSX (SEQ ID NO:37);
На фиг.43 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из 300 а.к. Corynebacterium\effciens\GDSX (SEQ ID NO:38);
На фиг.44 показан полипептид фермента липидацилтрансферазы из 284 а.к. Novosphingobium\aromaticivorans\GDSX (SEQ ID NO:39);
На фиг.45 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из 284 а.к. Novosphingobium\aromaticivorans\GDSX (SEQ ID NO:40);
На фиг.46 показан полипептид фермента липидацилтрансферазы из 268 а.к. Streptomyces coelicolor\GDSX (SEQ ID NO:41);
На фиг.47 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из 268 а.к. Streptomyces coelicolor\GDSX (SEQ ID NO:42);
На фиг.48 показан полипептид фермента липидацилтрансферазы из 269 а.к. Streptomyces avermitilis\GDSX (SEQ ID NO:43);
На фиг.49 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из 269 а.к. Streptomyces avermitilis\GDSX (SEQ ID NO:44);
На фиг.50 показан полипептид фермента липидацилтрансферазы из Streptomyces (SEQ ID NO:45);
На фиг.51 показана нуклеотидная последовательность, кодирующая фермент липидацилтрансферазу из Streptomyces (SEQ ID NO:46);
На фиг.52 показано ленточное представление кристаллической структуры 1IVN.PDB, обладающей глицерином в активном центре. Фигура сделана с применением программы просмотра Deep View Swiss-PDB;
На фиг.53 показана кристаллическая структура 1IVN.PDB -горизонтальная проекция с применением программы просмотра Deep View Swiss-PDB, с глицерином в активном центре - остатки в пределах 10 Å от глицерина активного центра закрашены черным;
На фиг.54 показана кристаллическая структура 1IVN.PDB - вид сверху с применением программы просмотра Deep View Swiss-PDB, с глицерином в активном центре - остатки в пределах 10 Å от глицерина активного центра закрашены черным;
На фиг.55 показано сравнение 1;
На фиг.56 показано сравнение 2;
На фиг.57 и 58 показано сравнение 1IVN по P10480 (P10480 представляет собой последовательность базы данных для фермента A. hydrophila), данное сравнение получено из базы данных PFAM и его применяют для процесса построения модели;
На фиг.59 показано сравнение, где P10480 представляет собой последовательность базы данных для Aeromonas hydrophila. Данную последовательность применяют для конструирования модели и выбора участка. Необходимо отметить, что изображен полноразмерный белок, зрелый белок (эквивалентный SEQ ID NO:2) начинается с остатка 19. A. sal представляет собой GDSX-липазу Aeromonas salmonicida (SEQ ID NO:28), A. hyd представляет собой GDSX-липазу Aeromonas hydrophila (SEQ ID NO:26). Консенсусная последовательность содержит * в положении различий между перечисленными последовательностями;
На фиг.60 показан типичный набор из 384 клонов, контроль дикого типа лежит на пересечении 0.9PC, 0.8DGDG; и
На фиг.61 показаны три представляющих интерес области. Область 1 содержит мутантов с повышенным отношением R, но с пониженной активностью в отношении DGDG. Область 2 содержит мутантов с повышенным отношением R и повышенной активностью в отношении DGDG. Область 3 содержит клоны с повышенной активностью в отношении PC или DGDG, но с повышенным отношением R.
ПРИМЕР 1
Моделирование GDSX-липазы Aeromonas hydrophila 1IVN
Сравнение аминокислотной последовательности GDSX-липазы Aeromonas hydrophila (P10480) c аминокислотной последовательностью тиоэстеразы Escherichia coli (1IVN) и аминокислотной последовательностью рамногалактуронанацетилэстеразы Aspergillus aculeatus (1DEO) получали из базы данных PFAM в формате FASTA. Сравнение P10480 и 1IVN вводили в автоматическое средство моделирования 3D-структур (сервер SWISS-MODELLER на www.expasy.org) вместе с файлом координат кристаллической структуры 1IVN.PDB фиг.52). Полученную модель для P10480 структурно сравнивали по координатам кристаллических структур 1IVN.PDB и 1DEO.PDB с применением "программы просмотра Deep View Swiss-PDB" (полученной с www.expasy.org/spdbv/) (фиг.53). Сравнение аминокислоты, полученной из базы данных PFAM (сравнение 1 - (фиг.55)), модифицировали на основе структурного сравнения 1DEO.PDB и 1IVN.PDB. Данное альтернативное сравнение аминокислот называют сравнение 2 (фиг.56).
Структура 1IVN.PDB содержит молекулу глицерина. Полагают, что эта молекула находится в активном центре, так как она находится вблизи входящих в состав каталитического центра остатков. Таким образом, можно провести выбор остатков, находящихся недалеко от активного центра, которые вследствие их близости, вероятно оказывают влияние на связывание субстрата, высвобождение продукта и/или катализ. В структуре 1IVN.PDB выбирали все аминокислоты в пределах сферы размером 10 Å, с центром в центральном углеродном атоме молекулы глицерина в активном центре (набор аминокислот 1) (см. фиг.53 и фиг.54).
Из последовательности P10480 выбирали следующие ниже аминокислоты: (1) все аминокислоты в P10480, соответствующие набору аминокислот 1 в сравнении 1; (2) все аминокислоты в P10480, соответствующие набору аминокислот 1 в сравнении 2; (3) из перекрывания модели P10480 и 1IVN все аминокислоты в модели P10480 в пределах 12 Å от молекулы глицерина в 1IVN. Все три группы совместно составляют набор аминокислот 2.
Последовательность P10480 сравнивали по "глицерофосфолипидхолестеринацилтрансферазе AAG09804.1 GI:9964017 [Aeromonas salmonicida]", а остатки в AAG09804, соответствующие набору аминокислот 2, выбирали с получением набора аминокислот 3.
Наборы 1, 2, и 3
Набор аминокислот 1:
Набор аминокислот 1 (необходимо отметить, что они представляют собой аминокислоты в 1IVN - фиг.57 и фиг.58).
Gly8, Asp9, Leu11, Ser12, Tyr15, Gly44, Asp45, Thr46, Glu69, Leu70, Gly71, Gly72, Asn73, Asp74, Gly75, Leu76, Gln106, Ile107, Arg108, Leu109, Pro110, Tyr113, Phe121, Phe139, Phe140, Met141, Tyr145, Met151, Asp154, His157, Gly155, Ile156, Pro158.
Высококонсервативные мотивы, такие как GDSX и входящие в состав каталитического центра остатки, исключены из набора 1 (подчеркнутые остатки). Во избежание неопределенности, набор 1 определяет аминокислотные остатки в пределах 10 Å от центрального углеродного атома глицерина в активном центре модели 1IVN.
Набор аминокислот 2:
Набор аминокислот 2 (необходимо отметить, что нумерация аминокислот относится к аминокислотам в зрелой последовательности P10480)
Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289 и Val290.
Таблица выбранных остатков в наборе 1 по сравнению с набором 2:

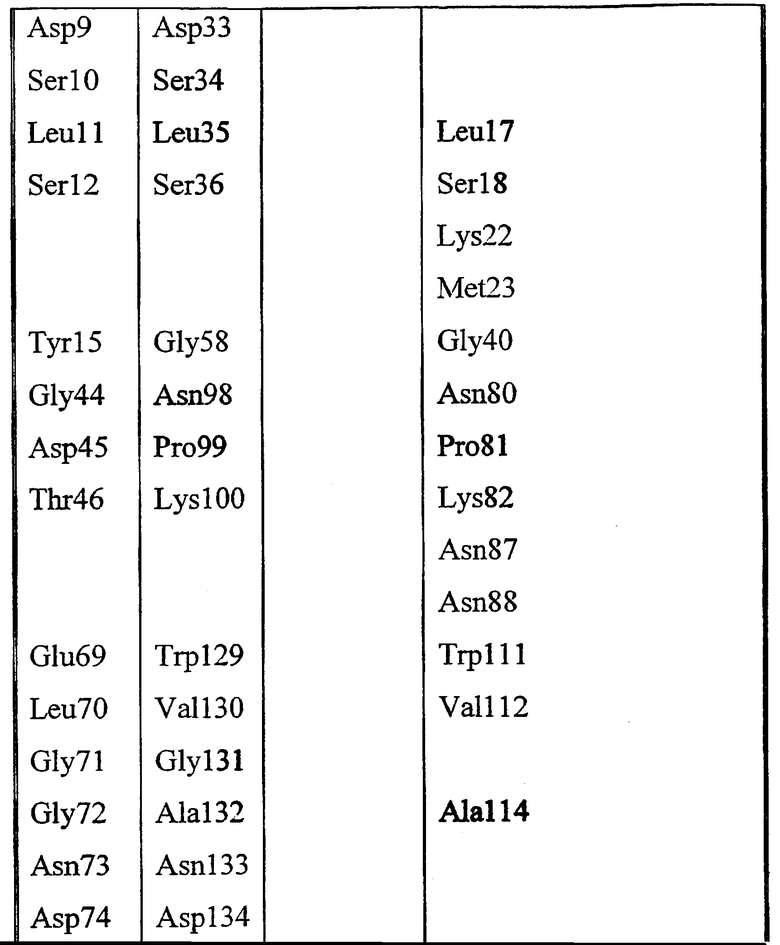
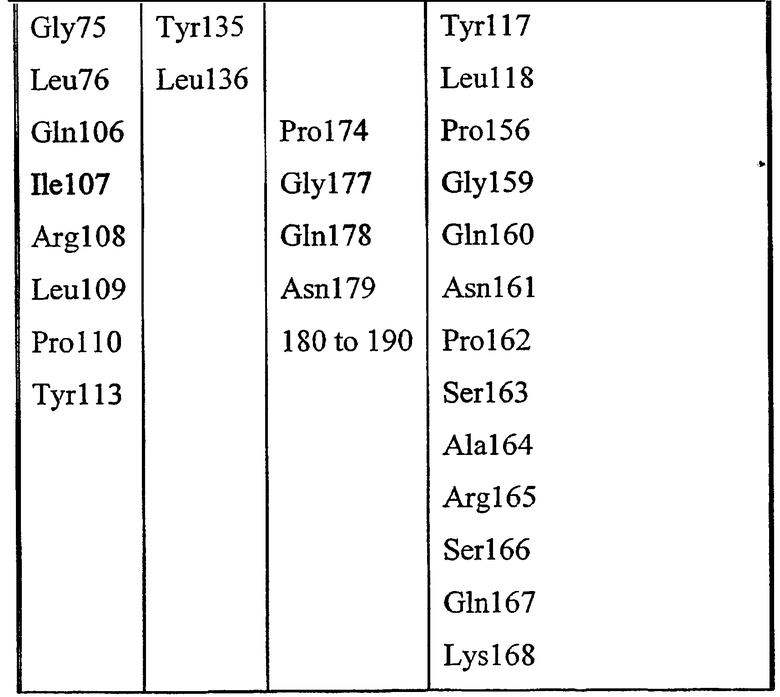
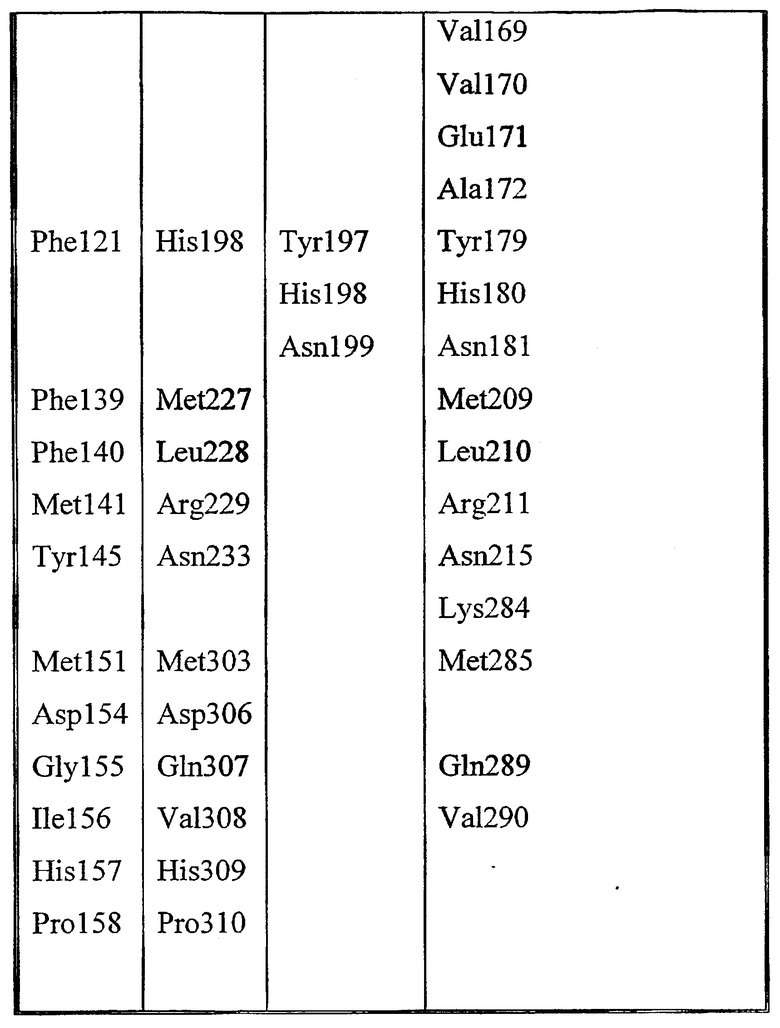
Набор аминокислот 3:
Набор аминокислот 3 идентичен набору 2, но относится к кодирующей последовательности Aeromonas salmonicida (SEQ ID NO:28), т.е. номера аминокислотных остатков в наборе 3 на 18 больше, так как это отражает разницу между нумерацией аминокислот в зрелом белке (SEQ ID NO:2) по сравнению с белком, включающим в себя сигнальную последовательность (SEQ ID NO:28).
