Область техники, к которой относится изобретение, и преимущественная область использования изобретения
Изобретение относится к генной инженерии, биохимии, биотехнологии и иммунологии и может быть использовано для получения рекомбинантного (химерного) белка Ag85A-DBD и иммуногенной композиции, содержащей его, направленной на индукцию иммунитета против туберкулезной инфекции. Изобретение позволяет получать штамм-продуцент, обеспечивающий высокий уровень продукции иммуногенного рекомбинантного белка, который может быть очищен и иммобилизован в одну стадию, а также получать эффективные иммуногенные композиции против туберкулеза.
Описание аналогов изобретения
В настоящее время единственной применяемой в мире вакциной против туберкулеза является БЦЖ (BCG, Bacillus Calmette-Guerin, 1921 г.), которая представляет собой живой аттенуированный штамм Mycobacterium bovis. Она безопасна, недорога и достаточно эффективно защищает детей как от заражения туберкулезом, так и от милиарного туберкулеза и туберкулезного менингита. Недостатком вакцины BCG является то, что она не защищает взрослых от легочного туберкулеза, а именно эта форма болезни приводит к распространению инфекции и тяжелой эпидемической ситуации во многих странах мира. Отсюда следует высокая актуальность разработки новых вакцин против туберкулеза.
Существует несколько направлений разработки новых противотуберкулезных вакцин, альтернативных БЦЖ: субъединичные вакцины, ДНК-вакцины и живые вакцины с улучшенными свойствами. Наиболее полно в экспериментальных моделях на животных охарактеризованы субъединичные вакцины, в том числе препараты, состоящие из нескольких секретируемых белков/пептидов М. tuberculosis. Идентифицировано много антигенов М. tuberculosis, потенциально важных в качестве компонентов новых вакцин. К ним относятся, в частности, иммунодоминантные секретируемые антигены, присутствующие в культуральном фильтрате М. tuberculosis. Среди секретируемых антигенов М. tuberculosis лучше всего изучены антигены ESAT-6 (Early Secret Antigen Target-6), CFP-10 (culture-filtrate protein-10) и белки комплекса 85 (Ag85A, В, С).
Известно изобретение US 2011/0206713 А1, которое описывает иммуногенную композицию или лекарственные средства, состоящие из одного антигена ESAT-6, Ag85A, Ag85B или ТВ 10.4 М. tuberculosis; сочетания различных химерных белков на основе этих антигенов; использование гибридных (fusion) белков в разных сочетаниях с вакциной BCG.
Изобретение US 2011/0027349 А1 касается иммуногенной композиции, включающей один из антигенов Ag85A, Ag85B, МРТ-64, Pst-S1, Ара, GroES, GroEL, Dnak, CFP-10, Rv083 1c, Rv1324 М. tuberculosis или их различные сочетания; использование полипептидов как компонентов для проведения prime-boost вакцинации в сочетании с вакциной BCG; использование различных адъювантов.
Также известно изобретение US 2010/0015171 А1, касающееся иммуногенной композиции, включающей антигены семейства Ag85 и ТВ 10.4 М. tuberculosis; различные сочетания этих антигенов; использование различных адъювантов (DDA, поликатионные пептиды, олигонуклеотиды, IC31® адъювант) для введения антигенов.
Общим недостатком изобретений является сложный многостадийный метод очистки рекомбинантных белков и формулирования иммуногенной композиции.
Изобретение RU (11)97110087 (13)А касается полинуклеотида, состоящего из отдельных антигенов семейства Ag85 М. tuberculosis или их различных сочетаний, который после введения в организм индуцирует ряд иммунных реакций; способ введения иммуногенной композиции в ткани позвоночных.
Недостатком изобретения является использование только антигенов микобактерий семейства Ag85, что ставит под сомнение протективность вакцины. ДНК-вакцина также имеет низкую иммуногенность и требует применения совместно с адъювантами, что усложняет формулирование готового препарата.
Изобретение US 6174700 относится к способу очистки полипептидов, несущих в своем составе карбогидратсвязывающий домен. В качестве карбогидратсвязывающего домена описывается целлюлозосвязывающий домен.
Недостатком изобретения является то, что метод очистки белков, приведенный в качестве примера, не может быть использован для фармацевтического применения вследствие загрязнения конечного продукта эндотоксинами Е. coli и сложности удаления карбогидратсвязывающего домена из белковой молекулы.
Патент (WO 2009/143413) описывает использование гибридного белка pneumolysin и микобактериальных белков ESAT6, mtb или Ag85, конъюгированных с полисахаридами, в том числе декстраном. Однако в данном патенте для иммобилизации белков не используются карбогидратсвязывающие белковые домены, специфически взаимодействующие с полисахаридом. Препараты, полученные описанным в изобретении способом, относятся к классическим конъюгированным вакцинам.
Известны патенты, в которых описывается использование декстрансвязывающего домена из микроорганизма Leuconostoc mesenteroides в качестве домена, взаимодействующего с декстраном. В патенте RU 2428477 приводится описание химерной белковой конструкции ДСД-сп-β-ГАЛ, содержащей декстрансвязывающий домен из микроорганизма Leuconostoc mesenteroides, который генно-инженерным путем объединен через глицин-сериновый спейсер с термостабильной бета-галактозидазой из термофильного микроорганизма Thermoanaerobacter ethanolicus. Однако в патенте не приводится способ очистки рекомбинантного белка с использованием декстрана в качестве сорбента, а приводимый в патенте пример по определению декстрансвязывающих свойств у химерного белка можно использовать только для очистки термостабильных рекомбинантных белков. Использовать этот метод для получения иммуногенных компонентов медицинских иммунобиологических препаратов невозможно по причине термолабильности антигенов.
