Ссылка на родственные заявки
В описании настоящего изобретения сделаны ссылки на нижеследующие родственные заявки: заявка на патент США № 09/750990, поданная 20 июля 1999 г.; заявка на патент США № 10/409391; заявка на патент США № 60/489441, поданная 23 июля 2003 г.; заявка на патент Соединенного Королевства № GB 0301117.8, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0301118.6, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0301119.4, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0301120.2, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0301121.0, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0301122.8, поданная 17 января 2003 г.; заявка на патент Соединенного Королевства № GB 0330016.7, поданная 24 декабря 2003 г., и международная заявка на патент РСТ/IB 2004/000655, поданная 15 января 2004 г. Все вышеуказанные заявки, все документы, приведенные в указанных заявках (“документы, на которые имеется ссылка в материалах заявки”), и все документы, приведенные в документах, на которые имеется ссылка в материалах заявки, указанные в тексте или использованные при рассмотрении дела по заявке на патент, а также доводы, подтверждающие патентоспособность изобретения, приведенные в процессе рассмотрения дела, включены в настоящее описание изобретения в качестве ссылки. В настоящем описании изобретения также приведены ссылки на различные документы (“документы, на которые имеется ссылка в материалах данной заявки”). Все документы, приведенные в данной заявке, и документы, на которые имеется ссылка в указанных документах, включены в настоящее описание изобретения в качестве ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение относится к новому способу ферментативного удаления и/или уменьшения содержания диглицерида (предпочтительно 1,2-диацилглицерида) в пищевом масле.
Уровень техники
Масла и жиры представляют собой сложные смеси триацилглицеринов (TAG), диацилглицеринов (DAG), свободных жирных кислот и других второстепенных компонентов. Кристаллизация указанных смесей зависит от характеристик TAG (структуры, длины цепи, насыщенности по сравнению с ненасыщенностью и тому подобных) и взаимодействия указанных TAG друг с другом. Что касается присутствия DAG, то ранее проведенные исследования показали, что диацилглицерины оказывают существенное влияние на физические свойства масел и жиров. Указанные свойства изменяются в зависимости от скорости кристаллизации, разной степени полиморфизма, температуры плавления, размеров и формы кристаллов (Siew, 2001).
В большинстве масел, экстрагированных из семян масличных, влияние DAG является менее выраженным, так как диацилглицерины присутствуют в таких маслах в небольших количествах. От присутствия DАG страдает главным образом качество пальмового и оливкового масел, которые характеризуются высоким природным содержанием DAG.
Пальмовое масло, получаемое из гвинейской масличной пальмы (Elaeis guineensis), является важным в коммерческом отношении пищевым маслом. Пальмовое масло является важным источником жира и масла для пищевой промышленности благодаря нескольким благоприятным свойствам, таким как высокая продуктивность, низкая стоимость, высокая тепловая стойкость, устойчивость к окислению и пластичность при комнатной температуре. Кроме того, по сравнению с другими растительными маслами пальмовое масло является богатым источником витамина Е, являющегося антиоксидантом.
Типичный химический состав рафинированного пальмового масла включает примерно 93% триглицеридов, 6% диглицеридов и 1% моноглицеридов (MAG) (Okiy, 1977).
В результате кристаллизации пальмового масла образуется сложная трехмерная сетка, состоящая из указанных компонентов. Из теории следует, что чем больше разнообразие строительных блоков (TAG, DAG и MAG) в указанной сетке, тем сложнее является данная сетка и тем медленнее происходит кристаллизация (Jacobsberg & Ho, 1976). Данная теория была подтверждена Дроздовским (1994). Кроме того, исследования, выполненные данным ученым, показывают, что чем разнообразнее является состав жирных кислот в молекуле триацилглицерина, тем труднее происходит переход между разными кристаллическими фазами.
Как было указано выше, высокое содержание диглицеридов в пальмовом масле влияет на его свойства кристаллизации (Okiy et al., 1978, Okiy, 1978).
Наличие диглицеридов в таких маслах является неблагоприятным фактором. Диглицериды в пищевых маслах (особенно в пальмовом масле), в частности, являются причиной низкого качества масла.
Проблемы, связанные с содержанием диглицеридов в пальмовом масле и других пищевых маслах и жирах, были предметом многих исследований и в научной литературе были описаны разные способы преодоления проблемы высокого содержания диглицеридов.
Японский производитель ферментов, компания Амано на своей информационной home-странице (Amano Enzyme Inc., 2004) рекомендует применение ферментативного процесса для удаления диглицерида из жиров и масел. Данный процесс основан на использовании фермента ЛИПАЗА G “AMANO” 50, который способен расщеплять диглицериды с образованием свободных жирных кислот и глицерина. Указанный фермент гидролизует диглицерид (DAG) и/или моноглицерид (MAG). Образовавшиеся свободные жирные кислоты удаляют путем вакуумной перегонки или фракционированной кристаллизации.
В европейском патенте № 0558112 описан способ ферментативного гидролиза остаточных диглицеридов в триглицеридных препаратах в эмульсиях. Данный способ основан на гидролизе диглицерида при помощи липазы G компании Amano, Япония (см. выше). Указанный способ был усовершенствован в результате выполнения ферментативной реакции в эмульсии для расщепления диглицерида с образованием жирных кислот и глицерина. Водную фазу отделяют после выполнения реакции и указанный фермент частично используют вторично.
В патенте Японии № 62061590 описано масло твердой консистенции с низким содержанием диглицерида, которое получают путем обработки масел или жиров частично глицеридспецифическим ферментом (например, липазой) в присутствии каталитического количества воды или липазой, которая является ферментом, специфичным к 1,3-диглицериду, в присутствии жирных кислот, сложных эфиров жирных кислот или других глицеридов масел или жиров. Полученный продукт представляет собой масло твердой консистенции, которое особенно пригодно для использования в качестве заменителя масла какао. Таким образом, липазу G и липазу Rhizopus deremer (фермент, специфичный к 1,3-диглицериду) смешивали с диатомитом и гранулировали. Гранулы смешивали с фракцией, имеющей температуру плавления пальмового масла (5,7% диглицерида, кислотное число 0,25), и водой (10% в расчете на частично глицеридспецифический фермент). Смесь перемешивали при комнатной температуре в течение 1 часа, после чего ферменты и воду удаляли с образованием масла твердой консистенции, содержащего 1,2% диглицерида (кислотное число 10,5).
Таким образом, ранее были описаны способы удаления или уменьшения содержания диглицерида в пальмовом масле и других пищевых маслах при помощи ферментативных реакций. В основе указанных способов лежит гидролиз диглицерида специфической диглицеридгидролизующей липазой с образованием свободных жирных кислот и глицерина. Затем свободные жирные кислоты могут быть удалены разными методами, такими как вакуумная перегонка или фракционирование.
Недостатком применения специфического диглицеридгидролизующего фермента является вредное образование свободных жирных кислот. Указанные свободные жирные кислоты должны быть удалены из пальмового масла. Таким образом, образование свободных жирных кислот часто рассматривается как потеря продукта.
Для преодоления проблем, связанных с удалением свободных жирных кислот и потерей продукта, вызываемого образованием свободных жирных кислот, авторы настоящего изобретения разработали новый способ, позволяющий снизить содержание диглицеридов в пальмовом масле и других растительных маслах.
В научной литературе было описано ферментативное удаление диглицеридов из пальмового масла при помощи липаз, которые обычно являются триацилглицерингидролизующими ферментами, специфичными к 1,3-диглицериду (Е.С. 3.1.1.3) (см., например, патент Японии № 62061590 или европейский патент № 0652289). В публикации WO 00/05396 наряду с прочим описана обработка липазой пищевого продукта, который может содержать глицерин, для осуществления глицеролиза в среде с низким содержанием воды.
Однако как триацилглицерингидролизующие ферменты, специфичные к 1,3-диглицериду (липазы), так и ферменты, гидролизующие DAG/MAG, вызывают значительное увеличение содержания свободных жирных кислот в масле, а также гидролиз моноглицеридов.
Однако в некоторых растительных маслах, предназначенных для определенных применений, может быть желательно увеличить содержание моноглицеридов, так как благодаря этому такое масло приобретает свойства эмульгатора. Таким образом, с одной стороны, желательно уменьшить содержание диглицеридов, не уменьшая содержания моноглицеридов. С другой стороны, может быть желательно уменьшить содержание как диглицеридов, так и моноглицеридов.
Кроме того, такие ферменты как липазы могут в значительной степени увеличивать содержание DAG вследствие гидролиза триацилглицерина (TAG), основного липида, присутствующего в пищевых маслах.
В публикациях WO 2000/36114, США 2003/0028923 и США 2003/0074695 описана трансформация растения нуклеиновой кислотой, которая имеет последовательность, кодирующую фермент диацилглицерин-ацилтрансферазу (DGAT), или его антисмысловую последовательность. В приведенных выше документах указано, что фермент DGAT катализирует конечную стадию “пути Кеннеди”, на которой диацилглицерин (DAG) объединяется с ацильными группами ацил-СоА с образованием триглицерида (TAG). Таким образом, в указанных документах описано получение трансгенных растений с модифицированным составом и/или содержанием TAG. Ферменты DGAT, как описано в вышеуказанных документах, не являются липид-ацилтрансферазами и/или диглицерид:глицерин-ацилтрансферазами по настоящему изобретению.
В частности, ферменты DGAT осуществляют воздействие только в присутствии ацил-СоА или СоА жирной кислоты. Ацил-СоА не пригоден для промышленного применения с целью обработки пищевых масел из-за слишком высокой стоимости. Однако указанные ферменты не действуют при отсутствии ацил-СоА. Кроме того, ферментативные реакции, осуществляемые в присутствии СоА жирной кислоты, трудно контролировать в промышленном масштабе.
Во всех приведенных выше документах указано, что часто желательно уменьшить экспрессию ферментов DGAT в растении и таким образом уменьшить содержание TAG. Такой подход резко отличается от настоящего изобретения, которое в конечном счете требует сохранения и/или продуцирования TAG при одновременном уменьшении содержания диглицеридов (DAG) в пищевом масле.
В публикации WO 03/100044 описана фосфолипид:диацилглицерин-ацилтрансфераза (PDAT), которая катализирует образование триглицеридов (TAG) в результате переноса ацильных групп из фосфолипидов (лецитин) в диацилглицерины (DAG). Ферменты PDAT требуют присутствия фосфолипида в качестве донора ацильных групп. Такой подход резко отличается от настоящего изобретения, в котором донором ацильных групп является DAG. В вышеуказанном документе не описано удаление DAG из пищевых масел с использованием липид-ацилтрансферазы по настоящему изобретению.
Сущность изобретения
Установлено, что использование липид-ацилтрансфераз, не зависящих от СоА жирной кислоты, в соответствии с определением, приведенным в настоящем описании изобретения, в частности диглицерид:глицерин-ацилтрансфераз, не зависящих от СоА жирной кислоты, вызывает избирательное уменьшение содержания и/или удаление диглицеридов (предпочтительно 1,2-диглицеридов) из пищевых масел.
Термин “избирательный” в используемом здесь значении означает, что в среде пищевого масла данный фермент использует диглицериды (DAG), предпочтительно 1,2-диглицериды, в качестве субстрата главным образом для триацилглицеридов (ТAG) или моноглицеридов (MAG). Таким образом, можно уменьшить содержание и/или удалить диглицериды из пищевого масла без изменения (или, по существу, без изменения) содержания триглицерида в масле. Количество моноглицеридов в масле остается неизмененным (или, по существу, неизмененным) или увеличивается. В некоторых применениях количество моноглицерида в масле может быть уменьшено.
Одним объектом настоящего изобретения является способ уменьшения содержания и/или удаления диглицерида из пищевого продукта, который включает смешивание пищевого продукта или его части с субстратом акцептора ацильных групп и диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, причем диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле.
Другим объектом настоящего изобретения является способ уменьшения содержания и/или удаления диглицерида из пищевого масла, который включает смешивание пищевого масла с субстратом акцептора ацильных групп и диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, причем диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле.
Способ по настоящему изобретению может далее включать добавление подвергнутого обработке пищевого масла или его части к одному или нескольким пищевым компонентам для получения пищевого продукта, такого как, например, маргарин или паста.
Настоящее изобретение далее относится к применению диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле, для производства пищевого продукта с пониженным содержанием и/или полностью удаленным (предпочтительно с избирательно пониженным содержанием и/или удаленным) диглицеридом.
Другим объектом настоящего изобретения является применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле, при производстве пищевого продукта для улучшения свойств кристаллизации указанного пищевого продукта.
Другим объектом настоящего изобретения является применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле, при производстве пищевого масла для уменьшения содержания и/или полного удаления (предпочтительно для избирательного уменьшения содержания и/или удаления) диглицерида из указанного пищевого масла.
Другим объектом настоящего изобретения является применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, способным переносить ацильную группу из диглицерида в глицерин в пищевом масле, при производстве пищевого масла для улучшения свойств кристаллизации указанного пищевого масла.
Подробное описание изобретения
Термины “липид-ацилтрансфераза, не зависящая от СоА жирной кислоты” и “диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты” в используемом здесь значении означает фермент, обладающий активностью ацилтрансферазы (обычно классифицируемый как Е.С. 2.3.1.х в соответствии с публикацией “Рекомендации по номенклатуре ферментов” (1992) Комитета по номенклатуре Международного союза биохимии и молекулярной биологии), причем указанный фермент способен переносить ацильную группу из диглицерида в один или несколько акцепторных субстратов).
Таким образом, “липид-ацилтрансфераза, не зависящая от СоА жирной кислоты” или “диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты”, по настоящему изобретению означает фермент, который обладает активностью ацилтрансферазы (обычно классифицируемый как Е.С. 2.3.1.х), но не является диацилглицерин-ацилтрансферазой (DGAT) или фосфолипид:диацилглицерин-ацилтрансферазой (PDAT). Ферменты DGAT обычно классифицируются как Е.С. 2.3.1.20. Ферменты PDAT обычно классифицируются как Е.С. 2.3.1.158. Таким образом, липид-ацилтрансфераза, не зависящая от СоА жирной кислоты или диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению представляет собой фермент, который обладает активностью ацилтрансферазы, но не является ни одним из ферментов, классифицируемых как Е.С. 2.3.1.20 или Е.С. 2.3.1.158.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, или фермент диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению является ферментом, способным переносить ацильную группу из DAG в глицерин в пищевом масле. Таким образом, реакцию, катализируемую ферментом по настоящему изобретению, можно представить следующим образом: диглицерид (DAG) + глицерин → 2 моноглицерида (MAG).
Указанный фермент резко отличается от ферментов, известных как диацилглицерин-ацилтрансферазы (или диацилглицерин-О-ацилтрансфераза) (DGAT) (такие ферменты классифицируются как Е.С. 2.3.1.20), которые катализируют конечную стадию процесса Кеннеди, а именно: 1,2-DAG + ацил-СоА → СоА + триацилглицерин (TAG).
Во избежание сомнений следует отметить, что ферменты DGAT не являются липид-ацилтрансферазами, не зависящими от СоА жирной кислоты, или диглицерид:глицерин-ацилтрансферазами, не зависящими от СоА жирной кислоты, по настоящему изобретению.
Реакция, катализируемая ферментом по настоящему изобретению, также отличается от реакции, катализируемой ферментами, известными как фосфолипиды:диацилглицерин-ацилтрансфераза (PDAT), а именно:
фосфолипиды (такие как лецитин) + 1,2-DAG → триацилглицерин (TAG) + лизофосфолипид
Во избежание сомнений следует отметить, что ферменты PDAT не являются липид-ацилтрансферазами, не зависящими от СоА жирной кислоты, или диглицерид:глицерин-ацилтрансферазами, не зависящими от СоА жирной кислоты, по настоящему изобретению.
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, или диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению является ферментом, классифицируемым как Е.С. 2.3.1.73.
Диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, может быть не связана от мембраны, то есть может быть белком, который в естественном окружении не связан с мембраной при помощи мембраносвязывающего фрагмента или трансмембранного домена.
Во избежание сомнений следует отметить, что липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению не является ферментом, описанным в любой из вышеуказанных публикаций WO 03/100044, WO 2000/36114, США 2003/0028923 или США 2003/0074695.
Помимо активности ацилтрансферазы указанный фермент может также обладать активностью липазы, например активностью фосфолипазы (обычно классифицируемый как Е.С. 3.1.1.х).
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению является диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты. Указанные термины могут иметь взаимозаменяемые значения в настоящем описании изобретения.
Диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению является ацилтрансферазой, которая содержит фрагмент аминокислотной последовательности GDSX, в котором Х означает один или несколько нижеследующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S.
Термин “диглицерид:глицерин-ацилтрансфераза” в значении, используемом в настоящем описании изобретения, синонимичен термину “диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты”.
Во избежание сомнений следует отметить, что термин “СоА жирной кислоты” аналогичен терминам “ацил-СоА”, “кофермент А жирной кислоты” и “ацил-кофермент А”. Указанные термины могут иметь взаимозаменяемые значения в настоящем описании изобретения.
Пищевое масло, используемое в способе или применении по настоящему изобретению, может представлять собой нерафинированное или рафинированное масло.
В одном варианте осуществления изобретения количество диглицерида предпочтительно уменьшают, а не удаляют полностью.
Термин “уменьшить” в используемом здесь значении означает, что количество диглицерида в пищевом масле, подвергнутом обработке ферментом по настоящему изобретению, меньше количества диглицерида в пищевом масле до обработки ферментом.
Диглицериды предпочтительно не удаляют полностью из пищевого масла.
В некоторых применениях, например, связанных с использованием подвергнутого обработке масла в маргаринах и/или шортенинге, количество диглицеридов, в частности 1,2-диглицеридов, должно быть уменьшено до такого количества, при котором скорость кристаллизации жировой смеси соответствует образованию мелких бета-кристаллов. Количество 1,2-диглицерида по отношению к количеству всего диглицерида в пальмовом масле зависит от возраста и условий хранения масла. Для коммерчески доступных масел соотношение 1,3-диглицерида и 1,2-диглицерида равно 1,8:3,3. Удаление 1,2-диглицеридов оказывает наибольшее влияние на свойства кристаллизации.
Как должно быть очевидно квалифицированному специалисту, уменьшение количества диглицерида можно контролировать по времени и температуре реакции. В проточном реакторе с использованием иммобилизованного фермента ход реакции можно контролировать по скорости потока.
Липид-ацилтрансфераза, предназначенная для использования в способах и/или применениях по настоящему изобретению, предпочтительно способна переносить ацильную группу из диглицерида в акцептор ацильных групп, причем акцептором ацильных групп является любое соединение, содержащее гидроксильную группу (-ОН).
Термин “диглицерид” в используемом здесь значении означает один или несколько 1,2-диглицеридов или 1,3-диглицеридов. Диглицеридом предпочтительно является 1,2-диглицерид.
Термины “диглицерид” и “диацилглицерин” имеют в данном описании изобретения взаимозаменяемые значения.
В определение термина “диглицерид” не входит дигалактозилдиглицерид (DGDG) и/или лецитин, например фосфатидилхолин.
Акцептором ацильных групп предпочтительно является вещество, растворимое в пищевом масле.
Акцептором ацильных групп может быть спирт, такой как, например, этанол или многоатомные спирты, в том числе глицерин, их смеси и производные. Акцептором ацильных групп может быть одно или несколько веществ, выбираемых из стерола, станола, оксикислоты, сорбита, сорбитана или других углеводов.
В одном предпочтительном варианте осуществления изобретения акцептором ацильных групп является глицерин.
Таким образом, один вариант осуществления настоящего изобретения относится к способу уменьшения содержания и/или удаления диглицерида из пищевого масла, который включает смешивание пищевого масла с глицерином и диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, причем диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, является ферментом, обладающим активностью ацилтрансферазы и содержащим фрагмент аминокислотной последовательности GDSX, где Х означает один или несколько нижеследующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S.
Акцептор ацильных групп по настоящему изобретению предпочтительно не является водой.
Акцептор ацильных групп предпочтительно не является моноглицеридом и/или диглицеридом.
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению способна расщеплять ацильную связь между одним или несколькими остатками жирной кислоты и глицериновым остовом липидного субстрата, причем липидный субстрат предпочтительно является диглицеридом, в частности 1,2-диглицеридом.
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению предпочтительно не воздействует на триглицериды и/или моноглицериды. Другими словами, липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, избирательно воздействует на диглицериды, предпочтительно на 1,2-диглицериды. Липидный субстрат может быть определен в настоящем изобретении как “липидный донор ацильных групп”.
Таким образом, настоящее изобретение обеспечивает достижение одного или нескольких благоприятных качеств: уменьшение содержания диглицеридов в пищевом масле; уменьшение содержания диглицеридов в пищевом масле без уменьшения содержания триглицеридов; уменьшение содержания диглицеридов в пищевом масле без увеличения содержания моноглицеридов; уменьшение содержания диглицеридов в пищевом масле с увеличением содержания моноглицеридов; уменьшение содержания диглицеридов и уменьшение содержания моноглицеридов в пищевом масле; уменьшение содержания диглицеридов в пищевом масле без значительного увеличения содержания жирных кислот.
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению выполняет реакцию алкоголиза (предпочтительно глицеролиза), осуществляя перенос ацильной группы жирной кислоты из диглицерида (предпочтительно 1,2-DAG) в спирт (предпочтительно глицерин), в результате чего образуются две молекулы моноглицерида, то есть одна молекула образуется из диглицерида и другая молекула образуется из глицерина вместе с захваченной ацильной группой.
Х во фрагменте DGSX предпочтительно означает L. Таким образом, фермент по настоящему изобретению предпочтительно содержит фрагмент аминокислотной последовательности GSDL.
Фрагмент GDSX состоит из четырех консервативных аминокислот. Серин в указанном фрагменте предпочтительно является каталитическим серином фермента липид-ацилтрансферазы. Серин во фрагменте GDSX может находиться в положении, соответствующем Ser-16 в липолитическом ферменте Aeromonas hydrophila, описанном в публикации Brumlik & Buckley (Journal of Bacteriology Apr. 1996, Vol.178, № 7, pр.2060-2064).
Чтобы определить наличие в белке фрагмента GDSX по настоящему изобретению, данную последовательность предпочтительно сравнивают с профилями скрытых моделей Маркова (профили НММ) в базе данных Рfam.
Pfam представляет собой базу данных семейств доменов белков. Pfam содержит множество проверенных последовательностей для каждого семейства, подвергнутых сравнительному анализу, а также профили скрытых моделей Маркова (профили НММ), которые предназначены для идентификации указанных доменов в новых последовательностях. С введением к базе данных Pfam можно ознакомиться в публикации Bateman A. et al. (2002) Nucleic Acids Res. 30; 276-280. Скрытые модели Маркова использованы в ряде баз данных, предназначенных для классифицикации белков; для ознакомления с указанными базами данных см. публикацию Bateman A. and Haft D.H. (2002) Brief Bioinform. 3; 236-245:
http://www.ncbi.nlm.nih.gov/entrez/guery.fcgi?cmd=Retrieve&db=PubMed&list- uids=12230032&dopt=Abstract
http://www.ncbi.nlm.nih.gov/entrez/guery.fcgi?cmd=Retrieve&db=PubMed&list- uids=11752314&dopt=Abstract
Для подробного ознакомления со скрытыми моделями Маркова и их применением в базе данных Pfam см. публикацию Durbin R., Eddy S., and Krogh A. (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4. Пакет программ Хаммера можно приобрести в Вашингтонском университете, Сент-Луис, США.
Альтернативно фрагмент GDSX можно идентифицировать при помощи пакета программ Хаммера, инструкции по использованию которого представлены в публикации Durbin R., Eddy S., and Krogh A. (1998) Biological sequence analysis; probabilistic models of proteins and nucleic acids. Cambridge University Press, ISBN 0-521-62041-4, и в приведенных в ней ссылках, и с помощью профиля НММЕR2, рассмотренного в настоящем описании изобретения.
Доступ к базе данных PFAM можно получить, например, с нескольких серверов, которые в настоящее время расположены на нижеследующих web-сайтах.
http://www.sanger.ac.uk/Software/Pfam/index.shtml
http://pfam.wustl.edu/
http://pfam.jouy.inra.fr/
http://pfam.cgb.ki.se/
Указанная база данных предоставляет средства поиска, с помощью которых можно войти в белковую последовательность. Используя параметры по умолчанию указанной базы данных, можно произвести анализ белковой последовательности на наличие доменов Pfam. Домен GDSX является установленным доментом в базе данных, поэтому его присутствие может быть обнаружено в любой запрашиваемой последовательности. Указанная база данных производит сравнительный анализ консенсусной последовательности Pfam00657 с запрашиваемой последовательностью.
Множественный сравнительный анализ, в том числе Aeromonas salmonicida или Aeromonas hydrophila, можно произвести
а) вручную
вышеописанным способом выполняют сравнительный анализ представляющего интерес белка с консенсусной последовательностью Pfam00657 и затем выполняют сравнительный анализ Р10480 с консенсусной последовательностью Pfam00657;
или
b) при помощи базы данных
после идентификации консенсусной последовательности Pfam00657 база данных предлагает вариант, показывающий сравнительный анализ запрашиваемой последовательности с отборочным сравнительным анализом консенсусной последовательности Pfam00657. Р10480 является частью указанного отборочного сравнительного анализа и обозначена GCAT_AERHY. Запрашиваемая последовательность и Р10480 будут отображены в одном окне.
Эталонная последовательность Aeromonas hydrophila:
Остатки липазы GDSX Aeromonas hydrophila пронумерованы в файле NCBI P10480, причем номера в настоящем описании изобретения соответствуют номерам, приведенным в указанном файле, который в настоящем изобретении использован для определения специфических аминокислотных остатков, присутствующих в предпочтительном варианте осуществления изобретения в ферментах липид-ацилтрансферазы по настоящему изобретению.
Выполнение сравнительного анализа при помощи базы данных Рfam (фиг.33 и 34):
Могут быть обнаружены нижеследующие консервативные остатки, которые могут присутствовать в предпочтительном варианте осуществления изобретения в ферментах, предназначенных для использования в композициях и способах по настоящему изобретению.
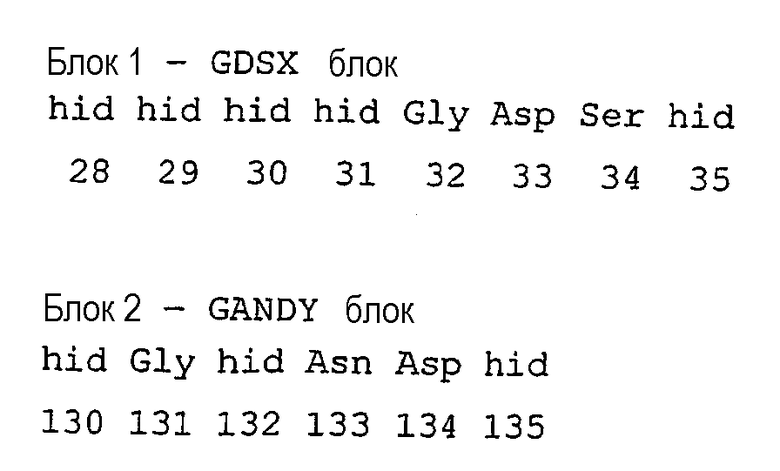