Зрелые белки Aeromonas salmonicida GDSX (SEQ ID NO:28) и Aeromonas hydrophila GDSX (SEQ ID NO:26) отличаются по пяти аминокислотам. Они представляют собой Thr3Ser, Gln182Lys, Glu309Ala, Ser310Asn, Gly318-, где остаток salmonicida представлен первым, а остаток hydrophila представлен последним (фиг.59). Белок hydrophila состоит в длину только из 317 аминокислот и у него остаток в положении 318 отсутствует. Aeromonas salmonicidae GDSX обладает значительно более высокой активностью в отношении полярных липидов, таких как галактолипидные субстраты, чем белок Aeromonas hydrophila. Сканирование участков проводили для всех пяти положений аминокислот.
Набор аминокислот 4:
Набор аминокислот 4 представляет собой S3, Q182, E309, S310, и -318.
Набор аминокислот 5:
F13S, D15N, S18G, S18V, Y30F, D116N, D116E, D157N, Y226F, D228N, Y230F.
Набор аминокислот 6:
Набор аминокислот 6 представляет собой Ser3, Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Gln182, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289, Val290, Glu309, Ser310, -318.
Нумерация аминокислот в наборе 6 относится к аминокислотным остаткам в P10480 (SEQ ID NO:2) - соответствующие аминокислоты в основных цепях других последовательностей можно определить посредством гомологичного сравнения и/или структурного сравнения с P10480 и/или 1IVN.
Набор аминокислот 7:
Набор аминокислот 7 представляет собой Ser3, Leu17, Lys22, Met23, Gly40, Asn80, Pro81, Lys82, Asn87, Asn88, Trp111, Val112, Ala114, Tyr117, Leu118, Pro156, Gly159, Gln160, Asn161, Pro162, Ser163, Ala164, Arg165, Ser166, Gln167, Lys168, Val169, Val170, Glu171, Ala172, Tyr179, His180, Asn181, Gln182, Met209, Leu210, Arg211, Asn215, Lys284, Met285, Gln289, Val290, Glu309, Ser310, -318, Y30X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Y226X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), Y230X (где X выбран из A, C, D, E, G, H, I, K, L, M, N, P, Q, R, S, T, V или W), S18X (где X выбран из A, C, D, E, F, H, I, K, L, M, N, P, Q, R, T, W или Y), D157X (где X выбран из A, C, E, F, G, H, I, K, L, M, P, Q, R, S, T, V, W или Y).
Нумерация аминокислот в наборе 7 относится к аминокислотным остаткам в P10480 (SEQ ID NO:2) - соответствующие аминокислоты в основных цепях других последовательностей можно определить посредством гомологичного сравнения и/или структурного сравнения с P10480 и/или 1IVN.
Из кристаллической структуры можно получить классификацию вторичной структуры. Это означает, что можно классифицировать каждую аминокислоту как являющуюся частью альфа-спирали или бета-слоя. На фиг.57 показано сравнение PFAM 1DEO, 1IVN и P10480 (база данных Aeromonas hydrophila). Добавленная ниже каждая строка последовательности представляет собой структурную классификацию.
База данных PFAM содержит сравнения белков с низкой идентичностью последовательностей. Таким образом, эти сравнения не являются очень хорошими. Хотя алгоритмы сравнения (профили HAMMER) хорошо подходят для распознавания консервативных мотивов, алгоритм не очень хорош на подробном уровне. Таким образом, нахождение несоответствия между сравнением PFAM и структурным сравнением не является неожиданным. Как легко поймут специалисты, сравнение PFAM можно модифицировать на основе структурных данных. В смысле того, что можно сравнивать те структурные элементы, которые перекрываются.
На фиг.55 показано исходное сравнение PFAM 1DEO, 1IVN и P10480. К сравнению добавлена информация о вторичной структуре из кристаллических структур 1DEO и 1IVN. Сравнение 2 на фиг.56 демонстрирует модифицированное вручную сравнение, где увеличено совпадение между элементами вторичной структуры. На основе консервативных остатков у 1DEO и P10480 или у 1IVN и P10480 сравнение для P10480 также модифицировали. Для легкого различения участков последовательностей идентификаторы последовательностей в сравнении 2 имеют дополнительную m (1DEOm, 1IVNm, P10480m).
Сравнение 3 является смесью 1 и 2, оно дает сравнение по участкам.
ПРИМЕР 2: Конструирование библиотек сканирования участков
Применяли набор Quick Change Multi Site-Directed Mutagenesis Kit из Stratagene по инструкциям производителя. Для каждой библиотеки разрабатывали вырожденный праймер с одним кодоном NNK или NNS (нуклеотидные сокращения). Разработку праймеров проводили с применением средств, доступных на веб-сайте Stratagene. Контроль качества праймера дополнительно проводили с применением стандартных средств анализа, анализирующих праймер на способность к формированию шпилек или к формированию димеров праймеров.
Основные концепции способа представляют собой следующее: с применением невытесняющей цепи ДНК-полимеразы с высокой точностью, такой как Pfu-Turbo, и одного праймера линейно амплифицировали матрицу ДНК. Это отличается от обычного процесса экспоненциальной амплификации в реакции ПЦР. Данный процесс линейной амплификации обеспечивает низкую частоту ошибок. Продукт представляет собой одноцепочечную неметилированную ДНК и двухцепочечную полуметилированную ДНК. Если матрица получена из природного организма-хозяина, затем матрица становится двухцепочечной метилированной ДНК. Это означает, что матричная ДНК может быть расщеплена эндонуклеазой Dpn I без расщепления ДНК продукта. Таким образом, при трансформации ДНК в подходящего хозяина только очень низкая частота трансформантов с немутированной плазмидой.
ПРИМЕР 3: Отбор лучших вариантов из библиотеки сканирования участков
Описаны два альтернативных подхода: определение последовательности библиотеки с последующим анализом уникальных аминокислот или анализ библиотеки с определением последовательности лучших вариантов.
Отбор лучших вариантов способом 1: определение последовательности библиотеки с последующим анализом уникальных аминокислот.
Для получения библиотек/вариантов сканирования участков у E. coli применяли трансформацию/экспрессирующий челночный вектор, а экспрессию вариантов в B. subtilis получали с pDP66S, Penninaga et al., (Biochemistry (1995), 3368-3376), посредством замещения кассеты селекции на кассету селекции канамицином. Вектор применяли для вставки гена варианта ацилтрансферазы вместо гена cgt вниз от промотора P32. Вектор использует промотор P32 для управления экспрессией гена варианта ацилтрансферазы в B. subtilis.
Экспрессирующий вектор трансформировали в nprE-, aprA- Bacillus subtillis DB104 (Kawamura и Doi, J. of Bacteriology Oct 1984, p442-444) с применением способов трансформации, как описано в Chapter 3, Molecular Biological Methods for Bacillus (Ed. C. R. Harwood и S. M. Cutting), 1990. John Wiley & Sons Ltd. Chichester, UK).