Описание прототипа изобретения
Прототипом заявленного изобретения является патент RU 2429292, в котором излагается способ получения рекомбинантного белка Ag85A-CBD, который содержит в своем составе целлюлозосвязывающий домен. Получают синтетический ген Ag85A М. tuberculosis, оптимизированный для гетерологичной экспрессии в непатогенных лабораторных штаммах. На его основе получают рекомбинантную плазмиду pAg85A-CBD, состоящую из искусственного бактериального оперона химерного белка, включающего ранний промотор бактериофага Т5, ген химерного белка и терминатор транскрипции; бактериального оперона бета-лактамазы и бактериального участка инициации репликации типа ColE1. Изобретение также включает штамм Е. coli - продуцент химерного белка Ag85A-CBD, а также способ иммобилизации, концентрирования и очистки полученного белка на целлюлозе. Изобретение относится к самому рекомбинантному белку Ag85A-CBD и иммуногенной композиции, содержащей его, направленной на индукцию иммунитета против туберкулезной инфекции. Изобретение позволяет получать штамм-продуцент, обеспечивающий высокий уровень продукции устойчивых иммуногенных белков, которые могут быть получены, иммобилизованы и очищены в одну стадию, а также получать эффективные иммуногенные композиции против туберкулеза.
Недостатком данного изобретения является низкая биодеградируемость целлюлозы, и, вследствие этого, невозможность использования иммуногенной композиции «рекомбинантный белок Ag85A-CBD - целлюлоза» в фармацевтической практике.
Раскрытие изобретения
Задачей изобретения является разработка вакцинных препаратов на основе отдельных белковых компонентов с высоким уровнем экспрессии и стабильности этих белков в бактериальной системе. Для решения этой задачи используется декстрансвязывающий домен из микроорганизма Leuconostoc citreum для получения стабильной в течение продолжительного времени (не менее года) иммуногенной композиции путем клонирования гена, включающего последовательность, кодирующую DBD, и последовательность микобактериального белка (антигена), его экспрессии в клетках Е. coli и иммобилизации полученного химерного антигена на полисахариде декстран, который применяется в качестве адъюванта.
Были созданы рекомбинантные белки, состоящие из двух компонентов: антигена М. tuberculosis и декстрансвязывающего домена декстрансукразы Leuconostoc citreum KM20. Рекомбинантные белки, имеющие в своем составе DBD, можно в одну стадию иммобилизовать на декстрановом носителе. В результате происходит их одновременная очистка, концентрирование и стабилизация, а также обеспечивается защита от протеаз бактерий-продуцентов. При этом исходные антигенные свойства функционально активного белкового домена не нарушаются. Использование декстрансвязывающего домена декстрансукразы Leuconostoc citreum определяется тем, что он обеспечивает большую стабильность рекомбинантных белков, чем описанный в литературе декстрансвязывающий домен из микроорганизмов Leuconostoc mesenteroides и Streptococcus sobrinus (S. Suwannarangsee, С. Moulis, G. Potocki-Veronese, P. Monsan, М. Remaud-Simeon, and W. Chulalaksananukul, "Search for a dextransucrase minimal motif involved in dextran binding", FEBS letters, vol. 581, №24, pp.4675-80, Oct. 2007).
Кроме того, для обеспечения высокого уровня экспрессии гена в гетерологичной системе, проводили оптимизацию кодонов гена Ag85A. В результате был спланирован синтетический ген Ag85A, который отличался от нативного микобактериального гена по нуклеотидному составу при сохраненной аминокислотной последовательности кодируемого им белка Ag85A. Этот подход позволил получить высокоэффективный штамм-продуцент, синтезирующий около 25-30% рекомбинантного антигена Ag85A от суммарного белка клетки.
При использовании этих подходов был получен двухкомпонентный рекомбинантный белок. Последовательность гена М. tuberculosis Ag85A кодирует полноразмерный белок, который формирует первый белковый домен, определяющий функциональные иммунологические свойства комплексной молекулы. Далее следует соединяющая (спейсерная) аминокислотная последовательность из нескольких чередующихся остатков глицина и серина (Gly-Ser). Второй белковый домен кодируется последовательностью фрагмента гена декстрансукразы Leuconostoc citreum KM20; он определяет способность продукта взаимодействовать с декстрановым сорбентом. Нековалентное взаимодействие декстрансвязывающего домена с декстраном обеспечивает постоянный, но медленный выход антигена из комплекса с субстратом за счет естественной диссоциации, тем самым обеспечивая пролонгированное взаимодействие компонентов вакцины с иммунной системой организма (депонирование), достаточное для индукции сильного и продолжительного иммунного ответа.
Белковые продукты нарабатываются в непатогенных лабораторных штаммах Е. coli. Иммобилизированные на декстране антигены представляют собой раствор декстрана с адсорбированными на нем белками. Декстран выступает в роли адъюванта при иммунизации, обеспечивающего укрупнение и полимеризацию антигена, без которого развивается более слабый иммунный ответ на белковые моноантигены.
Таким образом, заявляемая группа изобретений позволяет создать штамм Е. coli, обеспечивающий высокий уровень продукции рекомбинантного белка, содержащего белок М. tuberculosis Ag85A; получить простую и эффективную схему очистки рекомбинантного белка, специфически и эффективно индуцирующего иммунный ответ на антигены микобактерий, в том числе синтез ИФН-γ, необходимый для формирования протективного иммунитета против туберкулеза; создать иммуногенные композиции на основе рекомбинантного белка на декстране.
Указанный результат достигается за счет синтеза рекомбинатного белка Ag85A-DBD в клетках рекомбинантного штамма Е. coli M15 [pREP4, pAg85A-DBD], несущего рекомбинантную плазмиду pAg85A-DBD. Также за счет создания комплексного препарата Ag85A-DBD-декстран, в котором рекомбинантный белок Ag85A-DBD (концентрация 4 мг/мл) иммобилизован на декстране (50 мг/мл) и ресуспендирован в 1×PBS буфере рН 7,2-7,4.