где “hid” означает гидрофобный остаток, выбираемый из Met, Ile, Leu, Val, Ala, Gly, Cys, His, Lys, Trp, Tyr, Phe.
Сравнительный анализ фермента липид-ацилтрансферазы, предназначенного для использования в композициях/способах по настоящему изобретению, предпочтительно можно произвести, используя консенсусную последовательность Pfam00657.
Положительное совпадение с профилем скрытой модели Маркова (профиль НММ) семейства доменов pfam00657 указывает на присутствие домента GDSL или GDSX по настоящему изобретению.
При сравнении с консенсусной последовательностью Pfam00657 липид-ацилтрансфераза, предназначенная для использования в композициях/способах по настоящему изобретению, предпочтительно содержит по крайней мере один, предпочтительно более одного, предпочтительно более двух нижеследующих блоков: блок GDSx, блок GANDY, блок НРТ. Липид-ацилтрансфераза может иметь блок GDSx и блок GANDY. Альтернативно указанный фермент может иметь блок GDSx и блок НРТ. Указанный фермент предпочтительно содержит по крайней мере блок GDSx.
При сравнении с консенсусной последовательностью Pfam00657 фермент, предназначенный для использования в композициях/способах по настоящему изобретению, содержит по крайней мере один, предпочтительно более одного, предпочтительно более двух, предпочтительно более трех, предпочтительно более четырех, предпочтительно более пяти, предпочтительно более шести, предпочтительно более семи, предпочтительно более восьми, предпочтительно более девяти, предпочтительно более десяти, предпочтительно более одиннадцати, предпочтительно более двенадцати, предпочтительно более тринадцати, предпочтительно более четырнадцати нижеследующих аминокислотных остатков при сравнении с эталонной полипептидной последовательностью A. hydrophilia, а именно с SEQ ID NO:32: 28hid, 29hid, 30hid, 31hid, 32gly, 33Asp, 34Ser, 35hid, 130hid, 131Gly, 132Hid, 133Asn, 134Asp, 135hid, 309His.
Домен GDSX pfam00657 является однозначным идентификатором, который позволяет отличить белки, имеющие данный домен, от других ферментов.
Консенсусная последовательность pfam00657 представлена на фиг.1 как SEQ ID NO:1. Указанная последовательность получена в результате идентификации семейства 00657 при помощи базы данных pfam версии 6, которая в данном описании изобретения может быть также определена как pfam00657.6.
Консенсусная последовательность может быть модифицирована в результате использования последующих версий базы данных pfam.
Например, на фиг.33 и 34 показан сравнительный анализ семейства 00657 с использованием базы данных pfam версии 11, которая в данном описании изобретения может быть также определена как pfam00657.11.
Присутствие блоков GDSx, GANDY и НРТ обнаружено в семействе 00657 с использованием обеих версий базы данных pfam. Для идентификации семейства 00657 могут быть использованы последующие версии базы данных pfam.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению можно предпочтительно охарактеризовать на основании нижеследующих критериев:
(i) данный фермент обладает активностью ацилтрансферазы, которая может быть определена как активность переноса сложного эфира, в результате которого ацильная часть исходной сложноэфирной связи липидного донора ацильных групп переносится в акцептор ацильных групп с образованием нового сложного эфира;
(ii) данный фермент содержит фрагмент аминокислотной последовательности GDSX, в котором Х означает один или несколько нижеследующих аминокислотных остатков L, A, V, I, F, Y, H, Q, T, N, M или S;
(iii) данный фермент содержит остаток His-309 или остаток гистидина в положении, соответствующем остатку His-309 в липолитическом ферменте Aeromonas hydrophila, показанном на фиг.2 (SEQ ID NO:2 или SEQ ID NO:32).
Аминокислотным остатком фрагмента GDSХ предпочтительно является L.
В SEQ ID NO:2 или SEQ ID NO:32 первые 18 аминокислотных остатков образуют сигнальную последовательность. His-309 непроцессированной последовательности, которая представляет собой белок, включающий сигнальную последовательность, соответствует His-291 зрелой части белка, т.е. данной последовательности без сигнальной последовательности.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению включает три каталитических остатка: Ser-34, Asp-134 и His-309 или соответственно остаток серина, остаток аспарагиновой кислоты и остаток гистидина в положениях, соответствующих Sеr-34, Asp-134 и His-309 в липолитическом ферменте Aeromonas hydrophila, показанном на фиг.2 (SEQ ID NO:2) или на фиг.28 (SEQ ID NO:32). Как было указано выше, в последовательности, показанной в SEQ ID NO:2 или SEQ ID NO:32, первые 18 аминокислотных остатков образуют сигнальную последовательность. Ser-34, Asp-134 и His-309 непроцессированной последовательности, которая представляет собой белок, включающий сигнальную последовательность, соответствуют Ser-16, Asp-116 и Нis-291 зрелой части белка, то есть данной последовательности без сигнальной последовательности. В консенсусной последовательности pfam00657, как показано на фиг.1 (SEQ ID NO:1), остатки активного сайта соответствуют Ser-7, Asp-157 и His-348.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению можно предпочтительно охарактеризовать на основании нижеследующих критериев:
(i) данный фермент обладает активностью ацилтрансферазы, которая может быть определена как активность переноса сложного эфира, в результате которого ацильная часть исходной сложноэфирной связи первого липидного донора ацильных групп переносится в акцептор ацильных групп с образованием нового сложного эфира; и
(ii) данный фермент содержит по крайней мере Gly-32, Asp-33, Ser-34, Asp-134 и His-309 или остатки глицина, аспарагиновой кислоты, серина, аспарагиновой кислоты и гистидина в положениях, соответствующих Gly-32, Asp-33, Ser-34, Asp-134 и His-309 в липолитическом ферменте Aeromonas hydrophila, показанном на фиг.2 (SEQ ID NO:2) или на фиг.28 (SEQ ID NO:32).
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может быть предпочтительно получен из организмов, относящихся к одному или нескольким нижеследующим родам: Aeromonas, Streptomyces, Saccharomyces, Lactococcus, Mycobacterium, Streptococcus, Lactobacillus, Desulfitobacterium, Bacillus, Campylobacter, Vibrionaceae, Xylella, Sulfolobus, Aspergillus, Schizosaccharomyces, Listeria, Neisseria, Mesorhizobium, Ralstonia, Xanthomonas и Candida.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может быть предпочтительно получен из одного или нескольких нижеследующих организмов: Aeromonas hydrophila, Aeromonas salmonicida, Streptomyces coelicolor, Streptomyces rimosus, Mycobacterium, Streptococcus pyogenes, Lactococcus lactis, Streptococcus pyogenes, Streptococcus thermophilus, Lactobacillus helveticus, Desulfitobacterium dehalogenans, Bacillus sp., Campylobacter jejuni, Vibrionaceae, Xylella fastidiosa, Sulfolobus solfataricus, Saccharomyces cerevisiae, Aspergillus terreus, Schizosaccharomyces pombe, Listeria innocua, Listeria monocytogenes, Neisseria meningitides, Mesorhizobium loti, Ralstonia solanacearum, Xanthomonas campestris, Xanthomonas axonopodis и Candida parapsilosis.
В соответствии с одним объектом изобретения фермент липид-ацилтрансферазу, не зависящую от СоА жирной кислоты, по настоящему изобретению предпочтительно получают из одного или нескольких организмов Aeromonas hydrophila или Aeromonas salmonicida.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению содержит одну или несколько нижеследующих аминокислотных последовательностей:
(i) аминокислотная последовательность SEQ ID NO:2 (см. фиг.2)
(ii) аминокислотная последовательность SEQ ID NO:3 (см. фиг.3)
(iii) аминокислотная последовательность SEQ ID NO:4 (см. фиг.4)
(iv) аминокислотная последовательность SEQ ID NO:5 (см. фиг.5)
(v) аминокислотная последовательность SEQ ID NO:6 (см. фиг.6)
(vi) аминокислотная последовательность SEQ ID NO:12 (см. фиг.14)
(vii) аминокислотная последовательность SEQ ID NO:20 (фиг.16)
(viii) аминокислотная последовательность SEQ ID NO:22 (фиг.18)
(ix) аминокислотная последовательность SEQ ID NO:24 (фиг.20)
(х) аминокислотная последовательность SEQ ID NO:26 (фиг.22)
(xi) аминокислотная последовательность SEQ ID NO:28 (фиг.24)
(xii) аминокислотная последовательность SEQ ID NO:30 (фиг.26)
(xiii) аминокислотная последовательность SEQ ID NO:32 (фиг.28)
(xiv) аминокислотная последовательность SEQ ID NO:34 (фиг.30)
(xv) аминокислотная последовательность SEQ ID NO:55 (фиг.52)
(xvi) аминокислотная последовательность SEQ ID NO:58
(xvii) аминокислотная последовательность SEQ ID NO:60
(xviii) аминокислотная последовательность SEQ ID NO:61
(xix) аминокислотная последовательность SEQ ID NO:63
(хх) аминокислотная последовательность SEQ ID NO:65
(xxi) аминокислотная последовательность SEQ ID NO:67
(xxii) аминокислотная последовательность SEQ ID NO:70 или
(xxiii) аминокислотная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:55, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67 или SEQ ID NO:70.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению содержит аминокислотную последовательность SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:32 или SEQ ID NO:34, или аминокислотную последовательность, которая на 75% или более, предпочтительно на 80% или более, предпочтительно на 85% или более, предпочтительно на 90% или более, предпочтительно на 95% или более идентична аминокислотной последовательности SEQ ID NO:2, аминокислотной последовательности SEQ ID NO:3, аминокислотной последовательности SEQ ID NO:32, или аминокислотной последовательности SEQ ID NO:34.
В соответствии с целями настоящего изобретения степень идентичности определяется на основе числа одинаковых элементов последовательности. Степень идентичности по настоящему изобретению можно определить при помощи компьютерных программ, известных в данной области, таких как GAP в пакете программ GCG (Program Manual for the Wisconsin Package, Version 8, August 1994, Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, US53711) (Needleman & Wunsch (1970), J. of Molecular Biology 48, 443-45), с использованием нижеследующих установок для сравнения полипептидных последовательностей: штраф за создание пробела (GAP) равен 3,0 и штраф за удлинение пробела (GAP) равен 0,1.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению содержит аминокислотную последовательность, которая на 80% или более, предпочтительно на 85% или более, предпочтительнее на 90% или более или еще предпочтительнее на 95% или более идентична любой последовательности SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:55, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67 или SEQ ID NO:70.
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению содержит одну или несколько нижеследующих аминокислотных последовательностей:
(а) аминокислотная последовательность, представленная аминокислотными остатками 1-100 SEQ ID NO:2 или SEQ ID NO:32;
(b) аминокислотная последовательность, представленная аминокислотными остатками 101-200 SEQ ID NO:2 или SEQ ID NO:32;
(с) аминокислотная последовательность, представленная аминокислотными остатками 201-300 SEQ ID NO:2 или SEQ ID NO:32; или
(d) аминокислотная последовательность, которая на 75% или более, предпочтительно на 85% или более, предпочтительнее на 90% или более, еще предпочтительнее на 95% или более идентична любой из аминокислотных последовательностей, указанных в приведенных выше пп.(а) - (с).
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению содержит одну или несколько нижеследующих аминокислотных последовательностей:
(а) аминокислотная последовательность, представленная аминокислотными остатками 28-39 SEQ ID NO:2 или SEQ ID NO:32;
(b) аминокислотная последовательность, представленная аминокислотными остатками 77-88 SEQ ID NO:2 или SEQ ID NO:32;
(с) аминокислотная последовательность, представленная аминокислотными остатками 126-136 SEQ ID NO:2 или SEQ ID NO:32;
(d) аминокислотная последовательность, представленная аминокислотными остатками 163-175 SEQ ID NO:2 или SEQ ID NO:32;
(е) аминокислотная последовательность, представленная аминокислотными остатками 304-311 SEQ ID NO:2 или SEQ ID NO:32; или
(f) аминокислотная последовательность, которая на 75% или более, предпочтительно на 85% или более, предпочтительнее на 90% или более, еще предпочтительнее на 95% или более идентична любой из аминокислотных последовательностей, указанных в приведенных выше пп.(а) - (е).
Фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может содержать аминокислотную последовательность, полученную в результате экспрессии одной или нескольких нижеследующих нуклеотидных последовательностей:
(а) нуклеотидная последовательность SEQ ID NO:7 (см. фиг.9);
(b) нуклеотидная последовательность SEQ ID NO:8 (см. фиг.10);
(c) нуклеотидная последовательность SEQ ID NO:9 (см. фиг.11);
(d) нуклеотидная последовательность SEQ ID NO:10 (см. фиг.12);
(e) нуклеотидная последовательность SEQ ID NO:11 (см. фиг.13);
(f) нуклеотидная последовательность SEQ ID NO:13 (см. фиг.15);
(g) нуклеотидная последовательность SEQ ID NO:21 (см. фиг.17);
(h) нуклеотидная последовательность SEQ ID NO:23 (см. фиг.19);
(i) нуклеотидная последовательность SEQ ID NO:25 (см. фиг.21);
(j) нуклеотидная последовательность SEQ ID NO:27 (см. фиг.23);
(k) нуклеотидная последовательность SEQ ID NO:29 (см. фиг.25);
(l) нуклеотидная последовательность SEQ ID NO:31 (см. фиг.27);
(m) нуклеотидная последовательность SEQ ID NO:33 (см. фиг.29);
(n) нуклеотидная последовательность SEQ ID NO:35 (см. фиг.31);
(o) нуклеотидная последовательность SEQ ID NO:54 (см. фиг.51);
(p) нуклеотидная последовательность SEQ ID NO:59;
(q) нуклеотидная последовательность SEQ ID NO:62;
(r) нуклеотидная последовательность SEQ ID NO:64;
(s) нуклеотидная последовательность SEQ ID NO:66;
(t) нуклеотидная последовательность SEQ ID NO:68;
(u) нуклеотидная последовательность SEQ ID NO:69; или
(v) нуклеотидная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:54, SEQ ID NO:59, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68 или SEQ ID NO:69.
Нуклеотидная последовательность может быть на 80% или более, предпочтительно на 85% или более, предпочтительнее на 90% или более и еще предпочтительнее на 95% или более идентична любой последовательности SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:54, SEQ ID NO:59, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68 или SEQ ID NO:69.
В соответствии с одним объектом изобретения липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может представлять собой лецитин:холестерин-ацилтрансферазы (LCAT) или их вариант (например, вариант, полученный в результате молекулярной эволюции).
В данной области известны приемлемые LCAT, которые могут быть получены из одного или нескольких нижеследующих организмов, таких как, например, млекопитающие, крысы, мыши, цыплята, Drosophila melanogaster, растения, включающие Arabidopsis и Oryza sativa, нематоды, грибы и дрожжи.
В одном варианте осуществления изобретения фермент липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может представлять собой липид-ацилтрансферазу, предпочтительно полученную из штаммов ТОР 10 E. coli, включающих pPet12aAhydro и pPet12aASalmo, депонированных компанией Danisco A/S of Langebrogade 1, DK-1001 Copenhagen K, Denmark, на основании Будапештского соглашения о международном признании депонирования микроорганизмов в целях делопроизводства по выдаче патентов в Национальной коллекции промышленных, морских и пищевых бактерий (NCIMB), по адресу 23 St. Maсhar Street, Aberdeen Scotland, GB, 22 декабря 2003 г. под номерами доступа NICMB 41204 и NCIMB 41205.
При осуществлении способа по настоящему изобретению продукт предпочтительно получают без увеличения или значительного увеличения содержания свободных жирных кислот в пищевом продукте.
Термин “трансфераза” в настоящем описании изобретения имеет взаимозаменяемое значение с термином “липид-ацилтрансфераза”.
Липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, в используемом здесь значении катализирует одну или несколько нижеследующих реакций: переэтерификацию, трансэтерификацию, алкоголиз, гидролиз.
Термин “переэтерификация” означает катализируемый ферментом перенос ацильных групп между липидным донором и липидным акцептором, причем липидный донор не является свободной ацильной группой.
Термин “трансэтерификация” в используемом здесь значении означает катализируемый ферментом перенос ацильной группы из липидного донора (не являющегося свободной жирной кислотой) в акцептор ацильных групп (не являющийся водой).
В используемом здесь значении термин “алкоголиз” означает ферментативное расщепление ковалентной связи производного кислоты в результате взаимодействия со спиртом RОН, при этом один из продуктов связывается с атомом Н спирта и другой продукт связывается с группой OR спирта.
В используемом здесь значении термин “спирт” означает алкильное соединение, содержащее гидроксильную группу.
В используемом здесь значении термин “гидролиз” означает катализируемый ферментом перенос ацильной группы из липида в группу ОН молекулы воды. Перенос ацильной группы, осуществляемый в результате гидролиза, требует разделения молекулы воды.
Термин “без увеличения или без значительного увеличения количества свободных жирных кислот” в используемом здесь значении означает, что липид-ацилтрансфераза по настоящему изобретению предпочтительно обладает 100% активностью трансферазы (то есть переносит 100% ацильных групп из донора ацильных групп в акцептор ацильных групп при отсутствии гидролитической активности) в среде пищевого масла; однако фермент может переносить менее 100% ацильных групп, присутствующих в липидном доноре ацильных групп в акцептор ацильных групп. В данном случае активность ацилтрансферазы предпочтительно составляет по крайней мере 30%, предпочтительнее по крайней мере 40%, предпочтительнее по крайней мере 50%, предпочтительнее по крайней мере 70%, предпочтительнее по крайней мере 80%, предпочтительнее по крайней мере 90% и еще предпочтительнее по крайней мере 98% от общей ферментативной активности. Процентное значение активности трансферазы (то есть активности трансферазы в виде процентного значения от общей ферментативной активности) можно определить нижеследующим методом.
Метод определения процентного значения активности ацилтрансферазы
Пищевое масло, в которое была добавлена липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, по настоящему изобретению может быть экстрагировано в результате выполнения ферментативной реакции с использованием СНСl3:CH3OH в отношении 2:1, после чего органическую фазу, содержащую липид, выделяют и анализируют газожидкостной хроматографией (ГЖХ, GLC) методом, который подробно описан ниже. По результатам анализов ГЖХ определяют количество свободных жирных кислот и диглицеридов. Аналогичным образом анализируют контрольное пищевое масло, в которое не был добавлен фермент по настоящему изобретению.
Вычисление:
На основании результатов анализов ГЖХ можно вычислить увеличение количества свободных жирных кислот и уменьшение количества диглицеридов:
∆% жирной кислоты = % жирной кислоты (фермент) - % жирной кислоты (контрольный образец);
Mv Fa = средняя молекулярная масса жирных кислот;
∆% диглицерида = % диглицерида (контрольный образец) - % диглицерида (фермент);
Mv Di = средняя молекулярная масса диглицерида.
Активность трансферазы вычисляют в виде процентного значения от общей ферментативной активности:
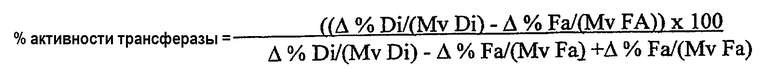
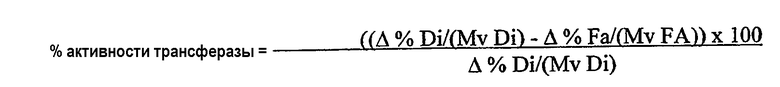
В случае увеличения количества свободных жирных кислот в пищевом масле такое увеличение предпочтительно является незначительным, то есть происходит в незначительной степени. Из вышеизложенного следует, что подобное увеличение свободных жирных кислот не оказывает вредного влияния на качество пищевого масла.
В соответствии с некоторыми объектами настоящего изобретения термин “без значительного увеличения количества свободных жирных кислот” в используемом здесь значении означает, что количество свободных жирных кислот в пищевом масле, подвергнутом обработке липид-ацилтрансферазой по настоящему изобретению, меньше количества свободных жирных кислот в пищевом масле или составе, образующихся при использовании фермента, не являющегося липид-ацилтрансферазой по настоящему изобретению, например, по сравнению с количеством свободных жирных кислот, образующихся при использовании обычной липазы, например липазы Pseudomonas cepacia (липаза PS, Amano, Japan) или липазы Rhizopus oryzae (липаза F, Amano, Japan).
Любой глицерин, оставшийся в пищевом масле после выполнения вышеуказанной реакции, можно удалить, например, центрифугированием или вакуумной перегонкой.
Фермент может быть необязательно удален из пищевого масла после осуществления ферментативной реакции. Альтернативно фермент может быть просто дезактивирован и оставлен в пищевом масле. Например, фермент может быть дезактивирован путем нагрева.
В одном варианте осуществления изобретения липид-ацилтрансфераза, не зависящая от СоА жирной кислоты, используемая при осуществлении способов по настоящему изобретению, может быть иммобилизована. При использовании иммобилизованного фермента смесь, включающая акцептор ацильных групп и пищевое масло, может быть пропущена через колонку, содержащую иммобилизованный фермент. Иммобилизованный фермент можно легко использовать вторично.
Иммобилизованный фермент можно использовать в проточном реакторе или в реакторе периодического действия, содержащем реакционную смесь, которая состоит из акцептора ацильных групп и пищевого масла в виде двухфазной системы. Реакционная смесь может быть необязательно подвергнута перемешиванию или воздействию ультразвука. После уравновешивания реакции реакционная смесь и иммобилизованный фермент могут быть разделены. После окончания реакции избыток акцептора ацильных групп (например, избыток глицерина) может быть удален центрифугированием или вакуумной перегонкой.
Иммобилизованную липид-ацилтрансферазу можно получить методами иммобилизации, известными в данной области. Существует много методов получения иммобилизованных ферментов, которые должны быть известны специалисту в данной области (например, методы, описанные в европейском патенте № 0746608 или в публикациях Balcao V.M., Paiva A.L., Malcata F.X., Enzyme Microb Technol. 1996 May 1; 18(6):392-416, или Retz M.T., Jaeger K.E. Chem. Phys. Lipids. 1998 Jun; 93(1-2):3-14; Bornscheuer U.T., Bessler C, Srinivas R, Krishna S.H. Trends Biotechnol. 2002 Oct; 20(10):433-7; Plou et al., J. Biotechnology 92 (2002) 55-66; Warmuth et al., 1992. Bio Forum 9, 282-283; Ferrer et al., 2000. J. Chem. Technol. Biotechnol. 75, 1-8; или Christensen et al., 1998. Nachwachsende Rohstoff 10, 98-105; Petersen and Christenen, 2000, Applied Biocatalysis. Harwood Academic Publishers, Amsterdam (которые включены в настоящее описание изобретения в качестве ссылки. Методы, которые могут быть использованы при осуществлении настоящего изобретения, включают, например, ковалентное связывание с эупергитом С, адсорбцию на полипропилене и гранулирование с диоксидом кремния.
Пищевое масло предпочтительно является любым пищевым маслом, содержащим диглицерид, предпочтительно значительное количество диглицерида.
Предпочтительно пищевое масло представляет собой одно или несколько из нижеследующих масел: масла, экстрагированные или выделенные из пальмового масла, олеин пальмового масла, стеарин пальмового масла, среднемолекулярную или любую фракцию пальмового масла или оливковое масло.
Более предпочтительно пищевым маслом является пальмовое масло, олеин пальмового масла и/или стеарин пальмового масла.
Что касается смешивания пищевого масла, субстрата акцептора ацильных групп и липид-ацилтрансферазы, то, как должно быть известно квалифицированному специалисту в данной области, указанное смешивание можно производить в любой комбинации и/или порядке. Только в качестве примера можно отметить, что пищевое масло может быть смешано с субстратом акцептора ацильных групп с последующим добавлением липид-ацилтрансферазы. Альтернативно пищевое масло может быть смешано с липид-ацилтрансферазой с последующим добавлением субстрата акцептора ацильных групп. Альтернативно сначала можно смешать липид-ацилтрансферазу и субстрат акцептора ацильных групп и затем смешать смесь фермента/субстрата с пищевым маслом. Альтернативно можно произвести одновременно смешивание всех трех веществ (а именно пищевого масла, субстрата акцептора ацильных групп и липид-ацилтрансферазы).
Данный способ предпочтительно осуществляют при температуре выше температуры плавления пищевого масла.
Указанный способ можно осуществлять при температуре в пределах 30-50°С, предпочтительно в пределах 35-45°С, предпочтительно в пределах 40-45°С.
Фермент предпочтительно добавляют к нерафинированному или рафинированному пищевому маслу. В объем настоящего изобретения предпочтительно не входит обработка ферментом пищевого масла, смешанного с водосодержащими компонентами. Таким образом, настоящее изобретение относится к обработке пищевого масла per se, то есть в среде с низким содержанием воды.
Хотя вода может быть добавлена к маслу до или во время осуществления способа по настоящему изобретению, в наиболее предпочтительном варианте осуществления изобретения воду не добавляют. При наличии воды в пищевом масле ее содержание предпочтительно составляет менее 10%, предпочтительнее менее 7,5%, менее 5%, менее 1%, менее 0,5%, менее 0,4%, менее 0,3%, менее 0,2% или менее 0,1%. Содержание воды в пищевом масле может составлять от 0,1% до 1%.
В одном варианте осуществления изобретения в пищевое масло, используемое при осуществлении способа по настоящему изобретению, может быть добавлен один или несколько доноров ацильных групп, один или несколько акцепторов ацильных групп и/или один или несколько акцепторов ацильных групп и один или несколько доноров ацильных групп. Термин “добавлен” означает, что донор ацильных групп и/или акцептор ацильных групп в естественном состоянии отсутствует в пищевом масле и добавлен одновременно, непосредственно после и/или непосредственно перед контактированием пищевого масла с липид-ацилтрансферазой по настоящему изобретению.
Дополнительным донором ацильных групп может не быть DAG. Дополнительным донором ацильных групп может быть, например, фосфолипид (в частности, лецитин).
Дополнительным акцептором ацильных групп может быть глицерин. Однако акцептором ацильных групп может быть не глицерин, а, например, растительный стерол и/или станол. Дополнительным акцептором ацильных групп может быть комбинация глицерина и одного или нескольких других акцепторов ацильных групп.
Использование масла, в которое добавлен фосфолипид, позволяет получить в масле дополнительный эмульгатор (лизофосфолипид). Использование масла, в которое добавлен растительный стерол и/или станол, позволяет получить в пищевом масле растительный сложный эфир стерола и/или сложный эфир станола. В научной литературе указано, что при введении в пищевой рацион растительные сложные эфиры стерола и сложные эфиры станола снижают содержание холестерина в сыворотке крови.
Ферментативную переэтерификацию с использованием иммобилизованных липаз, таких как Lipozyme® TL IM, 1,3-специфическая липаза (Novozymes, Denmark), применяют для уменьшения количества жирных транскислот в маслах, предназначенных для питания, и/или для модификации характеристик плавления пищевых масел и жиров.
Во время переэтерификации одна или две полиненасыщенные жирные кислоты в триглицеридах пищевого масла могут быть заменены жирной кислотой из другого масла с низким содержанием жирных транскислот, такого как пальмовое масло. Подобный перенос жирных кислот из пальмового масла позволяет изменить температуру плавления пищевого масла без введения жирных транскислот. Иммобилизация липаз описана в патентах США №№ 5776741, 4798793 и 5156963. В патенте США № 6284501 описана переэтерификация фосфолипидов.
Иммобилизованная трансфераза может быть получена методом, применяемым в отношении липаз.
В одном варианте осуществления изобретения липид-ацилтрансферазу, не зависящую от СоА жирной кислоты, описанную в настоящем изобретении, можно использовать в комбинации с липазой, используемой для переэтерификации. Стадии переэтерификации с помощью липазы и ацил-трансферазы предпочтительно осуществляют отдельно.
В соответствии с другим объектом изобретения липид-ацилтрансферазу, не зависящую от СоА жирной кислоты, описанную в настоящем изобретении, можно использовать в комбинации с известными ингибиторами кристаллизации.
Другим объектом настоящего изобретения является способ улучшения свойств кристаллизации пищевого продукта, содержащего пищевое масло, который включает смешивание пищевого масла с субстратом акцептора ацильных групп, диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, описанной в настоящем изобретении, и необязательно с ингибитором кристаллизации.
Один вариант осуществления настоящего изобретения относится к диглицерид:глицерин-ацилтрансферазе, не зависящей от СоА жирной кислоты, используемой при получении пищевого продукта, содержащего пищевое масло, для улучшения свойств кристаллизации указанного пищевого продукта.
В пищевой продукт могут быть необязательно добавлены один или несколько ингибиторов кристаллизации.
Настоящее изобретение далее относится к применению диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, описанной в настоящем изобретении, при производстве пищевого продукта, содержащего пищевое масло, для улучшения свойств кристаллизации указанного пищевого продукта.
Другим объектом настоящего изобретения является применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, в комбинации с дополнительным ингибитором кристаллизации при производстве пищевого продукта, содержащего пищевое масло, для улучшения свойств кристаллизации указанного пищевого продукта.
Дополнительным ингибитором кристаллизации может быть любой известный ингибитор кристаллизации, такой как, например, один или несколько тристеаратов сорбитана, лицетинов, PGE или полисорбатов.
Преимущества изобретения
При использовании способа по настоящему изобретению и, в частности, при выборе ферментов, описанных в настоящем изобретении, для удаления или уменьшения содержания диглицерида в пищевом масле может быть достигнуто избирательное уменьшение содержания диглицерида по сравнению с моно- и диглицеридами без уменьшения содержания триглицеридов и/или значительного увеличения количества свободных жирных кислот (FFA).
То, что способ по настоящему изобретению может быть осуществлен без значительного увеличения содержания свободных жирных кислот в пищевом масле, позволяет решить проблему потери продукта.
При обработке нерафинированного пальмового масла обычной липазой необходимо удалять свободные жирные кислоты в процессе рафинации растительного масла, а при обработке рафинированного пальмового масла липид-ацилтрансферазой по настоящему изобретению (благодаря чему устраняется необходимость обработки нерафинированного пальмового масла обычной липазой) не нужно удалять свободные кислоты, так как в данном случае не происходит значительного увеличения содержания свободных жирных кислот.
Кроме того, или альтернативно настоящее изобретение позволяет удалить и/или уменьшить содержание диглицеридов в пищевом масле без значительного снижения уровней моноглицеридов.
Кроме того, или альтернативно настоящее изобретение позволяет удалить и/или уменьшить содержание диглицеридов в пищевом масле при увеличении уровней моноглицеридов. В этом состоит отличие от ранее применявшихся ферментов, которые используют моноглицерид в качестве субстрата и поэтому уменьшают количество моноглицерида в пищевом масле.
Преимуществом настоящего изобретения является то, что оно не требует добавления СоА жирной кислоты. Такой подход резко отличается от использования ацил-СоА-зависимых ферментов DGAT. Действие фермента по настоящему изобретению происходит в присутствии (и при необходимости добавлении) глицерина, который является дешевым продуктом.
Коммерческая значимость удаления/уменьшения содержания диглицерида в пальмовом масле
Диглицериды замедляют кристаллизацию маргаринов и шортенингов на основе пальмового масла.
В течение многих лет авторы настоящего изобретения были свидетелями постоянного увеличения потребления пальмового масла частично благодаря его доступности, частично благодаря экономичности и главным образом благодаря продуктивности. Установлено, что присутствие пальмового масла в продуктах типа маргарина/шортенинга повышает качество β'-кристаллов масляной смеси. Однако применение пальмового масла ранее ограничивалось использованием стеарина и олеина пальмового масла при некотором использовании ядер кокосовых орехов и их компонентов. В настоящее время применение пальмового масла является более разнообразным вследствие того, что технологии производства кондитерских изделий и пищевых продуктов требуют весьма специфических характеристик плавления.
По мере увеличения использования пальмового масла в пищевых продуктах возникает все больше проблем, связанных с кристаллизацией. При необходимости в пищевые продукты добавляют стимуляторы образования “прокристаллов”, которые инициируют кристаллизацию в составах с низким содержанием жирных транскислот или без жирных транскислот, что позволяет предотвратить или отсрочить образование вторичных кристаллов до потребления пищевого продукта. Кроме того, наблюдается повышенный интерес к применению антикристаллизаторов, таких как тристеараты сорбитана, лецитины, РGE, полисорбаты, которые позволяют замедлить рост вторичных кристаллов. Рост вторичных кристаллов является признаком высокого содержания диглицеридов.
Настоящее изобретение обеспечивает достижение одной или нескольких нижеследующих коммерческих выгод:
а) уменьшает потребность к добавлению дополнительного моноглицерида к пищевому маслу во время обработки и/или последующего применения. Моноглицерид является широко используемым эмульгатором в пищевых системах;
b) уменьшает содержание диглицерида от около 6-8% до около 4-5% и даже больше;
с) увеличивает производительность вследствие сокращения общезаводских накладных расходов;
d) позволяет устранить основные проблемы, связанные с вторичной кристаллизацией;
е) позволяет удалить некоторые полностью насыщенные триглицериды масла в некоторых составах благодаря меньшей необходимости стимулировать кристаллизацию. Производители обычно добавляют полностью насыщенные триглицериды в составы для стимуляции образования кристаллов и устранения нежелательного образования кристаллов в продукте после его продажи. Такое образование кристаллов является признаком наличия значительного количества пальмового масла или его компонентов в составе, который должен содержать диглицериды в концентрациях, достаточных для замедления роста кристаллов до требуемого типа (то есть предпочтительно первичных бета-кристаллов в случае маргаринов и шортенингов) в процессе производства. Однако при использовании пальмового масла, подвергнутого обработке по настоящему изобретению, содержание диглицеридов в данной смеси или составе значительно уменьшается, благодаря чему добавление дополнительных полностью насыщенных триглицеридов для стимуляции образования кристаллов приобретает гораздо меньшее значение;
f) увеличивает разнообразие смесей на основе пальмового масла с учетом высокой стоимости жидкого масла и без значительных изменений технологического процесса;
g) позволяет полностью и/или частично заменить антикристаллизаторы.
В. Жирные транскислоты
Действующее в настоящее время законодательство и общественное мнение направлены против использования жирных транскислот в пищевых продуктах (Fodevareministeriet, Dеnmark, 2003). Вследствие этого возникают проблемы, связанные с производственным процессом. Там, где это возможно, производители хотели бы перейти на использование растворов на основе пальмового масла благодаря высокому содержанию изомеров С:16:0 и С:18:0, обнаруженному в масле данного типа. Результатом этого является более высокое потребление пальмового масла даже в тех отраслях промышленности, в которых пальмовое масло ранее не применялось.
С. Ингибирование кристаллизации диглицеридов в кондитерских изделиях
Эквиваленты масла какао (СВЕ) получают из специальных жиров, таких как фракционированное пальмовое масло, в частности среднемолекулярные фракции пальмового масла (PMF), фракционированный жир масляного дерева, а также жиры многих других экзотических растений. С учетом стоимости в эквиваленты масла какао желательно вводить как можно больше PMF. Многие эквиваленты масла какао могут содержать только фракции пальмового масла. Наиболее важным аспектом при производстве шоколада является кристаллизация жира. Диглицериды отрицательно влияют на кристаллизацию, например серьезную проблему может представлять извлечение из формы (Siew, 2001).
D. Качество кристаллизации
Качество триглицеридов оказывает существенное влияние на производительность, что явствует из примеров типичного состава масла, содержащего транскислоты, применяемого для изготовления стандартного столового маргарина с 82% содержанием жира, по сравнению с вариантом без жирных транскислот, содержащим обычную переэтерифицированную фракцию стеарина пальмового масла и ядра кокосовых орехов в качестве твердого сырья, которые имеют одинаковый состав SFC. Более медленная скорость и качество кристаллизации могут вызывать явление, известное как “вымасливание”. В свою очередь данное явление делает необходимым использование стимуляторов кристаллизации, таких как твердые MAG. Кроме того, замедление кристаллизации стабилизированного кристаллического состава вызывает явление, известное как “песчанистость”, в результате которого кристаллы требуемого типа трансформируются в более устойчивую бета-форму, которая образует песчанистую структуру. Вышеуказанное явление создает определенную проблему в процессе промышленного производства продуктов.
Некоторые применения
Модификация жиров
В процессе химического производства жиров для использования в твердых пищевых продуктах с высоким содержанием жира, таких как маргарины, пасты, кондитерские изделия/шоколад, жирные транскислоты вводят в пищевое масло на стадии гидрогенизации (отверждения). Такой подход позволяет изменить температуру плавления пищевого масла для достижения приемлемой консистенции конечного пищевого продукта, например столового маргарина, который является твердым, но пригоден для намазывания при комнатной температуре, или шоколада, который является твердым во время хранения, но тает во рту.
Однако химическое производство требует больших количеств растворителей, таких как гексан, которые считаются вредными для окружающей среды и здоровья и должны быть полностью удалены до использования модифицированного масла/жира в качестве компонента пищевого продукта.
Во многих странах применение частичной гидрогенизации для производства пищевых продуктов ограничено законодательством и контролирующими органами, поэтому многие производители пищевых продуктов в настоящее время переходят на альтернативное сырье с низким содержанием жирных транскислот.
Масло, полученное способом по настоящему изобретению, можно использовать в качестве компонента маргарина и/или паст.
Заменители/эквиваленты масла какао
Масло какао имеет уникальный состав, который позволяет получать твердые кондитерские изделия, которые тают во рту. Для замены масла какао более дешевым сырьем растительные масла гидрогенизируют, получая при этом жирные транскислоты, в результате чего повышается температура плавления пищевого масла, что позволяет изготовить подобные твердые кондитерские изделия, тающие при температуре тела. Такие модифицированные растительные масла известны как эквиваленты масла какао (СВЕ) или в том случае, когда модифицированные жиры улучшают характеристики шоколадного продукта, как заменители масла какао (CBR). Важной проблемой при использовании СВЕ и CBR является то, что в результате гидрогенизации растительных масел образуются жиры, содержащие жирные транскислоты, которые, как известно, являются вредными для здоровья и плохо влияют на температуру плавления. Это привело к использованию растительных масел с низким содержанием жирных транскислот или их фракций, которые характеризуются более высокой температурой плавления. Пальмовое масло и фракции пальмового масла считаются весьма перспективными для использования в качестве главного компонента CBR/CBE с низким содержанием жирных транскислот.
Жиры, являющиеся заменителями масла какао (CBR), используют для улучшения характеристик шоколадных продуктов, таких как отсутствие темперирования, менее продолжительное отверждение, стойкость, устойчивость к поседению. Особенно желательно использовать жиры, не содержащие лауриновой кислоты и жирных транскислот, такие как пальмовое масло. Свойства кристаллизации жиров, используемых в заменителях масла какао, играют важную роль в обеспечении приемлемого баланса между таянием продукта во рту и сохранением вышеуказанных предпочтительных характеристик. Использование жиров с низким содержанием жирных транскислот, таких как пальмовое масло, в смесях CBR является особенно желательным по соображениям здоровья. Применение пальмового масла в заменителях масла какао описано в европейском патенте № 0293194.
Масло, полученное способом по настоящему изобретению, можно использовать в качестве компонента шоколада, например в жировой смеси, применяемой в качестве заменителя и/или эквивалента масла какао.
Последовательности некоторых ферментов диглицерид:глицерин-ацилтрансфераз, предназначенных для использования в соответствии с настоящим изобретением
Приемлемые ферменты диглицерид:глицерин-ацилтрансферазы, предназначенные для использования в соответствии с настоящим изобретением и/или при осуществлении способов по настоящему изобретению, могут включать любые нижеследующие аминокислотные последовательности и/или аминокислотные последовательности, кодированные нижеследующими нуклеотидными последовательностями.
Termobifida\fusca GDSx 548 аминокислот
SEQ ID NO:58