Библиотеки сканирования участков конструировали с применением вырожденного олигонуклеотида, содержащего один кодон NNK, где K означает G или T, а N означает A, C, G или T. Это означает, что набор клонов, конструируемых на основании реакции амплификации с применением праймера NNK (также известные как "библиотека сканирования участков"), в принципе содержит 32 уникальных кодона (4×4×2 = 32 возможных комбинации). Не предполагая смещения ожидаемого, количество клонов, которые необходимо выбрать, чтобы иметь 95% шанс отбора каждого из 32 кодонов, по меньшей мере, один раз составляет 95. Это можно подсчитать с применением следующей формулы:
формула 1: n = {log (1-c)} / {log (1-f)}
где n представляет собой количество клонов, c представляет собой относительное значение доверительного интервала, например, 95% доверительный интервал имеет значение 0,95, а 99% доверительный интервал имеет относительное значение 0,99, и f представляет собой частоту, с которой появляется каждый кодон, которая для праймера NNK составляет 1/32 или 0,03125. Решение формулы для n дает 94,36 или 95 клонов. Если полагают, что 95% доверительный интервал является слишком маленьким или если невозможно избежать смещения на одной или нескольких стадий процесса конструирования библиотеки, можно принять решение анализировать или определять последовательность большего количества клонов. Например, в формуле 1, если n составляет 384, f составляет 1/32 или 0,03125, тогда доверительный интервал c будет значительно больше, чем 99%. Даже если 60% клонов содержат одинаковую мутацию или кодон дикого типа, то 363 клона дадут 99% уверенность получения всех 32 кодонов. Из этого можно заключить, что 384 клона дадут 99% уверенность содержания каждого из 32 кодонов, по крайней мере, один раз.
Для амплификации фрагмента из плазмиды внутри бактерии проводили ПЦР колоний (реакция ПЦР на бактериальной колонии или на жидкой бактериальной культуре), а последующее определение последовательности той части фрагмента, которую подвергали мутированию, является разработанным способом. ПЦР колоний можно проводить обычным способом для наборов из 96 образцов вследствие доступности заранее изготовленного материала (также известного как наборы) для ПЦР колоний, определения последовательностей и очистки последовательностей. Данная полная процедура предлагается как услуга несколькими коммерческими компаниями, такими как AGOWA GmbH, Glienicker weg 185, D-12489 Berlin, Germany.
После анализа 96 реакций определения последовательности, выбирали отдельные клоны, представляющие собой каждый кодон, доступный в наборе из 96 последовательностей. Затем каждый из клонов, представляющих собой мутантов, инокулировали в 5 мл бульона LB (ферментативно расщепленный казеин, 10 г/л; дрожжевой экстракт с низким содержанием натрия, 5 г/л; хлорид натрия, 5 г/л; инертные таблетированные вспомогательные средства, 2 г/л) дополненные 50 мг/л канамицина и инкубировали при 33°C в течение 6 часов при 205 об./мин. 0,7 мл этой культуры использовали для инокуляции в 50 мл субстрата SAS (K2HPO4, 10 г/л; MOPS (3-морфолинопропансульфоновая кислота), 40 г/л; хлорида натрия, 5 г/л; пеногасителя (Sin 260), 5 капель/л; обезжиренной соевой муки, 20 г/л; Biospringer 106 (100% dw YE), 20г/л), дополненного 50 мг/л канамицина и раствором высокомолекулярных гидролизатов мальтозного крахмала (60 г/л). Инкубацию продолжали в течение 40 часов при 33°C и 180 об/мин после чего супернатант культуры отделяли центрифугированием при 19000 об/мин в течение 30 мин. Супернатант переносили в чистую пробирку и непосредственно применяли для анализа.
Отбор лучших вариантов способом 2: скрининг библиотек и затем секвенирование лучших вариантов.
Хотя можно выбрать определение последовательностей 384 клонов, их также можно анализировать и перед определением последовательностей выбрать лучшие варианты.
При скрининге такого ряда образцов необходимо принять в рассмотрение ряд проблем. Не являясь исчерпывающим, несмотря на то, что можно выбрать варианты с измененной активностью для одного субстрата, различия в уровне экспрессии между 384 культурами могут быть существенными, даже если применять 384-луночный планшет для титрования, приводящий к высокому фоновому уровню. Таким образом, измерение двух видов активности и отбор лучших вариантов на основе изменения отношения является предпочтительным способом. В качестве иллюстрации, если два вида активности обладают определенным отношением R, тогда вне зависимости от присутствующего абсолютного количества фермента, отношение между двумя видами активности всегда будет R. Изменение значения R указывает на мутацию, изменяющую один вид активности относительно второго вида активности.
На фиг.60 показан набор данных, полученный из библиотеки сканирования участков. Все клоны тестировали на активность в отношении фосфатидилхолина (PC) и дигалактозилдиглицерида (DGDG). Все клоны, которые могут быть мутантными или нет, не демонстрирующие изменения значения R, лежат на прямой линии с определенной величиной ошибки. Не учитывая эти клоны на фиг.61, существуют три представляющих интерес группы.
Область 1 на фиг.61 содержит все те клоны, которые обладают значительно большим R, чем дикий тип (без мутации), но меньшей общей активностью для DGDG. Область 2 содержит те клоны, которые обладают и большим значением R, и большей активностью для DGDG, чем у дикого типа. Область 3 содержит клоны, которые не обладают повышенным значением R, но обладающие значительно большей активностью для DGDG или PC.
Если интерес представляют варианты с повышенной активностью для DGDG, тогда область 2 содержит наиболее интересные варианты и область 3 также содержит представляющие интерес варианты. Варианты в области 3, демонстрирующие большое увеличение гидролитической активности, могут сочетать уменьшение трансферазной активности.
Необходимо отметить один факт, если затронут определяющий специфичность остаток, большинство из 20 возможных аминокислот могут давать очень различные значения R. Однако, если в библиотеке присутствует большое смещение к одной аминокислоте (например, 60% представляет собой тирозин), тогда все такие варианты все еще будут лежать на прямой линии.
ПРИМЕР 4 : Анализы на активность для PC и DGDG в 384-луночном планшете для микротитрования
Исходный материал
Среды EM
Планшет для трансформантов
Планшет для дикого типа
384-луночные планшеты
сборщик колоний
Набор Waco NEFA-C
Растворы PC и DGDG в 384-луночном планшете
Часть 1 - сбор колоний
Сбор колоний в 384-луночный планшет, заполненный средой EM
Пропуск 4 лунок и инокуляция их колониями, содержащими основную цепь без мутации
Рост в течение ночи при 30°C при скорости перемешивания 200 об./мин.
Часть 2 - Инкубация на субстрате
Центрифугирование планшетов после роста в течение ночи; 2500 об/мин, 20 мин.
Перенос 10 мкл супернатанта из каждой лунки в 2 пустых 384-луночных планшета
Добавление 5 мкл 12,5 мМ DGDG в один из планшетов, добавление 5 мкл 12,5 мМ PC в другой планшет
Инкубация обоих планшетов в течение 2 часов при 37°C, перемешивание на старте для смешивания, а затем остановка перемешивания
Продолжение с применением процедуры NEFA-C
Часть 3 - процедура NEFA-C
Добавление 10 мкл раствора A
Инкубация 10 мин при 37°C, 300 об/мин
Добавление 20 мкл раствора B
Инкубация 10 мин при 37°C, 300 об/мин
Чтение планшета при 550 нм
Состав субстрата - в мМ
25 мМ PC или DGDG
10 мМ CaCl2
60 мМ Triton X 100
15 мМ NaN3
20 мМ Briton Robinson pH 5,0
ПРИМЕР 5 Выбранные варианты
Определение ферментативной активности
Для определения ферментативной активности для различных субстратов 4 мкл раствора фермента инкубировали с 11 мкл субстрата в течение 60 минут при 37°C. Затем с применением набора WACO NEFA-C определяли количество свободных жирных кислот. К 15 мкл смеси фермента и субстрата добавляли 75 мкл раствора A NEFA и инкубировали в течение 15 минут при 37°C. Затем добавляли 150 мкл раствора B NEFA и инкубировали в течение 15 минут. Затем измеряли оптическую плотность (OD) образца при 550 нм.
В качестве контроля из каждого варианта 4 мкл раствора фермента инкубировали с 11 мкл буфера HEPES в течение 60 мин при 37°C. Затем определяли количество свободных жирных кислот, как описано выше. Значения OD данного контрольного образца вычитали из наблюдаемого для каждого субстрата OD для получения скорректированной активности.
Применяли четыре различных субстрата, где состав в целом составлял 30 мг липида, 4,75 мл 50 мМ буфера HEPES pH 7, 42,5 мкл 0,6 М CaCl2, 200 мкл 10% Triton X-100 без H2O2. 30 мг липида представляли собой или фосфатидилхолин (PC), PC с холестерином в отношении 9 к 1, дигалактозилдиглицерид (DGDG), или DGDG с холестерином в отношении 9 к 1.
Отбор улучшенных вариантов
Варианты с повышенной активностью для PC
Те варианты, которые продемонстрировали увеличение OD по отношению к дикому типу фермента при инкубации на PC, выбирали как варианты с повышенной фосфолипазной активностью.
Варианты с повышенной активностью для DGDG
Те варианты, которые продемонстрировали увеличение OD по отношению к дикому типу фермента при инкубации на DGDG выбирали как варианты с повышенной активностью для DGDG.
Варианты с повышенной специфичностью для DGDG
Специфичность для DGDG представляет собой отношение активности для DGDG к активности для фосфатидилхолина (PC). Те варианты, которые продемонстрировали повышенное отношение DGDG к PC, по сравнению с диким типом отбирали как варианты с улучшенной специфичностью для DGDG.
Варианты с повышенной трансферазной активностью для PC в качестве донора ацила
Различие в количестве свободных жирных кислот, образуемое при инкубации фермента на PC и на PC с холестерином, представляет собой указание на степень трансферазной активности относительно степени гидролитической активности. Трансферазная активность не приводит к образованию свободных жирных кислот. Преимущество трансферазы представляет собой отношение свободных жирных кислот, образуемых, когда в качестве субстрата используют PC к свободным жирным кислотам, образуемым, когда в качестве субстрата используют PC с холестерином. Те варианты, которые демонстрируют увеличение преимущества трансферазы и демонстрируют большую, чем у дикого типа активность для PC, отбирают в качестве обладающих повышенной трансферазной активностью.