Сущность изобретения
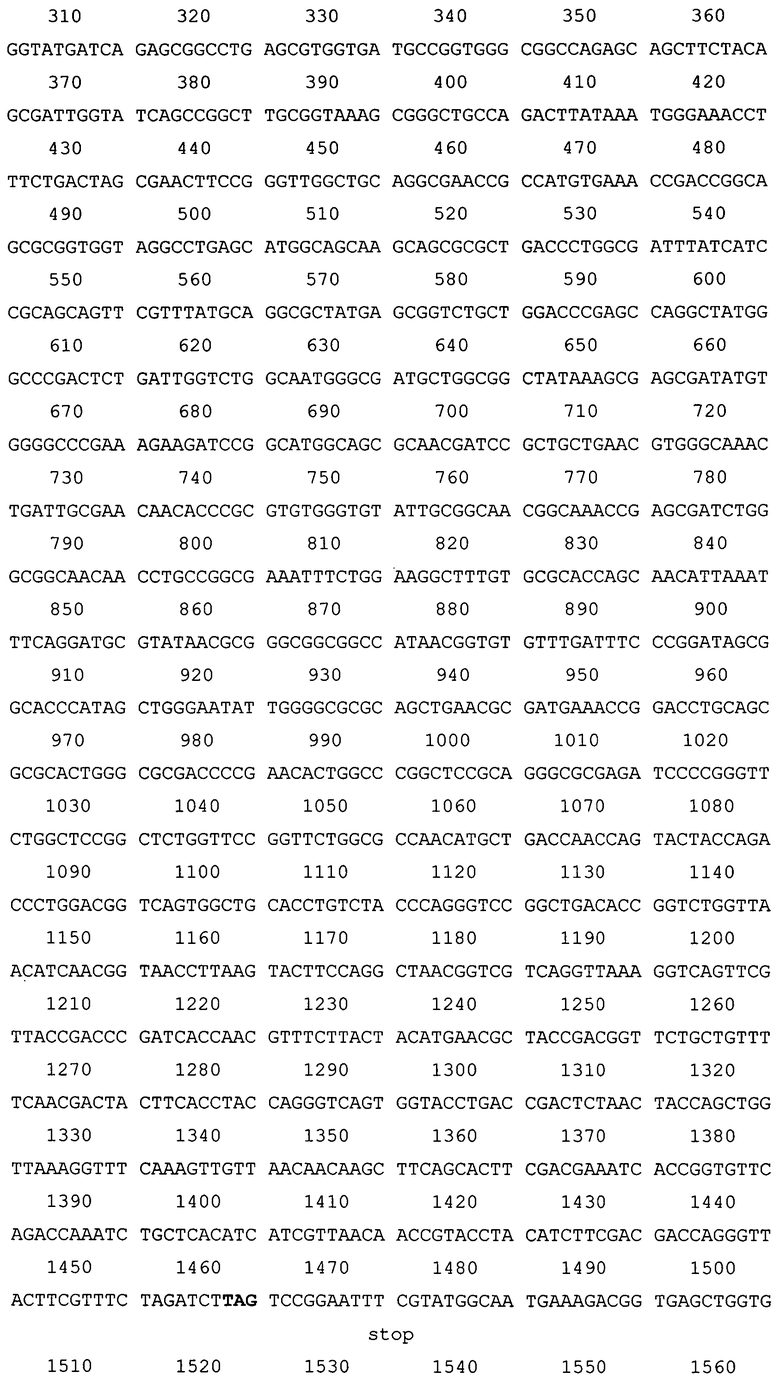
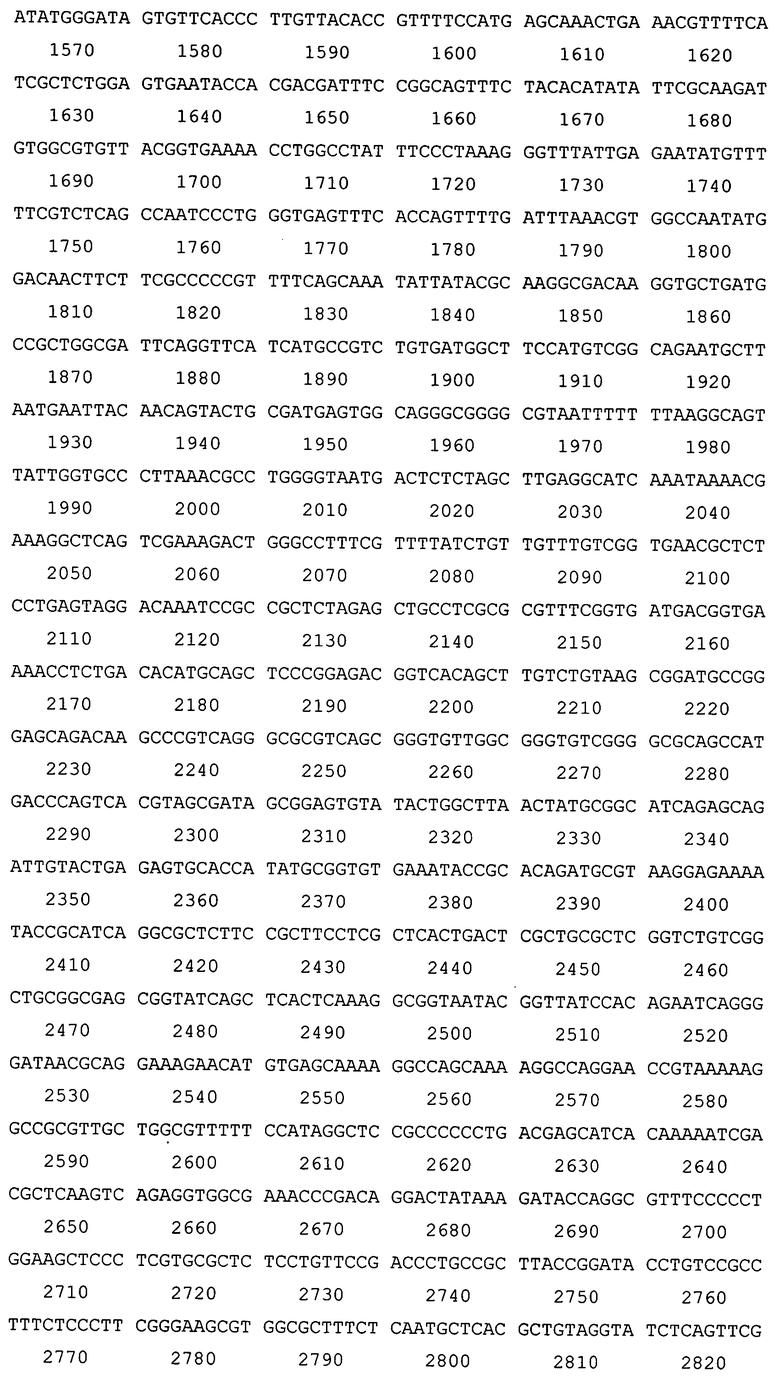
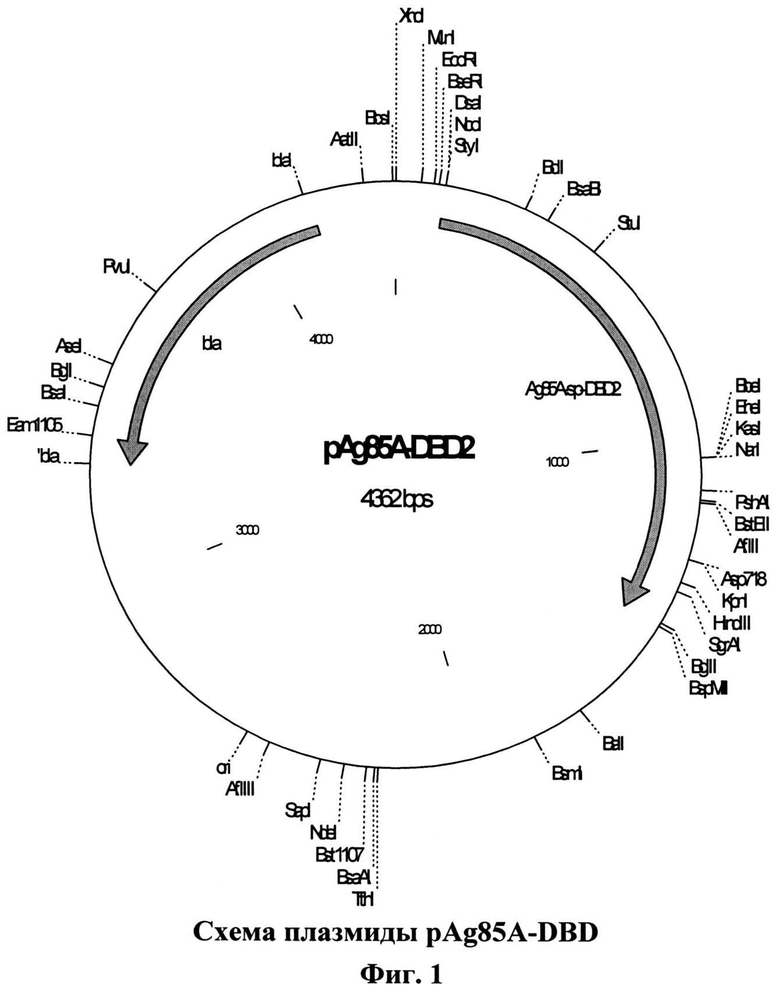
Создана рекомбинантная плазмида pAg85A-DBD (4362 п.н.), кодирующая бифункциональный рекомбинантный белок Ag85A-DBD (последовательность №1), обладающий способностью самопроизвольно связываться с декстрансодержащим сорбентом. Плазмида pAg85A-DBD (последовательность №2) содержит следующие существенные для ее функционирования структурные элементы:
а) синтетический ген, кодирующий последовательность белка Ag85A М. tuberculosis (900 п.н.);
б) искусственный бактериальный оперон химерного белка, состоящий из:
- промоторной области раннего промотора бактериофага Т5 (7-87 п.н.), обеспечивающей эффективную транскрипцию мРНК;
- рекомбинантного гена, кодирующего химерный белок «Ag85A М. tuberculosis - спейсер - декстран-связывающий домен декстрансукразы Leuconostoc citreum КМ20» - Ag85A-DBD (117-1460 п.н.);
- нетранслируемой области терминации транскрипции бактериального оперона, обеспечивающей эффективное окончание транскрипции pAg85A-DBD (1500-1596 п.н.);
в) бактериальный оперон bla, кодирующий белок бета-лактамазу, селективный маркер для трансформации штамма Е. coli pAg85A-DBD (3287-4147 п.н.) комплементарной цепи;
г) бактериальный участок инициации репликации типа ColE1, обеспечивающий репликацию плазмиды в штамме Е. coli: pAg85A-DBD (2529-2539 п.н.).
Штамм-продуцент рекомбинантного белка Ag85A-DBD (последовательность №3) получают трансформацией клеток Е. coli штамма M15 [pREP4] плазмидой pAg85A-DBD. Штамм обеспечивает продукцию рекомбинантного белка Ag85A-DBD после проведения процедуры индукции изопропил-β-D-тио-галактопиранозидом (ИПТГ) до 12-15% от тотального белка клетки.