SEQ ID NO:59
nt


Termobifida\fusca - GDSx
SEQ ID NO:60

Corynebacterium\effciens\GDSx 300 аминокислот
SEQ ID NO:61

SEQ ID NO:62
nt


Novosphingobium\aromaticivorans\GDSx 284 аминокислоты
SEQ ID NO:63
ZP 00094165

SEQ ID NO:64

S.coelicolor\GDSx 268 аминокислот
SEQ ID NO:65
NP 625998

SEQ ID NO:66


S.avermitilis\GDSx 269 аминокислот
SEQ ID NO:67

SEQ ID NO:68


Диглицерид:глицерин-ацилтрансфераза Streptomyces
SEQ ID NO:69

Диглицерид:глицерин-ацилтрансфераза Streptomyces
SEQ ID NO:70
Идентификация диглицерид:глицерин-ацилтрансферазы по настоящему изобретению
Анализ ферментативного уменьшения содержания диглицерида
в пальмовом масле
В стакан с крышкой отмеряют 1 г пальмового масла, содержащего 7% диглицерида. Добавляют 50 мг глицерина и 10 мкл раствора фермента. Реакционную смесь перемешивают при помощи магнитной мешалки в нагревательной камере при 40°С в течение 20 часов. Ферментативную реакцию прекращают, нагревая реакционную смесь до 100°С в течение 10 минут. Контрольный образец, полученный в результате добавления 10 мкл воды вместо раствора фермента, обрабатывают аналогичным образом. Образцы анализируют газожидкостной хроматографией (ГЖХ) стандартными методами (см. ниже) и высчитывают количество жирных кислот, моноглицерида и диглицерида.
Вычисление:
На основании результатов анализов ГЖХ можно вычислить увеличение содержания свободных жирных кислот и уменьшение количества диглицеридов:
∆% жирной кислоты = % жирной кислоты (фермент) - % жирной кислоты (контрольный образец);
Mv Fa = средняя молекулярная масса жирных кислот;
∆% диглицерида = % диглицерида (контрольный образец) - % диглицерида (фермент);
Mv Di = средняя молекулярная масса диглицерида.
Активность трансферазы вычисляют в виде процентного значения от общей ферментативной активности:
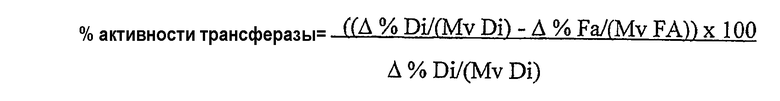
Анализ ГЖХ
Капиллярный газовый хроматограф Perkin Elmer Autosystem 9000, оснащенный колонкой из кварцевого стекла 12,5 м × внутренний диаметр 0,25 мм × 0,1 мкм толщину пленки из 5% фенилметилсиликона (СР Sil 8 CB компании Chrompack).
Газ-носитель: гелий.
Дозатор: раздельный ввод в холодном состоянии PSSI (начальная температура 50°С с нагревом до 385°С), объем 1,0 мкл.
Детектор FID: 395°С.
Получение образца: 30 мг образца растворяли в 9 мл смеси гептан:пиридин с соотношением 2:1, содержащей внутренний эталон гептадекан в количестве 0,5 мг/мл. 300 мкл раствора образца переносили в гофрированный сосуд, добавляли 300 мкл MSTFA (N-метил-N-триметилсилилтрифторацетамид) и подвергали взаимодействию в течение 20 минут при 60°С.
Выделение
В соответствии с одним объектом изобретения полипептид или белок, предназначенный для использования в настоящем изобретении, предпочтительно является выделенным. Термин “выделенный” означает, что данная последовательность по крайней мере, по существу, не содержит других компонентов, с которыми указанная последовательность взаимосвязана в естественных условиях и может быть обнаружена в природе.
Очистка
В соответствии с одним объектом изобретения полипептид или белок, предназначенный для использования в настоящем изобретении, предпочтительно является очищенным. Термин “очищенный” означает, что данная последовательность находится в относительно чистом состоянии, например является чистой по крайней мере примерно на 51%, по крайней мере примерно на 75%, по крайней мере примерно на 80%, по крайней мере примерно на 90%, по крайней мере примерно на 95% или по крайней мере примерно на 98%.
Клонирование нуклеотидной последовательности, кодирующей полипептид по настоящему изобретению
Нуклеотидная последовательность, кодирующая полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, или полипептид, пригодный для модификации, может быть выделена из любой клетки или организма, продуцирующего указанный полипептид. В данной области хорошо известны разные методы выделения нуклеотидных последовательностей.
Например, можно создать библиотеку геномных ДНК и/или кДНК, используя хромосомную ДНК или матричную РНК из организма, продуцирующего данный полипептид. Если известна аминокислотная последовательность данного полипептида, могут быть синтезированы меченые олигонуклеотидные зонды и использованы для идентификации полипептидкодирующих клонов из геномной библиотеки, полученной из данного организма. Альтернативно для идентификации полипептидкодирующих клонов можно использовать меченый олигонуклеотидный зонд, содержащий последовательности, гомологичные другому известному полипептидному гену. В последнем случае используют менее строгие условия гибридизации и промывки.
Альтернативно полипептидкодирующие клоны могут быть идентифицированы путем введения фрагментов геномной ДНК в экспрессирующий вектор, такой как плазмида, трансформирующий фермент-отрицательные бактерии полученной библиотекой геномных ДНК, и последующего культивирования трансформированных бактерий на агаре, содержащем фермент, ингибируемый данным полипептидом, что позволяет идентифицировать клоны, экспрессирующие указанный полипептид.
Кроме того, нуклеотидная последовательность, кодирующая данный полипептид, может быть получена путем синтеза стандартными методами, например методом на основе фосфорамидита, описанным в публикации Beucage S.L. et al. (1981) Tetrahedron Letters 22, рp.1859-1869, или методом, описанным в публикации Matthes et al. (1984) EMBO J., 3, рр.801-805. В методе на основе фосфорамидита синтезируют олигонуклеотиды, например, в автоматическом синтезаторе ДНК, которые очищают, гибридизируют, лигируют и клонируют в соответствующих векторах.
Нуклеотидная последовательность может иметь смешанное геномное и синтетическое происхождение, смешанное синтетическое и кДНК происхождение или смешанное геномное и кДНК происхождение, может быть получена лигированием фрагментов, имеющих синтетическое, геномное или кДНК происхождение (по потребности) стандартными методами. Все лигированные фрагменты соответствуют разным частям всей нуклеотидной последовательности. Последовательность ДНК может быть также получена в результате полимеразной реакции синтеза цепи (PCR) с использованием специфических праймеров, например, описанных в патенте США № 4683202 или в публикации Saiki R.K. et al. (Science (1988) 239, pp.487-491).
Нуклеотидные последовательности
Настоящее изобретение относится также к нуклеотидным последовательностям, кодирующим полипептиды, обладающие специфическими свойствами, как описано в настоящем изобретении. Термин “нуклеотидная последовательность” в используемом здесь значении означает олигонуклеотидную последовательность или полинуклеотидную последовательность и ее варианты, гомологи, фрагменты и производные (в частности, ее части). Нуклеотидная последовательность может быть геномной, синтетической или рекомбинантной, а также двухцепочечной или одноцепочечной, представляя собой смысловую и антисмысловую цепь.
Термин “нуклеотидная последовательность” применительно к настоящему изобретению включает геномную ДНК, кДНК, синтетическую ДНК и РНК. Указанный термин предпочтительно означает ДНК, более предпочтительно кДНК для кодирующей последовательности.
В предпочтительном варианте осуществления изобретения нуклеотидная последовательность, per se кодирующая полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, не является нативной нуклеотидной последовательностью в своем естественном окружении, где она связана с одной или несколькими естественно ассоциированными последовательностями, которые также находятся в своем естественном окружении. Для простоты понимания нуклеотидную последовательность, соответствующую данному предпочтительному варианту осуществления изобретения, можно определить как “ненативную нуклеотидную последовательность”. В данной связи термин “нативная нуклеотидная последовательность” означает всю нуклеотидную последовательность, которая находится в своем естественном окружении и функционально связана со всем промотором, с которым она ассоциирована в естественных условиях, причем указанный промотор также находится в своем естественном окружении. Таким образом, полипептид по настоящему изобретению может быть экспрессирован в своем естественном организме нуклеотидной последовательностью, не находящейся под контролем промотора, с которым она ассоциирована в естественных условиях в данном организме.
Полипептид предпочтительно не является нативным полипептидом. В данной связи термин “нативный полипептид” означает весь полипептид, находящийся в естественном окружении и экспрессированный своей нативной нуклеотидной последовательностью.
Нуклеотидную последовательность, кодирующую полипептиды, обладающие специфическими свойствами, как описано в настоящем изобретении, получают методами рекомбинантных ДНК (то есть при помощи рекомбинантной ДНК). Однако в альтернативном варианте осуществления изобретения нуклеотидная последовательность может быть синтезирована полностью или частично химическими методами, хорошо известными в данной области (см. публикации Caruthers M.H. et al. (1980) Nuc. Acids Res. Symp. Ser. 215-23 и Horn T. et al. (1980) Nuc. Acids Res. Symp. Ser. 225-232).
Молекулярная эволюция
После выделения фермент-кодирующей нуклеотидной последовательности или идентификации предполагаемой фермент-кодирующей нуклеотидной последовательности может быть желательно модифицировать выбранную нуклеотидную последовательность, например, может быть желательно мутировать последовательность для получения фермента по настоящему изобретению.
Мутации могут быть введены при помощи синтетических олигонуклеотидов. Указанные олигонуклеотиды содержат нуклеотидные последовательности, фланкирующие требуемые сайты мутации.
Приемлемый метод описан в публикации Morinaga et al. (Biotechnology (1984) 2, pр.646-649). Другой метод введения мутаций в фермент-кодирующие нуклеотидные последовательности описан в публикации Nelson and Long (Analytical Biochemistry (1989), 180, pр.147-151).
Вместо вышеописанного сайт-направленного мутагенеза мутации можно вводить произвольно, например, при помощи коммерчески доступного набора, такого как набор для мутагенеза методом РCR GeneMorph компании Stratagene или набора для произвольного мутагенеза методом PCR Diversify компании Clontech. В европейском патенте № 0583265 описаны методы оптимизации мутагенеза на основе РCR, которые могут быть также объединены с использованием мутагенных аналогов ДНК, описанных в европейском патенте № 0866796. Методы PCR, подверженные ошибкам, пригодны для получения вариантов липид-ацилтрансфераз с предпочтительными характеристиками. В публикации WO 0206457 описана молекулярная эволюция липаз.
Третьим методом получения новых последовательностей является фрагментация неидентичных нуклеотидных последовательностей с использованием любого числа рестрикционных ферментов или такого фермента как Dnase I, и последующая сборка полных нуклеотидных последовательностей, кодирующих функциональные белки. Альтернативно можно использовать одну или несколько неидентичных нуклеотидных последовательностей и ввести мутации во время вторичной сборки полной нуклеотидной последовательности. Методы перестановки ДНК или семейства пригодны для получения вариантов липид-ацилтрансфераз с предпочтительными характеристиками. Приемлемые методы выполнения “перестановки” описаны в европейских патентах №№ 0752008, 1138763, 1103606. Метод перестановки может быть объединен с другими формами мутагенеза ДНК, описанными в патенте США № 6180406 и в публикации WO 01/34835.
Таким образом, можно ввести многочисленные сайт-направленные или произвольные мутации в нуклеотидную последовательность in vivo или in vitro и затем произвести скрининг разными методами с целью выявления улучшенных функциональных свойств кодированного полипептида. Используя внутренне и внешне опосредуемые методы рекомбинации (см. WO 00/58517, патенты США №№ 6344328 и 6361974), например, можно произвести молекулярную эволюцию, в результате которой полученный вариант будет характеризоваться очень низкой гомологией с известными ферментами или белками. Такие варианты могут обладать значительной структурной аналогией с известными ферментами трансферазы и при этом отличаться очень низкой гомологией аминокислотных последовательностей.
Кроме того, мутации или естественные варианты полинуклеотидной последовательности могут быть объединены с мутациями дикого типа, другими мутациями или естественными вариантами для получения новых вариантов. Такие новые варианты могут быть также подвергнуты скринингу с целью обнаружения улучшенных функциональных свойств кодированного полипептида.
Применение вышеуказанных и подобных методов молекулярной эволюции позволяет идентифицировать и отобрать варианты ферментов по настоящему изобретению, которые обладают предпочтительными характеристиками, отличными от ранее известной структуры или функции белка, и получить непрогнозируемые, но благоприятные мутации или варианты. Существует много примеров применения молекулярной эволюции в данной области для оптимизации или изменения активности фермента, и такие примеры включают, не ограничиваясь ими, одно или несколько нижеследующих свойств: оптимизированную экспрессию и/или активность в клетке-хозяине или in vitro, повышенную ферментативную активность, измененную субстратную специфичность и/или специфичность продукта, повышенную или пониженную ферментативную или структурную стойкость, измененную ферментативную активность/специфичность в предпочтительных окружающих условиях, таких как температура, показатель рН, субстрат.
Как должно быть понятно специалисту в данной области, при помощи методов молекулярной эволюции можно изменить фермент с целью улучшения его функциональных свойств.
Подходящим образом липид-ацилтрансфераза, используемая в настоящем изобретении, может быть ее вариантом, то есть может содержать замену, делецию или добавление по крайней мере одной аминокислоты по сравнению с исходным ферментом. Вариантные ферменты сохраняют по крайней мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97%, 99% гомологию с исходным ферментом. Приемлемые исходные ферменты могут включать любой фермент, обладающий активностью эстеразы или липазы. Исходный фермент предпочтительно соответствует консенсусной последовательности pfam00657.
В предпочтительном варианте осуществления изобретения вариантный фермент липид-ацилтрансфераза сохраняет или включает по крайней мере один или несколько аминокислотных остатков консенсусной последовательности pfam00657, обнаруженных в блоках GDSх, GANDY и НРТ.
Ферменты, такие как липазы, не обладающие активностью липид-ацилтрансферазы или обладающие низкой активностью указанного фермента в водной среде, могут быть подвергнуты мутации методами молекулярной эволюции с целью введения или усиления активности трансферазы, в результате чего образуется фермент липид-ацилтрансфераза, обладающий значительной активностью трансферазы, который пригоден для использования в композициях по настоящему изобретению и при осуществлении способов по настоящему изобретению.
Подходящим образом липид-ацилтрансфераза, предназначенная для использования в настоящем изобретении, может быть вариантом с повышенной ферментативной активностью в отношении полярных липидов, предпочтительно фосфолипидов и/или гликолипидов, при сравнении с исходным ферментом. Такие варианты предпочтительно обладают также низкой активностью в отношении полярных лизолипидов или вообще не обладают такой активностью. Повышенная активность в отношении полярных липидов, фосфолипидов и/или гликолипидов может быть результатом гидролиза и/или активности трансферазы, или комбинации того и другого.
Вариантные липид-ацилтрансферазы, предназначенные для использования в настоящем изобретении, могут обладать пониженной активностью в отношении триглицеридов, моноглицеридов и/или диглицеридов по сравнению с исходным ферментом.
Вариантный фермент может не обладать активностью в отношении триглицеридов, моноглицеридов и/или диглицеридов.
Альтернативно вариантный фермент, предназначенный для использования в настоящем изобретении, может обладать повышенной активностью в отношении триглицеридов и/или может также обладать повышенной активностью в отношении одного или нескольких нижеследующих веществ, таких как полярные липиды, фосфолипиды, лецитин, фосфатидилхолин, гликолипиды, дигалактозилмоноглицерид, моногалактозилмоноглицерид.
Известны разные варианты липид-ацилтрансфераз, и один или несколько таких вариантов могут быть пригодны для использования в способах и применениях по настоящему изобретению и/или в композициях, содержащих фермент по настоящему изобретению. Только в качестве примера можно привести варианты липид-ацилтрансфераз, описанные в нижеследующих ссылках, которые могут быть использованы в настоящем изобретении: Hilton & Buckley J. Biol. Chem. 1991 Jan. 15: 266 (2): 997-1000; Robertson et al., J. Biol. Chem. 1994 Jan. 21; 269(3):2146-50; Brumlik et al., J. Bacteriol. 1996 Apr; 178(7): 2060-4; Peelman et al. Protein Sci. 1998 Mar; 7(3):587-99.
Аминокислотные последовательности
Настоящее изобретение относится также к аминокислотным последовательностям полипептидов, обладающих специфическими свойствами, описанных в настоящем изобретении.
В используемом здесь значении термин “аминокислотная последовательность” синонимичен термину “полипептид” и/или термину “белок”. В некоторых случаях термин “аминокислотная последовательность” синонимичен термину “пептид”.
Аминокислотная последовательность может быть получена/выделена из приемлемого источника, синтезирована или создана методами рекомбинантных ДНК.
Подходящим образом аминокислотные последовательности могут быть получены стандартными методами из выделенных полипептидов, описанных в настоящем изобретении.
Одним приемлемым методом получения аминокислотных последовательностей из выделенных полипептидов является нижеследующий метод.
Очищенный полипептид может быть подвергнут сублимационной сушке, после чего 100 мкг высушенного сублимацией вещества растворяют в 50 мкл смеси 8 М раствора мочевины и 0,4 М раствора гидрокарбоната аммония, рН 8,4. Растворенный белок денатурируют и восстанавливают в течение 15 минут при 50°С, затем реакционную смесь помещают в атмосферу азота и добавляют 5 мкл 45 мМ дитиотреитола. Реакционную смесь охлаждают до комнатной температуры и добавляют 5 мкл 100 мМ иодацетамида для получения производных соединений остатков цистеина в атмосфере азота в темном помещении в течение 15 минут при комнатной температуре.
К вышеуказанной реакционной смеси добавляют 135 мкл воды и 5 мкг эндопротеиназы Lys-C в 5 мкл воды и подвергают гидролизу при 37°С в атмосфере азота в течение 24 часов.
Полученные пептиды разделяют при помощи ВЭЖХ с обращенной фазой в колонке VYDAC C18 (0,46×15 см; 10 мкм; The Separation Group, California, USA) с использованием растворителя А: 0,1% ТФУ в воде и растворителя В: 0,1% ТФУ в ацетонитриле. Отобранные пептиды вторично хроматографируют в колонке Develosil C18 с использованием такой же системы растворителей с последующим секвенированием N-конца. Секвенирование может быть выполнено в секвенаторе Applied Biosystems 476A с использованием быстрых циклов пульсируемой жидкости в соответствии с инструкциями изготовителя (Applied Biosystems, California, USA).
Идентичность или гомология последовательностей
Настоящее изобретение относится также к использованию последовательностей, характеризующихся некоторой идентичностью или гомологией с аминокислотной(ыми) последовательностью(ями) полипептида, обладающего специфическими свойствами, описанного в настоящем изобретении, или с любой нуклеотидной последовательностью, кодирующей такой полипептид (далее определяемые как “гомологичная(ые) последовательность(и)”). Термин “гомолог” означает последовательность, обладающую определенной гомологией с аминокислотными и нуклеотидными последовательностями по настоящему изобретению. Термин “гомология” может быть эквивалентен термину “идентичность”.
Гомологичная аминокислотная последовательность и/или нуклеотидная последовательность должна определять и/или кодировать полипептид, который сохраняет функциональную активность и/или усиливает активность данного фермента.
В контексте настоящего изобретения предполагается, что гомологичная последовательность представляет собой аминокислотную последовательность, которая может быть по крайней мере на 75, 85 или 90% идентична, предпочтительно по крайней мере на 95 или 98% идентична последовательности по настоящему изобретению. Гомологи обычно имеют такие же активные сайты и т.д., что и аминокислотная последовательность по настоящему изобретению. Хотя гомология может быть также рассмотрена с точки зрения сходства (то есть с учетом аминокислотных остатков, обладающих подобными химическими свойствами/функциями), в контексте настоящего изобретения гомология предпочтительно выражена в виде идентичности последовательностей.
В контексте настоящего изобретения предполагается, что гомологичная последовательность представляет собой нуклеотидную последовательность, которая может быть по крайней мере на 75, 85 или 90% идентична, предпочтительно по крайней мере на 95 или 98% идентична нуклеотидной последовательности, кодирующей полипептид по настоящему изобретению (последовательность по настоящему изобретению). Гомологи включают такие же последовательности, кодирующие активные сайты и т.д., что и последовательность по настоящему изобретению. Хотя гомология может быть также рассмотрена с точки зрения сходства (то есть с учетом аминокислотных остатков, обладающих подобными химическими свойствами/функциями), в контексте настоящего изобретения гомология предпочтительно выражена в виде идентичности последовательностей.
Сравнение гомологии может быть произведено приблизительно или, как правило, при помощи существующих программ сравнения последовательностей. Указанные коммерчески доступные компьютерные программы позволяют вычислить % гомологии у двух или большего числа последовательностей.
Процентное значение (%) гомологии можно вычислить для смежных последовательностей, то есть одну последовательность сравнивают с другой последовательностью, причем каждую аминокислоту в одной последовательности непосредственно сравнивают с соответствующей аминокислотой в другой последовательности по одному остатку. Такое сравнение именуется сравнением “без пробелов”. Обычно такое сравнение без пробелов выполняют только в отношении относительно небольшого числа остатков.
Хотя такой метод является очень простым и сопоставимым, данный метод не позволяет принимать во внимание, например, то, что, по существу, в идентичной паре последовательностей в результате одной инсерции или делеции появляются аминокислотные остатки, выпадающие из сравнения, вследствие чего значительно сокращается % гомологии при выполнении глобального сравнительного анализа. Поэтому большинство методов сравнения последовательностей предполагает достижение оптимального сравнения, при котором учитываются все возможные инсерции и делеции без необоснованного начисления штрафов на общую оценку гомологии. Это достигается путем введения “пробелов” в сравниваемые последовательности с целью максимизации локальной гомологии.
Однако более сложные методы предполагают “штрафы за пробелы”, которые начисляются за каждый пробел, выявляемый в процессе сравнения, благодаря чему при одинаковом числе идентичных аминокислот сравниваемые последовательности, имеющие наименьшее количество пробелов, что отражает более высокую степень родства между двумя сравниваемыми последовательностями, получают более высокую оценку по сравнению с последовательностями, имеющими много пробелов. “Стоимость родственных пробелов” обычно используют для начисления относительно высокой стоимости за существование пробела и меньшего штрафа для каждого последующего остатка в пробеле. Указанная система является наиболее широко используемой системой оценки пробелов. Высокие штрафы, начисляемые за пробелы, позволяют произвести оптимизированное сравнение при наличии меньшего числа пробелов. Большинство программ сравнительного анализа позволяют модифицировать штрафы, начисляемые за пробелы. Однако желательно использовать значения по умолчанию при использовании такого программного обеспечения для сравнения последовательностей. Например, при использовании пакета программ GCG Wisconsin Bestfit штраф за пробелы по умолчанию для аминокислотных последовательностей равен -12 за каждый пробел и -4 за каждое удлинение пробела.