Варианты с повышенной трансферазной активностью для DGDG в качестве донора ацила
Различие в количестве свободных жирных кислот, образуемое при инкубации фермента на DGDG и на DGDG с холестерином, представляет собой указание на степень трансферазной активности относительно степени гидролитической активности. Трансферазная активность не приводит к образованию свободных жирных кислот. Преимущество трансферазы представляет собой отношение свободных жирных кислот, образуемых, когда в качестве субстрата используют DGDG к свободным жирным кислотам, образуемым, когда в качестве субстрата используют DGDG с холестерином. Те варианты, которые демонстрируют увеличение преимущества трансферазы и демонстрируют большую, чем у дикого типа активность для DGDG, отбирают в качестве обладающих повышенной трансферазной активностью.
Выбранные варианты
Для каждого из четырех критериев отбора, приведенных выше, отбирали ряд вариантов.
Фермент "дикого типа" в данном примере представляет собой фермент A. salmonicida (SEQ ID NO:28).
Варианты с повышенной активностью для PC
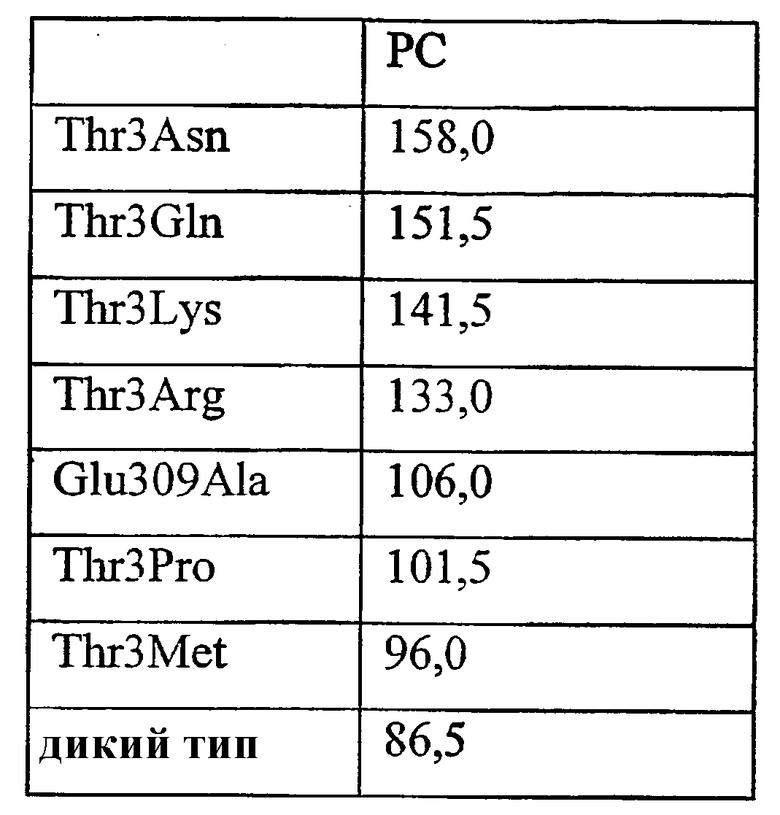
Варианты с повышенной активностью для DGDG
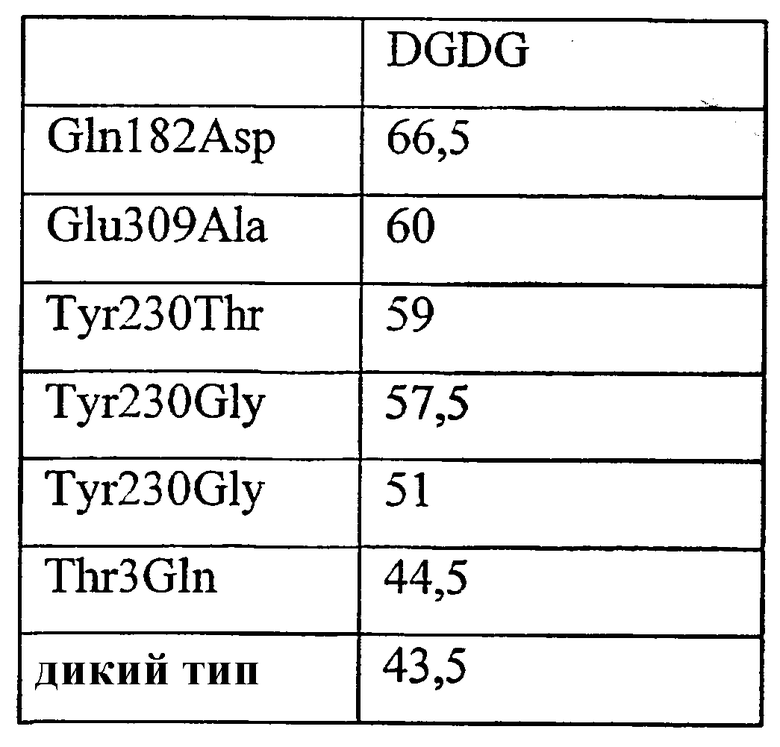
Варианты с повышенной специфичностью для DGDG
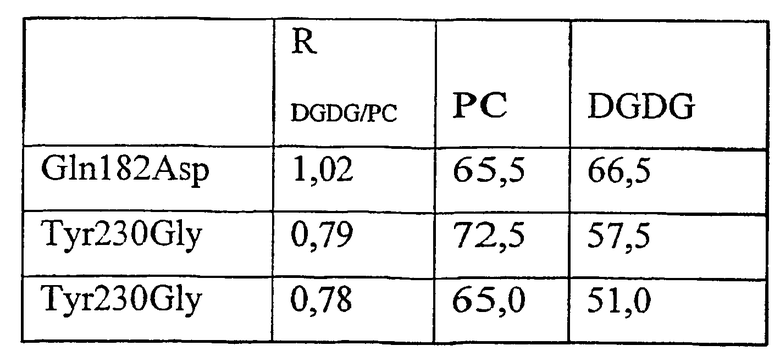
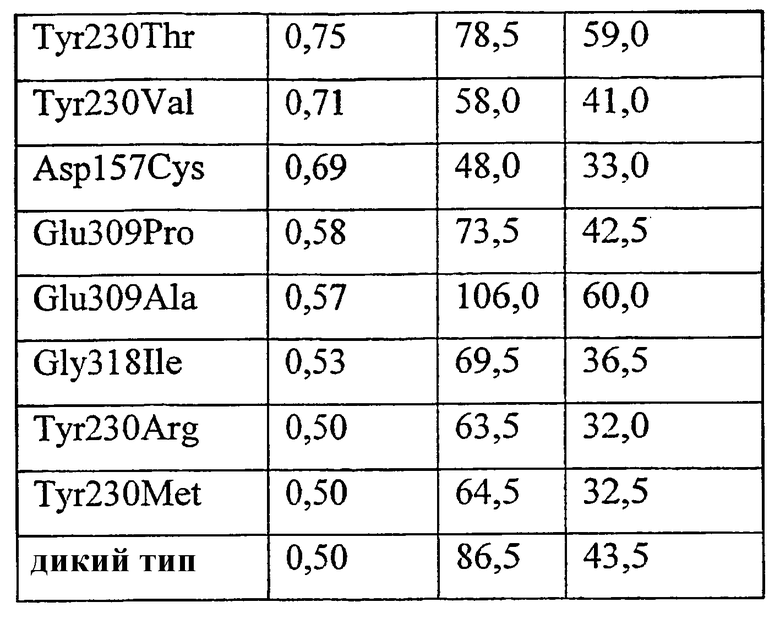
Варианты с повышенной трансферазной активностью для PC в качестве донора ацила
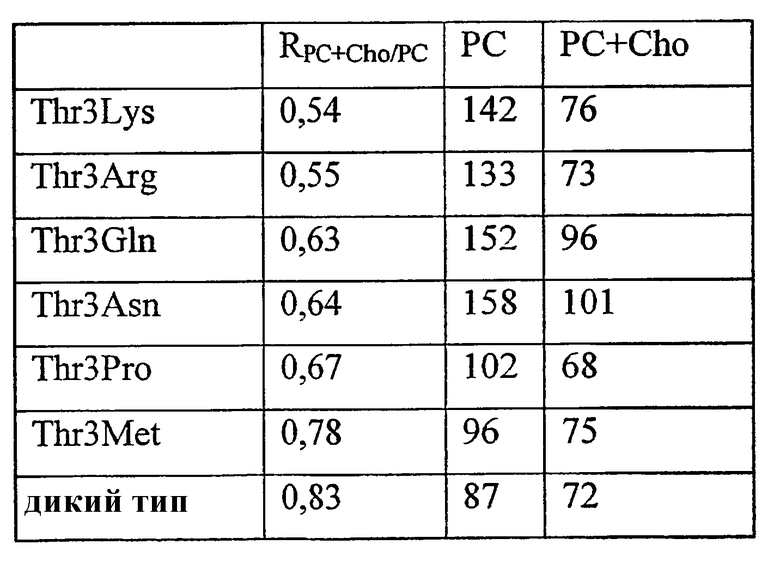
Варианты с повышенной трансферазной активностью для DGDG в качестве донора ацила
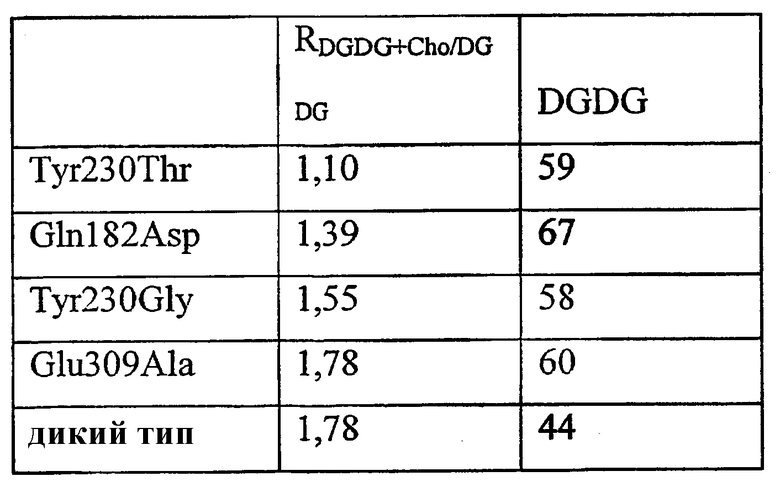
ПРИМЕР 6: Анализ трансферазной активности для фосфолипида:холестерина
Фосфолипид можно заменить на DGDG для обеспечения анализа трансферазы для галактолипида. Также в этом же анализе можно использовать другие акцепторы, например, глицерин, глюкозу, гидроксикислоты, белки или мальтозу.