Штамм Е. coli M15 [pREP4], несущий плазмиду pAg85A-DBD, - продуцент бифункционального рекомбинантного белка Ag85A-DBD характеризуется следующими признаками.
Культурально-морфологические признаки. Клетки прямые, палочковидные, неподвижные, грамотрицательные. При рассеве на чашке с 2,0% агаризованной средой LB рост в виде отдельных колоний, иногда в R-форме с неровными краями. Хорошо растет на плотных и жидких питательных средах (LB-бульон, LB-агар, МПА, МПБ).
Физиолого-биохимические признаки. Клетки растут при температуре от +4°С до +42°С, при оптимуме рН 6,8-7,5. Штамм разлагает глюкозу, маннит с образованием кислоты, не разлагает сахарозу, арабинозу, галактозу, сбраживает мальтозу, ксилозу, сорбит, рамнозу. Существенным при использовании данного штамма является его чувствительность к налидиксовой кислоте (25 мг/мл), стрептомицину (20 мг/мл) и рифампицину (25 мг/мл). Проявляет устойчивость к ампициллину (до 300 мкг/мл), обусловленную наличием плазмиды pAg85A-DBD, и к канамицину (до 50 мкг/мл), обусловленную наличием плазмиды pREP4.
По созданию комплексного препарата рекомбинантного белка и декстрана технический результат достигается за счет создания двухкомпонентного рекомбинатного белка Ag85A-DBD, состоящего из белка Ag85A M. tuberculosis и гена, кодирующего декстрансвязывающий домен декстрансукразы L. citreum KM20. А также за счет получения комплексного препарата (иммунологической композиции), в котором рекомбинатный белок (Ag85A-DBD) иммобилизован на декстране (композиция Ag85A-DBD-декстран).