Поэтому вычисление максимальной % гомологии сначала требует выполнения оптимального сравнения с учетом штрафов за пробелы. Приемлемой компьютерной программой, используемой для выполнения такого сравнительного анализа, является пакет программ GCG Wisconsin Bestfit (Devereux et al., 1984 Nuc. Acids Research 12 p.387). Примеры других программ, которые могут производить сравнения последовательностей, включают, не ограничиваясь ими, пакет программ BLAST (см. публикацию Ausubel et al. 1999 Short Protocols in Molecular Biology, 4th Ed - Chapter 18), FASTA (Altschul et al., 1990, J. Mol. Biol. 403-410) и пакет вспомогательных программ сравнения GENEWORKS. Программы BLAST и FASTA пригодны для автономного и оперативного поиска (см. публикацию Ausubel et al., 1999, pages 7-58 - 7-60). Однако в некоторых применениях желательно использовать программу GCG Bestfit. Для сравнения белковых и нуклеотидных последовательностей также создана новая вспомогательная программа, получившая название BLAST 2 Sequences (см. FEMS Microbiol. Lett. 1999 174(2): 247-50; FEMS Microbiol. Lett. 1999 177(1): 187-8 and mailto:tatiana@ncbi.nlm.nih.gov).
Хотя конечное значение % гомологии можно определить с точки зрения идентичности, сам по себе процесс сравнения обычно не основывается на сравнении всех пар. Вместо этого обычно используют масштабированную матрицу оценок сходства, которая позволяет присвоить оценки каждой паре в попарном сравнении на основании химического сходства или эволюционной дистанции. Примером такой обычно используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для пакета программ BLAST. В программах GCG Wisconsin обычно используются общепризнанные значения по умолчанию или заказная таблица сравнения символов в случае ее наличия (см. руководство пользователя для более подробного ознакомления). В некоторых применениях желательно использовать общепризнанные значения по умолчанию для пакета программ GCG или в случае другого программного обеспечения матрицу по умолчанию, такую как BLOSUM62.
Альтернативно процентное значение гомологии можно вычислить, используя множественный сравнительный анализ в программе DNASIS™ (Hitachi Software), выполняемый на основании алгоритма, аналогичного программе CLUSTAL (Higgins D.G. & Sharp P.M. (1988), Gene 73(1), 237-244).
После выполнения программой оптимального сравнительного анализа можно вычислить процентное значение гомологии, предпочтительно процентное значение идентичности последовательностей. Программа обычно выполняет данную операцию в виде части сравнения последовательностей и выводит числовой результат.
Последовательности могут также иметь делеции, инсерции или замены аминокислотных остатков, которые образуют молчащую замену и позволяют получить функционально эквивалентное вещество. Намеренные замены аминокислот могут быть произведены на основании сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатического характера остатков при сохранении вторичной связующей активности данного вещества. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными головными группами, характеризующиеся подобными значениями гидрофильности, включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин.
Консервативные замены могут быть произведены, например, в соответствии с приведенной ниже таблицей. Аминокислоты, приведенные в одном блоке второго столбца и предпочтительно в одной строке третьего столбца, могут быть заменены друг другом.
Настоящее изобретение относится также к гомологичному замещению (термины “замещение” и “замена” в используемом здесь значении означают замену существующего аминокислотного остатка альтернативным остатком), которое может иметь место, то есть к замещению подобными остатками, в частности основного остатка основным, кислотного остатка кислотным, полярного остатка полярным и т.д. Кроме того, может иметь место негомологичное замещение, то есть замена остатка одного класса остатком другого класса или альтернативно введение неприродных аминокислот, таких как орнитин (далее обозначаемый символом Z), орнитин диаминомасляной кислоты (далее обозначаемый символом В), орнитин норлейцина (далее обозначаемый символом О), пириилаланин, тиенилаланин, нафтилаланин и фенилглицин.
Кроме того, замены могут быть произведены неприродными аминокислотами.
Вариантные аминокислотные последовательности могут включать приемлемые спейсерные группы, которые могут быть введены между любыми двумя аминокислотными остатками последовательности, в частности алкильные группы, такие как метильные, этильные или пропильные группы, помимо аминокислотных спейсеров, таких как остатки глицина или β-аланина. Другая форма вариации, предполагающая наличие одного или нескольких аминокислотных остатков в пептоидной форме, должна быть хорошо известна специалистам в данной области. Во избежание сомнений термин “пептоидная форма” означает вариантные аминокислотные остатки, в которых группа, замещающая α-атом углерода, находится в положении атома азота остатка, а не α-атома углерода. Способы получения пептидов в пептоидной форме хорошо известны в данной области и описаны, например, в публикациях Simon R.J. et al., PNAS (1992) 89(20), 9367-9371 и Horwell D.C., Trends Biotechnol. (1995) 13(4), 132-134.
Нуклеотидные последовательности, предназначенные для использования в настоящем изобретении или кодирующие полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, могут включать синтетические или модифицированные нуклеотиды. В данной области известен целый ряд разных типов модификации олигонуклеотидов. Указанные модификации включают метилфосфонатные и фосфортиоатные остовы и/или добавление цепей акридина или полилизина по 3'- и/или 5'-концам молекулы. В соответствии с целями настоящего изобретения должно быть понятно, что нуклеотидные последовательности, как описано в настоящем изобретении, могут быть модифицированы любым методом, известным в данной области. Такие модификации могут быть выполнены для усиления активности in vivo или продолжительности жизни нуклеотидных последовательностей.
Настоящее изобретение относится также к использованию нуклеотидных последовательностей, которые являются комплементарными последовательностям, описанным в настоящем изобретении, или их производным, фрагментам или их производным. Последовательность, комплементарную ее фрагменту, можно использовать в качестве зонда для идентификации подобных кодирующих последовательностей в других организмах и т.д.
Полинуклеотиды, которые не являются на 100% гомологичными последовательностям по настоящему изобретению, но входят в объем изобретения, можно получить целым рядом способов. Другие варианты последовательностей, описанных в настоящем изобретении, можно получить, например, путем зондирования библиотек ДНК, созданных из ряда особей, например из особей, относящихся к разным популяциям. Кроме того, могут быть получены другие вирусные/бактериальные или клеточные гомологи, в частности клеточные гомологи, обнаруженные в клетках млекопитающих (например, в клетках крыс, мышей, крупного рогатого скота и приматов); такие гомологи и их фрагменты могут избирательно гибридизировать с последовательностями, представленными в списке последовательностей, прилагаемом к данному описанию изобретения. Такие последовательности могут быть получены путем зондирования библиотек кДНК, созданных из библиотек геномных ДНК других видов животных, при помощи зондов, содержащих всю последовательность или часть любой из последовательностей в прилагаемом списке последовательностей, в условиях от средней до высокой строгости. Аналогичные соображения относятся к получению видовых гомологов и аллельных вариантов полипептидных или нуклеотидных последовательностей по настоящему изобретению.
Варианты и гомологи штамма/вида могут быть также получены при помощи PCR вырожденной последовательности с использованием праймеров, созданных для целевых последовательностей внутри вариантов и гомологов, кодирующих консервативные аминокислотные последовательности в последовательностях по настоящему изобретению. Консервативные последовательности можно прогнозировать, например, путем сравнения аминокислотных последовательностей из нескольких вариантов/гомологов. Сравнительный анализ последовательностей может быть произведен при помощи компьютерных программ, известных в данной области. Например, широко используется программа GCG Wisconsin PileUp.
Праймеры, используемые при осуществлении PCR вырожденной последовательности, содержат одно или несколько вырожденных положений и могут быть использованы в условиях более низкой строгости по сравнению с применяемыми для клонирования последовательностей при помощи праймеров для одной последовательности против известных последовательностей.
Альтернативно такие полинуклеотиды могут быть получены при помощи сайт-направленного мутагенеза описанных последовательностей. Указанный метод может быть полезен при необходимости молчащего изменения последовательности кодона для оптимизации преференций кодона в отношении определенной клетки-хозяина, в которой предполагается экспрессировать данные полинуклеотидные последовательности. Другие изменения последовательности могут быть желательны для введения сайтов узнавания рестрикционного полипептида или для изменения свойства или функции полипептидов, кодированных указанными полинуклеотидами.
Полинуклеотиды (нуклеотидные последовательности) по настоящему изобретению могут быть использованы для создания праймера, например праймера для РCR, праймера для альтернативной реакции амплификации, зонда, например, меченного обнаруживаемой меткой известными методами с использованием радиоактивных или нерадиоактивных меток, либо указанные полинуклеотиды могут быть клонированы в векторах. Такие праймеры, зонды и другие фрагменты могут иметь длину, равную по крайней мере 15, предпочтительно по крайней мере 20, например, по крайней мере 25, 30 или 40 нуклеотидам, и входят в определение полинуклеотидов по настоящему изобретению, используемое в данном описании изобретения.
Полинуклеотиды, такие как полинуклеотиды ДНК и зонды по настоящему изобретению, могут быть получены рекомбинантными, синтетическими или любыми другими методами, известными специалистам в данной области. Указанные полинуклеотиды могут быть также клонированы стандартными методами.
Праймеры, как правило, получают синтетическими методами, включающими поэтапное создание требуемой последовательности нуклеиновой кислоты путем синтеза одного нуклеотида за другим. В данной области хорошо известны автоматические методы создания последовательностей нуклеиновых кислот.
Более длинные полинуклеотиды обычно получают рекомбинантными методами, например, используя методы клонирования при помощи PCR (полимеразная реакция синтеза цепи). Указанный метод включает создание пары праймеров (например, длиной около 15-30 нуклеотидов), фланкирующих область последовательности, направленно воздействующей на липид, которую желательно клонировать, осуществление контактирования указанных праймеров с мРНК или кДНК, полученными из животной клетки или клетки человека, при помощи полимеразной реакции синтеза цепи в условиях, вызывающих амплификацию требуемой области, выделение амплифицированного фрагмента (например, путем очистки реакционной смеси на агарозном геле) и получение амплифицированной ДНК. Созданные праймеры могут содержать приемлемые сайты узнавания рестрикционных ферментов, благодаря чему амплифицированная ДНК может быть клонирована в приемлемом клонирующем векторе.
Гибридизация
Настоящее изобретение относится также к последовательностям, которые являются комплементарными последовательностями по настоящему изобретению или последовательностям, способным гибридизировать с последовательностями по настоящему изобретению или с последовательностями, комплементарными указанным последовательностям.
Термин “гибридизация” в используемом здесь значении означает “процесс, в соответствии с которым цепь нуклеиновой кислоты соединяется с комплементарной цепью в результате спаривания оснований”, а также процесс амплификации, осуществляемый методами полимеразной реакции синтеза цепи (PCR).
Настоящее изобретение относится также к использованию нуклеотидных последовательностей, способных гибридизировать с последовательностями, комплементарными последовательностям по настоящему изобретению, или любому их производному, фрагменту или его производному.
Настоящее изобретение относится также к последовательностям, которые являются комплементарными последовательностям, способным гибридизировать с нуклеотидными последовательностями, описанными в настоящем изобретении.
Условия гибридизации выбирают с учетом температуры плавления (Тm) нуклеотид-связывающего комплекса, как это описано в публикации Berger and Kimmel (1987, Guide to Molecular Cloning Techniques, Methods in Enzymology, Vol.152, Academic Press, San Diego, CA), при этом условия гибридизации соответствуют определенной степени “строгости”, рассмотренной ниже.
Максимальная строгость обычно имеет место при температуре около Tm-5°С (на 5°С ниже Тm зонда); высокая строгость имеет место при температуре, которая примерно на 5°С-10°С ниже Tm; средняя строгость имеет место при температуре, которая примерно на 10°С-20°С ниже Тm; и низкая строгость имеет место при температуре, которая примерно на 20°С-25°С ниже Тm. Как должно быть понятно специалистам в данной области, гибридизация в условиях максимальной строгости может быть использована для идентификации или обнаружения идентичных нуклеотидных последовательностей, в то время как гибридизация в условиях средней (или низкой) строгости может быть использована для идентификации или обнаружения подобных или родственных полинуклеотидных последовательностей.
Настоящее изобретение предпочтительно относится к последовательностям, которые являются комплементарными последовательностям, способным гибридизировать в условиях высокой или средней строгости с нуклеотидными последовательностями, кодирующими полипептиды, обладающие специфическими свойствами, как описано в настоящем изобретении.
Более предпочтительно настоящее изобретение относится к последовательностям, которые являются комплементарными последовательностям, способным гибридизировать в условиях высокой строгости (например, при 65°С и 0,1хSSC {1xSSC = 0,15 M NaCl, 0,015 M цитрата натрия, рН 7,0}) с нуклеотидными последовательностями, кодирующими полипептиды, обладающие специфическими свойствами, как описано в настоящем изобретении.
Настоящее изобретение относится также к нуклеотидным последовательностям, способным гибридизировать с нуклеотидными последовательностями, описанными в данном изобретении (включая комплементарные последовательности описанных здесь последовательностей).
Настоящее изобретение относится также к нуклеотидным последовательностям, которые являются комплементарными последовательностям, способным гибридизировать с нуклеотидными последовательностями, описанными в данном изобретении (включая комплементарные последовательности описанных здесь последовательностей).
Кроме того, в объем настоящего изобретения входят полинуклеотидные последовательности, способные гибридизировать с нуклеотидными последовательностями, описанными в данном изобретении, в условиях от средней до максимальной строгости.
Предпочтительным объектом настоящего изобретения являются нуклеотидные последовательности, способные гибридизировать с нуклеотидными последовательностями, описанными в данном изобретении, или с их комплементом в строгих условиях (например, 50°С и 0,2хSSC).
Более предпочтительным объектом настоящего изобретения являются нуклеотидные последовательности, способные гибридизировать с нуклеотидными последовательностями, описанными в данном изобретении, или с их комплементом в условиях высокой строгости (например, 65°С и 0,1хSSC).
Экспрессия полипептидов
Нуклеотидная последовательность, предназначенная для использования в настоящем изобретении или кодирующая полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, может быть введена в рекомбинантный реплицируемый вектор. Указанный вектор может быть использован для репликации и экспрессии нуклеотидной последовательности в форме полипептида в совместимой клетке-хозяине и/или из совместимой клетки-хозяина. Экспрессию можно контролировать при помощи управляющих последовательностей, которые включают промоторы/энхансеры и другие сигналы регулирования экспрессии. Могут быть использованы прокариотические промоторы и промоторы, функционирующие в эукариотических клетках. Можно использовать тканеспецифичные или стимулоспецифичные промоторы. Кроме того, можно использовать химерные промоторы, включающие элементы последовательности из двух или более разных вышеописанных промоторов.
Полипептид, продуцированный рекомбинантной клеткой-хозяином в результате экспрессии нуклеотидной последовательности, может быть секретирован из клетки или может находиться внутри клетки в зависимости от используемой последовательности и/или вектора. Кодирующие последовательности могут иметь сигнальные последовательности, которые направляют секрецию кодирующих последовательностей вещества через мембрану определенной прокариотической или эукариотической клетки.
Экспрессирующий вектор
Термин “экспрессирующий вектор” означает конструкцию, способную экспрессировать последовательность in vivo или in vitro.
Экспрессирующий вектор предпочтительно встраивают в геном организма. Термин “встроенный” предпочтительно означает устойчивое введение в геном.
Нуклеотидная последовательность по настоящему изобретению или кодирующая полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, может находиться в векторе, в котором данная нуклеотидная последовательность функционально связана с регуляторными последовательностями, которые способны вызывать экспрессию данной нуклеотидной последовательности приемлемым организмом-хозяином, то есть такой вектор является экспрессирующим вектором.
Векторы по настоящему изобретению могут быть введены в приемлемую нижеописанную клетку-хозяин для экспрессии полипептида, обладающего специфическими свойствами, описанного в настоящем изобретении.
Выбор вектора, например плазмидного, космидного, вирусного или фагового вектора, часто зависит от клетки-хозяина, предназначенной для ввода данного вектора.
Векторы могут содержать один или несколько селектируемых генов-маркеров, таких как ген, который сообщает устойчивость к антибиотикам, например ампициллину, канамицину, хлорамфениколу или тетрациклину. Альтернативно выбор можно произвести методом котрансформации (описанным в WО 91/17243).
Векторы могут быть использованы in vitro, например, для продуцирования РНК или для трансфекции или трансформации клетки-хозяина.
Таким образом, еще один вариант осуществления изобретения относится к способу получения нуклеотидных последовательностей по настоящему изобретению или нуклеотидных последовательностей, кодирующих полипептиды, обладающие специфическими свойствами, как описано в настоящем изобретении, путем введения нуклеотидной последовательности в реплицируемый вектор, введения вектора в совместимую клетку-хозяин и выращивания клетки-хозяина в условиях, вызывающих репликацию вектора.
Вектор может далее содержать нуклеотидную последовательность, вызывающую репликацию вектора в клетке-хозяине. Примеры таких последовательностей включают ориджины репликации плазмид pUC19, pACYC177 pUB110, pE194, pAMB1 и pIJ702.
Регуляторные последовательности
В некоторых применениях нуклеотидная последовательность, предназначенная для использования в настоящем изобретении, или нуклеотидная последовательность, кодирующая полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, может быть функционально связана с регуляторной последовательностью, которая способна вызывать экспрессию нуклеотидной последовательности выбранной клеткой-хозяином. В качестве примера можно отметить, что настоящее изобретение относится к вектору, содержащему нуклеотидную последовательность по настоящему изобретению, функционально связанную с такой регуляторной последовательностью, то есть данный вектор является экспрессирующим вектором.
Термин “функционально связанный” означает смежное положение, в котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать требуемым образом. Регуляторная последовательность, “функционально связанная” с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с управляющими последовательностями.
В определение термина “регуляторные последовательности” входят промоторы, энхансеры и другие сигнальные последовательности, регулирующие экспрессию.
Термин “промотор” имеет обычное значение, используемое в данной области, например сайт связывания РНК-полимеразы.
Повышенная экспрессия нуклеотидной последовательности, кодирующей фермент, обладающий специфическими свойствами, описанными в настоящем изобретении, может быть также достигнута путем выбора гетерологичных регуляторных областей, например промоторных, лидерных и терминирующих областей, определяющих секрецию.
Нуклеотидная последовательность по настоящему изобретению может быть предпочтительно функционально связана по крайней мере с промотором.
Примеры приемлемых промоторов, направляющих транскрипцию нуклеотидной последовательности, в бактериальной, грибной или дрожжевой клетке-хозяине, хорошо известны в данной области.
Конструкции
Термин “конструкция”, который синонимичен таким терминам как “конъюгат”, “кластер” и “гибрид”, означает нуклеотидную последовательность, присоединенную к промотору прямой или непрямой связью, которая кодирует описанный здесь полипептид, обладающий специфическими свойствами и предназначенный для использования в настоящем изобретении. Примером непрямого присоединения является использование спейсерной группы, такой как интронная последовательность, в частности Sh1-интрон или интрон ADH, которая расположена между промотором и нуклеотидной последовательностью по настоящему изобретению. То же самое определение является верным для термина “слитый”, используемого в настоящем изобретении, который предполагает прямое или непрямое присоединение. В некоторых случаях указанные термины не относятся к природной комбинации нуклеотидной последовательности, кодирующей белок, обычно ассоциированный с промотором гена дикого типа, и к ситуациям, когда оба вышеуказанных элемента находятся в своем естественном окружении.
Указанная конструкция может даже содержать или экспрессировать маркер, позволяющий выбрать генетическую конструкцию.
В некоторых применениях указанная конструкция предпочтительно содержит по крайней мере одну нуклеотидную последовательность по настоящему изобретению или нуклеотидную последовательность, кодирующую описанный здесь полипептид, обладающий специфическими свойствами, который функционально связан с промотором.
Клетки-хозяева
Термин “клетка-хозяин”, используемый в настоящем изобретении, означает любую клетку, которая содержит нуклеотидную последовательность, кодирующую полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, или вышеуказанный экспрессирующий вектор и используется для рекомбинантного продуцирования описанного здесь полипептида, обладающего специфическими свойствами.
Таким образом, еще один вариант осуществления настоящего изобретения относится к клеткам-хозяевам, трансформированным или трансфецированным нуклеотидной последовательностью по настоящему изобретению или нуклеотидной последовательностью, экспрессирующей полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении. Выбираемые клетки должны быть совместимы с указанным вектором и могут быть, например, прокариотическими (например, бактериальными), грибными, дрожжевыми или растительными клетками. Клетки-хозяева предпочтительно не являются клетками человека.
Примерами приемлемых бактериальных организмов-хозяев являются грамотрицательные или грамположительные бактерии.
В зависимости от природы нуклеотидной последовательности, кодирующей полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, и/или желания дальнейшей обработки экспрессированного белка предпочтительными могут быть эукариотические клетки-хозяева, такие как дрожжи или другие грибы.
Дрожжевые клетки обычно являются более предпочтительными по сравнению с клетками грибов, так как ими легче манипулировать. Однако некоторые белки плохо секретируются из дрожжевой клетки и в некоторых случаях плохо процессируются (например, гипергликозилирование в дрожжах). В таких случаях необходимо выбрать организм-хозяин из других грибов.
Использование приемлемых клеток-хозяев, таких как дрожжевые, грибные и растительные клетки-хозяева, позволяет произвести пост-трансляционные модификации (например, миристоилирование, гликозилирование, усечение, лапидирование и фосфорилирование тирозина, серина или треонина), которые могут быть необходимы для сообщения оптимальной биологической активности рекомбинантным экспрессированным продуктам по настоящему изобретению.
Клетка-хозяин может не иметь протеазы или может относиться к штамму, лишенному протеазы.
Организм