300 мг фосфатидилхолина (Avanti №441601):холестерина (Sigma C8503) 9:1 помещали в пробирку Уитона. Добавляли 10 мл 50 мМ буфера HEPES pH 7,0 и перемешиванием при 40°C распределяли субстрат.
0,5 мл субстрата переносят в 4 мл флакон и помещают в нагревательное устройство при 40°C. Добавляют 0,05 мл раствора трансферазы, а также тем же способом анализируют контроль с 0,050 мл воды. Реакционную смесь перемешивают в течение 4 часов при 40°C. Затем образец замораживают и лиофилизируют, и анализируют посредством GLC.
Подсчет:
На основании анализа GLC подсчитывают содержание свободных жирных кислот и сложного эфира холестерина.
Ферментативную активность подсчитывают как:
% Трансферазной активности =
Δ % сложного эфира холестерина/(Mv сложного эфир стерина) × 100
Δ % сложного эфира холестерина/(Mv сложного эфира холестерина) + Δ % жирной кислоты/(Mv жирной кислоты)
% Гидролитической активности =
Δ % жирной кислоты/(Mv жирной кислоты) × 100
Δ % сложного эфира холестерина/(Mv сложного эфира холестерина)+ Δ %жирной кислоты/(Mv жирной кислоты)
Отношение трансферазной активности/гидролиза = % трансферазной активности/% гидролитической активности,
где
Δ % сложного эфира холестерина = % сложного эфира холестерина (образец) - % сложного эфира холестерина (контроль).
Δ % жирной кислоты = % жирной кислоты (образец) - % жирной кислоты (контроль).
Анализ трансферазной активности для галактолипида:холестерина.
300 мг дигалактозилдиглицерида (DGDG) (чистота > 95% галактолипида, применяемый DGDG очищали из пшеничных липидов. DGDG из Sigma D4651 также пригоден для применения):холестерин (Sigma) 9:1 помещали в пробирку Уитона. Добавляли 10 мл 50 мМ буфера HEPES pH 7,0 и перемешиванием при 40°C распределяли субстрат.
0,5 мл субстрата переносят в 4 мл флакон и помещают в нагревательное устройство при 40°C. Добавляют 0,05 мл раствора трансферазы, а также тем же способом анализируют контроль с 0,050 мл воды. Реакционную смесь перемешивают в течение 4 часов при 40°C. Затем образец замораживают и лиофилизируют, и анализируют посредством GLC.
Подсчет:
На основании анализа GLC подсчитывают содержание свободных жирных кислот и сложного эфира холестерина.
Ферментативную активность подсчитывают как:
% Трансферазной активности =
Δ % сложного эфира холестерина/(Mv сложного эфир стерина) × 100
Δ % сложного эфира холестерина/(Mv сложного эфира холестерина) + Δ % жирной кислоты/(Mv жирной кислоты)
% Гидролитической активности =
Δ % жирной кислоты/(Mv жирной кислоты) × 100
Δ % сложного эфира холестерина/(Mv сложного эфира холестерина) + Δ % жирной кислоты/(Mv жирной кислоты)
Отношение трансферазной активности/гидролиза = % трансферазной активности/% гидролитической активности
где
Δ % сложного эфира холестерина = % сложного эфира холестерина (образец) - % сложного эфира холестерина (контроль).
Δ % жирной кислоты = % жирной кислоты (образец) - % жирной кислоты (контроль).
ПРИМЕР 7 : Варианты липидацилтрансферазы для Aeromonas hydrophila (SEQ ID NO:26)
Мутации вносили с применением набора QuikChange™ Multi-Site Directed Mutagenesis из Stratagene, La Jolla, CA92037, USA по инструкциям, предоставленным Stratagene.
Варианты с Tyr256 продемонстрировали повышенную активность для фосфолипидов.
Варианты с Tyr256 и с Tyr260 продемонстрировали повышенную активность для галактолипидов.
Варианты с Tyr265 продемонстрировали повышенную трансферазную активность с галактолипидами в качестве донора ацила.
Номера указывают положения на следующей последовательности: фермент из Aeromonas hydrophila, аминокислотная последовательность которого представлена как SEQ ID NO:26. Нуклеотидная последовательность является такой, как представленная как SEQ ID NO:27.
ПРИМЕР 8 : Скрининг мутантов глицерофосфолипид:холестеринацилтрансферазы GCAT из Aeromonas salmonicida.
Мутанты с точковыми мутациями глицерофосфолипид:холестеринацилтрансферазы GCAT из Aeromonas salmonicida скринировали на трансферазную активность с применением фосфатидилхолина или дигалактозилдиглицерида в качестве донора и холестерина в качестве акцептора с целью отбора мутанта с улучшенной активностью для дигалактозилдиглицерида по сравнению с фосфатидилхолином.
Мутанты GCAT скринировали на трансферазную активность с применением дигалактозилдиглицерида (DG) и фосфатидилхолина (PC) в качестве донора и холестерин в качестве акцептора.
DG (чистота > 95% дигалактозилдиглицерида (применяемый DGDG очищали из липидов пшеницы. DGDG из Sigma D4651 также пригоден для применения из Signma D4651) и холестерин (Sigma C8503) помещали в отношении 9:1 и растворяли в хлороформе и выпаривали до сухости.
Субстрат получали растворением 3% DG:холестерина 9:1 в 50 мМ буфере HEPES pH 7.
0,250 мл субстрата переносили в 3 мл пробирку с закручивающейся крышкой. Добавляли 0,025 мл супернатанта из культуры мутанта GCAT и инкубировали при 40°C в течение 2 часов. Также получали эталонный образец с водой вместо фермента. Ферментативную реакцию останавливали нагреванием реакционной смеси в бане с кипящей водой в течение 10 минут.
Добавляли 2 мл 99% этанола и подвергали анализу на холестерин, а также анализу на свободную жирную кислоту.
Анализ на холестерин.
100 мкл субстрата, содержащего: 1,4 Ед/мл холестериноксидазы (SERVA Electrophoresis GmbH кат. № 17109), 0,4 мг/мл ABTS (Sigma A-1888), 6 Ед/мл пероксидазы (Sigma 6782) в 0,1 М буфере TRIS, HCl буфер pH 6,6 и 0,5% Triton X 100 (Sigma X-100) инкубировали при 37°C в течение 5 минут. Добавляли 5 мкл образца холестерина и перемешивали. Реакционную смесь инкубировали в течение дополнительных 5 минут и измеряли OD при 405 нм. Содержание холестерина подсчитывали на основании анализа стандартных растворов холестерина, содержащих 0,4 мг/мл, 0,3 мг/мл, 0,20 мг/мл, 0,1 мг/мл, 0,05 мг/мл и 0 мг/мл.
Анализ на свободную жирную кислоту.
Свободные жирные кислоты в образце измеряли с применением набора NEFA-C (WAKO Chemicals GmbH)
75 мкл реагента A NEFA инкубировали в течение 10 минут при 37°C. Добавляли 15 мкл образца фермента и перемешивали. Реакционную смесь инкубировали в течение 10 минут. Добавляли 150 мкл реагента B NEFA, перемешивали и инкубировали в течение дополнительных 10 минут и измеряли OD при 540 нм. Свободную жирную кислоту подсчитывали на основании стандартных растворов 0,4, 0,3, 0,2, 0,1, 0,05 и 0 мМ жирной кислоты.
Анализ трансферазной активности с применением фосфатидилхолина в качестве донора проводили тем же способом, но с применением фосфатидилхолина (Avanti №441601) вместо DG (DGDG).
Трансферазную активность выражали как % этерифицированного холестерина, подсчитанный на основании разницы свободного холестерина в эталонном образце и свободного холестерина в образце с ферментом.
Гидролитическую активность выражали как % образуемой свободной жирной кислоты, подсчитанный на основании разницы свободной жирной кислоты в образце с ферментом и свободной жирной кислоты в эталонном образце.
Относительную трансферазную активность для DG и PC подсчитывали как %TDG/TPC.
Для мутантов подсчитывали трансферазную активность TDG относительно гидролитической активности HDG для DG

где 386 = MW холестерина, а 280 = MW жирной кислоты.
Мутанты с TDG > 50% и TDG/TPC > 3 и
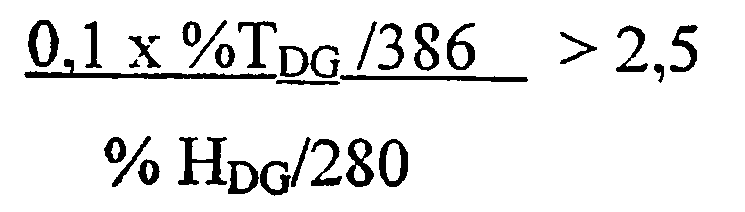
выбирали как улучшенные мутанты.
Данные, полученные из указанного выше примера, можно анализировать статистическими методами для идентификации и определения ключевых участков и/или конкретных аминокислотных замен, обеспечивающих желательный профиль активности, такой как повышенное отношение TDG к TPC. Например, предложена следующая надежная модель:
G40 L
N87 R, D, E, M
Y117 A, N, E, H,
Т
Q182K, T
M209K, M
L210 N
R211 G
N215 H
Y230 I
-318Y, H или S
N215 H
L210D, Q или T
E 309S, Q или R
H180K или Q
N 80 N, R или D
L210 G, I, H, E,
M, S, W, V, A, R,
N,
S310A, P, T, H, M,
K или G
V112C
Y30 G, I, L, S, E,
M, A или R
V290R, E, H или A
Q289R или N
K22E
G40L
Y179V
M209 L, К, М
L211 G, Q, K или D
Y230 V
S310 P
Y179 R
H180 T
Q289 T или D
G40 Q, L или V
N88 W
N87 R или D
S310Q, H или S
S3 E, A, G, K, M, Y,
R, P, N, T, или Q
-318 R, S, E, H, Q,
N или D
N215 L, G, V, R или
Y
K82S
N215 G
L210 D, H, R, E, A, Q, P, N, K, G, R,T, W, I, V или S
N80 G
Y30 L
N87 G
H180 I, T
M209Y
R211D, T или G
S18G, M или T
G40 R или M
N88 W
N87 С, D, R, E или G
ПРИМЕР 9 : Отбор улучшенных мутантов глицерофосфолипид:холестеринацилтрансферазы GCAT из Aeromonas salmonicida
"Исходный" фермент в данном примере представляет собой фермент A. salmonicida (SEQ ID NO:28).
