Область, к которой относится изобретение
Настоящее изобретение относится к новым грибковым липолитическим ферментам и к одному или нескольким полинуклеотидам, кодирующим один или несколько новых грибковых липолитических ферментов. Настоящее изобретение также относится к способам получения грибковых липолитических ферментов и к их применению. Кроме того, настоящее изобретение относится к получению продуктов питания улучшенного качества, и, в частности, к получению хлебобулочных изделий улучшенного качества. Более конкретно, настоящее изобретение относится к новым грибковым липолитическим ферментам, которые способны придавать улучшенные качества пищевым продуктам, включая хлебобулочные изделия.
Предшествующий уровень техники
Многие годы известно липолитические ферментов (Е.С.3.1.1.х) успешно применяют в промышленном производстве пищевых продуктов или кормов.
Так, например, в патенте ЕР 0585988 было заявлено, что добавление липазы в тесто обеспечивает положительный эффект против черствения. Было высказано предположение, что липаза, полученная из Rhizopus arrhizus, при ее добавлении в тесто, может повышать качество выпеченного хлеба, если она используется в сочетании с шортенингом/жиром. В заявке WO94/04035 сообщается, что повышение мягкости хлеба может быть достигнуто путем добавления липазы в тесто без добавления в это тесто дополнительного количества жира/масла. В публикации Castello, P.ESEGP 89-10 Dec. 1999 Helsenki, было показано, что экзогенные липазы могут модифицировать объем хлеба.
Субстратом для липазы в пшеничной муке является 1,5-3% эндогенных липидов пшеницы, которые представляют собой комплексную смесь из полярных и неполярных липидов. Полярные липиды можно разделить на гликолипиды и фосфолипиды. Эти липиды состоят из глицерина, этерифицированного двумя жирными кислотами и полярной группой. Такая полярная группа придает поверхностную активность этим липидам. Ферментативное расщепление одной из таких жирных кислот в этих липидах позволяют получить липиды с гораздо более высокой поверхностной активностью. Хорошо известно, что эмульгаторы, такие как DАТЕМ с высокой поверхностной активностью, являются в высокой степени функциональными при их добавлении в тесто.
Липолитические ферменты гидролизуют одну или несколько жирных кислот липидов, присутствующих в пищевом продукте, что может приводить к образованию молекул, действующих как сильные эмульгаторы в пищевом продукте, который представляет коммерческую ценность. Молекулами, придающими наилучшие эмульгирующие свойства, являются продукты частичного гидролиза, такие как молекулы лизофосфолипидов, лизогликолипидов и моноглицеридов. Особенно предпочтительными являются продукты гидролиза полярных липидов, а именно, лизофосфолипиды и лизогликолипиды. При выпечке хлеба, такие in situ эмульгаторы могут обладать функциональными свойствами, эквивалентными свойствам добавленных эмульгаторов, таких как DATEM.
Однако было также обнаружено, что активность липолитических ферментов также приводит к накоплению свободных жирных кислот, что может негативно влиять на свойства пищевого продукта. Такая природная активность липолитических ферментов ограничивает их применение.
Негативное влияние этих ферментов на объем хлеба часть объясняют их передозировкой. Передозировка может приводить к снижению эластичности клейковины, которая приводит к получению слишком жесткого теста, а поэтому и к снижению объема. Дополнительно или альтернативно такие липазы могут разлагать шортенинг, масло или молочный жир, добавляемые в тесто, что придает тесту и хлебобулочному изделию не свойственный им привкус. Избыточные уровни фермента и привкус являются следствием накопления в тесте свободных жирных кислот, а в частности, короткоцепочечных жирных кислот.
Присутствие высоких уровней свободных жирных кислот (СЖК) в исходных продуктах или в готовых пищевых продуктах является, вообще говоря, дефектом качества, но обычно производители пищевых продуктов и заказчики включают в технические условия на пищевые продукты максимальные уровни СЖК. Избыточные уровни СЖК могут приводить к ухудшению органолептических и/или функциональных свойств.
В ЕР 1193314 авторами изобретения было обнаружено, что использование липолитических ферментов, активных по отношению к гликолипидам, дает особенно благоприятный эффект при выпечке хлеба, поскольку было установлено, что продукты частичного гидролиза, а именно лизогликолипиды, действуют как сильные эмульгаторы, что, очевидно, приводит к более высокому уровню позитивного функционального эффекта эмульгирования по сравнению с негативным накоплением свободных жирных кислот. Однако было также обнаружено, что ферменты обладают значительной неселективной активностью по отношению триглицеридам, что приводит к чрезмерно высокому уровню свободных жирных кислот.
Эта проблема, связанная с высокой активностью по отношению к триглицеридам, обсуждается в заявке WO 02/094123, авторами которой было обнаружено, что улучшение функциональных свойств может быть достигнуто путем выбора липолитических ферментов, являющихся активными по отношению к полярным липидам (гликолипидам и фосфолипидам), присутствующих в тесте, но, в основном, не активными по отношению к триглицеридам или 1-моноглицеридам.
Предпочтительным коммерчески доступными источниками липазных ферментов являются гифомицеты, такие как Aspergillus spp. и Fusarium spp. Было обнаружено, что липазы, выделенные из гифомицетов, обладают ценными с промышленной точки зрения свойствами, а также было обнаружено, что они является наиболее подходящими для экспрессии в системах гетерологичного продуцирования, таких как Aspergillus oryzae, Fusarium и дрожжи.
Липаза, выделенная из Fusarium oxysporum, была идентифицирована в ЕР 0130064, а в работе Hoshino et al. (1992) Biosci. Biotech. Biochem. 56:660-664, было высказано предположение о возможном применении липаз F. oxysporum в пищевых продуктах.
В ЕР 0869167 описано клонирование и экспрессия липазы Fusarium oxysporum и ее применение в хлебопекарной промышленности. Этот фермент описан как обладающий фосфолипазной активностью. В настоящее время этот фермент имеется в продаже под торговым знаком как Lipopan FTM Novozymes A/S (Denmark).
В WO 02/00852 описаны пять липазных ферментов и кодирующих эти ферменты полинуклеотидов, выделенных из F.venenatum, F.sulphureum, A. berkeleyanum, F. culmorum и F. solani. Все эти пять ферментов описаны как ферменты, обладающие триацилглицерин-гидролизующей активностью, фосфолипазной активностью и галактолипазной активностью. Из них, три фермента обладают активностью, эквивалентной активности фермента F. oxysporum, описанного в ЕР 0869167: F. venenantum, F. sulphureum, F. culmorum.
Кроме того, было обнаружено, что некоторые липазы Fusarium, включая Lipopan FTM, обладают побочной активностью по отношению к полярным липидам, включая фосфолипиды и гликолипиды. Хотя в ЕР 0869167 было указано, что этот фермент обладает фосфолипазной активностью, однако, было установлено, что такой фермент Fusarium oxysporum обладает высокой липазной активностью. Этот фермент также обладает гликолипазной активностью. Однако, несмотря на значительную активность по отношению к полярным липидам, эффект, достигаемый с применением такого фермента, ограничен высокой липазной активностью (т.е. активностью по отношению к триглицеридам).
В публикации Nagao et al. (J. Biochem. 116(1994)536-540) описана липаза, выделенная из F.heterosporum, и такой фермент преимущественно функционирует как липаза (Е.С. 3.1.1.3), гидролизующая триглицериды. Этот фермент очень отличается от ферментов согласно изобретению.
Были получены варианты липолитических ферментов, имеющие специфические аминокислотные замены и вставки, некоторые из которых, по сравнению с исходными ферментами дикого типа, обладают повышенной активностью по отношению к полярным липидам. В WO 01/39602 описан такой вариант, обозначенный SP979, который представляет собой гибрид липазы Thermomyces lanuginosus и липазы Fusarium oxysporum, описанной в ЕР 0869167. Было обнаружено, что этот вариант обладает высоким отношением активности, направленной на фосфолипиды и гликолипиды, к активности, направленной на триглицериды.
Однако до подачи настоящей заявки, нигде не указывалось, что природные грибковые липолитические ферменты, а в частности, ферменты, происходящие от Fusarium spp., имеют высокое отношение активности, направленной на полярные липиды, к активности, направленной на триглицериды.
Описание сущности изобретения
В основном аспекте, настоящее изобретение относится к грибковому липолитическому ферменту, обладающему более высоким отношением активности, направленной на полярные липиды (фосфолипиды и/или гликолипиды) к активности, направленной на триглицериды, а в частности, более высоким отношением активности, направленной на гликолипиды, к активности, направленной на триглицериды.
В другом аспекте, настоящее изобретение относится к грибковому липолитическому ферменту дикого типа, имеющему более высокое отношение активности, направленной на полярные липиды (фосфолипиды и/или гликолипиды), к активности, направленной на триглицериды, а в частности, более высоким отношением активности, направленной на гликолипиды, к активности, направленной на триглицериды.
В другом своем аспекте, настоящее изобретение относится к нуклеиновой кислоте, кодирующей новый грибковый липолитический фермент, описанный в настоящей заявке.
В еще одном аспекте, настоящее изобретение относится к способу приготовления пищевого продукта, предпочтительно, продуктов с добавлением яиц, где указанный способ предусматривает добавление грибкового липолитического фермента по изобретению к одному или нескольким ингредиентам пищевого продукта.
Настоящее изобретение относится к способу получения теста, где указанный способ предусматривает добавление грибкового липолитического фермента по изобретению к одному или нескольким ингредиентам теста и смешивание этих ингредиентов для приготовления теста.
В другом своем аспекте, настоящее изобретение относится к способу приготовления хлебобулочного изделия из теста, где указанный способ предусматривает добавление грибкового липолитического фермента по изобретению в указанное тесто.
Настоящее изобретение также относится к способу получения грибкового липолитического фермента по изобретению, где указанный способ предусматривает трансформацию клетки-хозяина рекомбинантной нуклеиновой кислотой, содержащей нуклеиновую кислоту, кодирующую грибковый липолитический фермент, и где указанная клетка-хозяин обладает способностью экспрессировать нуклеотидную последовательность, кодирующую полипептид грибкового липолитического фермента; культивирование трансформированной клетки-хозяина в условиях, при которых экспрессируется указанная нуклеиновая кислота; и сбор полученного грибкового липолитического фермента.
В другом своем аспекте, настоящее изобретение относится к липолитическому ферменту, который сохраняет активность при низких температурах, то есть к низкотемпературному липолитическому ферменту.
Аспекты настоящего изобретения описаны в формуле изобретения и в нижеследующем описании.
Другие аспекты изобретения, относящиеся к нуклеотидным последовательностям, которые могут быть использованы в настоящем изобретении, включают конструкцию, содержащую последовательности согласно изобретению; вектор, содержащий последовательности, используемые в настоящем изобретении; плазмиду, содержащую последовательности, используемые в настоящем изобретении; трансформированную клетку, содержащую последовательности, используемые в настоящем изобретении; трансформированную ткань, содержащую последовательности, используемые в настоящем изобретении; трансформированный орган, содержащий последовательности, используемые в настоящем изобретении; трансформированного хозяина, содержащего последовательности, используемые в настоящем изобретении; трансформированный организм, содержащий последовательности, используемые в настоящем изобретении. Настоящее изобретение также относится к способам экспрессии нуклеотидной последовательности, используемой в настоящем изобретении, например, экспрессии в клетке-хозяине, включая способы переноса этой последовательности. Настоящее изобретение также относится к способам выделения нуклеотидной последовательности, таким как выделение последовательности из клетки-хозяина.
Другие аспекты настоящего изобретения, относящиеся к аминокислотной последовательности, используемой в настоящем изобретении, включают конструкцию, содержащую аминокислотные последовательности, используемые в настоящем изобретении; вектор, кодирующий аминокислотные последовательности, используемые в настоящем изобретении; плазмиду, кодирующую аминокислотные последовательности, используемые в настоящем изобретении; трансформированную клетку, экспрессирующую аминокислотные последовательности, используемые в настоящем изобретении; трансформированную ткань, экспрессирующую аминокислотные последовательности, используемые в настоящем изобретении; трансформированный орган, экспрессирующий аминокислотные последовательности, используемые в настоящем изобретении; трансформированный хозяин, экспрессирующий аминокислотные последовательности, используемые в настоящем изобретении; трансформированный организм, экспрессирующий аминокислотные последовательности, используемые в настоящем изобретении. Настоящее изобретение также охватывает способы очистки аминокислотной последовательности, используемой в настоящем изобретении с применением способа экспрессии этой последовательности, такой как экспрессия в клетке-хозяине, включая способы переноса этой последовательности с ее последующей очисткой.
Для лучшего понимания изобретения, эти и другие аспекты настоящего изобретения обсуждаются в настоящем описании в соответствующих разделах. Однако обсуждения, представленные в каждом разделе, не должны ограничиваться каким-либо конкретным разделом.
Подробное описание изобретения
В одном из своих аспектов, настоящее изобретение относится к грибковым липолитическим ферментам, имеющим более высокое отношение активности, направленной на полярные липиды, к активности, направленной на триглицериды.
В одном из своих аспектов, настоящее изобретение относится к грибковым липолитическим ферментам, содержащим аминокислотную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:4 или SEQ ID NO:6, или аминокислотную последовательность, которая по меньшей мере на 90% идентична указанной аминокислотной последовательности.
В другом своем аспекте, настоящее изобретение относится к нуклеиновой кислоте, кодирующей грибковый липолитический фермент, содержащий аминокислотную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:4 или SEQ ID NO:6, или аминокислотную последовательность, которая по меньшей мере на 90% идентична указанной аминокислотной последовательности.
SEQ ID NO:1 представлена на фиг.37, SEQ ID NO:2 представлена на фиг.38, SEQ ID NO:4 представлена на фиг.40, а SEQ ID NO:6 представлена на фиг.42.
В другом своем аспекте, настоящее изобретение относится к нуклеиновой кислоте, кодирующей грибковый липолитический фермент, где указанная нуклеиновая кислота выбрана из группы, состоящей из:
а) нуклеиновой кислоты, содержащей нуклеотидную последовательность, представленную в SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7;
b) нуклеиновой кислоты, которая соответствует нуклеотидной последовательности SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7 в соответствии с вырожденностью генетического кода; и
с) нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая по меньшей мере на 90% идентична нуклеотидной последовательности, представленной в SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
SEQ ID NO:3 представлена на фиг.39, SEQ ID NO:5 представлена на фиг.41, а SEQ ID NO:7 представлена на фиг.43.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению для приготовления пищевого продукта, такого как, например, тесто, хлебобулочные изделия, продукты с добавлением или основу которых составляют яица, макаронные изделия, сырные продукты, лепешки, корм для животных, растительное масло или пищевое масло. Добавление фермента по изобретению в пищевой продукт, преимущественно, приводит к улучшению эмульгирования с одновременным уменьшением содержания свободных жирных кислот.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению для приготовления теста и/или хлебобулочного изделия, где указанное применение включает добавление указанного липолитического фермента в тесто, и (необязательно) выпекание теста с получением хлебобулочного изделия, обладающего одним или несколькими свойствами, такими как: пониженная липкость теста: повышенная обрабатываемость теста; пониженный уровень вздутия во время выпекания хлебобулочного изделия; повышенный объем и/или мягкость хлеба; длительный срок хранения указанного хлебобулочного изделия и/или теста; повышенная устойчивость к черствению такого хлебобулочного изделия и/или теста; улучшенная структура мякиша такого хлебобулочного изделия; пониженная гетерогенность пор такого хлебобулочного изделия; повышенная гомогенность пор хлебобулочного изделия; уменьшенный средний размер пор хлебобулочного изделия; высокий глютеновый индекс теста; улучшенный вкус и/или запах хлебобулочного изделия, и более ярко выраженный цвет корочки такого хлебобулочного изделия.
В преимущественном варианте, фермент по изобретению может иметь более высокую активность, чем стандартные липофильные ферменты при низких рН, и таким образом, он может быть более подходящим для применения в кислом тесте с низким рН, чем стандартные липофильные ферменты.
В другом своем аспекте, настоящее изобретение относится к способу получения теста и/или хлебобулочного продукта, предусматривающему добавление в тесто грибкового липолитического фермента по изобретению и (необязательно) выпекание теста с получением хлебобулочного изделия.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению для приготовления продуктов с добавлением яиц в целях улучшения их консистенции, снижения среднего размера частиц, снижения среднего распределения частиц, повышения термостабильности и повышения производительности при выпечке в микроволновой печи, и/или повышения стабильности.
В другом своем аспекте, настоящее изобретение относится к способу обработки яиц или продуктов с добавлением яиц, где указанный способ предусматривает добавление к яйцам и яичным продуктам грибкового липолитического фермента согласно изобретению.
В другом своем аспекте, настоящее изобретение относится к способу приготовления лапши или теста для лапши, где указанный способ включает добавление в лапшу или в тесто для лапши или в продукт из лапши грибкового липолитического фермента по изобретению.
В одном из аспектов изобретения, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в приготовлении лапши или макаронных изделий для улучшения их цвета/придания им желтизны, а также для стабилизации цветовых характеристик, снижения белизны, снижения содержания жира, улучшения консистенции и уменьшения нежелательного привкуса (при разжевывании), снижения смачивающей активности, повышения устойчивости к разлому, повышения плотности изделия и повышения способности к сохранению формы в процессе обработки.
В другом своем аспекте, настоящее изобретение относится к способу приготовления лепешек или теста для лепешек, где указанный способ предусматривает добавление в эту лепешку или в тесто для лепешек грибкового липолитического фермента по изобретению.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению для изготовления лепешек или теста для лепешек в целях улучшения раскатываемости лепешки, повышения ее пластичности, повышения устойчивости против черствения лепешки и/или теста для лепешек, повышения мягкости и/или устранения привкуса данной лепешки и/или теста для лепешек.
Функциональные свойства липолитического фермента в лепешках и/или в лапше могут быть улучшены благодаря использованию этого фермента в сочетании с эмульгаторами, такими как DАТЕМ.
В другом своем аспекте, настоящее изобретение относится к способу обработки молока, молока для сыроделия, сыра или сырного продукта, где указанный способ предусматривает добавление в сыр или сырный продукт грибкового липолитического фермента по изобретению.
Кроме того, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в изготовлении сыра или сырного продукта для придания этому продукту одного или нескольких свойств, а именно, для улучшения вкуса и консистенции продукта и/или повышения его стабильности, а также для повышения маслянистости сыра и/или увеличения выхода сыра в сыроделии.
В другом своем аспекте, настоящее изобретение относится к способу обработки корма для животных, где указанный способ предусматривает добавление в корм для животных грибкового липолитического фермента согласно изобретению.
Настоящее изобретение также относится к применению грибкового липолитического фермента по изобретению в изготовлении корма для животных в целях достижения одной или нескольких целей, а именно, повышения эффективности утилизации и/или переработки кормов, увеличения прироста массы тела, улучшения усвояемости азота, усиления метаболизма сухих веществ и улучшения их вкусовых качеств.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в способе получения лизофосфолипида, например, лизолецитина путем обработки фосфолипида (например, лецитина) ферментом с продуцированием продукта частичного гидролиза, то есть лизофосфолипида.
В другом своем аспекте, настоящее изобретение относится к способу получения лизофосфолипида, например, лизолецитина, где указанный способ предусматривает обработку фосфолипида (например, лецитина) грибковым липолитическим ферментом по изобретению.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в способе получения лизогликолипида (например, дигалактозилмоноглицерида (ДГМГ) или моногалактозилмоноглицерида (МГМГ)) путем обработки гликолипида (например, дигалактозилдиглицерида (ДГДГ) или моногалактозилдиглицерида (МГДГ)) липолитическим ферментом по изобретению с получением продукта частичного гидролиза, то есть, лизогликолипида.
В другом своем аспекте, настоящее изобретение относится к способу получения лизогликолипида (например, дигалактозилмоноглицерида (ДГМГ) или моногалактозилмоноглицерида (МГМГ)), где указанный способ предусматривает обработку гликолипида (например, дигалактозилдиглицерида (ДГДГ) или моногалактозилдиглицерида (МГДГ)) грибковым липолитическим ферментом по изобретению.
Настоящее изобретение также относится к способу ферментативного рафинирования растительных или пищевых масел, предусматривающему обработку пищевого или растительного масла грибковым липолитическим ферментом по изобретению так, чтобы большая часть полярных липидов (например, фосфолипида и/или гликолипида) была гидролизована.
Для большей ясности, специалисту в данной области следует ознакомиться с соответствующей методикой проведения ферментативной обработки пищевых масел (см., например, ЕР0869167). При осуществлении настоящего изобретения может быть использован любой подходящий известный метод, при условии, что известный фермент будет заменен ферментом по изобретению.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента согласно изобретению для изготовления растительного или пищевого масла в целях снижения количества фосфолипида в растительном или пищевом масле при сохранении содержания триглицеридов в этом масле и/или в целях предотвращения или снижения накопления свободных жирных кислот.
В еще одном своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в способе, предусматривающем обработку фосфолипида в целях гидролиза ацильных групп жирных кислот.
В другом своем аспекте, настоящее изобретение относится к применению грибкового липолитического фермента по изобретению в способе снижения содержания фосфолипида в пищевом масле, где указанный способ предусматривает обработку указанного масла грибковым липолитическим ферментом по изобретению в целях гидролиза большей части фосфолипида, и удаление из этого масла водной фазы, содержащей гидролизованный фосфолипид.
В другом своем аспекте, настоящее изобретение относится к липолитическому ферменту, сохраняющему активность при низких температурах, то есть к низкотемпературному ферменту. В других своих аспектах, настоящее изобретение относится к применению низкотемпературного фермента, а именно грибкового липолитического фермента согласно изобретению, в описанных здесь способах.
Предпочтительно, грибковый липолитический фермент по изобретению имеет более высокое отношение активности, направленной на полярные липиды (например, гликолипидов и/или фосфолипидов), по сравнению с активностью, направленной на триглицериды.
Предпочтительно, грибковый липолитический фермент по изобретению имеет более высокое отношение фосфолипид-гидролизующей активности к триглицерид-гидролизующей активности.
Предпочтительно, грибковый липолитический фермент по изобретению имеет более высокое отношение гликолипид-гидролизующей активности по сравнению с триглицерид-гидролизующей активностью.
В соответствии с этим грибковый липолитический фермент по изобретению может иметь более высокое отношение гликолипид- и фосфолипид-гидролизующей активности к триглицерид-гидролизующей активности.
Более предпочтительно, грибковый липолитический фермент по изобретению имеет более высокое отношение дигалоктозилдиглицерид (ДГДГ)-гидролизующей активности к триглицерид-гидролизующей активности.
Предпочтительно, грибковый липолитический фермент по изобретению гидролизует ДГДГ или МГДГ с образованием ДГМГ или МГМГ, соответственно.
Используемый здесь термин “более высокое отношение активности, направленной на полярные липиды” означает, что грибковый липолитический фермент по изобретению имеет более высокое отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности, чем коммерчески доступный фермент Lipopan FTM (Novozymes A/S, Denmark).
Используемый здесь термин “полярные липиды” означает фосфолипиды и/или гликолипиды. Используемый здесь термин “полярные липиды”, предпочтительно, означает как фосфолипиды, так и гликолипиды.
Используемый здесь термин “более высокое отношение активности, направленной на гликолипиды” или “более высокое отношение активности, направленной на фосфолипиды” означает, что грибковый липолитический фермент по изобретению имеет более высокое отношение гликолипид-гидролизующей активности к триглицерид-гидролизующей активности, соответственно, чем коммерчески доступный фермент Lipopan FTM (Novozymes A/S, Denmark).
Предпочтительно, липолитический фермент по изобретению может иметь отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности, равное, по меньшей мере 4. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 5. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 8, предпочтительно, более 9, более предпочтительно, более 10, а еще более предпочтительно, более 15.
Предпочтительно, грибковый липолитический фермент по изобретению может иметь отношение фосфолипид-гидролизующей активности к триглицерид-гидролизующей активности, равное, по меньшей мере 4. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности, может составлять более 5. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 8, предпочтительно, более 9, более предпочтительно, более 10, а еще более предпочтительно, более 15.
Предпочтительно, грибковый липолитический фермент по изобретению может иметь отношение гликолипид-гидролизующей активности к триглицерид-гидролизующей активности, равное, по меньшей мере, 1,5, предпочтительно, по меньшей мере, 1,8, предпочтительно, по меньшей мере, 2, предпочтительно, по меньшей мере, 3, а предпочтительно, по меньшей мере, 4. В основном, отношение гликолипид-гидролизующей активности к триглицерид-гидролизующей активности, может составлять более 4. В основном, отношение гликолипид-гидролизующей активности к триглицерид-гидролизующей активности, может составлять более 5.
В другом своем аспекте, настоящее изобретение относится к грибковому липолитическому ферменту, имеющему отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности, равное, по меньшей мере, 4. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 5. В основном, отношение отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 8, предпочтительно, более 9, более предпочтительно, более 10, а еще более предпочтительно, более 15.
В другом своем аспекте, настоящее изобретение относится к грибковому липолитическому ферменту, имеющему отношение фосфолипид-гидролизующей активности к триглицерид-гидролизующей активности, равное, по меньшей мере, 4. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 5. В основном, отношение активности, направленной на гидролиз полярного липида, к триглицерид-гидролизующей активности может составлять более 8, предпочтительно, более 9, более предпочтительно, более 10, а еще более предпочтительно, более 15.
В еще одном своем аспекте, настоящее изобретение относится к грибковому липолитическому ферменту, имеющему отношение гликолипид-гидролизующей активности к триглицерид-гидролизующей активности, активностей, равное, по меньшей мере, 1,5, предпочтительно, по меньшей мере, 1,8, предпочтительно, по меньшей мере, 2, предпочтительно, по меньшей мере, 3, а предпочтительно, по меньшей мере, 4, предпочтительно, более 5, предпочтительно, более 10 и предпочтительно, более 15.
Предпочтительно, грибковый липолитический фермент по изобретению имеет в 1,5 раза, а предпочтительно, по меньшей мере, в 2 раза, более предпочтительно, по меньшей мере, в 3 раза, а еще более предпочтительно, по меньшей мере, в 4 раза большую активность по отношению к полярным липидам (например, фосфолипазную А2 (Е.С. 3.1.1.4) активность и/или фосфолипазную А1 (Е.С. 3.1.1.32) активность и/или гликолипазную (Е.С. 3.1.1.26) активность) по сравнению с триглицерид-липазной (Е.С. 3.1.1.3) активностью.
Предпочтительно, грибковый липолитический фермент по изобретению имеет в 1,5 раза, а предпочтительно, по меньшей мере, в 2 раза, более предпочтительно, по меньшей мере, в 3 раза, а еще более предпочтительно, по меньшей мере, в 4 раза большую гликолипазную (Е.С. 3.1.1.26) активность по сравнению с триглицерид-липазной (Е.С. 3.1.1.3) активностью.
При дозе, которая дает оптимальный объем хлеба, как было определено в анализе на выпечку в лабораторном масштабе, подробно описанном в примере 3, отношение ДГДГ-гидролизующей активности к триглицерид (ТГ)-гидролизующей активности, предпочтительно, составляет по меньшей мере 1,7%, предпочтительно по меньшей мере 1,8%, предпочтительно по меньшей мере 2%, предпочтительно по меньшей мере 3%, предпочтительно по меньшей мере 4%, предпочтительно, по меньшей мере 5%, предпочтительно, по меньшей мере 10%, предпочтительно, по меньшей мере 20%, предпочтительно, по меньшей мере 40%, а более предпочтительно, по меньшей мере 50%.
Используемый здесь термин “гликолипазная активность” охватывает “галактолипазную активность”.
Гликолипазная активность, фосфолипазная активность и триацилглицерид-липазная активность фермента могут быть определены в анализах, представленных ниже.
Определение галактолипазной активности (анализ на гликолипазную активность)
Субстрат:
0,6% дигалактозилдиглицерид (Sigma D 4651), 0,4% Тритона-Х 100 (Sigma X-100) и 5 мМ СаCl2 растворяют в 0,05М буфере HEPES при рН 7.
Процедура анализа:
400 мкл субстрата добавляют в 1,5 мл-пробирку Eppendorf и помещают в термомиксер Eppendorf на 5 минут при 37°С. В момент времени t=0 минут добавляют 50 мкл раствора фермента. В качестве контроля, вместо фермента использовали воду. Образец смешивали при 10 × 100 об/мин в термомиксере Eppendorf при 37°С в течение 10 минут. Через 10 минут пробирку Eppendorf помещают на 10 минут в другой термомиксер при 99°С для прекращения реакции.
Свободную жирную кислоту в образцах анализировали с использованием набора NEFA C, поставляемого WAKO GmbH.
Ферментативную активность GLU при рН 7 выражают в микромолях жирной кислоты, полученной за 1 минуту в условиях анализа.
Определение фосфолипазной активности (анализ на фосфолипазную активность)
Фосфолипазную активность измеряли двумя различными методами, которые давали сравнимые результаты. В соответствии с настоящим изобретением, для определения фосфолипазной активности может быть использован любой из этих методов. Для определения фосфолипазной активности любого фермента, предпочтительно, проводить анализ на PLU.
“Анализ на PLU-” для определения фосфолипазной активности
Субстрат:
0,6% L-α-фосфатидилхолин, 95% растительного экстракта (Avanti #441601), 0,4% Triton-Х 100 (Sigma X-100) и 5 мМ СаCl2 растворяют в 0,05М буфере HEPES при рН 7.
Процедура анализа:
400 мкл субстрата добавляли в 1,5 мл-пробирку Eppendorf и помещают на 5 минут в термомиксер Eppendorf при 37°С. На время t=0 минут добавляли 50 мкл раствора фермента. В качестве контроля, вместо фермента используют воду. Образец смешивают при 10 × 100 об/мин в термомиксере Eppendorf при 37°С в течение 10 минут. Через 10 минут пробирку Eppendorf помещают в другой термомиксер на 10 минут при 99°С для прекращения реакции.
Свободную жирную кислоту в образцах анализируют с использованием набора NEFA C, поставляемого WAKO GmbH.
Ферментативную активность PLU-7 при рН 7 выражают в микромолях жирной кислоты, продуцированной за 1 минуту в условиях анализа.
“Анализ на TIPU” для определения фосфолипазной активности
1 TIPU (титрационную единицу фосфолипазной активности) определяют как количество фермента, которое высвобождает 1 мкмоль свободной жирной кислоты в одну минуту в условиях данного анализа.
Фосфолипазы А1 и А2 катализируют превращение лецитина в лизолецитин с высвобождением свободной жирной кислоты из положения 1 и 2, соответственно. Фосфолипазная активность может быть определена путем непрерывного титрования жирных кислот, высвобождаемых из лецитина в процессе ферментации, поскольку количество поглощаемой щелочи равно количеству высвобождаемой жирной кислоты.
Субстрат:
4% лецитин, 4% Triton-Х 100 и 6 мМ СаCl2: 12 г лецитинового порошка (полярные липиды Avanti # 44160) и 12 г Triton-Х 100 (Merck 108643) диспергируют приблизительно в 200 мл деминерализованной воды при перемешивании с помощью магнитной мешалки. Затем добавляют 3,0 мл 0,6 М СаCl2 (р.а. Merck 1.02382). Объем доводят до 300 мл добавлением деминерализованной воды и эмульсию гомогенизируют с помощью устройства Ultra Thurax. Этот субстрат получают в свежем виде каждый день.
Процедура анализа:
Раствор фермента приготавливают с добавлением 300 мкл фермента и получают кривую титрования с наклоном от 0,06 до 0,18 мл/мин. В эту процедуру также включают контрольный образец с известной активностью.
Образцы растворяют в деминерализованной воде и перемешивают в течение 15 минут при 300 об/мин. 25,00 мл субстрата выдерживают в термостате в течение 10-15 минут при 37°С до тех пор, пока рН не будет составлять 7,0 после добавления 0,05М NaOH. К субстрату добавляют 300 мкл раствора фермента и осуществляют непрерывное титрование с добавлением 0,05М NaOH в устройстве для титрования, поддерживая постоянный рН с помощью рН-стата (Phm 290, Mettler Toledo). Два определения каждой активности проводят в каждом выбранном масштабе. Через 8 минут титрование прекращают и в течение 5-7 минут вычисляют наклон кривой титрования. Предел детекции составляет 3 TIPU/мл раствора фермента.
Вычисления:
Фосфолипазную активность (TIPU/г фермента) вычисляют по следующей формуле:
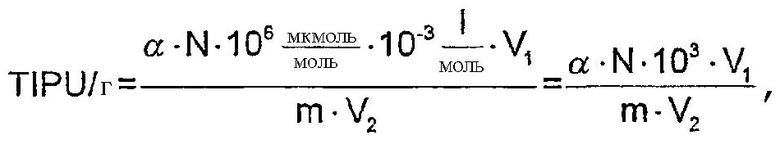
где: α означает наклон кривой титрования за время реакции от 5 до 7 минут (мл/мин.),
N означает нормальность используемого NaOH (моль/л),
V1 означает объем, в котором растворяется данный фермент (мл),
m означает количество фермента, добавляемого к V1 (г),
V2 означает объем раствора фермента, добавленного к субстрату (мл).
Определение триацилглицерид-липазной активности: анализ, проводимый с использованием триглицерида (трибутирина) в качестве субстрата (LIPU):
Липазную активность фермента на основе трибутирина измеряли методом, описанным в руководстве Food Chemical Codex, Forth Edition, National Academy Press, 1996, стр.803, за исключением того, что образец растворяли не в глициновом буфере, а в деионизованной воде, а постоянная величина рН, поддерживаемая с помощью рН-стата, составляла 5,5 вместо 7.
1 LIPU определяли как количество фермента, который может высвобождать 1 моль масляной кислоты за 1 минуту в условиях проведения анализа.
Исходя из анализов на активность по отношению к галактолипиду (GLU), фосфолипиду (PLU) и триглицериду (LIPU), можно вычислить отношения PLU/LIPU и GLU/LIPU.
Результаты анализа Lipopan FTM и липолитического фермента по изобретению, полученного из Fusarium heterosporum (образец 209)(см. пример 3), представлены ниже.
Отношения активности для Lipopan FTM и образца 209 имеют следующие значения:
В основном, используемые здесь термины “синергизм” или “синергический эффект” означают, что сочетание компонентов дает лучший эффект, чем каждый из этих компонентов (т.е. ферментов), используемый отдельно. Синергизм может быть определен путем приготовления продукта, например, теста и/или хлебобулочного изделия, при добавлении каждого компонента (т.е. фермента) по отдельности и комбинации этих компонентов, и сравнения этих эффектов.
Используемый здесь термин “грибковый липолитический фермент” означает, что природным источником фермента является грибок. Однако во избежание неясностей, следует отметить, что этот термин может включать грибковый фермент, который выделен из грибка и который экспрессируется в грибке-хозяине (либо в природном, либо в синтетическом), или который экспрессируется в не-грибковом хозяине (например, в бактериях или в дрожжах).
Предпочтительным грибковым липолитическим ферментом согласно изобретению является фермент дикого типа.
Используемые здесь термины “природный” и “дикого типа” относятся к природному ферменту. Следует отметить, что таким ферментом является фермент, экспрессируемый эндогенным геном и выделенный из эндогенного организма-хозяина и/или гетерологично продуцируемый фермент, который является немутированным (т.е. не содержит аминокислотных делеций, добавлений или замен) по сравнению с эндогенно продуцированной последовательностью зрелого белка (после событий ко- и пост-трансляционного расщепления). Природные белки и белки дикого типа по изобретению могут кодироваться оптимизированными по кодонам полинуклеотидами для гетерологичной экспрессии и могут также содержать не-эндогенный сигнальный пептид, выбранный для экспрессии в данном хозяине.
Используемый здесь термин “не-эндогенный сигнальный пептид” означает сигнальный пептид, который обычно отсутствует в растущей полипептидной цепи липолитического фермента до котрансляционного отщепления. В липолитическом ферменте согласно изобретению часть не-эндогенного сигнального пептида или весь этот пептид, например, про-пептид, может оставаться связанным со зрелым полипептидом, и такой пептид также входит в объем используемого здесь термина “фермент дикого типа”.
Как было указано выше, используемые здесь термины “природный” и “дикого типа” относятся к природному ферменту. Однако это не исключает возможности использования синтетического или химически синтезированного полипептида, содержащего такую же полипептидную последовательность, как и последовательность природного зрелого липолитического фермента.
Используемый здесь термин “вариант” означает белок, экспрессируемый не-эндогенным геном с образованием одной или нескольких аминокислотных модификаций (т.е. аминокислотных делеций, добавлений или замен) по сравнению с природной последовательностью или с последовательностью дикого типа, присутствующей в последовательности зрелого белка.
Предпочтительным грибковым липолитическим ферментом согласно изобретению является липолитический фермент, сохраняющий активность при низкой температуре, т.е. низкотемпературный липолитический фермент.
Термин “низкотемпературный липолитический фермент” означает фермент, который обладает значительной активностью при температуре 5-15°С, и, предпочтительно, фермент, который обладает значительной активностью при 10°С.
В одном из вариантов изобретения, указанным низкотемпературным липолитическим ферментом по изобретению не является липолитический фермент, содержащий мотив аминокислотной последовательности GDSX, как описано в WO 2004/064987, где Х представляет собой один или несколько из следующих аминокислотных остатков: L, A, V, I, F, Y, H, Q, T, N, M или S.
Низкотемпературным липолитическим ферментом по изобретению может быть фермент, который имеет относительную активность по меньшей мере 5%, предпочтительно по меньшей мере 7%, более предпочтительно по меньшей мере 10%, по отношению к лецитиновому субстрату при 10°С, при рН, составляющем 20% от оптимального рН для липолитического фермента, что было вычислено путем определения свободных жирных кислот методом NEFA C (см. пример 5, где процедуру проводили при рН 7). В примере 6 описан метод определения оптимального рН для липолитического фермента.
Низкотемпературным липолитическим ферментом по изобретению может быть фермент, который имеет относительную активность, по меньшей мере 10%, предпочтительно, по меньшей мере 15%, более предпочтительно, по меньшей мере 20%, еще более предпочтительно, по меньшей мере 25%, а наиболее предпочтительно, по меньшей мере 30%, по отношению к лецитиновому субстрату при 10°С, при рН, составляющем 20% от оптимального рН для липолитического фермента, что было вычислено путем определения свободных жирных кислот методом NEFA C (см. пример 5, в котором процедуру проводили при рН 7). В примере 6 описан метод определения оптимального рН для липолитического фермента.
Низкотемпературный липолитический фермент по изобретению может также обладать значительной активностью по отношению к лецитину яичного желтка при 5°С, отличающуюся тем, что она индуцирует высвобождение, по меньшей мере 1%, предпочтительно, по меньшей мере 1,5%, а более предпочтительно, по меньшей мере 2% свободной жирной кислоты после прохождения реакции в течение 480 минут при дозе фермента, эквивалентной дозе 20 единиц/г яичного желтка, в анализе, описанном в примере 9 и проиллюстрированном на фиг.24 и 25.
Предпочтительно, грибковый липолитический фермент по изобретению может быть получен (предпочтительно, получают) из гифомицетов. Более предпочтительно, такой грибковый липолитический фермент может быть получен (предпочтительно, получают) из Fusarium spp. Еще более предпочтительно, такой грибковый липолитический фермент по изобретению может быть получен (предпочтительно, получают) из Fusarium heterosporum или Fusarium semitectum. В основном, такой грибковый липолитический фермент по изобретению может быть получен (предпочтительно, получают) из Fusarium heterosporum (CBS 782.83) или Fusarium semitectum (IBT 9507).
Таким образом, в одном из аспектов, указанным липолитическим ферментом по изобретению, предпочтительно, является липолитический фермент гифомицетов, а более предпочтительно, липолитический фермент дикого типа, выделенный из гифомицетов.
Грибковый липолитический фермент по изобретению предпочтительно содержит аминокислотную последовательность, которая, по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, а наиболее предпочтительно, по меньшей мере на 99% идентична аминокислотной последовательности, представленной в SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:4 или SEQ ID NO:6.
Нуклеиновая кислота, кодирующая грибковый липолитический фермент согласно изобретению, предпочтительно, содержит нуклеотидную последовательность, которая, по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, а наиболее предпочтительно, по меньшей мере на 99% идентична нуклеотидной последовательности, представленной в SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
Предпочтительный грибковый липолитический фермент по изобретению не является гибридным белком, содержащим аминокислотную последовательность белка Thermomyces или его части, присоединенную к аминокислотной последовательности белка Fusarium или его части. В частности, предпочтительно, грибковый липолитический фермент по изобретению не является гибридным белком, содержащим аминокислотную последовательность белка Thermomyces lanuginosa или его части, присоединенную к аминокислотной последовательности белка Fusarium oxysporum или его части.
Предпочтительно, грибковый липолитический фермент по изобретению не является ферментом, выделенным из Thermomyces lanuginosa, и/или не является вариантом фермента, выделенного из Thermomyces lanuginosa.
Предпочтительно, грибковый липолитический фермент по изобретению выделяют из сбраживаемого бульона Fusarium heterosporum СBS 782.83 или Fusarium semitectum (IBT 9507).
В основном, указанный фермент может быть очищен с помощью жидкостной хроматографии.
Аминокислотная последовательность очищенного грибкового липолитического фермента может быть определена методом деградации по Эдману и с помощью анализа MALDI-TOF.
Частично очищенный липолитический фермент, полученный из Fusarium heterosporum СВS 782.83, был протестирован в лабораторных анализах на пробную выпечку и в экспериментальных анализах на пробную выпечку с очень хорошими результатами.
Было обнаружено, что результаты выпечки с использованием грибкового липолитического фермента, полученного из F. heterosporum СВS 782.83, превосходят результаты, полученные с использованием фермента Lipopan FTM, и эти результаты коррелируют с увеличением отношения активности, направленной на полярные липиды, а в частности, гликолипидов, таких как галактозильный диглицерид (ДГДГ), к активности, направленной на триглицериды.
Кроме того, липолитический фермент, полученный из Fusarium semitectum IBT 9507, был проанализирован на активность по отношению к липидам муки, присутствующим в тестовой суспензии, и этот анализ показал очень хорошие результаты.
Было показано, что липолитический фермент, полученный из F. semitectum IBT 9507, обладает значительной активностью, направленной на галактолипид в тесте, и относительно меньшей активностью, направленной на триглицерид, по сравнению с Lipopan FTM.
В общих чертах, используемый здесь термин “пищевой продукт” означает вещество, которое является пригодным для употребления в пищу человеком и/или животным.
В общих чертах, используемый здесь термин “пищевой продукт” обозначает продукт питания, приготовленный в форме, готовой для употребления. Однако альтернативно или дополнительно, используемый здесь термин “пищевой продукт” может означать один или несколько пищевых продуктов, которые используются для получения продукта питания. Термин “пищевой продукт”, но только лишь в качестве примера, охватывает хлебобулочные изделия, изготавливаемые из теста, а также тесто, используемое для изготовления указанных хлебобулочных изделий.
В своем предпочтительном аспекте, настоящее изобретение относится к пищевому продукту, определенному выше, где указанный пищевой продукт выбран из следующих продуктов: яиц, продуктов с добавлением яиц, включая, но не ограничиваясь ими, майонез, заправки для салатов, соусы, мороженное, яичный порошок, модифицированный яичный желток и изготавливаемые из них продукты; хлебобулочных изделий, включая хлеб, торты, изделия из сдобного теста, изделия из слоеного теста, пирожки из жидкого теста, круглые пышки, пончики, бисквиты, крекеры и печенье; кондитерских изделий, включая шоколад, конфеты, карамель, халву, жевательные резинки, включая жевательные резинки без сахара и с сахаром, жевательные резинки, которые при жевании образуют пузыри, мягкие жевательные резинки, которые при жевании образуют пузыри, стандартные жевательные резинки и пудинги; замороженных продуктов, включая шербеты, а предпочтительно, замороженные молочные продукты, включая мороженное и молочное мороженное; молочных продуктов, включая сыр, масло, молоко, кофе со сливками, взбитые сливки, сладкий крем, молочные напитки и йогурты; муссов, взбитых сливок на основе растительного масла; пищевых масел и жиров, газированных и негазированных прохладительных напитков, эмульсий типа “масло в воде”, эмульсий типа “вода в масле”, маргарина, шортенинга и паст, включая пасты с низким и с очень низким содержанием жира; заправок, майонеза, подливок, соусов-пюре, супов-пюре, напитков, пряных эмульсий и соусов.
В одном из аспектов изобретения, пищевым продуктом может быть продукт, выпекаемый из теста, или хлебобулочное изделие, такие как хлеб, обжаренный продукт, закусочные пищевые продукты, торты, пироги, шоколадные пирожные, печенье, лапша, лапша быстрого приготовления, лепешки, продукты для легкого завтрака, такие как крекеры, крекеры из муки грубого помола, крендели, картофельные чипсы и макаронные изделия.
В другом аспекте, пищевым продуктом по изобретению может быть корм для животных.
В одном из аспектов изобретения, пищевой продукт, предпочтительно, выбран из следующих продуктов, а именно, из яиц, продуктов с добавлением яиц, включая майонез, заправки для салатов, соусы, мороженное, яичный порошок, модифицированный яичный желток и изготавливаемые из них продукты.
В некоторых из упомянутых здесь применений, а в частности, для приготовления пищевых продуктов, таких как хлебобулочные изделия, липолитический фермент по изобретению может быть использован с одним или несколькими стандартными эмульгаторами, включая, например, моноглицериды, сложные эфиры диацетилвинной кислоты и моно- и диглицеридов жирных кислот, стеароиллактилат натрия (SSL) и лецитины.
Липолитический фермент по изобретению является особенно предпочтительным для выпекания хлеба с добавлением жира, и это может быть обусловлено низкой активностью указанного липолитического фермента по отношению к триглицеридам, которая приводит к снижению накопления свободных жирных кислот, а в случае короткоцепочечных триглицеридов, такая активность липолитического фермента позволяет уменьшить уровень постороннего запаха или устранить его.
В контексте настоящего изобретения, используемый термин “добавленный жир” не означает, что липид или жир добавляется в тесто, приготовленное из муки.
Дополнительно или альтернативно, фермент по изобретению может быть использован вместе с одним или несколькими другими подходящими пищевыми ферментами. Таким образом, помимо липолитического фермента по изобретению, в объем настоящего изобретения входит по меньшей мере один дополнительный фермент, который может быть добавлен в хлебобулочное изделие и/или в тесто. Такими дополнительными ферментами являются крахмал-разлагающие ферменты, такие как эндо- или экзоамилазы, пуллуланазы, деветвящие ферменты, гемицеллюлазы, включая ксиланазы, целлюлазы, оксидоредуктазы, например, глюкозооксидаза, пиранозооксидаза, сульфгидрилоксидаза или углевод-оксидаза, а именно, такие ферменты, которые окисляют мальтозу, например, гексозооксидаза (НОХ), липазы, фосфолипазы и гексозооксидазы, протеазы и ацилтрансферазы (такие как ферменты, описанные, например, в WO 04/064987).
Особенно предпочтительно, чтобы при приготовлении пищевых продуктов, липолитический фермент по изобретению использовался в комбинации с альфа-амилазами. В частности, такой амилазой может быть амилаза, не продуцирующая солод, такая как полипептид, обладающий не продуцирующей солод экзоамилазной активностью, а в частности, глюкан-1,4-альфа-мальтотетрагидролазной (ЕС.3.21.60) активностью (описанной в WO 05/003339). Подходящей амилазой, не продуцирующей солод, является коммерчески доступная амилаза PowersoftTM (поставляемая Danisco A/S, Denmark). При этом, могут быть также использованы и солод-образующие амилазы, такие как NovamylTM (Novozymes A/S, Denmark). В одном из вариантов изобретения, сочетание альфа-амилаз и липолитического фермента согласно изобретению может быть использовано для приготовления теста и/или для изготовления хлебобулочных изделий, таких как хлеб, торты, пончики, пирожки или бублики. Сочетание альфа-амилаз и липолитического фермента по изобретению также рассматривается как предпочтительная при ее использовании в методах выпекания лепешек, таких как пшеничные и/или кукурузные лепешки.
В другом предпочтительном варианте, при изготовлении пищевых продуктов, липолитический фермент по изобретению может быть использован в сочетании с ксиланазой. Примерами коммерчески доступных ксиланаз, поставляемых Danisco A/S, являются ксиланазы GRINDAMYLTM и POWERBake 7000. Другие примеры ферментов ксиланаз можно найти в WO 03/020923 и в WO 01/42433.
Липолитический фермент по изобретению, предпочтительно, может быть использован в сочетании с ксиланазой и альфа-амилазой. Обычно, альфа-амилазой может быть альфа-амилаза образующая солод или необразующая солод (такая как GRINDAMYLTM или POWERSoft, поставляемая Danisco A/S) или их сочетание.
Липолитический фермент по изобретению может быть, предпочтительно, также использован в сочетании с окисляющим ферментом, таким как окисляющий мальтозу фермент (МОХ), например, гексозооксидаза (НОХ). Подходящие методы описаны в WO 03/099016. Коммерчески доступные окисляющие мальтозу ферменты GRINDAMYLTM и SUPERBake поставляются фирмой Danisco A/S.
Альфа-амилаза, такая как экзоамилаза, непродуцирующая солод и/или амилаза, образующая солод, и/или окисляющий мальтозу фермент (МОХ), необязательно, в сочетании с ферментом по изобретению могут быть использованы в методах приготовления теста, хлебобулочных изделий, лепешек, тортов, лапши быстрого приготовления/обжаренных продуктов для закусок или молочного продукта, такого как сыр.
Липолитический фермент по изобретению обычно вводят в пищевой продукт или в другую композицию методами, известными специалистам. Такие методы предусматривают добавление липолитического фермента непосредственно в пищевой продукт или композицию, добавление липолитического фермента в комбинации со стабилизатором и/или носителем, и добавление смеси, содержащей липолитический фермент и стабилизатор и/или носитель.
Стабилизаторами, подходящими для использования в настоящем изобретении, являются, но не ограничиваются ими, неорганические соли (такие как NаСl, сульфат аммония), сорбит, эмульгаторы и детергенты (такое как твин 20, твин 80, панодан АВ100 без триглицеридов, сложные эфиры полиглицерина, сорбитанмоноолеат), масло (такое как рапсовое масло, подсолнечное масло и соевое масло), пектин, трегалоза и глицерин.
Носителями, подходящими для использования в настоящем изобретении, являются, но не ограничиваются ими, крахмал, дробленое пшеничное зерно, пшеничная мука, NаСl и цитрат.
Глютеновый индекс может быть измерен на аппарате Glutomatic 2200, поставляемом от Perten Instrumens (Швеция). Глютеновый индекс измеряют следующим образом: сразу после расстойки теста, отвешивают 15 г теста и помещают в аппарат Glutomatic, а затем промывают 500 мл 2% раствором (NаСl) в течение 10 минут. Затем промытое тесто может быть перенесено в центрифугу Gluten Index Centrifuge 2015, после чего две глютеновые фракции взвешивают и вычисляют глютеновый индекс по следующей формуле:
Глютеновый индекс = (масса клейковины, оставшаяся на сите х 100)/общая масса клейковины.
При этом, предпочтительно, чтобы глютеновый индекс теста был выше по меньшей мере на 5% по сравнению с тестом, приготовленным без добавления полипептида, при этом, глютеновый индекс может быть определен на упомянутом выше аппарате Glutomatic 2200.
Другие предпочтительные аспекты приводятся в прилагаемой ниже формуле изобретения и представлены в нижеследующем описании и в примерах.
Преимущества:
Удивительным и неожиданным было обнаружение, что грибковые липолитические ферменты по изобретению имеют гораздо более высокое отношение активности, направленной на гидролиз полярных липидов (фосфолипидов и/или гликолипидов), к триглицерид-гидролизующей активности, чем ранее идентифицированные липолитические ферменты (в частности, Lipopan FTM), выделенные из грибков. Этот факт оказался особенно неожиданным, потому, что до появления настоящего изобретения не было известно ни одного грибкового липолитического фермента дикого типа, который обладал бы подобной активностью. Хотя были проведены исследования вариантов липолитического фермента (то есть, вариантов фермента, подвергнутого неприродному мутагенезу и/или модифицированного некоторыми другими путями), однако, не было высказано каких-либо предположений, что природный фермент дикого типа, полученный из грибков, может обладать такими в высокой степени ценными свойствами.
Было обнаружено, что идентифицированные ферменты обладают превосходными функциональными свойствами при их применении в хлебопекарной промышленности. Преимуществом использования грибкового липолитического фермента по изобретению является значительное улучшение свойств теста и/или хлебобулочных изделий по сравнению с использованием других грибковых липолитических ферментов, в частности, Lipopan FTM.
Преимущество липолитического фермента, который сохраняет активность при низких температурах, то есть низкотемпературного липолитического фермента, заключается в том, что он может быть использован при низких температурах, что позволяет избежать необходимости нагревания субстрата. Это может оказаться особенно предпочтительным при таких применениях, как ферментативная обработка яичного желтка, ферментативное рафинирование пищевых масел и обработка молока или молочных продуктов, например, обработка молока для сыроделия перед приготовлением сыра. Другим преимуществом использования низкотемпературного липолитического фермента является то, что такая ферментативная обработка пищевых продуктов и/или кормов для животных позволяет сохранить значительную активность фермента при низких температурах с пониженным риском микробного, а в частности, бактериального заражения. Кроме того, если стабильность фермента повышается при более низких температурах, то это позволяет вводить эффективные дозы фермента и обеспечивает более длительное действие фермента в промышленном производстве.
Технические эффекты
Для приготовления хлебобулочных изделий, таких как например, хлеб, булочки с паровой обработкой и белый формовой хлеб (США), добавление липолитического фермента по изобретению может приводить к приготовлению изделия, обладающего одним или несколькими свойствами, такими как: увеличенный объем и/или повышенная мягкость хлеба; более длительный срок хранения и/или устойчивость к черствению; улучшенная структура мякиша; пониженная гетерогенность пор; уменьшенный средний размер пор; более высокий глютеновый индекс; улучшенный вкус и/или запах, и более ярко выраженный цвет корочки хлебобулочного изделия.
Фермент по изобретению может преимущественно служить заменой эмульгаторам в пищевых продуктах, таких как тесто и/или хлебобулочные изделия.
Липолитический фермент по изобретению может обладать синергическим действием совместно с эмульгаторами, такими как DATEM, SSL, CSL, моноглицерид, полисорбаты и твин. Так, например, липолитический фермент по изобретению может быть использован в сочетании с одним или несколькими эмульгаторами. Использование липолитического фермента по изобретению преимущественно в сочетании с одним или несколькими эмульгаторами дает возможность снижать общее количество используемого эмульгатора по сравнению с количеством, необходимым для его использования в отсутствии фермента согласно изобретению.
Липолитический фермент по изобретению может также обладать синергическим действием вместе с гидроколлоидами, гуаровой камедью, ксантаном и пектином и с ферментами, окисляющими мальтозу, такими как гексозооксидаза.
Так, например, в случае приготовления пончиков, пирожков, бубликов, закусочных пищевых продуктов и круглых пышек введение липолитического фермента по изобретению может приводить к синергическому эффекту при его использовании в сочетании с одной или несколькими альфа-амилазами, солодобразующей альфа-амилазой и непродуцирующей солод альфа-амилазой.
В случае приготовления тортов, например, бисквитных тортов и кокосовых тортов, введение липолитического фермента по изобретению может приводить к синергическому эффекту при его использовании в комбинации с одним или несколькими гидроколлоидами, такими как гуаровая камедь, и/или с одним или несколькими эмульгаторами, такими как DАТЕМ.
Для приготовления, например, бисквитов, использование липолитического фермента по изобретению приводит к улучшению раскатываемости и обработки теста, в частности, при пониженной температуре (холодная раскатка).
Преимуществом использования липолитического фермента по изобретению, в случае приготовления майонеза и других продуктов с добавлением яиц, является улучшение консистенции, снижение среднего размера частиц и/или снижение среднего распределения частиц, повышение термостабильности, повышение качества изделия при обработке в микроволновой печи и/или повышение стабильности.
Преимуществом такого применения по изобретению в случае приготовления тортов является увеличение мягкости и объема продукта, улучшение его свойств при хранении и увеличению срока его хранения.
Так, например, в случае приготовления лапши или макаронных изделий, например, лапши быстрого приготовления, липолитический фермент по изобретению может придавать продукту одно или несколько свойств, таких как: улучшенный цвет/желтизна, более стабильные цветовые характеристики, менее ярко выраженная белизна, пониженное содержание жира, улучшенная консистенция и отсутствие привкуса (при разжевывании), пониженная смачивающая активность, пониженная ломкость, повышенная твердость макаронных изделий и более длительное сохранение формы во время обработки.
Липолитический фермент по изобретению, предпочтительно, может быть использован для уменьшения содержания жира в лапше или в макаронных изделиях, например, в лапше быстрого приготовления.
В случае приготовления лепешек, например, применение фермента согласно изобретению может приводить к получению продукта с одним или несколькими из нижеследующих свойств, таких как улучшенная раскатываемость лепешек, например, улучшенная пластичность, повышенная устойчивость к черствению, повышенная мягкость и/или отсутствие привкуса.
Улучшенная раскатываемость и/или повышенная пластичность может, преимущественно, приводить к снижению вероятности разлома лепешки при ее раскатывании.
В случае приготовления сыра и/или сырных продуктов, например, применение фермента согласно изобретению может приводить к получению изделия с одним или несколькими из нижеследующих свойств, таких как улучшенный вкус, улучшенная консистенция и/или повышенная стабильность, пониженный эффект вымасливания сыра и/или увеличение выхода сыра.
Используемый здесь термин “эффект вымасливания” обозначает выделение свободного масла при плавлении сыра.
Липолитический фермент по изобретению может быть использован для приготовления сыра с низким содержанием жира. Преимуществом фермента согласно изобретению является то, что он может стабилизировать уровень жира в молоке и/или может улучшать его вкус.
Одним из преимуществ настоящего изобретения является то, что данный фермент функционирует (и, действительно, обладает хорошими функциональными свойствами) при низкой температуре. Такое его свойство дает ряд преимуществ в зависимости от конкретно используемого фермента. Так, например, при производстве сыра, такое действие фермента может приводить к снижению риска микробного заражения и размножения микробов в процессе ферментативной обработки. Это может быть обусловлено тем, что такой сыр может оставаться охлажденным в процессе ферментативной обработки. Так, например, липолитический фермент согласно изобретению может быть особенно подходящим для созревания сыра при низкой температуре, что способствует улучшению его вкуса.
Так, например, в случае приготовления корма для животных, фермент по изобретению может придавать одно или несколько из нижеследующих свойств, таких как: повышенная эффективность утилизации/усваиваемости кормов у животного, увеличение прироста массы животного, улучшение переваривания корма животным, повышение поглощения азота у животного, например, из корма, усиление метаболизма сухих веществ в корме и улучшение вкусовых качеств корма.
При рафинировании пищевого масла, такого как растительное масло, липолитический фермент по изобретению обладает высокой активностью при низкой температуре. Это свойство имеет то преимущество, что оно позволяет избежать необходимости нагревать масло до или во время ферментативной обработки. Важным эффектом такого преимущества является уменьшение количества энергии, необходимое для обработки. Фермент по изобретению может повышать селективность в отношении снижения уровня фосфолипидов по сравнению с триглицеридами. В соответствии с настоящим изобретением фермент, присутствующий в пищевом масле (таком как растительное масло), может обладать более низкой триглицеридам-гидролизующей активностью, чем фосфолипид-гидролизующая активность. Такая активность (по сравнению с активностью стандартных/фосфолипазных ферментов) может приводить к снижению уровня гидролизованных триглицеридов и к снижению потерь выхода масла и/или к снижению накопления свободных жирных кислот в масле (по сравнению со стандартными липолитическими/фосфолипазными ферментами).
Применение
Ферменты по изобретению имеют множество применений.
В частности, грибковые липолитические ферменты по изобретению могут быть использованы для изготовления продуктов питания.
Так, например, грибковые липолитические ферменты по изобретению могут быть, в частности, использованы для обработки яиц или продуктов с добавлением яиц.
Фосфолипазы, а в частности, фосфолипаза А2 (Е.С.3.1.1.4), уже много лет используется для обработки яиц или продуктов с добавлением яиц (см. патент США № 4034124 и, например, публикацию Dutihl & Groger 1981, J. Sci. Food Agric., 32, 451-458). Фосфолипазная активность в процессе обработки яиц или продуктов с добавлением яиц может приводить к накоплению полярного лизолецитина, который может действовать как эмульгатор.
Обработка яиц или продуктов с добавлением яиц грибковым липолитическим ферментом по изобретению может способствовать повышению стабильности и термостабильности продукта при тепловой обработке, такой как пастеризация, и может приводить к значительному загущению. Продуктами с добавлением яиц могут быть, но не ограничиваются ими, торты, майонез, заправки для салатов, соусы, мороженное и тому подобное.
Грибковые липолитические ферменты по изобретению могут быть, в частности, использованы для приготовления хлебобулочных изделий, таких как продукты, приготавливаемые из теста, включая хлеб, торты, изделия из сдобного теста, изделия из слоеного теста, пирожки из жидкого теста, круглые пышки, пончики, бисквиты, крекеры и печенье.
Грибковые липолитические ферменты по изобретению могут быть также использованы в добавках-улучшителях качества хлеба, например, в рецептурах приготовления теста, в добавках для теста, в кондиционерах для теста, в премиксах и аналогичных препаратах, обычно добавляемых в муку и/или в тесто в процессе выпекания хлеба или в других хлебобулочных изделиях для улучшения свойств хлеба или других хлебобулочных изделий.
Таким образом, настоящее изобретение также относится к композициям для улучшения качества хлеба и/или к композициям для улучшения теста, содержащим грибковый липолитический фермент по изобретению, а также к тесту или хлебобулочному изделию, содержащему такую композицию для улучшения качества хлеба и/или качества теста.
Указанная композиция для улучшения качества хлеба и/или композиция для улучшения теста, может, помимо грибкового липолитического фермента по изобретению, содержать и другие вещества, которые обычно используются при выпекании теста и/или хлебобулочных изделий в целях улучшения их свойств.
Указанная композиция для улучшения качества хлеба и/или композиция для улучшения теста, может содержать одну или несколько стандартных хлебопекарных добавок, таких как один или несколько из следующих ингредиентов: сухое молоко, клейковина, эмульгатор, гранулированный жир, окислитель, аминокислота, сахар, соль, мука или крахмал.
Примерами подходящих эмульгаторов являются: моноглицериды, сложные эфиры диацетилвинной кислоты и моно- и диглицеридов жирных кислот, сложные эфиры сахаров, стеароиллактилат натрия (SSL) и лецитины.
Указанная композиция для улучшения качества хлеба и/или композиция для улучшения теста может также содержать и другой фермент, такой как один или несколько других подходящих пищевых ферментов высокой чистоты, включая крахмал-разлагающие ферменты, такие как эндо- или экзоамилазы, пуллуланазы, деветвящие ферменты, гемицеллюлазы, включая ксиланазы, целлюлазы, оксидоредуктазы, например, глюкозооксидазу, пиранозооксидазу, сульфогидрилоксидазу или углевод-оксидазу, такую как фермент, который окисляет мальтозу, например, гексозооксидаза (НОХ); липазы; фосфолипазы и гексозооксидазы; протеазы и ацилтрансферазы (такие как ферменты, описанные, например, в WO 04/064987).
Используемый здесь термин “улучшенные свойства” означает любое свойство, которое может быть улучшено под действием грибковых липолитических ферментов по изобретению. В частности, использование грибкового липолитического фермента по изобретению позволяет получить продукт, обладающий одним или несколькими из следующих свойств, таких как: повышенный объем хлебобулочного изделия; улучшенная структура мякиша хлебобулочного изделия; а также повышенная устойчивость хлебобулочного изделия к черствению; повышенная плотность, повышенная стабильность, пониженная слипаемость и/или повышенная машинабельность теста.
Такие улучшенные свойства оценивают путем сравнения со свойствами теста и/или хлебобулочнго изделия, полученного без добавления липолитического фермента по изобретению.
Используемый здесь термин “хлебобулочное изделие” включает продукт, полученный из теста. Примерами хлебобулочных изделий (пшеничного хлеба, хлеба из теста на опаре или хлеба из пшеничной муки второго сорта), которые могут быть, преимущественно, получены способом по изобретению, являются одно или несколько из нижеследующих изделий, таких как хлеб (включая пшеничный хлеб, пшеничный хлеб из цельного зерна и ржаной хлеб), обычно в виде батонов, хлебцев или тостов, хлеб типа французского багета, лаваш, лепешки, лепешки с начинкой, кексы, блины, бисквиты, хрустящие хлебцы, макаронные изделия (паста), лапша и тому подобное.
Тесто в соответствии с изобретением может быть тестом на закваске или тестом, поставленным на закваску. Тесто может быть поставлено на закваску различными методами, такими как добавление бикарбоната натрия или тому подобное, или добавление подходящей дрожжевой культуры, такой как культура Saccharomyces cerevisiae (пекарские дрожжи).
Настоящее изобретение также относится к применению грибковых липолитических ферментов по изобретению в целях приготовления теста для макаронных изделий, предпочтительно, изделий из муки пшеницы дурум или из муки аналогичного качества.
Грибковые липолитические ферменты по изобретению могут быть использованы для ферментативного рафинирования растительных или пищевых масел. При обработке растительных или пищевых масел указанное пищевое или растительное масло обрабатывают грибковым липолитическим ферментом по изобретению так, чтобы он гидролизовал значительную часть полярных липидов (например, фосфолипида и/или гликолипида). Ацильные группы жирной кислоты, предпочтительно, гидролизуются из полярных липидов. Процесс рафинирования обычно приводит к снижению содержания полярных липидов, а в частности, фосфолипидов в пищевом масле, что обусловлено гидролизом значительной части (то есть, более чем 50%) полярного липида, например, гликолипида и/или фосфолипида. Обычно водную фазу, содержащую гидролизованный полярный липид (например, фосфолипид и/или гликолипид), отделяют от масла. В основном, пищевое или растительное масло изначально может (перед обработкой ферментом согласно изобретению) иметь содержание фосфора 50-250 м.д.
Кроме того, настоящее изобретение относится к применению липолитического фермента по изобретению для обработки сырных продуктов.
Липолитический фермент по изобретению также является особенно подходящим для его применения в изготовлении корма для животных.
Как известно специалистам, используемый здесь термин “рафинирование” означает очистку масла путем превращения фосфатидов (таких как лецитин, фосфолипиды и окклюдированное масло) в гидратируемые фосфатиды. Масло, подвергнутое рафинированию, является более жидким, а поэтому оно лучше поддается обработке, чем нерафинированное масло.
Нижеследующая таблица приводится лишь как общее руководство и общее представление об уровнях доз для липолитического фермента по изобретению, которые могут оказаться необходимыми для различных применений. В этой таблице также указаны оптимальные дозы липолитического фермента по изобретению при его использовании, например, в комбинации с эмульгатором. Разумеется, что для каждого конкретного применения, средний специалист в данной области может легко определить оптимальную дозу фермента, температуру и время реакции путем рутинного экспериментирования.
Выделение
В одном из аспектов изобретения, предпочтительная последовательность присутствует в выделенной форме. Термин “выделенный” означает, что данная последовательность не содержит по крайней мере значительного количества, по крайней мере одного из других компонентов, с которыми данная последовательность обычно ассоциируется и обнаруживается в природе.
Очистка
В одном из аспектов изобретения данная последовательность, предпочтительно, присутствует в очищенной форме. Термин “очищенная” означает, что данная последовательность присутствует в относительно чистой форме, например, имеет по меньшей мере примерно 90%-ную чистоту, по меньшей мере примерно 95%-ную чистоту или по меньшей мере примерно 98%-ную чистоту.
Нуклеотидная последовательность
В объем настоящего изобретения входят нуклеотидные последовательности, кодирующие ферменты, обладающие определенными здесь специфическими свойствами.
Используемый здесь термин “нуклеотидная последовательность” обозначает олигонуклеотидную последовательность или полинуклеотидную последовательность и их варианты, гомологи, фрагменты и производные (например, их части). Нуклеотидная последовательность может быть геномной, синтетической или рекомбинантной последовательностью, которая может быть двухцепочечной или одноцепочечной, независимо от того, присутствует ли она в смысловой или антисмысловой ориентации.
Термин “нуклеотидная последовательность” по изобретению включает геномную ДНК, кДНК, синтетическую ДНК и РНК. Этот термин, предпочтительно, означает ДНК, и, более предпочтительно, кДНК-последовательность, кодирующую фермент по настоящему изобретению.
В предпочтительном варианте изобретения нуклеотидная последовательность, если она относится к настоящему изобретению или входит в объем per se настоящего изобретения, не включает нативную нуклеотидную последовательность по изобретению, если она присутствует в своем природном окружении и если она присоединена к связанным с ней природным последовательностям, которые также присутствуют в своем природном окружении. Для удобства, авторы настоящего изобретения называют этот предпочтительный вариант “неприродной нуклеотидной последовательностью”. В соответствии с этим, термин “природная нуклеотидная последовательность” означает полноразмерную нуклеотидную последовательность, которая присутствует в своем природном окружении и которая функционально присоединена к полноразмерному промотору, с которым она связана в природе и который также присутствует в своем природном окружении. Однако аминокислотная последовательность, входящая в объем настоящего изобретения, может быть выделена и/или очищена после экспрессии нуклеотидной последовательности в организме, в котором она обнаруживается в природе. Однако, предпочтительно, чтобы аминокислотная последовательность, входящая в объем настоящего изобретения, экспрессировалась нуклеотидной последовательностью в своем природном организме, но чтобы, при этом, указанная нуклеотидная последовательность не находилась под контролем промотора, с которым она обычно связана в данном организме.
Получение нуклеотидной последовательности
Обычно нуклеотидную последовательность, входящую в объем настоящего изобретения, получают методами рекомбинантных ДНК (то есть, с использованием рекомбинантных ДНК). Однако в альтернативном варианте изобретения нуклеотидная последовательность может быть синтезирована, полностью или частично, химическими методами, хорошо известными специалистам (см. Caruthers M.H. et al., (1980) Nuc. Acids. Res. Symp. Ser. 215-23 и Horn T. et al. (1980) Nuc. Acids. Res. Symp. Ser. 225-232).
Нуклеотидная последовательность, кодирующая фермент, который обладает определенными здесь специфическими свойствами, может быть идентифицирована и/или выделена, и/или очищена из любой клетки или организма, продуцирующих указанный фермент. Для идентификации и/или выделения, и/или очистки нуклеотидных последовательностей существуют различные методы, хорошо известные специалистам. Так, например, после идентификации и/или выделения, и/или очистки подходящей последовательности можно получить дополнительное количество этой последовательности методами ПЦР-амплификации.
В другом примере, может быть сконструирована библиотека геномных ДНК и/или кДНК, используя хромосомную ДНК или информационную РНК, происходящей от организма, продуцирующего указанный фермент. Если известна аминокислотная последовательность данного фермента или часть аминокислотной последовательности такого фермента, то могут быть синтезированы меченные олигонуклеотидные зонды, которые могут быть использованы для идентификации фермент-кодирующих клонов, выделенных из геномной библиотеки, полученной из указанного организма. Альтернативно, для идентификации фермент-кодирующих клонов может быть использован меченный олигонуклеотидный зонд, содержащий последовательности, гомологичные последовательностям другого гена известного фермента. В последнем случае проводят гибридизацию и промывку в условиях низкой жесткости.
Альтернативно, фермент-кодирующие клоны могут быть идентифицированы путем встраивания фрагментов геномной ДНК в вектор экспрессии, такой как плазмида, трансформации негативной по ферменту бактерии полученной библиотекой геномных ДНК, и последующего посева трансформированных бактерий на агаровые планшеты, содержащие субстрат для фермента (например, мальтозу для фермента, продуцирующего глюкозидазу (мальтазу)), для экспрессии идентифицируемого фермента в клонах.
В еще одном из альтернативных вариантов изобретения, нуклеотидная последовательность, кодирующая указанный фермент, может быть получена стандартными методами синтеза, например, фосфорамидитным методом, описанным Beucage S.L. et al. (1981) Tetrahedron Letters 22, p. 1859-1869, или методом, описанным Matthes et al. (1984) EMBO J. 3, p.801-805. В указанном фосфороамидитном методе, олигонуклеотиды синтезируют, например, на автоматическом ДНК-синтезаторе, а затем очищают, отжигают, лигируют и клонируют в соответствующие векторы.
Нуклеотидная последовательность может представлять собой смесь геномной последовательности и синтетической последовательности, смесь синтетической последовательности и кДНК-последовательности, или смесь геномной последовательности и кДНК-последовательности, полученной путем лигирования фрагментов синтетической последовательности, геномной последовательности или кДНК-последовательности (если это необходимо) в соответствии со стандартными методами. Каждый лигированный фрагмент соответствует различным частям полноразмерной нуклеотидной последовательности. ДНК-последовательность может быть также получена с помощью полимеразной цепной реакции (ПЦР) с использованием специфических праймеров, например, как описано в патенте США №4683202 или у Saiki R.K. et al. (Science (1988) 239, p.487-491).
Из-за вырожденности генетического кода могут быть легко получены нуклеотидные последовательности, в которых может быть модифицирована встречаемость триплетных кодонов, кодирующих некоторые или все аминокислоты в данной природной нуклеотидной последовательности, что позволяет продуцировать нуклеотидную последовательность, с низкой гомологией с исходной нуклеотидной последовательностью, но кодирующую такую же аминокислотную последовательность или ее вариант, как и исходная нуклеотидная последовательность. Так, например, для большинства аминокислот, вырожденность генетического кода проявляется в третьем положении триплетного кодона (положении неоднозначного соответствия)(более подробно см. Stryer, Lubert, Biochemistry, Third Edition, Freeman Press, ISBN 0-7167-1920-7), и поэтому нуклеотидная последовательность, в которой все триплетные кодоны имели “неоднозначное соответствие” в третьем положении, должна быть примерно на 66% идентична исходной нуклеотидной последовательности. Однако модифицированная нуклеотидная последовательность должна кодировать такую же первичную аминокислотную последовательность или ее вариант, как и аминокислотная последовательность, кодируемая исходной нуклеотидной последовательностью.
Поэтому, настоящее изобретение также относится к любой нуклеотидной последовательности, которая имеет альтернативную встречаемость триплетных кодонов, и по меньшей мере для одного кодирующего аминокислоту триплетного кодона, но которая кодирует такую же полипептидную последовательность или ее вариант, как и полипептидная последовательность, кодируемая исходной нуклеотидной последовательностью.
Кроме того, в каждом конкретном организме обычно наблюдается присущая ему предпочтительность в использовании кодонов, кодирующих данную аминокислоту. Таблицы предпочтительности кодонов широко известны и могут служить ориентиром для получения оптимизированных по кодонам генов. Такие методы оптимизации кодонов обычно применяются для оптимизации экспрессии трансгенов в гетерологичном хозяине.