Термин “организм” применительно к настоящему изобретению означает любой организм, который может содержать нуклеотидную последовательность по настоящему изобретению или нуклеотидную последовательность, кодирующую полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, и/или полученные из него продукты.
Приемлемые организмы могут включать прокариоты, грибы, дрожжи или растения.
Термин “трансгенный организм” применительно к настоящему изобретению означает любой организм, который содержит нуклеотидную последовательность, кодирующую полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, и/или полученные из него продукты, и/или в котором промотор может стимулировать экспрессию нуклеотидной последовательности, кодирующей полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении. Нуклеотидную последовательность предпочтительно встраивают в геном организма.
В определение термина “трансгенный организм” не входят нативные нуклеотидные кодирующие последовательности в их естественном окружении, где они контролируются нативным промотором, который также находится в своем естественном окружении.
Таким образом, трансгенный организм по настоящему изобретению представляет собой организм, который включает одну или несколько нуклеотидных последовательностей, кодирующих полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, а также конструкции, векторы, плазмиды, клетки или их продукты по настоящему изобретению. Например, трансгенный организм может также содержать нуклеотидную последовательность, кодирующую полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, которая находится под контролем гетерологичного промотора.
Трансформация клеток-хозяев/организма
Как было указано выше, организм-хозяин может быть прокариотическим или эукариотическим организмом. Примеры приемлемых прокариотических хозяев включают E. coli и Bacillus subtilis.
Методы трансформации прокариотических хозяев всесторонне описаны в данной области; см., например, публикацию Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd edition, 1989, Cold Spring Harbor Laboratory Press). При использовании прокариотического хозяина нуклеотидная последовательность может потребовать соответствующей модификации перед трансформацией, например удаления интронов.
В другом варианте осуществления изобретения трансгенным организмом могут быть дрожжи.
Клетки нитевидных грибов могут быть трансформированы разными методами, известными в данной области, такими как образование протопластов и трансформация полученных протопластов с последующей регенерацией клеточной оболочки известным методом. Использование Aspergillus в качестве микроорганизма-хозяина описано в европейском патенте № 0238023.
Другим организмом-хозяином может быть растение. Общие методы, используемые для трансформации растений, описаны в статьях Potrykus (Annu. Rev. Plant Physiol. Plant Mol. Biol. [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994 17-27). Другие методы трансформации растений описаны в заявке на европейский патент № 0449375.
Общие методы трансформации грибов, дрожжей и растений представлены в нижеследующих разделах.
Трансформированные грибы
Организмом-хозяином может быть гриб, в частности нитевидный гриб. Примеры приемлемых грибов-хозяев включают любые грибы, относящиеся к родам Thermomyces, Acremonium, Aspergillus, Penicillium, Mucor, Neurospora, Trichoderma и тому подобным.
Методы трансформации нитевидных грибов описаны в заявке на патент США № 5741665, в которой указано, что в данной области хорошо известны стандартные методы трансформации нитевидных грибов и культивирования грибов. Широкий обзор методов трансформации грибов N. crassa приведен, например, в публикации Davis and de Serres, Methods Enzymol. (1971) 17A:79-143.
Другие методы трансформации нитевидных грибов описаны в заявке на патент США № 5674707.
В cоответствии с одним объектом изобретения организм-хозяин может относиться к роду Aspergillus, такому как Aspergillus niger.
Трансгенные грибы рода Aspergillus по настоящему изобретению могут быть также получены методами, описанными, например, в публикации Turner G. 1994 (Vectors for genetic manipulation. In: Martinelli S.D., Kinghorn J.R. (Editors) Aspergillus: 50 years on. Progress in industrial microbiology vol.29. Elsevier Amsterdam 1994, pp.641-666).
Методы экспрессии генов в нитевидных грибах описаны в публикации Punt et al. (2002) Trends Biotechnol. 2002 May; 20(5):200-6, Archer & Peberdy Crit. Rev. Biotechnol. (1997) 17(4):273-306.
Трансформированные дрожжи
В другом варианте осуществления изобретения трансгенным организмом могут быть дрожжи.
Принципы экспрессии гетерологичного гена в дрожжах описаны, например, в публикациях Methods Mol. Biol. (1995), 49:341-54 и Curr. Opin. Biotechnol. (1997) Oct; 8(5):554-60.
В данной связи следует отметить, что в качестве носителя для экспрессии гетерологичного гена можно использовать такие дрожжи, как виды Saccharomyces cerevisiае или Pichia pastoris (см. FEMS Microbiol Rev.(2000 24(1):45-66).
Принципы экспрессии гетерологичного гена в дрожжах Saccharomyces cerevisiae и секреции генных продуктов описаны в публикациях E. Hinchcliffe E Kenny (1993, “Yeast as a vehiclе for the expression of heterologous genes”, Yeasts, Vol.5, Antony H. Rose and J. Stuart Harrison, eds, 2nd edition, Academic Press Ltd.).
Для трансформации дрожжей было разработано несколько методов. Например, трансгенные дрожжи Saccharomyces по настоящему изобретению можно получить методами, описанными в нижеследующих публикациях Hinnen et al. (1978, Proceedings of the National Academy of Sciences of the USA 75, 1929); Beggs, J.D. (1978, Nature, London, 275, 104) и Ito, H. et al. (1983, J. Bacteriology 153, 163-168).
Трансформированные дрожжевые клетки могут быть отобраны при помощи разных селектируемых маркеров, таких как ауксотрофные маркеры, которые являются главными маркерами устойчивости к антибиотикам.
Приемлемые дрожжи-хозяева могут быть выбраны из биотехнологически приемлемых видов дрожжей, которые включают, не ограничиваясь ими, дрожжи, выбираемые из видов Pichia, Hansenula, Kluyveromyces, Yarrowinia, Saccharomyces, включая S. cerevisiae, или Schizosaссharomyce, включая Schizosaccharomyce pombe.
В качестве организма-хозяина можно использовать штамм метилотрофических дрожжей вида Pichia pastoris.
В одном варианте осуществления изобретения организмом-хозяином может быть вид Hansenula, такой как H. polymorpha (описанный в WO 01/39544).
Трансформированные растения/растительные клетки
Организмом-хозяином, пригодным для использования в настоящем изобретении, может быть растение. Общие методы трансформации растений описаны в статьях Potrykus (Annu. Rev. Plant Physiol. Plant Mol. Biol. [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994 17-27) или в WO 01/16308. Трансгенное растение может продуцировать большие количества сложных эфиров фитостерола и сложных эфиров фитостанола.
Поэтому настоящее изобретение относится также к способу получения трансгенного растения с повышенным содержанием сложных эфиров фитостерола и сложных эфиров фитостанола, который включает стадии трансформации растительной клетки липид-ацилтрансферазой, описанной в настоящем изобретении (в частности, экспрессирующим вектором или конструкцией, включающей липид-ацилтрансферазу по настоящему изобретению), и выращивания растения из трансформированной растительной клетки.
Секреция
Часто бывает желательно секретировать полипептид из экспрессирующего хозяина в культуральную среду, из которой фермент можно гораздо легче выделить. В соответствии с настоящим изобретением секретируемая лидерная последовательность может быть выбрана с учетом желаемого экспрессирущего хозяина. В контексте настоящего изобретения могут быть также использованы гибридные сигнальные последовательности.
Типичными примерами гетерологичных секретируемых лидерных последовательностей являются последовательности гена амилоглюкозидазы (AG) грибов (glaA включает варианты, состоящие из 18 и 24 аминокислот, полученные, например, из вида Aspergillus), гена а-фактора (например, дрожжи Saccharomyces, Kluyveromyces и Hansenula) или гена α-амилазы (Bacillus).
Обнаружение
В данной области известен целый ряд методов обнаружения и измерения экспрессии аминокислотных последовательностей. Такие методы включают твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) и клеточный сортинг с возбуждением флуоресценции (FACS).
Специалистам в данной области известно широкое разнообразие меток и методов конъюгации, которые могут быть использованы в разных анализах нуклеиновых кислот и аминокислот.
Ряд компаний, таких как Pharmacia Biotech (Piscataway, NJ), Promega (Madison, WI) и US Biochemical Corp. (Cleveland, OH), производят коммерчески доступные наборы, содержащие инструкции по выполнению указанных анализов.
Приемлемые репортерные молекулы или метки включают радиоактивные изотопы, ферменты, люминесцентные, хемилюминесцентные или хромогенные агенты, а также субстраты, кофакторы, ингибиторы, магнитные частицы и тому подобные. Патенты, в которых описано применение таких меток, включают US-A-3817837, US-A-3850752, US-A-3939350, US-A-3996345, US-A-4277437, US-A-4275149 и US-A-4366241.
Кроме того, могут быть получены рекомбинантные иммуноглобулины, описанные в патенте US-A-4816567.
Слитые белки
Полипептид, обладающий специфическими свойствами, описанный в настоящем изобретении, может быть получен в виде слитого белка, например, для облегчения его экстракции и очистки. Примерами партнеров слитых белков являются глутатион-S-трансфераза (GST), 6xHis, GAL4 (ДНК-связывающие и/или активирующие транскрипцию домены) и β-галактозидаза. Кроме того, между партнером слитого белка и представляющей интерес белковой последовательностью может быть введен сайт протеолитического расщепления, позволяющий удалить последовательности слитого белка. Слитый белок предпочтительно не подавляет активность белковой последовательности.
Системы экспрессии генов слитого белка в E. coli описаны в публикации Curr. Opin. Biotechnol. (1995) 6(5):501-6.
В другом варианте осуществления изобретения аминокислотная последовательность полипептида, обладающего специфическими свойствами, описанного в настоящем изобретении, может быть лигирована с гетерологичной последовательностью для кодирования слитого белка. Например, для скрининга пептидных библиотек в отношении агентов, способных влиять на активность вещества, может быть полезно кодировать химерное вещество, экспрессирующее гетерологичный эпитоп, узнаваемый коммерчески доступным антителом.
Настоящее изобретение далее будет описано только в качестве примера со ссылкой на нижеследующие фигуры и примеры.
На фиг.1 показана консенсусная последовательность pfam00657 из базы данных версии 6 (SEQ ID NO:1).
На фиг.2 показана аминокислотная последовательность (SEQ ID NO:2), полученная из организма Aeromonas hydrophila (P10480; GI:121051).
На фиг.3 показана аминокислотная последовательность (SEQ ID NO:3), полученная из организма Aeromonas salmonicida (AAG098404; GI:9964017).
На фиг.4 показана аминокислотная последовательность (SEQ ID NO:4), полученная из организма Streptomyces coelicolor A3(2) (номер доступа NP_631558 в банке генов Genbank).
На фиг.5 показана аминокислотная последовательность (SEQ ID NO:5), полученная из организма Streptomyces coelicolor A3(2) (номер доступа САС42140 в банке генов Genbank).
На фиг.6 показана аминокислотная последовательность (SEQ ID NO:6), полученная из организма Saccharomyces cerevisiae (номер доступа Р41734 в банке генов Genbank).
На фиг.7 показан сравнительный анализ выбранных последовательностей с консенсусной последовательностью pfam00657.
На фиг.8 показан попарный сравнительный анализ SEQ ID NO:3 с SEQ ID NO:2, результаты которого свидетельствуют о 93% идентичности аминокислотных последовательностей. Сигнальная последовательность подчеркнута. Символом + отмечены различия. Фрагмент GDSX, содержащий серин в положении 16 активного сайта, аспарагиновую кислоту в положении 116 и гистидин в положении 291 активных сайтов, выделен (см. заштрихованные участки). Числовые обозначения после аминокислоты указаны без сигнальной последовательности.
На фиг.9 показана нуклеотидная последовательность (SEQ ID NO:7), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Aeromonas hydrophila.
На фиг.10 показана нуклеотидная последовательность (SEQ ID NO:8), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Aeromonas salmonicida.
На фиг.11 показана нуклеотидная последовательность (SEQ ID NO:9), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Streptomyces coelicolor A3(2) (номер доступа NC_003888.1:8327480..8328367 в банке генов Genbank).
На фиг.12 показана нуклеотидная последовательность (SEQ ID NO:10), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Streptomyces coelicolor A3(2) (номер доступа AL939131.1:265480..266367 в банке генов Genbank).
На фиг.13 показана нуклеотидная последовательность (SEQ ID NO:11), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Saccharomyces cerevisiae (номер доступа Z75034 в банке генов Genbank).
На фиг.14 показана аминокислотная последовательность (SEQ ID NO:12), полученная из организма Ralstonia (номер доступа AL646052 а банке генов Genbank).
На фиг.15 показана нуклеотидная последовательность (SEQ ID NO:13), кодирующая липид-ацилтрансферазу по настоящему изобретению, полученную из организма Ralstonia.
На фиг.16 показана SEQ ID NO:20. Белок Scoe1 NCBI с кодом доступа САВ396707.1 GI:4539178, который является консервативным гипотетическим белком [Streptomyces coelicolor A3(2)].
На фиг.17 показана нуклеотидная последовательность в виде SEQ ID NO:21, кодирующая белок NCBI с кодом доступа САВ39707.1 GI:4539178, который является консервативным гипотетическим белком [Streptomyces coelicolor A3(2)].
На фиг.18 показана аминокислотная последовательность в виде SEQ ID NO:22. Белок Scoe2 NCBI с кодом доступа САС01477.1 GI:9716139, который является консервативным гипотетическим белком [Streptomyces coelicolor A3(2)].
На фиг.19 показана нуклеотидная последовательность в виде SEQ ID NO:23, кодирующая белок Scoe2 NCBI с кодом доступа САС01477.1 GI:9716139, который является консервативным гипотетическим белком [Streptomyces coelicolor A3(2)].
На фиг.20 показана аминокислотная последовательность (SEQ ID NO:24). Белок Scoe3 NCBI с кодом доступа САВ88833.1 GI:7635996, который является предполагаемым секретированным белком [Streptomyces coelicolor A3(2)].
На фиг.21 показана нуклеотидная последовательность в виде SEQ ID NO:25, кодирующая белок Scoe3 NCBI с кодом доступа САВ88833.1 GI:7635996, который является предполагаемым секретированным белком [Streptomyces coelicolor A3(2)].
На фиг.22 показана аминокислотная последовательность (SEQ ID NO:26). Белок Scoe4 NCBI с кодом доступа САВ89450.1 GI:7672261, который является предполагаемым секретированным белком [Streptomyces coelicolor A3(2)].
На фиг.23 показана нуклеотидная последовательность в виде SEQ ID NO:27, кодирующая белок Scoe4 NCBI с кодом доступа САВ89450.1 GI:7672261, который является предполагаемым секретированным белком [Streptomyces coelicolor A3(2)].
На фиг.24 показана аминокислотная последовательность (SEQ ID NO:28). Белок Scoe5 NCBI с кодом доступа САВ62724.1 GI:6562793, который является предполагаемым липопротеином [Streptomyces coelicolor A3(2)].
На фиг.25 показана нуклеотидная последовательность в виде SEQ ID NO:29, кодирующая белок Scoe5 NCBI с кодом доступа САВ62724.1 GI:6562793, который является предполагаемым липопротеином [Streptomyces coelicolor A3(2)].
На фиг.26 показана аминокислотная последовательность (SEQ ID NO:30). Белок Srim1 NCBI с кодом доступа ААК84028.1 GI:15082088, который является GDSL-липазой [Strepromyces rimosus].
На фиг.27 показана нуклеотидная последовательность в виде SEQ ID NO:31, кодирующая белок Srim1 NCBI с кодом доступа ААК84028.1 GI:15082088, который является GDSL-липазой [Streptomyces rimosus].
На фиг.28 показана аминокислотная последовательность (SEQ ID NO:32). Липид-ацилтрансфераза, полученная из Aeromonas hydrophila (АТСС № 7965).
На фиг.29 показана нуклеотидная последовательность (SEQ ID NO:33), кодирующая липид-ацилтрансферазу, полученную из Aeromonas hydrophila (АТСС № 7965).
На фиг.30 показана аминокислотная последовательность (SEQ ID NO:34) липид-ацилтрансферазы, полученной из подвида Aeromonas salmoniсida вида Salmonicida (АТСС № 14174).
На фиг.31 показана нуклеотидная последовательность (SEQ ID NO:35), кодирующая липид-ацилтрансферазу, полученную из подвида Aeromonas salmonicida вида Salmonicida (АТСС № 14174).
На фиг.32 показано, что гомологи генов Aeromonas могут быть идентифицированы при помощи базовой локальной поисковой инструментальной службы выполнения сравнительного анализа в Национальном центре биотехнологической информации, NIH, MD, США, с использованием существующих баз данных геномов. Для поиска в базах данных был использован фрагмент GDSX и при этом был идентифицирован ряд последовательностей/генов, предположительно кодирующих ферменты, обладающие липолитической активностью. Гены были идентифицированы в роде Streptomyces, Xanthomоnas и Ralstonia. Как указано в нижеследующем примере, ген Ralstonia solanacearum сравнивали с геном Aeromonas salmonicida (satA). Попарное сравнение позволило выявить 23% идентичность последовательностей. При этом можно идентифицировать серин в активном сайте у аминоконца и каталитические остатки гистидина и аспарагиновой кислоты.
На фиг.33 показана консенсусная последовательность Pfam00657.11 [семейство 00657, версия 11 базы данных] (далее именуемая консенсусной последовательностью Pfam) и сравнительный анализ разных последовательностей с консенсусной последовательностью Pfam. Стрелками показаны остатки активных сайтов, подчеркнутые блоки являются тремя гомологичными блоками, описанными в публикации [Upton C. and Buckley J.T. (1995) Trends Biochem. Sci. 20; 179-179]. Заглавными буквами в консенсусной последовательности Pfam обозначены консервативные остатки, имеющиеся во многих членах семейства. Символ “-” обозначает положение, в котором при помощи скрытой модели Маркова консенсусной последовательности Pfam предполагалось найти остаток, но такой остаток отсутствовал, поэтому был введен пробел. Символ “-” обозначает остаток без соответствующего остатка в консенсусной последовательности Pfam. Указанные последовательности являются аминокислотными последовательностями, показанными на фигурах 16, 18, 20, 22, 24, 26, 28 и 30.
На фиг.34 показана консенсусная последовательность Pfam00657.11 [семейство 00657, версия 11 базы данных] (далее именуемая консенсусной последовательностью Pfam) и сравнительный анализ разных последовательностей с консенсусной последовательностью Pfam. Стрелками показаны остатки активных сайтов, подчеркнутые блоки являются тремя гомологичными блоками, описанными в публикации [Upton C. and Buckley J.T. (1995) Trends Biochem. Sci. 20; 179-179]. Заглавными буквами в консенсусной последовательности Pfam обозначены консервативные остатки, имеющиеся во многих членах семейства. Символ “-” обозначает положение, в котором при помощи скрытой модели Маркова консенсусной последовательности Pfam предполагалось найти остаток, но такой остаток отсутствовал, поэтому был введен пробел. Символ “.” обозначает остаток без соответствующего остатка в консенсусной последовательности Pfam. Указанные последовательности являются аминокислотными последовательностями, показанными на фиг.2, 16, 18, 20, 26, 28 и 30. Было установлено, что все указанные белки обладают активностью против липидных субстратов.
На фиг.35 показан экспрессирующий вектор pet12-AsalGCAT=pSM, содержащий С-концевой His-меченый ген липид-ацилтрансферазы Aeromonas salmonicida.
На фиг.36 представлены результаты исследования клеточных экстрактов при помощи анализа с использованием набора NEFA, которые показывают активность рекомбинантной липид-ацилтрансферазы A.salmоnicida по отношению к лецитину. Лунки слева направо содержат: положительный контрольный образец, отрицательный контрольный образец (то есть экстракты из пустой плазмиды) и образцы, полученные через 0, 1, 2 и 3 часа культивирования после индукции IPTG.
На фиг.37 представлены результаты оптимизации роста клеток BL21(DE3)pLysS, содержащих экспрессирующий вектор pet12-AsalGCAT=pSM, которые показывают, что при культивировании при 30ºС происходит образование фермента, обладающего высокой активностью в отношении лецитина. Клеточные экстракты анализировали с целью обнаружения активности фосфолипазы с использованием набора NEFA. Лунки слева направо содержат: положительный контрольный образец, отрицательный контрольный образец, образец при 20°С, образец при 30°С.
На фиг.38 показаны неочищенные клеточные экстракты из клеток BL21(DE3)pLysS, экспрессирующих активную липид-ацилтрансферазу, инкубированную с лецитиновым субстратом, и результаты анализа реакционной смеси при помощи тонкослойной хроматографии, указывающие на присутствие продуктов распада. Полосы: 1. Отсутствие фермента; 2. +A.sal - 10 мкл, 37°С; 3. +A.sal - 20 мкл, 37°С; 4. +A.sal - 10 мкл, 24°С; 5. +A.sal - 20 мкл, 24°С.
На фиг.39 показаны результаты частичной очистки ацил-трансферазы Aeromonas salmonicida, указывающие на наличие активности фосфолипазы, ассоциированной с очищенным His-меченым белком. SЕ = экстракты, обработанные ультразвуком, His = очищенные при помощи набора Ni-NTA компании Qiagen.
На фиг.40 показан экспрессирующий вектор pet12-A.h. GCAT=pSMa, содержащий С-концевой His-меченый ген глицеролипид-ацилтрансферазы (GCAT) Aeromonas hydrophila, который был использован для трансформации штамма BL21(DE3)pLysS E.coli.
На фиг.41 показаны результаты исследования активности неочищенных экстрактов (5 и 10 мкл), содержащих рекомбинантный фермент GCAT Aeromonas hydrophila, в отношении лицитина с использованием набора неэтерифицированных жирных кислот (NEFA) (Roche, Switzerland), которые указывают на присутствие фермента, обладающего активностью в отношении фосфолипида, лецитина.
На фиг.42 представлены результаты оптимизации роста клеток BL21(DE3)pLysS, содержащих экспрессирующий вектор pet12-AsalGCAT=pSM, которые показывают, что при культивировании при 30°С происходит образование фермента, обладающего высокой активностью в отношении лецитина. Клеточные экстракты анализировали в отношении активности фосфолипазы с использованием набора NEFA.
На фиг.43 показаны результаты частичной очистки ацил-трансфераз Aeromonas hydrophila и А. salmonicida, указывающие на наличие активности фосфолипазы, ассоциированной с очищенным His-меченым белком. SЕ = экстракты, обработанные ультразвуком, His = очищенные при помощи набора Ni-NTA компании Qiagen.
На фиг.44 показаны результаты экспрессии генов Aeromonas в Bacillus cubtilis 163, указывающие на продуцирование секретированного фермента, обладающего активностью в отношении как лецитина, так и DGDG. pUB-AH = конструкция, содержащая ген A. hydrophila, и pUB-AS = конструкция, содержащая ген A. salmonicida. Культуральный фильтрат инкубировали с субстратами в течение 60 минут.
На фиг.45 показана аминокислотная последовательность (SEQ ID NO:36) гибридной конструкции, используемой для мутагенеза гена липид-ацилтрансферазы Aeromonas hydrophila в примере 17. Подчеркнутые аминокислоты являются сигнальным пептидом ксиланазы.
На фиг.46 показана нуклеотидная последовательность(SEQ ID NO:45), кодирующая фермент, полученный из Aeromonas hydrophila, которая включает сигнальный пептид ксиланазы.
На фиг.47 показаны результаты анализа методом высокоэффективной тонкослойной хроматографии (ВЭТСХ) в эксперименте I.
На фиг.48 показаны результаты анализа методом высокоэффективной тонкослойной хроматографии (ВЭТСХ) в эксперименте II.
На фиг.49 показана калибровочная кривая для стандартных растворов моноглицеридов.
На фиг.50 показана калибровочная кривая для стандартных растворов диглицеридов.
На фиг.51 показана нуклеотидная последовательность, кодирующая фермент липид-ацилтрансферазу по настоящему изобретению, полученная из Streptomyces (SEQ ID NO:54).
На фиг.52 показана полипептидная последовательность фермента липид-ацилтрансферазы по настоящему изобретению, полученная из Streptomyces (SEQ ID NO:55).
Примеры
Во избежание сомнений в настоящем описании изобретения использованы нижеследующие аббревиатуры:
MONO = моноглицерид
MAG = моноацилглицерин = моноглицерид
MAG и MONO имеют взаимозаменяемые значения в настоящем изобретении
DAG = диацилглицерин
FFA = свободная жирная кислота
Пример 1
Клонирование, секвенирование и гетерологичная экспрессия трансферазы из подвида Aeromonas salmonicida вида Salmonicida
Использованные штаммы
Подвид Aeromonas salmonicida вида Salmonicida (АТСС 14174), полученный из Американской коллекции типовых культур (АТСС), выращивали в течение ночи при 30°С в среде Луриа-Бертани (LB). Клетки центрифугировали и геномную ДНК выделяли методами, применяемыми для выделения геномной ДНК в компании Qiagen Ltd. Набор буферов для выделения геномной ДНК (№ по каталогу 19060), протеаза К (№ по каталогу 19131) и РНКаза А (№ по каталогу 19101) были приобретены в компании Qiagen Ltd. (Boundary court Gatwick Court, West Sussex, RH10 2AX).
Для продуцирования рекомбинантных ферментов Aeromonas использовали бактериальный штамм-хозяин BL21(DE3)pLysS (Novagen). Компетентные клетки штамма BL21(DE3)pLysS использовали в качестве хозяина для трансформации экспрессирующим вектором pet12-AsalGCAT=pSM. Трансформанты, содержащие соответствующую плазмиду, выращивали при 37°С на агаровой среде LB, содержащей 100 мкг ампициллина/мл.
Создание экспрессирущего вектора pet12-AsalGCAT-pSM
Для всех амплификаций ДНК генов трансферазы из Aeromonas геномную ДНК (0,2-1 мкл) использовали в качестве матрицы и ДНК-полимеразу pfu (2,5 ед.) использовали с 10 мкл 10-кратного буфера для pfu, 1 мкл каждого праймера (50 пмоль/мкл), 200 мкМ dNTP в общем объеме реакционной смеси, равном 100 мкл. Реакции PCR выполняли в программируемом термоблоке в нижеследующих условиях: 95°С в течение 30 секунд, 30 циклов при 95°С в течение 30 секунд, 60°С в течение 1 минуты и 68°С в течение 2 минут. Дополнительное удлинение цепи производили при 72°С в течение 5 минут.
Амплификацию методом PCR гена трансферазы из A. salmonicida осуществляли в результате выполнения 2 отдельных реакций PCR. Первую реакцию РCR выполняли с использованием пар праймеров as1USNEW (5' AGCATATGAAAA AATGGTTTGT TTGTTTATTG GGG 3' [SEQ ID NO:56]) и asls950new (5' GTG ATG GTG GGC GAG GAA CTC GTA CTG 3' [SEQ ID NO:37]). Вторую реакцию PCR выполняли для введения С-концевой гистидиновой метки, используя продукт первой реакции PCR и праймеры: as1USNEW (5' AGCATATGAAAA AATGGTTTGT TTGTTTATTG GGG 3' [SEQ ID NO:38] и AHLS1001 (5' TTGGATCC GAATTCAT CAATG GTG ATG GTG ATG GTG GGC 3' [SEQ ID NO:39]). Продукт второй реакции PCR очищали и гидролизовали рестрикционными ферментами Nde1 и BamHI. 2 мкг ДНК вектора pET 12a также гидролизовали рестрикционными ферментами Nde1 и BamHI и обрабатывали фосфатазой. Обработанные рестрикционными ферментами вектор pet12a и продукт второй реакции PCR очищали и лигировали, используя экспресс-набор для лигирования (Roche, Switzerland). Лигированную смесь использовали для трансформации клеток ТОР10 E. coli. Трансформанты культивировали на агаровой среде LB, содержащей 100 мкг/мл ампициллина.
Промоторный праймер Т7 (5'TAATACGACTCACTATAG 3' [SEQ ID NO:40] и терминирующий праймер Т7 (5'CTAGTTATTGCTCAGCGG 3' [SEQ ID NO:41]) использовали для контроля последовательностей и ориентации клонированных генов трансферазы в векторе pET12a. Секвенирование ДНК выполняли при помощи набора для секвенирования ABI Prism® BigDye™ Terminators Cycle с использованием 500 нг плазмидной ДНК в качестве матрицы и 3,2 пмоль промоторных и терминирующих праймеров Т7.
Конструкцию, показанную на фиг.35, использовали для трансформации компетентного бактериального штамма-хозяина BL21(DE3)pLysS (Novagen), после чего трансформанты, устойчивые к ампициллину, собирали и использовали для анализа экспрессии.
Экспрессия рекомбинантной липид-ацилтрансферазы Aeromonas salmonicida
Количественное определение активности фермента в отношении лецитина производили с использованием клеточных экстрактов при помощи набора неэтерифицированных жирных кислот (NEFA) (Roche, Switzerland).
Как показано на фиг.36, клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12-AsalGCAT=pSM, выращивали на среде LB, содержащей 100 мкг/мл ампициллина, и инкубировали, встряхивая, при 37°С до достижения OD600=0,6-1,0. Затем культуры индуцировали, используя IPTG (0,4 мМ), и продолжали инкубировать в течение следующих 3 часов. Образцы отбирали в периоды времени, равные 0, 1, 2 и 3 часам после индукции IPTG. Активность фермента анализировали с использованием набора NEFA и лецитина в качестве субстрата.
Оптимизация роста для получения более активных ферментов
Клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12-AsalGCAT=pSM, выращивали на среде LB, содержащей 100 мкг/мл ампициллина, и инкубировали, встряхивая, при разных температурах выращивания (37°С, 30°С и 20°С). Оптимальные условия продуцирования активного фермента липид-ацилтрансферазы имели место при выращивании культур при 30°С, как показано на фиг.37.
Частичная очистка рекомбинантной трансферазы Aeromonas salmonicida
Клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12-AsalGCAT=pSM, выращивали при 37°С и в результате ультразвуковой обработки и получали неочищенные клеточные экстракты. Рекомбинантный фермент выделяли из обработанных ультразвуком неочищенных клеточных экстрактов при помощи набора Ni-NTA компании Qiagen. Активность фосфолипазы определяли с использованием набора NEFA и лецитина в качестве субстрата. Неочищенные клеточные экстракты из клеток штамма BL21(DE3)pLysS, экспрессирующих активную трансферазу, инкубировали с лецитиновым субстратом и реакционную смесь анализировали при помощи тонкослойной хроматографии, результаты которой указывали на присутствие продуктов распада (см. фиг.38).
Частичная очистка рекомбинантной трансферазы Aeromonas salmonicidae
Клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12-AsalGCAT=pSM, выращивали при 37°С и в результате ультразвуковой обработки получали неочищенные клеточные экстракты. Рекомбинантный фермент выделяли из обработанных ультразвуком неочищенных клеточных экстрактов при помощи набора Ni-NTA компании Qiagen. Активность фосфолипазы анализировали с использованием набора NEFA и лецитина в качестве субстрата (см. фиг.39).
Пример 2
Клонирование и экспрессия трансферазы Aeromonas hydrophila в E. coli
Организм вида Aeromonas hydrophila (АТСС № 7965), полученный из Американской коллекции типовых культур (АТСС), выращивали в течение ночи при 30°С на среде Луриа-Бертани (LB). Клетки центрифугировали и выделяли геномную ДНК методами, применяемыми для выделения геномной ДНК в компании Qiagen Ltd. Набор буферов для выделения геномной ДНК (№ по каталогу 19060), протеаза К (№ по каталогу 19131) и РНКаза А (№ по каталогу 19101) были приобретены в компании Qiagen Ltd. (Boundary court Gatwick Court, West Sussex, RH10 2AX).
Для продуцирования рекомбинантных ферментов Aeromonas использовали бактериальный штамм-хозяин BL21(DE3)pLysS (Novagen). Компетентные клетки штамма BL21(DE3)pLysS использовали в качестве хозяина для трансформации экспрессирующим вектором pet12а-A.h.GCAT=pSMa. Трансформанты, содержащие соответствующую плазмиду, выращивали при 37°С на агаровой среде LB, содержащей 100 мкг ампициллина/мл.
Создание экспрессирующего вектора pet12a-A.h.GCAT-pSMa
Для всех амплификаций ДНК гена трансферазы из Aeromonas геномную ДНК (0,2-1 мкл) использовали в качестве матрицы и ДНК-полимеразу pfu (2,5 ед.) использовали с 10 мкл 10-кратного буфера для pfu, 1 мкл каждого праймера (50 пмоль/мкл), 200 мкМ dNTP в общем объеме реакционной смеси, равном 100 мкл. Реакции PCR выполняли в программируемом термоблоке в нижеследующих условиях: 95°С в течение 30 секунд, 30 циклов при 95°С в течение 30 секунд, 60°С в течение 1 минуты и 68°С в течение 2 минут. Дополнительное удлинение цепи производили при 72°С в течение 5 минут.
Амплификацию методом PCR гена трансферазы из A. hydrophila (АТСС № 7965) осуществляли в результате выполнения двух отдельных реакций PCR.
Первую реакцию PCR выполняли с использованием пар праймеров AHUS1 (5' GTCATATGAAAAAATGGTTTGTGTGTTTATTGGGATTGGTC 3', SEQ ID NO:42) и ahls950 (5' ATGGTGATGGTGGGCGAGGAACTCGTACTG 3', SEQ ID NO:43).
Вторую реакцию PCR выполняли для введения С-концевой гистидиновой метки, используя продукт первой реакции PCR и пары праймеров: AHUS1 (5' GTCATATGAAAAAATGGTTTGTGTGTTTATTGGGATTGGTC 3', SEQ ID NO:44) и AHLS1001 (5' TTGGATCCGAATTCATCAATGGTGATGGTGATGGTGGGC 3', SEQ ID NO:57).
Продукт второй реакции PCR очищали и гидролизовали рестрикционными ферментами Nde1 и BamHI. 2 мкг ДНК вектора pET 12a также гидролизовали рестрикционными ферментами Nde1 и BamHI и обрабатывали фосфатазой. Обработанные рестрикционными ферментами вектор pet12a и продукт второй реакции PCR очищали и лигировали, используя экспресс-набор для лигирования (Roche, Switzerland). Лигированную смесь использовали для трансформации клеток ТОР10 E. coli. Трансформанты культивировали на агаровой среде LB, содержащей 100 мкг/мл ампициллина.
Промоторный праймер Т7 (5' TAATACGACTCACTATAG 3') и терминирующий праймер Т7 (5' CTAGTTATTGCTCAGCGG 3') использовали для контроля последовательностей и ориентации клонированных генов GCAT в векторе pET12a. Секвенирование ДНК выполняли при помощи набора для секвенирования ABI Prism® BigDye™ Terminators Cycle с использованием 500 нг плазмидной ДНК в качестве матрицы и 3,2 пмоль промоторных и терминирующих праймеров Т7.
Конструкцию, показанную на фиг.40, использовали для трансформации компетентных клеток бактериального штамма-хозяина BL21(DE3)pLysS (Novagen), после чего трансформанты, устойчивые к ампициллину, собирали и использовали для анализа экспрессии.
Экспрессия трансферазы Aeromonas hydrophila в BL21(DE3)pLysS
Клетки штамма BL21(DE3)pLysS E. coli, содержащие экспрессирующий вектор pet12a-A.h.GCAT=РSMa, выращивали на среде LB, содержащей 100 мкг/мл ампициллина, и инкубировали, встряхивая, при 37°С до достижения ОD600=0,6-1,0. Затем культуры индуцировали, используя IPTG (0,4 мМ), и продолжали инкубировать в течение следующих 3 часов. Образцы отбирали в периоды времени, равные 0, 1, 2 и 3 часам после индукции IPTG. Активность фермента анализировали, используя набор NEFA и лецитин в качестве субстрата (фиг.41).
Оптимизация роста для продуцирования более активных ферментов
Клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12а-A.h.GCAT=pSMа, выращивали на среде LB, содержащей 100 мкг/мл ампициллина, и инкубировали, встряхивая, при разных температурах выращивания (37°С, 30°С и 20°С). Оптимальные условия продуцирования активного фермента GCAT имели место при выращивании культур при 30°С, как показано на фиг.42.
Частичная очистка рекомбинантной трансферазы A.hydrophila (GCAT)
Клетки штамма BL21(DE3)pLysS, содержащие экспрессирующий вектор pet12a-A.h.GCAT=pSMa, выращивали при 37°С и в результате ультразвуковой обработки получали неочищенные клеточные экстракты. Рекомбинантный фермент выделяли из обработанных ультразвуком неочищенных клеточных экстрактов при помощи набора Ni-NTA компании Qiagen. Активность фосфолипазы анализировали с использованием набора NEFA и лецитина в качестве субстрата (см. фиг.43).
Пример 3
Экспрессия трансфераз Aeromonas в Bacillus subtilis 163
Создание плазмиды
Для гетерологичной экспрессии генов Aeromonas в Bacillus subtilis использовали два разных экспрессирующих вектора для Bacillus subtilis (pUB110 и pBE5). Вектор pUB110 содержит альфа-амилазный промотор, в то время как вектор рВЕ содержит промотор Р32 в виде регуляторной области для экспрессии слитых генов Aeromonas. В векторе pUB110 первая аминокислота зрелых генов GCAT Aeromonas была слита в рамке считывания с последней аминокислотой сигнальной пептидной последовательности ксиланазы из Bacillus subtilis на сайте рестрикции Nhe1, в результате чего были получены две дополнительные аминокислоты перед зрелыми белками. Вектор рВЕ5 содержит слитую сигнальную последовательность cgtase на сайте Nco1 для секреции рекомбинантных белков в культуральный фильтрат.
Реакции PCR выполняли для слияния генов Aeromonas в рамке считывания с сигнальными последовательностями векторов pUB110 и рВЕ5. Реакции PCR выполняли с использованием нижеследующих пар праймеров для гена A. hydrophila:
Первая реакция PCR: usAHncol (5'-ATGCCATGGCCGACAGCCGTCCCGCC 3', SEQ ID NO:46) и IsAH (5'-TTGGATCCGAATTCATCAATGGTGATG 3', SEQ ID NO:47).
Вторая реакция PCR: US-AhnheI (5' TTGCTAGCGCCGACAGCCGTCCCGCC 3', SEQ ID NO:48) и IsAH (5' TTGGATCCGAATTCATCAATGGTGATG 3', SEQ ID NO:49).
Реакции PCR выполняли с использованием нижеследующих пар праймеров для гена A. salmonicida:
Третья реакция PCR: US-Asncol (5' TTGCCATGGCCGACACTCGCCCCGCC 3', SEQ ID NO:50) и IsAH (5' TTGGATCCGAATTCATCAATGGTGATG 3', SEQ ID NO:51).
Четвертая реакция PCR: US-ASnhe1 (5' TTGCTAGCGCCGACACTCGCCCCGCC 3', SEQ ID NO:52) и IsAH (5' TTGGATCCGAATTCATCAATGGTGATG 3', SEQ ID NO:53).
Все продукты PCR клонировали в векторе для PCR blunt II (вектор TOPO) и секвенировали при помощи нижних и верхних праймеров для секвенирования.
Клоны, полученные в результате выполнения первой и третьей реакций PCR, разрезали с помощью Nco1 и BamHI и использовали в качестве вставок для лигирования с вектором рВЕ5, разрезанным с помощью Nco1/BamHI/фосфатазы. Клоны, полученные в результате выполнения второй и четвертой реакций PCR, разрезали с помощью Nhe1 и BamHI и использовали в качестве вставок для лигирования с вектором pUB, разрезанным с помощью Nhe1/BamHI/фосфатазы.
Экспрессия генов трансферазы Aeromonas в Bacillus subtilis и исследование активности фермента
Ацилтрансферазы из двух видов Aeromonas были успешно экспрессированы в E. coli (результаты приведены выше). Слитые конструкции генов с векторами pUB110 и pBE5, предназначенные для Bacillus, использовали для трансформации Bacillus subtilis и трансформанты отбирали путем культивирования на чашках с канамицином. Трансформанты, устойчивые к канамицину, которые выделяли и выращивали на двукратной среде YT, пригодны для гетерологичной экспрессии генов Aeromonаs в виде Bacillus. Культуральные фильтраты обладают активностью галактолипазы в отношении дигалактозилдиацилглицерина (DGDG) помимо активностей ацилтрансферазы и фосфолипазы. Активность в отношении дигалактозилдиацилглицерина (DGDG), полученного из пшеничной муки (компании Sigma), и активность в отношении лецитина измеряли после инкубации культурального супернатанта с субстратом в течение 60 минут так, как показано на фиг.44. Фермент был продуцирован клетками Bacillus после культивирования в культуральной среде в течение периода времени от 20-24 часов до 48 часов в виде секретированного белка. В некоторых случаях было установлено, что экспрессия генов Aeromonas ухудшает жизнеспособность и рост клеток Bacillus и E.coli, поэтому необходимо тщательно выбирать экспрессирующие штаммы и оптимизировать условия роста для обеспечения экспрессии. Несколько штаммов-хозяев Bacillus (B.s 163, DB104 и OS 21) были трансформированы экспрессирующими векторами для сравнения роста. Штамм B.s163, трансформированный двумя генами Aeromonas, способен экспрессировать активный белок. Штамм DB104, трансформированный всеми конструкциями, способен экспрессировать только трансферазу A.salmonicida.
Пример 4
Ферментация и очистки липид-ацилтрансфераз Aeromonas, продуцированных в E.coli
Ферментация в E.coli
Микроорганизмы
В данном исследовании были использованы два штамма Eschericia coli, из которых один штамм содержал липид-ацилтрансферазу Aeromonas hydrophila (пример 2) и второй штамм содержал липид-ацилтрансферазы Aeromonas salmоnicida (пример 1).
Штамм E.coli, содержащий ген A. hydrophila, получил название DIDK0124, и штамм E.coli, содержащий ген A.salmonicida, получил название DIDK0125. Продукт ферментации с использованием DIDK0124 получил название HYDRO0303, и продукт ферментации с использованием DIDK0125 получил название SAL0302. Очищенный белок из HYDRO025 получил название REF №138. Очищенный белок из HYDRO0303 получил название REF №135.
Питательная среда и условия культивирования
Агаровая среда LB
Чашки с агаровой средой LB, используемые для поддержания штаммов, содержали: 10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 15 г/л агара, 100 мг/л ампициллина и 35 мг/л хлорамфеникола. Чашки с агаровой средой инкубировали при 30°С.
Встряхиваемая колба со средой LB
Среда LB (50 мл в одной встряхиваемой колбе), используемая для продуцирования инокулята, предназначенного для культивирования в биореакторе, содержала: 10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 100 мг/л ампициллина и 35 мг/л хлорамфеникола. Встряхиваемые колбы инокулировали материалом из чашек с агаровой средой LB и инкубировали при 30°С со скоростью вращения 200 оборотов/мин.
Культивирование в биореакторе
Культивирование в биореакторе осуществляли в 6 л созданных в собственной лаборатории биореакторах, заполненных 4 л среды, содержащей: 10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 8 г/л КН2РО4, 0,9 г/л MgSO4·7Н2О, 40 г/л моногидрата глюкозы, 0,4 мл/ антивспенивателя ADD APT® Foamstop Sin 260 (ADD APT Chemicals AG, Helmond, The Netherlands), 10 мг/л (NH4)2Fe(SO4)2·6H2O, 0,7 мг/л CuSO4·5H2O, 3 мг/л ZnSO4·7H2O, 3 мг/л MnSO4·H2O, 10 мг/л EDTA, 0,1 мг/л NiSO4·6H2O, 0,1 мг/л СоCl2, 0,1 мг/л Н3ВО4, 0,1 мг/л KI, 0,1 мг/л Na2MoO4·2H2O, 1 г/л ампициллина и 35 мг/л хлорамфеникола.
Биореакторы инокулировали таким количеством культуры на среде LB, которое гарантировало окончание роста после культивирования в течение примерно 20 часов (вычисленное с учетом максимальной удельной скорости роста, равной 0,6 час-1, значения OD600 встряхиваемой колбы со средой LB и конечного значения OD600 в биореакторе, равного примерно 20).
SAL0302 инокулировали 10 мл культуры на среде LB и HYDRO0303 инокулировали 4 мл культуры на среде LB.
Биоректоры действовали в нижеследующих условиях: температура 30ºС, скорость перемешивания 800-1000 оборотов/мин (в зависимости от эксперимента), аэрация со скоростью 5 л/мин, показатель рН доводили до 6,9, добавляя 8,75% (масс./об.) смесь NH3-вода и 2 М раствор H2SO4. Индукцию осуществляли путем добавления изопропил-β-D-тиогалактозида до конечной концентрации 0,6 мМ, при этом было соответственно продуцировано 0,4 моль (HYDRO0303) и 0,7 моль СО2.
Сбор биомассы
Сбор и гомогенизацию биомассы производили нижеследующим способом:
1. Ферментационный бульон, полученный в результате ферментации, центрифугировали с ускорением 5000×g при 4°С в течение 10 минут и супернатант выбрасывали. Биомассу хранили при -20°С до использования. Биомассу оттаивали и вторично суспендировали в 500 мл 20 мМ NaH2PO4, рН 7,4, 500 мМ NaCl, 10 мМ имидазола и полного (без EDTA) ингибитора протеазы (Roche, Germany).
2. Суспендированную биомассу гомогенизировали под давлением 2 кбар при 4°С в клеточном дезинтеграторе компании Constant Systems Ltd (Warwick, UK).
3. Клеточный дебрис удаляли центрифугированием с ускорением 10000×g при 4°С в течение 30 минут и собирали супернатант.
4. Супернатант очищали центрифугированием с ускорением 13700×g при 4°С в течение 60 минут и собирали.
5. Супернатант фильтровали через 0,2 мкм вакуум-фильтры с крышкой (Pall Life Sciences, UK) и фильтрат собирали для выполнения немедленной хроматографической очистки.
Хроматографическая очистка трансфераз
В колонку (2,5×10 см), заполненную 50 мл геля хелатирующей сефарозы ff., вводили сульфат никеля (в соответствии с методом, описанным изготовителем, Amersham Biosciences). Колонку уравновешивали 200 мл 20 мМ NaH2PO4, рН 7,4, 500 мМ NaCl, 10 мМ имидазола. В колонку вводили 400 мл неочищенного продукта со скоростью потока 5 мл/мин. Затем колонку промывали 20 мМ NaH2PO4, рН 7,4, 500 мМ NaCl, 10 мМ имидазола до тех пор, пока значение UV280 не достигало базовой линии. GCAT элюировали 40 мл 20 мМ NaH2PO4, рН 7,4, 500 мМ NaCl и 500 мМ имидазола.
Пример 5
Ферментация и очистка липид-ацилтрансфераз Aeromonas, продуцированных в Bacillus subtilis
Ферментация
ВАС0318-19, ВАС0323-24
Микроорганизм
Микроорганизмы, использованные в данном исследовании, были получены в результате трансформации штамма-хозяина Bacillus subtilis № 163 плазмидой, содержащей ген, кодирующий трансферазу Aeromonas salmonicida, введенный в вектор pUB110OIS. Экспрессию данного гена контролировали альфа-амилазным промотором, при этом секреция трансферазы была опосредована сигнальной последовательностью ксиланазы B. subtilis (пример 3). Указанные штаммы получили названия DIDK0138 (ферментация ВАС0318-19) и DIDK0153 (ферментация ВАС0323-24).
Питательные среды и условия культивирования
Прекультуральная среда
В встряхиваемую колбу (общим объемом 500 мл с перегородками) добавляли 100 мл среды, содержащей:
Показатель рН доводили до 7,0 перед автоклавированием.
После автоклавирования в колбу добавляли 6 мл 50% (масс./масс.) нутриозы. Канамицин добавляли в концентрации 50 мг/л после автоклавирования.
Инокуляция
Встряхиваемую колбу с прекультуральной средой инокулировали замороженной культурой непосредственно из запаса в 25% (масс./об.) глицерине. Встряхиваемую колбу инкубировали при 33°С и вращении со скоростью 175 оборотов/мин в течение примерно 16 часов, после чего 50 мл использовали для инокуляции ферментера.
Ферментация
Ферментацию выполняли в 6 л ферментерах, изготовленных в собственной лаборатории.
Порция среды (3 л) содержала:
Исходная смесь содержала:
Подачу исходной смеси при выполнении ферментации ВАС0318 и ВАС0323 начинали с учетом аккумулированного СО2, высчитываемого из нижеследующих уравнений:
Подача - скорость потока [г/час]=0, АсСО2<0,15
Подача - скорость потока [г/час]=2,85+t·1,54, AcCO2≥0,15 и t<12
Подача - скорость потока [г/час]=21,3, t>12
t: время (часы) от момента времени, когда аккумулированный СО2 (АсСО2) достигал 0,15 моль.
Подачу исходной смеси при выполнении ферментации ВАС0319 и ВАС0324 начинали с учетом аккумулированного СО2, высчитываемого из нижеследующих уравнений:
Подача - скорость потока [г/час]=0, АсСО2<0,15
Подача - скорость потока [г/час]=2,0 + t·1,08, AcCO2≥0,15 и t<12
Подача - скорость потока [г/час]=15, t>12
t: время (часы) от момента времени, когда аккумулированный СО2 (АсСО2) достигал 0,15 моль.
Показатель рН доводили до 7,0, добавляя 12,5% (масс./об.) смесь NH3-вода или 2 М раствор фосфорной кислоты.
Аэрацию производили со скоростью 3 л/мин, соответствующей 1 объему на один объем в минуту.
Температура была равна 33°С.
Ферментер был оснащен двумя лопастями Rushton диаметром 8 см, расположенными на расстоянии 10 см.
Сбор биомассы
Биомассу удаляли центрифугированием с ускорением 16000×g в течение 10 минут при комнатной температуре. Супернатант стерилизовали через фильтр и фильтрат использовали для анализов степени очистки и возможностей применения.
Пример 6
Ферментативное удаление диглицеридов из пальмового масла, катализируемое липид-ацилтрансферазой из Aeromonas salmonicidae
Краткий обзор
Эксперименты по выполнению ферментативного глицеролиза начинали, добавляя раствор глицерина/трансферазы в реактор, содержащий пальмовое масло и глицерин/воду в разных концентрациях. Все реакции выполняли при 43°С, производя перемешивание магнитной мешалкой в течение 24 часов. После окончания реакции образцы анализировали при помощи высокоэффективной тонкослойной хроматографии (ВЭТСХ) и в некоторых экспериментах полученный результат подтверждали газовой хроматографией (ГХ). Результаты выполненных анализов показали, что существует хорошая корреляция между концентрацией воды и уровнями DAG и MAG; самый высокий уровень MAG и самый низкий уровень DAG были получены при самых низких концентрациях воды.
На основании результатов анализов можно сделать вывод о том, что количество диглицеридов в пальмовом масле можно уменьшить в результате выполнения реакции, катализируемой трансферазой, причем фермент использует диглицериды в качестве донорных молекул и глицерин в качестве акцепторной молекулы при осуществлении реакции глицеролиза, в процессе которой синтезируются моноглицериды. Подобный результат отличается от реакции глицеролиза с использованием известных триглицеридгидролизующих липаз, при выполнении которой количество DAG увеличивается.
Кроме того, можно сделать вывод о том, что концентрация воды оказывает значительное влияние на выход синтезированных моноглицеридов и количество DAG. Была обнаружена нижеследующая корреляция: использование воды в низкой концентрации (<1%) и 5% глицерина увеличивает выход MAG и уменьшает концентрацию DAG. Полученные результаты также показывают, что главным образом уменьшается количество 1,2-изомера диглицерида.
Известно, что диглицериды, в частности 1,2-изомеры, замедляют кристаллизацию жира. Такой эффект вызывает последующую кристаллизацию жировых продуктов, таких как маргарин и шортенинги, в результате которой образуются крупные кристаллы (песчанистость). Было установлено, что в некоторых жировых смесях диглицериды улучшают устойчивость β'-кристаллов. Поэтому желательно лишь уменьшить, а не полностью удалить диглицериды в пальмовом масле. Таким образом, можно сделать вывод о том, что уменьшение диглицерида позволяет получить более однородное масло лучшего качества; факт, который нельзя недооценить.
Введение
Проблемы, связанные с диглицеридами, в некоторой степени зависят от типа продукта, например, является ли такой продукт маргарином или шортенингом. Таким образом, в зависимости от продукта присутствие диглицеридов может иметь как отрицательное, так и положительное значение. При производстве маргарина можно использовать только жировой продукт с требуемой кристаллической структурой для достижения необходимой консистенции и пластичности. β'-кристаллическая форма является наиболее приемлемой структурой, которая характеризуется мелкими кристаллами и высокой способностью сохранения жидкой фракции. Наиболее устойчивая кристаллическая форма (β-форма) является нежелательной, так как наличие крупных кристаллов является причиной образования зернистой структуры маргарина. Как указано в публикациях Hernquist & Anjou (1993) и Wahnelt et al. (1991), присутствие диглицеридов в жире замедляет превращение β'-кристаллов в β-кристаллы. Но подобно тому, как диглицериды замедляют превращение β'-формы в β-форму, они также замедляют превращение α-формы в β'-форму (Walnet et al., 1991). На основании вышеизложенного можно прийти к выводу, что присутствие диглицеридов замедляет процесс кристаллизации в целом. Медленная кристаллизация имеет важное значение для применения маргарина. Эффект замедления кристаллизации в маргариновых смесях, содержащих высокую долю пальмового масла, может быть причиной того, что после обычной обработки продукт окажется недостаточно твердым, создавая трудности с упаковкой, при этом последующая кристаллизация может вызвать образование более твердой структуры по сравнению с желаемой (Berger, 1990). Несмотря на достоинства и недостатки, очень важно найти правильный баланс между уменьшением содержания и полным удалением диглицеридов.
Было установлено, что количество диглицерида в пальмовом масле можно уменьшить путем ферментативного глицеролиза пальмового масла с использованием липид-ацилтрансферазы, например липид-ацилтрансферазы с фрагментом GDSх из Aeromonas salmonicida. Не ограничиваясь какой-либо теорией, можно отметить, что при осуществлении данного способа фермент добавляют к пальмовому маслу вместе с небольшим количеством глицерида и данный фермент катализирует нижеследующую реакцию:
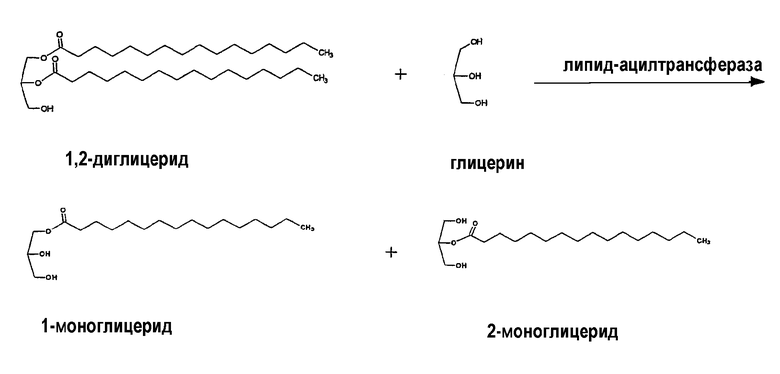
После окончания реакции избыток глицерина, в случае необходимости, можно легко отделить от реакционной смеси центрифугированием или другими методами.
Преимуществом данного способа является то, что количество диглицерида (предпочтительно 1,2-диглицерида) уменьшается в результате выполнения реакции глицеролиза, катализируемой ферментом, который является специфичным к диглицериду, без использования триглицерида в качестве донора для реакции, катализируемой трансферазой.
Такие продукты реакции как моноглицериды не требуют удаления из реакционной смеси и могут быть успешно использованы в качестве эффективного эмульгатора для производства пищевых продуктов, подобных маргарину и шортенингу. Таким образом, данный способ позволяет решить две проблемы. Во-первых, можно уменьшить количество диглицерида и предпочтительно 1,2-диглицерида, который оказывает отрицательное влияние на свойства кристаллизации триглицеридов. Во-вторых, такой продукт реакции как моноглицерид можно использовать в качестве эффективного эмульгатора при производстве пищевых продуктов, подобных маргарину и шортенингу.
В одном варианте осуществления изобретения желательно удалить моноглицериды, образовавшиеся (в случае их образования) в результате осуществления реакции, катализируемой трансферазой.
Подходящим образом при выполнении реакции, катализируемой трансферазой, в нерафинированном пальмовом масле моноглицерид, глицерин и/или остатки воды (в случае ее наличия) можно удалить методом дезодорации в процессе рафинирования пищевого масла.
Другим очень интересным преимуществом использования липид-ацилтрансферазы является то, что данный фермент в меньшей степени зависит от содержания воды в реакционной смеси, и, поскольку указанный фермент является трансферазой, имеет место низкая гидролитическая активность или полное отсутствие такой активности, из чего следует, что не происходит значительного увеличения количества свободных жирных кислот.
Хорошо известно, что содержание воды при осуществлении реакций глицеролиза имеет очень важное значение в случае использования липолитических ферментов, таких как липазы (Kristensen, 2004). Даже небольшое количество воды, которая необходима для осуществления большинства липолитических реакций, вызывает образование значительного количества свободных жирных кислот. Указанную проблему можно преодолеть благодаря использованию липид-ацилтрансферазы при осуществлении реакции глицеролиза.
Другой проблемой является то, что липазы, которые, как известно в данной области, катализируют реакцию глицеролиза, в качестве донора используют в основном триглицерид, что вызывает образование диглицерида вместо уменьшения его количества.
Вещества
Методы
Лиофилизация фермента
Перед лиофилизацией ферменты обессоливали (обессоливающие колонки PD-10, Аmersham Biosciences). Обессоленный фермент смешивали с глицерином (фермент:глицерин в соотношении 3,5:1). Образец лиофилизовали и добавляли 10% воды. Полученный образец содержал примерно 20 ед. (единицы фосфолипазы) на один грамм.
Определение активности фосфолипазы
(анализ активности фосфолипазы)
Субстрат
95% растительный субстрат, содержащий 0,6% L-α-фосфатидилхолина (Avanti, № 441601), 0,4% Triton-Х 100 (Sigma X-100) и 5 мМ CaCl2, растворяли в 0,05 М растворе буфера HEPES, рН 7.
Методика выполнения анализа
В пробирку Эппендорфа емкостью 1,5 мл добавляли 400 мкл субстрата и на 5 минут помещали в термосмеситель Эппендорфа при 37°С. В период времени t=0 минут добавляли 50 мкл раствора фермента. Кроме того, анализировали контрольный образец с использованием воды вместо фермента. Образец смешивали в термосмесителе Эппендорфа со скоростью 10*100 оборотов/мин при 37°С в течение 10 минут. В период времени t=10 минутам пробирку Эппендорфа на 10 минут помещали в другой термосмеситель при 99°С для прекращения реакции.
Содержание свободных жирных кислот в образцах анализировали, используя набор NEFA C компании WAKO GmbH.
Активность фермента PLU-7 при рН 7 высчитывали в микромолях жирной кислоты, продуцированной за одну минуту в условиях выполнения анализа.
Ферментативная реакция
Пальмовое масло подвергали взаимодействию с раствором глицерина/фермента с использованием составов, приведенных в таблице 1 и таблице 2.
Эксперимент I
Эксперимент II
Пальмовое масло отмеряли в 20 мл стакан Уитона и добавляли смесь глицерина/фермента и необязательно воду. Образец помещали в нагревательный блок (нагревательный блок Multitherm HP 15, нагреваемый в результате дифференциального перемешивания (15 камер), контролируемого термостатом VARIOMAG - Thermomodul 40 ST), и подвергали взаимодействию в нижеследующих условиях:
Фермент в реакционной смеси инактивировали при 97,5°С в течение 10 минут. После окончания реакции образец гомогенизировали (Ultra Turrax) в течение 20 секунд и гомогенный образец использовали для последующего анализа.
Высокоэффективная тонкослойная хроматография (ВЭТСХ)
Аппликатор: аппликатор LINOMAT 5, CAMAG.
Пластинка для ВЭТСХ: 10×10 см (Merck, № 1.05633)
Вышеуказанную пластинку активировали перед использованием путем сушки в термостате при 180°С в течение 20-30 минут.
Нанесение: 2,0 мкл 2,0% раствора, подвергнутого взаимодействию пальмового масла, растворенного в смеси хлороформ:метанол (2:1), наносили на пластинку для ВЭТСХ при помощи аппликатора LINOMAT 5.
Рабочий буфер: петролейный эфир:МТВЕ:уксусная кислота (50:50:1)
Время нанесения/элюирования: 8 минут.
Проявляющая жидкость: 6% куприацетата в 16% Н3РО4.
После элюирования пластинку сушили в термостате при 180°С в течение 10 минут, охлаждали и погружали в проявляющую жидкость, затем дополнительно сушили при 180°С в течение 20 минут. Производили визуальную оценку пластинки и прямое сканирование (Scan Wizard 5).
В эксперименте II производили количественное определение компонентов при помощи программы Adobe photoshоp 6.0 и количество MAG и DAG высчитывали из калибровочных кривых стандартных растворов DAG и MAG (см. фиг.49 и 50).
Газовая хроматография
Использовали капиллярный хроматограф Perkin Elmer 8420, оснащенный колонкой из кварцевого стекла WCOT 12,5 м с диаметром 0,25 мм, содержащей 0,1 мкм 5% фенилметилсиликона (СР Sil CB компании Crompack).
Газ-носитель: гелий
Ввод пробы: 1,5 мкл с делением пробы
Детектор: FID. 385°С
Получение образца: 50 мг липида растворяли в 12 мл смеси гептана/пиридина (2:1), содержащей внутренний эталон гептадекан в количестве 2 мг/мл. 500 мкл образца переносили в гофрированную пробирку. Добавляли 100 мкл MSTFA (N-метил-N-триметилсилилтрифторацетамид) и реакционную смесь инкубировали при 60°С в течение 15 минут.
Вычисление: факторы, влияющие на содержание моно-, ди-, триглицеридов и свободных жирных кислот, определяли на основании эталонных смесей указанных компонентов. Исходя из полученных значений вышеуказанных факторов, высчитывали содержание липидов в образцах.
Результаты
Эксперимент I
Целью данного эксперимента было исследование влияния диглицерид:глицерин-ацилтрансферазы из Aeromonas salmonicida на содержание DAG в пальмовом масле под действием реакции глицеролиза. Известно, что глицеролипид-ацилтрансфераза (GCAT) может переносить жирную кислоту из лецитина в холестерин в процессе образования сложного эфира холестерина и лизолецитина. В данном эксперименте была исследована способность фермента по настоящему изобретению использовать DAG в качестве донора и глицерин в качестве акцептора для уменьшения количества DAG в пальмовом масле и продуцирования моноглицерида.
В научной литературе всесторонне описано применение ферментов, таких как липазы, для катализа реакций глицеролиза. В указанных реакциях триглицериды являются основным субстратом, а моно- и диглицериды являются продуктами реакции. При осуществлении таких реакций происходит значительное увеличение количества диглицеридов, причем количество продуцированных диглицеридов соответствует количеству продуцированного моноглицерида или превышает указанное количество (Kristensen, 2004).
Четыре образца разного состава подвергали анализу и сравнивали с эталоном пальмового масла, смешанного с 5% глицерина без фермента, который обрабатывали аналогично экспериментальным образцам. На фиг.47 показаны результаты анализа ВЭТСХ образца, представленного в таблице 1.
Результаты, полученные в результате выполнения анализа ВЭТСХ, показывают, что количество диглицеридов изменяется в зависимости от условий реакции (в частности, от соотношения глицерина и Н2О). Одинаковым для всех реакций было то, что 1,3-изомеры диглицерида составляли большую часть, чем 1,2-(2,3-)изомеры диглицерида. Данное наблюдение может быть подтверждено теоретическими положениями, из которых следует, что соотношение 1,3-изомера и 1,2-(2,3-) изомера в нерафинированном пальмовом масле равно 7:3 (Siew & Ng, 1999; Timms, 2004). Кроме того, в результате анализа пластинки для ВЭТСХ было установлено, что трансфераза эффективно уменьшает количество диглицеридов. Достигнутое уменьшение в некоторой степени согласуется с количеством синтезированного моноглицерида (MONO).
Из сравнения 1,2-изомера с 1,3-изомером DAG на фиг.1 следует, что в результате катализируемой ферментом реакции прежде всего уменьшается количество 1,2-изомера DAG. Подобный результат согласуется с тем, что трансфераза по настоящему изобретению характеризуется предпочтительной специфичностью к жирным кислотам в положении sn2.
В выполненных экспериментах вода была использована в разных концентрациях в пределах 0,5-5,5%. Полученные результаты показывают, что концентрация воды оказывает значительное влияние на синтезированное количество моноглицерида наряду с уменьшением количества диглицеридов. При использовании трансферазы № 196 было установлено, что при низком содержании воды в системе образуется большее количество моноглицерида и соответственно уменьшается количество DAG. В данном эксперименте была также исследована возможность влияния количества глицерина на баланс в системе (полоса 5: 10% глицерина:1% Н2О). При использовании трансферазы № 196 было установлено, что более высокая концентрация глицерина, соответствующая двойной активности фермента, не вызывает увеличения содержания моноглицерида.
Настоящее изобретение предпочтительно осуществляют в среде с низким содержанием воды, то есть менее примерно 1%.
Одним преимуществом использования трансфераз вместо липаз является отсутствие корреляции между синтезом моноглицерида и чрезмерным увеличением концентрации свободных жирных кислот (FFA). Данный факт подтвержден настоящим экспериментом. На фиг.1 показано, что ни в одном из продуктов всех выполненных реакций не была обнаружена четкая полоса FFA (предполагалось, что полоса FFA должна находиться между 1,3-диглицеридами и триацилглицеридами).
На основании результатов данного эксперимента можно сделать вывод о том, что оптимальная концентрация глицерина и воды для трансферазы № 196 соответствует нижеследующему значению:
Трансфераза № 196: 5% глицерина:0,5% Н2О.
Процедура подготовки образцов для анализа ВЭТСХ не включала определение удельной массы, так как при этом нельзя было вычислить концентрации разных компонентов в реакционных смесях. По указанной причине исследование было основано только на визуальной оценке.
Результаты газовой хроматографии
На основании результатов ВЭТСХ образец № 2 был отобран для анализа газовой хроматографией. Данное исследование выполняли с целью подтверждения визуальной оценки пластинок для ВЭТСХ при помощи количественных анализов DAG. Перед выполнением анализа из образца удаляли глицерин центрифугированием и газовой хроматографии подвергали только липидную фазу. Результаты газовой хроматографии представлены в приведенной ниже таблице 3.
Результаты газовой хроматографии образца, отобранного по результатам эксперимента I
Анализ результатов газовой хроматографии показал, что отобранный образец отличается от эталона более высоким содержанием моноглицерида и более низким выходом DAG. Данное наблюдение подтверждает результаты, полученные при выполнении анализа ВЭТСХ.
Заключение. Эксперимент I
Что касается содержания моноглицеридов и диглицеридов в продукте глицеролиза, то результаты анализа ВЭТСХ показали, что существует корреляция между концентрацией воды и количествами моноглицерида и диглицеридов: чем меньше концентрация воды, тем ниже содержание диглицеридов и выше содержание моноглицерида.
Результаты газовой хроматографии подтвердили степень уменьшения DAG. В данном эксперименте уменьшение количества диглицеридов составляет 7,1% от общего количества диглицеридов в пальмовом масле (таблица 4).
Степень уменьшения диглицеридов
Эксперимент II
Целью данного эксперимента была дальнейшая оптимизация уменьшения DAG в пальмовом масле и анализ катализируемого трансферазой глицеролиза с использованием трансферазы № 196. Главной задачей было достижение хорошо сбалансированной реакции, при выполнении которой количество DAG должно уменьшаться одновременно с увеличением концентрации МONO.
В данной части экспериментальной работы было испытано шесть образцов разного состава, которые сравнивали с эталоном пальмового масла, смешенного с 5% глицерина (таблица 2). Профиль нагрева эталона был аналогичен профилю нагрева при использовании фермента, что позволило исследовать и определить степень теплового разложения в процессе уменьшения количества. Результаты анализа ВЭТСХ показаны на фиг.48.
Из результатов, приведенных на фиг.48, видно, что количество MAG является различным. Результаты анализа ВЭТСХ показывают, что в образцах, содержащих 5% глицерина (полосы 1-3), существует постоянная взаимосвязь между концентрацией воды и выходом МAG: уменьшение количества воды в реакционной смеси сопровождается увеличением выхода MAG. Такая же тенденция наблюдается при использовании реакционных смесей, содержащих 10% глицерина (полосы 5-7).
Что касается количества диглицеридов, то невозможно дифференцировать образцы только на основании визуальной оценки пластинки для ВЭТСХ. Поэтому для дифференцировки образцов производили количественное определение компонентов, анализируя стандартные вещества с известным составом МAG и DAG. Пластинки для ВЭТСХ сканировали и обрабатывали при помощи программы Adobe Photoshop 6.0 и дальнейшее вычисление производили с помощью специально созданной вычислительной макропрограммы (Microsoft Excel 2000). Результаты количественного анализа компонентов методом ВЭТСХ представлены в приведенной ниже таблице 5.
- = Вычисление не производилось
Результаты данного эксперимента подтверждают визуальную оценку и дают точную картину изменения уровня диглицеридов. Как видно из приведенных результатов, наибольшая степень уменьшения количества DAG была достигнута в образце № 1 (5% глицерина:1,5% Н2О). Далее было установлено, что данный образец содержит также самое высокое количество MAG.
Заключение. Эксперимент II
Результаты эксперимента II подтвердили ранее сделанное наблюдение, что липид-ацилтрансфераза GCAТ из Aeromonas salmonicida способна уменьшать количество диглицеридов в пальмовом масле. При выполнении экспериментов с использованием 5% глицерина была обнаружена корреляция между концентрацией воды в реакционной смеси и количествами MAG и DAG: чем ниже концентрация воды, тем меньше содержание DAG и выше содержание MAG.
Выполнение ВЭТСХ позволило произвести количественное определение степени уменьшения количества (см. таблицу 5). Из полученных результатов следует, что уменьшение количества диглицеридов составляет 13,29% от общего количества диглицеридов в пальмовом масле и количество синтезированного MONO увеличилось от 0,01% (эталон) до 0,38% (образец № 1).
На основании результатов данного эксперимента можно сделать вывод о том, что оптимальная концентрация глицерина и воды для трансферазы № 196 соответствует нижеследующему значению:
Трансфераза № 196: 5% глицерина:1,5% Н2О.
Сравнение указанных наблюдений с результатами, полученными в эксперименте I, свидетельствует о хорошей согласованности полученных результатов. Оптимальная концентрация воды составляет 0-1% в зависимости от активности фермента и содержания глицерина.
На основании результатов двух выполненных экспериментов можно сделать вывод о том, что количество диглицеридов в пальмовом масле можно уменьшить с помощью катализируемой трансферазой реакции глицеролиза, в которой фермент использует диглицериды в качестве донорных молекул и глицерин в качестве акцепторной молекулы, благодаря чему происходит синтез моноглицеридов. Вышеописанная реакция резко отличается от реакции глицеролиза, выполняемой с использованием известных липаз, гидролизующих триглицериды, при осуществлении которой количество DAG увеличивается вследствие частичной гидролизной активности липазы, гидролизующей триглицериды.
С учетом того, что диглицериды присутствуют в виде смеси 1,2- и 1,3-изомеров и трансфераза является специфичной к жирным кислотам в положении sn2, даже небольшое уменьшение количества диглицерида должно оказывать огромное физическое влияние на продукты на основе пальмового масла.
Пример 7
Иммобилизация диацилглицерин:глицерин-транферазы (липид-ацилтрансфераза по настоящему изобретению), полученной из Aeromonas salmonicida
Диацилглицерин:глицерин-трансферазу иммобилизуют на целите путем осаждения ацетоном. 10 мл раствора фермента в 20 мМ буфера с ТЕА, рН 7, медленно перемешивают с 0,1 г целита 535 (компании Fluka) в течение 2 часов при комнатной температуре.
При непрерывном перемешивании добавляют 50 мл холодного ацетона.
Осадок отделяют центрифугированием с ускорением 5000×g в течение 1 минуты.
Осадок дважды промывают 20 мл холодного ацетона.
Целит сушат при комнатной температуре в течение примерно 1 часа.
Иммобилизованную трансферазу испытывают в пальмовом масле (см. приведенную ниже таблицу 6).
Пальмовое масло и глицерин нагревают до 42°С. Добавляют иммобилизованную трансферазу.
Катализируемую трансферазой реакцию осуществляют при 42°С, осторожно перемешивая реакционную смесь магнитной мешалкой. Образцы отбирают для анализов через ½, 1, 3, 6 и 24 часа и анализируют ВЭТСХ. Реакцию прекращают через 24 часа и иммобилизованный фермент отфильтровывают.
Результаты анализа ВЭТСХ ясно свидетельствуют о влиянии иммобилизованной диацилглицерин:глицерин-трансферазы, полученной из A. salmonicida, на образование моноглицерида и уменьшение содержания диглицерида в пальмовом масле.
Все публикации, приведенные в описании изобретения, включены в данное описание изобретения в качестве ссылки. Специалистам в данной области должны быть очевидны разные модификации и варианты описанных способов и системы по настоящему изобретению, не выходящие за пределы объема и духа настоящего изобретения. Хотя настоящее изобретение было описано на основании конкретных предпочтительных вариантов осуществления, должно быть понятно, что данное изобретение не ограничивается такими конкретными вариантами осуществления. В объем нижеследующей формулы изобретения входят разные модификации описанных вариантов осуществления изобретения, которые должны быть очевидны специалистам в области биохимии и биотехнологии или родственных областях.
Источники информации
Amano Enzyme Inc. (2004). http://www.amano-enzyme.co.jp/english/productuse/oil_fat.html. Datо: 21.06.04.
Fødevareministeriet (2003). Bekendtgørelse om indhold af transfedtsyrer i olier og fedtstoffer. Bekendtgørelse nr. 160 af 11/03/2003.
Directive 2000/36/EC. http://europa.eu.int/scadplus/leg/en/lvb/121122b.htm. Datо: 16.06.04.
Karlshamns, (2004). Press information. http://www.karlshamns.com/investor/press_information-show.asp?ID=319. Dato: 16.06.04.
Berger, K. G. (1990). Recent developments in palm oil. In Oleagineux 45:437-443.
Drozdowski, B. (1994). General characteristics of eatable fats. WNT, Warszawa, Poland, pp.240-243 (In polish).
Hernquist, L. & Anjou, K. (1983). Diglycerides as a stabilizer of the β'-crystal Form in Margarines and Fats. In Fette Seifen Anstrichmittel. 2:64-66.
Hernquist, L. Herslof, B. Larsson, K. & Podlaha, O (1981). Polymorphism of rapeseed oil with low content of erucic acid and possibilities to stabilize the β' crystal form in fats. In Journal of Science and Food Agriculture. 32:1197-1202.
Jacobsberg, B. & Oh, C.H. (1976). Studies in Palm Oil Crystallisation. In Journal of the American Oil Chemist Society. 53:609-616.
Kristensen, A. C. J. (2004). Preparation of margarine and spreads by enzyme-generated emulsifiers. Master thesis, The Royal Veterinary and Agricultural University, Frederiksberg, Copenhagen.
McNeill, G. P. & Berger, R. G. (1993). Enzymatic glycerolysis of palm oil fractions and palm oil based model mixture: Relationship between fatty acid composition and monoglyceride yield. In Food Biotechnology 7:75-87.
Okiy, D. A. (1977). Partial glycerides and palm oil Crystallisation. In Journal of Science and Food Agriculture. 28:955.
Okiy, D. A. (1978). Interaction of triglycerides and diglycerides of palm oil. In Oleagineux. 33:625-628.
Okiy, D. A., Wright, W. В., Berger, K. G. & Morton, I. D. (1978). The physical properties of modified palm oil. In Journal of Science of Food and Agriculture. 29:625-628.
Walnett, S. V., Meusel, D. & Tülsner, M. (1991). Zur kenntnis des diglyceride influsses auf das kristallisationsverhalten von Fetten. In Fat Science Technology. 4:117-121.
Siew, N. L. (2001). Understanding the Interactions of Diacylglycerols with oil for better Product performance. Paper presented at the 2001 PIPOC International Palm Oil Congress - Chemistry and Technology Conference. 20-23 August 2001, Kuala Lumpur, Malaysia.
Siew, N. L. & Ng, W. L. (2000). Differential scanning thermograms of palm oil triglycerides in the presence of diglycerides. In Journal of Oil Palm Research. 12:1-7 Siew, N. L. & Ng, W. L. (1999). Influence of diglycerides on crystalisation of palm oil. In Journal of Science of Food and Agriculture. 79:722-726.
Sonntag, N. O. V. (1982a). Fat splitting, esterification and interesterification. Bailey's Industrial Oils and Fat products. Vol.2, 4th edn. John Wiley and Sons, New York pp.97-173.
Sonntag, N. O. V. (1982b). Glycerolysis of Fats and methyl esters - status, review and critique. In Journal of American Oil Chemist Society. 59:795A-802A.
Timms, R. (2004). Oral presentation (Lecture: Trends & delvlopment) at Danisco A/S, Brabrand, Denmark.
Будапештское соглашение о международном признании депонирования микроорганизмов в целях делопроизводства по выдаче патентов.






























































| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ | 2004 |
|
RU2376868C2 |
| БЕЛКИ | 2004 |
|
RU2518345C2 |
| СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА УГЛЕРОДА, СЛОЖНОГО ЭФИРА БЕЛКА, СЛОЖНОГО ЭФИРА БЕЛКОВОЙ СУБЪЕДИНИЦЫ ИЛИ СЛОЖНОГО ЭФИРА ГИДРОКСИКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЛИПИДАЦИЛТРАНСФЕРАЗЫ | 2004 |
|
RU2371474C2 |
| ВАРИАНТЫ ГЛИКОЛИПИДАЦИЛТРАНСФЕРАЗЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2004 |
|
RU2377300C2 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| БЕЛОК | 2005 |
|
RU2380414C2 |
| Способы получения липидов | 2011 |
|
RU2636344C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |








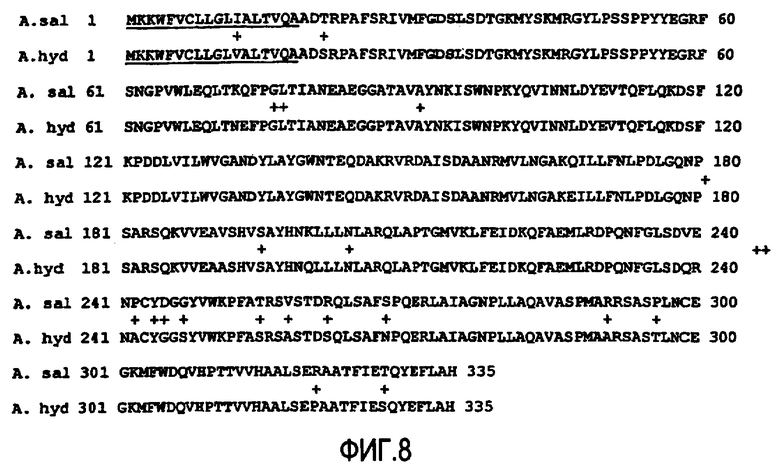


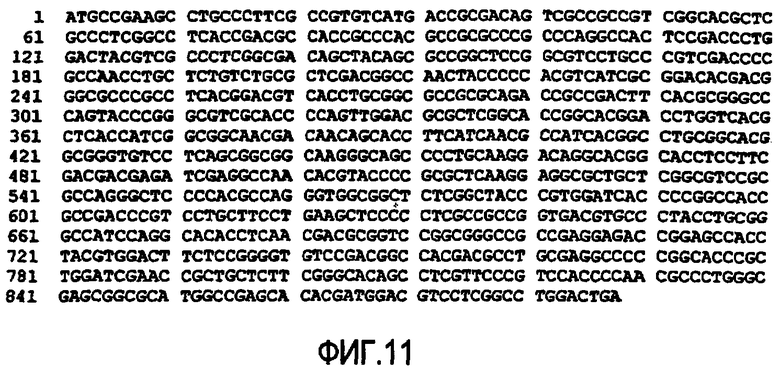





















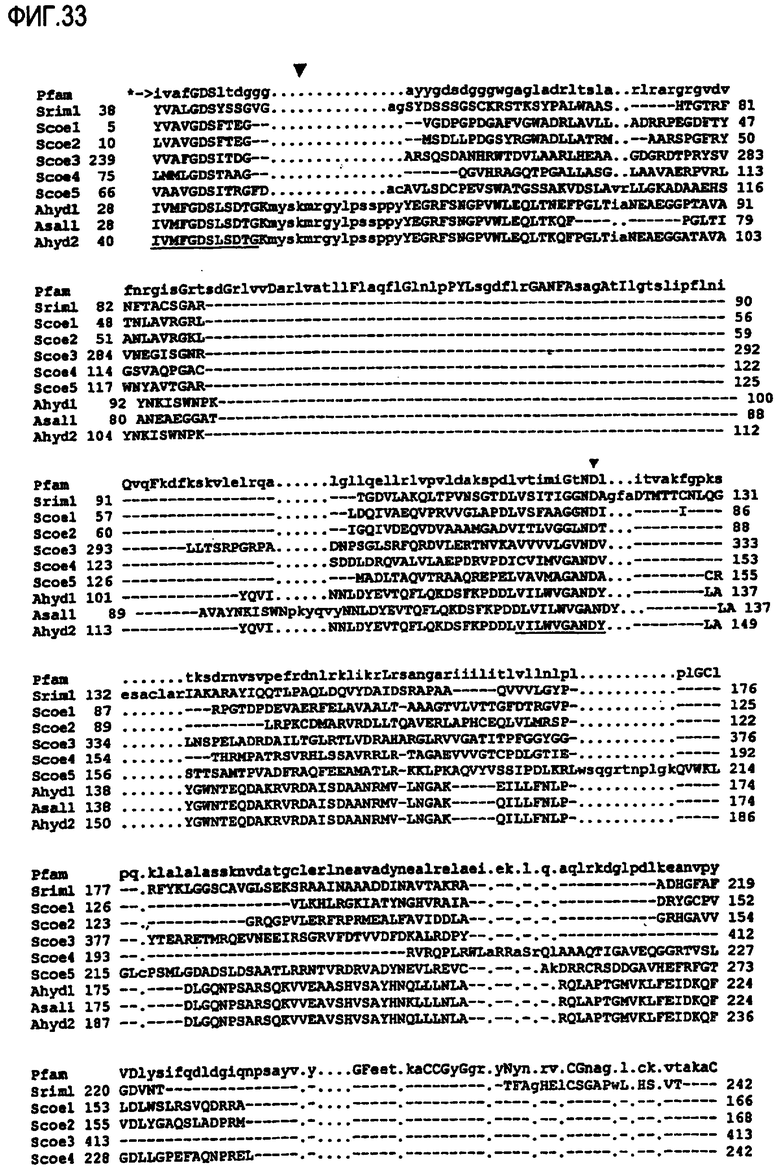

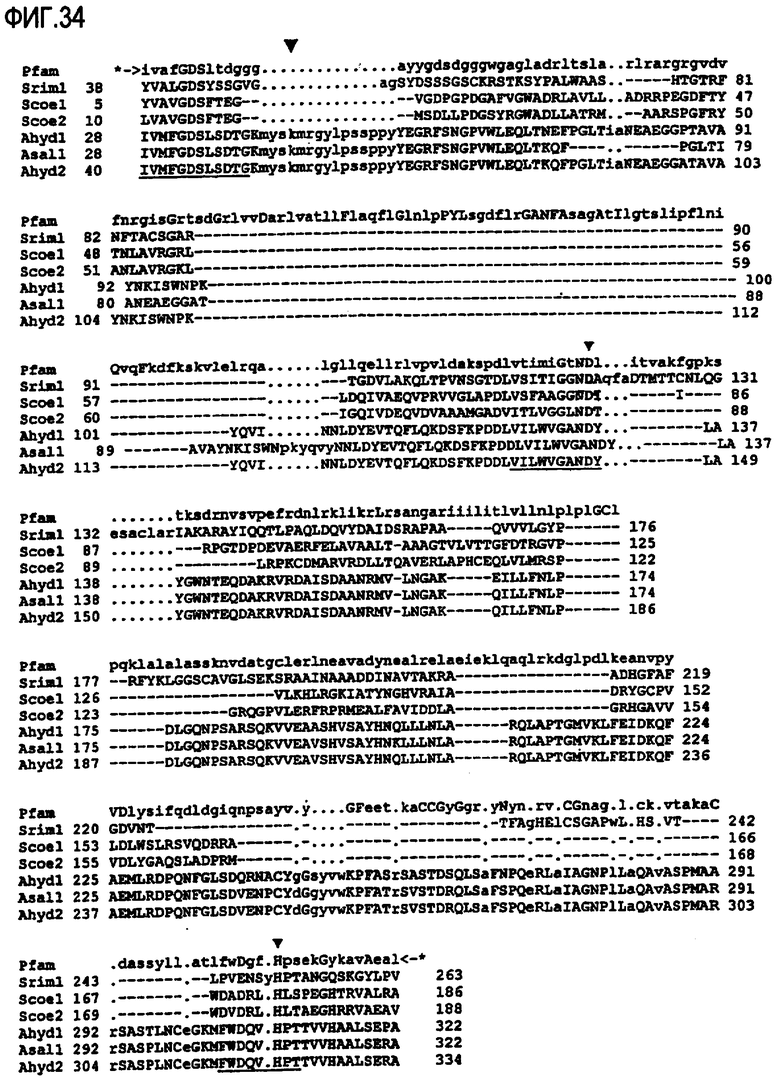


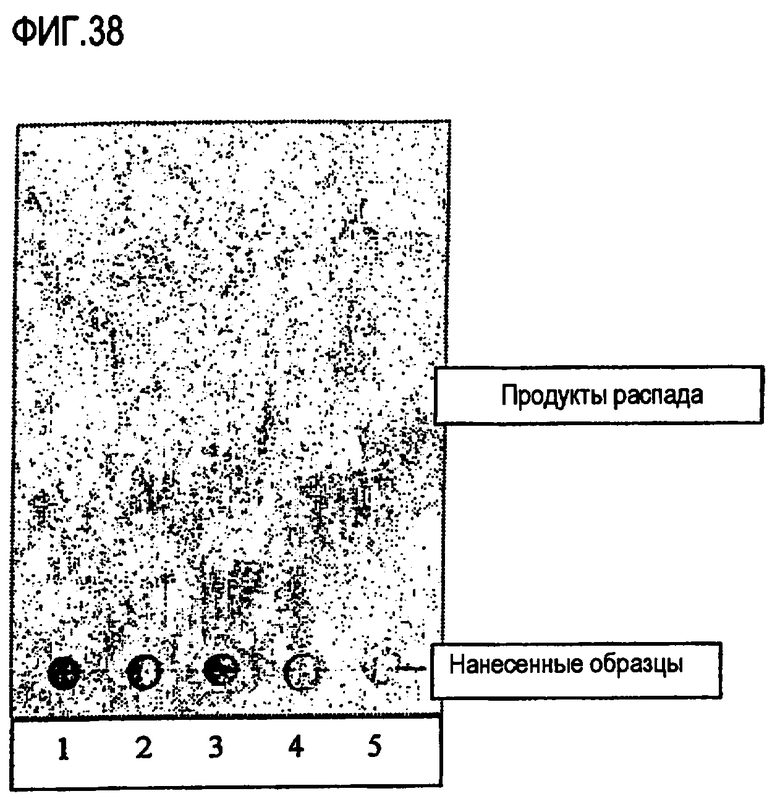
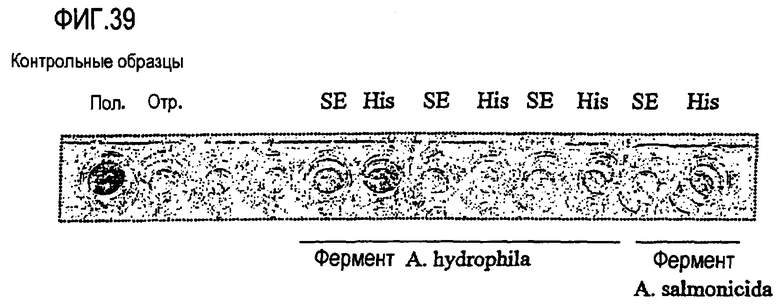

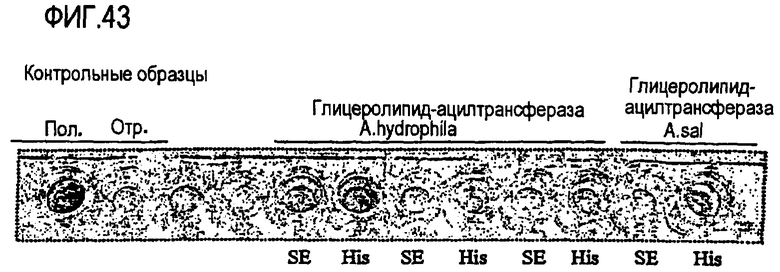
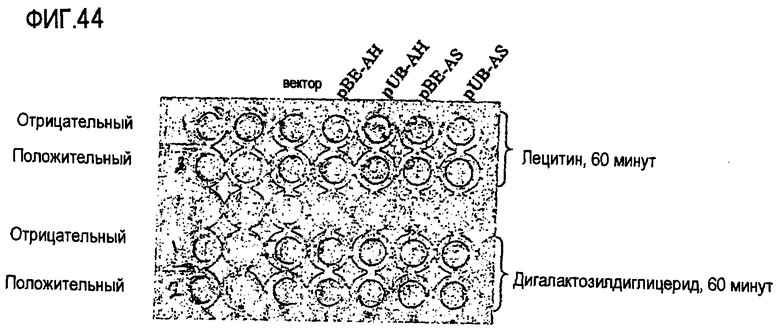


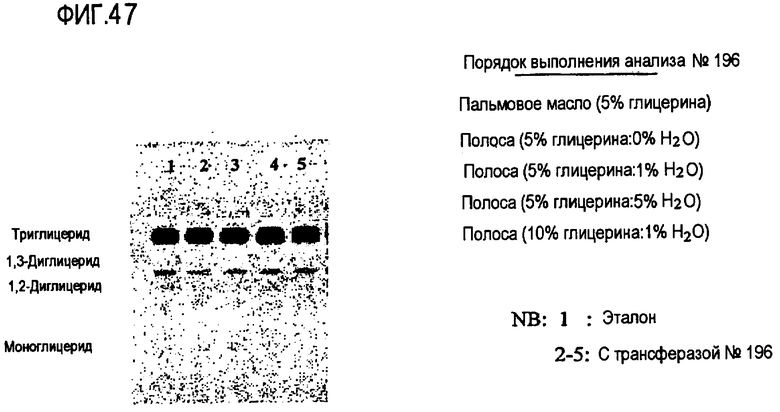
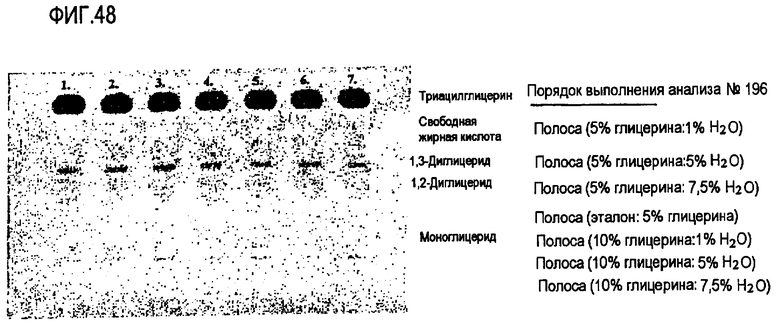
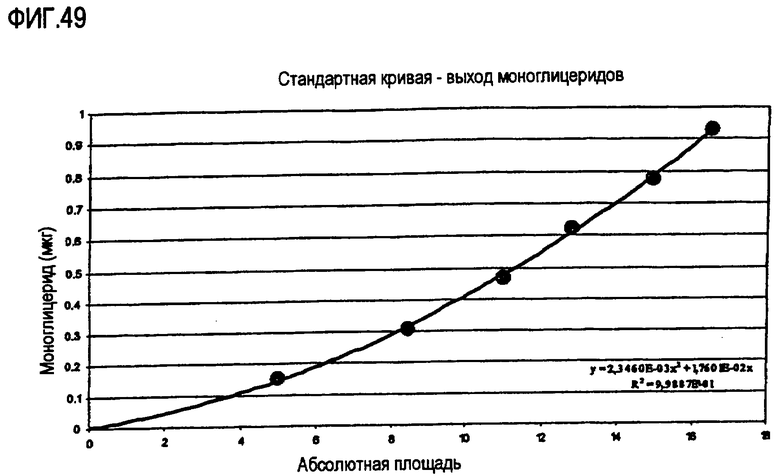
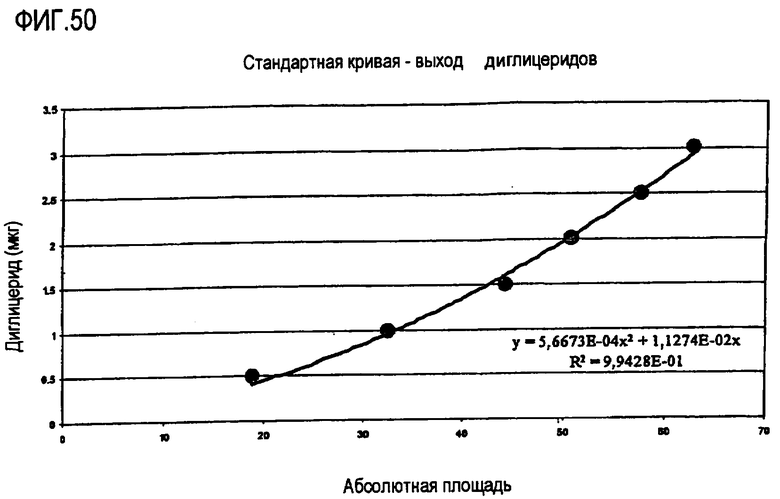


Изобретение относится к биотехнологии и касается способа уменьшения содержания и/или удаления диглицерида из пищевого масла, который включает смешивание пищевого масла с субстратом акцептора ацильных групп и диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, причем диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, является ферментом, способным переносить в пищевом масле ацильную группу из диглицерида в глицерин. Диглицерид:глицерин-ацилтрансфераза предпочтительно содержит фрагмент аминокислотной последовательности GDSX, где Х означает один или несколько нижеследующих аминокислотных остатков L, А, V, I, F, Y, H, Q, Т, N, М или S. Изобретение также касается применения диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, способным переносить в пищевом масле ацильную группу из диглицерида в глицерин, при производстве пищевого масла для уменьшения содержания и/или удаления диглицерида из указанного пищевого масла, и к применению указанного фермента при производстве пищевого продукта, содержащего пищевое масло, для улучшения свойств кристаллизации указанного пищевого продукта. Изобретение позволяет повысить эффективность удаления или уменьшения диглицерида из масла. 3 н. и 26 з.п. ф-лы, 52 ил., 6 табл.
1. Способ уменьшения содержания и/или удаления диглицерида из пищевого масла, который предусматривает смешивание пищевого масла с субстратом акцептора ацильных групп и диглицерид:глицерин-ацилтрансферазой, не зависящей от СоА жирной кислоты, причем диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, является ферментом, который в пищевом масле переносит ацильную группу из диглицерида в глицерин.
2. Способ по п.1, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит фрагмент аминокислотной последовательности GDSX, где Х означает один или несколько нижеследующих аминокислотных остатков: L, А, V, I, F, Y, H, Q, Т, N, М или S.
3. Способ по п.1, в котором происходит уменьшение количества диглицерида в пищевом масле.
4. Способ по п.1, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, переносит ацильную группу из диглицерида в акцептор ацильных групп, причем указанный акцептор ацильных групп является любым соединением, содержащим гидроксильную группу (-ОН).
5. Способ по любому из пп.1-4, в котором диглицерид является 1,2-диглицеридом.
6. Способ по любому из пп.1-4, в котором акцептор ацильных групп является веществом, растворимым в пищевом масле.
7. Способ по любому из пп.1-4, в котором акцептор ацильных групп является спиртом.
8. Способ по п.7, в котором акцептор ацильных групп является спиртом.
9. Способ по п.8, в котором акцептор ацильных групп является глицерином.
10. Способ по п.1, в котором фермент диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит остаток Н-309 или остаток гистидина в положении, соответствующем остатку His-309 в аминокислотной последовательности липолитического фермента Aeromonas hydrophila SEQ ID NO:2 или SEQ ID NO:32.
11. Способ по п.1, в котором диглицерид:глицерин-ацилтрансферазу, не зависящую от СоА жирной кислоты, получают из организма, относящегося к одному или нескольким нижеследующим родам: Aeromonas, Streptomyces, Saccharomyces, Lactococcus, Mycobacterium, Streptococcus, Lactobacillus, Desulfitobacterium, Bacillus, Campylobacter, Vibrionaceae, Xylella, Sulfolobus, Aspergillus, Schizosaccharomyces, Listeria, Neisseria, Mesorhizobium, Ralstonia, Xanthomonas и Candida.
12. Способ по п.1, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит одну или несколько нижеследующих аминокислотных последовательностей: (i) аминокислотная последовательность SEQ ID NO:2; (ii) аминокислотная последовательность SEQ ID NO:3; (iii) аминокислотная последовательность SEQ ID NO:4; (iv) аминокислотная последовательность SEQ ID NO:5; (v) аминокислотная последовательность SEQ ID NO:6; (vi) аминокислотная последовательность SEQ ID NO:12; (vii) аминокислотная последовательность SEQ ID NO:20; (viii) аминокислотная последовательность SEQ ID NO:22; (ix) аминокислотная последовательность SEQ ID NO:24; (x) аминокислотная последовательность SEQ ID NO:26; (xi) аминокислотная последовательность SEQ ID NO:28; (xii) аминокислотная последовательность, SEQ ID NO:30; (xiii) аминокислотная последовательность SEQ ID NO:32; (xiv) аминокислотная последовательность SEQ ID NO:34; (xv) аминокислотная последовательность SEQ ID NO:55; (xvi) аминокислотная последовательность SEQ ID NO:58; (xvii) аминокислотная последовательность SEQ ID NO:60; (xviii) аминокислотная последовательность SEQ ID NO:61; (xix) аминокислотная последовательность SEQ ID NO:63; (xx) аминокислотная последовательность SEQ ID NO:65; (xxi) аминокислотная последовательность SEQ ID NO:67; (xxii) аминокислотная последовательность SEQ ID NO:70 или (xxiii) аминокислотная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:55, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67 или SEQ ID NO:70.
13. Способ по п.1, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит аминокислотную последовательность, кодированную одной или несколькими нижеследующими нуклеотидными последовательностями:
(a) нуклеотидная последовательность SEQ ID NO:7;
(b) нуклеотидная последовательность SEQ ID NO:8;
(c) нуклеотидная последовательность SEQ ID NO:9;
(d) нуклеотидная последовательность SEQ ID NO:10;
(e) нуклеотидная последовательность SEQ ID NO:11;
(f) нуклеотидная последовательность SEQ ID NO:13;
(g) нуклеотидная последовательность SEQ ID NO:21;
(h) нуклеотидная последовательность SEQ ID NO:23;
(i) нуклеотидная последовательность SEQ ID NO:25;
(j) нуклеотидная последовательность SEQ ID NO:27;
(k) нуклеотидная последовательность SEQ ID NO:29;
(l) нуклеотидная последовательность SEQ ID NO:31;
(m) нуклеотидная последовательность SEQ ID NO:33;
(n) нуклеотидная последовательность SEQ ID NO:35;
(о) нуклеотидная последовательность SEQ ID NO:54;
(р) нуклеотидная последовательность SEQ ID NO:59;
(q) нуклеотидная последовательность SEQ ID NO:62;
(r) нуклеотидная последовательность SEQ ID NO:64;
(s) нуклеотидная последовательность SEQ ID NO:66;
(t) нуклеотидная последовательность SEQ ID NO:68;
(u) нуклеотидная последовательность SEQ ID NO:69 или
(v) нуклеотидная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:54, SEQ ID NO:59, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68 или SEQ ID NO:69.
14. Способ по п.1, который предусматривает смешивание подвергнутого обработке пищевого масла с одним или несколькими пищевыми компонентами с образованием пищевого продукта.
15. Применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, переносящим в пищевом масле ацильную группу из диглицерида в глицерин, при производстве пищевого масла для уменьшения содержания и/или удаления (предпочтительно избирательного уменьшения содержания и/или удаления) диглицерида из указанного пищевого масла.
16. Применение диглицерид:глицерин-ацилтрансферазы, не зависящей от СоА жирной кислоты, которая является ферментом, переносящим в пищевом масле ацильную группу из диглицерида в глицерин, при производстве пищевого продукта, содержащего пищевое масло, для улучшения свойств кристаллизации указанного пищевого продукта.
17. Применение по п.15 или 16, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит фрагмент аминокислотной последовательности GDSX, где Х означает один или несколько нижеследующих аминокислотных остатков: L, А, V, I, F, Y, Н, Q, Т, N, М или S.
18. Применение по п.15 или 16, в котором происходит уменьшение количества диглицерида в пищевом масле.
19. Применение по п.15 или 16, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, переносит ацильную группу из диглицерида в акцептор ацильных групп, причем акцептор ацильных групп является любым соединением, содержащим гидроксильную группу (-ОН).
20. Применение по п.15 или 16, в котором диглицерид является 1,2-диглицеридом.
21. Применение по п.15 или 16, в котором акцептор ацильных групп является веществом, растворимым в пищевом масле.
22. Применение по п.15 или 16, в котором акцептор ацильных групп является спиртом.
23. Применение по п.22, в котором акцептор ацильных групп является спиртом.
24. Применение по п.23, в котором акцептор ацильных групп является глицерином.
25. Применение по п.15 или 16, в котором фермент диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит остаток Н-309 или остаток гистидина в положении, соответствующем остатку His-309 в аминокислотной последовательности липолитического фермента Aeromonas hydrophila SEQ ID NO:2 или SEQ ID NO:32.
26. Применение по п.15 или п.16, в котором диглицерид:глицерин-ацилтрансферазу, не зависящую от СоА жирной кислоты, получают из организма, относящегося к одному или нескольким нижеследующим родам: Aeromonas, Streptomyces, Saccharomyces, Lactococcus, Mycobacterium, Streptococcus, Lactobacillus, Desulfitobacterium, Bacillus, Campylobacter, Vibrionaceae, Xylella, Sulfolobus, Aspergillus, Schizosaccharomyces, Listeria, Neisseria, Mesorhizobium, Ralstonia, Xanthomonas и Candida.
27. Применение по п.15 или 16, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит одну или несколько нижеследующих аминокислотных последовательностей:
(i)аминокислотная последовательность SEQ ID NO:2;
(ii)аминокислотная последовательность SEQ ID NO:3;
(iii)аминокислотная последовательность SEQ ID NO:4;
(iv)аминокислотная последовательность SEQ ID NO:5;
(v)аминокислотная последовательность SEQ ID NO:6; (vi)аминокислотная последовательность SEQ ID NO:12; (vii)аминокислотная последовательность SEQ ID NO:20; (viii)аминокислотная последовательность SEQ ID NO:22; (iх)аминокислотная последовательность SEQ ID NO:24; (х)аминокислотная последовательность SEQ ID NO:26; (хi)аминокислотная последовательность SEQ ID NO:28; (хii)аминокислотная последовательность SEQ ID NO:30; (хiii)аминокислотная последовательность SEQ ID NO:32; (хiv)аминокислотная последовательность SEQ ID NO:34; (хv)аминокислотная последовательность SEQ ID NO:55; (хvi)аминокислотная последовательность SEQ ID NO:58; (хvii)аминокислотная последовательность SEQ ID NO:60; (хviii)аминокислотная последовательность SEQ ID NO:61; (хiх)аминокислотная последовательность SEQ ID NO:63; (хх)аминокислотная последовательность SEQ ID NO:65; (ххi)аминокислотная последовательность SEQ ID NO:67; (ххii)аминокислотная последовательность SEQ ID NO:70 или (xxiii) аминокислотная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:12, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:55, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67 или SEQ ID NO:70.
28. Применение по п.15 или 16, в котором диглицерид:глицерин-ацилтрансфераза, не зависящая от СоА жирной кислоты, содержит аминокислотную последовательность, кодированную одной или несколькими нижеследующими нуклеотидными последовательностями:
(a) нуклеотидная последовательность SEQ ID NO:7;
(b) нуклеотидная последовательность SEQ ID NO:8;
(c) нуклеотидная последовательность SEQ ID NO:9;
(d) нуклеотидная последовательность SEQ ID NO:10;
(e) нуклеотидная последовательность SEQ ID NO:11;
(f) нуклеотидная последовательность SEQ ID NO:13;
(g) нуклеотидная последовательность SEQ ID NO:21;
(h) нуклеотидная последовательность SEQ ID NO:23;
(i) нуклеотидная последовательность SEQ ID NO:25;
(j) нуклеотидная последовательность SEQ ID NO:27;
(k) нуклеотидная последовательность SEQ ID NO:29;
(l) нуклеотидная последовательность SEQ ID NO:31;
(m) нуклеотидная последовательность SEQ ID NO:33;
(n) нуклеотидная последовательность SEQ ID NO:35;
(о) нуклеотидная последовательность SEQ ID NO:54;
(р) нуклеотидная последовательность SEQ ID NO:59;
(q) нуклеотидная последовательность SEQ ID NO:62;
(г) нуклеотидная последовательность SEQ ID NO:64;
(s) нуклеотидная последовательность SEQ ID NO:66;
(t) нуклеотидная последовательность SEQ ID NO:68;
(u) нуклеотидная последовательность SEQ ID NO:69 или
(v) нуклеотидная последовательность, которая на 75% или более идентична любой последовательности SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:54, SEQ ID NO:59, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68 или SEQ ID NO:69.
29. Применение по п.15 или 16, в котором диглицерид:глицерин-ацилтрансферазу, не зависящую от СоА жирной кислоты, используют в комбинации с ингибитором кристаллизации.
| HILTON S | |||
| et | |||
| al | |||
| Studies on the reaction mechanism of a microbial lipase/acyltransferase using chemical modification and site-directed mutagenesis | |||
| J | |||
| Biol | |||
| Chem | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| ROBERTSON D.L | |||
| et | |||
| al | |||
| Influence of active site and tyrosine modification on the secretion and activity of the Aeromonas hydrophila lipase/acyltransferase. | |||