32 положения GCAT из Aeromonas salmonicida (230 Tyr, 182 Lys, 3 Thr, 157 Asp, 310 Thr, 318 Gly, 309 Glu, 17 Leu, 111 Trp, 117 Tyr, 179 Tyr, 118 Leu, 215 Asn, 22 Lys, 290 Val, 289 Gln, 285 Met, 18 Ser, 23 Met, 180 His, 284 Lys, 181 Asn, 209 Met, 210 Leu, 211 Arg, 40 Gly, 81 Pro, 112 Val, 80 Asn, 82 Lys, 88 Asn, 87 Asn скринировали по схеме эксперимента из примера 8.
На основании результатов скринирования и трех критериев отбора отобрали следующих мутантов, перечисленных в таблице 1
Таблица 1
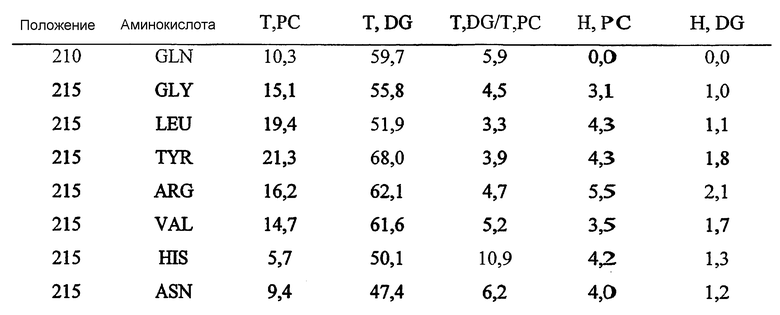
ПРИМЕР 10 : Отбор конкретных представляющих интерес аминокислотных областей для внесения мутаций глицерофосфолипид:холестеринацилтрансферазы GCAT из Aeromonas salmonicida
На основании сравнения pfam (сравнение 2; фиг.56) и перекрывания модель P10480 и 1IVN выбирали все аминокислоты в областях, окружающих молекулу глицерина в активном центре 1IVN, и использовали для определения областей особенного интереса (петли). (Номера относятся к аминокислотам в зрелой последовательности P10480 (SEQ ID NO:2)):
Thr 20-Arg 41 (Петля 1, L1)
Ile 77 - Leu 89 (Петля 2, L2)
Leu 118 - Asp 127 (Петля 3, L3)
Gly 146 - Val 176 (Петля 4, L4)
Glu 208 - Trp 287 (Петля 5, L5)
Промежуточные области (IVR) назвали следующим образом:
Ala 1 - Asp 19 (IVR1)
Phe 42 - Lys 76 (IVR2)
Asp 90 - Tyr 117 (IVR3)
Ala 128 - Asn 145 (IVR4)
Ser 177 - Ala 207 (IVR5)
Asp 288 - His 317 (IVR6)
Следующая таблица суммирует распределение предпочтительных положений для внесения мутаций глицерофосфолипид:холестеринацилтрансферазы GCAT из Aeromonas salmonicida. Результаты основаны на экспериментальных схемах, приведенных в примерах 8-10.
ные области для способов по изобретению
Y 30
V112,
W111,
A114
Q182,
H180,
Y179
L210,
R211,
N215,
Y230
R211, N215,
K284, M285,
Q289, V290
V290, E309, S310, -318
Все публикации, указанные в предоставленном выше описании, включены сюда в качестве ссылки. Специалистам в данной области будут очевидны различные модификации и вариации описанных способов и систем по настоящему изобретению без отклонения от области и сущности настоящего изобретения. Хотя настоящее изобретение описано применительно к конкретным предпочтительным вариантам изобретения, следует понимать, что заявленное изобретение, не следует излишне ограничивать такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления изобретения, которые очевидны специалистам в области биохимии и биотехнологии или связанных областей, должны находиться в пределах объема следующей ниже формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ГЛИКОЛИПИДАЦИЛТРАНСФЕРАЗЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2004 |