Аминокислотные последовательности
В объем настоящего изобретения также входят аминокислотные последовательности ферментов, обладающих определенными здесь специфическими свойствами.
Используемый здесь термин “аминокислотная последовательность” является синонимом термину “полипептид” и/или термину “белок”. В некоторых случаях термин “аминокислотная последовательность” является синонимом термину “пептид”. В некоторых случаях термин “аминокислотная последовательность” является синонимом термину “фермент”.
Аминокислотная последовательность может быть получена/выделена из подходящего источника, либо она может быть синтезирована или получена методами рекомбинантных ДНК.
Фермент, входящий в объем настоящего изобретения, может быть использован в комбинации с другими ферментами. Так, например, настоящее изобретение также относится к комбинации ферментов, где указанная комбинация содержит фермент согласно изобретению и другой фермент, которым может быть другой фермент по изобретению.
Предпочтительно, аминокислотной последовательностью, если она относится к настоящему изобретению и входит в объем per se настоящего изобретения, является неприродный фермент. В этой связи, термин “природный фермент” означает полноразмерный фермент, который присутствует в своем природном окружении и экспрессируется его природной нуклеотидной последовательностью.
Идентичность/гомология
Настоящее изобретение также относится к использованию гомологов любой аминокислотной последовательности фермента или любой нуклеотидной последовательности, кодирующей такой фермент.
Используемый здесь термин “гомолог” означает молекулу, имеющую определенную степень гомологии с аминокислотными последовательностями и с нуклеотидными последовательностями. Используемый здесь термин “гомология” может быть эквивалентным термину “идентичность”. Эти используемые здесь термины являются взаимозаменяемыми.
В контексте настоящего изобретения подразумевается, что гомологичная аминокислотная последовательность включает аминокислотную последовательность, которая может быть по меньшей мере на 92%, а предпочтительно, по меньшей мере на 95, 96, 97, 98 или 99% идентична исходной последовательности. В общих чертах, указанные гомологи содержат такие же активные центры, например, как и рассматриваемая аминокислотная последовательность. Хотя термин “гомология” последовательностей может также рассматриваться как “сходство” последовательностей (то есть, аминокислотные остатки этих последовательностей имеют аналогичные химические свойства/функции), однако, в контексте настоящего изобретения предпочтительно, чтобы под термином “гомология” последовательностей подразумевалась идентичность последовательностей.
Предпочтительно, гомологичная аминокислотная последовательность по изобретению представляет собой последовательность, которая имеет по меньшей мере 90%-ную идентичность, а более предпочтительно по меньшей мере 95%-, 96%-, 97%-, 98%- или 99%-ную идентичность в области, содержащей по меньшей мере 30, а более предпочтительно, 40 смежных аминокислот.
В контексте настоящего изобретения подразумевается, что гомологичная нуклеотидная последовательность включает нуклеотидную последовательность, которая может быть по меньшей мере на 92%, а предпочтительно по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична нуклеотидной последовательности, кодирующей фермент по изобретению (рассматриваемой последовательности). В общих чертах, указанные гомологи содержат последовательности, которые кодируют такие же активные центры и тому подобное, как и рассматриваемая последовательность. Хотя термин “гомология” последовательностей может также рассматриваться как “сходство” последовательностей (то есть, аминокислотные остатки этих последовательностей имеют аналогичные химические свойства/функции), однако, в контексте настоящего изобретения предпочтительно, чтобы термином “гомология” последовательностей подразумевалась идентичность аминокислотных последовательностей.
Предпочтительно, гомологичная нуклеотидная последовательность по изобретению представляет собой последовательность, которая имеет по меньшей мере 90%-ную идентичность, и, более предпочтительно, по меньшей мере 95%-, 96%-, 97%-, 98%- или 99%-ную идентичность в области, содержащей по меньшей мере 30, предпочтительно 40, и, более предпочтительно, 60 смежных нуклеотидов.
Для аминокислотных последовательностей и нуклеотидных последовательностей оценка гомологии может быть проведена визуально, или чаще всего, с помощью легко доступных компьютерных программ для сравнения последовательностей. Такие коммерчески доступные компьютерные программы позволяют вычислять процент гомологии между двумя или более последовательностями.
Процент гомологии может быть вычислен по всей длине последовательностей, то есть одну последовательность сравнивают с другой последовательностью путем выравнивания, и каждую аминокислоту в одной последовательности непосредственно сопоставляют с соответствующей аминокислотой в другой последовательности, по одному остатку за одно сравнение. Такое сравнение называют “выравнивание без пробелов”. Обычно такое выравнивание без пробелов осуществляют только при относительно небольшом числе остатков.
Хотя этот метод является очень простым и удобным, однако, он не дает удовлетворительных результатов при рассмотрении, например, двух более или менее идентичных последовательностей, в которых одна инсерция или делеция будет приводить к смещению аминокислотных остатков и нарушению их соответствия при выравнивании, а поэтому проведение такого глобального выравнивания, возможно, будет приводить к значительному снижению процента гомологии. Следовательно, большинство методов сравнения разработаны так, чтобы можно было проводить оптимальное выравнивание с учетов возможных инсерций и делеций без излишнего наложения “штрафов” при общей оценке гомологии. Это может быть достигнуто путем максимизации локальной гомологии, то есть путем введения “пробелов” при выравнивании последовательностей.
Однако в этих более сложных методах, каждому пробелу, который вводят при выравнивании, назначаются “штрафы за пробелы”, такие, чтобы для того же самого числа идентичных аминокислот, выравнивание последовательностей, по возможности, с небольшим числом пробелов, отражало наибольшее сходство между двумя сравниваемыми последовательностями, и чтобы, при этом, достигалась более высокая оценка, чем оценка, получаемая при использовании множества пробелов. Обычно используют такой параметр, как “цена аффинных пробелов”, который назначает относительно высокую “цену” за присутствие пробела и более низкий штраф за каждый последующий остаток в данном пробеле. В этом случае, чаще всего используется система оценки добавляемых пробелов. Очевидно, что высокие штрафы за пробелы позволяют оптимизировать выравнивание с меньшим числом пробелов. Большинство программ по выравниванию позволяют модифицировать штрафы за пробелы. Однако при применении такой компьютерной программы для сравнения последовательностей, предпочтительно, использовать параметры по умолчанию. Так, например, в пакете программ GCG Wisconsin Bestfit, такие параметры по умолчанию, как штраф за пробел в аминокислотных последовательностях составляет -12, а штраф за каждое удлинение составляет -4.
Поэтому, для вычисления максимального процента гомологии, прежде всего, требуется оптимальное выравнивание с учетом штрафов за пробел. Подходящей компьютерной программой для проведения такого выравнивания является пакет программ GCG Wisconsin Bestfit (Devereux et al. 1984 Nuc. Acids. Research 12, стp.387). Примерами другой программы, которая может быть использована для сравнения последовательностей, являются, но не ограничиваются ими, программа BLAST (см., Ausubel et al., 1999, Short Protocols in Molecular Biology, 4th Ed. - Chapter 18), FASTA (Altschul et al., 1990 J. Mol. Biol. 403-410) и пакет программ GENEWORKS для сравнения последовательностей. Программы BLAST и FASTA позволяют осуществлять поиск в автономном и в интерактивном режиме (см. Ausubel et al., 1999, Short Protocols in Molecular Biology, pages 7-58 - 7-60).
Однако для некоторых целей предпочтительно использовать программу GCG Bestfit. Для сравнения последовательностей белков и нуклеотидных последовательностей существует новая программа, называемая BLAST 2 Sequences (см. FEMS Microbiol. Lett. 1999 174(2):247-50; FEMS Microbiol Lett 1999 177(1):187-8 и tatiana@ncbi.nlm.nih.gov).
Хотя конечный процент гомологии может быть определен как идентичность, однако, сам процесс выравнивания обычно не основан на сравнении пары по типу “все или ничего”. Вместо этого, для оценки сходства обычно используется оценочная матрица, которая присваивает каждому попарному сравнению оценку исходя из химического сходства или эволюционной отдаленности. Примером такой обычно используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для пакета программ BLAST. В программах GCG Wisconsin обычно используются либо имеющиеся параметры по умолчанию, либо таблица специальных символов, если она имеется (более подробно смотри руководство для пользователя). В некоторых случаях, предпочтительно, использовать параметры по умолчанию, имеющиеся в пакете программ GСG, или, если используется другая программа, матрицу по умолчанию, такую как BLOSUM62.
Альтернативно, процент гомологии может быть вычислен с использованием множества параметров выравнивания в DNASISТМ (Hitachi Software), исходя из алгоритма, аналогичного алгоритму CLUSTAL (Higgins D.G. & Sharp P.M. (1988), Gene 73(1), 237-244).
После осуществления оптимального выравнивания с помощью компьютерной программы можно вычислить процент гомологии, а предпочтительно, процент идентичности последовательностей. Компьютерная программа обычно позволяет проводить такое вычисление в процессе сравнения последовательностей и дает численный результат.
Последовательности могут также иметь делеции, инсерции или замены аминокислотных остатков, которые продуцируют молчащие мутации и приводят к получению функционально эквивалентного вещества. Исходя из сходства свойств аминокислот (таких как полярность, заряд, растворимость, гидрофобность, гидрофильность и/или амфипатическая природа остатков) могут быть специально сделаны аминокислотные замены, а поэтому желательно, чтобы все аминокислоты были классифицированы по свойствам их функциональных групп. Аминокислоты могут быть подразделены на группы только по свойствам их боковых цепей. Однако, более предпочтительно, чтобы учитывались также данные об их мутациях. Классифицированные таким образом группы аминокислот будут, очевидно, консервативными по своим структурным признакам. Эти группы могут быть представлены в форме диаграммы Венна (Livingstone C.D. & Barton G.J. (1993) “Protein sequence alignments: a strategy for the hierarchical analysis of residue conversation” Comput. Appl. Biosci. 9:745-756)(Taylor W.R. (1986) “The classification of amino acid conservation” J. Theor. Biol. 119; 205-218. Консервативные замены могут быть сделаны, например, исходя из данных нижеследующей таблицы, в которой описаны обычно используемые группы аминокислот в соответствии с диаграммой Венна.
Настоящее изобретение также охватывает гомологичные замены (используемые здесь термины “замещения” и “замены” являются взаимозаменяемыми и означают замену присутствующего аминокислотного остатка альтернативным остатком), которые могут быть осуществлены путем замены одного остатка на подобный остаток, например, основного остатка на основный остаток, кислотного остатка на кислотный остаток, полярного остатка на полярный остаток и тому подобное. Может быть также сделана и не-гомологичная замена, то есть остаток одного класса может быть заменен остатком другого класса, или альтернативно он может быть заменен неприродными аминокислотами, такими как орнитин (далее называемый Z), диаминомасляная кислота-орнитин (далее называемый В), норлейцин-орнитин (далее называемый О), пиридилаланин, тиенилаланин, нафтилаланин и фенилглицин.
Могут быть также сделаны замены неприродных аминокислот.
Модифицированные аминокислотные последовательности, помимо аминокислотных спейсерных групп, таких как глициновый или β-аланиновый остатки, могут также включать подходящие спейсерные группы, которые могут быть введены между двумя аминокислотными остатками последовательности, включая алкильные группы, такие как метильная, этильная или пропильная группы. Другая форма модификации предусматривает присутствие одного или нескольких аминокислотных остатков в пептоидной форме, хорошо известной специалистам в данной области. Во избежание неясностей, следует указать, что используемый здесь термин “пептоидная форма” означает модифицированные аминокислотные остатки, где группа-заместитель α-углерода присутствует у атома азота данного остатка, а не у атома α-углерода. Способы получения пептидов в пептоидной форме известны специалистам и описаны, например, в работах Simon R.J. et al., PNAS (1992), 89(20), 9367-9371 и Horwell D.C. Trends Biotechnol. (1995), 13(4), 132-134.
Нуклеотидные последовательности, используемые в настоящем изобретении, могут включать в себя синтетические или модифицированные нуклеотиды. Специалистам известны различные типы модификаций олигонуклеотидов. Такие модифицированные последовательности имеют метилфосфонатный и фосфортиоатный остовы и/или добавленные акридиновые или полилизиновые цепи у 3'- и/или 5'-концов молекулы. При этом, следует отметить, что для осуществления настоящего изобретения, описанные здесь нуклеотидные последовательности могут быть модифицированы любым подходящим методом, известны специалистам. Такие модификации могут быть осуществлены для усиления in vivo активности или для увеличения времени жизни нуклеотидных последовательностей согласно изобретению.
Настоящее изобретение также охватывает применение нуклеотидных последовательностей, комплементарных представленным здесь последовательностям, или любого их производного, фрагмента или его производного. Если данная последовательность комплементарна фрагменту используемой здесь последовательности, то эта последовательность может быть использована в качестве зонда для идентификации аналогичных кодирующих последовательностей в других организмах и тому подобное.
Полинуклеотиды, которые не обладают 100%-ной гомологией с последовательностями по изобретению, но входят в объем настоящего изобретения, могут быть получены различными способами.
Другие варианты описанных здесь последовательностей могут быть получены, например, путем зондирования ДНК-библиотек, полученных от ряда индивидуумов, например, от индивидуумов, принадлежащих к различным популяциям. Кроме того, могут быть получены и другие гомологи, и такие гомологи и их фрагменты будут, по существу, способны селективно гибридизоваться с последовательностями, представленными в прилагаемом здесь списке последовательностей. Такие последовательности могут быть получены путем зондирования библиотек кДНК или геномных ДНК, полученных от других видов, зондами, содержащими всю последовательность или любую часть последовательности в прилагаемом списке последовательностей, в условиях средней или высокой жесткости. Аналогичные подходы могут быть применены для получения видовых гомологов и аллельных вариантов полипептидных или нуклеотидных последовательностей согласно изобретению.
Варианты и гомологи штаммов/видов могут быть также получены посредством ПЦР, в которой могут быть использованы вырожденные праймеры, сконструированные для последовательностей-мишеней, присутствующих в вариантах и гомологах, кодирующих консервативные аминокислотные последовательности, имеющиеся в последовательностях согласно изобретению. Консервативные последовательности могут быть предсказаны, например, путем выравнивания аминокислотных последовательностей нескольких вариантов/гомологов. Выравнивание таких последовательностей может быть осуществлено с помощью компьютерной программы, известной специалистам. Так, например, для этой цели широко используется программа GCG Wisconsin PileUp.
Праймеры, применяемые для “вырожденной” ПЦР, могут содержать одно или несколько положений вырожденности и могут быть использованы в условиях более низкой жесткости, чем условия, применяемые для клонирования последовательностей, осуществляемого с помощью одноцепочечных праймеров для известных последовательностей.
Альтернативно, такие полинуклеотиды могут быть получены с помощью сайт-направленного мутагенеза охарактеризованных последовательностей. Это может быть осуществлено, например, в том случае, когда в последовательность необходимо ввести молчащие замены кодонов в целях оптимизации предпочтительности кодонов для конкретной клетки-хозяина, в которой полинуклеотидные последовательности были экспрессированы. Другие замены в последовательностях могут оказаться желательными для введения сайтов распознавания для рестриктирующих ферментов либо для изменения свойства или функции полипептидов, кодируемых полинуклеотидами.
Полинуклеотиды (нуклеотидные последовательности) по изобретению могут быть использованы для продуцирования праймера, например, ПЦР-праймера; праймера для альтернативной реакции амплификации; зонда, например, зонда, меченного детектируемой меткой, в соответствии со стандартными способами, в которых используются радиоактивные или не радиоактивные метки; либо эти полинуклеотиды могут быть клонированы в векторы. Такие праймеры, зонды и другие фрагменты имеют длину, по меньшей мере, 15, предпочтительно, по меньшей мере, 20, например, по меньшей мере, 25, 30 или 40 нуклеотидов, и также подпадают под определение используемого здесь термина “полинуклеотиды согласно изобретению”.
Полинуклеотиды, такие как ДНК-полинуклеотиды и зонды по изобретению, могут быть получены рекомбинантным, синтетическим или любым другим способом, известным специалистам. Они могут быть также клонированы стандартными методами.
В общих чертах, праймеры могут быть получены методом синтеза, предусматривающим постадийный синтез нужной последовательности нуклеиновой кислоты путем последовательного присоединения по одному нуклеотиду. Методы осуществления такого синтеза с использованием автоматизированной технологии легко доступны специалистам в данной области.
Более длинные полинуклеотиды обычно получают методами рекомбинантных ДНК, например, методами клонирования посредством ПЦР (полимеразной цепной реакции). Могут быть сконструированы праймеры, содержащие сайты распознавания для рестрикционных ферментов, подходящие для клонирования амплифицированной ДНК в соответствующий клонирующий вектор.
Биологическая активность
Модифицированные последовательности, предпочтительно, обладают по меньшей мере такой же биологической активностью, как и представленные здесь последовательности.
Используемый здесь термин “биологически активный” относится к последовательностям, имеющим структурную функцию, и/или регуляторную функцию, и/или биохимическую функцию, аналогичную (но необязательно в той же степени) соответствующим структурным функциям природной последовательности.
Гибридизация
Настоящее изобретение также охватывает последовательности, которые являются комплементарными последовательностям нуклеиновой кислоты по изобретению или последовательностям, способным гибридизоваться либо с последовательностями согласно изобретению, либо с комплементарными им последовательностями.
Используемый здесь термин “гибридизация” включает понятие “процесс, посредством которого цепь нуклеиновой кислоты присоединяется к комплементарной цепи посредством спаривания оснований”, а также процесс амплификации, осуществляемый при полимеразной цепной реакции (ПЦР).
Настоящее изобретение также относится к применению нуклеотидных последовательностей, способных гибридизоваться с последовательностями, комплементарными представленным здесь последовательностям, либо с любым их производным, фрагментом или его производным.
Термин “вариант” также включает последовательности, комплементарные последовательностям, способным гибридизоваться с представленными здесь нуклеотидными последовательностями.
Предпочтительно, термин “вариант” включает последовательности, комплементарные последовательностям, способным гибридизоваться с представленными здесь нуклеотидными последовательностями в жестких условиях (например, при 50°С и 0,2 × SSС {1 × SSС = 0,15 М NаСl, 0,015 М Nа3-цитрат, рН 7,0}).
Более предпочтительно, термин “вариант” включает последовательности, комплементарные последовательностям, способным гибридизоваться с представленными здесь нуклеотидными последовательностями в условиях высокой жесткости (например, при 65°С и 0,1 × SSС {1 × SSС = 0,15 М NаСl, 0,015 М Nа3-цитрат, рН 7,0}).
Настоящее изобретение также относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями по изобретению (включая представленные здесь комплементарные последовательности).
Настоящее изобретение также относится к нуклеотидным последовательностям, комплементарным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями согласно изобретению (включая представленные здесь комплементарные последовательности).
В объем настоящего изобретения также входят полинуклеотидные последовательности, которые способны гибридизоваться с представленными здесь нуклеотидными последовательностями в условиях промежуточной или максимальной жесткости.
В предпочтительном аспекте, настоящее изобретение относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями согласно изобретению или с комплементарными им последовательностями в жестких условиях (например, при 50°С и 0,2 × SSС).
В более предпочтительном аспекте, настоящее изобретение относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями согласно изобретению или с комплементарными им последовательностями в условиях высокой жесткости (например, при 65°С и 0,1 × SSС).
Рекомбинантные последовательности
В одном из аспектов, последовательность, используемая в настоящем изобретении, является рекомбинантной последовательностью, то есть последовательностью, которая была получена методами рекомбинантных ДНК.
Эти методы рекомбинантных ДНК могут быть осуществлены специалистом в данной области. Такие методы описаны в литературе, см., например, Sambrook, J., Fritsh, E.F. & Maniatis T., 1989, Molecular Cloning: A Laboratory manual, Second Edition, Вooks 1-3, Cold Spring Harbor Laboratory Press.
Синтетические последовательности
В одном из аспектов последовательность, используемая в настоящем изобретении, является синтетической последовательностью, то есть последовательностью, которая была получена in vitro методами химического или ферментативного синтеза. Такими последовательностями являются, но не ограничиваются ими, последовательности, синтезированные с использованием оптимальной встречаемости кодонов для организмов-хозяинов, таких как метилотропные дрожжи Pichia и Hansenula.
Экспрессия ферментов
Нуклеотидная последовательность, используемая в настоящем изобретении, может быть введена в рекомбинантный реплицируемый вектор. Этот вектор может быть использован для репликации и экспрессии нуклеотидной последовательности с образованием фермента, продуцируемого в совместимой клетке-хозяине и/или секретируемого из этой клетки.
Такая экспрессия может регулироваться под контролем соответствующих последовательностей, например, регуляторных последовательностей.
Фермент, продуцируемый рекомбинантной клеткой-хозяином путем экспрессии нуклеотидной последовательности, может быть секретирован, либо он может содержаться внутри клетки, в зависимости от используемой последовательности или вектора. Кодирующие последовательности могут быть сконструированы так, чтобы они содержали сигнальную последовательность, которая направляет секрецию последовательности, кодирующей данное вещество, через мембрану соответствующей прокариотической или эукариотической клетки.
Вектор экспрессии
Термин “вектор экспрессии” означает конструкцию, способную к in vivo или in vitro экспрессии.
Такой вектор экспрессии, предпочтительно, встраивают к геном подходящего организма-хозяина. Термин “встроенный” относится к последовательности, стабильно встроенной в данный геном.
Нуклеотидная последовательность по изобретению может присутствовать в векторе, в котором эта нуклеотидная последовательность функционально присоединена к регуляторным последовательностям, способным обеспечивать экспрессию нуклеотидной последовательности в подходящем организме-хозяине.
Векторы, используемые в настоящем изобретении, могут быть трансформированы в подходящую клетку-хозяина, описанную ниже, для осуществления экспрессии полипептида по изобретению.
Выбор вектора, например, плазмиды, космиды или фага часто зависит от клетки-хозяина, в которую он должен быть встроен.
Векторы, используемые в настоящем изобретении, могут содержать один или несколько селектируемых маркерных генов, таких как ген, придающий резистентность к антибиотикам, например, ампициллину, канамицину, хлорамфениколу или тетрациклину. Альтернативно, такой отбор может быть осуществлен путем ко-трансформации (как описано в WO 91/17243).
Векторы могут быть использованы in vitro, например, для получения РНК или для трансфекции, трансформации, трансдукции или инфицирования клетки-хозяина.
Таким образом, в другом своем варианте, настоящее изобретение относится к способу получения нуклеотидных последовательностей по изобретению, предусматривающему встраивание нуклеотидной последовательности по изобретению в реплицируемый вектор; введение этого вектора в совместимую клетку-хозяина и культивирование указанной клетки-хозяина в условиях, способствующих репликации этого вектора.
Указанный вектор может также содержать нуклеотидную последовательность, обеспечивающую репликацию этого вектора в рассматриваемой клетке-хозяине. Примерами таких последовательностей является точка начала репликации плазмид pUC19, рАСYC177, pUВ110, рЕ194, рАМВ1 и pIJ702.
Регуляторные последовательности
В некоторых случаях, нуклеотидная последовательность, используемая в настоящем изобретении, функционально связана с регуляторной последовательностью, которая способна обеспечивать экспрессию нуклеотидной последовательности, например, в конкретно выбранной клетке-хозяине. Так, например, настоящее изобретение включает вектор, содержащий нуклеотидную последовательность по изобретению, функционально присоединенную к такой регуляторной последовательности, то есть этот вектор является экспрессионным вектором.
Термин “функционально связанный” относится к юкстаположению, где описанные компоненты связаны друг с другом так, чтобы это способствовало осуществлению их желательных функций. Регуляторная последовательность, “функционально связанная” с кодирующей последовательностью, должна быть связана так, чтобы экспрессия кодирующей последовательности осуществлялась в условиях, совместимых с условиями действия регуляторных последовательностей.
Термин “регуляторные последовательности” включает промоторы и энхансеры и другие сигналы регуляции экспрессии.
Термин “промотор” используется здесь в своем обычном смысле и означает, например, сайт связывания с РНК-полимеразой.
Повышенный уровень экспрессии нуклеотидной последовательности, кодирующей фермент по изобретению, может быть также достигнут путем выбора гетерологичных регуляторных областей, например, промотора, секретирующей лидерной последовательности и терминатора.
Предпочтительно, нуклеотидная последовательность по изобретению функционально связана по меньшей мере с одним промотором.
Примеры промоторов, подходящих для регуляции транскрипции нуклеотидной последовательности в бактериальном, грибковом или дрожжевом хозяине, хорошо известны специалистам.
Конструкции
Термин “конструкция”, который является синонимом таких терминов, как “конъюгат”, “кластер” и “гибрид”, включает нуклеотидную последовательность, используемую в соответствии с настоящим изобретением, которая непосредственно или опосредованно связана с промотором.
Примером опосредованного соединения является введение подходящей спейсерной группы, такой как интронная последовательность, например, Sh1-интрон или ADH-интрон, между промотором и нуклеотидной последовательности по изобретению. Это относится также и к термину “лигированный”, используемому в настоящем изобретении и включающему в себя понятие прямое или опосредуемое присоединение. В некоторых случаях, эти термины не относятся к природной комбинации белок-кодирующей нуклеотидной последовательности, обычно ассоциированной с промотором гена дикого типа, и к обеим этим последовательностям в их природном окружением.
Данная конструкция может даже содержать или экспрессировать маркер, который позволяет осуществлять отбор генетической конструкции.
В некоторых случаях, предпочтительно, чтобы конструкция по изобретению содержала по меньшей мере одну нуклеотидную последовательность по изобретению, функционально связанную с промотором.
Клетки-хозяева
Термин “клетка-хозяин”, используемый в настоящем изобретении, включает любую клетку, которая содержит либо нуклеотидную последовательность, либо вектор экспрессии, описанный выше, и которая используется в способе рекомбинантного получения фермента, обладающего специфическими свойствами, определенными в настоящей заявке.
Таким образом, в другом своем варианте, настоящее изобретение относится к клеткам-хозяевам, трансформированным или трансфецированным нуклеотидной последовательностью, которая экспрессирует фермент по изобретению. Эти клетки выбирают так, чтобы они были совместимыми с указанным вектором, и такими клетками могут быть прокариотические клетки (например, бактериальные), клетки грибов, дрожжей или растений. Предпочтительными клетками-хозяевами являются клетки, которые не являются клетками человека.
Подходящими примерами бактериальных организмов-хозяев являются грамположительные или грамотрицательные бактериальные клетки.
В зависимости от природы нуклеотидной последовательности, кодирующей фермент по изобретению, и/или от цели проведения последующего процессинга экспрессируемого белка, предпочтительными могут быть эукариотические хозяева, такие как дрожжи или другие грибки. В общих чертах, в отличие от грибковых клеток, дрожжевые клетки являются предпочтительными, поскольку они более легко поддаются модификации. Однако некоторые белки либо плохо секретируются из дрожжевых клеток, либо, в некоторых случаях, они не процессируются соответствующим образом (например, гипергликозилируются в дрожжах). В этих случаях, должен быть выбран другой грибковый организм-хозяин.
Использование подходящих клеток-хозяев, таких как дрожжевые, грибковые и растительные клетки-хозяева, может обеспечивать посттрансляционную модификацию (например, миристоилирование, гликозилирование, усечение, лапидирование и фосфорилирование по тирозину, серину или треонину), которая может оказаться необходимой для сообщения оптимальной биологической активности по отношению к рекомбинантно экспрессированным продуктам согласно изобретению.
Такой клеткой-хозяином может быть штамм, дефицитный по протеазе или не содержащий протеазы.
Для усиления экспрессии генотип клетки-хозяина может быть модифицирован.
Примерами модификаций клетки-хозяина являются дефицит по протеазе, добавление редких тРНК и модификация восстановительного потенциала в цитоплазме для стимуляции образования дисульфидных связей.
Так, например, клетка-хозяин E.coli может сверхэкспрессировать редкие тРНК, что обеспечивает усиление экспрессии гетерологичных белков, как было проиллюстрировано/описано в публикации Kane (Curr. Opin. Biotechnol (1995), 6, 494-500 “Effect of rare codon clusters on high-level expression of heterologous proteins in E.coli”). Клетка-хозяин может быть дефицитной по ряду редуцирующих ферментов, что благоприятствует образованию стабильных дисульфидных связей, как было проиллюстрировано/описано в публикации Bessette (Proc. Natl. Acad. Sci. USA (1999), 96, 13703-13708 “Efficient folding of proteins with multiple disulphide bonds in the Esсherichia coli cytoplasm”).
Организм
Используемый в настоящем изобретении термин “организм” включает любой организм, который может содержать нуклеотидную последовательность, кодирующую фермент согласно изобретению, и/или полученные из него продукты, и/или в котором промотор может регулировать экспрессию нуклеотидной последовательности по изобретению, если он присутствует в данном организме.
Подходящими организмами могут быть прокариоты, грибки, дрожжи или растения.
Используемый в настоящем изобретении термин “трансгенный организм” включает любой организм, который содержит нуклеотидную последовательность, кодирующую фермент по изобретению, и/или полученные из него продукты, и/или в котором промотор может регулировать экспрессию нуклеотидной последовательности по изобретению. Указанная нуклеотидная последовательность, предпочтительно, встроена в геном данного организма.
Термин “трансгенный организм” не относится к природным нуклеотидным кодирующим последовательностям в их природном окружении, если они находятся под контролем своего природного промотора, который также присутствует в их природном окружении.
Поэтому, трансгенный организм по изобретению включает организм, содержащий любые нуклеотидные последовательности, кодирующие фермент по изобретению; конструкции по изобретению; векторы по изобретению; плазмиды по изобретению; клетки по изобретению; ткани по изобретению или их продукты, или их сочетания.
Так, например, трансгенный организм может также включать нуклеотидную последовательность, кодирующую фермент согласно изобретению и находящуюся под контролем гетерологичного промотора.
Трансформация клеток-хозяев/организма
Как указывалось ранее, организмом-хозяином может быть прокариотический или эукариотический организм. Подходящими примерами прокариотических хозяев являются E.coli и Bacillus subtilis.
Методы трансформации прокариотических хозяев хорошо описаны в литературе, например, смотри Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd еdition, 1989, Cold Spring Harbor Laboratory Press). Если используется прокариотический хозяин, то нуклеотидная последовательность может быть подходящим образом модифицирована перед ее трансформацией, например, путем удаления интронов.
Клетки гифомицетов могут быть трансформированы различными методами, известными специалистам, такими как метод, предусматривающий образование протопластов и трансформацию протопластов с последующей регенерацией клеточной стенки известным способом. Использование Aspergillus в качестве микроорганизма-хозяина описано в ЕР 0238023.
Другим организмом-хозяином может быть растение. Описание общих методов трансформации растений можно найти в статьях Potrykus (Annu. Rev. Plant. Physiol. Plant Mol. Biol [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994, 17-27). Другие описания трансформации растений можно найти в ЕР-А-0449375.
Общие описания трансформации грибков, дрожжей и растений приводятся в нижеследующих разделах.
Трансформированные грибки
Организмом-хозяином могут быть грибки, такие как гифомицеты. Подходящими примерами таких хозяев являются любой хозяин, принадлежащий роду Thermomyces, Acremonium, Aspergillus, Penicillium, Mucor, Neurospora, Trichoderma и тому подобное.
Описания трансформации гифомицетов приводятся в патенте US-A-5741665, где указывается, что стандартные методы трансформации гифомицетов и культивирования грибков хорошо известны специалистам. Более широкое описание методов, применяемых к N. crassa, имеется, например, в публикации Davis и de Serres, Methods Enzymol. (1971) 17A:79-143.
Другие описания методов трансформации гифомицетов можно найти в патенте US-A-5674707.
В одном из аспектов изобретения, организмом-хозяином может быть микроорганизм рода Aspergillus, такой как Aspergillus niger.
Трансгенный Aspergillus по изобретению может быть также получен методом, описанным, например, Turner G. 1994 (Vectors for genetic manipulation. In: Martinelli S.D., Kinghorn J.R. (Editors) Aspergillus: 50 years on. Progress in industrial microbiology vol. 29. Elsevier Amsterdam 1994. pp.641-666).
Метод экспрессии генов в гифомицетах описан в публикации Punt et al. (2002) Trends Biotechnol. 2002 May; 20(5):200-6, Archer & Peberdy Crit. Rev. Biotechnol. (1997) 17(4):273-306.
Трансформированные дрожжи
В другом варианте изобретения, трансгенным организмом могут быть дрожжи.
Описание методов экспрессии гетерологичных генов в дрожжах можно найти, например, в публикации Methods Mol. Biol. (1995) 49:341-54 и Curr. Opin. Biotechnol. (1997) Oct.8(5):554-60.
В соответствии с этим, дрожжи, такие как дрожжи вида Saccharomyces cerevisiae или Pichia pastoris (см. FEMS Microbiol. Rev. (2000 24(1):45-66), могут быть использованы в качестве носителя для экспрессии гетерологичных генов.
Описание методов экспрессии гетерологичных генов в Saccharomyces cerevisiae и секреции генных продуктов можно найти в публикации E. Hinchcliffe E. Kenny (1993, “Yeast as a vehicle for the expression of heterologous genes”, Yeasts, Vol. 5, Anthony H. Rose & J. Stuart Harrison, Eds. 2nd edition, Academic Press Ltd).
Для трансформации дрожжей было разработано несколько протоколов трансформации. Так, например, трансгенные Saccharomyces по изобретению могут быть получены методами, описанными Hinnen et al. (1978, Proceedings of the National Academy Sciences of the USA 75, 1929); Beggs J.D. (1978, Nature, London, 275, 104) и Ito H. et al. (1983, J. Bacteriology 153, 163-168).
Трансформированные дрожжевые клетки могут быть отобраны с использованием различных селективных маркеров, таких как ауксотрофные маркеры, в основном, маркеры резистентности к антибиотикам.
Трансформированные растения/клетки растений
Организмом-хозяином, подходящим для его применения в настоящем изобретении, может быть растение. Описание общих методов можно найти в статьях Potrykus (Annu. Rev. Plant. Physiol. Plant Mol. Biol [1991] 42:205-225) и Christou (Agro-Food-Industry Hi-Tech March/April 1994, 17-27).
Культивирование и продуцирование
Клетки-хозяева, трансформированные нуклеотидной последовательностью по изобретению, могут быть культивированы в условиях, способствующих продуцированию кодируемого фермента и облегчающих выделение фермента из клеток и/или из культуральной среды.
Средой, используемой для культивирования клеток, может быть любая стандартная среда, подходящая для культивирования рассматриваемых клеток-хозяев и для осуществления экспрессии фермента.
Белок, продуцируемый рекомбинантной клеткой, может находиться на поверхности этой клетки.
Указанный фермент может секретироваться клетками-хозяевами и может быть соответствующим образом выделен из культуральной среды в соответствии с хорошо известными процедурами.
Секреция
В большинстве случаев желательно, чтобы фермент секретировался из экспрессионного хозяина в культуральную среду, из которой этот фермент может быть легко выделен. В соответствии с настоящим изобретением, секреторная лидерная последовательность может быть отобрана в зависимости от желательного экспрессионного хозяина. В целях настоящего изобретения могут быть также использованы гибридные сигнальные последовательности.
Типичными примерами гетерологичных секреторных лидерных последовательностей являются последовательности, которые являются производными гена амилоглюкозидазы (АГ) грибков (glaA - 18- и 24-аминокислотные варианты, полученные, например, из Aspergillus), гена α-фактора (дрожжей, например, Saccharomyces, Kluyveromyces и Hansenula) или гена α-амилазы (Bacillus).
Так, например, секреция гетерологичных белков в E.coli описана в публикации “Methods Enzymol (1990) 182:132-43”.
Детекция
Специалистам известны различные протоколы детекции и измерения экспрессии аминокислотных последовательностей. Примерами являются твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (РИА) и клеточный сортинг с активацией флуоресценции (FAСS).
Различные метки широкого ряда и методы конъюгирования известны специалистам и могут быть использованы в различных анализах нуклеиновых кислот и аминокислот.
Коммерчески доступные наборы и протоколы для проведения указанных анализов поставляются рядом компаний, таких как Pharmacia Biotech (Piscataway, NJ), Promega (Madison, WI) и US Biochemical Corp. (Cleveland, OH).
Подходящими репортерными молекулами или метками являются радионуклиды, ферменты, флуоресцентные, хемилюминесцентные или хромогенные агенты, а также субстраты, кофакторы, ингибиторы, магнитные частицы и тому подобное. Патентами, в которых описано использование таких меток, являются патенты США: US-A-3817837, US-A-3850752, US-A-3939350, US-A-3996345, US-A-4277437, US-A-4275149 и US-A-4366241.
Кроме того, могут быть получены рекомбинантные иммуноглобулины, как описано в US-A-4816567.
Гибридные белки
Аминокислотная последовательность, используемая в соответствии с настоящим изобретением, может быть получена в виде гибридного белка, например, для облегчения экстракции и очистки. Примерами партнеров гибридного белка являются глутатион-S-трансфераза (GST), 6хHis, GаL4 (ДНК-связывающие домены и/или домены активации транскрипции) и β-галактозидаза. Может также быть желательным введения сайт протеолитического расщепления между партнером гибридного белка и представляющей интерес последовательностью белка, что позволяет выделять последовательности гибридного белка.
Предпочтительно, чтобы гибридный белок не оказывал негативного влияния на активность последовательности белка.
Системы экспрессии гибридного гена в E.coli были описаны в Curr. Opin. Biotechnol. (1995) 6(5):501-6.
В другом варианте изобретения, аминокислотная последовательность может быть лигирована с гетерологичной последовательностью, кодирующей гибридный белок. Так, например, для скринига пептидных библиотек в целях поиска агентов, способных влиять на активность данного вещества, может оказаться полезным получение последовательности, кодирующей химерное вещество, экспрессирующее гетерологичный эпитоп, который распознается коммерчески доступным антителом.
Крупномасштабное производство
В одном из предпочтительных вариантов изобретения аминокислотная последовательность используется для крупномасштабного производства.
Предпочтительно, аминокислотную последовательность получают в количестве от 1 г до, примерно, 2 г на литр всего объема клеточной культуры после культивирования организма-хозяина.
Предпочтительно, аминокислотную последовательность получают в количестве от 100 мг до, примерно, 900 мг на литр всего объема клеточной культуры после культивирования организма-хозяина.
Предпочтительно, аминокислотную последовательность получают в количестве от 250 мг до, примерно, 500 мг на литр всего объема клеточной культуры после культивирования организма-хозяина.
Пищевой продукт
Композиция по изобретению может быть использована в качестве пищевого продукта или для его получения. Используемый здесь термин “пищевой продукт” имеет широкий смысл и охватывает как продукты питания для человека, так и продукты для кормления животных (то есть, корм для животных). В предпочтительном аспекте изобретения, таким продуктом питания является продукт, употребляемый в пищу человеком.
Указанный пищевой продукт может быть приготовлен в виде раствора или в виде твердого вещества, в зависимости от цели и/или способа его применения, и/или способа его введения.
Ингредиент пищевого продукта
Композиция по изобретению может быть использована в качестве ингредиента пищевого продукта.
Используемый здесь термин “ингредиент пищевого продукта” включает композицию, которая добавлена или может быть добавлена к функциональным (то есть, полезным для здоровья) пищевым продуктам или к обычным продуктам питания, а также включает композиции, которые могут быть использованы в небольших количествах в продуктах широкого ряда, требующих, например, подкисления или эмульгирования.
Указанный ингредиент пищевого продукта может быть приготовлен в виде раствора или в виде твердого вещества, в зависимости от цели и/или способа его применения, и/или способа его введения.
Пищевые продукты
Композиция по изобретению может быть использована для приготовления одного или нескольких пищевых продуктов, таких как кондитерские изделия, молочные продукты, птицепродукты, рыбные продукты и хлебобулочные изделия.
Настоящее изобретение также относится к способу приготовления пищевого продукта или ингредиента пищевого продукта, где указанный способ включает смешивание липолитического фермента согласно изобретению с другим ингредиентом пищевого продукта.
ПРИМЕРЫ
Настоящее изобретение описано на примерах, которые представлены лишь в целях иллюстрации, и в которых приводятся ссылки на нижеследующий графический материал.
На фиг.1 представлены профили липазной активности (заштрихованные участки, отмеченные как пул В) и белка (прерывистая линия), полученные после проведения ионообменной хроматографии.
На фиг.2 представлен очищенный грибковый липолитический фермент (дорожки 3-5), нанесенный на гель (NU-PAGE, 4-12%, Меs-буфер, полученный как описано производителем, Novex, США), который был затем окрашен кумасси.
На фиг.3 представлена хроматограмма #61.
На фиг.4 проиллюстрирован электрофорез в ДСН-ПААГ фракций из колонки с бутилсефарозой (Р: пул # 172-174, 100 ед/мл, разведенный 1:10; Std = набор стандартных белков).
На фиг.5 проиллюстрированы лабораторные эксперименты по пробной выпечке, где: 1) Chr #61, фракция 9, 2) пул #172-#174, 3) Chr.#61, фракция 14, 4) контроль, 5) липаза #3044.
На фиг.6 проиллюстрирован ГЖХ-анализ липидов теста, а именно, дигалактозилдиглицерида (ДГДГ) и дигалактозилмоноглицерида (ДГМГ) из BS8948-2.
На фиг.7 проиллюстрировано выравнивание аминокислотных последовательностей всех пептидов CBS путем сопоставления с последовательностями липазы японского штамма F. heterosporum (Nagao et al., 1994). Идентичные и аналогичные (в высокой степени консервативные) аминокислоты помечены ниже “*” и “·”, соответственно.
На фиг.8 показана нуклеотидная последовательность и транслированная аминокислотная последовательность гена синтезированного липолитического фермента F. heterosporum (СВS 782.83), присоединенная к синтетической альфа-сигнальной последовательности. Аминокислотная последовательность показана над нуклеотидной последовательностью. Нуклеотиды, кодирующие сайты рестриктирующих ферментов EcoRI и BamHI, подчеркнуты, а кодоны инициации и терминации трансляции подчеркнуты двойной линией. Острие стрелок указывает на положение стыка альфа-сигнальной последовательности и гена липолитического фермента. Стрелки указывают на праймеры, используемые для сборки гена.
На фиг.9 схематически представлен вектор экспрессии рВ14 Hansenula, содержащий ген синтезированного липолитического фермента (липазы) F. heterosporum (СВS 782.83), присоединенный к синтетической альфа-сигнальной последовательности (альфа-ss). URA3, ген оротидин-5'-фосфат-декарбоксилазы, обеспечивающий комплементацию урацила для отбора в Hansenula. HARS - автономно реплицирующаяся последовательность для репликации в Hansenula. FMD-P - промотор FMD для экспрессии в Hansenula.
На фиг.10 проиллюстрирована фосфолипазная активность отобранных клонов Hansenula polymorpha, содержащих ген синтезированного липолитического фермента F. heterosporum. Лецитин был использован в качестве субстрата, а свободная жирная кислота была определена с помощью набора NEFA (Roche).
На фиг.11 проиллюстрирована пробная лабораторная выпечка хлеба с повышенным содержанием (PLU) фосфолипазного образца 205 и Lipopan FTM.
На фиг.12 проиллюстрирован ГЖХ-анализ липидов теста. ДГДГ = дигалактозилдиглицерид. ДГМГ = дигалактозилмоноглицерид. Sum = ДГДГ + ДГМГ (пример 3).
На фиг.13 проиллюстрирована ВЭТСХ-хроматограмма А) стандартов: 1. фракционированные липиды муки; 2. гидролизованный ДГДГ; 3. ДГДГ; В) липидов, экстрагированных из теста: 4. контроль; 5. 2000 PLU-7/кг образца 205; 6. 40 м.д. Lipopan FTM.
На фиг.14 проиллюстрирован ГЖХ-анализ дигалактозилмоноглицеридного изомера в тесте, обработанном липолитическим ферментом, выделенным из Fusarium heterosporum.
На фиг.15 проиллюстрирована активность липолитического фермента, происходящего от Fusarium heterosporum, которая была определена путем 10-минутного воздействия фермента на лецитиновый субстрат, рН 7,0, при различных температурах, и последующего определения уровня свободных жирных кислот методом NEFA C.
На фиг.16 проиллюстрирована активность липолитического фермента, происходящего от Fusarium heterosporum, которая была определена после 30-минутного инкубирования в 50 мМ фосфатного буфера при 3 TIPU/мл при различных температурах (50 мМ фосфатный буфер, рН 7,0) и при 10-минутном воздействии фермента на лецитиновый субстрат (без СаCl2) в условиях 37°С и рН 7,0, с последующим определением свободных жирных кислот методом NEFA C.
На фиг.17 проиллюстрирована активность липолитического фермента, происходящего от Fusarium heterosporum, которая была определена путем 10-минутного воздействия фермента на лецитиновый субстрат (без СаCl2) при 37°С и при различных рН (50 мМ фосфатный буфер) с последующим определением свободных жирных кислот методом NEFA C.
На фиг.18 проиллюстрирована активность липолитического фермента, происходящего от Fusarium heterosporum, которая была определена после 30-минутного инкубирования в 50 мМ фосфатного буфера при 3 TIPU/мл и при различных рН (50 мМ фосфатный буфер) в условиях 10-минутного воздействия фермента на лецитиновый субстрат (без СаCl2) при 37°С и при рН 7,0, с последующим определением свободных жирных кислот методом NEFA C.
На фиг.19а и 19b проиллюстрировано определение молекулярной массы липолитического фермента, происходящего от Fusarium heterosporum, с помощью электрофореза в ДСН-ПААГ.
На фиг.20 указана оптимальная температура для липолитического фермента согласно изобретению. Ферментативную реакцию проводили при различных температурах.
На фиг.21 представлены кривые зависимости количества лецитина в модифицированном ферментом яичном желтке от времени реакции, А: 30°С, В: 40°С и С: 50°С. Количество лецитина было проанализировано с помощью ЖХ/МС-МС и выражено в процентах по массе яичного желтка.
На фиг.22 представлены кривые зависимости количества лизолецитина в модифицированном ферментом яичном желтке от времени реакции, А: 30°С, В: 40°С и С: 50°С. Количество лизолецитина было проанализировано с помощью ЖХ/МС-МС и выражено в процентах по массе яичного желтка.
На фиг.23 представлены кривые зависимости количества свободной жирной кислоты в модифицированном ферментом яичном желтке от времени реакции, А: 30°С, В: 40°С и С: 50°С. Количество свободной жирной кислоты было проанализировано с помощью ЖХ/МС-МС и выражено в процентах по массе яичного желтка.
На фиг.24 проиллюстрировано ферментативное превращение яичного желтка под действием липолитического фермента согласно изобретению (пример 4). Представлены кривые зависимости количества лизолецитина (А), свободной жирной кислоты (В) и лецитина (С) от времени реакции. Величина ошибки означает стандартное отклонение для двух определений (n=2). Количество лецитина и лизолецитина определяли с помощью ЖХ/МС-МС, а количество свободной жирной кислоты определяли методом NЕFA С. Результаты выражены в процентах по массе яичного желтка.
На фиг.25 проиллюстрировано ферментативное превращение яичного желтка под действием фосфолипазы Lecitase® Ultra, поставляемой от Novozymes A/S (пример 4). Представлены кривые зависимости количества лизолецитина (А), свободной жирной кислоты (В) и лецитина (С) от времени реакции. Величина ошибки означает стандартное отклонение для двух определений (n=2). Количество лецитина и лизолецитина определяли с помощью ЖХ/МС-МС, а количество свободной жирной кислоты определяли методом NЕFA С. Результаты выражены в процентах по массе яичного желтка.
На фиг.26 проиллюстрирован ТСХ-анализ (растворитель представлял собой смесь хлороформ:метанол:вода (65:24:4)) липидного экстракта из модифицированного яичного желтка (пример 4). 1: ФХ- и ЛФХ-стандарты. 2: Липолитический фермент согласно изобретению, 10°С, 240 мин. 3: Липолитический фермент согласно изобретению, 20°С, 240 мин. 4: Липолитический фермент согласно изобретению, 53°С, 240 мин. 5: Липолитический фермент согласно изобретению, 20°С, 1440 мин. 6: Lecitase® Ultra, 10°С, 4 ч. 7: Lecitase® Ultra, 20°С, 240 мин. 8: Lecitase® Ultra, 53°С, 4 ч. 9: Lecitase® Ultra, 20°С, 1440 мин. 10: Контрольный образец. Соединениями, указанными в левой части ТСХ-планшета, являются: холестерин (С), триацилглицерид (ТГ), диацилглицерид (ДГ), свободная жирная кислота (СЖК), моноацилглицерид (МГ), фосфатидилэтаноламин (ФЭ), фосфатидилхолин (ФX), лизофосфатидилэтаноламин (ЛФЭ) и лизофосфатидилхолин (ЛФX).
На фиг.27 показана взаимосвязь между изменением содержания лизолецитина и свободной жирной кислоты в процессе ферментативной обработки яичного желтка липолитическим ферментом согласно изобретению и фосфолипазами Lecitase® Ultra, соответственно (пример 4). Результаты получены исходя из молярной массы лизолецитина 523 и молярной массы свободных жирных кислот 283. Свободную жирную кислоту определяли методом NEFA C. Лизолецитин и лецитин определяли с помощью ЖХ/МС-МС.
На фиг.28 проиллюстрирован ВЭТСХ-анализ (растворитель представлял собой смесь петролейный эфир:МТВЕ:уксусная кислота (50:50:1)) липидного экстракта из модифицированного яичного желтка (пример 4). Соединениями, указанными в левой части ТСХ-планшета, являются: триацилглицерид (ТГ), свободная жирная кислота (СЖК), 1,3-диацилглицерид (1,3-ДГ), 1,2-диацилглицерид (1,2-ДГ), холестерин (С), моноацилглицерид (МГ), фосфатидилэтаноламин (ФЭ), фосфатидилхолин (ФX), лизофосфатидилэтаноламин (ЛФЭ) и лизофосфатидилхолин (ЛФX).
На фиг.29 проиллюстрирован ТСХ-анализ (растворитель IV) майонеза, приготовленного из модифицированного ферментом яичного желтка, поставляемого фирмой Sanofa A/S (пример 5).
На фиг.30 показан майонез, приготовленный из модифицированного ферментом яичного желтка, поставляемого фирмой Sanofa A/S, и прошедший термообработку в микроволновой печи (пример 5). Образец представлял собой контроль (вода), добавленный вместо ферментного раствора, образец 2 содержал 30 ед/г липолитического фермента согласно изобретению, а образец 3 содержал 30 ед/г Lecitase® Ultra.
На фиг.31 представлен удельный объем хлебобулочного изделия в виде хлебцев с твердой корочкой, изготовленных с различными концентрациями липолитического фермента согласно изобретению, добавленного отдельно или в комбинации с эмульгатором Panodan® M2020 DATEM, и протестированных с использованием комбинации Lipopan FTM и DАТЕМ, а также чистого Lipopan FTM или чистого DАТЕМ.
На фиг.32 представлен удельный объем хлебобулочного изделия в виде хлебцев с твердой корочкой, изготовленных с различными концентрациями липолитического фермента согласно изобретению, добавленного отдельно или в комбинации с эмульгатором Panodan® M2020 DATEM или SSL P 55, и протестированных с использованием комбинации Lipopan FTM/SSL P 55 или Lipopan FTM/DАТЕМ, а также чистого Lipopan FTM, чистого DАТЕМ и чистого SSL P 55.
На фиг.33 представлена нуклеотидная последовательность (SEQ ID NO:5) и выведенная аминокислотная последовательность (SEQ ID NO:4) кДНК липазы F. semitectum (IВТ 9507). Выведенная аминокислотная последовательность показана над нуклеотидной последовательностью. Стрелки указывают на праймеры, используемые для амплификации кДНК.
На фиг.34 схематически представлен вектор экспрессии рDВ14-alp-sem Hansenula, содержащий ген липазы (Lipase) F. semitectum, присоединенный к синтетической α-сигнальной последовательности (альфа-ss). АР(R), “URA3”, ген оротидин-5'-фосфат-декарбоксилазы, обеспечивающий комплементацию урацила для отбора. “HARS” означает автономно реплицирующуюся последовательность для репликации в Hansenula. “FMD-P” означает промотор FMD для экспрессии в Hansenula.
На фиг.35 представлена кривая зависимости фосфолипазной активности липолитического фермента Fusarium semitectum IВТ9507 от температуры.
На фиг.36 представлена кривая зависимости фосфолипазной активности липолитического фермента Fusarium semitectum IВТ9507 от рН.
На фиг.37 представлена аминокислотная последовательность (SEQ ID NO:1) грибкового липолитического фермента, происходящего от Fusarium heterosporum.
На фиг.38 представлена аминокислотная последовательность грибкового липолитического фермента, происходящего от Fusarium heterosporum, где указанная последовательность содержит N-концевую сигнальную последовательность (подчеркнута)(SEQ ID NO:2).
На фиг.39 представлена нуклеотидная последовательность (SEQ ID NO:3), кодирующая грибковый липолитический фермент согласно изобретению, происходящий от Fusarium heterosporum.
На фиг.40 представлена аминокислотная последовательность (SEQ ID NO:4) липолитического фермента, происходящего от Fusarium semitectum.
На фиг.41 представлена нуклеотидная последовательность (SEQ ID NO:5), кодирующая липолитический фермент, происходящий от Fusarium semitectum.
На фиг.42 представлена аминокислотная последовательность (SEQ ID NO:6) липолитического фермента, происходящего от Fusarium heterosporum (ЕАЕА представляет собой про-пептид, происходящий от сигнальной последовательности α-фактора).
На фиг.43 представлена нуклеотидная последовательность (SEQ ID NO:7) липолитического фермента, происходящего от Fusarium heterosporum, которая включает сигнальную последовательность α-фактора.
Пример 1. Экспрессия, очистка, секвенирование липолитического фермента Fusarium heterosporum и его испытание в условиях пробной выпечки
Ферментация
Штамм Fusarium heterosporum СВS 782.83 был получен от Centraalbureau voor Schimmelcultures (the Netherlands).
Среда для культивирования
Агар с глюкозодрожжевым экстрактом
Глюкозу добавляли после автоклавирования.
1.4. Среда для предварительного сбраживания
Среду приготавливали в шейкерных 500 мл-колбах с перегородками, и каждая шейкерная колба содержала 100 мл среды. Соевое масло добавляли отдельно в каждую колбу. Глюкозу добавляли после автоклавирования.
Среда для продуцирования
Среду приготавливали в шейкерных 500 мл-колбах с перегородками и каждая шейкерная колба содержала 100 мл среды. Твин ТМ-80 добавляли отдельно в каждую колбу. Перед автоклавированием, рН доводили до 6,0.
Условия культивирования
Планшет с агаром, содержащим глюкозодрожжевой экстракт, инокулировали Fusarium heterosporum СВS 782.83 и инкубировали при 24°С до появления спор. Шейкерную колбу, содержащую среду для предварительного сбраживания инокулировали агаровой пластиной размером 4 см2, содержащей культуру с хорошим спорообразованием. Эту шейкерную колбу инкубировали при 30°С и при вращении 200 об/мин. После трехдневного культивирования каждую из 30 шейкерных колб со средой для продуцирования инокулировали 5 мл сбраживающего бульона, взятого из шейкерной колбы для предварительного сбраживания. Шейкерные колбы со средой для продуцирования инкубировали при 30°С и при вращении 200 об/мин. Через 2, 3 и 4 дня после культивирования отбирали десять шейкерных колб с продуцирующей средой. Биомассу удаляли путем центрифугирования, а затем супернатант подвергали стерильной фильтрации через 0,2 мкм-фильтры (Фильтр VacuCap 90 Filter Unit масс./0,2 мкм - мембрана Supor), полученные из Gelman Laboratory. После фильтрации фильтрат замораживали при -80°С и хранили до проведения анализов.
Процедуры анализов
Фосфолипазную активность определяли с помощью “анализа на PLU”, описанного выше.
Применение
ТСХ-анализ
ТСХ-пластину активировали в нагревательном шкафу (110°С) в течение получаса. 100 мл рабочего буфера выливали в хроматографическую камеру с крышкой. Стенки этой камеры были покрыты фильтровальной бумагой (Whatman 2) для насыщения камеры парами растворителя. ТСХ-пластину помещали в рамку и на эту ТСХ-пластину наносили образец на уровне 2 см от дна. Затем ТСХ-пластину помещали в ТСХ-камеру с выбранным рабочим буфером. Когда уровень рабочего буфера достигал 14 см от дна пластины, ТСХ-пластину вынимали и сушили в дымовом шкафу, а затем помещали на 10 минут в нагревательный шкаф при 110°С. Затем ТСХ-пластину погружали в проявляющий реагент и сушили в нагревательном шкафу при 110°С в течение 15 минут.
Рабочий буфер:
№ IV: Хлороформ:метанол:H2O (65:25:4)
№ IV: Петролейный эфир:метил-трет-бутиловый эфир (МТВЕ):уксусная кислота (60:40:1).
Проявляющий буфер (ванадатный буфер):
32 г Na2СO3 и 300 мл H2O (1М)
Добавляли 18,2 г пентоксида ванадия (V2O5) и растворяли при легком нагревании, а затем содержимое подвергали горячей сушке в печи “BACO-LINE” в течение 6 минут.
Раствор охлаждали до комнатной температуры.
Затем осторожно добавляли 460 мл 2,5 М Н2SO4 (460 мл H2O + 61 мл Н2SO4).
Затем добавляли воду до 1000 мл.
Газовая хроматография
Капиллярную газовую хроматографию проводили на хроматографе Perkin-Elmer 8420, снабженном колонкой с WCOT-связанным силикагелем, 12,5 м × 0,25 мм внут. диам. × 0,1 мкм 5% фенилметилсиликона (СР Sil 8 CB от Crompack).
Носитель: гелий.
Инжекция: 1,5 мкл с перерывом.
Детектор: FID 385°С.
Получение образца: 50 мг липида пшеницы растворяли в 12 мл смеси гептан:пиридин, 2:1, содержащей гептадекан в качестве внутреннего стандарта, 2 мг/мл. 500 мкл образца переносили в гофрированный сосуд. Затем добавляли 100 мкл МSTFA (N-метил-N-триметилсилилтрифторацетамид) и реакционную смесь инкубировали в течение 15 минут при 90°С. Проводили вычисления коэффициентов отклика для моно-ди-триглицеридов, свободных жирных кислот и галактолипидов определяли с использованием контрольной смеси этих компонентов. Исходя из этих коэффициентов отклика вычисляли уровень липидов в тесте.
Пробная выпечка в лабораторном масштабе
В 50-граммовую дежу Брэбендера для перемешивания и замеса в течение 5 минут при 30°С добавляли следующие ингредиенты: 50 г муки, 10 г сухих дрожжей, 0,8 г сахара, 0,8 г соли, 70 м.д. аскорбиновой кислоты и воды (для доведения консистенции теста до 400 единиц Брэбендера). Время выдерживания составляло 10 минут при 34°С. Тесто отвешивали по 15 г. Затем тесто формовали на специальном устройстве, где его раскатывали между деревянной пластиной и рамой из плексигласа. Тесто оставляли на 45 минут в хлебопекарных формах для расстойки при 34°С, а затем выпекали в бытовой печи Voss в течение 8 минут при 225°С. После выпечки хлеб охлаждали до комнатной температуры, а затем оставляли на 20 минут. Хлеб взвешивали и его объем определяли методом вытеснения рапсовых семян. Затем хлеб разрезали и оценивали мякиш и корочку.
Экспериментальная пробная выпечка (хлебцы с твердой корочкой)
1500 г муки, соответствующей датскому стандарту, 90 г прессованных дрожжей, 24 г сахара, 24 г соли, 400 единиц воды по Брэбендеру + 2% замешивали в смесителе Хобарта с крюком в течение 2 минут при низкой скорости и в течение 9 минут при высокой скорости. Температура теста составляла 26°С. Тесто отвешивали по 1350 грамм. После этого тесто оставляли на 10 минут при 30°С и формовали на тестоформующей машине Fortuna. Затем тесто оставляли на 45 минут для расстойки при 34°С. Тесто выпекали в Bago-печи в течение 18 минут при 220°С и обрабатывали паром в течение 12 секунд. После охлаждения, хлебцы взвешивали и их объем измеряли методом вытеснения рапсовых семян.
Удельный объем хлеба:
Результаты и обсуждение
Ферментация
Образцы для ферментации анализировали на фосфолипазную активность и результаты представлены в таблице 1.
аСреда D. = среда-продуцент
Как видно из таблицы, фосфолипазная активность была почти идентичной на дни 2, 3 и 4, а поэтому все образцы объединяли и обозначали JBS-2254-97-3.
Очистка и секвенирование
Выделение фосфолипазы из сырого экстракта проводили с помощью анионообменной хроматографии.
Колонку (Q-сефароза FF, 1,5 × 2,8 см, 5 мл геля) приготавливали как рекомендовано производителями (Amersham Bio.), а затем уравновешивали в 20 мМ буфере Трис/HCl, 0,1 М NаСl, рН 7,5 (буфер А). К образцу (15 мл) добавляли 0,1 М NаСl и смесь наносили на колонку при скорости потока 3,5 мл/мин. Липолитический фермент элюировали линейным градиентом 0-0,6 М NаСl в буфере А (см. фиг.1). 3,5 мл-фракции собирали в процессе элюирования. 10 мкл каждой фракции подвергали анализу в многолуночном планшете. Липазную активность определяли с помощью анализа на трибутирин и лецитин в многолуночном планшете. (10 мкл каждой фракции переносили в свободную лунку и планшет инкубировали при 40°С. Образование гало на агарозных гелях происходило в зависимости от времени. Для сравнения, в одну из лунок также добавляли контроль без фермента). Затем фракции, содержащие липолитическую активность, подвергали электрофорезу в ДСН-ПААГ (см. фиг.2) и анализу N-концевых последовательностей.
Анализ методом ферментативного фингерпринтинга с помощью MALDI-TOF и секвенирование аминокислот
Белок восстанавливали дитиотреитолом и цистеиновые остатки защищали путем проведения реакции карбоксиметилирования с использованием иодацетамида. Затем белок гидролизовали трипсином и картину фрагментации триптических пептидов оценивали с помощью анализа MALDI-TOF. Пептиды разделяли хроматографией на обращенно-фазовой ВЭЖХ-колонке с С18 и проводили мониторинг степени очистки с помощью анализа MALDI-TOF. Аминокислотную последовательность определяли методом деградации по Эдману, как подробно описано ранее в публикации TR6452.
Определяли полноразмерную аминокислотную последовательность липолитического фермента Fusarium heterosporum. Гидролиз трипсином позволяет получить в высокой степени специфические пептиды, молекулярная масса которых может быть окончательно определена с помощью MALDI-TOF. Аминокислотные последовательности для всех пептидов также определяли методом деградации по Эдману. Аминокислотная последовательность, определенная методом деградации по Эдману, охватывает 99,64% полипептидной цепи липолитического фермента F. heterosporum.
Общие результаты исследований, проводимых с помощью MALDI-TOF и методом деградации по Эдману, представлены в Таблице 2.
Таблица 2
Результаты анализа, проводимого методом ферментативного фингерпринтинга; определение молекулярной массы (Мw) пептидов липолитического фермента Fusarium heterosporum; и определение полноразмерной аминокислотной последовательности методом деградации по Эдману.
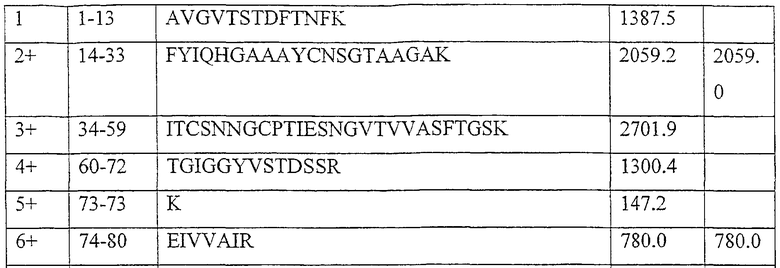
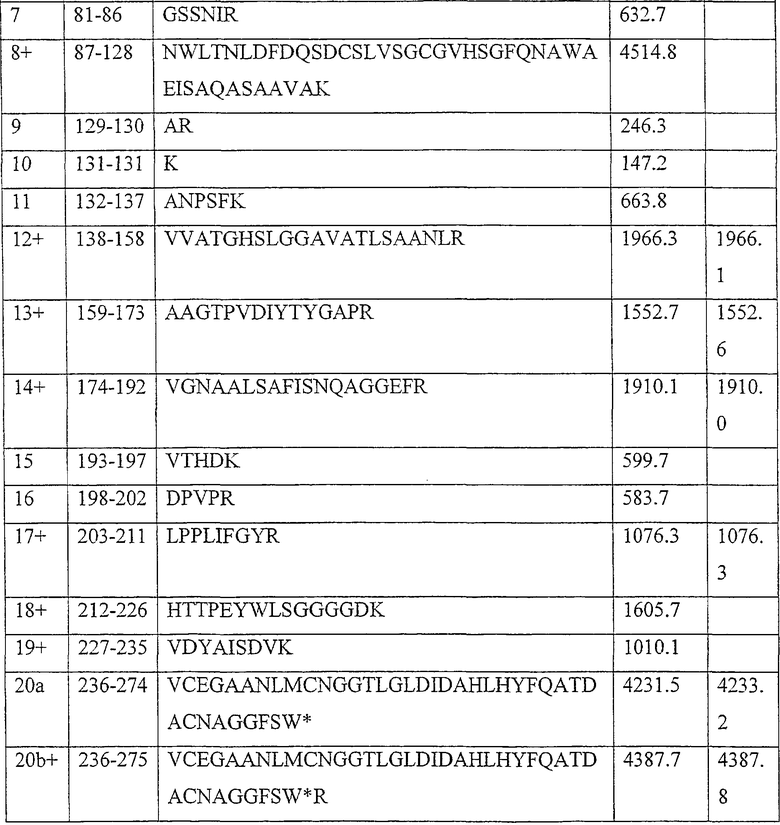
+ = подтверждено путем секвенирования по Эдману
* = окисленный триптофан
Доля последовательности = 99,64%.
Полноразмерная аминокислотная последовательность липолитического фермента Fusarium heterosporum представлена последовательностью SEQ ID NO:1 (см. Фиг.37).
Испытания в условиях практического применения
Пул из 2 литров, полученный от трех образцов F. heterosporum (таблица 1) и обозначенный как Пул #172-174, концентрировали путем ультрафильтрации (10 кДа-фильтр) на фильтрующем устройстве Amicon Ultrа. 230 мл ретентата содержали приблизительно 100 РLU-7/мл. Ретентат доводили до 1 М добавлением ацетата аммония, наносили на колонку (внут. диам. 2,5 см) с 27 мл бутилсефарозы и элюировали буфером А, содержащим 1М NН4-ацетат в 20 мМ ТЕА, рН 7,4, и буфером В, 20 мМ ТЕА, рН 7,4. Хроматограмма (#61), полученная после очистки, представлена на фиг.3.
Фракции для хроматограммы #61 анализировали с помощью электрофореза в ДСН-ПААГ, как показано на фиг.4.
Хроматографические 10 мл-фракции собирали и анализировали на фосфолипазную активность, указанную в таблице 3. Эти результаты показали, что самый высокий уровень фосфолипазной активности в этих фракциях элюировался в главном пике хроматограммы. Небольшое количество активности не было связано с колонкой и элюировалось в ранних фракциях. Хотя ДСН-гель не проходил достаточно хорошо, однако, было установлено, что эти фракции содержат несколько белков, а фракции 14 и 15 содержат одну главную полосу, которая, как предполагается, представляет собой грибковый липолитический фермент.
Фракция 9 и фракция 14 хроматограммы #61 были использованы в лабораторной пробной выпечке и в этом пробном испытании также оценивали неочищенный пул #172-174. Результаты этого эксперимента в условиях пробной выпечки приводятся в таблице 4. Эти результаты ясно показали, что очищенный липолитический фермент от F. heterosporum СВS 782.83 давал очень хорошие результаты по выпечке, а именно, способствовал увеличению объема хлеба. Неочищенный образец также значительно увеличивал объем хлеба. Структура мякиша хлеба также значительно улучшалась при добавлении липолитического фермента F. heterosporum, как показано на фиг.5, и, по оценкам, этот фермент был лучше, чем липаза #3044, выделенная из Pseudomonas cepacia.
Тесто, полученное в результате эксперимента по выпечке в лабораторном масштабе, экстрагировали бутанолом, насыщенным водой, и липиды анализировали с помощью ТСХ. ТСХ-анализ подтвердил, что липаза #3044 является более активной по отношению к триглицериду, чем липолитический фермент, выделенный из образцов F. heterosporum. Количество свободных жирных кислот (FFA) было также выше при использовании липазы #3044. ТСХ в растворителе IV указывала на присутствие компонента (ДГМГ), количество которого было заметно выше в образцах липидов теста, обработанных F. heterosporum, по сравнению с его количеством в образцах, обработанных триглицерид-гидролизующей липазой #3044.
Очищенные фракции от F. heterosporum также тестировали в эксперименте в условиях пробной выпечки и результаты этого эксперимента представлены в таблице 5.
Тесто, полученное в испытаниях с пробной выпечкой, экстрагировали насыщенным водой бутанолом, а затем липиды теста анализировали с помощью ГЖХ-анализа и полученные данные представлены в таблице 6.
Отношение уровня гидролиза ДГДГ к уровню гидролиза триглицеридов представлено в таблице 7.
ГЖХ-анализ галактолипидов также графически проиллюстрирован на фиг.6.
Результаты ГЖХ-анализа подтвердили, что количество ДГМГ, продуцированного в тесте под действием F. heterosporum, превышает количество, продуцируемое под действием 40 м.д. Lipopan F (#3016). Эти результаты также указывают на более высокий уровень гидролиза МГДГ по сравнению с гидролизом ДГДГ. Полученные результаты также показали, что количество гидролизованного триглицерида было низким по сравнению с нормальным количеством триглицерида, гидролизованным ферментом, подобным ферменту #3044, происходящему от P. cepacia. Результаты эксперимента по пробной выпечке и анализ на липиды подтвердили, что липолитический фермент, выделенный из F. heterosporum СВS 782.83, имел явно выраженную гидролитическую активность по отношению к дигалактозилдиглицериду (ДГДГ) и способствовал образованию дигалактозилмоноглицерида (ДГМГ) в тесте.
4. Выводы:
В этом исследовании, грибковый липолитический фермент, выделенный из F. heterosporum СВS 782.83, получали путем сбраживания в шейкерных колбах. Затем фермент очищали и определяли аминокислотную последовательность. Этот фермент имел примерно на 83%-ную гомологию с коммерчески доступной липазой, происходящей от F. oxysporum (Lipopan FTM). Указанный фермент давал очень хорошие результаты в испытании по пробной выпечке, а именно, в отношении увеличения объема хлеба и улучшения структуры мякиша. Анализ липидов теста подтвердил, что указанный фермент был активен по отношению к галактолипидам в процессе продуцирования галактомоноглицеридов. Результаты пробной выпечки, проводимой без какой-либо оптимизации дозы, показали, что грибковый липолитический фермент, выделенный из F. heterosporum CBS 782.83, является, по меньшей мере, эквивалентным коммерчески доступному ферменту Lipopan FTM, а сравнение активности по отношению к ДГДГ и к триглицеридам показало, что указанный фермент, присутствующий а тесте, обладает превосходной ферментативной активностью по сравнению с активностью фермента Lipopan FTM.
Пример 2: Конструирование и экспрессия синтетического гена, кодирующего липолитический фермент от Fusarium heterosporum (CBS 782.83), в Hansenula polymorpha.
Была определена аминокислотная последовательность грибкового липолитического фермента, выделенного из Fusarium heterosporum (CBS 782.83), и эту последовательность использовали для конструирования и клонирования синтетического гена липолитического фермента в целях его экспрессии в Hansenula polymorpha. Для стимуляции высокого уровня экспрессии кодоны синтетического гена оптимизировали в соответствии с предпочтительностью кодонов в Hansenula polymorpha. Оптимизированную по кодонам сигнальную последовательность альфа-фактора также синтезировали и клонировали в сайте, расположенном перед синтетическим геном липолитического фермента. Полученную конструкцию переносили в вектор экспрессии рВ14 и трансформировали в Hansenula polymorpha. рВ14 представляет собой плазмиду, в которой отсутствуют гены, сообщающие резистентность к антибиотикам, а поэтому она может быть использована в методах продуцирования.
Ряд штаммов Fusarium, продуцирующих липолитический фермент, скринировали на активность, и было обнаружено, что они имеют высокое отношение активности, направленной на галактолипиды и/или фосфолипиды, по сравнению с активностью, направленной на триглицериды.
В качестве штаммов-продуцентов липолитических ферментов было отобрано несколько штаммов, представляющих особый интерес. Из них, особый интерес представляет штамм Fusarium heterosporum (СВS 782.83). Поэтому, из этого штамма был выделен липолитический фермент и была определена его аминокислотная последовательность. Эту аминокислотную последовательность подвергали обратной трансляции с продуцированием последовательности нуклеиновой кислоты, которая была использована для моделирования и конструирования синтетического гена в целях его экспрессии в Hansenula polymorpha.
Эксперименты
Штамм Hansenula, используемый в этом исследовании, представляет собой ауксотрофный по урацилу штамм RВ11 (odcl) Hansenula polymorpha, полученный от Rhein Biotech GmbH (Дюссельдорф, Германия).
Анализ методом ферментативного фингерпринтинга с помощью MALDI-TOF и секвенирование аминокислот
Белок, обладающий липолитической ферментативной активностью, выделяли из Fusarium heterosporum (СВS 782.83). Этот белок восстанавливали дитиотреитолом и цистеиновые остатки защищали путем проведения реакции карбоксиметилирования с использованием иодацетамида. Затем белок гидролизовали трипсином и картину фрагментации триптических пептидов оценивали с помощью анализа MALDI-TOF. Пептиды разделяли хроматографией на обращенно-фазовой ВЭЖХ-колонке с С18 и проводили мониторинг степени очистки с помощью анализа MALDI-TOF. Аминокислотную последовательность определяли методом деградации по Эдману, как было подробно описано ранее в публикации TR6452.
Моделирование и конструирование синтетического гена липолитического фермента
Аминокислотные последовательности пептидных фрагментов сравнивали путем выравнивания с последовательностью японского штамма F. heterosporum (Nagao et al., 1994). Полученную таким образом полноразмерную аминокислотную последовательность подвергали обратной трансляции с продуцированием последовательности нуклеиновой кислоты для выявления всех возможных кодонов. Для каждой кодируемой кодоном аминокислоты выбирали кодон, наиболее предпочитаемый для экспрессии в Hansenula polymorpha, в соответствии с таблицей предпочтительности кодонов генов, экспрессируемых в Hansenula polymorpha. Были синтезированы синтетические олигонуклеотиды, каждый длиной примерно в 100 нуклеотидов, составляющие полноразмерный ген, и этот ген конструировали с помощью ПЦР. Для концевой амплификации этого гена использовали обратный праймер (alps.cbss), сконструированный так, чтобы большая часть его 5'-нуклеотидов находилась со стороны 3'-конца сигнальной последовательности альфа-фактора, для лигирования с сохранением рамки считывания, и прямой праймер (cbss.t), сконструированный с использованием рестрикционного BamHI-сайта для клонирования (таблица 8).
Нуклеотидную последовательность, кодирующую сигнальную последовательность дрожжевого альфа-фактора размножения дрожжей, синтезировали аналогичным образом с применением предпочтительных кодонов олигонуклеотидов и амплифицировали с помощью ПЦР. Для концевой амплификации этой альфа-сигнальной последовательности использовали обратный праймер (alpsynt), сконструированный с использованием рестрикционного EcoRI-сайта для клонирования, и прямой праймер (cbss.alps), сконструированный так, чтобы большая часть его 5'-нуклеотидов находилась со стороны 5'-конца синтетического гена липолитического фермента для лигирования с сохранением рамки считывания (таблица 8).
Для присоединения синтетической сигнальной последовательности альфа-фактора к синтетическому гену липолитического фермента два фрагмента смешивали и снова амплифицировали с использованием внешних праймеров alpsynt и cbss.t (таблица 8). ПЦР-продукт клонировали в вектор рСR 2.1-ТОРО (Invitrogen) и нуклеотидную последовательность вставок определяли с использованием набора для секвенирования (BigDye Terminator v3.0 cycle sequencing kit) (Apрlied Biosystems) и генного анализатора ABI Prism 3100 Genetic Analyzer (Apрlied Biosystems).
3'-cbss.t
5'-AGGATCCAATTCTCTCCATGGCCTATCTCCAGGAGAA
ACCTCCG
вует
BamHI
3'-cbss.alps
5'-AGGTCACTCCAACGGCTCTCTTGTCCAAGGAAACAC
CTTCC
отсутствует
Таблица 8. Праймерные последовательности использовались для амплификации и сборки синтетического гена липолитического фермента F. heterosporum (СВS 782.83) и синтетической альфа-сигнальной последовательности. Сайты рестриктирующих ферментов, введенные в каждый праймер в целях клонирования, подчеркнуты. Нуклеотиды, включенные для присоединения синтетического гена липолитического фермента к синтетической альфа-сигнальной последовательности, подчеркнуты двойной линией.
Экспрессия липолитического фермента в Hansenula polymorpha
Для экспрессии синтетического гена липолитического фермента F. heterosporum (СВS 782.83) в Hansenula гибридный ген “альфа-сигнальной последовательности/липолитического фермента” встраивали позади FMD-промотора в вектор экспрессии рВ14 Hansenula, то есть в плазмиду, не содержащую генов, сообщающих резистентность к антибиотикам. После определения конформации ожидаемой структуры собранной плазмиды в E.coli эту плазмиду вводили в компетентные клетки Hansenula polymorpha путем электропорации. Трансформанты отбирали на планшетах с YND и колонии снова отбирали на множественную интеграцию гена посредством 3 и 8 пассажей разведений 1:200 в жидких культурах YND. И наконец, отобранные культуры стабилизировали путем двухкратного переноса на среду YND. Для дополнительного отбора трансформантов с высокой степенью экспрессии каждую культуру, обнаруживающую высокий уровень экспрессии, засевали для образования моноколоний, каждую из которых анализировали на уровень экспрессии.
Для определения уровня экспрессии гена липолитического фермента отобранные клоны культивировали на среде YND, содержащей 1,8% глицерина и 0,2% глюкозы, в течение 2 дней при 24°С.
Ферментативная активность
Образцы культуральной среды анализировали на активность липолитического фермента с использованием лецитина или ДГДГ в качестве субстратов и с использованием набора NEFA (Roche) и эти образцы доводили до объемов, подходящих для микротитрационных планшетов и достаточных для определения уровня высвобождаемых свободных жирных кислот.
Результаты
Ферментативный фингерпринтинг с помощью MALDI-TOF и секвенирование аминокислот
Была определена полноразмерная аминокислотная последовательность липолитического фермента Fusarium heterosporum (см. SEQ ID NO:1 - фиг.37). Гидролиз трипсином приводил к образованию очень специфических пептидов, молекулярная масса (Мw) которых может быть окончательно определена с помощью MALDI-TOF. Аминокислотные последовательности для всех пептидов были также определены путем деградации по Эдману. Аминокислотные последовательности, определенные путем деградации по Эдману, охватывают 99,64% полипептидной цепи липолитического фермента F. heterosporum (СВS 782.83). Аминокислотные последовательности всех пептидов выравнивали путем сопоставления с последовательностями липазы японского штамма F. heterosporum (Nagao et al., 1994, J. Biochem. 116:536-540), в результате чего был выявлен порядок расположения пептидов, идентифицирующих аминокислотную последовательность зрелого белка. Выравнивание проиллюстрировано на фиг.7. Систематизированные результаты исследований с помощью MALDI-TOF и деградации по Эдману представлены в таблице 9, где указан порядок расположения пептидов в соответствии с выравниванием путем сопоставления с последовательностью Nagao.
Таблица 9
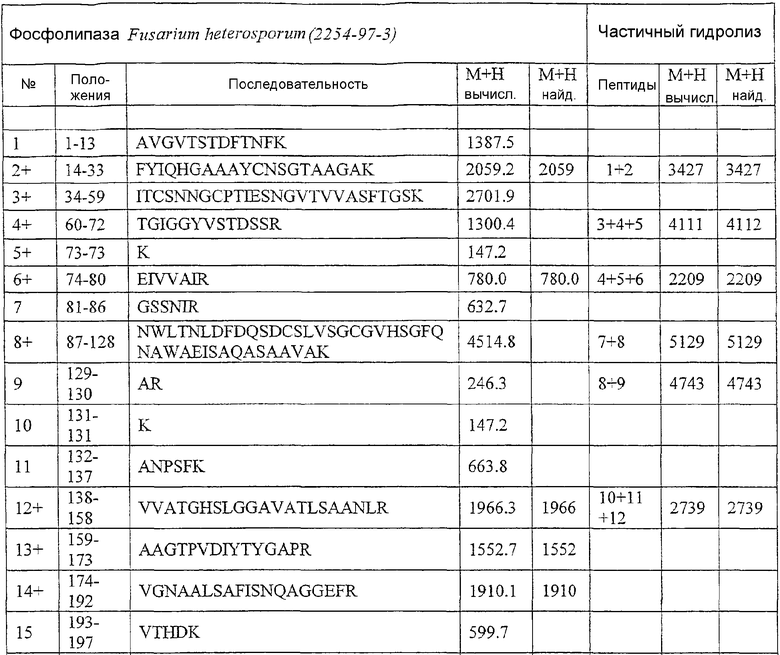

Таблица 9: Ферментативный фингерпринтинг, определение молекулярной массы (Мw) полноразмерной аминокислотной последовательности триптических пептидов липолитического фермента Fusarium heterosporum (СВS 782.83) путем деградации по Эдману. Пептидные последовательности, подтвержденные путем деградации по Эдману, обозначены “+”. Окисленный триптофан обозначен символом *. Доля последовательности = 99,64%.
Сходство с другими липазами Fusarium
В результате выравнивания аминокислотной и нуклеотидной последовательностей липолитического фермента F. heterosporum (СВS 782.83) с последовательностями других липаз Fusarium было выявлено родство между некоторыми липазами Fusarium (таблица 10).
Таблица 10. Определение идентичности липолитического фермента F. heterosporum (СВS 782.83) и других липаз Fusarium путем сравнения их аминокислотных и нуклеотидных последовательностей.
В результате смешивания и ПЦР-амплификации синтетических олигонуклеотидов гена липолитического фермента и альфа-сигнальной последовательности получали ДНК-фрагменты, которые клонировали и секвенировали. Фрагменты, содержащие “правильные” последовательности, использовали для сборки полноразмерного гена путем повторной амплификации с использованием праймеров, представленных в таблице 8. Собранная нуклеотидная последовательность представлена на фиг.8, где ее транслированная аминокислотная последовательность и используемые праймеры показаны стрелками.
ДНК-фрагмент, содержащий конструкцию собранного гена, переносили в вектор экспрессии 5рВ14 Hansenula с использованием встроенных рестрикционных сайтов. Полученная плазмида рВ14-alps.cbss схематически представлена на фиг.9.
Экспрессия активности грибкового липолитического фермента в выбранных клонах
Клоны, прошедшие процесс отбора, анализировали на экспрессию липолитического фермента. 10-микролитровые образцы супернатанта 2-дневных культур инкубировали либо с ДГДГ, либо с лецитином в течение 10 минут, и 10 микролитров этих реакционных смесей анализировали с использованием набора NEFA. Результаты, полученные после выделения одиночных колоний от 3 клонов, представлены на фиг.10.
Была определена аминокислотная последовательность липолитического фермента от штамма Fusarium heterosporum (СВS 782.83), а затем был сконструирован синтетический ген, кодирующий этот липолитический фермент и оптимизированный на экспрессию в Hansenula polymorpha. Ген, кодирующий зрелый фермент, был присоединен к синтетической сигнальной последовательности, происходящей от альфа-фактора размножения дрожжей. Ранее было показано, что комбинация альфа-сигнальной последовательности с промотором FMD вектора рВ14 Hansenula является подходящей для экспрессии липаз Fusarium.
Пример 3: Экспрессия липолитического фермента Fusarium heterosporum СВS 782.83 в Hansenula polymorpha и характеризация продукта в испытаниях с пробной выпечкой
Штамм В14:8-3,8 (DCDK0172) Hansenula polymorpha, содержащий ген, кодирующий липолитический фермент и происходящий от гифомицетов Fusarium heterosporum СВS 782.83, подвергали ферментации в культуре с подпиткой. После ферментации в течение 160 часов фосфолипазная активность достигала 1200 ед/мл. После ферментации получали три продукта, которые затем тестировали. Эти продукты обозначали следующим образом: образец 205, 206 и 209.
Образец 205 липолитического фермента от F. heterosporum, экспрессированного в H. polymorpha, тестировали в экспериментах с выпечкой в лабораторном масштабе. Тесто, полученное в этом эксперименте, анализировали с помощью ГЖХ и ВЭТСХ.
Результаты выпечки, проводимой в лабораторном масштабе, подтвердили явно выраженное улучшение свойств образца 205 липолитического фермента в отношении объема хлеба и структуры мякиша. Анализ липолитического фермента подтвердил высокую гидролитическую активность образца 205 липолитического фермента по отношению к дигалактозилдиглицерид (ДГДГ) с одновременным накоплением дигалактозилмоноглицерида (ДГМГ). При этом, указанный фермент обладал лишь незначительной активностью по отношению к триглицеридам, присутствующим в тесте.
Образцы 206 и 209 анализировали в испытаниях с пробной выпечкой в экспериментальном масштабе, и эти анализы подтвердили хороший хлебопекарный эффект липолитических ферментов, выражающийся в увеличении объема хлеба и улучшении структуры мякиша. Испытание с пробной выпечкой показало, что в процедуре приготовления безопарного теста, образец 206 давал несколько лучший результат, чем образец 209, однако, в этом испытании эти два продукта не сравнивали непосредственно друг с другом и для подтверждения этого результата необходимы дополнительные эксперименты по выпечке.
2. Эксперименты
Ферментация
Микроорганизм
В этом исследовании использовали штамм H. polymorpha, трансформированный плазмидой, содержащей липолитический фермент от F. heterosporum СВS 782.83 и описанный в примере 2. Промотором, используемым в этой конструкции, был промотор формиат-дегидрогеназы от H. polymorpha.
Среда и условия для культивирования
Среда с YNВ-глицерином
Среда, используемая для приготовления инокулята в целях проведения ферментации в биореакторе или для культивирования в шейкерных колбах, содержала 1,7 г/л азотистого основания дрожжей (DIFCO, Detroit, USA, 0335-15-9), 5 г/л (NH4)2SO4, 10 г/л глицерина и 0,1 М 2-[N-морфолино]этансульфоновой кислоты (MES) в качестве буфера. рН доводили до 6,1 (рКа MES) добавлением 4М NaOH (перед автоклавированием). Азотистое основание дрожжей и (NH4)2SO4 стерилизовали путем фильтрации в среду после автоклавирования. Эту среду использовали для культивирования в шейкерных колбах (250 мл среды в шейкерной колбе с полным объемом 500 мл).
Агаровая среда YNB
Среда, используемая для покрытия основных культур (при -80°С в 25% (об/масс.) глицирина), содержала 1,7 г/л азотистого основания дрожжей (DIFCO, Detroit, USA, 0335-15-9), 5 г/л (NH4)2SO4, 10 г/л глицерина и 20 г/л агара (DIFCO, Detroit, USA, 0140-01). Азотистое основание дрожжей и (NH4)2SO4 стерилизовали путем фильтрации в среду после автоклавирования.
Среда YPD
Для ограничения примесей в ферментерах использовали насыщенную среду. Среда содержала 10 г/л дрожжевого экстракта, 10 г/л пептона и 20 г/л глицерина.
Ферментация
В данном исследовании осуществляли три ферментизации: HET0401, HET0402 и HET0410, все эти штаммы описаны выше. Три ферментизации различались композицией формирующей среды и питательной среды. Все другие параметры были одинаковыми во всех трех ферментациях.
Формирующая среда (3 л), используемая для ферментации в 6 л ферментере, содержала: 13,3 г/л NH4H2PO4, 3,0 г/л MgSO4·H2O, 3,3 г/л KCl, 0,3 г/л NaCl, 15 г/л глицирина и 3 мл/л ADD APT® пеногасителя Foamstop Sin 260 (ADD APT Chemicals AG, Helmond, The Netherlands), 1,0 г/л СаСl2·2Н2О, 67 мг/л (NH4)2Fe(SO4)2·6Н2О, 5 мг/л CuSO4·5Н2О, 20 мг/л ZnSO4·7Н2О, 21 мг/л MnSO4·7Н2О и 67 мг/л EDTA), 0,65 мг/л NiSO4·6H2O, 0,65 мг/л CoCl2, 0,65 мг/л H3BO4, 0,65 мг/л KI, 0,65 мг/л Na2MoO4·2H2O), 2 мг/л D-биотина и 0,67 г/л тиаминхлорид-гидрохлорида.
Кроме описанной выше формирующей среды, ферментация HET0402 включала 10 г/л пептона в формирующей среде.
Кроме описанной выше формирующей среды, ферментация HET0410 включала 10 г/л триптона Bacto в формирующей среде.
Питательные среды HET0401 и HET0402:
Питательная среда содержала 635 г/кг глицерина и 130 г/кг муравьиной кислоты.
Питательная среда HET0410:
Питательная среда содержала 570 г/кг глицерина, 120 г/кг муравьиной кислоты и 95 г/кг бакто-триптона.
рН регулировали добавлением 25% (масс./об.) NH3-воды. Ферментацию осуществляли в режиме с подпиткой в бытовом 6-литровом ферментере. Эту ферментацию проводили при следующих условиях: рН 5, аэрация при 1 об.%/мин, температура 26°С и перемешивание при 400-700 об/мин. Ферментер инокулировали 2 × 250 мл культуры, содержащей YNB-глицерин и выращенной при 25°С с перемешиванием 180 об/мин, при этом OD600 составляла приблизительно 10.
Подпитка при ферментации регулировалась выделением аккумулированного CO2 и была вычислена по следующим уравнениям:
Подпитка - поток [г/ч] = 0, AcCO2 < 0,45
Подпитка - поток [г/ч] = 1,33 V AccCO 2 , 0,45 ≤ AccCO 2 ≤ 3,25
Подпитка - поток [г/ч] = 4,33 V, 3,25 ≤ AccCO 2
V: Объем сбраживаемой cреды [л]
AccCO 2 : Выделение аккумулированного СO2 [моль]
Cбор:
Сбраживаемую среду собирали путем центрифугирования в течение 10 минут при 16000 × g, а затем супернатант подвергали стерильной фильтрации через 0,2 мкм-фильтры (фильтрующее устройство VacuCap 90, 0,8/0,2 мкм-мембрана Supor), поставляемые Gelman Laboratory. Полученный продукт выдерживали при 4°С до его использования в экспериментах по пробной выпечке.
Процедуры анализов:
Определение липазной активности:
Образец для ферментации (10 мл) центрифугировали при 9000 × g в течение 10 минут и супернатант использовали для анализа фосфолипазной активности в соответствии с “анализом на PLU”, описанным выше.
Культивирование биомассы:
Концентрацию биомассы в культуральной жидкости определяли путем центрифугирования 10 мл культуральной жидкости при 9000 × g в течение 10 минут в предварительно взвешенном контейнере. После центрифугирования супернатант удаляли и биомассу сушили в течение 24 часов при 100°С, а затем взвешивали. Концентрацию биомассы вычисляли в граммах сухой массы клеток на один литр культуральной жидкости.
Характеризация фермента и выпечка в лабораторном масштабе:
Ферменты и мука:
Образец 205: Образец 7 (161-часовая ферментация) фосфолипазы Lipopan F HET0401, #2938.
Мука: в соответствии со стандартами 2003055.
Выпечка в лабораторном масштабе:
В 50-граммовую дежу Брэбендера для перемешивания и замеса в течение 5 минут при 30°С добавляли следующие ингредиенты: 50 г муки, 10 г сухих дрожжей, 0,8 г сахара, 0,8 г соли, 70 м.д. аскорбиновой кислоты и воду (в количестве, необходимом до доведения консистенции теста до 400 единиц Брэбендера). Время выдерживания составляло 10 минут при 34°С. Тесто отвешивали по 15 г. Затем тесто формовали на специальном устройстве, где его раскатывали между деревянной пластиной и рамой из плексигласа. После этого тесто оставляли на 45 минут в хлебопекарных формах для расстойки при 34°С, а затем выпекали в бытовой печи Voss в течение 8 минут при 225°С. После выпечки хлеб охлаждали до комнатной температуры, а затем оставляли на 20 минут. Хлеб взвешивали и объем определяли методом вытеснения рапсовых семян. Затем хлеб разрезали и оценивали мякиш и корочку.
Экстракция липида
10 г полностью расстоявшегося теста сразу замораживали и лиофилизовали. Лиофилизованное тесто измельчали в кофемолке и пропускали через сито 800 микрон. Лиофилизированное тесто делили по 1,5 г в центрифужной 15 мл-пробирке с завинчивающейся крышкой. Затем добавляли 7,5 мл насыщенного водой бутанола (WSB). Цетрифужную пробирку помещали на 10 минут в баню с кипящей водой. После этого пробирки помещали в роторный миксер Rotamix и вращали при 45 об/мин в течение 20 минут при комнатной температуре. Затем пробирки снова помещали на 10 минут в баню с кипящей водой и вращали в миксере Rotamix в течение 30 минут при комнатной температуре. После этого пробирки центрифугировали при 3500 × g в течение 5 минут. 5 мл супернатанта переносили в сосуд. WSB выпаривали досуха в потоке азота.
Газовая хроматография
Газовую хроматографию проводили как описано выше в процедурах анализа примера 1.
ВЭТСХ
Аппликатор: LINOMAT 5, CAMAG.
ВЭТСХ-пластина: 10 × 10 см, Merck № 1.05633.
Перед использованием пластину сушили в печи в течение 20-30 минут при 180°С.
Нанесение: 1,0 мкл 1% раствора в смеси CHCl3:МеОН, 85:15, наносили на ВЭТСХ-пластину с использованием аппликатора LINOMAT 5.
Рабочий буфер:
№ IV: хлороформ:метанол:H2O (65:25:4)
№ I: петролейный эфир:метил-трет-бутиловый эфир (МТВЕ):уксусная кислота (60:40:1).
Время нанесения/элюирования: 11 минут для рабочего буфера I и 18 минут для рабочего буфера IV.
Пластину сушили в печи при 180°С в течение 10 минут, охлаждали и проявляли в 6% ацетате меди в 16% H3PO4. Затем пластину сушили еще 10 минут при 180°С и сразу оценивали.
Эксперименты по пробной выпечке:
Тестируемые продукты: #3016 - Lipopan F, содержащий 8700 LIPU/г
НЕТ0401+НЕТ0402
НЕТ0410
Рецептура:
Хлебцы с твердой корочкой, приготовленные из муки, соответствующей стандарту: 2003159
Процедура выпекания:
Система размешивания Diosna:
- Медленное сухое размешивание в течение 1 минуты
- Медленное размешивание в течение 2 минут + быстрое размешивание в течение 4 минут
- Температура теста: 26°С
- Отвешивание теста: 1350 г
- Выдерживание: 10 мин при 30°С в нагревательной камере
- Формование: Fortuna 3/17/7
- Расстойка: 45 минут при 34°С, относительная влажность (ОВ) 85%
- Выпечка: Bago-печь: 13 минут, при 220°С, 13 сек - обработка паром + 5 минут - внешнее увлажнение
- Обработка в микроволновой печи с керамическим ярусом: прог. № 1
- После выпекания: охлаждение хлебцев в течение 25 минут, а затем взвешивание и измерение объема.
Результаты и обсуждение
Процесс ферментации и продуцирование фосфолипазы
Добавление триптона к экспериментальной и питательной среде НЕТ0410 приводило к более быстрому продуцированию биомассы и к большему конечному объему биомассы по сравнению с НЕТ0401-0402.
НЕТ0401 и НЕТ0402 были почти идентичными в отношении их фосфолипазной активности, а продуцируемость фосфолипазы в НЕТ0410 была значительно выше.
Сбор
Продукты ферментации собирали через 168 часов (НЕТ0401-0402) и через 161 часов (НЕТ0410) после ферментации. Продукт выдерживали при 4°С до его использования в экспериментах по пробной выпечке. Некоторые продукты НЕТ0401 были названы “образцом 205” и содержали приблизительно 700 PLU-7/мл. Некоторые продукты из НЕТ0401 и НЕТ0402 объединяли и называли “образцом 206”. Этот продукт содержал приблизительно 390 PLU-7/мл. Более низкая ферментативная активность образца 206 по сравнению с конечным продуктом НЕТ0401 и НЕТ0402 может быть обусловлена хранением и фильтрацией в стерильных условиях. Продукт НЕТ0410 был назван “образцом 209” и содержал приблизительно 950 PLU-7/мл.
Характеризация фермента и выпечка в лабораторном масштабе:
Образец 205 липолитического фермента, полученный после ферментации НЕТ0401, тестировали в эксперименте по пробной выпечке в лабораторном масштабе.
Затем добавляли различные дозы и сравнивали с контролем и Lipopan FTM. Удельный объем хлеба, полученный в этой пробной выпечке, указан в Таблице 11. Фотография хлеба представлена на фиг.11.
Результаты выпечки образца 205 подтвердили очень хорошее действие липолитического фермента с точки зрения увеличения объема хлеба и такое действие на объем хлеба было лучше, чем действие фермента Lipopan FTM, используемого в стандартной дозе 40 м.д.
Из фиг.11 также видно, что образец 205 значительно улучшает структуру мякиша и придает хороший цвет.
Полностью расстоявшееся тесто, полученное после проведения такого эксперимента по выпечке, лиофилизовали и экстрагировали насыщенным водой бутанолом, а затем выделенные липиды анализировали с помощью ГЖХ и ВЭТСХ.
ГЖХ-анализ липидов теста (таблица 12) подтвердил гидролитическое действие образца липолитического фермента 205 на дигалактозилдиглицерид (ДГДГ) с одновременным накоплением дигалактозилмоноглицерида (ДГМГ). Воздействие данного фермента на ДГМГ было совсем низким, поскольку при увеличении дозы фермента общее молярное количество ДГДГ (ммоль% = ммоль/100 г лиофилизованного теста) и ДГМГ (ммоль%) оставалось постоянным (фиг.12). Результаты ГЖХ также показали очень низкую активность образца 205 по отношению к триглицериду.
Таблица 12
ГЖХ-анализ липидов теста. СЖК = свободная жирная кислота. МГМГ = моногалактозилмоноглицерид. ДГМГ = дигалактозилмоноглицерид. МГДГ = моногалактозилдиглицерид. ДГДГ = дигалактозилдиглицерид. TГЛ = триглицерид.
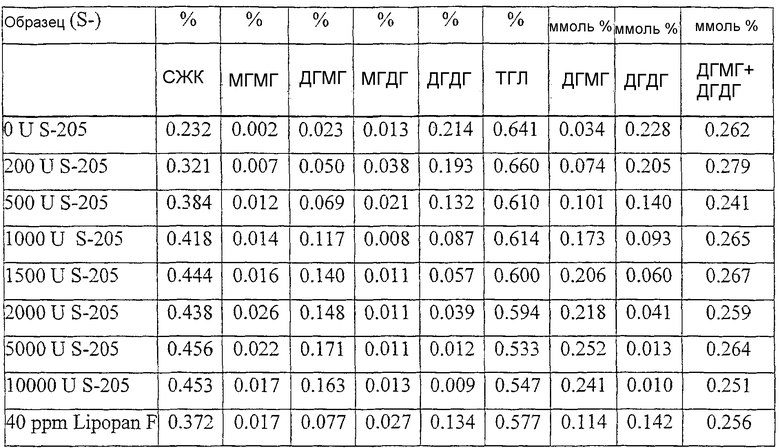
ммоль% = ммоль/100 г лиофилизованного теста.
При сравнении результатов выпечки и анализа липидов было сделано интересное наблюдение, свидетельствующее о том, что наилучший эффект выпечки не достигался при полном гидролизе ДГДГ в ДГМГ, а наоборот, как показали результаты, наилучшие хлебопекарные качества могут быть достигнуты при частичном гидролизе ДГДГ в ДГМГ.
При высокой дозе фермента продуцировалось большее количество ДГМГ, а также большее количество свободной жирной кислоты, которая, как предполагается, оказывает негативное влияние на хлебопекарные свойства, и это может служить другим объяснением, почему предпочтительным является только частичный гидролиз ДГДГ. В таблице 13 указано отношение уровня гидролиза ДГДГ к уровню гидролиза триглицеридов, вычисленное исходя из таблицы 12. Полученные результаты показали, что наилучшие хлебопекарные качества достигаются при дозе, которая дает самое высокое отношение уровня гидролиза ДГДГ к уровню гидролиза триглицеридов.
Некоторые образцы липидов также анализировали с помощью ВЭТСХ, как показано на фиг.13. Образцы 4, 5 и 6 представляют собой липиды теста, полученные в результате эксперимента по выпечке. ВЭТСХ-анализ подтвердил гидролиз ДГДГ и образование ДГМГ под действием образца липолитического фермента 205.
Отношение активности гидролиза полярных липидов к гидролизу триглицеридов для Lipopan F и образца 209, вычисленное с помощью описанных выше анализов, приводится ниже:
Образец 209 = 9
Образец 209 = 4
Липолитический фермент Fusarium heterosporum СВS 782.83 обнаруживал очень сильное действие в экспериментах по пробной выпечке в лабораторном масштабе, то есть способствовал значительному увеличению объема хлеба и улучшению структуры мякиша. Анализ липидов подтвердил его сильное гидролитическое действие на ДГДГ в тесте, сопровождающееся накоплением ДГМГ. При этом липолитический фермент Fusarium heterosporum CBS 782.83 обнаруживал низкую активность по отношению к триглицеридам в тесте.
Пример 4. Оценка активности по отношению к липидным субстратам и специфичности положения липолитического фермента Fusarium heterosporum CBS 782.83, экспрессируемого в Hansenula polymorpha.
Липолитический фермент согласно изобретению, происходящий от Fusarium heterosporum, был экспрессирован в Hansenula polymorpha, как описано в примере 3.
Процедуры анализа
Фосфолипазную активность определяли с помощью анализа на PLU как описано выше.
Галактолипазную активность определяли с помощью анализа на галактолипазу как описано выше.
Активность по отношению к триглицериду (трибутирину) определяли с помощью анализа на LIPU как описано выше.
Активность по отношению к подсолнечному маслу (LUSol, рН-стат, рН 6)
Реагенты:
8,4 г аравийской камеди растворяли в 100 мл деионизованной воды и добавляли 100 мл 30 мМ СаCl2. Затем медленно добавляли 36 г подсолнечного масла, перемешивая в миксере Turrax (20000 об/мин).
Анализ:
20 мл эмульсии подсолнечного масла в лабораторном сосуде уравновешивали при 30°С в течение 5 минут. рН доводили до 6,3-6,5 с использованием рН-стата. Затем добавляли 2 мл раствора фермента и непрерывно в течение 10 минут добавляли 0,05 н. NаОН, поддерживая рН при 6,5. После этого вычисляли угол наклона кривой зависимости добавления 0,05 н. NаОН от времени. 1 LUSol определяли как количество фермента, который может высвобождать 1 мкмоль жирной кислоты за 1 минуту в условиях анализа.
Липолитический фермент анализировали на его активность по отношению к различным субстратам в соответствии с вышеупомянутыми процедурами. Результаты представлены в таблице 14.
Липолитический фермент, происходящий от Fusarium heterosporum и экспрессируемый в Hansenula polymorpha, гидролизует жирные кислоты, главным образом, в положении sn-1 галактолипида и фосфолипидов в тесте. Специфичность фермента исследовали путем добавления различных концентраций фермента в тесто для выпекания хлеба. Полностью расстоявшееся тесто замораживали и лиофилизовали, а затем липиды теста экстрагировали насыщенным водой бутанолом.
Липиды теста анализировали с помощью ГЖХ- и ВЭЖХ-анализа.
С помощью ГЖХ можно проводить анализ на дигалактозилдиглицерид (ДГДГ) и дигалактозилмоноглицерид (ДГМГ). Кроме того, можно проанализировать изомеры положения дигалактозилмоноглицерида (1: дигалактозил-1-моноглицерида и 2: дигалактозил-2-моногилицерида, структурные формулы см. ниже). Эти компоненты разделяли и количественно оценивали с помощью ГЖХ.
1: R1 = Н и R2 = жирный ацил
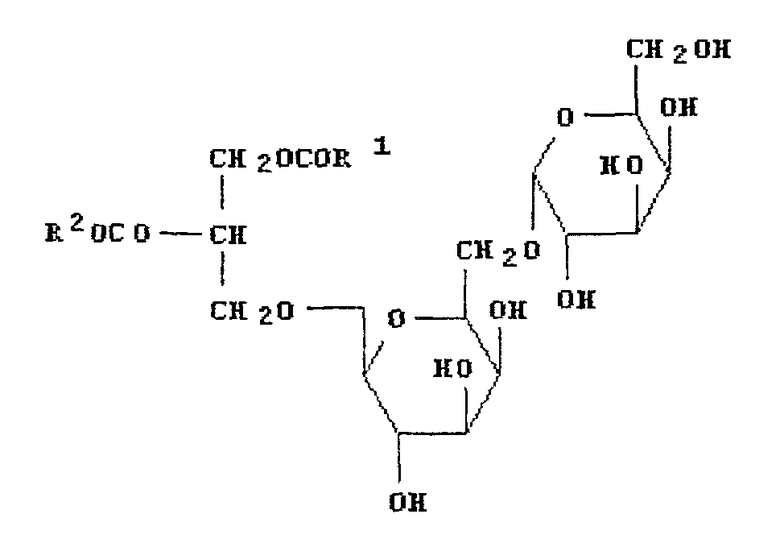
2: R1 = жирный ацил и R2 = Н
В пробной выпечке хлебцев с твердой корочкой добавляли различные дозы липолитического фермента и анализировали уровни галактолипидов в полностью расстоявшемся тесте. Количество изомеров дигалактозилмоноглицерида указано в таблице 15 и графически проиллюстрировано на фиг.14.
TIPU/кг муки
% по сухой массе теста
% по сухой массе теста
Выводы
Исходя из результатов, представленных в таблице 15 и на фиг.14, можно сделать вывод, что дигалактозилдиглицерид гидролизуется, главным образом, в 1-положении во время продуцирования дигалактозил-2-моноглицерида. Также наблюдалось увеличение, но в меньшей степени, количества дигалактозил-1-моноглицерида. Хорошо известно, что миграция ацила жирной кислоты в липидах происходит из 2-положения в 1-положение. Такая миграция ацила зависит от температуры, а равновесие между дигалактозил-2-моноглицеридом и дигалактозил-1-моноглицеридом зависит от времени. Этот феномен также объясняет наблюдаемое небольшое увеличение количества дигалактозил-1-моноглицерида.
Пример 5. Определение оптимальной температуры и стабильности липолитического фермента, происходящего от Fusarium heterosporum
Ферментативную активность осушенного распылением липолитического фермента, происходящего от F. heterosporum и экспрессированного в Hansenula polymorpha, определяли при различных температурах в соответствии с PLU-7 с модификациями, описанными ниже. Субстрат представлял собой эмульсию, состоящую из 0,6% фосфатидилхолина, 0,4% тритона-Х 100, 6 мМ СаCl2 и 50 мМ HEPES, рН 7,0. Осушенный распылением липолитический фермент разводили деминерализованной водой до 3 TIPU/мл. 400 мкл субстрата выдерживали в термостате в течение 5 минут при 10, 20, 30, 40, 45, 50 и 60°С и добавляли 50 мкл образца. Ровно через 10 минут ферментацию завершали путем инкубирования смеси при 99°С в течение еще 10 минут. И наконец, количество свободных жирных кислот определяли методом NЕFA С (Wako Chemicals GMbH, Neuss, Germany). Цветной реагент А и В получали в соответствии с протоколом, рекомендованным производителями. Затем 10 мкл повторно диспергированного экстрагированного липида и 100 мкл реагента А пипетировали в микротитрационный планшет и инкубировали при 37оС в течение 10 минут. После этого в микротитрационный планшет добавляли 200 мкл реагента В и инкубировали при 37оС в течение 10 минут. Затем измеряли оптическую плотность при 540 нм. Количество свободной жирной кислоты определяли путем считывания оптической плотности и строили стандартную кривую по данным, полученным для олеиновой кислоты. Результаты представлены на фиг.15.
Ферментативная стабильность осушенного распылением липолитического фермента определяли при различных температурах. Осушенный распылением липолитический фермент разводили 50 мМ фосфатным буфером, рН 7,0, 3 TIPU/мл. После 30-минутного инкубирования при 20, 30, 37, 40 и 45°С образец хранили на льду. Затем фосфолипазную активность определяли в соответствии с PLU-7 с модификациями, описанными ниже. Субстрат представлял собой эмульсию, состоящую из 0,6% фосфатидилхолина, 0,4% тритона-Х 100 и 50 мМ фосфатного буфера. СаCl2 был удален для предотвращения осаждения фосфата кальция и его влияния на ферментативную активность. 400 мкл субстрата выдерживали в термостате в течение 5 минут при 37°С и добавляли 50 мкл образца. Ровно через 10 минут ферментацию завершали путем инкубирования смеси при 99°С в течение еще 10 минут. И наконец, количество свободных жирных кислот определяли методом NЕFA С (Wako Chemicals GMbH, Neuss, Germany). Цветной реагент А и В получали в соответствии с протоколом, рекомендованным производителями. Затем 10 мкл повторно диспергированного экстрагированного липида и 100 мкл реагента А пипетировали в микротитрационный планшет и инкубировали при 37°С в течение 10 минут. После этого в микротитрационный планшет добавляли 200 мкл реагента В и инкубировали при 37°С в течение 10 минут. Затем измеряли оптическую плотность при 540 нм. Количество свободной жирной кислоты определяли путем считывания оптической плотности и строили стандартную кривую по данным, полученным для олеиновой кислоты. Результаты представлены на фиг.16.
Пример 6. Определение оптимального рН и стабильности липолитического фермента, происходящего от Fusarium heterosporum
Ферментативную активность осушенного распылением липолитического фермента, происходящего от F. heterosporum и экспрессированного в Hansenula polymorpha, определяли при различных рН. Субстрат представлял собой эмульсию, состоящую из 0,6% фосфатидилхолина, 0,4% тритона-Х 100 и 50 мМ фосфатного буфера, рН 4,0, 5,0, 6,0, 6,5, 7,0, 7,5, 8,0, 8,5, 9,0 и 10,0. СаCl2 был удален для предотвращения осаждения фосфата кальция и его влияния на ферментативную активность. Осушенный распылением липолитический фермент разводили деминерализованной водой до 3 TIPU/мл. 400 мкл субстрата выдерживали в термостате в течение 5 минут при 37°С и добавляли 50 мкл образца. Ровно через 10 минут ферментацию завершали путем инкубирования смеси при 99°С в течение еще 10 минут. И наконец, количество свободных жирных кислот определяли методом NЕFA С (Wako Chemicals GMbH, Neuss, Germany). Цветной реагент А и В получали в соответствии с протоколом, рекомендованным производителями. Затем 10 мкл повторно диспергированного экстрагированного липида и 100 мкл реагента А пипетировали в микротитрационный планшет и инкубировали при 37°С в течение 10 минут. После этого в микротитрационный планшет добавляли 200 мкл реагента В и инкубировали при 37°С в течение 10 минут. Затем измеряли оптическую плотность при 540 нм. Количество свободной жирной кислоты определяли путем считывания оптической плотности и строили стандартную кривую по данным, полученным для олеиновой кислоты. Результаты представлены на фиг.17.
Ферментативную стабильность осушенного распылением липолитического фермента определяли при различных рН. Осушенный распылением липолитический фермент разводили 50 мМ фосфатным буфером при рН 4,0, 5,0, 6,0, 6,5, 7,0, 7,5, 8,0, 8,5, 9,0 и 10,0 до 3 TIPU/мл. После 30-минутного инкубирования при 37°С образец хранили на льду. Затем фосфолипазную активность определяли в соответствии с PLU-7 с модификациями, описанными ниже. Субстрат представлял собой эмульсию, состоящую из 0,6% фосфатидилхолина, 0,4% тритона-Х 100 и 50 мМ фосфатного буфера, рН 7,0. СаCl2 был удален для предотвращения осаждения фосфата кальция и его влияния на ферментативную активность. 400 мкл субстрата выдерживали в термостате в течение 5 минут при 37°С и добавляли 50 мкл образца. Ровно через 10 минут ферментацию завершали путем инкубирования смеси при 99°С в течение еще 10 минут. И наконец, количество свободных жирных кислот определяли методом NЕFA С (Wako Chemicals GMbH, Neuss, Germany). Цветной реагент А и В получали в соответствии с протоколом, рекомендованным производителями. Затем 10 мкл повторно диспергированного экстрагированного липида и 100 мкл реагента А пипетировали в микротитрационный планшет и инкубировали при 37°С в течение 10 минут. После этого в микротитрационный планшет добавляли 200 мкл реагента В и инкубировали при 37°С в течение 10 минут. Затем измеряли оптическую плотность при 540 нм. Количество свободной жирной кислоты определяли путем считывания оптической плотности и строили стандартную кривую по данным, полученным для олеиновой кислоты. Результаты представлены на фиг.18.
Пример 7. Определение молекулярной массы очищенного липолитического фермента, происходящего от Fusarium heterosporum
Очищенный липолитический фермент согласно изобретению, происходящий от Fusarium heterosporum, подвергали электрофорезу в ДСН-ПААГ, фиг.19а и 19b, Молекулярную массу вычисляли исходя из стандартного маркера Novex, как показано в таблице 15 А.
Таблица 15 А
Определение молекулярной массы липолитического фермента согласно изобретению
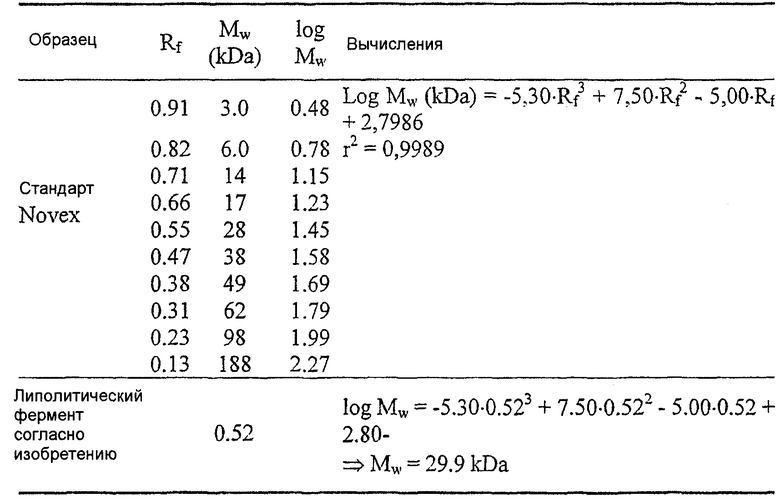
Вычисленная масса липолитического фермента составляла 29,9 кДа.
Пример 8. Определение изоэлектрической точки (рI) липолитического фермента, происходящего от Fusarium heterosporum
Изоэлектрическую точку (рI) липолитического фермента, происходящего от F. heterosporum, теоретически определяли исходя из аминокислотной последовательности SEQ ID NO:6.
Вычисление pI проводили с помощью компьютерной программы Vector NTI Suite 9, поставляемой от Informax (Invitrogen, CA, USA), и ее значение составляло 6,40.
Пример 9: Оценка ферментативного превращения лецитина в лизолецитин в яичном желтке при различных температурах под действием липолитического фермента Fusarium heterosporum CBS 782.83.
Липолитические ферменты могут превращать лецитин (фосфатидилхолин) в лизолецитин (лизофосфатидилхолин) с высвобождением свободной жирной кислоты. Ферментативное превращение лецитина в лизолецитин в яичном желтке приводит к улучшению эмульгирующих свойств продукта, поскольку лизолецитин является лучшим эмульгатором, чем лецитин. Хорошие эмульгирующие свойства яичного желтка имеют важное значение при приготовлении термостабильного майонеза и других пищевых продуктов; при промышленном изготовлении продуктов питания, таких как торты, но не ограничивающихся ими; и при созревании сыра.
Ферментный препарат:
Липолитический фермент, происходящий от Fusarium heterosporum, СВS 782.83, и экспрессированный в Hansenula polymorpha в результате ферментации в среде НЕТ0420, содержащей пшеничный крахмал, сушили распылением. Полученный ферментный препарат имел фосфолипазную активность 1265 Ед/г, определенную в анализе на TIPU, описанном выше. Маточный 10% (масс/об) или 20% (масс/об) раствор фермента получали путем растворения порошка фермента, осушенного распылением, в деминерализованной воде. После перемешивания в течение 15 минут раствор центрифугировали в течение пяти минут при 1370 × g. Супернатант использовали в качестве ферментного маточного раствора.
Ферментация
Для определения оптимальной комбинации дозы фермента, реакционной температуры и времени ферментативного превращения лецитина в лизолецитин яичного желтка проводили два различных эксперимента. В первом эксперименте, ферментацию осуществляли с использованием липолитического фермента согласно изобретению при следующих трех температурах: 30°С, 40°С и 50°С и при каждой температуре использовали четыре дозы фермента: 5 ед/г яичного желтка, 10 ед/г яичного желтка, 20 ед/г яичного желтка и 30 ед/г яичного желтка.
Во втором эксперименте ферментацию осуществляли с использованием липолитического фермента согласно изобретению и Lecitase® Ultra от Nоvozymes A/S (Denmark), соответственно, при следующих пяти температурах: 5°С, 10°С, 15°С, 20°С и 53°С и при дозе фермента 30 ед/г яичного желтка. При температуре 53°С фермент тестировали также при дозе 60 ед/г яичного желтка.
В обоих экспериментах, 10,0 г пастеризованного яичного желтка от Dan/Eg (Christiansfeld, Denmark) переносили в пробирку Уитона и помещали в нагревательный блок, снабженный термостатом для поддержания соответствующей температуры. Образцы непрерывно размешивали магнитной мешалкой. На время t=0 маточный раствор фермента добавляли в яичный желток, как указано в таблице 16. Каждый эксперимент проводили в дубликате. 1,0 г образцов брали из растворов яичного желтка/фермента как указано в таблице 17. После инкубирования в течение периода времени, указанного в таблице 17, ферментативную реакцию в этих образцах прекращали путем добавления 7,5 мл органического растворителя (CHCl3:МеОН, 2:1).
Таблица 17
Время реакции при экстракции образца в различных экспериментах
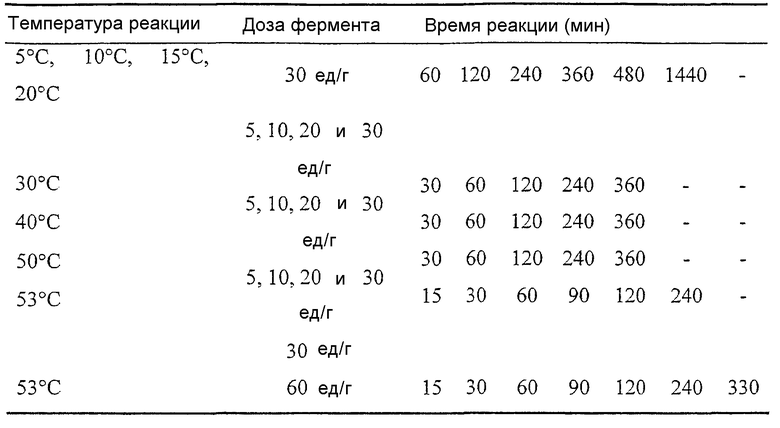
Экстракция липида:
Добавление 7,5 мл органического растворителя (CHCl3:МеОН, 2:1) к образцу не только приводило к прекращению ферментативной реакции, но также позволяло экстрагировать липиды. Кроме того, к образцу добавляли 0,2 мл деминерализованной H2O, а затем образец диспергировали в миксере Whirley в течение 1 минуты. Затем образец центрифугировали в течение десяти минут при 110 × g. Приблизительно 3 мл органической фазы переносили в другую пробирку и экстрагированный липид использовали в различных анализах. Образцы хранили при -18°С.
Определение свободных жирных кислот:
100 мкл раствора экстрагированного липида упаривали в атмосфере азота при 50°С. Затем добавляли 1,0 мл деминерализованной H2O и липид диспергировали в миксере Whirley. Количество свободных жирных кислот определяли с использованием набора NЕFA С от Wako Chemicals GMbH (Neuss, Germany). Цветной реагент А и В получали в соответствии с протоколом, рекомендованным производителями. Затем 10 мкл повторно диспергированного экстрагированного липида и 100 мкл раствора А пипетировали в микротитрационный планшет. Планшет инкубировали при 37°С в течение 15 минут. После этого в микротитрационный планшет добавляли 200 мкл раствора В и планшет инкубировали при 37°С в течение 10 минут. Оптическую плотность измеряли при 540 нм. Количество свободной жирной кислоты определяли путем считывания оптической плотности и строили стандартную кривую по данным, полученным для олеиновой кислоты.
Определение лецитина и лизолецитина с помощью ЖХ/МС-МС:
Материалы
Ацетон, метанол и хлороформ закупали у фирмы Lab Scan, Dublin, Ireland, 96%-ный этанол закупали у фирмы De Danske Spritfabrikker, а муравьиную кислоту закупали у AppliChem, Darmstadt, Germany.
Оборудование
ВЭЖХ-система состояла из четырехсекционного насоса (G1311А), капиллярного насоса (G1376А), устройства для автоматического сборника образцов (G1377А) и колоночной камеры (G1316А), которые были закуплены у Agilent Technologies (Waldbronn, Germany). Для разделения потока, выходящего из колонки в масс-спектрометр, и для введения полярного свежеприготовленного растворителя использовали устройство для разделения потока AcurateTM flowsplitter (ACM-CU-CR) от LC Packings (Amsterdam, Netherlands). Масс-спектрометр представлял собой ионную ловушку LCQ Deca, поставляемую фирмой Thermo Finnigan (San Jose, CA, USA).
Колонка представляла собой Hypersil SI, 100 × 4,6 см, внут. диам., 5 мкм, поставляемая от Thermo Hypersil-Keystone.
Хроматографические и МС-условия
Подвижные фазы:
А: - не использовали.
В: Хлороформ.
С: Смесь: метанол/муравьиная кислота (1000/0,190).
D: Смесь: хлороформ/метанол/вода/муравьиная кислота (300/550/150/0,190).
Свежеприготовленный раствор: 96%-ный этанол.
[мл/мин]
[мин]
[%]
[%]
[%]
Объем впрыска составлял 5 мкл, а температура колонки составляла 45°С.
Разделитель потока
Разделение составляет приблизительно 20:1
Условия МС
Установка параметров МС:
Установка параметров МС-детектора
Стандарт и получение образцов
Лизофосфатидилхолин (ЛФХ) (куриных яиц) (89865) и фосфатидилхолин (ФХ)(растений)(441601) были закуплены у Avanti Polar Lipids, Inc., Alabaster, AL, USA. Были приготовлены маточные растворы ФХ и ЛФХ (10 мг/20 мл CHCl3/МеОН). Эти растворы разводили для получения концентраций от 50 мкг/мл до 2,5 мкг/мл. 7,5 мкл липидного экстракта из 1 грамма яичного желтка разводили в 1,5 мл CHCl3:МеОН (1:1).
ТСХ-анализ:
ТСХ-анализ проводили как описано в примере 1. Для визуализации различных глицеридов 2 мкл липидного экстракта наносили в виде 3 мм-полос на ВЭТСХ-пластину 60 с двуокисью кремния (Merck) с помощью автоматического сборника образцов ТСХ 4 (CAMAG). Пластину с двуокисью кремния помещали в горизонтальную проявляющую камеру (CAMAG) с рабочим буфером I (смесь петролейного эфира:метил-трет-бутилового эфира:уксусной кислоты (50:50:1)). Для газовой фазы использовали 20 мл рабочего буфера, а для сквозного потока использовали 5 мл буфера и пластину элюировали примерно до 5 см от участка нанесения. Затем пластину сушили в нагревательном шкафу (160°С) в течение 5 минут. И наконец, ТСХ-пластину погружали в проявляющий реагент (6% Cu(СН3СОО)2 в 16% водном H3PO4) и обрабатывали двуокисью углерода в нагревательном шкафу (160°С) в течение 10 минут.
Результаты
Для определения оптимальной комбинации дозы фермента, реакционной температуры и времени ферментативного превращения лецитина в лизолецитин в яичном желтке проводили анализ с использованием четырех различных доз фермента при трех различных температурах и при пяти различных периодах времени реакции.
Используемые четыре дозы фермента: 5 ед/г, 10 ед/г, 20 ед/г и 30 ед/г, а также используемое время реакции: 30 минут, 60 минут, 120 минут, 240 минут и 360 минут были выбраны исходя из предварительных испытаний, описанных в настоящей заявке. Три температуры: 30°С, 40°С и 50°С были выбраны исходя из кривой оптимальных температур для липолитического фермента, см. фиг.20.
Количество лецитина и лизолецитина в модифицированном ферментом яичном желтке было определено с помощью ВЭЖХ и проиллюстрировано на фиг.21 и на фиг.22 в зависимости от времени реакции. На фиг.23 количество свободной жирной кислоты в модифицированном ферментом яичном желтке проиллюстрировано в зависимости от времени реакции.
Этот эксперимент показал, что превращение лецитина в лизолецитин под действием липолитического фермента согласно изобретению было оптимальным при дозе липолитического фермента 20 ед/г яичного желтка, при температуре 30°С и времени реакции 120 минут. Выбор дозы 20 ед/г яичного желтка был обусловлен наблюдаемым снижением уровней ЛФХ при дозе 30 ед/г яичного желтка в процессе реакции в период времени от 120 минут до 240 минут.
На основании этого результата был проведен анализ для определения способности липолитического фермента согласно изобретению и лецитазы (Lecitase® Ultra) воздействовать на липиды яичного желтка при температурах ниже 30°С, и для сравнения их активностей при 53°С, то есть температуры, которая в настоящее время применяется для промышленного получения лецитазы (Lecitase® Ultra).
Ферментативное превращение лецитина в лизолецитин яичного желтка тестировали при пяти различных температурах (5°С, 10°С, 15°С, 20°С и 53°С), и при шести различных периодах времени реакции. Фермент был также протестирован при дозе 30 ед/г яичного желтка, поскольку эта доза является самой высокой дозой, представляющей коммерческий интерес с точки зрения стоимости данного фермента, и поскольку предполагаемая скорость реакции при тестируемых температурах является низкой. Все вышеупомянутые количества ферментов были определены c помощью анализа на TIPU. Доза 30 ед/г яичного желтка также является рекомендованной дозой для лецитазы (Lecitase® Ultra). Кроме того, была протестирована доза 60 ед/г яичного желтка при 53°С. Время реакции составляло 60 минут, 120 минут, 240 минут, 360 минут, 480 минут и 1440 минут. Однако при 53°С реакцию проводили 15 минут, 30 минут, 60 минут, 90 минут, 120 минут и 240 минут. При 53°С и при дозе 60 ед/г образец подвергали реакции также в течение 330 минут.
На фиг.24 и 25 количества лизолецитина, свободной жирной кислоты и лецитина в модифицированном ферментом яичном желтке представлены в виде кривых зависимости от времени реакции, проводимой в присутствии липолитического фермента согласно изобретению и фосфлипаз Lecitase® Ultra, соответственно. Содержание лецитина и лизолецитина в образцах определяли с помощью ЖХ-МС, а содержание свободной жирной кислоты определяли методом NЕFA С. Количество СЖК в контрольных образцах (результаты не приводятся) и общее количество лизолецитина и лецитина оставалось постоянным во время проведения экспериментов, проиллюстрированных на фиг.24 и 25.
На фиг.24 приводятся результаты ферментативной обработки яичного желтка липолитическим ферментом согласно изобретению. При дозе фермента 30 ед/г яичного желтка (фиг.24b) и при 53°С активность липолитического фермента прекращалась через 30 минут после начала реакции, при достижении уровня ЛФХ 1,7% (масс/масс). Уровни СЖК составляли 1,0% (масс/масс) и 1,3% (масс/масс) при дозе фермента 30 ед/г и 60 ед/г яичного желтка, соответственно. При использовании фермента Lecitase® Ultra, количества ЛФХ и СЖК увеличивались за период времени 15-240 минут (фиг.19), и через 240 минут после начала реакции, при дозе 30 ед/г яичного желтка, выход ЛФХ составлял 2,7% (масс/масс). При дозе фермента 30 ед/г и 60 ед/г яичного желтка, уровни СЖК через 240 минут после начала реакции составляли 1,4% (масс/масс) и 2,1% (масс/масс), соответственно. Активность Lecitase® Ultra, при дозе фермента 60 ед/г яичного желтка, прекращалась через 330 минут после начала реакции. Липолитический фермент согласно изобретению имел более высокую начальную скорость реакции, чем фермент Lecitase® Ultra.
При 20°С и 53°С начальные скорости этого фермента были аналогичны скорости реакции липолитического фермента согласно изобретению (фиг.24). Во время эксперимента, количество ЛФХ и СЖК повышалось при температурах 5-20°С. При температурах ниже 20°С начальная скорость заметно снижалась по мере снижения температур. Через 60 минут после начала реакции, при 20°С и при дозе фермента 30 ед/г яичного желтка, уровень ЛФХ составлял 3,3% (масс/масс), а уровень СЖК составлял 1,6% (масс/масс). Через 240 минут после начала реакции при 53°С этот уровень был аналогичен уровню, достигаемому при использовании фермента Lecitase® Ultra. При анализе СЖК, через 1440 минут после начала реакции при 20°С было невозможно ресуспендировать липидный экстракт, не содержащий растворителя. Через 1440 минут после начала реакции и при температурах 5°С и 10°С, образцы с липолитическим ферментом имели высокую вязкость, что препятствовало их размешиванию. Наиболее вероятно, что это обусловлено кристаллизацией СЖК. В любом из этих экспериментов, проводимых при 30°С, через 120-240 минут после начала реакции, не наблюдалось снижения уровня ЛФХ, которое наблюдалось в образце ТN 6642 при дозе 30 ед/г яичного желтка.
Ферментация яичного желтка под действием фосфолипазы Lecitase® Ultra приводила к значительному снижению начальной скорости реакции при температуре 20°С и ниже, по сравнению с начальной скоростью реакции фосфолипазы Lecitase® Ultra при 53°С (фиг.25). При 20°С, через 1440 минут после начала реакции при дозе фермента 30 ед/г яичного желтка, уровень ЛФХ составлял 3,0% (масс/масс), а уровень СЖК составлял 1,5% (масс/масс). Этот уровень был аналогичен уровню, достигаемому через 240 минут после начала реакции при 53°С.
На фиг.26 проиллюстрирован ТСХ-анализ липида, экстрагированного из модифицированного ферментом яичного желтка. Этот анализ подтвердил результаты, полученные с помощью ЖХ-МС, и показал, что липолитический фермент согласно изобретению и фосфолипаза Lecitase® Ultra способствовали увеличению количества лизолецитина.
Ферментативная реакция, катализируемая липолитическими ферментами, продуцировала эквивалентные количества лизолецитина и свободных жирных кислот. Возможной и нежелательной побочной реакцией является гидролиз триацилглицеридов. Соотношения между количествами лизолецитина и свободных жирных кислот в процессе ферментативной реакции проиллюстрировано на фиг.27.
При использовании фермента Lecitase® Ultra наблюдалась хорошая корреляция между образованием лизолецитина и эквивалентным образованием свободных жирных кислот (фиг.27). Однако в большинстве образцов, обработанных ферментом Lecitase® Ultra, наблюдалась очень незначительная реакция. Ферментация яичного желтка под действием липолитического фермента согласно изобретению приводила к продуцированию более чем одной свободной жирной кислоты на лизолецитин, при этом уровень образования которого составлял выше 40 мМ, а уровень свободной жирной кислоты составлял выше 60 мМ. Максимальное превращение под действием фермента Lecitase® Ultra составляло 30 мМ для лизолецитина и 25 мМ для свободной жирной кислоты. Образцы, имеющие отношение уровней свободной жирной кислоты и лизолецитина ниже 0,8 или выше 1,2 (n/n) и содержание ЛФХ выше 1,0% (масс/масс), описаны в таблице 18.
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
фермент
Образцы, имеющие отношение уровней свободной жирной кислоты и лизолецитина выше 1,2 (n/n) и содержание ЛФХ выше 1,0% (масс/масс) (фиг.27), или образцы, содержащие менее чем 1,0% ФХ (масс/масс), обычно наблюдались при длительным времени реакции. Липолитический фермент при 15°С давал повышенное отношение уровня свободной жирной кислоты к уровню лизолецитина во всех образцах. Это указывало на то, при более длительной реакции или при низком содержании ФХ, изменялась специфичность фермента к субстрату. Это может быть обусловлено гидролизом фосфатидилэтаноламина, дигалактозилдиацилглицерида или триацилглицеридов, присутствующих в яичном желтке. Отношение уровней свободной жирной кислоты и лизолецитина ниже 0,8 (n/n) и содержание ЛФХ выше 1,0% (масс/масс) в образцах, в основном, наблюдалось при температуре реакции 53°С (таблица 18). Это может быть обусловлено реакцией переэтерификации.
На фиг.28 показано, что липолитический фермент согласно изобретению обладает гидролитической активностью по отношению к триглицеридам и к 1,3-диацилглицеридам при более длительной реакции или при низких концентрациях ФХ. Накопление 1,2-диацилглицеридов указывало на то, что липазная активность является 1,3-специфической. Образование моноглицеридов указывало на то, что фермент Lecitase® Ultra обладал гидролитическим действием на три- или диацилглицериды при 20°С. Поскольку уровни образования ЛФХ значительно отличались, то невозможно было определить, который из ферментов, а именно, липолитический фермент согласно изобретению или фосфолипаза Lecitase® Ultra, обладает наибольшей способностью к гидролизу триацилглицеридов. Снижение дозы фермента и уменьшение времени реакции липолитического фермента может приводить к снижению уровня гидролиза. В таблице 18А указаны время реакции, температура и доза, нанесенная на дорожки 1-30 на фиг.28.
Таблица 18А
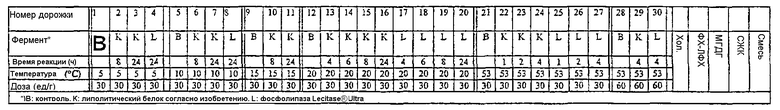
Специалистам в данной области известно, что для любого применения пищевого продукта, оптимальные значения дозы фермента, температуры реакции и времени реакции могут быть легко определены путем рутинного экспериментирования.
Выводы
Ферментацию яичного желтка Dan/Eg А/S липолитического фермента согласно изобретению и фосфолипазы Lecitase® Ultra осуществляли для оценки уровня превращения лецитина в лизолецитин. Эта процедура была проведена в целях определения ферментативной активности с использованием фермента в дозе 30 ед./г яичного желтка при пяти температурах (5-20°С и 53°С) и при шести различных периодах времени реакции (60-1440 минут, а при 53°С, 15-240 минут). В современной промышленности, температуру 53°С обычно используют для модификации яичного желтка ферментом Lecitase® Ultra.
Липолитический фермент согласно изобретению имел более высокую начальную скорость реакции при всех тестируемых температурах, чем фермент Lecitase® Ultra. При 53°С реакция под действием липолитического фермента прекращалась только через 30 минут после начала реакции. При дозе 30 ед/г яичного желтка и при 53°С уровень ЛФX для липолитического фермента и фермента Lecitase® Ultra составлял 1,7 и 2,7% (масс./масс.), соответственно. Уровень ЛФX 3,3% (масс./масс.) под действием липолитического фермента достигался только через 60 минут после начала реакции при температуре 20°С.
При низких температурах (5-20°С) превращение лецитина в лизолецитин под действием липолитического фермента было значительно более эффективным, чем под действием фермента Lecitase® Ultra. Скорость реакции под действием липолитического фермента была значительно меньше при 10°С и ниже по сравнению со скоростью реакции при температуре 15°С и выше. Липолитический фермент был активным при 5°С и через 24 часа после начала реакции наблюдалось образование более чем 2% (масс./масс.) лизолецитина. Кроме того, образцы с липолитическим ферментом были более вязкими при 10°С и ниже по сравнению с образцами, которые были получены при более высоких температурах.
Было обнаружено, что при более длительной реакции или при низком содержании ФX липолитический фермент изменяет свою специфичность к субстрату и, помимо фосфолипидов, гидролизует фосфатидилэтаноламин, дигалактозилдиацилглицерид или триацилглицериды. Такого изменения специфичности можно избежать путем введения меньшей дозы фермента и путем уменьшения времени реакции, что еще раз доказывает необходимость тщательной оптимизации условий обработки каждого рассматриваемого продукта. Реакции переэтерификации, проводимые при 53°С, позволяют установить, что под действием липолитического фермента и фермента Lecitase® Ultra продуцируется менее чем один эквивалент свободной жирной кислоты на лизолецитин.
В итоге, следует отметить, что липолитический фермент согласно изобретению является потенциальным кандидатом для использования при ферментации яичного желтка при низких температурах. Наблюдаемая активность этого фермента при низких температурах также представляет определенный интерес и для других применений.
Пример 10. Приготовление майонеза с использованием липолитического фермента Fusarium heterosporum CBS 782.83.
Приготовление майонеза:
6,25 г липолитического фермента, полученного как описано в примере 4, растворяли в 50 мл деминерализованной H2O, что соответствовало фосфолипазной активности при 150 ед/мл. После перемешивания в течение 15 минут, раствор центрифугировали в течение пяти минут при 1370 × g. Супернатант использовали для ферментации 150 г яичного желтка от Sanofa A/S, как показано в таблице 19. Другие 150 г яичного желтка от Sanofa A/S обрабатывали ферментом Lecitase® Ultra (Novozymes A/S, Denmark), как показано в таблице. Ферментацию проводили при 30°С в течение 180 минут при медленном помешивании. Экстракцию липидов проводили как описано в примере 4.
Таблица 19
Ферментация яичного желтка от Sanofa A/S под действием липолитического фермента согласно изобретению и фермента Lecitase® Ultra, соответственно. Активность раствора липолитического фермента составляла 150 ед/мл, а фосфолипазная активность фермента Lecitase® Ultra составляла 34500 ед/мл.
Майонез, приготавливаемый с использованием модифицированного ферментом яичного желтка от Sanofa A/S, получали путем перемешивания в миксере Koruma (Disho V60/10). В процессе обработки, майонез нагревали до 95°С в течение пяти минут.
контроль
Lecitase® Ultra
ТСХ-анализ:
ТСХ-анализ осуществляли как описано выше.
Определение размера частиц в майонезе:
2,0 г образца майонеза растворяли в 22,5 г 0,2% ДСН и перемешивали минимум 30 минут при 300 об/мин. Затем определяли распределение частиц по размерам на аппарате Mastersizer Malvern.
Результаты:
Для приготовления майонеза с использованием модифицированного ферментом яичного желтка брали яичный желток, поставляемый фирмой Sanofa A/S. Этот яичный желток содержал 8% соли (в отличие от яичного желтка, поставляемого фирмой Dan/Eg, который вообще не содержал соли). Начальные испытания (не приводятся) показали, что более высокая концентрация соли в яичном желтке, поставляемом Sanofa A/S, влияет на липолитическую активность, а поэтому фермент вводили в дозе 30 ед./г, а не 20 ед./г.
ТСХ-анализ липида, экстрагированного из модифицированного ферментом яичного желтка, поставляемого Sanofa A/S (фиг.29), показал, что липолитический фермент согласно изобретению способствовал снижению количества лецитина с одновременным увеличением количества лизолецитина (фиг.29). В отличие от этого, превращение лецитина в лизолецитин под действием фермента Lecitase® Ultra было крайне низким. Высокий уровень превращения лецитина в лизолецитин, обнаруженный с помощью ТСХ, хорошо коррелировал с высоким уровнем свободной жирной кислоты, определенным после экстракции липида из модифицированного ферментом яичного желтка, поставляемого Sanofa A/S (таблица 21). Количество свободной жирной кислоты, высвобождаемое под действием липолитического фермента, в 3,5 раз превышало количество свободной жирной кислоты, высвобождаемой под действием фермента Lecitase® Ultra.
Для оценки эмульгирующих свойств модифицированного различными ферментами яичного желтка, поставляемого Sanofa A/S, проводили гранулометрический анализ капелек масла в майонезе. В результате этого анализа было выявлено, что майонез, приготовленный с использованием яичного желтка, обработанного липолитическим ферментом согласно изобретению, имел наименьший средний размер частиц, и что, в нем также наблюдалось наиболее узкое распределение частиц по размеру по сравнению с майонезом, приготовленным с использованием яичного желтка, обработанного ферментом Lecitase® Ultra, или яичного желтка, не обработанного этим ферментом. Небольшой средний размер частиц, а также узкое распределение частиц по размеру указывали на хорошие эмульгирующие свойства продукта, а следовательно, яичный желток, модифицированный липолитическим ферментом, имел наилучшие эмульгирующие свойства.
Для оценки термостабильности эмульсий, приготовленных с использованием модифицированного ферментом яичного желтка от Sanofa A/S, майонез нагревали в микроволновой печи в течение 4 секунд. Как видно на фиг.30, майонез, содержащий модифицированный ферментом яичный желток, продуцировал термостабильные эмульсии, а контрольная эмульсия, содержащая необработанный яичный желток, разделялась на фазы после ее термообработки в микроволновой печи, а следовательно, эта эмульсия не является термостабильной.
Выводы:
Наблюдалась хорошая корреляция результатов ТСХ-анализа, результатов определения свободных жирных кислот модифицированного ферментом яичного желтка от Sanofa A/S, и результатов анализа на гранулометрический состав и анализа на термостабильность майонеза, приготовленного с использованием модифицированного ферментом яичного желтка Sanofa A/S. В яичном желтке, модифицированном с использованием липолитического фермента согласно изобретению, наблюдалась наибольшая скорость превращения лецитина в лизолецитин и наибольшее количество свободной жирной кислоты. Как и ожидалось, такое изменение отношения лецитин:лизолецитин приводило к получению майонеза, который был термостабильным, и в котором достигалось наиболее оптимальное распределение частиц по размеру.
Использование фермента Lecitase® Ultra для модификации яичного желтка Sanofa A/S не приводило к очень большому изменению отношения лецитин:лизолецитин или к продуцированию большого количества свободных жирных кислот. Такое менее выраженное превращение лецитина в лизолецитин проявлялось в гранулометрическом составе майонеза, который был аналогичен гранулометрическому составу немодифицированного яичного желтка. Изменение отношения лецитин:лизолецитин, которое наблюдалось при использовании фермента Lecitase® Ultra, было вполне достаточным для получения термостабильного майонеза.
Яичный желток от Sanofa A/S, модифицированный липолитическим ферментом в дозе 30 ед/г при 30°С в течение 120 минут, обнаруживал высокий уровень превращения лецитина в лизолецитин, и майонез, приготовленный с использованием такого яичного желтка, был термостабильным и имел оптимальный гранулометрический состав. По сравнению с этим, яичный желток Sanofa A/S, обработанный 30 ед/г фермента Lecitase® Ultra при 30°С в течение 120 минут, обнаруживал лишь небольшое изменение отношения лецитин:лизолецитин, и полученный майонез имел гранулометрический состав, аналогичный гранулометрическому составу майонеза, приготовленного с использованием необработанного яичного желтка, и фактически, был термостабильным. Следовательно, при изготовлении майонеза, липолитический фермент согласно изобретению по своим качествам превосходил фермент Lecitase® Ultra.
Пример 11. Тест на применение липолитического фермента, происходящего от Fusarium heterosporum, в комбинации с эмульгатором для приготовления хлебцев с твердой корочкой
В этом тесте, липолитический фермент согласно изобретению, происходящий от Fusarium heterosporum, использовали отдельно или в комбинации с эмульгаторами Panodan® F2020 DATEM и GRINDSTED® SSL Р55, поставляемыми Danisco A/S, для выпечки хлебцев с толстой корочкой. Влияние этого фермента на удельный объем хлеба сравнивали с влиянием на удельный объем хлеба фермента Lipopan FTM от NovoZymes, взятого отдельно или в комбинации с эмульгаторами.
Применение:
Хлебцы с твердой корочкой выпекали в соответствии с нижеследующей рецептурой и процедурой выпечки.
Процедура выпекания
Размешивание в системе Diosna
1. Медленное сухое размешивание в течение 1 минуты
2. Медленное размешивание в течение 2 минут + быстрое размешивание в течение 4 минут
3. Температура теста: 26°С
4. Отвешивание теста: 1350 г
5. Выдерживание: 10 мин при 30°С в нагревательной камере
6. Формование: в тестоформующей машине Fortuna 3/17/7
7. Расстойка: 45 минут при 34°С, относительная влажность 85%
8. Выпечка: в Bago-печи: 13 минут, при 220°С, 13 сек - обработка паром + 5 минут - внешнее увлажнение
9. Обработка в микроволновой печи с керамическим ярусом: прог. № 1
10. После выпекания: охлаждение хлебцев в течение 25 минут, а затем взвешивание. Объем хлебцев измеряли методом вытеснения рапсовых семян.
Удельный объем хлеба:
Удельный объем хлеба = объем хлеба, см3/масса хлеба, г.
Осушенный распылением липолитический фермент добавляли в муку. Этот фермент добавляли в муку после предварительного смешивания с водой, аскорбиновой кислотой и с прессованными дрожжами. Все другие сухие ингредиенты смешивали в стадии 1.
Результаты
Осушенный распылением липолитический фермент, происходящий от Fusarium heterosporum, использовали в комбинации с эмульгатором Panodan® М2020 DATEM от Danisco A/S и тестировали по сравнению с комбинацией Lipopan FTM/DATEM, а также с чистым Lipopan FTM или чистым DATEM. Результаты приводятся в таблице 23 и на фиг.31.
Таблица 23
Лиофилизованный липолитический фермент, происходящий от Fusarium heterosporum, использовали в комбинации с эмульгаторами Panodan® А2020 DATEM и GRINDSTED® SSL Р55, и тестировали по сравнению с комбинацией Lipopan FTM/SSL или Lipopan FTM/DATEM, а также с чистым Lipopan FTM, чистым DATEM и чистым SSL.
Результаты приводятся в таблице 24 и на фиг.32.
Таблица 24
Выводы:
Исходя из данных, представленных в таблице 23 и на фиг.31, можно сделать вывод, что оптимальная доза осушенного распылением липолитического фермента согласно изобретению составляет приблизительно 383 м.д. и что этот продукт может быть использован в низких дозах в комбинации с низкими дозами эмульгатора DАТЕМ. В параллельном эксперименте было показано, что при дозах от 574 м.д. до 1912 м.д. липолитического фермента наблюдается снижение удельного объема хлеба (данные не приводятся). Продуктивность дозы 383 м.д. липолитического фермента согласно изобретению аналогична продуктивности дозы 40 м.д. Lipopan FTM. Использование липолитического фермента согласно изобретению в комбинации с эмульгатором также дает уровень эффективности, достигаемой с использованием Lipopan FTM. В соответствии с определением фосфолипазной активности, достигаемой в анализе на TIPU, описанном выше, 10 м.д. Lipopan FTM соответствует приблизительно 120 TIPU на кг муки, а 96 м.д. липолитического фермента согласно изобретению соответствует 117 TIPU на кг муки.
Кроме того, на основании результатов испытаний, представленных в таблице 24 и на фиг.32, авторы пришли к выводу, что липолитический фермент согласно изобретению может быть использован в комбинации с SSL и с DАТЕМ, а поэтому он может усиливать действие эмульгаторов, используемых в небольших количествах. Такая функциональность липолитического фермента согласно изобретению может быть сравнимой с функциональностью Lipopan FTM при введении равных доз. Как было определено, оптимальный уровень фосфолипазной активности в комбинации с эмульгатором составляет приблизительно 100-150 TIPU/кг муки.
Таким образом, оптимальная доза чистого липолитического фермента согласно изобретению составляет примерно 500 TIPU на кг муки, а в комбинации с эмульгатором уровень липолитического фермента должен составлять 1/5 - 1/4 от оптимального уровня липолитического фермента, что примерно соответствует 120 TIPU на кг муки.
Пример 12. Тест на применение липолитического фермента, происходящего от Fusarium heterosporum, для приготовления пшеничных лепешек
Влияние липолитического фермента согласно изобретению, происходящего от Fusarium heterosporum, на раскатываемость пшеничной лепешки, изготавливаемой с использованием фумаровой кислоты (методика США), оценивали как описано в нижеследующем примере.
Пшеничные лепешки выпекали с использованием ингредиентов, указанных в таблице 25.
Процедура приготовления теста для пшеничных лепешек:
1. Желательная температура теста: 32°С.
2. Замешивание теста проводят при комнатной температуре в миксере Кемпера.
3. Все сухие ингредиенты помещают в дежу миксера (включая, но необязательно, липолитические ферменты и/или эмульгаторы).
4. Смесь сушат в течение 1 минуты.
5. Добавляют воду.
6. Смешивание: 11 минут при скорости 1.
7. Отвешивание теста: 1350 г × 3.
8. Придание формы: получают шарики теста на тестоделительной/тесто-округлительной машине Glimek.
9. Выдерживание в течение 10 минут при 32°С.
10. Выпекание: в печи для выпечки лепешек CFO 40, при следующих температурных режимах: в верхней части печи: 230°С, в средней части: 228°С и в нижней части: 160°С.
11. Охлаждение: 12 минут при 20°С, относительная влажность - 80%.
Липолитический фермент согласно изобретению добавляли в тесто при повышенных концентрациях (испытания № 3-7). Для сравнения включали контроль (испытание № 1) и проводили испытание с использованием эмульгатора Panodan® 205 Danisco A/S (испытание № 2). См. таблицу 26.
Липолитический фермент, Panodan® 205 и L-цистеин, если его использовали, добавляли в первой процедуре смешивания (стадии 3 и 4, описанные выше). L-цистеин может быть добавлен для повышения растяжимости приготовленного теста и, тем самым, для облегчения прессования теста перед его выпеканием.
Испытание №
Лепешки оценивали в испытании на холодное раскатывание при комнатной температуре, где эти лепешки раскатывали различными деревянными скалками различных диаметров, начиная с деревянной скалки самого большого диаметра. Раскатываемость соответствует номеру деревянной скалки, которая позволяет раскатывать тесто лепешки без его разлома. Чем выше номер, тем лучше раскатываемость.
Исходя из полученных результатов, авторы пришли к выводу, что доза липолитического фермента согласно изобретению, составляющая 200 м.д. или более, улучшает раскатываемость теста по сравнению с контрольной системой. С помощью анализа на TIPU, описанного выше, было определено, что уровень активности, необходимой для улучшения раскатываемости теста (в дозе 200 м.д.), соответствует приблизительно 650 единицам TIPU на кг муки. Исходя из этих результатов можно также сделать вывод, что нагрузка для проведения теста на продавливание повышается с увеличением уровня липолитического фермента, а это означает, что повышается устойчивость этой лепешки. Тест на продавливание осуществляют с использованием анализатора консистенции ТАХТ2, изготовленного Stable Micro System, с помощью которого измеряют нагрузку, необходимую для продавливания/разлома лепешки.
Это оборудование имеет следующие параметрические настройки:
Нагрузку измеряют путем сжатия
Пример 13. Молекулярное клонирование, анализ последовательностей и гетерологичная экспрессия синтетического гена, кодирующего липолитический фермент от Fusarium semitectum (IBT9507) в Hansenula polymorpha
Фрагмент гена липолитического фермента F. semitectum клонировали из геномной ДНК посредством ПЦР с использованием праймеров, сконструированных из консервативных блоков аминокислот, в выравниваемых последовательностях белка липолитических ферментов, происходящих от различных штаммов Fusarium. Вырожденные ПЦР-праймеры конструировали с помощью компьютерных программ CODEHOP (Rose et al., 2003 (Nucleic Acid Res., 18:3763-3766).
Для клонирования концов гена применяли методы конструирования 5'-и 3'-RACE (Frohman et al. 1988, Proc. Natl. Acad. Sci. USA 85:8998-9002). Полноразмерную РНК выделяли из культуры штамма F. semitectum, индуцированной 1%-ным подсолнечным маслом, и используемые праймеры конструировали на основе последовательности генного фрагмента, полученной с использованием праймеров CODEHOP.
Для идентификации полноразмерной кДНК-последовательности конструировали три фрагмента in silico, полученных в соответствии с вышеописанными процедурами. Анализ кДНК-последовательности длиной в 1236 нуклеотидов выявил открытую рамку считывания, содержащую 352 аминокислоты (фиг.33).
Для экспрессии гена липолитического фермента F. semitectum в Hansenula, к этому гену присоединяли сигнальную последовательность α-фактора размножения дрожжей и встраивали позади FMD-промотора в вектор экспрессии рB14 Hansenula. Полученную плазмиду, рВ14-alp.sem (схематически представленную на фиг.34) трансформировали в компетентные клетки Hansenula polymorpha путем электропорации. Трансформанты отбирали на планшетах с YND и колонии снова отбирали на множественную интеграцию гена путем проведения 10 пассажей при разведениях 1:200 в жидких культурах с YND. И наконец, отобранные культуры два раза переносили в среду с YPD.
Для определения уровня экспрессии гена липолитического фермента, отобранные клоны культивировали в YPD с 1,8% глицерином и 0,2% глюкозой в течение 2 дней при 37°С.
Пример 14. Определение рН и температуры, оптимальных для достижения активности липолитического фермента Fusarium semitectum.
Липолитический фермент согласно изобретению, выделенный из Fusarium semitectum IBT9507 и экспрессированный в Hansenula polymorpha, как описано в примере 8, использовали в функциональных анализах в тестовой взвеси для определения фосфолипазной и галактолипазной активности и проводили исследование активности этого фермента с точки зрения изменения рН и температуры.
Процедуры анализа
Газовая хроматография
Пшеничную муку отвешивали в количестве 0,8 грамма в центрифужную 12 мл-пробирку с крышкой. Затем добавляли 1,5 мл воды, содержащей фермент. Образец перемешивали в миксере Whirley и помещали на 60 минут в нагревательную камеру при 30°С. Затем добавляли 6 мл смеси н-бутанол:этанол, 9:1 и образец снова размешивали до тех пор, пока мука не образовывала тонкодисперсную суспензию в растворителе. Затем пробирки помещали на 10 минут в водяную баню при 95°С. После этого смесь снова перемешивали и помещали в роторный смеситель на 45 минут при 45 об/мин. Затем образец центрифугировали при 2000 × g в течение 10 минут, и 2 мл супернатанта переносили в 10 мл-сосуд из драмского стекла. Растворитель выпаривали при 70°С в потоке азота. Выделенные липиды анализировали с помощью ГЖХ.
Газовую хроматографию и анализ на галактолипазную активность осуществляли как описано в примере 1.
Оптимальная температура
Фосфолипазная активность
Для определения активности в зависимости от температуры анализ на фосфолипазную активность проводили, как описано в примере 1, но при температурах 30°С, 37оС, 45°С, 52°С или 60°С.
Оптимальный рН
Фосфолипазная активность
Для определения активности в зависимости от рН анализ на фосфолипазную активность проводили, как описано в примере 1, за исключением того, что 0,6% L-α-фосфатидилхолина, 95% растительного экстракта (Avanti #441601) и 0,4% тритона-Х 100 (Sigma X-100) растворяли в 0,05М фосфатном буфере HEPES при рН 5, рН 6, рН 7, рН 8 или рН 9.
Результаты
Липолитический фермент согласно изобретению, происходящий от Fusarium semitectum IBT9507, анализировали на фосфолипазную активность PLU-7 и галактолипазную активность GLU и результаты этого анализа приводятся в таблице 27.
Таблица 27
Ферментативная активность Fusarium semitectum
Fusarium semitectum IBT9507 анализировали в экспериментах, проводимых на тестовой взвеси путем добавления 1 PLU-7 к 0,8 грамма муки в соответствии с вышеупомянутой процедурой. Также получали контрольный образец, содержащий воду вместо фермента, и образец, содержащий Lipopan FTM. Липиды, экстрагированные из теста, анализировали с помощью ГЖХ и результаты этого анализа приводятся в таблице 28.
Результаты, представленные в таблице 28, показали, что, по сравнению с ферментом Lipopan FTM, липаза, выделенная из F.semitectum, обладает значительной активностью по отношению к галактолипидам и относительно небольшой активностью по отношению триглицеридам.
Fusarium semitectum IBT9507 также анализировали на активность в зависимости от температуры (таблица 29) и рН (таблица 30).
Активности, представленные в таблицах 29 и 30, также графически проиллюстрированы на фиг.35 и 36.
Выводы:
Было показано, что липолитический фермент согласно изобретению, происходящий от Fusarium semitectum, обладает очень сильной активностью по отношению к галактолипидам теста, а его активность по отношению к триглицериду меньше, чем активность по отношению к триглицериду у Lipopan FTM. Оптимальная температура для достижения активности этого фермента составляет приблизительно 45°С, а оптимальный рН составляет 7.
Пример 15: Использование липолитического фермента согласно изобретению при изготовлении корма для животных
Эффективность липолитического фермента согласно изобретению оценивали при различных дозах, используемых для приготовления стандартного корма, применяемого в течение всего периода выращивания цыплят-бройлеров.
Выводы:
Предварительные результаты позволяют предположить, что добавление липолитического фермента согласно изобретению в корм для цыплят-бройлеров представляет собой эффективный метод откармливания, способствующий улучшению качественных показателей птицы, более длительному сохранению микроэлементов и снижению выделения азота. Более конкретно, результаты предварительных исследований позволяют предположить, что добавление липолитического фермента согласно изобретению в корм для животных способствует увеличению массы тела, эффективному усвоению пищи и метаболизму сухих веществ и азота у животного.
22-42 дня, 9 обработок
32-42
Состав корма и режим кормления
Корм приготавливали в виде комбикорма в присутствии или в отсутствии липолитического фермента согласно изобретению.
Корм и воду давали по желанию (ad libitum). Контрольный корм давали в течение всего периода испытания. В образцы корма добавляли, но необязательно, липолитический фермент согласно изобретению в количестве 330 г/тонну. Этот фермент может быть добавлен при размешивании корма в виде сухого вещества.
Наблюдение проводили при следующих параметрах:
(в расчете на массу клетки):
Определяли общую массу клетки с кормом и птицей, а также общую массу погибших особей и число птиц в каждой клетке за период анализа. Потребление корма на одну клетку определяли без учета смертности. Данные эффективности конверсии корма определяли как общее потребление корма на живую массу и общую массу (включая массу погибших особей).
До начала исследования животных оценивали на наличие признаков заболевания и повреждения. Любую особь, у которой наблюдались признаки заболевания, исключали из исследования.
Исследуемых животных распределяли по группам обработки произвольным образом. Перед началом введения тестируемого корма животных и клетки для их содержания индивидуально помечали.
Данные, полученные для групп обработки, сравнивали с данными для релевантной контрольной группы путем проведения соответствующих статистических анализов, где уровень вероятности менее чем 0,05 принимался как показатель значимости.
Массу тела, потребление корма и конверсию корма анализировали путем оценки дисперсии и критериев минимального значимого различия.
Корм/содержание
Проводимые измерения основных параметров
фермент (1)
Пример 16: Оценка влияния липолитического фермента, происходящего от Fusarium heterosporum СВS 782.83, на качество лапши быстрого приготовления, изготовленной из китайской муки
Введение
За последние 5-8 лет в странах Юго-Восточной Азии и, до некоторой степени, в странах Европы и в США рынок продаж лапши быстрого приготовления (БП) достиг феноменального роста. Этот рост очевиден даже в регионах традиционного употребления продуктов из риса и/или пасты (Food Naviganor, 2000). Такая популярность лапши БП в последнее время, главным образом, обусловлена очень доступной ценой, а также удобством приготовления и легкостью процедуры ее приготовления.
Мука со средним содержанием белка (9-11%), низкой величиной зольности (~0,50%), высокой белизной L* (85) и желтизной b* (>8,0) и высокой вязкостью крахмального клейстера (<750BU) позволяет получить лапшу быстрого приготовления (БП), имеющую кремовый/желтый цвет и желательные органолептические свойства. Существует несколько различных типов имеющейся в продаже лапши, каждый из которых изготавливается из муки определенного сорта, что влияет на качество готового продукта.
Хотя в настоящее время рынок насыщен готовыми продуктами и может удовлетворять желания большого числа потребителей, однако, мукомольная промышленность пока не может в полной мере удовлетворить требования каждого конкретного потребителя. Специально разработанные ингредиенты и добавки, при их введении в соответствующих дозах, играют очень важную роль в улучшении вкуса, консистенции и внешнего вида, а также в увеличении срока хранения и/или улучшения питательной ценности готового продукта.
Хотя важность цвета и консистенции приготавливаемой лапши БП не может быть недооценена, однако, в последнее время, потребители становятся все более разборчивыми и больше заботятся о своем здоровье, а поэтому необходимо изготавливать альтернативные продукты с низким содержанием жира, которое не влияло бы на качество такого продукта.
Был проведен анализ липолитического фермента согласно изобретению, содержащегося в китайской муке, для оценки его влияния на содержание жира в лапше БП и для исследования изменения консистенции и цвета продукта в процессе его приготовления.
Материалы и методы
В этой программе исследований используется стандартная процедура изготовления лапши БП (Agrifood Technology) и обобщенный метод оценки. В качестве контроля используют китайскую муку, и новую партию муки получают каждый день. Содержание белка, влажность, зольность, цвет, сырую клейковину и диастатическую активность китайской муки измеряли с применением апробированных методов ААСС (Американская Ассоциация химиков-клиницистов). Реологические свойства теста определяют по фаринограмме, экстензограмме (45-минутному растяжению теста), альвеограмме и амилограмме.
Ниже приводится краткое описание изготовления лапши БП.
Каждую партию лапши БП изготавливают из 350 г муки и смешивают при низкой скорости, постепенно добавляя 33 части водного солевого раствора, содержащего 1% хлорид натрия и 0,2% соли щелочных металлов (карбонат калия:карбонат натрия в отношении 6:4). Для взятия доз образцов муку тщательно смешивают с измеренным количеством ингредиента, а затем добавляют водный солевой раствор.
Крошковатое тесто перемешивают еще 4 минуты при средней скорости и раскатывают 8 раз. Раскатывание начинают с помощью стального уплотнителя, а затем с помощью двух пластиковых рифленых вальцов, и наконец, с помощью пяти гладких вальцов из нержавеющей стали, при этом расстояние между вальцами снижалось на 30%. Толщина конечной тестовой ленты составляет 1,35 мм. Перед разрезанием тестовую ленту раскатывают еще один раз. При этом создается дифференциал скоростей между режущими вальцами и конвейерной лентой, что приводит к образованию тугих прядей лапши. Тугие пряди лапши обрабатывают паром в течение двух минут и обжаривают в пальмовом масле с обеих сторон при 180°С в течение 1 минуты. Блоки лапши охлаждают и упаковывают в герметично закрывающиеся пакеты для последующих анализов.
Образцы для анализа собирают в различных стадиях процесса изготовления. Цвет и размер частиц мякиша определяют с помощью хромометра Minolta и штангенциркуля с нониусом, соответственно. Цвет тестовой ленты и конечного продукта регистрируют с помощью хромометра Minolta и цифровой фотокамеры (фото не приводится). Измерения смачивающей активности проводят на обработанной паром лапше. Смачивающая активность может быть измерена путем определения массы обработанной паром лапши непосредственно после обработки паром и после полного удаления воды путем сушки в печи при 90°С, при этом содержание воды может быть определено путем деления разности масс до и после сушки на массу после сушки.
Оптимальное время варки, выход продукта, потери при варке (гравиметрический метод), цвет и консистенцию (твердость) сваренной лапши определяют стандартными методами Agrifood Technology, известными специалистам в данной области. Сваренную лапшу также анализируют на профиль консистенции (TPA) для определения слипаемости, упругости и разжевываемости.
Параметр “слипаемость” позволяет оценить степень устойчивости данного продукта ко второй деформации по сравнению с его поведением после первой деформации. Слипаемость измеряют как рабочую площадь во время второго уплотнения, деленную на рабочую площадь во время первого уплотнения, а поэтому эта величина не имеет единицы измерения. В данном случае, термин “слипаемость” означает степень прилипания продукта к зубам, “al-dente”, которое является нежелательным показателем для лапши БП.
Упругость означает физическую способность продукта принимать свою прежнюю форму после того, как он был подвергнут деформации в процессе первого уплотнения. Упругость измеряют несколькими способами, но чаще всего ее измеряют как высоту продукта после второго уплотнения.
Разжевываемость относится только к твердым продуктам и вычисляется как величина клейкости, умноженная на величину упругости. Термины “разжевываемость” и “клейкость” являются взаимоисключающими понятиями.
Один блок лапши, представляющий каждую дозу, измельчают в кофемолке и гомогенный субобразец используют для анализа на содержание жира методом кислотного гидролиза (могут быть использованы и альтернативные стандартные методы определения содержания жира).
Результаты и обсуждение
Содержание белка и цвет (с точки зрения белизны, L*) муки имели допустимые пределы, обычно принятые в производстве лапши быстрого приготовления. Поглощение воды слегка превышало верхний предел, принятый в производстве лапши БП, однако, поскольку тесто для лапши было крошащимся, то это не влияло на машинабельность теста.
Мука имела хорошую одноосную (экстензограмма) и двухосную (альвеограмма) растяжимость, которая оказывала положительное влияние на пищевые качества лапши. Пик вязкости на амилограмме составлял 879 единиц вязкости по Брукфилду (ед/Б), которая является хорошим показателем для лапши БП.
Потери при варке лапши БП, содержащей вторую самую высокую дозу липолитического фермента, были несколько выше, чем у контроля и у лапши БП, содержащей наименьшее количество липолитического фермента согласно изобретению. Содержание жира в лапше БП с наибольшим количеством липолитического фермента было значительно меньше, чем в контрольной лапше и в экспериментальной лапше БП с наименьшим количеством липолитического фермента. Упругость и разжевываемость некоторых экспериментальных образцов БП были лучше, чем у контрольных образцов. Исходя из этих данных, очевидно, что необходимы дополнительные исследования липолитического фермента при различных дозах.
Выводы
На основании проведенных исследований могут быть сделаны некоторые основополагающие выводы:
добавление липолитического фермента согласно изобретению в лапшу БП не оказывало значительного влияния на гранулометрический состав мякиша, липкость теста, его машинабельность или технологические свойства. Важно отметить, что увеличение доз липолитического фермента приводит к снижению содержания жира в лапше БП. Увеличение дозы липолитического фермента приводит к увеличению плотности лапши по сравнению с контролем, но не влияет на слипаемость лапши. Липолитический фермент оказывает положительное действие в отношении желтизны приготовленной лапши.
Таким образом, липолитический фермент снижает содержание жира в лапше БП, улучшает консистенцию и усиливает желтизну приготовленной лапши.
Все упомянутые выше публикации вводятся в настоящее описание посредством ссылки. Следует отметить, что в описанные методы и системы согласно изобретению могут быть внесены различные модификации и изменения, не выходящие за рамки и объем изобретения. Хотя настоящее изобретение описано на конкретных предпочтительных вариантах его осуществления, однако, следует иметь ввиду, что эти варианты не должны рассматриваться как ограничение заявленного изобретения. Само собой разумеется, что различные модификации описанных способов осуществления изобретения, которые являются очевидными для специалистов в области биохимии и биотехнологии или специалистов в родственных областях науки, входят в объем прилагаемой формулы изобретения.








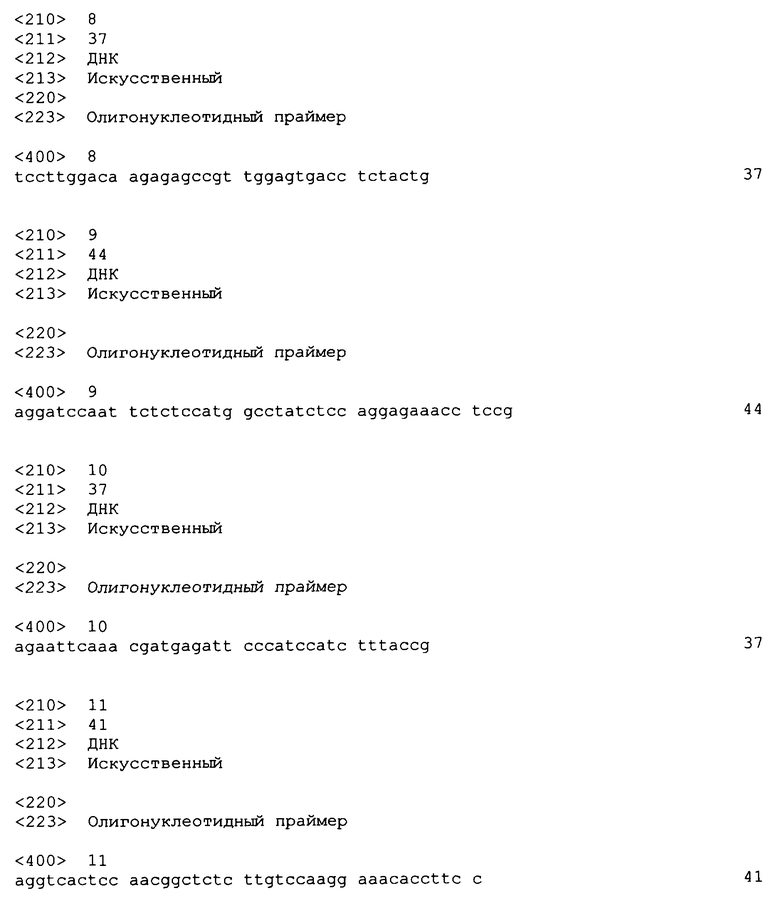
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ | 2004 |
|
RU2376868C2 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАРИАНТА ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА И ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ (ВАРИАНТЫ) | 1999 |
|
RU2235775C2 |
| СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА УГЛЕРОДА, СЛОЖНОГО ЭФИРА БЕЛКА, СЛОЖНОГО ЭФИРА БЕЛКОВОЙ СУБЪЕДИНИЦЫ ИЛИ СЛОЖНОГО ЭФИРА ГИДРОКСИКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЛИПИДАЦИЛТРАНСФЕРАЗЫ | 2004 |
|
RU2371474C2 |
| БЕЛКИ | 2004 |
|
RU2518345C2 |
| ВАРИАНТЫ ГЛИКОЛИПИДАЦИЛТРАНСФЕРАЗЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 2004 |
|
RU2377300C2 |
| ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ ДЛЯ ПРИМЕНЕНИЯ В ХЛЕБОПЕЧЕНИИ | 2018 |
|
RU2763469C2 |
| ФЕРМЕНТАТИВНАЯ ОБРАБОТКА МАСЕЛ | 2004 |
|
RU2377307C2 |

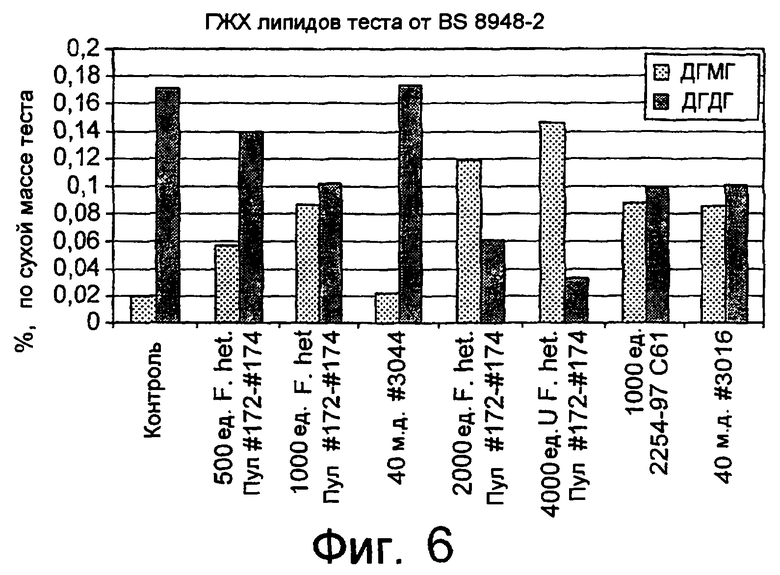
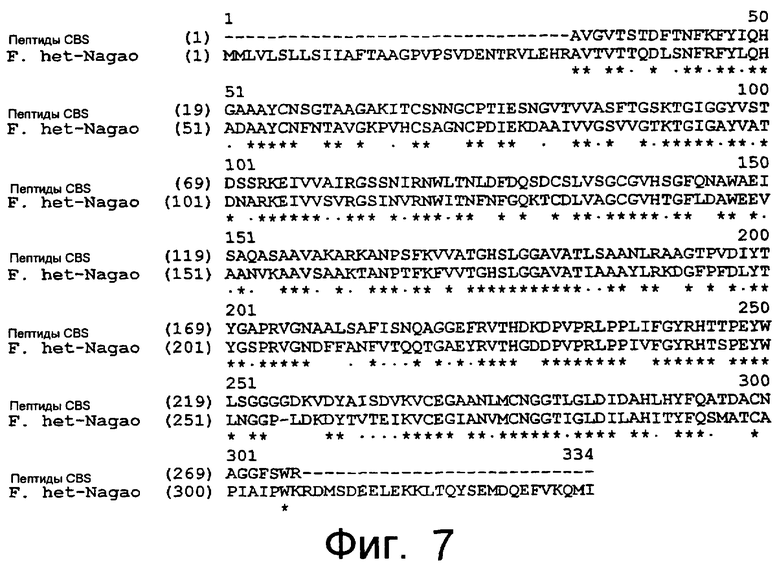

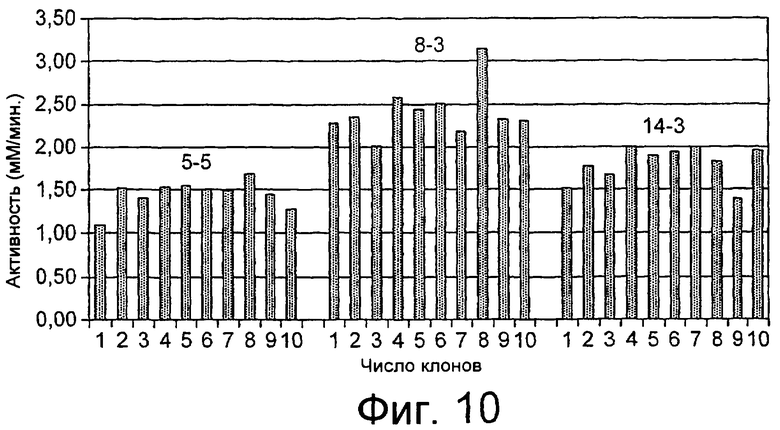
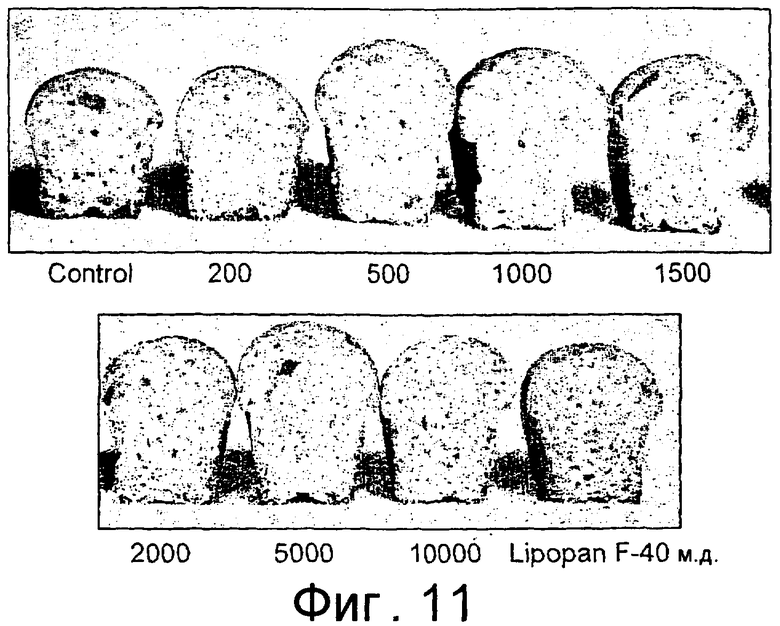
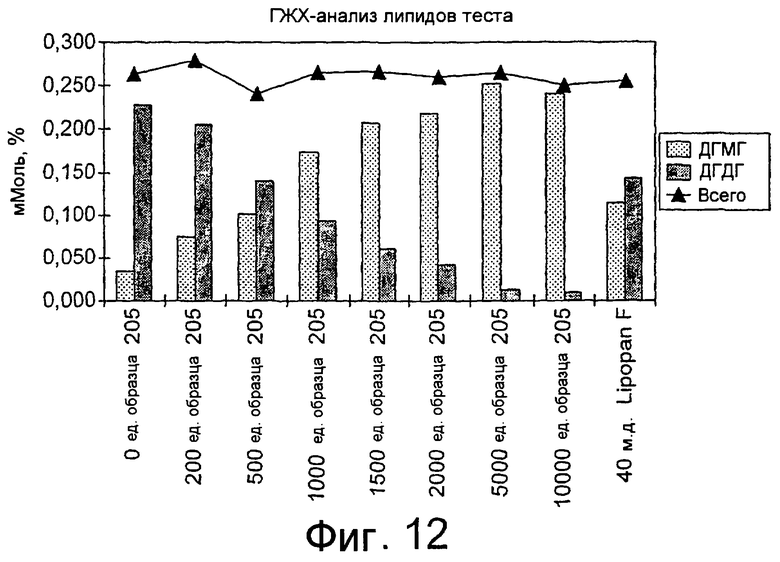
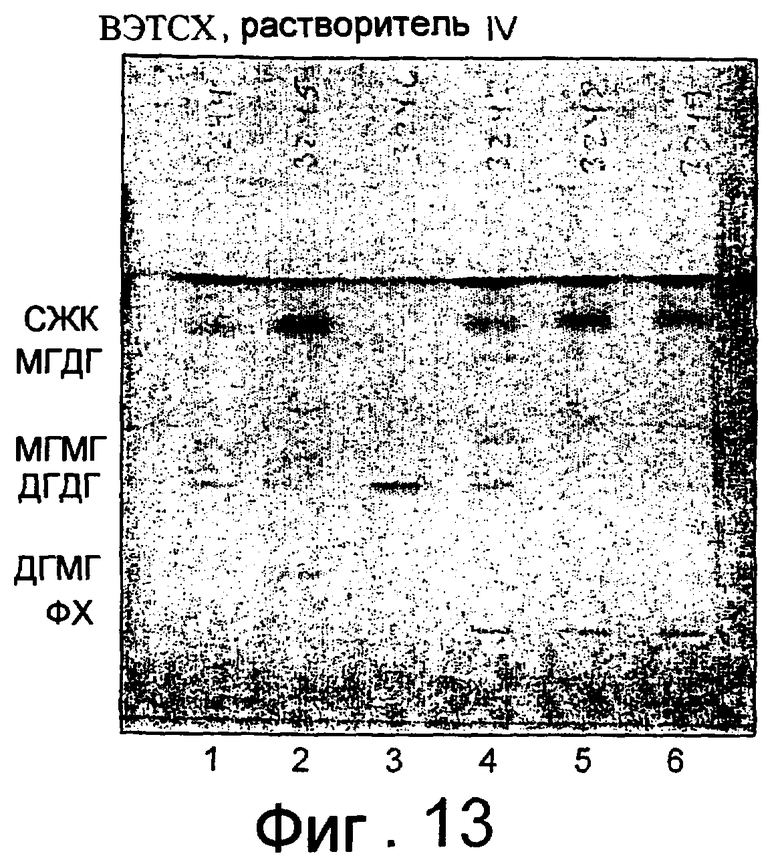
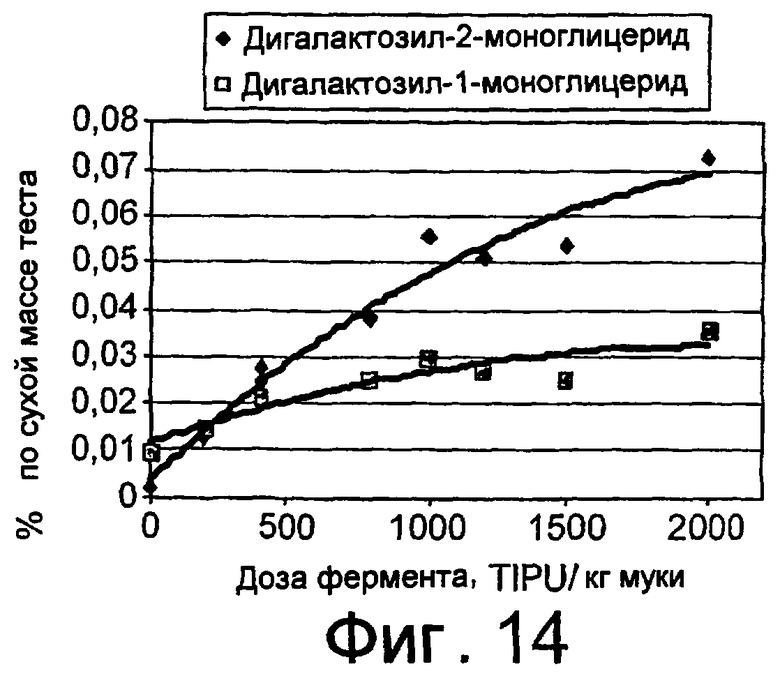
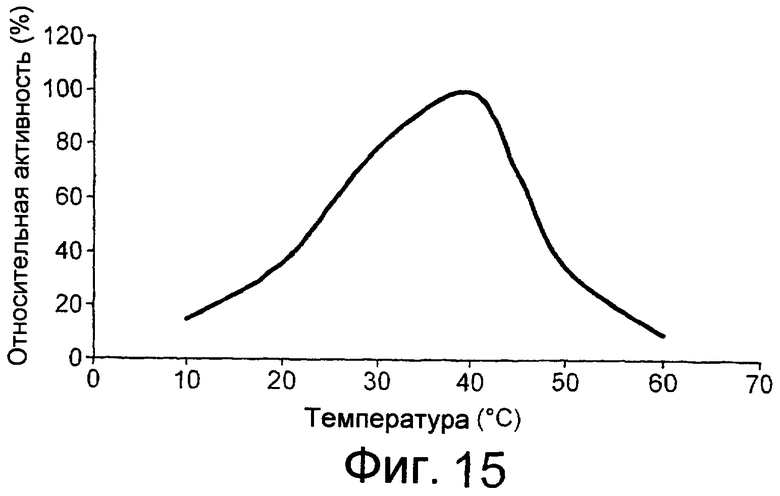
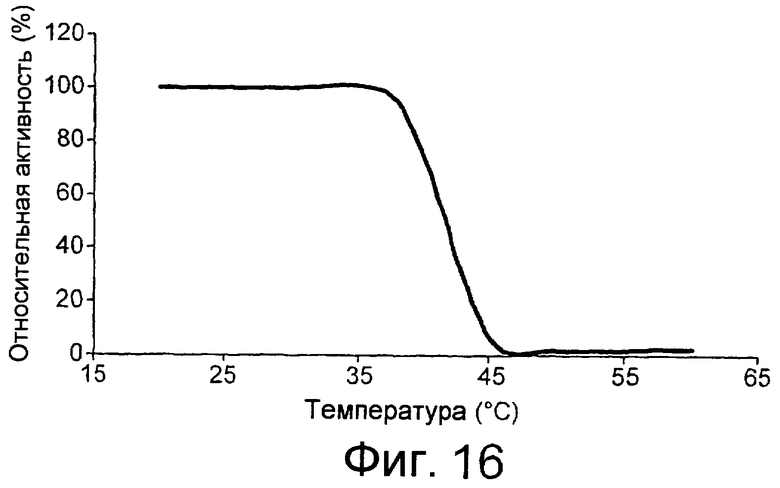

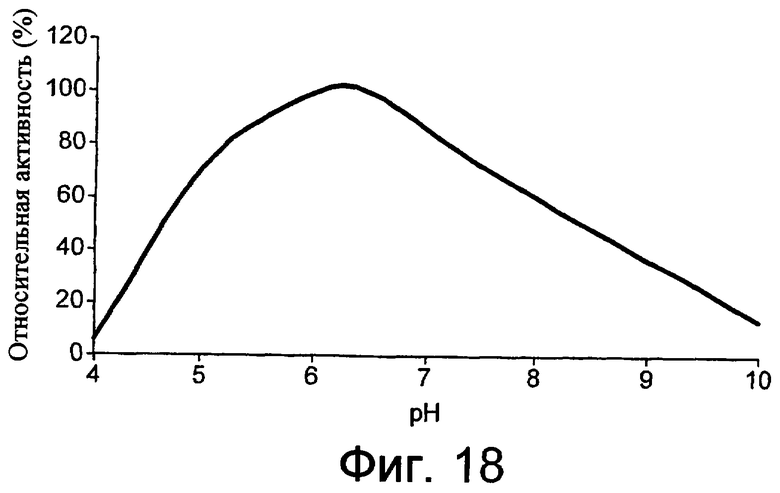

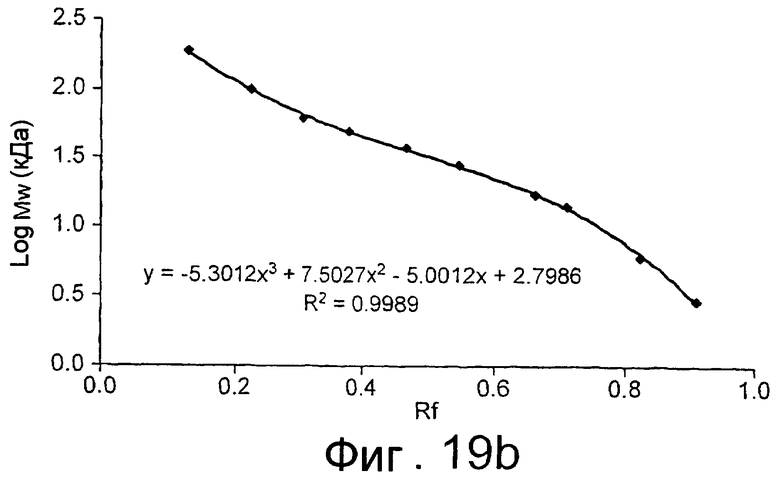
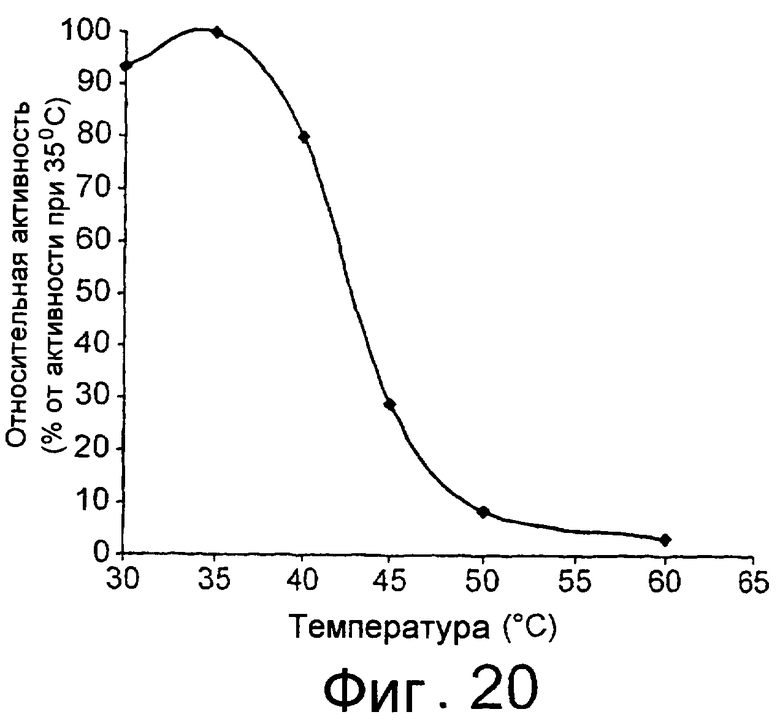
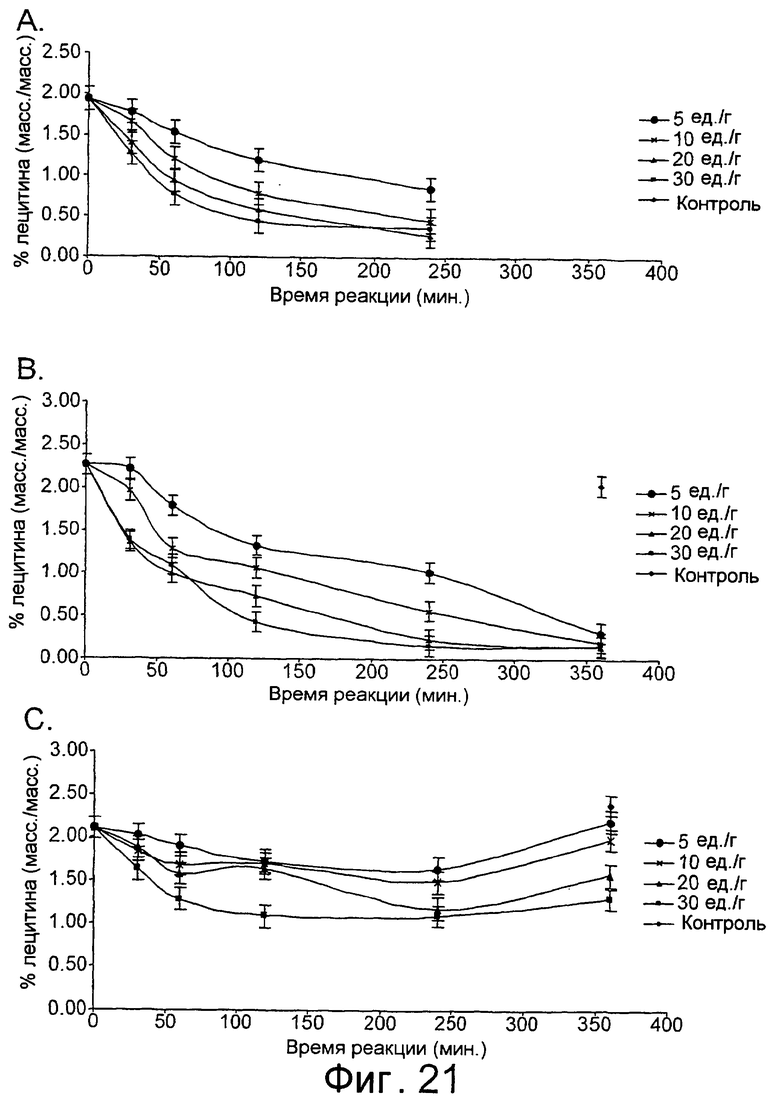
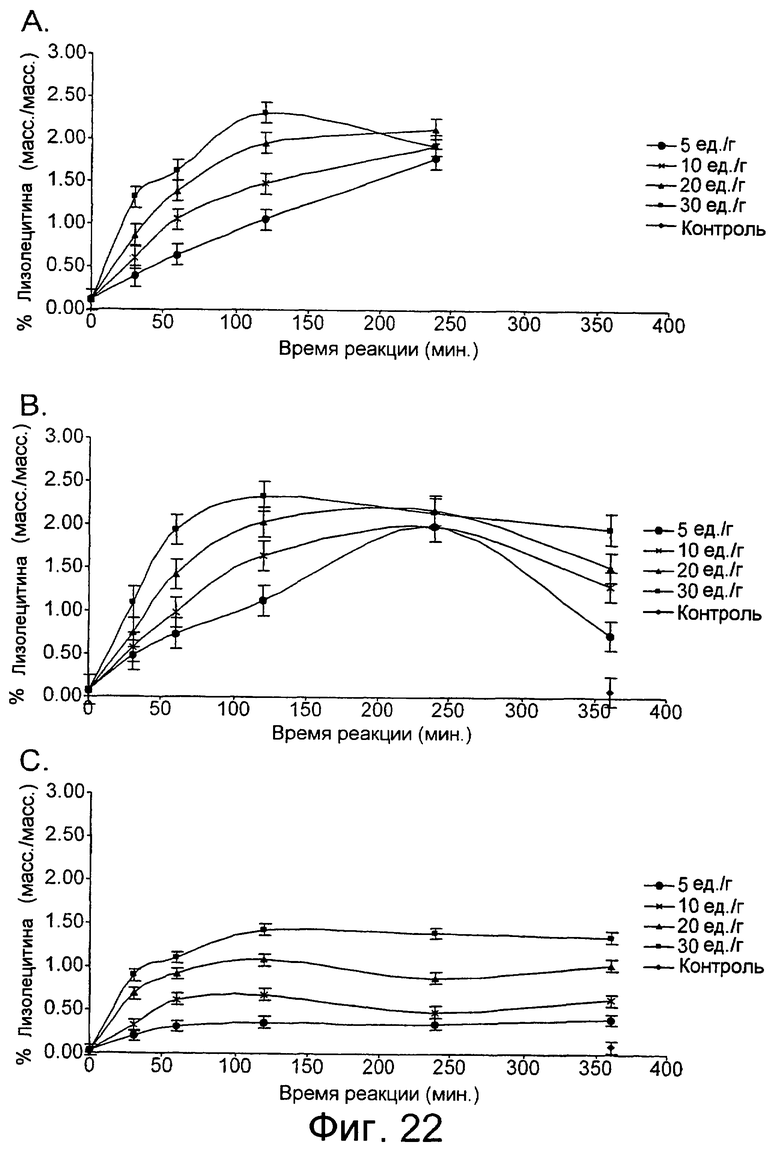
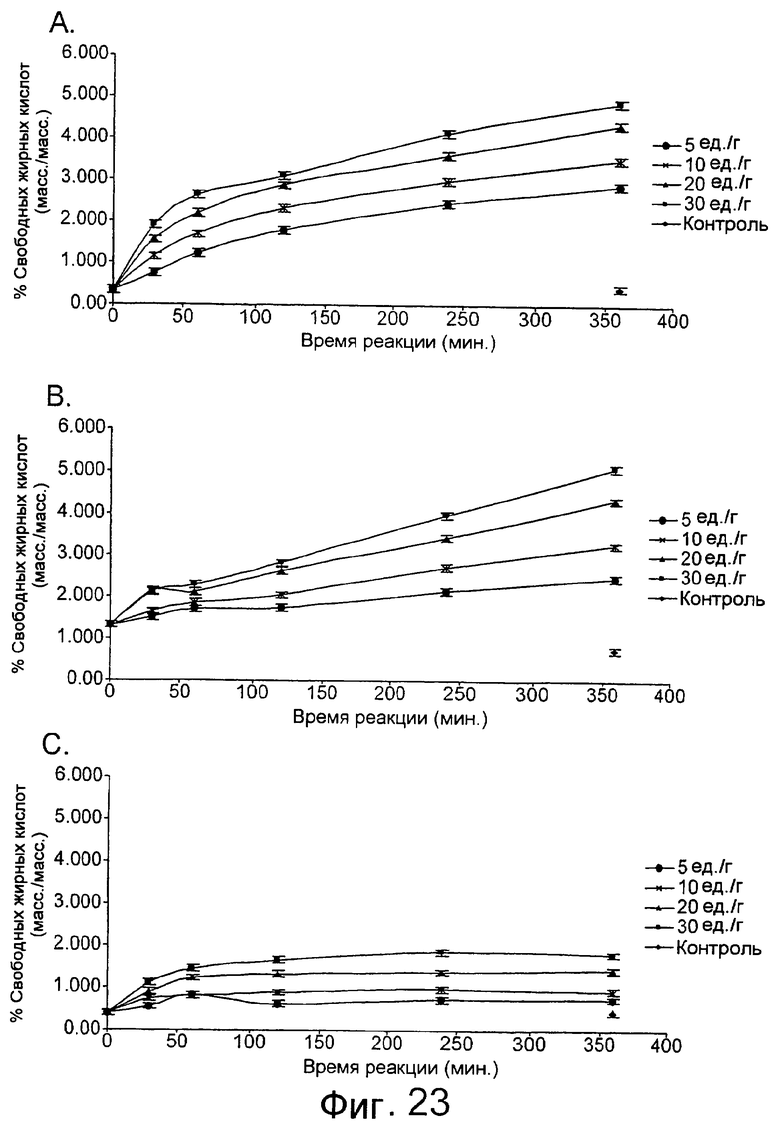
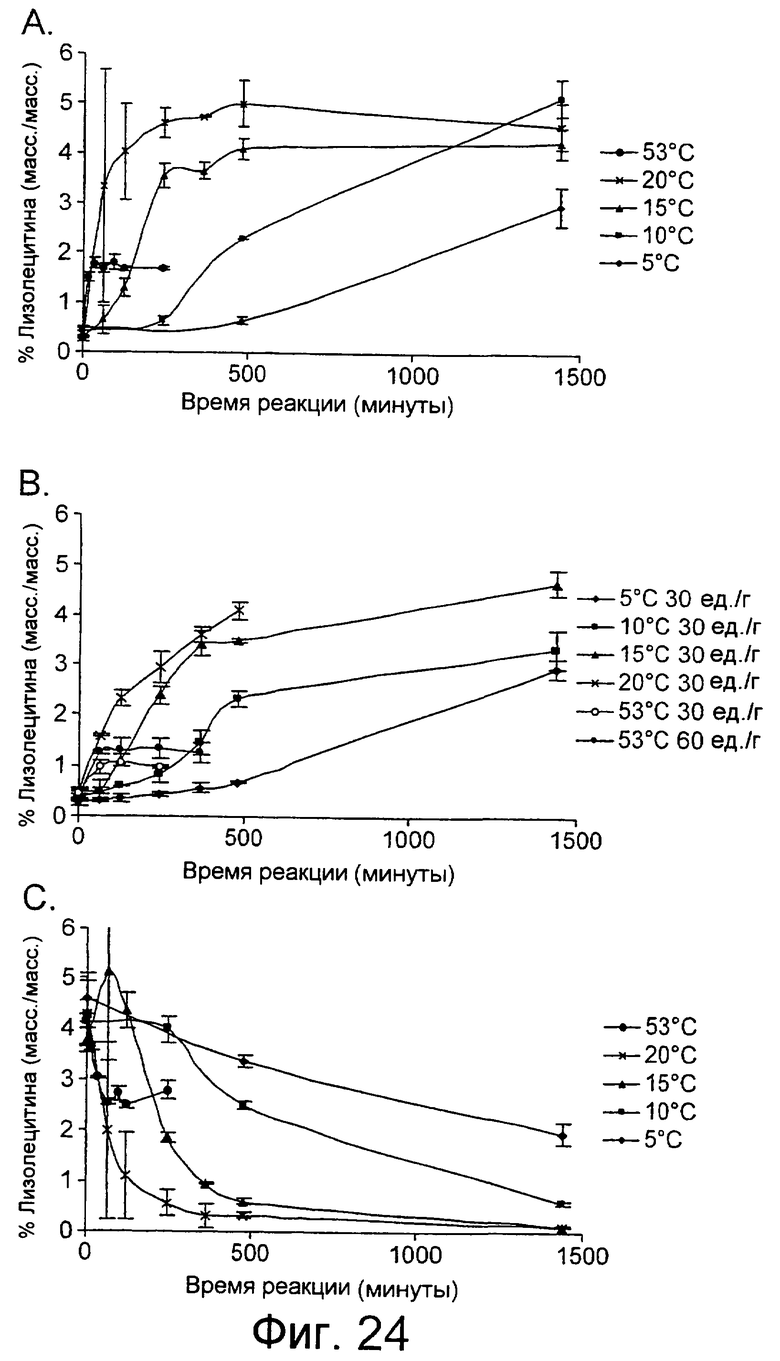
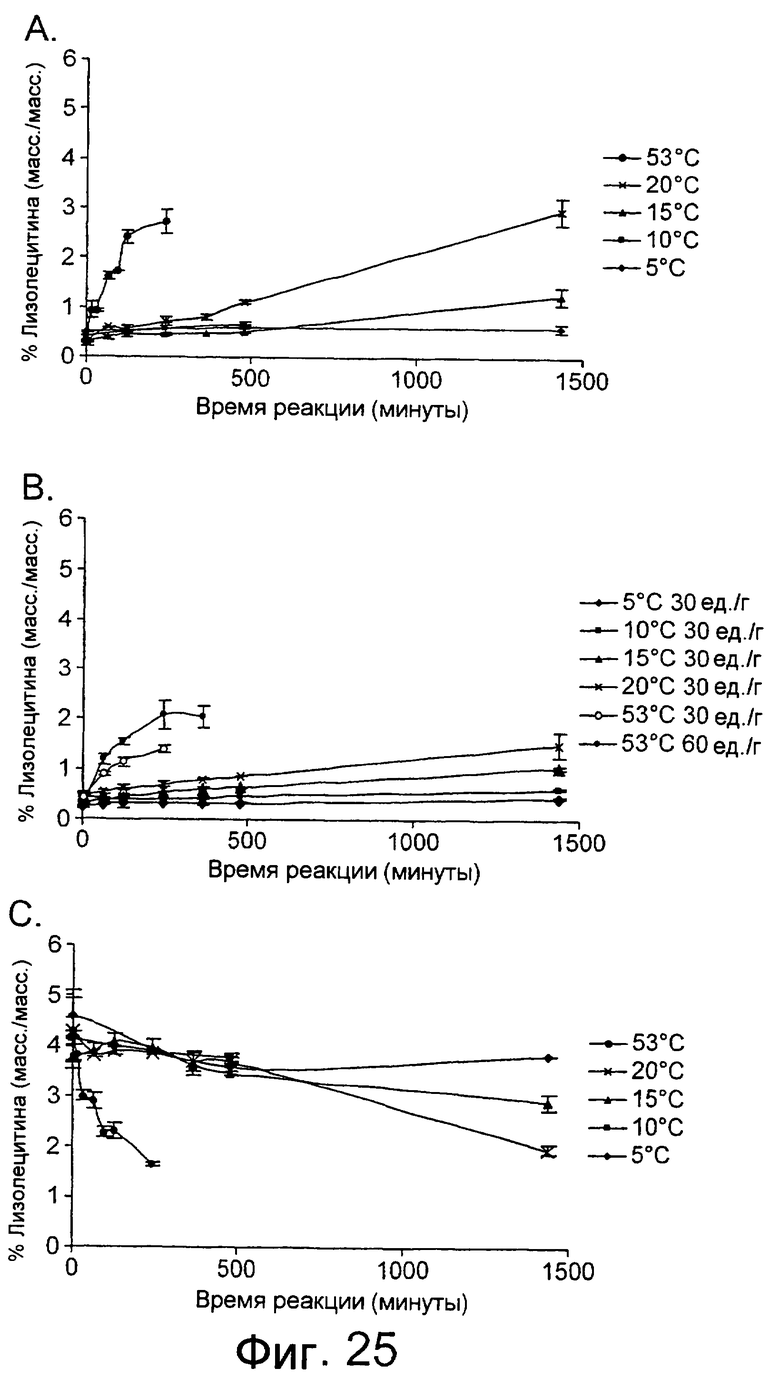
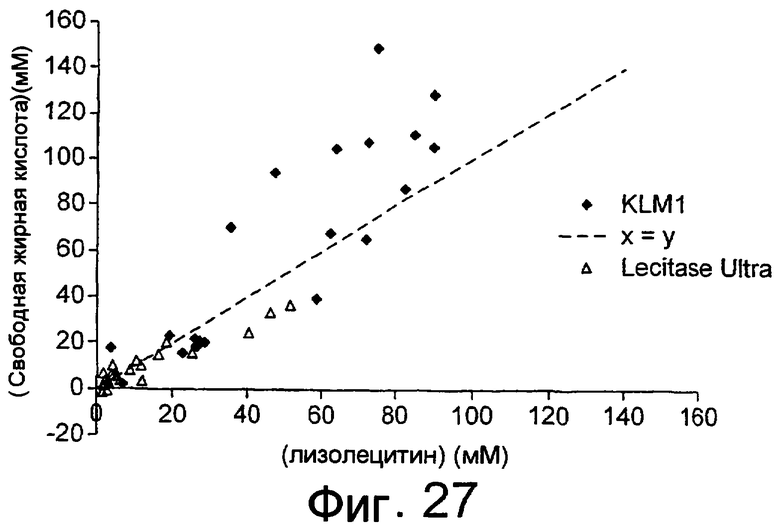

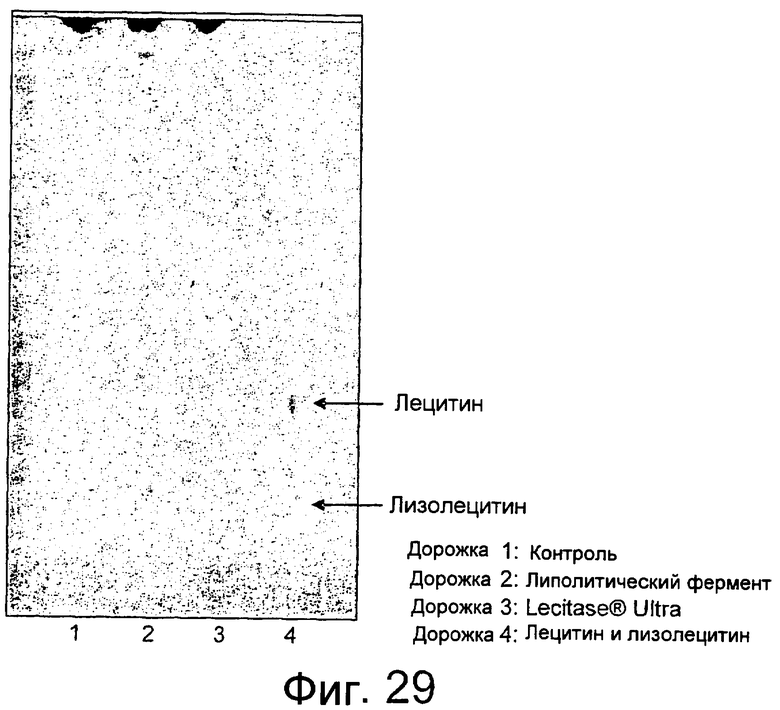
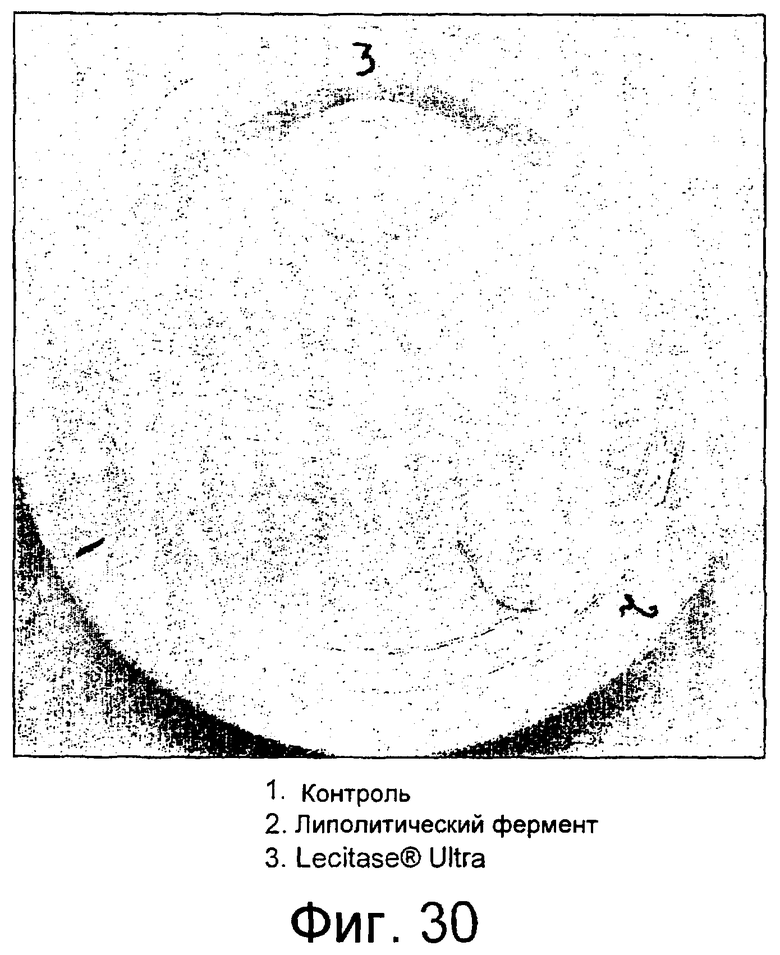
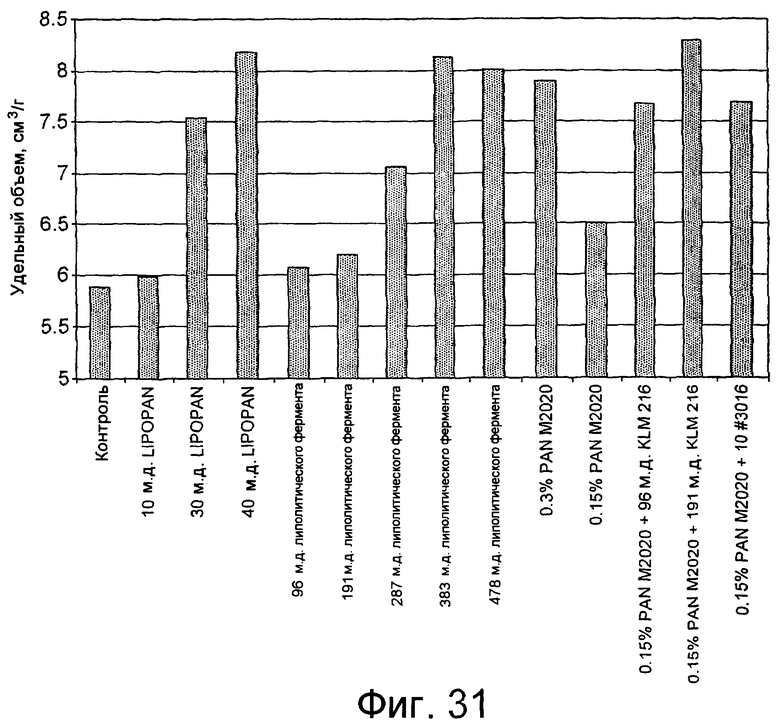
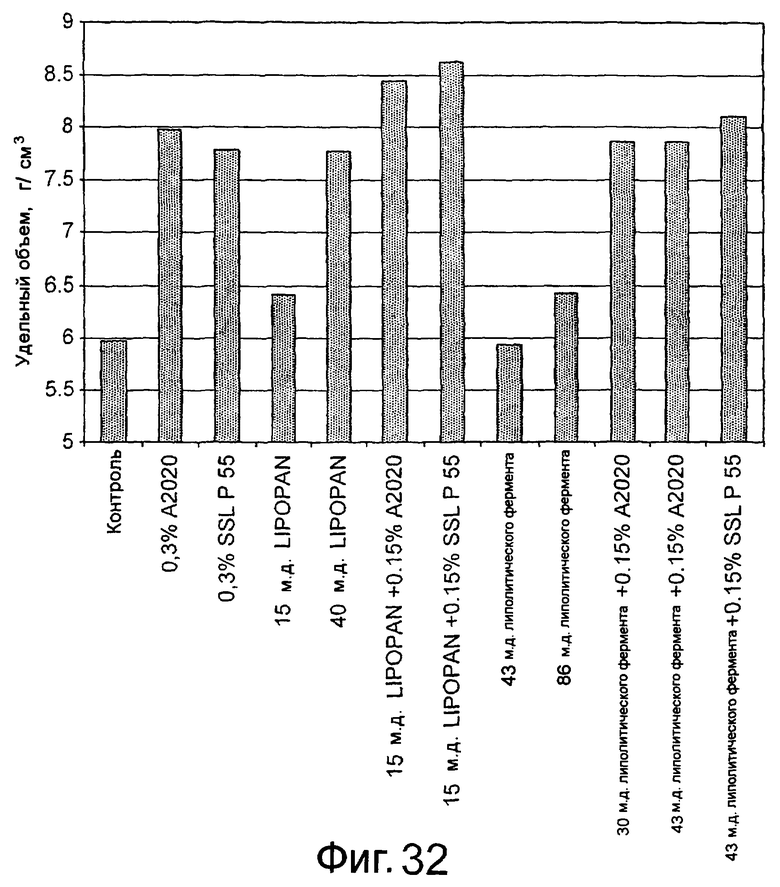
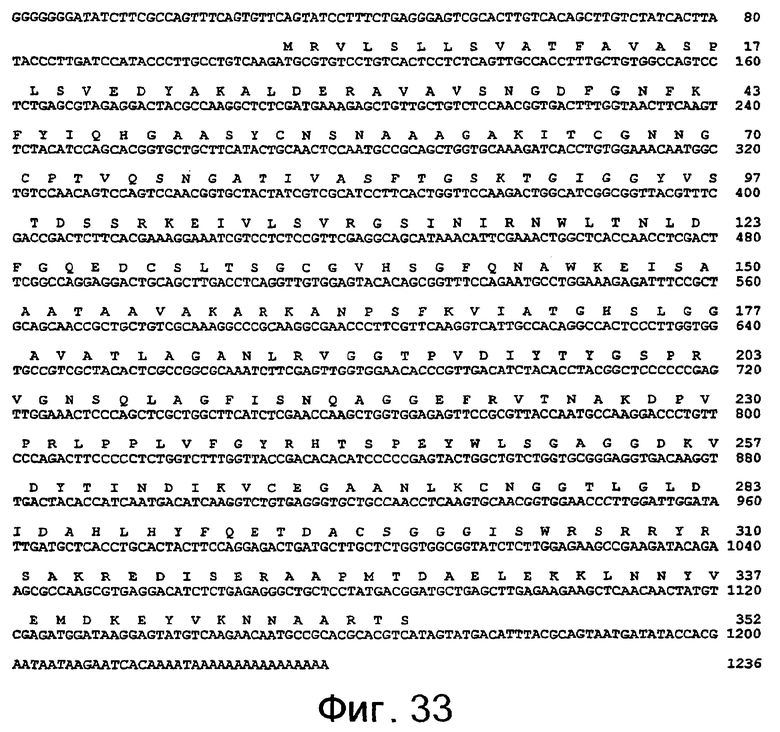
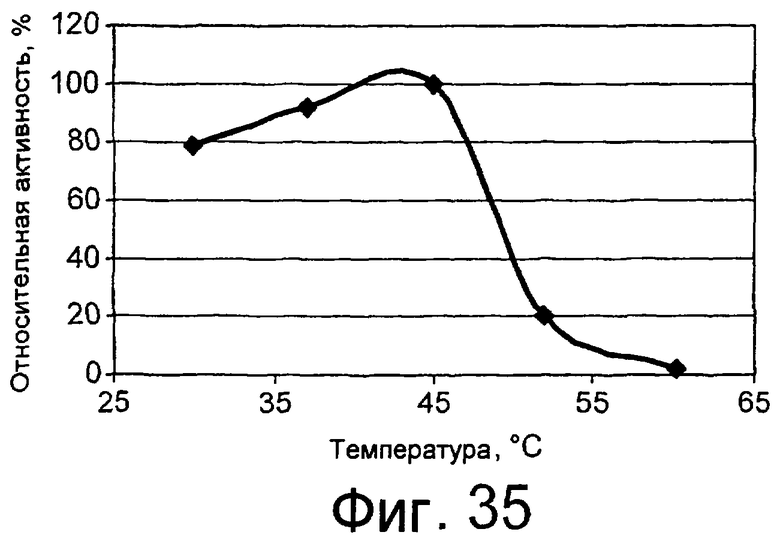
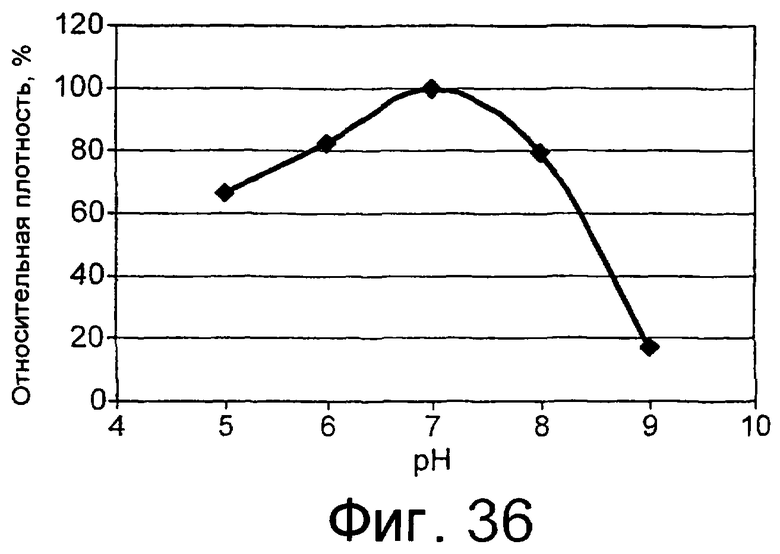


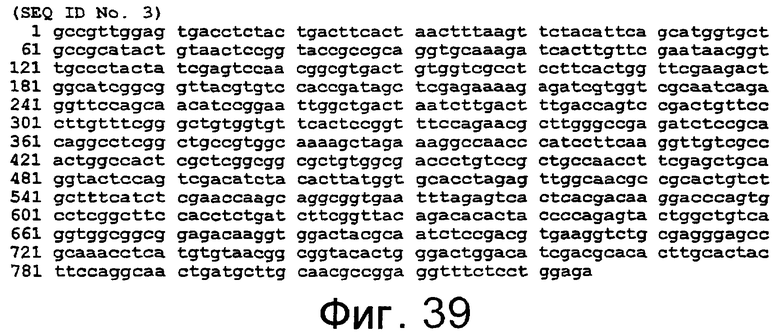

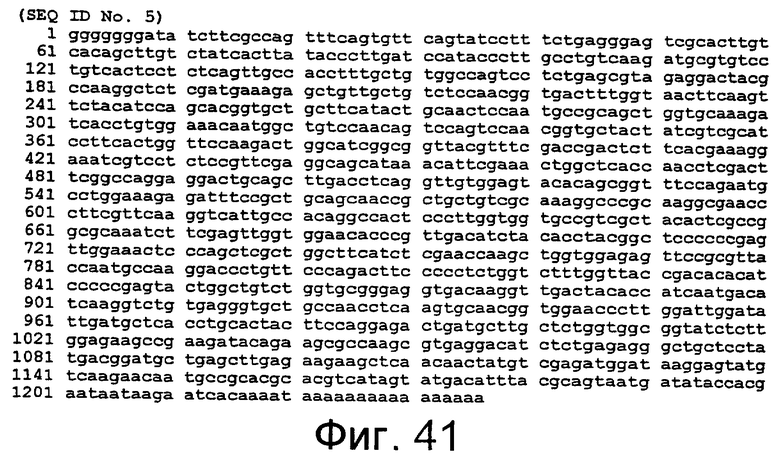


Изобретение относится к области биотехнологии, конкретно к новым грибковым липолитическим ферментам, имеющим более высокую активность, направленную на полярные липиды, по сравнению с активностью, направленной на триглицериды. Настоящее изобретение также относится к последовательностям нуклеиновых кислот, кодирующим указанные грибковые липолитические ферменты. Настоящее изобретение также относится пищевому продукту и к способу получения пищевого продукта, который включает в себя добавление грибкового липолитического фермента по настоящему изобретению к одному или нескольким ингредиентам приготовляемого продукта. Настоящее изобретения также включает в себя способ получения лизофосфолипида и способ получения лизогликолипида, где применяется грибковый липолитический фермент по настоящему изобретению. Использование грибковых липолитических ферментов по настоящему изобретению позволит получить пищевые продукты, включая хлебобулочные изделия, с улучшенным качеством. 11 н. и 11 з.п. ф-лы, 43 ил., 31 табл.
1. Грибковый липолитический фермент, где указанный фермент содержит аминокислотную последовательность, представленную в SEQ ID NO:1, или SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или аминокислотную последовательность, которая по меньшей мере на 90% идентична этим последовательностям.
2. Грибковый липолитический фермент по п.1, где указанный фермент получают из гифомицетов.
3. Грибковый липолитический фермент по п.2, где указанный фермент получают из Fusarium spp.
4. Грибковый липолитический фермент по п.3, где указанный фермент получают из Fusarium heterosporum.
5. Грибковый липолитический фермент по п.4, где указанный фермент получают из Fusarium heterosporum CBS 782.83.
6. Нуклеотидная последовательность, кодирующая грибковый липолитический фермент по любому из пп.1-5.
7. Нуклеиновая кислота, кодирующая грибковый липолитический фермент, где указанная нуклеиновая кислота выбрана из группы, состоящей из:
а) нуклеиновой кислоты, содержащей нуклеотидную последовательность, представленную в SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7;
b) нуклеиновой кислоты, которая соответствует нуклеотидной последовательности SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, с учетом вырожденности генетического кода; и
c) нуклеиновой кислоты, содержащей нуклеотидную последовательность, которая по меньшей мере на 90% идентична нуклеотидной последовательности, представленной в SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7.
8. Способ получения пищевого продукта, включающий добавление грибкового липолитического фермента по любому из пп.1-5 к одному или нескольким ингредиентам указанного пищевого продукта.
9. Способ по п.8, где указанным пищевым продуктом является один или несколько продуктов, выбранных из: яйцепродуктов или продуктов с добавлением яиц; хлебобулочного изделия; кондитерского изделия; замороженного продукта; молочного продукта, включая сыр; муссов; взбитых сливок на основе растительного масла; пищевого масла и жира; газированных и негазированных прохладительных напитков; эмульсий типа "масло в воде" и эмульсий типа "вода в масле"; маргарина; шортенинга; пасты, включая пасту с низким и с очень низким содержанием жира; заправок; майонеза; подливок; соусов-пюре; супов-пюре; напитков; пряных эмульсий и соусов.
10. Способ по любому из пп.8 или 9, включающий дополнительное добавление альфа-амилазы.
11. Способ приготовления хлебобулочного изделия, включающий добавление грибкового липолитического фермента по любому из пп.1-5 в тесто и выпекание из этого теста указанного хлебобулочного изделия.
12. Способ по п.11, включающий дополнительное добавление альфа-амилазы.
13. Способ получения лизофосфолипида, включающий обработку фосфолипида грибковым липолитическим ферментом по любому из пп.1-5 для получения указанного лизофосфолипида.
14. Способ по п.13, включающий дополнительное добавление альфа-амилазы.
15. Способ получения лизогликолипида, включающий обработку гликолипида грибковым липолитическим ферментом по любому из пп.1-5 для получения указанного лизогликолипида.
16. Способ по п.15, включающий дополнительное добавление альфа-амилазы.
17. Способ ферментативного рафинирования растительных или пищевых масел, включающий обработку пищевого или растительного масла грибковым липолитическим ферментом по любому из пп.1-5, так, чтобы большая часть описанных здесь полярных липидов была гидролизована.
18. Способ по п.17, включающий дополнительное добавление альфа-амилазы.
19. Пищевой продукт, полученный способом по п.8.
20. Хлебобулочное изделие, полученное способом по п.11.
21. Хлеб с улучшенным составом или тесто с улучшенным составом, содержащие грибковый липолиитический фермент по любому из пп.1-5.
22. Хлеб с улучшенным составом или тесто с улучшенным составом по п.21, дополнительно содержащие альфа-амилазу.
| US 6506588, 14.01.2003 | |||
| ЦЕНТРОБЕЖНЫЙ СЕПАРАТОР | 1996 |
|
RU2094123C1 |
| EP 0869167 A, 07.10.1998 | |||
| US 2003185939 A, 02.10.2003 | |||
| ЭЛЕКТРОДУГОВОЙ ПОДОГРЕВАТЕЛЬ ГАЗОИ | 0 |
|
SU203805A1 |
| HOSHINO T | |||
| ET AL.: "Purification and characterization of a lipolytic enzyme active at low temperature from Norwegian Typhula ishikariensis group III strain", European Journal of Plant Pathology, 1997, v.103, p.357-361. | |||