Рекомбинантный белок Ag85A-DBD имеет в своем составе домен, определяющий его способность связываться с декстраном, что позволяет проводить в одну стадию концентрирование, очистку и иммобилизацию белкового продукта на декстране. Иммобилизация на декстране обеспечивается за счет присутствия в рекомбинантном белке декстрансвязывающего домена декстрансукразы L. citreum KM20. Поскольку в Е. coli отсутствуют белки, связывающиеся с декстраном, то синтезируемый в клетках Е. coli, трансформированных плазмидой pAg85A-DBD, рекомбинантный белок Ag85A-DBD является единственным белком штамма-продуцента, прочно связывающимся с декстраном. Это обеспечивает возможность одностадийного получения высокоочищенного препарата белка, иммобилизованного на сорбенте. На фиг.1 приведена схема плазмиды pAg85A-DBD.
Сравнение стабильности рекомбинантных микобактериальных белков, слитых с декстрансвязывающими доменами из L. mesenteroides и L. Citreum, выявили, что белки, слитые с декстрансвязывающим доменом из L. mesenteroides являются нестабильными и склонны к деградации при хранении свыше полугода, что не позволяет использовать вышеупомянутый домен для очистки этих рекомбинантных белков. Использование DBD L. citreum в качестве субъединицы, которая связывается с декстраном, позволяет получать более стабильные иммуногенные композиции.
Была исследована иммуногенность комплексного препарата Ag85A-DBD-декстран на мышах. Мышей иммунизировали под кожу спины 2-кратно с 2-недельным интервалом иммуногенным и контрольным препаратом. Через 5 недель был исследован пролиферативный ответ Т-клеток и количество Т-клеток, продуцирующих ИФН-гамма. Был выявлен прирост ИФН-гамма-положительных Т-клеток и усиление пролиферации в присутствии антигена в культуре клеток по сравнению с контрольными животными.
Техническим результатом, достигаемым при осуществлении изобретения, является получение штамма-продуцента, обеспечивающего высокий уровень продукции устойчивого иммуногенного белка, который может быть получен, иммобилизован и очищен в одну стадию. Получена эффективная иммуногенная композиция против туберкулеза.
Изобретение проиллюстрировано следующими примерами, приведенными ниже. Генно-инженерные и микробиологические манипуляции, амплификацию и секвенирование ДНК проводили по стандартным методикам (T.Parish, N.G.Stoker. Mycobacterium tuberculosis protocols., Humana Press Inc., 2001, Methods in molecular Medicine, 54; Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д.Гловера, Пер. с англ., М.: Мир, 1988; Saiki R.K., Gelfand D.H., Stoffel S. et al. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 1988, Vol.239, No.4839, pp.487-491; Sanger F., Nicklen S., Coulson A.R. DNAsequencing with chain-terminating inhibitors (DNA polymerase/nucleotide sequences/bacteriophage 4Х174). Proc. Nat. Acad. Sci. USA, 1977, Vol.74, №12, pp.5463-5467).
Пример 1. Получение экспрессионной генетической конструкции.
а) Получение фрагмента гена белка sp-DBD с последующим его клонированном в вектор pQE6.
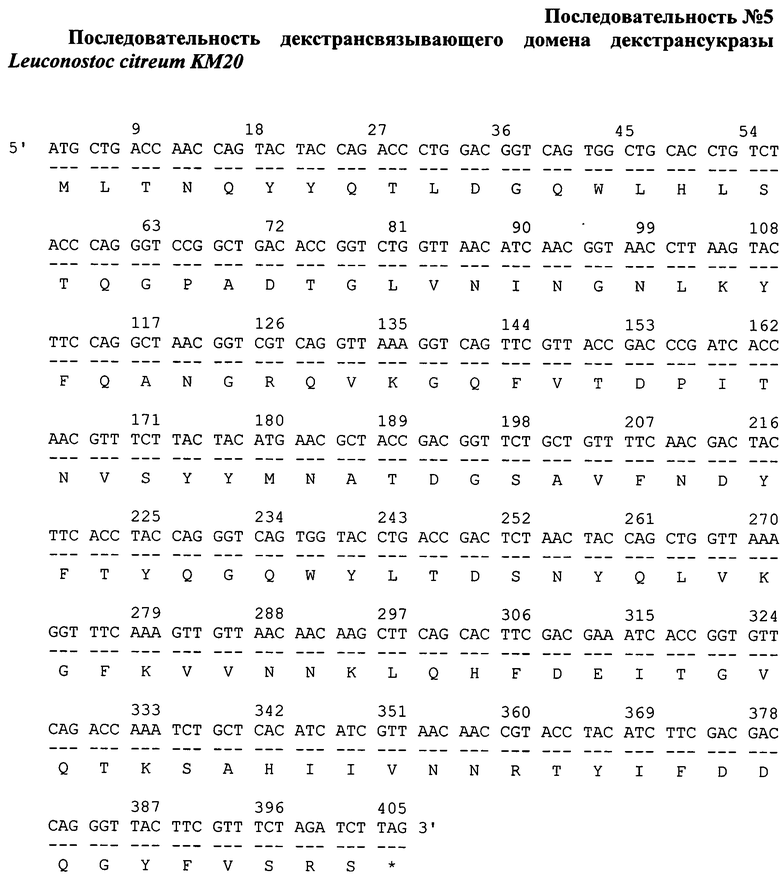
Фрагмент гена белка sp-DBD, содержащий Gly-Ser спейсер (последовательность №4) и последовательность декстрансвязывающего домена DBD из L. citreum KM20 (последовательность №5), получали синтетическим путем в фирме Евроген. Нуклеотидная последовательность, кодирующая белок sp-DBD, была спланирована с использованием оптимальных кодонов для экспрессии в Е. coli с учетом отсутствия выраженной вторичной структуры мРНК.
Для клонирования гена белка sp-DBD плазмидный вектор pQE6 и синтезированную в фирме Евроген плазмиду pAT-sp-DBD гидролизовали эндонуклеазами рестрикции NcoI и Kpn21 при 37°С в буфере, содержащем 33 мМ Трис-ацетата (рН 7,9 при 37°С), 10 мМ ацетата магния, 66 мМ ацетата калия и 0,1 мг/мл BSA в течение 1 ч. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенный из агарозного геля фрагмент вектора pQE6 (размером 3016 п.н.) объединяли с фрагментом плазмиды pAT-sp-DBD (464 п.н.). Далее фрагменты лигировали при 25°С в буфере, содержащем 40 мМ Трис-HCl, 10 мМ MgCl2, 10 мМ DTT, 0,5 мМ АТФ (рН 7,8 при 37°С) с помощью ДНК-лигазы бактериофага Т4 в течение 1 ч. Продукты реакции переосаждали при
-20°С 96% этиловым спиртом с добавлением ацетата аммония в течение 1,5 ч. После переосаждения смесь растворяли в деионизированной воде. Методом электропорации полученной лигазной смесью трансформировали компетентные клетки Е. coli штамма M15 [pREP4] (NalS, StrS, rifS, lac-, ara-, gal-, mtl-, F-, recA+, uvr+). После трансформации клоны отбирали на агаризованной среде LB, содержащей антибиотики ампициллин (50 мг/мл) и канамицин (25 мг/мл). Плазмидную ДНК из колоний выделяли методом щелочного лизиса; анализировали методом рестрикционного картирования эндонуклеазами рестрикции BglI, BglII, BspMII и NcoI. Отбирали клоны плазмидной ДНК psp-DBD (3480 п.н.), содержащие в своем составе последовательность гена белка sp-DBD. Первичную структуру полученных конструкций подтверждали секвенированием.
б) Получение фрагмента гена белка Ag85A с последующим его клонированном в плазмиду psp-DBD.
Фрагмент гена белка Ag85A из М. tuberculosis получали синтетическим путем в фирме Евроген. Нуклеотидная последовательность, кодирующая белок Ag85A, была спланирована с использованием оптимальных кодонов для экспрессии в Е. coli с учетом отсутствия выраженной вторичной структуры мРНК.
Для клонирования гена белка Ag85A-sp-DBD плазмиду psp-DBD и синтезированную в фирме Евроген плазмиду pAT-Ag85A гидролизовали эндонуклеазами рестрикции NcoI-BamHI и NcoI-BglII, соответственно, при 37°С в буфере, содержащем 33 мМ Трис-ацетата (рН 7,9 при 37°С), 10 мМ ацетата магния, 66 мМ ацетата калия и 0,1 мг/мл BSA в течение 1 ч. В 1,2% агарозном геле проводили разделение продуктов рестрикции. Выделенный из агарозного геля фрагмент вектора psp-DBD (размером 3469 п.н.) объединяли с фрагментом плазмиды pAT-Ag85A (893 п.н.). Далее фрагменты дотировали при 25°С в буфере, содержащем 40 мМ Трис-HCl, 10 мМ MgCl2, 10 мМ DTT, 0,5 мМ АТФ (рН 7,8 при 37°С) с помощью ДНК-лигазы бактериофага Т4 в течение 1 ч. Продукты реакции переосаждали при -20°С 96% этиловым спиртом с добавлением ацетата аммония в течение 1,5 ч. После переосаждения смесь растворяли в деионизированной воде. Методом электропорации полученной лигазной смесью трансформировали компетентные клетки Е. coli штамма M15 [pREP4] (NalS, StrS, rifS, lac-, ara-, gal-, mtl-, F-, recA+, uvr+). После трансформации клоны отбирали на агаризованной среде LB, содержащей антибиотики ампициллин (50 мг/мл) и канамицин (25 мг/мл). Плазмидную ДНК из колоний выделяли методом щелочного лизиса; анализировали методом рестрикционного картирования эндонуклеазами рестрикции BglI, BglII, BspMII и NcoI. Отбирали клоны плазмидной ДНК pAg85A-sp-DBD (4362 п.н.), содержащие в своем составе последовательность гена белка Ag85A-sp-DBD. Первичную структуру полученных конструкций подтверждали секвенированием.
Пример 2. Конструирование штамма Е. coli - продуцента антигена М. tuberculosis, соединенного с декстрансвязывающим доменом.
Для получения штамма Е. coli - продуцента рекомбинантного белка Ag85A-DBD, клетки штамма Е. coli M15 [pREP4] трансформировали плазмидой pAg85A-DBD. Трансформированные клетки выращивали в 3,5 мл среды LB с ампициллином и канамицином при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 550 нм (около 2,5 ч). В культуру добавляли 3 мкл 0,1 М раствора ИПТГ и выращивали в течение 3 ч. Для контроля продукции рекомбинантных белков в штамме Е. coli M15 [pREP4, pAg85A-DBD] применяли метод электрофореза по Лэммли в присутствии додецилсульфата натрия (ДСН). Разделение белков проводили в 12% полиакриламидном геле (ПААГ) в стандартной системе буферов (электродный буфер: 25 мМ Tris-HCl, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ Трис-HCl, рН 8,8). По окончании электрофореза гели окрашивали 0,15% раствором Кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывали в 10% уксусной кислоте.
При сравнении спектра белков в штаммах Е. coli M15 [pREP4] и Е. coli M15 [pREP4, pAg85A-DBD] обнаруживали появление дополнительной белковой полосы. Молекулярная масса дополнительной полосы 48,5 кДа соответствовала расчетной для белка Ag85A-DBD. Уровень синтеза белков в Е. coli определяли, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой соответствующего белка-стандарта Unstained Protein Molecular Weight Marker («Fermentas», Литва).
Для проверки растворимости синтезируемого белка биомассу выращенного штамма (Е. coli M15 [pREP4, pAg85A-DBD]) подвергали разрушению в 1% TES буфере (25 мМ Трис-HCl, 5 мМ ЭДТА, 50 мМ NaCl, 1% Тритон Х-100, 5 мг/мл лизоцима) в соотношении биомасса-буфер 1:2 и гомогенизировали до однородной массы. После центрифугирования пробы разделяли на 2 фракции: растворимых клеточных белков (надосадочная жидкость) и нерастворимых (осадок). Обе фракции подвергали разрушению в лизирующем буфере (0,05 М Трис-HCl, рН 6.8, 0,5% Triton Х-100, 0,5 М NaCl, 0,25 М MgCl2 5 мг/мл лизоцим, 10 мг/мл PMSF, 20 мг/мл ДНКаза) в соотношении 1:1; гомогенизировали до однородной массы; прогревали при +95°С 15 мин. Результаты контролировали с помощью электрофореза по Лэммли. Было показано, что рекомбинантный белок Ag85A-DBD синтезируется в клетках Е. coli как в растворимой фракции, так и виде телец-включений.
Пример 3. Получение рекомбинантного белка Ag85A-DBD в свободном состоянии и в виде белка, иммобилизованного на декстране.
Для получения рекомбинантного белка культуру штамма Е. coli M15 [pREP4, pAg85A-DBD] выращивали в 1000 мл среды LB с ампициллином и канамицином при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 550 нм. В среду добавляли 100 мкл 0,1 М раствора ИПТГ и выращивали в течение 3 ч. Клетки осаждали центрифугированием при 5500 об./мин в течение 15 мин.
Осадок ресуспендировали в лизирующем буфере, содержащем 50 мМ Трис-HCl (рН 8,0), 0,25 мМ NaCl, 5 мМ MgCl2, 0,15 мМ PMSF, 0,1% Тритон-Х100, 0,5 мг/мл лизоцима, 20 мг/мл ДНКазы; из расчета 1 г биомассы на 4 мл буфера. Дополнительно суспензию обрабатывали ультразвуком 3 раза по 20 с. После центрифугирования при 16000 об./мин в течение 30 мин фракция нерастворимых клеточных белков оставалась в осадке, растворимых - в надосадочной жидкости.
Осадок (нерастворимая фракция) суспендировали в лизирующем буфере (50 мМ Трис-HCl (рН 8,0), 10 мМ ЭДТА, 0,002 объема β-меркаптанола) и добавляли равный (по массе) объем мочевины. После перемешивания выдерживали на водяной бане при +37°С до полного растворения мочевины. Центрифугировали при 12000 об./мин 30 мин и отбирали надосадочную жидкость. Для очистки рекомбинантного белка надосадочную жидкость переносили на колонку с Sephadex G200 (декстран со сшивками). Элюцию белков с декстрана проводили градиентным раствором мочевины (8-2 М). Для удаления мочевины образцы диализовали против PBS буфера при 25°С в течение ночи.
Растворимую фракцию белков использовали для иммобилизации на декстране. К надосадочной жидкости добавляли декстран с молекулярной массой 500000 с концентрацией 50 мг/мл в буфере PBS; инкубировали при 25°С 1 ч. Концентрацию белков в свободной и иммобилизованной форме определяли по Бредфорду при длине волны 595 нм. Образцы подвергали лиофилизации. Таким образом, были получена иммуногенная композиция Ag85A-DBD-декстран. Степень чистоты рекомбинантного белка составила не менее 95%.
Пример 4. Оценка стабильности очищенного рекомбинантного белка.
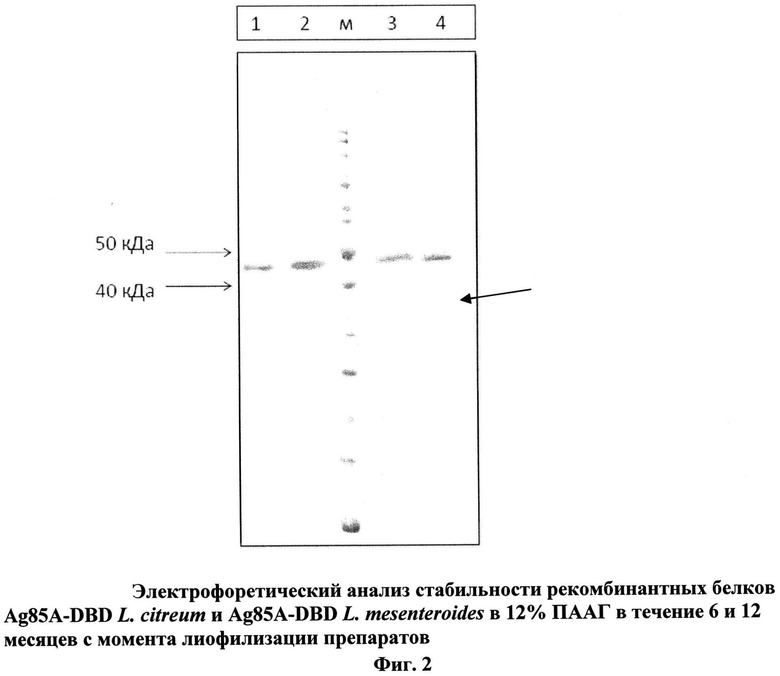
Оценка стабильности рекомбинантных белков, полученных путем слияния последовательности микобактериального белка Ag85A и DBD Leuconostoc citreum или DBD Leuconostoc mesenteroides, проводилась с помощью электрофореза в 12% ПААГ. Препараты химерных белков Ag85A-DBD Leuconostoc citreum и Ag85A-DBD Leuconostoc mesenteroides хранились при температуре 0-4°С в течение 6 и 12 месяцев с даты лиофилизации. Исследования выявили, что белок Ag85A-DBD Leuconostoc mesenteroides менее стабилен, чем Ag85A-DBD Leuconostoc citreum и после 6 месяцев хранения начинается его денатурация (см. фиг.2, где 1. Рекомбинантный белок Ag85A-DBD L. citreum после 6 месяцев хранения; 2. Рекомбинантный белок Ag85A-DBD L. citreum после 12 месяцев хранения; М - маркер молекулярных масс; 3. Рекомбинантный белок Ag85A-DBD L. mesenteroides после 6 месяцев хранения; 4. Рекомбинантный белок Ag85A-DBD L. mesenteroides после 12 месяцев хранения; Стрелкой отмечена деградация рекомбинантного белка).
Пример 5. Способ проверки иммуногенности комплексных препаратов Ag-DBD-декстран на мышах.
Продукция Т-клетками ИФН-γ в ответ на стимуляцию секретируемыми антигенами микобактерий считается самым важным элементом адаптивного защитного иммунного ответа при туберкулезной инфекции, поэтому вакцинация прежде всего должна стимулировать именно эту функцию.
Иммуногенные свойства сорбента, содержащего рекомбинантный антиген М. tuberculosis - Ag85A-DBD, проверяли на мышах инбредной линии C57BI/6. Положительным контролем была группа мышей, вакцинированная BCG. Количество Т-лимфоцитов, продуцирующих ИФН-γ, определяли в лимфатических узлах и селезенке методом ELISPOT.
У иммунизированных мышей исследовали селезенку и паховые лимфоузлы; готовили суспензию клеток. В работе использовали связывающие моноклональные антитела к ИФН-γ мыши клон R4-6A2, биотинилированные антитела к ИФН-γ мыши клон XMG1.2. Связывающие моноклональные антитела разводили в буфере PBS (рабочая концентрация 4 мкг/мл), вносили в лунки планшета и инкубировали 18 ч при 4°С. После этого содержимое лунок удаляли, вносили суспензию клеток (2,5-3,0×105 клеток/лунка) в культуральной среде и инкубировали 2-4 ч при 37°С. На этом этапе и далее перед внесением каждого следующего компонента планшет отмывали PBS. Содержимое лунок удаляли, в часть из них вносили образец рекомбинантного белка, разведенный в PBS (10 мкг/мл), в отрицательный контроль - PBS, инкубировали 48 ч при 37°С в 5% CO2. В лунки вносили биотинилированные проявляющие антитела в концентрации 1 мкг/мл, инкубировали 2-4 ч при 25°С. Добавляли конъюгат щелочной фосфатазы со стрептавидином, разведенный в PBS до концентрации 1:1000 (100 мкл/лунка), инкубировали 1-2 ч при 25°С. Добавляли субстрат щелочной фосфатазы, разведенный в PBS. Через 5-60 мин реакцию останавливали, планшет высушивали, оценивали количество пятен в лунках. Для выявления достоверности различий использовали t-критерий Стьюдента (Р<0,01). Результаты представлены в таблице 1.
Результаты показывают, что вакцинация моноантигенами, связанными с декстраном через DBD-домен, приводит к специфической активации продуцентов ИФН-гамма. Вакцинация теми же антигенами, в тех же количествах, но без адсорбции на декстране, не вызывает иммунного ответа.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА pESAT6-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-DBD, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК ESAT6-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2520737C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ТРЕХ ГИБРИДНЫХ БЕЛКОВ ОБОЛОЧКИ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОПРЕДЕЛЯЮЩИХ ПРИНАДЛЕЖНОСТЬ К СИБИРСКОМУ (DBD2-D3S), ЕВРОПЕЙСКОМУ (DBD2-D3E) И ДАЛЬНЕВОСТОЧНОМУ (DBD2-D3D) ПОДТИПАМ ВИРУСА; РЕКОМБИНАНТНЫЕ ПЛАЗМИДЫ pDBD2-D3S, pDBD2-D3E И pDBD2-D3D; ШТАММЫ-ПРОДУЦЕНТЫ Escherichia coli M15 [pREP4]; ХИМЕРНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2560588C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-CBD, ШТАММ Escherichia coli [pREP4, pAg85A-CBD], ХИМЕРНЫЙ БЕЛОК Ag85A-CBD И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2429292C1 |
| Иммуногенная композиция на основе рекомбинантных псевдоаденовирусных частиц, а также на основе белковых антигенов и способ получения иммуногенной композиции | 2018 |
|
RU2691302C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422525C1 |
| ВАКЦИНА РЕКОМБИНАНТНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ И АДЪЮВАНТ ДЛЯ НЕЕ | 2017 |
|
RU2665817C1 |
| ПОЛИАНТИГЕННАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ И ВСПОМОГАТЕЛЬНОГО ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА | 2019 |
|
RU2724896C1 |



Изобретение относится к биохимии и биотехнологии и представляет собой штамм Escherichia coli M15 [pREP4, pAg85A-DBD] - продуцент химерного белка Ag85A-DBD, а также способ иммобилизации, концентрирования и очистки полученного белка на декстране. Изобретение относится к способу получения иммуногенной композиции на основе рекомбинантного белка Ag85A-DBD в смеси с декстраном, самому рекомбинантному белку Ag85A-DBD. Изобретение позволяет получать штамм-продуцент, обеспечивающий высокий уровень продукции устойчивых иммуногенных белков, которые могут быть получены, иммобилизованы и очищены в одну стадию, а также получать эффективные иммуногенные композиции против туберкулеза. 4 н.п. ф-лы, 2 ил., 1 табл., 5 пр.
1. Способ получения очищенного иммобилизованного рекомбинантного белка Ag85A-DBD на декстране, включающий:
- выращивание клеток штамма Е. coli M15 [pREP4, pAg85A-DBD];
- иммобилизацию белка Ag85A-DBD в составе клеточных экстрактов штамма Е. coli M15 [pREP4, pAg85A-DBD] на декстрановом сорбенте за счет аффинного взаимодействия при процедуре инкубации с последующей отмывкой от несвязавшихся бактериальных белков, при этом происходит концентрирование целевого продукта и его очистка;
- выделение целевого продукта.
2. Рекомбинантный штамм Е. coli M15 [pREP4, pAg85A-DBD], полученный трансформацией плазмидой pAg85A-DBD с последовательностью №2, продуцент белка Ag85A-DBD.
3. Рекомбинантный белок Ag85A-DBD с молекулярной массой 48,5 кДа, включающий полноразмерный белок Ag85A с последовательностью №3, Gly-Ser спейсер с последовательностью №4, декстрансвязывающий домен гена декстрансукразы L. citreum KM20 с последовательностью №5.
4. Иммуногенная композиция, содержащая рекомбинантный белок по п.3 Ag85A-DBD, иммобилизованный на декстране в эффективном количестве, специфически активирующая Т-лимфоциты, синтезирующие ИФН-γ при стимуляции антигенами микобактерий.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-CBD, ШТАММ Escherichia coli [pREP4, pAg85A-CBD], ХИМЕРНЫЙ БЕЛОК Ag85A-CBD И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2429292C1 |
| РЕКОМБИНАНТНАЯ БЕЛКОВАЯ КОНСТРУКЦИЯ ДСД-сп-β-ГАЛ, ОБЛАДАЮЩАЯ АКТИВНОСТЬЮ ФЕРМЕНТА ТЕРМОСТАБИЛЬНОЙ β-ГАЛАКТОЗИДАЗЫ (ЛАКТАЗЫ) И СПОСОБНОСТЬЮ АФФИННО СВЯЗЫВАТЬСЯ С ДЕКСТРАНОМ, ПЛАЗМИДНАЯ ДНК pGD-10, ОПРЕДЕЛЯЮЩАЯ БИОСИНТЕЗ ДСД-сп-β-ГАЛ, И ШТАММ-ПРОДУЦЕНТ Escherichia coli DH5α/PGD-10 | 2009 |
|
RU2428477C2 |
| US 20110206713 A1, 25.08.2011 | |||
| RU 97110087 A, 27.06.1999 | |||
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА С УЛУЧШЕННОЙ ЭФФЕКТИВНОСТЬЮ | 2004 |
|
RU2342400C2 |