|
RU2377300C2 |
| СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА УГЛЕРОДА, СЛОЖНОГО ЭФИРА БЕЛКА, СЛОЖНОГО ЭФИРА БЕЛКОВОЙ СУБЪЕДИНИЦЫ ИЛИ СЛОЖНОГО ЭФИРА ГИДРОКСИКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЛИПИДАЦИЛТРАНСФЕРАЗЫ | 2004 |
|
RU2371474C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| СПОСОБ | 2004 |
|
RU2376868C2 |
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |
| БЕЛОК | 2005 |
|
RU2380414C2 |
| ВАРИАНТНЫЕ ФИТАЗНЫЕ ФЕРМЕНТЫ | 2005 |
|
RU2421520C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |

























































































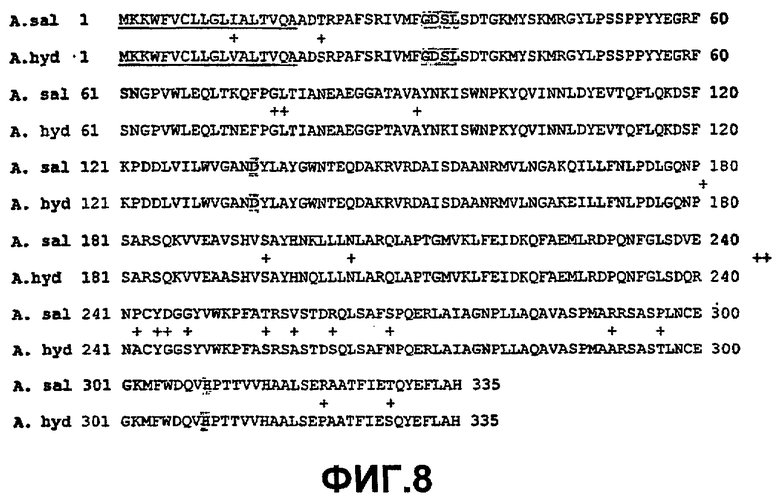
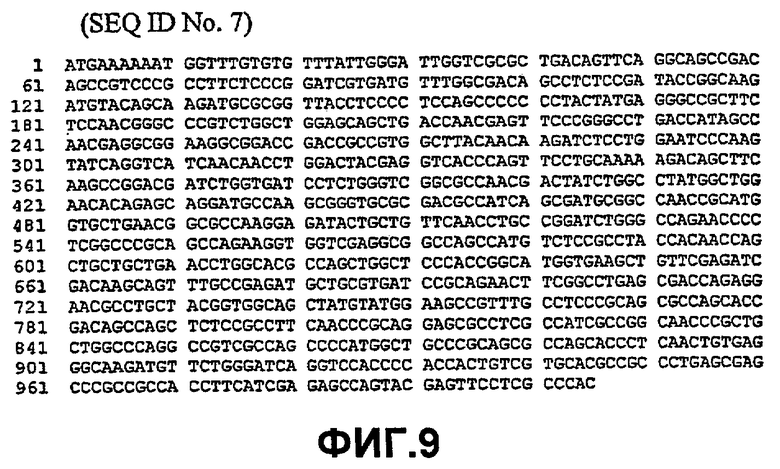
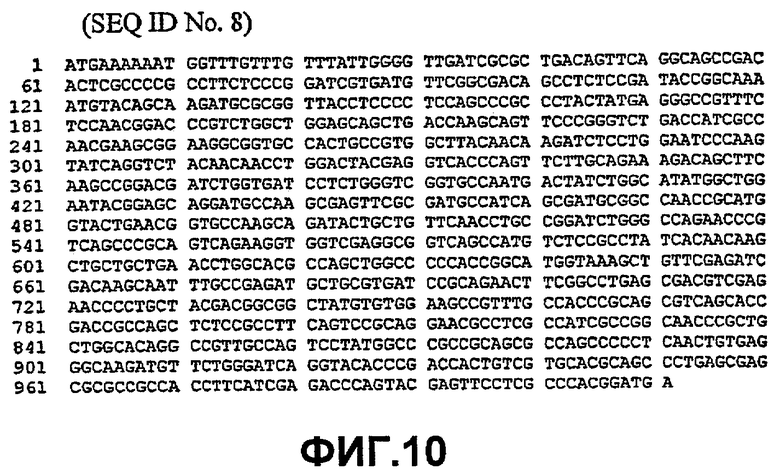
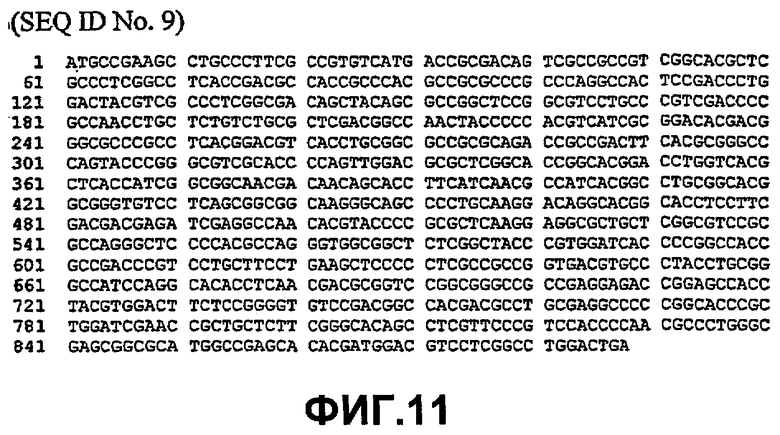
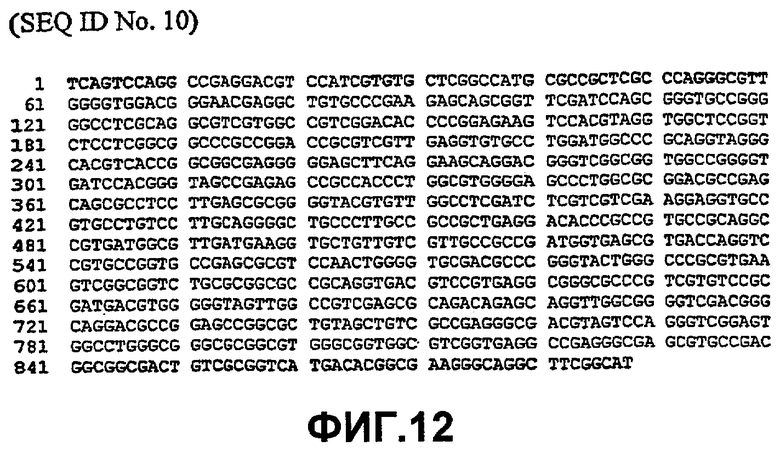
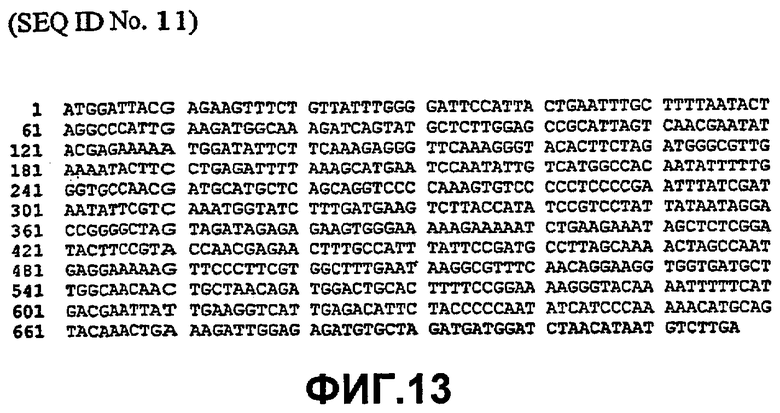









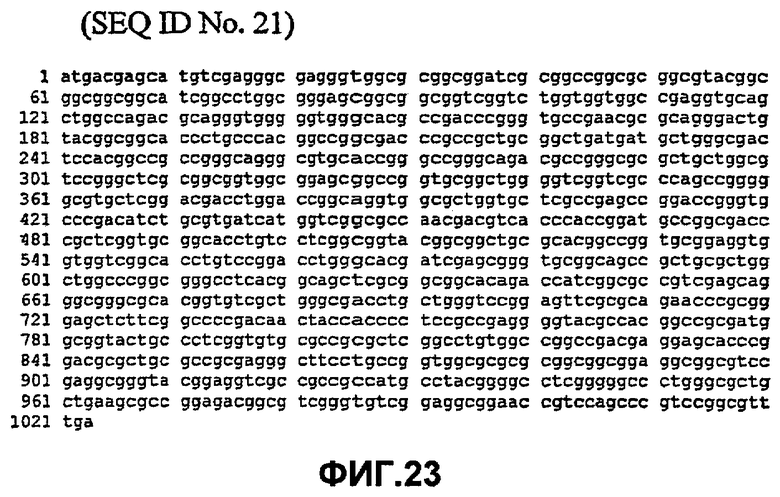









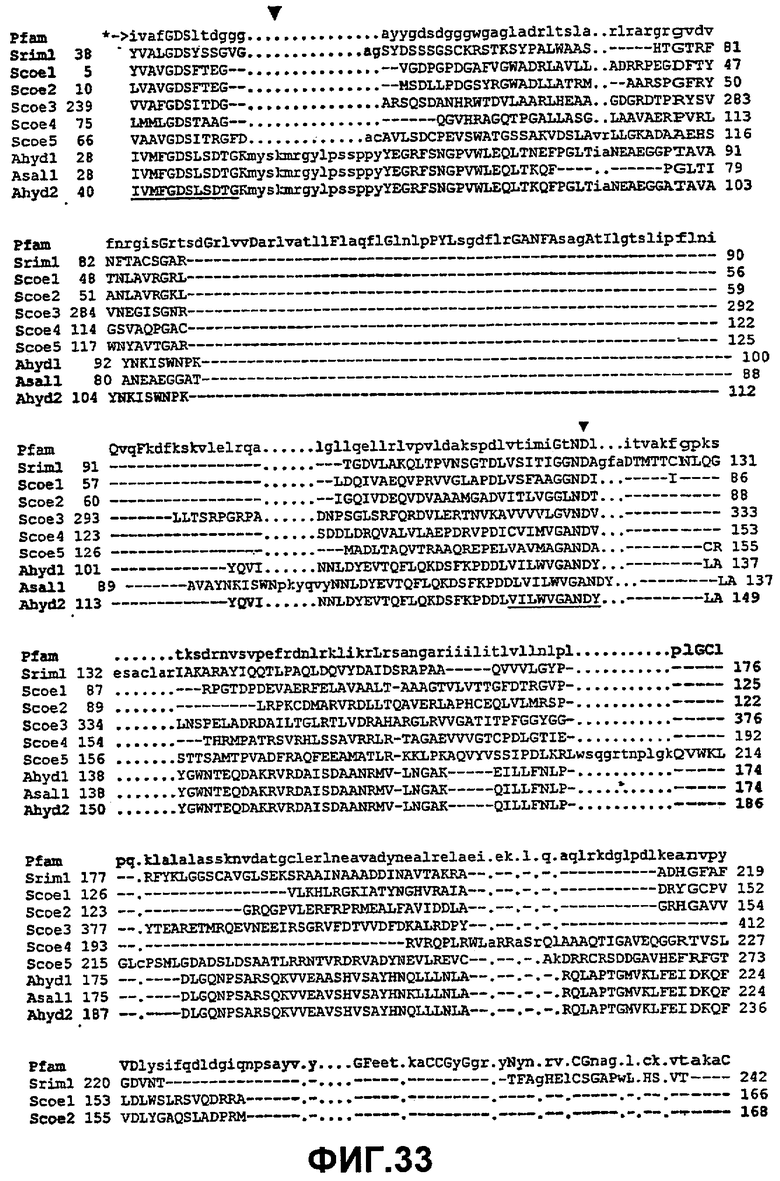




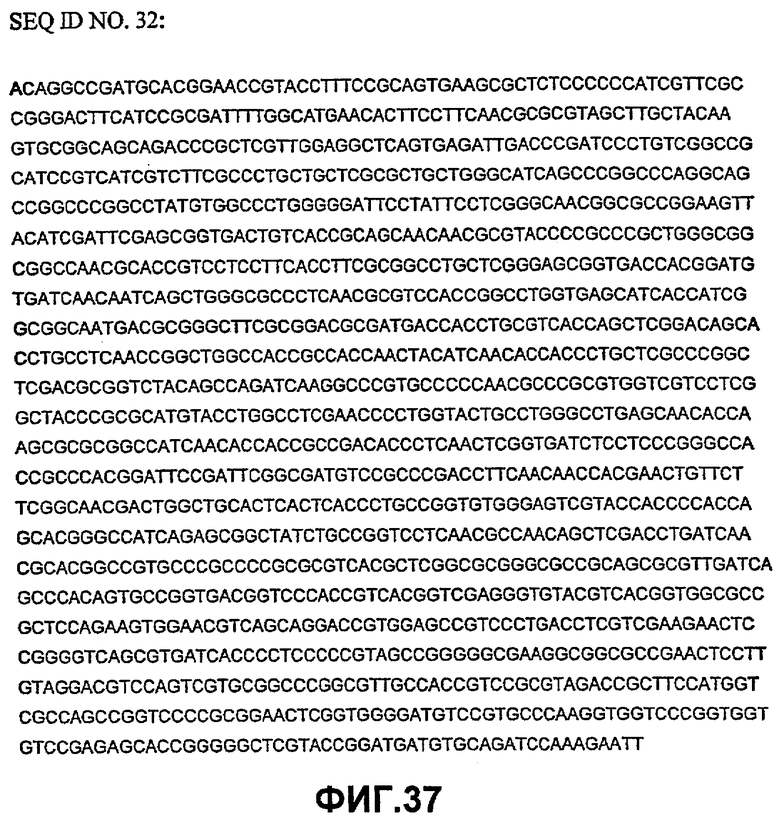














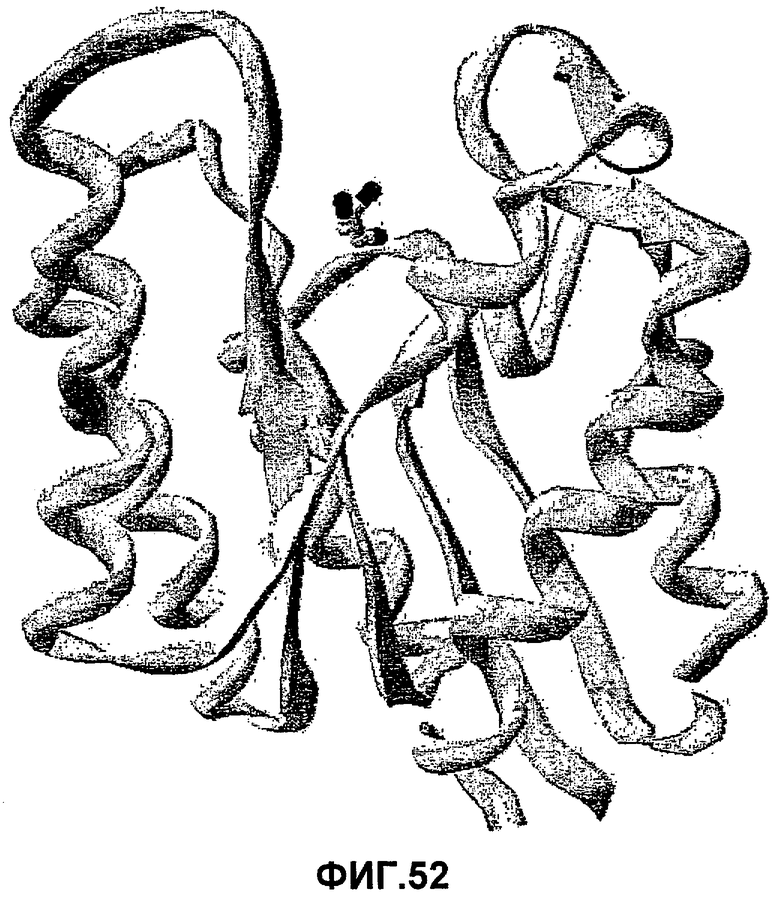
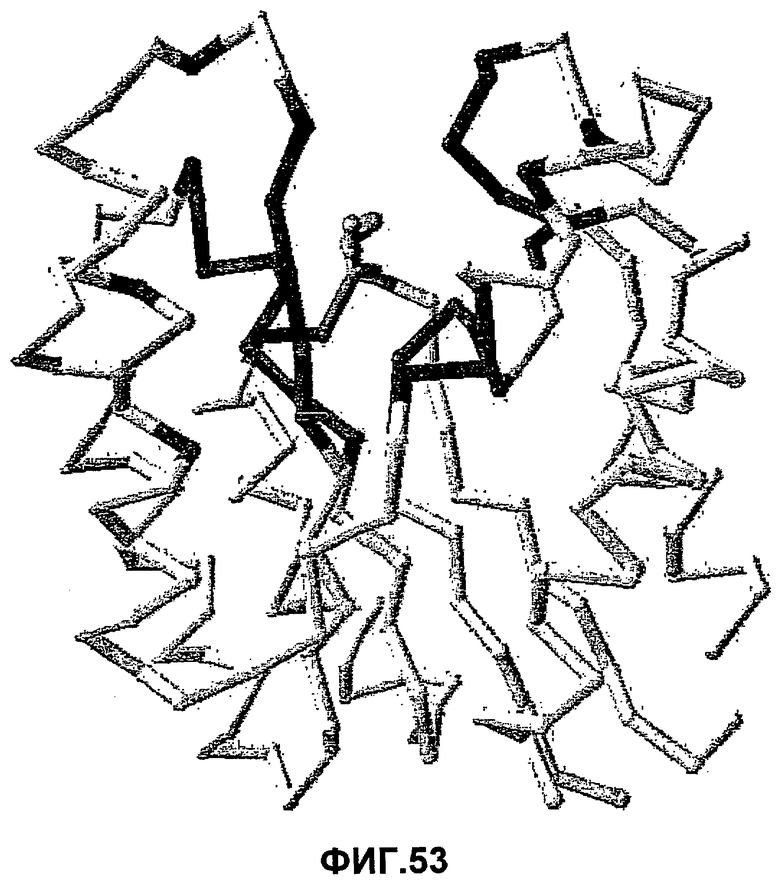




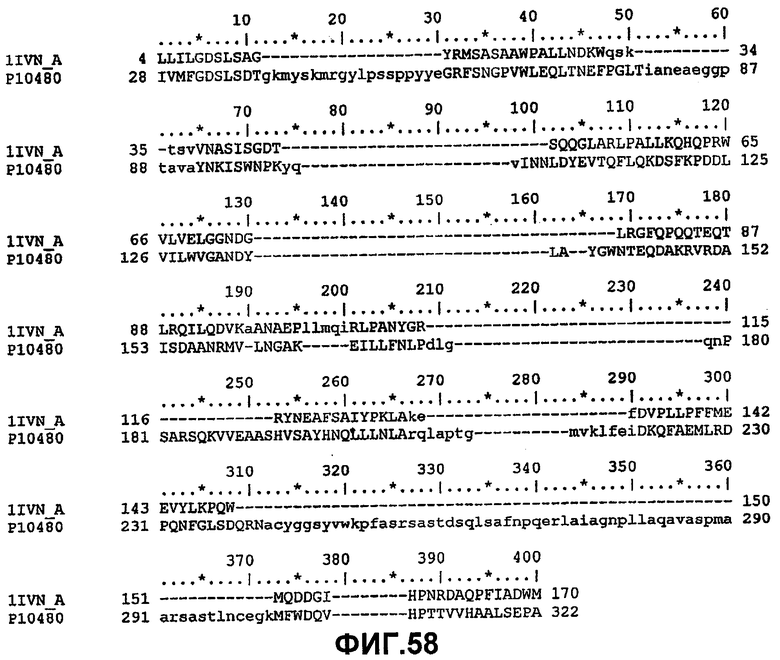


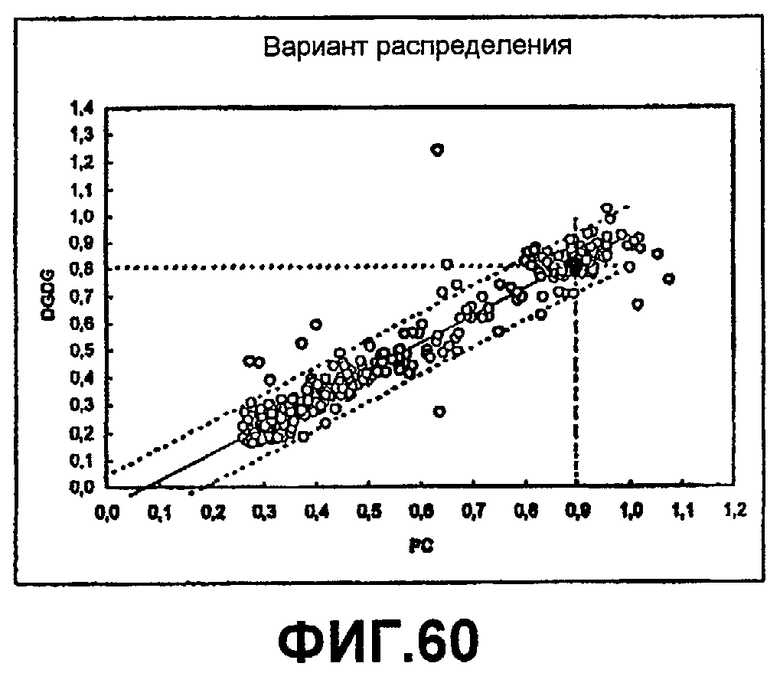
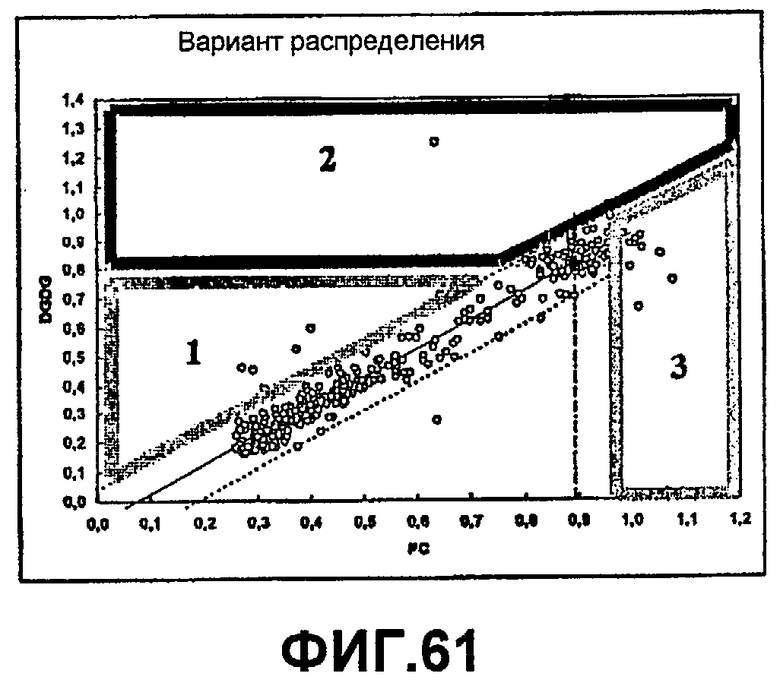
Изобретение относится к биотехнологии, в частности к варианту фермента липидацилтрансферазы, с повышенной фосфолипидтрансферазной активностью. Полученный фермент содержит аминокислотный мотив GDSX, где Х представляет собой один или несколько из следующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S и модификацию одной или нескольких аминокислот по сравнению с исходным ферментом. Данный фермент используют для получения пищевых продуктов, а также для уменьшения содержания фосфолипидов в пищевом масле. Изобретение позволяет получить эффективный вариант фермента с повышенной фосфолипидтранферазной активностью. 8 н.п. ф-лы, 61 ил., 1 табл., 10 пр.
1. Вариант фермента липидацилтрансферазы с повышенной фосфолипидтрансферазной активностью, отличающийся тем, что фермент содержит аминокислотный мотив GDSX, где X представляет собой один или несколько следующих аминокислотных остатков L, А, V, I, F, Y, Н, Q, Т, N, М или S, где вариант фермента содержит одну или несколько аминокислотных модификаций по сравнению с родительской последовательностью в любом, одном или нескольких, аминокислотном остатке:
S3N, Е, К, R, А, Р или М;
D157S, R, Е, G, Т, V, Q, К или С;
318Е;
E309R, Е, L, R или А;
Y179D, Т, Е, R, N, V, К, Q или S;
N215S, L, R или Y;
К22Е, R, С или А
Q289R, Е, G, Р или N;
М23К, Q, L, G, Т или S;
H180Q, R или К;
M209Q, S, R, A, N, Y, Е, V или L;
L210R, А, V, S, Т, I, W или М;
R211T;
P81G;
V112C;
N80R, G, N, D, Р, Т, Е, V, А или G;
L82N, S или Е;
N88C;
N87M или G,
где нумерация получена путем выравнивания последовательности варианта и эталонной последовательности SEQ ID NO:2, и где родительская последовательность получена из Aeromonas salmonicida.
2. Способ получения пищевого продукта, включающий добавление варианта фермента липидацилтрансферазы по п.1 к одному или нескольким ингредиентам пищевого продукта.
3. Способ получения выпеченного из теста продукта, включающий добавление варианта фермента липидацилтрансферазы по п.1 в тесто.
4. Применение варианта фермента липидацилтрансферазы по п.1 в способе обработки яичных продуктов или продуктов, основным ингредиентом которых являются яйца, для получения лизофосфолипидов.
5. Способ ферментативного рафинирования растительных или пищевых масел, включающий обработку пригодного в пищу или растительного масла вариантом фермента липидацилтрансферазы по п.1 так, чтобы гидролизовать основную часть полярных липидов (например, фосфолипид и/или гликолипид).
6. Применение варианта фермента липидацилтрансферазы по п.1 в способе уменьшения содержания фосфолипидов в пищевом масле, предусматривающее обработку масла указанным вариантом фермента липидацилтрасферазы так, чтобы гидролизовать большую часть фосфолипидов и отделить водную фазу, содержащую гидролизованные фосфолипиды от масла.
7. Применение варианта фермента липидацилтрансферазы по п.1 в биоконверсии полярных липидов (предпочтительно, гликолипидов) для получения высокоценных продуктов, таких как сложные эфиры углеводов, и/или сложные эфиры белков, и/или сложные эфиры белковых субъединиц, и/или сложные эфиры оксикислот.
8. Иммобилизованный вариант фермента липидацилтрансферазы по п.1.
| Преобразователь частоты | 1984 |
|
SU1275711A1 |
| RU 2001117497 A, 27.07.2003 | |||
| BRUMLIK M J Identification of the catalytic triad of the lipase/acyltransferase from Aeromonas hydrophila, J Bacteriol | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |

